RU2729383C2 - Сконструированные высокоаффинные t-клеточные рецепторы человека - Google Patents
Сконструированные высокоаффинные t-клеточные рецепторы человека Download PDFInfo
- Publication number
- RU2729383C2 RU2729383C2 RU2016124163A RU2016124163A RU2729383C2 RU 2729383 C2 RU2729383 C2 RU 2729383C2 RU 2016124163 A RU2016124163 A RU 2016124163A RU 2016124163 A RU2016124163 A RU 2016124163A RU 2729383 C2 RU2729383 C2 RU 2729383C2
- Authority
- RU
- Russia
- Prior art keywords
- tcr
- cell receptor
- cell
- modified
- cells
- Prior art date
Links
Images








Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K35/00—Medicinal preparations containing materials or reaction products thereof with undetermined constitution
- A61K35/12—Materials from mammals; Compositions comprising non-specified tissues or cells; Compositions comprising non-embryonic stem cells; Genetically modified cells
- A61K35/14—Blood; Artificial blood
- A61K35/17—Lymphocytes; B-cells; T-cells; Natural killer cells; Interferon-activated or cytokine-activated lymphocytes
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
- A61K38/16—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- A61K38/17—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- A61K38/177—Receptors; Cell surface antigens; Cell surface determinants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
- A61K38/16—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- A61K38/17—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- A61K38/177—Receptors; Cell surface antigens; Cell surface determinants
- A61K38/1774—Immunoglobulin superfamily (e.g. CD2, CD4, CD8, ICAM molecules, B7 molecules, Fc-receptors, MHC-molecules)
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P35/00—Antineoplastic agents
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/46—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates
- C07K14/47—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates from mammals
- C07K14/4701—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates from mammals not used
- C07K14/4747—Apoptosis related proteins
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/705—Receptors; Cell surface antigens; Cell surface determinants
- C07K14/70503—Immunoglobulin superfamily
- C07K14/7051—T-cell receptor (TcR)-CD3 complex
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/06—Animal cells or tissues; Human cells or tissues
- C12N5/0602—Vertebrate cells
- C12N5/0634—Cells from the blood or the immune system
- C12N5/0636—T lymphocytes
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/56—Immunoglobulins specific features characterized by immunoglobulin fragments variable (Fv) region, i.e. VH and/or VL
- C07K2317/565—Complementarity determining region [CDR]
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/60—Immunoglobulins specific features characterized by non-natural combinations of immunoglobulin fragments
- C07K2317/62—Immunoglobulins specific features characterized by non-natural combinations of immunoglobulin fragments comprising only variable region components
- C07K2317/622—Single chain antibody (scFv)
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/73—Inducing cell death, e.g. apoptosis, necrosis or inhibition of cell proliferation
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/90—Immunoglobulins specific features characterized by (pharmaco)kinetic aspects or by stability of the immunoglobulin
- C07K2317/92—Affinity (KD), association rate (Ka), dissociation rate (Kd) or EC50 value
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Immunology (AREA)
- Organic Chemistry (AREA)
- General Health & Medical Sciences (AREA)
- Zoology (AREA)
- Medicinal Chemistry (AREA)
- Cell Biology (AREA)
- Engineering & Computer Science (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Gastroenterology & Hepatology (AREA)
- Genetics & Genomics (AREA)
- Animal Behavior & Ethology (AREA)
- Pharmacology & Pharmacy (AREA)
- Public Health (AREA)
- Veterinary Medicine (AREA)
- Biochemistry (AREA)
- Biomedical Technology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Epidemiology (AREA)
- Toxicology (AREA)
- Biophysics (AREA)
- Molecular Biology (AREA)
- Biotechnology (AREA)
- Hematology (AREA)
- Wood Science & Technology (AREA)
- Developmental Biology & Embryology (AREA)
- Virology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Microbiology (AREA)
- General Engineering & Computer Science (AREA)
- Peptides Or Proteins (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
Abstract
Предложенная группа изобретений относится к иммунологии. Предложен модифицированный T-клеточный рецептор (TCR) и его антигенсвязывающий фрагмент, способные связывать сурвивин. Кроме того, представлены композиции, нацеленные на клетки рака, и способ лечения субъекта, имеющего рак. Данная группа изобретений может найти дальнейшее применение в терапии рака. 6 н. и 5 з.п. ф-лы, 5 табл., 9 пр., 9 ил.
Description
ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Настоящая заявка испрашивает приоритет в соответствии со статьей 35 § 119(e) Свода законов США на основании предварительной заявки на патент США № 61/907887, поданной 22 ноября 2013 г. и содержание данной предварител.ной заявки полностью включено в данную заявку посредством ссылки.
СВЕДЕНИЯ О ФИНАНСИРУЕМЫХ ИЗ ФЕДЕРАЛЬНОГО БЮДЖЕТА НАУЧНО-ИССЛЕДОВАТЕЛЬСКИХ И ОПЫТНО-КОНСТРУКТОРСКИХ РАБОТАХ
[0002] Настоящее изобретение было выполнено при государственной поддержке США грантами №№ R01 GM55767 и T32 GM070421, присужденными Национальными институтами здравоохранения. Правительство США имеет определенные права на настоящее изобретение.
СВЕДЕНИЯ, КАСАЮЩИЕСЯ ПЕРЕЧНЯ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0003] Перечень последовательностей, относящийся к настоящей заявке, предоставлен в текстовом формате вместо бумажной копии и настоящим включен посредством ссылки в описание настоящего изобретения. Название текстового файла, содержащего Перечень последовательностей, следующее: IMMU_003_01WO_ST25.txt. Размер указанного текстового файла составляет 12 килобайт, файл был создан 21 ноября 2014 г. и подан в электронном виде через EFS-Web.
ОБЛАСТЬ ТЕХНИКИ
[0004] В настоящем описании предложены высокоаффинные T-клеточные рецепторы (TCR), сконструированные с помощью способов in vitro, направленные против антигена сурвивина, а также способы получения модифицированных TCR и одноцепочечных TCR и соответствующие применения TCR для терапевтических, диагностических способов и способов визуализации.
УРОВЕНЬ ТЕХНИКИ
[0005] T-клеточные рецепторы (TCR) и антитела представляют собой молекулы, которые образовались для распознавания различных классов антигенов (лигандов) ((Murphy (2012), xix, стр. 868)). TCR представляют собой антигенспецифические молекулы, которые отвечают за распознавание антигенных пептидов, презентированных в составе главного комплекса гистосовместимости (MHC) на поверхности антигенпрезентирующих клеток (APC) или любой содержащей ядро клетки (например, всех клеток в организме человека, за исключением эритроцитов). Напротив, антитела обычно распознают растворимые антигены или антигены клеточной поверхности и не требуют презентации антигена с помощью MHC. Данная система наделяет T-клетки, посредством их TCR, потенциальной способностью распознавать весь набор внутриклеточных антигенов, экспрессируемых клеткой (включая белки вируса), которые процессируются внутри клетки с образованием коротких пептидов, связанных с внутриклеточной молекулой MHC, и доставляются на поверхность в виде комплекса пептид-MHC (pepMHC). Данная система позволяет фактически любому чужеродному белку (например, мутированному антигену рака или белку вируса) или аберрантно экспрессированному белку служить мишенью для T-клеток (обзор приведен в (Davis и Bjorkman (1988) Nature, 334, 395-402; Davis и др. (1998) Annu Rev Immunol, 16, 523-544; Murphy (2012), xix, стр. 868).
[0006] Взаимодействие TCR и pepMHC может запускать различные состояния активации T-клеток, в зависимости от аффинности (или скорости диссоциации) связывания. Процесс распознавания TCR позволяет T-клетке отличить нормальную здоровую клетку, например, от клетки, которая трансформировалась под воздействием вируса или в результате малигнизации, путем обеспечения разнообразного репертуара TCR, при этом существует высокая вероятность того, что будет присутствовать один или более TCR с аффинностью связывания с чужеродным пептидом, связанным с молекулой MHC, выше порога стимуляции активности T-клеток (Manning и Kranz (1999) Immunology Today, 20, 417-422).
[0007] На сегодняшний день было показано, что у TCR дикого типа, выделенных из клонов T-клеток либо человека, либо мыши, которые были обнаружены путем культивирования in vitro, наблюдают относительно низкие аффинности связывания (Kd = 1 - 300 мкМ) (Davis и др. (1998) Annu Rev Immunol, 16, 523-544). Часть объяснения этому, похоже, состоит в том, что T-клетки, которые развиваются в тимусе, подвергаются отрицательной селекции (индукция толерантности) по отношению к лигандам собственных пептидов-MHC, таким образом, что T-клетки со слишком высокой аффинностью удаляются (Starr и др. (2003) Annu Rev Immunol, 21, 139-76). Для того, чтобы компенсировать данные относительно низкие аффинности, T-клетки развили систему корецепторов, в которой молекулы клеточной поверхности CD4 и CD8 связываются с молекулами MHC (II класса и I класса, соответственно) и синергичным образом совместно с TCR опосредуют активность передачи сигналов. CD8 особенно эффективен в данном процессе, позволяя TCR с очень низкой аффинностью (например, Kd = 300 мкМ) опосредовать сильную антигенспецифическую активность.
[0008] Чтобы получить TCR с более высокой аффинностью к определенным pepMHC, использовали направленное развитие in vitro. Применяли три различных способа дисплея, описанных далее: дрожжевой дисплей (Holler и др. (2003) Nat Immunol, 4, 55-62; Holler и др. (2000) Proc Natl Acad Sci U S A, 97, 5387-92), фаговый дисплей (Li и др. (2005) Nat Biotechnol, 23, 349-54) и T-клеточный дисплей (Chervin и др. (2008) J Immunol Methods, 339, 175-84). Во всех трех подходах процесс включал конструирование или модифицирование TCR, который проявляет нормальную низкую аффинность, характерную для TCR дикого типа, таким образом, что мутантные TCR имеют аффинность, повышенную по отношению к узнаваемому pepMHC (исходному антигену, к которому специфичны T-клетки). Таким образом TCR дикого типа использовали в качестве матрицы для получения библиотек с мутациями в одном или более участках, определяющих комплементарность (CDR), и мутантов с более высокой аффинностью подвергали селекции по связыванию с антигеном, представляющим собой узнаваемый пептид-MHC.
[0009] В настоящей заявке описаны высокоаффинные T-клеточные рецепторы, специфичные к антигену рака сурвивину, сконструированные путем дрожжевого дисплея. Белок сурвивин стимулирует онкогенез путем ингибирования передачи сигналов, которые приводят апоптозу в норме (Dohi и др. (2004) Journal of Clinical Investigation 114, 1117-1127). Уровень экспрессии сурвивина в раковой ткани повышен (Ambrosini и др. (1997) Nat Med 3, 917-921). Он являлся мишенью при попытках создания вакцин и при различных адоптивных T-клеточных подходах, в которых применяют T-клетки с T-клеточными рецепторами дикого типа.
[0010] Пептидный антиген сурвивин занимает 21 место в списке 75 важнейших антигенов рака, составленном в зависимости от их приоритета Национальным институтом рака (Cheever и др. (2009) Clin Cancer Res, 15, 5323-5337). Соответственно, существует потребность в идентификации агентов, например, терапевтических агентов, которые специфично нацелены на данный антиген рака. Согласно настоящему изобретению предложены сконструированные in vitro TCR с более высокой афинностью, которые можно применять, например, в растворимой форме, для направленной доставки in vivo или в виде генов, которые внедряют в T-клетки, в адоптивном T-клеточном подходе.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0011] Настоящее изобретение относится к сконструированным in vitro T-клеточным рецепторам (TCR), которые связываются с антигеном сурвивином с улучшенной аффинностью. В частности, в настоящем описании предложены такие стабилизирующие и изменяющие аффинность мутации, которые выбрали посредством дисплея библиотек на поверхности дрожжей, фага или клеток млекопитающих; белки TCR, выбранные из данных библиотек благодаря связыванию с антигеном с повышенной аффинностью; и использование прошедших селекцию in vitro производных TCR для терапевтических, диагностических применений или применений для визуализации.
[0012] Один аспект настоящего изобретения относится к модифицированному T-клеточному рецептору, или к его антигенсвязывающему фрагменту, содержащему Vα и Vβ, полученные из T-клеточного рецептора дикого типа, при этом Vα, Vβ или обе из них содержат мутацию в одном или более участках, определяющих комплементарность (CDR), относительно T-клеточного рецептора дикого типа, при этом модифицированный T-клеточный рецептор связывается с антигеном пептид/MHC, известным как сурвивин/HLA-A2 (пептид сурвивина LMLGEFLKL (SEQ ID NO: 5), связанный с продуктом MHC, известным как HLA-A2).
[0013] В одном варианте реализации модифицированный T-клеточный рецептор содержит модифицированную Vα, включающую последовательность аминокислот, идентичную по меньшей мере на 80% последовательности аминокислот Vα SEQ ID NO: 3, при этом указанный модифицированный T-клеточный рецептор связывается с комплексом сурвивин/HLA-A2 с аффинностью (значением KA) 106 М и выше.
[0014] В другом варианте реализации модифицированный T-клеточный рецептор содержит модифицированную Vα, включающую последовательность аминокислот, идентичную по меньшей мере на 80% последовательности аминокислот Vα SEQ ID NO: 4, при этом указанный модифицированный T-клеточный рецептор связывается с комплексом сурвивин/HLA-A2 с аффинностью (значением KA) 106 М и выше.
[0015] В другом варианте реализации T-клеточный рецептор представляет собой одноцепочечный T-клеточный рецептор, включающий последовательность аминокислот SEQ ID NO: 6.
[0016] В другом варианте реализации T-клеточный рецептор представляет собой одноцепочечный T-клеточный рецептор, включающий последовательность аминокислот SEQ ID NO: 7.
[0017] В другом варианте реализации T-клеточный рецептор содержит по меньшей мере одну из мутаций в CDR3α, выбранных из N92S, N100K, A101G, R102Y и L103K, в последовательности аминокислот SEQ ID NO: 3.
[0018] В другом варианте реализации T-клеточный рецептор содержит по меньшей мере одну из мутаций в CDR3α, выбранных из N92H, N100G, A101W, R102Y и L103T, в последовательности аминокислот SEQ ID NO: 4.
[0019] В одном варианте реализации модифицированный T-клеточный рецептор получают путем селекции in vitro из библиотеки дрожжевого дисплея мутантных T-клеточных рецепторов.
[0020] В другом варианте реализации модифицированный T-клеточный рецептор экспрессируется в виде растворимого белка, который связывается с его целевым антигеном.
[0021] В другом варианте реализации модифицированный T-клеточный рецептор экспрессируется на поверхности T-клеток для того, чтобы опосредовать активность либо CD4+, либо CD8+ T-клеток.
[0022] Один аспект настоящего изобретения относится к терапевтическому агенту, который нацелен на клетки рака, экспрессирующие антиген сурвивин, при этом указанный терапевтический агент включает модифицированный T-клеточный рецептор, описанный в данной заявке. В одном варианте реализации предложен терапевтический агент, который нацелен на клетки рака, экспрессирующие антиген сурвивин, при этом указанный терапевтический агент включает T-клетку человека, которая экспрессирует модифицированный T-клеточный рецептор, описанный в данной заявке.
[0023] В одном варианте реализации предложен способ лечения субъекта, имеющего рак, который экспрессирует антиген сурвивин, включающий введение терапевтического агента, описанного в данной заявке.
КРАТКОЕ ОПИСАНИЕ ФИГУР
[0024] На фигуре 1 представлена диаграмма, на которой показан способ конструирования одноцепочечных TCR для улучшения их аффинности к комплексу пептид:HLA.A2. Показан обычный процесс, применяемый для конструирования высокоаффинных TCR.
[0025] На фигуре 2A представлена трехмерная диаграмма, на которой показан вид сбоку комплекса TCR:pepMHC (A6; PDB:1AO7). Указаны вариабельные (V) и константные (C) области α-цепи и β-цепи. Представленная структура не содержит область Cα TCR. HLA-A2 (α1, α2, α3 и β2m) показан серым цветом и пептид Tax (LLFGYPVYV, SEQ ID NO: 6) показан черным цветом. Во всех TCR, специфичных к A6, и TCR, специфичных к сурвивину, исследованных в настоящем изобретении, используется фрагмент Vα2 (также называемый TRAV12 на основании номенклатуры IMGT).
[0026] На фигуре 2B представлена трехмерная диаграмма, на которой показан вид сверху вниз следа TCR (CDR) на комплексе пептид-MHC (Tax/HLA-A2). Хотя не было описано кристаллических структур для специфичного к сурвивину TCR, используемого в настоящем описании, такую диагональную ориентацию стыковки с областью Vα, расположенной над спиралью α2 MHC и N-концом пептида, и областью Vβ, расположенной над спиралью α1 MHC и C-концом пептида, на сегодняшний день наблюдали фактически во всех комплексах.
[0027] На фигуре 3 представлено схематическое изображение системы дрожжевого дисплея для конструирования одноцепочечных фрагментов T-клеточного рецептора (Vα-линкер-Vβ или Vβ-линкер-Vα).
[0028] На фигурах 4A и 4B показаны гистограммы проточной цитометрии библиотеки специфичных к сурвивину TCR, полученной с помощью допускающей ошибки полимеразы, после сортировки с антителом, которое узнает конформационный эпитоп на Vβ20. Библиотеку специфичных к сурвивину TCR, полученную с помощью допускающей ошибки полимеразы, подвергали последовательной сортировке с разведенным 1:10 BC hVβ20 IgG, меченым FITC, а затем с меченым AlexaFluor® 488 вторичным антителом козы против IgG мыши (1:100), всего проводили 3 сортировки. Аликвоты дрожжевых клеток после каждой сортировки инкубировали с разведенным 1:10 BC hVβ20 (фигура 4A). Серым цветом показаны дрожжевые клетки, окрашенные только вторичным антителом. Стабильные клоны K2 окрашивали разведенным 1:20 hVβ20 IgG, меченым FITC, а затем меченым AlexaFlour 647 вторичным антителом козы против IgG мыши (1:100) (фигура 4B).
[0029] На фигурах 5A и 5B показаны гистограммы проточной цитометрии библиотеки специфичных к сурвивину CDR3α после сортировки с BC hVβ20 и SurvT2M:HLA-A2 и связывания двух высокоаффинных TCR с SurvT2M:HLA-A2. Библиотеку специфичных к сурвивину CDR3α подвергали сортировке сначала с помощью BC hVβ20 (1:10), а затем с помощью вторичного антитела к IgG мыши на микрогранулах MB (1:25), применяя магнитные колонки. Библиотеки специфичных к сурвивину CDR3α затем подвергали сортировке с помощью 100 нМ димера SurvT2M:HLA-A2 (DimerX; полученного от BD Pharmingen), а затем с помощью вторичного антитела к IgG мыши на микрогранулах MB (1:25), всего проводили три магнитных сортировки. Выделенные дрожжи впоследствии сортировали, применяя сортировку клеток с возбуждением флуоресценции (FACS), с помощью 100 нМ димера SurvT2M:HLA-A2 (DimerX; полученного от BD Pharmingen), а затем с помощью меченого AlexaFluor® 647 вторичного антитела козы против IgG мыши (1:100). Аликвоты дрожжевых клеток после каждой сортировки затем инкубировали со 100 нМ димера SurvT2M:HLA-A2 (DimerX; полученного от BD Pharmingen), а после с меченым AlexaFluor® 647 вторичным антителом козы против IgG мыши (1:100) (фигура 5A). Серым цветом показаны дрожжевые клетки, окрашенные только вторичным антителом. Клоны с улучшенным связыванием K2.4.1 (фигура 5B, левая панель) и K2.4.6 (фигура 5B, правая панель), выделенные после 4ой сортировки с применением FACS, окрашивали с помощью 100 нМ димера SurvT2M:HLA-A2 (DimerX; полученного от BD Pharmingen), а после с меченым AlexaFluor® 647 вторичным антителом козы против IgG мыши (1:100) (фигура 5B).
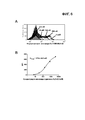
[0030] На фигурах 6A и 6B показаны свойства связывания высокоаффинного TCR K2.4.1 с мономерами SurvT2M:HLA-A2. На фигуре 6A представлена гистограмма проточной цитометрии, на которой показан высокоаффинный одноцепочечный TCR (scTCR) K2.4.1, окрашенный различными концентрациями мономера SurvT2M:HLA-A2, а затем вторичным антителом, конъюгированным со стрептавидином (SA) и меченым PE (1:100). На фигуре 6B представлена линейная диаграмма, на которой показаны значения средних интенсивностей флуоресценции (MFI) из гистограмм, представленных на фигуре 6A, нанесенные на график в зависимости от концентрации мономера SurvT2M:HLA-A2.
[0031] На фигуре 7 изображены последовательности специфичных к сурвивину высокоаффинных TCR (K2.4.1 и K2.4.6). Высокоаффинные одноцепочечные варианты выделяли из библиотек участков, определяющих комплементарность, которые затем подвергали скринингу для выявления созревшей аффинности. Мутации, выявленные в результате скрининга библиотек стабильности, подчеркнуты и выделены жирным шрифтом; мутации, выявленные в результате скрининга библиотек созревания аффинности, заключены в рамку и выделены жирным шрифтом. Также показаны последовательности дикого типа V-областей со «стабилизирующими» мутациями в клоне K2, представленном на поверхности дрожжей. Последовательности аминокислот, показанные для цепи Vβ, соответствуют последовательности SEQ ID NO: 12, и изображенная последовательность линкера представляет собой SEQ ID NO: 7. Последовательности аминокислот, показанные для цепи Vα, соответствуют последовательностям SEQ ID NO: 13, 1 и 2, сверху вниз.
[0032] На фигурах 8A - 8C показаны результаты анализа T-клеток, в котором T-клетки трансдуцировали TCR K2.4.1. T-клетки выделяли из трансгенных мышей AAD (это мыши, которые содержат гибридный ген I класса, состоящий из доменов α1 и α2 HLA-A2 и домена α3 Db мыши; данные мыши AAD доступны для приобретения у Jackson Laboratories). Клетки активировли с помощью гранул, к которым присоединены антитела к CD3 и к CD28, в течение 24 часов. T-клетки трансдуцировали с помощью ретровирусного вектора pMP71, содержащего домены Vα и β TCR K2.4.1, соединенные с доменами Cα и Cβ TCR 2C мыши (фигура 8A). Ложно трансдуцированные (серый) и трансдуцированные K2.4.1 (черная линия) T-клетки затем окрашивали с помощью тетрамера SurvT2M:HLA-A2 при концентрации 20 нМ (фигура 8B). T-клетки затем инкубировали при соотношении эффектор: мишень (E:T), равном 1:1, с клетками T2 человека, которые экспрессируют HLA-A2, и различными концентрациями пептида сурвивина в течение 24 часов. Супернатанты собирали и измеряли высвобождение IFN-γ, применяя ELISA (фигура 8C).
[0033] На фигурах 9A и 9B представлены диаграммы, на которых проиллюстрированы примеры терапевтических применений высокоаффинных одноцепочечных TCR, выделенных из библиотек каркасов. На фигуре 9A изображено пять примеров форматов TCR для применения в качестве растворимых терапевтических продуктов: 1) одноцепочечный TCR либо в ориентации Vα-Vβ, либо в ориентации Vβ-Vα (мутированные высокоаффинные V-домены показаны звездочками); 2) одноцепочечный TCR, слитый в одной рамке считывания с доменами константной области антитела; 3) иммуноглобулин, слитый в одной рамке считывания с константной областью либо легкой цепи, либо тяжелой цепи; 4) одноцепочечный TCR (или слитые иммуноглобулины, показанные в 2 и 3), непосредственно соединенный с лекарственным средством; и 5) одноцепочечный TCR, соединенный в одной рамке считывания с одноцепочечным Fv (VL-линкер-VH) с получением биспецифического агента. На фигуре 9B изображено два примера методов лечения на основе клеток, в которых будут применять высокоаффинные вариабельные домены (V), выделенные с помощью дрожжевого дисплея, клонированные в векторы для экспрессии в клетках млекопитающих, для экспрессии T-клетками в адоптивной T-клеточной терапии в виде: 1) одноцепочечных рецепторов в химерных антигенных рецепторах (CAR) и 2) полноразмерных TCR α и β.
КРАТКОЕ ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0034] SEQ ID NO: 1 представляет собой последовательность аминокислот модифицированной области Vα TCR (сурвивин-K2.4.1), которая связывается с высокой аффинностью с комплексом сурвивин/HLA-A2.
[0035] SEQ ID NO: 2 представляет собой последовательность аминокислот другой модифицированной области Vα TCR (сурвивин-K2.4.6), которая связывается с высокой аффинностью с комплексом сурвивин/HLA-A2.
[0036] SEQ ID NO: 3 представляет собой последовательность аминокислот одноцепочечного TCR (сурвивин-K2.4.1), который связывается с высокой аффинностью с комплексом сурвивин/HLA-A2.
[0037] SEQ ID NO: 4 представляет собой последовательность аминокислот другого одноцепочечного TCR (сурвивин-K2.4.6), который связывается с высокой аффинностью с комплексом сурвивин/HLA-A2.
[0038] SEQ ID NO: 5 представляет собой последовательность аминокислот антигена сурвивина.
[0039] SEQ ID NO: 6 представляет собой последовательность аминокислот антигена Tax.
[0040] SEQ ID NO: 7 представляет собой последовательность аминокислот линкера.
[0041] SEQ ID NO: 8 представляет собой полинуклеотидную последовательность праймера Splice 4L.
[0042] SEQ ID NO: 9 представляет собой полинуклеотидную последовательность праймера T7.
[0043] SEQ ID NO: 10 представляет собой полинуклеотидную последовательность обратного праймера, применяемого для получения перед SOE продуктов ПЦР №1 из библиотеки Surv CDR3α.
[0044] SEQ ID NO: 11 представляет собой полинуклеотидную последовательность прямого праймера, применяемого для получения перед SOE продуктов ПЦР №2 из библиотеки Surv CDR3α.
[0045] SEQ ID NO: 12 представляет собой последовательность аминокислот области Vb TCR (сурвивин-K2), которая связывается с комплексом сурвивин/HLA-A2.
[0046] SEQ ID NO: 13 представляет собой последовательность аминокислот области Va TCR (сурвивин-K2), которая связывается с комплексом сурвивин/HLA-A2.
[0047] SEQ ID NO: 14 представляет собой последовательность аминокислот антигена WT1.
[0048] SEQ ID NO: 15 представляет собой последовательность аминокислот пептида вируса гриппа типа A.
[0049] SEQ ID NO: 16 представляет собой последовательность аминокислот варианта пептида вируса гриппа типа A.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0050] Следующее описание предназначено для того, чтобы способствовать пониманию описания настоящего изобретения, но не предназначено для его ограничения.
[0051] Как правило, термины и формулировки, используемые в данной заявке, имеют известное в данной области техники значение, которое можно найти, обратившись к стандартным текстам, ссылочным материалам в журналах и контекстам, известным специалистам в данной области. Следующие определения предложены, чтобы разъяснить конкретное их применение в контексте настоящего описания.
[0052] В данной заявке «соединенный» относится к связи между двумя группами, которая может представлять собой ковалентную или нековалентную связь. Группы можно соединить, применяя пептидную цепь различной длины, не относящуюся к аминокислотам химическую группу или другие средства, известные в данной области. Линкерный участок может представлять собой последовательность аминокислот, которая функционально связывает два функциональных или структурных домена белка или пептида.
[0053] В данной заявке термин «химиотерапевтический агент» относится к любому веществу, способному уменьшать или предотвращать рост, пролиферацию или распространение клетки рака, популяции клеток рака, опухоли или другой злокачественной ткани. Предполагается, что в объем данного термина также входит любой противоопухолевый или противораковый агент.
[0054] В данной заявке предполагается, что в объем термина «эффективное количество» входят такие контексты, как фармацевтически эффективное количество или терапевтически эффективное количество. Например, в некоторых вариантах реализации эффективное количество позволяет добиться благоприятного состояния, благоприятного исхода, функциональной активности в скрининговом анализе или улучшения клинического состояния.
[0055] В данной заявке предполагается, что в объем термина «клетка рака» («раковая клетка») входят определения, понимаемые в широком смысле в данной области. В одном варианте реализации термин относится к клетке с аномальной регуляцией, которая может способствовать клиническому состоянию рака у человека или животного. В одном варианте реализации термин может относиться к культивируемой линии клеток, или к клетке внутри организма человека или животного, или к клетке, полученной из него. Клетка рака может принадлежать к любому из большого разнообразия типов дифференцированной клетки, ткани или органа, что очевидно в данной области. Конкретные примеры клеток рака включают клетки рака молочной железы, рака толстого кишечника, рака кожи, рака яичников, лейкемии, рака легких, рака печени, рака яичек, рака пищевода и других типов рака.
[0056] В данной заявке «лечить» или «лечение» относится к подходу для получения полезных или желательных результатов, включая и предпочтительно клинические результаты. Лечение может относиться либо к снижению выраженности симптомов заболевания или состояния, либо к отсрочке прогрессирования заболевания или состояния.
[0057] В данной заявке «предотвращение» или «предотвратить» относится к подходу для предотвращения, ингибирования или уменьшения вероятности начала или рецидива заболевания или состояния. Данный термин также относится к предотвращению, ингибированию или уменьшению вероятности возникновения или рецидива симптомов заболевания или состояния, и он также включает снижение интенсивности, эффекта, симптомов и/или нагрузки от заболевания или состояния перед началом или рецидивом заболевания или состояния.
[0058] В данной заявке «ингибирование роста клеток» или «ингибирование пролиферации клеток» относится к уменьшению скорости или остановке роста клеток. Например, при ингибировании роста опухолевых клеток скорость увеличения размера опухоли может замедляться. В других вариантах реализации опухоль может оставаться такого же размера или уменьшенного размера, т.е. регрессировать. В конкретных вариантах реализации скорость роста клеток или пролиферация клеток ингибирована по меньшей мере на 20%, по меньшей мере на 30%, по меньшей мере на 40%, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 80% или по меньшей мере на 90%.
[0059] Термины «дикого типа» и «ДТ» используют взаимозаменяемо в данной заявке и применяют по отношению к TCR, последовательность аминокислот или полинуклеотид которого кодирует вариабельные области, выделенные из встречающегося в природе или немодифицированного TCR, например, первоначального или исходного клона T-клетки, обладающего специфичностью к антигену.
[0060] На фигурах и в таблицах, в которых представлены последовательности аминокислот, дикий тип обозначен «ДТ». В последовательностях, представленных ниже верхней последовательности, прочерк указывает на то, что данная аминокислота такая же, как и присутствующая в ДТ или в верхней последовательности данного выравнивания. Буква указывает на то, что в данном положении была сделана замена относительно верхней последовательности.
[0061] В данной заявке термины «модифицированный», «вариант», «мутант», «мутированный» и «производный» T-клеточный рецептор относятся к последовательностям вариабельных областей TCR, содержащим одну или более мутаций по сравнению с исходным клоном T-клетки или клоном T-клетки дикого типа. Примеры модифицированных TCR включают высокоаффинные TCR.
[0062] «Кодирующая последовательность» представляет собой часть гена или кДНК, которая кодирует последовательность аминокислот белка или функциональную РНК, такую как тРНК или рРНК.
[0063] «Комплемент» или «комплементарная последовательность» означает последовательность нуклеотидов, которая образует связанный водородными связями дуплекс с другой последовательностью нуклеотидов по правилам спаривания оснований по Уотсону-Крику.
[0064] «По ходу транскрипции» относится к относительному положению в ДНК или РНК и описывает область, расположенную ближе к 3'-концу нити.
[0065] «Экспрессия» относится к транскрипции гена в структурную РНК (рРНК, тРНК) или информационную РНК (мРНК) и последующей трансляции мРНК в белок.
[0066] Две последовательности нуклеиновых кислот гетерологичны по отношению друг к другу, если указанные последовательности получены из отдельных организмов, независимо от того, принадлежат такие организмы к различным видам или нет, при условии, что указанные последовательности не встречаются вместе в природе в одном и том же расположении в одном и том же организме.
[0067] Гомология относится к степени идентичности между двумя последовательностями нуклеотидов или аминокислот.
[0068] Последовательность аминокислот, которая функционально эквивалентна приведенной в качестве конкретного примера последовательности TCR, представляет собой последовательность аминокислот, которую модифицировали путем одной замены или множества замен аминокислот, путем вставки и/или делеции аминокислот, или последовательность, в которой одну или более аминокислот модифицировали химическим способом, но которая, тем не менее, сохраняет специфичность связывания и активность высокоаффинного связывания связанного с клеткой или растворимого белка TCR согласно настоящему описанию. Функционально эквивалентные последовательности нуклеотидов представляют собой такие последовательности, которые кодируют полипептиды, обладающие по существу такой же биологической активностью, как и у приведенного в качестве конкретного примера связанного с клеткой или растворимого белка TCR. В контексте настоящего описания в растворимом белке TCR отсутствуют части нативного связанного с клеткой TCR, и он стабилен в растворе (т.е. он, как правило, не агрегирует в растворе, если с ним обращаться, как описано в данной заявке и при стандартных условиях для белковых растворов).
[0069] Термин «изолированный» относится к композиции, соединению, веществу или молекуле, измененной рукой человека относительно природного состояния. Например, композиция или вещество, которое встречается в природе, является изолированным, если оно было изменено или удалено из исходного окружения, или и то, и другое. Например, полинуклеотид или полипептид, присутствующий в природе в живом животном, не является изолированным, но тот же самый полинуклеотид или полипептид, отделенный от материалов, присутствующих совместно с ним в его природном состоянии, является изолированным, в этом смысле данный термин используется в данной заявке.
[0070] Конструкция нуклеиновой кислоты представляет собой молекулу нуклеиновой кислоты, которую выделили из встречающегося в природе гена или которая была модифицирована таким образом, чтобы в ней содержались фрагменты нуклеиновой кислоты, которые объединяют и соединяют таким образом, который в противном случае не существовал бы в природе.
[0071] Молекула нуклеиновой кислоты означает одно- или двунитевой линейный полинуклеотид, содержащий либо дезоксирибонуклеотиды, либо рибонуклеотиды, которые соединены 3'-5'-фосфодиэфирными связями.
[0072] Две последовательности ДНК являются функционально связанными, если природа связи не препятствует способности указанных последовательностей влиять на их обычные функции по отношению друг к другу. Например, промоторная область будет функционально связана с кодирующей последовательностью, если промотор будет способен осуществлять транскрипцию данной кодирующей последовательности.
[0073] Полипептид представляет собой линейный полимер аминокислот, которые соединены пептидными связями.
[0074] Термин «промотор» относится к цис-действующей последовательности ДНК, длина которой, как правило, составляет 80 - 120 пар оснований, и которая расположена против хода транскрипции от сайта инициации транскрипции гена, с которой может связываться РНК-полимераза и инициировать правильную транскрипцию. С ней могут быть ассоциированы дополнительные регулирующие транскрипцию последовательности, которые обеспечивают регуляцию включения/выключения транскрипции и/или которые увеличивают (повышают) экспрессию расположенной по ходу транскрипции кодирующей последовательности.
[0075] Рекомбинантная молекула нуклеиновой кислоты, например, рекомбинантная молекула ДНК, представляет собой новую последовательность нуклеиновой кислоты, образованную in vitro путем лигирования двух или более негомологичных молекул ДНК (например, рекомбинантную плазмиду, содержащую одну или более вставок чужеродной ДНК, клонированной в по меньшей мере один сайт клонирования).
[0076] Термины «трансформация» и «трансфекция» относятся к направленной модификации генома клетки путем нанесения снаружи очищенной рекомбинантной ДНК из другой клетки отличного генотипа, что приводит к ее поглощению и встраиванию в геном клетки субъекта. В бактериях рекомбинантная ДНК обычно не встраивается в бактериальную хромосому, но вместо этого автономно реплицируется в виде плазмиды. Термины «трансформированный» и «трансфицированный» используют взаимозаменяемо в данной заявке. Например, T-клетку можно трансфицировать последовательностью ДНК, кодирующей модифицированный или высокоаффинный TCR, описанный в данной заявке, перед адоптивным лечением T-клетками.
[0077] «Против хода транскрипции» означает с 5'-стороны от любого сайта в ДНК или РНК.
[0078] Вектор представляет собой молекулу нуклеиновой кислоты, которая способна автономно реплицироваться в клетке-хозяине и может принимать чужеродную ДНК. Вектор содержит точку начала собственной репликации, один или более уникальных сайтов распознавания для эндонуклеаз рестрикции, которые можно использовать для встраивания чужеродной ДНК, и, обычно, маркеры селекции, такие как гены, кодирующие устойчивость к антибиотику, и часто последовательности распознавания (например, промотор) для экспрессии встроенной ДНК. Обычные векторы включают плазмидные векторы и фаговые векторы.
[0079] Высокоаффинный T-клеточный рецептор (TCR) представляет собой сконструированный TCR с более сильным связыванием с целевым лигандом, чем у TCR дикого типа. Некоторые примеры высокой аффинности включают равновесную константу связывания с целевым лигандом в диапазоне приблизительно от 10-6 М до 10-12 М и все отдельные значения и диапазоны в указанных пределах. В данный диапазон входят аффинности в пределах описанных для дикого типа аффинностей (от 10-4 до 10-6 M) и такие, которые наблюдают у TCR, выделенных с помощью направленного развития (приблизительно 10-12 М).
[0080] Цитокин представляет собой белок, пептид или гликопротеин, производимый клетками, который влияет на другие клетки.
[0081] Млекопитающее включает как человека, так и не относящихся к человеку млекопитающих.
[0082] Для специалистов в данной области должно быть очевидно, что вследствие вырожденности генетического кода множество функционально эквивалентных последовательностей нуклеотидов кодируют одну и ту же последовательность аминокислот.
T-клеточные рецепторы.
[0083] T-клеточный рецептор (TCR) состоит из двух цепей (αβ или γδ), которые соединяются друг с другом на поверхности T-клетки с образованием гетеродимерного рецептора. TCR αβ экспрессируется на большинстве T-клеток в организме, и известно, что он участвует в узнавании рестриктированных (ограниченных) по MHC антигенов. Молекулярная генетика, структура и биохимия TCR αβ на данный момент полностью исследована. Каждая из α и β цепей состоит из двух доменов: константных доменов (C), которые заякоривают белок в мембране клетки и которые связаны с инвариантными субъединицами сигнального аппарата CD3, и вариабельных доменов (V), которые дают способность распознавать антиген посредством шести петель, называемых участками, определяющими комплементарность (CDR). В каждом из V-доменов содержится три участка, определяющих комплементарность. Данные участки, определяющие комплементарность, взаимодействуют с комплексом между антигенным пептидом и белком, кодируемым главным комплексом гистосовместимости (pepMHC) (Davis и Bjorkman (1988) Nature, 334, 395-402; Davis и др. (1998) Annu Rev Immunol, 16, 523-544; Murphy (2012), xix, стр. 868).
[0084] Молекулярно-генетический анализ TCR выявил процесс генетической рекомбинации между множеством генов, которые объединяются с образованием кодирующей области V-доменов. Указанный процесс аналогичен процессу развития антител, при котором гены тяжелой и легкой цепей перестраиваются (реаранжируются) с образованием колоссального разнообразия, проявляемого продуцируемыми B-клетками антителами (Tonegawa (1988) In Vitro Cell Dev Biol, 24, 253-65). В случае T-клеток V-домен α-цепи образуется путем перестройки одной V-области (из приблизительно 75 у людей) с одним фрагментом гена слияния (J) (из приблизительно 61 у людей) (фигура 5.8, Janeway, 8ое издание). V-домен β-цепи образуется путем перестройки одной V-области (из приблизительно 52 у людей) с одним геном разнообразия (D) (из 2 у людей) и с одним фрагментом гена слияния (J) (из 13 у людей) (фигура 5.8, (Murphy (2012), xix, стр. 868)). Соединенные перестроенные гены VαJα и VβDβJβ кодируют петли CDR3 каждой цепи, и они способствуют колоссальному разнообразию TCR αβ с теоретическим пределом свыше 1015 различных TCR (Davis и Bjorkman (1988) Nature, 334, 395-402), что значительно выше достижимого разнообразия у человека, так как существует всего лишь приблизительно 1011 T-клеток (Mason (1998) Immunol Today, 19, 395-404). Возможное разнообразие CDR1 и CDR2 каждой цепи представлено количеством V-генов, так как данные петли кодируются V-геном и TCR не подвержены соматическим мутациям in vivo. Хотя разнообразие петель CDR1 и CDR2 относительно ограничено по сравнению с петлями CDR3, было показано некоторое количество примеров, в которых осуществляли селекцию определенных V-областей на основе пептидного антигена и/или продукта MHC.
[0085] Продукты MHC I класса связываются с пептидами длиной от 8 до 10 аминокислот, и они экспрессируются на всех содержащих ядро клетках в организме (обзор приведен в Rock и Goldberg (1999) Annu Rev Immunol, 17, 739-79). Тогда как вся энергия связывания взаимодействия антитело-антиген сфокусирована чужеродном антигене, существенная часть энергии связывания комплекса TCR-пептид:MHC направлена на собственную молекулу MHC (Manning и Kranz (1999) Immunology Today, 20, 417-422). В действительности, результаты недавних исследований позволили предположить, что определенные остатки петель CDR1 и/или CDR2 эволюционировали, чтобы взаимодействовать с определенными остатками на спиралях MHC, таким образом обеспечивая базовую аффинность к MHC, учитывая процесс MHC-рестрикции (Garcia и др. (2009) Nat Immunol, 10, 143-7; Marrack и др. (2008) Annu Rev Immunol, 26, 171-203).
[0086] Было интересно использовать TCR, которые обладают аффинностью к антигену пептид-MHC (I класса) выше нормального диапазона (так называемые высокоаффинные TCR), для того, чтобы: 1) запускать активность CD4+ хелперных T-клеток (в которых отутствует корецептор CD8) или 2) разработать растворимые TCR, которые можно будет применять для непосредственного нацеливания на клетку путем присоединения «эффекторной» молекулы (например, Fc-областей антитела, токсичного вещества или scFv антитела, такого как антитело к CD3, с получением биспецифического белка) (Ashfield и Jakobsen (2006) IDrugs, 9, 554-9; Foote и Eisen (2000) Proc Natl Acad Sci USA, 97, 10679-81; Holler и др. (2000) Proc Natl Acad Sci U S A, 97, 5387-92; Molloy и др. (2005) Curr Opin Pharmacol, 5, 438-43; Richman и Kranz (2007) Biomol Eng, 24, 361-73). Данный подход также позволит преодолеть проблему, с которой встречаются ракаимеющие рак пациенты, состоящую в том, что их T-клетки не экспрессируют TCR с достаточной специфичностью и аффинностью связывания с потенциальными опухолевыми антигенами (отчасти обусловленную происходящими в тимусе процессами и процессами периферической толерантности). Например, на данный момент обнаружили свыше 300 рестриктированных по MHC распознаваемых T-клетками опухолевых антигенов (cancerimmunity.org/peptide/) (Boon и Old (1997) Curr Opin Immunol, 9, 681-3; Cheever и др. (2009) Clin Cancer Res, 15, 5323-37). Данные опухолевые антигены включают мутированные пептиды, антигены дифференциации и сверхэкспрессированные антигены, каждый из которых может служить в качестве мишени для терапии. Так как большинство антигенов рака, описанных на данный момент, было получено из внутриклеточных белков, которые могут служить мишенью только на клеточной поверхности в контексте молекулы MHC, TCR представляют собой идеальных кандидатов для получения лекарственных средств, так как они эволюционировали для распознавания данного класса антигенов.
[0087] Аналогично, TCR могут обнаруживать пептиды, полученные из вирусных белков, которые естественным образом были процессированы в инфицированных клетках и выставлены молекулой MHC на поверхности клетки. За последние 25 лет обнаружили множество целевых вирусных антигенов, включая пептиды, полученные из геномов вируса иммунодефицита человека (ВИЧ) и лимфотропного Т-клеточного вируса человека (ТЛВЧ) (например, Addo и др. (2007) PLoS ONE, 2, e321; Tsomides и др. (1994) J Exp Med, 180, 1283-93; Utz и др. (1996) J Virol, 70, 843-51). Тем не менее, у пациентов с данными заболеваниями могут отсутствовать оптимальные TCR для связывания и разрушения инфицированных клеток. Наконец, возможно, что TCR можно применять в качестве рецепторных антагонистов для аутоиммунных мишеней, или в качестве агентов доставки для подавления локального клеточного иммунного ответа, в процессе, который будет высоко специфичным, позволяя избежать общего подавления иммунного ответа ((Molloy и др. (2005) Curr Opin Pharmacol, 5, 438-43; Stone и др. (2012) Protein Construction)).
Модифицированные T-клеточные рецепторы.
[0088] Направленное развитие применяли для получения TCR с более высокой аффинностью к определенным pepMHC. Применяли три различных способа дисплея, описанных далее: дрожжевой дисплей (Holler и др. (2003) Nat Immunol, 4, 55-62; Holler и др. (2000) Proc Natl Acad Sci U S A, 97, 5387-92), фаговый дисплей (Li и др. (2005) Nat Biotechnol, 23, 349-54) и T-клеточный дисплей (Chervin и др. (2008) J Immunol Methods, 339, 175-84). Во всех трех подходах процесс включал конструирование TCR, который проявляет нормальную низкую аффинность, характерную для TCR дикого типа, таким образом, чтобы мутанты TCR обладали повышенной аффинностью к определенным pepMHC (т.е. к исходному антигену, к которому специфичны T-клетки). Таким образом TCR дикого типа использовали в качестве матрицы для получения библиотек с мутациями в одном или более участков, определяющих комплементарность, после чего осуществляли селекцию мутантов с более высокой аффинностью по связыванию с узнаваемым антигеном пептид-MHC. В данной области хорошо известно, что такое направленное развитие in vitro необходимо для того, чтобы сконструировать аффинности, которые выше, чем аффинность дикого типа, более чем лишь в несколько раз.
[0089] Дрожжевой дисплей позволяет экспрессировать интересующий белок в виде слитого белка с Aga2 на поверхности клетки (Boder и Wittrup (1997) Nat. Biotech., 15, 553-557; Boder и Wittrup (2000) Methods Enzymol, 328, 430-44). Данную систему успешно применяли для конструирования высокоаффинных TCR, одноцепочечных антител, фибронектина и других белков. В системе дрожжевого дисплея TCR воспроизведен в виде стабилизированного одноцепочечного белка в формах Vβ-линкер-Vα или Vα-линкер-Vβ (Aggen и др. (2011) Protein Engineering, Design, & Selection, 24, 361-72; Holler и др. (2000) Proc Natl Acad Sci U S A, 97, 5387-92; Kieke и др. (1999) Proc Natl Acad Sci U S A, 96, 5651-6; Richman и др. (2009) Mol Immunol, 46, 902-16; Weber и др. (2005) Proc Natl Acad Sci U S A, 102, 19033-8) или в виде двухцепочечного гетеродимера (Aggen и др. (2011) Protein Engineering, Design, & Selection, 24, 361-72; Richman и др. (2009) Mol Immunol, 46, 902-16). С применением данной системы сконструировали два TCR мыши с более высокой аффинностью: 2C (рестриктированный по MHC I класса) и 3.L2 (рестриктированный по MHC II класса) (Holler и др. (2000) Proc Natl Acad Sci U S A, 97, 5387-92; Weber и др. (2005) Proc Natl Acad Sci U S A, 102, 19033-8). Также недавно были разработаны одноцепочечные фрагменты VαVβ TCR человека (называемые scTv или scTCR) с использованием преимущества исключительной стабильности области Vα человека, называемой Vα2, также известной как TCRA12 по номенклатуре IMGT (Aggen и др. (2011) Protein Engineering, Design, & Selection, 24, 361-72). В данном случае сконструированные in vitro высокоаффинные T-клеточные рецепторы в одноцепочечном формате использовали для выделения стабилизированных scTv-фрагментов человека (Vβ-линкер-Vα), которые могли экспрессироваться в виде стабильных белков как на поверхности дрожжей, так и в растворимой форме в E. coli. Указанные TCR включали два стабилизированных scTv-фрагмента человека: scTv A6, который специфичен к пептиду, полученному из белка Tax Т-лимфотропного вируса человека, и scTv 868, который специфичен к пептиду, полученному из белка Gag вируса иммунодефицита человека (пептид: SL977-85). В обоих из данных TCR использовался ген Vα2 (IMGT: семейство TRAV12), но они содержали остатки CDR3α, CDR1β, CDR2β и CDR3β, полученные из исходного клона T-клетки, из которого были выделены указанные TCR. Таким образом, каждый из мутантов данных scTCR с более высокой аффинностью был получен из исходного (первичного) TCR против узнаваемых антигенов пептид-MHC.
[0090] Во второй системе, при фаговом дисплее, интересующий белок сливали с N-концом белка вирусной оболочки (Scott и Smith (1990) Science, 249, 386-90). Различные TCR, включая TCR, называемые A6, 868 и 1G4 (рестриктированные по MHC I класса), были сконструированы для получения более высокой аффинности с применением данного способа (Li и др. (2005) Nat Biotechnol, 23, 349-54; Sami и др. (2007) Protein Eng Des Sel, 20, 397-403; Varela-Rohena и др. (2008) Nat Med, 14, 1390-5). Фаговый дисплей данных TCR стал возможен благодаря введению ненативной дисульфидной связи между двумя C доменами для того, чтобы вызвать соединение цепей α и β. Таким образом, в данной системе используются полноразмерные (VαCα/VβCβ) гетеродимерные белки, полученные из исходных клонов T-клеток для конструирования таким образом, чтобы они были направлены против узнаваемого пептида-MHC.
[0091] Третья система для конструирования TCR, которую описывали, представляла собой дисплей в клетке млекопитающего (Chervin и др. (2008) J Immunol Methods, 339, 175-84; Kessels и др. (2000) Proc Natl Acad Sci U S A, 97, 14578-83). В данной системе используют ретровирусный вектор, чтобы ввести α- и β-цепи TCR в T-клеточную гибридому, не экспрессирующую TCR. В одном исследовании (Kessels и др. (2000) Proc Natl Acad Sci U S A, 97, 14578-83) было показано, что выбранный мутантный TCR связывается с пептидом, который был очень близок по структуре к узнаваемому пептиду (ASNENMDAM по сравнению с ASNENMETM, последовательности SEQ ID NO: 15 и 16, соответственно). В другом исследовании было показано, что аффинность мутантного TCR по отношению к узнаваемому pepMHC была выше (Chervin и др. (2008) J Immunol Methods, 339, 175-84). Во многих исследованиях показали, что такие высокоаффинные TCR также проявляют более высокие аффинности по отношению к близким по структуре вариантам узнаваемого пептида (например, Holler и др. (2003) Nat Immunol, 4, 55-62). В системе дисплея в клетке млекопитающего внедренные TCR экспрессировались на поверхности в нативной конформации в комплексе с субъединицами CD3, позволяя получить полностью функциональную T-клетку (способную осуществлять передачу сигналов). Таким образом, применяя данный способ сконструировали полноразмерные гетеродимерные TCR в их природном хозяине.
Высокоаффинные TCR, которые связываются с комплексом сурвивин/HLA-A2.
[0092] Согласно настоящему изобретению предложены высокоаффинные TCR, направленные против хорошо известного антигена рака сурвивин/HLA-A2. В некоторых вариантах реализации сконструированные TCR можно применять в растворимой форме для направленной доставки in vivo или в виде рекомбинантно экспрессируемых T-клетками TCR в способе адоптивного переноса или лечении адоптивным переносом. В некотором варианте реализации одноцепочечную форму VαVβ каркаса TCR (scTCR) можно получить и применять с нагрузкой, такой как цитокин, токсин, радиоактивный изотоп, химиотерапевтический агент или лекарственное средство (аналогично конъюгатам антитела с лекарственным средством), чтобы доставить эффекторную молекулу в локализацию, с которой связывается TCR (например, в опухоль). TCR также можно применять в клеточной терапии, такой как адоптивный перенос CD4+ T-клеток, CD8+ T-клеток и/или естественных клеток-киллеров (NK), чтобы опосредовать ответ против клеток рака, которые экспрессируют сурвивин. Каркасы scTCR, предложенные в данной заявке, также можно применять для диагностики, например, злокачественных или инфицированных вирусом клеток, посредством идентификации, например, неопластических или ассоциированных с вирусом антигенов клеточной поверхности путем ковалентного связывания, например, через аминореактивные или сульфгидрил-реактивные боковые цепи аминокислот TCR, с детектируемой группой, такой как радиоактивный изотоп или флуоресцентная молекула.
[0093] В одном варианте реализации белки scTCR, описанные в данной заявке, могут быть представлены на поверхности дрожжей, фага или клеток млекопитающих, и их можно применять для конструирования TCR с еще более высокой аффинностью к антигену сурвивину. В одном варианте реализации белки scTCR, описанные в данной заявке, можно экспрессировать в прокариотической клетке, такой как Escherichia coli, Aspergillus niger, Aspergillus ficuum, Aspergillus awamori, Aspergillus oryzae, Trichoderma reesei, Mucor miehei, Kluyveromyces lactis, Pichia pastoris, Saccharomyces cerevisiae, Bacillus subtilis или Bacillus licheniformis, в клетках насекомого (например, Drosophila melanogaster), в клетках млекопитающих, включая такие линии клеток, как линии клеток яичника китайского хомячка (CHO), или в видах растений (таких как канола, соя, кукуруза, картофель, ячмень, рожь, пшеница), например, или в других известных в данной области техники источниках экспрессии белка, и получить в больших количествах. TCR также можно применять, например и исключительно в качестве примера, чтобы обнаружить специфический комплекс пептид/MHC на поверхности клетки. В одном варианте реализации описанные гены scTCR можно соединить путем применения подходящих пептидных последовательностей, кодируемых конструкцией ДНК, с генами сигнальных доменов и внедрить в T-клетки, которые могут уничтожить целевые клетки. Данные конструкции назвали химерными антигенными рецепторами (CAR), которые теперь широко применяются в данной области, включая применение CAR, которые содержат scTCR.
[0094] В предложенных одноцепочечных белках VαVβ TCR вариабельная цепь альфа и вариабельная цепь бета соединены с применением любого подходящего пептидного линкера, включая линкеры, известные в данной области, например, с помощью связей одноцепочечного Fv антитела (Bird и др. (1988) Science, 242, 423-426; Holliger и др. (1993) Proc Natl Acad Sci U S A, 90, 6444-8; Hoogenboom (2005) Nat Biotechnol, 23, 1105-16; Turner и др. (1997) J Immunol Methods, 205, 43-54). В одном варианте реализации предложен растворимый одноцепочечный TCR человека, обладающий структурой Vα-L-Vβ или Vβ-L-Vα, где L представляет собой пептидный линкер, который соединяет Vβ с Vα, Vβ представляет собой вариабельную область β TCR и Vα представляет собой вариабельную область α TCR.
[0095] В одном варианте реализации VβVα TCR называют специфичным к сурвивину K2.4.1, где Vβ представляет собой вариабельную область β TCR из группы 20 и Vα2 представляет собой вариабельную область α TCR из группы 2 (Utz, U. и др., 1996) (Aggen, D.A. и др., 2011). В одном варианте реализации VβVα TCR называют специфичным к сурвивину K2.4.6, где Vβ представляет собой вариабельную область β TCR из группы 20 и Vα2 представляет собой вариабельную область α TCR из группы 2.
[0096] В одном варианте реализации пептидный линкер содержит более чем 5 остатков лизина. В одном варианте реализации пептидный линкер содержит от 5 до 30 аминокислот. В одном варианте реализации последовательность аминокислот пептидного линкера представляет собой GSADDAKKDAAKKDGKS (SEQ ID NO: 7). В одном варианте реализации предложенный одноцепочечный VβVα TCR (sc VβVα TCR) не содержит константную область. Если термин sc VβVα TCR используется в данной заявке, то должно быть очевидно, что sc VβVα TCR также включен как термин, понимаемый и используемый в данной области. Таким образом, цепи Vβ и Vα можно соединить друг с другом в любой конфигурации посредством линкера.
[0097] В некотором аспекте настоящего описания VβVα TCR согласно настоящему описанию специфично связывается с лигандом с равновесной константой связывания KD, равной приблизительно от 10-6 М до 10-12 M. В одном варианте реализации данного аспекта настоящего описания лиганд представляет собой лиганд пептид/MHC. В одном варианте реализации VβVα TCR согласно настоящему описанию проявляет повышенную аффинность по отношению к лиганду по сравнению с аффинностями нормальных TCR дикого типа.
Биологически активные группы.
[0098] Также предложены белки VβVα TCR, описанные в данной заявке, которые содержат биологически активную группу. В данной заявке «биологически активная группа» представляет собой группу, которая вызывает измеримый или детектируемый эффект в биологической системе. В одном варианте реализации биологически активную группу выбирают из: противоопухолевого агента, такого как, но не ограничиваясь перечисленными, ингибиторы ангиогенеза, ингибиторы ферментов, ингибиторы микротрубочек, ДНК-интеркаляторы или перекрестносшивающие агенты, ингибиторы синтеза ДНК; цитокина, такого как, но не ограничиваясь перечисленными, IL-2, IL-15, GM-CSF, IL-12, TNF-α, IFN-γ или LT-α (Schrama и др. (2006) Nat Rev Drug Discov, 5, 147-59; Wong и др. (2011) Protein Eng Des Sel, 24, 373-83); противовоспалительной группы, такой как, но не ограничиваясь перечисленными, TGF-β, IL-37, IL-10 (Nold и др. (2010) Nat Immunol, 11, 1014-22; Stone и др. (2012) Protein Construction), радиоактивного изотопа, такого как, но не ограничиваясь перечисленными, 90Y или 131I (Reichert и Valge-Archer (2007) Nat Rev Drug Discov, 6, 349-56); токсина, такого как, но не ограничиваясь перечисленными, экзотоксин A Pseudomonas, дифтерийный токсин или A-цепь рицина (Pastan и др. (2006) Nat Rev Cancer, 6, 559-65; Schrama и др. (2006) Nat Rev Drug Discov, 5, 147-59); лекарственного средства или антитела, такого как одноцепочечный Fv.
[0099] В одном варианте реализации данного аспекта настоящего описания биологически активная группа представляет собой цитотоксическую молекулу, иногда называемую лекарственным средством (например, в термине «конъюгат антитела с лекарственным средством»). В данной заявке «цитотоксический» означает токсический для клеток. Примеры цитотоксических молекул включают, но не ограничены перечисленными: доксорубицин, метотрексат, митомицин, 5-фторурацил, дуокармицин, ауристатины, майтанзины, калихеамицины и аналоги перечисленных выше молекул (Jarvis (2012) Chemical and Engineering News, 90, 12-18; Litvak-Greenfeld и Benhar (2012) Adv Drug Deliv Rev; Ricart и Tolcher (2007) Nat Clin Pract Oncol, 4, 245-55). Цитотоксические молекулы не обязательно должны вызывать полную гибель клеток, а, предпочтительнее, измеримое или детектируемое ингибирование роста или снижение активности клеток.
[00100] В одном варианте реализации TCR, описанный в данной заявке, соединен с ферментом, способным превращать пролекарство в лекарство. Это полезно, например, тем, что позволяет получить активную форму лекарственного средства в локализации, на которую нацелен TCR (например, в очаге опухоли).
[00101] В одном варианте реализации биологически активная группа связана с одноцепочечным TCR посредством линкера, что можно осуществить с помощью стандартных химических реакций, например, со свободными аминогруппами или сульфгидрильными группами TCR.
[00102] В другом варианте реализации TCR присоединен к одноцепочечному фрагменту антитела (scFv) с получением биспецифического агента. Биспецифические антитела, которые содержат один scFv против опухолевого антигена и один против молекулы CD3 T-клетки, на данный момент успешно применяются в клинической практике (Bargou и др. (2008) Science, 321, 974-7). Кроме того, также был описан биспецифический агент, содержащий TCR и scFv против CD3 (Liddy и др. (2012) Nat Med, 18, 980-7).
[00103] Также предложен одноцепочечный VβVα TCR, описанный в данной заявке, который содержит детектируемую группу. В одном варианте реализации детектируемая группа представляет собой такую группу, которую можно обнаружить с помощью спектроскопических или ферментативных способов. В одном варианте реализации детектируемая группа представляет собой флуоресцентную группу, такую как, но не ограничиваясь перечисленными: флуоресцеин, R-фикоэритрин (PE), PE-Cy5, PE-Cy7, техасский красный или аллофикоцианин (APC); меченую радиоактивной меткой группу, такую как, но не ограничиваясь перечисленными: 125I, 32P, 99mTc; поглощающую группу или фермент со свойствами, которые позволяют получить детектируемые продукты, такой как, но не ограничиваясь перечисленными: пероксидаза хрена или щелочная фосфатаза.
[00104] В данной области известно, что биологически активную группу, детектируемую группу или другую группу, присоединенную к TCR, можно присоединить, применяя гибкий пептидный линкер, или путем химического соединения, и она может быть ковалентно или нековалентно присоединена к TCR.
[00105] Также в настоящей заявке предложен TCR человека для применения в способе лечения или предотвращения рака у млекопитающего, включающий введение млекопитающему эффективного количества модифицированного TCR, соединенного с терапевтически эффективной молекулой. В некотором варианте реализации млекопитающее представляет собой человека. В другом варианте реализации млекопитающее представляет собой домашнее животное (например, собаку, кошку, кролика, грызуна, лошадь) или сельскохозяйственное животное (например, корову, лошадь, свинью).
[00106] Также предложен изолированный одноцепочечный TCR (scTCR), описанный в данной заявке, и способ получения одноцепочечного TCR в E. coli. Также предложена фармацевтическая композиция, содержащая scTCR, описанный в данной заявке, и фармацевтически приемлемый носитель.
[00107] Также предложены sc VαVβ TCR, описанные в данной заявке, которые соединяли с сигнальными доменами, позволяющими получить активный TCR на поверхности T-клетки. В одном варианте реализации такой scTCR можно применять в способе лечения рака у млекопитающего, включающем: клонирование TCR в вектор, внедрение вектора в T-клетки пациента и адоптивный перенос T-клеток обратно в пациента.
Модифицированные полипептиды и полинуклеотиды TCR.
[00108] В настоящем описании предложен ДНК-вектор, который содержит по меньшей мере один фрагмент ДНК, кодирующий одноцепочечный T-клеточный рецептор (scTCR).
[00109] Специалисты в данной области посредством стандартных методик мутагенеза вместе с анализами, описанными в данной заявке, могут получить измененные последовательности TCR и проверить их на определенную аффинность и/или специфичность связывания. Полезные методики мутагенеза, известные в данной области, включают, без ограничения, синтез гена de novo, олигонуклеотид-направленный мутагенез, область-специфический мутагенез, линкер-сканирующий мутагенез и сайт-направленный мутагенез посредством ПЦР (см., например, Sambrook и др. (1989) и Ausubel и др. (1999)).
[00110] При получении модифицированных последовательностей, кодирующих TCR, для средних специалистов в данной области будет очевидно, что полученные из TCR белки можно модифицировать путем замены, вставки, делеции некоторых аминокислот и посттрансляционных модификаций, без утраты или снижения биологической активности. В частности, хорошо известно, что консервативные замены аминокислот, то есть, замена одной аминокислоты на другую аминокислоту аналогичного размера, заряда, полярности и конформации, маловероятно могут значительно изменить функцию белка. 20 стандартных аминокислот, из которых состоят белки, можно в широком смысле разделить на четыре группы консервативных аминокислот, описанных далее: неполярная (гидрофобная) группа включает аланин, изолейцин, лейцин, метионин, фенилаланин, пролин, триптофан и валин; полярная (незаряженная, нейтральная) группа включает аспарагин, цистеин, глутамин, глицин, серин, треонин и тирозин; положительно заряженная (основная) группа включает аргинин, гистидин и лизин; и отрицательно заряженная (кислотная) группа включает аспарагиновую кислоту и глутаминовую кислоту. Замена в белке одной аминокислоты на другую в пределах одной и той же группы маловероятно окажет отрицательное влияние на биологическую активность белка.
[00111] В одном варианте реализации scTCR согласно настоящему описанию может содержать дополнительные мутации в любой области или областях вариабельного домена, которые приводят к стабильности белка. В одном варианте реализации одна или более дополнительных мутаций находится в одном или более из CDR1, CDR2, HV4, CDR3, FR2 и FR3. Области, используемые для мутагенеза, можно определить с помощью направленного развития, при котором кристаллические структуры или молекулярные модели используют для получения областей TCR, которые взаимодействуют с интересующим лигандом (например, антигеном). В других примерах вариабельной области можно придать новую форму путем вставки или делеции аминокислот, чтобы создать желательное взаимодействие между scTCR и лигандом.
[00112] Полипептиды согласно настоящему изобретению включают модифицированные TCR и их антигенсвязывающие фрагменты (например, scTCR) и химерные антигенные рецепторы (CAR). Термины «полипептид», «белок», «пептид» и «гликопротеин» используют взаимозаменяемо, и они означают полимер из аминокислот, не ограниченный какой-либо конкретной длиной. Термин не исключает модификации, такие как миристилирование, сульфатирование, гликозилирование, фосфорилирование и добавление или делецию сигнальных последовательностей. Термины «полипептид» или «белок» означают одну или более цепей аминокислот, при этом каждая цепь содержит аминокислоты, ковалентно соединенные пептидными связями, и при этом указанный полипептид или белок может содержать множество цепей, нековалентно и/или ковалентно соединенных друг с другом пептидными связями, последовательность которых соответствует последовательности нативных белков, то есть, белков, продуцированных встречающимися в природе и, в частности, нерекомбинантными клетками, или сконструированными генетически или рекомбинантными клетками и включают молекулы, последовательность аминокислот которых соответствует таковой у нативного белка, или молекулы, содержащие делеции, вставки и/или замены одной или более аминокислот нативной последовательности. В объем терминов «полипептид» и «белок», в особенности, входят модифицированные TCR, или их антигенсвязывающие фрагменты, согласно настоящему описанию, или последовательности содержащие делеции, вставки и/или замены одной или более аминокислот модифицированного TCR, или его антигенсвязывающего фрагмента. Таким образом, «полипептид» или «белок» может содержать одну (называется «мономер») или множество (называется «мультимер») цепей аминокислот.
[00113] Термин «изолированный белок», используемый в данной заявке, означает, что обсуждаемый белок (1) свободен от по меньшей мере некоторых других белков, с которыми он обычно будет встречаться в природе, (2) по существу свободен от других белков из того же источника, например, из того же вида, (3) экспрессируется клеткой из другого вида, (4) отделен от по меньшей мере приблизительно 50 процентов полинуклеотидов, липидов, углеводов или других материалов, с которыми он связан в природе, (5) не связан (посредством ковалентного или нековалентного взаимодействия) с частями белка, с которым «изолированный белок» связан в природе, (6) функционально связан (посредством ковалентного или нековалентного взаимодействия) с полипептидом, с которым он не связан в природе, или (7) не встречается в природе. Такой изолированный белок может кодироваться геномной ДНК, кДНК, мРНК или другой РНК, или может быть синтетического происхождения, или подходит любая комбинация перечисленных вариантов. В некоторых вариантах реализации изолированный белок по существу свободен от белков, или полипептидов, или других примесей, которые находятся в его естественном окружении и которые будут мешать его применению (терапевтическому, диагностическому, профилактическому, исследовательскому или другому).
[00114] В конкретных вариантах реализации обсуждаемый модифицированный TCR может содержать: a) вариабельную область альфа-цепи TCR, последовательность аминокислот которой по меньшей мере на 80% идентична, по меньшей мере на 85% идентична, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 98% или 99% идентична последовательности аминокислот вариабельной области альфа-цепи модифицированного TCR, описанного в данной заявке; и b) вариабельную область бета-цепи, последовательность аминокислот которой по меньшей мере на 80% идентична, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 98% или 99% идентична последовательности аминокислот вариабельной области бета-цепи модифицированного TCR, описанного в данной заявке.
[00115] В конкретных вариантах реализации модифицированный TCR может содержать: a) вариабельную область альфа-цепи TCR, включающую: i. участок 1, определяющий комплементарность (CDR1), который идентичен по последовательности аминокислот участку CDR1 альфа-цепи прошедшего процесс селекции TCR, описанного в данной заявке; ii. участок CDR2, который идентичен по последовательности аминокислот участку CDR2 альфа-цепи прошедшего процесс селекции TCR; и iii. участок CDR3, который идентичен по последовательности аминокислот участку CDR3 альфа-цепи прошедшего процесс селекции TCR; и b) вариабельную область бета-цепи, включающую: i. участок CDR1, который идентичен по последовательности аминокислот участку CDR1 бета-цепи прошедшего процесс селекции TCR; ii. участок CDR2, который идентичен по последовательности аминокислот участку CDR2 бета-цепи прошедшего процесс селекции TCR; и iii. участок CDR3, который идентичен по последовательности аминокислот участку CDR3 бета-цепи прошедшего процесс селекции TCR; при этом указанный TCR специфично связывает прошедший процесс селекции неузнаваемый антиген. В дополнительном варианте реализации модифицированный TCR, или его антигенсвязывающий фрагмент, представляет собой вариант модифицированного TCR, при этом указанный вариант содержит альфа-цепь и бета-цепь, идентичные таковым у подвергнутого селекции модифицированного TCR за исключением вплоть до 8, 9, 10, 11, 12, 13, 14, 15 или более замен аминокислот в участках, определяющих комплементарность, областей V-альфа и V-бета. В этом отношении, может быть 1, 2, 3, 4, 5, 6, 7, 8 или, в некоторых вариантах реализации, 9, 10, 11, 12, 13, 14, 15 или более замен аминокислот в участках, определяющих комплементарность, варианта подвергнутого селекции модифицированного TCR. Замены могут быть в участках, определяющих комплементарность, в областях V-альфа и/или V-бета (см., например, Muller, 1998, Structure 6:1153-1167).
[00116] В одном варианте реализации предложен полинуклеотид, кодирующий модифицированный TCR или его антигенсвязывающий фрагмент. В других близких вариантах реализации полинуклеотид может представлять собой вариант полинуклеотида, кодирующего модифицированный TCR. Варианты полинуклеотида могут быть в значительной степени идентичны полинуклеотидной последовательности, кодирующей модифицированный TCR, описанный в данной заявке. Например, полинуклеотид может представлять собой полинуклеотид, последовательность которого по меньшей мере на 70% идентична, предпочтительно по меньшей мере на 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% или более идентична эталонной полинуклеотидной последовательности, такой как последовательность, кодирующая TCR, описанная в данной заявке, что выявили, применяя способы, описанные в данной заявке (например, применяя анализ BLAST со стандартными параметрами, описанный ниже). Для специалиста в данной области будет очевидно, что данные значения можно подходящим образом скорректировать, чтобы определить соответствующую идентичность белков, кодируемых двумя последовательностями нуклеотидов, учитывая вырожденность кодонов, подобие аминокислот, расположение рамки считывания и тому подобные параметры.
[00117] Как правило, варианты полинуклеотида будут включать одну или более замен, добавлений, делеций и/или вставок, предпочтительно таким образом, что аффинность связывания TCR, кодируемого вариантом полинуклеотида, по существу не уменьшается по сравнению с антителом, кодируемым полинуклеотидной последовательностью, конкретно описанной в данной заявке.
[00118] При сравнении полинуклеотидных последовательностей две последовательности называют «идентичными», если последовательность нуклеотидов в двух указанных последовательностях одинакова при выравнивании для достижения максимального соответствия, как описано ниже. Сравнение двух последовательностей обычно осуществляют путем сравнения последовательностей в окне сравнения, чтобы определить и сравнить локальные области подобия последовательностей. В данной заявке термин «окно сравнения» относится к фрагменту, состоящему по меньшей мере приблизительно из 20 соседних положений, обычно от 30 до приблизительно 75, от 40 до приблизительно 50, в котором последовательность можно сравнить с эталонной последовательностью такого же количества соседних положений после оптимального выравнивания двух указанных последовательностей.
[00119] Оптимальное выравнивание последовательностей для сравнения можно осуществить, применяя программу Megalign в комплекте программного обеспечения для биоинформатики Lasergene (DNASTAR, Inc., Мэдисон, Висконсин) с использованием параметров по умолчанию. Данная программа осуществляет несколько схем выравнивания, описанных в следующих ссылках: Dayhoff, M.O. (1978) A model of evolutionary change in proteins - Matrices for detecting distant relationships. In Dayhoff, M.O. (ред.) Atlas of Protein Sequence and Structure, National Biomedical Research Foundation, Вашингтон, федеральный округ Колумбия, том 5, приложение 3, стр. 345 - 358; Hein J., Unified Approach to Alignment and Phylogenes, стр. 626-645 (1990); Methods in Enzymology, том 183, Academic Press, Inc., Сан-Диего, Калифорния; Higgins, D.G. и Sharp, P.M., CABIOS 5:151-153 (1989); Myers, E.W. и Muller W., CABIOS 4:11-17 (1988); Robinson, E.D., Comb. Theor 11:105 (1971); Santou, N. Nes, M., Mol. Biol. Evol. 4:406-425 (1987); Sneath, P.H.A. и Sokal, R.R., Numerical Taxonomy - the Principles and Practice of Numerical Taxonomy, Freeman Press, Сан-Франциско, Калифорния (1973); Wilbur, W.J. и Lipman, D.J., Proc. Natl. Acad., Sci. USA 80:726-730 (1983).
[00120] В качестве альтернативы, оптимальное выравнивание последовательностей для сравнения можно осуществить, применяя алгоритм локальной идентичности Smith и Waterman, Add. APL. Math 2:482 (1981), применяя алгоритм выравнивания идентичности Needleman и Wunsch, J. Mol. Biol. 48:443 (1970), с помощью способов поиска подобия Pearson и Lipman, Proc. Natl. Acad. Sci. USA 85: 2444 (1988), с помощью компьютеризированных реализаций данных алгоритмов (GAP, BESTFIT, BLAST, FASTA и TFASTA в пакете программного обеспечения Wisconsin Genetics Software Package, Genetics Computer Group (GCG), 575 Science Dr., Мэдисон, Висконсин), или путем проверки вручную.
[00121] Один предпочтительный пример алгоритмов, которые подходят для определения процента идентичности последовательностей и подобия последовательностей, представляет собой алгоритмы BLAST и BLAST 2.0, которые описаны в Altschul и др., Nucl. Acids Res. 25:3389-3402 (1977) и Altschul и др., J. Mol. Biol. 215:403-410 (1990), соответственно. BLAST и BLAST 2.0 можно применять, например, с параметрами, описанными в данной заявке, чтобы определить процент идентичности последовательностей среди двух или более полинуклеотидов. Программное обеспечение для выполнения анализов BLAST общедоступно через National Center for Biotechnology Information. В одном пояснительном примере совокупные балльные показатели можно рассчитать, применяя, для последовательностей нуклеотидов, параметры M (балл, присваиваемый за пару совпадающих остатков; всегда > 0) и N (штрафной балл за несовпадающие остатки; всегда < 0). Совпадения слов прекращают продлевать в каждом направлении, если: суммарный балл выравнивания снижается на величину X от максимально достигнутого им значения; суммарный балл опускается до нуля или ниже, вследствие накопления при выравнивании одного или более остатков с отрицательными баллами; или достигается конец любой последовательности. Параметры W, T и X алгоритма BLAST определяют чувствительность и скорость выравнивания. В программе BLASTN (для последовательностей нуклеотидов) используются в качестве параметров по умолчанию длина слова (W), равная 11, ожидание (E), равное 10, и выравнивания матрицы подсчета баллов BLOSUM62 (см. Henikoff и Henikoff, Proc. Natl. Acad. Sci. USA 89:10915 (1989)), (B), равное 50, ожидание (E), равное 10, M = 5, N = -4 и сравнение обеих нитей.
[00122] В некоторых вариантах реализации «процент идентичности последовательностей» определяют путем сравнения двух оптимально выровненных последовательностей в окне сравнения из по меньшей мере 20 положений, при этом часть полинуклеотидной последовательности в окне сравнения может включать добавления или делеции (т.е. гэпы) в количестве 20 процентов или менее, обычно от 5 до 15 процентов или от 10 до 12 процентов по сравнению с эталонными последовательностями (которые не содержат добавления или делеции) для оптимального выравнивания двух последовательностей. Указанный процент рассчитывают путем определения количества положений, в которых идентичные основания нуклеиновой кислоты встречаются в обеих последовательностях, с получением количества совпадающих положений, деления указанного количества совпадающих положений на общее количество положений в эталонной последовательности (т.е. размер окна) и умножения полученных результатов на 100 с получением процента идентичности последовательностей.
[00123] Для средних специалистов в данной области должно быть очевидно, что в результате вырожденности генетического кода существует множество последовательностей нуклеотидов, которые кодируют TCR, описанный в данной заявке. Последовательности некоторых из данных полинуклеотидов минимально идентичны последовательности нуклеотидов нативной или исходной полинуклеотидной последовательности, которая кодирует модифицированные TCR, которые связываются, например, с тем же самым антигеном. Тем не менее, полинуклеотиды, которые отличаются вследствие различий в использовании кодонов, явно предусмотрены в настоящем описании. В некоторых вариантах реализации, в частности, предусмотрены последовательности, которые были оптимизированы по использованию кодонов для экспрессии в млекопитающем.
[00124]Стандартные методики клонирования, выделения, амплификации и очистки ДНК для ферментативных реакций с участием ДНК-лигазы, ДНК-полимеразы, эндонуклеаз рестрикции и тому подобных ферментов, и различные методики разделения представляют собой методики, известные и широко применяемые специалистами в данной области. Множество стандартных методик описано в Sambrook и др. (1989) Molecular Cloning, второе издание, Cold Spring Harbor Laboratory, Плейнвью, Нью-Йорк; Maniatis и др. (1982) Molecular Cloning, Cold Spring Harbor Laboratory, Плейнвью, Нью-Йорк; Wu (ред.) (1993) Meth. Enzymol. 218, Часть I; Wu (ред.) (1979) Meth Enzymol. 68; Wu и др. (ред.) (1983) Meth. Enzymol. 100 и 101; Grossman and Moldave (ред.) Meth. Enzymol. 65; Miller (ред.) (1972) Experiments in Molecular Genetics, Cold Spring Harbor Laboratory, Колд-Спринг-Харбор, Нью-Йорк; Old и Primrose (1981) Principles of Gene Manipulation, University of California Press, Berkeley; Schleif и Wensink (1982) Practical Methods in Molecular Biology; Glover (ред.) (1985) DNA Cloning, том I и II, IRL Press, Оксфорд, Великобритания; Hames и Higgins (ред.) (1985) Nucleic Acid Hybridization, IRL Press, Оксфорд, Великобритания; и Setlow и Hollaender (1979) Genetic Engineering: Principles and Methods, тома 1-4, Plenum Press, Нью-Йорк. Сокращения и номенклатуру, когда они используются, считают стандартными в данной области техники и общепринятыми в профессиональных журналах, таких как цитируемые в данной заявке.
[00125] Гомологию между последовательностями нуклеотидов можно определить с помощью анализа гибридизации ДНК, в котором стабильность двунитевого ДНК-гибрида зависит от наблюдаемой степени спаривания оснований. Условия высокой температуры и/или низкого содержания соли уменьшают стабильность гибрида, и их можно изменить, чтобы предотвратить отжиг последовательностей со степенью гомологии, меньшей чем выбранная. Например, для последовательностей с GC-составом, приблизительно равным 55%, условия гибридизации и промывки 40 - 50°C, 6 X SSC (буфер из хлорида натрия/цитрата натрия) и 0,1% ДСН (додецилсульфат натрия) обозначают гомологию приблизительно на 60 - 70%, условия гибридизации и промывки 50 - 65°C, 1 X SSC и 0,1% ДСН обозначают гомологию приблизительно на 82 - 97% и условия гибридизации и промывки 52°C, 0,1 X SSC и 0,1% ДСН обозначают гомологию приблизительно на 99 - 100%. Также доступен широкий диапазон компьютерных программ для сравнения последовательностей нуклеотидов и аминокислот (и измерения степени гомологии), и перечень, в котором представлены источники как доступного для приобретения, так и бесплатного программного обеспечения можно найти в Ausubel и др. (1999). Легко доступные алгоритмы сравнения последовательностей и выравнивания множества последовательностей, соответственно, представляют собой программы BLAST (основное средство поиска локального выравнивания) (Altschul и др., 1997) и ClustalW. Программа BLAST доступна в сети Интернет на сайте ncbi.nlm.nih.gov и версия ClustalW доступна на сайте www2.ebi.ac.uk.
[00126] Промышленные штаммы микроорганизмов (например, Aspergillus niger, Aspergillus ficuum, Aspergillus awamori, Aspergillus oryzae, Trichoderma reesei, Mucor miehei, Kluyveromyces lactis, Pichia pastoris, Saccharomyces cerevisiae, Escherichia coli, Bacillus subtilis или Bacillus licheniformis), виды насекомого (Drosophila), млекопитающего (например, линии клеток яичника китайского хомячка, CHO) или растений (таких как канола, соя, кукуруза, картофель, ячмень, рожь, пшеница) можно применять в качестве клеток-хозяев для рекомбинантного получения белков TCR. В некоторых вариантах реализации на первом этапе гетерологичной экспрессии высокоаффинного белка TCR или растворимого белка получают экспрессионную конструкцию, содержащую последовательность, кодирующую TCR или растворимый TCR, и контролирующие экспрессию последовательности, такие как промоторы, энхансеры и терминаторы. Также можно включить в ее состав другие последовательности, такие как сигнальные последовательности и маркеры селекции. Для достижения внеклеточной экспрессии TCR экспрессионная конструкция может содержать секреторную сигнальную последовательность. В вариантах реализации указанная сигнальная последовательность не содержится на экспрессионной конструкции, если желательна цитоплазматическая экспрессия. В вариантах реализации промотор и сигнальная последовательность способны функционировать в клетке-хозяине и обеспечивают экспрессию и секрецию TCR или экспрессию растворимого белка TCR. Можно включить терминаторы транскрипции, чтобы обеспечить эффективную транскрипцию. Вспомогательные последовательности, улучшающие экспрессию или очистку белка, также можно включить в состав экспрессионной конструкции.
[00127] Согласно настоящему описанию можно применять различные промоторы (регуляторные области инициации транскрипции). Выбор подходящего промотора может зависеть от предложенного для экспрессии хозяина. Можно применять промоторы из гетерологичных источников, при условии, что они способны функционировать в выбранном хозяине.
[00128] Выбор промотора также зависит от желательной эффективности и уровня продукции пептида или белка. Индуцируемые промоторы, такие как tac, часто используют для того, чтобы резко увеличить уровень экспрессии белка в E. coli. Сверхэкспрессия белков может быть губительной для клеток-хозяев. Следовательно, рост клеток-хозяев может быть ограничен. Применение системы индуцируемого промотора позволяет культивировать клетки-хозяева до приемлемой плотности перед индукцией экспрессии гена, тем самым способствуя более высоким выходам продукта.
[00129] Согласно настоящему описанию можно применять различные сигнальные последовательности. Можно применять сигнальную последовательность, которая гомологична последовательности, кодирующей TCR. В качестве альтернативы, также можно применять сигнальную последовательность, которую выбрали или разработали для эффективной секреции и процессинга в хозяине, в котором осуществляют экспрессию. Например, подходящие пары сигнальная последовательность/клетка-хозяин включают B. subtilis/сигнальная последовательность sacB для секреции в B. subtilis и α-фактор спаривания Saccharomyces cerevisiae или сигнальные последовательности кислой фосфатазы phoI P. pastoris для секреции в P. pastoris. Сигнальную последовательность можно соединить напрямую через последовательность, кодирующую сайт расщепления сигнальной пептидазой, с кодирующей белок последовательностью, или через короткий нуклеотидный мостик, состоящий, как правило, менее чем из десяти кодонов, при этом указанный мостик обеспечивает правильную рамку считывания расположенной по ходу транскрипции последовательности TCR.
[00130] Элементы для улучшения транскрипции и трансляции были обнаружены для системы экспрессии эукариотических белков. Например, расположение промотора вируса мозаики цветной капусты (CaMV) в 1000 п.о. с любой стороны от гетерологичного промотора может повысить уровни транскрипции в 10 - 400 раз в клетках растения. Экспрессионная конструкция также должна содержать подходящие последовательности инициации трансляции. Модификация экспрессионной конструкции таким образом, чтобы она включала консенсусную последовательность Козак для правильной инициации трансляции, может повысить уровень трансляции в 10 раз.
[00131] Часто используют селективный маркер, который может быть частью экспрессионной конструкции или может быть отдельным от нее (например, его может нести вектор экспрессии), так что указанный маркер может встраиваться в сайт, отличный от сайта встраивания интересующего гена. Примеры включают маркеры, которые придают устойчивость к антибиотикам (например, bla придает устойчивость к ампициллину клеткам-хозяевам E. coli, nptII придает устойчивость к канамицину большому разнообразию прокариотических и эукариотических клеток), или которые позволяют хозяевам расти на минимальной среде (например, HIS4 позволяет P. pastoris или His- S. cerevisiae расти в отсутствие гистидина). У селектируемого маркера есть собственные регуляторные области инициации и терминации транскрипции и трансляции, чтобы обеспечить возможность независимой экспрессии маркера. Если в качестве маркера используется устойчивость к антибиотику, то концентрация антибиотика для селекции будет изменяться в зависимости от конкретного антибиотика, как правило в диапазоне от 10 до 600 мкг антибиотика/мл среды.
[00132] Сборку экспрессионной конструкции осуществляют с применением известных технологий рекомбинантных ДНК (Sambrook и др., 1989; Ausubel и др., 1999). Расщепление ферментами рестрикции и лигирование представляют собой основные этапы, используемые для соединения двух фрагментов ДНК. Может потребоваться модификация концов фрагмента ДНК перед лигированием, и этого можно добиться путем заполнения «липких» концов, удаления концевых участков фрагмента (фрагментов) с помощью нуклеаз (например, ExoIII), путем сайт-направленного мутагенеза или путем добавления новой пары оснований с помощью ПЦР. Полилинкеры и адапторы можно использовать для более легкого соединения выбранных фрагментов. Сборку экспрессионной конструкции обычно осуществляют на стадиях с применением раундов рестрикции, лигирования и трансформации E. coli. Множество клонирующих векторов, подходящих для конструирования экспрессионной конструкции, известно в данной области (λZAP и pBLUESCRIPT SK-1, Stratagene, Ла-Холья, Калифорния; pET, Novagen Inc., Мэдисон, Висконсин - цитированные в Ausubel и др., 1999), и конкретный их выбор не важен для настоящего описания. На выбор клонирующего вектора будет влиять система переноса генов, выбранная для внедрения экспрессионной конструкции в клетку-хозяина. По окончании каждой стадии полученную конструкцию можно исследовать с помощью анализов рестрикции, последовательности ДНК, гибридизации и ПЦР.
[00133] Хозяина можно трансформировать экспрессионной конструкцией в виде конструкции клонирующего вектора, либо линейной, либо кольцевой, или можно удалить из клонирующего вектора и использовать как есть, или вставить в вектор доставки. Вектор доставки облегчает внедрение и сохранение экспрессионной конструкции в выбранном типе клетки-хозяина. Экспрессионную конструкцию внедряют в клетки-хозяева с помощью любой из множества известных систем переноса генов (например, естественной компетенции, трансформации, опосредованной химическим соединением, трансформации протопластов, электропорации, биолистической трансформации, трансфекции или конъюгации) (Ausubel и др., 1999; Sambrook и др., 1989). Выбранная система переноса генов зависит от используемых клеток-хозяев и векторных систем.
[00134] Например, экспрессионную конструкцию можно внедрить в клетки S. cerevisiae путем трансформации протопластов или электропорации. Электропорацию S. cerevisiae легко осуществить, и она позволяет добиться эффективности трансформации, сравнимой с таковой для трансформации сферопластов.
[00135] Моноклональные или поликлональные антитела, предпочтительно моноклональные, специфически реагирующие с белком TCR в сайте, отличном от сайта связывания лиганда, можно получить с помощью способов, известных в данной области, и многие из них доступны для приобретения. См., например, Harlow и Lane (1988) Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratories; Goding (1986) Monoclonal Antibodies: Principles and Practice, 2ое изд., Academic Press, Нью-Йорк; и Ausubel и др. (1999) Current Protocols in Molecular Biology, John Wiley & Sons, Inc., Нью-Йорк.
[00136] TCR в связанной с клеткой или растворимой форме, которые специфичны к определенному целевому лиганду, полезны, например, в качестве диагностических зондов для скрининга биологических образцов (таких как клетки, образцы ткани, материал биопсии, физиологические жидкости и тому подобные образцы) или для обнаружения присутствия целевого лиганда в тестируемом образце. Часто TCR метят путем присоединения, либо ковалентно, либо нековалентно, вещества, которое дает детектируемый сигнал. Подходящие метки включают, но не ограничены перечисленными: радионуклиды, ферменты, субстраты, кофакторы, ингибиторы, флуоресцентные агенты, хемилюминестцентные агенты, магнитные частицы и тому подобные метки. Дополнительно TCR может быть соединен с лигандом для второй связывающей молекулы: например, TCR может быть биотинилирован. Обнаружение TCR, связанного с целевой клеткой или молекулой, затем можно осуществить путем связывания с детектируемым стрептавидином (стрептавидином, к которому присоединена флуоресцентная, радиоактивная, хемилюминестцентная или другая детектируемая молекула, или к которому присоединен фермент, для которого есть доступный хромофорный субстрат). Патенты Соединенных Штатов, описывающие применение таких меток и/или токсичных соединений для ковалентного связывания с scTCR, включают, но не ограничены перечисленными патентами с номерами: 3817837; 3850752; 3927193; 3939350; 3996345; 4277437; 4275149; 4331647; 4348376; 4361544; 4468457; 4444744; 4640561; 4366241; RE 35500; 5299253; 5101827; 5059413.
[00137] Меченные TCR можно обнаружить, применяя контролирующее устройство или способ, подходящие для используемой метки. Можно применять флуоресцентную микроскопию или сортировку клеток с возбуждением флуоресценции, если метка представляет собой флуоресцентную молекулу, и если метка представляет собой радионуклид, то, например, можно применять гамма-радиометрию, ауторадиографию или жидкостной сцинтилляционный счет, при условии, что данный способ подходит для анализируемого образца и используемого радионуклида. Кроме того, можно использовать вторичные детектируемые молекулы или частицы в тех случаях, когда детектируемая молекула или частица узнает часть TCR, которая не является частью сайта связывания целевого лиганда в отсутствие компонента MHC, что отмечали в данной заявке. В данной области известны полезные соединения для диагностической визуализации in situ; см., например, патенты США № 5101827; 5059413. Радионуклиды, полезные для терапии и/или визуализации in vivo, включают 111индий, 97рубидий, 125йод, 131йод, 123йод, 67галлий, 99технеций. Токсины включают дифтерийный токсин, рицин и токсин клещевины обыкновенной, среди прочих, при условии, что после того, как комплекс TCR-токсин связался с клеткой, токсичная молекула интернализируется, чтобы она могла оказывать цитотоксическое действие. Методика иммунотоксинов хорошо известна в данной области, и подходящие токсичные молекулы включают, без ограничения: химиотерапевтические средства, такие как виндезин, антифолаты, например, метотрексат, цисплатин, митомицин, антрациклины, такие как дауномицин, даунорубицин или адриамицин, и цитотоксические белки, такие как рибосом-инактивирующие белки (например, дифтерийный токсин, противовирусный белок лаконоса, абрин, рицин, экзотоксин A Pseudomonas), или рекомбинантные производные перечисленных токсинов. См., как правило, например, Olsnes и Pihl (1982) Pharmac. Ther. 25:355 - 381 и Monoclonal Antibodies for Cancer Detection and Therapy, ред. Baldwin и Byers, стр. 159 - 179, Academic Press, 1985.
[00138] Общая структура молекул TCR и способы их получения и применения, включая связывание с комплексом пептид:главный комплекс гистосовместимости, были описаны. См., например, PCT/US98/04274; PCT/US98/20263; WO99/60120.
Фармацевтические композиции и терапевтические агенты.
[00139] TCR, специфичные к конкретному целевому лиганду, полезны для лечения животных и млекопитающих, включая людей, которых считают страдающими от заболевания, связанного с конкретным антигеном, например, от неопластического заболевания или расстройства, такого как рак. Примеры типов рака, которые можно лечить согласно способам, описанным в данной заявке, включают, но не ограничены перечисленными: опухоль Вильмса, рак мочевого пузыря, рак молочной железы, рак толстого кишечника, колоректальный рак, карциномы пищевода, рак желудка, печеночно-клеточную карциному, рак почки, лейкемию, рак печени, рак легких, лимфому, меланому, нейробластому, немелкоклеточную карциному легкого, пероральный рак, остеосаркому, рак яичников, рак поджелудочной железы, рак предстательной железы, рак почки, рак кожи, мелкоклеточную карциному легкого и рак яичек.
[00140] Терапевтические продукты можно получить, применяя материалы, показанные в данной заявке. Эффективные количества терапевтических продуктов представляют собой минимальную дозу, которая вызывает измеримый эффект у субъекта. Терапевтические продукты легко сможет получить средний специалист в данной области. В одном варианте реализации TCR согласно настоящему описанию вводят непосредственно пациенту. В одном варианте реализации TCR согласно настоящему описанию соединяют с ПЭГ или с константными областями иммуноглобулина, как известно в данной области. Данный вариант реализации позволяет удлиннить клиренс из сыворотки. В одном варианте реализации TCR соединен с химиотерапевтическим агентом или лекарственным средством для того, чтобы доставить лекарственное средство в целевую клетку, такую как клетка рака. В одном варианте реализации TCR соединен с биологической эффекторной молекулой, такой как цитокин (Tayal и Kalra (2008) Eur J Pharmacol, 579, 1-12). В одном варианте реализации TCR соединен с цитокином, обладающим противоопухолевой активностью, таким как IL-2, IL-12 или TNFα (Wong и др. (2011) Protein Eng Des Sel, 24, 373-83). В одном варианте реализации scTCR соединен с иммуноингибирующим цитокином, таким как IL-10 или IL-13 (Stone и др. (2012) Protein Construction). В одном варианте реализации TCR соединен с другой антиген-связывающей молекулой с получением биспецифического агента (Miller и др. (2010) Protein Eng Des Sel, 23, 549-57; Thakur и Lum (2010) Curr Opin Mol Ther, 12, 340-9). В одном варианте реализации указанная биспецифическая молекула состоит из TCR, соединенного с одноцепочечным Fv, таким как направленный против CD3 ((Bargou и др. (2008) Science, 321, 974-7; Liddy и др. (2012) Nat Med, 18, 980-7), чтобы перекрестно связать T-клетки и больные клетки. В одном варианте реализации TCR соединен с сигнальными доменами TCR, такими как CD3, с получением химерного антигенного рецептора ((Porter и др. (2011) N Engl J Med, 365, 725 - 33; Sadelain и др. (2009) Curr Opin Immunol, 21, 215-23; Stroncek и др. (2012) J Transl Med, 10, 48). Данные способы и другие способы введения, такие как внутривенное введение, известны в данной области. Полезные дозировки может определить средний специалист в данной области.
[00141] Композиции TCR можно составить с помощью любых средств, известных в данной области. Их обычно можно получить в виде лекарственных форм для инъекций, особенно для внутривенного, интраперитонеального или синовиального введения (при этом путь определяют с учетом конкретного заболевания), или в виде лекарственных форм для интраназального или перорального введения, в виде либо жидких растворов, либо суспензий. Также можно получить твердые формы, подходящие для растворения или суспендирования в жидкости перед инъекцией или другим введением. Композиция также, например, может быть эмульгированной, или белок (белки)/пептид (пептиды) могут быть инкапсулированы в липосомы.
[00142] Активные ингредиенты часто смешивают с необязательными фармацевтическими вспомогательными веществами, такими как наполнители или носители, которые фармацевтически приемлемы и совместимы с активным ингредиентом. Подходящие вспомогательные вещества включают, но не ограничены перечисленными: воду, солевой раствор, декстрозу, глицерин, этанол или тому подобные вещества и комбинации перечисленных веществ. Концентрация scTCR в инъецируемых, аэрозольных или назальных лекарственных формах обычно находится в диапазоне от 0,05 до 5 мг/мл. Выбор конкретных эффективных дозировок известен, и его осуществляет без лишних экспериментов средний специалист в данной области. Аналогичные дозировки можно вводить в другие поверхности слизистых оболочек.
[00143] Кроме того, при желании, вакцины, которые могут включать scTCR, могут содержать незначительные количества фармацевтических вспомогательных веществ, таких как наполнители, такие как смачивающие или эмульгирующие агенты, pH-буферные вещества и/или адъюванты, которые повышают эффективность вакцины. Примеры адъювантов, которые могут быть эффективны, включают, но не ограничены перечисленными: гидроксид алюминия; N-ацетилмурамил-L-треонил-D-изоглутамин (thr-MDP); N-ацетил-нор-мурамил-L-аланил-D-изоглутамин (CGP 11637, называемый nor-MDP); N-ацетилмурамил-L-аланил-D-изоглутаминил-L-аланин-2-(1'-2'-дипальмитоил-sn-глицеро-3-гидроксифосфорилокси)-этиламин (CGP 19835A, называемый MTP-PE); и RIBI, который содержит три компонента, выделенных из бактерий: монофосфорил-липид А, димиколят трегалозы и каркас клеточной стенки (MPL+TDM+CWS) в 2% эмульсии сквален/Tween® 80. Такие дополнительные лекарственные формы и способы введения, известные в данной области, также можно применять.
[00144] TCR согласно настоящему описанию и/или связывающие фрагменты, первичная структура которых сходна (идентична более чем на 90%) с вариабельными областями TCR и которые сохраняют высокую аффинность к целевому лиганду, могут входить в состав вакцин в виде нейтральных или солевых форм. Фармацевтически приемлемые соли включают, но не ограничены перечисленными, соли присоединения кислоты (образованные со свободными аминогруппами пептида), которые образованы с неорганическими кислотами, например, соляной кислотой или фосфорными кислотами; и органическими кислотами, например, уксусной, щавелевой, винной или малеиновой кислотой. Соли, образованные со свободными карбоксильными группами, также можно получить из неорганических оснований, например, гидроксидов натрия, калия, аммония, кальция или железа, и органических оснований, например, изопропиламина, триметиламина, 2-этиламиноэтанола, гистидина и прокаина.
[00145] TCR для применения в терапии вводят способом, подходящим для дозированной лекарственной формы, и в таком количестве и таким способом, которые профилактически и/или терапевтически эффективны, в соответствии со знаниями в данной области. Количество, которое следует вводить, как правило, находится в диапазоне приблизительно от 100 до 20000 мкг белка на дозу, чаще в диапазоне приблизительно от 1000 до 10000 мкг белка на дозу. Сходные композиции можно вводить аналогичными путями, применяя меченые TCR для применения для визуализации, например, чтобы обнаружить клетки, с которыми связан целевой лиганд. Точные количества активного ингредиента, которые необходимо ввести, могут зависеть от решения врача или ветеринара, и могут быть особыми для каждого индивида, но такое определение находится в рамках компетенции такого практикующего врача.
[00146] Продукт TCR можно вводить в виде разовой дозы; в рамках режима введения двух доз, например, с интервалами от двух до восьми недель; или в рамках режима введения множества доз. Режим введения множества доз представляет собой такой режим, при котором первичный курс лечения может включать от 1 до 10 или более отдельных доз, после чего вводят другие дозы с последующими интервалами времени, при необходимости, для поддержания и/или усиления ответа.
[00147] Каждую лекарственную форму или комбинацию компонентов, описанную или приведенную в качестве примера, можно применять для осуществления настоящего изобретения, если не указано иное. Предполагается, что конкретные названия веществ приведены в качестве примера, так как известно, что средний специалист в данной области может назвать одно и то же вещество различными названиями. Если соединение описано в данной заявке таким образом, что конкретный изомер или энантиомер соединения не указан, например, в формуле или в химическом наименовании, то предполагается, что настоящее описание включает каждый изомер и энантиомер описанного соединения, отдельно или в любой комбинации. Для среднего специалиста в данной области должно быть очевидно, что способы, целевые лиганды, биологически активные группы, исходные материалы и способы синтеза, отличные от приведенных в качестве конкретного примера, можно применять для осуществления настоящего изобретения, не прибегая к лишним экспериментам. Предполагается, что все известные в данной области техники функциональные эквиваленты любых таких способов, целевых лигандов, биологически активных групп, исходных материалов и способов синтеза включены в настоящее описание. Всякий раз, когда в настоящем описании приведен диапазон, например, диапазон температуры, диапазон времени или диапазон композиций, предполагается, что все промежуточные диапазоны и поддиапазоны, а также все отдельные значения, включенные в указанные диапазоны, включены в настоящее описание.
[00148] Конкретную лекарственную форму, путь введения и дозировку может выбрать лечащий врач с учетом состояния пациента (см., например, Fingl и др., в The Pharmacological Basis of Therapeutics, 1975, глава 1, стр. 1).
[00149] Следует отметить, что лечащий врач будет знать, как и когда закончить, прервать или скорректировать введение вследствие токсичности или функциональной недостаточности органа. С другой стороны, лечащий врач будет знать, как скорректировать лечение путем введения более высоких уровней лекарственного средства, если клинический ответ был недостаточным (исключая токсичность). Размер вводимой дозы для лечения интересующего расстройства будет изменяться в зависимости от тяжести состояния, от которого лечат, и от пути введения. Тяжесть состояния можно, например, оценить отчасти с помощью стандартных прогностических способов оценки. Кроме того, доза и, возможно, частота введения доз также будет изменяться в зависимости от возраста, массы тела и реакции на лечение конкретного пациента. Программу, сравнимую с обсуждаемой выше, также можно применять в ветеринарии.
[00150] В зависимости от конкретных состояний, которые лечат, и выбранного способа нацеливания такие агенты можно включить в состав формы для системного или местного введения и вводить соответствующим способом. Методики получения и введения лекарственных форм можно найти в Alfonso и Gennaro (1995). Подходящие пути могут включать, например, пероральное, ректальное, трансдермальное, вагинальное, чреcслизистое введение или введение через кишечник; парентеральную доставку, включая внутримышечную, подкожную или интрамедуллярную инъекции, а также интратекальные, внутривенные или интраперитонеальные инъекции.
[00151] Для инъекции агенты согласно настоящему описанию можно включить в состав водных растворов, предпочтительно в физиологически совместимых буферах, таких как раствор Хенкса, раствор Рингера или физиологический раствор. Для введения через слизистую в лекарственной форме используют проникающие вещества, подходящие для барьера, через который нужно проникнуть. Такие проникающие вещества общеизвестны в данной области.
[00152] Применение фармацевтически приемлемых носителей для составления лекарственных форм соединений, описанных в данной заявке, для осуществления настоящего изобретения, в виде дозировок, подходящих для системного введения, входит в объем настоящего описания. При правильном выборе носителя и подходящей производственной практике композиции согласно настоящему описанию, в частности, составленные в виде растворов, можно вводить парентерально, например, путем внутривенной инъекции. Подходящие соединения можно легко включить в состав лекарственной формы, применяя фармацевтически приемлемые носители, хорошо известные в данной области, в виде дозировок, подходящих для перорального введения. Такие носители позволяют включить соединения согласно настоящему описанию в состав лекарственной формы в виде таблеток, пилюль, капсул, жидкостей, гелей, сиропов, взвесей, суспензий и тому подобных форм, для перорального приема пациентом, которого лечат.
[00153] Агенты, предназначенные для внутриклеточного введения, можно вводить, применяя методики, хорошо известные средним специалистам в данной области. Например, такие агенты можно инкапсулировать (заключить) в липосомы, а затем вводить, как описано выше. Липосомы представляют собой сферические липидные бислои с водным внутренним пространством. Все молекулы, присутствующие в водном растворе в момент образования липосомы, заключаются в водное внутреннее пространство. Содержимое липосомы защищено от внешнего микроокружения, а также, поскольку липосомы сливаются с мембранами клеток, эффективно доставляется в цитоплазму клетки. Кроме того, благодаря их гидрофобности, малые органические молекулы можно непосредственно вводить внутрь клетки.
[00154] Фармацевтические композиции, подходящие для применения в настоящем описании, включают композиции, в которые активные ингредиенты включены в количестве, эффективном для достижения предполагаемой цели. Определение эффективных количеств находится в рамках компетенции специалистов в данной области, особенно в свете подробного описания, предложенного в данной заявке.
[00155] Дополнительно к активным ингредиентам данные фармацевтические композиции могут содержать подходящие фармацевтически приемлемые носители, включающие наполнители и вспомогательные вещества, которые облегчают обработку активных соединений с получением композиций, которые можно применять в фармацевтике. Композиции, составленные для перорального введения, могут быть представлены в форме таблеток, драже, капсул или растворов, включая формы, составленные для отсроченного высвобождения, или для высвобождения, только когда фармацевтическая композиция достигнет тонкого или толстого кишечника.
[00156] Фармацевтические композиции согласно настоящему описанию можно производить способом, который сам по себе известен, например, с помощью процессов обычного смешивания, растворения, гранулирования, получения драже, растирания в порошок, эмульгирования, инкапсулирования, захвата или лиофилизации.
[00157] Фармацевтические композиции для парентерального введения включают водные растворы активных соединений в водорастворимой форме. Кроме того, суспензии активных соединений можно получить в виде подходящих маслянистых суспензий для инъекции. Подходящие липофильные растворители или носители включают жирные масла, такие как кунжутное масло, или синтетические сложные эфиры жирных кислот, такие как этилолеат или триглицериды, или липосомы. Водные суспензии для инъекций могут содержать вещества, которые повышают вязкость суспензии, такие как карбоксиметилцеллюлоза натрия, сорбит или декстран. Необязательно суспензия также может содержать подходящие стабилизаторы или агенты, которые повышают растворимость соединений, чтобы обеспечить возможность получения высококонцентрированных растворов.
[00158] Фармацевтические составы для перорального применения можно получить путем объединения активного соединения с твердым вспомогательным веществом, необязательно перемалывания полученной смеси и обработки смеси гранул, после добавления подходящих вспомогательных веществ, при желании, с получением таблеток или седцевин драже. Подходящие вспомогательные вещества, в частности, представляют собой наполнители, такие как сахара, включая лактозу, сахарозу, маннит или сорбит; препараты целлюлозы, такие как, например, кукурузный крахмал, пшеничный крахмал, рисовый крахмал, картофельный крахмал, желатин, трагакантовая камедь, метилцеллюлоза, гидроксипропилметилцеллюлоза, карбоксиметилцеллюлоза натрия и/или поливинилпирролидон (ПВП). При необходимости можно добавить дезинтегрирующие агенты, такие как перекрестно-сшитый поливинилпирролидон, агар или альгиновая кислота или ее соль, такая как альгинат натрия.
[00159] Седцевины драже предусмотрены с подходящими покрытиями. Для данной цели можно применять концентрированные сахарные растворы, которые могут необязательно содержать гуммиарабик, тальк, поливинилпирролидон, карбополовый гель, полиэтиленгликоль, и/или диоксид титана, растворы лака и подходящие органические растворители или смеси растворителей. В покрытия таблеток или драже можно добавить красители или пигменты для опознавания или для того, чтобы отличать различные комбинации доз активного соединения.
[00160] Фармацевтические составы, которые можно применять перорально, включают твердые капсулы, сделанные из желатина, а также мягкие запаянные капсулы, сделанные из желатина и пластификатора, такого как глицерин или сорбит. Твердые капсулы могут содержать активные ингредиенты в смеси с наполнителем, таким как лактоза, связующими веществами, такими как крахмалы, и/или лубрикантами, такими как тальк или стеарат магния, и необязательно стабилизаторами. В мягких капсулах активные соединения могут быть растворены или суспендированы в подходящих жидкостях, таких как жирные масла, жидкий парафин или жидкие полиэтиленгликоли. Кроме того, можно добавить стабилизаторы.
Способы лечения.
[00161] Высокоаффинные TCR и фармацевтические композиции, содержащие высокоаффинный TCR, можно применять, например, для лечения пациента, имеющего рак, опухоль, злокачественное новообразование или неопластическое заболевание или расстройство. В одном варианте реализации способ лечения пациента, имеющего рак, включает введение высокоаффинного TCR, описаного в данной заявке. В другом варианте реализации высокоаффинный TCR специфичен к сурвивину. В одном варианте реализации TCR содержит Vα, включающую последовательность аминокислот SEQ ID NO: 1. В другом варианте реализации TCR содержит Vα, включающую последовательность аминокислот SEQ ID NO: 2. В одном варианте реализации высокоаффинный TCR представляет собой одноцепочечный TCR, включающий последовательность аминокислот SEQ ID NO: 3. В другом варианте реализации высокоаффинный TCR представляет собой одноцепочечный TCR, включающий последовательность аминокислот SEQ ID NO: 4. В другом варианте реализации высокоаффинный TCR вводят в комбинации с терапевтическим агентом, например, химиотерапевтическим агентом. В еще одном варианте реализации высокоаффинный TCR конъюгирован с биологически активной группой.
[00162] В другом аспекте настоящего изобретения предложен способ адоптивного переноса T-клеток пациенту, нуждающемуся в этом, включающий введение T-клеток, которые экспрессируют высокоаффинный TCR, описанный в данной заявке. В одном варианте реализации T-клетки были трансфицированы полинуклеотидом, который кодирует высокоаффинный TCR, который специфичен к сурвивину. В одном варианте реализации TCR содержит Vα, включающую последовательность аминокислот SEQ ID NO: 1. В другом варианте реализации TCR содержит Vα, включающую последовательность аминокислот SEQ ID NO: 2. В одном варианте реализации высокоаффинный TCR представляет собой одноцепочечный TCR, включающий последовательность аминокислот SEQ ID NO: 3. В одном варианте реализации высокоаффинный TCR представляет собой одноцепочечный TCR, включающий последовательность аминокислот SEQ ID NO: 4.
ПРИМЕРЫ
[00163] В следующих примерах далее описаны лишь некоторые из примеров настоящего описания.
ПРИМЕР 1
Конструирование TCR с более высокой аффинностью к антигенам пептид/HLA-A2
[00164] Общая стратегия, используемая для обнаружения или получения одноцепочечных TCR с улучшенной аффинностью и стабильностью, показана на фигуре 1. Процесс включает шесть описанных далее этапов.
[00165] 1) Клонирование генов Vα и Vβ TCR из клона T-клетки, который узнает рестриктированный по MHC интересующий антигенный пептид, в одноцепочечный формат TCR для дисплея. В настоящем изобретении гены TCR из клона одной T-клетки человека, которая вступала в реакцию с антигеном сурвивином (от Delores Schendel, Thomas Blankenstein и Wolfgang Uckert; см., например, Leisegang и др. (2010) J Clin Invest. 120(11), 3869), клонировали в виде одноцепочечного формата (Vβ-линкер-Vα) и внедряли в вектор дрожжевого дисплея для экспрессии на поверхности дрожжей. Дополнительное описание TCR дикого типа, который вступал в реакцию с антигеном сурвивином, см. в US 2012/0128704.
[00166] 2) Создание библиотеки, полученной с помощью допускающей ошибки полимеразы, и селекция стабилизированных вариантов с применением FACS или магнитных гранул с помощью антитела к Vβ. Так как одноцепочечные Vα и Vβ TCR часто нестабильны вследствие утраты стабилизирующих константных областей, получают библиотеки путем мутагенеза с помощью допускающей ошибки полимеразы для селекции стабилизирующих мутаций, которые позволяют стабильную экспрессию на поверхности дрожжей, хотя можно применять другие форматы дисплея, включая, но не ограничиваясь перечисленными: фаговый дисплей и дисплей на клетках млекопитающих. Векторы фагового дисплея и клонирование позволили получить библиотеку размером 1011, тогда как векторы дрожжевого дисплея и этапы гомологичной рекомбинации позволили получить библиотеку размером 1010 ((Benatuil и др. (2010) Protein Eng Des Sel, 23, 155-9). Для селекции вариантов применяли различные способы, включая связывание на основе аффинности с иммобилизованными лигандами (фаговый дисплей), или селекцию на магнитных частицах с помощью антигенов (дрожжевой дисплей), или сортировку клеток с возбуждением флуоресценции с помощью антигенов меченый пептид-MHC (дрожжевой дисплей). Используя антитела против Vβ TCR, которые распознают свернутые эпитопы, в настоящем примере применяли сортировку клеток с возбуждением флуоресценции (FACS) или селекцию на магнитных гранулах для выделения вариантов с улучшенным связыванием антитела.
[00167] 3) Оценивали термическую стабильность клонов scTCR, выделенных в результате селекции из библиотеки, полученной с помощью допускающей ошибки полимеразы, и стабилизированный вариант выбирали в качестве матрицы для созревания аффинности и секвенировали. Обычно обнаруживают мутации в одном сайте, которые способствуют повышенным уровням на поверхности дрожжей и большей стабильности в растворе.
[00168] 4) Стабилизированные последовательности scTCR использовали в качестве матрицы для создания библиотек участков, определяющих комплементарность, обычно, в участках CDR1α, CDR3α, CDR3β, хотя также можно использовать другие участки, включая, но не ограничиваясь перечисленными: CDR1β, CDR2α, CDR2β и HV4. В настоящем описании воспроизведенные в дрожжах варианты подвергают селекции из библиотек участков, определяющих комплементарность, для выявления улучшенного связывания с комплексом пептид:MHC, применяя селекцию на магнитных гранулах и/или сортировку клеток с возбуждением флуоресценции (FACS), хотя можно применять селекцию с помощью других способов, включая, но не ограничиваясь перечисленными: пэннинг с фаговым дисплеем, или селекцию на магнитных гранулах, или FACS с дисплеем на клетках млекопитающего.
[00169] 5) Оценивали специфическое связывание клонов scTCR, выделенных путем селекции из библиотек участков, определяющих комплементарность, с комплексом пептид:MHC, против которого их сконструировали. Плазмиды высвобождали из дрожжевых клонов и секвенировали.
[00170] 6) Если потребуется дополнительное улучшение аффинности, то клон scTCR, выбранный на этапе 5, можно применять в качестве матрицы для создания дополнительных библиотек в других петлях или участках, в которых мутации не подвергали селекции, таких как CDR1α, CDR3α, CDR3β, хотя другие участки, включая, но не ограничиваясь перечисленными: CDR1β, CDR2α, CDR2β и HV4, также можно использовать. Примеры каждого из данных этапов дополнительно описаны ниже.
ПРИМЕР 2
Анализ TCR A6 человека, в котором используют Vα2 в комплексе с TAX:HLA.A2
[00171] Все TCR имеют сходную укладку Ig и сходный угол присоединения, и распознавание TCR pepMHC полностью опосредовано специфическими остатками на петлях CDR (Garcia и др. (2009) Nat Immunol, 10, 143-7; Marrack и др. (2008) Annu Rev Immunol, 26, 171-203; Rudolph и др. (2006) Annu Rev Immunol, 24, 419-66)). Хотя кристаллические структуры специфичных к сурвивину TCR не доступны на момент написания настоящей заявки, показана структура комплекса A6:Tax пептид:HLA-A2 (PDB: 1AO7) (Garboczi и др. (1996) Nature, 384, 134-141), в котором используется тот же самый домен Vα2, как и в специфичном к сурвивину TCR. Вид сбоку данного комплекса показал, что концы вариабельных доменов, которые содержат шесть участков, определяющих комплементарность, присоединились к молекуле Tax:HLA.A2, при этом центральная область сайта связывания расположена над пептидом Tax (фигура 2A). В данную кристаллическую структуру не включена константная область α, хотя константные области помогают стабилизировать полноразмерную конструкцию. Стабилизирующие мутации, выбранные на этапе 2, описанном выше, часто выбирают в каркасных областях, таких как поверхность контакта Vα/Vβ, или там, где происходит соединение поверхностей контакта Cα/Vα или Cβ/Vβ в полноразмерном TCR.
[00172] Показан вид сверху вниз комплекса Tax:HLA-A2, из которого «убрали» TCR, за исключением шести петель CDR (фигура 2B). На данном виде показано, что TCR занимает диагональное положение над комплексом пептид-MHC, это открытие теперь наблюдают для всех структур TCR:пептид-MHC. В данной ориентации две петли CDR3 расположены над пептидом, тогда как присутствуют различные остатки из петель CDR1 и CDR2, которые взаимодействуют преимущественно со спиралями молекулы MHC. С целью созревания аффинности на этапах 4 и 6 данные петли часто являются мишенями для создания библиотек созревания аффинности, хотя также можно использовать другие участки.
ПРИМЕР 3
Дрожжевой дисплей специфичных к сурвивину TCR
[00173] Для того, чтобы осуществить селекцию для выявления улучшенной стабильности (этап 2) или улучшенной аффинности (этап 5), необходимо использовать систему дисплея, в которой библиотеку мутантов TCR можно подвергнуть скринингу для выявления связывания с антителом, которое узнает конформационный эпитоп или лиганд пептид:MHC, соответственно. три системы дисплея применяли для конструирования TCR с более высокой аффинностью, и их можно применять для данного процесса: дрожжевой дисплей, фаговый дисплей и T-клеточный дисплей (на клетках млекопитающего). Альтернативные способы дисплея, такие как рибосомный, РНК-, ДНК- и цис-дисплей, также могут быть подходящими для данного процесса. Во всех данных случаях TCR дикого типа с низкой аффинностью к антигену клонировали в систему и использовали в качестве матрицы для конструирования TCR с повышенной стабильностью и аффинностью к лиганду пептид:MHC. Любую из данных систем можно применять в подходе, описанном в данной заявке, в котором один TCR используют в качестве матрицы для создания библиотек и селекции TCR с улучшенными свойствами связывания.
[00174] В настоящем примере дрожжевой дисплей использовали в качестве платформы (фигура 3). Специфичный к сурвивину TCR использовали в качестве матрицы для введения стабилизирующих мутаций с помощью допускающего ошибки мутагенеза и стабилизированные клоны, выделенные в результате селекции, использовали в качестве матриц для созревания аффинности.
ПРИМЕР 4
Конструирование библиотеки с помощью допускающей ошибки полимеразы и селекция стабилизированных специфичных к сурвивину TCR, Surv-K2
[00175] Специфичную к сурвивину библиотеку, созданную с помощью допускающей ошибки полимеразы, получали, как описано ранее (Richman и др. (2009) Methods Mol Biol, 504, 323-350), применяя в качестве матрицы реактивный по отношению к сурвивину TCR, полученный от коллаборатора, называемый «сурвивин 71». Специфичную к сурвивину человека библиотеку, полученную с помощью допускающей ошибки полимеразы, вставляли в вектор для дрожжевого дисплея путем объединения линеаризованного вектора pCT302, продукта допускающей ошибки ПЦР, специфичного к сурвивину, и компетентных дрожжевых клеток EBY 100. Полученную библиотеку оценили путем посева аликвот серийных разведений дрожжей после электропорации, и она содержала приблизительно 8,25 x 106 независимых клонов. Осуществляли селекцию клонов из библиотеки, которые связываются с антителом, которое узнает Vβ20 человека, - IgG к hVβ20, меченым FITC (Beckman Coulter), с помощью FACS согласно таблице 4.
Таблица 4. Условия сортировки
| Сортировка | Условия |
| 1 | BC hVβ20 FITC (1:10); меченые AlexaFluor® 488 антитела козы против IgG мыши (1:100) |
| 2 | BC hVβ20 FITC (1:10); меченые AlexaFluor® 488 антитела козы против IgG мыши (1:100) |
| 3 | BC hVβ20 FITC (1:10); меченые AlexaFluor® 488 антитела козы против IgG мыши (1:100) |
[00176] С помощью исследования методом термической денатурации мы обнаружили, что данное антитело узнает свернутые эпитопы на Vβ20 (результаты не представлены). Сигналы усиливали, применяя меченые AlexaFluor® 488 вторичные антитела козы против IgG мыши (Life Technologies). В процессе 3 повторных сортировок выявили положительно окрашенную на Vβ20 популяцию (фигура 4A). После 3ей сортировки выделили клон, названный Surv-K2, с улучшенной флуоресценцией Vβ20 (фигура 4B). Клон SurvK2 использовали в качестве матрицы для созревания аффинности.
ПРИМЕР 5
Конструирование библиотеки CDR 3α и селекция двух специфичных к сурвивину TCR с более сильным связыванием с комплексом сурвивин:HLA.A2: Surv-K2.4.1 и Surv-K2.4.6
[00177] Стабилизированный клон Surv-K2, выделенный в результате селекции из библиотек, полученных с помощью допускающей ошибки ПЦР, использовали в качестве матрицы для получения библиотеки CDR3α, охватывающей 5 остатков, путем сращивания генов перекрывающимися расширениями (SOE). Библиотеку Surv-K2 человека CDR3α scTCR, таким образом, вставляли в вектор для дрожжевого дисплея путем объединения линеаризованного вектора pCT302, продукта ПЦР библиотеки Surv-K2 CDR3β и компетентных дрожжевых клеток EBY100. Полученную библиотеку оценили путем посева аликвот серийных разведений дрожжей после электропорации, и она содержала 2,98 x 107 независимых клонов. Библиотеку Surv-K2 CDR3α подвергали сортировке три раза подряд, применяя магнитные колонки, а затем применяя FACS согласно таблице 5.
Таблица 5. Условия сортировки
| Сортировка | Условия |
| 1 | BC hVβ20, меченое FITC (1:20) Антитело к IgG мыши на микрогранулах MB (1:25) |
| 2 | 100 нМ димера SurvT2M:HLA.A2 Антитело к IgG мыши на микрогранулах MB (1:25) |
| 3 | 100 нМ димера SurvT2M:HLA.A2 Антитело к IgG мыши на микрогранулах MB (1:25) |
| 4 | 100 нМ димера SurvT2M:HLA.A2 Меченые AlexaFluor® 647 антитела козы против IgG мыши (1:100) |
[00178] После двух сортировок с применением магнитных гранул начала появляться умеренно положительно окрашенная популяция (фигура 5A). Клоны Surv-K2.4.1 и Surv-K2.4.6 выделяли после четвертой сортировки. Surv-K2.4.1 и SurvK2.4.6 проявили усиленное связывание с SurvT2M (LMLGEFLKL, SEQ ID NO: 5)/HLA-A2 (фигура 5B).
ПРИМЕР 6
Анализ связывания высокоаффинного специфичного к сурвивину TCR, Surv-K2.4.1
[00179] Для того, чтобы оценить связывание клона Surv-K2.4.1, выделенного в результате селекции из библиотек CDR3α, дрожжей, на которых выставлен Surv-K2.4.1, титровали с помощью мономеров SurvT2M (LMLGEFLKL, SEQ ID NO: 5)/HLA-A2 при концентрации 6,4 нМ, 32 нМ, 160 нМ, 800 нМ и 4 мкМ и анализировали с помощью проточной цитометрии (фигура 6A). Значения нормировали, применяя нелинейный регрессионный анализ и определили значения кажущейся константы диссоциации (KD,app), равные 279 ± 44,5 нМ (фигура 6B).
ПРИМЕР 7
Анализ последовательности выделенных TCR для улучшения аффинности к антигену сурвивину
[00180] Определяли последовательности стабилизированного клона scTCR K2 и специфичных к сурвивину (K2.4.1 и K2.4.6) высокоаффинных одноцепочечных вариантов, выделенных из библиотек созревания аффинности. На фигуре 7 показано, что обнаружили мутации в участках, определяющих комплементарность, двух высокоаффинных клонов, происходящие из библиотек дрожжевого дисплея. Подчеркнутые положения на фигуре 7 указывают на мутации, которые произошли в результате селекциии стабилизирующих мутаций из библиотеки, полученной с помощью допускающей ошибки полимеразы. Положения в рамках показывают улучшающие аффинность мутации, которые были выбраны из библиотек участков, определяющих комплементарность.
ПРИМЕР 8
Активность In Vitro TCR K2.4.1 в T-клетках
[00181] Для того, чтобы оценить активность TCR K2.4.1 в T-клетках, выделяли CD8+ T-клетки из трансгенных мышей AAD. Данные T-клетки затем активировали с помощью гранул, соединенных с антителами к CD3 и к CD28, в течение 24 часов. T-клетки трансдуцировали с помощью ретровирусного вектора pMP71, содержащего домены Vα и β TCR K2.4.1, соединенные с доменами Cα и Cβ TCR 2C мыши (фигура 8A). Для того, чтобы подтвердить экспрессию TCR K2.4.1, T-клетки окрашивали через 48 часов после трансдукции тетрамером SurvT2M:HLA-A2 при концентрации 20 нМ (фигура 8B). Трансдуцированные K2.4.1 T-клетки (черный) проявили повышенное связывание SurvT2M:HLA-A2 по сравнению с ложно трансдуцированными T-клетками (серый), что подтверждает поверхностную экспрессию высокоаффинного TCR. T-клетки затем инкубировали при соотношении E:T 1:1 с клетками T2, в которые экзогенно ввели титрованные концентрации пептида сурвивина. T-клетки, экспрессирующие TCR K2.4.1, активировались в присутствии пептида SurvT2M, но не тогда, когда им презентировали контрольный пептид, называемый WT1 (RMFPNAPYL, SEQ ID NO: 14), позволяя предположить, что данный TCR активен и специфичен в CD8+ T-клетках.
ПРИМЕР 9
Терапевтические форматы специфичных к сурвивину, Surv-K2.4.1 и Surv-K2.4.6, TCR
[00182] В настоящее время хорошо известно, что высокоаффинные TCR можно применять в различных форматах для нацеливания на клетки, которые экспрессируют соответствующий антиген. Таким образом, ясно, что TCR, полученные с помощью методик конструирования, описанных выше, можно применять либо в растворимой форме, либо в рамках генотерапии TCR для адоптивной Т-клеточной терапии, показанной на фигуре 9.
Материалы и методы
Антитела, пептид:HLA-A2, реагенты для магнитно-активированной сортировки клеток (MACS) и проточной цитометрии
[00183] Антитела, применяемые для обнаружения экспрессии на поверхности дрожжей, включают: антитело к эпитопу гемагглютининовой (HA) метки (клон HA.11; Covance), антитело к hVβ3, меченое FITC (клон CH92; Beckman-Coulter), антитело к hVβ3.1, меченое FITC (клон 8F10; Thermo Scientific), антитело к hVβ20 (клон ELL1.4; Beckman-Coulter), моноклональное антитело к Vα2, полученное в нашей лаборатории (результаты не представлены), антитело козы против IgM мыши, меченое APC (Life Technologies), вторичное антитело козы против IgG F(ab')2 мыши, меченое AlexaFluor® 647 (Invitrogen), стрептавидин-фикоэритрин (SA:PE, BD Pharmingen) и микрогранулы MACS (Miltenyl Biotec).
[00184] Пептид, который связывается с HLA-A2, SurvT2M: LMLGEFLKL (SEQ ID NO: 5), который заякоривается через остаток 2, модифицированный путем замены T на M для улучшения связывания с HLA-A2 (Andersen и др., 2001, Cancer Research 61, 5964-5968), синтезировали с помощью стандартной химии F-moc (N-(9-флуоренил)метоксикарбонил) в профильном центре Macromolecular Core Facility Медицинского колледжа университета штата Пенсильвания (Херши, Пенсильвания, США). Для анализа методом FACS и проточной цитометрии использовали рекомбинантный растворимый димерный слитый белок HLA-A2:Ig (BD™ dimerX). Кроме того, мономерный реагент HLA.A2-биотин, полученный путем замены расщепляемого УФ пептида на другой рестриктированный по HLA.A2 пептид в присутствии УФ света, использовали для проточной цитометрии и селекции методом MACS (Rodenko и др. (2006) Nat Protoc, 1, 1120-1132; Toebes и др. (2006) Nat Med, 12, 246-251).
Клонирование и экспрессия scTv в векторах дрожжевого дисплея
[00185] Фрагменты вариабельной области TCR (scTv) экспрессировали в плазмиде для дрожжевого дисплея pCT302 (Vβ-L-Vα) (Boder и Wittrup (2000) Methods Enzymol, 328, 430-444), которая содержит индуцируемый галактозой слитый ген AGA2, позволяющий рост в среде без Trp. Индукция гена scTv включает рост трансформированных дрожжевых клеток EBY100 до стационарной фазы в селективной среде, а затем их перенос в содержащую галактозу среду. Используемые в качестве матрицы гены специфичных к сурвивину одноцепочечных TCR синтезировали в Genscript (Пискатауэй, Нью-Джерси, США) с мутацией F49S в домене Vα2-указанной конструкции (Aggen и др. (2011) Protein Eng Des Sel, 24, 361-372).
[00186] Гены специфичных к сурвивину TCR выделяли из клонов CTL (гены TCR против сурвивина от Delores Schendel, Thomas Blankenstein и Wolfgang Uckert; например, Leisegang и др. (2010) J Clin Invest. 120(11), 3869), и указанные гены синтезировали в Genscript, клонировали в одноцепочечном формате (Vβ-линкер-Vα), внедряли в вектор дрожжевого дисплея для экспрессии на поверхности дрожжей. ScTv состояли из вариабельных доменов, соединенных линкерной областью GSADDAKKDAAKKDGKS (SEQ ID NO: 7) (Hoo и др. (1992) Proc Natl Acad Sci USA, 89, 4759-4763; Weber и др. (2005) Proc Natl Acad Sci USA, 102, 19033-19038; Aggen и др. (2011) Protein Eng Des Sel, 24, 361-372). ScTv встраивали в сайты рестрикции NheI и XhoI в плазмиде pCT302.
Получение, дисплей и селекция из мутированных библиотек дрожжевого дисплея scTv
[00187] Допускающую ошибки ПЦР использовали для получения случайных мутаций, как описано ранее (Richman и др. (2009) Mol Immunol, 46, 902-916). Библиотеки CDR1 и 3 получали, применяя ПЦР методом сращивания генов перекрывающимися расширениями (SOE), охватывающую 4 - 5 соседних кодонов одновременно (Horton и др. (1990) Biotechniques, 8, 528-535).
[00188] Для получения библиотеки Surv CDR3α продукты ПЦР перед SOE получали, применяя следующие пары праймеров: 5'-GGC AGC CCC ATA AAC ACA CAG TAT-3' (Splice 4L) (SEQ ID NO: 8) и 5'-CAC AGC GCA CAG ATA GGT AGC-3' (SEQ ID NO: 10) и 5'-CTG ATT CAG CTA CCT ATC TGT GCG CTG TGN NSN NSN NSN NSN NSA TGT TTG GCG ATG GTA CTC AGC TGG TTG TG-3' (SEQ ID NO: 11) и 5'-TAA TAC GAC TCA CTA TAG GG-3' (T7) (SEQ ID NO: 9). SOE ПЦР проводили с продуктами каждого соответствующего ПЦР перед SOE вместе с T7 и с Splice 4L.
[00189] Дрожжевые библиотеки получали путем гомологичной рекомбинации в дрожжах EBY100 посредством электропорации продуктов допускающей ошибки или SOE ПЦР вместе с расщепленной рестриктазами NheI и XhoI плазмидой pCT302 (Horton и др. (1990) Biotechniques, 8, 528-535). Полученные библиотеки индуцировали в содержащих галактозу средах (SG-CAA) в течение 48 ч, промывали 1 мл 1% ФБР/БСА и окрашивали антителами или реагентами пептид:MHC при концентрациях, указанных на фигурах 4A, 5A, 6A, 8A и 9A. Клетки промывали (1 мл, 1% ФБР/БСА) и клетки с наибольшей флуоресценцией выбирали, применяя высокоскоростной сортер FACS Aria (BD Bioscience), или с помощью LS-колонок MACS на сепараторе QuadroMACS™ (Miltenyl Biotec). Для того, чтобы проверить термическую стабильность выделенных клонов, дрожжи инкубировали при повышенной температуре в течение 30 мин перед выполнением протокола окрашивания (результаты не представлены).
Выделение и окрашивание высокоаффинных клонов
[00190] После селекции выделяли клоны из библиотеки путем посева серийных разведений. Колонии наращивали и индуцировали в содержащих галактозу средах (SG-CAA) в течение 48 часов, промывали 1 мл 1% ФБР/БСА и окрашивали различными концентрациями пептид/HLA.A2 dimerX, вторичного антитела козы против IgG мыши F(ab')2, меченого AlexaFluor® 647, или различными концентрациями комплекса УФ-замененный пептид/HLA.A2, SA-PE. Клетки промывали (1 мл, 1% ФБР/БСА) и анализировали на проточном цитометре Accuri C6.
[00191] Плазмиды выделяли, применяя Yeast Plasmid Miniprep II Zymoprep™ (Zymo Research), и вводили обратно в E. coli путем трансформации методом теплового шока в компетентные клетки Subcloning Efficiency™ DH5α™ (Invitrogen). Клетки E. coli наращивали и выделяли плазмиды, применяя набор QIAprep Spin Miniprep (Qiagen). Последовательности отдельных клонов определяли путем секвенирования по Сенгеру.
[00192] ПОЛОЖЕНИЯ, КАСАЮЩИЕСЯ ВКЛЮЧЕНИЯ ПОСРЕДСТВОМ ССЫЛКИ И ВАРИАНТОВ
[00193] Все ссылочные материалы, цитированные в данной заявке, например, патентные документы, включая опубликованные или выданные патенты или эквиваленты; публикации заявок на патент; и документы непатентной литературы или другие первоисточники; настоящим полностью включены в данную заявку посредством ссылки, как если бы они были индивидуально включены посредством ссылки, в той мере, насколько каждый ссылочный материал по меньшей мере частично не противоречит изложенному в настоящем описании (например, ссылочный материал, который частично противоречит, включен посредством ссылки за исключением частично противоречащей части данного ссылочного материала).
[00194] Все патенты и публикации, упомянутые в настоящем описании, указывают на уровни квалификации специалистов в области, к которой относится настоящее описание. Ссылочный материал, цитируемый в данной заявке, полностью включен в данную заявку посредством ссылки, чтобы указать на существующий уровень техники, в некоторых случаях, по его дате подачи, и предполагается, что данную информацию можно использовать в данной заявке, при необходимости, чтобы исключить конкретные варианты реализации (например, отказаться от них), которые принадлежат к известному уровню техники, или чтобы применять способы или материалы, которые принадлежат к известному уровню техники, без специального включения указанных способов или материалов в описание данной заявки. Например, если заявлено соединение, то должно быть очевидно, что соединения, известные в известном уровне техники, включая некоторые соединения, раскрытые в ссылочных материалах, описанных в данной заявке (особенно в ссылочных патентных документах), не предназначены для включения в формулу настоящего изобретения.
[00195] Если в данной заявке используются группы Маркуша или другие группы, то предполагается, что все отдельные представители данной группы и все возможные комбинации и подкомбинации представителей данной группы отдельно включены в настоящее описание.
[00196] Когда термины «включать», «включает», «состоит» или «включающий» используют в данной заявке, их следует истолковывать, как обозначающие присутствие указанных рассматриваемых свойств, целых чисел, этапов или компонентов, но не исключающие наличие или добавление одного или более других свойств, целых чисел, этапов, компонентов или их группы. Также предполагается, что включены отдельные варианты реализации согласно настоящему описанию, в которых термины «включающий» или «включают (включает)» или «состоит» возможно заменены на термины, грамматически аналогичные, например; «состоящий/состоят (состоит)» или «состоящий по существу из/состоят (состоит) по существу из», чтобы с их помощью описать дополнительные варианты реализации, которые не обязательно одинаковы по протяженности. Для ясности, в данной заявке термин «включающий» синонимичен терминам «обладающий», «заключающий в себе», «содержащий» или «характеризуемый» и является включительным или неограничивающим, и не исключает дополнительные, не перечисленные элементы или этапы способа. В данной заявке термин «состоящий из» исключает любой элемент, этап, компонент или ингредиент, не перечисленный в элементе пункта формулы изобретения. В данной заявке термин «состоящий по существу из» не исключает материалы или этапы, которые существенным образом не влияют на основные и новые свойства пункта формулы изобретения (например, не оказывают влияния на активный ингредиент). В каждом случае в данной заявке любой из терминов «включающий», «состоящий по существу из» и «состоящий из» можно заменить на любой из других двух терминов. Раскрытие, наглядно описанное в данной заявке, можно подходящим образом осуществить в отсутствие любого элемента или элементов, ограничения или ограничений, которые конкретно не описаны в данной заявке.
[00197] Раскрытие было описано на основании различных конкретных и предпочтительных вариантов реализации и методик. Тем не менее, должно быть очевидно, что можно осуществить множество вариаций и модификаций, оставаясь в рамках сущности и объема настоящего описания. Для среднего специалиста в данной области должно быть очевидно, что композиции, способы, устройства, элементы устройства, материалы, необязательные свойства, процедуры и методики, отличные от конкретно описанных в данной заявке, можно применять для осуществления настоящего изобретения, раскрытого в широком смысле в данной заявке, не прибегая к лишним экспериментам. Предполагается, что все известные в данной области техники функциональные эквиваленты композиций, способов, устройств, элементов устройств, материалов, процедур и методик, описанных в данной заявке; и их части; входят в объем настоящего описания. Всякий раз, когда описан диапазон, предполагается, что в объем входят все поддиапазоны и отдельные значения. Настоящее описание не должно быть ограничено описанными вариантами реализации, включая любые варианты реализации, показанные в рисунках или приведенные в качестве примера в настоящем описании, которые даны в качестве примера или иллюстрирования, но не ограничения. Некоторые ссылочные материалы, предложенные в данной заявке, включены в данную заявку посредством ссылки, чтобы предоставить подробности, касающиеся дополнительных исходных материалов, дополнительных способов синтеза, дополнительных способов анализа и дополнительных применений настоящего описания.
[00198] Специалист в данной области с легкостью поймет, что настоящее описание хорошо приспособлено для осуществления целей и достижения задач и преимуществ, как упомянутых, так и свойственных ему. Композиции и способы, а также и вспомогательные способы, описанные в данной заявке как, в настоящий момент, типичные предпочтительные варианты реализации, представляют собой примеры и не предназначены для ограничения объема настоящего изобретения. Изменения в них и другие применения, которые входят в объем сущности настоящего изобретения, придут в голову специалистам в данной области.
Перечень ссылочных материалов
1. Addo M. M., Draenert R., Rathod A., Verrill C. L., Davis B. T., Gandhi R. T., Robbins G. K., Basgoz N. O., Stone D. R., Cohen D. E., Johnston M. N., Flynn T., Wurcel A. G., Rosenberg E. S., Altfeld M. and Walker B. D. (2007) Fully Differentiated HIV-1 Specific CD8+ T Effector Cells Are More Frequently Detectable in Controlled than in Progressive HIV-1 Infection. PLoS ONE 2, e321.
2. Aggen D. H., Chervin A. S., Insaidoo F. K., Piepenbrink K., H., Baker B. M. and Kranz D. M. (2011) Identification and engineering of human variable regions that allow expression of stable single-chain T cell receptors. Protein Engineering, Design, & Selection 24, 361-72.
3. Anikeeva N., Mareeva T., Liu W. and Sykulev Y. (2009) Can oligomeric T-cell receptor be used as a tool to detect viral peptide epitopes on infected cells? Clin Immunol 130, 98-109.
4. Armstrong K. M., Piepenbrink K. H. and Baker B. M. (2008) Conformational changes and flexibility in T-cell receptor recognition of peptide-MHC complexes. Biochem J 415, 183-96.
5. Ashfield R. and Jakobsen B. K. (2006) Making high-affinity T-cell receptors: a new class of targeted therapeutics. IDrugs 9, 554-9.
6. Bargou R., Leo E., Zugmaier G., Klinger M., Goebeler M., Knop S., Noppeney R., Viardot A., Hess G., Schuler M., Einsele H., Brandl C., Wolf A., Kirchinger P., Klappers P., Schmidt M., Riethmuller G., Reinhardt C., Baeuerle P. A. and Kufer P. (2008) Tumor regression in cancer patients by very low doses of a T cell-engaging antibody. Science 321, 974-7.
7. Benatuil L., Perez J. M., Belk J. and Hsieh C. M. (2010) An improved yeast transformation method for the generation of very large human antibody libraries. Protein Eng Des Sel 23, 155-9.
8. Bird R. E., Hardman K. D., Jacobson J. W., Johnson S., Kaufman B. M., Lee S. M., Lee T., Pope S. H., Riordan G. S. and Whitlow M. (1988) Single-chain antigen-binding proteins. Science 242, 423-426.
9. Boder E. T. and Wittrup K. D. (1997) Yeast surface display for screening combinatorial polypeptide libraries. Nat. Biotech. 15, 553-557.
10. Boder E. T. and Wittrup K. D. (2000) Yeast surface display for directed evolution of protein expression, affinity, and stability. Methods Enzymol 328, 430-44.
11. Boon T. and Old L. J. (1997) Cancer tumor antigens. Curr Opin Immunol 9, 681-3.
12. Borbulevych O. Y., Santhanagopolan S. M., Hossain M. and Baker B. M. (2011) TCRs used in cancer gene therapy cross-react with MART-1/Melan-A tumor antigens via distinct mechanisms. J Immunol 187, 2453-63.
13. Brower V. (1997) Enbrel's phase III reinforces prospects in RA [news]. Nat Biotechnol 15, 1240.
14. Bulek A. M., Cole D. K., Skowera A., Dolton G., Gras S., Madura F., Fuller A., Miles J. J., Gostick E., Price D. A., Drijfhout J. W., Knight R. R., Huang G. C., Lissin N., Molloy P. E., Wooldridge L., Jakobsen B. K., Rossjohn J., Peakman M., Rizkallah P. J. and Sewell A. K. (2012) Structural basis for the killing of human beta cells by CD8(+) T cells in type 1 diabetes. Nat Immunol 13, 283-9.
15. Cheever M. A., Allison J. P., Ferris A. S., Finn O. J., Hastings B. M., Hecht T. T., Mellman I., Prindiville S. A., Viner J. L., Weiner L. M. and Matrisian L. M. (2009) The prioritization of cancer antigens: a national cancer institute pilot project for the acceleration of translational research. Clin Cancer Res 15, 5323-37.
16. Chervin A. S., Aggen D. H., Raseman J. M. and Kranz D. M. (2008) Engineering higher affinity T cell receptors using a T cell display system. J Immunol Methods 339, 175-84.
17. Chervin AS, Stone JD, Soto CM, Engels B, Schreiber H, Roy EJ, and Kranz DM. (2013) Design of T-cell receptor libraries with diverse binding properties to examine adoptive T-cell responses. Gene Ther. 20(6):634-44.
18. Colby D. W., Kellogg B. A., Graff C. P., Yeung Y. A., Swers J. S. and Wittrup K. D. (2004) Engineering antibody affinity by yeast surface display. Methods Enzymol 388, 348-58.
19. Davis M. M. and Bjorkman P. J. (1988) T-cell antigen receptor genes and T-cell recognition. Nature 334, 395-402.
20. Davis M. M., Boniface J. J., Reich Z., Lyons D., Hampl J., Arden B. and Chien Y. (1998) Ligand recognition by alpha beta T cell receptors. Annu Rev Immunol 16, 523-544.
21. Ding Y. H., Baker B. M., Garboczi D. N., Biddison W. E. and Wiley D. C. (1999) Four A6-TCR/peptide/HLA-A2 structures that generate very different T cell signals are nearly identical. Immunity 11, 45-56.
22. Foote J. and Eisen H. N. (2000) Breaking the affinity ceiling for antibodies and T cell receptors. Proc Natl Acad Sci U S A 97, 10679-81.
23. Garboczi D. N., Ghosh P., Utz U., Fan Q. R., Biddison W. E. and Wiley D. C. (1996) Structure of the complex between human T-cell receptor, viral peptide and HLA-A2. Nature 384, 134-141.
24. Garcia K. C., Adams J. J., Feng D. and Ely L. K. (2009) The molecular basis of TCR germline bias for MHC is surprisingly simple. Nat Immunol 10, 143-7.
25. Haidar J. N., Pierce B., Yu Y., Tong W., Li M. and Weng Z. (2009) Structure-based design of a T-cell receptor leads to nearly 100-fold improvement in binding affinity for pepMHC. Proteins 74, 948-60.
26. Harkiolaki M., Holmes S. L., Svendsen P., Gregersen J. W., Jensen L. T., McMahon R., Friese M. A., van Boxel G., Etzensperger R., Tzartos J. S., Kranc K., Sainsbury S., Harlos K., Mellins E. D., Palace J., Esiri M. M., van der Merwe P. A., Jones E. Y. and Fugger L. (2009) T cell-mediated autoimmune disease due to low-affinity crossreactivity to common microbial peptides. Immunity 30, 348-57.
27. Hawse W. F., Champion M. M., Joyce M. V., Hellman L. M., Hossain M., Ryan V., Pierce B. G., Weng Z. and Baker B. M. (2012) Cutting edge: evidence for a dynamically driven T cell signaling mechanism. J Immunol 188, 5819-23.
28. Holler P. D., Chlewicki L. K. and Kranz D. M. (2003) TCRs with high affinity for foreign pMHC show self-reactivity. Nat Immunol 4, 55-62.
29. Holler P. D., Holman P. O., Shusta E. V., O'Herrin S., Wittrup K. D. and Kranz D. M. (2000) In vitro evolution of a T cell receptor with high affinity for peptide/MHC. Proc Natl Acad Sci U S A 97, 5387-92.
30. Holliger P., Prospero T. and Winter G. (1993) "Diabodies": small bivalent and bispecific antibody fragments. Proc Natl Acad Sci U S A 90, 6444-8.
31. Hoogenboom H. R. (2005) Selecting and screening recombinant antibody libraries. Nat Biotechnol 23, 1105-16.
32. Jarvis L. M. (2012) Rethinking Antibody-Drug Conjugates. Chemical and Engineering News 90, 12-18.
33. Kessels H. W., van Den Boom M. D., Spits H., Hooijberg E. and Schumacher T. N. (2000) Changing T cell specificity by retroviral T cell receptor display. Proc Natl Acad Sci U S A 97, 14578-83.
34. Kieke M. C., Shusta E. V., Boder E. T., Teyton L., Wittrup K. D. and Kranz D. M. (1999) Selection of functional T cell receptor mutants from a yeast surface- display library. Proc Natl Acad Sci U S A 96, 5651-6.
35. Lauck F., Smith C. A., Friedland G. F., Humphris E. L. and Kortemme T. (2010) RosettaBackrub--a web server for flexible backbone protein structure modeling and design. Nucleic Acids Res 38, W569-75.
36. Li Y., Moysey R., Molloy P. E., Vuidepot A. L., Mahon T., Baston E., Dunn S., Liddy N., Jacob J., Jakobsen B. K. and Boulter J. M. (2005) Directed evolution of human T-cell receptors with picomolar affinities by phage display. Nat Biotechnol 23, 349-54.
37. Liddy N., Bossi G., Adams K. J., Lissina A., Mahon T. M., Hassan N. J., Gavarret J., Bianchi F. C., Pumphrey N. J., Ladell K., Gostick E., Sewell A. K., Lissin N. M., Harwood N. E., Molloy P. E., Li Y., Cameron B. J., Sami M., Baston E. E., Todorov P. T., Paston S. J., Dennis R. E., Harper J. V., Dunn S. M., Ashfield R., Johnson A., McGrath Y., Plesa G., June C. H., Kalos M., Price D. A., Vuidepot A., Williams D. D., Sutton D. H. and Jakobsen B. K. (2012) Monoclonal TCR-redirected tumor cell killing. Nat Med.
38. Litvak-Greenfeld D. and Benhar I. (2012) Risks and untoward toxicities of antibody-based immunoconjugates. Adv Drug Deliv Rev.
39. Manning T. C. and Kranz D. M. (1999) Binding energetics of T-cell receptors: correlation with immunological consequences. Immunology Today 20, 417-422.
40. Marrack P., Scott-Browne J. P., Dai S., Gapin L. and Kappler J. W. (2008) Evolutionarily conserved amino acids that control TCR-MHC interaction. Annu Rev Immunol 26, 171-203.
41. Marsh S. G. E., Parham P. and Barber L. D. (2000) The HLA Facts Book. Academic Press, London.
42. Mason D. (1998) A very high level of crossreactivity is an essential feature of the T- cell receptor. Immunol Today 19, 395-404.
43. Miller B. R., Demarest S. J., Lugovskoy A., Huang F., Wu X., Snyder W. B., Croner L. J., Wang N., Amatucci A., Michaelson J. S. and Glaser S. M. (2010) Stability engineering of scFvs for the development of bispecific and multivalent antibodies. Protein Eng Des Sel 23, 549-57.
44. Molloy P. E., Sewell A. K. and Jakobsen B. K. (2005) Soluble T cell receptors: novel immunotherapies. Curr Opin Pharmacol 5, 438-43.
45. Murphy K. (2012) Janeway's immunobiology. Garland Science, New York.
46. Nold M. F., Nold-Petry C. A., Zepp J. A., Palmer B. E., Bufler P. and Dinarello C. A. (2010) IL-37 is a fundamental inhibitor of innate immunity. Nat Immunol 11, 1014-22.
47. Pastan I., Hassan R., Fitzgerald D. J. and Kreitman R. J. (2006) Immunotoxin therapy of cancer. Nat Rev Cancer 6, 559-65.
48. Pierce B. G., Haidar J. N., Yu Y. and Weng Z. (2010) Combinations of affinity-enhancing mutations in a T cell receptor reveal highly nonadditive effects within and between complementarity determining regions and chains. Biochemistry 49, 7050-9.
49. Porter D. L., Levine B. L., Kalos M., Bagg A. and June C. H. (2011) Chimeric antigen receptor-modified T cells in chronic lymphoid leukemia. N Engl J Med 365, 725-33.
50. Reichert J. M. and Valge-Archer V. E. (2007) Development trends for monoclonal antibody cancer therapeutics. Nat Rev Drug Discov 6, 349-56.
51. Ricart A. D. and Tolcher A. W. (2007) Technology insight: cytotoxic drug immunoconjugates for cancer therapy. Nat Clin Pract Oncol 4, 245-55.
52. Richman S. A., Aggen D. H., Dossett M. L., Donermeyer D. L., Allen P. M., Greenberg P. D. and Kranz D. M. (2009) Structural features of T cell receptor variable regions that enhance domain stability and enable expression as single-chain ValphaVbeta fragments. Mol Immunol 46, 902-16.
53. Richman S. A. and Kranz D. M. (2007) Display, engineering, and applications of antigen-specific T cell receptors. Biomol Eng 24, 361-73.
54. Rock K. L. and Goldberg A. L. (1999) Degradation of cell proteins and the generation of MHC class I- presented peptides. Annu Rev Immunol 17, 739-79.
55. Rudolph M. G., Stanfield R. L. and Wilson I. A. (2006) How TCRs bind MHCs, peptides, and coreceptors. Annu Rev Immunol 24, 419-66.
56. Sadelain M., Brentjens R. and Riviere I. (2009) The promise and potential pitfalls of chimeric antigen receptors. Curr Opin Immunol 21, 215-23.
57. Sami M., Rizkallah P. J., Dunn S., Molloy P., Moysey R., Vuidepot A., Baston E., Todorov P., Li Y., Gao F., Boulter J. M. and Jakobsen B. K. (2007) Crystal structures of high affinity human T-cell receptors bound to peptide major histocompatibility complex reveal native diagonal binding geometry. Protein Eng Des Sel 20, 397-403.
58. Schrama D., Reisfeld R. A. and Becker J. C. (2006) Antibody targeted drugs as cancer therapeutics. Nat Rev Drug Discov 5, 147-59.
59. Scott J. K. and Smith G. P. (1990) Searching for peptide ligands with an epitope library. Science 249, 386-90.
60. Skowera A., Ellis R. J., Varela-Calvino R., Arif S., Huang G. C., Van-Krinks C., Zaremba A., Rackham C., Allen J. S., Tree T. I., Zhao M., Dayan C. M., Sewell A. K., Unger W. W., Drijfhout J. W., Ossendorp F., Roep B. O. and Peakman M. (2008) CTLs are targeted to kill beta cells in patients with type 1 diabetes through recognition of a glucose-regulated preproinsulin epitope. J Clin Invest 118, 3390-402.
61. Smith C. A. and Kortemme T. (2008) Backrub-like backbone simulation recapitulates natural protein conformational variability and improves mutant side-chain prediction. J Mol Biol 380, 742-56.
62. Soo Hoo W. F., Lacy M. J., Denzin L. K., Voss E. W. J., Hardman K. D. and Kranz D. M. (1992) Characterization of a single-chain T cell receptor expressed in E. Coli. Proc. Natl. Acad. Sci. 89, 4759-4763.
63. Starr T. K., Jameson S. C. and Hogquist K. A. (2003) Positive and negative selection of T cells. Annu Rev Immunol 21, 139-76.
64. Starwalt S. E., Masteller E. L., Bluestone J. A. and Kranz D. M. (2003) Directed evolution of a single-chain class II MHC product by yeast display. Protein Eng 16, 147-56.
65. Stone J. D., Chervin A. S., Aggen D. H. and Kranz D. M. (2012) T cell receptor engineering. Methods Enzymol 503, 189-222.
66. Stone J. D., Yin Y., Mo M., Weber K. S., Donermeyer D. L., Allen P. M., Mariuzza R. A. and Kranz D. M. (2012) Engineering High-Affinity T Cell Receptor/ Cytokine Fusions for Therapeutic Targeting. In Protein Engineering (Edited by Kaumaya P.). InTech.
67. Stroncek D. F., Berger C., Cheever M. A., Childs R. W., Dudley M. E., Flynn P., Gattinoni L., Heath J. R., Kalos M., Marincola F. M., Miller J. S., Mostoslavsky G., Powell D. J., Jr., Rao M., Restifo N. P., Rosenberg S. A., O'Shea J. and Melief C. J. (2012) New directions in cellular therapy of cancer: a summary of the summit on cellular therapy for cancer. J Transl Med 10, 48.
68. Swers J. S., Kellogg B. A. and Wittrup K. D. (2004) Shuffled antibody libraries created by in vivo homologous recombination and yeast surface display. Nucleic Acids Res 32, e36.
69. Tayal V. and Kalra B. S. (2008) Cytokines and anti-cytokines as therapeutics--an update. Eur J Pharmacol 579, 1-12.
70. Thakur A. and Lum L. G. (2010) Cancer therapy with bispecific antibodies: Clinical experience. Curr Opin Mol Ther 12, 340-9.
71. Tonegawa S. (1988) Nobel lecture in physiology or medicine--1987. Somatic generation of immune diversity. In Vitro Cell Dev Biol 24, 253-65.
72. Tsomides T. J., Aldovini A., Johnson R. P., Walker B. D., Young R. A. and Eisen H. N. (1994) Naturally processed viral peptides recognized by cytotoxic T lymphocytes on cells chronically infected by human immunodeficiency virus type 1. J Exp Med 180, 1283-93.
73. Turner D. J., Ritter M. A. and George A. J. (1997) Importance of the linker in expression of single-chain Fv antibody fragments: optimisation of peptide sequence using phage display technology. J Immunol Methods 205, 43-54.
74. Utz U., Banks D., Jacobson S. and Biddison W. E. (1996) Analysis of the T-cell receptor repertoire of human T-cell leukemia virus type 1 (HTLV-1) Tax-specific CD8+ cytotoxic T lymphocytes from patients with HTLV-1-associated disease: evidence for oligoclonal expansion. J Virol 70, 843-51.
75. Varela-Rohena A., Molloy P. E., Dunn S. M., Li Y., Suhoski M. M., Carroll R. G., Milicic A., Mahon T., Sutton D. H., Laugel B., Moysey R., Cameron B. J., Vuidepot A., Purbhoo M. A., Cole D. K., Phillips R. E., June C. H., Jakobsen B. K., Sewell A. K. and Riley J. L. (2008) Control of HIV-1 immune escape by CD8 T cells expressing enhanced T-cell receptor. Nat Med 14, 1390-5.
76. Weber K. S., Donermeyer D. L., Allen P. M. and Kranz D. M. (2005) Class II-restricted T cell receptor engineered in vitro for higher affinity retains peptide specificity and function. Proc Natl Acad Sci U S A 102, 19033-8.
77. Wong R. L., Liu B., Zhu X., You L., Kong L., Han K. P., Lee H. I., Chavaillaz P. A., Jin M., Wang Y., Rhode P. R. and Wong H. C. (2011) Interleukin-15:Interleukin-15 receptor alpha scaffold for creation of multivalent targeted immune molecules. Protein Eng Des Sel 24, 373-83.
Патенты США.
7569357; поданный 20 февраля 2004 г.; опубликованный 4 августа 2009 г.; Board of Trustees University of Illinois. Высокоаффинные белки TCR и методы.
7465787; поданный 16 декабря 2003 г.; опубликованный 16 декабря 2008 г.;Board of Trustees University of Illinois. Дисплей белков на клеточной поверхности дрожжей и его применения.
6759243; поданный 6 декабря 2000 г.; опубликованный 6 июля 2004 г.; Board of Trustees University of Illinois. Высокоаффинные белки TCR и методы.
6699658; поданный 20 января 1998 г.; опубликованный 2 марта 2004 г.; Board of Trustees University of Illinois. Дисплей белков на клеточной поверхности дрожжей и его применения.
6696251; поданный 28 ноября 2000 г.; опубликованный 24 февраля 2004 г.; Board of Trustees University of Illinois. Дисплей белков на клеточной поверхности дрожжей и его применения.
6423538; поданный 28 ноября 2000 г.; опубликованный 23 июля 2002 г.; Board of Trustees University of Illinois. Дисплей белков на клеточной поверхности дрожжей и его применения.
6300065; поданный 26 августа 1998 г.; опубликованный 9 октября 2001 г.; Board of Trustees University of Illinois. Дисплей белков на клеточной поверхности дрожжей и его применения.
8143376; поданный 18 мая 2005 г.; опубликованный 27 марта 2012 г.; Immunocore Limited; Высокоаффинный T-клеточный рецептор NY-ESO.
8088379; поданный 26 сентября 2007 г.; опубликованный 3 января 2012 г.; Immunocore Limited; Модифицированные T-клеточные рецепторы и связанные с ними материалы и методы.
8017730; поданный 19 мая 2006 г.; опубликованный 13 сентября 2011 г.; Immunocore Limited; T-клеточные рецепторы, которые связываются с VYGFVRACL-HLA-A24.
7763718; поданный 29 октября 2007 г.; опубликованный 27 июля 2010 г.; Immunocore Limited; Растворимые T-клеточные рецепторы.
7666604; поданный 9 июля 2003 г.; опубликованный 23 февраля 2010 г.; Immunocore Limited; Модифицированный растворимый T-клеточный рецептор.
7608410; поданный 7 октября 2008 г.; опубликованный 27 октября 2009 г.; Immunocore Limited; Способ улучшения T-клеточных рецепторов.
7569664; поданный 3 октября 2003 г.; опубликованный 4 августа 2009 г.; Immunocore Limited; Одноцепочечные рекомбинантные T-клеточные рецепторы.
8105830; поданный 5 ноября 2002 г.; опубликованный 31 января 2012 г.; Altor Bioscience Corporation; Молекулы с полиспецифическим связыванием и их применения.
6534633; поданный 21 октября 1999 г.; 18 марта 2003 г.; Altor Bioscience Corporation; Молекулы с полиспецифическим связыванием и их применения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Board of Trustees of the University of Illinois
Smith, Sheena N.
Harris, Daniel T.
Kranz, David M.
<120> СКОНСТРУИРОВАННЫЕ ВЫСОКОАФФИННЫЕ T-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ ЧЕЛОВЕКА
<130> IMMU-003/01WO
<150> US 61/907887
<151> 2013-11-22
<160> 16
<170> PatentIn версии 3.5
<210> 1
<211> 110
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Модифицированная вариабельная область альфа-цепи TCR (сурвивин-K2.4.1)
с высокой аффинностью к комплексу сурвивин/HLA-A2
<400> 1
Gln Lys Glu Val Glu Gln Asn Ser Gly Pro Leu Ser Val Pro Glu Gly
1 5 10 15
Ala Ile Ala Ser Leu Asn Cys Thr Tyr Ser Asp Arg Gly Ser Gln Ser
20 25 30
Phe Phe Trp Tyr Arg Gln Tyr Ser Gly Lys Ser Pro Glu Leu Ile Met
35 40 45
Ser Ile Tyr Ser Asn Gly Asp Lys Glu Asp Gly Arg Phe Thr Ala Gln
50 55 60
Leu Asn Lys Ala Ser Gln Tyr Val Ser Leu Leu Ile Arg Asp Ser Gln
65 70 75 80
Pro Ser Asp Ser Ala Thr Tyr Leu Cys Ala Val Ser Lys Gly Tyr Lys
85 90 95
Met Phe Gly Asp Gly Thr Gln Leu Val Val Lys Pro Asn Ile
100 105 110
<210> 2
<211> 110
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Модифицированная вариабельная область альфа-цепи TCR
с высокой аффинностью к комплексу сурвивин/HLA-A2
<400> 2
Gln Lys Glu Val Glu Gln Asn Ser Gly Pro Leu Ser Val Pro Glu Gly
1 5 10 15
Ala Ile Ala Ser Leu Asn Cys Thr Tyr Ser Asp Arg Gly Ser Gln Ser
20 25 30
Phe Phe Trp Tyr Arg Gln Tyr Ser Gly Lys Ser Pro Glu Leu Ile Met
35 40 45
Ser Ile Tyr Ser Asn Gly Asp Lys Glu Asp Gly Arg Phe Thr Ala Gln
50 55 60
Leu Asn Lys Ala Ser Gln Tyr Val Ser Leu Leu Ile Arg Asp Ser Gln
65 70 75 80
Pro Ser Asp Ser Ala Thr Tyr Leu Cys Ala Val His Gly Trp Tyr Thr
85 90 95
Met Phe Gly Asp Gly Thr Gln Leu Val Val Lys Pro Asn Ile
100 105 110
<210> 3
<211> 242
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Одноцепочечный TCR (сурвивин-K2.4.1), который связывается с
высокой аффинностью с комплексом сурвивин/HLA-A2
<400> 3
Ser Gln Thr Ile His Gln Trp Pro Ala Thr Leu Val Gln Pro Val Gly
1 5 10 15
Ser Pro Leu Ser Leu Glu Cys Thr Val Glu Gly Thr Ser Asn Pro Asn
20 25 30
Leu Tyr Trp Tyr Arg Gln Ala Ala Gly Arg Gly Leu Glu Leu Leu Phe
35 40 45
Tyr Ser Val Gly Ile Gly Gln Ile Ser Ser Glu Val Pro Gln Asn Leu
50 55 60
Phe Ala Ser Arg Pro Gln Asp Arg Gln Phe Ile Leu Ser Ser Lys Lys
65 70 75 80
Leu Leu Leu Ser Asp Ser Gly Phe Tyr Leu Cys Ala Trp Ser Ile Gly
85 90 95
Ala Glu Gln Phe Phe Gly Pro Gly Thr Arg Leu Thr Val Leu Glu Asp
100 105 110
Leu Lys Asn Gly Ser Ala Asp Asp Ala Lys Lys Asp Ala Ala Lys Lys
115 120 125
Asp Gly Lys Ser Gln Lys Glu Val Glu Gln Asn Ser Gly Pro Leu Ser
130 135 140
Val Pro Glu Gly Ala Ile Ala Ser Leu Asn Cys Thr Tyr Ser Asp Arg
145 150 155 160
Gly Ser Gln Ser Phe Phe Trp Tyr Arg Gln Tyr Ser Gly Lys Ser Pro
165 170 175
Glu Leu Ile Met Ser Ile Tyr Ser Asn Gly Asp Lys Glu Asp Gly Arg
180 185 190
Phe Thr Ala Gln Leu Asn Lys Ala Ser Gln Tyr Val Ser Leu Leu Ile
195 200 205
Arg Asp Ser Gln Pro Ser Asp Ser Ala Thr Tyr Leu Cys Ala Val Ser
210 215 220
Lys Gly Tyr Lys Met Phe Gly Asp Gly Thr Gln Leu Val Val Lys Pro
225 230 235 240
Asn Ile
<210> 4
<211> 242
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Одноцепочечный TCR (сурвивин-K2.4.6), который связывается с
высокой аффинностью с комплексом сурвивин/HLA-A2
<400> 4
Ser Gln Thr Ile His Gln Trp Pro Ala Thr Leu Val Gln Pro Val Gly
1 5 10 15
Ser Pro Leu Ser Leu Glu Cys Thr Val Glu Gly Thr Ser Asn Pro Asn
20 25 30
Leu Tyr Trp Tyr Arg Gln Ala Ala Gly Arg Gly Leu Glu Leu Leu Phe
35 40 45
Tyr Ser Val Gly Ile Gly Gln Ile Ser Ser Glu Val Pro Gln Asn Leu
50 55 60
Phe Ala Ser Arg Pro Gln Asp Arg Gln Phe Ile Leu Ser Ser Lys Lys
65 70 75 80
Leu Leu Leu Ser Asp Ser Gly Phe Tyr Leu Cys Ala Trp Ser Ile Gly
85 90 95
Ala Glu Gln Phe Phe Gly Pro Gly Thr Arg Leu Thr Val Leu Glu Asp
100 105 110
Leu Lys Asn Gly Ser Ala Asp Asp Ala Lys Lys Asp Ala Ala Lys Lys
115 120 125
Asp Gly Lys Ser Gln Lys Glu Val Glu Gln Asn Ser Gly Pro Leu Ser
130 135 140
Val Pro Glu Gly Ala Ile Ala Ser Leu Asn Cys Thr Tyr Ser Asp Arg
145 150 155 160
Gly Ser Gln Ser Phe Phe Trp Tyr Arg Gln Tyr Ser Gly Lys Ser Pro
165 170 175
Glu Leu Ile Met Ser Ile Tyr Ser Asn Gly Asp Lys Glu Asp Gly Arg
180 185 190
Phe Thr Ala Gln Leu Asn Lys Ala Ser Gln Tyr Val Ser Leu Leu Ile
195 200 205
Arg Asp Ser Gln Pro Ser Asp Ser Ala Thr Tyr Leu Cys Ala Val His
210 215 220
Gly Trp Tyr Thr Met Phe Gly Asp Gly Thr Gln Leu Val Val Lys Pro
225 230 235 240
Asn Ile
<210> 5
<211> 9
<212> ПРТ
<213> Homo sapiens
<400> 5
Leu Met Leu Gly Glu Phe Leu Lys Leu
1 5
<210> 6
<211> 9
<212> ПРТ
<213> Лимфотропный Т-клеточный вирус человека
<400> 6
Leu Leu Phe Gly Tyr Pro Val Tyr Val
1 5
<210> 7
<211> 17
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность линкера
<400> 7
Gly Ser Ala Asp Asp Ala Lys Lys Asp Ala Ala Lys Lys Asp Gly Lys
1 5 10 15
Ser
<210> 8
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер Splice 4L
<400> 8
ggcagcccca taaacacaca gtat 24
<210> 9
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер T7
<400> 9
taatacgact cactataggg 20
<210> 10
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер, используемый для получения библиотеки Surv CDR3-альфа
<400> 10
cacagcgcac agataggtag c 21
<210> 11
<211> 74
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер, используемый для получения библиотеки Surv CDR3-альфа
<220>
<221> misc_feature
<222> (30)..(31)
<220>
<221> misc_feature
<222> (30)..(31)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (33)..(34)
<220>
<221> misc_feature
<222> (33)..(34)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (36)..(37)
<220>
<221> misc_feature
<222> (36)..(37)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (39)..(40)
<220>
<221> misc_feature
<222> (39)..(40)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (42)..(43)
<220>
<221> misc_feature
<222> (42)..(43)
<223> n представляет собой a, c, g или t
<400> 11
ctgattcagc tacctatctg tgcgctgtgn nsnnsnnsnn snnsatgttt ggcgatggta 60
ctcagctggt tgtg 74
<210> 12
<211> 114
<212> ПРТ
<213> Homo sapiens
<400> 12
Ser Gln Thr Ile His Gln Trp Pro Ala Thr Leu Val Gln Pro Val Gly
1 5 10 15
Ser Pro Leu Ser Leu Glu Cys Thr Val Glu Gly Thr Ser Asn Pro Asn
20 25 30
Leu Tyr Trp Tyr Arg Gln Ala Ala Gly Arg Gly Leu Glu Leu Leu Phe Tyr
35 40 45
Ser Val Gly Ile Gly Gln Ile Ser Ser Glu Val Pro Gln Asn Leu Phe
50 55 60 65
Ala Ser Arg Pro Gln Asp Arg Gln Phe Ile Leu Ser Ser Lys Lys Leu
70 75 80
Leu Leu Ser Asp Ser Gly Phe Tyr Leu Cys Ala Trp Ser Ile Gly Ala
85 90 95
Glu Gln Phe Phe Gly Pro Gly Thr Arg Leu Thr Val Leu Glu Asp Leu
100 105 110
Lys Asn
<210> 13
<211> 110
<212> ПРТ
<213> Homo sapiens
<400> 13
Gln Lys Glu Val Glu Gln Asn Ser Gly Pro Leu Ser Val Pro Glu Gly
1 5 10 15
Ala Ile Ala Ser Leu Asn Cys Thr Tyr Ser Asp Arg Gly Ser Gln Ser
20 25 30
Phe Phe Trp Tyr Arg Gln Tyr Ser Gly Lys Ser Pro Glu Leu Ile Met
35 40 45
Ser Ile Tyr Ser Asn Gly Asp Lys Glu Asp Gly Arg Phe Thr Ala Gln
50 55 60
Leu Asn Lys Ala Ser Gln Tyr Val Ser Leu Leu Ile Arg Asp Ser Gln
65 70 75 80
Pro Ser Asp Ser Ala Thr Tyr Leu Cys Ala Ala Asn Asn Ala Arg Leu
85 90 95
Met Phe Gly Asp Gly Thr Gln Leu Val Val Lys Pro Asn Ile
100 105 110
<210> 14
<211> 9
<212> ПРТ
<213> Homo sapiens
<400> 14
Arg Met Phe Pro Asn Ala Pro Tyr Leu
1 5
<210> 15
<211> 9
<212> ПРТ
<213> Вирус гриппа A
<400> 15
Ala Ser Asn Glu Asn Met Asp Ala Met
1 5
<210> 16
<211> 9
<212> ПРТ
<213> Вирус гриппа A
<400> 16
Ala Ser Asn Glu Asn Met Glu Thr Met
1 5
<---
Claims (11)
1. Модифицированный T-клеточный рецептор или его антигенсвязывающий фрагмент, содержащий Vα и Vβ, происходящие из T-клеточного рецептора дикого типа, при этом аминокислотная последовательность Vα дикого типа представлена в SEQ ID NO: 13, а аминокислотная последовательность Vβ дикого типа представлена в SEQ ID NO: 12; и при этом указанный Vα содержит следующие участки, определяющие комплементарность (CDR): CDR1: YSDRGSQSF, CDR2: IYSNGDK, и CDR3: VSKGYKM или VHGWYTM, и при этом указанный Vβ содержит следующие CDR: CDR1: GTSNP, CDR2: YSVGIG, и CDR3: AWSIGAEQFFG.
2. Модифицированный T-клеточный рецептор по п. 1, отличающийся тем, что указанный модифицированный T-клеточный рецептор включает последовательность аминокислот Vα SEQ ID NO: 1.
3. Модифицированный T-клеточный рецептор по п. 1, отличающийся тем, что указанный модифицированный T-клеточный рецептор включает последовательность аминокислот Vα SEQ ID NO: 2.
4. Модифицированный T-клеточный рецептор по п. 1, отличающийся тем, что указанный модифицированный T-клеточный рецептор включает одноцепочечный T-клеточный рецептор с последовательностью аминокислот SEQ ID NO: 3.
5. Модифицированный T-клеточный рецептор по п. 1, отличающийся тем, что указанный модифицированный T-клеточный рецептор включает одноцепочечный T-клеточный рецептор с последовательностью аминокислот SEQ ID NO: 4.
6. Модифицированный T-клеточный рецептор по п. 1, который находится в растворимой форме.
7. Композиция, которая нацелена на клетки рака, экспрессирующие антиген сурвивин, при этом указанная композиция включает модифицированный T-клеточный рецептор по п. 6, и при этом указанный T-клеточный рецептор связан с токсином, радиоактивным изотопом, химиотерапевтическим агентом, scFv антитела к CD3, ингибитором ангиогенеза, ингибитором микротрубочек, ДНК-интеркалятором или перекрестносшивающим агентом, ингибитором синтеза ДНК или цитокином, выбранным из IL-10, IL-12, IL-15, IL-37, GM-CSF, TNF-α, TGF-β, IFN-γ или LT-α.
8. Композиция, которая нацелена на клетки рака, экспрессирующие антиген сурвивин, при этом указанная композиция включает T-клетку человека, которая экспрессирует модифицированный T-клеточный рецептор по п. 1.
9. Композиция, которая нацелена на клетки рака, экспрессирующие антиген сурвивин, при этом указанная композиция включает модифицированный T-клеточный рецептор по п. 3, и при этом указанный T-клеточный рецептор связан с токсином, радиоактивным изотопом, химиотерапевтическим агентом, scFv антитела к CD3, ингибитором ангиогенеза, ингибитором микротрубочек, ДНК-интеркалятором или перекрестносшивающим агентом, ингибитором синтеза ДНК или цитокином, выбранным из IL-10, IL-12, IL-15, IL-37, GM-CSF, TNF-α, TGF-β, IFN-γ или LT-α.
10. Композиция, которая нацелена на клетки рака, экспрессирующие антиген сурвивин, при этом указанная композиция включает T-клетку человека, которая экспрессирует модифицированный T-клеточный рецептор по п. 3.
11. Способ лечения субъекта, имеющего рак, который экспрессирует антиген сурвивин, при этом указанный способ включает введение композиции по любому из пп. 7, 8, 9 и 10.
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201361907887P | 2013-11-22 | 2013-11-22 | |
| US61/907,887 | 2013-11-22 | ||
| PCT/US2014/066892 WO2015077607A1 (en) | 2013-11-22 | 2014-11-21 | Engineered high-affinity human t cell receptors |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| RU2016124163A RU2016124163A (ru) | 2017-12-27 |
| RU2729383C2 true RU2729383C2 (ru) | 2020-08-06 |
Family
ID=53180203
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| RU2016124163A RU2729383C2 (ru) | 2013-11-22 | 2014-11-21 | Сконструированные высокоаффинные t-клеточные рецепторы человека |
| RU2016124179A RU2740648C2 (ru) | 2013-11-22 | 2014-11-21 | Сконструированные высокоаффинные t-клеточные рецепторы человека |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| RU2016124179A RU2740648C2 (ru) | 2013-11-22 | 2014-11-21 | Сконструированные высокоаффинные t-клеточные рецепторы человека |
Country Status (20)
| Country | Link |
|---|---|
| US (4) | US10023625B2 (ru) |
| EP (2) | EP3071594A4 (ru) |
| JP (4) | JP6697386B2 (ru) |
| KR (2) | KR102415259B1 (ru) |
| CN (2) | CN105873945B (ru) |
| AU (4) | AU2014352826B2 (ru) |
| BR (1) | BR112016011567A2 (ru) |
| CA (2) | CA2930847A1 (ru) |
| DK (1) | DK3071593T3 (ru) |
| ES (1) | ES2729406T3 (ru) |
| IL (2) | IL245468B (ru) |
| MX (3) | MX371202B (ru) |
| PL (1) | PL3071593T3 (ru) |
| PT (1) | PT3071593T (ru) |
| RU (2) | RU2729383C2 (ru) |
| SA (1) | SA516371174B1 (ru) |
| SG (2) | SG10201804330YA (ru) |
| TR (1) | TR201908404T4 (ru) |
| WO (2) | WO2015077615A1 (ru) |
| ZA (2) | ZA201603116B (ru) |
Families Citing this family (24)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CA2626238C (en) | 2005-10-17 | 2015-10-06 | Sloan Kettering Institute For Cancer Research | Wt1 hla class ii-binding peptides and compositions and methods comprising same |
| EP2010209B1 (en) | 2006-04-10 | 2016-06-15 | Sloan Kettering Institute For Cancer Research | Immunogenic wt-1 peptides and methods of use thereof |
| WO2013106834A2 (en) | 2012-01-13 | 2013-07-18 | Memorial Sloan Kettering Cancer Center | Immunogenic wt-1 peptides and methods of use thereof |
| US10815273B2 (en) | 2013-01-15 | 2020-10-27 | Memorial Sloan Kettering Cancer Center | Immunogenic WT-1 peptides and methods of use thereof |
| SI2945647T1 (sl) | 2013-01-15 | 2021-01-29 | Memorial Sloan Kettering Cancer Center | Imunogeni peptidi WT-1 in postopki za uporabo le-teh |
| PT3071593T (pt) | 2013-11-22 | 2019-06-27 | The Board Of Trustees Of The Univ Of Illionis | Recetores das células t de humanos manipulados de alta-afinidade |
| CA2999608A1 (en) * | 2015-09-22 | 2017-03-30 | Julius-Maximilians-Universitat Wurzburg | A method for high level and stable gene transfer in lymphocytes |
| AU2016329542C1 (en) * | 2015-10-01 | 2023-11-23 | Fondazione Centro San Raffaele | TCR and uses thereof |
| GB201520570D0 (en) * | 2015-11-23 | 2016-01-06 | Immunocore Ltd & Adaptimmune Ltd | Peptides |
| EP3394092A1 (en) | 2015-12-23 | 2018-10-31 | Fred Hutchinson Cancer Research Center | High affinity t cell receptors and uses thereof |
| MA45491A (fr) * | 2016-06-27 | 2019-05-01 | Juno Therapeutics Inc | Épitopes à restriction cmh-e, molécules de liaison et procédés et utilisations associés |
| AU2017356322B2 (en) * | 2016-11-14 | 2023-09-21 | Fred Hutchinson Cancer Center | High affinity merkel cell polyomavirus T antigen-specific TCRs and uses thereof |
| SG11201908527SA (en) * | 2017-03-15 | 2019-10-30 | Hutchinson Fred Cancer Res | High affinity mage-a1-specific tcrs and uses thereof |
| CN110785432A (zh) * | 2017-04-24 | 2020-02-11 | 圣拉斐尔医院有限责任公司 | Tcr和肽 |
| JP7181517B2 (ja) * | 2018-01-23 | 2022-12-01 | 国立大学法人三重大学 | T細胞レセプター |
| WO2020023556A1 (en) * | 2018-07-23 | 2020-01-30 | Magenta Therapeutics, Inc. | Use of anti-cd137 antibody drug conjugate (adc) in allogeneic cell therapy |
| GB201911066D0 (en) | 2019-08-02 | 2019-09-18 | Achilles Therapeutics Ltd | T cell therapy |
| AU2020334064A1 (en) * | 2019-08-20 | 2022-03-03 | Fred Hutchinson Cancer Center | T-cell immunotherapy specific for WT-1 |
| WO2021163366A1 (en) * | 2020-02-12 | 2021-08-19 | Abbvie Inc. | Bispecific binding molecules |
| IL297594A (en) | 2020-04-28 | 2022-12-01 | Achilles Therapeutics Uk Ltd | t cell therapy |
| GB202109886D0 (en) | 2021-07-08 | 2021-08-25 | Achilles Therapeutics Uk Ltd | Assay |
| US20220323500A1 (en) | 2021-04-09 | 2022-10-13 | Achilles Therapeutics Uk Limited | Cancer immunotherapy |
| KR20240023426A (ko) | 2021-06-22 | 2024-02-21 | 아킬레스 테라퓨틱스 유케이 리미티드 | 항원-특이적 t 세포를 생산하는 방법 |
| EP4373516A1 (en) * | 2021-07-19 | 2024-05-29 | Board of Regents, The University of Texas System | Peptides and engineered t cell receptors targeting fanci, rad51, and pbk antigens and methods of use |
Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EA008026B1 (ru) * | 2003-01-30 | 2007-02-27 | СУРВАК АпС | Производные от сурвивина пептиды и их применение |
| US20080125369A1 (en) * | 2001-08-31 | 2008-05-29 | Avidex Limited | Soluble t cell receptors |
| WO2010075417A1 (en) * | 2008-12-23 | 2010-07-01 | The United States Of America, As Represented By The Secretary, Department Of Health And Human Services | Survivin specific t cell receptor for treating cancer |
| US20120128704A1 (en) * | 2009-02-09 | 2012-05-24 | Max-Delbrueck-Centrum Fuer Molekulare Medizin | Repertoire of allo-restricted peptide-specific t cell receptor sequences and use thereof |
Family Cites Families (45)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| NL154598B (nl) | 1970-11-10 | 1977-09-15 | Organon Nv | Werkwijze voor het aantonen en bepalen van laagmoleculire verbindingen en van eiwitten die deze verbindingen specifiek kunnen binden, alsmede testverpakking. |
| US3817837A (en) | 1971-05-14 | 1974-06-18 | Syva Corp | Enzyme amplification assay |
| US3927193A (en) | 1973-05-18 | 1975-12-16 | Hoffmann La Roche | Localization of tumors by radiolabelled antibodies |
| US3939350A (en) | 1974-04-29 | 1976-02-17 | Board Of Trustees Of The Leland Stanford Junior University | Fluorescent immunoassay employing total reflection for activation |
| US3996345A (en) | 1974-08-12 | 1976-12-07 | Syva Company | Fluorescence quenching with immunological pairs in immunoassays |
| US4277437A (en) | 1978-04-05 | 1981-07-07 | Syva Company | Kit for carrying out chemically induced fluorescence immunoassay |
| US4275149A (en) | 1978-11-24 | 1981-06-23 | Syva Company | Macromolecular environment control in specific receptor assays |
| US4331647A (en) | 1980-03-03 | 1982-05-25 | Goldenberg Milton David | Tumor localization and therapy with labeled antibody fragments specific to tumor-associated markers |
| US4348376A (en) | 1980-03-03 | 1982-09-07 | Goldenberg Milton David | Tumor localization and therapy with labeled anti-CEA antibody |
| US4444744A (en) | 1980-03-03 | 1984-04-24 | Goldenberg Milton David | Tumor localization and therapy with labeled antibodies to cell surface antigens |
| US4361544A (en) | 1980-03-03 | 1982-11-30 | Goldenberg Milton David | Tumor localization and therapy with labeled antibodies specific to intracellular tumor-associated markers |
| US4366241A (en) | 1980-08-07 | 1982-12-28 | Syva Company | Concentrating zone method in heterogeneous immunoassays |
| US4468457A (en) | 1981-06-01 | 1984-08-28 | David M. Goldenberg | Method for producing a CSAp tryptic peptide and anti-CSAp antibodies |
| US5101827A (en) | 1985-07-05 | 1992-04-07 | Immunomedics, Inc. | Lymphographic and organ imaging method and kit |
| US4640561A (en) | 1985-11-15 | 1987-02-03 | Ford Motor Company | Flexible printed circuit connector |
| US5059413A (en) | 1988-04-18 | 1991-10-22 | Xoma Corporation | Scintigraphic monitoring of immunotoxins using radionuclides and heterobifunctional chelators |
| ATE172879T1 (de) | 1989-08-09 | 1998-11-15 | Rhomed Inc | Direkte radioetikettierung von antikörpern und sonstigen proteinen mittels technetium oder rhenium |
| US5299253A (en) | 1992-04-10 | 1994-03-29 | Akzo N.V. | Alignment system to overlay abdominal computer aided tomography and magnetic resonance anatomy with single photon emission tomography |
| US6699658B1 (en) | 1996-05-31 | 2004-03-02 | Board Of Trustees Of The University Of Illinois | Yeast cell surface display of proteins and uses thereof |
| US6696251B1 (en) | 1996-05-31 | 2004-02-24 | Board Of Trustees Of The University Of Illinois | Yeast cell surface display of proteins and uses thereof |
| US6300065B1 (en) | 1996-05-31 | 2001-10-09 | Board Of Trustees Of The University Of Illinois | Yeast cell surface display of proteins and uses thereof |
| US6759243B2 (en) | 1998-01-20 | 2004-07-06 | Board Of Trustees Of The University Of Illinois | High affinity TCR proteins and methods |
| JP2002515244A (ja) | 1998-05-19 | 2002-05-28 | アヴィデックス リミテッド | 可溶性t細胞受容体 |
| WO2000023087A1 (en) | 1998-10-21 | 2000-04-27 | Sunol Molecular Corporation | Polyspecific binding molecules and uses thereof |
| AU2003271904B2 (en) | 2002-10-09 | 2009-03-05 | Adaptimmune Limited | Single chain recombinant T cell receptors |
| GB0304068D0 (en) | 2003-02-22 | 2003-03-26 | Avidex Ltd | Substances |
| GB0328363D0 (en) | 2003-12-06 | 2004-01-14 | Imp College Innovations Ltd | Therapeutically useful molecules |
| ATE417065T1 (de) | 2004-05-19 | 2008-12-15 | Medigene Ltd | Hochaffiner ny-eso-t-zellen-rezeptor |
| JP4972549B2 (ja) | 2004-05-19 | 2012-07-11 | イムノコア リミテッド | T細胞レセプターを改善する方法 |
| GB0524477D0 (en) * | 2005-11-30 | 2006-01-11 | Avidex Ltd | Isolated T cell receptors which specifically bind to vygfvracl-hla-24 |
| GB0511124D0 (en) | 2005-06-01 | 2005-07-06 | Avidex Ltd | High affinity melan-a t cell receptors |
| AT503861B1 (de) | 2006-07-05 | 2008-06-15 | F Star Biotech Forsch & Entw | Verfahren zur manipulation von t-zell-rezeptoren |
| WO2008039818A2 (en) | 2006-09-26 | 2008-04-03 | Government Of The United States Of America, Represented By The Secretary, Department Of Health And Human Services | Modified t cell receptors and related materials and methods |
| US9281917B2 (en) | 2007-01-03 | 2016-03-08 | Nokia Technologies Oy | Shared control channel structure |
| CA2743669C (en) * | 2008-11-24 | 2018-10-16 | Helmholtz Zentrum Muenchen Deutsches Forschungszentrum Fuer Gesundheit Und Umwelt (Gmbh) | High affinity t cell receptor and use thereof |
| GB0911566D0 (en) * | 2009-07-03 | 2009-08-12 | Immunocore Ltd | T cell receptors |
| US10464987B2 (en) * | 2009-10-06 | 2019-11-05 | Abbvie Inc. | Human single-chain T cell receptors |
| EA201390011A1 (ru) | 2010-07-28 | 2013-07-30 | Иммьюнокор Лтд. | Т-клеточные рецепторы |
| DK2618835T3 (en) | 2010-09-20 | 2017-08-28 | Biontech Cell & Gene Therapies Gmbh | ANTIGEN-SPECIFIC T-CELL RECEPTORS AND T-CELL EPITOPES |
| MX361760B (es) | 2012-05-03 | 2018-12-17 | Hutchinson Fred Cancer Res | Receptores de celulas t con afinidad aumentada y metodos para hacer los mismos. |
| WO2014191465A1 (en) * | 2013-05-28 | 2014-12-04 | Møller Niels Iversen | Peptides derived from lawsonia intracellularis and their use in vaccination |
| PL228457B1 (pl) | 2013-08-30 | 2018-03-30 | Univ Jagiellonski | Tomograf hybrydowy TOF-PET/CT |
| PT3071593T (pt) | 2013-11-22 | 2019-06-27 | The Board Of Trustees Of The Univ Of Illionis | Recetores das células t de humanos manipulados de alta-afinidade |
| WO2016022400A1 (en) | 2014-08-04 | 2016-02-11 | Fred Hutchinson Cancer Research Center | T cell immunotherapy specific for wt-1 |
| CN113105562B (zh) * | 2018-09-26 | 2023-12-01 | 安源医药科技(上海)有限公司 | 突变型单链人凝血因子viii在制备融合蛋白中的应用 |
-
2014
- 2014-11-21 PT PT14863490T patent/PT3071593T/pt unknown
- 2014-11-21 SG SG10201804330YA patent/SG10201804330YA/en unknown
- 2014-11-21 AU AU2014352826A patent/AU2014352826B2/en active Active
- 2014-11-21 CA CA2930847A patent/CA2930847A1/en not_active Abandoned
- 2014-11-21 ES ES14863490T patent/ES2729406T3/es active Active
- 2014-11-21 KR KR1020167016131A patent/KR102415259B1/ko active IP Right Grant
- 2014-11-21 AU AU2014352834A patent/AU2014352834B2/en not_active Ceased
- 2014-11-21 RU RU2016124163A patent/RU2729383C2/ru active
- 2014-11-21 DK DK14863490.0T patent/DK3071593T3/da active
- 2014-11-21 PL PL14863490T patent/PL3071593T3/pl unknown
- 2014-11-21 WO PCT/US2014/066903 patent/WO2015077615A1/en active Application Filing
- 2014-11-21 EP EP14864079.0A patent/EP3071594A4/en active Pending
- 2014-11-21 EP EP14863490.0A patent/EP3071593B1/en active Active
- 2014-11-21 US US15/037,476 patent/US10023625B2/en active Active
- 2014-11-21 RU RU2016124179A patent/RU2740648C2/ru active
- 2014-11-21 TR TR2019/08404T patent/TR201908404T4/tr unknown
- 2014-11-21 MX MX2016006620A patent/MX371202B/es active IP Right Grant
- 2014-11-21 MX MX2016006625A patent/MX2016006625A/es unknown
- 2014-11-21 BR BR112016011567A patent/BR112016011567A2/pt not_active Application Discontinuation
- 2014-11-21 JP JP2016533163A patent/JP6697386B2/ja not_active Expired - Fee Related
- 2014-11-21 KR KR1020167016132A patent/KR20160085345A/ko not_active Application Discontinuation
- 2014-11-21 CA CA2930852A patent/CA2930852C/en active Active
- 2014-11-21 CN CN201480071620.2A patent/CN105873945B/zh active Active
- 2014-11-21 SG SG10201804335QA patent/SG10201804335QA/en unknown
- 2014-11-21 WO PCT/US2014/066892 patent/WO2015077607A1/en active Application Filing
- 2014-11-21 US US15/037,485 patent/US10344075B2/en active Active
- 2014-11-21 JP JP2016533150A patent/JP6476182B2/ja active Active
- 2014-11-21 CN CN201480071635.9A patent/CN105899530B/zh not_active Expired - Fee Related
-
2016
- 2016-05-04 IL IL245468A patent/IL245468B/en active IP Right Grant
- 2016-05-04 IL IL245467A patent/IL245467B/en not_active IP Right Cessation
- 2016-05-10 ZA ZA2016/03116A patent/ZA201603116B/en unknown
- 2016-05-11 ZA ZA2016/03169A patent/ZA201603169B/en unknown
- 2016-05-20 MX MX2021006932A patent/MX2021006932A/es unknown
- 2016-05-21 SA SA516371174A patent/SA516371174B1/ar unknown
-
2018
- 2018-05-25 US US15/990,524 patent/US20190055298A1/en not_active Abandoned
-
2019
- 2019-10-17 JP JP2019190139A patent/JP6970724B2/ja active Active
- 2019-11-28 AU AU2019272003A patent/AU2019272003B2/en not_active Ceased
-
2021
- 2021-04-15 AU AU2021202274A patent/AU2021202274A1/en not_active Abandoned
- 2021-11-01 JP JP2021178468A patent/JP2022023196A/ja active Pending
-
2022
- 2022-01-27 US US17/586,652 patent/US20220396606A1/en active Pending
Patent Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20080125369A1 (en) * | 2001-08-31 | 2008-05-29 | Avidex Limited | Soluble t cell receptors |
| EA008026B1 (ru) * | 2003-01-30 | 2007-02-27 | СУРВАК АпС | Производные от сурвивина пептиды и их применение |
| WO2010075417A1 (en) * | 2008-12-23 | 2010-07-01 | The United States Of America, As Represented By The Secretary, Department Of Health And Human Services | Survivin specific t cell receptor for treating cancer |
| US20120128704A1 (en) * | 2009-02-09 | 2012-05-24 | Max-Delbrueck-Centrum Fuer Molekulare Medizin | Repertoire of allo-restricted peptide-specific t cell receptor sequences and use thereof |
Non-Patent Citations (4)
Also Published As
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| AU2019272003B2 (en) | Engineered high-affinity human T cell receptors | |
| AU2018200222B2 (en) | Engineering T-cell receptors | |
| NZ719707B2 (en) | Engineered human t cell receptors with high affinity for binding survivin | |
| NZ719720B2 (en) | Engineered human t cell receptors with high affinity for wt1 |