JP7376331B2 - 分岐脂肪酸の代謝制御方法 - Google Patents
分岐脂肪酸の代謝制御方法 Download PDFInfo
- Publication number
- JP7376331B2 JP7376331B2 JP2019212465A JP2019212465A JP7376331B2 JP 7376331 B2 JP7376331 B2 JP 7376331B2 JP 2019212465 A JP2019212465 A JP 2019212465A JP 2019212465 A JP2019212465 A JP 2019212465A JP 7376331 B2 JP7376331 B2 JP 7376331B2
- Authority
- JP
- Japan
- Prior art keywords
- component
- branched fatty
- metabolism
- fatty acids
- microorganisms
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
Landscapes
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Description
本発明は、例えば生物学、生化学、生理科学、消費者使用製品の分野において、分岐脂肪酸の代謝を制御する方法に関する。
例えば、特許文献1には、皮脂汚れ成分が生乾き臭原因物質の1種である4-メチル-3-ヘキセン酸への変換されるのを抑制する剤として、特定の構造を有するケトン化合物が開示されている。
また、特許文献2には、β-グルコニダーゼの活性を阻害し、尿臭の生成を抑制する剤として、特定の構造を有する大環状ケトン化合物が開示されている。
さらに、安息香酸などの特定の構造を有する有機カルボン酸を使用し、微生物を殺菌することで臭いの生成を抑制する技術も知られている。
例えば、消費者使用製品の分野において、本発明は、不快臭の原因となる微生物を殺菌することなく、微生物が分岐脂肪酸を基質とする代謝により不快臭の原因物質を生成することを制御できる、分岐脂肪酸の代謝制御方法を提供する。
RO-(C2H4O)n-H (1)
〔式中、Rは、炭素数9以上16以下の直鎖脂肪族炭化水素基であり、nは0又は1である。〕
分岐脂肪酸は、微生物の細胞膜を通過する工程と、細胞膜内に取り込まれた分岐脂肪酸がβ酸化により代謝され代謝生成物を生成する工程とを経て代謝されると考えられる。(a)成分である前記一般式(1)で表される化合物は、対応する脂肪酸よりも速く微生物内に取り込まれることが考えられる。取り込まれた前記一般式(1)で表される化合物は、菌体内でアルコールデヒドロゲナーゼによる酸化、次いでアシルCoAシンセターゼによる修飾を受け、前記一般式(1)に対応する脂肪酸-CoAへ変換される。生成した脂肪酸-CoAが、脂肪酸の代謝・合成酵素の転写調節タンパクであるFadRへ作用し、菌体外に存在する分岐脂肪酸の微生物内への取り込み、そして代謝を抑制していると考えている。一般式(1)で表される化合物の微生物内への取り込みと代謝の制御により、分岐脂肪酸の代謝が制御されることは、従来、当業界では知られていなかった。しかしながら、FadRが活性化されると、脂肪酸伸長酵素が活性化される為に、微生物内で長鎖脂肪酸が生成し、長鎖脂肪酸の量が増加する。生成した長鎖脂肪酸は、FadRの不活化因子となるため、FadRが不活化し、菌体外脂肪酸の取込・代謝を開始してしまう。本発明の(b)成分である、脂肪酸合成酵素の阻害剤が、脂肪酸合成酵素の反応を阻害し、FadRの不活化を防ぐため、本発明の(a)成分の効果が高まる。微生物の代謝を制御する方法として、例えば微生物の数を減少させて、分岐脂肪酸の代謝を低下させる方法が一般的に考えられる。しかしながら、本発明のように、微生物の数を実質維持しつつ、微生物内の分岐脂肪酸の代謝の度合いを調整できる方法は知られていなかった。
また、分岐の脂肪族アルコールのように直鎖の炭化水素基を持たない化合物は、直鎖の炭化水素基を持つ前記一般式(1)で表される化合物、例えば直鎖の脂肪族アルコールよりも、菌体内での酸化工程から脂肪酸-CoAの生成工程が起こりにくく、分岐脂肪酸の微生物内への取り込みや代謝を十分に制御できないものと推定している。
本発明の方法で用いる(a)成分は、下記一般式(1)で表される化合物から選ばれる1種以上の化合物である。
RO-(C2H4O)n-H (1)
〔式中、Rは、炭素数9以上16以下の直鎖脂肪族炭化水素基であり、nは0又は1である。〕
一般式(1)中、分岐脂肪酸の代謝をより制御する観点、とりわけ分岐脂肪酸の代謝を抑制する観点から、nは0が好ましい。
(a)成分の質量当たりの分岐脂肪酸の代謝制御効果、とりわけ分岐脂肪酸の代謝を抑制する観点から、前記(a)成分中の、Rが炭素数12又は13の直鎖飽和脂肪族炭化水素基である化合物の含有割合は、好ましくは50質量%以上であり、更に好ましくは60質量%以上であり、より更に好ましくは70質量%以上であり、より更に好ましくは80質量%以上であり、より更に好ましくは90質量%以上であり、より更に好ましくは95質量%以上であり、より更に好ましくは97質量%以上であり、そして、100質量%以下であり、100質量%であってよい。
(b)成分は脂肪酸合成酵素の阻害剤である。微生物の細胞内の脂肪酸は、脂肪酸合成酵素により生体内で合成される場合がある。脂肪酸合成酵素には一般的に、細胞質に存在するタイプ1(FAS-I)と、ミトコンドリアに存在するタイプ2(FAS-II)が知られている。様々な化合物が脂肪酸合成酵素を阻害することが分かっており、本発明での使用に関し好適である。脂肪酸合成酵素の阻害剤を選択することは当業者の技術範囲内である。脂肪酸合成酵素を阻害する化合物は、精製脂肪酸合成酵素を用いてその化合物の脂肪酸合成酵素活性阻害能力を試験することで同定できる。例えば,Dilsら(1975年)Meth.Enzymol.35巻、p.74~83、に記載の方法で同定できる。脂肪酸合成酵素の阻害剤は、例えば国際特許出願公開WO94/02108号に例示されている。
(b1)成分:ジフェニルエーテル基を有する化合物
(b2)成分:チオラクトマイシン又はチオラクトマイシン誘導体
(b3)成分:セルレニン又はセルレニン誘導体
(b4)成分:α-メチレン-γ-ブチロラクトン又はα-メチレン-γ-ブチロラクトン誘導体
より低い量で分岐脂肪酸の代謝抑制効果が得やすい観点から、(b1)成分が好ましい。
(b1)成分は、ジフェニルエーテル基を有する化合物である。脂肪酸合成酵素の阻害効果がより高い観点から、好ましい(b1)成分は、フェニル基の水素原子の少なくとも一つ以上が、水酸基を有する化合物であり、より好ましくは、フェニル基の水素原子の二つ以上が、水酸基及びクロロ基を有する化合物であることが好ましい。より具体的な(b1)成分としては、4-4’-ジクロロ-2-ヒドロキシジフェニルエーテル(ジクロサン)及び2,4,4’-トリクロロ-2’-ヒドロキシジフェニルエーテル(トリクロサン)から選ばれる1種以上の化合物であることが好ましい。
(b2)成分は、チオラクトマイシン又はチオラクトマイシン誘導体である。チオラクトマイシン誘導体はチオラクトマイシンと構造的に関連しており、そして少なくとも測定可能量の脂肪酸合成酵素の阻害活性を保持している化合物を意味する。チオラクトマイシン又はチオラクトマイシン誘導体の非限定的な例としては、Wangら(1984年)Tetrahdron Lett.25巻、5243~5246ページ、Oishiら(1982年)J.Antibiotics 35巻、391~395ページおよびKremerら(2000年)、J.Bio.Chem.275巻、16857~16864ページに記載されている。
(b3)成分は、セルレニン又はセルレニン誘導体である。構造的には[2R,3S]-2,3-エポキシ-4-オキソ-7,10-trans,trans-ドデカン酸アミドとして特徴づけられる。セルレニン誘導体は、セルレニンと構造的に関連しており、そして少なくとも測定可能量の脂肪酸合成酵素の阻害活性を保持している化合物を意味する。セルレニンおよびセルレニン誘導体の例としては、Morisakiら(1992年)Chem.Pharm.Bull.40巻、2945~2953ページ、Shimazawaら(1992年)Chem.Pharm.Bull.40巻、2954~2957ページ、および米国特許第5539132号に記載のものが挙げられる。
アルコール
〔(b4)成分〕
(b4)成分は、α-メチレン-γ-ブチロラクトン又はα-メチレン-γ-ブチロラクトン誘導体である。α-メチレン-γ-ブチロラクトン誘導体は、それぞれのα-メチレン-γ-ブチロラクトンと構造的に関連しており、そして少なくとも測定可能量の脂肪酸合成酵素の阻害活性を有している化合物を意味する。α-メチレン-γ-ブチロラクトン誘導体は、例えば、米国特許第5981575号に記載のものが挙げられる。
本発明の対象とする微生物は分岐脂肪酸の代謝能を有する微生物であり、それらは環境中に存在する一般的な微生物に見出すことができる。例えば、嫌気性細菌、好気性細菌及び酵母から選ばれる1種以上の微生物が挙げられる。
本発明の分岐脂肪酸の代謝制御方法は、微生物の代謝系において、脂肪酸合成系の代謝に適用することができる。例えば、本発明の分岐脂肪酸の代謝制御方法は、微生物による分岐脂肪酸のβ酸化を抑制する方法であってよい。生体内でのβ酸化は、脂肪酸代謝において脂肪酸を酸化して脂肪酸アシルCoAを生成し、アセチルCoAを取り出す代謝経路であり、脂肪酸代謝の3つのステージ(β酸化、クエン酸回路、電子伝達系)の最初の段階に該当する。
また、分岐脂肪酸としては、分岐鎖としてメチル基を1つ以上有する分岐脂肪酸が挙げられる。
分岐脂肪酸の代謝とは、微生物内において、別の化合物に変換される工程を意味する。代謝には、分岐脂肪酸が微生物に取り込まれる工程や、取り込まれた分岐脂肪酸が、例えばβ酸化により代謝され、各種化合物を生成する工程も含まれる。例えば、炭素数17のアンテイソ脂肪酸が微生物内に取り込まれ、代謝されると、系中の分岐脂肪酸の量は低減して、別の化合物に変換される。炭素数17のアンテイソ脂肪酸が微生物内で代謝されると、例えば、4-メチル-3-ヘキセン酸(以下、4M3Hともいう)、4-メチルヘキサン酸(以下、4MHともいう)、2-メチル酪酸(以下、2MBAともいう)などが代謝物として生成される。4M3Hには、下記に示すようにシス体、トランス体が存在するが、本発明においては、そのいずれの構造の化合物も包含する。4M3H、4MH及び2MBAは、いずれも不快臭の原因物質として知られている。
本発明の代謝制御方法は、微生物に、(a)成分及び(b)成分を接触させて、当該微生物の分岐脂肪酸の代謝を制御するものである。
(I)(a)成分、(b)成分及び水を含む組成物と、微生物とを接触させる方法、
(II)水及び微生物を含む混合物と、(a)成分及び(b)成分を含む混合物とを接触させる方法、
(III)(a)成分と(b)成分と微生物が存在する部位に水を接触させる方法、
(IV)水及び微生物を含む混合物と、(a)成分と(b)成分を別々又は同時に接触させる方法、
(V)(a)成分及び水を含む組成物と、微生物と(b)成分とを別々又は同時に接触させる方法、
(VI)(b)成分及び水を含む組成物と、微生物と(a)成分とを別々又は同時に接触させる方法、
などが挙げられる。
分岐脂肪酸の存在下で(a)成分及び(b)成分を微生物に接触させる代謝制御方法の例として、例えば、
(X1)分岐脂肪酸と(a)成分と(b)成分とが共存する箇所に、微生物を接触させる方法、
(X2)(a)成分と(b)成分と微生物が共存する箇所に、分岐脂肪酸を接触させる方法、
(X3)分岐脂肪酸と微生物が共存する箇所に、(a)成分と(b)成分とを接触させる方法
などが挙げられる。分岐脂肪酸の代謝をより制御する観点、とりわけ分岐脂肪酸の代謝をより抑制する観点から、好ましくは前記の(X1)及び(X2)から選ばれる1種以上の方法であり、より好ましくは、(X1)の方法である。
前記微生物は、モラクセラ(Moraxella)属細菌、マイクロコッカス(Micrococcus)属細菌、エシェリキア(Escherichia)属細菌、スタフィロコッカス(Staphylococcus)属細菌、コリネバクテリウム(Corynebacterium)属細菌及びロドトルラ(Rhodotorula)属酵母から選ばれる1種以上の微生物が好ましい。
(a)成分は、前記一般式(1)において、nが0である化合物が好ましい。
(b)成分は、下記(b1)成分、(b2)成分、(b3)成分及び(b4)成分から選ばれる1種以上の化合物、更に下記(b1)成分、及び(b3)成分から選ばれる1種以上の化合物、更に下記(b1)成分から選ばれる1種以上の化合物、更に4-4’-ジクロロ-2-ヒドロキシジフェニルエーテル(ジクロサン)、2,4,4’-トリクロロ-2’-ヒドロキシジフェニルエーテル(トリクロサン)及びセルレニンから選ばれる1種以上の化合物が好ましい。
(b1)成分:ジフェニルエーテル基を有する化合物
(b2)成分:チオラクトマイシン又はチオラクトマイシン誘導体
(b3)成分:セルレニン又はセルレニン誘導体
(b4)成分:α-メチレン-γ-ブチロラクトン又はα-メチレン-γ-ブチロラクトン誘導体
前記処理液は、(a)成分の含有量が10ppm以上10質量%以下が好ましい。
前記処理液は、(b)成分の含有量が0.01ppm以上10ppm以下が好ましい。
前記処理液は、前記分岐脂肪酸の含有量が1ppm以上1質量%以下が好ましい。
前記処理液は、有機溶媒が残部であることが好ましい。有機溶媒はメタノールが好ましい。
前記処理液の接触時間は、1分以上64時間以下が好ましい。前記の(a)成分と微生物が接触している間、本発明の効果を得ることができる。本発明を適用する技術分野に合わせて、接触時間を適宜変えることができる。
前記処理液の接触温度は、1℃以上45℃以下が好ましく、本発明の効果が得られやすい観点から、好ましくは5℃以上であり、より好ましくは10℃以上であり、そして同じ観点から、好ましくは40℃以下であり、より好ましくは35℃以下である。
前記微生物と水とを含む混合物に、前記処理液を接触させてもよい。
[代謝制御剤]
<(a)成分>
C12OH:1-ドデカノール
・ダイクロサン
・トリクロサン
・セルレニン
・CCCP:カルボニルシアニド-m-クロロフェニルヒドラゾン
CCCPは、脱共役剤として知られており、脂肪酸合成酵素は阻害しない化合物である。
14-メチルヘキサデカン酸:SIGMA-ALDRICH製
モラクセラ(Moraxella)属細菌による14-メチルヘキサデカン酸の代謝抑制及び代謝生成物の抑制について、以下の方法で試験を行った。結果を表1~4に示した。
SCD-LP寒天培地(日本製薬)にMoraxella osloensis(衣類分離株)を37℃で24時間前培養し、イオン交換水で1/20に希釈したNB液体培地(Difco)を用いて106(CFU/ml)となるよう菌懸濁液を調製した。
滅菌した木綿平織布3cm×3cmに対して、14-メチルヘキサデカン酸(SIGMA-ALDRICH)を溶解させたメタノール溶液を滴下し布へ付着させた。14-メチルヘキサデカン酸の付着量は、木綿平織布1枚当たり10μg(100μg/布g)となるように設定した。さらに、(a)成分を溶解させたメタノール溶液、次いで(b)成分又は(b’)成分を溶解させたメタノール溶液を14-メチルヘキサデカン酸が付着している布に滴下し、その後1時間乾燥させた。前記メタノール溶液は、(a)成分又は(b)成分の量が表に記載の通りとなるように滴下した。この布を滅菌済No3スクリュー管(マルエム)に入れ、前記の菌懸濁液100μlを植菌して、37℃条件下で21時間培養を行った。
・GC:Agilent 6890N
・GCカラム:Agilent DB-1 30m×250μm×0.25μm
・昇温条件:50℃(3分ホールド)-(10℃/分)-300℃(5分ホールド)
・サンプル注入量:1μl
・スプリット比:1:12
・検出器:FID
代謝抑制率(%)=〔1-(100-A)/(100-B)〕×100
A:基準、試験例、又は比較試験例の14-メチルヘキサデカン酸の残存量(μg/布g)
B:対照の14-メチルヘキサデカン酸の残存量(μg/布g)
対照は、(a)成分、(b)成分又は(b’)成分を添加しない例であり、基準1-1、基準2-1、基準3-1、基準4-1が該当する(以下同様)。これらの対照では、代謝抑制率は0%となる。
上記(1)と同じ方法で準備した布を培養後、メタノール3mlを添加して30分間超音波照射下で抽出を行った(20℃)。抽出液をフナコシ社のADAM試薬(1000ppmメタノール溶液)と1:1(体積比)で混合し、室温暗所条件下で一晩放置させた。その後にHPLCを用い、14-メチルヘキサデカン酸の代謝生成物として4-メチル-3-ヘキセン酸の定量を行った。定量条件を下記に示す。
代謝生成物の定量条件
LC:HITACHI ELITE LaChrom
カラム:Zorbax C8 4.6×250mm
溶離液:アセトニトリル61%(v/v)、水39%(v/v)
カラム温度:40℃
サンプル注入量:10μL
流速:1.0mL/min
検出:FLD Ex.365nm、Em.412nm
代謝生成物抑制率(%)=100×〔(D-C)/D〕
C:基準、試験例、又は比較試験例の代謝生成物の生成量(μg/布g)
D:対照の代謝生成物の生成量(μg/布g)
上記(1)と同じ方法で準備した布を培養後、9mLのLP希釈液(日本製薬社製)を加えて10分間超音波下で菌の抽出を行った。抽出液をSCD-LP寒天培地(日本製薬)にて混釈後、37℃にて培養を1日間行った。それぞれの布について得られたコロニー数を計測し、その常用対数値を生菌数として測定した。代謝制御剤を添加しない場合の微生物の生菌数の対数値(1)と代謝制御剤を添加した場合の微生物の生菌数の対数値(2)との差である、対数値(1)-対数値(2)が-1以上2未満であることが代謝制御剤の接触の有無で生菌数に変化がなく、微生物の生菌数が維持されていることを意味する。また、値が0により近いほど、微生物の生菌数がより維持されていることを意味する。
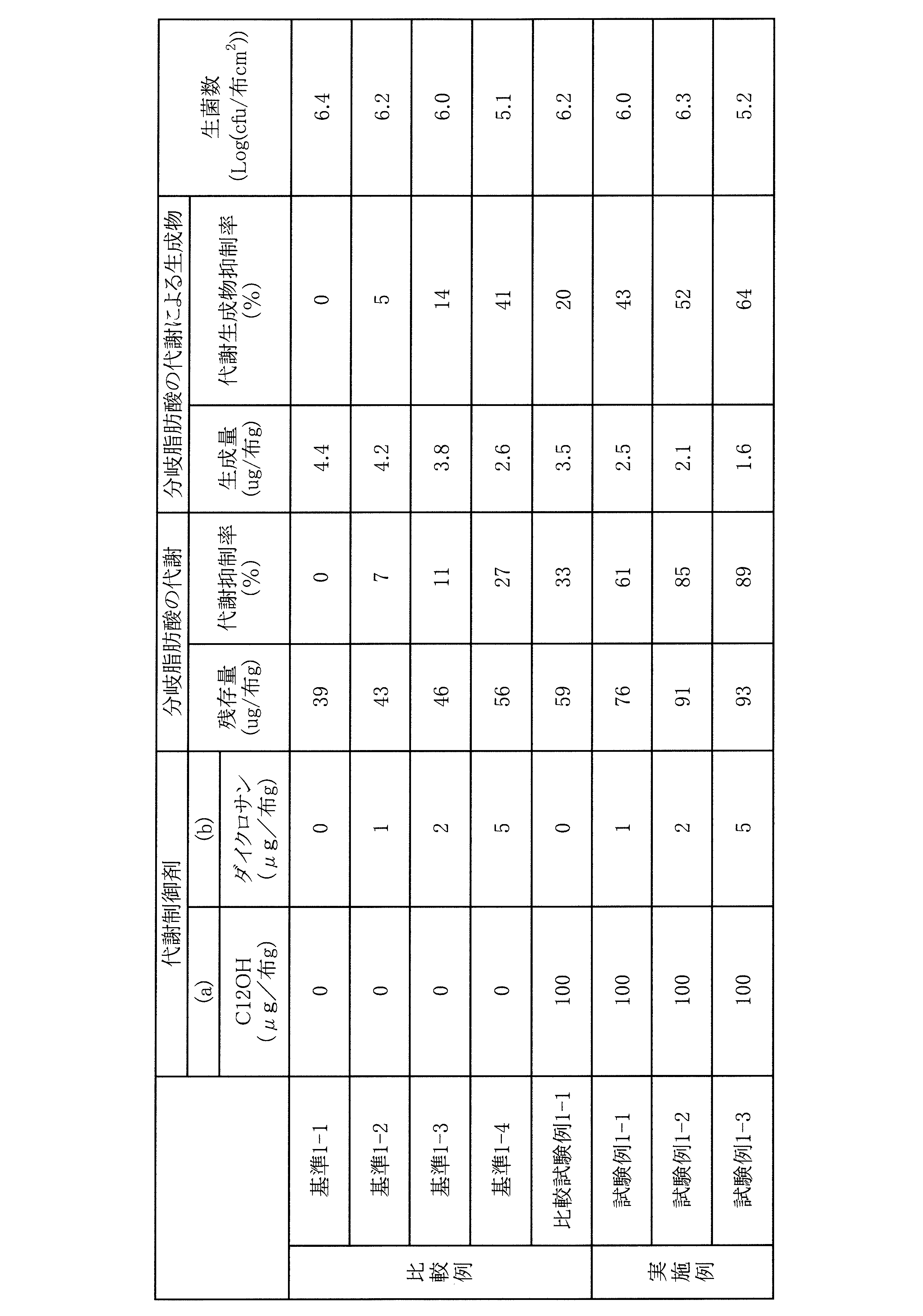



表1において、基準1-2は、ダイクロサンを共存させるが、C12OHの共存はない場合の値を記載している。比較試験例1-1は、ダイクロサンは共存しないが、C12OHが共存した場合の値を記載している。試験例1-1は、ダイクロサンとC12OHのいずれも共存する場合の値を記載している。ここで、分岐脂肪酸の代謝抑制率を見てみると、基準1-2((b)成分のみ)と比較試験例1-1((a)成分のみ)の値を合計すると、7%+33%で40%になるが、実施例である試験例1-1((a)成分と(b)成分)は61%であり、前記合計の40%よりも高く、ダイクロサンとC12OHの相乗効果により、より高い値となっている。試験例1-1の代謝生成物抑制率も、同様に相乗的な向上が確認できる。実施例である試験例1-2、試験例1-3も同様であり、代謝抑制率と代謝生成物抑制率の相乗的な向上が確認できる。
表2~3の実施例である各試験例についても、同様に、代謝抑制率と代謝生成物抑制率の相乗的な向上が確認できる。
Claims (6)
- 微生物に、(a)下記一般式(1)で表される化合物から選ばれる1種以上の化合物〔以下、(a)成分という〕と(b)脂肪酸合成酵素の阻害剤〔以下、(b)成分という〕を接触させて、当該微生物の分岐脂肪酸の代謝を抑制する、分岐脂肪酸の代謝抑制方法。
RO-(C2H4O)n-H (1)
〔式中、Rは、炭素数9以上16以下の直鎖脂肪族炭化水素基であり、nは0又は1である。〕 - (a)成分と微生物が接触しない場合の微生物の生菌数の対数値(1)と、(a)成分と微生物が接触した場合の微生物の生菌数の対数値(2)との差である、対数値(1)-対数値(2)が-1以上2未満である、請求項1に記載の分岐脂肪酸の代謝抑制方法。
- 分岐脂肪酸の存在下で(a)成分と(b)成分とを微生物に接触させる、請求項1又は2に記載の分岐脂肪酸の代謝抑制方法。
- 前記微生物の分岐脂肪酸の代謝がβ酸化である、請求項1~3の何れかに記載の分岐脂肪酸の代謝抑制方法。
- (a)下記一般式(1)で表される化合物から選ばれる1種以上の化合物及び(b)脂肪酸合成酵素の阻害剤を含有する、微生物における分岐脂肪酸の代謝抑制剤。
RO-(C2H4O)n-H (1)
〔式中、Rは、炭素数9以上16以下の直鎖脂肪族炭化水素基であり、nは0又は1である。〕 - 前記微生物の分岐脂肪酸の代謝がβ酸化である、請求項5に記載の分岐脂肪酸の代謝抑制剤。
Priority Applications (6)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2019212465A JP7376331B2 (ja) | 2019-11-25 | 2019-11-25 | 分岐脂肪酸の代謝制御方法 |
| EP20891690.8A EP4067475A4 (en) | 2019-11-25 | 2020-11-24 | Method for controlling branched fatty acid metabolism |
| CN202080081754.8A CN114761540A (zh) | 2019-11-25 | 2020-11-24 | 支链脂肪酸的代谢控制方法 |
| US17/776,393 US20220408724A1 (en) | 2019-11-25 | 2020-11-24 | Method for controlling metabolism of branched fatty acid |
| TW109141121A TWI865662B (zh) | 2019-11-25 | 2020-11-24 | 分枝脂肪酸之代謝控制方法 |
| PCT/JP2020/043557 WO2021106822A1 (ja) | 2019-11-25 | 2020-11-24 | 分岐脂肪酸の代謝制御方法 |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2019212465A JP7376331B2 (ja) | 2019-11-25 | 2019-11-25 | 分岐脂肪酸の代謝制御方法 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2021083326A JP2021083326A (ja) | 2021-06-03 |
| JP7376331B2 true JP7376331B2 (ja) | 2023-11-08 |
Family
ID=76084553
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2019212465A Active JP7376331B2 (ja) | 2019-11-25 | 2019-11-25 | 分岐脂肪酸の代謝制御方法 |
Country Status (1)
| Country | Link |
|---|---|
| JP (1) | JP7376331B2 (ja) |
Citations (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2009078987A (ja) | 2007-09-25 | 2009-04-16 | Kao Corp | バイオフィルム生成抑制方法 |
| JP2012127012A (ja) | 2010-12-13 | 2012-07-05 | Kao Corp | 生乾き臭抑制剤 |
| JP2018052980A (ja) | 2013-06-26 | 2018-04-05 | 花王株式会社 | 微生物の殺菌方法及び衣料の抑臭方法 |
Family Cites Families (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| ATE225167T1 (de) * | 1992-07-24 | 2002-10-15 | Univ Johns Hopkins | Verwendung von inhibitoren der synthese von fettsäuren zur behandlung von krebs |
-
2019
- 2019-11-25 JP JP2019212465A patent/JP7376331B2/ja active Active
Patent Citations (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2009078987A (ja) | 2007-09-25 | 2009-04-16 | Kao Corp | バイオフィルム生成抑制方法 |
| JP2012127012A (ja) | 2010-12-13 | 2012-07-05 | Kao Corp | 生乾き臭抑制剤 |
| JP2018052980A (ja) | 2013-06-26 | 2018-04-05 | 花王株式会社 | 微生物の殺菌方法及び衣料の抑臭方法 |
Also Published As
| Publication number | Publication date |
|---|---|
| JP2021083326A (ja) | 2021-06-03 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| Kerekes et al. | Anti‐biofilm forming and anti‐quorum sensing activity of selected essential oils and their main components on food‐related micro‐organisms | |
| Janek et al. | Antiadhesive activity of the biosurfactant pseudofactin II secreted by the Arctic bacterium Pseudomonas fluorescens BD5 | |
| FI109077B (fi) | Seokset ja niiden käyttö | |
| Ceresa et al. | Production of mannosylerythritol lipids (MELs) to be used as antimicrobial agents against S. aureus ATCC 6538 | |
| EP1221934B1 (en) | Method for producing antimicrobial perfuming compositions | |
| US20100158821A1 (en) | Antimicrobial agents, compositions and products containing the same, and methods of using the compositions and products | |
| JP7188558B2 (ja) | バイオフィルム処理剤及びバイオフィルム処理方法 | |
| Mimee et al. | In vitro antibacterial activity and antifungal mode of action of flocculosin, a membrane‐active cellobiose lipid | |
| Fernandes et al. | Effect of prolonged exposure to disinfectants in the antimicrobial resistance profile of relevant micro-organisms: a systematic review | |
| JP7376331B2 (ja) | 分岐脂肪酸の代謝制御方法 | |
| JP5767831B2 (ja) | 生乾き臭抑制剤 | |
| JP7376330B2 (ja) | 分岐脂肪酸の代謝制御方法 | |
| CN102427725A (zh) | 含有脂环族二醇抗微生物剂的组合物和产品以及使用组合物和产品的方法 | |
| TWI865662B (zh) | 分枝脂肪酸之代謝控制方法 | |
| JP5852313B2 (ja) | 生乾き臭抑制剤 | |
| Ge et al. | Benzalkonium chloride disinfection increases the difficulty of controlling foodborne pathogens identified in aquatic product processing | |
| EP1093355A1 (en) | Method of reducing or preventing malodour | |
| Haustein et al. | Growth inhibition of coryneform bacteria by a mixture of three natural products-Farnesol, glyceryl monolaurate, and phenoxyethanol: HGQ | |
| Ashby et al. | Antimicrobial potential of sophorolipids for anti-acne, anti-dental caries, hide preservation, and food safety applications | |
| Gao et al. | Design, synthesis and evaluation of quinolone quaternary ammonium antibacterial agent with killing ability to biofilm | |
| Aponte | Green tea polyphenol EGCG-S as an antimicrobial agent | |
| JP7321809B2 (ja) | 臭いの生成抑制方法 | |
| Obendorf et al. | Measurement of Odor Development Due to Bacterial Action on Antimicrobial Polyester Fabrics. | |
| Kart et al. | Assessment of the anti-quorum sensing effect of Lactobacillus sp. metabolites on expression levels of QS-related genes in Pseudomonas aeruginosa PAO1 | |
| KR102566917B1 (ko) | 쏘팔메토 오일 및 이의 불포화 지방산을 유효성분으로 함유하는 다중 병원성 미생물 바이오필름 형성 억제용 조성물 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20220913 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20230808 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20230915 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20231017 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20231026 |
|
| R151 | Written notification of patent or utility model registration |
Ref document number: 7376331 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R151 |