JP2004504603A - 抗微生物剤 - Google Patents
抗微生物剤 Download PDFInfo
- Publication number
- JP2004504603A JP2004504603A JP2002512679A JP2002512679A JP2004504603A JP 2004504603 A JP2004504603 A JP 2004504603A JP 2002512679 A JP2002512679 A JP 2002512679A JP 2002512679 A JP2002512679 A JP 2002512679A JP 2004504603 A JP2004504603 A JP 2004504603A
- Authority
- JP
- Japan
- Prior art keywords
- component
- bacterial
- substance
- cationic antimicrobial
- glycosaminoglycan
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
Images



Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q1/00—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions
- C12Q1/02—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions involving viable microorganisms
- C12Q1/18—Testing for antimicrobial activity of a material
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K31/00—Medicinal preparations containing organic active ingredients
- A61K31/70—Carbohydrates; Sugars; Derivatives thereof
- A61K31/715—Polysaccharides, i.e. having more than five saccharide radicals attached to each other by glycosidic linkages; Derivatives thereof, e.g. ethers, esters
- A61K31/716—Glucans
- A61K31/721—Dextrans
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K31/00—Medicinal preparations containing organic active ingredients
- A61K31/70—Carbohydrates; Sugars; Derivatives thereof
- A61K31/715—Polysaccharides, i.e. having more than five saccharide radicals attached to each other by glycosidic linkages; Derivatives thereof, e.g. ethers, esters
- A61K31/716—Glucans
- A61K31/722—Chitin, chitosan
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K31/00—Medicinal preparations containing organic active ingredients
- A61K31/74—Synthetic polymeric materials
- A61K31/785—Polymers containing nitrogen
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
- A61K38/02—Peptides of undefined number of amino acids; Derivatives thereof
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P1/00—Drugs for disorders of the alimentary tract or the digestive system
- A61P1/04—Drugs for disorders of the alimentary tract or the digestive system for ulcers, gastritis or reflux esophagitis, e.g. antacids, inhibitors of acid secretion, mucosal protectants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P17/00—Drugs for dermatological disorders
- A61P17/02—Drugs for dermatological disorders for treating wounds, ulcers, burns, scars, keloids, or the like
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/04—Antibacterial agents
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N2400/00—Assays, e.g. immunoassays or enzyme assays, involving carbohydrates
- G01N2400/10—Polysaccharides, i.e. having more than five saccharide radicals attached to each other by glycosidic linkages; Derivatives thereof, e.g. ethers, esters
- G01N2400/38—Heteroglycans, i.e. polysaccharides having more than one sugar residue in the main chain in either alternating or less regular sequence, e.g. gluco- or galactomannans, e.g. Konjac gum, Locust bean gum, Guar gum
- G01N2400/40—Glycosaminoglycans, i.e. GAG or mucopolysaccharides, e.g. chondroitin sulfate, dermatan sulfate, hyaluronic acid, heparin, heparan sulfate, and related sulfated polysaccharides
Abstract
プロテイナーゼ/グリコサミノグリカン経路の阻害作用を阻止することにより、陽イオン性抗微生物ペプチドの抗微生物活性を増強する物質の同定法であって:(i)第1の成分として、陽イオン性抗微生物ペプチドを提供し;(ii)第2の成分として、細菌を提供し;(iii)第3の成分として、プロテイナーゼ/グリコサミノグリカン経路の成分の一部またはすべてを提供し、こうして第3の成分が第1の成分(例えば、グリコサミノグリカン、または細菌、または細菌とプロテオグリカン、または細菌プロテイナーゼ、または細菌プロテイナーゼとプロテオグリカン)の抗微生物作用を低下させ;(iv)第1、第2、および第3の成分を、第3の成分の非存在下で抗微生物物質による細菌の死滅を可能にし、試験物質の非存在下で第3の成分による第1の成分の抗微生物活性の阻害を可能にする条件下で、試験物質に接触させ;
(v)細菌培養物の生存を監視して、試験物質が抗微生物活性を増強することができるかどうか(抗微生物活性を増強することができる試験物質は、細菌培養物の死滅を促進する)を決定する、ことを含む上記方法が提供される。そのような方法により同定される物質は、急性および慢性の感染症の治療、特に潰瘍の治療と創傷治癒の促進に有用である。
(v)細菌培養物の生存を監視して、試験物質が抗微生物活性を増強することができるかどうか(抗微生物活性を増強することができる試験物質は、細菌培養物の死滅を促進する)を決定する、ことを含む上記方法が提供される。そのような方法により同定される物質は、急性および慢性の感染症の治療、特に潰瘍の治療と創傷治癒の促進に有用である。
Description
【0001】
(発明の分野)
本発明は、慢性および急性の微生物感染症の治療に使用される物質に関する。本発明はまた、治療法、特に慢性および急性の微生物感染症の治療に有用な物質の同定方法に関する。
【0002】
(発明の背景)
感染の間の1つの基本的な宿主防御機構は、微生物の白血球介在死滅に依存する。最近、膜破壊により種々のグラム陰性菌ならびにグラム陽性細菌を死滅させることができる一群の陽イオン性抗微生物ペプチドが、多くの生物の先天的な宿主防御系の重要な成分として考えられている。これらのペプチドは、感染を受ける生物学的境界に局在する好中球や上皮細胞中で見いだされる。
【0003】
病原体は、この第一線の宿主防御を乗り越えて、急性ならびに慢性の一連の臨床的に重要な病状を発症させることができるようである。感染の間、細菌は、種々の方策を利用して宿主防御機構を乗り越える。細菌は、種々のプロテイナーゼを放出し、これらは、急性感染中に、カリクレイン、プラスミノーゲン、補体、サイトカイン、抗体、および抗プロテイナーゼを含む宿主の特定の経路を変化させる。細菌による外毒素の産生とタンパク質を変化させる細胞サイクルも報告されており、これは、宿主防御を易感染性にすることを目的とするさらなる経路である。
【0004】
化膿連鎖球菌(Streptococcus pyogenes)のような細菌は、容易に創傷感染や丹毒を生じる。緑膿菌(Pseudomonas aeruginosa)、エンテロコッカス・フェカーリス(Enterococcus faecalis)および霊菌(Proteus mirabilis)は、慢性の創傷中に存続することが知られている。これらの細菌はすべて、プロテオグリカン(PG)の豊富な結合組織中で増殖し繁殖する。
【0005】
(発明の要約)
本発明者らは、細菌により分泌される細胞外プロテイナーゼが、例えば結合組織中に存在するようなプロテオグリカンを分解することにより、グリコサミノグリカンを放出することを証明した。次にグリコサミノグリカンは、好中球と上皮細胞により分泌される陽イオン性抗微生物ペプチドに結合し、これらの抗微生物ペプチドが細菌を死滅させることを妨害する。特に本発明者らは、デルマタン硫酸含有プロテオグリカン(例えば、デコリン)が、緑膿菌(Pseudomonas aeruginosa)、エンテロコッカス・フェカーリス(Enterococcus faecalis)、化膿連鎖球菌(Streptococcus pyogenes)および霊菌(Proteus mirabilis)からの細菌プロテイナーゼにより分解されて、デルマタン硫酸を産生することを証明した。彼らは、デルマタン硫酸が好中球由来α−デフェンシンとLL−37に結合すること、そしてこの結合は、α−デフェンシンとLL−37の抗微生物活性を完全に中和することを証明した。従って、感染の間のグリコサミノグリカンの放出、例えばプロテオグリカン分解によるデルマタン硫酸の放出は、重要なかつ従来は知られていなかった細菌の防御機構であり、これは、新規な抗細菌法の標的となり得る。
【0006】
従って本発明は、陽イオン性抗微生物ペプチドの抗微生物活性を増強するのに使用するための、プロテイナーゼ/グリコサミノグリカン経路の阻害作用を阻止する物質を提供する。
【0007】
本発明はまた、プロテイナーゼ/グリコサミノグリカン経路の阻害作用を阻止することにより、陽イオン性抗微生物ペプチドの抗微生物活性を増強する物質の同定法であって:
(i)第1の成分として、陽イオン性抗微生物ペプチドを提供し;
(ii)第2の成分として、細菌を提供し;
(iii)第3の成分として、プロテイナーゼ/グリコサミノグリカン経路の成分の一部またはすべてを提供し、こうして第3の成分が第1の成分(例えば、グリコサミノグリカン、または細菌、または細菌とプロテオグリカン、または細菌プロテイナーゼ、または細菌プロテイナーゼとプロテオグリカン)の抗微生物作用を低下させ;
(iv)第1、第2、および第3の成分を、第3の成分の非存在下で抗微生物物質による細菌の死滅を可能にし、試験物質の非存在下で第3の成分による第1の成分の抗微生物活性の阻害を可能にする条件下で、試験物質に接触させ;
(v)細菌の生存を監視して、試験物質が抗微生物活性を増強することができるかどうか(抗微生物活性を増強することができる試験物質は、細菌の死滅を促進する)を決定する、ことを含む上記方法を提供する。
【0008】
陽イオン性抗微生物ペプチドの阻害に至るプロテイナーゼ/グリコサミノグリカン経路は、2つ以上の段階で阻害することができ、結果としての細菌への作用を監視する必要無く、各段階で試験物質の阻害作用を監視してもよいことが理解されるであろう。特に試験物質は、グリコサミノグリカンと陽イオン性抗微生物ペプチドとの相互作用、グリコサミノグリカンの放出、例えばプロテオグリカンからのグリコサミノグリカンの細菌プロテイナーゼ活性化放出、または細菌プロテイナーゼによる陽イオン性抗微生物ペプチドの直接の分解を阻害する。従って、プロテイナーゼ/グリコサミノグリカン経路の阻害作用を阻止することにより、陽イオン性抗微生物ペプチドの抗微生物活性を増強する物質は、本発明により提供される以下のいずれか1つの方法により同定される:
【0009】
− グリコサミノグリカンと陽イオン性抗微生物ペプチドとの相互作用を阻害することができる物質を同定する方法であって:
(i)第1の成分として、グリコサミノグリカンを提供し;
(ii)第2の成分として、陽イオン性抗微生物ペプチドを提供し;
(iii)第1および第2の成分を、試験物質の非存在下で第1および第2の成分が相互作用することを可能にする条件下で、試験物質に接触させ;そして
(iv)第1の成分と第2の成分の相互作用を監視して、こうして試験物質が第1の成分と第2の成分の相互作用を破壊することができるかどうかを決定する、ことを含む上記方法;
【0010】
− 陽イオン性抗微生物ペプチドのグリコサミノグリカン介在阻害を阻止することができる物質を同定する方法であって:
(i)第1の成分として、グリコサミノグリカンを提供し;
(ii)第2の成分として、陽イオン性抗微生物ペプチドを提供し;
(iii)第3の成分として、細菌を提供し;
(iv)第1、第2、および第3の成分を、第1の成分の非存在下で第2の成分による細菌の死滅を可能にし、試験物質の非存在下で第2の成分による細菌の死滅の、第1の成分による阻害を可能にする条件下で、試験物質に接触させ;そして
(v)細菌の生存を監視し、こうして試験物質が陽イオン性抗微生物ペプチドのグリコサミノグリカン介在阻害を妨害することができるかどうかを決定する、ことを含む上記方法;
【0011】
− 細菌プロテイナーゼによるプロテオグリカンの分解を阻害することができる物質を同定する方法であって:
(i)第1の成分として、細菌プロテイナーゼを提供し;
(ii)第2の成分として、プロテオグリカンを提供し;
(iii)第1および第2の成分を、試験物質の非存在下で第1の成分による第2の成分の分解を可能にする条件下で、試験物質に接触させ;そして
(iv)第2の成分の分解を監視して、こうして試験物質が、プロテイナーゼによるプロテオグリカンの分解を阻害することができるかどうかを決定する、ことを含む上記方法;
【0012】
− 細菌プロテイナーゼによる陽イオン性抗微生物ペプチドの分解を阻害することができる物質を同定する方法であって:
(i)第1の成分として、細菌プロテイナーゼを提供し;
(ii)第2の成分として、陽イオン性抗微生物ペプチドを提供し;
(iii)第1および第2の成分を、試験物質の非存在下で第1の成分による第2の成分の分解を可能にする条件下で、試験物質に接触させ;そして
(iv)第2の成分の分解を監視して、こうして試験物質が、プロテイナーゼによる陽イオン性抗微生物ペプチドの分解を阻害することができるかどうかを決定する、ことを含む上記方法。
【0013】
本発明はまた、以下を提供する:
− グリコサミノグリカンと陽イオン性抗微生物ペプチドとの相互作用を阻害することができる物質の同定に使用するためのキットであって:
(a)グリコサミノグリカン;と
(b)陽イオン性抗微生物ペプチド、とを含むキット;
【0014】
− 細菌プロテイナーゼによるプロテオグリカンの分解を阻害することができる物質の同定に使用するためのキットであって:
(a)プロテオグリカン;と
(b)細菌プロテイナーゼ、とを含むキット;
− 本発明の方法により同定される物質;
− 本発明の物質を含む医薬組成物;
− 治療法によりヒトまたは動物の体を治療する方法に使用するための本発明の物質または医薬組成物;
− 治療法によりヒトまたは動物の体を治療する方法において、同時にまたは連続的に使用するための本発明の物質と陽イオン性抗微生物ペプチドとを含む生成物;
【0015】
− 治療法によりヒトまたは動物の体を治療する方法において、同時にまたは連続的に使用するための、細菌の防御機構のインヒビターと陽イオン性抗微生物ペプチドとを含む生成物(ここで、該防御機構は、細菌プロテイナーゼによりプロテオグリカンを分解して、陽イオン性抗微生物ペプチドに結合して該ペプチドの抗微生物活性を阻害することができるグリコサミノグリカンを放出する);
− 慢性または急性の細菌感染を治療する方法で使用するための医薬の製造における、本発明の物質、医薬組成物または生成物の使用。
【0016】
(配列表の簡単な説明)
配列番号1は、陽イオン性抗微生物ペプチドLL−37のアミノ酸配列である。
配列番号2は、陽イオン性抗微生物ペプチドα−デフェンシンのアミノ酸配列である。
【0017】
(発明の詳細な説明)
本明細書と添付の請求の範囲を通して、「含んでなる」および「含む」という用語は、包括的であると理解されたい。すなわちこれらの用語は、文脈により、他の成分または整数を特に記載されていなくても含むことが可能であることを企図する。
【0018】
本発明は、細菌の防御機構を妨害することにより陽イオン性抗微生物ペプチドの作用を増強する物質の同定法を提供する。細菌の防御機構は、グリコサミノグリカンの放出と、放出されたグリコサミノグリカンの陽イオン性抗微生物ペプチドへの結合を含む。グリコサミノグリカンの陽イオン性抗微生物ペプチドへの結合は、ペプチドの抗微生物活性(本明細書において、プロテイナーゼ/グリコサミノグリカン経路と呼ぶ)を阻害する。グリコサミノグリカンは、プロテオグリカンの分解により放出される。好ましくはプロテオグリカンは、細菌プロテイナーゼにより分解される。グリコサミノグリカン放出の他の機構には、内因性金属プロテイナーゼにより仲介されるプロテオグリカンの放出の増強、プロテオグリカンの産生の増加、および結合組織中の結合部位からのプロテオグリカンの排除がある。例えば、細菌プロテイナーゼは、内因性マトリックス金属プロテイナーゼを直接活性化する。妨害は、例えば、例えばプロテイナーゼ結合または活性を阻害することによる、またはグリコサミノグリカンの陽イオン性抗微生物ペプチドへの結合を阻止することによる、プロテオグリカンの分解の阻止でもよい。
【0019】
陽イオン性抗微生物ペプチドは当該分野で公知である。例えば、レーラー(Lehrer, R.J.)とガンズ(Ganz, T.)(1999)、哺乳動物と昆虫宿主防御における抗微生物ペプチド、Curr Opin Immunol 11:23−27を参照されたい。
【0020】
陽イオン性抗微生物ペプチドは典型的には、細菌膜を破壊し従って細菌を死滅させることができる抗細菌性ペプチドである。陽イオン性抗微生物ペプチドは、外因性に加えてもよい。陽イオン性抗微生物ペプチドは好ましくは、感染部位で白血球および/または上皮細胞により産生される。好ましくは白血球は好中球である。陽イオン性抗微生物ペプチドは、好ましくはデフェンシンまたはカテリシジン(cathelicidin)である。さらに好ましくは陽イオン性抗微生物ペプチドは、α−デフェンシン(配列番号2)またはLL−37(配列番号1)である。陽イオン性抗微生物ペプチドは、配列番号1または配列番号2の変種または断片を含み、この変種または断片は、抗微生物活性、好ましくは抗細菌活性を示す。例えば、陽イオン性抗微生物ペプチドは、図7で下線を引いた配列を含む配列番号1の断片でもよい。
【0021】
本発明の方法で使用するために提供される陽イオン性抗微生物ペプチドは、任意の適当な供給源からのものである。例えば、陽イオン性抗微生物ペプチドは、組換えペプチドでも、陽イオン性抗微生物ペプチドは、白血球または上皮細胞から精製されても、または陽イオン性抗微生物ペプチドは、白血球、上皮細胞または培養物中で増殖させた組換えペプチドでトランスフェクションした細胞から抽出される増殖培地中に存在してもよい。ペプチドは、天然の配列を含んでも、人工的に変異させた配列を含んでもよい(ただし、変異は、ペプチドが細菌膜を破壊する能力またはグリコサミノグリカンと相互作用する能力に影響を与えないものとする)。
【0022】
ペプチドが抗微生物活性を示すかどうかを調べるための典型的な測定法は、細菌、例えば緑膿菌(Pseudomonas aeruginosa)、エンテロコッカス・フェカーリス(Enterococcus faecalis)、化膿連鎖球菌(Streptococcus pyogenes)、霊菌(Proteus mirabilis)または大腸菌(E. coli)を、ペプチドとインキュベートして、細菌の生存を監視することを含む。典型的には、抗微生物ペプチドは、4μg/mlからの濃度、例えば4μg/ml、5μg/ml、10μg/ml、20μg/ml、40μg/ml、100μg/ml、または500μg/mlの濃度で、細菌を有効に死滅させる。
【0023】
細菌の生存は、適当な方法により監視される。典型的にはコロニー形成単位の数が測定される。細菌の生存はまた、宿主動物または宿主細胞への細菌の作用を評価することにより、間接的に監視してもよい。
【0024】
細菌は任意の適当な株でよい。細菌は、グラム陰性でもグラム陽性でもよい。好ましくは細菌株は、ヒトまたは動物の感染部位に見いだされるものである。好ましくは感染部位は、皮膚または粘膜のようなヒトまたは動物への病原体の侵入の障壁になるようなものである。好ましくは細菌は、緑膿菌(Pseudomonas aeruginosa)、エンテロコッカス・フェカーリス(Enterococcus faecalis)、化膿連鎖球菌(Streptococcus pyogenes)、または霊菌(Proteus mirabilis)である。
【0025】
本発明の方法または本明細書に記載の測定法において、任意の適当な細菌調製物を使用することができる。例えば細菌は、TH培地またはC培地中で対数中期まで増殖してもよい。次に細菌を、例えば5mMグルコースを含有する10mMトリス−塩酸(pH7.5)中で洗浄し、希釈する。典型的には、1測定当たり5μl〜500μlの細菌、好ましくは10μl〜100μlの細菌(2×106 cfu/ml)を使用する。細菌と抗微生物ペプチド、抗微生物ペプチドとグリコサミノグリカン、抗微生物ペプチドと試験物質または抗微生物ペプチド、グリコサミノグリカンと試験物質のインキュベーションは、典型的には37℃で30分〜6時間、好ましくは2時間〜4時間行われる。抗微生物活性は、任意の適当な方法により定量され、例えばインキュベーション混合物の連続希釈物を作成し、希釈物を寒天(例えばTH寒天)上に蒔き、37℃でインキュベートし(好ましくは一晩)、産生されたコロニー形成単位(CFU)の数を測定することにより、定量される。
【0026】
本発明の方法での使用に適したグリコサミノグリカンは、ペプチドの抗微生物機能が阻害されるように、陽イオン性抗微生物ペプチドと相互作用することができる。グリコサミノグリカンは、典型的には内因性グリコサミノグリカンである。グリコサミノグリカンは、好ましくはヘパラン硫酸またはヘパリンであり、さらに好ましくはデルマタン硫酸である。好ましくは本発明の方法での使用に適したグリコサミノグリカンは、高度の硫酸化および/またはイズロン酸含量を示す。イズロン酸含量は、典型的には少なくとも50%、例えば少なくとも60%、少なくとも70%、少なくとも80%、または少なくとも90%である。硫酸化の程度は、少なくとも50%、例えば少なくとも60%、少なくとも70%、少なくとも80%、または少なくとも90%である。本発明の方法で使用するためのグリコサミノグリカンはまた、追加の修飾を含有してもよく、例えばイズロン酸は2−O−硫酸化されてもよい。
【0027】
グリコサミノグリカンが陽イオン性抗微生物ペプチドと相互作用することができるかどうかを決定するための典型的な測定法には、陽イオン性抗微生物ペプチドを例えばニトロセルロース上に固定化し、放射能標識グリコサミノグリカンとプローブ結合させる方法がある。好ましくはグリコサミノグリカンのペプチドへの結合は、過剰の非標識デルマタン硫酸で阻害される。
【0028】
グリコサミノグリカンペプチドが陽イオン性抗微生物ペプチドの抗微生物活性を阻害することができるかどうかを決定するための典型的な測定法には、細菌、例えば緑膿菌(Pseudomonas aeruginosa)、エンテロコッカス・フェカーリス(Enterococcus faecalis)、化膿連鎖球菌(Streptococcus pyogenes)または霊菌(Proteus mirabilis)の調製物または培養物を、グリコサミノグリカンおよびペプチドとインキュベートし、細菌の死滅を監視する方法がある。典型的には抗微生物ペプチドは、グリコサミノグリカンの非存在下で細菌を有効に死滅させる濃度で加えられるであろう。典型的にはグリコサミノグリカンは、細菌のペプチド介在死滅を、10μg/ml〜1mg/ml、さらに好ましくは10μg/ml〜500μg/ml、または最も好ましくは20μg/ml〜250μg/mlの濃度で阻害する。グリコサミノグリカンと陽イオン性抗微生物ペプチドとのモル比は、典型的には0.1:1から1000:1、好ましくは0.5:1から100:1、およびさらに好ましくは0.5:1から10:1である。グリコサミノグリカンは、ペプチドの抗微生物活性を部分的にのみ阻害してもよい。好ましくはグリコサミノグリカンは抗微生物活性を、少なくとも2%、例えば少なくとも6%、少なくとも16%、少なくとも30%、または少なくとも40%阻害する。さらに好ましくは抗微生物活性はグリコサミノグリカンにより、少なくとも50%、例えば少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、または少なくとも99%阻害される。
【0029】
グリコサミノグリカンは、任意の特定の供給源から得られる。好ましくはグリコサミノグリカンは、真皮(皮膚)または軟骨のような結合組織から得られる。グリコサミノグリカンは、任意の適当な方法により調製され、例えばローデン(L. Roden)、ベーカー(J. Baker)、シフォネッリ(J.A. Cifonelli)、マシューズ(M.B. Mathews)、Methods in Enzymology、ギンズバーグ(V. Ginsburg)編(アカデミックプレス(Academic Press)、ニューヨーク、1973年)、第28巻、73〜140頁、フランソン(L.−A. Fransson)、ニーヅスジンスキー(I.A. Nieduszynski)、フェルプス(C.F. Phelps)、シーハン(J.K. Sheehan)、Biochim. Biophys. Acta., 586, 179 (1979)、 またはフランソン(L.−A. Fransson)、シェーバーグ(I. Sjoberg)、ハブスマーク(B. Havsmark)、Eur. J. Biochem., 106, 59 (1980)に記載されている。
【0030】
典型的には、本発明の方法で使用するためのグリコサミノグリカンは、1つ以上のプロテイナーゼ、例えば細菌プロテイナーゼの、1つ以上のプロテオグリカンへの作用により産生される。グリコサミノグリカンは、適当な細菌プロテイナーゼと適当なプロテオグリカンを提供することにより、間接的に提供されてよい。プロテオグリカンは、繊維芽細胞培養物の細胞外生成物でもよい。好ましくはプロテオグリカンは、ヘパリン、ヘパラン硫酸、または好ましくはデルマタン硫酸を含む。好ましくはプロテオグリカンは、ビグリカン(biglycan)、シンデカン(syndecan)、グリピカン(glypican)またはCD44である。さらに好ましくはプロテオグリカンは、デコリン(decorin)またはベルシカン(versican)である。
【0031】
プロテオグリカンが細菌プロテイナーゼにより分解されるかどうかを決定するための典型的な測定法には、プロテオグリカンを35Sで放射能標識し、標識プロテオグリカンを細菌プロテイナーゼとともにインキュベートし、インキュベーション前に取った標識プロテオグリカンの試料とインキュベーション後に取った試料を、SDS−ポリアクリルアミドゲルに流し、標識プロテオグリカンと分解生成物の相対的サイズと量を測定する方法がある。例えば、もし細菌プロテイナーゼがシステインプロテイナーゼなら、還元剤(例えばDTT)をインキュベーション混合物中に含めることが必要かも知れない。プロテオグリカンは、任意の適当な供給源から取られ、例えば繊維芽細胞培養物の細胞外生成物でもよい。好ましくはプロテオグリカンは、80〜500kDa、好ましくは約100kDaまたは約400kDaの分子量を有する。分解産物は、好ましくは30〜50kDaの分子量を有し、これは遊離のまたはペプチド結合グリコサミノグリカン鎖に対応する。
【0032】
本明細書に記載の測定法に、任意の好適なプロテアーゼが使用できる。好ましくはプロテアーゼは、細菌プロテイナーゼである。任意の細菌調製物または細菌培養物が、細菌プロテイナーゼの供給源として使用される。細菌は、任意の適当な手段により刺激してプロテイナーゼを産生させてもよい。細菌の培養物からの増殖培地が使用できる。典型的には、細菌は、液体培地(例えば、TH培地またはC培地)中で静止期まで増殖される。本発明の方法で使用するのに適した増殖培地は、遠心分離により細菌をペレット化し、例えば、直径0.2μm〜0.4μmのフィルターを使用して上清をろ過することにより得られる。あるいは細菌プロテイナーゼは、増殖培地から単離される。細菌プロテイナーゼを単離するのに、任意の適当な方法が使用される。例えばプロテイナーゼは、硫酸アンモニウム沈殿、透析、および適当なカラム(例えば、ハイQ陰イオン交換クロマトグラフィー)で分離することにより調製される。あるいはプロテイナーゼは、ハーワルト(H. Herwald)、コリン(M. Collin)、ムラー−エスタール(Muller−Esterl)、ビヨルク(L. Bjorck)、J. Exp. Med. 184, 665 (1996)に記載のように調製される。本発明での使用に適した細菌プロテイナーゼには、緑膿菌(Pseudomonas aeruginosa)エラスターゼ、緑膿菌(Pseudomonas aeruginosa)アルカリ性プロテイナーゼ、エンテロコッカス・フェカーリス(Enterococcus faecalis)ゲラチナーゼ、および化膿連鎖球菌(Streptococcus pyogenes)システインプロテイナーゼがある。
【0033】
本発明により提供される陽イオン性抗微生物ペプチド活性を増強する物質の同定法は、グリコサミノグリカンと陽イオン性抗微生物ペプチドとの相互作用を阻害することができる物質の同定法を含む。そのような方法は基本的に以下からなる:
(i)第1の成分として、グリコサミノグリカンを提供し;
(ii)第2の成分として、陽イオン性抗微生物ペプチドを提供し;
(iii)第1および第2の成分を、試験物質の非存在下で第1および第2の成分が相互作用することを可能にする条件下で、試験物質に接触させ;そして
(iv)第1の成分と第2の成分の相互作用を監視して、こうして試験物質が第1の成分と第2の成分の相互作用を破壊することができるかどうかを決定する。
【0034】
この方法での使用に適したグリコサミノグリカンと陽イオン性抗微生物ペプチドは、既に本明細書に記載されている。第1および第2の成分を試験物質に接触させるために、任意の適当な条件が使用される。典型的には、陽イオン性ペプチドは、例えばニトロセルロース膜またはポリアクリルアミドゲル(これは好ましくは非変性性である)上に固定化され、固定化されたペプチドは、グリコサミノグリカンおよび試験物質とインキュベートされる。グリコサミノグリカンの固定化ペプチドへの結合、従って結合に対して試験物質が競合する能力は、任意の適当な手段により監視される。例えばグリコサミノグリカンは、例えば35Sで放射能標識され、固定化ペプチドに結合した放射能標識グリコサミノグリカンは、任意の適当な手段(例えば、シンチレーション計測、ホスホリメージャー(phosphorimager)、またはX線フィルム)を使用して検出される。好ましくは、同一の条件下で、好ましくは同時に、試験物質有りでおよび無しで実験を行い、実験の結果を比較して、試験物質がグリコサミノグリカンのペプチドへの結合の低下を引き起こすかどうかを決定する。一般に試験物質は、過剰に加えられる。好ましくは試験物質は、グリコサミノグリカンのペプチドへの結合を、少なくとも20%、少なくとも30%、少なくとも40%、少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも99%低下させるか、またはグリコサミノグリカンのペプチドへの結合を、物質の濃度0.1mg/ml、1mg/ml、10mg/ml、100mg/ml、500mg/ml、1mg/ml、10mg/ml、100mg/ml、または0.1g/mlで、実質的に妨害する。生成物の相互作用の程度と濃度の上記の低下パーセントの任意の組合せが、本発明の物質を規定するのに使用され、より低濃度でのより大きな分解が好ましい。本発明の好適な物質は、1mg/mlまたは0.5mg/mlの濃度で少なくとも50%レベルの阻害を示すものである。
【0035】
本発明により提供される陽イオン性抗微生物ペプチド活性を増強する物質の同定法は、陽イオン性抗微生物ペプチドの抗微生物活性へのグリコサミノグリカンの阻害作用を阻止することができる物質を同定する方法を含む。そのような方法は基本的に以下からなる:
(i)第1の成分として、グリコサミノグリカンを提供し;
(ii)第2の成分として、陽イオン性抗微生物ペプチドを提供し;
(iii)第3の成分として、細菌を提供し;
(iv)第1、第2、および第3の成分を、第1の成分の非存在下で第2の成分による細菌の死滅を可能にし、試験物質の非存在下で第2の成分による細菌の死滅の、第1の成分による阻害を可能にする条件下で、試験物質に接触させ;そして
(v)細菌の死滅を監視し、こうして試験物質が陽イオン性抗微生物ペプチドのグリコサミノグリカン介在阻害を妨害することができるかどうかを決定する。
【0036】
工程(iv)において第1および第2の成分は、まず試験物質と接触し、次に第3の成分と接触してもよい。第1および第2の成分を試験物質と接触させた後の適当な時間、例えば0〜4時間後、例えば30秒後、1分後、5分後、15分後、30分後、または1時間後に、第3の成分を、第1の成分、第2の成分、および試験物質の混合物に接触させてもよい。
【0037】
試験物質をまず、陽イオン性抗微生物ペプチドの活性に対する直接作用について試験してもよい。好ましくは、陽イオン性抗微生物ペプチド活性を阻害しない試験物質のみが、殺菌測定法で使用される。
【0038】
この方法での使用に適したグリコサミノグリカン、陽イオン性抗微生物ペプチドおよび細菌調製物は、既に記載されている。第1、第2、および第3の成分を試験物質に接触させるために、任意の適当な条件が使用される。典型的には、1測定について、5μl〜500μlの細菌、好ましくは10μl〜100μlの細菌(2×106 cfu/ml)が使用される。この3つの成分は典型的には、37℃で30分〜6時間、好ましくは2時間〜4時間インキュベートされる。細菌の死滅の監視は、公知の方法により行われ、例えばインキュベーション混合物の連続希釈物を作成し、希釈物を寒天(例えばTH寒天)上に蒔き、37℃で、好ましくは一晩インキュベートし、産生されたコロニー形成単位(CFU)の数を測定することにより定量される。好ましくは3つの測定[1つは、第2の成分(抗微生物ペプチド)のみを含有し、1つは、第1および第2の成分(グリコサミノグリカンと抗微生物ペプチド)のみを含有し、1つはすべての3つの成分を含有する]を、同一の条件下でかつ好ましくは同時に行う。3つの測定の結果を定量し、結果を比較して、グリコサミノグリカンによる抗微生物活性の阻害を阻止する試験物質の有効性を決定する。好ましくは試験物質は、ペプチドの抗微生物活性に対するグリコサミノグリカンの阻止作用を、少なくとも20%、少なくとも30%、少なくとも40%、少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも99%逆転させるか、または陽イオン性抗微生物ペプチド活性の阻害を、物質の濃度0.1mg/ml、1mg/ml、10mg/ml、100mg/ml、500mg/ml、1mg/ml、10mg/ml、100mg/ml、または0.1g/mlで、実質的に妨害する。阻害の程度と生成物の濃度の上記の低下パーセントの任意の組合せが、本発明の物質を規定するのに使用され、より低濃度でのより大きな分解が好ましい。本発明の好適な物質は、1mg/mlまたは0.5mg/mlの濃度で少なくとも50%レベルの阻害を示すものである。
【0039】
第2の成分の分解もまた、既に本明細書に記載の測定法を使用して、試験物質の存在下と非存在下で、陽イオン性抗微生物ペプチドによる細菌の死滅を監視することにより、間接的に監視される。
【0040】
本発明により提供される陽イオン性抗微生物ペプチド活性を増強する物質の同定法は、例えば細菌プロテイナーゼによるプロテオグリカンの分解を阻止することにより、グリコサミノグリカン(次にグリコサミノグリカンは、陽イオン性抗微生物ペプチドと自由に相互作用する)の放出を阻害することができる物質を同定する方法を含む。そのような方法は基本的に以下からなる:
(i)第1の成分として、細菌プロテイナーゼを提供し;
(ii)第2の成分として、プロテオグリカンを提供し;
(iii)第1および第2の成分を、試験物質の非存在下で第1の成分による第2の成分の分解を可能にする条件下で、試験物質に接触させ;そして
(iv)第2の成分の分解を監視して、こうして試験物質が、プロテイナーゼによるプロテオグリカンの分解を阻害することができるかどうかを決定する。
【0041】
適当な第1および第2の成分、およびプロテオグリカンが細菌プロテイナーゼにより分解されるかどうかを決定するための測定法は、既に本明細書に記載されている。好ましくは2つの測定(1つは、試験物質を含み、1つは試験物質を含まない)を、同一の条件下で行う。次に、2つの測定の間の分解生成物の相対量および/または非分解プロテオグリカンの相対量を比較して、試験物質による分解の阻害程度を決定する。各生成物の相対量は、ホスホリメージャー(phosphorimager)を使用して定量される。好ましくは、プロテイナーゼによるプロテオグリカンの分解を阻害する試験物質は、プロテオグリカンの分解を、少なくとも20%、少なくとも30%、少なくとも40%、少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも99%低下させるか、または物質の濃度0.1mg/ml、1mg/ml、10mg/ml、100mg/ml、500mg/ml、1mg/ml、10mg/ml、100mg/ml、または0.1g/mlで、実質的に妨害する。相互作用の程度と生成物の濃度の上記の低下パーセントの任意の組合せが、本発明の物質を規定するのに使用され、より低濃度でのより大きな分解が好ましい。本発明の好適な物質は、1mg/mlまたは0.5mg/mlの濃度で少なくとも50%レベルの阻害を示すものである。
【0042】
本発明により提供される陽イオン性抗微生物ペプチド活性を増強する物質の同定法は、細菌プロテイナーゼによる陽イオン性抗微生物ペプチドの分解を妨害することができる物質の同定法を含む。そのような方法は基本的に以下からなる:
(i)第1の成分として、細菌プロテイナーゼを提供し;
(ii)第2の成分として、陽イオン性抗微生物ペプチドを提供し;
(iii)第1および第2の成分を、試験物質の非存在下で第1の成分による第2の成分の分解を可能にする条件下で、試験物質に接触させ;そして
(iv)第2の成分の分解を監視して、こうして試験物質が、プロテイナーゼによる陽イオン性抗微生物ペプチドの分解を阻害することができるかどうかを決定する。
【0043】
適当な第1および第2の成分は、既に本明細書に記載されている。陽イオン性抗微生物ペプチドが細菌プロテイナーゼにより分解されるかどうかを決定する典型的な測定法は、ペプチドを35Sで放射能標識し、標識ペプチドを細菌プロテイナーゼとインキュベートし、インキュベーション前に取った標識ペプチドの試料とインキュベーション後に取った試料をSDS−ポリアクリルアミドゲルに流し、標識ペプチドと分解生成物の相対量を測定する方法がある。例えば、もし細菌プロテイナーゼがシステインプロテイナーゼなら、還元剤(例えばDTT)をインキュベーション混合物中に含めることが必要かも知れない。好ましくは2つの測定(1つは、試験物質を含み、1つは試験物質を含まない)を、同一の条件下で行う。次に、2つの測定の間の分解生成物の相対量および/または非分解ペプチドの相対量を比較して、試験物質による分解の阻害程度を決定する。各生成物の相対量は、ホスホリメージャー(phosphorimager)を使用して定量される。好ましくは、プロテイナーゼによるペプチドの分解を阻害する試験物質は、ペプチドの分解を、少なくとも20%、少なくとも30%、少なくとも40%、少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも99%低下させるか、またはペプチドの分解を、物質の濃度0.1mg/ml、1mg/ml、10mg/ml、100mg/ml、500mg/ml、1mg/ml、10mg/ml、100mg/ml、または0.1g/mlで、実質的に妨害する。相互作用の程度と生成物の濃度の上記の低下パーセントの任意の組合せが、本発明の物質を規定するのに使用され、より低濃度でのより大きな分解が好ましい。本発明の好適な物質は、1mg/mlまたは0.5mg/mlの濃度で少なくとも50%レベルの阻害を示すものである。
【0044】
ペプチド分解を監視するための他の適当な方法には、タンパク質染色、免疫ブロッティング、および放射性ヨード化がある。ペプチド分解はまた、機能的測定法、例えば分解産物に残存抗細菌活性があるかどうかを調べる方法により監視される。抗細菌活性は、殺菌測定法を使用して、コロニー形成単位(cfu)を測定することにより決定される。
【0045】
第2の成分の分解はまた、既に本明細書に記載の測定法を使用して、試験物質の存在下および非存在下で、陽イオン性抗微生物ペプチドによる細菌の死滅を監視することにより、間接的に監視される。
【0046】
本発明はまた、陽イオン性抗微生物ペプチド活性を増強する試験物質を同定するためのキットを提供し、このキットは以下を含む:
(i)グリコサミノグリカン;および
(ii)陽イオン性抗微生物ペプチド。
【0047】
キットはさらに以下を含む:
(iii)細菌試料;および/または
(iv)試験物質が、グリコサミノグリカンと陽イオン性抗微生物ペプチドとの相互作用を阻害するかどうかを決定するための手段。
【0048】
本発明はまた、陽イオン性抗微生物ペプチド活性を増強する試験物質を同定するためのキットを提供し、このキットは以下を含む:
(i)プロテオグリカン;
(ii)細菌プロテイナーゼ。
【0049】
キットはさらに以下を含んでよい:
(ii)試験物質が、細菌プロテイナーゼによるプロテオグリカンの分解を阻害するかどうかを決定するための手段。
【0050】
本発明により提供されるキットはまた以下を含む:
(v)または(iv)陽イオン性抗微生物ペプチド活性を増強する試験物質の能力を決定するための測定法を実施するための説明書。
【0051】
「物質」という用語は、単一の物質、および2、3またはそれ以上の物質の組合せを含むことを企図する。例えばこの用語は、単一のペプチド、2つ以上のペプチドの混合物、またはペプチドと規定される化学的物質との混合物を含んでよい。
【0052】
任意の適当な試験物質を本発明の方法で使用することができる。好ましくは試験物質は、真核細胞に対して非毒性である。上記測定法で試験される好適な試験物質には、炭水化物、化学的修飾(例えば、硫酸化および/またはカルボキシル化)を有する炭水化物骨格を含む分子、ペプチド、およびペプチド模倣物を含むことができる。特に好適な試験物質には、放出されたグリコサミノグリカンのスカベンジャーとして機能性する陽イオン性物質、ポリエチレンイミン、プロタミン、ポリブレン、ポリ−L−リジン、ポリ−L−アルギニン、キトサンおよびポリミキシンがある。他の適当な試験物質には、コンビナトリアルライブラリー、規定された化学的物質、オリゴヌクレオチドおよび例えばディスプレイライブラリー(例えば、ファージディスプレイライブラリー)のような天然産物ライブラリーがある。さらに抗体生成物(例えば、モノクローナル抗体およびポリクローナル抗体、1本鎖抗体、キメラ抗体、CDR移植抗体、およびヒト化抗体)、またはこれらの断片が使用される。適当な試験物質の例を表1に示す。
【0053】
【表1】
 【表2】
【表2】
【0054】
炭水化物試験物質は、単糖、二糖、オリゴ糖、または多糖でよい。例えばコンドロイチン硫酸のポリマー[GlcA−GalNAc]n またはデルマタン硫酸。好ましくは炭水化物試験物質は、陽イオン性抗微生物ペプチドに、その抗微生物活性に影響を与えることなく結合することができる。
【0055】
ペプチド試験物質は、陽イオン性抗微生物ペプチドの模倣物でもよい。好ましくは模倣物は、グリコサミノグリカンおよび/またはプロテアーゼに結合することができて、グリコサミノグリカンおよび/またはプロテアーゼを封鎖し、こうしてグリコサミノグリカンおよび/またはプロテアーゼが、陽イオン性抗微生物ペプチドと相互作用することを防ぐ。模倣物は、グリコサミノグリカンに結合する陽イオン性抗微生物ペプチドの領域のみでなく、抗微生物活性を有するペプチドの領域も模倣する。試験ペプチドはまた、任意の他のアミノ酸配列を含有してもよい。好ましくはペプチドは、グリコサミノグリカンおよび/またはプロテアーゼと相互作用して、これらが陽イオン性抗微生物ペプチドと結合するのを妨害することを可能にするアミノ酸配列を含む。
【0056】
試験物質は、例えば1回の反応につき10個の物質の最初のスクリーニングで使用され、阻害または活性化を示すこれらのバッチの物質が個々に試験される。試験物質は、1nM〜1000μM、好ましくは1μM〜100μM、または5μM〜50μM、さらに好ましくは1μM〜10μMの濃度で使用される。
【0057】
本発明はまた、治療法によりヒトまたは動物の体を治療する方法において同時にまたは連続的に使用するための、細菌プロテイナーゼ/グリコサミノグリカン防御経路のインヒビターと陽イオン性抗微生物ペプチドを含有する。
【0058】
インヒビターは、グリコサミノグリカンと陽イオン性抗微生物ペプチドとの相互作用を阻止することができ、細菌プロテイナーゼによるプロテオグリカンの分解を阻止することができ、または細菌プロテイナーゼによる陽イオン性抗微生物ペプチドの分解を阻止することができる。グリコサミノグリカンと陽イオン性抗微生物ペプチドとの相互作用を阻止することができるインヒビターは、グリコサミノグリカンに結合して、これが陽イオン性抗微生物ペプチドに結合することを妨害するか、または陽イオン性抗微生物ペプチドに結合して、ペプチドがその抗微生物活性を保持するが、阻害性グリコサミノグリカンには結合できないようにする。
【0059】
好適なインヒビターには、炭水化物、化学的修飾(例えば、硫酸化および/またはカルボキシル化)を有する炭水化物骨格を含む分子、ペプチド、およびペプチド模倣物を含む。他の適当なインヒビターには、コンビナトリアルライブラリー、規定された化学的物質、オリゴヌクレオチドおよび、例えば、ディスプレイライブラリー(例えば、ファージディスプレイライブラリー)のような天然産物ライブラリーがある。
【0060】
インヒビターは、陽イオン性抗微生物ペプチドの模倣物、例えばペプチドでもよい。好ましくは模倣物は、グリコサミノグリカンおよび/またはプロテアーゼに結合することができて、グリコサミノグリカンおよび/またはプロテアーゼを封鎖し、こうしてグリコサミノグリカンおよび/またはプロテアーゼが、陽イオン性抗微生物ペプチドと相互作用することを防ぐ。インヒビターは、グリコサミノグリカンに結合する陽イオン性抗微生物ペプチドの領域のみでなく、抗微生物活性を有するペプチドの領域も模倣する。好ましくは模倣物は、グリコサミノグリカンおよび/またはプロテアーゼと相互作用して、これらが陽イオン性抗微生物ペプチドと結合することを妨害することを可能にするアミノ酸配列を含む。
【0061】
インヒビターは好ましくは、単糖、二糖、オリゴ糖、または多糖である。さらに好ましくはインヒビターは、例えばポリエチレンイミン、プロタミン、ポリブレン、ポリ−L−リジン、ポリ−L−アルギニン、キトサンまたはポリミキシンのような陽イオン性物質である。好ましくはインヒビターは、陽イオン性抗微生物ペプチドに、その抗微生物活性に影響を与えることなく結合することができる。
【0062】
陽イオン性抗微生物ペプチドは、細菌膜を破壊し、こうして細菌を死滅させることができるペプチドである。好ましくは陽イオン性抗微生物ペプチドは、感染部位で白血球および/または上皮細胞により産生される。好ましくは白血球は好中球である。陽イオン性抗微生物ペプチドは好ましくは、デフェンシンまたはLL−37のような陽イオン性抗細菌性ペプチドである。さらに好ましくは陽イオン性抗微生物ペプチドはα−デフェンシンである。本発明の方法での使用のために提供される陽イオン性抗微生物ペプチドは、任意の適当な供給源から得られる。例えば、陽イオン性抗微生物ペプチドは、組換えペプチドでも、陽イオン性抗微生物ペプチドは、白血球または上皮細胞から精製されるか、または陽イオン性抗微生物ペプチドは、白血球、上皮細胞または培養物中で増殖させた組換えペプチドでトランスフェクションした細胞から抽出される増殖培地中に存在してもよい。ペプチドは、インビトロタンパク質合成により産生されてもよい。ペプチドは、天然の配列を含んでも、人工的に変異させた配列を含んでもよい(ただし、変異は、ペプチドが細菌膜を破壊する能力またはグリコサミノグリカンと相互作用する能力に影響を与えないものとする)。
【0063】
本発明はまた、治療法によりヒトまたは動物の体を治療する方法において同時にまたは連続的に使用するための、細菌プロテイナーゼ/グリコサミノグリカン防御経路の2つ以上のインヒビターを含有する生成物を提供する。さらに本発明は、治療法によりヒトまたは動物の体を治療する方法において同時にまたは連続的に使用するための、2つ以上の本発明の物質を含有する生成物を提供する。
【0064】
本発明により同定される物質と本発明の生成物は、治療法によりヒトまたは動物の体を治療する方法において使用される。特にそのような物質および生成物は、急性または慢性の細菌感染症の治療、潰瘍(例えば、慢性静脈潰瘍)の治療、および創傷の治療に使用される。潰瘍と他の創傷は、治癒を遅らせる細菌で感染されることがある。従って細菌の根絶により創傷治癒が促進される。本発明により同定される物質を使用して治療される他の創傷には、熱傷創、目、肺および尿路感染症、および無傷の皮膚の細菌感染症がある。任意の潰瘍(例えば、胃潰瘍)が、本発明の方法により同定される物質により治療される。本発明の物質と生成物はまた、無菌の創傷が細菌に感染されることを防ぐのに使用される。そのような物質と生成物はまた、急性もしくは慢性の感染の治療法、潰瘍の治療、または創傷治癒の促進法で使用される薬剤の製造のために使用できる。急性もしくは慢性の感染患者または潰瘍または創傷の患者の症状は、本発明の物質または生成物を投与することにより改善することができる。治療上有効量の本発明の物質または生成物は、必要とする宿主に投与される。物質は、例えば好中球により放出されるような天然に存在する(内因性)陽イオン性抗微生物ペプチドを増強してもよい。あるいは外因性陽イオン性抗微生物ペプチドと一緒に投与すると、物質は外因性ペプチドの抗微生物活性を増強することがある。
【0065】
本発明の方法により同定される物質は、治療法によりヒトまたは動物の体を治療する方法において、陽イオン性抗微生物ペプチドとともに使用してもよい。特にそのような併用療法は、急性または慢性の細菌感染の治療、潰瘍の治療、および創傷の治療で使用してもよい。物質と陽イオン性抗微生物ペプチドはまた、急性または慢性の感染の治療、潰瘍の治療、または創傷治癒の促進法で使用するための、薬剤の製造に使用される。急性もしくは慢性の感染患者または潰瘍または創傷の患者の症状は、本発明の物質と陽イオン性抗微生物ペプチドの組合せ投与により改善することができる。治療上有効量の本発明の物質と陽イオン性抗微生物ペプチドは、必要とする宿主に投与される。
【0066】
陽イオン性抗微生物ペプチドは、好中球中、生物学的境界(例えば、肺、皮膚、粘膜および尿管)で発現される。抗微生物ペプチド活性を調節する物質は、局所的治療、例えば皮膚または角膜、および内科的使用、例えば腸管、肺および尿管で使用される。
【0067】
上記で概説したスクリーニング法に従って同定され、本発明の投与のための物質は、標準的な薬剤学的に許容される担体および/または賦形剤、例えば薬剤学の分野で一般的なもの[例えばレミントンの薬剤科学(Remington’s Pharmaceutical Sciences)、マックパブリッシング社(Mack Publishing Company)、イースタンペンシルバニア、第17版、1985年(この開示内容は参照することによりその全体が本明細書に組み込まれる)に記載のようなもの]とともに調製される。医薬組成物は、本発明の物質を、陽イオン性抗微生物ペプチドおよび薬剤学的に許容される担体または賦形剤とともに含有してもよい。
【0068】
本発明はまた、細菌プロテアーゼ/グリコサミノグリカン防御経路のインヒビター、陽イオン性抗微生物ペプチド、および薬剤学的に許容される担体または賦形剤を含む医薬組成物を提供する。物質、組成物および生成物は、経腸的または腸管外(例えば、経口、頬、肛門、肺、静脈内、動脈内、筋肉内、腹腔内、局所的)、または他の適当な経路により投与される。
【0069】
物質の、物質と陽イオン性抗微生物ペプチドまたは生成物の治療上有効量のが、患者に投与される。物質の、物質と陽イオン性抗微生物ペプチドまたは生成物の用量は、特に使用される物質に従う種々のパラメータ(治療される患者の年齢、体重、症状;投与経路;および必要な処方)に従って決定される。医師が、特定の患者のための必要な投与経路と用量は決定できるであろう。典型的な1日用量は、物質、物質と陽イオン性抗微生物ペプチドまたは生成物の活性、治療される被験体の体重と症状、変性のタイプと重症度、および投与頻度と経路に従って、約0.1〜50mg/kg体重である。好ましくは1日用量は、5mg〜2gである。
【0070】
物質、インヒビター、およびペプチドは、同時にまたは逐次的に投与される。好ましくは物質またはインヒビターは、ペプチドの投与前に投与される。物質またはインヒビターは、ペプチドの投与前1分〜4時間、好ましくはペプチドの投与前5分〜1時間、ペプチドの投与前10分〜30分に投与される。さらに好ましくは物質またはインヒビターおよびペプチドは、同時に投与される。
【0071】
以下の例は、本発明を例示する。
【0072】
例1
35S−硫酸標識繊維芽細胞培養物の生成物と、細菌培養物からの増殖培地によるこれらの生成物の分解を、3〜12%SDS−ポリアクリルアミドゲル電気泳動(PAGE)により分析した。
【0073】
慢性静脈創傷から最初に得られた緑膿菌(Pseudomonas aeruginosa)とエンテロコッカス・フェカーリス(Enterococcus faecalis)分離株を、トッド−ヘウィット(Todd−Hewitt)培地中で37℃で18時間増殖させた。化膿連鎖球菌(Streptococcus pyogenes)AP1株(40/58)は、ストレプトコッカスの文献と研究のための世界保健機構協力センター(World Health Organization Collaborating Centre for References and Research on Streptococci)(プラハ、チェコ共和国)から得て、C培地(ハーワルト(H. Herwald)、コリン(M. Collin)、ムラー−エスタール(Muller−Esterl)、ビヨルク(L. Bjorck)、J. Exp. Med. 184, 665 (1996))で静止期まで増殖させた。遠心分離して細菌をペレットにし、上清を無菌ろ過した(0.3μm)。放射能標識プロテオグリカンの分解のために、10μlの35S−硫酸代謝標識された分泌された繊維芽細胞生成物(シュミチェン(A. Schmidtchen)、カールステット(I. Carlstedt)、マルムストローム(A. Malmstrom)、フランソン(L−A, Fransson)、Biochem. J. 265, 289 (1990))を、10μlの無菌ろ過細菌上清と、37℃で6時間インキュベートした。10μlのTH培地を対照中に加えた。ストレプトコッカスのシステインプロテイナーゼの活性化のために、10mMのDTTを加えた。
【0074】
結合組織プロテオグリカン(PG)デコリン/ビグリカンとベルシカンは、インビボならびに培養物中でヒト繊維芽細胞の主要な分泌生成物を構成する(イオッツォ(R.V. Iozzo)、Annu. Rev. Biochem., 67, 609 (1998))。3〜12%SDS−PAGEによる分析により、以前の結果が確認され、デコリン(〜100kDa)とベルシカン(〜400kDa)が35S−標識繊維芽細胞培養物の主要な細胞外生成物として同定された(シュミチェン(A. Schmidtchen)、カールステット(I. Carlstedt)、マルムストローム(A. Malmstrom)、フランソン(L−A, Fransson)、Biochem. J. 265, 289 (1990);シュミチェン(A. Schmidtchen)、フランソン(L−A, Fransson)、Biomed. Chrom., 7, 48 (1993))。緑膿菌(P. aeruginosa)とイー・フェカーリス(E. faecalis)培養物からの増殖培地を添加すると、デコリンが広範に分解された。また、還元剤ジチオスレイトール(DTT)を加えると、化膿連鎖球菌(S. pyogenes)細胞外生成物は、デコリンと高分子量プロテオグリカンを分解した。
【0075】
DTTの必要性は、化膿連鎖球菌(S. pyogenes)システインプロテイナーゼが分解に関与していることを示唆する。この仮定は、この作用を完全に阻止した特異的システインプロテイナーゼインヒビター(E64)の作用により確認された。
【0076】
分解された物質は、分子量30〜50kDaであり、既に記載されている(シュミチェン(A. Schmidtchen)、フランソン(L−A, Fransson)、Eur. J. Biochem. 208, 537 (1992))ように、遊離/またはペプチド結合GAG鎖のサイズに対応した。これらの実験は、細菌プロテイナーゼによる繊維芽細胞マトリックス金属プロテアーゼ(MMP)の活性化(バーンズ(E. H. Burns)、マーシエル(A.M. Marciel)、ムッサー(J.M. Musser)、Infect. Immun., 64, 4744 (1996);オカモト(T. Okamoto)ら、J. Biol. Chem., 272, 6059 (1997))の可能性を排除せず、分解されたPGを与えたため、特異的プロテアーゼの調製物を用いて一連の対照実験を行った。
【0077】
緑膿菌(P. aeruginosa)エラスターゼとアルカリ性プロテイナーゼタンパク質を、硫酸アンモニウム沈殿(70%飽和)により調製した。透析(10mMトリス、pH8.0)後、ハイQ陰イオン交換クロマトグラフィー(バイオラッド(Bio−Rad))で、10mMトリス(pH8.0)中0〜1.0M NaClの勾配を使用して、分離を行った。フロースルー画分中にエラスターゼが同定されたが、アルカリ性プロテイナーゼは、〜0.5M NaClで溶出された。
【0078】
化膿連鎖球菌(S. pyogenes)のシステインプロテイナーゼは、以前のプロトコール(ハーワルト(H. Herwald)、コリン(M. Collin)、ムラー−エスタール(Muller−Esterl)、ビヨルク(L. Bjorck)、J. Exp. Med. 184, 665 (1996))に従って精製された。
【0079】
イー・フェカーリス(E. faecalis)株はゲラチナーゼを含有し、上清を直接使用した。緑膿菌(P. aeruginosa)エラスターゼとアルカリ性プロテイナーゼ、化膿連鎖球菌(S. pyogenes)システインプロテイナーゼ、およびイー・フェカーリス(E. faecalis)ゲラチナーゼはすべて、精製したヒト子宮頸デコリンならびに調整培地中の35S−標識繊維芽細胞デコリンを分解した。段階的イオン交換クロマトグラフィー(シュミチェン(A. Schmidtchen)、カールステット(I. Carlstedt)、マルムストローム(A. Malmstrom)、フランソン(L−A, Fransson)、Biochem. J. 265, 289 (1990))(繊維芽細胞由来MMPの分離のため)そして次に放射能標識繊維芽細胞プロテオグリカンのPBS透析、または繊維芽細胞MMPを不活性化するための繊維芽細胞生成物の沸騰は、分解に影響を与えなかった。化膿連鎖球菌(S. pyogenes)システインプロテイナーゼをシステインプロテイナーゼインヒビターE64とプレインキュベーション(〜15分)すると、分解されなかった。
【0080】
別の創傷病原体である霊菌(Proteus mirabilis)もまた、デルマタン硫酸を放出する細胞外プロテアーゼを産生することが証明されている。
【0081】
例2
次に我々は、デルマタン硫酸(DS)、ヒトデコリンのグリコサミノグリカン(GAG)置換基(イオッツォ(R.V. Iozzo)、Annu. Rev. Biochem., 67, 609 (1998))、結合デフェンシン、および他の硫酸化多糖が、相互作用を阻止するかどうかを調べた。
【0082】
これらの実験で使用したグリコサミノグリカンは、デルマタン硫酸(DS)36、DS13、ヘパラン硫酸(HS)3−6、コンドロイチン硫酸(CS)−4およびCS−6である。これらの調製と性状解析は、既に記載されている(ローデン(L. Roden)、ベーカー(J. Baker)、シフォネッリ(J.A. Cifonelli)、マシューズ(M.B. Mathews)、Methods in Enzymology、ギンズバーグ(V. Ginsburg)編(アカデミックプレス(Academic Press)、ニューヨーク、1973年)、第28巻、73〜140頁;フランソン(L.−A. Fransson)、ニーヅスジンスキー(I.A. Nieduszynski)、フェルプス(C.F. Phelps)、シーハン(J.K. Sheehan)、Biochim. Biophys. Acta., 586, 179 (1979) ;フランソン(L.−A. Fransson)、シェーバーグ(I. Sjoberg)、ハブスマーク(B. Havsmark)、Eur. J. Biochem., 106, 59 (1980))。ヘパリンと追加の調製物CS−4(ウシ気管からのCS−A)と6(サメ軟骨からのCS−C)は、シグマ(Sigma)から購入した。デルマタン硫酸(分子量〜500kDa)は、ビーディーエィチ・バイオケミカル(BDH Biochemical)から得た。
【0083】
DS36の放射性ヨード化は、以前のプロトコール(チェング(F Cheng)、ヨシダ(K. Yoshida)、ヘイネガード(D. Heinegard)、フランソン(L.A. Fransson)、Glycobiology, 2, 553 (1992))に従って行った。スロット結合測定法を以下のように行った:α−デフェンシンペプチド1(バッケム(Bachem)、スイス)をニトロセルロース膜(ハイボンド(Hybond)、アマシャム(Amersham))にブロットした。膜を1時間ブロッキング(PBS、pH7.4、0.25%ツイーン20、3%BSA)し、同じ緩衝液中で放射能標識DS(〜20μg/ml)と1時間インキュベートした。結合を競合させるために、非標識多糖を加えた(2mg/ml)。膜を洗浄した(3×10分)(PBS,pH7.4、0.25%ツイーン20)。Bas2000放射性イメージングシステム(フジ(Fuji))を使用して、放射活性を視覚化した。
【0084】
0.2〜5μgのα−デフェンシン1をニトロセルロースにブロットし、放射性ヨード化デルマタン硫酸(DS)とプローブ結合させた。ペプチドに結合したDSと結合は、過剰(〜100倍)の非標識DS(DS36;75%イズロン酸塩[IdoA])ならびにDS13(>95% IdoA)により阻害された。コンドロイチン硫酸(CS)−4ならびにCS−6は、結合を破壊しなかった。CS−4とDSは、主にウロン酸(CS−4中のグルクロン酸とDS中のIdoA)のエピマー化が異なる(フランソン(L.−A. Fransson)、「多糖(The Polysaccharides)」中、アスピナール(G.O. Aspinall)編(アカデミックプレス(Academic Press)、ニューヨーク州、1985年)、第3巻、338〜406頁)。試験したヘパラン硫酸(HS)画分のうち(HS3〜6)、硫酸化とIdoA含量の高いもの(HS5;〜50%IdoA、および〜62%N−硫酸、HS6:65%IdoAおよび72%N−硫酸)のみが、DSを完全に排除することがわかった。さらにヘパリンとデキストラン硫酸(デキストランではない)は、放射能標識DSを排除した。まとめるとこの結果は、GAG−デキストラン相互作用が、多糖のIdoA残基ならびに硫酸化に依存することを示唆する。DSとHS/ヘパリン中のIdoAは、2−O−硫酸化されていることがあり(フランソン(L.−A. Fransson)、「多糖(The Polysaccharides)」中、アスピナール(G.O. Aspinall)編(アカデミックプレス(Academic Press)、ニューヨーク州、1985年)、第3巻、338〜406頁)、α−デフェンシン1への結合のために、DSとHSポリマーの追加の修飾が必要かも知れない。
【0085】
例3
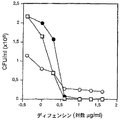
デフェンシン機能へのDSの作用を調べるために、試験微生物である化膿連鎖球菌(S. pyogenes)、イー・フェカーリス(E. faecalis)および緑膿菌(P. aeruginosa)に対するα−デフェンシン1の細菌活性を測定した。
【0086】
抗微生物測定法のために、化膿連鎖球菌(S. pyogenes)、イー・フェカーリス(E. faecalis)および緑膿菌(P. aeruginosa)を、TH培地中で対数中期まで増殖させた。細菌を洗浄し、5mMグルコース含有10mMトリス−塩酸(pH7.5)で希釈した。50μlの細菌(2×106 cfu/ml)を、0〜40μg/mlの濃度のα−デフェンシンと、2〜400μg/mlの濃度の種々のGAG鎖の添加有りまたは無しで、インキュベートした。インキュベーションは、37℃で2時間(化膿連鎖球菌(S. pyogenes)とイー・フェカーリス(E. faecalis))または4時間(緑膿菌(P. aeruginosa))行った。微生物活性を定量するために、このインキュベーション混合物の連続希釈物を、TH寒天に蒔き、37℃で一晩インキュベートし、CFUの数を測定した。
【0087】
細菌株を、0〜40μg/mlのα−デフェンシン1とインキュベートした。緑膿菌(P. aeruginosa)、イー・フェカーリス(E. faecalis)および化膿連鎖球菌(S. pyogenes)は、4μg/mlの濃度またはそれ以上のデフェンシンにより有効に死滅した。20〜40μg/ml(DS/デフェンシンモル比〜0.5〜1)の濃度のDSを添加すると、イー・フェカーリス(E. faecalis)のデフェンシン介在死滅がほとんど完全に逆転し、他の2つの種の死滅が部分的に阻害された(図1B)。400μg/mlのDS(モル比〜10:1)は、デフェンシンの抗微生物作用をほとんど完全に逆転させた。以前の実験は、DSがα−デフェンシン1に結合すること、および結合は〜100倍過剰のHS3ならびに2つのCS型により阻害されることを示したため、我々は、これらのGAGがデフェンシンを阻害する活性が小さいかどうかという問題を調べた。実際、測定法に含めると(試験成分として化膿連鎖球菌(S. pyogenes)を使用して)、デフェンシン介在細菌死滅は、ほんのわずかに阻害された;400μg/mlのHS3、CS−4およびCS−6(シグマ(Sigma))は、それぞれ6、16および2%阻害を示した。
【0088】
例4
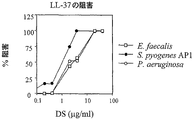
LL−37機能へのDSの作用を調べるために、試験微生物である化膿連鎖球菌(S. pyogenes)、イー・フェカーリス(E. faecalis)および緑膿菌(P. aeruginosa)に対するLL−37の細菌活性を測定した。
【0089】
抗微生物測定は、例3に記載のように行った。LL−37の抗細菌活性を例示する結果を図2に示す。0〜40μg/mlのDSの添加は、化膿連鎖球菌(S. pyogenes)、イー・フェカーリス(E. faecalis)および緑膿菌(P. aeruginosa)のLL−37死滅を阻害した。結果を図3に示す。
【0090】
例5
デルマタン硫酸(DS)がLL−37に結合し、および硫酸化多糖が相互作用をブロックするかどうかを調べるために、例2に示すように、スロット結合測定を行った。LL−37(0.2〜5μg)をニトロセルロースに適用し、ヨード化デルマタン硫酸(DS36)とインキュベートした。結合への競合のために、異なる多糖を加えた:DS36、DS18、DS13、CS−4、CS−6、CS−D(コンドロイチン硫酸D)およびCS−E(コンドロイチン硫酸E)。結果を図4に示す。CS−4、CS−6およびCS−DとCS−Dは、結合を破壊しなかった。しかし放射能標識DSは、DS−36、DS18、DS13、およびCS−Eにより排除された。
【0091】
例6
DS−LL−37相互作用への種々の陽イオン性分子の作用を、例2に記載のようにスロット−ブロット測定法を使用して監視した。結果を図5に示す。LL−37へのDSの結合は、ポリ−L−リジン、ポリ−L−アルギニン、DEAE−デキストラン、ポリブレン、キトサンおよびポリエチレンイミンによりブロックされた。結合レベルの低下は、プロタミンの存在下で観察された。ポリミキサンBおよびポリミキシンBノナペプチドは、LL−37へのDSの結合を阻止しなかった。
【0092】
例7
緑膿菌(P. aeruginosa)エラスターゼがLL−37を分解し不活性化することを証明するために、10μgのLL−37を50μlの10mMトリス−塩酸(pH7.5)中30mUの緑膿菌(P. aeruginosa)エラスターゼと、0、1、5、15もしくは30分または1、4もしくは20時間インキュベートした。インキュベーション物の等量のアリコートを、SDS−PAGE(16.5%トリス−トリシンゲル)で分析したかまたは細菌活性について測定した。細菌測定法のために、2×106 cfu/mlのイー・フェカーリス(E. faecalis)を0.1μgのLL−37と、10mMトリス−塩酸(pH7.5)、5mMグルコース中で37℃で2時間インキュベートした。図6に示す結果は、LL−37が緑膿菌(P. aeruginosa)エラスターゼにより分解され不活性化されることを示す。
【0093】
LL−37分解産物を、液体クロマトグラフィー質量スペクトル(LC−MS)、次に飛行時間(Time of flight)(TOF)MS−MSで測定した。断片質量が3708.859、3178.637、2822.450、2326.269および1401.748を与える主要なペプチド断片が検出され、さらに分析した。LL−37の切断点と提唱されている抗細菌領域を図7に示す。
【0094】
3mUのイー・フェカーリス(E. faecalis)ゲラチナーゼまたは3mUの化膿連鎖球菌(S. pyogenes)システインプロテイナーゼによるLL−37の分解を、1μgのLL−37を1時間と6時間インキュベートすることにより監視した。分解産物を16.5%のトリス−トリシンゲルのSDS−PAGEにより分析し、6mUの緑膿菌(P. aeruginosa)エラスターゼと1μgのLL−37を5分と1時間インキュベーション後の分解産物と比較した。結果を図8に示す。
【0095】
例8
1μgのLL−37を12mUの緑膿菌(P. aeruginosa)エラスターゼ、6mUのイー・フェカーリス(E. faecalis)ゲラチナーゼまたは6mUの化膿連鎖球菌(S. pyogenes)プロテアーゼと、インヒビターの存在下と非存在下で1時間インキュベートすることにより、LL−37の分解への種々の物質の作用を測定した。生じた物質をSDS−PAGE(16.5%トリス−トリシンゲル)により分析した。金属プロテアーゼインヒビターGM6001(100μM)と1,10−フェナントロリン(2mM)(これらは両方とも、緑膿菌(P. aeruginosa)エラスターゼとイー・フェカーリス(E. faecalis)ゲラチナーゼを阻害した)またはインヒビターE64(10μM)(これは、化膿連鎖球菌(S. pyogenes)システインプロテイナーゼを阻害した)により、分解は止まった(図9)。
【0096】
DS36とCS−E、および程度は低いがCS−AとCS−Cは、緑膿菌(P. aeruginosa)エラスターゼによるLL−37の分解を阻害した(図10)。構造[ΔUA(2S)−GalNAc(4,6S)](図11)の硫酸化二糖とシュクロースオクタ硫酸(図12)もまた、緑膿菌(P. aeruginosa)エラスターゼによるLL−37の分解を阻害した。
【図面の簡単な説明】
【図1】図1は、デルマタン硫酸(DS)によるα−デフェンシンの殺菌活性の阻止を例示する。図1Aは、α−デフェンシンによる細菌の死滅を例示する。2×106 CFU/mlの化膿連鎖球菌(S. pyogenes)(□)、イー・フェカーリス(E. faecalis)(●)および緑膿菌(P. aeruginosa)(○)を、α−デフェンシンと記載のペプチド濃度で37℃で2時間(化膿連鎖球菌(S. pyogenes))とイー・フェカーリス(E. faecalis))または4時間(緑膿菌(P. aeruginosa))インキュベートし、CFUを測定した。図1Bは、デルマタン硫酸による細菌のデフェンシン介在死滅の阻害を例示する。種々の濃度(0〜400μg/ml)のDSの添加は、化膿連鎖球菌(S. pyogenes)(□)、イー・フェカーリス(E. faecalis)(●)および緑膿菌(P. aeruginosa)(○)のデフェンシン死滅を阻害した。
【図2】図2は、LL−37の殺菌作用を例示する。2×106 コロニー形成単位(cfu)/mlのイー・フェカーリス(E. faecalis))(□)、化膿連鎖球菌(S. pyogenes)AP1(○)、および緑膿菌(P. aeruginosa)(●)を、LL−37と記載のペプチド濃度で37℃で、10mMトリス−塩酸(pH7.5)、5mMグルコース中で、2時間(化膿連鎖球菌(S. pyogenes)とイー・フェカーリス(E. faecalis))または4時間(緑膿菌(P. aeruginosa))インキュベートし、cfuを測定した。
【図3】図3は、デルマタン硫酸(DS)によるLL−37の細菌作用の阻止を例示する。種々の濃度(0〜40μg/ml)のDSの添加は、イー・フェカーリス(E. faecalis)(□)、化膿連鎖球菌(S. pyogenes)AP1(●)、および緑膿菌(P. aeruginosa)(○)のLL−37死滅を阻害した。種々の細菌株の2×106 CFU/mlを、10mMトリス−塩酸(pH7.5)、5mMグルコース中で、10μg/ml(イー・フェカーリス(E. faecalis)と緑膿菌(P. aeruginosa))または2μg/ml(化膿連鎖球菌(S. pyogenes)AP1)のLL−37、およびデルマタン硫酸と、2時間(AP1とイー・フェカーリス(E. faecalis))または4時間(緑膿菌(P. aeruginosa))インキュベートし、cfuを測定した。
【図4】図4は、LL−37へのデルマタン硫酸の結合を示す。LL−37(0.2〜5μg)をニトロセルロースに適用し、ヨード化したデルマタン硫酸(DS36)とインキュベートした。結合に競合させるために、異なる多糖を加えた。DS36、DS18、DS13(デルマタン硫酸36、18および13)、CS4とCS6(コンドロイチン硫酸4と6)、CS−D(コンドロイチン硫酸D)およびCS−E(コンドロイチン硫酸E)。
【図5】図5は、DS−LL−37相互作用に対する種々の陽イオン性分子の作用を示す。LL−37(0.2〜5μg)をニトロセルロースに適用し、ヨード化したデルマタン硫酸(DS36)とインキュベートした。結合に競合させるために、種々のパネルの上に示すように、異なる陽イオン性物質を加えた。(a)p−l−lys;ポリ−L−リジン、p−l−arg;ポリ−L−アルギニン、DEAE;DEAE−デキストラン、PB;ポリブレン。(b)CH;キトサン、PEI、ポリエチレンイミン、PM;プロタミン、PMX;ポリミキシンB、PMXnp;ポリマーミキシンBノナペプチド。
【図6】図6は、緑膿菌(Pseudomonas aeruginosa)エラスターゼがLL−37を分解し不活性化することを証明する実験の結果を示す。LL−37(10μg)を、50μlの10mMトリス−塩酸(pH7.5)中の緑膿菌エラスターゼ(P. aeruginosa)と、種々の時間(x軸上に記載)インキュベートした。次にインキュベート物の等量のアリコートを、SDS−PAGE(16.5%トリス−トリシンゲル;上の図)で分析したか、または殺菌活性(下の図)を測定した。2μgのLL−37に相当する容量を、ゲル上にのせた。分子量マーカーを左に示す(上の図)。殺菌測定(下の図)のために、2×106 cfu/mlのイー・フェカーリス(E. faecalis)を、分解の開始時に0.1μgの無傷のペプチドに相当する量のLL−37とインキュベートした。ペプチドを細菌と、10mMトリス−塩酸(pH7.5)、5mMグルコース中で、37℃で2時間インキュベートした。コロニー形成単位を測定した(左に記載)。
【図7】図7は、LL−37断片の性状解析を示す。LL−37(10μg)を、50μlの10mMトリス−塩酸(pH7.5)中で緑膿菌(P. aeruginosa)エラスターゼ(30mU)と、1、5、15、30分または1時間インキュベートした。生じた分解産物(図6参照)を、液体クロマトグラフィー質量スペクトル(LC−MS)、次に飛行時間(Time of flight)(TOF)MS−MSで測定した。主要なペプチド断片を検出し、さらに分析した。切断点を示し、LL−37の提唱されている抗細菌領域に下線を引いた。
【図8】図8は、細菌プロテイナーゼによるLL−37の分解を示す。LL−37(1μg)を、それぞれ緑膿菌(P. aeruginosa)エラスターゼ(ELA)と5分(1)または1時間(2)インキュベートし、イー・フェカーリス(E. faecalis)ゲラチナーゼ含有培地(GEL)または化膿連鎖球菌(S. pyogenes)システインプロテイナーゼ(CP)と1時間(1)または6時間(2)インキュベートした。加えたプロテイナーゼ活性(アゾカゼイン測定法により測定した)は、10μlの反応物当たり6mU(緑膿菌(P. aeruginosa)エラスターゼ)および3mU(他のプロテイナーゼ)に相当した。物質をSDS−PAGEで16.5%トリス−トリシンゲルで分析した。分子量マーカーを左に示す。
【図9】図9は、プロテイナーゼインヒビターによるLL−37分解の阻害を示す。LL−37(1μg)を、緑膿菌(P. aeruginosa)エラスターゼ(ELA、12mU)、イー・フェカーリス(E. faecalis)ゲラチナーゼ(GEL、6mU)または化膿連鎖球菌(S. pyogenes)システインプロテイナーゼ(CP、6mU)と、インヒビターの非存在下または存在下で1時間インキュベートした。GM6001;100μM GM6001、PHE;2mM 1,10−フェナトロリン、E64;10μM E64。物質をSDS−PAGE(16.5%トリス−トリシンゲル)で分析した。分子量マーカー(kDa)を左に示す。
【図10】図10は、LL−37の分解が、硫酸化グリコサミノグリカンにより阻害されることを示す。LL−37(1μg)を、緑膿菌(P. aeruginosa)エラスターゼ(12mU)と総量15μl(10mMトリス緩衝液)中で、記載のグリコサミノグリカンの非存在下(ELA)または存在下で、1時間インキュベートした。DS36、デルマタン硫酸36(CS−Bとも記載する)、CS−A;コンドロイチン硫酸A、CS−C、コンドロイチン硫酸C、CS−E;コンドロイチン硫酸E。物質をSDS−PAGEで16.5%トリス−トリシンゲルで分析した。分子量マーカー(kDa)を左に示す。
【図11】図11は、LL−37の分解が硫酸化二糖により阻害されることを示す。LL−37(1μg)を、緑膿菌(P. aeruginosa)エラスターゼ(12mU)と総量15μl(10mMトリス緩衝液)中で、硫酸化二糖の非存在下または存在下で、1時間インキュベートした。ELA;緑膿菌(P. aeruginosa)エラスターゼ、0S,UA−GalNAc;2S−4S;UA(2S)−GalNAc(4S)、2S−6S;UA(2S)−GalNAc(6S)、4,6diS;UA−GalNAc(4,6S)、2S−4,6diS;UA(2S)−GalNAc(4,6S)。物質をSDS−PAGEで16.5%トリス−トリシンゲルで分析した。分子量マーカー(kDa)を左に示す。
【図12】図12は、LL−37の分解がシュクロースオクタ硫酸により阻害されることを示す。IL−3(1μg)を、緑膿菌(P. aeruginosa)エラスターゼ(12mU;ELA)と総量15μl(10mMトリス緩衝液)中で、記載の量(ナノモル)のショ糖(S)またはシュクロースオクタ硫酸(SOS)の非存在下または存在下で、1時間インキュベートした。物質をSDS−PAGEで16.5%トリス−トリシンゲルで分析した。分子量マーカー(kDa)を左に示す。
(発明の分野)
本発明は、慢性および急性の微生物感染症の治療に使用される物質に関する。本発明はまた、治療法、特に慢性および急性の微生物感染症の治療に有用な物質の同定方法に関する。
【0002】
(発明の背景)
感染の間の1つの基本的な宿主防御機構は、微生物の白血球介在死滅に依存する。最近、膜破壊により種々のグラム陰性菌ならびにグラム陽性細菌を死滅させることができる一群の陽イオン性抗微生物ペプチドが、多くの生物の先天的な宿主防御系の重要な成分として考えられている。これらのペプチドは、感染を受ける生物学的境界に局在する好中球や上皮細胞中で見いだされる。
【0003】
病原体は、この第一線の宿主防御を乗り越えて、急性ならびに慢性の一連の臨床的に重要な病状を発症させることができるようである。感染の間、細菌は、種々の方策を利用して宿主防御機構を乗り越える。細菌は、種々のプロテイナーゼを放出し、これらは、急性感染中に、カリクレイン、プラスミノーゲン、補体、サイトカイン、抗体、および抗プロテイナーゼを含む宿主の特定の経路を変化させる。細菌による外毒素の産生とタンパク質を変化させる細胞サイクルも報告されており、これは、宿主防御を易感染性にすることを目的とするさらなる経路である。
【0004】
化膿連鎖球菌(Streptococcus pyogenes)のような細菌は、容易に創傷感染や丹毒を生じる。緑膿菌(Pseudomonas aeruginosa)、エンテロコッカス・フェカーリス(Enterococcus faecalis)および霊菌(Proteus mirabilis)は、慢性の創傷中に存続することが知られている。これらの細菌はすべて、プロテオグリカン(PG)の豊富な結合組織中で増殖し繁殖する。
【0005】
(発明の要約)
本発明者らは、細菌により分泌される細胞外プロテイナーゼが、例えば結合組織中に存在するようなプロテオグリカンを分解することにより、グリコサミノグリカンを放出することを証明した。次にグリコサミノグリカンは、好中球と上皮細胞により分泌される陽イオン性抗微生物ペプチドに結合し、これらの抗微生物ペプチドが細菌を死滅させることを妨害する。特に本発明者らは、デルマタン硫酸含有プロテオグリカン(例えば、デコリン)が、緑膿菌(Pseudomonas aeruginosa)、エンテロコッカス・フェカーリス(Enterococcus faecalis)、化膿連鎖球菌(Streptococcus pyogenes)および霊菌(Proteus mirabilis)からの細菌プロテイナーゼにより分解されて、デルマタン硫酸を産生することを証明した。彼らは、デルマタン硫酸が好中球由来α−デフェンシンとLL−37に結合すること、そしてこの結合は、α−デフェンシンとLL−37の抗微生物活性を完全に中和することを証明した。従って、感染の間のグリコサミノグリカンの放出、例えばプロテオグリカン分解によるデルマタン硫酸の放出は、重要なかつ従来は知られていなかった細菌の防御機構であり、これは、新規な抗細菌法の標的となり得る。
【0006】
従って本発明は、陽イオン性抗微生物ペプチドの抗微生物活性を増強するのに使用するための、プロテイナーゼ/グリコサミノグリカン経路の阻害作用を阻止する物質を提供する。
【0007】
本発明はまた、プロテイナーゼ/グリコサミノグリカン経路の阻害作用を阻止することにより、陽イオン性抗微生物ペプチドの抗微生物活性を増強する物質の同定法であって:
(i)第1の成分として、陽イオン性抗微生物ペプチドを提供し;
(ii)第2の成分として、細菌を提供し;
(iii)第3の成分として、プロテイナーゼ/グリコサミノグリカン経路の成分の一部またはすべてを提供し、こうして第3の成分が第1の成分(例えば、グリコサミノグリカン、または細菌、または細菌とプロテオグリカン、または細菌プロテイナーゼ、または細菌プロテイナーゼとプロテオグリカン)の抗微生物作用を低下させ;
(iv)第1、第2、および第3の成分を、第3の成分の非存在下で抗微生物物質による細菌の死滅を可能にし、試験物質の非存在下で第3の成分による第1の成分の抗微生物活性の阻害を可能にする条件下で、試験物質に接触させ;
(v)細菌の生存を監視して、試験物質が抗微生物活性を増強することができるかどうか(抗微生物活性を増強することができる試験物質は、細菌の死滅を促進する)を決定する、ことを含む上記方法を提供する。
【0008】
陽イオン性抗微生物ペプチドの阻害に至るプロテイナーゼ/グリコサミノグリカン経路は、2つ以上の段階で阻害することができ、結果としての細菌への作用を監視する必要無く、各段階で試験物質の阻害作用を監視してもよいことが理解されるであろう。特に試験物質は、グリコサミノグリカンと陽イオン性抗微生物ペプチドとの相互作用、グリコサミノグリカンの放出、例えばプロテオグリカンからのグリコサミノグリカンの細菌プロテイナーゼ活性化放出、または細菌プロテイナーゼによる陽イオン性抗微生物ペプチドの直接の分解を阻害する。従って、プロテイナーゼ/グリコサミノグリカン経路の阻害作用を阻止することにより、陽イオン性抗微生物ペプチドの抗微生物活性を増強する物質は、本発明により提供される以下のいずれか1つの方法により同定される:
【0009】
− グリコサミノグリカンと陽イオン性抗微生物ペプチドとの相互作用を阻害することができる物質を同定する方法であって:
(i)第1の成分として、グリコサミノグリカンを提供し;
(ii)第2の成分として、陽イオン性抗微生物ペプチドを提供し;
(iii)第1および第2の成分を、試験物質の非存在下で第1および第2の成分が相互作用することを可能にする条件下で、試験物質に接触させ;そして
(iv)第1の成分と第2の成分の相互作用を監視して、こうして試験物質が第1の成分と第2の成分の相互作用を破壊することができるかどうかを決定する、ことを含む上記方法;
【0010】
− 陽イオン性抗微生物ペプチドのグリコサミノグリカン介在阻害を阻止することができる物質を同定する方法であって:
(i)第1の成分として、グリコサミノグリカンを提供し;
(ii)第2の成分として、陽イオン性抗微生物ペプチドを提供し;
(iii)第3の成分として、細菌を提供し;
(iv)第1、第2、および第3の成分を、第1の成分の非存在下で第2の成分による細菌の死滅を可能にし、試験物質の非存在下で第2の成分による細菌の死滅の、第1の成分による阻害を可能にする条件下で、試験物質に接触させ;そして
(v)細菌の生存を監視し、こうして試験物質が陽イオン性抗微生物ペプチドのグリコサミノグリカン介在阻害を妨害することができるかどうかを決定する、ことを含む上記方法;
【0011】
− 細菌プロテイナーゼによるプロテオグリカンの分解を阻害することができる物質を同定する方法であって:
(i)第1の成分として、細菌プロテイナーゼを提供し;
(ii)第2の成分として、プロテオグリカンを提供し;
(iii)第1および第2の成分を、試験物質の非存在下で第1の成分による第2の成分の分解を可能にする条件下で、試験物質に接触させ;そして
(iv)第2の成分の分解を監視して、こうして試験物質が、プロテイナーゼによるプロテオグリカンの分解を阻害することができるかどうかを決定する、ことを含む上記方法;
【0012】
− 細菌プロテイナーゼによる陽イオン性抗微生物ペプチドの分解を阻害することができる物質を同定する方法であって:
(i)第1の成分として、細菌プロテイナーゼを提供し;
(ii)第2の成分として、陽イオン性抗微生物ペプチドを提供し;
(iii)第1および第2の成分を、試験物質の非存在下で第1の成分による第2の成分の分解を可能にする条件下で、試験物質に接触させ;そして
(iv)第2の成分の分解を監視して、こうして試験物質が、プロテイナーゼによる陽イオン性抗微生物ペプチドの分解を阻害することができるかどうかを決定する、ことを含む上記方法。
【0013】
本発明はまた、以下を提供する:
− グリコサミノグリカンと陽イオン性抗微生物ペプチドとの相互作用を阻害することができる物質の同定に使用するためのキットであって:
(a)グリコサミノグリカン;と
(b)陽イオン性抗微生物ペプチド、とを含むキット;
【0014】
− 細菌プロテイナーゼによるプロテオグリカンの分解を阻害することができる物質の同定に使用するためのキットであって:
(a)プロテオグリカン;と
(b)細菌プロテイナーゼ、とを含むキット;
− 本発明の方法により同定される物質;
− 本発明の物質を含む医薬組成物;
− 治療法によりヒトまたは動物の体を治療する方法に使用するための本発明の物質または医薬組成物;
− 治療法によりヒトまたは動物の体を治療する方法において、同時にまたは連続的に使用するための本発明の物質と陽イオン性抗微生物ペプチドとを含む生成物;
【0015】
− 治療法によりヒトまたは動物の体を治療する方法において、同時にまたは連続的に使用するための、細菌の防御機構のインヒビターと陽イオン性抗微生物ペプチドとを含む生成物(ここで、該防御機構は、細菌プロテイナーゼによりプロテオグリカンを分解して、陽イオン性抗微生物ペプチドに結合して該ペプチドの抗微生物活性を阻害することができるグリコサミノグリカンを放出する);
− 慢性または急性の細菌感染を治療する方法で使用するための医薬の製造における、本発明の物質、医薬組成物または生成物の使用。
【0016】
(配列表の簡単な説明)
配列番号1は、陽イオン性抗微生物ペプチドLL−37のアミノ酸配列である。
配列番号2は、陽イオン性抗微生物ペプチドα−デフェンシンのアミノ酸配列である。
【0017】
(発明の詳細な説明)
本明細書と添付の請求の範囲を通して、「含んでなる」および「含む」という用語は、包括的であると理解されたい。すなわちこれらの用語は、文脈により、他の成分または整数を特に記載されていなくても含むことが可能であることを企図する。
【0018】
本発明は、細菌の防御機構を妨害することにより陽イオン性抗微生物ペプチドの作用を増強する物質の同定法を提供する。細菌の防御機構は、グリコサミノグリカンの放出と、放出されたグリコサミノグリカンの陽イオン性抗微生物ペプチドへの結合を含む。グリコサミノグリカンの陽イオン性抗微生物ペプチドへの結合は、ペプチドの抗微生物活性(本明細書において、プロテイナーゼ/グリコサミノグリカン経路と呼ぶ)を阻害する。グリコサミノグリカンは、プロテオグリカンの分解により放出される。好ましくはプロテオグリカンは、細菌プロテイナーゼにより分解される。グリコサミノグリカン放出の他の機構には、内因性金属プロテイナーゼにより仲介されるプロテオグリカンの放出の増強、プロテオグリカンの産生の増加、および結合組織中の結合部位からのプロテオグリカンの排除がある。例えば、細菌プロテイナーゼは、内因性マトリックス金属プロテイナーゼを直接活性化する。妨害は、例えば、例えばプロテイナーゼ結合または活性を阻害することによる、またはグリコサミノグリカンの陽イオン性抗微生物ペプチドへの結合を阻止することによる、プロテオグリカンの分解の阻止でもよい。
【0019】
陽イオン性抗微生物ペプチドは当該分野で公知である。例えば、レーラー(Lehrer, R.J.)とガンズ(Ganz, T.)(1999)、哺乳動物と昆虫宿主防御における抗微生物ペプチド、Curr Opin Immunol 11:23−27を参照されたい。
【0020】
陽イオン性抗微生物ペプチドは典型的には、細菌膜を破壊し従って細菌を死滅させることができる抗細菌性ペプチドである。陽イオン性抗微生物ペプチドは、外因性に加えてもよい。陽イオン性抗微生物ペプチドは好ましくは、感染部位で白血球および/または上皮細胞により産生される。好ましくは白血球は好中球である。陽イオン性抗微生物ペプチドは、好ましくはデフェンシンまたはカテリシジン(cathelicidin)である。さらに好ましくは陽イオン性抗微生物ペプチドは、α−デフェンシン(配列番号2)またはLL−37(配列番号1)である。陽イオン性抗微生物ペプチドは、配列番号1または配列番号2の変種または断片を含み、この変種または断片は、抗微生物活性、好ましくは抗細菌活性を示す。例えば、陽イオン性抗微生物ペプチドは、図7で下線を引いた配列を含む配列番号1の断片でもよい。
【0021】
本発明の方法で使用するために提供される陽イオン性抗微生物ペプチドは、任意の適当な供給源からのものである。例えば、陽イオン性抗微生物ペプチドは、組換えペプチドでも、陽イオン性抗微生物ペプチドは、白血球または上皮細胞から精製されても、または陽イオン性抗微生物ペプチドは、白血球、上皮細胞または培養物中で増殖させた組換えペプチドでトランスフェクションした細胞から抽出される増殖培地中に存在してもよい。ペプチドは、天然の配列を含んでも、人工的に変異させた配列を含んでもよい(ただし、変異は、ペプチドが細菌膜を破壊する能力またはグリコサミノグリカンと相互作用する能力に影響を与えないものとする)。
【0022】
ペプチドが抗微生物活性を示すかどうかを調べるための典型的な測定法は、細菌、例えば緑膿菌(Pseudomonas aeruginosa)、エンテロコッカス・フェカーリス(Enterococcus faecalis)、化膿連鎖球菌(Streptococcus pyogenes)、霊菌(Proteus mirabilis)または大腸菌(E. coli)を、ペプチドとインキュベートして、細菌の生存を監視することを含む。典型的には、抗微生物ペプチドは、4μg/mlからの濃度、例えば4μg/ml、5μg/ml、10μg/ml、20μg/ml、40μg/ml、100μg/ml、または500μg/mlの濃度で、細菌を有効に死滅させる。
【0023】
細菌の生存は、適当な方法により監視される。典型的にはコロニー形成単位の数が測定される。細菌の生存はまた、宿主動物または宿主細胞への細菌の作用を評価することにより、間接的に監視してもよい。
【0024】
細菌は任意の適当な株でよい。細菌は、グラム陰性でもグラム陽性でもよい。好ましくは細菌株は、ヒトまたは動物の感染部位に見いだされるものである。好ましくは感染部位は、皮膚または粘膜のようなヒトまたは動物への病原体の侵入の障壁になるようなものである。好ましくは細菌は、緑膿菌(Pseudomonas aeruginosa)、エンテロコッカス・フェカーリス(Enterococcus faecalis)、化膿連鎖球菌(Streptococcus pyogenes)、または霊菌(Proteus mirabilis)である。
【0025】
本発明の方法または本明細書に記載の測定法において、任意の適当な細菌調製物を使用することができる。例えば細菌は、TH培地またはC培地中で対数中期まで増殖してもよい。次に細菌を、例えば5mMグルコースを含有する10mMトリス−塩酸(pH7.5)中で洗浄し、希釈する。典型的には、1測定当たり5μl〜500μlの細菌、好ましくは10μl〜100μlの細菌(2×106 cfu/ml)を使用する。細菌と抗微生物ペプチド、抗微生物ペプチドとグリコサミノグリカン、抗微生物ペプチドと試験物質または抗微生物ペプチド、グリコサミノグリカンと試験物質のインキュベーションは、典型的には37℃で30分〜6時間、好ましくは2時間〜4時間行われる。抗微生物活性は、任意の適当な方法により定量され、例えばインキュベーション混合物の連続希釈物を作成し、希釈物を寒天(例えばTH寒天)上に蒔き、37℃でインキュベートし(好ましくは一晩)、産生されたコロニー形成単位(CFU)の数を測定することにより、定量される。
【0026】
本発明の方法での使用に適したグリコサミノグリカンは、ペプチドの抗微生物機能が阻害されるように、陽イオン性抗微生物ペプチドと相互作用することができる。グリコサミノグリカンは、典型的には内因性グリコサミノグリカンである。グリコサミノグリカンは、好ましくはヘパラン硫酸またはヘパリンであり、さらに好ましくはデルマタン硫酸である。好ましくは本発明の方法での使用に適したグリコサミノグリカンは、高度の硫酸化および/またはイズロン酸含量を示す。イズロン酸含量は、典型的には少なくとも50%、例えば少なくとも60%、少なくとも70%、少なくとも80%、または少なくとも90%である。硫酸化の程度は、少なくとも50%、例えば少なくとも60%、少なくとも70%、少なくとも80%、または少なくとも90%である。本発明の方法で使用するためのグリコサミノグリカンはまた、追加の修飾を含有してもよく、例えばイズロン酸は2−O−硫酸化されてもよい。
【0027】
グリコサミノグリカンが陽イオン性抗微生物ペプチドと相互作用することができるかどうかを決定するための典型的な測定法には、陽イオン性抗微生物ペプチドを例えばニトロセルロース上に固定化し、放射能標識グリコサミノグリカンとプローブ結合させる方法がある。好ましくはグリコサミノグリカンのペプチドへの結合は、過剰の非標識デルマタン硫酸で阻害される。
【0028】
グリコサミノグリカンペプチドが陽イオン性抗微生物ペプチドの抗微生物活性を阻害することができるかどうかを決定するための典型的な測定法には、細菌、例えば緑膿菌(Pseudomonas aeruginosa)、エンテロコッカス・フェカーリス(Enterococcus faecalis)、化膿連鎖球菌(Streptococcus pyogenes)または霊菌(Proteus mirabilis)の調製物または培養物を、グリコサミノグリカンおよびペプチドとインキュベートし、細菌の死滅を監視する方法がある。典型的には抗微生物ペプチドは、グリコサミノグリカンの非存在下で細菌を有効に死滅させる濃度で加えられるであろう。典型的にはグリコサミノグリカンは、細菌のペプチド介在死滅を、10μg/ml〜1mg/ml、さらに好ましくは10μg/ml〜500μg/ml、または最も好ましくは20μg/ml〜250μg/mlの濃度で阻害する。グリコサミノグリカンと陽イオン性抗微生物ペプチドとのモル比は、典型的には0.1:1から1000:1、好ましくは0.5:1から100:1、およびさらに好ましくは0.5:1から10:1である。グリコサミノグリカンは、ペプチドの抗微生物活性を部分的にのみ阻害してもよい。好ましくはグリコサミノグリカンは抗微生物活性を、少なくとも2%、例えば少なくとも6%、少なくとも16%、少なくとも30%、または少なくとも40%阻害する。さらに好ましくは抗微生物活性はグリコサミノグリカンにより、少なくとも50%、例えば少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、または少なくとも99%阻害される。
【0029】
グリコサミノグリカンは、任意の特定の供給源から得られる。好ましくはグリコサミノグリカンは、真皮(皮膚)または軟骨のような結合組織から得られる。グリコサミノグリカンは、任意の適当な方法により調製され、例えばローデン(L. Roden)、ベーカー(J. Baker)、シフォネッリ(J.A. Cifonelli)、マシューズ(M.B. Mathews)、Methods in Enzymology、ギンズバーグ(V. Ginsburg)編(アカデミックプレス(Academic Press)、ニューヨーク、1973年)、第28巻、73〜140頁、フランソン(L.−A. Fransson)、ニーヅスジンスキー(I.A. Nieduszynski)、フェルプス(C.F. Phelps)、シーハン(J.K. Sheehan)、Biochim. Biophys. Acta., 586, 179 (1979)、 またはフランソン(L.−A. Fransson)、シェーバーグ(I. Sjoberg)、ハブスマーク(B. Havsmark)、Eur. J. Biochem., 106, 59 (1980)に記載されている。
【0030】
典型的には、本発明の方法で使用するためのグリコサミノグリカンは、1つ以上のプロテイナーゼ、例えば細菌プロテイナーゼの、1つ以上のプロテオグリカンへの作用により産生される。グリコサミノグリカンは、適当な細菌プロテイナーゼと適当なプロテオグリカンを提供することにより、間接的に提供されてよい。プロテオグリカンは、繊維芽細胞培養物の細胞外生成物でもよい。好ましくはプロテオグリカンは、ヘパリン、ヘパラン硫酸、または好ましくはデルマタン硫酸を含む。好ましくはプロテオグリカンは、ビグリカン(biglycan)、シンデカン(syndecan)、グリピカン(glypican)またはCD44である。さらに好ましくはプロテオグリカンは、デコリン(decorin)またはベルシカン(versican)である。
【0031】
プロテオグリカンが細菌プロテイナーゼにより分解されるかどうかを決定するための典型的な測定法には、プロテオグリカンを35Sで放射能標識し、標識プロテオグリカンを細菌プロテイナーゼとともにインキュベートし、インキュベーション前に取った標識プロテオグリカンの試料とインキュベーション後に取った試料を、SDS−ポリアクリルアミドゲルに流し、標識プロテオグリカンと分解生成物の相対的サイズと量を測定する方法がある。例えば、もし細菌プロテイナーゼがシステインプロテイナーゼなら、還元剤(例えばDTT)をインキュベーション混合物中に含めることが必要かも知れない。プロテオグリカンは、任意の適当な供給源から取られ、例えば繊維芽細胞培養物の細胞外生成物でもよい。好ましくはプロテオグリカンは、80〜500kDa、好ましくは約100kDaまたは約400kDaの分子量を有する。分解産物は、好ましくは30〜50kDaの分子量を有し、これは遊離のまたはペプチド結合グリコサミノグリカン鎖に対応する。
【0032】
本明細書に記載の測定法に、任意の好適なプロテアーゼが使用できる。好ましくはプロテアーゼは、細菌プロテイナーゼである。任意の細菌調製物または細菌培養物が、細菌プロテイナーゼの供給源として使用される。細菌は、任意の適当な手段により刺激してプロテイナーゼを産生させてもよい。細菌の培養物からの増殖培地が使用できる。典型的には、細菌は、液体培地(例えば、TH培地またはC培地)中で静止期まで増殖される。本発明の方法で使用するのに適した増殖培地は、遠心分離により細菌をペレット化し、例えば、直径0.2μm〜0.4μmのフィルターを使用して上清をろ過することにより得られる。あるいは細菌プロテイナーゼは、増殖培地から単離される。細菌プロテイナーゼを単離するのに、任意の適当な方法が使用される。例えばプロテイナーゼは、硫酸アンモニウム沈殿、透析、および適当なカラム(例えば、ハイQ陰イオン交換クロマトグラフィー)で分離することにより調製される。あるいはプロテイナーゼは、ハーワルト(H. Herwald)、コリン(M. Collin)、ムラー−エスタール(Muller−Esterl)、ビヨルク(L. Bjorck)、J. Exp. Med. 184, 665 (1996)に記載のように調製される。本発明での使用に適した細菌プロテイナーゼには、緑膿菌(Pseudomonas aeruginosa)エラスターゼ、緑膿菌(Pseudomonas aeruginosa)アルカリ性プロテイナーゼ、エンテロコッカス・フェカーリス(Enterococcus faecalis)ゲラチナーゼ、および化膿連鎖球菌(Streptococcus pyogenes)システインプロテイナーゼがある。
【0033】
本発明により提供される陽イオン性抗微生物ペプチド活性を増強する物質の同定法は、グリコサミノグリカンと陽イオン性抗微生物ペプチドとの相互作用を阻害することができる物質の同定法を含む。そのような方法は基本的に以下からなる:
(i)第1の成分として、グリコサミノグリカンを提供し;
(ii)第2の成分として、陽イオン性抗微生物ペプチドを提供し;
(iii)第1および第2の成分を、試験物質の非存在下で第1および第2の成分が相互作用することを可能にする条件下で、試験物質に接触させ;そして
(iv)第1の成分と第2の成分の相互作用を監視して、こうして試験物質が第1の成分と第2の成分の相互作用を破壊することができるかどうかを決定する。
【0034】
この方法での使用に適したグリコサミノグリカンと陽イオン性抗微生物ペプチドは、既に本明細書に記載されている。第1および第2の成分を試験物質に接触させるために、任意の適当な条件が使用される。典型的には、陽イオン性ペプチドは、例えばニトロセルロース膜またはポリアクリルアミドゲル(これは好ましくは非変性性である)上に固定化され、固定化されたペプチドは、グリコサミノグリカンおよび試験物質とインキュベートされる。グリコサミノグリカンの固定化ペプチドへの結合、従って結合に対して試験物質が競合する能力は、任意の適当な手段により監視される。例えばグリコサミノグリカンは、例えば35Sで放射能標識され、固定化ペプチドに結合した放射能標識グリコサミノグリカンは、任意の適当な手段(例えば、シンチレーション計測、ホスホリメージャー(phosphorimager)、またはX線フィルム)を使用して検出される。好ましくは、同一の条件下で、好ましくは同時に、試験物質有りでおよび無しで実験を行い、実験の結果を比較して、試験物質がグリコサミノグリカンのペプチドへの結合の低下を引き起こすかどうかを決定する。一般に試験物質は、過剰に加えられる。好ましくは試験物質は、グリコサミノグリカンのペプチドへの結合を、少なくとも20%、少なくとも30%、少なくとも40%、少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも99%低下させるか、またはグリコサミノグリカンのペプチドへの結合を、物質の濃度0.1mg/ml、1mg/ml、10mg/ml、100mg/ml、500mg/ml、1mg/ml、10mg/ml、100mg/ml、または0.1g/mlで、実質的に妨害する。生成物の相互作用の程度と濃度の上記の低下パーセントの任意の組合せが、本発明の物質を規定するのに使用され、より低濃度でのより大きな分解が好ましい。本発明の好適な物質は、1mg/mlまたは0.5mg/mlの濃度で少なくとも50%レベルの阻害を示すものである。
【0035】
本発明により提供される陽イオン性抗微生物ペプチド活性を増強する物質の同定法は、陽イオン性抗微生物ペプチドの抗微生物活性へのグリコサミノグリカンの阻害作用を阻止することができる物質を同定する方法を含む。そのような方法は基本的に以下からなる:
(i)第1の成分として、グリコサミノグリカンを提供し;
(ii)第2の成分として、陽イオン性抗微生物ペプチドを提供し;
(iii)第3の成分として、細菌を提供し;
(iv)第1、第2、および第3の成分を、第1の成分の非存在下で第2の成分による細菌の死滅を可能にし、試験物質の非存在下で第2の成分による細菌の死滅の、第1の成分による阻害を可能にする条件下で、試験物質に接触させ;そして
(v)細菌の死滅を監視し、こうして試験物質が陽イオン性抗微生物ペプチドのグリコサミノグリカン介在阻害を妨害することができるかどうかを決定する。
【0036】
工程(iv)において第1および第2の成分は、まず試験物質と接触し、次に第3の成分と接触してもよい。第1および第2の成分を試験物質と接触させた後の適当な時間、例えば0〜4時間後、例えば30秒後、1分後、5分後、15分後、30分後、または1時間後に、第3の成分を、第1の成分、第2の成分、および試験物質の混合物に接触させてもよい。
【0037】
試験物質をまず、陽イオン性抗微生物ペプチドの活性に対する直接作用について試験してもよい。好ましくは、陽イオン性抗微生物ペプチド活性を阻害しない試験物質のみが、殺菌測定法で使用される。
【0038】
この方法での使用に適したグリコサミノグリカン、陽イオン性抗微生物ペプチドおよび細菌調製物は、既に記載されている。第1、第2、および第3の成分を試験物質に接触させるために、任意の適当な条件が使用される。典型的には、1測定について、5μl〜500μlの細菌、好ましくは10μl〜100μlの細菌(2×106 cfu/ml)が使用される。この3つの成分は典型的には、37℃で30分〜6時間、好ましくは2時間〜4時間インキュベートされる。細菌の死滅の監視は、公知の方法により行われ、例えばインキュベーション混合物の連続希釈物を作成し、希釈物を寒天(例えばTH寒天)上に蒔き、37℃で、好ましくは一晩インキュベートし、産生されたコロニー形成単位(CFU)の数を測定することにより定量される。好ましくは3つの測定[1つは、第2の成分(抗微生物ペプチド)のみを含有し、1つは、第1および第2の成分(グリコサミノグリカンと抗微生物ペプチド)のみを含有し、1つはすべての3つの成分を含有する]を、同一の条件下でかつ好ましくは同時に行う。3つの測定の結果を定量し、結果を比較して、グリコサミノグリカンによる抗微生物活性の阻害を阻止する試験物質の有効性を決定する。好ましくは試験物質は、ペプチドの抗微生物活性に対するグリコサミノグリカンの阻止作用を、少なくとも20%、少なくとも30%、少なくとも40%、少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも99%逆転させるか、または陽イオン性抗微生物ペプチド活性の阻害を、物質の濃度0.1mg/ml、1mg/ml、10mg/ml、100mg/ml、500mg/ml、1mg/ml、10mg/ml、100mg/ml、または0.1g/mlで、実質的に妨害する。阻害の程度と生成物の濃度の上記の低下パーセントの任意の組合せが、本発明の物質を規定するのに使用され、より低濃度でのより大きな分解が好ましい。本発明の好適な物質は、1mg/mlまたは0.5mg/mlの濃度で少なくとも50%レベルの阻害を示すものである。
【0039】
第2の成分の分解もまた、既に本明細書に記載の測定法を使用して、試験物質の存在下と非存在下で、陽イオン性抗微生物ペプチドによる細菌の死滅を監視することにより、間接的に監視される。
【0040】
本発明により提供される陽イオン性抗微生物ペプチド活性を増強する物質の同定法は、例えば細菌プロテイナーゼによるプロテオグリカンの分解を阻止することにより、グリコサミノグリカン(次にグリコサミノグリカンは、陽イオン性抗微生物ペプチドと自由に相互作用する)の放出を阻害することができる物質を同定する方法を含む。そのような方法は基本的に以下からなる:
(i)第1の成分として、細菌プロテイナーゼを提供し;
(ii)第2の成分として、プロテオグリカンを提供し;
(iii)第1および第2の成分を、試験物質の非存在下で第1の成分による第2の成分の分解を可能にする条件下で、試験物質に接触させ;そして
(iv)第2の成分の分解を監視して、こうして試験物質が、プロテイナーゼによるプロテオグリカンの分解を阻害することができるかどうかを決定する。
【0041】
適当な第1および第2の成分、およびプロテオグリカンが細菌プロテイナーゼにより分解されるかどうかを決定するための測定法は、既に本明細書に記載されている。好ましくは2つの測定(1つは、試験物質を含み、1つは試験物質を含まない)を、同一の条件下で行う。次に、2つの測定の間の分解生成物の相対量および/または非分解プロテオグリカンの相対量を比較して、試験物質による分解の阻害程度を決定する。各生成物の相対量は、ホスホリメージャー(phosphorimager)を使用して定量される。好ましくは、プロテイナーゼによるプロテオグリカンの分解を阻害する試験物質は、プロテオグリカンの分解を、少なくとも20%、少なくとも30%、少なくとも40%、少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも99%低下させるか、または物質の濃度0.1mg/ml、1mg/ml、10mg/ml、100mg/ml、500mg/ml、1mg/ml、10mg/ml、100mg/ml、または0.1g/mlで、実質的に妨害する。相互作用の程度と生成物の濃度の上記の低下パーセントの任意の組合せが、本発明の物質を規定するのに使用され、より低濃度でのより大きな分解が好ましい。本発明の好適な物質は、1mg/mlまたは0.5mg/mlの濃度で少なくとも50%レベルの阻害を示すものである。
【0042】
本発明により提供される陽イオン性抗微生物ペプチド活性を増強する物質の同定法は、細菌プロテイナーゼによる陽イオン性抗微生物ペプチドの分解を妨害することができる物質の同定法を含む。そのような方法は基本的に以下からなる:
(i)第1の成分として、細菌プロテイナーゼを提供し;
(ii)第2の成分として、陽イオン性抗微生物ペプチドを提供し;
(iii)第1および第2の成分を、試験物質の非存在下で第1の成分による第2の成分の分解を可能にする条件下で、試験物質に接触させ;そして
(iv)第2の成分の分解を監視して、こうして試験物質が、プロテイナーゼによる陽イオン性抗微生物ペプチドの分解を阻害することができるかどうかを決定する。
【0043】
適当な第1および第2の成分は、既に本明細書に記載されている。陽イオン性抗微生物ペプチドが細菌プロテイナーゼにより分解されるかどうかを決定する典型的な測定法は、ペプチドを35Sで放射能標識し、標識ペプチドを細菌プロテイナーゼとインキュベートし、インキュベーション前に取った標識ペプチドの試料とインキュベーション後に取った試料をSDS−ポリアクリルアミドゲルに流し、標識ペプチドと分解生成物の相対量を測定する方法がある。例えば、もし細菌プロテイナーゼがシステインプロテイナーゼなら、還元剤(例えばDTT)をインキュベーション混合物中に含めることが必要かも知れない。好ましくは2つの測定(1つは、試験物質を含み、1つは試験物質を含まない)を、同一の条件下で行う。次に、2つの測定の間の分解生成物の相対量および/または非分解ペプチドの相対量を比較して、試験物質による分解の阻害程度を決定する。各生成物の相対量は、ホスホリメージャー(phosphorimager)を使用して定量される。好ましくは、プロテイナーゼによるペプチドの分解を阻害する試験物質は、ペプチドの分解を、少なくとも20%、少なくとも30%、少なくとも40%、少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも99%低下させるか、またはペプチドの分解を、物質の濃度0.1mg/ml、1mg/ml、10mg/ml、100mg/ml、500mg/ml、1mg/ml、10mg/ml、100mg/ml、または0.1g/mlで、実質的に妨害する。相互作用の程度と生成物の濃度の上記の低下パーセントの任意の組合せが、本発明の物質を規定するのに使用され、より低濃度でのより大きな分解が好ましい。本発明の好適な物質は、1mg/mlまたは0.5mg/mlの濃度で少なくとも50%レベルの阻害を示すものである。
【0044】
ペプチド分解を監視するための他の適当な方法には、タンパク質染色、免疫ブロッティング、および放射性ヨード化がある。ペプチド分解はまた、機能的測定法、例えば分解産物に残存抗細菌活性があるかどうかを調べる方法により監視される。抗細菌活性は、殺菌測定法を使用して、コロニー形成単位(cfu)を測定することにより決定される。
【0045】
第2の成分の分解はまた、既に本明細書に記載の測定法を使用して、試験物質の存在下および非存在下で、陽イオン性抗微生物ペプチドによる細菌の死滅を監視することにより、間接的に監視される。
【0046】
本発明はまた、陽イオン性抗微生物ペプチド活性を増強する試験物質を同定するためのキットを提供し、このキットは以下を含む:
(i)グリコサミノグリカン;および
(ii)陽イオン性抗微生物ペプチド。
【0047】
キットはさらに以下を含む:
(iii)細菌試料;および/または
(iv)試験物質が、グリコサミノグリカンと陽イオン性抗微生物ペプチドとの相互作用を阻害するかどうかを決定するための手段。
【0048】
本発明はまた、陽イオン性抗微生物ペプチド活性を増強する試験物質を同定するためのキットを提供し、このキットは以下を含む:
(i)プロテオグリカン;
(ii)細菌プロテイナーゼ。
【0049】
キットはさらに以下を含んでよい:
(ii)試験物質が、細菌プロテイナーゼによるプロテオグリカンの分解を阻害するかどうかを決定するための手段。
【0050】
本発明により提供されるキットはまた以下を含む:
(v)または(iv)陽イオン性抗微生物ペプチド活性を増強する試験物質の能力を決定するための測定法を実施するための説明書。
【0051】
「物質」という用語は、単一の物質、および2、3またはそれ以上の物質の組合せを含むことを企図する。例えばこの用語は、単一のペプチド、2つ以上のペプチドの混合物、またはペプチドと規定される化学的物質との混合物を含んでよい。
【0052】
任意の適当な試験物質を本発明の方法で使用することができる。好ましくは試験物質は、真核細胞に対して非毒性である。上記測定法で試験される好適な試験物質には、炭水化物、化学的修飾(例えば、硫酸化および/またはカルボキシル化)を有する炭水化物骨格を含む分子、ペプチド、およびペプチド模倣物を含むことができる。特に好適な試験物質には、放出されたグリコサミノグリカンのスカベンジャーとして機能性する陽イオン性物質、ポリエチレンイミン、プロタミン、ポリブレン、ポリ−L−リジン、ポリ−L−アルギニン、キトサンおよびポリミキシンがある。他の適当な試験物質には、コンビナトリアルライブラリー、規定された化学的物質、オリゴヌクレオチドおよび例えばディスプレイライブラリー(例えば、ファージディスプレイライブラリー)のような天然産物ライブラリーがある。さらに抗体生成物(例えば、モノクローナル抗体およびポリクローナル抗体、1本鎖抗体、キメラ抗体、CDR移植抗体、およびヒト化抗体)、またはこれらの断片が使用される。適当な試験物質の例を表1に示す。
【0053】
【表1】
炭水化物試験物質は、単糖、二糖、オリゴ糖、または多糖でよい。例えばコンドロイチン硫酸のポリマー[GlcA−GalNAc]n またはデルマタン硫酸。好ましくは炭水化物試験物質は、陽イオン性抗微生物ペプチドに、その抗微生物活性に影響を与えることなく結合することができる。
【0055】
ペプチド試験物質は、陽イオン性抗微生物ペプチドの模倣物でもよい。好ましくは模倣物は、グリコサミノグリカンおよび/またはプロテアーゼに結合することができて、グリコサミノグリカンおよび/またはプロテアーゼを封鎖し、こうしてグリコサミノグリカンおよび/またはプロテアーゼが、陽イオン性抗微生物ペプチドと相互作用することを防ぐ。模倣物は、グリコサミノグリカンに結合する陽イオン性抗微生物ペプチドの領域のみでなく、抗微生物活性を有するペプチドの領域も模倣する。試験ペプチドはまた、任意の他のアミノ酸配列を含有してもよい。好ましくはペプチドは、グリコサミノグリカンおよび/またはプロテアーゼと相互作用して、これらが陽イオン性抗微生物ペプチドと結合するのを妨害することを可能にするアミノ酸配列を含む。
【0056】
試験物質は、例えば1回の反応につき10個の物質の最初のスクリーニングで使用され、阻害または活性化を示すこれらのバッチの物質が個々に試験される。試験物質は、1nM〜1000μM、好ましくは1μM〜100μM、または5μM〜50μM、さらに好ましくは1μM〜10μMの濃度で使用される。
【0057】
本発明はまた、治療法によりヒトまたは動物の体を治療する方法において同時にまたは連続的に使用するための、細菌プロテイナーゼ/グリコサミノグリカン防御経路のインヒビターと陽イオン性抗微生物ペプチドを含有する。
【0058】
インヒビターは、グリコサミノグリカンと陽イオン性抗微生物ペプチドとの相互作用を阻止することができ、細菌プロテイナーゼによるプロテオグリカンの分解を阻止することができ、または細菌プロテイナーゼによる陽イオン性抗微生物ペプチドの分解を阻止することができる。グリコサミノグリカンと陽イオン性抗微生物ペプチドとの相互作用を阻止することができるインヒビターは、グリコサミノグリカンに結合して、これが陽イオン性抗微生物ペプチドに結合することを妨害するか、または陽イオン性抗微生物ペプチドに結合して、ペプチドがその抗微生物活性を保持するが、阻害性グリコサミノグリカンには結合できないようにする。
【0059】
好適なインヒビターには、炭水化物、化学的修飾(例えば、硫酸化および/またはカルボキシル化)を有する炭水化物骨格を含む分子、ペプチド、およびペプチド模倣物を含む。他の適当なインヒビターには、コンビナトリアルライブラリー、規定された化学的物質、オリゴヌクレオチドおよび、例えば、ディスプレイライブラリー(例えば、ファージディスプレイライブラリー)のような天然産物ライブラリーがある。
【0060】
インヒビターは、陽イオン性抗微生物ペプチドの模倣物、例えばペプチドでもよい。好ましくは模倣物は、グリコサミノグリカンおよび/またはプロテアーゼに結合することができて、グリコサミノグリカンおよび/またはプロテアーゼを封鎖し、こうしてグリコサミノグリカンおよび/またはプロテアーゼが、陽イオン性抗微生物ペプチドと相互作用することを防ぐ。インヒビターは、グリコサミノグリカンに結合する陽イオン性抗微生物ペプチドの領域のみでなく、抗微生物活性を有するペプチドの領域も模倣する。好ましくは模倣物は、グリコサミノグリカンおよび/またはプロテアーゼと相互作用して、これらが陽イオン性抗微生物ペプチドと結合することを妨害することを可能にするアミノ酸配列を含む。
【0061】
インヒビターは好ましくは、単糖、二糖、オリゴ糖、または多糖である。さらに好ましくはインヒビターは、例えばポリエチレンイミン、プロタミン、ポリブレン、ポリ−L−リジン、ポリ−L−アルギニン、キトサンまたはポリミキシンのような陽イオン性物質である。好ましくはインヒビターは、陽イオン性抗微生物ペプチドに、その抗微生物活性に影響を与えることなく結合することができる。
【0062】
陽イオン性抗微生物ペプチドは、細菌膜を破壊し、こうして細菌を死滅させることができるペプチドである。好ましくは陽イオン性抗微生物ペプチドは、感染部位で白血球および/または上皮細胞により産生される。好ましくは白血球は好中球である。陽イオン性抗微生物ペプチドは好ましくは、デフェンシンまたはLL−37のような陽イオン性抗細菌性ペプチドである。さらに好ましくは陽イオン性抗微生物ペプチドはα−デフェンシンである。本発明の方法での使用のために提供される陽イオン性抗微生物ペプチドは、任意の適当な供給源から得られる。例えば、陽イオン性抗微生物ペプチドは、組換えペプチドでも、陽イオン性抗微生物ペプチドは、白血球または上皮細胞から精製されるか、または陽イオン性抗微生物ペプチドは、白血球、上皮細胞または培養物中で増殖させた組換えペプチドでトランスフェクションした細胞から抽出される増殖培地中に存在してもよい。ペプチドは、インビトロタンパク質合成により産生されてもよい。ペプチドは、天然の配列を含んでも、人工的に変異させた配列を含んでもよい(ただし、変異は、ペプチドが細菌膜を破壊する能力またはグリコサミノグリカンと相互作用する能力に影響を与えないものとする)。
【0063】
本発明はまた、治療法によりヒトまたは動物の体を治療する方法において同時にまたは連続的に使用するための、細菌プロテイナーゼ/グリコサミノグリカン防御経路の2つ以上のインヒビターを含有する生成物を提供する。さらに本発明は、治療法によりヒトまたは動物の体を治療する方法において同時にまたは連続的に使用するための、2つ以上の本発明の物質を含有する生成物を提供する。
【0064】
本発明により同定される物質と本発明の生成物は、治療法によりヒトまたは動物の体を治療する方法において使用される。特にそのような物質および生成物は、急性または慢性の細菌感染症の治療、潰瘍(例えば、慢性静脈潰瘍)の治療、および創傷の治療に使用される。潰瘍と他の創傷は、治癒を遅らせる細菌で感染されることがある。従って細菌の根絶により創傷治癒が促進される。本発明により同定される物質を使用して治療される他の創傷には、熱傷創、目、肺および尿路感染症、および無傷の皮膚の細菌感染症がある。任意の潰瘍(例えば、胃潰瘍)が、本発明の方法により同定される物質により治療される。本発明の物質と生成物はまた、無菌の創傷が細菌に感染されることを防ぐのに使用される。そのような物質と生成物はまた、急性もしくは慢性の感染の治療法、潰瘍の治療、または創傷治癒の促進法で使用される薬剤の製造のために使用できる。急性もしくは慢性の感染患者または潰瘍または創傷の患者の症状は、本発明の物質または生成物を投与することにより改善することができる。治療上有効量の本発明の物質または生成物は、必要とする宿主に投与される。物質は、例えば好中球により放出されるような天然に存在する(内因性)陽イオン性抗微生物ペプチドを増強してもよい。あるいは外因性陽イオン性抗微生物ペプチドと一緒に投与すると、物質は外因性ペプチドの抗微生物活性を増強することがある。
【0065】
本発明の方法により同定される物質は、治療法によりヒトまたは動物の体を治療する方法において、陽イオン性抗微生物ペプチドとともに使用してもよい。特にそのような併用療法は、急性または慢性の細菌感染の治療、潰瘍の治療、および創傷の治療で使用してもよい。物質と陽イオン性抗微生物ペプチドはまた、急性または慢性の感染の治療、潰瘍の治療、または創傷治癒の促進法で使用するための、薬剤の製造に使用される。急性もしくは慢性の感染患者または潰瘍または創傷の患者の症状は、本発明の物質と陽イオン性抗微生物ペプチドの組合せ投与により改善することができる。治療上有効量の本発明の物質と陽イオン性抗微生物ペプチドは、必要とする宿主に投与される。
【0066】
陽イオン性抗微生物ペプチドは、好中球中、生物学的境界(例えば、肺、皮膚、粘膜および尿管)で発現される。抗微生物ペプチド活性を調節する物質は、局所的治療、例えば皮膚または角膜、および内科的使用、例えば腸管、肺および尿管で使用される。
【0067】
上記で概説したスクリーニング法に従って同定され、本発明の投与のための物質は、標準的な薬剤学的に許容される担体および/または賦形剤、例えば薬剤学の分野で一般的なもの[例えばレミントンの薬剤科学(Remington’s Pharmaceutical Sciences)、マックパブリッシング社(Mack Publishing Company)、イースタンペンシルバニア、第17版、1985年(この開示内容は参照することによりその全体が本明細書に組み込まれる)に記載のようなもの]とともに調製される。医薬組成物は、本発明の物質を、陽イオン性抗微生物ペプチドおよび薬剤学的に許容される担体または賦形剤とともに含有してもよい。
【0068】
本発明はまた、細菌プロテアーゼ/グリコサミノグリカン防御経路のインヒビター、陽イオン性抗微生物ペプチド、および薬剤学的に許容される担体または賦形剤を含む医薬組成物を提供する。物質、組成物および生成物は、経腸的または腸管外(例えば、経口、頬、肛門、肺、静脈内、動脈内、筋肉内、腹腔内、局所的)、または他の適当な経路により投与される。
【0069】
物質の、物質と陽イオン性抗微生物ペプチドまたは生成物の治療上有効量のが、患者に投与される。物質の、物質と陽イオン性抗微生物ペプチドまたは生成物の用量は、特に使用される物質に従う種々のパラメータ(治療される患者の年齢、体重、症状;投与経路;および必要な処方)に従って決定される。医師が、特定の患者のための必要な投与経路と用量は決定できるであろう。典型的な1日用量は、物質、物質と陽イオン性抗微生物ペプチドまたは生成物の活性、治療される被験体の体重と症状、変性のタイプと重症度、および投与頻度と経路に従って、約0.1〜50mg/kg体重である。好ましくは1日用量は、5mg〜2gである。
【0070】
物質、インヒビター、およびペプチドは、同時にまたは逐次的に投与される。好ましくは物質またはインヒビターは、ペプチドの投与前に投与される。物質またはインヒビターは、ペプチドの投与前1分〜4時間、好ましくはペプチドの投与前5分〜1時間、ペプチドの投与前10分〜30分に投与される。さらに好ましくは物質またはインヒビターおよびペプチドは、同時に投与される。
【0071】
以下の例は、本発明を例示する。
【0072】
例1
35S−硫酸標識繊維芽細胞培養物の生成物と、細菌培養物からの増殖培地によるこれらの生成物の分解を、3〜12%SDS−ポリアクリルアミドゲル電気泳動(PAGE)により分析した。
【0073】
慢性静脈創傷から最初に得られた緑膿菌(Pseudomonas aeruginosa)とエンテロコッカス・フェカーリス(Enterococcus faecalis)分離株を、トッド−ヘウィット(Todd−Hewitt)培地中で37℃で18時間増殖させた。化膿連鎖球菌(Streptococcus pyogenes)AP1株(40/58)は、ストレプトコッカスの文献と研究のための世界保健機構協力センター(World Health Organization Collaborating Centre for References and Research on Streptococci)(プラハ、チェコ共和国)から得て、C培地(ハーワルト(H. Herwald)、コリン(M. Collin)、ムラー−エスタール(Muller−Esterl)、ビヨルク(L. Bjorck)、J. Exp. Med. 184, 665 (1996))で静止期まで増殖させた。遠心分離して細菌をペレットにし、上清を無菌ろ過した(0.3μm)。放射能標識プロテオグリカンの分解のために、10μlの35S−硫酸代謝標識された分泌された繊維芽細胞生成物(シュミチェン(A. Schmidtchen)、カールステット(I. Carlstedt)、マルムストローム(A. Malmstrom)、フランソン(L−A, Fransson)、Biochem. J. 265, 289 (1990))を、10μlの無菌ろ過細菌上清と、37℃で6時間インキュベートした。10μlのTH培地を対照中に加えた。ストレプトコッカスのシステインプロテイナーゼの活性化のために、10mMのDTTを加えた。
【0074】
結合組織プロテオグリカン(PG)デコリン/ビグリカンとベルシカンは、インビボならびに培養物中でヒト繊維芽細胞の主要な分泌生成物を構成する(イオッツォ(R.V. Iozzo)、Annu. Rev. Biochem., 67, 609 (1998))。3〜12%SDS−PAGEによる分析により、以前の結果が確認され、デコリン(〜100kDa)とベルシカン(〜400kDa)が35S−標識繊維芽細胞培養物の主要な細胞外生成物として同定された(シュミチェン(A. Schmidtchen)、カールステット(I. Carlstedt)、マルムストローム(A. Malmstrom)、フランソン(L−A, Fransson)、Biochem. J. 265, 289 (1990);シュミチェン(A. Schmidtchen)、フランソン(L−A, Fransson)、Biomed. Chrom., 7, 48 (1993))。緑膿菌(P. aeruginosa)とイー・フェカーリス(E. faecalis)培養物からの増殖培地を添加すると、デコリンが広範に分解された。また、還元剤ジチオスレイトール(DTT)を加えると、化膿連鎖球菌(S. pyogenes)細胞外生成物は、デコリンと高分子量プロテオグリカンを分解した。
【0075】
DTTの必要性は、化膿連鎖球菌(S. pyogenes)システインプロテイナーゼが分解に関与していることを示唆する。この仮定は、この作用を完全に阻止した特異的システインプロテイナーゼインヒビター(E64)の作用により確認された。
【0076】
分解された物質は、分子量30〜50kDaであり、既に記載されている(シュミチェン(A. Schmidtchen)、フランソン(L−A, Fransson)、Eur. J. Biochem. 208, 537 (1992))ように、遊離/またはペプチド結合GAG鎖のサイズに対応した。これらの実験は、細菌プロテイナーゼによる繊維芽細胞マトリックス金属プロテアーゼ(MMP)の活性化(バーンズ(E. H. Burns)、マーシエル(A.M. Marciel)、ムッサー(J.M. Musser)、Infect. Immun., 64, 4744 (1996);オカモト(T. Okamoto)ら、J. Biol. Chem., 272, 6059 (1997))の可能性を排除せず、分解されたPGを与えたため、特異的プロテアーゼの調製物を用いて一連の対照実験を行った。
【0077】
緑膿菌(P. aeruginosa)エラスターゼとアルカリ性プロテイナーゼタンパク質を、硫酸アンモニウム沈殿(70%飽和)により調製した。透析(10mMトリス、pH8.0)後、ハイQ陰イオン交換クロマトグラフィー(バイオラッド(Bio−Rad))で、10mMトリス(pH8.0)中0〜1.0M NaClの勾配を使用して、分離を行った。フロースルー画分中にエラスターゼが同定されたが、アルカリ性プロテイナーゼは、〜0.5M NaClで溶出された。
【0078】
化膿連鎖球菌(S. pyogenes)のシステインプロテイナーゼは、以前のプロトコール(ハーワルト(H. Herwald)、コリン(M. Collin)、ムラー−エスタール(Muller−Esterl)、ビヨルク(L. Bjorck)、J. Exp. Med. 184, 665 (1996))に従って精製された。
【0079】
イー・フェカーリス(E. faecalis)株はゲラチナーゼを含有し、上清を直接使用した。緑膿菌(P. aeruginosa)エラスターゼとアルカリ性プロテイナーゼ、化膿連鎖球菌(S. pyogenes)システインプロテイナーゼ、およびイー・フェカーリス(E. faecalis)ゲラチナーゼはすべて、精製したヒト子宮頸デコリンならびに調整培地中の35S−標識繊維芽細胞デコリンを分解した。段階的イオン交換クロマトグラフィー(シュミチェン(A. Schmidtchen)、カールステット(I. Carlstedt)、マルムストローム(A. Malmstrom)、フランソン(L−A, Fransson)、Biochem. J. 265, 289 (1990))(繊維芽細胞由来MMPの分離のため)そして次に放射能標識繊維芽細胞プロテオグリカンのPBS透析、または繊維芽細胞MMPを不活性化するための繊維芽細胞生成物の沸騰は、分解に影響を与えなかった。化膿連鎖球菌(S. pyogenes)システインプロテイナーゼをシステインプロテイナーゼインヒビターE64とプレインキュベーション(〜15分)すると、分解されなかった。
【0080】
別の創傷病原体である霊菌(Proteus mirabilis)もまた、デルマタン硫酸を放出する細胞外プロテアーゼを産生することが証明されている。
【0081】
例2
次に我々は、デルマタン硫酸(DS)、ヒトデコリンのグリコサミノグリカン(GAG)置換基(イオッツォ(R.V. Iozzo)、Annu. Rev. Biochem., 67, 609 (1998))、結合デフェンシン、および他の硫酸化多糖が、相互作用を阻止するかどうかを調べた。
【0082】
これらの実験で使用したグリコサミノグリカンは、デルマタン硫酸(DS)36、DS13、ヘパラン硫酸(HS)3−6、コンドロイチン硫酸(CS)−4およびCS−6である。これらの調製と性状解析は、既に記載されている(ローデン(L. Roden)、ベーカー(J. Baker)、シフォネッリ(J.A. Cifonelli)、マシューズ(M.B. Mathews)、Methods in Enzymology、ギンズバーグ(V. Ginsburg)編(アカデミックプレス(Academic Press)、ニューヨーク、1973年)、第28巻、73〜140頁;フランソン(L.−A. Fransson)、ニーヅスジンスキー(I.A. Nieduszynski)、フェルプス(C.F. Phelps)、シーハン(J.K. Sheehan)、Biochim. Biophys. Acta., 586, 179 (1979) ;フランソン(L.−A. Fransson)、シェーバーグ(I. Sjoberg)、ハブスマーク(B. Havsmark)、Eur. J. Biochem., 106, 59 (1980))。ヘパリンと追加の調製物CS−4(ウシ気管からのCS−A)と6(サメ軟骨からのCS−C)は、シグマ(Sigma)から購入した。デルマタン硫酸(分子量〜500kDa)は、ビーディーエィチ・バイオケミカル(BDH Biochemical)から得た。
【0083】
DS36の放射性ヨード化は、以前のプロトコール(チェング(F Cheng)、ヨシダ(K. Yoshida)、ヘイネガード(D. Heinegard)、フランソン(L.A. Fransson)、Glycobiology, 2, 553 (1992))に従って行った。スロット結合測定法を以下のように行った:α−デフェンシンペプチド1(バッケム(Bachem)、スイス)をニトロセルロース膜(ハイボンド(Hybond)、アマシャム(Amersham))にブロットした。膜を1時間ブロッキング(PBS、pH7.4、0.25%ツイーン20、3%BSA)し、同じ緩衝液中で放射能標識DS(〜20μg/ml)と1時間インキュベートした。結合を競合させるために、非標識多糖を加えた(2mg/ml)。膜を洗浄した(3×10分)(PBS,pH7.4、0.25%ツイーン20)。Bas2000放射性イメージングシステム(フジ(Fuji))を使用して、放射活性を視覚化した。
【0084】
0.2〜5μgのα−デフェンシン1をニトロセルロースにブロットし、放射性ヨード化デルマタン硫酸(DS)とプローブ結合させた。ペプチドに結合したDSと結合は、過剰(〜100倍)の非標識DS(DS36;75%イズロン酸塩[IdoA])ならびにDS13(>95% IdoA)により阻害された。コンドロイチン硫酸(CS)−4ならびにCS−6は、結合を破壊しなかった。CS−4とDSは、主にウロン酸(CS−4中のグルクロン酸とDS中のIdoA)のエピマー化が異なる(フランソン(L.−A. Fransson)、「多糖(The Polysaccharides)」中、アスピナール(G.O. Aspinall)編(アカデミックプレス(Academic Press)、ニューヨーク州、1985年)、第3巻、338〜406頁)。試験したヘパラン硫酸(HS)画分のうち(HS3〜6)、硫酸化とIdoA含量の高いもの(HS5;〜50%IdoA、および〜62%N−硫酸、HS6:65%IdoAおよび72%N−硫酸)のみが、DSを完全に排除することがわかった。さらにヘパリンとデキストラン硫酸(デキストランではない)は、放射能標識DSを排除した。まとめるとこの結果は、GAG−デキストラン相互作用が、多糖のIdoA残基ならびに硫酸化に依存することを示唆する。DSとHS/ヘパリン中のIdoAは、2−O−硫酸化されていることがあり(フランソン(L.−A. Fransson)、「多糖(The Polysaccharides)」中、アスピナール(G.O. Aspinall)編(アカデミックプレス(Academic Press)、ニューヨーク州、1985年)、第3巻、338〜406頁)、α−デフェンシン1への結合のために、DSとHSポリマーの追加の修飾が必要かも知れない。
【0085】
例3
デフェンシン機能へのDSの作用を調べるために、試験微生物である化膿連鎖球菌(S. pyogenes)、イー・フェカーリス(E. faecalis)および緑膿菌(P. aeruginosa)に対するα−デフェンシン1の細菌活性を測定した。
【0086】
抗微生物測定法のために、化膿連鎖球菌(S. pyogenes)、イー・フェカーリス(E. faecalis)および緑膿菌(P. aeruginosa)を、TH培地中で対数中期まで増殖させた。細菌を洗浄し、5mMグルコース含有10mMトリス−塩酸(pH7.5)で希釈した。50μlの細菌(2×106 cfu/ml)を、0〜40μg/mlの濃度のα−デフェンシンと、2〜400μg/mlの濃度の種々のGAG鎖の添加有りまたは無しで、インキュベートした。インキュベーションは、37℃で2時間(化膿連鎖球菌(S. pyogenes)とイー・フェカーリス(E. faecalis))または4時間(緑膿菌(P. aeruginosa))行った。微生物活性を定量するために、このインキュベーション混合物の連続希釈物を、TH寒天に蒔き、37℃で一晩インキュベートし、CFUの数を測定した。
【0087】
細菌株を、0〜40μg/mlのα−デフェンシン1とインキュベートした。緑膿菌(P. aeruginosa)、イー・フェカーリス(E. faecalis)および化膿連鎖球菌(S. pyogenes)は、4μg/mlの濃度またはそれ以上のデフェンシンにより有効に死滅した。20〜40μg/ml(DS/デフェンシンモル比〜0.5〜1)の濃度のDSを添加すると、イー・フェカーリス(E. faecalis)のデフェンシン介在死滅がほとんど完全に逆転し、他の2つの種の死滅が部分的に阻害された(図1B)。400μg/mlのDS(モル比〜10:1)は、デフェンシンの抗微生物作用をほとんど完全に逆転させた。以前の実験は、DSがα−デフェンシン1に結合すること、および結合は〜100倍過剰のHS3ならびに2つのCS型により阻害されることを示したため、我々は、これらのGAGがデフェンシンを阻害する活性が小さいかどうかという問題を調べた。実際、測定法に含めると(試験成分として化膿連鎖球菌(S. pyogenes)を使用して)、デフェンシン介在細菌死滅は、ほんのわずかに阻害された;400μg/mlのHS3、CS−4およびCS−6(シグマ(Sigma))は、それぞれ6、16および2%阻害を示した。
【0088】
例4
LL−37機能へのDSの作用を調べるために、試験微生物である化膿連鎖球菌(S. pyogenes)、イー・フェカーリス(E. faecalis)および緑膿菌(P. aeruginosa)に対するLL−37の細菌活性を測定した。
【0089】
抗微生物測定は、例3に記載のように行った。LL−37の抗細菌活性を例示する結果を図2に示す。0〜40μg/mlのDSの添加は、化膿連鎖球菌(S. pyogenes)、イー・フェカーリス(E. faecalis)および緑膿菌(P. aeruginosa)のLL−37死滅を阻害した。結果を図3に示す。
【0090】
例5
デルマタン硫酸(DS)がLL−37に結合し、および硫酸化多糖が相互作用をブロックするかどうかを調べるために、例2に示すように、スロット結合測定を行った。LL−37(0.2〜5μg)をニトロセルロースに適用し、ヨード化デルマタン硫酸(DS36)とインキュベートした。結合への競合のために、異なる多糖を加えた:DS36、DS18、DS13、CS−4、CS−6、CS−D(コンドロイチン硫酸D)およびCS−E(コンドロイチン硫酸E)。結果を図4に示す。CS−4、CS−6およびCS−DとCS−Dは、結合を破壊しなかった。しかし放射能標識DSは、DS−36、DS18、DS13、およびCS−Eにより排除された。
【0091】
例6
DS−LL−37相互作用への種々の陽イオン性分子の作用を、例2に記載のようにスロット−ブロット測定法を使用して監視した。結果を図5に示す。LL−37へのDSの結合は、ポリ−L−リジン、ポリ−L−アルギニン、DEAE−デキストラン、ポリブレン、キトサンおよびポリエチレンイミンによりブロックされた。結合レベルの低下は、プロタミンの存在下で観察された。ポリミキサンBおよびポリミキシンBノナペプチドは、LL−37へのDSの結合を阻止しなかった。
【0092】
例7
緑膿菌(P. aeruginosa)エラスターゼがLL−37を分解し不活性化することを証明するために、10μgのLL−37を50μlの10mMトリス−塩酸(pH7.5)中30mUの緑膿菌(P. aeruginosa)エラスターゼと、0、1、5、15もしくは30分または1、4もしくは20時間インキュベートした。インキュベーション物の等量のアリコートを、SDS−PAGE(16.5%トリス−トリシンゲル)で分析したかまたは細菌活性について測定した。細菌測定法のために、2×106 cfu/mlのイー・フェカーリス(E. faecalis)を0.1μgのLL−37と、10mMトリス−塩酸(pH7.5)、5mMグルコース中で37℃で2時間インキュベートした。図6に示す結果は、LL−37が緑膿菌(P. aeruginosa)エラスターゼにより分解され不活性化されることを示す。
【0093】
LL−37分解産物を、液体クロマトグラフィー質量スペクトル(LC−MS)、次に飛行時間(Time of flight)(TOF)MS−MSで測定した。断片質量が3708.859、3178.637、2822.450、2326.269および1401.748を与える主要なペプチド断片が検出され、さらに分析した。LL−37の切断点と提唱されている抗細菌領域を図7に示す。
【0094】
3mUのイー・フェカーリス(E. faecalis)ゲラチナーゼまたは3mUの化膿連鎖球菌(S. pyogenes)システインプロテイナーゼによるLL−37の分解を、1μgのLL−37を1時間と6時間インキュベートすることにより監視した。分解産物を16.5%のトリス−トリシンゲルのSDS−PAGEにより分析し、6mUの緑膿菌(P. aeruginosa)エラスターゼと1μgのLL−37を5分と1時間インキュベーション後の分解産物と比較した。結果を図8に示す。
【0095】
例8
1μgのLL−37を12mUの緑膿菌(P. aeruginosa)エラスターゼ、6mUのイー・フェカーリス(E. faecalis)ゲラチナーゼまたは6mUの化膿連鎖球菌(S. pyogenes)プロテアーゼと、インヒビターの存在下と非存在下で1時間インキュベートすることにより、LL−37の分解への種々の物質の作用を測定した。生じた物質をSDS−PAGE(16.5%トリス−トリシンゲル)により分析した。金属プロテアーゼインヒビターGM6001(100μM)と1,10−フェナントロリン(2mM)(これらは両方とも、緑膿菌(P. aeruginosa)エラスターゼとイー・フェカーリス(E. faecalis)ゲラチナーゼを阻害した)またはインヒビターE64(10μM)(これは、化膿連鎖球菌(S. pyogenes)システインプロテイナーゼを阻害した)により、分解は止まった(図9)。
【0096】
DS36とCS−E、および程度は低いがCS−AとCS−Cは、緑膿菌(P. aeruginosa)エラスターゼによるLL−37の分解を阻害した(図10)。構造[ΔUA(2S)−GalNAc(4,6S)](図11)の硫酸化二糖とシュクロースオクタ硫酸(図12)もまた、緑膿菌(P. aeruginosa)エラスターゼによるLL−37の分解を阻害した。
【図面の簡単な説明】
【図1】図1は、デルマタン硫酸(DS)によるα−デフェンシンの殺菌活性の阻止を例示する。図1Aは、α−デフェンシンによる細菌の死滅を例示する。2×106 CFU/mlの化膿連鎖球菌(S. pyogenes)(□)、イー・フェカーリス(E. faecalis)(●)および緑膿菌(P. aeruginosa)(○)を、α−デフェンシンと記載のペプチド濃度で37℃で2時間(化膿連鎖球菌(S. pyogenes))とイー・フェカーリス(E. faecalis))または4時間(緑膿菌(P. aeruginosa))インキュベートし、CFUを測定した。図1Bは、デルマタン硫酸による細菌のデフェンシン介在死滅の阻害を例示する。種々の濃度(0〜400μg/ml)のDSの添加は、化膿連鎖球菌(S. pyogenes)(□)、イー・フェカーリス(E. faecalis)(●)および緑膿菌(P. aeruginosa)(○)のデフェンシン死滅を阻害した。
【図2】図2は、LL−37の殺菌作用を例示する。2×106 コロニー形成単位(cfu)/mlのイー・フェカーリス(E. faecalis))(□)、化膿連鎖球菌(S. pyogenes)AP1(○)、および緑膿菌(P. aeruginosa)(●)を、LL−37と記載のペプチド濃度で37℃で、10mMトリス−塩酸(pH7.5)、5mMグルコース中で、2時間(化膿連鎖球菌(S. pyogenes)とイー・フェカーリス(E. faecalis))または4時間(緑膿菌(P. aeruginosa))インキュベートし、cfuを測定した。
【図3】図3は、デルマタン硫酸(DS)によるLL−37の細菌作用の阻止を例示する。種々の濃度(0〜40μg/ml)のDSの添加は、イー・フェカーリス(E. faecalis)(□)、化膿連鎖球菌(S. pyogenes)AP1(●)、および緑膿菌(P. aeruginosa)(○)のLL−37死滅を阻害した。種々の細菌株の2×106 CFU/mlを、10mMトリス−塩酸(pH7.5)、5mMグルコース中で、10μg/ml(イー・フェカーリス(E. faecalis)と緑膿菌(P. aeruginosa))または2μg/ml(化膿連鎖球菌(S. pyogenes)AP1)のLL−37、およびデルマタン硫酸と、2時間(AP1とイー・フェカーリス(E. faecalis))または4時間(緑膿菌(P. aeruginosa))インキュベートし、cfuを測定した。
【図4】図4は、LL−37へのデルマタン硫酸の結合を示す。LL−37(0.2〜5μg)をニトロセルロースに適用し、ヨード化したデルマタン硫酸(DS36)とインキュベートした。結合に競合させるために、異なる多糖を加えた。DS36、DS18、DS13(デルマタン硫酸36、18および13)、CS4とCS6(コンドロイチン硫酸4と6)、CS−D(コンドロイチン硫酸D)およびCS−E(コンドロイチン硫酸E)。
【図5】図5は、DS−LL−37相互作用に対する種々の陽イオン性分子の作用を示す。LL−37(0.2〜5μg)をニトロセルロースに適用し、ヨード化したデルマタン硫酸(DS36)とインキュベートした。結合に競合させるために、種々のパネルの上に示すように、異なる陽イオン性物質を加えた。(a)p−l−lys;ポリ−L−リジン、p−l−arg;ポリ−L−アルギニン、DEAE;DEAE−デキストラン、PB;ポリブレン。(b)CH;キトサン、PEI、ポリエチレンイミン、PM;プロタミン、PMX;ポリミキシンB、PMXnp;ポリマーミキシンBノナペプチド。
【図6】図6は、緑膿菌(Pseudomonas aeruginosa)エラスターゼがLL−37を分解し不活性化することを証明する実験の結果を示す。LL−37(10μg)を、50μlの10mMトリス−塩酸(pH7.5)中の緑膿菌エラスターゼ(P. aeruginosa)と、種々の時間(x軸上に記載)インキュベートした。次にインキュベート物の等量のアリコートを、SDS−PAGE(16.5%トリス−トリシンゲル;上の図)で分析したか、または殺菌活性(下の図)を測定した。2μgのLL−37に相当する容量を、ゲル上にのせた。分子量マーカーを左に示す(上の図)。殺菌測定(下の図)のために、2×106 cfu/mlのイー・フェカーリス(E. faecalis)を、分解の開始時に0.1μgの無傷のペプチドに相当する量のLL−37とインキュベートした。ペプチドを細菌と、10mMトリス−塩酸(pH7.5)、5mMグルコース中で、37℃で2時間インキュベートした。コロニー形成単位を測定した(左に記載)。
【図7】図7は、LL−37断片の性状解析を示す。LL−37(10μg)を、50μlの10mMトリス−塩酸(pH7.5)中で緑膿菌(P. aeruginosa)エラスターゼ(30mU)と、1、5、15、30分または1時間インキュベートした。生じた分解産物(図6参照)を、液体クロマトグラフィー質量スペクトル(LC−MS)、次に飛行時間(Time of flight)(TOF)MS−MSで測定した。主要なペプチド断片を検出し、さらに分析した。切断点を示し、LL−37の提唱されている抗細菌領域に下線を引いた。
【図8】図8は、細菌プロテイナーゼによるLL−37の分解を示す。LL−37(1μg)を、それぞれ緑膿菌(P. aeruginosa)エラスターゼ(ELA)と5分(1)または1時間(2)インキュベートし、イー・フェカーリス(E. faecalis)ゲラチナーゼ含有培地(GEL)または化膿連鎖球菌(S. pyogenes)システインプロテイナーゼ(CP)と1時間(1)または6時間(2)インキュベートした。加えたプロテイナーゼ活性(アゾカゼイン測定法により測定した)は、10μlの反応物当たり6mU(緑膿菌(P. aeruginosa)エラスターゼ)および3mU(他のプロテイナーゼ)に相当した。物質をSDS−PAGEで16.5%トリス−トリシンゲルで分析した。分子量マーカーを左に示す。
【図9】図9は、プロテイナーゼインヒビターによるLL−37分解の阻害を示す。LL−37(1μg)を、緑膿菌(P. aeruginosa)エラスターゼ(ELA、12mU)、イー・フェカーリス(E. faecalis)ゲラチナーゼ(GEL、6mU)または化膿連鎖球菌(S. pyogenes)システインプロテイナーゼ(CP、6mU)と、インヒビターの非存在下または存在下で1時間インキュベートした。GM6001;100μM GM6001、PHE;2mM 1,10−フェナトロリン、E64;10μM E64。物質をSDS−PAGE(16.5%トリス−トリシンゲル)で分析した。分子量マーカー(kDa)を左に示す。
【図10】図10は、LL−37の分解が、硫酸化グリコサミノグリカンにより阻害されることを示す。LL−37(1μg)を、緑膿菌(P. aeruginosa)エラスターゼ(12mU)と総量15μl(10mMトリス緩衝液)中で、記載のグリコサミノグリカンの非存在下(ELA)または存在下で、1時間インキュベートした。DS36、デルマタン硫酸36(CS−Bとも記載する)、CS−A;コンドロイチン硫酸A、CS−C、コンドロイチン硫酸C、CS−E;コンドロイチン硫酸E。物質をSDS−PAGEで16.5%トリス−トリシンゲルで分析した。分子量マーカー(kDa)を左に示す。
【図11】図11は、LL−37の分解が硫酸化二糖により阻害されることを示す。LL−37(1μg)を、緑膿菌(P. aeruginosa)エラスターゼ(12mU)と総量15μl(10mMトリス緩衝液)中で、硫酸化二糖の非存在下または存在下で、1時間インキュベートした。ELA;緑膿菌(P. aeruginosa)エラスターゼ、0S,UA−GalNAc;2S−4S;UA(2S)−GalNAc(4S)、2S−6S;UA(2S)−GalNAc(6S)、4,6diS;UA−GalNAc(4,6S)、2S−4,6diS;UA(2S)−GalNAc(4,6S)。物質をSDS−PAGEで16.5%トリス−トリシンゲルで分析した。分子量マーカー(kDa)を左に示す。
【図12】図12は、LL−37の分解がシュクロースオクタ硫酸により阻害されることを示す。IL−3(1μg)を、緑膿菌(P. aeruginosa)エラスターゼ(12mU;ELA)と総量15μl(10mMトリス緩衝液)中で、記載の量(ナノモル)のショ糖(S)またはシュクロースオクタ硫酸(SOS)の非存在下または存在下で、1時間インキュベートした。物質をSDS−PAGEで16.5%トリス−トリシンゲルで分析した。分子量マーカー(kDa)を左に示す。
Claims (41)
- グリコサミノグリカンと陽イオン性抗微生物ペプチドとの相互作用を阻害することができる物質を同定する方法であって:
(i)第1の成分として、グリコサミノグリカンを提供し;
(ii)第2の成分として、陽イオン性抗微生物ペプチドを提供し;
(iii)第1および第2の成分を、試験物質の非存在下で第1および第2の成分が相互作用することを可能にする条件下で、試験物質に接触させ;そして
(iv)第1の成分と第2の成分の相互作用を監視して、こうして試験物質が第1の成分と第2の成分の相互作用を破壊することができるかどうかを決定する、ことを含む上記方法。 - 陽イオン性抗微生物ペプチドのグリコサミノグリカン介在阻害を阻止することができる物質を同定する方法であって:
(i)第1の成分として、グリコサミノグリカンを提供し;
(ii)第2の成分として、陽イオン性抗微生物ペプチドを提供し;
(iii)第3の成分として、細菌培養物を提供し;
(iv)第1、第2、および第3の成分を、第1の成分の非存在下で第2の成分による細菌培養物の死滅を可能にし、試験物質の非存在下で第2の成分による細菌培養物の死滅の、第1の成分による阻害を可能にする条件下で、試験物質に接触させ;そして
(v)細菌培養物の生存を監視し、こうして試験物質が陽イオン性抗微生物ペプチドのグリコサミノグリカン介在阻害を妨害することができるかどうかを決定する、ことを含む上記方法。 - 陽イオン性抗微生物ペプチドはデフェンシンである、請求項1または2の方法。
- デフェンシンはα−デフェンシンである、請求項3の方法。
- 陽イオン性抗微生物ペプチドはLL−37である、請求項1または2の方法。
- グリコサミノグリカンはイズロン酸塩含量が多い、前記請求項のいずれかの方法。
- グリコサミノグリカンは高度の硫酸化を有する、前記請求項のいずれかの方法。
- グリコサミノグリカンはデルマタン硫酸である、前記請求項のいずれかの方法。
- 細菌培養物は、プロテオグリカンの豊富な結合組織中で増殖することができる細菌株を含む、請求項2の方法。
- 細菌株は、緑膿菌(Pseudomonas aeruginosa)、エンテロコッカス・フェカーリス(Enterococcus faecalis)、化膿連鎖球菌(Streptococcus pyogenes)または霊菌(Proteus mirabilis)から選択される、請求項9の方法。
- 細菌プロテイナーゼによるプロテオグリカンの分解を阻害することができる物質を同定する方法であって:
(i)第1の成分として、細菌プロテイナーゼを提供し;
(ii)第2の成分として、プロテオグリカンを提供し;
(iii)第1および第2の成分を、試験物質の非存在下で第1の成分による第2の成分の分解を可能にする条件下で、試験物質に接触させ;そして
(iv)第2の成分の分解を監視して、こうして試験物質が、プロテイナーゼによるプロテオグリカンの分解を阻害することができるかどうかを決定する、ことを含む上記方法。 - プロテオグリカンはデルマタン硫酸を含む、請求項11の方法。
- プロテオグリカンはデコリンまたはベルシカンである、請求項11または12の方法。
- 工程(iv)においてプロテオグリカンからのデルマタン硫酸の放出が監視される、請求項12または13の方法。
- 細菌プロテイナーゼによる陽イオン性抗微生物ペプチドの分解を阻害することができる物質を同定する方法であって:
(i)第1の成分として、細菌プロテイナーゼを提供し;
(ii)第2の成分として、陽イオン性抗微生物ペプチドを提供し;
(iii)第1および第2の成分を、試験物質の非存在下で第1の成分による第2の成分の分解を可能にする条件下で、試験物質に接触させ;そして
(iv)第2の成分の分解を監視して、こうして試験物質が、プロテイナーゼによる陽イオン性抗微生物ペプチドの分解を阻害することができるかどうかを決定する、ことを含む上記方法。 - 陽イオン性抗微生物ペプチドは請求項3〜5のいずれか1項に記載に定義されたものである、請求項15の方法。
- 細菌プロテイナーゼは、請求項9または10で定義された細菌株から得られる、請求項11〜16のいずれか1項に記載の方法。
- 細菌プロテイナーゼは、緑膿菌(Pseudomonas aeruginosa)エラスターゼ、緑膿菌(Pseudomonas aeruginosa)アルカリ性プロテイナーゼ、エンテロコッカス・フェカーリス(Enterococcus faecalis)ゲラチナーゼ、および化膿連鎖球菌(Streptococcus pyogenes)システインプロテイナーゼから選択される、請求項11〜17のいずれか1項に記載の方法。
- 細菌プロテイナーゼは、細菌培養物から抽出される増殖培地中に存在する、請求項11〜18のいずれか1項に記載の方法。
- グリコサミノグリカンと陽イオン性抗微生物ペプチドとの相互作用を阻害することができる物質の同定に使用するためのキットであって:
(a)グリコサミノグリカン;と
(b)陽イオン性抗微生物ペプチド、
とを含むキット。 - (c)試験物質が、グリコサミノグリカンと陽イオン性抗微生物ペプチドとの相互作用を阻害するかどうかを決定するための手段、
をさらに含む請求項20のキット。 - (d)細菌試料、
をさらに含む請求項20または21のキット。 - 細菌プロテイナーゼによるプロテオグリカンの分解を阻害することができる物質の同定に使用するためのキットであって:
(a)プロテオグリカン;と
(b)細菌プロテイナーゼ、
とを含むキット; - (c)試験物質が、細菌プロテイナーゼによるプロテオグリカンの分解を阻害するかどうかを決定するための手段、
をさらに含む請求項23のキット。 - 前記請求項のいずれか1項に記載の方法により同定される物質。
- 単糖、二糖、オリゴ糖、または多糖である、請求項25の物質。
- 陽イオン性物質である、請求項25の物質。
- 陽イオン性物質は、ポリ−L−リジン、ポリ−L−アルギニン、DEAE−デキストラン、ポリブレン、キトサン、ポリエチレンイミンまたはプロタミンである、請求項27の物質。
- 請求項25〜28のいずれか1項に記載の物質と薬剤学的に許容される担体または希釈剤とを含む、医薬組成物。
- 陽イオン性抗微生物ペプチドをさらに含む、請求項29の医薬組成物。
- 治療法によりヒトまたは動物の体を治療する方法において使用するための、請求項25〜28のいずれか1項に記載の物質または請求項29または30の医薬組成物。
- 治療法によりヒトまたは動物の体を治療する方法において、別々に、同時に、または連続的に使用するための、請求項25〜28のいずれか1項に記載の物質および陽イオン性抗微生物ペプチドとを含む、生成物。
- 治療法によりヒトまたは動物の体を治療する方法において、同時にまたは連続的に使用するための、細菌の防御機構と陽イオン性抗微生物ペプチドのインヒビターとを含む生成物であって、ここで該防御機構は、細菌プロテイナーゼによりプロテオグリカンを分解して、陽イオン性抗微生物ペプチドに結合して該ペプチドの抗微生物活性を阻害することができるグリコサミノグリカンを放出する、上記生成物。
- グリコサミノグリカンは、細菌プロテイナーゼによるプロテオグリカンの分解により放出される、請求項33の生成物。
- インヒビターは、グリコサミノグリカンと陽イオン性抗微生物ペプチドとの相互作用を阻止することができる、請求項33または34の生成物。
- インヒビターは、細菌プロテイナーゼによるプロテオグリカンの分解を阻止することができる、請求項33または34の生成物。
- インヒビターは、細菌プロテイナーゼによる陽イオン性抗微生物ペプチドの分解を阻止することができる、請求項33または34の生成物。
- 慢性または急性の細菌感染を治療する方法で使用するための医薬の製造における、請求項25〜28のいずれか1項に記載の物質、請求項29または30に記載の医薬組成物、または請求項32〜36のいずれか1項に記載の生成物の使用。
- 方法は、潰瘍を治療する方法である、請求項38の物質の使用。
- 方法は、創傷治癒を促進する方法である、請求項38の使用。
- 慢性または急性の細菌感染を治療する方法であって:
(i)細菌による抗微生物ペプチドの不活性化を阻害する物質の同定、
(ii)必要な患者への治療上有効量の物質の投与、
を含む上記方法。
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| EP00306074 | 2000-07-17 | ||
| PCT/EP2001/008208 WO2002006821A2 (en) | 2000-07-17 | 2001-07-17 | Antimicrobial agent |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| JP2004504603A true JP2004504603A (ja) | 2004-02-12 |
Family
ID=8173129
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2002512679A Pending JP2004504603A (ja) | 2000-07-17 | 2001-07-17 | 抗微生物剤 |
Country Status (7)
| Country | Link |
|---|---|
| US (1) | US7335355B2 (ja) |
| EP (1) | EP1301797A2 (ja) |
| JP (1) | JP2004504603A (ja) |
| AU (1) | AU2001287628A1 (ja) |
| CA (1) | CA2418214A1 (ja) |
| WO (1) | WO2002006821A2 (ja) |
| ZA (1) | ZA200300013B (ja) |
Cited By (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2006502395A (ja) * | 2002-10-10 | 2006-01-19 | マブテック アーベー | 診断方法 |
| JP2015225036A (ja) * | 2014-05-29 | 2015-12-14 | 花王株式会社 | 抗菌性向上物質のスクリーニング方法 |
Families Citing this family (8)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| SE0300207D0 (sv) | 2003-01-29 | 2003-01-29 | Karolinska Innovations Ab | New use and composition |
| US8124370B2 (en) | 2003-01-31 | 2012-02-28 | Systagenix Wound Management (Us), Inc. | Cationic anti-microbial peptides and methods of use thereof |
| ATE432358T1 (de) * | 2003-11-03 | 2009-06-15 | Ethicon Inc | Kolorimetrische substrate, kolorimetrische sensoren und verwendungsverfahren |
| CA2848653A1 (en) | 2006-06-02 | 2007-12-13 | Synedgen, Inc. | Chitosan-lysine derivative compounds |
| US20080248508A1 (en) * | 2006-08-17 | 2008-10-09 | Shenda Baker | Methods of making a chitosan product having an ultra-low endotoxin concentration and the ultra-low endotoxin chitosan product derived therefrom and method of accurately determining inflammatory and anti-inflammatory cellular response to such materials |
| WO2009046851A1 (en) * | 2007-09-11 | 2009-04-16 | Mondobiotech Laboratories Ag | Cgrp as a therapeutic agent |
| EP2637682A4 (en) * | 2010-11-10 | 2014-05-14 | Univ Western Ontario | METHOD AND COMPOSITIONS WITH CYCLIC ANALOGUE OF HISTATIN-5 FOR THE TREATMENT OF WOUNDS |
| DE102011077052A1 (de) | 2011-06-07 | 2012-12-13 | Beiersdorf Ag | Verwendung von epsilon-Poly-L-Lysin zur Erhöhung der Bioverfügbarkeit antimikrobieller Peptide in menschlicher Haut. |
Family Cites Families (8)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| AU561066B2 (en) | 1982-03-22 | 1987-04-30 | Biocarb Ab | Antibacterial composition containing an oligosaccharide |
| US5166331A (en) * | 1983-10-10 | 1992-11-24 | Fidia, S.P.A. | Hyaluronics acid fractions, methods for the preparation thereof, and pharmaceutical compositions containing same |
| US4956350A (en) | 1988-08-18 | 1990-09-11 | Minnesota Mining And Manufacturing Company | Wound filling compositions |
| US5194585A (en) * | 1989-04-25 | 1993-03-16 | Igen, Inc. | Inhibitors of catalytic antibodies |
| US5338543A (en) * | 1992-02-27 | 1994-08-16 | Ambico, Inc. | Thimerosal inactivated mycoplasma hyopneumoniae vaccine |
| AU4993893A (en) * | 1992-07-31 | 1994-03-03 | Neose Pharmaceuticals, Inc. | Compositions for treating and inhibiting gastric and duodenal ulcers |
| JP3186607B2 (ja) * | 1996-11-08 | 2001-07-11 | 株式会社村田製作所 | 分布定数線路型フィルタ |
| US6017545A (en) * | 1998-02-10 | 2000-01-25 | Modi; Pankaj | Mixed micellar delivery system and method of preparation |
-
2001
- 2001-07-17 CA CA002418214A patent/CA2418214A1/en not_active Abandoned
- 2001-07-17 WO PCT/EP2001/008208 patent/WO2002006821A2/en active Application Filing
- 2001-07-17 AU AU2001287628A patent/AU2001287628A1/en not_active Abandoned
- 2001-07-17 JP JP2002512679A patent/JP2004504603A/ja active Pending
- 2001-07-17 US US10/333,319 patent/US7335355B2/en not_active Expired - Fee Related
- 2001-07-17 EP EP01967187A patent/EP1301797A2/en not_active Withdrawn
-
2003
- 2003-01-02 ZA ZA200300013A patent/ZA200300013B/en unknown
Cited By (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2006502395A (ja) * | 2002-10-10 | 2006-01-19 | マブテック アーベー | 診断方法 |
| JP4874545B2 (ja) * | 2002-10-10 | 2012-02-15 | マブテック アーベー | 口腔感染に対する被検体の感受性を決定する方法 |
| JP2015225036A (ja) * | 2014-05-29 | 2015-12-14 | 花王株式会社 | 抗菌性向上物質のスクリーニング方法 |
Also Published As
| Publication number | Publication date |
|---|---|
| WO2002006821A3 (en) | 2002-06-27 |
| US7335355B2 (en) | 2008-02-26 |
| US20040028672A1 (en) | 2004-02-12 |
| AU2001287628A1 (en) | 2002-01-30 |
| ZA200300013B (en) | 2004-03-24 |
| CA2418214A1 (en) | 2002-01-24 |
| EP1301797A2 (en) | 2003-04-16 |
| WO2002006821A2 (en) | 2002-01-24 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| Yamada et al. | Potential therapeutic application of chondroitin sulfate/dermatan sulfate | |
| JPH09508359A (ja) | 抗グラム陽性細菌学的方法および物質 | |
| US5837678A (en) | Therapeutic uses of bactericidal/permeability increasing protein products | |
| Petrosillo et al. | Colistin monotherapy vs. combination therapy: evidence from microbiological, animal and clinical studies | |
| US5807818A (en) | Therapeutic uses of bactericidal/permeability increasing protein (BPI) protein products | |
| JPH09502987A (ja) | 殺菌性/浸透性が向上した(bpi)タンパク質生成物および抗生物質の投与によるグラム陰性細菌感染の治療方法 | |
| AU4393600A (en) | Novel indications of mannan-binding lectin (mbl) in the treatment of immunocompromised individuals | |
| CN115920000A (zh) | 补体活性的调节剂 | |
| US7335355B2 (en) | Antimicrobial agent | |
| NZ237202A (en) | Composition containing beta-lactam antibiotic and cationic oligopeptide | |
| Hayashida et al. | 2-O-sulfated domains in syndecan-1 heparan sulfate inhibit neutrophil cathelicidin and promote Staphylococcus aureus corneal infection | |
| Żelechowska et al. | Endogenous antimicrobial factors in the treatment of infectious diseases | |
| JP2014510907A (ja) | 診断方法及び治療方法 | |
| US6245751B1 (en) | Methods for the treatment of apolipoprotein E related diseases | |
| Gervais et al. | Proteoglycans and amyloidogenic proteins in peripheral amyloidosis | |
| Weiss III et al. | Bactericidal and endotoxin neutralizing activity of a peptide derived from Limulus antilipopolysaccharide factor | |
| Henneke et al. | SIRS and group-B streptococcal sepsis in newborns: pathogenesis and perspectives in adjunctive therapy | |
| JP6713148B2 (ja) | 腹膜炎の治療のための組成物 | |
| Miklossy | Chronic inflammation and amyloidogenesis in Alzheimer’s disease: Putative role of bacterial peptidoglycan, a potent inflammatory and amyloidogenic factor | |
| CN114025783A (zh) | 治疗和预防骨和关节感染的方法 | |
| US20150072922A1 (en) | Rnase 7 antimicrobial peptides | |
| JP7237085B2 (ja) | 凝固異常を伴う敗血症の治療及び/又は改善のための医薬 | |
| CA2368937A1 (en) | Prevention of brain damage in stroke | |
| CN1899603B (zh) | 甘露聚糖-结合型凝集素(mbl)在免疫受损个体的治疗中的新意义 | |
| Popovych et al. | A Pragmatic, Individually Randomised, Open-Label, Retrospective Study of The Efficacy of Topical Therapy with Framycetin in The Treatment of Exacerbation of Chronic Nasopharyngitis |