JP2021515588A - ペプチド交換蛋白質 - Google Patents
ペプチド交換蛋白質 Download PDFInfo
- Publication number
- JP2021515588A JP2021515588A JP2020560595A JP2020560595A JP2021515588A JP 2021515588 A JP2021515588 A JP 2021515588A JP 2020560595 A JP2020560595 A JP 2020560595A JP 2020560595 A JP2020560595 A JP 2020560595A JP 2021515588 A JP2021515588 A JP 2021515588A
- Authority
- JP
- Japan
- Prior art keywords
- peptide
- cells
- cell
- mhc class
- target
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
- 108090000765 processed proteins & peptides Proteins 0.000 title claims abstract description 613
- 108090000623 proteins and genes Proteins 0.000 title claims abstract description 254
- 102000004169 proteins and genes Human genes 0.000 title claims abstract description 245
- 210000004027 cell Anatomy 0.000 claims abstract description 507
- 206010028980 Neoplasm Diseases 0.000 claims abstract description 104
- 230000002163 immunogen Effects 0.000 claims abstract description 90
- 210000001744 T-lymphocyte Anatomy 0.000 claims abstract description 69
- 238000011068 loading method Methods 0.000 claims abstract description 39
- 230000028993 immune response Effects 0.000 claims abstract description 34
- 210000004962 mammalian cell Anatomy 0.000 claims abstract description 33
- 230000001717 pathogenic effect Effects 0.000 claims abstract description 27
- 108091054437 MHC class I family Proteins 0.000 claims description 160
- 238000000034 method Methods 0.000 claims description 73
- 102000043129 MHC class I family Human genes 0.000 claims description 68
- 108010088652 Histocompatibility Antigens Class I Proteins 0.000 claims description 64
- 239000000427 antigen Substances 0.000 claims description 61
- 108091007433 antigens Proteins 0.000 claims description 61
- 102000036639 antigens Human genes 0.000 claims description 61
- 239000012634 fragment Substances 0.000 claims description 57
- 201000011510 cancer Diseases 0.000 claims description 56
- 210000000612 antigen-presenting cell Anatomy 0.000 claims description 46
- 230000014509 gene expression Effects 0.000 claims description 45
- 230000005847 immunogenicity Effects 0.000 claims description 36
- 102100028082 Tapasin Human genes 0.000 claims description 35
- 108010059434 tapasin Proteins 0.000 claims description 35
- 102000039446 nucleic acids Human genes 0.000 claims description 29
- 108020004707 nucleic acids Proteins 0.000 claims description 29
- 150000007523 nucleic acids Chemical class 0.000 claims description 29
- 102100028976 HLA class I histocompatibility antigen, B alpha chain Human genes 0.000 claims description 21
- 239000013598 vector Substances 0.000 claims description 19
- 244000052769 pathogen Species 0.000 claims description 17
- 210000004443 dendritic cell Anatomy 0.000 claims description 14
- 238000000338 in vitro Methods 0.000 claims description 14
- 230000002708 enhancing effect Effects 0.000 claims description 13
- 208000015181 infectious disease Diseases 0.000 claims description 9
- 239000003446 ligand Substances 0.000 claims description 8
- 206010022000 influenza Diseases 0.000 claims description 7
- 239000007787 solid Substances 0.000 claims description 7
- 102100022005 B-lymphocyte antigen CD20 Human genes 0.000 claims description 5
- 101000897405 Homo sapiens B-lymphocyte antigen CD20 Proteins 0.000 claims description 5
- 108010090804 Streptavidin Proteins 0.000 claims description 5
- 102000005962 receptors Human genes 0.000 claims description 5
- 101001117317 Homo sapiens Programmed cell death 1 ligand 1 Proteins 0.000 claims description 4
- 102100024216 Programmed cell death 1 ligand 1 Human genes 0.000 claims description 4
- 230000001086 cytosolic effect Effects 0.000 claims description 4
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 claims description 3
- 101710197241 Accessory gland-specific peptide 70A Proteins 0.000 claims description 3
- 208000006454 hepatitis Diseases 0.000 claims description 3
- 231100000283 hepatitis Toxicity 0.000 claims description 3
- 229960005486 vaccine Drugs 0.000 claims description 3
- 101150064935 HELI gene Proteins 0.000 claims description 2
- 101100449157 Human cytomegalovirus (strain AD169) GO gene Proteins 0.000 claims description 2
- 101100371784 Human cytomegalovirus (strain Merlin) UL142 gene Proteins 0.000 claims description 2
- 101100449158 Human cytomegalovirus (strain Merlin) UL74 gene Proteins 0.000 claims description 2
- 101800001690 Transmembrane protein gp41 Proteins 0.000 claims description 2
- 101150111135 UL1 gene Proteins 0.000 claims description 2
- 101150087840 UL11 gene Proteins 0.000 claims description 2
- 101150042088 UL16 gene Proteins 0.000 claims description 2
- 101150099617 UL5 gene Proteins 0.000 claims description 2
- 101150081415 UL55 gene Proteins 0.000 claims description 2
- 101150026859 UL9 gene Proteins 0.000 claims description 2
- 101150029683 gB gene Proteins 0.000 claims description 2
- 101150055782 gH gene Proteins 0.000 claims description 2
- 101150015940 gL gene Proteins 0.000 claims description 2
- 230000004936 stimulating effect Effects 0.000 claims description 2
- 125000003275 alpha amino acid group Chemical group 0.000 claims 3
- 208000022361 Human papillomavirus infectious disease Diseases 0.000 claims 1
- 230000003213 activating effect Effects 0.000 claims 1
- 230000008105 immune reaction Effects 0.000 claims 1
- 102000004196 processed proteins & peptides Human genes 0.000 abstract description 158
- 230000006870 function Effects 0.000 abstract description 19
- 239000003054 catalyst Substances 0.000 abstract description 14
- 239000012528 membrane Substances 0.000 abstract description 9
- 230000001939 inductive effect Effects 0.000 abstract description 8
- 230000001105 regulatory effect Effects 0.000 abstract description 7
- 102000018697 Membrane Proteins Human genes 0.000 abstract description 3
- 108010052285 Membrane Proteins Proteins 0.000 abstract description 3
- 108050001049 Extracellular proteins Proteins 0.000 abstract description 2
- 238000010586 diagram Methods 0.000 abstract 1
- 235000018102 proteins Nutrition 0.000 description 200
- 102000008949 Histocompatibility Antigens Class I Human genes 0.000 description 141
- 230000027455 binding Effects 0.000 description 100
- 108010075704 HLA-A Antigens Proteins 0.000 description 47
- 102000011786 HLA-A Antigens Human genes 0.000 description 47
- 101150029707 ERBB2 gene Proteins 0.000 description 38
- 102000037865 fusion proteins Human genes 0.000 description 38
- 108020001507 fusion proteins Proteins 0.000 description 38
- 150000001413 amino acids Chemical class 0.000 description 37
- 238000011282 treatment Methods 0.000 description 36
- 108010074032 HLA-A2 Antigen Proteins 0.000 description 30
- 102000025850 HLA-A2 Antigen Human genes 0.000 description 30
- 230000004927 fusion Effects 0.000 description 27
- 125000005647 linker group Chemical group 0.000 description 27
- 238000002474 experimental method Methods 0.000 description 24
- 102100037850 Interferon gamma Human genes 0.000 description 22
- 108010074328 Interferon-gamma Proteins 0.000 description 22
- 210000005220 cytoplasmic tail Anatomy 0.000 description 21
- 210000004881 tumor cell Anatomy 0.000 description 21
- 229920001184 polypeptide Polymers 0.000 description 20
- 108091008874 T cell receptors Proteins 0.000 description 19
- 102000016266 T-Cell Antigen Receptors Human genes 0.000 description 19
- 210000002472 endoplasmic reticulum Anatomy 0.000 description 17
- 108091006116 chimeric peptides Proteins 0.000 description 16
- 235000001014 amino acid Nutrition 0.000 description 15
- 229940024606 amino acid Drugs 0.000 description 15
- 210000000170 cell membrane Anatomy 0.000 description 15
- 230000001419 dependent effect Effects 0.000 description 14
- 239000002773 nucleotide Substances 0.000 description 14
- 241000699666 Mus <mouse, genus> Species 0.000 description 13
- LOKCTEFSRHRXRJ-UHFFFAOYSA-I dipotassium trisodium dihydrogen phosphate hydrogen phosphate dichloride Chemical compound P(=O)(O)(O)[O-].[K+].P(=O)(O)([O-])[O-].[Na+].[Na+].[Cl-].[K+].[Cl-].[Na+] LOKCTEFSRHRXRJ-UHFFFAOYSA-I 0.000 description 13
- 125000003729 nucleotide group Chemical group 0.000 description 13
- 239000002953 phosphate buffered saline Substances 0.000 description 13
- 230000003612 virological effect Effects 0.000 description 13
- 241000700605 Viruses Species 0.000 description 12
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 12
- 238000000684 flow cytometry Methods 0.000 description 12
- 230000001965 increasing effect Effects 0.000 description 12
- 230000003834 intracellular effect Effects 0.000 description 12
- 230000001225 therapeutic effect Effects 0.000 description 12
- 241000701022 Cytomegalovirus Species 0.000 description 11
- 101710192602 Latent membrane protein 1 Proteins 0.000 description 11
- 230000000890 antigenic effect Effects 0.000 description 11
- 201000010099 disease Diseases 0.000 description 11
- 230000001404 mediated effect Effects 0.000 description 11
- WGTODYJZXSJIAG-UHFFFAOYSA-N tetramethylrhodamine chloride Chemical compound [Cl-].C=12C=CC(N(C)C)=CC2=[O+]C2=CC(N(C)C)=CC=C2C=1C1=CC=CC=C1C(O)=O WGTODYJZXSJIAG-UHFFFAOYSA-N 0.000 description 11
- 206010006187 Breast cancer Diseases 0.000 description 10
- 208000026310 Breast neoplasm Diseases 0.000 description 10
- 125000000539 amino acid group Chemical group 0.000 description 10
- 238000003556 assay Methods 0.000 description 9
- 230000000694 effects Effects 0.000 description 9
- 238000001114 immunoprecipitation Methods 0.000 description 9
- 238000001727 in vivo Methods 0.000 description 9
- 238000011534 incubation Methods 0.000 description 9
- 239000000047 product Substances 0.000 description 9
- 238000010361 transduction Methods 0.000 description 9
- 230000026683 transduction Effects 0.000 description 9
- 108010074708 B7-H1 Antigen Proteins 0.000 description 8
- 102000008096 B7-H1 Antigen Human genes 0.000 description 8
- 108091003079 Bovine Serum Albumin Proteins 0.000 description 8
- 101000762808 Homo sapiens Tapasin-related protein Proteins 0.000 description 8
- 108010003723 Single-Domain Antibodies Proteins 0.000 description 8
- 230000005867 T cell response Effects 0.000 description 8
- 230000002950 deficient Effects 0.000 description 8
- 239000008194 pharmaceutical composition Substances 0.000 description 8
- 230000008685 targeting Effects 0.000 description 8
- 108010021625 Immunoglobulin Fragments Proteins 0.000 description 7
- 102000008394 Immunoglobulin Fragments Human genes 0.000 description 7
- 238000004458 analytical method Methods 0.000 description 7
- 239000003814 drug Substances 0.000 description 7
- 102000056944 human TAPBPL Human genes 0.000 description 7
- 239000000203 mixture Substances 0.000 description 7
- 238000012360 testing method Methods 0.000 description 7
- 238000001890 transfection Methods 0.000 description 7
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 6
- 102100028389 Melanoma antigen recognized by T-cells 1 Human genes 0.000 description 6
- 239000004365 Protease Substances 0.000 description 6
- 230000005907 cancer growth Effects 0.000 description 6
- 238000001514 detection method Methods 0.000 description 6
- YBYRMVIVWMBXKQ-UHFFFAOYSA-N phenylmethanesulfonyl fluoride Chemical compound FS(=O)(=O)CC1=CC=CC=C1 YBYRMVIVWMBXKQ-UHFFFAOYSA-N 0.000 description 6
- 210000001519 tissue Anatomy 0.000 description 6
- 238000005406 washing Methods 0.000 description 6
- 108010058607 HLA-B Antigens Proteins 0.000 description 5
- 241000699670 Mus sp. Species 0.000 description 5
- 108091005804 Peptidases Proteins 0.000 description 5
- 108010076504 Protein Sorting Signals Proteins 0.000 description 5
- 238000000692 Student's t-test Methods 0.000 description 5
- 101710148271 UDP-glucose:glycoprotein glucosyltransferase 1 Proteins 0.000 description 5
- 102100029151 UDP-glucuronosyltransferase 1A10 Human genes 0.000 description 5
- -1 VSVG Proteins 0.000 description 5
- 230000001363 autoimmune Effects 0.000 description 5
- 238000010494 dissociation reaction Methods 0.000 description 5
- 230000005593 dissociations Effects 0.000 description 5
- 238000004519 manufacturing process Methods 0.000 description 5
- 239000003550 marker Substances 0.000 description 5
- 239000000463 material Substances 0.000 description 5
- 230000007246 mechanism Effects 0.000 description 5
- 230000002018 overexpression Effects 0.000 description 5
- 230000037361 pathway Effects 0.000 description 5
- 230000008569 process Effects 0.000 description 5
- 238000000746 purification Methods 0.000 description 5
- 238000010186 staining Methods 0.000 description 5
- 208000024891 symptom Diseases 0.000 description 5
- 238000012353 t test Methods 0.000 description 5
- JVJGCCBAOOWGEO-RUTPOYCXSA-N (2s)-2-[[(2s)-2-[[(2s)-2-[[(2s)-2-[[(2s)-4-amino-2-[[(2s,3s)-2-[[(2s,3s)-2-[[(2s)-2-azaniumyl-3-hydroxypropanoyl]amino]-3-methylpentanoyl]amino]-3-methylpentanoyl]amino]-4-oxobutanoyl]amino]-3-phenylpropanoyl]amino]-4-carboxylatobutanoyl]amino]-6-azaniumy Chemical compound OC[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@H](C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(O)=O)CC1=CC=CC=C1 JVJGCCBAOOWGEO-RUTPOYCXSA-N 0.000 description 4
- 102100026559 Filamin-B Human genes 0.000 description 4
- 101000913551 Homo sapiens Filamin-B Proteins 0.000 description 4
- 101000578784 Homo sapiens Melanoma antigen recognized by T-cells 1 Proteins 0.000 description 4
- 241000713666 Lentivirus Species 0.000 description 4
- 230000004913 activation Effects 0.000 description 4
- 230000030741 antigen processing and presentation Effects 0.000 description 4
- 239000011324 bead Substances 0.000 description 4
- 230000009286 beneficial effect Effects 0.000 description 4
- 229940098773 bovine serum albumin Drugs 0.000 description 4
- 239000000969 carrier Substances 0.000 description 4
- 239000002771 cell marker Substances 0.000 description 4
- 125000002534 ethynyl group Chemical group [H]C#C* 0.000 description 4
- 239000012091 fetal bovine serum Substances 0.000 description 4
- 208000027866 inflammatory disease Diseases 0.000 description 4
- 238000003780 insertion Methods 0.000 description 4
- 230000037431 insertion Effects 0.000 description 4
- 230000035772 mutation Effects 0.000 description 4
- 210000000056 organ Anatomy 0.000 description 4
- 239000002243 precursor Substances 0.000 description 4
- 108020003175 receptors Proteins 0.000 description 4
- 230000002829 reductive effect Effects 0.000 description 4
- 239000000523 sample Substances 0.000 description 4
- 230000003248 secreting effect Effects 0.000 description 4
- 210000001082 somatic cell Anatomy 0.000 description 4
- 241000894007 species Species 0.000 description 4
- 238000006467 substitution reaction Methods 0.000 description 4
- 229940124597 therapeutic agent Drugs 0.000 description 4
- 238000002255 vaccination Methods 0.000 description 4
- POVNCJSPYFCWJR-USZUGGBUSA-N (4s)-4-[[(2s)-2-[[(2s)-2-amino-3-(4-hydroxyphenyl)propanoyl]amino]-4-methylpentanoyl]amino]-5-[(2s)-2-[[2-[(2s)-2-[[(2s)-1-[[(2s,3r)-1-[[(1s)-1-carboxy-2-methylpropyl]amino]-3-hydroxy-1-oxobutan-2-yl]amino]-3-methyl-1-oxobutan-2-yl]carbamoyl]pyrrolidin-1- Chemical compound C([C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(=O)N1[C@@H](CCC1)C(=O)NCC(=O)N1[C@@H](CCC1)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(O)=O)C1=CC=C(O)C=C1 POVNCJSPYFCWJR-USZUGGBUSA-N 0.000 description 3
- 239000012114 Alexa Fluor 647 Substances 0.000 description 3
- 108700028369 Alleles Proteins 0.000 description 3
- WVDDGKGOMKODPV-UHFFFAOYSA-N Benzyl alcohol Chemical compound OCC1=CC=CC=C1 WVDDGKGOMKODPV-UHFFFAOYSA-N 0.000 description 3
- 102000000905 Cadherin Human genes 0.000 description 3
- 108050007957 Cadherin Proteins 0.000 description 3
- 101000879203 Caenorhabditis elegans Small ubiquitin-related modifier Proteins 0.000 description 3
- 241000282472 Canis lupus familiaris Species 0.000 description 3
- 102100025064 Cellular tumor antigen p53 Human genes 0.000 description 3
- 241000282693 Cercopithecidae Species 0.000 description 3
- 102100035294 Chemokine XC receptor 1 Human genes 0.000 description 3
- 239000006144 Dulbecco’s modified Eagle's medium Substances 0.000 description 3
- PEDCQBHIVMGVHV-UHFFFAOYSA-N Glycerine Chemical compound OCC(O)CO PEDCQBHIVMGVHV-UHFFFAOYSA-N 0.000 description 3
- 239000004471 Glycine Substances 0.000 description 3
- 101000721661 Homo sapiens Cellular tumor antigen p53 Proteins 0.000 description 3
- 101000804783 Homo sapiens Chemokine XC receptor 1 Proteins 0.000 description 3
- 241000701806 Human papillomavirus Species 0.000 description 3
- 201000005505 Measles Diseases 0.000 description 3
- 102100022430 Melanocyte protein PMEL Human genes 0.000 description 3
- 241001465754 Metazoa Species 0.000 description 3
- 108010075205 OVA-8 Proteins 0.000 description 3
- 241000283973 Oryctolagus cuniculus Species 0.000 description 3
- 102100026450 POU domain, class 3, transcription factor 4 Human genes 0.000 description 3
- 101710133389 POU domain, class 3, transcription factor 4 Proteins 0.000 description 3
- 102000035195 Peptidases Human genes 0.000 description 3
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 description 3
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 description 3
- 102000051619 SUMO-1 Human genes 0.000 description 3
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 3
- 101800001271 Surface protein Proteins 0.000 description 3
- 239000002253 acid Substances 0.000 description 3
- IVRMZWNICZWHMI-UHFFFAOYSA-N azide group Chemical group [N-]=[N+]=[N-] IVRMZWNICZWHMI-UHFFFAOYSA-N 0.000 description 3
- 230000001580 bacterial effect Effects 0.000 description 3
- 230000008901 benefit Effects 0.000 description 3
- 239000000872 buffer Substances 0.000 description 3
- 239000003153 chemical reaction reagent Substances 0.000 description 3
- 150000001875 compounds Chemical class 0.000 description 3
- 125000000151 cysteine group Chemical class N[C@@H](CS)C(=O)* 0.000 description 3
- 230000003247 decreasing effect Effects 0.000 description 3
- 229940079593 drug Drugs 0.000 description 3
- RAXXELZNTBOGNW-UHFFFAOYSA-N imidazole Natural products C1=CNC=N1 RAXXELZNTBOGNW-UHFFFAOYSA-N 0.000 description 3
- 238000009169 immunotherapy Methods 0.000 description 3
- 230000003993 interaction Effects 0.000 description 3
- 230000002147 killing effect Effects 0.000 description 3
- 239000000546 pharmaceutical excipient Substances 0.000 description 3
- 239000013612 plasmid Substances 0.000 description 3
- 230000000069 prophylactic effect Effects 0.000 description 3
- 238000010188 recombinant method Methods 0.000 description 3
- 238000011160 research Methods 0.000 description 3
- 230000028327 secretion Effects 0.000 description 3
- 239000000243 solution Substances 0.000 description 3
- 230000000638 stimulation Effects 0.000 description 3
- UCSJYZPVAKXKNQ-HZYVHMACSA-N streptomycin Chemical compound CN[C@H]1[C@H](O)[C@@H](O)[C@H](CO)O[C@H]1O[C@@H]1[C@](C=O)(O)[C@H](C)O[C@H]1O[C@@H]1[C@@H](NC(N)=N)[C@H](O)[C@@H](NC(N)=N)[C@H](O)[C@H]1O UCSJYZPVAKXKNQ-HZYVHMACSA-N 0.000 description 3
- 238000002560 therapeutic procedure Methods 0.000 description 3
- 229960000575 trastuzumab Drugs 0.000 description 3
- 239000003656 tris buffered saline Substances 0.000 description 3
- 230000005909 tumor killing Effects 0.000 description 3
- 230000009385 viral infection Effects 0.000 description 3
- 238000001262 western blot Methods 0.000 description 3
- YBJHBAHKTGYVGT-ZKWXMUAHSA-N (+)-Biotin Chemical compound N1C(=O)N[C@@H]2[C@H](CCCCC(=O)O)SC[C@@H]21 YBJHBAHKTGYVGT-ZKWXMUAHSA-N 0.000 description 2
- QKNYBSVHEMOAJP-UHFFFAOYSA-N 2-amino-2-(hydroxymethyl)propane-1,3-diol;hydron;chloride Chemical compound Cl.OCC(N)(CO)CO QKNYBSVHEMOAJP-UHFFFAOYSA-N 0.000 description 2
- CIWBSHSKHKDKBQ-JLAZNSOCSA-N Ascorbic acid Chemical compound OC[C@H](O)[C@H]1OC(=O)C(O)=C1O CIWBSHSKHKDKBQ-JLAZNSOCSA-N 0.000 description 2
- 208000023275 Autoimmune disease Diseases 0.000 description 2
- 102100028681 C-type lectin domain family 4 member K Human genes 0.000 description 2
- 101100314454 Caenorhabditis elegans tra-1 gene Proteins 0.000 description 2
- UXVMQQNJUSDDNG-UHFFFAOYSA-L Calcium chloride Chemical compound [Cl-].[Cl-].[Ca+2] UXVMQQNJUSDDNG-UHFFFAOYSA-L 0.000 description 2
- 241000282465 Canis Species 0.000 description 2
- 241000283707 Capra Species 0.000 description 2
- 102000014914 Carrier Proteins Human genes 0.000 description 2
- 206010057248 Cell death Diseases 0.000 description 2
- 206010008342 Cervix carcinoma Diseases 0.000 description 2
- 241000588724 Escherichia coli Species 0.000 description 2
- 241000282326 Felis catus Species 0.000 description 2
- 102100040578 G antigen 7 Human genes 0.000 description 2
- 101710113436 GTPase KRas Proteins 0.000 description 2
- 102000003886 Glycoproteins Human genes 0.000 description 2
- 108090000288 Glycoproteins Proteins 0.000 description 2
- 102100036242 HLA class II histocompatibility antigen, DQ alpha 2 chain Human genes 0.000 description 2
- 108010075326 HLA-B51 Antigen Proteins 0.000 description 2
- 241000282412 Homo Species 0.000 description 2
- 101000893968 Homo sapiens G antigen 7 Proteins 0.000 description 2
- 101000767151 Homo sapiens General vesicular transport factor p115 Proteins 0.000 description 2
- 101000930801 Homo sapiens HLA class II histocompatibility antigen, DQ alpha 2 chain Proteins 0.000 description 2
- 101000934372 Homo sapiens Macrosialin Proteins 0.000 description 2
- 101001133056 Homo sapiens Mucin-1 Proteins 0.000 description 2
- 101001093919 Homo sapiens SEC14-like protein 2 Proteins 0.000 description 2
- 229940076838 Immune checkpoint inhibitor Drugs 0.000 description 2
- 108010054477 Immunoglobulin Fab Fragments Proteins 0.000 description 2
- 102000001706 Immunoglobulin Fab Fragments Human genes 0.000 description 2
- 102000037984 Inhibitory immune checkpoint proteins Human genes 0.000 description 2
- 108091008026 Inhibitory immune checkpoint proteins Proteins 0.000 description 2
- 108010050904 Interferons Proteins 0.000 description 2
- 102000014150 Interferons Human genes 0.000 description 2
- 206010025323 Lymphomas Diseases 0.000 description 2
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 2
- 239000004472 Lysine Substances 0.000 description 2
- 108010010995 MART-1 Antigen Proteins 0.000 description 2
- 102100025136 Macrosialin Human genes 0.000 description 2
- 241000124008 Mammalia Species 0.000 description 2
- 102100034256 Mucin-1 Human genes 0.000 description 2
- 239000012124 Opti-MEM Substances 0.000 description 2
- 229930182555 Penicillin Natural products 0.000 description 2
- JGSARLDLIJGVTE-MBNYWOFBSA-N Penicillin G Chemical compound N([C@H]1[C@H]2SC([C@@H](N2C1=O)C(O)=O)(C)C)C(=O)CC1=CC=CC=C1 JGSARLDLIJGVTE-MBNYWOFBSA-N 0.000 description 2
- ISWSIDIOOBJBQZ-UHFFFAOYSA-N Phenol Natural products OC1=CC=CC=C1 ISWSIDIOOBJBQZ-UHFFFAOYSA-N 0.000 description 2
- 102100021768 Phosphoserine aminotransferase Human genes 0.000 description 2
- 102000010780 Platelet-Derived Growth Factor Human genes 0.000 description 2
- 108010038512 Platelet-Derived Growth Factor Proteins 0.000 description 2
- 241000288906 Primates Species 0.000 description 2
- 108010072866 Prostate-Specific Antigen Proteins 0.000 description 2
- 241000700159 Rattus Species 0.000 description 2
- 102100030086 Receptor tyrosine-protein kinase erbB-2 Human genes 0.000 description 2
- 102100037486 Reverse transcriptase/ribonuclease H Human genes 0.000 description 2
- 239000008156 Ringer's lactate solution Substances 0.000 description 2
- 229920002684 Sepharose Polymers 0.000 description 2
- 241000723792 Tobacco etch virus Species 0.000 description 2
- 108090000631 Trypsin Proteins 0.000 description 2
- 102000004142 Trypsin Human genes 0.000 description 2
- 208000006105 Uterine Cervical Neoplasms Diseases 0.000 description 2
- 108010067390 Viral Proteins Proteins 0.000 description 2
- 208000036142 Viral infection Diseases 0.000 description 2
- 208000020329 Zika virus infectious disease Diseases 0.000 description 2
- 238000007792 addition Methods 0.000 description 2
- 230000001464 adherent effect Effects 0.000 description 2
- HSFWRNGVRCDJHI-UHFFFAOYSA-N alpha-acetylene Natural products C#C HSFWRNGVRCDJHI-UHFFFAOYSA-N 0.000 description 2
- 239000003963 antioxidant agent Substances 0.000 description 2
- 235000006708 antioxidants Nutrition 0.000 description 2
- 230000005975 antitumor immune response Effects 0.000 description 2
- 230000007416 antiviral immune response Effects 0.000 description 2
- 230000006472 autoimmune response Effects 0.000 description 2
- 150000001540 azides Chemical class 0.000 description 2
- 108091008324 binding proteins Proteins 0.000 description 2
- 230000000981 bystander Effects 0.000 description 2
- 239000001110 calcium chloride Substances 0.000 description 2
- 229910001628 calcium chloride Inorganic materials 0.000 description 2
- 235000011148 calcium chloride Nutrition 0.000 description 2
- 238000002619 cancer immunotherapy Methods 0.000 description 2
- YCIMNLLNPGFGHC-UHFFFAOYSA-N catechol Chemical compound OC1=CC=CC=C1O YCIMNLLNPGFGHC-UHFFFAOYSA-N 0.000 description 2
- 230000022534 cell killing Effects 0.000 description 2
- 230000005859 cell recognition Effects 0.000 description 2
- 230000001413 cellular effect Effects 0.000 description 2
- 201000010881 cervical cancer Diseases 0.000 description 2
- 230000008859 change Effects 0.000 description 2
- 238000007796 conventional method Methods 0.000 description 2
- 230000008878 coupling Effects 0.000 description 2
- 238000010168 coupling process Methods 0.000 description 2
- 238000005859 coupling reaction Methods 0.000 description 2
- 230000009089 cytolysis Effects 0.000 description 2
- 210000001151 cytotoxic T lymphocyte Anatomy 0.000 description 2
- 230000003111 delayed effect Effects 0.000 description 2
- 238000012217 deletion Methods 0.000 description 2
- 230000037430 deletion Effects 0.000 description 2
- 230000004069 differentiation Effects 0.000 description 2
- 239000000539 dimer Substances 0.000 description 2
- 231100000673 dose–response relationship Toxicity 0.000 description 2
- 238000004520 electroporation Methods 0.000 description 2
- 210000003527 eukaryotic cell Anatomy 0.000 description 2
- 239000007850 fluorescent dye Substances 0.000 description 2
- 125000000524 functional group Chemical group 0.000 description 2
- 238000001502 gel electrophoresis Methods 0.000 description 2
- 239000012274 immune-checkpoint protein inhibitor Substances 0.000 description 2
- 230000003053 immunization Effects 0.000 description 2
- 238000002649 immunization Methods 0.000 description 2
- 229940127121 immunoconjugate Drugs 0.000 description 2
- 230000001024 immunotherapeutic effect Effects 0.000 description 2
- 238000002347 injection Methods 0.000 description 2
- 239000007924 injection Substances 0.000 description 2
- 229940079322 interferon Drugs 0.000 description 2
- 238000002372 labelling Methods 0.000 description 2
- 210000004698 lymphocyte Anatomy 0.000 description 2
- RLSSMJSEOOYNOY-UHFFFAOYSA-N m-cresol Chemical compound CC1=CC=CC(O)=C1 RLSSMJSEOOYNOY-UHFFFAOYSA-N 0.000 description 2
- 210000002540 macrophage Anatomy 0.000 description 2
- 229910052751 metal Inorganic materials 0.000 description 2
- 239000002184 metal Substances 0.000 description 2
- 125000002496 methyl group Chemical group [H]C([H])([H])* 0.000 description 2
- 230000004048 modification Effects 0.000 description 2
- 238000012986 modification Methods 0.000 description 2
- 239000003068 molecular probe Substances 0.000 description 2
- 210000001616 monocyte Anatomy 0.000 description 2
- 238000002703 mutagenesis Methods 0.000 description 2
- 231100000350 mutagenesis Toxicity 0.000 description 2
- 239000013642 negative control Substances 0.000 description 2
- 229940049954 penicillin Drugs 0.000 description 2
- 230000000144 pharmacologic effect Effects 0.000 description 2
- 238000002360 preparation method Methods 0.000 description 2
- 238000011321 prophylaxis Methods 0.000 description 2
- QELSKZZBTMNZEB-UHFFFAOYSA-N propylparaben Chemical compound CCCOC(=O)C1=CC=C(O)C=C1 QELSKZZBTMNZEB-UHFFFAOYSA-N 0.000 description 2
- 235000019419 proteases Nutrition 0.000 description 2
- RXWNCPJZOCPEPQ-NVWDDTSBSA-N puromycin Chemical compound C1=CC(OC)=CC=C1C[C@H](N)C(=O)N[C@H]1[C@@H](O)[C@H](N2C3=NC=NC(=C3N=C2)N(C)C)O[C@@H]1CO RXWNCPJZOCPEPQ-NVWDDTSBSA-N 0.000 description 2
- 230000010076 replication Effects 0.000 description 2
- GHMLBKRAJCXXBS-UHFFFAOYSA-N resorcinol Chemical compound OC1=CC=CC(O)=C1 GHMLBKRAJCXXBS-UHFFFAOYSA-N 0.000 description 2
- 150000003839 salts Chemical class 0.000 description 2
- 239000011780 sodium chloride Substances 0.000 description 2
- 230000009870 specific binding Effects 0.000 description 2
- 239000003381 stabilizer Substances 0.000 description 2
- 235000000346 sugar Nutrition 0.000 description 2
- 150000008163 sugars Chemical class 0.000 description 2
- 239000006228 supernatant Substances 0.000 description 2
- 230000004083 survival effect Effects 0.000 description 2
- 230000009885 systemic effect Effects 0.000 description 2
- 238000002054 transplantation Methods 0.000 description 2
- 239000012588 trypsin Substances 0.000 description 2
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 2
- 230000003442 weekly effect Effects 0.000 description 2
- ORQXBVXKBGUSBA-QMMMGPOBSA-N β-cyclohexyl-alanine Chemical compound OC(=O)[C@@H](N)CC1CCCCC1 ORQXBVXKBGUSBA-QMMMGPOBSA-N 0.000 description 2
- HDTRYLNUVZCQOY-UHFFFAOYSA-N α-D-glucopyranosyl-α-D-glucopyranoside Natural products OC1C(O)C(O)C(CO)OC1OC1C(O)C(O)C(O)C(CO)O1 HDTRYLNUVZCQOY-UHFFFAOYSA-N 0.000 description 1
- SSOORFWOBGFTHL-OTEJMHTDSA-N (4S)-5-[[(2S)-1-[[(2S)-1-[[(2S)-1-[[(2S)-1-[[(2S)-1-[[(2S)-1-[[(2S)-1-[[(2S)-6-amino-1-[[(2S)-1-[[(2S)-1-[[(2S)-1-[[(2S)-1-[[2-[(2S)-2-[[(2S)-1-[[(2S)-1-[[(2S)-1-[[(2S)-1-[[(2S)-1-[[(2S)-1-[[(2S)-6-amino-1-[[(2S)-1-[[(2S)-1-[[(2S,3S)-1-[[(2S)-1-[[(2S)-1-[[(2S)-6-amino-1-[[(2S)-1-[[(2S)-1-[[(2S)-1-[[(2S)-1-[[(2S)-1-[[(2S)-5-amino-1-[[(2S)-1-[[(2S)-1-[[(2S)-6-amino-1-[[(2S)-6-amino-1-[[(2S)-1-[[(2S)-1-[[(2S)-5-amino-1-[[(2S)-5-carbamimidamido-1-[[(2S)-5-carbamimidamido-1-[[(1S)-4-carbamimidamido-1-carboxybutyl]amino]-1-oxopentan-2-yl]amino]-1-oxopentan-2-yl]amino]-1,5-dioxopentan-2-yl]amino]-5-carbamimidamido-1-oxopentan-2-yl]amino]-5-carbamimidamido-1-oxopentan-2-yl]amino]-1-oxohexan-2-yl]amino]-1-oxohexan-2-yl]amino]-5-carbamimidamido-1-oxopentan-2-yl]amino]-4-methyl-1-oxopentan-2-yl]amino]-1,5-dioxopentan-2-yl]amino]-4-methyl-1-oxopentan-2-yl]amino]-3-hydroxy-1-oxopropan-2-yl]amino]-3-hydroxy-1-oxopropan-2-yl]amino]-3-hydroxy-1-oxopropan-2-yl]amino]-1-oxopropan-2-yl]amino]-1-oxohexan-2-yl]amino]-3-hydroxy-1-oxopropan-2-yl]amino]-1-oxo-3-phenylpropan-2-yl]amino]-3-methyl-1-oxopentan-2-yl]amino]-3-methyl-1-oxobutan-2-yl]amino]-5-carbamimidamido-1-oxopentan-2-yl]amino]-1-oxohexan-2-yl]amino]-3-methyl-1-oxobutan-2-yl]amino]-5-carbamimidamido-1-oxopentan-2-yl]amino]-3-methyl-1-oxobutan-2-yl]amino]-4-methyl-1-oxopentan-2-yl]amino]-1-oxopropan-2-yl]amino]-5-carbamimidamido-1-oxopentan-2-yl]carbamoyl]pyrrolidin-1-yl]-2-oxoethyl]amino]-3-(1H-indol-3-yl)-1-oxopropan-2-yl]amino]-4-methyl-1-oxopentan-2-yl]amino]-1-oxo-3-phenylpropan-2-yl]amino]-5-carbamimidamido-1-oxopentan-2-yl]amino]-1-oxohexan-2-yl]amino]-3-methyl-1-oxobutan-2-yl]amino]-5-carbamimidamido-1-oxopentan-2-yl]amino]-4-methyl-1-oxopentan-2-yl]amino]-1-oxo-3-phenylpropan-2-yl]amino]-3-(1H-imidazol-4-yl)-1-oxopropan-2-yl]amino]-3-methyl-1-oxobutan-2-yl]amino]-4-methyl-1-oxopentan-2-yl]amino]-4-[[(2S)-2-[[(2S)-2-[[(2S)-2,6-diaminohexanoyl]amino]-3-methylbutanoyl]amino]propanoyl]amino]-5-oxopentanoic acid Chemical compound CC[C@H](C)[C@H](NC(=O)[C@@H](NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CCCCN)NC(=O)[C@@H](NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@@H](NC(=O)[C@H](CC(C)C)NC(=O)[C@H](C)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@@H]1CCCN1C(=O)CNC(=O)[C@H](Cc1c[nH]c2ccccc12)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](Cc1ccccc1)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CCCCN)NC(=O)[C@@H](NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](Cc1ccccc1)NC(=O)[C@H](Cc1c[nH]cn1)NC(=O)[C@@H](NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](NC(=O)[C@@H](N)CCCCN)C(C)C)C(C)C)C(C)C)C(C)C)C(C)C)C(C)C)C(=O)N[C@@H](Cc1ccccc1)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C)C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O SSOORFWOBGFTHL-OTEJMHTDSA-N 0.000 description 1
- OWEGMIWEEQEYGQ-UHFFFAOYSA-N 100676-05-9 Natural products OC1C(O)C(O)C(CO)OC1OCC1C(O)C(O)C(O)C(OC2C(OC(O)C(O)C2O)CO)O1 OWEGMIWEEQEYGQ-UHFFFAOYSA-N 0.000 description 1
- LKKMLIBUAXYLOY-UHFFFAOYSA-N 3-Amino-1-methyl-5H-pyrido[4,3-b]indole Chemical compound N1C2=CC=CC=C2C2=C1C=C(N)N=C2C LKKMLIBUAXYLOY-UHFFFAOYSA-N 0.000 description 1
- 102100030310 5,6-dihydroxyindole-2-carboxylic acid oxidase Human genes 0.000 description 1
- 101710163881 5,6-dihydroxyindole-2-carboxylic acid oxidase Proteins 0.000 description 1
- YMZMTOFQCVHHFB-UHFFFAOYSA-N 5-carboxytetramethylrhodamine Chemical compound C=12C=CC(N(C)C)=CC2=[O+]C2=CC(N(C)C)=CC=C2C=1C1=CC=C(C(O)=O)C=C1C([O-])=O YMZMTOFQCVHHFB-UHFFFAOYSA-N 0.000 description 1
- 101710163573 5-hydroxyisourate hydrolase Proteins 0.000 description 1
- 102100031585 ADP-ribosyl cyclase/cyclic ADP-ribose hydrolase 1 Human genes 0.000 description 1
- 102100030840 AT-rich interactive domain-containing protein 4B Human genes 0.000 description 1
- 108010006533 ATP-Binding Cassette Transporters Proteins 0.000 description 1
- 102000005416 ATP-Binding Cassette Transporters Human genes 0.000 description 1
- 102100033350 ATP-dependent translocase ABCB1 Human genes 0.000 description 1
- RZVAJINKPMORJF-UHFFFAOYSA-N Acetaminophen Chemical class CC(=O)NC1=CC=C(O)C=C1 RZVAJINKPMORJF-UHFFFAOYSA-N 0.000 description 1
- 102100023635 Alpha-fetoprotein Human genes 0.000 description 1
- 101100181128 Arabidopsis thaliana KIN14F gene Proteins 0.000 description 1
- 239000004475 Arginine Substances 0.000 description 1
- DCXYFEDJOCDNAF-UHFFFAOYSA-N Asparagine Natural products OC(=O)C(N)CC(N)=O DCXYFEDJOCDNAF-UHFFFAOYSA-N 0.000 description 1
- 102100024222 B-lymphocyte antigen CD19 Human genes 0.000 description 1
- 241000894006 Bacteria Species 0.000 description 1
- 102100032412 Basigin Human genes 0.000 description 1
- 208000009137 Behcet syndrome Diseases 0.000 description 1
- 102100027314 Beta-2-microglobulin Human genes 0.000 description 1
- 108060000903 Beta-catenin Proteins 0.000 description 1
- 102000015735 Beta-catenin Human genes 0.000 description 1
- 206010004593 Bile duct cancer Diseases 0.000 description 1
- 206010005003 Bladder cancer Diseases 0.000 description 1
- 206010005949 Bone cancer Diseases 0.000 description 1
- 208000018084 Bone neoplasm Diseases 0.000 description 1
- 241000283690 Bos taurus Species 0.000 description 1
- 208000003174 Brain Neoplasms Diseases 0.000 description 1
- 102100022595 Broad substrate specificity ATP-binding cassette transporter ABCG2 Human genes 0.000 description 1
- 102100028668 C-type lectin domain family 4 member C Human genes 0.000 description 1
- 101710183165 C-type lectin domain family 4 member K Proteins 0.000 description 1
- 102100039521 C-type lectin domain family 9 member A Human genes 0.000 description 1
- 102100032912 CD44 antigen Human genes 0.000 description 1
- 101100005789 Caenorhabditis elegans cdk-4 gene Proteins 0.000 description 1
- 102100024423 Carbonic anhydrase 9 Human genes 0.000 description 1
- 102100026548 Caspase-8 Human genes 0.000 description 1
- 241000700198 Cavia Species 0.000 description 1
- 201000006082 Chickenpox Diseases 0.000 description 1
- VEXZGXHMUGYJMC-UHFFFAOYSA-M Chloride anion Chemical compound [Cl-] VEXZGXHMUGYJMC-UHFFFAOYSA-M 0.000 description 1
- 241000579895 Chlorostilbon Species 0.000 description 1
- 108090000317 Chymotrypsin Proteins 0.000 description 1
- 102100035167 Coiled-coil domain-containing protein 54 Human genes 0.000 description 1
- 241000699800 Cricetinae Species 0.000 description 1
- 108010072210 Cyclophilin C Proteins 0.000 description 1
- FBPFZTCFMRRESA-FSIIMWSLSA-N D-Glucitol Natural products OC[C@H](O)[C@H](O)[C@@H](O)[C@H](O)CO FBPFZTCFMRRESA-FSIIMWSLSA-N 0.000 description 1
- FBPFZTCFMRRESA-KVTDHHQDSA-N D-Mannitol Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-KVTDHHQDSA-N 0.000 description 1
- FBPFZTCFMRRESA-JGWLITMVSA-N D-glucitol Chemical compound OC[C@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-JGWLITMVSA-N 0.000 description 1
- WQZGKKKJIJFFOK-QTVWNMPRSA-N D-mannopyranose Chemical compound OC[C@H]1OC(O)[C@@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-QTVWNMPRSA-N 0.000 description 1
- 108020004414 DNA Proteins 0.000 description 1
- 241000702421 Dependoparvovirus Species 0.000 description 1
- 229920002307 Dextran Polymers 0.000 description 1
- 239000004375 Dextrin Substances 0.000 description 1
- 229920001353 Dextrin Polymers 0.000 description 1
- 101100216227 Dictyostelium discoideum anapc3 gene Proteins 0.000 description 1
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 1
- 201000011001 Ebola Hemorrhagic Fever Diseases 0.000 description 1
- 241001115402 Ebolavirus Species 0.000 description 1
- 102000004190 Enzymes Human genes 0.000 description 1
- 108090000790 Enzymes Proteins 0.000 description 1
- 102000018651 Epithelial Cell Adhesion Molecule Human genes 0.000 description 1
- 108010066687 Epithelial Cell Adhesion Molecule Proteins 0.000 description 1
- 108010069621 Epstein-Barr virus EBV-associated membrane antigen Proteins 0.000 description 1
- 208000000461 Esophageal Neoplasms Diseases 0.000 description 1
- XZWYTXMRWQJBGX-VXBMVYAYSA-N FLAG peptide Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CCCCN)NC(=O)[C@@H](NC(=O)[C@@H](N)CC(O)=O)CC1=CC=C(O)C=C1 XZWYTXMRWQJBGX-VXBMVYAYSA-N 0.000 description 1
- 108010074860 Factor Xa Proteins 0.000 description 1
- 108010087819 Fc receptors Proteins 0.000 description 1
- 102000009109 Fc receptors Human genes 0.000 description 1
- 102100028073 Fibroblast growth factor 5 Human genes 0.000 description 1
- 108090000380 Fibroblast growth factor 5 Proteins 0.000 description 1
- 102100039699 G antigen 4 Human genes 0.000 description 1
- 101710092267 G antigen 5 Proteins 0.000 description 1
- 102100039698 G antigen 5 Human genes 0.000 description 1
- 101710092269 G antigen 6 Proteins 0.000 description 1
- 102100039713 G antigen 6 Human genes 0.000 description 1
- 108091005973 GFP derivatives Proteins 0.000 description 1
- 102100039788 GTPase NRas Human genes 0.000 description 1
- 108010012015 GVYDGREHTV Proteins 0.000 description 1
- 102100040510 Galectin-3-binding protein Human genes 0.000 description 1
- 101710197901 Galectin-3-binding protein Proteins 0.000 description 1
- 108010010803 Gelatin Proteins 0.000 description 1
- 108700028146 Genetic Enhancer Elements Proteins 0.000 description 1
- 108700039691 Genetic Promoter Regions Proteins 0.000 description 1
- 108700007698 Genetic Terminator Regions Proteins 0.000 description 1
- YLJHCWNDBKKOEB-IHRRRGAJSA-N Glu-Glu-Phe Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O YLJHCWNDBKKOEB-IHRRRGAJSA-N 0.000 description 1
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 1
- 108010070675 Glutathione transferase Proteins 0.000 description 1
- 241000282575 Gorilla Species 0.000 description 1
- 206010053759 Growth retardation Diseases 0.000 description 1
- 102100028972 HLA class I histocompatibility antigen, A alpha chain Human genes 0.000 description 1
- 102100028971 HLA class I histocompatibility antigen, C alpha chain Human genes 0.000 description 1
- 108010036972 HLA-A11 Antigen Proteins 0.000 description 1
- 108010034115 HLA-A29 antigen Proteins 0.000 description 1
- 108010021736 HLA-B15 Antigen Proteins 0.000 description 1
- 108010061486 HLA-B27 Antigen Proteins 0.000 description 1
- 102000012153 HLA-B27 Antigen Human genes 0.000 description 1
- 108010086707 HLA-B38 Antigen Proteins 0.000 description 1
- 108010033369 HLA-B57 antigen Proteins 0.000 description 1
- 108010052199 HLA-C Antigens Proteins 0.000 description 1
- 108010059045 HLA-C*06 antigen Proteins 0.000 description 1
- 102100029100 Hematopoietic prostaglandin D synthase Human genes 0.000 description 1
- 241000238631 Hexapoda Species 0.000 description 1
- 208000017604 Hodgkin disease Diseases 0.000 description 1
- 208000021519 Hodgkin lymphoma Diseases 0.000 description 1
- 208000010747 Hodgkins lymphoma Diseases 0.000 description 1
- 241001272567 Hominoidea Species 0.000 description 1
- 101000777636 Homo sapiens ADP-ribosyl cyclase/cyclic ADP-ribose hydrolase 1 Proteins 0.000 description 1
- 101000792935 Homo sapiens AT-rich interactive domain-containing protein 4B Proteins 0.000 description 1
- 101001017818 Homo sapiens ATP-dependent translocase ABCB1 Proteins 0.000 description 1
- 101000980825 Homo sapiens B-lymphocyte antigen CD19 Proteins 0.000 description 1
- 101000798441 Homo sapiens Basigin Proteins 0.000 description 1
- 101000823298 Homo sapiens Broad substrate specificity ATP-binding cassette transporter ABCG2 Proteins 0.000 description 1
- 101000766907 Homo sapiens C-type lectin domain family 4 member C Proteins 0.000 description 1
- 101000888548 Homo sapiens C-type lectin domain family 9 member A Proteins 0.000 description 1
- 101100165850 Homo sapiens CA9 gene Proteins 0.000 description 1
- 101000868273 Homo sapiens CD44 antigen Proteins 0.000 description 1
- 101000914324 Homo sapiens Carcinoembryonic antigen-related cell adhesion molecule 5 Proteins 0.000 description 1
- 101000914321 Homo sapiens Carcinoembryonic antigen-related cell adhesion molecule 7 Proteins 0.000 description 1
- 101000737052 Homo sapiens Coiled-coil domain-containing protein 54 Proteins 0.000 description 1
- 101000866749 Homo sapiens Elongation factor 2 Proteins 0.000 description 1
- 101000886678 Homo sapiens G antigen 2D Proteins 0.000 description 1
- 101000886136 Homo sapiens G antigen 4 Proteins 0.000 description 1
- 101000744505 Homo sapiens GTPase NRas Proteins 0.000 description 1
- 101100395310 Homo sapiens HLA-A gene Proteins 0.000 description 1
- 101100395311 Homo sapiens HLA-B gene Proteins 0.000 description 1
- 101001014223 Homo sapiens MAPK/MAK/MRK overlapping kinase Proteins 0.000 description 1
- 101001057156 Homo sapiens Melanoma-associated antigen C2 Proteins 0.000 description 1
- 101000946889 Homo sapiens Monocyte differentiation antigen CD14 Proteins 0.000 description 1
- 101000610208 Homo sapiens Poly(A) polymerase gamma Proteins 0.000 description 1
- 101000617725 Homo sapiens Pregnancy-specific beta-1-glycoprotein 2 Proteins 0.000 description 1
- 101000880770 Homo sapiens Protein SSX2 Proteins 0.000 description 1
- 101001012157 Homo sapiens Receptor tyrosine-protein kinase erbB-2 Proteins 0.000 description 1
- 101001062222 Homo sapiens Receptor-binding cancer antigen expressed on SiSo cells Proteins 0.000 description 1
- 101000591201 Homo sapiens Receptor-type tyrosine-protein phosphatase kappa Proteins 0.000 description 1
- 101001073409 Homo sapiens Retrotransposon-derived protein PEG10 Proteins 0.000 description 1
- 101001094545 Homo sapiens Retrotransposon-like protein 1 Proteins 0.000 description 1
- 101000973629 Homo sapiens Ribosome quality control complex subunit NEMF Proteins 0.000 description 1
- 101000824971 Homo sapiens Sperm surface protein Sp17 Proteins 0.000 description 1
- 101000946843 Homo sapiens T-cell surface glycoprotein CD8 alpha chain Proteins 0.000 description 1
- 101000847107 Homo sapiens Tetraspanin-8 Proteins 0.000 description 1
- 101000848653 Homo sapiens Tripartite motif-containing protein 26 Proteins 0.000 description 1
- 101000671653 Homo sapiens U3 small nucleolar RNA-associated protein 14 homolog A Proteins 0.000 description 1
- 241000701044 Human gammaherpesvirus 4 Species 0.000 description 1
- 108010052919 Hydroxyethylthiazole kinase Proteins 0.000 description 1
- 108010027436 Hydroxymethylpyrimidine kinase Proteins 0.000 description 1
- 241000282596 Hylobatidae Species 0.000 description 1
- 206010020751 Hypersensitivity Diseases 0.000 description 1
- 102000009438 IgE Receptors Human genes 0.000 description 1
- 108010073816 IgE Receptors Proteins 0.000 description 1
- DGAQECJNVWCQMB-PUAWFVPOSA-M Ilexoside XXIX Chemical class C[C@@H]1CC[C@@]2(CC[C@@]3(C(=CC[C@H]4[C@]3(CC[C@@H]5[C@@]4(CC[C@@H](C5(C)C)OS(=O)(=O)[O-])C)C)[C@@H]2[C@]1(C)O)C)C(=O)O[C@H]6[C@@H]([C@H]([C@@H]([C@H](O6)CO)O)O)O.[Na+] DGAQECJNVWCQMB-PUAWFVPOSA-M 0.000 description 1
- 108060003951 Immunoglobulin Proteins 0.000 description 1
- 108010067060 Immunoglobulin Variable Region Proteins 0.000 description 1
- 102000017727 Immunoglobulin Variable Region Human genes 0.000 description 1
- 206010061218 Inflammation Diseases 0.000 description 1
- 108010030506 Integrin alpha6beta4 Proteins 0.000 description 1
- 108010014726 Interferon Type I Proteins 0.000 description 1
- 102000002227 Interferon Type I Human genes 0.000 description 1
- 208000008839 Kidney Neoplasms Diseases 0.000 description 1
- 241000235058 Komagataella pastoris Species 0.000 description 1
- ODKSFYDXXFIFQN-BYPYZUCNSA-P L-argininium(2+) Chemical compound NC(=[NH2+])NCCC[C@H]([NH3+])C(O)=O ODKSFYDXXFIFQN-BYPYZUCNSA-P 0.000 description 1
- DCXYFEDJOCDNAF-REOHCLBHSA-N L-asparagine Chemical compound OC(=O)[C@@H](N)CC(N)=O DCXYFEDJOCDNAF-REOHCLBHSA-N 0.000 description 1
- 102100031413 L-dopachrome tautomerase Human genes 0.000 description 1
- 101710093778 L-dopachrome tautomerase Proteins 0.000 description 1
- WHUUTDBJXJRKMK-VKHMYHEASA-N L-glutamic acid Chemical compound OC(=O)[C@@H](N)CCC(O)=O WHUUTDBJXJRKMK-VKHMYHEASA-N 0.000 description 1
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 1
- HNDVDQJCIGZPNO-YFKPBYRVSA-N L-histidine Chemical compound OC(=O)[C@@H](N)CC1=CN=CN1 HNDVDQJCIGZPNO-YFKPBYRVSA-N 0.000 description 1
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 1
- KDXKERNSBIXSRK-YFKPBYRVSA-N L-lysine Chemical compound NCCCC[C@H](N)C(O)=O KDXKERNSBIXSRK-YFKPBYRVSA-N 0.000 description 1
- FFEARJCKVFRZRR-BYPYZUCNSA-N L-methionine Chemical compound CSCC[C@H](N)C(O)=O FFEARJCKVFRZRR-BYPYZUCNSA-N 0.000 description 1
- 208000032420 Latent Infection Diseases 0.000 description 1
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 1
- 206010058467 Lung neoplasm malignant Diseases 0.000 description 1
- 108010059255 MAGE-A10 antigen Proteins 0.000 description 1
- 102100031520 MAPK/MAK/MRK overlapping kinase Human genes 0.000 description 1
- PEEHTFAAVSWFBL-UHFFFAOYSA-N Maleimide Chemical compound O=C1NC(=O)C=C1 PEEHTFAAVSWFBL-UHFFFAOYSA-N 0.000 description 1
- GUBGYTABKSRVRQ-PICCSMPSSA-N Maltose Natural products O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@@H](CO)OC(O)[C@H](O)[C@H]1O GUBGYTABKSRVRQ-PICCSMPSSA-N 0.000 description 1
- 102100027252 Melanoma-associated antigen C2 Human genes 0.000 description 1
- 102000003939 Membrane transport proteins Human genes 0.000 description 1
- 108090000301 Membrane transport proteins Proteins 0.000 description 1
- 206010027476 Metastases Diseases 0.000 description 1
- 102100024193 Mitogen-activated protein kinase 1 Human genes 0.000 description 1
- 101710166115 Mitogen-activated protein kinase 2 Proteins 0.000 description 1
- 101710166076 Mitogen-activated protein kinase 5 Proteins 0.000 description 1
- 108010006519 Molecular Chaperones Proteins 0.000 description 1
- 102100035877 Monocyte differentiation antigen CD14 Human genes 0.000 description 1
- 102100021339 Multidrug resistance-associated protein 1 Human genes 0.000 description 1
- 208000034578 Multiple myelomas Diseases 0.000 description 1
- 208000005647 Mumps Diseases 0.000 description 1
- 101100328099 Mus musculus Clec9a gene Proteins 0.000 description 1
- 101710135898 Myc proto-oncogene protein Proteins 0.000 description 1
- 102100038895 Myc proto-oncogene protein Human genes 0.000 description 1
- 102000003505 Myosin Human genes 0.000 description 1
- 108060008487 Myosin Chemical class 0.000 description 1
- 108010069196 Neural Cell Adhesion Molecules Proteins 0.000 description 1
- 102100027347 Neural cell adhesion molecule 1 Human genes 0.000 description 1
- 102000005348 Neuraminidase Human genes 0.000 description 1
- 108010006232 Neuraminidase Proteins 0.000 description 1
- 206010030155 Oesophageal carcinoma Diseases 0.000 description 1
- 206010033128 Ovarian cancer Diseases 0.000 description 1
- 206010061535 Ovarian neoplasm Diseases 0.000 description 1
- 208000012868 Overgrowth Diseases 0.000 description 1
- 229910019142 PO4 Inorganic materials 0.000 description 1
- 102100034640 PWWP domain-containing DNA repair factor 3A Human genes 0.000 description 1
- 108050007154 PWWP domain-containing DNA repair factor 3A Proteins 0.000 description 1
- 241000282579 Pan Species 0.000 description 1
- 206010061902 Pancreatic neoplasm Diseases 0.000 description 1
- 108090000526 Papain Proteins 0.000 description 1
- 102100024968 Peptidyl-prolyl cis-trans isomerase C Human genes 0.000 description 1
- 108010004729 Phycoerythrin Proteins 0.000 description 1
- 206010035226 Plasma cell myeloma Diseases 0.000 description 1
- 208000000474 Poliomyelitis Diseases 0.000 description 1
- 102100040153 Poly(A) polymerase gamma Human genes 0.000 description 1
- 239000002202 Polyethylene glycol Substances 0.000 description 1
- 229920001213 Polysorbate 20 Polymers 0.000 description 1
- 102100022019 Pregnancy-specific beta-1-glycoprotein 2 Human genes 0.000 description 1
- 206010060862 Prostate cancer Diseases 0.000 description 1
- 208000000236 Prostatic Neoplasms Diseases 0.000 description 1
- 101710118538 Protease Proteins 0.000 description 1
- 229940124158 Protease/peptidase inhibitor Drugs 0.000 description 1
- 108010029485 Protein Isoforms Proteins 0.000 description 1
- 102000001708 Protein Isoforms Human genes 0.000 description 1
- 102100037686 Protein SSX2 Human genes 0.000 description 1
- 201000004681 Psoriasis Diseases 0.000 description 1
- 201000001263 Psoriatic Arthritis Diseases 0.000 description 1
- 208000036824 Psoriatic arthropathy Diseases 0.000 description 1
- 101710100968 Receptor tyrosine-protein kinase erbB-2 Proteins 0.000 description 1
- 102100029165 Receptor-binding cancer antigen expressed on SiSo cells Human genes 0.000 description 1
- 102100034089 Receptor-type tyrosine-protein phosphatase kappa Human genes 0.000 description 1
- 208000015634 Rectal Neoplasms Diseases 0.000 description 1
- 206010038389 Renal cancer Diseases 0.000 description 1
- 102100035844 Retrotransposon-derived protein PEG10 Human genes 0.000 description 1
- 102100022213 Ribosome quality control complex subunit NEMF Human genes 0.000 description 1
- 241000283984 Rodentia Species 0.000 description 1
- 206010039491 Sarcoma Diseases 0.000 description 1
- 108010071390 Serum Albumin Proteins 0.000 description 1
- 102000007562 Serum Albumin Human genes 0.000 description 1
- 101710173693 Short transient receptor potential channel 1 Proteins 0.000 description 1
- 101710173694 Short transient receptor potential channel 2 Proteins 0.000 description 1
- 208000000453 Skin Neoplasms Diseases 0.000 description 1
- 241000713880 Spleen focus-forming virus Species 0.000 description 1
- 208000006045 Spondylarthropathies Diseases 0.000 description 1
- 208000005718 Stomach Neoplasms Diseases 0.000 description 1
- 244000057717 Streptococcus lactis Species 0.000 description 1
- 235000014897 Streptococcus lactis Nutrition 0.000 description 1
- 241000282887 Suidae Species 0.000 description 1
- 230000006044 T cell activation Effects 0.000 description 1
- 102100034922 T-cell surface glycoprotein CD8 alpha chain Human genes 0.000 description 1
- 108700019889 TEL-AML1 fusion Proteins 0.000 description 1
- 101150031162 TM4SF1 gene Proteins 0.000 description 1
- 102100032802 Tetraspanin-8 Human genes 0.000 description 1
- 108090000190 Thrombin Proteins 0.000 description 1
- 208000024770 Thyroid neoplasm Diseases 0.000 description 1
- 101710150448 Transcriptional regulator Myc Proteins 0.000 description 1
- 108010033576 Transferrin Receptors Proteins 0.000 description 1
- 102100026144 Transferrin receptor protein 1 Human genes 0.000 description 1
- 102100034902 Transmembrane 4 L6 family member 1 Human genes 0.000 description 1
- HDTRYLNUVZCQOY-WSWWMNSNSA-N Trehalose Natural products O[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@H](O)[C@@H](O)[C@@H](O)[C@@H](CO)O1 HDTRYLNUVZCQOY-WSWWMNSNSA-N 0.000 description 1
- 241000255993 Trichoplusia ni Species 0.000 description 1
- 108700015934 Triose-phosphate isomerases Proteins 0.000 description 1
- 102100033598 Triosephosphate isomerase Human genes 0.000 description 1
- 239000013504 Triton X-100 Substances 0.000 description 1
- 229920004890 Triton X-100 Polymers 0.000 description 1
- LVTKHGUGBGNBPL-UHFFFAOYSA-N Trp-P-1 Chemical compound N1C2=CC=CC=C2C2=C1C(C)=C(N)N=C2C LVTKHGUGBGNBPL-UHFFFAOYSA-N 0.000 description 1
- 108010028230 Trp-Ser- His-Pro-Gln-Phe-Glu-Lys Proteins 0.000 description 1
- 206010053613 Type IV hypersensitivity reaction Diseases 0.000 description 1
- 102100039094 Tyrosinase Human genes 0.000 description 1
- 108060008724 Tyrosinase Proteins 0.000 description 1
- 102100040099 U3 small nucleolar RNA-associated protein 14 homolog A Human genes 0.000 description 1
- HSCJRCZFDFQWRP-JZMIEXBBSA-N UDP-alpha-D-glucose Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1OP(O)(=O)OP(O)(=O)OC[C@@H]1[C@@H](O)[C@@H](O)[C@H](N2C(NC(=O)C=C2)=O)O1 HSCJRCZFDFQWRP-JZMIEXBBSA-N 0.000 description 1
- 108090000848 Ubiquitin Proteins 0.000 description 1
- 102000044159 Ubiquitin Human genes 0.000 description 1
- HSCJRCZFDFQWRP-UHFFFAOYSA-N Uridindiphosphoglukose Natural products OC1C(O)C(O)C(CO)OC1OP(O)(=O)OP(O)(=O)OCC1C(O)C(O)C(N2C(NC(=O)C=C2)=O)O1 HSCJRCZFDFQWRP-UHFFFAOYSA-N 0.000 description 1
- 208000007097 Urinary Bladder Neoplasms Diseases 0.000 description 1
- 208000002495 Uterine Neoplasms Diseases 0.000 description 1
- 206010046980 Varicella Diseases 0.000 description 1
- IXKSXJFAGXLQOQ-XISFHERQSA-N WHWLQLKPGQPMY Chemical compound C([C@@H](C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N1CCC[C@H]1C(=O)NCC(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(O)=O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(O)=O)NC(=O)[C@@H](N)CC=1C2=CC=CC=C2NC=1)C1=CNC=N1 IXKSXJFAGXLQOQ-XISFHERQSA-N 0.000 description 1
- 241000269370 Xenopus <genus> Species 0.000 description 1
- 230000005856 abnormality Effects 0.000 description 1
- QPMSXSBEVQLBIL-CZRHPSIPSA-N ac1mix0p Chemical compound C1=CC=C2N(C[C@H](C)CN(C)C)C3=CC(OC)=CC=C3SC2=C1.O([C@H]1[C@]2(OC)C=CC34C[C@@H]2[C@](C)(O)CCC)C2=C5[C@]41CCN(C)[C@@H]3CC5=CC=C2O QPMSXSBEVQLBIL-CZRHPSIPSA-N 0.000 description 1
- 150000007513 acids Chemical class 0.000 description 1
- 239000000654 additive Substances 0.000 description 1
- 230000000996 additive effect Effects 0.000 description 1
- 208000009956 adenocarcinoma Diseases 0.000 description 1
- 201000005188 adrenal gland cancer Diseases 0.000 description 1
- 208000024447 adrenal gland neoplasm Diseases 0.000 description 1
- 238000001042 affinity chromatography Methods 0.000 description 1
- 230000000735 allogeneic effect Effects 0.000 description 1
- HDTRYLNUVZCQOY-LIZSDCNHSA-N alpha,alpha-trehalose Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 HDTRYLNUVZCQOY-LIZSDCNHSA-N 0.000 description 1
- 108010026331 alpha-Fetoproteins Proteins 0.000 description 1
- 238000010171 animal model Methods 0.000 description 1
- 230000014102 antigen processing and presentation of exogenous peptide antigen via MHC class I Effects 0.000 description 1
- 238000013459 approach Methods 0.000 description 1
- 239000007864 aqueous solution Substances 0.000 description 1
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 1
- 238000003491 array Methods 0.000 description 1
- 238000010420 art technique Methods 0.000 description 1
- 229960005070 ascorbic acid Drugs 0.000 description 1
- 235000010323 ascorbic acid Nutrition 0.000 description 1
- 239000011668 ascorbic acid Substances 0.000 description 1
- 229960001230 asparagine Drugs 0.000 description 1
- 235000009582 asparagine Nutrition 0.000 description 1
- 229960003852 atezolizumab Drugs 0.000 description 1
- 230000002238 attenuated effect Effects 0.000 description 1
- 230000004888 barrier function Effects 0.000 description 1
- 108010056708 bcr-abl Fusion Proteins Proteins 0.000 description 1
- 102000004441 bcr-abl Fusion Proteins Human genes 0.000 description 1
- 229960000686 benzalkonium chloride Drugs 0.000 description 1
- 229960001950 benzethonium chloride Drugs 0.000 description 1
- UREZNYTWGJKWBI-UHFFFAOYSA-M benzethonium chloride Chemical compound [Cl-].C1=CC(C(C)(C)CC(C)(C)C)=CC=C1OCCOCC[N+](C)(C)CC1=CC=CC=C1 UREZNYTWGJKWBI-UHFFFAOYSA-M 0.000 description 1
- 235000019445 benzyl alcohol Nutrition 0.000 description 1
- CADWTSSKOVRVJC-UHFFFAOYSA-N benzyl(dimethyl)azanium;chloride Chemical compound [Cl-].C[NH+](C)CC1=CC=CC=C1 CADWTSSKOVRVJC-UHFFFAOYSA-N 0.000 description 1
- 108010081355 beta 2-Microglobulin Proteins 0.000 description 1
- WQZGKKKJIJFFOK-VFUOTHLCSA-N beta-D-glucose Chemical compound OC[C@H]1O[C@@H](O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-VFUOTHLCSA-N 0.000 description 1
- 230000001588 bifunctional effect Effects 0.000 description 1
- 210000000941 bile Anatomy 0.000 description 1
- 208000026900 bile duct neoplasm Diseases 0.000 description 1
- 230000003115 biocidal effect Effects 0.000 description 1
- 229960002685 biotin Drugs 0.000 description 1
- 235000020958 biotin Nutrition 0.000 description 1
- 239000011616 biotin Substances 0.000 description 1
- 210000001124 body fluid Anatomy 0.000 description 1
- 239000010839 body fluid Substances 0.000 description 1
- UDSAIICHUKSCKT-UHFFFAOYSA-N bromophenol blue Chemical compound C1=C(Br)C(O)=C(Br)C=C1C1(C=2C=C(Br)C(O)=C(Br)C=2)C2=CC=CC=C2S(=O)(=O)O1 UDSAIICHUKSCKT-UHFFFAOYSA-N 0.000 description 1
- LRHPLDYGYMQRHN-UHFFFAOYSA-N butyl alcohol Substances CCCCO LRHPLDYGYMQRHN-UHFFFAOYSA-N 0.000 description 1
- 125000000484 butyl group Chemical group [H]C([*])([H])C([H])([H])C([H])([H])C([H])([H])[H] 0.000 description 1
- 210000004899 c-terminal region Anatomy 0.000 description 1
- 239000001506 calcium phosphate Substances 0.000 description 1
- 229910000389 calcium phosphate Inorganic materials 0.000 description 1
- 235000011010 calcium phosphates Nutrition 0.000 description 1
- 238000009566 cancer vaccine Methods 0.000 description 1
- 229940022399 cancer vaccine Drugs 0.000 description 1
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 description 1
- 230000003197 catalytic effect Effects 0.000 description 1
- 238000004113 cell culture Methods 0.000 description 1
- 230000030570 cellular localization Effects 0.000 description 1
- 208000013557 cerebral hemisphere cancer Diseases 0.000 description 1
- 201000008860 cerebrum cancer Diseases 0.000 description 1
- 239000013522 chelant Substances 0.000 description 1
- 239000002738 chelating agent Substances 0.000 description 1
- 125000003636 chemical group Chemical group 0.000 description 1
- 230000014564 chemokine production Effects 0.000 description 1
- 238000002512 chemotherapy Methods 0.000 description 1
- 208000006990 cholangiocarcinoma Diseases 0.000 description 1
- 229960002376 chymotrypsin Drugs 0.000 description 1
- 150000001860 citric acid derivatives Chemical class 0.000 description 1
- 238000010367 cloning Methods 0.000 description 1
- 230000019771 cognition Effects 0.000 description 1
- 230000000112 colonic effect Effects 0.000 description 1
- 238000002591 computed tomography Methods 0.000 description 1
- 230000021615 conjugation Effects 0.000 description 1
- 239000000356 contaminant Substances 0.000 description 1
- 230000001276 controlling effect Effects 0.000 description 1
- 239000013078 crystal Substances 0.000 description 1
- 208000035250 cutaneous malignant susceptibility to 1 melanoma Diseases 0.000 description 1
- HPXRVTGHNJAIIH-UHFFFAOYSA-N cyclohexanol Chemical compound OC1CCCCC1 HPXRVTGHNJAIIH-UHFFFAOYSA-N 0.000 description 1
- 235000018417 cysteine Nutrition 0.000 description 1
- 238000004163 cytometry Methods 0.000 description 1
- 231100000433 cytotoxic Toxicity 0.000 description 1
- 230000001472 cytotoxic effect Effects 0.000 description 1
- 230000006378 damage Effects 0.000 description 1
- 230000007547 defect Effects 0.000 description 1
- 230000001934 delay Effects 0.000 description 1
- 238000013461 design Methods 0.000 description 1
- 235000019425 dextrin Nutrition 0.000 description 1
- 238000003745 diagnosis Methods 0.000 description 1
- 150000002016 disaccharides Chemical class 0.000 description 1
- 208000037771 disease arising from reactivation of latent virus Diseases 0.000 description 1
- 208000035475 disorder Diseases 0.000 description 1
- 239000002552 dosage form Substances 0.000 description 1
- 229960003722 doxycycline Drugs 0.000 description 1
- XQTWDDCIUJNLTR-CVHRZJFOSA-N doxycycline monohydrate Chemical compound O.O=C1C2=C(O)C=CC=C2[C@H](C)[C@@H]2C1=C(O)[C@]1(O)C(=O)C(C(N)=O)=C(O)[C@@H](N(C)C)[C@@H]1[C@H]2O XQTWDDCIUJNLTR-CVHRZJFOSA-N 0.000 description 1
- 229950009791 durvalumab Drugs 0.000 description 1
- 239000012636 effector Substances 0.000 description 1
- 238000001378 electrochemiluminescence detection Methods 0.000 description 1
- 229910052876 emerald Inorganic materials 0.000 description 1
- 239000010976 emerald Substances 0.000 description 1
- 230000012202 endocytosis Effects 0.000 description 1
- 238000005516 engineering process Methods 0.000 description 1
- 229940088598 enzyme Drugs 0.000 description 1
- 230000008029 eradication Effects 0.000 description 1
- 201000004101 esophageal cancer Diseases 0.000 description 1
- 238000009472 formulation Methods 0.000 description 1
- 206010017758 gastric cancer Diseases 0.000 description 1
- 239000008273 gelatin Substances 0.000 description 1
- 229920000159 gelatin Polymers 0.000 description 1
- 235000019322 gelatine Nutrition 0.000 description 1
- 235000011852 gelatine desserts Nutrition 0.000 description 1
- 239000008103 glucose Substances 0.000 description 1
- 229930195712 glutamate Natural products 0.000 description 1
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Natural products OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 description 1
- 102000035122 glycosylated proteins Human genes 0.000 description 1
- 108091005608 glycosylated proteins Proteins 0.000 description 1
- 108010072094 gp100(280-288) melanoma antigen peptide Proteins 0.000 description 1
- 230000012010 growth Effects 0.000 description 1
- 239000001963 growth medium Substances 0.000 description 1
- 229940093915 gynecological organic acid Drugs 0.000 description 1
- 229910052736 halogen Inorganic materials 0.000 description 1
- 150000002367 halogens Chemical class 0.000 description 1
- 201000010536 head and neck cancer Diseases 0.000 description 1
- 208000014829 head and neck neoplasm Diseases 0.000 description 1
- 208000002672 hepatitis B Diseases 0.000 description 1
- 239000000833 heterodimer Substances 0.000 description 1
- 238000004128 high performance liquid chromatography Methods 0.000 description 1
- HNDVDQJCIGZPNO-UHFFFAOYSA-N histidine Natural products OC(=O)C(N)CC1=CN=CN1 HNDVDQJCIGZPNO-UHFFFAOYSA-N 0.000 description 1
- 102000046101 human AFP Human genes 0.000 description 1
- 108010007811 human immunodeficiency virus p17 gag peptide Proteins 0.000 description 1
- 230000028996 humoral immune response Effects 0.000 description 1
- 238000003384 imaging method Methods 0.000 description 1
- 230000005746 immune checkpoint blockade Effects 0.000 description 1
- 230000007256 immune mediated inflammatory response Effects 0.000 description 1
- 230000008073 immune recognition Effects 0.000 description 1
- 210000000987 immune system Anatomy 0.000 description 1
- 230000036039 immunity Effects 0.000 description 1
- 102000018358 immunoglobulin Human genes 0.000 description 1
- 230000001976 improved effect Effects 0.000 description 1
- 230000006872 improvement Effects 0.000 description 1
- 230000004054 inflammatory process Effects 0.000 description 1
- 108010061181 influenza matrix peptide (58-66) Proteins 0.000 description 1
- 238000001802 infusion Methods 0.000 description 1
- 239000004615 ingredient Substances 0.000 description 1
- 230000002401 inhibitory effect Effects 0.000 description 1
- 210000004020 intracellular membrane Anatomy 0.000 description 1
- 238000007918 intramuscular administration Methods 0.000 description 1
- 230000002601 intratumoral effect Effects 0.000 description 1
- 238000001990 intravenous administration Methods 0.000 description 1
- 229940029329 intrinsic factor Drugs 0.000 description 1
- PGLTVOMIXTUURA-UHFFFAOYSA-N iodoacetamide Chemical compound NC(=O)CI PGLTVOMIXTUURA-UHFFFAOYSA-N 0.000 description 1
- 230000007794 irritation Effects 0.000 description 1
- 238000002955 isolation Methods 0.000 description 1
- 201000010982 kidney cancer Diseases 0.000 description 1
- 230000003902 lesion Effects 0.000 description 1
- 208000032839 leukemia Diseases 0.000 description 1
- 210000000265 leukocyte Anatomy 0.000 description 1
- 230000021633 leukocyte mediated immunity Effects 0.000 description 1
- 239000002502 liposome Substances 0.000 description 1
- 239000007788 liquid Substances 0.000 description 1
- 201000007270 liver cancer Diseases 0.000 description 1
- 208000014018 liver neoplasm Diseases 0.000 description 1
- 230000004807 localization Effects 0.000 description 1
- 201000005202 lung cancer Diseases 0.000 description 1
- 208000020816 lung neoplasm Diseases 0.000 description 1
- 230000014759 maintenance of location Effects 0.000 description 1
- 230000003211 malignant effect Effects 0.000 description 1
- 208000015486 malignant pancreatic neoplasm Diseases 0.000 description 1
- 210000004216 mammary stem cell Anatomy 0.000 description 1
- 241001515942 marmosets Species 0.000 description 1
- 239000002609 medium Substances 0.000 description 1
- 201000001441 melanoma Diseases 0.000 description 1
- 230000009061 membrane transport Effects 0.000 description 1
- 230000009401 metastasis Effects 0.000 description 1
- 229930182817 methionine Natural products 0.000 description 1
- 235000010270 methyl p-hydroxybenzoate Nutrition 0.000 description 1
- 239000004292 methyl p-hydroxybenzoate Substances 0.000 description 1
- 229960002216 methylparaben Drugs 0.000 description 1
- 235000013336 milk Nutrition 0.000 description 1
- 239000008267 milk Substances 0.000 description 1
- 210000004080 milk Anatomy 0.000 description 1
- 239000007758 minimum essential medium Substances 0.000 description 1
- 238000010369 molecular cloning Methods 0.000 description 1
- 150000002772 monosaccharides Chemical class 0.000 description 1
- 238000000465 moulding Methods 0.000 description 1
- 229940126619 mouse monoclonal antibody Drugs 0.000 description 1
- 108010066052 multidrug resistance-associated protein 1 Proteins 0.000 description 1
- 208000010805 mumps infectious disease Diseases 0.000 description 1
- AEMBWNDIEFEPTH-UHFFFAOYSA-N n-tert-butyl-n-ethylnitrous amide Chemical compound CCN(N=O)C(C)(C)C AEMBWNDIEFEPTH-UHFFFAOYSA-N 0.000 description 1
- 230000001613 neoplastic effect Effects 0.000 description 1
- 239000002736 nonionic surfactant Substances 0.000 description 1
- 150000007524 organic acids Chemical class 0.000 description 1
- 235000005985 organic acids Nutrition 0.000 description 1
- LXCFILQKKLGQFO-UHFFFAOYSA-N p-hydroxybenzoic acid methyl ester Natural products COC(=O)C1=CC=C(O)C=C1 LXCFILQKKLGQFO-UHFFFAOYSA-N 0.000 description 1
- 238000004806 packaging method and process Methods 0.000 description 1
- 201000002528 pancreatic cancer Diseases 0.000 description 1
- 208000008443 pancreatic carcinoma Diseases 0.000 description 1
- 229940055729 papain Drugs 0.000 description 1
- 235000019834 papain Nutrition 0.000 description 1
- 229960005489 paracetamol Drugs 0.000 description 1
- 230000036961 partial effect Effects 0.000 description 1
- 239000008188 pellet Substances 0.000 description 1
- 239000000137 peptide hydrolase inhibitor Substances 0.000 description 1
- 229940023041 peptide vaccine Drugs 0.000 description 1
- 230000002085 persistent effect Effects 0.000 description 1
- 230000003285 pharmacodynamic effect Effects 0.000 description 1
- 235000021317 phosphate Nutrition 0.000 description 1
- 150000003013 phosphoric acid derivatives Chemical class 0.000 description 1
- 230000008488 polyadenylation Effects 0.000 description 1
- 229920001223 polyethylene glycol Polymers 0.000 description 1
- 229920000642 polymer Polymers 0.000 description 1
- 102000054765 polymorphisms of proteins Human genes 0.000 description 1
- 239000000256 polyoxyethylene sorbitan monolaurate Substances 0.000 description 1
- 235000010486 polyoxyethylene sorbitan monolaurate Nutrition 0.000 description 1
- 239000001267 polyvinylpyrrolidone Substances 0.000 description 1
- 229920000036 polyvinylpyrrolidone Polymers 0.000 description 1
- 235000013855 polyvinylpyrrolidone Nutrition 0.000 description 1
- 230000001323 posttranslational effect Effects 0.000 description 1
- 244000144977 poultry Species 0.000 description 1
- 239000000843 powder Substances 0.000 description 1
- 238000009021 pre-vaccination Methods 0.000 description 1
- 239000003755 preservative agent Substances 0.000 description 1
- 230000003449 preventive effect Effects 0.000 description 1
- 238000012545 processing Methods 0.000 description 1
- 210000001236 prokaryotic cell Anatomy 0.000 description 1
- 230000001737 promoting effect Effects 0.000 description 1
- 235000010232 propyl p-hydroxybenzoate Nutrition 0.000 description 1
- 239000004405 propyl p-hydroxybenzoate Substances 0.000 description 1
- 229960003415 propylparaben Drugs 0.000 description 1
- 229950010131 puromycin Drugs 0.000 description 1
- 239000002510 pyrogen Substances 0.000 description 1
- JFINOWIINSTUNY-UHFFFAOYSA-N pyrrolidin-3-ylmethanesulfonamide Chemical compound NS(=O)(=O)CC1CCNC1 JFINOWIINSTUNY-UHFFFAOYSA-N 0.000 description 1
- 238000003908 quality control method Methods 0.000 description 1
- 230000005855 radiation Effects 0.000 description 1
- 239000012217 radiopharmaceutical Substances 0.000 description 1
- 229940121896 radiopharmaceutical Drugs 0.000 description 1
- 230000002799 radiopharmaceutical effect Effects 0.000 description 1
- 238000001959 radiotherapy Methods 0.000 description 1
- 108010014186 ras Proteins Proteins 0.000 description 1
- 102000016914 ras Proteins Human genes 0.000 description 1
- 238000003259 recombinant expression Methods 0.000 description 1
- 238000011084 recovery Methods 0.000 description 1
- 206010038038 rectal cancer Diseases 0.000 description 1
- 201000001275 rectum cancer Diseases 0.000 description 1
- 230000012121 regulation of immune response Effects 0.000 description 1
- 230000004044 response Effects 0.000 description 1
- 230000000717 retained effect Effects 0.000 description 1
- 229960004641 rituximab Drugs 0.000 description 1
- 201000005404 rubella Diseases 0.000 description 1
- 239000012723 sample buffer Substances 0.000 description 1
- 238000000926 separation method Methods 0.000 description 1
- 238000012163 sequencing technique Methods 0.000 description 1
- 201000000849 skin cancer Diseases 0.000 description 1
- 229910052708 sodium Inorganic materials 0.000 description 1
- 239000011734 sodium Substances 0.000 description 1
- 239000008354 sodium chloride injection Substances 0.000 description 1
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 1
- 238000010532 solid phase synthesis reaction Methods 0.000 description 1
- 239000000600 sorbitol Substances 0.000 description 1
- 230000003393 splenic effect Effects 0.000 description 1
- 201000005671 spondyloarthropathy Diseases 0.000 description 1
- 238000003153 stable transfection Methods 0.000 description 1
- 238000010561 standard procedure Methods 0.000 description 1
- SFVFIFLLYFPGHH-UHFFFAOYSA-M stearalkonium chloride Chemical compound [Cl-].CCCCCCCCCCCCCCCCCC[N+](C)(C)CC1=CC=CC=C1 SFVFIFLLYFPGHH-UHFFFAOYSA-M 0.000 description 1
- 210000000130 stem cell Anatomy 0.000 description 1
- 239000008223 sterile water Substances 0.000 description 1
- 201000011549 stomach cancer Diseases 0.000 description 1
- 229960005322 streptomycin Drugs 0.000 description 1
- 238000007920 subcutaneous administration Methods 0.000 description 1
- 230000008093 supporting effect Effects 0.000 description 1
- 230000001629 suppression Effects 0.000 description 1
- 101150047061 tag-72 gene Proteins 0.000 description 1
- 210000001550 testis Anatomy 0.000 description 1
- 125000003396 thiol group Chemical group [H]S* 0.000 description 1
- 150000003573 thiols Chemical class 0.000 description 1
- 210000000115 thoracic cavity Anatomy 0.000 description 1
- 229960004072 thrombin Drugs 0.000 description 1
- 201000002510 thyroid cancer Diseases 0.000 description 1
- 230000030968 tissue homeostasis Effects 0.000 description 1
- 231100000419 toxicity Toxicity 0.000 description 1
- 230000001988 toxicity Effects 0.000 description 1
- 230000002103 transcriptional effect Effects 0.000 description 1
- 238000012546 transfer Methods 0.000 description 1
- 230000009466 transformation Effects 0.000 description 1
- 230000014616 translation Effects 0.000 description 1
- 230000014621 translational initiation Effects 0.000 description 1
- 108010020589 trehalose-6-phosphate synthase Proteins 0.000 description 1
- 125000001425 triazolyl group Chemical group 0.000 description 1
- QORWJWZARLRLPR-UHFFFAOYSA-H tricalcium bis(phosphate) Chemical compound [Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O QORWJWZARLRLPR-UHFFFAOYSA-H 0.000 description 1
- 230000004614 tumor growth Effects 0.000 description 1
- 230000005951 type IV hypersensitivity Effects 0.000 description 1
- 208000027930 type IV hypersensitivity disease Diseases 0.000 description 1
- 238000002604 ultrasonography Methods 0.000 description 1
- 241000701161 unidentified adenovirus Species 0.000 description 1
- 241001529453 unidentified herpesvirus Species 0.000 description 1
- 241001515965 unidentified phage Species 0.000 description 1
- 241001430294 unidentified retrovirus Species 0.000 description 1
- 201000005112 urinary bladder cancer Diseases 0.000 description 1
- 206010046766 uterine cancer Diseases 0.000 description 1
- 210000005166 vasculature Anatomy 0.000 description 1
- 210000005253 yeast cell Anatomy 0.000 description 1
- DGVVWUTYPXICAM-UHFFFAOYSA-N β‐Mercaptoethanol Chemical compound OCCS DGVVWUTYPXICAM-UHFFFAOYSA-N 0.000 description 1
Images




















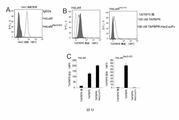

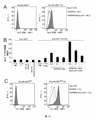
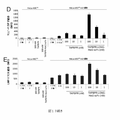





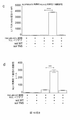






Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
- A61K38/16—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/0005—Vertebrate antigens
- A61K39/0011—Cancer antigens
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/46—Cellular immunotherapy
- A61K39/461—Cellular immunotherapy characterised by the cell type used
- A61K39/4611—T-cells, e.g. tumor infiltrating lymphocytes [TIL], lymphokine-activated killer cells [LAK] or regulatory T cells [Treg]
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/46—Cellular immunotherapy
- A61K39/463—Cellular immunotherapy characterised by recombinant expression
- A61K39/4632—T-cell receptors [TCR]; antibody T-cell receptor constructs
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/46—Cellular immunotherapy
- A61K39/464—Cellular immunotherapy characterised by the antigen targeted or presented
- A61K39/4643—Vertebrate antigens
- A61K39/4644—Cancer antigens
- A61K39/464402—Receptors, cell surface antigens or cell surface determinants
- A61K39/464403—Receptors for growth factors
- A61K39/464406—Her-2/neu/ErbB2, Her-3/ErbB3 or Her 4/ ErbB4
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/705—Receptors; Cell surface antigens; Cell surface determinants
- C07K14/70503—Immunoglobulin superfamily
- C07K14/70539—MHC-molecules, e.g. HLA-molecules
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/2803—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the immunoglobulin superfamily
- C07K16/2827—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the immunoglobulin superfamily against B7 molecules, e.g. CD80, CD86
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/32—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against translation products of oncogenes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/06—Animal cells or tissues; Human cells or tissues
- C12N5/0602—Vertebrate cells
- C12N5/0634—Cells from the blood or the immune system
- C12N5/0636—T lymphocytes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/06—Animal cells or tissues; Human cells or tissues
- C12N5/0602—Vertebrate cells
- C12N5/0634—Cells from the blood or the immune system
- C12N5/0639—Dendritic cells, e.g. Langherhans cells in the epidermis
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/56—Immunoglobulins specific features characterized by immunoglobulin fragments variable (Fv) region, i.e. VH and/or VL
- C07K2317/569—Single domain, e.g. dAb, sdAb, VHH, VNAR or nanobody®
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/60—Immunoglobulins specific features characterized by non-natural combinations of immunoglobulin fragments
- C07K2317/62—Immunoglobulins specific features characterized by non-natural combinations of immunoglobulin fragments comprising only variable region components
- C07K2317/622—Single chain antibody (scFv)
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/70—Fusion polypeptide containing domain for protein-protein interaction
- C07K2319/74—Fusion polypeptide containing domain for protein-protein interaction containing a fusion for binding to a cell surface receptor
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Immunology (AREA)
- Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Organic Chemistry (AREA)
- Biomedical Technology (AREA)
- Medicinal Chemistry (AREA)
- Microbiology (AREA)
- Genetics & Genomics (AREA)
- Cell Biology (AREA)
- Zoology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Pharmacology & Pharmacy (AREA)
- Veterinary Medicine (AREA)
- Public Health (AREA)
- Animal Behavior & Ethology (AREA)
- Epidemiology (AREA)
- Biotechnology (AREA)
- Wood Science & Technology (AREA)
- Mycology (AREA)
- Biochemistry (AREA)
- Oncology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- General Engineering & Computer Science (AREA)
- Hematology (AREA)
- Biophysics (AREA)
- Molecular Biology (AREA)
- Gastroenterology & Hepatology (AREA)
- Toxicology (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Peptides Or Proteins (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Medicinal Preparation (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
Abstract
Description
本発明は、ペプチド交換触媒および哺乳類細胞の表面のMHCクラスI分子により提示されるペプチドレパートリーを調節する上におけるその使用に関するものである。
癌の免疫療法は最近大きな成長を見たが、現在の治療に対する腫瘍耐性の出現とともに、患者は新たな治療法を切実に必要としている1。抗PD1抗体や抗CTLA4抗体のような免疫チェックポイント阻害剤の使用により、現在、Tリンパ球が腫瘍1、2を認識、破壊する機能を活用できる可能性が出てきた。しかし、このような治療は、現在一部の患者、特に高い突然変異3-6荷重を持つ腫瘍患者にのみ有効である。腫瘍は自然の免疫コントロールあるいは免疫療法による免疫コントロールを数多くのメカニズムにより逃れており、例えば免疫編集プロセスが免疫原性7の低い腫瘍細胞を選択的に残す場合などである。従って、腫瘍の免疫原性を増加させることができれば、より多くの患者集団に有効な治療が提供できる可能性があり、これには突然変異負荷の低い腫瘍も含まれる。
本発明の発明者は、TAPBPRの内腔ドメインは、可用性細胞外蛋白質あるいは膜結合性細胞表面蛋白質として哺乳類細胞に提示された場合、MHCクラスI分子交換触媒として機能する能力を保持していることを期せずして発見した。可溶性もしくは細胞外ペプチド交換触媒は、例えば免疫原性ペプチドを腫瘍細胞あるいは他の病的細胞に負荷し、T細胞による認識を誘導するといったように、免疫反応の調節を介する多くの治療面での応用上有用である可能性がある。
図1はペプチド受容MHCクラスIが、表面TAPRBRを発現している細胞上にあることを示す。(a)TAPRBRが過剰発現すると細胞表面にTAPRBRが現れる。IFN-γ処理HeLaM細胞とHeLaM-TAPBPRKO( -/+TAPBPRWT形質導入)は、TAPBPR特異的mAb PeTe- 4で染色した。(b-e) TAPBPRWTを過剰発現している細胞は、対照細胞に比べ外来性ペプチドとの結合増加を示す。IFN-γ処理細胞は、HLA-A*68:02 特異的蛍光性ペプチド ETVSK*QSNVもしくはその非結合型変異体EGVSK*QSNG (アンカー残基に突然変異を有している)とインキュベートし、フローサイトメトリ―で分析した。(b、c)細胞を37℃で15分間10 nM (b) ETVSK*QSNV もしくは(c) EGVSK*QSNGとインキュベートした場合の典型的なペプチド結合のヒストグラム。細胞を(d) ETVSK*QSNV濃度を増加させ15分間処理した場合、または(e)10 nM ETVSK*QSNVで37℃、0-180分間処理した場合の(d)用量反応曲線および(e)時間経過で、HeLaM、TAPBPR欠損(TAPBPRKO)あるいはHLA-A、-B、-C 欠損(HLA-ABCKO)変異型と比べTAPRBRWTを過剰発現している細胞には外来性ペプチドの結合が増加していることを示す。(e)において、10 nM EGVSK*QSNGで観察された結合は対照である。線グラフは3つの異なる実験からの平均蛍光強度(MFI)-/+ s.e.mを示す。
標識されていない競合ペプチドYVVPFVAKV (YVV)、ETVSEQSNV(ETV) もしくは ETVSEQSNG (ETVΔ2/9)のある状態、あるいは無い状態でのIFN-γ処理HeLaKOTAPBPRWT細胞からの蛍光性ペプチド(a、b) YVVPKVAK*V (YVV*) もしくは (c、d) ETVSK*QSNV(ETV*)の解離。細胞は37℃、15分間10 nM蛍光性ペプチドとインキュベート後、洗浄し、次いで未標識の競合ペプチドの濃度を上げ37℃で15分間インキュベートした。(a、c)ヒストグラムは100 nM競合ペプチドとのインキュベーション後に観察された蛍光性ペプチドの典型的解離を示す。(b、d)線グラフは、未標識ペプチドの濃度を上げ処理した場合の残存蛍光性ペプチドのパーセンテージ-/+ s.e.mで、(b)4つの、(d)3つの独立した実験から得られた結果を示す。
この発明は、TAP結合蛋白関連(TAPBPR)のフラグメントを含む組み換えペプチド交換蛋白質に関するものである。TAPBPRの内腔ドメインは、ここにおいてペプチドエディターとして機能することが示され、このドメインを含むペプチド交換蛋白質は、外来性ペプチドを細胞表面のMHCクラスI分子に負荷することができる細胞外あるいは細胞表面MHCクラスIペプチド交換触媒として活動することが示される。
好適なリンカーは当技術分野で周知のように、科学的、およびペプチジアルリンカーを含む。例えば、ペプチジアルリンカーはアミノ酸残基配列、例えば5から30もしくは5から22のアミノ酸残基を含む場合があり、10から20のアミノ酸残基を含むことが望ましく、約12のアミノ酸残基がより望ましい。どのようなリンカー配列も使用可能である。リンカー配列は異種配列であることが望ましい。好適なリンカーアミノ酸配列は、当技術分野で周知のように、アミノ酸配列GGGGS、(GGGGS)3 もしくは GSTVAAPSTVAAPSTVAAPSGS、HVGGGGSGGGGSGGGGSTSもしくはその変異体を含む場合がある。
表面MHCクラスI分子を持つ哺乳類細胞集団を提供する、
哺乳類細胞集団を、外来性ペプチドおよびTAPBPRの内腔ドメインから成るTAPBPRフラグメントを含むペプチド交換蛋白質と接触させる、
こうしてペプチド交換蛋白質が外来性ペプチドを集団の細胞上のMHCクラスI分子に負荷するようにし、これにより哺乳類細胞の免疫原性を調整する。
前もって個体から得られた抗原提示細胞の集団を提供し、そして抗原提示細胞にペプチド交換蛋白質および免疫原性ペプチドを接触させ、ペプチド交換蛋白質が抗原提示細胞の表面MHCクラスI分子に免疫原性ペプチドを負荷するようにし、抗原提示細胞がT細胞を刺激し免疫反応を惹起することができるようにする。
前記個体における癌細胞と結合する標的ドメインを含む上に記載されたペプチド交換蛋白質を個体に投与し、そして免疫原性ペプチドを個体に投与し、ペプチド交換蛋白質が、免疫原性ペプチドを前記個体の癌細胞の表面 MHCクラスI分子に負荷するようにし、これにより前記個体における癌細胞に対する免疫反応を惹起もしくは増強する。
1.材料と方法
1.1 構成体
以前に記載されているように、全長性TAPBPRWTおよびTAPBPRTN5の生成は、レンチウィルスベクターpHRSIN-C56W-UbEM を用いて行われ、これは脾臓フォーカス形成ウイルス(SFFV)プロモーターのコントロール下でTAPBPRを、そしてユビキチンプロモーターのコントロール下でGFP誘導体エメラルドを作り出す11,15。キメラ構成体TAPBPRPM および tapasinPMのクローニングは、同じレンチウィルスベクターで、二段階PCR 手順により行われ、ここでTAPBPRもしくはタパシンのいずれかの細胞外ドメインおよび膜貫通性ドメインは増幅され、次いでCD8の細胞質尾部と融合させられた。TAPBPRERは同様の手順で作成され、ここでTAPBPRの細胞外ドメインはタパシンの膜貫通性および細胞質ドメインと融合させられた。TAPBPRWTもしくはTAPBPRTN5の分泌型を作成するために、Li et al 22に記載されているように、両者の内腔ドメインがpiggyBacトランスポゾンに基づく哺乳類細胞発現系にクローニングされた。
IFN 23に対してより反応性の高いHeLa細胞株の変異型 であるHeLaM細胞、修飾を受けたその変異型であるHEK-293T 細胞とMCF7細胞が10%ウシ胎仔血清(FBS)(Gibco, Thermo Fisher Scientific)、100 U/ml ペニシリンおよび100 μg/ml ストレプトマイシン(Gibco, Thermo Fisher Scientific)を添加されたダルベッコ改変培地(DMEM; Sigma-Aldrich, UK)で37℃、5% CO2環境下で維持された。内因性に発現しているTAPBPRの発現を誘導し、MHCクラスI抗原処理および提示経路の他の構成要素を上方制御するため、指定されているところでHeLaM および MCF7細胞が200 U/ml IFN-γ(Peprotech、UK)で 48-72時間処理された。
次のTAPBPR特異抗体が使用された。TAPBPR の未変性コンフォーメーションに特異的なマウスモノクローナル抗体(mAb)で、ヒトTAPBPR11のアミノ酸22-406に対して作成され、タパシン15とは交差反応しないPeTe4および TAPBPRのアミノ酸23-122に対して作成され、変性TAPBPRに反応するマウスmAb、 ab57411 (Abcam、UK)。次のMHCクラスI特異的抗体が使用された;MHC class I α2ドメイン上のコンフォーメーション特異的エピトープを認識する汎MHC class I mAb で、β2mおよびペプチド24の存在からは独立しているW6/32; α1ドメイン25,26のアミノ酸 57-62にPxxWDR モチーフを含むHLA-A、 -Bおよび-C 分子を認識するMHC class I特異的mAb、HC10; HLA-A2 および - A68重鎖/β2mヘテロ二量体に特異的なビオチン化抗 HLA-A68反応性mAb(One Lambda, Thermo Fisher Scientific、 Canoga Park、 CA); HLA-A2重鎖/β2mヘテロ二量体特異的抗体、 BB7.2。使用された他の抗体には次のものがある:Pasta-1,タパシン特異的mAb (Dick et al., 2002); ウサギ抗カルネキシン抗体(Enzo Life Sciences、 UK); UGT1(ab124879, Abcam)に対するウサギmAb; 陰性対照としてIgG2aアイソタイプコントロール(Sigma-Aldrich)。
レンチウィルスはFugene(Promega、 UK)を使用して、HEK-293T細胞にレンチウィルスベクターを パッケージングベクターpCMVΔR8.91および エンベロープベクターpMD.Gとともにトランスフェクションすることにより作成された。ウィルス上清は48時間で収集され、以前に記載されたTAPBPR-ノックアウトHeLaM細胞株(HeLaM-TAPBPRKO) (Neerincx et al., 2017)の形質導入に使用された。TAPBPRWT, TAPBPRTN5、 TAPBPRPPM、 TAPBPRER、 tapasinWTおよび tapasinPMがHeLaM-TAPBPRKO細胞株で再構成された。
次のMHCクラスI特異的ペプチドが使用された:HLA-A*68:02結合ペプチドETVSEQSNV、 そのアンカー残基(2位および9位アミノ酸)をグリシンで置換することによって得られたその誘導体EGVSEQSNG, また、その蛍光標識型ETVSKTAMRAQSNVおよびそれぞれ5位グルタミン酸塩をリジンで置換することにより得られたEGVSKTAMRAQSNG、 5-カルボキシルテトラメチルローダミン [TAMRA] (Peptide Synthetics、 UKより)で標識された標識型; HLA-A*02:01結合ペプチドNLVPMVATV、 YLLEMLWRL、 YLLEMLWRL、 CLGGLLTMV およびYVVPFVAKV、加えてこれらの蛍光標識変異型NLVPKTAMRAVATV、 FMVFKTAMRAQTHI、 CLGGKTAMRALTMV、 YLLEKTAMRALWRLおよびそれぞれYVVPFVAKTAMRAV(Peptide Synthetics、 UKから);HLA-B*27:05特異ペプチドSRYWAIRTRおよびその蛍光標識変異型SRYWKTAMRAIRTR (Peptide Synthetics, UKより)。
トリプシン処理後、細胞を1% 牛血清アルブミン(BSA)で洗い, 4℃ で1x PBSの中に浮遊させ、次いで次の抗体の一つを含む 1% BSA内で 30分間4℃で染色した。W6/32, pete4, pasta-1、 抗HLA-A68反応性mAb、 BB7.2もしくはアイソタイプコントロール抗体。過剰の未結合の抗体を除くため細胞を洗浄した後、細胞と結合した1次抗体は、ビオチン化抗HLA-A68 mAbに対するヤギ抗マウスAlexa-Fluor 647 IgG (Invitrogen Molecular Probes、 Thermo Fisher Scientific)あるいはAlexa-Fluor 647結合 ストレプトアビジン (Invitrogen Molecular Probes、 Thermo Fisher Scientific) で4℃、25 分間インキュベートすることにより検出された。次いで3回洗浄後、蛍光標識はCytek 修正機能付きBD FACScanアナライザーを使い検出され、FlowJo(FlowJo、 LLC、 Ashland、 OR)を用いて分析された。
細胞を採取し、次いでリン酸緩衝生食水(PBS)で洗浄した。表面TAPBPR免疫沈降実験のため、細胞を4℃で、回転をかけつつ1時間1x PBSに溶かした1% BSA中で2 μg Pete4抗体とインキュベートした。過剰の抗体は細胞を4℃の1x PBSで5回洗浄することにより除去した。次いで細胞を溶解し、全細胞溶解液からのTAPBPR免疫沈降と同様に細胞内TAPBPR免疫沈降がさらに行われた。
TAPBPRWT もしくは TAPBPRTN5の一方の選択された形が、22に記載されているようにPiggyBac発現システムを用いて 293T細胞に発現された。このため、いずれかの蛋白質のC末端Hisタグ付き細胞外ドメインをNheI と NotI(Thermo Fisher Scientific)を用いプロテインAを欠くPB-T-PAFベクターの修飾版へ組み込ませた。簡潔に述べると、2x105 293T細胞に100 ng PB-RNプラスミッド、100 ng PBaseプラスミッド、および800ng PB-T-TAPBPRあるいはPB-Tタパシンをトランスフェクトさせた。トランスフェクションの48時間後、細胞は10 %ウシ胎仔血清(FBS)(Gibco, Thermo Fisher Scientific)、1xP/S、 3 μg/ml ピューロマイシン(Invivogen)および700 μg G418 (Gibco, Thermo Scientific)を添加したダルベッコ改変培地 (DMEM; Sigma-Aldrich、 UK)を用い7日間安定したトランスフェクションのために選択された。蛋白質生成のため、6x107の細胞が5% FCSと100U/mlペニシリンを添加した200 ml DMEMの中で5-7日間2 μg/mlドキシサイクリンで誘導を受けた。7日後、培養液を収集し、NiSepharoseTMexcelビーズ(GE Lifesciences)を用いてTAPBPRを精製した。蛋白質はPBS (Sigma)に溶かした250 mMイミダゾールで溶出し、次いで48時間PBS(Sigma)で透析した。純度の評価のため、溶出画分をSDSPAGEで分析し、次いでクマシー染色を行った。
標的細胞株を25,000−30,000 細胞/ウェルで12ウェルプレートに播種し、IFN-γで刺激した。
刺激期間に続き、細胞は1x PBSで3回洗浄し、前もって温められた300 μL のopti-MEM (Thermo Fisher Scientific、 UK)でインキュベートした。ペプチド結合が組み換えTAPBPRの存在下でなされた場合、細胞は次いで組み換えTAPBPR (HLA-A*68:02は100 nM あるいは HLA-A* 02:01は1 μM)の存在下もしくは不在下で処理された。15分後、望みのTAMRA標識ペプチドが細胞に加えられ、37℃(HLA-A*68:02は15分間、あるいは HLA-A*02:01は60分間)でインキュベートされた。ペプチドの結合が過剰発現したTAPBPRにより促進される場合には、標識ペプチドは組み換えTAPBPRを使用することなく、直接に細胞に加えられた。ペプチド処理に続き、細胞は3回1x PBSで洗浄し、収穫した。結合ペプチド/細胞のレベルを、YelFL1 channel(Cytek)を用い、フローサイトメトリーにより決定した。
細胞は25,000細胞/ウェルで播種し、48時間IFN-γで刺激後洗浄し、opti-MEMで希釈した関心の対象である10 nM TAMRA標識ペプチドで、上に記載されているように37℃で15分間処理した。結合ステップに続き、ペプチドを含む培養液を除去し、細胞を洗浄後、培養液単独、もしくは様々な濃度の非標識ペプチドで37℃、さらに15分間処理した。次いで細胞を洗浄、収穫、細胞当たりの結合ペプチドのレベルをYelFL1 channel(Cytek)を用い、フローサイトメトリーにより決定した。
組み換えTAPBPRの存在下で、対応するペプチド10nMで処理後、HLA-A*02:01(Sim et al., 2013)と関連する エプスタイン・バー・ウィルス (EBV)潜伏感染遺伝子産物潜伏膜蛋白質 2A(LMP2A426-434: CLGGLLTMV)および潜伏膜蛋白質1(LMP1125-133: YLLEMLWRL)由来のペプチドに特異的なTCR様mAb が標的細胞を染色するために使用された。洗浄後、結合したTCR様mAbのレベルをヤギ抗マウスAlexa-Fluor 647 IgGを使用することにより検出し、次いでフローサイトメトリーで測定した。
標的細胞(MCF-7 細胞あるいはHLA heavy chain A、 B および Cを欠き、HLA-A*02:01 重鎖で再構成された HeLaM細胞)を6ウェルプレートに80,000細胞/ウェルに播種し、72時間、200単位の IFN-γ(Peprotech、 UK)で刺激した。次いで細胞を3回1x PBS(Sigma-Aldrich、UK)で洗浄し、組み換えWT TAPBPR、 TN5 TAPBPR 突然変異体のいずれかを含む状態で、あるいはTAPBPRなしの状態で600 μLのあらかじめ温められたopti-MEMとインキュベートした。15分後、100 pM NLVPMVATVペプチドを望む試料に加え、さらに60分間インキュベートした。ペプチド処理に続き、細胞を3回1x PBSで洗浄し収穫した。各試料は次いで再度2回1x PBSで洗浄し、X-VIVO 15培養液に1 mil 細胞/ mLの濃度で再浮遊させた標的細胞には最初放射線照射を行い、次いで一晩あらかじめブロックされた96ウェルプレートに8,000 T細胞/ウェルとともに50,000細胞/ウェル加えた。プレートは37℃で一晩インキュベートし、翌日展開した。
2.1 ペプチド受容性MHC class Iは表面 TAPBPRを発現している細胞上に存在する
表面TAPBPRが表面MHCクラスI分子上のペプチドエディターとして機能する可能性があるかどうかを探求するために、私たちはまず、蛍光性ペプチドETVSK*QSNV、すなわちHLA-A*68:02に高い親和性で結合する新生抗原ETVSEQSNVの変異型が、表面TAPBPRを欠損する細胞に比べて表面TAPBPRを発現している細胞へより多く結合するかどうかを検索した。IFN-γ 処理を受けた HeLaMTAPBPRKO (HeLaMKO)および HeLaM細胞は表面になんらTAPBPRを発現することはないが、HeLaM-TAPBPRKOにおけるTAPBPRWTの過剰発現の結果、表面発現したTAPBPRはかなりの量となった(図 1a)。表面TAPBPRを発現している細胞を10 nM外来性ペプチドと37℃で15分間インキュベートし、我々はこれら細胞へのペプチド結合を示す蛍光の大幅な増強を観察した(図 1 b)。対照的に、我々は同一の条件下で表面TAPBPRを欠如する細胞株へのETVSK*QSNV結合は観察できなかった(図 1 b)。観察された細胞性蛍光がMHCクラスIへのペプチド結合の直接の結果であることを確認するため、我々はまた、細胞を蛍光ペプチド(EGVSK*QSNG)の変異型とともにインキュベートした。ここでHLAA* 68:02に結合を許しているアンカー残基は突然変異を起こしたものである。これは細胞株のいずれに対しても結合を起こさなかった(図1c)。HeLaMもしくは HeLaMKO細胞と比べてIFN-γ 処理を受けた HeLaKOTAPBPRWTへのペプチド結合の増加は広い範囲のペプチド濃度にわたり観察できた(図1d)。ペプチド濃度がより高い場合、表面TAPBPRを欠如している細胞への外来性ペプチド結合が観察された(図1d)。しかし、このTAPBPR非依存性の細胞へのペプチド結合は約120倍効率が低かった。それはTAPBPRで促進される様式で必要な10 nMのペプチドで観察されるのと同じ蛍光を得るためには約1.2 μMが必要であったからである(図1d)。HLA-A、B、C 欠損HeLaM細胞は、外来性ペプチド濃度が10 μMとなるまでペプチドを結合できなかったことを考えると、細胞へのペプチド結合はMHCクラスIに依存性であった。時間経過に伴うTAPBPR促進性ペプチド結合の動力学を探求した際、私たちは10 nMペプチドとインキュベートすると、TAPBPR非依存性条件の場合に比べ、細胞が表面MHCクラスIへ外来性ペプチドを負荷する能力の顕著な増加を観察した(図1e)。例えば、60分後、私たちはHeLaMおよび HeLaMKO対照に比べてHeLaKO-TAPBPRWTに結合した蛍光ペプチドのレベルが21倍に増加したのを観察した図1e)。これらの所見は、表面MHCクラスI分子に抗原ペプチドを負荷するうえでの表面TAPBPRの役割を支持している。さらに、私たちは4℃でHeLaMKOTAPBPRWT細胞への高レベルの外来性ペプチド負荷を観察したが、この温度は膜輸送を阻害するため、さらに、ペプチド負荷はエンドサイト―シス小胞内というよりむしろ細胞表面で直接起きていることが示唆される。
過剰発現した蛋白質というよりむしろTAPBPRの表面プールが外来性ペプチドのMHCクラスIへの負荷に責任があるという決定的証拠を提供するため、私たちは、TAPBPRの標的を異なる細胞内部位とするキメラTAPBPR構成体を2つ作成した。TAPBPRの内腔部分が細胞膜(PM)を標的とすることは、TAPBPRの細胞質尾部をCD8 (TAPBPRPM)12で置換することにより達成し、一方TAPBPRは、その膜貫通性ドメインおよび細胞質尾部をタパシンのそれらで置換(TAPBPRER)13,14することにより小胞体(ER)内に保持された。非常に高レベルに細胞表面に発現しているTAPBPRPMとは対照的に、TAPBPRERは細胞膜上には検出されなかった(図2a)。TAPBPRの表面プールの免疫沈降が示すことは、MHCクラスIはTAPBPRWTおよび TAPBPRPM で形質導入された細胞上に表面発現したTAPBPRと会合したが、TAPBPR の突然変異型でMHCクラスI15とは結合しないTAPBPRTN5で形質導入された細胞からのTAPBPRとは会合しなかったということである(図2b)。単離された表面TAPBPRの量(図 2b)は、TAPBPRERを発現している細胞から単離されたかろうじて検出可能な量でフローサイトメトリーを使用して観察された表面TAPBPR発現(図 2a)と密接な相関があり、UGT1は表面プルダウンでは検出されなかった(図 2b)ため、私たちはTAPBPRの表面プールのみを単離し、細胞内プールは単離しなかったたようであった。表面TAPBPRをあらかじめ除去した細胞からの細胞内TAPBPRプールの単離により、すべてのTAPBPR変異型が発現されており、またTAPBPRWT, TAPBPRPM およびTAPBPRER分子はMHCクラスIと強い会合を示したことが確認された(図2b)。TAPBPRPMとは対照的に, UGT1の TAPBPRWTおよび TAPBPRER両者との強い会合が観察され、
キメラ蛋白質の予想されていた細胞内局在が支持された(図2b)。2つの外来性HLA-A*68:02特異的蛍光ペプチド、 ETVSK*QSNV およびYVVPFVAK*Vに結合する細胞株の能力が検査された際、表面TAPBPRを発現している細胞のみが、37℃で15分間、10 nMの外来性ペプチド存在下で有意のペプチド会合を示した(図2c & d)。TAPBPRERを発現している細胞上では蛍光ペプチド結合は観察されなかった(図2c & d)。これらの結果は、TAPBPRの過剰発現というよりむしろ、表面TAPBPRが外来性ペプチドをMHCクラスIに負荷することに責任があることを示している。
タパシンもまたMHCクラスI分子エディターであり、私たちは、この分子が細胞表面に発現した場合、同様に外来性ペプチドをMHCクラスIに負荷することができるのか検索した。TAPBPRWTの過剰発現とは対照的に, タパシンWTの過剰発現は、おそらくその細胞質尾部に見いだされるER保持モチーフのため、この蛋白質の細胞表面への発現を生じなかった(図2e)13, 14。従って、私たちはタパシンの細胞質尾部をCD8 (タパシンPM)のそれで置換し、細胞表面にその発現を見た(図2e)。タパシンを過剰発現する細胞のETVSK*QSNV および YVVPFVAK*Vへの結合能力を検査した際、細胞表面にタパシンを発現している細胞で、HLA-A*68:02に結合する外来性蛍光ペプチドのわずかながら有意の増加が認められた(図 2f & g)。こうした結果は、表面タパシンも外来性ペプチドのMHCクラスIへの会合を増強することができるということを示す。しかし、タパシンPMで観察された外来性ペプチドの結合は、TAPBPRPMで観察されたそれよりも約10 倍少なかった。
表面発現したTAPBPRがMHCクラスIに外来性ペプチド負荷を促進し得るメカニズムとして考えられるものが2つある;TAPBPRがペプチド受容性MHCクラスI分子を一緒に分泌経路を通して細胞表面に移動させるか、および/またはそれがこの非典型的局在部位でペプチド交換触媒として機能する能力を保持するか、である。この問題をさらに探求するために、私たちは表面発現TAPBPRがMHCクラスI分子上でペプチド交換を促進することができるかどうかを決定するアッセイを開発した。まず、細胞を37℃で15分間、10 nM蛍光標識ペプチドでインキュベートし、表面MHCクラスI分子を標識ペプチドと結合させた。未結合の蛍光ペプチドを除去するために、十分洗浄後、標識ペプチドを交換する細胞の能力を、細胞を様々な非標識競合ペプチドと37℃で15分間インキュベートすることにより評価した。この方法を使用することにより、私たちは高親和性非標識競合ペプチド(ETVSEQSNVもしくは YVVPFVAKV)のいずれかの存在下で、YVVPKVAK*V (図3a & b) およびETVSK*QSNV(図3c & d)両者の解離を認めた。HLA-A*68:02と結合できないEGVAK*QSNGの存在下では、HeLaKOTAPBPRWT上におけるHLA-A*68:02からの蛍光ペプチドの解離は見られなかった(図 3)。我々の結果は、表面TAPBPRは(YVVPFVAKV> ETVSEQSNV>EGVSEQSNQ)のペプチド親和性順で、またペプチド濃度依存性に、MHCクラスI分子上のペプチド交換を促進することができることを示す(図 3b & d)。これらの所見は、TAPBPRが、細胞表面に発現した場合でも依然としてペプチド交換触媒として機能する能力を保持しており、またMHCクラスI分子上でペプチド交換を行うことができることを証明している。
細胞膜結合TAPBPRはHLA-A*68:02上でペプチド交換触媒として機能するので、私たちは細胞に加えられた外来性可溶性TAPBPRが同じ触媒として機能することができるかどうか、あるいは正しい方向性のために膜アンカーを必要とするかどうかに興味を持った。まず、私たちはN末端のIgVおよびIgCドメインからなり、膜貫通および細胞質尾部を欠如する外来性TAPBPRが表面MHCクラスI分子と結合できるかどうかを検査した。HeLaM細胞を37℃で15分間、100 nMの外来性TAPBPRWTとインキュベートした際、TAPBPR特異的mAb PeTe-4を使用することにより、TAPBPRは明瞭に細胞表面に検出可能であった(図4a)。以下の理由により、TAPBPRの細胞への結合は、そのMHCクラスI分子との会合に完全に依存していると思われた: 1)外来性TAPBPRTN5 (MHC クラス I15と結合できない突然変異型)は細胞に結合しなかった(図 4a), 2)外来性TAPBPRWTは古典的MHCクラスI発現を欠如するHeLaM (HeLaMHLA- ABCKO)とはもはや結合できなかった(図4b)および3) 外来性TAPBPRWTのHeLaMHLA-ABCKO細胞への結合は、 HLA-A*A68:02発現が再構成されると回復した(図4b)。これらの結果は、外来性可溶性TAPBPRが細胞表面に発現しているHLA-A*68:02と結合できること証明している。
私たちは次に、表面MHCクラスI分子に結合した外来性TAPBPRが細胞膜にアンカーしたそれと同様にペプチド交換を行うことができるかを調べた。100 nMの外来性TAPBPRの存在下もしくは不在下で、37℃、15分間、細胞をインキュベート後、さらに15分間10 nM蛍光ペプチドとともに、あるいはこれなしで処理した。TAPBPRWT不在下で、あるいはTAPBPRTN5で処理された場合にHeLaM細胞上に観察された外来性ペプチド結合の著しく低いレベルとは対照的に、細胞が外来性TAPBPRWTで処理された場合、外来性蛍光性ペプチドETVSK*QSNV およびYVVPKVAK*VのHeLaMへの結合の有意の増強が観察された (図4c &d)。EGVAK*QSNGとの会合は、検査したどの条件でも見られなかった(図 4c & d)。TAPBPRを介する外来性ペプチドの細胞への結合はHLA-A*68:02を介して仲介されることが示されたと言え、これはMHCクラスI発現を欠く細胞にはETVSK*QSNV (図4e & f) もしくはYVVPKVAK*Vの結合が見られず、またHLA-A*68:02 発現が再構成されるとペプチド会合の回復が見られたことによる(図 4e & f)。可溶性TAPBPRWTは広い範囲のペプチド濃度にわたり細胞への外来性ペプチド結合を増強した(図4g)。こうした結果は、外来性TAPBPRがペプチドをHLA-A*68:02に負荷することができ、またTAPBPRの内腔ドメインがMHCクラスI分子へのTAPBPR負荷の場として十分であることを明瞭に証明している。
観察された結果はHLA-A*68:02の異常でないことを確認するため、私たちは分析をさらに広げ、TAPBPRが他のヒトMHCクラスI分子、HLA-A*02:01へ種々の外来性ペプチドを負荷する能力を検査した。外来性TAPBPRWTは、NLVPMVATV(CMV蛋白質pp6516由来免疫原性ペプチド)、 YVVPFVAKV (ヒトCCR4-NOT転写複合体サブユニット18)およびYLLEMLWRL (EBV 潜伏膜蛋白質1 (LMP1)17由来免疫原性ペプチド17(図 4h & i)の蛍光性変異型の結合を有意に促進した。TAPBPRが促進するこれらの外来性ペプチドの負荷はHLA-A2に依存しており、これはHLA-A2 陰性細胞には蛍光性ペプチドの結合が観察されないことによる。外来性TAPBPRWTはまたCLGGLLTMV (EBV蛋白質潜伏膜蛋白質1由来の免疫原性ペプチド)の蛍光性変異型の結合を有意なレベルではないが、わずかに促進するように思われたが、(図4h & i) 他のMHCクラスI分子に特異的な他のペプチドのHLA-A2への結合は促進しなかった (図 4i)。MHCクラスIに結合できない外来性TAPBPRTN5はどのペプチドにおいてもHLA-A2への結合を促進することはなかった (図4i)。総合すると、私たちのデータは、外来性TAPBPRが親和性に基づく様式で、表面MHCクラスIに対し外来性ペプチドの負荷を促進できることを強く示唆している。
私たちは次いで、TAPBPRを介して負荷されたペプチドがT細胞レセプター(TCR)検出に利用可能であるか決定した。希望の持てることに、可溶性TAPBPRは高親和性ペプチドが表面MHCクラスI分子に結合すると細胞から解離することが見いだされており(図 17)、TAPBPR負荷されたペプチド:MHC複合体はT細胞レセプター(TCR)による検出のために完全にアクセス可能である可能性があった。
腫瘍細胞に免疫原性ペプチドを負荷する能力は癌免疫療法に非常に有益であると思われるため、私たちは、腫瘍またはウィルスペプチドを乳癌細胞株MCF-7に負荷するTAPBPRの能力を検査した。
TAPBPRリンカー-GFPナノボディ融合産物(種々のリンカーと)が生成された(図7)。これらの産物は、TAPBPR-抗体融合産物が完全な機能を保持していることを証明するために使用された。TAPBPRと抗体フラグメントの間に様々な長さのリンカーを持つTAPBPR融合蛋白質が効果的に機能することが示された(図21)。
私たちは、GFP特異的ナノボディと融合したTAPBPRが表面にGFPを発現しているHeLaM細胞に結合するが、表面GFP,すなわちバイスタンダー細胞なしにはペプチドと結合、もしくはペプチドをHeLaM細胞上に負荷できないことを見出した(図 8)。組み換え蛋白質100 nMで、HLA-A*68:02との相互作用のため、可溶性TAPBPRのみでも低レベルのHeLaM細胞への結合が見られたが、TAPBPR-GFPNB融合蛋白質は表面GFPを発現しているHeLaM 細胞への著しく高レベルの結合を示した(図8B & 8C)。しかし、100 nMでも、TAPBPR-GFPNB融合蛋白質はGFP を欠如するHeLaM細胞には結合を示さなかった(図 8Bおよび8C)。組み換え蛋白質10 nMでは、可溶性TAPBPR単独ではHeLaM細胞株のいずれにも結合を示さなかったが(図8C)、TAPBPR-GFPNB融合蛋白質は高レベルの結合を、特にGFPを発現している細胞に結合を示した(図8C)。これは、このTAPBPR融合蛋白質の結合の大部分が抗体タグから由来することの証明である。
TAPBPR-GFP融合蛋白質は、表面GFPの不在下では、HeLaM 細胞上に外来性ペプチドを効果的に負荷できなかったが、表面GFP 陽性細胞にペプチドを負荷するのは極めて効率的であった(図 8D &8E)。1 nMでもTAPBPR-GFPNB融合は外来性ペプチドを抗体-標的発現細胞に負荷することができた(図 8E)。可溶性TAPBPRは同様の蛋白質濃度を使用した場合、ペプチド負荷は示さなかった(図 8E)。示されたデータは長リンカーを持つTAPBPR融合産物に関するものであるが、3つの異なるリンカーすべて(短リンカー、長リンカー、および極長リンカー)に対して同様の結果が見いだされた。総合すると、これらの結果はTAPBPRのMHCI結合部位は、非表面結合状態では抗体フラグメントによりマスクされていることを示唆する。しかし、抗体フラグメントがその標的に結合すると、TAPBPR上のMHCクラスI結合部位が露出し、TAPBPRは細胞表面に発現したMHCクラスI分子上でペプチド交換触媒として機能することができるようになる。これはTAPBPRの機能が特異的マーカーを発現している望みの細胞型に向けられ得るという本質的な概念実証データを提供する。
GFPナノボディと融合させたTAPBPRの乳癌細胞株MCF-7への効果を決定した。MCF-7は極めて一般的なMHCクラスI分子であるHLA-A2を発現している。その細胞表面にGFPを発現させ(図9a)、TAPBPR-GFPナノボディ融合の前記細胞への結合能力を検査した(図9b)。繰り返しになるが、これは、TAPBPR-GFPナノボディ融合が表面にGFPを発現している細胞と結合するが、GFP陰性細胞とは結合しない(図9b)ことを明らかにし、TAPBPRの標的を、特定の細胞表面マーカーを発現している特異的細胞集団に絞る能力を証明した。次いでTAPBPR-GFP-ナノボディ融合が免疫原性ウィルスペプチド(CMV 由来NLVPMVATVペプチド)をMCF-7細胞に負荷する能力を検査した (図 9c)。これは、TAPBPR融合産物が的を絞ったやり方で(結合している抗体の特性に依存)、乳癌細胞株上にウィルスペプチドを負荷することができることを証明した。
TAPBPR-GFP-ナノボディに関する上記データは、選択的にTAPBPRを標的に向ける能力に関係する概念実証データを提供するが、私たちは、次にTAPBPRが腫瘍細胞株に自然に見いだされるマーカーを標的にすることができるTAPBPR-抗体結合物を設計した。従って、私たちは腫瘍細胞マーカーErbB2 (Her2)に対する特異性を有するscFvにリンクしたTAPBPRを作成した (図 10)。
私たちは、TAPBPR-抗体融合蛋白質がMHCクラスIに結合することができず、その結果、抗体に対するリガンド不在下では、ペプチド交換を仲介できないように見えることを観察した。私たちの所見は、TAPBPR上のMHCクラスI結合部位は細胞膜に結合していない状態の抗体フラグメント(ナノボディあるいはscFV)によりマスクされているが、その後、抗体が細胞表面のそのリガンドに結合すると露出することを示唆している。非結合状態の抗体融合フラグメントによる立体障害を支持するデータは、図8のTAPBPR-GFPナノボディ融合蛋白質(HeLa細胞上、すなわち表面GFPを発現していない)および図12のTAPBPR-Her2-scFv(Her2がノックアウトされたHeLa細胞上)に関して見いだされる。図8および図12の両者において、私たちは、可溶性TAPBPR単独でもHeLa/HeLa-Her2KO細胞上に発現しているMHCクラスIによく結合するが、TAPBPR-抗体融合はこれら細胞(HeLa上は図8B & C、 HeLa-Her2KO上は図12B & C)上に発現しているHLA分子に結合できないことを観察した。しかし、抗体に対するリガンドが細胞上に存在する場合、TAPBPR融合蛋白質は極めてよく細胞に結合できた(HeLa+GFP上は図8B & C、 HeLa上は図12B & C)。同様に、私たちがTAPBPR-抗体融合蛋白質の外来性ペプチド負荷仲介能力を検査したとき、図8および図12の両者において可溶性TAPBPRが単独で効率よくHeLa/HeLa- Her2KO細胞上に発現しているMHCクラスIに外来性ペプチドを負荷することができ、TAPBPR-抗体融合はこれら細胞にペプチドを負荷することができないことを観察した(HeLa細胞は図8B & C、 HeLa-Her2KO細胞は図12B & C)。繰り返しになるが、抗体に対するリガンドが細胞上に存在している場合、TAPBPR融合蛋白質は効率的に外来性ペプチド負荷を仲介することができた(HeLa+GFPは図8B & C、 HeLaは図12B & C)。この興味ある観察結果は、こうした生成物は選ばれた標的に対し高度に選択的であり、MHCクラスIのみを発現している健常細胞に対する効果が限定的であろうと思われることより、TAPBPR融合蛋白質の治療的応用を考える場合、有望であるといえる。
TAPBPR-Her2-svFcは、HLA-A2 を発現しているHeLa細胞上にウィルスペプチドを負荷することが示された(図 13a-d)。さらに、T細胞レセプター(TCR)は負荷を受けた細胞を認識することが示された(図 13e)。これはTAPBPR-Her2-svFcが機能し、生じるペプチド/MHCクラスI複合体がT細胞結合のためにアクセス可能であることを示している。
マウスTAPBPRの内腔ドメインはHeLa細胞上に発現しているHLA-A68(MHCクラスI)に蛍光性ヒト新生抗原ETVSK*QSNV(ETV*)を負荷することが示された。こうした結果は、外来性マウスTAPBPRをヒトTAPBPRと同様の方法で、ヒトMHCクラスI分子に免疫原性ペプチドを負荷するために使用することもできることを証明している。ヒトTAPBPRよりも効率は落ちるが、マウスTAPBPRは依然としてT細胞反応を惹起する十分な免疫原性ペプチドを負荷することができる。
TAPBPRWT、TAPBPRのTMDを有するが細胞質尾部を欠く TAPBPRtailless および細胞質尾部がCD8で置換されたTAPBPRCD8tailで形質導入された細胞は、TAPBPRで形質導入されていない細胞に比べた場合、細胞表面にTAPBPRを発現すること(図 16A) および蛍光性ペプチドETVSK*QSNV (ETV*)を結合すること (図16B)が示された。図16の結果は、細胞質尾部を欠くTAPBPRは効率的に細胞表面に発現されることを証明している。TAPBPRCD8tailを発現している細胞の形質導入は低レベルであるが、CD8細胞質尾部をTAPBPRに追加すると低い形質導入効率でより効率的な細胞発現が可能である。TAPBPR発現細胞が同様の細胞集団でゲーティングされると(Aのゲート参照)、すべてのTAPBPR変異型は同程度のペプチド編集が可能である(B)。
上記の結果は、可溶性TAPBPRは標的細胞に免疫原性ペプチドを結合させ、腫瘍に対するT細胞反応を増強するために使用できることを示すが、私たちは次に、これが腫瘍細胞の殺滅を増強するかどうかを決定した。私たちは、ヒトTAPBPRおよび極めて低い濃度のSIINFEKLペプチド存在下で、OT1T細胞によるマウスEL4細胞の殺滅を評価した。可溶性ヒトTAPBPRWTはEL4細胞に結合し、(図19a) EL4上に発現しているH-2Kb へのSIINFEKL の負荷を有意に増強した(図19b, 19c & 19d)。私たちがH-2Kbの存在下で SIINFEKLを認識する OT1細胞傷害性T細胞の、ペプチドをパルスされた EL4標的細胞を溶解する能力を検査したとき、私たちは可溶性ヒト TAPBPRWT 存在下で殺滅の有意な増強を観察したが、TAPBPRTN5の存在下では、これは見られなかった(図 19e)。こうした結果は、TAPBPRがペプチド−特異的CD8+ Tリンパ球による腫瘍の殺滅増強に使用可能であることを証明している。
1.Sharma, P et al Cell 168, 707-723, doi:10.1016/j.cell.2017.01.017 (2017).
2.Pardoll, D. M. Cancer 12, 252-264, doi: 10.1038/nrc3239 (2012).
3.Rizvi, N. A. et al.Science 348, 124-128, doi:10.1126/science.aaa1348 (2015).
4.Snyder, A. et al.The New England journal of medicine 371, 2189-2199, doi:10.1056/NEJMoa1406498 (2014).
5.Van Allen, E. M. et al.Science 350, 207-211, doi:10.1126/science.aad0095 (2015).
6.Lauss, M. et al..Nature Communications 8, 1738, doi: 10.1038/s41467-017-01460-0 (2017).
7.Mittal, D et al.Curr Opin Immunol 27, 16-25, doi:10.1016/j.coi.2014.01.004 (2014).
8.Hermann, C. et al. eLife 4, e09617, doi:10.7554/eLife.09617 (2015).
9.Morozov, G. I. et al.Proc Natl Acad Sci U S A 113, E1006-1015, doi:10.1073/pnas.1519894113 (2016).
10.Neerincx, A. et al. eLife 6, doi:10.7554/eLife.23049 (2017).
11.Boyle, L. H. et al.Proc Natl Acad Sci U S A 110, 3465-3470, doi:10.1073/pnas.1222342110 1222342110 [pii] (2013).
12.Nilsson, T et al.Cell 58, 707-718 (1989).
13.Li, S., et al.Proc Natl Acad Sci U S A 94, 8708-8713 (1997).
14.Ortmann, B. et al.Science 277, 1306-1309 (1997).
15.Hermann, C., et al J Immunol 191, 5743-5750, doi:10.4049/jimmunol.1300929 (2013).
16.Wills, M. R. et al.J Virol 70, 7569-7579 (1996).
17.Sim, A. C. et al.Scientific reports 3, 3232, doi:10.1038/srep03232 (2013).
18.Wearsch, P. A. et al Nat Immunol 8, 873-881, doi: ni1485 [pii] 10.1038/ni1485 (2007).
19.Chen, M. et al EMBO J 26, 1681-1690, doi: 7601624 [pii] 10.1038/sj.emboj.7601624 (2007).
20.Jiang, J. et al.Science 358, 1064-1068, doi:10.1126/science.aao5154 (2017).
21.Thomas, C. et al.Science 358, 1060-1064, doi:10.1126/science.aao6001 (2017).
22.Li, Z et al.Proc Natl Acad Sci U S A 110, 5004-5009, doi:10.1073/pnas.1218620110 (2013).
23.Tiwari, R. K. et al EMBO J 6, 3373-3378 (1987).
24.Barnstable, C. J. et al.Cell 14, 9-20 (1978).
25.Stam, N. J. et al Journal of Immunology 137, 2299-2306 (1986).
26.Perosa, F. et al.J Immunol 171, 1918-1926 (2003).
27.Hogan, K. T. et al.Cancer research 58, 5144-5150 (1998).
28.Parkhurst MR, et al.(1996) J Immunol 157(6):2539-2548.
29.Valmori D, et al.(1998) J Immunol 160(4):1750-1758.
30.Theobald M, et al.(1995) Proc Natl Acad Sci U S A 92(26):11993-11997.
31.Kawashima I, et al.(1998) Hum Immunol 59(1):1-14.
Aagccccacccagcagaggggcagtggcgggcagtggacgtggtcctagactgtttcctggtgaaggacggtgcgcaccgtggagctctcgccagcagtgaggacagggcaagggcctcccttgtgctgaagcaggtgccagtgctggacgatggctccctggaggacttcaccgatttccaagggggcacactggcccaagatgacccacctattatctttgaggcctcagtggacctggtccagattccccaggccgaggccttgctccatgctgactgcagtgggaaggaggtgacctgtgagatctcccgctactttctccagatgacagagaccactgttaagacagcagcttggttcatggccaacgtgcaggtctctggagggggacctagcatctccttggtgatgaagactcccagggtcgccaagaatgaggtgctctggcacccaacgctgaacttgccactgagcccccaggggactgtgcgaactgcagtggagttccaggtgatgacacagacccaatccctgagcttcctgctggggtcctcagcctccttggactgtggcttctccatggcaccgggcttggacctcatcagtgtggagtggcgactgcagcacaagggcaggggtcagttggtgtacagctggaccgcagggcaggggcaggctgtgcggaagggcgctaccctggagcctgcacaactgggcatggccagggatgcctccctcaccctgcccggcctcactatacaggacgaggggacctacatttgccagatcaccacctctctgtaccgagctcagcagatcatccagctcaacatccaagcttcccctaaagtacgactgagcttggcaaacgaagctctgctgcccaccctcatctgcgacattgctggctattaccctctggatgtggtggtgacgtggacccgagaggagctgggtgggtccccagcccaagtctctggtgcctccttctccagcctcaggcaaagcgtggcaggcacctacagcatctcctcctctctcaccgcagaacctggctctgcaggtgccacttacacctgccaggtcacacacatctctctggaggagccccttggggccagcacccaggttgtcccaccagagcggaga
SEQ ID NO: 1 -nucleotide sequence encoding TAPBPR luminal domain (mature sequence without 21 aa leader sequence)
KPHPAEGQWRAVDVVLDCFLVKDGAHRGALASSEDRARASLVLKQVPVLDDGSLEDFTDFQGGTLAQDDPPIIFEASVDLVQIPQAEALLHADCSGKEVTCEISRYFLQMTETTVKTAAWFMANVQVSGGGPSISLVMKTPRVAKNEVLWHPTLNLPLSPQGTVRTAVEFQVMTQTQSLSFLLGSSASLDCGFSMAPGLDLISVEWRLQHKGRGQLVYSWTAGQGQAVRKGATLEPAQLGMARDASLTLPGLTIQDEGTYICQITTSLYRAQQIIQLNIQASPKVRLSLANEALLPTLICDIAGYYPLDVVVTWTREELGGSPAQVSGASFSSLRQSVAGTYSISSSLTAEPGSAGATYTCQVTHISLEEPLGASTQVVPPERR
25 SEQ ID NO: 2 -TAPBPR luminal domain (mature sequence without 21 aa leader sequence)

atgggcacacaggagggctggtgcctgctgctctgcctggctctatctggagcagcagaaaccaagccccacccagcagaggggcagtggcgggcagtggacgtggtcctagactgtttcctggtgaaggacggtgcgcaccgtggagctctcgccagcagtgaggacagggcaagggcctcccttgtgctgaagcaggtgccagtgctggacgatggctccctggaggacttcaccgatttccaagggggcacactggcccaagatgacccacctattatctttgaggcctcagtggacctggtccagattccccaggccgaggccttgctccatgctgactgcagtgggaaggaggtgacctgtgagatctcccgctactttctccagatgacagagaccactgttaagacagcagcttggttcatggccaacgtgcaggtctctggagggggacctagcatctccttggtgatgaagactcccagggtcgccaagaatgaggtgctctggcacccaacgctgaacttgccactgagcccccaggggactgtgcgaactgcagtggagttccaggtgatgacacagacccaatccctgagcttcctgctggggtcctcagcctccttggactgtggcttctccatggcaccgggcttggacctcatcagtgtggagtggcgactgcagcacaagggcaggggtcagttggtgtacagctggaccgcagggcaggggcaggctgtgcggaagggcgctaccctggagcctgcacaactgggcatggccagggatgcctccctcaccctgcccggcctcactatacaggacgaggggacctacatttgccagatcaccacctctctgtaccgagctcagcagatcatccagctcaacatccaagcttcccctaaagtacgactgagcttggcaaacgaagctctgctgcccaccctcatctgcgacattgctggctattaccctctggatgtggtggtgacgtggacccgagaggagctgggtgggtccccagcccaagtctctggtgcctccttctccagcctcaggcaaagcgtggcaggcacctacagcatctcctcctctctcaccgcagaacctggctctgcaggtgccacttacacctgccaggtcacacacatctctctggaggagccccttggggccagcacccaggttgtcccaccagagcggagacacgtgggtggcggtggctccggtggcggtggctccggtggcggtggctccactagtgactccccagacaggccctggaacccccccaccttcttcccagccctgctcgtggtgaccgaaggggacaacgccaccttcacctgcagcttctccaacacatcggagagcttccacgtgatctggcaccgcgagagccccagcggccagacggacaccctggccgccttccccgaggaccgcagccagcccggccaggactgccgcttccgtgtcacacaactgcccaacgggcgtgacttccacatgagcgtggtcagggcccggcgcaatgacagcggcacctacgtgtgtggggtgatctccctggcccccaagatccagatcaaagagagcctg
SEQ ID NO: 5 nucleotide sequence encoding sTAPBPR-sPD1
atgggcacacaggagggctggtgcctgctgctctgcctggctctatctggagcagcagaaaccaagccccacccagcagaggggcagtggcgggcagtggacgtggtcctagactgtttcctggtgaaggacggtgcgcaccgtggagctctcgccagcagtgaggacagggcaagggcctcccttgtgctgaagcaggtgccagtgctggacgatggctccctggaggacttcaccgatttccaagggggcacactggcccaagatgacccacctattatctttgaggcctcagtggacctggtccagattccccaggccgaggccttgctccatgctgactgcagtgggaaggaggtgacctgtgagatctcccgctactttctccagatgacagagaccactgttaagacagcagcttggttcatggccaacgtgcaggtctctggagggggacctagcatctccttggtgatgaagactcccagggtcgccaagaatgaggtgctctggcacccaacgctgaacttgccactgagcccccaggggactgtgcgaactgcagtggagttccaggtgatgacacagacccaatccctgagcttcctgctggggtcctcagcctccttggactgtggcttctccatggcaccgggcttggacctcatcagtgtggagtggcgactgcagcacaagggcaggggtcagttggtgtacagctggaccgcagggcaggggcaggctgtgcggaagggcgctaccctggagcctgcacaactgggcatggccagggatgcctccctcaccctgcccggcctcactatacaggacgaggggacctacatttgccagatcaccacctctctgtaccgagctcagcagatcatccagctcaacatccaagcttcccctaaagtacgactgagcttggcaaacgaagctctgctgcccaccctcatctgcgacattgctggctattaccctctggatgtggtggtgacgtggacccgagaggagctgggtgggtccccagcccaagtctctggtgcctccttctccagcctcaggcaaagcgtggcaggcacctacagcatctcctcctctctcaccgcagaacctggctctgcaggtgccacttacacctgccaggtcacacacatctctctggaggagccccttggggccagcacccaggttgtcccaccagagcggagacacgtgggtggcggtggctccggtggcggtggctccggtggcggtggctccactagtgcggatcttggatcccgggccatggcccaggtgcagctggtgcagtctggggcagaggtgaaaaagcccggggagtctctgaagatctcctgtaagggttctggatacagctttaccagctactggatcgcctgggtgcgccagatgcccgggaaaggcctggagtacatggggctcatctatcctggtgactctgacaccaaatacagcccgtccttccaaggccaggtcaccatctcagtcgacaagtccgtcagcactgcctacttgcaatggagcagtctgaagccctcggacagcgccgtgtatttttgtgcgagacatgacgtgggatattgcagtagttccaactgcgcaaagtggcctgaatacttccagcattggggccagggcaccctggtcaccgtctcctcaggtggaggcggttcaggcggaggtggctctggcggtggcggatcgcagtctgtgttgacgcagccgccctcagtgtctgcggccccaggacagaaggtcaccatctcctgctctggaagcagctccaacattgggaataattatgtatcctggtaccagcagctcccaggaacagcccccaaactcctcatctatgatcacaccaatcggcccgcaggggtccctgaccgattctctggctccaagtctggcacctcagcctccctggccatcagtgggttccggtccgaggatgaggctgattattactgtgcctcctgggactacaccctctcgggctgggtgttcggcggaggaaccaagctgaccgtcctaggtgcggccgccggcggaggaggatct
SEQ ID NO: 7 nucleotide sequence encoding TAPBPR-Her2scFv
atgggcacacaggagggctggtgcctgctgctctgcctggctctatctggagcagcagaaaccaagccccacccagcagaggggcagtggcgggcagtggacgtggtcctagactgtttcctggtgaaggacggtgcgcaccgtggagctctcgccagcagtgaggacagggcaagggcctcccttgtgctgaagcaggtgccagtgctggacgatggctccctggaggacttcaccgatttccaagggggcacactggcccaagatgacccacctattatctttgaggcctcagtggacctggtccagattccccaggccgaggccttgctccatgctgactgcagtgggaaggaggtgacctgtgagatctcccgctactttctccagatgacagagaccactgttaagacagcagcttggttcatggccaacgtgcaggtctctggagggggacctagcatctccttggtgatgaagactcccagggtcgccaagaatgaggtgctctggcacccaacgctgaacttgccactgagcccccaggggactgtgcgaactgcagtggagttccaggtgatgacacagacccaatccctgagcttcctgctggggtcctcagcctccttggactgtggcttctccatggcaccgggcttggacctcatcagtgtggagtggcgactgcagcacaagggcaggggtcagttggtgtacagctggaccgcagggcaggggcaggctgtgcggaagggcgctaccctggagcctgcacaactgggcatggccagggatgcctccctcaccctgcccggcctcactatacaggacgaggggacctacatttgccagatcaccacctctctgtaccgagctcagcagatcatccagctcaacatccaagcttcccctaaagtacgactgagcttggcaaacgaagctctgctgcccaccctcatctgcgacattgctggctattaccctctggatgtggtggtgacgtggacccgagaggagctgggtgggtccccagcccaagtctctggtgcctccttctccagcctcaggcaaagcgtggcaggcacctacagcatctcctcctctctcaccgcagaacctggctctgcaggtgccacttacacctgccaggtcacacacatctctctggaggagccccttggggccagcacccaggttgtcccaccagagcggagacacgtgggtggcggtggctccggtggcggtggctccggtggcggtggctccactagtcaggttcagctggttgaaagcggtggtgcactggttcagcctggtggtagcctgcgtctgagctgtgcagcaagcggttttccggttaatcgttatagcatgcgttggtatcgtcaggcaccgggtaaagaacgtgaatgggttgcaggtatgagcagtgccggtgatcgtagcagctatgaagatagcgttaaaggtcgttttaccatcagccgtgatgatgcacgtaataccgtttatctgcaaatgaatagcctgaaaccggaagataccgcagtgtattattgcaatgttaacgtgggctttgaatattggggtcagggcacccaggttaccgttagcagcaaa
SEQ ID NO: 9 nucleotide sequence encoding TAPBPR-GFP-NB
atgggcacacaggagggctggtgcctgctgctctgcctggctctatctggagcagcagaaaccaagccccacccagcagaggggcagtggcgggcagtggacgtggtcctagactgtttcctggtgaaggacggtgcgcaccgtggagctctcgccagcagtgaggacagggcaagggcctcccttgtgctgaagcaggtgccagtgctggacgatggctccctggaggacttcaccgatttccaagggggcacactggcccaagatgacccacctattatctttgaggcctcagtggacctggtccagattccccaggccgaggccttgctccatgctgactgcagtgggaaggaggtgacctgtgagatctcccgctactttctccagatgacagagaccactgttaagacagcagcttggttcatggccaacgtgcaggtctctggagggggacctagcatctccttggtgatgaagactcccagggtcgccaagaatgaggtgctctggcacccaacgctgaacttgccactgagcccccaggggactgtgcgaactgcagtggagttccaggtgatgacacagacccaatccctgagcttcctgctggggtcctcagcctccttggactgtggcttctccatggcaccgggcttggacctcatcagtgtggagtggcgactgcagcacaagggcaggggtcagttggtgtacagctggaccgcagggcaggggcaggctgtgcggaagggcgctaccctggagcctgcacaactgggcatggccagggatgcctccctcaccctgcccggcctcactatacaggacgaggggacctacatttgccagatcaccacctctctgtaccgagctcagcagatcatccagctcaacatccaagcttcccctaaagtacgactgagcttggcaaacgaagctctgctgcccaccctcatctgcgacattgctggctattaccctctggatgtggtggtgacgtggacccgagaggagctgggtgggtccccagcccaagtctctggtgcctccttctccagcctcaggcaaagcgtggcaggcacctacagcatctcctcctctctcaccgcagaacctggctctgcaggtgccacttacacctgccaggtcacacacatctctctggaggagccccttggggccagcacccaggttgtcccaccagagcggagacacgtgggtggcggtggctccggtggcggtggctccggtggcggtggctccactagtgaggtgcagctggtggagagcggcggcggcctggtgcagcccggcggcagcctgagactgagctgcgccgccagcggcttcaccctggactactacgccatcggctggttcagacaggcccccggcaaggagagagagtgggccagcagcatcagcagcagcgacggcagcacctactacgccgacagcgtgaagggcagattcaccatcagcagagacaacgccaagaacaccgtgttcctgcagatgaacagcctgaagcccgaggacaccgccgtgtacagctgcgccgccagccaggcccccatcaccatcgccaccatgatgaagcccttctacgactactggggccagggcacccaggtgaccgtgagcagcggcggaggaggatct
SEQ ID NO: 11 nucleotide sequence encoding sTAPBPR-LONG-PD-L1-NB1
atgggcacacaggagggctggtgcctgctgctctgcctggctctatctggagcagcagaaaccaagccccacccagcagaggggcagtggcgggcagtggacgtggtcctagactgtttcctggtgaaggacggtgcgcaccgtggagctctcgccagcagtgaggacagggcaagggcctcccttgtgctgaagcaggtgccagtgctggacgatggctccctggaggacttcaccgatttccaagggggcacactggcccaagatgacccacctattatctttgaggcctcagtggacctggtccagattccccaggccgaggccttgctccatgctgactgcagtgggaaggaggtgacctgtgagatctcccgctactttctccagatgacagagaccactgttaagacagcagcttggttcatggccaacgtgcaggtctctggagggggacctagcatctccttggtgatgaagactcccagggtcgccaagaatgaggtgctctggcacccaacgctgaacttgccactgagcccccaggggactgtgcgaactgcagtggagttccaggtgatgacacagacccaatccctgagcttcctgctggggtcctcagcctccttggactgtggcttctccatggcaccgggcttggacctcatcagtgtggagtggcgactgcagcacaagggcaggggtcagttggtgtacagctggaccgcagggcaggggcaggctgtgcggaagggcgctaccctggagcctgcacaactgggcatggccagggatgcctccctcaccctgcccggcctcactatacaggacgaggggacctacatttgccagatcaccacctctctgtaccgagctcagcagatcatccagctcaacatccaagcttcccctaaagtacgactgagcttggcaaacgaagctctgctgcccaccctcatctgcgacattgctggctattaccctctggatgtggtggtgacgtggacccgagaggagctgggtgggtccccagcccaagtctctggtgcctccttctccagcctcaggcaaagcgtggcaggcacctacagcatctcctcctctctcaccgcagaacctggctctgcaggtgccacttacacctgccaggtcacacacatctctctggaggagccccttggggccagcacccaggttgtcccaccagagcggagacacgtgggtggcggtggctccggtggcggtggctccggtggcggtggctccactagtgaggtgcagctggtggagagcggcggcggcctggtgcagcccggcggcagcctgagactgagctgcgccgccagcggcttcaccctggactactacgccaagtgctggttcagacaggcccccggcaaggagagagagtgggtgagctgcatcagcagcagcgacggcagcacctactacgccgacagcgtgaagggcagattcaccatcagcagagacaacgccaagaacaccgtgtacctgcagatgaacagcctgaagcccgaggacaccgccgtgtacttctgcgccgccagacacggcggccccctgaccgtggagtacttcttcgactactggggccagggcacccaggtgaccgtgagcagcggcggaggaggatct
SEQ ID NO: 13 nucleotide sequence encoding sTAPBPR-LONG-PD-L1-NB2
atgggcacacaggagggctggtgcctgctgctctgcctggctctatctggagcagcagaaaccaagccccacccagcagaggggcagtggcgggcagtggacgtggtcctagactgtttcctggtgaaggacggtgcgcaccgtggagctctcgccagcagtgaggacagggcaagggcctcccttgtgctgaagcaggtgccagtgctggacgatggctccctggaggacttcaccgatttccaagggggcacactggcccaagatgacccacctattatctttgaggcctcagtggacctggtccagattccccaggccgaggccttgctccatgctgactgcagtgggaaggaggtgacctgtgagatctcccgctactttctccagatgacagagaccactgttaagacagcagcttggttcatggccaacgtgcaggtctctggagggggacctagcatctccttggtgatgaagactcccagggtcgccaagaatgaggtgctctggcacccaacgctgaacttgccactgagcccccaggggactgtgcgaactgcagtggagttccaggtgatgacacagacccaatccctgagcttcctgctggggtcctcagcctccttggactgtggcttctccatggcaccgggcttggacctcatcagtgtggagtggcgactgcagcacaagggcaggggtcagttggtgtacagctggaccgcagggcaggggcaggctgtgcggaagggcgctaccctggagcctgcacaactgggcatggccagggatgcctccctcaccctgcccggcctcactatacaggacgaggggacctacatttgccagatcaccacctctctgtaccgagctcagcagatcatccagctcaacatccaagcttcccctaaagtacgactgagcttggcaaacgaagctctgctgcccaccctcatctgcgacattgctggctattaccctctggatgtggtggtgacgtggacccgagaggagctgggtgggtccccagcccaagtctctggtgcctccttctccagcctcaggcaaagcgtggcaggcacctacagcatctcctcctctctcaccgcagaacctggctctgcaggtgccacttacacctgccaggtcacacacatctctctggaggagccccttggggccagcacccaggttgtcccaccagagcggagacacgtgggtggcggtggctccggtggcggtggctccggtggcggtggctccactagtgaggtgcagctggtggagagcggcggcggcctggtgcaggccggcggcagcctgagactgagctgcgccgccagcggcagcaccttcagccagtacgacgtgggctggtacagacaggcccccggcaagcagagagagctggtggccttcagcagcagcggcggcagaaccatctaccccgacagcgtgaagggcagattcaccttcagcagagacaacaccaagaacaccgtgtacctgcagatgaccagcctgaagcccgaggacaccgccgtgtactactgcaagatcgactggtacctgaacagctactggggccagggcacccaggtgaccgtgagcagcggcggaggaggatct
SEQ ID NO: 15 nucleotide sequence encoding sTAPBPR-LONG-PD-L1-NB4
atgggcacacaggagggctggtgcctgctgctctgcctggctctatctggagcagcagaaaccaagccccacccagcagaggggcagtggcgggcagtggacgtggtcctagactgtttcctggtgaaggacggtgcgcaccgtggagctctcgccagcagtgaggacagggcaagggcctcccttgtgctgaagcaggtgccagtgctggacgatggctccctggaggacttcaccgatttccaagggggcacactggcccaagatgacccacctattatctttgaggcctcagtggacctggtccagattccccaggccgaggccttgctccatgctgactgcagtgggaaggaggtgacctgtgagatctcccgctactttctccagatgacagagaccactgttaagacagcagcttggttcatggccaacgtgcaggtctctggagggggacctagcatctccttggtgatgaagactcccagggtcgccaagaatgaggtgctctggcacccaacgctgaacttgccactgagcccccaggggactgtgcgaactgcagtggagttccaggtgatgacacagacccaatccctgagcttcctgctggggtcctcagcctccttggactgtggcttctccatggcaccgggcttggacctcatcagtgtggagtggcgactgcagcacaagggcaggggtcagttggtgtacagctggaccgcagggcaggggcaggctgtgcggaagggcgctaccctggagcctgcacaactgggcatggccagggatgcctccctcaccctgcccggcctcactatacaggacgaggggacctacatttgccagatcaccacctctctgtaccgagctcagcagatcatccagctcaacatccaagcttcccctaaagtacgactgagcttggcaaacgaagctctgctgcccaccctcatctgcgacattgctggctattaccctctggatgtggtggtgacgtggacccgagaggagctgggtgggtccccagcccaagtctctggtgcctccttctccagcctcaggcaaagcgtggcaggcacctacagcatctcctcctctctcaccgcagaacctggctctgcaggtgccacttacacctgccaggtcacacacatctctctggaggagccccttggggccagcacccaggttgtcccaccagagcggagaacagccttgggagtcatctttgccagcagtctcttccttcttgcactgatgttcctggggcttcagagacggcaagcacctacaggacttgggctgcttcaggctgaacgctgggagaccacttcctgtgctgacacacagagctcccatctccatgaagaccgcacagcgcgtgtaagccagcccagctga
SEQ ID NO: 17 nucleotide sequence encoding hTAPBPR full length (with TMD and cytoplasmic tail)
(V41_G151_V169 polymorphisms as used in our studies)
atgggcttggagcccagctggtatctgctgctctgtttggctgtctctggggcagcagggactgaccctcccacagcgcccaccacagcagaaagacagcggcagcccacggacatcatcttagactgcttcttggtgacagaagacaggcaccgcggggcttttgccagcagtggggacagggagagggccttgcttgtgctgaagcaggtaccagtgctggatgatggctccctggaaggcatcacagatttccaggggagcactgagaccaaacaggattcacctgttatctttgaggcctcagtggacttggtacagattccccaggcagaggcgttgctccatgctgactgcagcgggaaggcagtgacctgcgagatctccaagtatttcctccaggccagacaagaggccacttttgagaaagcacattggttcatcagcaacatgcaggtttctagaggtggccccagtgtctccatggtgatgaagactctaagagatgctgaagttggagctgtccggcaccctacactgaacctacctctgagtgcccagggcacagtgaagactcaagtggagttccaggtgacatcagagacccaaaccctgaaccacctgctggggtcctctgtctccctgcactgcagtttctccatggcaccagacctggacctcactggcgtggagtggcggctgcagcataaaggcagcggccagctggtgtacagctggaagacagggcaggggcaggccaagcgcaagggcgctacactggagcctgaggagctactcagggctggaaacgcctctctcaccttacccaacctcactctaaaggatgaggggacctacatctgccagatctccacctctctgtatcaagctcaacagatcatgccacttaacatcctggctccccccaaagtacaactgcacttggcaaacaaggaccctctgccttccctcgtctgcagcattgccggctactatcctctggatgtgggagtgacgtggattcgagaggagctgggtggaattccagcccaagtctctggtgcctccttctccagcctcaggcagagcacgatgggaacctacagcatttcttccacggtgatggctgacccaggccccacaggtgccacttatacctgccaagtcgcccacgtctccctggaggagccccttacaaccagcatgagggttttgccaaatccagagcagagaggaaccttgggagtcatctttgccagcatcatcttcctttctgcgctgttgttgtttctgggacttcacagacagcaagcttcttcgtcaaggtccaccaggcctatgaggcattctgggtga
SEQ ID NO: 19 nucleotide sequence encoding mouse TAPBPR full length
MGLEPSWYLLLCLAVSGAAGTDPPTAPTTAERQRQPTDIILDCFLVTEDRHRGAFASSGDRERALLVLKQVPVLDDGSLEGITDFQGSTETKQDSPVIFEASVDLVQIPQAEALLHADCSGKAVTCEISKYFLQARQEATFEKAHWFISNMQVSRGGPSVSMVMKTLRDAEVGAVRHPTLNLPLSAQGTVKTQVEFQVTSETQTLNHLLGSSVSLHCSFSMAPDLDLTGVEWRLQHKGSGQLVYSWKTGQGQAKRKGATLEPEELLRAGNASLTLPNLTLKDEGTYICQISTSLYQAQQIMPLNILAPPKVQLHLANKDPLPSLVCSIAGYYPLDVGVTWIREELGGIPAQVSGASFSSLRQSTMGTYSISSTVMADPGPTGATYTCQVAHVSLEEPLTTSMRVLPNPEQRGTLGVIFASIIFLSALLLFLGLHRQQASSSRSTRPMRHSG*
SEQ ID NO: 20 mouse TAPBPR full length
gaccctcccacagcgcccaccacagcagaaagacagcggcagcccacggacatcatcttagactgcttcttggtgacagaagacaggcaccgcggggcttttgccagcagtggggacagggagagggccttgcttgtgctgaagcaggtaccagtgctggatgatggctccctggaaggcatcacagatttccaggggagcactgagaccaaacaggattcacctgttatctttgaggcctcagtggacttggtacagattccccaggcagaggcgttgctccatgctgactgcagcgggaaggcagtgacctgcgagatctccaagtatttcctccaggccagacaagaggccacttttgagaaagcacattggttcatcagcaacatgcaggtttctagaggtggccccagtgtctccatggtgatgaagactctaagagatgctgaagttggagctgtccggcaccctacactgaacctacctctgagtgcccagggcacagtgaagactcaagtggagttccaggtgacatcagagacccaaaccctgaaccacctgctggggtcctctgtctccctgcactgcagtttctccatggcaccagacctggacctcactggcgtggagtggcggctgcagcataaaggcagcggccagctggtgtacagctggaagacagggcaggggcaggccaagcgcaagggcgctacactggagcctgaggagctactcagggctggaaacgcctctctcaccttacccaacctcactctaaaggatgaggggacctacatctgccagatctccacctctctgtatcaagctcaacagatcatgccacttaacatcctggctccccccaaagtacaactgcacttggcaaacaaggaccctctgccttccctcgtctgcagcattgccggctactatcctctggatgtgggagtgacgtggattcgagaggagctgggtggaattccagcccaagtctctggtgcctccttctccagcctcaggcagagcacgatgggaacctacagcatttcttccacggtgatggctgacccaggccccacaggtgccacttatacctgccaagtcgcccacgtctccctggaggagccccttacaaccagcatgagggttttgccaaatccagagcagagaggaacc
SEQ ID NO: 21 nucleotide sequence encoding luminal domains of mouse TAPBPR
DPPTAPTTAERQRQPTDIILDCFLVTEDRHRGAFASSGDRERALLVLKQVPVLDDGSLEGITDFQGSTETKQDSPVIFEASVDLVQIPQAEALLHADCSGKAVTCEISKYFLQARQEATFEKAHWFISNMQVSRGGPSVSMVMKTLRDAEVGAVRHPTLNLPLSAQGTVKTQVEFQVTSETQTLNHLLGSSVSLHCSFSMAPDLDLTGVEWRLQHKGSGQLVYSWKTGQGQAKRKGATLEPEELLRAGNASLTLPNLTLKDEGTYICQISTSLYQAQQIMPLNILAPPKVQLHLANKDPLPSLVCSIAGYYPLDVGVTWIREELGGIPAQVSGASFSSLRQSTMGTYSISSTVMADPGPTGATYTCQVAHVSLEEPLTTSMRVLPNPEQRGT*
SEQ ID NO: 22 luminal domains of mouse TAPBPR
ADLGSRAMAQVQLVQSGAEVKKPGESLKISCKGSGYSFTSYWIAWVRQMPGKGLEYMGLIYPGDSDTKYSPSFQGQVTISVDKSVSTAYLQWSSLKPSDSAVYFCARHDVGYCSSSNCAKWPEYFQHWGQGTLVTVSSGGGGSGGGGSGGGGSQSVLTQPPSVSAAPGQKVTISCSGSSSNIGNNYVSWYQQLPGTAPKLLIYDHTNRPAGVPDRFSGSKSGTSASLAISGFRSEDEADYYCASWDYTLSGWVFGGGTKLTVLGAAA
SEQ ID NO: 23 Anti-HER2 svFc
DIQMTQSPSSLSASVGDRVTITCRASQDVNTAVAWYQQKPGKAPKLLIYSASFLYSGVPSRFSGSRSGTDFTLTISSLQPEDFATYYCQQHYTTPPTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
SEQ ID NO: 24 Anti-HER2 (trastuzumab) Light chain
EVQLVESGGGLVQPGGSLRLSCAASGFNIKDTYIHWVRQAPGKGLEWVARIYPTNGYTRYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCSRWGGDGFYAMDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
SEQ ID NO: 25 Anti-HER2 (trastuzumab) Heavy chain
EVQLVESGGGLVQPGGSLRLSCAASGFTLDYYAIGWFRQAPGKEREWASSISSSDGSTYYADSVKGRFTISRDNAKNTVFLQMNSLKPEDTAVYSCAASQAPITIATMMKPFYDYWGQGTQVTVSS
SEQ ID NO: 26 Anti-PDL1 nanobody
EVQLVESGGGLVQPGGSLRLSCAASGFTLDYYAKCWFRQAPGKEREWVSCISSSDGSTYYADSVKGRFTISRDNAKNTVYLQMNSLKPEDTAVYFCAARHGGPLTVEYFFDYWGQGTQVTVSS
SEQ ID NO: 27 Anti-PDL1 nanobody
EVQLVESGGGLVQPGGSLRLSCAASGFTFDYYAIGWFRQAPGKAREGVSCISGGDNSTYYADSVKGRFTISRDNAKNTVYLQMNSLKPEDTAVYYCATGGWKYCSGYDPEYIYWGQGTQVTVSS
SEQ ID NO: 28 Anti-PDL1 nanobody
EVQLVESGGGLVQAGGSLRLSCAASGSTFSQYDVGWYRQAPGKQRELVAFSSSGGRTIYPDSVKGRFTFSRDNTKNTVYLQMTSLKPEDTAVYYCKIDWYLNSYWGQGTQVTVSS
SEQ ID NO: 29 Anti-PDL1 nanobody
EVQLVESGGGLVQAGGSLRLSCAASGVDASNSAMGWYRQAPGKQREWVARITGGGLIAYTDSVKGRFTISRDNAKSTVYLQMWSLEPEDTAVYYCNTINSRDGWGQGTQVTVSS
SEQ ID NO: 30 Anti-PDL1 nanobody
EVQLVESGGGLVQAGGSLTISCAASGITFSDSIVSWYRRARGKQREWVAGISNGGTTKYAESVLGRFTISRDNAKNMVYLQMWGLNPEDTAVYLCKVRQYWGQGTQVTVSS
SEQ ID NO: 31 Anti-PDL1 nanobody
TALGVIFASSLFLLALMFLGL
SEQ ID NO: 32 Human TAPBPR TMD
atgggcacacaggagggctggtgcctgctgctctgcctggctctatctggagcagcagaaaccaagccccacccagcagaggggcagtggcgggcagtggacgtggtcctagactgtttcctggtgaaggacggtgcgcaccgtggagctctcgccagcagtgaggacagggcaagggcctcccttgtgctgaagcaggtgccagtgctggacgatggctccctggaggacttcaccgatttccaagggggcacactggcccaagatgacccacctattatctttgaggcctcagtggacctggtccagattccccaggccgaggccttgctccatgctgactgcagtgggaaggaggtgacctgtgagatctcccgctactttctccagatgacagagaccactgttaagacagcagcttggttcatggccaacgtgcaggtctctggagggggacctagcatctccttggtgatgaagactcccagggtcgccaagaatgaggtgctctggcacccaacgctgaacttgccactgagcccccaggggactgtgcgaactgcagtggagttccaggtgatgacacagacccaatccctgagcttcctgctggggtcctcagcctccttggactgtggcttctccatggcaccgggcttggacctcatcagtgtggagtggcgactgcagcacaagggcaggggtcagttggtgtacagctggaccgcagggcaggggcaggctgtgcggaagggcgctaccctggagcctgcacaactgggcatggccagggatgcctccctcaccctgcccggcctcactatacaggacgaggggacctacatttgccagatcaccacctctctgtaccgagctcagcagatcatccagctcaacatccaagcttcccctaaagtacgactgagcttggcaaacgaagctctgctgcccaccctcatctgcgacattgctggctattaccctctggatgtggtggtgacgtggacccgagaggagctgggtgggtccccagcccaagtctctggtgcctccttctccagcctcaggcaaagcgtggcaggcacctacagcatctcctcctctctcaccgcagaacctggctctgcaggtgccacttacacctgccaggtcacacacatctctctggaggagccccttggggccagcacccaggttgtcccaccagagcggagaacagccttgggagtcatctttgccagcagtctcttccttcttgcactgatgttcctggggcttcagagacgaagacgtgtttgcaaatgtccccggcctgtggtcaaatcgggagacaagcccagcctttcggcgagatacgtctaa
SEQ ID NO: 33 nucleotide sequence encoding hTAPBPR-CD8 cytoplasmic tail
MGTQEGWCLLLCLALSGAAETKPHPAEGQWRAVDVVLDCFLVKDGAHRGALASSEDRARASLVLKQVPVLDDGSLEDFTDFQGGTLAQDDPPIIFEASVDLVQIPQAEALLHADCSGKEVTCEISRYFLQMTETTVKTAAWFMANVQVSGGGPSISLVMKTPRVAKNEVLWHPTLNLPLSPQGTVRTAVEFQVMTQTQSLSFLLGSSASLDCGFSMAPGLDLISVEWRLQHKGRGQLVYSWTAGQGQAVRKGATLEPAQLGMARDASLTLPGLTIQDEGTYICQITTSLYRAQQIIQLNIQASPKVRLSLANEALLPTLICDIAGYYPLDVVVTWTREELGGSPAQVSGASFSSLRQSVAGTYSISSSLTAEPGSAGATYTCQVTHISLEEPLGASTQVVPPERRTALGVIFASSLFLLALMFLGLQRRRRVCKCPRPVVKSGDKPSLSARYV*
SEQ ID NO: 34 TAPBPR-CD8 cytoplasmic tail
Claims (46)
- TAP結合蛋白質関連(TAPBPR)のフラグメントを含む単離ペプチド交換蛋白質であって、TAPBPR内腔ドメインから成る前記フラグメント。
- SEQ ID NO: 2もしくはSEQ ID NO:22と少なくとも50%の配列同一性を持つアミノ酸配列から成るTAPBPRフラグメントである請求項1に記載の単離ペプチド交換蛋白質。
- さらに標的ドメインを含む請求項1もしくは請求項2に記載の単離ペプチド交換蛋白質。
- 標的ドメインがリンカーを介してTAPBPRフラグメントと結合している請求項3に記載の単離ペプチド交換蛋白質。
- 標的ドメインが標的細胞に特異的に結合する請求項3もしくは請求項4に記載の単離ペプチド交換蛋白質。
- 標的細胞が癌細胞である請求項5に記載の単離ペプチド交換蛋白質。
- 標的ドメインが癌細胞表面の標的分子に特異的に結合する請求項6に記載の単離ペプチド交換蛋白質。
- 標的分子が、ErbB2, PDL1またはCD20である請求項7に記載の単離ペプチド交換蛋白質。
- 標的細胞が感染を受けた細胞である、請求項5に記載の単離ペプチド交換蛋白質。
- 病原体感染細胞がHIV、CMV、EBV、HPV、インフルエンザあるいは肝炎である、請求項9に記載の単離ペプチド交換蛋白質。
- 標的ドメインが、感染を受けた細胞表面の病原体抗原もしくは病原体感染により発現亢進した分子と特異的に結合する、請求項9もしくは請求項10に記載の単離ペプチド交換蛋白質。
- 病原体抗原がgp120およびgp41から選ばれたHIV抗原;UL11、UL142、UL9、UL1、UL5、UL16、UL55、UL74、UL75およびUL155 (gL)から選ばれたCMV抗原もしくはヘモアグルチニンおよびノリラミニダーゼから選択されたインフルエンザ抗原である、請求項10に記載の単離ペプチド交換蛋白質。
- 標的細胞が抗原提示細胞である、請求項3もしくは請求項4に記載の単離ペプチド交換蛋白質。
- 抗原提示細胞が樹状細胞である、請求項13に記載の単離ペプチド交換蛋白質。
- 標的ドメインが抗体分子、場合によりscFvあるいはナノボディである、請求項3から14のいずれかに記載の単離ペプチド交換蛋白質。
- 標的ドメインが標的細胞の表面レセプターに対するリガンドである、請求項3から14までのいずれかに記載の単離ペプチド交換蛋白質。
- 標的ドメインがCD4、CD20、PD1もしくは抗体のFcドメインである、請求項16に記載の単離ペプチド交換蛋白質。
- SEQ ID NO: 4、SEQ ID NO: 6、SEQ ID NO: 8、SEQ ID NO: 10、SEQ ID NO: 12、SEQ ID NO: 14、もしくは SEQ ID NO:16と少なくとも50%の配列同一性を持つアミノ酸配列から成る、請求項3から17のいずれかに記載の単離ペプチド交換蛋白質。
- ペプチド交換蛋白質であり、
(i)TAP結合蛋白質関連(TAPBPR)フラグメント、TAPBPR内腔ドメインを含む前記フラグメントおよびTAPBPR膜貫通性ドメイン;もしくは
(ii)TAPBPR内腔ドメインと非相同膜貫通性ドメインを含むTAP結合蛋白質関連(TAPBPR)のフラグメントを含むペプチド交換蛋白質。 - 前記フラグメントがTAPBPR細胞質ドメインを欠除している、請求項19に記載の単離ペプチド交換蛋白質。
- さらに異種細胞の表面標的配列を含む、請求項19もしくは請求項20に記載の単離ペプチド交換蛋白質。
- 異種細胞表面の標的配列がCD8の細胞質ドメインを含む、請求項21に記載の単離ペプチド交換蛋白質。
- SEQ ID NO:34と少なくとも50%の配列同一性を持つアミノ酸配列から成る、請求項22に記載の単離ペプチド交換蛋白質。
- 請求項1から23のいずれかに記載の単離ペプチド交換蛋白質をコードする核酸。
- 請求項24に記載の核酸を含むベクター。
- 請求項24に記載の核酸もしくは請求項25に記載のベクターを含む哺乳類細胞。
- その表面に請求項19から26のいずれかに記載のペプチド交換蛋白質を含む哺乳類細胞。
- 哺乳類細胞の免疫原性を高める方法であり、表面MHCクラスI分子を持つ哺乳類細胞集団を提供し、前記哺乳類細胞集団を免疫原性ペプチドおよび請求項1から23のいずれかに記載のペプチド交換蛋白質もしくは請求項27に記載の哺乳類細胞と接触させ、ペプチド交換蛋白質が免疫原性ペプチドを前記集団の細胞の表面MHCクラスI分子に負荷するようにし、これにより哺乳類細胞の免疫原性を高めることを含む方法。
- 抗原提示細胞を生成し、個体における免疫反応を刺激する方法であり、あらかじめ個体から得られた抗原提示細胞集団を提供し、in vitroで抗原提示細胞を請求項1から23のいずれかに記載のペプチド交換蛋白質もしくは請求項27に記載の哺乳類細胞および免疫原性ペプチドと接触させ、ペプチド交換蛋白質が免疫原性ペプチドを抗原提示細胞の表面MHCクラスI分子に負荷するようにすることを含む方法。
- 抗原提示細胞が樹状細胞である、請求項29に記載の方法。
- さらに抗原提示細胞でT細胞集団を活性化することを含む請求項29から30のいずれかに記載の方法。
- in vitroもしくはex vivoで行われる請求項29から31までのいずれかに記載の方法。
- さらに個体に哺乳類細胞、抗原提示細胞もしくは活性化T細胞を投与することを含む請求項28から32のいずれかに記載の方法。
- 個体の標的細胞の免疫原性を高める方法であり、請求項3から18までのいずれかに記載の、個体の標的細胞と結合する標的ドメインを含むぺプチド交換蛋白質を個体に投与すること、個体に免疫原性ペプチドを投与することを含み、そうしてペプチド交換蛋白質が免疫原性ペプチドを標的細胞の表面MHCクラスI分子に負荷するようにし、これにより、前記標的細胞の免疫原性を高める方法。
- 請求項34に記載の個体の標的細胞の免疫原性を高める方法で使用するための、請求項3から18までのいずれかに記載のペプチド交換蛋白質。
- 標的細胞が病的細胞である、請求項34もしくは35の記載のように使用される方法あるいは蛋白質。
- 病的細胞が癌細胞あるいは病原体感染細胞である、請求項36の記載のように使用される方法あるいは蛋白質。
- 個体の免疫反応を刺激する方法であり、個体に、請求項3から18までのいずれかに記載の、標的ドメインが個体の抗原提示細胞に結合するペプチド交換蛋白質を投与すること、個体に免疫原性ペプチドを投与することを含み、ペプチド交換蛋白質が免疫原性ペプチドを抗原提示細胞の表面MHCクラスI分子に負荷するようにし、前記抗原提示細胞が個体の免疫反応を刺激するようにする方法。
- 抗原提示細胞が樹状細胞である、請求項38に記載の方法。
- 免疫原性ペプチドがワクチンである、請求項26から39までのいずれかの記載のように使用される方法もしくは蛋白質。
- 標的ペプチドを提示しているMHCクラスI分子を生成する方法であり、MHCクラスI分子を請求項1または請求項2のペプチド交換蛋白質および標的ペプチドと接触させることを含み、ペプチド交換蛋白質が標的ペプチドをMHCクラスI分子に負荷するようにし、これにより標的ペプチドを提示するMHCクラスI分子を生成する方法。
- MHCクラスI分子が、ペプチド交換蛋白質との接触後に標的ペプチドに置換される初期ペプチドを提示する請求項41に記載の方法。
- MHCクラスI分子が固体支持体に固定化される、請求項41もしくは42に記載の方法。
- MHCクラスI分子が、複数のMHCクラスI分子を含む多量体のサブユニットである請求項41もしくは42に記載の方法。
- 前記多量体がストレプトアビジンで結合したビオチン化MHCクラスI分子を含む四両体である、請求項44に記載の方法。
- 標的ペプチドを提示するMHCクラスI分子をT細胞の集団と接触させること、およびMHCクラスI分子が前記集団内のT細胞への結合することを決定することをさらに含む請求項41から45までのいずれかに記載の方法。
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2023213107A JP2024029012A (ja) | 2018-01-26 | 2023-12-18 | ペプチド交換蛋白質 |
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| GB1801323.5 | 2018-01-26 | ||
| GBGB1801323.5A GB201801323D0 (en) | 2018-01-26 | 2018-01-26 | Peptide exchange protein |
| GB1813737.2 | 2018-08-23 | ||
| GBGB1813737.2A GB201813737D0 (en) | 2018-08-23 | 2018-08-23 | Peptide exchange protein |
| PCT/EP2019/051907 WO2019145509A1 (en) | 2018-01-26 | 2019-01-25 | Peptide exchange protein |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2023213107A Division JP2024029012A (ja) | 2018-01-26 | 2023-12-18 | ペプチド交換蛋白質 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2021515588A true JP2021515588A (ja) | 2021-06-24 |
| JPWO2019145509A5 JPWO2019145509A5 (ja) | 2022-01-19 |
Family
ID=65234583
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2020560595A Pending JP2021515588A (ja) | 2018-01-26 | 2019-01-25 | ペプチド交換蛋白質 |
| JP2023213107A Pending JP2024029012A (ja) | 2018-01-26 | 2023-12-18 | ペプチド交換蛋白質 |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2023213107A Pending JP2024029012A (ja) | 2018-01-26 | 2023-12-18 | ペプチド交換蛋白質 |
Country Status (7)
| Country | Link |
|---|---|
| US (1) | US20210052695A1 (ja) |
| EP (1) | EP3743093A1 (ja) |
| JP (2) | JP2021515588A (ja) |
| CN (1) | CN111836635A (ja) |
| AU (1) | AU2019211121B2 (ja) |
| CA (1) | CA3089154A1 (ja) |
| WO (1) | WO2019145509A1 (ja) |
Families Citing this family (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US11814420B2 (en) | 2018-07-06 | 2023-11-14 | The Regents Of The University Of California | Peptide deficient-MHC class I/chaperone compositions and methods |
| WO2021138688A1 (en) * | 2020-01-03 | 2021-07-08 | The Regents Of The University Of California | Systems and methods for identification of mhc-i peptide epitopes |
| WO2023163980A1 (en) * | 2022-02-22 | 2023-08-31 | The Children's Hospital Of Philadelphia | Systems and methods for chaperone-mediated ligand exchange on mhc-i and mhc-related molecules using chicken tapbpr |
Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2017509317A (ja) * | 2014-01-27 | 2017-04-06 | モレキュラー テンプレーツ, インク.Molecular Templates, Inc. | Mhcクラスiエピトープ送達ポリペプチド |
Family Cites Families (15)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US5342774A (en) | 1991-05-23 | 1994-08-30 | Ludwig Institute For Cancer Research | Nucleotide sequence encoding the tumor rejection antigen precursor, MAGE-1 |
| US6235525B1 (en) | 1991-05-23 | 2001-05-22 | Ludwig Institute For Cancer Research | Isolated nucleic acid molecules coding for tumor rejection antigen precursor MAGE-3 and uses thereof |
| US5541104A (en) | 1991-05-23 | 1996-07-30 | Ludwig Institute For Cancer Research | Monoclonal antibodies which bind to tumor rejection antigen precursor mage-1 |
| DK0658113T3 (da) | 1992-08-31 | 2005-02-14 | Ludwig Inst Cancer Res | Isoleret nonapeptid afledt af MAGE-3-gen og præsenteret af HLA-A1, og anvendelse deraf |
| US6222012B1 (en) | 1992-08-31 | 2001-04-24 | Ludwig Institute For Cancer Research | Isolated nonapeptides presented by HLA molecules, and uses thereof |
| EP0672142B1 (en) | 1992-12-04 | 2001-02-28 | Medical Research Council | Multivalent and multispecific binding proteins, their manufacture and use |
| CA2184482A1 (en) | 1994-03-01 | 1995-09-08 | Etienne De Plaen | Determination of cancerous conditions by mage gene expression |
| US6291430B1 (en) | 1997-09-12 | 2001-09-18 | Ludwig Institute For Cancer Research | Mage-3 peptides presented by HLA class II molecules |
| US7060479B2 (en) * | 1999-12-08 | 2006-06-13 | Serono Genetics Institute, S.A. | Full-length human cDNAs encoding potentially secreted proteins |
| US6692923B2 (en) * | 1999-04-14 | 2004-02-17 | Incyte Corporation | Tapasin-like protein |
| AU2001260847A1 (en) * | 2000-05-24 | 2001-12-03 | Genesis Research And Development Corporation Limited | Compositions isolated from skin cells and methods for their use |
| AU2005316169B2 (en) * | 2004-12-13 | 2011-11-17 | Alethia Biotherapeutics Inc. | Polynucleotides and polypeptide sequences involved in the process of bone remodeling |
| CN102159241A (zh) * | 2008-01-28 | 2011-08-17 | 塔皮穆内公司 | 用于增强免疫应答的泰帕森(Tapasin)增加 |
| US9209965B2 (en) | 2014-01-14 | 2015-12-08 | Microsemi Semiconductor Ulc | Network interface with clock recovery module on line card |
| CN107857819A (zh) * | 2017-07-03 | 2018-03-30 | 江苏西迪尔生物技术有限公司 | 多功能融合蛋白及其应用 |
-
2019
- 2019-01-25 AU AU2019211121A patent/AU2019211121B2/en active Active
- 2019-01-25 JP JP2020560595A patent/JP2021515588A/ja active Pending
- 2019-01-25 CN CN201980018422.2A patent/CN111836635A/zh active Pending
- 2019-01-25 WO PCT/EP2019/051907 patent/WO2019145509A1/en unknown
- 2019-01-25 US US16/964,695 patent/US20210052695A1/en active Pending
- 2019-01-25 CA CA3089154A patent/CA3089154A1/en active Pending
- 2019-01-25 EP EP19701859.1A patent/EP3743093A1/en active Pending
-
2023
- 2023-12-18 JP JP2023213107A patent/JP2024029012A/ja active Pending
Patent Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2017509317A (ja) * | 2014-01-27 | 2017-04-06 | モレキュラー テンプレーツ, インク.Molecular Templates, Inc. | Mhcクラスiエピトープ送達ポリペプチド |
Non-Patent Citations (4)
| Title |
|---|
| ""RecName: Full=Tapasin-related protein; Short=TAPASIN-R; AltName: Full=TAP-binding protein-like; Al", UNIPROT: Q8VD31 [ONLINE], vol. entry version 116, JPN7022005941, 2017, ISSN: 0005133116 * |
| ""RecName: Full=Tapasin-related protein; Short=TAPASIN-R; AltName: Full=TAP-binding protein-like; Al", UNIPROT: Q9BX59 [ONLINE], vol. entry version 135, JPN7022005944, 2017, ISSN: 0005133115 * |
| CHEN X ET AL: "The modification of tapasin enhances cytotoxic T lymphocyte activity of intracellularly delivered CT", ACTA BIOCHIM BIOPHYS SIN, vol. 45, no. 3, JPN7022005943, 2013, pages 203 - 212, ISSN: 0005133118 * |
| HERMANN C ET AL: "The binding of TAPBPR and tapasin to MHC Class I is mutually exclusive", JOURNAL OF IMMUNOLOGY, vol. 191, no. 11, JPN7022005942, 2013, pages 5743 - 5750, XP055669294, ISSN: 0005133117, DOI: 10.4049/jimmunol.1300929 * |
Also Published As
| Publication number | Publication date |
|---|---|
| JP2024029012A (ja) | 2024-03-05 |
| US20210052695A1 (en) | 2021-02-25 |
| CN111836635A (zh) | 2020-10-27 |
| AU2019211121A1 (en) | 2020-08-27 |
| CA3089154A1 (en) | 2019-08-01 |
| WO2019145509A1 (en) | 2019-08-01 |
| EP3743093A1 (en) | 2020-12-02 |
| AU2019211121B2 (en) | 2024-09-12 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| Kato et al. | Targeting antigen to Clec9A primes follicular Th cell memory responses capable of robust recall | |
| Ossendorp et al. | Specific T helper cell requirement for optimal induction of cytotoxic T lymphocytes against major histocompatibility complex class II negative tumors | |
| KR100457647B1 (ko) | 펩티드및애쥬번트에기초한면역조절용약제학적조성물 | |
| JP2024029012A (ja) | ペプチド交換蛋白質 | |
| JP2014509841A (ja) | 癌を治療するための組成物および方法 | |
| JP2000510689A (ja) | T細胞ペプチド・エピトープの選択と産生方法および選択したエピトープを組込むワクチン | |
| US11746138B2 (en) | Artificial synapses | |
| KR20200039765A (ko) | T 세포 변형 | |
| SA519410007B1 (ar) | مركبات ببتيد وطرق لمعالجة السكر | |
| US20230074462A1 (en) | Methods and compositions for stimulating immune response | |
| US20200385441A1 (en) | Immunotherapeutic composition for the treatment of cancer | |
| JP4587566B2 (ja) | Hlaクラスi分子と付着手段とを含む複合体を用いた、標的細胞に対するt細胞応答を作出又は強化する方法 | |
| Botelho et al. | Combination of synthetic long peptides and XCL1 fusion proteins results in superior tumor control | |
| US20040096429A1 (en) | Method for producing cytotoxic T-cells | |
| Zhao et al. | TAT-mediated gp96 transduction to APCs enhances gp96-induced antiviral and antitumor T cell responses | |
| Peng et al. | Novel vaccines for the treatment of chronic HBV infection based on mycobacterial heat shock protein 70 | |
| Deloizy et al. | The anti-influenza M2e antibody response is promoted by XCR1 targeting in pig skin | |
| WO2020145222A1 (ja) | 新規ネオアンチゲン及びそれらを用いたがん免疫治療薬 | |
| US20030044415A1 (en) | Method for producing or enhancing a T-cell response against a target cell using a complex comprising an HLA class I molecule and an attaching means | |
| KR20220083773A (ko) | 암 항원을 포함하는 개선된 lamp 구축물 | |
| Brosi et al. | Processing in the endoplasmic reticulum generates an epitope on the insulin A chain that stimulates diabetogenic CD8 T cell responses | |
| Padula et al. | Secreted heat shock protein gp96-Ig and OX40L-Fc combination vaccine enhances SARS-CoV-2 Spike (S) protein-specific B and T cell immune responses | |
| Wang et al. | Gene conjugation of molecular adjuvant C3d3 to hCGβ increased the anti‐hCGβ Th2 and humoral immune response in DNA immunization | |
| Guo et al. | Antigen-display exosomes provide adjuvant-free protection against SARS-CoV-2 disease at nanogram levels of spike protein | |
| Babon et al. | Cross-presentation of a CMV pp65 epitope by human dendritic cells using bee venom PLA2 as a membrane-binding vector |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20220111 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20220111 |
|
| A977 | Report on retrieval |
Free format text: JAPANESE INTERMEDIATE CODE: A971007 Effective date: 20221206 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20230110 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20230407 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20230606 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A821 Effective date: 20230606 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20230822 |