JP4776216B2 - 新規な植物細胞死誘導因子NbCD1 - Google Patents
新規な植物細胞死誘導因子NbCD1 Download PDFInfo
- Publication number
- JP4776216B2 JP4776216B2 JP2004348749A JP2004348749A JP4776216B2 JP 4776216 B2 JP4776216 B2 JP 4776216B2 JP 2004348749 A JP2004348749 A JP 2004348749A JP 2004348749 A JP2004348749 A JP 2004348749A JP 4776216 B2 JP4776216 B2 JP 4776216B2
- Authority
- JP
- Japan
- Prior art keywords
- plant
- cell death
- gene
- promoter
- seq
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Expired - Fee Related
Links
Images

Landscapes
- Breeding Of Plants And Reproduction By Means Of Culturing (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Peptides Or Proteins (AREA)
Description
(a)配列番号1で表される塩基配列を含むDNA
(b)配列番号1で表される塩基配列からなるDNAと相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ植物細胞死誘導活性を有するタンパク質をコードするベンサミアナタバコ由来のDNA
前記NbCD1遺伝子は、以下の(c)、(d)または(e)のタンパク質をコードする。
(c)配列番号2で表されるアミノ酸配列からなるタンパク質
(d)配列番号2で表されるアミノ酸配列において1または数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列からなり、かつ植物細胞死誘導活性を有するタンパク質
(e)配列番号2で表されるアミノ酸配列と80%以上の相同性を有し、かつ植物細胞死誘導活性を有するタンパク質
但し、上記(d)および(e)において、NbCD1活性に不可欠なAP2ドメイン(配列番号2のアミノ酸番号20〜84)、AD(同200〜219)、EARモチーフ(同220〜226)の配列は保存されるものとする。
本発明は、上記(c)、(d)または(e)に示すNbCD1組換えタンパク質も提供する。
本発明は、過剰発現によって植物に細胞死を誘導する因子NbCD1に関する。
1.1 NbCD1のクローニング
植物にストレスやエリシター処理を施すと、時間をおいて細胞死が誘導されることが知られている。そこで、ストレスあるいはエリシター処理後、細胞死にいたる過程で植物において発現する遺伝子を対象として、目的とする植物細胞死誘導因子のスクリーニングを行った。
発明者らは、上記のようにして細胞死誘導活性が確認された複数のクローンから、最も早く細胞死を誘導するクローンを1つ選択した。そして、このクローンのcDNAをベクターより単離し、常法にしたがって塩基配列を決定したところ、配列番号1に示される塩基配列を有することが確認された。この塩基配列は、全長981塩基対からなり、配列番号2に示される231アミノ酸からなるタンパク質をコードする696 塩基対のORF(Open Reading Frame)を含むことが確認された。確認されたDNA配列および予想アミノ酸配列は公開されているDNAおよびタンパク質データベース上には報告のない新規な遺伝子、およびタンパク質であることが確認された。そこで、発明者らはこの新規な植物細胞死誘導因子をNbCD1と命名した。
本発明のベクターは、プラスミド等の公知のベクターに本発明のNbCD1遺伝子を連結(挿入)して得ることができる。前記ベクターは宿主中で複製可能なものであれば特に限定されず、例えば、プラスミドDNA、ファージDNA等が挙げられる。
本発明の形質転換体は、本発明のベクターを、目的遺伝子が発現し得るように宿主中に導入することにより得ることができる。宿主は、本発明の遺伝子を発現できるものであれば特に限定されない。例えば、エッシェリヒア・コリ(Escherichia coli)等のエッシェリヒア属、バチルス・ズブチリス(Bacillus subtilis)等のバチルス属、シュードモナス・プチダ(Pseudomonas putida)等のシュードモナス属、リゾビウム・メリロティ(Rhizobium meliloti)等のリゾビウム属に属する細菌、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、シゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)等の酵母、シロイヌナズナ、タバコ、トウモロコシ、イネ、ニンジン等から株化した植物細胞やプロトプラスト、COS細胞、CHO細胞等の動物細胞、あるいはSf9、Sf21等の昆虫細胞等が挙げられる。
(1) 植物導入用組換えベクターの作製およびアグロバクテリウムの形質転換
植物細胞導入用ベクターは、本発明の遺伝子を含むDNAを適当な制限酵素で切断後、必要に応じて適切なリンカーを連結し、植物細胞用のクローニングベクターに挿入することにより得ることができる。クローニング用ベクターとしては、pBI2113Not、pBI2113、pBI101、pBI121、pGA482、pGAH、pBIG等のバイナリーベクター系のプラスミドやpLGV23Neo、pNCAT、pMON200等の中間ベクター系のプラスミドを用いることができる。
本発明の形質転換体の宿主は特に限定されないが、植物であることが好ましい。該植物は、植物培養細胞、栽培植物の植物体全体、植物器官(例えば、葉、花弁、茎、根、根茎、種子等)、または植物組織(例えば、表皮、師部、柔組織、木部、維管束等)のいずれであってもよい。植物はイネ、トウモロコシ、コムギ等の単子葉植物であることがより好ましい。植物培養細胞、植物体、植物器官または植物組織を宿主とする場合、本発明のタンパク質をコードするDNAは、採取した植物切片にベクターをアグロバクテリウム感染法、パーティクルガン法またはポリエチレングリコール法等で導入し、植物宿主を形質転換することができる。あるいはプロトプラストにエレクトロポレーション法で導入して形質転換植物を作製することもできる。
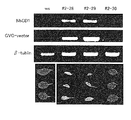
本発明の遺伝子を導入したトランスジェニック植物では、該遺伝子の発現が活性化(過剰発現)されることにより、当該発現部位特異的に細胞死が観察される。当該部位における遺伝子の発現レベルは、これらの細胞および組織から常法に従ってRNAを抽出し、RT-PCR法またはノーザン分析を用いてNbCD1遺伝子のmRNAを検出することにより確認することができる。あるいは、NbCD1タンパク質に特異的な抗体を用いたウエスタン分析等により直接、分析することによっても行うことができる。
本発明のNbCD1タンパク質は、前述の形質転換体(宿主細胞)を適当な培地で培養し、その培養物からNbCD1活性を有するタンパク質を採取することによって得ることができる。本発明の形質転換体の培養は、常法に従って行えばよい。例えば、大腸菌や酵母等の微生物を宿主とする形質転換体の場合は、微生物が資化しうる炭素源、窒素源、無機塩類等を含有し、形質転換体を効率的に培養しうる天然培地、あるいは合成培地で培養すればよい。また、植物細胞を宿主として用いている場合には、チアミン、ピリドキシン等のビタミン類を添加した植物細胞用の培地で培養すればよい。
本発明のNbCD1は、過剰発現することにより、植物に細胞死を誘導する。したがって、条件特異的(例えば、組織特異的あるいは環境特異的)に活性化されるプロモーターに本発明のNbCD1遺伝子を連結して宿主植物に導入することにより、当該植物に条件特異的細胞死を誘導することが可能になる。
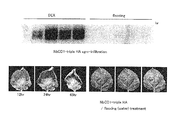
ベンサミアナタバコ(Nicotiana benthamiana)植物葉身にジャガイモ疫病菌(Phytophthora infestans)由来のエリシターINF1処理を施すと、約24時間で過敏感細胞死が誘導される(Plant Cell 10:1413-1426)。そこで、INF1誘導性細胞死に関わるベンサミアナタバコ遺伝子を同定する目的で、INF1処理後1、2、4時間後のベンサミアナ植物葉からSDSフェノール法(Biochemistry 13:3606-3615)により全RNAを抽出した。この全RNAよりオリゴdTカラム(Amersham Co. UK)を利用してmRNAを抽出し、次いでSuperScriptTM Plasmid System with Gateway Technology for cDNA Synthesis and Cloningキット (Life Technology Co.)を用いて、2本鎖cDNAを合成した。このキットにより、各cDNAの5’末端はSalI、3’末端は、NotIと相補的なcohesive末端となっている。
pSfinxベクター(Plant Journal 24:275-283)は、植物形質転換用のバイナリーベクターのRight Border(RB)配列とLeft Border (LB)配列間に、ジャガイモウイルスX (Potato virus X: PVX)の転写ユニットが挿入された構造を有する。本ベクター中のPVX 転写ユニットのコートプロテイン(CP)遺伝子プロモーターは、重複しており、その内の一つのCPプロモーターの下流に、目的遺伝子の断片を挿入するためのmultiple cloning site (MCS)が設けられている。このMCSの制限酵素サイトAsc1 サイトとSalIサイトをそれぞれの制限酵素で切断した後、SalI-Not1リンカー断片をライゲートして、pSfinx-SalI-NotI ベクターを作製した。なお、SalI-Not1リンカーは、リンカー1:P-5’-CGCGGTCGACGGGATATCACGCGGCCGCG-3’(配列番号3); リンカー2:P-5’-TCGACGCGGCCGCGTGATATCCCGTCGAC-3’(配列番号4)を互いにアニールして作製した。pSfinX-SalI-NotIベクターを制限酵素SalIとNotIによって消化後、ベンサミアナタバコ由来のcDNAをライゲートした。ライゲート産物を用いて、エレクトロポレーション法(GenePulser, BioRad社使用)によって大腸菌DH10Bのコンピテントセル(ElectroMAX DH10B Cells, Invitrogen)を形質転換した。しかるのち形質転換細胞を10 ppmカナマイシンを含むLB寒天培地上で選抜した。その結果、約50万個のクローンからなるcDNAライブラリーが作製された。
上記約50万個の大腸菌クローンから、プラスミド単離キット(QIAprep Spin Miniprep Kit, Quiagen)を用いて、プラスミドをまとめて回収した。この混合プラスミド液によって、Agrobacterium tumefaciens MOG101 株をエレクトロポレーション法(GenePulser, BioRad社使用)を用いて形質転換した。形質転換細胞は、カナマイシン50 ppm、リファンピシン 50 ppmを含むYEB寒天培地(培地1Lあたり5 g beef extract, 1 g yeast extract, 5 g tryptone, 5 g sucrose, 2 mL 1M MgSO4, 15 g Bactoagarを含有)上で選抜した。これにより、計約20万個の形質転換アグロバクテリウムクローンが得られた。これらのクローンは、滅菌した爪楊枝を用いて、15%グリセリン、カナマイシン50 ppm、リファンピシン 50 ppmを含むYEB培地を満たした384穴のマイクロタイタープレートに移し、整列させて-80度で保存した。
上記アグロバクテリウムクローンを96個ずつ96穴マイクロタイタープレートに移し、カナマイシン50 ppm、リファンピシン 50 ppmを含むYEB液体培地で28℃1日間振とう培養した。液体培養後のクローンを塗布した爪楊枝を、ベンサミアナタバコ葉身に突き刺すことにより、アグロバクテリウムを葉に接種した。1枚の葉に50クローンを整列して接種した。
アグロバクテリウム接種後の植物を閉鎖系温室で栽培し、1週間後以降接種部位の変化を観察した。接種部位周辺が黒褐色に変色し、細胞死が認められるクローンを同定した。これらのクローンについては、再度新しい植物体の葉に接種し、細胞死誘導性を追試した。
候補クローン(Agrobacterium株)を、カナマイシン50 ppm、リファンピシン 50 ppmを含むYEB液体培地で28℃1日間振とう培養した。バクテリアを、遠心操作によって沈澱後、これを注射液(10mM MgCl2, 10mM MES, 100μM acetosyringone)に溶解し、波長600nmにおける吸光度が1.0になるように調整した。この溶液を、室温で2〜3時間静置した後、針なし注射器(1mL)によって、ベンサミアナタバコの葉身に注入した。3〜4日後に注入箇所が枯死することを確認した。
NbCD1遺伝子全長の内、植物細胞死誘導にとって必須の領域を明らかにするために、以下に示す欠失実験をおこなった(図2)。NbCD1アミノ酸配列中には、3つの特徴的なタンパク質領域が存在する。すなわち、AP2ドメイン、酸性ドメイン(Acidic Domain:AD)およびERF-associated amphiphilic repression (EAR)モチーフである。AP2ドメインは、配列番号2の20〜84番目のアミノ酸(配列番号1の186〜380番目の塩基)に対応し、ADは配列番号2の200〜219番目のアミノ酸(配列番号1の727〜785番目の塩基)に対応し、EARモチーフは配列番号2の220〜226番目のアミノ酸(配列番号1の786〜806番目の塩基)に対応する。そこで、これらの領域を一つずつ欠失させたcDNAを作製し、pSfinXベクターに導入して、Agroinfiltration法によってベンサミアナタバコの葉身に接種して表現型を観察したところ、細胞死誘導にとって、これら3領域:AP2ドメイン、AD、EARモチーフが必須であることが確認された。AP2ドメインはGCC-boxと呼ばれるシスエレメントに結合するためのDNA結合ドメイン、EARモチーフは転写抑制(active repression)部位として知られているが、3領域のいずれも細胞死との関連についてはこれまでに報告されたことはない。したがって、NbCD1の細胞死誘導活性は、その構造からは予測がつかないものであった。
NbCD1はタバコ属以外の双子葉植物でも細胞死誘導効果をもつことを確認するため、NbCD1のシロイヌナズナに対する細胞死誘導効果について検討した。
酸性ドメイン
EAR モチーフ
配列番号2−AP2ドメイン
酸性ドメイン
EAR モチーフ
配列番号3−人工配列の説明:合成DNA(リンカー1)
配列番号4−人工配列の説明:合成DNA(リンカー2)
配列番号5−人工配列の説明:合成アミノ酸(HA エピトープ)
配列番号6−人工配列の説明:合成アミノ酸(triple HA エピトープ)
配列番号7−人工配列の説明:プライマー(NbCD1)
配列番号8−人工配列の説明:プライマー(NbCD1)
配列番号9−人工配列の説明:プライマー(GVG-vector)
配列番号10−人工配列の説明:プライマー(GVG-vector)
配列番号11−人工配列の説明:プライマー(β-tublin)
配列番号12−人工配列の説明:プライマー(β-tublin)
Claims (10)
- 配列番号1で表される塩基配列を含むDNAからなる遺伝子。
- 以下の(c)、(d)または(e)のタンパク質をコードする遺伝子。
(c)配列番号2で表されるアミノ酸配列からなるタンパク質;
(d)配列番号2で表されるアミノ酸配列において1または数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列からなり、かつ植物細胞死誘導活性を有するタンパク質;
(e)配列番号2で表されるアミノ酸配列と90%以上の同一性を有し、かつ植物細胞死誘導活性を有するタンパク質;
但し、上記(d)および(e)において、配列番号2のアミノ酸番号20〜84、200〜219、および220〜226における配列は保存されるものとする。 - 以下の(c)、(d)または(e)の組換えタンパク質。
(c)配列番号2で表されるアミノ酸配列からなるタンパク質;
(d)配列番号2で表されるアミノ酸配列において1または数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列からなり、かつ植物細胞死誘導活性を有するタンパク質;
(e)配列番号2で表されるアミノ酸配列と90%以上の同一性を有し、かつ植物細胞死誘導活性を有するタンパク質;
但し、上記(d)および(e)において、配列番号2のアミノ酸番号20〜84、200〜219、および220〜226における配列は保存されるものとする。 - 請求項1または2記載の遺伝子を機能しうる態様でプロモーターに連結させて含む、ベクター。
- 前記プロモーターが、条件特異的に活性化されるプロモーターである、請求項4記載のベクター。
- 前記条件特異的に活性化されるプロモーターが、キャベツのBcp1遺伝子プロモーターを含む葯特異的プロモーター、およびパセリのPR1遺伝子プロモーターを含む病原菌エリシター応答性プロモーターから選ばれるいずれかのプロモーターである、請求項5記載のベクター。
- 請求項4〜6のいずれか1項に記載のベクターを宿主に導入して得られる、形質転換体。
- 請求項4〜6のいずれか1項に記載のベクターを植物に導入して得られる、トランスジェニック植物。
- 請求項5または6記載のベクターを植物に導入して得られる、条件特異的に細胞死を引き起こすトランスジェニック植物。
- 請求項5または6記載のベクターを植物に導入することにより、該植物に条件特異的細
胞死を引き起こす方法。
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2004348749A JP4776216B2 (ja) | 2004-03-02 | 2004-12-01 | 新規な植物細胞死誘導因子NbCD1 |
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2004057653 | 2004-03-02 | ||
| JP2004057653 | 2004-03-02 | ||
| JP2004348749A JP4776216B2 (ja) | 2004-03-02 | 2004-12-01 | 新規な植物細胞死誘導因子NbCD1 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2005278634A JP2005278634A (ja) | 2005-10-13 |
| JP4776216B2 true JP4776216B2 (ja) | 2011-09-21 |
Family
ID=35177684
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2004348749A Expired - Fee Related JP4776216B2 (ja) | 2004-03-02 | 2004-12-01 | 新規な植物細胞死誘導因子NbCD1 |
Country Status (1)
| Country | Link |
|---|---|
| JP (1) | JP4776216B2 (ja) |
Families Citing this family (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN110468136B (zh) * | 2019-09-04 | 2022-11-01 | 中国农业大学 | 一种花特异性启动子及其应用 |
| CN116120412B (zh) * | 2023-02-24 | 2023-07-18 | 南京林业大学 | 樟疫霉效应子蛋白scr83830及其应用 |
Family Cites Families (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| AU5796099A (en) * | 1998-08-31 | 2000-03-21 | Monsanto Company | A new method of identifying non-host plant disease resistance genes |
| JP2004506415A (ja) * | 2000-07-03 | 2004-03-04 | シンジェンタ リミテッド | クラドスポリウム菌に由来するエリシター |
-
2004
- 2004-12-01 JP JP2004348749A patent/JP4776216B2/ja not_active Expired - Fee Related
Also Published As
| Publication number | Publication date |
|---|---|
| JP2005278634A (ja) | 2005-10-13 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| Checker et al. | Molecular and functional characterization of mulberry EST encoding remorin (MiREM) involved in abiotic stress | |
| CN101130785B (zh) | 一个与耐旱相关的水稻wrky基因的克隆及应用 | |
| CN100526465C (zh) | 利用水稻转录因子基因snac2提高植物耐冷耐盐能力 | |
| US9809827B2 (en) | Transgenic maize | |
| CN106047889A (zh) | 大豆myb转录因子基因提高大豆异黄酮生物合成的应用 | |
| CN111218470B (zh) | 一种调控植物抗逆性的方法 | |
| CN110734482A (zh) | 一种岷江百合WRKY转录因子基因LrWRKY4及应用 | |
| US20120278948A1 (en) | Transgenic plant having increased biomass and improved environmental stress resistance, and process for production thereof | |
| CN103819549B (zh) | 一种水稻蛋白及其编码基因在调控植物抗旱性中的应用 | |
| CN110295175B (zh) | 一个大豆NAC转录因子家族基因Glyma08g41995的应用 | |
| CN113666993B (zh) | 紫花苜蓿MsSPL12蛋白及其相关生物材料与它们在提高植物抗逆性中的应用 | |
| CN111118036B (zh) | 刚毛柽柳phd3转录因子编码基因及其应用 | |
| CN113372423A (zh) | 一个野生大豆Bet_v_1家族基因GsMLP328的应用 | |
| CN103421835B (zh) | 转录因子基因RrMYB11在调控玫瑰花型中的应用 | |
| CN101139385B (zh) | 一种植物抗逆性相关蛋白及其编码基因与应用 | |
| CN1328383C (zh) | 野生稻的一个抗旱基因及其编码蛋白与应用 | |
| CN103320448B (zh) | 一种岷江百合bZIP转录因子基因LrbZIP1及应用 | |
| JP2013212105A (ja) | 高い種子収量性を有する環境ストレス耐性植物及びその作製方法 | |
| CN102336826A (zh) | 一种大豆逆境相关转录因子erf及其编码基因与应用 | |
| CN105175522B (zh) | 百脉根ap2/erf转录因子及其编码基因和应用 | |
| CN104004073A (zh) | 来源于小麦的抗病性相关蛋白TaCPK7-R及其相关生物材料与应用 | |
| JP4776216B2 (ja) | 新規な植物細胞死誘導因子NbCD1 | |
| CN115786358B (zh) | 抗旱相关蛋白IbbHLH118及其编码基因与应用 | |
| CN101942448A (zh) | 棉花nodulin-like基因的一个耐盐、耐旱新功能及其应用 | |
| KR102674984B1 (ko) | CaSIRF1 유전자 및 이를 이용한 식물체의 건조 스트레스 저항성 증진 방법 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20071120 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20100803 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20100930 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20110531 |
|
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20110628 |
|
| R150 | Certificate of patent or registration of utility model |
Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20140708 Year of fee payment: 3 |
|
| LAPS | Cancellation because of no payment of annual fees |