JP6068336B2 - 植物種子へのオメガ−7脂肪酸の蓄積 - Google Patents
植物種子へのオメガ−7脂肪酸の蓄積 Download PDFInfo
- Publication number
- JP6068336B2 JP6068336B2 JP2013516790A JP2013516790A JP6068336B2 JP 6068336 B2 JP6068336 B2 JP 6068336B2 JP 2013516790 A JP2013516790 A JP 2013516790A JP 2013516790 A JP2013516790 A JP 2013516790A JP 6068336 B2 JP6068336 B2 JP 6068336B2
- Authority
- JP
- Japan
- Prior art keywords
- plant
- seq
- amino acid
- acid sequence
- plant material
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Expired - Fee Related
Links
Images





Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8241—Phenotypically and genetically modified plants via recombinant DNA technology
- C12N15/8242—Phenotypically and genetically modified plants via recombinant DNA technology with non-agronomic quality (output) traits, e.g. for industrial processing; Value added, non-agronomic traits
- C12N15/8243—Phenotypically and genetically modified plants via recombinant DNA technology with non-agronomic quality (output) traits, e.g. for industrial processing; Value added, non-agronomic traits involving biosynthetic or metabolic pathways, i.e. metabolic engineering, e.g. nicotine, caffeine
- C12N15/8247—Phenotypically and genetically modified plants via recombinant DNA technology with non-agronomic quality (output) traits, e.g. for industrial processing; Value added, non-agronomic traits involving biosynthetic or metabolic pathways, i.e. metabolic engineering, e.g. nicotine, caffeine involving modified lipid metabolism, e.g. seed oil composition
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8216—Methods for controlling, regulating or enhancing expression of transgenes in plant cells
- C12N15/8222—Developmentally regulated expression systems, tissue, organ specific, temporal or spatial regulation
- C12N15/823—Reproductive tissue-specific promoters
- C12N15/8234—Seed-specific, e.g. embryo, endosperm
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/0004—Oxidoreductases (1.)
- C12N9/0071—Oxidoreductases (1.) acting on paired donors with incorporation of molecular oxygen (1.14)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/0004—Oxidoreductases (1.)
- C12N9/0071—Oxidoreductases (1.) acting on paired donors with incorporation of molecular oxygen (1.14)
- C12N9/0083—Miscellaneous (1.14.99)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y114/00—Oxidoreductases acting on paired donors, with incorporation or reduction of molecular oxygen (1.14)
- C12Y114/19—Oxidoreductases acting on paired donors, with incorporation or reduction of molecular oxygen (1.14) with oxidation of a pair of donors resulting in the reduction of molecular oxygen to two molecules of water (1.14.19)
- C12Y114/19001—Stearoyl-CoA 9-desaturase (1.14.19.1), i.e. DELTA9-desaturase
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y02—TECHNOLOGIES OR APPLICATIONS FOR MITIGATION OR ADAPTATION AGAINST CLIMATE CHANGE
- Y02A—TECHNOLOGIES FOR ADAPTATION TO CLIMATE CHANGE
- Y02A40/00—Adaptation technologies in agriculture, forestry, livestock or agroalimentary production
- Y02A40/10—Adaptation technologies in agriculture, forestry, livestock or agroalimentary production in agriculture
- Y02A40/146—Genetically Modified [GMO] plants, e.g. transgenic plants
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Genetics & Genomics (AREA)
- Chemical & Material Sciences (AREA)
- Engineering & Computer Science (AREA)
- Organic Chemistry (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Biotechnology (AREA)
- Biomedical Technology (AREA)
- General Engineering & Computer Science (AREA)
- Molecular Biology (AREA)
- General Health & Medical Sciences (AREA)
- Biochemistry (AREA)
- Microbiology (AREA)
- Medicinal Chemistry (AREA)
- Physics & Mathematics (AREA)
- Biophysics (AREA)
- Cell Biology (AREA)
- Plant Pathology (AREA)
- Nutrition Science (AREA)
- Oil, Petroleum & Natural Gas (AREA)
- Developmental Biology & Embryology (AREA)
- Pregnancy & Childbirth (AREA)
- Reproductive Health (AREA)
- Breeding Of Plants And Reproduction By Means Of Culturing (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Enzymes And Modification Thereof (AREA)
Description
優先権主張
本願は、「ACCUMULATION OF N-7 FATTY ACIDS IN PLANT SEEDS」について2010年6月24日に出願された米国仮特許出願第61/358,318号の出願日の利益を主張する。
[1]配列番号1と少なくとも60%同一であるヌクレオチド配列を含むΔ 9 デサチュラーゼ酵素をコードする核酸分子。
[2]遺伝子調節要素をさらに含む、上記[1]に記載の核酸分子。
[3]上記遺伝子調節要素がファゼオリンプロモータまたはLTP170プロモータである、上記[2]に記載の核酸分子。
[4]配列番号2と少なくとも80%同一であるアミノ酸配列を含むΔ 9 デサチュラーゼ酵素。
[5]配列番号2と少なくとも80%同一である上記アミノ酸が配列番号2中の位置114に類似した位置のセリン、配列番号2中の位置117に類似した位置のアルギニン、配列番号2中の位置118に類似した位置のシステイン、配列番号2中の位置179に類似した位置のロイシン、または配列番号2中の位置188に類似した位置のトレオニンを含む、上記[4]に記載のΔ 9 デサチュラーゼ酵素。
[6]配列番号2と少なくとも80%同一である上記アミノ酸が配列番号2中の位置114に類似した位置のセリン、配列番号2中の位置117に類似した位置のアルギニン、配列番号2中の位置118に類似した位置のシステイン、配列番号2中の位置179に類似した位置のロイシン、および配列番号2中の位置188に類似した位置のトレオニンを含む、上記[4]に記載のΔ 9 デサチュラーゼ酵素。
[7]植物物質を上記[1]に記載の核酸分子によって、上記植物物質中の異常脂肪酸の量が増加されるように形質転換することを含む、植物物質中の異常脂肪酸の量を増加させる方法。
[8]植物物質を上記[1]に記載の追加の核酸分子によって形質転換することをさらに含む、上記[7]に記載の方法。
[9]上記植物物質が植物物質中の16:0−ACPのレベルを上昇させる手段を含む、上記[7]に記載の方法。
[10]上記植物物質中の16:0−ACPのレベルを上昇させる手段がプラスチド外デサチュラーゼの発現である、上記[9]に記載の方法。
[11]植物物質中の16:0−ACPのレベルを上昇させる手段がKASIIの抑制である、上記[9]に記載の方法。
[12]KASIIの抑制がfab1遺伝子中に突然変異を導入することによって達成される、上記[11]に記載の方法。
[13]上記植物物質中の16:0−ACPのレベルを上昇させる手段が植物物質中の16:0脂肪酸の伸長を減少させることである、上記[9]に記載の方法。
[14]植物物質中の16:0脂肪酸の伸長を減少させることが、fae1遺伝子中に突然変異を導入することによって達成される、上記[13]に記載の方法。
[15]植物物質がアラビドプシス、ボラゴ、キャノーラ、リシヌス、テオブロマ、ジーア、ゴシピウム、クランべ、クフェア、リヌム、レスクエレラ、リムナンテス、リノーラ、トロパエオルム、オエノテラ、オレア、エラエイス、アラキス、アブラナ、カルタムス、グリシン、ソヤ、ヘリアンタス、ニコチアナ、ベルノニア、トリチカム、ホルデウム、オリザ、アベナ、ソルガム、セカレまたはグラミネアエの他のメンバーを含む群より選択される属より選択される植物から得られる、上記[7]に記載の方法。
[16]上記植物物質が植物物質中の16:0−ACPのレベルを上昇させる2つの手段を含む、上記[7]に記載の方法。
[17]植物物質中の16:0−ACPのレベルを上昇させる第1の手段がKASIIの抑制であり、植物物質中の16:0−ACPのレベルを上昇させる第2の手段が植物物質中の16:0脂肪酸の伸長を減少させることである、上記[16]に記載の方法。
[18]上記植物物質が植物物質中の16:0−ACPのレベルを上昇させる3つの手段を含む、上記[7]に記載の方法。
[19]植物物質中の16:0−ACPのレベルを上昇させる第1の手段がKASIIの抑制であり、植物物質中の16:0−ACPのレベルを上昇させる第2の手段が植物物質中の16:0脂肪酸の伸長を減少させることであり、植物物質中の16:0−ACPのレベルを上昇させる第3の手段がプラスチド外デサチュラーゼの発現である、上記[18]に記載の方法。
[20]上記プラスチド外デサチュラーゼがLnΔ9DまたはAnΔ9デサチュラーゼである、上記[10]に記載の方法。
[21]野生型植物と比較して植物中に増加した量の異常脂肪酸を含む遺伝子操作植物を生成する方法であって:植物物質を上記[1]に記載の核酸分子によって形質転換すること、および植物を得るために形質転換された植物物質を培養することを含む方法。
[22]上記植物がアラビドプシス、ボラゴ、キャノーラ、リシヌス、テオブロマ、ジーア、ゴシピウム、クランべ、クフェア、リヌム、レスクエレラ、リムナンテス、リノーラ、トロパエオルム、オエノテラ、オレア、エラエイス、アラキス、アブラナ、カルタムス、グリシン、ソヤ、ヘリアンタス、ニコチアナ、ベルノニア、トリチカム、ホルデウム、オリザ、アベナ、ソルガム、セカレまたはグラミネアエの他のメンバーを含む群より選択される属より選択される、上記[21]に記載の方法。
[23]上記[21]に記載の方法によって得られた植物。
[24]上記[23]に記載の植物から得られた植物物質。
[25]上記植物物質が種子である、上記[24]に記載の植物物質。
また、植物種子中の異常脂肪酸のパーセント組成が上昇するように、WTトウゴマΔ9−18:0デサチュラーゼと比べてCom25酵素の向上したデサチュラーゼ活性を利用するために、植物細胞中でCom25を発現する方法も開示される。いくつかの実施形態において、方法はアラビドプシス中でCom25を発現させることを包含する。ある実施形態において、植物種子中で増加した異常脂肪酸はω−7脂肪酸である。これらの実施形態において、ω−7脂肪酸は16:1Δ19および/または18:1Δ11であり得る。
本明細書では、配列番号1と少なくとも60%同一であるヌクレオチド配列を含むΔ9デサチュラーゼ酵素をコードするヌクレオチド酸分子を開示する。核酸分子は、遺伝子調節要素をさらに含み得る。いくつかの実施形態において、遺伝子調節要素は、ファゼオリンプロモータであり得る。
II.略語
x:yΔz x個の炭素およびカルボキシル端から数えた位置zにおけるy個の二重結合を含有する脂肪酸
ACP アシル担体タンパク質
COA 補酵素A
KASII β−ケトアシル−ACPシンターゼII
FA 脂肪酸
FAS 脂肪酸合成
FAME 脂肪酸メチルエステル
WT 野生型
III.用語
脂肪酸:本明細書で使用する場合、「脂肪酸」という用語は、約C12からC22の鎖長の長鎖脂肪族酸(アルカン酸)を指すが、より長いまたはより短い鎖長の酸のどちらも公知である。脂肪酸の構造は、表記x:yΔzによって表され、式中、「x」は特定の脂肪酸中の炭素(C)原子の総数であり、「y」は、酸のカルボキシル端からカウントした位置「z」における炭素鎖中の二重結合の数である。
A.概要
本発明の実施形態は、例えば植物種子中のパルミトレイン酸(16:1Δ9)およびバクセン酸(18:1Δ11)からなるω−7脂肪酸(FA)の蓄積を代謝操作することへの系統的アプローチを包含する。プラスチドにおいて新たに合成された脂肪酸の流れを遮断する方法を例示するために、トウゴマΔ9−18:0−デサチュラーゼの16:0−デサチュラーゼ活性を向上させる指向性進化プログラムから生じる16:0−ACPデサチュラーゼであるCom25を、種子特異的ファゼオリンプロモータの制御下で発現させた。本明細書で開示する実施形態では、いずれの種子特異的プロモータも使用され得る。このアプローチは、ω−7 FAの蓄積を野生型(WT)における2%未満からCom25トランスフォーマントにおける約14%まで上昇させた。
本発明のいくつかの実施形態における核酸配列は、配列番号1と整列させたときに増加するパーセンテージ同一性を示す。これらおよび他の実施形態の中の特異的核酸配列は、配列番号2と例えば少なくとも60%、65%、70%、75%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%または100%の同一性を有する配列を含み得る。例えばコドン縮重による許容されるヌクレオチド置換によって、コードされたポリペプチドのアミノ酸配列が実質的に変更されることなく、核酸分子が修飾され得ることが当業者によって理解される。
本発明のいくつかの実施形態によるアミノ酸配列は、配列番号2と整列させたときに増加するパーセンテージ同一性を示す。これらおよび他の実施形態の中の特異的アミノ酸配列は、配列番号2と例えば少なくとも70%、75%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%または100%の同一性を有する配列を含み得る。多くの実施形態において、配列番号2と整列させたときに上記の配列同一性を有するアミノ酸配列は、酵素Δ9−18:0−ACPデサチュラーゼ活性を持つペプチドをコードする。
本発明の態様は、親トウゴマデサチュラーゼから誘導された新規の遺伝子操作デサチュラーゼに関する。詳細な実施形態において、遺伝子操作デサチュラーゼはCom25である。Com25は、親トウゴマデサチュラーゼとは以下の5つのアミノ酸位置:M114S、T117R、L118C、P179L、およびG188T(成熟トウゴマデサチュラーゼPDBエントリ1AFRに従ってナンバリング)が異なる。さらなる実施形態において、遺伝子操作デサチュラーゼは、Com25におけるこれらの5つの突然変異の1つ以上を含み得る。例えば遺伝子操作デサチュラーゼは、親トウゴマデサチュラーゼとは以下の位置が異なり得る:M114S;T117R;L118C;P179L;G188T;M114SおよびT117R;M114SおよびL118C;M114SおよびP179L;M114SおよびG188T;T117RおよびL118C;T117RおよびP179L;T117RおよびG188T;L118CおよびP179L;L118CおよびG188T;P179LおよびG188T;M114S、T117R、およびL118C;M114S、T117R、およびP179L;M114S、T1117R、およびG188T;M114S、L118C、およびP179L;M114S、L118C、およびG188T;M114S、P179L、およびG188T;T117R、L118C、およびP179L;T117R、L118C、およびG188T;T117R、P179L、およびG188T;またはL118C、P179L、およびG188T。
好ましい実施形態において、Com25によって形質転換された宿主細胞または物質は、16:0脂肪酸の上昇したレベルを呈し得る。宿主細胞は、例えばこれらの宿主細胞において16:0−ACPの代謝を低下させることによって、16:0脂肪酸の上昇したレベルを呈し得る。宿主細胞において16:0脂肪酸のレベルを上昇させる他の方法が使用され得て、このような方法は、当業者の裁量の行使によって選ばれ得る。宿主細胞における16:0脂肪酸のレベルを上昇させる方法の例は、これに限定されるわけではないが、1)宿主細胞におけるプラスチド外デサチュラーゼの発現、2)例えばfab1遺伝子に突然変異を導入することによる、宿主細胞におけるKASIIの抑制、および3)例えばfae1遺伝子に突然変異を導入することによる、16:0脂肪酸の伸長を減少させることを包含する。
本発明は、配列番号1に少なくとも60%同一である核酸配列を含む1つ以上の核酸分子を含む形質転換細胞を産生する方法にも関する。このような核酸分子は、例えば非コード調節要素、例えばプロモータも含み得る。他の配列も非コード調節要素および転写可能核酸分子配列と共に、細胞中に導入され得る。このような他の配列は、3’転写ターミネータ、3’ポリアデニル化シグナル、他の非翻訳配列、トランジットまたはターゲティング配列、選択可能マーカー、エンハンサ、およびオペレータを包含し得る。
本発明のいくつかの実施形態において、トランスジェニック種子は、配列番号2と少なくとも90%同一であるアミノ酸配列を含むポリペプチドを含む。これらおよび他の実施形態において、トランスジェニック種子は、配列番号1と少なくとも60%同一である核酸配列を含む。ある実施形態において、本発明の種子は異常脂肪酸、例えばω−7脂肪酸、例えば16:1Δ 9 -および/または18:1Δ11の上昇したレベルを呈する。種子は、受精トランスジェニック植物から収穫されて、本発明による核酸配列および興味のある別の遺伝子または核酸コンストラクトを含むハイブリッド植物系統を包含する、本発明の形質転換された植物の子孫世代を成長させるために使用することができる。
物質および方法
植物成長および形質転換
Com25は、長さが18C未満のアシル鎖に対して改善された活性を持つバリアントを同定するように設計されたコンビナトリアル飽和変異誘発/選択のプログラムから生じた、リシヌス・コムニス(Ricinus communis)Δ9−18:0−ACPデサチュラーゼのバリアントである。Whittle and Shanklin (2001) J. Biol. Chem. 276(24):21500-5。Com25は、親トウゴマデサチュラーゼとは以下の5つのアミノ酸位置:M114S、T117R、L118C、P179L、およびG188T(成熟トウゴマデサチュラーゼPDBエントリ1AFRに従ってナンバリング)が異なる。
Phas:Com25。その標準(authentic)輸送ペプチドを含有するように操作され、5’PacIおよび3’XhoI制限部位が隣接したトウゴマバリアントCom25のオープン・リーディング・フレーム全体がプラスミドpDs−Red−Phas、Pidkowich et al., (2007) Proc. Natl. Acad. Sci. USA 104(11): 4742-7、(Ds−Redマーカーあり)の対応する部位にクローニングされてPhas:Com25(図5)が生成された。
単一の種子の脂肪酸を分析するために、脂肪酸メチルエステル(FAME)が、メタノール中の0.2Mトリメチルスルホニウムヒドロキシドによって種子をインキュベートすることによって調製された。Butte et al., (1982) Anal. Lett. 15(10):841-50。バルク種子を同様に分析するために、FAMEは、0.5mL BCl3中で種子を80℃にて1時間インキュベートすること、それを1mLヘキサンによって抽出すること、および次にN2下で乾燥させることによって調製された。FAMEは、60m×250μM SP−2390キャピラリーカラム(スペルコ)を装着した、HP6890(商標)ガスクロマトグラフ−水素炎イオン化型検出器(アジレントテクノロジー)、またはHP5890(商標)ガスクロマトグラフ−質量分析計(ヒューレット・パッカード)のどちらかで分析された。分析中に炉温度は100℃から240℃まで15℃/分の速度で上昇され、流速は1.1mL/分であった。質量分析はHP5973(商標)質量選択検出器(ヒューレット−パッカード)によって行われた。本発明者らは、一不飽和FAMEの二重結合位置をジメチルスルフィド誘導体化によって決定した。Yamamoto et al., (1991) Chem. and Phys. Lipids 60(1):39-50。
実施例II
WTアラビドプシスにおけるCom25の発現
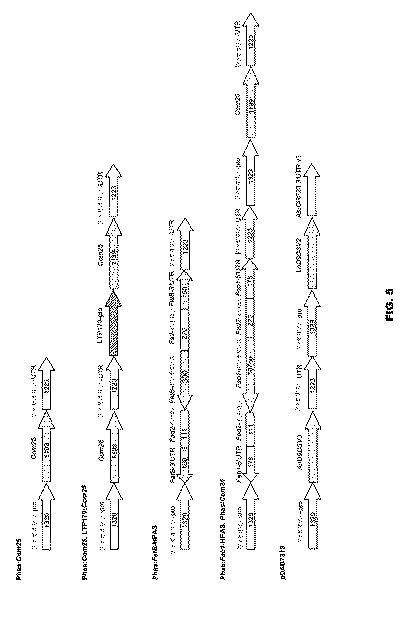
実施例IV
16:0の上昇したレベルを含有する宿主におけるCom25の発現
実施例V
増加するCom25遺伝子の用量は増加したω−7蓄積を生じる
実施例VI
プラスチド外Δ9−16:0デサチュラーゼの発現はω−7 FA蓄積を上昇させる
Claims (11)
- 異種プラスチド外LnΔ9DSまたはAnΔ9DSデサチュラーゼを発現する植物物質を、配列番号2のアミノ酸配列に少なくとも90%同一であるアミノ酸配列を含むΔ9デサチュラーゼ酵素をコードする配列番号1のヌクレオチド配列に少なくとも80%同一であるヌクレオチド配列を含む核酸分子によって、前記植物物質中の異常脂肪酸の量が増加されるように形質転換することを含む、植物物質中の異常脂肪酸の量を増加させる方法であって、前記Δ9デサチュラーゼ酵素のアミノ酸配列が、配列番号2のアミノ酸配列中の114位に類似する位置にセリンを、配列番号2のアミノ酸配列中の117位に類似する位置にアルギニンを、配列番号2のアミノ酸配列の118位に類似する位置にシステインを、配列番号2のアミノ酸配列中の179位に類似する位置にロイシンを、または配列番号2のアミノ酸配列中の188位に類似する位置にトレオニンを含む、方法。
- 植物物質を、配列番号2のアミノ酸配列に少なくとも90%同一であるアミノ酸配列を含むΔ9デサチュラーゼ酵素をコードする配列番号1のヌクレオチド配列に少なくとも80%同一であるヌクレオチド配列を含む追加の核酸分子によって形質転換することをさらに含む、請求項1に記載の方法であって、前記Δ9デサチュラーゼ酵素のアミノ酸配列が、配列番号2のアミノ酸配列中の114位に類似する位置にセリンを、配列番号2のアミノ酸配列中の117位に類似する位置にアルギニンを、配列番号2のアミノ酸配列の118位に類似する位置にシステインを、配列番号2のアミノ酸配列中の179位に類似する位置にロイシンを、または配列番号2のアミノ酸配列中の188位に類似する位置にトレオニンを含む、方法。
- 前記植物物質がfab1遺伝子中に突然変異を含む、請求項1に記載の方法。
- 前記植物物質が、fae1遺伝子中に突然変異を含む、請求項1に記載の方法。
- 植物物質がアラビドプシス、ボラゴ、キャノーラ、リシヌス、テオブロマ、ジーア、ゴシピウム、クランべ、クフェア、リヌム、レスクエレラ、リムナンテス、リノーラ、トロパエオルム、オエノテラ、オレア、エラエイス、アラキス、アブラナ、カルタムス、グリシン、ソヤ、ヘリアンタス、ニコチアナ、ベルノニア、トリチカム、ホルデウム、オリザ、アベナ、ソルガム、セカレまたはグラミネアエの他のメンバーを含む群より選択される属より選択される植物から得られる、請求項1に記載の方法。
- 前記植物物質がfab1遺伝子中に突然変異、およびfae1遺伝子中に突然変異を含む、請求項1に記載の方法。
- 植物を得るために前記形質転換された植物物質を培養することをさらに含む、請求項1に記載の方法。
- 前記植物がアラビドプシス、ボラゴ、キャノーラ、リシヌス、テオブロマ、ジーア、ゴシピウム、クランべ、クフェア、リヌム、レスクエレラ、リムナンテス、リノーラ、トロパエオルム、オエノテラ、オレア、エラエイス、アラキス、アブラナ、カルタムス、グリシン、ソヤ、ヘリアンタス、ニコチアナ、ベルノニア、トリチカム、ホルデウム、オリザ、アベナ、ソルガム、セカレまたはグラミネアエの他のメンバーを含む群より選択される属より選択される、請求項7に記載の方法。
- 請求項7に記載の方法によって得られた植物。
- 請求項9に記載の植物から得られた植物物質。
- 前記植物物質が種子である、請求項10に記載の植物物質。
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US35831810P | 2010-06-24 | 2010-06-24 | |
| US61/358,318 | 2010-06-24 | ||
| PCT/US2011/041759 WO2011163557A2 (en) | 2010-06-24 | 2011-06-24 | Accumulation of omega-7 fatty acids in plant seeds |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2013532975A JP2013532975A (ja) | 2013-08-22 |
| JP6068336B2 true JP6068336B2 (ja) | 2017-01-25 |
Family
ID=45353924
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2013516790A Expired - Fee Related JP6068336B2 (ja) | 2010-06-24 | 2011-06-24 | 植物種子へのオメガ−7脂肪酸の蓄積 |
Country Status (21)
| Country | Link |
|---|---|
| US (2) | US9340776B2 (ja) |
| EP (1) | EP2585599B1 (ja) |
| JP (1) | JP6068336B2 (ja) |
| KR (1) | KR20130026487A (ja) |
| CN (1) | CN103429741B (ja) |
| AR (1) | AR081302A1 (ja) |
| AU (1) | AU2011270751B2 (ja) |
| BR (1) | BR112012033037A2 (ja) |
| CA (1) | CA2803599C (ja) |
| CL (1) | CL2012003663A1 (ja) |
| EA (1) | EA022985B1 (ja) |
| ES (1) | ES2645239T3 (ja) |
| HU (1) | HUE037067T2 (ja) |
| IL (1) | IL223440A (ja) |
| MX (2) | MX355329B (ja) |
| NZ (1) | NZ603968A (ja) |
| PL (1) | PL2585599T3 (ja) |
| TW (1) | TWI570239B (ja) |
| UA (1) | UA113834C2 (ja) |
| WO (1) | WO2011163557A2 (ja) |
| ZA (1) | ZA201209082B (ja) |
Families Citing this family (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP3232770A4 (en) * | 2014-12-19 | 2018-06-06 | Dow AgroSciences LLC | Generation of transgenic canola with low or no saturated fatty acids |
| CN105053256A (zh) * | 2015-07-29 | 2015-11-18 | 广州金酮医疗科技有限公司 | 一种风味型营养均衡调和油 |
| CN114431101B (zh) * | 2022-01-28 | 2023-02-28 | 广西壮族自治区农业科学院 | 一种甘蔗13c同位素标记的方法及装置 |
Family Cites Families (27)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US5188958A (en) | 1986-05-29 | 1993-02-23 | Calgene, Inc. | Transformation and foreign gene expression in brassica species |
| NZ221259A (en) | 1986-07-31 | 1990-05-28 | Calgene Inc | Seed specific transcriptional regulation |
| US5004863B2 (en) | 1986-12-03 | 2000-10-17 | Agracetus | Genetic engineering of cotton plants and lines |
| ATE87032T1 (de) | 1986-12-05 | 1993-04-15 | Ciba Geigy Ag | Verbessertes verfahren zur transformation von pflanzlichen protoplasten. |
| US5416011A (en) | 1988-07-22 | 1995-05-16 | Monsanto Company | Method for soybean transformation and regeneration |
| US7705215B1 (en) | 1990-04-17 | 2010-04-27 | Dekalb Genetics Corporation | Methods and compositions for the production of stably transformed, fertile monocot plants and cells thereof |
| US5550318A (en) | 1990-04-17 | 1996-08-27 | Dekalb Genetics Corporation | Methods and compositions for the production of stably transformed, fertile monocot plants and cells thereof |
| US5484956A (en) | 1990-01-22 | 1996-01-16 | Dekalb Genetics Corporation | Fertile transgenic Zea mays plant comprising heterologous DNA encoding Bacillus thuringiensis endotoxin |
| WO1991010725A1 (en) | 1990-01-22 | 1991-07-25 | Dekalb Plant Genetics | Fertile transgenic corn plants |
| US6403865B1 (en) | 1990-08-24 | 2002-06-11 | Syngenta Investment Corp. | Method of producing transgenic maize using direct transformation of commercially important genotypes |
| US5384253A (en) | 1990-12-28 | 1995-01-24 | Dekalb Genetics Corporation | Genetic transformation of maize cells by electroporation of cells pretreated with pectin degrading enzymes |
| US5518908A (en) | 1991-09-23 | 1996-05-21 | Monsanto Company | Method of controlling insects |
| CA2084348A1 (en) * | 1991-12-31 | 1993-07-01 | David F. Hildebrand | Fatty acid alteration by a d9 desaturase in transgenic plant tissue |
| ATE398679T1 (de) | 1992-07-07 | 2008-07-15 | Japan Tobacco Inc | Verfahren zur transformation einer monokotyledon pflanze |
| US7109392B1 (en) * | 1996-10-09 | 2006-09-19 | Cargill, Incorporated | Methods for increasing oleic acid content in seeds from transgenic plants containing a mutant delta 12 desaturase |
| US5981840A (en) | 1997-01-24 | 1999-11-09 | Pioneer Hi-Bred International, Inc. | Methods for agrobacterium-mediated transformation |
| WO2000042207A2 (en) | 1999-01-14 | 2000-07-20 | Monsanto Technology Llc | Soybean transformation method |
| US7323335B2 (en) * | 1999-01-19 | 2008-01-29 | Brookhaven Science Associates, Llc | Mutant fatty acid desaturase and methods for directed mutagenesis |
| ATE342362T1 (de) | 1999-06-09 | 2006-11-15 | Brookhaven Science Ass Llc | Mutierte fettsäuredesaturase |
| US7214859B2 (en) * | 2002-08-16 | 2007-05-08 | National Research Council Of Canada | Brassica pyruvate dehydrogenase kinase gene |
| US7655833B2 (en) * | 2003-05-29 | 2010-02-02 | Brookhaven Science Associates, Llc | ADS genes for reducing saturated fatty acid levels in seed oils |
| US7122367B2 (en) * | 2003-06-03 | 2006-10-17 | Board Of Trustees Operating Michigan State University | Diacylglycerol acyltransferase genes, proteins, and uses thereof |
| EP3318121A1 (en) * | 2004-10-08 | 2018-05-09 | Dow AgroSciences LLC | Certain plants with "no saturate" or reduced saturate levels of fatty acids in seeds, and oil derived from the seeds |
| US20080260933A1 (en) | 2004-10-08 | 2008-10-23 | Dow Agroscience Llc | Certain Plants with "No Saturate" or Reduced Saturate Levels of Fatty Acids in Seeds, and Oil Derived from the Seeds |
| AU2006312126A1 (en) * | 2005-11-04 | 2007-05-18 | Dow Agrosciences Llc | Preparation of vaccine master cell lines using recombinant plant suspension cultures |
| US7943823B2 (en) * | 2006-04-28 | 2011-05-17 | E.I. Du Pont De Nemours And Company | Delta-8 desaturase and its use in making polyunsaturated fatty acids |
| US8957280B2 (en) * | 2007-05-03 | 2015-02-17 | E. I. Du Pont De Nemours And Company | Delta-5 desaturases and their use in making polyunsaturated fatty acids |
-
2011
- 2011-06-23 AR ARP110102187A patent/AR081302A1/es unknown
- 2011-06-23 TW TW100122017A patent/TWI570239B/zh not_active IP Right Cessation
- 2011-06-24 EA EA201291242A patent/EA022985B1/ru not_active IP Right Cessation
- 2011-06-24 HU HUE11798970A patent/HUE037067T2/hu unknown
- 2011-06-24 ES ES11798970.7T patent/ES2645239T3/es active Active
- 2011-06-24 AU AU2011270751A patent/AU2011270751B2/en active Active
- 2011-06-24 PL PL11798970T patent/PL2585599T3/pl unknown
- 2011-06-24 CA CA2803599A patent/CA2803599C/en active Active
- 2011-06-24 KR KR1020137001833A patent/KR20130026487A/ko not_active Abandoned
- 2011-06-24 US US13/168,320 patent/US9340776B2/en not_active Expired - Fee Related
- 2011-06-24 JP JP2013516790A patent/JP6068336B2/ja not_active Expired - Fee Related
- 2011-06-24 CN CN201180031133.XA patent/CN103429741B/zh not_active Expired - Fee Related
- 2011-06-24 MX MX2016010612A patent/MX355329B/es unknown
- 2011-06-24 WO PCT/US2011/041759 patent/WO2011163557A2/en not_active Ceased
- 2011-06-24 BR BR112012033037A patent/BR112012033037A2/pt not_active IP Right Cessation
- 2011-06-24 UA UAA201300815A patent/UA113834C2/uk unknown
- 2011-06-24 NZ NZ603968A patent/NZ603968A/en not_active IP Right Cessation
- 2011-06-24 MX MX2012015291A patent/MX343272B/es active IP Right Grant
- 2011-06-24 EP EP11798970.7A patent/EP2585599B1/en not_active Not-in-force
-
2012
- 2012-11-30 ZA ZA2012/09082A patent/ZA201209082B/en unknown
- 2012-12-04 IL IL223440A patent/IL223440A/en active IP Right Grant
- 2012-12-21 CL CL2012003663A patent/CL2012003663A1/es unknown
-
2016
- 2016-04-15 US US15/130,788 patent/US9976155B2/en active Active
Also Published As
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US20240150780A1 (en) | Enzymes and methods for producing omega-3 fatty acids | |
| CN103068986B (zh) | 降低植物种子的饱和脂肪酸含量 | |
| WO2016099568A1 (en) | Generation of transgenic canola with low or no saturated fatty acids | |
| AU2023241326A1 (en) | Enzymes and methods for producing omega-3 fatty acids | |
| US9976155B2 (en) | Accumulation of omega-7 fatty acids in plant seeds | |
| US10370674B2 (en) | Generation of transgenic canola with low or no saturated fatty acids | |
| AU2020277266B2 (en) | Enzymes and methods for producing omega-3 fatty acids | |
| HK1191975A (en) | Accumulation of omega-7 fatty acids in plant seeds | |
| HK1191975B (en) | Accumulation of omega-7 fatty acids in plant seeds |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20130521 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20140618 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20150811 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20151008 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20160108 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20160329 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20160729 |
|
| A911 | Transfer to examiner for re-examination before appeal (zenchi) |
Free format text: JAPANESE INTERMEDIATE CODE: A911 Effective date: 20160921 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20161122 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20161222 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 6068336 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| LAPS | Cancellation because of no payment of annual fees |