JP2004254697A - 殺菌/浸透性が向上した安定なタンパク質生成物およびそれを含む薬剤組成物 - Google Patents
殺菌/浸透性が向上した安定なタンパク質生成物およびそれを含む薬剤組成物 Download PDFInfo
- Publication number
- JP2004254697A JP2004254697A JP2004108131A JP2004108131A JP2004254697A JP 2004254697 A JP2004254697 A JP 2004254697A JP 2004108131 A JP2004108131 A JP 2004108131A JP 2004108131 A JP2004108131 A JP 2004108131A JP 2004254697 A JP2004254697 A JP 2004254697A
- Authority
- JP
- Japan
- Prior art keywords
- rbpi
- product
- bpi
- cells
- fragment
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Withdrawn
Links
- 239000008194 pharmaceutical composition Substances 0.000 title description 10
- 102100033735 Bactericidal permeability-increasing protein Human genes 0.000 title 1
- 101000871785 Homo sapiens Bactericidal permeability-increasing protein Proteins 0.000 title 1
- 239000012634 fragment Substances 0.000 claims abstract description 87
- 235000001014 amino acid Nutrition 0.000 claims description 37
- 150000001413 amino acids Chemical class 0.000 claims description 27
- 125000000151 cysteine group Chemical group N[C@@H](CS)C(=O)* 0.000 claims description 26
- 230000000844 anti-bacterial effect Effects 0.000 claims description 23
- 108090000765 processed proteins & peptides Proteins 0.000 claims description 20
- 229920001184 polypeptide Polymers 0.000 claims description 19
- 102000004196 processed proteins & peptides Human genes 0.000 claims description 19
- 230000000149 penetrating effect Effects 0.000 claims description 2
- 108020004414 DNA Proteins 0.000 abstract description 73
- 108090000623 proteins and genes Proteins 0.000 abstract description 71
- 102000004169 proteins and genes Human genes 0.000 abstract description 55
- 235000018102 proteins Nutrition 0.000 abstract description 52
- 239000013612 plasmid Substances 0.000 abstract description 44
- 235000018417 cysteine Nutrition 0.000 abstract description 28
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 abstract description 26
- 239000000539 dimer Substances 0.000 abstract description 20
- 230000015572 biosynthetic process Effects 0.000 abstract description 19
- 238000000034 method Methods 0.000 abstract description 16
- 239000000203 mixture Substances 0.000 abstract description 11
- 230000035699 permeability Effects 0.000 abstract description 10
- 238000002360 preparation method Methods 0.000 abstract description 7
- 108091028043 Nucleic acid sequence Proteins 0.000 abstract description 6
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 abstract description 5
- 125000001909 leucine group Chemical group [H]N(*)C(C(*)=O)C([H])([H])C(C([H])([H])[H])C([H])([H])[H] 0.000 abstract description 5
- 239000013600 plasmid vector Substances 0.000 abstract description 4
- 230000002401 inhibitory effect Effects 0.000 abstract 1
- 230000001954 sterilising effect Effects 0.000 abstract 1
- 238000004659 sterilization and disinfection Methods 0.000 abstract 1
- 239000000047 product Substances 0.000 description 144
- 210000004027 cell Anatomy 0.000 description 130
- 125000003295 alanine group Chemical group N[C@@H](C)C(=O)* 0.000 description 70
- 239000002609 medium Substances 0.000 description 37
- 229940024606 amino acid Drugs 0.000 description 34
- 239000013598 vector Substances 0.000 description 34
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 32
- 230000014509 gene expression Effects 0.000 description 28
- 108020004705 Codon Proteins 0.000 description 25
- 238000004458 analytical method Methods 0.000 description 25
- 239000002158 endotoxin Substances 0.000 description 24
- 238000004519 manufacturing process Methods 0.000 description 24
- 239000003550 marker Substances 0.000 description 24
- 229920006008 lipopolysaccharide Polymers 0.000 description 20
- 238000002965 ELISA Methods 0.000 description 18
- LFQSCWFLJHTTHZ-UHFFFAOYSA-N Ethanol Chemical compound CCO LFQSCWFLJHTTHZ-UHFFFAOYSA-N 0.000 description 18
- 230000009466 transformation Effects 0.000 description 18
- 102000052586 bactericidal permeability increasing protein Human genes 0.000 description 17
- 108010032816 bactericidal permeability increasing protein Proteins 0.000 description 17
- 239000000523 sample Substances 0.000 description 16
- 239000011780 sodium chloride Substances 0.000 description 16
- 238000004520 electroporation Methods 0.000 description 15
- HPNSFSBZBAHARI-RUDMXATFSA-N mycophenolic acid Chemical compound OC1=C(C\C=C(/C)CCC(O)=O)C(OC)=C(C)C2=C1C(=O)OC2 HPNSFSBZBAHARI-RUDMXATFSA-N 0.000 description 15
- 239000006228 supernatant Substances 0.000 description 15
- LRFVTYWOQMYALW-UHFFFAOYSA-N 9H-xanthine Chemical compound O=C1NC(=O)NC2=C1NC=N2 LRFVTYWOQMYALW-UHFFFAOYSA-N 0.000 description 14
- 108091003079 Bovine Serum Albumin Proteins 0.000 description 14
- 235000004279 alanine Nutrition 0.000 description 14
- 230000027455 binding Effects 0.000 description 14
- 238000010276 construction Methods 0.000 description 14
- HPNSFSBZBAHARI-UHFFFAOYSA-N micophenolic acid Natural products OC1=C(CC=C(C)CCC(O)=O)C(OC)=C(C)C2=C1C(=O)OC2 HPNSFSBZBAHARI-UHFFFAOYSA-N 0.000 description 14
- 229960000951 mycophenolic acid Drugs 0.000 description 14
- 241000588724 Escherichia coli Species 0.000 description 13
- 125000003275 alpha amino acid group Chemical group 0.000 description 13
- 241000699666 Mus <mouse, genus> Species 0.000 description 12
- 230000014621 translational initiation Effects 0.000 description 12
- MTCFGRXMJLQNBG-REOHCLBHSA-N (2S)-2-Amino-3-hydroxypropansäure Chemical compound OC[C@H](N)C(O)=O MTCFGRXMJLQNBG-REOHCLBHSA-N 0.000 description 11
- 239000006144 Dulbecco’s modified Eagle's medium Substances 0.000 description 11
- 238000000338 in vitro Methods 0.000 description 11
- 238000013519 translation Methods 0.000 description 11
- 229920002684 Sepharose Polymers 0.000 description 10
- 108010022394 Threonine synthase Proteins 0.000 description 10
- 239000002299 complementary DNA Substances 0.000 description 10
- 102000004419 dihydrofolate reductase Human genes 0.000 description 10
- 229940079593 drug Drugs 0.000 description 10
- 239000003814 drug Substances 0.000 description 10
- 239000012091 fetal bovine serum Substances 0.000 description 10
- 239000002245 particle Substances 0.000 description 10
- 238000003860 storage Methods 0.000 description 10
- 239000004094 surface-active agent Substances 0.000 description 10
- 230000014616 translation Effects 0.000 description 10
- 241000701024 Human betaherpesvirus 5 Species 0.000 description 9
- RVGRUAULSDPKGF-UHFFFAOYSA-N Poloxamer Chemical compound C1CO1.CC1CO1 RVGRUAULSDPKGF-UHFFFAOYSA-N 0.000 description 9
- 108091036066 Three prime untranslated region Proteins 0.000 description 9
- 210000004899 c-terminal region Anatomy 0.000 description 9
- 230000012010 growth Effects 0.000 description 9
- 125000000729 N-terminal amino-acid group Chemical group 0.000 description 8
- 108010076504 Protein Sorting Signals Proteins 0.000 description 8
- 230000029087 digestion Effects 0.000 description 8
- 102000037865 fusion proteins Human genes 0.000 description 8
- 108020001507 fusion proteins Proteins 0.000 description 8
- 239000001963 growth medium Substances 0.000 description 8
- 238000000746 purification Methods 0.000 description 8
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 8
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 description 7
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 7
- 230000010261 cell growth Effects 0.000 description 7
- 230000000694 effects Effects 0.000 description 7
- 238000002330 electrospray ionisation mass spectrometry Methods 0.000 description 7
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Natural products OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 description 7
- 238000004128 high performance liquid chromatography Methods 0.000 description 7
- 230000008488 polyadenylation Effects 0.000 description 7
- 229920000136 polysorbate Polymers 0.000 description 7
- 238000004007 reversed phase HPLC Methods 0.000 description 7
- 230000035897 transcription Effects 0.000 description 7
- 238000013518 transcription Methods 0.000 description 7
- 230000005030 transcription termination Effects 0.000 description 7
- 229940075420 xanthine Drugs 0.000 description 7
- WEVYAHXRMPXWCK-UHFFFAOYSA-N Acetonitrile Chemical compound CC#N WEVYAHXRMPXWCK-UHFFFAOYSA-N 0.000 description 6
- 108091026890 Coding region Proteins 0.000 description 6
- IAZDPXIOMUYVGZ-UHFFFAOYSA-N Dimethylsulphoxide Chemical compound CS(C)=O IAZDPXIOMUYVGZ-UHFFFAOYSA-N 0.000 description 6
- ZQISRDCJNBUVMM-UHFFFAOYSA-N L-Histidinol Natural products OCC(N)CC1=CN=CN1 ZQISRDCJNBUVMM-UHFFFAOYSA-N 0.000 description 6
- ZQISRDCJNBUVMM-YFKPBYRVSA-N L-histidinol Chemical compound OC[C@@H](N)CC1=CNC=N1 ZQISRDCJNBUVMM-YFKPBYRVSA-N 0.000 description 6
- 238000002835 absorbance Methods 0.000 description 6
- 239000011324 bead Substances 0.000 description 6
- YTRQFSDWAXHJCC-UHFFFAOYSA-N chloroform;phenol Chemical compound ClC(Cl)Cl.OC1=CC=CC=C1 YTRQFSDWAXHJCC-UHFFFAOYSA-N 0.000 description 6
- 230000005764 inhibitory process Effects 0.000 description 6
- 239000012528 membrane Substances 0.000 description 6
- 229920001983 poloxamer Polymers 0.000 description 6
- 229960000502 poloxamer Drugs 0.000 description 6
- UCSJYZPVAKXKNQ-HZYVHMACSA-N streptomycin Chemical compound CN[C@H]1[C@H](O)[C@@H](O)[C@H](CO)O[C@H]1O[C@@H]1[C@](C=O)(O)[C@H](C)O[C@H]1O[C@@H]1[C@@H](NC(N)=N)[C@H](O)[C@@H](NC(N)=N)[C@H](O)[C@H]1O UCSJYZPVAKXKNQ-HZYVHMACSA-N 0.000 description 6
- 238000006467 substitution reaction Methods 0.000 description 6
- VMHLLURERBWHNL-UHFFFAOYSA-M Sodium acetate Chemical compound [Na+].CC([O-])=O VMHLLURERBWHNL-UHFFFAOYSA-M 0.000 description 5
- 239000000872 buffer Substances 0.000 description 5
- 230000021615 conjugation Effects 0.000 description 5
- 239000013604 expression vector Substances 0.000 description 5
- 238000000855 fermentation Methods 0.000 description 5
- 230000004151 fermentation Effects 0.000 description 5
- 239000002773 nucleotide Substances 0.000 description 5
- 125000003729 nucleotide group Chemical group 0.000 description 5
- -1 oxygen side-chain polysaccharide Chemical class 0.000 description 5
- 229950008882 polysorbate Drugs 0.000 description 5
- 239000001632 sodium acetate Substances 0.000 description 5
- 235000017281 sodium acetate Nutrition 0.000 description 5
- KIUMMUBSPKGMOY-UHFFFAOYSA-N 3,3'-Dithiobis(6-nitrobenzoic acid) Chemical compound C1=C([N+]([O-])=O)C(C(=O)O)=CC(SSC=2C=C(C(=CC=2)[N+]([O-])=O)C(O)=O)=C1 KIUMMUBSPKGMOY-UHFFFAOYSA-N 0.000 description 4
- 102000006496 Immunoglobulin Heavy Chains Human genes 0.000 description 4
- 108010019476 Immunoglobulin Heavy Chains Proteins 0.000 description 4
- TWRXJAOTZQYOKJ-UHFFFAOYSA-L Magnesium chloride Chemical compound [Mg+2].[Cl-].[Cl-] TWRXJAOTZQYOKJ-UHFFFAOYSA-L 0.000 description 4
- 125000001429 N-terminal alpha-amino-acid group Chemical group 0.000 description 4
- 229940098773 bovine serum albumin Drugs 0.000 description 4
- 238000005277 cation exchange chromatography Methods 0.000 description 4
- 239000007979 citrate buffer Substances 0.000 description 4
- 239000000499 gel Substances 0.000 description 4
- 238000011534 incubation Methods 0.000 description 4
- 230000014759 maintenance of location Effects 0.000 description 4
- 210000004962 mammalian cell Anatomy 0.000 description 4
- 229960004452 methionine Drugs 0.000 description 4
- 230000035772 mutation Effects 0.000 description 4
- 210000004898 n-terminal fragment Anatomy 0.000 description 4
- 239000002777 nucleoside Substances 0.000 description 4
- 125000003835 nucleoside group Chemical group 0.000 description 4
- 235000010482 polyoxyethylene sorbitan monooleate Nutrition 0.000 description 4
- 229920000053 polysorbate 80 Polymers 0.000 description 4
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 4
- 230000003362 replicative effect Effects 0.000 description 4
- 230000028327 secretion Effects 0.000 description 4
- 239000000243 solution Substances 0.000 description 4
- 241000894007 species Species 0.000 description 4
- 241000894006 Bacteria Species 0.000 description 3
- 102000016928 DNA-directed DNA polymerase Human genes 0.000 description 3
- 108010014303 DNA-directed DNA polymerase Proteins 0.000 description 3
- 241000238631 Hexapoda Species 0.000 description 3
- 241000282412 Homo Species 0.000 description 3
- FFEARJCKVFRZRR-BYPYZUCNSA-N L-methionine Chemical compound CSCC[C@H](N)C(O)=O FFEARJCKVFRZRR-BYPYZUCNSA-N 0.000 description 3
- FBOZXECLQNJBKD-ZDUSSCGKSA-N L-methotrexate Chemical compound C=1N=C2N=C(N)N=C(N)C2=NC=1CN(C)C1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 FBOZXECLQNJBKD-ZDUSSCGKSA-N 0.000 description 3
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 3
- OKKJLVBELUTLKV-UHFFFAOYSA-N Methanol Chemical compound OC OKKJLVBELUTLKV-UHFFFAOYSA-N 0.000 description 3
- 241000699670 Mus sp. Species 0.000 description 3
- 241000283973 Oryctolagus cuniculus Species 0.000 description 3
- 238000012408 PCR amplification Methods 0.000 description 3
- 229930182555 Penicillin Natural products 0.000 description 3
- JGSARLDLIJGVTE-MBNYWOFBSA-N Penicillin G Chemical compound N([C@H]1[C@H]2SC([C@@H](N2C1=O)C(O)=O)(C)C)C(=O)CC1=CC=CC=C1 JGSARLDLIJGVTE-MBNYWOFBSA-N 0.000 description 3
- 239000007983 Tris buffer Substances 0.000 description 3
- KZSNJWFQEVHDMF-UHFFFAOYSA-N Valine Natural products CC(C)C(N)C(O)=O KZSNJWFQEVHDMF-UHFFFAOYSA-N 0.000 description 3
- 241000700605 Viruses Species 0.000 description 3
- 239000002671 adjuvant Substances 0.000 description 3
- 125000000539 amino acid group Chemical group 0.000 description 3
- 238000003556 assay Methods 0.000 description 3
- 230000037429 base substitution Effects 0.000 description 3
- KRKNYBCHXYNGOX-UHFFFAOYSA-N citric acid Chemical compound OC(=O)CC(O)(C(O)=O)CC(O)=O KRKNYBCHXYNGOX-UHFFFAOYSA-N 0.000 description 3
- 239000003085 diluting agent Substances 0.000 description 3
- 239000003623 enhancer Substances 0.000 description 3
- 230000009036 growth inhibition Effects 0.000 description 3
- 208000015181 infectious disease Diseases 0.000 description 3
- 239000006166 lysate Substances 0.000 description 3
- 229930182817 methionine Natural products 0.000 description 3
- 229960000485 methotrexate Drugs 0.000 description 3
- 239000000178 monomer Substances 0.000 description 3
- 108091027963 non-coding RNA Proteins 0.000 description 3
- 102000042567 non-coding RNA Human genes 0.000 description 3
- 229940049954 penicillin Drugs 0.000 description 3
- 229920001993 poloxamer 188 Polymers 0.000 description 3
- 239000000244 polyoxyethylene sorbitan monooleate Substances 0.000 description 3
- 229940068968 polysorbate 80 Drugs 0.000 description 3
- 230000008929 regeneration Effects 0.000 description 3
- 238000011069 regeneration method Methods 0.000 description 3
- 229960005322 streptomycin Drugs 0.000 description 3
- LENZDBCJOHFCAS-UHFFFAOYSA-N tris Chemical compound OCC(N)(CO)CO LENZDBCJOHFCAS-UHFFFAOYSA-N 0.000 description 3
- 241000701447 unidentified baculovirus Species 0.000 description 3
- 239000004474 valine Substances 0.000 description 3
- QKNYBSVHEMOAJP-UHFFFAOYSA-N 2-amino-2-(hydroxymethyl)propane-1,3-diol;hydron;chloride Chemical compound Cl.OCC(N)(CO)CO QKNYBSVHEMOAJP-UHFFFAOYSA-N 0.000 description 2
- FERIUCNNQQJTOY-UHFFFAOYSA-M Butyrate Chemical compound CCCC([O-])=O FERIUCNNQQJTOY-UHFFFAOYSA-M 0.000 description 2
- FERIUCNNQQJTOY-UHFFFAOYSA-N Butyric acid Natural products CCCC(O)=O FERIUCNNQQJTOY-UHFFFAOYSA-N 0.000 description 2
- 230000004544 DNA amplification Effects 0.000 description 2
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 2
- 208000037487 Endotoxemia Diseases 0.000 description 2
- SIKJAQJRHWYJAI-UHFFFAOYSA-N Indole Chemical compound C1=CC=C2NC=CC2=C1 SIKJAQJRHWYJAI-UHFFFAOYSA-N 0.000 description 2
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 2
- 102000052508 Lipopolysaccharide-binding protein Human genes 0.000 description 2
- 108010053632 Lipopolysaccharide-binding protein Proteins 0.000 description 2
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 2
- 239000004472 Lysine Substances 0.000 description 2
- 101100370002 Mus musculus Tnfsf14 gene Proteins 0.000 description 2
- 239000000020 Nitrocellulose Substances 0.000 description 2
- 108091034117 Oligonucleotide Proteins 0.000 description 2
- 229920003171 Poly (ethylene oxide) Polymers 0.000 description 2
- 229920001214 Polysorbate 60 Polymers 0.000 description 2
- 206010040047 Sepsis Diseases 0.000 description 2
- 206010040070 Septic Shock Diseases 0.000 description 2
- XSQUKJJJFZCRTK-UHFFFAOYSA-N Urea Chemical compound NC(N)=O XSQUKJJJFZCRTK-UHFFFAOYSA-N 0.000 description 2
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 2
- 239000002253 acid Substances 0.000 description 2
- RJURFGZVJUQBHK-UHFFFAOYSA-N actinomycin D Natural products CC1OC(=O)C(C(C)C)N(C)C(=O)CN(C)C(=O)C2CCCN2C(=O)C(C(C)C)NC(=O)C1NC(=O)C1=C(N)C(=O)C(C)=C2OC(C(C)=CC=C3C(=O)NC4C(=O)NC(C(N5CCCC5C(=O)N(C)CC(=O)N(C)C(C(C)C)C(=O)OC4C)=O)C(C)C)=C3N=C21 RJURFGZVJUQBHK-UHFFFAOYSA-N 0.000 description 2
- 230000003321 amplification Effects 0.000 description 2
- 238000010171 animal model Methods 0.000 description 2
- 238000000376 autoradiography Methods 0.000 description 2
- 230000004071 biological effect Effects 0.000 description 2
- 229920001400 block copolymer Polymers 0.000 description 2
- 230000008859 change Effects 0.000 description 2
- 238000012512 characterization method Methods 0.000 description 2
- 210000004978 chinese hamster ovary cell Anatomy 0.000 description 2
- 238000003776 cleavage reaction Methods 0.000 description 2
- 230000000295 complement effect Effects 0.000 description 2
- 239000012228 culture supernatant Substances 0.000 description 2
- 210000000805 cytoplasm Anatomy 0.000 description 2
- 230000034994 death Effects 0.000 description 2
- 230000002950 deficient Effects 0.000 description 2
- 235000014113 dietary fatty acids Nutrition 0.000 description 2
- 238000010790 dilution Methods 0.000 description 2
- 239000012895 dilution Substances 0.000 description 2
- 210000003527 eukaryotic cell Anatomy 0.000 description 2
- 239000013613 expression plasmid Substances 0.000 description 2
- 229930195729 fatty acid Natural products 0.000 description 2
- 239000000194 fatty acid Substances 0.000 description 2
- 125000000291 glutamic acid group Chemical group N[C@@H](CCC(O)=O)C(=O)* 0.000 description 2
- 238000002347 injection Methods 0.000 description 2
- 239000007924 injection Substances 0.000 description 2
- 229910001629 magnesium chloride Inorganic materials 0.000 description 2
- 238000005259 measurement Methods 0.000 description 2
- 125000001360 methionine group Chemical group N[C@@H](CCSC)C(=O)* 0.000 description 2
- SXTAYKAGBXMACB-UHFFFAOYSA-N methionine sulfoximine Chemical compound CS(=N)(=O)CCC(N)C(O)=O SXTAYKAGBXMACB-UHFFFAOYSA-N 0.000 description 2
- 229920001220 nitrocellulos Polymers 0.000 description 2
- 238000003199 nucleic acid amplification method Methods 0.000 description 2
- 229940044519 poloxamer 188 Drugs 0.000 description 2
- 229920002401 polyacrylamide Polymers 0.000 description 2
- 238000001556 precipitation Methods 0.000 description 2
- 230000008569 process Effects 0.000 description 2
- 230000006824 pyrimidine synthesis Effects 0.000 description 2
- 230000009467 reduction Effects 0.000 description 2
- 150000003839 salts Chemical class 0.000 description 2
- 230000007017 scission Effects 0.000 description 2
- 238000012216 screening Methods 0.000 description 2
- 230000003248 secreting effect Effects 0.000 description 2
- 125000003607 serino group Chemical group [H]N([H])[C@]([H])(C(=O)[*])C(O[H])([H])[H] 0.000 description 2
- 210000002966 serum Anatomy 0.000 description 2
- 239000004017 serum-free culture medium Substances 0.000 description 2
- 239000001509 sodium citrate Substances 0.000 description 2
- NLJMYIDDQXHKNR-UHFFFAOYSA-K sodium citrate Chemical compound O.O.[Na+].[Na+].[Na+].[O-]C(=O)CC(O)(CC([O-])=O)C([O-])=O NLJMYIDDQXHKNR-UHFFFAOYSA-K 0.000 description 2
- 239000000126 substance Substances 0.000 description 2
- 125000003396 thiol group Chemical group [H]S* 0.000 description 2
- LWIHDJKSTIGBAC-UHFFFAOYSA-K tripotassium phosphate Chemical compound [K+].[K+].[K+].[O-]P([O-])([O-])=O LWIHDJKSTIGBAC-UHFFFAOYSA-K 0.000 description 2
- 125000002987 valine group Chemical group [H]N([H])C([H])(C(*)=O)C([H])(C([H])([H])[H])C([H])([H])[H] 0.000 description 2
- 230000003612 virological effect Effects 0.000 description 2
- 238000012800 visualization Methods 0.000 description 2
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 2
- 101710088758 23kDa protein Proteins 0.000 description 1
- DGZSVBBLLGZHSF-UHFFFAOYSA-N 4,4-diethylpiperidine Chemical compound CCC1(CC)CCNCC1 DGZSVBBLLGZHSF-UHFFFAOYSA-N 0.000 description 1
- QFVHZQCOUORWEI-UHFFFAOYSA-N 4-[(4-anilino-5-sulfonaphthalen-1-yl)diazenyl]-5-hydroxynaphthalene-2,7-disulfonic acid Chemical compound C=12C(O)=CC(S(O)(=O)=O)=CC2=CC(S(O)(=O)=O)=CC=1N=NC(C1=CC=CC(=C11)S(O)(=O)=O)=CC=C1NC1=CC=CC=C1 QFVHZQCOUORWEI-UHFFFAOYSA-N 0.000 description 1
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 description 1
- 241000714175 Abelson murine leukemia virus Species 0.000 description 1
- 208000013824 Acidemia Diseases 0.000 description 1
- 208000010444 Acidosis Diseases 0.000 description 1
- 206010001052 Acute respiratory distress syndrome Diseases 0.000 description 1
- 208000002109 Argyria Diseases 0.000 description 1
- 241001367049 Autographa Species 0.000 description 1
- 108090001008 Avidin Proteins 0.000 description 1
- 101150043508 BPI gene Proteins 0.000 description 1
- 208000035143 Bacterial infection Diseases 0.000 description 1
- OYPRJOBELJOOCE-UHFFFAOYSA-N Calcium Chemical compound [Ca] OYPRJOBELJOOCE-UHFFFAOYSA-N 0.000 description 1
- 241000589877 Campylobacter coli Species 0.000 description 1
- 102000005367 Carboxypeptidases Human genes 0.000 description 1
- 108010006303 Carboxypeptidases Proteins 0.000 description 1
- 241000178041 Ceropegia media Species 0.000 description 1
- KRKNYBCHXYNGOX-UHFFFAOYSA-K Citrate Chemical compound [O-]C(=O)CC(O)(CC([O-])=O)C([O-])=O KRKNYBCHXYNGOX-UHFFFAOYSA-K 0.000 description 1
- 108020004635 Complementary DNA Proteins 0.000 description 1
- 241000699800 Cricetinae Species 0.000 description 1
- 101150074155 DHFR gene Proteins 0.000 description 1
- 108010092160 Dactinomycin Proteins 0.000 description 1
- BWGNESOTFCXPMA-UHFFFAOYSA-N Dihydrogen disulfide Chemical compound SS BWGNESOTFCXPMA-UHFFFAOYSA-N 0.000 description 1
- 241000192125 Firmicutes Species 0.000 description 1
- 108010010803 Gelatin Proteins 0.000 description 1
- 108700028146 Genetic Enhancer Elements Proteins 0.000 description 1
- 108700039691 Genetic Promoter Regions Proteins 0.000 description 1
- 108090000079 Glucocorticoid Receptors Proteins 0.000 description 1
- 102100033417 Glucocorticoid receptor Human genes 0.000 description 1
- WHUUTDBJXJRKMK-UHFFFAOYSA-N Glutamic acid Natural products OC(=O)C(N)CCC(O)=O WHUUTDBJXJRKMK-UHFFFAOYSA-N 0.000 description 1
- 239000012981 Hank's balanced salt solution Substances 0.000 description 1
- 101000573199 Homo sapiens Protein PML Proteins 0.000 description 1
- 101100273566 Humulus lupulus CCL10 gene Proteins 0.000 description 1
- 208000001953 Hypotension Diseases 0.000 description 1
- 102000018251 Hypoxanthine Phosphoribosyltransferase Human genes 0.000 description 1
- 108010091358 Hypoxanthine Phosphoribosyltransferase Proteins 0.000 description 1
- 108060003951 Immunoglobulin Proteins 0.000 description 1
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 description 1
- KZSNJWFQEVHDMF-BYPYZUCNSA-N L-valine Chemical compound CC(C)[C@H](N)C(O)=O KZSNJWFQEVHDMF-BYPYZUCNSA-N 0.000 description 1
- 108091026898 Leader sequence (mRNA) Proteins 0.000 description 1
- 241001124569 Lycaenidae Species 0.000 description 1
- 101100278853 Mus musculus Dhfr gene Proteins 0.000 description 1
- 101000966481 Mus musculus Dihydrofolate reductase Proteins 0.000 description 1
- 241001045988 Neogene Species 0.000 description 1
- 102000004316 Oxidoreductases Human genes 0.000 description 1
- 108090000854 Oxidoreductases Proteins 0.000 description 1
- 238000009004 PCR Kit Methods 0.000 description 1
- 102000003992 Peroxidases Human genes 0.000 description 1
- 206010035226 Plasma cell myeloma Diseases 0.000 description 1
- 229920001213 Polysorbate 20 Polymers 0.000 description 1
- 101710093543 Probable non-specific lipid-transfer protein Proteins 0.000 description 1
- 101100534621 Psychromonas ingrahamii (strain 37) sucC gene Proteins 0.000 description 1
- 206010037660 Pyrexia Diseases 0.000 description 1
- 208000001647 Renal Insufficiency Diseases 0.000 description 1
- 208000013616 Respiratory Distress Syndrome Diseases 0.000 description 1
- 241001352312 Salivirus Species 0.000 description 1
- 241000607142 Salmonella Species 0.000 description 1
- BQCADISMDOOEFD-UHFFFAOYSA-N Silver Chemical compound [Ag] BQCADISMDOOEFD-UHFFFAOYSA-N 0.000 description 1
- 108091081024 Start codon Proteins 0.000 description 1
- GSEJCLTVZPLZKY-UHFFFAOYSA-N Triethanolamine Chemical compound OCCN(CCO)CCO GSEJCLTVZPLZKY-UHFFFAOYSA-N 0.000 description 1
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 1
- 108091023045 Untranslated Region Proteins 0.000 description 1
- RJURFGZVJUQBHK-IIXSONLDSA-N actinomycin D Chemical compound C[C@H]1OC(=O)[C@H](C(C)C)N(C)C(=O)CN(C)C(=O)[C@@H]2CCCN2C(=O)[C@@H](C(C)C)NC(=O)[C@H]1NC(=O)C1=C(N)C(=O)C(C)=C2OC(C(C)=CC=C3C(=O)N[C@@H]4C(=O)N[C@@H](C(N5CCC[C@H]5C(=O)N(C)CC(=O)N(C)[C@@H](C(C)C)C(=O)O[C@@H]4C)=O)C(C)C)=C3N=C21 RJURFGZVJUQBHK-IIXSONLDSA-N 0.000 description 1
- 208000011341 adult acute respiratory distress syndrome Diseases 0.000 description 1
- 201000000028 adult respiratory distress syndrome Diseases 0.000 description 1
- 238000001042 affinity chromatography Methods 0.000 description 1
- 229940126575 aminoglycoside Drugs 0.000 description 1
- 208000007502 anemia Diseases 0.000 description 1
- 230000000845 anti-microbial effect Effects 0.000 description 1
- 208000022362 bacterial infectious disease Diseases 0.000 description 1
- 239000003899 bactericide agent Substances 0.000 description 1
- 230000008901 benefit Effects 0.000 description 1
- 230000000903 blocking effect Effects 0.000 description 1
- 210000004369 blood Anatomy 0.000 description 1
- 239000008280 blood Substances 0.000 description 1
- 239000007853 buffer solution Substances 0.000 description 1
- 239000007975 buffered saline Substances 0.000 description 1
- 239000011575 calcium Substances 0.000 description 1
- 229910052791 calcium Inorganic materials 0.000 description 1
- 239000001506 calcium phosphate Substances 0.000 description 1
- 229910000389 calcium phosphate Inorganic materials 0.000 description 1
- 235000011010 calcium phosphates Nutrition 0.000 description 1
- 239000004202 carbamide Substances 0.000 description 1
- 238000005341 cation exchange Methods 0.000 description 1
- 238000004113 cell culture Methods 0.000 description 1
- 239000013592 cell lysate Substances 0.000 description 1
- 238000006243 chemical reaction Methods 0.000 description 1
- 238000002038 chemiluminescence detection Methods 0.000 description 1
- 238000010367 cloning Methods 0.000 description 1
- 230000015271 coagulation Effects 0.000 description 1
- 238000005345 coagulation Methods 0.000 description 1
- 150000001875 compounds Chemical class 0.000 description 1
- 230000009089 cytolysis Effects 0.000 description 1
- 231100000433 cytotoxic Toxicity 0.000 description 1
- 230000001472 cytotoxic effect Effects 0.000 description 1
- 229960000640 dactinomycin Drugs 0.000 description 1
- 238000001514 detection method Methods 0.000 description 1
- UREBDLICKHMUKA-CXSFZGCWSA-N dexamethasone Chemical compound C1CC2=CC(=O)C=C[C@]2(C)[C@]2(F)[C@@H]1[C@@H]1C[C@@H](C)[C@@](C(=O)CO)(O)[C@@]1(C)C[C@@H]2O UREBDLICKHMUKA-CXSFZGCWSA-N 0.000 description 1
- 229960003957 dexamethasone Drugs 0.000 description 1
- OZRNSSUDZOLUSN-LBPRGKRZSA-N dihydrofolic acid Chemical compound N=1C=2C(=O)NC(N)=NC=2NCC=1CNC1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 OZRNSSUDZOLUSN-LBPRGKRZSA-N 0.000 description 1
- 239000012470 diluted sample Substances 0.000 description 1
- 238000007865 diluting Methods 0.000 description 1
- 238000006471 dimerization reaction Methods 0.000 description 1
- 238000009826 distribution Methods 0.000 description 1
- VHJLVAABSRFDPM-QWWZWVQMSA-N dithiothreitol Chemical compound SC[C@@H](O)[C@H](O)CS VHJLVAABSRFDPM-QWWZWVQMSA-N 0.000 description 1
- 238000001035 drying Methods 0.000 description 1
- 230000005672 electromagnetic field Effects 0.000 description 1
- 230000008030 elimination Effects 0.000 description 1
- 238000003379 elimination reaction Methods 0.000 description 1
- 238000010828 elution Methods 0.000 description 1
- 238000005516 engineering process Methods 0.000 description 1
- 238000011156 evaluation Methods 0.000 description 1
- 238000002474 experimental method Methods 0.000 description 1
- 210000002950 fibroblast Anatomy 0.000 description 1
- 239000008273 gelatin Substances 0.000 description 1
- 229920000159 gelatin Polymers 0.000 description 1
- 235000019322 gelatine Nutrition 0.000 description 1
- 235000011852 gelatine desserts Nutrition 0.000 description 1
- 238000007429 general method Methods 0.000 description 1
- 235000013922 glutamic acid Nutrition 0.000 description 1
- 239000004220 glutamic acid Substances 0.000 description 1
- 101150106093 gpt gene Proteins 0.000 description 1
- 208000027096 gram-negative bacterial infections Diseases 0.000 description 1
- 210000000224 granular leucocyte Anatomy 0.000 description 1
- 239000008187 granular material Substances 0.000 description 1
- 238000003306 harvesting Methods 0.000 description 1
- 101150113423 hisD gene Proteins 0.000 description 1
- 102000054896 human PML Human genes 0.000 description 1
- 210000004408 hybridoma Anatomy 0.000 description 1
- 230000036543 hypotension Effects 0.000 description 1
- 238000007654 immersion Methods 0.000 description 1
- 102000018358 immunoglobulin Human genes 0.000 description 1
- 238000012744 immunostaining Methods 0.000 description 1
- PZOUSPYUWWUPPK-UHFFFAOYSA-N indole Natural products CC1=CC=CC2=C1C=CN2 PZOUSPYUWWUPPK-UHFFFAOYSA-N 0.000 description 1
- RKJUIXBNRJVNHR-UHFFFAOYSA-N indolenine Natural products C1=CC=C2CC=NC2=C1 RKJUIXBNRJVNHR-UHFFFAOYSA-N 0.000 description 1
- 229910052500 inorganic mineral Inorganic materials 0.000 description 1
- 230000003993 interaction Effects 0.000 description 1
- 238000004255 ion exchange chromatography Methods 0.000 description 1
- 238000002955 isolation Methods 0.000 description 1
- 229960000310 isoleucine Drugs 0.000 description 1
- AGPKZVBTJJNPAG-UHFFFAOYSA-N isoleucine Natural products CCC(C)C(N)C(O)=O AGPKZVBTJJNPAG-UHFFFAOYSA-N 0.000 description 1
- 125000000741 isoleucyl group Chemical group [H]N([H])C(C(C([H])([H])[H])C([H])([H])C([H])([H])[H])C(=O)O* 0.000 description 1
- 238000005304 joining Methods 0.000 description 1
- 210000003734 kidney Anatomy 0.000 description 1
- 201000006370 kidney failure Diseases 0.000 description 1
- 238000011031 large-scale manufacturing process Methods 0.000 description 1
- 230000001665 lethal effect Effects 0.000 description 1
- 208000032839 leukemia Diseases 0.000 description 1
- 201000002364 leukopenia Diseases 0.000 description 1
- 231100001022 leukopenia Toxicity 0.000 description 1
- 238000012454 limulus amebocyte lysate test Methods 0.000 description 1
- GZQKNULLWNGMCW-PWQABINMSA-N lipid A (E. coli) Chemical compound O1[C@H](CO)[C@@H](OP(O)(O)=O)[C@H](OC(=O)C[C@@H](CCCCCCCCCCC)OC(=O)CCCCCCCCCCCCC)[C@@H](NC(=O)C[C@@H](CCCCCCCCCCC)OC(=O)CCCCCCCCCCC)[C@@H]1OC[C@@H]1[C@@H](O)[C@H](OC(=O)C[C@H](O)CCCCCCCCCCC)[C@@H](NC(=O)C[C@H](O)CCCCCCCCCCC)[C@@H](OP(O)(O)=O)O1 GZQKNULLWNGMCW-PWQABINMSA-N 0.000 description 1
- 239000007788 liquid Substances 0.000 description 1
- 210000004698 lymphocyte Anatomy 0.000 description 1
- 239000012139 lysis buffer Substances 0.000 description 1
- 238000004949 mass spectrometry Methods 0.000 description 1
- 210000003622 mature neutrocyte Anatomy 0.000 description 1
- 230000002503 metabolic effect Effects 0.000 description 1
- 238000013508 migration Methods 0.000 description 1
- 230000005012 migration Effects 0.000 description 1
- 239000011707 mineral Substances 0.000 description 1
- 230000004048 modification Effects 0.000 description 1
- 238000012986 modification Methods 0.000 description 1
- 238000010369 molecular cloning Methods 0.000 description 1
- 201000000050 myeloid neoplasm Diseases 0.000 description 1
- 101150091879 neo gene Proteins 0.000 description 1
- 231100001083 no cytotoxicity Toxicity 0.000 description 1
- 235000015097 nutrients Nutrition 0.000 description 1
- 229920001542 oligosaccharide Polymers 0.000 description 1
- 150000002482 oligosaccharides Chemical class 0.000 description 1
- 230000007170 pathology Effects 0.000 description 1
- 238000012510 peptide mapping method Methods 0.000 description 1
- 108040007629 peroxidase activity proteins Proteins 0.000 description 1
- 239000000546 pharmaceutical excipient Substances 0.000 description 1
- 239000000256 polyoxyethylene sorbitan monolaurate Substances 0.000 description 1
- 235000010486 polyoxyethylene sorbitan monolaurate Nutrition 0.000 description 1
- 229920001282 polysaccharide Polymers 0.000 description 1
- 239000005017 polysaccharide Substances 0.000 description 1
- 239000011148 porous material Substances 0.000 description 1
- 230000004481 post-translational protein modification Effects 0.000 description 1
- 229910000160 potassium phosphate Inorganic materials 0.000 description 1
- 235000011009 potassium phosphates Nutrition 0.000 description 1
- 238000012545 processing Methods 0.000 description 1
- 230000006916 protein interaction Effects 0.000 description 1
- 230000002797 proteolythic effect Effects 0.000 description 1
- 239000010453 quartz Substances 0.000 description 1
- 230000009257 reactivity Effects 0.000 description 1
- 238000003259 recombinant expression Methods 0.000 description 1
- 230000006798 recombination Effects 0.000 description 1
- 238000005215 recombination Methods 0.000 description 1
- 230000001172 regenerating effect Effects 0.000 description 1
- 210000001995 reticulocyte Anatomy 0.000 description 1
- 239000012723 sample buffer Substances 0.000 description 1
- 230000035945 sensitivity Effects 0.000 description 1
- 230000036303 septic shock Effects 0.000 description 1
- 238000012163 sequencing technique Methods 0.000 description 1
- 239000012679 serum free medium Substances 0.000 description 1
- 206010040560 shock Diseases 0.000 description 1
- VYPSYNLAJGMNEJ-UHFFFAOYSA-N silicon dioxide Inorganic materials O=[Si]=O VYPSYNLAJGMNEJ-UHFFFAOYSA-N 0.000 description 1
- 229910052709 silver Inorganic materials 0.000 description 1
- 239000004332 silver Substances 0.000 description 1
- 239000011734 sodium Substances 0.000 description 1
- MFBOGIVSZKQAPD-UHFFFAOYSA-M sodium butyrate Chemical compound [Na+].CCCC([O-])=O MFBOGIVSZKQAPD-UHFFFAOYSA-M 0.000 description 1
- 239000012475 sodium chloride buffer Substances 0.000 description 1
- 230000002269 spontaneous effect Effects 0.000 description 1
- 230000000087 stabilizing effect Effects 0.000 description 1
- 238000010186 staining Methods 0.000 description 1
- 239000013595 supernatant sample Substances 0.000 description 1
- 208000024891 symptom Diseases 0.000 description 1
- 230000002195 synergetic effect Effects 0.000 description 1
- 238000003786 synthesis reaction Methods 0.000 description 1
- 238000012360 testing method Methods 0.000 description 1
- 238000002560 therapeutic procedure Methods 0.000 description 1
- 206010043554 thrombocytopenia Diseases 0.000 description 1
- 210000001519 tissue Anatomy 0.000 description 1
- 238000012546 transfer Methods 0.000 description 1
- 230000007704 transition Effects 0.000 description 1
- QORWJWZARLRLPR-UHFFFAOYSA-H tricalcium bis(phosphate) Chemical compound [Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O QORWJWZARLRLPR-UHFFFAOYSA-H 0.000 description 1
- 238000011144 upstream manufacturing Methods 0.000 description 1
- 230000002792 vascular Effects 0.000 description 1
- 238000005406 washing Methods 0.000 description 1
- 238000001262 western blot Methods 0.000 description 1
Images







Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K19/00—Hybrid peptides, i.e. peptides covalently bound to nucleic acids, or non-covalently bound protein-protein complexes
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
- A61K38/16—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P11/00—Drugs for disorders of the respiratory system
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P13/00—Drugs for disorders of the urinary system
- A61P13/12—Drugs for disorders of the urinary system of the kidneys
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P29/00—Non-central analgesic, antipyretic or antiinflammatory agents, e.g. antirheumatic agents; Non-steroidal antiinflammatory drugs [NSAID]
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/04—Antibacterial agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P39/00—General protective or antinoxious agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P39/00—General protective or antinoxious agents
- A61P39/02—Antidotes
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P7/00—Drugs for disorders of the blood or the extracellular fluid
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P7/00—Drugs for disorders of the blood or the extracellular fluid
- A61P7/02—Antithrombotic agents; Anticoagulants; Platelet aggregation inhibitors
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P7/00—Drugs for disorders of the blood or the extracellular fluid
- A61P7/06—Antianaemics
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P9/00—Drugs for disorders of the cardiovascular system
- A61P9/02—Non-specific cardiovascular stimulants, e.g. drugs for syncope, antihypotensives
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/46—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates
- C07K14/47—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates from mammals
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/46—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates
- C07K14/47—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates from mammals
- C07K14/4701—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates from mammals not used
- C07K14/4742—Bactericidal/Permeability-increasing protein [BPI]
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/85—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P21/00—Preparation of peptides or proteins
- C12P21/02—Preparation of peptides or proteins having a known sequence of two or more amino acids, e.g. glutathione
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2830/00—Vector systems having a special element relevant for transcription
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2830/00—Vector systems having a special element relevant for transcription
- C12N2830/15—Vector systems having a special element relevant for transcription chimeric enhancer/promoter combination
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2830/00—Vector systems having a special element relevant for transcription
- C12N2830/30—Vector systems having a special element relevant for transcription being an enhancer not forming part of the promoter region
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2830/00—Vector systems having a special element relevant for transcription
- C12N2830/60—Vector systems having a special element relevant for transcription from viruses
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2830/00—Vector systems having a special element relevant for transcription
- C12N2830/80—Vector systems having a special element relevant for transcription from vertebrates
- C12N2830/85—Vector systems having a special element relevant for transcription from vertebrates mammalian
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2840/00—Vectors comprising a special translation-regulating system
- C12N2840/44—Vectors comprising a special translation-regulating system being a specific part of the splice mechanism, e.g. donor, acceptor
Abstract
【解決手段】
本発明は、カルボキシ末端残基として第193位にロイシン残基を有する殺菌性/浸透性が向上したタンパク質の生物学的に活性な断片をコードするDNA。
【選択図】 なし
Description
本発明は、殺菌性/浸透性が向上した新規なタンパク質生成物およびそれを含む薬剤組成物に関する。
−4761(1992)。
活性なポリペプチドを安定化せしめる相乗効果を得る目的で、共同所有に係る、係属中で、同時に出願された米国特許出願No.08/012,360にて教示されている。rBPI生成物が、0.1重量%のポロキサマー188(Pluronic F−68,BASF Wyandotte,パルシパニィー、ニュージャージー州)および0.002重量%のポリソルベート80(Tween 80,ICI Americas Inc.,ウィルミントン、デラウェア州)を含むクエン酸緩衝化生理食塩水(0.02Mクエン酸、0.15M塩化ナトリウム、pH5.0)に、1mg/mlの濃度で存在させることが最も好ましい。
本発明は、以下の項を提供する:
項1.第132位あるいは第135位のシステイン残基が異なるアミノ酸で置換されている、殺菌性/浸透性が向上したタンパク質のポリペプチド類似体あるいはその生物学的に活性な断片;
項2.前記システイン残基を置換するアミノ酸が、アラニンおよびセリンからなるグループから選択された非極性アミノ酸である請求の範囲第1項に記載のポリペプチド類似体;項3.第132位のシステイン残基が、アラニンで置換される請求の範囲第1項に記載のポリペプチド類似体;
項4.第135位のシステイン残基が、セリンで置換される請求の範囲第1項に記載のポリペプチド類似体;
項5.殺菌性/浸透性が向上したタンパク質の、第132位あるいは第135位のシステイン置換を除く、第176位から第199位の開始アミノ末端アミノ酸残基を含む請求の範囲第1項、第2項、第3項もしくは第4項に記載のポリペプチド類似体;
項6.殺菌性/浸透性が向上したタンパク質の第193位のアミノ末端アミノ酸残基を含む請求の範囲第5項に記載のポリペプチド類似体;
項7.第132位あるいは第135位のシステイン残基が異なるアミノ酸で置換されている、殺菌性/浸透性が向上したタンパク質のポリペプチド類似体あるいはその生物学的に活性な断片をコードするDNA;
項8.31個のアミノ酸リーダー配列ならびに殺菌性/浸透性が向上したタンパク質の最初の193位のN−末端残基をコードし、第193位のロイシン残基のためのコドンの直後に続くコドンに続く停止コドンを有する請求の範囲第7項に記載のDNA;
項9.31個のアミノ酸リーダー配列ならびに殺菌性/浸透性が向上したタンパク質の最初の199位のN−末端残基をコードし、第199位のイソロイシンのためのコドンの直後に続くコドンに続く停止コドンを有する請求の範囲第7項に記載のDNA;
項10.請求の範囲第7項に記載のDNAを含む自律複製DNA;
項11.前記ポリペプチド類似体の宿主細胞での発現を許容する方法にて、請求の範囲第7項に記載のDNAで安定的に形質変換あるいは形質転換された宿主細胞;
項12.請求の範囲第11項に記載の真核細胞;
項13.ATCC CRL 11246およびATCC HB 11247からなるグループから選択されたATCC受託番号を有する請求の範囲第12項に記載の宿主細胞;
項14.適切な培養培地で請求の範囲第11項に記載の宿主細胞を生長させ、および前記宿主細胞あるいは前記培養培地からポリペプチド類似体を単離することを含む、殺菌性/
浸透性が向上したタンパク質のポリペプチド類似体あるいはその生物学的に活性な断片を製造する方法;
項15.アミノ末端に請求の範囲第1項に記載の類似体ポリペプチドを、および、カルボキシ末端に免疫グロブリン重鎖あるいはその対立性変異体の少なくとも一つの定常領域を含むハイブリッド融合タンパク質;
項16.請求の範囲第15項に記載のハイブリッド融合タンパク質をコードするDNA;項17.請求の範囲第16項に記載のDNAを含む自律複製DNAベクター;
項18.前記融合タンパク質の宿主細胞での発現を許容する方法にて、請求の範囲第16項に記載のDNAで安定的に形質変換あるいは形質転換された宿主細胞;
項19.適切な培養培地で請求の範囲第18項に記載の宿主細胞を生長させ、および前記宿主細胞あるいは前記培養培地から前記類似体を単離することを含む、請求の範囲第15項に記載のハイブリッド融合タンパク質を製造する方法;
項20.請求の範囲第1項に記載のポリペプチド類似体および薬学的に許容される希釈剤、補助剤、あるいは担体を含む薬学的組成物;
項21.請求の範囲第15項に記載のハイブリッド融合タンパク質および薬学的に許容される希釈剤、補助剤、あるいは担体を含む薬学的組成物;
項22.請求の範囲第20項もしくは第21項に記載の薬学的組成物の有効量を投与することを含む、細菌性疾患あるいはその後遺症を治療するための方法;
項23.カルボキシ末端残基の第193位にロイシン残基を有する殺菌性/浸透性が向上したタンパク質の生物学的に活性な断片をコードするDNA;
項24.31個のアミノ酸リーダー配列ならびに殺菌性/浸透性が向上したタンパク質の最初の193位のN−末端アミノ酸をコードし、第193位のロイシン残基のためのコドンの直後に続くコドンに続く停止コドンを有する請求の範囲第23項に記載のDNA;
項25.請求の範囲第23項に記載のDNAを含む自律複製DNAベクター;
項26.宿主細胞での発現を許容する方法にて、請求の範囲第23項に記載のDNAで安定的に形質変換あるいは形質転換された宿主細胞;
項27.請求の範囲第23項に記載のDNAで安定的に形質変換あるいは形質転換された宿主細胞を適切な培養培地で生長させ、および前記宿主細胞あるいは前記培養培地から前記断片を単離することを含む、生物学的に活性な殺菌性/浸透性が向上した断片を製造する方法。
イシンのためのコドンの直後に停止コドンを有するポリペプチドが教示されている。
(BPIのシステイン置換類似体を含むベクターの構築)
(A.プラスミドpING4519とpING4520の構築)
発現ベクターpING4503を、rBPI(1−199)と称する組換え発現生成物をコードするDNA源として、すなわち、第151位のバリンがGTCよりもむしろGTGで特定され、また第185位の残基が(AAGで特定される)リシンよりむしろ(GAGで特定される)グルタミン酸であることを除いて、配列番号:1および2に説明したような、31残基のシグナル配列と成熟したヒトBPIのN末端の最初の199個のアミノ酸を含むポリペプチドをコードするDNA源として用いた。
るために三片接合した。
pING4519のようにアラニンからシステインへの置換を含み、pING4503からpING4519に運ばれたDHFRマーカーに代えて(マイコフェノール酸耐性を許容する)gpt選択可能なマーカーを含む他のベクター、pING4530を構築した。pING4530を構築するために、1629bpのSalI−DraIII制限断片が、pING4519から単離された。この断片は、rBPI(1−199)ala132のコード領域をすべて、ならびにこのコード領域の3’末端での約895bpのベクター配列を含んでいる。この断片は、pING4530を生成するために、pING4513から単離された大きな(約7230bp)DraIII−SalIベクター断片に接合した。
BPIシグナル配列の5番目のアミノ酸である、−27位のメチオニン(ATG)を特定するコドンを、Kozakの定常翻訳開始配列 GCCACCRCCATGG(配列番号:7)[Kozak,Nucl.Acid.Res.,15:8125(1987)]との関連で置換し、BPIシグナルの最初の四つのアミノ酸をコードするDNA配列を削除して、rBPI(1−199)ala132の発現のためのプラスミドpING4533を構築した。BPIシグナルの−27位のATG(メチオニン)の前にSalI制限部位およびヌクレオチドGCCACCを組み込んだPCRプライマーBPI−23:ACTGTCGACGCCACCATGGCCAGGGGC(配列番号:8)、およびrBPI(1−199)コード配列の3’端に対応するプライマーBPI−2:CCGCGGCTCGAGCTATATTTTGGTCAT(配列番号:9)を用いて、[pGEM−7zf (+)中の]ヒトBPI cDNAの全長を含むプラスミドからBPI配列のPCR増幅によって構築を行った。約700bpのPCR増幅したDNAをSalIおよびEcoRIで消化し、BPI(1−199)コード配列の約1/3を含む270bpの得られた断片を精製した。このSalI−EcoRI断片を、gptマーカーを含む pING4533を生成するために、(1)132位のシステインをアラニンで置換したBPI(1−199)の残部をコードするpING4519からの 420bpのEcoRI−S
stII断片、および(2)pING4502からの約8000bpのSstII−SalIベクター断片(30bpの5’非翻訳配列を含まず、DHFRよりもむしろgPtマーカーを有することを除けば実質的にpING4503と同様のベクター)の二つの断片と接合した。
シグナル配列の−27位のメチオニン残基に対応する至適なKozakの翻訳開始部位と、システインからアラニンへの132位での置換を含む挿入体を有する pING4533と同様のベクターを構築した。しかしながら、これら構築体では193位残基にて、BPI断片がコードする配列は終了している。上記したように、このDNAの宿主発現から得られた組換え生成物は、「rBPI(1−193)ala132」と称した。これら挿入体を含むベクターは、BPIのDNA挿入体の5’端を切断する、SalIでまずpING4533を消化し、192位残基の3’側に覆い被さる3bpを残すAlwNI で消化することで作製される。そして、得られた約700bpの断片は精製された。この断片は、二つのアニールした相補性オリゴヌクレオチド、BPI− 30:CTGTAGCTCGAGCCGC(配列番号:10)およびBPI−31:GGCTCGAGCTACAGAGT(配列番号:11)によって、 SstII−SalIでのpING4533消化から得られた大きな断片へ再接合された。これは、AlwNIとSstII部位の間の領域を、193位の残基(ロイシン)のためのコドン、停止コドン、およびXhoI制限5’部位と共にSstII部位に置換し、AlwNIとSstII部位双方を再生し、また、193位のアミノ酸(ロイシン)のためのコドン(CTG)の直後に停止コドンとTAGを置くことになる。得られたプラスミドをpING4223と称し、gPtマーカーを有していた。異なる選択マーカーを用いてベクターを生成するために、異なるSstII−SalIベクター断片が用いられていることを除けば、pING4223のために記載された方法と同じくして同様の構築体が作製される。例えば、gptマーカーの代わりに(ヒスチジノールに耐性を付与する)hisマーカーを含むことを除けば、pING4221はpING4223と同一であり、また、gptの代わりにDHFRマーカーを含むことを除けば、pING4222はpING4223と同一である。
rBPI(1−193)ala132をコードする挿入体、至適化されたKozakの翻訳開始部位、SstII部位の3’端のヒトゲノミックガンマ1重鎖のポリアデニル化および転写終了領域が、ヒト軽鎖ポリアデニアル化配列、続いてマウス軽鎖(カッパ)ゲノミック転写終了配列で置換されていることを除いて実質的にpING4221、 pING4222およびpING4223と同一である異なる選択マーカーを含んだ一連のベクターが構築された。並行した遺伝子発現の研究にて、軽鎖ポリアデニアル化シグナルおよび転写終止領域は、Sp2/0およびCH0−K1細胞でのBPI発現レベルの2.5〜5倍の増大に寄与しているものと思われる。
39:3521(1987)にてΔ8と称されている構築体にて記載されているBam HIリンカーが続くポリアデニル化シグナルから構成されている。約1350bp断片の残りの部分は、転写終止配列を供給するBglII−Bam HIマウスカッパ3’ゲノミック断片〔Xu et al.,J.Biol.Chem.261:3838(1986)の断片”D”〕から構成されている。この断片は、BPI挿入体の全部とSstIIでの消化、T4ポリメラーゼ処理、および(BPI挿入体の全部とベクターの一部を含む)NotI消化により得られたベクターの一部を含む3044bp断片であるpING4533と、約4574bpのBam HI−NotI断片の二つの断片による三片接合に用いた。得られたベクター、pING4537は、上記したゲノミック3’非翻訳領域の相違点を除いて pING4533と同一である。
rBPIコード配列の発現が、Abelsonマウス白血病ウィルス(A−MuLv)LTRプロモーターに代えてヒトサイトメガロウィルス(hCMV)の直接の初期エンハンサー/プロモーターの制御下にあることを除けば、pING4143とpING4150のそれぞれと同一であるpING4144と pING4151の二つのプラスミドを構築した。よって、pING4144とpING4151の双方は、第132位でのシステインからアラニンへの変異、至適化されたKozak翻訳開始配列、およびヒト軽鎖ポリ−A/マウスカッパゲノミック転写終止領域を含んでいる。当初のベクター(pING4143と pING4150)の第879位から第1708位のヌクレオチド領域を、本明細書に参考までに採り入れた、Boshart et al.,Cell41:521(1985)の図3に見られるように、第−598位から第+174位のヌクレオチドに対応するhCMVエンハンサー/プロモーターの領域と置換した。BPI発現ベクターにhCMVプロモーター領域に導入するために、抗体軽鎖挿入体の発現を進めるhCMVプロモーターを含むプラスミドpING2250からの断片を含む約1054bpのEc
oRI−SalI/hCMVプロモーターで、pING4222の断片を含む約1117bpのEcoRI−SalI/A−MuLv プロモーターでまず置換することで、プラスミドpING4538を構築した。pING4144を構築するために、三つの断片を接合した。すなわち、(1) pING4538のNotI−XhoI断片を含む約2955bpのrBPI(1−193)、(2)pING4537からの約1360bpのXhoI− Bam HI、および(3)pING4221からのhis遺伝子を含む約4770bpのBam HI−NotI断片である。
第132位のアラニン置換に代えて野性型(自然配列)のシステインを含むことを除けば、pING4143、pING4146、およびpING4150と同一である、プラスミドpING4145、 pING4148、およびpING4152を構築した。よって、これら三つのプラスミドは、rBPI(1−193)挿入体、至適化したKozakの翻訳開始配列、およびヒト軽鎖ポリA/マウスカッパゲノミック転写終止領域を含んでいた。これら三つのプラスミドは、以下のようにして構築した。 pING4145を構築するために、三つの断片を接合した。すなわち、(1)(第132位に野性型システインが含まれていることを除けば、 pING4221と同一である)pING4140からの断片を含む約3000bpのNot I−XhoIBPI(1−193)、(2)pING4537からの約1360bpのXhoI− Bam HI断片、および(3)pING4222からのDHFR遺伝子を含む約4150bpの Bam HI− Not I断片である。pING4152を構築するために、三つの断片を接合した。すなわち、(1)(第132位に野性型システインが含まれていることを除けば、pING4223と同一である)pING4142からの断片を含む約3000bpのNotI− XhoI断片、(2)pING4537からのXhoI−BamHI断片、および(3)pING4221からのhis遺伝子を含む約4770bpのBam HI−Not I断片である。

(rBPIのシステイン置換類似体の発現のための細胞の形質変換)
哺乳類細胞は、その好適な分泌、保持力、および発現タンパク質の翻訳後の修飾を許容
するので、哺乳類細胞は、本発明のrBPIタンパク質類似体の生成のための好適な宿主である。目下のところ、本発明の類似体の生成のための好適な哺乳類宿主細胞には、CH0−K1細胞(ATCC CCL61);CHO−DG44細胞、コロンビア大学のLawrence Chasin博士よりいただいたCHO Torontoのジヒドロ葉酸レダクターゼ欠損[DHFR−]変異株、Lawrence Chasin博士よりいただいたCHO−K1のDHFR−変異株であるCHO−DBX−11、Velo細胞(ATCC CRL81)、幼ハムスター腎臓(BHK)細胞(ATCC CCL10)、Sp2/0−Ag14ハイブリドーマ細胞(ATCC CRL1581)およびNSO骨髄腫(ECACC No.85110503)のような、繊維芽細胞およびリンパ球由来の細胞が含まれる。
al.,Proc.Natl.Acad.Sci.USA,78:2072−2076(1981)]、(3)ヌクレオシドの欠乏下でのDHFR−細胞の生長と、メトトレキセートの増大した濃度の下での遺伝子増幅を許容するジヒドロ葉酸レダクターゼ(DHFR)、(4)ヒスチジノルの存在下での生長を許容する Salmonella ryphimuriumの hisD遺伝子[Hartman,et al.,Proc.Natl.Acad.Sci.USA,85:8047−8051(1988)]、(5)インドールの存在下で(トリプトファン無しに)の生長を許容するE.coliのtrPB遺伝子[Hartman,et al.,Proc.Natl.Acad.Sci.USA,85:8047−8051(1988)]、および(6)グルタミンを欠いた培地での生長を許容するグルタミンシンターゼ遺伝子などがある。
プラスミドpING4533は、A−MuLvプロモーターに融合したrBPI(1−199)ala132をコードする遺伝子配列、至適化したKozakの翻訳開始配列、ヒトガンマ−1重鎖3’非翻訳領域、およびMPA−耐性細胞の選択のためのgptマーカーを含む。
DNAの40μgでエレクトロポレーションして形質変換した。エレクトロポレーションに続いて、細胞を非選択性HamのF12培地で24時間かけて再生した。そして、この細胞をトリプシン処理し、MPA(25μg/ml)とキサンチン(250μg/ml
)を補充したHamのF12培地で5×104細胞/mlの濃度で再懸濁し、そして、96ウェルプレートにて、104細胞/ウェルの割合で置いた。形質変換していないCHO−K1細胞は、MPAによるピリミジン合成の阻害のために、この培地では生長しなかった。
1.5L醗酵にて生長した。S−セファロースビーズ(50ml分画)から溶出した生成物のC4 HPLC分析により、生産効率を、120から140時間にわたって分析した。醗酵のこの段階での生産効率は、15から25μg/Lであった。
プラスミドpING4222は、A−MuLvプロモーターに融合したrBPI(1−193)ala132をコードするDNA、至適化したKozakの翻訳開始配列、ヒトガンマ−1重鎖3’非翻訳領域、およびヌクレオシドを含まない培地での形質転換した細胞の選択のためのマウスDHFR遺伝子を含む。
所望のrBPI生成物の至適な発現を行うために適用する手法は、第一マーカーでの第一発現プラスミドを有する細胞の形質変換、最高の生成体のスクリーニング、および異なる第二マーカーを有する第二の発現プラスミドを用いたスクリーニングした生成体の形質変換を含む。この手法を、Sp2/O細胞を用いた場合について、下記する。
ン、および8mMヒスチジノールを足したDMEM/FBS中に、約104個の細胞/ウェルになるように、96ウェルプレートに細胞を置いた。形質変換していない細胞は、MPAおよびヒスチジノールの存在下では生長しなかった。 1.5から2週間後に、形質転換した細胞が、96ウェルプレートで観察された。単一コロニーを含むウェルからの上清を、ELISAによって、rBPI反応性タンパク質の存在に関して分析した。
プラスミドpING4143は、A−MuLvプロモーターに融合したrBPI(1−193)ala132をコードするDNA、至適化したKozakの翻訳開始配列、ヒトガンマ−1重鎖3’非翻訳領域、およびMPA−耐性細胞の選択のためのgptマーカーに沿ったマウスカッパ軽鎖3’非翻訳領域を含む。Sp2/O細胞は、NotIで消化し、フェノール−クロロホルムで抽出し、そしてエタノール沈澱したpING4143 DNAの40μgを用いたエレクトロポレーションで形質変換した。エレクトロポレーションに続いて、非選択性DMEM培地で48時間置いて再生した。細胞を遠心分離し、MPA(6μg/ml)とキサンチン(250μg/ml)を足したDMEM培地中に5×104細胞/mlの濃度にまで再懸濁し、そして、104細胞/ウェルの割合で96ウェルプレートに置いた。約2週間後に、形質転換した細胞を含むコロニーが、96ウェルプレートで観察された。単一コロニーを含むウェルの上清を、ELISAによって、BPI−反応性タンパク質の存在に関して分析した。最も良好な生成体を24ウェルプレートに移した。10−7Mのデキサメトサンの有無により、24ウェルプレートでの停止した細胞生長から生産効率を測定した。最高の生成体であるクローン134は、デキサメトサンの有無により、それぞれ、約12μg/mlと28μg/mlの生成物を分泌した。
CHO−K1細胞系を、pING4533によるCHO−K1細胞の形質変換のための、上記A節に記載した方法により、pING4143で形質変換した。約2週間後に、単一コロニーを含む約800個のウェルからの上清を、ELISAによって、BPI−反応性タンパク質の存在に関して分析した。約9から 13μg/mlの生成物を分泌する最高の生成体は、醗酵時の生長のための血清を含まない培地に適用されうる。rBPI生成物を高レベルで生成する細胞系を提供するために、選択マーカーとしてhisあるいはneoを有するpING4150あるいはpING4154のようなベクターを用いて再度形質変換する。
A−MuLvプロモーターの代わりにヒトサイトメガロウィルス(hCMV)プロモーターを含むことを除けば、プラスミドpING4144は、pING4143と同様である。CHO−K1細胞系を、上記A節に記載した方法により、pING4144で形質変換した。約2週間後に、単一コロニーを含む約200個のウェルからの上清を、ELISAによって、BPI−反応性タンパク質の存在に関して分析した。最高の生成体を、24ウェルプレートに移し、酪酸ナトリウムを入れた24ウェルプレートにてrBPI発現を決定した。この分析にて、最高の生成体(クローン174)は、酪酸塩の欠乏下で約3から5μg/mlの生成物を、また5mM酪酸塩の存在下で約15から18μg/mlの生成物を分泌した。クローンC1771と改めて命名したこのクローンは、受託番号CRL
11246として、メリーランド州、20852、ロックヴィル、12301パークロウンドライブに所在のアメリカンタイプカルチャーコレクションに寄託されている。最高の生成体は、醗酵時の生長のための血清を含まない培地に適用されうる。 BPI生成物を高レベルで生成する細胞系を提供するために、選択マーカーとしてhisあるいはneoを有する、hCMVプロモーターの制御下でのBPI遺伝子を含む、pING4151あるいはpING4155のようなベクターを用いて再度形質変換されうる。
NSO細胞を、pING4143を用いたエレクトロポレーションで形質変換した。
約3週間後に、形質変換した細胞からなるコロニーが、96ウェルプレートにて観察された。単一コロニーを含むウェルからの上清を、ELISAによって、 BPI−反応性タンパク質の存在に関して分析した。最高の生成体を、24ウェルプレートに移した。生産効率を、細胞の生長が停止した24ウェル培養にて測定した。最高の生成体は、15から16μg/mlの生成物を分泌した。最高の生成体は、より生成能の優れた生成体を得るために、上記したようにして、pING4150のようなベクターで再度形質変換されうる。
ベクターpEE13[Bebbington,et al.,Biotechnology,10:169−175(1992)]にクローニングされた至適化したKozakの翻訳開始配列に融合したrBPI(1−193)ala132をコードするDNAを含むpING4132を用いたエレクトロポレーションによってNSO細胞を形質変換した。ベクターpEE13は、グルタミンが欠乏した培地でも生長できる形質変換体を選択するためのグルタミン合成遺伝子を含んでいる。
最高の生成体を、24ウェルプレートに移した。生産効率を、細胞の生長が停止した24ウェル培養にて測定した。7から15μg/mlの生成物を24ウェル培養で分泌した最
高の生成体は、様々な濃度のメチオニンスルホキシミンの存在下での増幅に適用されうる。
プラスミドpING4145は、A−MuLvプロモーターに融合したrBPI(1−193)ala132をコードするDNA、至適化したKozakの翻訳開始配列、マウスカッパ軽鎖3’非翻訳領域、およびMPA−耐性細胞の選択のためのgpt遺伝子を含む。 Sp2/O細胞は、Not Iで消化し、フェノール−クロロホルムで抽出し、そしてエタノール沈澱したpING4145 DNAの40μgを用いたエレクトロポレーションで形質変換した。エレクトロポレーションに続いて、非選択性DMEM培地で48時間置いて再生し、細胞を遠心分離し、MPA(6μg/ml)とキサンチン(250μg/ml)を足したDMEM培地中に5×104細胞/mlの濃度にまで再懸濁した。そして、104細胞/ウェルの割合で96ウェルプレートに置いた。約2週間後に、形質転換した細胞を含むコロニーが、96ウェルプレートで観察された。単一コロニーを含むウェルの上清を、ELISAによって、BPI−反応性タンパク質の存在に関して分析した。最も良好な生成体を24ウェルプレートに移し、10−7Mのデキサメトサンの有無により、24ウェルプレートでの停止した細胞生長から生産効率を測定した。BPIの発現を最高ならしめるために、最高の生成体であるSp2/O形質転換体は、A−MuLvプロモーターに融合したrBPI(1−193)ala132をコードする遺伝子配列、および形質転換体の選択のためのhis遺伝子を有するマウスカッパ軽鎖3’非翻訳領域を含むベクターを用いたエレクトロポレーションによって形質変換されうる。
CHO−K1細胞系を、上記A節に記載した方法により、pING4145DNAで形質変換した。約2週間後に、単一コロニーを含む約500から800個のウェルからの上清を、ELISAによって、BPI−反応性タンパク質の存在に関して分析した。最高の生成体を24ウェルプレートに移し、S−セファロースを含む24ウェルプレートにてBPI発現を決定した。この最高の生成体は、次に、醗酵時の生長のための血清を含まない培地に適用されうる。rBPI生成物を高レベルで生成する細胞系を提供するために、異なる選択マーカーを含むベクターを用いて、これらは再度形質変換されうる。
rBPI生成物が発現しうる他の真核細胞系は、rBPI生成物をコードするDNAを含む組換えバキュロウィルスで感染した昆虫細胞である。組換えウィルスを生成し、またそこからの組換え生成物の発現を達成するシステム(Invitrogen社、サンディエゴ、カリフォルニア州)が、通常利用されている。 31個のアミノ酸シグナル配列を含むrBPI(1−199)をコードするDNAは、pB1ueBac転移ベクター(Invitrogen社)のNheI部位にクロ−ニングされる。Sf9昆虫細胞(BRL社、ATCC CRL 1711)を、このベクターと、野性型AcMNPV(Autographa california多核ポリヒドロシスウィルス、Invitrogen社)を用いて共形質変換した。組換えウィルスプラークを同定し、精製し、そして、 BRL社から入手した実験説明書に従って、より高い力価の組換えウィルス性株を生成するために用いた。
0、150mM NaCl、50mM Tris−HCl、pH 8.0)で溶菌した。そして細胞を、細胞掻き取り器で、エッペンドルフ管に回収した。細胞溶解物を、2分間、マイクロ遠心分離した。溶解物の上清を、新しい管に移し、そして、−20℃で保管した。毎日の採取時間に得られた培地試料は、ELISAによってBPI含量が分析され、そして、溶解物を抗−BPI抗体を用いたウェスターン法によって解析した。
(rBPI(1−199)ala132およびrBPI(1−199)Ser135のin vitro での転写および翻訳のためのプラスミドの構築)
rBPI(1−199)をコードするDNAの源としてのプラスミドpIC127を用いてin vitroでの転写/翻訳の研究を行った。pIC127の構築は、以下のようにして行った。31個のアミノ酸シグナル配列を含むrBPI(1−199)をコードするDNAを、pGEM−72f(+)にあるBPIをコードするcDNAの全長を含んだプラスミドからPCR増幅した。プライマーBPI−3:GTCGACGCATGCGAGAGAACATGGC(配列番号:12)およびBPI−2: CCGCGGCTCGAGCTATATTTTGGTCAT(配列番号:9)を用いて、rBPIをコードする配列の3’端にXhoIとSstII部位を、また、5’端にSalI部位が組み込まれるようにして増幅を行った。得られたPCR増幅した断片は、pIC102を生成するために、プラスミドpT7T3 18u(Pharmacia LKB Technology社、ピスカタウェイ、ニュージャージー州)のマルチプルクローニング領域のSmaI部位にその切断端をクローニングした。
Bam HI部位は、pIC102のSalI部位を、また、AsP 718I部位はpIC102のSstII部位をそれぞれ切断した。切り出した断片の末端は、T4 DNAポリメラーゼで平滑化され、そして、PstIとEcoRIでまず消化され、T4 DNAポリメラーゼで平滑化したプラスミドpGEM1(ProMega社、マジソン、ウィスコンシン州)へその平滑化断片はクローニングされる。得られた構築体はpIC124と命名され、5’端がpGEM1のSp6プロモーターに近接するよう置かれた挿入体をコードするrBPI(1−199)を有している。
二つのプラスミド、pML101およびpML102を、rBPI(1−199)ala132およびrBPI(1−199)Ser135のin vitroでの転写/翻訳のためにpIC127を用いて構築した。
生成できる。rBPI(1−199)cys135およびrBPI(1−199)ala132における二量体生成物の発現が、細胞を含まないin vitroでの転写/翻訳システムを用いた結果によるものと考えられる。
上記したin vitro発現システムにて生成した標識付けしたタンパク質を、LPS結合活性について試験した。マイクロタイタープレートのウェルを、pH 9.4の0.1M Na2CO3/20mM EDTA(エチレンジアミン四酢酸)中のSalmonrlla minnesota R7(Rd変異体)(メタノール保管培養基中の5mg/ml)からのLPS(50μlウェルの全量で2μgのLPS)で被覆した。4℃で、一晩インキュベートした後、ウェルを水で洗浄し、37℃で乾燥した。次に、ウェルを215μlの Dulbecco’s−PBS/0.1%BSAを用いて、37℃で、3時間ブロックした。ブロック溶液を廃棄し、ウェルをPBS/0.2%Tween− 20で洗浄した。rBPI試料を、PBS/0.2%Tweenに全量が50μl(翻訳反応物の2μl)となるよう添加した。
(生成物の特徴)
(A.物理的特徴)
rBPIの生成物の特徴付けを、逆相(C4)HPLC、陽イオン交換(MA7C)HPLC、SDS−PAGE、および電子噴射イオン化質量分光測定法(ESI−MS)を用いて実施した。
酸ナトリウムおよび1000mM塩化ナトリウムを用いて、rBPI生成物を溶出した。そして、精製したrBPI生成物を、その物理的特徴を決定するために分析した。
rBPI生成物のSDS−PAGE分析を、14%ポリアクリルアミドゲルおよび、還元あるいは非還元条件下でのtris−グリシン緩衝液システムを用いて実施した。視覚化のためにクーマシーブルーあるいは銀染色のいずれかによって、タンパク質バンドを染色した。
MA7Cカラムを用いた陽イオン交換HPLCも、rBPI生成物の二量体の程度を測定するために使用した。40%緩衝液B(20mMMES、1M塩化ナトリウム、pH 5.5)で、1.0ml/分で平衡化したBio−Rad MA7Cカートリッジ(4.6×30mm、Bio−RadカタログNo.125−00556)を使用した。rBPI(1−199)生成物を、1mlの試料を、100μg/mlにまで希釈し、200μlの希釈試料をカラムに注入して分析した。rBPIを、40から100%の緩衝液Bの勾配で、6分間にわたって溶出した。緩衝液Aは、pH 5.5にて20mM MESを含んでいた。229nmにて吸収度を測定した。
イオン化質量分光測定法による分析 rBPI生成物の微小な不均一性が、逆相HPLCおよび電子噴射イオン化質量分光測定法(ESI−MS)によって示された。HPLC分析のために、Brownlee BU−300 C4カラムを、流速0.5ml/分で、37%移動相B(80%アセトニトリル/ 0.65%TFA)で平衡化した。rBP
I(1−199)の試料(各1ml)を、100μl/mlにまで希釈し、その50mlを注入した。カラムを、 2.5分間にわたって37%移動相Bで洗浄し、37から50%の移動相Bの勾配で、20分間にわたって溶出した。移動相Aは、5%アセトニトリル
/0.1%TFAであり、220nmで吸収度を測定した。
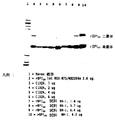
rBPI(1−199)ala132生成物を分析するために、同じ手順を踏んだ。図6に示したように、上記した第二(主要)ピークに対応する単一ピークとして、rBPI(1−199)ala132は溶出した。
rBPI(1−199)ala132に関してHPLCから取得された単一ピークを生成する溶出液の分析は、上記した第二(主要)ピークを生成する溶出液から取得したデータと同様のものであった。これらデータは、rBPI(1−199)生成物の切り詰めたカルボキシ末端を示すペプチドマッピングのデータと符合する。
ESI−MSデータも、rBPI生成物のアミノ末端での微小な不均一性の存在を示した。アミノ末端にアラニン残基を有するrBPI生成物の形態が知見され、そして、トリプシンペプチドの配列決定によって確認された。
MS研究は、各質量値が約119−120kDa大きいことを除けば、それが、第二(主要)
ピークを形成するタンパク質と同様の質量分布を有するタンパク質であることを示した。これらデータは、上記した第一(小さな)逆相HPLCピークを生成した溶出液が、質量値の均質な移動に寄与する、ジスルフィド結合したシステイン付加物を含むことを示唆している。
20mMクエン酸ナトリウム、0.15M塩化ナトリウム緩衝液、0.1%ポロキサマー、および0.002%ポリソルベート80を含むpH5.0の緩衝液中のrBPI(1−199)の試料(1mg/ml)を、好ましいとされる保管温度2から8℃と、高めの保管温度22℃と37℃で、8週間にわたってその保存安定性を決定すべく分析を行った。
表4に示した、2から8℃での保存の結果は、二量体の割合の(1%から4%への)増大が示されているが、システイン付加物あるいは試料中の粒子形成において顕著な増大は認められなかった。
薬剤組成物を含む様々なrBPIの濁度を決定するための実験を行った。この実施例にて、濁度とは薬剤組成物の崩れ(すなわち、タンパク質の三次元構造の喪失)、および/または粒子形成(大きな(>10μm)粒子を形成する各タンパク質間の相互作用)に関係する程度を指す。試験した薬剤組成物は、クエン酸緩衝液(20mMクエン酸ナトリウム/150mM塩化ナトリウム、pH5.0)、あるいは0.1%ポロキサマー188(ポリオキシプロピレン−ポリオキシエチレンブロックコポリマーを含むポロキサマー界面活性剤)および0.002%ポリソルベート80(ポリオキシエチレンソルビタン脂肪酸エステルを含むポリソルベート界面活性剤)を含むクエン酸緩衝液中のいずれかにある、rBPI(1−199)、rBPI(1−199)ala132あるいはrBPI(1−193)ala132のいずれかを含んでいた。上述したように、ポロキサマー/ポリソルベート界面活性剤系の組み合わせの使用は、本明細書に参考までに取り入れた、共同所有に係る、McGregor,et alによる、係属中の、1993年2月1日に出願された米国特許出願No. (代理人整理番号27129/31162)に教示されたような薬剤組成物を安定ならしめるのである。
収度を測定した。これに続いて、吸収度の経時的な変化を決定するために、350nmでの試料の吸収度を2分ごとに1時間にわたって計測した。
rBPI(1−199)ala132生成物のin vitro活性を、LPSへ生成物を結合することにより、LAL阻害分析によって、また、生成物の殺菌性強度により決定した。
E.coli(0111−B4株)あるいはS.minnesota(Rd変異体)リポ多糖(Sigma Chemical社、セントルイス、モンタナ州)の試料(それぞれ、20μgから60μg)を、LPSへのrBPI(1−199)ala132生成物の結合能力を決定するために用いた。
rBPI生成物の殺菌性強度を決定するために、E.coliをrBPI(1−199)またはrBPI(1−199)ala132類似体で処理し、殺菌性活性の測定として培地での成長阻害を観察することで、E.coliの培地での成長阻害の分析を行った。J5(E.coli株 0111:B4のUDP−4−エピメラーゼの少ない自然変異体)と命名された、短鎖LPSを有するE.coliの「自然」株をこの分析にて用いた。BPI に対する感受性をE.coliに付与する、トリエタノールアミンで緩衝化した鉱物塩培地(Simon,et al.,Proc.Natl.Acad.Scz,51:877(1964))にて細胞を成長させた。 細胞を洗浄し、約5×108細胞/mlの密度になるまで、0.9%塩化ナトリウム中で再懸濁を行った。
rBPI(1−199)ala132の LPSへの結合性を決定するために、LAL阻害検定を行った。LAL阻害検定は、Gazzano−Santoro,Infect.Immun.,60: 4754−4761(1992)に記載した方法を使用した。LAL検定の結果は、rBPI(1−199)と同等の10というIC−50値を、rBPI(1 −199)ala132が有していたことを実証するものであった。
これらデータは、LPSに結合するための野性型rBPI生成物に類似体が匹敵することを示すものである。
内毒素血症の動物モデルを、内毒素ショックに対するrBPI(1−199)ala132類似体とrBPI(1−199)の効果の比較を行うために用いた。
Claims (1)
- 第132位あるいは第135位のシステイン残基が異なるアミノ酸で置換されている、殺菌性/浸透性が向上したタンパク質のポリペプチド類似体あるいはその生物学的に活性な断片。
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US08/013,801 US5420019A (en) | 1993-02-02 | 1993-02-02 | Stable bactericidal/permeability-increasing protein muteins |
Related Parent Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP51821094A Division JP4139867B2 (ja) | 1993-02-02 | 1994-02-02 | 殺菌性/浸透性が向上した安定なタンパク質生成物およびそれを含む薬剤組成物 |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2006204005A Division JP2006321813A (ja) | 1993-02-02 | 2006-07-26 | 殺菌/浸透性が向上した安定なタンパク質生成物およびそれを含む薬剤組成物 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| JP2004254697A true JP2004254697A (ja) | 2004-09-16 |
Family
ID=21761817
Family Applications (4)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP51821094A Expired - Fee Related JP4139867B2 (ja) | 1993-02-02 | 1994-02-02 | 殺菌性/浸透性が向上した安定なタンパク質生成物およびそれを含む薬剤組成物 |
| JP2003355706A Withdrawn JP2004024276A (ja) | 1993-02-02 | 2003-10-15 | 殺菌/浸透性が向上した安定なタンパク質生成物およびそれを含む薬剤組成物 |
| JP2004108131A Withdrawn JP2004254697A (ja) | 1993-02-02 | 2004-03-31 | 殺菌/浸透性が向上した安定なタンパク質生成物およびそれを含む薬剤組成物 |
| JP2006204005A Withdrawn JP2006321813A (ja) | 1993-02-02 | 2006-07-26 | 殺菌/浸透性が向上した安定なタンパク質生成物およびそれを含む薬剤組成物 |
Family Applications Before (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP51821094A Expired - Fee Related JP4139867B2 (ja) | 1993-02-02 | 1994-02-02 | 殺菌性/浸透性が向上した安定なタンパク質生成物およびそれを含む薬剤組成物 |
| JP2003355706A Withdrawn JP2004024276A (ja) | 1993-02-02 | 2003-10-15 | 殺菌/浸透性が向上した安定なタンパク質生成物およびそれを含む薬剤組成物 |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2006204005A Withdrawn JP2006321813A (ja) | 1993-02-02 | 2006-07-26 | 殺菌/浸透性が向上した安定なタンパク質生成物およびそれを含む薬剤組成物 |
Country Status (19)
| Country | Link |
|---|---|
| US (6) | US5420019A (ja) |
| EP (3) | EP1310558B1 (ja) |
| JP (4) | JP4139867B2 (ja) |
| KR (1) | KR100361997B1 (ja) |
| CN (1) | CN1142996C (ja) |
| AT (3) | ATE196650T1 (ja) |
| AU (1) | AU693089B2 (ja) |
| CA (1) | CA2155004C (ja) |
| DE (3) | DE69434980T2 (ja) |
| DK (2) | DK0689592T3 (ja) |
| ES (2) | ES2150983T3 (ja) |
| FI (2) | FI112367B (ja) |
| GR (1) | GR3034492T3 (ja) |
| HK (1) | HK1014548A1 (ja) |
| NO (3) | NO315705B1 (ja) |
| NZ (1) | NZ262284A (ja) |
| PT (2) | PT689592E (ja) |
| WO (1) | WO1994018323A1 (ja) |
| ZA (1) | ZA94703B (ja) |
Families Citing this family (59)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US5198541A (en) * | 1987-08-11 | 1993-03-30 | New York University | Dna encoding bactericidal/permeability-increasing proteins |
| US6265187B1 (en) * | 1989-02-14 | 2001-07-24 | Incyte Pharmaceuticals, Inc. | Recombinant endotoxin-neutralizing proteins |
| US5840836A (en) * | 1989-02-17 | 1998-11-24 | Bayer Corporation | Pancreatic islet cell antigens obtained by molecular cloning |
| US6811989B1 (en) * | 1989-02-17 | 2004-11-02 | Bayer Corporation | Pancreatic islet cell antigens obtained by molecular cloning |
| US6274348B1 (en) | 1992-05-19 | 2001-08-14 | Xoma Corporation | Methods for the preparation of positively charged proteins |
| US5643570A (en) * | 1992-05-19 | 1997-07-01 | Xoma Corporation | BPI-immunoglobulin fusion proteins |
| US5420019A (en) * | 1993-02-02 | 1995-05-30 | Xoma Corporation | Stable bactericidal/permeability-increasing protein muteins |
| US5858974A (en) * | 1995-07-20 | 1999-01-12 | Xoma Corporation | Anti-fungal peptides |
| JPH08509703A (ja) * | 1993-03-12 | 1996-10-15 | ゾーマ コーポレイション | 殺菌/透過性増加タンパク質産物の治療用途 |
| US5652332A (en) * | 1993-03-12 | 1997-07-29 | Xoma | Biologically active peptides from functional domains of bactericidal/permeability-increasing protein and uses thereof |
| US6214789B1 (en) | 1993-03-12 | 2001-04-10 | Xoma Corporation | Treatment of mycobacterial diseases by administration of bactericidal/permeability-increasing protein products |
| AU7175694A (en) * | 1993-06-17 | 1995-01-17 | Xoma Corporation | Lipopolysaccharide binding protein derivatives |
| AU7330994A (en) * | 1993-07-14 | 1995-02-13 | Xoma Corporation | Method for potentiating bpi protein product bactericidal activity by administration of lbp protein products |
| US5770561A (en) * | 1993-07-14 | 1998-06-23 | Xoma Corporation | Method for potentiating BPI protein product bactericidal activity by administration of LBP protein products |
| WO1995008344A1 (en) * | 1993-09-22 | 1995-03-30 | Xoma Corporation | Method of treating gram-negative bacterial infection by administration of bactericidal/permeability-increasing (bpi) protein product and antibiotic |
| CN1133634A (zh) * | 1993-09-22 | 1996-10-16 | 爱克斯欧玛公司 | 定量体液中的bpi的方法 |
| US6759203B1 (en) | 1993-09-22 | 2004-07-06 | Xoma Corporation | Method for quantifying BPI in body fluids |
| AU703192B2 (en) * | 1994-01-14 | 1999-03-18 | Xoma Corporation | Anti-gram-positive bacterial methods and materials |
| CA2181165C (en) * | 1994-01-14 | 2007-06-05 | Roger G. Ii Little | Anti-fungal methods and materials |
| US5447913A (en) * | 1994-03-11 | 1995-09-05 | Xoma Corporation | Therapeutic uses of bactericidal/permeability-increasing protein dimer products |
| US5830860A (en) * | 1994-03-24 | 1998-11-03 | Regents Of The University Of Minnesota | Peptides with bactericidal activity and endotoxin neutralizing activity for gram negative bacteria and methods for their use |
| US5786324A (en) * | 1994-03-24 | 1998-07-28 | Regents Of The University Of Minnesota | Synthetic peptides with bactericidal activity and endotoxin neutralizing activity for gram negative bacteria and methods for their use |
| US5578568A (en) * | 1994-04-22 | 1996-11-26 | Xoma Corporation | Method of treating conditions associated with intestinal ischemia/reperfusion |
| US6271203B1 (en) | 1994-07-07 | 2001-08-07 | Xoma Corporation | Anti-protozoan methods and materials |
| US5646114A (en) * | 1994-07-11 | 1997-07-08 | Xoma Corporation | Anti-protozoan methods |
| AU4230396A (en) * | 1994-11-08 | 1996-05-31 | Novagen, Inc. | Method for in vitro protein synthesis |
| US5494896A (en) * | 1995-03-31 | 1996-02-27 | Xoma Corporation | Method of treating conditions associated with burn injuries |
| EP0861088A1 (en) * | 1995-11-14 | 1998-09-02 | Xoma Corporation | Bactericidal permeability increasing protein (bpi) for treating conditions associated with corneal injury |
| US5686414A (en) * | 1995-11-14 | 1997-11-11 | Xoma Corporation | Methods of treating conditions associated with corneal transplantation |
| US5856159A (en) * | 1996-03-27 | 1999-01-05 | Cytel Corporation | Production of galactosyltransferase |
| US5741779A (en) | 1996-05-10 | 1998-04-21 | Xoma Corporation | Antithrombotic materials and methods |
| CA2253837C (en) | 1996-05-10 | 2002-07-02 | Xoma Corporation | Therapeutic uses of bpi protein products for human meningococcemia |
| CA2255539C (en) * | 1996-05-23 | 2005-09-27 | Xoma Corporation | Therapeutic uses of bpi protein products in humans with hemorrhage due to trauma |
| CA2255865C (en) | 1996-05-24 | 2008-10-14 | The Regents Of The University Of Minnesota | Synthesis of soluble beta-sheet forming peptides |
| US5888973A (en) * | 1996-08-09 | 1999-03-30 | Xoma Corporation | Anti-chlamydial uses of BPI protein products |
| US6482796B2 (en) * | 1996-11-01 | 2002-11-19 | Xoma Corporation | Therapeutic uses of N-terminal BPI protein products in ANCA-positive patients |
| CN1062564C (zh) * | 1996-12-20 | 2001-02-28 | 周红 | 猪血中提纯杀菌性/通透性增加蛋白的方法 |
| CA2278924A1 (en) * | 1997-01-24 | 1998-07-30 | Avi Biopharma, Inc. | Method and conjugate for treating h. pylori infection |
| US6093573A (en) * | 1997-06-20 | 2000-07-25 | Xoma | Three-dimensional structure of bactericidal/permeability-increasing protein (BPI) |
| FR2767323B1 (fr) * | 1997-08-12 | 2001-01-05 | Synt Em | Peptides lineaires derives de peptides antibiotiques, leur preparation et leur utilisation pour vectoriser des substances actives |
| US6013631A (en) * | 1998-06-19 | 2000-01-11 | Xoma Corporation | Bactericidal/permeability-increasing protein (BPI) deletion analogs |
| US6156514A (en) | 1998-12-03 | 2000-12-05 | Sunol Molecular Corporation | Methods for making recombinant cells |
| US6242219B1 (en) * | 1999-03-18 | 2001-06-05 | Xoma (Us) Llc | Methods for recombinant peptide production |
| US7041644B2 (en) * | 1999-04-01 | 2006-05-09 | Xoma Technology | Therapeutic uses of BPI protein products in BPI-deficient humans |
| US6153584A (en) * | 1999-04-01 | 2000-11-28 | Levy; Ofer | Therapeutic uses of BPI protein products in BPI-deficient humans |
| AU1431101A (en) | 1999-10-04 | 2001-05-10 | University Of Maryland Biotechnology Institute | Novel adjuvant comprising a lipopolysaccharide antagonist |
| CA2430588A1 (en) * | 2000-12-01 | 2002-07-18 | Xoma Technology Ltd. | Modulation of pericyte proliferation using bpi protein products or bpi inhibitors |
| US20030049616A1 (en) * | 2001-01-08 | 2003-03-13 | Sydney Brenner | Enzymatic synthesis of oligonucleotide tags |
| AU2002248557A1 (en) * | 2001-03-05 | 2002-09-19 | Transtech Pharma, Inc. | High level insect expression of rage proteins |
| EP1572719A4 (en) | 2002-02-20 | 2007-11-14 | Univ Minnesota | PARTIAL PEPTIDE MIMETICS AND METHOD |
| ES2557741T3 (es) * | 2002-03-27 | 2016-01-28 | Immunex Corporation | Procedimientos para incrementar la producción de polipéptidos |
| DE60335794D1 (de) | 2002-03-29 | 2011-03-03 | Xoma Technology Ltd | Multigene vectoren und methoden für eine erhöhte expression von rekombinanten polypeptiden |
| US7906722B2 (en) * | 2005-04-19 | 2011-03-15 | Palo Alto Research Center Incorporated | Concentrating solar collector with solid optical element |
| EP1741440A1 (en) | 2005-07-08 | 2007-01-10 | Mellitus S.L. | Use of BPI protein for the treatment of disorders of the metabolism and cardiovascular disorders |
| MX2010003622A (es) | 2007-10-12 | 2010-04-14 | Hoffmann La Roche | Expresion proteinica a partir de acidos nucleicos multiples. |
| CN103648515B (zh) | 2011-04-05 | 2018-01-02 | 达纳-法伯癌症研究所有限公司 | 作为辐射缓和剂和辐射防护剂的bpi和其同源物 |
| GB201319621D0 (en) | 2013-11-06 | 2013-12-18 | Norwegian University Of Science And Technology | Antimicrobial agents and their use in therapy |
| GB201319620D0 (en) | 2013-11-06 | 2013-12-18 | Norwegian University Of Science And Technology | Immunosuppressive agents and their use in therapy |
| MA48723A (fr) | 2017-03-10 | 2020-04-08 | Hoffmann La Roche | Procédé de production d'anticorps multispécifiques |
Family Cites Families (31)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US4588585A (en) * | 1982-10-19 | 1986-05-13 | Cetus Corporation | Human recombinant cysteine depleted interferon-β muteins |
| US4853332A (en) * | 1982-10-19 | 1989-08-01 | Cetus Corporation | Structural genes, plasmids and transformed cells for producing cysteine depleted muteins of biologically active proteins |
| US4518584A (en) * | 1983-04-15 | 1985-05-21 | Cetus Corporation | Human recombinant interleukin-2 muteins |
| US4816566A (en) * | 1983-06-01 | 1989-03-28 | Hoffmann-La Roche, Inc. | Polypeptides having interferon activity |
| US4879226A (en) * | 1984-04-06 | 1989-11-07 | Asahi Kasei Kogyo Kabushiki Kaisha | Novel human physiologically active polypeptide |
| US4677063A (en) * | 1985-05-02 | 1987-06-30 | Cetus Corporation | Human tumor necrosis factor |
| US4677064A (en) * | 1984-11-09 | 1987-06-30 | Cetus Corporation | Human tumor necrosis factor |
| US4959314A (en) * | 1984-11-09 | 1990-09-25 | Cetus Corporation | Cysteine-depleted muteins of biologically active proteins |
| US5206344A (en) * | 1985-06-26 | 1993-04-27 | Cetus Oncology Corporation | Interleukin-2 muteins and polymer conjugation thereof |
| US5198541A (en) * | 1987-08-11 | 1993-03-30 | New York University | Dna encoding bactericidal/permeability-increasing proteins |
| US5576292A (en) * | 1987-08-11 | 1996-11-19 | New York University | Biologically active bactericidal/permeability-increasing protein fragments |
| WO1989001486A1 (en) * | 1987-08-11 | 1989-02-23 | New York University | Biologically active bactericidal/permeability-increasing protein fragments |
| US5308834A (en) * | 1989-02-14 | 1994-05-03 | Incyte Pharmaceuticals, Inc. | Treatment of endotoxin-associated shock and prevention thereof using a BPI protein |
| US5089274A (en) * | 1989-02-14 | 1992-02-18 | Incyte Pharmaceuticals, Inc. | Use of bactericidal/permeability increasing protein or biologically active analogs thereof to treat endotoxin-related disorders |
| US5234912A (en) * | 1989-02-14 | 1993-08-10 | Incyte Pharmaceuticals, Inc. | Pharmaceutical compositions comprising recombinant BPI proteins and a lipid carrier and uses thereof |
| US5770694A (en) * | 1990-08-13 | 1998-06-23 | Incyte Pharmaceuticals, Inc. | Genetically engineered BPI variant proteins |
| US5334584A (en) * | 1989-02-14 | 1994-08-02 | Incyte Pharamaceuticals, Inc. | Recombinant, non-glycosylated bpi protein and uses thereof |
| US5171739A (en) * | 1989-02-14 | 1992-12-15 | Incyte Pharmaceuticals, Inc. | Treatment of endotoxin-associated shock and preventation thereof using a BPI protein |
| US5166322A (en) * | 1989-04-21 | 1992-11-24 | Genetics Institute | Cysteine added variants of interleukin-3 and chemical modifications thereof |
| DE69128968T2 (de) * | 1990-12-03 | 1998-10-08 | Univ New York | Biologisch aktive bakterizide/permeabilitätserhöhende proteinbruchstücke |
| JPH07502642A (ja) * | 1991-09-26 | 1995-03-23 | インサイト ファーマシューティカルズ,インコーポレイテッド | 新規な形態のリポ糖結合性タンパク質(lbp) |
| ATE153202T1 (de) * | 1991-10-31 | 1997-05-15 | Kwang Sil Lee | Elektronisches identifizierungssystem mit automatischer fernantwort und identifizierungsverfahren hierzu |
| US5643570A (en) * | 1992-05-19 | 1997-07-01 | Xoma Corporation | BPI-immunoglobulin fusion proteins |
| ES2132239T3 (es) * | 1992-05-19 | 1999-08-16 | Xoma Corp | Metodos mejorados para la preparacion de proteinas de union a endotoxinas. |
| US5420019A (en) * | 1993-02-02 | 1995-05-30 | Xoma Corporation | Stable bactericidal/permeability-increasing protein muteins |
| AU695125B2 (en) * | 1993-02-02 | 1998-08-06 | Xoma Corporation | Pharmaceutical compositions containing bactericidal permeability increasing protein and a surfactant |
| ATE169304T1 (de) * | 1993-03-12 | 1998-08-15 | Xoma Corp | Biologisch aktive peptide aus funktionellen domaenen des bakteriziden/ die permeabilitaet erhoehenden proteins und ihre verwendung |
| US5348942A (en) * | 1993-03-12 | 1994-09-20 | Xoma Corporation | Therapeutic uses of bactericidal/permeability increasing protein products |
| CA2157925C (en) * | 1993-03-12 | 2000-06-06 | Lewis H. Lambert, Jr. | Treatment of mycobacterial diseases by administration of bactericidal/permeability-increasing protein products |
| JPH08509703A (ja) * | 1993-03-12 | 1996-10-15 | ゾーマ コーポレイション | 殺菌/透過性増加タンパク質産物の治療用途 |
| AU6942994A (en) * | 1993-04-30 | 1994-11-21 | Incyte Pharmaceuticals, Inc. | Recombinant bpi-based and lbp-based proteins, nucleic acid molecules encoding same, methods of producing same, and uses thereof |
-
1993
- 1993-02-02 US US08/013,801 patent/US5420019A/en not_active Expired - Lifetime
-
1994
- 1994-02-02 WO PCT/US1994/001235 patent/WO1994018323A1/en active IP Right Grant
- 1994-02-02 AT AT94908704T patent/ATE196650T1/de not_active IP Right Cessation
- 1994-02-02 NZ NZ262284A patent/NZ262284A/en not_active IP Right Cessation
- 1994-02-02 AT AT03000252T patent/ATE363535T1/de not_active IP Right Cessation
- 1994-02-02 JP JP51821094A patent/JP4139867B2/ja not_active Expired - Fee Related
- 1994-02-02 EP EP03000252A patent/EP1310558B1/en not_active Expired - Lifetime
- 1994-02-02 AU AU61702/94A patent/AU693089B2/en not_active Ceased
- 1994-02-02 KR KR1019950703197A patent/KR100361997B1/ko not_active IP Right Cessation
- 1994-02-02 CA CA2155004A patent/CA2155004C/en not_active Expired - Fee Related
- 1994-02-02 CN CNB941916723A patent/CN1142996C/zh not_active Expired - Fee Related
- 1994-02-02 PT PT94908704T patent/PT689592E/pt unknown
- 1994-02-02 EP EP00103901A patent/EP1013760B1/en not_active Expired - Lifetime
- 1994-02-02 DE DE69434980T patent/DE69434980T2/de not_active Expired - Fee Related
- 1994-02-02 DE DE69426019T patent/DE69426019T2/de not_active Expired - Lifetime
- 1994-02-02 AT AT00103901T patent/ATE230797T1/de not_active IP Right Cessation
- 1994-02-02 DK DK94908704T patent/DK0689592T3/da active
- 1994-02-02 ES ES94908704T patent/ES2150983T3/es not_active Expired - Lifetime
- 1994-02-02 DK DK03000252T patent/DK1310558T3/da active
- 1994-02-02 EP EP94908704A patent/EP0689592B1/en not_active Expired - Lifetime
- 1994-02-02 ES ES03000252T patent/ES2288199T3/es not_active Expired - Lifetime
- 1994-02-02 ZA ZA94703A patent/ZA94703B/xx unknown
- 1994-02-02 DE DE69431995T patent/DE69431995T2/de not_active Expired - Fee Related
- 1994-02-02 PT PT03000252T patent/PT1310558E/pt unknown
-
1995
- 1995-04-28 US US08/430,417 patent/US5674834A/en not_active Expired - Lifetime
- 1995-06-06 US US08/466,822 patent/US5827816A/en not_active Expired - Lifetime
- 1995-08-01 FI FI953658A patent/FI112367B/fi active
- 1995-08-01 NO NO19953033A patent/NO315705B1/no not_active IP Right Cessation
-
1998
- 1998-12-28 HK HK98115811A patent/HK1014548A1/xx not_active IP Right Cessation
-
1999
- 1999-10-19 US US09/425,034 patent/US6433140B1/en not_active Expired - Fee Related
-
2000
- 2000-09-28 GR GR20000402074T patent/GR3034492T3/el not_active IP Right Cessation
-
2002
- 2002-07-03 NO NO20023224A patent/NO319156B1/no unknown
- 2002-07-16 US US10/196,460 patent/US6828418B2/en not_active Expired - Fee Related
-
2003
- 2003-08-26 FI FI20031199A patent/FI115633B/fi active IP Right Grant
- 2003-10-15 JP JP2003355706A patent/JP2004024276A/ja not_active Withdrawn
-
2004
- 2004-03-31 JP JP2004108131A patent/JP2004254697A/ja not_active Withdrawn
- 2004-11-03 US US10/980,722 patent/US20060009383A1/en not_active Abandoned
-
2005
- 2005-02-24 NO NO20050998A patent/NO20050998D0/no not_active Application Discontinuation
-
2006
- 2006-07-26 JP JP2006204005A patent/JP2006321813A/ja not_active Withdrawn
Also Published As
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP2004254697A (ja) | 殺菌/浸透性が向上した安定なタンパク質生成物およびそれを含む薬剤組成物 | |
| US5643570A (en) | BPI-immunoglobulin fusion proteins | |
| US5703038A (en) | Therapeutic uses of bactericidal-permeability-increasing protein dimer products | |
| JPH01501283A (ja) | 新規な型のコロニー刺激因子―1 | |
| HUT50870A (en) | Process for producing new recombinant and chimeric antibodies against human adenocarcinoma antigene | |
| JP2007510403A (ja) | ケモカイン変異体の治療への使用 | |
| US5185431A (en) | Recombinant natural killer cell activator | |
| US8932601B2 (en) | Hybrid and chimeric polypeptides that regulate activation of complement | |
| JPH02255699A (ja) | 新規血液抗凝固物質及びその製法 | |
| JP2012511309A (ja) | Ec−sodのカルボキシル末端のアポプチンタンパク質導入ドメインの融合蛋白質 | |
| WO2021143733A1 (zh) | 一种融合蛋白及其制法和用途 | |
| US6274348B1 (en) | Methods for the preparation of positively charged proteins | |
| JP2839837B2 (ja) | 顆粒球コロニー刺激因子受容体のリガンド結合領域蛋白質をコードしているdna |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20050907 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20051117 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20051124 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20060306 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20060331 |
|
| A761 | Written withdrawal of application |
Free format text: JAPANESE INTERMEDIATE CODE: A761 Effective date: 20080219 |