KR20200047176A - 투라노스의 제조방법 - Google Patents
투라노스의 제조방법 Download PDFInfo
- Publication number
- KR20200047176A KR20200047176A KR1020180129298A KR20180129298A KR20200047176A KR 20200047176 A KR20200047176 A KR 20200047176A KR 1020180129298 A KR1020180129298 A KR 1020180129298A KR 20180129298 A KR20180129298 A KR 20180129298A KR 20200047176 A KR20200047176 A KR 20200047176A
- Authority
- KR
- South Korea
- Prior art keywords
- leu
- asp
- ala
- arg
- gly
- Prior art date
Links
Images





Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P19/00—Preparation of compounds containing saccharide radicals
- C12P19/12—Disaccharides
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P19/00—Preparation of compounds containing saccharide radicals
- C12P19/14—Preparation of compounds containing saccharide radicals produced by the action of a carbohydrase (EC 3.2.x), e.g. by alpha-amylase, e.g. by cellulase, hemicellulase
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y204/00—Glycosyltransferases (2.4)
- C12Y204/01—Hexosyltransferases (2.4.1)
- C12Y204/01004—Amylosucrase (2.4.1.4)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y302/00—Hydrolases acting on glycosyl compounds, i.e. glycosylases (3.2)
- C12Y302/01—Glycosidases, i.e. enzymes hydrolysing O- and S-glycosyl compounds (3.2.1)
- C12Y302/01003—Glucan 1,4-alpha-glucosidase (3.2.1.3), i.e. glucoamylase
Landscapes
- Organic Chemistry (AREA)
- Chemical & Material Sciences (AREA)
- Engineering & Computer Science (AREA)
- Life Sciences & Earth Sciences (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Health & Medical Sciences (AREA)
- Biochemistry (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Microbiology (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Biotechnology (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Enzymes And Modification Thereof (AREA)
Abstract
본 발명은 투라노스 생산 효소 및 이를 이용하여 투라노스를 효율적으로 생산하는 방법에 관한 것이다.
Description
본 발명은 투라노스 생산 효소 및 이를 이용하여 투라노스를 효율적으로 생산하는 방법에 관한 것이다.
투라노스는 자연적으로 벌에서 생기는 환원성 이당류로서 설탕 단맛의 약 50%에 해당하는 단맛을 나타내는 설탕의 유사체이며 3-O-α-D-글루코피라노실-D-프락토즈의 화학적 구조를 지닌다. 투라노스는 치아 우식 유발 미생물에 의해 발효되지 않기 때문에 충치 예방에 도움이 되며, 칼로리 없는 감미료로서 사용될 수 있으며, 따라서 음식, 화장품 및 약학 산업계에서 중요한 역할을 수행할 수 있다.
한국특허 제10-11772184호는 아밀로수크라제를 이용하여 설탕과 프럭토스를 기질로 투라노스를 제조하는 방법을 포함한다. 그러나, 기존 효소들은 고열에서 활성을 나타내지 못하거나 유지하지 못하여, 공정에 적합한 최적 온도인 50℃에서 투라노스 전환반응 시 수크로스가 모두 소모되기 전에 활성을 잃는다. 결과적으로 아밀로수크라제에 의한 투라노스 전환 반응 시 최종 전환액에 설탕이 남게 되어 추후 투라노스 고순도 분리를 하는 데 결정적 방해 요인이 된다. 또한 기존의 효소에 의한 투라노스 제조는 반응 속도가 매우 느려서 상업화에 적합하지 않다. 따라서 50℃이상의 온도에서 투라노스 전환반응 시에 수크로스를 모두 소모하며, 반응 속도가 개선된 아밀로수크라제를 개발하는 것이 매우 중요하다.
투라노스 생산 활성을 갖는 아밀로수크라제는 미생물 Neisseria polysaccharea, Deinococcus geothermalis 및 Alteromonas macleodii 등에서 유래된 것이 알려져 있다.
투라노스 생산 활성을 갖는 아밀로수크라제를 이용하여, 수크로스 함유 당류를 기질로 사용하여 투라노스를 제조하는 경우, 반응 생성물에는 투라노스 뿐만 아니라 다양한 당류를 포함하며, 고순도 및 고수율로 투라노스를 생산하기 위해서는, 높은 전환율로 투라노스를 생산하는 효소를 포함하는 촉매 조성물이 필요하지만, 또한 분리 및 정제 단계에서 반응생성물에 포함된 투라노스 이외의 당류의 존재로 인한 영향을 최소화하고 고순도로 투라노스를 얻을 필요가 있다.
본 발명은 높은 전환율로 투라노스를 생산하고 기타 당류의 생산을 최소화하는, 비피도박테리움 터모필럼 유래의 아밀로수크라제 효소 단백질 또는 이의 변이체를 제공하는 것을 목적으로 한다.
본 발명의 또 다른 목적은 상기 아밀로수크라제 효소 단백질 또는 이의 변이체를 이용하여 투라노스를 생산하는 방법에 관한 것이다.
본 발명의 추가 목적은, 상기 아밀로수크라제 효소 단백질 또는 이의 변이체를 이용하여 투라노스를 생산하는 단계 및 모사이동층 크로마토그래피를 이용하여 고순도의 투라노스를 얻은 단계를 포함하는 고순도 투라노스를 생산하는 방법에 관한 것이다.
본 발명의 일 예는 높은 전환율로 투라노스를 생산하고 기타 당류의 생산을 최소화하는, 비피도박테리움 터모필럼 유래의 아밀로수크라제 효소 단백질 또는 이의 변이체, 및 상기 효소 단백질 또는 이의 변이체를 이용하여 투라노스를 생산하는 방법에 관한 것이다.
본 발명의 일 예는 비피도박테리움 터모필럼에서 유래되며 투라노스의 전환 활성을 갖는 효소 단백질 또는 변이형 효소 단백질, 이를 발현하는 재조합 미생물, 상기 재조합 미생물의 배양물 또는 상기 재조합 미생물의 파쇄물(또는 파쇄물의 상등액)로 이루어진 군에서 선택된 1 이상을 포함하는 투라노스 생산용 조성물을 제공한다.
본 발명의 또 다른 일 예로서, 투라노스를 생성하는 것을 특징으로 하는 비피도박테리움 터모필럼 유래의 효소 단백질, 이를 발현하는 재조합 미생물, 상기 재조합 미생물의 배양물 또는 상기 재조합 미생물의 파쇄물(또는 파쇄물의 상등액)로 이루어진 군에서 선택된 1 이상을 포함하는 투라노스 생산용 조성물을 기질과 반응시키는 단계를 포함하는 투라노스 생산 방법을 제공한다. 상기 효소 단백질에 관한 사항은 상기 투라노스 생산용 조성물 및 제조 방법에 동일하게 적용될 수 있다.
또한, 본 발명의 일 예는, 상기 아밀로수크라제 효소 단백질 또는 이의 변이체를 이용하여 투라노스를 생산하는 단계 및 모사이동층 크로마토그래피를 이용하여 고순도의 투라노스를 얻은 단계를 포함하는 고순도 투라노스를 생산하는 방법에 관한 것이다.
본 발명에 따른 투라노스 생산 활성을 갖는 아밀로수크라제는 수크로스 함유 당류를 기질로 사용하여 투라노스를 제조하는 경우, 높은 전환율로 투라노스를 생산할 뿐만 아니라, 반응 생성물에는 투라노스 이외의 당류들, 예를 들면 트레할룰로스 및 올리고당의 함량이 낮아 분리 및 정제 단계에서 반응생성물에 포함된 투라노스 이외의 당류의 존재로 인한 영향을 최소화하고 고순도로 투라노스를 얻을 수 있다.
이하, 본 발명을 더욱 자세히 설명하고자 한다.
본 발명의 일 예는 비피도박테리움 터모필럼에서 유래되며 투라노스의 전환 활성을 갖는 효소 단백질 또는 변이형 효소 단백질, 이를 발현하는 재조합 미생물, 상기 재조합 미생물의 배양물 또는 상기 재조합 미생물의 파쇄물(또는 파쇄물의 상등액)로 이루어진 군에서 선택된 1 이상을 포함하는 투라노스 생산용 조성물을 제공한다.
본 발명의 일 예는 비피도박테리움 터모필럼에서 유래되며 투라노스의 전환 활성을 갖는 효소 단백질 또는 변이형 효소 단백질, 이를 발현하는 재조합 미생물, 상기 재조합 미생물의 배양물 또는 상기 재조합 미생물의 파쇄물(또는 파쇄물의 상등액)로 이루어진 군에서 선택된 1 이상을 포함하는 투라노스 생산용 조성물과 수크로스 또는 수크로스와 프럭토스를 포함하는 기질과 반응하여 투라노스를 생산하는 방법에 관한 것이다.
또한, 본 발명은 비피도박테리움 터모필럼 유래의 아밀로수크라제 효소 단백질 또는 이의 변이체를 이용하여 투라노스를 생산하는 단계를 포함하는 투라노스 제조방법에 관한 것으로서, 또한, 투라노스를 생산하는 단계에 더하여 모사이동층 크로마토그래피를 이용하여 분리하는 단계를 포함한다. 생물학적 방법으로 설탕을 이용하여 투라노스를 제조하는 경우, 반응산물에는 분리컬럼 상에서 투라노스를 기준으로 앞뒤로 밀접하게 붙어있는 올리고머와 트레할룰로스 때문에 투라노스만 선택적으로 분리하기 어려운 단점이 있다. 특히 올리고머, 투라노스, 트레할룰로스가 크로마토그램 상에서 연속적으로 인접하여 피크를 형성하게 되어 분리가 어렵다는 문제점이 있다.
본 발명에 적용되는 투라노스 생산용 조성물은 비피도박테리움 터모필럼에서 유래되며 투라노스의 전환 활성을 갖는 효소 단백질 또는 변이형 효소 단백질, 이를 발현하는 재조합 미생물, 상기 재조합 미생물의 배양물 또는 상기 재조합 미생물의 파쇄물(또는 파쇄물의 상등액)로 이루어진 군에서 선택된 1 이상을 포함할 수 있다. 상기 조성물은, 비피도박테리움 터모필럼 유래의 효소 단백질을 발현하는 재조합 미생물의 배양물을 원심분리하는 단계; 및 상기 원심분리된 농축물을 포함할 수 침전물을 파쇄하는 단계; 및 상기 파쇄물의 상등액을 얻는 단계를 포함하여 제조된 세포일 수 있다.
본 발명의 효소는 비피도박테리움 터모필럼(Bifidobacterium thermophilum) 유래의 효소 단백질일 수 있다. 본 발명의 효소 단백질은 투라노스를 생산하는 아밀로수크라제(amylosucrase)로서, 수크로스 단독 또는 수크로스 및 프럭토스를 포함하는 기질 용액으로부터 투라노스를 생성할 수 있다.
본 발명에 따른 투라노스 제조방법에 적용 가능한 투라노스 생성 특성을 갖는 아밀로수크라제 효소 및 이의 변이 효소 단백질은 생산조건을 만족하는 한 사용될 수 있다. 상기 비피도박테리움 터모필럼 유래의 효소 단백질은 서열번호 1의 아미노산 서열을 갖는 단백질일 수 있으나, 투라노스를 생성할 수 있는 활성이 있는 한 이에 한정되지 않고 상기 서열번호 1의 아미노산 서열 중에서 1 이상의 아미노산이 치환, 삽입 또는 결실된 변이 단백질을 모두 포함할 수 있다. 예를 들어, 상기 효소 단백질 또는 이의 변이형 효소는, 서열번호 1의 아미노산 서열과 70% 이상, 바람직하게는 80% 이상, 더욱 바람직하게는 90% 이상, 93% 이상 또는 95% 이상, 더욱 바람직하게는 98% 이상, 99%이상 또는 99.5%이상의 상동성 있는 아미노산 서열을 갖는 효소 단백질일 수 있다.
본 발명에서 적용되는 투라노스 생산용 조성물은 비피도박테리움 터모필럼에서 유래되며 투라노스의 전환 활성을 가지는 치환 변이에 의한 변이형 효소 단백질의 예는, 서열번호 1를 갖는 아미노산 서열의 200번째, 202번째, 265번째, 305번째, 393번째, 414번째, 420번째 및 542번째 위치로 이루어진 군에서 선택된 1 이상의 위치의 아미노산이 치환된 것일 수 있다. 예를 들어, 서열번호 1를 갖는 아미노산 서열의 200번째, 202번째, 265번째, 305번째, 393번째, 414번째, 420번째 및 542번째 위치로 이루어진 군에서 선택된 1 이상의 위치의 아미노산이, 아르기닌, 히스티딘, 아스파르트산, 글루탐산, 이소루신, 페닐알라닌 및 트레오닌으로 이루어진 군에서 선택된 적어도 1종의 아미노산으로 치환된 것일 수 있다.
구체적인 일예에서, 상기 치환 변이에 의한 변이형 효소 단백질의 예는, 상기 서열번호 1를 갖는 아미노산 서열의 200번째, 202번째, 265번째, 305번째, 393번째, 414번째 및 420번째 위치로 이루어진 군에서 선택된 1 이상의 위치의 아미노산이 아르기닌, 이소루신, 페닐알라닌 및 트레오닌으로 이루어진 군에서 선택된 1종의 아미노산으로 치환된 것일 수 있다. 서열번호 1를 갖는 아미노산 서열의 아미노산이 아래의 아미노산 치환 중 하나 이상을 포함할 수 있다:
200번째 아미노산이 아르기닌으로 치환,
202번째 아미노산이 이소루신으로 치환,
265번째 아미노산이 페닐알라닌으로 치환,
305번째 아미노산이 이소루신으로 치환,
393번째 아미노산이 아르기닌으로 치환,
414번째 아미노산이 페닐알라닌으로 치환, 및
420번째 아미노산이 트레오닌으로 치환.
예를 들어, 상기 변이형 효소 단백질은 아미노산 변이 1개를 갖는 서열번호 2 내지 10의 아미노산 서열 중 어느 하나의 서열을 갖는 것일 수 있으며, 또는 서열번호 1를 갖는 아미노산 서열의 200번째, 202번째, 265번째, 305번째, 393번째, 414번째 및 420번째 위치로 이루어진 군에서 선택된 2 이상의 위치의 아미노산이 아르기닌, 이소루신, 페닐알라닌 및 트레오닌으로 이루어진 군에서 선택된 적어도 1종 이상의 아미노산으로 치환된 아미노산 변이 2개 이상을 포함하는 변이형 효소일 수 있으며, 예를 들면 서열번호 1를 갖는 아미노산 서열의 200번째, 202번째, 265번째, 305번째, 393번째, 414번째 및 420번째 위치로 이루어진 군에서 선택된 2 개 위치에서, 아미노산이 아르기닌, 이소루신, 페닐알라닌 및 트레오닌으로 이루어진 군에서 선택된 적어도 1종 이상의 아미노산으로 치환된 2개 변이를 포함하는 변이형 효소, 구체적으로 서열번호 11 내지 13의 아미노산 서열 중에서 선택될 수 있다.
구체적으로, 본 발명의 효소 단백질은 서열번호 2 (변이효소 V542K) 또는 서열번호 3 (변이효소 V542D)의 아미노산 서열을 포함하는 아미노산 서열 중 선택된 어느 하나의 아미노산 서열을 포함하는 효소 단백질일 수 있다. 서열번호 1의 효소는 야생형 효소, 서열번호 2는 서열번호 1의 야생형 효소의 아미노산 서열에서 V542D 변이를 갖는 효소, 서열번호 3은 서열번호 1의 야행형 효소의 아미노산 서열에서 V542K 변이를 갖는 효소를 의미한다. 상기 V542D 변이 또는 V542K 변이를 갖는 변이형 효소 단백질은 활성이 2배 이상 증가하였기 때문에 원가절감과 직결되는 생산성 문제를 개선하는 효과를 기대할 수 있다.
본 발명에 따른 야생형 효소 및 변이 효소의 일 예를 하기 표 1에 나타낸다.
| 구분 | 서열 종류 | 서열번호 |
| 야생형 효소 | 아미노산 서열 | 1 |
| 변이효소 V542K | 아미노산 서열 | 2 |
| 변이효소 V542D | 아미노산 서열 | 3 |
| 변이효소 P200R | 아미노산 서열 | 4 |
| 변이효소 V202I | 아미노산 서열 | 5 |
| 변이효소 Y265F | 아미노산 서열 | 6 |
| 변이효소 V305I | 아미노산 서열 | 7 |
| 변이효소 K393R | 아미노산 서열 | 8 |
| 변이효소 S420T | 아미노산 서열 | 9 |
| 변이효소 Y414F | 아미노산 서열 | 10 |
| 변이효소 Y414F*P200R | 아미노산 서열 | 11 |
| 변이효소 Y414F*V202I | 아미노산 서열 | 12 |
| 변이효소 Y414F*Y265F | 아미노산 서열 | 13 |
| 야생형 효소 | 뉴클레오타이드 서열 | 14 |
| 변이효소 V542K | 뉴클레오타이드 서열 | 15 |
| 변이효소 V542D | 뉴클레오타이드 서열 | 16 |
| 변이효소 P200R | 뉴클레오타이드 서열 | 17 |
| 변이효소 V202I | 뉴클레오타이드 서열 | 18 |
| 변이효소 Y265F | 뉴클레오타이드 서열 | 19 |
| 변이효소 V305I | 뉴클레오타이드 서열 | 20 |
| 변이효소 K393R | 뉴클레오타이드 서열 | 21 |
| 변이효소 S420T | 뉴클레오타이드 서열 | 22 |
| 변이효소 Y414F | 뉴클레오타이드 서열 | 23 |
| 변이효소 Y414F*P200R | 뉴클레오타이드 서열 | 24 |
| 변이효소 Y414F*V202I | 뉴클레오타이드 서열 | 25 |
| 변이효소 Y414F*Y265F | 뉴클레오타이드 서열 | 26 |
종래 알려진 아밀로수크라제는 최적 활성 온도가 35℃ 내외이고, 50℃에서 전환반응을 실행할 경우 낮은 활성과 내열성의 부재로 인하여 전환액에 수크로스를 모두 소진하고 투라노스를 포함한 생성물만 남을 때 까지 효소의 활성이 유지가 되지 않았기 때문에 산업화에 어려움이 있었다. 그러나 본 발명의 효소 단백질은 50℃에서 수크로스가 모두 분해가 될 때까지 반응이 유지가 가능하기 때문에 산업화의 조건을 만족한다고 할 수 있다.
본 발명의 효소 단백질은 최적 온도가 30 내지 65℃, 최적 pH가 5.0 내지 9.0 및 설탕 함유 기질로부터 투라노스 생성하는 활성을 가지며, 바람직하게는 (i) pH 6.0 및 50℃의 온도에서 기질 2M 수크로스를 반응 시, 기질 농도 100%에 대한 생성된 투라노스 농도로 산출되는 투라노스 전환율이 10 내지 60% 및/또는 (ii) pH 6.0 및 50℃의 온도에서 기질 2M 수크로스를 반응 시 시간 당 생성되는 투라노스가 3 내지 80g/L/h로 생산하는 특성을 가질 수 있다.
상기 최적 온도는 효소의 투라노스 생산 활성이 최대가 되는 온도로서, 30 내지 65℃, 바람직하게는 40 내지 65℃, 41 내지 65℃, 42 내지 65℃, 또는 41 내지 60℃, 더욱 바람직하게는 45 내지 55℃, 더욱 바람직하게는 50℃일 수 있다. 종래 알려진 아밀로수크라제는 최적 활성 온도가 35℃내외이고, 40℃ 이상, 예를 들면 45℃ 또는 50℃ 의 온도에서 전환반응을 실행 할 경우 낮은 활성과 내열성의 부재로 인하여 전환액에 수크로스가 남지 않고 투라노스를 포함한 생성물만 남을 때 까지 효소의 활성이 유지가 되지 않았기 때문에 산업화에 어려움이 있었다. 그러나 본 발명의 효소 단백질은 40℃ 이상 또는 40℃ 초과하는 온도, 예를 들면 41℃, 42℃?, 43℃?, 45℃? 또는 50℃ 의 온도에서 수크로스가 모두 분해가 될 때까지 반응이 유지되기 때문에 내열성이 우수하여 산업화에 바람직하다.
변이형 효소 P200R, Y265F, V305I, K393R, S420T, Y414F, Y414F*P200R, Y414F*V202I 또는 Y414F*Y265F는 상기 최적 pH는 효소의 투라노스 생산 활성이 최대가 되는 pH로서, pH 5.0 내지 9.0 또는 pH 5.0 내지 7.0, 바람직하게는 pH 5.5 내지 6.5, 더욱 바람직하게는 pH 6.0일 수 있다.
본 발명의 효소 단백질의 투라노스 전환율은 pH 6.0 및 50℃의 온도에서 기질 2M 수크로스를 반응 시, 기질 농도 100%에 대한 생성된 투라노스 농도로 산출되는 투라노스 전환율이 10 내지 70%, 바람직하게는 20 내지 40% 또는 35 내지 60%, 더욱 바람직하게는 45% 내지 55%일 수 있다. 본 발명의 효소 단백질은, pH6 및 50℃의 온도에서 기질 2M 수크로스를 반응 시, 시간 당 생성되는 투라노스가 3 내지 80g/L/h, 바람직하게는 10 내지 30 g/L/h 또는 20 내지 60 g/L/h, 더욱 바람직하게는 50 g/L/h 내지 60 g/L/h 일 수 있다.
기존에 많은 연구가 선행된 아밀로수크라제 효소 중 투라노스 생산율이 높은 효소로 NpAS(Neisseria polysaccharea) 가 알려져 있다. NpAS는 최적 pH는 pH 7.0, 최적 온도는 35℃, 기질 2 M 수크로스 기준으로 투라노스 전환율은 50.9%으로 알려져 있다. 본 발명의 신규 아밀로수크라제 야생형 효소는 내열성이 높아 보다 더 안정적이고 투라노스의 전환 수율이 높아 산업적으로 유용하다.
본 명세서에서 효소 단백질에 의해 설탕을 포함하는 기질로부터 생성되는 당 시럽 조성물 내 투라노스 함량은 50℃의 온도 및 pH 6.0에서 기질과 1 내지 48시간, 바람직하게는 3 내지 48시간, 더욱 바람직하게는 12 내지 48시간 반응하여 수크로스가 반응액에 남아있지 않게 되는 시점의 생성물에 포함된 투라노스 양, 즉 상기 기질로부터 상대적으로 전환된 투라노스의 중량%로 정의된다. 예를 들어, 서열번호 1의 아미노산 서열을 갖는 효소 단백질의 투라노스 전환율은 35 내지 45 중량%, 바람직하게는 40 내지 43 중량%일 수 있으며, 서열번호 2 또는 서열번호 3의 아미노산 서열을 갖는 효소 단백질의 투라노스 전환율은 40 내지 50 중량%, 바람직하게는 43 내지 49 중량%일 수 있다.
또한 본 발명의 효소 단백질은 GRAS(Generally Recognized as Safe) 균주에서 발현하여 활성을 나타내며, 예를 들어 서열번호 1의 아미노산 서열을 갖는 야생형 효소 단백질뿐 아니라, 변이된 효소 단백질, 예를 들어 서열번호 2 또는 서열번호 3의 아미노산 서열을 갖는 변이 효소 단백질로의 변형에 따른 활성 증가 효과가 GRAS 균주에서도 재현된다. 따라서 본 발명은 투라노스를 GRAS 균주에서 발현하여 안전한 식품원료로 제공할 수 있을 뿐만 아니라, 효소 개량을 통해 높은 반응 안정성과 활성을 가지고 있는 효소를 확보함으로써 산업에 적용하기에 적합하다.
본 발명의 또 다른 예는, 상기 효소 단백질을 암호화하는 폴리뉴클레오티드, 상기 단백질을 암호화하는 폴리뉴클레오티드를 포함하는 재조합 벡터, 또는 상기 단백질을 발현하는 재조합 미생물을 제공한다.
상기 효소 단백질에 관한 사항은 효소 단백질을 암호화하는 폴리뉴클레오티드, 이를 포함하는 재조합 벡터 또는 이를 발현하는 재조합 미생물에 동일하게 적용될 수 있다.
본 발명의 효소 단백질을 암호화하는 폴리뉴클레오티드는, 서열번호 1 내지 서열번호 13의 아미노산 서열 중 선택된 어느 하나의 아미노산 서열을 암호화하는 폴리뉴클레오티드일 수 있으며, 바람직하게는 상기 아미노산 서열을 암호화하는 폴리뉴클레오티드는 서열번호 14 내지 서열번호 26 중 어느 하나일 수 있으나, 상기한 염기 서열에 대하여 실질적인 동일성을 나타내는 서열도 포함하는 것이다. 상기의 실질적인 동일성은, 상기한 본 발명의 염기 서열과 임의의 다른 서열을 최대한 대응되도록 배열하고, 그 서열을 분석한 경우, 서열번호 14 내지 서열번호 26의 염기 서열과 70% 이상, 보다 바람직하게는 80% 이상, 더욱 바람직하게는 90% 이상, 가장 바람직하게는 98% 이상 상동성이 있는 염기 서열을 포함한다.
예를 들어 본 발명의 효소 단백질을 암호화하는 폴리뉴클레오티드는 서열번호 1 내지 서열번호 13의 아미노산 서열 중 선택된 어느 하나의 아미노산 서열을 암호화하는 폴리뉴클레오티드일 수 있으며, 상기 폴리뉴클레오티드는 서열번호 14 내지 26의 염기서열로 이루어진 군에서 선택된 1 이상일 수 있다.
본 발명의 재조합 발현 벡터는 예를 들어 서열번호 1 내지 13의 아미노산 서열 중 선택된 어느 하나의 아미노산 서열을 암호화하는 폴리뉴클레오티드를 포함할 수 있으며, 상기 서열번호 1의 아미노산 서열을 암호화하는 폴리뉴클레오티드를 포함하는 재조합 발현 벡터는 도 1의 개열지도를 갖는 것일 수 있다. 본 발명의 재조합 미생물은 예를 들어 서열번호 1 내지 13의 아미노산 서열 중 선택된 어느 하나의 아미노산 서열을 암호화하는 폴리뉴클레오티드를 포함하는 재조합 발현벡터로 형질전환된 아밀로수크라제 효소 단백질을 발현하는 재조합 균주일 수 있다.
상기 투라노스 생산 방법에 있어서, 효율적인 투라노스 생산을 위하여, 사용되는 기질은 수크로스 단독 또는 상기 수크로스에 프럭토스를 추가적으로 사용할 수 있다.
본 발명에 따른 투라노스 생산 방법에서 프럭토스를 기질에 첨가하여 수크로스 및 프럭토스의 혼합 기질로 사용하는 경우에, 기질 중에서 설탕 대비 과당의 중량을 높이면 올리고머의 생성이 줄어들고 상대적으로 설탕의 비율이 적어지므로 투라노스의 생성양은 감소한다. 본 발명의 구체적인 예에서, 반응 생성물에 포함된 트레할룰로스는 설탕 대비 과당 비율에 크게 영향을 받지 않으며 비교적 낮은 수준으로 유지한다. 따라서, 본 발명에 따른 투라노스 생산 방법에서 수크로스 및 프럭토스의 혼합 기질을 사용하는 경우에, 반응 생성물에 포함된 전체 당류 중에서 목적 생성물인 투라노스 함량은 높이고, 바람직하지 않는 부산물인 트레할룰로스 함량과 올리고당 함량을 낮추도록 기질의 조성을 적절히 조절할 수 있다. 본 발명에 따른 투라노스 생산에서 얻어지는 반응 생성물의 전체 당류 고형분 함량을 기준으로 바람직한 당조성은, 올리고당 함량이 20 중량%이하, 18중량%이하, 17중량%이하, 15중량%이하, 13중량%이하, 11중량%이하, 또는 10 중량%이하이고 트레할룰로스 함량이 7중량%이하, 6중량%이하, 5 중량%, 4.5중량%이하, 또는 4.0 중량%이하인 조건을 만족하고, 투라노스 함량은 높을수록 좋으며 예를 들면, 30중량%이상, 35중량%이상, 40중량%이상, 41중량%이상, 43중량%이상, 45중량%이상, 47중량%이상, 49중량%이상 또는 50 중량%이상일 수 있다. 상기 올리고당의 대부분은 포도당이 알파 1-4 결합으로 연결된 DP3 및 DP4이상의 중합도를 가지며, DP3 구성당이 주성분이다. 상기 올리고당의 80~85 중량% 정도는 glucoamylase 효소 처리하면 포도당으로 분해될 수 있다.
본 발명의 일예에서, 반응 생성물에서 투라노스 생성량 대비 올리고당 함량이 줄어드는 기질의 조성은, 설탕 100중량부를 기준으로 과당 5 내지 250중량부, 5 내지 200중량부, 5 내지 175중량부, 5 내지 150중량부, 5 내지 125중량부, 5 내지 100중량부, 5 내지 90중량부, 5 내지 80중량부, 5 내지 75중량부, 5 내지 70중량부, 10 내지 250중량부, 10 내지 200중량부, 10 내지 175중량부, 10 내지 150중량부, 10 내지 125중량부, 10 내지 100중량부, 10내지 90중량부, 10 내지 80중량부, 10 내지 75중량부, 10 내지 70중량부, 15 내지 250중량부, 15 내지 200중량부, 15 내지 175중량부, 15 내지 150중량부, 15 내지 125중량부, 15 내지 100중량부, 15 내지 90중량부, 15 내지 80중량부, 15 내지 75중량부, 15 내지 70중량부, 20 내지 250중량부, 20 내지 200중량부, 20 내지 175중량부, 20 내지 150중량부, 20 내지 125중량부, 20 내지 100중량부, 20 내지 90중량부, 20 내지 80중량부, 20 내지 75중량부, 20 내지 70중량부, 30 내지 250중량부, 30 내지 200중량부, 30 내지 175중량부, 30 내지 150중량부, 30 내지 125중량부, 30 내지 100중량부, 30 내지 90중량부, 30 내지 80중량부, 또는 30 내지 75중량부, 30 내지 70중량부, 35 내지 250중량부, 35 내지 200중량부, 35 내지 175중량부, 35 내지 150중량부, 35 내지 125중량부, 35 내지 100중량부, 35 내지 90중량부, 35 내지 80중량부, 35 내지 75중량부, 또는 35 내지 70중량부 일 수 있으며, 예를 들면 50 중량부이다.
본 발명에 따른 투라노스 제조방법은 투라노즈 제조 반응의 반응 생성물에서 투라노스 생성량 대비 DP3이상의 올리고당 함량이 감소하며, 이에 SMB 크로마토그래피에 적합하다. 설탕에 첨가하는 과당 함량이 지나치게 높으면 반응 생성물의 올리고당 함량은 낮으나 투라노스 함량이 낮아 수율도 낮고 분리 및 정제도 용이하지 않다. 따라서, 반응 생성물에 포함된 투라노스 함량이 높고, 올리고당, 트레할룰로스 및 과당으로 이루어지는 군에서 선택된 1종 이상의 함량이 낮으며, 바람직하게는 올리고당 함량이 낮도록 혼합 기질의 당조성을 적절히 설정할 수 있다.
예를 들면, 본 발명의 투라노스 제조를 위한 기질 반응의 수크로스(설탕) 농도는 0.1M 내지 2.5M, 바람직하게는 0.5M 내지 2.0M을 포함할 수 있다. 또한 본 발명의 투라노스 제조를 위한 기질이 수크로스와 프럭토스를 포함하는 경우, 기질 용액 중 프럭토스 농도는 0.1 내지 4M, 또는 0.75 내지 3M을 추가로 포함할 수 있다.
본 발명의 효소 단백질은 기질 반응의 수크로스 농도는 0.1 내지 2.5M, 0.25 내지 2.5M, 0.75 내지 2.5M, 1.0 내지 2.5M, 0.1 내지 2.25M, 0.25 내지 2.25M, 0.75 내지 2.25M, 1.0 내지 2.25M, 0.1 내지 2.0M, 0.25 내지 2.0M, 0.75 내지 2.0M, 1.0 내지 2.0M, 0.1 내지 1.5M, 0.25 내지 1.5M, 0.75 내지 1.5M, 1.0 내지 1.5M, 0.1 내지 1.25M, 0.25 내지 1.25M, 0.75 내지 1.25M, 또는 1.0 내지 1.25M일 수 있다.
상기 효소 단백질을 기질과 반응시키는 단계는 상기 단백질을 기질과 접촉시키는 단계에 의하여 수행될 수 있다. 상기 효소 단백질을 기질과 접촉시키는 단계는, 예컨대, 상기 효소 단백질 등을 기질과 혼합하는 단계 또는 상기 효소 단백질 등이 고정화된 담체에 기질을 접촉시키는 단계에 의하여 수행될 수 있다. 또 다른 예에서 상기 효소 단백질 등을 기질과 반응시키는 단계는 상기 재조합 균주의 균체를 기질이 포함된 배양 배지에서 배양하는 단계에 의하여 수행될 수 있다. 이와 같이 상기 효소 단백질은 수크로스 또는 프럭토스를 투라노스로 전환하여 수크로스 또는 프럭토스로부터 투라노스를 생산할 수 있다.
상기 생산 방법은 서열번호 1 내지 서열번호 13 중에서 선택된 1종 이상의 아미노산 서열을 갖는 효소 단백질을 발현하는 재조합 미생물을 배양하는 단계; 및 상기 재조합 미생물 또는 상기 재조합 미생물로부터 분리된 효소 단백질을 기질과 반응시키는 단계를 포함할 수 있다.
상기 재조합 미생물의 배양 단계는, 사용되는 미생물, 예를 들어 균주의 특성에 따라 본 발명이 속하는 기술 분야의 당업자에 의해 용이하게 선택되는 배지 및 배양 조건 하에서 이루어질 수 있다. 예를 들어, 상기 배지로서는 대장균을 비롯한 임의의 숙주 세포, 및 세포 내용물을 지지하거나 또는 함유할 수 있는 임의의 배양 배지, 용액, 고체, 반고체 또는 강성 지지체를 포함하며, 바람직하게는 2YT 배지, LB 배지, SOB 배지 또는 TB 배지 등이 될 수 있다. 상기 배양은 연속, 반연속, 또는 회분식 배양일 수 있다.
상기 균체는 상기 미생물의 배양물에 대해 원심분리, 여과 등을 수행하여 얻을 수 있으며, 또한 상기 얻어진 균체를 균질화시키고 원심분리하여 수득된 상층액 또는 상기 상층액을 분획화하거나, 크로마토그래피 등을 통해 분리 정제하여 효소 단백질을 얻을 수 있다. 예컨대, 회수된 균체를 50mM 인산 완충용액으로 현탁한 후 파쇄하여 원심분리 한 후, 상등액만 Ni-NTA 컬럼 (Qiagen) 에서 흡착시킨 후 20mM, 200mM 이미다졸의 농도로 목적 단백질을 회수할 수 있다.
상기 투라노스 생산 방법에 있어서, 효율적인 투라노스 생산을 위하여, 사용되는 효소 단백질의 양은 전체 반응물(기질 및 단백질 모두을 포함) 기준으로 50 U/ml 내지 1500 U/ml 일 수 있다. 효소의 사용량이 상기 농도보다 낮으면 투라노스 전환 효율이 낮아질 수 있고, 상기 농도보다 높으면 산업에서의 경제성이 낮아지므로 상기 범위가 적당하다.
또한, 상기 기질과 반응시키는 단계는 바람직하게는 본 발명의 효소 단백질의 최적 활성화 조건 하에 이루어질 수 있다. 즉, 상기 반응은 30 내지 65℃, 바람직하게는 40 내지 60℃, 더욱 바람직하게는 45 내지 55℃, 더욱 바람직하게는 50℃의 온도에서 이루어질 수 있다. 상기 기질과 반응시키는 단계는 pH 5.0 내지 9.0, 바람직하게는 pH 5.5 내지 8.0, 또는 pH 5.5 내지 6.5에서 이루어질 수 있다.
본 발명의 방법에 의하여 수득된 투라노스는 모사이동층 크로마토그래피를 이용하여 투라노스를 고순도로 분리할 수 있다. 상기 분리방법에서. 예를 들어 원심분리, 여과, 이온정제 및 이들의 조합으로 이루어진 군으로부터 선택된 하나 이상의 단계를 추가로 수행할 수 있다.
본 발명에 따른 투라노스 제조방법에 있어서, 투라노스를 함유하는 당 조성물을 모사 이동층(simulated moving bed, SMB) 크로마토그래피로 분리하여 투라노스 분획과 라피네이트를 얻는 단계를 포함하는 고순도 투라노스의 제조방법에 관한 것이다.
본 발명에 따른 투라노스의 제조 공정은 연속식과 배치식 모두 사용 가능하며, 바람직하게는 연속식 공정이다.
본 명세서에서, 용어 "라피네이트(raffinate)"라 함은 추잔액이라고도 하며, 분리공정에 투입된 원료가 분리공정을 통과하여 얻어지는 산물에는 분리공정으로 함량을 높이고자 하는 목적 물질을 포함하는 목적 분획과, 분리공정에서 제거 또는 함량을 감소하고자 하는 물질등을 포함하는 잔류액을 포함하며, 상기 잔류액 라피네이트라고 한다. 본 발명의 일 예에서 투라노스 전환 공정에서 얻어지는 산물은 원료 기질인 설탕 및/또는 과당과 생산물인 투라노스를 포함하는 혼합물이며, 고순도 분리공정을 거치면서 목적 물질인 투라노스의 함량이 증가된 투라노스 분획과 잔류액을 얻으며, 잔류액에는 투라노스 전환 반응의 기질 또는 산물인 과당이 다량 포함되므로 과당 라피네이트를 의미할 수 있다.
본 발명의 일 예에서, SMB 크로마토그래피를 이용한 고순도 분리 공정은 분리과정에서 상 변화가 없어 물질의 안정성 확보에 용이한 분리방법이다. 이러한 흡착 분리방법 중에서 액상 흡착 분리방법으로는 크로마토그래피 분리방법이 많이 사용되고 있다. 이중, 모사 이동층 흡착 분리 방법(simulated moving bed, SMB)은 1961년 미국특허 제2,985,589호 에서 제안된 분리 기술로, 다수의 컬럼을 이용하여 연속적으로 분리함으로써 기존의 회분식 크로마토그라피에 비해 순도 및 생산성이 우수하고, 적은 용매의 사용이 가능하다는 장점을 지닌다. 상기 모사 이동층(SMB) 흡착 분리 공정은 분리대상 혼합물의 주입과 라피네이트 및 추출물의 생산이 연속적으로 이루어지는 공정이다.
SMB 분리 후 이온 정제 공정으로서, 이온교환수지가 충진된 분리탑을 1개 또는 3개 사용하며, 이온교환수지는 강산성 양이온교환수지와 약염기성 음이온교환수지를 사용하여 35~50℃에서 수행할 수 있다.
본 발명의 또 다른 일 예로서, 상기 투라노스 생산 방법으로 생산된 투라노스 함유 당 조성물을 제공한다. 상기 효소 단백질 및 투라노스 생산 방법에 관한 사항은 상기 투라노스 함유 당 조성물에 동일하게 적용될 수 있다.
상기 효소 단백질을 수크로스 단독 또는 수크로스 및 프럭토스를 포함하는 기질과 반응하여 생성된 투라노스를 함유하는 당 조성물, 예를 들어, 투라노스, 수크로스, 프럭토스, 트레할룰로스, 포도당 및 올리고당, 희소당(알룰로스, 알로스 등)으로 이루어진 군에서 선택된 1종 이상의 당류를 포함하는 당 조성물을 제조할 수 있다.
상기 반응 생성물을 목적 산물인 투라토스를 높은 함량으로 포함하고, 불순물인 올리고당 및 트레할룰로스를 낮은 함량을 포함하는 것이 바람직하다. 예를 들면, 반응생성물에 포함된 당류 고형분 함량 100중량%를 기준으로, 투라노스는 30중량%이상, 35중량%이상, 40중량%이상, 41중량%이상, 43중량%이상, 45중량%이상, 47중량%이상, 49중량%이상 또는 50 중량%이상일 수 있다. 상기 본 발명에 따른 투라노스 생산에서 얻어지는 반응 생성물의 전체 당류 고형분 함량을 기준으로 바람직한 당조성은, 올리고당 함량이 20 중량%이하, 18중량%이하, 17중량%이하, 15중량%이하, 13중량%이하, 11중량%이하, 또는 10 중량%이하이고 트레할룰로스 함량이 7중량%이하, 6중량%이하, 5 중량%, 4.5중량%이하, 또는 4.0 중량%이하인 조건을 만족할 수 있다.
구체적인 일예에서, 본 발명에 따른 아밀로수크라제 효소를, 반응 기질과 50℃의 온도 및 pH 6.0에서 24시간 동안 반응하여 얻어진 반응생성물 고형분 100중량%를 기준으로 투라노스 30 중량%이상, 올리고당 20중량%이하, 및 트레할룰로스 7중량%이하로 포함하는 반응생성물을 제조할 수 있다.
본 발명에 따른 비피도박테리움 터모필럼(Bifidobacterium thermophilum) 유래 효소 단백질은 수크로스로부터 투라노스를 고수율로 생성할 수 있어 투라노스의 산업적 이용에 유용하다.
도 1은 본 발명의 일 예에 따라 재조합 투라노스 3-에피머화 효소 발현용 재조합 DNA를 포함하는 발현벡터 pRSet(A)를 나타내는 도면이다.
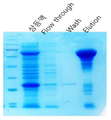
도 2는 본 발명의 일 예에 따라 비피도박테리움 터모필럼 아밀로수크라제 발현을 나타내는 SDS-PAGE 도면이다.
도 3는 야생형 아밀로수크라제의 온도에 따른 상대 활성을 나타낸 결과이다.
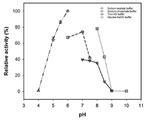
도 4은 야생형 아밀로수크라제의 pH에 따른 상대 효소 활성을 나타낸 결과이다.
도 5는 야생형 아밀로수크라제의 온도에 따른 상대 효소 활성을 나타낸 결과이다.
도 6는 본 발명의 일 예에 따라 기질인 수크로스 및 프럭토스 농도에 따른 투라노스 전환율을 나타낸 결과이다.
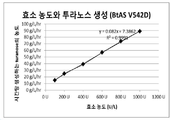
도 7은 본 발명의 일 예에 따라 정제된 비피도박테리움 터모필럼 아밀로수크라제의 돌연변이체 V542D(서열번호6)의 효소 농도에 따른 시간당 생성하는 투라노스의 농도 측정량을 나타내는 도면이다.
도 8는 본 발명의 일 예에 따라 정제된 비피도박테리움 터모필럼 아밀로수크라제의 돌연변이체 V542K(서열번호5)의 효소 농도에 따른 시간당 생성하는 투라노스의 농도 측정량을 나타내는 도면이다.
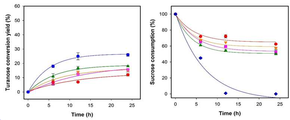
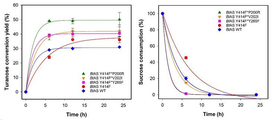
도 9는 본 발명의 일 예에 따라 pH 6 및 50℃의 온도에서, 기질 2 M 수크로스와 400 U/L 및 1600 U/L로 반응시킨 후 산출된 투라노스 전환율을 나타낸 결과이다.
도 10은 본 발명의 일 예에 따라 pH 6 및 50℃의 온도에서, 기질 2 M 수크로스와 400 U/L 및 1600 U/L로 반응시킨 후 산출된 기질 수크로스 소비량을 나타낸 결과이다.
도 2는 본 발명의 일 예에 따라 비피도박테리움 터모필럼 아밀로수크라제 발현을 나타내는 SDS-PAGE 도면이다.
도 3는 야생형 아밀로수크라제의 온도에 따른 상대 활성을 나타낸 결과이다.
도 4은 야생형 아밀로수크라제의 pH에 따른 상대 효소 활성을 나타낸 결과이다.
도 5는 야생형 아밀로수크라제의 온도에 따른 상대 효소 활성을 나타낸 결과이다.
도 6는 본 발명의 일 예에 따라 기질인 수크로스 및 프럭토스 농도에 따른 투라노스 전환율을 나타낸 결과이다.
도 7은 본 발명의 일 예에 따라 정제된 비피도박테리움 터모필럼 아밀로수크라제의 돌연변이체 V542D(서열번호6)의 효소 농도에 따른 시간당 생성하는 투라노스의 농도 측정량을 나타내는 도면이다.
도 8는 본 발명의 일 예에 따라 정제된 비피도박테리움 터모필럼 아밀로수크라제의 돌연변이체 V542K(서열번호5)의 효소 농도에 따른 시간당 생성하는 투라노스의 농도 측정량을 나타내는 도면이다.
도 9는 본 발명의 일 예에 따라 pH 6 및 50℃의 온도에서, 기질 2 M 수크로스와 400 U/L 및 1600 U/L로 반응시킨 후 산출된 투라노스 전환율을 나타낸 결과이다.
도 10은 본 발명의 일 예에 따라 pH 6 및 50℃의 온도에서, 기질 2 M 수크로스와 400 U/L 및 1600 U/L로 반응시킨 후 산출된 기질 수크로스 소비량을 나타낸 결과이다.
이하 본 발명을 구체적인 실시예에 의해 더 상세히 설명하고자 한다. 하지만 본 발명은 하기 실시예에 한정된 것이 아니다.
실시예 1. 야생형 아밀로수크라제 제조
발현균주로 사용될 대장균에 최적화되도록 비피도박테리움 터모필럼으로부터 유래된 아밀로수크라제(서열번호 4)를 암호화하는 폴리뉴클레오타이드(서열번호 1)를 바이오니아(Bioneer.Co.Korea)에 의뢰하여 합성하였다. 합성된 서열번호 1의 폴리뉴클레오타이드를 제한효소 NheI과 XhoI(NEB)을 사용하여 발현 벡터인 pRSet(A)(Invitrogen)의 동일한 제한효소 부위에 삽입하여 재조합 벡터 pRSet(A)-아밀로수크라제(pRSet(A)-BtAS)를 제조하였다(도 1).
이후 제조된 재조합 벡터로 heat shock 방법(Sambrook and Russell: Molecular Cloning)에 의하여 대장균 BL21(DE3)(Invitrogen)를 형질전환하여 서열번호 4의 아미노산 서열을 암호화하는 폴리뉴클레오타이드를 포함하는 재조합 균주를 제조하였다.
형질전환된 재조합 균주를 5ml LB-ampicillin 배지(Difco)에 접종한 후 600nm에서의 흡광도(OD)가 1.5에 도달할 때까지 37℃의 온도 및 200rpm에서 진탕배양 한 후, 이 배양액을 500ml LB-ampicillin 배지에 접종, 37 ℃의 온도 및 200rpm에서 진탕배양하였다. 이 배양액의 600nm에서 흡광도가 0.5일 때 0.1mM의 IPTG를 첨가하여 목적 효소의 과발현을 유도하였다. (이때 과발현 유도시점부터 배양조건은 16℃의 온도 및 150rpm으로 전환하여 16시간 동안 유지하는 것이 바람직하나, 30℃의 온도에서 실험을 해도 무관하다.
이후, 원심분리기 6000rpm에서 20분간 원심분리하여 균체를 회수하였다. 회수한 균체는 0.85%(w/v) NaCl로 2회 세척 후 lysis buffer(PBS buffer, 10mM imidazole)에 혼탁시킨 후 Bead beater를 20초 3회 회전으로 작동으로 파쇄하였다. 파쇄액을 13000rpm에서 20분 동안 원심분리하여 상등액만을 모은 후, 미리 lysis buffer로 평형시킨 Ni-NTA컬럼(Ni-NTA Superflow. Qiagen)에 적용시킨 다음 PBS 버퍼에 40 mM imidazole이 함유된 완충용액을 순차적으로 흘려주었다. 마지막 과정인 PBS 버퍼에 200 mM imidazole을 흘려줌으로써 목적 단백질을 용출하였다. 용출된 단백질은 효소 활성 측정용 완충용액(50 mM Tris-HCl pH 7.0)으로 전환하여 다음 실험에 사용하였다.
이 방법으로 부분 정제된 아밀로수크라제를 얻었으며, SDS-PAGE를 통하여 단량체의 크기가 약 68 킬로달톤(kDa)인 것을 확인하였다(도 2).
실시예 2. 변이형 아밀로수크라제 제조
2.1 PCR에 의한 돌연변이 작제 및 클로닝
아밀로수크라제 돌연변이체는 Stratagene사의 Quikchange 부위-지향성 돌연변이 유발법을 사용하여 제작되었다. 구체적으로 insert만을 PCR을 통해 증폭하고 플라스미드 DNA로 subcloning하는 방식이 아닌, plasmid DNA 전체를 PCR을 통해 증폭하고, template DNA는 DpnI 효소로 분해하는 방식을 사용하였다. Quikchange 부위-지향성 돌연변이 유발법은 보다 빠르게 돌연변이를 유발할 수 있다는 장점이 있다.
야생형 아밀로수크라제 효소를 암호화하는 DNA pET21a의 벡터 DNA에 클로닝되어 있는 플라스미드 DNA를 복제를 하는 주형으로 사용하였다. 제조된 변이형 아밀로수크라제 발현용 재조합 DNA를 포함하는 발현벡터는, 도 1에 도식화된 백터에서 insert에 해당하는 서열, 즉 각각 하나의 뉴클레오타이드가 치환된 pRSet(A)-BtAS V542D, pRSet(A)-BtAS V542K이다.
V542D 돌연변이체를 제작하기 위하여 주형 DNA 250ng, V542D 정방향 프라이머 125ng, V542D 역방향 프라이머 125ng, pfu ultra high fidelity DNA polymerase 2.5ul, 10X 반응 버퍼 5ul, 10mM dNTP 1ul를 포함하는 최종부피 50ul에 PCR을 세팅하였다. 상기 반응은 GeneAmp PCR system 9700 에서 95 ℃(1분)에서 1회의 사이클, [95℃(50초), 60 ℃(50초), 68 ℃(6분)]의 18회의 사이클, 68 ℃(7분)에서 1회의 사이클을 포함한다. 돌연변이가 유발되지 않은 주형 DNA를 인식하여 분해하는 DpnI 효소를 PCR이 끝난 50ul 부피의 반응액에 500U(10U/ul) 넣고 37 ℃에서 1 시간 동안 처리하였다.
또한, V5425K 돌연변이체를 제작하기 위하여 주형 DNA 250ng, V5425K 정방향 프라이머 125ng, V5425K 역방향 프라이머 125ng, pfu ultra high fidelity DNA polymerase 2.5ul, 10X 반응 버퍼 5ul, 10mM dNTP 1ul를 포함하는 최종부피 50ul에 PCR을 세팅하였다. 상기 반응에 필요한 프라이머 염기 서열은 표1과 같다. 상기 반응은 GeneAmp PCR system 9700 에서 95℃(1분)에서 1회의 사이클, [95℃(50초), 60℃(50초), 68℃(7분)]의 18회의 사이클, 68℃(7분)에서 1회의 사이클을 포함한다. 돌연변이가 유발되지 않은 주형 DNA를 인식하여 분해하는 DpnI 효소를 PCR이 끝난 50ul 부피의 반응액에 500U(10U/ul) 넣고 37℃에서 1 시간 동안 처리하였다.
돌연변이체 작제에 사용된 프라이머의 서열은 아래 표에 나타내었으며, PCR 과정을 아래 표에 정리하였다.
| 명칭 | 서열 | Tm |
| V542D_FWD (서열번호 27) | 5'- CGCCCCTGACGCTTGGGACACGACGTGGGACGCGC -3' | 73℃ |
| V542D_RVS (서열번호 28) | 5'- GCGCGTCCCACGTCGTGTCCCAAGCGTCAGGGGCG -3' | 72℃ |
| V542K_FWD (서열번호 29) | 5'- CGCCCCTGACGCTTGGAAGACGACGTGGGACGCGC -3' | 73℃ |
| V542K_RVS (서열번호 30) | 5'- GCGCGTCCCACGTCGTCTTCCAAGCGTCAGGGGCG -3' | 72℃ |
| segment | step | cycles | temperature(℃) | time |
| 1 | initialization | 1 | 95 | 1 min |
| 2 | denaturation | 18 | 95 | 50 sec |
| annealing | 118 | 60 | 50 sec | |
| elongation | 18 | 68 | 7 min | |
| 3 | final elongation | 1 | 68 | ∞ |
2.2 형질 전환 및 발현
E.coli DH10B를 최초 숙주로 사용하였다. 실시예 2.1 에서 기술한 PCR 반응액에 DpnI이 첨가 된 혼합액을 바로 DH10B에 형질 전환한다. PCR 반응액과 DpnI의 혼합 용액을 5ul 피펫팅 하여 초저온 냉동고에 보관되어 있던 CaCl2법으로 제작된 competent cell인 DH10B 50ul에 heat shock 방법으로 형질 전환한다. LB-ampicillin 고체 배지에 도말하고 37 ℃에서 배양한다. 고체 배지 상에 생성된 콜로니 중에서 3개를 선별하여 3ml LB-ampicillin 액체 배지에서 배양한 후 miniprep kit(Quiagen) 을 이용하여 plasmid DNA를 추출한다. 시퀀싱은 외부기관(마크로젠, 한국)에서 실시하였으며, 그 결과 투라노스 생성용 아밀로수크라제 효소를 암호화 하는 DNA 부위에 돌연변이가 유발되었으며, 구체적으로 아미노산 서열 V542D 변이(서열번호 2) 및 아미노산 서열 V542K 변이(서열번호 3)가 유발되었음을 확인하였다. 돌연변이 유발이 확인된 plasmid DNA는 냉동고에 보관 되어있던 CaCl2법으로 제작된 competent cell인 E.coli BL21 solu (DE3) 50ul에 heat shock 방법으로 형질전환 하였다. LB-ampicillin 고체 배지에 도말하고 37℃에서 배양하였다.
형질전환으로 획득한 아밀로수크라제 돌연변이를 암호화 하는 plasmid DNA를 포함하고 있는 E.coli BL21 solu (DE3)의 콜로니를 피킹하여 3ml LB-ampicillin 액체 배지에 접종하고 37 ℃에서 12시간 배양하였다. 뿌옇게 자란 3ml 배양볼륨의 미생물을 1ml 피펫팅 하여 250ml 삼각 플라스크에 담긴 50ml의 LB-ampicillin 액체배지에 접종하여 37 ℃에서 배양하였다. 600nm에서 흡광도가 0.6이 될 때 IPTG가 0.1mM 되도록 induction을 한 후 16℃에서 배양하였다. 모든 배양은 200 rpm 교반 인큐베이터에서 이루어졌다. 16℃에서 배양한 지 12시간 후에 3500gX의 원심분리기에서 30분 동안 원심분리 하여 미생물 균체를 회수하였다.
상기 회수된 균체를 lysis buffer(PBS buffer, 10 mM imidazole)에 혼탁시킨후 Bead beater를 20초 3회 회전으로 작동으로 파쇄한다. 파쇄액을 13000rpm에서 20분 동안 원심분리하여 상등액만을 모은 후, 미리 lysis buffer로 평형시킨 Ni-NTA컬럼(Ni-NTA Superflow. Qiagen)에 적용시킨 다음 PBS 버퍼에 40 mM imidazole이함유된 완충용액을 순차적으로 흘려주었다. 마지막 과정인 PBS 버퍼에 200 mM imidazole을 흘려줌으로써 목적 단백질을 용출하였다. 용출된 단백질은 효소 활성 측정용 완충용액(50 mM Tris-HCl pH 7.0)으로 전환하여 이후 실험에 사용하였다.
실시예 3. 변이형 아밀로수크라제 제조
단일 점(single point) 돌연변이 효소를 제작하기 위하여 아래 표의 프라이머와 Quickchange site-directed mutagenesis kit를 이용하여 돌연변이를 유발하여 재조합 벡터 pBT7-N-His/아밀로수크라제를 제작하였다.
| 유전자 | 방향 | 서열번호 | 서열 |
| BtAS P200R | Forward | 31 | 5'-GCCGTCGTCCGCCAAGTCTTC-3' |
| Reverse | 32 | 5'-GAAGACTTGGCGGACGACGGC-3' | |
| BtAS V202I | Forward | 33 | 5'-GTCCCGCAAATCTTCCCGACC-3' |
| Reverse | 34 | 5'-GGTCGGGAAGATTTGCGGGAC-3' | |
| BtAS Y265F | Forward | 35 | 5'-GACGCGGTGCCGTTCATCTGGAAGCAA-3' |
| Reverse | 36 | 5'-TTGCTTCCAGATGAACGGCACCGCGTC-3' | |
| BtAS V305I | Forward | 37 | 5'-AAAGGTGAAGTCATCATGGCTCCCAAG-3' |
| Reverse | 38 | 5'-CTTGGGAGCCATGATGACTTCACCTTT-3' | |
| BtAS K393R | Forward | 39 | 5'-CCGCTCAAGCACAGGGAATTCCTCTAC-3' |
| Reverse | 40 | 5'-GTAGAGGAATTCCCTGTGCTTGAGCGG-3' | |
| BtAS S420T | Forward | 41 | 5'-TATGATCCGGCGACGGGTGACGCGCGC -3' |
| Reverse | 42 | 5'-GCGCGCGTCACCCGTCGCCGGATCATA-3' | |
| BtAS Y414F | Forward | 43 | 5'-ATGGGCGAGCTGTTCAACTATGATCCG-3' |
| Reverse | 44 | 5'-CGGATCATAGTTGAACAGCTCGCCCAT-3' |
상기 유발된 점 돌연변이는, 야생형 아밀로수크라제 효소의 200번째 프롤린을 아르기닌으로(서열번호 4), 202번째 발린은 이소루신으로(서열번호 5), 265번째 티로신은 페닐알라닌으로(서열번호 6) 305번째 발린은 이소루신으로(서열번호 7) 393번째 리신은 아르기닌으로(서열번호 8), 414번째 티로신은 페닐알라닌으로(서열번호 9), 420번째 세린은 트레오닌으로(서열번호 10) 치환시킨 것이다.
상기 플라스미드를 형질전환 방법에 의하여 대장균 BL21(DE3)에 형질전환 하였고, 상기 실시예 2.2와 동일한 방법으로 변이형 아밀로수크라제 효소를 분리하였다.
실시예 4. 변이형 아밀로수크라제 효소 제조
이중 점(double point) 돌연변이를 갖는 변이형 효소를 제작하기 위하여 Y414F 변이체와 야생형 효소보다 반응종결 시간이 빨라진 변이체의 이중 점 변이 (P200R를 Y414F에 적용)를 접목시켰다.
pBT7-N-His/아밀로수크라제 Y414F 돌연변이 효소 벡터에 P200R 프라이머를 이용하여 이중 점 변이형 아밀로수크라제 효소, Y414F*P200R(서열번호9)를 제작하였다. 상기 플라스미드를 형질전환 방법에 의하여 대장균 BL21(DE3)에 형질전환하였고, 아밀로수크라제를 대량생산하기 위하여 상기 재조합 E.coli BL21의 스탁 배양물을 LB배지 5mL에 접종하고 온도 37℃ 12시간 동안 180rpm에서 전 배양시킨 후, 전 배양물을 120℃에서 15분 동안 멸균시킨 2 L 플라스크에 들어있는 1 L LB배지에 접종한 후 0.01%(w/v) 암피실린의 존재 하 배양온도는 37℃, 교반 속도는 180rpm에서 배양하였다. 600nm에서의 흡광도가 0.6에 도달했을 때, 최종 농도 0.2mM의 이소프로필-β-D-치오갈락토피라노사이드를 첨가하여 효소의 대량 발현을 유도하였다. 이소프로필-β-D-치오갈락토피라노사이드를 첨가한 후에는 배양온도를 16℃로 낮춰 20시간 배양하였다. 과발현 된 아밀로수크라제 효소 배양물을 4℃에서 10분 동안 3,000 x g에서 원심분리하여 침전물을 수확하였다. 상기 침전물을 50 mM Tris-HCl 완충 용액(pH 7) 45mL에서 볼텍싱하여 완전하게 재현탁 시킨 후에 세포 용액을 초음파 분쇄기(Sonic Dismembrator 550, Fisher Scientific Co.) Model D100을 이용하여 세포를 파괴하였다. 세포 파쇄물은 4℃에서 20분 동안 11,000 x g에서 원심분리하여 침전물인 세포 잔해물은 제외하고 상등액 만을 얻는다. 이 과정에서 얻은 반응물을 세포 추출물(Cell extract)라고 명명하였다.
수확한 세포 추출물은 효소 정제 수율을 알기 위하여 소량 채취해 보관한다. 이 세포 상등액을 멸균된 0.45 μm 시린지 필터를 통해 여과시킨 후, 45 mL의 여과액을 니켈-니트릴로트리아세트산(Ni-NTA) 친화성 칼럼을 통해 통과시켜 재조합 히스 텍(His-tag)된 아밀로수크라제를 통과시켜 정제된 아밀로수크라제 효소를 얻었다. 컬럼을 40mL 세정 완충액(Lysis buffer) (50mM Tris-HCl, 300mM 염화나트륨, 20mM 이미다졸, pH 7.0)으로 먼저 세정한 후, 40mL의 용출 완충액 (Elution buffer) (50mM Tris-HCl, 300mM 염화나트륨, 250mM 이미다졸, pH 7.0)을 이용하여 원하는 단백질을 얻는다. 정제한 단백질은 4℃에서 Amicon Centrifugal Filter Devices (Millipore Corporation, Billerica, USA)를 이용하여 농축하여 투라노스 생산에 사용되는 아밀로수크라제 효소로서 분리하였다.
비교예 1. 네이세리아 서브플라바 유래 아밀로수크라제 제조
발현균주로 사용될 대장균에 최적화되도록 네이세리아 서브플라바로부터 유래된 아밀로수크라제(서열번호 1)를 암호화하는 폴리뉴클레오타이드(서열번호 4)를 바이오니아(Bioneer.Co.Korea)에 의뢰하여 합성하였다.
합성된 서열번호 4의 폴리뉴클레오타이드를 제한효소 NheI과 XhoI(NEB)을 사용하여 발현 벡터인 pRSet(A)(invitrogen)의 동일한 제한효소 부위에 삽입하여 재조합 벡터 pRSet(A)-아밀로수크라제(pRSet(A)-NsAS)를 제조하였다. 이후 제조된 재조합 벡터로 heat shock 방법(Sambrook and Russell MolecularCloning.)에 의하여 대장균 BL21(DE3)(invitrogen)를 형질전환하여 서열번호 1의 아미노산 서열을 암호화하는 폴리뉴클레오타이드를 포함하는 재조합 균주를 제조하였다.
형질전환된 재조합 균주를 5ml LB-ampicilline 배지(Difco)에 접종한 후 600nm에서의 흡광도(OD)가 1.5에 도달할 때까지 37℃의 온도 및 200rpm에서 진탕 배양한 후, 이 배양액을 500ml LB-ampicilline 배지에 접종, 37℃ 의 온도 및 200rpm에서 진탕 배양하였다. 이 배양액의 600nm에서 흡광도가 0.5일 때 0.1mM의 IPTG를 첨가하여 목적 효소의 과발현을 유도하였다. (이때 과발현 유도시점부터 배양조건은 16℃ 의 온도 및 150rpm으로 전환하여 16시간 동안 유지하였다. 이후 원심분리기 6000rpm에서 20분간 원심분리하여 균체를 회수하였다. 회수한 균체는 0.85%(w/v) NaCl로 2회 세척 후 균체를 이용한 투라노스 생산 및 효소정제에 사용하였다.
실시예 5. 야생형 아밀로수크라제의 활성 분석
5.1 온도에 따른 효소 활성
실시예 1에서 얻어진 아밀로수크라제 효소의 효소 활성에 대한 온도의 영향은 25 ~ 65℃ 의 범위에서 결정되었다. 0.1M 수크로스, 50mM Tris-HCl 완충액 (pH 7.0) 및 희석된 효소를 함유하는 총 부피가 0.5mL 인 반응 혼합물을 25, 30, 35, 40, 45, 50, 55, 60 및 65℃ 의 온도에서 배양 하였다. BtAS 활성의 온도 의존성은 다양한 온도에서 환원당의 방출량을 결정하기 위한 DNS 방법으로서, 즉, DNS 용액을 이용한 환원당량 측정법을 통해 온도별 효소활성을 구하고 상대활성도로 표시하였으며 그 결과를 도 3에 나타내었다. 구체적으로, 상기 DNS 방법에 의한 효소 활성 측정은, 기질인 100mM 수크로스에 상기 아밀로수크라제 0.02mg/ml을 처리하고 상기 pH 및 온도 조건에서 30분간 반응하고, DNS 시약을 넣어 반응을 중지하고, 100℃ 의 온도에서 5분간 가열하여 DNS의 환원당 발색 반응을 유도하고 얼음 위에서 식힌 후, 575nm에서 ABS를 측정 한 후 프럭토스 스탠다드와 비교하여 계산하였다.
도 3의 결과에서 확인할 수 있듯이, 기질인 수크로스를 가수분해하는 능력은 50℃에서 최대로 나타냈다. 55℃에서는 98%의 상대활성을 보이며, 60℃에서 약 85 %의 잔여활성을 보인다. 65℃에서는 대부분 활성이 감소하는 것을 확인 하였다. 따라서 야생형 아밀로수크라제 BtAS는 50℃에서도 안정하면서도 최대활성을 보이므로 50℃를 최적온도로 확인하였다.
5.2 pH에 따른 활성 분석
실시예 1에서 정제된 아밀로수크라제 효소에 대한 최적 pH를 확인하기 위하여, 효소 활성에 대한 최적 pH는 4.0 내지 10.0의 범위에서 측정하였다. 효소 활성에 대한 pH 효과를 분석하기 위해 4 개의 완충 시스템을 사용하였다: 50 mM 소듐 아세테이트 완충액 (pH 4.0 내지 6.0); 50 mM 인산 나트륨 완충액 (pH6.0 - 7.5); 50 mM Tris - HCl 완충액 (pH 7.0 - 9.0); 50 mM 글리신 -수산화나트륨 (pH 8.0 - 10.0) 0.1M 의 기질인 수크로스, pH가 다른 50mM의 각 완충액 및 희석된 효소를 함유한 반응 혼합물을 50℃ 에서 30 분 동안 수행 하였다. 효소 활성은 실시예 5-1에 제시된 DNS 법을 이용하여 측정하고 결과를 도 4에 나타내었다.
그 결과, 야생형 효소의 가장 큰 상대 활성은 pH 6.0의 50 mM 소듐 아세테이트 완충용액에서 관찰되었다. pH가 최적 pH 6.0에서 산성 (pH 5.5-4.0) 및 알칼리 (pH 7.0-9.0) 범위로 이동하면, 그 상대 활성은 현저하게 감소하였다. 완충 시스템은 효소 활성에 영향을 주어서는 안 되지만, 본 발명에서는 동일한 pH에서도 완충 성분의 유형에 따라 효소 활성이 유의하게 영향을 받았다. 따라서, 50mM 소듐아세테이트 완충용액 중 pH 6.0 를 이후의 실험에 사용하였다.
최적 온도와 최적 pH인 6.0에서 비피도박테리움 터모필럼 아밀로수크라제 변이주 V542D 유래 아밀로수크라제는 1.7U/mg, 비피도박테리움 터모필럼 아밀로수크라제 변이주 V542K 유래 아밀로수크라제는 1.4U/mg, 비피도박테리움 터모필럼 아밀로수크라제 변이주 Y414F*P200R 유래 아밀로수크라제는 2.6U/mg의 최적 활성을 지닌다.
5.3. 아밀로수크라제의 내열성 및 효소실활상수(Kd) 평가
야생형 아밀로수크라제의 내열성, 효소실활상수를 평가하기 위하여 상기 실시예 1에서 정제된 야생형 아밀로수크라제를 항온 진탕 수조에서 최대 120시간동안 열처리하였다. 45, 50, 55 및 60℃ 의 온도에서 열처리 후, 잔류 효소 활성을 측정하기 위하여 0.1 M 수크로스가 함유된 50 mM 완충액 490 ㎕에 상기 열처리한 효소를 적당히 희석한 희석액 10 ㎕를 넣고 35℃에서 30분간 반응시킨 후 반응 종결액에 500 ㎕의 DNS 용액을 첨가하고 100℃에서 5분간 끓인 뒤 5분간 얼음물에 방랭 시켰다. Abs575에서 흡광도를 측정해 Standard curve로부터 환원 당량을 측정하여 온도에 따른 효소활성의 상대 값을 도출하고 그 결과를 도 5(A, 45℃; B, 50℃?; C, 55℃?; D, 60℃?) 에 나타내었다. 이 값을 바탕으로 온도 별 시간에 따른 상대 활성도를 함수로 표기하고, 이를 다시 자연로그 함수로 변환시켜 kd 및 온도 반감기를 구하여 아래 표에 나타내었다.
| 온도(℃) | Kd(h-1) | t1/2(h) |
| 45 | 0.00003 | 23104.9 |
| 50 | 0.0012 | 577.6 |
| 55 | 0.0098 | 70.7 |
| 60 | 0.762 | 0.9 |
상기 표에서 확인할 수 있는 바와 같이, 야생형 아밀로수크라제 BtAS는 45℃에서 온도 반감기가 약 23,104시간, 50℃에서 온도반감기가 약 577시간으로 최대 120시간 동안의 효소 반응 시 매우 우수한 온도 안정성을 보였다. 이 온도 보다 5℃가 높은 55℃에서는 온도 반감기가 약 70시간으로 급격히 감소하는 경향을 보였으나, 전체적으로 열에 대한 안정성이 우수하였다.
실시예 6. 아밀로수크라제의 투라노스 전환 반응
6.1 비피도박테리움 터모필럼 유래 아밀로수크라제 투라노스 전환율
실시예 1 내지 4의 아밀로수크라제의 투라노스 전환율을 검토하기 위하여, 설탕과 과당을 1:0.2의 중량 비율로 혼합하여 농도가 63.5bx가 되도록 용해시켜 기질을 준비하고 pH는 6.0으로 조정하였다. 실시예 1의 비피도박테리움 터모필럼 유래 투라노스 전환 효소 (BtAS)의 야생형을 700U/L 투입하였고 50℃에서 24시간 반응을 진행하였다. 실시예 2의 BtAS의 변이형1(V542D) 및 변이형2(V542K), 실시예 4에 따른 변이형3(Y414F*P200R)을 야생형과 같이 동일한 조건에서 반응시켰다. 각 효소의 전환반응 후 당조성 분석은 아래 표의 조건으로 HPLC 분석하였다.
| 컬럼 | Bio-rad Aminex HPX-87C |
| 주입량 | 10㎕ |
| 유속 | 0.6ml/min |
| 컬럼온도 | 80℃ |
| 이동상 | Water 100% |
상기 비피도박테리움 터모필럼 유래 BtAS 효소에 따른 전환반응 후 당조성 결과를 아래 표에 나타내었다. 상기 표에서 전환결과 얻어진 산물의 당조성 분석은, 반응 생성물에 포함된 전체 당류 고형분 함량(중량)을 기준으로, 각 구성 당류의 고형분 함량(중량)을 백분율로 나타낸 것이다(하기 표의 단위는 중량%이다). 상기 올리고당은 주로 포도당이 알파 1-4 결합으로 연결된 포도당 올리고머이며, 대부분은 DP3 및 DP4이상의 당류로 구성된다.
| 구분 | 올리고당 | 설탕 | 투라노스 | 트레할룰로스 | 과당 |
| BtAS 야생형 | 16.1 | 0 | 46.7 | 3.3 | 33.9 |
| BtAS V542D | 18.5 | 0 | 46.9 | 3.4 | 31.2 |
| BtAS V542K | 18.1 | 0 | 46.1 | 4.2 | 31.6 |
| BtAS Y414F*P200R | 15.8 | 0 | 50.0 | 3.2 | 31.0 |
상기 표에서 확인한 바와 같이, 비피도박테리움 유래의 아밀로수크라제는 반응온도가 50℃로 높아 반응속도가 빠르며 24시간 내 700U/L의 효소로 설탕 기질을 모두 소모하고 투라노스를 최대 생성하며 반응을 종결시킬 수 있었다.
6.2 나이세리아 서브플라바 유래 아밀로수크라제 투라노스 전환율
비교예 1의 나이세리아 서브플라바 유래 아밀로수크라제의 투라노스 전환율을 검토하기 위하여, 상기 실시예 6.1과 동일한 조건 및 방법으로 분석하되 나이세리아 서브플라바 유래 투라노스 전환효소(NsAS)는 500, 700, 1000, 1400, 1900U/L를 투입하였고 35℃에서 48시간 반응을 진행하고 그 결과를 아래 표에 나타내었다.
| 구분 | 올리고머 | 설탕 | 투라노스 | 트레할룰로스 | 과당 |
| NsAS 500U/L | 1.9 | 41.8 | 25.2 | 8.8 | 22.3 |
| NsAS 700U/L | 3.5 | 29.7 | 35.3 | 8.8 | 22.7 |
| NsAS 1000U/L | 6.5 | 16.6 | 47.7 | 7.7 | 21.5 |
| NsAS 1400U/L | 13.0 | 0 | 55.9 | 7.7 | 23.4 |
| NsAS 1900U/L | 12.6 | 0 | 56.7 | 7.9 | 22.8 |
상기 표에서 확인한 바와 같이, NsAS는 반응온도가 35℃로 낮아 반응속도가 느리며 1000U/L의 효소 투입으로는 48시간 내 반응을 완료시키기 어려웠으며, 48시간 내에 설탕 기질을 모두 소진하고 투라노스를 최대로 생산하기 위해서는 1400U/L이상의 많은 효소가 필요한 것을 확인하였다. 나이세리아 서브플라바 유래 아밀로수크라제를 이용한 투라노스 생산 결과를 보면, 트레할룰로스 함량이 높아 투라노스 분리 정제에 바람직하지 않음을 확인하였다.
따라서, 비피도박테리움 유래의 아밀로수크라제가 나이세리아 서브플라바 유래의 아밀로수크라제 대비 적은 양의 효소 투입량으로 기질을 빠르게 소진하여 전체 전환 반응을 빠르게 종결시킬 수 있어, 효소 투입량 절감 및 반응속도를 단축함을 확인하였다.
실시예 7. 변이형 아밀로수크라제의 전환반응
7.1 야생형 아밀로수크라제의 투라노스 전환반응
50 mM 소듐아세테이트 완충용액 (pH 6.0)에서 수크로스를 1.0, 1.5, 2.0 및 2.5M의 농도(각각 1.0S, 1.5S, 2.0S 및 2.5S)로, 프럭토즈를 0.25, 0.5, 0.75 및 1M의 농도(각각 0.25F, 0.5F, 0.75F 및 1F)로 포함하는 기질로부터 투라노스를 합성시켰다. 효소 반응은 400 U/L의 야생형 효소로 48 시간 동안 수행되었다. 배양 후, 반응 혼합물을 끓는 물에서 10 분 동안 열처리하여 효소를 불 활성화시켰다. 불 활성화된 반응 용액을 0.2 ㎛ 주사기 필터로 여과하고 가용성 분획을 HPAEC로 분석 하였다. 가용성 분획은 CarboPacTM PA1 분석 칼럼 (4 x 250mm, DIONEX, SUNNYVALE, CA, USA)에서 pulsed amperometric detection (HPAEC-PAD)과 결합 된 고성능 음이온 교환 크로마토그램으로 분석하여 적은 양의 포도당, 과당, 자당 및 투라노스를 정량화하였다. 반응물의 상층 액을 증류수로 적절히 희석하고 0.2 μm 주사기 필터로 여과하였다. 주입 부피는 50 μl이고 100 mM NaOH 용액으로 1.0 mL/min의 유속으로 분석했다. 당류의 함량 조성은 표준 물질과의 면적 비교에 의해 결정되었다.
투라노스 전환율에 대한 기질 조성의 영향은 400 U/L의 BtAS로 50℃ 의 온도 및 50 mM 소듐아세테이트 완충액으로 pH 6.0에서 수행되었었으며, 분석된 결과를 도 6에 나타내었다. 투라노스 전환율은 기질(수크로스)로부터 전환된 투라노스의 생성량을 의미하는 것으로, 기질 농도 대비 생성된 투라노스 농도의 비(%)로 나타낸다.
실험 결과에서 확인할 수 있듯이, 48 시간 동안의 효소 반응 후, 최종 투라노스 전환율은 프럭토스 농도가 1.0 M까지 증가함에 따라 유의하게 증가 하였다. 이는 co-substrate로서 외인성 프럭토스이 투라노스 생산을 촉진함을 시사했다. 반응 혼합물 중에 1.0 M의 과당을 첨가함으로써, 투라노스 전환율은 1.0 M의 수크로스로 22.7 %에서 43.3 %로 증가하였고, 1.5 M의 수크로스에서 23.7 %에서 39.4 %로 증가 하였다. 따라서, 투라노스 수확량은 BtAS 생체 전환 시스템의 초기 프럭토스 농도에 크게 의존하는 것을 확인하였다.
즉, 수크로스로부터 유래된 과당 부분이 효소로부터 방출되지 않고서 투라노스를 생산하기 위한 수용체로 직접 이용되고 외인과적인 과당 첨가가 투라노스 생산 수율을 더욱 향상 시키는 것으로 보인다. 최적의 기질 조건은 수크로스와 과당 각각 1.0 M이었고, 이 때 투라노스 전환율은 43.3 %이었다.
7.2. 실시예 2의 변이형 효소의 투라노스 생산성
실시예 2의 2종의 변이형 아밀로수크라제의 투라노스 생산성을 살펴보기 위하여, 2M 설탕, 0.75M 과당의 기질에 아밀로수크라제 100U/L~500U/L(여기서 Unit은 가수분해 활성으로, 100mM의 설탕에 1mg의 아밀로수크라제를 처리하여 50℃에서 30분 간 반응했을 때 분당 생성되는 과당의 양 μmole을 의미한다.)을 넣고 투라노스 전환반응을 시작했다.
도 7 및 도 8은 효소의 농도와 시간당 생성하는 투라노스의 농도의 상관 관계를 나타내는 그래프이다. 도 7 및 도 8 의 수식 모두 x값은 넣어준 효소의 농도 (U/L)를 의미하며, y값은 시간당 생성되는 투라노스의 농도를 g/L로 나타낸 것으로서 단위는 g/L/hr이다. 수식은 y = 0.082x + 7.3862 이고, 변이 아밀로수크라제 V542D를 2M 설탕과 0.75M의 과당에 첨가 했을 때 시간당 생성되는 투라노스를 계산하였다. 그 결과, 비피도박테리움 터모필럼 유래의 아밀로수크라제 V542D의 투라노스 생산성은 8.9 U/mg임을 확인하였다.
동일한 방법으로 아밀로수크라제 V542K를 2M설탕과 0.75M의 과당에 100U/L~500U/L만큼 넣고 시간대 별로 시료를 채취하여 분석한 결과, 도 8에서 보는 효소 농도와 투라노스 생성에 관한 상관 관계는 y = 0.1879x + 8.0043이었다. 이를 통하여 비피도박테리움 터모필럼 유래의 변이 아밀로수크라제 V542K의 투라노스 생산성은 9.9U/mg임을 확인하였다.
2M 설탕 또는 0.75M 과당을 기질로 하여 50℃에서 반응 할 때, 변이 아밀로수크라제 V542D는 8.9U/mg, 변이 아밀로수크라제 V542K는 9.9U/mg의 활성을 지녀, 야생형 아밀로수크라제 3.0U/mg의 두 배 이상에 해당하는 수치로, DNA돌연변이법으로 효소 활성을 상당한 수준으로 개선한 것이라고 할 수 있다.
7.3 실시예 3의 변이형 효소의 활성도 및 생산성
실시예 3의 단일 점 변이형 아밀로수크라제 효소의 효소 활성도를 실시예 5.1과 동일한 방법으로 측정하고, 상기 단일 점 변이형 아밀로수크라제 효소의 투라노스 전환율을 pH 6 및 50℃의 온도에서, 기질 2M 수크로스와 400U/L 반응시킨 후 기질의 농도 대비 생성된 투라노스 농도의 비(%)를 산출하였다. 또한 기질로부터 생성된 투라노스의 농도를 반응 종결 시간으로 나눈, 기질로부터 투라노스 전환 시 시간 당 생성되는 투라노스(농도)를 산출하고 아래 표에 나타내었다.
Y414F는 400 U/L로 반응시켰을 때, 수크로스를 완전하게 소비하지 못하고 속도가 느려졌으며, 이는 상대적으로 단백질 양이 적게 들어가서 반응을 못한 것이라 예측하여 동일 조건에서 단백질 양을 4배 늘려 1600U/L로 반응시켰다. 상기 측정 및 산출된 결과를 아래 표에 나타내었다.
| 정제후 | 반응 효소 양 (unit/L) |
단백질량 (mg/10ml) |
효소 비활성 (unit/mg) |
투라노스 전환율 (%) |
반응 종결 시간(h) | 시간 당 생성되는 투라노스 (g/L/h) |
| 야생형 효소(BtAS) | 400 | 3 | 1.3±0.1 | 25±2.5 | 12 | 14.25 |
| P200R | 400 | 8 | 0.5±0.0 | 27±1.5 | 12 | 15.39 |
| V202I | 400 | 7 | 0.9±0.0 | 29±2.0 | 12 | 16.53 |
| Y265F | 400 | 6 | 0.6±0.2 | 23±1.0 | 6 | 26.22 |
| V305I | 400 | 3 | 1.6±0.1 | 21±1.0 | 24 | 6 |
| K393R | 400 | 4 | 1.0±0.0 | 22±0.0 | 12 | 12.54 |
| S420T | 400 | 4 | 1.5±0.3 | 20 | 24 | 5.7 |
| Y414F | 400 | 1 | 4.1±0.8 | 12.5±2.5 | >24 | - |
| Y414F | 1600 | 4 | 4.1±0.8 | 35±1.4 | 12 | 19.95 |
상기 결과에 나타낸 바와 같이, 야생형 효소와 비교하면 비활성은 414번째 티로신을 페닐알라닌으로 치환시킨 변이체가 특이적으로 4배가 높아졌다. 모든 변이체들의 투라노스 전환율을 확인하기 위하여 스크리닝 개념으로 400 U/L로 효소 양을 맞춰 반응을 시킨 결과, P200R, V202I가 야생형 효소보다 투라노스 전환율이 높게 나왔으며, 반응속도는 Y265F가 빨라진 것을 확인 할 수 있었다. 또한 1600 U/L로 Y414F를 반응시킨 결과, 수크로스를 완전하게 소비하고 12시간 만에 반응이 종결되었으며 투라노스 전환율이 약 35%로, 변이체 중에서 1600 U/L로 반응시킨 Y414F 돌연변이 효소가 투라노스 전환율이 가장 높다는 것을 확인 할 수 있었다.
7.4. 이중 점 변이형 효소의 활성도 및 생산성
실시예 4의 이중 점 변이형 아밀로수크라제 효소를 상기 7.3의 실험과 동일한 방법으로 pH 6 및 50℃의 온도에서, 기질 2 M 수크로스와 400 U/L 및 1600 U/L로 반응시킨 후 투라노스 전환율 및 수크로스 소비량을 산출하여 도 9 및 도 10에 나타내었다.
| 정제후 | 반응 효소 양 (unit/L) |
단백질량 (mg/10ml) |
효소 비활성 (unit/mg) |
투라노스 전환율(%) | 반응 종결 시간(h) | 시간 당 생성되는 투라노스 (g/L/h) |
| 야생형 효소(BtAS) | 400 | 3 | 1.3±0.1 | 25±2.5 | 12 | 14.25 |
| Y414F*P200R | 400 | 1.5 | 2.6±0.5 | 17±0.5 | >24 | - |
| Y414F*V202I | 400 | 1 | 4.0±0.2 | 14±2.5 | >24 | - |
| Y414F*Y265F | 400 | 1 | 4.2±0.1 | 13±2.0 | >24 | - |
| Y414F*P200R | 1600 | 6 | 2.6±0.5 | 50±4 | 6 | 57 |
| Y414F*V202I | 1600 | 4 | 4.0±0.2 | 41.5±4.9 | 6 | 47 |
| Y414F*Y265F | 1600 | 4 | 4.2±0.1 | 40.5±2.1 | 6 | 46 |
Y414F 기반의 더블포인트 돌연변이 변이체도 Y414F 싱글포인트 돌연변이 변이체와 마찬가지로 400 U/L로 반응시켰을 때 수크로스 소비를 완전하게 하지 못하여 최대 투라노스 전환율을 확인하지 못했다. 다만 1600U/L로 반응시킨 결과, 투라노스 전환율이 높게 나왔으며 그 중에서도 Y414F 변이체와 P200R을 접목시킨 Y414F*P200R 더블 포인트 돌연변이 변이체가 투라노스 전환율이 50%로 가장 높은 투라노스 전환율을 보였다.
실시예 8. 이중 점 변이형 효소를 함유하는 (세포 추출물) 투라노스 생성
2M 수크로스와, 2M 수크로스와 0.75M 프럭토스의 혼합 기질이 각각 함유된 50mM 소듐 아세테이트 완충액(pH 6.0) 10 ml을 10분간 pre-heat해준 뒤 실시예 4의 Y414F*P200R 변이체 효소 Cell extract 상태에서 16 unit/10ml의 농도가 되도록 앞서 정제한 신규 아밀로수크라제 16 unit을 수크로스 완충용액에 첨가하였다. 실시예 4에서 수확한 침전물을 50mM Tris-HCl 완충 용액(pH 7) 45mL에서 볼텍싱하여 완전하게 재 현탁 시킨 후에 세포 용액을 초음파 분쇄기(Sonic Dismembrator 550, Fisher Scientific Co.) Model D100을 이용하여 세포를 파괴하였다. 세포 파쇄물을 4에서 20분 동안 11,000 x g에서 원심분리하여 침전물인 세포 잔해물은 제외하고 상등액, 세포 추출물을 얻었다.
50℃에서 12시간 동안 효소반응을 수행하고 10분간 끓는 물에 가열하여 효소반응을 종결시킨 후 얻어진 반응물을 10000배 희석하여 UltiMate 3000 HPLC system(Dionex, USA)을 이용하여 분석하였다. 이때, 글루코스, 프락토스, 수크로스, 투라노스, 말토오스(Sigma-Aldrich Chemical Co., St. Louis, MO), 트레할룰로스(Samyang, Seongnam, Korea)를 표준물질로 이용하여 효소반응물을 분석하였으며, 투라노스의 전환율은 여러 가지 농도의 투라노스 표준물질을 분석하여 얻은 Standard curve를 이용하여 계산하였다.
투라노스를 분석하기 전에 샘플은 수크로스를 제거하기 위하여 Invertase 로 전처리한 후에 분석한다. 수크로스를 제거하는 이유는 투라노스와 수크로스의 피크가 겹쳐 투라노스의 정량에 영향을 미치기 때문이다. 투라노스 분석조건으로 HPLC 칼럼은 Bio-rad Aminex HPX-87C를 사용하였고, 샘플 주입량(Injection volume)은 10ul로 Column oven 온도는 80℃에서 이동상 용매는 100% 물로 분석하였다.
2M의 수크로스를 기질로 사용한 반응 후 6시간이 경과한 샘플의 당 분석 결과 및, 2M 수크로스 및 0.75M 프럭토스를 기질로 사용한 반응 후 6시간이 경과한 샘플의 당 분석 결과를 아래 표에 정리하였다.
| 기질 | 올리고머 (중량%) |
투라노스 (중량%) |
글루코스+트레할룰로스 (중량%) |
| 2M 수크로스 | 30.02 | 42.01 | 0.16 |
| 2M 수크로스 +0.75M 프럭토스 | 17.05 | 51.12 | 0.95 |
상기 결과에 나타낸 바와 같이, 기질을 2M 수크로스만 반응시키면 투라노스는 42.01%이고 기질을 2M 수크로스 뿐만 아니라 0.75M의 프럭토스를 첨가하여 반응시킨 결과 51.1%로 더 높은 수준으로 투라노스를 생성하는 것을 확인하였다.
실시예 9. 기질 조성에 따른 아밀로수크라제의 전환반응
아밀로수크라제의 전환 반응에서 기질 조성에 따른 반응성을 검토하기 위하여, 기질로 쓰이는 설탕과 과당의 고형분 중량비를 1:0, 1:0.2, 1:0.5, 1:0.75, 1:1, 1:1.5로 하여 물을 첨가하여 농도가 63.5bx가 되도록 용해시켜 기질을 준비하고 pH는 6로 조정하였다. 상기 설탕 고형분 함량은 2M에 해당되며 구체적인 설탕, 및 과당의 중량은 하기 표 12에 나타냈다.
| 설탕:과당의 혼합비율 | 설탕(g/L) | 설탕 몰농도(M) | 과당(g/L) | 과당 몰농도(M) |
| 1:0.0 | 685 | 2 | 0 | 0 |
| 1:0.2 | 685 | 2 | 135 | 0.75 |
| 1:0.5 | 685 | 2 | 342 | 1.9 |
| 1:0.7 | 685 | 2 | 513 | 2.8 |
| 1:1.0 | 685 | 2 | 685 | 3.8 |
| 1:1.5 | 685 | 2 | 1027 | 5.7 |
설탕과 과당의 조성이 다른 기질에 실시예 4의 BtAS(Y414F*P200R) 효소를 700U/L 투입하였고 50℃에서 24시간 반응을 진행하였다. 각 기질에서의 효소 전환반응 후 당조성 분석은 상기 실시예 6과 같은 조건으로 수행하였다.
기질 조성에 따른 BtAS(Y414F*P200R) 효소의 전환반응 후 당조성 결과는 아래 표에 나타내었다. 하기 표 13에서 함량은 반응산물 내 전체 당류 고형분 100중량%에 포함된 각각 당류의 함량을 중량%로 표시한 것이다.
| 설탕:과당의 혼합비율 | 올리고당 | 올리고당/투라노스 비율 | 설탕 | 투라노스 | 트레할룰로스 | 과당 |
| 1:0.0 | 32.4 | 82.2% | 0 | 39.4 | 2.9 | 25.3 |
| 1:0.2 | 16.5 | 33.0% | 0 | 50.0 | 3.5 | 30.0 |
| 1:0.5 | 8.2 | 16.5% | 0 | 49.7 | 3.5 | 38.6 |
| 1:0.7 | 5.8 | 12.8% | 0 | 45.2 | 3.3 | 45.7 |
| 1:1.0 | 4.0 | 9.7% | 0 | 41.3 | 3.1 | 51.6 |
| 1:1.5 | 2.4 | 7.0% | 0 | 34.4 | 2.5 | 60.7 |
상기 결과에서 확인한 바와 같이, 기질에서 설탕 대비 과당의 중량을 높이면 올리고당의 생성이 감소하고 상대적으로 설탕의 비율이 적어지므로 투라노스의 생성양은 줄어드는 것을 확인하였다. 트레할룰로스는 과당 비율에 크게 영향을 받지 않고 3~4 중량% 수준을 유지하였다. 투라노스 생성량 대비 올리고당이 현저히 줄어드는 기질의 조성은 설탕:과당=1:0.2 부터 시작하여 1:0.5에서 크게 감소하며 이후 과당함이 증가할수록 점진적으로 감소하였다. 설탕과 과당의 혼합비를 살펴보면, 동일 설탕 고형분 함량 대비 혼합되는 과당의 함량이 증가할수록, 투라노스 생성량 대비 올리고당 함량이 적어지며, SMB 크로마토그래피에 적합한 당조성으로 판단되었다. 다만, 설탕 기질 대비 첨가되는 과당 함량이 지나치게 높은 경우, 투라노스 생성량이 감소하므로, 투라노스의 생성량을 특정 ?량이상으로 확보하면서, 투라노스 함량 대비 불순물인 올리고당의 함량의 비율이 낮은 생성물을 얻을 수 있도록 설탕과 과당의 혼합 비율을 적절히 선택하는 것이 바람직하다.
실시예 10. 아밀로수크라제의 전환반응 조성물의 투라노스 고순도 분리
10.1 비피도박테리움 터모필럼 유래 아밀로수크라제의 전환반응 조성물
비피도박테리움 터모필럼 유래 아밀로수크라제 효소의 전환반응 조성물은 기질의 조성을 설탕:과당=1:0.5로 하고 실시예 4의 BtAS(Y414F*P200R) 효소 700U/L 투입하여 실시예 9와 동일한 방법으로 전환 반응하여 얻었다.
상기 아밀로수크라제에 의한 전환반응 조성물은 SMB 크로마토그래피를 원활하게 진행하게 위해 Glucoamlyase(노보자임 사)를 투라노스 전환 반응 후에 얻어진 반응 생성물의 고형분 함량(중량) 대비 0.3 중량%, PH 4.5, 60℃에서 반응하여 올리고당을 포도당으로 분해하였다. 그 후 Na+ 타입의 양이온 교환수지를 사용하여 투라노스 분획을 분리하였다. SMB 크로마토그래피에 의한 투라노스 분획 분리 전 후 조성물의 당조성을 아래 표에 나타내었다.
| 구분 | 올리고당 | 설탕 | 투라노스 | 트레할룰로스 | 포도당 | 과당 |
| 분리 전 | 1.5 | 0 | 52.2 | 2.8 | 7.0 | 36.5 |
| 분리 후 | 4.0 | 0 | 91.6 | 3.8 | 0.6 | 0 |
SMB 크로마토그래피를 활용한 투라노스 고순도 분리 결과, 91.6% 함량의 투라노스 조성물을 수율 65%로 얻을 수 있었다. SMB 크로마토그래피는 2상 분리에 적합하게 설계되어 있어 글루코아밀라제 효소 처리로 올리고당을 포도당으로 분해시키고, SMB 크로마토그램 상에서 투라노스 피크와 트레할룰로스 피크 사이를 기점으로 양쪽 두 개의 분획을 얻고자 투라노스와 트레할룰로스 사이를 분리하였으며, 그 결과 좌측 분획은 투라노스와 일부 올리고머만 존재하기 때문에 좌측 분획에서 투라노스의 순도는 상승하게 되어 투라노스 분획을 얻고, 우측 분획은 투라노스가 거의 없기 때문에 라피네이트가 되어 결국, 투라노스 고순도 분획을 얻었다.
BtAS 효소의 전환반응 조성물은 SMB 분리 전 조성물의 트레할룰로스 함량이 적어 SMB 크로마토그래피에 의한 분리 후의 투라노스 순도 및 수율을 높일 수 있었다. 즉, 투라노스 피크와 트레할룰로스 피크는 약간 겹쳐 있어 둘 사이를 기점으로 양쪽 분획을 얻을 때, 완벽하게 두 물질이 분리되지 않고 양쪽 분획에 각각 조금씩 딸려가게 된다. 그렇기 때문에 트레할룰로스 함량이 적으면 투라노스 분획쪽으로 분리되는 트레할룰로스가 그만큼 적어지기 때문에 투라노스쪽 분획에서 좀 더 높은 함량 및 순도로 투라노스를 얻을 수 있다.
10.2 나이세리아 서브플라바 유래 아밀로수크라제의 전환반응 조성물
나이세리아 서브플라바 유래 효소의 전환반응 조성물은 비교예 1의 NsAS 효소를 1400U/L 투입하고 실시예 10.1과 동일한 방법으로 전환 반응하여 얻었다. 전환반응 조성물은 SBM 크로마토그래피를 원활하게 진행하게 위해 Glucoamlyase(노보자임 사)를 투라노스 전환 반응 후에 얻어진 반응 생성물의 고형분 함량(중량) 대비 0.3 중량%, pH4.5, 60℃에서 반응하여 올리고머를 포도당으로 분해하였다.
그 후 Na+ 타입의 양이온 교환수지를 사용하여 투라노스 분획을 분리하였다. SMB 크로마토그래피에 의한 투라노스 분획 분리 전과 후 조성물의 당조성은 아래 표에 나타내었다.
| 구분 | 올리고당 | 설탕 | 투라노스 | 트레할룰로스 | 포도당 | 과당 |
| 분리 전 | 1.6 | 0 | 60.6 | 8.0 | 7.4 | 22.4 |
| 분리 후 | 4.8 | 0 | 87.2 | 7.3 | 0.7 | 0 |
나이세리아 서브플라바 유래 아밀로수크라제의 전환 반응 생성물의 경우, SMB 크로마토그래피를 이용한 고순도 분리 후 87.2% 함량의 투라노스 조성물을 수율 38%로 얻을 수 있었다. 투라노스와 근접하여 함께하는 트레할룰로스의 함량이 높아 투라노스 분리 순도는 90%이상 도달하지 못하였다.
<110> SAMYANG CORPORATION
INDUSTRY ACADEMY COOPERATION FOUNDATION OF SEJONG UNIVERSITY
<120> Method ofproducing turanose
<130> DPP20173326KR
<160> 44
<170> KoPatentIn 3.0
<210> 1
<211> 603
<212> PRT
<213> Artificial Sequence
<220>
<223> amylosucrase BtAS wild type
<400> 1
Met Glu Ala Thr Tyr Arg Asp Ser Val Phe Ala Glu Arg Leu Ala Pro
1 5 10 15
Arg Cys Ala Glu Leu Glu Gln Leu Phe Arg Ser Leu Tyr Gly Asp Ser
20 25 30
Pro Glu Phe Asp His Phe Glu Gln Val Met Ala Lys Ala His Ala Asp
35 40 45
Arg Pro Ala Asp Leu Lys Arg Leu Asp Ala Ala Arg Glu His Asp Pro
50 55 60
Gln Trp Tyr Arg Arg Gly Asp Met Phe Gly Met Thr Met Tyr Thr Asp
65 70 75 80
Leu Phe Ala Gly Lys Leu Thr Asp Leu Ala Lys His Ile Asp Tyr Leu
85 90 95
Lys Glu Gln His Leu Thr Tyr Leu His Leu Met Pro Leu Leu Thr Met
100 105 110
Pro His Pro Asp Asn Asp Gly Gly Tyr Ala Ile Glu Asp Phe Asp Thr
115 120 125
Val Asp Pro Thr Ile Gly Thr Asn Glu Asp Leu Ala Asp Leu Thr Ala
130 135 140
Lys Leu Arg Glu Ala Gly Ile Ser Leu Cys Leu Asp Phe Val Met Asn
145 150 155 160
His Thr Ala Ser Thr His Arg Trp Ala Lys Ala Ala Gln Ala Gly Asp
165 170 175
Pro Glu Tyr Gln Asp Tyr Tyr Phe Cys Tyr Asp Asp Arg Thr Ile Pro
180 185 190
Asp Gln Tyr Asp Ala Val Val Pro Gln Val Phe Pro Thr Ala Ala Pro
195 200 205
Gly Asn Phe Thr Trp Asn Glu Gln Met Gly Lys Trp Val Met Thr Gln
210 215 220
Phe Tyr Pro Phe Gln Trp Asp Leu Asn Tyr Arg Asn Pro Lys Val Phe
225 230 235 240
Val Val Met Met Ser Ser Leu Leu His Leu Ala Asn Leu Gly Val Glu
245 250 255
Val Phe Arg Ile Asp Ala Val Pro Tyr Ile Trp Lys Gln Leu Gly Thr
260 265 270
Asn Cys Arg Asn Leu Pro Gln Val His Thr Ile Val Arg Met Met Arg
275 280 285
Ile Met Ser Glu Ile Val Cys Pro Ala Val Val Phe Lys Gly Glu Val
290 295 300
Val Met Ala Pro Lys Glu Leu Ala Ala Tyr Phe Gly Thr Pro Glu Lys
305 310 315 320
Pro Glu Cys His Met Leu Tyr Asn Val Ser Val Met Val Asn Leu Trp
325 330 335
Ser Ala Leu Ala Asn Gly Asp Thr Arg Leu Leu Lys Thr Gln Ile Asp
340 345 350
Lys Leu Asp Ala Leu Pro Asp Asn Cys Trp Phe Val Asn Tyr Leu Arg
355 360 365
Cys His Asp Asp Ile Gly Trp Gly Leu Asp Glu Asp Val Glu Arg Gln
370 375 380
Leu Gly Ile Asp Pro Leu Lys His Lys Glu Phe Leu Tyr His Phe Tyr
385 390 395 400
Glu Gly Met Val Pro Gly Ser Trp Ala Met Gly Glu Leu Tyr Asn Tyr
405 410 415
Asp Pro Ala Ser Gly Asp Ala Arg Ser Cys Gly Thr Thr Ala Ser Leu
420 425 430
Cys Gly Ile Glu Arg Ala Leu Ile Thr His Asp Arg Pro Leu Tyr Glu
435 440 445
Arg Ser Ile Gln Arg Asp Leu Leu Met His His Ala Met Gly Phe Leu
450 455 460
Arg Gly Phe Pro Met Leu Asn Cys Gly Asp Glu Ile Gly Gln Leu Asn
465 470 475 480
Gly Trp Asp Tyr Lys Glu Asp Pro Asp Arg Val Ala Asp Ser Arg Asn
485 490 495
Leu His Arg Ser Lys Phe Asn Trp Lys Asn Ala Ala Lys Arg Asp Val
500 505 510
Pro Gly Thr Leu Pro Asn Arg Leu Trp Glu Gly Met Ala Asp Val Arg
515 520 525
Gln Met Arg Ser Asp Pro Cys Phe Ala Pro Asp Ala Trp Val Thr Thr
530 535 540
Trp Asp Ala His Asp Asp Gly Ile Leu Ala Met Val Arg Gln Ser Gly
545 550 555 560
Gly Arg Thr Leu Leu Gly Val Phe Asn Phe Ala Asn Arg Asp Ala Thr
565 570 575
Ala Thr Leu Asp Ser Ile Glu Gly Val Ser Leu Pro Arg Thr Val Ala
580 585 590
Leu Lys Pro Tyr Glu Trp Lys Ile Glu Ala Cys
595 600
<210> 2
<211> 603
<212> PRT
<213> Artificial Sequence
<220>
<223> amylosucrase BtAS V542K
<400> 2
Met Glu Ala Thr Tyr Arg Asp Ser Val Phe Ala Glu Arg Leu Ala Pro
1 5 10 15
Arg Cys Ala Glu Leu Glu Gln Leu Phe Arg Ser Leu Tyr Gly Asp Ser
20 25 30
Pro Glu Phe Asp His Phe Glu Gln Val Met Ala Lys Ala His Ala Asp
35 40 45
Arg Pro Ala Asp Leu Lys Arg Leu Asp Ala Ala Arg Glu His Asp Pro
50 55 60
Gln Trp Tyr Arg Arg Gly Asp Met Phe Gly Met Thr Met Tyr Thr Asp
65 70 75 80
Leu Phe Ala Gly Lys Leu Thr Asp Leu Ala Lys His Ile Asp Tyr Leu
85 90 95
Lys Glu Gln His Leu Thr Tyr Leu His Leu Met Pro Leu Leu Thr Met
100 105 110
Pro His Pro Asp Asn Asp Gly Gly Tyr Ala Ile Glu Asp Phe Asp Thr
115 120 125
Val Asp Pro Thr Ile Gly Thr Asn Glu Asp Leu Ala Asp Leu Thr Ala
130 135 140
Lys Leu Arg Glu Ala Gly Ile Ser Leu Cys Leu Asp Phe Val Met Asn
145 150 155 160
His Thr Ala Ser Thr His Arg Trp Ala Lys Ala Ala Gln Ala Gly Asp
165 170 175
Pro Glu Tyr Gln Asp Tyr Tyr Phe Cys Tyr Asp Asp Arg Thr Ile Pro
180 185 190
Asp Gln Tyr Asp Ala Val Val Pro Gln Val Phe Pro Thr Ala Ala Pro
195 200 205
Gly Asn Phe Thr Trp Asn Glu Gln Met Gly Lys Trp Val Met Thr Gln
210 215 220
Phe Tyr Pro Phe Gln Trp Asp Leu Asn Tyr Arg Asn Pro Lys Val Phe
225 230 235 240
Val Val Met Met Ser Ser Leu Leu His Leu Ala Asn Leu Gly Val Glu
245 250 255
Val Phe Arg Ile Asp Ala Val Pro Tyr Ile Trp Lys Gln Leu Gly Thr
260 265 270
Asn Cys Arg Asn Leu Pro Gln Val His Thr Ile Val Arg Met Met Arg
275 280 285
Ile Met Ser Glu Ile Val Cys Pro Ala Val Val Phe Lys Gly Glu Val
290 295 300
Val Met Ala Pro Lys Glu Leu Ala Ala Tyr Phe Gly Thr Pro Glu Lys
305 310 315 320
Pro Glu Cys His Met Leu Tyr Asn Val Ser Val Met Val Asn Leu Trp
325 330 335
Ser Ala Leu Ala Asn Gly Asp Thr Arg Leu Leu Lys Thr Gln Ile Asp
340 345 350
Lys Leu Asp Ala Leu Pro Asp Asn Cys Trp Phe Val Asn Tyr Leu Arg
355 360 365
Cys His Asp Asp Ile Gly Trp Gly Leu Asp Glu Asp Val Glu Arg Gln
370 375 380
Leu Gly Ile Asp Pro Leu Lys His Lys Glu Phe Leu Tyr His Phe Tyr
385 390 395 400
Glu Gly Met Val Pro Gly Ser Trp Ala Met Gly Glu Leu Tyr Asn Tyr
405 410 415
Asp Pro Ala Ser Gly Asp Ala Arg Ser Cys Gly Thr Thr Ala Ser Leu
420 425 430
Cys Gly Ile Glu Arg Ala Leu Ile Thr His Asp Arg Pro Leu Tyr Glu
435 440 445
Arg Ser Ile Gln Arg Asp Leu Leu Met His His Ala Met Gly Phe Leu
450 455 460
Arg Gly Phe Pro Met Leu Asn Cys Gly Asp Glu Ile Gly Gln Leu Asn
465 470 475 480
Gly Trp Asp Tyr Lys Glu Asp Pro Asp Arg Val Ala Asp Ser Arg Asn
485 490 495
Leu His Arg Ser Lys Phe Asn Trp Lys Asn Ala Ala Lys Arg Asp Val
500 505 510
Pro Gly Thr Leu Pro Asn Arg Leu Trp Glu Gly Met Ala Asp Val Arg
515 520 525
Gln Met Arg Ser Asp Pro Cys Phe Ala Pro Asp Ala Trp Lys Thr Thr
530 535 540
Trp Asp Ala His Asp Asp Gly Ile Leu Ala Met Val Arg Gln Ser Gly
545 550 555 560
Gly Arg Thr Leu Leu Gly Val Phe Asn Phe Ala Asn Arg Asp Ala Thr
565 570 575
Ala Thr Leu Asp Ser Ile Glu Gly Val Ser Leu Pro Arg Thr Val Ala
580 585 590
Leu Lys Pro Tyr Glu Trp Lys Ile Glu Ala Cys
595 600
<210> 3
<211> 603
<212> PRT
<213> Artificial Sequence
<220>
<223> amylosucrase BtAS V542D
<400> 3
Met Glu Ala Thr Tyr Arg Asp Ser Val Phe Ala Glu Arg Leu Ala Pro
1 5 10 15
Arg Cys Ala Glu Leu Glu Gln Leu Phe Arg Ser Leu Tyr Gly Asp Ser
20 25 30
Pro Glu Phe Asp His Phe Glu Gln Val Met Ala Lys Ala His Ala Asp
35 40 45
Arg Pro Ala Asp Leu Lys Arg Leu Asp Ala Ala Arg Glu His Asp Pro
50 55 60
Gln Trp Tyr Arg Arg Gly Asp Met Phe Gly Met Thr Met Tyr Thr Asp
65 70 75 80
Leu Phe Ala Gly Lys Leu Thr Asp Leu Ala Lys His Ile Asp Tyr Leu
85 90 95
Lys Glu Gln His Leu Thr Tyr Leu His Leu Met Pro Leu Leu Thr Met
100 105 110
Pro His Pro Asp Asn Asp Gly Gly Tyr Ala Ile Glu Asp Phe Asp Thr
115 120 125
Val Asp Pro Thr Ile Gly Thr Asn Glu Asp Leu Ala Asp Leu Thr Ala
130 135 140
Lys Leu Arg Glu Ala Gly Ile Ser Leu Cys Leu Asp Phe Val Met Asn
145 150 155 160
His Thr Ala Ser Thr His Arg Trp Ala Lys Ala Ala Gln Ala Gly Asp
165 170 175
Pro Glu Tyr Gln Asp Tyr Tyr Phe Cys Tyr Asp Asp Arg Thr Ile Pro
180 185 190
Asp Gln Tyr Asp Ala Val Val Pro Gln Val Phe Pro Thr Ala Ala Pro
195 200 205
Gly Asn Phe Thr Trp Asn Glu Gln Met Gly Lys Trp Val Met Thr Gln
210 215 220
Phe Tyr Pro Phe Gln Trp Asp Leu Asn Tyr Arg Asn Pro Lys Val Phe
225 230 235 240
Val Val Met Met Ser Ser Leu Leu His Leu Ala Asn Leu Gly Val Glu
245 250 255
Val Phe Arg Ile Asp Ala Val Pro Tyr Ile Trp Lys Gln Leu Gly Thr
260 265 270
Asn Cys Arg Asn Leu Pro Gln Val His Thr Ile Val Arg Met Met Arg
275 280 285
Ile Met Ser Glu Ile Val Cys Pro Ala Val Val Phe Lys Gly Glu Val
290 295 300
Val Met Ala Pro Lys Glu Leu Ala Ala Tyr Phe Gly Thr Pro Glu Lys
305 310 315 320
Pro Glu Cys His Met Leu Tyr Asn Val Ser Val Met Val Asn Leu Trp
325 330 335
Ser Ala Leu Ala Asn Gly Asp Thr Arg Leu Leu Lys Thr Gln Ile Asp
340 345 350
Lys Leu Asp Ala Leu Pro Asp Asn Cys Trp Phe Val Asn Tyr Leu Arg
355 360 365
Cys His Asp Asp Ile Gly Trp Gly Leu Asp Glu Asp Val Glu Arg Gln
370 375 380
Leu Gly Ile Asp Pro Leu Lys His Lys Glu Phe Leu Tyr His Phe Tyr
385 390 395 400
Glu Gly Met Val Pro Gly Ser Trp Ala Met Gly Glu Leu Tyr Asn Tyr
405 410 415
Asp Pro Ala Ser Gly Asp Ala Arg Ser Cys Gly Thr Thr Ala Ser Leu
420 425 430
Cys Gly Ile Glu Arg Ala Leu Ile Thr His Asp Arg Pro Leu Tyr Glu
435 440 445
Arg Ser Ile Gln Arg Asp Leu Leu Met His His Ala Met Gly Phe Leu
450 455 460
Arg Gly Phe Pro Met Leu Asn Cys Gly Asp Glu Ile Gly Gln Leu Asn
465 470 475 480
Gly Trp Asp Tyr Lys Glu Asp Pro Asp Arg Val Ala Asp Ser Arg Asn
485 490 495
Leu His Arg Ser Lys Phe Asn Trp Lys Asn Ala Ala Lys Arg Asp Val
500 505 510
Pro Gly Thr Leu Pro Asn Arg Leu Trp Glu Gly Met Ala Asp Val Arg
515 520 525
Gln Met Arg Ser Asp Pro Cys Phe Ala Pro Asp Ala Trp Asp Thr Thr
530 535 540
Trp Asp Ala His Asp Asp Gly Ile Leu Ala Met Val Arg Gln Ser Gly
545 550 555 560
Gly Arg Thr Leu Leu Gly Val Phe Asn Phe Ala Asn Arg Asp Ala Thr
565 570 575
Ala Thr Leu Asp Ser Ile Glu Gly Val Ser Leu Pro Arg Thr Val Ala
580 585 590
Leu Lys Pro Tyr Glu Trp Lys Ile Glu Ala Cys
595 600
<210> 4
<211> 603
<212> PRT
<213> Artificial Sequence
<220>
<223> BtAS P200R
<400> 4
Met Glu Ala Thr Tyr Arg Asp Ser Val Phe Ala Glu Arg Leu Ala Pro
1 5 10 15
Arg Cys Ala Glu Leu Glu Gln Leu Phe Arg Ser Leu Tyr Gly Asp Ser
20 25 30
Pro Glu Phe Asp His Phe Glu Gln Val Met Ala Lys Ala His Ala Asp
35 40 45
Arg Pro Ala Asp Leu Lys Arg Leu Asp Ala Ala Arg Glu His Asp Pro
50 55 60
Gln Trp Tyr Arg Arg Gly Asp Met Phe Gly Met Thr Met Tyr Thr Asp
65 70 75 80
Leu Phe Ala Gly Lys Leu Thr Asp Leu Ala Lys His Ile Asp Tyr Leu
85 90 95
Lys Glu Gln His Leu Thr Tyr Leu His Leu Met Pro Leu Leu Thr Met
100 105 110
Pro His Pro Asp Asn Asp Gly Gly Tyr Ala Ile Glu Asp Phe Asp Thr
115 120 125
Val Asp Pro Thr Ile Gly Thr Asn Glu Asp Leu Ala Asp Leu Thr Ala
130 135 140
Lys Leu Arg Glu Ala Gly Ile Ser Leu Cys Leu Asp Phe Val Met Asn
145 150 155 160
His Thr Ala Ser Thr His Arg Trp Ala Lys Ala Ala Gln Ala Gly Asp
165 170 175
Pro Glu Tyr Gln Asp Tyr Tyr Phe Cys Tyr Asp Asp Arg Thr Ile Pro
180 185 190
Asp Gln Tyr Asp Ala Val Val Arg Gln Val Phe Pro Thr Ala Ala Pro
195 200 205
Gly Asn Phe Thr Trp Asn Glu Gln Met Gly Lys Trp Val Met Thr Gln
210 215 220
Phe Tyr Pro Phe Gln Trp Asp Leu Asn Tyr Arg Asn Pro Lys Val Phe
225 230 235 240
Val Val Met Met Ser Ser Leu Leu His Leu Ala Asn Leu Gly Val Glu
245 250 255
Val Phe Arg Ile Asp Ala Val Pro Tyr Ile Trp Lys Gln Leu Gly Thr
260 265 270
Asn Cys Arg Asn Leu Pro Gln Val His Thr Ile Val Arg Met Met Arg
275 280 285
Ile Met Ser Glu Ile Val Cys Pro Ala Val Val Phe Lys Gly Glu Val
290 295 300
Val Met Ala Pro Lys Glu Leu Ala Ala Tyr Phe Gly Thr Pro Glu Lys
305 310 315 320
Pro Glu Cys His Met Leu Tyr Asn Val Ser Val Met Val Asn Leu Trp
325 330 335
Ser Ala Leu Ala Asn Gly Asp Thr Arg Leu Leu Lys Thr Gln Ile Asp
340 345 350
Lys Leu Asp Ala Leu Pro Asp Asn Cys Trp Phe Val Asn Tyr Leu Arg
355 360 365
Cys His Asp Asp Ile Gly Trp Gly Leu Asp Glu Asp Val Glu Arg Gln
370 375 380
Leu Gly Ile Asp Pro Leu Lys His Lys Glu Phe Leu Tyr His Phe Tyr
385 390 395 400
Glu Gly Met Val Pro Gly Ser Trp Ala Met Gly Glu Leu Tyr Asn Tyr
405 410 415
Asp Pro Ala Ser Gly Asp Ala Arg Ser Cys Gly Thr Thr Ala Ser Leu
420 425 430
Cys Gly Ile Glu Arg Ala Leu Ile Thr His Asp Arg Pro Leu Tyr Glu
435 440 445
Arg Ser Ile Gln Arg Asp Leu Leu Met His His Ala Met Gly Phe Leu
450 455 460
Arg Gly Phe Pro Met Leu Asn Cys Gly Asp Glu Ile Gly Gln Leu Asn
465 470 475 480
Gly Trp Asp Tyr Lys Glu Asp Pro Asp Arg Val Ala Asp Ser Arg Asn
485 490 495
Leu His Arg Ser Lys Phe Asn Trp Lys Asn Ala Ala Lys Arg Asp Val
500 505 510
Pro Gly Thr Leu Pro Asn Arg Leu Trp Glu Gly Met Ala Asp Val Arg
515 520 525
Gln Met Arg Ser Asp Pro Cys Phe Ala Pro Asp Ala Trp Val Thr Thr
530 535 540
Trp Asp Ala His Asp Asp Gly Ile Leu Ala Met Val Arg Gln Ser Gly
545 550 555 560
Gly Arg Thr Leu Leu Gly Val Phe Asn Phe Ala Asn Arg Asp Ala Thr
565 570 575
Ala Thr Leu Asp Ser Ile Glu Gly Val Ser Leu Pro Arg Thr Val Ala
580 585 590
Leu Lys Pro Tyr Glu Trp Lys Ile Glu Ala Cys
595 600
<210> 5
<211> 603
<212> PRT
<213> Artificial Sequence
<220>
<223> BtAS V202I
<400> 5
Met Glu Ala Thr Tyr Arg Asp Ser Val Phe Ala Glu Arg Leu Ala Pro
1 5 10 15
Arg Cys Ala Glu Leu Glu Gln Leu Phe Arg Ser Leu Tyr Gly Asp Ser
20 25 30
Pro Glu Phe Asp His Phe Glu Gln Val Met Ala Lys Ala His Ala Asp
35 40 45
Arg Pro Ala Asp Leu Lys Arg Leu Asp Ala Ala Arg Glu His Asp Pro
50 55 60
Gln Trp Tyr Arg Arg Gly Asp Met Phe Gly Met Thr Met Tyr Thr Asp
65 70 75 80
Leu Phe Ala Gly Lys Leu Thr Asp Leu Ala Lys His Ile Asp Tyr Leu
85 90 95
Lys Glu Gln His Leu Thr Tyr Leu His Leu Met Pro Leu Leu Thr Met
100 105 110
Pro His Pro Asp Asn Asp Gly Gly Tyr Ala Ile Glu Asp Phe Asp Thr
115 120 125
Val Asp Pro Thr Ile Gly Thr Asn Glu Asp Leu Ala Asp Leu Thr Ala
130 135 140
Lys Leu Arg Glu Ala Gly Ile Ser Leu Cys Leu Asp Phe Val Met Asn
145 150 155 160
His Thr Ala Ser Thr His Arg Trp Ala Lys Ala Ala Gln Ala Gly Asp
165 170 175
Pro Glu Tyr Gln Asp Tyr Tyr Phe Cys Tyr Asp Asp Arg Thr Ile Pro
180 185 190
Asp Gln Tyr Asp Ala Val Val Pro Gln Ile Phe Pro Thr Ala Ala Pro
195 200 205
Gly Asn Phe Thr Trp Asn Glu Gln Met Gly Lys Trp Val Met Thr Gln
210 215 220
Phe Tyr Pro Phe Gln Trp Asp Leu Asn Tyr Arg Asn Pro Lys Val Phe
225 230 235 240
Val Val Met Met Ser Ser Leu Leu His Leu Ala Asn Leu Gly Val Glu
245 250 255
Val Phe Arg Ile Asp Ala Val Pro Tyr Ile Trp Lys Gln Leu Gly Thr
260 265 270
Asn Cys Arg Asn Leu Pro Gln Val His Thr Ile Val Arg Met Met Arg
275 280 285
Ile Met Ser Glu Ile Val Cys Pro Ala Val Val Phe Lys Gly Glu Val
290 295 300
Val Met Ala Pro Lys Glu Leu Ala Ala Tyr Phe Gly Thr Pro Glu Lys
305 310 315 320
Pro Glu Cys His Met Leu Tyr Asn Val Ser Val Met Val Asn Leu Trp
325 330 335
Ser Ala Leu Ala Asn Gly Asp Thr Arg Leu Leu Lys Thr Gln Ile Asp
340 345 350
Lys Leu Asp Ala Leu Pro Asp Asn Cys Trp Phe Val Asn Tyr Leu Arg
355 360 365
Cys His Asp Asp Ile Gly Trp Gly Leu Asp Glu Asp Val Glu Arg Gln
370 375 380
Leu Gly Ile Asp Pro Leu Lys His Lys Glu Phe Leu Tyr His Phe Tyr
385 390 395 400
Glu Gly Met Val Pro Gly Ser Trp Ala Met Gly Glu Leu Tyr Asn Tyr
405 410 415
Asp Pro Ala Ser Gly Asp Ala Arg Ser Cys Gly Thr Thr Ala Ser Leu
420 425 430
Cys Gly Ile Glu Arg Ala Leu Ile Thr His Asp Arg Pro Leu Tyr Glu
435 440 445
Arg Ser Ile Gln Arg Asp Leu Leu Met His His Ala Met Gly Phe Leu
450 455 460
Arg Gly Phe Pro Met Leu Asn Cys Gly Asp Glu Ile Gly Gln Leu Asn
465 470 475 480
Gly Trp Asp Tyr Lys Glu Asp Pro Asp Arg Val Ala Asp Ser Arg Asn
485 490 495
Leu His Arg Ser Lys Phe Asn Trp Lys Asn Ala Ala Lys Arg Asp Val
500 505 510
Pro Gly Thr Leu Pro Asn Arg Leu Trp Glu Gly Met Ala Asp Val Arg
515 520 525
Gln Met Arg Ser Asp Pro Cys Phe Ala Pro Asp Ala Trp Val Thr Thr
530 535 540
Trp Asp Ala His Asp Asp Gly Ile Leu Ala Met Val Arg Gln Ser Gly
545 550 555 560
Gly Arg Thr Leu Leu Gly Val Phe Asn Phe Ala Asn Arg Asp Ala Thr
565 570 575
Ala Thr Leu Asp Ser Ile Glu Gly Val Ser Leu Pro Arg Thr Val Ala
580 585 590
Leu Lys Pro Tyr Glu Trp Lys Ile Glu Ala Cys
595 600
<210> 6
<211> 603
<212> PRT
<213> Artificial Sequence
<220>
<223> BtAS Y265F
<400> 6
Met Glu Ala Thr Tyr Arg Asp Ser Val Phe Ala Glu Arg Leu Ala Pro
1 5 10 15
Arg Cys Ala Glu Leu Glu Gln Leu Phe Arg Ser Leu Tyr Gly Asp Ser
20 25 30
Pro Glu Phe Asp His Phe Glu Gln Val Met Ala Lys Ala His Ala Asp
35 40 45
Arg Pro Ala Asp Leu Lys Arg Leu Asp Ala Ala Arg Glu His Asp Pro
50 55 60
Gln Trp Tyr Arg Arg Gly Asp Met Phe Gly Met Thr Met Tyr Thr Asp
65 70 75 80
Leu Phe Ala Gly Lys Leu Thr Asp Leu Ala Lys His Ile Asp Tyr Leu
85 90 95
Lys Glu Gln His Leu Thr Tyr Leu His Leu Met Pro Leu Leu Thr Met
100 105 110
Pro His Pro Asp Asn Asp Gly Gly Tyr Ala Ile Glu Asp Phe Asp Thr
115 120 125
Val Asp Pro Thr Ile Gly Thr Asn Glu Asp Leu Ala Asp Leu Thr Ala
130 135 140
Lys Leu Arg Glu Ala Gly Ile Ser Leu Cys Leu Asp Phe Val Met Asn
145 150 155 160
His Thr Ala Ser Thr His Arg Trp Ala Lys Ala Ala Gln Ala Gly Asp
165 170 175
Pro Glu Tyr Gln Asp Tyr Tyr Phe Cys Tyr Asp Asp Arg Thr Ile Pro
180 185 190
Asp Gln Tyr Asp Ala Val Val Pro Gln Val Phe Pro Thr Ala Ala Pro
195 200 205
Gly Asn Phe Thr Trp Asn Glu Gln Met Gly Lys Trp Val Met Thr Gln
210 215 220
Phe Tyr Pro Phe Gln Trp Asp Leu Asn Tyr Arg Asn Pro Lys Val Phe
225 230 235 240
Val Val Met Met Ser Ser Leu Leu His Leu Ala Asn Leu Gly Val Glu
245 250 255
Val Phe Arg Ile Asp Ala Val Pro Phe Ile Trp Lys Gln Leu Gly Thr
260 265 270
Asn Cys Arg Asn Leu Pro Gln Val His Thr Ile Val Arg Met Met Arg
275 280 285
Ile Met Ser Glu Ile Val Cys Pro Ala Val Val Phe Lys Gly Glu Val
290 295 300
Val Met Ala Pro Lys Glu Leu Ala Ala Tyr Phe Gly Thr Pro Glu Lys
305 310 315 320
Pro Glu Cys His Met Leu Tyr Asn Val Ser Val Met Val Asn Leu Trp
325 330 335
Ser Ala Leu Ala Asn Gly Asp Thr Arg Leu Leu Lys Thr Gln Ile Asp
340 345 350
Lys Leu Asp Ala Leu Pro Asp Asn Cys Trp Phe Val Asn Tyr Leu Arg
355 360 365
Cys His Asp Asp Ile Gly Trp Gly Leu Asp Glu Asp Val Glu Arg Gln
370 375 380
Leu Gly Ile Asp Pro Leu Lys His Lys Glu Phe Leu Tyr His Phe Tyr
385 390 395 400
Glu Gly Met Val Pro Gly Ser Trp Ala Met Gly Glu Leu Tyr Asn Tyr
405 410 415
Asp Pro Ala Ser Gly Asp Ala Arg Ser Cys Gly Thr Thr Ala Ser Leu
420 425 430
Cys Gly Ile Glu Arg Ala Leu Ile Thr His Asp Arg Pro Leu Tyr Glu
435 440 445
Arg Ser Ile Gln Arg Asp Leu Leu Met His His Ala Met Gly Phe Leu
450 455 460
Arg Gly Phe Pro Met Leu Asn Cys Gly Asp Glu Ile Gly Gln Leu Asn
465 470 475 480
Gly Trp Asp Tyr Lys Glu Asp Pro Asp Arg Val Ala Asp Ser Arg Asn
485 490 495
Leu His Arg Ser Lys Phe Asn Trp Lys Asn Ala Ala Lys Arg Asp Val
500 505 510
Pro Gly Thr Leu Pro Asn Arg Leu Trp Glu Gly Met Ala Asp Val Arg
515 520 525
Gln Met Arg Ser Asp Pro Cys Phe Ala Pro Asp Ala Trp Val Thr Thr
530 535 540
Trp Asp Ala His Asp Asp Gly Ile Leu Ala Met Val Arg Gln Ser Gly
545 550 555 560
Gly Arg Thr Leu Leu Gly Val Phe Asn Phe Ala Asn Arg Asp Ala Thr
565 570 575
Ala Thr Leu Asp Ser Ile Glu Gly Val Ser Leu Pro Arg Thr Val Ala
580 585 590
Leu Lys Pro Tyr Glu Trp Lys Ile Glu Ala Cys
595 600
<210> 7
<211> 603
<212> PRT
<213> Artificial Sequence
<220>
<223> BtAS V305I
<400> 7
Met Glu Ala Thr Tyr Arg Asp Ser Val Phe Ala Glu Arg Leu Ala Pro
1 5 10 15
Arg Cys Ala Glu Leu Glu Gln Leu Phe Arg Ser Leu Tyr Gly Asp Ser
20 25 30
Pro Glu Phe Asp His Phe Glu Gln Val Met Ala Lys Ala His Ala Asp
35 40 45
Arg Pro Ala Asp Leu Lys Arg Leu Asp Ala Ala Arg Glu His Asp Pro
50 55 60
Gln Trp Tyr Arg Arg Gly Asp Met Phe Gly Met Thr Met Tyr Thr Asp
65 70 75 80
Leu Phe Ala Gly Lys Leu Thr Asp Leu Ala Lys His Ile Asp Tyr Leu
85 90 95
Lys Glu Gln His Leu Thr Tyr Leu His Leu Met Pro Leu Leu Thr Met
100 105 110
Pro His Pro Asp Asn Asp Gly Gly Tyr Ala Ile Glu Asp Phe Asp Thr
115 120 125
Val Asp Pro Thr Ile Gly Thr Asn Glu Asp Leu Ala Asp Leu Thr Ala
130 135 140
Lys Leu Arg Glu Ala Gly Ile Ser Leu Cys Leu Asp Phe Val Met Asn
145 150 155 160
His Thr Ala Ser Thr His Arg Trp Ala Lys Ala Ala Gln Ala Gly Asp
165 170 175
Pro Glu Tyr Gln Asp Tyr Tyr Phe Cys Tyr Asp Asp Arg Thr Ile Pro
180 185 190
Asp Gln Tyr Asp Ala Val Val Pro Gln Val Phe Pro Thr Ala Ala Pro
195 200 205
Gly Asn Phe Thr Trp Asn Glu Gln Met Gly Lys Trp Val Met Thr Gln
210 215 220
Phe Tyr Pro Phe Gln Trp Asp Leu Asn Tyr Arg Asn Pro Lys Val Phe
225 230 235 240
Val Val Met Met Ser Ser Leu Leu His Leu Ala Asn Leu Gly Val Glu
245 250 255
Val Phe Arg Ile Asp Ala Val Pro Tyr Ile Trp Lys Gln Leu Gly Thr
260 265 270
Asn Cys Arg Asn Leu Pro Gln Val His Thr Ile Val Arg Met Met Arg
275 280 285
Ile Met Ser Glu Ile Val Cys Pro Ala Val Val Phe Lys Gly Glu Val
290 295 300
Ile Met Ala Pro Lys Glu Leu Ala Ala Tyr Phe Gly Thr Pro Glu Lys
305 310 315 320
Pro Glu Cys His Met Leu Tyr Asn Val Ser Val Met Val Asn Leu Trp
325 330 335
Ser Ala Leu Ala Asn Gly Asp Thr Arg Leu Leu Lys Thr Gln Ile Asp
340 345 350
Lys Leu Asp Ala Leu Pro Asp Asn Cys Trp Phe Val Asn Tyr Leu Arg
355 360 365
Cys His Asp Asp Ile Gly Trp Gly Leu Asp Glu Asp Val Glu Arg Gln
370 375 380
Leu Gly Ile Asp Pro Leu Lys His Lys Glu Phe Leu Tyr His Phe Tyr
385 390 395 400
Glu Gly Met Val Pro Gly Ser Trp Ala Met Gly Glu Leu Tyr Asn Tyr
405 410 415
Asp Pro Ala Ser Gly Asp Ala Arg Ser Cys Gly Thr Thr Ala Ser Leu
420 425 430
Cys Gly Ile Glu Arg Ala Leu Ile Thr His Asp Arg Pro Leu Tyr Glu
435 440 445
Arg Ser Ile Gln Arg Asp Leu Leu Met His His Ala Met Gly Phe Leu
450 455 460
Arg Gly Phe Pro Met Leu Asn Cys Gly Asp Glu Ile Gly Gln Leu Asn
465 470 475 480
Gly Trp Asp Tyr Lys Glu Asp Pro Asp Arg Val Ala Asp Ser Arg Asn
485 490 495
Leu His Arg Ser Lys Phe Asn Trp Lys Asn Ala Ala Lys Arg Asp Val
500 505 510
Pro Gly Thr Leu Pro Asn Arg Leu Trp Glu Gly Met Ala Asp Val Arg
515 520 525
Gln Met Arg Ser Asp Pro Cys Phe Ala Pro Asp Ala Trp Val Thr Thr
530 535 540
Trp Asp Ala His Asp Asp Gly Ile Leu Ala Met Val Arg Gln Ser Gly
545 550 555 560
Gly Arg Thr Leu Leu Gly Val Phe Asn Phe Ala Asn Arg Asp Ala Thr
565 570 575
Ala Thr Leu Asp Ser Ile Glu Gly Val Ser Leu Pro Arg Thr Val Ala
580 585 590
Leu Lys Pro Tyr Glu Trp Lys Ile Glu Ala Cys
595 600
<210> 8
<211> 603
<212> PRT
<213> Artificial Sequence
<220>
<223> BtAS K393R
<400> 8
Met Glu Ala Thr Tyr Arg Asp Ser Val Phe Ala Glu Arg Leu Ala Pro
1 5 10 15
Arg Cys Ala Glu Leu Glu Gln Leu Phe Arg Ser Leu Tyr Gly Asp Ser
20 25 30
Pro Glu Phe Asp His Phe Glu Gln Val Met Ala Lys Ala His Ala Asp
35 40 45
Arg Pro Ala Asp Leu Lys Arg Leu Asp Ala Ala Arg Glu His Asp Pro
50 55 60
Gln Trp Tyr Arg Arg Gly Asp Met Phe Gly Met Thr Met Tyr Thr Asp
65 70 75 80
Leu Phe Ala Gly Lys Leu Thr Asp Leu Ala Lys His Ile Asp Tyr Leu
85 90 95
Lys Glu Gln His Leu Thr Tyr Leu His Leu Met Pro Leu Leu Thr Met
100 105 110
Pro His Pro Asp Asn Asp Gly Gly Tyr Ala Ile Glu Asp Phe Asp Thr
115 120 125
Val Asp Pro Thr Ile Gly Thr Asn Glu Asp Leu Ala Asp Leu Thr Ala
130 135 140
Lys Leu Arg Glu Ala Gly Ile Ser Leu Cys Leu Asp Phe Val Met Asn
145 150 155 160
His Thr Ala Ser Thr His Arg Trp Ala Lys Ala Ala Gln Ala Gly Asp
165 170 175
Pro Glu Tyr Gln Asp Tyr Tyr Phe Cys Tyr Asp Asp Arg Thr Ile Pro
180 185 190
Asp Gln Tyr Asp Ala Val Val Pro Gln Val Phe Pro Thr Ala Ala Pro
195 200 205
Gly Asn Phe Thr Trp Asn Glu Gln Met Gly Lys Trp Val Met Thr Gln
210 215 220
Phe Tyr Pro Phe Gln Trp Asp Leu Asn Tyr Arg Asn Pro Lys Val Phe
225 230 235 240
Val Val Met Met Ser Ser Leu Leu His Leu Ala Asn Leu Gly Val Glu
245 250 255
Val Phe Arg Ile Asp Ala Val Pro Tyr Ile Trp Lys Gln Leu Gly Thr
260 265 270
Asn Cys Arg Asn Leu Pro Gln Val His Thr Ile Val Arg Met Met Arg
275 280 285
Ile Met Ser Glu Ile Val Cys Pro Ala Val Val Phe Lys Gly Glu Val
290 295 300
Val Met Ala Pro Lys Glu Leu Ala Ala Tyr Phe Gly Thr Pro Glu Lys
305 310 315 320
Pro Glu Cys His Met Leu Tyr Asn Val Ser Val Met Val Asn Leu Trp
325 330 335
Ser Ala Leu Ala Asn Gly Asp Thr Arg Leu Leu Lys Thr Gln Ile Asp
340 345 350
Lys Leu Asp Ala Leu Pro Asp Asn Cys Trp Phe Val Asn Tyr Leu Arg
355 360 365
Cys His Asp Asp Ile Gly Trp Gly Leu Asp Glu Asp Val Glu Arg Gln
370 375 380
Leu Gly Ile Asp Pro Leu Lys His Arg Glu Phe Leu Tyr His Phe Tyr
385 390 395 400
Glu Gly Met Val Pro Gly Ser Trp Ala Met Gly Glu Leu Tyr Asn Tyr
405 410 415
Asp Pro Ala Ser Gly Asp Ala Arg Ser Cys Gly Thr Thr Ala Ser Leu
420 425 430
Cys Gly Ile Glu Arg Ala Leu Ile Thr His Asp Arg Pro Leu Tyr Glu
435 440 445
Arg Ser Ile Gln Arg Asp Leu Leu Met His His Ala Met Gly Phe Leu
450 455 460
Arg Gly Phe Pro Met Leu Asn Cys Gly Asp Glu Ile Gly Gln Leu Asn
465 470 475 480
Gly Trp Asp Tyr Lys Glu Asp Pro Asp Arg Val Ala Asp Ser Arg Asn
485 490 495
Leu His Arg Ser Lys Phe Asn Trp Lys Asn Ala Ala Lys Arg Asp Val
500 505 510
Pro Gly Thr Leu Pro Asn Arg Leu Trp Glu Gly Met Ala Asp Val Arg
515 520 525
Gln Met Arg Ser Asp Pro Cys Phe Ala Pro Asp Ala Trp Val Thr Thr
530 535 540
Trp Asp Ala His Asp Asp Gly Ile Leu Ala Met Val Arg Gln Ser Gly
545 550 555 560
Gly Arg Thr Leu Leu Gly Val Phe Asn Phe Ala Asn Arg Asp Ala Thr
565 570 575
Ala Thr Leu Asp Ser Ile Glu Gly Val Ser Leu Pro Arg Thr Val Ala
580 585 590
Leu Lys Pro Tyr Glu Trp Lys Ile Glu Ala Cys
595 600
<210> 9
<211> 603
<212> PRT
<213> Artificial Sequence
<220>
<223> BtAS S420T
<400> 9
Met Glu Ala Thr Tyr Arg Asp Ser Val Phe Ala Glu Arg Leu Ala Pro
1 5 10 15
Arg Cys Ala Glu Leu Glu Gln Leu Phe Arg Ser Leu Tyr Gly Asp Ser
20 25 30
Pro Glu Phe Asp His Phe Glu Gln Val Met Ala Lys Ala His Ala Asp
35 40 45
Arg Pro Ala Asp Leu Lys Arg Leu Asp Ala Ala Arg Glu His Asp Pro
50 55 60
Gln Trp Tyr Arg Arg Gly Asp Met Phe Gly Met Thr Met Tyr Thr Asp
65 70 75 80
Leu Phe Ala Gly Lys Leu Thr Asp Leu Ala Lys His Ile Asp Tyr Leu
85 90 95
Lys Glu Gln His Leu Thr Tyr Leu His Leu Met Pro Leu Leu Thr Met
100 105 110
Pro His Pro Asp Asn Asp Gly Gly Tyr Ala Ile Glu Asp Phe Asp Thr
115 120 125
Val Asp Pro Thr Ile Gly Thr Asn Glu Asp Leu Ala Asp Leu Thr Ala
130 135 140
Lys Leu Arg Glu Ala Gly Ile Ser Leu Cys Leu Asp Phe Val Met Asn
145 150 155 160
His Thr Ala Ser Thr His Arg Trp Ala Lys Ala Ala Gln Ala Gly Asp
165 170 175
Pro Glu Tyr Gln Asp Tyr Tyr Phe Cys Tyr Asp Asp Arg Thr Ile Pro
180 185 190
Asp Gln Tyr Asp Ala Val Val Pro Gln Val Phe Pro Thr Ala Ala Pro
195 200 205
Gly Asn Phe Thr Trp Asn Glu Gln Met Gly Lys Trp Val Met Thr Gln
210 215 220
Phe Tyr Pro Phe Gln Trp Asp Leu Asn Tyr Arg Asn Pro Lys Val Phe
225 230 235 240
Val Val Met Met Ser Ser Leu Leu His Leu Ala Asn Leu Gly Val Glu
245 250 255
Val Phe Arg Ile Asp Ala Val Pro Tyr Ile Trp Lys Gln Leu Gly Thr
260 265 270
Asn Cys Arg Asn Leu Pro Gln Val His Thr Ile Val Arg Met Met Arg
275 280 285
Ile Met Ser Glu Ile Val Cys Pro Ala Val Val Phe Lys Gly Glu Val
290 295 300
Val Met Ala Pro Lys Glu Leu Ala Ala Tyr Phe Gly Thr Pro Glu Lys
305 310 315 320
Pro Glu Cys His Met Leu Tyr Asn Val Ser Val Met Val Asn Leu Trp
325 330 335
Ser Ala Leu Ala Asn Gly Asp Thr Arg Leu Leu Lys Thr Gln Ile Asp
340 345 350
Lys Leu Asp Ala Leu Pro Asp Asn Cys Trp Phe Val Asn Tyr Leu Arg
355 360 365
Cys His Asp Asp Ile Gly Trp Gly Leu Asp Glu Asp Val Glu Arg Gln
370 375 380
Leu Gly Ile Asp Pro Leu Lys His Lys Glu Phe Leu Tyr His Phe Tyr
385 390 395 400
Glu Gly Met Val Pro Gly Ser Trp Ala Met Gly Glu Leu Tyr Asn Tyr
405 410 415
Asp Pro Ala Thr Gly Asp Ala Arg Ser Cys Gly Thr Thr Ala Ser Leu
420 425 430
Cys Gly Ile Glu Arg Ala Leu Ile Thr His Asp Arg Pro Leu Tyr Glu
435 440 445
Arg Ser Ile Gln Arg Asp Leu Leu Met His His Ala Met Gly Phe Leu
450 455 460
Arg Gly Phe Pro Met Leu Asn Cys Gly Asp Glu Ile Gly Gln Leu Asn
465 470 475 480
Gly Trp Asp Tyr Lys Glu Asp Pro Asp Arg Val Ala Asp Ser Arg Asn
485 490 495
Leu His Arg Ser Lys Phe Asn Trp Lys Asn Ala Ala Lys Arg Asp Val
500 505 510
Pro Gly Thr Leu Pro Asn Arg Leu Trp Glu Gly Met Ala Asp Val Arg
515 520 525
Gln Met Arg Ser Asp Pro Cys Phe Ala Pro Asp Ala Trp Val Thr Thr
530 535 540
Trp Asp Ala His Asp Asp Gly Ile Leu Ala Met Val Arg Gln Ser Gly
545 550 555 560
Gly Arg Thr Leu Leu Gly Val Phe Asn Phe Ala Asn Arg Asp Ala Thr
565 570 575
Ala Thr Leu Asp Ser Ile Glu Gly Val Ser Leu Pro Arg Thr Val Ala
580 585 590
Leu Lys Pro Tyr Glu Trp Lys Ile Glu Ala Cys
595 600
<210> 10
<211> 603
<212> PRT
<213> Artificial Sequence
<220>
<223> BtAS Y414F
<400> 10
Met Glu Ala Thr Tyr Arg Asp Ser Val Phe Ala Glu Arg Leu Ala Pro
1 5 10 15
Arg Cys Ala Glu Leu Glu Gln Leu Phe Arg Ser Leu Tyr Gly Asp Ser
20 25 30
Pro Glu Phe Asp His Phe Glu Gln Val Met Ala Lys Ala His Ala Asp
35 40 45
Arg Pro Ala Asp Leu Lys Arg Leu Asp Ala Ala Arg Glu His Asp Pro
50 55 60
Gln Trp Tyr Arg Arg Gly Asp Met Phe Gly Met Thr Met Tyr Thr Asp
65 70 75 80
Leu Phe Ala Gly Lys Leu Thr Asp Leu Ala Lys His Ile Asp Tyr Leu
85 90 95
Lys Glu Gln His Leu Thr Tyr Leu His Leu Met Pro Leu Leu Thr Met
100 105 110
Pro His Pro Asp Asn Asp Gly Gly Tyr Ala Ile Glu Asp Phe Asp Thr
115 120 125
Val Asp Pro Thr Ile Gly Thr Asn Glu Asp Leu Ala Asp Leu Thr Ala
130 135 140
Lys Leu Arg Glu Ala Gly Ile Ser Leu Cys Leu Asp Phe Val Met Asn
145 150 155 160
His Thr Ala Ser Thr His Arg Trp Ala Lys Ala Ala Gln Ala Gly Asp
165 170 175
Pro Glu Tyr Gln Asp Tyr Tyr Phe Cys Tyr Asp Asp Arg Thr Ile Pro
180 185 190
Asp Gln Tyr Asp Ala Val Val Pro Gln Val Phe Pro Thr Ala Ala Pro
195 200 205
Gly Asn Phe Thr Trp Asn Glu Gln Met Gly Lys Trp Val Met Thr Gln
210 215 220
Phe Tyr Pro Phe Gln Trp Asp Leu Asn Tyr Arg Asn Pro Lys Val Phe
225 230 235 240
Val Val Met Met Ser Ser Leu Leu His Leu Ala Asn Leu Gly Val Glu
245 250 255
Val Phe Arg Ile Asp Ala Val Pro Tyr Ile Trp Lys Gln Leu Gly Thr
260 265 270
Asn Cys Arg Asn Leu Pro Gln Val His Thr Ile Val Arg Met Met Arg
275 280 285
Ile Met Ser Glu Ile Val Cys Pro Ala Val Val Phe Lys Gly Glu Val
290 295 300
Val Met Ala Pro Lys Glu Leu Ala Ala Tyr Phe Gly Thr Pro Glu Lys
305 310 315 320
Pro Glu Cys His Met Leu Tyr Asn Val Ser Val Met Val Asn Leu Trp
325 330 335
Ser Ala Leu Ala Asn Gly Asp Thr Arg Leu Leu Lys Thr Gln Ile Asp
340 345 350
Lys Leu Asp Ala Leu Pro Asp Asn Cys Trp Phe Val Asn Tyr Leu Arg
355 360 365
Cys His Asp Asp Ile Gly Trp Gly Leu Asp Glu Asp Val Glu Arg Gln
370 375 380
Leu Gly Ile Asp Pro Leu Lys His Lys Glu Phe Leu Tyr His Phe Tyr
385 390 395 400
Glu Gly Met Val Pro Gly Ser Trp Ala Met Gly Glu Leu Phe Asn Tyr
405 410 415
Asp Pro Ala Ser Gly Asp Ala Arg Ser Cys Gly Thr Thr Ala Ser Leu
420 425 430
Cys Gly Ile Glu Arg Ala Leu Ile Thr His Asp Arg Pro Leu Tyr Glu
435 440 445
Arg Ser Ile Gln Arg Asp Leu Leu Met His His Ala Met Gly Phe Leu
450 455 460
Arg Gly Phe Pro Met Leu Asn Cys Gly Asp Glu Ile Gly Gln Leu Asn
465 470 475 480
Gly Trp Asp Tyr Lys Glu Asp Pro Asp Arg Val Ala Asp Ser Arg Asn
485 490 495
Leu His Arg Ser Lys Phe Asn Trp Lys Asn Ala Ala Lys Arg Asp Val
500 505 510
Pro Gly Thr Leu Pro Asn Arg Leu Trp Glu Gly Met Ala Asp Val Arg
515 520 525
Gln Met Arg Ser Asp Pro Cys Phe Ala Pro Asp Ala Trp Val Thr Thr
530 535 540
Trp Asp Ala His Asp Asp Gly Ile Leu Ala Met Val Arg Gln Ser Gly
545 550 555 560
Gly Arg Thr Leu Leu Gly Val Phe Asn Phe Ala Asn Arg Asp Ala Thr
565 570 575
Ala Thr Leu Asp Ser Ile Glu Gly Val Ser Leu Pro Arg Thr Val Ala
580 585 590
Leu Lys Pro Tyr Glu Trp Lys Ile Glu Ala Cys
595 600
<210> 11
<211> 603
<212> PRT
<213> Artificial Sequence
<220>
<223> BtAS Y414F*P200R
<400> 11
Met Glu Ala Thr Tyr Arg Asp Ser Val Phe Ala Glu Arg Leu Ala Pro
1 5 10 15
Arg Cys Ala Glu Leu Glu Gln Leu Phe Arg Ser Leu Tyr Gly Asp Ser
20 25 30
Pro Glu Phe Asp His Phe Glu Gln Val Met Ala Lys Ala His Ala Asp
35 40 45
Arg Pro Ala Asp Leu Lys Arg Leu Asp Ala Ala Arg Glu His Asp Pro
50 55 60
Gln Trp Tyr Arg Arg Gly Asp Met Phe Gly Met Thr Met Tyr Thr Asp
65 70 75 80
Leu Phe Ala Gly Lys Leu Thr Asp Leu Ala Lys His Ile Asp Tyr Leu
85 90 95
Lys Glu Gln His Leu Thr Tyr Leu His Leu Met Pro Leu Leu Thr Met
100 105 110
Pro His Pro Asp Asn Asp Gly Gly Tyr Ala Ile Glu Asp Phe Asp Thr
115 120 125
Val Asp Pro Thr Ile Gly Thr Asn Glu Asp Leu Ala Asp Leu Thr Ala
130 135 140
Lys Leu Arg Glu Ala Gly Ile Ser Leu Cys Leu Asp Phe Val Met Asn
145 150 155 160
His Thr Ala Ser Thr His Arg Trp Ala Lys Ala Ala Gln Ala Gly Asp
165 170 175
Pro Glu Tyr Gln Asp Tyr Tyr Phe Cys Tyr Asp Asp Arg Thr Ile Pro
180 185 190
Asp Gln Tyr Asp Ala Val Val Arg Gln Val Phe Pro Thr Ala Ala Pro
195 200 205
Gly Asn Phe Thr Trp Asn Glu Gln Met Gly Lys Trp Val Met Thr Gln
210 215 220
Phe Tyr Pro Phe Gln Trp Asp Leu Asn Tyr Arg Asn Pro Lys Val Phe
225 230 235 240
Val Val Met Met Ser Ser Leu Leu His Leu Ala Asn Leu Gly Val Glu
245 250 255
Val Phe Arg Ile Asp Ala Val Pro Tyr Ile Trp Lys Gln Leu Gly Thr
260 265 270
Asn Cys Arg Asn Leu Pro Gln Val His Thr Ile Val Arg Met Met Arg
275 280 285
Ile Met Ser Glu Ile Val Cys Pro Ala Val Val Phe Lys Gly Glu Val
290 295 300
Val Met Ala Pro Lys Glu Leu Ala Ala Tyr Phe Gly Thr Pro Glu Lys
305 310 315 320
Pro Glu Cys His Met Leu Tyr Asn Val Ser Val Met Val Asn Leu Trp
325 330 335
Ser Ala Leu Ala Asn Gly Asp Thr Arg Leu Leu Lys Thr Gln Ile Asp
340 345 350
Lys Leu Asp Ala Leu Pro Asp Asn Cys Trp Phe Val Asn Tyr Leu Arg
355 360 365
Cys His Asp Asp Ile Gly Trp Gly Leu Asp Glu Asp Val Glu Arg Gln
370 375 380
Leu Gly Ile Asp Pro Leu Lys His Lys Glu Phe Leu Tyr His Phe Tyr
385 390 395 400
Glu Gly Met Val Pro Gly Ser Trp Ala Met Gly Glu Leu Phe Asn Tyr
405 410 415
Asp Pro Ala Ser Gly Asp Ala Arg Ser Cys Gly Thr Thr Ala Ser Leu
420 425 430
Cys Gly Ile Glu Arg Ala Leu Ile Thr His Asp Arg Pro Leu Tyr Glu
435 440 445
Arg Ser Ile Gln Arg Asp Leu Leu Met His His Ala Met Gly Phe Leu
450 455 460
Arg Gly Phe Pro Met Leu Asn Cys Gly Asp Glu Ile Gly Gln Leu Asn
465 470 475 480
Gly Trp Asp Tyr Lys Glu Asp Pro Asp Arg Val Ala Asp Ser Arg Asn
485 490 495
Leu His Arg Ser Lys Phe Asn Trp Lys Asn Ala Ala Lys Arg Asp Val
500 505 510
Pro Gly Thr Leu Pro Asn Arg Leu Trp Glu Gly Met Ala Asp Val Arg
515 520 525
Gln Met Arg Ser Asp Pro Cys Phe Ala Pro Asp Ala Trp Val Thr Thr
530 535 540
Trp Asp Ala His Asp Asp Gly Ile Leu Ala Met Val Arg Gln Ser Gly
545 550 555 560
Gly Arg Thr Leu Leu Gly Val Phe Asn Phe Ala Asn Arg Asp Ala Thr
565 570 575
Ala Thr Leu Asp Ser Ile Glu Gly Val Ser Leu Pro Arg Thr Val Ala
580 585 590
Leu Lys Pro Tyr Glu Trp Lys Ile Glu Ala Cys
595 600
<210> 12
<211> 603
<212> PRT
<213> Artificial Sequence
<220>
<223> BtAS Y414F*V202I
<400> 12
Met Glu Ala Thr Tyr Arg Asp Ser Val Phe Ala Glu Arg Leu Ala Pro
1 5 10 15
Arg Cys Ala Glu Leu Glu Gln Leu Phe Arg Ser Leu Tyr Gly Asp Ser
20 25 30
Pro Glu Phe Asp His Phe Glu Gln Val Met Ala Lys Ala His Ala Asp
35 40 45
Arg Pro Ala Asp Leu Lys Arg Leu Asp Ala Ala Arg Glu His Asp Pro
50 55 60
Gln Trp Tyr Arg Arg Gly Asp Met Phe Gly Met Thr Met Tyr Thr Asp
65 70 75 80
Leu Phe Ala Gly Lys Leu Thr Asp Leu Ala Lys His Ile Asp Tyr Leu
85 90 95
Lys Glu Gln His Leu Thr Tyr Leu His Leu Met Pro Leu Leu Thr Met
100 105 110
Pro His Pro Asp Asn Asp Gly Gly Tyr Ala Ile Glu Asp Phe Asp Thr
115 120 125
Val Asp Pro Thr Ile Gly Thr Asn Glu Asp Leu Ala Asp Leu Thr Ala
130 135 140
Lys Leu Arg Glu Ala Gly Ile Ser Leu Cys Leu Asp Phe Val Met Asn
145 150 155 160
His Thr Ala Ser Thr His Arg Trp Ala Lys Ala Ala Gln Ala Gly Asp
165 170 175
Pro Glu Tyr Gln Asp Tyr Tyr Phe Cys Tyr Asp Asp Arg Thr Ile Pro
180 185 190
Asp Gln Tyr Asp Ala Val Val Pro Gln Ile Phe Pro Thr Ala Ala Pro
195 200 205
Gly Asn Phe Thr Trp Asn Glu Gln Met Gly Lys Trp Val Met Thr Gln
210 215 220
Phe Tyr Pro Phe Gln Trp Asp Leu Asn Tyr Arg Asn Pro Lys Val Phe
225 230 235 240
Val Val Met Met Ser Ser Leu Leu His Leu Ala Asn Leu Gly Val Glu
245 250 255
Val Phe Arg Ile Asp Ala Val Pro Tyr Ile Trp Lys Gln Leu Gly Thr
260 265 270
Asn Cys Arg Asn Leu Pro Gln Val His Thr Ile Val Arg Met Met Arg
275 280 285
Ile Met Ser Glu Ile Val Cys Pro Ala Val Val Phe Lys Gly Glu Val
290 295 300
Val Met Ala Pro Lys Glu Leu Ala Ala Tyr Phe Gly Thr Pro Glu Lys
305 310 315 320
Pro Glu Cys His Met Leu Tyr Asn Val Ser Val Met Val Asn Leu Trp
325 330 335
Ser Ala Leu Ala Asn Gly Asp Thr Arg Leu Leu Lys Thr Gln Ile Asp
340 345 350
Lys Leu Asp Ala Leu Pro Asp Asn Cys Trp Phe Val Asn Tyr Leu Arg
355 360 365
Cys His Asp Asp Ile Gly Trp Gly Leu Asp Glu Asp Val Glu Arg Gln
370 375 380
Leu Gly Ile Asp Pro Leu Lys His Lys Glu Phe Leu Tyr His Phe Tyr
385 390 395 400
Glu Gly Met Val Pro Gly Ser Trp Ala Met Gly Glu Leu Phe Asn Tyr
405 410 415
Asp Pro Ala Ser Gly Asp Ala Arg Ser Cys Gly Thr Thr Ala Ser Leu
420 425 430
Cys Gly Ile Glu Arg Ala Leu Ile Thr His Asp Arg Pro Leu Tyr Glu
435 440 445
Arg Ser Ile Gln Arg Asp Leu Leu Met His His Ala Met Gly Phe Leu
450 455 460
Arg Gly Phe Pro Met Leu Asn Cys Gly Asp Glu Ile Gly Gln Leu Asn
465 470 475 480
Gly Trp Asp Tyr Lys Glu Asp Pro Asp Arg Val Ala Asp Ser Arg Asn
485 490 495
Leu His Arg Ser Lys Phe Asn Trp Lys Asn Ala Ala Lys Arg Asp Val
500 505 510
Pro Gly Thr Leu Pro Asn Arg Leu Trp Glu Gly Met Ala Asp Val Arg
515 520 525
Gln Met Arg Ser Asp Pro Cys Phe Ala Pro Asp Ala Trp Val Thr Thr
530 535 540
Trp Asp Ala His Asp Asp Gly Ile Leu Ala Met Val Arg Gln Ser Gly
545 550 555 560
Gly Arg Thr Leu Leu Gly Val Phe Asn Phe Ala Asn Arg Asp Ala Thr
565 570 575
Ala Thr Leu Asp Ser Ile Glu Gly Val Ser Leu Pro Arg Thr Val Ala
580 585 590
Leu Lys Pro Tyr Glu Trp Lys Ile Glu Ala Cys
595 600
<210> 13
<211> 603
<212> PRT
<213> Artificial Sequence
<220>
<223> BtAS YY414F*Y265F
<400> 13
Met Glu Ala Thr Tyr Arg Asp Ser Val Phe Ala Glu Arg Leu Ala Pro
1 5 10 15
Arg Cys Ala Glu Leu Glu Gln Leu Phe Arg Ser Leu Tyr Gly Asp Ser
20 25 30
Pro Glu Phe Asp His Phe Glu Gln Val Met Ala Lys Ala His Ala Asp
35 40 45
Arg Pro Ala Asp Leu Lys Arg Leu Asp Ala Ala Arg Glu His Asp Pro
50 55 60
Gln Trp Tyr Arg Arg Gly Asp Met Phe Gly Met Thr Met Tyr Thr Asp
65 70 75 80
Leu Phe Ala Gly Lys Leu Thr Asp Leu Ala Lys His Ile Asp Tyr Leu
85 90 95
Lys Glu Gln His Leu Thr Tyr Leu His Leu Met Pro Leu Leu Thr Met
100 105 110
Pro His Pro Asp Asn Asp Gly Gly Tyr Ala Ile Glu Asp Phe Asp Thr
115 120 125
Val Asp Pro Thr Ile Gly Thr Asn Glu Asp Leu Ala Asp Leu Thr Ala
130 135 140
Lys Leu Arg Glu Ala Gly Ile Ser Leu Cys Leu Asp Phe Val Met Asn
145 150 155 160
His Thr Ala Ser Thr His Arg Trp Ala Lys Ala Ala Gln Ala Gly Asp
165 170 175
Pro Glu Tyr Gln Asp Tyr Tyr Phe Cys Tyr Asp Asp Arg Thr Ile Pro
180 185 190
Asp Gln Tyr Asp Ala Val Val Pro Gln Val Phe Pro Thr Ala Ala Pro
195 200 205
Gly Asn Phe Thr Trp Asn Glu Gln Met Gly Lys Trp Val Met Thr Gln
210 215 220
Phe Tyr Pro Phe Gln Trp Asp Leu Asn Tyr Arg Asn Pro Lys Val Phe
225 230 235 240
Val Val Met Met Ser Ser Leu Leu His Leu Ala Asn Leu Gly Val Glu
245 250 255
Val Phe Arg Ile Asp Ala Val Pro Phe Ile Trp Lys Gln Leu Gly Thr
260 265 270
Asn Cys Arg Asn Leu Pro Gln Val His Thr Ile Val Arg Met Met Arg
275 280 285
Ile Met Ser Glu Ile Val Cys Pro Ala Val Val Phe Lys Gly Glu Val
290 295 300
Val Met Ala Pro Lys Glu Leu Ala Ala Tyr Phe Gly Thr Pro Glu Lys
305 310 315 320
Pro Glu Cys His Met Leu Tyr Asn Val Ser Val Met Val Asn Leu Trp
325 330 335
Ser Ala Leu Ala Asn Gly Asp Thr Arg Leu Leu Lys Thr Gln Ile Asp
340 345 350
Lys Leu Asp Ala Leu Pro Asp Asn Cys Trp Phe Val Asn Tyr Leu Arg
355 360 365
Cys His Asp Asp Ile Gly Trp Gly Leu Asp Glu Asp Val Glu Arg Gln
370 375 380
Leu Gly Ile Asp Pro Leu Lys His Lys Glu Phe Leu Tyr His Phe Tyr
385 390 395 400
Glu Gly Met Val Pro Gly Ser Trp Ala Met Gly Glu Leu Phe Asn Tyr
405 410 415
Asp Pro Ala Ser Gly Asp Ala Arg Ser Cys Gly Thr Thr Ala Ser Leu
420 425 430
Cys Gly Ile Glu Arg Ala Leu Ile Thr His Asp Arg Pro Leu Tyr Glu
435 440 445
Arg Ser Ile Gln Arg Asp Leu Leu Met His His Ala Met Gly Phe Leu
450 455 460
Arg Gly Phe Pro Met Leu Asn Cys Gly Asp Glu Ile Gly Gln Leu Asn
465 470 475 480
Gly Trp Asp Tyr Lys Glu Asp Pro Asp Arg Val Ala Asp Ser Arg Asn
485 490 495
Leu His Arg Ser Lys Phe Asn Trp Lys Asn Ala Ala Lys Arg Asp Val
500 505 510
Pro Gly Thr Leu Pro Asn Arg Leu Trp Glu Gly Met Ala Asp Val Arg
515 520 525
Gln Met Arg Ser Asp Pro Cys Phe Ala Pro Asp Ala Trp Val Thr Thr
530 535 540
Trp Asp Ala His Asp Asp Gly Ile Leu Ala Met Val Arg Gln Ser Gly
545 550 555 560
Gly Arg Thr Leu Leu Gly Val Phe Asn Phe Ala Asn Arg Asp Ala Thr
565 570 575
Ala Thr Leu Asp Ser Ile Glu Gly Val Ser Leu Pro Arg Thr Val Ala
580 585 590
Leu Lys Pro Tyr Glu Trp Lys Ile Glu Ala Cys
595 600
<210> 14
<211> 1812
<212> DNA
<213> Artificial Sequence
<220>
<223> amylosucrase BtAS wild type
<400> 14
atggaagcca catatcgcga ttccgtattc gccgaacggc tcgccccgcg ctgcgcagaa 60
cttgaacaac ttttccgctc gttgtacggg gattcccctg aattcgacca cttcgaacag 120
gtcatggcca aggcccacgc cgaccggcca gccgacctca aacgcctcga cgccgcccgt 180
gaacacgatc cgcaatggta ccgtcgcggc gacatgttcg gcatgaccat gtacaccgac 240
ctgttcgccg gcaaactcac cgatctcgcc aagcatatcg actatctcaa agagcagcat 300
ctgacctacc tgcacctcat gccgctgctg accatgcccc accccgacaa cgacggcggc 360
tacgccatcg aggatttcga caccgtcgac ccgactatcg gcaccaatga ggacctcgcc 420
gacctcaccg cgaaactgcg cgaagccggc atcagcctgt gccttgattt cgtcatgaac 480
cacaccgcat ccacccaccg gtgggcgaaa gccgcacaag ccggcgaccc cgaataccag 540
gactactact tctgctatga cgaccgcacc atccccgacc aatatgacgc cgtcgtcccg 600
caagtcttcc cgaccgccgc ccccggcaac ttcacatgga atgagcagat gggcaaatgg 660
gtcatgaccc agttctaccc gttccaatgg gacctcaact accgcaatcc caaggtcttc 720
gtcgtcatga tgtccagcct gctgcacctg gccaacctcg gcgtcgaagt cttccgcatc 780
gacgcggtgc cgtacatctg gaagcaactc ggcaccaact gccgcaacct gccgcaagtc 840
cacaccatcg tgcgcatgat gcgcatcatg tccgaaatcg tctgcccggc cgtcgtgttc 900
aaaggtgaag tcgtcatggc tcccaaggag ctcgccgcct acttcggcac ccccgagaag 960
cccgaatgcc acatgctgta caacgtgtcc gtcatggtca acttgtggag cgcgctcgcc 1020
aacggcgaca cccgcctgct taaaacccag atcgacaagc tcgacgccct gcccgacaac 1080
tgctggttcg tcaactatct gcgctgccat gacgatatcg gctggggtct ggacgaggat 1140
gtcgaacgcc agttgggcat cgacccgctc aagcacaagg aattcctcta ccacttctac 1200
gagggcatgg tgcccggcag ctgggcgatg ggcgagctgt acaactatga tccggcgtcc 1260
ggtgacgcgc gcagctgcgg caccacggcg agcttgtgcg gtattgagcg tgcgctgatc 1320
acgcatgacc ggccgctgta tgagcgttcc atccagcgtg atctgctcat gcaccacgct 1380
atgggcttcc tgcgtgggtt cccgatgctc aactgcggcg acgagatcgg ccagctcaac 1440
ggctgggatt ataaggaaga cccggaccgt gtcgctgaca gccgcaatct gcaccgcagc 1500
aagttcaact ggaagaacgc cgcgaagcgc gatgtccccg gaaccttgcc aaaccggctg 1560
tgggaaggca tggcggatgt gcggcagatg cgctcggacc catgcttcgc ccctgacgct 1620
tgggtgacga cgtgggacgc gcatgatgac ggtattctcg cgatggtccg gcagtcaggt 1680
gggcgcacac tgctcggcgt gttcaatttc gcgaaccgtg acgccacggc gacgcttgac 1740
agcatcgagg gcgtgagcct gccgcgtacg gtggcgctca agccatacga gtggaagatc 1800
gaggcctgct ga 1812
<210> 15
<211> 1812
<212> DNA
<213> Artificial Sequence
<220>
<223> amylosucrase BtAS V542K
<400> 15
atggaagcca catatcgcga ttccgtattc gccgaacggc tcgccccgcg ctgcgcagaa 60
cttgaacaac ttttccgctc gttgtacggg gattcccctg aattcgacca cttcgaacag 120
gtcatggcca aggcccacgc cgaccggcca gccgacctca aacgcctcga cgccgcccgt 180
gaacacgatc cgcaatggta ccgtcgcggc gacatgttcg gcatgaccat gtacaccgac 240
ctgttcgccg gcaaactcac cgatctcgcc aagcatatcg actatctcaa agagcagcat 300
ctgacctacc tgcacctcat gccgctgctg accatgcccc accccgacaa cgacggcggc 360
tacgccatcg aggatttcga caccgtcgac ccgactatcg gcaccaatga ggacctcgcc 420
gacctcaccg cgaaactgcg cgaagccggc atcagcctgt gccttgattt cgtcatgaac 480
cacaccgcat ccacccaccg gtgggcgaaa gccgcacaag ccggcgaccc cgaataccag 540
gactactact tctgctatga cgaccgcacc atccccgacc aatatgacgc cgtcgtcccg 600
caagtcttcc cgaccgccgc ccccggcaac ttcacatgga atgagcagat gggcaaatgg 660
gtcatgaccc agttctaccc gttccaatgg gacctcaact accgcaatcc caaggtcttc 720
gtcgtcatga tgtccagcct gctgcacctg gccaacctcg gcgtcgaagt cttccgcatc 780
gacgcggtgc cgtacatctg gaagcaactc ggcaccaact gccgcaacct gccgcaagtc 840
cacaccatcg tgcgcatgat gcgcatcatg tccgaaatcg tctgcccggc cgtcgtgttc 900
aaaggtgaag tcgtcatggc tcccaaggag ctcgccgcct acttcggcac ccccgagaag 960
cccgaatgcc acatgctgta caacgtgtcc gtcatggtca acttgtggag cgcgctcgcc 1020
aacggcgaca cccgcctgct taaaacccag atcgacaagc tcgacgccct gcccgacaac 1080
tgctggttcg tcaactatct gcgctgccat gacgatatcg gctggggtct ggacgaggat 1140
gtcgaacgcc agttgggcat cgacccgctc aagcacaagg aattcctcta ccacttctac 1200
gagggcatgg tgcccggcag ctgggcgatg ggcgagctgt acaactatga tccggcgtcc 1260
ggtgacgcgc gcagctgcgg caccacggcg agcttgtgcg gtattgagcg tgcgctgatc 1320
acgcatgacc ggccgctgta tgagcgttcc atccagcgtg atctgctcat gcaccacgct 1380
atgggcttcc tgcgtgggtt cccgatgctc aactgcggcg acgagatcgg ccagctcaac 1440
ggctgggatt ataaggaaga cccggaccgt gtcgctgaca gccgcaatct gcaccgcagc 1500
aagttcaact ggaagaacgc cgcgaagcgc gatgtccccg gaaccttgcc aaaccggctg 1560
tgggaaggca tggcggatgt gcggcagatg cgctcggacc catgcttcgc ccctgacgct 1620
tggaagacga cgtgggacgc gcatgatgac ggtattctcg cgatggtccg gcagtcaggt 1680
gggcgcacac tgctcggcgt gttcaatttc gcgaaccgtg acgccacggc gacgcttgac 1740
agcatcgagg gcgtgagcct gccgcgtacg gtggcgctca agccatacga gtggaagatc 1800
gaggcctgct ga 1812
<210> 16
<211> 1812
<212> DNA
<213> Artificial Sequence
<220>
<223> amylosucrase BtAS V542D
<400> 16
atggaagcca catatcgcga ttccgtattc gccgaacggc tcgccccgcg ctgcgcagaa 60
cttgaacaac ttttccgctc gttgtacggg gattcccctg aattcgacca cttcgaacag 120
gtcatggcca aggcccacgc cgaccggcca gccgacctca aacgcctcga cgccgcccgt 180
gaacacgatc cgcaatggta ccgtcgcggc gacatgttcg gcatgaccat gtacaccgac 240
ctgttcgccg gcaaactcac cgatctcgcc aagcatatcg actatctcaa agagcagcat 300
ctgacctacc tgcacctcat gccgctgctg accatgcccc accccgacaa cgacggcggc 360
tacgccatcg aggatttcga caccgtcgac ccgactatcg gcaccaatga ggacctcgcc 420
gacctcaccg cgaaactgcg cgaagccggc atcagcctgt gccttgattt cgtcatgaac 480
cacaccgcat ccacccaccg gtgggcgaaa gccgcacaag ccggcgaccc cgaataccag 540
gactactact tctgctatga cgaccgcacc atccccgacc aatatgacgc cgtcgtcccg 600
caagtcttcc cgaccgccgc ccccggcaac ttcacatgga atgagcagat gggcaaatgg 660
gtcatgaccc agttctaccc gttccaatgg gacctcaact accgcaatcc caaggtcttc 720
gtcgtcatga tgtccagcct gctgcacctg gccaacctcg gcgtcgaagt cttccgcatc 780
gacgcggtgc cgtacatctg gaagcaactc ggcaccaact gccgcaacct gccgcaagtc 840
cacaccatcg tgcgcatgat gcgcatcatg tccgaaatcg tctgcccggc cgtcgtgttc 900
aaaggtgaag tcgtcatggc tcccaaggag ctcgccgcct acttcggcac ccccgagaag 960
cccgaatgcc acatgctgta caacgtgtcc gtcatggtca acttgtggag cgcgctcgcc 1020
aacggcgaca cccgcctgct taaaacccag atcgacaagc tcgacgccct gcccgacaac 1080
tgctggttcg tcaactatct gcgctgccat gacgatatcg gctggggtct ggacgaggat 1140
gtcgaacgcc agttgggcat cgacccgctc aagcacaagg aattcctcta ccacttctac 1200
gagggcatgg tgcccggcag ctgggcgatg ggcgagctgt acaactatga tccggcgtcc 1260
ggtgacgcgc gcagctgcgg caccacggcg agcttgtgcg gtattgagcg tgcgctgatc 1320
acgcatgacc ggccgctgta tgagcgttcc atccagcgtg atctgctcat gcaccacgct 1380
atgggcttcc tgcgtgggtt cccgatgctc aactgcggcg acgagatcgg ccagctcaac 1440
ggctgggatt ataaggaaga cccggaccgt gtcgctgaca gccgcaatct gcaccgcagc 1500
aagttcaact ggaagaacgc cgcgaagcgc gatgtccccg gaaccttgcc aaaccggctg 1560
tgggaaggca tggcggatgt gcggcagatg cgctcggacc catgcttcgc ccctgacgct 1620
tgggacacga cgtgggacgc gcatgatgac ggtattctcg cgatggtccg gcagtcaggt 1680
gggcgcacac tgctcggcgt gttcaatttc gcgaaccgtg acgccacggc gacgcttgac 1740
agcatcgagg gcgtgagcct gccgcgtacg gtggcgctca agccatacga gtggaagatc 1800
gaggcctgct ga 1812
<210> 17
<211> 1812
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS P200R
<400> 17
atggaagcca catatcgcga ttccgtattc gccgaacggc tcgccccgcg ctgcgcagaa 60
cttgaacaac ttttccgctc gttgtacggg gattcccctg aattcgacca cttcgaacag 120
gtcatggcca aggcccacgc cgaccggcca gccgacctca aacgcctcga cgccgcccgt 180
gaacacgatc cgcaatggta ccgtcgcggc gacatgttcg gcatgaccat gtacaccgac 240
ctgttcgccg gcaaactcac cgatctcgcc aagcatatcg actatctcaa agagcagcat 300
ctgacctacc tgcacctcat gccgctgctg accatgcccc accccgacaa cgacggcggc 360
tacgccatcg aggatttcga caccgtcgac ccgactatcg gcaccaatga ggacctcgcc 420
gacctcaccg cgaaactgcg cgaagccggc atcagcctgt gccttgattt cgtcatgaac 480
cacaccgcat ccacccaccg gtgggcgaaa gccgcacaag ccggcgaccc cgaataccag 540
gactactact tctgctatga cgaccgcacc atccccgacc aatatgacgc cgtcgtccgc 600
caagtcttcc cgaccgccgc ccccggcaac ttcacatgga atgagcagat gggcaaatgg 660
gtcatgaccc agttctaccc gttccaatgg gacctcaact accgcaatcc caaggtcttc 720
gtcgtcatga tgtccagcct gctgcacctg gccaacctcg gcgtcgaagt cttccgcatc 780
gacgcggtgc cgtacatctg gaagcaactc ggcaccaact gccgcaacct gccgcaagtc 840
cacaccatcg tgcgcatgat gcgcatcatg tccgaaatcg tctgcccggc cgtcgtgttc 900
aaaggtgaag tcgtcatggc tcccaaggag ctcgccgcct acttcggcac ccccgagaag 960
cccgaatgcc acatgctgta caacgtgtcc gtcatggtca acttgtggag cgcgctcgcc 1020
aacggcgaca cccgcctgct taaaacccag atcgacaagc tcgacgccct gcccgacaac 1080
tgctggttcg tcaactatct gcgctgccat gacgatatcg gctggggtct ggacgaggat 1140
gtcgaacgcc agttgggcat cgacccgctc aagcacaagg aattcctcta ccacttctac 1200
gagggcatgg tgcccggcag ctgggcgatg ggcgagctgt acaactatga tccggcgtcc 1260
ggtgacgcgc gcagctgcgg caccacggcg agcttgtgcg gtattgagcg tgcgctgatc 1320
acgcatgacc ggccgctgta tgagcgttcc atccagcgtg atctgctcat gcaccacgct 1380
atgggcttcc tgcgtgggtt cccgatgctc aactgcggcg acgagatcgg ccagctcaac 1440
ggctgggatt ataaggaaga cccggaccgt gtcgctgaca gccgcaatct gcaccgcagc 1500
aagttcaact ggaagaacgc cgcgaagcgc gatgtccccg gaaccttgcc aaaccggctg 1560
tgggaaggca tggcggatgt gcggcagatg cgctcggacc catgcttcgc ccctgacgct 1620
tgggtgacga cgtgggacgc gcatgatgac ggtattctcg cgatggtccg gcagtcaggt 1680
gggcgcacac tgctcggcgt gttcaatttc gcgaaccgtg acgccacggc gacgcttgac 1740
agcatcgagg gcgtgagcct gccgcgtacg gtggcgctca agccatacga gtggaagatc 1800
gaggcctgct ga 1812
<210> 18
<211> 1812
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS V202I
<400> 18
atggaagcca catatcgcga ttccgtattc gccgaacggc tcgccccgcg ctgcgcagaa 60
cttgaacaac ttttccgctc gttgtacggg gattcccctg aattcgacca cttcgaacag 120
gtcatggcca aggcccacgc cgaccggcca gccgacctca aacgcctcga cgccgcccgt 180
gaacacgatc cgcaatggta ccgtcgcggc gacatgttcg gcatgaccat gtacaccgac 240
ctgttcgccg gcaaactcac cgatctcgcc aagcatatcg actatctcaa agagcagcat 300
ctgacctacc tgcacctcat gccgctgctg accatgcccc accccgacaa cgacggcggc 360
tacgccatcg aggatttcga caccgtcgac ccgactatcg gcaccaatga ggacctcgcc 420
gacctcaccg cgaaactgcg cgaagccggc atcagcctgt gccttgattt cgtcatgaac 480
cacaccgcat ccacccaccg gtgggcgaaa gccgcacaag ccggcgaccc cgaataccag 540
gactactact tctgctatga cgaccgcacc atccccgacc aatatgacgc cgtcgtcccg 600
caaatcttcc cgaccgccgc ccccggcaac ttcacatgga atgagcagat gggcaaatgg 660
gtcatgaccc agttctaccc gttccaatgg gacctcaact accgcaatcc caaggtcttc 720
gtcgtcatga tgtccagcct gctgcacctg gccaacctcg gcgtcgaagt cttccgcatc 780
gacgcggtgc cgtacatctg gaagcaactc ggcaccaact gccgcaacct gccgcaagtc 840
cacaccatcg tgcgcatgat gcgcatcatg tccgaaatcg tctgcccggc cgtcgtgttc 900
aaaggtgaag tcgtcatggc tcccaaggag ctcgccgcct acttcggcac ccccgagaag 960
cccgaatgcc acatgctgta caacgtgtcc gtcatggtca acttgtggag cgcgctcgcc 1020
aacggcgaca cccgcctgct taaaacccag atcgacaagc tcgacgccct gcccgacaac 1080
tgctggttcg tcaactatct gcgctgccat gacgatatcg gctggggtct ggacgaggat 1140
gtcgaacgcc agttgggcat cgacccgctc aagcacaagg aattcctcta ccacttctac 1200
gagggcatgg tgcccggcag ctgggcgatg ggcgagctgt acaactatga tccggcgtcc 1260
ggtgacgcgc gcagctgcgg caccacggcg agcttgtgcg gtattgagcg tgcgctgatc 1320
acgcatgacc ggccgctgta tgagcgttcc atccagcgtg atctgctcat gcaccacgct 1380
atgggcttcc tgcgtgggtt cccgatgctc aactgcggcg acgagatcgg ccagctcaac 1440
ggctgggatt ataaggaaga cccggaccgt gtcgctgaca gccgcaatct gcaccgcagc 1500
aagttcaact ggaagaacgc cgcgaagcgc gatgtccccg gaaccttgcc aaaccggctg 1560
tgggaaggca tggcggatgt gcggcagatg cgctcggacc catgcttcgc ccctgacgct 1620
tgggtgacga cgtgggacgc gcatgatgac ggtattctcg cgatggtccg gcagtcaggt 1680
gggcgcacac tgctcggcgt gttcaatttc gcgaaccgtg acgccacggc gacgcttgac 1740
agcatcgagg gcgtgagcct gccgcgtacg gtggcgctca agccatacga gtggaagatc 1800
gaggcctgct ga 1812
<210> 19
<211> 1812
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS Y265F
<400> 19
atggaagcca catatcgcga ttccgtattc gccgaacggc tcgccccgcg ctgcgcagaa 60
cttgaacaac ttttccgctc gttgtacggg gattcccctg aattcgacca cttcgaacag 120
gtcatggcca aggcccacgc cgaccggcca gccgacctca aacgcctcga cgccgcccgt 180
gaacacgatc cgcaatggta ccgtcgcggc gacatgttcg gcatgaccat gtacaccgac 240
ctgttcgccg gcaaactcac cgatctcgcc aagcatatcg actatctcaa agagcagcat 300
ctgacctacc tgcacctcat gccgctgctg accatgcccc accccgacaa cgacggcggc 360
tacgccatcg aggatttcga caccgtcgac ccgactatcg gcaccaatga ggacctcgcc 420
gacctcaccg cgaaactgcg cgaagccggc atcagcctgt gccttgattt cgtcatgaac 480
cacaccgcat ccacccaccg gtgggcgaaa gccgcacaag ccggcgaccc cgaataccag 540
gactactact tctgctatga cgaccgcacc atccccgacc aatatgacgc cgtcgtcccg 600
caagtcttcc cgaccgccgc ccccggcaac ttcacatgga atgagcagat gggcaaatgg 660
gtcatgaccc agttctaccc gttccaatgg gacctcaact accgcaatcc caaggtcttc 720
gtcgtcatga tgtccagcct gctgcacctg gccaacctcg gcgtcgaagt cttccgcatc 780
gacgcggtgc cgttcatctg gaagcaactc ggcaccaact gccgcaacct gccgcaagtc 840
cacaccatcg tgcgcatgat gcgcatcatg tccgaaatcg tctgcccggc cgtcgtgttc 900
aaaggtgaag tcgtcatggc tcccaaggag ctcgccgcct acttcggcac ccccgagaag 960
cccgaatgcc acatgctgta caacgtgtcc gtcatggtca acttgtggag cgcgctcgcc 1020
aacggcgaca cccgcctgct taaaacccag atcgacaagc tcgacgccct gcccgacaac 1080
tgctggttcg tcaactatct gcgctgccat gacgatatcg gctggggtct ggacgaggat 1140
gtcgaacgcc agttgggcat cgacccgctc aagcacaagg aattcctcta ccacttctac 1200
gagggcatgg tgcccggcag ctgggcgatg ggcgagctgt acaactatga tccggcgtcc 1260
ggtgacgcgc gcagctgcgg caccacggcg agcttgtgcg gtattgagcg tgcgctgatc 1320
acgcatgacc ggccgctgta tgagcgttcc atccagcgtg atctgctcat gcaccacgct 1380
atgggcttcc tgcgtgggtt cccgatgctc aactgcggcg acgagatcgg ccagctcaac 1440
ggctgggatt ataaggaaga cccggaccgt gtcgctgaca gccgcaatct gcaccgcagc 1500
aagttcaact ggaagaacgc cgcgaagcgc gatgtccccg gaaccttgcc aaaccggctg 1560
tgggaaggca tggcggatgt gcggcagatg cgctcggacc catgcttcgc ccctgacgct 1620
tgggtgacga cgtgggacgc gcatgatgac ggtattctcg cgatggtccg gcagtcaggt 1680
gggcgcacac tgctcggcgt gttcaatttc gcgaaccgtg acgccacggc gacgcttgac 1740
agcatcgagg gcgtgagcct gccgcgtacg gtggcgctca agccatacga gtggaagatc 1800
gaggcctgct ga 1812
<210> 20
<211> 1812
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS V305I
<400> 20
atggaagcca catatcgcga ttccgtattc gccgaacggc tcgccccgcg ctgcgcagaa 60
cttgaacaac ttttccgctc gttgtacggg gattcccctg aattcgacca cttcgaacag 120
gtcatggcca aggcccacgc cgaccggcca gccgacctca aacgcctcga cgccgcccgt 180
gaacacgatc cgcaatggta ccgtcgcggc gacatgttcg gcatgaccat gtacaccgac 240
ctgttcgccg gcaaactcac cgatctcgcc aagcatatcg actatctcaa agagcagcat 300
ctgacctacc tgcacctcat gccgctgctg accatgcccc accccgacaa cgacggcggc 360
tacgccatcg aggatttcga caccgtcgac ccgactatcg gcaccaatga ggacctcgcc 420
gacctcaccg cgaaactgcg cgaagccggc atcagcctgt gccttgattt cgtcatgaac 480
cacaccgcat ccacccaccg gtgggcgaaa gccgcacaag ccggcgaccc cgaataccag 540
gactactact tctgctatga cgaccgcacc atccccgacc aatatgacgc cgtcgtcccg 600
caagtcttcc cgaccgccgc ccccggcaac ttcacatgga atgagcagat gggcaaatgg 660
gtcatgaccc agttctaccc gttccaatgg gacctcaact accgcaatcc caaggtcttc 720
gtcgtcatga tgtccagcct gctgcacctg gccaacctcg gcgtcgaagt cttccgcatc 780
gacgcggtgc cgtacatctg gaagcaactc ggcaccaact gccgcaacct gccgcaagtc 840
cacaccatcg tgcgcatgat gcgcatcatg tccgaaatcg tctgcccggc cgtcgtgttc 900
aaaggtgaag tcatcatggc tcccaaggag ctcgccgcct acttcggcac ccccgagaag 960
cccgaatgcc acatgctgta caacgtgtcc gtcatggtca acttgtggag cgcgctcgcc 1020
aacggcgaca cccgcctgct taaaacccag atcgacaagc tcgacgccct gcccgacaac 1080
tgctggttcg tcaactatct gcgctgccat gacgatatcg gctggggtct ggacgaggat 1140
gtcgaacgcc agttgggcat cgacccgctc aagcacaagg aattcctcta ccacttctac 1200
gagggcatgg tgcccggcag ctgggcgatg ggcgagctgt acaactatga tccggcgtcc 1260
ggtgacgcgc gcagctgcgg caccacggcg agcttgtgcg gtattgagcg tgcgctgatc 1320
acgcatgacc ggccgctgta tgagcgttcc atccagcgtg atctgctcat gcaccacgct 1380
atgggcttcc tgcgtgggtt cccgatgctc aactgcggcg acgagatcgg ccagctcaac 1440
ggctgggatt ataaggaaga cccggaccgt gtcgctgaca gccgcaatct gcaccgcagc 1500
aagttcaact ggaagaacgc cgcgaagcgc gatgtccccg gaaccttgcc aaaccggctg 1560
tgggaaggca tggcggatgt gcggcagatg cgctcggacc catgcttcgc ccctgacgct 1620
tgggtgacga cgtgggacgc gcatgatgac ggtattctcg cgatggtccg gcagtcaggt 1680
gggcgcacac tgctcggcgt gttcaatttc gcgaaccgtg acgccacggc gacgcttgac 1740
agcatcgagg gcgtgagcct gccgcgtacg gtggcgctca agccatacga gtggaagatc 1800
gaggcctgct ga 1812
<210> 21
<211> 1812
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS K393R
<400> 21
atggaagcca catatcgcga ttccgtattc gccgaacggc tcgccccgcg ctgcgcagaa 60
cttgaacaac ttttccgctc gttgtacggg gattcccctg aattcgacca cttcgaacag 120
gtcatggcca aggcccacgc cgaccggcca gccgacctca aacgcctcga cgccgcccgt 180
gaacacgatc cgcaatggta ccgtcgcggc gacatgttcg gcatgaccat gtacaccgac 240
ctgttcgccg gcaaactcac cgatctcgcc aagcatatcg actatctcaa agagcagcat 300
ctgacctacc tgcacctcat gccgctgctg accatgcccc accccgacaa cgacggcggc 360
tacgccatcg aggatttcga caccgtcgac ccgactatcg gcaccaatga ggacctcgcc 420
gacctcaccg cgaaactgcg cgaagccggc atcagcctgt gccttgattt cgtcatgaac 480
cacaccgcat ccacccaccg gtgggcgaaa gccgcacaag ccggcgaccc cgaataccag 540
gactactact tctgctatga cgaccgcacc atccccgacc aatatgacgc cgtcgtcccg 600
caagtcttcc cgaccgccgc ccccggcaac ttcacatgga atgagcagat gggcaaatgg 660
gtcatgaccc agttctaccc gttccaatgg gacctcaact accgcaatcc caaggtcttc 720
gtcgtcatga tgtccagcct gctgcacctg gccaacctcg gcgtcgaagt cttccgcatc 780
gacgcggtgc cgtacatctg gaagcaactc ggcaccaact gccgcaacct gccgcaagtc 840
cacaccatcg tgcgcatgat gcgcatcatg tccgaaatcg tctgcccggc cgtcgtgttc 900
aaaggtgaag tcgtcatggc tcccaaggag ctcgccgcct acttcggcac ccccgagaag 960
cccgaatgcc acatgctgta caacgtgtcc gtcatggtca acttgtggag cgcgctcgcc 1020
aacggcgaca cccgcctgct taaaacccag atcgacaagc tcgacgccct gcccgacaac 1080
tgctggttcg tcaactatct gcgctgccat gacgatatcg gctggggtct ggacgaggat 1140
gtcgaacgcc agttgggcat cgacccgctc aagcacaggg aattcctcta ccacttctac 1200
gagggcatgg tgcccggcag ctgggcgatg ggcgagctgt acaactatga tccggcgtcc 1260
ggtgacgcgc gcagctgcgg caccacggcg agcttgtgcg gtattgagcg tgcgctgatc 1320
acgcatgacc ggccgctgta tgagcgttcc atccagcgtg atctgctcat gcaccacgct 1380
atgggcttcc tgcgtgggtt cccgatgctc aactgcggcg acgagatcgg ccagctcaac 1440
ggctgggatt ataaggaaga cccggaccgt gtcgctgaca gccgcaatct gcaccgcagc 1500
aagttcaact ggaagaacgc cgcgaagcgc gatgtccccg gaaccttgcc aaaccggctg 1560
tgggaaggca tggcggatgt gcggcagatg cgctcggacc catgcttcgc ccctgacgct 1620
tgggtgacga cgtgggacgc gcatgatgac ggtattctcg cgatggtccg gcagtcaggt 1680
gggcgcacac tgctcggcgt gttcaatttc gcgaaccgtg acgccacggc gacgcttgac 1740
agcatcgagg gcgtgagcct gccgcgtacg gtggcgctca agccatacga gtggaagatc 1800
gaggcctgct ga 1812
<210> 22
<211> 1812
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS S420T
<400> 22
atggaagcca catatcgcga ttccgtattc gccgaacggc tcgccccgcg ctgcgcagaa 60
cttgaacaac ttttccgctc gttgtacggg gattcccctg aattcgacca cttcgaacag 120
gtcatggcca aggcccacgc cgaccggcca gccgacctca aacgcctcga cgccgcccgt 180
gaacacgatc cgcaatggta ccgtcgcggc gacatgttcg gcatgaccat gtacaccgac 240
ctgttcgccg gcaaactcac cgatctcgcc aagcatatcg actatctcaa agagcagcat 300
ctgacctacc tgcacctcat gccgctgctg accatgcccc accccgacaa cgacggcggc 360
tacgccatcg aggatttcga caccgtcgac ccgactatcg gcaccaatga ggacctcgcc 420
gacctcaccg cgaaactgcg cgaagccggc atcagcctgt gccttgattt cgtcatgaac 480
cacaccgcat ccacccaccg gtgggcgaaa gccgcacaag ccggcgaccc cgaataccag 540
gactactact tctgctatga cgaccgcacc atccccgacc aatatgacgc cgtcgtcccg 600
caagtcttcc cgaccgccgc ccccggcaac ttcacatgga atgagcagat gggcaaatgg 660
gtcatgaccc agttctaccc gttccaatgg gacctcaact accgcaatcc caaggtcttc 720
gtcgtcatga tgtccagcct gctgcacctg gccaacctcg gcgtcgaagt cttccgcatc 780
gacgcggtgc cgtacatctg gaagcaactc ggcaccaact gccgcaacct gccgcaagtc 840
cacaccatcg tgcgcatgat gcgcatcatg tccgaaatcg tctgcccggc cgtcgtgttc 900
aaaggtgaag tcgtcatggc tcccaaggag ctcgccgcct acttcggcac ccccgagaag 960
cccgaatgcc acatgctgta caacgtgtcc gtcatggtca acttgtggag cgcgctcgcc 1020
aacggcgaca cccgcctgct taaaacccag atcgacaagc tcgacgccct gcccgacaac 1080
tgctggttcg tcaactatct gcgctgccat gacgatatcg gctggggtct ggacgaggat 1140
gtcgaacgcc agttgggcat cgacccgctc aagcacaagg aattcctcta ccacttctac 1200
gagggcatgg tgcccggcag ctgggcgatg ggcgagctgt acaactatga tccggcgacg 1260
ggtgacgcgc gcagctgcgg caccacggcg agcttgtgcg gtattgagcg tgcgctgatc 1320
acgcatgacc ggccgctgta tgagcgttcc atccagcgtg atctgctcat gcaccacgct 1380
atgggcttcc tgcgtgggtt cccgatgctc aactgcggcg acgagatcgg ccagctcaac 1440
ggctgggatt ataaggaaga cccggaccgt gtcgctgaca gccgcaatct gcaccgcagc 1500
aagttcaact ggaagaacgc cgcgaagcgc gatgtccccg gaaccttgcc aaaccggctg 1560
tgggaaggca tggcggatgt gcggcagatg cgctcggacc catgcttcgc ccctgacgct 1620
tgggtgacga cgtgggacgc gcatgatgac ggtattctcg cgatggtccg gcagtcaggt 1680
gggcgcacac tgctcggcgt gttcaatttc gcgaaccgtg acgccacggc gacgcttgac 1740
agcatcgagg gcgtgagcct gccgcgtacg gtggcgctca agccatacga gtggaagatc 1800
gaggcctgct ga 1812
<210> 23
<211> 1812
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS Y414F
<400> 23
atggaagcca catatcgcga ttccgtattc gccgaacggc tcgccccgcg ctgcgcagaa 60
cttgaacaac ttttccgctc gttgtacggg gattcccctg aattcgacca cttcgaacag 120
gtcatggcca aggcccacgc cgaccggcca gccgacctca aacgcctcga cgccgcccgt 180
gaacacgatc cgcaatggta ccgtcgcggc gacatgttcg gcatgaccat gtacaccgac 240
ctgttcgccg gcaaactcac cgatctcgcc aagcatatcg actatctcaa agagcagcat 300
ctgacctacc tgcacctcat gccgctgctg accatgcccc accccgacaa cgacggcggc 360
tacgccatcg aggatttcga caccgtcgac ccgactatcg gcaccaatga ggacctcgcc 420
gacctcaccg cgaaactgcg cgaagccggc atcagcctgt gccttgattt cgtcatgaac 480
cacaccgcat ccacccaccg gtgggcgaaa gccgcacaag ccggcgaccc cgaataccag 540
gactactact tctgctatga cgaccgcacc atccccgacc aatatgacgc cgtcgtcccg 600
caagtcttcc cgaccgccgc ccccggcaac ttcacatgga atgagcagat gggcaaatgg 660
gtcatgaccc agttctaccc gttccaatgg gacctcaact accgcaatcc caaggtcttc 720
gtcgtcatga tgtccagcct gctgcacctg gccaacctcg gcgtcgaagt cttccgcatc 780
gacgcggtgc cgtacatctg gaagcaactc ggcaccaact gccgcaacct gccgcaagtc 840
cacaccatcg tgcgcatgat gcgcatcatg tccgaaatcg tctgcccggc cgtcgtgttc 900
aaaggtgaag tcgtcatggc tcccaaggag ctcgccgcct acttcggcac ccccgagaag 960
cccgaatgcc acatgctgta caacgtgtcc gtcatggtca acttgtggag cgcgctcgcc 1020
aacggcgaca cccgcctgct taaaacccag atcgacaagc tcgacgccct gcccgacaac 1080
tgctggttcg tcaactatct gcgctgccat gacgatatcg gctggggtct ggacgaggat 1140
gtcgaacgcc agttgggcat cgacccgctc aagcacaagg aattcctcta ccacttctac 1200
gagggcatgg tgcccggcag ctgggcgatg ggcgagctgt tcaactatga tccggcgtcc 1260
ggtgacgcgc gcagctgcgg caccacggcg agcttgtgcg gtattgagcg tgcgctgatc 1320
acgcatgacc ggccgctgta tgagcgttcc atccagcgtg atctgctcat gcaccacgct 1380
atgggcttcc tgcgtgggtt cccgatgctc aactgcggcg acgagatcgg ccagctcaac 1440
ggctgggatt ataaggaaga cccggaccgt gtcgctgaca gccgcaatct gcaccgcagc 1500
aagttcaact ggaagaacgc cgcgaagcgc gatgtccccg gaaccttgcc aaaccggctg 1560
tgggaaggca tggcggatgt gcggcagatg cgctcggacc catgcttcgc ccctgacgct 1620
tgggtgacga cgtgggacgc gcatgatgac ggtattctcg cgatggtccg gcagtcaggt 1680
gggcgcacac tgctcggcgt gttcaatttc gcgaaccgtg acgccacggc gacgcttgac 1740
agcatcgagg gcgtgagcct gccgcgtacg gtggcgctca agccatacga gtggaagatc 1800
gaggcctgct ga 1812
<210> 24
<211> 1812
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS Y414F*P200R
<400> 24
atggaagcca catatcgcga ttccgtattc gccgaacggc tcgccccgcg ctgcgcagaa 60
cttgaacaac ttttccgctc gttgtacggg gattcccctg aattcgacca cttcgaacag 120
gtcatggcca aggcccacgc cgaccggcca gccgacctca aacgcctcga cgccgcccgt 180
gaacacgatc cgcaatggta ccgtcgcggc gacatgttcg gcatgaccat gtacaccgac 240
ctgttcgccg gcaaactcac cgatctcgcc aagcatatcg actatctcaa agagcagcat 300
ctgacctacc tgcacctcat gccgctgctg accatgcccc accccgacaa cgacggcggc 360
tacgccatcg aggatttcga caccgtcgac ccgactatcg gcaccaatga ggacctcgcc 420
gacctcaccg cgaaactgcg cgaagccggc atcagcctgt gccttgattt cgtcatgaac 480
cacaccgcat ccacccaccg gtgggcgaaa gccgcacaag ccggcgaccc cgaataccag 540
gactactact tctgctatga cgaccgcacc atccccgacc aatatgacgc cgtcgtccgc 600
caagtcttcc cgaccgccgc ccccggcaac ttcacatgga atgagcagat gggcaaatgg 660
gtcatgaccc agttctaccc gttccaatgg gacctcaact accgcaatcc caaggtcttc 720
gtcgtcatga tgtccagcct gctgcacctg gccaacctcg gcgtcgaagt cttccgcatc 780
gacgcggtgc cgtacatctg gaagcaactc ggcaccaact gccgcaacct gccgcaagtc 840
cacaccatcg tgcgcatgat gcgcatcatg tccgaaatcg tctgcccggc cgtcgtgttc 900
aaaggtgaag tcgtcatggc tcccaaggag ctcgccgcct acttcggcac ccccgagaag 960
cccgaatgcc acatgctgta caacgtgtcc gtcatggtca acttgtggag cgcgctcgcc 1020
aacggcgaca cccgcctgct taaaacccag atcgacaagc tcgacgccct gcccgacaac 1080
tgctggttcg tcaactatct gcgctgccat gacgatatcg gctggggtct ggacgaggat 1140
gtcgaacgcc agttgggcat cgacccgctc aagcacaagg aattcctcta ccacttctac 1200
gagggcatgg tgcccggcag ctgggcgatg ggcgagctgt tcaactatga tccggcgtcc 1260
ggtgacgcgc gcagctgcgg caccacggcg agcttgtgcg gtattgagcg tgcgctgatc 1320
acgcatgacc ggccgctgta tgagcgttcc atccagcgtg atctgctcat gcaccacgct 1380
atgggcttcc tgcgtgggtt cccgatgctc aactgcggcg acgagatcgg ccagctcaac 1440
ggctgggatt ataaggaaga cccggaccgt gtcgctgaca gccgcaatct gcaccgcagc 1500
aagttcaact ggaagaacgc cgcgaagcgc gatgtccccg gaaccttgcc aaaccggctg 1560
tgggaaggca tggcggatgt gcggcagatg cgctcggacc catgcttcgc ccctgacgct 1620
tgggtgacga cgtgggacgc gcatgatgac ggtattctcg cgatggtccg gcagtcaggt 1680
gggcgcacac tgctcggcgt gttcaatttc gcgaaccgtg acgccacggc gacgcttgac 1740
agcatcgagg gcgtgagcct gccgcgtacg gtggcgctca agccatacga gtggaagatc 1800
gaggcctgct ga 1812
<210> 25
<211> 1812
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS Y414F*V202I
<400> 25
atggaagcca catatcgcga ttccgtattc gccgaacggc tcgccccgcg ctgcgcagaa 60
cttgaacaac ttttccgctc gttgtacggg gattcccctg aattcgacca cttcgaacag 120
gtcatggcca aggcccacgc cgaccggcca gccgacctca aacgcctcga cgccgcccgt 180
gaacacgatc cgcaatggta ccgtcgcggc gacatgttcg gcatgaccat gtacaccgac 240
ctgttcgccg gcaaactcac cgatctcgcc aagcatatcg actatctcaa agagcagcat 300
ctgacctacc tgcacctcat gccgctgctg accatgcccc accccgacaa cgacggcggc 360
tacgccatcg aggatttcga caccgtcgac ccgactatcg gcaccaatga ggacctcgcc 420
gacctcaccg cgaaactgcg cgaagccggc atcagcctgt gccttgattt cgtcatgaac 480
cacaccgcat ccacccaccg gtgggcgaaa gccgcacaag ccggcgaccc cgaataccag 540
gactactact tctgctatga cgaccgcacc atccccgacc aatatgacgc cgtcgtcccg 600
caaatcttcc cgaccgccgc ccccggcaac ttcacatgga atgagcagat gggcaaatgg 660
gtcatgaccc agttctaccc gttccaatgg gacctcaact accgcaatcc caaggtcttc 720
gtcgtcatga tgtccagcct gctgcacctg gccaacctcg gcgtcgaagt cttccgcatc 780
gacgcggtgc cgtacatctg gaagcaactc ggcaccaact gccgcaacct gccgcaagtc 840
cacaccatcg tgcgcatgat gcgcatcatg tccgaaatcg tctgcccggc cgtcgtgttc 900
aaaggtgaag tcgtcatggc tcccaaggag ctcgccgcct acttcggcac ccccgagaag 960
cccgaatgcc acatgctgta caacgtgtcc gtcatggtca acttgtggag cgcgctcgcc 1020
aacggcgaca cccgcctgct taaaacccag atcgacaagc tcgacgccct gcccgacaac 1080
tgctggttcg tcaactatct gcgctgccat gacgatatcg gctggggtct ggacgaggat 1140
gtcgaacgcc agttgggcat cgacccgctc aagcacaagg aattcctcta ccacttctac 1200
gagggcatgg tgcccggcag ctgggcgatg ggcgagctgt tcaactatga tccggcgtcc 1260
ggtgacgcgc gcagctgcgg caccacggcg agcttgtgcg gtattgagcg tgcgctgatc 1320
acgcatgacc ggccgctgta tgagcgttcc atccagcgtg atctgctcat gcaccacgct 1380
atgggcttcc tgcgtgggtt cccgatgctc aactgcggcg acgagatcgg ccagctcaac 1440
ggctgggatt ataaggaaga cccggaccgt gtcgctgaca gccgcaatct gcaccgcagc 1500
aagttcaact ggaagaacgc cgcgaagcgc gatgtccccg gaaccttgcc aaaccggctg 1560
tgggaaggca tggcggatgt gcggcagatg cgctcggacc catgcttcgc ccctgacgct 1620
tgggtgacga cgtgggacgc gcatgatgac ggtattctcg cgatggtccg gcagtcaggt 1680
gggcgcacac tgctcggcgt gttcaatttc gcgaaccgtg acgccacggc gacgcttgac 1740
agcatcgagg gcgtgagcct gccgcgtacg gtggcgctca agccatacga gtggaagatc 1800
gaggcctgct ga 1812
<210> 26
<211> 1812
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS Y414F*Y265F
<400> 26
atggaagcca catatcgcga ttccgtattc gccgaacggc tcgccccgcg ctgcgcagaa 60
cttgaacaac ttttccgctc gttgtacggg gattcccctg aattcgacca cttcgaacag 120
gtcatggcca aggcccacgc cgaccggcca gccgacctca aacgcctcga cgccgcccgt 180
gaacacgatc cgcaatggta ccgtcgcggc gacatgttcg gcatgaccat gtacaccgac 240
ctgttcgccg gcaaactcac cgatctcgcc aagcatatcg actatctcaa agagcagcat 300
ctgacctacc tgcacctcat gccgctgctg accatgcccc accccgacaa cgacggcggc 360
tacgccatcg aggatttcga caccgtcgac ccgactatcg gcaccaatga ggacctcgcc 420
gacctcaccg cgaaactgcg cgaagccggc atcagcctgt gccttgattt cgtcatgaac 480
cacaccgcat ccacccaccg gtgggcgaaa gccgcacaag ccggcgaccc cgaataccag 540
gactactact tctgctatga cgaccgcacc atccccgacc aatatgacgc cgtcgtcccg 600
caagtcttcc cgaccgccgc ccccggcaac ttcacatgga atgagcagat gggcaaatgg 660
gtcatgaccc agttctaccc gttccaatgg gacctcaact accgcaatcc caaggtcttc 720
gtcgtcatga tgtccagcct gctgcacctg gccaacctcg gcgtcgaagt cttccgcatc 780
gacgcggtgc cgttcatctg gaagcaactc ggcaccaact gccgcaacct gccgcaagtc 840
cacaccatcg tgcgcatgat gcgcatcatg tccgaaatcg tctgcccggc cgtcgtgttc 900
aaaggtgaag tcgtcatggc tcccaaggag ctcgccgcct acttcggcac ccccgagaag 960
cccgaatgcc acatgctgta caacgtgtcc gtcatggtca acttgtggag cgcgctcgcc 1020
aacggcgaca cccgcctgct taaaacccag atcgacaagc tcgacgccct gcccgacaac 1080
tgctggttcg tcaactatct gcgctgccat gacgatatcg gctggggtct ggacgaggat 1140
gtcgaacgcc agttgggcat cgacccgctc aagcacaagg aattcctcta ccacttctac 1200
gagggcatgg tgcccggcag ctgggcgatg ggcgagctgt tcaactatga tccggcgtcc 1260
ggtgacgcgc gcagctgcgg caccacggcg agcttgtgcg gtattgagcg tgcgctgatc 1320
acgcatgacc ggccgctgta tgagcgttcc atccagcgtg atctgctcat gcaccacgct 1380
atgggcttcc tgcgtgggtt cccgatgctc aactgcggcg acgagatcgg ccagctcaac 1440
ggctgggatt ataaggaaga cccggaccgt gtcgctgaca gccgcaatct gcaccgcagc 1500
aagttcaact ggaagaacgc cgcgaagcgc gatgtccccg gaaccttgcc aaaccggctg 1560
tgggaaggca tggcggatgt gcggcagatg cgctcggacc catgcttcgc ccctgacgct 1620
tgggtgacga cgtgggacgc gcatgatgac ggtattctcg cgatggtccg gcagtcaggt 1680
gggcgcacac tgctcggcgt gttcaatttc gcgaaccgtg acgccacggc gacgcttgac 1740
agcatcgagg gcgtgagcct gccgcgtacg gtggcgctca agccatacga gtggaagatc 1800
gaggcctgct ga 1812
<210> 27
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS V542D forward primer
<400> 27
cgcccctgac gcttgggaca cgacgtggga cgcgc 35
<210> 28
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS V542D reverse primer
<400> 28
gcgcgtccca cgtcgtgtcc caagcgtcag gggcg 35
<210> 29
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS V542K forward primer
<400> 29
cgcccctgac gcttggaaga cgacgtggga cgcgc 35
<210> 30
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS V542K reverse primer
<400> 30
gcgcgtccca cgtcgtcttc caagcgtcag gggcg 35
<210> 31
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS P200R forward primer
<400> 31
gccgtcgtcc gccaagtctt c 21
<210> 32
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS P200R reverse primer
<400> 32
gaagacttgg cggacgacgg c 21
<210> 33
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS V202I forward primer
<400> 33
gtcccgcaaa tcttcccgac c 21
<210> 34
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS V202I reverse primer
<400> 34
ggtcgggaag atttgcggga c 21
<210> 35
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS Y265F forward primer
<400> 35
gacgcggtgc cgttcatctg gaagcaa 27
<210> 36
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS Y265F reverse primer
<400> 36
ttgcttccag atgaacggca ccgcgtc 27
<210> 37
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS Y305I forward primer
<400> 37
aaaggtgaag tcatcatggc tcccaag 27
<210> 38
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS Y305I reverse primer
<400> 38
cttgggagcc atgatgactt caccttt 27
<210> 39
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS K393R forward primer
<400> 39
ccgctcaagc acagggaatt cctctac 27
<210> 40
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS K393R reverse primer
<400> 40
gtagaggaat tccctgtgct tgagcgg 27
<210> 41
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS S420T forward primer
<400> 41
tatgatccgg cgacgggtga cgcgcgc 27
<210> 42
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS S420T reverse primer
<400> 42
gcgcgcgtca cccgtcgccg gatcata 27
<210> 43
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS Y414F forward primer
<400> 43
atgggcgagc tgttcaacta tgatccg 27
<210> 44
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> BtAS Y414F reverse primer
<400> 44
cggatcatag ttgaacagct cgcccat 27
Claims (15)
- 설탕 및 과당으로 이루어진 군에서 선택된 1종 이상의 반응 기질에 아밀로수크라제 효소를 포함하는 촉매 조성물을 반응시켜 반응생성물을 제조하는 단계; 및
상기 반응 생성물을 모사 이동층(SMB) 크로마토그래피를 이용한 투라노스 분획과 라피네이트로 분리하는 단계를 포함하는,
투라노스의 제조방법. - 제1항에 있어서, 상기 반응생성물에 글루코아밀라제 효소를 반응시키는 단계를 추가로 포함하는, 제조방법.
- 제1항에 있어서, 상기 반응생성물은 투라노스 및 트레할룰로스로 이루어진 군에서 선택된 1종 이상을 포함하는 것인, 제조방법.
- 제3항에 있어서, 상기 반응생성물에 포함된 전체 당류 고형분 함량 100중량%를 기준으로, 트레할룰로스 함량이 7.0 중량%이하인 것인 제조방법.
- 제1항에 있어서, 상기 투라노스 분획에 포함된 당류 고형분 함량 100중량%를 기준으로, 트레할룰로스 함량이 7.0 중량%이하인 것인 제조방법.
- 제1항에 있어서, 상기 투라노스 분획에 포함된 당류 고형분 함량 100중량%를 기준으로, 투라노스 함량이 70 중량%이상인 것인 제조방법.
- 제1항에 있어서, 상기 반응 기질은 설탕 고형분 함량 100중량부를 기준으로 과당 고형분 함량이 5 내지 250중량부로 포함하는 것인 제조방법.
- 제1항에 있어서, 상기 반응생성물 고형분 100중량%를 기준으로, 반응 생성물은 DP3이상의 올리고당 함량이 20중량%이고, 투라노스 30 중량%이상으로 포함하는 것인 제조방법.
- 제1항에 있어서, 상기 촉매 조성물은 아밀로수크라제 효소, 아밀로 수크라제 효소를 생산하는 재조합 미생물, 상기 재조합 미생물의 배양물 및 상기 재조합 미생물의 파쇄물로 이루어진 군에서 선택된 1종 이상을 포함하는 것인, 제조방법.
- 제1항에 있어서, 탈염, 탈색, 농축, 결정화, 원심분리 및 여과로 이루어진 군에서 선택된 1종 이상의 단계를 추가로 포함하는, 제조방법.
- 제1항에 있어서, 상기 아밀로수크라제 효소는 아래 특성으로 이루어진 군에서 선택된 1종 이상의 특성을 갖는 것인, 제조방법:
(1) 단량체의 분자량이 65 내지 75 kDa
(2) 최적 온도가 30 내지 65℃
(3) 최적 pH가 6.0 내지 9.0. - 제1항에 있어서, 상기 아밀로수크라제 효소는, 상기 반응 기질과 50℃의 온도 및 pH 6.0에서 24시간 동안 반응하여 얻어진 반응생성물 고형분 100중량%를 기준으로 투라노스 30 중량%이상, 올리고당 20중량%이하 및 트레할룰로스 7중량%이하로 포함하는 반응생성물을 제조하는 것을 특징으로 하는, 제조방법.
- 제1항에 있어서, 상기 아밀로수크라제 효소는 비피도박테리움 터모필럼(Bifidobacterium thermophilum) 유래 효소인, 제조방법.
- 제1항에 있어서, 상기 아밀로수크라제 효소는 서열번호 1의 아미노산 서열, 상기 서열번호 1의 아미노산 서열과 90% 이상의 서열 상동성이 있는 아미노산 서열을 갖는 것인, 제조방법.
- 제14항에 있어서, 상기 아밀로수크라제 효소는 서열번호 1 내지 서열번호 11의 아미노산 서열들로 이루어지는 군에서 선택된 1종 이상의 효소인 제조방법.
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| KR1020180129298A KR102466543B1 (ko) | 2018-10-26 | 2018-10-26 | 투라노스의 제조방법 |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| KR1020180129298A KR102466543B1 (ko) | 2018-10-26 | 2018-10-26 | 투라노스의 제조방법 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| KR20200047176A true KR20200047176A (ko) | 2020-05-07 |
| KR102466543B1 KR102466543B1 (ko) | 2022-11-11 |
Family
ID=70733777
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020180129298A KR102466543B1 (ko) | 2018-10-26 | 2018-10-26 | 투라노스의 제조방법 |
Country Status (1)
| Country | Link |
|---|---|
| KR (1) | KR102466543B1 (ko) |
Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| KR20120049421A (ko) * | 2010-11-02 | 2012-05-17 | 세종대학교산학협력단 | 아밀로수크라제를 이용한 투라노즈의 제조방법 및 상기 투라노즈를 이용한 감미료 |
-
2018
- 2018-10-26 KR KR1020180129298A patent/KR102466543B1/ko active IP Right Grant
Patent Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| KR20120049421A (ko) * | 2010-11-02 | 2012-05-17 | 세종대학교산학협력단 | 아밀로수크라제를 이용한 투라노즈의 제조방법 및 상기 투라노즈를 이용한 감미료 |
Non-Patent Citations (3)
| Title |
|---|
| Chromatography Research International, Vol. 2013, pp. 1-7 (2013.)* * |
| Food Chemistry, Vol. 132, pp. 773-779 (2011.11.17.)* * |
| NCBI GenBank Accession No. KFJ08006.1 (2014.08.18.)* * |
Also Published As
| Publication number | Publication date |
|---|---|
| KR102466543B1 (ko) | 2022-11-11 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US20210198648A1 (en) | 3-Epimerase | |
| AU2011356210B8 (en) | Novel fucosyltransferases and their applications | |
| RU2642307C2 (ru) | Новые фукозилтрансферазы и их применение | |
| CN105821027B (zh) | 一种3-差向异构酶的用途 | |
| US11866758B2 (en) | Methods and compositions for preparing tagatose from fructose | |
| CN112063666B (zh) | 一种重组蔗糖异构酶在转化蔗糖制备异麦芽酮糖中的应用 | |
| KR101787331B1 (ko) | 내열성 한천분해효소 및 이를 이용한 단당류의 생산방법 | |
| CA2454277C (en) | Process for manufacturing of tagatose | |
| TW201823456A (zh) | D-阿洛酮糖表異構酶、使用其製備d-阿洛酮糖的方法、編碼其的聚何苷酸、重組載體、微生物以及組成物 | |
| US20180216086A1 (en) | Cellobiose Phosphorylase | |
| CN109576239B (zh) | 耐热磷酸化酶及其应用 | |
| JP2023506154A (ja) | アルロースエピマー化酵素変異体、その製造方法及びそれを利用したアルロースの製造方法 | |
| KR102004945B1 (ko) | 아밀로수크라제의 발현 시스템 및 이를 이용한 투라노스의 생산 | |
| KR102466543B1 (ko) | 투라노스의 제조방법 | |
| KR102141174B1 (ko) | 아밀로수크라제의 발현 시스템 및 이를 이용한 투라노스의 생산 | |
| CN108034649B (zh) | 一种葡萄糖异构酶突变体及其应用 | |
| CN111057698B (zh) | L-阿拉伯糖异构酶、突变体及其应用 | |
| JP7445947B2 (ja) | アラビノースイソメラーゼ変異体 | |
| US20240052389A1 (en) | Allulose epimerase variant with excellent thermal stability, preparation method therefor, and preparation method for allulose using same | |
| EP3757209A1 (en) | Enzymatic production of levan-based, prebiotic fructooligosaccharides | |
| CN117327684A (zh) | 用于制备塔格糖的酶、其组合物及其应用 | |
| KR101294598B1 (ko) | 4-알파-글루카노트랜스퍼레이즈와 아밀로오스의 복합체 및 이의 이용 | |
| KR20200087064A (ko) | 가야도모나스 주비니에게 g7 유래 알파-네오아가로올리고당 가수분해효소의 이용 | |
| NO320145B1 (no) | Fremgangsmate for fremstilling av tagatose |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| E902 | Notification of reason for refusal | ||
| E701 | Decision to grant or registration of patent right | ||
| GRNT | Written decision to grant |