JP7206424B2 - mRNAを増幅する方法及び完全長mRNAライブラリを調製する方法 - Google Patents
mRNAを増幅する方法及び完全長mRNAライブラリを調製する方法 Download PDFInfo
- Publication number
- JP7206424B2 JP7206424B2 JP2021569082A JP2021569082A JP7206424B2 JP 7206424 B2 JP7206424 B2 JP 7206424B2 JP 2021569082 A JP2021569082 A JP 2021569082A JP 2021569082 A JP2021569082 A JP 2021569082A JP 7206424 B2 JP7206424 B2 JP 7206424B2
- Authority
- JP
- Japan
- Prior art keywords
- primer
- mrna
- molecule
- cdna
- sample
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
- 238000000034 method Methods 0.000 title claims description 122
- 108020004999 messenger RNA Proteins 0.000 title claims description 97
- 239000002773 nucleotide Substances 0.000 claims description 89
- 125000003729 nucleotide group Chemical group 0.000 claims description 89
- 239000002299 complementary DNA Substances 0.000 claims description 81
- 108091032973 (ribonucleotides)n+m Proteins 0.000 claims description 79
- 230000000295 complement effect Effects 0.000 claims description 45
- 239000005546 dideoxynucleotide Substances 0.000 claims description 39
- 238000012163 sequencing technique Methods 0.000 claims description 37
- 108020004414 DNA Proteins 0.000 claims description 31
- 150000001540 azides Chemical class 0.000 claims description 29
- 238000012650 click reaction Methods 0.000 claims description 20
- 102100034343 Integrase Human genes 0.000 claims description 19
- 238000006243 chemical reaction Methods 0.000 claims description 18
- 108010092799 RNA-directed DNA polymerase Proteins 0.000 claims description 17
- 108010014303 DNA-directed DNA polymerase Proteins 0.000 claims description 16
- 102000016928 DNA-directed DNA polymerase Human genes 0.000 claims description 16
- GNFTZDOKVXKIBK-UHFFFAOYSA-N 3-(2-methoxyethoxy)benzohydrazide Chemical compound COCCOC1=CC=CC(C(=O)NN)=C1 GNFTZDOKVXKIBK-UHFFFAOYSA-N 0.000 claims description 13
- 102000053602 DNA Human genes 0.000 claims description 13
- 150000001345 alkine derivatives Chemical class 0.000 claims description 13
- 108010008286 DNA nucleotidylexotransferase Proteins 0.000 claims description 12
- 102100033215 DNA nucleotidylexotransferase Human genes 0.000 claims description 12
- 230000015572 biosynthetic process Effects 0.000 claims description 12
- 238000012986 modification Methods 0.000 claims description 11
- 230000004048 modification Effects 0.000 claims description 11
- 239000000872 buffer Substances 0.000 claims description 10
- FGUUSXIOTUKUDN-IBGZPJMESA-N C1(=CC=CC=C1)N1C2=C(NC([C@H](C1)NC=1OC(=NN=1)C1=CC=CC=C1)=O)C=CC=C2 Chemical compound C1(=CC=CC=C1)N1C2=C(NC([C@H](C1)NC=1OC(=NN=1)C1=CC=CC=C1)=O)C=CC=C2 FGUUSXIOTUKUDN-IBGZPJMESA-N 0.000 claims description 9
- 102000040430 polynucleotide Human genes 0.000 claims description 9
- 108091033319 polynucleotide Proteins 0.000 claims description 9
- 239000002157 polynucleotide Substances 0.000 claims description 9
- 108020004682 Single-Stranded DNA Proteins 0.000 claims description 4
- 230000001413 cellular effect Effects 0.000 claims description 4
- 201000010099 disease Diseases 0.000 claims description 4
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 claims description 4
- 238000010461 azide-alkyne cycloaddition reaction Methods 0.000 claims description 2
- 238000011835 investigation Methods 0.000 claims description 2
- 238000013507 mapping Methods 0.000 claims description 2
- 239000002904 solvent Substances 0.000 claims description 2
- 230000004077 genetic alteration Effects 0.000 claims 1
- 238000007671 third-generation sequencing Methods 0.000 claims 1
- 239000000523 sample Substances 0.000 description 44
- 238000003199 nucleic acid amplification method Methods 0.000 description 26
- 230000003321 amplification Effects 0.000 description 25
- 210000004027 cell Anatomy 0.000 description 17
- 239000000203 mixture Substances 0.000 description 16
- 239000000047 product Substances 0.000 description 16
- 239000012634 fragment Substances 0.000 description 14
- 108090000623 proteins and genes Proteins 0.000 description 14
- QTBSBXVTEAMEQO-UHFFFAOYSA-N Acetic acid Chemical compound CC(O)=O QTBSBXVTEAMEQO-UHFFFAOYSA-N 0.000 description 12
- 102000039446 nucleic acids Human genes 0.000 description 11
- 108020004707 nucleic acids Proteins 0.000 description 11
- 150000007523 nucleic acids Chemical class 0.000 description 11
- 238000010839 reverse transcription Methods 0.000 description 11
- 239000007787 solid Substances 0.000 description 11
- 102000040650 (ribonucleotides)n+m Human genes 0.000 description 9
- IAZDPXIOMUYVGZ-UHFFFAOYSA-N Dimethylsulphoxide Chemical compound CS(C)=O IAZDPXIOMUYVGZ-UHFFFAOYSA-N 0.000 description 9
- 108091034117 Oligonucleotide Proteins 0.000 description 9
- 239000003153 chemical reaction reagent Substances 0.000 description 9
- 239000000499 gel Substances 0.000 description 8
- 238000002360 preparation method Methods 0.000 description 8
- 239000011541 reaction mixture Substances 0.000 description 8
- 238000012408 PCR amplification Methods 0.000 description 7
- IVRMZWNICZWHMI-UHFFFAOYSA-N azide group Chemical group [N-]=[N+]=[N-] IVRMZWNICZWHMI-UHFFFAOYSA-N 0.000 description 7
- 102000004169 proteins and genes Human genes 0.000 description 6
- 108020004635 Complementary DNA Proteins 0.000 description 5
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 5
- 239000008049 TAE buffer Substances 0.000 description 5
- 239000007983 Tris buffer Substances 0.000 description 5
- HGEVZDLYZYVYHD-UHFFFAOYSA-N acetic acid;2-amino-2-(hydroxymethyl)propane-1,3-diol;2-[2-[bis(carboxymethyl)amino]ethyl-(carboxymethyl)amino]acetic acid Chemical compound CC(O)=O.OCC(N)(CO)CO.OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O HGEVZDLYZYVYHD-UHFFFAOYSA-N 0.000 description 5
- 238000004458 analytical method Methods 0.000 description 5
- 238000009739 binding Methods 0.000 description 5
- 238000001514 detection method Methods 0.000 description 5
- 230000002068 genetic effect Effects 0.000 description 5
- 230000037452 priming Effects 0.000 description 5
- 238000011160 research Methods 0.000 description 5
- 238000007480 sanger sequencing Methods 0.000 description 5
- 239000006228 supernatant Substances 0.000 description 5
- 238000003786 synthesis reaction Methods 0.000 description 5
- 102000004190 Enzymes Human genes 0.000 description 4
- 108090000790 Enzymes Proteins 0.000 description 4
- 108091028043 Nucleic acid sequence Proteins 0.000 description 4
- ISAKRJDGNUQOIC-UHFFFAOYSA-N Uracil Chemical compound O=C1C=CNC(=O)N1 ISAKRJDGNUQOIC-UHFFFAOYSA-N 0.000 description 4
- XSQUKJJJFZCRTK-UHFFFAOYSA-N Urea Chemical compound NC(N)=O XSQUKJJJFZCRTK-UHFFFAOYSA-N 0.000 description 4
- 108020000999 Viral RNA Proteins 0.000 description 4
- 239000011543 agarose gel Substances 0.000 description 4
- 125000004429 atom Chemical group 0.000 description 4
- OPTASPLRGRRNAP-UHFFFAOYSA-N cytosine Chemical compound NC=1C=CNC(=O)N=1 OPTASPLRGRRNAP-UHFFFAOYSA-N 0.000 description 4
- 230000002255 enzymatic effect Effects 0.000 description 4
- UYTPUPDQBNUYGX-UHFFFAOYSA-N guanine Chemical compound O=C1NC(N)=NC2=C1N=CN2 UYTPUPDQBNUYGX-UHFFFAOYSA-N 0.000 description 4
- 238000009396 hybridization Methods 0.000 description 4
- 239000008188 pellet Substances 0.000 description 4
- 230000008569 process Effects 0.000 description 4
- 238000000746 purification Methods 0.000 description 4
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 4
- OSBLTNPMIGYQGY-UHFFFAOYSA-N 2-amino-2-(hydroxymethyl)propane-1,3-diol;2-[2-[bis(carboxymethyl)amino]ethyl-(carboxymethyl)amino]acetic acid;boric acid Chemical compound OB(O)O.OCC(N)(CO)CO.OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O OSBLTNPMIGYQGY-UHFFFAOYSA-N 0.000 description 3
- VAKXPQHQQNOUEZ-UHFFFAOYSA-N 3-[4-[[bis[[1-(3-hydroxypropyl)triazol-4-yl]methyl]amino]methyl]triazol-1-yl]propan-1-ol Chemical compound N1=NN(CCCO)C=C1CN(CC=1N=NN(CCCO)C=1)CC1=CN(CCCO)N=N1 VAKXPQHQQNOUEZ-UHFFFAOYSA-N 0.000 description 3
- 208000024556 Mendelian disease Diseases 0.000 description 3
- 238000003559 RNA-seq method Methods 0.000 description 3
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 3
- 230000008901 benefit Effects 0.000 description 3
- 108010005774 beta-Galactosidase Proteins 0.000 description 3
- 238000010804 cDNA synthesis Methods 0.000 description 3
- NHVNXKFIZYSCEB-XLPZGREQSA-N dTTP Chemical compound O=C1NC(=O)C(C)=CN1[C@@H]1O[C@H](COP(O)(=O)OP(O)(=O)OP(O)(O)=O)[C@@H](O)C1 NHVNXKFIZYSCEB-XLPZGREQSA-N 0.000 description 3
- 238000003745 diagnosis Methods 0.000 description 3
- 238000010790 dilution Methods 0.000 description 3
- 239000012895 dilution Substances 0.000 description 3
- 238000011534 incubation Methods 0.000 description 3
- 239000000463 material Substances 0.000 description 3
- 230000001404 mediated effect Effects 0.000 description 3
- 238000007481 next generation sequencing Methods 0.000 description 3
- 239000000243 solution Substances 0.000 description 3
- 125000006850 spacer group Chemical group 0.000 description 3
- 239000000126 substance Substances 0.000 description 3
- 150000003852 triazoles Chemical group 0.000 description 3
- 229930024421 Adenine Natural products 0.000 description 2
- GFFGJBXGBJISGV-UHFFFAOYSA-N Adenine Chemical compound NC1=NC=NC2=C1N=CN2 GFFGJBXGBJISGV-UHFFFAOYSA-N 0.000 description 2
- 108091033409 CRISPR Proteins 0.000 description 2
- 108020004638 Circular DNA Proteins 0.000 description 2
- 108091026890 Coding region Proteins 0.000 description 2
- RYGMFSIKBFXOCR-UHFFFAOYSA-N Copper Chemical compound [Cu] RYGMFSIKBFXOCR-UHFFFAOYSA-N 0.000 description 2
- 108700039887 Essential Genes Proteins 0.000 description 2
- 108090000331 Firefly luciferases Proteins 0.000 description 2
- 102100031181 Glyceraldehyde-3-phosphate dehydrogenase Human genes 0.000 description 2
- 101710203526 Integrase Proteins 0.000 description 2
- 108091034057 RNA (poly(A)) Proteins 0.000 description 2
- 239000013614 RNA sample Substances 0.000 description 2
- HDRRAMINWIWTNU-NTSWFWBYSA-N [[(2s,5r)-5-(2-amino-6-oxo-3h-purin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl] phosphono hydrogen phosphate Chemical class C1=2NC(N)=NC(=O)C=2N=CN1[C@H]1CC[C@@H](COP(O)(=O)OP(O)(=O)OP(O)(O)=O)O1 HDRRAMINWIWTNU-NTSWFWBYSA-N 0.000 description 2
- 229960000643 adenine Drugs 0.000 description 2
- 125000002355 alkine group Chemical group 0.000 description 2
- ROOXNKNUYICQNP-UHFFFAOYSA-N ammonium persulfate Chemical compound [NH4+].[NH4+].[O-]S(=O)(=O)OOS([O-])(=O)=O ROOXNKNUYICQNP-UHFFFAOYSA-N 0.000 description 2
- -1 azide modified oligonucleotides Chemical class 0.000 description 2
- WQZGKKKJIJFFOK-FPRJBGLDSA-N beta-D-galactose Chemical compound OC[C@H]1O[C@@H](O)[C@H](O)[C@@H](O)[C@H]1O WQZGKKKJIJFFOK-FPRJBGLDSA-N 0.000 description 2
- 239000012472 biological sample Substances 0.000 description 2
- 239000004202 carbamide Substances 0.000 description 2
- 239000012141 concentrate Substances 0.000 description 2
- 229910052802 copper Inorganic materials 0.000 description 2
- 239000010949 copper Substances 0.000 description 2
- 230000001351 cycling effect Effects 0.000 description 2
- 229940104302 cytosine Drugs 0.000 description 2
- RGWHQCVHVJXOKC-SHYZEUOFSA-J dCTP(4-) Chemical compound O=C1N=C(N)C=CN1[C@@H]1O[C@H](COP([O-])(=O)OP([O-])(=O)OP([O-])([O-])=O)[C@@H](O)C1 RGWHQCVHVJXOKC-SHYZEUOFSA-J 0.000 description 2
- HAAZLUGHYHWQIW-KVQBGUIXSA-N dGTP Chemical compound C1=NC=2C(=O)NC(N)=NC=2N1[C@H]1C[C@H](O)[C@@H](COP(O)(=O)OP(O)(=O)OP(O)(O)=O)O1 HAAZLUGHYHWQIW-KVQBGUIXSA-N 0.000 description 2
- 238000010586 diagram Methods 0.000 description 2
- 238000009826 distribution Methods 0.000 description 2
- ZMMJGEGLRURXTF-UHFFFAOYSA-N ethidium bromide Chemical compound [Br-].C12=CC(N)=CC=C2C2=CC=C(N)C=C2[N+](CC)=C1C1=CC=CC=C1 ZMMJGEGLRURXTF-UHFFFAOYSA-N 0.000 description 2
- 229960005542 ethidium bromide Drugs 0.000 description 2
- 238000013467 fragmentation Methods 0.000 description 2
- 238000006062 fragmentation reaction Methods 0.000 description 2
- 108020004445 glyceraldehyde-3-phosphate dehydrogenase Proteins 0.000 description 2
- 238000002372 labelling Methods 0.000 description 2
- 238000011068 loading method Methods 0.000 description 2
- 230000035772 mutation Effects 0.000 description 2
- 229920002401 polyacrylamide Polymers 0.000 description 2
- 239000011535 reaction buffer Substances 0.000 description 2
- 229920002477 rna polymer Polymers 0.000 description 2
- RWQNBRDOKXIBIV-UHFFFAOYSA-N thymine Chemical compound CC1=CNC(=O)NC1=O RWQNBRDOKXIBIV-UHFFFAOYSA-N 0.000 description 2
- 238000013518 transcription Methods 0.000 description 2
- 230000035897 transcription Effects 0.000 description 2
- 229940035893 uracil Drugs 0.000 description 2
- 238000007482 whole exome sequencing Methods 0.000 description 2
- 150000000177 1,2,3-triazoles Chemical class 0.000 description 1
- ZSNNBSPEFVIUDS-SHYZEUOFSA-N 1-[(2r,4s,5s)-4-azido-5-(hydroxymethyl)oxolan-2-yl]pyrimidine-2,4-dione Chemical compound C1[C@H](N=[N+]=[N-])[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C=C1 ZSNNBSPEFVIUDS-SHYZEUOFSA-N 0.000 description 1
- QKNYBSVHEMOAJP-UHFFFAOYSA-N 2-amino-2-(hydroxymethyl)propane-1,3-diol;hydron;chloride Chemical compound Cl.OCC(N)(CO)CO QKNYBSVHEMOAJP-UHFFFAOYSA-N 0.000 description 1
- ASJSAQIRZKANQN-CRCLSJGQSA-N 2-deoxy-D-ribose Chemical group OC[C@@H](O)[C@@H](O)CC=O ASJSAQIRZKANQN-CRCLSJGQSA-N 0.000 description 1
- STRZQWQNZQMHQR-UAKXSSHOSA-N 5-fluorocytidine Chemical compound C1=C(F)C(N)=NC(=O)N1[C@H]1[C@H](O)[C@H](O)[C@@H](CO)O1 STRZQWQNZQMHQR-UAKXSSHOSA-N 0.000 description 1
- OIRDTQYFTABQOQ-KQYNXXCUSA-N Adenosine Natural products C1=NC=2C(N)=NC=NC=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O OIRDTQYFTABQOQ-KQYNXXCUSA-N 0.000 description 1
- 102000002260 Alkaline Phosphatase Human genes 0.000 description 1
- 108020004774 Alkaline Phosphatase Proteins 0.000 description 1
- 239000002126 C01EB10 - Adenosine Substances 0.000 description 1
- 208000005623 Carcinogenesis Diseases 0.000 description 1
- 108010076499 DNA polymerase X Proteins 0.000 description 1
- 238000001712 DNA sequencing Methods 0.000 description 1
- 241000238557 Decapoda Species 0.000 description 1
- 108700024394 Exon Proteins 0.000 description 1
- 108091092195 Intron Proteins 0.000 description 1
- 102000003960 Ligases Human genes 0.000 description 1
- 108090000364 Ligases Proteins 0.000 description 1
- 241001465754 Metazoa Species 0.000 description 1
- KWYHDKDOAIKMQN-UHFFFAOYSA-N N,N,N',N'-tetramethylethylenediamine Chemical compound CN(C)CCN(C)C KWYHDKDOAIKMQN-UHFFFAOYSA-N 0.000 description 1
- 206010028980 Neoplasm Diseases 0.000 description 1
- 102000003832 Nucleotidyltransferases Human genes 0.000 description 1
- 108090000119 Nucleotidyltransferases Proteins 0.000 description 1
- 108020002230 Pancreatic Ribonuclease Proteins 0.000 description 1
- 102000005891 Pancreatic ribonuclease Human genes 0.000 description 1
- 108010002747 Pfu DNA polymerase Proteins 0.000 description 1
- 208000035977 Rare disease Diseases 0.000 description 1
- 239000008051 TBE buffer Substances 0.000 description 1
- 102000008579 Transposases Human genes 0.000 description 1
- 108010020764 Transposases Proteins 0.000 description 1
- 229920004890 Triton X-100 Polymers 0.000 description 1
- 239000013504 Triton X-100 Substances 0.000 description 1
- JZBFFCANYXZLES-KVQBGUIXSA-N [[(2s,3s,5r)-5-(2-amino-6-oxo-3h-purin-9-yl)-3-azidooxolan-2-yl]methoxy-hydroxyphosphoryl] phosphono hydrogen phosphate Chemical compound C1=2NC(N)=NC(=O)C=2N=CN1[C@H]1C[C@H](N=[N+]=[N-])[C@@H](COP(O)(=O)OP(O)(=O)OP(O)(O)=O)O1 JZBFFCANYXZLES-KVQBGUIXSA-N 0.000 description 1
- ARLKCWCREKRROD-POYBYMJQSA-N [[(2s,5r)-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl] phosphono hydrogen phosphate Chemical class O=C1N=C(N)C=CN1[C@@H]1O[C@H](COP(O)(=O)OP(O)(=O)OP(O)(O)=O)CC1 ARLKCWCREKRROD-POYBYMJQSA-N 0.000 description 1
- 230000005856 abnormality Effects 0.000 description 1
- 239000012190 activator Substances 0.000 description 1
- 229960005305 adenosine Drugs 0.000 description 1
- 229910001870 ammonium persulfate Inorganic materials 0.000 description 1
- 239000012491 analyte Substances 0.000 description 1
- 238000010462 azide-alkyne Huisgen cycloaddition reaction Methods 0.000 description 1
- 125000000852 azido group Chemical group *N=[N+]=[N-] 0.000 description 1
- 239000011324 bead Substances 0.000 description 1
- 239000012620 biological material Substances 0.000 description 1
- 239000000090 biomarker Substances 0.000 description 1
- 239000008280 blood Substances 0.000 description 1
- 210000004369 blood Anatomy 0.000 description 1
- 201000011510 cancer Diseases 0.000 description 1
- 230000036952 cancer formation Effects 0.000 description 1
- 238000005251 capillar electrophoresis Methods 0.000 description 1
- 125000004432 carbon atom Chemical group C* 0.000 description 1
- 231100000504 carcinogenesis Toxicity 0.000 description 1
- 238000006555 catalytic reaction Methods 0.000 description 1
- 230000007248 cellular mechanism Effects 0.000 description 1
- 230000004640 cellular pathway Effects 0.000 description 1
- 238000006352 cycloaddition reaction Methods 0.000 description 1
- SUYVUBYJARFZHO-RRKCRQDMSA-N dATP Chemical compound C1=NC=2C(N)=NC=NC=2N1[C@H]1C[C@H](O)[C@@H](COP(O)(=O)OP(O)(=O)OP(O)(O)=O)O1 SUYVUBYJARFZHO-RRKCRQDMSA-N 0.000 description 1
- SUYVUBYJARFZHO-UHFFFAOYSA-N dATP Natural products C1=NC=2C(N)=NC=NC=2N1C1CC(O)C(COP(O)(=O)OP(O)(=O)OP(O)(O)=O)O1 SUYVUBYJARFZHO-UHFFFAOYSA-N 0.000 description 1
- 239000000539 dimer Substances 0.000 description 1
- VHJLVAABSRFDPM-QWWZWVQMSA-N dithiothreitol Chemical compound SC[C@@H](O)[C@H](O)CS VHJLVAABSRFDPM-QWWZWVQMSA-N 0.000 description 1
- 239000012154 double-distilled water Substances 0.000 description 1
- 230000002349 favourable effect Effects 0.000 description 1
- 238000001502 gel electrophoresis Methods 0.000 description 1
- 238000010353 genetic engineering Methods 0.000 description 1
- 238000010438 heat treatment Methods 0.000 description 1
- 125000005842 heteroatom Chemical group 0.000 description 1
- 230000006872 improvement Effects 0.000 description 1
- 238000010348 incorporation Methods 0.000 description 1
- 238000005304 joining Methods 0.000 description 1
- 239000003550 marker Substances 0.000 description 1
- 238000005259 measurement Methods 0.000 description 1
- 230000009456 molecular mechanism Effects 0.000 description 1
- 150000004713 phosphodiesters Chemical class 0.000 description 1
- 230000008488 polyadenylation Effects 0.000 description 1
- HJRIWDYVYNNCFY-UHFFFAOYSA-M potassium;dimethylarsinate Chemical compound [K+].C[As](C)([O-])=O HJRIWDYVYNNCFY-UHFFFAOYSA-M 0.000 description 1
- 230000002265 prevention Effects 0.000 description 1
- 238000005215 recombination Methods 0.000 description 1
- 230000006798 recombination Effects 0.000 description 1
- 230000004044 response Effects 0.000 description 1
- 238000005096 rolling process Methods 0.000 description 1
- 230000035945 sensitivity Effects 0.000 description 1
- 238000010532 solid phase synthesis reaction Methods 0.000 description 1
- 210000000130 stem cell Anatomy 0.000 description 1
- 239000000758 substrate Substances 0.000 description 1
- 238000001308 synthesis method Methods 0.000 description 1
- 230000008685 targeting Effects 0.000 description 1
- 238000012360 testing method Methods 0.000 description 1
- 229940113082 thymine Drugs 0.000 description 1
- 238000013519 translation Methods 0.000 description 1
- LENZDBCJOHFCAS-UHFFFAOYSA-N tris Chemical compound OCC(N)(CO)CO LENZDBCJOHFCAS-UHFFFAOYSA-N 0.000 description 1
- 238000012800 visualization Methods 0.000 description 1
Images











Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q1/00—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions
- C12Q1/68—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions involving nucleic acids
- C12Q1/6844—Nucleic acid amplification reactions
- C12Q1/6853—Nucleic acid amplification reactions using modified primers or templates
- C12Q1/6855—Ligating adaptors
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/10—Processes for the isolation, preparation or purification of DNA or RNA
- C12N15/1096—Processes for the isolation, preparation or purification of DNA or RNA cDNA Synthesis; Subtracted cDNA library construction, e.g. RT, RT-PCR
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/12—Transferases (2.) transferring phosphorus containing groups, e.g. kinases (2.7)
- C12N9/1241—Nucleotidyltransferases (2.7.7)
- C12N9/1276—RNA-directed DNA polymerase (2.7.7.49), i.e. reverse transcriptase or telomerase
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q1/00—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions
- C12Q1/68—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions involving nucleic acids
- C12Q1/6806—Preparing nucleic acids for analysis, e.g. for polymerase chain reaction [PCR] assay
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q2521/00—Reaction characterised by the enzymatic activity
- C12Q2521/10—Nucleotidyl transfering
- C12Q2521/107—RNA dependent DNA polymerase,(i.e. reverse transcriptase)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q2521/00—Reaction characterised by the enzymatic activity
- C12Q2521/10—Nucleotidyl transfering
- C12Q2521/131—Terminal transferase
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q2523/00—Reactions characterised by treatment of reaction samples
- C12Q2523/10—Characterised by chemical treatment
- C12Q2523/109—Characterised by chemical treatment chemical ligation between nucleic acids
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q2525/00—Reactions involving modified oligonucleotides, nucleic acids, or nucleotides
- C12Q2525/10—Modifications characterised by
- C12Q2525/155—Modifications characterised by incorporating/generating a new priming site
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q2525/00—Reactions involving modified oligonucleotides, nucleic acids, or nucleotides
- C12Q2525/10—Modifications characterised by
- C12Q2525/191—Modifications characterised by incorporating an adaptor
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q2531/00—Reactions of nucleic acids characterised by
- C12Q2531/10—Reactions of nucleic acids characterised by the purpose being amplify/increase the copy number of target nucleic acid
- C12Q2531/113—PCR
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q2535/00—Reactions characterised by the assay type for determining the identity of a nucleotide base or a sequence of oligonucleotides
- C12Q2535/122—Massive parallel sequencing
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y207/00—Transferases transferring phosphorus-containing groups (2.7)
- C12Y207/07—Nucleotidyltransferases (2.7.7)
- C12Y207/07049—RNA-directed DNA polymerase (2.7.7.49), i.e. telomerase or reverse-transcriptase
Description
a)増幅対象である少なくとも1つのRNAの3’末端又はその近傍に位置する配列と相補的な配列を含む少なくとも1つの第1プライマーを用意し、少なくとも1つの第1プライマーを少なくとも1つのRNAにハイブリダイズできる条件下で上記少なくとも1つの第1プライマーを上記サンプルに添加する工程;
b)逆転写酵素及びヌクレオチドを上記サンプルに添加することを含む条件下で上記少なくとも1つのRNAを逆転写して、上記少なくとも1つのRNAの完全長cDNAを得る工程;
c)少なくとも過剰なヌクレオチドから上記サンプルを精製する工程;
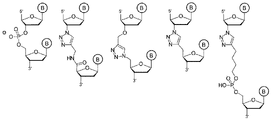
d)アジド分子-アルキン分子ペアの第1パートナーを含むように3’位で修飾されたジデオキシヌクレオチドと、テンプレート非依存的ポリメラーゼとを添加して、工程b)で得られたcDNAの3’末端に単一の3’-アジド又は3’-アルキン修飾ジデオキシヌクレオチドを結合させる工程;
e)少なくとも、工程d)で添加した過剰な修飾ジデオキシヌクレオチドから上記サンプルを精製する工程;
f)ポリヌクレオチド配列と、その5’末端に結合した上記アジド分子-アルキン分子ペアの第2パートナーとを含むアダプター分子を、クリック反応を実施し、且つトリアゾール結合の形成下、工程d)で得られた3’修飾cDNAに上記アダプター分子をライゲーションするための条件下で添加する工程;
g)上記アダプター分子の5’末端のヌクレオチドのうち少なくとも6個と相補的であり、且つその3’末端に、上記cDNAの3’末端のジデオキシヌクレオチドと相補的なヌクレオチドを含むヌクレオチド配列を有する第2プライマーを添加して、上記トリアゾール結合と重なる位置で、工程f)で得られたライゲーションされたアダプター/cDNA分子に上記第2プライマーをハイブリダイズ及び結合させる工程;
あるいは、
h)DNAポリメラーゼを添加して上記第2プライマーを鎖伸長させることで、工程b)で得られた全cDNAと相補的な配列を含む第二鎖DNAを生成する工程;及び
i)工程h)の上記第二鎖DNAを配列決定する工程;若しくは
j)上記第1プライマーの少なくとも一部と同一である第3プライマーを添加し、上記完全長cDNAを増幅して完全長RNAライブラリを得る工程;
又は
h’)DNAポリメラーゼを添加して上記第2プライマーを鎖伸長させると共に、上記第二鎖DNA及び上記全cDNAの配列を決定する工程
を有する方法に関する。
a)増幅対象である少なくとも1つのmRNAの3’末端又はその近傍に位置する配列と相補的な配列を含み、且つその5’末端にアジド分子-アルキン分子ペアの第1パートナーによる修飾を含む少なくとも1つの第1プライマーを用意し、少なくとも1つの第1プライマーを少なくとも1つのmRNAにハイブリダイズできる条件下で上記少なくとも1つの第1プライマーを上記サンプルに添加する工程;
b)逆転写酵素及びヌクレオチドを上記サンプルに添加することを含む条件下で上記少なくとも1つのmRNAを逆転写して、上記少なくとも1つのmRNAの完全長cDNAを得る工程;
c)少なくとも過剰なヌクレオチドから上記サンプルを精製する工程;
d)アジド分子-アルキン分子ペアの第2パートナーを含むように3’位で修飾されたジデオキシヌクレオチドと、テンプレート非依存的ポリメラーゼとを添加して、工程b)で得られたcDNAの3’末端に単一の3’-アジド又は3’-アルキン修飾ジデオキシヌクレオチドを結合させる工程;
e)少なくとも、工程d)で添加した過剰な修飾ジデオキシヌクレオチドから上記サンプルを精製する工程;
f)クリックライゲーション反応を実施して、トリアゾール結合の形成下、環状一本鎖cDNAを生成する工程;
g)上記第1プライマーの5’末端のヌクレオチドのうち少なくとも6個と相補的であり、且つその3’末端に、上記cDNAの3’末端のジデオキシヌクレオチドと相補的なヌクレオチドを含むヌクレオチド配列を有する第2プライマーを添加して、上記トリアゾール結合と重なる位置で、工程f)で得られたライゲーションされた環状第1プライマー/cDNA分子に上記第2プライマーをハイブリダイズ及び結合させる工程;
及び、
h)DNAポリメラーゼを添加して上記第2プライマーを鎖伸長させた後、上記完全長cDNAを増幅する工程;又は
h’)DNAポリメラーゼを添加して上記第2プライマーを鎖伸長させると共に、上記全cDNAを含む上記環状一本鎖DNAの配列を決定する工程
を有する方法に関する。
a)増幅対象である少なくとも1つのRNAの3’末端又はその近傍に位置する配列と相補的な配列を含む第1プライマー;
b)逆転写酵素;
c)アジド分子-アルキン分子ペアの第1パートナーを含むように3’位で修飾されたジデオキシヌクレオチド;
d)テンプレート非依存的ポリメラーゼ;
e)ポリヌクレオチド配列と、その5’末端に結合した上記アジド分子-アルキン分子ペアの第2パートナーとを含むアダプター分子;
f)上記アダプター分子の5’部分のヌクレオチドのうち少なくとも6個と相補的であり、且つその3’末端にc)の上記ジデオキシヌクレオチドと相補的なヌクレオチドを含むヌクレオチド配列を有する第2プライマー;
g)上記第1プライマーの少なくとも一部と同一である第3プライマー
を含むキットに関する。
a)増幅対象である少なくとも1つのmRNAの3’末端又はその近傍に位置する配列と相補的な配列を含み、且つその5’末端にアジド分子-アルキン分子ペアの第1パートナーによる修飾を含む第1プライマー;
b)逆転写酵素;
c)アジド分子-アルキン分子ペアの第2パートナーを含むように3’位で修飾されたジデオキシヌクレオチド;
d)テンプレート非依存的ポリメラーゼ;
e)上記第1プライマーの5’部分のヌクレオチドのうち少なくとも6個と相補的であり、且つその3’末端にc)の上記ジデオキシヌクレオチドと相補的なヌクレオチドを含むヌクレオチド配列を有する第2プライマー
を含むキットである。
a)増幅対象である少なくとも1つの(m)RNAの3’末端又はその近傍に位置する配列と相補的な配列を含む第1プライマー;
b)逆転写酵素;
c)アジド分子-アルキン分子ペアの第1パートナーを含むように3’位で修飾されたジデオキシヌクレオチド;
d)テンプレート非依存的ポリメラーゼ;
e)ポリヌクレオチド配列と、その5’末端に結合した上記アジド分子-アルキン分子ペアの第2パートナーとを含むアダプター分子;
f)上記アダプター分子の5’部分のヌクレオチドのうち少なくとも6個と相補的であり、且つその3’末端にc)の上記ジデオキシヌクレオチドと相補的なヌクレオチドを含むヌクレオチド配列を有する第2プライマー;
g)必要に応じて、上記第1プライマーの少なくとも一部と同一である第3プライマー
を含む。
a)増幅対象である少なくとも1つのmRNAの3’末端又はその近傍に位置する配列と相補的な配列を含み、且つその5’末端にアジド分子-アルキン分子ペアの第1パートナーによる修飾を含む第1プライマー;
b)逆転写酵素;
c)アジド分子-アルキン分子ペアの第2パートナーを含むように3’位で修飾されたジデオキシヌクレオチド;
d)テンプレート非依存的ポリメラーゼ;
e)上記第1プライマーの5’部分のヌクレオチドのうち少なくとも6個と相補的であり、且つその3’末端にc)の上記ジデオキシヌクレオチドと相補的なヌクレオチドを含むヌクレオチド配列を有する第2プライマー
を含む。
h)RNase;
i)全4種の天然由来ヌクレオチド;
j)クリック反応を実施するための試薬;
k)バッファ及び溶媒;
l)cDNA増幅を実施するための試薬;
m)精製用試薬
のうち少なくとも1つを含んでもよい。
内部コントロールとして、GAPDH等のハウスキーピング遺伝子の次に、eGFP(Baseclick)、CleanCap(登録商標)Cas9(Trilink)、CleanCap(登録商標)β-Gal(Trilink)、及びCleanCap(登録商標)Fluc(Trilink)のmRNA(0.1μg)を2μgのJurkat細胞全RNAプールにスパイクした。このRNAプールに、1μLのdNTPミックス(10mM)及び2μLのポリ(dT)プライマー(100μM)を合わせて、RNaseを含まないH2Oで総体積を13μLにした。混合物を65℃で5分間インキュベートし、0℃まで3分間冷却してハイブリダイズした。cDNA合成では、4μLの5×SuperScript IVバッファ、1μLのジチオスレイトール(100mM)、200ユニットのSuperscript IV逆転写酵素を添加し、RNaseを含まない水でフィルアップして総体積を20μLにした。混合物を50℃で20分間、80℃で10分間インキュベートし、4℃まで3分間冷却した。cDNA合成後、3μLの10×RNase Hバッファ、1μLのRNase A(10mg/mL)、1.4μLのRNase H(5U/μL)、及び4μLのシュリンプアルカリフォスファターゼ(1U/μL)、及び0.6μLのdH2Oを添加して、RNA及び過剰のヌクレオチドを除去した。混合物を37℃で25分間、65℃で15分間インキュベートし、0℃まで3分間冷却した後、PCR産物用のメーカー説明書に従ってスピンカラム法(BaseClean Kit)で浄化し、17μLのdH2Oで溶出した。



クリックライゲーション(実施例3に記載の手順)後、特定のPCRフラグメントが得られる標的プライマー(各種の内部リバース及びフォワードプライマー、下記表を参照)を用いて、cDNAプールから特定の遺伝子を増幅した。200μLの反応バイアル中で、11.5μLのdH2O、4μLの5×OneTaq標準反応バッファ、1μLの内部リバースプライマー(10μM)、1μLの内部フォワードプライマー(10μM)、0.4μLのdNTPミックス(10mM)、2μLの精製クリックライゲーションcDNA混合物、及び0.13μLのOneTaq DNAポリメラーゼ(5000U/μL)(New England Biolabs)を合わせた。このサンプルをサーモサイクラー(BIORAD)で熱サイクルプログラムにかけた。


クリックライゲーション(図3の例に記載の手順)後、特定のPCRフラグメントが得られる遺伝子特異的プライマー及び一般的なアダプタープライマー標的プライマー(各種の内部リバースプライマー及びアダプタープライマー1、下記表を参照)を用いて、cDNAプールから特定の遺伝子を増幅した。200μLの反応バイアル中で、11.5μLのdH2O、4μLの5×OneTaq標準反応バッファ、1μLの内部リバースプライマー(10μM)、1μLのアダプタープライマー1(10μM)、0.4μLのdNTPミックス(10mM)、2μLの精製クリックライゲーションcDNA混合物、及び0.13μLのOneTaq DNAポリメラーゼ(5000U/μL)(New England Biolabs)を合わせた。このサンプルをサーモサイクラー(BIORAD)で熱サイクルプログラムにかけた。


トリアゾール読み通しの実現可能性を、モデルRNA配列の逆転写について例示した。RNAをプライマー1にハイブリダイズさせた後、200μMのdTTP、dGTP、dCTP、及び3’-アジド-ddATPの存在下、MuLV逆転写酵素を用いて逆転写した。メーカー説明書に従い、ヌクレオチド除去キット(QIAGEN)を用いてcDNAを精製して、ヌクレオチド及び酵素を除去した。







最終体積25μLのサンプルは、1×末端ヌクレオチジルトランスフェラーゼバッファ(25mM Tris-HCl(pH7.2)、200mMカコジル酸カリウム、0.01%(v/v)Triton X-100、1mM CoCl2)中、1μMのオリゴ2、1mMの3’-アジド-2’,3’-ジデオキシグアノシン-5’-トリフォスフェート又は3’-アジド-2’,3’-ジデオキシウリジン、及びTdT-酵素(2U/μl)を含み、TdT-反応を実施した。反応物を37℃で一晩(12~15時間)インキュベートし、70℃まで10分間加熱して停止した。QIAquick Nucleotide Removal Kitを用いて混合物を精製し、30μLの水で溶出した。


Claims (24)
- サンプルに含まれる少なくとも1つのRNAを増幅及び/又は配列決定する方法であって、
a)増幅対象である少なくとも1つのRNAの3’末端又はその近傍に位置する配列と相補的な配列を含む少なくとも1つの第1プライマーを用意し、少なくとも1つの第1プライマーを少なくとも1つのRNAにハイブリダイズできる条件下で上記少なくとも1つの第1プライマーを上記サンプルに添加する工程;
b)逆転写酵素及びヌクレオチドを上記サンプルに添加することを含む条件下で上記少なくとも1つのRNAを逆転写して、上記少なくとも1つのRNAの完全長cDNAを得る工程;
c)少なくとも過剰なヌクレオチドから上記サンプルを精製する工程;
d)アジド分子-アルキン分子ペアの第1パートナーを含むように3’位で修飾されたジデオキシヌクレオチドと、テンプレート非依存的ポリメラーゼとを添加して、工程b)で得られたcDNAの3’末端に単一の3’-アジド又は3’-アルキン修飾ジデオキシヌクレオチドを結合させる工程;
e)少なくとも、工程d)で添加した過剰な修飾ジデオキシヌクレオチドから上記サンプルを精製する工程;
f)ポリヌクレオチド配列と、その5’末端に結合した上記アジド分子-アルキン分子ペアの第2パートナーとを含むアダプター分子を、クリック反応を実施し、且つトリアゾール結合の形成下、工程d)で得られた3’修飾cDNAに上記アダプター分子をライゲーションするための条件下で添加する工程;及び
g)上記アダプター分子の5’末端のヌクレオチドのうち少なくとも6個と相補的であり、且つその3’末端に、上記cDNAの3’末端のジデオキシヌクレオチドと相補的なヌクレオチドを含むヌクレオチド配列を有する第2プライマーを添加して、上記トリアゾール結合と重なる位置で、工程f)で得られたライゲーションされたアダプター/cDNA分子に上記第2プライマーをハイブリダイズ及び結合させる工程;
を有し、さらに、
工程g)に続いて、
h)DNAポリメラーゼを添加して上記第2プライマーを鎖伸長させることで、工程b)で得られた全cDNAと相補的な配列を含む第二鎖DNAを生成する工程;及び
i)工程h)に続いて、工程h)の上記第二鎖DNAを配列決定する工程;若しくは
j)工程h)に続いて、上記第1プライマーの少なくとも一部と同一である第3プライマーを添加し、上記完全長cDNAを増幅して完全長RNAライブラリを得る工程;
を有するか、又は
工程g)に続いて、
h’)DNAポリメラーゼを添加して上記第2プライマーを鎖伸長させると共に、上記第二鎖DNA及び上記全cDNAの配列を決定する工程
を有する方法。 - サンプルに含まれる少なくとも1つのRNAを増幅及び/又は配列決定する方法であって、
a)増幅対象である少なくとも1つのmRNAの3’末端又はその近傍に位置する配列と相補的な配列を含み、且つその5’末端にアジド分子-アルキン分子ペアの第1パートナーによる修飾を含む少なくとも1つの第1プライマーを用意し、少なくとも1つの第1プライマーを少なくとも1つのmRNAにハイブリダイズできる条件下で上記少なくとも1つの第1プライマーを上記サンプルに添加する工程;
b)逆転写酵素及びヌクレオチドを上記サンプルに添加することを含む条件下で上記少なくとも1つのmRNAを逆転写して、上記少なくとも1つのmRNAの完全長cDNAを得る工程;
c)少なくとも過剰なヌクレオチドから上記サンプルを精製する工程;
d)アジド分子-アルキン分子ペアの第2パートナーを含むように3’位で修飾されたジデオキシヌクレオチドと、テンプレート非依存的ポリメラーゼとを添加して、工程b)で得られたcDNAの3’末端に単一の3’-アジド又は3’-アルキン修飾ジデオキシヌクレオチドを結合させる工程;
e)少なくとも、工程d)で添加した過剰な修飾ジデオキシヌクレオチドから上記サンプルを精製する工程;
f)クリックライゲーション反応を実施して、トリアゾール結合の形成下、環状一本鎖cDNAを生成する工程;
g)上記第1プライマーの5’末端のヌクレオチドのうち少なくとも6個と相補的であり、且つその3’末端に、上記cDNAの3’末端のジデオキシヌクレオチドと相補的なヌクレオチドを含むヌクレオチド配列を有する第2プライマーを添加して、上記トリアゾール結合と重なる位置で、工程f)で得られたライゲーションされた環状第1プライマー/cDNA分子に上記第2プライマーをハイブリダイズ及び結合させる工程;
及び、
h)DNAポリメラーゼを添加して上記第2プライマーを鎖伸長させた後、上記完全長cDNAを増幅する工程、又は
h’)DNAポリメラーゼを添加して上記第2プライマーを鎖伸長させると共に、上記全cDNAを含む上記環状一本鎖DNAの配列を決定する工程
を有する方法。 - 前記RNAがmRNAである、請求項1または2に記載の方法。
- 工程a)において、上記少なくとも1つの第1プライマーとして2~50個のヌクレオチドの5’伸長を含むポリ(dT)プライマーを用意し、上記サンプル中に存在する上記少なくとも1つのmRNAのポリA鎖にハイブリダイズできる条件下で上記サンプルに添加する、請求項1または3に記載の方法。
- 第1プライマーがアンカー配列である、請求項4に記載の方法。
- 工程d)において、上記ジデオキシヌクレオチドとして、3’-アルキン若しくは3’-アジド修飾ddGTP、又は3’-アルキン若しくは3’-アジド修飾ddCTPを使用する、請求項1~5のいずれかに記載の方法。
- 工程d)において、上記ジデオキシヌクレオチドとして3’-アジド修飾ddGTPを使用する、請求項1~6のいずれかに記載の方法。
- 工程d)において、上記ジデオキシヌクレオチドとして3’-アジド-2’,3’-ジデオキシGTP(AzddGTP)を使用する、請求項1~6のいずれかに記載の方法。
- 工程g)において、3’末端にdC又はdGを含む第2プライマーを添加する、請求項1~8のいずれか1項に記載の方法。
- 工程g)において、3’末端にdCを含む第2プライマーを添加する、請求項1~9のいずれか1項に記載の方法。
- 工程g)において、上記ヌクレオチド配列の3’末端の2位にdC又はdGを含む第2プライマーを添加する、請求項1~10のいずれか1項に記載の方法。
- 工程g)において、上記ヌクレオチド配列の3’末端の2位にdCを含む第2プライマーを添加する、請求項1~11のいずれか1項に記載の方法。
- 工程d)において、上記テンプレート非依存的ポリメラーゼとして、末端デオキシヌクレオチジルトランスフェラーゼ(TdT)を使用する、請求項1~12のいずれかに記載の方法。
- 工程f)において、上記アダプター分子の5’末端にアルキンを結合させる、請求項1及び3~13のいずれか1項に記載の方法。
- 工程f)において、上記クリック反応は、銅触媒によるアジド-アルキン環化付加(CuAAC)又は歪み促進型銅フリークリックライゲーション(SPAAC)を含む、請求項1~14のいずれか1項に記載の方法。
- 複数のmRNA分子を含むサンプルから完全長mRNAライブラリを調製するための、請求項1~15のいずれか1項に記載の方法の使用。
- 上記複数のmRNA分子を含むサンプルは、1種以上の細胞の全mRNA又は生物の全エクソームを含む、請求項16に記載の使用。
- サンプルに含まれる複数のmRNA、1種以上の細胞の全mRNA、又は個体の全エクソームを配列決定する方法であって、上記複数のmRNA、上記全細胞mRNA、又は個体のエクソームを含むサンプルを用意する工程;請求項1~15のいずれか1項に記載の方法を実施することで、又は請求項16に記載の使用に従って、完全長mRNAのライブラリを調製する工程;及び増幅されたmRNA又は得られたmRNAライブラリの配列を決定する工程を有する方法。
- ロングリードシーケンシング技術を適用する、請求項18に記載の方法。
- 複合疾患のバリアントマッピング、遺伝子異常の調査のための、請求項18又は19に記載の方法。
- サンプルに含まれる少なくとも1つのRNAを増幅するためのキットであって、
a)増幅対象である少なくとも1つのmRNAの3’末端又はその近傍に位置する配列と相補的な配列を含む第1プライマー;
b)逆転写酵素;
c)アジド分子-アルキン分子ペアの第1パートナーを含むように3’位で修飾されたジデオキシヌクレオチド;
d)テンプレート非依存的ポリメラーゼ;
e)ポリヌクレオチド配列と、その5’末端に結合した上記アジド分子-アルキン分子ペアの第2パートナーとを含むアダプター分子;
f)上記アダプター分子の5’部分のヌクレオチドのうち少なくとも6個と相補的であり、且つその3’末端にc)の上記ジデオキシヌクレオチドと相補的なヌクレオチドを含むヌクレオチド配列を有する第2プライマー;
g)上記第1プライマーの少なくとも一部と同一である第3プライマー
を含むキット。 - サンプルに含まれる少なくとも1つのRNAを増幅及び/又は配列決定するためのキットであって、
a)増幅対象である少なくとも1つのmRNAの3’末端又はその近傍に位置する配列と相補的な配列を含み、且つその5’末端にアジド分子-アルキン分子ペアの第1パートナーによる修飾を含む第1プライマー;
b)逆転写酵素;
c)アジド分子-アルキン分子ペアの第2パートナーを含むように3’位で修飾されたジデオキシヌクレオチド;
d)テンプレート非依存的ポリメラーゼ;
e)上記第1プライマーの5’部分のヌクレオチドのうち少なくとも6個と相補的であり、且つその3’末端にc)の上記ジデオキシヌクレオチドと相補的なヌクレオチドを含むヌクレオチド配列を有する第2プライマー
を含むキット。 - 前記RNAがmRNAである、請求項21または22に記載のキット。
- h)RNase;
i)全4種の天然由来ヌクレオチド;
k)バッファ及び溶媒;
のうち少なくとも1つを更に含む、請求項21~23のいずれか1項に記載のキット。
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| EP19219326.6A EP3842532A1 (en) | 2019-12-23 | 2019-12-23 | Method of amplifying mrnas and for preparing full length mrna libraries |
| EP19219326.6 | 2019-12-23 | ||
| EP20158810 | 2020-02-21 | ||
| EP20158810.0 | 2020-02-21 | ||
| PCT/EP2020/087393 WO2021130151A1 (en) | 2019-12-23 | 2020-12-21 | Method of amplifying mrnas and for preparing full length mrna libraries |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2022534213A JP2022534213A (ja) | 2022-07-28 |
| JP7206424B2 true JP7206424B2 (ja) | 2023-01-17 |
Family
ID=74175803
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2021569082A Active JP7206424B2 (ja) | 2019-12-23 | 2020-12-21 | mRNAを増幅する方法及び完全長mRNAライブラリを調製する方法 |
Country Status (17)
| Country | Link |
|---|---|
| US (1) | US20220333184A1 (ja) |
| EP (1) | EP3947675B1 (ja) |
| JP (1) | JP7206424B2 (ja) |
| KR (1) | KR102445790B1 (ja) |
| CN (1) | CN114341353B (ja) |
| AU (1) | AU2020410970B2 (ja) |
| BR (1) | BR112021021491A2 (ja) |
| CA (1) | CA3137993C (ja) |
| CL (1) | CL2021003338A1 (ja) |
| ES (1) | ES2953918T3 (ja) |
| HU (1) | HUE062776T2 (ja) |
| IL (1) | IL290043B2 (ja) |
| MX (1) | MX2021014430A (ja) |
| PL (1) | PL3947675T3 (ja) |
| SG (1) | SG11202111700UA (ja) |
| WO (1) | WO2021130151A1 (ja) |
| ZA (1) | ZA202108516B (ja) |
Families Citing this family (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2023280156A1 (zh) * | 2021-07-08 | 2023-01-12 | 深圳华大生命科学研究院 | 修饰的核苷或核苷酸 |
| WO2023194331A1 (en) | 2022-04-04 | 2023-10-12 | Ecole Polytechnique Federale De Lausanne (Epfl) | CONSTRUCTION OF SEQUENCING LIBRARIES FROM A RIBONUCLEIC ACID (RNA) USING TAILING AND LIGATION OF cDNA (TLC) |
Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2015026853A2 (en) | 2013-08-19 | 2015-02-26 | Abbott Molecular Inc. | Next-generation sequencing libraries |
| WO2018075785A1 (en) | 2016-10-19 | 2018-04-26 | Illumina, Inc. | Methods for chemical ligation of nucleic acids |
Family Cites Families (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20200361984A1 (en) | 2017-09-29 | 2020-11-19 | Baseclick Gmbh | Click based ligation |
| EP3461832A1 (en) * | 2017-09-29 | 2019-04-03 | baseclick GmbH | Click based ligation |
-
2020
- 2020-12-21 AU AU2020410970A patent/AU2020410970B2/en active Active
- 2020-12-21 BR BR112021021491A patent/BR112021021491A2/pt active Search and Examination
- 2020-12-21 PL PL20838999.9T patent/PL3947675T3/pl unknown
- 2020-12-21 CN CN202080061992.2A patent/CN114341353B/zh active Active
- 2020-12-21 US US17/639,743 patent/US20220333184A1/en active Pending
- 2020-12-21 MX MX2021014430A patent/MX2021014430A/es unknown
- 2020-12-21 HU HUE20838999A patent/HUE062776T2/hu unknown
- 2020-12-21 KR KR1020217037391A patent/KR102445790B1/ko active IP Right Grant
- 2020-12-21 WO PCT/EP2020/087393 patent/WO2021130151A1/en unknown
- 2020-12-21 JP JP2021569082A patent/JP7206424B2/ja active Active
- 2020-12-21 SG SG11202111700UA patent/SG11202111700UA/en unknown
- 2020-12-21 ES ES20838999T patent/ES2953918T3/es active Active
- 2020-12-21 EP EP20838999.9A patent/EP3947675B1/en active Active
- 2020-12-21 IL IL290043A patent/IL290043B2/en unknown
- 2020-12-21 CA CA3137993A patent/CA3137993C/en active Active
-
2021
- 2021-11-02 ZA ZA2021/08516A patent/ZA202108516B/en unknown
- 2021-12-14 CL CL2021003338A patent/CL2021003338A1/es unknown
Patent Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2015026853A2 (en) | 2013-08-19 | 2015-02-26 | Abbott Molecular Inc. | Next-generation sequencing libraries |
| WO2018075785A1 (en) | 2016-10-19 | 2018-04-26 | Illumina, Inc. | Methods for chemical ligation of nucleic acids |
Non-Patent Citations (1)
| Title |
|---|
| Journal of molecular biology,2015年,Vol.427,p.2610-2616 |
Also Published As
| Publication number | Publication date |
|---|---|
| MX2021014430A (es) | 2023-01-05 |
| CL2021003338A1 (es) | 2022-09-02 |
| IL290043B1 (en) | 2023-07-01 |
| ES2953918T3 (es) | 2023-11-17 |
| CN114341353A (zh) | 2022-04-12 |
| EP3947675C0 (en) | 2023-08-09 |
| BR112021021491A2 (pt) | 2022-11-16 |
| PL3947675T3 (pl) | 2023-11-20 |
| EP3947675A1 (en) | 2022-02-09 |
| WO2021130151A1 (en) | 2021-07-01 |
| IL290043A (en) | 2022-03-01 |
| AU2020410970A1 (en) | 2021-11-18 |
| SG11202111700UA (en) | 2021-11-29 |
| HUE062776T2 (hu) | 2023-12-28 |
| JP2022534213A (ja) | 2022-07-28 |
| CA3137993C (en) | 2022-10-11 |
| US20220333184A1 (en) | 2022-10-20 |
| CA3137993A1 (en) | 2021-07-01 |
| AU2020410970B2 (en) | 2022-09-15 |
| KR102445790B1 (ko) | 2022-09-21 |
| KR20210152551A (ko) | 2021-12-15 |
| IL290043B2 (en) | 2023-11-01 |
| CN114341353B (zh) | 2023-01-06 |
| ZA202108516B (en) | 2023-05-31 |
| EP3947675B1 (en) | 2023-08-09 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US20210071171A1 (en) | Compositions and methods for targeted nucleic acid sequence enrichment and high efficiency library generation | |
| US8999677B1 (en) | Method for differentiation of polynucleotide strands | |
| EP3913066A1 (en) | Compositions for rapid nucleic acid library preparation | |
| WO2011032053A1 (en) | Compositions and methods for whole transcriptome analysis | |
| JP2011500092A (ja) | 非ランダムプライマーを用いたcDNA合成の方法 | |
| JP7206424B2 (ja) | mRNAを増幅する方法及び完全長mRNAライブラリを調製する方法 | |
| EP2785865A1 (en) | Method and kit for characterizing rna in a composition | |
| JP2024515076A (ja) | 一本鎖dnaの増幅 | |
| KR20230124636A (ko) | 멀티플렉스 반응에서 표적 서열의 고 감응성 검출을위한 조성물 및 방법 | |
| WO2021202403A1 (en) | Methods and compositions for preparing nucleic acid libraries | |
| EP3842532A1 (en) | Method of amplifying mrnas and for preparing full length mrna libraries | |
| WO2008127901A1 (en) | Region-specific hyperbranched amplification |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20220131 |
|
| A871 | Explanation of circumstances concerning accelerated examination |
Free format text: JAPANESE INTERMEDIATE CODE: A871 Effective date: 20220131 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20220602 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20220816 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20221116 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20221213 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20230104 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 7206424 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |