JP5284554B2 - 前立腺特異的抗原の形態およびその検出方法 - Google Patents
前立腺特異的抗原の形態およびその検出方法 Download PDFInfo
- Publication number
- JP5284554B2 JP5284554B2 JP54726998A JP54726998A JP5284554B2 JP 5284554 B2 JP5284554 B2 JP 5284554B2 JP 54726998 A JP54726998 A JP 54726998A JP 54726998 A JP54726998 A JP 54726998A JP 5284554 B2 JP5284554 B2 JP 5284554B2
- Authority
- JP
- Japan
- Prior art keywords
- ppsa
- psa
- antibody
- sample
- determining
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Expired - Lifetime
Links
- 108010072866 Prostate-Specific Antigen Proteins 0.000 title description 224
- 238000001514 detection method Methods 0.000 title description 7
- 102000007066 Prostate-Specific Antigen Human genes 0.000 title 1
- 210000002966 serum Anatomy 0.000 claims abstract description 48
- 238000000034 method Methods 0.000 claims abstract description 34
- 239000012530 fluid Substances 0.000 claims abstract description 10
- 108090000765 processed proteins & peptides Proteins 0.000 claims description 38
- 102000004196 processed proteins & peptides Human genes 0.000 claims description 35
- 229920001184 polypeptide Polymers 0.000 claims description 31
- 150000001413 amino acids Chemical class 0.000 claims description 14
- 239000012634 fragment Substances 0.000 claims description 12
- 210000001519 tissue Anatomy 0.000 claims description 8
- 239000007790 solid phase Substances 0.000 claims description 6
- 239000003153 chemical reaction reagent Substances 0.000 claims description 5
- 230000009918 complex formation Effects 0.000 claims description 3
- 230000008105 immune reaction Effects 0.000 claims description 3
- 238000002156 mixing Methods 0.000 claims description 3
- 230000002860 competitive effect Effects 0.000 claims description 2
- 210000005267 prostate cell Anatomy 0.000 claims description 2
- 206010060862 Prostate cancer Diseases 0.000 abstract description 26
- 208000000236 Prostatic Neoplasms Diseases 0.000 abstract description 22
- 239000002243 precursor Substances 0.000 abstract description 10
- 239000013604 expression vector Substances 0.000 abstract description 9
- 238000003745 diagnosis Methods 0.000 abstract description 4
- 238000002405 diagnostic procedure Methods 0.000 abstract description 3
- 238000009007 Diagnostic Kit Methods 0.000 abstract description 2
- 102100038358 Prostate-specific antigen Human genes 0.000 description 223
- 101001050577 Homo sapiens Kinesin-like protein KIF2A Proteins 0.000 description 58
- 210000004027 cell Anatomy 0.000 description 53
- 239000002609 medium Substances 0.000 description 31
- 102000004169 proteins and genes Human genes 0.000 description 25
- 108090000623 proteins and genes Proteins 0.000 description 25
- 239000000523 sample Substances 0.000 description 25
- 235000018102 proteins Nutrition 0.000 description 23
- 210000000582 semen Anatomy 0.000 description 22
- 238000003556 assay Methods 0.000 description 20
- 230000000694 effects Effects 0.000 description 17
- 210000004962 mammalian cell Anatomy 0.000 description 14
- 238000006243 chemical reaction Methods 0.000 description 13
- 210000002307 prostate Anatomy 0.000 description 12
- 238000001262 western blot Methods 0.000 description 12
- 108090000631 Trypsin Proteins 0.000 description 11
- 102000004142 Trypsin Human genes 0.000 description 11
- 239000000203 mixture Substances 0.000 description 11
- 239000012588 trypsin Substances 0.000 description 11
- 238000004191 hydrophobic interaction chromatography Methods 0.000 description 10
- 108020004414 DNA Proteins 0.000 description 9
- 206010028980 Neoplasm Diseases 0.000 description 9
- 239000000427 antigen Substances 0.000 description 9
- 102000036639 antigens Human genes 0.000 description 9
- 108091007433 antigens Proteins 0.000 description 9
- 239000000872 buffer Substances 0.000 description 8
- 238000011534 incubation Methods 0.000 description 8
- 230000014759 maintenance of location Effects 0.000 description 8
- 102000004190 Enzymes Human genes 0.000 description 7
- 108090000790 Enzymes Proteins 0.000 description 7
- 201000011510 cancer Diseases 0.000 description 7
- 229940088598 enzyme Drugs 0.000 description 7
- 239000003550 marker Substances 0.000 description 7
- KFZMGEQAYNKOFK-UHFFFAOYSA-N Isopropanol Chemical compound CC(C)O KFZMGEQAYNKOFK-UHFFFAOYSA-N 0.000 description 6
- 150000007523 nucleic acids Chemical class 0.000 description 6
- 238000012163 sequencing technique Methods 0.000 description 6
- 206010004446 Benign prostatic hyperplasia Diseases 0.000 description 5
- 102000001399 Kallikrein Human genes 0.000 description 5
- 108060005987 Kallikrein Proteins 0.000 description 5
- 208000004403 Prostatic Hyperplasia Diseases 0.000 description 5
- 239000000499 gel Substances 0.000 description 5
- 239000012528 membrane Substances 0.000 description 5
- 108020004707 nucleic acids Proteins 0.000 description 5
- 102000039446 nucleic acids Human genes 0.000 description 5
- 239000000137 peptide hydrolase inhibitor Substances 0.000 description 5
- 239000001488 sodium phosphate Substances 0.000 description 5
- 229910000162 sodium phosphate Inorganic materials 0.000 description 5
- 241000894007 species Species 0.000 description 5
- RYFMWSXOAZQYPI-UHFFFAOYSA-K trisodium phosphate Chemical compound [Na+].[Na+].[Na+].[O-]P([O-])([O-])=O RYFMWSXOAZQYPI-UHFFFAOYSA-K 0.000 description 5
- 239000013598 vector Substances 0.000 description 5
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 4
- 239000000020 Nitrocellulose Substances 0.000 description 4
- 108091028043 Nucleic acid sequence Proteins 0.000 description 4
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 4
- 102000057032 Tissue Kallikreins Human genes 0.000 description 4
- 239000007983 Tris buffer Substances 0.000 description 4
- 235000001014 amino acid Nutrition 0.000 description 4
- 229940024606 amino acid Drugs 0.000 description 4
- BFNBIHQBYMNNAN-UHFFFAOYSA-N ammonium sulfate Chemical compound N.N.OS(O)(=O)=O BFNBIHQBYMNNAN-UHFFFAOYSA-N 0.000 description 4
- 229910052921 ammonium sulfate Inorganic materials 0.000 description 4
- 235000011130 ammonium sulphate Nutrition 0.000 description 4
- 238000004458 analytical method Methods 0.000 description 4
- 239000002299 complementary DNA Substances 0.000 description 4
- 239000007857 degradation product Substances 0.000 description 4
- 229940042399 direct acting antivirals protease inhibitors Drugs 0.000 description 4
- 210000000981 epithelium Anatomy 0.000 description 4
- 229920001220 nitrocellulos Polymers 0.000 description 4
- 201000001514 prostate carcinoma Diseases 0.000 description 4
- 238000000746 purification Methods 0.000 description 4
- 230000009257 reactivity Effects 0.000 description 4
- 239000011347 resin Substances 0.000 description 4
- 229920005989 resin Polymers 0.000 description 4
- 239000000758 substrate Substances 0.000 description 4
- 238000012360 testing method Methods 0.000 description 4
- LENZDBCJOHFCAS-UHFFFAOYSA-N tris Chemical compound OCC(N)(CO)CO LENZDBCJOHFCAS-UHFFFAOYSA-N 0.000 description 4
- 239000003656 tris buffered saline Substances 0.000 description 4
- 108010039627 Aprotinin Proteins 0.000 description 3
- 108091026890 Coding region Proteins 0.000 description 3
- 108010076504 Protein Sorting Signals Proteins 0.000 description 3
- 230000004913 activation Effects 0.000 description 3
- 229960004405 aprotinin Drugs 0.000 description 3
- 239000013060 biological fluid Substances 0.000 description 3
- 230000015572 biosynthetic process Effects 0.000 description 3
- 238000009835 boiling Methods 0.000 description 3
- 238000004587 chromatography analysis Methods 0.000 description 3
- 239000003593 chromogenic compound Substances 0.000 description 3
- 238000001378 electrochemiluminescence detection Methods 0.000 description 3
- 238000005516 engineering process Methods 0.000 description 3
- 239000003623 enhancer Substances 0.000 description 3
- 230000007062 hydrolysis Effects 0.000 description 3
- 238000006460 hydrolysis reaction Methods 0.000 description 3
- ZPNFWUPYTFPOJU-LPYSRVMUSA-N iniprol Chemical compound C([C@H]1C(=O)NCC(=O)NCC(=O)N[C@H]2CSSC[C@H]3C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@H](C(N[C@H](C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC=4C=CC(O)=CC=4)C(=O)N[C@@H](CC=4C=CC=CC=4)C(=O)N[C@@H](CC=4C=CC(O)=CC=4)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C)C(=O)NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CSSC[C@H](NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](C)NC(=O)[C@H](CO)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CC=4C=CC=CC=4)NC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CCCCN)NC(=O)[C@H](C)NC(=O)[C@H](CCCNC(N)=N)NC2=O)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CSSC[C@H](NC(=O)[C@H](CC=2C=CC=CC=2)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H]2N(CCC2)C(=O)[C@@H](N)CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(=O)N2[C@@H](CCC2)C(=O)N2[C@@H](CCC2)C(=O)N[C@@H](CC=2C=CC(O)=CC=2)C(=O)N[C@@H]([C@@H](C)O)C(=O)NCC(=O)N2[C@@H](CCC2)C(=O)N3)C(=O)NCC(=O)NCC(=O)N[C@@H](C)C(O)=O)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@H](C(=O)N[C@@H](CC=2C=CC=CC=2)C(=O)N[C@H](C(=O)N1)C(C)C)[C@@H](C)O)[C@@H](C)CC)=O)[C@@H](C)CC)C1=CC=C(O)C=C1 ZPNFWUPYTFPOJU-LPYSRVMUSA-N 0.000 description 3
- 238000002360 preparation method Methods 0.000 description 3
- 230000001105 regulatory effect Effects 0.000 description 3
- 239000012723 sample buffer Substances 0.000 description 3
- 230000003248 secreting effect Effects 0.000 description 3
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 3
- 239000007787 solid Substances 0.000 description 3
- 238000005406 washing Methods 0.000 description 3
- YBJHBAHKTGYVGT-ZKWXMUAHSA-N (+)-Biotin Chemical compound N1C(=O)N[C@@H]2[C@H](CCCCC(=O)O)SC[C@@H]21 YBJHBAHKTGYVGT-ZKWXMUAHSA-N 0.000 description 2
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 2
- BFSVOASYOCHEOV-UHFFFAOYSA-N 2-diethylaminoethanol Chemical compound CCN(CC)CCO BFSVOASYOCHEOV-UHFFFAOYSA-N 0.000 description 2
- 241000894006 Bacteria Species 0.000 description 2
- 241000283707 Capra Species 0.000 description 2
- 108090000317 Chymotrypsin Proteins 0.000 description 2
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 2
- 108010067306 Fibronectins Proteins 0.000 description 2
- 102000016359 Fibronectins Human genes 0.000 description 2
- 101710103262 Glandular kallikrein Proteins 0.000 description 2
- 239000004471 Glycine Substances 0.000 description 2
- 241000238631 Hexapoda Species 0.000 description 2
- FBOZXECLQNJBKD-ZDUSSCGKSA-N L-methotrexate Chemical compound C=1N=C2N=C(N)N=C(N)C2=NC=1CN(C)C1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 FBOZXECLQNJBKD-ZDUSSCGKSA-N 0.000 description 2
- 241001465754 Metazoa Species 0.000 description 2
- 108091034117 Oligonucleotide Proteins 0.000 description 2
- 102000035195 Peptidases Human genes 0.000 description 2
- 108091005804 Peptidases Proteins 0.000 description 2
- 206010035226 Plasma cell myeloma Diseases 0.000 description 2
- 239000004365 Protease Substances 0.000 description 2
- 108020004511 Recombinant DNA Proteins 0.000 description 2
- 102000012479 Serine Proteases Human genes 0.000 description 2
- 108010022999 Serine Proteases Proteins 0.000 description 2
- VYPSYNLAJGMNEJ-UHFFFAOYSA-N Silicium dioxide Chemical compound O=[Si]=O VYPSYNLAJGMNEJ-UHFFFAOYSA-N 0.000 description 2
- PMZURENOXWZQFD-UHFFFAOYSA-L Sodium Sulfate Chemical compound [Na+].[Na+].[O-]S([O-])(=O)=O PMZURENOXWZQFD-UHFFFAOYSA-L 0.000 description 2
- 238000001042 affinity chromatography Methods 0.000 description 2
- 238000001261 affinity purification Methods 0.000 description 2
- 239000003098 androgen Substances 0.000 description 2
- 238000013459 approach Methods 0.000 description 2
- 238000004113 cell culture Methods 0.000 description 2
- 230000010261 cell growth Effects 0.000 description 2
- 229960002376 chymotrypsin Drugs 0.000 description 2
- 238000012875 competitive assay Methods 0.000 description 2
- 238000002967 competitive immunoassay Methods 0.000 description 2
- 238000010276 construction Methods 0.000 description 2
- 230000009260 cross reactivity Effects 0.000 description 2
- 238000010828 elution Methods 0.000 description 2
- 230000002255 enzymatic effect Effects 0.000 description 2
- 238000011067 equilibration Methods 0.000 description 2
- 210000003527 eukaryotic cell Anatomy 0.000 description 2
- 238000003384 imaging method Methods 0.000 description 2
- 238000003018 immunoassay Methods 0.000 description 2
- 230000002163 immunogen Effects 0.000 description 2
- 239000006166 lysate Substances 0.000 description 2
- 238000005259 measurement Methods 0.000 description 2
- 108020004999 messenger RNA Proteins 0.000 description 2
- 229960000485 methotrexate Drugs 0.000 description 2
- 235000013336 milk Nutrition 0.000 description 2
- 239000008267 milk Substances 0.000 description 2
- 210000004080 milk Anatomy 0.000 description 2
- 230000004048 modification Effects 0.000 description 2
- 238000012986 modification Methods 0.000 description 2
- 238000010369 molecular cloning Methods 0.000 description 2
- 238000012544 monitoring process Methods 0.000 description 2
- 201000000050 myeloid neoplasm Diseases 0.000 description 2
- 239000013642 negative control Substances 0.000 description 2
- 239000012071 phase Substances 0.000 description 2
- 229920002401 polyacrylamide Polymers 0.000 description 2
- 229920002981 polyvinylidene fluoride Polymers 0.000 description 2
- 238000007639 printing Methods 0.000 description 2
- 238000012545 processing Methods 0.000 description 2
- 208000017497 prostate disease Diseases 0.000 description 2
- 239000011780 sodium chloride Substances 0.000 description 2
- 229910052938 sodium sulfate Inorganic materials 0.000 description 2
- 235000011152 sodium sulphate Nutrition 0.000 description 2
- 239000000126 substance Substances 0.000 description 2
- 238000001890 transfection Methods 0.000 description 2
- 230000001810 trypsinlike Effects 0.000 description 2
- 239000000439 tumor marker Substances 0.000 description 2
- 102100022524 Alpha-1-antichymotrypsin Human genes 0.000 description 1
- 239000004475 Arginine Substances 0.000 description 1
- DCXYFEDJOCDNAF-UHFFFAOYSA-N Asparagine Natural products OC(=O)C(N)CC(N)=O DCXYFEDJOCDNAF-UHFFFAOYSA-N 0.000 description 1
- 108090000227 Chymases Proteins 0.000 description 1
- 102000003858 Chymases Human genes 0.000 description 1
- 102100031673 Corneodesmosin Human genes 0.000 description 1
- 101710139375 Corneodesmosin Proteins 0.000 description 1
- -1 DEAE Chemical compound 0.000 description 1
- 239000006144 Dulbecco’s modified Eagle's medium Substances 0.000 description 1
- 238000002965 ELISA Methods 0.000 description 1
- 101710108755 Extracellular serine protease Proteins 0.000 description 1
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 1
- 108090000288 Glycoproteins Proteins 0.000 description 1
- 102000003886 Glycoproteins Human genes 0.000 description 1
- 102100037852 Insulin-like growth factor I Human genes 0.000 description 1
- 102000004374 Insulin-like growth factor binding protein 3 Human genes 0.000 description 1
- 108090000965 Insulin-like growth factor binding protein 3 Proteins 0.000 description 1
- 102100027612 Kallikrein-11 Human genes 0.000 description 1
- 101710176220 Kallikrein-2 Proteins 0.000 description 1
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 description 1
- DCXYFEDJOCDNAF-REOHCLBHSA-N L-asparagine Chemical compound OC(=O)[C@@H](N)CC(N)=O DCXYFEDJOCDNAF-REOHCLBHSA-N 0.000 description 1
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 1
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 1
- 239000004472 Lysine Substances 0.000 description 1
- 239000002033 PVDF binder Substances 0.000 description 1
- 102000003992 Peroxidases Human genes 0.000 description 1
- 229940124158 Protease/peptidase inhibitor Drugs 0.000 description 1
- 101800004937 Protein C Proteins 0.000 description 1
- 102000017975 Protein C Human genes 0.000 description 1
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 description 1
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 description 1
- 108090000783 Renin Proteins 0.000 description 1
- 102100028255 Renin Human genes 0.000 description 1
- 101800001700 Saposin-D Proteins 0.000 description 1
- 229920005654 Sephadex Polymers 0.000 description 1
- 239000012507 Sephadex™ Substances 0.000 description 1
- 238000012300 Sequence Analysis Methods 0.000 description 1
- 108700022175 Tissue Kallikreins Proteins 0.000 description 1
- 102000003978 Tissue Plasminogen Activator Human genes 0.000 description 1
- 108090000373 Tissue Plasminogen Activator Proteins 0.000 description 1
- GLNADSQYFUSGOU-GPTZEZBUSA-J Trypan blue Chemical compound [Na+].[Na+].[Na+].[Na+].C1=C(S([O-])(=O)=O)C=C2C=C(S([O-])(=O)=O)C(/N=N/C3=CC=C(C=C3C)C=3C=C(C(=CC=3)\N=N\C=3C(=CC4=CC(=CC(N)=C4C=3O)S([O-])(=O)=O)S([O-])(=O)=O)C)=C(O)C2=C1N GLNADSQYFUSGOU-GPTZEZBUSA-J 0.000 description 1
- 101710152431 Trypsin-like protease Proteins 0.000 description 1
- 108060008682 Tumor Necrosis Factor Proteins 0.000 description 1
- 102000000852 Tumor Necrosis Factor-alpha Human genes 0.000 description 1
- 241000251539 Vertebrata <Metazoa> Species 0.000 description 1
- 238000002835 absorbance Methods 0.000 description 1
- 239000002253 acid Substances 0.000 description 1
- 150000007513 acids Chemical class 0.000 description 1
- 230000009471 action Effects 0.000 description 1
- 208000009956 adenocarcinoma Diseases 0.000 description 1
- 230000002776 aggregation Effects 0.000 description 1
- 238000004220 aggregation Methods 0.000 description 1
- 235000004279 alanine Nutrition 0.000 description 1
- 108010091628 alpha 1-Antichymotrypsin Proteins 0.000 description 1
- 150000001408 amides Chemical class 0.000 description 1
- 238000012870 ammonium sulfate precipitation Methods 0.000 description 1
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 1
- 229960001230 asparagine Drugs 0.000 description 1
- 235000009582 asparagine Nutrition 0.000 description 1
- 230000008901 benefit Effects 0.000 description 1
- 239000012620 biological material Substances 0.000 description 1
- 230000001851 biosynthetic effect Effects 0.000 description 1
- 229960002685 biotin Drugs 0.000 description 1
- 235000020958 biotin Nutrition 0.000 description 1
- 239000011616 biotin Substances 0.000 description 1
- 210000004369 blood Anatomy 0.000 description 1
- 239000008280 blood Substances 0.000 description 1
- 239000001506 calcium phosphate Substances 0.000 description 1
- 229910000389 calcium phosphate Inorganic materials 0.000 description 1
- 235000011010 calcium phosphates Nutrition 0.000 description 1
- 238000007707 calorimetry Methods 0.000 description 1
- 230000015556 catabolic process Effects 0.000 description 1
- 230000003197 catalytic effect Effects 0.000 description 1
- 230000030833 cell death Effects 0.000 description 1
- 230000006037 cell lysis Effects 0.000 description 1
- 230000003833 cell viability Effects 0.000 description 1
- 108091092328 cellular RNA Proteins 0.000 description 1
- 238000005119 centrifugation Methods 0.000 description 1
- 238000012512 characterization method Methods 0.000 description 1
- 238000011098 chromatofocusing Methods 0.000 description 1
- 230000037012 chymotrypsin-like activity Effects 0.000 description 1
- 238000010367 cloning Methods 0.000 description 1
- 201000010897 colon adenocarcinoma Diseases 0.000 description 1
- 208000029742 colonic neoplasm Diseases 0.000 description 1
- 150000001875 compounds Chemical class 0.000 description 1
- 238000007796 conventional method Methods 0.000 description 1
- 238000006731 degradation reaction Methods 0.000 description 1
- 238000013461 design Methods 0.000 description 1
- 238000010586 diagram Methods 0.000 description 1
- 238000010790 dilution Methods 0.000 description 1
- 239000012895 dilution Substances 0.000 description 1
- 238000001962 electrophoresis Methods 0.000 description 1
- 210000002919 epithelial cell Anatomy 0.000 description 1
- 238000012869 ethanol precipitation Methods 0.000 description 1
- 238000011156 evaluation Methods 0.000 description 1
- 238000002474 experimental method Methods 0.000 description 1
- 235000013861 fat-free Nutrition 0.000 description 1
- 230000001605 fetal effect Effects 0.000 description 1
- 238000005194 fractionation Methods 0.000 description 1
- 230000004927 fusion Effects 0.000 description 1
- 238000002523 gelfiltration Methods 0.000 description 1
- 238000007429 general method Methods 0.000 description 1
- 210000004907 gland Anatomy 0.000 description 1
- 239000008103 glucose Substances 0.000 description 1
- 230000013595 glycosylation Effects 0.000 description 1
- 238000006206 glycosylation reaction Methods 0.000 description 1
- 239000011544 gradient gel Substances 0.000 description 1
- 238000004128 high performance liquid chromatography Methods 0.000 description 1
- 210000004408 hybridoma Anatomy 0.000 description 1
- 238000001727 in vivo Methods 0.000 description 1
- 230000002779 inactivation Effects 0.000 description 1
- 238000011081 inoculation Methods 0.000 description 1
- 230000003993 interaction Effects 0.000 description 1
- PNDPGZBMCMUPRI-UHFFFAOYSA-N iodine Chemical compound II PNDPGZBMCMUPRI-UHFFFAOYSA-N 0.000 description 1
- 238000005342 ion exchange Methods 0.000 description 1
- 238000002955 isolation Methods 0.000 description 1
- 210000003734 kidney Anatomy 0.000 description 1
- 238000002372 labelling Methods 0.000 description 1
- 239000003446 ligand Substances 0.000 description 1
- 238000001638 lipofection Methods 0.000 description 1
- 238000011068 loading method Methods 0.000 description 1
- 238000004519 manufacturing process Methods 0.000 description 1
- 230000009826 neoplastic cell growth Effects 0.000 description 1
- 229920001542 oligosaccharide Polymers 0.000 description 1
- 150000002482 oligosaccharides Chemical class 0.000 description 1
- 210000000496 pancreas Anatomy 0.000 description 1
- 230000037361 pathway Effects 0.000 description 1
- 125000001151 peptidyl group Chemical group 0.000 description 1
- 108040007629 peroxidase activity proteins Proteins 0.000 description 1
- 239000013612 plasmid Substances 0.000 description 1
- 230000008488 polyadenylation Effects 0.000 description 1
- 238000003752 polymerase chain reaction Methods 0.000 description 1
- 229920000136 polysorbate Polymers 0.000 description 1
- 239000013641 positive control Substances 0.000 description 1
- 238000001556 precipitation Methods 0.000 description 1
- 230000008569 process Effects 0.000 description 1
- 239000000047 product Substances 0.000 description 1
- 229960000856 protein c Drugs 0.000 description 1
- 239000011546 protein dye Substances 0.000 description 1
- 238000012207 quantitative assay Methods 0.000 description 1
- 230000005855 radiation Effects 0.000 description 1
- 230000002285 radioactive effect Effects 0.000 description 1
- 230000009467 reduction Effects 0.000 description 1
- 238000011160 research Methods 0.000 description 1
- 230000004044 response Effects 0.000 description 1
- 238000004007 reversed phase HPLC Methods 0.000 description 1
- 210000003079 salivary gland Anatomy 0.000 description 1
- 230000028327 secretion Effects 0.000 description 1
- 230000035945 sensitivity Effects 0.000 description 1
- 239000003001 serine protease inhibitor Substances 0.000 description 1
- 239000000377 silicon dioxide Substances 0.000 description 1
- 230000019100 sperm motility Effects 0.000 description 1
- 210000004989 spleen cell Anatomy 0.000 description 1
- 238000010186 staining Methods 0.000 description 1
- 238000003756 stirring Methods 0.000 description 1
- 239000006228 supernatant Substances 0.000 description 1
- 239000013589 supplement Substances 0.000 description 1
- 239000012085 test solution Substances 0.000 description 1
- 229960000187 tissue plasminogen activator Drugs 0.000 description 1
- 238000013518 transcription Methods 0.000 description 1
- 230000035897 transcription Effects 0.000 description 1
- 238000012546 transfer Methods 0.000 description 1
- 230000001131 transforming effect Effects 0.000 description 1
- 238000013519 translation Methods 0.000 description 1
- QORWJWZARLRLPR-UHFFFAOYSA-H tricalcium bis(phosphate) Chemical compound [Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O QORWJWZARLRLPR-UHFFFAOYSA-H 0.000 description 1
- 238000003211 trypan blue cell staining Methods 0.000 description 1
- 241000701447 unidentified baculovirus Species 0.000 description 1
- 241001515965 unidentified phage Species 0.000 description 1
Images














Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/48—Hydrolases (3) acting on peptide bonds (3.4)
- C12N9/50—Proteinases, e.g. Endopeptidases (3.4.21-3.4.25)
- C12N9/64—Proteinases, e.g. Endopeptidases (3.4.21-3.4.25) derived from animal tissue
- C12N9/6421—Proteinases, e.g. Endopeptidases (3.4.21-3.4.25) derived from animal tissue from mammals
- C12N9/6424—Serine endopeptidases (3.4.21)
- C12N9/6445—Kallikreins (3.4.21.34; 3.4.21.35)
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/40—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against enzymes
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Organic Chemistry (AREA)
- Engineering & Computer Science (AREA)
- Genetics & Genomics (AREA)
- General Health & Medical Sciences (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Medicinal Chemistry (AREA)
- Molecular Biology (AREA)
- Zoology (AREA)
- Biochemistry (AREA)
- Wood Science & Technology (AREA)
- Biomedical Technology (AREA)
- Biotechnology (AREA)
- Biophysics (AREA)
- Microbiology (AREA)
- Immunology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- General Engineering & Computer Science (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Peptides Or Proteins (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Description
本発明は、一般的に、タンパク質の検出および同定に関し、加えて、診断マーカーとして潜在的な有用性を有するタンパク質の種々な形態およびサブユニットに関する。より詳細には、本発明は前立腺に特異的な抗原の不活性前駆体形態の検出に関する。
発明の背景
前立腺癌(PCa)は米国人男性に最も頻繁に診断される癌である。パーカー(Parker, S.L)ら、CA Cancer J. Clin., 46:5-27, 1996。前立腺特異的抗原、又はPSAは前立腺癌患者の管理において信頼できる予後マーカーとして広く用いられてきた。カタロナ(Catalona, W. J.)ら、N. Engl. J. Med., 324:1156-1161, 1991;エステルリング(Oesterling, J. E.), J. Urol., 145:907-923, 1991;ラブリー(Labrie, F.)ら、J. Urol., 147:846-852, 1992。PSA検定における主な制約の一つは良性前立腺肥大(BPH)とPCaとを区別する特異性の欠如である。マッコーマック(McCormack, R. T.)ら、Urology, 45:729-744, 1995。血清PSAの診断精度を改良するために、PSA密度、PSA速度、遊離PSAと全PSAとの比率、または全PSAに対する複合体の比率等の様々なアプローチが導入されてきた。ベンソン(Benson, M. C.)ら、J. Urol., 147:815-816, 1992:カーター(Carter, H. B.)ら、J. Am. Med. Assoc., 267:2215-2220, 1992:エステルリングら、J. Am. Med. Assoc., 270:860-864, 1993。
セリンプロテアーゼのヒトカリクレインファミリーのメンバーであるPSA(hK3としても知られる)は、アスパラギン45に付着結合したN-連鎖オリゴ糖を1個有する30-34kDaの単鎖糖タンパク質である。ベランジャー(Belanger, A.)ら、Prostate 27:187-197, 1995。cDNAの分子クローニングにおいて、PSAのmRNAは、17aaの親水性シグナル配列(プレプロ領域)と7 aaのプロペプチドとが237aaの成熟タンパク質に先行する261 aaプレプロタンパク質をコードすることが明らかにされている。ランドウォール(Lundwall, A.)ら、FEBS Lett., 214:317-322, 1987;リーグマン(Riegman, P. H. J.)ら、Biochem. Biophys. Res. Commun., 155:181-188, 1988;ヘンツ(Henttu, P.)ら、Biochem. Biophys. Res. Commun., 160:903-910, 1989。PSA mRNA発現は前立腺上皮に支配的であり(クイ(Qui, S.)ら、J. Urol. 144:1550-1556, 1990)、アンドロゲンで制御される(ヤング(Young, C. Y. F.)ら、Cancer Res., 51:3748-3752, 1991)。PSAはキモトリプシン様活性を有することが見出された。ワット(Watt, K. W. K.)ら、Proc. Natl. Acad. Sci. USA, 83:3166-3170, 1986:バン(Ban, Y.)ら、Biochem. Biophys. Res. Commun., 123:482-488, 1984。PSAの提起された第一義的な生物学的役割は、主要ゲル生成タンパク質であるゼーメノゲリンI、IIおよび精液中のフィブロネクチンを切り離し、精子の運動性を増強させることである。リリャ(Lilja, H.)、J. Clin. Invest., 76:1899-1903, 1985。更に最近では、PSAはIGF結合タンパク質3を切り離し、前立腺での上皮細胞成長の制御に重要と思われる機能であるIGFの活性増加をもたらすことが報告された。カネティ(Kanety, H.)ら、J. Clin. Endocrin. Metab., 77:229-233, 1992。上記の研究はPSAの生理的基質の可能性を強調するが、その生合成、活性化および活性制御に関する基本的な疑問には答えていない。
ヒトカリクレインファミリーはhK1、hK2およびhK3(PSA)と表わされる3つのメンバーからなる。クレメンツ(Clements, J. A.)ら、Endocr. Rev., 10:393-419, 1989;カルビニ(Carbini, L. A.)ら、J. Hypertens., 11:893-898, 1993。hK1は主として腎臓、膵臓および顎下唾液腺で産生される。フクシマ(Fukushima, D.)ら、Biochemistry, 24:8037-8043, 1985。hK2はPSA同様、前立腺上皮で主に産生され(モリス(Morris, B. J.)、Clin. Exp. Pharm. Phys., 16:345-351, 1989;チャプデライン(Chapdelaine, P.)ら、FEBS Lett., 236:205-208, 1988)、アンドロゲンで制御され(ヤングら、Biochemistry, 31:818-824, 1992;グラウェル(Grauer, L.)ら、J. Androl., 17:353-359, 1966)、PSAと78%のaa相同性を有する(シェドリッヒ(Schedlich, L. J.)ら、DNA, 6:429-437, 1987;リリャら、World J. Urol., 11:188-191, 1993)。潜在的な前立腺癌マーカーとしてのhK2の寄与が最近調べられている。ヤングら、The Prostate Supplement、7:17-24, 1996;ダルソン(Darson, M. F.)ら、「前立腺上皮内新形成および腺癌腫におけるヒト腺カリクレイン2(hK2)発現:新規な前立腺癌マーカー」、Urology(印刷中);リッテンハウス(Rittenhouse, H. G)ら、「hK2の特徴づけと評価:PSAに緊密に関連した潜在的前立腺癌マーカー」、前立腺癌に関する第1回国際会議議事録(proceedings)(印刷中)。PSAはキモトリプシン様プロテアーゼであるが、hK2はトリプシン様プロテアーゼであり(ミコラジャクツィク(Mikolajczyk, S. D.)ら、「ヒト腺カリクレイン(hK2)はアルギニン制限特異性を示し、血漿プロテアーゼ阻害剤と複合体を形成する」、Prostate(印刷中))、このことはこれら二つの酵素が異なる生理学的役割を持つことを示す。hK2の生理学的役割はほとんど知られていない。デパーテス(Deperthes)らは最近、射精液中に存在するフィブロネクチンがhK2によりPSAによるよりも効率的に加水分解されたことを示した(J. Androl. 17:659-665, 1996)。一方、ゼーメノゲリン類のhK2による加水分解はPSAによる加水分解と同程度であった。PSAおよびhK2は同じ環境に存在すること明らかにされているが、これらの相互作用はまだ研究されていない。
PSAの生合成および活性化制御を研究し、hK2とPSAとの間の生理学的関係を調べるには、哺乳動物細胞中でPSAを発現させることが絶対的に必要である。PSAおよびhK2の両方を哺乳動物細胞に発現させるためのいくつかの試みがなされた。哺乳動物細胞におけるhK2の発現および精製は以前に報告されている。クマール(Kumar, A.)ら、Cancer Res., 56:5397-5402, 1996。ロヴグレンらはSFV系を用いてBHK21細胞にPSAを発現させた(ロヴグレン(Lovgren, J.)ら、Biochem. Biophys. Res. Commun., 231:888-895, 1995)。しかし、この系では、PSAの生合成プロセッシングの研究に必要な安定細胞系は得られなかった。カールらは抗-PSA治療の標的として使用する目的で、PSAを発現する安定なマウス結腸腺癌腫細胞系を産出したが、PSAに関する生化学的研究は報告されなかった。カール(Karr)ら、Cancer Res., 55:2455-2462, 1995。
従って、哺乳動物系においてPSAを安定して発現させるための方法が必要とされている。更に、PSA前駆体および誘導体を含め、かかる系の発現生成物を検出する必要もある。更に、前立腺癌の診断および病期分類を改良し、前立腺癌と良性前立腺肥大をよりよく区別する必要もある。
発明の概要
本発明により、哺乳動物細胞においてPSAを安定に発現し、クローニングする発現ベクターが提供される。PSAは哺乳動物細胞によってプロPSAとして使用培地に分泌されることが本明細書中で初めて実証されている。このように分泌されたプロPSAは培地中では酵素的に不活性、かつ安定である。プロPSAがhK2によって細胞外培地において酵素的に活性なPSAに変換され得ることも実証されている。これはhK2とPSAとの間の生理的関係の可能性、およびhK2がインビボでPSA活性を制御しているであろうことを示唆する。
本発明の実施態様は核酸分子を含んでなるキメラ発現ベクターである。核酸分子はpPSAポリペプチドをコード化する。核酸分子は、好ましくは、前記発現ベクターで形質変換されるホスト細胞によって認識される制御配列に使用可能に(operably)連結している。ホスト細胞は哺乳動物源に由来することが好ましい。
本発明はヒトの生理学的液中のプロPSAを検出する方法をも包含する。本発明のこの局面は、プロPSAが生物学的液中に存在し、前立腺癌の有用なマーカーとして働くだろうという発見に基づく。具体的には、いくつかのPSAの不活性な前駆体の形態が同定され、少なくとも1つが血清中に検出された。これらPSAの不活性な前駆体の形態の測定は前立腺癌の診断および病期分類について重要な情報を提供するだろう。
従って、本発明の方法によって産生されたプロPSAポリペプチドは、その変異体およびサブユニットと同様に、抗体集団の産生に用いることができる。その抗体集団は、次いで、精液、血液または血清などの生理学的液、前立腺癌腫などの組織、あるいは前立腺細胞などの細胞に由来する試料中のプロPSAポリペプチド(または「タンパク質」)を検出、定量する直接検定または競合検定の基礎として用いられる。
従って、本発明の別の実施態様は、抗体、および好ましくはプロPSAに特異的に結合するモノクローナル抗体を包含する。[-4]pPSA、[-5]pPSAおよび[-7]pPSA(但し、これらに限定されない)を含む種々のプロPSAの不活性な前駆体の形態に対する抗体をも意図するものである。
プロPSAを検出するための直接検定および競合検定も本発明の範囲内に包含される。ヒト生理学的液試料中のプロPSAを検出する方法につき説明するが、この方法は、pPSAに対する精製された抗体を用意し、抗体と試料とを接触させて抗体とpPSAとの間に複合体を生成させ、そして抗体と複合体形成したpPSAの存在または量を決定することを含む。診断方法およびキットも本発明の実施態様として含まれる。
【図面の簡単な説明】
図1はPSA発現ベクター、pGTD-PSAを示す模式図である。
図2はAV12-PSA#8細胞によるPSAの発現を示す。AV12-PSA#8細胞の使用培地を含む血清を6日間連続して毎日採取した。タンデム(Tandem▲R▼)-MP PSA検定を用いてPSA濃度を測定した。トリパンブルーを用いて生存する菌数を毎日計測した。
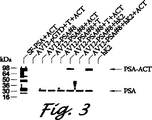
図3は、トリプシンおよびhK2で処理してACTと複合体を形成したAV12-PSA#8使用培地を示す。AV12-PSA#8細胞の7日使用培地およびAV12-PGTD細胞を適切に濃縮した。濃縮した使用培地(タンデム▲R▼-MP PSA検定で測定したPSA(1μg)を含む)を精製トリプシン(0.02μg)または精製hK2(1μg)と37℃で60分間混合した。アプロチニン(0.05μg)を加えて反応を停止させた。いくつかの試料はACT(5μg)で更に4時間インキュベートした。試料緩衝液+βMEと煮沸により反応を停止させた。反応物量の約1/10を4-20%勾配ゲル上で電気泳動した。タンパク質をニトロセルロースフィルター上で電気ブロットし、PSA特異的mAb PSM773でプローブした。分子量マーカーを左側に表示する。SF-PSA=精液から精製したPSA、AV12-PSA#8=AV12-PSA#8細胞からの使用培地、T=トリプシン、ACT=α1-抗キモトリプシン。
図4は精製プロPSAと共にインキュベートしたhK2のHICクロマトグラフィーを示す。パネルA:pPSAと混合して直ちに注入したhK2。ピーク1はhK2、ピーク2は精製pPSAである。パネルB:37℃で2時間、pPSAと共にインキュベートしたhK2。ピーク3はN-末端配列決定で確認された成熟PSA。
図5はPSAおよびACT標準のHICクロマトグラフプロフィールを示す。図5Aは活性ACTの保持時間を示す。図5Bは精液から精製したPSAと共にインキュベートした活性ACTを示す。図5Cは精液PSAの保持時間を示す。
図6は異なる形態のPSAのHICクロマトグラフプロフィールを示す。図6Aは成熟PSAおよびpPSAの保持時間を示し、[-4]pPSA、[-5]pPSAおよび[-7]pPSAを含む。図6BはPSM773アフィニティカラムに結合した血清試料からのPSA形態の保持時間を示す。
図7は精製した成熟PSAとpPSAの混合物のクロマトグラフプロフィールを示す。図7AはACTを添加していないタンパク質混合物のクロマトグラフプロフィールを示す。図7Bは、同じタンパク質混合物を37℃で2時間、ACTと共にインキュベートした後のクロマトグラフプロフィールを示す。
図8はウェスタンブロットを示し、前立腺癌腫血清中のPSAの33kDaの形態が同定されている。
図9は血清および精液中のPSAのウェスタンブロット分析を示す。レーン1:前立腺癌腫血清(レーン当たりの担持:1μl、28ng PSA)、レーン2:精液から精製したPSA(28ng)、レーン3:雌性血清(1μl)中にスパイクした精液から精製したPSA(28ng)、レーン4:分子量マーカー。
好ましい実施態様の詳細な説明
血清中のPSAの不活性な前駆体の形態が同定されることは、プロPSAの血清濃度の測定が前立腺癌の診断およびモニターに有用であり得ることを示唆する。PSAの生合成およびプロPSAの成熟PSAへの活性化に関与するステップを見分けるために、哺乳動物細胞におけるPSA発現が必要である。
本明細書及び請求の範囲で使用する用語、「PSA」および「PSAポリペプチド」は区別することなく用いられ、組換体プレプロ、プロおよび成熟PSAポリペプチドを包含する。用語、「プロPSA」、「pPSA」、「プロPSAポリペプチド」および「pPSAポリペプチド」は区別することなく用いられ、好ましくはPSAの不活性な前駆体の形態の全てを包含し、[-4]プロPSA、[-7]プロPSAおよび[-5]プロPSAを包含するが、これに制限されるものではない。
本明細書及び請求の範囲で用いる「キメラ」とはベクターが少なくとも2つの異なる種のDNAを含むか、或いは同一種のDNAを含み、「自然」または野生型の種では起こらない方法で連結または会合していることを意味する。
「制御配列」とは、特定のホスト生物における使用可能に連結したコーディング配列の発現に必要なDNA配列を意味するものと定義する。原核細胞に適した制御配列は、例えばプロモーター、任意にオペレーター配列、リボソーム結合部位を含む。真核細胞では、プロモーター、ポリアデニル化シグナルおよびエンハンサーが用いられることが知られている。
「使用可能に連結した」とは核酸が別の核酸配列と機能的関係に置かれていることを意味する。例えば、プレ配列または分泌リーダーのDNAは、ポリペプチドの分泌に関与するプレタンパク質として発現すれば、そのポリペプチドのDNAに使用可能に連結している。プロモーターまたはエンハンサーは、配列の転写に影響を及ぼせば、コーディング配列に使用可能に連結している。また、リボソーム結合部位は翻訳を促進するように位置していれば、コーディング配列に使用可能に連結している。一般的に、「使用可能に連結した」とは連結した複数のDNA配列が連続していることを意味し、分泌リーダーの場合には連続して、かつ読み取り相にある。しかしながら、エンハンサーは連続している必要はない。連結は好都合な制限部位での連結反応によって達成される。かかる部位が存在しなければ、合成オリゴヌクレオチドアダプターまたはリンカーが通常の方法に従って用いられる。
標的細胞を形質変換することができる組換えDNAを構築する一般的な方法は当業者に周知であり、同じ組成および構築方法を本明細書で有用なDNAの産生に用いてもよい。例えば、サンブルーク(J. Sambrook)ら、「分子クローニング:実験室マニュアル」、(コールドスプリングハーバープレス(Cold Spring Harbor Press), 2d ed. 1989)は適当な構築方法を提供する。
組換えDNAは、PSAをコード化するcDNAを含む発現ベクターでトランスフェクトすることによって、例えばチェン(C. Chen)らのリン酸カルシウム沈殿法の変法(Mol. Cell. Biol. 7, 2745, 1987)によって、容易に標的細胞に導入することができる。トランスフェクションは市販キット(例えば、BRLライフ・テクノロジーズ社(BRL Life Technologies, Inc.)提供)を用いるリポフェクションによっても達成することができる。
PSA発現に適当なホスト細胞は多細胞生物に由来する。かかるホスト細胞は複合体プロセッシングおよび糖鎖形成活性の能力がある。しかしながら、哺乳動物の細胞は、タンパク質の自然ホストに密接に関連した方法で組換えタンパク質を改変し、プロセッシングするので、哺乳動物細胞は哺乳動物のタンパク質発現に好適なホストである。原則的に、脊椎動物または非脊椎動物の培養であろうと、いかなる高度な真核細胞の培養も本発明の実施に用いることができる。非脊椎動物細胞の例には植物および昆虫細胞が含まれる。多数のバキュロウィルス菌株および変異体、対応する許容昆虫ホスト細胞が同定されている。
「ポリメラーゼ連鎖反応」、或いは「PCR」とは、予め選択された核酸、RNAおよび/またはDNAの断片の量が米国特許第4,683,195号記載のように増幅される操作または方法をいう。一般的に、目的の領域の末端またはそれ以外の配列情報がオリゴヌクレオチドプライマーの設計に用いられる。これらプライマーは増幅される鋳型の反対鎖と配列が同一か類似であろう。PCRは特異的RNA配列、全ゲノムDNAからの特異的DNA配列、全細胞RNAから転写されたcDNA、バクテリオファージまたはプラスミド配列などの増幅に用いられてもよい。一般的には、ムリス(Mullis)らの「Cold Spring Harbor Symp.」[Quant. Biol., 51, 263(1987)]、エーリッヒ(Erlich)編:「PCR Technology」(ストックトンプレス(Stockton Press), NY, 1989)を参照。
PSAポリペプチドがヒト起源以外の組換え細胞に発現すると、PSAポリペプチドはヒト起源のタンパク質またはポリペプチドを全く含んでいない。しかしながら、PSAポリペプチドに実質的に均質な調製物を得る為には、組換え細胞タンパク質またはポリペプチドからのPSAポリペプチドを精製する必要がある。例えば、培地または溶解質を遠心して顆粒細胞の破片を除去する。こうして膜および可溶性タンパク質画分を分離する。PSAポリペプチドを可溶性タンパク質画分から、必要によっては培養溶解質の膜画分から精製してもよい。次に、PSAポリペルチドを、混入する可溶性タンパク質およびポリペプチドから、免疫アフィニティ分画またはイオン交換カラム、エタノール沈殿、逆相HPLC、シリカ上またはDEAEなどのアニオン交換樹脂上でのクロマトグラフィ、クロマトフォーカシング、SDS-PAGE、硫酸アンモニウム沈殿、例えばセファデックス(Sephadex)G-75を用いるゲルろ過、或いはリガンドアフィニティクロマトグラフィーで精製することができる。
本発明の一実施態様に従って、単離後に、プロPSA領域に対応するプロPSAポリペプチドまたはペプチドを用いて、抗-pPSA抗体を産生することができる。本発明に従って抗体を産生するために用いられたプロPSAポリペプチドには、-7、-5および-4プロPSAがこれに限定されることなく含まれる。プロPSA領域に対応するペプチドも抗-pPSA抗体の産生に用いてもよく、pPSAポリペプチドのプロ領域のいずれかの部分を含む全てのペプチドが包含される。これらペプチドは、好ましくは、約8-15のアミノ酸を含み、免疫原生エピトープを含む。精製pPSA(全タンパク質)または上記ペプチドに対するモノクローナル抗体は、公知なハイブリドーマ細胞培養法、例えばハーロー(E. Harlow)らの「抗体:実験室マニュアル」(コールドスプリングハーバーラボラトリーズ(Cold Spring Harbor Laboratory), 1988)に記載の方法で調製することができる。一般的に、この方法は、(例えば、骨髄腫細胞の和合性連続系と融合した始原脾臓細胞の)抗体産生融合細胞系を調製すること、集団培養または用いた骨髄腫細胞系由来または和合性の動物種のいずれかで融合細胞を成長させることを含む。かかる抗体は高度に特異的で感受性があり、かつ免疫化学的に比較的「純粋」であるので、動物接種によって産生した抗体に比べ多くの利点がある。抗体の免疫学的に活性な断片(例えばf(ab)断片)も部分的にヒト化されたモノクローナル抗体であるので本発明の範囲内である。
この抗-pPSA抗体を産生する抗原として有用であることに加え、本発明の方法に従って産生された単離pPSAポリペプチドおよびその抗原的に活性な変異体、誘導体およびその断片は、プロPSAまたは抗-プロPSA抗体を含む疑いのある生物学的物質に由来する試料中のプロPSA検定に用いることができる。
本発明の実施態様に従って実施できる有用な免疫検定は二抗体サンドイッチ法である。これらの検定は主に未知試料中の抗原濃度の決定に用いられる。二抗体検定は迅速かつ正確であり、純粋な抗原源(この場合プロPSA)が利用できれば、検定は未知試料中における抗原の絶対量の決定に利用できる。検定には抗原上で重複しないエピトープに結合する二つの抗体が必要である。別々の部位を認識する二つのモノクローナル抗体または1バッチのアフィニティ精製ポリクローナル抗体のいずれかを用いることができる。
二抗体検定では、1つの抗体は精製されて固相に結合する。いかなる固相も利用できるが、ほとんどの用途ではPVCマイクロタイタープレートが好適である。例えば、マイクロタイタープレートのウェルに結合した抗体は標識されておらず、「捕獲された抗体」と呼ばれる。用いられる抗体量は個々の検定に依存するが、一般的に約1μg/ウェルの量が最大結合を与える。より高用量または低用量の捕獲抗体も用いられる。次に、ウェルを洗浄して、ウェルに試料を加えて試験液中の抗原(この場合はpPSA)を固相に結合させることができる。非結合タンパク質を洗浄して除去し、標識された第2の抗体を加えることができる。別法として、試料および第2の標識された抗体を同時に加えてもよい。洗浄後、固相に結合した標識された第2の抗体の量を測定することによって定量検定を行ってもよい。本発明の最も好適な実施態様では、第1の非標識抗体としてモノクローナル抗体を、第2の標識抗体としてモノクローナル抗体を用いる。結合した標識抗体の量を定量するのに用いる検出方法は使用した標識に依存する。抗体の標識は通常ヨウ素、酵素またはビオチンで行われる。熱量測定またはその他の検出方法を用いてもよい。
本発明のプロPSAポリペプチドは、固定化されて、抗-pPSA抗体の検定試料からの抗-pPSA抗体を結合して固定化する「捕獲抗原」として用いることができる。プロPSAポリペプチドおよび抗-pPSA抗体の二価複合体を次に検出する。例えば、ヒトの生理学的物質の場合には、検出可能な標識または検出可能な標識の結合部位を含む抗-ヒトIgG抗体と反応させることによって検出される。後者の場合、結合部位は結合部位に特異的な化合物と反応したそれ自体であり、それ自体が検出可能な標識を含む。有用な検出可能標識には、酵素、放射性標識、或いは蛍光標識が包含される。次に、得られた三元複合体または四元複合体を検出可能標識を介して、すなわち酵素-基質呈色反応、放射線放出、凝集などを介して検出し/または定量してもよい。
別法として、プロPSAポリペプチドを、1つ以上の放射能標識されたペプチジル残基を介するなどして検出可能標識で標識することもでき、抗-プロPSA抗体への結合について内性プロPSAと競合させるために用いることができる。すなわち、種々の競合免疫検定フォーマットを介して生理学的液の試料中の抗-プロPSA抗体に結合する捕獲抗原として競合させることができる。例えば、固定化された抗-プロPSA抗体を用いるプロPSAの競合免疫検定は次のように行われる。
(a) 固体表面に付着した所定量の抗-プロPSA抗体を用意する、
(b) 検定すべき生理学的液の試料を、検出可能な標識を含む既知量のプロPSAポリペプチドと混合して混合試料を調製する、
(c) 上記の抗体と混合試料とを十分な時間接触させて、抗体とプロPSAとの間で免疫学的な反応を起こさせて、抗体-プロPSA複合体を生成し、および、抗体と標識ポリペプチドとの間で抗体-標識ポリペプチド複合体を生成させる、
(d) 混合試料からプロPSAに結合した抗体と標識ポリペプチドに結合した抗体とを分離する、
(e) 混合試料中に残存するかまたは固体表面上の抗体に結合する、標識ポリペプチドの存在または量を検出または決定する、および
(f) ステップ(e)の結果から試料中のプロPSAの存在または量を決定する。
ここに詳述した免疫検定は、前立腺癌の診断およびモニターを目的とする、血清および組織などの、ヒト生理学的試料中のpPSA検出方法としても利用できる。本発明の検定は、前立腺癌を良性前立腺肥大から区別する方法にも利用可能である。本発明の好ましい実施態様では、診断方法は、試料中のpPSA量を決定し、試料中の成熟PSA量を決定し、そして数値を数学的に結合して診療に有用な結果、すなわち前立腺癌の予測となるカットオフ値を得ることなどを含む。試料中の全PSAに対するプロPSAの量の比は、例えば、本発明に拠って決定することができる。
以下に詳細に記述する例を参照して本発明を更に説明する。
例1
pPSAの発現とhK2によるPSAへの変換
試料および方法
発現ベクター、細胞系およびトランスフェクション
全ppPSAをコードする0.8kb DNA断片をGBMTプロモーターの制御のもとでpGT-dのBc11部位にクローニングし(ベルグ(Berg, D. T.)ら、「分泌タンパク質の安定、高レベル発現のためのE1a-応答哺乳動物ホスト/ベクター系」,Nucl. Acids Res., 20:5485-5486, 1992)、発現ベクターpGTD-PSAを得た(図1)。挿入の配向および配列を確認した。DMEM(高グルコース)および10%胎児クローン[ハイクローン(Hyclone),ローガン(Logan), UT]中で培養したAV12-664(ATCC CRL 9595)をリポフェクタミン(Lipofectamine)(ライフテクノロジーズ社)を用いてpGTD-PSAでトランスフェクトした。トランスフェクトしたAV12-664細胞(AV12-PSA)を400nMメトトレキセート[シグマケミカルカンパニー(Sigma Chemical Company)]中で選択した。エンプティーベクター(AV12-PGTD)でトランスフェクトしたAV12-664も負のコントロール(対照)として用いるために、同様の方法で選択した。単細胞クローンを単離した。細胞の生存をトリパンブルー色素排除法によって評価した。
抗体
マウスmAbs PSM773およびHK1G586.1を用いた。PSM773はタンデム▲R▼-MP PSAキット[ハイブリテック(Hybritech Incorporated),サンディエゴ(San Diego), CA)]の一成分であり、PSAに特異的であることが判明した[ワン(Wang, T. J.)ら、「抗-PSAモノクローナル抗体と組換えヒト腺カリクレインとの交差反応性分析」,J. Urol., 155:695A, 1996]。HK1G586.1は抗-hK2抗体である[クマール(Kumar A.)ら、Cancer Research, 5397-5402, 1996]。
ELISAおよびウェスタンブロット分析
AV12-PSA細胞からの使用培地を含む血清を特定日に採取した。タンデム▲R▼-MP PSAおよび遊離(free)PSA検定(両者ともハイブリテック,サンディエゴ, CA)を用い、製造者の指示に従ってPSAを測定した。負のコントロールとしてAV12およびAV12-PGTD細胞からの使用培地を用いた。
ウェスタンブロット分析のために、AV12-PSA細胞を含む組織培養フラスコを約60-70%密集成長させ、PBSで洗浄して血清を含まないHH4培養液を加えた。特定日に採取した使用培地をセントリコン10(Centricon 10)[アミコン社(Amicon, Inc.)]を用いて濃縮し、精製トリプシンまたは精製hK2のいずれかでインキュベートしてからACTと共にインキュベートし、4-20%ゲル[バイオ-ラド社(Bio-Rad, Inc.)]上でSDS-PAGEに付した。電気泳動の後、タンパク質をニトロセルロース膜上で電気ブロットした。第1次抗体(1-10μg/ml)および第2次抗体(ヤギ抗-マウスIgG-ホースラディッシュペルオキシダーゼ、1:500;[ジャクソンイムノサーチラボラトリーズ社(Jackson Immunosearch Laboratories, Inc.)]を用いてブロットをプローブした。増強化学発光[アマーシャム(Amersham), バッキンガムシャー(Buckinghamshire),イギリス]システムを用い、製造者の指示に従って免疫反応シグナルを検出した。
PSAの精製
製造者の指示に従ってmAb PSM773をアミノリンク(AminoLink)[ピアース(Pierce),ロックフォード(Rockford), IL]にカップリングした。AV12-PSA細胞を含む組織培養フラスコを約60-70%密集成長させ、PBSで洗浄してから血清を含まないHH4培養液を加えた。特定日に使用培地を採取し、4℃にて一晩、撹拌しながら上記の樹脂で濃縮、インキュベートした。次に、樹脂をカラム内に入れ、PBSで洗浄し、PSAを100mMグリシン、0.5M NaCl、pH2.5で溶出させた。試料を直ちに1Mトリス(Tris)、pH8.0で中和した。
PSA活性測定のための検定
PSAの酵素活性をクリステンソン(Christensson, A.)らが発表した手順[「前立腺特異的抗原の酵素活性およびその細胞外セリンプロテアーゼインヒビターとの反応」,Eur. J. Biochem., 194:755-763, 1990]に従って測定した。簡単に説明すると、PSA調製物(精液からの精製物またはAV12-PSA#8細胞の7日使用培地のいずれか)を、200mMトリス/5mM EDTA(pH8.0)中37℃で、1mM pNA-誘導ペプチド色素原基質(メトキシスクシニル-Arg-Pro-Tyr-pNA, S2586;[ファルマシアヘパー社(Pharmacia Hepar, Inc.)]と共にインキュベートした。PSAの酵素活性をペプチド色素原基質の加水分解によって決定し、405nmでの吸光度の増加が認められた。
精製hK2による精製プロPSAのPSAへの変換
精製pPSA(4.7μM)を100mMトリス、2mM EDTA中、pH8、37℃で精製hK2(0.56μM)と共にインキュベートした。hK2のアフィニティ精製はクマールら[「哺乳動物細胞におけるヒト腺カリクレイン、hK2、の発現」,Cancer Res. 56: 5397-5402, 1996]およびミコラジャクツィクら[「ヒト腺カリクレイン(hK2)はアルギニン制限特異性を示し、血漿プロテアーゼンヒビターと複合体を形成する」、Prostate(印刷中)]が記述している。最初の混合は4℃で行い、アリコートを直ちに疎水性相互作用クロマトグラフィ(HIC)で分解した。試料を37℃に保持し、別のアリコートを2時間まで分析した。HICカラムの仕様および緩衝液組成は次の通りであった。ポリプロピルアスパラギン酸アミドカラム4.6×250mm、ポリLC(PolyLC)[ウェスタンアナリティカル社(Western Analytical), テメキュラ(Temecula), CAより販売]、緩衝液A:1.2M硫酸ナトリウム、50mMリン酸ナトリウム、pH6.3、緩衝液B:50mMリン酸ナトリウム、5%v/v 2-プロパノール、pH7.3。試料を1.5M硫酸アンモニウム中で調製し、次いで以下に示す勾配でHICカラム上に注入した。0-35%緩衝液Bを0-1分、30-80%緩衝液Bを1-12分、そして、緩衝液Aでの再平衡より前に12-14分80%緩衝液Bで無勾配。プロソーブカートリッジ(アプライドバイオシステムズインスツルメンツ(Applied Biosystems Instruments))を用いて、カラムから採取したピークをPVDF(ポリビニリデンジフルオライド)膜に吸着させ、ABIモデル492プロサイス(Procise)シーケンサーを用いてN-末端配列決定を行った。ピーク試料の酵素活性を上述したように決定した。
結果
クマールらがhK2について記述したアプローチに類似な方法を用いて、GBMTプロモーター制御の下でPSAのcDNAをpGT-dベクター中にクローニングした。PSAの発現を調べるために、AV12細胞をpGTD-PSA発現ベクターでトランスフェクトした。細胞を400nMのメトトレキセート中で2-3週間選択し、単細胞クローンをPSA発現につきタンデム▲R▼-MP PSA検定を用いて分析し、mAb PSM773を用いてウェスタンブロット分析した。クローンAV12-PSA#8をその〜32kDaでの高発現レベルのPSA免疫反応性バンドに基づいて選択した。
哺乳動物細胞中のPSA発現パターンを決定するために、AV12-PSA#8細胞からの使用培地試料を連続6日間採取し、タンデム▲R▼-MP PSA検定を用いて分析した。図2はPSAが使用培地中において1日目に検出され、6日目までに>9μg/mlまで蓄積したことを示す。PSAの発現は細胞成長の対数期で高く、これはPSAの安定な形態が、細胞死および溶菌に続いて放出されるのとは対照的に、細胞によって分泌されていることを示す。同じ試料を遊離PSAについて分析すると、類似な数値が得られ(データは示さず)、これはAV12-PSA#8細胞が非複合体または遊離PSAを発現することを示す。
1日目に分泌されたタンパク質を同定するために、AV12-PSA#8細胞からの使用培地を採取して濃縮した。倍地中のPSAをPSM773、PSA特異的mAbを用いてアフィニティクロマトグラフィで精製した。精製タンパク質のN-末端配列決定で配列、APLILSRIVGG、が明らかになった。この配列は、-7aaで始まるプロPSAの末端について予測された配列に対応する。-5 aa(ロイシン)で始まる他の種も同定された。エドマン分解法によって逐次放出されたaa(アミノ酸)プロフィールからPSAの成熟形態に対応する競合配列は明らかでなかった。同じ精製プロトコルを用いてAV12-PSA#8細胞の7日目の使用培地からタンパク質を精製すると、プロPSAだけが検出され、成熟PSAは検出されなかった。この結果はPSAがプロPSAとしてAV12-PSA#8細胞によって1日目から分泌され始め、プロPSAは7日後でも使用培地中で安定であることを示す。
AV12-PSA#8によって発現したプロPSAは酵素的に不活性であり、緩和なトリプシン処理で酵素的に活性なPSAに変換され得ることを実証するために、市販の色素原基質、メトキシスクシニル-Arg-Pro-Tyr-pNA(S-2586)を用いて分光光度検定を行った。AV12-PSA#8(タンデム▲R▼-MP PSA検定で測定されたPSA(14μg)と等量を含む)からの使用培地をトリプシン(2%w/w)で37℃、60分間処理してから4%w/wアプロチニンで反応停止したときに、活性が10倍(活性単位/分で2.38×10-4から2.27×10-3)に増加したことが観察された。同様にトリプシンで処理してからアプロチニンで反応停止したAV12-PGTD細胞(エンプティpGT-dベクターでトランスフェクトされたAV12細胞)からの使用培地は、検出可能な活性を示さず、このことはAV12細胞によって内性PSA様プロテアーゼが分泌されないことを示す。正のコントロール、精液血漿から精製され、HH4中で同様にトリプシン処理されたPSA(14μg)は活性単位/分9.49×10-3の活性を示した。
PSAは血清中のACTと複合体形成することは従来から判明していた。我々は、酵素的に不活性なプロPSAがACTなどのプロテアーゼインヒビターと複合体化しないのであろうと推論した。トリプシン変換PSAがACTと複合体化するかどうかを試験するために、AV12-PSA#8細胞の使用培地にACTを加え、トリプシンと共にプレインキュベートした。反応を、4時間後に試料緩衝液+βMEの添加および煮沸で停止させた。試料を電気泳動、電気ブロットしてPSM 773mAbでプローブした(図3)。PSA-ACT(レーン1)と共泳動する〜94kDaの複合体をAV12-PSA#8細胞のトリプシン処理使用培地中で検出した(レーン6)。対照的に、未処理AV12-PSA#8使用培地をACTと共にインキュベートした場合には、PSA-ACT複合体は観察されなかった(レーン4)。トリプシンプレインキュベートAV12-PGTDの7日目使用培地をACTと共にインキュベートした場合には、免疫反応性バンドは観察されなかった(レーン2)。これらの結果により、AV12-PSA#8細胞の7日目使用培地中のプロPSAが酵素的に不活性であり、プロテアーゼインヒビター、ACTと複合体形成しないことを確認した。更に、これらの結果はプロPSAが緩和なトリプシン処理でPSAに変換され、このPSAが酵素的に活性であり、ACTと共有結合複合体を形成できたことを示した。PSA分解生成物がPF1D215mAbを用いるウェスタンブロットによって検出されないので(データは示さず)、プロPSAの-1から+1領域はトリプシン反応性の主要部位であると思われる。PF1D215mAbは従来から精液中に存在するPSA分解生成物を検出することが判明していた[ワン(Wang, T. J.)ら、「SDS-PAGEウェスタンブロット分析によるPSAおよびPSA断片の抗体特異性」,J. Tumor Biology, 1997(印刷中)]。
hK2はヒトカリクレインファミリーの別のメンバーであり、そのほとんどは、PSAもまた大量に発現している部位である前立腺上皮に発現する。hK2はアルギニン制限トリプシン様活性を示す。プロPSAがhK2によってPSAに変換され得るかを試験するために、精製hK2をAV12-PSA#8細胞の使用培地に加えた。37℃で60分間、インキュベートした後に、この試料にACTを加えた。更に、4時間、37℃でインキュベートした後に、反応を試料緩衝液+bMEの添加および煮沸で停止させた。次に、試料を電気泳動、電気ブロットしてPSM773でプローブした(図3)。精製PSA-ACTに共泳動する〜94kDaのバンド(レーン1)がAV12-PSA#8のhK2処理使用培地中に検出された(レーン8)。hK2とPSAはaa類似度〜78%を共有するが、hK2はPSM773mAbによって免疫検出されず(レーン9)、このmAbのPSA特異性が確認された。これらの結果はAV12-PSA#8使用培地では、hK2が酵素的に不活性なプロPSAをPSAの活性形態に変換し得ることを示す。
以上の結果を確認するために、hK2およびpPSAのアフィニティ精製調製物をインキュベートし、pPSAのPSAへの変換をHICおよびアミノ酸配列決定法で分析した。すでに説明したように、混合物をHICカラム上で分解した(図4)。これらの類似性にもかかわらず、hK2、pPSAおよび成熟PSAの保持時間は明らかに異なっていた。図4Aは時間ゼロでのhK2とpPSAの混合物を示し、図4Bは37℃で2時間、インキュベートした後の同じ混合物を示す。hK2(ピーク1)とpPSA(ピーク2)の保持時間は、それぞれ別々に注入すると、精製hK2および精製pPSAとの保持時間と同じであった(データは示さず)。hK2変換PSAのピーク(ピーク3)は、精液から精製した成熟PSAと同じ位置に溶出した(データは示さず)。図4Bは、pPSAピークが、精製hK2と共に2時間、37℃でインキュベートした後に、ほとんど完全に成熟PSAに変換されたことを示す。20分および1時間における別の時間での点も分析したが、PSAは中間レベルを示した(データは示さず)。プロPSAから成熟PSAへの変換をピークのaa配列決定法で確認し、それにより、ピーク2がN-末端プロPSAリーダーペプチド(-7aaで始まる)を含み、一方ピーク3が成熟配列(+1aaで始まる)で始まることが判明した。更に、hK2変換PSAはS-2586基質上で58nmol/分/mgの酵素活性を含んでいた。この値は同じ基質上で精液から精製されたPSAの酵素活性に相当した。pPSAと対照的に、hK2処理前のpPSA試料はPSA酵素活性の<5%を含んでいた。hK2を加えずに精製PSAをインキュベートした場合、PSAへの変換を示さなかった(データは示さず)。
他のセリンプロテアーゼのように、PSAは不活性プレプロPSA前駆体として翻訳される。分泌経路を通過した後に、シグナルペプチドが切断されてタンパク質のプロ形態を生じる。一般的に、プロペプチドは細胞内で切断されるが(例えば、組織プラスミノーゲン活性化因子、プロテインC、および腫瘍壊死因子)、プロプロテインとして分泌され、細胞外で切断される例外もある(例えば、レニン、トリプシン、キモトリプシン、およびhK2)[クマールら、Cancer Res. 56:5397-5402, 1996]。以下に示す証拠はPSAが哺乳動物細胞からプロPSAとして分泌されることを明白に実証する。すなわち、(a)AV12-PSA#8細胞の使用培地はPSA様酵素活性を有しなかった、(b)AV12-PSA#8細胞の使用培地でのPSA形態は、ウェスタンブロット分析で実証されたように、ACTと複合体化しなかった、(c)AV12-PSA#8の使用培地をトリプシン処理すると、酵素的に活性な成熟PSAを生じ、かつACTと複合体を形成した、および(d)AV12-PSA#8の使用培地に存在するPSA形態の精製および配列分析でプロPSAとしての同一性が確認された。
PSAはSFおよび血清中に多くの形態で存在する。これらの形態にはACTおよびPCIなどのプロテアーゼインヒビターとPSAとの複合体が含まれる。PSAはPSA-MG複合体および遊離PSAとしても存在する。血清およびSF中の遊離PSAの一部は、実際にプロPSAであることも可能である。我々のデータはプロPSAが生物学的液中に存在し、前立腺疾患の有用なマーカーとなり得ることを示唆する。
これまでのところ、プロPSAからプロペプチドを切断して活性分子を生成する原因となるプロテアーゼは同定されていない。ヒトカリクレインファミリーの他のメンバーであるhK2はアルギニン制限トリプシン様活性を示し、PSAに類似であり、前立腺上皮に主に発現する[モリス(Morris, B. J.)、前掲;チャプデライン(Chapdelaine, P.)、前掲]。hK2は哺乳動物細胞によってプロhK2として分泌される。プロhK2は細胞外で酵素的に活性なhK2に変換される。hK2は-1-+1残基間でそれ自身を切り出すことができる自家触媒活性を有することが判明している[ミコラジャクツィキら、「アラニン217は前立腺特異的ヒトカリクレイン、hK2の触媒機能および自己活性化に重要である」、Eur. J. Biochem. (印刷中)]。
本明細書に掲載したデータはhK2がプロPSAからプロペプチドを切り出してそれを酵素的に活性な成熟PSAに変換することを明白に示す(図4)。成熟PSAはACTと複合体化できるが、プロPSAは複合体化することができない(図3)。この結果は、ヒトカリクレインファミリーのこれら二つのメンバー間における生理学的制御関係の可能性を示唆する。さらなる研究によって、PSAの-1-+1がhK2による作用の好ましい部位であるか否かが決定されるものと予期される。典型的に、精液から単離された30-40%PSAは、85-86、145-146または182-183残基の間で切り出される[クリステンソンら、Eur. J. Biochem., 194:755-763, 1990]。PSAの145-146(リジン-リジン)間の切り出しはPSAを不活性化させることが知られている[クリステンソンら、前掲]。85-86および182-183間の切り出しも不活性化を導くと思われる。hK2は上述した或る部位での切り出しによって、PSA活性およびプロテアーゼインヒビターとの複合体生成を制御しているかもしれない。同様に、PSAはキモトリプシン感受性部位でhK2を切り出すことによってhK2活性を制御しているかもしれない。本明細書に記載した試薬に加えて、哺乳動物細胞に発現したhK2はこれらの実験を容易にするはずである。
上述した結果は包括的に、PSAが哺乳動物細胞中にプロ形で発現し、hK2によって細胞外的に酵素的に活性な成熟形態に変換され得ることを示す。これらの結果はまた、プロPSAが生物学的液中に存在し、それゆえに前立腺疾患のマーカーとして有用であり得ることを示唆する。本明細書に記述した細胞系は免疫原として利用されるプロPSAの貴重な源であり、かつプロPSAおよびPSAの生物学的役割の詳細な研究に利用される。
例2
ヒト血清中におけるpPSAの検出
ヒト血清中にpPSAが存在することは次のことを示すだろう。第1に、PSAがヒト組織中にpPSAの形態として分泌され、細胞外的に成熟PSAに変換されること。第2に、pPSAはヒト血清中で安定であり、pCaまたはBPHの有用な診断マーカーであり得ること。我々はヒト血清中のpPSAの存在を、最初にアフィニティ精製を用いてヒト血清プールに存在する全形態のPSAを精製することによって評価した。次に、HPLC上で溶出PSA形態を分画して、カラムからの溶出プロフィールに基づき、それぞれのPSA形態を同定した。この分析はpPSAがヒト血清中に存在することを示した。
方法
PSAが高められた前立腺癌患者からプールしたヒト血清(75ml)を得た。固体の硫酸アンモニウムを血清に加え、最終濃度を2Mにした後、試料を2M硫酸アンモニウムに対し4℃で16時間透析した。血清を遠心分離で浄化し、上清液を20mMリン酸ナトリウム(2リットル)、pH7、に対し3回(各1時間)で透析した。次に、試料を0.2μメンブランフィルターを通して濾過し、1ml/分で0.5mlアフィニティカラムに通した。アフィニティカラムはアミノリンク(Aminolink)[ピアース(Pierce)]樹脂に、樹脂1mlあたりmAbが5mgの濃度で共有結合したmAb PSM773からなるものであった。
アフィニティカラムをPBS(50ml)で洗浄し、PSAを100mMグリシン、0.5M塩化ナトリウム、pH2.5の1ml容積で3回で溶出した。溶出液(3ml)を1Mトリス(300μl)、pH8で中和した。硫酸アンモニウムを溶出液に加えて最終濃度を2Mとし、この試料をHPLCカラムにかけて、疎水性相互作用クロマトグラフィ(PolyLC、ポリプロピルアスパラギン酸アミドカラム、孔径1000Å、4.6mm×200mm(製造元、ウェスタンアナリティカル(Western Analytical), テメキュラ(Temecula), CA)によって分解した。緩衝液Aは20mMリン酸ナトリウム、1.2M硫酸ナトリウム、pH6.3であり、緩衝液Bは50mMリン酸ナトリウム、5% 2-プロパノール、pH7.4であった。溶出勾配は、緩衝液Aにおける再平衡前、0-1分0-35%B、1-14分35-80%Bであり、流速は1ml/分であった。
結果
図5は上述したHICクロマトグラフィ条件で分解した標準の保持時間(RT)を示す。図5Aは活性ACTのRTを示す。図5Bは精液から精製したPSAと共にインキュベートした活性ACTのRTを示す。精液PSAは約50%不活性なPSAを含有することが知られているので(ACTと複合体化しないので「遊離」PSAとして知られる)、PSAのACTとのインキュベーションの結果、PSA-ACTに加え遊離PSAが得られる。また、PSAはACTを切り出して不活性化させるので、不活性ACT(iACT)も見出される。図5Cは精液PSAのRTを示す。これらのクロマトグラフィ条件下、活性PSAと不活性PSAとの間の保持時間に違いはない。
図6Aは形態が異なるPSAの標準のRTを示す。全ての形態はアミノ酸配列法で照合された。[-7,-5]pPSAピークは互いに分離されないほぼ等しいレベルの形態の[-7]pPSAおよび[-5]pPSAを含む。図6Bは上述のPSM 773アフィニティカラムに結合した血清からのPSA形態のプロフィールを示す。試料を0.5mlの画分に採取し、タンデム▲R▼-MP遊離PSA検定[fPSA検定、ハイブリテック社(Hybritech Incorporated)]によって検定した。fPSA検定はpPSAと遊離(不活性)PSAの両方を検出する。7分における小ピークはアフィニティカラムから溶出したPSA-ACTへのfPSA検定の僅かな交差反応性に由来する。この試料中のPSA-ACTの実際レベルは遊離PSAのレベルよりも約10倍の高さである(データは示さず)。10分および12分でのピークはそれぞれ成熟PSAおよび[-4]pPSAに対応する。これらのデータはpPSAの少なくとも一形態([-4]pPSA)がヒト血清中に存在し、相対的ピーク面積から判断して、血清中の約25%の遊離または非複合PSAを占めることを示す。
例3
PSA形態のACTとの反応性
PSAのプロPSA形態はACTと反応性を有さないことを確認するために、精製した成熟PSA、[-4]pPSAおよび[-7,-5]pPSAの混合物をACTと共にインキュベートした。図7A(W/O ACT)はACTを添加しないタンパク質混合物のクロマトグラフィプロフィールを示す。図7B(+ACT)は、ACTと共に2時間、37℃でインキュベートした後における等量の同じ混合物のクロマトグラフィプロフィールを示す。成熟PSAだけがACT複合体を形成する。pPSAの[-4]、[-5]、および[-7]形態はピーク面積の減少が見られなかったので、ACTと複合体を形成しなかった。このデータはACTと複合体形成していない血清中の[-4]pPSAと一致する。
例4
ヒト血清における不活性非複合PSAのウェスタンブロットによる検出
方法
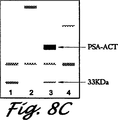
異なる量のPSA(971ng/ml-5,330ng/ml)を含む前立腺癌腫血清(2μl/レーン)の異なる3つの試料を、還元し、変性して4%-20%ポリアクリルアミドゲルに担持した。ゲルをニトロセルロースにブロットし、抗-PSAモノクローナル抗体PSM773と共に2μl/mL、1時間インキュベートした。よく洗浄した後、HRPに複合した第2の抗体を加え、ブロットとともに1時間インキュベートしてから再度PBSで洗浄した。抗体反応性を増幅化学発光(ECL)試薬[アマーシャム(Amersham),アーリントンハイツ(Arlington Heights),イリノイ州]を用い、製造者の指示に従って検出した。
結果
抗-PSAモノクローナル抗体は、図8に示すように、3つの全ての血清試料中に、PSA-ACTを表わす約90kDaに主バンドを検出する。約33kDaの小バンドも3つの血清試料中2つでも検出される。PSAのこの低分子量の形態は、活性PSAが容易にACTと複合化するので、PSAの不活性な非複合形態を表す。PSAレベルの高い血清試料中に33kDa形態のPSAが存在することは、この方法および抗体の感度を反映していると思われる。
例5
PSAの切り取られた形態はヒト(前立腺癌)血清中に検出されないこと
方法:
血清のウェスタンブロット
還元および変性されたタンパク質を4-20%ポリアクリルアミドゲル上で電気泳動し、ニトロセルロース上にブロットした(タウビン(Towbin), 1979)。十分に転移されことをポンソー(Ponceau)Sタンパク質染料の染色ブロットでチェックした。ブロットをトリス緩衝生理食塩水(TBS)中の5%無脂肪乾燥乳中でブロックし、抗-PSAモノクローナル抗体PF1D215.1、1μg/mlとともに、4℃で振とうしながら1時間インキュベートした。TBSでよく洗浄した後、0.05%トゥイーン(tween)、5%ミルク中のヤギ抗-マウスIgG-HRPの1:2000希釈液(ジャクソンラボラトリーズ(Jackson Laboratories),ウェストグローヴ(Westgrove), PA)を振とうしながら、ブロットとともに30分間、インキュベートした。ブロットをTBSで5回洗浄し、抗体反応性を増幅化学発光(ECL)試薬[アマーシャム,アーリントンハイツ,IL]を用い、製造者の指示に従って検出した。ブロットをケミイメージャー低光画像システム[ChemiImager Low Light Imaging System(アルファイノテックコーポレーション(Alpha Innotech Corporation), サンレンドロ(San Leandro), CA)]で走査した。
精液中に切り出された形態のPSAが存在することはよく記録されているが、ヒト血清中におけるこの切り出された形態の存在は実証されておらず、複合体形成していない遊離形態で存在するPSAの割合に寄与していると推論されていたにすぎない。PSAの切り出された分解生成物、PFID215を検出する抗体を用い、我々はウェスタンブロット分析によって分解生成物の存在に関し前立腺癌血清を試験した。図9で前立腺癌血清が精液と比較されており、それぞれの試料は等量のPSAを含有する。精液から精製したPSAをゲルに直接加えたものと正常雌性血清にスパイクさせたものとを試験した。我々は、切り出され/分解した形態のPSAは精液試料中に明らかに存在するが(レーン3:スパイクなし、レーン4:雌性血清中にスパイクした)、ヒト前立腺癌血清に切り取られた形態の証拠は存在しない(レーン2)ことを実証した。更に、血清のウェスタンブロット分析で見出された優勢なバンドはPSA-α2-マクログロブリン複合体、PSA-ACT複合体、および32kdでの完全(切り出されていない)であるが不活性な形態の遊離PSAに一致した。図6はこの形態を一部プロPSAとして、一部は別の不活性なPSAの形態(まだ特徴付けられていない)として示す。これらのデータは血清中の遊離PSAおよび精液が異なる分子種を含んでなることを実証する。
全ての文献、特許および特許文献は、それぞれ個別に参照したが、参照によって本明細書の記載の一部を構成する。本発明を種々の特定の実施態様および技術方法を参照して説明してきた。しかし、多くの変形および変更を本発明の精神および範囲内において行うことができるということが理解されるべきである。
Claims (16)
- (a)[−4]pPSAおよび[−5]pPSAからなる群より選択されるpPSAを特異的に結合するが、PSAの成熟形態には有意な結合を提示しない、精製された抗体を所定量用意し、
(b)前記抗体と検定される試料とを十分な時間接触させて、少なくとも前記抗体の一部と前記pPSAの一部との間に二元複合体を形成させ、そして
(c)前記抗体と複合体化したpPSAの存在または量を検出または決定する各工程を含み、ここで、[−4]pPSAは、−4アミノ酸(Ile)で始まり、[−5]pPSAは、−5アミノ酸(Leu)で始まる、pPSAを含むヒトの生理学的液試料中でpPSAを検出または決定する方法。 - 前記ヒトの生理学的液が血清である請求項1に記載の方法。
- 前記抗体がモノクローナル抗体である請求項1に記載の方法。
- 工程(c)において、前記pPSAをpPSAと特異的に結合する所定量の抗体と反応させ、前記抗体が検出可能な標識を含むかまたは検出可能な標識に結合して、検出可能な三元複合体を形成する請求項1に記載の方法。
- 前記抗体がモノクローナル抗体である請求項4に記載の方法。
- 前記抗体が固相に付着結合している請求項1に記載の方法。
- (a)[−4]pPSAおよび[−5]pPSAからなる群より選択されるpPSAを特異的に結合するが、PSAの成熟形態には有意な結合を提示しない、精製された抗体を所定量用意し、
(b)検定される試料を、既知量のpPSA、または検出可能な標識を含みpPSAに特異的に結合する抗体に結合するpPSAの免疫反応サブユニットと混合して混合試料を作成し、
(c)前記抗体を前記混合試料と十分な時間接触させて、前記抗体と前記試料中のpPSAと
の間および前記抗体と前記標識されたpPSAとの間で免疫反応を起こさせ、
(d)前記混合試料から前記抗体を分離し、
(e)前記抗体に結合しているかまたは前記混合試料中に残存する標識pPSAの存在または
量を検出または決定し、そして
(f)工程(e)の結果から、前記試料中の前記pPSAの存在または量を決定する各工程を含み、ここで、[−4]pPSAは、−4アミノ酸(Ile)で始まり、[−5]pPSAは、−5アミノ酸(Leu)で始まる、pPSAを含むヒトの生理学的液試料中のpPSAを検出または決定する競合方法。 - (a)[−4]pPSAおよび[−5]pPSAからなる群より選択されるpPSAポリペプチドに特異的に結合するが、PSAの成熟形態には有意な結合を提示しない、所定量の抗体またはその免疫学的に活性な断片と哺乳動物組織試料の細胞とを混合して、前記 抗体またはその免疫学的に活性な断片と前記細胞とを含む二元複合体を生成させ、そして
(b)前記試料中の複合体生成の存在または量を決定または検出する各工程を含み、ここで、[−4]pPSAは、−4アミノ酸(Ile)で始まり、[−5]pPSAは、−5アミノ酸(Leu)で始まる、哺乳動物の組織試料中のpPSAを検出または決定する方法。 - 前記抗体がモノクローナル抗体である請求項8に記載の方法。
- 前記細胞が前立腺細胞である請求項8に記載の方法。
- 複合体生成が、検出可能な標識を含むかまたは検出可能な標識と結合して、検出可能な三元複合体を形成する第2の試薬によって検出される請求項8に記載の方法。
- 前記第2の試薬が抗体である請求項11に記載の方法。
- [−4]pPSAおよび[−5]pPSAからなる群より選択されるpPSAを特異的に結合するが、PSAの成熟形態には有意な結合を提示しない、既知量の少なくとも一つの抗体またはその免疫学的に活性な断片を、前記抗体またはその免疫学的に活性な断片が検出可能なように標識されているかまたは検出可能な標識に結合するように含み、ここで、[−4]pPSAは、−4アミノ酸(Ile)で始まり、[−5]pPSAは、−5アミノ酸(Leu)で始まる、ヒトの生理学的液中のpPSAを検出または決定するためのキット。
- 前記抗体を付着結合させることができる固相を更に含む請求項13に記載のキット。
- [−4]pPSAおよび[−5]pPSAからなる群より選択されるpPSAポリペプチドに特異的に結合するが、PSAの成熟形態には有意な結合を提示しない、既知量の少なくとも一つの抗体またはその免疫学的に活性な断片を含み、ここで、[−4]pPSAは、−4アミノ酸(Ile)で始まり、[−5]pPSAは、−5アミノ酸(Leu)で始まる、哺乳動物の組織試料の細胞中のpPSAを検出または決定するためのキットであって、前記少なくとも一つの抗体またはその免疫学的に活性な断片が検出可能なように標識されているかまたは検出可能な標識に結合する、キット。
- 前記抗体がモノクローナル抗体である請求項15に記載のキット。
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US84640897A | 1997-04-30 | 1997-04-30 | |
| US08/846,408 | 1997-04-30 | ||
| PCT/US1998/008498 WO1998049323A1 (en) | 1997-04-30 | 1998-04-28 | Forms of prostate specific antigen and methods for their detection |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2002501384A JP2002501384A (ja) | 2002-01-15 |
| JP5284554B2 true JP5284554B2 (ja) | 2013-09-11 |
Family
ID=25297849
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP54726998A Expired - Lifetime JP5284554B2 (ja) | 1997-04-30 | 1998-04-28 | 前立腺特異的抗原の形態およびその検出方法 |
Country Status (8)
| Country | Link |
|---|---|
| EP (1) | EP0981629B1 (ja) |
| JP (1) | JP5284554B2 (ja) |
| AT (1) | ATE404676T1 (ja) |
| AU (1) | AU739546B2 (ja) |
| CA (1) | CA2286090C (ja) |
| DE (1) | DE69839876D1 (ja) |
| ES (1) | ES2312186T3 (ja) |
| WO (1) | WO1998049323A1 (ja) |
Families Citing this family (11)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US7288636B2 (en) * | 1997-04-30 | 2007-10-30 | Hybritech Incorporated | Forms of prostate specific antigens and methods for their detection |
| US7659073B2 (en) | 1997-04-30 | 2010-02-09 | Hybritech Incorporated | Forms of prostate specific antigens and methods for their detection |
| US6300088B1 (en) * | 1997-11-24 | 2001-10-09 | Duke University | Method of detecting prostate specific antigen |
| CA2360073C (en) | 1999-01-28 | 2011-01-18 | Gen-Probe Incorporated | Probes and primers for detecting prostate specific antigen (psa) in a biological sample |
| US6423503B1 (en) * | 1999-04-30 | 2002-07-23 | Hybritech Incorporated | Forms of free prostate-specific antigen (PSA) and their association with prostate tissues from prostate peripheral zone and transition zone |
| US20030219840A1 (en) * | 2002-05-24 | 2003-11-27 | Mikolajczyk Stephen D. | Method of analyzing proenzyme forms of prostate specific antigen in serum to improve prostate cancer detection |
| AU783356B2 (en) * | 1999-07-29 | 2005-10-20 | Medarex, Inc. | Human monoclonal antibodies to prostate specific membrane antigen |
| FR2807438B1 (fr) * | 2000-04-10 | 2004-09-17 | Bio Merieux | Nouveaux anticorps reconnaissant specifiquement le psa libre inactif, et leurs applications |
| WO2001090194A2 (de) * | 2000-05-24 | 2001-11-29 | Roche Diagnostics Gmbh | Antikörper gegen spezielle formen des propsa und deren verwendung in immunoassays |
| FI20002127A0 (fi) | 2000-09-27 | 2000-09-27 | Artic Partners Oy Ab | Uusi vasta-aine, immunomääritys ja menetelmä eturauhassyövän havaitsemiseksi |
| CN113237162B (zh) * | 2021-05-28 | 2022-12-27 | 西安建筑科技大学 | 一种冷冻水循环系统控制优化方法、系统及设备 |
Family Cites Families (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CA2148350A1 (en) * | 1992-10-29 | 1994-05-11 | Carlo Croce | Methods of detecting micrometastasis of prostate cancer |
| US6103237A (en) * | 1993-07-22 | 2000-08-15 | Hybritech Incorporated | Stable variant hK2 polypeptide |
| EP0725139A3 (en) * | 1995-02-03 | 1997-11-26 | Bayer Corporation | Recombinant prostate-specific antigen and its use as an immunoassay calibrator |
| CA2218754A1 (en) * | 1996-11-15 | 1998-05-15 | Eli Lilly And Company | Recombinant psa expression vectors and assays using the same |
-
1998
- 1998-04-28 WO PCT/US1998/008498 patent/WO1998049323A1/en active IP Right Grant
- 1998-04-28 JP JP54726998A patent/JP5284554B2/ja not_active Expired - Lifetime
- 1998-04-28 CA CA2286090A patent/CA2286090C/en not_active Expired - Lifetime
- 1998-04-28 ES ES98918814T patent/ES2312186T3/es not_active Expired - Lifetime
- 1998-04-28 AT AT98918814T patent/ATE404676T1/de not_active IP Right Cessation
- 1998-04-28 EP EP98918814A patent/EP0981629B1/en not_active Expired - Lifetime
- 1998-04-28 DE DE69839876T patent/DE69839876D1/de not_active Expired - Lifetime
- 1998-04-28 AU AU71668/98A patent/AU739546B2/en not_active Expired
Also Published As
| Publication number | Publication date |
|---|---|
| AU739546B2 (en) | 2001-10-18 |
| AU7166898A (en) | 1998-11-24 |
| ES2312186T3 (es) | 2009-02-16 |
| ATE404676T1 (de) | 2008-08-15 |
| CA2286090C (en) | 2013-01-08 |
| EP0981629A1 (en) | 2000-03-01 |
| WO1998049323A1 (en) | 1998-11-05 |
| JP2002501384A (ja) | 2002-01-15 |
| EP0981629B1 (en) | 2008-08-13 |
| DE69839876D1 (de) | 2008-09-25 |
| CA2286090A1 (en) | 1998-11-05 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| Kumar et al. | Expression of pro form of prostate-specific antigen by mammalian cells and its conversion to mature, active form by human kallikrein 2 | |
| Christensson et al. | Complex formation between protein C inhibitor and prostate‐specific antigen in vitro and in human semen | |
| Lilja et al. | Prostate specific antigen predominantly forms a complex with alpha1‐antichymotrypsin in blood. Implications for procedures to measure prostate specific antigen in serum | |
| Mikolajczyk et al. | Ala217 is important for the catalytic function and autoactivation of prostate‐specific human kallikrein 2 | |
| JP4205738B2 (ja) | 安定hk2ポリペプチド変異体 | |
| Piironen et al. | Determination and analysis of antigenic epitopes of prostate specific antigen (PSA) and human glandular kallikrein 2 (hK2) using synthetic peptides and computer modeling | |
| JP5284554B2 (ja) | 前立腺特異的抗原の形態およびその検出方法 | |
| US20100151501A1 (en) | Forms of Prostate Specific Antigens and Methods for their Detection | |
| Becker et al. | Individual prostate-specific antigen (PSA) forms as prostate tumor markers | |
| US6482599B1 (en) | Forms of prostate specific antigen (PSA) specific for benign prostatic hyperplasia (BPH) and methods of using such | |
| EP0837930B1 (en) | A TRP-226 MUTANT HUMAN GLANDULAR KALLIKREIN-1 (hK2) | |
| Herrala et al. | Androgen‐sensitive human prostate cancer cells, LNCaP, produce both N‐terminally mature and truncated prostate‐specific antigen isoforms | |
| Kurkela et al. | Expression of active, secreted human prostate-specific antigen by recombinant baculovirus-infected insect cells on a pilot-scale | |
| US6140468A (en) | Recombinant human prostate specific antigen | |
| US7288636B2 (en) | Forms of prostate specific antigens and methods for their detection | |
| EP0974057B1 (en) | Methods to detect hk2 polypeptides | |
| JP2002538818A (ja) | 前立腺腫瘍組織におけるヒトカリクレイン2とプロテアーゼインヒビター−6の新規な複合体および該複合体の利用方法 | |
| WO1998002748A9 (en) | Methods to detect hk2 polypeptides | |
| JP2001503991A (ja) | 転移性前立腺癌の検出方法 | |
| Kallikrein | Expression of Pro Form of Prostate-specific Antigen by | |
| CHRISTENSSON et al. | in zyxwvutsrqponmlkji | |
| US20030166036A1 (en) | Protease and an aminopeptidase associated with development of benign prostatic hyperplasia (BPH) | |
| HOLLINNGSWORTH et al. | Copyright zyxwvutsrqponmlkjihg |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20050427 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20080507 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20080807 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20081014 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20090107 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20090216 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20090213 |
|
| RD02 | Notification of acceptance of power of attorney |
Free format text: JAPANESE INTERMEDIATE CODE: A7422 Effective date: 20090213 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20090323 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20090313 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20090427 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20090409 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20090609 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20091006 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A821 Effective date: 20091006 |
|
| A911 | Transfer to examiner for re-examination before appeal (zenchi) |
Free format text: JAPANESE INTERMEDIATE CODE: A911 Effective date: 20091112 |
|
| A912 | Re-examination (zenchi) completed and case transferred to appeal board |
Free format text: JAPANESE INTERMEDIATE CODE: A912 Effective date: 20091126 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20111104 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20111111 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20111202 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20111207 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20111228 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20120111 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20120820 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20120823 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20121121 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A821 Effective date: 20121121 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20130415 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20130530 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| EXPY | Cancellation because of completion of term |