KR20200044015A - 유전자조작 미생물을 개선하기 위한 방법 및 조성물 - Google Patents
유전자조작 미생물을 개선하기 위한 방법 및 조성물 Download PDFInfo
- Publication number
- KR20200044015A KR20200044015A KR1020207006825A KR20207006825A KR20200044015A KR 20200044015 A KR20200044015 A KR 20200044015A KR 1020207006825 A KR1020207006825 A KR 1020207006825A KR 20207006825 A KR20207006825 A KR 20207006825A KR 20200044015 A KR20200044015 A KR 20200044015A
- Authority
- KR
- South Korea
- Prior art keywords
- nitrogen
- plant
- bacteria
- bacterial
- microorganisms
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Abandoned
Links
- 238000000034 method Methods 0.000 title claims abstract description 157
- 239000000203 mixture Substances 0.000 title claims abstract description 132
- 244000005700 microbiome Species 0.000 title description 249
- IJGRMHOSHXDMSA-UHFFFAOYSA-N Atomic nitrogen Chemical compound N#N IJGRMHOSHXDMSA-UHFFFAOYSA-N 0.000 claims abstract description 505
- 229910052757 nitrogen Inorganic materials 0.000 claims abstract description 252
- 108090000623 proteins and genes Proteins 0.000 claims abstract description 178
- 230000001580 bacterial effect Effects 0.000 claims abstract description 140
- 230000001965 increasing effect Effects 0.000 claims abstract description 60
- 230000008635 plant growth Effects 0.000 claims abstract description 27
- 230000001737 promoting effect Effects 0.000 claims abstract description 18
- 101100498062 Escherichia coli (strain K12) cysK gene Proteins 0.000 claims abstract description 8
- 101150111924 cysZ gene Proteins 0.000 claims abstract description 8
- 101150116936 fhaB gene Proteins 0.000 claims abstract description 8
- 101150009166 pehA gene Proteins 0.000 claims abstract description 8
- 101150019926 glgA gene Proteins 0.000 claims abstract description 7
- 101150065899 glgA1 gene Proteins 0.000 claims abstract description 7
- 101150037310 glgM gene Proteins 0.000 claims abstract description 7
- 101150095512 otsB gene Proteins 0.000 claims abstract description 7
- 101150056091 yjbE gene Proteins 0.000 claims abstract description 7
- 101150117042 treZ gene Proteins 0.000 claims abstract description 6
- 241000196324 Embryophyta Species 0.000 claims description 374
- 241000894006 Bacteria Species 0.000 claims description 235
- 230000014509 gene expression Effects 0.000 claims description 70
- 241000894007 species Species 0.000 claims description 43
- 238000004519 manufacturing process Methods 0.000 claims description 41
- 240000008042 Zea mays Species 0.000 claims description 40
- 102000004190 Enzymes Human genes 0.000 claims description 39
- 108090000790 Enzymes Proteins 0.000 claims description 39
- 235000002017 Zea mays subsp mays Nutrition 0.000 claims description 39
- 235000005824 Zea mays ssp. parviglumis Nutrition 0.000 claims description 36
- 235000005822 corn Nutrition 0.000 claims description 36
- 230000037361 pathway Effects 0.000 claims description 34
- 238000011282 treatment Methods 0.000 claims description 31
- 238000012239 gene modification Methods 0.000 claims description 21
- 230000005017 genetic modification Effects 0.000 claims description 21
- 235000013617 genetically modified food Nutrition 0.000 claims description 21
- 235000015097 nutrients Nutrition 0.000 claims description 19
- 239000000969 carrier Substances 0.000 claims description 15
- 108010020943 Nitrogenase Proteins 0.000 claims description 14
- 240000007594 Oryza sativa Species 0.000 claims description 13
- 235000007164 Oryza sativa Nutrition 0.000 claims description 13
- 235000021307 Triticum Nutrition 0.000 claims description 13
- 235000010469 Glycine max Nutrition 0.000 claims description 12
- 235000009566 rice Nutrition 0.000 claims description 12
- 244000068988 Glycine max Species 0.000 claims description 11
- NINIDFKCEFEMDL-UHFFFAOYSA-N Sulfur Chemical group [S] NINIDFKCEFEMDL-UHFFFAOYSA-N 0.000 claims description 11
- 230000002829 reductive effect Effects 0.000 claims description 11
- 229910052717 sulfur Inorganic materials 0.000 claims description 11
- 239000011593 sulfur Substances 0.000 claims description 11
- 240000005979 Hordeum vulgare Species 0.000 claims description 9
- 235000007340 Hordeum vulgare Nutrition 0.000 claims description 9
- 235000011684 Sorghum saccharatum Nutrition 0.000 claims description 9
- 150000004676 glycans Chemical class 0.000 claims description 8
- 229920001282 polysaccharide Polymers 0.000 claims description 8
- 239000005017 polysaccharide Substances 0.000 claims description 8
- HDTRYLNUVZCQOY-UHFFFAOYSA-N α-D-glucopyranosyl-α-D-glucopyranoside Natural products OC1C(O)C(O)C(CO)OC1OC1C(O)C(O)C(O)C(CO)O1 HDTRYLNUVZCQOY-UHFFFAOYSA-N 0.000 claims description 7
- HDTRYLNUVZCQOY-WSWWMNSNSA-N Trehalose Natural products O[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@H](O)[C@@H](O)[C@@H](O)[C@@H](CO)O1 HDTRYLNUVZCQOY-WSWWMNSNSA-N 0.000 claims description 7
- HDTRYLNUVZCQOY-LIZSDCNHSA-N alpha,alpha-trehalose Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 HDTRYLNUVZCQOY-LIZSDCNHSA-N 0.000 claims description 7
- 108010059820 Polygalacturonase Proteins 0.000 claims description 4
- 230000004048 modification Effects 0.000 claims description 4
- 238000012986 modification Methods 0.000 claims description 4
- 101710154643 Filamentous hemagglutinin Proteins 0.000 claims description 3
- 244000098338 Triticum aestivum Species 0.000 claims description 2
- 240000006394 Sorghum bicolor Species 0.000 claims 1
- 230000003100 immobilizing effect Effects 0.000 abstract 1
- QGZKDVFQNNGYKY-UHFFFAOYSA-N Ammonia Chemical compound N QGZKDVFQNNGYKY-UHFFFAOYSA-N 0.000 description 87
- 230000000813 microbial effect Effects 0.000 description 80
- 210000001519 tissue Anatomy 0.000 description 80
- 241000193830 Bacillus <bacterium> Species 0.000 description 68
- 230000035772 mutation Effects 0.000 description 67
- 230000000694 effects Effects 0.000 description 65
- 239000002689 soil Substances 0.000 description 62
- 108020004414 DNA Proteins 0.000 description 52
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 51
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Natural products OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 description 49
- 210000004027 cell Anatomy 0.000 description 47
- 230000001105 regulatory effect Effects 0.000 description 47
- 102000004169 proteins and genes Human genes 0.000 description 45
- 229910021529 ammonia Inorganic materials 0.000 description 42
- 230000001976 improved effect Effects 0.000 description 39
- 235000018102 proteins Nutrition 0.000 description 38
- 239000003337 fertilizer Substances 0.000 description 37
- 229940088598 enzyme Drugs 0.000 description 35
- 230000007614 genetic variation Effects 0.000 description 35
- 239000013612 plasmid Substances 0.000 description 35
- 239000000126 substance Substances 0.000 description 35
- 108091033409 CRISPR Proteins 0.000 description 32
- 125000003729 nucleotide group Chemical group 0.000 description 32
- 238000004458 analytical method Methods 0.000 description 31
- 230000012010 growth Effects 0.000 description 30
- 238000002703 mutagenesis Methods 0.000 description 30
- 231100000350 mutagenesis Toxicity 0.000 description 30
- 239000013615 primer Substances 0.000 description 30
- QGZKDVFQNNGYKY-UHFFFAOYSA-O Ammonium Chemical compound [NH4+] QGZKDVFQNNGYKY-UHFFFAOYSA-O 0.000 description 29
- -1 NifA Proteins 0.000 description 29
- 239000002773 nucleotide Substances 0.000 description 29
- 108091033319 polynucleotide Proteins 0.000 description 28
- 102000040430 polynucleotide Human genes 0.000 description 28
- 239000002157 polynucleotide Substances 0.000 description 28
- 239000002609 medium Substances 0.000 description 27
- 238000003753 real-time PCR Methods 0.000 description 27
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 27
- 102000005396 glutamine synthetase Human genes 0.000 description 26
- 108020002326 glutamine synthetase Proteins 0.000 description 26
- 238000003752 polymerase chain reaction Methods 0.000 description 25
- 238000013518 transcription Methods 0.000 description 25
- 230000035897 transcription Effects 0.000 description 25
- 230000008569 process Effects 0.000 description 24
- 150000001875 compounds Chemical class 0.000 description 23
- 239000007788 liquid Substances 0.000 description 23
- 230000029142 excretion Effects 0.000 description 22
- 239000000463 material Substances 0.000 description 22
- 239000000047 product Substances 0.000 description 22
- 239000000523 sample Substances 0.000 description 22
- 230000027455 binding Effects 0.000 description 21
- 150000007523 nucleic acids Chemical class 0.000 description 21
- 239000002028 Biomass Substances 0.000 description 20
- XSQUKJJJFZCRTK-UHFFFAOYSA-N Urea Chemical compound NC(N)=O XSQUKJJJFZCRTK-UHFFFAOYSA-N 0.000 description 20
- 241000179039 Paenibacillus Species 0.000 description 19
- 230000002068 genetic effect Effects 0.000 description 19
- 230000003993 interaction Effects 0.000 description 19
- 230000015572 biosynthetic process Effects 0.000 description 18
- 238000012217 deletion Methods 0.000 description 18
- 230000037430 deletion Effects 0.000 description 18
- 235000013399 edible fruits Nutrition 0.000 description 18
- 239000000417 fungicide Substances 0.000 description 18
- 230000006872 improvement Effects 0.000 description 18
- 230000003321 amplification Effects 0.000 description 17
- 230000007613 environmental effect Effects 0.000 description 17
- 238000003199 nucleic acid amplification method Methods 0.000 description 17
- PEDCQBHIVMGVHV-UHFFFAOYSA-N Glycerine Chemical compound OCC(O)CO PEDCQBHIVMGVHV-UHFFFAOYSA-N 0.000 description 16
- XEEYBQQBJWHFJM-UHFFFAOYSA-N Iron Chemical compound [Fe] XEEYBQQBJWHFJM-UHFFFAOYSA-N 0.000 description 16
- 101100007970 Mus musculus Ctdspl gene Proteins 0.000 description 16
- 235000013339 cereals Nutrition 0.000 description 16
- 230000035882 stress Effects 0.000 description 16
- 241000233866 Fungi Species 0.000 description 15
- 101710163270 Nuclease Proteins 0.000 description 15
- QVGXLLKOCUKJST-UHFFFAOYSA-N atomic oxygen Chemical compound [O] QVGXLLKOCUKJST-UHFFFAOYSA-N 0.000 description 15
- 238000002955 isolation Methods 0.000 description 15
- 239000001301 oxygen Substances 0.000 description 15
- 229910052760 oxygen Inorganic materials 0.000 description 15
- 238000012216 screening Methods 0.000 description 15
- 238000009395 breeding Methods 0.000 description 14
- 230000001488 breeding effect Effects 0.000 description 14
- 108091008053 gene clusters Proteins 0.000 description 14
- 230000005764 inhibitory process Effects 0.000 description 14
- 230000003834 intracellular effect Effects 0.000 description 14
- 239000002207 metabolite Substances 0.000 description 14
- 101150001904 amtB gene Proteins 0.000 description 13
- 238000003556 assay Methods 0.000 description 13
- 230000003115 biocidal effect Effects 0.000 description 13
- 230000033228 biological regulation Effects 0.000 description 13
- 239000003795 chemical substances by application Substances 0.000 description 13
- 238000009472 formulation Methods 0.000 description 13
- 210000004215 spore Anatomy 0.000 description 13
- 101100323184 Aquifex aeolicus (strain VF5) amt gene Proteins 0.000 description 12
- 101100162669 Bacillus subtilis (strain 168) nrgA gene Proteins 0.000 description 12
- 241000209140 Triticum Species 0.000 description 12
- 239000000853 adhesive Substances 0.000 description 12
- 230000001070 adhesive effect Effects 0.000 description 12
- 238000006243 chemical reaction Methods 0.000 description 12
- 230000001939 inductive effect Effects 0.000 description 12
- 238000003780 insertion Methods 0.000 description 12
- 230000037431 insertion Effects 0.000 description 12
- 101150004139 nifL gene Proteins 0.000 description 12
- 102000039446 nucleic acids Human genes 0.000 description 12
- 108020004707 nucleic acids Proteins 0.000 description 12
- 229920000642 polymer Polymers 0.000 description 12
- 238000010354 CRISPR gene editing Methods 0.000 description 11
- 101100448732 Rhizobium etli (strain CFN 42 / ATCC 51251) glnB gene Proteins 0.000 description 11
- 238000013459 approach Methods 0.000 description 11
- 230000006870 function Effects 0.000 description 11
- 101150106336 glnK gene Proteins 0.000 description 11
- 238000000338 in vitro Methods 0.000 description 11
- 238000005259 measurement Methods 0.000 description 11
- 108010015857 nitrogenase reductase Proteins 0.000 description 11
- 230000009467 reduction Effects 0.000 description 11
- 238000012163 sequencing technique Methods 0.000 description 11
- 239000013598 vector Substances 0.000 description 11
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 10
- 101100229207 Dictyostelium discoideum glnA3 gene Proteins 0.000 description 10
- 108020005004 Guide RNA Proteins 0.000 description 10
- 230000010757 Reduction Activity Effects 0.000 description 10
- VYPSYNLAJGMNEJ-UHFFFAOYSA-N Silicium dioxide Chemical compound O=[Si]=O VYPSYNLAJGMNEJ-UHFFFAOYSA-N 0.000 description 10
- 244000062793 Sorghum vulgare Species 0.000 description 10
- 239000004202 carbamide Substances 0.000 description 10
- 239000012634 fragment Substances 0.000 description 10
- 101150019449 glnB gene Proteins 0.000 description 10
- 108010064177 glutamine synthetase I Proteins 0.000 description 10
- 230000006801 homologous recombination Effects 0.000 description 10
- 238000002744 homologous recombination Methods 0.000 description 10
- 108020004999 messenger RNA Proteins 0.000 description 10
- 101150035327 nifA gene Proteins 0.000 description 10
- 101150108916 nifA1 gene Proteins 0.000 description 10
- 239000000575 pesticide Substances 0.000 description 10
- 230000000243 photosynthetic effect Effects 0.000 description 10
- 239000000843 powder Substances 0.000 description 10
- 230000019491 signal transduction Effects 0.000 description 10
- 108020004465 16S ribosomal RNA Proteins 0.000 description 9
- 108010039224 Amidophosphoribosyltransferase Proteins 0.000 description 9
- 108091026890 Coding region Proteins 0.000 description 9
- 241000588914 Enterobacter Species 0.000 description 9
- 108091028043 Nucleic acid sequence Proteins 0.000 description 9
- 150000001413 amino acids Chemical class 0.000 description 9
- 230000021615 conjugation Effects 0.000 description 9
- 230000000670 limiting effect Effects 0.000 description 9
- 230000004060 metabolic process Effects 0.000 description 9
- 239000003921 oil Substances 0.000 description 9
- 230000004044 response Effects 0.000 description 9
- 239000004094 surface-active agent Substances 0.000 description 9
- 241000203069 Archaea Species 0.000 description 8
- 101100134102 Escherichia coli (strain K12) glnL gene Proteins 0.000 description 8
- 206010064571 Gene mutation Diseases 0.000 description 8
- 229910002651 NO3 Inorganic materials 0.000 description 8
- NHNBFGGVMKEFGY-UHFFFAOYSA-N Nitrate Chemical compound [O-][N+]([O-])=O NHNBFGGVMKEFGY-UHFFFAOYSA-N 0.000 description 8
- 240000000111 Saccharum officinarum Species 0.000 description 8
- 238000010521 absorption reaction Methods 0.000 description 8
- HSFWRNGVRCDJHI-UHFFFAOYSA-N alpha-acetylene Natural products C#C HSFWRNGVRCDJHI-UHFFFAOYSA-N 0.000 description 8
- 229940024606 amino acid Drugs 0.000 description 8
- 235000001014 amino acid Nutrition 0.000 description 8
- 238000003776 cleavage reaction Methods 0.000 description 8
- 238000000576 coating method Methods 0.000 description 8
- 125000005677 ethinylene group Chemical group [*:2]C#C[*:1] 0.000 description 8
- 230000000855 fungicidal effect Effects 0.000 description 8
- 230000004077 genetic alteration Effects 0.000 description 8
- 101150035089 glnR gene Proteins 0.000 description 8
- 230000007017 scission Effects 0.000 description 8
- 235000002639 sodium chloride Nutrition 0.000 description 8
- 108020004705 Codon Proteins 0.000 description 7
- 241000588748 Klebsiella Species 0.000 description 7
- WHUUTDBJXJRKMK-VKHMYHEASA-N L-glutamic acid Chemical compound OC(=O)[C@@H](N)CCC(O)=O WHUUTDBJXJRKMK-VKHMYHEASA-N 0.000 description 7
- 101150062830 NIFK gene Proteins 0.000 description 7
- OAICVXFJPJFONN-UHFFFAOYSA-N Phosphorus Chemical compound [P] OAICVXFJPJFONN-UHFFFAOYSA-N 0.000 description 7
- 241000589516 Pseudomonas Species 0.000 description 7
- 235000007201 Saccharum officinarum Nutrition 0.000 description 7
- 230000009286 beneficial effect Effects 0.000 description 7
- 239000002274 desiccant Substances 0.000 description 7
- 238000001514 detection method Methods 0.000 description 7
- 235000013305 food Nutrition 0.000 description 7
- 239000001963 growth medium Substances 0.000 description 7
- 238000009396 hybridization Methods 0.000 description 7
- 229910052742 iron Inorganic materials 0.000 description 7
- 230000001404 mediated effect Effects 0.000 description 7
- 235000019198 oils Nutrition 0.000 description 7
- 229910052698 phosphorus Inorganic materials 0.000 description 7
- 239000011574 phosphorus Substances 0.000 description 7
- 108090000765 processed proteins & peptides Proteins 0.000 description 7
- 230000035945 sensitivity Effects 0.000 description 7
- 230000004083 survival effect Effects 0.000 description 7
- 235000014698 Brassica juncea var multisecta Nutrition 0.000 description 6
- 235000004977 Brassica sinapistrum Nutrition 0.000 description 6
- 241001465754 Metazoa Species 0.000 description 6
- 241000244206 Nematoda Species 0.000 description 6
- 244000082988 Secale cereale Species 0.000 description 6
- 235000007238 Secale cereale Nutrition 0.000 description 6
- 108010017070 Zinc Finger Nucleases Proteins 0.000 description 6
- 239000003242 anti bacterial agent Substances 0.000 description 6
- 239000002585 base Substances 0.000 description 6
- 244000275904 brauner Senf Species 0.000 description 6
- 229930002875 chlorophyll Natural products 0.000 description 6
- 235000019804 chlorophyll Nutrition 0.000 description 6
- ATNHDLDRLWWWCB-AENOIHSZSA-M chlorophyll a Chemical compound C1([C@@H](C(=O)OC)C(=O)C2=C3C)=C2N2C3=CC(C(CC)=C3C)=[N+]4C3=CC3=C(C=C)C(C)=C5N3[Mg-2]42[N+]2=C1[C@@H](CCC(=O)OC\C=C(/C)CCC[C@H](C)CCC[C@H](C)CCCC(C)C)[C@H](C)C2=C5 ATNHDLDRLWWWCB-AENOIHSZSA-M 0.000 description 6
- 239000004927 clay Substances 0.000 description 6
- 239000011248 coating agent Substances 0.000 description 6
- 230000003247 decreasing effect Effects 0.000 description 6
- 238000005516 engineering process Methods 0.000 description 6
- 230000002538 fungal effect Effects 0.000 description 6
- 235000003869 genetically modified organism Nutrition 0.000 description 6
- 101150062928 glnE gene Proteins 0.000 description 6
- 101150051351 glnL gene Proteins 0.000 description 6
- 229930195712 glutamate Natural products 0.000 description 6
- SEOVTRFCIGRIMH-UHFFFAOYSA-N indole-3-acetic acid Chemical compound C1=CC=C2C(CC(=O)O)=CNC2=C1 SEOVTRFCIGRIMH-UHFFFAOYSA-N 0.000 description 6
- 238000011081 inoculation Methods 0.000 description 6
- 235000021374 legumes Nutrition 0.000 description 6
- 239000003550 marker Substances 0.000 description 6
- 101150101270 nifD gene Proteins 0.000 description 6
- 101150112302 nifH gene Proteins 0.000 description 6
- 101150046736 ntrB gene Proteins 0.000 description 6
- 239000003415 peat Substances 0.000 description 6
- 230000006798 recombination Effects 0.000 description 6
- 238000005215 recombination Methods 0.000 description 6
- 238000007634 remodeling Methods 0.000 description 6
- 150000003839 salts Chemical class 0.000 description 6
- 238000002741 site-directed mutagenesis Methods 0.000 description 6
- 239000000243 solution Substances 0.000 description 6
- 230000002103 transcriptional effect Effects 0.000 description 6
- 230000009466 transformation Effects 0.000 description 6
- 239000001993 wax Substances 0.000 description 6
- 229920001817 Agar Polymers 0.000 description 5
- 108091093088 Amplicon Proteins 0.000 description 5
- 241000193755 Bacillus cereus Species 0.000 description 5
- 241000193417 Brevibacillus laterosporus Species 0.000 description 5
- 241000193403 Clostridium Species 0.000 description 5
- 241001057636 Dracaena deremensis Species 0.000 description 5
- 244000299507 Gossypium hirsutum Species 0.000 description 5
- 101710130691 HTH-type transcriptional regulator TnrA Proteins 0.000 description 5
- 241000033245 Kosakonia Species 0.000 description 5
- 235000007688 Lycopersicon esculentum Nutrition 0.000 description 5
- 101100327510 Methanosarcina acetivorans (strain ATCC 35395 / DSM 2834 / JCM 12185 / C2A) cfbD gene Proteins 0.000 description 5
- 235000018290 Musa x paradisiaca Nutrition 0.000 description 5
- 241000193411 Paenibacillus amylolyticus Species 0.000 description 5
- 241001478280 Rahnella Species 0.000 description 5
- 101100278502 Rhodospirillum rubrum draT gene Proteins 0.000 description 5
- 240000003768 Solanum lycopersicum Species 0.000 description 5
- 235000002595 Solanum tuberosum Nutrition 0.000 description 5
- 244000061456 Solanum tuberosum Species 0.000 description 5
- 241000607479 Yersinia pestis Species 0.000 description 5
- 239000008272 agar Substances 0.000 description 5
- 239000012298 atmosphere Substances 0.000 description 5
- 230000000903 blocking effect Effects 0.000 description 5
- 230000008859 change Effects 0.000 description 5
- 230000000295 complement effect Effects 0.000 description 5
- 229920001577 copolymer Polymers 0.000 description 5
- 238000011161 development Methods 0.000 description 5
- 230000018109 developmental process Effects 0.000 description 5
- 201000010099 disease Diseases 0.000 description 5
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 5
- 238000002474 experimental method Methods 0.000 description 5
- 231100000118 genetic alteration Toxicity 0.000 description 5
- 230000035784 germination Effects 0.000 description 5
- 101150065098 glnG gene Proteins 0.000 description 5
- 235000011187 glycerol Nutrition 0.000 description 5
- 229910052739 hydrogen Inorganic materials 0.000 description 5
- 239000002054 inoculum Substances 0.000 description 5
- 101150012415 ntrC gene Proteins 0.000 description 5
- 244000052769 pathogen Species 0.000 description 5
- 229920001184 polypeptide Polymers 0.000 description 5
- 230000001323 posttranslational effect Effects 0.000 description 5
- 102000004196 processed proteins & peptides Human genes 0.000 description 5
- 239000003223 protective agent Substances 0.000 description 5
- 239000004576 sand Substances 0.000 description 5
- 239000007787 solid Substances 0.000 description 5
- PAWQVTBBRAZDMG-UHFFFAOYSA-N 2-(3-bromo-2-fluorophenyl)acetic acid Chemical compound OC(=O)CC1=CC=CC(Br)=C1F PAWQVTBBRAZDMG-UHFFFAOYSA-N 0.000 description 4
- 244000291564 Allium cepa Species 0.000 description 4
- 101100204370 Bacillus subtilis (strain 168) sufU gene Proteins 0.000 description 4
- 235000006008 Brassica napus var napus Nutrition 0.000 description 4
- 240000007124 Brassica oleracea Species 0.000 description 4
- 235000006618 Brassica rapa subsp oleifera Nutrition 0.000 description 4
- 241001453380 Burkholderia Species 0.000 description 4
- 229920000742 Cotton Polymers 0.000 description 4
- 241000195493 Cryptophyta Species 0.000 description 4
- 230000007018 DNA scission Effects 0.000 description 4
- 241000134884 Ericales Species 0.000 description 4
- 241000588722 Escherichia Species 0.000 description 4
- PLUBXMRUUVWRLT-UHFFFAOYSA-N Ethyl methanesulfonate Chemical compound CCOS(C)(=O)=O PLUBXMRUUVWRLT-UHFFFAOYSA-N 0.000 description 4
- 241000219427 Fagales Species 0.000 description 4
- UFHFLCQGNIYNRP-UHFFFAOYSA-N Hydrogen Chemical compound [H][H] UFHFLCQGNIYNRP-UHFFFAOYSA-N 0.000 description 4
- 241001477369 Kosakonia sacchari Species 0.000 description 4
- 241000186660 Lactobacillus Species 0.000 description 4
- 240000004658 Medicago sativa Species 0.000 description 4
- 240000008790 Musa x paradisiaca Species 0.000 description 4
- 101100294147 Nostoc sp. (strain PCC 7120 / SAG 25.82 / UTEX 2576) nifV1 gene Proteins 0.000 description 4
- 108091034117 Oligonucleotide Proteins 0.000 description 4
- ZLMJMSJWJFRBEC-UHFFFAOYSA-N Potassium Chemical compound [K] ZLMJMSJWJFRBEC-UHFFFAOYSA-N 0.000 description 4
- 241000191025 Rhodobacter Species 0.000 description 4
- 229920002472 Starch Polymers 0.000 description 4
- 241000187747 Streptomyces Species 0.000 description 4
- 108700019146 Transgenes Proteins 0.000 description 4
- 101100187081 Trichormus variabilis (strain ATCC 29413 / PCC 7937) nifS1 gene Proteins 0.000 description 4
- HCHKCACWOHOZIP-UHFFFAOYSA-N Zinc Chemical compound [Zn] HCHKCACWOHOZIP-UHFFFAOYSA-N 0.000 description 4
- 239000002253 acid Substances 0.000 description 4
- 230000006154 adenylylation Effects 0.000 description 4
- 230000008901 benefit Effects 0.000 description 4
- 230000015556 catabolic process Effects 0.000 description 4
- 244000038559 crop plants Species 0.000 description 4
- 230000007423 decrease Effects 0.000 description 4
- 238000006731 degradation reaction Methods 0.000 description 4
- 230000001419 dependent effect Effects 0.000 description 4
- 230000003828 downregulation Effects 0.000 description 4
- 238000004520 electroporation Methods 0.000 description 4
- 244000053095 fungal pathogen Species 0.000 description 4
- 238000010353 genetic engineering Methods 0.000 description 4
- 101150013801 glnD gene Proteins 0.000 description 4
- 239000004009 herbicide Substances 0.000 description 4
- 239000001257 hydrogen Substances 0.000 description 4
- 230000002779 inactivation Effects 0.000 description 4
- 230000002401 inhibitory effect Effects 0.000 description 4
- 101150021879 iscS gene Proteins 0.000 description 4
- 101150027367 iscU gene Proteins 0.000 description 4
- 229940039696 lactobacillus Drugs 0.000 description 4
- UMFJAHHVKNCGLG-UHFFFAOYSA-N n-Nitrosodimethylamine Chemical compound CN(C)N=O UMFJAHHVKNCGLG-UHFFFAOYSA-N 0.000 description 4
- 101150095858 nifB gene Proteins 0.000 description 4
- 101150046169 nifE gene Proteins 0.000 description 4
- 101150009446 nifN gene Proteins 0.000 description 4
- 101150082753 nifS gene Proteins 0.000 description 4
- 101150087952 nifU gene Proteins 0.000 description 4
- 101150033171 nifV gene Proteins 0.000 description 4
- 101150093734 nifW gene Proteins 0.000 description 4
- 101150043856 nifZ gene Proteins 0.000 description 4
- 210000001672 ovary Anatomy 0.000 description 4
- 230000002018 overexpression Effects 0.000 description 4
- 239000002245 particle Substances 0.000 description 4
- 230000026731 phosphorylation Effects 0.000 description 4
- 238000006366 phosphorylation reaction Methods 0.000 description 4
- 229920000036 polyvinylpyrrolidone Polymers 0.000 description 4
- 239000001267 polyvinylpyrrolidone Substances 0.000 description 4
- 235000013855 polyvinylpyrrolidone Nutrition 0.000 description 4
- 239000011591 potassium Substances 0.000 description 4
- 229910052700 potassium Inorganic materials 0.000 description 4
- FGIUAXJPYTZDNR-UHFFFAOYSA-N potassium nitrate Chemical compound [K+].[O-][N+]([O-])=O FGIUAXJPYTZDNR-UHFFFAOYSA-N 0.000 description 4
- 239000002987 primer (paints) Substances 0.000 description 4
- 238000012545 processing Methods 0.000 description 4
- 238000000746 purification Methods 0.000 description 4
- 230000008707 rearrangement Effects 0.000 description 4
- 230000001850 reproductive effect Effects 0.000 description 4
- 238000011160 research Methods 0.000 description 4
- 108091008146 restriction endonucleases Proteins 0.000 description 4
- 101150090202 rpoB gene Proteins 0.000 description 4
- 238000005070 sampling Methods 0.000 description 4
- 239000004055 small Interfering RNA Substances 0.000 description 4
- 238000005507 spraying Methods 0.000 description 4
- 239000008107 starch Substances 0.000 description 4
- 235000019698 starch Nutrition 0.000 description 4
- 235000000346 sugar Nutrition 0.000 description 4
- 239000000725 suspension Substances 0.000 description 4
- 238000012360 testing method Methods 0.000 description 4
- 235000015112 vegetable and seed oil Nutrition 0.000 description 4
- FHVDTGUDJYJELY-UHFFFAOYSA-N 6-{[2-carboxy-4,5-dihydroxy-6-(phosphanyloxy)oxan-3-yl]oxy}-4,5-dihydroxy-3-phosphanyloxane-2-carboxylic acid Chemical compound O1C(C(O)=O)C(P)C(O)C(O)C1OC1C(C(O)=O)OC(OP)C(O)C1O FHVDTGUDJYJELY-UHFFFAOYSA-N 0.000 description 3
- 241000726119 Acidovorax Species 0.000 description 3
- 241000756998 Alismatales Species 0.000 description 3
- 235000002732 Allium cepa var. cepa Nutrition 0.000 description 3
- 235000007319 Avena orientalis Nutrition 0.000 description 3
- 244000075850 Avena orientalis Species 0.000 description 3
- 241000589941 Azospirillum Species 0.000 description 3
- 241000589149 Azotobacter vinelandii Species 0.000 description 3
- 241000194103 Bacillus pumilus Species 0.000 description 3
- 244000063299 Bacillus subtilis Species 0.000 description 3
- 235000014469 Bacillus subtilis Nutrition 0.000 description 3
- WVDDGKGOMKODPV-UHFFFAOYSA-N Benzyl alcohol Chemical compound OCC1=CC=CC=C1 WVDDGKGOMKODPV-UHFFFAOYSA-N 0.000 description 3
- 240000002791 Brassica napus Species 0.000 description 3
- 235000003899 Brassica oleracea var acephala Nutrition 0.000 description 3
- 241000589513 Burkholderia cepacia Species 0.000 description 3
- 108091079001 CRISPR RNA Proteins 0.000 description 3
- 244000025254 Cannabis sativa Species 0.000 description 3
- 229920002134 Carboxymethyl cellulose Polymers 0.000 description 3
- 108010078791 Carrier Proteins Proteins 0.000 description 3
- 229920001661 Chitosan Polymers 0.000 description 3
- 241000193468 Clostridium perfringens Species 0.000 description 3
- 235000009854 Cucurbita moschata Nutrition 0.000 description 3
- 240000001980 Cucurbita pepo Species 0.000 description 3
- 235000009852 Cucurbita pepo Nutrition 0.000 description 3
- 241000203813 Curtobacterium Species 0.000 description 3
- 102000053602 DNA Human genes 0.000 description 3
- 108010076525 DNA Repair Enzymes Proteins 0.000 description 3
- 102000011724 DNA Repair Enzymes Human genes 0.000 description 3
- 238000007400 DNA extraction Methods 0.000 description 3
- 108010014303 DNA-directed DNA polymerase Proteins 0.000 description 3
- 102000016928 DNA-directed DNA polymerase Human genes 0.000 description 3
- LYCAIKOWRPUZTN-UHFFFAOYSA-N Ethylene glycol Chemical compound OCCO LYCAIKOWRPUZTN-UHFFFAOYSA-N 0.000 description 3
- 240000009088 Fragaria x ananassa Species 0.000 description 3
- 241000896533 Gliocladium Species 0.000 description 3
- 102000009127 Glutaminase Human genes 0.000 description 3
- 108010073324 Glutaminase Proteins 0.000 description 3
- 244000020551 Helianthus annuus Species 0.000 description 3
- 235000003222 Helianthus annuus Nutrition 0.000 description 3
- 241000605016 Herbaspirillum Species 0.000 description 3
- 241000238631 Hexapoda Species 0.000 description 3
- 239000005909 Kieselgur Substances 0.000 description 3
- 235000003228 Lactuca sativa Nutrition 0.000 description 3
- 240000008415 Lactuca sativa Species 0.000 description 3
- 241000253389 Methylobacteriaceae Species 0.000 description 3
- 241001467578 Microbacterium Species 0.000 description 3
- NWBJYWHLCVSVIJ-UHFFFAOYSA-N N-benzyladenine Chemical compound N=1C=NC=2NC=NC=2C=1NCC1=CC=CC=C1 NWBJYWHLCVSVIJ-UHFFFAOYSA-N 0.000 description 3
- VZUNGTLZRAYYDE-UHFFFAOYSA-N N-methyl-N'-nitro-N-nitrosoguanidine Chemical compound O=NN(C)C(=N)N[N+]([O-])=O VZUNGTLZRAYYDE-UHFFFAOYSA-N 0.000 description 3
- 244000061176 Nicotiana tabacum Species 0.000 description 3
- 235000002637 Nicotiana tabacum Nutrition 0.000 description 3
- 108700026244 Open Reading Frames Proteins 0.000 description 3
- 241000178961 Paenibacillus alvei Species 0.000 description 3
- 241000611801 Paenibacillus illinoisensis Species 0.000 description 3
- 241000178960 Paenibacillus macerans Species 0.000 description 3
- 241000194105 Paenibacillus polymyxa Species 0.000 description 3
- 241000592795 Paenibacillus sp. Species 0.000 description 3
- 241000520272 Pantoea Species 0.000 description 3
- 241001668579 Pasteuria Species 0.000 description 3
- 241001668578 Pasteuria penetrans Species 0.000 description 3
- 239000002202 Polyethylene glycol Substances 0.000 description 3
- 239000004372 Polyvinyl alcohol Substances 0.000 description 3
- DNIAPMSPPWPWGF-UHFFFAOYSA-N Propylene glycol Chemical compound CC(O)CO DNIAPMSPPWPWGF-UHFFFAOYSA-N 0.000 description 3
- 108010026552 Proteome Proteins 0.000 description 3
- 241000220221 Rosales Species 0.000 description 3
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 3
- 241000533281 Stagonospora Species 0.000 description 3
- 108091081024 Start codon Proteins 0.000 description 3
- 241000122971 Stenotrophomonas Species 0.000 description 3
- 108700005078 Synthetic Genes Proteins 0.000 description 3
- ZMANZCXQSJIPKH-UHFFFAOYSA-N Triethylamine Chemical compound CCN(CC)CC ZMANZCXQSJIPKH-UHFFFAOYSA-N 0.000 description 3
- 229920001807 Urea-formaldehyde Polymers 0.000 description 3
- 240000006365 Vitis vinifera Species 0.000 description 3
- 235000014787 Vitis vinifera Nutrition 0.000 description 3
- 235000007244 Zea mays Nutrition 0.000 description 3
- DPKHZNPWBDQZCN-UHFFFAOYSA-N acridine orange free base Chemical compound C1=CC(N(C)C)=CC2=NC3=CC(N(C)C)=CC=C3C=C21 DPKHZNPWBDQZCN-UHFFFAOYSA-N 0.000 description 3
- 230000003044 adaptive effect Effects 0.000 description 3
- 229940072056 alginate Drugs 0.000 description 3
- 235000010443 alginic acid Nutrition 0.000 description 3
- 229920000615 alginic acid Polymers 0.000 description 3
- 229940088710 antibiotic agent Drugs 0.000 description 3
- PYMYPHUHKUWMLA-UHFFFAOYSA-N arabinose Natural products OCC(O)C(O)C(O)C=O PYMYPHUHKUWMLA-UHFFFAOYSA-N 0.000 description 3
- 244000052616 bacterial pathogen Species 0.000 description 3
- 239000000440 bentonite Substances 0.000 description 3
- 229910000278 bentonite Inorganic materials 0.000 description 3
- SVPXDRXYRYOSEX-UHFFFAOYSA-N bentoquatam Chemical compound O.O=[Si]=O.O=[Al]O[Al]=O SVPXDRXYRYOSEX-UHFFFAOYSA-N 0.000 description 3
- DZBUGLKDJFMEHC-UHFFFAOYSA-N benzoquinolinylidene Natural products C1=CC=CC2=CC3=CC=CC=C3N=C21 DZBUGLKDJFMEHC-UHFFFAOYSA-N 0.000 description 3
- SRBFZHDQGSBBOR-UHFFFAOYSA-N beta-D-Pyranose-Lyxose Natural products OC1COC(O)C(O)C1O SRBFZHDQGSBBOR-UHFFFAOYSA-N 0.000 description 3
- 150000001720 carbohydrates Chemical class 0.000 description 3
- 235000014633 carbohydrates Nutrition 0.000 description 3
- KRKNYBCHXYNGOX-UHFFFAOYSA-N citric acid Chemical compound OC(=O)CC(O)(C(O)=O)CC(O)=O KRKNYBCHXYNGOX-UHFFFAOYSA-N 0.000 description 3
- 229910052570 clay Inorganic materials 0.000 description 3
- 239000008199 coating composition Substances 0.000 description 3
- 230000000875 corresponding effect Effects 0.000 description 3
- 230000006378 damage Effects 0.000 description 3
- MNNHAPBLZZVQHP-UHFFFAOYSA-N diammonium hydrogen phosphate Chemical compound [NH4+].[NH4+].OP([O-])([O-])=O MNNHAPBLZZVQHP-UHFFFAOYSA-N 0.000 description 3
- 229910001873 dinitrogen Inorganic materials 0.000 description 3
- 239000002270 dispersing agent Substances 0.000 description 3
- 238000009826 distribution Methods 0.000 description 3
- 238000003379 elimination reaction Methods 0.000 description 3
- 239000000839 emulsion Substances 0.000 description 3
- 238000000855 fermentation Methods 0.000 description 3
- 230000004151 fermentation Effects 0.000 description 3
- 239000000835 fiber Substances 0.000 description 3
- 235000013312 flour Nutrition 0.000 description 3
- 239000007789 gas Substances 0.000 description 3
- 238000010362 genome editing Methods 0.000 description 3
- IXORZMNAPKEEDV-UHFFFAOYSA-N gibberellic acid GA3 Natural products OC(=O)C1C2(C3)CC(=C)C3(O)CCC2C2(C=CC3O)C1C3(C)C(=O)O2 IXORZMNAPKEEDV-UHFFFAOYSA-N 0.000 description 3
- 239000003630 growth substance Substances 0.000 description 3
- 210000004209 hair Anatomy 0.000 description 3
- 150000002500 ions Chemical class 0.000 description 3
- 238000002307 isotope ratio mass spectrometry Methods 0.000 description 3
- 238000007726 management method Methods 0.000 description 3
- 238000013507 mapping Methods 0.000 description 3
- 239000012528 membrane Substances 0.000 description 3
- 230000002503 metabolic effect Effects 0.000 description 3
- 230000037353 metabolic pathway Effects 0.000 description 3
- MBABOKRGFJTBAE-UHFFFAOYSA-N methyl methanesulfonate Chemical compound COS(C)(=O)=O MBABOKRGFJTBAE-UHFFFAOYSA-N 0.000 description 3
- 239000013586 microbial product Substances 0.000 description 3
- 239000011733 molybdenum Substances 0.000 description 3
- 229910052750 molybdenum Inorganic materials 0.000 description 3
- 239000003471 mutagenic agent Substances 0.000 description 3
- 231100000707 mutagenic chemical Toxicity 0.000 description 3
- 230000003505 mutagenic effect Effects 0.000 description 3
- 125000000740 n-pentyl group Chemical group [H]C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])* 0.000 description 3
- 101150106615 nifF gene Proteins 0.000 description 3
- 101150117431 nifJ gene Proteins 0.000 description 3
- 101150077076 nifQ gene Proteins 0.000 description 3
- 101150038920 nifX gene Proteins 0.000 description 3
- 101150098909 nifY gene Proteins 0.000 description 3
- 235000016709 nutrition Nutrition 0.000 description 3
- 230000008520 organization Effects 0.000 description 3
- 230000001717 pathogenic effect Effects 0.000 description 3
- 239000008188 pellet Substances 0.000 description 3
- 230000029553 photosynthesis Effects 0.000 description 3
- 238000010672 photosynthesis Methods 0.000 description 3
- 229920001223 polyethylene glycol Polymers 0.000 description 3
- 229920002689 polyvinyl acetate Polymers 0.000 description 3
- 239000011118 polyvinyl acetate Substances 0.000 description 3
- 229920002451 polyvinyl alcohol Polymers 0.000 description 3
- 108700022487 rRNA Genes Proteins 0.000 description 3
- 238000011084 recovery Methods 0.000 description 3
- 230000010076 replication Effects 0.000 description 3
- 230000002441 reversible effect Effects 0.000 description 3
- JQXXHWHPUNPDRT-WLSIYKJHSA-N rifampicin Chemical compound O([C@](C1=O)(C)O/C=C/[C@@H]([C@H]([C@@H](OC(C)=O)[C@H](C)[C@H](O)[C@H](C)[C@@H](O)[C@@H](C)\C=C\C=C(C)/C(=O)NC=2C(O)=C3C([O-])=C4C)C)OC)C4=C1C3=C(O)C=2\C=N\N1CC[NH+](C)CC1 JQXXHWHPUNPDRT-WLSIYKJHSA-N 0.000 description 3
- 239000013049 sediment Substances 0.000 description 3
- 230000007226 seed germination Effects 0.000 description 3
- 238000000926 separation method Methods 0.000 description 3
- 239000002002 slurry Substances 0.000 description 3
- 239000011734 sodium Substances 0.000 description 3
- 239000003381 stabilizer Substances 0.000 description 3
- 239000006228 supernatant Substances 0.000 description 3
- 238000003786 synthesis reaction Methods 0.000 description 3
- 239000000454 talc Substances 0.000 description 3
- 229910052623 talc Inorganic materials 0.000 description 3
- 230000008685 targeting Effects 0.000 description 3
- 238000013519 translation Methods 0.000 description 3
- 230000032258 transport Effects 0.000 description 3
- 238000011144 upstream manufacturing Methods 0.000 description 3
- 239000010455 vermiculite Substances 0.000 description 3
- 229910052902 vermiculite Inorganic materials 0.000 description 3
- 235000019354 vermiculite Nutrition 0.000 description 3
- JLIDBLDQVAYHNE-YKALOCIXSA-N (+)-Abscisic acid Chemical compound OC(=O)/C=C(/C)\C=C\[C@@]1(O)C(C)=CC(=O)CC1(C)C JLIDBLDQVAYHNE-YKALOCIXSA-N 0.000 description 2
- LNAZSHAWQACDHT-XIYTZBAFSA-N (2r,3r,4s,5r,6s)-4,5-dimethoxy-2-(methoxymethyl)-3-[(2s,3r,4s,5r,6r)-3,4,5-trimethoxy-6-(methoxymethyl)oxan-2-yl]oxy-6-[(2r,3r,4s,5r,6r)-4,5,6-trimethoxy-2-(methoxymethyl)oxan-3-yl]oxyoxane Chemical compound CO[C@@H]1[C@@H](OC)[C@H](OC)[C@@H](COC)O[C@H]1O[C@H]1[C@H](OC)[C@@H](OC)[C@H](O[C@H]2[C@@H]([C@@H](OC)[C@H](OC)O[C@@H]2COC)OC)O[C@@H]1COC LNAZSHAWQACDHT-XIYTZBAFSA-N 0.000 description 2
- IIZPXYDJLKNOIY-JXPKJXOSSA-N 1-palmitoyl-2-arachidonoyl-sn-glycero-3-phosphocholine Chemical compound CCCCCCCCCCCCCCCC(=O)OC[C@H](COP([O-])(=O)OCC[N+](C)(C)C)OC(=O)CCC\C=C/C\C=C/C\C=C/C\C=C/CCCCC IIZPXYDJLKNOIY-JXPKJXOSSA-N 0.000 description 2
- 108020004463 18S ribosomal RNA Proteins 0.000 description 2
- UHPMCKVQTMMPCG-UHFFFAOYSA-N 5,8-dihydroxy-2-methoxy-6-methyl-7-(2-oxopropyl)naphthalene-1,4-dione Chemical compound CC1=C(CC(C)=O)C(O)=C2C(=O)C(OC)=CC(=O)C2=C1O UHPMCKVQTMMPCG-UHFFFAOYSA-N 0.000 description 2
- ARSRBNBHOADGJU-UHFFFAOYSA-N 7,12-dimethyltetraphene Chemical compound C1=CC2=CC=CC=C2C2=C1C(C)=C(C=CC=C1)C1=C2C ARSRBNBHOADGJU-UHFFFAOYSA-N 0.000 description 2
- PWJFNRJRHXWEPT-UHFFFAOYSA-N ADP ribose Natural products C1=NC=2C(N)=NC=NC=2N1C1OC(COP(O)(=O)OP(O)(=O)OCC(O)C(O)C(O)C=O)C(O)C1O PWJFNRJRHXWEPT-UHFFFAOYSA-N 0.000 description 2
- 230000005730 ADP ribosylation Effects 0.000 description 2
- SRNWOUGRCWSEMX-KEOHHSTQSA-N ADP-beta-D-ribose Chemical compound C([C@H]1O[C@H]([C@@H]([C@@H]1O)O)N1C=2N=CN=C(C=2N=C1)N)OP(O)(=O)OP(O)(=O)OC[C@H]1O[C@@H](O)[C@H](O)[C@@H]1O SRNWOUGRCWSEMX-KEOHHSTQSA-N 0.000 description 2
- 101150067778 ARF18 gene Proteins 0.000 description 2
- 240000004507 Abelmoschus esculentus Species 0.000 description 2
- NIXOWILDQLNWCW-UHFFFAOYSA-M Acrylate Chemical compound [O-]C(=O)C=C NIXOWILDQLNWCW-UHFFFAOYSA-M 0.000 description 2
- 241000251468 Actinopterygii Species 0.000 description 2
- 241000589158 Agrobacterium Species 0.000 description 2
- 239000005995 Aluminium silicate Substances 0.000 description 2
- NLXLAEXVIDQMFP-UHFFFAOYSA-N Ammonia chloride Chemical compound [NH4+].[Cl-] NLXLAEXVIDQMFP-UHFFFAOYSA-N 0.000 description 2
- 244000099147 Ananas comosus Species 0.000 description 2
- 235000007119 Ananas comosus Nutrition 0.000 description 2
- 241000219194 Arabidopsis Species 0.000 description 2
- 108010070255 Aspartate-ammonia ligase Proteins 0.000 description 2
- 241000208837 Asterales Species 0.000 description 2
- PYIXHKGTJKCVBJ-UHFFFAOYSA-N Astraciceran Natural products C1OC2=CC(O)=CC=C2CC1C1=CC(OCO2)=C2C=C1OC PYIXHKGTJKCVBJ-UHFFFAOYSA-N 0.000 description 2
- 229930192334 Auxin Natural products 0.000 description 2
- 241000589152 Azotobacter chroococcum Species 0.000 description 2
- 241000194107 Bacillus megaterium Species 0.000 description 2
- 108020000946 Bacterial DNA Proteins 0.000 description 2
- 208000034309 Bacterial disease carrier Diseases 0.000 description 2
- NDVRQFZUJRMKKP-UHFFFAOYSA-N Betavulgarin Natural products O=C1C=2C(OC)=C3OCOC3=CC=2OC=C1C1=CC=CC=C1O NDVRQFZUJRMKKP-UHFFFAOYSA-N 0.000 description 2
- 241000589173 Bradyrhizobium Species 0.000 description 2
- 241000589174 Bradyrhizobium japonicum Species 0.000 description 2
- 235000011299 Brassica oleracea var botrytis Nutrition 0.000 description 2
- 235000011301 Brassica oleracea var capitata Nutrition 0.000 description 2
- 235000001169 Brassica oleracea var oleracea Nutrition 0.000 description 2
- 240000003259 Brassica oleracea var. botrytis Species 0.000 description 2
- 241000555281 Brevibacillus Species 0.000 description 2
- 241000498637 Brevibacillus agri Species 0.000 description 2
- 241000193764 Brevibacillus brevis Species 0.000 description 2
- 241000253402 Burkholderiaceae Species 0.000 description 2
- VTYYLEPIZMXCLO-UHFFFAOYSA-L Calcium carbonate Chemical compound [Ca+2].[O-]C([O-])=O VTYYLEPIZMXCLO-UHFFFAOYSA-L 0.000 description 2
- 241001164374 Calyx Species 0.000 description 2
- 235000002566 Capsicum Nutrition 0.000 description 2
- 235000003255 Carthamus tinctorius Nutrition 0.000 description 2
- 244000020518 Carthamus tinctorius Species 0.000 description 2
- 241000219504 Caryophyllales Species 0.000 description 2
- 240000001829 Catharanthus roseus Species 0.000 description 2
- VEXZGXHMUGYJMC-UHFFFAOYSA-M Chloride anion Chemical compound [Cl-] VEXZGXHMUGYJMC-UHFFFAOYSA-M 0.000 description 2
- 241000611330 Chryseobacterium Species 0.000 description 2
- 244000241235 Citrullus lanatus Species 0.000 description 2
- 241001672694 Citrus reticulata Species 0.000 description 2
- 240000000560 Citrus x paradisi Species 0.000 description 2
- 241000763622 Cladosporiaceae Species 0.000 description 2
- 241000193401 Clostridium acetobutylicum Species 0.000 description 2
- 241000193454 Clostridium beijerinckii Species 0.000 description 2
- 241000193469 Clostridium pasteurianum Species 0.000 description 2
- 241000193449 Clostridium tetani Species 0.000 description 2
- 108700010070 Codon Usage Proteins 0.000 description 2
- 241001600130 Comamonadaceae Species 0.000 description 2
- 241000186216 Corynebacterium Species 0.000 description 2
- 241000219112 Cucumis Species 0.000 description 2
- 235000015510 Cucumis melo subsp melo Nutrition 0.000 description 2
- 240000008067 Cucumis sativus Species 0.000 description 2
- 241000192700 Cyanobacteria Species 0.000 description 2
- CMSMOCZEIVJLDB-UHFFFAOYSA-N Cyclophosphamide Chemical compound ClCCN(CCCl)P1(=O)NCCCO1 CMSMOCZEIVJLDB-UHFFFAOYSA-N 0.000 description 2
- SRBFZHDQGSBBOR-IOVATXLUSA-N D-xylopyranose Chemical compound O[C@@H]1COC(O)[C@H](O)[C@H]1O SRBFZHDQGSBBOR-IOVATXLUSA-N 0.000 description 2
- 230000004568 DNA-binding Effects 0.000 description 2
- 241001600129 Delftia Species 0.000 description 2
- 108010008532 Deoxyribonuclease I Proteins 0.000 description 2
- 102000007260 Deoxyribonuclease I Human genes 0.000 description 2
- 239000005696 Diammonium phosphate Substances 0.000 description 2
- 235000009355 Dianthus caryophyllus Nutrition 0.000 description 2
- 240000006497 Dianthus caryophyllus Species 0.000 description 2
- RUPBZQFQVRMKDG-UHFFFAOYSA-M Didecyldimethylammonium chloride Chemical compound [Cl-].CCCCCCCCCC[N+](C)(C)CCCCCCCCCC RUPBZQFQVRMKDG-UHFFFAOYSA-M 0.000 description 2
- ZFIVKAOQEXOYFY-UHFFFAOYSA-N Diepoxybutane Chemical compound C1OC1C1OC1 ZFIVKAOQEXOYFY-UHFFFAOYSA-N 0.000 description 2
- 235000001950 Elaeis guineensis Nutrition 0.000 description 2
- 108010042407 Endonucleases Proteins 0.000 description 2
- 102000004533 Endonucleases Human genes 0.000 description 2
- 241000588921 Enterobacteriaceae Species 0.000 description 2
- 241000588724 Escherichia coli Species 0.000 description 2
- VGGSQFUCUMXWEO-UHFFFAOYSA-N Ethene Chemical compound C=C VGGSQFUCUMXWEO-UHFFFAOYSA-N 0.000 description 2
- 239000005977 Ethylene Substances 0.000 description 2
- IAYPIBMASNFSPL-UHFFFAOYSA-N Ethylene oxide Chemical compound C1CO1 IAYPIBMASNFSPL-UHFFFAOYSA-N 0.000 description 2
- 244000166124 Eucalyptus globulus Species 0.000 description 2
- 240000002989 Euphorbia neriifolia Species 0.000 description 2
- 240000002395 Euphorbia pulcherrima Species 0.000 description 2
- 240000008620 Fagopyrum esculentum Species 0.000 description 2
- 235000009419 Fagopyrum esculentum Nutrition 0.000 description 2
- 241000192125 Firmicutes Species 0.000 description 2
- 235000016623 Fragaria vesca Nutrition 0.000 description 2
- 235000011363 Fragaria x ananassa Nutrition 0.000 description 2
- 241000223218 Fusarium Species 0.000 description 2
- 241000427940 Fusarium solani Species 0.000 description 2
- 229930191978 Gibberellin Natural products 0.000 description 2
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 2
- 241001626381 Gnomoniaceae Species 0.000 description 2
- 241000206596 Halomonas Species 0.000 description 2
- 241001014264 Klebsiella variicola Species 0.000 description 2
- 229930195714 L-glutamate Natural products 0.000 description 2
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 2
- 241000207832 Lamiales Species 0.000 description 2
- 241000143673 Lasiosphaeriaceae Species 0.000 description 2
- 241000234269 Liliales Species 0.000 description 2
- 240000006240 Linum usitatissimum Species 0.000 description 2
- 235000004431 Linum usitatissimum Nutrition 0.000 description 2
- 240000003183 Manihot esculenta Species 0.000 description 2
- 235000010624 Medicago sativa Nutrition 0.000 description 2
- 235000017587 Medicago sativa ssp. sativa Nutrition 0.000 description 2
- 235000006679 Mentha X verticillata Nutrition 0.000 description 2
- 235000002899 Mentha suaveolens Nutrition 0.000 description 2
- 235000001636 Mentha x rotundifolia Nutrition 0.000 description 2
- 239000005807 Metalaxyl Substances 0.000 description 2
- 101100015014 Methanothermobacter marburgensis (strain ATCC BAA-927 / DSM 2133 / JCM 14651 / NBRC 100331 / OCM 82 / Marburg) glnBA gene Proteins 0.000 description 2
- 101100175820 Methanothermobacter marburgensis (strain ATCC BAA-927 / DSM 2133 / JCM 14651 / NBRC 100331 / OCM 82 / Marburg) glnBB gene Proteins 0.000 description 2
- 241000589323 Methylobacterium Species 0.000 description 2
- 241000736262 Microbiota Species 0.000 description 2
- 241001112070 Microvirga Species 0.000 description 2
- 240000003433 Miscanthus floridulus Species 0.000 description 2
- ZOKXTWBITQBERF-UHFFFAOYSA-N Molybdenum Chemical compound [Mo] ZOKXTWBITQBERF-UHFFFAOYSA-N 0.000 description 2
- 108010038629 Molybdoferredoxin Proteins 0.000 description 2
- 241000529935 Monacrosporium Species 0.000 description 2
- 241000186359 Mycobacterium Species 0.000 description 2
- 241000187488 Mycobacterium sp. Species 0.000 description 2
- IOVCWXUNBOPUCH-UHFFFAOYSA-M Nitrite anion Chemical compound [O-]N=O IOVCWXUNBOPUCH-UHFFFAOYSA-M 0.000 description 2
- GQPLMRYTRLFLPF-UHFFFAOYSA-N Nitrous Oxide Chemical compound [O-][N+]#N GQPLMRYTRLFLPF-UHFFFAOYSA-N 0.000 description 2
- 238000012408 PCR amplification Methods 0.000 description 2
- 229910019142 PO4 Inorganic materials 0.000 description 2
- 241000178959 Paenibacillus durus Species 0.000 description 2
- 241000194109 Paenibacillus lautus Species 0.000 description 2
- 241000689135 Paenibacillus macquariensis Species 0.000 description 2
- 241000193397 Paenibacillus pabuli Species 0.000 description 2
- 241001310339 Paenibacillus popilliae Species 0.000 description 2
- 241000123637 Pandanales Species 0.000 description 2
- 240000001090 Papaver somniferum Species 0.000 description 2
- 235000008753 Papaver somniferum Nutrition 0.000 description 2
- 241000588701 Pectobacterium carotovorum Species 0.000 description 2
- 239000006002 Pepper Substances 0.000 description 2
- 240000007377 Petunia x hybrida Species 0.000 description 2
- 241000233805 Phoenix Species 0.000 description 2
- XYFCBTPGUUZFHI-UHFFFAOYSA-N Phosphine Chemical compound P XYFCBTPGUUZFHI-UHFFFAOYSA-N 0.000 description 2
- IHPVFYLOGNNZLA-UHFFFAOYSA-N Phytoalexin Natural products COC1=CC=CC=C1C1OC(C=C2C(OCO2)=C2OC)=C2C(=O)C1 IHPVFYLOGNNZLA-UHFFFAOYSA-N 0.000 description 2
- 235000016761 Piper aduncum Nutrition 0.000 description 2
- 240000003889 Piper guineense Species 0.000 description 2
- 235000017804 Piper guineense Nutrition 0.000 description 2
- 235000008184 Piper nigrum Nutrition 0.000 description 2
- 241000532838 Platypus Species 0.000 description 2
- 241001136502 Pleosporaceae Species 0.000 description 2
- 241000209504 Poaceae Species 0.000 description 2
- 241001536628 Poales Species 0.000 description 2
- 241000219000 Populus Species 0.000 description 2
- WCUXLLCKKVVCTQ-UHFFFAOYSA-M Potassium chloride Chemical compound [Cl-].[K+] WCUXLLCKKVVCTQ-UHFFFAOYSA-M 0.000 description 2
- 241000192142 Proteobacteria Species 0.000 description 2
- 241001378513 Pseudoclavibacter Species 0.000 description 2
- 241000589540 Pseudomonas fluorescens Species 0.000 description 2
- 241000589615 Pseudomonas syringae Species 0.000 description 2
- 241000187603 Pseudonocardia Species 0.000 description 2
- 238000003559 RNA-seq method Methods 0.000 description 2
- 238000011529 RT qPCR Methods 0.000 description 2
- 241000232299 Ralstonia Species 0.000 description 2
- 244000061121 Rauvolfia serpentina Species 0.000 description 2
- 241001633102 Rhizobiaceae Species 0.000 description 2
- 241000589157 Rhizobiales Species 0.000 description 2
- 241000589180 Rhizobium Species 0.000 description 2
- 241000316848 Rhodococcus <scale insect> Species 0.000 description 2
- 241000209051 Saccharum Species 0.000 description 2
- 101000888131 Schizosaccharomyces pombe (strain 972 / ATCC 24843) Glutamate synthase [NADH] Proteins 0.000 description 2
- 241000607720 Serratia Species 0.000 description 2
- 235000005775 Setaria Nutrition 0.000 description 2
- 241000232088 Setaria <nematode> Species 0.000 description 2
- 241000607768 Shigella Species 0.000 description 2
- 241001647968 Shinella Species 0.000 description 2
- 241001135312 Sinorhizobium Species 0.000 description 2
- 108091027967 Small hairpin RNA Proteins 0.000 description 2
- 108020004459 Small interfering RNA Proteins 0.000 description 2
- DBMJMQXJHONAFJ-UHFFFAOYSA-M Sodium laurylsulphate Chemical compound [Na+].CCCCCCCCCCCCOS([O-])(=O)=O DBMJMQXJHONAFJ-UHFFFAOYSA-M 0.000 description 2
- 241000131972 Sphingomonadaceae Species 0.000 description 2
- 241000736131 Sphingomonas Species 0.000 description 2
- 241000383873 Sphingopyxis Species 0.000 description 2
- 235000009337 Spinacia oleracea Nutrition 0.000 description 2
- 244000300264 Spinacia oleracea Species 0.000 description 2
- 241000187389 Streptomyces lavendulae Species 0.000 description 2
- 229930006000 Sucrose Natural products 0.000 description 2
- CZMRCDWAGMRECN-UGDNZRGBSA-N Sucrose Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 CZMRCDWAGMRECN-UGDNZRGBSA-N 0.000 description 2
- QAOWNCQODCNURD-UHFFFAOYSA-L Sulfate Chemical compound [O-]S([O-])(=O)=O QAOWNCQODCNURD-UHFFFAOYSA-L 0.000 description 2
- 108010006785 Taq Polymerase Proteins 0.000 description 2
- 244000269722 Thea sinensis Species 0.000 description 2
- 244000299461 Theobroma cacao Species 0.000 description 2
- 235000009470 Theobroma cacao Nutrition 0.000 description 2
- 108091023040 Transcription factor Proteins 0.000 description 2
- 102000040945 Transcription factor Human genes 0.000 description 2
- 108020004566 Transfer RNA Proteins 0.000 description 2
- 241000489523 Veratrum Species 0.000 description 2
- XTXRWKRVRITETP-UHFFFAOYSA-N Vinyl acetate Chemical compound CC(=O)OC=C XTXRWKRVRITETP-UHFFFAOYSA-N 0.000 description 2
- 241000700605 Viruses Species 0.000 description 2
- 235000009754 Vitis X bourquina Nutrition 0.000 description 2
- 235000012333 Vitis X labruscana Nutrition 0.000 description 2
- 241001453327 Xanthomonadaceae Species 0.000 description 2
- 241000209149 Zea Species 0.000 description 2
- 235000007241 Zea diploperennis Nutrition 0.000 description 2
- 244000083398 Zea diploperennis Species 0.000 description 2
- 241000482268 Zea mays subsp. mays Species 0.000 description 2
- MCMNRKCIXSYSNV-UHFFFAOYSA-N Zirconium dioxide Chemical compound O=[Zr]=O MCMNRKCIXSYSNV-UHFFFAOYSA-N 0.000 description 2
- FJJCIZWZNKZHII-UHFFFAOYSA-N [4,6-bis(cyanoamino)-1,3,5-triazin-2-yl]cyanamide Chemical compound N#CNC1=NC(NC#N)=NC(NC#N)=N1 FJJCIZWZNKZHII-UHFFFAOYSA-N 0.000 description 2
- 230000036579 abiotic stress Effects 0.000 description 2
- PEJLNXHANOHNSU-UHFFFAOYSA-N acridine-3,6-diamine;10-methylacridin-10-ium-3,6-diamine;chloride Chemical compound [Cl-].C1=CC(N)=CC2=NC3=CC(N)=CC=C3C=C21.C1=C(N)C=C2[N+](C)=C(C=C(N)C=C3)C3=CC2=C1 PEJLNXHANOHNSU-UHFFFAOYSA-N 0.000 description 2
- 239000013543 active substance Substances 0.000 description 2
- 239000000654 additive Substances 0.000 description 2
- 239000002671 adjuvant Substances 0.000 description 2
- UCTWMZQNUQWSLP-UHFFFAOYSA-N adrenaline Chemical compound CNCC(O)C1=CC=C(O)C(O)=C1 UCTWMZQNUQWSLP-UHFFFAOYSA-N 0.000 description 2
- 230000009418 agronomic effect Effects 0.000 description 2
- 230000004075 alteration Effects 0.000 description 2
- 235000012211 aluminium silicate Nutrition 0.000 description 2
- LFVGISIMTYGQHF-UHFFFAOYSA-N ammonium dihydrogen phosphate Chemical compound [NH4+].OP(O)([O-])=O LFVGISIMTYGQHF-UHFFFAOYSA-N 0.000 description 2
- 229910000387 ammonium dihydrogen phosphate Inorganic materials 0.000 description 2
- BFNBIHQBYMNNAN-UHFFFAOYSA-N ammonium sulfate Chemical compound N.N.OS(O)(=O)=O BFNBIHQBYMNNAN-UHFFFAOYSA-N 0.000 description 2
- 229910052921 ammonium sulfate Inorganic materials 0.000 description 2
- 235000011130 ammonium sulphate Nutrition 0.000 description 2
- 238000004873 anchoring Methods 0.000 description 2
- 239000005557 antagonist Substances 0.000 description 2
- 230000000845 anti-microbial effect Effects 0.000 description 2
- 239000007864 aqueous solution Substances 0.000 description 2
- PYMYPHUHKUWMLA-WDCZJNDASA-N arabinose Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)C=O PYMYPHUHKUWMLA-WDCZJNDASA-N 0.000 description 2
- 230000006793 arrhythmia Effects 0.000 description 2
- 206010003119 arrhythmia Diseases 0.000 description 2
- 239000002363 auxin Substances 0.000 description 2
- SKKMWRVAJNPLFY-UHFFFAOYSA-N azanylidynevanadium Chemical compound [V]#N SKKMWRVAJNPLFY-UHFFFAOYSA-N 0.000 description 2
- 239000011230 binding agent Substances 0.000 description 2
- 239000002551 biofuel Substances 0.000 description 2
- 239000012620 biological material Substances 0.000 description 2
- 229920001222 biopolymer Polymers 0.000 description 2
- GZUXJHMPEANEGY-UHFFFAOYSA-N bromomethane Chemical compound BrC GZUXJHMPEANEGY-UHFFFAOYSA-N 0.000 description 2
- 235000015115 caffè latte Nutrition 0.000 description 2
- ZCCIPPOKBCJFDN-UHFFFAOYSA-N calcium nitrate Chemical compound [Ca+2].[O-][N+]([O-])=O.[O-][N+]([O-])=O ZCCIPPOKBCJFDN-UHFFFAOYSA-N 0.000 description 2
- YYRMJZQKEFZXMX-UHFFFAOYSA-N calcium;phosphoric acid Chemical compound [Ca+2].OP(O)(O)=O.OP(O)(O)=O YYRMJZQKEFZXMX-UHFFFAOYSA-N 0.000 description 2
- 235000010948 carboxy methyl cellulose Nutrition 0.000 description 2
- 239000001768 carboxy methyl cellulose Substances 0.000 description 2
- 239000008112 carboxymethyl-cellulose Substances 0.000 description 2
- 229940105329 carboxymethylcellulose Drugs 0.000 description 2
- 238000004113 cell culture Methods 0.000 description 2
- JCKYGMPEJWAADB-UHFFFAOYSA-N chlorambucil Chemical compound OC(=O)CCCC1=CC=C(N(CCCl)CCCl)C=C1 JCKYGMPEJWAADB-UHFFFAOYSA-N 0.000 description 2
- 229960004630 chlorambucil Drugs 0.000 description 2
- 210000000349 chromosome Anatomy 0.000 description 2
- 230000002860 competitive effect Effects 0.000 description 2
- 238000007796 conventional method Methods 0.000 description 2
- 230000002596 correlated effect Effects 0.000 description 2
- 238000012258 culturing Methods 0.000 description 2
- 238000005520 cutting process Methods 0.000 description 2
- 125000004122 cyclic group Chemical group 0.000 description 2
- 229960004397 cyclophosphamide Drugs 0.000 description 2
- 239000004062 cytokinin Substances 0.000 description 2
- UQHKFADEQIVWID-UHFFFAOYSA-N cytokinin Natural products C1=NC=2C(NCC=C(CO)C)=NC=NC=2N1C1CC(O)C(CO)O1 UQHKFADEQIVWID-UHFFFAOYSA-N 0.000 description 2
- 230000004665 defense response Effects 0.000 description 2
- 230000007812 deficiency Effects 0.000 description 2
- 238000010586 diagram Methods 0.000 description 2
- 229910000388 diammonium phosphate Inorganic materials 0.000 description 2
- 235000019838 diammonium phosphate Nutrition 0.000 description 2
- DENRZWYUOJLTMF-UHFFFAOYSA-N diethyl sulfate Chemical compound CCOS(=O)(=O)OCC DENRZWYUOJLTMF-UHFFFAOYSA-N 0.000 description 2
- 229940008406 diethyl sulfate Drugs 0.000 description 2
- 230000029087 digestion Effects 0.000 description 2
- 239000003085 diluting agent Substances 0.000 description 2
- 238000010790 dilution Methods 0.000 description 2
- 239000012895 dilution Substances 0.000 description 2
- XBDQKXXYIPTUBI-UHFFFAOYSA-N dimethylselenoniopropionate Natural products CCC(O)=O XBDQKXXYIPTUBI-UHFFFAOYSA-N 0.000 description 2
- 230000024346 drought recovery Effects 0.000 description 2
- 238000001035 drying Methods 0.000 description 2
- 239000000975 dye Substances 0.000 description 2
- 239000012636 effector Substances 0.000 description 2
- 239000003623 enhancer Substances 0.000 description 2
- 238000011156 evaluation Methods 0.000 description 2
- 239000000284 extract Substances 0.000 description 2
- 238000000605 extraction Methods 0.000 description 2
- 230000002349 favourable effect Effects 0.000 description 2
- 238000000684 flow cytometry Methods 0.000 description 2
- 230000009969 flowable effect Effects 0.000 description 2
- 239000012530 fluid Substances 0.000 description 2
- 238000000799 fluorescence microscopy Methods 0.000 description 2
- 239000006260 foam Substances 0.000 description 2
- HKQYGTCOTHHOMP-UHFFFAOYSA-N formononetin Chemical compound C1=CC(OC)=CC=C1C1=COC2=CC(O)=CC=C2C1=O HKQYGTCOTHHOMP-UHFFFAOYSA-N 0.000 description 2
- 108091006104 gene-regulatory proteins Proteins 0.000 description 2
- 102000034356 gene-regulatory proteins Human genes 0.000 description 2
- 238000003205 genotyping method Methods 0.000 description 2
- 239000003448 gibberellin Substances 0.000 description 2
- 101150092620 glnBI gene Proteins 0.000 description 2
- 101150010000 glnBII gene Proteins 0.000 description 2
- RWSXRVCMGQZWBV-WDSKDSINSA-N glutathione Chemical compound OC(=O)[C@@H](N)CCC(=O)N[C@@H](CS)C(=O)NCC(O)=O RWSXRVCMGQZWBV-WDSKDSINSA-N 0.000 description 2
- 239000008187 granular material Substances 0.000 description 2
- 229920000591 gum Polymers 0.000 description 2
- 238000003306 harvesting Methods 0.000 description 2
- 230000036541 health Effects 0.000 description 2
- 238000010438 heat treatment Methods 0.000 description 2
- 101150028208 hesA gene Proteins 0.000 description 2
- GNOIPBMMFNIUFM-UHFFFAOYSA-N hexamethylphosphoric triamide Chemical compound CN(C)P(=O)(N(C)C)N(C)C GNOIPBMMFNIUFM-UHFFFAOYSA-N 0.000 description 2
- LELOWRISYMNNSU-UHFFFAOYSA-N hydrogen cyanide Chemical compound N#C LELOWRISYMNNSU-UHFFFAOYSA-N 0.000 description 2
- 238000011065 in-situ storage Methods 0.000 description 2
- 239000003617 indole-3-acetic acid Substances 0.000 description 2
- 230000004941 influx Effects 0.000 description 2
- 229910052500 inorganic mineral Inorganic materials 0.000 description 2
- 230000037041 intracellular level Effects 0.000 description 2
- NLYAJNPCOHFWQQ-UHFFFAOYSA-N kaolin Chemical compound O.O.O=[Al]O[Si](=O)O[Si](=O)O[Al]=O NLYAJNPCOHFWQQ-UHFFFAOYSA-N 0.000 description 2
- 238000002372 labelling Methods 0.000 description 2
- JVTAAEKCZFNVCJ-UHFFFAOYSA-N lactic acid Chemical compound CC(O)C(O)=O JVTAAEKCZFNVCJ-UHFFFAOYSA-N 0.000 description 2
- 235000010445 lecithin Nutrition 0.000 description 2
- 239000000787 lecithin Substances 0.000 description 2
- 229940067606 lecithin Drugs 0.000 description 2
- 230000004807 localization Effects 0.000 description 2
- 230000008774 maternal effect Effects 0.000 description 2
- 230000007246 mechanism Effects 0.000 description 2
- 229960004961 mechlorethamine Drugs 0.000 description 2
- ZQEIXNIJLIKNTD-GFCCVEGCSA-N metalaxyl-M Chemical compound COCC(=O)N([C@H](C)C(=O)OC)C1=C(C)C=CC=C1C ZQEIXNIJLIKNTD-GFCCVEGCSA-N 0.000 description 2
- CKFGINPQOCXMAZ-UHFFFAOYSA-N methanediol Chemical compound OCO CKFGINPQOCXMAZ-UHFFFAOYSA-N 0.000 description 2
- WSFSSNUMVMOOMR-NJFSPNSNSA-N methanone Chemical compound O=[14CH2] WSFSSNUMVMOOMR-NJFSPNSNSA-N 0.000 description 2
- ZQEIXNIJLIKNTD-UHFFFAOYSA-N methyl N-(2,6-dimethylphenyl)-N-(methoxyacetyl)alaninate Chemical compound COCC(=O)N(C(C)C(=O)OC)C1=C(C)C=CC=C1C ZQEIXNIJLIKNTD-UHFFFAOYSA-N 0.000 description 2
- 229920000609 methyl cellulose Polymers 0.000 description 2
- 230000011987 methylation Effects 0.000 description 2
- 238000007069 methylation reaction Methods 0.000 description 2
- 235000010981 methylcellulose Nutrition 0.000 description 2
- 239000001923 methylcellulose Substances 0.000 description 2
- 229960002900 methylcellulose Drugs 0.000 description 2
- 108091070501 miRNA Proteins 0.000 description 2
- 239000002679 microRNA Substances 0.000 description 2
- 239000011785 micronutrient Substances 0.000 description 2
- 235000013369 micronutrients Nutrition 0.000 description 2
- 235000010755 mineral Nutrition 0.000 description 2
- 239000011707 mineral Substances 0.000 description 2
- 239000007003 mineral medium Substances 0.000 description 2
- 230000000394 mitotic effect Effects 0.000 description 2
- 238000002156 mixing Methods 0.000 description 2
- 239000006012 monoammonium phosphate Substances 0.000 description 2
- 235000019837 monoammonium phosphate Nutrition 0.000 description 2
- SQMWSBKSHWARHU-SDBHATRESA-N n6-cyclopentyladenosine Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C2=NC=NC(NC3CCCC3)=C2N=C1 SQMWSBKSHWARHU-SDBHATRESA-N 0.000 description 2
- 229910017464 nitrogen compound Inorganic materials 0.000 description 2
- 150000002830 nitrogen compounds Chemical class 0.000 description 2
- 230000014075 nitrogen utilization Effects 0.000 description 2
- 235000021231 nutrient uptake Nutrition 0.000 description 2
- 230000035764 nutrition Effects 0.000 description 2
- 230000003287 optical effect Effects 0.000 description 2
- 150000007524 organic acids Chemical class 0.000 description 2
- 239000000123 paper Substances 0.000 description 2
- 239000010452 phosphate Substances 0.000 description 2
- NBIIXXVUZAFLBC-UHFFFAOYSA-K phosphate Chemical compound [O-]P([O-])([O-])=O NBIIXXVUZAFLBC-UHFFFAOYSA-K 0.000 description 2
- 238000013081 phylogenetic analysis Methods 0.000 description 2
- 239000000280 phytoalexin Substances 0.000 description 2
- 150000001857 phytoalexin derivatives Chemical class 0.000 description 2
- 239000004069 plant analysis Substances 0.000 description 2
- 239000005648 plant growth regulator Substances 0.000 description 2
- 239000004323 potassium nitrate Substances 0.000 description 2
- 235000010333 potassium nitrate Nutrition 0.000 description 2
- 238000004382 potting Methods 0.000 description 2
- 230000000644 propagated effect Effects 0.000 description 2
- 108020001580 protein domains Proteins 0.000 description 2
- 230000004850 protein–protein interaction Effects 0.000 description 2
- 238000011002 quantification Methods 0.000 description 2
- 230000005855 radiation Effects 0.000 description 2
- 230000033458 reproduction Effects 0.000 description 2
- 238000007894 restriction fragment length polymorphism technique Methods 0.000 description 2
- 239000011435 rock Substances 0.000 description 2
- 230000028327 secretion Effects 0.000 description 2
- 239000011780 sodium chloride Substances 0.000 description 2
- VWDWKYIASSYTQR-UHFFFAOYSA-N sodium nitrate Chemical compound [Na+].[O-][N+]([O-])=O VWDWKYIASSYTQR-UHFFFAOYSA-N 0.000 description 2
- 125000006850 spacer group Chemical group 0.000 description 2
- 235000020354 squash Nutrition 0.000 description 2
- UCSJYZPVAKXKNQ-HZYVHMACSA-N streptomycin Chemical compound CN[C@H]1[C@H](O)[C@@H](O)[C@H](CO)O[C@H]1O[C@@H]1[C@](C=O)(O)[C@H](C)O[C@H]1O[C@@H]1[C@@H](NC(N)=N)[C@H](O)[C@@H](NC(N)=N)[C@H](O)[C@H]1O UCSJYZPVAKXKNQ-HZYVHMACSA-N 0.000 description 2
- 239000005720 sucrose Substances 0.000 description 2
- 239000002426 superphosphate Substances 0.000 description 2
- 230000009469 supplementation Effects 0.000 description 2
- 230000031068 symbiosis, encompassing mutualism through parasitism Effects 0.000 description 2
- 208000024891 symptom Diseases 0.000 description 2
- 230000009885 systemic effect Effects 0.000 description 2
- 230000002792 vascular Effects 0.000 description 2
- 239000008158 vegetable oil Substances 0.000 description 2
- 235000013311 vegetables Nutrition 0.000 description 2
- 229940057613 veratrum Drugs 0.000 description 2
- OGWKCGZFUXNPDA-XQKSVPLYSA-N vincristine Chemical compound C([N@]1C[C@@H](C[C@]2(C(=O)OC)C=3C(=CC4=C([C@]56[C@H]([C@@]([C@H](OC(C)=O)[C@]7(CC)C=CCN([C@H]67)CC5)(O)C(=O)OC)N4C=O)C=3)OC)C[C@@](C1)(O)CC)CC1=C2NC2=CC=CC=C12 OGWKCGZFUXNPDA-XQKSVPLYSA-N 0.000 description 2
- 229960004528 vincristine Drugs 0.000 description 2
- OGWKCGZFUXNPDA-UHFFFAOYSA-N vincristine Natural products C1C(CC)(O)CC(CC2(C(=O)OC)C=3C(=CC4=C(C56C(C(C(OC(C)=O)C7(CC)C=CCN(C67)CC5)(O)C(=O)OC)N4C=O)C=3)OC)CN1CCC1=C2NC2=CC=CC=C12 OGWKCGZFUXNPDA-UHFFFAOYSA-N 0.000 description 2
- 229920002554 vinyl polymer Polymers 0.000 description 2
- 244000052613 viral pathogen Species 0.000 description 2
- 238000005406 washing Methods 0.000 description 2
- 239000000080 wetting agent Substances 0.000 description 2
- 239000002023 wood Substances 0.000 description 2
- 241000228158 x Triticosecale Species 0.000 description 2
- 239000011701 zinc Substances 0.000 description 2
- 229910052725 zinc Inorganic materials 0.000 description 2
- YNWVFADWVLCOPU-MDWZMJQESA-N (1E)-1-(4-chlorophenyl)-4,4-dimethyl-2-(1H-1,2,4-triazol-1-yl)pent-1-en-3-ol Chemical compound C1=NC=NN1/C(C(O)C(C)(C)C)=C/C1=CC=C(Cl)C=C1 YNWVFADWVLCOPU-MDWZMJQESA-N 0.000 description 1
- 239000001100 (2S)-5,7-dihydroxy-2-(3-hydroxy-4-methoxyphenyl)chroman-4-one Substances 0.000 description 1
- FYGDTMLNYKFZSV-URKRLVJHSA-N (2s,3r,4s,5s,6r)-2-[(2r,4r,5r,6s)-4,5-dihydroxy-2-(hydroxymethyl)-6-[(2r,4r,5r,6s)-4,5,6-trihydroxy-2-(hydroxymethyl)oxan-3-yl]oxyoxan-3-yl]oxy-6-(hydroxymethyl)oxane-3,4,5-triol Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@H]1OC1[C@@H](CO)O[C@@H](OC2[C@H](O[C@H](O)[C@H](O)[C@H]2O)CO)[C@H](O)[C@H]1O FYGDTMLNYKFZSV-URKRLVJHSA-N 0.000 description 1
- LDVVMCZRFWMZSG-OLQVQODUSA-N (3ar,7as)-2-(trichloromethylsulfanyl)-3a,4,7,7a-tetrahydroisoindole-1,3-dione Chemical compound C1C=CC[C@H]2C(=O)N(SC(Cl)(Cl)Cl)C(=O)[C@H]21 LDVVMCZRFWMZSG-OLQVQODUSA-N 0.000 description 1
- PPDBOQMNKNNODG-NTEUORMPSA-N (5E)-5-(4-chlorobenzylidene)-2,2-dimethyl-1-(1,2,4-triazol-1-ylmethyl)cyclopentanol Chemical compound C1=NC=NN1CC1(O)C(C)(C)CC\C1=C/C1=CC=C(Cl)C=C1 PPDBOQMNKNNODG-NTEUORMPSA-N 0.000 description 1
- BJEPYKJPYRNKOW-REOHCLBHSA-N (S)-malic acid Chemical compound OC(=O)[C@@H](O)CC(O)=O BJEPYKJPYRNKOW-REOHCLBHSA-N 0.000 description 1
- RMOGWMIKYWRTKW-UONOGXRCSA-N (S,S)-paclobutrazol Chemical compound C([C@@H]([C@@H](O)C(C)(C)C)N1N=CN=C1)C1=CC=C(Cl)C=C1 RMOGWMIKYWRTKW-UONOGXRCSA-N 0.000 description 1
- KANAPVJGZDNSCZ-UHFFFAOYSA-N 1,2-benzothiazole 1-oxide Chemical class C1=CC=C2S(=O)N=CC2=C1 KANAPVJGZDNSCZ-UHFFFAOYSA-N 0.000 description 1
- VUWCWMOCWKCZTA-UHFFFAOYSA-N 1,2-thiazol-4-one Chemical class O=C1CSN=C1 VUWCWMOCWKCZTA-UHFFFAOYSA-N 0.000 description 1
- IHGSAQHSAGRWNI-UHFFFAOYSA-N 1-(4-bromophenyl)-2,2,2-trifluoroethanone Chemical compound FC(F)(F)C(=O)C1=CC=C(Br)C=C1 IHGSAQHSAGRWNI-UHFFFAOYSA-N 0.000 description 1
- PXMNMQRDXWABCY-UHFFFAOYSA-N 1-(4-chlorophenyl)-4,4-dimethyl-3-(1H-1,2,4-triazol-1-ylmethyl)pentan-3-ol Chemical compound C1=NC=NN1CC(O)(C(C)(C)C)CCC1=CC=C(Cl)C=C1 PXMNMQRDXWABCY-UHFFFAOYSA-N 0.000 description 1
- 108091000130 1-aminocyclopropane-1-carboxylate deaminase Proteins 0.000 description 1
- UKIDUMMXBQMTKO-UHFFFAOYSA-N 1-methyl-1-nitro-2-nitrosoguanidine Chemical compound [O-][N+](=O)N(C)C(=N)NN=O UKIDUMMXBQMTKO-UHFFFAOYSA-N 0.000 description 1
- PRPINYUDVPFIRX-UHFFFAOYSA-N 1-naphthaleneacetic acid Chemical compound C1=CC=C2C(CC(=O)O)=CC=CC2=C1 PRPINYUDVPFIRX-UHFFFAOYSA-N 0.000 description 1
- ZMZGFLUUZLELNE-UHFFFAOYSA-N 2,3,5-triiodobenzoic acid Chemical compound OC(=O)C1=CC(I)=CC(I)=C1I ZMZGFLUUZLELNE-UHFFFAOYSA-N 0.000 description 1
- WCOXQTXVACYMLM-UHFFFAOYSA-N 2,3-bis(12-hydroxyoctadecanoyloxy)propyl 12-hydroxyoctadecanoate Chemical compound CCCCCCC(O)CCCCCCCCCCC(=O)OCC(OC(=O)CCCCCCCCCCC(O)CCCCCC)COC(=O)CCCCCCCCCCC(O)CCCCCC WCOXQTXVACYMLM-UHFFFAOYSA-N 0.000 description 1
- VEPOHXYIFQMVHW-XOZOLZJESA-N 2,3-dihydroxybutanedioic acid (2S,3S)-3,4-dimethyl-2-phenylmorpholine Chemical compound OC(C(O)C(O)=O)C(O)=O.C[C@H]1[C@@H](OCCN1C)c1ccccc1 VEPOHXYIFQMVHW-XOZOLZJESA-N 0.000 description 1
- PVWMAOPFDINGAY-UHFFFAOYSA-N 2-(3-methylbutanoyl)indene-1,3-dione Chemical compound C1=CC=C2C(=O)C(C(=O)CC(C)C)C(=O)C2=C1 PVWMAOPFDINGAY-UHFFFAOYSA-N 0.000 description 1
- MWBWWFOAEOYUST-UHFFFAOYSA-N 2-aminopurine Chemical compound NC1=NC=C2N=CNC2=N1 MWBWWFOAEOYUST-UHFFFAOYSA-N 0.000 description 1
- UDPGUMQDCGORJQ-UHFFFAOYSA-M 2-chloroethyl(hydroxy)phosphinate Chemical compound OP([O-])(=O)CCCl UDPGUMQDCGORJQ-UHFFFAOYSA-M 0.000 description 1
- JLIDBLDQVAYHNE-LXGGSRJLSA-N 2-cis-abscisic acid Chemical compound OC(=O)/C=C(/C)\C=C\C1(O)C(C)=CC(=O)CC1(C)C JLIDBLDQVAYHNE-LXGGSRJLSA-N 0.000 description 1
- FVTWJXMFYOXOKK-UHFFFAOYSA-N 2-fluoroacetamide Chemical compound NC(=O)CF FVTWJXMFYOXOKK-UHFFFAOYSA-N 0.000 description 1
- 239000001763 2-hydroxyethyl(trimethyl)azanium Substances 0.000 description 1
- LNCCBHFAHILMCT-UHFFFAOYSA-N 2-n,4-n,6-n-triethyl-1,3,5-triazine-2,4,6-triamine Chemical compound CCNC1=NC(NCC)=NC(NCC)=N1 LNCCBHFAHILMCT-UHFFFAOYSA-N 0.000 description 1
- 125000000094 2-phenylethyl group Chemical group [H]C1=C([H])C([H])=C(C([H])=C1[H])C([H])([H])C([H])([H])* 0.000 description 1
- HBEMYXWYRXKRQI-UHFFFAOYSA-N 3-(8-methoxyoctoxy)propyl-methyl-bis(trimethylsilyloxy)silane Chemical compound COCCCCCCCCOCCC[Si](C)(O[Si](C)(C)C)O[Si](C)(C)C HBEMYXWYRXKRQI-UHFFFAOYSA-N 0.000 description 1
- SSZWWUDQMAHNAQ-UHFFFAOYSA-N 3-chloropropane-1,2-diol Chemical compound OCC(O)CCl SSZWWUDQMAHNAQ-UHFFFAOYSA-N 0.000 description 1
- HEGWNIMGIDYRAU-UHFFFAOYSA-N 3-hexyl-2,4-dioxabicyclo[1.1.0]butane Chemical compound O1C2OC21CCCCCC HEGWNIMGIDYRAU-UHFFFAOYSA-N 0.000 description 1
- 102000034279 3-hydroxybutyrate dehydrogenases Human genes 0.000 description 1
- 108090000124 3-hydroxybutyrate dehydrogenases Proteins 0.000 description 1
- CCXYYNHQDVOMEP-UHFFFAOYSA-N 4-(quinoxalin-2-ylamino)benzenesulfonamide Chemical compound C1=CC(S(=O)(=O)N)=CC=C1NC1=CN=C(C=CC=C2)C2=N1 CCXYYNHQDVOMEP-UHFFFAOYSA-N 0.000 description 1
- SATHPVQTSSUFFW-UHFFFAOYSA-N 4-[6-[(3,5-dihydroxy-4-methoxyoxan-2-yl)oxymethyl]-3,5-dihydroxy-4-methoxyoxan-2-yl]oxy-2-(hydroxymethyl)-6-methyloxane-3,5-diol Chemical compound OC1C(OC)C(O)COC1OCC1C(O)C(OC)C(O)C(OC2C(C(CO)OC(C)C2O)O)O1 SATHPVQTSSUFFW-UHFFFAOYSA-N 0.000 description 1
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 description 1
- YHQDZJICGQWFHK-UHFFFAOYSA-N 4-nitroquinoline N-oxide Chemical compound C1=CC=C2C([N+](=O)[O-])=CC=[N+]([O-])C2=C1 YHQDZJICGQWFHK-UHFFFAOYSA-N 0.000 description 1
- 125000001572 5'-adenylyl group Chemical group C=12N=C([H])N=C(N([H])[H])C=1N=C([H])N2[C@@]1([H])[C@@](O[H])([H])[C@@](O[H])([H])[C@](C(OP(=O)(O[H])[*])([H])[H])([H])O1 0.000 description 1
- 108020004565 5.8S Ribosomal RNA Proteins 0.000 description 1
- 108020005075 5S Ribosomal RNA Proteins 0.000 description 1
- 235000003934 Abelmoschus esculentus Nutrition 0.000 description 1
- 244000215068 Acacia senegal Species 0.000 description 1
- 241000590020 Achromobacter Species 0.000 description 1
- 241000266272 Acidithiobacillus Species 0.000 description 1
- 241000589291 Acinetobacter Species 0.000 description 1
- HRPVXLWXLXDGHG-UHFFFAOYSA-N Acrylamide Chemical compound NC(=O)C=C HRPVXLWXLXDGHG-UHFFFAOYSA-N 0.000 description 1
- 241001156739 Actinobacteria <phylum> Species 0.000 description 1
- 241000187844 Actinoplanes Species 0.000 description 1
- 241000466670 Adlercreutzia Species 0.000 description 1
- 241000193798 Aerococcus Species 0.000 description 1
- 241000607534 Aeromonas Species 0.000 description 1
- 229920000936 Agarose Polymers 0.000 description 1
- 241000589155 Agrobacterium tumefaciens Species 0.000 description 1
- 241001467490 Agromyces Species 0.000 description 1
- 241000500137 Agromyces sp. Species 0.000 description 1
- 102000007698 Alcohol dehydrogenase Human genes 0.000 description 1
- 108010021809 Alcohol dehydrogenase Proteins 0.000 description 1
- 241000640374 Alicyclobacillus acidocaldarius Species 0.000 description 1
- 241000193412 Alicyclobacillus acidoterrestris Species 0.000 description 1
- 235000005255 Allium cepa Nutrition 0.000 description 1
- 241000556588 Alstroemeria Species 0.000 description 1
- 102000006941 Amino Acid Transport System X-AG Human genes 0.000 description 1
- 102100039338 Aminomethyltransferase, mitochondrial Human genes 0.000 description 1
- 108091006987 Ammonia transporters Proteins 0.000 description 1
- VHUUQVKOLVNVRT-UHFFFAOYSA-N Ammonium hydroxide Chemical compound [NH4+].[OH-] VHUUQVKOLVNVRT-UHFFFAOYSA-N 0.000 description 1
- 239000004254 Ammonium phosphate Substances 0.000 description 1
- 239000004114 Ammonium polyphosphate Substances 0.000 description 1
- 108050001492 Ammonium transporters Proteins 0.000 description 1
- 241000192542 Anabaena Species 0.000 description 1
- 241000272522 Anas Species 0.000 description 1
- 241000862972 Ancylobacter Species 0.000 description 1
- 244000118350 Andrographis paniculata Species 0.000 description 1
- 241000193741 Aneurinibacillus aneurinilyticus Species 0.000 description 1
- 241001312221 Anthurium Species 0.000 description 1
- 239000001904 Arabinogalactan Substances 0.000 description 1
- 241001014371 Archontophoenix cunninghamiana Species 0.000 description 1
- 241000123640 Arecales Species 0.000 description 1
- 235000001405 Artemisia annua Nutrition 0.000 description 1
- 240000000011 Artemisia annua Species 0.000 description 1
- 241000186063 Arthrobacter Species 0.000 description 1
- 241000186073 Arthrobacter sp. Species 0.000 description 1
- 241000215383 Arthrobotrys gephyropaga Species 0.000 description 1
- 241000722807 Arthrobotrys oligospora Species 0.000 description 1
- 102000015790 Asparaginase Human genes 0.000 description 1
- 108010024976 Asparaginase Proteins 0.000 description 1
- 244000003416 Asparagus officinalis Species 0.000 description 1
- 235000005340 Asparagus officinalis Nutrition 0.000 description 1
- 241001136151 Atopostipes Species 0.000 description 1
- 241001465356 Atropa belladonna Species 0.000 description 1
- 241000894008 Azorhizobium Species 0.000 description 1
- 241000589151 Azotobacter Species 0.000 description 1
- 241000099686 Azotobacter sp. Species 0.000 description 1
- 101100269818 Azotobacter vinelandii anfA gene Proteins 0.000 description 1
- 101100484827 Azotobacter vinelandii vnfA gene Proteins 0.000 description 1
- 239000005730 Azoxystrobin Substances 0.000 description 1
- 241000193375 Bacillus alcalophilus Species 0.000 description 1
- 241000193744 Bacillus amyloliquefaciens Species 0.000 description 1
- 241000193410 Bacillus atrophaeus Species 0.000 description 1
- 241000193409 Bacillus azotoformans Species 0.000 description 1
- 241000193408 Bacillus badius Species 0.000 description 1
- 241000193752 Bacillus circulans Species 0.000 description 1
- 241000193749 Bacillus coagulans Species 0.000 description 1
- 241000193422 Bacillus lentus Species 0.000 description 1
- 241000194108 Bacillus licheniformis Species 0.000 description 1
- 241000194106 Bacillus mycoides Species 0.000 description 1
- 241000893637 Bacillus nematocida Species 0.000 description 1
- 241000194104 Bacillus psychrosaccharolyticus Species 0.000 description 1
- 241000278457 Bacillus siamensis Species 0.000 description 1
- 241000193400 Bacillus simplex Species 0.000 description 1
- 241000193399 Bacillus smithii Species 0.000 description 1
- 101000950981 Bacillus subtilis (strain 168) Catabolic NAD-specific glutamate dehydrogenase RocG Proteins 0.000 description 1
- 101100269722 Bacillus subtilis (strain 168) alsT gene Proteins 0.000 description 1
- 101100271045 Bacillus subtilis (strain 168) ansZ gene Proteins 0.000 description 1
- 101100267082 Bacillus subtilis (strain 168) yflA gene Proteins 0.000 description 1
- 101100489082 Bacillus subtilis (strain 168) yrbD gene Proteins 0.000 description 1
- 241000193388 Bacillus thuringiensis Species 0.000 description 1
- 108091032955 Bacterial small RNA Proteins 0.000 description 1
- 235000017166 Bambusa arundinacea Nutrition 0.000 description 1
- 235000017491 Bambusa tulda Nutrition 0.000 description 1
- 241000604933 Bdellovibrio Species 0.000 description 1
- 235000021537 Beetroot Nutrition 0.000 description 1
- 241000588882 Beijerinckia Species 0.000 description 1
- FMMWHPNWAFZXNH-UHFFFAOYSA-N Benz[a]pyrene Chemical compound C1=C2C3=CC=CC=C3C=C(C=C3)C2=C2C3=CC=CC2=C1 FMMWHPNWAFZXNH-UHFFFAOYSA-N 0.000 description 1
- 240000000724 Berberis vulgaris Species 0.000 description 1
- 235000021533 Beta vulgaris Nutrition 0.000 description 1
- 241000335053 Beta vulgaris Species 0.000 description 1
- 229920002498 Beta-glucan Polymers 0.000 description 1
- JKSKMVKDSOOOFA-UHFFFAOYSA-N Bisthiosemi Chemical compound NC(=S)NNCNNC(N)=S JKSKMVKDSOOOFA-UHFFFAOYSA-N 0.000 description 1
- 235000006010 Bixa orellana Nutrition 0.000 description 1
- 244000017106 Bixa orellana Species 0.000 description 1
- 108010006654 Bleomycin Proteins 0.000 description 1
- ZOXJGFHDIHLPTG-UHFFFAOYSA-N Boron Chemical compound [B] ZOXJGFHDIHLPTG-UHFFFAOYSA-N 0.000 description 1
- 241000427199 Bosea <angiosperm> Species 0.000 description 1
- 241000743774 Brachypodium Species 0.000 description 1
- 241000589171 Bradyrhizobium sp. Species 0.000 description 1
- 235000011331 Brassica Nutrition 0.000 description 1
- 241000219198 Brassica Species 0.000 description 1
- 235000011303 Brassica alboglabra Nutrition 0.000 description 1
- 244000178993 Brassica juncea Species 0.000 description 1
- 235000011332 Brassica juncea Nutrition 0.000 description 1
- 235000014700 Brassica juncea var napiformis Nutrition 0.000 description 1
- 235000011293 Brassica napus Nutrition 0.000 description 1
- 235000011302 Brassica oleracea Nutrition 0.000 description 1
- 235000017647 Brassica oleracea var italica Nutrition 0.000 description 1
- 235000012905 Brassica oleracea var viridis Nutrition 0.000 description 1
- 241000218980 Brassicales Species 0.000 description 1
- IXVMHGVQKLDRKH-VRESXRICSA-N Brassinolide Natural products O=C1OC[C@@H]2[C@@H]3[C@@](C)([C@H]([C@@H]([C@@H](O)[C@H](O)[C@H](C(C)C)C)C)CC3)CC[C@@H]2[C@]2(C)[C@@H]1C[C@H](O)[C@H](O)C2 IXVMHGVQKLDRKH-VRESXRICSA-N 0.000 description 1
- 241000131407 Brevundimonas Species 0.000 description 1
- 239000005966 Bromadiolone Substances 0.000 description 1
- OWNRRUFOJXFKCU-UHFFFAOYSA-N Bromadiolone Chemical compound C=1C=C(C=2C=CC(Br)=CC=2)C=CC=1C(O)CC(C=1C(OC2=CC=CC=C2C=1O)=O)C1=CC=CC=C1 OWNRRUFOJXFKCU-UHFFFAOYSA-N 0.000 description 1
- OYPRJOBELJOOCE-UHFFFAOYSA-N Calcium Chemical compound [Ca] OYPRJOBELJOOCE-UHFFFAOYSA-N 0.000 description 1
- 240000001432 Calendula officinalis Species 0.000 description 1
- 235000005881 Calendula officinalis Nutrition 0.000 description 1
- 241000759909 Camptotheca Species 0.000 description 1
- 241000572575 Candidatus Pasteuria usgae Species 0.000 description 1
- 241000218236 Cannabis Species 0.000 description 1
- 240000004160 Capsicum annuum Species 0.000 description 1
- 239000005745 Captan Substances 0.000 description 1
- 108090000994 Catalytic RNA Proteins 0.000 description 1
- 102000053642 Catalytic RNA Human genes 0.000 description 1
- 241000863012 Caulobacter Species 0.000 description 1
- 241000632385 Celastrales Species 0.000 description 1
- 241000186321 Cellulomonas Species 0.000 description 1
- 241000863387 Cellvibrio Species 0.000 description 1
- DMKDRSIXSDKEFQ-KDXMTYKHSA-N Cephaline Natural products CCCCCCCCCCCCCCCCCOC(=O)C[C@@H](COP(=O)(O)CCN)C(=O)OCCCCCCCC=CCCCCCCCC DMKDRSIXSDKEFQ-KDXMTYKHSA-N 0.000 description 1
- 241000488899 Cephalotaxus Species 0.000 description 1
- 240000006162 Chenopodium quinoa Species 0.000 description 1
- 229920002101 Chitin Polymers 0.000 description 1
- 108010022172 Chitinases Proteins 0.000 description 1
- 102000012286 Chitinases Human genes 0.000 description 1
- ZAMOUSCENKQFHK-UHFFFAOYSA-N Chlorine atom Chemical compound [Cl] ZAMOUSCENKQFHK-UHFFFAOYSA-N 0.000 description 1
- 241000191382 Chlorobaculum tepidum Species 0.000 description 1
- 235000019743 Choline chloride Nutrition 0.000 description 1
- 241001332334 Chromobacterium subtsugae Species 0.000 description 1
- 235000000604 Chrysanthemum parthenium Nutrition 0.000 description 1
- 235000010523 Cicer arietinum Nutrition 0.000 description 1
- 244000045195 Cicer arietinum Species 0.000 description 1
- 235000007542 Cichorium intybus Nutrition 0.000 description 1
- 244000298479 Cichorium intybus Species 0.000 description 1
- 235000021516 Cinchona officinalis Nutrition 0.000 description 1
- 244000182633 Cinchona succirubra Species 0.000 description 1
- 241000132536 Cirsium Species 0.000 description 1
- 241000588923 Citrobacter Species 0.000 description 1
- 235000009831 Citrullus lanatus Nutrition 0.000 description 1
- 235000012828 Citrullus lanatus var citroides Nutrition 0.000 description 1
- 235000008733 Citrus aurantifolia Nutrition 0.000 description 1
- 235000005979 Citrus limon Nutrition 0.000 description 1
- 244000175448 Citrus madurensis Species 0.000 description 1
- 240000004307 Citrus medica Species 0.000 description 1
- 244000131522 Citrus pyriformis Species 0.000 description 1
- 241000333459 Citrus x tangelo Species 0.000 description 1
- 241001149472 Clonostachys rosea Species 0.000 description 1
- 241000896542 Clonostachys rosea f. catenulata Species 0.000 description 1
- 241000193464 Clostridium sp. Species 0.000 description 1
- 235000013162 Cocos nucifera Nutrition 0.000 description 1
- 244000060011 Cocos nucifera Species 0.000 description 1
- 235000005320 Coleus barbatus Nutrition 0.000 description 1
- 241000233971 Commelinales Species 0.000 description 1
- 108020004394 Complementary RNA Proteins 0.000 description 1
- VPAXJOUATWLOPR-UHFFFAOYSA-N Conferone Chemical compound C1=CC(=O)OC2=CC(OCC3C4(C)CCC(=O)C(C)(C)C4CC=C3C)=CC=C21 VPAXJOUATWLOPR-UHFFFAOYSA-N 0.000 description 1
- RYGMFSIKBFXOCR-UHFFFAOYSA-N Copper Chemical compound [Cu] RYGMFSIKBFXOCR-UHFFFAOYSA-N 0.000 description 1
- 241001266001 Cordyceps confragosa Species 0.000 description 1
- 241000134970 Cornales Species 0.000 description 1
- 241000186249 Corynebacterium sp. Species 0.000 description 1
- 235000005956 Cosmos caudatus Nutrition 0.000 description 1
- 244000293323 Cosmos caudatus Species 0.000 description 1
- JFIXKFSJCQNGEK-UHFFFAOYSA-N Coumafuryl Chemical group OC=1C2=CC=CC=C2OC(=O)C=1C(CC(=O)C)C1=CC=CO1 JFIXKFSJCQNGEK-UHFFFAOYSA-N 0.000 description 1
- 244000241257 Cucumis melo Species 0.000 description 1
- 235000009842 Cucumis melo Nutrition 0.000 description 1
- 235000009849 Cucumis sativus Nutrition 0.000 description 1
- 235000010799 Cucumis sativus var sativus Nutrition 0.000 description 1
- 240000004244 Cucurbita moschata Species 0.000 description 1
- 241001528480 Cupriavidus Species 0.000 description 1
- 241000770189 Curvibacter Species 0.000 description 1
- 244000007835 Cyamopsis tetragonoloba Species 0.000 description 1
- 241000196114 Cycadales Species 0.000 description 1
- 241000723247 Cylindrocarpon Species 0.000 description 1
- 206010011732 Cyst Diseases 0.000 description 1
- 102000018832 Cytochromes Human genes 0.000 description 1
- 108010052832 Cytochromes Proteins 0.000 description 1
- 102100028717 Cytosolic 5'-nucleotidase 3A Human genes 0.000 description 1
- FBPFZTCFMRRESA-FSIIMWSLSA-N D-Glucitol Natural products OC[C@H](O)[C@H](O)[C@@H](O)[C@H](O)CO FBPFZTCFMRRESA-FSIIMWSLSA-N 0.000 description 1
- FBPFZTCFMRRESA-KVTDHHQDSA-N D-Mannitol Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-KVTDHHQDSA-N 0.000 description 1
- FBPFZTCFMRRESA-JGWLITMVSA-N D-glucitol Chemical compound OC[C@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-JGWLITMVSA-N 0.000 description 1
- 108020001019 DNA Primers Proteins 0.000 description 1
- 108020003215 DNA Probes Proteins 0.000 description 1
- 101710099946 DNA mismatch repair protein Msh6 Proteins 0.000 description 1
- 102100021147 DNA mismatch repair protein Msh6 Human genes 0.000 description 1
- 239000003155 DNA primer Substances 0.000 description 1
- 239000003298 DNA probe Substances 0.000 description 1
- 238000001712 DNA sequencing Methods 0.000 description 1
- 241000858015 Dactylellina drechsleri Species 0.000 description 1
- 239000005975 Daminozide Substances 0.000 description 1
- NOQGZXFMHARMLW-UHFFFAOYSA-N Daminozide Chemical compound CN(C)NC(=O)CCC(O)=O NOQGZXFMHARMLW-UHFFFAOYSA-N 0.000 description 1
- 241000208296 Datura Species 0.000 description 1
- 241000192093 Deinococcus Species 0.000 description 1
- 241001600125 Delftia acidovorans Species 0.000 description 1
- 241000205646 Devosia Species 0.000 description 1
- 229920001353 Dextrin Polymers 0.000 description 1
- 239000004375 Dextrin Substances 0.000 description 1
- FEWJPZIEWOKRBE-JCYAYHJZSA-N Dextrotartaric acid Chemical compound OC(=O)[C@H](O)[C@@H](O)C(O)=O FEWJPZIEWOKRBE-JCYAYHJZSA-N 0.000 description 1
- MDNWOSOZYLHTCG-UHFFFAOYSA-N Dichlorophen Chemical compound OC1=CC=C(Cl)C=C1CC1=CC(Cl)=CC=C1O MDNWOSOZYLHTCG-UHFFFAOYSA-N 0.000 description 1
- 101100380328 Dictyostelium discoideum asns gene Proteins 0.000 description 1
- 241000618813 Dilleniales Species 0.000 description 1
- 235000005903 Dioscorea Nutrition 0.000 description 1
- 244000281702 Dioscorea villosa Species 0.000 description 1
- 235000000504 Dioscorea villosa Nutrition 0.000 description 1
- 241000207977 Dipsacales Species 0.000 description 1
- 208000035240 Disease Resistance Diseases 0.000 description 1
- 241001647153 Dokdonella Species 0.000 description 1
- 241001262066 Drechslerella dactyloides Species 0.000 description 1
- 241001277594 Duganella Species 0.000 description 1
- 241000600050 Dyella Species 0.000 description 1
- 240000003133 Elaeis guineensis Species 0.000 description 1
- 244000127993 Elaeis melanococca Species 0.000 description 1
- 241001552883 Enhydrobacter Species 0.000 description 1
- 101000889905 Enterobacteria phage RB3 Intron-associated endonuclease 3 Proteins 0.000 description 1
- 101000889904 Enterobacteria phage T4 Defective intron-associated endonuclease 3 Proteins 0.000 description 1
- 101000889900 Enterobacteria phage T4 Intron-associated endonuclease 1 Proteins 0.000 description 1
- 101000889899 Enterobacteria phage T4 Intron-associated endonuclease 2 Proteins 0.000 description 1
- 241000194033 Enterococcus Species 0.000 description 1
- 241000218671 Ephedra Species 0.000 description 1
- 241001465251 Ephedra sinica Species 0.000 description 1
- 241000588698 Erwinia Species 0.000 description 1
- 240000006890 Erythroxylum coca Species 0.000 description 1
- 101100134117 Escherichia coli (strain K12) glnG gene Proteins 0.000 description 1
- 101100184192 Escherichia coli (strain K12) mlaD gene Proteins 0.000 description 1
- 241001302160 Escherichia coli str. K-12 substr. DH10B Species 0.000 description 1
- LFQSCWFLJHTTHZ-UHFFFAOYSA-N Ethanol Chemical compound CCO LFQSCWFLJHTTHZ-UHFFFAOYSA-N 0.000 description 1
- 239000001856 Ethyl cellulose Substances 0.000 description 1
- ZZSNKZQZMQGXPY-UHFFFAOYSA-N Ethyl cellulose Chemical compound CCOCC1OC(OC)C(OCC)C(OCC)C1OC1C(O)C(O)C(OC)C(CO)O1 ZZSNKZQZMQGXPY-UHFFFAOYSA-N 0.000 description 1
- 241001468125 Exiguobacterium Species 0.000 description 1
- 108091029865 Exogenous DNA Proteins 0.000 description 1
- 241001247262 Fabales Species 0.000 description 1
- 241000531184 Ferroglobus Species 0.000 description 1
- 241001585410 Filimonas Species 0.000 description 1
- 241001617393 Finegoldia Species 0.000 description 1
- 241000589565 Flavobacterium Species 0.000 description 1
- 239000005781 Fludioxonil Substances 0.000 description 1
- PWNAWOCHVWERAR-UHFFFAOYSA-N Flumetralin Chemical compound [O-][N+](=O)C=1C=C(C(F)(F)F)C=C([N+]([O-])=O)C=1N(CC)CC1=C(F)C=CC=C1Cl PWNAWOCHVWERAR-UHFFFAOYSA-N 0.000 description 1
- 235000017317 Fortunella Nutrition 0.000 description 1
- 241000187809 Frankia Species 0.000 description 1
- 241000187808 Frankia sp. Species 0.000 description 1
- 241000382489 Frigoribacterium Species 0.000 description 1
- 206010017533 Fungal infection Diseases 0.000 description 1
- 241001423728 Fusarium neocosmosporiellum Species 0.000 description 1
- 230000005526 G1 to G0 transition Effects 0.000 description 1
- 101150065368 GLTP gene Proteins 0.000 description 1
- 241000234271 Galanthus Species 0.000 description 1
- 108010010803 Gelatin Proteins 0.000 description 1
- 229920002148 Gellan gum Polymers 0.000 description 1
- 229930182566 Gentamicin Natural products 0.000 description 1
- CEAZRRDELHUEMR-URQXQFDESA-N Gentamicin Chemical compound O1[C@H](C(C)NC)CC[C@@H](N)[C@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](NC)[C@@](C)(O)CO2)O)[C@H](N)C[C@@H]1N CEAZRRDELHUEMR-URQXQFDESA-N 0.000 description 1
- 241000208326 Gentianales Species 0.000 description 1
- 241000134874 Geraniales Species 0.000 description 1
- 239000005980 Gibberellic acid Substances 0.000 description 1
- 241000218790 Ginkgoales Species 0.000 description 1
- 241000032681 Gluconacetobacter Species 0.000 description 1
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 1
- 108020000311 Glutamate Synthase Proteins 0.000 description 1
- 102000016901 Glutamate dehydrogenase Human genes 0.000 description 1
- 108091006151 Glutamate transporters Proteins 0.000 description 1
- 108010024636 Glutathione Proteins 0.000 description 1
- 239000004471 Glycine Substances 0.000 description 1
- 229920002527 Glycogen Polymers 0.000 description 1
- 108010001483 Glycogen Synthase Proteins 0.000 description 1
- 241000218664 Gnetales Species 0.000 description 1
- 235000009432 Gossypium hirsutum Nutrition 0.000 description 1
- 229920000084 Gum arabic Polymers 0.000 description 1
- 101100238555 Haemophilus influenzae (strain ATCC 51907 / DSM 11121 / KW20 / Rd) msbA gene Proteins 0.000 description 1
- 241000588731 Hafnia Species 0.000 description 1
- 241000159657 Halobaculum Species 0.000 description 1
- 241001134726 Heliobacillus Species 0.000 description 1
- 241000207153 Heliobacterium chlorum Species 0.000 description 1
- 241001494519 Heliobacterium sp. Species 0.000 description 1
- 241001628319 Heliophilum Species 0.000 description 1
- 240000001194 Heliotropium europaeum Species 0.000 description 1
- 102000001554 Hemoglobins Human genes 0.000 description 1
- 108010054147 Hemoglobins Proteins 0.000 description 1
- 244000043261 Hevea brasiliensis Species 0.000 description 1
- 241000143459 Hirsutella Species 0.000 description 1
- 241000339365 Hirsutella minnesotensis Species 0.000 description 1
- 241000161908 Hirsutella rhossiliensis Species 0.000 description 1
- 241000209219 Hordeum Species 0.000 description 1
- 241001090156 Huperzia serrata Species 0.000 description 1
- VEXZGXHMUGYJMC-UHFFFAOYSA-N Hydrochloric acid Chemical compound Cl VEXZGXHMUGYJMC-UHFFFAOYSA-N 0.000 description 1
- 229920002153 Hydroxypropyl cellulose Polymers 0.000 description 1
- 241000033356 Hymenobacter Species 0.000 description 1
- 241000208278 Hyoscyamus Species 0.000 description 1
- PWGOWIIEVDAYTC-UHFFFAOYSA-N ICR-170 Chemical compound Cl.Cl.C1=C(OC)C=C2C(NCCCN(CCCl)CC)=C(C=CC(Cl)=C3)C3=NC2=C1 PWGOWIIEVDAYTC-UHFFFAOYSA-N 0.000 description 1
- DGAQECJNVWCQMB-PUAWFVPOSA-M Ilexoside XXIX Chemical compound C[C@@H]1CC[C@@]2(CC[C@@]3(C(=CC[C@H]4[C@]3(CC[C@@H]5[C@@]4(CC[C@@H](C5(C)C)OS(=O)(=O)[O-])C)C)[C@@H]2[C@]1(C)O)C)C(=O)O[C@H]6[C@@H]([C@H]([C@@H]([C@H](O6)CO)O)O)O.[Na+] DGAQECJNVWCQMB-PUAWFVPOSA-M 0.000 description 1
- 108010093096 Immobilized Enzymes Proteins 0.000 description 1
- IMQLKJBTEOYOSI-GPIVLXJGSA-N Inositol-hexakisphosphate Chemical compound OP(O)(=O)O[C@H]1[C@H](OP(O)(O)=O)[C@@H](OP(O)(O)=O)[C@H](OP(O)(O)=O)[C@H](OP(O)(O)=O)[C@@H]1OP(O)(O)=O IMQLKJBTEOYOSI-GPIVLXJGSA-N 0.000 description 1
- 239000005796 Ipconazole Substances 0.000 description 1
- 235000002678 Ipomoea batatas Nutrition 0.000 description 1
- 244000017020 Ipomoea batatas Species 0.000 description 1
- 108010044467 Isoenzymes Proteins 0.000 description 1
- 241000221089 Jatropha Species 0.000 description 1
- 241001048891 Jatropha curcas Species 0.000 description 1
- 241001110439 Klebsiella michiganensis M5al Species 0.000 description 1
- 241000588749 Klebsiella oxytoca Species 0.000 description 1
- 241000588747 Klebsiella pneumoniae Species 0.000 description 1
- 241000588754 Klebsiella sp. Species 0.000 description 1
- 241000579722 Kocuria Species 0.000 description 1
- 241001386813 Kraken Species 0.000 description 1
- 239000004395 L-leucine Substances 0.000 description 1
- 235000019454 L-leucine Nutrition 0.000 description 1
- KZSNJWFQEVHDMF-BYPYZUCNSA-N L-valine Chemical compound CC(C)[C@H](N)C(O)=O KZSNJWFQEVHDMF-BYPYZUCNSA-N 0.000 description 1
- 240000001046 Lactobacillus acidophilus Species 0.000 description 1
- 235000013956 Lactobacillus acidophilus Nutrition 0.000 description 1
- 235000010671 Lathyrus sativus Nutrition 0.000 description 1
- 240000005783 Lathyrus sativus Species 0.000 description 1
- 241000218194 Laurales Species 0.000 description 1
- 241001121967 Lecanicillium Species 0.000 description 1
- 241001647840 Leclercia Species 0.000 description 1
- 235000014647 Lens culinaris subsp culinaris Nutrition 0.000 description 1
- 244000043158 Lens esculenta Species 0.000 description 1
- 240000003271 Leonurus japonicus Species 0.000 description 1
- 241000589902 Leptospira Species 0.000 description 1
- 102000003960 Ligases Human genes 0.000 description 1
- 108090000364 Ligases Proteins 0.000 description 1
- 241000234435 Lilium Species 0.000 description 1
- 108010028921 Lipopeptides Proteins 0.000 description 1
- 241000209082 Lolium Species 0.000 description 1
- 241000219745 Lupinus Species 0.000 description 1
- 235000010649 Lupinus albus Nutrition 0.000 description 1
- 240000000894 Lupinus albus Species 0.000 description 1
- 241000986841 Luteibacter Species 0.000 description 1
- 241001647883 Luteimonas Species 0.000 description 1
- 241000195947 Lycopodium Species 0.000 description 1
- 241000193386 Lysinibacillus sphaericus Species 0.000 description 1
- 241001660189 Lysobacter antibioticus Species 0.000 description 1
- 241000863030 Lysobacter enzymogenes Species 0.000 description 1
- 108700036248 MT-RNR1 Proteins 0.000 description 1
- FYYHWMGAXLPEAU-UHFFFAOYSA-N Magnesium Chemical compound [Mg] FYYHWMGAXLPEAU-UHFFFAOYSA-N 0.000 description 1
- 239000005953 Magnesium phosphide Substances 0.000 description 1
- 239000005983 Maleic hydrazide Substances 0.000 description 1
- BGRDGMRNKXEXQD-UHFFFAOYSA-N Maleic hydrazide Chemical compound OC1=CC=C(O)N=N1 BGRDGMRNKXEXQD-UHFFFAOYSA-N 0.000 description 1
- 241000219171 Malpighiales Species 0.000 description 1
- 239000005913 Maltodextrin Substances 0.000 description 1
- 229920002774 Maltodextrin Polymers 0.000 description 1
- 244000081841 Malus domestica Species 0.000 description 1
- 235000011430 Malus pumila Nutrition 0.000 description 1
- 235000015103 Malus silvestris Nutrition 0.000 description 1
- 241000134966 Malvales Species 0.000 description 1
- 235000004456 Manihot esculenta Nutrition 0.000 description 1
- 235000016735 Manihot esculenta subsp esculenta Nutrition 0.000 description 1
- 101100507576 Mannheimia haemolytica hslU gene Proteins 0.000 description 1
- 229930195725 Mannitol Natural products 0.000 description 1
- 241001358049 Massilia Species 0.000 description 1
- OKIBNKKYNPBDRS-UHFFFAOYSA-N Mefluidide Chemical compound CC(=O)NC1=CC(NS(=O)(=O)C(F)(F)F)=C(C)C=C1C OKIBNKKYNPBDRS-UHFFFAOYSA-N 0.000 description 1
- 108090000301 Membrane transport proteins Proteins 0.000 description 1
- 102000003939 Membrane transport proteins Human genes 0.000 description 1
- 235000014435 Mentha Nutrition 0.000 description 1
- 241001072983 Mentha Species 0.000 description 1
- 244000024873 Mentha crispa Species 0.000 description 1
- 235000014749 Mentha crispa Nutrition 0.000 description 1
- 241000970829 Mesorhizobium Species 0.000 description 1
- 206010027476 Metastases Diseases 0.000 description 1
- 101100038261 Methanococcus vannielii (strain ATCC 35089 / DSM 1224 / JCM 13029 / OCM 148 / SB) rpo2C gene Proteins 0.000 description 1
- 241000500375 Microbacterium sp. Species 0.000 description 1
- 108700005443 Microbial Genes Proteins 0.000 description 1
- 241001074116 Miscanthus x giganteus Species 0.000 description 1
- 241000699666 Mus <mouse, genus> Species 0.000 description 1
- 101100226013 Mus musculus Ercc1 gene Proteins 0.000 description 1
- 241000234295 Musa Species 0.000 description 1
- 241000204031 Mycoplasma Species 0.000 description 1
- 208000031888 Mycoses Diseases 0.000 description 1
- 241000223251 Myrothecium Species 0.000 description 1
- 241000134886 Myrtales Species 0.000 description 1
- FUSGACRLAFQQRL-UHFFFAOYSA-N N-Ethyl-N-nitrosourea Chemical compound CCN(N=O)C(N)=O FUSGACRLAFQQRL-UHFFFAOYSA-N 0.000 description 1
- ZRKWMRDKSOPRRS-UHFFFAOYSA-N N-Methyl-N-nitrosourea Chemical compound O=NN(C)C(N)=O ZRKWMRDKSOPRRS-UHFFFAOYSA-N 0.000 description 1
- 241001230286 Narenga Species 0.000 description 1
- 241000588653 Neisseria Species 0.000 description 1
- 241000124176 Neocosmospora Species 0.000 description 1
- 241000556984 Neonectria galligena Species 0.000 description 1
- 101710096620 Nitrogen regulatory protein Proteins 0.000 description 1
- 241000187654 Nocardia Species 0.000 description 1
- 241000192656 Nostoc Species 0.000 description 1
- 241000424623 Nostoc punctiforme Species 0.000 description 1
- 108020004711 Nucleic Acid Probes Proteins 0.000 description 1
- 241000039470 Nymphaeales Species 0.000 description 1
- 241000588843 Ochrobactrum Species 0.000 description 1
- 241001486832 Okibacterium Species 0.000 description 1
- 108020005187 Oligonucleotide Probes Proteins 0.000 description 1
- 241000121201 Oligotropha Species 0.000 description 1
- 241000209094 Oryza Species 0.000 description 1
- 241000712830 Oryzihumus Species 0.000 description 1
- 241000178984 Oxalophagus Species 0.000 description 1
- KYGZCKSPAKDVKC-UHFFFAOYSA-N Oxolinic acid Chemical compound C1=C2N(CC)C=C(C(O)=O)C(=O)C2=CC2=C1OCO2 KYGZCKSPAKDVKC-UHFFFAOYSA-N 0.000 description 1
- 239000004100 Oxytetracycline Substances 0.000 description 1
- 238000010222 PCR analysis Methods 0.000 description 1
- 238000002944 PCR assay Methods 0.000 description 1
- 239000005985 Paclobutrazol Substances 0.000 description 1
- 241001236817 Paecilomyces <Clavicipitaceae> Species 0.000 description 1
- 241000321594 Paenibacillus borealis Species 0.000 description 1
- 241000933952 Paenibacillus campinasensis Species 0.000 description 1
- 241000611799 Paenibacillus chibensis Species 0.000 description 1
- 241000611786 Paenibacillus glucanolyticus Species 0.000 description 1
- 241001310335 Paenibacillus lentimorbus Species 0.000 description 1
- 241000611735 Paenibacillus peoriae Species 0.000 description 1
- 241000220897 Paenibacillus riograndensis Species 0.000 description 1
- 241000775032 Paenibacillus riograndensis SBR5 Species 0.000 description 1
- 241000209117 Panicum Species 0.000 description 1
- 235000006443 Panicum miliaceum subsp. miliaceum Nutrition 0.000 description 1
- 235000009037 Panicum miliaceum subsp. ruderale Nutrition 0.000 description 1
- 241000588912 Pantoea agglomerans Species 0.000 description 1
- 235000014370 Papaver orientale Nutrition 0.000 description 1
- 244000293991 Papaver orientale Species 0.000 description 1
- 241001495454 Parthenium Species 0.000 description 1
- AVFIYMSJDDGDBQ-UHFFFAOYSA-N Parthenium Chemical compound C1C=C(CCC(C)=O)C(C)CC2OC(=O)C(=C)C21 AVFIYMSJDDGDBQ-UHFFFAOYSA-N 0.000 description 1
- 241001495453 Parthenium argentatum Species 0.000 description 1
- 241001242657 Pasteuria nishizawae Species 0.000 description 1
- 241000291055 Pasteuria ramosa Species 0.000 description 1
- 206010034133 Pathogen resistance Diseases 0.000 description 1
- 241000372441 Pelomonas Species 0.000 description 1
- 244000115721 Pennisetum typhoides Species 0.000 description 1
- 235000007195 Pennisetum typhoides Nutrition 0.000 description 1
- 241000542733 Perlucidibaca Species 0.000 description 1
- PCNDJXKNXGMECE-UHFFFAOYSA-N Phenazine Natural products C1=CC=CC2=NC3=CC=CC=C3N=C21 PCNDJXKNXGMECE-UHFFFAOYSA-N 0.000 description 1
- 108090000608 Phosphoric Monoester Hydrolases Proteins 0.000 description 1
- 102000004160 Phosphoric Monoester Hydrolases Human genes 0.000 description 1
- 101710155159 Phosphotransferase enzyme IIA component Proteins 0.000 description 1
- 244000082204 Phyllostachys viridis Species 0.000 description 1
- 235000015334 Phyllostachys viridis Nutrition 0.000 description 1
- 235000002489 Physalis philadelphica Nutrition 0.000 description 1
- 240000009134 Physalis philadelphica Species 0.000 description 1
- IMQLKJBTEOYOSI-UHFFFAOYSA-N Phytic acid Natural products OP(O)(=O)OC1C(OP(O)(O)=O)C(OP(O)(O)=O)C(OP(O)(O)=O)C(OP(O)(O)=O)C1OP(O)(O)=O IMQLKJBTEOYOSI-UHFFFAOYSA-N 0.000 description 1
- 241000233614 Phytophthora Species 0.000 description 1
- RZKYEQDPDZUERB-UHFFFAOYSA-N Pindone Chemical compound C1=CC=C2C(=O)C(C(=O)C(C)(C)C)C(=O)C2=C1 RZKYEQDPDZUERB-UHFFFAOYSA-N 0.000 description 1
- 241000218633 Pinidae Species 0.000 description 1
- 241000758713 Piperales Species 0.000 description 1
- 235000010582 Pisum sativum Nutrition 0.000 description 1
- 240000004713 Pisum sativum Species 0.000 description 1
- 108020005120 Plant DNA Proteins 0.000 description 1
- 108700001094 Plant Genes Proteins 0.000 description 1
- 108010064851 Plant Proteins Proteins 0.000 description 1
- 241001408562 Plantibacter Species 0.000 description 1
- 241000131459 Plectranthus barbatus Species 0.000 description 1
- 241001002196 Pochonia Species 0.000 description 1
- 241000500034 Podostemaceae Species 0.000 description 1
- 208000000474 Poliomyelitis Diseases 0.000 description 1
- 229920003171 Poly (ethylene oxide) Polymers 0.000 description 1
- 229920002565 Polyethylene Glycol 400 Polymers 0.000 description 1
- 241000178315 Polynucleobacter Species 0.000 description 1
- 239000004793 Polystyrene Substances 0.000 description 1
- 241000161288 Populus candicans Species 0.000 description 1
- 241000243142 Porifera Species 0.000 description 1
- 101710093543 Probable non-specific lipid-transfer protein Proteins 0.000 description 1
- 239000005986 Prohexadione Substances 0.000 description 1
- 241000186429 Propionibacterium Species 0.000 description 1
- 241001156046 Propioniciclava Species 0.000 description 1
- 241000617410 Proteales Species 0.000 description 1
- 241000589517 Pseudomonas aeruginosa Species 0.000 description 1
- 241001646398 Pseudomonas chlororaphis Species 0.000 description 1
- 241001645955 Pseudomonas chlororaphis subsp. aureofaciens Species 0.000 description 1
- 241000589776 Pseudomonas putida Species 0.000 description 1
- 241001647875 Pseudoxanthomonas Species 0.000 description 1
- 241000588671 Psychrobacter Species 0.000 description 1
- 241001465752 Purpureocillium lilacinum Species 0.000 description 1
- 241000233639 Pythium Species 0.000 description 1
- 238000002123 RNA extraction Methods 0.000 description 1
- 108090000951 RNA polymerase sigma 70 Proteins 0.000 description 1
- 101710097340 RNA polymerase sigma factor RpoD Proteins 0.000 description 1
- 101710198277 RNA polymerase sigma factor sigA Proteins 0.000 description 1
- 239000013614 RNA sample Substances 0.000 description 1
- 238000011530 RNeasy Mini Kit Methods 0.000 description 1
- 230000002292 Radical scavenging effect Effects 0.000 description 1
- 241001128129 Rafflesiaceae Species 0.000 description 1
- 241001478271 Rahnella aquatilis Species 0.000 description 1
- 238000001530 Raman microscopy Methods 0.000 description 1
- 241000133533 Ranunculales Species 0.000 description 1
- 244000088415 Raphanus sativus Species 0.000 description 1
- 235000006140 Raphanus sativus var sativus Nutrition 0.000 description 1
- 108700005075 Regulator Genes Proteins 0.000 description 1
- 108091081062 Repeated sequence (DNA) Proteins 0.000 description 1
- 241000948188 Rheinheimera Species 0.000 description 1
- 241001148115 Rhizobium etli Species 0.000 description 1
- 241000589194 Rhizobium leguminosarum Species 0.000 description 1
- 241000191023 Rhodobacter capsulatus Species 0.000 description 1
- 241001464989 Rhodococcus globerulus Species 0.000 description 1
- 241000190932 Rhodopseudomonas Species 0.000 description 1
- 108091028664 Ribonucleotide Proteins 0.000 description 1
- 240000000528 Ricinus communis Species 0.000 description 1
- 235000004443 Ricinus communis Nutrition 0.000 description 1
- 201000001718 Roberts syndrome Diseases 0.000 description 1
- 208000012474 Roberts-SC phocomelia syndrome Diseases 0.000 description 1
- 235000011449 Rosa Nutrition 0.000 description 1
- 241000220317 Rosa Species 0.000 description 1
- 241000332814 Roseateles Species 0.000 description 1
- 240000007651 Rubus glaucus Species 0.000 description 1
- 235000011034 Rubus glaucus Nutrition 0.000 description 1
- 235000009122 Rubus idaeus Nutrition 0.000 description 1
- 108091006629 SLC13A2 Proteins 0.000 description 1
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 1
- 241000124033 Salix Species 0.000 description 1
- 244000001385 Sanguinaria canadensis Species 0.000 description 1
- 241000134968 Sapindales Species 0.000 description 1
- 241000208437 Sarraceniaceae Species 0.000 description 1
- 241000134890 Saxifragales Species 0.000 description 1
- 241000242873 Scopolia Species 0.000 description 1
- 241001453170 Sebaldella Species 0.000 description 1
- 241000206467 Sediminibacillus Species 0.000 description 1
- 241000346882 Sediminibacterium Species 0.000 description 1
- BUGBHKTXTAQXES-UHFFFAOYSA-N Selenium Chemical compound [Se] BUGBHKTXTAQXES-UHFFFAOYSA-N 0.000 description 1
- 241000270295 Serpentes Species 0.000 description 1
- 241000147799 Serratia entomophila Species 0.000 description 1
- 241000607715 Serratia marcescens Species 0.000 description 1
- 244000000231 Sesamum indicum Species 0.000 description 1
- 235000003434 Sesamum indicum Nutrition 0.000 description 1
- 235000008515 Setaria glauca Nutrition 0.000 description 1
- 229920001800 Shellac Polymers 0.000 description 1
- 239000000589 Siderophore Substances 0.000 description 1
- 241000589196 Sinorhizobium meliloti Species 0.000 description 1
- 241000144249 Sinorhizobium sp. Species 0.000 description 1
- 241001227511 Sinosporangium Species 0.000 description 1
- 239000005708 Sodium hypochlorite Substances 0.000 description 1
- 235000002597 Solanum melongena Nutrition 0.000 description 1
- 244000061458 Solanum melongena Species 0.000 description 1
- 235000007230 Sorghum bicolor Nutrition 0.000 description 1
- 241001136275 Sphingobacterium Species 0.000 description 1
- 241000371136 Sphingosinicella Species 0.000 description 1
- 241000191940 Staphylococcus Species 0.000 description 1
- 241000194017 Streptococcus Species 0.000 description 1
- 101100309436 Streptococcus mutans serotype c (strain ATCC 700610 / UA159) ftf gene Proteins 0.000 description 1
- 239000005838 Streptomyces K61 (formerly S. griseoviridis) Substances 0.000 description 1
- 241000970268 Streptomyces colombiensis Species 0.000 description 1
- 241001467541 Streptomyces galbus Species 0.000 description 1
- 241000187218 Streptomyces goshikiensis Species 0.000 description 1
- 241000191251 Streptomyces griseoviridis Species 0.000 description 1
- 241000946809 Streptomyces prasinus Species 0.000 description 1
- 241000970858 Streptomyces saraceticus Species 0.000 description 1
- 241000187180 Streptomyces sp. Species 0.000 description 1
- 241000531819 Streptomyces venezuelae Species 0.000 description 1
- 241000132988 Stygiolobus Species 0.000 description 1
- 108091027544 Subgenomic mRNA Proteins 0.000 description 1
- KDYFGRWQOYBRFD-UHFFFAOYSA-N Succinic acid Natural products OC(=O)CCC(O)=O KDYFGRWQOYBRFD-UHFFFAOYSA-N 0.000 description 1
- 241000985077 Sulfurisphaera Species 0.000 description 1
- 241001110323 Syagrus romanzoffiana Species 0.000 description 1
- 241000192584 Synechocystis Species 0.000 description 1
- 244000103423 Tanacetum pathenium Species 0.000 description 1
- 241001622829 Tatumella Species 0.000 description 1
- 241000202349 Taxus brevifolia Species 0.000 description 1
- 244000162450 Taxus cuspidata Species 0.000 description 1
- 235000009065 Taxus cuspidata Nutrition 0.000 description 1
- 239000005839 Tebuconazole Substances 0.000 description 1
- 241000189180 Tepidimonas Species 0.000 description 1
- 241000623377 Terminalia elliptica Species 0.000 description 1
- 235000006468 Thea sinensis Nutrition 0.000 description 1
- 241000473945 Theria <moth genus> Species 0.000 description 1
- 241001249784 Thermomonas Species 0.000 description 1
- FZWLAAWBMGSTSO-UHFFFAOYSA-N Thiazole Chemical compound C1=CSC=N1 FZWLAAWBMGSTSO-UHFFFAOYSA-N 0.000 description 1
- 241000605118 Thiobacillus Species 0.000 description 1
- 239000005843 Thiram Substances 0.000 description 1
- 235000007303 Thymus vulgaris Nutrition 0.000 description 1
- 240000002657 Thymus vulgaris Species 0.000 description 1
- 235000011941 Tilia x europaea Nutrition 0.000 description 1
- 240000006909 Tilia x europaea Species 0.000 description 1
- 108091028113 Trans-activating crRNA Proteins 0.000 description 1
- 241000219793 Trifolium Species 0.000 description 1
- OKJPEAGHQZHRQV-UHFFFAOYSA-N Triiodomethane Natural products IC(I)I OKJPEAGHQZHRQV-UHFFFAOYSA-N 0.000 description 1
- 239000007983 Tris buffer Substances 0.000 description 1
- 239000005859 Triticonazole Substances 0.000 description 1
- 108090000848 Ubiquitin Proteins 0.000 description 1
- 102000044159 Ubiquitin Human genes 0.000 description 1
- 235000013419 Uniola paniculata Nutrition 0.000 description 1
- 240000007492 Uniola paniculata Species 0.000 description 1
- LEHOTFFKMJEONL-UHFFFAOYSA-N Uric Acid Chemical compound N1C(=O)NC(=O)C2=C1NC(=O)N2 LEHOTFFKMJEONL-UHFFFAOYSA-N 0.000 description 1
- TVWHNULVHGKJHS-UHFFFAOYSA-N Uric acid Natural products N1C(=O)NC(=O)C2NC(=O)NC21 TVWHNULVHGKJHS-UHFFFAOYSA-N 0.000 description 1
- KZSNJWFQEVHDMF-UHFFFAOYSA-N Valine Natural products CC(C)C(N)C(O)=O KZSNJWFQEVHDMF-UHFFFAOYSA-N 0.000 description 1
- 241001478283 Variovorax Species 0.000 description 1
- 241000251539 Vertebrata <Metazoa> Species 0.000 description 1
- BZHJMEDXRYGGRV-UHFFFAOYSA-N Vinyl chloride Chemical compound ClC=C BZHJMEDXRYGGRV-UHFFFAOYSA-N 0.000 description 1
- 229930003779 Vitamin B12 Natural products 0.000 description 1
- MECHNRXZTMCUDQ-UHFFFAOYSA-N Vitamin D2 Natural products C1CCC2(C)C(C(C)C=CC(C)C(C)C)CCC2C1=CC=C1CC(O)CCC1=C MECHNRXZTMCUDQ-UHFFFAOYSA-N 0.000 description 1
- QYSXJUFSXHHAJI-XFEUOLMDSA-N Vitamin D3 Natural products C1(/[C@@H]2CC[C@@H]([C@]2(CCC1)C)[C@H](C)CCCC(C)C)=C/C=C1\C[C@@H](O)CCC1=C QYSXJUFSXHHAJI-XFEUOLMDSA-N 0.000 description 1
- 241000589634 Xanthomonas Species 0.000 description 1
- 241000589636 Xanthomonas campestris Species 0.000 description 1
- 241000607735 Xenorhabdus nematophila Species 0.000 description 1
- 235000016383 Zea mays subsp huehuetenangensis Nutrition 0.000 description 1
- 229920002494 Zein Polymers 0.000 description 1
- 239000006011 Zinc phosphide Substances 0.000 description 1
- 241000234675 Zingiberales Species 0.000 description 1
- QFHMNFAUXJAINK-UHFFFAOYSA-N [1-(carbamoylamino)-2-methylpropyl]urea Chemical compound NC(=O)NC(C(C)C)NC(N)=O QFHMNFAUXJAINK-UHFFFAOYSA-N 0.000 description 1
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 1
- JPHMPWQBESMBJL-UHFFFAOYSA-N [N].[Ca] Chemical compound [N].[Ca] JPHMPWQBESMBJL-UHFFFAOYSA-N 0.000 description 1
- OLBVUFHMDRJKTK-UHFFFAOYSA-N [N].[O] Chemical compound [N].[O] OLBVUFHMDRJKTK-UHFFFAOYSA-N 0.000 description 1
- CWVZGJORVTZXFW-UHFFFAOYSA-N [benzyl(dimethyl)silyl]methyl carbamate Chemical compound NC(=O)OC[Si](C)(C)CC1=CC=CC=C1 CWVZGJORVTZXFW-UHFFFAOYSA-N 0.000 description 1
- 238000002679 ablation Methods 0.000 description 1
- 235000010489 acacia gum Nutrition 0.000 description 1
- 239000000205 acacia gum Substances 0.000 description 1
- 238000009825 accumulation Methods 0.000 description 1
- 230000021736 acetylation Effects 0.000 description 1
- 238000006640 acetylation reaction Methods 0.000 description 1
- 230000002378 acidificating effect Effects 0.000 description 1
- 229920006243 acrylic copolymer Polymers 0.000 description 1
- 230000004913 activation Effects 0.000 description 1
- 239000012190 activator Substances 0.000 description 1
- 230000001154 acute effect Effects 0.000 description 1
- 230000004721 adaptive immunity Effects 0.000 description 1
- 230000004520 agglutination Effects 0.000 description 1
- 239000003513 alkali Substances 0.000 description 1
- 125000002947 alkylene group Chemical group 0.000 description 1
- BJEPYKJPYRNKOW-UHFFFAOYSA-N alpha-hydroxysuccinic acid Natural products OC(=O)C(O)CC(O)=O BJEPYKJPYRNKOW-UHFFFAOYSA-N 0.000 description 1
- BIGPRXCJEDHCLP-UHFFFAOYSA-N ammonium bisulfate Chemical compound [NH4+].OS([O-])(=O)=O BIGPRXCJEDHCLP-UHFFFAOYSA-N 0.000 description 1
- 235000019270 ammonium chloride Nutrition 0.000 description 1
- 229910000148 ammonium phosphate Inorganic materials 0.000 description 1
- 235000019289 ammonium phosphates Nutrition 0.000 description 1
- 235000019826 ammonium polyphosphate Nutrition 0.000 description 1
- 229920001276 ammonium polyphosphate Polymers 0.000 description 1
- XYXNTHIYBIDHGM-UHFFFAOYSA-N ammonium thiosulfate Chemical compound [NH4+].[NH4+].[O-]S([O-])(=O)=S XYXNTHIYBIDHGM-UHFFFAOYSA-N 0.000 description 1
- 230000003698 anagen phase Effects 0.000 description 1
- 101150104862 anfD gene Proteins 0.000 description 1
- 101150040823 anfG gene Proteins 0.000 description 1
- 101150101246 anfH gene Proteins 0.000 description 1
- 101150036396 anfK gene Proteins 0.000 description 1
- 235000020244 animal milk Nutrition 0.000 description 1
- 125000000129 anionic group Chemical group 0.000 description 1
- 238000000137 annealing Methods 0.000 description 1
- 230000000844 anti-bacterial effect Effects 0.000 description 1
- 230000003466 anti-cipated effect Effects 0.000 description 1
- 230000000692 anti-sense effect Effects 0.000 description 1
- 239000003429 antifungal agent Substances 0.000 description 1
- 229940121375 antifungal agent Drugs 0.000 description 1
- 230000006907 apoptotic process Effects 0.000 description 1
- 239000007900 aqueous suspension Substances 0.000 description 1
- 235000019312 arabinogalactan Nutrition 0.000 description 1
- 238000003491 array Methods 0.000 description 1
- 230000011681 asexual reproduction Effects 0.000 description 1
- 238000013465 asexual reproduction Methods 0.000 description 1
- 101150062095 asnA gene Proteins 0.000 description 1
- 101150057409 asnB gene Proteins 0.000 description 1
- 229960003272 asparaginase Drugs 0.000 description 1
- DCXYFEDJOCDNAF-UHFFFAOYSA-M asparaginate Chemical compound [O-]C(=O)C(N)CC(N)=O DCXYFEDJOCDNAF-UHFFFAOYSA-M 0.000 description 1
- 229940009098 aspartate Drugs 0.000 description 1
- 238000003149 assay kit Methods 0.000 description 1
- WFDXOXNFNRHQEC-GHRIWEEISA-N azoxystrobin Chemical compound CO\C=C(\C(=O)OC)C1=CC=CC=C1OC1=CC(OC=2C(=CC=CC=2)C#N)=NC=N1 WFDXOXNFNRHQEC-GHRIWEEISA-N 0.000 description 1
- 229940054340 bacillus coagulans Drugs 0.000 description 1
- 229940097012 bacillus thuringiensis Drugs 0.000 description 1
- 210000004666 bacterial spore Anatomy 0.000 description 1
- 239000011425 bamboo Substances 0.000 description 1
- AYJRCSIUFZENHW-DEQYMQKBSA-L barium(2+);oxomethanediolate Chemical compound [Ba+2].[O-][14C]([O-])=O AYJRCSIUFZENHW-DEQYMQKBSA-L 0.000 description 1
- 239000011324 bead Substances 0.000 description 1
- 235000013871 bee wax Nutrition 0.000 description 1
- 239000012166 beeswax Substances 0.000 description 1
- DMSMPAJRVJJAGA-UHFFFAOYSA-N benzo[d]isothiazol-3-one Chemical compound C1=CC=C2C(=O)NSC2=C1 DMSMPAJRVJJAGA-UHFFFAOYSA-N 0.000 description 1
- MMIMIFULGMZVPO-UHFFFAOYSA-N benzyl 3-bromo-2,6-dinitro-5-phenylmethoxybenzoate Chemical compound [O-][N+](=O)C1=C(C(=O)OCC=2C=CC=CC=2)C([N+](=O)[O-])=C(Br)C=C1OCC1=CC=CC=C1 MMIMIFULGMZVPO-UHFFFAOYSA-N 0.000 description 1
- 235000019445 benzyl alcohol Nutrition 0.000 description 1
- WQZGKKKJIJFFOK-VFUOTHLCSA-N beta-D-glucose Chemical compound OC[C@H]1O[C@@H](O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-VFUOTHLCSA-N 0.000 description 1
- 230000001588 bifunctional effect Effects 0.000 description 1
- 102000023732 binding proteins Human genes 0.000 description 1
- 108091008324 binding proteins Proteins 0.000 description 1
- 239000000227 bioadhesive Substances 0.000 description 1
- 230000008238 biochemical pathway Effects 0.000 description 1
- 239000012681 biocontrol agent Substances 0.000 description 1
- 230000004071 biological effect Effects 0.000 description 1
- 238000004178 biological nitrogen fixation Methods 0.000 description 1
- 239000012472 biological sample Substances 0.000 description 1
- 235000012978 bixa orellana Nutrition 0.000 description 1
- 229960001561 bleomycin Drugs 0.000 description 1
- OYVAGSVQBOHSSS-UAPAGMARSA-O bleomycin A2 Chemical compound N([C@H](C(=O)N[C@H](C)[C@@H](O)[C@H](C)C(=O)N[C@@H]([C@H](O)C)C(=O)NCCC=1SC=C(N=1)C=1SC=C(N=1)C(=O)NCCC[S+](C)C)[C@@H](O[C@H]1[C@H]([C@@H](O)[C@H](O)[C@H](CO)O1)O[C@@H]1[C@H]([C@@H](OC(N)=O)[C@H](O)[C@@H](CO)O1)O)C=1N=CNC=1)C(=O)C1=NC([C@H](CC(N)=O)NC[C@H](N)C(N)=O)=NC(N)=C1C OYVAGSVQBOHSSS-UAPAGMARSA-O 0.000 description 1
- 229920001400 block copolymer Polymers 0.000 description 1
- 238000006664 bond formation reaction Methods 0.000 description 1
- 229910052796 boron Inorganic materials 0.000 description 1
- IXVMHGVQKLDRKH-KNBKMWSGSA-N brassinolide Chemical compound C1OC(=O)[C@H]2C[C@H](O)[C@H](O)C[C@]2(C)[C@H]2CC[C@]3(C)[C@@H]([C@H](C)[C@@H](O)[C@H](O)[C@@H](C)C(C)C)CC[C@H]3[C@@H]21 IXVMHGVQKLDRKH-KNBKMWSGSA-N 0.000 description 1
- 150000001647 brassinosteroids Chemical class 0.000 description 1
- KDYFGRWQOYBRFD-NUQCWPJISA-N butanedioic acid Chemical compound O[14C](=O)CC[14C](O)=O KDYFGRWQOYBRFD-NUQCWPJISA-N 0.000 description 1
- 229910052791 calcium Inorganic materials 0.000 description 1
- 239000011575 calcium Substances 0.000 description 1
- 229910000019 calcium carbonate Inorganic materials 0.000 description 1
- 229920005551 calcium lignosulfonate Polymers 0.000 description 1
- RYAGRZNBULDMBW-UHFFFAOYSA-L calcium;3-(2-hydroxy-3-methoxyphenyl)-2-[2-methoxy-4-(3-sulfonatopropyl)phenoxy]propane-1-sulfonate Chemical compound [Ca+2].COC1=CC=CC(CC(CS([O-])(=O)=O)OC=2C(=CC(CCCS([O-])(=O)=O)=CC=2)OC)=C1O RYAGRZNBULDMBW-UHFFFAOYSA-L 0.000 description 1
- 238000004364 calculation method Methods 0.000 description 1
- 239000004204 candelilla wax Substances 0.000 description 1
- 235000013868 candelilla wax Nutrition 0.000 description 1
- 229940073532 candelilla wax Drugs 0.000 description 1
- 229940041514 candida albicans extract Drugs 0.000 description 1
- 229940117949 captan Drugs 0.000 description 1
- 229910052799 carbon Inorganic materials 0.000 description 1
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 description 1
- 239000004203 carnauba wax Substances 0.000 description 1
- 235000013869 carnauba wax Nutrition 0.000 description 1
- 238000006555 catalytic reaction Methods 0.000 description 1
- 125000002091 cationic group Chemical group 0.000 description 1
- 230000030833 cell death Effects 0.000 description 1
- 210000000170 cell membrane Anatomy 0.000 description 1
- 230000003833 cell viability Effects 0.000 description 1
- 210000002421 cell wall Anatomy 0.000 description 1
- 230000001413 cellular effect Effects 0.000 description 1
- 230000005754 cellular signaling Effects 0.000 description 1
- 229920002678 cellulose Polymers 0.000 description 1
- 239000001913 cellulose Substances 0.000 description 1
- 238000005119 centrifugation Methods 0.000 description 1
- KXZJHVJKXJLBKO-UHFFFAOYSA-N chembl1408157 Chemical compound N=1C2=CC=CC=C2C(C(=O)O)=CC=1C1=CC=C(O)C=C1 KXZJHVJKXJLBKO-UHFFFAOYSA-N 0.000 description 1
- 239000002962 chemical mutagen Substances 0.000 description 1
- 239000007795 chemical reaction product Substances 0.000 description 1
- 230000035605 chemotaxis Effects 0.000 description 1
- 239000012174 chinese wax Substances 0.000 description 1
- 229940045110 chitosan Drugs 0.000 description 1
- OJYGBLRPYBAHRT-IPQSZEQASA-N chloralose Chemical compound O1[C@H](C(Cl)(Cl)Cl)O[C@@H]2[C@@H](O)[C@@H]([C@H](O)CO)O[C@@H]21 OJYGBLRPYBAHRT-IPQSZEQASA-N 0.000 description 1
- 229950009941 chloralose Drugs 0.000 description 1
- 239000000460 chlorine Substances 0.000 description 1
- 229910052801 chlorine Inorganic materials 0.000 description 1
- 210000003763 chloroplast Anatomy 0.000 description 1
- SGMZJAMFUVOLNK-UHFFFAOYSA-M choline chloride Chemical compound [Cl-].C[N+](C)(C)CCO SGMZJAMFUVOLNK-UHFFFAOYSA-M 0.000 description 1
- 229960003178 choline chloride Drugs 0.000 description 1
- MYSWGUAQZAJSOK-UHFFFAOYSA-N ciprofloxacin Chemical compound C12=CC(N3CCNCC3)=C(F)C=C2C(=O)C(C(=O)O)=CN1C1CC1 MYSWGUAQZAJSOK-UHFFFAOYSA-N 0.000 description 1
- 229920003211 cis-1,4-polyisoprene Polymers 0.000 description 1
- 235000015165 citric acid Nutrition 0.000 description 1
- 235000020971 citrus fruits Nutrition 0.000 description 1
- 239000013599 cloning vector Substances 0.000 description 1
- AGVAZMGAQJOSFJ-WZHZPDAFSA-M cobalt(2+);[(2r,3s,4r,5s)-5-(5,6-dimethylbenzimidazol-1-yl)-4-hydroxy-2-(hydroxymethyl)oxolan-3-yl] [(2r)-1-[3-[(1r,2r,3r,4z,7s,9z,12s,13s,14z,17s,18s,19r)-2,13,18-tris(2-amino-2-oxoethyl)-7,12,17-tris(3-amino-3-oxopropyl)-3,5,8,8,13,15,18,19-octamethyl-2 Chemical compound [Co+2].N#[C-].[N-]([C@@H]1[C@H](CC(N)=O)[C@@]2(C)CCC(=O)NC[C@@H](C)OP(O)(=O)O[C@H]3[C@H]([C@H](O[C@@H]3CO)N3C4=CC(C)=C(C)C=C4N=C3)O)\C2=C(C)/C([C@H](C\2(C)C)CCC(N)=O)=N/C/2=C\C([C@H]([C@@]/2(CC(N)=O)C)CCC(N)=O)=N\C\2=C(C)/C2=N[C@]1(C)[C@@](C)(CC(N)=O)[C@@H]2CCC(N)=O AGVAZMGAQJOSFJ-WZHZPDAFSA-M 0.000 description 1
- 235000013353 coffee beverage Nutrition 0.000 description 1
- 239000003086 colorant Substances 0.000 description 1
- 239000003283 colorimetric indicator Substances 0.000 description 1
- 239000002299 complementary DNA Substances 0.000 description 1
- 239000003184 complementary RNA Substances 0.000 description 1
- JECGPMYZUFFYJW-UHFFFAOYSA-N conferone Natural products CC1=CCC2C(C)(C)C(=O)CCC2(C)C1COc3cccc4C=CC(=O)Oc34 JECGPMYZUFFYJW-UHFFFAOYSA-N 0.000 description 1
- 238000012790 confirmation Methods 0.000 description 1
- 238000010276 construction Methods 0.000 description 1
- 230000001276 controlling effect Effects 0.000 description 1
- 239000010949 copper Substances 0.000 description 1
- 229910052802 copper Inorganic materials 0.000 description 1
- 235000012343 cottonseed oil Nutrition 0.000 description 1
- 239000002385 cottonseed oil Substances 0.000 description 1
- HENZOLWOIZODCT-UHFFFAOYSA-N coumachlor Chemical compound OC=1OC2=CC=CC=C2C(=O)C=1C(CC(=O)C)C1=CC=C(Cl)C=C1 HENZOLWOIZODCT-UHFFFAOYSA-N 0.000 description 1
- 238000012272 crop production Methods 0.000 description 1
- 238000003967 crop rotation Methods 0.000 description 1
- 208000031513 cyst Diseases 0.000 description 1
- 230000009089 cytolysis Effects 0.000 description 1
- RGWHQCVHVJXOKC-SHYZEUOFSA-J dCTP(4-) Chemical group O=C1N=C(N)C=CN1[C@@H]1O[C@H](COP([O-])(=O)OP([O-])(=O)OP([O-])([O-])=O)[C@@H](O)C1 RGWHQCVHVJXOKC-SHYZEUOFSA-J 0.000 description 1
- 230000000593 degrading effect Effects 0.000 description 1
- 239000008367 deionised water Substances 0.000 description 1
- 229910021641 deionized water Inorganic materials 0.000 description 1
- 238000004925 denaturation Methods 0.000 description 1
- 230000036425 denaturation Effects 0.000 description 1
- 239000005547 deoxyribonucleotide Substances 0.000 description 1
- 125000002637 deoxyribonucleotide group Chemical group 0.000 description 1
- 230000030609 dephosphorylation Effects 0.000 description 1
- 238000006209 dephosphorylation reaction Methods 0.000 description 1
- 230000008021 deposition Effects 0.000 description 1
- 238000013461 design Methods 0.000 description 1
- 239000000645 desinfectant Substances 0.000 description 1
- FCRACOPGPMPSHN-UHFFFAOYSA-N desoxyabscisic acid Natural products OC(=O)C=C(C)C=CC1C(C)=CC(=O)CC1(C)C FCRACOPGPMPSHN-UHFFFAOYSA-N 0.000 description 1
- 230000004407 detection of glutamine Effects 0.000 description 1
- 230000001627 detrimental effect Effects 0.000 description 1
- 235000019425 dextrin Nutrition 0.000 description 1
- GUJOJGAPFQRJSV-UHFFFAOYSA-N dialuminum;dioxosilane;oxygen(2-);hydrate Chemical compound O.[O-2].[O-2].[O-2].[Al+3].[Al+3].O=[Si]=O.O=[Si]=O.O=[Si]=O.O=[Si]=O GUJOJGAPFQRJSV-UHFFFAOYSA-N 0.000 description 1
- KTTMEOWBIWLMSE-UHFFFAOYSA-N diarsenic trioxide Chemical compound O1[As](O2)O[As]3O[As]1O[As]2O3 KTTMEOWBIWLMSE-UHFFFAOYSA-N 0.000 description 1
- 229960003887 dichlorophen Drugs 0.000 description 1
- VSVAQRUUFVBBFS-UHFFFAOYSA-N difethialone Chemical compound OC=1SC2=CC=CC=C2C(=O)C=1C(C1=CC=CC=C1C1)CC1C(C=C1)=CC=C1C1=CC=C(Br)C=C1 VSVAQRUUFVBBFS-UHFFFAOYSA-N 0.000 description 1
- 238000009792 diffusion process Methods 0.000 description 1
- FWCBATIDXGJRMF-UHFFFAOYSA-N dikegulac Natural products C12OC(C)(C)OCC2OC2(C(O)=O)C1OC(C)(C)O2 FWCBATIDXGJRMF-UHFFFAOYSA-N 0.000 description 1
- 238000007865 diluting Methods 0.000 description 1
- WZISDKTXHMETKG-UHFFFAOYSA-H dimagnesium;dipotassium;trisulfate Chemical compound [Mg+2].[Mg+2].[K+].[K+].[O-]S([O-])(=O)=O.[O-]S([O-])(=O)=O.[O-]S([O-])(=O)=O WZISDKTXHMETKG-UHFFFAOYSA-H 0.000 description 1
- 239000000539 dimer Substances 0.000 description 1
- 235000004879 dioscorea Nutrition 0.000 description 1
- 238000007598 dipping method Methods 0.000 description 1
- 239000006185 dispersion Substances 0.000 description 1
- 230000005782 double-strand break Effects 0.000 description 1
- 101150005728 draT gene Proteins 0.000 description 1
- 230000008641 drought stress Effects 0.000 description 1
- 239000003814 drug Substances 0.000 description 1
- 229940079593 drug Drugs 0.000 description 1
- 239000000428 dust Substances 0.000 description 1
- 210000005069 ears Anatomy 0.000 description 1
- 229920001971 elastomer Polymers 0.000 description 1
- 230000008030 elimination Effects 0.000 description 1
- 239000003995 emulsifying agent Substances 0.000 description 1
- 230000002708 enhancing effect Effects 0.000 description 1
- 230000009088 enzymatic function Effects 0.000 description 1
- 229960002061 ergocalciferol Drugs 0.000 description 1
- 239000003797 essential amino acid Substances 0.000 description 1
- 235000020776 essential amino acid Nutrition 0.000 description 1
- 235000019441 ethanol Nutrition 0.000 description 1
- ZMMJGEGLRURXTF-UHFFFAOYSA-N ethidium bromide Chemical compound [Br-].C12=CC(N)=CC=C2C2=CC=C(N)C=C2[N+](CC)=C1C1=CC=CC=C1 ZMMJGEGLRURXTF-UHFFFAOYSA-N 0.000 description 1
- 229960005542 ethidium bromide Drugs 0.000 description 1
- 235000019325 ethyl cellulose Nutrition 0.000 description 1
- 229920001249 ethyl cellulose Polymers 0.000 description 1
- 229960004667 ethyl cellulose Drugs 0.000 description 1
- 239000005038 ethylene vinyl acetate Substances 0.000 description 1
- 210000003527 eukaryotic cell Anatomy 0.000 description 1
- 239000013604 expression vector Substances 0.000 description 1
- 239000003925 fat Substances 0.000 description 1
- 235000019197 fats Nutrition 0.000 description 1
- 101150100173 fdx gene Proteins 0.000 description 1
- 230000035558 fertility Effects 0.000 description 1
- 235000008384 feverfew Nutrition 0.000 description 1
- 239000000945 filler Substances 0.000 description 1
- 239000010408 film Substances 0.000 description 1
- 239000000834 fixative Substances 0.000 description 1
- 229930003935 flavonoid Natural products 0.000 description 1
- 235000017173 flavonoids Nutrition 0.000 description 1
- 150000002215 flavonoids Chemical class 0.000 description 1
- 235000004426 flaxseed Nutrition 0.000 description 1
- OWUZCVPRFKSBRG-UHFFFAOYSA-N flocoumafen Chemical compound OC=1OC2=CC=CC=C2C(=O)C=1C(C1=CC=CC=C1C1)CC1C(C=C1)=CC=C1OCC1=CC=C(C(F)(F)F)C=C1 OWUZCVPRFKSBRG-UHFFFAOYSA-N 0.000 description 1
- MUJOIMFVNIBMKC-UHFFFAOYSA-N fludioxonil Chemical compound C=12OC(F)(F)OC2=CC=CC=1C1=CNC=C1C#N MUJOIMFVNIBMKC-UHFFFAOYSA-N 0.000 description 1
- 239000007850 fluorescent dye Substances 0.000 description 1
- 239000003269 fluorescent indicator Substances 0.000 description 1
- XWROTTLWMHCFEC-LGMDPLHJSA-N fluthiacet Chemical compound C1=C(Cl)C(SCC(=O)O)=CC(\N=C/2N3CCCCN3C(=O)S\2)=C1F XWROTTLWMHCFEC-LGMDPLHJSA-N 0.000 description 1
- 230000002431 foraging effect Effects 0.000 description 1
- 210000003953 foreskin Anatomy 0.000 description 1
- 235000019256 formaldehyde Nutrition 0.000 description 1
- RIKPNWPEMPODJD-UHFFFAOYSA-N formononetin Natural products C1=CC(OC)=CC=C1C1=COC2=CC=CC=C2C1=O RIKPNWPEMPODJD-UHFFFAOYSA-N 0.000 description 1
- 239000013505 freshwater Substances 0.000 description 1
- 230000005078 fruit development Effects 0.000 description 1
- 235000013376 functional food Nutrition 0.000 description 1
- 101150055609 fusA gene Proteins 0.000 description 1
- 101150022010 gam gene Proteins 0.000 description 1
- JLYXXMFPNIAWKQ-GNIYUCBRSA-N gamma-hexachlorocyclohexane Chemical compound Cl[C@H]1[C@H](Cl)[C@@H](Cl)[C@@H](Cl)[C@H](Cl)[C@H]1Cl JLYXXMFPNIAWKQ-GNIYUCBRSA-N 0.000 description 1
- 238000004817 gas chromatography Methods 0.000 description 1
- 239000000499 gel Substances 0.000 description 1
- 238000001502 gel electrophoresis Methods 0.000 description 1
- 239000008273 gelatin Substances 0.000 description 1
- 229920000159 gelatin Polymers 0.000 description 1
- 235000019322 gelatine Nutrition 0.000 description 1
- 235000011852 gelatine desserts Nutrition 0.000 description 1
- 235000010492 gellan gum Nutrition 0.000 description 1
- 239000000216 gellan gum Substances 0.000 description 1
- 238000012246 gene addition Methods 0.000 description 1
- 238000012224 gene deletion Methods 0.000 description 1
- 102000054767 gene variant Human genes 0.000 description 1
- 238000012252 genetic analysis Methods 0.000 description 1
- 230000037442 genomic alteration Effects 0.000 description 1
- 229960002518 gentamicin Drugs 0.000 description 1
- IXORZMNAPKEEDV-OBDJNFEBSA-N gibberellin A3 Chemical compound C([C@@]1(O)C(=C)C[C@@]2(C1)[C@H]1C(O)=O)C[C@H]2[C@]2(C=C[C@@H]3O)[C@H]1[C@]3(C)C(=O)O2 IXORZMNAPKEEDV-OBDJNFEBSA-N 0.000 description 1
- 101150024902 glnS gene Proteins 0.000 description 1
- 101150088916 glnT gene Proteins 0.000 description 1
- 101150041871 gltB gene Proteins 0.000 description 1
- 101150058504 gltS gene Proteins 0.000 description 1
- 101150115352 gltT gene Proteins 0.000 description 1
- 101150103988 gltX gene Proteins 0.000 description 1
- 239000008103 glucose Substances 0.000 description 1
- 235000001727 glucose Nutrition 0.000 description 1
- 239000003292 glue Substances 0.000 description 1
- 229960003180 glutathione Drugs 0.000 description 1
- 229940096919 glycogen Drugs 0.000 description 1
- 230000013595 glycosylation Effects 0.000 description 1
- 238000006206 glycosylation reaction Methods 0.000 description 1
- 235000011868 grain product Nutrition 0.000 description 1
- 235000002532 grape seed extract Nutrition 0.000 description 1
- 239000005431 greenhouse gas Substances 0.000 description 1
- 101150013736 gyrB gene Proteins 0.000 description 1
- CJNBYAVZURUTKZ-UHFFFAOYSA-N hafnium(IV) oxide Inorganic materials O=[Hf]=O CJNBYAVZURUTKZ-UHFFFAOYSA-N 0.000 description 1
- 125000001475 halogen functional group Chemical group 0.000 description 1
- 230000009931 harmful effect Effects 0.000 description 1
- 230000008642 heat stress Effects 0.000 description 1
- IUJAMGNYPWYUPM-UHFFFAOYSA-N hentriacontane Chemical compound CCCCCCCCCCCCCCCCCCCCCCCCCCCCCCC IUJAMGNYPWYUPM-UHFFFAOYSA-N 0.000 description 1
- 230000002363 herbicidal effect Effects 0.000 description 1
- AIONOLUJZLIMTK-UHFFFAOYSA-N hesperetin Natural products C1=C(O)C(OC)=CC=C1C1OC2=CC(O)=CC(O)=C2C(=O)C1 AIONOLUJZLIMTK-UHFFFAOYSA-N 0.000 description 1
- AIONOLUJZLIMTK-AWEZNQCLSA-N hesperetin Chemical compound C1=C(O)C(OC)=CC=C1[C@H]1OC2=CC(O)=CC(O)=C2C(=O)C1 AIONOLUJZLIMTK-AWEZNQCLSA-N 0.000 description 1
- 229960001587 hesperetin Drugs 0.000 description 1
- 235000010209 hesperetin Nutrition 0.000 description 1
- 238000013537 high throughput screening Methods 0.000 description 1
- 238000012165 high-throughput sequencing Methods 0.000 description 1
- FTODBIPDTXRIGS-UHFFFAOYSA-N homoeriodictyol Natural products C1=C(O)C(OC)=CC(C2OC3=CC(O)=CC(O)=C3C(=O)C2)=C1 FTODBIPDTXRIGS-UHFFFAOYSA-N 0.000 description 1
- 238000003898 horticulture Methods 0.000 description 1
- VDTIMXCBOXBHER-UHFFFAOYSA-N hydroxy-bis(sulfanyl)-sulfanylidene-$l^{5}-phosphane Chemical compound OP(S)(S)=S VDTIMXCBOXBHER-UHFFFAOYSA-N 0.000 description 1
- 229920003063 hydroxymethyl cellulose Polymers 0.000 description 1
- 229940031574 hydroxymethyl cellulose Drugs 0.000 description 1
- 235000010977 hydroxypropyl cellulose Nutrition 0.000 description 1
- 239000001863 hydroxypropyl cellulose Substances 0.000 description 1
- 229940071676 hydroxypropylcellulose Drugs 0.000 description 1
- 230000028993 immune response Effects 0.000 description 1
- 238000001727 in vivo Methods 0.000 description 1
- 238000010348 incorporation Methods 0.000 description 1
- 239000003317 industrial substance Substances 0.000 description 1
- 239000003112 inhibitor Substances 0.000 description 1
- 238000013101 initial test Methods 0.000 description 1
- 238000002347 injection Methods 0.000 description 1
- 239000007924 injection Substances 0.000 description 1
- 239000002917 insecticide Substances 0.000 description 1
- 230000010354 integration Effects 0.000 description 1
- 239000000138 intercalating agent Substances 0.000 description 1
- 230000009545 invasion Effects 0.000 description 1
- 238000011835 investigation Methods 0.000 description 1
- INQOMBQAUSQDDS-UHFFFAOYSA-N iodomethane Chemical compound IC INQOMBQAUSQDDS-UHFFFAOYSA-N 0.000 description 1
- 239000002563 ionic surfactant Substances 0.000 description 1
- QTYCMDBMOLSEAM-UHFFFAOYSA-N ipconazole Chemical compound C1=NC=NN1CC1(O)C(C(C)C)CCC1CC1=CC=C(Cl)C=C1 QTYCMDBMOLSEAM-UHFFFAOYSA-N 0.000 description 1
- UWKQSNNFCGGAFS-XIFFEERXSA-N irinotecan Chemical compound C1=C2C(CC)=C3CN(C(C4=C([C@@](C(=O)OC4)(O)CC)C=4)=O)C=4C3=NC2=CC=C1OC(=O)N(CC1)CCC1N1CCCCC1 UWKQSNNFCGGAFS-XIFFEERXSA-N 0.000 description 1
- 229930013032 isoflavonoid Natural products 0.000 description 1
- 150000003817 isoflavonoid derivatives Chemical class 0.000 description 1
- 235000012891 isoflavonoids Nutrition 0.000 description 1
- BPHPUYQFMNQIOC-NXRLNHOXSA-N isopropyl beta-D-thiogalactopyranoside Chemical compound CC(C)S[C@@H]1O[C@H](CO)[C@H](O)[C@H](O)[C@H]1O BPHPUYQFMNQIOC-NXRLNHOXSA-N 0.000 description 1
- 230000000155 isotopic effect Effects 0.000 description 1
- 229960000318 kanamycin Drugs 0.000 description 1
- 229930027917 kanamycin Natural products 0.000 description 1
- SBUJHOSQTJFQJX-NOAMYHISSA-N kanamycin Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CN)O[C@@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](N)[C@H](O)[C@@H](CO)O2)O)[C@H](N)C[C@@H]1N SBUJHOSQTJFQJX-NOAMYHISSA-N 0.000 description 1
- 229930182823 kanamycin A Natural products 0.000 description 1
- 230000002147 killing effect Effects 0.000 description 1
- 239000004310 lactic acid Substances 0.000 description 1
- 235000014655 lactic acid Nutrition 0.000 description 1
- 229940039695 lactobacillus acidophilus Drugs 0.000 description 1
- 101150053222 lapA gene Proteins 0.000 description 1
- 231100000518 lethal Toxicity 0.000 description 1
- 230000001665 lethal effect Effects 0.000 description 1
- 229960003136 leucine Drugs 0.000 description 1
- 239000004571 lime Substances 0.000 description 1
- 230000029226 lipidation Effects 0.000 description 1
- 239000012669 liquid formulation Substances 0.000 description 1
- 239000006194 liquid suspension Substances 0.000 description 1
- 230000002132 lysosomal effect Effects 0.000 description 1
- 210000003712 lysosome Anatomy 0.000 description 1
- 230000001868 lysosomic effect Effects 0.000 description 1
- 238000010801 machine learning Methods 0.000 description 1
- 235000021073 macronutrients Nutrition 0.000 description 1
- 239000011777 magnesium Substances 0.000 description 1
- 229910052749 magnesium Inorganic materials 0.000 description 1
- 235000009973 maize Nutrition 0.000 description 1
- 239000001630 malic acid Substances 0.000 description 1
- 235000011090 malic acid Nutrition 0.000 description 1
- 229940035034 maltodextrin Drugs 0.000 description 1
- YKSNLCVSTHTHJA-UHFFFAOYSA-L maneb Chemical compound [Mn+2].[S-]C(=S)NCCNC([S-])=S YKSNLCVSTHTHJA-UHFFFAOYSA-L 0.000 description 1
- 229920000940 maneb Polymers 0.000 description 1
- 229910001437 manganese ion Inorganic materials 0.000 description 1
- WPBNNNQJVZRUHP-UHFFFAOYSA-L manganese(2+);methyl n-[[2-(methoxycarbonylcarbamothioylamino)phenyl]carbamothioyl]carbamate;n-[2-(sulfidocarbothioylamino)ethyl]carbamodithioate Chemical compound [Mn+2].[S-]C(=S)NCCNC([S-])=S.COC(=O)NC(=S)NC1=CC=CC=C1NC(=S)NC(=O)OC WPBNNNQJVZRUHP-UHFFFAOYSA-L 0.000 description 1
- 239000000594 mannitol Substances 0.000 description 1
- 235000010355 mannitol Nutrition 0.000 description 1
- 239000006224 matting agent Substances 0.000 description 1
- 235000012054 meals Nutrition 0.000 description 1
- HAWPXGHAZFHHAD-UHFFFAOYSA-N mechlorethamine Chemical class ClCCN(C)CCCl HAWPXGHAZFHHAD-UHFFFAOYSA-N 0.000 description 1
- SGDBTWWWUNNDEQ-LBPRGKRZSA-N melphalan Chemical compound OC(=O)[C@@H](N)CC1=CC=C(N(CCCl)CCCl)C=C1 SGDBTWWWUNNDEQ-LBPRGKRZSA-N 0.000 description 1
- 229960001924 melphalan Drugs 0.000 description 1
- 239000001220 mentha spicata Substances 0.000 description 1
- 229910052751 metal Inorganic materials 0.000 description 1
- 239000002184 metal Substances 0.000 description 1
- FQPSGWSUVKBHSU-UHFFFAOYSA-N methacrylamide Chemical compound CC(=C)C(N)=O FQPSGWSUVKBHSU-UHFFFAOYSA-N 0.000 description 1
- 229940102396 methyl bromide Drugs 0.000 description 1
- DSKJXGYAJJHDOE-UHFFFAOYSA-N methylideneurea Chemical compound NC(=O)N=C DSKJXGYAJJHDOE-UHFFFAOYSA-N 0.000 description 1
- 238000002493 microarray Methods 0.000 description 1
- 238000012775 microarray technology Methods 0.000 description 1
- 238000009629 microbiological culture Methods 0.000 description 1
- 244000005706 microflora Species 0.000 description 1
- 238000000386 microscopy Methods 0.000 description 1
- 239000004005 microsphere Substances 0.000 description 1
- 230000005012 migration Effects 0.000 description 1
- 238000013508 migration Methods 0.000 description 1
- 235000019713 millet Nutrition 0.000 description 1
- 239000002480 mineral oil Substances 0.000 description 1
- 235000010446 mineral oil Nutrition 0.000 description 1
- 239000003147 molecular marker Substances 0.000 description 1
- 239000000178 monomer Substances 0.000 description 1
- 229910052901 montmorillonite Inorganic materials 0.000 description 1
- 230000004899 motility Effects 0.000 description 1
- 238000002887 multiple sequence alignment Methods 0.000 description 1
- QTGVGIVRLSGTJJ-UHFFFAOYSA-N n-(acetamidomethyl)-2-chloro-n-(2,6-diethylphenyl)acetamide Chemical compound CCC1=CC=CC(CC)=C1N(CNC(C)=O)C(=O)CCl QTGVGIVRLSGTJJ-UHFFFAOYSA-N 0.000 description 1
- 239000013642 negative control Substances 0.000 description 1
- 238000007481 next generation sequencing Methods 0.000 description 1
- 101150027931 nifT gene Proteins 0.000 description 1
- 239000000618 nitrogen fertilizer Substances 0.000 description 1
- QJGQUHMNIGDVPM-UHFFFAOYSA-N nitrogen group Chemical group [N] QJGQUHMNIGDVPM-UHFFFAOYSA-N 0.000 description 1
- OSTGTTZJOCZWJG-UHFFFAOYSA-N nitrosourea Chemical compound NC(=O)N=NO OSTGTTZJOCZWJG-UHFFFAOYSA-N 0.000 description 1
- 239000001272 nitrous oxide Substances 0.000 description 1
- 239000002736 nonionic surfactant Substances 0.000 description 1
- 231100001160 nonlethal Toxicity 0.000 description 1
- DNTHHIVFNQZZRD-CYYJNZCTSA-N norbormide Chemical compound C=1C=CC=CC=1C(C=1N=CC=CC=1)(O)C1=CC2C(C(NC3=O)=O)C3C1\C2=C(C=1N=CC=CC=1)/C1=CC=CC=C1 DNTHHIVFNQZZRD-CYYJNZCTSA-N 0.000 description 1
- 238000010899 nucleation Methods 0.000 description 1
- 239000002853 nucleic acid probe Substances 0.000 description 1
- 235000006180 nutrition needs Nutrition 0.000 description 1
- 230000000050 nutritive effect Effects 0.000 description 1
- JPMIIZHYYWMHDT-UHFFFAOYSA-N octhilinone Chemical compound CCCCCCCCN1SC=CC1=O JPMIIZHYYWMHDT-UHFFFAOYSA-N 0.000 description 1
- 239000002751 oligonucleotide probe Substances 0.000 description 1
- 229920001542 oligosaccharide Polymers 0.000 description 1
- 150000002482 oligosaccharides Chemical class 0.000 description 1
- 238000005457 optimization Methods 0.000 description 1
- 230000008723 osmotic stress Effects 0.000 description 1
- 230000001590 oxidative effect Effects 0.000 description 1
- 229960000321 oxolinic acid Drugs 0.000 description 1
- 229960000625 oxytetracycline Drugs 0.000 description 1
- IWVCMVBTMGNXQD-PXOLEDIWSA-N oxytetracycline Chemical compound C1=CC=C2[C@](O)(C)[C@H]3[C@H](O)[C@H]4[C@H](N(C)C)C(O)=C(C(N)=O)C(=O)[C@@]4(O)C(O)=C3C(=O)C2=C1O IWVCMVBTMGNXQD-PXOLEDIWSA-N 0.000 description 1
- 235000019366 oxytetracycline Nutrition 0.000 description 1
- 229940094443 oxytocics prostaglandins Drugs 0.000 description 1
- 239000005022 packaging material Substances 0.000 description 1
- 230000003071 parasitic effect Effects 0.000 description 1
- 229910001562 pearlite Inorganic materials 0.000 description 1
- 235000010987 pectin Nutrition 0.000 description 1
- 239000001814 pectin Substances 0.000 description 1
- 229920001277 pectin Polymers 0.000 description 1
- 239000000816 peptidomimetic Substances 0.000 description 1
- 239000003209 petroleum derivative Substances 0.000 description 1
- 239000000546 pharmaceutical excipient Substances 0.000 description 1
- HOKBIQDJCNTWST-UHFFFAOYSA-N phosphanylidenezinc;zinc Chemical compound [Zn].[Zn]=P.[Zn]=P HOKBIQDJCNTWST-UHFFFAOYSA-N 0.000 description 1
- 229910000073 phosphorus hydride Inorganic materials 0.000 description 1
- 230000035479 physiological effects, processes and functions Effects 0.000 description 1
- 230000035790 physiological processes and functions Effects 0.000 description 1
- 235000002949 phytic acid Nutrition 0.000 description 1
- 239000000467 phytic acid Substances 0.000 description 1
- 229940068041 phytic acid Drugs 0.000 description 1
- 239000003375 plant hormone Substances 0.000 description 1
- 230000001863 plant nutrition Effects 0.000 description 1
- 235000021118 plant-derived protein Nutrition 0.000 description 1
- 238000007747 plating Methods 0.000 description 1
- 229920001084 poly(chloroprene) Polymers 0.000 description 1
- 229920002401 polyacrylamide Polymers 0.000 description 1
- 229920000058 polyacrylate Polymers 0.000 description 1
- 229920002239 polyacrylonitrile Polymers 0.000 description 1
- 235000010958 polyglycerol polyricinoleate Nutrition 0.000 description 1
- 239000002861 polymer material Substances 0.000 description 1
- 238000006116 polymerization reaction Methods 0.000 description 1
- ODGAOXROABLFNM-UHFFFAOYSA-N polynoxylin Chemical compound O=C.NC(N)=O ODGAOXROABLFNM-UHFFFAOYSA-N 0.000 description 1
- 229920001451 polypropylene glycol Polymers 0.000 description 1
- 229920001296 polysiloxane Polymers 0.000 description 1
- 229920002223 polystyrene Polymers 0.000 description 1
- 239000013641 positive control Substances 0.000 description 1
- 230000029279 positive regulation of transcription, DNA-dependent Effects 0.000 description 1
- 230000004481 post-translational protein modification Effects 0.000 description 1
- 229940097322 potassium arsenite Drugs 0.000 description 1
- 239000001103 potassium chloride Substances 0.000 description 1
- 235000011164 potassium chloride Nutrition 0.000 description 1
- OTYBMLCTZGSZBG-UHFFFAOYSA-L potassium sulfate Chemical compound [K+].[K+].[O-]S([O-])(=O)=O OTYBMLCTZGSZBG-UHFFFAOYSA-L 0.000 description 1
- 229910052939 potassium sulfate Inorganic materials 0.000 description 1
- 235000011151 potassium sulphates Nutrition 0.000 description 1
- HEQWEGCSZXMIJQ-UHFFFAOYSA-M potassium;oxoarsinite Chemical compound [K+].[O-][As]=O HEQWEGCSZXMIJQ-UHFFFAOYSA-M 0.000 description 1
- 230000002028 premature Effects 0.000 description 1
- 239000003755 preservative agent Substances 0.000 description 1
- 230000037452 priming Effects 0.000 description 1
- CPTBDICYNRMXFX-UHFFFAOYSA-N procarbazine Chemical compound CNNCC1=CC=C(C(=O)NC(C)C)C=C1 CPTBDICYNRMXFX-UHFFFAOYSA-N 0.000 description 1
- 229960000624 procarbazine Drugs 0.000 description 1
- 229960001586 procarbazine hydrochloride Drugs 0.000 description 1
- BUCOQPHDYUOJSI-UHFFFAOYSA-N prohexadione Chemical compound CCC(=O)C1C(=O)CC(C(O)=O)CC1=O BUCOQPHDYUOJSI-UHFFFAOYSA-N 0.000 description 1
- 235000019260 propionic acid Nutrition 0.000 description 1
- 150000003180 prostaglandins Chemical class 0.000 description 1
- 239000011814 protection agent Substances 0.000 description 1
- 229960000856 protein c Drugs 0.000 description 1
- 108010076078 protein kinase-phosphatase NTRB Proteins 0.000 description 1
- 235000021251 pulses Nutrition 0.000 description 1
- CLKZWXHKFXZIMA-UHFFFAOYSA-N pyrinuron Chemical compound C1=CC([N+](=O)[O-])=CC=C1NC(=O)NCC1=CC=CN=C1 CLKZWXHKFXZIMA-UHFFFAOYSA-N 0.000 description 1
- 229910052903 pyrophyllite Inorganic materials 0.000 description 1
- 108010005284 pyruvate-flavodoxin oxidoreductase Proteins 0.000 description 1
- 238000004445 quantitative analysis Methods 0.000 description 1
- 238000010791 quenching Methods 0.000 description 1
- 230000000171 quenching effect Effects 0.000 description 1
- JETDZFFCRPFPDH-UHFFFAOYSA-N quinacrine mustard dihydrochloride Chemical compound [H+].[H+].[Cl-].[Cl-].C1=C(Cl)C=CC2=C(NC(C)CCCN(CCCl)CCCl)C3=CC(OC)=CC=C3N=C21 JETDZFFCRPFPDH-UHFFFAOYSA-N 0.000 description 1
- IUVKMZGDUIUOCP-BTNSXGMBSA-N quinbolone Chemical compound O([C@H]1CC[C@H]2[C@H]3[C@@H]([C@]4(C=CC(=O)C=C4CC3)C)CC[C@@]21C)C1=CCCC1 IUVKMZGDUIUOCP-BTNSXGMBSA-N 0.000 description 1
- 230000018612 quorum sensing Effects 0.000 description 1
- 239000000700 radioactive tracer Substances 0.000 description 1
- 238000002708 random mutagenesis Methods 0.000 description 1
- 230000007420 reactivation Effects 0.000 description 1
- 101150079601 recA gene Proteins 0.000 description 1
- 238000010188 recombinant method Methods 0.000 description 1
- 238000004064 recycling Methods 0.000 description 1
- 230000022532 regulation of transcription, DNA-dependent Effects 0.000 description 1
- 238000002271 resection Methods 0.000 description 1
- 230000012423 response to bacterium Effects 0.000 description 1
- 230000000717 retained effect Effects 0.000 description 1
- 238000003757 reverse transcription PCR Methods 0.000 description 1
- 239000002336 ribonucleotide Substances 0.000 description 1
- 125000002652 ribonucleotide group Chemical group 0.000 description 1
- 108020004418 ribosomal RNA Proteins 0.000 description 1
- 108091092562 ribozyme Proteins 0.000 description 1
- 239000004170 rice bran wax Substances 0.000 description 1
- 235000019384 rice bran wax Nutrition 0.000 description 1
- 229960001225 rifampicin Drugs 0.000 description 1
- 230000002786 root growth Effects 0.000 description 1
- 101150085857 rpo2 gene Proteins 0.000 description 1
- 238000005001 rutherford backscattering spectroscopy Methods 0.000 description 1
- 101150025220 sacB gene Proteins 0.000 description 1
- 210000003296 saliva Anatomy 0.000 description 1
- 238000002804 saturated mutagenesis Methods 0.000 description 1
- 239000013535 sea water Substances 0.000 description 1
- 238000001004 secondary ion mass spectrometry Methods 0.000 description 1
- 239000011669 selenium Substances 0.000 description 1
- 229910052711 selenium Inorganic materials 0.000 description 1
- 230000009919 sequestration Effects 0.000 description 1
- 239000004208 shellac Substances 0.000 description 1
- 235000013874 shellac Nutrition 0.000 description 1
- 229940113147 shellac Drugs 0.000 description 1
- ZLGIYFNHBLSMPS-ATJNOEHPSA-N shellac Chemical compound OCCCCCC(O)C(O)CCCCCCCC(O)=O.C1C23[C@H](C(O)=O)CCC2[C@](C)(CO)[C@@H]1C(C(O)=O)=C[C@@H]3O ZLGIYFNHBLSMPS-ATJNOEHPSA-N 0.000 description 1
- 239000012176 shellac wax Substances 0.000 description 1
- 108091006024 signal transducing proteins Proteins 0.000 description 1
- 102000034285 signal transducing proteins Human genes 0.000 description 1
- 230000011664 signaling Effects 0.000 description 1
- 239000000377 silicon dioxide Substances 0.000 description 1
- 229910052708 sodium Inorganic materials 0.000 description 1
- PTLRDCMBXHILCL-UHFFFAOYSA-M sodium arsenite Chemical compound [Na+].[O-][As]=O PTLRDCMBXHILCL-UHFFFAOYSA-M 0.000 description 1
- JGFYQVQAXANWJU-UHFFFAOYSA-M sodium fluoroacetate Chemical compound [Na+].[O-]C(=O)CF JGFYQVQAXANWJU-UHFFFAOYSA-M 0.000 description 1
- SUKJFIGYRHOWBL-UHFFFAOYSA-N sodium hypochlorite Chemical compound [Na+].Cl[O-] SUKJFIGYRHOWBL-UHFFFAOYSA-N 0.000 description 1
- 239000004317 sodium nitrate Substances 0.000 description 1
- 235000010344 sodium nitrate Nutrition 0.000 description 1
- DEWVPZYHFVYXMZ-QCILGFJPSA-M sodium;(3ar,4as,8ar,8bs)-2,2,7,7-tetramethyl-4a,5,8a,8b-tetrahydro-[1,3]dioxolo[3,4]furo[1,3-d][1,3]dioxine-3a-carboxylate Chemical compound [Na+].O([C@H]12)C(C)(C)OC[C@@H]1O[C@]1(C([O-])=O)[C@H]2OC(C)(C)O1 DEWVPZYHFVYXMZ-QCILGFJPSA-M 0.000 description 1
- 244000000000 soil microbiome Species 0.000 description 1
- 239000008247 solid mixture Substances 0.000 description 1
- 238000005063 solubilization Methods 0.000 description 1
- 230000007928 solubilization Effects 0.000 description 1
- 239000000600 sorbitol Substances 0.000 description 1
- 235000012424 soybean oil Nutrition 0.000 description 1
- 239000003549 soybean oil Substances 0.000 description 1
- 108010068698 spleen exonuclease Proteins 0.000 description 1
- 230000002269 spontaneous effect Effects 0.000 description 1
- 239000007921 spray Substances 0.000 description 1
- 229910001220 stainless steel Inorganic materials 0.000 description 1
- 239000010935 stainless steel Substances 0.000 description 1
- 238000010561 standard procedure Methods 0.000 description 1
- 239000008223 sterile water Substances 0.000 description 1
- 230000001954 sterilising effect Effects 0.000 description 1
- 238000004659 sterilization and disinfection Methods 0.000 description 1
- 235000021012 strawberries Nutrition 0.000 description 1
- 229960005322 streptomycin Drugs 0.000 description 1
- 238000006467 substitution reaction Methods 0.000 description 1
- 150000005846 sugar alcohols Chemical class 0.000 description 1
- 150000008163 sugars Chemical class 0.000 description 1
- 239000013589 supplement Substances 0.000 description 1
- 238000001356 surgical procedure Methods 0.000 description 1
- 238000013268 sustained release Methods 0.000 description 1
- 239000012730 sustained-release form Substances 0.000 description 1
- 230000009044 synergistic interaction Effects 0.000 description 1
- 235000020357 syrup Nutrition 0.000 description 1
- 239000006188 syrup Substances 0.000 description 1
- 230000009897 systematic effect Effects 0.000 description 1
- 239000008399 tap water Substances 0.000 description 1
- 235000020679 tap water Nutrition 0.000 description 1
- 229940095064 tartrate Drugs 0.000 description 1
- 230000002123 temporal effect Effects 0.000 description 1
- IWVCMVBTMGNXQD-UHFFFAOYSA-N terramycin dehydrate Natural products C1=CC=C2C(O)(C)C3C(O)C4C(N(C)C)C(O)=C(C(N)=O)C(=O)C4(O)C(O)=C3C(=O)C2=C1O IWVCMVBTMGNXQD-UHFFFAOYSA-N 0.000 description 1
- 239000004753 textile Substances 0.000 description 1
- YTQVHRVITVLIRD-UHFFFAOYSA-L thallium sulfate Chemical compound [Tl+].[Tl+].[O-]S([O-])(=O)=O YTQVHRVITVLIRD-UHFFFAOYSA-L 0.000 description 1
- 229940119523 thallium sulfate Drugs 0.000 description 1
- 229910000374 thallium(I) sulfate Inorganic materials 0.000 description 1
- 239000002562 thickening agent Substances 0.000 description 1
- RYYWUUFWQRZTIU-UHFFFAOYSA-K thiophosphate Chemical compound [O-]P([O-])([O-])=S RYYWUUFWQRZTIU-UHFFFAOYSA-K 0.000 description 1
- 229960002447 thiram Drugs 0.000 description 1
- KUAZQDVKQLNFPE-UHFFFAOYSA-N thiram Chemical compound CN(C)C(=S)SSC(=S)N(C)C KUAZQDVKQLNFPE-UHFFFAOYSA-N 0.000 description 1
- 239000001585 thymus vulgaris Substances 0.000 description 1
- 238000003971 tillage Methods 0.000 description 1
- 229940098465 tincture Drugs 0.000 description 1
- 101150031871 tnrA gene Proteins 0.000 description 1
- 238000003325 tomography Methods 0.000 description 1
- 239000003053 toxin Substances 0.000 description 1
- 231100000765 toxin Toxicity 0.000 description 1
- 108700012359 toxins Proteins 0.000 description 1
- 229910021655 trace metal ion Inorganic materials 0.000 description 1
- UZKQTCBAMSWPJD-UQCOIBPSSA-N trans-Zeatin Natural products OCC(/C)=C\CNC1=NC=NC2=C1N=CN2 UZKQTCBAMSWPJD-UQCOIBPSSA-N 0.000 description 1
- UZKQTCBAMSWPJD-FARCUNLSSA-N trans-zeatin Chemical compound OCC(/C)=C/CNC1=NC=NC2=C1N=CN2 UZKQTCBAMSWPJD-FARCUNLSSA-N 0.000 description 1
- 108091006106 transcriptional activators Proteins 0.000 description 1
- 238000012085 transcriptional profiling Methods 0.000 description 1
- 238000012546 transfer Methods 0.000 description 1
- 230000009261 transgenic effect Effects 0.000 description 1
- 238000003146 transient transfection Methods 0.000 description 1
- 230000014621 translational initiation Effects 0.000 description 1
- 230000005945 translocation Effects 0.000 description 1
- 238000002054 transplantation Methods 0.000 description 1
- 150000003852 triazoles Chemical class 0.000 description 1
- ZIBGPFATKBEMQZ-UHFFFAOYSA-N triethylene glycol Chemical compound OCCOCCOCCO ZIBGPFATKBEMQZ-UHFFFAOYSA-N 0.000 description 1
- 230000001228 trophic effect Effects 0.000 description 1
- 229940116269 uric acid Drugs 0.000 description 1
- 239000004474 valine Substances 0.000 description 1
- 235000014393 valine Nutrition 0.000 description 1
- 230000017260 vegetative to reproductive phase transition of meristem Effects 0.000 description 1
- 239000003981 vehicle Substances 0.000 description 1
- 108700026220 vif Genes Proteins 0.000 description 1
- 239000011782 vitamin Substances 0.000 description 1
- 235000013343 vitamin Nutrition 0.000 description 1
- 229930003231 vitamin Natural products 0.000 description 1
- 229940088594 vitamin Drugs 0.000 description 1
- 235000019163 vitamin B12 Nutrition 0.000 description 1
- 239000011715 vitamin B12 Substances 0.000 description 1
- 235000001892 vitamin D2 Nutrition 0.000 description 1
- 239000011653 vitamin D2 Substances 0.000 description 1
- MECHNRXZTMCUDQ-RKHKHRCZSA-N vitamin D2 Chemical compound C1(/[C@@H]2CC[C@@H]([C@]2(CCC1)C)[C@H](C)/C=C/[C@H](C)C(C)C)=C\C=C1\C[C@@H](O)CCC1=C MECHNRXZTMCUDQ-RKHKHRCZSA-N 0.000 description 1
- 235000005282 vitamin D3 Nutrition 0.000 description 1
- 239000011647 vitamin D3 Substances 0.000 description 1
- QYSXJUFSXHHAJI-YRZJJWOYSA-N vitamin D3 Chemical compound C1(/[C@@H]2CC[C@@H]([C@]2(CCC1)C)[C@H](C)CCCC(C)C)=C\C=C1\C[C@@H](O)CCC1=C QYSXJUFSXHHAJI-YRZJJWOYSA-N 0.000 description 1
- 229940021056 vitamin d3 Drugs 0.000 description 1
- 101150001289 vnfH gene Proteins 0.000 description 1
- 101150087820 vnfK gene Proteins 0.000 description 1
- 229960005080 warfarin Drugs 0.000 description 1
- PJVWKTKQMONHTI-UHFFFAOYSA-N warfarin Chemical compound OC=1C2=CC=CC=C2OC(=O)C=1C(CC(=O)C)C1=CC=CC=C1 PJVWKTKQMONHTI-UHFFFAOYSA-N 0.000 description 1
- 239000002699 waste material Substances 0.000 description 1
- 235000015099 wheat brans Nutrition 0.000 description 1
- 238000012070 whole genome sequencing analysis Methods 0.000 description 1
- 229920001285 xanthan gum Polymers 0.000 description 1
- 239000000230 xanthan gum Substances 0.000 description 1
- 235000010493 xanthan gum Nutrition 0.000 description 1
- 229940082509 xanthan gum Drugs 0.000 description 1
- 239000012138 yeast extract Substances 0.000 description 1
- 229940023877 zeatin Drugs 0.000 description 1
- 239000005019 zein Substances 0.000 description 1
- 229940093612 zein Drugs 0.000 description 1
- 229940048462 zinc phosphide Drugs 0.000 description 1
Images


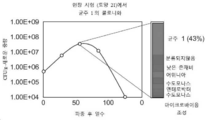
















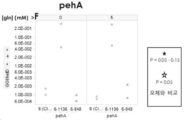






























Classifications
-
- C—CHEMISTRY; METALLURGY
- C05—FERTILISERS; MANUFACTURE THEREOF
- C05F—ORGANIC FERTILISERS NOT COVERED BY SUBCLASSES C05B, C05C, e.g. FERTILISERS FROM WASTE OR REFUSE
- C05F11/00—Other organic fertilisers
- C05F11/08—Organic fertilisers containing added bacterial cultures, mycelia or the like
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/02—Preparation of hybrid cells by fusion of two or more cells, e.g. protoplast fusion
- C12N15/03—Bacteria
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01C—PLANTING; SOWING; FERTILISING
- A01C21/00—Methods of fertilising, sowing or planting
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01N—PRESERVATION OF BODIES OF HUMANS OR ANIMALS OR PLANTS OR PARTS THEREOF; BIOCIDES, e.g. AS DISINFECTANTS, AS PESTICIDES OR AS HERBICIDES; PEST REPELLANTS OR ATTRACTANTS; PLANT GROWTH REGULATORS
- A01N63/00—Biocides, pest repellants or attractants, or plant growth regulators containing microorganisms, viruses, microbial fungi, animals or substances produced by, or obtained from, microorganisms, viruses, microbial fungi or animals, e.g. enzymes or fermentates
- A01N63/20—Bacteria; Substances produced thereby or obtained therefrom
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/195—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/195—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria
- C07K14/235—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria from Bordetella (G)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/52—Genes encoding for enzymes or proenzymes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/74—Vectors or expression systems specially adapted for prokaryotic hosts other than E. coli, e.g. Lactobacillus, Micromonospora
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/1048—Glycosyltransferases (2.4)
- C12N9/1051—Hexosyltransferases (2.4.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/16—Hydrolases (3) acting on ester bonds (3.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/24—Hydrolases (3) acting on glycosyl compounds (3.2)
- C12N9/2402—Hydrolases (3) acting on glycosyl compounds (3.2) hydrolysing O- and S- glycosyl compounds (3.2.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y204/00—Glycosyltransferases (2.4)
- C12Y204/01—Hexosyltransferases (2.4.1)
- C12Y204/01021—Starch synthase (2.4.1.21)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y301/00—Hydrolases acting on ester bonds (3.1)
- C12Y301/03—Phosphoric monoester hydrolases (3.1.3)
- C12Y301/03012—Trehalose-phosphatase (3.1.3.12)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y302/00—Hydrolases acting on glycosyl compounds, i.e. glycosylases (3.2)
- C12Y302/01—Glycosidases, i.e. enzymes hydrolysing O- and S-glycosyl compounds (3.2.1)
- C12Y302/01015—Polygalacturonase (3.2.1.15)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y302/00—Hydrolases acting on glycosyl compounds, i.e. glycosylases (3.2)
- C12Y302/01—Glycosidases, i.e. enzymes hydrolysing O- and S-glycosyl compounds (3.2.1)
- C12Y302/01141—4-Alpha-D-{(1->4)-alpha-D-glucano} trehalose trehalohydrolase (3.2.1.141)
Landscapes
- Chemical & Material Sciences (AREA)
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Organic Chemistry (AREA)
- Genetics & Genomics (AREA)
- Engineering & Computer Science (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biochemistry (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Biomedical Technology (AREA)
- Biotechnology (AREA)
- Molecular Biology (AREA)
- Microbiology (AREA)
- Medicinal Chemistry (AREA)
- Biophysics (AREA)
- Plant Pathology (AREA)
- Physics & Mathematics (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Gastroenterology & Hepatology (AREA)
- Soil Sciences (AREA)
- Environmental Sciences (AREA)
- Cell Biology (AREA)
- Agronomy & Crop Science (AREA)
- Pest Control & Pesticides (AREA)
- Virology (AREA)
- Dentistry (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Breeding Of Plants And Reproduction By Means Of Culturing (AREA)
Abstract
본 발명은 다음을 포함하는 박테리아 조성물을 제공한다: 농업 시스템에서 대기 질소를 고정시키는 하나 이상의 유전자 조작된 박테리아 균주, 이 때 상기 박테리아 균주는 bcsII, bcsIII, yjbE, fhaB, pehA, glgA, otsB, treZ 및 cysZ로 이루어진 군에서 선택된 하나 이상의 유전자에서의 변형을 포함한다. 본 발명은 식물에서 식물 성장 촉진 박테리아 균주의 콜로니화를 증가시키는 박테리아 조성물 및 방법을 추가로 제공하며, 이 때 상기 식물 성장 촉진 박테리아 균주는 상기 식물의 콜로니화를 증가시키도록 리모델링 되어있다. 또 다른 양상에서, 본 발명은 식물에 이용가능한 질소 또는 질소 고정화를 증가시키는 방법을 제공한다.
Description
상호 참조
본 출원은 2017년 8월 9일 출원된 미국 가특허출원 제 62/543,288호의 이익을 주장하며, 이 출원은 본 명세서에 참고문헌으로 포함된다.
발명의 배경
2050년까지 유엔 식량 농업기구는 증가하는 인구의 수요를 충족시키기 위해 총 식량 생산이 70% 증가해야 한다고 예상하는데, 이러한 과제는 담수 자원 감소, 경작지 경쟁 증가, 에너지 가격 상승, 투입 비용 증가, 그리고 작물이 보다 건조하고, 보다 뜨겁고 보다 극단적인 지구 기후의 압력에 적응해야 할 필요 가능성을 비롯한 수많은 요인들에 의해 악화된다.
관심 분야 중 하나는 질소 고정의 개선에 있다. 질소 기체 (N2) 는 지구 대기의 주요 성분이다. 또한, 원소 질소 (N) 는 살아있는 유기체를 구성하는 많은 화합물의 중요한 성분이다. 그러나 많은 유기체는 성장과 번식과 같은 생리학적 과정에 사용되는 화학 물질을 합성하기 위해 N2를 직접 사용할 수 없다. N2를 이용하기 위해서는 N2를 수소와 결합시켜야 한다. 수소와 N2의 결합을 질소 고정이라 지칭한다. 화학적으로든 생물학적으로든 질소 고정에는 많은 양의 에너지 투자가 필요하다. 생물학적 시스템에서, 질소고정효소로 알려진 효소는 질소 고정을 가져오는 반응을 촉매한다. 질소 고정 연구의 중요한 목표는 이 표현형을 비-콩과 식물, 특히, 중요한 작물학적 풀, 가령, 밀, 쌀 및 옥수수로 확장하는 것이다. 리조비아 (rhizobia)와 콩과 식물 사이의 질소 고정 공생의 발달을 이해하는데 엄청난 진전이 있었음에도 불구하고 비-콩과 작물에 질소 고정 결절을 유도하기 위해 그 지식을 사용하는 경로는 여전히 명확하지 않다. 한편, 비료와 같이 충분한 질소의 보충 공급원을 제공해야 하는 과제는 식품 생산 증가에 대한 수요가 증가함에 따라 계속 증가 할 것이다.
발명의 요약
한 양상에서, 본 발명은 농업 시스템에서 대기 질소를 고정시키는 하나 이상의 유전자 조작된 박테리아 균주를 포함하는 박테리아 조성물을 제공하며, 여기서 박테리아 균주는 bcsII, bcsIII, yjbE, fhaB, pehA, glgA, otsB, treZ 및 cysZ로 이루어진 군에서 선택된 하나 이상의 유전자에서의 변형을 포함한다.
또 다른 양상에서, 본 발명은 식물 성장 촉진 박테리아 균주를 포함하는 박테리아 조성물을 제공하며, 여기서 상기 균주는 식물에서 상기 식물 성장 촉진 박테리아 균주의 콜로니화를 증가시키도록 리모델링되었다. 일부 경우에, 상기 식물 성장 촉진 박테리아 균주의 콜로니화는 상기 식물의 뿌리에서 발생한다. 일부 경우에, 상기 박테리아 균주는 세포외 다당류의 생성에 관여하는 효소 또는 경로에서의 유전자 변형을 포함한다. 일부 경우에, 상기 유전자 변형은 bcsII, bcsIII 및 yjbE로 구성된 그룹에서 선택된 유전자에 존재한다. 일부 경우에, 상기 박테리아 균주는 사상성 헤마글루티닌의 생성에 관여하는 효소 또는 경로에서의 유전자 변형을 포함한다. 일부 경우에, 상기 유전자 변형은 fhaB 유전자에 존재한다. 일부 경우에, 상기 박테리아 균주는 엔도-폴리갈락투로나제 생성에 관여하는 효소 또는 경로에서의 유전자 변형을 포함한다. 일부 경우에, 상기 유전자 변형은 pehA 유전자에 존재한다. 일부 경우에, 상기 박테리아 균주는 트레할로스의 생성에 관여하는 효소 또는 경로에서의 유전자 변형을 포함한다. 일부 경우에, 상기 유전자 변형은 otsB 및 treZ로 구성된 그룹에서 선택된 유전자에 존재한다. 일부 경우에, 상기 박테리아 조성물은 현장(field)에 처리하기 위해 제형화된다. 일부 경우에, 상기 식물 성장 촉진 박테리아는 상기 식물에 영양소를 제공한다. 일부 경우에, 상기 식물 성장 촉진 박테리아는 상기 식물에 고정 질소를 제공한다. 일부 경우에, 상기 식물은 옥수수, 보리, 밀, 수수, 대두 및 쌀로 구성된 군에서 선택된다.
또 다른 양상에서, 본 발명은 식물에서 식물 성장 촉진 박테리아 균주의 콜로니화를 증가시키는 방법을 제공하며, 이 방법은 다음을 포함한다: 세포외 다당류 생성, 엔도-폴리갈락투로나제 생성, 및 트레할로스 생성으로 구성된 그룹에서 선택된 경로에 관여하는 유전자에서의 유전자 변형을 상기 식물 성장 촉진 박테리아 균주에 도입하는 단계.
또 다른 양상에서, 본 발명은 식물에 이용 가능한 질소를 증가시키는 방법을 제공하며, 상기 방법은 식물에 복수의 리모델링된 박테리아를 처리하는 단계를 포함하고, 상기 복수의 리모델링된 박테리아는 glgA의 감소된 발현을 가진다.
일부 경우에, 상기 리모델링된 박테리아는 상기 리모델링된 박테리아와 동일한 종의 비-리모델링된 박테리아의 동화도에 비해 상기 리모델링된 박테리아 내에서 보다 낮은 질소 동화도를 가진다.
또 다른 양상에서, 본 발명은 식물에 이용가능한 질소 고정을 증가시키는 방법을 제공하며, 상기 방법은 식물에 복수의 리모델링된 박테리아를 처리하는 단계를 포함하고, 상기 복수의 리모델링된 박테리아는 최소한 하나의 질소분해효소 보조인자의 증가된 발현을 가진다. 일부 경우에서, 상기 증가된 질소분해효소 보조인자는 황이다. 일부 경우에서, 상기 리모델링된 박테리아는 cysZ의 증가된 발현을 가진다. 일부 경우에서, cysZ는 황 운반체이다.
참고문헌으로 포함
본 명세서에 언급된 모든 간행물 및 특허 출원은 각각의 개별 간행물 또는 특허 출원이 구체적으로 및 개별적으로 참조로 포함되도록 지시된 것과 동일한 정도로 본 명세서에 참조 문헌으로 인용된다.
본 발명의 새로운 특징들은 첨부된 청구범위에 구체적으로 제시된다. 본 발명의 특징 및 이점들은 본 발명의 원리를 이용하는 예시적 구체예들을 제시하는 하기 상세한 설명 그리고 다음과 같은 첨부도면을 참고하여 보다 잘 이해될 것이다:
도 1A는 콜로니화에 대해 시험한 다양한 현장 토양의 토양 조직도를 도시한다. 몇 가지 미생물들을 본래 얻은 토양들은 별표로 표시된다.
도 1B는 4가지 상이한 토양 유형 (원)에 걸쳐 시험된 균주 1 및 균주 5의 콜로니화율을 도시한다. 두 균주는 다양한 토양 유형에 걸쳐 비교적 강력한 콜로니화 프로파일을 나타냈다.
도 1C는 성장기의 기간에 걸쳐 현장 시험에서 시험된 균주 1의 콜로니화를 도시한다. 균주 1은 식재 후 12주까지 옥수수 조직에서 유지되고, 그 시간 이후에 콜로니화 감소를 나타내기 시작한다.
도 2A는 구체예들에 따른 미생물 육종의 개략도를 도시한다.
도 2B는 도 2A에 도시된 마이크로바이옴 조성의 측정에 관한 확대도를 도시한다.
도 2C는 실시예 7에서 이용된 뿌리의 샘플링을 도시한다.
도 3A-3C는 질산염이 많은 농경 토양과 유사한 조건에서 체외에서 질소를 고정하고 배설하는 유도체 미생물을 보여준다. 도 3A는 2-도메인 ATase-AR 효소 및 AmtB로 도시된 주요 노드 NifL, NifA, GS, GlnE를 포함하여 PBC6.1에서 질소 고정 및 동화를 제어하는 조절 네트워크를 도시한다. 도 3B는 코사코니아 사카리 (Kosakonia sacchari) 단리주 PBC6.1의 게놈을 도시함을 나타낸다. 게놈을 둘러싸는 3개의 트랙은 각각 PBC6.1, PBC6.38로부터의 전사 데이터 및 균주 사이의 차등 발현을 전달한다. 도 3C는 질소 고정 유전자 클러스터를 도시하고 전사 데이터는 보다 상세히 확대된다.
도 4는 옥수수 뿌리의 뿌리-관련 미생물군 중 PBC6.1 콜로니화가 대략 21% 존재비임을 보여준다. 존재비 데이터는 PBC6.1을 접종하고 온실 조건에서 성장한 옥수수 식물의 뿌리줄기 및 내생포자의 16S 앰플리콘 시퀀싱에 기초한다.
도 5는 K. 사카리의 리모델링된 균주에서 qPCR로 나타낸 3가지 관심 유전자의 증가된 발현을 도시한다. 균주들을 5 mM 글루타민을 함유하는 배지에서 배양하였다.
도 6은 글루타민의 존재 또는 부재하에 몇몇 리모델링된 균주의 질소 환원 활성을 도시한다.
도 7은 본 발명에 기재된 몇몇 균주를 사용한 시험관내 부착 분석의 결과를 도시한다.
도 8A는 본 명세서에 기재된 바와 같이 fhaB의 발현이 증가된 리모델링된 균주를 사용한 시험관내 부착 분석의 결과를 도시한다. 이 균주는 모 균주 6-848에 비해 유의하게 증가한 폴드 회수를 나타내었다. (2.3x, p = 0.013).
도 8B는 본 발명에 기재된 몇몇 균주를 사용한 시험관내 부착 분석의 결과를 도시한다.
도 9는 상향조절된 yjbE2 유전자 및 bcsIII 오페론을 갖는 균주에 의한 부착 증가를 도시한다. 2 개의 리모델링된 균주는 6-848에 비해 약간의 증가를 보여주었다: 6-112 (6-848 + yjbE2-prm1): 1.9x (p = 0.07), 및 6-1126 (6-848 + bcsIII-prm2): 1.7x (p = 0.06). 균주 6-1127 (6-848 + bcsIII-prm9)은 6-848에 비해 유의한 2.5x 증가를 보여주었다 (p = 0.0005)
도 10은 시험관내 부착 분석 결과를 도시한다. 0은 물에 미생물이 첨가되지 않았고, 462는 물에 추가된 DH10B 대장균 (식물과 관련이 없는 대조군)이며, 6-848 및 6-881은 본 발명의 분석에 사용된 다른 균주들의 모체 균주 (양성 대조군)이다. 접종되지 않은 대장균 대조군은 일부 배경을 보여주지만, 야생형 균주 6 및 모체 대조군 6-848 및 6-881은 음성 대조군에 비해 유의한 부착을 나타낸다 (p <0.05).
도 11은 pehA 유전자, yjbE2 유전자 및 bcsII 오페론에서 변형을 갖는 균주의 부착 증가를 도시한다.
도 12는 글루타민의 존재 및 부재하에 리모델링된 균주에서 변형된 유전자의 과발현을 도시한다.
도 13A는 글루타민의 존재 및 부재하에 리모델링된 균주에서 변형된 유전자의 과발현을 도시한다.
도 13B는 글루타민의 존재 및 부재하에 리모델링된 균주에서 변형된 유전자의 과발현을 도시한다.
도 13C는 글루타민의 존재 및 부재하에 리모델링된 균주에서 변형된 유전자의 과발현을 도시한다.
도 14는 본 명세서에 기재된 3가지 균주에 대한 혐기성 조건 하에서의 암모늄 배설을 도시한다.
도 15는 본 명세서에 기재된 몇몇 균주에 대한 암모늄 배설을 도시한다.
도 16은 본 명세서에 기재된 또 다른 균주에 대한 암모늄 배설을 도시한다.
도 17은 본 명세서에 기재된 또 다른 균주에 대한 암모늄 배설을 도시한다.
도 18은 본 명세서에 기재된 몇몇 균주에 대한 질소 환원 활성을 도시한다.
도 19는 본 명세서에 기재된 또 다른 균주에 대한 암모늄 배설을 도시한다.
도 20은 본 명세서에 기재된 몇몇 균주에 대한 질소 환원 활성을 도시한다.
도 21은 본 명세서에 기재된 또 다른 균주에 대한 암모늄 배설을 도시한다.
도 22A는 질소의 존재 또는 부재시 본 명세서에 기재된 몇몇 균주에 대한 질소 환원 활성을 도시한다.
도 22B는 질소의 존재 또는 부재시 본 명세서에 기재된 몇몇 균주에 대한 질소 환원 활성을 도시한다.
도 23A는 본 명세서에 기재된 균주들에 대한 암모늄 배설을 도시한다.
도 23B는 본 명세서에 기재된 균주들에 대한 암모늄 배설을 도시한다.
도 24는 질소의 존재 또는 부재시 본 명세서에 기재된 몇몇 균주에 대한 질소 환원 활성을 도시한다.
도 25는 본 명세서에 기재된 균주들에 대한 암모늄 배설을 도시한다.
도 26A는 질소의 존재 또는 부재시 본 명세서에 기재된 몇몇 균주에 대한 질소 환원 활성을 도시한다.
도 26B는 질소의 존재 또는 부재시 본 명세서에 기재된 몇몇 균주에 대한 질소 환원 활성을 도시한다.
도 27A는 본 명세서에 기재된 균주들에 대한 암모늄 배설을 도시한다.
도 27B는 본 명세서에 기재된 균주들에 대한 암모늄 배설을 도시한다.
도 28은 온실 분석에서 본 명세서에 기재된 균주들의 콜로니화를 도시한다. 콜로니화에 대한 온실 분석은 p < 0.05의 차이를 구분할 검정력이 없었다.
도 29는 온실 분석에서 본 명세서에 기재된 또 다른 균주들의 콜로니화를 도시한다.
도 30은 온실 분석에서 본 명세서에 기재된 균주들의 콜로니화를 도시한다.
도 31은 온실 분석에서 본 명세서에 기재된 균주들의 콜로니화를 도시한다.
도 32는 온실 분석에서 본 명세서에 기재된 균주들의 콜로니화를 도시한다.
도 33은 본 명세서에 기재된 균주들에 대한 암모늄 배설을 도시한다.
도 34는 질소의 존재 또는 부재시 본 명세서에 기재된 몇몇 균주에 대한 질소 환원 활성을 도시한다.
도 35는 본 명세서에 기재된 균주들에 대한 암모늄 배설을 도시한다.
도 36은 질소의 존재 또는 부재시 본 명세서에 기재된 몇몇 균주에 대한 질소 환원 활성을 도시한다.
도 37은 본 명세서에 기재된 균주들의 콜로니화를 도시한다.
도 38은 본 명세서에 기재된 균주들의 콜로니화를 도시한다.
도 1A는 콜로니화에 대해 시험한 다양한 현장 토양의 토양 조직도를 도시한다. 몇 가지 미생물들을 본래 얻은 토양들은 별표로 표시된다.
도 1B는 4가지 상이한 토양 유형 (원)에 걸쳐 시험된 균주 1 및 균주 5의 콜로니화율을 도시한다. 두 균주는 다양한 토양 유형에 걸쳐 비교적 강력한 콜로니화 프로파일을 나타냈다.
도 1C는 성장기의 기간에 걸쳐 현장 시험에서 시험된 균주 1의 콜로니화를 도시한다. 균주 1은 식재 후 12주까지 옥수수 조직에서 유지되고, 그 시간 이후에 콜로니화 감소를 나타내기 시작한다.
도 2A는 구체예들에 따른 미생물 육종의 개략도를 도시한다.
도 2B는 도 2A에 도시된 마이크로바이옴 조성의 측정에 관한 확대도를 도시한다.
도 2C는 실시예 7에서 이용된 뿌리의 샘플링을 도시한다.
도 3A-3C는 질산염이 많은 농경 토양과 유사한 조건에서 체외에서 질소를 고정하고 배설하는 유도체 미생물을 보여준다. 도 3A는 2-도메인 ATase-AR 효소 및 AmtB로 도시된 주요 노드 NifL, NifA, GS, GlnE를 포함하여 PBC6.1에서 질소 고정 및 동화를 제어하는 조절 네트워크를 도시한다. 도 3B는 코사코니아 사카리 (Kosakonia sacchari) 단리주 PBC6.1의 게놈을 도시함을 나타낸다. 게놈을 둘러싸는 3개의 트랙은 각각 PBC6.1, PBC6.38로부터의 전사 데이터 및 균주 사이의 차등 발현을 전달한다. 도 3C는 질소 고정 유전자 클러스터를 도시하고 전사 데이터는 보다 상세히 확대된다.
도 4는 옥수수 뿌리의 뿌리-관련 미생물군 중 PBC6.1 콜로니화가 대략 21% 존재비임을 보여준다. 존재비 데이터는 PBC6.1을 접종하고 온실 조건에서 성장한 옥수수 식물의 뿌리줄기 및 내생포자의 16S 앰플리콘 시퀀싱에 기초한다.
도 5는 K. 사카리의 리모델링된 균주에서 qPCR로 나타낸 3가지 관심 유전자의 증가된 발현을 도시한다. 균주들을 5 mM 글루타민을 함유하는 배지에서 배양하였다.
도 6은 글루타민의 존재 또는 부재하에 몇몇 리모델링된 균주의 질소 환원 활성을 도시한다.
도 7은 본 발명에 기재된 몇몇 균주를 사용한 시험관내 부착 분석의 결과를 도시한다.
도 8A는 본 명세서에 기재된 바와 같이 fhaB의 발현이 증가된 리모델링된 균주를 사용한 시험관내 부착 분석의 결과를 도시한다. 이 균주는 모 균주 6-848에 비해 유의하게 증가한 폴드 회수를 나타내었다. (2.3x, p = 0.013).
도 8B는 본 발명에 기재된 몇몇 균주를 사용한 시험관내 부착 분석의 결과를 도시한다.
도 9는 상향조절된 yjbE2 유전자 및 bcsIII 오페론을 갖는 균주에 의한 부착 증가를 도시한다. 2 개의 리모델링된 균주는 6-848에 비해 약간의 증가를 보여주었다: 6-112 (6-848 + yjbE2-prm1): 1.9x (p = 0.07), 및 6-1126 (6-848 + bcsIII-prm2): 1.7x (p = 0.06). 균주 6-1127 (6-848 + bcsIII-prm9)은 6-848에 비해 유의한 2.5x 증가를 보여주었다 (p = 0.0005)
도 10은 시험관내 부착 분석 결과를 도시한다. 0은 물에 미생물이 첨가되지 않았고, 462는 물에 추가된 DH10B 대장균 (식물과 관련이 없는 대조군)이며, 6-848 및 6-881은 본 발명의 분석에 사용된 다른 균주들의 모체 균주 (양성 대조군)이다. 접종되지 않은 대장균 대조군은 일부 배경을 보여주지만, 야생형 균주 6 및 모체 대조군 6-848 및 6-881은 음성 대조군에 비해 유의한 부착을 나타낸다 (p <0.05).
도 11은 pehA 유전자, yjbE2 유전자 및 bcsII 오페론에서 변형을 갖는 균주의 부착 증가를 도시한다.
도 12는 글루타민의 존재 및 부재하에 리모델링된 균주에서 변형된 유전자의 과발현을 도시한다.
도 13A는 글루타민의 존재 및 부재하에 리모델링된 균주에서 변형된 유전자의 과발현을 도시한다.
도 13B는 글루타민의 존재 및 부재하에 리모델링된 균주에서 변형된 유전자의 과발현을 도시한다.
도 13C는 글루타민의 존재 및 부재하에 리모델링된 균주에서 변형된 유전자의 과발현을 도시한다.
도 14는 본 명세서에 기재된 3가지 균주에 대한 혐기성 조건 하에서의 암모늄 배설을 도시한다.
도 15는 본 명세서에 기재된 몇몇 균주에 대한 암모늄 배설을 도시한다.
도 16은 본 명세서에 기재된 또 다른 균주에 대한 암모늄 배설을 도시한다.
도 17은 본 명세서에 기재된 또 다른 균주에 대한 암모늄 배설을 도시한다.
도 18은 본 명세서에 기재된 몇몇 균주에 대한 질소 환원 활성을 도시한다.
도 19는 본 명세서에 기재된 또 다른 균주에 대한 암모늄 배설을 도시한다.
도 20은 본 명세서에 기재된 몇몇 균주에 대한 질소 환원 활성을 도시한다.
도 21은 본 명세서에 기재된 또 다른 균주에 대한 암모늄 배설을 도시한다.
도 22A는 질소의 존재 또는 부재시 본 명세서에 기재된 몇몇 균주에 대한 질소 환원 활성을 도시한다.
도 22B는 질소의 존재 또는 부재시 본 명세서에 기재된 몇몇 균주에 대한 질소 환원 활성을 도시한다.
도 23A는 본 명세서에 기재된 균주들에 대한 암모늄 배설을 도시한다.
도 23B는 본 명세서에 기재된 균주들에 대한 암모늄 배설을 도시한다.
도 24는 질소의 존재 또는 부재시 본 명세서에 기재된 몇몇 균주에 대한 질소 환원 활성을 도시한다.
도 25는 본 명세서에 기재된 균주들에 대한 암모늄 배설을 도시한다.
도 26A는 질소의 존재 또는 부재시 본 명세서에 기재된 몇몇 균주에 대한 질소 환원 활성을 도시한다.
도 26B는 질소의 존재 또는 부재시 본 명세서에 기재된 몇몇 균주에 대한 질소 환원 활성을 도시한다.
도 27A는 본 명세서에 기재된 균주들에 대한 암모늄 배설을 도시한다.
도 27B는 본 명세서에 기재된 균주들에 대한 암모늄 배설을 도시한다.
도 28은 온실 분석에서 본 명세서에 기재된 균주들의 콜로니화를 도시한다. 콜로니화에 대한 온실 분석은 p < 0.05의 차이를 구분할 검정력이 없었다.
도 29는 온실 분석에서 본 명세서에 기재된 또 다른 균주들의 콜로니화를 도시한다.
도 30은 온실 분석에서 본 명세서에 기재된 균주들의 콜로니화를 도시한다.
도 31은 온실 분석에서 본 명세서에 기재된 균주들의 콜로니화를 도시한다.
도 32는 온실 분석에서 본 명세서에 기재된 균주들의 콜로니화를 도시한다.
도 33은 본 명세서에 기재된 균주들에 대한 암모늄 배설을 도시한다.
도 34는 질소의 존재 또는 부재시 본 명세서에 기재된 몇몇 균주에 대한 질소 환원 활성을 도시한다.
도 35는 본 명세서에 기재된 균주들에 대한 암모늄 배설을 도시한다.
도 36은 질소의 존재 또는 부재시 본 명세서에 기재된 몇몇 균주에 대한 질소 환원 활성을 도시한다.
도 37은 본 명세서에 기재된 균주들의 콜로니화를 도시한다.
도 38은 본 명세서에 기재된 균주들의 콜로니화를 도시한다.
발명의 상세한 설명
본 발명의 다양한 구체예들을 본 출원에 나타내고 설명하였으나, 해당 분야의 숙련된 기술자들에게 이러한 구체예들이 단지 예로서 제공되는 것임은 자명할 것이다. 본 발명의 범위에서 벗어나지 않는 수많은 변형들, 변화들, 및 치환들이 해당 분야의 숙련된 기술자들에 의해 이루어질 수 있다. 본 출원에 기재된 본 발명의 구체예들에 대한 다양한 대체예들이 사용될 수 있음을 이해하여야 한다.
비료 이용률이 증가하면 환경 문제가 발생하는데 이는 세계의 많은 경제적 스트레스 지역들에서는 그렇지 않을 수도 있다. 또한, 미생물 분야의 많은 통상의 기술자들은 속간 미생물 생성에 초점을 맞추고 있다. 그러나, 속간 미생물로 특성화/분류되는 유전자 조작된 미생물에 가해지는 중대한 규제 부담이 존재한다. 이러한 속간 미생물들은 광범위한 채택 및 실시를 어렵게 하는 보다 높은 규제 부담에 직면할 뿐만 아니라, 방대한 대중 인식 조사에도 직면하게 된다.
현재 시중에는 비-속간인 그리고 비-콩과 작물에서 질소 고정을 증가시킬 수 있는 유전자 조작 미생물이 없다. 이러한 미생물의 부족은 진정으로 환경 친화적이고 보다 지속가능한 21세기 농업 시스템을 도입하는데 도움이 되지 않는 요소이다.
본 발명은 전술한 문제점을 해결하고 작물에서 질소를 용이하게 고정 시키도록 유전자 조작된 비-속간 미생물을 제공한다. 이들 미생물은 속간 미생물로 특성화/분류되지 않기 때문에 이러한 미생물의 가혹한 규제 부담에 직면하지 않을 것이다. 또한, 개시된 비-속간 미생물은 21 세기 농민이 점점 더 많은 양의 외인성 질소 비료를 이용하는 것에 덜 의존하게 하는데 도움이 될 것이다.
유도 미생물 리모델링 (Guided microbial remodeling)은 작물 뿌리를 콜로니화하고 영양소 흡수를 증가시키는 그 최대 잠재력을 구현하기 위해 미생물을 식별, 특성화 및 미세 조정하는 최첨단 프로세스이다. 일부 경우에, 유도 미생물 리모델링은 각 미생물에 자연적으로 존재하는 유전 물질을 사용하여, 작물에 의한 질소 흡수를 증가시킬 수 있다. 더 나은 영양 공급은 작물 품질과 수확량을 향상시킨다.
대기 질소를 암모니아로 환원시키는 것을 담당하는 미생물에는 질소고정효소라 불리는 효소의 복합체가 있으며, 미생물과 식물 모두 글루타민 합성효소 (GS)에 의해 암모니아를 글루타민으로 전환하여 미생물 및 식물 모두의 성장 및 발달에 필수적인 다른 아미노산으로 전환한다. nif 유전자는 질소고정효소의 복합체를 인코드하는 유전자이다. 질소고정효소 이외에도, nif 유전자는 또한 질소 고정과 관련된 많은 조절 단백질을 인코드한다. nif 유전자의 발현은 뿌리줄기에서 낮은 수준의 고정 질소 및 산소 농도에 대한 반응으로 유도된다. 대부분의 질소-고정 박테리아에서 nif 유전자 전사의 조절은 질소에 민감한 NifA 단백질에 의해 이루어진다. 유기체에 사용할 수 있는 고정 질소가 충분하지 않으면 NifA 발현이 활성화되고 NifA는 나머지 nif 유전자를 활성화시킨다. 뿌리줄기에 충분한 양의 환원 질소 또는 산소가 존재하는 경우, 또 다른 단백질 인 NifL이 활성화된다. NifL은 NifA 활성을 억제하여 질소고정효소 형성을 억제한다.
곡물 작물에 대한 질소 이용률을 증가시키기 위해 변경 될 수 있는 미생물 질소 고정의 양상들의 예는 다음을 포함한다:
뿌리줄기에서 미생물과 작물 뿌리 사이의 상호작용을 향상시킨다.
NifA 단백질의 음성 조절을 제거하여 미생물 질소 고정을 향상시킨다. 예를 들어, nifL 유전자의 결실은 NifA 활성의 억제를 제거하고 산소 및 외인성 고정 질소 둘 모두의 존재하에서 nif 발현을 개선시킨다. 또한, 질소-독립적 프로모터의 제어하에 nifA를 발현시키는 것은 질소고정효소 생합성을 환경 질소 및 산소의 감지로부터 단리시킬 수 있다.
미생물의 글루타민에 대한 암모니아 동화를 억제하고 미생물의 암모니아 배출을 증가시켜 식물에 대한 암모니아 가용성을 향상시킨다. 예를 들면, 미생물에 의한 고정 질소의, GS에 의한 글루타민으로의 신속한 동화는 2-도메인 아데닐릴트랜스퍼라제 (ATase) 효소 GlnE에 의하여, 활성을 각각 약화 및 회복시키는 GS의 아데닐릴화 및 탈아데닐릴화를 통해 가역적으로 조절된다. GlnE 단백질을 절두하여 그 아데닐릴제거 (AR) 도메인을 제거하면 항시적으로 아데닐릴화된 글루타민 합성효소가 생성되어, 미생물에 의한 암모니아 동화를 제한하고 세포간 및 세포외 암모니아를 증가시킨다. 마지막으로, 미생물에 의한 암모니아 흡수를 담당하는 운반체를 인코드하는 amtB 유전자를 결실시키면, 암모니아 배설이 더 많아진다.
일부 경우에, 본 명세서에 기재된 방법에 의해 생성된 최종 미생물은 외래 (형질전환) 유전 물질을 함유하지 않으며, 현장에 배포하기에 적합 할 수 있다. 미생물의 유전자 변화는 유기체 자체의 게놈 내에서의 서열 결실 또는 작은 서열 재배열, 가령, 미생물의 게놈 내에서 프로모터 서열의 카피본을 생성하고 다른 유전자 앞에 그 카피본을 삽입하여 이루어질 수 있다. 따라서, 미생물의 게놈에 도입되는 임의의 유전자 요소는 동일한 모체 미생물로부터 유래 될 수 있다. 또한, 미생물 게놈에 도입된 유전자 요소는 비-코딩 유전자 요소 일 수 있다. 생성된 돌연변이는 "마커리스" 일 수 있는데, 이는 박테리아에서 돌연변이를 생성하는데 종종 사용되는 도입되는 항생제 저항성 또는 다른 선별 마커 유전자를 함유하지 않음을 의미한다. 일부 경우에, 유도 미생물 리모델링 과정은 비-형질전환 돌연변이의 생성을 용이하게 하기 위해 일부 보조 DNA 분자를 박테리아 세포 내로 도입하는 것을 포함 할 수 있다. 그러나, 모든 보조 DNA는 상기 균주들로부터 완전히 제거될 수 있다. 예를 들면, 일부 경우 모든 외래 DNA는 상기 균주가 현장 배포에 고려되기 전에 균주로부터 제거될 수 있다. 모든 보조 DNA의 제거는 예를 들어 보조 DNA 서열들의 단일 분자조차도 검출 할 수 있는 민감한 방법인 일루미나 시퀀싱을 통해 확인할 수 있다. 따라서, 유도 미생물 리모델링을 통해 생성된 미생물은 트랜스진 또는 외래 DNA를 포함하지 않을 수 있으며, 균주의 유전자 변화는 서열 결실 및 프로모터 재배열로만 이루어지는데, 이는 자연에서 발생하는 것으로 밝혀졌다.
정의
용어 "폴리뉴클레오티드", "뉴클레오티드", "뉴클레오티드 서열", "핵산" 및 "올리고뉴클레오티드"는 호환적으로 사용된다. 이들은 데옥시리보뉴클레오티드 또는 리보뉴클레오티드, 또는 이의 유사체인, 임의의 길이의 뉴클레오티드들의 폴리머 형태를 지칭한다. 폴리뉴클레오티드는 임의의 3차원 구조를 가질 수 있고, 공지되거나 알려지지 않은 임의의 기능을 수행 할 수 있다. 다음은 폴리뉴클레오티드의 비-제한적 예이다: 유전자 또는 유전자 단편의 코딩 또는 비-코딩 영역, 링키지 분석으로 결정된 유전자좌 (locus), 엑손, 인트론, 메신저 RNA (mRNA), 전달 RNA (tRNA), 리보솜 RNA (rRNA), 짧은 간섭 RNA (siRNA), 짧은 헤어핀 RNA (shRNA), 마이크로 RNA (miRNA), 리보자임, cDNA, 재조합 폴리뉴클레오티드, 분지형 폴리뉴클레오티드, 플라스미드, 벡터, 임의의 서열의 단리된 DNA, 임의의 서열의 단리된 RNA, 핵산 프로브 및 프라이머. 폴리뉴클레오티드는 하나 이상의 변형된 뉴클레오티드, 가령, 메틸화된 뉴클레오티드 및 뉴클레오티드 유사체를 포함할 수 있다. 존재하는 경우, 뉴클레오티드 구조에 대한 변형들은 폴리머 어셈블리 전 또는 이후에 부여될 수 있다. 뉴클레오티드들의 서열은 비-뉴클레오티드 성분들에 의해 중단될 수 있다. 폴리뉴클레오티드는 중합 후, 가령, 표지 성분과의 결합에 의해 추가로 변형될 수 있다.
본 명세서에서 사용되는 "발현"은 폴리뉴클레오티드가 DNA 주형으로부터 (예를 들어, mRNA 또는 다른 RNA 전사체로) 전사되는 과정 및/또는 전사된 mRNA가 후속적으로 펩티드, 폴리펩티드, 또는 단백질로 번역되는 과정을 지칭한다. 전사체 및 암호화된 폴리펩티드는 통칭하여 "유전자 산물"로 지칭 될 수 있다. 폴리뉴클레오티드가 게놈 DNA로부터 유래된 경우, 발현은 진핵 세포에서 mRNA의 스플라이싱을 포함 할 수 있다.
용어 "폴리펩티드", "펩티드" 및 "단백질"은 임의의 길이의 아미노산 폴리머들을 지칭하기 위해 본 명세서에서 호환적으로 사용된다. 상기 중합체는 선형 또는 분지형일 수 있고, 변형된 아미노산을 포함할 수 있으며, 비-아미노산에 의해 불연속성을 가질 수 있다. 상기 용어는 또한 변형된 아미노산 중합체를 포함하는데; 예를 들어, 이황화물 결합 형성, 글리코실화, 지질화, 아세틸화, 인산화 또는 라벨링 성분과의 접합(conjugation)과 같은 임의의 다른 조작을 포함할 수 있다. 본 명세서에서 사용되는 용어 “아미노산”은 글리신 및 D 또는 L 모두의 광학 이성질체를 비롯한 자연 및/또는 비자연 또는 합성 아미노산, 및 아미노산 유사체, 및 펩티도모방체를 지칭한다.
본 명세서에서 사용되는 용어 "약"은 용어 "대략"과 동의어로 사용된다. 예시적으로, 양과 관련된 용어 "약"의 사용은 인용된 값의 약간 밖에 있는 값, 예를 들어, 0.1% 내지 10%의 플러스 또는 마이너스를 나타낸다.
식량 작물 안팎의 미생물은 그 작물의 특성에 영향을 줄 수 있다. 미생물에 의해 영향을 받을 수 있는 식물 특성은 다음을 포함한다: 수율 (예컨대, 곡물 생산, 바이오매스 생성, 과일 발육, 꽃 세트); 영양 (예컨대, 질소, 인, 칼륨, 철, 미량 영양소 획득); 비 생물적 스트레스 관리 (예컨대, 가뭄 내성, 내염성, 내열성); 생물학적 스트레스 관리 (예컨대, 해충, 잡초, 곤충, 곰팡이 및 박테리아). 작물 특성을 변경하기 위한 전략에는 다음이 포함된다: 주요 대사산물 농도 증가; 주요 대사산물에 대한 미생물 영향의 시간적 역학 변화; 미생물 대사산물 생성/분해를 새로운 환경 신호에 연결; 부정적 대사산물 감소; 및 대사산물 또는 기초 단백질의 균형을 개선.
일부 구체예에서, 본 발명의 유전자의 고유 또는 내인성 제어 서열은 하나 이상의 유전자내 제어 서열로 대체된다.
본 명세서에서 사용되는 "도입된"은 자연 발생 도입이 아니라 현대 생명 공학에 의한 도입을 지칭한다.
일부 구체예들에서, 본 발명의 박테리아는 변형되었으므로, 이들은 자연 발생 박테리아가 아니다.
일부 구체예들에서, 본 발명의 박테리아는 식물의 신선 또는 건조 중량 그램 당 최소한 103 cfu, 104 cfu, 105 cfu, 106 cfu, 107 cfu, 108 cfu, 109 cfu, 1010 cfu, 1011 cfu, 또는 1012 cfu의 양으로 식물에 존재한다. 일부 구체예들에서, 본 발명의 박테리아는 식물의 신선 또는 건조 중량 그램 당 최소한 약 103 cfu, 약 104 cfu, 약 105 cfu, 약 106 cfu, 약 107 cfu, 약 108 cfu, 약 109 cfu, 약 1010 cfu, 약 1011 cfu, 또는 약 1012 cfu의 양으로 식물에 존재한다. 일부 구체예들에서, 본 발명의 박테리아는 식물의 신선 또는 건조 중량 그램 당 최소한 103 내지 109, 103 내지 107, 103 내지 105, 105 내지 109, 105 내지 107, 106 내지 1010, 106 내지 107 cfu의 양으로 식물에 존재한다.
본 발명의 비료 및 외인성 질소는 다음과 같은 질소-함유 분자를 포함 할 수 있다: 암모늄, 질산염, 아질산염, 암모니아, 글루타민 등. 본 발명의의 질소 공급원은 무수 암모니아, 암모니아 설페이트, 우레아, 다이암모늄 포스페이트, 우레아-형태, 모노암모늄 포스페이트, 암모늄 니트레이트, 질소 용액, 칼슘 니트레이트, 포타슘 니트레이트, 소듐 니트레이트, 등을 포함할 수 있다.
본 명세서에서 사용되는 "외인성 질소"는 암모니아, 암모늄, 질산염, 아질산염, 우레아, 요산, 암모늄 산 등을 포함하여 비-질소 제한 조건하에 존재하는 토양, 현장 또는 성장 배지에서 쉽게 이용할 수 있는 비-대기성 질소를 지칭한다.
본 명세서에서 사용된 "비-질소 제한 조건"은 Kant 외의 문헌 (2010. J. Exp. Biol. 62(4):1499-1509)에 개시된 바와 같이 약 4 mM 질소보다 큰 농도에서 토양, 현장, 매질에서 이용가능한 비-대기 질소를 지칭하며, 이는 본 명세서에 참고문헌으로 포함된다.
본 명세서에서 사용되는 "속간 미생물"은 처음에 상이한 분류학적 기원의 유기체로부터 단리된 유전 물질의 의도적인 조합에 의해 형성되는 미생물이다. “속간 돌연변이체”는 “속간 미생물”과 호환적으로 사용될 수 있다. 예시적인 "속간 미생물"은 수용자 미생물과 상이한 속의 미생물에서 먼저 확인된 이동성 유전 요소를 함유하는 미생물을 포함한다.
본 명세서에서 사용되는 "속내 미생물"은 처음에 동일한 분류학적 기원의 유기체로부터 단리된 유전 물질의 의도적인 조합에 의해 형성되는 미생물이다. “속내 돌연변이체”는 “속내 미생물”과 호환적으로 사용될 수 있다.
본 명세서에서 사용되는 "도입된 유전 물질"은 수용자의 게놈에 첨가되고 그 성분으로서 남아있는 유전 물질을 의미한다.
일부 구체예에서, 질소 고정 및 동화 유전자 조절 네트워크는 미생물 질소 고정 및/또는 동화를 지시, 조절 및/또는 조절하는 유전자 및 비-코딩 서열을 인코드하는 폴리뉴클레오티드를 포함하고, nif 클러스터 (예컨대, nifA, nifB, nifC,……nifZ)의 폴리 뉴클레오티드 서열, 질소 조절 단백질 C를 인코드하는 폴리뉴클레오티드, 질소 조절 단백질 B를 인코드하는 폴리뉴클레오티드, gln 클러스터 (예컨대, glnA 및 glnD)의 폴리뉴클레오티드 서열, draT, 및 암모니아 운반체/투과효소를 포함 할 수 있다. 일부 경우에, Nif 클러스터는 NifB, NifH, NifD, NifK, NifE, NifN, NifX, hesa, 및 NifV를 포함할 수 있다. 일부 경우에, Nif 클러스터는 NifB, NifH, NifD, NifK, NifE, NifN, NifX, hesa, 및 NifV의 부분집합을 포함할 수 있다.
일부 구체예들에서, 본 발명의 비료는 중량으로 최소한 5%, 6%, 7%, 8%, 9%, 10%, 11%, 12%, 13%, 14%, 15%, 16%, 17%, 18%, 19%, 20%, 21%, 22%, 23%, 24%, 25%, 26%, 27%, 28%, 29%, 30%, 31%, 32%, 33%, 34%, 35%, 36%, 37%, 38%, 39%, 40%, 41%, 42%, 43%, 44%, 45%, 46%, 47%, 48%, 49%, 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%의 질소를 포함한다.
일부 구체예들에서, 본 발명의 비료는 중량으로 최소한 약 5%, 약 6%, 약 7%, 약 8%, 약 9%, 약 10%, 약 11%, 약 12%, 약 13%, 약 14%, 약 15%, 약 16%, 약 17%, 약 18%, 약 19%, 약 20%, 약 21%, 약 22%, 약 23%, 약 24%, 약 25%, 약 26%, 약 27%, 약 28%, 약 29%, 약 30%, 약 31%, 약 32%, 약 33%, 약 34%, 약 35%, 약 36%, 약 37%, 약 38%, 약 39%, 약 40%, 약 41%, 약 42%, 약 43%, 약 44%, 약 45%, 약 46%, 약 47%, 약 48%, 약 49%, 약 50%, 약 51%, 약 52%, 약 53%, 약 54%, 약 55%, 약 56%, 약 57%, 약 58%, 약 59%, 약 60%, 약 61%, 약 62%, 약 63%, 약 64%, 약 65%, 약 66%, 약 67%, 약 68%, 약 69%, 약 70%, 약 71%, 약 72%, 약 73%, 약 74%, 약 75%, 약 76%, 약 77%, 약 78%, 약 79%, 약 80%, 약 81%, 약 82%, 약 83%, 약 84%, 약 85%, 약 86%, 약 87%, 약 88%, 약 89%, 약 90%, 약 91%, 약 92%, 약 93%, 약 94%, 약 95%, 약 96%, 약 97%, 약 98%, 또는 약 99%의 질소를 포함한다.
일부 구체예들에서, 본 발명의 비료는 중량으로 약 5% 내지 50%, 약 5% 내지 75%, 약 10% 내지 50%, 약 10% 내지 75%, 약 15% 내지 50%, 약 15% 내지 75%, 약 20% 내지 50%, 약 20% 내지 75%, 약 25% 내지 50%, 약 25% 내지 75%, 약 30% 내지 50%, 약 30% 내지 75%, 약 35% 내지 50%, 약 35% 내지 75%, 약 40% 내지 50%, 약 40% 내지 75%, 약 45% 내지 50%, 약 45% 내지 75%, 또는 약 50% 내지 75%의 질소를 포함한다.
일부 구체예에서, 식물에서 질소 고정 및/또는 질소 생성의 증가는 본 발명의 박테리아에 노출되지 않은 대조 식물에 대해 측정된다. 박테리아의 모든 증가 또는 감소는 대조 박테리아에 대해 측정된다. 식물의 모든 증가 또는 감소는 대조 식물에 대해 측정된다.
본 명세서에서 사용되는 "항시적 프로모터"는 대부분의 조건 및/또는 대부분의 발달 단계에서 활성인 프로모터이다. 생명 공학에 사용되는 발현 벡터에서 항시적 프로모터를 사용하는 것에는 다음과 같은 몇 가지 장점이 있다: 형질전환 세포 또는 유기체를 선택하는데 사용되는 단백질의 높은 수준의 생산; 쉽게 검출 및 정량화 할 수 있는 리포터 단백질 또는 측정가능한 마커의 높은 수준의 발현; 조절 전사 시스템의 일부인 전사 인자의 높은 수준의 생산; 유기체에서 편재된 활성을 요구하는 화합물의 생산; 그리고 모든 말달 단계에서 필요한 화합물의 생산. 비-제한적인 예시적인 항시적 프로모터는 CaMV 35S 프로모터, 오핀 프로모터, 유비퀴틴 프로모터, 알코올 탈수소효소 프로모터 등을 포함한다.
본 명세서에서 사용되는 "비-항시적 프로모터"는 특정 조건, 특정 유형의 세포 및/또는 특정 발달 단계 동안 활성인 프로모터이다. 예를 들어, 조직 특이적, 조직 선호적, 세포 유형 특이적, 세포 유형 선호적, 유도성 프로모터 및 발달 제어하의 프로모터는 비-항시적 프로모터이다. 발달 제어하에 있는 프로모터의 예는 특정 조직에서 전사를 우선적으로 개시하는 프로모터를 포함한다.
본 명세서에서 사용되는 "유도성" 또는 "억제성" 프로모터는 화학적 또는 환경적 요인 제어하에 있는 프로모터이다. 유도성 프로모터에 의해 전사에 영향을 줄 수 있는 환경 조건의 예는 혐기성 조건, 특정 화학 물질, 빛의 존재, 산성 또는 염기성 조건 등을 포함한다.
본 명세서에서 사용되는 "조직 특이적"프로모터는 특정 조직에서만 전사를 개시하는 프로모터이다. 유전자의 항시적 발현과는 달리, 조직-특이적 발현은 몇몇 상호작용 수준의 유전자 조절의 결과이다. 이와 같이, 해당 분야에서 때때로 특정 조직에서 트랜스진의 효율적이고 신뢰할 수 있는 발현을 구현하기 위해 상동성 또는 밀접하게 관련된 종으로부터의 프로모터를 사용하는 것이 바람직하다. 이것은 다량의 조직-특이적 프로모터가 과학 및 특허 문헌 모두에서 발견되는 특정 조직으로부터 단리되는 주된 이유 중 하나이다.
본 명세서에서 사용되는 용어 "작동가능하게 연결된"은 하나의 기능이 다른 하나에 의해 조절되도록 단일 핵산 단편상의 핵산 서열들이 연결되는 것을 지칭한다. 예를 들어, 프로모터는 코딩 서열의 발현을 조절할 수 있을 때 (즉, 코딩 서열이 프로모터의 전사 제어하에 있음) 코딩 서열과 작동가능하게 연결된다. 코딩 서열은 센스 또는 안티센스 배향으로 조절 서열에 작동가능하게 연결될 수 있다. 또 다른 예에서, 본 발명의 상보적 RNA 영역은, 직접적으로 또는 간접적으로, 5'을 표적 mRNA에, 또는 3'을 표적 mRNA에, 또는 표적 mRNA 내부에 작동가능하게 연결시킬 수 있거나, 또는 제 1 상보성 영역은 5'이고 그의 보체는 표적 mRNA에 대해 3'이다.
양상들에서, “식물에 복수의 비-속간 박테리아를 처리하는 것”은 식물 (식물 부분, 가령, 종자, 뿌리, 줄기, 조직 등을 포함)을 식물의 임의의 수명 사이클 단계에서 상기 박테리아와 접촉 (즉, 노출)되게 하는 임의의 수단을 포함한다. 결과적으로, "식물에 복수의 비-속간 박테리아를 처리하는 것"은 식물 (식물 부분, 가령, 종자, 뿌리, 줄기, 조직 등을 포함)을 상기 박테리아에 노출시키는 다음과 같은 임의의 수단들을 포함한다: 식물에 분사, 식물에 드립핑(dripping), 종자 코트로서 처리, 현장에 처리한 다음 여기에 종자를 식재, 종자를 이미 식재한 현장에 처리, 성체 식물이 있는 현장에 처리, 등.
용어 mmol은 밀리몰에 대한 약자로, 본 명세서에서 mol로 약칭되는 1몰의 1000분의 1 (103 )이다.
본 명세서에서 사용되는 용어 “미생물 (microorganism)”또는 “미생물(microbe)”은 광범위하게 사용되어야 한다. 호환적으로 사용되는 이들 용어는 2개의 원핵 도메인, 박테리아 및 고세균을 포함하지만 이에 제한되지는 않는다. 이 용어는 또한 진핵생물 진균 및 원생생물을 포함 할 수 있다.
용어 "미생물 컨소시아" 또는 "미생물 컨소시엄"은 개별 미생물 종 또는 종의 균주들의 미생물 커뮤니티의 부분집합을 지칭하며, 이는 공통 기능을 수행하는 것으로 기재될 수 있거나 또는 인식가능한 매개 변수, 가령, 관심 표현형 형질에 관여, 또는 이를 초래 또는 이와 상관관계가 있는 것으로 기재될 수 있다.
"미생물 커뮤니티 (microbial community)"라는 용어는 둘 이상의 종 또는 균주를 포함하는 미생물 군을 의미한다. 미생물 컨소시아와 달리, 미생물 커뮤니티는 공통 기능을 수행할 필요는 없으며 또는 인식가능한 매개 변수, 가령, 관심 표현형 형질에 관여, 또는 이를 초래 또는 이와 상관관계가 있을 필요는 없다.
본 명세서에서 사용되는, "단리하다", "단리된", "단리된 미생물"등의 용어는 하나 이상의 미생물이 특정 환경 (예를 들면, 토양, 물, 식물 조직 등)에서 그와 연결된 물질 중 하나 이상으로부터 단리되었음을 의미하는 것으로 한다. 따라서 “단리된 미생물”은 자연 발생 환경에 존재하지 않으며; 오히려, 본 명세서에 기재된 다양한 기술을 통해 미생물이 자연 환경에서 제거되어 자연적으로 발생하지 않는 존재 상태에 놓이게 된다. 따라서, 단리된 균주 또는 단리된 미생물은, 예를 들어, 생물학적으로 순수한 배양물 또는 포자 (또는 다른 형태의 균주)로서 존재할 수 있다. 양상들에서, 단리된 미생물은 허용가능한 담체와 결합될 수 있으며, 이는 농업 적으로 허용가능한 담체 일 수 있다.
본 발명의 특정 양상에서, 단리된 미생물은 "단리된 생물학적으로 순수한 배양물"로서 존재한다. 특정 미생물의 단리된 생물학적으로 순수한 배양물은 상기 배양물에 다른 살아있는 유기체가 실질적으로 없으며 문제의 개별 미생물만을 함유한다는 것을 숙련된 기술자는 이해할 것이다. 배양물은 다양한 농도의 상기 미생물을 함유 할 수 있다. 본 발명은 단리된 생물학적으로 순수한 미생물이 종종 "덜 순수하거나 불순한 물질과는 필연적으로 다름"을 나타낸다. 예컨대, In re Bergstrom, 427 F.2d 1394, (CCPA 1970)(정제된 프로스타글란딘을 논의)를 참고하고, 또한 In re Bergy, 596 F.2d 952 (CCPA 1979)(정제된 미생물을 논의)를 참고하며, 또한 Parke-Davis & Co. v. H.K. Mulford & Co., 189 F. 95 (S.D.N.Y. 1911) (정제된 아드레날린을 논의한 러니드 핸드), aff'd in part, rev'd in part, 196 F. 496 (2d Cir. 1912)를 참고하며, 각 문헌은 본 명세서에 참고문헌으로 포함된다. 또한, 일부 양상들에서, 본 발명은 단리된 생물학적으로 순수한 미생물 배양물에서 발견되어야 하는 농도 또는 순도 한계에 관한 특정 정량적 측정법을 제공한다. 특정 구체예들에서, 이러한 순도 값의 제공은 본 발명에 개시된 미생물을 자연 상태에 존재하는 미생물과 구별시키는 또 다른 속성이다. 예컨대, Merck & Co. v. Olin Mathieson Chemical Corp., 253 F.2d 156 (4th Cir. 1958) (미생물에 의해 생성된 비타민 B12에 대한 순도 한계를 논의)를 참고하고, 이는 본 명세서에 참고문헌으로 포함된다.
본 명세서에서 사용되는 "개별 단리물"은 하나 이상의 다른 미생물로부터 단리된 후, 미생물의 단일 속, 종 또는 균주 우세를 포함하는 조성물 또는 배양 물을 의미하는 것으로 여겨져야 한다.
본 발명의 미생물은 포자 및/또는 식물 세포를 포함 할 수 있다. 일부 구체예들에서, 본 발명의 미생물은 생존 가능하지만 배양 불가능한 (VBNC) 상태의 미생물을 포함한다. 본 명세서에서 사용된 "포자" 또는 "포자들"은 생존 및 분산에 적합한 박테리아 및 진균에 의해 생성되는 구조를 지칭한다. 포자는 일반적으로 휴면 구조로 특징되지만; 포자는 발아 과정을 통해 분화 할 수 있다. 발아는 포자가 대사 활동, 성장 및 번식이 가능한 식물 세포로 분화하는 것이다. 단일 포자의 발아는 단일 곰팡이 또는 박테리아 식물 세포를 생성한다. 곰팡이 포자는 무성 생식의 단위이며, 일부 경우, 곰팡이 수명주기에 필요한 구조이다. 박테리아 포자는 일반적으로 식물 세포의 생존 또는 성장을 촉진하지 않을 수 있는 생존 조건을 위한 구조이다.
본 명세서에서 사용되는, "미생물 조성물"은 본 발명의 하나 이상의 미생물을 포함하는 조성물을 지칭한다. 일부 구체 예에서, 미생물 조성물은 식물 (다양한 식물 부분 포함) 및/또는 농경지에 투여된다.
본 명세서에서 사용되는, "담체", "허용 가능한 담체" 또는 "농업적으로 허용가능한 담체"는 미생물과 함께 투여 될 수 있는 희석제, 보조제, 부형제 또는 비히클을 말하며, 이는 미생물에 해로운 영향을 미치지 않는다.
본 발명을 기재하는 내용에서 (특히 청구범위의 내용에서) 용어 “하나” 및 “그것” 그리고 이와 유사한 관사들의 사용은, 본 출원에서 달리 언급이 없는 한 또는 내용상 명확하게 반대가 아닌 한, 단수형 및 복수형 모두를 포함하는 것으로 해석되어야 한다. "포함하는", "갖는", "비롯한" 및 "함유하는" 이라는 용어는 달리 언급되지 않는 한 개방형 용어로 해석되어야 한다 (즉, "포함하지만 이에 제한되지 않는"을 의미). 본 명세서에서 값의 범위의 언급은 달리 지시되지 않는 한, 단지 범위 내에 속하는 각각의 개별 값을 개별적으로 지칭하는 속기 방법으로서 제공되는 것으로 하며, 각각의 개별 값은 마치 본 명세서에서 개별적으로 언급된 것처럼 명세서에 포함된다. 예를 들어, 범위 10-15가 개시되면, 11, 12, 13 및 14도 개시된다. 본 출원에 달리 언급이 없는 한 또는 내용상 달리 명확하게 반대의 언급이 없는 한, 본 출원에 기재된 모든 방법들은 임의의 적합한 순서로 실시될 수 있다. 본 출원에서 제공되는 임의의 그리고 모든 예시들, 그리고 예시적인 언어 (예컨대, “가령”)의 사용은 본 발명을 보다 잘 설명하기 위한 것이며 청구범위에 기재된 발명의 범위를 달리 제한하고자 하는 것이 아니다. 명세서의 어떠한 언어도 본 발명의 실시에 필수적인 비-청구범위 기재 요소를 나타내는 것으로 해석되어서는 안된다.
박테리아 콜로니화
일부 구체예에서, 미생물은 식물로 질소의 전달을 증가시키기 위해 변형 될 수 있다. 예를 들어, 미생물은 식물 뿌리의 콜로니화를 증가시키거나 질소 고정에 필요한 보조 인자를 증가시키기 위해 변형 될 수 있다.
일부 경우에, 세포외 다당류는 식물의 박테리아 콜로니화에 관여할 수 있다. 일부 경우, 식물 콜로니화 미생물은 생물막을 생성 할 수 있다. 일부 경우에, 식물 콜로니화 미생물은 식물에 대한 접착 또는 식물 면역 반응의 회피를 도울 수 있는 분자를 분비한다. 일부 경우에, 식물 콜로니화 미생물은 미생물에 대한 식물 반응을 변경시키는 신호 전달 분자를 배설 할 수 있다. 일부 경우에, 식물 콜로니화 미생물은 국소 미세 환경을 변화시키는 분자를 분비 할 수 있다. 일부 경우에, 식물 콜로니화 미생물은 상기 미생물이 근접한 식물에 적응하도록 하는 유전자의 발현을 변경시킬 수 있다. 일부 경우에, 식물 콜로니화 미생물은 국소 환경에서 식물의 존재를 탐지 할 수 있고 반응에 따라 유전자의 발현을 변화시킬 수 있다.
질소 고정의 조절
일부 경우에, 질소 고정 경로가 유전 공학 및 최적화의 표적으로 작용할 수 있다. 본 명세서에 기재된 방법에 의한 조절을 위하여 표적 될 수 있는 한 가지 특성은 질소 고정이다. 질소 비료는 농장에서 가장 큰 운영비이며 옥수수나 밀과 같은 줄뿌림 작물에서 더 높은 수확량을 가져오는 가장 큰 원동력이다. 비-콩과 작물에서 재생 가능한 형태의 질소를 전달할 수 있는 미생물 산물이 본 명세서에 기재되어 있다. 일부 내생미생물은 순수한 배양믈에서 질소를 고정시킴에 필요한 유전학을 가지고 있지만, 근본적인 기술적 과제는 곡물과 풀의 야생형 내생미생물이 비료처리된 현장에서 질소 고정을 멈춘다는 것이다. 현장 토양에 화학 비료와 잔류 질소 수준을 적용하면 미생물은 질소 고정을 위한 생화학적 경로를 차단하는 신호를 생성한다.
질소 고정 조절 네트워크의 성분의 전사 및 번역 후 수준의 변화는 비료의 존재 하에서 질소를 옥수수로 고정 및 전달할 수 있는 미생물의 개발에 유리할 수 있다. 이를 위해, 규제 네트워크를 정확하게 진화시키고 신규한 표현형을 도출하기 위한 숙주-미생물 진화 (HoME) 기술이 본 명세서에 기재되어 있다. 또한 질소 스트레스 및 과잉과 같은 다른 환경 조건 하에서 미생물과 숙주 식물의 상호 작용을 둘러싼 광범위한 오믹스 데이터와 결합된, 옥수수에서 분리된 질소 고정 내생미생물의 독특하고 독점적인 라이브러리가 본 명세서에 기재되어 있다. 일부 구체예들에서, 이 기술은 내생미생물의 유전자 조절 네트워크의 정밀 진화를 통해 현장에서 비료의 존재 하에서도 질소를 능동적으로 고정시키는 미생물을 생성 할 수 있다. 또한, 옥수수 뿌리 조직을 콜로니화하고 비료화된 식물에 대한 질소를 생성하는 미생물을 진화시키는 기술적 잠재력에 대한 평가 및 미생물을 현대 질소 관리 전략에 통합시켜야 하는 타당성을 결정하기 위한 표준 배합 관행 및 다양한 토양과 내생미생물의 상용성에 대한 평가가 본 명세서에 기재되어 있다.
화학적 합성을 위해 원소 질소 (N)를 이용하기 위해, 생명 형태는 질소 고정으로 알려진 과정에서 대기 중 이용가능한 질소 기체 (N2)와 수소를 결합시킨다. 생물학적 질소 고정의 에너지 집약적 특성으로 인해, 이조류 (대기 질소 기체를 고정시키는 박테리아 및 고세균)는 환경 산소 및 이용가능한 질소에 반응하여 nif 유전자 클러스터의 정교하고 엄격한 조절을 진화시켰다. Nif 유전자는 질소 고정에 관여하는 효소 (가령, 질소고정효소 복합체) 및 질소 고정을 조절하는 단백질을 인코드한다. Shamseldin의 문헌 (2013. Global J. Biotechnol. Biochem. 8(4):84-94)은 nif 유전자 및 이들의 생성물에 대한 상세한 설명을 개시하고, 이 문헌은 본 명세서에 참조로 포함된다. 본 명세서에는 제 1 식물로부터 박테리아를 단리하는 단계, 단리된 박테리아의 유전자에 질소 고정을 증가시키는 유전자 변이를 도입하는 단계, 제 2 식물을 변이체 박테리아에 노출시키는 단계, 제 1 식물에 비해 개선된 특성을 가지는 제 2 식물로부터 박테리아를 단리하는 단계, 그리고 제 2 식물로부터 단리된 박테리아를 이용하여 상기 단계들을 반복하는 단계를 포함하는, 개선된 특성을 가진 식물의 제조 방법이 기재되어 있다.
프로테오박테리아에서, 질소 고정의 조절은 nif 클러스터의 양성 전사 조절 인자인, σ54-의존적 인핸서-결합 단백질 NifA를 중심으로 한다. 활성 NifA의 세포내 수준은 두 가지 주요 인자: nifLA 오페론의 전사 및 NifL과의 단백질-단백질 상호작용에 의한 NifA 활성의 억제에 의해 제어된다. 이들 두 과정 모두는 PII 단백질 신호전달 캐스케이드를 통한 세포내 글루타민 수준에 반응한다. 이 캐스케이드는 GlnD에 의해 매개되는데, 이는 글루타민을 직접 감지하고, 결합 된 글루타민의 부재 또는 존재에 반응하여 2개의 PII 조절 단백질 - GlnB 및 GlnK-의 유리딜릴화 또는 탈유리딜릴화를 촉매한다. 질소 과잉 조건 하에서, 변형되지 않은 GlnB는 nifLA 프로모터의 비활성화를 신호전달한다. 그러나, 질소 제한 조건 하에서, GlnB는 번역 후 변형되어, 그의 활성을 억제하고 nifLA 오페론의 전사를 초래한다. 이러한 방식으로, nifLA 전사는 PII 단백질 신호전달 캐스케이드를 통해 환경 질소에 반응하여 엄격하게 제어된다. 번역 후의 NifA 조절 수준에서, GlnK는 세포내 유리 GlnK의 전체 수준에 의존하는 물질에서 NifL/NifA 상호작용을 억제한다.
NifA는 nifLA 오페론으로부터 전사되며, 그의 프로모터는 또 다른 σ54- 의존적 조절제인 인산화된 NtrC에 의해 활성화된다. NtrC의 인산화 상태는 히스티딘 키나제 NtrB에 의해 매개되는데, 이것은 탈유리딜릴화된 GlnB와 상호작용하지만 유리딜릴화된 GlnB와는 상호작용하지 않는다. 질소 과잉 조건 하에서, 높은 세포내 수준의 글루타민은 GlnB의 탈유리딜릴화를 초래하고, 이어서 NtrB와 상호작용하여 그 인산화 활성을 비활성화시키고 그 포스파타제 활성을 활성화시켜 NtrC의 탈인산화 및 nifLA 프로모터의 비활성화를 초래한다. 그러나, 질소 제한 조건 하에서, 낮은 수준의 세포내 글루타민은 GlnB의 유리딜릴화를 초래하여, NtrB와의 상호작용을 억제하고 NtrC의 인산화 및 nifLA 오페론의 전사를 가능하게 한다. 이러한 방식으로, nifLA 발현은 PII 단백질 신호전달 캐스케이드를 통해 환경 질소에 반응하여 엄격하게 제어된다. nifA, ntrB, ntrC 및 glnB는 본 명세서에 기재된 방법에서 돌연변이 될 수 있는 모든 유전자이다. 이들 과정은 또한 세포내 또는 세포외 수준의 암모니아, 우레아 또는 질산염에 반응 할 수도 있다.
NifA의 활성은 또한 환경 질소에 반응하여 번역 후, 가장 전형적으로는 NifA 활성의 NifL- 매개 억제를 통해 조절된다. 일반적으로, NifL과 NifA의 상호 작용은 GlnK를 통한 PII 단백질 신호 전달 캐스케이드에 의해 영향을 받지만, GlnK와 NifL/NifA 사이의 상호작용의 특성은 질소자급영양생물들 간에 상당히 다르다. 클레브시엘라 뉴모니아에 (klebsiella pneumoniae)에서, GlnK의 두 형태는 NifL/NifA 상호작용을 억제하고, GlnK와 NifL/NifA 사이의 상호작용은 세포내 유리 GlnK의 전체 수준에 의해 결정된다. 질소 과잉 조건 하에서, 탈유리딜릴화된 GlnK는 AmtB에 의한 암모늄 흡수와 막에 대한 GlnK의 격리를 막는 역할을 하는 암모늄 운반체 AmtB와 상호작용하여 NifL에 의한 NifA의 억제를 가능하게 한다. 한편, 아조토박터 비넬란디 (azotobacter vinelandii)에서, NifL/NifA 상호작용 및 NifA 억제를 위해서는 탈유리딜릴화된 GlnK와의 상호 작용이 요구되는 반면, GlnK의 유리딜릴화는 NifL과의 상호 작용을 억제한다. nifL 유전자가 결여된 질소자급영양생물에서, 질소 과잉 조건 하에서 GlnK 및 GlnB모두의 탈유리딜릴화된 형태와의 상호 작용에 의해 NifA 활성이 직접적으로 억제된다는 증거가 있다. 일부 박테리아에서 Nif 클러스터는 glnR에 의해 조절 될 수 있으며, 일부 경우에는 음성 조절을 포함 할 수 있다. 메커니즘에 관계없이, NifA의 번역 후 억제는 대부분의 공지된 질소자급영양생물에서 nif 클러스터의 중요한 조절자이다. 추가로, nifL, amtB, glnK, 및 glnR은 본 명세서에 기재된 방법에서 돌연변이 될 수 있는 유전자이다.
nif 유전자 클러스터의 전사를 조절하는 것 외에도, 많은 질소자급영양생물은 번역 후 변형 및 질소고정효소 차단으로 공지된 질소고정효소 효소 자체의 억제를 위한 메커니즘을 진화시켰다. 이것은 질소 과잉 조건 하에서 Fe 단백질 (NifH)의 ADP-리보실화에 의해 매개되며, 이는 MoFe 단백질 복합체 (NifDK)와의 상호 작용을 방해하고 질소고정효소 활성을 제거한다. DraT는 Fe 단백질의 ADP-리보실화 및 질소고정효소 차단을 촉매하는 반면, DraG는 ADP-리보스 제거 및 질소고정효소 재활성화를 촉매한다. nifLA 전사 및 NifA 억제에서와 같이, 질소고정효소 차단 또한 PII 단백질 신호 전달 캐스케이드를 통해 조절된다. 질소 과잉 조건에서 탈유리딜릴화된 GlnB는 DraT와 상호작용하여 활성화하는 반면 탈유리딜릴화된 GlnK는 DraG 및 AmtB 모두와 상호작용하여 DraG를 막에 대해 격리시키는 복합체를 형성한다. 질소-제한 조건 하에서, 유리딜릴화된 형태의 GlnB 및 GlnK는 각각 DraT 및 DraG와 상호작용하지 않아, DraT의 불활성화 및 DraG의 Fe 단백질로의 확산을 가져오며, 이 때 ADP-리보스를 제거하고 질소고정효소를 활성화시킨다. 본 명세서에 기재된 방법은 또한 nifH, nifD, nifK 및 draT 유전자 내로 유전자 변이를 도입하는 것을 고려한다.
일부 내생미생물는 시험관내에서 질소를 고정시키는 능력을 갖지만, 종종 높은 수준의 외인성 화학 비료에 의해 유전학은 현장에서 침묵된다. 현장 기반 질소 고정을 용이하게 하기 위해 외인성 질소의 감지와 질소고정효소 효소의 발현을 분리시킬 수 있다. 시간에 따른 질소고정효소 활성의 적분을 개선하는 것은 작물이 이용하는 질소의 생산을 증대시키는 역할을 한다. 본 명세서에 기재된 방법을 사용하여 현장-기반 질소 고정을 용이하게 하기 위한 유전자 변이를 위한 구체적인 표적은 nifA, nifL, ntrB, ntrC, glnA, glnB, glnK, draT, amtB, glnD, glnE, nifJ, nifH, nifD, nifK , nifY, nifE, nifN, nifU, nifS, nifV, nifW, nifZ, nifM, nifF, nifB, 및 nifQ로 구성된 군에서 선택된 하나 이상의 유전자들을 포함한다.
본 명세서에 기재된 방법을 사용하여 현장-기반 질소 고정을 용이하게 하는 유전자 변이를 위한 또 다른 표적은 NifA 단백질이다. NifA 단백질은 전형적으로 질소 고정 유전자의 발현을 위한 활성화제이다. (항시적으로 또는 높은 암모니아 조건 동안) NifA의 생성을 증가시키는 것은 고유 암모니아-감지 경로를 우회한다. 또한, NifA의 공지된 억제제인 NifL 단백질의 생성을 감소시키는 것은 또한 자유 활성 NifA의 수준을 증가시킨다. 또한, (항시적으로 또는 높은 암모니아 조건 동안) nifAL 오페론의 전사 수준을 증가시키는 것은 또한 전체적으로 높은 수준의 NifA 단백질을 초래한다. 상승된 nifAL 발현 수준은 프로모터 자체를 변경하거나 NtrB의 발현을 감소시킴으로써 달성된다 (ntrB 및 ntrC 신호 전달 캐스케이드의 일부는 원래 높은 질소 조건에서 nifAL 오페론의 차단을 초래할 것이다). 본 명세서에 기재된 이들 또는 임의의 다른 방법에 의해 구현된 높은 수준의 NifA는 내생미생물의 질소 고정 활성을 증가시킨다.
본 명세서에 기재된 방법을 사용하여 현장-기반 질소 고정을 용이하게하는 유전자 변이에 대한 또 다른 표적은 GlnD/GlnB/GlnK PII 신호 전달 캐스케이드이다. 세포 내 글루타민 수준은 GlnD/GlnB/GlnK PII 신호 전달 캐스케이드를 통해 감지된다. GlnD의 유리딜릴 제거 활성을 없애는 GlnD의 활성 부위 돌연변이는 질소 감지 캐스케이드를 파괴한다. 또한, GlnB 농도의 감소는 글루타민-감지 캐스케이드를 단락시킨다. 이들 돌연변이는 세포를 질소 제한 상태를 인지하도록 "트릭 (trick)"함으로써, 질소 고정 레벨 활성을 증가시킨다. 이들 과정은 또한 세포내 또는 세포외 수준의 암모니아, 우레아 또는 질산염에 반응 할 수도 있다.
amtB 단백질은 또한 본 명세서에 기재된 방법을 사용하여 현장-기반 질소 고정을 용이하게하는 유전적 변이의 표적이다. amtB 단백질의 발현 수준을 감소시킴으로써 환경으로부터의 암모니아 흡수를 감소시킬 수 있다. 세포 내 암모니아가 없으면, 내생미생물은 높은 수준의 암모니아를 감지 할 수 없어 질소 고정 유전자의 하향 조절을 방지한다. 세포 내 구획으로 들어가는 암모니아는 글루타민으로 전환된다. 세포 내 글루타민 수준은 질소 감지의 주요 흐름이다. 세포 내 글루타민 수준을 감소시키면 세포가 환경에서 높은 암모늄 수치를 감지하지 못한다. 이 효과는 글루타민을 글루타메이트로 전환시키는 효소인 글루타미나제의 발현 수준을 증가시킴으로써 구현될 수있다. 또한, 글루타민 합성효소 (암모니아를 글루타민으로 전환시키는 효소)를 감소시킴으로써 세포 내 글루타민을 감소시킬 수도 있다. 질소자급영양생물에서, 고정된 암모니아는 글루타민 및 글루타메이트로 빠르게 동화되어 세포 공정에 사용된다. 암모니아 동화로의 중단은 세포로부터 배출될 고정된 질소의 암모니아로서의 전환을 가능하게 할 수 있다. 고정된 암모니아는 글루타민 합성효소 (GS)에 의해 글루타민으로 주로 동화되고, glnA에 의해 인코드되고,이어서 글루타민 옥소글루타레이트 아미노트랜스퍼라제 (GOGAT)에 의해 글루타민으로 동화된다. 일부 예에서, glnS는 글루타민 합성효소를 인코드한다. GS는 GS 아데닐릴 트랜스퍼라제 (GlnE)에 의해 번역 후 조절되는데, 이는 아데닐릴 트랜스퍼라제 (AT) 및 아데닐릴 제거 (AR) 도메인의 활성을 통해 GS의 아데닐릴화 및 탈아데닐릴화를 촉매하는 glnE에 의해 인코드되는 이작용성 효소이다. 질소 제한 조건 하에서 glnA가 발현되고 GlnE의 AR 도메인은 GS를 탈아데닐릴화시켜 이를 활성화시킨다. 질소 과잉 조건에서 glnA 발현이 꺼지고 GlnE의 AT 도메인은 글루타민에 의해 동종이계로 활성화되어 GS의 아데닐릴화 및 비활성화를 유발한다.
또한, draT 유전자 또한 본원에 기재된 방법을 사용하여 현장-기반 질소 고정을 용이하게 하기 위한 "유전자 변이의 표적" 일 수 있다. 세포에 의해 질소 고정 효소가 생성되면, 질소고정효소 차단은 세포가 높은 질소 조건에서 고정 활성을 하향조절하는 또 다른 수준을 나타낸다. 이 차단은 DraT의 발현 수준을 감소시켜 제거 할 수 있다.
새로운 미생물 표현형을 부여하는 방법은 전사, 번역 및 번역 후 수준에서 수행 될 수 있다. 전사 수준은 프로모터에서의 변화 (가령, 시그마 인자 친화성 변화 또는 프로모터의 전부 또는 일부의 결실을 포함하는 전사 인자에 대한 결합 부위 변화) 또는 전사 종결자 및 감쇠제를 변화시키는 것을 포함한다. 번역 수준은 리보솜 결합 부위에서의 변화 및 mRNA 분해 신호 변화를 포함한다. 번역 후 수준은 효소의 활성 부위 돌연변이 및 단백질-단백질 상호작용의 변화를 포함한다. 이러한 변화는 다양한 방법으로 구현 될 수 있다. 발현 수준의 감소 (또는 완전한 폐지)는 고유 리보솜 결합 부위 (RBS) 또는 프로모터를 더 낮은 강도/효율을 갖는 다른 것과 교환함으로써 구현 될 수 있다. ATG 시작 부위는 GTG, TTG 또는 CTG 시작 코돈으로 교환 될 수 있으며, 이는 코딩 영역의 번역 활성을 감소시킨다. 유전자의 코딩 영역을 녹아웃 (결실)함으로써 발현의 완전한 제거가 이루어질 수 있다. 오픈 리딩 프레임 (ORF)을 틀이동시키면 ORF를 따라 조기 정지 코돈이 생성되고, 이로써 비-기능적 절두 산물이 생성될 수 있을 것이다. 인-프레임 정지 코돈의 삽입 또한 유사하게 비-기능적 절두 생성물을 생성할 것이다. N 또는 C 말단에 분해 태그를 추가하여 특정 유전자의 유효 농도를 감소시킬 수도 있다.
반대로, 본 명세서에 기재된 유전자의 발현 수준은 더 강한 프로모터를 사용함으로써 구현 될 수 있다. 높은 질소 수준 조건 (또는 다른 조건) 동안 높은 프로모터 활성을 보장하기 위해, 높은 질소 수준 조건에서 전체 게놈의 전사 프로파일을 얻을 수 있고 원하는 전사 수준의 활성 프로모터를 해당 데이터 세트에서 선택하여 약한 프로모터를 대체할 수 있다. 약한 시작 코돈을 ATG 시작 코돈으로 교체하여 번역 개시 효율을 높일 수 있다. 약한 리보솜 결합 부위 (RBS)는 또한 높은 번역 개시 효율을 갖는 다른 RBS와 교환 될 수 있다. 또한, 효소의 활성을 변경시키기 위해 부위 특이적 돌연변이유발이 또한 수행 될 수 있다.
질소고정효소 유전자 및 질소 동화 유전자의 조절에 더하여, 배출된 질소의 수준은 또한 고정 및/또는 동화에 관여하는 보조 인자의 이용가능성에 의해 영향을 받을 수 있다. 예를 들면, 황은 일부 질소고정효소의 성분이다. 황은 일반적으로 세포막을 가로 질러 이동되어야 한다. 로투스 자포니쿠스 (L. japonicus)에서 설페이트 운반체 LjSST1은 질소 고정에 필수적이며; 녹아웃 돌연변이체는 기능적 노듈을 발달시킬 수 없다. 일부 구체 예에서, 황과 같은 보조인자의 이용가능성을 증가시키는 운반체를 상향조절함으로써 질소고정효소 활성이 증가 될 수 있다. 예를 들어, 황 운반체, cysZ의 발현 또는 활성이 증가 될 수있다.
식물에서 발생하는 질소 고정 수준을 높이면 작물 생산에 필요한 화학 비료의 양을 줄이고 온실 가스 배출량 (예컨대, 아산화 질소)을 감소시킬 수 있다.
박테리아 집단의 생성
박테리아의 단리
본원에 개시된 방법 및 조성물에 유용한 미생물은 천연 식물의 표면 또는 조직으로부터 미생물을 추출함으로써 얻을 수 있다. 미생물은 종자를 분쇄하여 미생물을 단리하여 얻을 수 있다. 미생물은 다양한 토양 샘플에 종자를 심고 조직에서 미생물을 회수하여 얻을 수 있다. 또한, 미생물은 식물에 외인성 미생물을 접종하고 식물 조직에 어떤 미생물이 나타나는지를 결정함으로써 얻을 수 있다. 식물 조직의 비-제한적인 예는 종자, 묘목, 잎, 절단, 식물, 구근 또는 괴경을 포함 할 수 있다.
미생물을 얻는 방법은 토양에서 박테리아를 단리하는 것이다. 박테리아는 다양한 토양 유형에서 수집 될 수 있다. 일부 예에서, 토양은 높은 또는 낮은 생식 능력, 수분 수준, 미네랄 수준 및 다양한 경작 관행과 같은 특성을 특징으로 할 수 있다. 예를 들면, 토양은 연속 재배기에서 동일한 토양에 상이한 작물들이 식재되는 작물 회전과 관련될 수 있다. 같은 토양에서 다른 작물의 순차적인 성장은 특정 미네랄의 불균형 고갈을 막을 수 있다. 박테리아는 선택된 토양에서 자라는 식물로부터 단리 될 수 있다. 묘목 식물은 2-6주 성장시에 수확될 수 있다. 예를 들면, 최소한 400개 단리물들이 1회 수확시 수집될 수 있다. 토양 및 식물 유형은 식물 표현형, 뿐만 아니라 특정 표현형의 하류 농축을 허용하는 조건을 나타낸다.
미생물 특성을 평가하기 위해 식물 조직으로부터 미생물을 단리 할 수 있다. 조직 샘플을 처리하기 위한 파라미터는 다양한 유형의 회합성 미생물, 가령, 뿌리줄기 박테리아, 착생식물, 또는 내생미생물을 단리하기 위해 변화될 수 있다. 단리물은 질소 고정을 수행하는 박테리아를 풍부하게 하기 위해 무-질소 배지에서 배양할 수 있다. 대안적으로, 미생물은 세계 균주 은행들로부터 얻을 수 있다.
식물에서 미생물 특성을 평가하기 위한 분석이 실시된다. 일부 구체예에서, 식물 조직은 DNA 및 RNA에 대한 높은 처리량 처리에 의한 스크리닝을 위해 처리 될 수 있다. 또한 비-침습적 측정을 사용하여 콜로니화와 같은 식물 특성을 평가할 수 있다. 야생 미생물에 대한 측정값은 식물별로 얻을 수 있다. 야생 미생물에 대한 측정은 또한 중간 처리량 방법을 사용하여 현장에서 얻을 수 있다. 측정은 시간 경과에 따라 연속적으로 이루어질 수 있다. 세타리아를 비롯한 (그러나 이에 제한되는 것은 아님) 모델 식물 시스템이 사용될 수 있다.
식물 시스템에서 미생물은 식물 시스템 내 미생물의 전사 프로파일링을 통해 스크리닝 될 수 있다. 전사 프로파일링을 통한 스크리닝의 예는 정량적 중합효소 연쇄반응 (qPCR), 전사체 검출을 위한 분자 바코드, 차세대 시퀀싱 및 형광 마커를 이용한 미생물 태깅 방법을 사용하는 것이다. 충격인자를 측정하여, 마이크로바이옴, 비생물적 요인, 토양 조건, 산소, 수분, 온도, 접종원 조건 및 뿌리 국소화를 포함한 (그러나 이에 제한되는 것은 아님) 온실에서의 콜로니화를 평가 할 수 있다. 본 명세서에 기재된 IRMS 또는 NanoSIMS로 15N 가스/비료 (희석)를 측정함으로써 박테리아에서 질소 고정을 평가할 수 있다. NanoSIMS는 고분해능 2차 이온 질량 분석법이다. NanoSIMS 기술은 생물학적 시료의 화학적 활성을 조사하는 방법이다. 미생물의 대사를 유발하는 산화 반응의 환원의 촉매작용은 세포, 아세포, 분자 및 원소 수준에서 조사 될 수 있다. NanoSIMS는 0.1 μm 보다 큰 높은 공간 분해능을 제공할 수 있다. NanoSIMS는 동위원소 추적자, 가령, 13C, 15N, 및 18O의 이용을 검출할 수 있다. 그러므로, NanoSIMS는 세포에서 화학적 활성 질소의 검출에 사용될 수 있다.
식물 분석에 자동화 온실이 사용될 수 있다. 미생물 노출에 반응하는 식물 측정항목에는 바이오매스, 엽록체 분석, CCD 카메라, 체적 단층촬영 측정이 포함되나 이에 제한되지 않는다.
미생물 집단을 풍부하게 하는 한 가지 방법은 유전자형에 따른 것이다. 예를 들어, 표적된 프라이머 또는 특정 프라이머를 사용한 중합효소 연쇄 반응 (PCR) 분석. 질소자급영양생물은 질소 고정 과정에서 nifH 유전자를 발현하기 때문에 nifH 유전자 용으로 설계된 프라이머를 사용하여 질소자급영양생물을 식별 할 수 있다. 미생물 집단은 단일 세포 배양 독립 접근법과 화학주성-유도 단리 접근법을 통해 풍부해질 수 있다. 대안적으로, 미생물의 표적된 단리는 선별 배지상에서 미생물을 배양함으로써 실시될 수 있다. 원하는 형질에 대한 미생물 집단을 풍부하게 하기 위한 사전계획된 접근법은 생물정보학 데이터에 의해 안내 될 수 있으며 본 명세서에 기재되어 있다.본 명세서에 기재된
미생물의 순화
자연으로부터 단리된 미생물은 순화 과정을 거칠 수 있는데, 여기서 미생물은 유전자 추적 및 식별 가능한 형태로 전환된다. 미생물을 순화시키는 한 가지 방법은 항생제 내성을 가지고 이를 조작하는 것이다. 항생제 내성을 조작하는 과정은 야생형 미생물 균주에서 항생제 감수성을 결정함으로써 시작할 수 있다. 박테리아가 항생제에 민감하다면 항생제는 항생제 내성 조작을 위한 좋은 후보가 될 수 있다. 이어서, 항생제 내성 유전자 또는 역선별 자살 벡터를 재조합 방법을 사용하여 미생물의 게놈에 도입 할 수 있다. 역선별 자살 벡터는 관심 유전자의 결실, 선별 마커 및 역선별 마커 sacB로 구성 될 수 있다. 역선별은 고유 미생물 DNA 서열을 항생제 내성 유전자와 교환하는데 사용될 수 있다. 병렬 순화가 가능한 다수의 미생물을 동시에 평가하기 위해 중간 처리량 방법을 사용할 수 있다. 대안적인 순화 방법들에는, 자살 벡터 서열들이 순환하는 것 또는 개재 벡터 서열들을 얻는 것을 방지하기 위해 호밍 뉴클레아제를 사용하는 것이 포함된다.
DNA 벡터는 전기천공 및 화학적 형질전환을 비롯한 여러 방법을 통해 박테리아에 도입 될 수 있다. 표준 벡터 라이브러리들을 형질전환에 사용할 수 있다. 유전자 편집 방법의 예는 CRISPR 후 Cas9 테스트하여 미생물에서 Cas9의 활성을 확인하는 것이다.
이러한 리모델링 과정은 특정 플라스미드를 미생물로 일시적으로 형질감염시키는 것을 포함할 수 있다. 이어서 이들 플라스미드는 이를 제거하기 위해 미생물로부터 큐어링(curing) 될 수 있다. 플라스미드를 제거하기 위해 화학적 및 물리적 제제를 포함하는 다양한 방법이 개발되었다. 플라스미드 큐어링 프로토콜은 종종 몇몇 화학 물질들, 예컨대, 아크리딘 오렌지, 아크리플라빈 및 소듐 도데실 설페이트의 하위-억제 농도 또는 초-최적 온도에 배양물을 노출한 후, 큐어링된 유도체 선택으로 구성된다.
플라스미드가 안정하거나 성질 소실을 결정하기 어려운 경우, 박테리아는 큐어링제로 처리 될 수 있다. 큐어링제는 화학 및 물리적 물질들을 포함하며, 이들 중 일부는 DNA를 돌연변이 시키거나 구체적으로 그 복제를 방해하거나 박테리아 세포의 특정 구조 성분이나 효소에 영향을 줄 수 있다. 플라스미드 큐어링 프로토콜은 종종 몇몇 화학 물질들, 예컨대, 아크리딘 오렌지, 아크리플라빈 및 소듐 도데실 설페이트의 하위-억제 농도 또는 초-최적 온도에 배양물을 노출한 후, 큐어링된 유도체 선택으로 구성된다. 아크리딘 오렌지와 에티듐 브로마이드와 같은 DNA 삽입 물질(DNA intercalating agent)은 다양한 종류의 플라스미드에 대해 효과적이기 때문에 가장 일반적으로 사용된다. 이들 제제는 모두 플라스미드가 적은 다양한 박테리아 유도체의 회수를 향상시키기 위해 사용되었지만, 이들은 일부 플라스미드에 대해서만 개별적으로 효과적이며, 그들의 반응은 예측할 수 없다. 큐어링 효율은 또한 플라스미드 및 이를 운반하는 특정 박테리아 숙주에 따라 크게 달라질 수 있다. 일부 경우에, 큐어링하기 어려운 플라스미드는 큐어링하기 어려운 플라스미드를 표적하고 분해 할 수 있는 단백질 또는 시스템을 인코드하는 또 다른 큐어링하기 쉬운 플라스미드를 도입함으로써 제거 될 수 있다. 예를 들어, 큐어링하기 어려운 플라스미드는 큐어링하기 어려운 플라스미드를 표적으로하는 CRIPSR 시스템을 인코드하는 플라스미드를 도입함으로써 큐어링 될 수 있다. 박테리아 균주가 큐어링되면, 플라스미드 서열의 완전한 제거를 확인하기 위해 샘플을 시퀀싱 할 수 있다.
미생물의 비-형질전환 유전자조작
유리한 특성을 가진 미생물 집단은 직접 진화를 통해 얻을 수 있다. 직접 진화는 자연 선택 과정을 모방하여 사용자 정의 목표를 향해 단백질 또는 핵산을 진화시키는 접근법이다. 직접 진화의 예는 무작위 돌연변이가 미생물 집단에 도입되고, 가장 유리한 특성을 갖는 미생물이 선택되고, 선택된 미생물의 성장이 계속되는 경우이다. 성장 촉진 뿌리박테리아 (PGPRs)에서 가장 유리한 특성은 질소 고정에 있을 수 있다. 직접 진화 방법은 각 반복 후의 선택 프로세스에 기초하여 반복적이고 적응적일 수 있다.
질소 고정 능력이 높은 식물 성장 촉진 뿌리박테리아 (PGPR)가 생성 될 수 있다. PGPR의 진화는 유전자 변이의 도입을 통해 수행 될 수 있다. 유전자 변이는 중합효소 연쇄반응 돌연변이유발, 올리고뉴클레오티드-지정 돌연변이유발, 포화 돌연변이 유발, 단편 셔플링 돌연변이유발, 상동 재조합, CRISPR/Cas9 시스템, 화학적 돌연변이유발 및 이들의 조합을 통해 도입 될 수 있다. 이러한 접근법은 미생물 집단에 무작위 돌연변이를 도입 할 수 있다. 예를 들어, 돌연변이체는 올리고 뉴클레오티드-지정 돌연변이유발을 통한 합성 DNA 또는 RNA를 사용하여 생성 될 수 있다. 돌연변이체는 플라스미드에 포함된 툴을 사용하여 생성 할 수 있으며, 나중에 치유된다. 관심 유전자는, 개선된 PGPR 특성, 개선된 곡물의 콜로니화, 증가된 산소 민감성, 증가된 질소 고정 및 증가된 암모니아 배설을 비롯한 (그러나 이에 제한되는 것은 아님) 개선된 특성을 갖는 다른 종의 라이브러리를 사용하여 식별 될 수 있다. 이러한 라이브러리들에 기초하여 Geneious 또는 Platypus 설계 소프트웨어와 같은 소프트웨어를 사용하여 속내 유전자들을 설계할 수 있다. 기계 학습을 통해 돌연변이를 설계 할 수 있다. 대사 모델을 통해 돌연변이를 설계 할 수 있다. la Platypus를 사용하여 돌연변이의 자동 설계를 수행 할 수 있으며 Cas-지정 돌연변이유발을 위한 RNA를 안내 할 것이다.
속내 유전자는 숙주 미생물 내로 전달 될 수 있다. 또한, 리포터 시스템이 미생물에 전달될 수도 있다. 리포터 시스템은 프로모터의 특성을 규명하고 형질전환 성공 여부를 결정하며 돌연변이체를 선별하고 음성 선별 툴로 작용한다.
돌연변이를 수반하는 미생물은 연속 계대배양을 통해 배양 될 수 있다. 미생물 콜로니는 미생물의 단일 변이체를 함유한다. 미생물 콜로니는 자동 콜로니 피커 및 액체 처리기를 사용하여 스크리닝된다. 유전자 복제 및 증가된 카피수를 갖는 돌연변이체는 원하는 형질의 유전자형을 더 많이 발현한다.
질소 고정을 기반으로 한 식물 성장 촉진 미생물 선택
미생물 콜로니는 질소 고정을 평가하기 위해 다양한 분석을 사용하여 스크리닝될 수 있다. 질소 고정을 측정하는 한 가지 방법은 질소 배설을 측정하는 단일 발효 분석법에 의한 것이다. 다른 방법은 시간에 따른 인라인 샘플링을 통한 아세틸렌 환원 분석 (ARA)이다. ARA는 마이크로튜브 어레이의 고 처리량 플레이트에서 수행 될 수 있다. ARA는 살아있는 식물 및 식물 조직으로 수행 될 수 있다. 배지 배합 및 배지 산소 농도는 ARA 분석에서 변화될 수 있다. 미생물 변이체를 선별하는 또 다른 방법은 바이오센서를 사용하는 것이다. NanoSIMS 및 Raman 현미경을 사용하여 미생물의 활성을 조사 할 수 있다. 일부 경우에, 생물반응기에서 발효 방법을 사용하여 박테리아를 배양 및 확장시킬 수도 있다. 생물반응기는 박테리아 성장의 견고성을 개선하고 산소에 대한 박테리아의 민감성을 감소시키도록 설계된다. 중간 내지 높은 TP 플레이트 기반 마이크로발효기를 사용하여 산소 감수성, 영양 요구, 질소 고정 및 질소 배설을 평가한다. 박테리아는 또한 숨어있는 경로를 밝히기 위해 경쟁적이거나 유리한 미생물과 공동-배양 될 수 있다. 유세포 분석법은 화학, 비색 또는 형광 표시기를 사용하여 높은 수준의 질소를 생성하는 박테리아를 선별하는 데 사용될 수 있다. 박테리아는 질소원의 존재 또는 부재하에서 배양 될 수 있다. 예를 들어, 박테리아는 글루타민, 암모니아, 우레아 또는 질산염과 함께 배양 될 수 있다.
미생물 육종
미생물 육종은 작물 마이크로바이옴 내에서 종의 역할을 체계적으로 식별하고 개선하는 방법이다. 이 방법은 다음 세 단계를 포함한다: 1) 식물-미생물 상호작용을 매핑하고 특정 표현형에 연결된 조절 네트워크를 예측하여 후보 종 선택, 2) 조절 네트워크와 유전자 클러스터의 종내 교잡을 통한 미생물 표현형의 실용적이고 예측가능한 개선 3) 원하는 작물 표현형을 생산하는 새로운 미생물 유전자형의 스크리닝 및 선택. 균주의 개선을 체계적으로 평가하기 위해, 미생물 커뮤니티의 콜로니화 역학을 주요 종별의 유전적 활동과 연결하는 모델이 생성된다. 이 모델을 사용하여 유전자 표적 육종을 예측하고 농업학적 관련성이 있는 마이크로바이옴-인코드된 형질에 있어서의 개선을 선택하는 빈도를 개선시킨다. 상기 과정의 한 구체예의 도식적 표현은 도 2A를 참조한다. 특히, 도 2A는 구체예들에 따른 미생물 육종의 개략도를 도시한다. 도 2A에 도시된 바와 같이, 작물 마이크로바이옴의 합리적인 개선은 토양 생물다양성을 증가시키고, 핵심종의 영향을 조정하며, 및/또는 중요한 대사 경로의 타이밍 및 발현을 변경하는데 사용될 수 있다. 이를 위해, 본 발명자들은 작물 마이크로바이옴 내에서 균주의 역할을 확인하고 개선하기 위한 미생물 육종 파이프 라인을 개발하였다. 이 방법은 다음 세 단계를 포함한다: 1) 식물-미생물 상호작용을 매핑하고 특정 표현형에 연결된 조절 네트워크를 예측하여 후보 종 선택, 2) 유전자 조절 네트워크와 유전자 클러스터의 종내 교잡을 통한 미생물 표현형의 실용적이고 예측가능한 개선 3) 원하는 작물 표현형을 생산하는 새로운 미생물 유전자형의 스크리닝 및 선택. 균주의 개선을 체계적으로 평가하기 위해, 발명자들은 미생물 커뮤니티의 콜로니화 역학을 주요 종별의 유전적 활동과 연결하는 모델을 이용한다. 이 과정은 육종 및 농업적으로 관련있는 마이크로바이옴-인코드된 특성들의 개선을 선택하는 방법론을 나타낸다.
식물 특성(예컨대, 질소 고정)을 개선하기 위한 박테리아의 생산은 연속 계대를 통해 구현될 수 있다. 이 박테리아의 생산은 하나 이상의 식물에 하나 이상의 개선된 특성을 부여할 수 있는 박테리아 및/또는 조성물을 확인하는 것 외에, 미생물 무리에 의해 영향을 받는 특정한 개선된 특성을 갖는 식물을 선택함으로써 수행 될 수 있다. 식물 특성을 개선하는 박테리아를 생산하는 한 가지 방법은 다음 단계들을 포함한다: (a) 제 1 식물의 조직 또는 토양으로부터 박테리아를 단리하는 단계; (b) 하나 이상의 변이체 박테리아를 생성하기 위해 하나 이상의 박테리아에 유전자 변이를 도입하는 단계; (c) 복수의 식물을 변이체 박테리아에 노출시키는 단계; (d) 복수의 식물 중 하나의 조직 또는 토양으로부터 박테리아를 단리하는 단계, 이 때 박테리아가 단리되는 식물은 복수의 식물 내 다른 식물에 비해 개선된 특성을 갖는 단계; 및 (e) 개선된 특성을 가지는 식물로부터 단리된 (단계 (d)) 박테리아로 단계 (b) 내지 (d)를 반복하는 단계. 단계 (b) 내지 (d)는 식물의 개선된 특성이 원하는 수준에 도달 할 때까지 임의의 횟수 (예를 들어, 1 회, 2 회, 3 회, 4 회, 5 회, 10 회 또는 그 이상)로 반복 될 수 있다. 또한, 복수의 식물은 2이상의 식물, 가령, 10 개 내지 20 개 식물, 또는 20 개 이상, 50 개 이상, 100 개 이상, 300 개 이상, 500 개 이상 또는 1000 개 이상의 식물 일 수 있다.
특성이 개선된 식물을 얻는 것 이외에도, 하나 이상의 유전자 (예를 들어, 질소 고정을 조절하는 유전자)에 도입된 하나 이상의 유전자 변이체를 포함하는 박테리아를 포함하는 박테리아 집단이 수득된다. 상기 단계를 반복함으로써, 관심 식물 특성과 상관관계가 있는 집단 중 가장 적절한 구성원을 포함하는 박테리아 집단을 얻을 수 있다. 이 집단 내 박테리아는, 가령, 유전자 및/또는 표현형 분석에 의해 식별되고 그 유리한 특성들이 결정될 수 있다. 단계 (a)에서 단리된 박테리아의 유전자 분석을 할 수 있다. 표현형 및/ 또는 유전자형 정보는 다음을 포함하는 기술들을 사용하여 얻을 수 있다: 식물 기원의 화학 성분의 고 처리량 스크리닝, 유전 물질의 고 처리량 시퀀싱을 포함하는 시퀀싱 기술, 차등 디스플레이 기술 (DDR-PCR 및 DD-PCR 포함), 핵산 마이크로 어레이 기술, RNA-시퀀싱 (전장 전사체 샷건 시퀀싱) 및 qRT-PCR (정량 실시간 PCR). 얻은 정보를 사용하여, 존재하는 박테리아의 식별 및 활성에 대한 커뮤니티 프로파일링 정보, 가령, 계통발생학적 분석 또는 rRNA 오페론의 구성요소 또는 기타 분류학적으로 유익한 유전자좌의 구성요소를 코딩하는 핵산의 마이크로어레이-기반 스크리닝을 얻을 수 있다. 분류 학적으로 유익한 유전자좌의 예는 16S rRNA 유전자, 23S rRNA 유전자, 5S rRNA 유전자, 5.8S rRNA 유전자, 12S rRNA 유전자, 18S rRNA 유전자, 28S rRNA 유전자, gyrB 유전자, rpoB 유전자, fusA 유전자, recA 유전자, coxl 유전자, nifD 유전자를 포함한다. 집단에 존재하는 분류군을 결정하기 위한 분류학적 프로파일링 공정의 예는 US20140155283에 기재되어 있다. 박테리아 식별은 하나 이상의 유전자 또는 하나 이상의 신호전달 경로, 가령, 질소 고정 경로와 관련된 유전자의 활성을 특성규명하는 것을 포함 할 수 있다. 상이한 박테리아 종들 사이의 상승적인 상호작용 (두 성분이 이들의 조합에 의해 원하는 효과를 추가량 이상 증가시키는 경우) 또한 박테리아 집단에 존재할 수 있다.
유전자 변이 - 게놈 변경의 위치와 출처
유전자 변이는 다음으로 구성된 군에서 선택된 유전자일 수 있다: nifA, nifL, ntrB, ntrC, glnA, glnB, glnK, draT, amtB, glnD, glnE, nifJ, nifH, nifD, nifK , nifY, nifE, nifN, nifU, nifS, nifV, nifW, nifZ, nifM, nifF, nifB, nifQ, bcsII, bcsIII, yjbE, fhaB, pehA, glgA, otsB, treZ, 및 cysZ. 유전자 변이는 다음으로 구성된 군에서 선택된 기능성을 가지는 단백질을 인코드하는 유전자에서의 변이일 수 있다: 글루타민 합성효소, 글루타미나제, 글루타민 합성효소 아데닐릴트랜스퍼라제, 전사 활성화제, 항-전사 활성화제, 피루베이트 플라보독신 산화환원효소, 플라보독신, NAD +-이질소-환원효소 aDP-D- 리보실트랜스퍼라제, 세포외 다당류 생성, 사상 헤마글루티닌, 글리코겐 합성효소, 트레할로스 합성, 또는 설페이트 운반체. 유전자 변이는 다음 중 하나 이상에서 생성되는 돌연변이 일 수 있다: NifA 글루타미나제, bcsII, bcsIII, yjbE, fhaB, pehA, otsB, treZ 또는 CysZ의 증가된 발현 또는 활성; NifL, NtrB, 글루타민 합성효소, GlnB, GlnK, DraT, AmtB 또는 glgA의 감소된 발현 또는 활성; GlnE의 감소된 아데닐릴-제거 활성; 또는 GlnD의 감소된 유리딜릴-제거 활성. 유전자 변이를 도입하는 것은 표적 부위에서 하나 이상의 뉴클레오티드, 가령, 1, 2, 3, 4, 5, 10, 25, 50, 100, 250, 500개 이상의 뉴클레오티드의 삽입 및/또는 결실을 포함 할 수 있다. 본 명세서에 개시된 방법의 하나 이상의 박테리아에 도입된 유전자 변이는 녹아웃 돌연변이 (예를 들어, 프로모터의 결실, 조기 정지 코돈을 생성하기 위한 삽입 또는 결실, 전체 유전자의 결실) 일 수 있거나, 또는 단백질 도메인의 활성의 제거 또는 폐지 (예를 들어, 활성 부위에 영향을 미치는 점 돌연변이, 또는 단백질 산물의 관련 부분을 인코드하는 유전자의 일부 결실)일 수 있거나, 또는 표적 유전자의 조절 서열을 변경 또는 폐지할 수 있다. 유전적 변이가 도입되는 박테리아에 상응하는 박테리아 종 또는 속의 게놈내에서 발견되는 이종조절 서열 및 조절 서열을 포함하는 하나 이상의 조절 서열이 또한 삽입 될 수 있다. 더욱이, 조절 서열은 박테리아 배양물 또는 식물 조직 내에서 유전자의 발현 수준에 기초하여 선택 될 수있다. 유전자 변이는 표적 부위에 특이적으로 도입된 미리 결정된 유전자 변이 일 수 있다. 유전자 변이는 표적 부위 내의 무작위 돌연변이 일 수 있다. 유전자 변이는 하나 이상의 뉴클레오티드의 삽입 또는 결실 일 수 있다. 몇몇 경우에, 특성 개선을 평가하기 위해 박테리아를 식물에 노출시키기 전에 복수의 상이한 유전자 변이 (예를 들어, 2, 3, 4, 5, 10 또는 그 이상)가 하나 이상의 단리된 박테리아에 도입된다. 복수의 유전자 변이는 상기 유형 중 임의의 유형, 동일하거나 상이한 유형, 및 임의의 조합 일 수 있다. 일부 경우에, 연관된 식물들에 점진적으로 개선된 특성을 부여하는 박테리아에서 복수의 유전자 변이를 축적하기 위해, 복수의 상이한 유전자 변화는 순차적으로, 제 1 단리 단계 후 제 1 유전자 변이, 제 2 단리 단계 후 제 2 유전자 변이 등을 도입함으로써 도입된다.
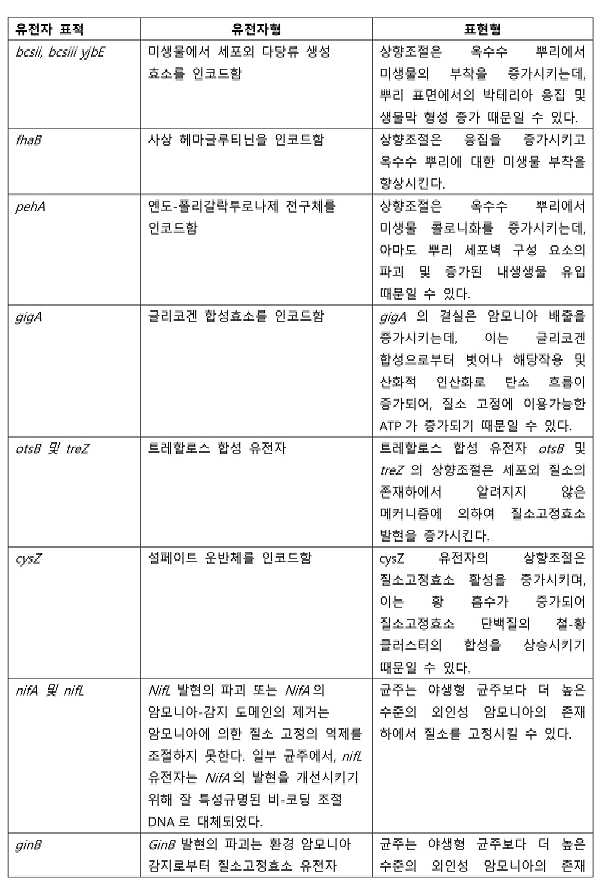
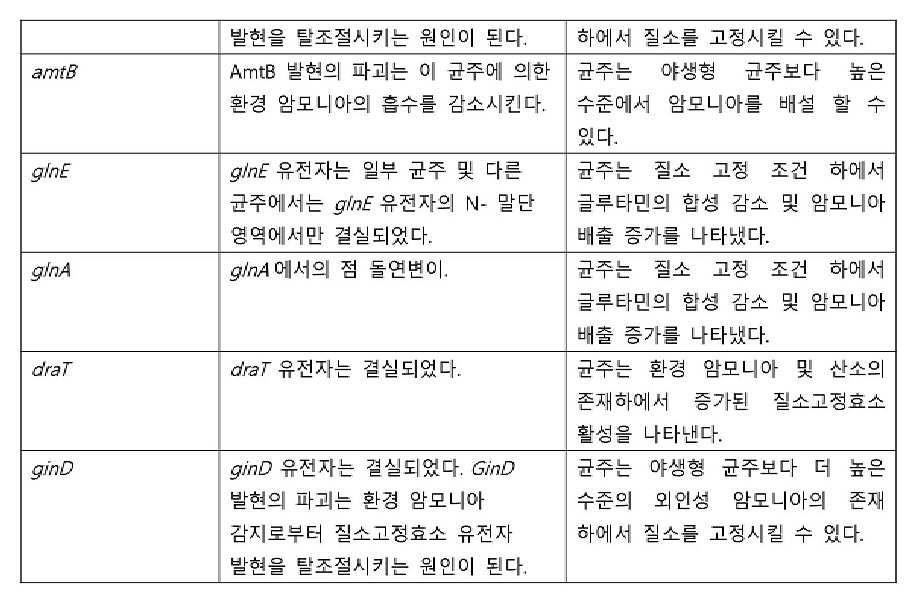
유전자 변이 - 게놈 변경 도입 방법
일반적으로, "유전자 변이"라는 용어는 참조 폴리뉴클레오티드, 가령, 참조 게놈 또는 이의 일부, 또는 참조 유전자 또는 이의 일부와 관련하여 폴리뉴클레오티드 서열에 도입된 임의의 변화를 지칭한다. 유전자 변이는 "돌연변이"로 지칭 될 수 있고, 유전자 변이를 포함하는 서열 또는 유기체는 "유전자 변이체"또는 "돌연변이체"로 지칭 될 수 있다. 유전자 변이는 유전자 발현, 대사 및 세포 신호 전달을 포함한 일부 생물학적 활성의 증가 또는 감소와 같은 많은 효과를 가질 수 있다. 유전자 변이는 구체적으로 표적 부위에 도입되거나 무작위로 도입 될 수 있다. 유전자 변이를 도입하기 위한 다양한 분자 툴 및 방법이 이용 가능하다. 예를 들면, 유전자 변이는 중합효소 연쇄반응 돌연변이유발, 올리고뉴클레오티드-지정 돌연변이유발, 포화 돌연변이 유발, 단편 셔플링 돌연변이유발, 상동 재조합, 재조합매개 유전자공학 (recombineering), 람다 레드 매개 재조합, CRISPR/Cas9 시스템, 화학적 돌연변이유발 및 이들의 조합을 통해 도입 될 수 있다. 유전자 변이를 도입하는 화학적 방법은 화학적 돌연변이원, 예를 들어 에틸 메탄설포네이트 (EMS), 메틸 메탄설포네이트 (MMS), N-니트로소우레아 (EN U), N-메틸-N-니트로-N'-니트로소구아니딘, 4-니트로퀴놀린 N- 옥사이드, 디 에틸 설페이트, 벤조피렌, 사이클로포스파미드, 블레오마이신, 트리에틸멜라민, 아크릴아미드 모노머, 질소 머스타드, 빈크리스틴, 다이에폭시알칸 (예를 들면, 다이에폭시부탄), ICR-170, 포름알데히드, 프로카르바진 하이드로클로라이드, 에틸렌옥사이드, 디메틸니트로사민, 7,12 다이메틸벤즈(아)안트라센, 클로람부실, 헥사메틸포스포르아미드, 바이설판 등에 대한 DNA의 노출을 포함한다. 방사선 돌연변이-유도 물질은 자외선, γ조사, X-선 및 빠른 중성자 충격을 포함한다. 유전자 변이는 또한 예컨대, 자외선과 함께 트리메틸소랄렌을 사용하여 핵산에 도입 될 수 있다. 이동성 DNA 요소, 예를 들어, 전위 요소의 무작위 또는 표적화된 삽입은 유전자 변이를 생성하는 또 다른 적합한 방법이다. 유전자 변이는 예를 들어, 중합효소 연쇄 반응 (PCR) 기술, 가령, 에러-프론 PCR을 사용하여 무세포 시험관내 시스템에서 증폭하는 동안 핵산에 도입 될 수 있다. 유전자 변이는 DNA 셔플링 기술 (예컨대, 엑손 셔플링, 도메인 스와핑 등)을 사용하여 시험관내 핵산에 도입 될 수 있다. 유전자 변이는 또한 세포에서 DNA 복구 효소 결핍의 결과로서 핵산에 도입될 수도 있으며, 예컨대, 돌연변이체 DNA 복구 효소를 인코드하는 돌연변이체 유전자의 세포에서의 존재는 세포 게놈에서 높은 빈도의 돌연변이 (즉, 약 1 돌연변이/100 유전자 -1 돌연변이/10,000 유전자)를 생성하는 것으로 예상된다. DNA 복구 효소를 인코드하는 유전자의 예는 Mut H, Mut S, Mut L 및 Mut U 및 다른 종에서의 상동체 (예컨대, MSH 1 6, PMS 1 2, MLH 1, GTBP, ERCC-1 등)를 포함하지만 이에 제한되지는 않는다. 유전자 변이를 도입하기 위한 다양한 방법들의 예시적 설명은 예컨대, Stemple (2004) Nature 5:1-7; Chiang 외. (1993) PCR Methods Appl 2(3): 210-217; Stemmer (1994) Proc. Natl. Acad. Sci. USA 91:10747-10751; 및 미국 특허 제 6,033,861, 및 6,773,900에 제공되어 있다.
미생물에 도입된 유전자 변이는 형질전환, 시스제닉 (cisgenic), 유전체내 (intragenomic), 속내, 속간, 합성, 진화, 재배열, 또는 SNP로서 분류될 수 있다.
유전자 변이는 상기 기재된 특성의 개선을 유도하기 위해 미생물 내의 수많은 대사 경로에 도입 될 수 있다. 대표적인 경로는 황 흡수 경로, 글리코겐 생합성, 글루타민 조절 경로, 몰리브덴 흡수 경로, 질소 고정 경로, 암모니아 동화, 암모니아 배설 또는 분비, n 질소 흡수, 글루타민 생합성, 아나목스 (Annamox), 인산 가용화, 유기산 수송, 유기산 생성, 응집소 생성, 반응성 산소 라디칼 소거 유전자, 인돌 아세트산 생합성, 트레할로스 생합성, 식물 세포벽 분해 효소 또는 경로, 뿌리 부착 유전자, 세포외 다당류 분비, 글루타메이트 합성효소 경로, 철 흡수 경로, 시데로포어 (siderophore) 경로, 키틴분해효소 경로, ACC 탈아미노효소, 글루타티온 생합성, 인 신호전달 유전자, 쿼럼 ?칭 경로, 시토크롬 경로, 헤모글로빈 경로, 박테리아 헤모글로빈-유사 경로, 작은 RNA rsmZ, 리조비톡신 생합성, lapA 접착 단백질, AHL 쿼럼 센싱 경로, 페나진 생합성, 사이클릭 리포펩티드 생합성, 및 항생제 생성을 포함한다.
CRISPR/Cas9 (클러스터된 규칙적으로 간격을 둔 짧은 회문 반복서열)/CRISPR 관련 (Cas) 시스템을 사용하여 원하는 돌연변이를 도입 할 수 있다. CRISPR/Cas9는 침입 핵산의 침묵을 유도하기 위해 CRISPR RNA (crRNA)를 사용함으로써 바이러스 및 플라스미드에 대한 적응 면역을 박테리아 및 고세균에 제공한다. Cas9 단백질 (또는 이의 기능적 균등물 및/또는 그의 변이체, 즉, Cas9-유사 단백질)은 crRNA 및 tracrRNA라 불리는 2개의 자연 발생 또는 합성 RNA 분자 (가이드 RNA라고도 함)와 단백질의 결합에 의존하는 DNA 엔도뉴클레아제 활성을 자연적으로 함유한다. 일부 경우에, 2개의 분자는 공유 결합하여 단일 분자 (단일 가이드 RNA ( "sgRNA")로도 불림)를 형성한다. 따라서, Cas9 또는 Cas9-유사 단백질은 DNA-표적화 RNA (이 용어는 2-분자 가이드 RNA 구조 및 단일-분자 가이드 RNA 구조 둘 모두를 포함함)와 결합되며, 이는 Cas9 또는 Cas9-유사 단백질을 활성화시키고 표적 핵산 서열로 단백질을 유도한다. Cas9 또는 Cas9-유사 단백질이 자연적인 효소 기능을 유지하는 경우, 표적 DNA를 절단하여, 게놈 변경 (즉, 편집: 결실, 삽입 (공여체 폴리뉴클레오티드가 존재하는 경우), 대체 등)을 가져올 수 있는 이중 가닥 파괴를 생성함으로써 유전자 발현이 변경될 것이다. Cas9의 일부 변이체들 (이러한 변이체들은 용어 Cas9-유사에 포함됨)은 감소된 DNA 절단 활성을 가지도록 변형되었다 (일부 경우에서, 이들은 표적 DNA의 두 가닥 모두 대신에 단일 가닥을 절단하는 반면, 다른 경우에, 이들은 DNA 절단 활성이 없는 것으로 심각하게 감소되었다). 유전자 변이를 도입하기 위한 CRISPR 시스템의 추가의 예시적인 설명은 예컨대, US8795965에서 찾을 수 있다.
주기적 증폭 기술로서, 중합효소 연쇄 반응 (PCR) 돌연변이유발은 돌연변이유발 프라이머를 사용하여 원하는 돌연변이를 도입한다. PCR은 변성, 어닐링, 및 확장의 주기에 의해 실시된다. PCR에 의한 증폭 후, 돌연변이된 DNA의 선택 및 모체 플라스미드 DNA의 제거는 다음에 의해 구현될 수 있다: 1) PCR 동안 하이드록시메틸화-dCTP로 dCTP를 대체한 후, 비-하이드록시메틸화 모체 DNA만을 제거하는 제한 효소로 소화; 2) 플라스미드를 상이한 항생제 내성으로 변화시키는, 항생제 내성 유전자 및 연구대상 유전자 모두의 동시 돌연변이유발, 이러한 새로운 항생제 내성은 차후 원하는 돌연변이의 선택을 용이하게 하며; 3) 원하는 돌연변이 도입 후, 모체 메틸화 템플릿 DNA by 메틸화 DNA만을 절단하는 제한 효소 Dpnl에 의한 모체 메틸화 주형 DNA 소화, 이로써 돌연변이유발된 비메틸화 사슬들이 회수되고; 또는 4) 돌연변이된 DNA의 형질전환 효율을 증가시키기 위하여 추가 결찰 반응에서 돌연변이된 PCR 산물들을 순환. 예시적인 방법들에 대한 또 다른 설명은 예컨대, US7132265, US6713285, US6673610, US6391548, US5789166, US5780270, US5354670, US5071743, 및 US20100267147에서 찾을 수 있다.
부위-지정 돌연변이유발이라고도 하는 올리고뉴클레오티드-지정 돌연변이유발은 전형적으로 합성 DNA 프라이머를 이용한다. 이 합성 프라이머는 원하는 돌연변이를 함유하고, 돌연변이 부위 주위의 주형 DNA에 상보적이어서, 관심 유전자의 DNA와 혼성화 할 수 있다. 돌연변이는 단일 염기 변화 (점 돌연변이), 다중 염기 변화, 결실 또는 삽입, 또는 이들의 조합 일 수 있다. 이어서 단일-가닥 프라이머는 DNA 중합효소를 사용하여 연장되며, 이는 나머지 유전자를 복제한다. 이와 같이 복제된 유전자는 돌연변이 부위를 포함하고, 벡터로서 숙주 세포에 도입되어 클로닝 될 수 있다. 마지막으로, 돌연변이체는 원하는 돌연변이를 함유하는지 확인하기 위해 DNA 시퀀싱에 의해 선택될 수 있다.
유전자 변이는 에러-프론 PCR을 사용하여 도입될 수 있다. 이 기술에서, 관심 유전자는 서열 복제의 충실도가 부족한 조건하에서 DNA 중합효소를 사용하여 증폭된다. 결과적으로 증폭 산물은 시퀀스에서 최소한 하나의 에러를 포함한다. 유전자가 증폭되고 생성된 반응 산물(들)이 주형 분자와 비교하여 서열에서 하나 이상의 변경을 포함하는 경우, 생성된 산물은 주형과 비교하여 돌연변이유발된다. 무작위 돌연변이를 도입하는 또 다른 수단은 세포를 니트로소구아니딘 또는 에틸 메탄설포네이트와 같은 화학적 돌연변이원에 노출시키는 것 (Nestmann, Mutat Res 1975 June; 28 (3):323-30)이며, 이어서 유전자를 함유하는 벡터는 숙주로부터 분리된다.
포화 돌연변이유발은 무작위 돌연변이유발의 또 다른 형태이며, 특정 부위 또는 유전자의 좁은 영역에서 모든 또는 거의 모든 가능한 돌연변이를 생성하려고 시도한다. 일반적인 의미로, 포화 돌연변이유발은 돌연변이유발될 정의된 폴리뉴클레오티드 서열 (이 때 돌연변이될 서열은, 예를 들면, 15 내지 100,000 염기 길이임)에서 전체 돌연변이유발 카세트 세트 (이 때 각 카세트는, 예를 들면, 1-500 염기 길이임)를 돌연변이유발하는 것을 포함한다. 따라서, 돌연변이유발 시키기 위해 돌연변이 그룹 (예를 들어, 1 내지 100 돌연변이 범위)이 각 카세트에 도입된다. 하나의 카세트에 도입 될 돌연변이 그룹화는 1 라운드의 포화 돌연변이유발 동안 제 2 카세트에 도입 될 제 2 돌연변이 그룹화와 상이하거나 동일 할 수 있다. 이러한 그룹화는 특정 코돈의 결실, 부가, 그룹화, 및 특정 뉴클레오티드 카세트들의 그룹화로 예시된다.
DNA 셔플링으로도 불리는 단편 셔플링 돌연변이유발은, 유익한 돌연변이를 신속하게 전파시키는 한 방법이다. 셔플링 과정의 예에서, DNAse를 사용하여 한 세트의 모체 유전자를, 예컨대, 약 50-100 bp 길이의 조각으로 단편화한다. 이후 프라이머 없이 중합효소 연쇄 반응 (PCR)이 후속되며 상동성 서열이 충분히 중첩된 DNA 단편들은 서로에 대해 어닐링된 다음 DNA 중합효소에 의해 확장될 것이다. 이러한 PCR 확장을 수회 라운드 발생하도록 하면, 이후 DNA 분자들 중 일부는 모체 유전자의 크기에 도달한다. 이러한 유전자는 또 다른 PCR을 사용하여 증폭 될 수 있으며 이번에는 가닥의 단부를 보완하도록 설계된 프라이머를 추가한다. 프라이머는 5' 단부에 또 다른 서열, 가령, 클로닝 벡터 내로의 결찰에 필요한 제한 효소 인식 부위에 대한 서열을 가질 수 있다. 셔플링 기술의 또 다른 예들은 US20050266541에 제공되어 있다.
상동 재조합 돌연변이 유발은 외인성 DNA 단편과 표적화된 폴리뉴클레오티드 서열 사이의 재조합을 포함한다. 이중-가닥 파열이 발생한 후, 파열부의 5' 단부 주위의 DNA 섹션이 절제술 (resection)이라는 과정에서 절단된다. 이어지는 가닥 침습 단계에서, 파열된 DNA 분자의 오버행 3' 단부는 이후 파열되지 않은 유사 또는 동일한 DNA 분자를 "침습한다". 이 방법은 유전자 결실, 엑손 제거, 유전자 부가 및 점 돌연변이 도입에 사용할 수 있다. 상동 재조합 돌연변이유발은 영구적이거나 조건부 일 수 있다. 일반적으로, 재조합 주형 또한 제공된다. 재조합 주형은 다른 벡터의 성분이거나, 별도의 벡터에 포함되거나, 또는 별도의 폴리뉴클레오티드로서 제공 될 수 있다. 일부 구체예에서, 재조합 주형은, 가령, 부위-특이적 뉴클레아제에 의해 닉킹되거나 (nicked) 절단되는 표적 서열 내에서 또는 근처에서 상동 재조합에서 주형으로서 기능하도록 설계된다. 주형 폴리뉴클레오티드 임의의 적합한 길이, 가령, 약 또는 약 10, 15, 20, 25, 50, 75, 100, 150, 200, 500, 1000 이상, 또는 그 이상의 뉴클레오티드 길이 일 수 있다. 일부 구체예들에서, 주형 폴리뉴클레오티드는 표적 서열을 포함하는 폴리뉴클레오티드의 일부에 상보적이다. 최적으로 정렬시, 주형 폴리뉴클레오티드는 표적 서열의 하나 이상의 뉴클레오티드 (예컨대, 약 또는 약 1, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80, 90, 100 이상 또는 그 이상의 뉴클레오티드)와 중첩될 수 있다. 일부 구체예에서, 주형 서열 및 표적 서열을 포함하는 폴리뉴클레오티드가 최적으로 정렬 될 때, 주형 폴리뉴클레오티드의 가장 가까운 뉴클레오티드는 표적 서열로부터 약 1, 5, 10, 15, 20, 25, 50, 75, 100, 200, 300, 400, 500, 1000, 5000, 10000, 또는 그 이상의 뉴클레오티드 이내에 존재한다. 상동 재조합 방법에 유용한 부위-지정 뉴클레아제의 비-제한적 예는 아연 핑거 뉴클레아제, CRISPR 뉴클레아제, TALE 뉴클레아제 및 메가뉴클레아제를 포함한다. 이러한 뉴클레아제의 사용에 관한 또 다른 설명은 예컨대, US8795965 및 US20140301990을 참고하라.
화학적 돌연변이원 또는 방사선을 비롯하여, 주로 점 돌연변이를 생성하고 짧은 결실, 삽입, 전환 및/또는 전이를 생성하는 돌연변이원을 사용하여 유전자 변이를 생성할 수 있다. 돌연변이원에는, 에틸 메탄설포네이트, 메틸메탄 설포네이트, N-에틸-N-니트로소우레아, 트라이에틸아민, N-메틸-N-니트로소우레아, 프로카르바진, 클로람부실, 사이클로포스파미드, 다이에틸 설페이트, 아크릴아미드 모노머, 멜팔란, 질소 머스터드, 빈크리스틴, 다이메틸니트로사민, N-메틸-N'-니트로-니트로소구아니딘, 니트로소구아니딘, 2-아미노퓨린, 7,12 다이메틸-벤즈(아)안트라센, 에틸렌 옥사이드, 헥사메틸포스포라미드, 바이설판, 다이에폭시알칸 (다이에폭시옥탄, 다이에폭시부탄, 등), 2-메톡시-6-클로로-9[3-(에틸-2-클로로- 에틸)아미노프로필아미노]아크리딘 다이하이드로클로라이드 및 포름알데하이드가 포함되나 이에 제한되는 것은 아니다.
유전자 변이를 도입하는 것은 불완전한 과정일 수 있으므로, 처리된 박테리아 집단에서 일부 박테리아는 원하는 돌연변이를 보유하지만 다른 박테리아는 그렇지 않다. 일부 경우에, 원하는 유전자 변이를 보유하는 박테리아를 풍부하게 하기 위해 선별 압력을 적용하는 것이 바람직하다. 전통적으로, 성공적인 유전자 변이체의 선택은 항생제 내성 유전자를 삽입하거나 치명적이지 않은 화합물을 치명적인 대사산물로 전환시킬 수 있는 대사 활성을 제거하는 경우와 같이, 유전자 변이에 의해 부여되거나 제거되는 몇몇 기능에 대한 또는 기능에 맞서는 선별을 포함하였다. 폴리뉴클레오티드 서열 그 자체를 기준으로 한 선별 압력을 적용하여, 원하는 유전자 변이만이 도입될 필요가 있게 하는 것도 가능하다 (예컨대, 선별 마커를 필요로 하지 않고). 이 경우에, 선별 압력은 표적 부위에 유전자 변이가 도입되지 않은 절단 게놈을 포함 할 수 있어서, 선별은 유전자 변이를 도입하고자 하는 참조 서열에 대해 효과적으로 지시된다. 전형적으로, 절단은 표적 부위의 100개 뉴클레오티드 이내에서 (예컨대, 표적 부위에서 또는 그 내부에서의 절단을 포함하여 표적 부위로부터 75, 50, 25, 10, 또는 그 미만의 뉴클레오티드 내에서) 발생한다. 절단은 아연 핑거 뉴클레아제, CRISPR 뉴클레아제, TALE 뉴클레아제 (TALEN) 또는 메가뉴클레아제로 이루어진 군으로부터 선택된 부위-특이적 뉴클레아제에 의해 지시 될 수 있다. 이러한 과정은 상동 재조합을 위한 주형이 제공되지 않는다는 점을 제외하고는 표적 부위에서 상동 재조합을 향상시키는 과정과 유사하다. 결과적으로, 원하는 유전자 변이가 없는 박테리아는, 복구되지 않은 채, 세포 사멸을 가져오는 절단을 거칠 가능성이 더 크다. 이어서, 개선된 특성의 부여를 평가하기 위한 식물에 대한 노출에 사용하기 위하여 선별에 살아남은 박테리아를 단리 할 수 있다.
CRISPR 뉴클레아제는 표적 부위에 대한 절단을 지시하는 부위-특이적 뉴클레아제로서 사용될 수 있다. 돌연변이되지 않은 세포를 사멸시키는 Cas9를 사용하여 돌연변이된 미생물의 선별을 개선할 수 있다. 이어서, 돌연변이된 미생물을 식물에 접종하여 공생을 재확인하고 효율적인 공생 제를 선별하기 위한 진화 압력을 생성한다. 그 후 식물 조직에서 미생물을 다시 단리 할 수 있다. 비-변이체에 대한 선별에 사용된 CRISPR 뉴클레아제 시스템은 상동 재조합을 위한 주형이 제공되지 않는 것을 제외하고는 유전자 변이 도입에 관해 상기 기재된 것과 유사한 요소를 사용할 수 있다. 표적 부위로 지시된 절단은 영향을 받는 세포의 사멸을 향상시킨다.
아연 핑거 뉴클레아제, TALE 뉴클레아제 (TALEN) 시스템 및 메가뉴클레아제와 같이, 표적 부위에서의 절단을 구체적으로 유도하기 위한 다른 방안이 이용가능하다. 아연 핑거 뉴클레아제 (ZFN)는 아연 핑거 DNA 결합 도메인을 DNA 절단 도메인에 융합시켜 생성된 인공 DNA 엔도뉴클레아제이다. ZFN은 원하는 DNA 서열을 표적하도록 조작 될 수 있으며, 이는 아연-핑거 뉴클레아제가 독특한 표적 서열을 절단 할 수 있게 한다. 세포 내로 도입 될 때, ZFN은 이중 가닥 파괴를 유도함으로써 세포 (예컨대, 세포의 게놈)에서 표적 DNA를 편집하는데 사용될 수 있다. 전사 활성화인자-유사 이펙터 뉴클레아제 (TALEN)는 TAL (전사 활성화인자-유사) 이펙터 DNA 결합 도메인을 DNA 절단 도메인에 융합시켜 생성된 인공 DNA 엔도뉴클레아제이다. TALENS는 실질적으로 임의의 원하는 DNA 서열에 결합하도록 신속하게 조작될 수 있고, 세포에 도입 될 때, TALEN은 이중 가닥 파열을 유도함으로써 세포 (예를 들어, 세포의 게놈)에서 표적 DNA를 편집하는데 사용될 수 있다. 메가뉴클레아제 (호밍 엔도뉴클레아제)는 큰 인식 부위 (12 내지 40개 염기쌍의 이중-가닥 DNA 서열)를 특징으로 하는 엔도데옥시리보뉴클레아제이다. 메가뉴클레아제는 고도로 표적화된 방식으로 서열을 대체, 제거 또는 변형하는데 사용될 수 있다. 단백질 유전공학을 통해 인식 서열을 변형함으로써 표적 서열은 변경될 수 있다. 메가뉴클레아제는 박테리아, 식물 또는 동물 등 모든 게놈 유형을 변형시킴에 사용될 수 있으며, 일반적으로 다음 네 가지 패밀리로 분류된다: LAGLIDADG 패밀리 (서열 번호: 1), GIY-YIG 패밀리, His-Cyst 박스 패밀리 및 HNH 패밀리. 예시적인 호밍 엔도뉴클레아제는 I-SceI, I-CeuI, PI-PspI, PI-Sce, I-SceIV, I-CsmI, I-PanI, I-SceII, I-PpoI, I-SceIII, I-CreI, I-TevI, I-TevII 및 I-TevIII을 포함한다.
유전자 변이 - 식별 방법
본 발명의 미생물은 상기 미생물에 도입된 하나 이상의 유전자 변형 또는 변경에 의해 식별 될 수 있다. 상기 유전자 변형 또는 변경을 식별할 수 있는 한 가지 방법은 유전자 변형 또는 변경을 식별하기에 충분한 미생물 게놈 서열의 일부를 함유하는 서열 번호를 참조하는 것이다.
또한, 게놈에 도입된 유전자 변형 또는 변경을 가지지 않은 미생물 (예를 들어, 야생형, WT)의 경우, 본 발명은 16S 핵산 서열을 이용하여 상기 미생물을 식별할 수 있다. 16S 핵산 서열은 "분자 마커" 또는 "유전자 마커"의 예이며, 이는 핵산 서열의 특성의 차이를 시각화하는 방법에 사용되는 지표를 지칭한다. 이러한 지표의 다른 예는 제한 단편 길이 다형성 (RFLP) 마커, 증폭된 단편 길이 다형성 (AFLP) 마커, 단일 뉴클레오티드 다형성 (SNP), 삽입 돌연변이, 초위성체 마커 (SSR), 서열-특성화된 증폭 영역 (SCAR), 절단된 증폭 다형성 서열 (CAPS) 마커 또는 아이소자임 마커 또는 특정 유전자 및 염색체 위치를 정의하는 본 명세서에 기재된 마커들의 조합이다. 마커들은 또한 16S 또는 18S rRNA를 인코드하는 폴리뉴클레오티드 서열, 및 내부 전사 스페이서 (ITS) 서열을 추가로 포함하는데, 이는 서로 비교시 관계 또는 구별을 명확히 밝힘에 있어 특히 유용한 것으로 밝혀진, 소형-소단위 및 대형-소단위 rRNA 유전자들 사이에서 발견되는 서열들이다. 또한, 본 발명은 본 명세서에 개시된 미생물을 식별하기 위해 관심 유전자에서 발견되는 독특한 서열 (예컨대, nif H, D, K, L, A, glnE, amtB 등)을 이용한다.
주요 rRNA 소단위 (16S)의 일차 구조는 상이한 속도로 진화하는 도메인과 같은 매우 오래된 계통 및 속과 같은 보다 현대적인 계통의 분리를 가능하게 하는 보존된, 가변 및 초가변 영역의 특정 조합을 포함한다. 16S 소단위의 이차 구조는 대략 50%의 나선을 포함하며, 이는 잔기의 약 67%의 염기 페어링을 가져온다. 이러한 고도로 보존된 2차 구조 특징은 기능적으로 매우 중요하며 다중 서열 정렬 및 계통발생학적 분석에서 위치상 상동성을 확인함에 사용될 수 있다. 지난 수십년 동안 16S rRNA 유전자는 가장 잘 시퀀싱된 분류학적 마커가 되었으며 현재 체계적인 박테리아 및 고세균 분류의 초석이다 (Yarza 외. 2014. Nature Rev. Micro. 12 : 635-45).
유전자 변이 - 검출 방법: 프라이머, 프로브 및 분석
본 발명은 본 명세서에 개시된 미생물을 검출하는데 유용한 프라이머, 프로브 및 분석법을 개시한다. 일부 양상에서, 본 발명은 WT 모체 균주를 검출하는 방법을 제공한다. 다른 양상에서, 본 발명은 WT 균주로부터 유래된 비-속간 유전자조작 미생물을 검출하는 방법을 제공한다. 측면에서, 본 발명은 미생물에서 비-속간 유전자 변경을 식별하는 방법을 제공한다.
측면에서, 본 발명의 유전자 조작 방법은 유도된 비-속간 미생물에서 비-천연 뉴클레오티드 "접합" 서열의 생성을 가져온다. 이러한 비-자연 발생 뉴클레오티드 접합은 본 명세서에 개시된 미생물에서 특정 유전자 변경의 존재를 나타내는 진단 유형으로 사용될 수 있다.
본 기술은 고유하게 설계된 프라이머 및 프로브를 포함하는 특수한 정량적 PCR 방법을 이용하여 이러한 비-자연 발생 뉴클레오티드 접합을 검출 할 수 있다. 일부 양상에서, 본 발명의 프로브는 비-자연 발생 뉴클레오티드 접합 서열에 결합한다. 일부 양상들에서, 전통적인 PCR이 이용된다. 다른 양상들에서, 실시간 PCR이 이용된다. 일부 양상들에서, 정량적 PCR (qPCR)이 이용된다.
그러므로 본 발명은 실시간으로 PCR 산물을 검출하는 다음 2가지 통상적인 방법들의 사용을 포함할 수 있다: (1) 임의의 이중-가닥 DNA에 개재 (intercalate)되는 비특이적 형광 염료, 및 (2) 프로브와 이의 상보적 서열과의 혼성화 후에만 검출이 가능한 형광 리포터로 표지된 올리고뉴클레오티드들로 구성된 서열-특이적 DNA 프로브. 일부 양상들에서, 비-자연 발생 뉴클레오티드 접합만이 개시된 프라이머를 통해 증폭될 것이며, 그 결과 비-특이적 염료 또는 특이적 혼성화 프로브를 이용하여 검출 될 수 있다. 다른 양상들에서, 본 발명의 프라이머는 접합 서열의 한 쪽에 연접하도록 선택되므로, 그 결과, 증폭 반응이 일어나는 경우, 상기 접합 서열이 존재하게 된다.
본 발명의 양상들은 온화한 내지 엄격한 혼성화 조건하에서 상기 비-자연 발생 뉴클레오티드 접합 서열에 결합 할 수 있는 다른 뉴클레오티드 분자와 함께, 비-자연 발생 뉴클레오티드 접합 서열 분자 그 자체를 포함한다. 일부 양상에서, 온화한 내지 엄격한 혼성화 조건하에서 상기 비-자연 발생 뉴클레오티드 접합 서열에 결합 할 수 있는 뉴클레오티드 분자는 "뉴클레오티드 프로브"로 지칭된다.
양상들에서, 게놈 DNA를 샘플들로부터 추출하고 이를 qPCR을 사용하여 본 발명의 미생물의 존재를 정량화하기 위해 사용할 수 있다. qPCR 반응에 이용된 프라이머는 프라이머 Blast (https://www.ncbi.nlm.nih.gov/tools/프라이머-blast/)에 의해 설계된 프라이머 일 수 있는데, 이는 야생형 게놈의 독특한 영역 또는 유전자조작된 비-속간 돌연변이체 균주의 독특한 영역을 증폭시킨다. qPCR 반응은 정방향 및 역방향 증폭 프라이머만을 사용하는 SYBR GreenER qPCR SuperMix Universal (Thermo Fisher P/N 11762100) 키트를 사용하여 수행 될 수 있고; 대안적으로, Kapa Probe Force 키트 (Kapa Biosystems P/N KK4301)가 증폭 프라이머 및 5' 단부에 FAM 염료 라벨, 내부 ZEN 소광제 및 마이너 그루브 결합제 및 3' 단부에 형광 소광제를 함유하는 TaqMan 프로브 (Integrated DNA Technologies)와 함께 사용될 수 있다.
qPCR 반응 효율은 표적 게놈으로부터 알려진 양의 gDNA로부터 생성된 표준 곡선을 사용하여 측정 될 수 있다. 데이터는 조직 중량 및 추출 부피를 사용하여 새로운 중량 g 당 게놈 카피에 대해 정규화될 수 있다.
정량적 중합효소 연쇄 반응 (qPCR)은 하나 이상의 핵산 서열의 증폭을 실시간으로 정량하는 방법이다. PCR 분석의 실시간 정량은 증폭중인 관심 핵산과 보정 표준으로 기능할 수 있는 적절한 대조 핵산 서열을 비교함으로써 PCR 증폭 단계에 의해 생성되는 핵산의 양을 측정 할 수 있게 한다.
TaqMan 프로브는 종종 표적 핵산 서열을 정량화함에 증가된 특이성을 필요로 하는 qPCR 분석에 이용된다. TaqMan 프로브는 5' 단부에 형광단이 부착되고 프로브의 3' 단부에 소광제가 부착된 올리고뉴클레오티드 프로브를 포함한다. TaqMan 프로브가 프로브의 5' 및 3' 단부가 서로 밀착된 상태로 유지되면 소광제는 형광단에서 형광 신호가 전송되는 것을 방지한다. TaqMan 프로브는 특정 프라이머 세트에 의해 증폭된 핵산 영역 내에서 어닐링하도록 설계된다. Taq 중합효소가 프라이머를 확장시키고 신생 가닥을 합성함에 따라, Taq 중합효소의 5' -> 3' 엑소뉴클레아제 활성은 주형에 어닐링된 프로브를 분해시킨다. 이 프로브 분해는 형광단을 방출하여, 소광제에 대한 근접성을 파괴하고 형광단의 형광을 가능하게 한다. qPCR 분석에서 검출되는 형광은 방출된 형광단 및 반응에 존재하는 DNA 주형의 양에 정비례한다.
qPCR의 특징은 진료의로 하여금 겔 전기영동 제재의 노동집약적 증폭후 단계를 제거할 수 있게 하는데, 이 단계는 일반적으로 전통적인 PCR 분석의 증폭 산물의 관찰에 필요하다. 종래의 PCR에 비해 qPCR의 이점은 상당하며, 속도, 사용 편의성, 재현성 및 정량 능력의 증가를 포함한다.
형질의 개선
본 발명의 방법은 하나 이상의 다양한 바람직한 형질을 도입하거나 개선하기 위해 사용될 수 있다. 개선될 형질은 박테리아의 형질 일 수 있으며, 또는 박테리아는 관련 식물에서 형질을 개선하기 위해 변형 될 수 있다. 도입되거나 개선 될 수 있는 형질의 예는 다음을 포함한다: 뿌리바이오매스, 뿌리 길이, 높이, 싹 길이, 잎 수, 물 사용 효율, 전체 바이오매스, 수율, 과실 크기, 입자 크기, 광합성율, 가뭄에 대한 내성, 내열성, 내염성, 선충 스트레스에 대한 내성, 곰팡이 병원체에 대한 내성, 세균성 병원체에 대한 내성, 바이러스 병원체에 대한 내성, 대사 산물의 수준 및 프로테옴 발현. 키, 전체 바이오매스, 뿌리 및/또는 싹 바이오매스, 종자 발아, 묘목 생존, 광합성 효율, 증산율, 종자/과실 수 또는 질량, 식물 곡물 또는 과일 수확량, 잎 엽록소 함량, 광합성 속도, 뿌리 길이, 또는 이들의 임의의 조합을 비롯한 원하는 형질을 사용하여 성장을 측정하고 동일한 조건하에서 성장한 참조 농업 식물 (예를 들어, 형질이 개선되지 않은 식물)의 성장 속도와 비교할 수 있다.
개선 될 수 있는 형질의 또 다른 예는 박테리아가 식물에 부착하여 콜로니화하는 능력을 포함한다. 예를 들어, 박테리아는 식물의 뿌리에 더 잘 접착되거나 콜로니화에 유리한 화합물을 생산 또는 분비하도록 변형 될 수 있다.
도입되거나 개선되는 바람직한 형질은 본 발명에 기재된 질소 고정이다. 일부 경우에서, 본 발명에 기재된 방법에 의해 생성된 식물은 형질에 있어서, 동일한 토양 조건하에서 성장한 참조 농업 식물 보다 최소한 약 5% 더 큰, 예를 들면, 최소한 약 5%, 최소한 약 8%, 최소한 약 10%, 최소한 약 15%, 최소한 약 20%, 최소한 약 25%, 최소한 약 30%, 최소한 약 40%, 최소한 약 50%, 최소한 약 60%, 최소한 약 75%, 최소한 약 80%, 최소한 약 80%, 최소한 약 90%, 또는 최소한 100%, 최소한 약 200%, 최소한 약 300%, 최소한 약 400% 또는 그 이상의 차이를 나타낸다. 또 다른 예들에서, 본 발명에 기재된 방법에 의해 생성된 식물은 형질에 있어서, 유사한 토양 조건하에서 성장한 참조 농업 식물 보다 최소한 약 5% 더 큰, 예를 들면, 최소한 약 5%, 최소한 약 8%, 최소한 약 10%, 최소한 약 15%, 최소한 약 20%, 최소한 약 25%, 최소한 약 30%, 최소한 약 40%, 최소한 약 50%, 최소한 약 60%, 최소한 약 75%, 최소한 약 80%, 최소한 약 80%, 최소한 약 90%, 또는 최소한 100%, 최소한 약 200%, 최소한 약 300%, 최소한 약 400% 또는 그 이상의 차이를 나타낸다.
개선될 형질은 하나 이상의 생물학적 또는 비생물학적 스트레스요인의 적용을 포함하는 조건하에서 평가 될 수 있다. 스트레스요인의 예는 비생물적 스트레스 (가령, 열 스트레스, 염 스트레스, 가뭄 스트레스, 저온 스트레스 및 낮은 영양소 스트레스) 및 생물 스트레스 (가령, 선충 스트레스, 초식 곤충 스트레스, 곰팡이 병원체 스트레스, 박테리아 병원체 스트레스 및 바이러스 병원체 스트레스)를 포함한다.
본 발명의 방법 및 조성물에 의해 개선되는 형질은 이전에 질소 고정 할 수 없었던 식물에서를 포함하여, 질소 고정 일 수 있다. 일부 경우들에서, 본 발명에 기재된 방법에 따라 단리된 박테리아는 식물의 질소의 1% 이상 (예컨대, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 15%, 20%, 또는 그 이상)을 생산하며, 이는 임의의 유전자 변이를 도입하기 전에 제 1 식물로부터 단리된 박테리아와 비교하여 최소한 2-배 (예컨대, 3-배, 4-배, 5-배, 6-배, 7-배, 8-배, 9-배, 10-배, 20-배, 50-배, 100-배, 1000-배, 또는 그 이상)의 질소 고정능의 증가를 나타낼 수 있다. 일부 경우들에서, 박테리아는 식물 질소의 5% 이상을 생산한다. 원하는 질소 고정 수준은 유전자 변이 도입, 복수의 식물들에 대한 노출, 및 개선된 형질을 가지는 식물들로부터 박테리아를 단리하는 단계를 1회 이상 (예컨대, 1, 2, 3, 4, 5, 10, 15, 25, 또는 그 이상 회수) 반복한 후 구현될 수 있다. 일부 경우에서, 향상된 질소 고정 수준은 글루타민, 암모니아, 또는 다른 화학적 질소원들이 보충된 비료의 존재하에서 구현된다. 질소 고정도를 평가하는 방법은 공지되어 있으며, 이들의 예가 본 명세서에 기재되어 있다.
미생물 육종은 작물 마이크로바이옴 내에서 종의 역할을 체계적으로 식별하고 개선하는 방법이다. 이 방법은 다음 세 단계를 포함한다: 1) 식물-미생물 상호작용을 매핑하고 특정 표현형에 연결된 조절 네트워크를 예측하여 후보 종 선택, 2) 조절 네트워크와 유전자 클러스터의 종내 교잡을 통한 미생물 표현형의 실용적이고 예측가능한 개선 3) 원하는 작물 표현형을 생산하는 새로운 미생물 유전자형의 스크리닝 및 선택. 균주의 개선을 체계적으로 평가하기 위해, 미생물 커뮤니티의 콜로니화 역학을 주요 종별의 유전적 활동과 연결하는 모델이 생성된다. 이 모델을 사용하여 유전자 표적 육종을 예측하고 농업학적 관련성이 있는 마이크로바이옴-인코드된 형질에 있어서의 개선을 선택하는 빈도를 개선시킨다.
농업상 관련된 현장에서 전달된 질소 측정
현장에서, 전달되는 질소의 양은 콜로니화에 활성을 곱한 함수에 의해 결정된다.

상기 방정식은 (1) 식물 조직 단위 당 평균 콜로니화, 및 (2) 각 미생물 세포에 의해 배설된 암모니아의 양 또는 고정된 질소의 양으로서의 활성을 필요로 한다. 에이커 당 질소 파운드로 변환하기 위해, 옥수수 성장 생리학을 성숙 시기 전반에 걸쳐 시간 경과에 따라, 예를 들어, 식물의 크기 및 관련 뿌리 시스템을 추적한다.
에이커-작기 당 작물에 전달되는 질소 파운드는 다음 방정식으로 계산할 수 있다:
식물 조직 (t)는 성장 시간 (t)에 따른 옥수수 식물 조직의 새로운 중량이다. 합리적으로 계산하기 위한 수치들은 Roots, Growth and Nutrient Uptake (Mengel. Dept. of Agronomy Pub. # AGRY-95-08 (Rev. May-95. p. 1-8.) 라는 제목의 간행물에 자세히 설명되어 있다.
콜로니화 (t)는 성장시기 동안 임의의 특정 시간, t에서 식물 조직의 새로운 중량 그램 당 식물 조직 내에서 발견되는 관심 미생물의 양이다. 단일 시점만 이용가능한 경우 단일 시점은 해당 시기 동안 피크 콜로니화 비율로서 정규화되고 나머지 시점의 콜로니화 비율은 그에 따라 조정된다.
활성(t)는 성장 시기 동안, 임의의 특정 시간, t에서 단위 시간 당 관심 미생물에 의해 고정되는 N의 비율이다. 본 명세서에 개시된 구체예들에서, 이러한 활성도는 5 mM 글루타민의 존재하의 ARA 배지에서 시험관내 아세틸렌 환원 분석 (ARA) 또는 5mM 암모늄 이온 존재하의 ARA 배지에서 암모늄 배설 분석에 의해 근사된다.
이어서 전달되는 질소 양을 상기 함수를 수치적으로 적분하여 계산한다. 상기 기재한 변수의 값이 설정된 시점에서 이산적으로 측정되는 경우, 이들 시점 사이의 값은 선형 보간을 수행함으로써 근사된다.
질소 고정
질소 고정을 조절하는 하나 이상의 유전자에 도입된 하나 이상의 유전자 변이를 포함하는 박테리아에 식물을 노출시키는 것을 포함하는, 식물에서 질소 고정을 증가시키는 방법이 본 발명에 기재되어 있으며, 여기서 박테리아는 식물에서 1% 이상의 질소 (예컨대, 2%, 5 %, 10 % 또는 그 이상)를 생성하며, 이는 박테리아가 없는 식물과 비교하여 최소한 2-배의 질소 고정 능력을 나타낼 수 있다. 이러한 박테리아는 글루타민, 우레아, 질산염 또는 암모니아를 보충한 비료의 존재시 질소를 생산할 수 있다. 유전자 변이는 상기 제공되 예들을 비롯하여 본 발명에 기재된, 임의의 수 및 임의의 조합의 임의의 유전자 변이일 수 있다. 유전자 변이는 nifA, nifL, ntrB, ntrC, 글루타민 합성효소, glnA, glnB, glnK, draT, amtB, 글루탐산분해효소, glnD, glnE, nifJ, nifH, nifD, nifK , nifY, nifE, nifN, nifU, nifS, nifV, nifW, nifZ, nifM, nifF, nifB, 및 nifQ로 구성된 그룹에서 선택된 유전자에 도입될 수 있다. 유전자 변이는 다음 중 하나 이상을 가져오는 돌연변이 일 수 있다: nifA 또는 글루탐산분해효소의 발현 또는 활성 증가; nifL, ntrB, 글루타민 합성효소, glnB, glnK, draT, amtB의 발현 또는 활성 감소; GlnE의 아데닐릴-제거 활성 감소; 또는 GlnD의 유리딜릴-제거 활성 감소. 본 발명에 개시된 방법의 하나 이상의 박테리아에 도입되는 유전자 변이는 녹아웃 돌연변이 일 수 있거나, 또는 표적 유전자의 조절 서열을 제거할 수 있거나, 또는 이종 조절 서열의 삽입, 예를 들어, 동일한 박테리아 종 또는 속의 게놈 내에서 발견되는 조절 서열의 삽입을 포함할 수 있다. 조절 서열은 박테리아 배양물 또는 식물 조직 내에서 유전자의 발현 수준에 기초하여 선택 될 수 있다. 유전자 변이는 화학적 돌연변이유발에 의해 생성될 수 있다. 단계 (c)에서 성장한 식물은 생물적 또는 비생물적 스트레스요인에 노출될 수 있다.
본 명세서에 기재된 식물에서 발생하는 질소 고정의 양은 여러 방식, 예를 들어, 아세틸렌-환원 (AR) 분석에 의해 측정 될 수 있다. 아세틸렌-환원 분석은 시험관내 또는 생체내에서 수행 될 수 있다. 특정 박테리아가 식물에 고정 질소를 제공한다는 증거는 다음을 포함할 수 있다: 1) 총 식물 N은 접종시 유의하게 증가하며, 바람직하게는 식물 내 N 농도가 동시에 증가함; 2) 접종시 N-제한 조건하에서 질소 결핍 증상이 경감됨 (이는 건조 물질의 증가를 포함하여야 함); 3) N2 고정은 15N 접근법을 사용하여 기록됨 (이는 동위원소 희석 실험, 15N2 환원 분석, 또는 15N 자연 존재비 분석일 수 있음); 4) 고정된 N은 식물 단백질 또는 대사산물에 혼입됨; 그리고 5) 이 효과들 모두는 접종물 균주의 돌연변이체가 접종된 식물 또는 비-접종 식물에서는 관찰되지 않음.
야생형 질소 고정 조절 캐스케이드는 디지털 논리 회로로 나타낼 수 있으며, 여기서 투입 O2 및 NH4 +는 NOR 게이트를 통과하고, 그 산출물은 ATP 외에 AND 게이트로 들어간다. 일부 구체예들에서, 본 명세서에 개시된 방법은 조절 캐스케이드의 다수의 지점들에서 이 회로에 대한 NH4 +의 영향을 방해하고, 그 결과 미생물들이 비료화된 현장에서 조차도 질소를 생산할 수 있게 된다. 그러나, 본 명세서에 개시된 방법은 또한 회로에 대한 ATP 또는 O2의 영향을 변경하거나, 회로를 세포 내 다른 조절 캐스케이드로 대체하거나, 질소 고정 이외의 유전자 회로를 변경하는 것을 고려한다. 이종 조절 시스템의 제어하에 기능적 생성물을 생성하도록 유전자 클러스터를 재 설계 할 수 있다. 유전자 클러스터의 코딩 서열의 외부 및 내부의 고유 조절 요소들을 제거하고 이를 대안적인 조절 시스템으로 대체함으로써, 복잡한 유전자 오페론 및 다른 유전자 클러스터의 기능적 생성물은 제어되고/되거나 고유 유전자들이 유래하였던 종들 이외의 상이한 종들의 세포를 비롯한 이종 세포로 이동될 수 있다. 일단 재설계된 후, 합성 유전자 클러스터는 유전자 회로 또는 다른 유도성 조절 시스템에 의해 제어되고, 이로써, 산물의 발현을 필요에 따라 제어 할 수 있다. 발현 카세트는 논리 게이트, 펄스 발생기, 발진기, 스위치 또는 메모리 장치로 작동하도록 설계 될 수 있다. 제어 발현 카세트는 발현 카세트가 산소, 온도, 접촉, 삼투 스트레스, 막 스트레스 또는 산화 환원 센서와 같은 환경 센서로서 기능하도록 프로모터에 연결될 수있다.
한 예로서, nifL, nifA, nifT, 및 nifX 유전자는 nif 유전자 클러스터로부터 제거될 수 있다. 합성 유전자는 각 아미노산 서열을 인코드하는 DNA를 코돈 랜덤화함으로써 설계될 수 있다. 코돈 선택이 수행되며, 코돈 사용이 고유 유전자에서의 코돈 사용과 다를 가능성이 있음을 명시한다. 제안된 서열은 임의의 원하지 않는 특성들, 가령, 제한 효소 인식 부위, 트랜스포존 인식 부위, 반복 서열, 시그마 54 및 시그마 70 프로모터, 크립틱 리보솜 결합 부위 (cryptic ribosome binding sites) 및 rho 독립 종결인자에 대해 스캔된다. 합성 리보솜 결합 부위는 각각의 상응하는 고유 리보솜 결합 부위의 강도와 일치하도록, 가령, 유전자의 시작 코돈을 둘러싸는 150 bp (-60 내지 +90)가 형광 유전자에 융합되어 있는 형광 리포터 플라스미드를 작제함으로써 선택된다. 이 키메라는 Ptac 프로모터의 제어하에 발현 될 수 있고, 형광은 유세포 분석을 통해 측정된다. 합성 리보솜 결합 부위를 생성하기 위해, 150 bp (-60 내지 +90)의 합성 발현 카세트를 사용하는 리포터 플라스미드 라이브러리가 생성된다. 간략하게, 합성 발현 카세트는 랜덤 DNA 스페이서, RBS 라이브러리를 인코드하는 축퇴 서열, 및 각각의 합성 유전자에 대한 코딩 서열로 구성 될 수 있다. 고유 리보솜 결합 부위와 가장 일치하는 합성 리보솜 결합 부위를 확인하기 위해 다수의 클론을 스크리닝 하였다. 그리하여 고유 오페론과 동일한 유전자로 구성된 합성 오페론을 작제하고 기능적 상보성에 대해 시험한다. 합성 오페론에 관한 또 다른 예의 설명은 US20140329326에 제공되어 있다.
박테리아 종
본 명세서에 개시된 방법 및 조성물에 유용한 미생물은 임의의 출처로부터 얻을 수 있다. 일부 경우, 미생물은 박테리아, 고세균, 원생동물 또는 진균일 수 있다. 본 발명의 미생물은 질소 고정 미생물, 예를 들면, 질소 고정 박테리아, 질소 고정 고세균, 질소 고정 진균, 질소 고정 효모, 또는 질소 고정 원생동물일 수 있다. 본 명세서에 개시된 방법 및 조성물에 유용한 미생물은 포자 형성 미생물, 예를 들어 포자 형성 박테리아 일 수 있다. 일부 경우에, 본 명세서에 개시된 방법 및 조성물에 유용한 박테리아는 그람 양성 박테리아 또는 그람 음성 박테리아 일 수 있다. 일부 경우에, 박테리아는 퍼미큐트 (Firmicute) 문의 내생포자 형성 박테리아 일 수 있다. 일부 경우에, 박테리아는 질소자급영양생물일 수 있다. 일부 경우에, 박테리아는 질소자급영양생물이 아닐 수 있다.
클레브시엘라 바리이콜라 (Klebsiella variicola)
클레브시엘라 바리이콜라는 자유 생존 질소 고정 토양 박테리아이며 바나나, 쌀, 사탕수수 및 옥수수 뿌리줄기에서 단리되었다. 137 균주는 원래 세인트 찰스 카운티의 미주리에 있는 현장에서 옥수수 뿌리의 뿌리줄기에서 수집한 토양 샘플에서 단리되었다. 동일한 균주가 캘리포니아와 푸에르토 리코의 현장에서 또한 발견되었으며, 이는 캘리포니아와 푸에르토 리코의 토양에서 137 균주 16S rRNA와 자연적으로 존재하는 K. 바리이콜라 유기체의 16S rRNA 사이의 정렬에 의해 확인되었다. 클레브시엘라 바리이콜라는 어떤 식물 해충 특성도 나타내지 않으나; 한 연구 그룹은 K. 바리이콜라의 균주가 중국에서 Banana Soft Rot1을 유발할 수 있다고 보고한 바 있다. K. 바리이콜라의 137 균주의 게놈에서는 발병 요인이 발견되지 않았다.
코사코니아 사카리 (Kosakonia sacchari)
코사코니아 사카리 는 엔테로박터2 속에 포함되어 있는 새로운 코사코니아 속에 속하는 새로운 종이다. K 사카리는 사탕수수와의 결합을 이유로 명명된 자유-생존 질소 고정 박테리아 (사카룸 오피시나룸 L., Saccharum officinarum L.)이다. K 사카리 박테리아는 그람 음성, 호기성, 비-포자 형성의, 운동성 막대이며 사탕수수 식물과 연계하여 콜로니화하고 질소를 고정시킬 수 있어 식물 성장을 촉진한다. 균주 CI006은 캘리포니아주 산 호아킨 카운티 (San Joaquin County)에서 채취한 토양 샘플에서 단리되었다. 코사코니아 사카리는 식물 해충 특성을 나타내는 것으로 알려져 있지 않다.
상기 기재된 2가지 야생형 미생물 (균주 137 및 CI006)의 동일성은 원핵생물 계통발생학적 연구에 확립된 방법인 16S rRNA 유전자의 서열 분석에 의해 확인되었다. 이들 두 균주들에 대한 생물학적 기탁 정보가 본 명세서에 포함되어 있다.
본 발명의 방법 및 조성물은 고세균, 가령, 예를 들면, 메타노써모박터 써모오토트로피쿠스 (Methanothermobacter thermoautotrophicus)와 함께 사용될 수 있다.
일부 경우에, 유용할 수 있는 박테리아는, 아그로박테리움 라디오박터 (Agrobacterium radiobacter), 바실루스 아시도칼다리우스 (Bacillus acidocaldarius), 바실루스 아시도테레스트리스 (Bacillus acidoterrestris), 바실루스 아그리 (Bacillus agri), 바실루스 아이자와이 (Bacillus aizawai), 바실루스 알보락티스 (Bacillus albolactis), 바실루스 알칼로필루스 (Bacillus alcalophilus), 바실루스 알베이 (Bacillus alvei), 바실루스 아미노글루코시디쿠스 (Bacillus aminoglucosidicus), 바실루스 아미노보란스 (Bacillus aminovorans), 바실루스 아밀로리티쿠스 (Bacillus amylolyticus) (또한 파에니바실루스 아밀로리티쿠스 (Paenibacillus amylolyticus)로도 공지됨) 바실루스 아밀로리쿠에파시엔스 (Bacillus amyloliquefaciens), 바실루스 아뉴리놀리티쿠스 (Bacillus aneurinolyticus), 바실루스 아트로파에수스 (Bacillus atrophaeus), 바실루스 아조토포르만스 (Bacillus azotoformans), 바실루스 바디우스 (Bacillus badius), 바실루스 세레우스 (Bacillus cereus) (동의어: 바실루스 엔도리쓰모스 (Bacillus endorhythmos), 바실루스 메두사 (Bacillus medusa)), 바실루스 키티노스포루스 (Bacillus chitinosporus), 바실루스 서큘란 (Bacillus circulans), 바실러스 코아굴란스 (Bacillus coagulans), 바실루스 엔도파라시티쿠스 (Bacillus endoparasiticus), 바실루스 파스티디오수스 (Bacillus fastidiosus), 바실루스 퍼르무스 (Bacillus firmus), 바실루스 쿠르스타키 (Bacillus kurstaki), 바실루스 락티콜라 (Bacillus lacticola), 바실루스 락티모르부스 (Bacillus lactimorbus), 바실루스 락티스 (Bacillus lactis), 바실루스 라테로스포루스 (Bacillus laterosporus) (또한 브레비바실루스 라테로스포루스 (Brevibacillus laterosporus)로도 공지됨), 바실루스 라우투스 (Bacillus lautus), 바실루스 렌티모르부스 (Bacillus lentimorbus), 바실루스 렌투스 (Bacillus lentus), 바실루스 리체니포르미스 (Bacillus licheniformis), 바실루스 마로카누스 (Bacillus maroccanus), 바실루스 메가테리움 (Bacillus megaterium), 바실루스 메티엔스 (Bacillus metiens), 바실루스 마이코이데스 (Bacillus mycoides), 바실루스 나토 (Bacillus natto), 바실루스 네마토시다 (Bacillus nematocida), 바실루스 니그리피칸스 (Bacillus nigrificans), 바실루스 니그룸 (Bacillus nigrum), 바실루스 판토텐티쿠스 (Bacillus pantothenticus), 바실루스 포필라에 (Bacillus popillae), 바실루스 사이코로사카롤리티쿠스 (Bacillus psychrosaccharolyticus), 바실루스 푸밀루스 (Bacillus pumilus), 바실루스 시아멘시스 (Bacillus siamensis), 바실루스 스미티이 (Bacillus smithii), 바실루스 스파에리쿠스 (Bacillus sphaericus), 바실루스 서브틸리스 (Bacillus subtilis), 바실루스 투링기엔시스 (Bacillus thuringiensis), 바실루스 유니플라겔라투스 (Bacillus uniflagellatus), 브래디리조비움 자포니쿰 (Bradyrhizobium japonicum), 브레비바실루스 브레비스 (Brevibacillus brevis) 브레비바실루스 라테로스포루스 (Brevibacillus laterosporus) (이전에, 바실루스 라테로스포루스 (Bacillus laterosporus)), 크로모박테리움 서브추가에 (Chromobacterium subtsugae), 델프티아 아시도보란스 (Delftia acidovorans), 락토바실루스 아시도필루스 (Lactobacillus acidophilus), 라이소박터 안티비오티쿠스 (Lysobacter antibioticus), 라이소박터 엔지모게네스 (Lysobacter enzymogenes), 파에니바실루스 알베이 (Paenibacillus alvei), 파에니바실루스 폴리믹사 (Paenibacillus polymyxa), 파에니바실루스 포필리아에 (Paenibacillus popilliae) (이전에 바실루스 포필리아에 (Bacillus popilliae)), 판토에아 아글로머란스 (Pantoea agglomerans), 파스테우리아 페네트란스 (Pasteuria penetrans) (이전에 바실루스 페네트란스 (Bacillus penetrans)), 파스테우리아 우스가에 (Pasteuria usgae), 펙토박테리움 카로토보룸 (Pectobacterium carotovorum) (이전에 어위니아 카로토보라 (Erwinia carotovora)), 수도모나스 아에루기노사 (Pseudomonas aeruginosa), 수도모나스 아우레오파시엔스 (Pseudomonas aureofaciens), 수도모나스 세파시아 (Pseudomonas cepacia) (이전에 버크홀데리아 세파시아 (Burkholderia cepacia)로 공지), 수도모나스 클로로라피스 (Pseudomonas chlororaphis), 수도모나스 플루오레센스 (Pseudomonas fluorescens), 수도모나스 프로라딕스 (Pseudomonas proradix), 수도모나스 푸티다 (Pseudomonas putida), 수도모나스 시링가에 (Pseudomonas syringae), 세라티아 엔토모필라 (Serratia entomophila), 세라티아 마르세센스 (Serratia marcescens), 스트렙토마이세스 콜롬비엔시스 (Streptomyces colombiensis), 스트렙토마이세스 갈부스 (Streptomyces galbus), 스트렙토마이세스 고시키엔시스 (Streptomyces goshikiensis), 스트렙토마이세스 그리세오비리디스 (Streptomyces griseoviridis), 스트렙토마이세스 라벤둘라에 (Streptomyces lavendulae), 스트렙토마이세스 프라시누스 (Streptomyces prasinus), 스트렙토마이세스 사라세티쿠스 (Streptomyces saraceticus), 스트렙토마이세스 베네주엘라에 (Streptomyces venezuelae), 잔토모나스 캄페스트리스 (Xanthomonas campestris), 제노르합두스 루미네센스 (Xenorhabdus luminescens), 제노르합두스 네마토필라 (Xenorhabdus nematophila), 로도코쿠스 글로베룰루스 AQ719 (Rhodococcus globerulus AQ719) (NRRL 수탁 번호 B-21663), 바실루스 종. AQ175 (ATCC 수탁 번호 55608), 바실루스 종. AQ 177 (ATCC 수탁 번호 55609), 바실루스 종. AQ178 (ATCC 수탁 번호 53522), 및 스트렙토마이세스 종. 균주 NRRL 수탁 번호 B-30145를 포함하나 이에 제한되는 것은 아니다. 일부 경우에서 상기 박테리아는 아조토박터 크루코쿰 (Azotobacter chroococcum), 메타노사르시나 바르케리 (Methanosarcina barkeri), 클레브시엘라 뉴모니아에 (Klesiella pneumoniae), 아조토박터 비넬란디 (azotobacter vinelandii), 로도박터 스파로이데스 (Rhodobacter spharoides), 로도박터 캡술라투스 (Rhodobacter capsulatus), 로도박터 팔루스트리스 (Rhodobcter palustris), 로도스포릴룸 루브룸 (Rhodosporillum rubrum), 리조비움 레구미노사룸 (Rhizobium leguminosarum) 또는 리조비움 에틀리 (Rhizobium etli) 일 수 있다.
일부 경우에, 상기 박테리아는 클로스트리디움 (Clostridium) 종, 예를 들면, 클로스트리디움 파스테우리아눔 (Clostridium pasteurianum), 클로스트리디움 베이제린키이 (Clostridium beijerinckii), 클로스트리디움 퍼프린겐스 (Clostridium perfringens), 클로스트리디움 테타니 (Clostridium tetani), 클로스트리디움 아세토부틸리쿰 (Clostridium acetobutylicum) 일 수 있다.
일부 경우에, 본 발명의 방법 및 조성물과 함께 사용되는 박테리아는 시아노박테리아 일 수 있다. 시아노박테리아 속의 예들은 아나바에나 (Anabaena) (예를 들면, 아나가에나 종 (Anagaena sp.) PCC7120), 노스톡 (Nostoc) (예를 들면, 노스톡 펑크티포르메 (Nostoc punctiforme)), 또는 시네코시스티스 (Synechocystis) (예를 들면, 시네코시스티스 종 PCC6803)를 포함한다.
일부 경우에, 본 발명의 방법 및 조성물과 함께 사용되는 박테리아는 클로로비 문, 예를 들어, 클로로비움 테피둠 (Chlorobium tepidum)에 속할 수 있다.
일부 경우에, 본 발명의 방법 및 조성물과 함께 사용되는 미생물은 공지된 NifH 유전자와 상동성인 유전자를 포함 할 수 있다. 공지된 NifH 유전자들의 서열은, 예를 들면, Zehr lab NifH 데이터베이스, (https://wwwzehr.pmc.ucsc.edu/nifH_Database_Public/, April 4, 2014), 또는 Buckley lab NifH 데이터베이스 (http://www.css.cornell.edu/faculty/buckley/nifh.htm), 및 Gaby, John Christian, 및 Daniel H. Buckley "A comprehensive aligned nifH gene database: a multipurpose tool for studies of nitrogen-fixing bacteria." Database 2014 (2014): bau001.)에서 찾을 수 있다. 일부 경우에, 본 발명의 방법 및 조성물과 함께 사용되는 미생물은 Zehr lab NifH 데이터베이스, (https://wwwzehr.pmc.ucsc.edu/nifH_Database_Public/, April 4, 2014)의 서열에 대해 최소한 60%, 70%, 80%, 85%, 90%, 95%, 96%, 96%, 98%, 99% 또는 99% 이상의 서열 동일성을 가지는 폴리펩티드를 인코드하는 서열을 포함할 수 있다. 일부 경우에, 본 발명의 방법 및 조성물과 함께 사용되는 미생물은 Buckley lab NifH 데이터베이스, (Gaby, John Christian, and Daniel H. Buckley. "A comprehensive aligned nifH gene database: a multipurpose tool for studies of nitrogen-fixing bacteria."Database 2014 (2014): bau001.)의 서열에 대해 최소한 60%, 70%, 80%, 85%, 90%, 95%, 96%, 96%, 98%, 99% 또는 99% 이상의 서열 동일성을 가지는 폴리펩티드를 인코드하는 서열을 포함할 수 있다.
본 명세서에 개시된 방법 및 조성물에서 유용한 미생물들은 고유 식물의 표면 또는 조직으로부터 미생물을 추출하는 단계; 종자를 분쇄하여 미생물을 단리하는 단계; 다양한 토양 샘플에 종자를 식재하고 조직들로부터 미생물을 회수하는 단계; 또는 외인성 미생물을 식물에 접종하는 단계 및 식물 조직에서 어떤 미생물이 나타나는지 결정하는 단계에 의해 얻을 수 있다. 식물 조직의 비-제한적 예들에는 종자, 묘목, 잎, 절단, 식물, 구근 또는 괴경이 포함된다. 일부 경우에서, 박테리아는 종자로부터 단리된다. 샘플을 처리하기 위한 파라미터는 다양한 유형의 회합성 미생물, 가령, 뿌리줄기 박테리아, 착생식물, 또는 내생미생물을 단리하기 위해 변화될 수 있다. 박테리아는 처음에 제 1 식물로부터 단리하는 대신, 환경 균주 콜렉션과 같은 저장소로부터 얻을 수도 있다. 미생물들은 단리된 미생물의 게놈들을 시퀀싱하고; 식물체 내 커뮤니티의 조성을 프로파일링하고; 커뮤니티 또는 단리된 미생물들의 전사체 기능성을 특성규명하고; 또는 선택 또는 표현형 배지 (예컨대, 질소 고정 또는 포스페이트 가용화 표현형)를 사용하여 미생물 특징을 스크리닝함을 통해, 유전자형과 표현형이 결정될 수 있다. 선별된 후보 균주들 또는 집단들은 서열 데이터; 표현형 데이터; 식물 데이터 (예컨대, 게놈, 표현형, 및/또는 수율 데이터); 토양 데이터 (예컨대, pH, N/P/K 함량, 및/또는 벌크 토양 생물학적 커뮤니티); 또는 이의 이의의 조합을 통해 얻을 수 있다.
본 명세서에 기재된 박테리아 및 박테리아의 제조 방법은 식물 방어 반응을 손상시키지 않으면서 잎 표면, 뿌리 표면 또는 식물 조직 내부에서 효율적으로 자가-증식 할 수 있는 박테리아, 또는 식물 방어 반응에 내성인 박테리아에 적용될 수 있다. 본 명세서에 기재된 박테리아는 질소가 첨가되지 않은 배지에서 식물 조직 추출물을 배양함으로써 또는 잎 표면 세척에 의해 단리 될 수 있다. 그러나, 상기 박테리아는 배양불가능할 수 있다, 즉, 해당 분야에 공지된 표준 방법들을 사용하여 배양가능한 것으로 알려져있지 않거나 배양하기 어려울 수 있다. 본 명세서에 기재된 박테리아는 내생미생물 또는 착생생물 또는 식물 뿌리줄기에 서식하는 박테리아 (뿌리줄기 박테리아) 일 수 있다. 유전자 변이 도입, 복수의 식물들에 대한 노출, 및 개선된 형질을 가지는 식물들로부터 박테리아를 단리하는 단계들을 1회 이상 (예컨대, 1, 2, 3, 4, 5, 10, 15, 25회, 또는 그 이상) 반복한 후 얻어진 박테리아는 내생미생물, 착생생물, 또는 뿌리줄기 일 수 있다. 내생미생물은 질병 증상을 일으키거나 공생 구조의 형성을 유도하지 않고 식물의 내부로 들어가는 유기체로서, 식물 성장을 향상시키고 식물의 영양을 향상(예컨대, 질소 고정)시킬 수 있기 때문에 농업에서 관심 대상이 된다. 이 박테리아는 종자-유래 내생미생물 일 수 있다. 종자-유래 내생미생물은 잔디 또는 식물의 종자와 관련되거나 그로부터 유래된 박테리아, 가령, 성숙, 건조, 손상되지 않은 (예컨대, 균열, 진균 감염 또는 조기 발아가 없는) 종자에서 발견되는 종자-유래 박테리아 내생미생물을 포함한다. 종자-유래 박테리아 내생미생물은 종자의 표면과 결합되거나 종자 표면으로부터 유래할 수 있으며; 대안적으로, 또는 부가적으로, 내부 종자 구획 (예컨대, 표면-살균 종자)와 결합되거나 이로부터 유래될 수 있다. 일부 경우에, 종자-유래 박테리아 내생미생물은 식물 조직 내부에서, 예를 들면, 종자 내부에서 복제할 수 있다. 또한, 일부 경우에, 종자-유래 박테리아 내생미생물은 생존 건조 가능하다.
본 발명의 방법에 따라 단리되거나 본 발명의 방법 또는 조성물에 사용된, 복수의 상이한 박테리아 분류군을 조합하여 포함 할 수 있다. 한 예로서, 박테리아는 프로테오박테리아 (가령, 수도모나스 (Pseudomonas), 엔테로박터 (Enterobacter), 스테노트로포모나스 (Stenotrophomonas), 버크홀데리아 (Burkholderia), 리조비움 (Rhizobium), 허바스피릴룸 (Herbaspirillum), 판토에아 (Pantoea), 세라티아 (Serratia), 라넬라 (Rahnella), 아조스피릴룸 (Azospirillum), 아조리조비움 (Azorhizobium), 아조토박터 (Azotobacter), 두가넬라 (Duganella), 델프티아 (Delftia), 브래디리조비움 (Bradyrhizobium), 시노리조비움 (Sinorhizobium) 및 할로모나스 (Halomonas)), 퍼미큐트 (가령, 바실루스 (Bacillus), 파에니바실루스 (Paenibacillus), 락토바실루스 (Lactobacillus), 마이코플라즈마 (Mycoplasma), 및 아세타박테리움 (Acetabacterium)), 및 악티노박테리아 (Actinobacteria) (가령, 스트렙토마이세스 (Streptomyces), 로다코쿠스 (Rhodacoccus), 마이크로박테리움 (Microbacterium), 및 쿠르토박테리움 (Curtobacterium))을 포함할 수 있다. 본 발명의 방법 및 조성물에서 사용되는 박테리아는 2종 이상의 질소 고정 박테리아 컨소시엄을 포함할 수 있다. 일부 경우에, 상기 박테리아 컨소시엄 중 1종 이상의 박테리아는 질소를 고정시킬 수 있다. 일부 경우에, 1종 이상의 박테리아 컨소시엄은 다른 박테리아의 질소 고정 능력을 촉진시키거나 향상시킬 수 있다. 질소를 고정시키는 박테리아와 다른 박테리아의 질소 고정 능력을 향상시키는 박테리아는 동일하거나 상이 할 수 있다. 일부 예에서, 박테리아 균주는 다른 박테리아 균주와 조합하여, 또는 특정 박테리아 컨소시엄에서 존재할 때 질소를 고정시킬 수 있으나, 단일배양물에서는 질소를 고정시키지 못할 수 있다. 질소 고정 박테리아 컨소시엄에서 발견될 수 있는 박테리아 속의 예들에는, 허바스피릴룸 (Herbaspirillum), 아조스피릴룸 (Azospirillum), 엔테로박터 (Enterobacter), 및 바실루스 (Bacillus)가 포함되나 이에 제한되는 것은 아니다.
본 발명에 개시된 방법에 의해 생성될 수 있는 박테리아는 아조토박터 종 (Azotobacter sp.), 브래디리조비움 종 (Bradyrhizobium sp.), 클레브시엘라 종 (Klebsiella sp.), 및 시노리조비움 종 (Sinorhizobium sp.)을 포함한다. 일부 경우에, 박테리아는 다음으로 구성된 군에서 선택될 수 있다: 아조토박터 비넬란디 (azotobacter vinelandii), 브래디리조비움 자포니쿰 (Bradyrhizobium japonicum), 클레브시엘라 뉴모니아에 (klebsiella pneumoniae), 및 시노리조비움 메릴로티 (Sinorhizobium meliloti). 일부 경우에, 박테리아는 엔테로박터 (Enterobacter) 또는 라넬라 (Rahnella) 속 일 수 있다. 일부 경우에, 박테리아는 프랭키아 (Frankia), 또는 클로스트리디움 (Clostridium) 속 일 수 있다. 클로스트리디움 속의 박테리아의 예들은, 클로스트리디움 아세토부틸리쿰 (Clostridium acetobutilicum), 클로스트리디움 파스테우리아눔 (Clostridium pasteurianum), 클로스트리디움 베이제린키이 (Clostridium beijerinckii), 클로스트리디움 퍼프린겐스 (Clostridium perfringens), 및 클로스트리디움 테타니 (Clostridium tetani)를 포함하나 이에 제한되는 것은 아니다. 일부 경우에, 박테리아는 파에니바실루스 (Paenibacillus) 속일 수 있으며, 예를 들면, 파에니바실루스 아조토픽산스 (Paenibacillus azotofixans), 파에니바실루스 보레알리스 (Paenibacillus borealis), 파에니바실루스 두루스 (Paenibacillus durus), 파에니바실루스 마세란스 (Paenibacillus macerans), 파에니바실루스 폴리믹사 (Paenibacillus polymyxa), 파에니바실루스 알베이 (Paenibacillus alvei), 파에니바실루스 아밀로리티쿠스 (Paenibacillus amylolyticus), 파에니바실루스 캄피나센시스 (Paenibacillus campinasensis), 파에니바실루스 키벤시스 (Paenibacillus chibensis), 파에니바실루스 글루카놀리티쿠스 (Paenibacillus glucanolyticus), 파에니바실루스 일리노이센시스 (Paenibacillus illinoisensis), 파에니바실루스 라바에 아종 라바에 (Paenibacillus larvae subsp. Larvae), 파에니바실루스 라바에 아종 풀비파시엔스 (Paenibacillus larvae subsp. Pulvifaciens), 파에니바실루스 라우투스 (Paenibacillus lautus), 파에니바실루스 마세란스 (Paenibacillus macerans), 파에니바실루스 마쿠아리엔시스 (Paenibacillus macquariensis), 파에니바실루스 마쿠아리엔시스 (Paenibacillus macquariensis), 파에니바실루스 파불리 (Paenibacillus pabuli), 파에니바실루스 페오리아에 (Paenibacillus peoriae), 또는 파에니바실루스 폴리믹사 (Paenibacillus polymyxa)이다.
일부 예들에서, 본 발명의 방법에 따라 단리된 박테리아는 다음 분류군 중 하나 이상의 구성원일 수 있다: 아크로모박터 (Achromobacter), 아시디티오바실루스 (Acidithiobacillus), 아시도보락스 (Acidovorax), 아시도보라즈 (Acidovoraz), 아시네토박터 (Acinetobacter), 악티노플라네스 (Actinoplanes), 아들러크레우치아 (Adlercreutzia), 아에로코쿠스 (Aerococcus), 아에로모나스 (Aeromonas), 아피피아 (Afipia), 아그로마이세스 (Agromyces), 안사일로박터 (Ancylobacter), 아르트로박터 (Arthrobacter), 아토포스티페스 (Atopostipes), 아조스피릴룸 (Azospirillum), 바실루스 (Bacillus), 브델로비브리오 (Bdellovibrio), 베이제린키아 (Beijerinckia), 보세아 (Bosea), 브래디리조비움 (Bradyrhizobium), 브레비바실루스 (Brevibacillus), 브레분디모나스 (Brevundimonas), 버크홀데리아 (Burkholderia), 칸디다투스 할로레디비부스 (Candidatus Haloredivivus), 카울로박터 (Caulobacter), 셀룰로모나스 (Cellulomonas), 셀비브리오 (Cellvibrio), 크리세오박테리움 (Chryseobacterium), 시트로박터 (Citrobacter), 클로스트리디움 (Clostridium), 코랄리오마르가리타 (Coraliomargarita), 코리네박테리움 (Corynebacterium), 쿠프리아비두스 (Cupriavidus), 쿠르토박테리움 (Curtobacterium), 쿠르비박터 (Curvibacter), 데이노코쿠스 (Deinococcus), 델프티아 (Delftia), 데셈지아 (Desemzia), 데보시아 (Devosia), 독도넬라 (Dokdonella), 디엘라 (Dyella), 엔하이드로박터 (Enhydrobacter), 엔테로박터 (Enterobacter), 엔테로코쿠스 (Enterococcus), 어위니아 (Erwinia), 에스체리키아 (Escherichia), 에스체리키아/시겔라 (Escherichia/Shigella), 엑시구오박테리움 (Exiguobacterium), 페로글로부스 (Ferroglobus), 필리모나스 (Filimonas), 피네골디아 (Finegoldia), 플라비솔리박터 (Flavisolibacter), 플라보박테리움 (Flavobacterium), 프리고리박테리움 (Frigoribacterium), 글루코나세토박터 (Gluconacetobacter), 하프니아 (Hafnia), 할로배큘럼 (Halobaculum), 할로모나스 (Halomonas), 할로심플렉스 (Halosimplex), 허바스피릴룸 (Herbaspirillum), 하이메노박터 (Hymenobacter), 클레브시엘라 (Klebsiella), 코쿠리아 (Kocuria), 코사코니아 (Kosakonia), 락토바실루스 (Lactobacillus), 레클레르시아 (Leclercia), 렌체아 (Lentzea), 루테이박터 (Luteibacter), 루테이모나스 (Luteimonas), 마실리아 (Massilia), 메조리조비움 (Mesorhizobium), 메틸로박테리움 (Methylobacterium), 마이크로박테리움 (Microbacterium), 마이크로코쿠스 (Micrococcus), 마이크로비르가 (Microvirga), 마이코박테리움 (Mycobacterium), 네이세리아 (Neisseria), 노카르디아 (Nocardia), 오세아니바쿨룸 (Oceanibaculum), 오크로박트룸 (Ochrobactrum), 오키박테리움 (Okibacterium), 올리고트로파 (Oligotropha), 오리지후무스 (Oryzihumus), 옥살로파구스 (Oxalophagus), 파에니바실루스 (Paenibacillus), 판테오아 (Panteoa), 판토에아 (Pantoea), 펠로모나스 (Pelomonas), 퍼루시디바카 (Perlucidibaca), 플랜티박터 (Plantibacter), 폴리뉴클레오박터 (Polynucleobacter), 프로피오니박테리움 (Propionibacterium), 프로피오니시클라바 (Propioniciclava), 수도클라비박터 (Pseudoclavibacter), 수도모나스 (Pseudomonas), 수도노카르디아 (Pseudonocardia), 수도잔토모나스 (Pseudoxanthomonas), 사이크로박터 (Psychrobacter), 라넬라 (Rahnella), 랄스토니아 (Ralstonia), 레인헤이메라 (Rheinheimera), 리조비움 (Rhizobium), 로도코쿠스 (Rhodococcus), 로도수도모나스 (Rhodopseudomonas), 로세아텔레스 (Roseateles), 루미노코쿠스 (Ruminococcus), 세발델라 (Sebaldella), 세디미니바실루스 (Sediminibacillus), 세디미니박테리움 (Sediminibacterium), 세라티아 (Serratia), 시겔라 (Shigella), 시넬라 (Shinella), 시노리조비움 (Sinorhizobium), 시노스포란지움 (Sinosporangium), 스핑고박테리움 (Sphingobacterium), 스핑고모나스 (Sphingomonas), 스핑고픽시스 (Sphingopyxis), 스핑고시니셀라 (Sphingosinicella), 스타필로코쿠스 (Staphylococcus), 25 스테노트로포모나스 (Stenotrophomonas), 스트레노트로포모나스 (Strenotrophomonas), 스트렙토코쿠스 (Streptococcus), 스트렙토마이세스 (Streptomyces), 스티지오로부스 (Stygiolobus), 술푸리스파에라 (Sulfurisphaera), 타투멜라 (Tatumella), 테피디모나스 (Tepidimonas), 테르모모나스 (Thermomonas), 티오바실루스 (Thiobacillus), 바리오보락스 (Variovorax), WPS-2 속 인서타에 세디스 (genera incertae sedis), 잔토모나스 (Xanthomonas), 및 짐머만넬라 (Zimmermannella).
일부 경우에, 다음 속들 중 최소한 하나에서 선택된 박테리아 종들이 이용된다: 엔테로박터 (Enterobacter), 클레브시엘라 (Klebsiella), 코사코니아 (Kosakonia), 및 라넬라 (Rahnella). 일부 경우에, 다음 속의 박테리아 종들의 조합이 이용된다: 엔테로박터 (Enterobacter), 클레브시엘라 (Klebsiella), 코사코니아 (Kosakonia), 및 라넬라 (Rahnella). 일부 경우에, 이용되는 종들은 다음 중 하나 이상일 수 있다: 엔테로박터 사카리 (Enterobacter sacchari), 클레브시엘라 바리이콜라 (Klebsiella variicola), 코사코니아 사카리 (Kosakonia sacchari), 및 라넬라 아쿠아틸리스 (Rahnella aquatilis).
일부 경우에, 그람 양성 미생물은 다음을 포함하는 몰리브덴-철 질소고정효소 시스템을 가질 수 있다: nifH, nifD, nifK, nifB, nifE, nifN, nifX, hesA, nifV, nifW, nifU, nifS, nifI1, 및 nifI2. 일부 경우에, 그람 양성 미생물은 다음을 포함하는 바나듐 질소고정효소 시스템을 가질 수 있다: vnfDG, vnfK, vnfE, vnfN, vupC, vupB, vupA, vnfV, vnfR1, vnfH, vnfR2, vnfA (전사 조절인자). 일부 경우에, 그람 양성 미생물은 다음을 포함하는 철-단독 질소고정효소 시스템을 가질 수 있다: anfK, anfG, anfD, anfH, anfA (전사 조절인자). 일부 경우에 그람 양성 미생물은 glnB, 및 glnK (질소 신호전달 단백질)를 포함하는 질소고정효소 시스템을 가질 수 있다. 그람 양성 미생물의 질소 대사에 관여하는 효소들의 일부 예들에는 glnA (글루타민 합성효소), gdh (글루타메이트 탈수소효소), bdh (3-하이드록시부티레이트 탈수소효소), 글루탐산분해효소, gltAB/gltB/gltS (글루타메이트 합성효소), asnA/asnB (아스파테이트- 암모니아 연결효소/아스파라긴 합성효소), 및 ansA/ansZ (아스파라기나제)가 포함된다. 그람 양성 미생물에서 질소 운반에 관여하는 단백질들의 일부 예들에는 amtB (암모늄 운반체), glnK (암모늄 운반 조절인자), glnPHQ/ glnQHMP (ATP-의존성 글루타민/글루타메이트 운반체), glnT/alsT/yrbD/yflA (글루타민-유사 양성자 운반체), 및 gltP/gltT/yhcl/nqt (글루타메이트-유사 양성자 심포트 운반체)가 포함된다.
특히 관심 대상이 될 수 있는 그람 양성 미생물들의 예들에는, 파에니바실루스 폴리믹사 (Paenibacillus polymixa), 파에니바실루스 리오그란덴시스 (Paenibacillus riograndensis), 파에니바실루스 종 (Paenibacillus sp.), 프란키아 종 (Frankia sp.), 헬리오박테리움 종 (Heliobacterium sp.), 헬리오박테리움 클로룸 (Heliobacterium chlorum), 헬리오바실루스 종 (Heliobacillus sp.), 헬리오필룸 종 (Heliophilum sp.), 헬리오레스티스 종 (Heliorestis sp.), 클로스트리디움 아세토부틸리쿰 (Clostridium acetobutylicum), 클로스트리디움 종 (Clostridium sp.), 마이코박테리움 플라움 (Mycobacterium flaum), 마이코박테리움 종 (Mycobacterium sp.), 아트로박터 종 (Arthrobacter sp.), 아그로마이세스 종 (Agromyces sp.), 코리네박테리움 아우티트로피쿰 (Corynebacterium autitrophicum), 코리네박테리움 종 (Corynebacterium sp.), 마이크로몬스포라 종 (Micromonspora sp.), 프로피오니박테리아 종 (Propionibacteria sp.), 스트렙토마이세스 종 (Streptomyces sp.), 및 마이크로박테리움 종 (Microbacterium sp.)이 포함된다.
그람 양성 미생물에서 이루어질 수 있는 유전자 변경의 일부 예들에는 다음이 포함된다: 환경 질소의 존재시 BNF의 음성 조절을 제거하기 위해 glnR을 결실, 환경 질소에 반응하는 GlnR에 의한 조절을 제거하기 위해 nif 클러스터의 상류에 직접 상이한 프로모터를 삽입, GS-GOGAT 경로에 의한 암모늄 동화 속도를 감소시키기 위해 glnA를 돌연변이, 배지로부터의 암모늄 흡수를 감소시키기 위해 amtB 를 결실, GS-GOGAT 경로에 의한 암모늄 동화를 감소시키기 위해 glnA를 돌연변이시켜 항시적으로 피드백-억제된 (FBI-GS) 상태로 존재하게 하는 것.
일부 경우에, glnR은 파에니바실루스 종 (Paenibacillus species)에서 N 대사 및 고정의 주요 조절인자이다. 일부 경우에, 파에니바실루스 종 (Paenibacillus species)의 게놈은 glnR을 생성하는 유전자를 함유하지 않을 수 있다. 일부 경우에, 파에니바실루스 종 (Paenibacillus species)의 게놈은 glnE 또는 glnD을 생성하는 유전자를 함유하지 않을 수 있다. 일부 경우에, 파에니바실루스 종 (Paenibacillus species)의 게놈은 glnB 또는 glnK를 생성하는 유전자를 함유할 수 있다. 예를 들면, 파에니바실루스 종 (Paenibacillus sp.) WLY78은 고세균 메타노코쿠스 마리팔루디스 (Methanococcus maripaludis)에서 발견되는 glnB에 대한 유전자, 또는 이의 동족체 nifI1 및 nifI2를 함유하지 않는다. 일부 경우에, 파에니바실루스 종 (Paenibacillus species)의 게놈은 가변적일 수 있다. 예를 들면, 파에니바실루스 폴리믹사 (Paenibacillus polymixa) E681은 glnK 및 gdh가 없으며, 몇 가지 질소 화합물 운반체를 가지지만, amtB만이 GlnR에 의해 제어되는 것으로 보인다. 또 다른 예로, 파에니바실루스 종 (Paenibacillus species) JDR2는 glnK, gdh 및 대부분의 다른 중심 질소 대사 유전자를 가지며, 보다 적은 수의 많은 질소 화합물 운반체들을 가지지만, GlnR에 의해 제어되는 glnPHQ는 가지지 않는다. 파에니바실루스 리오그란덴시스 (Paenibacillus riograndensis) SBR5는 표준 glnRA 오페론, fdx 유전자, 주요 nif 오페론, 제 2 nif 오페론, 및 anf 오페론 (철-단독 질소고정효소를 인코드함)을 함유한다. 추정 glnR/tnrA 부위는 이들 오페론 각각의 상류에서 발견되었다. GlnR은 anf 오페론을 제외한 상기 모든 오페론을 조절할 수 있다. GlnR은 이들 조절 서열 각각에 이량체로서 결합 할 수 있다.
파에니바실루스 (Paenibacillus) N-고정 균주는 다음 2개 하위그룹에 속할 수 있다: 최소 nif 유전자 클러스터만을 함유하는 하위그룹 I 및 최소 클러스터, 그리고 nifX와 hesA 사이의 특성화되지 않은 유전자, 그리고 종종 nif 유전자들의 일부, 가령, nifH, nifHDK, nifBEN을 복제하는 다른 클러스터 또는 바나듐 질소고정효소 (vnf) 또는 철-단독 질소고정효소 (anf) 유전자를 인코드하는 클러스터를 함유하는 하위그룹 II.
일부 경우에, 파에니바실루스 종 (paenibacillus species)의 게놈은 glnB 또는 glnK를 생성하는 유전자를 함유하지 않을 수 있다. 일부 경우에, 파에니바실루스 종 (paenibacillus species)의 게놈은 시그마-70 프로모터로부터 전사된 9개 유전자들을 가지는 최소 nif 클러스터를 함유할 수 있다. 일부 경우에 파에니바실루스 (Paenibacillus) nif 클러스터는 질소 또는 산소에 의해 음성 조절될 수 있다. 일부 경우에, 파에니바실루스 종 (Paenibacillus species)의 게놈은 시그마-54를 생성하는 유전자를 함유하지 않을 수 있다. 예를 들면, 파에니바실루스 종 (Paenibacillus sp.) WLY78은 시그마-54에 관한 유전자를 함유하지 않는다. 일부 경우에, nif 클러스터는 glnR, 및/또는 TnrA에 의해 조절될 수 있다. 일부 경우에, nif 클러스터의 활성은 glnR, 및/또는 TnrA의 활성을 변경시킴으로써 변경될 수 있다.
바실루스에서, 글루타민 합성효소 (GS)는 높은 세포내 글루타민 농도에 의해 피드백 억제되어, 확인의 이동을 유발한다 (FBI-GS로 지칭됨). Nif 클러스터는 몇몇 바실루스 종에서 조절인자 GlnR 및 TnrA에 대한 서로 다른 결합 부위들을 함유한다. GlnR은 과량의 세포내 글루타민 및 AMP의 존재하에서 결합하여 유전자 발현을 억제한다. GlnR의 역할은 높은 질소 가용성 조건하에서 글루타민과 암모늄의 유입 및 세포내 생성을 저해하는 것일 수 있다. TnrA는 제한적인 세포내 글루타민의 존재 및/또는 FBI-GS의 존재하에서 결합하고 및/또는 유전자 발현을 활성화 (또는 억제) 할 수 있다. 일부 경우에 바실루스 nif 클러스터의 활성은 GlnR의 활성을 변경함으로써 변경될 수 있다.
피드백-억제된 글루타민 합성효소 (FBI-GS)는 GlnR에 결합하여 인식 서열에 대한 GlnR의결합을 안정화시킬 수 있다. 몇몇 박테리아 종들은 nif 클러스터의 상류에 GlnR/TnrA 결합 부위를 가진다. FBI-GS 및 GlnR의 결합을 변경시키는 것은 nif 경로의 활성을 변경시킬 수 있다.
미생물의 출처
박테리아 (또는 본 발명에 따른 임의의 미생물)는 토양, 식물, 진균, 동물 (무척추 동물 포함) 그리고 호수 및 강의 퇴적물, 물 및 생물상을 비롯한 기타 생물상을 포함하는 임의의 일반 지상 환경으로부터; 해양 환경, 그 생물상 및 퇴적물 (예를 들어, 해수, 해양 진흙, 해양 식물, 해양 무척추 동물 (예를 들어, 해면), 해양 척추 동물 (예를 들어, 물고기)); 지상 및 해양 지구 (표토 및 암석, 예를 들어, 쇄석된 지하암, 모래 및 점토); 빙권 및 그 용융수; 대기 (예를 들어, 여과된 공중 먼지, 구름 및 빗방울); 도시, 산업 및 기타 인공 환경 (예를 들어, 콘크리트, 노변 배수로, 지붕 표면 및 노면에 축적된 유기 및 무기물)으로부터 얻을 수 있다.
박테리아 (또는 본 발명에 따른 임의의 미생물)를 얻는 식물은 하나 이상의 바람직한 특성을 갖는 식물, 예를 들어, 특정 환경 또는 특정 관심 조건 하에서 자연적으로 성장하는 식물 일 수 있다. 예로서, 특정 식물은 모래 토양이나 염분이 높은 모래 또는 극한의 온도에서 또는 물이 거의 없는 상태에서 자연적으로 성장할 수 있거나 환경에 존재하는 특정 해충이나 질병에 내성을 가질 수 있으며, 상업적 작물이 이러한 조건에서, 특히 이들이 특정 지리적 위치에서 이용가능한 유일한 조건인 경우에 그러한 조건에서 재배 될 수 있다. 추가 예로서, 박테리아는 이러한 환경에서 재배된 상업용 작물로부터, 또는 보다 구체적으로는 임의의 특정 환경에서 재배된 작물 중 관심 특성을 가장 잘 나타내는 개별 작물 식물: 예를 들어, 식염수 제한 토양에서 재배된 작물 중 가장 빠르게 성장하는 식물, 또는 심각한 곤충 손상 또는 질병 전염병에 노출된 작물에서 가장 손상이 적은 식물, 또는 섬유소 함량, 오일 함량 등을 비롯하여 원하는 양의 특정 대사 산물 및 기타 화합물을 가진 식물 또는 바람직한 색, 맛 또는 냄새를 나타내는 식물로부터 수집 될 수 있다. 박테리아는 상기 언급된 진균 및 다른 동물 및 식물 생물상, 토양, 물, 퇴적물 및 다른 환경 요소를 비롯하여, 관심 식물 또는 관심 환경에서 발생하는 임의의 물질로부터 수집 될 수 있다.
박테리아 (또는 본 발명에 따른 임의의 미생물)는 식물 조직으로부터 단리 될 수 있다. 이러한 단리는 예를 들어 뿌리, 줄기 및 잎을 비롯한 임의의 적절한 식물 조직, 그리고 식물 생식 조직으로부터 발생 할 수 있다. 예로서, 식물로부터 단리하는 통상적인 방법은 전형적으로 관심 식물 물질의 멸균 절제 (예를 들어, 뿌리 또는 줄기 길이, 잎), 적절한 용액으로 표면 멸균 (예컨대, 2% 소듐 하이포클로라이트)을 포함하며, 그 후 식물 물질을 미생물 성장을 위한 영양 배지에 넣는다. 대안적으로, 표면-멸균된 식물 물질은 멸균 액체 (보통 물) 중에서 분쇄될 수 있고, 적합한 고체 한천 배지 또는 선별 또는 비선별적일 수 있는 매질 (예컨대, 인의 공급원으로서 피트산만을 함유)의 표면에 도포된 분쇄된 식물 물질의 작은 조각을 포함하는 액체 현탁액 일 수 있다. 이러한 접근법은 단리된 콜로니를 형성하는 박테리아에 특히 유용한데, 이 방법으로 박테리아는 영양 배지 플레이트를 분리하기 위해 개별적으로 선택될 수 있고, 널리 공지된 방법에 의해 단일 종들로 추가 정제될 수 있다. 대안적으로, 식물 뿌리 또는 잎 샘플은 표면 멸균되지 않고 단지 부드럽게 세척만 되므로 단리 과정에서 표면-서식 착생생물 미생물을 포함하게 될 수 있으며, 또는 착생생물 미생물은, 식물 뿌리, 줄기 또는 잎 조각들을 각인하고 한천 배지 표면 위로 제거한 다음 상기와 같은 개별적인 콜로니들로 단리함으로써, 별도로 단리될 수 있다. 이러한 접근법은, 예를 들면, 박테리아에 특히 유용하다. 대안적으로, 뿌리에 붙은 소량의 토양을 세척하지 않고 뿌리가 가공될 수 있으므로, 식물 뿌리줄기를 콜로니화하는 미생물이 포함될 수 있다. 그렇지 않으면, 뿌리에 붙은 토양을 제거하고, 희석하여, 적합한 선별 및 비-선별 배지 위에 도포하여 개별적인 뿌리줄기 박테리아 콜로니들을 단리할 수 있다.
특허 절차를 위한 미생물의 기탁에 관한 국제적 승인에 대한 부다페스트 조약
본 발명의 미생물 기탁은 특허 절차를 위한 미생물의 기탁에 관한 국제적 승인에 대한 부다페스트 조약 (부다페스트 조약) 규정하에 이루어졌다.
출원인은, 37 C.F.R. §1.808(a)(2)에 따라 출원인은 “기탁된 물질의 공중 이용에 대하여 기탁자가 가한 모든 제한사항은 특허 등록시 최종적으로 제거될 것임”을 진술한다. 이러한 진술은 이 장 (즉, 37 C.F.R. §1.808(b))의 (b)항에 따른 것이다.
생물학적으로 순수한 코사코니아 사카리 배양물 (WT)은 2017년 1월 6일에, 미국 메인 04544, 이스트 부스베이, 비겔로우 드라이브 60에 위치한 Bigelow National Center for Marine Algae and Microbiota (NCMA)에 기탁되었으며, NCMA 특허 기탁 지정 번호 201701001을 받았다. 기탁 정보는 아래 표 1에 제공된다.
생물학적으로 순수한 클레브시엘라 바리이콜라 배양물 (WT)은 2017년 8월 11일에, 미국 메인 04544, 이스트 부스베이, 비겔로우 드라이브 60에 위치한 Bigelow National Center for Marine Algae and Microbiota (NCMA)에 기탁되었으며, NCMA 특허 기탁 지정 번호 201708001을 받았다. 기탁 정보는 아래 표 1에 제공된다.

단리된 생물학적으로 순수한 미생물
특정 구체예에서, 본 발명은 특히 농업에 적용되는 단리되어 생물학적으로 순수한 미생물을 제공한다. 개시된 미생물은 단리되어 생물학적으로 순수한 상태로 사용될 수 있을 뿐만 아니라 조성물로 제제화 될 수 있다 (예시적인 조성물 설명은 하기 섹션 참조). 또한, 본 발명은 개시된 단리되어 생물학적으로 순수한 미생물 중 최소한 2개의 구성원을 함유하는 미생물 조성물, 뿐만 아니라 상기 미생물 조성물의 이용 방법을 제공한다. 또한, 본 발명은 개시된 단리되어 생물학적으로 순수한 미생물의 이용을 통해 식물에서 질소 고정을 조절하는 방법을 제공한다.
일부 양상들에서, 본 발명의 단리되어 생물학적으로 순수한 미생물은 표 1의 것들이다. 다른 양상들에서, 본 발명의 단리되어 생물학적으로 순수한 미생물은 표 1의 미생물로부터 유래된다. 예를 들면, 표 1의 미생물의 균주, 차일드 (child), 돌연변이체, 또는 유도체가 본 발명에 제공되어 있다. 본 발명은 표 1에 열거된 미생물들의 모든 가능한 조합을 고려하며, 상기 조합은 때때로 미생물 컨소시엄을 형성한다. 표 1의 미생물은, 개별적으로 또는 임의의 조합으로, 본 발명에서 언급된 임의의 식물, 활성제 (합성, 유기 등), 보강제, 담체, 보충제, 또는 생물학적 물질과 조합될 수 있다.
조성물
본 명세서에 기재된 방법에 따라 생성 및/또는 본 명세서에 기재된 특징을 갖는 박테리아 또는 박테리아 집단을 포함하는 조성물은 액체, 발포체 또는 건조 생성물의 형태 일 수 있다. 본 명세서에 기재된 방법에 따라 생성 및/또는 본 명세서에 기재된 특징을 갖는 박테리아 또는 박테리아 집단을 포함하는 조성물은 또한 식물 형질을 개선하기 위해 사용될 수 있다. 일부 예들에서, 박테리아 집단을 포함하는 조성물은 건조 분말, 분말과 물의 슬러리, 또는 유동가능한 종자 처리제 형태일 수 있다. 박테리아 집단을 포함하는 조성물은 종자 표면에 코팅될 수 있고, 액체 형태로 존재할 수 있다.
조성물은 연속 교반 탱크 반응기, 배치 반응기 및 농장과 같은 생물 반응기에서 제조 될 수 있다. 일부 예에서, 조성물은 용기, 가령, 항아리에 또는 미니 벌크에 보관될 수 있다. 일부 예에서, 조성물은 병, 통, 앰플, 패키지, 용기, 백, 박스, 빈, 봉투, 판지 (carton), 용기, 사일로 (silo), 선적 컨테이너, 트럭 베드, 및/또는 케이스로 구성된 그룹에서 선택된 물체 내부에 보관될 수 있다.
조성물은 또한 식물 형질을 개선하기 위해 사용될 수 있다. 일부 예들에서, 하나 이상의 조성물은 종자에 코팅될 수 있다. 일부 예들에서, 하나 이상의 조성물은 묘목에 코팅될 수 있다. 일부 예들에서, 하나 이상의 조성물은 종자 표면에 코팅될 수 있다. 일부 예들에서, 하나 이상의 조성물은 종자 표면 위의 층으로 코팅될 수 있다. 일부 예에서, 종자에 코팅된 조성물은 액체 형태, 건조 생성물 형태, 기포 형태, 분말과 물의 슬러리 형태 또는 유동가능한 종자 처리제로 존재할 수 있다. 일부 예에서, 하나 이상의 조성물은 종자 및/또는 묘목에 분사, 함침, 코팅, 캡슐화함으로써, 및/또는 종자 및/또는 묘목을 하나 이상의 조성물로 비산시킴으로써 처리될 수 있다. 일부 예에서, 복수의 박테리아 또는 박테리아 집단은 식물의 종자 및/또는 묘목에 코팅될 수 있다. 일부 예에서, 박테리아 조합의 최소한 2, 최소한 3, 최소한 4, 최소한 5, 최소한 6, 최소한 7, 최소한 8, 최소한 9, 최소한 10, 또는 10종 이상의 박테리아는 다음 속들 중 하나에서 선택될 수 있다: 아시도보락스 (Acidovorax), 아그로박테리움 (Agrobacterium), 바실루스 (Bacillus), 버크홀데리아 (Burkholderia), 크리세오박테리움 (Chryseobacterium), 쿠르토박테리움 (Curtobacterium), 엔테로박터 (Enterobacter), 에셰리키아 (Escherichia), 메틸로박테리움 (Methylobacterium), 파에니바실루스 (Paenibacillus), 판토에아 (Pantoea), 수도모나스 (Pseudomonas), 랄스토니아 (Ralstonia), 사카리바실루스 (Saccharibacillus), 스핑고모나스 (Sphingomonas), 및 스테노트로포모나스 (Stenotrophomonas).
일부 예에서, 내생미생물 조합의 최소한 2, 최소한 3, 최소한 4, 최소한 5, 최소한 6, 최소한 7, 최소한 8, 최소한 9, 최소한 10, 또는 10종 이상의 박테리아 및 박테리아 집단은 다음 패밀리 중 하나에서 선택된다: 바실라세아에 (Bacillaceae), 버크홀데리아세아에 (Burkholderiaceae), 코마모나다세아에 (Comamonadaceae), 엔테로박테리아세아에 (Enterobacteriaceae), 플라보박테리아세아에 (Flavobacteriaceae), 메틸로박테리아세아에 (Methylobacteriaceae), 마이브로박테리아세아에 (Microbacteriaceae), 파에노바실리레아에 (Paenibacillileae), 수도모나세아에 (Pseudomonnaceae), 리조비아세아에 (Rhizobiaceae), 스핑고모나다세아에 (Sphingomonadaceae), 잔토모나다세아에 (Xanthomonadaceae), 클라도스포리아세아에 (Cladosporiaceae), 그노모니아세아에 (Gnomoniaceae), 인서타에 세디스 (Incertae sedis), 라시오스파에리아세아에 (Lasiosphaeriaceae), 네트리아세아에 (Netriaceae), 및 플레오스포라세아에 (Pleosporaceae).
일부 예에서, 내생미생물 조합의 박테리아 및 박테리아 집단의 최소한 2, 최소한 3, 최소한 4, 최소한 5, 최소한 6, 최소한 7, 최소한 8, 최소한 9, 최소한 10, 또는 10종 이상의 박테리아는 다음 패밀리 중 하나에서 선택된다: 바실라세아에 (Bacillaceae), 버크홀데리아세아에 (Burkholderiaceae), 코마모나다세아에 (Comamonadaceae), 엔테로박테리아세아에 (Enterobacteriaceae), 플라보박테리아세아에 (Flavobacteriaceae), 메틸로박테리아세아에 (Methylobacteriaceae), 마이브로박테리아세아에 (Microbacteriaceae), 파에노바실리레아에 (Paenibacillileae), 수도모나세아에 (Pseudomonnaceae), 리조비아세아에 (Rhizobiaceae), 스핑고모나다세아에 (Sphingomonadaceae), 잔토모나다세아에 (Xanthomonadaceae), 클라도스포리아세아에 (Cladosporiaceae), 그노모니아세아에 (Gnomoniaceae), 인서타에 세디스 (Incertae sedis), 라시오스파에리아세아에 (Lasiosphaeriaceae), 네트리아세아에 (Netriaceae), 및 플레오스포라세아에 (Pleosporaceae).
조성물의 예는 상업적으로 중요한 농작물, 예를 들어, 수수, 카놀라, 토마토, 딸기, 보리, 쌀, 옥수수 및 밀에 대한 종자 코팅을 포함 할 수 있다. 조성물의 예는 또한 옥수수, 대두, 카놀라, 수수, 감자, 쌀, 야채, 시리얼 및 유지 종자용 종자 코팅을 포함 할 수 있다. 본 발명에서 제공되는 종자는 유전자 변형 유기체 (GMO), 비-GMO, 유기 또는 전통적인 것일 수 있다. 일부 예에서, 조성물은 식물의 공중부에 분사되거나, 식물 종자를 심은 고랑에 삽입하거나, 토양에 물을 주거나, 조성물의 현탁액에 뿌리를 담그어 뿌리에 처리될 수 있다. 일부 예에서, 조성물은 세포 생존력 및 숙주 식물을 인공적으로 접종하여 콜로니화하는 능력을 유지하는 적절한 방식으로 탈수 될 수 있다. 박테리아 종은 108 내지 1010 CFU/ml의 농도로 조성물에 존재할 수 있다. 일부 예에서, 조성물에 미량 금속 이온, 가령, 몰리브덴 이온, 철 이온, 망간 이온, 또는 이들 이온의 조합을 보충할 수 있다. 본 발명에 기재된 조성물의 예에서 이온 농도는 약 0.1 mM 내지 약 50 mM 일 수 있다. 조성물의 일부 예는 또한 담체, 가령, 베타-글루칸, 카르복실메틸 셀룰로오스 (CMC), 박테리아 세포외 중합체 물질 (EPS), 설탕, 동물성 우유 또는 다른 적합한 담체와 함께 제형화 될 수 있다. 일부 예에서, 이탄 또는 식재 재료가 담체로서 사용될 수 있거나, 조성물이 생체고분자에 포획되어 있는 생체고분자가 담체로서 사용될 수 있다. 본 발명에 기재된 박테리아 집단을 포함하는 조성물은 식물 형질을 개선, 가령, 식물 성장 촉진, 잎에서 높은 엽록소 함량 유지, 과실 또는 종자 수 증가, 과실 또는 종자 단위 중량 증가시킬 수 있다.
본 발명에 기재된 박테리아 집단을 포함하는 조성물은 종자의 표면에 코팅될 수 있다. 이에 따라, 본 발명에 기재된 하나 이상의 박테리아로 코팅된 종자를 포함하는 조성물이 또한 고려된다. 종자 코팅은 박테리아 집단을 화학적으로 불활성인 과립상의 다공성 담체와 혼합하여 형성 될 수 있다. 대안적ㅈ으로, 조성물은 종자를 심는 고랑에 직접 삽입되거나 식물 잎에 분사되거나 조성물의 현탁액에 뿌리를 침지시켜 처리될 수 있다. 유효량의 조성물을 사용하여 식물의 뿌리에 인접한 서브-토양 영역을 생존가능한 박테리아 성장으로 채우거나 식물의 잎을 생존가능한 박테리아 성장으로 채울 수 있다. 일반적으로, 유효량은 개선된 형질 (예컨대, 원하는 수준의 질소 고정)을 가지는 식물을 생성하기에 충분한 양이다.
본 발명에 기재된 박테리아 조성물은 농업상 허용가능한 담체를 사용하여 제형화 될 수 있다. 이들 구체예에 유용한 제제는 점착제, 미생물 안정화제, 살진균제, 항박테리아제, 보존제, 안정화제, 계면활성제, 항-복합제, 제초제, 살선충제, 살충제, 식물 성장 조절제, 비료, 살서제, 건조제, 살균제, 영양소 또는 이들의 임의의 조합으로 구성된 그룹으로부터 선택된 하나 이상의 구성원을 포함 할 수 있다. 일부 예에서, 조성물은 상온에서 안정 할 수 있다. 예를 들어, 본 발명에 기재된 임의의 조성물은 농업상 허용가능한 담체 (예를 들어, 비-자연 발생 비료와 같은 비료, 비-자연 발생 접착제와 같은 접착제, 및 비-자연 발생 살충제와 같은 살충제)를 포함 할 수 있다. 비-자연 접착제는, 예를 들어, 중합체, 공중합체 또는 합성 왁스 일 수 있다. 예를 들어, 본 발명에 기재된 임의의 코팅된 종자, 묘목 또는 식물은 종자 코팅에 이러한 농업상 허용가능한 담체를 함유 할 수 있다. 본 명세서에 기재된 임의의 조성물 또는 방법에서, 농업상 허용가능한 담체는 비-자연 발생 화합물 (예컨대, 비-자연 발생 비료, 비-자연 발생 접착제, 가령, 중합체, 공중합체, 또는 합성 왁스 또는 비-자연 발생 살충제) 일 수 있거나 이를 포함할 수 있다. 농업상 허용가능한 담체의 비-제한적인 예는 이하에 기재되어 있다. 농업상 허용가능한 담체의 추가예는 해당 분야에 공지되어있다.
일부 경우에, 박테리아는 농업상 허용가능한 담체와 혼합된다. 담체는 고체 담체 또는 액체 담체 일 수 있으며, 미소구체, 분말, 에멀전 등을 포함하는 다양한 형태 일 수 있다. 담체는 다양한 성질들, 가령, 증가된 안정성, 습윤성, 또는 분산성을 부여하는 수많은 담체 중 하나 이상일 수 있다. 비이온성 또는 이온성 계면활성제 일 수 있는 천연 또는 합성 계면 활성제와 같은 습윤제 또는 이들의 조합이 조성물에 포함될 수 있다. 유중수 에멀전은 또한 단리된 박테리아를 포함하는 조성물을 제형화하는데 사용될 수 있다 (예를 들어, 미국 특허 제 7,485,451 참조). 제조 될 수 있는 적합한 제형은 습윤성 분말, 과립, 겔, 한천 스트립 또는 펠렛, 증점제 등, 미세 캡슐화된 입자 등, 수성 유동성, 수성 현탁액, 유중수 에멀전 등과 같은 액체를 포함한다. 제제는 곡물 또는 콩과 제품, 예를 들어, 분쇄된 곡물 또는 콩, 곡물 또는 콩, 전분, 설탕 또는 오일로부터 유래된 액체배지 또는 가루를 포함 할 수 있다.
일부 구체예에서, 농업용 담체는 토양 또는 식물 성장 배지 일 수 있다. 사용될 수 있는 다른 농업용 담체에는 물, 비료, 식물계 오일, 습윤제 또는 이들의 조합이 포함된다. 대안적으로, 농업용 담체는 고체, 가령, 규조토, 양토, 실리카, 알기네이트, 점토, 벤토나이트, 질석, 종자 케이스, 다른 식물 및 동물 제품, 또는 과립, 펠릿 또는 현탁액을 포함한 조합물 일 수 있다. 상기 언급된 성분들 중 임의의 성분들의 혼합물 또한 담체, 가령, 페스타 (가루 및 카올린 점토), 한천 또는 양토, 모래 또는 점토 중의 가루-계 펠렛 등 (그러나 이에 제한되는 것은 아님)으로 고려된다. 제제들은 박테리아를 위한 식품 공급원, 가령, 보리, 쌀, 또는 다른 생물학적 재료들, 가령, 종자, 식물 부분, 사탕 수수 찌꺼기, 곡물 가공에서 생긴 깍지 또는 줄기, 분쇄된 식물 재료 또는 건축 현장 쓰레기에서 얻은 목재, 종이, 섬유, 또는 목재 재활용으로부터 생기는 톱밥 또는 작은 섬유들을 포함할 수 있다.
예를 들면, 비료는 종자, 묘목, 또는 식물의 성장을 촉진하고 이들에게 영양소를 제공함에 도움을 주기 위해 사용될 수 있다. 비료의 비-제한적인 예는 질소, 인, 칼륨, 칼슘, 황, 마그네슘, 붕소, 염소, 망간, 철, 아연, 구리, 몰리브덴 및 셀레늄 (또는 그의 염)을 포함한다. 비료의 추가 예는 하나 이상의 아미노산, 염, 탄수화물, 비타민, 포도당, NaCl, 효모 추출물, NH4H2PO4, (NH4)2SO4, 글리세롤, 발린, L-류신, 락트산, 프로피온산, 숙신산, 말산, 시트르산, KH 타르트레이트, 자일로스, 릭소스 및 레시틴을 포함한다. 한 구체예에서, 제제는 다른 활성제를 물질 (예를 들어, 종자의 표면)에 결합시키는 것을 돕기 위해 점착제 또는 부착제 (접착제로 지칭됨)를 포함 할 수 있다. 이러한 제제는 박테리아를 다른 화합물 (예컨대, 생물학적 물질이 아닌 대조 물질)을 함유 할 수 있는 담체와 조합하여 코팅 조성물을 생성하는데 유용하다. 이러한 조성물은 식물 또는 종자 주위에 코팅을 생성하여 미생물 및 다른 물질과 식물 또는 식물 부분과의 접촉을 유지시킴에 도움을 준다. 한 구체예에서, 접착제는 알기네이트, 검, 전분, 레시틴, 포르모노네틴, 폴리비닐 알코올, 알칼리 포르모노네티네이트, 헤스페레틴, 폴리비닐 아세테이트, 세팔린, 검 아라빅, 잔탄 검, 미네랄 오일, 폴리에틸렌 글리콜 (PEG), 폴리비닐 피롤리돈 (PVP), 아라비노-갈락탄, 메틸 셀룰로오스, PEG 400, 키토산, 폴리아크릴아미드, 폴리아크릴레이트, 폴리아크릴로니트릴, 글리세롤, 트리에틸렌 글리콜, 비닐 아세테이트, 겔란 검, 폴리스티렌, 폴리비닐, 카르복시메틸 셀룰로오스, 검 가티, 및 폴리옥시에틸렌-폴리옥시부틸렌 블록 공중합체로 이루어진 군으로부터 선택된다.
한 구체예에서, 접착제는 예컨대, 왁스, 가령, 카르나우바 왁스, 밀랍, 중국 왁스, 셸락 왁스, 경납 왁스, 칸데릴라 왁스, 피마자 왁스, 엉겅퀴 왁스 및 쌀겨 왁스, 다당류 (예컨대, 전분, 덱스트린, 말토덱스트린, 알기네이트 및 키토산), 지방, 오일, 단백질 (예컨대, 젤라틴 및 제인), 경작용 검 및 셸락 일 수 있다. 접착제는 비-자연 발생 화합물, 예컨대, 중합체, 공중합체 및 왁스 일 수 있다. 예를 들면, 접착제로서 사용될 수 있는 비-제한적 중합체의 예들에는 다음이 포함된다: 폴리비닐 아세테이트, 폴리비닐 아세테이트 공중합체, 에틸렌비닐 아세테이트 (EVA) 공중합체, 폴리비닐 알코올, 폴리비닐 알코올 공중합체, 셀룰로오스 (예컨대, 에틸 셀룰로오스, 메틸 셀룰로오스, 하이드록시 메틸셀룰로오스, 하이드록시 프로필셀룰로오스 및 카르복시 메틸셀룰로오스), 폴리비닐 피롤리돈, 비닐 클로라이드, 비닐리덴 클로라이드 공중합체, 칼슘 리그노설포네이트, 아크릴 공중합체, 폴리비닐 아크릴레이트, 폴리에틸렌 옥사이드, 아실아미드 중합체 및 공중합체, 폴리히드록시에틸 아크릴레이트, 메틸아크릴아미드 단량체 및 폴리클로로프렌.
일부 예에서, 하나 이상의 접착제, 항진균제, 성장 조절제 및 살충제 (예컨대, 살곤충제)는 비-자연 발생 화합물 (예컨대, 임의의 조합으로)이다. 농업상 허용가능한 담체의 추가 예는 분산제 (예컨대, 폴리비닐피롤리돈/비닐 아세테이트 PVPIVA S-630), 계면활성제, 결합제 및 충전제를 포함한다.
제제는 또한 계면활성제를 함유할 수도 있다. 계면활성제의 비-제한적 예들에는, 질소-계면활성제 블렌드, 가령, Prefer 28 (Cenex), Surf-N(US), Inhance (Brandt), P-28 (Wilfarm) 및 Patrol (Helena)이 포함되고; 에스터화 종자유에는 Sun-It II (AmCy), MSO (UAP), Scoil (Agsco), Hasten (Wilfarm) 및 Mes-100 (Drexel)이 포함되며; 유기-실리콘 계면활성제에는 Silwet L77 (UAP), Silikin (Terra), Dyne-Amic (Helena), Kinetic (Helena), Sylgard 309 (Wilbur-Ellis) 및 Century (Precision)가 포함된다. 한 구체예에서, 계면활성제는 0.01% v/v 내지 10% v/v의 농도로 존재한다. 또 다른 구체예에서, 계면활성제는 0.1% v/v 내지 1% v/v의 농도로 존재한다.
특정 경우에서, 제제는 미생물 안정화제를 포함한다. 이러한 물질은 건조제를 포함할 수 있으며, 이 건조제는 해당 화합물 또는 화합물들이 사실상 액체 접종제에 대한 건조 효과를 가지는 농도로 사용되는지 여부와 관계없이 건조제로서 분류될 수 있는 임의의 화합물 또는 화합물의 혼합물을 포함할 수 있다. 이러한 건조제는 사용된 박테리아 집단과 이상적으로 상용가능하며, 종자에 처리시 생존하고 건조시 생존하는 미생물 집단의 능력을 촉진시켜야 한다. 적합한 건조제의 예는 트레할로스, 수크로스, 글리세롤 및 메틸렌 글리콜 중 하나 이상을 포함한다. 다른 적합한 건조제는 비 환원당 및 당 알코올 (예컨대, 만니톨 또는 소르비톨)을 포함하지만, 이에 제한되지는 않는다. 제제에 도입되는 건조제의 양은 중량/부피로 약 5% 내지 약 50%, 예를 들어, 약 10% 내지 약 40%, 약 15% 내지 약 35%, 또는 약 20% 내지 약 30%의 범위 일 수 있다. 일부 경우에, 제제는 살진균제, 항균제, 제초제, 살선충제, 살충제, 식물 성장 조절제, 살서제, 살균제 또는 영양소와 같은 물질을 함유하는 것이 유리하다. 일부 예에서, 물질은 종자 표면-유래 병원체에 대한 보호를 제공하는 보호제를 포함 할 수 있다. 일부 예에서, 보호제는 일정 수준의 토양-유래 병원균 제어를 제공 할 수 있다. 일부 예에서, 보호제는 종자 표면에서 우세하게 효과적 일 수 있다.
일부 예에서, 살진균제는 화학적인 것이든 또는 생물학적인 것이든 곰팡이의 성장을 억제하거나 곰팡이를 사멸시킬 수 있는 화합물 또는 물질을 포함 할 수 있다. 일부 예에서, 살진균제는 정진균성 또는 살진균성일 수 있는 화합물을 포함할 수 있다. 일부 예에서, 살진균제는 보호제, 또는 종자 표면에서 우세하게 효과적인 물질일 수 있으며, 이는 종자 표면-유래 병원체에 대한 보호를 제공하고 토양-유래 병원체의 일정 수준의 제어를 제공한다. 보호제 살진균제의 비-제한적 예에는 캡탄, 마네브, 티람 또는 플루디옥소닐을 포함한다.
일부 예에서, 살진균제는 전신 살진균제 일 수 있으며, 이는 신생 묘목에 흡수되어 숙주 식물 조직 내부의 진균을 억제 또는 사멸시킬 수 있다. 종자 처리에 사용되는 전신 살진균제는 아족시스트로빈, 카르복신, 메페녹삼, 메탈락실, 티아벤다졸, 트라이플루옥시스트로빈, 및, 디페노코나졸, 입코나졸, 테부코나졸 및 트라이티코나졸을 비롯한 다양한 트라이아졸 살진균제를 포함하지만 이에 제한되지는 않는다. 메페녹삼 및 메탈락실은 주로 물곰팡이균 피티움 (Pythium) 및 피토프토라 (Phytophthora)를 표적하기 위해 사용된다. 병원성 진균 종의 민감도의 미묘한 차이 또는 식물의 살진균제 분포 또는 민감도의 차이로 인해 식물 종에 따라 일부 살진균제가 다른 것보다 바람직하다. 일부 예에서, 살진균제는 박테리아 또는 진균과 같은 생물학적 제어 물질일 수 있다. 이러한 유기체는 병원성 진균에 기생하거나, 독소 또는 진균을 사멸 또는 진균의 성장을 방지 할 수 있는 다른 물질들을 분비 할 수 있다. 임의의 유형의 살진균제, 특히, 식물에 일반적으로 사용되는 살진균제는 종자 조성물에서 제어물질로서 사용될 수 있다.
일부 예들에서, 종자 코팅 조성물은 항균 성질을 가지는 제어 물질을 포함한다. 한 구체예에서, 항균 성질을 갖는 제어 물질은 본 명세서의 다른 부분에 기재된 화합물로부터 선택된다. 또 다른 예에서, 화합물은 스트렙토마이신, 옥시테트라사이클린, 옥솔린산 또는 젠타마이신이다. 종자 코팅 조성물의 일부로서 사용될 수 있는 항균 화합물의 다른 예들에는 디클로로펜 및 벤질알코올 헤미 포말 (ICI의 Proxel®또는 Thor Chemie의 Acticide®RS 및 Rohm & Haas의 Kathon®MK 25) 및 이소티아졸리논 유도체, 가령, 알킬이소티아졸리논 및 벤즈이소티아졸리논 (Thor Chemie의 Acticide®MBS)을 기초로 한 것을 포함한다.
일부 예들에서, 성장 조절제는 다음으로 구성된 그룹에서 선택된다: 앱시스산, 아미도클로르, 안시미돌, 6-벤질아미노퓨린, 브래시놀라이드, 부트랄린, 클로르메? (클로르메? 클로라이드), 콜린 클로라이드, 사이클라닐리드, 다미노자이드, 디케굴락, 다이메티핀, 2,6-다이메틸퓨리딘, 에테폰, 플루메트랄린, 플루르프리미돌, 플루티아셋, 포클로르페누론, 지베렐린산, 이나베니피드, 인돌-3-아세트산, 말레익 하이드라지드, 메플루이다이드, 메피? (메피? 클로라이드), 나프탈렌아세틱 애시드, N-6-벤질아데닌, 파클로부트라졸, 프로헥사다이온 포스포로트라이티오에이트, 2,3,5-트라이-아이오도벤조익 애시드, 트라이넥사팍-에틸 및 유니코나졸. 성장 조절제의 또 다른 비-제한적 예들에는, 브래시노스테로이드, 사이토키닌 (예컨대, 키네틴 및 제아틴), 옥신 (예컨대, 인돌릴아세틱 애시드 및 인돌릴아세틸 아스파테이트), 플라보노이드 및 이소플라보노이드 (예컨대, 포르모노네틴 및 다이오스메틴), 피토알렉신 (예컨대, 글리세올린), 및 피토알렉신-유도 올리고당 (예컨대, 펙틴, 키틴, 키토산, 폴리갈라쿠로닉 애시드, 및 올리고갈락투로닉 애시드), 및 지베렐린이 포함된다. 이러한 물질들은 이상적으로 해당 제제가 처리되는 농업용 종자 또는 묘목과 상용가능하다 (예컨대, 이는 식물의 성장 또는 건강에 유해하지 않아야 한다). 더욱이, 인간, 동물 또는 산업 용도에서 안전성 문제를 일으키지 않는 물질이 이상적이다 (안전성 문제가 없거나, 또는 화합물이 충분히 불안정하여, 식물로부터 유래된 상품 식물 제품이 무시할만한 양의 화합물을 무시할만한 양으로 함유한다).
선충-길항제 생물제어 물질의 일부 예들에는 ARF18; 30 아스로보트리스 종 (Arthrobotrys spp.); 카에토뮴 종 (Chaetomium spp.); 실린드로카르폰 종 (Cylindrocarpon spp.); 엑소필리아 종 (Exophilia spp.); 푸사리움 종 (Fusarium spp.); 글리오클라디움 종 (Gliocladium spp.); 히르수텔라 종 (Hirsutella spp.); 레카니실리움 종 (Lecanicillium spp.); 모나크로스포리움 종 (Monacrosporium spp.); 미로테시움 종 (Myrothecium spp.); 네오코스모스포라 종 (Neocosmospora spp.); 파에실로마이세스 종 (Paecilomyces spp.); 포코니아 종 (Pochonia spp.); 스타고노스포라 종 (Stagonospora spp.); 소포상-수지상 마이코리자 진균 (vesicular- arbuscular mycorrhizal fungi), 버크홀데리아 종 (Burkholderia spp.); 파스테우리아 종 (Pasteuria spp.), 브레비바실루스 종 (Brevibacillus spp.); 수도모나스 종 (Pseudomonas spp.); 및 리조박테리아 (Rhizobacteria)가 포함된다. 특히 바람직한 선충-길항제 생물제어 물질의 예에는 ARF18, 아스로보트리스 올리고스포라 (Arthrobotrys oligospora), 아스로보트리스 닥틸로이데스 (Arthrobotrys dactyloides), 카에토뮴 글로보숨 (Chaetomium globosum), 실린드로카르폰 헤테로네마 (Cylindrocarpon heteronema), 엑소필리아 진셀메이 (Exophilia jeanselmei), 엑소필리아 피시필라 (Exophilia pisciphila), 푸사리움 아르퍼길루스 (Fusarium aspergilus), 푸사리움 솔라니 (Fusarium solani), 글리오클라디움 카테눌라툼 (Gliocladium catenulatum), 글리오클라디움 로세움 (Gliocladium roseum), 글리오클라디움 빅센스 (Gliocladium vixens), 히르수텔라 포실리엔시스 (Hirsutella rhossiliensis), 히르수텔라 미네소텐시스 (Hirsutella minnesotensis), 레카니실리움 레카니이 (Lecanicillium lecanii), 모나크로스포리움 드레드레슬레리 (Monacrosporium drechsleri), 모나크로스포리움 게피로파굼 (Monacrosporium gephyropagum), 마이로테시움 베루카리아 (Myrotehcium verrucaria), 네오코스모스포라 바신펙타 (Neocosmospora vasinfecta), 파에실로마이세스 릴라시누스 (Paecilomyces lilacinus), 포코니아 클라마이도스포리아 (Pochonia chlamydosporia), 스타고노스포라 헤테로데라에 (Stagonospora heteroderae), 스타고노스포라 파세올리 (Stagonospora phaseoli), 소포상-수지상 마이코리자 진균 (vesicular- arbuscular mycorrhizal fungi), 버크홀데리아 세파시아 (Burkholderia cepacia), 파스테우리아 페네트란스 (Pasteuria penetrans), 파스테우리아 토르네이 (Pasteuria thornei), 파스테우리아 니시자와에 (Pasteuria nishizawae), 파스테우리아 라모사 (Pasteuria ramosa), 파스테우리아 우스가에 (Pastrueia usage), 브레비바실루스 라테로스포루스 (Brevibacillus laterosporus) 균주 G4, 수도모나스 플루오레센스 (Pseudomonas fluorescens) 및 리조박테리아 (Rhizobacteria)가 포함된다.
영양소들의 일부 예들은 우레아, 암모늄 니트레이트, 암모늄 설페이트, 비-압력 질소 용액, 수성 암모니아, 무수 암모니아, 암모늄 티오설페이트, 황-코팅된 우레아, 우레아-포름알데하이드, IBDU, 중합체-코팅된 우레아, 칼슘 니트레이트, 우레아포름, 및 메틸렌 우레아를 비롯한 (그러나 이에 제한되는 것은 아님) 질소 비료, 인 비료, 가령, 다이암모늄 포스페이트, 모노암모늄 포스페이트, 암모늄 폴리포스페이트, 농축 수퍼포스페이트 및 트리플 수퍼포스페이트, 및 칼륨 비료, 가령, 포타슘 클로라이드, 포타슘 설페이트, 포타슘-마그네슘 설페이트, 포타슘 니트레이트로 구성된 그룹에서 선택될 수 있다. 이러한 조성물은 종자 코트 조성물 내에서 유리 염 또는 이온으로 존재할 수 있다. 대안적으로, 영양소/비료는 시간 경과에 따른 지속형 방출을 제공하도록 복합체형성 또는 킬레이트 될 수 있다.
살서제의 일부 예들은 2-아이소발러릴인단-1,3-다이온, 4-(퀴녹살린-2-일아미노) 벤젠설폰아미드, 알파-클로로하이드린, 알루미늄 포스파이드, 안투, 아르세너스 옥사이드 (arsenous oxide), 바륨 카보네이트, 비스티오세미, 브로디파쿰, 브로마디올론, 브로메탈린, 칼슘 시아나이드, 클로랄로스, 클로로파시논, 콜레칼시페롤, 쿠마클로르, 쿠마푸릴, 쿠마테트랄릴, 크리미딘, 디페나쿰, 디페티알론, 디파시논, 에르고칼시페롤, 플로쿠마펜, 플루오로아세트아미드, 플루프로파딘, 플루프로파딘 하이드로클로라이드, 하이드로겐 시아나이드, 아이오도메탄, 린단, 마그네슘 포스파이드, 메틸 브로마이드, 노르보르마이드, 포사세팀, 포스핀, 인, 핀돈, 포타슘 아르세나이트, 피리누론, 실리로사이드, 소듐 아르세나이트, 소듐 시아나이드, 소듐 플루오로아세테이트, 스트리크닌, 탈륨 설페이트, 와파린 및 징크 포스파이드로 구성된 물질들의 그룹에서 선택된 것들을 포함할 수 있다.
액체 형태에서, 예를 들어, 용액 또는 현탁액, 박테리아 집단은 물 또는 수성 용액에서 혼합 또는 현탁될 수 있다. 적합한 액체 희석제 또는 담체에는 물, 수용액, 석유 증류액 또는 다른 액체 담체가 포함된다.
고형 조성물은 이탄, 밀, 밀기울, 질석, 점토, 활석, 벤토나이트, 규조토, 풀러 어스 (fuller's earth), 저온 살균 토양 등과 같은 적절하게 분할된 고형 담체에 박테리아 집단을 분산시킴으로써 제조 될 수 있다. 이러한 제형이 습윤성 분말로서 사용되는 경우, 비이온성, 음이온성, 양쪽성 또는 양이온성 분산제 및 유화제와 같은 생물학적으로 상용가능한 분산제가 사용될 수 있다.
제제화시 사용되는 고체 분말은, 예를 들면, 무기질 담체, 가령, 카올린 점토, 파이로필라이트, 벤토나이트, 몬트모릴로나이트, 규조토, 산성 백 토양, 질석, 및 펄라이트, 및 무기염, 가령, 암모늄 설페이트, 암모늄 포스페이트, 암모늄 니트레이트, 우레아, 암모늄 클로라이드, 및 칼슘 카보네이트를 포함한다. 또한, 유기 미세분말, 가령, 밀가루, 밀겨, 및 쌀겨가 사용될 수 있다. 액체 담체에는 식물성 오일, 가령, 대두유 및 면실유, 글리세롤, 에틸렌 글리콜, 폴리에틸렌 글리콜, 프로필렌 글리콜, 폴리프로필렌 글리콜, 등이 포함된다.
작물에 대한 박테리아 집단의 처리
본 발명에 기재된 박테리아 또는 박테리아 집단의 조성물은 고랑에서, 활석으로 또는 종자 처리로 적용될 수 있다. 조성물은 벌크, 미니 벌크, 백, 또는 활석으로 종자 패키지에 처리될 수 있다.
재배자는 처리된 종자를 심고 기존 방식, 쌍줄 또는 경작이 필요 없는 방식에 따라 작물을 재배 할 수 있다. 종자는 제어 호퍼 또는 개별 호퍼를 사용하여 분배 될 수 있다. 종자는 압축 대기를 사용하여 또는 수작업으로 분배될 수 있다. 종자 배치는 가변 속도 기술을 사용하여 수행될 수 있다. 추가적으로, 본 명세서에 기재된 박테리아 또는 박테리아 집단의 처리는 가변 속도 기술을 사용하여 처리될 수 있다. 일부 예에서, 박테리아는 옥수수, 대두, 카놀라, 수수, 감자, 쌀, 야채, 시리얼, 수도시리얼 (pseudocereals) 및 유지종자의 종자에 처리될 수 있다. 곡물의 예에는 보리, 포니오, 귀리, 팔머 잔디, 호밀, 진주 기장, 사탕수수, 스펠트밀, 테프, 라이밀 및 밀이 포함될 수 있다. 수도시리얼의 예에는 브래드넛, 메밀, 부들, 치아, 아마, 곡물 아마란스, 한자, 퀴노아, 및 참깨가 포함될 수 있다. 일부 예에서, 종자는 유전자 변형 유기체 (GMO), 비-GMO, 유기 또는 전통적인 것일 수 있다.
미세 비료, PGR, 제초제, 살충제 및 살진균제와 같은 첨가제를 추가로 사용하여 작물을 처리 할 수 있다. 첨가제의 예들에는, 작물 보호제, 가령, 살충제, 살선충제, 진균제, 증강제, 가령, 착색제, 중합체, 펠릿화제, 프라이밍제, 및 소독제, 그리고 다른 물질들, 가령, 접종 물질, PGR, 연화제, 및 미량영양소가 포함된다. PGR은 뿌리 성장, 개화, 또는 줄기 신장에 영향을 주는 천연 또는 합성 식물 호르몬 일 수 있다. PGR은 옥신, 지베렐린, 사이토키닌, 에틸렌, 및 앱시스산 (ABA)을 포함할 수 있다.
상기 조성물은 액체 비료와 조합하여 고랑에 처리될 수 있다. 일부 예들에서, 액체 비료는 탱크에 보유될 수 있다. NPK 비료는 나트륨, 인, 및 칼륨의 다량영양소를 함유한다.
상기 조성물은 식물 형질을 개선, 가령, 식물 성장 촉진, 잎에서 높은 엽록소 함량 유지, 과실 또는 종자 수 증가, 과실 또는 종자 단위 중량 증가시킬 수 있다. 본 발명의 방법은 하나 이상의 다양한 바람직한 형질을 도입하거나 개선하기 위해 사용될 수 있다. 도입되거나 개선 될 수 있는 형질의 예는 다음을 포함한다: 뿌리바이오매스, 뿌리 길이, 높이, 싹 길이, 잎 수, 물 사용 효율, 전체 바이오매스, 수율, 과실 크기, 입자 크기, 광합성율, 가뭄에 대한 내성, 내열성, 내염성, 낮은 질소 스트레스에 대한 내성, 질소 사용 효율성, 선충 스트레스에 대한 내성, 곰팡이 병원체에 대한 내성, 세균성 병원체에 대한 내성, 바이러스 병원체에 대한 내성, 대사 산물의 수준, 대사산물의 수준 조절 및 프로테옴 발현. 키, 전체 바이오매스, 뿌리 및/또는 싹 바이오매스, 종자 발아, 묘목 생존, 광합성 효율, 증산율, 종자/과실 수 또는 질량, 식물 곡물 또는 과일 수확량, 잎 엽록소 함량, 광합성 속도, 뿌리 길이, 또는 이들의 임의의 조합을 비롯한 원하는 형질을 사용하여 성장을 측정하고 동일한 조건하에서 성장한 참조 농업 식물 (예를 들어, 형질이 도입되지 않은 및/또는 개선되지 않은 식물)의 성장 속도와 비교할 수 있다. 일부 예들에서, 키, 전체 바이오매스, 뿌리 및/또는 싹 바이오매스, 종자 발아, 묘목 생존, 광합성 효율, 증산율, 종자/과실 수 또는 질량, 식물 곡물 또는 과일 수확량, 잎 엽록소 함량, 광합성 속도, 뿌리 길이, 또는 이들의 임의의 조합을 비롯한 원하는 형질을 사용하여 성장을 측정하고 유사한 조건하에서 성장한 참조 농업 식물 (예를 들어, 형질이 도입되지 않은 및/또는 개선되지 않은 식물)의 성장 속도와 비교할 수 있다.
숙주 식물의 작물학적 형질에는 다음이 포함될 수 있으나 이에 제한되는 것은 아니다: 상기 종자 처리 제제 없이 종자로부터 성장한 모본 식물과 비교하여, 변경된 오일 함량, 변경된 단백질 함량, 변경된 종자 탄수화물 조성, 변경된 종자 오일 조성, 및 변경된 종자 단백질 조성, 화학적 내성, 저온 내성, 노쇠 지연, 질병 내성, 가뭄 내성, 이삭 무게, 성장 개선, 건강 향상, 열 내성, 제초제 내성, 초식동물 내성, 질소 고정 개선, 질소 이용 개선, 뿌리 구조 개선, 물 이용 효율성 개선, 바이오매스 증가, 뿌리 길이 증가, 종자 중량 증가, 지상부 길이 증가, 수율 증가, 물-제한 조건하에서 수율 증가, 속씨 질량, 속씨 수분 함량, 금속 내성, 이삭 수, 이삭 당 속씨 수, 꼬투리 수, 영양 증강, 병원체 내성, 해충 내성, 광합성 능력 개선, 염분 내성, 녹기-연장성, 활력 개선, 성숙 종자의 건조 중량 개선, 성숙 종자의 신생 중량 개선, 식물 당 성숙 종자수 증가, 엽록소 함량 증가, 식물 당 꼬투리 수 증가, 식물 당 꼬투리 길이 증가, 식물 당 시든 잎의 수 감소, 식물 당 심각하게 시든 잎의 수 감소, 및 식물 당 시들지 않은 잎의 수 증가, 대사산물 수준의 탐지가능한 조절, 대사체 수준의 탐지가능한 조절, 및 프로테옴에서 탐지가능한 조절.
일부 경우에, 접종되는 식물의 식물 요소와 동일한 식물 종으로부터 단리된 박테리아 또는 박테리아 집단을 식물에 접종한다. 예를 들면, 한 제아 마이 (Zea mays, 옥수수) 품종에서 일반적으로 발견되는 박테리아 또는 박테리아 집단은 그 자연 상태에서 상기 박테리아 및 박테리아 집단이 없는 또 다른 제아 마이 품종 식물의 식물 요소와 결합된다. 한 구체예에서, 박테리아 및 박테리아 집단은 접종된 식물의 식물 요소와 관련된 식물 종의 식물로부터 유래된다. 예를 들면, Zea diplo perennis Iltis 외, (diploperennial teosinte)에서 일반적으로 발견되는 박테리아 및 박테리아 집단이 제아 메이 (Zea mays, 옥수수)에 처리되거나, 또는 그 역으로 처리된다. 일부 경우에, 접종되는 식물의 식물 요소에 이종인 박테리아 및 박테리아 집단을 식물에 접종한다. 한 구체예에서, 박테리아 및 박테리아 집단은 또 다른 종의 식물로부터 유래된다. 예를 들면, 다이코트에서 일반적으로 발견되는 박테리아 및 박테리아 집단이 모노코트 식물에 처리되거나 (예컨대, 대두-유래 박테리아 및 박테리아 집단을 옥수수에 접종), 또는 그 역으로 처리된다. 다른 경우, 식물에 접종될 박테리아 및 박테리아 집단은 접종 중인 식물과 관련된 종로부터 유래된다. 한 구체예에서, 박테리아 및 박테리아 집단은 관련 분류군, 예를 들면, 관련된 종들로부터 유래된다. 또 다른 종들의 식물은 농업상의 식물일 수 있다. 또 다른 구체예에서, 박테리아 및 박테리아 집단은 임의의 숙주 식물 요소에 접종되도록 설계된 조성의 일부이다.
일부 예들에서, 박테리아 또는 박테리아 집단은 외인성이며, 이 때 박테리아 및 박테리아 집단은 접종된 식물과 상이한 식물에서 단리된다. 예를 들면, 한 구체예에서, 박테리아 또는 박테리아 집단은 접종되는 식물과 동일한 종의 상이한 식물로부터 단리될 수 있다. 일부 경우에, 박테리아 또는 박테리아 집단은 접종되는 식물과 관련된 종들로부터 단리될 수 있다.
일부 예들에서, 본 명세서에 기재된 박테리아 및 박테리아 집단은 하나의 조직 유형에서 또 다른 조직 유형으로 이동할 수 있다. 예를 들면, 종자 외부에 코팅한 후 식물의 성숙한 조직 내에서 본 발명의 박테리아 및 박테리아 집단의 탐지 및 단리는 종자 외부로부터 성숙 식물의 영양 조직으로 이동하는 능력을 나타낸다. 그러므로, 한 구체예에서, 박테리아 및 박테리아 집단은 종자 외부로부터 식물의 영양 조직으로 이동할 수 있다. 한 구체예에서, 식물의 종자에 코팅되는 박테리아 및 박테리아 집단은 종자가 영양 상태로 발아될 때, 상이한 식물 조직으로 국소화할 수 있다. 예를 들면, 박테리아 및 박테리아 집단은 다음을 비롯한 식물 조직들 중 어느 하나에 국소화될 수 있다: 뿌리, 부정근, 종자근, 뿌리털, 지상부, 잎, 꽃, 싹, 수염, 분열조직, 꽃가루, 암술, 씨방, 수술, 과실, 기는 줄기, 리좀, 뿌리혹, 덩이줄기, 모용, 공변 세포, 수공, 꽃잎, 꽃받침, 포영, 총엽병, 유관속 형성층, 체관부, 및 물관부. 한 구체예에서, 상기 박테리아 및 박테리아 집단은 식물의 뿌리 및/또는 뿌리털에 국소화할 수 있다. 또 다른 구체예에서, 박테리아 및 박테리아 집단은 광합성 조직, 예를 들면, 식물의 잎 및 지상부에 국소화할 수 있다. 다른 경우에, 박테리아 및 박테리아 집단은 식물의 유관 조직, 예를 들면, 물관부 및 체관부에 국소화된다. 또 다른 구체예에서, 박테리아 및 박테리아 집단은 식물의 번식 조직 (꽃, 꽃가루, 암술, 씨방, 수술, 과실)에 국소화할 수 있다. 또 다른 구체예에서, 박테리아 및 박테리아 집단은 식물의 뿌리, 지상부, 잎 및 생식 조직에 국소화할 수 있다. 또 다른 구체예에서, 박테리아 및 박테리아 집단은 식물의 과실 또는 종자 조직을 콜로니화한다. 또 다른 구체예에서, 박테리아 및 박테리아 집단은 식물 표면에 존재하도록 식물을 콜로니화할 수 있다 (즉, 이의 존재는 식물 외부에서 또는 식물의 외권에서 탐지가능하게 존재한다). 또 다른 구체예에서, 박테리아 및 박테리아 집단은 식물의 실질적으로 모든, 또는 모든 조직에 국소화 될 수 있다. 특정 구체예에서, 박테리아 및 박테리아 집단은 식물 뿌리에 국소화되지 않는다. 다른 경우에, 박테리아 및 박테리아 집단은 식물의 광합성 조직에 국소화되지 않는다.
본 조성물의 효율성은 또한 작물의 상대 성숙도 또는 작물 가열 유닛 (CHU)을 측정함으로써 평가될 수 있다. 예를 들면, 박테리아 집단은 옥수수에 처리될 수 있고, 옥수수 성장은 옥수수 속씨의 상대 성숙도 또는 옥수수 속씨가 최대 중량이 되는 시간에 따라 평가될 수 있다. 작물 가열 유닛 (CHU) 또한 옥수수 작물의 성숙도를 예측하기 위해 사용될 수 있다. CHU는 매일 최대 온도를 측정함으로써 작물 성장에 대한 열 축적의 양을 결정한다.
예에서, 박테리아는 다음을 비롯한 임의의 식물 조직들 중 하나에 국소화할 수 있다: 뿌리, 부정근, 종자근, 뿌리털, 지상부, 잎, 꽃, 싹, 수염, 분열조직, 꽃가루, 암술, 씨방, 수술, 과실, 기는 줄기, 리좀, 뿌리혹, 덩이줄기, 모용, 공변 세포, 수공, 꽃잎, 꽃받침, 포영, 총엽병, 유관속 형성층, 체관부, 및 물관부. 또 다른 구체예에서, 박테리아 또는 박테리아 집단은 광합성 조직, 예를 들면, 식물의 잎 및 지상부에 국소화할 수 있다. 다른 경우에, 박테리아 및 박테리아 집단은 식물의 유관 조직, 예를 들면, 물관부 및 체관부에 국소화된다. 또 다른 구체예에서, 박테리아 또는 박테리아 집단은 식물의 생식 조직 (꽃, 꽃가루, 암술, 씨방, 수술, 또는 과실)에 국소화할 수 있다. 또 다른 구체예에서, 박테리아 및 박테리아 집단은 식물의 뿌리, 지상부, 잎 및 생식 조직에 국소화할 수 있다. 또 다른 구체예에서, 박테리아 또는 박테리아 집단은 식물의 과실 또는 종자 조직을 콜로니화한다. 또 다른 구체예에서, 박테리아 또는 박테리아 집단은 식물 표면에 존재하도록 식물을 콜로니화할 수 있다. 또 다른 구체예에서, 박테리아 또는 박테리아 집단은 식물의 실질적으로 모든, 또는 모든 조직에 국소화 될 수 있다. 특정 구체예에서, 박테리아 또는 박테리아 집단은 식물 뿌리에 국소화되지 않는다. 다른 경우에, 박테리아 및 박테리아 집단은 식물의 광합성 조직에 국소화되지 않는다.
작물에 처리되는 박테리아 조성물의 유효성은 재배 속도, 묘목 활력, 뿌리 강도, 가뭄 내성, 식물 높이, 드라이 다운, 및 테스트 중량을 비롯한 (그러나 이에 제한되는 것은 아님) 다양한 작물 성장 특징들을 측정함으로써 평가될 수 있다.
식물 종
본 명세서에 기재된 방법 및 박테리아는 호르데움 (Hordeum), 오리자 (Oryza), 제아 (Zea) 및 트리티케아 (Trichea) 속의 식물과 같은 다양한 식물에 적합하다. 적합한 식물에 대한 다른 비-제한적 예들에는, 이끼, 태선, 및 조류가 포함된다. 일부 경우에, 식물, 가령, 식품 작물, 섬유 작물, 오일 작물, 임업 또는 펄프 및 제지 산업의 식물, 바이오 연료 생산을위한 공급 원료 및/또는 관상용 식물은 경제적, 사회적 및/또는 환경적 가치를 가진다. 일부 예들에서, 식물들은 경제적으로 가치있는 제품들, 가령, 곡물, 밀가루, 전분, 시럽, 식사, 오일, 필름, 포장재, 기능성 식품, 펄프, 동물 사료, 어류 사료, 산업 화학 물질을위한 벌크 재료, 곡물 제품, 가공 된 인간 식품, 설탕, 알코올 및 / 또는 단백질을 생산하기 위해 사용될 수 있다. 작물 식물의 비-제한적인 예는 옥수수, 쌀, 밀, 보리, 수수, 기장, 귀리, 호밀 삼피, 메밀, 단 옥수수, 사탕 수수, 양파, 토마토, 딸기 및 아스파라거스를 포함한다.
일부 예에서, 본 명세서에 개시된 방법 및 조성물을 사용하여 수득 또는 개선될 수 있는 식물은 농업, 원예, 바이오연료 분자 및 다른 화학 물질 생성을 위한 바이오매스 및/또는 임업에 중요하거나 흥미로운 식물을 포함 할 수 있다. 이러한 식물들의 일부 예들은 파인애플, 바나나, 코코넛, 백합, 그라스완두 및 잔디; 및 쌍떡잎식물, 가령, 예를 들면, 완두류, 알팔파, 토마틸로, 멜론, 병아리콩, 치커리, 클로버, 케일, 렌틸, 대두, 담배, 감자, 고구마, 무, 양배추, 유채, 사과 나무, 포도, 목화, 해바라기, 애기 장대, 카놀라, 감귤류 (오렌지, 만다린, 금귤, 레몬, 라임, 자몽, 감귤, 탄젤로, 시트론, 및 포멜로 포함), 후추, 콩, 상추, 지팽이풀 (스위치), 소르검 바이컬러 (Sorghum bicolor) (수수, 수단), 미스칸투스 기간테우스 (Miscanthus giganteus) (미스칸투스), 사카룸 종 (에너지수수), 포퓰러스 발사미페라 (Populus balsamifera) (포플러), 제아 메이스 (옥수수), 글리신 맥스 (대두), 브라시카 나푸스 (Brassica napus) (유채), 트리티쿰 에스티붐 (Triticum aestivum) (밀), 고시퓸 히르수툼 (Gossypium hirsutum) (면) 오리자 사티바 ( Oryza sativa) (쌀), 헬리안투스 아누스 (Helianthus annuus) (해바라기), 메디카고 사티바 (Medicago sativa) (알팔파), 베타 불가리스 (Beta vulgaris) (사탕무), 페니세툼 글라우쿰 (Pennisetum glaucum) (펄 밀렛), 파니쿰 종 (Panicum spp.), 소르검 종 (Sorghum spp.), 미스칸투스 종 (Miscanthus spp.), 사카룸 종 (Saccharum spp.), 에리안투스 종 (Erianthus spp.), 포퓰루스 종 (Populus spp.), 세칼 세레알 (Secale cereale) (호밀), 살릭스 종 (Salix spp.) (버드나무), 유칼립투스 종 (Eucalyptus spp.) (유칼립투스), 트리티코세칼 종 (Triticosecale spp.) (트리티쿰- 25 밀 X 호밀), 대나무, 카르타무스 팅크토리우스 (Carthamus tinctorius) (잇꽃), 자트로파 커카스 (Jatropha curcas) (자트로파), 리시누스 코뮤니스 (Ricinus communis) (피마자), 엘라에이스 귀닌시스 (Elaeis guineensis) (오일 팜), 피닉스 닥틸리페라 (Phoenix dactylifera) (데이트 팜), 아르콘토피닉스 쿤닝가미아나 (Archontophoenix cunninghamiana) (킹 팜), 시아그루스 로만조피아나 (Syagrus romanzoffiana) (퀸 팜), 리눔 우시타티시뭄 (Linum usitatissimum) (아마), 브라시카 준세아 (Brassica juncea), 마니홋 에스쿨렌타 (Manihot esculenta) (카사바), 라이코페르시콘 에스큘렌튬(Lycopersicon esculentum) (토마토), 락투카 사티바 (Lactuca sativa) (상추), 무사 파라디시아카 (Musa paradisiaca) (바나나), 솔라눔 투베로숨 (Solanum tuberosum) (감자), 브라시카 올레라세아 (Brassica oleracea) (브로콜리, 컬리플라워, 방울다다기 양배추), 카멜리아 시넨시스 (Camellia sinensis) (차), 프라가리아 아나나사 (Fragaria ananassa) (딸기), 테오브로마 카카오 (Theobroma cacao) (코코아), 코페아 아라비카 (Coffea arabica) (커피), 비티스 비니페라 (Vitis vinifera) (포도), 아나나스 코모수스 (Ananas comosus) (파인애플), 캡시쿰 안눔 (Capsicum annum) (핫 & 스윗 페퍼), 알리움 세파(Allium cepa) (양파), 큐큐미스 멜로 (멜론), 큐큐미스 사티부스 (Cucumis sativus) (오이), 쿠커비타 맥시마 (Cucurbita maxima) (스쿼시), 쿠커비타 모스차타 (Cucurbita moschata) (스쿼시), 스피나세아 올레라세아 (Spinacea oleracea) (시금치), 시트룰루스 라나투스 (Citrullus lanatus) (수박), 아벨모추스 에스쿨렌투스 (Abelmoschus esculentus) (오크라), 솔라눔 멜론게나 (Solanum melongena) (가지), 파파베르 솜니페룸 (Papaver somniferum) (양귀비), 파파베르 오리엔탈레 (Papaver orientale), 탁수스 바카타 (Taxus baccata), 탁수스 브레비폴리아 (Taxus brevifolia), 아르테미시아 안누아 (Artemisia annua), 카나비스 살리바 (Cannabis saliva), 캄프토테카 아큐미네이트 (Camptotheca acuminate), 카타란투스 로세우스 (Catharanthus roseus), 빈카 로세아 (Vinca rosea), 신코나 오피시날리스 (Cinchona officinalis), 코이키쿰 아우툼날레 (Coichicum autumnale), 베라트룸 캘리포르니카 (Veratrum californica), 디지탈리스 라나타 (Digitalis lanata), 디기탈리스 푸푸레아 (Digitalis purpurea), 디오스코레아 5 종 (Dioscorea 5 spp.), 안드로그라피스 파니쿨라타 (Andrographis paniculata), 아트로파 벨라돈나 (Atropa belladonna), 다투라 스토모니움 (Datura stomonium), 베르베리스 종 (Berberis spp.), 세팔로탁수스 종 (Cephalotaxus spp.), 에페드라 시니카 (Ephedra sinica), 에페드라 종 (Ephedra spp.), 에리트록실룸 코카 (Erythroxylum coca), 갈란투스 워르노리이 (Galanthus wornorii), 스코폴리아 종 (Scopolia spp.), 라이코포디움 세라툼 (Lycopodium serratum) (뱀톱), 라이코포디움 종 (Lycopodium spp.), 라우월피아 세르펜티나 (Rauwolfia serpentina), 라우월피아 종 (Rauwolfia spp.), 산귀나리아 카나덴시스 (Sanguinaria canadensis), 히오스시아무스 종 (Hyoscyamus spp.), 카렌둘라 오피시날리스 (Calendula officinalis), 크리산테뭄 파르테늄 (Chrysanthemum parthenium), 콜레우스 포르스콜리이 (Coleus forskohlii), 타나세툼 파르테늄 (Tanacetum parthenium), 파르테늄 아르젠타눔 (Parthenium argentatum) (구아율), 헤베아 종 (Hevea spp.) (고무), 멘타 스피카타 (Mentha spicata) (민트), 멘타 피페리타 (Mentha piperita) (민트), 빅사 오렐라나 (Bixa orellana), 알스트로에메리아 종 (Alstroemeria spp.), 로사 종 (Rosa spp.) (장미), 디안투스 카리오필루스 (Dianthus caryophyllus) (카네이션), 페투니아 종 (Petunia spp.) (페투니아), 포인세티아 풀케리마 (Poinsettia pulcherrima) (포인세티아), 니코티아나 타바쿰 (Nicotiana tabacum) (담배), 루피누스 알부스 (Lupinus albus) (루핀), 유니올라 파리쿨라타 (Uniola paniculata) (귀리), 호르데움 불가레 (Hordeum vulgare) (보리), 및 롤리움 종 (Lolium spp.) (호밀)을 포함할 수 있다.
일부 예에서, 외떡잎식물이 사용될 수 있다. 외떡잎식물은 알리스마탈레스 (Alismatales), 아랄레스 (Arales), 아레칼레스 (Arecales), 브로멜리알레스 (Bromeliales), 코멜리날레스 (Commelinales), 사이클란탈레스 (Cyclanthales), 사이페랄레스 (Cyperales), 에리오카울랄레스 (Eriocaulales), 하이드로카리탈레스 (Hydrocharitales), 준칼레스 (Juncales), 릴리알레스 (Lilliales), 나자달레스 (Najadales), 오르키달레스 (Orchidales), 판다날레스 (Pandanales), 포알레스 (Poales), 레스티오날레스 (Restionales), 트리우리달레스 (Triuridales), 티팔레스 (Typhales), 및 진기베랄레스 (Zingiberales) 목에 속한다. 겉씨식물 (Gymnospermae) 강에 속하는 식물들은 사이카달레스 (Cycadales), 진크고알레스 (Ginkgoales), 그네탈레스 (Gnetales), 및 피날레스 (Pinales)이다. 일부 예들에서, 외떡잎식물은 옥수수, 쌀, 밀, 보리, 및 사탕수수로 구성된 군에서 선택될 수 있다.
일부 예들에서, 아리스토키알레스 (Aristochiales), 아스테랄레스 (Asterales), 바탈레스 (Batales), 캄파누랄레스 (Campanulales), 카파랄레스 (Capparales), 카리오필랄레스 (Caryophyllales), 카수아리날레스 (Casuarinales), 셀라스트랄레스 (Celastrales), 코르날레스 (Cornales), 디아펜살레스 (Diapensales), 딜레니알레스 (Dilleniales), 디프사칼레스 (Dipsacales), 에베날레스 (Ebenales), 에리칼레스 (Ericales), 유코미알레스 (Eucomiales), 유포르비알레스 (Euphorbiales), 파발레스 (Fabales), 파갈레스 (Fagales), 겐티아날레스 (Gentianales), 게라니알레스 (Geraniales), 할로라갈레스 (Haloragales), 하마멜리달레스 (Hamamelidales), 미들레스 (Middles), 주글란달레스 (Juglandales), 라미알레스 (Lamiales), 라우랄레스 (Laurales), 레시티달레스 (Lecythidales), 레이트네리알레스 (Leitneriales), 마그니오랄레스 (Magniolales), 말발레스 (Malvales), 미리칼레스 (Myricales), 미르탈레스 (Myrtales), 님파에알레스 (Nymphaeales), 파페베랄레스 (Papeverales), 피페랄레스 (Piperales), 플란타기날레스 (Plantaginales), 플룸 아기날레스 (Plumb aginales), 포도스테말레스 (Podostemales), 폴레모니알레스 (Polemoniales), 폴리가랄레스 (Polygalales), 폴리고날레스 (Polygonales), 프리뮬라레스 (Primulales), 프로테알레스 (Proteales), 라플레시알레스 (Rafflesiales), 라눈쿠랄레스 (Ranunculales), 람날레스 (Rhamnales), 로잘레스 (Rosales), 루비알레스 (Rubiales), 살리칼레스 (Salicales), 산탈레스 (Santales), 사핀달레스 (Sapindales), 사라세피아세아에 (Sarraceniaceae), 스크로풀라리알레스 (Scrophulariales), 테알레스 (Theales), 트로코덴드랄레스 (Trochodendrales), 움벨랄레스 (Umbellales), 우르티칼레스 (Urticales), 및 비올라테스 (Violates) 목에 속하는 것들을 비롯한 쌍떡잎식물이 사용될 수 있다. 일부 예들에서, 쌍떡잎식물은 목화, 대두, 페퍼, 및 토마토로 구성된 그룹에서 선택될 수 있다.
일부 경우에, 개선될 식물은 실험 조건에 맞기가 쉽지 않을 수 있다. 예를 들면, 작물 식물은 여러번의 반복에 걸쳐 연속적으로 개선된 형질을 실제적으로 평가할 수 있게 충분히 성장시키기에 너무 오랜 시간이 걸릴 수 있다. 따라서, 박테리아가 초기에 단리되는 제 1 식물 및/또는 유전자 조작된 박테리아가 처리되는 복수의 식물은 모형 식물, 가령, 원하는 조건하에서 평가하기에 더 적합한 식물 일 수 있다. 모형 식물의 비-제한적 예들은 세타리아 (Setaria), 브래키포디움 (Brachypodium), 및 아라비돕시스 (Arabidopsis)를 포함한다. 이어서 모델 식물을 사용하여 본 발명의 방법에 따라 분리 된 박테리아의 능력을 개선 된 특성의 부여를 확인하기 위해 다른 유형의 식물 (예컨대, 작물 식물)에 처리 할 수 있다.
본 명세서에 개시된 방법에 의해 개선 될 수 있는 형질은, 관찰가능한 임의의 식물의 특성, 예를 들어, 성장 속도, 높이, 중량, 색, 맛, 냄새, 식물에 의한 하나 이상의 화합물 (예를 들어, 대사산물, 단백질, 약물, 탄수화물, 오일 및 임의의 다른 화합물)의 생성에서의 변화를 포함한다. 유전자형 정보에 기초한 식물의 선택 또한 예상된다 (예를 들어, 박테리아에 반응한 식물 유전자 발현의 패턴, 또는 유전자 마커, 가령, 질소 고정 증가와 관련된 것들의 존재를 식별하는 것을 포함). 식물은 또한 특정 특징 또는 형질 (가령, 바람직한 특징 또는 형질)의 존재와 대조되는 특정 특징 또는 형질 (가령, 바람직하지 않은 특징 또는 형질)의 부재, 억제 또는 저해에 기초하여 선택 될 수 있다.
농업상 조성물의 처리 농도 및 비율
전술한 바와 같이, 개시된 미생물을 포함하는 본 발명의 농업상 조성물은 다양한 방식으로 식물에 처리될 수 있다. 두 가지 특정 양상들에서, 본 발명은 고랑-내 처리 또는 종자 처리를 고려한다.
종자 처리 구체예에서, 본 발명의 미생물은 종자에 다양한 농도로 존재할 수 있다. 예를 들면, 종자 처리에서 미생물은 각 종자 당 cfu 농도로 발견될 수 있다: 1 × 101, 1 × 102, 1 × 103, 1 × 104, 1 × 105, 1 × 106, 1 × 107, 1 × 108, 1 × 109, 1 × 1010, 또는 그 이상. 특정 양상들에서, 종자 처리 조성물은 종자 당 약 1 × 104 내지 약 1 × 108 cfu를 포함한다. 다른 특정 양상들에서, 종자 처리 조성물은 종자 당 약 1 × 105 내지 약 1 × 107 cfu를 포함한다. 다른 양상들에서, 종자 처리 조성물은 종자 당 약 1 × 106 cfu를 포함한다.
미국에서는 약 10%의 옥수수 면적이 에이커 당 약 36,000개 종자 이상의 종자 밀도로 식재되는데; 옥수수 면적의 1/3은 에이커 당 약 33,000 내지 36,000 개 종자의 종자 밀도로 식재되고; 옥수수 면적의 1/3은 에이커 당 약 30,000 내지 33,000개 종자의 종자 밀도로 식재되며, 나머지 면적은 가변적이다. 다음 사이트에서 볼 수 있는 Steve Butzen 저, “Seeding Rate Considerations”를 참고하라: https://www.pioneer.com/home/site/us/agronomy/library/corn-seeding-rate-considerations/
아래 표 2는 고려되는 종자 처리 구체예 (열) 및 종자 면적 식재 밀도 (제 1 열: 15K-41K)에서 종자 당 다양한 cfu 농도를 이용하여, 에이커 당 총 cfu의 양을 계산한 것이며, 이는 다양한 농업 시나리오에 이용 될 수 있다 (즉, 종자 당 종자 처리 농도 × 에이커 당 식재된 종자 밀도). 그러므로, 종자 당 1 × 106 cfu의 종자 처리 및 에이커 당 30,000개 종자 식재를 이용하고자 하는 경우, 에이커 당 총 cfu 함량은 3 × 1010 (즉, 30K * 1 × 106)이 될 것이다.

고랑-내 구체예들에서, 본 발명의 미생물은 다음과 같은 에이커 당 cfu 농도로 처리될 수 있다: 1 × 106, 3.20 × 1010, 1.60 × 1011, 3.20 × 1011, 8.0 × 1011, 1.6 × 1012, 3.20 × 1012, 또는 그 이상. 그러므로, 양상들에서, 고랑-내 액체 조성물은 에이커 당 약 1 × 106 내지 약 3 × 1012 cfu의 농도로 처리될 수 있다.
일부 양상들에서, 고랑-내 조성물은 액체 제제에 함유된다. 액체 고랑-내 구체예에서, 미생물은 다음과 같은 밀리리터 당 cfu 농도로 존재할 수 있다: 1 × 101, 1 × 102, 1 × 103, 1 × 104, 1 × 105, 1 × 106, 1 × 107, 1 × 108, 1 × 109, 1 × 1010, 1 × 1011, 1 × 1012, 1 × 1013, 또는 그 이상. 특정 양상들에서, 액체 고랑-내 조성물은 밀리리터 당 약 1 × 106 내지 약 1 × 1011 cfu의 농도로 미생물을 포함한다. 다른 양상들에서, 액체 고랑-내 조성물은 밀리리터 당 약 1 × 107 내지 약 1 × 1010 cfu의 농도로 미생물을 포함한다. 다른 양상들에서, 액체 고랑-내 조성물은 밀리리터 당 약 1 × 108 내지 약 1 × 109 cfu의 농도로 미생물을 포함한다. 다른 양상들에서, 액체 고랑-내 조성물은 밀리리터 당 최대 약 1 × 1013 cfu의 농도로 미생물을 포함한다.
실시예
본 발명에서 제공되는 실시예는 박테리아 단리, 박테리아 및 식물 분석, 및 식물 형질 개선 방법을 설명한다. 이러한 실시예들은 단지 설명을 위한 것이며 어떤 방식으로든 제한으로 해석되어서는 안된다.
실시예 1: 후보 미생물의 전사체 프로파일링
환경 질소의 존재하에서 활성인 프로모터들을 식별하기 위해 균주 CI010의 전사체 프로파일링을 실시하였다. 균주 CI010을 10 mM 글루타민으로 보충된 정해진, 무-질소 배지에서 배양하였다. 이러한 배양물들로부터 전체 RNA를 추출하여 (QIAGEN RNeasy 키트) Illumina HiSeq (SeqMatic, Fremont CA)로 RNAseq 시퀀싱 하였다. 시퀀싱 리드를 Geneious를 사용하여 CI010 게놈 데이터에 맵핑하고, 근위 전사 프로모터의 제어하에 고도로 발현된 유전자들을 식별하였다.
표 3 및 4는 전체 RNA의 RNASeq 시퀀싱을 통해 측정된 유전자 및 이의 상대 발현 수준을 열거한다. nif 경로, 질소 이용 관련 경로, 콜로니화 관련 경로의 돌연변이유발에서 사용하기 위한 근위 프로모터의 서열들, 또는 원하는 발현 수준의 다른 유전자들을 기록하였다.
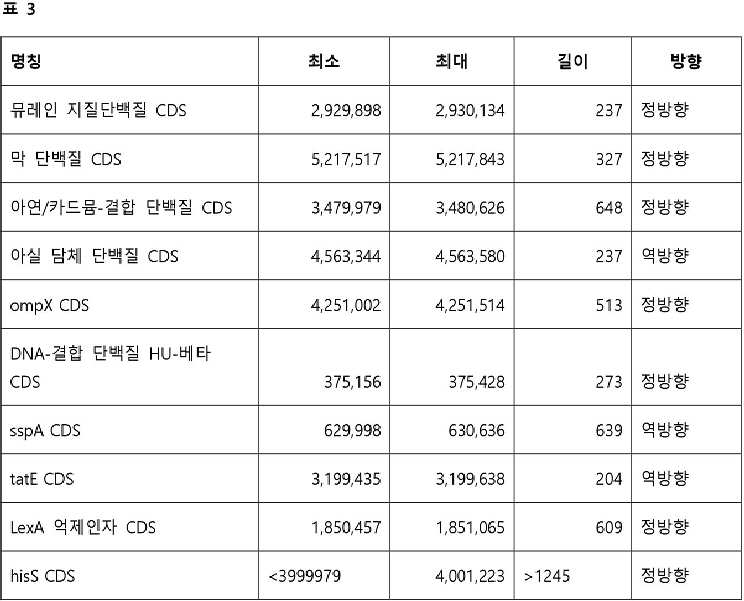
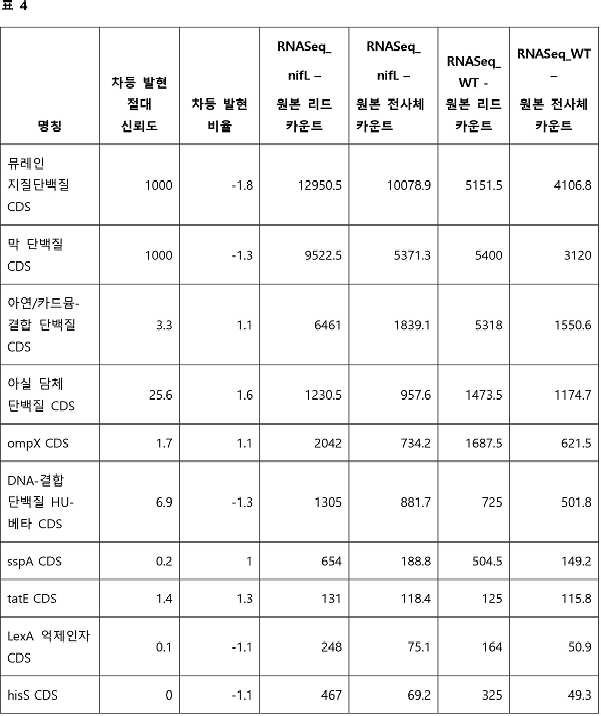
실시예 2: 후보 미생물의 돌연변이유발
람다-레드 매개 녹아웃
플라스미드 pKD46 또는 카나마이신 내성 마커를 함유하는 유도체를 사용하여 후보 미생물의 여러 돌연변이체가 생성되었다 (Datsenko 외. 2000; PNAS 97 (12) : 6640-6645). 표적 유전자에 연접하는 250bp 상동성을 가지는 녹아웃 카세트가 설계되었고 중첩 확장 PCR을 통해 생성되었다. 후보 미생물을 pKD46으로 형질전환시키고, 아라비노스의 존재하에 배양하여 람다-레드 기계식 발현을 유도하고, 전기천공을 위해 프렙하고, 녹아웃 카세트로 형질전환하여 후보 돌연변이체 균주를 생성하였다. 표 5에서 보는 바와 같이, 4개의 후보 미생물 및 1개의 실험실 균주 클레브시엘라 옥시토카 (Klebsiella oxytoca) M5A1을 사용하여 질소 고정 조절 유전자 nifL, glnB 및 amtB의 13개의 후보 돌연변이체를 생성하였다.

올리고-지정 돌연변이유발과 Cas9 선택
올리고-지정 돌연변이유발을 사용하여 대장균 DH10B에서 rpoB 유전자에 대한 게놈 변화를 표적하고, CRISPR-Cas 시스템을 사용하여 돌연변이체들을 선택하였다. rpoB 유전자에 대한 4-bp 돌연변이를 통해 리팜피신 내성을 부여하기 위하여 돌연변이원 올리고 (ss1283: “TTCcaagGTTTCGATCGGACATACGCGACCGTAGTGGGTCGGGTGTACGTCTCGAACTTCAAAGCC” (서열 번호: 2), 이 때 *는 포스포로티오에이트 결합을 표시함)가 설계되었다. Cas9를 인코드하는 플라스미드를 내포하는 세포들을 Cas9 발현을 위해 유도하고, 전기천공을 위해 프렙한 다음, WT rpoB 서열의 Cas9 절단을 표적하는 가이드 RNA (gRNA)의 항시적 발현을 인코드하는 플라스미드 및 돌연변이원 올리고 모두로 전기천공하였다. 전기천공된 세포들을 비선별 배지에서 하룻밤 회수하여, 생성된 돌연변이체 염색체가 충분히 분리되도록 하였다. gRNA-인코딩 플라스미드에 대한 도말 또는 선별 후, 스크리닝된 10개 콜로니들 중 2개가 원하는 돌연변이를 함유하는 것으로 나타났던 반면, 나머지는 gRNA 플라스미드 또는 Cas9 플라스미드 손실에서 프로토스페이서 돌연변이를 통해 생성된 돌연변이체들을 벗어나는 것으로 나타났다.
람다-레드 돌연변이유발과 Cas9 선택
람다-레드 돌연변이유발을 통해 후보 미생물 CI006 및 CI010의 돌연변이체들을 생성하고 CRISPR-Cas로 선별하였다. 녹아웃 카세트는 전사체 프로파일링을 통해 확인된 내인성 프로모터 (표 3-4에 도시됨) 및 결실 표적에 연접하는 ~250bp 상동성 영역을 함유하였다. CI006 및 CI010을 아라비노스 유도성 프로모터의 제어하에 람다-레드 재조합 시스템 (엑소, 베타, gam 유전자)을 인코드하는 플라스미드 및 IPTG 유도성 프로모터의 제어하에 Cas9로 형질전환시켰다. 생성된 형질전환체에서 적색 재조합 및 Cas9 시스템을 유도하고, 전기천공을 위한 균주를 제조하였다. 녹아웃 카세트 및 플라스미드-인코드된 선별 gRNA들이 후속하여 적격 세포에 형질전환되었다. Cas9 플라스미드 및 gRNA 플라스미드 모두에 대하여 선별적인 항생제에서 도말 후, 스크리닝된 10개 콜로니 중 7개는 의도한 녹아웃 돌연변이를 보여주었다.
실시예 3: 후보 미생물의 식물체 내 표현형분석
후보 미생물에 의한 식물들의 콜로니화
후보 미생물에 의한 원하는 숙주 식물들의 콜로니화를 단기 식물 성장 실험들을 통하여 정량하였다. 옥수수 식물들을 플라스미드로부터 또는 Tn5-통합된 RFP 발현 카세트로부터 RFP를 발현시키는 균주들로 접종시켰다. 식물들을 무균 모래 및 미무균 이탄 배지 모두에서 성장시키고, 1 mL의 세포 배양물을 피펫팅하여 발아 후 3일에 출현하는 식물 초엽 위에 직접 접종하였다. 적절한 항생제를 함유하는 용액을 식물에 주어 플라스미드를 유지시켰다. 3주 후, 식물 뿌리들을 수집하여, 무균수로 3회 세척하여 눈에 보이는 토양을 제거하고, 2개 샘플들로 분할하였다. 후보 미생물의 국소화 패턴을 식별하기 위해 하나의 뿌리 샘플을 형광 현미경검사법으로 분석하였다. 가장 양호한 무손상 식물 뿌리들 10 mm 길이에 대해 현미경검사를 실시하였다.
콜로니화를 평가하기 위한 두 번째 정량법이 개발되었다. 내생미생물을 접종한 식물들의 뿌리들로부터 얻은 DNA 표본들 전부에 대하여 정량 PCR 분석법을 실시하였다. 옥수수 종자들 (Dekalb DKC-66-40)을 2.5 인치 X 2.5 인치 X 10 인치 포트의 미리 오토클레이브처리된 모래에서 발아시켰다. 식재한 다음날에, 하룻밤 배양한 (SOB 배지) 1ml의 내생미생물로 종자가 위치하였던 바로 그 지점을 흠뻑 적셨다. 이러한 하룻밤 배양물 1mL는 약 10^9 cfu와 대략 균등하며, 어떠한 균주가 사용되는지에 따라 서로 3-배 이내에서 달라진다. 각 묘목에 2.5mM 또는 0.25mM 암모늄 니트레이트를 보충한 개질된 호글랜드 용액 50mL로 매주 3회 비료를 주었다. 식재 후 4주차에, DNA 추출을 위해 뿌리 샘플들을 수집하였다. 가압 물 스프레이를 사용하여 토양 파편들을 세척하였다. 권고된 프로토콜에 따라 이후 이들 조직 샘플들을 QIAGEN 조직분쇄기를 사용하여 균질화시킨 다음, DNA를 QIAmp DNA 미니 키트 (QIAGEN)를 사용하여 추출하였다. 각 내생미생물의 게놈에 있는 좌위에 특이적이도록 설계된 프라이머를 사용하여 (NCBI의 프라이머 BLAST를 사용하여) 이들 DNA 추출물들에 대해 Stratagene Mx3005P RT-PCR을 이용한 qPCR 분석을 실시하였다. 내생미생물의 게놈 카피들의 존재를 정량하였다. 내생미생물의 동일성을 추가로 확인하기 위해, PCR 증폭 산물들을 시퀀싱하고 정확한 서열을 가지는지 확인한다. 후보 미생물의 균주 CI006 및 CI008의 콜로니화 프로파일 요약이 표 6에 제시된다. 균주 CI008에서 10^7x cfu/뿌리 g fw 만큼 높은 콜로니화율을 나타내었다.
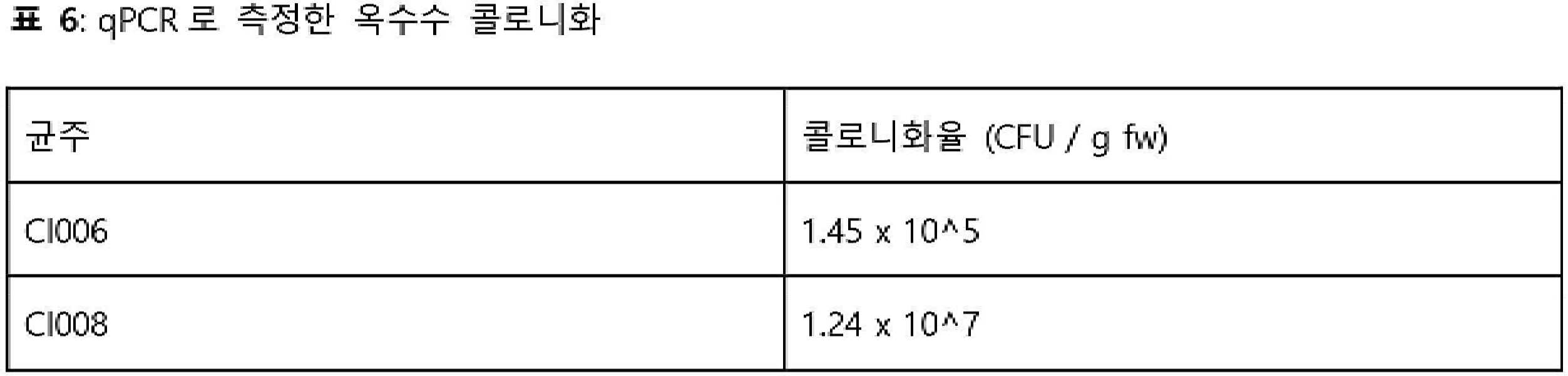
식물체내
RNA 프로파일링
식물체내 nif 경로 성분의 생합성은 nif 유전자의 전사를 측정함으로써 추정되었다. CI006 접종된 (전술한 바와 같은 이식 방법) 식물의 뿌리 식물 조직으로부터 전체 RNA를 얻었다. 권고된 프로토콜 (QIAGEN)에 따라 RNEasy 미니 키트를 사용하여 RNA 추출을 실시하였다. 이후 이들 식물 조직으로부터 얻은 전체 RNA를 균주 CI006의 게놈에서 nif 유전자에 특이적이었던 프로브들을 사용하여 Nanostring Elements 키트 (NanoString Technologies, Inc.)를 이용해 분석하였다. 식물체내 nif 유전자 발현 데이터를 표 7에 요약한다. nifH 유전자 발현은 CM013 균주를 접종한 식물에서 검출되었으며 반면 nifH 발현은 CI006 접종된 식물에서 검출불가능하였다. 균주 CM013은 nifL 유전자가 녹아웃 되어있는 균주 CI006의 유도체이다.
고도로 발현된 CM011 유전자들을 백만 킬로베이스 당 전사체 (transcripts per kilobase million, TPM)로 순위를 정하고, 이는 비료 조건하에서 식물체내 측정되었다. 이들 고도로 발현된 유전자 중 일부의 발현을 제어하는 프로모터를 표적된 질소 고정 및 동화 좌위로의 상동 재조합을 위한 주형으로서 사용하였다. 온실 성장 CM011 접종 식물로부터 RNA 샘플을 추출하고, rRNA를 Ribo-Zero 키트를 사용하여 제거하고, Illumina의 Truseq 플랫폼을 사용하여 시퀀싱하고 CM011의 게놈에 다시 맵핑하였다. CM011로부터 고도로 발현된 유전자들을 표 8에 열거한다.
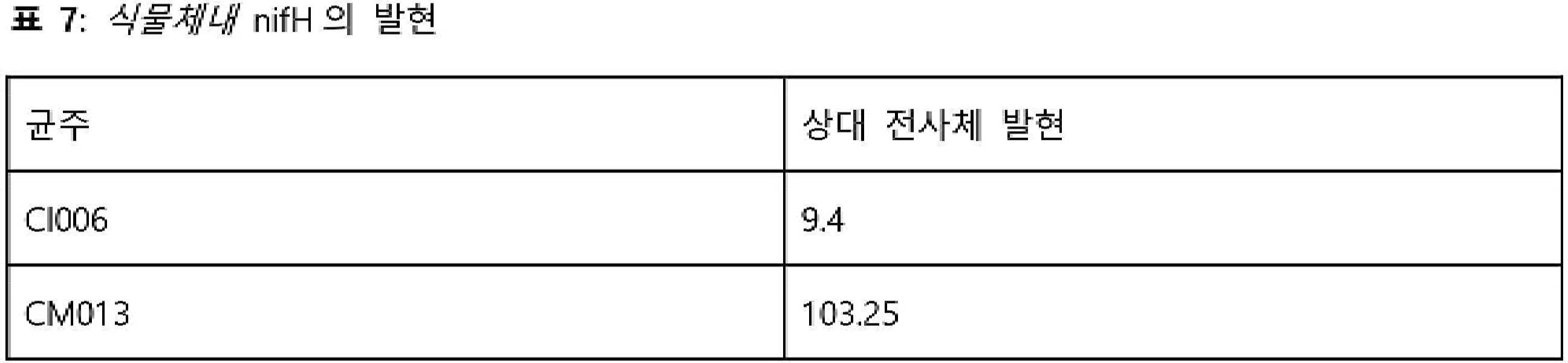

실시예 4: 지리학적 상용성
접종된 식물을 콜로니화시키는 개선된 미생물의 능력은 현장 조건에서 식물의 성공에 중요하다. 기재된 단리 방법은 옥수수와 같은 작물과 밀접한 관계를 가질 수 있는 토양 미생물로부터 선택하도록 설계되지만, 많은 균주는 다양한 식물 유전자형, 환경, 토양 유형 또는 접종 조건에 걸쳐 유효하게 콜로니화하지 않을 수 있다. 콜로니화는 미생물 균주와 숙주 식물 사이의 다양한 상호 작용을 요구하는 복잡한 과정이기 때문에, 콜로니화 능력에 대한 스크리닝은 추가 개발을 위해 우선순위 균주를 선택하는 중심적인 방법이 되었다. 콜로니화를 평가하기 위한 초기의 노력은 균주의 형광 태그처리를 사용하였는데, 이는 효과적이지만 시간 소모적이고 균주 별로 측정할 수 없었다. 콜로니화 활성은 직접적인 개선이 불가능하기 때문에, 잠재적인 생성물 후보가 자연 콜로니화제인 균주로부터 선택되는 것이 필수적이다.
qPCR 및 커뮤니티 샘플에서 균주 특이적으로 설계된 프라이머를 사용하여 임의의 주어진 숙주 식물에서 야생형 균주의 강력한 콜로니화를 시험하기 위한 분석을 설계하였다. 이러한 분석은 옥수수 조직 샘플들로부터 얻은 미생물의 콜로니화율을 신속하게 측정하고자 하는 것이다. 형광 현미경법 및 플레이트-기반 기술을 사용하여 가능한 콜로니화제로 평가된 균주를 사용한 초기 테스트는 qPCR 접근법이 정량적이며 측정가능함을 나타내었다.
전형적인 분석은 다음과 같이 실시된다: 대부분의 옥수수와 밀 품종은 온실의 이탄 포팅 믹스에서 균주 당 6회 반복하여 재배된다. 식재 후 4일 또는 5일에, 0.6-1.0 (대략 5E+08 CFU/mL)의 OD590까지 희석된 박테리아의 초기 고정상 배양액 1mL 드렌치를 신생 초엽 위에 피펫팅한다. 식물에 수돗물 만으로 물을 주고 샘플링 전 4주 동안 성장시키고, 샘플링시, 식물을 뿌리뽑아 뿌리를 철저히 세척하여 대부분의 이탄 잔류물을 제거한다. 깨끗한 뿌리 샘플들을 절제하고 균질화하여 식물 세포 파편 및 관련 박테리아 세포들의 슬러리를 생성한다. 우리는 qPCR을 위한 주형으로 사용하기 위한 식물과 박테리아 DNA의 혼합물을 효과적으로 제조하는 고-처리량 DNA 추출 프로토콜을 개발하였다. 박테리아 세포 스파이크-인 (spike-in) 실험에 기초하여, 이러한 DNA 추출 과정은 새로운 뿌리 중량에 대하여 정량적인 박테리아 DNA 샘플을 제공한다. 프라이머 BLAST (Ye 2012)를 사용하여 설계된 균주 특이적 프라이머를 사용하여 각 균주를 평가하고 접종되지 않은 식물의 배경 증폭과 비교한다. 일부 프라이머는 접종되지 않은 식물에서 오프타겟 증폭을 나타내므로, 콜로니화는 배경 수준과 비교하여 정확한 산물의 증폭의 존재 또는 상승된 증폭에 의해 결정된다.
이 분석은 상이한 토양 지리학에 따른 미생물 산물의 상용성을 측정하기 위해 사용되었다. 현장 토양 품질 및 현장 조건은 미생물 산물의 효과에 큰 영향을 미칠 수 있다. 토양 pH, 물 보유능, 및 경쟁적 미생물들은 접종물 생존 및 콜로니화 능력에 영향을 줄 수 있는 토양 내 요인들 중 단지 몇 가지 예이다. 콜로니화 분석은 캘리포니아의 농업 현장으로부터 샘플링된 3가지 서로 다른 토양 유형들을 식물 성장 배지로서 사용하여 실시되었다 (도 1A). 중간 접종 밀도를 사용하여 현실적인 농업 조건들에 가깝게 하였다. 3주 이내, 균주 5는 1E+06 내지 1E+07 CFU/g FW에서 모든 식물들을 콜로니화하였다. 식물 성장 7주 후, 균주 1의 진화된 버전은 모든 토양 유형에서 높은 콜로니화율 (1E+06 CFU/g FW)을 나타내었다. (도 1B).
추가적으로, 현장 조건의 복잡성에서 콜로니화를 평가하기 위하여, 2015년 6월에 San Luis Obispo에서 1-에이커 현장 시험을 시작하여, 2가지 현장 옥수수 품종들에서 야생형 균주들 중 7개의 콜로니화 및 영향을 평가하였다. 계약 현장 연구 기관인 Pacific Ag Research가 농업 설계 및 시행을 수행했다. 접종을 위해, 접종 방법 실험에서 시험했던 동일한 이탄 배양 종자 코팅 기술을 사용하였다. 성장기 동안, 뿌리와 줄기 내부의 콜로니화를 평가하기 위해 식물 샘플을 수집했다. 식재 후 4주 및 8주에 각 처리를 3회 반복 플롯, 및 16주에 수확 직전에 각 처리를 모두 6회 반복한 것으로부터 샘플을 수집하였다. 12주차에, 미처리 대조군 뿐만 아니라, 균주 1 및 균주 2로 접종한 6회 반복 처리 플롯 모두로부터 추가 샘플을 수집하였다. 새로운 세척 뿌리 그램 중량 당 세포의 수를 qPCR 및 균주-특이적 프라이머를 이용한 다른 콜로니화 분석으로 평가하였다. 균주 1과 균주 2의 두 균주는 12주에 정점에 도달 한 후 급격히 감소하는 일관되고 광범위한 뿌리 콜로니화를 보여주었다 (도 1C). 균주 2는 균주 1보다 수십 배 낮은 수로 존재하는 것으로 나타났지만, 식물별로 더 일관된 수로 발견되었다. 줄기 내부를 효과적으로 콜로니화하는 것으로 나타난 균주는 없었다. qPCR 콜로니화 데이터를 지원하기 위해, 매칭 서열의 단리물을 식별하기 위하여 도말 및 16S 시퀀싱을 사용하여 뿌리 샘플들로부터 두 균주들 모두를 성공적으로 재단리하였다.
실시예 5: 미생물 육종
미생물 육종의 예들은 도 2A에 도식에 요약될 수 있다. 도 2A는 미생물 육종을 나타내며, 여기서 마이크로바이옴의 조성이 먼저 측정 될 수 있고 관심 종이 식별된다. 마이크로바이옴의 대사를 매핑하고 유전학에 연결시킬 수 있다. 그 후, 접합 및 재조합, 화학적 돌연변이유발, 적응 진화 및 유전자 편집을 포함한 (그러나 이에 제한되지 않는) 표적된 유전자 변이가 도입 될 수 있다. 유도 미생물을 사용하여 작물들을 접종한다. 일부 예에서, 가장 우수한 표현형을 갖는 작물이 선택된다.
도 2A에서 제공된 바와 같이, 마이크로바이옴의 조성을 먼저 측정 할 수 있고 관심 종이 확인된다. 도 2B는 마이크로바이옴 단계 측정의 확대도를 도시한다. 마이크로바이옴의 대사를 매핑하고 유전학에 연결시킬 수 있다. 질소 대사는 AmtB 운반체를 통해 뿌리줄기로부터 박테리아의 세포액으로 암모니아 (NH4 +)의 유입을 포함할 수 있다. 암모니아 및 L-글루타메이트 (L-Glu)는 글루타민 합성효소 및 ATP에 의해 글루타민으로 촉매화된다. 글루타민은 바이오매스의 형성 (식물 성장)을 야기 할 수 있고, 또한 nif 오페론의 발현을 억제 할 수 있다. 그 후, 접합 및 재조합, 화학적 돌연변이유발, 적응 진화 및 유전자 편집을 포함한 (그러나 이에 제한되지 않는) 표적된 유전자 변이가 도입 될 수 있다. 유도 미생물을 사용하여 작물들을 접종한다. 가장 우수한 표현형을 갖는 작물이 선택된다.
실시예 6: 본 발명의 미생물을 이용한 현장 시험
농업상 토양을 비롯한 자연에서 다양한 질소 고정 박테리아가 발견 될 수 있다. 그러나, 미생물이 작물에 충분한 질소를 제공하여 비료 사용을 감소시킬 수 있는 가능성은 비료처리 토양에서 질소고정효소 유전자의 억제 뿐만 아니라 작물 뿌리와 밀접하게 연관된 낮은 존재비에 의해 제한 될 수 있다. 주요 상업용 작물과 밀접한 관련이 있는 미생물의 식별, 단리 및 육종은 질소 감지와 질소 고정을 연결하는 조절 네트워크를 방해하고 개선 할 수 있으며 작물 관련 미생물에 의한 상당한 질소 기여를 해제시킬 수 있다. 이를 위해, 옥수수 뿌리 시스템과 관련이 있고 이를 콜로니화하는 질소 고정 미생물이 식별되었다.
농업상 적절한 토양에서 자란 옥수수 식물로부터 뿌리 샘플을 수집하고, 뿌리줄기 및 내생포자에서 미생물 집단을 추출했다. 이들 샘플로부터의 게놈 DNA를 추출한 후, 커뮤니티 조성을 프로파일링하기 위해 16S 앰플리콘 시퀀싱을 수행하였다. 코사코니아 사카리 미생물 (균주 PBC6.1)을 단리하고 16S rRNA 및 전체 게놈 시퀀싱을 통해 분류하였다. 이는 뿌리-관련 미생물총의 거의 21% 존재비까지 콜로니화 할 수 있는 특히 흥미로운 질소 고정제이다 (도 4). 외인성 질소에 대한 균주 민감성을 평가하기 위해, 순수한 배양물에서 질소 고정율을 고전적인 아세틸렌 환원 분석 (ARA) 및 다양한 수준의 글루타민 보충으로 측정 하였다. 이 종들은 무-질소 배지에서 높은 수준의 질소 고정 활성을 보였지만, 외인성 고정된 질소는 nif 유전자 발현 및 질소고정효소 활성을 억제하였다 (균주 PBC6.1, 도 3C). 또한, 질소-고정 조건에서 성장된 PBC6.1의 상청액에서 방출된 암모니아를 측정할 때, 고정 질소의 방출이 거의 검출되지 않을 수 있다.
우리는 질소 대사를 제어하는 조절 네트워크가 고정 질소의 존재하에서 최적의 질소고정효소 발현 및 암모니아 방출을 가능하도록 하기 위해 다시 연결된다면, PBC6.1이 비료처리 현장에서 고정 질소의 중요한 기여자가 될 수 있다고 가정했다. 트랜스진 또는 합성 조절 요소의 삽입없이 광범위한 표현형 리모델링을 가능하게 하기 위해 PBC6.1 게놈 내에 충분한 유전자 다양성이 존재해야 한다. 단리된 균주는 적어도 5.4 Mbp의 게놈 및 표준 질소 고정 유전자 클러스터를 갖는다. PBC6.1의 관련 질소 대사 경로는 질소 고정을 위한 모델 유기체인, 클레브시엘라 옥시토카 (Klebsiella oxytoca) m5al의 경로와 유사합니다.
특히 높은 외인성 고정 질소 농도에서, 질소 고정을 증대시키고 후속하여 숙주 식물로 이동시킬 수 있는 몇 가지 유전자 조절 네트워크 노드들이 확인되었다 (도 3A). nifLA 오페론은 NifA에 의한 전사 활성화 및 NifL에 의한 NifA의 질소-및 산소-의존적 억제를 통해 나머지 nif 클러스터를 직접 조절한다. nifL의 파괴는 산소 및 외인성 고정 질소 둘 모두의 존재하에서 NifA의 억제를 없애고 nif 발현을 개선시킬 수있다. 또한, 질소-독립적 프로모터의 제어하에 nifA를 발현시키는 것은 질소고정효소 생합성을 NtrB/NtrC 질소 감지 복합체에 의한 조절로부터 분리시킬 수 있다. 미생물에 의한 고정 질소의, 글루타민 합성효소 (GS)에 의한 글루타민으로의 동화는 2-도메인 아데닐릴트랜스퍼라제 (ATase) 효소 GlnE에 의하여, 활성을 각각 약화 및 회복시키는 GS의 아데닐릴화 및 탈아데닐릴화를 통해 가역적으로 조절된다. GlnE 단백질을 절두하여 그 아데닐릴제거 (AR) 도메인을 제거하면 항시적으로 아데닐릴화된 글루타민 합성효소가 생성되어, 미생물에 의한 암모니아 동화를 제한하고 세포간 및 세포외 암모니아를 증가시킬 수 있다. 마지막으로, 암모니아 흡수를 담당하는 운반체인 AmtB의 발현 감소는 보다 많은 세포외 암모니아를 유발할 수 있다. 트랜스지을 사용하지 않고 합리적으로 설계된 미생물 표현형을 생성하기 위해, 단백질 도메인 또는 전체 유전자를 인코드하는 게놈 서열의 마커리스 결실을 생성하는 것, 그리고 게놈 내 프로모터 재배열에 의해 조절 네트워크를 다시 연결하는 것의 두 가지 접근법이 사용되었다. 반복 돌연변이유발 과정을 통해, PBC6.1의 여러 비-형질전환 유도체 균주가 생성되었다 (표 9).
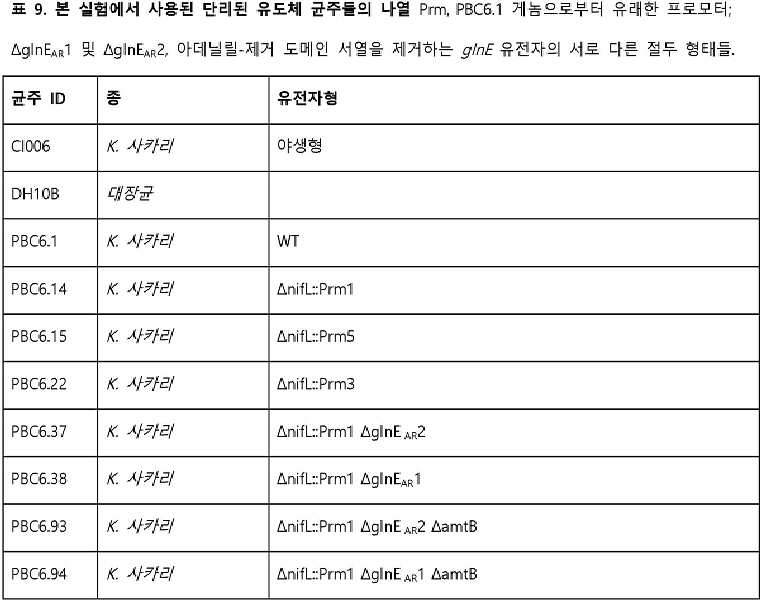
유도체 균주의 특정 표현형을 특성화하기 위해 몇몇 시험관내 분석을 수행 하였다. ARA를 사용하여 외인성 질소에 대한 균주 민감성을 평가하였고, 여기서 PBC6.1은 높은 글루타민 농도에서 질소고정효소 활성의 억제를 나타냈다. 대조적으로, 대부분의 유도체 균주는 높은 글루타민 농도에서 관찰되는 다양한 수준의 아세틸렌 감소와 함께 탈억제 (derepressed) 표현형을 나타냈다. qPCR에 의해 분석된 샘플에서 nifA의 전사 속도는 아세틸렌 환원 속도와 잘 상관되어 있으며, 질소-독립 프로모터의 nifL 파괴 및 삽입으로 nifA를 구동시키는 것이 nif 클러스터 탈억제을 유발할 수 있다는 가설을 뒷받침한다. 변경된 GlnE 또는 AmtB 활성을 갖는 균주는 이러한 돌연변이가 없는 야생형 또는 유도체 균주에 비해 암모늄 배설 속도가 현저하게 증가함을 보여주었으며, 이는 암모니아 동화 및 재흡수에 대한 이들 유전자형의 효과를 보여주는 것이다.
옥수수 식물과 PBC6.1 유도체의 상호작용을 연구하고 고정 질소의 식물 조직으로의 혼입을 정량화하기 위해 2개의 실험을 수행 하였다. 먼저, 동위원소 추적자를 사용하여 온실 연구에서 미생물 질소 고정 속도를 정량화하였다. 요약하면, 식물은 15N 표지된 비료로 재배되며, 식물 조직에서 15N의 희석된 농도는 미생물로부터의 고정 질소의 기여를 나타낸다. 옥수수 묘목에 선택된 미생물 균주를 접종하고, 식물을 V6 성장 단계까지 성장시켰다. 미생물 콜로니화 및 유전자 발현의 측정, 뿐만 아니라 동위원소 비율 질량 분석법 (IRMS)에 의한 식물 조직에서 15N/14N 비율 측정을 가능하게 하기 위해 식물을 후속하여 분해하였다. 공중 조직의 분석은 식물 질소 수준에 대한 PBC6.38에 의한 작은, 유의하지 않은 기여 및 PBC6.94에 의한 유의한 기여를 보여주었다 (p = 0.011). 지상 옥수수 잎에서 발견되는 질소의 약 20%는 PBC6.94에 의해 생성되었으며, 나머지는 다른 토양유래 미생물에 의한 종자, 포팅 믹스 또는 "배경" 고정에서 온 것이다. 이는 우리의 미생물 육종 파이프라인이 질소 비료의 존재하에서 식물에 상당한 질소 기여를 할 수있는 균주를 생성 할 수 있음을 보여준다. 식물 조직 내 미생물 전사를 측정하고, 야생형 균주가 아닌 유도체 균주에서 nif 유전자 클러스터의 발현이 관찰되었는데, 이는 비료처리 조건에서 작물에 대한 BNF의 기여에 대한 nif 탈억제의 중요성을 보여주는 것이다. qPCR에 의해 측정된 뿌리 콜로니화는 콜로니화 밀도가 시험된 각각의 균주에 대해 다르다는 것을 입증하였다. PBC6.38과 PBC6.94 사이에서 50배 콜로니화 차이가 관찰되었다. 이러한 차이는 PBC6.94가 높은 수준의 고정 및 배설의 결과로서 PBC6.38에 비해 뿌리줄기에서 감소된 적합성을 가짐을 나타내는 것일 수 있다.
방법
배지
미네랄 배지는 (리터 당) 25 g Na2HPO4, 0.1g CaCL2-2H2O, 3 g KH2PO4, 0.25 g MgSO4·7H2O, 1 g NaC1, 2.9 mg FeCl3, 0.25 mg Na2MoO4·2H2O, 및 20 g 수크로스를 함유한다. 성장 배지는 리터 당 50 ml의 200 mM 글루타민이 보충된 미네랄 배지로 정의된다.
질소자급영양생물의 단리
옥수수 묘목을 22°C (밤) 내지 26°C (낮)로 제어되는 온실 환경에서 2주 동안 종자(DKC 66-40, DeKalb, IL)로부터 성장시키고 캘리포니아주 San Joaquin County에서 수집한 토양에서 16시간 명 주기에 노출시켰다. 뿌리를 수확하고 무균 탈이온수로 세척하여 벌크 토양을 제거하였다. 뿌리 조직을 설정 30에서 3 분 동안 조직 용해장치 (TissueLyser II, Qiagen P/N 85300)에서 2mm 스테인레스 스틸 비드로 균질화하고, 샘플을 13,000rpm에서 1 분 동안 원심분리하여 뿌리-관련 박테리아로부터 조직을 분리시켰다. 상청액을 2개의 분획으로 나누고, 1개를 사용하여 16S rRNA 앰플리콘 시퀀싱을 통해 마이크로바이옴을 특성화하고, 나머지 분획을 1.5% 한천이 보충된 무-질소 액체배지 (NfB) 배지에 희석 및 도말하였다. 플레이트를 30°C에서 5-7일 동안 배양하였다. 나타난 콜로니들을 nifH 유전자의 존재에 대해 프라이머 Ueda19f 및 Ueda406r을 이용하여 콜로니 PCR로 시험하였다. 양성 nifH 콜로니 PCR을 갖는 균주로부터의 게놈 DNA를 단리하고 (QIAamp DNA 미니 키트, 카탈로그 번호 51306, QIAGEN, 독일) 시퀀싱하였다 (Illumina MiSeq v3, SeqMatic, Fremont, CA). 서열 어셈블리 및 주석달기 후, 질소 고정 유전자 클러스터를 함유하는 분리주를 하위 연구에 사용하였다.
단리 묘목들의 마이크로바이옴 프로파일링
게놈 DNA를 ZR-96 게놈 DNA I 키트 (Zymo Research P/N D3011)를 사용하여 뿌리-관련 박테리아로부터 단리하고, 799f 및 1114r을 표적으로하는 nextera-바코드 프라이머를 사용하여 16S rRNA 앰플리콘을 생성하였다. 앰플리콘 라이브러리를 정제하고 Illumina MiSeq v3 플랫폼 (SeqMatic, Fremont, CA)으로 시퀀싱하였다. minikraken 데이터베이스를 사용한 Kraken을 사용하여 리드를 분류학적으로 분류하였다.
아세틸렌 환원 분석 (ARA)
순수한 배양 조건에서 아세틸렌 환원 분석의 변형된 버전을 사용하여 질소고정효소 활성을 측정하였다. 균주들을 30 °C의 SOB (RPI, P/N S25040-1000)에서 200 RPM에서 24시간 동안 진탕하면서 단일 콜로니로부터 번식시킨 다음, 1:25로 성장 배지에 계대배양하고 24 시간 (30 °C, 200 RPM) 동안 호기적으로 성장시켰다. 이어서, 1 ml의 최소 배지 배양물을 기밀 Hungate 튜브에서 0 내지 10 mM 글루타민이 보충된 4ml의 최소 배지에 첨가하고 4 시간 (30 ℃, 200 RPM) 동안 혐기적으로 성장시켰다. 이어서 10% 헤드스페이스를 제거한 후, 주입에 의해 동일한 부피의 아세틸렌으로 대체하고, 1시간 동안 배양을 계속하였다. 이어서, 불꽃 이온화 검출기 (FID)가 장착된 Agilent 6850 기체 크로마토그래피를 사용하여 에틸렌 생성을 정량화하기 위해 가스 기밀 주사기를 통해 2 ml의 헤드스페이스를 제거하였다.
암모늄 배설 분석
혐기성 생물 반응기에서 회분식 발효를 사용하여 암모니아 형태의 고정 질소의 배출을 측정 하였다. 균주를 96 웰 DeepWell 플레이트에서 1 ml/웰의 SOB 중 단일 콜로니로부터 번식시켰다. 플레이트를 200 RPM에서 24시간 동안 진탕시키면서 30 ℃에서 배양한 다음, 1 ml/웰의 성장 배지를 함유하는 새로운 플레이트에 1:25로 희석시켰다. 세포를 24 시간 동안 (30 ℃, 200 RPM) 배양한 후, 최소 배지를 함유한 새로운 플레이트에 1:10으로 희석시켰다. 플레이트를 >98.5% 질소, 1.2-1.5% 수소 및 <30 ppM 산소의 가스 혼합물이 있는 혐기성 챔버로 옮기고, 1350 RPM, 실온에서 66-70 시간 동안 배양하였다. 초기 배양 바이오매스를 590 nm에서 광학 밀도를 측정함으로써 종료 바이오매스와 비교하였다. 이어서, 세포를 원심 분리에 의해 분리하고, 각 시점에서 바이오 매스에 대해 정규화된 Megazyme 암모니아 분석 키트 (P/N K-AMIAR)를 사용하여 반응기 액체배지로부터의 상청액을 유리 암모니아에 대해 분석하였다.
뿌리-관련 마이크로바이옴의 추출
뿌리를 부드럽게 흔들어 느슨한 입자를 제거하고, 뿌리 시스템을 분리하고 RNA 안정화 용액 (Thermo Fisher P/N AM7021)에 30분 동안 담갔다. 이어서 뿌리를 무균 탈이온수로 간단히 헹구었다. 2ml의 용해 완충액 (Qiagen P/N 79216)에서 조직 용해장치 (TissueLyser II, Qiagen P/N 85300)에서 ½-인치 스테인레스 스틸 볼 베어링으로 비드 비팅하여 샘플을 균질화하였다. 게놈 DNA 추출은 ZR-96 Quick-gDNA 키트 (Zymo Research P/N D3010) 및 RNeasy 키트 (Qiagen P/N 74104)를 사용한 RNA 추출로 수행되었다.
뿌리 콜로니화 분석
식재 4일 후, 1 ml의 박테리아 하룻밤 배양물 (약 109 cfu)을 식재된 종자 위 토양에 처리하였다. 묘목은 0.5 mM 암모늄 니트레이트로 보충된 25 ml 개질 Hoagland 용액으로 매주 3회 비료처리되었다. 식재 4주 후, 뿌리 샘플을 수집하고 전체 게놈 DNA (gDNA)를 추출하였다. 야생형 또는 유도체 균주 게놈의 독특한 영역을 증폭시키도록 설계된 프라이머를 이용하는 qPCR을 사용하여 뿌리 콜로니화를 정량하였다. qPCR 반응 효율은 표적 게놈으로부터 알려진 양의 gDNA로부터 생성된 표준 곡선을 사용하여 측정되었다. 데이터는 조직 중량 및 추출 부피를 사용하여 새로운 중량 g 당 게놈 카피에 대해 정규화되었다. 각 실험에 대해, 콜로니화 수를 비처리 대조 묘목들과 비교하였다.
식물체내
전사체
뿌리-관련 미생물의 전사 프로파일링은 뿌리 콜로니화 분석에 기재된 바와 같이 성장 및 가공된 묘목에서 측정되었다. 정제된 RNA는 Illumina NextSeq 플랫폼 (SeqMatic, Fremont, CA)을 사용하여 시퀀싱되었다. '-매우 민감한 국소' 파라미터 및 최소 정렬 점수 30을 사용하는 bowtie2를 사용하여 접종된 균주의 게놈에 리드를 맵핑하였다. 게놈 전반에 걸친 커버리지를 샘툴(samtools)을 사용하여 계산하였다. 차등적 커버리지는 하우스키핑 유전자 발현에 대해 정규화되었고 Circos를 사용하는 게놈 및 DNAplotlib을 사용하는 nif 유전자 클러스터 전반에 걸쳐 시각화되었다. 또한, 식물체 내 전사 프로파일을 표적화된 Nanostring 분석을 통해 정량화하였다 정제된 RNA를 nCounter Sprint (Core Diagnostics, Hayward, CA)에서 처리하였다.
15N 희석 온실 연구
식물체 내 최적화된 균주 활성을 평가하기 위해 15N 비료 희석 실험을 수행하였다. 질석과 세척된 모래의 혼합물 (DI H2O에서 5회 헹굼)을 사용하여 최소의 배경 N을 함유한 식재 배지를 제조하였다. 모래 혼합물을 122 ℃에서 1시간 동안 오토클레이브하고 약 600g을 40 입방 인치 (656mL) 포트로 측정하였고, 무균 DI H2O로 포화시키고 식재 전 24시간 동안 배수시켰다. 옥수수 종자 (DKC 66-40)를 0.625% 소듐 하이포클로라이트에서 10분 동안 표면 무균한 다음 무균 증류수에서 5회 헹구고 1cm 깊이로 심었다. 식물은 평균 22 ℃(밤) 내지 26 ℃(일)의 실온에서 형광 램프하에 16-시간 하루 길이의 4주 동안 유지되었다.
식재 후 5일에, 묘목들에 1 ml의 세포 현탁액을 로 나타난 초엽 위에 직접 적셔 접종하였다. 접종물을 SOB 중 5 ml 하룻밤 배양물로부터 제조하고, 이를 스핀다운시키고 5 ml PBS 중에 2회 재현탁시켜 잔류 SOB를 제거한 다음, 1.0의 OD로 최종 희석하였다 (약 109 CFU/ml). 대조 식물을 무균 PBS로 처리하고, 각 처리를 10개의 복제 식물에 처리하였다.
식물은 식재 후 5, 9, 14 및 19일에 2% 15N-풍부 2mM KNO3를 함유하는 25ml 비료 용액으로 그리고 식재 후 7, 12, 16 및 18 일에 KNO3가 없는 동일한 용액으로 비료처리되었다. 비료 용액은 (리터 당) 3 mmol CaCl2, 0.5 mmol KH2PO4, 2 mmol MgSO4, 17.9 μmol FeSO4, 2.86 mg H3BO3, 1.81 mg MnCl2·4H2O, 0.22 mg ZnSO4·7H2O, 51 μg CuSO4·5H2O, 0.12 mg Na2MoO4·2H2O, 및 0.14nmol NiCl2를 함유하였다. 유출없이 일관된 토양 수분을 유지하기 위해 필요에 따라 모든 포트에 무균 DI H2O로 물을 주었다.
4주에, 식물을 수확하고 최하위 노드에서 뿌리 gDNA 및 RNA 추출을 위한 샘플 및 IRMS를 위한 공중 조직으로 분리하였다. 모래를 제거하기 위해 필요에 따라 공중 조직을 닦아내고, 종이백에 전체를 넣고 60 ℃에서 72시간 이상 동안 건조시켰다. 완전히 건조되면, 전체 공중 조직을 비드 비팅에 의해 균질화하고, MBL 안정성 동위원소 실험실 (The Ecosystems Center, Woods Hole, MA)에 의해 δ에 대한 동위원소 비율 질량 분광분석법 (IRMS)에 의해 5-7 mg 샘플을 분석하였다. 퍼센트 NDFA를 다음 식을 사용하여 계산하였다: %NDFA = (UTC 평균의 δ-샘플의 δ/ (UTC 평균의 δx 100.
실시예 7: 비-속간 유전자조작 미생물을 검출하기 위한 방법 및 분석
본 발명은 전술한 다양한 실시에들에서 이용된 미생물을 검출하는데 유용한 프라이머, 프로브 및 분석법을 개시한다. 이 분석은 유도된/ 돌연변이체 비-속간 미생물에서 비-천연 뉴클레오티드 "접합"서열을 검출 할 수 있다. 이러한 비-자연 발생 뉴클레오티드 접합은 미생물에서 특정 유전자 변경의 존재를 나타내는 진단 유형으로 사용될 수 있다.
본 기술은 고유하게 설계된 프라이머 및 프로브를 포함하는 특수한 정량적 PCR 방법을 이용하여 이러한 비-자연 발생 뉴클레오티드 접합을 검출 할 수 있다. 상기 프로브는 비-자연 발생 뉴클레오티드 접합 서열에 결합할 수 있다. 즉, 프로브와 이의 상보적 서열과의 혼성화 후에만 검출이 가능한 형광 리포터로 표지된 올리고뉴클레오티드들로 구성된 서열-특이적 DNA 프로브가 사용될 수 있다. 정량 방법들은 비-자연 발생 뉴클레오티드 접합만이 개시된 프라이머를 통해 증폭될 것이며, 그 결과 비-특이적 염료 또는 특이적 혼성화 프로브를 이용하여 검출 될 수 있음을 확인시켜 줄 수 있다. 상기 방법의 또 다른 양상은 접합 서열 중 한 쪽에 프라이머가 연접하도록 프라이머를 선택하는 것이며, 그 결과 증폭 반응이 발생하면, 상기 접합 서열에 존재하게 된다.
결과적으로, 게놈 DNA를 샘플들로부터 추출하고 이를 qPCR을 사용하여 본 발명의 미생물의 존재를 정량화하기 위해 사용할 수 있다. qPCR 반응에 이용된 프라이머는 프라이머 Blast (https://www.ncbi.nlm.nih.gov/tools/프라이머-blast/)에 의해 설계된 프라이머 일 수 있는데, 이는 야생형 게놈의 독특한 영역 또는 유전자조작된 비-속간 돌연변이체 균주의 독특한 영역을 증폭시킨다. qPCR 반응은 정방향 및 역방향 증폭 프라이머만을 사용하는 SYBR GreenER qPCR SuperMix Universal (Thermo Fisher P/N 11762100) 키트를 사용하여 수행 될 수 있고; 대안적으로, Kapa Probe Force 키트 (Kapa Biosystems P/N KK4301)가 증폭 프라이머 및 5' 단부에 FAM 염료 라벨, 내부 ZEN 소광제 및 마이너 그루브 결합제 및 3' 단부에 형광 소광제를 함유하는 TaqMan 프로브 (Integrated DNA Technologies)와 함께 사용될 수 있다.

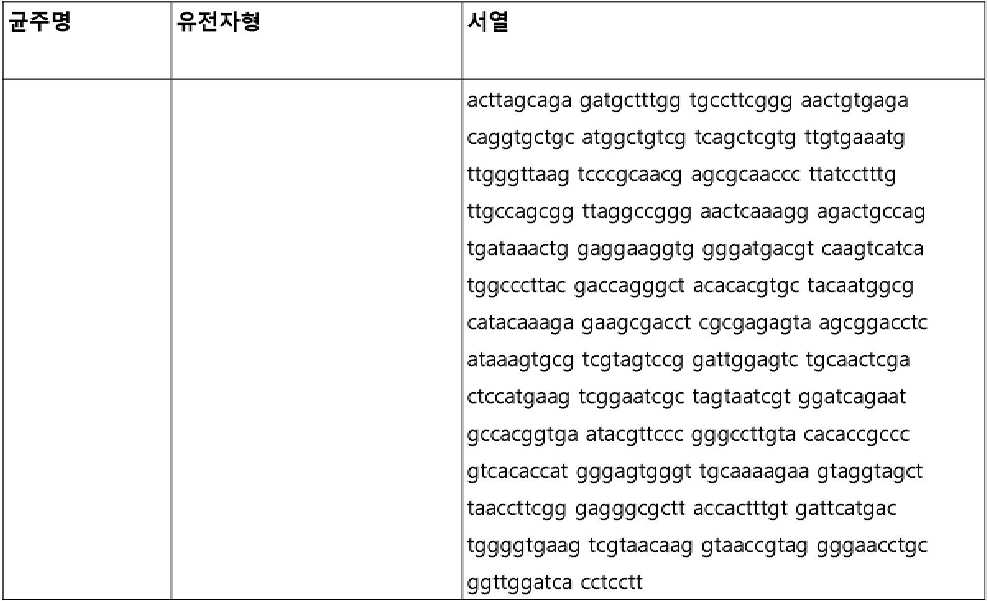
실시예 8:
클레브시엘라 바리이콜라 및 코사코니아 사카리
의 식별
.
박테리아 계통발생 및 분류학을 연구하기 위한 16S rRNA 유전자 서열은 여러 가지 이유로 지금까지 가장 일반적으로 사용된 하우스키핑 유전자 마커였다. 이러한 이유들에는 (i) 종종 다유전자 패밀리로 존재하는 거의 모든 박테리아, 또는 오페론에서의 이의 존재; (ii) 시간 경과에 따라 16S rRNA 유전자의 기능이 변화하지 않았으며, 이는 무작위 서열 변화가 더욱 정확한 시간 (진화)의 측정치임을 제시하는 것이며; 그리고 (iii) 16S rRNA 유전자 (1.500 bp)는 정보학 목적에 충분히 크다는 것이 포함된다.
클레브시엘라 바리이콜라 및 코사코니아 사카리의 16S rRNA 유전자의 뉴클레오티드 서열은 PCR 분석에 의해 결정되었다. 사용된 프라이머는 범용 16S 프라이머로서, 다음 뉴클레오티드 서열을 가진다:
정방향 16S 프라이머 27f (AGAGTTTGATCMTGGCTCAG)
역방향 165 프라이머 1492r (GGTTACCTTGTTACGACTT).
생성된 PCR 산물을 시퀀싱하고, 생성된 서열을 최초 단리 동안 종 식별을 위해 미국 국립생물공학 정보센터 (National Center for Biotechnology Information, NCBI) 데이터베이스와 비교 하였다. 임의의 후속 미생물 생성에 있어서, 오염물이 없는 순수한 검증된 미생물임을 확인하기 위해 동일한 PCR 분석을 수행하였다.
두 개의 모체 균주들이 이 방법으로 식별되었다. 첫 번째 모체 균주의 학명은 클레브시엘라 바리이콜라이다. 종 동일성은 유기체의 16S rRNA를 복수회 시퀀싱하고 속 및 종 검증을 위해 NCBI 데이터베이스에서 BLAST를 사용하여 결정되었다. 또한, 유기체의 전체 게놈을 시퀀싱하고 추출된 16S rRNA를 (게놈 서열로부터) BLAST 분석하였다. 이 서열을 이전에 결정된 16S rRNA 서열에 대해 정렬하여 유기체가 순수한 배양물로 단리되었음을 확인하였다.
두 번째 유기체의 학명은 코사코니아 사카리이다. 종 동일성은 유기체의 16S rRNA를 복수회 시퀀싱하고 속 및 종 검증을 위해 NCBI 데이터베이스에서 BLAST를 사용하여 결정되었다. 또한, 유기체의 전체 게놈을 시퀀싱하고 추출된 16S rRNA를 (게놈 서열로부터) BLAST 분석하였다. 이 서열을 이전에 결정된 16S rRNA 서열에 대해 정렬하여 유기체가 순수한 배양물로 단리되었음을 확인하였다.
실시예 9: 증가된 콜로니화 또는 질소 고정 활성을 가지는 리모델링된 균주
콜로니화 능력, 질소 고정 활성, 또는 질소 배설을 증가시킬 수 있는 새로운 변형을 조사하기 위하여 몇가지 돌연변이체 균주들을 개발하였다. 이들 균주들은 표 11에 요약되어 있다. 프로모터와 유전자 서열들은 표 12에 열거되어 있다. 리모델링된 균주들을 야생형 또는 모체 균주와 비교 평가하여, 변형된 유전자의 발현 변화 (도 5, 12, 및 13), 질소 환원 활성 (도 6, 18, 20, 22A, 22B, 24, 26A, 26B, 34, 및 36), 질소 배설 (도 14, 15, 16, 17, 19, 21, 23A, 23B, 25, 27A, 27B, 33, 및 35), 시험관내 콜로니화 (도 7, 8, 9, 10, 11), 및 온실 콜로니화 (도 28, 29, 30, 31, 32, 37 및 38)에 대해 확인하였다. 암모늄 배설 분석을 아래 설명한 바와 같이 수행하였다. 데이터 중 일부를 표 13 및 14에 추가로 요약한다.
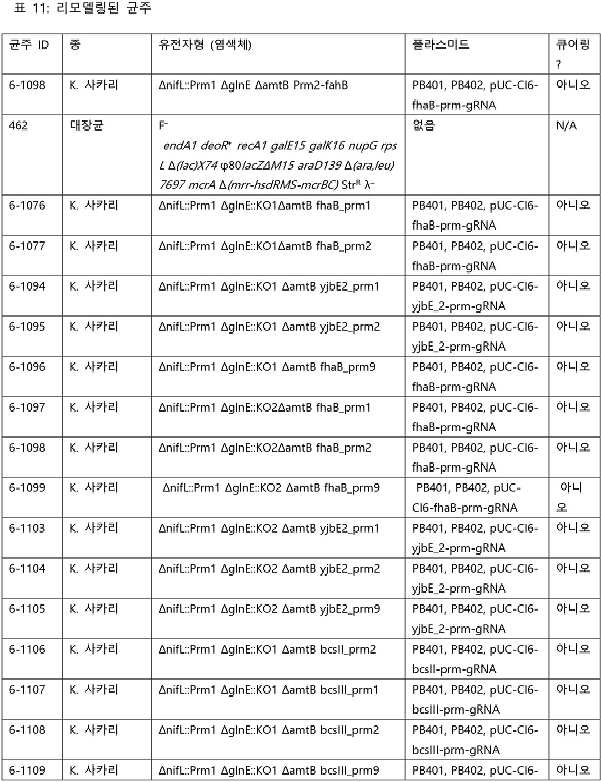

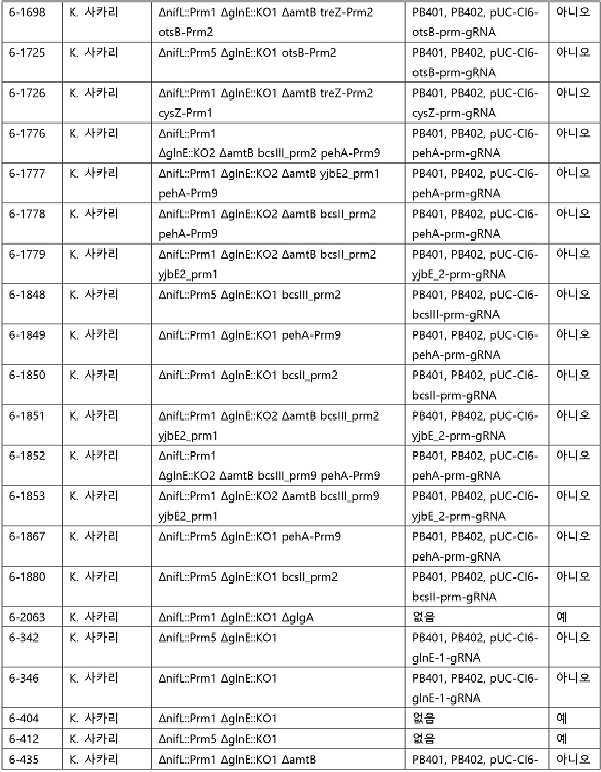
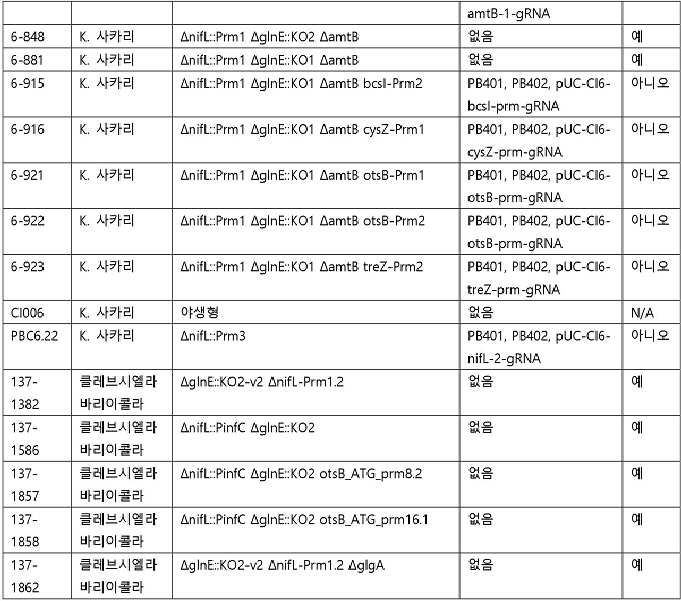
방법: 암모늄 배설 분석
1. 콜로니들을 선택하여 96-웰 딥 웰 플레이트 풍부 배지에서 30C에서 하룻밤동안 배양한다
2. 상기 하룻밤 배양물을 사용하여 10mM 글루타민 (gln)으로 보충된 ARA 배지 (N-없는 배지) (또한 96-웰 플레이트에서)의 배양물을 접종한다
3. ARA+gln에서의 배양물을 사용하여 gln이 없는 ARA의 배양물을 접종한다
4. 혐기성 챔버에 옮기고 3 일 동안 실온에서 진탕하면서 배양한다
5. 96-웰 플레이트를 세포와 함께 스핀다운하여 상청액 샘플을 제거한다
6. Megazyme 암모늄 분석 키트를 사용하여 배지의 암모늄 농도를 분석한다


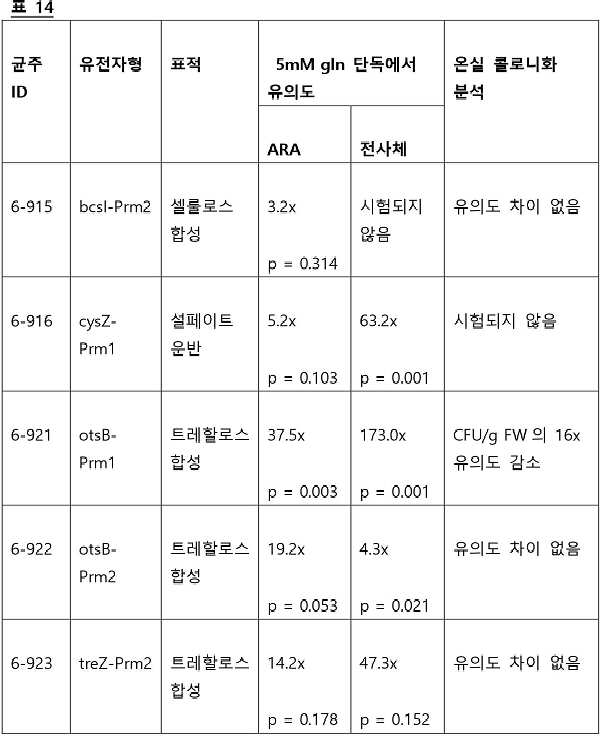
실시예 10: 독성학 연구
클레브시엘라 바리이콜라 및 코사코니아 사카리에 대한 인간 또는 포유동물 독성이 없음을 확인하기 위하여, 이들 유기체들을 사용하여 독성 연구를 수행하였다. 계약자는 독성 연구에 광범위한 경험을 가진 회사인 Smithers Avanza Toxicology Services였다.
이 연구의 목적은 단일 피하 주사 후 마우스에서 박테리아 단리물의 독성을 결정하는 것이었다. CD-1 마우스를 사용하여 체중 kg 당 박테리아 단리물의 수를 최대화하였다. 피하 경로는 박테리아 단리물을 동물에게 접종하기에 적합한 경로이기 때문에 선택되었으며, 이는 미생물에 대한 가장 가능성이 높은 인간 노출을 가장 잘 모사한다.
클레브시엘라 바리이콜라 또는 코사코니아 사카리 박테리아 단리물을 사용한 처리는 마우스의 사망율에 영향을 미치지 않았으며, 모든 동물은 연구 기간 종료시 예정된 종결까지 생존하였다. 연구 과정 내내 클레브시엘라 바리이콜라 및 코사코니아 사카리 치료군에서 동물의 임상 관찰에는 어떠한 이상도 없었다. 또한, 케이지 관찰 동안에도 어떠한 이상도 나타나지 않았다. 또한, 클레브시엘라 바리이콜라 및 코사코니아 사카리 치료군에 대한 피부 관찰에서도 이상이 없었다. 마우스 체온은 모든 그룹에서 정상적이며 평균 온도 범위는 36.66 " 내지 37.62”였다. 클레브시엘라 바리이콜라 및 코사코니아 사카리 처리 그룹의 마우스에 있어서 약간의 체중 감소가 존재하였으나, 이는 본 연구의 마우스들에 있어 그리고 이러한 유형의 연구에 대한 과거 데이터로부터 정상 범위에 속하였다. 이 연구의 결과는 클레브시엘라 바리이콜라 및 코사코니아 사카리 박테리아 단리물이 인간이나 다른 포유류에 독성이나 병원성을 유발할 위험이 없음을 확인시켜 준다.
이 연구의 목적은 단일 피하 주사 후 마우스에서 박테리아 단리물의 독성을 결정하는 것이었다. CD-1 마우스를 사용하여 체중 kg 당 박테리아 단리물의 수를 최대화하였다. 피하 경로는 박테리아 단리물을 동물에게 접종하기에 적합한 경로이기 때문에 선택되었다.
대략 8주령의 미감작 암컷 CD-1 마우스 45 마리를 2017년 6월 8일 Charles River Breeding Labs으로부터 입수하였다. 경재 침대 (hardwood bedding)를 포함하고 있는 폴리카보네이트 케이지에 동물들을 그룹 수용시켰다 (케이지 당 5마리 동물). 동물에게 Certified Global Teklad Laboratory Diet 2018 (펠릿) 먹이를 수시 제공하였으며 물병을 통해 식수를 제공했다. 동물실 환경 제어는 온도 20-26℃ 상대 습도 30-70%, 시간당 최소 10회의 공기 교환 및 12-시간 명/12-시간 암주기를 유지하도록 설정되었다.
40마리의 마우스를 각각 5마리의 마우스로 구성된 8개의 그룹으로 무작위 배당하였다. 남은 동물들을 훈련/스톡 콜로니로 이동시켰다. 동물들을 표 15에 나타낸 바와 같이 처리하였다.

시험 품목들은 스폰서에 의해 투여 준비 형태로 공급되었다. 모든 동물은 1 연구일 (SD)에 견갑골 전방 영역에 피하 주사를 통해 1회 투약되었다.
신체 검사를 매일 수행하고 케이지측 관찰을 매일 2회 수행했다. 피부 관찰 또한 표 16에 따라 매일 수행되었다. 추가 관찰내용들을 표 17-27에 열거한다.

용량 투여 전 (SD 1) 및 종료 전 (SD 8) 체중을 기록하였다. 종료 전 (SD 8)에 체온을 측정하였다. SD 8에서 최종 데이터 수집 후, 동물을 CO2 흡입 및 경추 탈골을 통해 안락사시키고 부검없이 폐기하였다.
어떠한 박테리아 단리물의 처리도 사망율에 영향을 주지 않았다. 모든 동물들은 예정된 종료시까지 생존하였다.
신체 검사 동안 임상 관찰 내용을 나타내었다. 연구 과정 내내 어떤 동물에서도 이상이 관찰되지 않았다.
케이지측 관찰 동안에도 어떠한 이상도 발견되지 않았다.
피부 드레이즈 관찰을 수행하였다. 그룹 6의 한 동물은 SD 7에서 시작하여 관찰 기간이 끝날 때까지 (SD 8) 최소 부종 및 홍반을 나타냈다. 그룹 7의 한 동물은 SD 5에서 시작하여 관찰 기간이 끝날 때까지 (SD 8) 경도 홍반을 나타냈다. 그룹 8에서, 1일에서 4일까지 2마리 동물에서 최소 또는 경도 부종이 관찰되었다. SD 7부터 SD 8까지 동물 22029 (8f)에서 경도 홍반이 관찰되었고, SD 5 및 6에서 동물 22032 (8f)에서 중등도 홍반이 나타났으며, 이는 SD 7 및 8에서 최소 홍반으로 가라앉았다.
체중 및 체중 변화를 기록하였다. 그룹 1, 2, 3, 5, 6 및 8은 연구 전반에 걸쳐 체중이 최소로 감소되었으며, 평균 퍼센트 변화는 -1.91% 내지 -6.87% 범위였다. 그룹 4 및 7은 연구 전반에 걸쳐 체중이 최소 증가되었으며, 평균 퍼센트 변화는 각각 0.31% 및 2.73%였다.
체온을 추적하였다. 체온은 모든 그룹에서 정상이었이며 평균 체온은 36.66℃ 내지 37.62℃ 범위였다.
실시예 10 현장 시험
다양한 질소 상황하에서 옥수수 성장 및 생산성에 대한 Pivot 균주의 효능을 평가하기 위해 현장 시험이 수행 될 수 있다.
재료 및 방법
실험 설계:
N 비율을 메인 구획(Main plots)으로 처리를 서브 구획 (Subplots)으로 하는 분할 구획 (Split Plot) 설계.
위치:
아르헨티나의 3개의 지리적으로 다양한 위치.
데이터
식재 1주 이내에 제공될 지리 참조 TAG 좌표, 현장 구획, 처리 위치 및 디코더 차트를 제공.
처리: 5개 메인 구획-100%, 100% - 25 lbs., 100%- 50 lbs., 100% - 75 lbs., 0%
14개 서브 구획 종자 처리 (AGT에 의해 공급된 처리된 종자)
질소 타이밍: 식재전 (Pre-plant) 우레아 또는 다른 승인된 최상의 관리 관행.
반복:
6
플롯
위치 당 420
하이브리드:
AgriThority에 의해 공급됨, 하나는 북부 위치 & 하나는 남부 위치.
어떠한 생물학제제도 종자에 처리되지 않음.
구획 크기 최소:
30' 길이의 4개 이랑 (총 ~3 에이커)
관찰:
모든 관찰 내용은 (달리 언급이 없는 한) 구획의 중심 2개 이랑으로부터 가져옴. 모든 파괴적 샘플링은 이랑 외부에서 가져옴.
샘플 보관: 처리된 종자 샘플은 사용시까지 냉장 보관. 샘플 처리 1.5 내지 2시간 전에 냉장고에서 샘플을 제거.
비료 처리
협력자에게 권장 비료 비율을 제공하고 처리 전 Pivot Bio사의 동의를 얻을 것.
현지 농업: 관행
● 종자: (제공된) 시판 옥수수는 생물학제제없이 상업적으로 종자 처리됨. 파종 비율 - 현지 관행.
● 식재일: 현지 관행.
● 현지 표준 생산 관행 - 잡초, 곤충 관리, 등.
● 살진균제 처리를 제외하고 표준 관리 관행을 준수해야 함. 살진균제 처리는 고객과의 상담을 필요로 함.
● 농작물 재배에 관하여 준수한 모든 농업 관행을 기록할 것.
토양 특성화
● 토양 텍스쳐
● 토양 비옥도 분석
o 질소 비료 수준을 측정하기 위해 식재 전 각 현지로부터 토양 샘플을 채취하여, 0-12”및 잠재적으로 12”- 24”질산염 질소가 50 lbs,/ Ac 미만인지를 확인할 것. 표준 토양 시험 그리고 질산염 질소, 암모늄 질소, 총 질소, 유기물 및 CEC를 완료할 것. 곧바로 실험실로 보낼 것,
o 허용가능한 질소 반응성. 0 & 100% 질소 비옥화 간의 높은 델타 수율.
o pH, CEC, 총 K 및 P,
표준 토양 샘플링 절차, 예컨대 토양 중심 0 cm 내지 30cm 및 30 cm - 60 cm
o 식재 및 비료처리 전 Pivot 토양 샘플을 채취할 것- UTC로부터 0 내지 6-12 인치에서 2 ml 토양 샘플. 이랑의 중간부를 사용하여 N 체계 당 각 반복에 대해 하나의 샘플을 채취. (5개 비료 체계* 6회 반복 = 30개 토양 샘플). Pivot Bio.사로 보낼 것
o 식재 (V4-V6) 후 Pivot 토양 샘플을 채취할 것 - UTC로부터 0 내지 6-12 인치에서 2 ml 토양 샘플. 이랑의 중간부를 사용하여 N 체계 당 각 반복에 대해 하나의 샘플을 채취. (5개 비료 체계* 6회 반복 = 30개 토양 샘플).
o 수확 후 Pivot 토양 샘플을 채취할 것- UTCV로부터 0 내지 6-12 인치에서 2 ml 토양 샘플. 이랑의 중간부를 사용하여 N 체계 당 각 반복에 대해 하나의 샘플을 채취. (5개 비료 체계* 6회 반복 = 30개 토양 샘플).
o 0-12”에서 그리고 잠재적으로 12”- 24”에서 각 위치에서 얻은 수확 후 토양 샘플. 표준 토양 시험 그리고 질산염 질소, 암모늄 질소, 총 질소, 유기물 및 CEC를 완료할 것. 곧바로 실험실로 보낼 것.
o 각 비료 체계 (모든 비료 체계에 대해 100% 및 100% + 25 lbs. [100% 블록에서]의 처리를 제외함]로부터 0-12”및 12”- 24”에서 얻은 V6-V10 토양 샘플.
표준 토양 시험 그리고 질산염 질소, 암모늄 질소, 총 질소, 유기물 및 CEC를 완료할 것. 곧바로 실험실로 보낼 것. (5개 체계 곱하기 2개 깊이 = 위치 당 10개 샘플).
o 각 비료 체계 (모든 비료 체계에 대해 100% 및 100% + 25 lbs. [100% 블록에서]의 처리를 제외함)로부터 0-12”- 24”에서 얻은 수확 후 토양 샘플.
표준 토양 시험 그리고 질산염 질소, 암모늄 질소, 총 질소, 유기물 및 CEC를 완료할 것. 곧바로 실험실로 보낼 것. {5개 체계 곱하기 2개 깊이 = 위치 당 10개 샘플}.
평가
● 온도 프로브 (모니터링 진행중)
● UTC의 @~50%에서 초기 식물 집단
● 수확 전 최종 식물 집단
● 활력 (1 내지 10 스케일, w/ 10 = 탁월) V4 & V8 - V10
● 식물 높이 V8-V10 & V14
● 수율 (Bu/에이커), 표준 수분 pct에 대해 보정됨.
● 시험 중량
● 입자 수분 %
● 흑색층에서 줄기 질산염 시험 (420개 구획 X 7개 위치)
● 콜로니화. V4 - V6에서 0% 및 100% 비료에서 1개 집 락 백에 구획당 1개 식물 (1개 식물 * 14개 처리 * 6 반복 * 2개 비료 체계 = 168개 식물)
● 전사체학. V4-V6에서 0% 및 100% 비료에서 1개 튜브에 구획 당 1개 식물 (1개 식물 * 14개 처리 * 6 반복 * 2개 비료 체계 - 168개 식물)
● 콜로니화. V10 - V12에서 0% 및 100% 비료에서 1개 집 락 백에 구획당 1개 식물 (1개 식물 * 14개 처리 * 6 반복 * 2개 비료 체계 = 168개 식물)
● NDVI 또는 NDRE 결정 2개 시점 (V4-V6, & VT)에서 Greenseeker 기기를 사용
o 모두 7개 위치들에서 각 구획을 평가 (420개 구획 X 2개 시점 X 7개 위치)
● 줄기 특성규명 R2 & R5 사이의 모든 7개 위치에서 측정함
o 6인치 높이에서 10개 식물/구획의 줄기 직경 기록
o 6 인치 마크 위 첫번째 인터노드 (internode) 길이를 기록
o 2개 내부 이랑들에서 중심으로부터 5개 연속 식물들, 10개 식물을 모니터할 것.
o 7개 위치 (420개 구획 X 7개 위치)
a. 모니터링 일정
·
AgriThority 전문가는 V3-V4 단계에서 모든 시험을 방문하여 처리에 대한 초기 시즌 반응을 평가하고 번식기 성장 동안 성숙도를 모니터링
·
조건에 따라 그리고 고객이 원하는 경우 Pivot사와 함께 공동 방문.
b. 날씨 정보
식재시부터 수확시까지 날씨 데이터는 다음으로 이루어짐:
● 일일 최고 및 최저 온도,
● 파종시 토양 온도
● 일일 강수 및 관류 (적용가능한 경우)
● 비정상적인 날씨 사건 (예컨대, 과도한 비, 바람, 추위, 열)
c. 데이터 분석 및 보고
● AgriThority는 8월 1일까지 Pivot 중간-시즌 보고서, 11월 30일까지 기능적 초안 및 12월 31일까지 최종 보고서를 제공한다.
● 추가 보고서는 협상할 수 있다.
● 협력자는 시험이 완료되면 AgriThority에 2016년 11월 15일까지 식재 레이아웃 맵을 포함하여 복제 데이터 (원본 데이터)를 Excel 형식 (제공됨)으로 제공한다.
● 보고서에는 아래에 언급된 정보를 포함한 모든 관련 정보가 포함되어야 한다:
● 토양 텍스쳐 & 시험 보고서
● 이랑 간격, 구획 크기, 관류 및 처리된 물의 기록, 경작지, 선행 작물
● 파종 비율 & 식물 개체수
● 시기 당 비료 투입량 - 출처, 비율, 시기, 배치
● 농경법상 투입물 (예컨대, 관류, 사용된 제초제 등)
● 수확 면적 크기
● 수확 방법 - 수작업, 기기, 측정 툴 (스케일, 수율 모니터, 등).
표 17

표 18
표 19

표 20
표 21
표 22

표 23
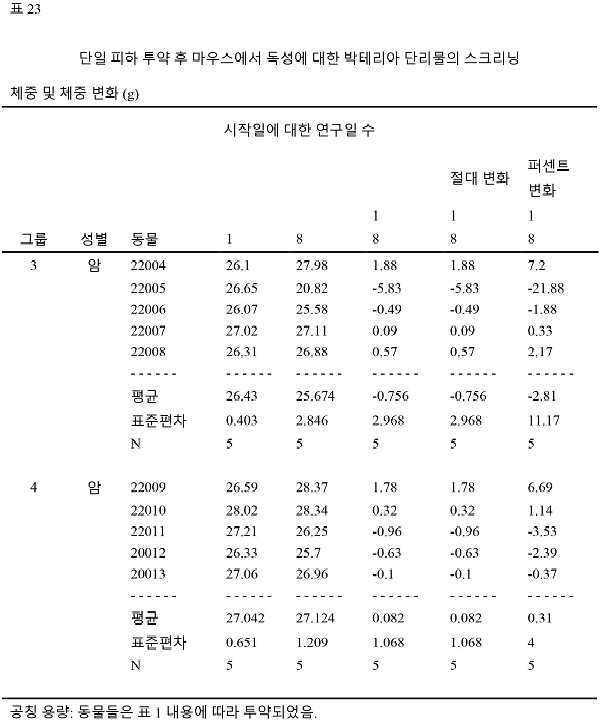
표 24

표 25

표 26
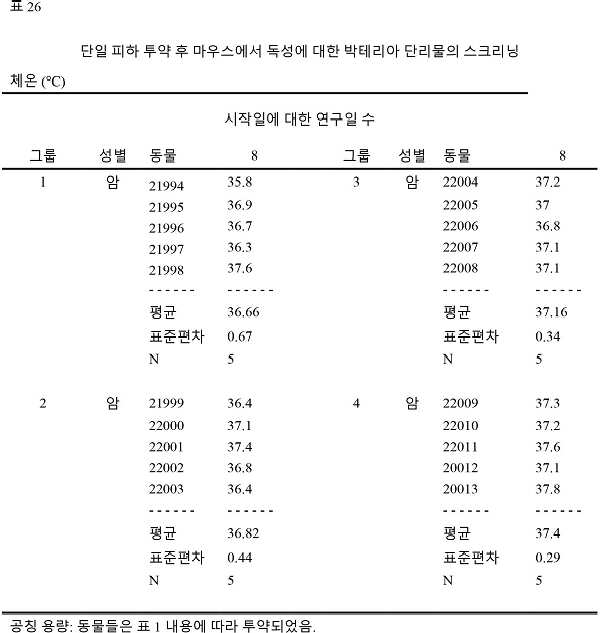
표 27

본 발명의 바람직한 구체예들을 본 출원에 나타내고 설명하였으나, 해당 분야의 숙련된 기술자들에게 이러한 구체예들이 단지 예로서 제공되는 것임은 자명할 것이다. 본 발명에서 벗어나지 않고 수많은 변형들, 변화들, 및 치환들이 해당 분야의 숙련된 기술자들에 의해 이루어질 것이다. 본 출원에 기재된 본 발명의 구체예들에 대한 다양한 대체예들이 본 발명을 실시함에 사용될 수 있음을 이해하여야 한다. 하기 청구범위는 본 발명의 범위를 정의하며 이러한 청구범위에 속하는 방법들 및 구성들 및 이들의 균등예들은 청구범위에 의해 뒷받침되는 것으로 본다.
SEQUENCE LISTING
<110> PIVOT BIO, INC.
<120> METHODS AND COMPOSITIONS FOR IMPROVING ENGINEERED MICROBES
<130> 47736-712.601
<140> PCT/US2018/046148
<141> 2018-08-09
<150> 62/543,288
<151> 2017-08-09
<160> 28
<170> PatentIn version 3.5
<210> 1
<211> 9
<212> PRT
<213> Unknown
<220>
<223> Description of Unknown:
"LAGLIDADG" family peptide motif sequence
<400> 1
Leu Ala Gly Leu Ile Asp Ala Asp Gly
1 5
<210> 2
<211> 90
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 2
gttgatcaga ccgatgttcg gaccttccaa ggtttcgatc ggacatacgc gaccgtagtg 60
ggtcgggtgt acgtctcgaa cttcaaagcc 90
<210> 3
<211> 1536
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 3
ttgaagagtt tgatcatggc tcagattgaa cgctggcggc aggcctaaca catgcaagtc 60
gaacggtagc acagagagct tgctctcggg tgacgagtgg cggacgggtg agtaatgtct 120
gggaaactgc ctgatggagg gggataacta ctggaaacgg tagctaatac cgcataacgt 180
cgcaagacca aagaggggga ccttcgggcc tcttgccatc agatgtgccc agatgggatt 240
agctagtagg tggggtaacg gctcacctag gcgacgatcc ctagctggtc tgagaggatg 300
accagccaca ctggaactga gacacggtcc agactcctac gggaggcagc agtggggaat 360
attgcacaat gggcgcaagc ctgatgcagc catgccgcgt gtgtgaagaa ggccttcggg 420
ttgtaaagca ctttcagcgg ggaggaaggg agtaaggtta ataaccttat tcattgacgt 480
tacccgcaga agaagcaccg gctaactccg tgccagcagc cgcggtaata cggagggtgc 540
aagcgttaat cggaattact gggcgtaaag cgcacgcagg cggtctgtca agtcggatgt 600
gaaatccccg ggctcaacct gggaactgca tccgaaactg gcaggcttga gtctcgtaga 660
gggaggtaga attccaggtg tagcggtgaa atgcgtagag atctggagga ataccggtgg 720
cgaaggcggc ctcctggacg aagactgacg ctcaggtgcg aaagcgtggg gagcaaacag 780
gattagatac cctggtagtc cacgccgtaa acgatgtcta tttggaggtt gtgcccttga 840
ggcgtggctt ccggagctaa cgcgttaaat agaccgcctg gggagtacgg ccgcaaggtt 900
aaaactcaaa tgaattgacg ggggcccgca caagcggtgg agcatgtggt ttaattcgat 960
gcaacgcgaa gaaccttacc tggtcttgac atccacagaa ctttccagag atggattggt 1020
gccttcggga actgtgagac aggtgctgca tggctgtcgt cagctcgtgt tgtgaaatgt 1080
tgggttaagt cccgcaacga gcgcaaccct tatcctttgt tgccagcggt ccggccggga 1140
actcaaagga gactgccagt gataaactgg aggaaggtgg ggatgacgtc aagtcatcat 1200
ggcccttacg accagggcta cacacgtgct acaatggcgc atacaaagag aagcgacctc 1260
gcgagagtaa gcggacctca taaagtgcgt cgtagtccgg attggagtct gcaactcgac 1320
tccatgaagt cggaatcgct agtaatcgtg gatcagaatg ccacggtgaa tacgttcccg 1380
ggccttgtac acaccgcccg tcacaccatg ggagtgggtt gcaaaagaag taggtagctt 1440
aaccttcggg agggcgctta ccactttgtg attcatgact ggggtgaagt cgtaacaagg 1500
taaccgtagg ggaacctgcg gttggatcac ctcctt 1536
<210> 4
<211> 1537
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<220>
<221> modified_base
<222> (450)..(450)
<223> a, c, t, g, unknown or other
<220>
<221> modified_base
<222> (452)..(452)
<223> a, c, t, g, unknown or other
<220>
<221> modified_base
<222> (455)..(455)
<223> a, c, t, g, unknown or other
<220>
<221> modified_base
<222> (473)..(473)
<223> a, c, t, g, unknown or other
<400> 4
ttgaagagtt tgatcatggc tcagattgaa cgctggcggc aggcctaaca catgcaagtc 60
gaacggtagc acagagagct tgctctcggg tgacgagtgg cggacgggtg agtaatgtct 120
gggaaactgc ctgatggagg gggataacta ctggaaacgg tagctaatac cgcataacgt 180
cgcaagacca aagaggggga ccttcgggcc tcttgccatc agatgtgccc agatgggatt 240
agctagtagg tggggtaacg gctcacctag gcgacgatcc ctagctggtc tgagaggatg 300
accagccaca ctggaactga gacacggtcc agactcctac gggaggcagc agtggggaat 360
attgcacaat gggcgcaagc ctgatgcagc catgccgcgt gtgtgaagaa ggccttcggg 420
ttgtaaagca ctttcagcgg ggaggaaggn antanggtta ataacctgtg ttnattgacg 480
ttacccgcag aagaagcacc ggctaactcc gtgccagcag ccgcggtaat acggagggtg 540
caagcgttaa tcggaattac tgggcgtaaa gcgcacgcag gcggtctgtc aagtcggatg 600
tgaaatcccc gggctcaacc tgggaactgc atccgaaact ggcaggcttg agtctcgtag 660
agggaggtag aattccaggt gtagcggtga aatgcgtaga gatctggagg aataccggtg 720
gcgaaggcgg cctcctggac gaagactgac gctcaggtgc gaaagcgtgg ggagcaaaca 780
ggattagata ccctggtagt ccacgccgta aacgatgtct atttggaggt tgtgcccttg 840
aggcgtggct tccggagcta acgcgttaaa tagaccgcct ggggagtacg gccgcaaggt 900
taaaactcaa atgaattgac gggggcccgc acaagcggtg gagcatgtgg tttaattcga 960
tgcaacgcga agaaccttac ctggtcttga catccacaga acttagcaga gatgctttgg 1020
tgccttcggg aactgtgaga caggtgctgc atggctgtcg tcagctcgtg ttgtgaaatg 1080
ttgggttaag tcccgcaacg agcgcaaccc ttatcctttg ttgccagcgg ttaggccggg 1140
aactcaaagg agactgccag tgataaactg gaggaaggtg gggatgacgt caagtcatca 1200
tggcccttac gaccagggct acacacgtgc tacaatggcg catacaaaga gaagcgacct 1260
cgcgagagta agcggacctc ataaagtgcg tcgtagtccg gattggagtc tgcaactcga 1320
ctccatgaag tcggaatcgc tagtaatcgt ggatcagaat gccacggtga atacgttccc 1380
gggccttgta cacaccgccc gtcacaccat gggagtgggt tgcaaaagaa gtaggtagct 1440
taaccttcgg gagggcgctt accactttgt gattcatgac tggggtgaag tcgtaacaag 1500
gtaaccgtag gggaacctgc ggttggatca cctcctt 1537
<210> 5
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
primer
<400> 5
agagtttgat cmtggctcag 20
<210> 6
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
primer
<400> 6
ggttaccttg ttacgactt 19
<210> 7
<211> 449
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 7
tcaccacggc gataaccata ggttttcggc gtggccacat ccatggtgaa tcccactttt 60
tccagcacgc gcgccacttc atcgggtctt aaatacatag attttcctcg tcatctttcc 120
aaagcctcgc caccttacat gactgagcat ggaccgtgac tcagaaaatt ccacaaacga 180
acctgaaagg cgtgattgcc gtctggcctt aaaaattatg gtctaaacta aaatttacat 240
cgaaaacgag ggaggatcct atgcataaca atgaacccaa aacgcagtca gaccctactc 300
tgggctatac attccaaaat gattttctgg cgttaagtaa ggcattttca ttgcctgaaa 360
ttgattacgc cgatatttcc cagcgagaac agttggcggc ggcatttaag cgctggccgc 420
tgctggcaga attagcccgt caacaataa 449
<210> 8
<211> 1425
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 8
cgtcctgtaa taataaccgg acaattcgga ctgattaaaa aagcgccctt gtggcgcttt 60
ttttatattc ccgcctccat ttaaaataaa aaatccaatc ggatttcact atttaaactg 120
gccattatct aagatgaatc cgatggaagc tcgctgtttt aacacgcgtt ttttaacctt 180
ttattgaaag tcggtgcttc tttgagcgaa cgatcaaatt taagtggatt cccatcaaaa 240
aaatattctc aacctaaaaa agtttgtgta atacttgtaa cgctacatgg agattaactc 300
aatctagagg gtattaataa tgaatcgtac taaactggta ctgggcatgc ctttattctt 360
ttctgtgcaa gatgataaca acatgaaaaa aatcatcgga attattttca cgctgatgct 420
gctgttatta tcgctgctgg tagtgattac tcccatgagc agcgataaac aatacctctt 480
tggccttagc gtgatggcag cggtatttat cctgggaaga gtgaaaagca aaaaagccgt 540
actggcaatg ctttccctct ccgtattgat gtcgacacgt tatatttggt ggcgcgccac 600
gacaacgctg cattttgatt cgacggtaga gatgctgctc ggcagtttac tttttgctgc 660
ggaaatttac tcgtggacga ttttactgct gggttatatt caaatggcgt ggccattaga 720
acgcccgatt gcgcctttac cggcagataa atcaacctgg ccgaccgtcg atatttatgt 780
cccctcttat aatgaaagcc ttgaggttgt tcgcgatacg gttctggcgg cgcagtgtat 840
cgaatatccg caagataaag tgaaggttta tattcttgac gacggaaagc gcgatgaatt 900
ccgcgatttc gccgctgaag cgggcgttgg gtatattacc cgcccggata acagccacgc 960
caaagccggt aatctcaacc acgcgatgac catcacccat ggtgagctta tctgcgtgtt 1020
cgactgcgac cacgtggcaa cgcgcgtgtt cctgcaagcc acgattggcg agtttttccg 1080
cgacgataaa ctggcgctga tccagacgcc gcaccacttc tattcgctcg atccatttga 1140
gcgcaacctg acagcagcaa agcgcgttcc ccacgaaggg gcgctgttct acggcccggt 1200
gcagcaaggc aatgacaact ggaacgccac gttcttctgc ggctcctgcg cggtgatccg 1260
tcgtagcgcg ctcaatgaag tcggcggctt tgccgtggaa accgtgaccg aagatgcgca 1320
taccgcgctg aaattacagc gtcgcggctg gaacaccgcg tttctggata ttccgctggc 1380
ggcggggctg gcaaccgagc gtctggcgct gcatgttaac cagcg 1425
<210> 9
<211> 1339
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 9
tcaccacggc gataaccata ggttttcggc gtggccacat ccatggtgaa tcccactttt 60
tccagcacgc gcgccacttc atcgggtctt aaatacatag attttcctcg tcatctttcc 120
aaagcctcgc caccttacat gactgagcat ggaccgtgac tcagaaaatt ccacaaacga 180
acctgaaagg cgtgattgcc gtctggcctt aaaaattatg gtctaaacta aaatttacat 240
cgaaaacgag ggaggatcct atgcctttat tcttttctgt gcaagatgat aacaacatga 300
aaaaaatcat cggaattatt ttcacgctga tgctgctgtt attatcgctg ctggtagtga 360
ttactcccat gagcagcgat aaacaatacc tctttggcct tagcgtgatg gcagcggtat 420
ttatcctggg aagagtgaaa agcaaaaaag ccgtactggc aatgctttcc ctctccgtat 480
tgatgtcgac acgttatatt tggtggcgcg ccacgacaac gctgcatttt gattcgacgg 540
tagagatgct gctcggcagt ttactttttg ctgcggaaat ttactcgtgg acgattttac 600
tgctgggtta tattcaaatg gcgtggccat tagaacgccc gattgcgcct ttaccggcag 660
ataaatcaac ctggccgacc gtcgatattt atgtcccctc ttataatgaa agccttgagg 720
ttgttcgcga tacggttctg gcggcgcagt gtatcgaata tccgcaagat aaagtgaagg 780
tttatattct tgacgacgga aagcgcgatg aattccgcga tttcgccgct gaagcgggcg 840
ttgggtatat tacccgcccg gataacagcc acgccaaagc cggtaatctc aaccacgcga 900
tgaccatcac ccatggtgag cttatctgcg tgttcgactg cgaccacgtg gcaacgcgcg 960
tgttcctgca agccacgatt ggcgagtttt tccgcgacga taaactggcg ctgatccaga 1020
cgccgcacca cttctattcg ctcgatccat ttgagcgcaa cctgacagca gcaaagcgcg 1080
ttccccacga aggggcgctg ttctacggcc cggtgcagca aggcaatgac aactggaacg 1140
ccacgttctt ctgcggctcc tgcgcggtga tccgtcgtag cgcgctcaat gaagtcggcg 1200
gctttgccgt ggaaaccgtg accgaagatg cgcataccgc gctgaaatta cagcgtcgcg 1260
gctggaacac cgcgtttctg gatattccgc tggcggcggg gctggcaacc gagcgtctgg 1320
cgctgcatgt taaccagcg 1339
<210> 10
<211> 1408
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 10
atattgacac catgacgcgc gtaatgctga ttggttctgt gacgctggta atgattgtcg 60
aaattctgaa cagtgccatc gaagccgtag tagaccgtat tggtgcagaa ttccatgaac 120
tttccgggcg ggcgaaggat atggggtcgg cggcggtgct gatgtccatc ctgctggcga 180
tgtttacctg gatcgcatta ctctggtcac attttcgata acgcttccag aattcgataa 240
cgccctggtt ttttgcttaa atttggttcc aaaatcgcct ttagctgtat atactcacag 300
cataactgta tatacaccca gggggcggga tgcctttatt cttttctgtg caagatgata 360
acaacatgaa aaaaatcatc ggaattattt tcacgctgat gctgctgtta ttatcgctgc 420
tggtagtgat tactcccatg agcagcgata aacaatacct ctttggcctt agcgtgatgg 480
cagcggtatt tatcctggga agagtgaaaa gcaaaaaagc cgtactggca atgctttccc 540
tctccgtatt gatgtcgaca cgttatattt ggtggcgcgc cacgacaacg ctgcattttg 600
attcgacggt agagatgctg ctcggcagtt tactttttgc tgcggaaatt tactcgtgga 660
cgattttact gctgggttat attcaaatgg cgtggccatt agaacgcccg attgcgcctt 720
taccggcaga taaatcaacc tggccgaccg tcgatattta tgtcccctct tataatgaaa 780
gccttgaggt tgttcgcgat acggttctgg cggcgcagtg tatcgaatat ccgcaagata 840
aagtgaaggt ttatattctt gacgacggaa agcgcgatga attccgcgat ttcgccgctg 900
aagcgggcgt tgggtatatt acccgcccgg ataacagcca cgccaaagcc ggtaatctca 960
accacgcgat gaccatcacc catggtgagc ttatctgcgt gttcgactgc gaccacgtgg 1020
caacgcgcgt gttcctgcaa gccacgattg gcgagttttt ccgcgacgat aaactggcgc 1080
tgatccagac gccgcaccac ttctattcgc tcgatccatt tgagcgcaac ctgacagcag 1140
caaagcgcgt tccccacgaa ggggcgctgt tctacggccc ggtgcagcaa ggcaatgaca 1200
actggaacgc cacgttcttc tgcggctcct gcgcggtgat ccgtcgtagc gcgctcaatg 1260
aagtcggcgg ctttgccgtg gaaaccgtga ccgaagatgc gcataccgcg ctgaaattac 1320
agcgtcgcgg ctggaacacc gcgtttctgg atattccgct ggcggcgggg ctggcaaccg 1380
agcgtctggc gctgcatgtt aaccagcg 1408
<210> 11
<211> 773
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 11
tcaccacggc gataaccata ggttttcggc gtggccacat ccatggtgaa tcccactttt 60
tccagcacgc gcgccacttc atcgggtctt aaatacatag attttcctcg tcatctttcc 120
aaagcctcgc caccttacat gactgagcat ggaccgtgac tcagaaaatt ccacaaacga 180
acctgaaagg cgtgattgcc gtctggcctt aaaaattatg gtctaaacta aaatttacat 240
cgaaaacgag ggaggatcct atgggtcatt acgatgattt gcagaggttt aaggacaaga 300
ctcgcaatca gaagcacgat tttaaggatc tctccgcaca gaacctcgca caagaccaga 360
gcaattgggc aatcattaac cagctttctc ctgccacgga tgaatcaatg ctggcaatgg 420
gcggccatgt gtcactgcct gtcccgcaat ccgtcgatcc tgcgcttttt gccgcgcaag 480
agccggctgc gttggtggat gatattgcac ctgcgcctgc gtcaacaact tctctgctaa 540
aagacgtcgc cagccagctt gatgcagtgg ctccggcatt caacgccgca gcctacgcgc 600
ctgcgcctgc aagcgttgca ccggttatgc agatgccatc gcctgcaaat ccttcttcgg 660
ctcctgtcag ctatgcgcgc ctgttcgcag cgaaagcgac ggaggagaag ccccgcgcgg 720
aaaaaaatca gcccttacac tcgttgttag aaaggatcgc ttcatgccgt tga 773
<210> 12
<211> 1108
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 12
cgtcctgtaa taataaccgg acaattcgga ctgattaaaa aagcgccctt gtggcgcttt 60
ttttatattc ccgcctccat ttaaaataaa aaatccaatc ggatttcact atttaaactg 120
gccattatct aagatgaatc cgatggaagc tcgctgtttt aacacgcgtt ttttaacctt 180
ttattgaaag tcggtgcttc tttgagcgaa cgatcaaatt taagtggatt cccatcaaaa 240
aaatattctc aacctaaaaa agtttgtgta atacttgtaa cgctacatgg agattaactc 300
aatctagagg gtattaataa tgaatcgtac taaactggta ctgggcatgg tttcatcatc 360
cccgtctgtt gctcgcagcg gcatgtttta cttttcccag ggctggaaac tggtcacact 420
gccaggcatt cgtcgctttg tgatcttgcc gttaatggtg aatattttgc tgatgggcgg 480
cgcgttctgg tggctgttta cccgcctgga tagttggatc ccttcgctga tgagccatgt 540
gccggactgg ctgcaatggc tgagctattt actgtggcca attgccgttg tctcaattct 600
gttggtcttc ggctacttct tctcaaccct tgcgaactgg attgccgctc cctttaatgg 660
cttgctggca gaacagctgg aagcacgcct taccggcgcg acaccacccg acgtcggcgt 720
ttttgggctg ctcaaagatg taccgcgcat catgaagcgc gaatggcaaa aactggcctg 780
gtatctgccc cgcgccttgc tgctgctggt gctttatttt attcccggta ttgggcaaac 840
cgtcgccccg gtgctgtggt tcctgttcag tgcgtggatg ttggctatcc agtattgcga 900
ctatccgttt gataaccata aggtgccgtt taaagcgatg cgcgtttcct tgcgcgaacg 960
caaggttatt aatatgcaat tcggtgcgct caccagcctg tttacgatga ttccggtgtt 1020
gaatttggtc gtgatgcccg tagcggtgtg tggcgcaacg gcgatgtggg tcgattgcta 1080
ccgggaaaaa cacgcggtct ggcgataa 1108
<210> 13
<211> 4222
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 13
cgtcctgtaa taataaccgg acaattcgga ctgattaaaa aagcgccctt gtggcgcttt 60
ttttatattc ccgcctccat ttaaaataaa aaatccaatc ggatttcact atttaaactg 120
gccattatct aagatgaatc cgatggaagc tcgctgtttt aacacgcgtt ttttaacctt 180
ttattgaaag tcggtgcttc tttgagcgaa cgatcaaatt taagtggatt cccatcaaaa 240
aaatattctc aacctaaaaa agtttgtgta atacttgtaa cgctacatgg agattaactc 300
aatctagagg gtattaataa tgaatcgtac taaactggta ctgggcgtga ttaattcgga 360
aaagaaagtc caacgccttg taagaactca gctggccccg ttgacggtaa caatgatatt 420
cgcattacct gctgttgcgc attccgctgg tttaagtatg aaaaatggca cgctatataa 480
tgcgaatggc gtgccggtcg tggatattaa taaacctaac agcaatggct tgtcacataa 540
cgtctgggac aacctgaacg tcgataaaaa cggtgttatt tttaataaca gcggctcagc 600
atccgatacg gtattgggcg gcaccatcca gggtaacagc aatctgacat ccgggtcggc 660
aaaagttatt ctcaacgaag ttacctccaa aaatgcgtcg acgatcaacg gtatgatgga 720
agttgcgggt gataaagccg cgttgattat cgccaacccc aatggcattg cggtaaatgg 780
tggcggtacg atcaataccg ataaactcac tctgaccact ggcacaccag atattcagaa 840
cgacaaactc gctggttatt ccgtcaacgg cggtaccatc acgatcagca aactggataa 900
cgccagccca actgaaattt tgtcacgcaa cgtagtgctc agcgggaaag ttaccgctga 960
cgaactgaat attgttgcgg gtaataacta cgttaataca gacggccagg tgaccggaac 1020
tgtcactgcg tcaggctcgc gtaacggcta tagcgttgac gttgcgcaag ttggcggcat 1080
gtacgcgaat aaaattaatc ttatcagtac cgaaagcggc gtgggcgtgc gtaatcaggg 1140
cgtaattgcc ggtggcactg gtgggatcag cattgatgct aatggccagc tactgaacag 1200
taatgcgcag attcaatcga ccggttcgac aaagattaat accaacggca aattggacaa 1260
tacaacgggt cttattactt ctgctggtgc tatttatctc gatactaata agaacgctat 1320
tgttaatgcc cgttcaggta cgatttcgac gacggcagat atctacgtta atagcggcgc 1380
agttgataat accaacggca aaattgccgc cagcggcgtg ctggctgtcg ataccaataa 1440
caatactttt accaacgccg gtaaagggaa cgatgtcggt atcgaagcgg gtattgtcac 1500
cttaaaaacc ggtacgctaa ataatagtaa cggccagatc cacggcggct atctggggct 1560
tgaatccggc gcgattaaca ataacagcgg tattctggaa acgtcaggcg acatggacat 1620
tgtcagcctt ggaaatgtcg acaataacaa aggcctgatc cgatctgcca gcggccatat 1680
caacatcctg tctggcggca ctatcaacaa cggcagtacc aaaacagcgg ataccgtcag 1740
ttccgactcc ttaggcatcg cagccgatac cggtgtagta atcaacgcaa acagcatcaa 1800
taatagcggt ggtcagatcg catcgaacgg cgatgtgtcg ctaaacagta agggcctggt 1860
caataacgat tccggtaatc ttctttccaa tagcaaagtg attgtgaagg gcggttctta 1920
tcgtaatgac tacggtggca tcggcggcaa gaagggcgtg gaggtgtcgg ttaacggcga 1980
cctgagcaac tatatcggcg ttgtgagtgc ggaagaaggt gatatctcac tgagtgcaaa 2040
tgcagtggat acccacggcg gtttcatgat ggggcaaaat atctccatcg acgcgaaagc 2100
cggcgtaaat accacccgat cgctggtatt agccagtgaa aagctggcta tccatgcagg 2160
cggaaatgtt gataaccgtg atggtaatag ctttggcaat gacttcggcc tctatttcgg 2220
catgccgcaa cagaccggtg gcttgatcgg taaaggtggc gttgagattt ccgggcagaa 2280
tatttacaac aacaacagcc gcatcattgc agaaaatggg ccaatgagcc tgcaagcgaa 2340
agggaacatt gataacacgc gtgcgctgat ggtcagcggg gccgataccg caatcaaaac 2400
cggcggggct ttctacaata actacgcaac gatttccagc ggaggcaacc tcacactcga 2460
taccgtttcc ctggaaaact ccagcagcgg cacgcttatc gataataacg ccacaggttt 2520
catcaactca gataaagata tcacgctgaa tgtggacaac aatttcacga actacggctg 2580
gatcagtggt aaaggcgcag tggcgctgaa tgtcgtgaaa ggggcgtttt ataaccgtaa 2640
taccattgcg gctgataagt cacttgcgat cagtgccctg aacggtatcg agaactttaa 2700
agatatctct gcaggcggca gcctgacgct ggatacgcaa cgtcatgtga caaataacag 2760
caacagcaat atggtcggta acgggattac tattaacgcg gtaaacgaca ttaataaccg 2820
cggcaatatt gtcagtgatg gcgatatgag tgttgtgacc aaaggtaacc tgtataacta 2880
cctctatatg ctgggcgctg gtgatgtcgc gattaccgca aacaacgtca acaacaataa 2940
ttcacttatt gaggctggtg gtgatctgac gattacctct actggcaatg tgggtaataa 3000
ccgtggcacg ctgcgctccc tgacgggcga tgtaaatgtg gcggcaaaag gaacggttga 3060
taactatagc ggtaagatta cctctgctgg cgatgtcgga gtcagtggaa actggttgct 3120
caatgattat ggtcagattg ctggcttaag caatattaac ctgattctga cgggcaactt 3180
cgacagttac aaaggttctg tgacctctca aatgggcgat atcaagctgg ttgccaactc 3240
ggtggataat aacaacgggt taatcgcagg tgaaaatatc tccattgatt ccaaatcagg 3300
tgtctataac aacacggcgc tgattgccgc gaataaaaca cttaccgtaa atgctgtcgg 3360
caatattgaa aacagagacg gtaataactt cagttattac aacggcgagt attttggtgt 3420
cgcaggcgat gtgggtggca tggtgggtaa cgcgggcatc tcgctttccg cacagaatat 3480
ctacagcaca aacagccgta ttctggcaga aaatggccca cttaacgtgc tgtcccgggg 3540
cgtattcgat aacacccgca gcatgctgat gagcggtgcc aacgccatta ttaaagcggc 3600
cggtgttttc tataataact atgcgatgac atatagcgca ggtaacctcg acatcacttc 3660
tgcttacctt gagaactaca gcaacggtaa gctggaagat ggaacggcaa caggcgttat 3720
cgcttcggat aagaacctga acctgagcgt ggataatagc accaccaact acggctggat 3780
cagcggtaag ggcgatgtga atttcagcgt gttaaaaggt gtgctgtata accgcaacac 3840
catcgccgct aataacgcgc tggcgattaa tgccctgaac ggtatcgaga acttcaaaga 3900
tatcctggcg ggaaccagcc ttacccttac tacccaacgg cacgttacca acaacagcaa 3960
cagtaatatg cttgggcaaa gcattgggat taatgcggta aacgatatta ataaccgcgg 4020
caatattgtc agcgattatg cattaaccgt gaaaacagac gggaacgtgt ataactacct 4080
caatatgctg agttatggca ccgcaaaagt aaccgcgaat aaagtcacga acagcggcaa 4140
ggacgcggtg ctgggcggtt tttacggcct tgaccttgaa tcgaatgcga tcaccaatac 4200
cggcaccatt gtaggtctgt aa 4222
<210> 14
<211> 4136
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 14
tcaccacggc gataaccata ggttttcggc gtggccacat ccatggtgaa tcccactttt 60
tccagcacgc gcgccacttc atcgggtctt aaatacatag attttcctcg tcatctttcc 120
aaagcctcgc caccttacat gactgagcat ggaccgtgac tcagaaaatt ccacaaacga 180
acctgaaagg cgtgattgcc gtctggcctt aaaaattatg gtctaaacta aaatttacat 240
cgaaaacgag ggaggatcct gtgattaatt cggaaaagaa agtccaacgc cttgtaagaa 300
ctcagctggc cccgttgacg gtaacaatga tattcgcatt acctgctgtt gcgcattccg 360
ctggtttaag tatgaaaaat ggcacgctat ataatgcgaa tggcgtgccg gtcgtggata 420
ttaataaacc taacagcaat ggcttgtcac ataacgtctg ggacaacctg aacgtcgata 480
aaaacggtgt tatttttaat aacagcggct cagcatccga tacggtattg ggcggcacca 540
tccagggtaa cagcaatctg acatccgggt cggcaaaagt tattctcaac gaagttacct 600
ccaaaaatgc gtcgacgatc aacggtatga tggaagttgc gggtgataaa gccgcgttga 660
ttatcgccaa ccccaatggc attgcggtaa atggtggcgg tacgatcaat accgataaac 720
tcactctgac cactggcaca ccagatattc agaacgacaa actcgctggt tattccgtca 780
acggcggtac catcacgatc agcaaactgg ataacgccag cccaactgaa attttgtcac 840
gcaacgtagt gctcagcggg aaagttaccg ctgacgaact gaatattgtt gcgggtaata 900
actacgttaa tacagacggc caggtgaccg gaactgtcac tgcgtcaggc tcgcgtaacg 960
gctatagcgt tgacgttgcg caagttggcg gcatgtacgc gaataaaatt aatcttatca 1020
gtaccgaaag cggcgtgggc gtgcgtaatc agggcgtaat tgccggtggc actggtggga 1080
tcagcattga tgctaatggc cagctactga acagtaatgc gcagattcaa tcgaccggtt 1140
cgacaaagat taataccaac ggcaaattgg acaatacaac gggtcttatt acttctgctg 1200
gtgctattta tctcgatact aataagaacg ctattgttaa tgcccgttca ggtacgattt 1260
cgacgacggc agatatctac gttaatagcg gcgcagttga taataccaac ggcaaaattg 1320
ccgccagcgg cgtgctggct gtcgatacca ataacaatac ttttaccaac gccggtaaag 1380
ggaacgatgt cggtatcgaa gcgggtattg tcaccttaaa aaccggtacg ctaaataata 1440
gtaacggcca gatccacggc ggctatctgg ggcttgaatc cggcgcgatt aacaataaca 1500
gcggtattct ggaaacgtca ggcgacatgg acattgtcag ccttggaaat gtcgacaata 1560
acaaaggcct gatccgatct gccagcggcc atatcaacat cctgtctggc ggcactatca 1620
acaacggcag taccaaaaca gcggataccg tcagttccga ctccttaggc atcgcagccg 1680
ataccggtgt agtaatcaac gcaaacagca tcaataatag cggtggtcag atcgcatcga 1740
acggcgatgt gtcgctaaac agtaagggcc tggtcaataa cgattccggt aatcttcttt 1800
ccaatagcaa agtgattgtg aagggcggtt cttatcgtaa tgactacggt ggcatcggcg 1860
gcaagaaggg cgtggaggtg tcggttaacg gcgacctgag caactatatc ggcgttgtga 1920
gtgcggaaga aggtgatatc tcactgagtg caaatgcagt ggatacccac ggcggtttca 1980
tgatggggca aaatatctcc atcgacgcga aagccggcgt aaataccacc cgatcgctgg 2040
tattagccag tgaaaagctg gctatccatg caggcggaaa tgttgataac cgtgatggta 2100
atagctttgg caatgacttc ggcctctatt tcggcatgcc gcaacagacc ggtggcttga 2160
tcggtaaagg tggcgttgag atttccgggc agaatattta caacaacaac agccgcatca 2220
ttgcagaaaa tgggccaatg agcctgcaag cgaaagggaa cattgataac acgcgtgcgc 2280
tgatggtcag cggggccgat accgcaatca aaaccggcgg ggctttctac aataactacg 2340
caacgatttc cagcggaggc aacctcacac tcgataccgt ttccctggaa aactccagca 2400
gcggcacgct tatcgataat aacgccacag gtttcatcaa ctcagataaa gatatcacgc 2460
tgaatgtgga caacaatttc acgaactacg gctggatcag tggtaaaggc gcagtggcgc 2520
tgaatgtcgt gaaaggggcg ttttataacc gtaataccat tgcggctgat aagtcacttg 2580
cgatcagtgc cctgaacggt atcgagaact ttaaagatat ctctgcaggc ggcagcctga 2640
cgctggatac gcaacgtcat gtgacaaata acagcaacag caatatggtc ggtaacggga 2700
ttactattaa cgcggtaaac gacattaata accgcggcaa tattgtcagt gatggcgata 2760
tgagtgttgt gaccaaaggt aacctgtata actacctcta tatgctgggc gctggtgatg 2820
tcgcgattac cgcaaacaac gtcaacaaca ataattcact tattgaggct ggtggtgatc 2880
tgacgattac ctctactggc aatgtgggta ataaccgtgg cacgctgcgc tccctgacgg 2940
gcgatgtaaa tgtggcggca aaaggaacgg ttgataacta tagcggtaag attacctctg 3000
ctggcgatgt cggagtcagt ggaaactggt tgctcaatga ttatggtcag attgctggct 3060
taagcaatat taacctgatt ctgacgggca acttcgacag ttacaaaggt tctgtgacct 3120
ctcaaatggg cgatatcaag ctggttgcca actcggtgga taataacaac gggttaatcg 3180
caggtgaaaa tatctccatt gattccaaat caggtgtcta taacaacacg gcgctgattg 3240
ccgcgaataa aacacttacc gtaaatgctg tcggcaatat tgaaaacaga gacggtaata 3300
acttcagtta ttacaacggc gagtattttg gtgtcgcagg cgatgtgggt ggcatggtgg 3360
gtaacgcggg catctcgctt tccgcacaga atatctacag cacaaacagc cgtattctgg 3420
cagaaaatgg cccacttaac gtgctgtccc ggggcgtatt cgataacacc cgcagcatgc 3480
tgatgagcgg tgccaacgcc attattaaag cggccggtgt tttctataat aactatgcga 3540
tgacatatag cgcaggtaac ctcgacatca cttctgctta ccttgagaac tacagcaacg 3600
gtaagctgga agatggaacg gcaacaggcg ttatcgcttc ggataagaac ctgaacctga 3660
gcgtggataa tagcaccacc aactacggct ggatcagcgg taagggcgat gtgaatttca 3720
gcgtgttaaa aggtgtgctg tataaccgca acaccatcgc cgctaataac gcgctggcga 3780
ttaatgccct gaacggtatc gagaacttca aagatatcct ggcgggaacc agccttaccc 3840
ttactaccca acggcacgtt accaacaaca gcaacagtaa tatgcttggg caaagcattg 3900
ggattaatgc ggtaaacgat attaataacc gcggcaatat tgtcagcgat tatgcattaa 3960
ccgtgaaaac agacgggaac gtgtataact acctcaatat gctgagttat ggcaccgcaa 4020
aagtaaccgc gaataaagtc acgaacagcg gcaaggacgc ggtgctgggc ggtttttacg 4080
gccttgacct tgaatcgaat gcgatcacca ataccggcac cattgtaggt ctgtaa 4136
<210> 15
<211> 4205
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 15
atattgacac catgacgcgc gtaatgctga ttggttctgt gacgctggta atgattgtcg 60
aaattctgaa cagtgccatc gaagccgtag tagaccgtat tggtgcagaa ttccatgaac 120
tttccgggcg ggcgaaggat atggggtcgg cggcggtgct gatgtccatc ctgctggcga 180
tgtttacctg gatcgcatta ctctggtcac attttcgata acgcttccag aattcgataa 240
cgccctggtt ttttgcttaa atttggttcc aaaatcgcct ttagctgtat atactcacag 300
cataactgta tatacaccca gggggcgggg tgattaattc ggaaaagaaa gtccaacgcc 360
ttgtaagaac tcagctggcc ccgttgacgg taacaatgat attcgcatta cctgctgttg 420
cgcattccgc tggtttaagt atgaaaaatg gcacgctata taatgcgaat ggcgtgccgg 480
tcgtggatat taataaacct aacagcaatg gcttgtcaca taacgtctgg gacaacctga 540
acgtcgataa aaacggtgtt atttttaata acagcggctc agcatccgat acggtattgg 600
gcggcaccat ccagggtaac agcaatctga catccgggtc ggcaaaagtt attctcaacg 660
aagttacctc caaaaatgcg tcgacgatca acggtatgat ggaagttgcg ggtgataaag 720
ccgcgttgat tatcgccaac cccaatggca ttgcggtaaa tggtggcggt acgatcaata 780
ccgataaact cactctgacc actggcacac cagatattca gaacgacaaa ctcgctggtt 840
attccgtcaa cggcggtacc atcacgatca gcaaactgga taacgccagc ccaactgaaa 900
ttttgtcacg caacgtagtg ctcagcggga aagttaccgc tgacgaactg aatattgttg 960
cgggtaataa ctacgttaat acagacggcc aggtgaccgg aactgtcact gcgtcaggct 1020
cgcgtaacgg ctatagcgtt gacgttgcgc aagttggcgg catgtacgcg aataaaatta 1080
atcttatcag taccgaaagc ggcgtgggcg tgcgtaatca gggcgtaatt gccggtggca 1140
ctggtgggat cagcattgat gctaatggcc agctactgaa cagtaatgcg cagattcaat 1200
cgaccggttc gacaaagatt aataccaacg gcaaattgga caatacaacg ggtcttatta 1260
cttctgctgg tgctatttat ctcgatacta ataagaacgc tattgttaat gcccgttcag 1320
gtacgatttc gacgacggca gatatctacg ttaatagcgg cgcagttgat aataccaacg 1380
gcaaaattgc cgccagcggc gtgctggctg tcgataccaa taacaatact tttaccaacg 1440
ccggtaaagg gaacgatgtc ggtatcgaag cgggtattgt caccttaaaa accggtacgc 1500
taaataatag taacggccag atccacggcg gctatctggg gcttgaatcc ggcgcgatta 1560
acaataacag cggtattctg gaaacgtcag gcgacatgga cattgtcagc cttggaaatg 1620
tcgacaataa caaaggcctg atccgatctg ccagcggcca tatcaacatc ctgtctggcg 1680
gcactatcaa caacggcagt accaaaacag cggataccgt cagttccgac tccttaggca 1740
tcgcagccga taccggtgta gtaatcaacg caaacagcat caataatagc ggtggtcaga 1800
tcgcatcgaa cggcgatgtg tcgctaaaca gtaagggcct ggtcaataac gattccggta 1860
atcttctttc caatagcaaa gtgattgtga agggcggttc ttatcgtaat gactacggtg 1920
gcatcggcgg caagaagggc gtggaggtgt cggttaacgg cgacctgagc aactatatcg 1980
gcgttgtgag tgcggaagaa ggtgatatct cactgagtgc aaatgcagtg gatacccacg 2040
gcggtttcat gatggggcaa aatatctcca tcgacgcgaa agccggcgta aataccaccc 2100
gatcgctggt attagccagt gaaaagctgg ctatccatgc aggcggaaat gttgataacc 2160
gtgatggtaa tagctttggc aatgacttcg gcctctattt cggcatgccg caacagaccg 2220
gtggcttgat cggtaaaggt ggcgttgaga tttccgggca gaatatttac aacaacaaca 2280
gccgcatcat tgcagaaaat gggccaatga gcctgcaagc gaaagggaac attgataaca 2340
cgcgtgcgct gatggtcagc ggggccgata ccgcaatcaa aaccggcggg gctttctaca 2400
ataactacgc aacgatttcc agcggaggca acctcacact cgataccgtt tccctggaaa 2460
actccagcag cggcacgctt atcgataata acgccacagg tttcatcaac tcagataaag 2520
atatcacgct gaatgtggac aacaatttca cgaactacgg ctggatcagt ggtaaaggcg 2580
cagtggcgct gaatgtcgtg aaaggggcgt tttataaccg taataccatt gcggctgata 2640
agtcacttgc gatcagtgcc ctgaacggta tcgagaactt taaagatatc tctgcaggcg 2700
gcagcctgac gctggatacg caacgtcatg tgacaaataa cagcaacagc aatatggtcg 2760
gtaacgggat tactattaac gcggtaaacg acattaataa ccgcggcaat attgtcagtg 2820
atggcgatat gagtgttgtg accaaaggta acctgtataa ctacctctat atgctgggcg 2880
ctggtgatgt cgcgattacc gcaaacaacg tcaacaacaa taattcactt attgaggctg 2940
gtggtgatct gacgattacc tctactggca atgtgggtaa taaccgtggc acgctgcgct 3000
ccctgacggg cgatgtaaat gtggcggcaa aaggaacggt tgataactat agcggtaaga 3060
ttacctctgc tggcgatgtc ggagtcagtg gaaactggtt gctcaatgat tatggtcaga 3120
ttgctggctt aagcaatatt aacctgattc tgacgggcaa cttcgacagt tacaaaggtt 3180
ctgtgacctc tcaaatgggc gatatcaagc tggttgccaa ctcggtggat aataacaacg 3240
ggttaatcgc aggtgaaaat atctccattg attccaaatc aggtgtctat aacaacacgg 3300
cgctgattgc cgcgaataaa acacttaccg taaatgctgt cggcaatatt gaaaacagag 3360
acggtaataa cttcagttat tacaacggcg agtattttgg tgtcgcaggc gatgtgggtg 3420
gcatggtggg taacgcgggc atctcgcttt ccgcacagaa tatctacagc acaaacagcc 3480
gtattctggc agaaaatggc ccacttaacg tgctgtcccg gggcgtattc gataacaccc 3540
gcagcatgct gatgagcggt gccaacgcca ttattaaagc ggccggtgtt ttctataata 3600
actatgcgat gacatatagc gcaggtaacc tcgacatcac ttctgcttac cttgagaact 3660
acagcaacgg taagctggaa gatggaacgg caacaggcgt tatcgcttcg gataagaacc 3720
tgaacctgag cgtggataat agcaccacca actacggctg gatcagcggt aagggcgatg 3780
tgaatttcag cgtgttaaaa ggtgtgctgt ataaccgcaa caccatcgcc gctaataacg 3840
cgctggcgat taatgccctg aacggtatcg agaacttcaa agatatcctg gcgggaacca 3900
gccttaccct tactacccaa cggcacgtta ccaacaacag caacagtaat atgcttgggc 3960
aaagcattgg gattaatgcg gtaaacgata ttaataaccg cggcaatatt gtcagcgatt 4020
atgcattaac cgtgaaaaca gacgggaacg tgtataacta cctcaatatg ctgagttatg 4080
gcaccgcaaa agtaaccgcg aataaagtca cgaacagcgg caaggacgcg gtgctgggcg 4140
gtttttacgg ccttgacctt gaatcgaatg cgatcaccaa taccggcacc attgtaggtc 4200
tgtaa 4205
<210> 16
<211> 860
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 16
gaatttactt acattaaggc ggcgaggggc gcctatactt gatagttctg ataccagaag 60
aaggaagaac tatggataac cagatatctg taccgcctgc gctaaccgga aactacgctt 120
ttttctttga cctcgacggt actcttgccg atatccaacc gcaccccgat caggtggtga 180
tacccgacaa cacgttgcag gcgctcaacg cgctggccca gcagcagggt ggggcggtgg 240
cattgatttc agggcgctca atggctgaac ttgacgcgct aacccatccc tggcggctac 300
cgctggccgg cgttcacggg gcagaacgcc gtgatatcaa tggtaaaacc tatatcgttt 360
ctctgccgtc ggcgctgcgc gatgaaatcg cgactgagct gacctcggcg ctgcaggggc 420
tctccggctg cgagctggag agtaaggaga tggccttcgc gctgcactac cgccaggctc 480
cgcagcagca gagcgcagta ctggagctgg ctcagcgcat cgttcagcgc tacccgctgc 540
tggcactgca gctgggtaaa tgtgtggtgg agattaaacc ccgcggagtg aacaaagggg 600
aggcgatcag cgccttcatg caggaggcgc cgtttgccgg ccgcgaaccg gtttttgtcg 660
gcgatgattt gaccgatgag gcgggattta gcgtggttaa tcaactgcag ggaatgtcgg 720
taaaagtcgg cgccggggaa actcaggcgc actggcggtt ggcggatgcc gccgctgtaa 780
ggacgtggtt gcaacatctg gcgtatgacg cgcaaaccga aaggagggat gaccatgagt 840
cgtttagtcg tagtctctaa 860
<210> 17
<211> 1088
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 17
cgccgtcctc gcagtaccat tgcaaccgac tttacagcaa gaagtgattc tggcacgcat 60
ggaacaaatt cttgccagtc gggctttatc cgatgacgaa cgcgcacagc ttttatatga 120
gcgcggagtg ttgtatgata gtctcggtct gagggcatta gcgcgaaatg atttttcaca 180
agcgctggca atccgacccg atatgcctga agtattcaat tacttaggca tttacttaac 240
gcaggcaggc aattttgatg ctgcctatga agcgtttgat tctgtacttg agcttgatca 300
tggataacca gatatctgta ccgcctgcgc taaccggaaa ctacgctttt ttctttgacc 360
tcgacggtac tcttgccgat atccaaccgc accccgatca ggtggtgata cccgacaaca 420
cgttgcaggc gctcaacgcg ctggcccagc agcagggtgg ggcggtggca ttgatttcag 480
ggcgctcaat ggctgaactt gacgcgctaa cccatccctg gcggctaccg ctggccggcg 540
ttcacggggc agaacgccgt gatatcaatg gtaaaaccta tatcgtttct ctgccgtcgg 600
cgctgcgcga tgaaatcgcg actgagctga cctcggcgct gcaggggctc tccggctgcg 660
agctggagag taaggagatg gccttcgcgc tgcactaccg ccaggctccg cagcagcaga 720
gcgcagtact ggagctggct cagcgcatcg ttcagcgcta cccgctgctg gcactgcagc 780
tgggtaaatg tgtggtggag attaaacccc gcggagtgaa caaaggggag gcgatcagcg 840
ccttcatgca ggaggcgccg tttgccggcc gcgaaccggt ttttgtcggc gatgatttga 900
ccgatgaggc gggatttagc gtggttaatc aactgcaggg aatgtcggta aaagtcggcg 960
ccggggaaac tcaggcgcac tggcggttgg cggatgccgc cgctgtaagg acgtggttgc 1020
aacatctggc gtatgacgcg caaaccgaaa ggagggatga ccatgagtcg tttagtcgta 1080
gtctctaa 1088
<210> 18
<211> 1153
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 18
cgtcctgtaa taataaccgg acaattcgga ctgattaaaa aagcgccctt gtggcgcttt 60
ttttatattc ccgcctccat ttaaaataaa aaatccaatc ggatttcact atttaaactg 120
gccattatct aagatgaatc cgatggaagc tcgctgtttt aacacgcgtt ttttaacctt 180
ttattgaaag tcggtgcttc tttgagcgaa cgatcaaatt taagtggatt cccatcaaaa 240
aaatattctc aacctaaaaa agtttgtgta atacttgtaa cgctacatgg agattaactc 300
aatctagagg gtattaataa tgaatcgtac taaactggta ctgggcgtgg aagacgcact 360
atctataccg cctctcattt ccggaaattt tgcctttttt ttcgatttgg atggcaccct 420
tgcggacatc aaaccgcacc cggatgatgt ttttatccct gaagatgtgc tgaccagact 480
ttcattgctt gctgagctaa atgacagggc gttggcattg atttcagggc gttcaattgc 540
tgagctggaa cagctcgcca aaccgtatcg tttcccgctg gccggggtac atggggcgga 600
gcgccgcgat atcaatggtc gaaccgaacg aatgaccttg ccggaagcgg ttttacaacc 660
gctggaacgt gaactccggc atggcattgc gccgctgacc ggcgttgaac ttgaaaccaa 720
agggatggca tttgcactgc attatcgcca ggcgccgcag catgaagagg cggtctttgc 780
cctggcaaaa acactcatcg cgcactaccc acaactggca atgcagccgg gaaaatgtgt 840
ggtcgagctg aaaccctctg gcattcataa aggcgccgct attggcgcgt ttatgcaaac 900
gccgcccttt ctcggcaaga aaccggtgtt tcttggcgac gatctgactg acgaacatgg 960
atttaaaacc gttaataaac ttggcggcat atcgataaaa gttggccctg gcgcaacaca 1020
ggcaatgtgg cgtctggctg gcgtaaatga tgtctatcaa tggcttgaaa aacttgtcga 1080
ccatcaacaa caagaacaac gggcgctaac aaacaggaga gatggctatg agtcgcttag 1140
tcgtagtatc taa 1153
<210> 19
<211> 1067
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 19
tcaccacggc gataaccata ggttttcggc gtggccacat ccatggtgaa tcccactttt 60
tccagcacgc gcgccacttc atcgggtctt aaatacatag attttcctcg tcatctttcc 120
aaagcctcgc caccttacat gactgagcat ggaccgtgac tcagaaaatt ccacaaacga 180
acctgaaagg cgtgattgcc gtctggcctt aaaaattatg gtctaaacta aaatttacat 240
cgaaaacgag ggaggatcct gtggaagacg cactatctat accgcctctc atttccggaa 300
attttgcctt ttttttcgat ttggatggca cccttgcgga catcaaaccg cacccggatg 360
atgtttttat ccctgaagat gtgctgacca gactttcatt gcttgctgag ctaaatgaca 420
gggcgttggc attgatttca gggcgttcaa ttgctgagct ggaacagctc gccaaaccgt 480
atcgtttccc gctggccggg gtacatgggg cggagcgccg cgatatcaat ggtcgaaccg 540
aacgaatgac cttgccggaa gcggttttac aaccgctgga acgtgaactc cggcatggca 600
ttgcgccgct gaccggcgtt gaacttgaaa ccaaagggat ggcatttgca ctgcattatc 660
gccaggcgcc gcagcatgaa gaggcggtct ttgccctggc aaaaacactc atcgcgcact 720
acccacaact ggcaatgcag ccgggaaaat gtgtggtcga gctgaaaccc tctggcattc 780
ataaaggcgc cgctattggc gcgtttatgc aaacgccgcc ctttctcggc aagaaaccgg 840
tgtttcttgg cgacgatctg actgacgaac atggatttaa aaccgttaat aaacttggcg 900
gcatatcgat aaaagttggc cctggcgcaa cacaggcaat gtggcgtctg gctggcgtaa 960
atgatgtcta tcaatggctt gaaaaacttg tcgaccatca acaacaagaa caacgggcgc 1020
taacaaacag gagagatggc tatgagtcgc ttagtcgtag tatctaa 1067
<210> 20
<211> 730
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 20
cgtcctgtaa taataaccgg acaattcgga ctgattaaaa aagcgccctt gtggcgcttt 60
ttttatattc ccgcctccat ttaaaataaa aaatccaatc ggatttcact atttaaactg 120
gccattatct aagatgaatc cgatggaagc tcgctgtttt aacacgcgtt ttttaacctt 180
ttattgaaag tcggtgcttc tttgagcgaa cgatcaaatt taagtggatt cccatcaaaa 240
aaatattctc aacctaaaaa agtttgtgta atacttgtaa cgctacatgg agattaactc 300
aatctagagg gtattaataa tgaatcgtac taaactggta ctgggcatgg gatatgacgc 360
gtgtaatttt attaactcga tcgttaaaaa ttttaaagaa tccattggaa taatgaacgt 420
gttgttactg gaacctgatg agtccgacgc tttgcgctta ttgtatctaa aactggatgt 480
tatcagggct ggcattgagc aggatgtccg catgttaacc aaagcccgtc tggacagcaa 540
cgccgcactt tgcctttacc aggtacataa tgcgttaaat gctttgaact attgcattgg 600
gattgtcaga aatacgccgt taaatcagca ggtttattac gatttcgaag atctaaagcg 660
tattgtgaac actcttgatg caacatccgg acgtgttttt aataaactgg atggtaatga 720
gtacacataa 730
<210> 21
<211> 644
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 21
tcaccacggc gataaccata ggttttcggc gtggccacat ccatggtgaa tcccactttt 60
tccagcacgc gcgccacttc atcgggtctt aaatacatag attttcctcg tcatctttcc 120
aaagcctcgc caccttacat gactgagcat ggaccgtgac tcagaaaatt ccacaaacga 180
acctgaaagg cgtgattgcc gtctggcctt aaaaattatg gtctaaacta aaatttacat 240
cgaaaacgag ggaggatcct atgggatatg acgcgtgtaa ttttattaac tcgatcgtta 300
aaaattttaa agaatccatt ggaataatga acgtgttgtt actggaacct gatgagtccg 360
acgctttgcg cttattgtat ctaaaactgg atgttatcag ggctggcatt gagcaggatg 420
tccgcatgtt aaccaaagcc cgtctggaca gcaacgccgc actttgcctt taccaggtac 480
ataatgcgtt aaatgctttg aactattgca ttgggattgt cagaaatacg ccgttaaatc 540
agcaggttta ttacgatttc gaagatctaa agcgtattgt gaacactctt gatgcaacat 600
ccggacgtgt ttttaataaa ctggatggta atgagtacac ataa 644
<210> 22
<211> 713
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 22
atattgacac catgacgcgc gtaatgctga ttggttctgt gacgctggta atgattgtcg 60
aaattctgaa cagtgccatc gaagccgtag tagaccgtat tggtgcagaa ttccatgaac 120
tttccgggcg ggcgaaggat atggggtcgg cggcggtgct gatgtccatc ctgctggcga 180
tgtttacctg gatcgcatta ctctggtcac attttcgata acgcttccag aattcgataa 240
cgccctggtt ttttgcttaa atttggttcc aaaatcgcct ttagctgtat atactcacag 300
cataactgta tatacaccca gggggcggga tgggatatga cgcgtgtaat tttattaact 360
cgatcgttaa aaattttaaa gaatccattg gaataatgaa cgtgttgtta ctggaacctg 420
atgagtccga cgctttgcgc ttattgtatc taaaactgga tgttatcagg gctggcattg 480
agcaggatgt ccgcatgtta accaaagccc gtctggacag caacgccgca ctttgccttt 540
accaggtaca taatgcgtta aatgctttga actattgcat tgggattgtc agaaatacgc 600
cgttaaatca gcaggtttat tacgatttcg aagatctaaa gcgtattgtg aacactcttg 660
atgcaacatc cggacgtgtt tttaataaac tggatggtaa tgagtacaca taa 713
<210> 23
<211> 2048
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 23
tcaccacggc gataaccata ggttttcggc gtggccacat ccatggtgaa tcccactttt 60
tccagcacgc gcgccacttc atcgggtctt aaatacatag attttcctcg tcatctttcc 120
aaagcctcgc caccttacat gactgagcat ggaccgtgac tcagaaaatt ccacaaacga 180
acctgaaagg cgtgattgcc gtctggcctt aaaaattatg gtctaaacta aaatttacat 240
cgaaaacgag ggaggatcct atggcgtcga aatcgttttg taaaagttgg ggagccgaat 300
acctcgcgaa tggtcgcgtg cgttttcgct tatgggccac cgggcagcag caggtttcac 360
tgcgtttacc ggctggcgaa ttgccaatgc aacgccttga tgacggctgg tttgaactga 420
ccgtcgataa tatcgccgca ggtacggcgt accagtttgt cctgagcaac ggtttggcgg 480
ttcccgatcc cgcctcgcgc gcgcaggccg ccgatgttaa cggcccttcg ctggttgtcg 540
acccggatca ttacgtctgg caacacccgc agtggcaggg tcgcccgtgg caagaatccg 600
tggtctatga actgcatatt gggacattta ccccgcgggg cacttttcgc gcggcaatgg 660
aaaaactgcc gtacctcgcg caactcggca tcaccatgat agaggtgatg cccgtctccc 720
agtttggcgg caatcgcggc tggggctatg acggcgtgct gctctatgcg ccgcactcgg 780
cgtatggcac gccggatgat tttaaagcct ttatcgacgc cgctcacggc tacggccttt 840
cagtggtgct ggatattgtg ctcaaccact ttggcccgga aggtaactac ttacccgcgc 900
tgtcaccgga ttttttccac gccgaacggc agacgccgtg ggggccgggc atcgcgtatg 960
acgttgacgc ggtgcggcgt tacatcgttg aagcaccgct ctactggctc aacgaataca 1020
acctggacgg gctgcgattt gatgccatcg atcagattga agacagcgcc gaaccgccgg 1080
tgctggtgga gattgcccgc cgtattcgcg aggagatcac cgaccggccg gtgcatctga 1140
ccaccgaaga ttgccgcaat attattgcgc tgcatccgcg cgagccggat gggcgtgcgc 1200
cgctgtttac cgccgagtgg aatgacgatc tgcacaatgc cgcgcatgtg tttgccaccg 1260
gtgaaaccca tgcctattac caggattttg cggataagcc cgaagtgtgg ctggcgcgca 1320
cgttaactga aggattcgcg tttcaggggg agattgcccc gtcaaccggt gaaccacgcg 1380
gagtggacag ccgcaaccag ccgccggtag cgtttgtcga ttttatccag aaccacgatc 1440
agaccggcaa ccgcgcccag ggcgagcggc ttatcacgct ggcgggcgag gagagaacgc 1500
gcgtgctgct cggcgcgctg ctgttttccc cgcatattcc gctgctgttt atgggcgaag 1560
agtatggcga aaccaatcct ttcctgttct ttaccgattt tcacggcgat ctggcgcgcg 1620
ccgtgcgtga agggcgcgcg cgggaattcc agggccatgc cggacatgaa ggggaagcgg 1680
ttcccgatcc gaatgcgcct gacactttta cccgatcaca actggactgg gcaaaaacgc 1740
agcaggagag cggccagcgc tggctgacgc aaacccgcga ctggctgacg ctgcgccaga 1800
aatatgtggt tccgctgctg acaaatgcgc agcggcaaaa tggggaagtg atcgccaccg 1860
cgccagggtt tgtcgcggtc cgctggcgct tcccgcaagg gacattatcg ctggcgctca 1920
atattggcga ggcgacgcaa ccgttgccgg atcttcccgg caaaacgctg gcagcctggc 1980
cgcgcggaca agatgacttg ccgccgaatg cgttcattgt tcgtctggct ttgggagagg 2040
gaatgtga 2048
<210> 24
<211> 589
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 24
cgtcctgtaa taataaccgg acaattcgga ctgattaaaa aagcgccctt gtggcgcttt 60
ttttatattc ccgcctccat ttaaaataaa aaatccaatc ggatttcact atttaaactg 120
gccattatct aagatgaatc cgatggaagc tcgctgtttt aacacgcgtt ttttaacctt 180
ttattgaaag tcggtgcttc tttgagcgaa cgatcaaatt taagtggatt cccatcaaaa 240
aaatattctc aacctaaaaa agtttgtgta atacttgtaa cgctacatgg agattaactc 300
aatctagagg gtattaataa tgaatcgtac taaactggta ctgggcatga aaaaagtcct 360
gtatggcatt tttgccatat ccgcgcttgc ggcgacttct gcttttgctg caccggtgca 420
aattggtgaa gcggcagggt cggcagcgac ctctgtttct gcgggtagct ccgcagcaac 480
gagcgttagc accgtagggt ccgcggtggg tgtcgcgctg gcggcaaccg gtggcggtga 540
tggctccaac acgggaacca cgaccaccac gaccacaagt acccagtaa 589
<210> 25
<211> 503
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 25
tcaccacggc gataaccata ggttttcggc gtggccacat ccatggtgaa tcccactttt 60
tccagcacgc gcgccacttc atcgggtctt aaatacatag attttcctcg tcatctttcc 120
aaagcctcgc caccttacat gactgagcat ggaccgtgac tcagaaaatt ccacaaacga 180
acctgaaagg cgtgattgcc gtctggcctt aaaaattatg gtctaaacta aaatttacat 240
cgaaaacgag ggaggatcct atgaaaaaag tcctgtatgg catttttgcc atatccgcgc 300
ttgcggcgac ttctgctttt gctgcaccgg tgcaaattgg tgaagcggca gggtcggcag 360
cgacctctgt ttctgcgggt agctccgcag caacgagcgt tagcaccgta gggtccgcgg 420
tgggtgtcgc gctggcggca accggtggcg gtgatggctc caacacggga accacgacca 480
ccacgaccac aagtacccag taa 503
<210> 26
<211> 572
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 26
atattgacac catgacgcgc gtaatgctga ttggttctgt gacgctggta atgattgtcg 60
aaattctgaa cagtgccatc gaagccgtag tagaccgtat tggtgcagaa ttccatgaac 120
tttccgggcg ggcgaaggat atggggtcgg cggcggtgct gatgtccatc ctgctggcga 180
tgtttacctg gatcgcatta ctctggtcac attttcgata acgcttccag aattcgataa 240
cgccctggtt ttttgcttaa atttggttcc aaaatcgcct ttagctgtat atactcacag 300
cataactgta tatacaccca gggggcggga tgaaaaaagt cctgtatggc atttttgcca 360
tatccgcgct tgcggcgact tctgcttttg ctgcaccggt gcaaattggt gaagcggcag 420
ggtcggcagc gacctctgtt tctgcgggta gctccgcagc aacgagcgtt agcaccgtag 480
ggtccgcggt gggtgtcgcg ctggcggcaa ccggtggcgg tgatggctcc aacacgggaa 540
ccacgaccac cacgaccaca agtacccagt aa 572
<210> 27
<211> 600
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 27
gataccgagt gagagcaagg cgttagaaag ggtgcgacca atcagaaact ccatcgataa 60
gtagtaaacc tggcgcactt cctgagagag ctgtgcacgg gtagaacgca gccagcgctc 120
caccagccga tcgcgcacgg caaacagcgt ggcgttcagc cattcgtggc gattggcgac 180
agccggatct ttaccgatgg tgaacatcag tttgtaggca atcgaatgtt ttaacgcctc 240
aatacttaac gtaggtgaag cataagtaaa aggtgcattc atataatgat tcctggaaaa 300
tatcgctcct gttcaaaacc cgttggcctt gatgccagaa atggggaaac agcgcgcgga 360
ccccttccgc gcaaggttat aatcagatgc tttcagcttt taatttgcgc agcatttcac 420
gtgtgaccaa cacaatgcct tcttcggaac ggtaaaagcg acgcgcatct tcttccgcat 480
tttcaccaat caccgttcct tccggtatca cgcagcctct gtcgataatg cagcgatgga 540
ggcgacagga gcgccccacc cacacttccg gcagcaagac cgaagagtcg atattgcaga 600
<210> 28
<211> 600
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 28
cggcggctgc attatctccg ggtcggtggt ggtgcagtcg gtgctgttcc cgcgcgtgcg 60
ggtgaactca ttctgtaaca ttgattcggc agtcttgttg ccagatgtct gggtggggcg 120
ctcatgccgc ctgcgtcgct gcgtgattga ccgcgcctgc gtcatcccgg aaggtatggt 180
gattggtgag aacgcggaag aggatgcacg ccgcttctat cgctcagagg aaggtatcgt 240
gctggtgacc cgcgacatgc tgcggaaatt ggggcacaaa caggagcgct aatgcaggtc 300
ttgatgtaac cactggagtc attgttatga atgtaccctt tagctatgca tcaccgacgc 360
tgagcgttga ggcgttaaag cactctattg cctataagct gatgtttatc atcggtaaag 420
atccggccat cgccaacaag catgaatggc ttaacgccac gctgtttgcc gtccgcgatc 480
gcatggtgga gcgctggctg cgctccaacc gcgcgcagct ctcgcaggaa gtgcgccagg 540
tatactatct gtcgatggag ttcctgattg gccgtaccct ctctaacgcg ctgctgtcgc 600
Claims (22)
- 다음을 포함하는 박테리아 조성물:
bcsII, bcsIII, yjbE, fhaB, pehA, glgA, otsB, treZ 및 cysZ로 이루어진 군에서 선택된 하나 이상의 유전자에서의 변형을 포함하며, 농업 시스템에서 대기 질소를 고정시키는, 하나 이상의 유전자 조작된 박테리아 균주. - 다음을 포함하는 박테리아 조성물:
식물에서 식물 성장 촉진 박테리아 균주의 콜로니화를 증가시키도록 리모델링되어 있는, 식물 성장 촉진 박테리아 균주. - 청구항 2에 있어서, 상기 식물 성장 촉진 박테리아 균주의 콜로니화는 상기 식물의 뿌리에서 발생함을 특징으로 하는 박테리아 조성물.
- 청구항 2에 있어서, 상기 박테리아 균주는 세포외 다당류의 생성에 관여하는 효소 또는 경로에서의 유전자 변형을 포함함을 특징으로 하는 박테리아 조성물.
- 청구항 4에 있어서, 상기 유전자 변형은 bcsII, bcsIII 및 yjbE로 구성된 그룹에서 선택된 유전자에 존재함을 특징으로 하는 박테리아 조성물.
- 청구항 2에 있어서, 상기 박테리아 균주는 사상성 헤마글루티닌의 생성에 관여하는 효소 또는 경로에서의 유전자 변형을 포함함을 특징으로 하는 박테리아 조성물.
- 청구항 6에 있어서, 상기 유전자 변형은 fhaB 유전자에 존재함을 특징으로 하는 박테리아 조성물.
- 청구항 2에 있어서, 상기 박테리아 균주는 엔도-폴리갈락투로나제 생성에 관여하는 효소 또는 경로에서의 유전자 변형을 포함함을 특징으로 하는 박테리아 조성물.
- 청구항 8에 있어서, 상기 유전자 변형은 pehA 유전자에 존재함을 특징으로 하는 박테리아 조성물.
- 청구항 2에 있어서, 상기 박테리아 균주는 트레할로스의 생성에 관여하는 효소 또는 경로에서의 유전자 변형을 포함함을 특징으로 하는 박테리아 조성물.
- 청구항 6에 있어서, 상기 유전자 변형은 otsB 및 treZ로 구성된 그룹에서 선택된 유전자에 존재함을 특징으로 하는 박테리아 조성물.
- 청구항 2에 있어서, 상기 박테리아 조성물은 현장(field)에 처리용으로 제형화됨을 특징으로 하는 박테리아 조성물.
- 청구항 2-12 중 어느 한 항에 있어서, 상기 식물 성장 촉진 박테리아는 상기 식물에 영양소를 제공함을 특징으로 하는 박테리아 조성물.
- 청구항 2-13 중 어느 한 항에 있어서, 상기 식물 성장 촉진 박테리아는 상기 식물에 고정된 질소를 제공함을 특징으로 하는 박테리아 조성물.
- 청구항 2-14 중 어느 한 항에 있어서, 상기 식물은 옥수수, 보리, 밀, 수수, 대두 및 쌀로 구성된 그룹에서 선택됨을 특징으로 하는 박테리아 조성물.
- 다음 단계를 포함하는, 식물에서 식물 성장 촉진 박테리아 균주의 콜로니화를 증가시키는 방법:
세포외 다당류 생성, 엔도-폴리갈락투로나제 생성, 및 트레할로스 생성으로 구성된 그룹에서 선택된 경로에 관여하는 유전자에서의 유전자 변형을 상기 식물 성장 촉진 박테리아 균주에 도입하는 단계. - 다음 단계를 포함하는, 식물에 이용가능한 질소를 증가시키는 방법:
glgA의 발현이 감소되어 있는 복수의 리모델링된 박테리아를 식물에 처리하는 단계. - 청구항 17에 있어서, 상기 리모델링된 박테리아는 상기 리모델링된 박테리아와 동일한 종의 동일한 양의 비-리모델링된 박테리아의 질소 동화도와 비교하여 상기 리모델링된 박테리아 내에서 보다 적은 질소 동화를 가짐을 특징으로 하는 방법.
- 다음 단계를 포함하는, 식물에 이용가능한 질소를 증가시키는 방법:
복수의 리모델링된 박테리아를 식물에 처리하는 단계, 상기 복수의 리모델링된 박테리아는 상기 리모델링된 박테리아 내부에 증가된 양의 최소한 하나의 질소분해효소 보조인자를 가짐. - 청구항 19에 있어서, 상기 보조인자는 황임을 특징으로 하는 방법.
- 청구항 19에 있어서, 상기 리모델링된 박테리아는 cysZ의 증가된 발현을 가짐을 특징으로 하는 방법.
- 청구항 19에 있어서, 상기 리모델링된 박테리아는 황 운반체의 증가된 발현을 가짐을 특징으로 하는 방법.
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201762543288P | 2017-08-09 | 2017-08-09 | |
| US62/543,288 | 2017-08-09 | ||
| PCT/US2018/046148 WO2019032926A1 (en) | 2017-08-09 | 2018-08-09 | METHODS AND COMPOSITIONS FOR ENHANCING MODIFIED MICROBES |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| KR20200044015A true KR20200044015A (ko) | 2020-04-28 |
Family
ID=65271714
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020207006825A Abandoned KR20200044015A (ko) | 2017-08-09 | 2018-08-09 | 유전자조작 미생물을 개선하기 위한 방법 및 조성물 |
Country Status (10)
| Country | Link |
|---|---|
| US (1) | US20210163374A1 (ko) |
| EP (1) | EP3665141A4 (ko) |
| KR (1) | KR20200044015A (ko) |
| CN (1) | CN111542507A (ko) |
| AU (1) | AU2018313949A1 (ko) |
| BR (1) | BR112020002654A2 (ko) |
| CA (1) | CA3072466A1 (ko) |
| MX (1) | MX2020001599A (ko) |
| RU (1) | RU2020109871A (ko) |
| WO (1) | WO2019032926A1 (ko) |
Families Citing this family (32)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2014071182A1 (en) | 2012-11-01 | 2014-05-08 | Massachusetts Institute Of Technology | Directed evolution of synthetic gene cluster |
| MX377432B (es) | 2015-07-13 | 2025-03-10 | Pivot Bio Inc | Metodos y composiciones para mejorar atributos de plantas. |
| EP3359664A4 (en) | 2015-10-05 | 2019-03-20 | Massachusetts Institute Of Technology | NITROGEN FIXATION WITH REINFORCED NIF CLUSTERS |
| MX2019008285A (es) | 2017-01-12 | 2019-11-11 | Pivot Bio Inc | Métodos y composiciones para mejorar atributos de plantas. |
| EP3700879A4 (en) | 2017-10-25 | 2022-01-12 | Pivot Bio, Inc. | GEN TARGETS FOR NITROGEN FIXATION TARGETING TO IMPROVE PLANT CHARACTERISTICS |
| MX2020004343A (es) | 2017-10-25 | 2021-01-08 | Pivot Bio Inc | Métodos y composiciones para mejorar microbios modificados que fijan nitrógeno. |
| US11963530B2 (en) | 2018-06-27 | 2024-04-23 | Pivot Bio, Inc. | Agricultural compositions comprising remodeled nitrogen fixing microbes |
| CN109536431A (zh) * | 2018-12-04 | 2019-03-29 | 江苏省农业科学院 | 复合生化复苏因子组合物及其应用 |
| EP3897105A2 (en) | 2018-12-21 | 2021-10-27 | Pivot Bio, Inc. | Methods, compositions, and media for improving plant traits |
| WO2020185590A1 (en) | 2019-03-08 | 2020-09-17 | Zymergen Inc. | Iterative genome editing in microbes |
| US11053515B2 (en) | 2019-03-08 | 2021-07-06 | Zymergen Inc. | Pooled genome editing in microbes |
| JP2022524044A (ja) * | 2019-03-08 | 2022-04-27 | ザイマージェン インコーポレイテッド | 微生物のプール型ゲノム編集 |
| WO2020190363A1 (en) | 2019-03-19 | 2020-09-24 | Massachusetts Institute Of Technology | Control of nitrogen fixation in rhizobia that associate with cereals |
| AU2020263508A1 (en) * | 2019-04-25 | 2021-10-14 | Pivot Bio, Inc. | High-throughput methods for isolating and characterizing ammonium-excreting mutant libraries generated by chemical mutagenesis |
| WO2021221690A1 (en) | 2020-05-01 | 2021-11-04 | Pivot Bio, Inc. | Modified bacterial strains for improved fixation of nitrogen |
| EP4143293A1 (en) | 2020-05-01 | 2023-03-08 | Pivot Bio, Inc. | Measurement of nitrogen fixation and incorporation |
| WO2021222567A2 (en) | 2020-05-01 | 2021-11-04 | Pivot Bio, Inc. | Modified bacterial strains for improved fixation of nitrogen |
| CN112047490B (zh) * | 2020-08-24 | 2022-11-18 | 上海理工大学 | 生物炭协同复合菌去除城市河道水体中氨氮的方法及装置 |
| TWI793554B (zh) * | 2021-03-19 | 2023-02-21 | 蜻蜓創意有限公司 | 塔形連續自動化生物飼養器及其飼養槽體裝置 |
| CN112877263A (zh) * | 2021-04-09 | 2021-06-01 | 黑龙江省科学院高技术研究院 | 一种汉麻内生菌的分离方法 |
| CN113149768A (zh) * | 2021-05-18 | 2021-07-23 | 保定瑞沃斯环境科技有限公司 | 一种菌酶复合微生物肥料及制备方法 |
| AU2022301301A1 (en) | 2021-07-02 | 2023-12-14 | Pivot Bio, Inc. | Genetically-engineered bacterial strains for improved fixation of nitrogen |
| CN113322212B (zh) * | 2021-07-09 | 2022-05-20 | 华中农业大学 | 菌株Dyella sp.LX-1及其在氧化有机硒或单质硒中的应用 |
| CN113402327B (zh) * | 2021-07-28 | 2023-02-03 | 甘肃省农业科学院植物保护研究所 | 一种蔬菜专用的微生物复合肥及其制备方法 |
| CN113913327B (zh) * | 2021-09-30 | 2024-04-16 | 中国科学院东北地理与农业生态研究所农业技术中心 | 一株可分泌iaa的耐盐碱促生菌ys-at1及其应用 |
| CN114317584B (zh) * | 2021-11-23 | 2023-06-23 | 海南医学院 | 新型转座子突变株文库的构建系统、新型转座子突变文库和应用 |
| KR20250035505A (ko) * | 2022-04-14 | 2025-03-12 | 인트린식스 바이오 인코포레이티드 | 식물 영양, 성장 및 성능을 개선하기 위한 내생균을 포함하는 조성물 및 이의 이용 방법 |
| EP4526277A1 (en) * | 2022-05-17 | 2025-03-26 | BioConsortia, Inc. | Methods and compositions for refactoring nitrogen fixation clusters |
| CN115747358B (zh) * | 2022-11-19 | 2023-08-18 | 河南农业大学 | 一种鉴定pgpr趋化根系分泌物到达根际的趋化速率的方法及其应用 |
| CN115852001A (zh) * | 2022-11-23 | 2023-03-28 | 深圳海关动植物检验检疫技术中心 | 一种小麦病原菌检测方法及其应用 |
| WO2024121871A1 (en) * | 2022-12-09 | 2024-06-13 | Fertis India Pvt. Ltd. | Genetic modification of microbes for production of nitrogen and carbon-containing compounds |
| CN117004540B (zh) * | 2023-09-28 | 2023-12-15 | 北京量维生物科技研究院有限公司 | 变栖克雷伯氏固氮菌的筛选及其应用 |
Family Cites Families (13)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| FR2718750B1 (fr) * | 1994-04-19 | 1996-06-14 | Pasteur Institut | Protéines recombinantes de l'hémagglutinine filamenteuse de Bordetella, notamment, B. Pertussis, production et application à la production de protéines étrangères ou de principes actifs vaccinants. |
| US8389250B2 (en) * | 2006-01-04 | 2013-03-05 | Metabolic Explorer | Methods for producing methionine by culturing a microorganism modified to enhance production of cysteine |
| US8481026B1 (en) * | 2009-04-17 | 2013-07-09 | Peter J. Woodruff | Bacteria with increased trehalose production and method for using the same in bioremediation |
| MX377432B (es) * | 2015-07-13 | 2025-03-10 | Pivot Bio Inc | Metodos y composiciones para mejorar atributos de plantas. |
| MX2019008285A (es) * | 2017-01-12 | 2019-11-11 | Pivot Bio Inc | Métodos y composiciones para mejorar atributos de plantas. |
| MX2020004343A (es) * | 2017-10-25 | 2021-01-08 | Pivot Bio Inc | Métodos y composiciones para mejorar microbios modificados que fijan nitrógeno. |
| EP3700879A4 (en) * | 2017-10-25 | 2022-01-12 | Pivot Bio, Inc. | GEN TARGETS FOR NITROGEN FIXATION TARGETING TO IMPROVE PLANT CHARACTERISTICS |
| WO2020006246A1 (en) * | 2018-06-27 | 2020-01-02 | Pivot Bio, Inc. | Guided microbial remodeling, a platform for the rational improvement of microbial species for agriculture |
| US11963530B2 (en) * | 2018-06-27 | 2024-04-23 | Pivot Bio, Inc. | Agricultural compositions comprising remodeled nitrogen fixing microbes |
| MX2021000187A (es) * | 2018-07-11 | 2021-06-15 | Pivot Bio Inc | Suministro de nitrógeno dinámico temporal y espacialmente dirigido mediante microbios remodelados. |
| EP3826646A4 (en) * | 2018-07-25 | 2022-07-27 | Convergent Genomics, Inc. | URINARY MICROBIOMIC PROFILING |
| AR116964A1 (es) * | 2018-11-01 | 2021-06-30 | Pivot Bio Inc | Composiciones de biopelículas con estabilidad mejorada para fijación del nitrógeno en productos microbianos |
| KR20210133967A (ko) * | 2019-02-05 | 2021-11-08 | 피벗 바이오, 인크. | 생물적 질소 고정에 의한 작물 수확량 일관성의 개선 |
-
2018
- 2018-08-09 CA CA3072466A patent/CA3072466A1/en active Pending
- 2018-08-09 RU RU2020109871A patent/RU2020109871A/ru unknown
- 2018-08-09 MX MX2020001599A patent/MX2020001599A/es unknown
- 2018-08-09 KR KR1020207006825A patent/KR20200044015A/ko not_active Abandoned
- 2018-08-09 CN CN201880065665.7A patent/CN111542507A/zh active Pending
- 2018-08-09 US US16/637,565 patent/US20210163374A1/en not_active Abandoned
- 2018-08-09 EP EP18843845.1A patent/EP3665141A4/en not_active Withdrawn
- 2018-08-09 WO PCT/US2018/046148 patent/WO2019032926A1/en unknown
- 2018-08-09 BR BR112020002654-9A patent/BR112020002654A2/pt not_active Application Discontinuation
- 2018-08-09 AU AU2018313949A patent/AU2018313949A1/en not_active Abandoned
Also Published As
| Publication number | Publication date |
|---|---|
| RU2020109871A (ru) | 2021-09-14 |
| AU2018313949A1 (en) | 2020-02-27 |
| EP3665141A4 (en) | 2021-08-25 |
| BR112020002654A2 (pt) | 2020-08-18 |
| US20210163374A1 (en) | 2021-06-03 |
| CA3072466A1 (en) | 2019-02-14 |
| RU2020109871A3 (ko) | 2022-04-22 |
| MX2020001599A (es) | 2020-08-17 |
| EP3665141A1 (en) | 2020-06-17 |
| WO2019032926A1 (en) | 2019-02-14 |
| CN111542507A (zh) | 2020-08-14 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| CN111587287B (zh) | 用于改良固氮的工程微生物的方法和组合物 | |
| US20250115529A1 (en) | Gene targets for nitrogen fixation targeting for improving plant traits | |
| US11565979B2 (en) | Methods and compositions for improving plant traits | |
| KR20200044015A (ko) | 유전자조작 미생물을 개선하기 위한 방법 및 조성물 | |
| US20220017911A1 (en) | Methods, compositions, and media for improving plant traits | |
| US20220211048A1 (en) | Gene targets for nitrogen fixation targeting for improving plant traits | |
| RU2805085C2 (ru) | Гены-мишени для направленного воздействия на азотфиксацию для улучшения качеств растений | |
| BR122024009210A2 (pt) | Método para identificar uma cepa microbiana, composição e método para aumentar a fixação de nitrogênio em uma planta não leguminosa |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| PA0105 | International application |
Patent event date: 20200309 Patent event code: PA01051R01D Comment text: International Patent Application |
|
| PG1501 | Laying open of application | ||
| PA0201 | Request for examination |
Patent event code: PA02012R01D Patent event date: 20210723 Comment text: Request for Examination of Application |
|
| E902 | Notification of reason for refusal | ||
| PE0902 | Notice of grounds for rejection |
Comment text: Notification of reason for refusal Patent event date: 20230801 Patent event code: PE09021S01D |
|
| PC1902 | Submission of document of abandonment before decision of registration |