JP6445434B2 - スーパーカインおよびシンセカイン:新規かつ増強されたシグナル伝達活性を有する再利用されるサイトカイン - Google Patents
スーパーカインおよびシンセカイン:新規かつ増強されたシグナル伝達活性を有する再利用されるサイトカイン Download PDFInfo
- Publication number
- JP6445434B2 JP6445434B2 JP2015526711A JP2015526711A JP6445434B2 JP 6445434 B2 JP6445434 B2 JP 6445434B2 JP 2015526711 A JP2015526711 A JP 2015526711A JP 2015526711 A JP2015526711 A JP 2015526711A JP 6445434 B2 JP6445434 B2 JP 6445434B2
- Authority
- JP
- Japan
- Prior art keywords
- receptor
- shared
- cells
- mutein
- ligand
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
- 102000004127 Cytokines Human genes 0.000 title description 94
- 108090000695 Cytokines Proteins 0.000 title description 94
- 230000007781 signaling event Effects 0.000 title 1
- 102000004388 Interleukin-4 Human genes 0.000 claims description 342
- 108090000978 Interleukin-4 Proteins 0.000 claims description 342
- 102000005962 receptors Human genes 0.000 claims description 121
- 108020003175 receptors Proteins 0.000 claims description 121
- 102100020791 Interleukin-13 receptor subunit alpha-1 Human genes 0.000 claims description 78
- 101710112663 Interleukin-13 receptor subunit alpha-1 Proteins 0.000 claims description 78
- 108090000765 processed proteins & peptides Proteins 0.000 claims description 65
- 230000027455 binding Effects 0.000 claims description 60
- 229920001184 polypeptide Polymers 0.000 claims description 59
- 102000004196 processed proteins & peptides Human genes 0.000 claims description 59
- 239000000203 mixture Substances 0.000 claims description 51
- 239000003446 ligand Substances 0.000 claims description 48
- 238000000034 method Methods 0.000 claims description 37
- 102000003675 cytokine receptors Human genes 0.000 claims description 34
- 108010057085 cytokine receptors Proteins 0.000 claims description 34
- 150000001413 amino acids Chemical class 0.000 claims description 28
- 238000006467 substitution reaction Methods 0.000 claims description 23
- 230000002829 reductive effect Effects 0.000 claims description 16
- 238000011282 treatment Methods 0.000 claims description 16
- 125000003275 alpha amino acid group Chemical group 0.000 claims description 15
- 230000009467 reduction Effects 0.000 claims description 11
- 239000003795 chemical substances by application Substances 0.000 claims description 10
- 101001002709 Homo sapiens Interleukin-4 Proteins 0.000 claims description 9
- 102000055229 human IL4 Human genes 0.000 claims description 9
- 239000008194 pharmaceutical composition Substances 0.000 claims description 9
- 230000036755 cellular response Effects 0.000 claims description 6
- 102000018682 Interleukin Receptor Common gamma Subunit Human genes 0.000 claims description 5
- 108010066719 Interleukin Receptor Common gamma Subunit Proteins 0.000 claims description 5
- 108010017511 Interleukin-13 Receptors Proteins 0.000 claims description 4
- 208000027866 inflammatory disease Diseases 0.000 claims description 4
- 102100039078 Interleukin-4 receptor subunit alpha Human genes 0.000 claims 2
- ROMPPAWVATWIKR-UHFFFAOYSA-N 4-[3-(4-chlorophenyl)-1,2,4-oxadiazol-5-yl]butanoic acid Chemical compound O1C(CCCC(=O)O)=NC(C=2C=CC(Cl)=CC=2)=N1 ROMPPAWVATWIKR-UHFFFAOYSA-N 0.000 claims 1
- 229940028885 interleukin-4 Drugs 0.000 description 285
- 210000004027 cell Anatomy 0.000 description 149
- 230000014509 gene expression Effects 0.000 description 61
- 108090000623 proteins and genes Proteins 0.000 description 46
- 210000001616 monocyte Anatomy 0.000 description 41
- 230000026731 phosphorylation Effects 0.000 description 38
- 238000006366 phosphorylation reaction Methods 0.000 description 38
- 102000003816 Interleukin-13 Human genes 0.000 description 32
- 108090000176 Interleukin-13 Proteins 0.000 description 32
- 210000001744 T-lymphocyte Anatomy 0.000 description 32
- 102000004169 proteins and genes Human genes 0.000 description 31
- 235000001014 amino acid Nutrition 0.000 description 29
- 238000002474 experimental method Methods 0.000 description 29
- 235000018102 proteins Nutrition 0.000 description 29
- 230000004044 response Effects 0.000 description 29
- 229940024606 amino acid Drugs 0.000 description 27
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 27
- 108010017213 Granulocyte-Macrophage Colony-Stimulating Factor Proteins 0.000 description 25
- 102100039620 Granulocyte-macrophage colony-stimulating factor Human genes 0.000 description 25
- 210000004443 dendritic cell Anatomy 0.000 description 25
- 230000000694 effects Effects 0.000 description 25
- YBJHBAHKTGYVGT-ZKWXMUAHSA-N (+)-Biotin Chemical group N1C(=O)N[C@@H]2[C@H](CCCCC(=O)O)SC[C@@H]21 YBJHBAHKTGYVGT-ZKWXMUAHSA-N 0.000 description 22
- 230000004069 differentiation Effects 0.000 description 20
- 238000004458 analytical method Methods 0.000 description 19
- 230000001965 increasing effect Effects 0.000 description 18
- 230000004913 activation Effects 0.000 description 17
- 230000000638 stimulation Effects 0.000 description 17
- 102000008300 Mutant Proteins Human genes 0.000 description 16
- 108010021466 Mutant Proteins Proteins 0.000 description 16
- 210000003719 b-lymphocyte Anatomy 0.000 description 16
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 15
- 208000035475 disorder Diseases 0.000 description 15
- 239000000523 sample Substances 0.000 description 15
- 101000946889 Homo sapiens Monocyte differentiation antigen CD14 Proteins 0.000 description 14
- 102100035877 Monocyte differentiation antigen CD14 Human genes 0.000 description 14
- 208000006673 asthma Diseases 0.000 description 14
- 230000011664 signaling Effects 0.000 description 13
- 125000000539 amino acid group Chemical group 0.000 description 12
- 230000008901 benefit Effects 0.000 description 12
- 201000010099 disease Diseases 0.000 description 12
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 11
- 102000000535 Type II Interleukin-4 Receptors Human genes 0.000 description 11
- 108010002120 Type II Interleukin-4 Receptors Proteins 0.000 description 11
- 229960002685 biotin Drugs 0.000 description 11
- 235000020958 biotin Nutrition 0.000 description 11
- 239000011616 biotin Substances 0.000 description 11
- 230000006698 induction Effects 0.000 description 11
- -1 lipidoids Substances 0.000 description 11
- 238000000684 flow cytometry Methods 0.000 description 10
- 108020004999 messenger RNA Proteins 0.000 description 10
- 150000003839 salts Chemical class 0.000 description 10
- OKKJLVBELUTLKV-UHFFFAOYSA-N Methanol Chemical compound OC OKKJLVBELUTLKV-UHFFFAOYSA-N 0.000 description 9
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 9
- 230000008859 change Effects 0.000 description 9
- 239000013078 crystal Substances 0.000 description 9
- 230000006870 function Effects 0.000 description 9
- 230000003993 interaction Effects 0.000 description 9
- 210000002540 macrophage Anatomy 0.000 description 9
- 238000007427 paired t-test Methods 0.000 description 9
- 102000010787 Interleukin-4 Receptors Human genes 0.000 description 8
- 108010038486 Interleukin-4 Receptors Proteins 0.000 description 8
- 206010028980 Neoplasm Diseases 0.000 description 8
- 201000011510 cancer Diseases 0.000 description 8
- 230000003828 downregulation Effects 0.000 description 8
- 210000000265 leukocyte Anatomy 0.000 description 8
- 239000002609 medium Substances 0.000 description 8
- 210000000440 neutrophil Anatomy 0.000 description 8
- 229920000642 polymer Polymers 0.000 description 8
- 102100020793 Interleukin-13 receptor subunit alpha-2 Human genes 0.000 description 7
- 102000009509 Type I Interleukin-4 Receptors Human genes 0.000 description 7
- 108010009386 Type I Interleukin-4 Receptors Proteins 0.000 description 7
- 239000003814 drug Substances 0.000 description 7
- 238000009472 formulation Methods 0.000 description 7
- 238000000338 in vitro Methods 0.000 description 7
- 108040003607 interleukin-13 receptor activity proteins Proteins 0.000 description 7
- 108040006852 interleukin-4 receptor activity proteins Proteins 0.000 description 7
- 238000003032 molecular docking Methods 0.000 description 7
- 239000002245 particle Substances 0.000 description 7
- 230000035755 proliferation Effects 0.000 description 7
- 230000001105 regulatory effect Effects 0.000 description 7
- 239000000243 solution Substances 0.000 description 7
- QTBSBXVTEAMEQO-UHFFFAOYSA-N Acetic acid Chemical compound CC(O)=O QTBSBXVTEAMEQO-UHFFFAOYSA-N 0.000 description 6
- 206010020649 Hyperkeratosis Diseases 0.000 description 6
- 208000001126 Keratosis Diseases 0.000 description 6
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 6
- 239000004698 Polyethylene Substances 0.000 description 6
- 230000000903 blocking effect Effects 0.000 description 6
- 230000037396 body weight Effects 0.000 description 6
- 230000024245 cell differentiation Effects 0.000 description 6
- KRKNYBCHXYNGOX-UHFFFAOYSA-N citric acid Chemical compound OC(=O)CC(O)(C(O)=O)CC(O)=O KRKNYBCHXYNGOX-UHFFFAOYSA-N 0.000 description 6
- 239000007788 liquid Substances 0.000 description 6
- 210000004698 lymphocyte Anatomy 0.000 description 6
- 239000000463 material Substances 0.000 description 6
- 238000005259 measurement Methods 0.000 description 6
- 239000000047 product Substances 0.000 description 6
- 230000019491 signal transduction Effects 0.000 description 6
- 208000017520 skin disease Diseases 0.000 description 6
- 238000010186 staining Methods 0.000 description 6
- UCSJYZPVAKXKNQ-HZYVHMACSA-N streptomycin Chemical compound CN[C@H]1[C@H](O)[C@@H](O)[C@H](CO)O[C@H]1O[C@@H]1[C@](C=O)(O)[C@H](C)O[C@H]1O[C@@H]1[C@@H](NC(N)=N)[C@H](O)[C@@H](NC(N)=N)[C@H](O)[C@H]1O UCSJYZPVAKXKNQ-HZYVHMACSA-N 0.000 description 6
- 238000002198 surface plasmon resonance spectroscopy Methods 0.000 description 6
- 238000012360 testing method Methods 0.000 description 6
- 210000001519 tissue Anatomy 0.000 description 6
- QWPXBEHQFHACTK-KZVYIGENSA-N (10e,12e)-86-chloro-12,14,4-trihydroxy-85,14-dimethoxy-33,2,7,10-tetramethyl-15,16-dihydro-14h-7-aza-1(6,4)-oxazina-3(2,3)-oxirana-8(1,3)-benzenacyclotetradecaphane-10,12-dien-6-one Chemical compound CN1C(=O)CC(O)C2(C)OC2C(C)C(OC(=O)N2)CC2(O)C(OC)\C=C\C=C(C)\CC2=CC(OC)=C(Cl)C1=C2 QWPXBEHQFHACTK-KZVYIGENSA-N 0.000 description 5
- 201000004624 Dermatitis Diseases 0.000 description 5
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 5
- 102100025092 Insulin receptor substrate 2 Human genes 0.000 description 5
- 108010002335 Interleukin-9 Proteins 0.000 description 5
- 102000000585 Interleukin-9 Human genes 0.000 description 5
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 5
- 238000007792 addition Methods 0.000 description 5
- 238000013459 approach Methods 0.000 description 5
- 239000011324 bead Substances 0.000 description 5
- 230000003111 delayed effect Effects 0.000 description 5
- 238000001514 detection method Methods 0.000 description 5
- 210000002919 epithelial cell Anatomy 0.000 description 5
- 239000000499 gel Substances 0.000 description 5
- 230000003834 intracellular effect Effects 0.000 description 5
- 230000001404 mediated effect Effects 0.000 description 5
- 230000035772 mutation Effects 0.000 description 5
- 210000004967 non-hematopoietic stem cell Anatomy 0.000 description 5
- 239000000546 pharmaceutical excipient Substances 0.000 description 5
- 230000003389 potentiating effect Effects 0.000 description 5
- 239000000843 powder Substances 0.000 description 5
- 230000008569 process Effects 0.000 description 5
- 230000028327 secretion Effects 0.000 description 5
- 239000011780 sodium chloride Substances 0.000 description 5
- 208000024891 symptom Diseases 0.000 description 5
- 231100000419 toxicity Toxicity 0.000 description 5
- 230000001988 toxicity Effects 0.000 description 5
- 108020004414 DNA Proteins 0.000 description 4
- LYCAIKOWRPUZTN-UHFFFAOYSA-N Ethylene glycol Chemical compound OCCO LYCAIKOWRPUZTN-UHFFFAOYSA-N 0.000 description 4
- 101001077604 Homo sapiens Insulin receptor substrate 1 Proteins 0.000 description 4
- 102100025087 Insulin receptor substrate 1 Human genes 0.000 description 4
- 101710201820 Insulin receptor substrate 2 Proteins 0.000 description 4
- PXHVJJICTQNCMI-UHFFFAOYSA-N Nickel Chemical compound [Ni] PXHVJJICTQNCMI-UHFFFAOYSA-N 0.000 description 4
- NBIIXXVUZAFLBC-UHFFFAOYSA-N Phosphoric acid Chemical compound OP(O)(O)=O NBIIXXVUZAFLBC-UHFFFAOYSA-N 0.000 description 4
- 229920002319 Poly(methyl acrylate) Polymers 0.000 description 4
- 208000002200 Respiratory Hypersensitivity Diseases 0.000 description 4
- 230000002159 abnormal effect Effects 0.000 description 4
- 239000000443 aerosol Substances 0.000 description 4
- 239000000556 agonist Substances 0.000 description 4
- 230000010085 airway hyperresponsiveness Effects 0.000 description 4
- 238000003556 assay Methods 0.000 description 4
- 230000015572 biosynthetic process Effects 0.000 description 4
- 210000004369 blood Anatomy 0.000 description 4
- 239000008280 blood Substances 0.000 description 4
- 210000004899 c-terminal region Anatomy 0.000 description 4
- 230000001413 cellular effect Effects 0.000 description 4
- 229920002678 cellulose Chemical class 0.000 description 4
- 229920001577 copolymer Polymers 0.000 description 4
- 238000003745 diagnosis Methods 0.000 description 4
- 229940079593 drug Drugs 0.000 description 4
- 239000012636 effector Substances 0.000 description 4
- 239000003102 growth factor Substances 0.000 description 4
- 230000002209 hydrophobic effect Effects 0.000 description 4
- 230000005764 inhibitory process Effects 0.000 description 4
- 238000002826 magnetic-activated cell sorting Methods 0.000 description 4
- 238000004519 manufacturing process Methods 0.000 description 4
- 108020004707 nucleic acids Proteins 0.000 description 4
- 102000039446 nucleic acids Human genes 0.000 description 4
- 150000007523 nucleic acids Chemical class 0.000 description 4
- 230000037361 pathway Effects 0.000 description 4
- 229920001223 polyethylene glycol Polymers 0.000 description 4
- 239000000126 substance Substances 0.000 description 4
- 230000001225 therapeutic effect Effects 0.000 description 4
- 102000042286 type I cytokine receptor family Human genes 0.000 description 4
- 108091052247 type I cytokine receptor family Proteins 0.000 description 4
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Chemical compound O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 4
- MTCFGRXMJLQNBG-REOHCLBHSA-N (2S)-2-Amino-3-hydroxypropansäure Chemical compound OC[C@H](N)C(O)=O MTCFGRXMJLQNBG-REOHCLBHSA-N 0.000 description 3
- FYGDTMLNYKFZSV-URKRLVJHSA-N (2s,3r,4s,5s,6r)-2-[(2r,4r,5r,6s)-4,5-dihydroxy-2-(hydroxymethyl)-6-[(2r,4r,5r,6s)-4,5,6-trihydroxy-2-(hydroxymethyl)oxan-3-yl]oxyoxan-3-yl]oxy-6-(hydroxymethyl)oxane-3,4,5-triol Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@H]1OC1[C@@H](CO)O[C@@H](OC2[C@H](O[C@H](O)[C@H](O)[C@H]2O)CO)[C@H](O)[C@H]1O FYGDTMLNYKFZSV-URKRLVJHSA-N 0.000 description 3
- 208000024893 Acute lymphoblastic leukemia Diseases 0.000 description 3
- 102100024222 B-lymphocyte antigen CD19 Human genes 0.000 description 3
- 229920002498 Beta-glucan Polymers 0.000 description 3
- 102000004190 Enzymes Human genes 0.000 description 3
- 108090000790 Enzymes Proteins 0.000 description 3
- AEMRFAOFKBGASW-UHFFFAOYSA-N Glycolic acid Polymers OCC(O)=O AEMRFAOFKBGASW-UHFFFAOYSA-N 0.000 description 3
- 101000980825 Homo sapiens B-lymphocyte antigen CD19 Proteins 0.000 description 3
- 101001057504 Homo sapiens Interferon-stimulated gene 20 kDa protein Proteins 0.000 description 3
- 101001055144 Homo sapiens Interleukin-2 receptor subunit alpha Proteins 0.000 description 3
- 101000878605 Homo sapiens Low affinity immunoglobulin epsilon Fc receptor Proteins 0.000 description 3
- 206010020751 Hypersensitivity Diseases 0.000 description 3
- 206010061218 Inflammation Diseases 0.000 description 3
- 102100027268 Interferon-stimulated gene 20 kDa protein Human genes 0.000 description 3
- 108010050904 Interferons Proteins 0.000 description 3
- 102000014150 Interferons Human genes 0.000 description 3
- 102000003814 Interleukin-10 Human genes 0.000 description 3
- 108090000174 Interleukin-10 Proteins 0.000 description 3
- 108010002350 Interleukin-2 Proteins 0.000 description 3
- 102000000588 Interleukin-2 Human genes 0.000 description 3
- 102100030703 Interleukin-22 Human genes 0.000 description 3
- 108010002386 Interleukin-3 Proteins 0.000 description 3
- 102000000646 Interleukin-3 Human genes 0.000 description 3
- 108090001005 Interleukin-6 Proteins 0.000 description 3
- 108010019421 Janus Kinase 3 Proteins 0.000 description 3
- AYFVYJQAPQTCCC-GBXIJSLDSA-N L-threonine Chemical compound C[C@@H](O)[C@H](N)C(O)=O AYFVYJQAPQTCCC-GBXIJSLDSA-N 0.000 description 3
- 102100038007 Low affinity immunoglobulin epsilon Fc receptor Human genes 0.000 description 3
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 3
- QWPXBEHQFHACTK-UHFFFAOYSA-N Maytansinol Natural products CN1C(=O)CC(O)C2(C)OC2C(C)C(OC(=O)N2)CC2(O)C(OC)C=CC=C(C)CC2=CC(OC)=C(Cl)C1=C2 QWPXBEHQFHACTK-UHFFFAOYSA-N 0.000 description 3
- 241000699670 Mus sp. Species 0.000 description 3
- 108091007960 PI3Ks Proteins 0.000 description 3
- 206010034277 Pemphigoid Diseases 0.000 description 3
- 229930182555 Penicillin Natural products 0.000 description 3
- JGSARLDLIJGVTE-MBNYWOFBSA-N Penicillin G Chemical compound N([C@H]1[C@H]2SC([C@@H](N2C1=O)C(O)=O)(C)C)C(=O)CC1=CC=CC=C1 JGSARLDLIJGVTE-MBNYWOFBSA-N 0.000 description 3
- 102000003993 Phosphatidylinositol 3-kinases Human genes 0.000 description 3
- 108090000430 Phosphatidylinositol 3-kinases Proteins 0.000 description 3
- 229920002732 Polyanhydride Polymers 0.000 description 3
- 239000004793 Polystyrene Substances 0.000 description 3
- OFOBLEOULBTSOW-UHFFFAOYSA-N Propanedioic acid Natural products OC(=O)CC(O)=O OFOBLEOULBTSOW-UHFFFAOYSA-N 0.000 description 3
- 108010076504 Protein Sorting Signals Proteins 0.000 description 3
- 201000004681 Psoriasis Diseases 0.000 description 3
- 239000002253 acid Substances 0.000 description 3
- 208000026935 allergic disease Diseases 0.000 description 3
- 239000000427 antigen Substances 0.000 description 3
- 102000036639 antigens Human genes 0.000 description 3
- 108091007433 antigens Proteins 0.000 description 3
- 230000006907 apoptotic process Effects 0.000 description 3
- 201000008937 atopic dermatitis Diseases 0.000 description 3
- 208000010668 atopic eczema Diseases 0.000 description 3
- 239000011230 binding agent Substances 0.000 description 3
- 229920002988 biodegradable polymer Polymers 0.000 description 3
- 239000004621 biodegradable polymer Substances 0.000 description 3
- 239000000872 buffer Substances 0.000 description 3
- 239000001913 cellulose Chemical class 0.000 description 3
- 235000010980 cellulose Nutrition 0.000 description 3
- 239000003153 chemical reaction reagent Substances 0.000 description 3
- 238000012217 deletion Methods 0.000 description 3
- 230000037430 deletion Effects 0.000 description 3
- 230000001419 dependent effect Effects 0.000 description 3
- 239000000539 dimer Substances 0.000 description 3
- 231100000673 dose–response relationship Toxicity 0.000 description 3
- 239000003937 drug carrier Substances 0.000 description 3
- 239000000839 emulsion Substances 0.000 description 3
- 230000002349 favourable effect Effects 0.000 description 3
- 238000001943 fluorescence-activated cell sorting Methods 0.000 description 3
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Natural products OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 description 3
- 239000001963 growth medium Substances 0.000 description 3
- 230000003394 haemopoietic effect Effects 0.000 description 3
- 210000003958 hematopoietic stem cell Anatomy 0.000 description 3
- 210000003630 histaminocyte Anatomy 0.000 description 3
- 210000005260 human cell Anatomy 0.000 description 3
- 239000001257 hydrogen Substances 0.000 description 3
- 229910052739 hydrogen Inorganic materials 0.000 description 3
- 230000001976 improved effect Effects 0.000 description 3
- 230000004054 inflammatory process Effects 0.000 description 3
- 238000001990 intravenous administration Methods 0.000 description 3
- PGHMRUGBZOYCAA-ADZNBVRBSA-N ionomycin Chemical compound O1[C@H](C[C@H](O)[C@H](C)[C@H](O)[C@H](C)/C=C/C[C@@H](C)C[C@@H](C)C(/O)=C/C(=O)[C@@H](C)C[C@@H](C)C[C@@H](CCC(O)=O)C)CC[C@@]1(C)[C@@H]1O[C@](C)([C@@H](C)O)CC1 PGHMRUGBZOYCAA-ADZNBVRBSA-N 0.000 description 3
- PGHMRUGBZOYCAA-UHFFFAOYSA-N ionomycin Natural products O1C(CC(O)C(C)C(O)C(C)C=CCC(C)CC(C)C(O)=CC(=O)C(C)CC(C)CC(CCC(O)=O)C)CCC1(C)C1OC(C)(C(C)O)CC1 PGHMRUGBZOYCAA-UHFFFAOYSA-N 0.000 description 3
- 208000032839 leukemia Diseases 0.000 description 3
- 210000004072 lung Anatomy 0.000 description 3
- 230000035800 maturation Effects 0.000 description 3
- 238000002493 microarray Methods 0.000 description 3
- 230000004048 modification Effects 0.000 description 3
- 238000012986 modification Methods 0.000 description 3
- 239000000178 monomer Substances 0.000 description 3
- 230000001613 neoplastic effect Effects 0.000 description 3
- 239000002674 ointment Substances 0.000 description 3
- 230000001575 pathological effect Effects 0.000 description 3
- 229940049954 penicillin Drugs 0.000 description 3
- 210000003819 peripheral blood mononuclear cell Anatomy 0.000 description 3
- 229920001606 poly(lactic acid-co-glycolic acid) Polymers 0.000 description 3
- 229920002223 polystyrene Polymers 0.000 description 3
- 238000002360 preparation method Methods 0.000 description 3
- 239000003755 preservative agent Substances 0.000 description 3
- 238000002818 protein evolution Methods 0.000 description 3
- 238000003753 real-time PCR Methods 0.000 description 3
- 230000007115 recruitment Effects 0.000 description 3
- 230000004043 responsiveness Effects 0.000 description 3
- 210000002966 serum Anatomy 0.000 description 3
- 239000007921 spray Substances 0.000 description 3
- 238000010561 standard procedure Methods 0.000 description 3
- 210000000130 stem cell Anatomy 0.000 description 3
- 229960005322 streptomycin Drugs 0.000 description 3
- 239000000829 suppository Substances 0.000 description 3
- 239000000725 suspension Substances 0.000 description 3
- 208000011580 syndromic disease Diseases 0.000 description 3
- 238000012546 transfer Methods 0.000 description 3
- 235000002374 tyrosine Nutrition 0.000 description 3
- 125000001493 tyrosinyl group Chemical group [H]OC1=C([H])C([H])=C(C([H])=C1[H])C([H])([H])C([H])(N([H])[H])C(*)=O 0.000 description 3
- 241000701447 unidentified baculovirus Species 0.000 description 3
- 230000003827 upregulation Effects 0.000 description 3
- 230000002792 vascular Effects 0.000 description 3
- 210000005253 yeast cell Anatomy 0.000 description 3
- VQZYZXLBKBUOHE-UHFFFAOYSA-N (2,5-dioxopyrrolidin-1-yl) 3-(pyridin-2-yldisulfanyl)butanoate Chemical compound C=1C=CC=NC=1SSC(C)CC(=O)ON1C(=O)CCC1=O VQZYZXLBKBUOHE-UHFFFAOYSA-N 0.000 description 2
- GTBCXYYVWHFQRS-UHFFFAOYSA-N (2,5-dioxopyrrolidin-1-yl) 4-(pyridin-2-yldisulfanyl)pentanoate Chemical compound C=1C=CC=NC=1SSC(C)CCC(=O)ON1C(=O)CCC1=O GTBCXYYVWHFQRS-UHFFFAOYSA-N 0.000 description 2
- JKMHFZQWWAIEOD-UHFFFAOYSA-N 2-[4-(2-hydroxyethyl)piperazin-1-yl]ethanesulfonic acid Chemical compound OCC[NH+]1CCN(CCS([O-])(=O)=O)CC1 JKMHFZQWWAIEOD-UHFFFAOYSA-N 0.000 description 2
- FWBHETKCLVMNFS-UHFFFAOYSA-N 4',6-Diamino-2-phenylindol Chemical compound C1=CC(C(=N)N)=CC=C1C1=CC2=CC=C(C(N)=N)C=C2N1 FWBHETKCLVMNFS-UHFFFAOYSA-N 0.000 description 2
- 208000014697 Acute lymphocytic leukaemia Diseases 0.000 description 2
- 208000036762 Acute promyelocytic leukaemia Diseases 0.000 description 2
- 208000016683 Adult T-cell leukemia/lymphoma Diseases 0.000 description 2
- 206010059313 Anogenital warts Diseases 0.000 description 2
- 239000004475 Arginine Substances 0.000 description 2
- IJGRMHOSHXDMSA-UHFFFAOYSA-N Atomic nitrogen Chemical compound N#N IJGRMHOSHXDMSA-UHFFFAOYSA-N 0.000 description 2
- VTYYLEPIZMXCLO-UHFFFAOYSA-L Calcium carbonate Chemical compound [Ca+2].[O-]C([O-])=O VTYYLEPIZMXCLO-UHFFFAOYSA-L 0.000 description 2
- CURLTUGMZLYLDI-UHFFFAOYSA-N Carbon dioxide Chemical compound O=C=O CURLTUGMZLYLDI-UHFFFAOYSA-N 0.000 description 2
- 201000009030 Carcinoma Diseases 0.000 description 2
- 108010012236 Chemokines Proteins 0.000 description 2
- 102000019034 Chemokines Human genes 0.000 description 2
- 206010012438 Dermatitis atopic Diseases 0.000 description 2
- 108010010803 Gelatin Proteins 0.000 description 2
- WHUUTDBJXJRKMK-UHFFFAOYSA-N Glutamic acid Natural products OC(=O)C(N)CCC(O)=O WHUUTDBJXJRKMK-UHFFFAOYSA-N 0.000 description 2
- 239000004471 Glycine Substances 0.000 description 2
- 239000007995 HEPES buffer Substances 0.000 description 2
- 102000006354 HLA-DR Antigens Human genes 0.000 description 2
- 108010058597 HLA-DR Antigens Proteins 0.000 description 2
- 108010093488 His-His-His-His-His-His Proteins 0.000 description 2
- 101100005713 Homo sapiens CD4 gene Proteins 0.000 description 2
- 101000763314 Homo sapiens Thrombomodulin Proteins 0.000 description 2
- 241000701806 Human papillomavirus Species 0.000 description 2
- CPELXLSAUQHCOX-UHFFFAOYSA-N Hydrogen bromide Chemical compound Br CPELXLSAUQHCOX-UHFFFAOYSA-N 0.000 description 2
- 108090000177 Interleukin-11 Proteins 0.000 description 2
- 102000004559 Interleukin-13 Receptors Human genes 0.000 description 2
- 102000003812 Interleukin-15 Human genes 0.000 description 2
- 108090000172 Interleukin-15 Proteins 0.000 description 2
- 108010002616 Interleukin-5 Proteins 0.000 description 2
- 102000000743 Interleukin-5 Human genes 0.000 description 2
- 108010002586 Interleukin-7 Proteins 0.000 description 2
- 102000000704 Interleukin-7 Human genes 0.000 description 2
- 125000000998 L-alanino group Chemical group [H]N([*])[C@](C([H])([H])[H])([H])C(=O)O[H] 0.000 description 2
- DCXYFEDJOCDNAF-REOHCLBHSA-N L-asparagine Chemical compound OC(=O)[C@@H](N)CC(N)=O DCXYFEDJOCDNAF-REOHCLBHSA-N 0.000 description 2
- CKLJMWTZIZZHCS-REOHCLBHSA-N L-aspartic acid Chemical compound OC(=O)[C@@H](N)CC(O)=O CKLJMWTZIZZHCS-REOHCLBHSA-N 0.000 description 2
- 229930182816 L-glutamine Natural products 0.000 description 2
- AGPKZVBTJJNPAG-WHFBIAKZSA-N L-isoleucine Chemical compound CC[C@H](C)[C@H](N)C(O)=O AGPKZVBTJJNPAG-WHFBIAKZSA-N 0.000 description 2
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 2
- COLNVLDHVKWLRT-QMMMGPOBSA-N L-phenylalanine Chemical compound OC(=O)[C@@H](N)CC1=CC=CC=C1 COLNVLDHVKWLRT-QMMMGPOBSA-N 0.000 description 2
- 208000019693 Lung disease Diseases 0.000 description 2
- AFVFQIVMOAPDHO-UHFFFAOYSA-N Methanesulfonic acid Chemical compound CS(O)(=O)=O AFVFQIVMOAPDHO-UHFFFAOYSA-N 0.000 description 2
- 108700011259 MicroRNAs Proteins 0.000 description 2
- 108091028043 Nucleic acid sequence Proteins 0.000 description 2
- 241000101040 Pityriasis Species 0.000 description 2
- 239000004952 Polyamide Substances 0.000 description 2
- 229920000954 Polyglycolide Polymers 0.000 description 2
- 208000006664 Precursor Cell Lymphoblastic Leukemia-Lymphoma Diseases 0.000 description 2
- 208000033766 Prolymphocytic Leukemia Diseases 0.000 description 2
- 208000033826 Promyelocytic Acute Leukemia Diseases 0.000 description 2
- 229920002472 Starch Chemical class 0.000 description 2
- QAOWNCQODCNURD-UHFFFAOYSA-N Sulfuric acid Chemical compound OS(O)(=O)=O QAOWNCQODCNURD-UHFFFAOYSA-N 0.000 description 2
- 208000031673 T-Cell Cutaneous Lymphoma Diseases 0.000 description 2
- 102100026966 Thrombomodulin Human genes 0.000 description 2
- 102000004887 Transforming Growth Factor beta Human genes 0.000 description 2
- 108090001012 Transforming Growth Factor beta Proteins 0.000 description 2
- 102100025387 Tyrosine-protein kinase JAK3 Human genes 0.000 description 2
- KZSNJWFQEVHDMF-UHFFFAOYSA-N Valine Chemical compound CC(C)C(N)C(O)=O KZSNJWFQEVHDMF-UHFFFAOYSA-N 0.000 description 2
- 208000016025 Waldenstroem macroglobulinemia Diseases 0.000 description 2
- 208000033559 Waldenström macroglobulinemia Diseases 0.000 description 2
- 150000007513 acids Chemical class 0.000 description 2
- 239000012190 activator Substances 0.000 description 2
- 239000004480 active ingredient Substances 0.000 description 2
- 239000013543 active substance Substances 0.000 description 2
- 201000006966 adult T-cell leukemia Diseases 0.000 description 2
- 208000037883 airway inflammation Diseases 0.000 description 2
- 239000013566 allergen Substances 0.000 description 2
- 229910000147 aluminium phosphate Inorganic materials 0.000 description 2
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 2
- 238000007845 assembly PCR Methods 0.000 description 2
- 229920001400 block copolymer Polymers 0.000 description 2
- 210000001185 bone marrow Anatomy 0.000 description 2
- KQNZDYYTLMIZCT-KQPMLPITSA-N brefeldin A Chemical compound O[C@@H]1\C=C\C(=O)O[C@@H](C)CCC\C=C\[C@@H]2C[C@H](O)C[C@H]21 KQNZDYYTLMIZCT-KQPMLPITSA-N 0.000 description 2
- JUMGSHROWPPKFX-UHFFFAOYSA-N brefeldin-A Natural products CC1CCCC=CC2(C)CC(O)CC2(C)C(O)C=CC(=O)O1 JUMGSHROWPPKFX-UHFFFAOYSA-N 0.000 description 2
- 239000002775 capsule Substances 0.000 description 2
- 239000000969 carrier Substances 0.000 description 2
- OSASVXMJTNOKOY-UHFFFAOYSA-N chlorobutanol Chemical compound CC(C)(O)C(Cl)(Cl)Cl OSASVXMJTNOKOY-UHFFFAOYSA-N 0.000 description 2
- HVYWMOMLDIMFJA-DPAQBDIFSA-N cholesterol Chemical compound C1C=C2C[C@@H](O)CC[C@]2(C)[C@@H]2[C@@H]1[C@@H]1CC[C@H]([C@H](C)CCCC(C)C)[C@@]1(C)CC2 HVYWMOMLDIMFJA-DPAQBDIFSA-N 0.000 description 2
- 238000000576 coating method Methods 0.000 description 2
- 239000002299 complementary DNA Substances 0.000 description 2
- 239000006071 cream Substances 0.000 description 2
- 238000002425 crystallisation Methods 0.000 description 2
- 230000008025 crystallization Effects 0.000 description 2
- 239000012228 culture supernatant Substances 0.000 description 2
- 201000007241 cutaneous T cell lymphoma Diseases 0.000 description 2
- 230000003247 decreasing effect Effects 0.000 description 2
- 210000004207 dermis Anatomy 0.000 description 2
- 238000010586 diagram Methods 0.000 description 2
- 239000002552 dosage form Substances 0.000 description 2
- 231100000371 dose-limiting toxicity Toxicity 0.000 description 2
- 239000006196 drop Substances 0.000 description 2
- 238000012377 drug delivery Methods 0.000 description 2
- 230000002500 effect on skin Effects 0.000 description 2
- 238000005516 engineering process Methods 0.000 description 2
- 210000002615 epidermis Anatomy 0.000 description 2
- 230000005714 functional activity Effects 0.000 description 2
- 239000007789 gas Substances 0.000 description 2
- 229920000159 gelatin Polymers 0.000 description 2
- 239000008273 gelatin Substances 0.000 description 2
- 235000019322 gelatine Nutrition 0.000 description 2
- 235000011852 gelatine desserts Nutrition 0.000 description 2
- 239000008187 granular material Substances 0.000 description 2
- 201000009277 hairy cell leukemia Diseases 0.000 description 2
- 210000002216 heart Anatomy 0.000 description 2
- 230000009610 hypersensitivity Effects 0.000 description 2
- 206010021198 ichthyosis Diseases 0.000 description 2
- 210000002865 immune cell Anatomy 0.000 description 2
- 238000002513 implantation Methods 0.000 description 2
- 230000006872 improvement Effects 0.000 description 2
- 238000011534 incubation Methods 0.000 description 2
- 230000001939 inductive effect Effects 0.000 description 2
- 230000028709 inflammatory response Effects 0.000 description 2
- 238000001802 infusion Methods 0.000 description 2
- 238000002347 injection Methods 0.000 description 2
- 239000007924 injection Substances 0.000 description 2
- NOESYZHRGYRDHS-UHFFFAOYSA-N insulin Chemical compound N1C(=O)C(NC(=O)C(CCC(N)=O)NC(=O)C(CCC(O)=O)NC(=O)C(C(C)C)NC(=O)C(NC(=O)CN)C(C)CC)CSSCC(C(NC(CO)C(=O)NC(CC(C)C)C(=O)NC(CC=2C=CC(O)=CC=2)C(=O)NC(CCC(N)=O)C(=O)NC(CC(C)C)C(=O)NC(CCC(O)=O)C(=O)NC(CC(N)=O)C(=O)NC(CC=2C=CC(O)=CC=2)C(=O)NC(CSSCC(NC(=O)C(C(C)C)NC(=O)C(CC(C)C)NC(=O)C(CC=2C=CC(O)=CC=2)NC(=O)C(CC(C)C)NC(=O)C(C)NC(=O)C(CCC(O)=O)NC(=O)C(C(C)C)NC(=O)C(CC(C)C)NC(=O)C(CC=2NC=NC=2)NC(=O)C(CO)NC(=O)CNC2=O)C(=O)NCC(=O)NC(CCC(O)=O)C(=O)NC(CCCNC(N)=N)C(=O)NCC(=O)NC(CC=3C=CC=CC=3)C(=O)NC(CC=3C=CC=CC=3)C(=O)NC(CC=3C=CC(O)=CC=3)C(=O)NC(C(C)O)C(=O)N3C(CCC3)C(=O)NC(CCCCN)C(=O)NC(C)C(O)=O)C(=O)NC(CC(N)=O)C(O)=O)=O)NC(=O)C(C(C)CC)NC(=O)C(CO)NC(=O)C(C(C)O)NC(=O)C1CSSCC2NC(=O)C(CC(C)C)NC(=O)C(NC(=O)C(CCC(N)=O)NC(=O)C(CC(N)=O)NC(=O)C(NC(=O)C(N)CC=1C=CC=CC=1)C(C)C)CC1=CN=CN1 NOESYZHRGYRDHS-UHFFFAOYSA-N 0.000 description 2
- 229940079322 interferon Drugs 0.000 description 2
- 108010074108 interleukin-21 Proteins 0.000 description 2
- 238000007918 intramuscular administration Methods 0.000 description 2
- 238000007912 intraperitoneal administration Methods 0.000 description 2
- 238000002955 isolation Methods 0.000 description 2
- 210000002510 keratinocyte Anatomy 0.000 description 2
- JVTAAEKCZFNVCJ-UHFFFAOYSA-N lactic acid Chemical compound CC(O)C(O)=O JVTAAEKCZFNVCJ-UHFFFAOYSA-N 0.000 description 2
- 238000007726 management method Methods 0.000 description 2
- 239000003550 marker Substances 0.000 description 2
- BDAGIHXWWSANSR-UHFFFAOYSA-N methanoic acid Natural products OC=O BDAGIHXWWSANSR-UHFFFAOYSA-N 0.000 description 2
- 239000002679 microRNA Substances 0.000 description 2
- 239000011325 microbead Substances 0.000 description 2
- 201000005962 mycosis fungoides Diseases 0.000 description 2
- 239000013642 negative control Substances 0.000 description 2
- 229910052759 nickel Inorganic materials 0.000 description 2
- 239000002773 nucleotide Chemical group 0.000 description 2
- 125000003729 nucleotide group Chemical group 0.000 description 2
- 238000007254 oxidation reaction Methods 0.000 description 2
- FJKROLUGYXJWQN-UHFFFAOYSA-N papa-hydroxy-benzoic acid Natural products OC(=O)C1=CC=C(O)C=C1 FJKROLUGYXJWQN-UHFFFAOYSA-N 0.000 description 2
- 230000008823 permeabilization Effects 0.000 description 2
- 229920001281 polyalkylene Polymers 0.000 description 2
- 229920002647 polyamide Polymers 0.000 description 2
- 229920001610 polycaprolactone Polymers 0.000 description 2
- 229920002635 polyurethane Polymers 0.000 description 2
- 239000004814 polyurethane Substances 0.000 description 2
- 229920002451 polyvinyl alcohol Polymers 0.000 description 2
- 239000002243 precursor Substances 0.000 description 2
- 230000002265 prevention Effects 0.000 description 2
- 208000025638 primary cutaneous T-cell non-Hodgkin lymphoma Diseases 0.000 description 2
- 230000002062 proliferating effect Effects 0.000 description 2
- 238000003127 radioimmunoassay Methods 0.000 description 2
- 230000033300 receptor internalization Effects 0.000 description 2
- 230000002441 reversible effect Effects 0.000 description 2
- YGSDEFSMJLZEOE-UHFFFAOYSA-N salicylic acid Chemical compound OC(=O)C1=CC=CC=C1O YGSDEFSMJLZEOE-UHFFFAOYSA-N 0.000 description 2
- 230000035945 sensitivity Effects 0.000 description 2
- 210000003491 skin Anatomy 0.000 description 2
- DAEPDZWVDSPTHF-UHFFFAOYSA-M sodium pyruvate Chemical compound [Na+].CC(=O)C([O-])=O DAEPDZWVDSPTHF-UHFFFAOYSA-M 0.000 description 2
- 239000007787 solid Substances 0.000 description 2
- 235000019698 starch Nutrition 0.000 description 2
- 238000007619 statistical method Methods 0.000 description 2
- 230000004936 stimulating effect Effects 0.000 description 2
- 239000006190 sub-lingual tablet Substances 0.000 description 2
- 238000007920 subcutaneous administration Methods 0.000 description 2
- 239000006228 supernatant Substances 0.000 description 2
- 230000001629 suppression Effects 0.000 description 2
- 238000013268 sustained release Methods 0.000 description 2
- 239000012730 sustained-release form Substances 0.000 description 2
- 238000003786 synthesis reaction Methods 0.000 description 2
- 239000003826 tablet Substances 0.000 description 2
- ZRKFYGHZFMAOKI-QMGMOQQFSA-N tgfbeta Chemical compound C([C@H](NC(=O)[C@H](C(C)C)NC(=O)CNC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@H](CC(C)C)NC(=O)CNC(=O)[C@H](C)NC(=O)[C@H](CO)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@@H](NC(=O)[C@H](C)NC(=O)[C@H](C)NC(=O)[C@@H](NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CCSC)C(C)C)[C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)N[C@@H](C)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(=O)N1[C@@H](CCC1)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O)C1=CC=C(O)C=C1 ZRKFYGHZFMAOKI-QMGMOQQFSA-N 0.000 description 2
- 229940124597 therapeutic agent Drugs 0.000 description 2
- JOXIMZWYDAKGHI-UHFFFAOYSA-N toluene-4-sulfonic acid Chemical compound CC1=CC=C(S(O)(=O)=O)C=C1 JOXIMZWYDAKGHI-UHFFFAOYSA-N 0.000 description 2
- 230000000699 topical effect Effects 0.000 description 2
- OUYCCCASQSFEME-UHFFFAOYSA-N tyrosine Chemical group OC(=O)C(N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-UHFFFAOYSA-N 0.000 description 2
- NQPDZGIKBAWPEJ-UHFFFAOYSA-N valeric acid Chemical compound CCCCC(O)=O NQPDZGIKBAWPEJ-UHFFFAOYSA-N 0.000 description 2
- 239000003981 vehicle Substances 0.000 description 2
- 230000003612 virological effect Effects 0.000 description 2
- DGVVWUTYPXICAM-UHFFFAOYSA-N β‐Mercaptoethanol Chemical compound OCCS DGVVWUTYPXICAM-UHFFFAOYSA-N 0.000 description 2
- JWDFQMWEFLOOED-UHFFFAOYSA-N (2,5-dioxopyrrolidin-1-yl) 3-(pyridin-2-yldisulfanyl)propanoate Chemical compound O=C1CCC(=O)N1OC(=O)CCSSC1=CC=CC=N1 JWDFQMWEFLOOED-UHFFFAOYSA-N 0.000 description 1
- KIUKXJAPPMFGSW-DNGZLQJQSA-N (2S,3S,4S,5R,6R)-6-[(2S,3R,4R,5S,6R)-3-Acetamido-2-[(2S,3S,4R,5R,6R)-6-[(2R,3R,4R,5S,6R)-3-acetamido-2,5-dihydroxy-6-(hydroxymethyl)oxan-4-yl]oxy-2-carboxy-4,5-dihydroxyoxan-3-yl]oxy-5-hydroxy-6-(hydroxymethyl)oxan-4-yl]oxy-3,4,5-trihydroxyoxane-2-carboxylic acid Chemical compound CC(=O)N[C@H]1[C@H](O)O[C@H](CO)[C@@H](O)[C@@H]1O[C@H]1[C@H](O)[C@@H](O)[C@H](O[C@H]2[C@@H]([C@@H](O[C@H]3[C@@H]([C@@H](O)[C@H](O)[C@H](O3)C(O)=O)O)[C@H](O)[C@@H](CO)O2)NC(C)=O)[C@@H](C(O)=O)O1 KIUKXJAPPMFGSW-DNGZLQJQSA-N 0.000 description 1
- DDMOUSALMHHKOS-UHFFFAOYSA-N 1,2-dichloro-1,1,2,2-tetrafluoroethane Chemical compound FC(F)(Cl)C(F)(F)Cl DDMOUSALMHHKOS-UHFFFAOYSA-N 0.000 description 1
- 108020004463 18S ribosomal RNA Proteins 0.000 description 1
- OSWFIVFLDKOXQC-UHFFFAOYSA-N 4-(3-methoxyphenyl)aniline Chemical compound COC1=CC=CC(C=2C=CC(N)=CC=2)=C1 OSWFIVFLDKOXQC-UHFFFAOYSA-N 0.000 description 1
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 description 1
- UZOVYGYOLBIAJR-UHFFFAOYSA-N 4-isocyanato-4'-methyldiphenylmethane Chemical compound C1=CC(C)=CC=C1CC1=CC=C(N=C=O)C=C1 UZOVYGYOLBIAJR-UHFFFAOYSA-N 0.000 description 1
- LLLVZDVNHNWSDS-UHFFFAOYSA-N 4-methylidene-3,5-dioxabicyclo[5.2.2]undeca-1(9),7,10-triene-2,6-dione Chemical compound C1(C2=CC=C(C(=O)OC(=C)O1)C=C2)=O LLLVZDVNHNWSDS-UHFFFAOYSA-N 0.000 description 1
- SQDAZGGFXASXDW-UHFFFAOYSA-N 5-bromo-2-(trifluoromethoxy)pyridine Chemical compound FC(F)(F)OC1=CC=C(Br)C=N1 SQDAZGGFXASXDW-UHFFFAOYSA-N 0.000 description 1
- ZCYVEMRRCGMTRW-UHFFFAOYSA-N 7553-56-2 Chemical compound [I] ZCYVEMRRCGMTRW-UHFFFAOYSA-N 0.000 description 1
- 101150106774 9 gene Proteins 0.000 description 1
- 208000002874 Acne Vulgaris Diseases 0.000 description 1
- 206010000830 Acute leukaemia Diseases 0.000 description 1
- 208000031261 Acute myeloid leukaemia Diseases 0.000 description 1
- 229920001817 Agar Polymers 0.000 description 1
- 102000009027 Albumins Human genes 0.000 description 1
- 108010088751 Albumins Proteins 0.000 description 1
- DCXYFEDJOCDNAF-UHFFFAOYSA-N Asparagine Natural products OC(=O)C(N)CC(N)=O DCXYFEDJOCDNAF-UHFFFAOYSA-N 0.000 description 1
- 208000010839 B-cell chronic lymphocytic leukemia Diseases 0.000 description 1
- 208000032791 BCR-ABL1 positive chronic myelogenous leukemia Diseases 0.000 description 1
- 102100032367 C-C motif chemokine 5 Human genes 0.000 description 1
- 102100032366 C-C motif chemokine 7 Human genes 0.000 description 1
- 102000000496 Carboxypeptidases A Human genes 0.000 description 1
- 108010080937 Carboxypeptidases A Proteins 0.000 description 1
- 201000000274 Carcinosarcoma Diseases 0.000 description 1
- 102000014914 Carrier Proteins Human genes 0.000 description 1
- 229920000623 Cellulose acetate phthalate Polymers 0.000 description 1
- DQEFEBPAPFSJLV-UHFFFAOYSA-N Cellulose propionate Chemical compound CCC(=O)OCC1OC(OC(=O)CC)C(OC(=O)CC)C(OC(=O)CC)C1OC1C(OC(=O)CC)C(OC(=O)CC)C(OC(=O)CC)C(COC(=O)CC)O1 DQEFEBPAPFSJLV-UHFFFAOYSA-N 0.000 description 1
- 229920002284 Cellulose triacetate Polymers 0.000 description 1
- 229920001287 Chondroitin sulfate Polymers 0.000 description 1
- 108020004705 Codon Proteins 0.000 description 1
- 102000008186 Collagen Human genes 0.000 description 1
- 108010035532 Collagen Proteins 0.000 description 1
- 102100032218 Cytokine-inducible SH2-containing protein Human genes 0.000 description 1
- IGXWBGJHJZYPQS-SSDOTTSWSA-N D-Luciferin Chemical compound OC(=O)[C@H]1CSC(C=2SC3=CC=C(O)C=C3N=2)=N1 IGXWBGJHJZYPQS-SSDOTTSWSA-N 0.000 description 1
- CYCGRDQQIOGCKX-UHFFFAOYSA-N Dehydro-luciferin Natural products OC(=O)C1=CSC(C=2SC3=CC(O)=CC=C3N=2)=N1 CYCGRDQQIOGCKX-UHFFFAOYSA-N 0.000 description 1
- 206010012434 Dermatitis allergic Diseases 0.000 description 1
- 206010012455 Dermatitis exfoliative Diseases 0.000 description 1
- 229920002307 Dextran Polymers 0.000 description 1
- FEWJPZIEWOKRBE-JCYAYHJZSA-N Dextrotartaric acid Chemical compound OC(=O)[C@H](O)[C@@H](O)C(O)=O FEWJPZIEWOKRBE-JCYAYHJZSA-N 0.000 description 1
- 239000004338 Dichlorodifluoromethane Substances 0.000 description 1
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 1
- 241000196324 Embryophyta Species 0.000 description 1
- 241000792859 Enema Species 0.000 description 1
- 206010015150 Erythema Diseases 0.000 description 1
- 239000001856 Ethyl cellulose Substances 0.000 description 1
- ZZSNKZQZMQGXPY-UHFFFAOYSA-N Ethyl cellulose Chemical compound CCOCC1OC(OC)C(OCC)C(OCC)C1OC1C(O)C(O)C(OC)C(CO)O1 ZZSNKZQZMQGXPY-UHFFFAOYSA-N 0.000 description 1
- 108010087819 Fc receptors Proteins 0.000 description 1
- 102000009109 Fc receptors Human genes 0.000 description 1
- 206010016654 Fibrosis Diseases 0.000 description 1
- BJGNCJDXODQBOB-UHFFFAOYSA-N Fivefly Luciferin Natural products OC(=O)C1CSC(C=2SC3=CC(O)=CC=C3N=2)=N1 BJGNCJDXODQBOB-UHFFFAOYSA-N 0.000 description 1
- 101150074790 G3 gene Proteins 0.000 description 1
- 206010059024 Gastrointestinal toxicity Diseases 0.000 description 1
- 108010017080 Granulocyte Colony-Stimulating Factor Proteins 0.000 description 1
- 102000004269 Granulocyte Colony-Stimulating Factor Human genes 0.000 description 1
- 108010043121 Green Fluorescent Proteins Proteins 0.000 description 1
- 102000004144 Green Fluorescent Proteins Human genes 0.000 description 1
- 102100034221 Growth-regulated alpha protein Human genes 0.000 description 1
- 108010041384 HLA-DPA antigen Proteins 0.000 description 1
- 208000036066 Hemophagocytic Lymphohistiocytosis Diseases 0.000 description 1
- 208000032672 Histiocytosis haematophagic Diseases 0.000 description 1
- 208000017604 Hodgkin disease Diseases 0.000 description 1
- 208000010747 Hodgkins lymphoma Diseases 0.000 description 1
- 101000797762 Homo sapiens C-C motif chemokine 5 Proteins 0.000 description 1
- 101000797758 Homo sapiens C-C motif chemokine 7 Proteins 0.000 description 1
- 101000943420 Homo sapiens Cytokine-inducible SH2-containing protein Proteins 0.000 description 1
- 101001069921 Homo sapiens Growth-regulated alpha protein Proteins 0.000 description 1
- 101001077600 Homo sapiens Insulin receptor substrate 2 Proteins 0.000 description 1
- 101000982628 Homo sapiens Prolyl 3-hydroxylase OGFOD1 Proteins 0.000 description 1
- 101001059454 Homo sapiens Serine/threonine-protein kinase MARK2 Proteins 0.000 description 1
- 101000914514 Homo sapiens T-cell-specific surface glycoprotein CD28 Proteins 0.000 description 1
- 101000997835 Homo sapiens Tyrosine-protein kinase JAK1 Proteins 0.000 description 1
- 101000997832 Homo sapiens Tyrosine-protein kinase JAK2 Proteins 0.000 description 1
- VEXZGXHMUGYJMC-UHFFFAOYSA-N Hydrochloric acid Chemical compound Cl VEXZGXHMUGYJMC-UHFFFAOYSA-N 0.000 description 1
- 229920002153 Hydroxypropyl cellulose Polymers 0.000 description 1
- DGAQECJNVWCQMB-PUAWFVPOSA-M Ilexoside XXIX Chemical compound C[C@@H]1CC[C@@]2(CC[C@@]3(C(=CC[C@H]4[C@]3(CC[C@@H]5[C@@]4(CC[C@@H](C5(C)C)OS(=O)(=O)[O-])C)C)[C@@H]2[C@]1(C)O)C)C(=O)O[C@H]6[C@@H]([C@H]([C@@H]([C@H](O6)CO)O)O)O.[Na+] DGAQECJNVWCQMB-PUAWFVPOSA-M 0.000 description 1
- 108060003951 Immunoglobulin Proteins 0.000 description 1
- 102000008394 Immunoglobulin Fragments Human genes 0.000 description 1
- 108010021625 Immunoglobulin Fragments Proteins 0.000 description 1
- 102100023915 Insulin Human genes 0.000 description 1
- 108090001061 Insulin Proteins 0.000 description 1
- 102100037850 Interferon gamma Human genes 0.000 description 1
- 108010074328 Interferon-gamma Proteins 0.000 description 1
- 108010065805 Interleukin-12 Proteins 0.000 description 1
- 102000013462 Interleukin-12 Human genes 0.000 description 1
- 102000014158 Interleukin-12 Subunit p40 Human genes 0.000 description 1
- 108010011429 Interleukin-12 Subunit p40 Proteins 0.000 description 1
- 102000015696 Interleukins Human genes 0.000 description 1
- 108010063738 Interleukins Proteins 0.000 description 1
- 230000004163 JAK-STAT signaling pathway Effects 0.000 description 1
- 208000002260 Keloid Diseases 0.000 description 1
- 208000008839 Kidney Neoplasms Diseases 0.000 description 1
- XUJNEKJLAYXESH-REOHCLBHSA-N L-Cysteine Chemical compound SC[C@H](N)C(O)=O XUJNEKJLAYXESH-REOHCLBHSA-N 0.000 description 1
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 description 1
- WHUUTDBJXJRKMK-VKHMYHEASA-N L-glutamic acid Chemical compound OC(=O)[C@@H](N)CCC(O)=O WHUUTDBJXJRKMK-VKHMYHEASA-N 0.000 description 1
- 125000000393 L-methionino group Chemical group [H]OC(=O)[C@@]([H])(N([H])[*])C([H])([H])C(SC([H])([H])[H])([H])[H] 0.000 description 1
- 125000000174 L-prolyl group Chemical group [H]N1C([H])([H])C([H])([H])C([H])([H])[C@@]1([H])C(*)=O 0.000 description 1
- KZSNJWFQEVHDMF-BYPYZUCNSA-N L-valine Chemical compound CC(C)[C@H](N)C(O)=O KZSNJWFQEVHDMF-BYPYZUCNSA-N 0.000 description 1
- GUBGYTABKSRVRQ-QKKXKWKRSA-N Lactose Natural products OC[C@H]1O[C@@H](O[C@H]2[C@H](O)[C@@H](O)C(O)O[C@@H]2CO)[C@H](O)[C@@H](O)[C@H]1O GUBGYTABKSRVRQ-QKKXKWKRSA-N 0.000 description 1
- 208000006404 Large Granular Lymphocytic Leukemia Diseases 0.000 description 1
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 1
- 102100032352 Leukemia inhibitory factor Human genes 0.000 description 1
- 108090000581 Leukemia inhibitory factor Proteins 0.000 description 1
- 102000003960 Ligases Human genes 0.000 description 1
- 108090000364 Ligases Proteins 0.000 description 1
- 108091007460 Long intergenic noncoding RNA Proteins 0.000 description 1
- DDWFXDSYGUXRAY-UHFFFAOYSA-N Luciferin Natural products CCc1c(C)c(CC2NC(=O)C(=C2C=C)C)[nH]c1Cc3[nH]c4C(=C5/NC(CC(=O)O)C(C)C5CC(=O)O)CC(=O)c4c3C DDWFXDSYGUXRAY-UHFFFAOYSA-N 0.000 description 1
- 229910052765 Lutetium Inorganic materials 0.000 description 1
- 206010025323 Lymphomas Diseases 0.000 description 1
- 239000004472 Lysine Substances 0.000 description 1
- 101710141347 Major envelope glycoprotein Proteins 0.000 description 1
- 241000124008 Mammalia Species 0.000 description 1
- 208000035490 Megakaryoblastic Acute Leukemia Diseases 0.000 description 1
- 102000018697 Membrane Proteins Human genes 0.000 description 1
- 108010052285 Membrane Proteins Proteins 0.000 description 1
- 206010054949 Metaplasia Diseases 0.000 description 1
- CERQOIWHTDAKMF-UHFFFAOYSA-M Methacrylate Chemical compound CC(=C)C([O-])=O CERQOIWHTDAKMF-UHFFFAOYSA-M 0.000 description 1
- 241001529936 Murinae Species 0.000 description 1
- 241000699666 Mus <mouse, genus> Species 0.000 description 1
- 102100025243 Myeloid cell surface antigen CD33 Human genes 0.000 description 1
- ZDZOTLJHXYCWBA-VCVYQWHSSA-N N-debenzoyl-N-(tert-butoxycarbonyl)-10-deacetyltaxol Chemical compound O([C@H]1[C@H]2[C@@](C([C@H](O)C3=C(C)[C@@H](OC(=O)[C@H](O)[C@@H](NC(=O)OC(C)(C)C)C=4C=CC=CC=4)C[C@]1(O)C3(C)C)=O)(C)[C@@H](O)C[C@H]1OC[C@]12OC(=O)C)C(=O)C1=CC=CC=C1 ZDZOTLJHXYCWBA-VCVYQWHSSA-N 0.000 description 1
- 230000004988 N-glycosylation Effects 0.000 description 1
- 238000005481 NMR spectroscopy Methods 0.000 description 1
- GRYLNZFGIOXLOG-UHFFFAOYSA-N Nitric acid Chemical compound O[N+]([O-])=O GRYLNZFGIOXLOG-UHFFFAOYSA-N 0.000 description 1
- 239000000020 Nitrocellulose Substances 0.000 description 1
- 208000015914 Non-Hodgkin lymphomas Diseases 0.000 description 1
- 241000238413 Octopus Species 0.000 description 1
- 229930012538 Paclitaxel Natural products 0.000 description 1
- 208000027190 Peripheral T-cell lymphomas Diseases 0.000 description 1
- 102000004160 Phosphoric Monoester Hydrolases Human genes 0.000 description 1
- 108090000608 Phosphoric Monoester Hydrolases Proteins 0.000 description 1
- 108091000080 Phosphotransferase Proteins 0.000 description 1
- 229920003171 Poly (ethylene oxide) Polymers 0.000 description 1
- 229920001305 Poly(isodecyl(meth)acrylate) Polymers 0.000 description 1
- 229920002845 Poly(methacrylic acid) Polymers 0.000 description 1
- 229920001030 Polyethylene Glycol 4000 Polymers 0.000 description 1
- 239000002202 Polyethylene glycol Substances 0.000 description 1
- 229920000331 Polyhydroxybutyrate Polymers 0.000 description 1
- 239000004743 Polypropylene Substances 0.000 description 1
- ZLMJMSJWJFRBEC-UHFFFAOYSA-N Potassium Chemical compound [K] ZLMJMSJWJFRBEC-UHFFFAOYSA-N 0.000 description 1
- 229910052777 Praseodymium Inorganic materials 0.000 description 1
- 102100026942 Prolyl 3-hydroxylase OGFOD1 Human genes 0.000 description 1
- 201000001263 Psoriatic Arthritis Diseases 0.000 description 1
- 208000036824 Psoriatic arthropathy Diseases 0.000 description 1
- 208000006311 Pyoderma Diseases 0.000 description 1
- 206010038389 Renal cancer Diseases 0.000 description 1
- 241001303601 Rosacea Species 0.000 description 1
- 239000006146 Roswell Park Memorial Institute medium Substances 0.000 description 1
- 102000014400 SH2 domains Human genes 0.000 description 1
- 108050003452 SH2 domains Proteins 0.000 description 1
- 206010039491 Sarcoma Diseases 0.000 description 1
- 206010039793 Seborrhoeic dermatitis Diseases 0.000 description 1
- 206010039796 Seborrhoeic keratosis Diseases 0.000 description 1
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 1
- 102100028904 Serine/threonine-protein kinase MARK2 Human genes 0.000 description 1
- 108020004459 Small interfering RNA Proteins 0.000 description 1
- PMZURENOXWZQFD-UHFFFAOYSA-L Sodium Sulfate Chemical compound [Na+].[Na+].[O-]S([O-])(=O)=O PMZURENOXWZQFD-UHFFFAOYSA-L 0.000 description 1
- 241000256251 Spodoptera frugiperda Species 0.000 description 1
- 229930182558 Sterol Natural products 0.000 description 1
- 108010090804 Streptavidin Proteins 0.000 description 1
- KDYFGRWQOYBRFD-UHFFFAOYSA-N Succinic acid Natural products OC(=O)CCC(O)=O KDYFGRWQOYBRFD-UHFFFAOYSA-N 0.000 description 1
- 239000012505 Superdex™ Substances 0.000 description 1
- 241000950638 Symphysodon discus Species 0.000 description 1
- 230000005867 T cell response Effects 0.000 description 1
- 208000031672 T-Cell Peripheral Lymphoma Diseases 0.000 description 1
- 201000008717 T-cell large granular lymphocyte leukemia Diseases 0.000 description 1
- 102100027213 T-cell-specific surface glycoprotein CD28 Human genes 0.000 description 1
- FEWJPZIEWOKRBE-UHFFFAOYSA-N Tartaric acid Natural products [H+].[H+].[O-]C(=O)C(O)C(O)C([O-])=O FEWJPZIEWOKRBE-UHFFFAOYSA-N 0.000 description 1
- 229940123237 Taxane Drugs 0.000 description 1
- AYFVYJQAPQTCCC-UHFFFAOYSA-N Threonine Natural products CC(O)C(N)C(O)=O AYFVYJQAPQTCCC-UHFFFAOYSA-N 0.000 description 1
- 239000004473 Threonine Substances 0.000 description 1
- 108091023040 Transcription factor Proteins 0.000 description 1
- 102000040945 Transcription factor Human genes 0.000 description 1
- 102000006747 Transforming Growth Factor alpha Human genes 0.000 description 1
- 102000009618 Transforming Growth Factors Human genes 0.000 description 1
- 108010009583 Transforming Growth Factors Proteins 0.000 description 1
- 101800004564 Transforming growth factor alpha Proteins 0.000 description 1
- 241000255993 Trichoplusia ni Species 0.000 description 1
- 108060008682 Tumor Necrosis Factor Proteins 0.000 description 1
- 102000000852 Tumor Necrosis Factor-alpha Human genes 0.000 description 1
- 102100033438 Tyrosine-protein kinase JAK1 Human genes 0.000 description 1
- 102100033444 Tyrosine-protein kinase JAK2 Human genes 0.000 description 1
- 208000024780 Urticaria Diseases 0.000 description 1
- XTXRWKRVRITETP-UHFFFAOYSA-N Vinyl acetate Chemical compound CC(=O)OC=C XTXRWKRVRITETP-UHFFFAOYSA-N 0.000 description 1
- 206010047642 Vitiligo Diseases 0.000 description 1
- VWQVUPCCIRVNHF-OUBTZVSYSA-N Yttrium-90 Chemical compound [90Y] VWQVUPCCIRVNHF-OUBTZVSYSA-N 0.000 description 1
- 229920002494 Zein Polymers 0.000 description 1
- SMEGJBVQLJJKKX-HOTMZDKISA-N [(2R,3S,4S,5R,6R)-5-acetyloxy-3,4,6-trihydroxyoxan-2-yl]methyl acetate Chemical compound CC(=O)OC[C@@H]1[C@H]([C@@H]([C@H]([C@@H](O1)O)OC(=O)C)O)O SMEGJBVQLJJKKX-HOTMZDKISA-N 0.000 description 1
- NNLVGZFZQQXQNW-ADJNRHBOSA-N [(2r,3r,4s,5r,6s)-4,5-diacetyloxy-3-[(2s,3r,4s,5r,6r)-3,4,5-triacetyloxy-6-(acetyloxymethyl)oxan-2-yl]oxy-6-[(2r,3r,4s,5r,6s)-4,5,6-triacetyloxy-2-(acetyloxymethyl)oxan-3-yl]oxyoxan-2-yl]methyl acetate Chemical compound O([C@@H]1O[C@@H]([C@H]([C@H](OC(C)=O)[C@H]1OC(C)=O)O[C@H]1[C@@H]([C@@H](OC(C)=O)[C@H](OC(C)=O)[C@@H](COC(C)=O)O1)OC(C)=O)COC(=O)C)[C@@H]1[C@@H](COC(C)=O)O[C@@H](OC(C)=O)[C@H](OC(C)=O)[C@H]1OC(C)=O NNLVGZFZQQXQNW-ADJNRHBOSA-N 0.000 description 1
- FJWGYAHXMCUOOM-QHOUIDNNSA-N [(2s,3r,4s,5r,6r)-2-[(2r,3r,4s,5r,6s)-4,5-dinitrooxy-2-(nitrooxymethyl)-6-[(2r,3r,4s,5r,6s)-4,5,6-trinitrooxy-2-(nitrooxymethyl)oxan-3-yl]oxyoxan-3-yl]oxy-3,5-dinitrooxy-6-(nitrooxymethyl)oxan-4-yl] nitrate Chemical compound O([C@@H]1O[C@@H]([C@H]([C@H](O[N+]([O-])=O)[C@H]1O[N+]([O-])=O)O[C@H]1[C@@H]([C@@H](O[N+]([O-])=O)[C@H](O[N+]([O-])=O)[C@@H](CO[N+]([O-])=O)O1)O[N+]([O-])=O)CO[N+](=O)[O-])[C@@H]1[C@@H](CO[N+]([O-])=O)O[C@@H](O[N+]([O-])=O)[C@H](O[N+]([O-])=O)[C@H]1O[N+]([O-])=O FJWGYAHXMCUOOM-QHOUIDNNSA-N 0.000 description 1
- 230000001594 aberrant effect Effects 0.000 description 1
- 230000005856 abnormality Effects 0.000 description 1
- 229940081735 acetylcellulose Drugs 0.000 description 1
- 206010000496 acne Diseases 0.000 description 1
- NIXOWILDQLNWCW-UHFFFAOYSA-N acrylic acid group Chemical group C(C=C)(=O)O NIXOWILDQLNWCW-UHFFFAOYSA-N 0.000 description 1
- 229910052767 actinium Inorganic materials 0.000 description 1
- QQINRWTZWGJFDB-UHFFFAOYSA-N actinium atom Chemical compound [Ac] QQINRWTZWGJFDB-UHFFFAOYSA-N 0.000 description 1
- 208000021841 acute erythroid leukemia Diseases 0.000 description 1
- 208000013593 acute megakaryoblastic leukemia Diseases 0.000 description 1
- 208000020700 acute megakaryocytic leukemia Diseases 0.000 description 1
- 125000004423 acyloxy group Chemical group 0.000 description 1
- 239000000654 additive Substances 0.000 description 1
- 239000002671 adjuvant Substances 0.000 description 1
- 230000009824 affinity maturation Effects 0.000 description 1
- 239000008272 agar Substances 0.000 description 1
- 235000004279 alanine Nutrition 0.000 description 1
- 239000000783 alginic acid Substances 0.000 description 1
- 229920000615 alginic acid Polymers 0.000 description 1
- 235000010443 alginic acid Nutrition 0.000 description 1
- 229960001126 alginic acid Drugs 0.000 description 1
- 150000004781 alginic acids Chemical class 0.000 description 1
- 229910052783 alkali metal Inorganic materials 0.000 description 1
- 150000001340 alkali metals Chemical class 0.000 description 1
- 229920013820 alkyl cellulose Polymers 0.000 description 1
- 125000000217 alkyl group Chemical group 0.000 description 1
- 125000002947 alkylene group Chemical group 0.000 description 1
- 230000000172 allergic effect Effects 0.000 description 1
- 230000007815 allergy Effects 0.000 description 1
- 208000004631 alopecia areata Diseases 0.000 description 1
- 230000001668 ameliorated effect Effects 0.000 description 1
- 230000003321 amplification Effects 0.000 description 1
- 239000003708 ampul Substances 0.000 description 1
- 230000033115 angiogenesis Effects 0.000 description 1
- 150000008064 anhydrides Chemical class 0.000 description 1
- 239000005557 antagonist Substances 0.000 description 1
- 230000006909 anti-apoptosis Effects 0.000 description 1
- 230000000692 anti-sense effect Effects 0.000 description 1
- 239000008135 aqueous vehicle Substances 0.000 description 1
- 125000000637 arginyl group Chemical group N[C@@H](CCCNC(N)=N)C(=O)* 0.000 description 1
- 238000003491 array Methods 0.000 description 1
- 125000003118 aryl group Chemical group 0.000 description 1
- 235000009582 asparagine Nutrition 0.000 description 1
- 229960001230 asparagine Drugs 0.000 description 1
- 235000003704 aspartic acid Nutrition 0.000 description 1
- 238000000429 assembly Methods 0.000 description 1
- 230000000712 assembly Effects 0.000 description 1
- 230000001580 bacterial effect Effects 0.000 description 1
- 210000000270 basal cell Anatomy 0.000 description 1
- 239000002585 base Substances 0.000 description 1
- 210000003651 basophil Anatomy 0.000 description 1
- 229960000686 benzalkonium chloride Drugs 0.000 description 1
- SRSXLGNVWSONIS-UHFFFAOYSA-N benzenesulfonic acid Chemical compound OS(=O)(=O)C1=CC=CC=C1 SRSXLGNVWSONIS-UHFFFAOYSA-N 0.000 description 1
- 229940092714 benzenesulfonic acid Drugs 0.000 description 1
- CADWTSSKOVRVJC-UHFFFAOYSA-N benzyl(dimethyl)azanium;chloride Chemical compound [Cl-].C[NH+](C)CC1=CC=CC=C1 CADWTSSKOVRVJC-UHFFFAOYSA-N 0.000 description 1
- OQFSQFPPLPISGP-UHFFFAOYSA-N beta-carboxyaspartic acid Natural products OC(=O)C(N)C(C(O)=O)C(O)=O OQFSQFPPLPISGP-UHFFFAOYSA-N 0.000 description 1
- 108091008324 binding proteins Proteins 0.000 description 1
- 230000004071 biological effect Effects 0.000 description 1
- 230000031018 biological processes and functions Effects 0.000 description 1
- 230000008512 biological response Effects 0.000 description 1
- 238000001574 biopsy Methods 0.000 description 1
- 229910052797 bismuth Inorganic materials 0.000 description 1
- JCXGWMGPZLAOME-UHFFFAOYSA-N bismuth atom Chemical compound [Bi] JCXGWMGPZLAOME-UHFFFAOYSA-N 0.000 description 1
- KGBXLFKZBHKPEV-UHFFFAOYSA-N boric acid Chemical compound OB(O)O KGBXLFKZBHKPEV-UHFFFAOYSA-N 0.000 description 1
- 239000004327 boric acid Substances 0.000 description 1
- 210000000481 breast Anatomy 0.000 description 1
- 238000013276 bronchoscopy Methods 0.000 description 1
- 239000007853 buffer solution Substances 0.000 description 1
- 208000000594 bullous pemphigoid Diseases 0.000 description 1
- DQXBYHZEEUGOBF-UHFFFAOYSA-N but-3-enoic acid;ethene Chemical compound C=C.OC(=O)CC=C DQXBYHZEEUGOBF-UHFFFAOYSA-N 0.000 description 1
- KDYFGRWQOYBRFD-NUQCWPJISA-N butanedioic acid Chemical compound O[14C](=O)CC[14C](O)=O KDYFGRWQOYBRFD-NUQCWPJISA-N 0.000 description 1
- 229910000019 calcium carbonate Inorganic materials 0.000 description 1
- 239000001506 calcium phosphate Substances 0.000 description 1
- 229910000389 calcium phosphate Inorganic materials 0.000 description 1
- 235000011010 calcium phosphates Nutrition 0.000 description 1
- 159000000007 calcium salts Chemical class 0.000 description 1
- HXCHCVDVKSCDHU-LULTVBGHSA-N calicheamicin Chemical compound C1[C@H](OC)[C@@H](NCC)CO[C@H]1O[C@H]1[C@H](O[C@@H]2C\3=C(NC(=O)OC)C(=O)C[C@](C/3=C/CSSSC)(O)C#C\C=C/C#C2)O[C@H](C)[C@@H](NO[C@@H]2O[C@H](C)[C@@H](SC(=O)C=3C(=C(OC)C(O[C@H]4[C@@H]([C@H](OC)[C@@H](O)[C@H](C)O4)O)=C(I)C=3C)OC)[C@@H](O)C2)[C@@H]1O HXCHCVDVKSCDHU-LULTVBGHSA-N 0.000 description 1
- 229930195731 calicheamicin Natural products 0.000 description 1
- 230000005907 cancer growth Effects 0.000 description 1
- 208000035269 cancer or benign tumor Diseases 0.000 description 1
- 230000000711 cancerogenic effect Effects 0.000 description 1
- 239000001569 carbon dioxide Substances 0.000 description 1
- 229910002092 carbon dioxide Inorganic materials 0.000 description 1
- 229960004424 carbon dioxide Drugs 0.000 description 1
- 229910002091 carbon monoxide Inorganic materials 0.000 description 1
- 125000002915 carbonyl group Chemical group [*:2]C([*:1])=O 0.000 description 1
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 description 1
- 125000002843 carboxylic acid group Chemical class 0.000 description 1
- 231100000315 carcinogenic Toxicity 0.000 description 1
- 230000015556 catabolic process Effects 0.000 description 1
- 230000020411 cell activation Effects 0.000 description 1
- 238000004113 cell culture Methods 0.000 description 1
- 230000003915 cell function Effects 0.000 description 1
- 230000010261 cell growth Effects 0.000 description 1
- 230000004663 cell proliferation Effects 0.000 description 1
- 239000002458 cell surface marker Substances 0.000 description 1
- 230000005754 cellular signaling Effects 0.000 description 1
- 229920002301 cellulose acetate Polymers 0.000 description 1
- 229920006217 cellulose acetate butyrate Polymers 0.000 description 1
- 229940081734 cellulose acetate phthalate Drugs 0.000 description 1
- 229920003086 cellulose ether Polymers 0.000 description 1
- 229920006218 cellulose propionate Polymers 0.000 description 1
- 238000005119 centrifugation Methods 0.000 description 1
- 125000003636 chemical group Chemical group 0.000 description 1
- 238000002512 chemotherapy Methods 0.000 description 1
- 210000000038 chest Anatomy 0.000 description 1
- 229960004926 chlorobutanol Drugs 0.000 description 1
- 235000012000 cholesterol Nutrition 0.000 description 1
- 150000001840 cholesterol esters Chemical class 0.000 description 1
- 229940059329 chondroitin sulfate Drugs 0.000 description 1
- 238000004587 chromatography analysis Methods 0.000 description 1
- 230000001684 chronic effect Effects 0.000 description 1
- 238000004140 cleaning Methods 0.000 description 1
- 239000011248 coating agent Substances 0.000 description 1
- 229940110456 cocoa butter Drugs 0.000 description 1
- 235000019868 cocoa butter Nutrition 0.000 description 1
- 229920001436 collagen Polymers 0.000 description 1
- 210000001072 colon Anatomy 0.000 description 1
- 239000003086 colorant Substances 0.000 description 1
- 238000004440 column chromatography Methods 0.000 description 1
- 239000002131 composite material Substances 0.000 description 1
- 239000007891 compressed tablet Substances 0.000 description 1
- 238000004590 computer program Methods 0.000 description 1
- 235000009508 confectionery Nutrition 0.000 description 1
- 239000002872 contrast media Substances 0.000 description 1
- 238000013270 controlled release Methods 0.000 description 1
- 238000007796 conventional method Methods 0.000 description 1
- 239000002577 cryoprotective agent Substances 0.000 description 1
- 238000002447 crystallographic data Methods 0.000 description 1
- 210000004748 cultured cell Anatomy 0.000 description 1
- 238000012258 culturing Methods 0.000 description 1
- 210000005220 cytoplasmic tail Anatomy 0.000 description 1
- 238000013480 data collection Methods 0.000 description 1
- 230000007423 decrease Effects 0.000 description 1
- 238000006731 degradation reaction Methods 0.000 description 1
- 230000004041 dendritic cell maturation Effects 0.000 description 1
- 238000000432 density-gradient centrifugation Methods 0.000 description 1
- 238000011161 development Methods 0.000 description 1
- 230000018109 developmental process Effects 0.000 description 1
- PXBRQCKWGAHEHS-UHFFFAOYSA-N dichlorodifluoromethane Chemical compound FC(F)(Cl)Cl PXBRQCKWGAHEHS-UHFFFAOYSA-N 0.000 description 1
- 235000019404 dichlorodifluoromethane Nutrition 0.000 description 1
- 229940042935 dichlorodifluoromethane Drugs 0.000 description 1
- 229940087091 dichlorotetrafluoroethane Drugs 0.000 description 1
- 235000014113 dietary fatty acids Nutrition 0.000 description 1
- 239000003085 diluting agent Substances 0.000 description 1
- 238000006471 dimerization reaction Methods 0.000 description 1
- 239000007884 disintegrant Substances 0.000 description 1
- 239000002270 dispersing agent Substances 0.000 description 1
- 239000012153 distilled water Substances 0.000 description 1
- 230000009977 dual effect Effects 0.000 description 1
- 238000004520 electroporation Methods 0.000 description 1
- 210000003038 endothelium Anatomy 0.000 description 1
- 239000007920 enema Substances 0.000 description 1
- 229940079360 enema for constipation Drugs 0.000 description 1
- 230000007613 environmental effect Effects 0.000 description 1
- 230000007071 enzymatic hydrolysis Effects 0.000 description 1
- 238000006047 enzymatic hydrolysis reaction Methods 0.000 description 1
- 210000003979 eosinophil Anatomy 0.000 description 1
- 231100000321 erythema Toxicity 0.000 description 1
- 230000000925 erythroid effect Effects 0.000 description 1
- 239000003797 essential amino acid Substances 0.000 description 1
- 235000020776 essential amino acid Nutrition 0.000 description 1
- 150000002148 esters Chemical class 0.000 description 1
- 229920001249 ethyl cellulose Polymers 0.000 description 1
- 235000019325 ethyl cellulose Nutrition 0.000 description 1
- 239000005038 ethylene vinyl acetate Substances 0.000 description 1
- 230000007717 exclusion Effects 0.000 description 1
- 208000004526 exfoliative dermatitis Diseases 0.000 description 1
- 238000010195 expression analysis Methods 0.000 description 1
- 239000003925 fat Substances 0.000 description 1
- 235000019197 fats Nutrition 0.000 description 1
- 229930195729 fatty acid Natural products 0.000 description 1
- 239000000194 fatty acid Substances 0.000 description 1
- 150000004665 fatty acids Chemical class 0.000 description 1
- 230000005294 ferromagnetic effect Effects 0.000 description 1
- 210000002950 fibroblast Anatomy 0.000 description 1
- 230000004761 fibrosis Effects 0.000 description 1
- 239000012997 ficoll-paque Substances 0.000 description 1
- 239000000945 filler Substances 0.000 description 1
- 239000012530 fluid Substances 0.000 description 1
- GNBHRKFJIUUOQI-UHFFFAOYSA-N fluorescein Chemical compound O1C(=O)C2=CC=CC=C2C21C1=CC=C(O)C=C1OC1=CC(O)=CC=C21 GNBHRKFJIUUOQI-UHFFFAOYSA-N 0.000 description 1
- 108091006047 fluorescent proteins Proteins 0.000 description 1
- 102000034287 fluorescent proteins Human genes 0.000 description 1
- 230000003325 follicular Effects 0.000 description 1
- 235000003599 food sweetener Nutrition 0.000 description 1
- 235000019253 formic acid Nutrition 0.000 description 1
- 239000012458 free base Substances 0.000 description 1
- 231100000414 gastrointestinal toxicity Toxicity 0.000 description 1
- 210000001654 germ layer Anatomy 0.000 description 1
- 235000013922 glutamic acid Nutrition 0.000 description 1
- 239000004220 glutamic acid Substances 0.000 description 1
- 125000000404 glutamine group Chemical group N[C@@H](CCC(N)=O)C(=O)* 0.000 description 1
- 125000005456 glyceride group Chemical group 0.000 description 1
- 230000013595 glycosylation Effects 0.000 description 1
- 238000006206 glycosylation reaction Methods 0.000 description 1
- 210000002175 goblet cell Anatomy 0.000 description 1
- PCHJSUWPFVWCPO-UHFFFAOYSA-N gold Chemical compound [Au] PCHJSUWPFVWCPO-UHFFFAOYSA-N 0.000 description 1
- 229910052737 gold Inorganic materials 0.000 description 1
- 239000010931 gold Substances 0.000 description 1
- 239000005090 green fluorescent protein Substances 0.000 description 1
- 230000012010 growth Effects 0.000 description 1
- 210000003128 head Anatomy 0.000 description 1
- 230000003862 health status Effects 0.000 description 1
- 208000014752 hemophagocytic syndrome Diseases 0.000 description 1
- 231100000304 hepatotoxicity Toxicity 0.000 description 1
- 238000004128 high performance liquid chromatography Methods 0.000 description 1
- 230000006801 homologous recombination Effects 0.000 description 1
- 238000002744 homologous recombination Methods 0.000 description 1
- 229920002674 hyaluronan Polymers 0.000 description 1
- 229960003160 hyaluronic acid Drugs 0.000 description 1
- 150000004678 hydrides Chemical class 0.000 description 1
- BHEPBYXIRTUNPN-UHFFFAOYSA-N hydridophosphorus(.) (triplet) Chemical compound [PH] BHEPBYXIRTUNPN-UHFFFAOYSA-N 0.000 description 1
- 239000000017 hydrogel Substances 0.000 description 1
- 229910000042 hydrogen bromide Inorganic materials 0.000 description 1
- IXCSERBJSXMMFS-UHFFFAOYSA-N hydrogen chloride Substances Cl.Cl IXCSERBJSXMMFS-UHFFFAOYSA-N 0.000 description 1
- 229910000041 hydrogen chloride Inorganic materials 0.000 description 1
- 229920013821 hydroxy alkyl cellulose Polymers 0.000 description 1
- 230000033444 hydroxylation Effects 0.000 description 1
- 238000005805 hydroxylation reaction Methods 0.000 description 1
- 239000001863 hydroxypropyl cellulose Substances 0.000 description 1
- 235000010977 hydroxypropyl cellulose Nutrition 0.000 description 1
- 239000001866 hydroxypropyl methyl cellulose Substances 0.000 description 1
- 229920003088 hydroxypropyl methyl cellulose Polymers 0.000 description 1
- 235000010979 hydroxypropyl methyl cellulose Nutrition 0.000 description 1
- UFVKGYZPFZQRLF-UHFFFAOYSA-N hydroxypropyl methyl cellulose Chemical compound OC1C(O)C(OC)OC(CO)C1OC1C(O)C(O)C(OC2C(C(O)C(OC3C(C(O)C(O)C(CO)O3)O)C(CO)O2)O)C(CO)O1 UFVKGYZPFZQRLF-UHFFFAOYSA-N 0.000 description 1
- 230000003463 hyperproliferative effect Effects 0.000 description 1
- 230000004047 hyperresponsiveness Effects 0.000 description 1
- 230000002519 immonomodulatory effect Effects 0.000 description 1
- 230000036039 immunity Effects 0.000 description 1
- 238000003018 immunoassay Methods 0.000 description 1
- 238000003119 immunoblot Methods 0.000 description 1
- 238000010166 immunofluorescence Methods 0.000 description 1
- 102000018358 immunoglobulin Human genes 0.000 description 1
- 239000007943 implant Substances 0.000 description 1
- 238000001727 in vivo Methods 0.000 description 1
- 239000005414 inactive ingredient Substances 0.000 description 1
- 208000015181 infectious disease Diseases 0.000 description 1
- 239000004615 ingredient Substances 0.000 description 1
- 210000005007 innate immune system Anatomy 0.000 description 1
- 238000003780 insertion Methods 0.000 description 1
- 230000037431 insertion Effects 0.000 description 1
- 229940125396 insulin Drugs 0.000 description 1
- 230000035992 intercellular communication Effects 0.000 description 1
- 229940047124 interferons Drugs 0.000 description 1
- 229940047122 interleukins Drugs 0.000 description 1
- 230000031146 intracellular signal transduction Effects 0.000 description 1
- 230000004068 intracellular signaling Effects 0.000 description 1
- 238000010212 intracellular staining Methods 0.000 description 1
- 238000007916 intrasternal administration Methods 0.000 description 1
- 229910052740 iodine Inorganic materials 0.000 description 1
- 239000011630 iodine Substances 0.000 description 1
- 239000003456 ion exchange resin Substances 0.000 description 1
- 229920003303 ion-exchange polymer Polymers 0.000 description 1
- 230000000622 irritating effect Effects 0.000 description 1
- HOQADATXFBOEGG-UHFFFAOYSA-N isofenphos Chemical compound CCOP(=S)(NC(C)C)OC1=CC=CC=C1C(=O)OC(C)C HOQADATXFBOEGG-UHFFFAOYSA-N 0.000 description 1
- 229960000310 isoleucine Drugs 0.000 description 1
- AGPKZVBTJJNPAG-UHFFFAOYSA-N isoleucine Natural products CCC(C)C(N)C(O)=O AGPKZVBTJJNPAG-UHFFFAOYSA-N 0.000 description 1
- 210000000281 joint capsule Anatomy 0.000 description 1
- 210000001117 keloid Anatomy 0.000 description 1
- 208000011379 keloid formation Diseases 0.000 description 1
- 206010023332 keratitis Diseases 0.000 description 1
- 201000010982 kidney cancer Diseases 0.000 description 1
- 238000002372 labelling Methods 0.000 description 1
- 239000004310 lactic acid Substances 0.000 description 1
- 235000014655 lactic acid Nutrition 0.000 description 1
- 239000008101 lactose Substances 0.000 description 1
- 210000001821 langerhans cell Anatomy 0.000 description 1
- 208000002741 leukoplakia Diseases 0.000 description 1
- 201000011486 lichen planus Diseases 0.000 description 1
- 150000002632 lipids Chemical class 0.000 description 1
- 239000002502 liposome Substances 0.000 description 1
- 239000012669 liquid formulation Substances 0.000 description 1
- 210000004185 liver Anatomy 0.000 description 1
- 239000006210 lotion Substances 0.000 description 1
- 239000000314 lubricant Substances 0.000 description 1
- 239000002932 luster Substances 0.000 description 1
- OHSVLFRHMCKCQY-UHFFFAOYSA-N lutetium atom Chemical compound [Lu] OHSVLFRHMCKCQY-UHFFFAOYSA-N 0.000 description 1
- 208000019420 lymphoid neoplasm Diseases 0.000 description 1
- 230000005291 magnetic effect Effects 0.000 description 1
- 238000007885 magnetic separation Methods 0.000 description 1
- 230000014759 maintenance of location Effects 0.000 description 1
- VZCYOOQTPOCHFL-UPHRSURJSA-N maleic acid Chemical compound OC(=O)\C=C/C(O)=O VZCYOOQTPOCHFL-UPHRSURJSA-N 0.000 description 1
- 239000011976 maleic acid Substances 0.000 description 1
- 230000003211 malignant effect Effects 0.000 description 1
- 208000020968 mature T-cell and NK-cell non-Hodgkin lymphoma Diseases 0.000 description 1
- 230000007246 mechanism Effects 0.000 description 1
- 210000002752 melanocyte Anatomy 0.000 description 1
- 201000001441 melanoma Diseases 0.000 description 1
- 210000000716 merkel cell Anatomy 0.000 description 1
- 230000015689 metaplastic ossification Effects 0.000 description 1
- 230000001394 metastastic effect Effects 0.000 description 1
- 206010061289 metastatic neoplasm Diseases 0.000 description 1
- 229940071648 metered dose inhaler Drugs 0.000 description 1
- 229940098779 methanesulfonic acid Drugs 0.000 description 1
- 229920000609 methyl cellulose Polymers 0.000 description 1
- 239000001923 methylcellulose Substances 0.000 description 1
- 235000010981 methylcellulose Nutrition 0.000 description 1
- 238000013508 migration Methods 0.000 description 1
- 230000005012 migration Effects 0.000 description 1
- 230000003278 mimic effect Effects 0.000 description 1
- 238000002156 mixing Methods 0.000 description 1
- 238000010369 molecular cloning Methods 0.000 description 1
- 230000000877 morphologic effect Effects 0.000 description 1
- 239000012452 mother liquor Substances 0.000 description 1
- 210000000214 mouth Anatomy 0.000 description 1
- 239000002324 mouth wash Substances 0.000 description 1
- 229940051866 mouthwash Drugs 0.000 description 1
- 230000003232 mucoadhesive effect Effects 0.000 description 1
- PSZYNBSKGUBXEH-UHFFFAOYSA-N naphthalene-1-sulfonic acid Chemical compound C1=CC=C2C(S(=O)(=O)O)=CC=CC2=C1 PSZYNBSKGUBXEH-UHFFFAOYSA-N 0.000 description 1
- 210000000822 natural killer cell Anatomy 0.000 description 1
- 229920005615 natural polymer Polymers 0.000 description 1
- 239000006199 nebulizer Substances 0.000 description 1
- 210000003739 neck Anatomy 0.000 description 1
- 210000005170 neoplastic cell Anatomy 0.000 description 1
- 231100000417 nephrotoxicity Toxicity 0.000 description 1
- 230000000926 neurological effect Effects 0.000 description 1
- 231100000228 neurotoxicity Toxicity 0.000 description 1
- 230000007935 neutral effect Effects 0.000 description 1
- 230000003472 neutralizing effect Effects 0.000 description 1
- 229910017604 nitric acid Inorganic materials 0.000 description 1
- 229920001220 nitrocellulos Polymers 0.000 description 1
- 229910052757 nitrogen Inorganic materials 0.000 description 1
- 238000003199 nucleic acid amplification method Methods 0.000 description 1
- 238000001668 nucleic acid synthesis Methods 0.000 description 1
- 239000003921 oil Substances 0.000 description 1
- 210000000056 organ Anatomy 0.000 description 1
- 230000002611 ovarian Effects 0.000 description 1
- 230000003647 oxidation Effects 0.000 description 1
- 238000012856 packing Methods 0.000 description 1
- 229960001592 paclitaxel Drugs 0.000 description 1
- 206010033675 panniculitis Diseases 0.000 description 1
- 230000005298 paramagnetic effect Effects 0.000 description 1
- 244000045947 parasite Species 0.000 description 1
- 239000008188 pellet Substances 0.000 description 1
- 230000000149 penetrating effect Effects 0.000 description 1
- 238000010647 peptide synthesis reaction Methods 0.000 description 1
- 239000002304 perfume Substances 0.000 description 1
- 210000005259 peripheral blood Anatomy 0.000 description 1
- 239000011886 peripheral blood Substances 0.000 description 1
- 210000004976 peripheral blood cell Anatomy 0.000 description 1
- 210000005105 peripheral blood lymphocyte Anatomy 0.000 description 1
- 102000013415 peroxidase activity proteins Human genes 0.000 description 1
- 108040007629 peroxidase activity proteins Proteins 0.000 description 1
- 230000002085 persistent effect Effects 0.000 description 1
- 239000000825 pharmaceutical preparation Substances 0.000 description 1
- 230000003285 pharmacodynamic effect Effects 0.000 description 1
- COLNVLDHVKWLRT-UHFFFAOYSA-N phenylalanine Natural products OC(=O)C(N)CC1=CC=CC=C1 COLNVLDHVKWLRT-UHFFFAOYSA-N 0.000 description 1
- 230000000865 phosphorylative effect Effects 0.000 description 1
- 102000020233 phosphotransferase Human genes 0.000 description 1
- 239000006069 physical mixture Substances 0.000 description 1
- 230000004983 pleiotropic effect Effects 0.000 description 1
- 229920000233 poly(alkylene oxides) Polymers 0.000 description 1
- 229920001490 poly(butyl methacrylate) polymer Polymers 0.000 description 1
- 229920001483 poly(ethyl methacrylate) polymer Polymers 0.000 description 1
- 229920001200 poly(ethylene-vinyl acetate) Polymers 0.000 description 1
- 239000005015 poly(hydroxybutyrate) Substances 0.000 description 1
- 229920000212 poly(isobutyl acrylate) Polymers 0.000 description 1
- 229920000205 poly(isobutyl methacrylate) Polymers 0.000 description 1
- 229920000747 poly(lactic acid) Polymers 0.000 description 1
- 229920001306 poly(lactide-co-caprolactone) Polymers 0.000 description 1
- 229920000196 poly(lauryl methacrylate) Polymers 0.000 description 1
- 229920003229 poly(methyl methacrylate) Polymers 0.000 description 1
- 229920000184 poly(octadecyl acrylate) Polymers 0.000 description 1
- 238000002264 polyacrylamide gel electrophoresis Methods 0.000 description 1
- 229920001515 polyalkylene glycol Polymers 0.000 description 1
- 229920000515 polycarbonate Polymers 0.000 description 1
- 239000004417 polycarbonate Substances 0.000 description 1
- 229920000728 polyester Polymers 0.000 description 1
- 229920000573 polyethylene Polymers 0.000 description 1
- 239000004633 polyglycolic acid Substances 0.000 description 1
- 229920000129 polyhexylmethacrylate Polymers 0.000 description 1
- 229920000197 polyisopropyl acrylate Polymers 0.000 description 1
- 239000004626 polylactic acid Substances 0.000 description 1
- 239000004926 polymethyl methacrylate Substances 0.000 description 1
- 108091033319 polynucleotide Proteins 0.000 description 1
- 102000040430 polynucleotide Human genes 0.000 description 1
- 239000002157 polynucleotide Substances 0.000 description 1
- 229920000182 polyphenyl methacrylate Polymers 0.000 description 1
- 229920001155 polypropylene Polymers 0.000 description 1
- 229920001282 polysaccharide Polymers 0.000 description 1
- 239000005017 polysaccharide Substances 0.000 description 1
- 150000004804 polysaccharides Chemical class 0.000 description 1
- 229920001296 polysiloxane Polymers 0.000 description 1
- 235000019422 polyvinyl alcohol Nutrition 0.000 description 1
- 229920000915 polyvinyl chloride Polymers 0.000 description 1
- 239000004800 polyvinyl chloride Substances 0.000 description 1
- 229920001290 polyvinyl ester Polymers 0.000 description 1
- 229920001289 polyvinyl ether Polymers 0.000 description 1
- 229920001291 polyvinyl halide Polymers 0.000 description 1
- 229920000036 polyvinylpyrrolidone Polymers 0.000 description 1
- 239000001267 polyvinylpyrrolidone Substances 0.000 description 1
- 235000013855 polyvinylpyrrolidone Nutrition 0.000 description 1
- 239000011148 porous material Substances 0.000 description 1
- 230000035409 positive regulation of cell proliferation Effects 0.000 description 1
- 238000002600 positron emission tomography Methods 0.000 description 1
- 230000001323 posttranslational effect Effects 0.000 description 1
- 229910052700 potassium Inorganic materials 0.000 description 1
- 239000011591 potassium Substances 0.000 description 1
- PUDIUYLPXJFUGB-UHFFFAOYSA-N praseodymium atom Chemical compound [Pr] PUDIUYLPXJFUGB-UHFFFAOYSA-N 0.000 description 1
- 230000002335 preservative effect Effects 0.000 description 1
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 1
- 238000004393 prognosis Methods 0.000 description 1
- 108060006613 prolamin Proteins 0.000 description 1
- XXRYFVCIMARHRS-UHFFFAOYSA-N propan-2-yl n-dimethoxyphosphorylcarbamate Chemical compound COP(=O)(OC)NC(=O)OC(C)C XXRYFVCIMARHRS-UHFFFAOYSA-N 0.000 description 1
- 239000003380 propellant Substances 0.000 description 1
- 230000000069 prophylactic effect Effects 0.000 description 1
- 210000002307 prostate Anatomy 0.000 description 1
- 238000000746 purification Methods 0.000 description 1
- 238000003908 quality control method Methods 0.000 description 1
- 238000011002 quantification Methods 0.000 description 1
- 230000000306 recurrent effect Effects 0.000 description 1
- 230000009711 regulatory function Effects 0.000 description 1
- 230000008439 repair process Effects 0.000 description 1
- 238000011160 research Methods 0.000 description 1
- 230000000241 respiratory effect Effects 0.000 description 1
- 230000000717 retained effect Effects 0.000 description 1
- 238000003757 reverse transcription PCR Methods 0.000 description 1
- 238000012552 review Methods 0.000 description 1
- 206010039073 rheumatoid arthritis Diseases 0.000 description 1
- PYWVYCXTNDRMGF-UHFFFAOYSA-N rhodamine B Chemical compound [Cl-].C=12C=CC(=[N+](CC)CC)C=C2OC2=CC(N(CC)CC)=CC=C2C=1C1=CC=CC=C1C(O)=O PYWVYCXTNDRMGF-UHFFFAOYSA-N 0.000 description 1
- 108020004418 ribosomal RNA Proteins 0.000 description 1
- 201000004700 rosacea Diseases 0.000 description 1
- 239000012146 running buffer Substances 0.000 description 1
- 229960004889 salicylic acid Drugs 0.000 description 1
- 231100000241 scar Toxicity 0.000 description 1
- 238000006748 scratching Methods 0.000 description 1
- 230000002393 scratching effect Effects 0.000 description 1
- 208000008742 seborrheic dermatitis Diseases 0.000 description 1
- 201000003385 seborrheic keratosis Diseases 0.000 description 1
- 238000000926 separation method Methods 0.000 description 1
- 238000012163 sequencing technique Methods 0.000 description 1
- 238000013207 serial dilution Methods 0.000 description 1
- 229920000260 silastic Polymers 0.000 description 1
- 210000004927 skin cell Anatomy 0.000 description 1
- 150000003384 small molecules Chemical class 0.000 description 1
- 210000002460 smooth muscle Anatomy 0.000 description 1
- 229910052708 sodium Inorganic materials 0.000 description 1
- 239000011734 sodium Substances 0.000 description 1
- 229940054269 sodium pyruvate Drugs 0.000 description 1
- ZNJHFNUEQDVFCJ-UHFFFAOYSA-M sodium;2-[4-(2-hydroxyethyl)piperazin-1-yl]ethanesulfonic acid;hydroxide Chemical compound [OH-].[Na+].OCCN1CCN(CCS(O)(=O)=O)CC1 ZNJHFNUEQDVFCJ-UHFFFAOYSA-M 0.000 description 1
- 239000002904 solvent Substances 0.000 description 1
- 125000006850 spacer group Chemical group 0.000 description 1
- 230000000087 stabilizing effect Effects 0.000 description 1
- 239000008107 starch Substances 0.000 description 1
- 235000003702 sterols Nutrition 0.000 description 1
- 150000003432 sterols Chemical class 0.000 description 1
- 210000000434 stratum corneum Anatomy 0.000 description 1
- 210000004304 subcutaneous tissue Anatomy 0.000 description 1
- 229940098466 sublingual tablet Drugs 0.000 description 1
- 235000000346 sugar Nutrition 0.000 description 1
- 150000008163 sugars Chemical class 0.000 description 1
- 239000002511 suppository base Substances 0.000 description 1
- 230000004083 survival effect Effects 0.000 description 1
- 239000003765 sweetening agent Substances 0.000 description 1
- 230000008961 swelling Effects 0.000 description 1
- 229920001059 synthetic polymer Polymers 0.000 description 1
- 239000006188 syrup Substances 0.000 description 1
- 235000020357 syrup Nutrition 0.000 description 1
- 239000011975 tartaric acid Substances 0.000 description 1
- 235000002906 tartaric acid Nutrition 0.000 description 1
- RCINICONZNJXQF-MZXODVADSA-N taxol Chemical compound O([C@@H]1[C@@]2(C[C@@H](C(C)=C(C2(C)C)[C@H](C([C@]2(C)[C@@H](O)C[C@H]3OC[C@]3([C@H]21)OC(C)=O)=O)OC(=O)C)OC(=O)[C@H](O)[C@@H](NC(=O)C=1C=CC=CC=1)C=1C=CC=CC=1)O)C(=O)C1=CC=CC=C1 RCINICONZNJXQF-MZXODVADSA-N 0.000 description 1
- 229940063683 taxotere Drugs 0.000 description 1
- 238000002560 therapeutic procedure Methods 0.000 description 1
- RTKIYNMVFMVABJ-UHFFFAOYSA-L thimerosal Chemical compound [Na+].CC[Hg]SC1=CC=CC=C1C([O-])=O RTKIYNMVFMVABJ-UHFFFAOYSA-L 0.000 description 1
- 229940033663 thimerosal Drugs 0.000 description 1
- CNHYKKNIIGEXAY-UHFFFAOYSA-N thiolan-2-imine Chemical compound N=C1CCCS1 CNHYKKNIIGEXAY-UHFFFAOYSA-N 0.000 description 1
- 229940034610 toothpaste Drugs 0.000 description 1
- 239000000606 toothpaste Substances 0.000 description 1
- 231100000331 toxic Toxicity 0.000 description 1
- 230000002588 toxic effect Effects 0.000 description 1
- VZCYOOQTPOCHFL-UHFFFAOYSA-N trans-butenedioic acid Natural products OC(=O)C=CC(O)=O VZCYOOQTPOCHFL-UHFFFAOYSA-N 0.000 description 1
- 238000013518 transcription Methods 0.000 description 1
- 230000035897 transcription Effects 0.000 description 1
- 230000002103 transcriptional effect Effects 0.000 description 1
- 230000002463 transducing effect Effects 0.000 description 1
- 238000001890 transfection Methods 0.000 description 1
- 238000013519 translation Methods 0.000 description 1
- 238000002054 transplantation Methods 0.000 description 1
- 150000003626 triacylglycerols Chemical class 0.000 description 1
- QORWJWZARLRLPR-UHFFFAOYSA-H tricalcium bis(phosphate) Chemical compound [Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O QORWJWZARLRLPR-UHFFFAOYSA-H 0.000 description 1
- CYRMSUTZVYGINF-UHFFFAOYSA-N trichlorofluoromethane Chemical compound FC(Cl)(Cl)Cl CYRMSUTZVYGINF-UHFFFAOYSA-N 0.000 description 1
- 229940029284 trichlorofluoromethane Drugs 0.000 description 1
- 230000001960 triggered effect Effects 0.000 description 1
- 102000042287 type II cytokine receptor family Human genes 0.000 description 1
- 108091052254 type II cytokine receptor family Proteins 0.000 description 1
- 150000003668 tyrosines Chemical class 0.000 description 1
- 238000000870 ultraviolet spectroscopy Methods 0.000 description 1
- 238000002255 vaccination Methods 0.000 description 1
- 229940005605 valeric acid Drugs 0.000 description 1
- 239000004474 valine Substances 0.000 description 1
- 239000011534 wash buffer Substances 0.000 description 1
- 238000001262 western blot Methods 0.000 description 1
- 239000005019 zein Substances 0.000 description 1
- 229940093612 zein Drugs 0.000 description 1
Images









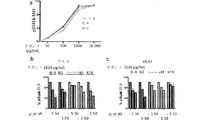













Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/52—Cytokines; Lymphokines; Interferons
- C07K14/54—Interleukins [IL]
- C07K14/5406—IL-4
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P29/00—Non-central analgesic, antipyretic or antiinflammatory agents, e.g. antirheumatic agents; Non-steroidal antiinflammatory drugs [NSAID]
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P35/00—Antineoplastic agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/52—Cytokines; Lymphokines; Interferons
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Organic Chemistry (AREA)
- Medicinal Chemistry (AREA)
- General Health & Medical Sciences (AREA)
- Biochemistry (AREA)
- Biophysics (AREA)
- Gastroenterology & Hepatology (AREA)
- Genetics & Genomics (AREA)
- Zoology (AREA)
- Molecular Biology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Toxicology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Pharmacology & Pharmacy (AREA)
- Animal Behavior & Ethology (AREA)
- Public Health (AREA)
- Veterinary Medicine (AREA)
- Immunology (AREA)
- Engineering & Computer Science (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Pain & Pain Management (AREA)
- Rheumatology (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Peptides Or Proteins (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Description
サイトカインは、1つの細胞により作られて、他に作用する分子として広く定義される。他の命名の中でも、インターロイキン、インターフェロン、増殖因子、およびTNFとよばれるこれらの分子は、また、細胞の増殖から炎症、免疫、遊走、線維症、修復、および血管新生まで、あらゆる本質的に重要な生物学的過程に関わる。これらの特性に起因して、治療法としてのサイトカインの使用が探究されてきた。残念ながら、実例の大多数において、治療法としての野生型サイトカインの使用は、用量制限の毒性または不十分な効能を伴うことが多い。
別様に定義されない限り、本明細書で使用される全ての技術および科学用語は、概して、本発明が属する技術分野の当業者により共通に理解されるものと同じ意味を有する。一般的に、本明細書で使用される命名法および細胞培養、分子遺伝学、有機化学における実験手順は、当該技術分野でよく知られたものであり、共通に採用される。標準的な技術が、核酸およびペプチド合成のために使用される。技術および手順は、概して、当該技術分野における従来の方法および種々の一般的な参照文献に従って行われ(概して、参照により本明細書に組み込まれる、Sambrook et al. MOLECULAR CLONING: A LABORATORY MANUAL, 2nd ed. (1989) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.を参照)、それはこの文献の全体にわたって提供される。本明細書で使用される命名法および以下に記載される分析および有機合成化学の実験手順は、よく知られるものであり、当該技術分野で共通に採用される。標準的な技術またはその改変形態が、化学合成および化学分析のために使用される。
IL‐4は、古典的な4つのα‐ヘリックスバンドルサイトカインである。その一次結合鎖は、IL‐4Rαであり、細胞外ドメイン、膜貫通ドメイン、および長い細胞内テイル(intracellular tail)からなり、Box1ドメイン、PTBドメインタンパク質および3つのSTAT6ドッキングについての結合部位、並びに活性化部位を含む、II型サイトカインレセプター鎖である。
種々の実施形態では、本開示は、必ずではないが、実質的に精製され、野生型IL‐4のアゴニストとして機能し得るIL‐4変異体ポリペプチドを提供し、それらは1つまたはそれ以上のIL‐4の生物活性を保持する(例えば、細胞増殖の刺激)。
増殖および分化の障害
いくつかの実施形態では、変異体サイトカインポリペプチドおよび/またはそれらを発現する核酸が、異常なアポトーシスまたは分化のプロセスに関連する障害(例えば、細胞増殖性障害または細胞分化性障害、例えば癌)を治療するために対象に投与され得る。
開示されるサイトカイン変異タンパク質の組成物およびそれらの使用の方法による寛解ができる病的症状は、例えば、喘息およびアレルギー性炎症応答のような肺障害を含む。
1つもしくはそれ以上の治療薬が単独で、または1つもしくはそれ以上の化学療法と組み合わせて、対象への投与のために薬学的に許容可能な担体とともに処方され得る。活性成分は、逐次投与のために単独で(個々に)処方され得、または同時投与のために一緒に処方されることとしてもよい。
投与の様式は、経口投与、舌下投与、鼻腔内投与、気管内投与、吸入、点眼、局所投与、経皮投与、皮内投与、直腸投与、腟内投与、皮下投与、静脈内投与、筋肉内投与、腹腔内投与、胸骨内の、投与、経粘膜的投与を介するもの、を含む、あらゆる医学的に許容可能な様式であることとしてもよい。また、投与の様式は、体外装置および/または組織貫通電磁気装置を介することとしてもよい。
本明細書に記載される変異タンパク質は、例えば、酵素、放射標識、エピトープ、または蛍光タンパク質(例えば、緑色蛍光タンパク質)のような、シグナルを産生するまたは標識である部分で直接または間接的に標識され得る。本明細書に記載される変異タンパク質は、例えば、標準的なイムノブロッティング、免疫蛍光法、酵素免疫測定法(EIA)、ラジオイムノアッセイ(RIA)、蛍光エネルギー転移、ウエスタンブロット、並びに他の診断および検出技術を用いて、レセプターがサンプルにまたは細胞上に存在するかどうかを測定するために、サンプルにまたは細胞に接触され得る。
IL−4/IL−13系を用いて、共有サイトカインレセプターの相対親和性の操作が特徴的な応答性のパターンを有するサイトカインをもたらし得るという考えを試験した。2つのアプローチをとって、γc(図1b)またはIL−13Rα1(図1c)に結合するより高い親和性のためにIL−4を操作した。
得られたIL−4変異タンパク質の結合の特徴を、特徴づけした。表面プラズモン共鳴(SPR)によってそれらの部位2結合親和性および動態をIL−13Rα1およびγcに関して測定するために、バキュロウイルスおよびIL−4Rαと形成された複合体を用いた組み換えIL−4および変異体KF、KFR、RQおよびRGAである(表1および図2を参照)。

理想的には、スーパー4といったIL−4変異タンパク質の増大した親和性は、野生型IL−4/γc結合の配向の最小限の妨害によって実現された。第2の鎖、γcとのスーパー4ドッキング様式が野生型IL−4に比べて乱されたかどうかを決定するために、結晶解析を実施した。遊離(たとえば、IL−4Rαに結合していない)スーパー4の、γcに対するより高い部位2親和性(KD約300nM)を考慮すると、IL−4Rαの不在下で、スーパー4/γc二元複合体を3.25Åの分解能まで結晶化させることが可能であった(図3aおよび4、表2)
様々な細胞型に対する、開示されるIL−4変異タンパク質の影響を試験した。スーパーカインの第2の鎖の動員の平衡定数がIL−4のものよりも劇的に異なったため、主にI型もしくはII型IL−4レセプターまたは顕著な量の双方を発現する細胞型のスーパーカインの挙動を試験した。
スーパー4が約3〜10倍より強力な、STAT6リン酸化の活性剤であった一方で、γcに関する三次元平衡はIL−4のものよりも約3700倍高かったという考えは、シグナル誘導レセプター形成が、どのように第2の鎖の発現によって決定されるかという分析を促した。マクロファージの約40のレセプターは、ほとんど最大限のSTAT6活性化をもたらすであろうと計算されていた。従って、細胞レベルでスーパー4が有した驚くほどに控えめな利点は、形成されたレセプター複合体の比較的低い数と何か関係があり得、および/または、第2の鎖の数が低い、たとえば、リガンドが利用可能な第2の鎖と競合する場合として観測され得る。
γc発現での増加はIL−4に対してスーパー4が有した利点を低下させることが予測され得るため、ラモス細胞よりもはるかに高いγc発現を有したHH細胞系統で、IL−4およびスーパー4に対する感受性を実験した(図6aおよびb)。スーパー4は、10pg/mlから10,000pg/mlの濃度範囲にわたって、HH細胞でのSTAT6のリン酸化においてIL−4よりも優れてはいなかった(図10a)。
様々なスーパーカインに対する応答は、どの第2の鎖をそれらが第1に使用するか次第であったことを確認するために、ヒト末梢血液白血球(PBL)のStat6リン酸化応答を試験した(図11)。PBLは、刺激されなかったか、15分間、IL−4または様々なスーパーカインで刺激されたかのいずれかであった。STAT6 Y641リン酸化をフローサイトメトリーで測定し、様々な末梢血液細胞型の応答性を試験した。スーパー4に対するリンパ球の応答は、低いリガンド濃度で著しくより良好であった。
IL−4およびスーパーカインの機能上の特異性および免疫調節性能力を試験するために、CD4 T細胞および単球を含む一連の実験を実施した。TGF−βおよびIL−4の組み合わせは、未処置のヒトCD4 T細胞の、TH9細胞への分化を促進する。スーパー4が野生型IL−4よりも強力にTH9分化を誘導するかどうかを試験するために、未処置のCD4+CD45RA+CD45RO−CD25−T細胞を単離した。
GM−CSFと組み合わせたIL−4は、ヒト単球から、樹状細胞(DC)のインビトロ分化を誘導する。このプロセスに関するI型またはII型レセプターの相対的な役割はまだ明らかになっていない。スーパー4がγcに優先的に結合し、KFRはIL−13Rα1ライゲーションに向かってそれることを考慮し、ヒトDC分化の間のγcおよびIL−13Rα1経路のために必要な要件を解明した。
IL−4および2つのスーパーカインが単球において同じ程度までSTAT6を活性化したため、スーパー4のDC分化を誘導する能力を決定した(図11(e)、図12および図19)。精製した単球を2つの用量のサイトカインで処理し、1つの用量は、P−STAT6 EC50値(30pM)(図15)に対応しており、もう一方の用量は、飽和状態(50nM)(図20)に対応していた。STAT6およびIRS1リン酸化の量ならびにγcおよびIL−13Rα1レセプターの下方制御を示した時間で分析した。低用量では、スーパー4およびKFRはIL−4と比較して遅れたSTAT6およびIRS1の活性化を示した。γcまたはIL−13Rα1のいずれの顕著な内部移行も一切観察されなかった(図15)。高用量では、3つのサイトカインは、STAT6およびIRS1活性化の同一の動態プロファイルを誘導した。KFRは、刺激の終わりに近い時間で、IL−13Rα1のより強力な内部移行を示した(図20)。概して、これらの結果によって、表面レセプター内部移行とシグナル伝達活性化との間の相関関係の欠如が示される。さらに、シグナル伝達活性化の遅れた動態だけでは、スーパー4のDC分化を誘導する非効率性を説明できず、II型レセプター特異性シグナル伝達がDC分化に必要であることを示唆する。
DCの分化において、IL−4およびスーパーカインによって誘導される遺伝子プログラムの重複性の程度に対して定性的な見識を得るために、GM−CSFで同時に処理した単球の野生型IL−4および2つのスーパーカインに応答した遺伝子発現のゲノム全体の分析を実施した。CD14+単球を、5人の健康なドナーから単離し、6時間、50ng/mlのGM−CSF単独で、または、20ng/mlのIL−4、スーパー4、またはKFRで刺激した。細胞をPBS中で洗浄し、1mlのTRIzol試薬(シグマ社)に溶解した。組み合わせたTRIzol/RNeasy Micro(キアゲン社)プロトコルを用いて全RNAを単離した。RNAの品質を2100Bioanalyzer(アジレント社)で確認した。cDNAを作製し、Two−color Low Input Quick Amp Labelingキット(アジレント社)を用いて、Cy3で標識される単一処理試料(GM−CSF単独)およびCy5で標識される二重処理試料で標識した。各ドナーに関し、各Cy5標識化二重処理(IL−4、Super−4またはKFRのいずれかを伴うGM−CSF)の試料を、8×60K SurePrint G3 Gene発現マイクロアレイ(アジレント社)上にGM−CSFを伴う対応するCy3標識化単一処理と同時ハイブリダイズし、製造業者の説明書に従って処理した。Feature Extractionソフトウェア(アジレント社)を用いて生のデータを正規化した。二重処理と単一処理の間、および変異形の間の変化の統計的分析のために、対応のあるt検定分析を用いた。遺伝子発現分析をGeneSpring GX11.5およびExcel上で実施した。6個未満のアレイでしか検知されなかった実体、品質管理プローブおよび長い遺伝子間非コードRNAのためのプローブは除外した。GM−CSF単独と比較して(Cy5対Cy3)少なくとも1つの二重処理群で有意な変化(対応のあるt検定)があった実体を保持した。2つの所与のサイトカインの間の倍変化統計的分析を、対応のあるt検定によって決定した。ミクロアレイデータをNCBI(GEOシリーズ受託番号:GSE40200)のGene Expression Omnibus(GEO)Databaseに提出した。
操作されたサイトカインによって誘導されるDCの機能性をさらに評価するため、8日間、最後の24時間はLPS刺激を行う、または行わないで、培養した細胞の上澄み上でLuminexアッセイを実施することで、サイトカイン、ケモカインおよび増殖因子の分泌パターンを比較した。
Claims (25)
- 2つまたはそれ以上の共有レセプターポリペプチドにより認識される野生型IL‐4リガンドに対して、標的細胞における細胞応答を選択的に操作するための方法であって、IL‐4変異タンパク質リガンドを提供することを含み、前記IL‐4変異タンパク質リガンドが、前記野生型IL‐4リガンドよりも高い親和性で前記2つまたはそれ以上の共有レセプターポリペプチドの1つと結合することにより、前記細胞応答を選択的に操作し、前記IL‐4変異タンパク質が以下のアミノ酸置換:117R、118V、121Q、122S、124W、125F、128G、および129Aを含み、前記アミノ酸の番号付けは野生型ヒトIL‐4に従っている、方法。
- 前記IL‐4変異タンパク質リガンドが、前記野生型IL‐4リガンドよりも高い親和性で前記2つまたはそれ以上の共有レセプターポリペプチドのうちの2つと結合することにより、前記細胞応答を選択的に操作する、請求項1の方法。
- 前記IL‐4変異タンパク質リガンドが、前記野生型IL‐4リガンドよりも高い親和性で前記2つまたはそれ以上の共有レセプターポリペプチドのうちの1つと結合し、前記野生型IL‐4リガンドよりも低い親和性で前記2つまたはそれ以上のうちの共有レセプターポリペプチドの別の1つと結合することにより、前記細胞応答を選択的に操作する、請求項1の方法。
- 前記2つまたはそれ以上の共有レセプターポリペプチドが、前記標的細胞の表面に等しく存在しない、請求項1の方法。
- 前記IL‐4変異タンパク質リガンドにより結合される前記共有レセプターが、前記IL‐4変異タンパク質リガンドにより結合されていない前記共有レセプターよりも、前記標的細胞の表面に、より低いレベルで存在する、請求項4の方法。
- 前記IL‐4変異タンパク質リガンドにより結合される前記共有レセプターが、前記IL‐4変異タンパク質リガンドにより結合されていない前記共有レセプターよりも、前記標的細胞の表面に、より高いレベルで存在する、請求項4の方法。
- 前記IL‐4変異タンパク質リガンドにより結合されていない前記共有レセプターが、前記IL‐4変異タンパク質リガンドへの低減された到達性を有する、請求項1の方法。
- 前記IL‐4変異タンパク質リガンドにより結合されていない前記共有レセプターへの到達性が、前記IL‐4変異タンパク質リガンドにより結合されていない前記共有レセプターの前記到達性に干渉する反応剤を提供することにより低減される、請求項7の方法。
- 前記反応剤は、前記IL‐4変異タンパク質リガンドにより結合されていない前記共有レセプターを認識する抗体である、請求項8の方法。
- 前記共有レセプターは、共通γ鎖(γc)、またはインターロイキン‐13レセプターアルファ1(IL‐13Rα1)である、請求項1の方法。
- 前記より高い親和性は少なくとも10倍である、請求項1の方法。
- 前記より低い親和性は少なくとも5倍である、請求項3の方法。
- IL‐4変異タンパク質を含み、前記IL‐4変異タンパク質が、野生型IL‐4と比較して、共有サイトカインレセプターに結合する、より高い親和性をもたらし、前記IL‐4変異タンパク質が以下のアミノ酸置換:117R、118V、121Q、122S、124W、125F、128G、および129Aを含み、前記アミノ酸の番号付けは野生型ヒトIL‐4に従っている、組成物。
- 前記IL‐4変異タンパク質が、野生型IL‐4と比較して、第1の共有サイトカインレセプターに結合するより高い親和性、および野生型IL‐4vと比較して、第2の共有サイトカインレセプターへの低減された親和性をもたらす請求項13に記載の組成物。
- 前記第1の共有サイトカインレセプターは、前記第2の共有サイトカインレセプターよりも低いレベルで発現される、請求項14に記載の組成物。
- 前記第1の共有サイトカインレセプターは、前記第2の共有サイトカインレセプターよりも高いレベルで発現される、請求項14に記載の組成物。
- 前記IL‐4変異タンパク質が、野生型IL‐4と比較して、第1および第2の共有サイトカインレセプターに結合するより高い親和性をもたらす、請求項13に記載の組成物。
- 前記親和性における増加は少なくとも10倍である、請求項13から17のいずれか1項に記載の組成物。
- 前記親和性における低減は少なくとも5倍である、請求項14に記載の組成物。
- 前記共有サイトカインレセプターは、共通γ鎖(γc)またはインターロイキン‐13レセプターアルファ1(IL‐13Rα1)である、請求項13に記載の組成物。
- 前記第1の共有サイトカインレセプターはγcであり、前記第2の共有サイトカインレセプターはIL‐13Rα1である、請求項14に記載の組成物。
- 前記第1の共有サイトカインレセプターはIL‐13Rα1であり、前記第2の共有サイトカインレセプターはγcである、請求項14に記載の組成物。
- 前記第1および第2の共有サイトカインレセプターはγcおよびIL‐13Rα1である、請求項14に記載の組成物。
- 必要とする対象の治療において使用するための医薬組成物であって、治療上有効量の請求項13に記載に記載の組成物を含み、必要とする対象に投与される、医薬組成物。
- 前記対象が炎症性疾患に罹っている、請求項24に記載の医薬組成物。
Applications Claiming Priority (7)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201261681490P | 2012-08-09 | 2012-08-09 | |
| US61/681,490 | 2012-08-09 | ||
| US201261725791P | 2012-11-13 | 2012-11-13 | |
| US61/725,791 | 2012-11-13 | ||
| US201361825980P | 2013-05-21 | 2013-05-21 | |
| US61/825,980 | 2013-05-21 | ||
| PCT/US2013/054164 WO2014074186A2 (en) | 2012-08-09 | 2013-08-08 | Superkines and synthekines: repurposed cytokines with new and enhanced signaling activities |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2018118931A Division JP2018161143A (ja) | 2012-08-09 | 2018-06-22 | スーパーカインおよびシンセカイン:新規かつ増強されたシグナル伝達活性を有する再利用されるサイトカイン |
Publications (3)
| Publication Number | Publication Date |
|---|---|
| JP2015525574A JP2015525574A (ja) | 2015-09-07 |
| JP2015525574A5 JP2015525574A5 (ja) | 2016-03-10 |
| JP6445434B2 true JP6445434B2 (ja) | 2018-12-26 |
Family
ID=50685291
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2015526711A Active JP6445434B2 (ja) | 2012-08-09 | 2013-08-08 | スーパーカインおよびシンセカイン:新規かつ増強されたシグナル伝達活性を有する再利用されるサイトカイン |
| JP2018118931A Pending JP2018161143A (ja) | 2012-08-09 | 2018-06-22 | スーパーカインおよびシンセカイン:新規かつ増強されたシグナル伝達活性を有する再利用されるサイトカイン |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2018118931A Pending JP2018161143A (ja) | 2012-08-09 | 2018-06-22 | スーパーカインおよびシンセカイン:新規かつ増強されたシグナル伝達活性を有する再利用されるサイトカイン |
Country Status (6)
| Country | Link |
|---|---|
| US (3) | US9738696B2 (ja) |
| EP (1) | EP2882449B1 (ja) |
| JP (2) | JP6445434B2 (ja) |
| CN (2) | CN110615835A (ja) |
| IN (1) | IN2015KN00329A (ja) |
| WO (1) | WO2014074186A2 (ja) |
Families Citing this family (10)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CA3038533A1 (en) * | 2016-09-30 | 2018-04-05 | The Board Of Trustees Of The Leland Stanford Junior University | Variant type iii interferons and synthekines |
| JP2018157812A (ja) * | 2017-03-23 | 2018-10-11 | 東ソー株式会社 | 試料中に含まれる細胞の検出方法 |
| CN110582293A (zh) * | 2017-03-31 | 2019-12-17 | 小利兰·斯坦福大学托管委员会 | 合成因子组合物及使用方法 |
| CN113840832A (zh) | 2018-05-14 | 2021-12-24 | 狼人治疗公司 | 可活化白介素-2多肽及其使用方法 |
| WO2019222294A1 (en) | 2018-05-14 | 2019-11-21 | Werewolf Therapeutics, Inc. | Activatable cytokine polypeptides and methods of use thereof |
| WO2019232523A1 (en) | 2018-06-01 | 2019-12-05 | The Board Of Trustees Of The Leland Stanford Junior University | Il-13/il-4 superkines: immune cell targeting constructs and methods of use thereof |
| CA3137058A1 (en) | 2019-04-19 | 2020-10-22 | Synerkine Pharma B.V. | A fusion protein comprising il13 |
| SG11202112541RA (en) | 2019-05-14 | 2021-12-30 | Werewolf Therapeutics Inc | Separation moieties and methods and use thereof |
| KR20220101147A (ko) | 2019-11-14 | 2022-07-19 | 웨어울프 세라퓨틱스, 인크. | 활성화가능한 사이토카인 폴리펩티드 및 이의 사용 방법 |
| WO2023230163A1 (en) * | 2022-05-24 | 2023-11-30 | University Of Pittsburgh - Of The Commonwealth System Of Higher Education | Interleukin 4 and/or interleukin 13-based eye compositions |
Family Cites Families (6)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| DE4423131A1 (de) * | 1994-07-01 | 1996-01-04 | Bayer Ag | Neue hIL-4-Mutantenproteine als Antagonisten oder partielle Agonisten des humanen Interleukin 4 |
| US6335426B1 (en) * | 1996-06-14 | 2002-01-01 | Bayer Corporation | T-cell selective interleukin-4 agonists |
| US6028176A (en) * | 1996-07-19 | 2000-02-22 | Bayer Corporation | High-affinity interleukin-4 muteins |
| US6333410B1 (en) | 2000-08-18 | 2001-12-25 | Immunogen, Inc. | Process for the preparation and purification of thiol-containing maytansinoids |
| CA2423469A1 (en) | 2000-10-18 | 2002-04-25 | Massachusetts Institute Of Technology | Methods and products related to pulmonary delivery of polysaccharides |
| WO2011106779A1 (en) * | 2010-02-26 | 2011-09-01 | Aerovance Inc. | Use of modified il-4 mutien receptor antagonists to treat dermatitis |
-
2013
- 2013-08-08 CN CN201910717548.0A patent/CN110615835A/zh active Pending
- 2013-08-08 EP EP13852778.3A patent/EP2882449B1/en active Active
- 2013-08-08 IN IN329KON2015 patent/IN2015KN00329A/en unknown
- 2013-08-08 JP JP2015526711A patent/JP6445434B2/ja active Active
- 2013-08-08 US US14/419,873 patent/US9738696B2/en active Active
- 2013-08-08 CN CN201380047839.4A patent/CN104619334A/zh active Pending
- 2013-08-08 WO PCT/US2013/054164 patent/WO2014074186A2/en active Application Filing
-
2017
- 2017-07-27 US US15/662,180 patent/US10738096B2/en active Active
-
2018
- 2018-06-22 JP JP2018118931A patent/JP2018161143A/ja active Pending
-
2020
- 2020-08-07 US US16/988,460 patent/US20210188932A1/en active Pending
Also Published As
| Publication number | Publication date |
|---|---|
| US20180016316A1 (en) | 2018-01-18 |
| US20210188932A1 (en) | 2021-06-24 |
| US20160244499A1 (en) | 2016-08-25 |
| US9738696B2 (en) | 2017-08-22 |
| WO2014074186A2 (en) | 2014-05-15 |
| JP2018161143A (ja) | 2018-10-18 |
| EP2882449A4 (en) | 2016-03-09 |
| CN104619334A (zh) | 2015-05-13 |
| WO2014074186A3 (en) | 2014-06-19 |
| US10738096B2 (en) | 2020-08-11 |
| EP2882449A2 (en) | 2015-06-17 |
| IN2015KN00329A (ja) | 2015-07-10 |
| EP2882449B1 (en) | 2019-05-15 |
| CN110615835A (zh) | 2019-12-27 |
| JP2015525574A (ja) | 2015-09-07 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP6445434B2 (ja) | スーパーカインおよびシンセカイン:新規かつ増強されたシグナル伝達活性を有する再利用されるサイトカイン | |
| JP7503794B2 (ja) | インターロイキン-2のスーパーアンタゴニスト、パーシャルアゴニスト及びアンタゴニスト | |
| JP6441867B2 (ja) | インターロイキン2のスーパーアゴニストおよびアンタゴニスト | |
| Udalova et al. | Macrophage heterogeneity in the context of rheumatoid arthritis | |
| TWI357336B (en) | Uses of il-23 agonists and antagonists; related re | |
| CN112843222B (zh) | Ankrd22蛋白在制备治疗或延缓自身免疫性疾病的产品中的应用 | |
| US20110311475A1 (en) | Means and method for the treatment of antibody deficiency diseases based on il-21 and il-21 variants | |
| CN114380919B (zh) | 经修饰的il-2分子及其用途 | |
| Rusiñol et al. | Review of the Il-18 and IL37 Implications in the Pathogenesis of Atopic Dermatitis and Psoriasis: Prospective Treatment Targets | |
| CN116854821A (zh) | IL-15突变体-Fc/IL-15Rα亚基-Fc异源二聚体及其用途 | |
| Mori et al. | Eosinophil lineage-committed progenitors | |
| JP2000510684A (ja) | インターロイキン−3(il−3)受容体アゴニスト |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20160715 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20170704 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20171004 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20180223 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20180622 |
|
| A911 | Transfer to examiner for re-examination before appeal (zenchi) |
Free format text: JAPANESE INTERMEDIATE CODE: A911 Effective date: 20180802 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20181018 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20181023 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20181102 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20181129 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 6445434 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |