JP6861422B2 - エストロゲン様作用組成物と、これを含む医薬、飲食品、エストロゲン様作用組成物の製造方法およびハナビラタケのdna塩基配列の利用法 - Google Patents
エストロゲン様作用組成物と、これを含む医薬、飲食品、エストロゲン様作用組成物の製造方法およびハナビラタケのdna塩基配列の利用法 Download PDFInfo
- Publication number
- JP6861422B2 JP6861422B2 JP2019506060A JP2019506060A JP6861422B2 JP 6861422 B2 JP6861422 B2 JP 6861422B2 JP 2019506060 A JP2019506060 A JP 2019506060A JP 2019506060 A JP2019506060 A JP 2019506060A JP 6861422 B2 JP6861422 B2 JP 6861422B2
- Authority
- JP
- Japan
- Prior art keywords
- estrogen
- sparassis crispa
- mycelium
- crispa
- action composition
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
Images
















Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K36/00—Medicinal preparations of undetermined constitution containing material from algae, lichens, fungi or plants, or derivatives thereof, e.g. traditional herbal medicines
- A61K36/06—Fungi, e.g. yeasts
- A61K36/07—Basidiomycota, e.g. Cryptococcus
-
- A—HUMAN NECESSITIES
- A23—FOODS OR FOODSTUFFS; TREATMENT THEREOF, NOT COVERED BY OTHER CLASSES
- A23L—FOODS, FOODSTUFFS OR NON-ALCOHOLIC BEVERAGES, NOT OTHERWISE PROVIDED FOR; PREPARATION OR TREATMENT THEREOF
- A23L33/00—Modifying nutritive qualities of foods; Dietetic products; Preparation or treatment thereof
- A23L33/10—Modifying nutritive qualities of foods; Dietetic products; Preparation or treatment thereof using additives
- A23L33/105—Plant extracts, their artificial duplicates or their derivatives
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K45/00—Medicinal preparations containing active ingredients not provided for in groups A61K31/00 - A61K41/00
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P3/00—Drugs for disorders of the metabolism
- A61P3/06—Antihyperlipidemics
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P43/00—Drugs for specific purposes, not provided for in groups A61P1/00-A61P41/00
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P5/00—Drugs for disorders of the endocrine system
- A61P5/24—Drugs for disorders of the endocrine system of the sex hormones
- A61P5/30—Oestrogens
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P9/00—Drugs for disorders of the cardiovascular system
- A61P9/10—Drugs for disorders of the cardiovascular system for treating ischaemic or atherosclerotic diseases, e.g. antianginal drugs, coronary vasodilators, drugs for myocardial infarction, retinopathy, cerebrovascula insufficiency, renal arteriosclerosis
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N1/00—Microorganisms; Compositions thereof; Processes of propagating, maintaining or preserving microorganisms or compositions thereof; Processes of preparing or isolating a composition containing a microorganism; Culture media therefor
- C12N1/14—Fungi; Culture media therefor
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Medicinal Chemistry (AREA)
- Organic Chemistry (AREA)
- Public Health (AREA)
- Veterinary Medicine (AREA)
- Animal Behavior & Ethology (AREA)
- Pharmacology & Pharmacy (AREA)
- Mycology (AREA)
- Natural Medicines & Medicinal Plants (AREA)
- Biotechnology (AREA)
- Genetics & Genomics (AREA)
- Botany (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Microbiology (AREA)
- Epidemiology (AREA)
- Biomedical Technology (AREA)
- Biochemistry (AREA)
- Medical Informatics (AREA)
- Diabetes (AREA)
- Alternative & Traditional Medicine (AREA)
- General Engineering & Computer Science (AREA)
- Obesity (AREA)
- Polymers & Plastics (AREA)
- Molecular Biology (AREA)
- Urology & Nephrology (AREA)
- Vascular Medicine (AREA)
- Cardiology (AREA)
- Heart & Thoracic Surgery (AREA)
- Nutrition Science (AREA)
- Food Science & Technology (AREA)
- Virology (AREA)
Description
ハナビラタケは未だ全ゲノム塩基配列の決定がされていない。ハナビラタケゲノムの標準化を図るためにハナビラタケの全ゲノム塩基配列の決定を行った。
ハナビラタケ(S.crispa)の種菌株(Scrmy26株)の菌糸体の液体培養物はインタートレード社より提供を受けた。この菌糸体培養物よりゲノム抽出キット(Macherey-Nagel社:Nucleo Bond buffer set III)を用いてゲノムDNA を抽出した。S. crispa菌糸体(Scrmy26株)のゲノムは全ゲノムショットガン配列決定法を用いて配列決定した。
RNA-Seqリードは、最初にHISAT(非特許文献2)の参照ゲノムに整列させ、次いで転写配列を構築するためにStringTie(非特許文献3)によってアセンブルした。得られた15,184転写物を用いてハナビラタケ(S. crispa)の遺伝子モデルのパラメーターについてAUGUSTUS(非特許文献4)を使用し、RNA-seqマッピングデータから得られたエクソンおよびイントロンヒントを入力データとして実行した。
約21.3Gbpのリード(> 500Xカバレッジ)をアセンブルすることによって、39.0Mbのゲノム配列が得られた(表1)。

予測されたハナビラタケ(S. crispa)のプロテオームを、配列決定されている25の糸状菌と比較した。
ハナビラタケ(S. crispa)とともに担子菌類(Basidiomycota)または子嚢菌類(Ascomycota)に分類される26種の菌種を系統発生分析に用いた。 これらの26種の菌類のタンパク質配列を、BLASTPにより、e値<1e-5、ヒット数<500と比較した。次いで、オーソロガス遺伝子を得るためのデフォルトパラメータを用いてOrthoMCL(非特許文献10)によって、895のシングルコピーオーソロガス遺伝子を決定した。 これらの895の遺伝子のマルチ配列アラインメントをMAFFT v7 .309(非特許文献11)ソフトウェアによって計算し、それぞれの種についてひとつの長い配列にまとめた。 次に、アラインメントされた配列の中で保存されたブロック領域を、デフォルトパラメータを用いてGblock 0.91b(非特許文献12)により選別し、最終アラインメント長として221,266aaが得られた。
ハナビラタケ(S. crispa)の進化の歴史は、OrthoMCL分析によって得られたこれらの26の菌類で保存された895の単一コピーオーソロガス遺伝子を用いて構築された系統樹(図2)で調べた。
ハナビラタケ(S. crispa)のゲノム配列における2つの交配型遺伝子座A、Bが異なるコンティグ上に同定された。
A型交配型遺伝子座は、HD1およびHD2ホメオドメイン転写因子およびCoprinopsis cinereaおよびSchizophyllum communeのMIP遺伝子との相同性検索によって同定され、MIPについては相対的に高いタンパク質相同性(66〜67%同一性)が見い出され、HD1およびHD2については低い相同性(28〜34%同一性)が見出された(図3、表4)。
ハナビラタケ(S. crispa)のゲノムにおいて、合計246の糖質関連酵素(CAZymes)が同定された。
糖質関連酵素(CAZymes)は、dbCAN HMM 5.0(デフォルトカットオフ閾値の条件)によるHMM検索およびCAZyデータベースのBLASTP検索(非特許文献20)(e値≦1e-6; 分数比> = 0.2、最大ヒット数= 500)によってそれぞれ別々に分類された。
ハナビラタケ(S. crispa)のゲノム中に、131のグリコシド加水分解酵素(GH)、10の炭水化物エステラーゼ(CE)、61のグリコシルトランスフェラーゼ(GT)、4の多糖リアーゼ、19の炭水化物結合モジュール(CBM)および31の補助活性酵素(AA)を含む合計246個の糖質関連酵素遺伝子(CAZymes)候補が特定された。
報告されている遺伝子を用いたBLASTP検索によって、β-グルカン合成酵素遺伝子の同定を行った。
β-グルカン合成酵素遺伝子の同定は、I型およびII型遺伝子について報告されている遺伝子を用いたBLAST検索(非特許文献19)により行い、触媒ドメインおよび膜貫通領域のドメイン分析は、PfamまたはTMHMM (http://www.cbs.dtu.dk/services/TMHMM/) データベースによって行った。
(2)結果
真菌細胞壁の多糖類のうち、β-1,3-グルカン(β-グルカン)が最も多く、様々な薬理作用のために広く使用されている。β-グルカンは、ウリジン5'-二リン酸(UDP)-グルコースからβ-グルカン合成酵素により合成される。β-グルカン合成酵素は、触媒ドメインおよび膜貫通ドメインからなる膜タンパク質である。Saccharomyces cerevisiaeには、2つの独立したβ-グルカン合成酵素遺伝子(FKS1およびFKS2)が存在する。β-グルカン合成酵素遺伝子の同定は、報告されている遺伝子を用いたBLASTP検索によって実施した。ここで、2つの新規のβ-グルカン合成酵素遺伝子ScrFKS1およびScrFKS2が同定され、それぞれI型およびII型遺伝子に相同であることが判明した(表12、図6)。
ハナビラタケ有効成分の探索のために、菌体の抽出法の確立とその抽出法による抽出効率の評価を行った。
1)乾燥菌体(菌糸体)を水に溶かし、10分間煮沸した。
2)遠心機で分離し、上清をフィルター濾過した。
3)濃縮・乾固し、一部凍結保存した。
4)乾固されたハナビラタケ菌糸体の熱水抽出物(以下、「SCE」と記載する)を滅菌水
で10 mg/ml に調整した。
5)調製した SCE と酢酸エチルを混合した。
6)遠心分離し、酢酸エチル層 (上清)を回収した。
7)5)〜6)の操作を2回行った。
8)酢酸エチル層を濃縮乾固し、SCEの酢酸エチル抽出物(以下、「SCE-EtOAc」と記載する)を得た。
9)SCE-EtOAc をメタノールに溶解した(10 mg/ml)。
10)リン酸化アッセイとHPLC 分析(資料2、3)。
乾燥菌体100%に対し、SCEの抽出効率(回収率)は30%、SCE-EtOAcの抽出効率(回収率)は2.5%であった。
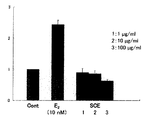
ハナビラタケ菌糸体抽出物のHPLC分画による細胞増殖効果を評価した。方法は(「Brefeldin A is an estrogenic, Erk1/2-activating component in the extract of Agaricusblazei mycelia.」(J.Agri.Food Chem.,2013)に従った。
1)細胞と刺激物質
細胞:MCF-7 細胞
刺激物質:ポジティブコントロール(10 nM エストロゲン(E2))、ネガティブコントロール(0.1 % DMSO)、 SCE(1μg/ml、10μg/ml、100 μg/ml)を使用した。
2)培養:MCF-7 細胞を、1ウェル当たり 0.5×104細胞数になるように計測し、24 wellプレートに接種した。培地は10 %のチャコールデキストラン処理をしたFBS(CD-FBS)を補充したフェノールレッドを含まないRPMI培地を使用した。細胞はCO2インキュベーター内で37℃で3日間培養した。
3)刺激:細胞の接着、増殖を確認した後に、培地を除去し、各刺激物質の入った培地を各ウェルに添加し、細胞をさらに72 時間培養した。
4)固定:培地を除去し、各ウェルをPBSで洗浄した。 10 %トリクロル酢酸を加えて4℃で30分間静置し、蒸留水で洗浄後、乾燥させた。
5)染色: 各ウェ ルに0.4 % SRB/1%酢酸溶液 を加えて細胞を20分間染色した。
6)洗浄: 1 %酢酸で洗浄し、SRB を取り除き、一晩乾燥させた。
7)測定: 各ウェルに10 mMトリス塩酸緩衝液(pH 10.4)を加え、細胞に取り込まれたSRB を溶出させた後に、細胞を96 ウェルプレートに移し、マイクロプレートリーダー (SPECTRA MAX190 Molecular Devices社)を用いて吸光度(OD490)を測定した。
結果を図9に示す。細胞増殖アッセイは再現性を評価するため3 回行った結果である。エストロゲン(E2)がコントロールに対して2倍以上の細胞増殖促進効果を示したのに対して、SCEは増殖活性を有していないことが確認された。この結果は、SCEを体内に摂取しても安全であること根拠になり得る。
エストロゲン応答遺伝子を用いて、化学物質のエストロゲン活性評価用オリゴDNAマイクロアレイ(DNAチップ)を作製し、エストロゲン活性の評価を行った。アッセイシステム(「DNA microarray-based gene expression profiling of estrogenic chemicals」 (Cell. Mol. Life Sci., 2014))を利用して、E2とSCEサンプル(SCE10、SCE100、SCE-EtOAc)の間で細胞刺激時のエストロゲン応答遺伝子の発現のプロファイルをそれぞれ比較し、ハナビラタケ抽出物のエストロゲン活性を評価した。
1)細胞と刺激物質
細胞:ヒト由来MCF-7 細胞
刺激物質:ポジティブコントロール(10 nM エストロゲン(E2))、ネガティブコントロール(0.1 % DMSO)、SCE10 (SCE 10μg/ml)、SCE100 (SCE 100μg/ml)、SCE-EtOAc (ハナビラタケ酢酸エチル抽出分画、10μg/ml)を用いた。
マイクロアレイスキャナーFLA8000(FUJI社)を使用し、測定を行った。
得られたマイクロアレイデータの統計解析は相関解析とクラスター解析により行った。相関解析は以下のように行った。まず、それぞれのエストロゲン応答遺伝子(172 遺伝子)について、各化合物を添加したサンプル(E2+あるいはSCE+)の蛍光強度と化合物を添加しないサンプル(E2−あるいはSCE−)の蛍光強度の比を求めた。次いで、28個の内部コントロール遺伝子を用いて蛍光強度の比を補正した後((E2+/E2−)あるいは(SCE+/SCE−) )、各補正値をlog2値に変換して、それぞれのサンプルについて172 遺伝子の発現の全体像(遺伝子発現プロファイル)を得た。2つのプロファイル間の相関は散布図により示し、相関係数(R値)は直線回帰に基づいて計算した。
DNAマイクロアレイアッセイを用いてE2とSCEサンプルの間で細胞を刺激した時のエストロゲン応答遺伝子の発現のプロファイルをそれぞれ比較したところ、SCE10のR値が0.76、SCE100がR値0.62と、高いことが分かった(図10)。
DNAマイクロアレイアッセイの結果からSCEはE2と似たエストロゲン活性を有していることが分かり、さらにその活性は水分画に溶出されることが分かった。このことから、水分画に溶出されたハナビラタケ菌糸体抽出物分画を逆相カラムを使用して分離し、分離された各分画についてはリン酸化アッセイを用いて活性評価を行った。
1)逆相カラムによる分離
以下の条件で逆相カラムによる分離を行った。
a: 100%H2O → 20% AcN: 0分→ 60分(60分間)
b: 20% AcN → 100% AcN: 60分→ 75分(15分間)
c: 100% AcN → 100% H2O: 75分→ 80分(5分間)
カラム:CAPCELL PAK C18 MG II S5(20 mm×250 mm)
流速:10 ml/min
分離したサンプルは、濃縮・乾固し、重さを量り、滅菌したH2Oで10 mg/mlに調製後、ウェスタンブロット法によるリン酸化アッセイに用いた。
細胞:MCF-7 細胞
刺激物質:ポジティブコントロール(10 nM E2)、SCE(10 μg/ml)、逆相カラムで分離した26サンプル(10 μg/ml)を用いた。
リン酸化タンパク質を検出する抗体:Phospho-p44/42 MAPK(ERK) 、Phospho-Akt(Ser473)を用いた。
1)逆相カラムを用いて上記の溶出条件により、図12に示す溶出パターンを得た。それぞれ図に示すアセトニトリルの濃度の分画を回収した。
2)逆相カラムを用いて2回の分離精製を行ったところ、計26の分画を得ることができた。それらを10 mg/mlに調製後、ウェスタンブロット法によるリン酸化アッセイにより活性評価を行ったところ、図13に示すように、P-ERKにおいては、ポジティブコントロールであるE2やSCEのように刺激後5分間から15分間で活性の上昇が認められた。顕著な例は6B3や6B4、6B6、6B7、6B10である。一方、6B5、6B8のように低い活性を示す分画も確認された。
生物学的安全性評価として、ハナビラタケ菌糸体培養乾燥物およびハナビラタケ菌糸体熱水抽出物SCE(抽出乾燥物)をマウスに単回経口投与し、その急性毒性を評価した。
以下の表7に示す群構成を行い、ハナビラタケ菌糸体培養乾燥物およびハナビラタケ菌糸体抽出乾燥物を雌雄各5匹のICR系マウスに2000 mg/kgでそれぞれ単回経口投与を行い、投与後14日間観察し、14日目の観察終了後、剖検(肉眼的観察)により急性毒性を評価した。
投与後の一般状態、体重推移(図14、図15)および投与後14日の剖検所見においてハナビラタケ菌糸体培養乾燥物およびハナビラタケ菌糸体熱水抽出物SCE(抽出乾燥物)に起因する変化は認められなかった。
高脂肪食を与えたBALB/c.KOR/Stm Slc-Apoeshlマウス(以下、ApoE 欠損マウス)に、ハナビラタケ菌糸体培養乾燥物を180 mg/kg/day、ハナビラタケ菌糸体抽出乾燥物を1000 mg/kg/dayで、それぞれ6週間連日経口投与した。投与期間終了時に血液生化学的検査と内臓脂肪蓄積量の測定を実施した。
(1)方法
高脂肪食を与えたApoE 欠損マウスに、ハナビラタケ菌糸体培養乾燥物を180 mg/kg/day、ハナビラタケ菌糸体抽出乾燥物を1000 mg/kg/dayで、それぞれ6週間連日経口投与した。投与期間終了時に麻酔下で採血後、血液生化学的検査を行った。また内臓脂肪(精巣周囲脂肪、腎臓周囲脂肪)を摘出し、生理食塩水で各臓器に付着している体液を取り除いた後、重量を測定した。
総コレステロール値は、6週間連日経口投与後において、ハナビラタケ菌糸体培養乾燥物、ハナビラタケ菌糸体抽出乾燥物をそれぞれ投与したApoE欠損マウスは非投与群ApoE欠損マウスに比べ、それぞれ統計的有意に低値傾向を示した。中性脂質値も、6週間連日経口投与後において、ハナビラタケ菌糸体培養乾燥物、ハナビラタケ菌糸体抽出乾燥物をそれぞれ投与したApoE欠損マウスは非投与群ApoE欠損マウスに比べ、それぞれ統計的有意に低値傾向を示した。また遊離脂肪酸値も、6週間連日経口投与後において、ハナビラタケ菌糸体培養乾燥物、ハナビラタケ菌糸体抽出乾燥物をそれぞれ投与したApoE欠損マウスは非投与群ApoE欠損マウスに比べ、それぞれ統計的有意に低値傾向を示した(図16)。また内臓脂肪の蓄積量は、6週間連日経口投与後において、ハナビラタケ菌糸体培養乾燥物、ハナビラタケ菌糸体抽出乾燥物をそれぞれ投与したApoE欠損マウスは非投与群ApoE欠損マウスに比べ、それぞれ統計的有意に低値傾向を示した(図17)。
Claims (3)
- エストロゲンの欠乏による障害または疾患を治療または予防するための医薬であって、ハナビラタケ菌糸体の熱水抽出物を含むエストロゲン様作用組成物を含み、
前記障害および前記疾患は、更年期障害、泌尿生殖器障害、前立腺癌、前立腺肥大症、骨粗鬆症、動脈硬化、記憶障害、アルツハイマー病、血栓性疾患、内臓脂肪蓄積のうちの1種または2種以上であることを特徴とする医薬。 - エストロゲンの欠乏による障害または疾患を治療または予防するための飲食品であって、ハナビラタケ菌糸体の熱水抽出物を含むエストロゲン様作用組成物を含み、
前記障害および前記疾患は、更年期障害、泌尿生殖器障害、前立腺癌、前立腺肥大症、骨粗鬆症、動脈硬化、記憶障害、アルツハイマー病、血栓性疾患、内臓脂肪蓄積のうちの1種または2種以上であること特徴とする飲食品。 - 請求項1の医薬または請求項2の飲食品の製造方法であって、
ハナビラタケ菌糸体を熱水に接触させて熱水抽出物を得る工程を含むことを特徴とする製造方法。
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2017047723 | 2017-03-13 | ||
| JP2017047723 | 2017-03-13 | ||
| PCT/JP2018/009817 WO2018168879A1 (ja) | 2017-03-13 | 2018-03-13 | エストロゲン様作用組成物と、これを含む医薬、飲食品、エストロゲン様作用組成物の製造方法およびハナビラタケのdna塩基配列の利用法 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JPWO2018168879A1 JPWO2018168879A1 (ja) | 2020-02-20 |
| JP6861422B2 true JP6861422B2 (ja) | 2021-04-21 |
Family
ID=63523070
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2019506060A Active JP6861422B2 (ja) | 2017-03-13 | 2018-03-13 | エストロゲン様作用組成物と、これを含む医薬、飲食品、エストロゲン様作用組成物の製造方法およびハナビラタケのdna塩基配列の利用法 |
Country Status (2)
| Country | Link |
|---|---|
| JP (1) | JP6861422B2 (ja) |
| WO (1) | WO2018168879A1 (ja) |
Families Citing this family (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP4547184A1 (en) | 2022-06-30 | 2025-05-07 | The Procter & Gamble Company | Absorbent articles and methods for making absorbent articles with frangible pathways |
| JP7440838B1 (ja) | 2023-07-06 | 2024-02-29 | 株式会社インタートレードヘルスケア | 卵巣機能欠落生体の内臓脂肪蓄積抑制飲食品、及び卵巣機能欠落生体の内臓脂肪蓄積抑制薬 |
| KR20250067506A (ko) * | 2023-11-08 | 2025-05-15 | 이희찬 | 침출차 가공공법을 기반으로 한 꽃송이버섯 티백 제조방법 |
Family Cites Families (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2006126488A1 (ja) * | 2005-05-25 | 2006-11-30 | Unitika Ltd. | ハナビラタケ抽出物 |
-
2018
- 2018-03-13 JP JP2019506060A patent/JP6861422B2/ja active Active
- 2018-03-13 WO PCT/JP2018/009817 patent/WO2018168879A1/ja not_active Ceased
Also Published As
| Publication number | Publication date |
|---|---|
| JPWO2018168879A1 (ja) | 2020-02-20 |
| WO2018168879A1 (ja) | 2018-09-20 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| WoldemariamYohannes et al. | Prebiotic, probiotic, antimicrobial, and functional food applications of Bacillus amyloliquefaciens | |
| Wu et al. | Isolation and characterization of three antihypertension peptides from the mycelia of Ganoderma lucidum (Agaricomycetes) | |
| Marcotuli et al. | Non-starch polysaccharides in durum wheat: a review | |
| Jiang et al. | Detection of active mammalian GH31 α-glucosidases in health and disease using in-class, broad-spectrum activity-based probes | |
| Hu et al. | Structure and characteristic of β‐glucan in cereal: A review | |
| Xia et al. | Preparation of saltiness-enhancing enzymatic hydrolyzed pea protein and identification of the functional small peptides of salt reduction | |
| JP6861422B2 (ja) | エストロゲン様作用組成物と、これを含む医薬、飲食品、エストロゲン様作用組成物の製造方法およびハナビラタケのdna塩基配列の利用法 | |
| Mohapatra et al. | In silico investigation of black tea components on α-amylase, α-glucosidase and lipase | |
| Ermawar et al. | Distribution, structure and biosynthetic gene families of (1, 3; 1, 4)‐β‐glucan in Sorghum bicolor | |
| Corzo-Martínez et al. | Tofu whey permeate is an efficient source to enzymatically produce prebiotic fructooligosaccharides and novel fructosylated α-galactosides | |
| CN111568821B (zh) | 包括用冠突曲霉菌株固体发酵的发酵人参提取物的用于改善皮肤美容的组合物 | |
| Wu et al. | The toxicology mechanism of endophytic fungus and swainsonine in locoweed | |
| Lu et al. | Genome survey and transcriptome analysis on mycelia and primordia of Agaricus blazei | |
| JP6679202B2 (ja) | Iv型アレルギー用組成物 | |
| Lee et al. | Metabolomics reveal optimal grain preprocessing (milling) toward rice koji fermentation | |
| KR102309911B1 (ko) | 신규한 아커만시아 뮤시니필라 ak32 균주의 발견 및 장 손상의 예방 또는 치료를 위한 응용 | |
| Salwan et al. | Recent developments in shiitake mushrooms and their nutraceutical importance | |
| Ren et al. | Research progress of α-glucosidase inhibitors produced by microorganisms and their applications | |
| Pan et al. | Structural characteristics and hypoglycemic activity of a polysaccharide from an edible bolete Phlebopus portentosus | |
| US20090227001A1 (en) | Fungal mixture and preparation thereof | |
| TWI590766B (zh) | Thrombotic disease prevention preparation | |
| JP2008212137A (ja) | 生物転換法を利用したペリヌスリンテウス菌糸体の液体培養による人参からの新規のジンセノサイド製造方法 | |
| EP3855934A2 (en) | A consumable product comprising malted dehulled oat | |
| CN105166387B (zh) | 一种银杏叶生物饲料添加剂的制备方法 | |
| JP5684989B2 (ja) | トゲドコロ根茎の酵素処理物及び麹菌発酵処理物 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| AA64 | Notification of invalidation of claim of internal priority (with term) |
Free format text: JAPANESE INTERMEDIATE CODE: A241764 Effective date: 20191126 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20191206 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20191210 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20201027 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20201216 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20210105 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20210302 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20210322 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 6861422 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |