CN111406111A - 一种改进的生物技术生产l-色氨酸的方法 - Google Patents
一种改进的生物技术生产l-色氨酸的方法 Download PDFInfo
- Publication number
- CN111406111A CN111406111A CN201880073668.5A CN201880073668A CN111406111A CN 111406111 A CN111406111 A CN 111406111A CN 201880073668 A CN201880073668 A CN 201880073668A CN 111406111 A CN111406111 A CN 111406111A
- Authority
- CN
- China
- Prior art keywords
- ala
- leu
- val
- gly
- ser
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 title claims description 60
- 229960004799 tryptophan Drugs 0.000 title claims description 21
- 238000004519 manufacturing process Methods 0.000 title claims description 19
- RWZYAGGXGHYGMB-UHFFFAOYSA-N anthranilic acid Chemical compound NC1=CC=CC=C1C(O)=O RWZYAGGXGHYGMB-UHFFFAOYSA-N 0.000 claims abstract description 219
- 230000001580 bacterial effect Effects 0.000 claims abstract description 36
- 230000002829 reductive effect Effects 0.000 claims abstract description 22
- 230000035945 sensitivity Effects 0.000 claims abstract description 20
- NQEQTYPJSIEPHW-UHFFFAOYSA-N 1-C-(indol-3-yl)glycerol 3-phosphate Chemical compound C1=CC=C2C(C(O)C(COP(O)(O)=O)O)=CNC2=C1 NQEQTYPJSIEPHW-UHFFFAOYSA-N 0.000 claims abstract description 13
- ALYNCZNDIQEVRV-UHFFFAOYSA-N 4-aminobenzoic acid Chemical compound NC1=CC=C(C(O)=O)C=C1 ALYNCZNDIQEVRV-UHFFFAOYSA-N 0.000 claims abstract description 5
- 229960004050 aminobenzoic acid Drugs 0.000 claims abstract description 4
- 230000000694 effects Effects 0.000 claims description 65
- 102000004190 Enzymes Human genes 0.000 claims description 42
- 108090000790 Enzymes Proteins 0.000 claims description 42
- 241000588724 Escherichia coli Species 0.000 claims description 21
- 230000005764 inhibitory process Effects 0.000 claims description 20
- COLNVLDHVKWLRT-QMMMGPOBSA-N L-phenylalanine Chemical compound OC(=O)[C@@H](N)CC1=CC=CC=C1 COLNVLDHVKWLRT-QMMMGPOBSA-N 0.000 claims description 17
- 108010037870 Anthranilate Synthase Proteins 0.000 claims description 14
- 150000001413 amino acids Chemical class 0.000 claims description 13
- 230000035772 mutation Effects 0.000 claims description 13
- 229940024606 amino acid Drugs 0.000 claims description 10
- 235000001014 amino acid Nutrition 0.000 claims description 10
- 240000004808 Saccharomyces cerevisiae Species 0.000 claims description 9
- 235000014680 Saccharomyces cerevisiae Nutrition 0.000 claims description 9
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 claims description 8
- 230000002401 inhibitory effect Effects 0.000 claims description 8
- 238000000034 method Methods 0.000 claims description 8
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 claims description 7
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Natural products OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 claims description 6
- KZSNJWFQEVHDMF-UHFFFAOYSA-N Valine Natural products CC(C)C(N)C(O)=O KZSNJWFQEVHDMF-UHFFFAOYSA-N 0.000 claims description 5
- 239000004474 valine Substances 0.000 claims description 5
- 241000228212 Aspergillus Species 0.000 claims description 4
- 241000228245 Aspergillus niger Species 0.000 claims description 4
- COLNVLDHVKWLRT-UHFFFAOYSA-N phenylalanine Natural products OC(=O)C(N)CC1=CC=CC=C1 COLNVLDHVKWLRT-UHFFFAOYSA-N 0.000 claims description 4
- 241000894006 Bacteria Species 0.000 claims description 3
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 claims description 3
- KZSNJWFQEVHDMF-BYPYZUCNSA-N L-valine Chemical compound CC(C)[C@H](N)C(O)=O KZSNJWFQEVHDMF-BYPYZUCNSA-N 0.000 claims description 3
- 239000004472 Lysine Substances 0.000 claims description 3
- ONIBWKKTOPOVIA-UHFFFAOYSA-N Proline Natural products OC(=O)C1CCCN1 ONIBWKKTOPOVIA-UHFFFAOYSA-N 0.000 claims description 3
- 239000004471 Glycine Substances 0.000 claims description 2
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 claims description 2
- AGPKZVBTJJNPAG-WHFBIAKZSA-N L-isoleucine Chemical compound CC[C@H](C)[C@H](N)C(O)=O AGPKZVBTJJNPAG-WHFBIAKZSA-N 0.000 claims description 2
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 claims description 2
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 claims description 2
- 235000004279 alanine Nutrition 0.000 claims description 2
- 238000012258 culturing Methods 0.000 claims description 2
- 229960000310 isoleucine Drugs 0.000 claims description 2
- AGPKZVBTJJNPAG-UHFFFAOYSA-N isoleucine Natural products CCC(C)C(N)C(O)=O AGPKZVBTJJNPAG-UHFFFAOYSA-N 0.000 claims description 2
- 241000235070 Saccharomyces Species 0.000 claims 2
- 239000003112 inhibitor Substances 0.000 abstract description 8
- 239000013612 plasmid Substances 0.000 description 44
- 108020004414 DNA Proteins 0.000 description 31
- 108090000623 proteins and genes Proteins 0.000 description 25
- 210000004027 cell Anatomy 0.000 description 24
- NQEQTYPJSIEPHW-MNOVXSKESA-N (1S,2R)-1-C-(indol-3-yl)glycerol 3-phosphate Chemical compound C1=CC=C2C([C@H](O)[C@@H](COP(O)(O)=O)O)=CNC2=C1 NQEQTYPJSIEPHW-MNOVXSKESA-N 0.000 description 23
- 230000015572 biosynthetic process Effects 0.000 description 20
- 238000000855 fermentation Methods 0.000 description 19
- 230000004151 fermentation Effects 0.000 description 19
- 230000035508 accumulation Effects 0.000 description 17
- 238000009825 accumulation Methods 0.000 description 17
- SITLTJHOQZFJGG-UHFFFAOYSA-N N-L-alpha-glutamyl-L-valine Natural products CC(C)C(C(O)=O)NC(=O)C(N)CCC(O)=O SITLTJHOQZFJGG-UHFFFAOYSA-N 0.000 description 15
- VPZXBVLAVMBEQI-UHFFFAOYSA-N glycyl-DL-alpha-alanine Natural products OC(=O)C(C)NC(=O)CN VPZXBVLAVMBEQI-UHFFFAOYSA-N 0.000 description 15
- QQACQIHVWCVBBR-GVARAGBVSA-N Ala-Ile-Tyr Chemical compound [H]N[C@@H](C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O QQACQIHVWCVBBR-GVARAGBVSA-N 0.000 description 12
- 108091028043 Nucleic acid sequence Proteins 0.000 description 12
- 108010010147 glycylglutamine Proteins 0.000 description 12
- 235000018102 proteins Nutrition 0.000 description 12
- 102000004169 proteins and genes Human genes 0.000 description 12
- OTCJMMRQBVDQRK-DCAQKATOSA-N Arg-Asp-Leu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O OTCJMMRQBVDQRK-DCAQKATOSA-N 0.000 description 11
- -1 phosphoribosylaminobenzoic acid Chemical compound 0.000 description 11
- 108010026333 seryl-proline Proteins 0.000 description 11
- 238000003786 synthesis reaction Methods 0.000 description 11
- WTFXTQVDAKGDEY-UHFFFAOYSA-N (-)-chorismic acid Natural products OC1C=CC(C(O)=O)=CC1OC(=C)C(O)=O WTFXTQVDAKGDEY-UHFFFAOYSA-N 0.000 description 10
- IJGRMHOSHXDMSA-UHFFFAOYSA-N Atomic nitrogen Chemical compound N#N IJGRMHOSHXDMSA-UHFFFAOYSA-N 0.000 description 10
- WTFXTQVDAKGDEY-HTQZYQBOSA-N Chorismic acid Natural products O[C@@H]1C=CC(C(O)=O)=C[C@H]1OC(=C)C(O)=O WTFXTQVDAKGDEY-HTQZYQBOSA-N 0.000 description 10
- 230000014509 gene expression Effects 0.000 description 10
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 9
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 9
- 108010047495 alanylglycine Proteins 0.000 description 9
- 239000008103 glucose Substances 0.000 description 9
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 8
- LFSQWRSVPNKJGP-WDCWCFNPSA-N Leu-Thr-Glu Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@H](C(O)=O)CCC(O)=O LFSQWRSVPNKJGP-WDCWCFNPSA-N 0.000 description 8
- 108010050848 glycylleucine Proteins 0.000 description 8
- 108010061238 threonyl-glycine Proteins 0.000 description 8
- RUXQNKVQSKOOBS-JURCDPSOSA-N Ala-Phe-Ile Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O RUXQNKVQSKOOBS-JURCDPSOSA-N 0.000 description 7
- DCVYRWFAMZFSDA-ZLUOBGJFSA-N Ala-Ser-Ala Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CO)C(=O)N[C@@H](C)C(O)=O DCVYRWFAMZFSDA-ZLUOBGJFSA-N 0.000 description 7
- NTAZNGWBXRVEDJ-FXQIFTODSA-N Arg-Asp-Asp Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O NTAZNGWBXRVEDJ-FXQIFTODSA-N 0.000 description 7
- XQDGOJPVMSWZSO-SRVKXCTJSA-N Gln-Pro-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)[C@@H]1CCCN1C(=O)[C@H](CCC(=O)N)N XQDGOJPVMSWZSO-SRVKXCTJSA-N 0.000 description 7
- LPIKVBWNNVFHCQ-GUBZILKMSA-N Gln-Ser-Leu Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(O)=O LPIKVBWNNVFHCQ-GUBZILKMSA-N 0.000 description 7
- HXIDVIFHRYRXLZ-NAKRPEOUSA-N Ile-Ser-Val Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(=O)O)N HXIDVIFHRYRXLZ-NAKRPEOUSA-N 0.000 description 7
- CFZZDVMBRYFFNU-QWRGUYRKSA-N Leu-His-Gly Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CNC=N1)C(=O)NCC(O)=O CFZZDVMBRYFFNU-QWRGUYRKSA-N 0.000 description 7
- QJXHMYMRGDOHRU-NHCYSSNCSA-N Leu-Ile-Gly Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)NCC(O)=O QJXHMYMRGDOHRU-NHCYSSNCSA-N 0.000 description 7
- 108090000364 Ligases Proteins 0.000 description 7
- 102000003960 Ligases Human genes 0.000 description 7
- FUMGHWDRRFCKEP-CIUDSAMLSA-N Ser-Leu-Ala Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C)C(O)=O FUMGHWDRRFCKEP-CIUDSAMLSA-N 0.000 description 7
- AEFJNECXZCODJM-UWVGGRQHSA-N Val-Val-Gly Chemical compound CC(C)[C@H]([NH3+])C(=O)N[C@@H](C(C)C)C(=O)NCC([O-])=O AEFJNECXZCODJM-UWVGGRQHSA-N 0.000 description 7
- 125000000539 amino acid group Chemical group 0.000 description 7
- 108010018691 arginyl-threonyl-arginine Proteins 0.000 description 7
- 230000003197 catalytic effect Effects 0.000 description 7
- 238000010276 construction Methods 0.000 description 7
- 229940023064 escherichia coli Drugs 0.000 description 7
- 108010049041 glutamylalanine Proteins 0.000 description 7
- 108010092114 histidylphenylalanine Proteins 0.000 description 7
- 239000000543 intermediate Substances 0.000 description 7
- 108010057821 leucylproline Proteins 0.000 description 7
- 108010048818 seryl-histidine Proteins 0.000 description 7
- 108010044087 AS-I toxin Proteins 0.000 description 6
- CXRCVCURMBFFOL-FXQIFTODSA-N Ala-Ala-Pro Chemical compound C[C@H](N)C(=O)N[C@@H](C)C(=O)N1CCC[C@H]1C(O)=O CXRCVCURMBFFOL-FXQIFTODSA-N 0.000 description 6
- SKHCUBQVZJHOFM-NAKRPEOUSA-N Ala-Arg-Ile Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O SKHCUBQVZJHOFM-NAKRPEOUSA-N 0.000 description 6
- GSCLWXDNIMNIJE-ZLUOBGJFSA-N Ala-Asp-Asn Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O GSCLWXDNIMNIJE-ZLUOBGJFSA-N 0.000 description 6
- SFNFGFDRYJKZKN-XQXXSGGOSA-N Ala-Gln-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)N)NC(=O)[C@H](C)N)O SFNFGFDRYJKZKN-XQXXSGGOSA-N 0.000 description 6
- YIGLXQRFQVWFEY-NRPADANISA-N Ala-Gln-Val Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](C(C)C)C(O)=O YIGLXQRFQVWFEY-NRPADANISA-N 0.000 description 6
- PNALXAODQKTNLV-JBDRJPRFSA-N Ala-Ile-Ala Chemical compound C[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](C)C(O)=O PNALXAODQKTNLV-JBDRJPRFSA-N 0.000 description 6
- QJABSQFUHKHTNP-SYWGBEHUSA-N Ala-Ile-Trp Chemical compound [H]N[C@@H](C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(O)=O QJABSQFUHKHTNP-SYWGBEHUSA-N 0.000 description 6
- SUMYEVXWCAYLLJ-GUBZILKMSA-N Ala-Leu-Gln Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(O)=O SUMYEVXWCAYLLJ-GUBZILKMSA-N 0.000 description 6
- MDNAVFBZPROEHO-UHFFFAOYSA-N Ala-Lys-Val Natural products CC(C)C(C(O)=O)NC(=O)C(NC(=O)C(C)N)CCCCN MDNAVFBZPROEHO-UHFFFAOYSA-N 0.000 description 6
- BTRULDJUUVGRNE-DCAQKATOSA-N Ala-Pro-Lys Chemical compound C[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCCCN)C(O)=O BTRULDJUUVGRNE-DCAQKATOSA-N 0.000 description 6
- PEEYDECOOVQKRZ-DLOVCJGASA-N Ala-Ser-Phe Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O PEEYDECOOVQKRZ-DLOVCJGASA-N 0.000 description 6
- KTXKIYXZQFWJKB-VZFHVOOUSA-N Ala-Thr-Ser Chemical compound [H]N[C@@H](C)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CO)C(O)=O KTXKIYXZQFWJKB-VZFHVOOUSA-N 0.000 description 6
- ZCUFMRIQCPNOHZ-NRPADANISA-N Ala-Val-Gln Chemical compound C[C@@H](C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCC(=O)N)C(=O)O)N ZCUFMRIQCPNOHZ-NRPADANISA-N 0.000 description 6
- YJHKTAMKPGFJCT-NRPADANISA-N Ala-Val-Glu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O YJHKTAMKPGFJCT-NRPADANISA-N 0.000 description 6
- RWDVGVPHEWOZMO-GUBZILKMSA-N Arg-Cys-Val Chemical compound CC(C)[C@H](NC(=O)[C@H](CS)NC(=O)[C@@H](N)CCCNC(N)=N)C(O)=O RWDVGVPHEWOZMO-GUBZILKMSA-N 0.000 description 6
- HPKSHFSEXICTLI-CIUDSAMLSA-N Arg-Glu-Ala Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(O)=O HPKSHFSEXICTLI-CIUDSAMLSA-N 0.000 description 6
- NKBQZKVMKJJDLX-SRVKXCTJSA-N Arg-Glu-Leu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O NKBQZKVMKJJDLX-SRVKXCTJSA-N 0.000 description 6
- FKQITMVNILRUCQ-IHRRRGAJSA-N Arg-Phe-Asp Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(O)=O)C(O)=O FKQITMVNILRUCQ-IHRRRGAJSA-N 0.000 description 6
- GOVUDFOGXOONFT-VEVYYDQMSA-N Asn-Arg-Thr Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)O)C(O)=O GOVUDFOGXOONFT-VEVYYDQMSA-N 0.000 description 6
- JQSWHKKUZMTOIH-QWRGUYRKSA-N Asn-Gly-Phe Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)O)NC(=O)CNC(=O)[C@H](CC(=O)N)N JQSWHKKUZMTOIH-QWRGUYRKSA-N 0.000 description 6
- NLDNNZKUSLAYFW-NHCYSSNCSA-N Asn-Lys-Val Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C(C)C)C(O)=O NLDNNZKUSLAYFW-NHCYSSNCSA-N 0.000 description 6
- KZYSHAMXEBPJBD-JRQIVUDYSA-N Asn-Thr-Tyr Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O KZYSHAMXEBPJBD-JRQIVUDYSA-N 0.000 description 6
- MYRLSKYSMXNLLA-LAEOZQHASA-N Asn-Val-Glu Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O MYRLSKYSMXNLLA-LAEOZQHASA-N 0.000 description 6
- CBHVAFXKOYAHOY-NHCYSSNCSA-N Asn-Val-Leu Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O CBHVAFXKOYAHOY-NHCYSSNCSA-N 0.000 description 6
- HPNDBHLITCHRSO-WHFBIAKZSA-N Asp-Ala-Gly Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](C)C(=O)NCC(O)=O HPNDBHLITCHRSO-WHFBIAKZSA-N 0.000 description 6
- LIVXPXUVXFRWNY-CIUDSAMLSA-N Asp-Lys-Ala Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C)C(O)=O LIVXPXUVXFRWNY-CIUDSAMLSA-N 0.000 description 6
- GGRSYTUJHAZTFN-IHRRRGAJSA-N Asp-Pro-Tyr Chemical compound C1C[C@H](N(C1)C(=O)[C@H](CC(=O)O)N)C(=O)N[C@@H](CC2=CC=C(C=C2)O)C(=O)O GGRSYTUJHAZTFN-IHRRRGAJSA-N 0.000 description 6
- JSNWZMFSLIWAHS-HJGDQZAQSA-N Asp-Thr-Leu Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC(C)C)C(=O)O)NC(=O)[C@H](CC(=O)O)N)O JSNWZMFSLIWAHS-HJGDQZAQSA-N 0.000 description 6
- LBOLGUYQEPZSKM-YUMQZZPRSA-N Cys-Gly-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)CNC(=O)[C@H](CS)N LBOLGUYQEPZSKM-YUMQZZPRSA-N 0.000 description 6
- VXLXATVURDNDCG-CIUDSAMLSA-N Cys-Lys-Asp Chemical compound C(CCN)C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)O)NC(=O)[C@H](CS)N VXLXATVURDNDCG-CIUDSAMLSA-N 0.000 description 6
- VIOQRFNAZDMVLO-NRPADANISA-N Cys-Val-Glu Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O VIOQRFNAZDMVLO-NRPADANISA-N 0.000 description 6
- 102220504690 Cytochrome P450 11B1, mitochondrial_S60G_mutation Human genes 0.000 description 6
- NNQHEEQNPQYPGL-FXQIFTODSA-N Gln-Ala-Gln Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CCC(N)=O)C(O)=O NNQHEEQNPQYPGL-FXQIFTODSA-N 0.000 description 6
- PHZYLYASFWHLHJ-FXQIFTODSA-N Gln-Asn-Glu Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O PHZYLYASFWHLHJ-FXQIFTODSA-N 0.000 description 6
- CYTSBCIIEHUPDU-ACZMJKKPSA-N Gln-Asp-Ala Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](C)C(O)=O CYTSBCIIEHUPDU-ACZMJKKPSA-N 0.000 description 6
- BLOXULLYFRGYKZ-GUBZILKMSA-N Gln-Glu-Arg Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O BLOXULLYFRGYKZ-GUBZILKMSA-N 0.000 description 6
- JKGHMESJHRTHIC-SIUGBPQLSA-N Gln-Ile-Tyr Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)O)NC(=O)[C@H](CCC(=O)N)N JKGHMESJHRTHIC-SIUGBPQLSA-N 0.000 description 6
- VNTGPISAOMAXRK-CIUDSAMLSA-N Gln-Pro-Ser Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CO)C(O)=O VNTGPISAOMAXRK-CIUDSAMLSA-N 0.000 description 6
- FHPXTPQBODWBIY-CIUDSAMLSA-N Glu-Ala-Arg Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O FHPXTPQBODWBIY-CIUDSAMLSA-N 0.000 description 6
- HNVFSTLPVJWIDV-CIUDSAMLSA-N Glu-Glu-Gln Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(N)=O)C(O)=O HNVFSTLPVJWIDV-CIUDSAMLSA-N 0.000 description 6
- BUZMZDDKFCSKOT-CIUDSAMLSA-N Glu-Glu-Glu Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O BUZMZDDKFCSKOT-CIUDSAMLSA-N 0.000 description 6
- FBEJIDRSQCGFJI-GUBZILKMSA-N Glu-Leu-Ser Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(O)=O FBEJIDRSQCGFJI-GUBZILKMSA-N 0.000 description 6
- AQNYKMCFCCZEEL-JYJNAYRXSA-N Glu-Lys-Tyr Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 AQNYKMCFCCZEEL-JYJNAYRXSA-N 0.000 description 6
- FQFWFZWOHOEVMZ-IHRRRGAJSA-N Glu-Phe-Gln Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCC(N)=O)C(O)=O FQFWFZWOHOEVMZ-IHRRRGAJSA-N 0.000 description 6
- YQAQQKPWFOBSMU-WDCWCFNPSA-N Glu-Thr-Leu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(O)=O YQAQQKPWFOBSMU-WDCWCFNPSA-N 0.000 description 6
- NTNUEBVGKMVANB-NHCYSSNCSA-N Glu-Val-Met Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCSC)C(O)=O NTNUEBVGKMVANB-NHCYSSNCSA-N 0.000 description 6
- PUUYVMYCMIWHFE-BQBZGAKWSA-N Gly-Ala-Arg Chemical compound NCC(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CCCN=C(N)N PUUYVMYCMIWHFE-BQBZGAKWSA-N 0.000 description 6
- JBRBACJPBZNFMF-YUMQZZPRSA-N Gly-Ala-Lys Chemical compound NCC(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CCCCN JBRBACJPBZNFMF-YUMQZZPRSA-N 0.000 description 6
- GVVKYKCOFMMTKZ-WHFBIAKZSA-N Gly-Cys-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](CS)NC(=O)CN GVVKYKCOFMMTKZ-WHFBIAKZSA-N 0.000 description 6
- BYYNJRSNDARRBX-YFKPBYRVSA-N Gly-Gln-Gly Chemical compound NCC(=O)N[C@@H](CCC(N)=O)C(=O)NCC(O)=O BYYNJRSNDARRBX-YFKPBYRVSA-N 0.000 description 6
- XPJBQTCXPJNIFE-ZETCQYMHSA-N Gly-Gly-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)CNC(=O)CN XPJBQTCXPJNIFE-ZETCQYMHSA-N 0.000 description 6
- YTSVAIMKVLZUDU-YUMQZZPRSA-N Gly-Leu-Asp Chemical compound [H]NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(O)=O)C(O)=O YTSVAIMKVLZUDU-YUMQZZPRSA-N 0.000 description 6
- CCBIBMKQNXHNIN-ZETCQYMHSA-N Gly-Leu-Gly Chemical compound NCC(=O)N[C@@H](CC(C)C)C(=O)NCC(O)=O CCBIBMKQNXHNIN-ZETCQYMHSA-N 0.000 description 6
- SAPLASXFNUYUFE-CQDKDKBSSA-N His-Phe-Ala Chemical compound C[C@@H](C(=O)O)NC(=O)[C@H](CC1=CC=CC=C1)NC(=O)[C@H](CC2=CN=CN2)N SAPLASXFNUYUFE-CQDKDKBSSA-N 0.000 description 6
- KDDKJKKQODQQBR-NHCYSSNCSA-N His-Val-Asp Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)O)NC(=O)[C@H](CC1=CN=CN1)N KDDKJKKQODQQBR-NHCYSSNCSA-N 0.000 description 6
- QYZYJFXHXYUZMZ-UGYAYLCHSA-N Ile-Asn-Asn Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CC(=O)N)C(=O)O)N QYZYJFXHXYUZMZ-UGYAYLCHSA-N 0.000 description 6
- RGSOCXHDOPQREB-ZPFDUUQYSA-N Ile-Asp-Leu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CC(C)C)C(=O)O)N RGSOCXHDOPQREB-ZPFDUUQYSA-N 0.000 description 6
- XQLGNKLSPYCRMZ-HJWJTTGWSA-N Ile-Phe-Val Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](C(C)C)C(=O)O)N XQLGNKLSPYCRMZ-HJWJTTGWSA-N 0.000 description 6
- ZNOBVZFCHNHKHA-KBIXCLLPSA-N Ile-Ser-Glu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N ZNOBVZFCHNHKHA-KBIXCLLPSA-N 0.000 description 6
- LJHGALIOHLRRQN-DCAQKATOSA-N Leu-Ala-Arg Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CCCN=C(N)N LJHGALIOHLRRQN-DCAQKATOSA-N 0.000 description 6
- WNGVUZWBXZKQES-YUMQZZPRSA-N Leu-Ala-Gly Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C)C(=O)NCC(O)=O WNGVUZWBXZKQES-YUMQZZPRSA-N 0.000 description 6
- OIARJGNVARWKFP-YUMQZZPRSA-N Leu-Asn-Gly Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(=O)NCC(O)=O OIARJGNVARWKFP-YUMQZZPRSA-N 0.000 description 6
- ZURHXHNAEJJRNU-CIUDSAMLSA-N Leu-Asp-Asn Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O ZURHXHNAEJJRNU-CIUDSAMLSA-N 0.000 description 6
- WMTOVWLLDGQGCV-GUBZILKMSA-N Leu-Glu-Cys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CS)C(=O)O)N WMTOVWLLDGQGCV-GUBZILKMSA-N 0.000 description 6
- VBZOAGIPCULURB-QWRGUYRKSA-N Leu-Gly-His Chemical compound CC(C)C[C@@H](C(=O)NCC(=O)N[C@@H](CC1=CN=CN1)C(=O)O)N VBZOAGIPCULURB-QWRGUYRKSA-N 0.000 description 6
- WMIOEVKKYIMVKI-DCAQKATOSA-N Leu-Pro-Ala Chemical compound [H]N[C@@H](CC(C)C)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C)C(O)=O WMIOEVKKYIMVKI-DCAQKATOSA-N 0.000 description 6
- ICYRCNICGBJLGM-HJGDQZAQSA-N Leu-Thr-Asp Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@H](C(O)=O)CC(O)=O ICYRCNICGBJLGM-HJGDQZAQSA-N 0.000 description 6
- ARNIBBOXIAWUOP-MGHWNKPDSA-N Leu-Tyr-Ile Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O ARNIBBOXIAWUOP-MGHWNKPDSA-N 0.000 description 6
- FZIJIFCXUCZHOL-CIUDSAMLSA-N Lys-Ala-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCCCN FZIJIFCXUCZHOL-CIUDSAMLSA-N 0.000 description 6
- KCXUCYYZNZFGLL-SRVKXCTJSA-N Lys-Ala-Leu Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(O)=O KCXUCYYZNZFGLL-SRVKXCTJSA-N 0.000 description 6
- MRWXLRGAFDOILG-DCAQKATOSA-N Lys-Gln-Gln Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(N)=O)C(O)=O MRWXLRGAFDOILG-DCAQKATOSA-N 0.000 description 6
- OIYWBDBHEGAVST-BZSNNMDCSA-N Lys-His-Tyr Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O OIYWBDBHEGAVST-BZSNNMDCSA-N 0.000 description 6
- XOQMURBBIXRRCR-SRVKXCTJSA-N Lys-Lys-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](CCCCN)NC(=O)[C@@H](N)CCCCN XOQMURBBIXRRCR-SRVKXCTJSA-N 0.000 description 6
- VVURYEVJJTXWNE-ULQDDVLXSA-N Lys-Tyr-Val Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](C(C)C)C(O)=O VVURYEVJJTXWNE-ULQDDVLXSA-N 0.000 description 6
- DRRXXZBXDMLGFC-IHRRRGAJSA-N Lys-Val-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](C(C)C)NC(=O)[C@@H](N)CCCCN DRRXXZBXDMLGFC-IHRRRGAJSA-N 0.000 description 6
- AUEJLPRZGVVDNU-UHFFFAOYSA-N N-L-tyrosyl-L-leucine Natural products CC(C)CC(C(O)=O)NC(=O)C(N)CC1=CC=C(O)C=C1 AUEJLPRZGVVDNU-UHFFFAOYSA-N 0.000 description 6
- CDNPIRSCAFMMBE-SRVKXCTJSA-N Phe-Asn-Ser Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(O)=O CDNPIRSCAFMMBE-SRVKXCTJSA-N 0.000 description 6
- MQVFHOPCKNTHGT-MELADBBJSA-N Phe-Asp-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC(=O)O)NC(=O)[C@H](CC2=CC=CC=C2)N)C(=O)O MQVFHOPCKNTHGT-MELADBBJSA-N 0.000 description 6
- LLGTYVHITPVGKR-RYUDHWBXSA-N Phe-Gln-Gly Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCC(N)=O)C(=O)NCC(O)=O LLGTYVHITPVGKR-RYUDHWBXSA-N 0.000 description 6
- GXDPQJUBLBZKDY-IAVJCBSLSA-N Phe-Ile-Ile Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O GXDPQJUBLBZKDY-IAVJCBSLSA-N 0.000 description 6
- BAONJAHBAUDJKA-BZSNNMDCSA-N Phe-Tyr-Asp Chemical compound C([C@H](N)C(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(=O)N[C@@H](CC(O)=O)C(O)=O)C1=CC=CC=C1 BAONJAHBAUDJKA-BZSNNMDCSA-N 0.000 description 6
- VXCHGLYSIOOZIS-GUBZILKMSA-N Pro-Ala-Arg Chemical compound NC(N)=NCCC[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H]1CCCN1 VXCHGLYSIOOZIS-GUBZILKMSA-N 0.000 description 6
- HRNQLKCLPVKZNE-CIUDSAMLSA-N Ser-Ala-Leu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(O)=O HRNQLKCLPVKZNE-CIUDSAMLSA-N 0.000 description 6
- SNVIOQXAHVORQM-WDSKDSINSA-N Ser-Gly-Gln Chemical compound [H]N[C@@H](CO)C(=O)NCC(=O)N[C@@H](CCC(N)=O)C(O)=O SNVIOQXAHVORQM-WDSKDSINSA-N 0.000 description 6
- IOVHBRCQOGWAQH-ZKWXMUAHSA-N Ser-Gly-Ile Chemical compound [H]N[C@@H](CO)C(=O)NCC(=O)N[C@@H]([C@@H](C)CC)C(O)=O IOVHBRCQOGWAQH-ZKWXMUAHSA-N 0.000 description 6
- TWLMXDWFVNEFFK-FJXKBIBVSA-N Thr-Arg-Gly Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)NCC(O)=O TWLMXDWFVNEFFK-FJXKBIBVSA-N 0.000 description 6
- PKXHGEXFMIZSER-QTKMDUPCSA-N Thr-Arg-His Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)N)O PKXHGEXFMIZSER-QTKMDUPCSA-N 0.000 description 6
- BOBZBMOTRORUPT-XIRDDKMYSA-N Trp-Ser-Leu Chemical compound C1=CC=C2C(C[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(O)=O)=CNC2=C1 BOBZBMOTRORUPT-XIRDDKMYSA-N 0.000 description 6
- KSFXWENSJABBFI-ZKWXMUAHSA-N Val-Ser-Asn Chemical compound [H]N[C@@H](C(C)C)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(O)=O KSFXWENSJABBFI-ZKWXMUAHSA-N 0.000 description 6
- HTONZBWRYUKUKC-RCWTZXSCSA-N Val-Thr-Val Chemical compound CC(C)[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(O)=O HTONZBWRYUKUKC-RCWTZXSCSA-N 0.000 description 6
- 108010076324 alanyl-glycyl-glycine Proteins 0.000 description 6
- 108010070944 alanylhistidine Proteins 0.000 description 6
- 108010062796 arginyllysine Proteins 0.000 description 6
- 108010069205 aspartyl-phenylalanine Proteins 0.000 description 6
- 108010038633 aspartylglutamate Proteins 0.000 description 6
- 108010092854 aspartyllysine Proteins 0.000 description 6
- 238000006243 chemical reaction Methods 0.000 description 6
- 108010078144 glutaminyl-glycine Proteins 0.000 description 6
- 108010037389 glutamyl-cysteinyl-lysine Proteins 0.000 description 6
- 108010026364 glycyl-glycyl-leucine Proteins 0.000 description 6
- 108010082286 glycyl-seryl-alanine Proteins 0.000 description 6
- 108010037850 glycylvaline Proteins 0.000 description 6
- 108010025153 lysyl-alanyl-alanine Proteins 0.000 description 6
- 108010017391 lysylvaline Proteins 0.000 description 6
- 101150016309 trpC gene Proteins 0.000 description 6
- HAUUXTXKJNVIFY-ONGXEEELSA-N Lys-Gly-Val Chemical compound [H]N[C@@H](CCCCN)C(=O)NCC(=O)N[C@@H](C(C)C)C(O)=O HAUUXTXKJNVIFY-ONGXEEELSA-N 0.000 description 5
- 238000004458 analytical method Methods 0.000 description 5
- 238000003032 molecular docking Methods 0.000 description 5
- 229910052757 nitrogen Inorganic materials 0.000 description 5
- 230000037361 pathway Effects 0.000 description 5
- QGZKDVFQNNGYKY-UHFFFAOYSA-N Ammonia Chemical compound N QGZKDVFQNNGYKY-UHFFFAOYSA-N 0.000 description 4
- 108090000769 Isomerases Proteins 0.000 description 4
- 102000004195 Isomerases Human genes 0.000 description 4
- AZWNCEBQZXELEZ-FXQIFTODSA-N Ser-Pro-Ser Chemical compound OC[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CO)C(O)=O AZWNCEBQZXELEZ-FXQIFTODSA-N 0.000 description 4
- 108090000992 Transferases Proteins 0.000 description 4
- 102000004357 Transferases Human genes 0.000 description 4
- 238000006555 catalytic reaction Methods 0.000 description 4
- 239000013078 crystal Substances 0.000 description 4
- 230000002255 enzymatic effect Effects 0.000 description 4
- 230000002068 genetic effect Effects 0.000 description 4
- 230000012010 growth Effects 0.000 description 4
- 230000001965 increasing effect Effects 0.000 description 4
- 230000006698 induction Effects 0.000 description 4
- 230000002018 overexpression Effects 0.000 description 4
- 230000001105 regulatory effect Effects 0.000 description 4
- 239000000758 substrate Substances 0.000 description 4
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 3
- 241000880493 Leptailurus serval Species 0.000 description 3
- 101100370749 Streptomyces coelicolor (strain ATCC BAA-471 / A3(2) / M145) trpC1 gene Proteins 0.000 description 3
- RAXXELZNTBOGNW-UHFFFAOYSA-N imidazole Natural products C1=CNC=N1 RAXXELZNTBOGNW-UHFFFAOYSA-N 0.000 description 3
- BPHPUYQFMNQIOC-NXRLNHOXSA-N isopropyl beta-D-thiogalactopyranoside Chemical compound CC(C)S[C@@H]1O[C@H](CO)[C@H](O)[C@H](O)[C@H]1O BPHPUYQFMNQIOC-NXRLNHOXSA-N 0.000 description 3
- 230000002503 metabolic effect Effects 0.000 description 3
- 229960005190 phenylalanine Drugs 0.000 description 3
- 238000012360 testing method Methods 0.000 description 3
- 101150044170 trpE gene Proteins 0.000 description 3
- LXCUAFVVTHZALS-UHFFFAOYSA-N 3-(3-methoxyphenyl)piperidine Chemical compound COC1=CC=CC(C2CNCCC2)=C1 LXCUAFVVTHZALS-UHFFFAOYSA-N 0.000 description 2
- LMFXXZPPZDCPTA-ZKWXMUAHSA-N Ala-Gly-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)CNC(=O)[C@H](C)N LMFXXZPPZDCPTA-ZKWXMUAHSA-N 0.000 description 2
- 108020001077 Anthranilate Phosphoribosyltransferase Proteins 0.000 description 2
- PQKSVQSMTHPRIB-ZKWXMUAHSA-N Asn-Val-Ser Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CO)C(O)=O PQKSVQSMTHPRIB-ZKWXMUAHSA-N 0.000 description 2
- AYFVRYXNDHBECD-YUMQZZPRSA-N Asp-Leu-Gly Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)NCC(O)=O AYFVRYXNDHBECD-YUMQZZPRSA-N 0.000 description 2
- 101000623895 Bos taurus Mucin-15 Proteins 0.000 description 2
- 101000981883 Brevibacillus parabrevis ATP-dependent tryptophan/phenylalanine/tyrosine adenylase Proteins 0.000 description 2
- 101000981889 Brevibacillus parabrevis Linear gramicidin-PCP reductase Proteins 0.000 description 2
- 101000906861 Chondromyces crocatus ATP-dependent tyrosine adenylase Proteins 0.000 description 2
- MUSGDMDGNGXULI-DCAQKATOSA-N Glu-Glu-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](N)CCC(O)=O MUSGDMDGNGXULI-DCAQKATOSA-N 0.000 description 2
- SYWCGQOIIARSIX-SRVKXCTJSA-N Glu-Pro-Leu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(C)C)C(O)=O SYWCGQOIIARSIX-SRVKXCTJSA-N 0.000 description 2
- UQJNXZSSGQIPIQ-FBCQKBJTSA-N Gly-Gly-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)CNC(=O)CN UQJNXZSSGQIPIQ-FBCQKBJTSA-N 0.000 description 2
- NBJAAWYRLGCJOF-UGYAYLCHSA-N Ile-Asp-Asn Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CC(=O)N)C(=O)O)N NBJAAWYRLGCJOF-UGYAYLCHSA-N 0.000 description 2
- SIKJAQJRHWYJAI-UHFFFAOYSA-N Indole Chemical compound C1=CC=C2NC=CC2=C1 SIKJAQJRHWYJAI-UHFFFAOYSA-N 0.000 description 2
- RCFDOSNHHZGBOY-UHFFFAOYSA-N L-isoleucyl-L-alanine Natural products CCC(C)C(N)C(=O)NC(C)C(O)=O RCFDOSNHHZGBOY-UHFFFAOYSA-N 0.000 description 2
- WSGXUIQTEZDVHJ-GARJFASQSA-N Leu-Ala-Pro Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C)C(=O)N1CCC[C@@H]1C(O)=O WSGXUIQTEZDVHJ-GARJFASQSA-N 0.000 description 2
- GKFNXYMAMKJSKD-NHCYSSNCSA-N Lys-Asp-Val Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O GKFNXYMAMKJSKD-NHCYSSNCSA-N 0.000 description 2
- 101100261636 Methanothermobacter marburgensis (strain ATCC BAA-927 / DSM 2133 / JCM 14651 / NBRC 100331 / OCM 82 / Marburg) trpB2 gene Proteins 0.000 description 2
- 241000187479 Mycobacterium tuberculosis Species 0.000 description 2
- 108010079364 N-glycylalanine Proteins 0.000 description 2
- WFIVLLFYUZZWOD-RHYQMDGZSA-N Pro-Lys-Thr Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)O)C(O)=O WFIVLLFYUZZWOD-RHYQMDGZSA-N 0.000 description 2
- 101100408135 Pseudomonas aeruginosa (strain ATCC 15692 / DSM 22644 / CIP 104116 / JCM 14847 / LMG 12228 / 1C / PRS 101 / PAO1) phnA gene Proteins 0.000 description 2
- RIAKPZVSNBBNRE-BJDJZHNGSA-N Ser-Ile-Leu Chemical compound OC[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(C)C)C(O)=O RIAKPZVSNBBNRE-BJDJZHNGSA-N 0.000 description 2
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 2
- ZMYCLHFLHRVOEA-HEIBUPTGSA-N Thr-Thr-Ser Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CO)C(O)=O ZMYCLHFLHRVOEA-HEIBUPTGSA-N 0.000 description 2
- 108060008539 Transglutaminase Proteins 0.000 description 2
- 108010075344 Tryptophan synthase Proteins 0.000 description 2
- OVBMCNDKCWAXMZ-NAKRPEOUSA-N Val-Ile-Ser Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CO)C(=O)O)NC(=O)[C@H](C(C)C)N OVBMCNDKCWAXMZ-NAKRPEOUSA-N 0.000 description 2
- 101100386506 Xenopus laevis dazap1 gene Proteins 0.000 description 2
- 230000004913 activation Effects 0.000 description 2
- 108010087924 alanylproline Proteins 0.000 description 2
- 229940064734 aminobenzoate Drugs 0.000 description 2
- 108010040443 aspartyl-aspartic acid Proteins 0.000 description 2
- 238000003556 assay Methods 0.000 description 2
- WQZGKKKJIJFFOK-VFUOTHLCSA-N beta-D-glucose Chemical compound OC[C@H]1O[C@@H](O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-VFUOTHLCSA-N 0.000 description 2
- 239000012148 binding buffer Substances 0.000 description 2
- 230000008033 biological extinction Effects 0.000 description 2
- 230000033228 biological regulation Effects 0.000 description 2
- 238000011138 biotechnological process Methods 0.000 description 2
- 230000010261 cell growth Effects 0.000 description 2
- 238000004440 column chromatography Methods 0.000 description 2
- 230000029087 digestion Effects 0.000 description 2
- 125000000404 glutamine group Chemical group N[C@@H](CCC(N)=O)C(=O)* 0.000 description 2
- 108010018006 histidylserine Proteins 0.000 description 2
- 238000001727 in vivo Methods 0.000 description 2
- 238000002955 isolation Methods 0.000 description 2
- 239000003446 ligand Substances 0.000 description 2
- 108010009298 lysylglutamic acid Proteins 0.000 description 2
- 108010064235 lysylglycine Proteins 0.000 description 2
- 239000003550 marker Substances 0.000 description 2
- 108010005942 methionylglycine Proteins 0.000 description 2
- 230000000813 microbial effect Effects 0.000 description 2
- 244000005700 microbiome Species 0.000 description 2
- 238000002703 mutagenesis Methods 0.000 description 2
- 231100000350 mutagenesis Toxicity 0.000 description 2
- 230000006959 non-competitive inhibition Effects 0.000 description 2
- 230000036963 noncompetitive effect Effects 0.000 description 2
- 108010070409 phenylalanyl-glycyl-glycine Proteins 0.000 description 2
- 239000002243 precursor Substances 0.000 description 2
- 239000000047 product Substances 0.000 description 2
- 238000000746 purification Methods 0.000 description 2
- 230000009467 reduction Effects 0.000 description 2
- 229930000044 secondary metabolite Natural products 0.000 description 2
- 125000003607 serino group Chemical group [H]N([H])[C@]([H])(C(=O)[*])C(O[H])([H])[H] 0.000 description 2
- JXOHGGNKMLTUBP-HSUXUTPPSA-N shikimic acid Chemical compound O[C@@H]1CC(C(O)=O)=C[C@@H](O)[C@H]1O JXOHGGNKMLTUBP-HSUXUTPPSA-N 0.000 description 2
- JXOHGGNKMLTUBP-JKUQZMGJSA-N shikimic acid Natural products O[C@@H]1CC(C(O)=O)=C[C@H](O)[C@@H]1O JXOHGGNKMLTUBP-JKUQZMGJSA-N 0.000 description 2
- 238000012916 structural analysis Methods 0.000 description 2
- 239000000126 substance Substances 0.000 description 2
- 102000003601 transglutaminase Human genes 0.000 description 2
- 101150019416 trpA gene Proteins 0.000 description 2
- 101150081616 trpB gene Proteins 0.000 description 2
- 101150111232 trpB-1 gene Proteins 0.000 description 2
- 101150100816 trpD gene Proteins 0.000 description 2
- 125000002987 valine group Chemical group [H]N([H])C([H])(C(*)=O)C([H])(C([H])([H])[H])C([H])([H])[H] 0.000 description 2
- BRPMXFSTKXXNHF-IUCAKERBSA-N (2s)-1-[2-[[(2s)-pyrrolidine-2-carbonyl]amino]acetyl]pyrrolidine-2-carboxylic acid Chemical compound OC(=O)[C@@H]1CCCN1C(=O)CNC(=O)[C@H]1NCCC1 BRPMXFSTKXXNHF-IUCAKERBSA-N 0.000 description 1
- MEYZIGGCNFHINA-UHFFFAOYSA-N (6-aminoquinolin-2-yl) n-(2,5-dioxopyrrolidin-1-yl)-n-hydroxycarbamate Chemical compound C1=CC2=CC(N)=CC=C2N=C1OC(=O)N(O)N1C(=O)CCC1=O MEYZIGGCNFHINA-UHFFFAOYSA-N 0.000 description 1
- XBOJYPMLUAMOQB-ZBCCYFLUSA-N 2-amino-3-[(3R,4S,5R)-3,4-dihydroxy-5-(hydroxymethyl)oxolan-2-yl]-4-phosphonobenzoic acid Chemical compound P(=O)(O)(O)C1=C(C(=C(C(=O)O)C=C1)N)C1[C@H](O)[C@H](O)[C@H](O1)CO XBOJYPMLUAMOQB-ZBCCYFLUSA-N 0.000 description 1
- ALYNCZNDIQEVRV-UHFFFAOYSA-M 4-aminobenzoate Chemical compound NC1=CC=C(C([O-])=O)C=C1 ALYNCZNDIQEVRV-UHFFFAOYSA-M 0.000 description 1
- BUANFPRKJKJSRR-ACZMJKKPSA-N Ala-Ala-Gln Chemical compound C[C@H]([NH3+])C(=O)N[C@@H](C)C(=O)N[C@H](C([O-])=O)CCC(N)=O BUANFPRKJKJSRR-ACZMJKKPSA-N 0.000 description 1
- YLTKNGYYPIWKHZ-ACZMJKKPSA-N Ala-Ala-Glu Chemical compound C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CCC(O)=O YLTKNGYYPIWKHZ-ACZMJKKPSA-N 0.000 description 1
- YYSWCHMLFJLLBJ-ZLUOBGJFSA-N Ala-Ala-Ser Chemical compound C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CO)C(O)=O YYSWCHMLFJLLBJ-ZLUOBGJFSA-N 0.000 description 1
- QDRGPQWIVZNJQD-CIUDSAMLSA-N Ala-Arg-Gln Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(N)=O)C(O)=O QDRGPQWIVZNJQD-CIUDSAMLSA-N 0.000 description 1
- ZIWWTZWAKYBUOB-CIUDSAMLSA-N Ala-Asp-Leu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O ZIWWTZWAKYBUOB-CIUDSAMLSA-N 0.000 description 1
- DECCMEWNXSNSDO-ZLUOBGJFSA-N Ala-Cys-Ala Chemical compound C[C@H](N)C(=O)N[C@@H](CS)C(=O)N[C@@H](C)C(O)=O DECCMEWNXSNSDO-ZLUOBGJFSA-N 0.000 description 1
- NJPMYXWVWQWCSR-ACZMJKKPSA-N Ala-Glu-Asn Chemical compound C[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O NJPMYXWVWQWCSR-ACZMJKKPSA-N 0.000 description 1
- GGNHBHYDMUDXQB-KBIXCLLPSA-N Ala-Glu-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](C)N GGNHBHYDMUDXQB-KBIXCLLPSA-N 0.000 description 1
- HXNNRBHASOSVPG-GUBZILKMSA-N Ala-Glu-Leu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O HXNNRBHASOSVPG-GUBZILKMSA-N 0.000 description 1
- HMRWQTHUDVXMGH-GUBZILKMSA-N Ala-Glu-Lys Chemical compound C[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@H](C(O)=O)CCCCN HMRWQTHUDVXMGH-GUBZILKMSA-N 0.000 description 1
- WMYJZJRILUVVRG-WDSKDSINSA-N Ala-Gly-Gln Chemical compound C[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CCC(N)=O WMYJZJRILUVVRG-WDSKDSINSA-N 0.000 description 1
- BEMGNWZECGIJOI-WDSKDSINSA-N Ala-Gly-Glu Chemical compound [H]N[C@@H](C)C(=O)NCC(=O)N[C@@H](CCC(O)=O)C(O)=O BEMGNWZECGIJOI-WDSKDSINSA-N 0.000 description 1
- GSHKMNKPMLXSQW-KBIXCLLPSA-N Ala-Ile-Gln Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)O)NC(=O)[C@H](C)N GSHKMNKPMLXSQW-KBIXCLLPSA-N 0.000 description 1
- TZDNWXDLYFIFPT-BJDJZHNGSA-N Ala-Ile-Leu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(C)C)C(O)=O TZDNWXDLYFIFPT-BJDJZHNGSA-N 0.000 description 1
- LXAARTARZJJCMB-CIQUZCHMSA-N Ala-Ile-Thr Chemical compound [H]N[C@@H](C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)O)C(O)=O LXAARTARZJJCMB-CIQUZCHMSA-N 0.000 description 1
- HHRAXZAYZFFRAM-CIUDSAMLSA-N Ala-Leu-Asn Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(O)=O HHRAXZAYZFFRAM-CIUDSAMLSA-N 0.000 description 1
- CCDFBRZVTDDJNM-GUBZILKMSA-N Ala-Leu-Glu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O CCDFBRZVTDDJNM-GUBZILKMSA-N 0.000 description 1
- SOBIAADAMRHGKH-CIUDSAMLSA-N Ala-Leu-Ser Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(O)=O SOBIAADAMRHGKH-CIUDSAMLSA-N 0.000 description 1
- QUIGLPSHIFPEOV-CIUDSAMLSA-N Ala-Lys-Ala Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C)C(O)=O QUIGLPSHIFPEOV-CIUDSAMLSA-N 0.000 description 1
- VCSABYLVNWQYQE-SRVKXCTJSA-N Ala-Lys-Lys Chemical compound NCCCC[C@H](NC(=O)[C@@H](N)C)C(=O)N[C@@H](CCCCN)C(O)=O VCSABYLVNWQYQE-SRVKXCTJSA-N 0.000 description 1
- VCSABYLVNWQYQE-UHFFFAOYSA-N Ala-Lys-Lys Natural products NCCCCC(NC(=O)C(N)C)C(=O)NC(CCCCN)C(O)=O VCSABYLVNWQYQE-UHFFFAOYSA-N 0.000 description 1
- MDNAVFBZPROEHO-DCAQKATOSA-N Ala-Lys-Val Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C(C)C)C(O)=O MDNAVFBZPROEHO-DCAQKATOSA-N 0.000 description 1
- KLALXKYLOMZDQT-ZLUOBGJFSA-N Ala-Ser-Asn Chemical compound C[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CC(N)=O KLALXKYLOMZDQT-ZLUOBGJFSA-N 0.000 description 1
- XAXMJQUMRJAFCH-CQDKDKBSSA-N Ala-Tyr-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)C)CC1=CC=C(O)C=C1 XAXMJQUMRJAFCH-CQDKDKBSSA-N 0.000 description 1
- IYKVSFNGSWTTNZ-GUBZILKMSA-N Ala-Val-Arg Chemical compound C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CCCN=C(N)N IYKVSFNGSWTTNZ-GUBZILKMSA-N 0.000 description 1
- OMSKGWFGWCQFBD-KZVJFYERSA-N Ala-Val-Thr Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O OMSKGWFGWCQFBD-KZVJFYERSA-N 0.000 description 1
- QGZKDVFQNNGYKY-UHFFFAOYSA-O Ammonium Chemical compound [NH4+] QGZKDVFQNNGYKY-UHFFFAOYSA-O 0.000 description 1
- OOBVTWHLKYJFJH-FXQIFTODSA-N Arg-Ala-Ala Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C)C(=O)N[C@@H](C)C(O)=O OOBVTWHLKYJFJH-FXQIFTODSA-N 0.000 description 1
- GIVATXIGCXFQQA-FXQIFTODSA-N Arg-Ala-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCCN=C(N)N GIVATXIGCXFQQA-FXQIFTODSA-N 0.000 description 1
- WESHVRNMNFMVBE-FXQIFTODSA-N Arg-Asn-Asp Chemical compound C(C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CC(=O)O)C(=O)O)N)CN=C(N)N WESHVRNMNFMVBE-FXQIFTODSA-N 0.000 description 1
- TTXYKSADPSNOIF-IHRRRGAJSA-N Arg-Asp-Phe Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O TTXYKSADPSNOIF-IHRRRGAJSA-N 0.000 description 1
- GXXWTNKNFFKTJB-NAKRPEOUSA-N Arg-Ile-Ser Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CO)C(O)=O GXXWTNKNFFKTJB-NAKRPEOUSA-N 0.000 description 1
- LVMUGODRNHFGRA-AVGNSLFASA-N Arg-Leu-Arg Chemical compound NC(N)=NCCC[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O LVMUGODRNHFGRA-AVGNSLFASA-N 0.000 description 1
- NOZYDJOPOGKUSR-AVGNSLFASA-N Arg-Leu-Met Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCSC)C(O)=O NOZYDJOPOGKUSR-AVGNSLFASA-N 0.000 description 1
- CVXXSWQORBZAAA-SRVKXCTJSA-N Arg-Lys-Glu Chemical compound OC(=O)CC[C@@H](C(O)=O)NC(=O)[C@H](CCCCN)NC(=O)[C@@H](N)CCCN=C(N)N CVXXSWQORBZAAA-SRVKXCTJSA-N 0.000 description 1
- FOQFHANLUJDQEE-GUBZILKMSA-N Arg-Pro-Cys Chemical compound C1C[C@H](N(C1)C(=O)[C@H](CCCN=C(N)N)N)C(=O)N[C@@H](CS)C(=O)O FOQFHANLUJDQEE-GUBZILKMSA-N 0.000 description 1
- JKRPBTQDPJSQIT-RCWTZXSCSA-N Arg-Thr-Met Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCSC)C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N)O JKRPBTQDPJSQIT-RCWTZXSCSA-N 0.000 description 1
- XRLOBFSLPCHYLQ-ULQDDVLXSA-N Arg-Tyr-His Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)N[C@@H](CC2=CN=CN2)C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N)O XRLOBFSLPCHYLQ-ULQDDVLXSA-N 0.000 description 1
- 239000004475 Arginine Substances 0.000 description 1
- HOIFSHOLNKQCSA-FXQIFTODSA-N Asn-Arg-Asp Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(O)=O HOIFSHOLNKQCSA-FXQIFTODSA-N 0.000 description 1
- JJGRJMKUOYXZRA-LPEHRKFASA-N Asn-Arg-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CC(=O)N)N)C(=O)O JJGRJMKUOYXZRA-LPEHRKFASA-N 0.000 description 1
- HAJWYALLJIATCX-FXQIFTODSA-N Asn-Asn-Arg Chemical compound C(C[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)N)NC(=O)[C@H](CC(=O)N)N)CN=C(N)N HAJWYALLJIATCX-FXQIFTODSA-N 0.000 description 1
- CUQUEHYSSFETRD-ACZMJKKPSA-N Asn-Asp-Gln Chemical compound C(CC(=O)N)[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)O)NC(=O)[C@H](CC(=O)N)N CUQUEHYSSFETRD-ACZMJKKPSA-N 0.000 description 1
- GQRDIVQPSMPQME-ZPFDUUQYSA-N Asn-Ile-Leu Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(C)C)C(O)=O GQRDIVQPSMPQME-ZPFDUUQYSA-N 0.000 description 1
- NCFJQJRLQJEECD-NHCYSSNCSA-N Asn-Leu-Val Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C(C)C)C(O)=O NCFJQJRLQJEECD-NHCYSSNCSA-N 0.000 description 1
- UGXYFDQFLVCDFC-CIUDSAMLSA-N Asn-Ser-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CO)NC(=O)[C@@H](N)CC(N)=O UGXYFDQFLVCDFC-CIUDSAMLSA-N 0.000 description 1
- WLVLIYYBPPONRJ-GCJQMDKQSA-N Asn-Thr-Ala Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C)C(O)=O WLVLIYYBPPONRJ-GCJQMDKQSA-N 0.000 description 1
- HBUJSDCLZCXXCW-YDHLFZDLSA-N Asn-Val-Tyr Chemical compound NC(=O)C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 HBUJSDCLZCXXCW-YDHLFZDLSA-N 0.000 description 1
- HMQDRBKQMLRCCG-GMOBBJLQSA-N Asp-Arg-Ile Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O HMQDRBKQMLRCCG-GMOBBJLQSA-N 0.000 description 1
- WJHYGGVCWREQMO-GHCJXIJMSA-N Asp-Cys-Ile Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CS)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O WJHYGGVCWREQMO-GHCJXIJMSA-N 0.000 description 1
- FTNVLGCFIJEMQT-CIUDSAMLSA-N Asp-Cys-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)[C@H](CS)NC(=O)[C@H](CC(=O)O)N FTNVLGCFIJEMQT-CIUDSAMLSA-N 0.000 description 1
- ZSJFGGSPCCHMNE-LAEOZQHASA-N Asp-Gln-Val Chemical compound CC(C)[C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)N)NC(=O)[C@H](CC(=O)O)N ZSJFGGSPCCHMNE-LAEOZQHASA-N 0.000 description 1
- HSWYMWGDMPLTTH-FXQIFTODSA-N Asp-Glu-Gln Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(N)=O)C(O)=O HSWYMWGDMPLTTH-FXQIFTODSA-N 0.000 description 1
- PZXPWHFYZXTFBI-YUMQZZPRSA-N Asp-Gly-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)CNC(=O)[C@@H](N)CC(O)=O PZXPWHFYZXTFBI-YUMQZZPRSA-N 0.000 description 1
- SCQIQCWLOMOEFP-DCAQKATOSA-N Asp-Leu-Arg Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O SCQIQCWLOMOEFP-DCAQKATOSA-N 0.000 description 1
- CLUMZOKVGUWUFD-CIUDSAMLSA-N Asp-Leu-Asn Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(O)=O CLUMZOKVGUWUFD-CIUDSAMLSA-N 0.000 description 1
- XLILXFRAKOYEJX-GUBZILKMSA-N Asp-Leu-Gln Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(O)=O XLILXFRAKOYEJX-GUBZILKMSA-N 0.000 description 1
- ZRUBWRCKIVDCFS-XPCJQDJLSA-N Asp-Leu-Thr-Ser Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CO)C(O)=O ZRUBWRCKIVDCFS-XPCJQDJLSA-N 0.000 description 1
- NVFSJIXJZCDICF-SRVKXCTJSA-N Asp-Lys-Lys Chemical compound C(CCN)C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CC(=O)O)N NVFSJIXJZCDICF-SRVKXCTJSA-N 0.000 description 1
- XXAMCEGRCZQGEM-ZLUOBGJFSA-N Asp-Ser-Asn Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(O)=O XXAMCEGRCZQGEM-ZLUOBGJFSA-N 0.000 description 1
- KGHLGJAXYSVNJP-WHFBIAKZSA-N Asp-Ser-Gly Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](CO)C(=O)NCC(O)=O KGHLGJAXYSVNJP-WHFBIAKZSA-N 0.000 description 1
- QSFHZPQUAAQHAQ-CIUDSAMLSA-N Asp-Ser-Leu Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(O)=O QSFHZPQUAAQHAQ-CIUDSAMLSA-N 0.000 description 1
- ZQFRDAZBTSFGGW-SRVKXCTJSA-N Asp-Ser-Phe Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O ZQFRDAZBTSFGGW-SRVKXCTJSA-N 0.000 description 1
- XWKBWZXGNXTDKY-ZKWXMUAHSA-N Asp-Val-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](C(C)C)NC(=O)[C@@H](N)CC(O)=O XWKBWZXGNXTDKY-ZKWXMUAHSA-N 0.000 description 1
- XMKXONRMGJXCJV-LAEOZQHASA-N Asp-Val-Glu Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O XMKXONRMGJXCJV-LAEOZQHASA-N 0.000 description 1
- SFJUYBCDQBAYAJ-YDHLFZDLSA-N Asp-Val-Phe Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 SFJUYBCDQBAYAJ-YDHLFZDLSA-N 0.000 description 1
- RKXVTTIQNKPCHU-KKHAAJSZSA-N Asp-Val-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@H](C(C)C)NC(=O)[C@@H](N)CC(O)=O RKXVTTIQNKPCHU-KKHAAJSZSA-N 0.000 description 1
- 101000765308 Aspergillus niger N-(5'-phosphoribosyl)anthranilate isomerase Proteins 0.000 description 1
- 239000002028 Biomass Substances 0.000 description 1
- NOCCABSVTRONIN-CIUDSAMLSA-N Cys-Ala-Leu Chemical compound C[C@@H](C(=O)N[C@@H](CC(C)C)C(=O)O)NC(=O)[C@H](CS)N NOCCABSVTRONIN-CIUDSAMLSA-N 0.000 description 1
- UPURLDIGQGTUPJ-ZKWXMUAHSA-N Cys-Gly-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)CNC(=O)[C@H](CS)N UPURLDIGQGTUPJ-ZKWXMUAHSA-N 0.000 description 1
- CUXIOFHFFXNUGG-HTFCKZLJSA-N Cys-Ile-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)O)NC(=O)[C@H](CS)N CUXIOFHFFXNUGG-HTFCKZLJSA-N 0.000 description 1
- UCSXXFRXHGUXCQ-SRVKXCTJSA-N Cys-Leu-Lys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CS)N UCSXXFRXHGUXCQ-SRVKXCTJSA-N 0.000 description 1
- CNBIWHCVAZHRBI-IHRRRGAJSA-N Cys-Met-Phe Chemical compound CSCC[C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)O)NC(=O)[C@H](CS)N CNBIWHCVAZHRBI-IHRRRGAJSA-N 0.000 description 1
- HEPLXMBVMCXTBP-QWRGUYRKSA-N Cys-Phe-Gly Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)NCC(O)=O HEPLXMBVMCXTBP-QWRGUYRKSA-N 0.000 description 1
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 1
- 241001522878 Escherichia coli B Species 0.000 description 1
- YJIUYQKQBBQYHZ-ACZMJKKPSA-N Gln-Ala-Ala Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](C)C(O)=O YJIUYQKQBBQYHZ-ACZMJKKPSA-N 0.000 description 1
- KVYVOGYEMPEXBT-GUBZILKMSA-N Gln-Ala-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCC(N)=O KVYVOGYEMPEXBT-GUBZILKMSA-N 0.000 description 1
- XOKGKOQWADCLFQ-GARJFASQSA-N Gln-Arg-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CCC(=O)N)N)C(=O)O XOKGKOQWADCLFQ-GARJFASQSA-N 0.000 description 1
- TWHDOEYLXXQYOZ-FXQIFTODSA-N Gln-Asn-Gln Chemical compound C(CC(=O)N)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CCC(=O)N)C(=O)O)N TWHDOEYLXXQYOZ-FXQIFTODSA-N 0.000 description 1
- KCJJFESQRXGTGC-BQBZGAKWSA-N Gln-Glu-Gly Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)NCC(O)=O KCJJFESQRXGTGC-BQBZGAKWSA-N 0.000 description 1
- IKFZXRLDMYWNBU-YUMQZZPRSA-N Gln-Gly-Arg Chemical compound NC(=O)CC[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CCCN=C(N)N IKFZXRLDMYWNBU-YUMQZZPRSA-N 0.000 description 1
- ORYMMTRPKVTGSJ-XVKPBYJWSA-N Gln-Gly-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)CNC(=O)[C@@H](N)CCC(N)=O ORYMMTRPKVTGSJ-XVKPBYJWSA-N 0.000 description 1
- MLSKFHLRFVGNLL-WDCWCFNPSA-N Gln-Leu-Thr Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O MLSKFHLRFVGNLL-WDCWCFNPSA-N 0.000 description 1
- KPNWAJMEMRCLAL-GUBZILKMSA-N Gln-Ser-Lys Chemical compound C(CCN)C[C@@H](C(=O)O)NC(=O)[C@H](CO)NC(=O)[C@H](CCC(=O)N)N KPNWAJMEMRCLAL-GUBZILKMSA-N 0.000 description 1
- SYZZMPFLOLSMHL-XHNCKOQMSA-N Gln-Ser-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CO)NC(=O)[C@H](CCC(=O)N)N)C(=O)O SYZZMPFLOLSMHL-XHNCKOQMSA-N 0.000 description 1
- ZMXZGYLINVNTKH-DZKIICNBSA-N Gln-Val-Phe Chemical compound NC(=O)CC[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 ZMXZGYLINVNTKH-DZKIICNBSA-N 0.000 description 1
- SOEXCCGNHQBFPV-DLOVCJGASA-N Gln-Val-Val Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](C(C)C)C(O)=O SOEXCCGNHQBFPV-DLOVCJGASA-N 0.000 description 1
- RUFHOVYUYSNDNY-ACZMJKKPSA-N Glu-Ala-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCC(O)=O RUFHOVYUYSNDNY-ACZMJKKPSA-N 0.000 description 1
- UTKUTMJSWKKHEM-WDSKDSINSA-N Glu-Ala-Gly Chemical compound OC(=O)CNC(=O)[C@H](C)NC(=O)[C@@H](N)CCC(O)=O UTKUTMJSWKKHEM-WDSKDSINSA-N 0.000 description 1
- CKRUHITYRFNUKW-WDSKDSINSA-N Glu-Asn-Gly Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(N)=O)C(=O)NCC(O)=O CKRUHITYRFNUKW-WDSKDSINSA-N 0.000 description 1
- KASDBWKLWJKTLJ-GUBZILKMSA-N Glu-Glu-Met Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCSC)C(O)=O KASDBWKLWJKTLJ-GUBZILKMSA-N 0.000 description 1
- CAVMESABQIKFKT-IUCAKERBSA-N Glu-Gly-His Chemical compound C1=C(NC=N1)C[C@@H](C(=O)O)NC(=O)CNC(=O)[C@H](CCC(=O)O)N CAVMESABQIKFKT-IUCAKERBSA-N 0.000 description 1
- LRPXYSGPOBVBEH-IUCAKERBSA-N Glu-Gly-Leu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)NCC(=O)N[C@@H](CC(C)C)C(O)=O LRPXYSGPOBVBEH-IUCAKERBSA-N 0.000 description 1
- HILMIYALTUQTRC-XVKPBYJWSA-N Glu-Gly-Val Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)NCC(=O)N[C@@H](C(C)C)C(O)=O HILMIYALTUQTRC-XVKPBYJWSA-N 0.000 description 1
- LZMQSTPFYJLVJB-GUBZILKMSA-N Glu-Leu-Cys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CCC(=O)O)N LZMQSTPFYJLVJB-GUBZILKMSA-N 0.000 description 1
- DXVOKNVIKORTHQ-GUBZILKMSA-N Glu-Pro-Glu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCC(O)=O)C(O)=O DXVOKNVIKORTHQ-GUBZILKMSA-N 0.000 description 1
- WIKMTDVSCUJIPJ-CIUDSAMLSA-N Glu-Ser-Arg Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CCCN=C(N)N WIKMTDVSCUJIPJ-CIUDSAMLSA-N 0.000 description 1
- SYAYROHMAIHWFB-KBIXCLLPSA-N Glu-Ser-Ile Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O SYAYROHMAIHWFB-KBIXCLLPSA-N 0.000 description 1
- BDISFWMLMNBTGP-NUMRIWBASA-N Glu-Thr-Asp Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(O)=O)C(O)=O BDISFWMLMNBTGP-NUMRIWBASA-N 0.000 description 1
- UUTGYDAKPISJAO-JYJNAYRXSA-N Glu-Tyr-Leu Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](CC(C)C)C(O)=O)CC1=CC=C(O)C=C1 UUTGYDAKPISJAO-JYJNAYRXSA-N 0.000 description 1
- ZYRXTRTUCAVNBQ-GVXVVHGQSA-N Glu-Val-Lys Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CCC(=O)O)N ZYRXTRTUCAVNBQ-GVXVVHGQSA-N 0.000 description 1
- QXUPRMQJDWJDFR-NRPADANISA-N Glu-Val-Ser Chemical compound CC(C)[C@H](NC(=O)[C@@H](N)CCC(O)=O)C(=O)N[C@@H](CO)C(O)=O QXUPRMQJDWJDFR-NRPADANISA-N 0.000 description 1
- RLFSBAPJTYKSLG-WHFBIAKZSA-N Gly-Ala-Asp Chemical compound NCC(=O)N[C@@H](C)C(=O)N[C@@H](CC(O)=O)C(O)=O RLFSBAPJTYKSLG-WHFBIAKZSA-N 0.000 description 1
- MFVQGXGQRIXBPK-WDSKDSINSA-N Gly-Ala-Glu Chemical compound NCC(=O)N[C@@H](C)C(=O)N[C@@H](CCC(O)=O)C(O)=O MFVQGXGQRIXBPK-WDSKDSINSA-N 0.000 description 1
- LJPIRKICOISLKN-WHFBIAKZSA-N Gly-Ala-Ser Chemical compound NCC(=O)N[C@@H](C)C(=O)N[C@@H](CO)C(O)=O LJPIRKICOISLKN-WHFBIAKZSA-N 0.000 description 1
- QIZJOTQTCAGKPU-KWQFWETISA-N Gly-Ala-Tyr Chemical compound [NH3+]CC(=O)N[C@@H](C)C(=O)N[C@H](C([O-])=O)CC1=CC=C(O)C=C1 QIZJOTQTCAGKPU-KWQFWETISA-N 0.000 description 1
- KFMBRBPXHVMDFN-UWVGGRQHSA-N Gly-Arg-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)CN)CCCNC(N)=N KFMBRBPXHVMDFN-UWVGGRQHSA-N 0.000 description 1
- GGEJHJIXRBTJPD-BYPYZUCNSA-N Gly-Asn-Gly Chemical compound NCC(=O)N[C@@H](CC(N)=O)C(=O)NCC(O)=O GGEJHJIXRBTJPD-BYPYZUCNSA-N 0.000 description 1
- XLFHCWHXKSFVIB-BQBZGAKWSA-N Gly-Gln-Gln Chemical compound NCC(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(N)=O)C(O)=O XLFHCWHXKSFVIB-BQBZGAKWSA-N 0.000 description 1
- STVHDEHTKFXBJQ-LAEOZQHASA-N Gly-Glu-Ile Chemical compound [H]NCC(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O STVHDEHTKFXBJQ-LAEOZQHASA-N 0.000 description 1
- YYPFZVIXAVDHIK-IUCAKERBSA-N Gly-Glu-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)CN YYPFZVIXAVDHIK-IUCAKERBSA-N 0.000 description 1
- GDOZQTNZPCUARW-YFKPBYRVSA-N Gly-Gly-Glu Chemical compound NCC(=O)NCC(=O)N[C@H](C(O)=O)CCC(O)=O GDOZQTNZPCUARW-YFKPBYRVSA-N 0.000 description 1
- YFGONBOFGGWKKY-VHSXEESVSA-N Gly-His-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC2=CN=CN2)NC(=O)CN)C(=O)O YFGONBOFGGWKKY-VHSXEESVSA-N 0.000 description 1
- SWQALSGKVLYKDT-UHFFFAOYSA-N Gly-Ile-Ala Natural products NCC(=O)NC(C(C)CC)C(=O)NC(C)C(O)=O SWQALSGKVLYKDT-UHFFFAOYSA-N 0.000 description 1
- LUJVWKKYHSLULQ-ZKWXMUAHSA-N Gly-Ile-Cys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)CN LUJVWKKYHSLULQ-ZKWXMUAHSA-N 0.000 description 1
- HAXARWKYFIIHKD-ZKWXMUAHSA-N Gly-Ile-Ser Chemical compound NCC(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CO)C(O)=O HAXARWKYFIIHKD-ZKWXMUAHSA-N 0.000 description 1
- SCWYHUQOOFRVHP-MBLNEYKQSA-N Gly-Ile-Thr Chemical compound NCC(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)O)C(O)=O SCWYHUQOOFRVHP-MBLNEYKQSA-N 0.000 description 1
- DKEXFJVMVGETOO-LURJTMIESA-N Gly-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)CN DKEXFJVMVGETOO-LURJTMIESA-N 0.000 description 1
- LHYJCVCQPWRMKZ-WEDXCCLWSA-N Gly-Leu-Thr Chemical compound [H]NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O LHYJCVCQPWRMKZ-WEDXCCLWSA-N 0.000 description 1
- PTIIBFKSLCYQBO-NHCYSSNCSA-N Gly-Lys-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](CCCCN)NC(=O)CN PTIIBFKSLCYQBO-NHCYSSNCSA-N 0.000 description 1
- OQQKUTVULYLCDG-ONGXEEELSA-N Gly-Lys-Val Chemical compound CC(C)[C@H](NC(=O)[C@H](CCCCN)NC(=O)CN)C(O)=O OQQKUTVULYLCDG-ONGXEEELSA-N 0.000 description 1
- LPHQAFLNEHWKFF-QXEWZRGKSA-N Gly-Met-Ile Chemical compound [H]NCC(=O)N[C@@H](CCSC)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O LPHQAFLNEHWKFF-QXEWZRGKSA-N 0.000 description 1
- SSFWXSNOKDZNHY-QXEWZRGKSA-N Gly-Pro-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)CN SSFWXSNOKDZNHY-QXEWZRGKSA-N 0.000 description 1
- GAAHQHNCMIAYEX-UWVGGRQHSA-N Gly-Pro-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)CN GAAHQHNCMIAYEX-UWVGGRQHSA-N 0.000 description 1
- DBUNZBWUWCIELX-JHEQGTHGSA-N Gly-Thr-Glu Chemical compound [H]NCC(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(O)=O)C(O)=O DBUNZBWUWCIELX-JHEQGTHGSA-N 0.000 description 1
- RHRLHXQWHCNJKR-PMVVWTBXSA-N Gly-Thr-His Chemical compound NCC(=O)N[C@@H]([C@H](O)C)C(=O)N[C@H](C(O)=O)CC1=CN=CN1 RHRLHXQWHCNJKR-PMVVWTBXSA-N 0.000 description 1
- WSWWTQYHFCBKBT-DVJZZOLTSA-N Gly-Thr-Trp Chemical compound C[C@@H](O)[C@H](NC(=O)CN)C(=O)N[C@@H](Cc1c[nH]c2ccccc12)C(O)=O WSWWTQYHFCBKBT-DVJZZOLTSA-N 0.000 description 1
- GJHWILMUOANXTG-WPRPVWTQSA-N Gly-Val-Arg Chemical compound [H]NCC(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O GJHWILMUOANXTG-WPRPVWTQSA-N 0.000 description 1
- YDIDLLVFCYSXNY-RCOVLWMOSA-N Gly-Val-Asn Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)CN YDIDLLVFCYSXNY-RCOVLWMOSA-N 0.000 description 1
- NGRPGJGKJMUGDM-XVKPBYJWSA-N Gly-Val-Gln Chemical compound NCC(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCC(N)=O)C(O)=O NGRPGJGKJMUGDM-XVKPBYJWSA-N 0.000 description 1
- BNMRSWQOHIQTFL-JSGCOSHPSA-N Gly-Val-Phe Chemical compound NCC(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 BNMRSWQOHIQTFL-JSGCOSHPSA-N 0.000 description 1
- JBCLFWXMTIKCCB-UHFFFAOYSA-N H-Gly-Phe-OH Natural products NCC(=O)NC(C(O)=O)CC1=CC=CC=C1 JBCLFWXMTIKCCB-UHFFFAOYSA-N 0.000 description 1
- OWXMKDGYPWMGEB-UHFFFAOYSA-N HEPPS Chemical compound OCCN1CCN(CCCS(O)(=O)=O)CC1 OWXMKDGYPWMGEB-UHFFFAOYSA-N 0.000 description 1
- 239000007996 HEPPS buffer Substances 0.000 description 1
- 102220609562 Hemoglobin subunit alpha_K61Q_mutation Human genes 0.000 description 1
- BIAKMWKJMQLZOJ-ZKWXMUAHSA-N His-Ala-Ala Chemical compound C[C@H](NC(=O)[C@H](C)NC(=O)[C@@H](N)Cc1cnc[nH]1)C(O)=O BIAKMWKJMQLZOJ-ZKWXMUAHSA-N 0.000 description 1
- WZOGEMJIZBNFBK-CIUDSAMLSA-N His-Asp-Asn Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O WZOGEMJIZBNFBK-CIUDSAMLSA-N 0.000 description 1
- RAVLQPXCMRCLKT-KBPBESRZSA-N His-Gly-Phe Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)NCC(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O RAVLQPXCMRCLKT-KBPBESRZSA-N 0.000 description 1
- FYTCLUIYTYFGPT-YUMQZZPRSA-N His-Gly-Ser Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)NCC(=O)N[C@@H](CO)C(O)=O FYTCLUIYTYFGPT-YUMQZZPRSA-N 0.000 description 1
- UMBKDWGQESDCTO-KKUMJFAQSA-N His-Lys-Lys Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCCN)C(O)=O UMBKDWGQESDCTO-KKUMJFAQSA-N 0.000 description 1
- PYNPBMCLAKTHJL-SRVKXCTJSA-N His-Pro-Glu Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCC(O)=O)C(O)=O PYNPBMCLAKTHJL-SRVKXCTJSA-N 0.000 description 1
- OWYIDJCNRWRSJY-QTKMDUPCSA-N His-Pro-Thr Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)O)C(O)=O OWYIDJCNRWRSJY-QTKMDUPCSA-N 0.000 description 1
- PZAJPILZRFPYJJ-SRVKXCTJSA-N His-Ser-Leu Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(O)=O PZAJPILZRFPYJJ-SRVKXCTJSA-N 0.000 description 1
- DRKZDEFADVYTLU-AVGNSLFASA-N His-Val-Val Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](C(C)C)C(O)=O DRKZDEFADVYTLU-AVGNSLFASA-N 0.000 description 1
- MKWSZEHGHSLNPF-NAKRPEOUSA-N Ile-Ala-Val Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](C)C(=O)N[C@@H](C(C)C)C(=O)O)N MKWSZEHGHSLNPF-NAKRPEOUSA-N 0.000 description 1
- YOTNPRLPIPHQSB-XUXIUFHCSA-N Ile-Arg-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CCCCN)C(=O)O)N YOTNPRLPIPHQSB-XUXIUFHCSA-N 0.000 description 1
- FJWYJQRCVNGEAQ-ZPFDUUQYSA-N Ile-Asn-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CCCCN)C(=O)O)N FJWYJQRCVNGEAQ-ZPFDUUQYSA-N 0.000 description 1
- PFTFEWHJSAXGED-ZKWXMUAHSA-N Ile-Cys-Gly Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CS)C(=O)NCC(=O)O)N PFTFEWHJSAXGED-ZKWXMUAHSA-N 0.000 description 1
- WZDCVAWMBUNDDY-KBIXCLLPSA-N Ile-Glu-Ala Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](C)C(=O)O)N WZDCVAWMBUNDDY-KBIXCLLPSA-N 0.000 description 1
- NZOCIWKZUVUNDW-ZKWXMUAHSA-N Ile-Gly-Ala Chemical compound CC[C@H](C)[C@H](N)C(=O)NCC(=O)N[C@@H](C)C(O)=O NZOCIWKZUVUNDW-ZKWXMUAHSA-N 0.000 description 1
- MTONDYJJCIBZTK-PEDHHIEDSA-N Ile-Ile-Met Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCSC)C(=O)O)N MTONDYJJCIBZTK-PEDHHIEDSA-N 0.000 description 1
- FJWALBCCVIHZBS-QXEWZRGKSA-N Ile-Met-Gly Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCSC)C(=O)NCC(=O)O)N FJWALBCCVIHZBS-QXEWZRGKSA-N 0.000 description 1
- SNHYFFQZRFIRHO-CYDGBPFRSA-N Ile-Met-Val Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCSC)C(=O)N[C@@H](C(C)C)C(=O)O)N SNHYFFQZRFIRHO-CYDGBPFRSA-N 0.000 description 1
- UAELWXJFLZBKQS-WHOFXGATSA-N Ile-Phe-Gly Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](Cc1ccccc1)C(=O)NCC(O)=O UAELWXJFLZBKQS-WHOFXGATSA-N 0.000 description 1
- FGBRXCZYVRFNKQ-MXAVVETBSA-N Ile-Phe-Ser Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CO)C(=O)O)N FGBRXCZYVRFNKQ-MXAVVETBSA-N 0.000 description 1
- KCTIFOCXAIUQQK-QXEWZRGKSA-N Ile-Pro-Gly Chemical compound CC[C@H](C)[C@H](N)C(=O)N1CCC[C@H]1C(=O)NCC(O)=O KCTIFOCXAIUQQK-QXEWZRGKSA-N 0.000 description 1
- CAHCWMVNBZJVAW-NAKRPEOUSA-N Ile-Pro-Ser Chemical compound CC[C@H](C)[C@@H](C(=O)N1CCC[C@H]1C(=O)N[C@@H](CO)C(=O)O)N CAHCWMVNBZJVAW-NAKRPEOUSA-N 0.000 description 1
- JHNJNTMTZHEDLJ-NAKRPEOUSA-N Ile-Ser-Arg Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O JHNJNTMTZHEDLJ-NAKRPEOUSA-N 0.000 description 1
- KBDIBHQICWDGDL-PPCPHDFISA-N Ile-Thr-Leu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(=O)O)N KBDIBHQICWDGDL-PPCPHDFISA-N 0.000 description 1
- 108010063678 Indole-3-Glycerol-Phosphate Synthase Proteins 0.000 description 1
- HGCNKOLVKRAVHD-UHFFFAOYSA-N L-Met-L-Phe Natural products CSCCC(N)C(=O)NC(C(O)=O)CC1=CC=CC=C1 HGCNKOLVKRAVHD-UHFFFAOYSA-N 0.000 description 1
- SITWEMZOJNKJCH-UHFFFAOYSA-N L-alanine-L-arginine Natural products CC(N)C(=O)NC(C(O)=O)CCCNC(N)=N SITWEMZOJNKJCH-UHFFFAOYSA-N 0.000 description 1
- ODKSFYDXXFIFQN-BYPYZUCNSA-P L-argininium(2+) Chemical group NC(=[NH2+])NCCC[C@H]([NH3+])C(O)=O ODKSFYDXXFIFQN-BYPYZUCNSA-P 0.000 description 1
- 125000000510 L-tryptophano group Chemical group [H]C1=C([H])C([H])=C2N([H])C([H])=C(C([H])([H])[C@@]([H])(C(O[H])=O)N([H])[*])C2=C1[H] 0.000 description 1
- RIMMMMYKGIBOSN-DCAQKATOSA-N Leu-Asn-Met Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCSC)C(O)=O RIMMMMYKGIBOSN-DCAQKATOSA-N 0.000 description 1
- WGNOPSQMIQERPK-UHFFFAOYSA-N Leu-Asn-Pro Natural products CC(C)CC(N)C(=O)NC(CC(=O)N)C(=O)N1CCCC1C(=O)O WGNOPSQMIQERPK-UHFFFAOYSA-N 0.000 description 1
- FIJMQLGQLBLBOL-HJGDQZAQSA-N Leu-Asn-Thr Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O FIJMQLGQLBLBOL-HJGDQZAQSA-N 0.000 description 1
- PJYSOYLLTJKZHC-GUBZILKMSA-N Leu-Asp-Gln Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@H](C(O)=O)CCC(N)=O PJYSOYLLTJKZHC-GUBZILKMSA-N 0.000 description 1
- MMEDVBWCMGRKKC-GARJFASQSA-N Leu-Asp-Pro Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N1CCC[C@@H]1C(=O)O)N MMEDVBWCMGRKKC-GARJFASQSA-N 0.000 description 1
- QCSFMCFHVGTLFF-NHCYSSNCSA-N Leu-Asp-Val Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O QCSFMCFHVGTLFF-NHCYSSNCSA-N 0.000 description 1
- NEEOBPIXKWSBRF-IUCAKERBSA-N Leu-Glu-Gly Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(=O)NCC(O)=O NEEOBPIXKWSBRF-IUCAKERBSA-N 0.000 description 1
- KGCLIYGPQXUNLO-IUCAKERBSA-N Leu-Gly-Glu Chemical compound CC(C)C[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CCC(O)=O KGCLIYGPQXUNLO-IUCAKERBSA-N 0.000 description 1
- VWHGTYCRDRBSFI-ZETCQYMHSA-N Leu-Gly-Gly Chemical compound CC(C)C[C@H](N)C(=O)NCC(=O)NCC(O)=O VWHGTYCRDRBSFI-ZETCQYMHSA-N 0.000 description 1
- OYQUOLRTJHWVSQ-SRVKXCTJSA-N Leu-His-Ser Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CO)C(O)=O OYQUOLRTJHWVSQ-SRVKXCTJSA-N 0.000 description 1
- OMHLATXVNQSALM-FQUUOJAGSA-N Leu-Ile-Pro Chemical compound CC[C@H](C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CC(C)C)N OMHLATXVNQSALM-FQUUOJAGSA-N 0.000 description 1
- HRTRLSRYZZKPCO-BJDJZHNGSA-N Leu-Ile-Ser Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CO)C(O)=O HRTRLSRYZZKPCO-BJDJZHNGSA-N 0.000 description 1
- FAELBUXXFQLUAX-AJNGGQMLSA-N Leu-Leu-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CC(C)C FAELBUXXFQLUAX-AJNGGQMLSA-N 0.000 description 1
- HVHRPWQEQHIQJF-AVGNSLFASA-N Leu-Lys-Glu Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(O)=O HVHRPWQEQHIQJF-AVGNSLFASA-N 0.000 description 1
- REPBGZHJKYWFMJ-KKUMJFAQSA-N Leu-Lys-His Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)N REPBGZHJKYWFMJ-KKUMJFAQSA-N 0.000 description 1
- LZHJZLHSRGWBBE-IHRRRGAJSA-N Leu-Lys-Val Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C(C)C)C(O)=O LZHJZLHSRGWBBE-IHRRRGAJSA-N 0.000 description 1
- IBSGMIPRBMPMHE-IHRRRGAJSA-N Leu-Met-Lys Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CCCCN)C(O)=O IBSGMIPRBMPMHE-IHRRRGAJSA-N 0.000 description 1
- MVHXGBZUJLWZOH-BJDJZHNGSA-N Leu-Ser-Ile Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O MVHXGBZUJLWZOH-BJDJZHNGSA-N 0.000 description 1
- ZJZNLRVCZWUONM-JXUBOQSCSA-N Leu-Thr-Ala Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C)C(O)=O ZJZNLRVCZWUONM-JXUBOQSCSA-N 0.000 description 1
- VUBIPAHVHMZHCM-KKUMJFAQSA-N Leu-Tyr-Ser Chemical compound CC(C)C[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](CO)C(O)=O)CC1=CC=C(O)C=C1 VUBIPAHVHMZHCM-KKUMJFAQSA-N 0.000 description 1
- AIMGJYMCTAABEN-GVXVVHGQSA-N Leu-Val-Glu Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O AIMGJYMCTAABEN-GVXVVHGQSA-N 0.000 description 1
- YQFZRHYZLARWDY-IHRRRGAJSA-N Leu-Val-Lys Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CCCCN YQFZRHYZLARWDY-IHRRRGAJSA-N 0.000 description 1
- WXJKFRMKJORORD-DCAQKATOSA-N Lys-Arg-Ala Chemical compound NC(=N)NCCC[C@@H](C(=O)N[C@@H](C)C(O)=O)NC(=O)[C@@H](N)CCCCN WXJKFRMKJORORD-DCAQKATOSA-N 0.000 description 1
- LZWNAOIMTLNMDW-NHCYSSNCSA-N Lys-Asn-Val Chemical compound CC(C)[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)N)NC(=O)[C@H](CCCCN)N LZWNAOIMTLNMDW-NHCYSSNCSA-N 0.000 description 1
- GCMWRRQAKQXDED-IUCAKERBSA-N Lys-Glu-Gly Chemical compound [NH3+]CCCC[C@H]([NH3+])C(=O)N[C@@H](CCC([O-])=O)C(=O)NCC([O-])=O GCMWRRQAKQXDED-IUCAKERBSA-N 0.000 description 1
- DUTMKEAPLLUGNO-JYJNAYRXSA-N Lys-Glu-Phe Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O DUTMKEAPLLUGNO-JYJNAYRXSA-N 0.000 description 1
- ZMMDPRTXLAEMOD-BZSNNMDCSA-N Lys-His-Phe Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O ZMMDPRTXLAEMOD-BZSNNMDCSA-N 0.000 description 1
- SLQJJFAVWSZLBL-BJDJZHNGSA-N Lys-Ile-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H]([C@@H](C)CC)NC(=O)[C@@H](N)CCCCN SLQJJFAVWSZLBL-BJDJZHNGSA-N 0.000 description 1
- IUWMQCZOTYRXPL-ZPFDUUQYSA-N Lys-Ile-Asp Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(O)=O)C(O)=O IUWMQCZOTYRXPL-ZPFDUUQYSA-N 0.000 description 1
- IZJGPPIGYTVXLB-FQUUOJAGSA-N Lys-Ile-Pro Chemical compound CC[C@H](C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CCCCN)N IZJGPPIGYTVXLB-FQUUOJAGSA-N 0.000 description 1
- VMTYLUGCXIEDMV-QWRGUYRKSA-N Lys-Leu-Gly Chemical compound OC(=O)CNC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CCCCN VMTYLUGCXIEDMV-QWRGUYRKSA-N 0.000 description 1
- HVAUKHLDSDDROB-KKUMJFAQSA-N Lys-Lys-Leu Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(O)=O HVAUKHLDSDDROB-KKUMJFAQSA-N 0.000 description 1
- DIBZLYZXTSVGLN-CIUDSAMLSA-N Lys-Ser-Ser Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(O)=O DIBZLYZXTSVGLN-CIUDSAMLSA-N 0.000 description 1
- MIFFFXHMAHFACR-KATARQTJSA-N Lys-Ser-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@H](CO)NC(=O)[C@@H](N)CCCCN MIFFFXHMAHFACR-KATARQTJSA-N 0.000 description 1
- RMOKGALPSPOYKE-KATARQTJSA-N Lys-Thr-Ser Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CO)C(O)=O RMOKGALPSPOYKE-KATARQTJSA-N 0.000 description 1
- SUZVLFWOCKHWET-CQDKDKBSSA-N Lys-Tyr-Ala Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](C)C(O)=O SUZVLFWOCKHWET-CQDKDKBSSA-N 0.000 description 1
- XATKLFSXFINPSB-JYJNAYRXSA-N Lys-Tyr-Gln Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CCC(N)=O)C(O)=O XATKLFSXFINPSB-JYJNAYRXSA-N 0.000 description 1
- IKXQOBUBZSOWDY-AVGNSLFASA-N Lys-Val-Val Chemical compound CC(C)[C@@H](C(=O)N[C@@H](C(C)C)C(=O)O)NC(=O)[C@H](CCCCN)N IKXQOBUBZSOWDY-AVGNSLFASA-N 0.000 description 1
- VHGIWFGJIHTASW-FXQIFTODSA-N Met-Ala-Asp Chemical compound CSCC[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CC(O)=O VHGIWFGJIHTASW-FXQIFTODSA-N 0.000 description 1
- OGAZPKJHHZPYFK-GARJFASQSA-N Met-Glu-Pro Chemical compound CSCC[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N1CCC[C@@H]1C(=O)O)N OGAZPKJHHZPYFK-GARJFASQSA-N 0.000 description 1
- GVIVXNFKJQFTCE-YUMQZZPRSA-N Met-Gly-Gln Chemical compound CSCC[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CCC(N)=O GVIVXNFKJQFTCE-YUMQZZPRSA-N 0.000 description 1
- AEQVPPGEJJBFEE-CYDGBPFRSA-N Met-Ile-Arg Chemical compound CSCC[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O AEQVPPGEJJBFEE-CYDGBPFRSA-N 0.000 description 1
- DBXMFHGGHMXYHY-DCAQKATOSA-N Met-Leu-Ser Chemical compound CSCC[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(O)=O DBXMFHGGHMXYHY-DCAQKATOSA-N 0.000 description 1
- GGXZOTSDJJTDGB-GUBZILKMSA-N Met-Ser-Val Chemical compound [H]N[C@@H](CCSC)C(=O)N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(O)=O GGXZOTSDJJTDGB-GUBZILKMSA-N 0.000 description 1
- YBAFDPFAUTYYRW-UHFFFAOYSA-N N-L-alpha-glutamyl-L-leucine Natural products CC(C)CC(C(O)=O)NC(=O)C(N)CCC(O)=O YBAFDPFAUTYYRW-UHFFFAOYSA-N 0.000 description 1
- XMBSYZWANAQXEV-UHFFFAOYSA-N N-alpha-L-glutamyl-L-phenylalanine Natural products OC(=O)CCC(N)C(=O)NC(C(O)=O)CC1=CC=CC=C1 XMBSYZWANAQXEV-UHFFFAOYSA-N 0.000 description 1
- AYPMIIKUMNADSU-IHRRRGAJSA-N Phe-Arg-Asn Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(N)=O)C(O)=O AYPMIIKUMNADSU-IHRRRGAJSA-N 0.000 description 1
- JIYJYFIXQTYDNF-YDHLFZDLSA-N Phe-Asn-Val Chemical compound CC(C)[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)N)NC(=O)[C@H](CC1=CC=CC=C1)N JIYJYFIXQTYDNF-YDHLFZDLSA-N 0.000 description 1
- LWPMGKSZPKFKJD-DZKIICNBSA-N Phe-Glu-Val Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O LWPMGKSZPKFKJD-DZKIICNBSA-N 0.000 description 1
- APJPXSFJBMMOLW-KBPBESRZSA-N Phe-Gly-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)CNC(=O)[C@@H](N)CC1=CC=CC=C1 APJPXSFJBMMOLW-KBPBESRZSA-N 0.000 description 1
- SFKOEHXABNPLRT-KBPBESRZSA-N Phe-His-Gly Chemical compound N[C@@H](Cc1ccccc1)C(=O)N[C@@H](Cc1cnc[nH]1)C(=O)NCC(O)=O SFKOEHXABNPLRT-KBPBESRZSA-N 0.000 description 1
- BEEVXUYVEHXWRQ-YESZJQIVSA-N Phe-His-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC2=CN=CN2)NC(=O)[C@H](CC3=CC=CC=C3)N)C(=O)O BEEVXUYVEHXWRQ-YESZJQIVSA-N 0.000 description 1
- HTXVATDVCRFORF-MGHWNKPDSA-N Phe-Ile-His Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)[C@H](CC2=CC=CC=C2)N HTXVATDVCRFORF-MGHWNKPDSA-N 0.000 description 1
- LRBSWBVUCLLRLU-BZSNNMDCSA-N Phe-Leu-Lys Chemical compound CC(C)C[C@H](NC(=O)[C@@H](N)Cc1ccccc1)C(=O)N[C@@H](CCCCN)C(O)=O LRBSWBVUCLLRLU-BZSNNMDCSA-N 0.000 description 1
- AUJWXNGCAQWLEI-KBPBESRZSA-N Phe-Lys-Gly Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCCCN)C(=O)NCC(O)=O AUJWXNGCAQWLEI-KBPBESRZSA-N 0.000 description 1
- BPIMVBKDLSBKIJ-FCLVOEFKSA-N Phe-Thr-Phe Chemical compound C([C@H](N)C(=O)N[C@@H]([C@H](O)C)C(=O)N[C@@H](CC=1C=CC=CC=1)C(O)=O)C1=CC=CC=C1 BPIMVBKDLSBKIJ-FCLVOEFKSA-N 0.000 description 1
- ABEFOXGAIIJDCL-SFJXLCSZSA-N Phe-Thr-Trp Chemical compound C([C@H](N)C(=O)N[C@@H]([C@H](O)C)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(O)=O)C1=CC=CC=C1 ABEFOXGAIIJDCL-SFJXLCSZSA-N 0.000 description 1
- VDTYRPWRWRCROL-UFYCRDLUSA-N Phe-Val-Phe Chemical compound C([C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC=1C=CC=CC=1)C(O)=O)C1=CC=CC=C1 VDTYRPWRWRCROL-UFYCRDLUSA-N 0.000 description 1
- 108010086950 Phosphoribosylanthranilate isomerase Proteins 0.000 description 1
- AJLVKXCNXIJHDV-CIUDSAMLSA-N Pro-Ala-Gln Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](C)C(=O)N[C@@H](CCC(N)=O)C(O)=O AJLVKXCNXIJHDV-CIUDSAMLSA-N 0.000 description 1
- SFECXGVELZFBFJ-VEVYYDQMSA-N Pro-Asp-Thr Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O SFECXGVELZFBFJ-VEVYYDQMSA-N 0.000 description 1
- ZPPVJIJMIKTERM-YUMQZZPRSA-N Pro-Gln-Gly Chemical compound OC(=O)CNC(=O)[C@H](CCC(=O)N)NC(=O)[C@@H]1CCCN1 ZPPVJIJMIKTERM-YUMQZZPRSA-N 0.000 description 1
- UAYHMOIGIQZLFR-NHCYSSNCSA-N Pro-Gln-Val Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](C(C)C)C(O)=O UAYHMOIGIQZLFR-NHCYSSNCSA-N 0.000 description 1
- WVOXLKUUVCCCSU-ZPFDUUQYSA-N Pro-Glu-Ile Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O WVOXLKUUVCCCSU-ZPFDUUQYSA-N 0.000 description 1
- WSRWHZRUOCACLJ-UWVGGRQHSA-N Pro-Gly-His Chemical compound C([C@@H](C(=O)O)NC(=O)CNC(=O)[C@H]1NCCC1)C1=CN=CN1 WSRWHZRUOCACLJ-UWVGGRQHSA-N 0.000 description 1
- GSPPWVHVBBSPSY-FHWLQOOXSA-N Pro-His-Trp Chemical compound OC(=O)[C@H](Cc1c[nH]c2ccccc12)NC(=O)[C@H](Cc1cnc[nH]1)NC(=O)[C@@H]1CCCN1 GSPPWVHVBBSPSY-FHWLQOOXSA-N 0.000 description 1
- UREQLMJCKFLLHM-NAKRPEOUSA-N Pro-Ile-Ser Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CO)C(O)=O UREQLMJCKFLLHM-NAKRPEOUSA-N 0.000 description 1
- CLJLVCYFABNTHP-DCAQKATOSA-N Pro-Leu-Asp Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(O)=O)C(O)=O CLJLVCYFABNTHP-DCAQKATOSA-N 0.000 description 1
- GURGCNUWVSDYTP-SRVKXCTJSA-N Pro-Leu-Gln Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(O)=O GURGCNUWVSDYTP-SRVKXCTJSA-N 0.000 description 1
- ULWBBFKQBDNGOY-RWMBFGLXSA-N Pro-Lys-Pro Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CCCCN)C(=O)N2CCC[C@@H]2C(=O)O ULWBBFKQBDNGOY-RWMBFGLXSA-N 0.000 description 1
- ZVEQWRWMRFIVSD-HRCADAONSA-N Pro-Phe-Pro Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CC2=CC=CC=C2)C(=O)N3CCC[C@@H]3C(=O)O ZVEQWRWMRFIVSD-HRCADAONSA-N 0.000 description 1
- RCYUBVHMVUHEBM-RCWTZXSCSA-N Pro-Pro-Thr Chemical compound [H]N1CCC[C@H]1C(=O)N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)O)C(O)=O RCYUBVHMVUHEBM-RCWTZXSCSA-N 0.000 description 1
- OWQXAJQZLWHPBH-FXQIFTODSA-N Pro-Ser-Asn Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(O)=O OWQXAJQZLWHPBH-FXQIFTODSA-N 0.000 description 1
- CZCCVJUUWBMISW-FXQIFTODSA-N Pro-Ser-Cys Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CO)C(=O)N[C@@H](CS)C(=O)O CZCCVJUUWBMISW-FXQIFTODSA-N 0.000 description 1
- SXJOPONICMGFCR-DCAQKATOSA-N Pro-Ser-Lys Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCCCN)C(=O)O SXJOPONICMGFCR-DCAQKATOSA-N 0.000 description 1
- MKGIILKDUGDRRO-FXQIFTODSA-N Pro-Ser-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H](CO)NC(=O)[C@@H]1CCCN1 MKGIILKDUGDRRO-FXQIFTODSA-N 0.000 description 1
- HRIXMVRZRGFKNQ-HJGDQZAQSA-N Pro-Thr-Gln Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(N)=O)C(O)=O HRIXMVRZRGFKNQ-HJGDQZAQSA-N 0.000 description 1
- VDHGTOHMHHQSKG-JYJNAYRXSA-N Pro-Val-Phe Chemical compound CC(C)[C@H](NC(=O)[C@@H]1CCCN1)C(=O)N[C@@H](Cc1ccccc1)C(O)=O VDHGTOHMHHQSKG-JYJNAYRXSA-N 0.000 description 1
- 108010019477 S-adenosyl-L-methionine-dependent N-methyltransferase Proteins 0.000 description 1
- HBOABDXGTMMDSE-GUBZILKMSA-N Ser-Arg-Val Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C(C)C)C(O)=O HBOABDXGTMMDSE-GUBZILKMSA-N 0.000 description 1
- UGJRQLURDVGULT-LKXGYXEUSA-N Ser-Asn-Thr Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O UGJRQLURDVGULT-LKXGYXEUSA-N 0.000 description 1
- ICHZYBVODUVUKN-SRVKXCTJSA-N Ser-Asn-Tyr Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O ICHZYBVODUVUKN-SRVKXCTJSA-N 0.000 description 1
- IXUGADGDCQDLSA-FXQIFTODSA-N Ser-Gln-Gln Chemical compound C(CC(=O)N)[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)O)NC(=O)[C@H](CO)N IXUGADGDCQDLSA-FXQIFTODSA-N 0.000 description 1
- XWCYBVBLJRWOFR-WDSKDSINSA-N Ser-Gln-Gly Chemical compound OC[C@H](N)C(=O)N[C@@H](CCC(N)=O)C(=O)NCC(O)=O XWCYBVBLJRWOFR-WDSKDSINSA-N 0.000 description 1
- GZFAWAQTEYDKII-YUMQZZPRSA-N Ser-Gly-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)CNC(=O)[C@@H](N)CO GZFAWAQTEYDKII-YUMQZZPRSA-N 0.000 description 1
- XXXAXOWMBOKTRN-XPUUQOCRSA-N Ser-Gly-Val Chemical compound [H]N[C@@H](CO)C(=O)NCC(=O)N[C@@H](C(C)C)C(O)=O XXXAXOWMBOKTRN-XPUUQOCRSA-N 0.000 description 1
- BKZYBLLIBOBOOW-GHCJXIJMSA-N Ser-Ile-Asp Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(O)=O)C(O)=O BKZYBLLIBOBOOW-GHCJXIJMSA-N 0.000 description 1
- MOINZPRHJGTCHZ-MMWGEVLESA-N Ser-Ile-Pro Chemical compound CC[C@H](C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CO)N MOINZPRHJGTCHZ-MMWGEVLESA-N 0.000 description 1
- GJFYFGOEWLDQGW-GUBZILKMSA-N Ser-Leu-Gln Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)O)NC(=O)[C@H](CO)N GJFYFGOEWLDQGW-GUBZILKMSA-N 0.000 description 1
- XNCUYZKGQOCOQH-YUMQZZPRSA-N Ser-Leu-Gly Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(=O)NCC(O)=O XNCUYZKGQOCOQH-YUMQZZPRSA-N 0.000 description 1
- XXNYYSXNXCJYKX-DCAQKATOSA-N Ser-Leu-Met Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCSC)C(O)=O XXNYYSXNXCJYKX-DCAQKATOSA-N 0.000 description 1
- ADJDNJCSPNFFPI-FXQIFTODSA-N Ser-Pro-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)CO ADJDNJCSPNFFPI-FXQIFTODSA-N 0.000 description 1
- RHAPJNVNWDBFQI-BQBZGAKWSA-N Ser-Pro-Gly Chemical compound OC[C@H](N)C(=O)N1CCC[C@H]1C(=O)NCC(O)=O RHAPJNVNWDBFQI-BQBZGAKWSA-N 0.000 description 1
- QPPYAWVLAVXISR-DCAQKATOSA-N Ser-Pro-His Chemical compound C1C[C@H](N(C1)C(=O)[C@H](CO)N)C(=O)N[C@@H](CC2=CN=CN2)C(=O)O QPPYAWVLAVXISR-DCAQKATOSA-N 0.000 description 1
- HHJFMHQYEAAOBM-ZLUOBGJFSA-N Ser-Ser-Ala Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)N[C@@H](C)C(O)=O HHJFMHQYEAAOBM-ZLUOBGJFSA-N 0.000 description 1
- FZXOPYUEQGDGMS-ACZMJKKPSA-N Ser-Ser-Gln Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCC(N)=O)C(O)=O FZXOPYUEQGDGMS-ACZMJKKPSA-N 0.000 description 1
- SRSPTFBENMJHMR-WHFBIAKZSA-N Ser-Ser-Gly Chemical compound OC[C@H](N)C(=O)N[C@@H](CO)C(=O)NCC(O)=O SRSPTFBENMJHMR-WHFBIAKZSA-N 0.000 description 1
- BMKNXTJLHFIAAH-CIUDSAMLSA-N Ser-Ser-Leu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(O)=O BMKNXTJLHFIAAH-CIUDSAMLSA-N 0.000 description 1
- CUXJENOFJXOSOZ-BIIVOSGPSA-N Ser-Ser-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CO)NC(=O)[C@H](CO)N)C(=O)O CUXJENOFJXOSOZ-BIIVOSGPSA-N 0.000 description 1
- VGQVAVQWKJLIRM-FXQIFTODSA-N Ser-Ser-Val Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(O)=O VGQVAVQWKJLIRM-FXQIFTODSA-N 0.000 description 1
- WUXCHQZLUHBSDJ-LKXGYXEUSA-N Ser-Thr-Asp Chemical compound OC[C@H](N)C(=O)N[C@@H]([C@H](O)C)C(=O)N[C@@H](CC(O)=O)C(O)=O WUXCHQZLUHBSDJ-LKXGYXEUSA-N 0.000 description 1
- ODRUTDLAONAVDV-IHRRRGAJSA-N Ser-Val-Tyr Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O ODRUTDLAONAVDV-IHRRRGAJSA-N 0.000 description 1
- 241000205091 Sulfolobus solfataricus Species 0.000 description 1
- DWYAUVCQDTZIJI-VZFHVOOUSA-N Thr-Ala-Ser Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CO)C(O)=O DWYAUVCQDTZIJI-VZFHVOOUSA-N 0.000 description 1
- JMQUAZXYFAEOIH-XGEHTFHBSA-N Thr-Arg-Cys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CS)C(=O)O)N)O JMQUAZXYFAEOIH-XGEHTFHBSA-N 0.000 description 1
- CEXFELBFVHLYDZ-XGEHTFHBSA-N Thr-Arg-Ser Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(O)=O CEXFELBFVHLYDZ-XGEHTFHBSA-N 0.000 description 1
- UNURFMVMXLENAZ-KJEVXHAQSA-N Thr-Arg-Tyr Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O UNURFMVMXLENAZ-KJEVXHAQSA-N 0.000 description 1
- LXWZOMSOUAMOIA-JIOCBJNQSA-N Thr-Asn-Pro Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N1CCC[C@@H]1C(=O)O)N)O LXWZOMSOUAMOIA-JIOCBJNQSA-N 0.000 description 1
- PZVGOVRNGKEFCB-KKHAAJSZSA-N Thr-Asn-Val Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](C(C)C)C(=O)O)N)O PZVGOVRNGKEFCB-KKHAAJSZSA-N 0.000 description 1
- LKEKWDJCJSPXNI-IRIUXVKKSA-N Thr-Glu-Tyr Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 LKEKWDJCJSPXNI-IRIUXVKKSA-N 0.000 description 1
- YJCVECXVYHZOBK-KNZXXDILSA-N Thr-Ile-Pro Chemical compound CC[C@H](C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H]([C@@H](C)O)N YJCVECXVYHZOBK-KNZXXDILSA-N 0.000 description 1
- XYFISNXATOERFZ-OSUNSFLBSA-N Thr-Ile-Val Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](C(C)C)C(=O)O)NC(=O)[C@H]([C@@H](C)O)N XYFISNXATOERFZ-OSUNSFLBSA-N 0.000 description 1
- KRDSCBLRHORMRK-JXUBOQSCSA-N Thr-Lys-Ala Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C)C(O)=O KRDSCBLRHORMRK-JXUBOQSCSA-N 0.000 description 1
- WTMPKZWHRCMMMT-KZVJFYERSA-N Thr-Pro-Ala Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C)C(O)=O WTMPKZWHRCMMMT-KZVJFYERSA-N 0.000 description 1
- FWTFAZKJORVTIR-VZFHVOOUSA-N Thr-Ser-Ala Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CO)C(=O)N[C@@H](C)C(O)=O FWTFAZKJORVTIR-VZFHVOOUSA-N 0.000 description 1
- BJJRNAVDQGREGC-HOUAVDHOSA-N Thr-Trp-Asn Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)N[C@@H](CC(=O)N)C(=O)O)N)O BJJRNAVDQGREGC-HOUAVDHOSA-N 0.000 description 1
- BKIOKSLLAAZYTC-KKHAAJSZSA-N Thr-Val-Asn Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(N)=O)C(O)=O BKIOKSLLAAZYTC-KKHAAJSZSA-N 0.000 description 1
- HYVLNORXQGKONN-NUTKFTJISA-N Trp-Ala-Lys Chemical compound C1=CC=C2C(C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCCN)C(O)=O)=CNC2=C1 HYVLNORXQGKONN-NUTKFTJISA-N 0.000 description 1
- CZWIHKFGHICAJX-BPUTZDHNSA-N Trp-Glu-Glu Chemical compound C1=CC=C2C(C[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O)=CNC2=C1 CZWIHKFGHICAJX-BPUTZDHNSA-N 0.000 description 1
- RCLOWEZASFJFEX-KKUMJFAQSA-N Tyr-Asp-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 RCLOWEZASFJFEX-KKUMJFAQSA-N 0.000 description 1
- VFJIWSJKZJTQII-SRVKXCTJSA-N Tyr-Asp-Ser Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(O)=O VFJIWSJKZJTQII-SRVKXCTJSA-N 0.000 description 1
- HVPPEXXUDXAPOM-MGHWNKPDSA-N Tyr-Ile-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H]([C@@H](C)CC)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 HVPPEXXUDXAPOM-MGHWNKPDSA-N 0.000 description 1
- CDKZJGMPZHPAJC-ULQDDVLXSA-N Tyr-Leu-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 CDKZJGMPZHPAJC-ULQDDVLXSA-N 0.000 description 1
- ZOBLBMGJKVJVEV-BZSNNMDCSA-N Tyr-Lys-Lys Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCCN)C(=O)O)N)O ZOBLBMGJKVJVEV-BZSNNMDCSA-N 0.000 description 1
- NVZVJIUDICCMHZ-BZSNNMDCSA-N Tyr-Phe-Ser Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CO)C(O)=O NVZVJIUDICCMHZ-BZSNNMDCSA-N 0.000 description 1
- VBFVQTPETKJCQW-RPTUDFQQSA-N Tyr-Phe-Thr Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H]([C@@H](C)O)C(O)=O VBFVQTPETKJCQW-RPTUDFQQSA-N 0.000 description 1
- BIWVVOHTKDLRMP-ULQDDVLXSA-N Tyr-Pro-Leu Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(C)C)C(O)=O BIWVVOHTKDLRMP-ULQDDVLXSA-N 0.000 description 1
- RWOKVQUCENPXGE-IHRRRGAJSA-N Tyr-Ser-Arg Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O RWOKVQUCENPXGE-IHRRRGAJSA-N 0.000 description 1
- NHOVZGFNTGMYMI-KKUMJFAQSA-N Tyr-Ser-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CO)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 NHOVZGFNTGMYMI-KKUMJFAQSA-N 0.000 description 1
- JHDZONWZTCKTJR-KJEVXHAQSA-N Tyr-Thr-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 JHDZONWZTCKTJR-KJEVXHAQSA-N 0.000 description 1
- LABUITCFCAABSV-BPNCWPANSA-N Val-Ala-Tyr Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 LABUITCFCAABSV-BPNCWPANSA-N 0.000 description 1
- HNWQUBBOBKSFQV-AVGNSLFASA-N Val-Arg-His Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)N HNWQUBBOBKSFQV-AVGNSLFASA-N 0.000 description 1
- ZMDCGGKHRKNWKD-LAEOZQHASA-N Val-Asn-Glu Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N ZMDCGGKHRKNWKD-LAEOZQHASA-N 0.000 description 1
- PVPAOIGJYHVWBT-KKHAAJSZSA-N Val-Asn-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CC(=O)N)NC(=O)[C@H](C(C)C)N)O PVPAOIGJYHVWBT-KKHAAJSZSA-N 0.000 description 1
- ZQGPWORGSNRQLN-NHCYSSNCSA-N Val-Asp-His Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)N ZQGPWORGSNRQLN-NHCYSSNCSA-N 0.000 description 1
- TZVUSFMQWPWHON-NHCYSSNCSA-N Val-Asp-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)O)NC(=O)[C@H](C(C)C)N TZVUSFMQWPWHON-NHCYSSNCSA-N 0.000 description 1
- SCBITHMBEJNRHC-LSJOCFKGSA-N Val-Asp-Val Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](C(C)C)C(=O)O)N SCBITHMBEJNRHC-LSJOCFKGSA-N 0.000 description 1
- WBUOKGBHGDPYMH-GUBZILKMSA-N Val-Cys-Met Chemical compound CSCC[C@@H](C(O)=O)NC(=O)[C@H](CS)NC(=O)[C@@H](N)C(C)C WBUOKGBHGDPYMH-GUBZILKMSA-N 0.000 description 1
- VFOHXOLPLACADK-GVXVVHGQSA-N Val-Gln-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)N)NC(=O)[C@H](C(C)C)N VFOHXOLPLACADK-GVXVVHGQSA-N 0.000 description 1
- ZEVNVXYRZRIRCH-GVXVVHGQSA-N Val-Gln-Lys Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H](CCCCN)C(=O)O)N ZEVNVXYRZRIRCH-GVXVVHGQSA-N 0.000 description 1
- JXGWQYWDUOWQHA-DZKIICNBSA-N Val-Gln-Phe Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)O)N JXGWQYWDUOWQHA-DZKIICNBSA-N 0.000 description 1
- VVZDBPBZHLQPPB-XVKPBYJWSA-N Val-Glu-Gly Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)NCC(O)=O VVZDBPBZHLQPPB-XVKPBYJWSA-N 0.000 description 1
- OQWNEUXPKHIEJO-NRPADANISA-N Val-Glu-Ser Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CO)C(=O)O)N OQWNEUXPKHIEJO-NRPADANISA-N 0.000 description 1
- PMXBARDFIAPBGK-DZKIICNBSA-N Val-Glu-Tyr Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 PMXBARDFIAPBGK-DZKIICNBSA-N 0.000 description 1
- UEHRGZCNLSWGHK-DLOVCJGASA-N Val-Glu-Val Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O UEHRGZCNLSWGHK-DLOVCJGASA-N 0.000 description 1
- WFENBJPLZMPVAX-XVKPBYJWSA-N Val-Gly-Glu Chemical compound CC(C)[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CCC(O)=O WFENBJPLZMPVAX-XVKPBYJWSA-N 0.000 description 1
- PTFPUAXGIKTVNN-ONGXEEELSA-N Val-His-Gly Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)NCC(=O)O)N PTFPUAXGIKTVNN-ONGXEEELSA-N 0.000 description 1
- UKEVLVBHRKWECS-LSJOCFKGSA-N Val-Ile-Gly Chemical compound CC[C@H](C)[C@@H](C(=O)NCC(=O)O)NC(=O)[C@H](C(C)C)N UKEVLVBHRKWECS-LSJOCFKGSA-N 0.000 description 1
- FTKXYXACXYOHND-XUXIUFHCSA-N Val-Ile-Leu Chemical compound CC(C)[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(C)C)C(O)=O FTKXYXACXYOHND-XUXIUFHCSA-N 0.000 description 1
- OTJMMKPMLUNTQT-AVGNSLFASA-N Val-Leu-Arg Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)NC(=O)[C@H](C(C)C)N OTJMMKPMLUNTQT-AVGNSLFASA-N 0.000 description 1
- SYSWVVCYSXBVJG-RHYQMDGZSA-N Val-Leu-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](C(C)C)N)O SYSWVVCYSXBVJG-RHYQMDGZSA-N 0.000 description 1
- YMTOEGGOCHVGEH-IHRRRGAJSA-N Val-Lys-Lys Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCCN)C(O)=O YMTOEGGOCHVGEH-IHRRRGAJSA-N 0.000 description 1
- VNGKMNPAENRGDC-JYJNAYRXSA-N Val-Phe-Arg Chemical compound NC(N)=NCCC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)C(C)C)CC1=CC=CC=C1 VNGKMNPAENRGDC-JYJNAYRXSA-N 0.000 description 1
- ZXYPHBKIZLAQTL-QXEWZRGKSA-N Val-Pro-Asp Chemical compound CC(C)[C@@H](C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(=O)O)C(=O)O)N ZXYPHBKIZLAQTL-QXEWZRGKSA-N 0.000 description 1
- QSPOLEBZTMESFY-SRVKXCTJSA-N Val-Pro-Val Chemical compound CC(C)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C(C)C)C(O)=O QSPOLEBZTMESFY-SRVKXCTJSA-N 0.000 description 1
- PZTZYZUTCPZWJH-FXQIFTODSA-N Val-Ser-Ser Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)O)N PZTZYZUTCPZWJH-FXQIFTODSA-N 0.000 description 1
- DOBHJKVVACOQTN-DZKIICNBSA-N Val-Tyr-Gln Chemical compound NC(=O)CC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)C(C)C)CC1=CC=C(O)C=C1 DOBHJKVVACOQTN-DZKIICNBSA-N 0.000 description 1
- 238000002835 absorbance Methods 0.000 description 1
- 230000009471 action Effects 0.000 description 1
- 108010045350 alanyl-tyrosyl-alanine Proteins 0.000 description 1
- 108010005233 alanylglutamic acid Proteins 0.000 description 1
- 108010044940 alanylglutamine Proteins 0.000 description 1
- 108010070783 alanyltyrosine Proteins 0.000 description 1
- KOSRFJWDECSPRO-UHFFFAOYSA-N alpha-L-glutamyl-L-glutamic acid Natural products OC(=O)CCC(N)C(=O)NC(CCC(O)=O)C(O)=O KOSRFJWDECSPRO-UHFFFAOYSA-N 0.000 description 1
- 229910021529 ammonia Inorganic materials 0.000 description 1
- AVKUERGKIZMTKX-NJBDSQKTSA-N ampicillin Chemical compound C1([C@@H](N)C(=O)N[C@H]2[C@H]3SC([C@@H](N3C2=O)C(O)=O)(C)C)=CC=CC=C1 AVKUERGKIZMTKX-NJBDSQKTSA-N 0.000 description 1
- 229960000723 ampicillin Drugs 0.000 description 1
- 239000002246 antineoplastic agent Substances 0.000 description 1
- 229940041181 antineoplastic drug Drugs 0.000 description 1
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 1
- 108010008355 arginyl-glutamine Proteins 0.000 description 1
- 108010038850 arginyl-isoleucyl-tyrosine Proteins 0.000 description 1
- 125000003118 aryl group Chemical group 0.000 description 1
- 108010068265 aspartyltyrosine Proteins 0.000 description 1
- 230000008901 benefit Effects 0.000 description 1
- WPYMKLBDIGXBTP-UHFFFAOYSA-N benzoic acid group Chemical group C(C1=CC=CC=C1)(=O)O WPYMKLBDIGXBTP-UHFFFAOYSA-N 0.000 description 1
- 230000001588 bifunctional effect Effects 0.000 description 1
- 230000001851 biosynthetic effect Effects 0.000 description 1
- 238000001815 biotherapy Methods 0.000 description 1
- 239000006227 byproduct Substances 0.000 description 1
- 239000007795 chemical reaction product Substances 0.000 description 1
- 210000000349 chromosome Anatomy 0.000 description 1
- 235000018417 cysteine Nutrition 0.000 description 1
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 1
- 230000002950 deficient Effects 0.000 description 1
- 238000010790 dilution Methods 0.000 description 1
- 239000012895 dilution Substances 0.000 description 1
- 108010054812 diprotin A Proteins 0.000 description 1
- 230000002708 enhancing effect Effects 0.000 description 1
- 238000009585 enzyme analysis Methods 0.000 description 1
- 238000002474 experimental method Methods 0.000 description 1
- 239000012467 final product Substances 0.000 description 1
- 230000004907 flux Effects 0.000 description 1
- 238000009472 formulation Methods 0.000 description 1
- 230000006870 function Effects 0.000 description 1
- 230000002538 fungal effect Effects 0.000 description 1
- 108010055341 glutamyl-glutamic acid Proteins 0.000 description 1
- 125000003630 glycyl group Chemical group [H]N([H])C([H])([H])C(*)=O 0.000 description 1
- 108010019832 glycyl-asparaginyl-glycine Proteins 0.000 description 1
- 108010089804 glycyl-threonine Proteins 0.000 description 1
- 108010015792 glycyllysine Proteins 0.000 description 1
- 108010040030 histidinoalanine Proteins 0.000 description 1
- 108010028295 histidylhistidine Proteins 0.000 description 1
- 108010085325 histidylproline Proteins 0.000 description 1
- 238000000338 in vitro Methods 0.000 description 1
- PZOUSPYUWWUPPK-UHFFFAOYSA-N indole Natural products CC1=CC=CC2=C1C=CN2 PZOUSPYUWWUPPK-UHFFFAOYSA-N 0.000 description 1
- RKJUIXBNRJVNHR-UHFFFAOYSA-N indolenine Natural products C1=CC=C2CC=NC2=C1 RKJUIXBNRJVNHR-UHFFFAOYSA-N 0.000 description 1
- 125000001041 indolyl group Chemical group 0.000 description 1
- 238000009776 industrial production Methods 0.000 description 1
- 230000007154 intracellular accumulation Effects 0.000 description 1
- 230000003834 intracellular effect Effects 0.000 description 1
- 108010053037 kyotorphin Proteins 0.000 description 1
- 108010083708 leucyl-aspartyl-valine Proteins 0.000 description 1
- 108010044311 leucyl-glycyl-glycine Proteins 0.000 description 1
- 108010047926 leucyl-lysyl-tyrosine Proteins 0.000 description 1
- 230000000670 limiting effect Effects 0.000 description 1
- 125000003588 lysine group Chemical group [H]N([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])(N([H])[H])C(*)=O 0.000 description 1
- 108010003700 lysyl aspartic acid Proteins 0.000 description 1
- 108010045397 lysyl-tyrosyl-lysine Proteins 0.000 description 1
- 108010038320 lysylphenylalanine Proteins 0.000 description 1
- 230000007246 mechanism Effects 0.000 description 1
- 108020004999 messenger RNA Proteins 0.000 description 1
- 108010056582 methionylglutamic acid Proteins 0.000 description 1
- 108010068488 methionylphenylalanine Proteins 0.000 description 1
- 239000000203 mixture Substances 0.000 description 1
- 108010000785 non-ribosomal peptide synthase Proteins 0.000 description 1
- 108020004707 nucleic acids Proteins 0.000 description 1
- 102000039446 nucleic acids Human genes 0.000 description 1
- 150000007523 nucleic acids Chemical class 0.000 description 1
- 230000036961 partial effect Effects 0.000 description 1
- 108010024654 phenylalanyl-prolyl-alanine Proteins 0.000 description 1
- 239000002244 precipitate Substances 0.000 description 1
- 230000008569 process Effects 0.000 description 1
- 108090000765 processed proteins & peptides Proteins 0.000 description 1
- 125000001500 prolyl group Chemical group [H]N1C([H])(C(=O)[*])C([H])([H])C([H])([H])C1([H])[H] 0.000 description 1
- 108010070643 prolylglutamic acid Proteins 0.000 description 1
- 108010090894 prolylleucine Proteins 0.000 description 1
- 108010053725 prolylvaline Proteins 0.000 description 1
- 238000011002 quantification Methods 0.000 description 1
- 108020003175 receptors Proteins 0.000 description 1
- 238000005215 recombination Methods 0.000 description 1
- 230000006798 recombination Effects 0.000 description 1
- 238000011160 research Methods 0.000 description 1
- 102200081479 rs121908458 Human genes 0.000 description 1
- 150000003839 salts Chemical class 0.000 description 1
- 238000012216 screening Methods 0.000 description 1
- 238000011218 seed culture Methods 0.000 description 1
- 238000000926 separation method Methods 0.000 description 1
- 238000012163 sequencing technique Methods 0.000 description 1
- 108010069117 seryl-lysyl-aspartic acid Proteins 0.000 description 1
- IFGCUJZIWBUILZ-UHFFFAOYSA-N sodium 2-[[2-[[hydroxy-(3,4,5-trihydroxy-6-methyloxan-2-yl)oxyphosphoryl]amino]-4-methylpentanoyl]amino]-3-(1H-indol-3-yl)propanoic acid Chemical compound [Na+].C=1NC2=CC=CC=C2C=1CC(C(O)=O)NC(=O)C(CC(C)C)NP(O)(=O)OC1OC(C)C(O)C(O)C1O IFGCUJZIWBUILZ-UHFFFAOYSA-N 0.000 description 1
- 239000011780 sodium chloride Substances 0.000 description 1
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 1
- 239000001488 sodium phosphate Substances 0.000 description 1
- 229910000162 sodium phosphate Inorganic materials 0.000 description 1
- 241000894007 species Species 0.000 description 1
- 238000002798 spectrophotometry method Methods 0.000 description 1
- 108010005652 splenotritin Proteins 0.000 description 1
- 230000000087 stabilizing effect Effects 0.000 description 1
- 230000002194 synthesizing effect Effects 0.000 description 1
- QJVXKWHHAMZTBY-GCPOEHJPSA-N syringin Chemical compound COC1=CC(\C=C\CO)=CC(OC)=C1O[C@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 QJVXKWHHAMZTBY-GCPOEHJPSA-N 0.000 description 1
- QJVXKWHHAMZTBY-KSXIZUIISA-N syringin Natural products COc1cc(C=CCO)cc(OC)c1O[C@H]2O[C@@H](CO)[C@H](O)[C@@H](O)[C@@H]2O QJVXKWHHAMZTBY-KSXIZUIISA-N 0.000 description 1
- 238000013518 transcription Methods 0.000 description 1
- 230000035897 transcription Effects 0.000 description 1
- 230000009466 transformation Effects 0.000 description 1
- RYFMWSXOAZQYPI-UHFFFAOYSA-K trisodium phosphate Chemical compound [Na+].[Na+].[Na+].[O-]P([O-])([O-])=O RYFMWSXOAZQYPI-UHFFFAOYSA-K 0.000 description 1
- 101150079930 trpGD gene Proteins 0.000 description 1
- 101150059846 trpS gene Proteins 0.000 description 1
- 101150004782 trpS2 gene Proteins 0.000 description 1
- 201000008827 tuberculosis Diseases 0.000 description 1
- 229960004441 tyrosine Drugs 0.000 description 1
- 108010073969 valyllysine Proteins 0.000 description 1
- 230000035899 viability Effects 0.000 description 1
Images




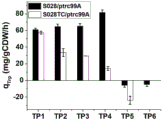







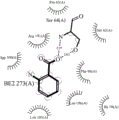
Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/88—Lyases (4.)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P13/00—Preparation of nitrogen-containing organic compounds
- C12P13/04—Alpha- or beta- amino acids
- C12P13/22—Tryptophan; Tyrosine; Phenylalanine; 3,4-Dihydroxyphenylalanine
- C12P13/227—Tryptophan
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y401/00—Carbon-carbon lyases (4.1)
- C12Y401/01—Carboxy-lyases (4.1.1)
- C12Y401/01048—Indole-3-glycerol-phosphate synthase (4.1.1.48)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y401/00—Carbon-carbon lyases (4.1)
- C12Y401/03—Oxo-acid-lyases (4.1.3)
- C12Y401/03027—Anthranilate synthase (4.1.3.27)
Abstract
本发明的目的之一在于通过基因突变细菌细胞,使之表达的吲哚‑3‑甘油磷酸合成酶IGPs相较于野生型IGPs,对抑制剂邻氨基苯甲酸的敏感度下降,甚至可以被氨基苯甲酸激活。
Description
技术领域
本发明涉及利用生物技术生产色氨酸及其衍生物的方法。
背景技术
L-色氨酸(L-trp)是一种营养必需的氨基酸,广泛用于食品和制药行业。L-trp还可以作为多种活性次级代谢产物和抗肿瘤药物的合成前体[1],例如紫丁香素和脱氧紫丁香素[2-4],这为基于L-trp的高价值生物疗法奠定了基础。目前,工业上通常通过微生物合成法等生物技术手段来生产L-trp。
在微生物中,色氨酸是由分支酸产生的,是莽草酸途径的代谢终产物。从分支酸开始,色氨酸通过邻氨基苯甲酸(ANT)、磷酸核糖氨基苯甲酸(PRA 或 PA)、羧苯氨基-脱氧核糖-5-磷酸(CdRP)、吲哚-3-甘油磷酸(IGP)和吲哚进行生物合成。合成过程所涉及的酶包括由trpE基因编码的邻氨基苯甲酸合成酶(EC 4.1.3.27),由trpD基因编码的邻氨基苯甲酸磷酸核糖基转移酶(EC 2.4.1.28),由trpC基因编码的磷酸核糖氨基苯甲酸异构酶(EC5.3.1.24,PRAi)和吲哚-3-甘油磷酸合成酶(EC 4.1.1.48,IGP),以及由trpB和trpA基因编码的色氨酸合成酶(EC 4.2.1.20)。 这些基因聚集在trp操纵子上。 TrpC(IGPs)具有磷酸核糖氨基苯甲酸异构酶(PRAi)和吲哚-3-甘油磷酸合酶(IGPs)的活性。
现有技术已经公开了多种提高微生物合成L-色氨酸的方法,过表达整个trp操纵子并不会提高色氨酸的产量,只会导致代谢链中第一个中间产物邻氨基苯甲酸的积累(Lee KHet al [12])。专利EP 2803720 A2中,发明人建议将trp操纵子进行部分过表达,即将trpD,trpC, trpB和trpA而不是trpE进行过表达。另一现有技术是在大肠杆菌中进行酵母磷酸核糖转移酶的表达(US 2016/0153014 A1)。
然而,利用生物技术生产L-色氨酸的方法仍需改进,因此,本发明提供了一种微生物高产L-色氨酸的方法。
发明内容
本发明的目的之一在于通过基因突变细菌细胞,使之表达的吲哚-3-甘油磷酸合成酶IGPs相较于野生型IGPs,对抑制剂邻氨基苯甲酸盐的敏感度下降。
本发明的研究是基于发现了微生物吲哚-3-甘油磷酸合成酶(IGPs),例如来源于大肠杆菌的EcIGPs 或eIGPs,对抑制剂邻氨基苯甲酸具有很高的敏感度。由于邻氨基苯甲酸是分支酸合成L-色氨酸途径中的中间产物,并且该中间产物是在IGPs之前合成的,因此这种类型的抑制也被称为“前馈抑制”。发明人认为,此前用于提高色氨酸生产率的方法都没有成功或者效果不佳正是因为没有考虑到这种前馈抑制。然而,本发明利用邻氨基苯甲酸对IGPs的负调节解决了这一难题。本发明利用IGPs或具有IGPs活性的酶,其相较于野生型IGPs,对邻氨基苯甲酸盐的抑制敏感度下降,从而提高色氨酸的产率。
本申请中术语“异源”为本领域技术人员公知含义,表示外源的元素,例如酶或者蛋白。“外源”表示该元素没有在靶细胞中存在,例如源于不同遗传组成的细胞或生物,如物种不同的生物体。
术语“同源”表示这个酶或蛋白为天然的酶或蛋白质,即该酶或蛋白在目的细胞内是天然存在的,与之相对应的是异源的酶或蛋白。
“表达”在这里的是指遗传信息的转化,例如基于遗传信息形成蛋白或核酸。尤其要指出,该术语的意思包括基于遗传信息合成蛋白质,包括合成蛋白质之前的步骤,例如基于DNA模板转录形成mRNA。
“经过基因突变、用于表达吲哚-3-甘油磷酸合成酶的细菌细胞”是指一种通过基因工程改造,使之能表达吲哚-3-甘油磷酸合成酶的细菌细胞。术语“吲哚-3-甘油磷酸合成酶”(IGPs)为一种具有IGPs活性的酶(EC 4.1.1.48),该酶具有催化羧苯氨基-脱氧核糖-5-磷酸(CdRP)转化为吲哚-3-甘油磷酸(IGP)的作用。该术语包括除IGPs活性外还具有一种或多种其它的活性,例如磷酸核糖氨基苯甲酸异构酶(PRAi)活性。
术语“对邻氨基苯甲酸盐的抑制敏感度下降”是指在相同邻氨基苯甲酸盐浓度以及相同试验条件(如温度、pH、盐浓度等)下,前者的酶活高于后者,此处因为相同的酶催化反应。
“突变体”是指一种蛋白质,例如一种酶,它的氨基酸序列与野生型不同。该术语涵盖由于编码该蛋白的基因序列发生突变,从而导致氨基酸序列与野生型相比存在不同。
“具有IGPs活性的异源酶”是指具有吲哚-3-甘油磷酸合成酶活性的异源酶,该酶还可以具有一种或多种酶活性,例如磷酸核糖氨基苯甲酸异构酶(PRAi)活性或邻氨基苯甲酸合成酶活性。
“邻氨基苯甲酸合成酶II结构域”或“AS II结构域”是指多功能邻氨基苯甲酸合成酶的组分II,该组分具有谷氨酰胺转移酶活性。邻氨基苯甲酸合成酶催化分支酸盐形成邻氨基苯甲酸的过程可以由邻氨基苯甲酸合成酶的组分I或者组分II催化完成,组分I利用的是氨而非谷氨酰胺,而组分II具有谷氨酰胺转移酶活性。
“邻氨基苯甲酸结合位点”是指酶的一个区域,在这里是指吲哚-3-甘油磷酸合成酶与邻氨基苯甲酸(2-氨基苯甲酸,CAS 118-92-3)结合的区域。在本发明中,“区域”不限于一部分连续的氨基酸,也包括位于酶中不同位置但通过空间折叠彼此靠近的几个氨基酸残基。“邻氨基苯甲酸结合位点”涵盖与邻氨基苯甲酸形成暂时性键的氨基酸残基。并且该术语还涵盖与邻氨基苯甲酸形成暂时性键的氨基酸残基相邻的氨基酸残基。“相邻”表示在于与邻氨基苯甲酸形成暂时性键的氨基酸残基的左侧和/或右侧包含的3个、优选两个、最优选1个的氨基酸残基。本文中“邻氨基苯甲酸结合位点”和“邻氨基苯甲酸结合区域”为相同的意思。
本发明优选实施方案:本发明基因突变的细菌细胞用于表达:
a)细菌IGPs突变体,该突变体相较于野生型细胞产生的IGPs,对邻氨基苯甲酸的抑制敏感度降低,或
b)具有IGPs活性的异源酶,该酶相较于野生型IGPs,对邻氨基苯甲酸的抑制敏感度降低。
本申请发明人发现,细菌IGPs具有邻氨基苯甲酸结合位点,邻氨基苯甲酸与IGPs结合后,非竞争性抑制了磷酸核糖氨基苯甲酸(PRA 或 PA)通过羧苯氨基-脱氧核糖-5-磷酸(CdRP)向吲哚-3-甘油磷酸(IGP)的酶促转化;本发明基因工程改造后的IGPs,可表现出对抑制剂邻氨基苯甲酸盐的敏感度下降。因此,本发明公开了一种细菌IGPs突变体,该突变体产生的IGPs与野生型细胞产生的IGPs相比,对氨基苯甲酸盐的抑制敏感度下降。
在一个优选的实施例中,本发明所述细菌IGPs突变体与基因突变的细菌细胞是同源的。
优选地。本发明基因突变的细菌细胞为用于表达突变IGPs的大肠杆菌细胞。
或者,本发明通过细菌细胞基因突变,使其异源表达产生具有IGPs活性的酶,该异源酶与野生型IGPs相比,对邻氨基苯甲酸的抑制敏感度下降。该异源酶可以是细菌或是其它来源的,例如利用酵母进行异源表达。发明人还发现来源于非细菌细胞(例如酿酒酵母和曲霉菌)的IGPs也具有活性,但是对邻氨基苯甲酸不敏感,甚至可以被激活。在本发明的一个实施例中,利用基因突变的细菌细胞表达具有活性的异源IGPs,但是该酶对邻氨基苯甲酸不敏感并且(或者)可以被邻氨基苯甲酸激活;例如来源于酿酒酵母或曲霉属的酶,优选为来源于酿酒酵母和黑曲霉,该酶具有邻氨基苯甲酸合成酶II结构域。
在本发明另一个优选实施例中,IGPs突变菌株为:
a)与SEQ ID NO: 1序列相比,突变位点为:用丝氨酸替代第60位的丙氨酸或甘氨酸,并且(或者)用异亮氨酸替代第8位的缬氨酸,并且(或者)用亮氨酸代替第188位的苯丙氨酸,或者用丝氨酸代替第58位的谷氨酰胺,用脯氨酸代替第59位的缬氨酸,用丝氨酸替代第60位的苯丙氨酸,用赖氨酸替代第61位的谷氨酰胺;或者
b)突变序列与SEQ ID NO: 1序列相比,第8-188位中至少有一个氨基酸被其它氨基酸取代,但突变体仍然具有IGPs活性,并且与野生型菌株表达的IGPs相比,突变后的IGPs对邻氨基苯甲酸的抑制敏感度降低。
在本发明优选实施例中,细菌IGPs突变序列为SEQ ID NO: 2 至 SEQ ID NO: 5,或SEQID NO:30其中之一。
优选地,突变细菌细胞是大肠杆菌细胞。
SEQ ID NO: 1为野生型大肠杆菌IGPs序列(EcTrpC),SEQ ID NO: 2为I8V突变序列,SEQ ID NO: 3为S60A突变序列,SEQ ID NO: 4为S60G突变序列,SEQ ID NO: 5为L188F突变序列,SEQ ID NO: 30为S58Q, P59V, S60F, K61Q突变序列,SEQ ID NO: 6为野生型酿酒酵母IGPs序列(ScTrpC),SEQ ID NO: 7为野生型黑曲霉IGPs序列(AgTrpC)。
第二方面,本发明还涉及通过分离法或合成法获得SEQ ID NO: 2 -5或 SEQ ID NO:30对应的酶。
第三方面,本发明还涉及一种利用生物技术法生产L-色氨酸的方法,包括根据第一方面所述的方法,利用生物反应器中培养突变的细菌细胞,
优选地,在本发明方法中使用的基因突变细菌为大肠杆菌。
另一方面,本发明还涉及利用上述方法获得的细菌细胞和酶来生产L-色氨酸,并且能以在生物反应器中进行工业化L-色氨酸生产。
本发明的优点在于:通过基因突变细菌细胞,使之表达的吲哚-3-甘油磷酸合成酶IGPs相较于野生型IGPs,对抑制剂邻氨基苯甲酸盐的敏感度下降,甚至可以被氨基苯甲酸酯激活。
附图说明
图1为大肠杆菌中分支酸生物合成L-trp的流程图。
图2为邻氨基苯甲酸对eIGPS活性的影响;其中(a)表示邻氨基苯甲酸对eIGPs活性的抑制作用;(b)表示非竞争性抑制的动力学曲线。
图3为邻氨基苯甲酸对野生型和突变eIGPs活性的影响。
图4为四株菌(S028/ptrpE(S40F), S028TC/ptrpE(S40F), S028/ptrc99A, S028TC/ptrc99A)的基因类型和发酵结果;其中(a)为生长曲线;(b)为葡萄糖消耗曲线;(c)为脱氢莽草酸酯积累曲线;(d)为莽草酸酯积累曲线;(e)为铵离子消耗曲线;(f)为L-trp生产曲线;(g)为L-tyr形成曲线;(h)为L-phe形成曲线;上述为在发酵3h时加入0.2mM IPTG进行诱导的试验结果。
图5为菌株S028/ptrpE(S40F) 和S028TC/ptrpE(S40F)在分批发酵过程中Trp,邻氨基苯甲酸,Phe和Tyr的形成速率;TP1至TP6分别对应于发酵时间3-8.5、8.5-14.5、14.5-22.5、22.5-27.5、27.5-33和33-37.5h。
图6为摇瓶发酵过程中,菌株S028/ptrc99A(黑色)和菌株S028TC/ptrc99A(白色)的L-trp产生速率;TP1至TP6分别对应于发酵时间3-8.5、8.5-14.5、14.5-22.5、22.5-27.5、27.5-33和33-37.5h。
图7为邻氨基苯甲酸对TrpC合成过程中吲哚甘油磷酸合酶活性的前馈调节作用。大肠杆菌的EcTrpC,TrpC受到邻氨基苯甲酸酯的负前馈调控,而酿酒酵母和黑曲霉的ScTrpC和AgTrpC,TrpC分别受到邻氨基苯甲酸正调控。
图8为邻氨基苯甲酸对ScTrpC和ScIGPs的活性的影响;其中ScIGPs为没有邻氨基苯甲酸合成酶II结构域的ScTrpC。
图9至图11分别为pAgTrpC、pEcTrpC和pScTrpC的质粒图谱。
图12为邻氨基苯甲酸浓度对EcTrpC WT和TrpC QVFQ活力的影响。
图13 A为来自大肠杆菌和结核分枝杆菌的吲哚磷酸合成酶晶体结构比对(PDB_ID:3T44, 浅灰色, MtIGPS 或 mIGPs;PDB_ID: 1PII, 黑色, EcIGPS 或 eIGPs);B为来自大肠杆菌和结核分枝杆菌的吲哚磷酸合成酶与邻氨基苯甲酸结合位点比对(浅灰色为mIGPs,黑色为eIGPs);标记为eIGPS与邻氨基苯甲酸结合的残基位点(黑色柱状);其中ANT为邻氨基苯甲酸。
图14为邻氨基苯甲酸和mIGPs PDB_ID: 3T44的结合位点;Igp 300(A) 和 BE2 273(A)分别代表IGPs产生的吲哚-3-甘油磷酸酯和邻氨基苯甲酸配体(或抑制剂;它们不是残基,也不属于IGPs酶本身,两者都与邻氨基苯甲酸结合位点无关。
具体实施方式
如图1所示,L-trp是由分支酸合成的,分支酸也是芳香族氨基酸L-苯丙氨酸和L-酪氨酸共同的前体。在大肠杆菌中,分支酸合成L-trp是在TrpE、TrpD、TrpC、TrpB、TrpA五个结构基因编码的酶的作用下完成的,这五个结构基因组成了trp操纵子。先前的研究表明,trp操纵子会受到终产物L-trp的严格调控,包括反馈抑制、衰减调控和弱化作用[7-10]。L-trp的生物合成分六个步骤完成,有五个酶参与催化反应,分别为 TrpE,TrpD,TrpC,TrpA和TrpB。其中TrpC为双功能基因,同时具有具有磷酸核糖氨基苯甲酸异构酶活性(PRAi)和吲哚-3-甘油磷酸合成酶活性(IGPs)。首先,在trpE基因编码的邻氨基苯甲酸合成酶(EC4.1.3.27)的作用下,将分支酸转化为邻氨基苯甲酸(ANT),随后在TrpD编码的邻氨基苯甲酸磷酸核糖基转移酶(EC 2.4.1.28)的作用下,将邻氨基苯甲酸转化为磷酸核糖氨基苯甲酸(PRA 或 PA)。接着PRA在磷酸核糖氨基苯甲酸异构酶活性((EC 5.3.1.24, PRAi)的催化下,转化为羧苯氨基-脱氧核糖-5-磷酸(CdRP),CdRP在吲哚-3-甘油磷酸合成酶(EC4.1.1.48, IGPs)催化下,转化为吲哚-3-甘油磷酸(IGP)。上述两个反应都是由TrpC编码的酶进行催化完成的。随后IGP转化为吲哚,进而合成L-色氨酸,这两个步骤都是在TrpA和TrpB编码的色氨酸合成酶(EC 4.2.1.20)的催化下完成的。
实施例1 质粒与菌株的构建方法
表1为本发明使用的菌株与质粒。表2列出了本发明所用到的引物序列。
表1 本发明菌株与质粒
质粒:

菌株:

表2 引物

用表2中的引物SmaI-trpE和trpE-XbaI扩增大肠杆菌S028(表1)中的色氨酸抗性基因trpES40F,并利用SmaI和XbaI双酶切,将trpES40F基因克隆至pTrc99A质粒中,获得ptrpE(S40F)质粒(表1)。用引物NdeI-eIGPs和eIGPs-XhoI(表2)从大肠杆菌S028的trpC基因中克隆出eIGPs的ORF序列(eIGPs, the IGPs in E. coli TrpC, EcTrpC),然后利用NdeI-eIGPs 和eIGPs-XhoI(表2)双酶切插入pET22(b)载体,获得pET-eIGPs(WT)质粒。克隆的eIGPs的理论肽序列包含TrpC的259个残基和C端用于纯化的标签LGHHHHHH。然后通过使用典型诱变引物对(表2)扩增整个pET-eIGPs(WT)质粒,以获得eIGPs突变质粒,突变质粒分别被命名为pET-eIGPs(I8V), pET-eIGPs(I8A), pET-eIGPs(S60A), pET-eIGPs(S60G),pET-eIGPs(L188A)和pET-eIGPs(L188F)。将引物EcoRI-6H-trpC和trpC-XbaI(表2)从大肠杆菌S028扩增得到的基因插入pTrc99A中以获得p6HTrpC质粒;同样地,利用EcoRI-6H-trpC和 eIGPs-XbaI引物构建获得p6HeIGPs质粒。在质粒p6HTrpC和p6HeIGPs中编码蛋白的N端带有6His标签。
本发明S028TC菌株(表1)的构建方法参照Lin 等人[11]公开的方法,利用选择/反选择标记进行染色体无缝编辑。具体为:利用表2中的trpC-O1 and trpC-O2 引物对从pJLK[11]质粒模板中扩增出选择/反选择标记盒,然后利用引物对trpC-INF/trpC-INR从pET-eIGPs(S60A)质粒中扩增得到突变点S60A,经重组、筛选、测序,最终获得突变菌株S028TC。
实施例2 培养条件
本发明分批发酵在摇瓶中进行,其中种子培养基和发酵培养基配方参照Lin 等人[11]文献。本发明发酵条件均为37℃、250rpm。具体方法为:将分离获得的菌落接种到含有5mLLB培养基的50mL 锥形瓶中,过夜预培养,然后将预培养物接种到含有10 mL种子培养基的100 mL摇瓶中,初始OD600 = 0.2,待培养8-10小时后,将种子培养物接种到含有30 mL发酵培养基的300mL挡板摇瓶中,控制初始OD600 = 0.1,三个平行。待生长3h(OD600约为0.9)后,加入0.2mM的IPTG进行诱导。上述所有培养均添加了100μg/mL的氨苄青霉素。
实施例3 分子对接试验
使用UCSF Chimera软件将结核分枝杆菌IGP(mIGPs 或or MtIGPs)的IGP构象复制到eIGPs中,构建eIGPs和mIGPs复合物[21],结合Chimera和AutoDock Vina[22]进行柔性配体与刚性受体对接。
实施例4 eIGPs的表达与纯化
将pET-eIGPs(wt)和含有eIGPs突变体的质粒分别转化到大肠杆菌BL21中。将质粒p6HTrpC 和 p6HeIGPs分别转化到大肠杆菌TOP10中。将37℃过夜培养的菌种分离后,接种至含有50mL LB培养基的300mL摇瓶中进行50倍稀释。在37℃、220rpm条件下培养至OD600大约为0.6时,添加0.5mM的IPTG进行诱导,然后在20℃、220rpm条件下继续培养12-16h。培养结束后取出置于冰上冷却30min后,4℃,5000rpm离心收集菌体细胞,并用30mL结合缓冲液(20mM磷酸钠,500mM NaCl,20mM咪唑,pH 7.4)洗涤一次,再将沉淀重悬于3mL结合缓冲液中,利用FastPrep®-24破碎细胞。接着将破碎后的样品置于4℃,13000rpm条件下离心20min,用预先安装好的His SpinTrap色谱柱(GE Healthcare)纯化目的蛋白,并利用400μl洗脱缓冲液(20 mM磷酸钠,500 mM NaCl,500 mM咪唑,pH 7.4)洗脱,随后将洗脱缓冲液更换为HEPPS缓冲液(50 mM HEPPS pH 7.5,4 mM EDTA),在4°C条件下,利用Amicon® Ultra-0.5 超滤管浓缩后进行酶活测定。
实施例5 IGPs酶活力测定
本发明通过测定278nm[23]处的吸光值(摩尔消光系数为5500 M-1 cm-1 [24])获得IGP的生成量,从而计算得到IGPs酶活。比色皿中的测定体系为:50 mM HEPPS pH 7.5, 4mM EDTA,20-25 μg/mL纯酶,测定温度为30℃。为了研究邻氨基苯甲酸对IGPs活性的影响,本发明加入在不同浓度的邻氨基苯甲酸,测定其对IGPs活性的影响。除非另有说明,上述反应都是在添加180μM底物CdRP条件下进行的。 CdRP的合成方法是参考Kirschner等人[24]的报道进行。通过用完全转化的反应产物IGP的浓度来确定储备溶液中合成的CdRP的浓度。
实施例6 分析方法
葡萄糖的定量通过HPLC法,利用DSA和SA进行定量[25, 26]。L-trp利用灵敏分光光度法进行定量[27]。其它氨基酸、铵离子、邻氨基苯甲酸用6-氨基喹啉基-N-羟基琥珀酰亚胺基氨基甲酸酯进行衍生化作用后,利用HPLC进行定量[25]。
实施例7潜在的邻氨基苯甲酸与eIGPs结合位点研究
包括IGP产物和邻氨基苯甲酸复合物形成的mIGPs的晶体结构,硫磺矿硫化叶菌产生的IGP(sIGPs, PDB ID: 1A53)[28]与大肠杆菌TrpC(PDB ID: 1PII) [29]形成复合物的晶体结构。IGP产物和邻氨基苯甲酸复合物形成的mIGPs的晶体结构表明,邻氨基苯甲酸结合位点的残基位于一个螺旋结构中,并形成三个环,组成一个“门”结构(图13、14)。邻氨基苯甲酸与IGP通过非化学键结合,阻止IGP从催化位点释放。sIGPs,mIGPs和eIGPs的序列之间的二级结构即使有一些区别,但还是非常保守,相似度大于70%。并且本发明还发现在mIGPs和eIGPs中,邻氨基苯甲酸结合位点的残基序列是100%保守的(图13),并且三个IGPs蛋白都为(β/α)8桶状蛋白,且(β/α)结构都能精确对齐,与eIGPs相比,sIGPs和mIGPs的第一条β链前面,分别有一个和两个螺旋结构,但是他们的活性位点都是高度保守的,邻氨基苯甲酸的结合可以使IGP在活性位点的结合更加紧密,因为吲哚环在mIGPs比在sIGPs中更加靠近桶状结构的位置。
为了确定eIGPs中是否存在邻氨基苯甲酸的潜在结合位点,本发明进行了分子对接研究。结果表明,邻氨基苯甲酸可以对接到eIGPs和IGP的复合物中。前三个对接的邻氨基苯甲酸与IGP是面对面结构(图中未显示),而前五个对接的邻氨基苯甲酸的苯甲酸环与参照对接的苯环都在平面上,该平面与IGP中的吲哚基团表面垂直(图中未显示)。其中,邻氨基苯甲酸对接mIGPs(IGP)和eIGPs(IGP)的最高得分分别为7.5和6.5,表明邻氨基苯甲酸与eIGPs结合的可能性很高。eIGPs、sIGPs和mIGPs中,与邻氨基苯甲酸结合位点的残基是100%保守的,这些结果表明,eIGPs和mIGPs与邻氨基苯甲酸结合的位点相同。然而,在与邻氨基苯甲酸结合时,结合位点残基的构型会进行调整。
实施例8 邻氨基苯甲酸非竞争性抑制eIGPs活性
本发明为了研究邻氨基苯甲酸对eIGPs活性的影响,测定了eIGPs (eIGPs-6His)在不同浓度邻氨基苯甲酸存在体系中的催化活性,由图2可知,eIGPs活性随着邻氨基苯甲酸浓度的增加,呈现下降趋势。结果表明0.5mM的邻氨基苯甲酸可以抑制46%的eIGPs活性,而在2mM的浓度下,eIGPs活性丧失70%。测定得到eIGPs-6His的抑制常数(IC50,半抑制浓度)约为0.70mM。
大肠杆菌TtrpC具有两个共价连接的结构域(PRAi结构域和IGPs结构域),如图1所示,每个结构域均具有催化活性。但是在很多其它生物中,IGPs是以单链酶的形式存在的[6]。此前的研究表明[30],PRAi结构域对IGPs的结构具有稳定作用。为了确定PRAi结构域是否对eIGPs的稳定性产生影响,本发明测定了邻氨基苯甲酸对eIGPs-PRAi(6His-TrpC)活性的影响,为了保证实验准确性,本发明将分离的6His-eIGPs作为对照。结果表明,邻氨基苯甲酸对6His-TrpC和6His-eIGPs的活性都具有显著抑制作用(图2a),6His-eIGPs对邻氨基苯甲酸的IC50约为0.4mM,而6His-TrpC的IC50位1.3mM,由此可知,PRAi结构域可以协助IGPs抵抗邻氨基苯甲酸的抑制作用。
为了确定邻氨基苯甲酸对eIGPs的抑制机理,本发明用6His-TrpC研究了eIGPs的米氏常数。本发明在无氨基苯甲酸和0.5mM氨基苯甲酸存在的体系中,利用不同浓度的CdRP(2.6- 260 μM)进行反应,绘制Lineweaver-Burk图(图2b)。由图可知,邻氨基苯甲酸抑制剂降低了Vmax,但对Km几乎没有影响,据线性拟合函数可知,无邻氨基苯甲酸酯和0.5mM邻氨基苯甲酸酯的Vmax和Km值分别为3.64、2.63μmol/ min / mg和1.57、1.53μM。 这表明邻氨基苯甲酸是eIGPS的非竞争性抑制剂。 根据晶体结构分析可知,邻氨基苯甲酸的催化位点与结合位点不同,这与本发明研究结果一致[19]。
实施例9 eIGPs邻氨基苯甲酸结合位点的突变分析
据推测,无法通过增强trp操纵子来提高L-trp产量的原因是因为邻氨基苯甲酸积累导致的IGP前馈抑制所致[12],为了检验该假设,本发明将trp操纵子中的野生型IGP替换为突变型,为此,本发明基于结构分析,来设计突变位点。由于N端的His标签离结合位点太近,可能会对试验造成影响,且6His-eIGPs的溶解度比eIGPs-6His差很多(数据未显示),因此本发明所有的突变分析都是基于eIGPs-6His进行的。
eIGPs和mIGPs与邻氨基苯甲酸结合位的残基之间高度保守。 表3列出了eIGPs和mIGPs的结合位点的残基及其所在位置。
表3. eIGPs和mIGPs与邻氨基苯甲酸酯结合位点中的保守残基及其所在的位置(见图13、14)。

其中,选择I8, S60, and L188三个残基,通过诱变引物(表4)分别对其进行单点突变。
表4 诱变引物

SDS-PAGE分析表明,所有的突变体均比野生型eIGPs的溶解度低,尤其是I8A(数据未显示)。突变体I8A对邻氨基苯甲酸的敏感性显著降低,但其活性和溶解度非常低(数据未显示)。而突变体L188A在相同的条件下没有活性,因此没有进行邻氨基苯甲酸活性研究。如图3所示,其它突变体与野生型eIGPs相比,对邻氨基苯甲酸的敏感性均有所降低,其中,突变体I8V和L188F对邻氨基苯甲酸的抗性有小幅度增加,而突变体S60A和S60G的活性则显著提高。在2mM的邻氨基苯甲酸存在下,突变体S60G和S60A的活性仅分别被抑制了20%和46%,而野生型的活性被损失了70%(数据未显示)。经测定,突变体S60A的IC50约为2.0 mM,约为野生型eIGPs的3倍。以上结果表明,残基I8,S60和L188都是eIGPs与邻氨基苯甲酸酯的潜在结合位点。
由表5可知,在没有邻氨基苯甲酸存在的体系中,突变体S60A的活性最高,但相比野生型还是有所降低。
表5 野生型和突变体的eIGPs比活力。从转化的IGP确定CdRP的浓度,其摩尔消光系数值为5500 M-1 cm-1[24]。 数据表示为平均值±标准偏差,-未进行酶分析。

在存在超过0.1 mM邻氨基苯甲酸和180μM CdRP的体系下,S60A和S60G的比活均高于野生型。据报道,由于底物的亲和力降低,突变体S60A的效率(Kcat / Km)仅为双结构域野生型酶的效率的30%[31]。如表5所示,在单结构域形式中也发现催化效率较低的情况。在含有18μM CdRP且不含邻氨基苯甲酸酯的体系中,突变体S60A和S60G的活性分别仅为野生型的74%和52%
实施例9 抗邻氨基苯甲酸的eIGPs对L-trp形成的影响
为了证明邻氨基苯甲酸对eIGPs的体内抑制作用,并探讨在邻氨基苯甲酸积累的情况下,抗邻氨基苯甲酸的eIGPs是否更适合L-trp的合成,因此本发明构建一种带有抗邻氨基苯甲酸的eIGPs的重组菌株。如上所述,突变体S60A具有最高的催化效率,型eIGPs相比,S60A对邻氨基苯甲酸的敏感性也大大降低。因此,我们用突变体基因trpCS60A替换了菌株S028中的野生型基因trpC,得到了重组菌株S028TC(表1)。为了使邻氨基苯甲酸在细胞内的积累量到可以达到显着抑制IGPs活性的水平,我们需要增强trp操纵子的第一反应,即将分支酸转化为邻氨基苯甲酸的反应。因此,本发明用质粒ptrpE(S40F)过表达trpES40F基因,增加了抗反馈抑制的邻氨基苯甲酸合成酶(TrpES40F)的有效性(表1)。本发明将质粒ptrpE(S40F)导入菌株S028和S028TC,构建菌株S028 / ptrpE(S40F)和S028TC / ptrpE(S40F)。同时,通过将空白载体ptrc99A导入宿主中,构建了参考菌株S028 / ptrc99A和S028TC /ptrc99A。这四种菌株之间的差异如图4所示。
如图4a所示,带有突变体TrpCS60A的菌株S028TC / ptrc99A与带有TrpC的野生型菌株S028 / ptrc99A相比,其IGPs活性降低,但生长速率、产量、生物量都有提高。高生长速率导致了葡萄糖消耗速率也提高了,但是降低了用于L-trp生物合成的代谢通量。经计算,在发酵时间8.5至27.5 h内,菌株S028 / ptrc99A和S028TC / ptrc99A的葡萄糖消耗率分别为1.06(R2 = 0.9764)和1.47 g / L / h(R2 = 0.9807)(图4b),且在发酵过程中,中间体DSA(图4c)和SA(图4d)在显著积累。由图4f可知,尽管菌株S028 / ptrc99A中的中间体积累高于突变菌株S028TC / ptrc99A中的积累,但S028TC / ptrc99A的L-trp产量显著提高,约为野生型的1.7倍。同时,与菌株S028TC / ptrc99A相比,菌株S028 / ptrc99A的副产物Tyr(图4g)和Phe(图4h)产量更少。在发酵结束时,菌株S028 / ptrc99A的所有可测量中间体(DSA和SA),副产物(Tyr和Phe)和L-trp之和约为20 mM,而S028TC / ptrc99A约为12 mM,这表明突变型的分支酸途径代谢通量更小,而更多的代谢通量被用于细胞生长,这是由于L-trp分支途径中IGPs的催化效率严重降低所致。这表明,高活性的IGPs对于实现高效率的trp操纵子具有至关重要的作用。
值得注意的是,发酵过程中氮的消耗要早于葡萄糖(图4b和4e),当葡萄糖尚未成为限制性因素是,L-trp的生产受到氮供应的限制。如图4e,f,g和h所示,氮限制可能会停止L-trp的生产并触发副产物(Phe和Tyr)的积累。由于NH4是L-gln(L-trp生产的底物)的生物合成的必需物质,因此,NH4的缺乏会阻断分支酸转化为L-trp分支途径的反应,使得用于合成Phe和Tyr的分支酸增加。从这个角度来看,如果提供足够的氮,L-trp的产量和产率将会提高。 因此,有必要在新设计的发酵培养基中添加更多的氮源。
有趣的是,发明人发现发现当基因trpES40F在S028或S028TC中过表达时,细胞生长被显着抑制,但原因尚不清楚。因此,这两个菌株的葡萄糖消耗率以及中间体(DSA和SA)的积累量相对较低(图4a,b,c和d)。
如图5a所示,菌株S028TC / ptrpE(S40F)的L-trp产率比对照菌株S028 / ptrpE(S40F)高得多,但两株菌在27.5 h后都几乎停止合成L-trp,但是在结束发酵时(37.5h),S028TC / ptrpE(S40F)菌株的L-trp产量(575±33 mg / L)显著高于对照菌株(366±22mg/L),产量提高了57%。不出所料,由于基因trpES40F的过表达,导致中间体邻氨基苯甲酸的积累,且在参考菌株中积累严重(图5b)。与L-trp不同的是,诱导后,邻氨基苯甲酸积累量持续增加,在发酵结束时,对照菌株的邻氨基苯甲酸积累量达到32.3±1.3 mg/L,是S028TC/ ptrpE(S40F)菌株(13.2±3.9 mg/L)的1.4倍,且对照菌株也产生了更多的副产物Phe(图5c) 和Tyr (图5d)。
值得注意的是,两种菌株的L-trp(qTrp)的比生产率保持下降趋势,而邻氨基苯甲酸、Phe和Tyr的比合成率在诱导后持续快速增长直到保持稳定。但是,菌株S028TC / ptrpE(S40F)的qTrp值比对照菌株高,其突变体TrpC(S60A)对邻氨基苯甲酸的敏感性较低。由于菌株S028 / ptrpE(S40F)和S028TC / ptrpE(S40F)的生长曲线几乎相同(图4a),因此可以认为,L-trp分支途径中的底物和从其他途径得到的底物供应量几乎相同。结合eIGPs的活性可在体外被邻氨基苯甲酸抑制的事实,以上结果表明该抑制作用也可以在体内发生。由试验结果可知,随着L-trp的合成,邻氨基苯甲酸,Phe和Tyr积累量增加,表明两个菌株中的IGPs活性在27.5 h后都可能被显着抑制。
如图6所示,当不存在氮和葡萄糖限制的条件下,诱导后菌株S028 / ptrc99A的qTrp量没有降低,当氮和葡萄糖充分供应时,菌株S028TC / ptrc99A的qTrp量在发酵8.5-14.5h时开始下降。表明菌株S028 / ptrc99A细胞内邻氨基苯甲酸的浓度没有达到可以显着抑制IGPs活性的水平。然而,尽管在菌株S028TC / ptrc99A四倍稀释的发酵液样品中未检测到胞外邻氨基苯甲酸,但它在细胞内可能已经达到了显着抑制IGPs的水平。
上述结构研究和分子对接结果表明邻氨基苯甲酸能够结合eIGPs。 酶分析表明,邻氨基苯甲酸以非竞争性方式前馈抑制eIGPs的酶活性。 对涉及的三个残基(I8,S60和L188)的eIGPs邻氨基苯甲酸结合位点的突变研究表明,单点突变体,尤其是S60A和S60G,会导致eIGPs的邻氨基苯甲酸敏感性显着降低,然而,突变体的酶催化效率却急剧下降。体内研究表明,即使发酵过程中邻氨基苯甲酸积累,导致突变体S60A催化效率降低,但是与野生型相比,L-trp的产量还是具有显著性增加。
实施例10 具有邻氨基苯甲酸合成酶II结构域的真菌IGPs不受邻氨基苯甲酸抑制
如图7所示,来自酿酒酵母和黑曲霉的TrpC,即ScTrpC和AgTrpC可被邻氨基苯甲酸激活。结构分析表明,正调控的TrpC(ScTrpC和AgTrpC)包含邻氨基苯甲酸合成酶II结构域(ASII结构域),而负调控的TrpC(EcTrpC)则不包含该结构域。 从ScTrpC移除邻氨基苯甲酸合成酶II结构域后,则未观察到ScIGP的激活(图8)。 这表明邻氨基苯甲酸合成酶II结构域对正调控至关重要。
实施例11 邻氨基苯甲酸活化TrpC对L-trp产量的影响
本发明通过从色氨酸产生菌株S028中敲除trpC基因构建trpC缺陷菌株S092,从而研究邻氨基苯甲酸活化TrpC对L-trp产量的影响。接着,在S092中引入EcTrpC,ScTrpC和AgTrpC以获得重组菌株S092 / pEcTrpC,S092 / pScTrpC和S092 / pAgTrpC。所用的质粒如图9至11所示。在生物反应器中用这三种菌株进行分批发酵, 结果如表6所示,菌株S092 /pScTrpC和菌株S092 / pAgTrpC均具有比对照菌株S092 / pEcTrpC更高的色氨酸产量和产率。这些结果表明,邻氨基苯甲酸激活的TrpC有利于色氨酸的产生。
表6.带有EcTrpC,ScTrpC和AgTrpC的菌株色氨酸生产率的比较。

实施例12 大肠杆菌IGPs多点突变的影响
进一步地,本发明在大肠杆菌IGPs的第58- 61位诱导多点突变,将多点突变菌株S092::TrpSen::pIBB24-trpCS58Q-P59V-S60F-K61Q(58位的丝氨酸突变为谷氨酰胺,59位的脯氨酸突变为缬氨酸,60位的丝氨酸突变为精氨酸,61位的赖氨酸突变为半胱氨酸)与S092::TrpSen::pIBB24-trpCSPSK进行比较。发酵结果如表7所示。
表7 EctrpCSPSK (TrpC WT), EctrpCS58Q-P59V-S60F-K61Q (TrpC QVFQ), and EctrpC S58Q -P59L-S60R-K61C (TrpC QLRC)摇瓶发酵20h结果比较。DCW = 细胞干重; con. = 浓度; qtrp =比产率(mg/gDCW/h)。

结果表明,在58-61位具有QVFQ序列的突变体菌体生长密度更高,并且就绝对产量而言可产生更多的色氨酸。
在邻氨基苯甲酸酯浓度增加的情况下对TrpCs比活进行测定,结果表明,与野生型TrpC相比,QVFQ变体的邻氨基苯甲酸的抑制程度明显降低,如表8所示(也参见图12)。
表8 TrpC WT 和 TrpC QVFQ比活力对比

表9为了野生型和QVFQ突变体的表观动力学常数。
表9.野生型和QVFQ TrpC变体的表观动力学常数

以下给出检索的相关文献 :
1.Wunsch C, Mundt K, Li S-M (2015) Targeted production of secondarymetabolites by coexpression of non-ribosomal peptide synthetase andprenyltransferase genes in As-pergillus. Appl Microbiol Biotechnol 99(10):4213–4223. doi: 10.1007/s00253-015-6490-8
2.Fang M-Y, Zhang C, Yang S et al. (2015) High crude violacein productionfrom glucose by Escherichia coli engineered with interactive control oftryptophan pathway and viola-cein biosynthetic pathway. Microb. Cell Fact.14: 8. doi: 10.1186/s12934-015-0192-x
3.Fang M, Wang T, Zhang C et al. (2016) Intermediate-sensor assistedpush-pull strategy and its application in heterologous deoxyviolaceinproduction in Escherichia coli. Metab Eng 33: 41–51. doi: 10.1016/j.ymben.2015.10.006
4.Rodrigues AL, Trachtmann N, Becker J et al. (2013) Systems metabolicengineering of Escherichia coli for production of the antitumor drugsviolacein and deoxyviolacein. Metab. Eng. 20: 29–41
6.Merino E, Jensen RA, Yanofsky C (2008) Evolution of bacterial trpoperons and their regulation. Curr Opin Microbiol 11(2): 78–86. doi: 10.1016/j.mib.2008.02.005
7.Bertrand K, Yanofsky C (1976) Regulation of Transcription Terminationin Leader Region of Tryptophan Operon of Escherichia-Coli Involves Tryptophanor Its Metabolic Product. J. Mol. Biol. 103(2): 339–349
8.Kwak JH, Hong KW, Lee SH et al. (1999) Identification of amino acidresidues involved in feedback inhibition of the anthranilate synthase inEscherichia coli. J. Biochem. Mol. Bi-ol. 32(1): 20–24
9.Platt T (1981) Termination of Transcription and Its Regulation in theTryptophan Operon of Escherichia-Coli. Cell 24(1): 10–23
10.Zurawski G, Elseviers D, Stauffer GV et al. (1978) Translationalcontrol of transcription termination at the attenuator of the Escherichiacoli tryptophan operon. Proc. Natl. Acad. Sci. U.S.A. 75(12): 5988–5992
11.Chen L, Zeng A-P (2017) Rational design and metabolic analysis ofEscherichia coli for effective production of L-tryptophan at highconcentration. Appl Microbiol Biotechnol 101(2): 559–568. doi: 10.1007/s00253-016-7772-5
12.Lee KH, Park HM, Lee HH et al. (2014) Microorganisms of Escherichiacoli having en-hanced L-tryptophan production and method for producing L-tryptophan by using same(EP2803720)
19.Reddy M, Bruning JB, Thurman C et al. (2012) Crystal structure ofMycobacterium tu-berculosis Indole Glycerol Phosphate Synthase (IGPS) incomplex with indole glycerol phosphate (IGP) amd anthranilate. doi: 10.2210/pdb3t44/pdb
20.Amann E, Ochs B, Abel KJ (1988) Tightly regulated tac promoter vectorsuseful for the expression of unfused and fused proteins in Escherichia coli.Gene 69(2): 301–315
21.Pettersen EF, Goddard TD, Huang CC et al. (2004) UCSF Chimera--avisualization system for exploratory research and analysis. J Comput Chem 25(13): 1605–1612. doi: 10.1002/jcc.20084
22.Trott O, Olson AJ (2010) AutoDock Vina: improving the speed andaccuracy of docking with a new scoring function, efficient optimization, andmultithreading. J Comput Chem 31(2): 455–461. doi: 10.1002/jcc.21334
23.Zaccardi MJ, Mannweiler O, Boehr DD (2012) Differences in thecatalytic mechanisms of mesophilic and thermophilic indole-3-glycerolphosphate synthase enzymes at their adaptive temperatures. Biochem. Biophys.Res. Commun. 418(2): 324–329. doi: 10.1016/j.bbrc.2012.01.020
24.Kirschner K, Szadkowski H, Jardetzky TS et al. (1987)Phosphoribosylanthranilate isomerase—indoleglycerol-phosphate synthase fromEscherichia coli. In: Metabolism of Aromatic Amino Acids and Amines, vol 142.Academic Press, pp 386–397
25.da Luz JA, Hans E, Zeng A-P (2014) Automated fast filtration and on-filter quenching improve the intracellular metabolite analysis ofmicroorganisms. Eng. Life Sci. 14(2): 135–142. doi: 10.1002/elsc.201300099
26.Rajesh Reddy Bommareddy, Zhen Chen, Sugima Rappert et al. (2014) A denovo NADPH generation pathway for improving lysine production ofCorynebacterium glutamicum by rational design of the coenzyme specificity ofglyceraldehyde 3-phosphate dehydrogen-ase. Metab. Eng. 25(0): 30–37. doi:10.1016/j.ymben.2014.06.005
27.Nagaraja P, Yathirajan HS, Vasantha RA (2003) Highly sensitivereaction of tryptophan with p-phenylenediamine. Anal. Biochem. 312(2): 157–161. doi: 10.1016/S0003-2697(02)00504-3
28.Hennig M, Darimont B, Jansonius J et al. (2002) The CatalyticMechanism of Indole-3-glycerol Phosphate Synthase: Crystal Structures ofComplexes of the Enzyme from Sul-folobus solfataricus with SubstrateAnalogue, Substrate, and Product. J. Mol. Biol. 319(3): 757–766. doi:10.1016/S0022-2836(02)00378-9
29.Wilmanns M, Priestle JP, Niermann T et al. (1992) Three-dimensionalstructure of the bifunctional enzyme phosphoribosylanthranilate isomerase:indoleglycerolphosphate synthase from Escherichia coli refined at 2.0 Aresolution. Journal of Molecular Biology 223(2): 477–507
30.Eberhard M, Tsai-Pflugfelder M, Bolewska K et al. (1995)Indoleglycerol phosphate syn-thase-phosphoribosyl anthranilate isomerase:comparison of the bifunctional enzyme from Escherichia coli with engineeredmonofunctional domains. Biochemistry 34(16): 5419–5428
31.Darimont B, Stehlin C, Szadkoski H et al. (1998) Mutational analysisof the active site of indoleglycerol phosphate synthase from Escherichiacoli. Protein Sci. 7(5): 1221–1232. doi: 10.1002/pro.5560070518。
序列表
<110> 汉堡工业大学
<120> 一种改进的生物技术生产L-色氨酸的方法
<130> 30
<160> 30
<170> SIPOSequenceListing 1.0
<210> 1
<211> 452
<212> PRT
<213> 人工序列()
<400> 1
Met Gln Thr Val Leu Ala Lys Ile Val Ala Asp Lys Ala Ile Trp Val
1 5 10 15
Glu Ala Arg Lys Gln Gln Gln Pro Leu Ala Ser Phe Gln Asn Glu Val
20 25 30
Gln Pro Ser Thr Arg His Phe Tyr Asp Ala Leu Gln Gly Ala Arg Thr
35 40 45
Ala Phe Ile Leu Glu Cys Lys Lys Ala Ser Pro Ser Lys Gly Val Ile
50 55 60
Arg Asp Asp Phe Asp Pro Ala Arg Ile Ala Ala Ile Tyr Lys His Tyr
65 70 75 80
Ala Ser Ala Ile Ser Val Leu Thr Asp Glu Lys Tyr Phe Gln Gly Ser
85 90 95
Phe Asn Phe Leu Pro Ile Val Ser Gln Ile Ala Pro Gln Pro Ile Leu
100 105 110
Cys Lys Asp Phe Ile Ile Asp Pro Tyr Gln Ile Tyr Leu Ala Arg Tyr
115 120 125
Tyr Gln Ala Asp Ala Cys Leu Leu Met Leu Ser Val Leu Asp Asp Asp
130 135 140
Gln Tyr Arg Gln Leu Ala Ala Val Ala His Ser Leu Glu Met Gly Val
145 150 155 160
Leu Thr Glu Val Ser Asn Glu Glu Glu Gln Glu Arg Ala Ile Ala Leu
165 170 175
Gly Ala Lys Val Val Gly Ile Asn Asn Arg Asp Leu Arg Asp Leu Ser
180 185 190
Ile Asp Leu Asn Arg Thr Arg Glu Leu Ala Pro Lys Leu Gly His Asn
195 200 205
Val Thr Val Ile Ser Glu Ser Gly Ile Asn Thr Tyr Ala Gln Val Arg
210 215 220
Glu Leu Ser His Phe Ala Asn Gly Phe Leu Ile Gly Ser Ala Leu Met
225 230 235 240
Ala His Asp Asp Leu His Ala Ala Val Arg Arg Val Leu Leu Gly Glu
245 250 255
Asn Lys Val Cys Gly Leu Thr Arg Gly Gln Asp Ala Lys Ala Ala Tyr
260 265 270
Asp Ala Gly Ala Ile Tyr Gly Gly Leu Ile Phe Val Ala Thr Ser Pro
275 280 285
Arg Cys Val Asn Val Glu Gln Ala Gln Glu Val Met Ala Ala Ala Pro
290 295 300
Leu Gln Tyr Val Gly Val Phe Arg Asn His Asp Ile Ala Asp Val Val
305 310 315 320
Asp Lys Ala Lys Val Leu Ser Leu Ala Ala Val Gln Leu His Gly Asn
325 330 335
Glu Glu Gln Leu Tyr Ile Asp Thr Leu Arg Glu Ala Leu Pro Ala His
340 345 350
Val Ala Ile Trp Lys Ala Leu Ser Val Gly Glu Thr Leu Pro Ala Arg
355 360 365
Glu Phe Gln His Val Asp Lys Tyr Val Leu Asp Asn Gly Gln Gly Gly
370 375 380
Ser Gly Gln Arg Phe Asp Trp Ser Leu Leu Asn Gly Gln Ser Leu Gly
385 390 395 400
Asn Val Leu Leu Ala Gly Gly Leu Gly Ala Asp Asn Cys Val Glu Ala
405 410 415
Ala Gln Thr Gly Cys Ala Gly Leu Asp Phe Asn Ser Ala Val Glu Ser
420 425 430
Gln Pro Gly Ile Lys Asp Ala Arg Leu Leu Ala Ser Val Phe Gln Thr
435 440 445
Leu Arg Ala Tyr
450
<210> 2
<211> 452
<212> PRT
<213> 人工序列()
<400> 2
Met Gln Thr Val Leu Ala Lys Ile Val Ala Asp Lys Ala Ile Trp Val
1 5 10 15
Glu Ala Arg Lys Gln Gln Gln Pro Leu Ala Ser Phe Gln Asn Glu Val
20 25 30
Gln Pro Ser Thr Arg His Phe Tyr Asp Ala Leu Gln Gly Ala Arg Thr
35 40 45
Ala Phe Ile Leu Glu Cys Lys Lys Ala Ser Pro Ser Lys Gly Val Ile
50 55 60
Arg Asp Asp Phe Asp Pro Ala Arg Ile Ala Ala Ile Tyr Lys His Tyr
65 70 75 80
Ala Ser Ala Ile Ser Val Leu Thr Asp Glu Lys Tyr Phe Gln Gly Ser
85 90 95
Phe Asn Phe Leu Pro Ile Val Ser Gln Ile Ala Pro Gln Pro Ile Leu
100 105 110
Cys Lys Asp Phe Ile Ile Asp Pro Tyr Gln Ile Tyr Leu Ala Arg Tyr
115 120 125
Tyr Gln Ala Asp Ala Cys Leu Leu Met Leu Ser Val Leu Asp Asp Asp
130 135 140
Gln Tyr Arg Gln Leu Ala Ala Val Ala His Ser Leu Glu Met Gly Val
145 150 155 160
Leu Thr Glu Val Ser Asn Glu Glu Glu Gln Glu Arg Ala Ile Ala Leu
165 170 175
Gly Ala Lys Val Val Gly Ile Asn Asn Arg Asp Leu Arg Asp Leu Ser
180 185 190
Ile Asp Leu Asn Arg Thr Arg Glu Leu Ala Pro Lys Leu Gly His Asn
195 200 205
Val Thr Val Ile Ser Glu Ser Gly Ile Asn Thr Tyr Ala Gln Val Arg
210 215 220
Glu Leu Ser His Phe Ala Asn Gly Phe Leu Ile Gly Ser Ala Leu Met
225 230 235 240
Ala His Asp Asp Leu His Ala Ala Val Arg Arg Val Leu Leu Gly Glu
245 250 255
Asn Lys Val Cys Gly Leu Thr Arg Gly Gln Asp Ala Lys Ala Ala Tyr
260 265 270
Asp Ala Gly Ala Ile Tyr Gly Gly Leu Ile Phe Val Ala Thr Ser Pro
275 280 285
Arg Cys Val Asn Val Glu Gln Ala Gln Glu Val Met Ala Ala Ala Pro
290 295 300
Leu Gln Tyr Val Gly Val Phe Arg Asn His Asp Ile Ala Asp Val Val
305 310 315 320
Asp Lys Ala Lys Val Leu Ser Leu Ala Ala Val Gln Leu His Gly Asn
325 330 335
Glu Glu Gln Leu Tyr Ile Asp Thr Leu Arg Glu Ala Leu Pro Ala His
340 345 350
Val Ala Ile Trp Lys Ala Leu Ser Val Gly Glu Thr Leu Pro Ala Arg
355 360 365
Glu Phe Gln His Val Asp Lys Tyr Val Leu Asp Asn Gly Gln Gly Gly
370 375 380
Ser Gly Gln Arg Phe Asp Trp Ser Leu Leu Asn Gly Gln Ser Leu Gly
385 390 395 400
Asn Val Leu Leu Ala Gly Gly Leu Gly Ala Asp Asn Cys Val Glu Ala
405 410 415
Ala Gln Thr Gly Cys Ala Gly Leu Asp Phe Asn Ser Ala Val Glu Ser
420 425 430
Gln Pro Gly Ile Lys Asp Ala Arg Leu Leu Ala Ser Val Phe Gln Thr
435 440 445
Leu Arg Ala Tyr
450
<210> 3
<211> 452
<212> PRT
<213> 人工序列()
<400> 3
Met Gln Thr Val Leu Ala Lys Ile Val Ala Asp Lys Ala Ile Trp Val
1 5 10 15
Glu Ala Arg Lys Gln Gln Gln Pro Leu Ala Ser Phe Gln Asn Glu Val
20 25 30
Gln Pro Ser Thr Arg His Phe Tyr Asp Ala Leu Gln Gly Ala Arg Thr
35 40 45
Ala Phe Ile Leu Glu Cys Lys Lys Ala Ser Pro Ala Lys Gly Val Ile
50 55 60
Arg Asp Asp Phe Asp Pro Ala Arg Ile Ala Ala Ile Tyr Lys His Tyr
65 70 75 80
Ala Ser Ala Ile Ser Val Leu Thr Asp Glu Lys Tyr Phe Gln Gly Ser
85 90 95
Phe Asn Phe Leu Pro Ile Val Ser Gln Ile Ala Pro Gln Pro Ile Leu
100 105 110
Cys Lys Asp Phe Ile Ile Asp Pro Tyr Gln Ile Tyr Leu Ala Arg Tyr
115 120 125
Tyr Gln Ala Asp Ala Cys Leu Leu Met Leu Ser Val Leu Asp Asp Asp
130 135 140
Gln Tyr Arg Gln Leu Ala Ala Val Ala His Ser Leu Glu Met Gly Val
145 150 155 160
Leu Thr Glu Val Ser Asn Glu Glu Glu Gln Glu Arg Ala Ile Ala Leu
165 170 175
Gly Ala Lys Val Val Gly Ile Asn Asn Arg Asp Leu Arg Asp Leu Ser
180 185 190
Ile Asp Leu Asn Arg Thr Arg Glu Leu Ala Pro Lys Leu Gly His Asn
195 200 205
Val Thr Val Ile Ser Glu Ser Gly Ile Asn Thr Tyr Ala Gln Val Arg
210 215 220
Glu Leu Ser His Phe Ala Asn Gly Phe Leu Ile Gly Ser Ala Leu Met
225 230 235 240
Ala His Asp Asp Leu His Ala Ala Val Arg Arg Val Leu Leu Gly Glu
245 250 255
Asn Lys Val Cys Gly Leu Thr Arg Gly Gln Asp Ala Lys Ala Ala Tyr
260 265 270
Asp Ala Gly Ala Ile Tyr Gly Gly Leu Ile Phe Val Ala Thr Ser Pro
275 280 285
Arg Cys Val Asn Val Glu Gln Ala Gln Glu Val Met Ala Ala Ala Pro
290 295 300
Leu Gln Tyr Val Gly Val Phe Arg Asn His Asp Ile Ala Asp Val Val
305 310 315 320
Asp Lys Ala Lys Val Leu Ser Leu Ala Ala Val Gln Leu His Gly Asn
325 330 335
Glu Glu Gln Leu Tyr Ile Asp Thr Leu Arg Glu Ala Leu Pro Ala His
340 345 350
Val Ala Ile Trp Lys Ala Leu Ser Val Gly Glu Thr Leu Pro Ala Arg
355 360 365
Glu Phe Gln His Val Asp Lys Tyr Val Leu Asp Asn Gly Gln Gly Gly
370 375 380
Ser Gly Gln Arg Phe Asp Trp Ser Leu Leu Asn Gly Gln Ser Leu Gly
385 390 395 400
Asn Val Leu Leu Ala Gly Gly Leu Gly Ala Asp Asn Cys Val Glu Ala
405 410 415
Ala Gln Thr Gly Cys Ala Gly Leu Asp Phe Asn Ser Ala Val Glu Ser
420 425 430
Gln Pro Gly Ile Lys Asp Ala Arg Leu Leu Ala Ser Val Phe Gln Thr
435 440 445
Leu Arg Ala Tyr
450
<210> 4
<211> 452
<212> PRT
<213> 人工序列()
<400> 4
Met Gln Thr Val Leu Ala Lys Ile Val Ala Asp Lys Ala Ile Trp Val
1 5 10 15
Glu Ala Arg Lys Gln Gln Gln Pro Leu Ala Ser Phe Gln Asn Glu Val
20 25 30
Gln Pro Ser Thr Arg His Phe Tyr Asp Ala Leu Gln Gly Ala Arg Thr
35 40 45
Ala Phe Ile Leu Glu Cys Lys Lys Ala Ser Pro Gly Lys Gly Val Ile
50 55 60
Arg Asp Asp Phe Asp Pro Ala Arg Ile Ala Ala Ile Tyr Lys His Tyr
65 70 75 80
Ala Ser Ala Ile Ser Val Leu Thr Asp Glu Lys Tyr Phe Gln Gly Ser
85 90 95
Phe Asn Phe Leu Pro Ile Val Ser Gln Ile Ala Pro Gln Pro Ile Leu
100 105 110
Cys Lys Asp Phe Ile Ile Asp Pro Tyr Gln Ile Tyr Leu Ala Arg Tyr
115 120 125
Tyr Gln Ala Asp Ala Cys Leu Leu Met Leu Ser Val Leu Asp Asp Asp
130 135 140
Gln Tyr Arg Gln Leu Ala Ala Val Ala His Ser Leu Glu Met Gly Val
145 150 155 160
Leu Thr Glu Val Ser Asn Glu Glu Glu Gln Glu Arg Ala Ile Ala Leu
165 170 175
Gly Ala Lys Val Val Gly Ile Asn Asn Arg Asp Leu Arg Asp Leu Ser
180 185 190
Ile Asp Leu Asn Arg Thr Arg Glu Leu Ala Pro Lys Leu Gly His Asn
195 200 205
Val Thr Val Ile Ser Glu Ser Gly Ile Asn Thr Tyr Ala Gln Val Arg
210 215 220
Glu Leu Ser His Phe Ala Asn Gly Phe Leu Ile Gly Ser Ala Leu Met
225 230 235 240
Ala His Asp Asp Leu His Ala Ala Val Arg Arg Val Leu Leu Gly Glu
245 250 255
Asn Lys Val Cys Gly Leu Thr Arg Gly Gln Asp Ala Lys Ala Ala Tyr
260 265 270
Asp Ala Gly Ala Ile Tyr Gly Gly Leu Ile Phe Val Ala Thr Ser Pro
275 280 285
Arg Cys Val Asn Val Glu Gln Ala Gln Glu Val Met Ala Ala Ala Pro
290 295 300
Leu Gln Tyr Val Gly Val Phe Arg Asn His Asp Ile Ala Asp Val Val
305 310 315 320
Asp Lys Ala Lys Val Leu Ser Leu Ala Ala Val Gln Leu His Gly Asn
325 330 335
Glu Glu Gln Leu Tyr Ile Asp Thr Leu Arg Glu Ala Leu Pro Ala His
340 345 350
Val Ala Ile Trp Lys Ala Leu Ser Val Gly Glu Thr Leu Pro Ala Arg
355 360 365
Glu Phe Gln His Val Asp Lys Tyr Val Leu Asp Asn Gly Gln Gly Gly
370 375 380
Ser Gly Gln Arg Phe Asp Trp Ser Leu Leu Asn Gly Gln Ser Leu Gly
385 390 395 400
Asn Val Leu Leu Ala Gly Gly Leu Gly Ala Asp Asn Cys Val Glu Ala
405 410 415
Ala Gln Thr Gly Cys Ala Gly Leu Asp Phe Asn Ser Ala Val Glu Ser
420 425 430
Gln Pro Gly Ile Lys Asp Ala Arg Leu Leu Ala Ser Val Phe Gln Thr
435 440 445
Leu Arg Ala Tyr
450
<210> 5
<211> 452
<212> PRT
<213> 人工序列()
<400> 5
Met Gln Thr Val Leu Ala Lys Ile Val Ala Asp Lys Ala Ile Trp Val
1 5 10 15
Glu Ala Arg Lys Gln Gln Gln Pro Leu Ala Ser Phe Gln Asn Glu Val
20 25 30
Gln Pro Ser Thr Arg His Phe Tyr Asp Ala Leu Gln Gly Ala Arg Thr
35 40 45
Ala Phe Ile Leu Glu Cys Lys Lys Ala Ser Pro Ser Lys Gly Val Ile
50 55 60
Arg Asp Asp Phe Asp Pro Ala Arg Ile Ala Ala Ile Tyr Lys His Tyr
65 70 75 80
Ala Ser Ala Ile Ser Val Leu Thr Asp Glu Lys Tyr Phe Gln Gly Ser
85 90 95
Phe Asn Phe Leu Pro Ile Val Ser Gln Ile Ala Pro Gln Pro Ile Leu
100 105 110
Cys Lys Asp Phe Ile Ile Asp Pro Tyr Gln Ile Tyr Leu Ala Arg Tyr
115 120 125
Tyr Gln Ala Asp Ala Cys Leu Leu Met Leu Ser Val Leu Asp Asp Asp
130 135 140
Gln Tyr Arg Gln Leu Ala Ala Val Ala His Ser Leu Glu Met Gly Val
145 150 155 160
Leu Thr Glu Val Ser Asn Glu Glu Glu Gln Glu Arg Ala Ile Ala Leu
165 170 175
Gly Ala Lys Val Val Gly Ile Asn Asn Arg Asp Phe Arg Asp Leu Ser
180 185 190
Ile Asp Leu Asn Arg Thr Arg Glu Leu Ala Pro Lys Leu Gly His Asn
195 200 205
Val Thr Val Ile Ser Glu Ser Gly Ile Asn Thr Tyr Ala Gln Val Arg
210 215 220
Glu Leu Ser His Phe Ala Asn Gly Phe Leu Ile Gly Ser Ala Leu Met
225 230 235 240
Ala His Asp Asp Leu His Ala Ala Val Arg Arg Val Leu Leu Gly Glu
245 250 255
Asn Lys Val Cys Gly Leu Thr Arg Gly Gln Asp Ala Lys Ala Ala Tyr
260 265 270
Asp Ala Gly Ala Ile Tyr Gly Gly Leu Ile Phe Val Ala Thr Ser Pro
275 280 285
Arg Cys Val Asn Val Glu Gln Ala Gln Glu Val Met Ala Ala Ala Pro
290 295 300
Leu Gln Tyr Val Gly Val Phe Arg Asn His Asp Ile Ala Asp Val Val
305 310 315 320
Asp Lys Ala Lys Val Leu Ser Leu Ala Ala Val Gln Leu His Gly Asn
325 330 335
Glu Glu Gln Leu Tyr Ile Asp Thr Leu Arg Glu Ala Leu Pro Ala His
340 345 350
Val Ala Ile Trp Lys Ala Leu Ser Val Gly Glu Thr Leu Pro Ala Arg
355 360 365
Glu Phe Gln His Val Asp Lys Tyr Val Leu Asp Asn Gly Gln Gly Gly
370 375 380
Ser Gly Gln Arg Phe Asp Trp Ser Leu Leu Asn Gly Gln Ser Leu Gly
385 390 395 400
Asn Val Leu Leu Ala Gly Gly Leu Gly Ala Asp Asn Cys Val Glu Ala
405 410 415
Ala Gln Thr Gly Cys Ala Gly Leu Asp Phe Asn Ser Ala Val Glu Ser
420 425 430
Gln Pro Gly Ile Lys Asp Ala Arg Leu Leu Ala Ser Val Phe Gln Thr
435 440 445
Leu Arg Ala Tyr
450
<210> 6
<211> 484
<212> PRT
<213> 人工序列()
<400> 6
Met Ser Val His Ala Ala Thr Asn Pro Ile Asn Lys His Val Val Leu
1 5 10 15
Ile Asp Asn Tyr Asp Ser Phe Thr Trp Asn Val Tyr Glu Tyr Leu Cys
20 25 30
Gln Glu Gly Ala Lys Val Ser Val Tyr Arg Asn Asp Ala Ile Thr Val
35 40 45
Pro Glu Ile Ala Ala Leu Asn Pro Asp Thr Leu Leu Ile Ser Pro Gly
50 55 60
Pro Gly His Pro Lys Thr Asp Ser Gly Ile Ser Arg Asp Cys Ile Arg
65 70 75 80
Tyr Phe Thr Gly Lys Ile Pro Val Phe Gly Ile Cys Met Gly Gln Gln
85 90 95
Cys Met Phe Asp Val Phe Gly Gly Glu Val Ala Tyr Ala Gly Glu Ile
100 105 110
Val His Gly Lys Thr Ser Pro Ile Ser His Asp Asn Cys Gly Ile Phe
115 120 125
Lys Asn Val Pro Gln Gly Ile Ala Val Thr Arg Tyr His Ser Leu Ala
130 135 140
Gly Thr Glu Ser Ser Leu Pro Ser Cys Leu Lys Val Thr Ala Ser Thr
145 150 155 160
Glu Asn Gly Ile Ile Met Gly Val Arg His Lys Lys Tyr Thr Val Glu
165 170 175
Gly Val Gln Phe His Pro Glu Ser Ile Leu Thr Glu Glu Gly His Leu
180 185 190
Met Ile Arg Asn Ile Leu Asn Val Ser Gly Gly Thr Trp Glu Glu Asn
195 200 205
Lys Ser Ser Pro Ser Asn Ser Ile Leu Asp Arg Ile Tyr Ala Arg Arg
210 215 220
Lys Ile Asp Val Asn Glu Gln Ser Lys Ile Pro Gly Phe Thr Phe Gln
225 230 235 240
Asp Leu Gln Ser Asn Tyr Asp Leu Gly Leu Ala Pro Pro Leu Gln Asp
245 250 255
Phe Tyr Thr Val Leu Ser Ser Ser His Lys Arg Ala Val Val Leu Ala
260 265 270
Glu Val Lys Arg Ala Ser Pro Ser Lys Gly Pro Ile Cys Leu Lys Ala
275 280 285
Val Ala Ala Glu Gln Ala Leu Lys Tyr Ala Glu Ala Gly Ala Ser Ala
290 295 300
Ile Ser Val Leu Thr Glu Pro His Trp Phe His Gly Ser Leu Gln Asp
305 310 315 320
Leu Val Asn Val Arg Lys Ile Leu Asp Leu Lys Phe Pro Pro Lys Glu
325 330 335
Arg Pro Cys Val Leu Arg Lys Glu Phe Ile Phe Ser Lys Tyr Gln Ile
340 345 350
Leu Glu Ala Arg Leu Ala Gly Ala Asp Thr Val Leu Leu Ile Val Lys
355 360 365
Met Leu Ser Gln Pro Leu Leu Lys Glu Leu Tyr Ser Tyr Ser Lys Asp
370 375 380
Leu Asn Met Glu Pro Leu Val Glu Val Asn Ser Lys Glu Glu Leu Gln
385 390 395 400
Arg Ala Leu Glu Ile Gly Ala Lys Val Val Gly Val Asn Asn Arg Asp
405 410 415
Leu His Ser Phe Asn Val Asp Leu Asn Thr Thr Ser Asn Leu Val Glu
420 425 430
Ser Ile Pro Lys Asp Val Leu Leu Ile Ala Leu Ser Gly Ile Thr Thr
435 440 445
Arg Asp Asp Ala Glu Lys Tyr Lys Lys Glu Gly Val His Gly Phe Leu
450 455 460
Val Gly Glu Ala Leu Met Lys Ser Thr Asp Val Lys Lys Phe Ile His
465 470 475 480
Glu Leu Cys Glu
<210> 7
<211> 770
<212> PRT
<213> 人工序列()
<400> 7
Met Ala Asp Ser Gly Leu Val Asp His Ser Pro His His Pro Thr Lys
1 5 10 15
Ala Ala Gln Leu Asn Thr Ala Ser Asn Val Ile Leu Ile Asp Asn Tyr
20 25 30
Asp Ser Phe Thr Trp Asn Val Tyr Gln Tyr Leu Val Leu Glu Gly Ala
35 40 45
Thr Val Asn Val Phe Arg Asn Asp Gln Ile Thr Leu Glu Glu Leu Ile
50 55 60
Ala Lys Lys Pro Thr Gln Leu Val Ile Ser Pro Gly Pro Gly His Pro
65 70 75 80
Glu Thr Asp Ala Gly Ile Ser Ser Ala Ala Ile Gln Tyr Phe Ser Gly
85 90 95
Lys Ile Pro Ile Phe Gly Val Cys Met Gly Gln Gln Cys Ile Ile Thr
100 105 110
Cys Phe Gly Gly Lys Val Asp Val Thr Gly Glu Ile Leu His Gly Lys
115 120 125
Thr Ser Ala Leu Lys His Asp Gly Lys Gly Ala Tyr Glu Gly Leu Pro
130 135 140
Asp Ser Leu Ala Val Thr Arg Tyr His Ser Leu Ala Gly Thr His Ala
145 150 155 160
Thr Ile Pro Asp Cys Leu Glu Val Ser Ser Ser Val Gln Leu Thr Asp
165 170 175
Asp Ser Asn Lys Asp Val Ile Met Gly Val Arg His Lys Lys Leu Ala
180 185 190
Val Glu Gly Val Gln Phe His Pro Glu Ser Ile Leu Thr Glu Tyr Gly
195 200 205
Arg Thr Met Phe Arg Asn Phe Leu Lys Leu Thr Ala Gly Thr Trp Glu
210 215 220
Gly Asn Gly Lys His Phe Asp Glu Gln Ser Asn Thr Thr Lys Ala Thr
225 230 235 240
Val Ser Ser Asn Thr Ala Pro Lys Thr Asp Lys Lys Leu Ser Ile Leu
245 250 255
Glu Arg Ile Tyr Asp His Arg Arg Ala Ala Val Ala Val Gln Lys Thr
260 265 270
Ile Pro Ser Gln Arg Pro Ala Asp Leu Gln Ala Ala Tyr Asp Leu Asn
275 280 285
Leu Ala Pro Pro Gln Val Pro Phe Pro Ala Arg Leu Arg Gln Ser Pro
290 295 300
Tyr Pro Leu Ser Leu Met Ala Glu Ile Lys Arg Ala Ser Pro Ser Lys
305 310 315 320
Gly Met Ile Ala Glu Asn Ala Cys Ala Pro Ala Gln Ala Arg Gln Tyr
325 330 335
Ala Lys Ala Gly Ala Ser Val Ile Ser Val Leu Thr Glu Pro Glu Trp
340 345 350
Phe Lys Gly Ser Ile Asp Asp Leu Arg Ala Val Arg Gln Ser Leu Glu
355 360 365
Gly Leu Thr Asn Arg Pro Ala Ile Leu Arg Lys Glu Phe Val Phe Asp
370 375 380
Glu Tyr Gln Ile Leu Glu Ala Arg Leu Ala Gly Ala Asp Thr Val Leu
385 390 395 400
Leu Ile Val Lys Met Leu Ser Val Glu Leu Leu Thr Arg Leu Tyr His
405 410 415
Tyr Ser Arg Ser Leu Gly Met Glu Pro Leu Val Glu Val Asn Thr Pro
420 425 430
Glu Glu Met Lys Ile Ala Val Asp Leu Gly Ala Glu Val Ile Gly Val
435 440 445
Asn Asn Arg Asp Leu Thr Ser Phe Glu Val Asp Leu Gly Thr Thr Ser
450 455 460
Arg Leu Met Asp Gln Val Pro Ser Ser Thr Ile Val Cys Ala Leu Ser
465 470 475 480
Gly Ile Ser Gly Pro Lys Asp Val Glu Ala Tyr Lys Lys Glu Gly Val
485 490 495
Lys Ala Ile Leu Val Gly Glu Ala Leu Met Arg Ala Ala Asp Thr Ala
500 505 510
Ala Phe Ile Ala Glu Leu Leu Gly Gly Ser Ser Gln Asn Val Ser Lys
515 520 525
Glu Ser Arg Ser Ser Pro Leu Val Lys Ile Cys Gly Thr Arg Ser Glu
530 535 540
Glu Ala Ala Arg Ala Ala Ile Glu Ala Gly Ala Asp Leu Ile Gly Ile
545 550 555 560
Ile Met Val Gln Gly Arg Thr Arg Cys Val Pro Asp Asp Val Ala Leu
565 570 575
Arg Ile Ser Gln Val Val Lys Ser Thr Pro Lys Pro Ala Gly Gln Thr
580 585 590
Pro Pro Thr Ser Gln Gly Thr Pro Ala Ala Ala Ser Val Glu Tyr Phe
595 600 605
Asp His Ser Ala Arg Ile Leu Arg His Pro Ser Arg Ala Leu Leu Val
610 615 620
Gly Val Phe Gln Asn Gln Pro Leu Asp Tyr Ile Leu Ser Gln Gln Gln
625 630 635 640
Lys Leu Gly Leu Asp Val Val Gln Leu His Gly Ser Glu Pro Leu Glu
645 650 655
Trp Ala Lys Leu Ile Pro Val Pro Val Ile Arg Lys Phe Gly Leu Asp
660 665 670
Glu Pro Ala Ile Ala Arg Arg Ala Tyr His Ser Leu Pro Leu Leu Asp
675 680 685
Ser Gly Val Gly Gly Thr Gly Glu Leu Leu Asp Gln Ser Arg Val Gln
690 695 700
Asn Val Leu Asp Lys Asp Ser Gly Leu Arg Val Ile Leu Ala Gly Gly
705 710 715 720
Leu Asp Pro Thr Asn Val Ala Gly Ile Val Gln Lys Leu Gly Glu Ser
725 730 735
Gly Arg Lys Val Val Gly Val Asp Val Ser Ser Gly Val Glu Ser Asp
740 745 750
Gly Ala Gln Asp Val Gly Lys Ile Arg Ala Phe Val Gln Ala Val Arg
755 760 765
Gly Leu
770
<210> 8
<211> 50
<212> DNA
<213> 人工序列()
<400> 8
ttgttcccgg gtataaagga ggccatccat gcaaacacaa aaaccgactc 50
<210> 9
<211> 36
<212> DNA
<213> 人工序列()
<400> 9
gcagaatcta gatcatcaga aagtctcctg tgcatg 36
<210> 10
<211> 70
<212> DNA
<213> 人工序列()
<400> 10
gcgctacagg gtgcgcgcac ggcgtttatt ctggagtgca agaaagcgtc gttgacagct 60
agctcagtcc 70
<210> 11
<211> 70
<212> DNA
<213> 人工序列()
<400> 11
gatgccggat tcgctgatta ccgtcacgtt gtgccccagt ttcggcgcaa atttgatgcc 60
tgggcatgcg 70
<210> 12
<211> 20
<212> DNA
<213> 人工序列()
<400> 12
atgcaaaccg ttttagcgaa 20
<210> 13
<211> 20
<212> DNA
<213> 人工序列()
<400> 13
caaatcgtca tgggccatca 20
<210> 14
<211> 37
<212> DNA
<213> 人工序列()
<400> 14
gcaacgcata tgcaaaccgt tttagcgaaa atcgtcg 37
<210> 15
<211> 37
<212> DNA
<213> 人工序列()
<400> 15
agtcgcctcg agtactttat tctcacccag caacacc 37
<210> 16
<211> 74
<212> DNA
<213> 人工序列()
<400> 16
cggcgcgaat tcagaaggag atatacatat gcaccaccac caccaccacc aaaccgtttt 60
agcgaaaatc gtcg 74
<210> 17
<211> 31
<212> DNA
<213> 人工序列()
<400> 17
agcgtctcta gacttaatat gcgcgcagcg t 31
<210> 18
<211> 41
<212> DNA
<213> 人工序列()
<400> 18
agcgtctcta gacttatact ttattctcac ccagcaacac c 41
<210> 19
<211> 22
<212> DNA
<213> 人工序列()
<400> 19
gcagacaagg cgatttgggt ag 22
<210> 20
<211> 27
<212> DNA
<213> 人工序列()
<400> 20
gacggctttc gctaaaacgg tttgcat 27
<210> 21
<211> 27
<212> DNA
<213> 人工序列()
<400> 21
gacgactttc gctaaaacgg tttgcat 27
<210> 22
<211> 21
<212> DNA
<213> 人工序列()
<400> 22
gcaaaaggcg tgatccgtga t 21
<210> 23
<211> 20
<212> DNA
<213> 人工序列()
<400> 23
cggcgacgct ttcttgcact 20
<210> 24
<211> 28
<212> DNA
<213> 人工序列()
<400> 24
tcgccgggaa aaggcgtgat ccgtgatg 28
<210> 25
<211> 19
<212> DNA
<213> 人工序列()
<400> 25
cgctttcttg cactccaga 19
<210> 26
<211> 21
<212> DNA
<213> 人工序列()
<400> 26
atcgcggttg ttgatgccaa c 21
<210> 27
<211> 20
<212> DNA
<213> 人工序列()
<400> 27
gcgcgtgatt tgtcgattga 20
<210> 28
<211> 19
<212> DNA
<213> 人工序列()
<400> 28
gttgttgatg ccaacgacc 19
<210> 29
<211> 34
<212> DNA
<213> 人工序列()
<400> 29
cgcgattttc gtgatttgtc gattgatctc aacc 34
<210> 30
<211> 452
<212> PRT
<213> 人工序列()
<400> 30
Met Gln Thr Val Leu Ala Lys Ile Val Ala Asp Lys Ala Ile Trp Val
1 5 10 15
Glu Ala Arg Lys Gln Gln Gln Pro Leu Ala Ser Phe Gln Asn Glu Val
20 25 30
Gln Pro Ser Thr Arg His Phe Tyr Asp Ala Leu Gln Gly Ala Arg Thr
35 40 45
Ala Phe Ile Leu Glu Cys Lys Lys Ala Gln Val Phe Gln Gly Val Ile
50 55 60
Arg Asp Asp Phe Asp Pro Ala Arg Ile Ala Ala Ile Tyr Lys His Tyr
65 70 75 80
Ala Ser Ala Ile Ser Val Leu Thr Asp Glu Lys Tyr Phe Gln Gly Ser
85 90 95
Phe Asn Phe Leu Pro Ile Val Ser Gln Ile Ala Pro Gln Pro Ile Leu
100 105 110
Cys Lys Asp Phe Ile Ile Asp Pro Tyr Gln Ile Tyr Leu Ala Arg Tyr
115 120 125
Tyr Gln Ala Asp Ala Cys Leu Leu Met Leu Ser Val Leu Asp Asp Asp
130 135 140
Gln Tyr Arg Gln Leu Ala Ala Val Ala His Ser Leu Glu Met Gly Val
145 150 155 160
Leu Thr Glu Val Ser Asn Glu Glu Glu Gln Glu Arg Ala Ile Ala Leu
165 170 175
Gly Ala Lys Val Val Gly Ile Asn Asn Arg Asp Leu Arg Asp Leu Ser
180 185 190
Ile Asp Leu Asn Arg Thr Arg Glu Leu Ala Pro Lys Leu Gly His Asn
195 200 205
Val Thr Val Ile Ser Glu Ser Gly Ile Asn Thr Tyr Ala Gln Val Arg
210 215 220
Glu Leu Ser His Phe Ala Asn Gly Phe Leu Ile Gly Ser Ala Leu Met
225 230 235 240
Ala His Asp Asp Leu His Ala Ala Val Arg Arg Val Leu Leu Gly Glu
245 250 255
Asn Lys Val Cys Gly Leu Thr Arg Gly Gln Asp Ala Lys Ala Ala Tyr
260 265 270
Asp Ala Gly Ala Ile Tyr Gly Gly Leu Ile Phe Val Ala Thr Ser Pro
275 280 285
Arg Cys Val Asn Val Glu Gln Ala Gln Glu Val Met Ala Ala Ala Pro
290 295 300
Leu Gln Tyr Val Gly Val Phe Arg Asn His Asp Ile Ala Asp Val Val
305 310 315 320
Asp Lys Ala Lys Val Leu Ser Leu Ala Ala Val Gln Leu His Gly Asn
325 330 335
Glu Glu Gln Leu Tyr Ile Asp Thr Leu Arg Glu Ala Leu Pro Ala His
340 345 350
Val Ala Ile Trp Lys Ala Leu Ser Val Gly Glu Thr Leu Pro Ala Arg
355 360 365
Glu Phe Gln His Val Asp Lys Tyr Val Leu Asp Asn Gly Gln Gly Gly
370 375 380
Ser Gly Gln Arg Phe Asp Trp Ser Leu Leu Asn Gly Gln Ser Leu Gly
385 390 395 400
Asn Val Leu Leu Ala Gly Gly Leu Gly Ala Asp Asn Cys Val Glu Ala
405 410 415
Ala Gln Thr Gly Cys Ala Gly Leu Asp Phe Asn Ser Ala Val Glu Ser
420 425 430
Gln Pro Gly Ile Lys Asp Ala Arg Leu Leu Ala Ser Val Phe Gln Thr
435 440 445
Leu Arg Ala Tyr
450
Claims (13)
1.一种基因突变以表达吲哚3-甘油磷酸合成酶IGPs的细菌细胞,其特征在于,该细菌细胞所产生的IGPs与野生型IGPs相比,对邻氨基苯甲酸的抑制敏感度降低。
2.根据权利要求1所述的细菌细胞,其特征在于,该细胞突变后产生:
a)细菌IGPs突变体,该突变体相较于野生型细胞产生的IGPs,对邻氨基苯甲酸的抑制敏感度降低,或
b)具有IGPs活性的异源酶,该酶相较于野生型IGPs,对邻氨基苯甲酸的抑制敏感度降低。
3.根据权利要求2所述的细菌细胞,其特征在于,所述细菌IGPs突变体与基因突变的细菌细胞是同源的。
4.根据权利要求2-3任一所述的细菌细胞,其特征在于,该细胞所产生的IGPs与野生型细胞IGPs相比,在邻氨基苯甲酸结合位点至少也有一个氨基酸发生突变,且突变后的IGPs具有活性并对氨基苯甲酸的抑制敏感度降低。
5.根据权利要求2-4任一所述的细菌细胞,其特征在于,所述细菌细胞突变后的IGPs具有以下特征:
a)与SEQ ID NO: 1序列相比,用丝氨酸替代第60位的丙氨酸或甘氨酸,并且/或者用异亮氨酸替代第8位的缬氨酸,并且/或者用亮氨酸代替第188位的苯丙氨酸,或者用丝氨酸代替第58位的谷氨酰胺,用脯氨酸代替第59位的缬氨酸,用丝氨酸替代第60位的苯丙氨酸,用赖氨酸替代第61位的谷氨酰胺;或者
b)突变序列与SEQ ID NO: 1序列相比,第8-188位中至少有一个氨基酸被其它氨基酸取代,但突变体仍然具有IGPs活性,并且与野生型菌株表达的IGPs相比,突变后的IGPs对邻氨基苯甲酸的抑制敏感度降低。
6.据权利要求2-5任一所述的细菌细胞,其特征在于,所述细菌细胞突变后的IGPs序列为SEQ ID NO: 2 至 SEQ ID NO: 5,或SEQ ID NO:30其中之一。
7.根据权利要求1-6任一所述的细菌细胞,其特征在于,所述细菌细胞为大肠杆菌细胞。
8.根据权利要求2所述的细菌细胞,其特征在于,所述具有IGPs活性的异源酶为具有邻氨基苯甲酸合成酶II结构域的酶,所述邻氨基苯甲酸合成酶II结构域的酶为来源于酿酒酵母属或曲霉属的酶,所述酿酒酵母属的菌为酿酒酵母,所述曲霉属的菌为黑曲霉。
9.一种分离或合成的酶,其特征在于,所述酶的氨基酸序列为SEQ ID NO: 2 至 SEQID NO: 5,或SEQ ID NO:30其中之一。
10.一种利用生物技术生产L-色氨酸的方法,其特征在于,包括以下步骤:在生物反应器中培养权利要求1-8任一所述的基因突变的细菌细胞。
11.根据权利要求10所述的方法,其特征在于所述基因突变的细菌细胞为大肠杆菌。
12.一种如权利要求1-8任一所述的细菌细胞或如权利要求9所述的酶在生产L-色氨酸中的应用。
13.根据权利要求12所述的应用,其特征在于,利用生物反应器进行规模化工业生产L-色氨酸。
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| DE102017126895.2 | 2017-11-15 | ||
| DE102017126895 | 2017-11-15 | ||
| LU100521 | 2017-11-15 | ||
| LU100521A LU100521B1 (en) | 2017-11-15 | 2017-11-15 | Improved biotechnological production of L-tryptophan |
| PCT/EP2018/080908 WO2019096727A1 (en) | 2017-11-15 | 2018-11-12 | Improved biotechnological production of l-tryptophan |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| CN111406111A true CN111406111A (zh) | 2020-07-10 |
Family
ID=64332280
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN201880073668.5A Pending CN111406111A (zh) | 2017-11-15 | 2018-11-12 | 一种改进的生物技术生产l-色氨酸的方法 |
Country Status (4)
| Country | Link |
|---|---|
| US (1) | US10947523B2 (zh) |
| EP (1) | EP3710593A1 (zh) |
| CN (1) | CN111406111A (zh) |
| WO (1) | WO2019096727A1 (zh) |
Family Cites Families (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| HUE057864T2 (hu) | 2012-01-10 | 2022-06-28 | Cj Cheiljedang Corp | Fokozott L-triptofán termeléssel rendelkezõ Escherichia coli mikroorganizmusok és eljárás L-triptofán elõállítására azok alkalmazásával |
| KR101530817B1 (ko) | 2013-04-16 | 2015-06-25 | 씨제이제일제당 (주) | L-트립토판 생산능을 갖는 미생물 및 이를 이용하여 l-트립토판을 생산하는 방법 |
| CN106520652B (zh) * | 2016-11-29 | 2019-07-05 | 天津科技大学 | 一株谷氨酸棒杆菌及其合成色氨酸的关键基因 |
-
2018
- 2018-11-12 EP EP18804259.2A patent/EP3710593A1/en not_active Withdrawn
- 2018-11-12 CN CN201880073668.5A patent/CN111406111A/zh active Pending
- 2018-11-12 WO PCT/EP2018/080908 patent/WO2019096727A1/en unknown
- 2018-11-12 US US16/763,017 patent/US10947523B2/en active Active
Non-Patent Citations (8)
Also Published As
| Publication number | Publication date |
|---|---|
| WO2019096727A1 (en) | 2019-05-23 |
| US20200277593A1 (en) | 2020-09-03 |
| EP3710593A1 (en) | 2020-09-23 |
| US10947523B2 (en) | 2021-03-16 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| Webb et al. | Biosynthesis of pantothenate | |
| EP1177311B1 (en) | In vitro macromolecule biosynthesis methods using exogenous amino acids and a novel atp regeneration system | |
| US6994986B2 (en) | In vitro synthesis of polypeptides by optimizing amino acid metabolism | |
| KR101944150B1 (ko) | 메티오닌의 발효적 생산을 위한 재조합 미생물 | |
| KR102116734B1 (ko) | 대사 공학을 통한 에르고티오네인 생성 | |
| Chen et al. | Discovery of feed-forward regulation in L-tryptophan biosynthesis and its use in metabolic engineering of E. coli for efficient tryptophan bioproduction | |
| CN112105734A (zh) | 生产色胺的方法 | |
| Ravikumar et al. | Incorporating unnatural amino acids to engineer biocatalysts for industrial bioprocess applications | |
| Zhang et al. | Tyrosine decarboxylase from Lactobacillus brevis: soluble expression and characterization | |
| Ramzi et al. | 5-Aminolevulinic acid production in engineered Corynebacterium glutamicum via C5 biosynthesis pathway | |
| RU2014104668A (ru) | Гены и белки для синтеза алканоил-соа | |
| US11306331B2 (en) | Processes for the production of hydroxycinnamic acids using polypeptides having tyrosine ammonia lyase activity | |
| US20220348974A1 (en) | Biotin synthases for efficient production of biotin | |
| JPWO2016017631A1 (ja) | γ−グルタミルシステイン及びグルタチオンの製造方法 | |
| JP2011500077A (ja) | 方法及び組成物 | |
| WO2016002884A1 (ja) | 酸化型γ-グルタミルシステイン及び酸化型グルタチオンの製造方法 | |
| KR20140135622A (ko) | 발효에 의한 감마-글루타밀시스테인 및 이 디펩티드의 유도체의 과다 생산을 위한 미생물 및 방법 | |
| Liu et al. | The 138th residue of acetohydroxyacid synthase in Corynebacterium glutamicum is important for the substrate binding specificity | |
| KR102149044B1 (ko) | 2-히드록시 감마 부티로락톤 또는 2,4-디히드록시-부티레이트 의 제조 방법 | |
| US10480003B2 (en) | Constructs and systems and methods for engineering a CO2 fixing photorespiratory by-pass pathway | |
| US20140370557A1 (en) | Genetically engineered microbes and methods for producing 4-hydroxycoumarin | |
| US10704036B2 (en) | Protein having synthetic activity for imidazole dipeptid and production method of imidazole dipeptide | |
| CN111406111A (zh) | 一种改进的生物技术生产l-色氨酸的方法 | |
| Zhai et al. | Mining of aminotransferase gene ota3 from Bacillus pumilus W3 via genome analysis, gene cloning and expressing for compound bioamination | |
| Ferrer et al. | L-serine biosensor-controlled fermentative production of l-tryptophan derivatives by Corynebacterium glutamicum. Biology. 2022; 11: 744 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| PB01 | Publication | ||
| PB01 | Publication | ||
| SE01 | Entry into force of request for substantive examination | ||
| SE01 | Entry into force of request for substantive examination | ||
| WD01 | Invention patent application deemed withdrawn after publication |
Application publication date: 20200710 |
|
| WD01 | Invention patent application deemed withdrawn after publication |