JP2004531225A - ペプチド、蛋白質及び疑似ペプチド合成のための方法及び組成物 - Google Patents
ペプチド、蛋白質及び疑似ペプチド合成のための方法及び組成物 Download PDFInfo
- Publication number
- JP2004531225A JP2004531225A JP2002559580A JP2002559580A JP2004531225A JP 2004531225 A JP2004531225 A JP 2004531225A JP 2002559580 A JP2002559580 A JP 2002559580A JP 2002559580 A JP2002559580 A JP 2002559580A JP 2004531225 A JP2004531225 A JP 2004531225A
- Authority
- JP
- Japan
- Prior art keywords
- peptide
- trna
- translation system
- translation
- product
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
Images















Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/67—General methods for enhancing the expression
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P35/00—Antineoplastic agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P43/00—Drugs for specific purposes, not provided for in groups A61P1/00-A61P41/00
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/10—Processes for the isolation, preparation or purification of DNA or RNA
- C12N15/1034—Isolating an individual clone by screening libraries
- C12N15/1062—Isolating an individual clone by screening libraries mRNA-Display, e.g. polypeptide and encoding template are connected covalently
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P21/00—Preparation of peptides or proteins
- C12P21/02—Preparation of peptides or proteins having a known sequence of two or more amino acids, e.g. glutathione
Landscapes
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Engineering & Computer Science (AREA)
- Organic Chemistry (AREA)
- Genetics & Genomics (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- Biomedical Technology (AREA)
- General Health & Medical Sciences (AREA)
- Biochemistry (AREA)
- Molecular Biology (AREA)
- Microbiology (AREA)
- Biophysics (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Physics & Mathematics (AREA)
- Plant Pathology (AREA)
- General Chemical & Material Sciences (AREA)
- Pharmacology & Pharmacy (AREA)
- Bioinformatics & Computational Biology (AREA)
- Medicinal Chemistry (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Crystallography & Structural Chemistry (AREA)
- Animal Behavior & Ethology (AREA)
- Public Health (AREA)
- Veterinary Medicine (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Peptides Or Proteins (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
Abstract
Description
【0001】
本発明は、翻訳因子EF−P、W、W2又はレスキュー(rescue)の添加を必要としない単純化され、高度に精製された前進的な翻訳系に関する。
【背景技術】
【0002】
リガンドの認識及び結合は、免疫認識、細胞シグナリング、細胞連絡、転写、翻訳、細胞内シグナリング及び触媒反応、即ち酵素反応のようなほとんど全ての生物学的プロセスを調節する。ホルモン、増殖因子及び神経伝達物質のようなアゴニストとして作用し又はリガンド活性を作動若しくは拮抗させることができる天然又は非天然のリガンド分子(このものは、B細胞(抗体仲介)免疫又はT細胞(細胞仲介)免疫を誘導し、化学反応を触媒でき、或いは遺伝子発現を転写又は翻訳レベルで調節できる)を同定及び合成するための技術には、長年にわたって関心が寄せられている。このようなリガンドの大部分は、蛋白質、ペプチド及び疑似ペプチド(peptidomimetic)である。
【0003】
リガンド及び薬剤発見への伝統的なアプローチは、運と激務の混ぜ合わせに大きく依存している。動物及び植物組織からの天然産物又は発酵培養基の産物をスクリーニングすること、或いは保管された合成分子を無作為スクリーニングすることが新規のリード化合物を同定するために最も生産的な手段であった。
【0004】
しかしながら、新薬の探索における最近の動向は、新薬発見に関して可能性のある新規のリード化合物源としてのコンビナトリアル・ライブラリーを調製することに集中している。「コンビナトリアル・ケミストリー」というこの新しい分野の核心は、非生合成又は生合成のいずれかで調製でき且つ生物活性ごとに様々な形式でスクリーニングできる異なった分子を集めることである。非生合成技術、例えば、コード化、空間的アドレッシング及び/又はデコンボリューションの使用によって、ペプチド、疑似ペプチド及び非ペプチドベースの分子のコンビナトリアル・ライブラリーがバッチプロセスで合成でき、そして重要なことに、このライブラリーの個々の要素の分子同一性は、薬剤スクリーニング形式で確認できる(例えば、「Lam他(1993)Gene 137,13−16」、Dooley他(1994)Science 266,2019−2022)。非生合成ライブラリーは、生物学的単量体(例えば天然のアミノ酸及びヌクレオチド)及びそれらの誘導体に制限されないという利点を有するが、このライブラリーは、数週間のうちにスクリーニングできる分子の数が制限されるという不利点を有する(通常せいぜい105〜108個であり、これは、対象の標的に対して高い親和性のリガンドを同定するには小数すぎる分子である)(「Roberts(1999)Curr.Op.Chem.Biol.3,268−273」、「Wilson他(2001)PNAS98,3750−3755」)。しかしながら、生合成ライブラリーは、多くの場合この制限をほとんど受けない。なぜならば、1015個の異なるペプチド、RNA又はDNA分子を数週間のうちにスクリーニングできるというこのようなライブラリーの例が存在するからである(Roberts,上記)。これは、個々の生合成ライブラリー要素を、多くの場合時として定向進化と呼ばれるダーウィンの進化論に類似したプロセスである関連した突然変異誘発段階(例えば、親和性成熟、突然変異誘発PCR又はDNAシャフリング(Roberts,上記))でもって反復選択し且つ増幅させることによって達成される。
【0005】
部分的に又は完全に無作為化されたプールからの蛋白質の単離を可能にした従来の方法の多くは、そのような in vivo 段階によるものであった。この種の方法には、モノクローナル抗体技術(「Milstein,Sci.Amer.243:66(1980)」及び「Schultz他,J.Chem.Engng.News 68:26(1990)」)、ファージディスプレイ(「Smith,Science 228:1315(1985)」、「Parmley及びSmith,Gene 73:305(1988)」並びに「McCafferty他,Nature 348:552(1990)」)、ペプチド・lacリプレッサー融合(Cull他、PNAS 89:1865(1992))及び古典的な遺伝学的選択が含まれる。これらの方法のそれぞれは、蛋白質と核酸の間のトポロジカルな結合に依拠する。なぜならば、核酸しか複製できないからである。しかして、蛋白質の情報は保持され且つ読み込み可能な核酸の形で回収できる。
【0006】
他の蛋白質選択技術は、in vivo 段階なしに実施される。多くの場合リボソームディスプレイと呼ばれる失速翻訳法は、リボソーム及びmRNAとさらに複合体を形成する新生蛋白質鎖のいくらかの特性ごとに選択を行う技術である(「川崎,米国特許第5658754号」、「Tuerk及びGold,Science 249:505(1990)」、「Irvine他,J.Mol.Biol.222:379(1991)」、「Korman他,PNAS 79:1844−1848(1982)」、「Mattheakis他,PNAS 91:9022−9026(1994)」、「Mattheakis他,Meth.Enzymol.267:195(1996)」並びに「Hanes及びPluckthun,PNAS 94:4937(1997)」)。mRNA・蛋白質融合法又はmRNAディスプレイ法(「根本他、FEBS Lett.414,405−408(1997)」、「柳川他,米国特許第6228994号」、「Szostak他,米国特許第6281344号、同6261804号、同6258558号、同6214553号及び同6207446号」並びに「Roberts及びSzostak,PNAS 94,12297−12302(1997)」)は、mRNAをDNA/プロマイシンリンカーによってその蛋白質産物に直接共有結合させる。終止コドンの存在によっては損なわれない「裸の」mRNA・ペプチド融合体を合成させるための方法は、ペプチドをミセルの状態でそれらのペプチドをリボソームから分離させ、次いでそれらの特異的mRNAに再結合できるような方法でもって合成させることである(例えば、ストレプトアビジン配列を含む蛋白質は、ビオチニル化mRNAに結合するであろう。土井及び柳川(1999)FEBS Lett.475,227−230)。
【0007】
しかしながら、この先行技術の「天然」(L−)ペプチドライブラリー技術は、多くの不都合がある。第1に、ほぼ完全にキラルな単量体(アミノ酸)からなるこのライブラリーは、このキラル単量体の鏡像異性体を欠失している。例えば、L−ペプチドライブラリーについて、この20種の天然型アミノ酸は、幅広い立体性、電子性及び官能性の基を与えるが、C−α炭素のキラリティーは、先行技術のディスプレイ技術によって達成できる三次元形状の空間を事実上制限する。また、L−ペプチドライブラリーは、標的物との非共有結合又は共有結合複合体を形成させるのに有用であり得る多数の一般的な有機化学官能基も欠失しており(例えば、アルケン、アルキル尿素、ハロゲン化アルキル及びケトン)、しかも「非天然」アミノ酸(予め合成されたものか又は理論上のものかのいずれか)で達成できる非常に大きい付加形状の多様性も欠如している。さらに、治療剤として、天然L−アミノ酸のペプチドは、通常それらの非天然鏡像異性体(D−ペプチド)又はアナログよりも好ましいわけではない。なぜならば、L−ペプチドは、in vivo プロセシングのため乏しい薬物動態学的プロファイルによって使用が制限され得るからである。例えば、L−ペプチドは、動物への投与後にプロテアーゼによって迅速に分解され得、しかしてより高い有効投与量を必要とする。さらに、薬用ペプチドは、患者の強力な免疫原性応答を誘導でき、さらにその迅速なクリアランスに寄与し、また有害であるかもしれない炎症反応も生じさせる。この薬用ペプチドの分解を回避するための一方法は、レトロインベルソアナログ(Sisto他の米国特許第4522752号を参照)、レトロエナンチオアナログ(Goissis他のJ.Med.Chem 19:1287−90(1976)を参照)のような非加水分解性のペプチドアナログ、トランス−オレフィン誘導体(Shue他のTetrahedron Letters 28:3225(1987)を参照)及びホスホネート誘導体(Loots他の「ケミストリーアンドバイオロジー」(エスコムサイエンス出版,ライデン,1988,p118)の「ペプチド」を参照)を生成させることであった。しかしながら、多くの場合、ペプチドの主鎖は、蛋白質分解に耐性のある疑似ペプチドにするために変質される。このようにすると、得られた疑似ペプチドは、天然ペプチド主鎖と標的受容体との間のある種の結合性接触の欠如並びに二面角などの変化による該ペプチドに対する立体空間の変化によって生物活性が低下する欠点を有し得る。別の問題は、ほとんど全てのL−ペプチドがその親水性のために生体膜を横断しないことである。対照的に、D−ペプチド及びN−メチルアミノ酸のようなその他の非天然アミノ酸を含むペプチド(疑似ペプチド)はプロテアーゼ耐性を増大させ、そして疑似ペプチド薬のシクロスポリンAは、膜を横断でき且つ経口的に利用できる。なぜならば、部分的にではあるが、このものは、天然のペプチド結合よりも疎水性であるいくつかのN−メチルペプチド結合を含有するからである(「Zawadzke及びBerg(1992)J.Am.Chem.Soc 114:4002」、Walsh他(1992)J.Biol.Chem.267,13115−13118)。残念ながら、D−ペプチドライブラリーのような化学的に合成された(非生合成の)疑似ペプチドライブラリー(Lam他(上記)、Dooly(上記))は上で論じたライブラリーサイズの制限を受け、鏡像ファージ又はリボソームディスプレイ(「Schumacher他(1996)Science 271,1854−1857」、「Eckert他(1999)Cell 99,103−115」及び「Forster他PCT公開WO97/35194」)のような疑似ペプチドライブラリーのサイズ制限を克服するための方法論的策略は、鏡像異性体の標的物を化学的に合成するという厄介な必要条件によって制限される。
【0008】
蛋白質、ペプチド及び疑似ペプチドは、一般に、3つの異なる方法で合成され、それぞれ固有の制限を有している。
【0009】
1.合成ペプチド化学は、長さが約30残基までの非常に多様な疑似ペプチドを高収率且つ高純度で合成するために日常的に使用され得る(Eckert他,上記)。
しかしながら、この方法は、非効率的なカップリング段階、精製の問題及び折りたたみの困難性のためにより長い産物については非効率的又は非実用的である。また、所望の生成物の反応性側鎖の全てについて相溶性のある保護基が必要であるため、合成が制限される。さらに合成疑似ペプチドは、反復選択、増幅及び変異(進化)のために遺伝学的にコードされ得ず、合成疑似ペプチドライブラリーの複雑性を最適な薬剤発見には少なすぎる約108種の分子に制限させる。
【0010】
2.生細胞を使用する in vitro 翻訳が、遺伝学的にコードされた天然又は組換えDNA配列からの短い又は長い蛋白質を効率的に合成し且つ翻訳後に修飾するために広く使用されている。
しかしながら、合成は、その遺伝子産物が有毒である場合には役に立たず、特にその蛋白質が封入体中で発現する場合には精製が困難であり得、しかも復元の問題もあり得る。最も重要なことには、この方法は、複数の非天然アミノ酸を選択的に取り込むことができない、或いは翻訳後修飾プロセス(例えば、プロテアーゼ触媒プロセシング又は分解)を制御することができない欠点がある。
【0011】
3.粗製細胞抽出物による in vitro 翻訳は、一般に、毒性の問題を克服し(しかし、翻訳後修飾を制御しない)、より容易な精製及び折りたたみを生じさせることができ、しかも人工のサプレッサーtRNAを使用して蛋白質当たり1個の非天然アミノ酸の選択的な取り込みを可能にさせる(Noren他(1989)Science 244,182−188)。
しかしながら、この方法による非天然アミノ酸の取り込みは、in vivo 系よりも収率が非常に低い難点がある。なぜならば、この方法は、終結因子と競合する本質的に非効率なサプレッサーtRNAに依存するからである。100個以上の異なる非天然アミノ酸が個々に取り込まれたが(例えば、Mendel他(1995)Annu.Rev.Biophys.Biomol.Struct.24,435−462)、この戦略は、3種の終止(ナンセンス)コドンの一つ(UAGコドン)のみでの蛋白質当たり1個だけの非天然アミノ酸の選択的な取り込みに制限された。これは、20種の異なるアミノアシルtRNAシンテターゼのtRNA結合活性及びプルーフリーディング活性によって触媒される天然アミノ酸によるアミノ酸(センス)コドンでの競合のため及びリボソームによるリードスルーのため第2の終止コドン(UGA)を使用しようとする試みが失敗したためである(Cload他(1996)Chem.and Biol.3,1033−1038)。
【0012】
また、非天然アミノ酸をセンスコドンに一般的な態様で選択的に取り込もうとする多くの試みも失敗した。例えば、非天然型アミノ酸を取り込むために最も一般的に使用される方法では、天然アミノ酸の高比活性放射性同位元素誘導体を in vitro 翻訳によって取り込んで放射性標識された蛋白質を合成させる場合に、この放射性アミノ酸の比活性が、通常、粗製翻訳系に存在するアミノ酸の非標識型による取り込みのための競合によってこの非標識型を添加された非標識アミノ酸プールから与えないにもかかわらず減少することは周知である。プロメガ社によって、類似するアナログの希釈の結果が別のレポーター群であるビオチン標識リシンの取り込み用の市販キットを使用して得られている(Trascend(商標)非放射性翻訳系の付属文献)。さらに、粗製の翻訳抽出液を濾過して天然アミノ酸を除去し、次いでリシンを除く天然アミノ酸の全てを追加し且つアミノ酸アナログが付加されたリシンtRNAを追加すると、リシンアナログ対リシンがわずか1:3〜1:4の比で取り込まれた(Crowley他(1993)Cell 73,1101−1115)。アミノ酸アナログの取り込みの低い選択性は、ある種の用途には十分であるが(Rothschild他,米国特許第5643722号)、遺伝学的にコードされた特異的な疑似ペプチド配列の増幅及び特徴付けを必要とするような多くの用途には明らかに適さない。2種の異なるフレームシフトサプレッサーtRNAを使用して2種の異なる非天然アミノ酸を取り込めることが証明され(芳坂他(1999)JACS 121,12194−12195)、多くの同一の非天然アミノ酸がPheアミノアシルtRNAシンテターゼに対して特異的な阻害剤を使用して取り込まれた(Baldini他(1988)Biochem.27,7951−7959)。しかしながら、これらの方法の両方は、多くの異なる非天然アミノ酸を単一の疑似ペプチドに取り込むのに必要な態様には一般化できない。粗製及び in vivo 翻訳に固有のこれらの制限を克服するために、遺伝暗号を直交tRNA及び直交非天然核酸塩基対に基づいて拡張させるための綿密な戦略が提案されたが、単一の試験管内改変終止コドン(Bain他(1992)Nature 356,537−539)を超える開発は、技術的に非常に困難であることが分かった(Service(2000)Science 289,232−235)。
【0013】
本発明者は、この問題が純粋な in vitro 翻訳系を使用して解決できるかもしれないと考えた。非天然アミノ酸と天然アミノ酸又は終結因子との間の競合は、ある種のアミノ酸、tRNA、アミノアシルtRNAシンテターゼ及び/又は終結因子のようなある種の成分を除外することによって回避できたかもしれない。不運にも、mRNA依存性ポリペプチド合成のための最低限の必要条件は、含まれる高分子が非常に多いために定義するのが困難であった。精製された成分からの翻訳の再構成がE.coliについて達成されたが、必要とされる翻訳因子の数は、いまだ議論の余地がある。
【0014】
ワイスバッハ研究所によって構築された最初の精製翻訳系は、4種のE.coli mRNAを、高度に塩洗浄されたリボソーム、開始因子(部分的なIF−1依存性)、伸長因子Tu(EF−TuH)及びアミノアシルtRNAシンテターゼの群に強く依存して、met−tRNAi fMetホルミルトランスフェラーゼ及び伸長因子G(EF−G)に部分的に依存して、伸長因子Ts(EF−Ts)又は終結因子に全く依存しないで効率的に翻訳した(Kung他(1978)Arch.Biochem.and Biophys.187,457−463)。非常に多くの精製成分を維持し且つ微量の汚染物を除去する際の困難性のために、追加の一般的な翻訳因子についてのサーチは、fMet−tRNAi fMet及び1又は2種の伸長体アミノアシルtRNAによるジペプチド又はトリペプチドの合成へとこの系を単純化させることによって促進され、それによってアミノアシルtRNAシンテターゼのための必要条件が回避された(Weissbach他(1984)Biotechniques2,16−22))。
【0015】
Ganozaが率いる第2グループは、後者の単純化された方法を試験管内で付加された全tRNA及び終結因子を使用してより長いペプチドに拡大させたときに、バクテリオファージMS2及びf1の翻訳がEF−P、W及びレスキュー(rescue)と呼ばれる3種の追加の因子に依存することを発見した(「Green他(1985)Biochem.Biophys.Res.Com.126,792−798」、「Ganoza他(1985)PNAS 82,1648−1652」)。これらの因子を存在させないと、連続移動性が非効率的になった。例えば、ヘキサペプチド合成反応ではジ、トリ、テトラ及びペンタペプチドの休止産物又は早期終結産物が支配的であった。deaD/W2(Wよりも数kD大きい)と呼ばれる恐らくは関連のある翻訳因子及びEF−Pもクローン化され、これらのものは最大限の成長にとって必要であり、そして真核細胞の開始因子に相同的である(「青木他(1991)Nucleic Acids Res.19,6215−6220」、「青木他(1997)J.Biol.Chem.272,32254−32259」、「Lu他(1999)Int.J.Biochem.Cell Biol.31,215−229」)。
【0016】
Ganozaの結果とは明らかに相反して、その他2つのグループが精製成分をEF−P、W、W2又はレスキューを加えることなく使用してアミノアシルtRNA基質から短いペプチドを合成することを報告したが、これら2つのグループは、それらの系の連続移動性又はそれらのリボソームの純度を直接記録していなかった(「Stade他(1995)Nucleic Acids Res.23,2371−2380」、「Pavlov他(1997)EMBO J.16,4134−4141」)。この矛盾が本当ならば、これは単に解釈に関して推測しているにすぎない。例えば、EF−P、W及びレスキューは、リボソーム調製物から精製できるため(Ganoza他(1996)Biochemie 78,51−61)、Ganozaのグループが使用したのとはかなり異なる手順によって調製された後者2つのグループが使用したリボソームは、EF−P、W、W2及び/又はレスキューで汚染されていた可能性がある。これは問題が多い。なぜならば、EF−P、W、W2及び/又はレスキューによる汚染は、アミノアシルtRNAシンテターゼ及び終結因子のような、望ましくない反応の原因となり得るより多数の蛋白質による汚染を暗示しかねないからである。一方、EF−P、W、W2及び/又はレスキューは、Ganozaの系では単に効率的な連続移動性のために必要とされ得るにすぎない。
【0017】
可能ならば、ペプチド又は蛋白質を純粋な翻訳系によってEF−P、W(多くの場合W2と呼ばれる)及びレスキューを添加することなく合成できることが望ましい。なぜならば、これらの蛋白質は、機能に関してあまりよく理解されておらず、結果的にそれらの活性を検査するのが困難となり、しかしてその前に活性な蛋白質を精製するのが困難になるからである。さらに、W(又はW2)の実際寸法及びWとW2が同一蛋白質の誘導体に相当するのかどうかについての議論があり、また、レスキューについての遺伝子をさらにクローン化しなければならない。
【発明の開示】
【課題を解決するための手段】
【0018】
本発明は、翻訳因子EF−P、W、W2又はレスキューの添加を必要としない単純化され、高度に精製された前進的な翻訳系である。新規な翻訳プロセスは、これまでのところ別経路によって合成されていた全てのペプチド及び蛋白質に新規でともすれば改善された経路を提供する。このプロセスは、蛋白質、ペプチド及び疑似ペプチド合成のために上記された方法1、2及び3に固有の制限を克服する。
【0019】
ある好ましい具体例では、この精製された系は、インスリン、成長ホルモン又はエリスロポエチンのようなペプチド又は蛋白質リガンド又は触媒の合成のために使用できる。
【0020】
別の好ましい具体例では、この精製系は、粗製の細胞抽出液に依存する既存のリボソーム及びmRNAディスプレイ系とは異なり、「純粋なリボソームディスプレイ」及び「純粋なmRNAディスプレイ」選択実験のために使用できる。ペプチドの翻訳後修飾の欠如が予期されること、しばしば蛋白質分解の問題を生じさせるプロテアーゼがないこと及び選択段階で汚染物による競合がないことのような、純粋系でペプチド及び蛋白質ディスプレイを実施することに関連するいくつかの利点がある。追加の利点として、以下のことが挙げられる。
(i)リボヌクレアーゼが存在しないこと(本発明の純粋なE.coli系で測定された放射性mRNAの長期間安定性によって立証される(結果は示さない))は、様々な粗製系、特にE.Coli(Roberts,上記)で観察されるmRNA分解に関わる問題を回避させる。mRNAが翻訳及び選択され得る前に分解されないことは明らかに重要である。
(ii)純粋な再構成系は翻訳終結因子によって汚染されないことが予期される。実際に、本発明の系は、終結因子の添加によって刺激される。最も一般的な粗製ディスプレイ系を使用する研究者が未完成ペプチドの急速な放出を回避するために選択前又は選択前のmRNA結合体への融合前にmRNAから終止コドンを除去することが必要であることを発見した(融合及び選択の両方は、緩慢なプロセスである)。この終止コドンの除去は、通常、個々のmRNAの場合には特別の突然変異誘発段階を必要とし、しかも特に全ての終止コドンを変異させることが不可能である天然mRNAライブラリー又は合成コンビナトリアルライブラリーについては多くの問題がある。このライブラリーにおける終止コドンに関連する問題は、蛋白質の小さなサブドメインのライブラリー(このものは、全長の蛋白質を欠いており、十分に現れないカルボキシ末端ドメインを有し、正確に折りたたむ能力に対して多くの不適切な境界をコードし、しかも非天然のオープンリーディングフレームからの多数の配列を含有する)を無作為に生成させることによって、或いは終止コドンを含有する無作為ライブラリーの要素を選択することによって回避されてきた(それによって、合成反応及び多様性の大部分を浪費する。「Cho他(2000)J.Mol.Biol.297,309−319」)。精製リボソーム及び精製mRNAディスプレイ系では、ペプチジルtRNAが1日以上にわたってリボソームと安定に結合したままであることができる(翻訳が完了した後に、温度を低下させ及び/又は塩濃度を増加させる場合に安定していることが特に有利である(Schaffitzel他(1999)J.Immunol.Methods 231,119−135)):リボソームは、tRNAが与えられないセンスコドンで伸長を停止させるか(それによって終結因子との競合を完全に回避させる)又は終止コドンで停止させるか又は終止コドンを欠失したmRNAの末端で停止させるかのいずれかである。しかして、天然mRNAからの全長翻訳産物のライブラリーが本発明により調製でき、そして、このような発現ライブラリーは、in vitro 選択に直接付され、或いはこれらのものは、特に、ハイブリダイゼーションによってゲノム及びプロテオミクス研究のためのDNAマイクロチップに対応できる。
(iii)この純粋な再構成系は、終止コドンを欠失したmRNAから合成されたペプチドを分解させるtmRNA系によって汚染されないことが予期される。粗製系を使用する研究者がこのtmRNA系を阻害するように試みることが重要であることを発見した(Hanes及びPluckthun(1997)PNAS 94,4937−4942)。
【0021】
他の好ましい具体例では、本発明は、特定の疑似ペプチド(ペプチドアナログ)のmRNA指令合成を一般的な態様でできるようにし、得られる疑似ペプチドの多様性及び長さを大いに増大させることができる。見込まれるものとしては、既存の疑似ペプチドリガンド及び疑似ペプチド薬(シクロスポリンAのようなリボソームによらないで生合成されたリガンドを含めて)並びにそれらの誘導体が挙げられる。
【0022】
他の具体例では、本発明は、in vitro 進化によって(例えば、上記の純粋なリボソームディスプレイ又は純粋なmRNAディスプレイを使用することによって)触媒、リガンド及び薬剤発見のために疑似ペプチド産物を遺伝学的にコードできるようにする。疑似ペプチドの特異的な合成は、既存の粗製リボソーム及びmRNAディスプレイの手法では不可能である。なぜならば、粗製翻訳系では、天然アミノ酸が非天然アミノ酸と取り込みのために競合するからである。
【0023】
本発明は、所望の特性を有するペプチド及び疑似ペプチドの単離を容易にさせる。一つの具体例では、この方法は、標的分子のためのリガンドを同定することに関する。例として、標的分子にはペプチド、核酸、炭水化物及び非重合性分子、例えば、ステロイド、イノシトール、脂溶性ビタミン、テルペン、アセトゲニム、神経伝達物質又は遷移状態のアナログが含まれる。好ましい具体例では、標的分子は蛋白質である。蛋白質の標的は、例示すると、受容体、酵素、DNA結合蛋白質又は蛋白質複合体又はスクリーニングできる活性を保持しているそれらの部分若しくはドメインであることができる。
【0024】
好ましい具体例では、主題の方法を使用して少なくとも103種の異なる配列、好ましくは少なくとも108種の異なる配列、最も好ましくは少なくとも1015種の異なる配列の試験ペプチド又は疑似ペプチドの多様な集団を生成させる。
【0025】
本発明のさらに別の態様は、主題の方法によって同定されたペプチド及び疑似ペプチドのような化合物及びそれらの使用に関する。また、これらのものには、このようなペプチド及び疑似ペプチドの結合体及び誘導体も含まれる(例えば、結合したペプチド又は疑似ペプチド配列を膜を横切って効率的に輸送できるようにする陽イオン性ペプチド配列への結合体)(Moore及びRosbash(2001)Science 294,1841−1842)。
【0026】
本発明の別の態様は、ペプチド又は疑似ペプチドの合成及び/又は進化用のキットに関する。
【0027】
図面の説明
図1:E.coliからの5種のhisタグE.coli翻訳因子の過剰発現及び精製。15%ゲルでのSDS−PAGE後に、試料をクマシーブルーで染色した。Uは誘導されなかった全細胞であり、IはIPTG誘導された全細胞であり、PはNi2+ビーズから溶出した精製蛋白質であり、Mは分子量マーカーの蛋白質(寸法はkDで示される)である。
【0028】
図2:リボソーム指令ペプチド合成段階の概略図である。矢印によって示された3種の酵素反応は、開始(上)、第1伸長段階(右)、次の転位及び伸長段階(下)である。ペプチド産物は、分析のために塩基触媒加水分解によってペプチジルtRNAから放出され得る。産物のGDP及びPiは示されていない。Eは出口部位であり、Pはペプチジル部位、Aはアミノアシル部位である。
【0029】
図3:短鎖mRNA鋳型。最長mRNAを合成するのに使用したDNAプライマーと鋳型の対を上部に示している。また、本発明の精製系からの予測される翻訳産物も示している(3’の終止コドンGAA(Glu)及びUUC(Phe)についてのアミノアシルtRNAは使用しなかった)。bKはビオチン標識されたリシンであり、S−Dはシャイン・ダルガーノリボソーム結合部位である。
【0030】
図4:精製hisタグ翻訳系におけるmRNAからのオリゴペプチド合成速度の特徴付け。fMTVは、IF1H、IF2H、IF3H、EF−TuH、EF−GH及び0.020A260/μlのリボソームを含有する翻訳系でペプチド産物に取り込まれた3H−バリンによって測定された。三角形:翻訳は、プレインキュベートされた開始成分をプレインキュベートされた伸長成分と混合することによって開始された。四角形:翻訳は、翻訳混合物を0℃から37℃に移すことによって開始された。アリコートを1分から始める所定の時間にNaOHで終了させた。ペプチド産物のd.p.m.は、37℃のインキュベーション前に終了されたアリコートで得られたd.p.m.を引き算することによって計算された。代表的な実験からの個々のデータのポイントがプロットされ、推定される偏差は20%未満であった。プレインキュベーション反応の曲線に対して接線を引いて定常状態の速度を推定する。
【0031】
図5:mRNAのMTVを有する精製翻訳系における翻訳因子の依存性。淡い棒:ペプチド産物に取り込まれた3H−バリン(fMTVの測定値)は、IF2H、IF3H、EF−TuH及びEF−GHへの強い依存性を示している(IF1Hは、これらの翻訳系から除外された。材料及び方法を参照されたい)。黒い棒:同一の産物に取り込まれた14C−トレオニン(fMTとfMTVを一緒に測定したもの)。0℃から37℃に移すことによって開始された30分の翻訳でのペプチド合成は、全d.p.m.からmRNAなしの対照反応で得られたd.p.m(最大のd.p.mの1.3%)を引き算することによって計算された。得られた合成産物の最大濃度は、T及びV両者について0.12μMであった。
【0032】
図6:MTTVのmRNA鋳型から産生された産物のHPLC分析。二重標識翻訳のペプチド産物は、まず、tRNAから塩基により放出され、次いで標識されていないマーカーペプチドと混合された。この混合物を酸性化させ、微量遠心分離して不溶性の物質を除去し、そして分析のために注入前に10kDフィルターによって微量遠心分離した(材料及び方法参照)。マーカーペプチドの溶出位置は、クロマトグラムの上に示されている。黒丸は14C−トレオニンの全d.p.m.である。白丸は同一画分の3H−バリンの全d.p.m.である。産物の合成量は、30μl当たり2.1pmol(70nM)であった。
【0033】
図7:ε配列の包含は、精製翻訳系でのオリゴペプチド産物の合成を向上させる。mRNAであるMVT(丸)、スクランブルされたεMTV(四角形)及びεを欠失したMVT(三角形)からのfMVT合成の速度の間の比較。IF1H、IF2H、IF3H、EF−TuH及びEF−GHを含有する二重標識翻訳系を図5に記載されるように分析した。30分で合成されたオリゴペプチドの最大濃度は、それぞれのアミノアシルtRNAのうち0.5μMを使用して0.25μMであった。
【0034】
図8:精製hisタグ翻訳系を使用した非天然アミノ酸を含むペプチドの合成及び選択。ビオチン標識lys−tRNAlys、fmet−tRNAi fmet、thr−tRNA3 thr及び3H−val−tRNA1 val基質並びにmRNAであるMTKV又はMTV(翻訳産物については図3を参照されたい)のいずれかを含有する翻訳混合物を37℃で30分間インキュベートした。ペプチド及びアミノ酸をtRNA及びリボソームから塩基により放出させ、中和し、そしてこの混合物を軟質のアビジンビーズと共にインキュベートしてビオチン含有分子を結合させた。結合された3H(黒い棒:3H−バリンに共有結合したビオチン標識リシンを含有する産物の測定値)をカウントする前にこのビーズを4回洗浄してビオチニル化していない分子を除去した。プールされた洗浄物をろ過し、酸性化させ、そして陽イオン交換カラムに通して結合していない3H(淡い棒:3H−バリンを含有し、ビオチン標識リシン又はリシンを含有しないホルミル化ペプチド産物の測定値)をカウントした。結合した及び結合していない3Hのd.p.m.は、mRNAなしの対照反応で得られたd.p.m.(最大の結合及び非結合のd.p.m.のそれぞれ23%及び15%)を引き算した後にプロットしている。ビオチン標識lys−tRNAlysがmRNAのMTKVの翻訳から除外されたときに、3Hのビーズへの結合は観察されない(図示しない)。
【0035】
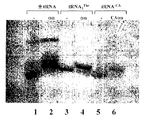
図9:Cys−tRNAcysの化学的ビオチニル化。
【0036】
図10:ビオチニル−Cys−tRNAcysの精製。ビオチニル−Cys−tRNAは、図16に示すように電気泳動され(以下参照)、その産物は、適切なバンド(即ち、レーン7又は9の大きい方のバンド)を切り出し、4℃で溶出させることによって精製された。tRNAにCysを付加させたものとさせていないもの、ビオチニル化させたもの(pH6.0又は6.9のいずれかで)とさせていないもの及びその後にアミノアシル結合をTris−HClpH8.8で加水分解させたものとさせていないものを含む対照(レーン1〜6、8、10)は、ビオチニル化がCysに特異的であり、しかもCys−tRNAがビオチニル−Cys−tRNAと共移動しなかったことを示した。
【0037】
図11:精製系を使用した精製ビオチニルCysのfM−T−bC−V疑似ペプチドへの取り込み。翻訳は、mMTCV、fMとTとVとをそれぞれ結合したtRNA及び非結合の、Cys結合の又は精製ビオチニル−Cys−結合のtRNAを含み、対照はmRNAがないものであった。選択は図8に示すような軟質アビジンによるものであった。3Hペプチドのビオチニル−Cys基質との不完全な結合は、軟質アビジン(ほぼ10-7Mの非常に高いKdを有するように誘導体化されたアビジン)の単一のビオチンを含有するペプチドに対する低い親和性のためであると思われた。
【0038】
図12:隣接する多数の非天然アミノ酸をfM−T−bC−bC−V疑似ペプチドに取り込むためのアッセイ。この実験は、図11に示すように実施された。軟質アビジンによる結合は、1個以上のビオチンを含有するペプチドに対する非常に低い親和性のために予想されるように、fM−T−bC−bC−Vについては完全であったが、fM−T−bC−Vについては不完全であったことに留意されたい。
【0039】
図13:全tRNAと15種の異なる14C標識アミノ酸の混合物(ニュー・イングランド・ニュークレア社)との結合活性(TCA沈殿法によって測定される)についての本発明の純粋な翻訳系のアッセイ(「蛋白質ポリメラーゼ」)。添加されたシンテターゼは、tRNAを含まない粗製のアミノアシルtRNAシンテターゼ細胞抽出物からなる。
【0040】
図14:選択のコドンに対して特異的な非天然アミノ酸を結合したアミノアシルtRNAを合成するために一般化できる方法。合成オリゴデオキシリボヌクレオチドから調製された本発明の組換えDNAクローンから試験管内で合成された伸長体tRNAAsn-CA(図16のレーン5)は、矢印で示される天然tRNAAsnからの実質的な塩基改変物を含有する。このtRNAのアンチコドンは、大きい文字で示されている。アミノNVOCで保護された天然アミノ酸は、pdCA上で化学的にアミノアシル化され(右上参照)、次いでtRNAAsn-CA(FokI切断鋳型のランオフ転写法によって形成された)にT4RNAリガーゼで連結された(図16、レーン6)。この方法は一般化できる。なぜならば、アミノアシルtRNAシンテターゼ又は天然tRNAが全く必要とされなかったからである。
【0041】
図15:左から右にtRNAAsn(T)tRNAAsn(S)tRNAAsn(V)と命名されたtRNAAsn(N)の3種のアンチコドン変異体。これらtRNAの新規なアンチコドンは、これらのtRNAが認識するコドンの上に大きい文字で示されている。この遺伝暗号は、コドンが、どちらのアミノ酸(天然又は非天然)がそれぞれのtRNAAsn-CAに連結するために選択されるのかを直ちに指定するように再設計された。
【0042】
図16:非結合tRNA(レーン1、3及び5)及びアミノアシルtRNA基質(レーン2、4及び6)の酸/尿素ポリアクリルアミドゲル電気泳動(Varshney他(1991)J.Biol.Chem.266,24712−24718)。このゲル系において、遊離のRNA種(レーン1、3及び5)の観察された移動度は、アミノアシル化(レーン2、4)によって、或いはアミノアシル化CAジヌクレオチド(レーン6)の連結によって抑制された。
【0043】
図17:隣接するaGアミノ酸を疑似ペプチドに選択的に取り込むための一般的な方法。この実験は、図8及び11と同様に実施したが、ただし選択段階は実施しなかった。ここで、tRNAAsn(N)はAsnをベースとしてtRNAAsnBと略記されていることに留意されたい。
【0044】
図18:純粋リボソームディスプレイ。これは、ペプチドと疑似ペプチドの反復合成及び選択(定向進化)のために使用できるMattheakis他(上記)の粗製 in vitro 系(生体物質なしの)の純粋なタイプである。疑似ペプチドは、選択されるためには非放出 de novo 合成ペプチドを取り囲むほぼ100Åの長さのリボソームトンネルを超える程度に長いことが必要である。mRNA(及びリボソーム)からのペプチドの分離は、終止コドン、終結因子又はある種のアミノアシルtRNAを除外し且つリボソームを立ち往生させる抗体を使用することによって防止できる。
【0045】
図19:本発明のスペーサーmRNA及びそれらのコードペプチド産物。
【0046】
図20:本発明の精製系を使用した図19のスペーサーmRNAの翻訳。ペプチド産物に取り込まれた伸長体バリン(3H標識)対開始体ホルミルメチオニン(35S標識)の比率は、二重標識d.p.m.計数プログラムを使用してTCA沈殿産物を分析することによって測定された。観察された直線のグラフは、高い連続移動性について予期されるものである。
【0047】
図21:純粋mRNAディスプレイ。これは、Mattheakis他(図18)の粗製 in vitro 系を基にした根本他(上記)並びにRoberts及びSzostak(上記)の粗製 in vitro 系の純粋なタイプであって、mRNAをプロマイシン(Pm)と結合させてそのmRNAをそれらのペプチド又は疑似ペプチド産物に共有結合的に融合させるようにできるという点で異なるものである。しかして、このmRNAと疑似ペプチドの融合体は、選択前にその他の翻訳成分から精製できるため、非常に短い疑似ペプチドをリボソームトンネルによって遮蔽することなく表示できる。この融合反応は緩やかであるため、終止コドンを含むmRNAを使用するときに終結因子を除外して終結因子が触媒するペプチドの放出を防止することが重要である。典型的には、リボソームをmRNAに結合したデオキシヌクレオチド配列によって立ち往生させる。本発明者は、mMTKV及びmMTCV(図8及び11)を、注文合成された「添え木」DNA(mRNA及びpdA27dCdCPmのいずれかの末端にハイブリダイズさせるように設計されたTTTTTTTTTTAATTCAAC)及びT4DNAリガーゼを使用してpdA27dCdCPmに効率的に連結させ(トリ連結)、次いで融合及び選択実験に使用するためにこの結合体をゲル精製した(Roberts及びSzostak(1997)PNAS 94,12297−12302)。ここで、図中のXは、C又はbC(図9)又はbK(図22)のような容易に選択できるアミノ酸である。
【0048】
図22:プロメガ社製のTranscendビオチニル−リシンtRNAlys(E.coli、材料及び方法を参照)。
【発明を実施するための最良の形態】
【0049】
発明の詳細な説明
(I)概説
本発明は、複数の異なる天然及び非天然アミノ酸残基を高度に制御され且つ一般化できる態様で特異的に取り込めるようにするための精製成分から再構成された in vitro 翻訳系に関する。ある種の野生型のアミノ酸及び終結因子による望ましくない競合を取り除くため、遺伝学的にコードされた小さい及び長い疑似ペプチドの分子及びライブラリーを効率的且つ特異的に合成することが可能である。
【0050】
本発明は、斯界の多数の研究者による長年にわたる相当な努力にもかかわらず未解決のままであった遺伝学的にコードされた疑似ペプチドを合成することのできる生合成方法に対する長く切望された必要性のために開発された。
【0051】
また、本発明は、効率的な連続移動性のためにEF−P、W、W2又はレスキューの添加を必要としない、単純化され、高度に精製された翻訳系に対する長く切望された必要性のためにも開発された。斯界におけるこのような系の最も高度に精製されたタイプでの翻訳の連続移動性の問題及び明らかな解答のないこの分野における10年間の研究を基にすれば、成功する見込みは小さいように思われる。
【0052】
(II)用語の定義
ここで、便宜上、明細書、実施例及び添付された特許請求の範囲で使用したある種の用語を収録する。
【0053】
「蛋白質」とは、1個以上のペプチド結合によって結合された任意の2個以上の天然型アミノ酸を意味する。ここでは、「蛋白質」及び「ペプチド」が互換性を持つように使用される。
【0054】
用語「ペプチド」とは、それらの単量体がアミド結合によって互いに結合された天然アミノ酸(α−アミノ酸)であるオリゴマーをいう。ペプチドは、2個以上のアミノ酸単量体の長さであるが、たいていは5〜10個のアミノ酸単量体の長さであり、さらにはそれよりも長い、即ち20個まで又はそれ以上のアミノ酸であることができるが、20個のアミノ酸よりも長いペプチドは、「ポリペプチド」と呼ぶのが適当である。用語「蛋白質」とは、斯界に周知のものであり、いくつかの生物学的機能を有する非常に大きなポリペプチド又は会合した相同若しくは異種ポリペプチドをいう。本発明の目的のために、用語「ペプチド」、「ポリペプチド」及び「蛋白質」は、3つのタイプ全てが翻訳系によって合成できるときにはたいてい置き換えることができ、そのため一括して蛋白質と呼ばれる。
【0055】
「疑似ペプチド」とは、非天然主鎖を有する産物及びさらにエステル、チオエステル若しくはその他の結合で部分的に又は全体的に置換されたアミド(ペプチド)結合を有するもの(Mendel、前述)を含めて、1種以上の非天然アミノ酸(例えば、非天然側鎖、非天然キラリティー、N−置換アミノ酸若しくはβ−アミノ酸)、非天然トポロジー(例えば、環状若しくは分岐)又は非天然化学誘導体(例えば、メチル化若しくは末端がブロックされたもの)を含むペプチドアナログ、或いは天然アミノ酸を含むペプチド以外の任意の分子であってリボソームによって合成されるものを意味する。
【0056】
用語「アミノ酸残基」及び「ペプチド残基」とは、アミノ酸又はペプチド分子であってそのカルボキシル基の−OHがないもの(C末端が結合されたもの)又はそのアミノ基のプロトンがないもの(N末端が結合されたもの)を意味する。一般的に、アミノ酸及び保護基を示すためにここで使用される略称は、IUPAC−IUB生化学命名法委員会の提言に基づくものである(Biochemistry(1972)11:1726−1732参照)。ペプチド中のアミノ酸残基は、次のように略記する。アラニンはAla又はA、システインはCys又はC、アスパラギン酸はAsp又はD、グルタミン酸はGlu又はE、フェニルアラニンはPhe又はF、グリシンはGly又はG、ヒスチジンはHis又はH、イソロイシンはIle又はI、リシンはLys又はK、ロイシンはLeu又はL、メチオニンはMet又はM、アスパラギンはAsn又はN、プロリンはPro又はP、グルタミンはGln又はQ、アルギニンはArg又はR、セリンはSer又はS、トレオニンはThr又はT、バリンはVal又はV、トリプトファンはTrp又はW及びチロシンはTyr又はYである。ホルミルメチオニンはfMet又はfMと略記する。用語「残基」とは、相当するα−アミノ酸からそのカルボシキル基のOH部分及びα−アミノ基のH部分を取り除くことによって誘導される基を意味する。用語「アミノ酸側鎖」とは、「K.D.Kopple,”ペプチド及びアミノ酸”,W.A.Benjamin社,ニューヨーク及びアムステルダム,1966年,2及び33ページ」によって定義されるように、−CH(NH2)COOH部分を除いたアミノ酸の一部分のことである。一般的なアミノ酸のこのような側鎖の例は、−CH2CH2SCH3(メチオニンの側鎖)、−CH2(CH3)−CH2CH3(イソロイシンの側鎖)、−CH2CH(CH3)2(ロイシンの側鎖)又は−H(グリシンの側鎖)である。
【0057】
ある具体例では、本発明の用途で使用されるアミノ酸は、蛋白質中に見いだされる天然型のアミノ酸である。特に好適なアミノ酸側鎖には、次の21種の天然アミノ酸:アラニン、アルギニン、アスパラギン、アスパラギン酸、システイン、グルタミン酸、グルタミン、グリシン、ヒスチジン、イソロイシン、ロイシン、リシン、メチオニン、フェニルアラニン、プロリン、セレノシステイン、セリン、トレオニン、トリプトファン、チロシン及びバリンの側鎖から選択されるものが含まれる。しかしながら、本発明は、ここで言及される任意の特定のアミノ酸のアナログ、誘導体及び同族体を使用することを意図する。例えば、本発明は、放射性アミノ酸アナログ、側鎖を長くし又は短くすると同時に重合のためのカルボキシル、アミノ又はその他の反応性の前駆体官能基をさらに与えるアミノ酸アナログ並びに異なる側鎖(適切な官能基を有する)を有するアミノ酸アナログを使用することを意図する。例えば、主題の疑似ペプチドは、例えばβ−シアノアラニン、カナバニン、ジエンコル酸、ノルロイシン、3−ホスホセリン、ホモセリン、ジヒドロキシフェニルアラニン、5−ヒドロキシトリプトファン、1−メチルヒスチジン、3−メチルヒスチジン、アリルグリシン(又はそのアルキン誘導体)、O−メチルセリン、ビオチニルリシン、ビオチニルシステイン(又はその他のビオチン標識アミノ酸)、シクロヘキシルアラニン、ホモグルタメート、D−アラニン(又はその他のD−アミノ酸)、N−メチルグリシン(又はその他のN−メチルアミノ酸)、ε−N−メチルリシンとしてのアミノ酸アナログ及び21種の天然アミノ酸又は非天然アミノ酸の放射性同位元素誘導体を包含できる。ここで好適なその他の天然型アミノ酸又は非天然型アミノ酸は、当業者であれば容易に認識され且つ本発明の範囲に含められる。
【0058】
用語「キラル」とは、鏡像の相手を重ね合わすことができないという特性を有する分子をいい、一方、用語「アキラル」とは、その鏡像の相手に重ね合わすことができる分子をいう。
【0059】
用語「立体異性体」とは、同一の化学構成を有する化合物であるが、原子又は基の空間配置に関して異なるものをいう。特に、「鏡像異性体」とは、互いの鏡像を重ね合わすことができない化合物の2種の立体異性体をいう。一方、「ジアステレオマー」とは、非対称である2つ以上の中心を有する立体異性体であってその分子が互いに鏡像でないものをいう。キラル中心の命名に関して、用語「D」及び「L」立体配置は、IUPACの推奨によって定義されるようなものである。この用語の使用に関して、ジアステレオマー、ラセミ化合物及び鏡像異性体は、ペプチド調製物の立体化学を説明するためにその一般的な状況で使用されるであろう。
【0060】
用語「D−アミノ酸」及び「L−アミノ酸」とは、それぞれ、グリセルアルデヒドの予想される立体異性体に関する規則による絶対配置を意味する。しかして、所定の異性体による偏光面の回転方向にかかわらず、立体化学的にL−グリセルアルデヒドに関わる全ての立体異性体はL−と表記され、D−グリセルアルデヒドに関わるものはD−と表記される。トレオニン及びイソロイシンの場合には、2つの立体化学中心、即ち、Cα及びCβ原子がある。ここで使用されるD−トレオニン及びD−イソロイシンは、好ましくは、両方のキラル部位においてこれらのアミノ酸のL−立体異性体の立体配置とは逆である(鏡像異性である)立体配置を有する。例えば、これらのものは完全に鏡像である。グリシンは一般的に生じる唯一のアキラルなアミノ酸である。グリシンのようなアキラルなアミノ酸残基の存在は、そのキラリティーの名称に影響を及ぼさない。
【0061】
自然界で de novo 合成される蛋白質の全てのキラルなアミノ酸(例えば、「天然型」)は、L−アミノ酸である。
【0062】
「D−鏡像異性体」又は「D−ペプチド鏡像異性体」とは、L−アミノ酸ではなく、D−アミノ酸残基からなるペプチドをいう。
【0063】
用語「合成」とは、試験管内での化学的又は酵素的合成による産生をいう。
【0064】
DNAとは、2個以上の共有結合された天然型又は修飾デオキシリボヌクレオチドの配列を意味する。
【0065】
「RNA」とは、2個以上の共有結合された天然型又は修飾リボヌクレオチドの配列を意味する。この用語に包含される修飾RNAの2つの例は、ホスホロチオエート型RNA及び天然の修飾塩基を含有する「トランスファーRNA」である。
【0066】
「トランスファーRNA」又は「tRNA」とは、ペプチド又は疑似ペプチド前駆体をmRNAへの部分的な塩基の対合によって特定される態様でリボソームに運搬できる任意のRNA分子を意味する。
【0067】
「翻訳開始配列」とは、機能性のリボソーム進入部位を与えることができる任意の配列を意味する。細菌の系では、この領域は、時としてリボソーム結合配列又はシャイン・ダルガーノ配列と呼ばれる。
【0068】
「メッセンジャーRNA」又は「mRNA」とは、「翻訳開始配列」を含有する任意の核酸を意味する。
【0069】
「再構成翻訳系」とは、in vitro 翻訳、例えばmRNA依存性蛋白質合成を行うことのできる反応混合物(a)及び細胞溶解物の翻訳系又は麦芽抽出物の翻訳系に見いだされる汚染性蛋白質の10%未満を有し、さらに好ましくはこのような汚染性蛋白質の5%未満又はさらに1%未満を有することを特徴とする反応混合物(b)をいう。ある好ましい具体例では、主題の再構成翻訳系は、組換えによって生産された及び/又は精製された蛋白質を混合することによって生成される。
【0070】
「開始コドン」とは、蛋白質コード配列の始まりを表す3種の塩基を意味する。一般的には、これらの塩基は、AUG(又はATG)であるが、この態様で使用できる任意のその他の塩基のトリプレットは置換されていてよい。
【0071】
「終止コドン」とは、蛋白質コード配列の終わりを表す3種の塩基を意味する。一般的には、これらの塩基は、UAG、UAA又はUGA(ここで、UはTで置換されていてよい)であるが、この態様で使用できる任意のその他の塩基のトリプレットは置換されていていよい。
【0072】
「休止配列」とは、リボソームにその翻訳速度の遅延及び停止を生じさせる核酸配列、例えばDNA配列を意味する。
【0073】
「ペプチドアクセプター」とは、リボソームのペプチジルトランスフェラーゼ機能の触媒活性によって、成長しつつある蛋白質鎖のカルボキシル末端に付加できる任意の分子を意味する。典型的には、このような分子は、(i)ヌクレオチド又はヌクレオチド様の部分(例えば、アデノシン又はアデノシンアナログ(N−6アミノ部位でのジメチル化が許容できる))、(ii)アミノ酸又はアミノ酸様の部分(例えば、20種のD−又はL−アミノ酸又はそれらの任意のアミノ酸アナログのいずれか(例えば、O−メチルチロシン又は「Ellman他,Meth.Enzymol.202:301,1991」に記載されたアナログのうち任意のもの))及び(iii)これら2種の間の結合(例えば、エステル、アミド又は3’の位置若しくはあまり好ましくはないが2’の位置でのケトン結合)(好ましくは、この結合は、天然リボヌクレオチドのコンホメーションによる環の縮みを有意に摂動させない)を含有する。また、ペプチドアクセプターは求核基も有し、このものは、アミノ基、ヒドロキシル基又はスルフヒドリル基であることができるが、これらに制限されるわけではない。さらに、ペプチドアクセプターは、ヌクレオチドの疑似体、アミノ酸の疑似体又はヌクレオチド・アミノ酸の結合構造の疑似体から構成されていてもよい。
【0074】
「それぞれのコドンでの高選択的な取り込み」とは、コドンに相当するペプチド又は疑似ペプチド中のある位置でのアミノ酸残基の少なくとも80%の選択的な取り込み、より好ましくは少なくとも90、95又はさらには98%の選択的な取り込みを意味する。
【0075】
ペプチドアクセプターが蛋白質コード配列の「3’末端」に配置されるとは、ペプチドアクセプター分子がその蛋白質コード配列の最後のコドンの後に配置されることを意味する。この用語には、蛋白質コード配列の3’末端に正確に配置されるペプチドアクセプター分子並びにコード配列又は非コード配列を介在させることによって最後のコドンから隔てられるもの(例えば、休止部位に相当する配列)が含まれるが、これに限定されるものではない。また、この用語には、コード又は非コード配列がペプチドアクセプター分子に追従する(即ち、3’からそれに)構築物も含まれる。さらに、この用語には、蛋白質コード配列に共有結合される(直接的に又は核酸配列を介在させることによって間接的に)ペプチドアクセプター分子並びに蛋白質コード配列にいくつかの非共有結合の手段によって、例えば、蛋白質コード配列の3’末端に又はその近傍に結合する第2の核酸配列を使用するハイブリダイゼーションによって結合し、しかもそれ自体がペプチドアクセプター分子に結合するものが含まれるが、これに限定されるわけではない。
【0076】
ペプチドアクセプターに「共有結合する」とは、ペプチドアクセプターが「蛋白質コード配列」に共有結合によって直接的に、或いは別の共有結合した配列(例えば、休止部位に相当するDNA)によって間接的に結合することを意味する。
【0077】
「mRNAディスプレイ」とは、共有又は非共有結合によって、mRNAの翻訳からのペプチド又は疑似ペプチド産物をそのペプチド又は疑似ペプチド産物に関わる同種の(コグネイト)mRNA又はcDNA又はヌクレオチド配列に結合させるための任意の方法を意味する。一般的に使用される結合はプロマイシンを含む。
【0078】
「変化した機能」とは、ある分子の機能における任意の定性的な又は定量的な変化を意味する。
【0079】
ここで使用するときに、「結合の相手」とは、所望のmRNA・ペプチド/疑似ペプチドのリボソーム複合体又は融合体の一部分に対する特異的で共有結合的又は非共有結合的な親和性を有する任意の分子を意味する。結合の相手の例として、抗原/抗体の対、蛋白質/阻害剤の対、受容体/リガンドの対(例えば、ホルモン受容体/ペプチドホルモンの対のような細胞表面受容体/リガンドの対)、酵素/基質の対(例えば、キナーゼ/基質の対)、レクチン/炭水化物の対、オリゴマー又はヘテロオリゴマー蛋白質集合体、DNA結合蛋白質/DNA結合部位の対、RNA/蛋白質の対及び核酸の二本鎖、ヘテロ二本鎖又は連結鎖並びにRNA・蛋白質融合体の任意の部分と1個以上の共有又は非共有結合(例えば、ジスルフィド結合)を形成することができる任意の分子よりなる構成要素が挙げられるが、これらに限定されない。
【0080】
用語「リガンド」とは、蛋白質、例えば受容体のような特定の標的によって認識される分子をいう。標的が結合する又はそれと反応する任意の薬剤が「リガンド」と呼ばれるため、この用語には、酵素の基質及び触媒反応の反応体が含まれる。用語「リガンド」とは、任意の特定の分子の大きさ又は問題の物質が標的と結合でき、さもなくばそれと相互作用できること以外のその他の構造的特徴又は組成的特徴を意味するものではない。「リガンド」は、標的が結合する天然のリガンド又はアゴニスト若しくはアンタゴニストとして作用できる機能性アナログとしての役割を果たし得る。
【0081】
用語「基質」とは、酵素のリガンドであって触媒として作用し且つ酵素によって生成物に化学的に変換されるものをいう。
【0082】
用語「受容体」とは、所定のリガンドに対して親和性を有する分子をいう。
【0083】
用語「固体担体」とは、硬質又は半硬質の表面を有する材料をいう。このような材料は、好ましくは、小型のビーズ、ペレット、ディスク、チップ、皿状物、マルチウェルプレート、ウエハーなどの形をとるであろうが、その他の形を使用してもよい。いくらかの具体例では、この材料の少なくとも一つの表面は、実質的に平坦である。用語「表面」とは、ある固形物上の任意の一般的には二次元構造をいい、表面が無くならなければ段差、隆起、屈曲、テラスなどがあってもよい。
【0084】
ここで使用するときに、「集団」とは、1種以上の分子(例えば、1種以上のRNA、DNA、RNA・蛋白質又はRNA・疑似ペプチド融合分子)を意味する。本発明の方法は、所望ならば多数の候補分子から始める選択を容易にするものであるから、本発明に従う「集団」とは、好ましくは、少なくとも103種の異なる配列、さらに好ましくは少なくとも108種の異なる配列、そして最も好ましくは少なくとも1015種の異なる配列を意味する。
【0085】
用語「無作為ペプチドライブラリー」とは、無作為の又は半無作為のペプチド集団並びにこれらの無作為ペプチドを含有する融合蛋白質の集団(応用できるように)を意味する。疑似ペプチドの1種以上の残基が非天然型アミノ酸様の部分であるということを認識した上で、同様の意味が用語「無作為疑似ペプチドライブラリー」に与えられる。
【0086】
「選択」とは、ある分子をある集団内のその他の分子から実質的に分離することを意味する。ここで使用されるときに、「選択」段階は、この選択段階の後に所望の分子を集団中の望ましくない分子に対して少なくとも2倍、好ましくは10倍、さらに好ましくは100倍、最も好ましくは1000倍以上富化させる。ここに示されるときに、選択段階を何度でも繰り返してよく、また、異なるタイプの選択段階を所定の方法に組み合わせてもよい。
【0087】
試験ペプチド又は疑似ペプチドを標的と結合させることに関して、句「個々に選択的な態様」及び「個々に選択的な結合」とは、標的の分子同一性に対して特異的な結合及びそれに依存した結合をいう。
【0088】
用語「差異結合手段」並びに「親和性選択」及び「親和性富化」とは、ペプチド又は疑似ペプチドディスプレイライブラリーの構成要素を、このライブラリーのディスプレイパッケージのそれぞれについて試験ペプチド又は疑似ペプチドを標的に結合する異なる能力に基づいて分離させることをいう。ディスプレイ分子による標的の差異結合は、標的と特に相互作用する分子を相互作用しない分子から親和性によって分離するのに使用できる。親和性選択の例として、アフィニティークロマトグラフィー、沈殿法、蛍光表示式細胞分取法及びプラークリフト法が挙げられる。以下に記載するように、アフィニティークロマトグラフィーには、例えば固定化された標的蛋白質を使用するパンニング技術が含まれる。
【0089】
用語「レポーター群」又は「タグ」とは、他の分子に結合したとき及び化学的な分離プロセスで利用されたときに容易に検出できる原子、化合物又は生体分子若しくは複合体をいう。レポーター群は、例えば、蛍光性若しくは放射性原子又は1個以上のこのような原子を含有する化合物であることができる。
【0090】
アミノアシルtRNAシンテターゼの「プルーフリーディング」活性とは、同種でない(非コグネイト)アミノ酸(天然又は非天然)を認識し、次いでこれをシンテターゼのアミノアシル化された同種のtRNAアイソアクセプターから除去するというこの酵素の加水分解触媒活性を意味する。
【0091】
(III)代表的な具体例
一般に、本発明の方法は、ペプチド又は疑似ペプチドを生成させる in vitro 又は in situ 転写/翻訳プロトコルからなる。これは、mRNA分子を天然型及び/又は非天然型アミノ酸又はアミノ酸アナログを結合した1種以上のtRNA分子で合成し且つ in vitro 又は in situ 翻訳することによって達成される。本発明者は、細菌の翻訳がEF−P、W、W2又はレスキューを添加することなく再構成できることを発見した。
【0092】
一般に、好ましい最小の翻訳系は、次の高分子成分:リボソーム、mRNA、アミノアシルtRNA及び翻訳因子であるIF2H、IF3H、EF−TuH、EF−GHを含む。IF1HとEF−Tsは促進性であるので、しばしば添加される。更なる別法を以下に詳述する。
【0093】
標識開始因子の高レベル過剰発現用のクローンが入手できないため、精製翻訳系を再構成するために大量の高度に精製された因子を再現可能に調製することが大きな技術的障害であった。実際に、多くの研究が主要成分の資質に依存していた。この問題を克服するために、本発明者は、(His)6タグ(hisタグと呼ばれる)を有する3種のE.coli開始因子全てをサブクローニングし且つ過剰産生させ、精製翻訳系でそれぞれの因子の活性を検査した。
【0094】
hisタグE.coli翻訳因子のサブクローニング、過剰発現及び精製。
公表されたE.coliのIF1、IF2及びIF3発現用クローンは、多数の初期研究には有用ではあるものの、親和精製できず且つ一つの例外を除いて熱誘導できる非標識因子のためのものである(「Calogero他(1987)Mol Gen Genet 208,63−9」、「Laalami他(1991)J Mol Biol 220,335−49」、「De Bellis及びSchwartz(1990)Nucleic Acids Res 18,1311」、「Mortensen他(1991)Biochimie 73,983−9」、「Brombach及びPon(1987)Mol Gen Genet 208,94−100」)。IF3の過剰発現について考慮すべき重要な事項は、希少なAUU開始コドンが存在することである(Brombach及びPon(1987)Mol Gen Genet 208,94−100)。fmet−tRNAi fmetと対合する開始コドンは、IF3によって直接校正され(Meinnel他(1999)J Mol Biol 290,825−37)、それによって生体内で自身の遺伝子の翻訳をIF3仲介フィードバック抑制することができる(Brombach及びPon(1987)Mol Gen Genet 208,94−100)。従って、発現レベルを増大させ且つ3種の開始因子全ての精製を単純化させるために、本発明者は、開始コドンをPCR法によってAUG(CAC)6配列で置き換え、そのコード配列をpET由来の発現用プラスミドにサブクローニングした(材料及び方法参照)。得られたhisタグクローンは、非毒性であり且つ溶解物の可溶性画分中でそれら因子(IF1H、IF2H及びIF3Hと呼ぶ)を非常に高いレベルで過剰産生した(図1、レーン1、2、5、6、8、9)。さらに、N末端his6タグを有するE.coliのEF−Tu及びEF−G(EF−TuH及びEF−GHと呼ぶ)を市販のクローンを使用して過剰発現させた(図1、レーン12、13、15、16(「Hwang他(1997)Arch Biochem Biphys 348,157−62」、「Semenkov他(1996)Proc Natl Acad Sci USA 93,12183−8」))。また、Ni2+精製開始因子及び伸長因子のゲル分析も図1に示す(レーン3,7、10、14及び17)。IF2H、IF3H、EF−TuH及びEF−GHは、真正のE.coli蛋白質と共に移動し(データは示さない)、IF1Hは、分子量マーカーとの比較に基づいて予期された電気泳動度を有していた(図1、レーン1〜4)。IF2のより短い形のIF2−2は、おそらくIF2Hと共に過剰発現したが(図1、レーン6)、内部翻訳開始機構によるその合成(Sacerdot他(1992)J Mol Biol 225,67−80)はhis6タグを取り込まないであろうから、IF2Hと共に精製しなかった(図1、レーン7)。
【0095】
開始アッセイにおけるhisタグ開始因子の依存性。
3種の開始因子の活性及び純度を、リボソーム:fmet−tRNAi fmet:ApUpGのトリヌクレオチド複合体構成によって測定した(図2、頂部線)。3×塩洗浄リボソームを有する開始複合体構成中でのhisタグ開始因子の依存性は、ネイティブ因子について報告された依存性に匹敵した(表1、「Kung他(1974)Arch Biochem Biphys 162,578−84」、「Dubnoff及びMaitra(1972)J Biol Chem 247,2876−83」)。IF3依存の変差は、異なる70Sリボソーム調製物中の異なる量の遊離30Sリボソームサブユニットに起因する可能性がある。なぜならば、IF3は、このアッセイで70Sリボソームの代わりに遊離30Sサブユニットを使用すると、開始複合体構成物を強くは刺激しないからである(Canonaco他(1987)Biochimie 69,957−63)。開始因子がリボソームと共に精製される傾向があれば(Kung他(1974)Arch Biochem Biophys 162,578−84)、この結果もリボソームの純粋さを裏付けるものである。AUGトリヌクレオチド鋳型をMTTV mRNA鋳型と非常に低濃度(0.3μM)で置き換えると(図3、以下に説明される)、同等の収率の複合体構成が可能になる(データは示さない)。
【0096】
トリペプチド合成におけるhisタグ因子の依存性。
本発明者は、次に、開始複合体が図2に示される成分を使用する精製翻訳系で伸長を受けることができたかどうかを試験した。本発明者が選択したmRNAのデザイン(Pavlov他(1997)Biochimie 79,415−22)は、最適なシャイン・ダルガーノ配列及びε翻訳エンハンサー(Olins及びRangwala(1989)J Biol Chem 264,16973−6)を含む。本発明のデザイン(図3)は、標準的な18merオリゴデオキシリボヌクレオチドにハイブリダイズされる一本鎖の長い合成DNAの鋳型から直接合成される程度に短いmRNAを、クローン化する必要なくコードするという利点を有する(Milligan及びUhlenbeck(1989)Methods in Enzymology 180,51−62)。この鋳型の翻訳は、リボソームがあるコドンに転位し、この場合に同種の(コグネイト)tRNAが全く供給されないとき完了する(Weissbach他(1984)BioTechniques 2,16−22)。
【0097】
図4に示すように、3×塩洗浄リボソーム及びMTV mRNA(図3)を使用して図2に示される成分の全てによりオリゴペプチド合成を再構成すると、3H標識バリンの単離ペプチド産物への取り込みによって判断されるように、fMTVの合成に帰着する。合成のタイムコースがプレインキュベートされた開始因子の混合物をプレインキュベートされた伸長因子の混合物と結合させることによって開始されるときには(図4の三角形、凡例参照)、最初の1分以内での産物合成の迅速な初期バースト及び定常状態での緩やかな合成速度が存在する。10分間のプレインキュベーションなしでは(図4の四角形、凡例参照)、最初の1分以内では産物がほとんど合成されず、また合成は、時間経過につれてほぼ直線である(図7も参照)。
【0098】
図5は、14C標識トレオニン及び3H標識バリンのペプチド産物への取り込みに基づいて、in vitro 翻訳に必要なそれぞれの因子(IF1はこの実験では除外された)への依存性を示している。この完全な系は、fMTV合成について予期されるように、1.0のトレオニン/バリン(T/V)比のペプチド産物を産生した。任意の1種の因子の除外は、fMTV合成を劇的に減少させ(図5)、ネイティブ因子について報告された依存性に匹敵する依存性を与える(「Robakis他(1981)Proc Natl Acad Sci USA 78,4261−4」、「Cenatiempo他(1982)Arch Biochem Biophys 218,572−8」)。翻訳因子EF−GHの除外は、転位におけるEF−Gの既知の主要な役割と一致して翻訳をトリペプチド合成からfMTジペプチド合成に転換させた(図5)。ジペプチドの合成はIF1の添加を必要とせず(Robakis他(1981)Proc Natl Acad Sci USA 78,4261−4)、しかも本発明の翻訳系にIF1Hを含めても全体的な収率の劇的な増加には至らないが、IF1Hは、前に報告されたネイティブIF1の研究(Robakis他(1981)Proc Natl Acad Sci USA 78,4261−4)と一致して、プレインキュベーションなしで実施される翻訳の最初の数分の間にfMTV合成の速度を2.5倍促進させる(データは示さない)。従って、この翻訳の結果は、開始アッセイからの知見を確認し且つ拡大するものである(表1)。翻訳系のその他の高分子成分を除外した追加の対照の翻訳は、予期された依存性を示す(材料及び方法参照)。1回だけ洗浄されたリボソームは、図5に示されるタイプの翻訳アッセイでは不十分な因子依存性を示した(データは示さない)。
【0099】
テトラペプチド合成。
次に、本発明者は、開始及び伸長の全ての段階が高度に精製された系で起こり得ることを示すために、テトラペプチド合成についてのこの系の適合性を調査した。トリペプチド合成とは対照的に、テトラペプチド合成は、脱アシル化tRNAのリボソーム出口部位からの解離なしにはあり得ない(Wilson及びNoller(1998)Cell 92,337−49)。テトラペプチドfMTTVをコードする鋳型を使用して(図3)、二重標識産物の合成を逆相HPLCで評価した(図6)。放射性ペプチドを化学合成された非放射性標準物との共移動に基づいて同定した。ペプチジル分離範囲の主要放射性ピークは、予期されるT/V比に近い比を有するfMTTVテトラペプチドに相当する(この範囲内の14C又は3H放射活性の80〜85%)。2つの副ピークは、予期されるように、3Hが全く取り込まれていない休止又は早期終結産物であるfMT及びfMTTに相当する。fMT及びfMTTは、共に、ペプチジル範囲内の14C放射活性の12%を含有し、これはモルベースでは結合産物fMTTV、fMTT及びfMTの20%に相当する。残りの2つの副ピーク(24及び50分での)は、おそらくfMTVの誘導体(例えば、メチオニン酸化産物又はホルミル化されていないペプチド)、その他のペプチジル産物及び/又はペプチドでなはない放射性汚染物に相当する。従って、精製されたhisタグテトラペプチド翻訳系は、大多数ではあるが完全に前進的ではなく、全長テトラペプチド産物の収率は、ペプチド産物の80%に等しい。この詳細な分析は、添加されたEF−P、W及びレスキューがない場合の連続移動性が以前に報告されたもの(Ganoza他(1985)Proc Natl Acad Sci USA 82,1648−52)よりも非常に高いことを示した。
【0100】
本発明の in vitro 翻訳系によって効率的に合成されるペプチドのさらなる例を表2に示す。予期される7−merの全長fMTTTTTVペプチド産物の合成は非常に前進的であった。これは、早期終結産物の圧倒性が、測定された6.4の値よりもさらに大きいT/V比を生じさせたためであろう。
【0101】
オリゴペプチド合成に及ぼす上流でのmRNA変異の影響。
翻訳開始に関して未解決の問題は、精製系におけるε配列の影響である。従って、εをスクランブリング又は欠失させる効果を、mRNAのεスクランブルMVTとε欠失MVT(図3)及びmRNAのMVTに対して飽和する鋳型濃度(1μM)を使用して、開始速度を制限する条件下(図4参照)で決定した。図7は、Uリッチなε配列をスクランブリングするとfMTV合成が低下し、この配列を欠失させると産物合成の初速度が5倍低下することを示している。mRNAε欠失MVTの濃度がそれよりも高い14倍までであると、fMVTの収率を実質的に増加させることができなかった(データは示さない)。
【0102】
非天然アミノ酸の取り込み及び選択。
本発明のテトラペプチド合成方式は、力学的実験又は選択実験用のリボソームによる非天然アミノ酸の取り込みについての試験を含めて、多くのタイプの開始及び伸長アッセイに直接適用できる。例えば、基質である3H−fmet−tRNAi fmet、14C−thr−tRNA3 thr、ビオチン標識lys−tRNAlys(プロメガ社)及び3H−val−tRNAl valを使用してmRNAのmMTKVを翻訳すると(図3)、mRNAのリシンコドンとビオチン標識lys−tRNAlysとに依存する態様で産物を軟質アビジンビーズ(プロメガ社)と共に精製することによって評価されるように、ビオチンと3H−バリンの両方を含有するペプチド産物(fM−T−bK−V)を産生した(図8)。この実験は、この精製翻訳系が容易に選択できる大きな非天然アミノ酸を取り込むことができることを示している。
【0103】
ビオチン・アビジン相互作用のKd(10-15M)が標的に非共有結合的に且つ1価として結合される任意のリガンドで知られている最も低いものの一つであれば、本発明者が合成したビオチン含有疑似ペプチドは、そのコグネイト標的(アビジン)の1種に対して親和性を有し、これはそれと同様の長さのどんな天然ペプチドのそのコグネイト標的に対する親和性よりも大きいことが予期される。非常に高い親和性のリガンドの同定がライブラリースクリーニング実験に最も望ましいならば、非天然アミノ酸を使用できるリガンドスクリーニング法の天然アミノ酸しか使用できない方法を超える見込まれる利点は、本発明のビオチニルリシン含有ペプチドについての上記の例によって例示される。
【0104】
また、図8の実験に類似する実験もビオチニルCys−tRNAcysで実施したが、ただし、この場合には、使用したビオチニル−アミノアシルtRNA種を他の手段で精製した。純粋なtRNAcysアイソアクセプター(Subriden RNA)にtRNAを含まない粗製のシンテターゼ抽出物を使用してCysを結合させ、得られたCys−tRNAcysを、公開された方法(大塚他(1997)Nucleic Acid Symp.37,125−126)を多少改変してビオチンで化学的に標識し(図9)、ビオチニル−Cys−tRNACysをゲル精製し(図10)、次いでビオチニル−Cys(bCと略記する)を疑似ペプチドに取り込ませ、そして軟質アビジンに結合させた(図11及び12)。図12の結果は、本発明の in vitro 系が特に2種の隣接する大きな非天然アミノ酸を取り込むことができる可能性があることを示唆している。
【0105】
重要なことには、図11の非結合tRNAcysによる翻訳は、3Hの取り込みを全く与えなかったので、この精製系は結合活性がなく且つ外因性の基質に特異的であったことを示している。また、これは、アミノアシルtRNAシンテターゼを含まない純粋な系について予期されるように、非結合tRNAThr又は非結合tRNAValによって「種をまかれる」翻訳のためであることも示している。このシンテターゼはE.coliの最も数の多い蛋白質の1種であり、しかもリボソーム調製物はそれらの精製のために使用できる(Ganoza他(1996)Biochemie 78,51−61)ので、本発明の純粋な翻訳系がシンテターゼ活性を欠損していたことを確かめるためにさらなる実験を行った。図13は、本発明の「蛋白質ポリメラーゼ」が汚染性のアミノアシルtRNAシンテターゼを欠損していたことを裏付けた。
【0106】
ペプチド及び疑似ペプチド産物の精製。
斯界において標準的ないくつかの別法(非変性法又は変性法のいずれか)がペプチジルtRNAから遊離のペプチド又は疑似ペプチドを放出させるために使用できる。この方法の例として、塩基による化学的加水分解(例えば、図4〜8)、終結因子(表3)又はペプチジルtRNA加水分解酵素(Karimi他(1998)J.Mol.Biol.281,241−252のように精製され且つ使用される)による酵素的触媒作用及びプロマイシン抗生物質又はその誘導体(図21)による求核攻撃が挙げられるが、これらに限定されない。ペプチド又は蛋白質精製のための斯界において標準的な多数の方法うち任意の方法(非変性法又は変性法)がペプチド及び疑似ペプチド産物の精製のために使用できる。これらの方法の例として、固形物担体を使用する親和性精製(例えば、図8、11及び12の軟質アビジンビーズとビオチンの間の相互作用を使用する)、クロマトグラフィー(例えば、図4及び5の陽イオン交換クロマトグラフィー若しくは図6の逆相HPLCを使用する)又は溶液からの沈殿(例えば、図20のTCAを使用する)が挙げられるが、これらに限定されない。
【0107】
高いことが示された本発明の翻訳系の純度(表1、図1、5、11及び13)並びにEF−P、W及びレスキューが前進型翻訳のために明らかに重要でないことにもかかわらず、これらの因子の1種以上がリボソームを汚染しているという議論(発明の背景参照)を除外するためにさらなる実験を行った。リボソームからEF−P、W及びレスキューを除去するための公開された方法(Green他(1985)Biochem.Biophys.Res.Comm.126,792−798)は、リボソームを5倍にまでそれぞれの遠心分離の間に高濃度塩(0.5〜1M)で一晩洗浄することによってペレット化することである(塩洗浄が多ければ多いほどリボソームの純度は高い)。本発明のリボソームは3回の高濃度塩洗浄で4倍にペレット化されたため、本発明のリボソームはGoanzaのものに匹敵する純度を有していよう。それにもかかわらず、追加の(4回目の)高濃度塩洗浄を取り入れた異なるバッチのリボソームを調製し(Goanzaの最も高度に洗浄されたリボソームと同一の総数)、さらに、最後の150000gの遠心分離物を、ペレット化物を容易に取り出すために2段階に分けた。第1段階では、遠心分離を最大速度で1分間にわたって行い、得られたペレットを処分した(この物質は260nmでの低い吸光度に基づく主にリボソームではないものであった)。第2段階では、上澄み液からリボソームを長期間にわたる遠心分離によってペレット化させた。これら4×洗浄リボソームの純度は、開始及び翻訳のための翻訳因子への大きな依存に基づいて非常に高いものであることが立証された(表1及び図5のようにアッセイされた)。追加の対照として、本発明の翻訳因子の全て(このものはNiアフィニティークロマトグラフィーによって予め精製された)をさらに追加のゲルろ過クロマトグラフィー段階によって別々に精製した。これら4×洗浄リボソームとNi/ゲル精製因子の組合せは、101個のアミノ酸(図20参照)程度に長いペプチドの合成では活性且つ前進的でり、しかもこの追加の精製段階は、3×洗浄リボソーム及びNi精製因子と比較して本発明に有意に影響を及ぼさないことが分かった。これらの研究は、本発明の翻訳系がEF−P、W及びレスキューに依存しないことを確認するものである。
【0108】
本発明の他の具体例は、EF−P、W及びレスキューを欠失した本発明の単純化された翻訳系のバージョンであって、その成分の1種以上が実質的に濃度調節され又は完全に除外されたものである。例えば、たいていの場合、PEGを除外すると収率がほぼ25%だけ減少し、効率的な翻訳はIF1なしで生じる(表2、図5)。本発明のその他の標準的な翻訳開始因子及び伸長因子を含めることは、一般的に使用される条件下では効率のために重要であるが、産物合成にとって必須ではない(図5)。実際に、モデル系での効率的な翻訳は、細菌の開始因子(即ち、IF1、IF2及びIF3)をより高濃度の陽イオンで置き換える(例えば、Mg2+又はポリアミン、Wagner他(1982)Eur.J.Biochem,122,193−197)ならば、この開始因子のいずれかなしに可能である。
【0109】
本発明のさらに別の具体例は、上に詳述した本発明の単純化された精製翻訳系のバージョンであって、翻訳に関与し又は翻訳を刺激することが知られているその他の精製高分子及び小分子のうち1種以上が添加されたものである。これらは、細胞性の全tRNA又はそれらの画分、細胞性の全アミノアシルtRNA又はそれらの画分、合成型の結合又は非結合tRNA、20種の天然アミノ酸のそれぞれに対するアミノアシルtRNAシンテターゼのうち1種以上、Met−tRNAi fMetホルミルトランスフェラーゼ(メチオニルtRNAトランスフェラーゼとも呼ばれる)、N10−ホルミルTHFシンテターゼ及びTHF誘導体、伸長因子Ts(材料及び方法参照)、終結因子(RF1、RF2、RF3及びRRF又はRF4)、転写と翻訳を対で行うためのDNA鋳型とRNAポリメラーゼ、RNAヘリカーゼ、シャペロン(Hardesty他(1999)Curr Opin Struct Biol 9,111−4)、ショ糖密度勾配遠心分離法のような異なる手順により精製されたリボソーム(サブユニットへの分離を含めて)、ポリアミンを含めて「ポリミックス緩衝液」の成分(Jelenc及びKurland(1979)PNAS 76,3174−3178)並びにクレアチンキナーゼ、ミオキナーゼ及び/又はピロホスファターゼ(清水他(2001)Nat.Biotech.19,751−755)を有する系のような本発明のピルビン酸キナーゼ系とは異なるエネルギー関連系を包含するが、これらに限定されるべきではない。また、非天然アミノ酸の取り込みを改善させる本発明の組換え非標識IF1、IF2及びIF3(材料及び方法参照)又はEF−Tu誘導体のような天然成分の非標識の又は変異されたタイプを添加し、或いはこれらで置き換えることも純粋な翻訳系では可能である。
【0110】
例えば、図6及び11のような本発明の単純化された精製翻訳系は、細菌の終結因子の添加によって刺激される(表3)。本発明の翻訳にシンテターゼ(例えば、Thr及びValに対するシンテターゼ)をアミノ酸ThrとVal及びATPと共に添加すると、それぞれの非結合tRNAからそれぞれのアミノアシルtRNAを生成及び再生成できる(示さない)。DNA鋳型、NTP及びRNAポリメラーゼを添加すると、転写と翻訳を対で行うことができた(表2)。また、ポリエチレングリコールのようなその他の分子の添加でも刺激されることが分かった(示さない)。また、細胞から酸・フェノール抽出によって単離された全アミノアシルtRNA(「Varshney他(1991)J.Biol.Chem.266,24712−24718」、以下の図16、レーン2も参照)も本発明の精製翻訳に有効である。基質調製のための別法は、脱アシル化された全tRNAを翻訳前に試験管内で結合させることである(Green他(1985)Biochem.Biophys.Res.Comm.126,792−798)。後者の方法は、適切なtRNA上に結合した放射性標識された又は非天然のアミノ酸の取り込みを可能にさせるように、ある種のアイソアクセプターtRNAを選択的に消耗させ又は不活性化させる(例えば、アイソアクセプターに特異的なDNAオリゴマーとRNアーゼHを使用して(神田他(1998)FEBS Lett.440,273−276))ことにさらに容易に適合できるという利点を有する。
【0111】
仮特許出願の6ヶ月後に、精製翻訳系におけるEF−P、W及びレスキューのこの可欠性に関してさらなる証拠が公開された(所望ならば清水他(2001)Nat.Biotech.19,751−755を参照されたい)。本発明の好ましい系とは対照的に、これらの本発明系の変形系は、アミノアシルtRNAシンテターゼを含有していた。翻訳は、よく特徴付けられた翻訳因子及びシンテターゼの全ての組換えタイプでもってEF−P、W、W2及びレスキューを添加することなく効率的に再構成されていた。しかしながら、リボソームは、Ganoza及び本発明の方法とは別の方法(より少ない塩洗浄を使用する)によって調製され、そして20種の異なるシンテターゼのうち3種への依存性が不十分であった。これは、EF−P、W、W2及びレスキューによる汚染の可能性を高める。それにもかかわらず、これらの因子の大部分への強い依存性が報告され、いくらかの蛋白質を効率的に合成した。
【0112】
シンテターゼの認識を回避するように変異された化学結合サプレッサーtRNAを使用してバリンを効率的に取り込むためにある種の終止コドンが補充されたが、この文献の主張に反して、非天然アミノ酸の取り込みは試験されなかった(清水他(2001)Nat.Biotech.19,751−755)。場合によっては、この本発明の系を変形した系でのサプレッサーtRNAによる非天然アミノ酸の取り込みは、ある種の終結因子との競合をこれらのものを単に除外することによって回避できるという利点を有するかもしれない。しかしながら、粗製抽出物を使用する非天然アミノ酸の取り込みのための既存の系(発明の背景参照)と同様に、この戦略は、おそらく3種の終止コドンのうち1種のみ(UAGコドン)での1蛋白質当たり1個だけのタイプの非天然アミノ酸の取り込みに制限される。これは、添加された20種の異なるアミノアシルtRNAシンテターゼのtRNA結合活性とプルーフリーディング活性によって触媒されるセンスコドンでの天然アミノ酸による競合のため及びリボソームによるリードスルーのため第2の終止コドン(UGA)を使用する試みが失敗したためである(Cload他(1996)Chem.and Biol.3,1033−1038)。場合によっては、このような系で複数の異なる非天然アミノ酸の取り込みを可能にさせるために、このサプレッサーアンチコドンは、ある種のセンスコドンを認識させるように変異できたかもしれず(塩基対合によって)、これらのコドンでの競合は、同種の(コグネイト)天然アミノ酸及びシンテターゼを除外することによって回避できたかもしれない。しかしながら、20種のシンテターゼの大部分にとっては、そのコグネイトtRNAアイソアクセプター基質のアンチコドンは最も重要な認識要素であるため、サプレッサーtRNAのアンチコドンが変化すると望ましくないシンテターゼの認識に至り(時として交差認識の予測できない特異性によって)、しかしてプルーフリーディング及び/又は天然アミノ酸の結合に至ると思われる(Giege他(1998)Nucleic Acid Res.26,5017−5035)。さらに、シンテターゼの認識を防止することを意図したtRNAの変異は、EF−Tu及び/又はリボソームの認識効率をも減少させ、それによって運ばれたアミノ酸の取り込み効率を減少させる可能性がある。これらのシンテターゼ認識の困難性及び20種類もの数の異なるシンテターゼ蛋白質を過剰発現させることに関わる困難性のために(Swartz(2001)Nat.Biotech.19,732−733)、本発明者は、非天然アミノ酸による本発明の試験のためにシンテターゼを含まない翻訳系を選んだ。
【0113】
本発明者は、化学的に結合されたtRNAを使用して数種の非天然アミノ酸の取り込みを試験した。第1段階は、非天然アミノ酸による化学的な誤アシル化を可能にさせるように、末端CAジヌクレオチドを欠失した合成伸長体tRNAを一般化できる態様で構築することであった。現在の技術は、シンテターゼのいずれかによる結合及びプルーフリーディングを避けるように特別に設計された人工サプレッサーtRNAに依存している。本発明の精製系では、本発明者は、tRNA塩基修飾活性の予期される欠如について見込まれる効果のみを問題にした。なぜならば、このような修飾は機能にとって重要であり得(Bjork他(1999)FEBS Lett.452,47−51)、粗製翻訳系は合成tRNAを修飾し得るからである(Claesson他(1990)FEBS Lett.273,173−6)。新規の試験の雛形として、本発明者は、E.ColiのtRNAAsn(図14、大橋他(1976)Nucleic Acids Res.3,3369−3376)及びtRNAAla(以下に議論する。Picking他(1991)Nucleic Acids Res.19,5749−5754)を選択した。tRNAAsnの5’末端配列をT7RNAポリメラーゼによる最適な転写のために変異させたが、M1RNA又はRNアーゼPを使用して合成非修飾tRNA前駆体を処理するような突然変異誘発とは別の戦略もある(Forster及びAltman(1990)Science 249,783−786)。また、両者のtRNAのアンチコドンも改変されたコドン認識特性を有するいくつかの変異体を生じさせるように変異させた(本発明のtRNAAsn変異体のうち3種を図15に示す。tRNAによって認識されるアミノ酸コドンはその括弧内に示されている)。非天然アミノ酸であるアリルグリシン(aG、或いは2Pと略記される)をNVOC基でアミノ保護し、そしてT4RNAリガーゼを使用して標準的且つ一般的な戦略で(材料及び方法参照。「Thorson他(1988)Methods in Molecular Biology 77,43−47」、「Steward及びChamberlin(1998)Methods in Molecular Biology 77,325−354」)tRNAAsn(N)に連結して(図14)ゲル上で予期される移動度でもって移動する種を得た(図16、レーン6)。
【0114】
NVOCaG−tRNAAsn(N)のアミノ基を紫外線光分解法によって脱保護し、そのaG−tRNAAsn(N)をmMTNV鋳型を含有する純粋な翻訳反応に添加した。aGは、下流の3H−Vの取り込みを可能にするようにNコドンにうまく取り込まれたが、mMTVの対照の翻訳よりも低い収率を示した(図17)。このより低い収率は、主にこの系を飽和させるのには不十分なaG−tRNAAsn(N)を使用するためであることが分かった。なぜならば、aG−tRNAAsnのRNA濃度を1μMにまで倍加させる(転写及び連結反応からのaG−tRNAAsn(N)の収率が100%であると見なして翻訳系中のそれぞれの天然アミノアシルtRNA濃度を倍加させる)と、収率が倍増し且つ約65%の連続移動率でアナログを取り込んだ(アナログの前の14C−T及びその後の3H−Vの取り込みに基づく)からである。mMTNNVの翻訳は、隣接したアナログの取り込みを示し、この第2のアナログの取り込みについての連続移動率(%)は、これらのアナログ基質制限条件下では第1のアナログと同様であった。一般的な偏在性CCA3’配列で終結することを除いて図14のtRNAAsn(N)と同一構造を有する非結合tRNAによるmMTNV及びmMTNNV(BstNIで切断された鋳型のランオフ転写によって生じる)の翻訳は、取り込みを全く与えなかったことに留意されたい。これは、この精製系がこの変異tRNAAsn(N)についての結合活性を欠損しており、しかもNコドンでの外因性基質の取り込みに対して特異的であったことを示している。追加の対照として、非結合tRNAAsn(N)は、mMTVの翻訳を抑制しなかった(図17)。
【0115】
センスコドンが選択的なアミノ酸の取り込みに一般的な態様で奪い取られたという確証を得たならば、次の段階は、aG及びその他の非天然アミノ酸の取り込み効率を最適化することなので、O−メチルセリン(mS)及びアラニンのNVOCアミノ保護型を調製した(材料及び方法参照)。aG、mS又はAlaで化学的にアミノアシル化された高濃度(7μMまで)の様々なtRNAAsn(図14及び15)を使用することによって、Thr−tRNAThrで得られた取り込み効率の90%以上の取り込み効率が得られた(示さない)。
【0116】
興味深いことに、非修飾aG−tRNAAla(N)であって、公表された合成tRNAAla(Picking他(1991)Nucleic Acids Res.19,5749−5754)と比較してそのアンチコドンにGUU置換を有するものは、翻訳が全体的に不活性であった。しかしながら、粗製のtRNAを含まない細胞抽出物を使用してAlaを結合させると、効率的に取り込まれるAla−tRNAAla(N)を生じた。この活性の獲得は、aGがtRNAAsn(N)に良好に作用したので恐らくアミノ酸の相違によるものではなく、粗製抽出物の塩基修飾活性によるものと思われる。この結果は、tRNAの塩基修飾がある種のtRNAの翻訳活性にとって極めて重要であり、しかも適切な原型tRNAの初期選択が試行錯誤を必要とする自明でないプロセスであるという本発明の仮説と一致する。別の問題は、ある種の非修飾アミノアシルtRNAが標準的な精製手順中に変性によって不活性化されることである(Harrington他(1993)Biochemistry 32,7617−7622)。それにもかかわらず、いったん本発明のtRNAAla(N)のような安定な活性の非修飾tRNAが同定されたならば、アンチコドン変異体であってその他のコドンを予測できる態様で認識するものを作るのは容易である(図15、下部参照)。
【0117】
表4は、5個の連続的な非天然アミノ酸(aG)の7mer疑似ペプチド産物内への添加されたaG−tRNAAsn(T)に完全に依存する態様での典型的な取り込みを示している。これは、長さの点だけでなく、より重要なことには一般化及び有用性の点でも、2個の連続的な非天然アミノ酸について公表された記録(芳坂他(1999)JACS 121,12194−12195)を向上させている。十分に異なるアミノ酸単量体を与えると、無作為の7mer疑似ペプチドライブラリーは、実質的な疑似ペプチド多様性をコードできる。4種の異なる非修飾tRNAAsn変異体であってそのTコドンについてmSで及びそのN、S及びVコドンについてaGで化学的にアミノアシル化されたものを使用することが図14及び15に示されており、全長産物がカルボキシ末端3H−Eの取り込みに基づいて合成された(表5)。これは、単一産物中に複数且つ異なる非天然アミノ酸が取り込まれた証拠を与える。付加されたmSへの完全な依存性は、mSが実際に取り込まれ、しかもこのTコドンがtRNAAsn(T)に対して特異的である(即ち、図14及び15に示される密接に関連したアンチコドン変異体のtRNAAsn(N)、tRNAAsn(S)及びtRNAAsn(V)によって誤読されない)ということを立証した。
【0118】
非天然アミノ酸によるセンスコドン置換という本発明の戦略は一般化できるものであり、しかして61種のセンスコドン全てに拡張すべきである。自然型(21種の天然アミノ酸のみ)によるコドン使用の有意な重複を与えるならば、単一のmRNAを実質的に21種以上の異なるアミノ酸に翻訳できるアミノアシルtRNAライブラリーを創作することが可能であろう。3種の終止コドンは本発明の系では必ずしも必要ではないが、これら全てはサプレッサーにより翻訳され得るかもしれないので、見込まれるリードスルーの問題(Cload他(1996)Chem.and Biol.3,1033−1038)は、リードスルーを引き起こす天然のアミノアシルtRNAを除外することによって克服される。AUG開始コドンとfMet開始体アミノ酸を効果的に置換すると、伸長から一連の異なる分子間接触が維持されるが、これは既知の許容できる置換に基づいて生体内及び試験管内でできる(「Picking他(1991)Nucleic Acids Res.19,5749−5754」、「Wu及びRajBhandary(1997)J.Biol.Chem.272,1891−1895」)。
【0119】
A.in vitro ライブラリー
リボソーム上で発現するペプチドアナログのコンビナトリアル・ライブラリーからのペプチド又は疑似ペプチドの試験管内での定向進化は、任意の選択の標的分子に結合するリガンド又は薬剤候補物の迅速な進化のために効果的な方法である。粗製抽出物で実施されるリボソームディスプレイ(Mattheakis及びDower(1995)PCT WO95/11922)に関連する本発明の「純粋なリボソームディスプレイ」(図18)の例は、次の段階及びそれらの別法からなる。
【0120】
DNAライブラリーの構築:
無作為化された又は部分的に無作為化されたコード配列を含む合成オリゴデオキシリボヌクレオチドを化学的に合成し、RNAポリメラーゼプロモーター(例えば、バクテリオファージRNAポリメラーゼ)、翻訳開始配列及び開始コドンを含むオリゴデオキシリボヌクレオチドにハイブリダイズさせ、DNAポリメラーゼで伸長させ、DNAリガーゼによってオープンリーディングフレームをコードするプラスミド制限断片(例えば、図19に示される本発明の長鎖配列のような反復スペーサーアミノ酸配列)に連結し、ゲルで精製し、そして定量する。
【0121】
段階1(図18):合成DNAライブラリーをRNAポリメラーゼでmRNAライブラリーに転写する。
【0122】
段階2:mRNAライブラリーを、本発明の精製されたリボソーム、翻訳因子及びある種の(全てではない)野生型アミノアシルtRNAとアミノ酸アナログを結合した特別に合成されたtRNAとの混合物を含有する再構成アミノアシルtRNAプールでペプチドアナログに翻訳する。好ましくは、それぞれのmRNAコドンは、該プール中のユニークな結合tRNAによって解読されるため、このmRNAの配列は、ユニークなペプチドアナログ配列を規定している。例えば、図19の本発明のスペーサー配列であってそのいくつかがリボソームトンネルを横断するほどに長いものは、良好な連続移動率で合成された(図20)。蛋白質合成を、例えば、終止コドンを欠失している鋳型の末端に到達させることによって、終結因子を除外することによって、供給されるアミノアシルtRNAが全くないコドン(図19及び20の最長産物の場合にはAsnコドンのような)に到達させることによって又は抗生物質で立ち往生させることによって停止させる。
【0123】
段階3:無作為ペプチドアナログを含有するリボソームを遠心分離によって単離し、次いで固定化された標的物と共にインキュベートし、そして結合していないリボソームを洗い流す。結合したリボソームを解離させ(例えばEDTAで)、放出された選択mRNAを精製する。
【0124】
段階4:cDNA合成及びPCR増幅。Error−prone PCRを使用して突然変異を導入する。
【0125】
段階1〜4の反復:これらの選択及び増幅段階を必要に応じて1回以上繰り返す(以下参照)。
【0126】
段階5(示さない):増幅されたDNAを、選択され且つ増幅された疑似ペプチドの構造を推定するために、DNAシーケンス法による分析用のプラスミドにクローン化させる。選択されたコンセンサス配列の同定は、反復した選択及び増幅(段階1〜4)の十分なラウンドが使用され、しかもリガンドがこの実験によって同定されたという証拠となる。さらに、サイクルを使用してより高い親和性を有するリガンドを生じさせることができる。
【0127】
リボゾームの非存在下での選択段階を可能にする方法の改変:
DNA鋳型を、タンパク合成中にペプチドアナログに導入されるときに、mRNA又はmRNAに結合する分子(例えば、ペプチドタグに対して高い親和性を有する抗体に共有結合するハイブリダイズした相補DNAプライマー)に対して高い親和性を有するペプチドタグをコードするように修飾する。このとき、このペプチド・mRNAを選択段階の前にリボソームから分離してもよい。これらの方法及びその他の可能な別法は今までに記載されている(Mattheakis及びDower,上記、土井及び柳川(1999)FEBS Lett.457,227−230)。別法として、mRNAは、プロマイシンで3’末端標識されていてもよく、そのため「純粋なmRNAディスプレイ」用のペプチドに直接結合できる(図21、以下参照)。
【0128】
Szostak他のPCT公開WO00/047775号及びWO98/31700号(所望ならば参照されたい)は、対象のペプチド及び疑似ペプチドをコードするRNA分子に共有結合される該ペプチド及び疑似ペプチドの形を生じさせるために、本発明に容易に適合できる方法を記載している。即ち、本発明は、疑似ペプチドであってその自身のmRNAの3’末端に共有結合したもの、即ち、RNA・疑似ペプチド融合体を生成させるプロトコルを提供する。
【0129】
これは、mRNA分子をその3’末端に結合したペプチドアクセプターと共に合成及び in vitro 又は in situ 翻訳することによって達成される。一つの好ましいペプチドアクセプターは、成長しつつあるペプチド鎖のC末端に付加し且つ翻訳を終結させるヌクレオシドアナログのプロマイシンである。一つの好ましい設計では、あるDNA配列が遺伝子情報の末端とリボソームをオープンリーディングフレームの末端で停止させるためのペプチドアクセプターとの間に含まれ、このペプチドアクセプター(例えば、プロマイシン)がペプチジルtRNA連結体の加水分解前に初期のペプチド鎖を受容する追加時間を与える(図21)。
【0130】
所望ならば、得られたRNA・疑似ペプチド融合体は、選択及び増幅のラウンドを繰り返してもよい。なぜならば、このコード配列の情報は、逆転写及び増幅(例えば、PCR増幅並びに3SR又はTSAのようなRNAベースの増幅技術を含めてその他の増幅技術によって)によって回復できるからである。次いで、増幅された核酸を転写させ、修飾させ、in vitro 又は in situ 翻訳させて次の選択ラウンド用のmRNA・疑似ペプチド融合体を生成させる。この選択及び増幅の複数ラウンドを実施する能力は、非常に希少な分子(例えば、105種の構成分子からの1種の所望の分子)を富化及び単離するのを可能にする。このことは、結果として、新規な又は改良された、特に任意の標的を実質的に認識し又は所望の化学反応を触媒するペプチド及び疑似ペプチドの単離を可能にする。
【0131】
従って、ある態様では、本発明は、(a)候補のRNA分子集団であって、これらのいずれかが翻訳開始配列及びある候補の蛋白質コード配列に作動可能に連結した開始コドンを含み、これらのいずれかが該候補蛋白質コード配列の3’末端でペプチドアクセプターに作動可能に連結したものを与え、(b)天然型及び/又は非天然型アミノ酸の存在下に該候補蛋白質コード配列を in vitro 又は in situ 翻訳させて候補のRNA・疑似ペプチド融合体の集団を産生させ、(c)所望のRNA・疑似ペプチド融合体を選択し、それによって所望の疑似ペプチドを選択する段階を含む、所望の蛋白質又は疑似ペプチドの選択方法に関する。
【0132】
上記方法の好ましい具体例では、候補RNA分子の集団は、少なくとも102種、好ましくは、少なくとも105種、より好ましくは1010種又は1015程度の異なるRNA分子を含む。重要なことには、in situ 翻訳反応は、好ましくは、粗製翻訳系ではなく再構成された精製混合物中で実施される。選択段階は、疑似ペプチドを固定化された結合の相手に結合させること又は疑似ペプチドの機能活性についてアッセイすることを含む。
【0133】
別の関連する態様では、本発明は、ここに記載される選択方法のいずれかを実施するためのキットに関する。
【0134】
最後の態様では、本発明は、固定化された本発明の疑似ペプチドの配置を含むマイクロチップに関する。
【0135】
B.標的分子
標的分子は、実質的には、本発明のペプチド又は疑似ペプチドとの相互作用が有用であり得る任意の分子であることができる。ある種の具体例では、標的分子は、核酸(DNA又はRNA)、蛋白質、脂質、炭水化物などのような生体高分子である。
【0136】
ポリペプチドスクリーニング用標的物を選択する際に考慮され得る因子には、溶解度、ペプチド鎖の長さ、翻訳後修飾の必要条件若しくはコファクターの付加及び/又はこの標的がベースとしている蛋白質のモノマー又はオリゴマーの性質が含まれる。一般に、ペプチドの標的は、可溶性で、部分的に精製され又は純粋であり、しかも斯界に標準的な方法によって固体担体上に固定化されていることが望ましいであろう。
【0137】
従って、本発明は、SH2ドメイン、SH3ドメイン、アンキリン様リピート、WD40モチーフ、Kunitz型阻害剤のドメイン、EFG様ドメインのような増殖因子様ドメイン、クリングルドメイン、フィブロネクチンンフィンガー様ドメイン、ヘパリン結合ドメイン、デスドメイン、TRAFドメイン、プレクストリン相同(PH)ドメイン、ITAM、キナーゼドメインのような触媒ドメイン、ホスファターゼドメイン、ホスホリパーゼドメイン、グアニンヌクレオチド交換因子(GEF)ドメイン及びヒドロラーゼドメイン(例えばプロテアーゼドメイン)又はDNA結合ドメイン、例えばロイシンジッパー、ジンクフィンガー及びへリックス・ループ・へリックスモチーフのようなドメイン構造に相当する(例えば、これらを含む)スクリーニング用標的物を意図する。
【0138】
対象の蛋白質が膜貫通蛋白質である場合には、スクリーニング用標的物は、可溶性の細胞外又は細胞質ドメインから得ることができる。例示のために、スクリーニング用標的物は、グアニリルシクラーゼ、シトキン受容体、チロシンキナーゼ受容体又はセリン/トレオニンキナーゼ受容体の細胞外ドメインに相当し得る。その他の具体例では、スクリーニング用標的物は、リガンド結合活性を保持しているG蛋白質共役型受容体(GCR)の可溶性部分に相当し得る。例えば、上記のように、GCRの膜貫通部分の間の細胞外ループのうちのある種のものは、溶液状でなくなったときでさえもリガンド結合活性を保持することが示された。さらに別の具体例では、スクリーニング用標的物は、リポソームのような脂質二重層又はその他の小胞中で再構成でき(例えば、Kalva Kolanu他(1990)Biotechniques 11:248並びにHuangの米国特許第4957735号及び同第4708933号を参照)、この脂質/蛋白質結合体をスクリーニング用標的物として使用できる。
【0139】
単なる例示の目的のために、以下の蛋白質標的物を対象の方法に使用するために記載する。
【0140】
一具体例では、見込まれる治療用標的物は、neu受容体ファミリーからの受容体である。米国の女性において、乳癌は最も一般的な癌であり且つ癌死亡率が肺ガンに次いで単独で第2位である。主な乳癌の標的は、185kDの膜貫通ホスホ糖タンパク質のチロシンキナーゼであるneu/erbB−2/HER−2である(Shih他、(1981)Nature 290:261)。neu癌遺伝子の増幅又は過剰発現は、乳房及び卵巣の約30%で発生し、これは一次治療のまずい対応に相関することが分かっている(Slamon他(1987)Science 235:177及びHayes他(1993)Annals of Oncology4:807)。NIH 3T3細胞にneu癌遺伝子をトランスフェクトすると形質転換を生じ(Shih他,上記)、活性化されたneu癌遺伝子をマウスに導入すると全乳房上皮組織の形質転換(Muller他,(1988)Cell 54:105)を生じ又は乳房腫瘍が確率的に出現する(Bouchard他(1989)Cell 57:931)。
【0141】
乳癌がneu又はヘレグリンファミリーの一員のようなneuのリガンドに結合する分子によって抑制できるという証拠が蓄積している(Holmes他、1992,Nature 256:1205)。neuの細胞外ドメインに結合するMAb及びそれらの放射性標識結合体は、neu陰性細胞クローンの選択なしに培養液中及びヌードマウス内で乳癌細胞の増殖を遅延させる(DeSantes他,(1992)Cancer Res.52:1916及びDrebin他(1986)PNAS 83:9129)。高度乳癌治療のための診療所で使用される組換えヒト化Mabであるヘルセプチン(Colomer他(2001)Cancer Investigation 19,49−56)のような当該MAb及びそれらの誘導体は、neuの情報伝達経路を変更し且ついくつかの異なる方法で腫瘍の増殖に影響を及ぼし得る。これらのものは、(i)neuを過剰に刺激し、それによって分化を生じさせ(Bacus他(1992)Cancer Res.52:2580)、(ii)neuのホモ又はヘテロ二量体化を妨げ、それによってneuを不活性化させ(Caraway他(1994)Cell 78:5)、(iii)neuの細胞性インターナリゼーション及びダウンレギュレーションを生じさせ(Tagliabue他(1991)Int.J.Cancer 47:93)、(iv)細胞表面又は細胞質に共役細胞傷害性放射性ヌクレオチド又は毒素を送達し、或いは(v)リガンドがneuに結合しないようにする。
【0142】
本発明によって誘導され得る疑似ペプチドは、例えばneuのそのリガンド又はその他の蛋白質との結合を競争的に開裂させ又はneuに関わる酵素活性のアロステリックな活性化を阻害することによってneuの生物学的機能を抑制するのに有用であり得る。別法として、これらのものは、neuの過剰刺激又はダウンレギュレーションを生じさせるアゴニストとして有用であり得る。
【0143】
さらに別の見込まれる標的は、インターロイキン8(IL−8)である。IL−8は、好中球の化学誘引物質及び活性化物質であり且つ様々な急性及び慢性の炎症性疾患に関与している(Murphy(1994)Annu.Rev.Immunol.12:593−633)。ヒトIL−8は、腫瘍壊死因子のような因子、インターロイキン1及びリポ多糖による誘導に基づいて単球、繊維芽細胞、角化細胞及び内皮細胞によって産生される72アミノ酸残基のポリペプチドである(Murphy,上記)。IL−8のある種のアナログは、試験管内で好中球の活性化(走化性、エキソサイトーシス及び呼吸バースト)を阻害することによってIL−8アンタゴニストとして作用するため、IL−8抑制剤が炎症性疾患に対する治療上の可能性を有し得ることを示唆する(Moser他(1993)J.Biol.Chem.268:7125−7128)。
【0144】
単量体のIL−8ペプチドは、試験管内で20MのKdでもって二量体を形成する(Paolini他(1994)J Immunology 153:2704及びBurrows他(1994)Biochemistry 33:12741−12745)ため、この単量体及び/又は二量体は生体内で活性であることが可能である。二量体化できない変異体は、試験管内での機能アッセイでは活性である(Rajarathnam他(1994)Science 264:90)。興味深いことに、IL−8二量体の三次元構造のNMR及びX線測定(Clore他(1990)Biochemistry 29:1689−1696及びBaldwin他(1991)PNAS 88:502)により、このものがMHCのクラスI及びII蛋白質のペプチド結合の溝に類似していることが明らかになった(Bjorkman他(1987)Nature 329:506)ため、IL−8二量体は、MHC分子と同様の態様で in vitro 選択ペプチド又は疑似ペプチド配列に結合できる可能性があると考えられる。従って、IL−8単量体に加えて、その二量体は、魅力的な標的である。
【0145】
IL−8経路の場合には、IL−8とは別の標的は、IL−8受容体の機能性断片である。それぞれ39個及び44個のアミノ酸のヒトIL−8タイプ1受容体及びウサギIL−8タイプ1受容体のさらに小さい断片は、IL−8結合アッセイでは機能的である(Gayle他(1993)J Biol Chem 268:7283−7289)。従って、最も大きな受容体のファミリーの一部である7種の膜貫通受容体が見込まれる標的である。
【0146】
細胞性の癌原遺伝子であるc−mycは、細胞増殖及び形質転換に関与するか、プログラム細胞死(アポトーシス)の誘導にも関わる。c−Myc蛋白質は、カルボキシル末端塩基性領域/ヘリックス・ループ・ヘリックス(HLH)/ロイシンジッパー(LZ)ドメインを有する転写活性化物質である。このものは、HLH/LZ蛋白質のMaxとヘテロ二量体を形成し、DNAのEボックス要素に結合した後に遺伝子発現をトランスに活性化する。Max蛋白質は、現在までに分析された多くのその生物学的機能にとって必須のc−Mycのパートナーである。例えば、Mycは、DNAに結合し且つその癌遺伝子活性を果たすためにはMaxと共にヘテロ二量体化しなければならない。
【0147】
本発明によれば、主題の方法は、MycとMaxのようなその他の蛋白質との間の複合体の形成を阻害できる及び/又はMyc複合体が遺伝子内のmyc応答配列に結合するのを阻害できるペプチド及び疑似ペプチドを誘導するために使用できる。Myc・Max及びMax・Max二量体の全合成は、Canne他(1995)J Am Chem Soc 117:2998−3007に記載されている。
【0148】
TGFの全合成は、Woo他(1989)Protein Eng 3:29−37に記載されており、これは別の見込まれる標的分子を与えるものである。
【0149】
主題の方法に使用するために誘導できるさらに別の標的は、細胞接着、組織器質化及び創傷治癒に関わる糖蛋白質のフィブロネクチンである。フィブロネクチンン分子の全合成は、例えば、Williams他(1994)J Am Chem Soc 116:10797−10798に記載されている。
【0150】
これまでに、ヒト免疫不全ウイルス1型(HIV−1)の主要gag蛋白質であるp24の発現は、HIV−1に感染した細胞の表面上で持続することが示された(西野他(1992)Vaccine 10:677−683)。p24のC末端の100アミノ酸断片の全合成は、Mascagni他(1990)Tetrahedron Lett 31:4637−4640)によって説明され、p24蛋白質のその部分を使用してスクリーニング用標的物を生成することができる。
【0151】
同様に、HIVプロテアーゼは、すべて化学合成手段によって合成され(kent他PCT公開WO93/20098)且つ触媒活性の阻害剤並びにこのプロテアーゼに関わる蛋白質・蛋白質相互作用の阻害剤を開発するためのユニークな標的となる。
【0152】
上に例示された標的物の多くは化学合成によって調製できるが、これは決して必要条件又はさらに優先的に選択されるものではない。本発明の利点は、生物学的供給源からの精製、生合成又は組換えDNAベースの発現のような任意の他の方法を使用して部分的に精製された又は純粋な標的物を調製できることである。
【0153】
材料及び方法
hisタグ及び非標識E.coli IF1、IF2及びIF3蛋白質の過剰発現用プラスミドの構築。
N末端メチオニンのすぐ後に6個のヒスチジン挿入物をそれぞれ含むE.coliの開始因子コード配列を公表されたプラスミドからPCRによって合成し、pET24a(Novagen社)由来のベクターにサブクローニングした。E.coli優先コドン(infAコドンの代わりに)の人工配列によってコードされるネイティブIF1配列を含むプラスミドpXR201を、R.Spurio及びC.Gualerzi両氏から快く提供してもらい(Calogero他(1987)Mol Gen Genet 208,63−9)、所定のpAF1Hにサブクローニングした。infBによってコードされるネイティブIF2配列を含むプラスミドpSL4を、S.Laalami及びM.Grunberg−manago両氏から快く提供してもらい(Laalami他(1991)J Mol Biol 220,335−49)、所定のpAF2Hにサブクローニングした。infCによってコードされるネイティブIF3配列を含むプラスミドpDD1を、N.brot及びI.Schwartz両氏から快く提供してもらい(De Bellis及びSchwartz(1990)Nucleic Acids Res 18,1311)、所定のpAF3Hにサブクローニングした。この3種のサブクローンの配列は、制限酵素消化と配列分析の組み合わせによって特徴付けられ、第2アミノ酸の前のTATACA/TATG(CAC)6から始まる(連結部位に下線を施した)。最後のアミノ酸の後にTAAG/AATTCGAGCTCCGTCGA/42bp欠失/AGATCC配列が続き、この配列の残余は、pET24aからのものである。また、IF1、IF2及びIF3の非標識バージョンをクローン化し且つ過剰発現させるために類似の方法も使用した。
【0154】
E.coli翻訳因子蛋白質の過剰発現及び精製。
TufAの最初の2つのコドン間に挿入されたhis6配列を含むhisタグE.coliEF−TuをコードするプラスミドpHTA7(E.coli BL21(DE3)内の)及びTsfの最初の2つのコドン間に挿入されたhis6配列を含むプラスミドpHTSは、Y−W.Hwang及びD.Miller両氏が提供してくれた(Hwang他(1997)Arch Biochem Biophys 348,157−62参照)。his6を含めて約30個のアミノ酸のN末端伸長を含む、hisタグE.coliEF−GをコードするプラスミドpRSET/EF−G(His)(pLysSプラスミドと共にE.coliBL21(DE3)細胞内の)は、A.Savelsbergh及びW.Wintermeyer両氏が提供してくれた(Semenkov他(1996)Proc Natl Acad Sci USA93,12183−8)。本発明の3種のhisタグ開始因子のサブクローン(E.coli BL21(DE3)pLysSの:Novagen社)及び提供された3種のクローンの発現をIPTGによって誘導した。これらの因子全てを主として可溶性細胞画分中で発現させ、標準的なプロトコルを使用してNiNTAアガロースカラム(Qiagen社)から段階溶出によって精製した(ただし、10μMのGDPをEF−TuHのための最後の透析段階まで含めた)。全ての因子を緩衝液A(100mMのTris−HClpH7.4,1mMのMgCl2,1mMのDTT)に透析した。沈殿したIF3Hを5Mの尿素に再溶解させることによって回収し(Hershey他(1977)Arch Biochem Biophys 182,626−38)、希釈し、次いで100mMのNH4Clを含有する緩衝液Aに透析した。ネイティブIF2の過剰発現及び精製中に他の者によって観察された広範囲にわたる蛋白質分解性の分解(Mortensen他(1991)Biochimie 73,983−9)とは異なり、組換えIF2Hは、蛋白質分解に不安定ではなかった。純粋なE.coli終結因子(RF)のRF1、RF2、RF3及びRRFを、記載されているように(Yu他(1998)J.Mol.Biol.284,579−590)調製した。全ての因子を−80℃で保存した。これらのものを、EF−TuHを除いて活性を損失させることなく何度も解凍し、解凍後には4℃で保存し且つ数週間以内に使用した。
【0155】
E.coliリボソームの精製(Kung他(1974)Arch Biochem Biophys 162,578−84)。
対数増殖中期にまで増殖したSOLR(商標)細胞(Stratagene社)を緩衝液B(60mMのKOAc,14mMのMg(OAc)2,10mMのTris−HOAc,1mMのDTT,pH7.9)に再懸濁し、超音波処理し、10000gで遠心分離した。上澄液を30000gで遠心分離し、次いで得られた上澄液を150000gで遠心分離した。このリボソームペレットを1MのNH4Clを含有する緩衝液B中で4℃で一晩攪拌し、次いで150000gで再ペレット化することによって洗浄した。この洗浄を2回以上繰り返して3×洗浄リボソームを与え、このものを緩衝液C(10mMのMg(OAc)2,1mMのTris−HClpH7.4,1mMのDTT)に再懸濁し、−80℃で保存した。
【0156】
mRNAの合成。
mRNAをオリゴデオキシリボヌクレオチド(Research Genetics社)から転写し(Milligan及びUhlenbeck(1989)Methods in Enzymology 180,51−62)、ゲル電気泳動により精製した(Forster及びSymons(1987)Cell 49,211−20)。鋳型が比較的長いため、最適な転写を可能にするにはブロックされたオリゴヌクレオチドの合成の後に長期の脱保護時間が必要であった(12時間)。
【0157】
アミノアシルtRNAの調製。
純粋なE.coli tRNAアイソアクセプターは、SubridenのRNAからのものであった。それぞれのアイソアクセプターは、E.coliの全tRNA(Plenum社)から3つのカラムクロマトグラフィー段階を使用してメーカーにより製造された。第1の分画はBDセルロースを使用し、第2はpH7でのDEAEセファデックスを使用し、第3はpH5でのDEAEセファデックス又はセファロースを使用した。天然のアミノアシルtRNAをこれらのアイソアクセプターから次のように調製した。高特異的活性3H−fmet−tRNAi fmet(表1において24000d.p.m./pmol)及び低特異的活性3H−又は35S−fmet−tRNAi fmet(数百d.p.m./pmol、全ての他の研究のために使用される)を、MetRS、met−tRNAi fmetホルミルトランスフェラーゼ及びN5,10−メテニルTHF(N10−ホルミルTHFシンテターゼを使用して合成された)を使用して記載されるように(Robakis他(1981)Proc Natl Acad Sci USA 78,4261−4)調製した。14C−thr−tRNA3 thr(510d.p.m./pmol)及び3H−val−tRNA1 val(図5〜7において21000d.p.m./pmol、図4及び8において28000d.p.m./pmol)を、記載されるように(Robakis他(1981)Proc Natl Acad Sci USA 78,4261−4)、ThrRS、ValRS又は、低特異的活性val−tRNAについては、150000gの上澄液から0.3MのKClでDEAEセファロースから段階溶出することによって部分的に精製された全E.coliシンテターゼ(Kung他(1975)J Biol Chem 250,1556−62参照)のtRNAを含まない調製物でもってアミノアシル化した。
【0158】
ビオチン標識lys−tRNAlys(Transcend(商標)tRNA、図22)をプロメガ社から購入した。この材料は、このメーカーによって、E.coliの全tRNAから全シンテターゼの粗製調製物を使用してリシンを結合させ、lys−tRNAlysをイオン交換クロマトグラフィーにより富化させ、そして非電荷の13個の炭素の長さのスペーサーを介してビオチンに化学的に結合させることによって調製された。
【0159】
NVOCアミノ保護アミノアシルtRNAを斯界に標準的な方法によって調製し、保存し、使用した(「Thorson他(1988)Methods in Molecular Biology 77,43−73」、「Steward及びChamberlin(1998)Methods in Molecular Biology 77,325−354」に詳述されている)。手短に言えば、pdCpAをアデノシンとデオキシシチジン誘導体から化学的に合成した。種々の非天然アミノ酸を標準的な業者(例えば、Sigma社、Aldrich社、Fluka社、Bachem社及びNovabiochem社)から保護されていない形で購入した。aG、mS及びAlaのような本発明者が最も一般的に使用したNVOCアミノ酸は、化学的な保護を必要とする反応性のアミノ酸側鎖を欠失していたため、これを保護されていないアミノ酸からNVOC−Clによるアミノ保護によって直接合成した。標準法によって合成された追加の側鎖保護を有するものを含めて、適切に保護された全てのアミノ酸をシアノメチルエステル合成によって活性化させ、次いでpdCpAに結合させた。得られたNVOC保護アミノアシル化pdCpA化合物(及びそれらのRNAとの結合体)は、水溶液中に−80℃の暗所でほぼ5のpHでもって保存したときに安定であった。NVOC保護アミノアシル化pdCpA化合物を、末端CAジヌクレオチドを欠失した様々なtRNA誘導体にT4RNAリガーゼで連結し、そして結合体を精製し、保存した(上記参照)。得られたNVOC保護アミノアシルtRNAを翻訳に使用する前に直ちにUV照射によって脱保護した。
【0160】
hisタグE.coli翻訳因子蛋白質のアッセイ。
開始因子アッセイ(Kung他(1974)Arch Biochem Biophys 162,578−84)は、受注合成されたApUpG RNA鋳型(TriLink Biotechnologies社)を使用した。0.95μMのIF1H、0.15μMのIF2H、0.78μMのIF3H、0.029A260/μlの3×洗浄リボソーム(33nMが翻訳において活性であると見積もられる。以下参照)、0.29μMの3H−fmet−tRNAi fmet、150μMのAUG及び0.4mMのGTPを含んでなる(50mMのTris−HClpH7.4,100mMのNH4Cl,5mMのMg(OAc)2,2mMのDTT)混合溶液を37℃で10分間インキュベートした。希釈後、この混合液を素早くニトロセルロースで濾過して開始複合体結合fmet−tRNAi fmetを非結合種から分離した。鋳型を欠いた反応を非特異的な結合のための対照として使用した(表1に関して最大のd.p.m.の29%、3H−fmet−tRNAi fmetの70%を結合したより高(飽和)濃度を使用して最大d.p.m.の9%)。EF−Tsに対するEF−Tuの高い親和性のため、EF−Tsの活性アッセイ(Weissbach及びPestka(1977)Molecuar Mechanisms of Protein Biosynthesis,Academic Press,ニューヨーク,NY)をEF−TuHと共に実施したところ、約2%の共精製レベルを示した。EF−TuH活性をGDPの結合によって測定し(Weissbach及びPestka(1977)Molecular Mechanisms of Protein Biosynthesis,Academic Press,ニューヨーク,NY)、EF−GH活性をリボソーム依存型GTPアーゼアッセイによって測定した(Weissbach及びPestka(1977)Molecular Mechanisms of Protein Biosynthesis,Academic Press,ニューヨーク,NY)。
【0161】
翻訳。
翻訳混合物の成分を公表された実験法(「Robakis他(1981)Proc Natl Acad Sci USA 78,4261−4」、「Cenatiempo他(1982)Arch Biochem Biophys 218,572−8」)から改変した。5×プレミックス緩衝液を、180mMのTris−HOAc(pH7.5)、50mMの3,3−ジメチルグルタル酸ナトリウム(pH6.0)、180mMのNH4OAc、10mMのDTT、140mMのホスホエノールピルビン酸カリウム(pH6.6)、195mMのKOAc及び4mMのスペルミジン・3HClを含有する溶液からこれをNaOHでpH6.8に調整することによって調製した。代表例として、MTTVmRNA鋳型(図6、30μlの総容量)を翻訳する際の混合物は、1×プレミックス緩衝液、9.5mMのMg(OAc)2、1mMのGTP、14ng/μlのピルベートキナーゼ、4.3%のPEG8000、0.95μMのIF1H、0.15μMのIF2H、0.78μMのIF3H、3.1μMのEF−TuH、0.88μMのEF−GH、0.029A260/μlの3×洗浄リボソーム(33nMが翻訳の際に活性であると見積もられる。図4参照)、0.29μMの3H−fmet−tRNAi fmet、0.58μMの14C−thr−tRNA3 thr、0.29μMの3H−val−tRNA1 val及び1μMのmRNAを包含した。この混合物を37℃で50分間インキュベートしたが、いくらかの翻訳は1分程であった(図4)。また、いくらかの翻訳のための設定は、開始混合物(GTP、IF1F、IF2H、IF3H、リボソーム、3H−fmet−tRNAi fmet及びmRNA)及び伸長因子混合物(GTP、EF−TuH、EF−GH、14C−thr−tRNA3 thr及び3H−val−tRNA1 val(図4))の37℃で10分間のプレインキュベーションを包含した。
【0162】
HPLC分析(図6)のために、ペプチド及びアミノ酸を1MのNaOH(6μl)の添加及び37℃で20分間のインキュベーションによりtRNAから放出させた。次いで、非標識マーカーのペプチド(Research Genetics社)を添加し(H2O中に10μg/μlの合わせたペプチド濃度で18μl)、この溶液を氷酢酸(5μl)で酸性化し、微量遠心分離し、次いで上澄液をMicrocon10限外濾過装置(10kDカットオフ、Amicon社)によって微量遠心分離した。濾過液の一部分(20μl)をC18逆相カラム(Vydac社)上に添加し、0.1%のTFAを含有する0〜31.5%のH2O/MeCN勾配でもって1ml/分で溶出させ、229nmの吸光度で検出し、二重標識d.p.m.プログラム(Packard社)を使用して42個の滴下画分をシンチレーション計数した。
【0163】
fMTV及びfMVT合成の分析のために、ホルミル化されたペプチド及びホルミルメチオニンを塩基加水分解、酸性化及び小型カラム上での陽イオン交換クロマトグラフィーによってホルミル化されていないアミノ酸から分離した(Peacock他(1984)Proc Natl Acad Sci USA81,6009−13)。
【0164】
対照の翻訳。
追加の翻訳を実施して翻訳因子以外の成分への依存性を評価した。3H−fmet−tRNAi fmet又は14C−thr−tRNA3 thrの除外は、3H−valの産物への取り込みに基づいてmRNAMTVからのfMTV合成を停止させる。14C−thr−tRNA3 thrを除外した実験において、非結合tRNA3 thrを添加すると精製系で予期されるような測定可能なfMTV合成を再構成しなことから、翻訳条件下ではThrRS活性がないことを示している(さらなる対照として、thr−tRNA3 thrの添加はfMTV合成を阻害しなかった)。3H−val−tRNA1 valの除外は、14C−thr−tRNA3 thrの産物への取り込みに基づいてmRNAのMVTからのfMTV合成(図4)を停止させる。mRNAMVT及びmRNAmMV(図3)産物中のT/V比は、予期されるように、それぞれ約1.0及び0である。リボソームを除外すると、最も低いバックグラウンド放射能測定値が得られ、mRNAを欠失した翻訳は、ホルミル化ペプチド産物を時間経過と共に蓄積しない。ウサギ筋肉のピルベートキナーゼ(シグマ社製のこの蛋白質(Jelenc及びKurland(1979)Proc Natl Acad Sci USA 76,3174−8)は、SDS−PAGEで単一の主要バンドとして移動する)を除外すると、産物の収率が50%だけ減少する。Mg2+の標準濃度(9.5mM)が最適であるが、翻訳はそれよりも高い及び低い濃度で効率的に起こり得る。
【0165】
本研究の一部は、NIH承認K08−CA80833によって裏付けられた。アメリカ合衆国政府は本発明に関して一定の権利を有する。
【0166】
【表1】

【表2】
【表3】
【表4】
【表5】
【図面の簡単な説明】
【0171】
【図1】E.coliからの5種のhisタグE.coli翻訳因子の過剰発現及び精製。
【図2】リボソーム指令ペプチド合成段階の概略図。
【図3】短鎖mRNA鋳型。
【図4】精製hisタグ翻訳系におけるmRNAからのオリゴペプチド合成速度の特徴付け。
【図5】mRNAのMTVを有する精製翻訳系における翻訳因子の依存性。
【図6】MTTVのmRNA鋳型から産生された産物のHPLC分析。
【図7】精製翻訳系でのオリゴペプチド産物の合成。
【図8】精製hisタグ翻訳系を使用した非天然アミノ酸を含むペプチドの合成及び選択。
【図9】Cys−tRNAcysの化学的ビオチニル化。
【図10】ビオチニル−Cys−tRNAcysの精製。
【図11】精製系を使用した精製ビオチニルCysのfM−T−bC−V疑似ペプチドへの取り込み。
【図12】隣接する多数の非天然アミノ酸をfM−T−bC−bC−V疑似ペプチドに取り込むためのアッセイ。
【図13】全tRNAと15種の異なる14C標識アミノ酸の混合物(ニュー・イングランド・ニュークレア社)との結合活性(TCA沈殿法によって測定される)についての本発明の純粋な翻訳系のアッセイ(「蛋白質ポリメラーゼ」)。
【図14】選択のコドンに対して特異的な非天然アミノ酸を結合したアミノアシルtRNAを合成するために一般化できる方法。
【図15】左から右にtRNAAsn(T)tRNAAsn(S)tRNAAsn(V)と命名されたtRNAAsn(N)の3種のアンチコドン変異体。
【図16】非結合tRNA(レーン1、3及び5)及びアミノアシルtRNA基質(レーン2、4及び6)の酸/尿素ポリアクリルアミドゲル電気泳動。
【図17】隣接するaGアミノ酸を疑似ペプチドに選択的に取り込むための一般的な方法。
【図18】純粋リボソームディスプレイ。
【図19】本発明のスペーサーmRNA及びそれらのコードペプチド産物。
【図20】本発明の精製系を使用した図19のスペーサーmRNAの翻訳。
【図21】純粋mRNAディスプレイ。
【図22】プロメガ社製のTranscendビオチニル−リシンtRNAlys。
Claims (22)
- 細胞を含まない再構成翻訳系において、翻訳因子及びtRNA種であって外因的に添加されたmRNAをそれぞれのコドンでの高選択的な取り込みでもって翻訳してペプチド産物又は該系が非天然型アミノ酸若しくはアミノ酸アナログを結合したtRNAの1種以上を含むときには疑似ペプチド産物を形成できるものを含む、細胞を含まない再構成翻訳系。
- 調製物が翻訳因子EF−P、W、W2又はレスキューを実質的に含まない、ペプチド産物生成用の請求項1に記載の翻訳系。
- 疑似ペプチド産物の主鎖に取り込まれる非天然型アミノ酸又はアミノ酸アナログを結合したtRNA種の1種以上を含む、疑似ペプチド産物生成用の請求項1に記載の翻訳系。
- 調製物が翻訳因子EF−P、W、W2又はレスキューを実質的に含まない、疑似ペプチド産物生成用の請求項2に記載の翻訳系。
- アミノ酸アナログがβ−シアノアラニン、カナバニン、ジエンコル酸、ノルロイシン、3−ホスホセリン、ホモセリン、ジヒドロキシフェニルアラニン、5−ヒドロキシトリプトファン、1−メチルヒスチジン、3−メチルヒスチジン、アリルグリシン(又はそのアルキン誘導体)、O−メチルセリン、ビオチニルリシン、ビオチニルシステイン(又はその他のビオチン標識アミノ酸)、シクロヘキシルアラニン、ホモグルタメート、D−アラニン(又はその他のD−アミノ酸)、N−メチルグリシン(又はその他のN−メチルアミノ酸)、ε−N−メチルリシン及び21種の天然アミノ酸又は非天然アミノ酸の放射性同位元素誘導体よりなる群から選択される請求項2に記載の翻訳系。
- 1種以上の外因的に添加されたmRNA種をさらに含む請求項1に記載の翻訳系。
- 翻訳因子及びtRNA種であって外因的に添加されたmRNAを翻訳して疑似ペプチド産物を形成できるものを含む細胞を含まない翻訳系において、その調製物が
(a)1種以上の活性な野生型アミノアシルtRNAと該野生型アミノアシルtRNAを合成する能力とを欠損しており、
(b)該活性のないtRNA種の代わりに非天然アミノ酸種又はアミノ酸アナログを結合した少なくとも1種の外因性アミノアシルtRNAを含む、
細胞を含まない翻訳系。 - 複数の異なるmRNA種を含む請求項7に記載の翻訳系。
- (a)外因的に添加されたRNAを翻訳してペプチド又は疑似ペプチドを形成できる細胞を含まない再構成翻訳系を形成させるために混合され得る請求項1に記載の翻訳系の成分及び(b)それに関する説明書からなるキット。
- (a)外因的に添加されたRNAを翻訳して疑似ペプチドを形成できる細胞を含まない翻訳系を形成させるために混合され得る請求項7に記載の翻訳系の成分及び(b)それに関する説明書からなるキット。
- 次の段階:
(a)請求項1に記載の細胞を含まない翻訳系を準備し、
(b)該翻訳系を1種以上の外因性mRNA種と接触させ、
(c)該翻訳系のペプチド若しくは疑似ペプチド産物を単離及び/又は同定すること
を含むペプチド又は疑似ペプチドの生成方法。 - 翻訳系のtRNA種の1種以上に疑似ペプチド産物の主鎖に取り込まれる非天然型アミノ酸又はアミノ酸アナログを結合させる請求項11に記載の方法。
- 前記方法を少なくとも100種の異なるRNA種のライブラリーについて実施し、しかもペプチド又は疑似ペプチド産物を翻訳系から触媒活性又は結合活性に基づいて同定又は単離する、請求項11に記載の方法。
- mRNAを前記翻訳系で in vitro 転写によって生成させる請求項11に記載の方法。
- ペプチド又は疑似ペプチド産物を、該産物をコードする外因性mRNAの共有結合付加物として形成させる、請求項11に記載の方法。
- 翻訳系を異なる外因性mRNA種のライブラリーと接触させて少なくとも103種の異なる配列のペプチド又は疑似ペプチド産物の多様な集団を生じさせる請求項11に記載の方法。
- 少なくとも108種の異なる配列を生じさせる請求項16に記載の方法。
- 請求項11に記載の方法によって同定若しくは合成されたペプチド又は疑似ペプチド。
- 次の段階:
(a)請求項7に記載の細胞を含まない翻訳系を準備し、
(b)該翻訳系を1種以上の外因性mRNA種と接触させ、
(c)該翻訳系の疑似ペプチド産物を単離及び/又は同定すること
を含む疑似ペプチドの生成方法。 - 次の段階:
(a)請求項11に記載の方法によって所望の生物学的活性を有する翻訳系のペプチド又は疑似ペプチド産物を同定し、
(b)段階(a)で同定されたペプチド又は疑似ペプチド産物の生物学的活性をさらに試験管内又は生体内アッセイによって確認し、
(c)段階(b)で確認された産物又はさらにそのアナログの動物における有効性及び毒性についての治療プロファイリングを行い、
(d)許容できる治療プロファイルを有する段階(c)で同定された1種以上の産物を含む医薬品を処方すること
を含む薬剤発見ビジネスを行う方法。 - 販売用の前記医薬品を流通させるための流通システムを確立する追加の段階を含み、随意として該医薬品を販売するための販売部門を設立することを含む、請求項20に記載の方法。
- 次の段階:
(a)請求項11に記載の方法によって所望の生物学的活性を有する翻訳系のペプチド又は疑似ペプチド産物を同定し、
(b)段階(a)で同定されたペプチド又は疑似ペプチド産物の生物学的活性をさらに試験管内又は生体内アッセイによって確認し、
(c)随意として段階(b)で確認された産物又はさらにそのアナログの動物における有効性及び毒性についての治療プロファイリングを行い、
(d)確認された産物のさらなる薬剤開発についての権利を第三者にライセンス供与すること
を含む薬剤発見ビジネスを行う方法。
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US26414701P | 2001-01-25 | 2001-01-25 | |
| PCT/US2002/002344 WO2002059293A2 (en) | 2001-01-25 | 2002-01-25 | Process and compositions for peptide, protein and peptidomimetic synthesis |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2010152192A Division JP4972191B2 (ja) | 2001-01-25 | 2010-07-02 | ペプチド、蛋白質及びペプチドミメティック合成のための方法及び組成物 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2004531225A true JP2004531225A (ja) | 2004-10-14 |
| JP2004531225A5 JP2004531225A5 (ja) | 2005-12-22 |
Family
ID=23004801
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2002559580A Pending JP2004531225A (ja) | 2001-01-25 | 2002-01-25 | ペプチド、蛋白質及び疑似ペプチド合成のための方法及び組成物 |
| JP2010152192A Expired - Lifetime JP4972191B2 (ja) | 2001-01-25 | 2010-07-02 | ペプチド、蛋白質及びペプチドミメティック合成のための方法及び組成物 |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2010152192A Expired - Lifetime JP4972191B2 (ja) | 2001-01-25 | 2010-07-02 | ペプチド、蛋白質及びペプチドミメティック合成のための方法及び組成物 |
Country Status (7)
| Country | Link |
|---|---|
| US (1) | US6977150B2 (ja) |
| EP (1) | EP1356036B1 (ja) |
| JP (2) | JP2004531225A (ja) |
| AT (1) | ATE394482T1 (ja) |
| CA (1) | CA2435812C (ja) |
| DE (1) | DE60226411D1 (ja) |
| WO (1) | WO2002059293A2 (ja) |
Families Citing this family (17)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP4061043B2 (ja) | 2000-12-28 | 2008-03-12 | 株式会社ポストゲノム研究所 | invitro転写/翻訳系によるペプチド等の製造方法 |
| AU2003225085A1 (en) * | 2002-04-19 | 2003-11-03 | California Institute Of Technology | Unnatural amino acid containing display libraries |
| WO2004110964A2 (en) * | 2003-06-16 | 2004-12-23 | Nuevolution A/S | Encoded molecules by translation (emt) |
| WO2005074524A2 (en) | 2004-02-02 | 2005-08-18 | Ambrx, Inc. | Modified human interferon polypeptides and their uses |
| WO2007139573A1 (en) * | 2006-05-31 | 2007-12-06 | The Trustees Of Columbia University In The City Of New York | Ribosome and rna display of biologically active small molecules |
| WO2007140471A2 (en) * | 2006-05-31 | 2007-12-06 | The Trustees Of Columbia University In The City Of New York | Ribosome and rna display of biologically active small molecules |
| WO2008005529A2 (en) | 2006-07-07 | 2008-01-10 | The Trustees Columbia University In The City Of New York | Cell-mediated directed evolution |
| US9182406B2 (en) * | 2008-08-04 | 2015-11-10 | Biodesy, Inc. | Nonlinear optical detection of molecules comprising an unnatural amino acid possessing a hyperpolarizability |
| CN102348807B (zh) * | 2009-01-12 | 2015-04-15 | 苏特罗生物制药公司 | 利用体外蛋白合成体系将非天然氨基酸引入蛋白的单装载体系 |
| EP2742153B1 (en) | 2011-08-01 | 2018-09-05 | The General Hospital Corporation | Protein and peptide libraries |
| DK2906592T3 (en) | 2012-10-12 | 2018-12-10 | Sutro Biopharma Inc | Proteolytic inactivation of selected proteins in bacterial extracts for enhanced expression |
| WO2014189768A1 (en) * | 2013-05-19 | 2014-11-27 | The Board Of Trustees Of The Leland | Devices and methods for display of encoded peptides, polypeptides, and proteins on dna |
| US10563193B2 (en) | 2013-12-02 | 2020-02-18 | Brandeis University | Multivalent glycopeptides that tightly bind to target proteins |
| EP3237906B8 (en) | 2014-12-23 | 2020-10-28 | Bluelight Therapeutics, Inc. | Attachment of proteins to interfaces for use in nonlinear optical detection |
| CN107614689A (zh) * | 2015-03-27 | 2018-01-19 | 昆士兰大学 | 用于将非天然氨基酸并入蛋白质中的平台 |
| US11174479B2 (en) | 2018-08-13 | 2021-11-16 | The Board Of Trustees Of The Leland Stanford Junior University | Devices and methods for display of encoded peptides, PolyPeptides, and proteins on DNA |
| MX2022001664A (es) | 2019-08-09 | 2022-07-19 | Nutcracker Therapeutics Inc | Aparato microfluídico y metodos de uso del mismo. |
Family Cites Families (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| AU8124694A (en) | 1993-10-29 | 1995-05-22 | Affymax Technologies N.V. | In vitro peptide and antibody display libraries |
| US5643722A (en) * | 1994-05-11 | 1997-07-01 | Trustees Of Boston University | Methods for the detection and isolation of proteins |
| JP4061043B2 (ja) * | 2000-12-28 | 2008-03-12 | 株式会社ポストゲノム研究所 | invitro転写/翻訳系によるペプチド等の製造方法 |
-
2002
- 2002-01-25 DE DE60226411T patent/DE60226411D1/de not_active Expired - Lifetime
- 2002-01-25 JP JP2002559580A patent/JP2004531225A/ja active Pending
- 2002-01-25 WO PCT/US2002/002344 patent/WO2002059293A2/en active IP Right Grant
- 2002-01-25 EP EP02724888A patent/EP1356036B1/en not_active Expired - Lifetime
- 2002-01-25 CA CA2435812A patent/CA2435812C/en not_active Expired - Lifetime
- 2002-01-25 US US10/057,783 patent/US6977150B2/en not_active Expired - Lifetime
- 2002-01-25 AT AT02724888T patent/ATE394482T1/de not_active IP Right Cessation
-
2010
- 2010-07-02 JP JP2010152192A patent/JP4972191B2/ja not_active Expired - Lifetime
Also Published As
| Publication number | Publication date |
|---|---|
| WO2002059293A3 (en) | 2003-05-30 |
| AU2002255488A2 (en) | 2002-08-06 |
| WO2002059293A2 (en) | 2002-08-01 |
| JP4972191B2 (ja) | 2012-07-11 |
| EP1356036B1 (en) | 2008-05-07 |
| JP2010259440A (ja) | 2010-11-18 |
| ATE394482T1 (de) | 2008-05-15 |
| EP1356036A2 (en) | 2003-10-29 |
| CA2435812C (en) | 2012-03-27 |
| DE60226411D1 (de) | 2008-06-19 |
| US6977150B2 (en) | 2005-12-20 |
| CA2435812A1 (en) | 2002-08-01 |
| US20040091955A1 (en) | 2004-05-13 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP4972191B2 (ja) | ペプチド、蛋白質及びペプチドミメティック合成のための方法及び組成物 | |
| JP6239888B2 (ja) | 直交tRNA−アミノアシルtRNAシンテターゼ対を生産するための方法及び組成物 | |
| Wang et al. | Expanding the genetic code | |
| JP2004531225A5 (ja) | ||
| Achenbach et al. | Outwitting EF-Tu and the ribosome: translation with d-amino acids | |
| WO2014181888A1 (ja) | ペプチドライブラリの製造方法、ペプチドライブラリ、及びスクリーニング方法 | |
| CN109072203B (zh) | 镜像核酸复制体系 | |
| US20160068835A1 (en) | Flexible display method | |
| JP7079018B6 (ja) | D-アミノ酸及びβ-アミノ酸の取り込みを増強するtRNAのD及びTアームの改変 | |
| Anthony-Cahill et al. | Expanding the natural repertoire of protein structure and function | |
| WO2003062417A1 (fr) | Produit de ligation arn-adn, et utilisation correspondante | |
| AU2002255488B2 (en) | Process and compositions for peptide, protein and peptidomimetic synthesis | |
| WO2015115661A1 (ja) | アゾール誘導体骨格を有するペプチドの製造方法 | |
| AU2002255488A1 (en) | Process and compositions for peptide, protein and peptidomimetic synthesis | |
| US20060210982A1 (en) | Cleavable assigned molecules and screening method using the same | |
| WO2021100833A1 (ja) | N-メチルアミノ酸の取り込みを増強するtRNAのTステムの改変 | |
| WO2023069816A2 (en) | Compositions and methods for multiplex decoding of quadruplet codons | |
| More et al. | Posttranslational modification of proteins Incorporating nonnatural amino acids | |
| Schultz et al. | In vivo incorporation of unnatural amino acids | |
| Young | Expanding the genetic lexicon | |
| JP2007097423A (ja) | 新規な非天然アミノ酸のタンパク質への導入法 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20050121 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20050121 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20080819 |
|
| A711 | Notification of change in applicant |
Free format text: JAPANESE INTERMEDIATE CODE: A711 Effective date: 20081107 |
|
| RD04 | Notification of resignation of power of attorney |
Free format text: JAPANESE INTERMEDIATE CODE: A7424 Effective date: 20081107 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20081119 |
|
| RD13 | Notification of appointment of power of sub attorney |
Free format text: JAPANESE INTERMEDIATE CODE: A7433 Effective date: 20081119 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A821 Effective date: 20081107 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A821 Effective date: 20081119 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20081225 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20090219 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20100302 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A821 Effective date: 20100702 Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20100702 |