JP3662021B2 - 補体関連蛋白質および炭水化物よりなる組成物、ならびに該組成物の製法および使用方法 - Google Patents
補体関連蛋白質および炭水化物よりなる組成物、ならびに該組成物の製法および使用方法 Download PDFInfo
- Publication number
- JP3662021B2 JP3662021B2 JP52569594A JP52569594A JP3662021B2 JP 3662021 B2 JP3662021 B2 JP 3662021B2 JP 52569594 A JP52569594 A JP 52569594A JP 52569594 A JP52569594 A JP 52569594A JP 3662021 B2 JP3662021 B2 JP 3662021B2
- Authority
- JP
- Japan
- Prior art keywords
- complement
- protein
- scr1
- cells
- selectin
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Expired - Fee Related
Links
Images







Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/705—Receptors; Cell surface antigens; Cell surface determinants
- C07K14/7056—Lectin superfamily, e.g. CD23, CD72
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P29/00—Non-central analgesic, antipyretic or antiinflammatory agents, e.g. antirheumatic agents; Non-steroidal antiinflammatory drugs [NSAID]
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/705—Receptors; Cell surface antigens; Cell surface determinants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Organic Chemistry (AREA)
- Medicinal Chemistry (AREA)
- Immunology (AREA)
- General Health & Medical Sciences (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Cell Biology (AREA)
- Molecular Biology (AREA)
- Genetics & Genomics (AREA)
- Biophysics (AREA)
- Biochemistry (AREA)
- Gastroenterology & Hepatology (AREA)
- Zoology (AREA)
- Toxicology (AREA)
- Public Health (AREA)
- General Chemical & Material Sciences (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Veterinary Medicine (AREA)
- Animal Behavior & Ethology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Pharmacology & Pharmacy (AREA)
- Pain & Pain Management (AREA)
- Engineering & Computer Science (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Rheumatology (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Peptides Or Proteins (AREA)
- Pharmaceuticals Containing Other Organic And Inorganic Compounds (AREA)
- Carbon And Carbon Compounds (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
Description
最も広い態様において、本発明は、少なくとも1つの補体部位および少なくとも1つの炭水化物部位よりなる組成物、ならびにかかる組成物の製法を提供する。特に、本発明の組成物は、補体受容体1型に関連する補体蛋白質よりなり、さらに、セレクチンのごとき細胞間接着分子についてのリガンドよりなる。好ましい態様において、該組成物は、ルイスX抗原またはシアリルルイスX抗原と組み合わせた、補体受容体1型、またはその断片もしくは誘導体よりなる。本発明の組成物は、補体活性および炎症が関与する疾患の診断または治療に用途を有する。また、不適当な補体活性および細胞間接着によって媒介される炎症を治療または緩和させるための医薬組成物も提供する。
2.発明の背景
2.1.補体系
補体系は、ヒトの正常血清中のグロブリン類の約10パーセントを構成する一群の蛋白質である(Hood,L.E.,et al.,1984,Immunology,第2版,The Benjamin/Cummings Publishing Co.,Menlo Park,California,第339頁)。補体(C)は免疫およびアレルギー反応の媒介において重要な役割を果たす(Rapp,H.J.and Borsos,T,1970,Molecular Basis of Complement Action,Appleton−Century−Crofts(Meredity),New York)。補体成分の活性化は、補体依存性の病気に関連する炎症を媒介する走化性ペプチドを含めた、一群の因子の生成に導く。補体カスケードの連続的活性化は、抗原−抗体複合体が関与する古典的経路を介して、あるいはある種の細胞壁多糖のごとき外来性構造の認識が関与する代替経路によって起こり得る。活性化補体蛋白質によって媒介される活性は、標的細胞の溶解、走化性、オプソニン化、血管および他の平滑筋細胞の刺激、および肥満細胞の脱顆粒反応のごとき機能的異常、小血管の浸透性の増大、白血球の定方向化移動、Bリンパ球およびマクロファージの活性化を含む(Eisen,H.N.,1974,Immunology,Harper & Row Publishers,Inc.Hagerstown,Maryland,第512頁)。
蛋白質分解カスケードステップの間に、生物学的活性ペプチド断片、アナフィラトキシンC3a、C4a、およびC5a(WHO Scientific Group,1977,WHO Tech Rep.Ser.606:5およびそこに引用された文献参照)が、第3(C3)、第4(C4)、および第5(C5)天然補体成分から放出される(Hugli,T.E.,1981,CRC Crit.Rev.Immunol.1:321;Bult,H.and Herman,A.G.,1983,Agents Actions 13:405)。
2.2 補体受容体
補体受容体1(CR1)。CR1またはCD35と呼ばれるヒトC3b/C4b受容体は、赤血球、単球/マクロファージ、顆粒球、B細胞、いくつかのT細胞、脾臓濾胞樹枝状細胞、および糸球体有足細胞上に存在する(Fearon D.T.,1980,J.Exp.Med.152:20,Wilson,J.G.,et al.,1983,J.Immunol.131:684;Reynes,M.,et al.,1976 N.Engl.J.Med.295:10;Kazatchkine,M.D.,et al.,1982,Clin.Immunol.Immunopathol.27:210)。CR1はC3b、C4bおよびiC3bに特異的に結合する。
CR1は、古典的経路および代替経路C3/C5転換酵素を阻害し、I因子によるC3bおよびC4bの切断についての助因子として作用し、これは、CR1は受容体として働くに加えて補体調節機能も有することを示す(Fearon,D.T.,1979,Proc.Natl.Acad.Sci.U.S.A.76:5867;Iida,K.I.and Nussenzweig,V.,1981,J.Exp.Med.153:1138)。補体活性化の代替経路において、二分子複合体C3b,BbはC3酵素(転換酵素)である。CR1(およびH因子、高濃度において)はC3bに結合でき、またC3b,Bbの解離を促進する。さらに、C3b、CR1(およびC3b、H)はC3bを、I因子による不可逆的蛋白質分解不活化に感受性とし、その結果、不活化C3b(iC3b)が形成される。補体活性化の古典的経路において、複合体C4b,2aはC3転換酵素である。
CR1(およびC4結合蛋白質、C4bp、高濃度において)はC4bに結合でき、C4b,2aの解離を促進する。結合はC4bを、C4cおよびC4d(不活化補体蛋白質)への切断を通じてのI因子による不可逆的蛋白質不活化に感受性とする。
CR1は補体受容体2型(CR2)に対して相同性を有することが示された(Weis,J.J.,et al.,1986,Proc.Natl.Acad.Sci.U.S.A.83:5639−5643)。CR1は4つの長鎖相同性反復(LHR)に配置された多重のショートコンセンサスリピート(SCR)よりなる糖蛋白質である。LHR−Dと呼ばれる最もC−末端のLHRに続いて、2つのさらなるSCR、膜貫通領域および細胞質領域がある(Klickstein,et al.,1987,J.Exp.Med.,165:1095:Klickstein,et al.,1988,J.Exp.Med.,168:1699−1717)。赤血球CR1は自己免疫患者における循環免疫複合体の除去に関与し、そのレベルはAIDSの進展に関係し得る(Inada,et al.,1986,AIDS Res.2:235;Inada,et al.,1989,Ann.Rheu.Dis.4:287)。
4つのCR1のアロタイプ形態が見い出されており、40,1000−50,000ダルトンの分子量の増加だけ異なる。AおよびBアロタイプとも呼ばれる2つの最も通常の形態であるFおよびSアロタイプは、各々、250,000および290,000ダルトンの分子量を有し(Dykman,T.R.,et al.,1983,Proc.Natl.Acad.Sci.U.S.A.80:1698;Wong,W.W.et al.,1983、J.Clin.Invest.72:685)、2つのより稀な形態は210,000および290,000ダルトンの分子量を有する(Dykman,T.R.,et al.,1984,J.Exp.Med.159:691;Dykman,T.R.,et al.,1985,J.Immunol.134:1787)。これらの違いは、糖鎖付加状態よりもむしろCR1のポリペプチド鎖における変異を表しているようである。何故ならば、それらはエンドグリコシダーゼFでの精製された受容体蛋白質の処理によって分解されなかったからであり(Wong,W.W.,et al.,1983,J.Clin,Invest.72:685)、それらは、糖鎖付加阻害剤ツニカマイシンの存在下で受容体アロタイプが生合成された場合に観察された(Lublin,D.M.,et al.,1986,J.Biol.Chem.261:5736)。すべての4つのCR1アロタイプはC3b−結合活性を有する(Dykman,T.R.,et al.,1983,Proc.Natl.Acad.Sci,U.S.A.80:1698;Wong,W.W.,et al.,1983,J.Clin.Invest.72:685;Dykman,T.R.,et al.,1984,J.Exp.Med.,159:691;Dykman,T.R.,et al.,1985,J.Immunol.134:1787)。〜250kDのF(またはA)アロタイプには、5'から3'にかけて、各々、LHR−A、−B、−C、および−Dと呼ばれる4つのLHRがある(Wong,et al.,1989,J.Exp.Med.169:847)。LHR−Aにおける最初の2つのSCRはC4bに結合するその能力を決定し、LHR−Bおよび−Cにおける対応するユニットはC3bについてのそのより高いアフィニティーを決定する。〜290kdのより大きいS(またはB)アロタイプは、LHR−Bの5'側半分とLHR−Aの3'側半分のキメラである第5のLHRを有し、第3のC3b結合部位を含有すると予測される(Wong,et al.,1989,J.Exp.Med.169:847)。全身性紅斑性狼瘡(SLE)を持つ患者で多く発生し、多発性狼瘡ファミリー(Dykman,et al.,1984,J.Exp.Med.,159:691;Van Dyne,et al.,1987,Clin.Exp.Immunol.68:570)における患者に関連して見い出される、〜210kDのCR1の最も小さいF'(またはC)アロタイプは、1つのLHRの欠失に由来するらしく、補体断片で被覆された免疫複合体に対して効果的に結合するその能力が損なわれているらしい。
CR1の天然に生じる可溶性形態が、正常個体およびSLEを持つある種の個体の血漿で同定されている(Yoon,et al.,1985 J.Immunol.134:3332−3338)。その構造的および機能的特徴は、構造的および機能的双方において、赤血球(細胞表面)CR1のそれと同様である。また、Hourcadeら(1988、J.Exp.Med.168:1255−1270)は、C4b結合ドメインを含有するCR1の分泌形態を産生することが予測されたヒトCR1転写ユニットにおいて別のポリアデニル化部位を観察した。
また、CR1のいくつかの可溶性断片が、発現されるべきDNAからの膜貫通領域を除去することによって、組換えDNA法により作製されている(Fearon,et al.,1989年10月5日に公開された国際公開WO89/09220;Fearon,et al.1991年4月18日に公開された国際公開WO91/05047)。可溶性CR1断片は機能的に活性であり、C3bおよび/またはC4bに結合し、それらが含有する領域に応じて、I因子助因子活性を示した。かかる構築体は、好中球酸化的バースト、補体媒介溶血、およびC3aおよびC5a産生のごとき補体活性化の結果をin vitroで阻害した。また、可溶性構築体sCR1/pBSCR1cは、逆の受動的アルサス反応(Fearon,et al.,1989,前掲;Fearon,et al.,1991,前掲;Yeh,et al.,1991,前掲)、抑制された虚血性心筋炎およびネクロシス(Fearon,et al.,1989,前掲;Fearon,et al.,1991,前掲;Weismann,et al.,1990,Science,249:146−151)および移植後の延長された生存率(Pruitt and Bollinger,1991,J.Surg.Res.50:350;Pruitt,et al.,1991,Transplantation 52:868)においてin vivo活性を示した[Mulligan et al.,1992,J.Immunol.148:3086−3092(免疫複合体沈積後の傷害)、Mulligan,et al.,1992,J.Immunol.148:1479−1485(好中球媒介組織傷害からの保護)、Lindsay,et al.,1992,Annals of Surg.216:677,Hill,et al.,1992,J.Immunol.149:1722−1728(組織虚血性再潅流傷害)]。
CR2。補体受容体2型(CR2、CD21)は、15もしくは16のCSR、24アミノ酸膜貫通領域、および34アミノ酸細胞質ドメインからなる細胞外ドメインよりなる膜貫通リン蛋白質である(Moore,et al.,1987,Proc.Natl.,Acad.Sci.U.S.A.84:9194−9198;Weis,et al.,1988,J.Exp.Med.167:1047−1066)。可溶性組換えCR2の電子顕微鏡観察は、CR1のように、それが、39.6ナノメーターの推定輪郭長を持つ高度にフレキシブルな分子で、3.2ナノメーターだけ伸長され、そこでは、各CSRが長さ2.4ナノメーターの小環として見えることを示した(Moore,et al.,1989,J.Biol.Chem.,34:20576−20582)。
COS細胞でCR2の欠失もしくは置換突然変異体を発現する真核生物発現ベクターでの組換えDNA実験によって、CR2のリガンド結合部位は当該分子の2つのN−末端SCRに位置付けられた(Lowell,et al.,1989,J.Exp.Med.170:1931−1946)。iC3bおよびC3dgのごときC3リガンドの多価形態の細胞表面CR2による結合は、B−細胞の活性化を引き起こす(Melchers,et al.,1985,Nature,317:264−267;Bohnsack,et al.,1988,J.Immunol.141:457−463;Carter,et al.,1988,J.Immunol.143:1755−1760)。
組換え可溶性CR2の形態が産生されている(Moore,et al.,1989,J.Biol.Chem.264:20576−20582)。可溶性CR1系と同様にして、受容体の全細胞外ドメインを含有するが膜貫通および細胞質ドメインを含有しないCR2が、発現ベクターから組換え系で産生された。この組換えCR2は、27.5mMと同等のKdと1:1複合体にてC3dgに結合し、3.2nMのKdと1:1複合体にてエプスタインバール蛋白質gp350/220に結合すると報告されている(Moore,et al.,1989,J.Biol.Chem.264:20576−20582)。
CR3。第3の補体受容体CR3もiC3bに結合する。iC3bのCR3への結合は、炎症の間において補体−活性化内皮細胞への好中球の接着を促進する(Marks,et al.,1989,Nature,339:314)。また、CR3はファゴサイトーシスに関与しており、そこでは、iC3bで被覆された粒子が好中球またはマクロファージによって飲み込まれる(Wright,et al.,1982,J.Exp.Med.156:1149;Wright,et al.,1983,J.Exp.Med.158:1338)。
CR4。また、CR4(CD11)は白血球接着に関与するらしい(Kishimoto,et al.,1989,Adv.Immunol.46:149−82)。
DAF。DAF、または崩壊促進因子は、C4bからC2bへの機能的解離を引き起こす点でC4Bpと同様の作用を有するらしい膜蛋白質である。DAFはホスファチジルイノシトール糖脂質を介して膜に結合しており、赤血球からその不存在は、発作性夜間血色素尿症における主要な原因因子であることが示されている(Encyclopedia of Human Biology,Academic Press,Inc.1991)。DAFはC3b/C4bならびにC3転換酵素に結合する(EP 0512 733 A2)。
DAFは、4つのSCR、続いてのO−結合糖鎖付加領域を含有し、糖脂質アンカーで終わる(EP 0512 733 A2)。DAFを発現する細胞は、補体媒介細胞溶解に対して耐性の実質的増加を示す(Lublin,D.M.et al.,1991,J.Exp.Med.174:35;Oglesby,T.J.,et al.,1991;Trans.Assoc.Am.Phys.CIV:164−172;White,D.J.G.et al.,1992;Transplant Proc.24:474−476)。
MCP。MCPまたは膜助因子蛋白質は、DAFと同様に、4つのSCR、続いてのO−結合糖鎖付加領域を含有する。MCPは細胞質外セグメント(その重要性は未知)、膜貫通領域および細胞内ドメインで終止する(EP 0512 733 A2)。また、DAFと同様に、MCPを発現する細胞は補体−媒介細胞溶解に対する耐性の実質的増加を与える(EP 0512 733 A2およびLublin,D.M.,et al.,J.Exp.Med.(19)174:35;Oglesby,T.J.et al.,Trans Assoc Am Phsy.(1991)CIV:164−172;White,D.J.G.,et al.,1992;Transplant Proc.24:474−476)。
H因子。H因子は、専ら、または支配的にSCRからなる血漿蛋白質である(Chung,L.P.,et al.,1985,Biochem.J.230:133;Kristensen,T.,et al.,1986,J.Immunol.136:3407)。H因子は代替経路のレギュレーターである。H因子はC3bおよびSC3転換酵素(C3b,Bb)のC3b部分に結合し(Encyclopedia of Human Biology,前掲)、これらの複合体からのBbの解離を加速し、それにより、それらを不活化する。また、H因子は、C3bに対する結合についてC5と競合し、かくして、C3/C5転換酵素の活性を不活化することによって、古典的経路におけるC5の使用を調節する(Encyclopedia of Human Biology,前掲)。
2.3.セレクチンおよびセレクチンリガンド
セレクチンは、動物レクチンについて記載されている炭水化物認識構造に関連するNH2末端レクチンドメイン、上皮細胞成長因子ドメイン、および補体活性のレギュレーター(RCA)と呼ばれ、第1染色体の領域にマップされる補体制御蛋白質で見い出されたものと類似の短鎖反復配列よりなるドメインとを特徴的に有する一群の細胞表面糖蛋白質である(Harlan & Liu,Adhesion;Its Role in Inflammatory Disease,W.H.Freeman & Co.,1992)。3つの独立して研究されたセレクチンが特徴付けられ、各々が同定された細胞型に応じて命名されている。現在の命名法の下では、サイトカイン−活性化内皮細胞で同定されたE−セレクチン(Bevilacque,M.P.et al.,(1985)J.Clin.Invest.76:2003−2011);活性化血小板で発見されたP−セレクチン(Hsu−Lin,P.E.,et al.,(1984)J.Biol.Chem.259:9121−9126);および最後に、リンパ球、好中球および単球を含めたほとんどの白血球上の細胞表面マーカーとして認識されるL−セレクチン(Kansas,G.S.et al.,(1985)J.Immunol.134:2995−3002)がある。各セレクチンは細胞接着および認識で重要な事象における鍵となる因子として示されている。それ自体、分子のNH2−末端部分におけるその炭水化物認識構造ならびにその炭水化物リガンドは広く研究されてきた。
従って、セレクチンは、炎症状況において、血小板および白血球の血管表面への付着および組織への引き続いてのその浸潤の原因となる細胞接着分子である。通常の炎症反応の間に、種々のシグナルに応答して、白血球は組織に入り、侵入する生物を貪食する。乾癬および慢性関節リウマチのごとき種々の病理学的炎症疾患において、この反応は重篤な器官組織損傷を導き得る。同様に、再潅流傷害において、侵入する白血球は組織損傷の原因である。また、炎症へのその関与とは別に、セレクチンに関する細胞接着分子は腫瘍転移のごとき他の病気で中心的な役割を演ずる。
炎症状況において、すべての3種のセレクチンは炎症部位への白血球の動員に関係している。炎症反応での初期の事象は、組織損傷部位への好中球の動員を含む。通常の状況において、循環リンパ球は低結合活性にて血管内皮細胞に結合する。しかしながら、身体が細菌病原体に侵された場合あるいは組織損傷が起こった場合のように窮迫の状況下では、白血球は他の様式で活性化内皮細胞と相互作用する。まず、内皮細胞および血小板でセレクチンの上昇(up)調節が起こって、炎症した内皮細胞に対する白血球の局所化を制御する。細静脈の内側をおおう内皮細胞への好中球の付着の初期段階はセレクチンによって制御され、好中球「ローリング(rolling)」として知られている(von Andrian,U.H.et al.,(1991)Proc.Natl.Acad.Sci.,U.S.A.88:7538−7542;Smith,C.W.,et al.,(1991)J.Clin.Invest.,87:609−618)。この「ローリング」は、白血球、特に好中球の、インテグリンとして知られている異なるクラスの受容体によって制御される内皮細胞への強固な接着に先行する(Lawrence,M.B.and Springer,T.S.(1991)Cell 65:859−873;von Andrain,U.H.et al.,(1991)Proc.Natl.Sci.U.S.A.88:7538−7542;Larson R.S.and Springer,T.A.(1990)Immunol.Res.114:181−217)。周囲の組織への細胞の管外遊出は、前記した付着プロセスが各々達成された後に進行する。
セレクチン類のうちの1つ、E−セレクチン(ELAM−1、内皮細胞接着分子、LECCAM−2)は、内皮細胞上で発現され、続いて、インターロイキン−1β、腫瘍壊死因子−α、リンホトキシン、細菌エンドトキシン、インターフェロン−γおよび神経ペプチドサブスタンス−Pのごときサイトカインによって誘導される(Harlan & Liu,前掲)。活性化内皮細胞上でのE−セレクチンの発現はde novo合成を必要とし、4〜6時間においてピークとなり、初期刺激の後に2〜48時間続く。ELAM−1受容体を発現する活性化内皮には、好中球(Bevilacque M.P.,et al.(1987)Proc.Natl.Acad.Sci.U.S.A.84:9238−9242);単球(Walz,G.et al.,(1990)Science 250:1132−1135);好酸球(Kyan−Aung(1991)J.Immunol.146:521−528)およびNK細胞(Goelz,S.E.(1990)Cell 63:1149−1356)が結合することが示されている。加えて、活性化内皮細胞はいくつかの癌腫細胞に結合し(Rice,G.E.and Bevilacqua M.P.(1989)Science 246:1303−1306;Walz,G.et al.,(1990)250 1132−1135)、血管壁に対する腫瘍細胞の付着におけるE−セレクチンの役割に関与する。
P−セレクチン(CD62、顆粒球膜蛋白質−140、GMP−140、血小板活性化依存性顆粒球外部膜、Padgem、LECCAM−3)は活性化血小板ならびに内皮細胞上で発現される。P−セレクチンの発現は、活性化後数分以内に細胞内貯蔵から動員され得る。P−セレクチンは好中球および単球、ならびに癌腫細胞に結合する(Walz,G.,et al.,(1990)250:1132−1135)。
P−セレクチン、またはCD62の発現はde novo合成を必要としない。何故ならば、このセレクチンは、血小板および内皮細胞双方において、ウェイベル−パラド(Weibel−Palade)体とも呼ばれる分泌顆粒に貯蔵されるからである。かくして、例えば、トロンビン、ヒスタミン、あるいはフォルボルエステルによるいずれかの細胞型の活性化から数分以内に、CD62は細胞表面に迅速に輸送され、そこで、それは好中球、単球および他の細胞上で見い出されるリガンドに結合できる。従って、これらのリガンドを担持する細胞はCD62受容体を発現する血小板または内皮細胞に接着する。
Patelらは、内皮細胞は、また、フリーラジカルの生産を通じて過酸化水素または他の酸化剤の低レベルに応答してCD62を発現することを見い出している(Patel et al.,1991,J.Cell Biol.112:749−759)。内皮細胞は通常は活性化から数分以内にCD62を再度内部に取り込むが、フリーラジカルによる誘導はセレクチンの発現の遅延を生じる。好中球は活性化の後に酸化剤およびフリーラジカルを放出するので、一時的に発現されたCD62による好中球の初期動員は、好中球によるフリーラジカル生成を通じてCD62の発現を効果的に遅らし得る(Harlan & Liu,Adhesion,前掲)。
L−セレクチン(白血球ホーミング(homing)受容体、LECCAM−1、Mel−14、Leu−8、TQ−1、Ly−22、LAM−1)は細胞表面で構成的に発現され、活性化の後に放出される(Jung,T.M.et al.,(1988)J.Immunol.,141:4110−4117)。
接着分子分野の最近の進歩は、蛋白質−炭水化物相互作用の役割の理解に導いた。特に、セレクチンについてのリガンドが最近研究されている(Bevilaque,M.P.and Nelson,R.M.(1993)J.Clin.Invest.91:379−387)。同定されたリガンドのうちには、ルイスX血液抗原(LeX)およびシアリルルイスX抗原がある。ルイスX抗原はある程度前から知られており、好中球および前骨髄球細胞系の細胞表面糖蛋白質および糖脂質上の末端構造として同定されている(Harlan & Liu,Adhesion,前掲)。
Loweらは、ルイス血液型フコシルトランスフェラーゼ(Galβ1,3/4GlcNacα1,3フコシルトランスフェラーゼ)についてのcDNAのチャイニーズハムスター卵巣細胞(CHO)へのトランスフェクションの結果、LeXおよびSLeX抗原が発現され、TNF−α−活性化ヒト臍静脈内皮細胞(HUVEC)に接着するトランスフェクト細胞の能力が同時に生じることを示している(Lowe et al.,1990,Cell 63:475−484)。細胞のシアリターゼ処理は、活性化HUVECへ接着するその能力を無くし、これは、シアリル化構造が接着に必要であることを示す。加えて、SLeXを発現した前骨髄球性白血病−60(HL−60)細胞クローンはHUVECに結合したが、SLeXを発現しなかった他のクローンはHUVECに結合しなかったことが観察されている。
Phillipsらは、CHO糖鎖付加突然変異体を産生したが、これは野生型細胞とは異なり、細胞表面糖蛋白質上の末端糖構造としてLeXおよびSLeX(LEC11)双方またはLeXのみ(LEC12)を合成するフコシルトランスフェラーゼ活性を発現した(Phillips et al.,1990 Science 250:1130−1132)。LEC11細胞のみが活性化HUVEC上のE−セレクチンに結合し、シアリターゼでのLEC11細胞の予備処理によって接着はなくなり、これはリガンドとしてのSLeXを意味する。
α1,3グリコシド結合を介して、ELAMのごとき適当な炭水化物へフコシル残基を付加することを担うα1,3−フコシルトランスフェラーゼの核酸配列が報告されている(国際公開WO91/16900)。この報告は、該トランスフェラーゼで形質転換された組換えCOSおよびCHO細胞も記載している。
セレクチンに結合する他のリガンドも開示されている。これらのリガンドは構造的にルイスX抗原に類似する(国際公開WO92/02527および国際公開WO91/19502)。
2.4.不適当な補体活性が関与する病気
全身性紅斑性狼瘡(SLE)を持つ患者の赤血球上のCR1発現の低下が、日本(Miyakawa,et al.,1981,Lancet 2:493−497;Minota,et al.,1984,Arthr.Rheum.27:1329−1335)、米国(Iida,et al.,1982,J.Exp.Med.155:1427−1438;Wilson,et al.,1982,N.Engl.J.Med.307:981−986)およびヨーロッパ(Walport,et al.,1985,Clin.Exp.Immunol.59:547;Jouvin,et al.,1986,Complement 3:88−96;Holme,et al.,1986,Clin.Exp.Immunol.63:41−48)を含めたいくつかの地理的領域からの研究者によって報告されている。CR1数は、免疫複合体の血清レベル、C3dの血清レベル、および赤血球結合C3dgの量に逆相関することも見い出されており、これは、恐らくは、補体活性化免疫複合体の摂取および「無害性バイスタンダー(innocent bystander)」としての赤血球上への沈積を反映する(Ross,et al.,1985,J.Immunol.135:2005−2014;Holme,et el.,1986,Clin.Exp.Immunol.63:41−48;Walport,et al.,1985,Clin.Exp.Immunol.59:547)。
SLEにおける補体受容体発現の異常性は赤血球CR1に限定されない。SLE患者の好中球の合計の細胞性CR1およびBリンパ球の形質膜CR1との相対的欠乏が起こることが示されている(Wilson,et al.,1986,Arthr.Rheum.29:739747)。
また、赤血球からのCR1の相対的喪失が、ヒト免疫不全症ウイルス(HIV)感染(Tausk,F.A.,et al.,1986,J.Clin.Invest.78:977−982)およびらい腫らい(Tausk,F.A.,et al.,1985,J.Invest.Dermat.85:58s−61s)を持つ患者で観察されている。
また、補体活性化は炎症が関与する病気状態に関連する。クローン氏病(限局性回腸炎)の腸炎症は単球および多形核白血球のリンパ系浸潤によって特徴付けられる。最近、クローン氏病(限局性回腸炎)患者の空腸液における補体C4濃度が正常な対照と比べて増加してことが見い出されている(Ahrenstedt,et al.,1990,New Engl.J.Med.322:1345−9)。炎症において補体系が関与する他の病気状態は、熱傷害(火傷、凍傷)(Gelfand,et al.,1982,J.Clin.Invest.70:1170;Demling,et al.,1989,Surgery 106:52−9)、血液透析(Deppisch,et al.,1990,Kidney Inst.37:696−706;Kojima,et al.,1989,Nippon Jenzo Gakkai Shi 31:91−7)、および心肺バイパスにおけるポストポンプ(post pump)症候群(Chenoweth,et al.,1981,Complement Inflamm.3:152−165;Chenoweth,et al.,1986,Complement 3:152−165;Salama,et al.,1988,N.Engl.J.Med.318:408−14)を含む。補体および白血球は共に成人呼吸困難症候群(Zilow,et al.,1990,Clin.Exp.Immunol.79:151−57;Langlois,et al.,1989,Heart Lung 18:71−84)の病因に関与することが報告されている。補体系の活性化は、敗血症における致命的な合併症の進展に関与することが示唆されており(Hack,et al.,1989,Am.J.Med.86:20−26)、免疫複合体誘導脈管炎(Cochrane,1984,Springer Seminar Immunopathol.7:263)、糸球体腎炎(Couser et al,1985,Kidney Inst.29:879)、溶血性貧血(Schreiber and Frank,1972,J.Clin.Invest.51:575)、重症筋無力症(Lennon,et al.,1978,J.Exp.Med.147:973;Biesecker and Gomez,1989,J.Immunol.142:2654)、II型コラーゲン誘導関節炎(Watson and Townes,1985,J.Exp.Med.162:1878)、および実験的アレルギーおよび激症異種移植片拒絶(Knechtle,et al.,1985,J.Heart Transplant 4(5):541;Guttman,1974,Transplantation 17:383;Adachi,et al.,1987,Trans.Proc.19(1):1145)のごとき自己免疫疾患の動物モデルにおいて組織傷害を引き起こす。組換えIL−2での免疫療法の間における補体活性化は、IL−2処置から観察される重篤な毒性および副作用を引き起こすようである(This,et al.,1990,J.Immunol.144:2419)。
また、補体は免疫複合体が関与する病気で役割を演じ得る。免疫複合体は多くの病理学的状態で見い出されており、慢性関節リウマチまたはSLEのごとき自己免疫疾患、AIDSのごとき血液学的悪性病(Taylor,et al.,1983,Arthritis Rheum.26:736−44;Inada,et al.,1986,AIDS Research 2:235−247)および自己抗体および/または補体の活性化関与の障害(Ross,et al.,1985,J.Immunol.135:2005−14)を含むがそれらに限定されるものではない。
多数の動物モデルにおいて補体活性化を阻害するのに可溶性CR1が成功して用いられている(Moat,B.P.,et al.,1992,Amer.Review of Respiratory disease 145:A845;Mulligan,M.S.et al.,1992,J.Immunol.148:1479−1485;Yeh,C.G.et al.,1991,J.Immunol.146 250−256;Weisman,et al.,1990,Science 249:146−51;Pruitt,et al.,1991,Transplantation 52(5):868−73;Pruitt and Bollinger,1991,J.Surg,Res.50:350−55;Rabinovici,et al.,1992,J.Immunol.149:1744−50;Mulligan,et al.,1992,J.Immunol.148:1479−1485;Lindsay,et al.,1992,Annals of Surg.216:677)。
Weismanらの研究(1990,Science 249:146−151)は、酵母壁成分ザイモサンによって活性化されたヒト血清において、sCR1はC3aおよびC5aの生成の90%を防止できることを示している。また、Weismanら(1990,前掲)は、補体活性化を阻害し、心筋梗塞による損傷を減少させるためにラットにおいてsCR1を利用した。また、可溶性CR1は逆アルサス反応(Yeh,et al.,1991,J.Immuno.146:250−256)、および激症異種移植片拒絶(Pruitt,et al.,1991,Transplantation 52:868−873)の補体依存性プロセスを阻害するようである。最近のデータ(Moat,et al.,1992,Amer.Rev.Respiratory Disease 145:A845)は、sCR1は、ブタにおける心肺バイパスの実験モデルで補体活性化を防止するに価値あることを示しており、そこでは補体活性化の状況が説明されている。
本出願のセクション2のいずれの文献の引用および確認も自認として解釈されるべきではなく、本発明の先行技術として利用されるべきである。
3.発明の概要
本発明により、その最も広い態様において、補体部位および炭水化物部位よりなる組成物が提供される。これらの組成物は補体関与の病気または疾患を治療するのに、ならびに細胞間接着分子およびそのリガンドの間の相互作用をブロックするごとき炎症反応における最初の事象を阻害するのに有用である。好ましい態様において、組成物が細胞間接着分子についてのリガンドよりなるのは本発明の利点である。補体部位は、補体成分に結合できるか、または、SCRモチーフを含有することによって補体受容体1型に関連する多数の蛋白質のうちいずれか1つであればよい。炭水化物部位は、細胞間接着分子に結合するかまたはそれとの相互作用を防止する多数の炭水化物のうちのいずれか1つであればよい。この構築体は、補体蛋白質の傷害部位への局所化を容易とし、有利には、とりわけ低用量の治療を可能とする。同組成物が炎症反応における初期事象に干渉できるのは、本発明のさらなる利点である。従って、細胞接着分子リガンドよりなる補体蛋白質も、細胞間接着によって媒介される炎症、ならびに補体関連病気または疾患を治療するのに有用である。
本発明の組成物の炭水化物部位は、化学的もしくは酵素的付着のごとき細胞外事象によって補体部位に付着しており、あるいは適当な酵素の発現によって達成される細胞内プロセッシング事象の結果であり得る。ある具体例において、炭水化物部位は細胞間接着分子に特異的に結合するであろう。1の具体例において、炭水化物はセレクチンとして知られている特別のクラスの接着分子に結合する。かくして、好ましい態様において、本発明は、少なくとも1つの補体部位および少なくとも1つの炭水化物部位よりなる組成物を提供し、該組成物は特定のセレクチンに優先的に結合する。セレクチンのうちには、E−セレクチン、L−セレクチンまたはP−セレクチンがある。特に好ましい具体例は、少なくとも1つの補体部位および少なくとも1つの炭水化物部位を備え、該炭水化物部位は、好ましくは複合体タイプの、より好ましくはフコシル化されたおよびシアリル化されたN−結合炭水化物よりなる。最も好ましい態様において、炭水化物はルイスX抗原、特にシアリルルイスX抗原に関係している。
1の具体例において、補体部位は、少なくとも1つのショートコンセンサスリピートを含有する蛋白質であり、より好ましくは、補体カスケードの成分に結合しおよび/または補体に関連する活性を阻害する。より好ましい具体例において、補体部位は補体受容体1型のすべてまたは一部よりなる。好ましくは、補体蛋白質は可溶性補体蛋白質である。最も好ましい具体例において、補体部位は可溶性補体受容体1型(sCR1)、またはその断片もしくは誘導体である。
さらに、本発明は、適当な医薬担体と混合した、少なくとも1つの補体蛋白質および少なくとも1つの炭水化物部位よりなる医薬組成物を提供する。好ましい具体例において、補体蛋白質は可溶性であり、特にsCR1またはその断片もしくは誘導体である。これらの好ましい具体例において、炭水化物はN−結合炭水化物であり、好ましくはフコシル化されており、より好ましくは、フコシル化されかつシアリル化されている。これらのうち、ルイスX(LeX)抗原またはシアリルルイスX(sLeX)抗原が特に好ましい。
また、本発明は、本明細書に記載する組成物の製法を提供する。1つの好ましい具体例において、本発明は、補体蛋白質にLeX抗原、好ましくはSLeX抗原の糖鎖付加する細胞において補体蛋白質を発現させ、該蛋白質を回収することを提供する。もう1つの具体例において、本発明は、炭水化物部位を蛋白質に化学的に結合させることによって補体蛋白質を修飾することを提供し、該炭水化物部位は好ましくはセレクチンリガンドである。
さらにもう1つの具体例において、本発明は、望ましくないまたは不適当な補体活性が関与する病気を持つ対象を治療することを提供する。かかる治療は、本発明の医薬組成物を、該望ましくない補体活性を調節するのに適した量および時間にて、治療を要する対象に投与することよりなる。好ましくは、かかる医薬組成物における炭水化物部位はLeXのごときセレクチンリガンドであり、より好ましくは該リガンドはSLeXである。セレクチンリガンドよりなる補体蛋白質での治療は、不適当な補体活性化の病気または疾患、特に炎症疾患を含むが、それらの限定されるものではない。かかる疾患は、虚血後再潅流疾患、感染症、敗血症、免疫複合体疾患および自己免疫疾患を含むがそれらに限定されるものではない。
本発明の組成物は、補体部位、好ましくはCR1、より好ましくはsCR1を、活性化内皮細胞上のセレクチンのごとき接着分子にホーミングするのに用いることができ、とりわけ、sCR1単独またはその既存の糖形態での使用と比較して低用量を可能とする。従って、本発明の組成物は、炎症部位に執拗に存在し、それにより、さらなる活性化を防止する。セレクチン/リガンド相互作用に依存する初期の好中球接着事象もブロックされ得る。加えて、sCR1のin vivo半減期が延長され得る。特別の具体例において、CR1部位は、古典的経路および代替経路双方において転換酵素C3およびC5をブロックし、かくして、C5aの放出を防止する。C5aの放出の防止は、さらに、とりわけ、好中球の活性化および化学誘引を阻害する。
本明細書で提示する組成物は抗原性が低いことはさらにもう1つの利点である。これは、ルイスX抗原に関係する炭水化物が、他の炭水化物構造、例えば、非ヒト宿主細胞等から得られたものと比較して、その糖鎖付加パターン化においてより「天然」であり得るので、本明細書に記載した好ましい具体例の意味で特に関連し得る。
3.1 略語
CR1−補体受容体1
CR2−補体受容体2
CR3−補体受容体3
CR4−補体受容体4
DAF−崩壊促進因子
ELAM−内皮細胞接着分子
LeX−ルイスX抗原
LHR−長鎖相同反復
MCP−膜助因子蛋白質
sCR1−可溶性補体受容体1
SLeX−シアリルルイスX抗原
SCR−ショートコンセンサスリピート
CD15−ルイスX抗原
4.図面の簡単な記載
図1Aは、クーマシー染色したSDS−PAGEゲルを示し;レーン1は分子量標準であり;レーン2は対照HL−60細胞溶解物であり;レーン3はDUKX.B11細胞から産生されたsCR1[des−A]を含有し;レーン4はLEC−11細胞から回収したsCR1[des−A]を含有する。
図1Bは、SLeX炭水化物抗原決定基と反応する抗体、CSLEX1でプローブした、Aに記載したのと同一ゲルの化学ルミネッセンス・ウェスタンブロットを示す。レーン4は(LHRのBCおよびDを含有するsCR1の可溶性形態(LEC−11細胞から回収したsCR1[des−A]sLeXを含有する)sCR1[des−A]sLeXの異なる糖鎖付加形態を表す2つの区別されるバッド(bad)を示す。
図1Cは、細片化し、CR1に対するアフィニティー精製したウサギポリクローナル抗体でプローブした、図Bに記載したのと同一ゲルである。該ブロットは、前記ゲルBからのsCR1[des−A]の2つの糖鎖付加形態も抗−補体受容体1型抗体と反応性であることを示す。
図2Aは、クーマシーブルー染色したポリアクリルアミドゲルのパターンである。レーン2、および4〜6におけるほぼ187kdにおける支配的なバンドはsCR1[des−A]蛋白質であり、レーン2はDUKX−B11細胞から得られ、レーン4〜6はLEC−11細胞から得られた。
図2Bは、ウェスタンブロットし、抗−sCR1[des−A]ポリクローナル血清でプローブした、図2Aと同一のゲルである。予測されるごとく、DUKX−B11細胞またはLEC−11細胞に由来するかを問わず、sCR1[des−A]を含有するすべてのレーンはsCR1[des−A]について陽性である。
図2Cは、細片化し、簡略表示NeuNAcα2−3Galβ1−4(Fucα1−3)GlcNacβ1−3Galβ1−4(Fucα1−3)GlcNAcによって表されるシアリ−ジルイスX抗原(FH6)について特異的な抗体で再プローブした、図2Bと同一のブロットである。予測されるごとく、LEC−11細胞から得られたsCR1[des−A]sLeXを含有するレーン4〜6のみが適当な炭水化物構造について陽性である。
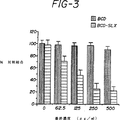
図3は、静的接着結合アッセイの結果を示す。黒い棒線はDUKX−B11細胞から得られたsCR1[des−A]物質を表す。水平線を持つ棒線はLEC−11細胞から得られたsCR1[des−A]sLeX物質を表す。該sCR1[des−A]sLeX物質は、濃度依存的に、活性化動脈内皮細胞へのU937細胞の結合を阻害した。
図4は、sCR1、sCR1[des−A]およびsCR1[des−A]sLeXの、CVFにより誘導された肺傷害からの保護効果を記載する。Aは浸透性、肺血管からの放射性同位体標識蛋白質の漏れの測定である。Bは、放射性同位体標識赤血球の血管から肺への漏れによって測定した、出血の対照よりも低い測定値である。Cはミエロペルオキシダーゼ活性の測定によって評価した、好中球の肺への蓄積の測定である。
5.詳細な記載
本発明は、少なくとも1つの補体部分および少なくとも1つの炭水化物部位よりなる組成物に指向される。本発明の組成物は、適当な受容体を発現する細胞と細胞レベルで相互作用する。ある好ましい具体例において、該組成物の炭水化物部位はセレクチンに結合する。
明確にするために、組成物の各種成分、かかる組成物の製法、該組成物の活性の測定のため機能的アッセイ、および該組成物を用いる診断、治療および予防方法に関するセクションで本発明を詳細に記載する。
5.1.補体蛋白質
本発明の範囲内にある「補体部位」は、補体カスケードに関連するいずれかの蛋白質、またはショートコンセンサスリピートの少なくとも一部を含有する蛋白質のすべてまたは一部を含有するいずれの蛋白質をも意味する。ある種の有用な補体蛋白質は発明の背景のセクション2.1および2.2に記載されており、好ましくは、補体成分C3bおよびC4bについての受容体である補体受容体1型(CR1);C3dについての受容体である補体受容体2型(CR2);iC3bについての受容体である補体受容体3型(CR3);iC3bに対して特異的である補体受容体4型(CR4);iC3b、C3dg、およびC3dのC3d部分について特異的である補体受容体5型(CR5);c5a受容体(C5a−R);およびC3aおよびC4aについての受容体:の完全な蛋白質もしくはいずれかの断片を含むが、それらの限定されるものではない。好ましい態様において、本発明は、保存されたショートコンセンサスリピート(SCR)モチーフを含有する補体制御蛋白質のファミリーのメンバーを含有することを意図する。SCRモチーフは、補体受容体1型、およびいくつかの他のC3/C4−結合蛋白質、最も顕著にはCR2、H因子、C4−結合蛋白質(C4−BP)、膜助因子蛋白質(MCP)、および崩壊促進因子(DAF)で見い出される。H因子、C4−BP、CR2、およびDAFについての遺伝子は、「補体活性化のレギュレーター」(RCA)と命名された第1染色体の領域にマップされる(Hourcade,D.,et al.,1989,Advances in Immunol.,45:381−416)。補体活性化のこれらのレギュレーターの特別の類似体は1992年11月11日に公開されたAtkinson,et al.のヨーロッパ特許出願公開番号0 512 733 A2に見い出される。かくして、好ましい具体例において、補体蛋白質は少なくとも1つのCSRを含有し、補体の成分に結合できる。かかる補体蛋白質は、1の具体例において、本明細書で記載された蛋白質のごとく、C3bもしくはC4bまたはC3もしくはC4の断片に結合するであろう。
CR1は広く研究されており、ショートコンセンサスリピート(SCR)と命名された60〜70アミノ酸の構造的モチーフが発見されている。該SCRモチーフは、CR1のF−アロタイプにおいて30回縦列反復されており、さらなる反復サイクルが他のアロタイプで起こる。SCRのコンセンサス配列は、すべてのSCRのうちで不変である4つのシステイン、グリシンおよびトリプトファンを包含する。16個の他の位置は保存されており、同一のアミノ酸または保存的置換が他の30のSCRの半分以上で見い出されている(Klickstein,et al.,1987,J.Exp.Med.165:1095−1112;Klickstein et al.,1988,J.Exp.Med.168:1699−1717;Hourcade et al.,1988,J.Exp.Med.168:1255−1270)。各SCRの寸法はほぼ2.5〜3.0nm×2nm×2nmと見積もられている。
SCRの縦列反復(同一の不変残基およびシステイン間の同様のスペーシング)が、補体系の12個のさらなる蛋白質で同定されている(Ahearn et al.,1989,Adv.Immunol.46:183−219)。これらの蛋白質は、各々、代替および古典的C3−C4転換酵素および膜侵襲複合体のサブユニットである同質の補体蛋白質のセット、C3、C4、またはC5と相互作用する能力を共に有する。SCRを含有する補体関連蛋白質は、活性化機能(Clr、Cls、B因子およびC2)、負の調節役割(H因子、C4−BP、DAF、MCP、およびCR1)を有し、ファゴサイトおよびリンパ球の機能を誘導できる細胞受容体(CR1およびCR2)として働くか、あるいは補体チャンネル形成膜侵襲複合体の形成を促進する(C6およびC7)。かくして、SCRは、補体系の最も特徴的な構造の1つである。インターロイキン−2受容体α鎖、β2−糖蛋白質1、および第XIII因子におけるごとく、非補体蛋白質におけるSCRの発見は、この可能性は排除されないが、補体関連機能を必ずしも示すものではない。
所望の結果を得るために適した組合せにて、組成物が前記したSCRの1以上よりなることは本発明の範囲内のものである。さらなる基準として、組織によって容易に吸収される、迅速な代謝から保護されるおよび/または延長された半減期を提供する補体蛋白質またはその断片の形態は、本発明の組成物の製造で優先的に選択される。当業者ならば、吸収を行うために、蛋白質処方の修飾を行うことができるであろう。これらの修飾は、プロドラッグや一次構造の化学的修飾の使用を含むが、これらに限定されるものではない(Wearley,L.L.,1991,Crit.Rev.in Ther.Drug Carrier Systems,8(4):333)。補体蛋白質の代謝を最小化し、それにより、蛋白質の有効量を増加させるにおいて、かかる修飾は化学的修飾およびポリマーへの共有結合付着を含むが、それらに限定されるものではない(Wearly,L.L.,1991,前掲)。
本発明の組成物は、リポソームのごとき送達系の一部であってもよい。リポソーム関連送達系は国際公開WO91/02805および国際公開WO91/19501、ならびにJanoffらに対する米国特許第4,880,635号に記載されている。これらの文献および特許は、リポソーム薬物送達についての技術の有用な記載を提供する。
補体関連蛋白質についての遺伝子は容易に入手可能であり、例えば、核酸配列および/または本発明の補体蛋白質をコードする遺伝子は、例えば、DAF(1989年2月9日に公開された国際公開WO89/01041);MCP(Lublin M.D.,et al.,1988,J.Exp.Med.168:181−194);およびCR2(Weis,J.J.,et al.,1988,J.Exp.Med.168:1047−1066)のごとく公知である。CR1遺伝子およびそれがコードする蛋白質は、1989年10月5日に公開された「ヒトC3b/C4b受容体(CR1)」なる発明の名称の国際出願WO89/09220に提供されている。一旦、遺伝子およびそれがコードする蛋白質が入手可能となれば、当該分野で知られているいずれの技術を用いても遺伝子自体またはそれがコードする蛋白質を修飾することができる。本発明は、かかる補体蛋白質関連断片、誘導体、および類似体を包含することを意図する。本発明の組成物および処方で用いる補体蛋白質関連断片、誘導体、および類似体は、当該分野で公知の種々の方法によって製造できる。結果としてその製造ができる操作は、遺伝子または蛋白質レベルで起こるか、あるいは化学的合成方法による。例えば、クローン化補体遺伝子は、当該分野で公知の多数の戦略のうちのいずれかによって修飾することができる(Maniatis,T.,1982,Molecular Cloning,A Laboratory Manual,Cold Spring Harbor Laboratory,Cold Spring Harbor,New York)。補体蛋白質遺伝子配列は制限エンドヌクレアーゼ(類)で適当な部位を切断することができ、続いて、所望ならば酵素修飾でき、単離し、in vitroで連結する。補体蛋白質に関連する誘導体、類似体、またはペプチドをコードする遺伝子の産生においては、所望の補体阻害性特異的活性がコードされた遺伝子領域において、修飾された遺伝子が、翻訳終結シグナルによって干渉されない、天然補体蛋白質遺伝子と同一の翻訳リーディングフレーム内に残存することを確認するよう注意すべきである。
加えて、補体蛋白質遺伝子はin vitroまたはin vivoにて突然変異させて、翻訳、開始、および終結配列を生じるかおよび/または破壊するか、あるいはコーディング領域に変異を生じさせおよび/または新しい制限エンドヌクレアーゼ部位を形成させ、あるいは予め存在していたものを破壊して、さらなるin vitro修飾を容易とすることができる。in vitroの部位特異的突然変異誘発(Hutchinson,C.,et al.,1978,J.Biol.Chem.235:6551)、TABXリンカー(Pharmacia)の使用等を含めた、当該分野で公知のいずれの突然変異誘発技術も使用できるが、それらに限定されるものではない。
また、補体蛋白質の操作を蛋白質レベルで行うこともできる。臭化シアン、トリプシン、キモトリプシン、パパイン、V8プロテアーゼ、NaBH4による特異的化学切断、アセチル化、ホルミル化、酸化、還元等を含めた、いずれの多数の化学的修飾も行うことができるが、これらに限定されるものではない。
補体蛋白質がCR1である特別の具体例において、例えば、CR1のヌクレオチド配列の特異的修飾を組換えDNA法によってなすことができ、その結果、多重LHR−B配列を有する蛋白質をコードする配列が得られる。例えば、1991年4月18日に公開された国際公開WO91/05047参照。かかる結合価修飾は、免疫もしくは炎症疾患のごときかかる機能と関連する疾患で重要であるC3b結合の程度を変更する。例えば、所望の活性を呈する全長CR1またはその断片および関連分子は、I因子助因子として働き、補体成分C3bまたはC4bの不可逆的不活化を促進する能力(Fearon,D.T.,1979,Proc.Natl.Acad.Sci.U.S.A.76:5867;Iida,K.and Nussenzweig,v.,1981,J.Exp.Med.153:1138)によって、および/または代替もしくは古典的C3もしくはC5転換酵素を阻害する能力によって補体を阻害し治療用途を有し得る。
もう1つの具体例において、特異的な十分に定義されたLHRまたはSCRの組合せを含有するCR1配列の特異的部分を生じさせることもできる。これらの化合物の活性は、特異的活性を含有する全長CR1分子の一部を選択することによって予測できる。得られた断片は、親分子の少なくとも1つの機能を含有すべきであるが、含有しなくともよい。かかる機能は、遊離もしくは複合体形態のC3bおよび/またはC4bの結合;ファゴサイトーシスの促進、補体調節、免疫刺激;I因子助因子として作用する能力;補体成分C3bまたはC4bの不可逆的不活化の促進(Fearon,D.T.,1979,Proc.Natl.Acad.Sci.U.S.A.76:5867;Iida,K.and Nussenweig.V.,1981,J.Exp.Med.153:1138);免疫複合体除去を行うことおよび/または代替もしくは古典的C3もしくはC5転換酵素を阻害する能力を含むが、これらに限定されるものではない。特別の具体例において、CR1は、LHR B、CおよびDを包含し、LHR Aを包含しない。
加えて、補体蛋白質に関連する類似体およびペプチドは化学的に合成することができる。例えば、所望の活性(例えば、C3bおよび/またはC4b結合、免疫刺激、補体調節等)を媒介する補体蛋白質の一部に対応するペプチドはペプチド合成機を使用することによって合成できる。
本発明の特別の具体例において、組換えDNA法または化学的合成法によるかを問わず、その誘導体、類似体または断片を含めたかかる補体蛋白質は、一次アミノ酸配列として、機能的に同等のアミノ酸残基が配列内の残基の代わりに置き換えられて、その結果サイレント変化が生じる改変配列を包含する天然補体蛋白質のアミノ酸配列のすべてまたは一部を含有するものを含むが、それらに限定されるものではない。例えば、配列内の1以上のアミノ酸残基を、機能的に同等物として作用し、その結果サイレント改変を生じるような同様の極性を有する別のアミノ酸によって置き換えることができる。また、非保存的置換によっても、機能的に同等な蛋白質が得られる。
1つの具体例において、補体蛋白質配列内のアミノ酸の置換は、アミノ酸が属するクラスの他のメンバーから選択することができる。例えば、非極性(疎水性)アミノ酸はアラニン、ロイシン、イソロイシン、バリン、プロリン、フェニルアラニン、トリプトファン、およびメチオニンを含む。極性中性アミノ酸はグリシン、セリン、スレオニン、システイン、チロシン、アスパラギン、およびグルタミンを含む。正に荷電した(塩基性)アミノ酸はアルギニン、リジンおよびヒスチジンを含む。負に荷電した(酸性)アミノ酸はアスパラギン酸およびグルタミン酸を含む。
特別の具体例において、補体蛋白質配列の一部と非補体蛋白質配列とを合わせた配列からなる分子よりなる融合蛋白質をコードする核酸配列を産生することができる。例えば、国際公開WO91/05047参照。例えば、LHRまたはSCRを含有する補体蛋白質のさらなる修飾は、得られたキメラに溶解性、薬理学もしくはクリアランスに影響を与えることを目的とした他の分子を付着したLHRおよび/またはSCR配列の一部を含有したキメラ分子の生成を包含する。かかるキメラは、融合蛋白質としての遺伝子レベルにおいて、または化学的に製造された誘導体としての蛋白質レベルにおいて製造できる。免疫グロブリン鎖の一部および補体蛋白質からなるキメラ蛋白質は、該キメラ蛋白質の免疫グロブリン部分における非補体活性化イソタイプのFc領域を除去するために重鎖のヒンジ領域の後を蛋白質分解するかまたはそこに停止コドンを導入して、補体活性化複合体のFc受容体媒介クリアランスを得ることによって産生されたFabまたは(Fab')2分子を含有できる。キメラを形成するのに使用できる他の分子は、他のSCR含有蛋白質、血清アルブミン、ヘパリンもしくは免疫グロブリンのごとき蛋白質、ポリエチレングリコールもしくはポリオキシエチル化ポリオールのごときポリマー、例えば、ポリエチレングリコールで誘導体化することによって抗原性を低下させるように修飾した蛋白質を包含するが、これらの限定されるものではない。適当な分子は当該分野で公知であり、例えば、米国特許第4,745,180号、第4,766,106号および第4,847,325号およびそこに引用された文献に記載されている。生物学的化合物またはその断片の誘導体を形成するのに使用できるさらなる分子はプロテインAまたはプロテインGを包含する(1987年9月24日に公開され、「Method and means for producing a protein having the same IgG specificity as protein G」なる発明の名称の国際公開WO87/05631;Bjorck,et al.,1987,Mol.Immunol.24:1113−1122;Guss,et al.,1986,EMBOJ.5:1567−1575;Nygren,et al.,1988,J.Molecular Recognition 1:69−74)。補体係合部位を有する複数のショートコンセンサスリピートよりなる構築体、免疫グロブリンまたは可溶性で生理学的に適合するマクロ分子担体に付着した該構築体も本明細書で教示する補体部位として適当である。これらの構築体の調製は、ここに出典明示して本明細書の一部とみなす国際公開WO91/16437に開示されている。
コードされた蛋白質の単離および回収は、当該分野で公知の技術によって行うことができる。補体蛋白質は、クロマトグラフィー(例えば、イオン交換、アフィニティー、およびサイズカラムクロマトグラフィー、高速液体クロマトグラフィー)、遠心、溶解度分別を含めた標準的な方法によって、あるいは蛋白質の精製についてのいずれかの他の標準的な技術によって単離し精製できる。補体蛋白質がそれを産生する細胞によって輸送される場合、蛋白質の精製について特別に効果的な方法は以下の通りである:蛋白質を含有する細胞培養培地をa)カチオン交換クロマトグラフィー、b)硫酸アンモニウム沈殿、c)疎水性相互作用クロマトグラフィー、d)アニオン交換クロマトグラフィー、e)さらなるカチオン交換クロマトグラフィー、およびf)サイズ排除クロマトグラフィーの連続的工程に付す。
より好ましい具体例において、本発明は、可溶性CR1分子に関する。本明細書で用いるごとく、可溶性CR1分子なる語は、発現に際し、膜蛋白質として細胞表面には位置しないCR1蛋白質の一部を意味する。特別の例として、膜貫通領域を実質的に欠くCR1分子は可溶性CR1分子である。本発明の特別の具体例において、(例えば、最もC−末端のSCRによってコードされたアスパルテートに対するカルボキシル−末端の削除によって)膜貫通領域を欠くCR1分子をコードする発現ベクターを構築でき、その結果、可溶性CR1断片が産生される。1の具体例において、かかる断片は、遊離または複合体形態で、C3bおよび/またはC4bに結合できる能力を保持することができる。特別の具体例において、かかる可溶性CR1蛋白質はもはやI因子助因子活性を示さないであろう。
また、CR1の結合部位のいくつかまたはすべてを担持する可溶性構築体も考えられる。かかる構築体は、ある好ましい具体例においては、補体の活性化および細胞の補体依存性活性化を阻害するであろう。例えば、特別の具体例において、例えば、古典的補体媒介溶血、古典的C5a産生、古典的C3a産生、またはin vitroでの好中球酸化的バーストを阻害するその能力によって示されるごとく、所望の機能的活性を保有する可溶性CR1分子を使用できる。1つの具体例において、かかる断片は、遊離または複合体形態にて、C3bおよび/またはC4bに結合する能力を保有できる。かく産生されたsCR1分子は、膜貫通領域の最初のアラニン残基を含み、また該残基までのLHR−A、LHR−B、LHR−C、LHR−D、SCR29、SCR30を含有できる。本発明の好ましい態様において、可溶性CR1蛋白質は、ATCCに寄託され、受託番号CRL 10052が与えられたプラスミドpBSCR1/pTCSgptを保有するチャイニーズハムスター卵巣細胞DUX B11によって発現される蛋白質の特徴を有する。
さらに特別の具体例において、当該CR1分子のLHR−A領域を欠くCR1分子を産生できる。この目的には、発現ベクターを構築して、膜貫通領域およびSCR1−7を欠くCR1分子をコードさせ、その結果、代替経路を優先的に阻害することが予測される可溶性CR1断片が産生される。かく構築された発現ベクターは第一義的にC3bに結合するための部位を含有することが予測される。従って、かかる構築体は、本明細書に記載したin vitro溶血アッセイによって評価して、代替補体経路を優先的に阻害することが予測される。
さらに別の具体例において、発現ベクターを構築して、補体受容体1型のSCR1−18のみを含有させることができる。かかる構築体は、C3bおよびC4bに結合する部位を含有することによって、補体受容体1型に関連する十分な機能を有することが期待される。かかる産物は、本明細書に記載したin bitroアッセイによって評価して、補体の古典的経路および代替経路を阻害することが予測される。さらにもう1つの具体例において、構築体はSCR15−18のみを含有させることができる。かかる構築体は、第一義的にC3bに結合し、補体の代替経路を優先的に阻害すると予測される。
これらの構築体、ならびに本出願の他の構築体は、糖鎖付加の差異に起因する利点を有し得る。かかる差異は、分子のin vivo半減期のごときパラメーターに影響すると予測される。当業者ならば、N−結合糖鎖付加についての可能な部位はかかる構築体の産物で変化するであろうことを認識するであろう。糖鎖付加の差異は、個々の細胞接着分子についての天然リガンドの結合をブロックするその能力につき本明細書に記載した機能的アッセイによって評価できる。
本発明の補体蛋白質は、その補体関連活性を証明するために、当該分野で公知の技術によってアッセイできる。かかるアッセイは、補体蛋白質と相互作用する、補体活性を阻害する、あるいは補体由来ペプチドの産生を選択的に阻害する能力についての以下のin vitroテストを包含するが、それらに限定されるものではない;
(i)細胞、例えば赤血球細胞の補体媒介溶解の阻害の測定(IH50アッセイ)(国際公開WO92/10096)
(ii)c5aおよびC5adesArgのごとき補体活性化産物の形成を阻害する能力の測定および/またはC3aまたはC3adesArgの形成を阻害する能力の測定、またはC5b−9またはsC5b−9の形成を阻害する能力の測定(国際公開WO92/10096)
(iii)例えば、C3bまたはC4bのI因子分解についての助因子として働く能力の測定(Makrides et al.,(1992)267:24754−24761,Wiesman,H.F.,et al.(1990)Science,244:146−151)
(iv)C3bまたは他のC4b由来蛋白質に結合する能力、またはC4bまたは他のC4b由来蛋白質の結合の測定(Makrides et al.,前掲,Weisman et al.,前掲)
(v)代替経路媒介溶血の阻害の測定(AH50アッセイ)(国際公開WO92/10096)
補体受容体に関連する活性のいずれか1つを有する補体蛋白質またはその断片、誘導体もしくは類似体、特にCR1蛋白質は、本明細書で提供する組成物の補体部位として本発明の範囲内のものである。
補体受容体1型に通常に関連する活性は当該分野にてよく記載されている。例えば、可溶性CR1蛋白質については、かかる活性は、好中球酸化的バーストを阻害する、補体媒介溶血を阻害する、C3aおよび/またはC5a産生を阻害する、C3bおよび/またはC4bに結合する、I因子助因子活性を呈する、およびC3および/またはC5転換酵素活性を阻害するin vitro能力を含む。活性およびアッセイの代表的な開示は、とりわけ、WO89/09220として1989年10月5日に公開されたPCT/US89/01358;およびWeissman,et al.,1990,Science 249:146−151;Fearon,D.T.and Wong,W.W.,1989,Ann.Rev.Immunol.1:243;Fearon,D.T.,1979,Proc.Natl.Acad.Sci.U.S.A.76:5867;Iida,K.and Nussenzweig,V.,1981,J.Exp.Med.153:1138;Klickstein et al.,1987,J.Exp.Med.,165:1095;Weiss,et al.,1988,J.Exp.Med.,167:1047−1066;Moore,et al.,1987,Proc.Natl.Acad.Sci.84:9194;Moore,et al.,1989,J.Biol.Chem.264:205−76に記載されている。
5.2.セレクチンリガンドよりなる炭水化物構造
本発明の組成物の炭水化物部位は種々の炭水化物構造から選択できる。好ましい具体例において、この部位はセレクチンのごとき特別の細胞接着分子に補体部位を結合させることを担う。発明の背景のセクション2.3は、炭水化物部位が適当に結合するいくつかのセレクチンを詳しく記載する。セレクチンを含めた、細胞間接着分子に結合する炭水化物部位は当該分野でよく知られている。例えば、1991年12月26日に公開され、「Intercellular Adhesion Mediators」なる発明の名称の国際出願公開番号WO91/19502;1992年2月20日に公開され、「New Carbohydrate−Based Anti−Inflammatory Agents」なる発明の名称の国際出願公開番号WO92/02527;1992年11月12日に公開され、「GLYCAM−1(Spg 50),A Selectin Ligand」なる発明の名称の国際出願公開番号WO92/19735;1992年2月6日に公開され、「Functionally Active Selectin−derived Peptides and Ligands for GMP−140」なる発明の名称の国際出願公開番号WO92/01718;1991年12月26日に公開され、「Intercellular Adhesion Mediators」なる発明の名称の国際出願公開番号WO91/19501はすべて本発明で有用な炭水化物分子の開示を提供し:公開された特許出願は、出典明示して本明細書の一部とみなす。また、炭水化物の合成および加工は当該分野でよく知られている(Hubbard,S.C.and Ivatt,R.J.(1981)Ann.Rev.Biochem.50:555−83およびそこで引用された文献;Goochee,C.F.,(1991)Biotechnology,9:1347−1355およびそこに引用された文献;Kobata,A.(1992)Eur.J.Bochem.209,483−501、およびそこに引用された文献)。従って、本発明の炭水化物部位は細胞接着分子と効果的に相互作用する。
また、セレクチンについての特別のリガンドが記載されている(Howard,D.R.,et al.,(1987)J.Biol.Chem.262:16830−16837,Phillips,M.L.,et al.,(1990)Science 250:1130−1132,Walz,G.et al.,(1990)Science,250:1132−1135,Stanley,P.,and Atkinson,P.,(1986)J.Biol.Chem.263:11374−11381;Butcher,E.,(1991)Cell,67:1033−1036)。ルイスXおよびシアリルルイスXオリゴ糖はセレクチン結合で特に重要であることが示されている。最近の研究は、セレクチンについてのリガンド構造をさらに特徴付け、ルイスXおよびシアリルルイスXオリゴ糖の修飾は該オリゴ糖とセレクチンとの間の相互作用を促進できることを記載している(Bevilacqua,M.P.and Nelson,R.M.(1993)J.Clin.Invest.91:379−387,Nelson,R.M.,et al.,(1993)J.Clin.Invest.91:1157−1166,Norgard,K.E.et al.,(1993)Proc.Natl.Acad.Sci.U.S.A.90:1068−1072;Imai,Y.et al.,(1993)Nature 361:555−557)。
本発明の炭水化物部位は、今や、オリゴ糖の記載のための通常に使用される命名法を参照して記載する。この命名法を用いる炭水化物化学の総括は、Hubbard and Ivatt(1981)、前掲に見い出される。この命名法は、例えば、マンノースを表すMan;2−N−アセチルグルコサミンを表すGlcNAc;フコースを表すFuc;ガラクトースを表すGal;およびグルコースをいうGlcを包含する。好ましい具体例において、炭水化物部位はシアル酸残基よりなる。2つの好ましいシアル酸残基は5−N−アセチルノイラミン酸につき「NeuNAc」によって、5−グルコリルノイラミン酸につき「NeuNGc」によって短縮表示にて記載される(J.Biol.Chem.,1982,257:3347;J.Biol.Chem.,1982,257:3352)。
当業者に容易に理解されるごとく、炭水化物を記載するこの方法は炭水化物のネーミングに関連する種々のグリコシド結合についての表示を含む。従って、オリゴ糖を形成する2以上の単糖を連結させる結合の記載において、ガラクトースのC−1およびグルコースのC−4の間のβグリコシド結合は通常Galβ1−4Glcによって表される。表示βおよびαは、グリコシド環構造に対する結合の向きを表すことを意図する。D−糖については、例えば、表示βは、C−1に付着したヒドロキシルは環面の上方にあることを意味する。逆に、D−糖については、表示αは、C−1に付着したヒドロキシル基は環面の下方にあることを意味する。炭水化物部位はこの短縮表示を参照して記載する。
その最も広い態様において、本発明で有用な炭水化物構造は広範囲の構造から選択できる。好ましくは、炭水化物はいくらかのレベルで接着分子と相互作用するであろう。例えば、かかる部位は細胞接着分子に結合し、あるいはそれへの天然リガンドの結合を防止する、あるいは内因的に起こるリガンドを置き換えさえする。当該分野で十分に理解されているごとく、個々のリガンドおよびその受容体の間の相互作用は、一般に、アフィニティー定数によって記載される。「結合アフィニティー」は、一般に、リガンドおよびその受容体の会合したおよび解離した形の平衡濃度についてのアフィニティー定数によって測定される。本発明は、炭水化物リガンドとその内皮細胞接着分子受容体の間のかかる相互作用を考慮する。一般に、炭水化物部位の結合は、本発明で有用である約Ka=10-4Mまたはそれ以上の親和性にて起こり、約10-8Mを超えるのがより好ましく、最も好ましくは約10-8Mと約10-10Mの間である。
特別の具体例において、本発明の炭水化物構造は、セレクチンとして知られている細胞接着分子のクラスについてのリガンドである。セレクチンは、広く3つの群に分類される種々の炭水化物構造に結合することが示されている。第1の群は、N−結合およびO−結合炭水化物を包含する。N−結合およびO−結合炭水化物は、そのコア構造が第一義的に異なる。N−結合炭水化物はすべて共通のManα1−6(Manα1−3)Manβ1−4GlcNAcβ1−4GlcNAcβ−Rコア構造を含有する。N−結合炭水化物のうち、本発明で最も重要なのは複合体N−結合炭水化物である。かかる複合体N−結合炭水化物はいくつかのアンテナ構造を含有するであろう。かくして、モノ−、ビ−、トリ−、テトラ−、およびペンタ−アンテナ外方鎖が重要である。かかる外方鎖構造は、本発明の炭水化物よりなる特異的糖および結合についてのさらなる部位を提供する。N−結合糖鎖付加とは、GlcNAcを介しての炭水化物部位のペプチド鎖中のアスパラギン残基への付着をいう。従って、記載したコア構造において、Rはアスパラギン残基を表す。補体部位のペプチド配列、アスパラギン−X−セリン、アスパラギン−X−スレオニン、およびアスパラギン−X−システイン(ここに、Xはプロリンを除くいずれかのアミノ酸)は本発明のN−結合炭水化物部位の酵素的付着のための可能な認識部位である。対照的に、O−結合炭水化物は、スレオニンまたはセリンのヒドロキシル基に付着したGalNAcである共通のコア構造によって特徴付けられる。
N−結合グリカンは、適当な糖を付加させる一連の酵素によって細胞内で起こる一連の複合体工程によって形成される。別法として、コア構造の酵素的合成は、適当な炭水化物を得るための化学的および酵素的工程によって細胞外で達成できる。これらの化学的および酵素的合成は当該分野で、例えば、出典明示して本明細書の一部とみなす、国際出願公開番号WO91/19502およびそこで引用された文献に記載されている。
特異的グリコシルトランスフェラーゼは複合体炭水化物の最終外方鎖構造で重要である。これらのグリコシルトランスフェラーゼは適当な単糖について高度に特異的である。本発明で特に重要なもののうち、N−結合およびO−結合オリゴ糖で見い出されるGalβ1−4GlcNAc基のシアリル化およびフコシル化に関与する酵素である。末端糖鎖付加配列は異なることが当業者に理解されよう。複合体オリゴ糖鎖の外方鎖部位で見い出される種々の構造のうちには、特定のセレクチンに結合することが知られている炭水化物部位がある。
本発明の範囲内にある特別に好ましいのは、特異的位置および結合におけるシアル酸およびフコース残基を共に有するシアリル化フコシル化N−アセチルグルコサミンである。従って、ルイスX(LeX)炭水化物に関連するオリゴ糖(Galβ1−4(Fucα1−3)GlcNAc)は特に有用である。一般式Iの構造は特に関連する:

によって表される。好ましい具体例において、シアル酸は5−N−アセチルノイラミン酸である。もう1つの具体例において、シアル酸は5−グリコリルノイラミン酸である。
本発明の組成物で有用な特異的炭水化物構造のさらなる例は国際出願公開番号WO92/02527に開示されており、以下の通りに表される:
引き続いてのセクションで詳細に記載するごとく、これらの構造は、限定されるものではないが特定の補体を発現する細胞を適当なフコシルトランスフェラーゼ酵素でトランスフェクションすることを含めた種々のメカニズムによって補体部位上に提供することができる。別法として、該構造は、適当なフコシルトランスフェラーゼおよびシアリルトランスフェラーゼを用いて化学的に合成でき、また、補体部位に化学的に結合させることができる。かかるトランスフェラーゼは、一般に、後記するごとく入手可能である。
前記したごとく、セレクチンリガンドの特異的修飾は、炭水化物抗原決定基と個々のセレクチンとの間の相互作用を増強し得る。Nelsonらは、SLeXおよびSlea(sLeaはある種の癌細胞上でのその重要な発現のため腫瘍転移で特に重要であり得る)構造に基づいて一連のオリゴ糖の結合相互作用を研究した(Nelson,et al.,(1993)J.Clin.Invest.91:1157−1166)。Nelsonは、特別な位置および結合におけるシアル酸およびフコースは共にE−セレクチン認識を増強することを示唆している(Nelson,前掲)。SLeXおよびSLeaはα2−3結合でガラクトース(Gal)に結合した末端シアル酸(Neu5Ac)を含有し、今度はこれがN−アセチルグルコサミンに結合している(GlcNAc)。また、両構造はサブ末端GlcNAcにカップリングしたフコースを含有する。この特徴的構造は一般により大きい糖蛋白質の一部である。従って、ある種の好ましい具体例において、炭水化物は、少なくとも1つのフコース残基と適合する少なくとも1つのシアル酸を含有するように修飾される。
また、E−セレクチンは、末端シアル酸を欠くがその代わりに硫酸基を有するSLeXおよびSLeaに関連するオリゴ糖に結合する(Yuen,C.T.et al.,(1992)Biochemistry 31:9126−9131)。この一次構造の修飾は、炭水化物部位を特定のセレクチンにホーミングさせる選択的利点を有し得る。従って、末端シアル酸を欠くがその代わりに硫酸基を有する炭水化物は本発明の範囲内のものである。加えて、糖蛋白質の硫酸化はL−セレクチンへのリガンド結合を増強する(Imai,Y,Lasky,L.A.,and Rosen,S.D.(1993)Science 361:555−557)。この点につき、Yuenら、(1992)Biochemistry 31:9126−91341は指導的であり、ここに出典を明示して本明細書の一部とみなす。
土台となるオリゴ糖に影響するか否かを問わず、シアル酸残基の選択的酸化は、出典を明示して本明細書の一部とみなすNorgardら((1993),Proc.Natl.Acad.Sci.U.S.A.90:1086−1072)によって記載されているごとく、L−セレクチンとの相互反応を増強する。炭水化物担持補体蛋白質の結合または選択的結合の増強の結果となり得る一次構造の他の修飾も本発明の範囲内のものである。
また、本発明の範囲内にある炭水化物部位は、構造修飾によって、対応する天然に生じる炭水化物部位よりも代謝分解に対して耐性な構造を有する安定した炭水化物部位を提供することが示されている炭水化物を含む。また、かかる修飾構造は特定の標的細胞接着分子について高アフィニティーを示し得る。かくして、本発明の範囲内にある炭水化物部位は、特定の細胞間接着分子につきアフィニティーを獲得するように特異的に設計された炭水化物も含む。かかる炭水化物は、構造変異体もしくは模擬体が対応する天然に生じる炭水化物構造とほぼ同等またはそれよりも良好なセレクチン結合活性、免疫原性および抗原性を有するように、構造的に修飾した炭水化物であるかまたは炭水化物構造の模擬体であり得る。従って、認識および接着についての炭水化物構造に応じて相互作用を増強する炭水化物構造に対するいずれの修飾も本発明の範囲内のものである。ある種の炭水化物および天然に生じる炭水化物の構造的および機能的変異体である炭水化物模擬体は、例えば、Toyokuniらによる1993年11月29日に公開された国際出願公開番号WO93/23031に見い出される。炭水化物化学および炭水化物模擬体の分野の当業者ならば、適当なそれらの構造は本明細書における教示に基づいて本発明の範囲内のものであることを認識するであろう。
セレクチンと相互反応し、本発明に含まれる第2の群の炭水化物はマンノース−6−リン酸のごときリン酸化された単糖または多糖である。このリン酸化単糖、ならびに高分子量の酵母由来ホスホマンノン(PPME)は、P−セレクチンおよびE−セレクチンがこれらの分子に結合しないごとく、L−セレクチンのパートナーに専ら結合するようである(Bevilacqua,MP and Nelson,R.M.(1993)J.Clin.Invest.91:379−387)。
最後に、ヘパリンのごときいくつかの硫酸化多糖はセレクチンに結合する(Nelson,R.M.et al.,(1993)J.Clin.Invest.91:1157−1166)。
本発明は、補体部位の一部に付着した少なくとも1つの区別される炭水化物単位を含む。当業者ならば、本発明の範囲内にある補体蛋白質は、糖部位の付着のためのN−結合およびO−結合糖鎖付加のいくつかの部位を含有し得ることを認識するであろう。従って、本発明はいずれかの所与の補体部位に付着した1つまたは多くの炭水化物単位を含むことを意図する。当該組成物の炭水化物部位の特定の炭水化物側鎖のなかには、しばしば、特定の一次構造が生じるためのいくつかの部位があるであろう。例えば、N−結合複合体炭水化物は、本発明の特異的炭水化物構造の補体部位への付着のための可能な位置である1以上のアンテナ構造を含有し、従って、特定の補体部位の糖鎖付加の量は、組成物全体として達成しようと試みる生物学的活性に応じて大いに変化するであろう。
補体部位の糖鎖付加パターンの差異は、そのin vivo活性に基づいて特定の組成物を評価するのを助力するにおいて有利である。従って、半減期および吸収のごとき種々の因子を評価でき、これらの特性に基づいて、特定の組成物が選択される。糖鎖付加に影響する条件は、培地処方、細胞密度、酸素付加、pH等のごときパラメーターを包含するが、これらに限定されるものではない。別法として、適当な位置および結合において炭水化物残基を付加するのに関与する特異的トランスフェラーゼのごとき特別の酵素を増幅することが希望されるであろう。
糖鎖付加分析のための当該分野で公知のいくつかの方法は、本発明で有用である。かかる方法は、ペプチドに付着したオリゴ糖の同一性および組成に関する情報を提供する。本発明で有用な糖鎖付加分析の方法は、レクチンクロマトグラフィー;電荷に基づいてオリゴ糖を分離するために高pHアニオン交換クロマトグラフィーを用いるHPAEC−PAD;NMR;質量分析;HPLC;GPC;単糖組成分析;連続的酵素分解を含むが、これらに限定されるものではない。加えて、3つの主要な方法を用いて、オリゴ糖を糖蛋白質から放出させることができる。これらの方法は、1)通常ペプチド−N−グリコシダーゼF/エンド−β−ガラクトシダーゼを用いて行われる酵素的なもの、2)主としてO−結合構造を放出させるために厳しいアルカリ性環境を用いるβ−脱離;および3)N−およびO−結合オリゴ糖を共に放出させるために無水ヒドラジンを用いる化学的方法である。
本明細書で提示するいくつかの方法は当該分野で公知であり、特定のセレクチンのための分子のアフィニティーを測定するのに有用である。一般に、多数の方法を用いて、セレクチンによって媒介される細胞間接着を阻害する本発明の組成物の能力をアッセイすることができる。例えば、実施例セクションに記載した競合アッセイは、特別の方法を開示する。例えば、天然の細胞リガンドを特定のセレクチンを発現する細胞へ接着するのを阻害する能力を用いることができる。典型的には、本発明の補体蛋白質を、天然リガンド担持細胞の存在下でセレクチン担持細胞と共にインキュベートするが、ここに、セレクチン担持細胞は固体支持体に固定されたものである。次いで、結合した補体部位の量を計算するか、あるいは置き換えられた細胞を評価することによって、細胞接着の阻害を評価する。この点において、HL−60細胞および活性化ヒト血小板および内皮細胞が特に有用である。
好ましい具体例において、補体部位は、補体受容体1型のすべてまたは一部、特別には、前記したセクション5.1で記載した補体受容体1型のいずれかの可溶性断片よりなる。特別に好ましい具体例において、補体部位はsCR1よりなる。その全長形態であるこの蛋白質はN−結合糖鎖付加のための25の部位を有する。この具体例において、炭水化物側鎖がsCR1分子上に提供され、その側鎖は内皮細胞受容体のための特定のリガンドに結合できるか、あるいはその結合を妨げることができる1以上の炭水化物構造よりなる。特に、これらの炭水化物部位はセレクチンについてのリガンドである。特別に好ましい具体例において、これらの炭水化物部位はルイスXオリゴ糖、シアリル化ルイスXオリゴ糖または双方の組合せである。当業者ならば、糖鎖付加の量は、利用可能な部位の完全な飽和から、やや少ない部位まで変わることを認識するであろう。
5.3 セレクチンリガンドよりなる補体蛋白質の産生
本発明は、開示し特許請求する組成物の種々の製法で、セレクチン結合活性を有する、すなわち、LeX、より好ましくはSLeXのごときセレクチンリガンドよりなる補体蛋白質の製法を提供する。
5.3.1 同時トランスフェクション
本明細書中で用いる「同時トランスフェクション」なる語は、少なくとも1つの補体部位をコードする核酸およびフコースをラクトサミン配列に移行させることができる酵素をコードする少なくとも1つの核酸との導入をいう。この結果、少なくとも1つの補体部位および該酵素が細胞で同時発現される。有用な酵素はα1,3フコシルトランスフェラーゼである。適当な結合に適当な糖を付加するのに有用なこれらの酵素は、α1,3フコシルトランスフェラーゼ、α2,3シアリルトランスフェラーゼ、α2,6シアリルトランスフェラーゼ、α2,6シアリルトランスフェラーゼ、β1,4ガラクトシルトランスフェラーゼ、β1,3ガラクトシルトランスフェラーゼ、およびβ1,4N−アセチルグルコシルトランスフェラーゼを含むが、これらに限定されるものではない。これらは、Genzyme,Inc.,Cambridge,MA,Sigma,St.Louis,MO,the Albert Einstein College of Medicine,New York,NY,Biogen,Inc.,Cambridge,MA等の入手先から容易に入手される。かかるトランスフェラーゼについての遺伝子は継続的にクローン化されており、さらに将来は容易に入手できることが期待されるであろう。
好ましい具体例において、a1,3−フコシルトランスフェラーゼはこの目的では特に有用であることが見い出されている。本明細書で用いる「α1,3−フコシルトランスフェラーゼ」なる語は、LeX抗原決定基を形成できる、例えば、ラクトサミン配列にフコースを移行させることができるいずれの酵素をもいう。特に、本発明のα1,3−フコシルトランスフェラーゼは、公知の基質特異性のいずれか1つを示すことができる(Harlan and Liu,Adhesion,前掲参照)。好ましくは、細胞はCOSまたはチャイニーズハムスター卵巣(CHO)細胞のごとき哺乳動物細胞である。
α1,3−フコシルトランスフェラーゼを発現する遺伝子は種々の入手源から得ることができる(Kukowska−Latallo et al.,1990,Genes Dev.4:1288−1303;国際出願公開番号WO91/16900;およびPaulson & Colley,1989,J.Biol.Chem.264:17615−17618参照)。
少なくとも1つの補体蛋白質をコードする核酸およびα1,3−フコシルトランスフェラーゼ蛋白質についてコードする核酸は、適当な発現ベクター、または2つのベクターに挿入できる。本明細書で用いる「発現ベクター」なる語は、挿入された蛋白質をコードする配列の転写および翻訳のために必要なエレメントを含有するベクターをいう。必要な転写および翻訳シグナルは天然遺伝子および/またはそのフランキング領域によって供給できる。
当該系が同時トランスフェクトされた酵素系を用いて補体部位の糖鎖付加を提供できる限り、種々の宿主−ベクター系を蛋白質をコードする配列を発現させるのに利用することができる。可能な宿主−ベクター系は、ウイルス(例えば、ワクシニアウイルス、アデノウイルス等)で感染させた哺乳動物細胞系;ウイルス(例えば、バクロウイルス)で感染させた昆虫細胞系;または酵母ベクターを含有する酵母のごとき微生物を含むが、これらに限定されるものではない。ベクターの発現エレメントはその強度および特異性が変動する。利用する宿主−ベクター系に応じて、多数の適当な転写および翻訳エレメントのうちのいずれか1つを用いることができる。
1つの具体例において、発現ベクターまたはベクター類は複製起点を含有する。別の具体例において、少なくとも1つの補体部位および少なくとも1つの酵素を含むベクターまたはベクター類は、補体蛋白質および酵素(例えば、α1,3−フコシルトランスフェラーゼ)をコードする配列を組換え法によって染色体に統合された後に染色体で発現される。当業者ならば、種々のトランスフェラーゼ酵素または他の酵素をコードする多数の遺伝子を挿入して、かく挿入された少なくとも1つの酵素がここに記載する目的で最適となることを確実とするのが望ましいことが理解されるだろう。かくして、糖鎖付加を行う異なる能力を示す酵素をコードする種々の遺伝子の挿入は、かかる遺伝子の1つのみを挿入することが望ましい。また、補体関連蛋白質をコードする1以上の遺伝子を同時トランスフェクトして、本発明の組成物のこの部分の構築体を変えるのが望まれるであろう。
ベクターにDNA断片を挿入するための当該分野で公知のいずれの方法を用いても、補体蛋白質の発現用の少なくとも1つの遺伝子および適当な酵素の発現用の少なくとも1つの遺伝子、ならびに適当な転写および/または翻訳制御シグナルを含有する発現ベクターまたはベクター類を構築することができる。これらの方法は、in vitro組換えDNA法および合成技術およびin vivo組換え(遺伝的組換え)を含む。
補体蛋白質またはペプチド断片をコードするさらなる核酸配列の発現は、該補体蛋白質またはペプチドならびに酵素用遺伝子が組換えDNA法で形質転換された宿主で発現されるように、さらなる核酸配列によって調節できる。例えば、補体蛋白質およびα1,3−フコシルトランスフェラーゼの発現は、当該分野で公知のいずれのプロモーター/エンハンサーによっても制御できるが、これらの調節エレメントは発現用に選択した宿主で機能するものでなければならない。遺伝子発現を制御するのに使用できるプロモーターは、SV40初期プロモーター領域(Benoist and Chambon,1981,Nature 290:304−310)、ラウス肉腫ウイルスの3'長鎖末端反復に含まれるプロモーター(Yamamoto,et al.,1980,Cell 22:787−797)、ヘルペスチミジンキナーゼプロモーター(Wagner et al.,1981,Proc.Natl.Acad.Sci.U.S.A.78:1441−1445)、メタロチオネイン遺伝子の調節配列(Brinster et al.,1982,Nature 296:39−42);ノパリンシンテターゼプロモーター領域よりなる植物発現ベクター(Herrera−Estrella et al.,Nature 303:209−213)またはカリフラワーモザイクウイルス35S RNAプロモーター(Gardner,et al.,1981,Nucl.Acids Res.9:2871)、および光合成酵素リブロース二リン酸カルボキシラーゼのプロモーター(Herrera−Estrella et al.,1984,Nature 310:115−120);Gal 4プロモーター、ADC(アルコールデヒドロゲナーゼ)プロモーター、PGK(ホスホグリセロールキナーゼ)プロモーター、アルカリホスファターゼプロモーターのごとき酵母または他の菌類からのプロモーターエレメント、ならびに、組織特異性を示し、トランスジェニック動物で利用されてきた以下の動物転写制御領域:膵臓腺房細胞で活性なエラスターゼI遺伝子制御領域(Swift et al.,1984,Cell 38:639−646;Ornitz et al.,1986,Cold Spring Harbor Symp.Quant.Biol.50:399−409;MacDonald,1987,Hepatology 7:425−515);膵臓ベータ細胞で活性なインスリン遺伝子制御領域(Hanahan,1985,Nature 315:115−122)、リンパ系細胞で活性な免疫グロブリン遺伝子制御領域(Grosschedl et al.,1984,Cell 38:647−658;Adames et al.,1985,Nature 318:533−538;Alexander et al.,1987,Mol.Cell.Biol.7:1436−1444)、精巣、乳房、リンパ系および肥満細胞で活性なマウス乳癌ウィルス制御領域(Leder et al.,1986,Cell 45:485−495)、肝臓で活性なアルブミン遺伝子制御領域(Pinkert et al.,1987,Genes and Devel.1:268−276)、肝臓で活性なアルファ−胎児性蛋白質遺伝子制御領域(Krumlauf et al.,1985,Mol.Cell.Biol.5:1639−1648;Hammer et al.,1987,Science 235:53−58)、肝臓で活性なアルファ1−抗トリプシン遺伝子制御領域(Kelsey et al.,1987,Genes and Devel.1:161−171)、骨髄性細胞で活性なベータ−グロビン遺伝子制御領域(Mogram et al.,1985,Nature 315:338−340;Kollias et al.,1986,Cell 46:89−94)、脳の乏突起膠細胞で活性なミエリン塩基性蛋白質遺伝子制御領域(Readhead et al.,1987,Cell 48:703−712)、骨格筋で活性なミオシン軽鎖−2遺伝子制御領域(Sani,1985,Nature 314:283−286)、および視床下部で活性なゴナドトロピン放出ホルモン遺伝子制御領域(Mason et al.,1986,Science 234:1372−1378)を含むが、これらに限定されるものではない。
好ましい具体例において、トランスフェラーゼのための少なくとも1つの遺伝子をもった少なくとも1つの補体蛋白質を哺乳動物細胞、より好ましくはチャイニーズハムスター卵巣(CHO)細胞で発現させる(例えば、Stanley et al.,1990,J.Biol.Chem.265:1615−1622)。
特別の具体例において、細胞からのゲノムDNAおよびプラスミドDNAは標準的な方法(Maniatis)によって調製し、トリス−EDTA(10:1)緩衝液に溶解させる。もしポリブレン トランスフェクションを用いるならば、細胞ゲノムDNAは剪断される。細胞はポリブレンまたはリン酸カルシウム法でトランスフェクトできる(例えば、Stanley et al.,1990,前掲参照)。
また、同時トランスフェクションは、DETA−デキストラン法を用いて達成できる(Lowe et al.,1990,Cell,63:475−484;Davis et al.,Basic Methods in Molecular Biology,Elsevier Publishing Co.,1986)。
少なくとも1つの補体部位およびいずれかの酵素用の少なくとも1つの核酸挿入断片を含有する発現ベクターまたはベクター類は、4つの一般的アプローチ:(a)所望のプラスミドDNAまたは特異的mRNAのPCR増幅、(b)核酸ハイブリダイゼーション、(c)「マーカー」遺伝子機能の存在または不存在、および(d)挿入された配列の発現によって同定される。第1のアプローチにおいて、放射性同位体標識ヌクレオチドを取り込むか臭化エチジウム染色して核酸をPCRによって増幅して、増幅された産物の検出ができる。第2のアプローチにおいて、発現ベクターに挿入された外来性遺伝子の存在は、挿入された補体蛋白質およびα1,3−フコシルトランスフェラーゼ遺伝子に相同な配列からなるプローブを用いて核酸ハイブリダイゼーションによって検出できる。第3のアプローチにおいて、組換えベクター/宿主系は、当該ベクターへの外来性遺伝子の挿入によって引き起こされたある種の「マーカー」遺伝子機能(例えば、β−ガラクトシダーゼ活性、チミジンキナーゼ活性、抗生物質に対する耐性、形質転換表現型、バクロウイルスにおける封入体形成等)の存在または不存在に基づいて同定し選択できる。特別の例において、もし補体蛋白質またはα1,3−フコシルトランスフェラーゼ遺伝子がベクターのマーカー遺伝子配列内に挿入されれば、該挿入断片を含有する組換体はマーカー遺伝子機能の不存在によって同定できる。第4のアプローチにおいて、組換え発現ベクターは、組換体によって発現された遺伝子産物の活性についてアッセイすることによって同定できる。かかるアッセイは、例えば、遺伝産物in vitroアッセイの物理的もしくは機能的特性、例えば、補体阻害性活性、または抗体またはセレクチンとの結合に基づくものであり得る(セクション5.1、前掲、およびセクション5.3参照)。
一旦特定の組換えDNA分子が同定され単離されたならば、当該分野で公知のいくつかの方法を用いてそれを増殖できる。一旦適当な宿主系および増殖条件が確立されたならば、組換え発現ベクターを増殖し、量産できる。前記したごとく、使用できる発現ベクターは以下のベクターおよびその誘導体:少し名前を挙げると、ワクシニアウイルスもしくはアデノウイルスのごときヒトもしくは動物ウイルス;バクロウイルスのごとき昆虫ウイルス;およびプラスミドおよびコスミドDNAベクターを含むが、それらに限定されるものではない。
加えて、炭水化物部位(例えば、セレクチンリガンド)を付加するように追加して挿入された配列の発現を変調しするか、あるいは遺伝子産物を所望により特異的に修飾し加工する宿主細胞系を選択できる。ある種のプロモーターからの発現は、ある種のインデューサーの存在下で上昇させることができ;遺伝的に操作した補体部位および酵素産物の発現を制御できる。さらに、異なる宿主細胞は、翻訳および翻訳後プロセッシングおよび蛋白質の修飾(例えば、糖鎖付加、切断[例えば、シグナル配列の切断]について、特徴的かつ特異的なメカニズムを有する。適当な細胞系または宿主系を選択して、発現された外来性蛋白質の所望の修飾およびプロセッシングを確実とすることができる。
少なくとも1つの補体蛋白質および適当な酵素をコードする少なくとも1つの核酸配列を含有するベクターを、当該分野で公知の方法、例えば、トランスフェクション、エレクトロポレーション、マイクロインジェクション、形質導入、細胞融合、DEAEデキストラン、リン酸カルシウム沈殿、リポフェクション(リポソーム融合)、遺伝子銃の使用、またはDNAベクターのトランスポーターによって所望の宿主細胞に導入する(例えば、Wu et al.,1992,J.Biol.Chem.267:963−967;Wu and Wu,1988,J.Bil.Chem.263:14621−14624;Hartmut et al.,1990年3月15日に出願されたカナダ特許出願第2,012,311号参照)。
cDNAおよびゲノム配列は共にクローン化し、発現させることができる。
一旦、適当な酵素遺伝子または遺伝子類と共に補体蛋白質遺伝子または遺伝子類を発現させる組換体を同定したら、遺伝子産物を分析すべきである。これは、産物の物理的、免疫学的、または機能的特性に基づいてアッセイすることができる。
本発明の組成物よりなる発現された蛋白質産物の回収は、クロマトグラフィー(例えば、イオン交換、アフィニティー、およびサイズ分画カラムクロマトグラフィー、高速液体クロマトグラフィー)、遠心、溶解度分別によって、あるいは蛋白質の精製のためのいずれかの他の標準的な技術を含めた標準的な方法によって達成できる。
いずれのヒト細胞も、補体部位遺伝子もしくは遺伝子類および酵素遺伝子もしくは遺伝子類の分子クローニングのための核酸源として潜在的に働く。遺伝子の単離は、補体蛋白質関連構造もしくは特性、例えば、C3bもしくはC4bまたは免疫複合体の結合、ファゴサイトーシスの変調、免疫刺激もしくは増殖、および補体の調節を示す蛋白質をコードするDNA配列の単離を含む。DNAはクローン化DNA(例えば、DNA「ライブラリー」)から当該分野で公知の標準的な手法によって、化学的合成によって、cDNAクローニングによって、あるいは所望のヒト細胞から精製したゲノミックDNA、もしくはその断片のクローニングによって得ることができる(例えば、Maniatis et al.,1982),Molecular Cloning,A Laboratory Manual,Cold Spring Harbor Laboratory,Cold Spring Harbor,New York;Glover,D.M.(編),1985,DNA Cloning:A Practical Approach,MRL Press,Ltd.,Oxford,U.K.Vol.I,II参照)。遺伝子のcDNAクローニング用の核酸の入手源として供される細胞は、単球/マクロファージ、顆粒球、B細胞、T細胞、脾臓濾胞樹枝状細胞および糸球体有足細胞を含むが、それらに限定されるものではない。ゲノムDNAに由来するクローンは、暗号配列に加えて調節およびイントロンDNA領域を含有し得;cDNAに由来するクローンは唯一のエクソン配列を含有するであろう。入手源を問わず、遺伝子は、遺伝子の増殖用の適当なベクターに分子的にクローン化させるべきである。
好ましい具体例において、C3b/C4b受容体(CR1)蛋白質およびルイスX抗原を用いる。より好ましい具体例において、CR1蛋白質およびシアリルルイスX抗原を用いる。CR1遺伝子およびそれがコードする蛋白質は、1989年10月5日に公開され、「The Human C3b/C4b Receptor(CR1)」なる発明の名称の国際出願公開番号WO89/09220に提供されている。適当な酵素はα1,3−フコシルトランスフェラーゼであり、その遺伝子およびコードする蛋白質はLowe et al.,1992,J.Biol.Chem.267:4152−4160に提供されている。α1,3−フコシルトランスフェラーゼは国際出願公開番号WO91/16900、Kukowska−Latallo et al.,1990,Genes Dev.4:1288−1303,and Paulson et al.,1989,J.Biol.Chem.264:17615−17618に記載されている。
この好ましい具体例において、適当な炭水化物分子で蛋白質に糖鎖付加できるα1,3−フコシルトランスフェラーゼで同時トランスフェクトした細胞の選択は、トロンビンで活性化した血小板上のCD15構造と共に細胞を加熱することによって進行させることができる。例えば、トロンビン、ADP、コラーゲン、またはエピネフリンで活性化した血小板は、セレクチン受容体CD62/PADGEM/GMP140を発現する。結合した細胞はEDTAのごときキレート化剤の存在下で除去できる。というのは、セレクチン/炭水化物の相互作用はCa++およびMg++に依存するからである。次いで、これらの放出された細胞をクローン化し、適当な活性についてスクリーニングする。もう1つの具体例において、考慮すべき細胞を、HL60またはU937細胞の活性化血小板への結合競合アッセイで、セレクチン結合活性についてアッセイすることができる。
α1,3−フコシルトランスフェラーゼ活性について直接的にスクリーニングするアッセイは、種々の手段によって達成することができる。例えば、アッセイは、放射性同位体標識フコースをアクセプター分子に結合させるα1,3−フコシルトランスフェラーゼの能力をテストすることができる(国際出願公開番号WO91/16900参照)。α1,3−フコシルトランスフェラーゼについてテストするアッセイも当該分野で公知である(Stanley et al.,J.Biol.Chem.1987,262:16830−16837,Lowe et al.,1992,J.Biol.Chem.267:4152−4160;Stanley et al.,1990,J.Biol.,Chem.265:1615−1622)。
5.3.2 突然変異誘発
また、本発明は、補体蛋白質を発現して、本発明による組成物(好ましくは、該組成物は補体蛋白質およびLeX抗原、より好ましくはSLeXのごときセレクチンリガンドよりなる)を産生できる細胞系を得るための適当な細胞系での、当該分野でよく知られた方法による化学的突然変異誘発の使用を含む。本発明で考えられる1つの適当な方法は、エチルメタンスルホネートを用いる、α1,3−フコシルトランスフェラーゼのごとき適当な酵素を発現する細胞系の産生である(Stanley et al.,1983,Somatic Cell Genetics 9:593−608)。
所望の補体蛋白質を発現するCHOのごとき親細胞系を、エチルメタンスルホネート(EMS:Eastman Chemical Co.,Rochester,New York)で、100μg/mlの濃度にて、34℃および38.5℃で突然変異誘発できる。所望の補体蛋白質を発現するいずれの細胞系、好ましくは哺乳動物細胞系を用いることもできるが、突然変異誘発はα1,3−フコシルトランスフェラーゼのごとき1以上の酵素を潜在的に誘導して補体蛋白質に糖鎖付加できることを条件とする。例えば、内因的に補体蛋白質を発現する細胞、ならびに所望の補体蛋白質を発現する能力をもつトランスフェクトした細胞を用いることができる。特別の具体例については、実施例セクション6.4.1参照。
突然変異誘発の他の方法は当該分野でよく知られており、使用することができる(Maniatis,Ad.,前掲)。加えて、細胞に放射線を照射し、突然変異を誘発した細胞を本明細書で記載する技術によって選択する。
本発明の組成物を発現する突然変異誘発細胞をスクリーニングする方法は当該分野で公知であり、セクション5.2およびこのセクションに続くセクションに記載される。
5.3.3.補体蛋白質と共に適当な酵素活性を有する細胞の トランスフェクション
トランスフェラーゼ酵素活性を発現する細胞は多くの入手源から得ることができる。例えば、α1,3−フコシルトランスフェラーゼのごとき適当な酵素を内因的に発現する細胞を用いるか、あるいは細胞を前記セクション5.3.1で教示した方法によってかかる酵素をコードする遺伝子でトランスフェクトして用いることができる。また、予め、α1,3−フコシルトランスフェラーゼのごとき、炭水化物部位での糖鎖付加に必要な酵素を発現させるために、突然変異を誘発した細胞を用いることができる(セクション5.3.2、前掲)。あるいは、予めトランスフェクトした細胞を用いることもできる。一旦適当な酵素を発現する細胞が得られたら、それを当該分野で公知の方法によって補体蛋白質遺伝子でトランスフェクトする。特に、補体蛋白質はセクション5.1(前掲)に記載され;既に適当な酵素活性を発現する適当な宿主細胞にかかる蛋白質をコードする核酸を導入する方法はセクション5.3.1(前掲)に記載されている。
特に、補体蛋白質をコードする核酸を、次いで、それを本発明の組成物を発現する細胞系、特にHL−60(ATCC ♯CCL−240)およびK562(ATCC ♯CCL−243)細胞系に導入することができる。
5.3.4.細胞融合
本発明の組成物を得るための別の方法は細胞融合による。前掲した5.3.3を通じて、セクション5.3.1.で記載したごとくに、適当な酵素および補体蛋白質を発現する能力を有する細胞系を、標準的な技術を用いて相互に融合させることができる(Current Protocols in Molecular Biology,Greene and Wiley−Interscience(1989)参照)。
特別の具体例において、補体蛋白質を発現する細胞を酵素活性を有する細胞と融合させる。この具体例の特別の例は後記する実施例のセクションに示す。
好ましくは、かかる細胞融合技術によりハイブリッド細胞を調製する場合、1の細胞を、例えば、突然変異誘発を介して、ヒポキサンチン−グアニンホスホリボシルトランスフェラーゼ遺伝子を欠くように選択し設計すべきである。これらの細胞はPRPPを利用するサルベージ経路を介してプリンをリサイクルする活性を欠くであろう。これらの細胞は、融合現象が突然変異体細胞を含有しないハイブリッド細胞系を生じないように、過剰に提供すべきである。細胞系、例えば、α1,3−フコシルトランスフェラーゼを発現する細胞系、あるいは補体蛋白質を発現する細胞系のいずれかを、突然変異誘発することができる。突然変異誘発されなかった細胞はサルベージ経路を利用する能力を維持するであろう。従って、突然変異誘発細胞系を含有しない少数のハイブリッド細胞系のみが(アミノプテリンの存在のため)HAT培地で生き残るであろう。突然変異誘発細胞を大量に用いて融合を行うことによって、非突然変異誘発細胞(「正常」細胞)のほとんどが、突然変異原と融合し、正常細胞のうち少数のみが突然変異原と融合しないことになるであろう。突然変異原の融合の結果となるすべての細胞;突然変異細胞はまもなく死滅するであろう、というのはこれらの細胞はプリンサルベージ経路を利用する手段を有しないからである。かくして、この負の選択により、α1,3−フコシルトランスフェラーゼ活性および補体蛋白質を発現するハイブリッド細胞系が得られるであろう。
5.3.5.IN VITRO修飾
前記した適当な補体蛋白質を発現する能力を有する細胞系を、引き続いての産生後修飾用の補体蛋白質源として使用する。存在する炭水化物構造の修飾は、例えばセクション5.3.1.に記載した適当な酵素のいずれかを用いて達成できる。特別の具体例において、産生後修飾は、GDP−フコースおよび適当なα1,3−フコシルトランスフェラーゼを用いて、適当な条件下、in vitroで起こる。かかるトランスフェラーゼは前記した。上述の修飾により、Galβ1−4GlcNAcのごとき存在するコア炭水化物構造上でフコシル化されたオリゴ糖を得ることが期待される。適当なシアル酸と共に適当なシアリルトランスフェラーゼは、末端シアル酸残基をGalβ1−4GlcNAcまたはGalβ1−4(Fucα1−4)GlcNAcのごとき適当なコア構造に付加することが予測される。得られた炭水化物は前記のものを含め当該分野で公知のいずれかの方法で分析することができる。
5.3.6.化学的修飾
本発明は、さらに、当該分野でよく知られた化学合成技術を用いて、炭水化物部位を補体部位に共有結合させることによって本発明の組成物を調製することを含む。
かくして、本発明の補体蛋白質は化学的修飾によって炭水化物リガンドで糖鎖付加される。この修飾の結果、糖蛋白質が得られ、そこでは、補体蛋白質が炭水化物リガンドに直接結合しているか、あるいは、別の具体例においては、結合活性を有する不活性蛋白質を補体蛋白質に共有結合にて架橋させ、それにより、不活性蛋白質は炭水化物にブリッジすることができる。もしかかる不活性蛋白質を用いるならば、ショートコンセンサスリピート(SCR)が好ましい。というのは、SCRは多くの補体蛋白質上で見い出される構造モチーフだからであり(セクション5.1.、前掲参照)、従って、補体蛋白質の構造および機能に最小限の影響しか与えないようである。
当業者に認識されるごとく、炭水化物部位は天然源から精製し、収集できる。このプロセスの例は、Stanely et al.,J.Biol.Chem.263:11374(1988)にLeXおよびsLeXについて開示されている(WO91/19502(PCT/US91/04284)およびWO92/02527(PCT/US91/05416)参照)。また、セクション5.1(前掲)に記載したごとく精製された補体蛋白質も得られる。別法として、炭水化物部位は合成により得られる(Wong et al.,1992,J.Am.Chem.Soc.114:9283,C.F.Borman,1992,C & En Dec.7;p25参照)。
いずれの入手源からの炭水化物部位も、いずれの入手源からの補体蛋白質に結合でき、化学合成技術を用いて本発明の組成物が得られる。特に好ましい具体例において、炭水化物部位はルイスXであり、より好ましくは、それはシリアルルイスXである。好ましくは補体蛋白質はCR1、より好ましくは可溶性CR1である。
セレクチンリガンドの補体タンパク質への化学的架橋は、ホルムアルデヒド(ホルムアルデヒドは発癌性のため使用が示されていないにも拘わらず)のごとき、続けて1度以上反応できる1の官能基を有する分子、ならびに1以上の反応性基を持つ分子のような従来の架橋剤を用いて進行させることができるが、これらに限定されるものではない。本明細書で用いる「反応性基」なる語は、補体蛋白質上の官能基と反応して、当該架橋剤と蛋白質との間に共有結合を形成する架橋剤上の官能基をいう。架橋剤は炭水化物部位と反応するための第2の官能基を有するべきである。「官能基」なる語は、有機化学におけるその標準的な意味を有する。好ましくは、本発明における架橋剤は多官能性分子である。すなわち、それは1以上の反応性基を含む。使用できる多官能性分子は生体適合性リンカーである、すなわち、それらはin vivoにて非発癌性、非毒性、かつ実質的に非免疫原性である。当該分野で知られており、かつ本明細書に記載するもののごとき多官能性架橋剤は、その生体適合性を測定するための動物モデルで容易にテストすることができる。
好ましくは、多官能性分子は二官能性である。本明細書で用いる「二官能性分子」なる語は、2個の反応性基を持つ分子をいう。二官能性分子はヘテロ二官能性またはホモ二官能性であってよい。好ましくは、二官能性分子は、炭水化物部位および補体部位の方向性のある結合を可能とするヘテロ二官能性である。典型的には、多官能性分子は、補体蛋白質上のスルフヒドリル基および炭水化物部位上のヒドロキシル基、アミノ基、アルデヒド基もしくはカルボン酸と共有結合する。しかしながら、カルボン酸またはヒドロキシル基のごとき、補体蛋白質上での他の官能基と反応する多官能性分子も本発明で使用できる。
ホモ二官能性分子は、同一である少なくとも2個の反応性官能基を有する。ホモ二官能性分子上の反応性官能基は、例えば、アルデヒド基および反応性エステル基を含む。アルデヒド基を有するホモ二官能性分子は、例えば、グルタルアルデヒド(Poznansky et al.,1984,Science 223:1304−1306)およびスバルアルデヒドを含む。少なくとも2個の反応性エステル単位を有するホモ二官能性分子は、ジカルボン酸およびN−ヒドロキシスクシンイミドのエステルを含む。かかるN−スクシンイミジルエステルのいくつかの例は、ジスクシンイミジルスベレート、ジチオ−ビス−(スクシンイミジルプロピオネート)、ならびにその可溶性−ビススルホン酸およびそのナトリウムおよびカリウム塩のごときビス−スルホン酸塩である。これらのホモ二官能性試薬はPierce Chemicals,Rockford,Illinoisから入手可能である。
ヘテロ二官能性分子の反応性基がアミノ基と共有結合を形成する場合、共有結合は通常アミド、より特別にはイミド結合である。アミノ基と共有結合を形成する反応性基は、例えば、活性化カルボキシレート基、ハロカルボニル基、またはエステル基である。好ましいハロカルボニル基はクロロカルボニル基である。エステル基は、好ましくは、例えば、N−ヒドロキシ−スクシンイミドエステル基のごとき反応性エステル基、または1−ヒドロキシ−2−ニトロベンゼン−4−スルホン酸ナトリウム塩のN−マレイミド−6−アミノカプロイルエステルのそれ(Mal−Sac−HNSA;Bachem Biosciences,Inc.;Philadelphia,PA)である。
典型的には、補体蛋白質上のもう1つの官能基はチオール基、チオール基に変換できる基、またはチオール基と共有結合を形成する基のうちのいずれかである。遊離スルフヒドリル基は、1以上のジスルフィドを含有する補体蛋白質(またはペプチド)のジスルフィド結合から生成し得る。これは、蛋白質分子の温和な還元によって達成される。温和な還元条件は、蛋白質機能を干渉しないように、当該蛋白質の2次および3次構造が有意に変化するのが好ましい。過剰に還元すると、蛋白質の変性が起こり得る。かかる反応性基は、ジスルフィド移行を介して遊離チオールと反応し得るジスルフィド、例えば、ピリジルジスルフィド、p−マーキュリーベンゾエート基およびミカエル型付加反応できる基(例えば、Mitra and Lawton,1979,J.Amer.Chem.Soc.101:3097−3110に記載されているタイプの基やマレイミドを含む)を含むが、これらに限定されるものではない。共有結合は、通常は、チオエステル結合またはジスルフィドであろう。チオール基と共有結合を形成する反応性基は、例えば、チオール基もしくは活性化ジスルフィドと反応する二重結合である。チオール基と反応できる二重結合を含有する反応性基はマレイミド基であり、アクリロニトリルのごとき他のものも可能である。反応性ジスルフィド基は、例えば、2−ピリジルジチオ基または5,5'−ジチオ−ビス−(2−ニトロ安息香酸)基であり得る。
本発明によると、還元された蛋白質のスルフヒドリル基への付着のためには、マレイミドもしくはジスルフィド基を当該リンカーの一端に付着させることによって基質リンカーを修飾することができる。該リンカー上の未修飾部位を炭水化物部位上の官能基に共有結合させる。例えば、前記した化合物に連結させたエステルまたはアミドである基質リンカー(Partis et al.,1983,J.Pro Chem.2:263;Means and Feeney,1990 Bioconjugate Chem 1:2−12)。
反応性ジスルフィド結合を含有するヘテロ二官能性試薬のいくつかの例は、N−スクシンイミジル3−(2−ピリジル−ジチオ)プロピオネート(Carlsson,et al.,1978,Biochem J.,173:723−737)、ナトリウムS−4−スクシンイミジルオキシカルボニル−アルファ−メチルベンジルチオスルフェート、および4−スクシンイミジルオキシカルボニル−アルファ−メチル−(2−ピリジルジチオ)トルエンを含む。チオール基と反応する二重結合を有する反応性基よりなるヘテロ二官能性試薬のいくつかの例は、スクシンイミジル4−(N−マレイミドメチル)シクロヘキサン−1−カルボキシレートおよびスクシンイミジルm−マレイミドベンゾエートを含む。
他のヘテロ二官能性分子は、スクシンイミジル3−(マレイミド)プロピオネート、スルホスクシンイミジル4−(p−マレイミド−フェニル)ブチレート、スルホスクシンイミジル4−(N−マレイミドメチルシクロヘキサン)−1−カルボキシレート、およびマレイミドベンゾイル−N−ヒドロキシ−スクシンイミドエステルを含む。前記したヘテロ二官能性薬剤およびそれらのスルホネート塩の多くはPierce Chemicals(前掲)から入手可能である。これらをいかにして製造するかまたは使用するか、ならびに得ることができる他の多官能性試薬についてのさらなる情報は当該分野でよく知られている。例えば、架橋の方法はMeans and Feeney,1990,Bioconjugate Chem.1:2−12に総括されている。
架橋剤の反応性基は、(飽和および不飽和を含めた)アルキル基、環状アルキル基、置換されたアルキルもしくは環状アルキル基、またはペプチド配列を含めた同等のスペーサ−基を介してスペースを入れることができる。特別の具体例において、架橋反応性基は、20原子を超えるスペースも考えられるが、0ないし約20原子だけ相互からスペースを設ける。
もう1つの具体例において、補体糖蛋白質の炭水化物側鎖を選択的に酸化してアルデヒドを生じさせることができる(例えば、Jackson,1944,Organic Reactions 2:341;Bunton 1965,Oxidation in Organic Chemistry,Vol.1(Wiberg編),Academic Press,New York,p367;(Cooper,et al.,1959,J.Biol.Chem.234:445−448)。これは、炭水化物側鎖がセレクチンリガンドでない場合に好ましい。次いで、得られたアルデヒドをアミン基(例えば、第一級アミンのごときアンモニア誘導体、ヒドロキシルアミン、ヒドラジン、ヒドラジド、チオヒドラジド、フェニルヒドラジン、セミカルバジドまたはチオセミカルバジド)と反応させてシッフ塩基または還元されたシッフ塩基(例えば、イミン、オキシム、ヒドラゾン、フェニルヒドラゾン、セミカルバゾンまたはチオセミカルバゾン、またはそれらの還元形態)を得ることができる。
エステルもしくはアミド結合または炭素−炭素結合を介して、セレクチンリガンド、例えばLeXまたはSLeXにヒドラジド架橋剤を付着させ、次いで、酸化された炭水化物を含有する酸化補体糖蛋白質と反応させることができる。この結果、ヒドラゾンが形成され、架橋基を介して当該化合物が糖蛋白質の炭水化物側鎖に共有結合する。
別法として、補体蛋白質の糖蛋白質形態をフコースの存在下でα1,3−フコシルトランスフェラーゼと反応させて、補体阻害性糖蛋白質のフコシル化形態を得ることができる。
5.4.機能的活性
本発明は、さらに、本発明の組成物の機能的活性を評価するためのアッセイを提供する。特に、本発明は、LeX、好ましくはSLeXのごときセレクチンリガンドよりなるCR1分子についてのある種の有用な機能的アッセイを提供する。本明細書で用いる「機能的活性」なる語は、分子の生物学的機能とは別に免疫学的結合をいう。また、本発明の組成物の補体部位および炭水化物部位の性質を測定するために物理化学的アッセイも考えられる。
1つの具体例において、補体部位および炭水化物部位の活性は別々に評価することができる。かくして、本明細書の教示による炭水化物部位よりなる補体蛋白質は、例えば、セクション5.1(前掲)に記載したごとき、同様もしくは同一の、電気泳動移動、等電点電気泳動挙動、蛋白分解消化地図、C3bおよび/またはC4bおよび/または免疫複合体結合活性、補体調節活性、ファゴサイトーシスもしくは免疫刺激に対する効果または補体阻害性蛋白質について公知の抗原性を有している。同様に、炭水化物部位の機能的活性は、例えば、セクション5.2(前掲)に記載され、国際出願公開番号WO/19502に記載されているごとくに直接アッセイすることができる。
多数の現在入手可能なモノクローナル抗体を本発明で用いて、セレクチンによって媒介される細胞間接着を阻害することができる。例えば、SLeX(Macher et al.,前掲)とわずかに異なる配列を認識する、CSLEX−1(Campbell et al.,J.Biol.Chem.259:11208−11214(1984))、VIM−2、FH6(米国特許第4,904,596号に記載)(すべての文献を出典明示して本明細書の一部とみなす)またはワシントン州のシアトルのBiomembrane InstituteのS.Hakomori博士によって生成されたSH3およびSH4。
もう1つの具体例において、補体部位および炭水化物部位の機能的活性または物理化学的特性は、同一のアッセイで評価される。例えば、特別の具体例において、セレクチンリガンドよりなる補体阻害性蛋白質の分子量は、PAGEによって評価することができる(LeX、好ましくはSLeXのごときセレクチンリガンドの蛋白質への結合を示す見掛け分子量の増加)。もう1つの具体例において、サンドイッチイムノアッセイを用いて機能的活性をアッセイすることができる。例えば、補体阻害性蛋白質およびセレクチンリガンドに対する抗体を用いることによって、組成物を同定することができる。特別の具体例において、CR1に特異的な抗体をアッセイプレートに吸着させる。セレクチンリガンドSLeXまたはLeXからなる推定される可溶性CR1を、抗体結合を可能とする条件下でプレートに添加する。SLeXまたはLeXよりなる結合した可溶性CR1の存在は、各々、FITCで標識したCSLEX抗体または抗−CD15抗体、続いて、ホースラディッシュペルオキシターゼで標識した抗−FITC抗体を添加することによって検出される。当業者に容易に理解されるごとく、かかるサンドイッチイムノアッセイは、固相上にCSLEX抗体または抗−CD15抗体を配置するか、あるいは間接アッセイよりも直接アッセイとすることができる。さらにもう1つの具体例において、ウェスタンブロットアッセイを用いて、産物がセクレチンリガンドよりなる補体蛋白質を含むことを示すことができる。1つの態様において、補体蛋白質に対する抗体で1のレーンにて検出された蛋白質およびセレクチンリガンドに対する抗体でもう1つのレーンにて検出された蛋白質の見掛けの分子量をを比較することができる。同一の分子量を示すという結果は、分子の明確な同一性を示す。もう1つの態様において、抗−補体蛋白質カラムまたは抗−セレクチンリガンドカラムのいずれかで、蛋白質をアフィニティークロマトグラフィーによって精製し、精製蛋白質を別の蛋白質を用いウェスタンブロッティングにて検出する。
当業者に容易に理解されるごとく、十分に高いアフィニティーの補体阻害性蛋白質またはセクレチンリガンドのいずれのアフィニティー結合パートナーも、特異的抗体分子の代わりにアッセイで用いることができる。
当業者ならば、組成物の個々の成分の活性をアッセイすることができ、また組成物全体としての総じての活性をアッセイすることができる他の方法があることを理解するであろう。これらのタイプのアッセイは、治療の場のごとき所望のセッティングにおいて組成物が所望の総じての機能を達成するかについての情報である。従って、このセクション、ならびに実施例セクションは、ある種の十分に受け入れられた技術の例であることを意図する。
5.5 治療用の組成物および使用
本発明の組成物の1つの主要な利点は、炭水化物部位が炎症内皮細胞に「ホーミングし」、かくして、組成物を組織損傷部位に局所化させ、それにより、その抗−補体活性を増強させ、好中球のローリングおよび管外遊出のごとき好中球−内皮細胞相互作用をブロックする点にある。補体蛋白質を損傷部位に止まらせることによって、それがそこに執拗に存在することとなり、特許を請求する組成物は、有利には、成分のいずれか単独で投与した場合に可能なよりも低い用量で治療することが可能となる。また、本発明の組成物はin vivo半減期の増大および/または大きな生物学的利用性を示す。
セレクチンの発現は、炎症の部位に細胞を動員する役割を果たす。多数の接着蛋白質およびそのリガンドが、白血球を内皮細胞に接着させ、また内皮細胞を横切って管外遊出するのに必要であることが文献でよく書かれている。例えば、ELAM−1(炎症サイトカイン、エンドトキシン)およびCD62(トロンビン、ヒスタミン等)の発現の公知アクチベーターで行った実験に基づき、その発現は組織傷害に対する炎症反応およびホメオスタティック反応を表すと考えられている。
血管壁を横切っての血管外組織への白血球の移動は、微生物または外来制抗原に対する宿主の防御および組織傷害の回復に必要である。しかしながら、ある状況下では、白血球−内皮細胞相互作用は宿主にとって悪い結果を与えかねない。接着および経内皮細胞移動の間に、白血球は、直接内皮細胞を損傷するかあるいは種々の炎症メディエーター(Harlan & Lie,前掲)を放出することによって内皮細胞の損傷を引き起こす酸化剤、プロテアーゼ、またはサイトカインを放出し得る。酸化剤のごときこれらのメディエーターのいくつかは、補体を直接活性化し、これは、次いで、フィードバックされてC3aおよびC4aを介して好中球をさらに活性化する。これは、さらなる組織損傷に導く。このセレクチン−相互作用によって内皮細胞微小環境に「ホーミングされた」補体阻害性蛋白質によるこのプロセスの介在は、このプロセスを停止するかあるいは速度を低下させ得る。
最後に、単一の白血球の毛細血管内腔への固着または白血球のより大きな血管内への集合は、微小血管閉塞に導きかねず、また、虚血を生じ得る。白血球−媒介の血管および組織の損傷は、広範囲の臨床疾患の病因に関与している。内皮細胞への白血球接着の阻害−「抗−接着」療法は、白血球が血管および組織の損傷に有意に寄与する炎症および免疫疾患の治療に対する新規なアプローチを表す。in vitro実験は、内皮細胞への白血球の密接な近接が、白血球と、血漿阻害剤に接近できない内皮細胞との界面保護された微小環境を形成することを示している。該界面において接着性白血球によって放出された高い反応性の酸化剤、プロテアーゼ、およびホスホリパーゼ産物は内皮細胞と反応し、それを損傷しかねない。かかる強い接着の阻害は、保護された微小環境の形成を妨げ、それにより、内皮細胞に対してこのタイプの「無害な傍観者」損傷を低下させる。また、内皮細胞への白血球接着の阻害は、組織への移動を妨げ、結果として、移動した白血球によって生じる組織損傷を減少させる。最後に、内皮細胞への白血球接着またはホモタイプの集合の阻害は、微小血管閉塞を妨げる。
本発明の医薬組成物を用いて、多数の疾患に関連する細胞接着をブロックまたは阻害することができる。例えば、多数の炎症性疾患は血管内皮細胞および血小板に発現されたセレクチンに関連する。本明細書で用いる「炎症」なる語は、特異的および非特異的防御系双方の反応をいうように用いる。特異的防御系反応は抗原に対する特異的免疫系反応である。特異的防御系反応の例は、ウイルスのごとき抗原に対する抗原反応、および遅延型過敏性反応を含む。非特異的防御系反応は一般に免疫学的記憶ができない白血球によって媒介される炎症性反応である。かかる細胞は、マクロファージ、好酸球および好中球を含む。非特異的反応の例は、蜂に刺された直後の腫れ、および細菌感染部位におけるPMN白血球の集合、例えば、細菌性肺炎における肺浸潤物および膿傷における膿形成を含む。
加えて、本発明の医薬組成物は、移植器官で起こる補体損傷を除去もしくはブロックするのに用いることができる。移植用に調製した器官を本発明の組成物を用いて潅流することができる。別法として、移植用器官を、本発明の組成物を含有する溶液中で貯蔵することもできる。かかる貯蔵は、例えば、輸送の間に行うことができる。さらなる具体例において、該組成物は、死体からのごとく、移植器官を取り出した領域をフラッシュするのに使用できる。引き続いての潅流および/または貯蔵も考えられる。
他の治療可能な疾患は、例えば、慢性関節リウマチ、虚血後白血球−媒介組織損傷(再潅流損傷)、凍傷損傷もしくはショック、急性白血球−媒介肺損傷(例えば、成人呼吸困難症候群、喘息、外傷性ショック、敗血症性ショック、腎炎、脈管炎ならびに急性および慢性炎症を包含し、アトピー性皮膚炎、乾癬、および炎症性腸疾患を含む。アテローム性動脈硬化症および凝固のごとき種々の血小板−媒介病理学も治療することができる。加えて、循環癌細胞の接着を阻害することによって腫瘍転移を阻害または防止することができる。その例は結腸の癌腫およびメラノーマを含む。これらの具体例において、組成物の補体部位の部分がほとんど担体蛋白質として作用する。
本発明の組成物は、敗血症性ショックまたは血管内凝固症候群(DIC)の二次効果を治療するという用途がある。敗血症性ショックまたはDICの間における組織への白血球の移動は、しばしば、病理学的組織破壊を生じる。さらに、これらの患者は広く分布した微小循環血栓およびび慢性炎症を有し得る。本明細書で提供する治療用組成物は、これらの部位における白血球移動を阻害し、組織損傷を緩和する。
また、抗−補体作用とカップリングした、セレクチン−リガンド相互作用の阻害剤は、外傷性ショックおよびそれに伴う急性組織損傷を治療するのに有用である。セレクチンは損傷部位への白血球の動員で役割を果たすので(特に、急性損傷および炎症の場合のELAM−1)、その阻害剤を局所または全身投与して、かかる傷害に関連する組織損傷を制御することができる。さらに、炎症部位、例えば、ELAM−1受容体が発現される部位での阻害剤の特異性のために、これらの組成物は、従来の抗炎症剤と比較して、より効果的で、合併症をより引き起こさないようである。
本発明の組成物は、それを必要とする対象に投与して、予防的に病気状態を処置するか、またはそれが開始した後に軽減することによって該対象を治療することができる。本発明の組成物は、非経口、局所、経口、または(エアロゾルもしくは経皮のごとき)局所投与またはそれらの組合せを含め、いずれかの適当な手法で投与することができる。好ましくは、該組成物は医薬上許容される担体と共に投与し、該担体の性質は投与法により異なり、例えば、経口投与では通常固体担体を用い、静脈内投与では液体塩溶液担体を用いる。
本発明の組成物は、患者ならびに本発明の組成物の蛋白質および炭水化物部位に適合する医薬上許容される成分を含む。これらは、一般に、懸濁液、溶液およびエリキシル剤、ならびにリン酸緩衝生理食塩水、生理食塩水のごとき最も特別な生物学的緩衝液、ダルベッコの培地等を含む。また、エアロゾルを用いることができ、また、スターチ、糖、微結晶性セルロース、希釈剤、顆粒剤、潤滑剤、結合剤、崩壊剤等(散剤、カプセル剤、および錠剤のごとき経口固体製剤の場合)のような担体を用いることができる。
本明細書で用いる「医薬上許容される」なる語は、好ましくは、動物、特にヒトに使用するものとして、連邦政府または州政府の管理当局によって認可されているか、米国薬局方または他の一般に認められている局方に収録されていることを意味する。
選択した処方は、前記した種々の緩衝液、または例えば医薬グレードのマンニトール、ラクトース、スターチ、ステアリン酸マグネシウム、サッカリンナトリウムセルロース、炭酸マグネシウム等を含めた賦形剤を用いて達成できる。組成物の「ペグレーション(peglation)」は、当該分野で公知の技術を用いて達成できる(例えば、国際出願公開番号WO92/16555、Enzonに対する米国特許第5,122,614号、および国際出願公開番号WO92/00748参照)。経口組成物は液剤、懸濁剤、錠剤、丸剤、カプセル剤、徐放性処方、または散剤の形態とすることができる。特に有用なのは、DMSOのごとき浸透促進剤を含む経皮処方で組成物を間接的に投与することである。他の局所処方を投与して皮膚炎症を治療することができる。
炎症を引き起こすことが予測されるかまたは炎症を現実に引き起こしているセレクチンリガンドの実質的部分を調節し、ならびに最適濃度の補体部位を当該部位に送達して不適当な補体関連活性と戦うことを確実にするためには、十分な量の本発明の組成物を患者に投与すべきである。このように、炎症は、防止するか軽減することができる。組成物の選択、投与の頻度および投与する組成物の量は、治療すべき個々の病気およびその重症度、患者の総じての症状、ならびに治療する医者の判断による。典型的な用量範囲は、抗体および他の生物医薬の使用によるこれらの病気状態の治療と同様である。典型的には、本発明の組成物は、有効成分約1%ないし約95%、好ましくは約10%ないし約50%含有する。好ましくは、投与量は約1〜10mg/kgである。子供には約1mgないし約50mgを投与し、成人には約25mgおよび約1000mgの間を投与する。他の効果的な投与量は、用量反応曲線を確立する日常的試行を通じて当業者が容易に決定できる。
投与すべき組成物の用量を決定するにおいて、セレクチン受容体のすべてを完全にブロックすることは望まれないであろうし、あるいはかかる受容体を限定した時間だけ(すなわち、虚血現象後の数時間)完全にブロックすることが望まれるであろうことを銘記すべきである。通常の治癒プロセスを進行させるためには、少なくともいくらかの白血球細胞および好中球を、負傷、感染または疾患状態が起こりつつある領域の組織に運ばなければならない。かくして、ブロッキング剤として投与すべき組成物の量は、治療されるべき病気のタイプのごとき種々の因子を考慮しつつ、患者の個々の必要性に基づいて注意深く調整しなければならない。例えば、関節炎の関節で好中球が再発生するのを望まないが、心筋梗塞、組織挫傷損傷等後のいくつかの部位ではかかる再発生を期待している。
好ましい具体例において、本発明は、セレクチンに結合できる補体阻害性蛋白質よりなる医薬組成物を含む。好ましくは、医薬組成物は、LeX、最も好ましくはSLeXのごときセレクチンリガンドからなる可溶性CR1分子よりなる。1つの態様において、かかる可溶性CR1分子は、LHR A、B、CおよびDを有する。もう1つの態様において、かかる可溶性CR1はLHR B、CおよびDを有する。
従って、本発明の医薬組成物は送達され、蛋白質の血漿レベルの上昇を達成して、炎症活性が関与するか否かを問わず、不適当な補体活性が関与する病気または疾患を治療するであろうと考えられる。補体調節蛋白質の全身または循環レベルを要する補体関与の病気または疾患は、前記したセクション2.2および以下の表Iで詳しく述べる。
表I
補体関与の全身性病気および疾患
神経学的疾患
多発性硬化症
卒中
ギラン・バレー症候群
外傷性脳傷害
パーキンソン病
不適当なもしくは望ましくない補体活性化の疾患
血液透析合併症
激症同種移植片拒絶
異種移植片拒絶
IL−2治療中のインターロイキン−2誘導毒性
炎症性疾患
自己免疫疾患の炎症
クローン氏病(限局性回腸炎)
成人呼吸困難症候群
火傷および凍傷を含めた熱傷害
虚血後再潅流疾患
心筋梗塞
バルーン血管形成
心肺バイパスまたは腎臓バイパスでのポストポンプ(post−pump)症候群
血液透析
腎臓虚血
動脈再構成後の腸間膜動脈再潅流
移植器官再潅流
感染症または敗血症
器官保存
免疫複合体疾患および自己免疫疾患
慢性関節リウナチ
全身性紅斑狼瘡(SLE)
SLE腎炎
増殖性腎炎
糸球体腎炎
溶血性貧血
重症筋無力症
特に、全身投与によって治療し得る疾患は前記セクション2.2に記載されている。特別の具体例において、ショック症候群としても知られている火傷または心筋梗塞−誘導外傷、成人呼吸困難症候群(ARDS)による、組織破壊の広がったゾーンに関連する疾患は、本明細書の教示に従いセレクチンリガンドよりなる補体阻害性蛋白質の有効量を非経口投与することによって治療することができる。
かくして、有効量の本発明組成物は、その他の効果に加えて、補体活性を阻害するのに効果的な量である。
好ましい具体例において、セレクチン結合活性を持つ補体阻害性蛋白質の使用は、抗−炎症療法で特に有用であるはずである。セレクチンリガンドは補体阻害性蛋白質を傷害部位にホーミングさせるので、これは好中球のローリングを妨げるであろう。これは、セレクチンリガンドが血管壁のセレクチンに結合し、かくして、白血球、特に好中球の接着を妨げるからである。
例えば、特に好ましい態様において、SLeX部位を付加することによって、sCR1活性を組織傷害部位に局所化させ、かくして、その抗−C活性が増強され、好中球のローリングおよび管外遊出のごとき好中球−内皮細胞相互作用がブロックされる。
加えて、当該組成物は、活性化された内皮細胞上のそのリガンド(セレクチン)にCR1をホーミングするのに用いることができ、sCR1単独またはその現行糖形態の単独投与と比べて、低用量でより効果的とすることができる。また、SLeX−sCR1を炎症部位に執拗に止まらせることが達成され、それにより、さらなる活性化を妨げる。セレクチン/リガンド相互作用に依存する初期の好中球接着事象もブロックされる。最後に、sCR1のin vivo半減期および/または生物学的利用性が延長される。
5.6.診断分野における適用
本発明の組成物を用いて、細胞接着分子からなる放出されまたは循環する複合体に結合できる検出試薬を構成することができる。かかる放出されまたは循環する接着分子は、細胞接着分子を有する特定の細胞の活性化の結果存在するらしい。例えば、L−セレクチンは、細胞表面に構成的に発現され、活性化に続いて迅速に放出されることがよく知られている(Bevilacqua,M.P.,and Nelson,R.N.(1983)前掲)。かくして、セレクチンは、細胞表面からの出現または消失によって制御されているようである。活性化に際して放出される循環受容体は、当業者によく知られた技術によってアッセイすることができる。かかるアッセイの例は、ここに出典明示して本明細書の一部とみなす1987年6月18日に公開された国際出願番号WO87/03600に見い出すことができる。かかる細胞接着分子は、例えば、細胞膜への付着に必要なある種のドメインを欠く受容体を生じる別のスプライシングの産物として、細胞表面に存在する受容体とは物理的に区別できる。また、かかる受容体は天然受容体の断片または一部であるか、あるいはより大きな膜断片と会合していてもよい。さらに、かかる受容体は無傷細胞上に存在し得る。
本発明の組成物は、例えば、受容体を発現しているかどうか疑われる患者からの血清試料または他の試料において、循環中の受容体の存在または不存在を検出するのに有用であり得る。別法として、当該組成物は検出可能に標識し、細胞接着受容体の存在についてのin vitroもしくはin vivo診断イメージングに用いることができる。
再潅流傷害、敗血症性ショック、および(例えば、乾癬および慢性関節リウマチのごとき)他の慢性炎症性疾患のようなある種の炎症性疾患において、炎症した内皮細胞は細胞の傷害部位への動員に寄与する。従って、本発明の組成物および方法は、活性化細胞に結合し、天然リガンドの結合を置き換えるかまたは妨げる能力により、かかる炎症性疾患の存在または不存在を検出するのに有用である。この具体例において、本発明の組成物は、当該分野でよく知られた技術によって検出可能に標識される。
さらなる具体例において、本発明の組成物は、固体支持体に固定化し、起こる結合の量を測定または計算することによって、ある種の細胞接着分子の存在または不存在が検出される。この具体例において、当該分野でよく知られたある種のモノクローナル抗体を組成物と共に用いることもできる。
また、当該組成物は、本明細書に記載した炎症のメディエーターとのその直接相互作用により、炎症および補体媒介の病気もしくは疾患を研究するのに使用できる。特に、当該組成物をin vitroまたはin vivo方法いずれかにて用いることができる。in vitro方法において、試料は液体検体または組織検体でよく、免疫ペルオキシダーゼアッセイまたは組織試料の染色のごとき酵素連結アッセイを包含し得る。
本発明の組成物はキット、特に診断キットの一部として使用できる。かかるキットは、例えば、本発明の組成物、ならびに検出可能に標識できる成分、例えば、特定の細胞接着分子に対するモノクローナル抗体を含むことができる。1の具体例において、該キットは、適当な希釈用およびインキュベーション用緩衝液と共に、1以上の組成物、サンドイッチアッセイのフォーマットで用いるのに適した検出可能に標識された結合パートナー、および基質薬剤を含む。
6.実施例1
6.1.補体受容体1の可溶性欠失突然変異体の産生
以下の実験は、本発明で有用な補体受容体1型のいくつかの可溶性欠失突然変異体の産生を詳細に記載する。
6.2.LHR−Aを欠くCR1の可溶性欠失突然変異体(SCR1 [des−A])の産生
プラスミドpBSABCDは、1989年10月5日に公開された、Fearon,D.T.,et al.による国際公開番号WO89/09220「The Human C3b/C4b Receptor(CR1)」に記載されている(また、Klickstein,L.B.,et al.,(1989)J.Exp.Med.168:1699−1717参照)。このプラスミドは、pBluescript KS+(Stratagene,La Jolla,CA)のEcoR I−Sma I部位に6.86−キロベース(kb)EcoR I−EcoR V片として挿入されたヒトCR1についての全長cDNAを保有し;かくして、EcoR VおよびSma I部位は再生されなかった。細胞外および膜貫通領域の結合点に翻訳停コドンを導入することによってpBSABCDをさらに修飾して、膜貫通および細胞質ドメインを欠く可溶性CR1蛋白質を発現できるpBL−sCR1を得た。(1989年10月5日に公開されたFearon,D.T.,et al.による国際出願公開番号WO89/09220「The Human C3b/C4b Receptor(CR1)」;Weisman,H.F.,et al.,(1990)Science 249:146−151)。
pBL−sCR1をCla IおよびBal Iで消化し、得られた断片(3.96および5.9kb)を低融点アガロースゲルから精製した。プラスミドpBR322をCla IおよびBal Iで消化し、2.9−kb断片をアガロースゲルから精製し、pBL−sCR1からの5.9−kb断片に連結した。連結ミックスを受容可能なイー・コリ(E.coli)DH5α細胞(GIBCO BRL)に形質転換し、得られたプラスミドpBR8.8を精製し、Xba Iで消化して、7.45および1.35kbの2つの断片を得た。7.45断片を精製し、環状形態に再連結した。得られたプラスミドpBR7.45をCla IおよびBal Iで消化し、CR1 cDNAを含有する4.5−kb断片をpBL−sCR1からの3.96−kb断片に連結して、LHR−Bを欠くpBL−sACDを得た。
pBL−sACD(Makrides et al.,(1992)J.Biol.Chem.267:24754−24761)とも呼ばれるpBL−CR1c2をNar IおよびNsi Iで消化して、リーダーの3'末端から76bp、全LHR−A、およびLHRCの5'末端からの57bpを除去し;7.07kb断片をアガロースゲルから精製し、以下の配列を有する長さが68および66bpの2つの合成二本鎖オリゴヌクレオチド(Operon Technologies,Alameda,CA)に連結した。
pBL−CR1c8をNsi Iで線状化し、製造業者の指示に従い細菌アルカリホスファターゼ(Gibco BRL)を用いて脱リン酸化した。pBL−sCR1とも呼ばれるpBL−CR1c[Weisman et al.,(1990)Science 249:146−151]をNsi Iで消化し、LHR−BのほとんどおよびLHR−Cからの最初の56ヌクレオチドを含有する1.35kb断片をアガロースゲルから精製し、線状化したpBL−CR1c8に連結した。これにより、LHR B、CおよびDを含有するpBL−CR1c6Aが組み立てられた。BCD挿入断片の正しい向きは、制限消化分析によって決定した。
該挿入断片をXho Iで消化することによって切り出し、アガロースゲルから精製した。発現プラスミドpTCSgpt(1989年10月5日に公開されたFearon,D.T.et al.による国際出願公開番号WO89/09220「The Human C3b/C4b Receptor(CR1)」;Carson et al.,(1991)J.Biol.Chem.266:7883−7887)をXho Iで消化し、細菌アルカリホスファターゼで脱リン酸化し、BCD断片に連結した。連結ミックスをイー・コリ(E.coli)DH1に形質転換し、プラスミドpT−CR1c6Aを得た。正しい挿入断片向きをBgl I制限消化によって決定し、pT−CR1c6Aを大規模に調製した。pT−CR1c6Aは、LHR−Aならびに膜貫通および細胞質ドメインを欠くCR1の可溶性欠失突然変異体についての暗号配列を保有するプラスミドである。得られた可溶性欠失突然変異体を、LHR B、CおよびDを含有するsCR1[des−A]と命名した。
7.実施例2
7.1 SCR15〜18を含有するCR1の可溶性欠失突然変異体 の構築
CR1リーダーおよびショートコンセンサスリピート(SCR)15ないし18からなるDNA断片を、鋳型としてpBL−CR1c8を用いてPCR−合成した[Makrides et al.(1992)J.Biol.Chem.267,24754−24761]。5'「センス」プライマーは、CR1リーダーの上流のpBluescriptポリリンカー領域にハイブリダイズし、下線を施したXho I制限部位を含有していた。
CR1リーダーおよびSCR15−18からなるDNA断片をXho Iでの消化によってpB−CR1(15−18A)から切り出し、Genecleanキット(BIO 101,La Jolla,CA)を用いてアガロースゲルから精製し、予めXho Iで制限酵素処理し、ウシ腸アルカリホスファターゼ(Boehringer Mannheim,Indianapolis,IN)で脱リン酸化した発現ベクターpTCSgpt[Carson et al.,(1991)J.Biol.Chem.266,7883−7887]に連結した。連結ミックスのイー・コリ(E.coli)DH1受容可能な細胞への形質転換により、プラスミドpT−CR1c12(8.52kb)を得た。
7.2 安定な細胞系のトランスフェクションおよび選択
pT−CR1c12をFsp I消化によって線状化し、フェノール抽出し、エタノール沈殿させ、滅菌水に再懸濁した。960μFおよび230VにてのGene Pulser(Bio−Rad)でのエレクトロポレーションを用い、組換えプラスミド30μgを、3μgのpTCSdhfr[Carson et al.,(1991)J.Biol.Chem.266,7883−7887]と共に、ジヒドロ葉酸レダクターゼを欠くCHO DUKX−B11細胞[Urlaub and Chasin(1980)Proc.Natl.Acad.Sci.U.S.A.77,4216−4220]に同時トランスフェクトした。トランスフェクトした細胞を、10%加熱不活化子ウシ血清、1%ペニシリン−ストレプトマイシン、50μg/mlゲンタマイシン、4mMグルタミン(Gibco BRL)、各10μg/mlのチミジン、アデノシンおよびデオキシグアノシン(Sigma,St.Louis.MO)を加えた非選択的α−最小培地(α−MEM)に移した。2日後、細胞を、10%透析子ウシ血清、1%ペニシリン−ストレプトマイシン、50μg/mlゲンタマイシン、4mMグルタミン、20mM HEPES pH7.0、6μg/mlミコフェノール酸、250μg/mlキサンチン、および15μg/mlヒポキサンチン(Sigma)を加えたα−MEM中で選択した。SCR15〜18を分泌するクローンを酵素免疫法(CellfreeR CD35;T Cell Sciences,Inc.)によって同定し、蛋白質の補体阻害性活性を溶血アッセイを用いて確認した[Yeh et al.,(1991)J.Immunol.146,250−256]。高発現クローンをメトトレキセート(Lederle,Pearl River,NY)を含有する増殖培地中で選択した。
8.実施例3
8.1 LHE Dを欠くCR1の可溶性欠失突然変異体(sCR1 [des−D])の構築
プラスミドpBSABCDはDouglas Fearon博士「Klickstein et al.,(1988)J.Exp.Med.168,1699−1717]から得た。このプラスミドは、pBluescript KS+(Stratagene,La Jolla,CA)のEcoR I−Sma I部位に6.86キロベース(kb)EcoR I−EcoR V部位として挿入されたヒトCR1についての全長cDNAを保有しており;かくして、EcoR VおよびSma I部位は再生されなかった。pBSABCDを記載されているごとくさらに修飾して[Weisman et al.,(1990)Science 249,146−151]、膜貫通および細胞質ドメインを欠く可溶性CR1蛋白質を発現できるpBL−sCR1を得た。
ユニークなNru I制限部位を、4200塩基対(bp)の位置、すなわち、LHR−Cおよび−Dの結合点において、pBL−sCR1に導入した。酵素部位は、Muta−gene Phagemidキット(Bio−Rad Laboratories,Melville,NY)を用い、部位特異的突然変異誘発[Kunkel(1985)Proc.Natl.Acad.Sci.U.S.A.82,488−492]によって作製した。40−塩基のリン酸化突然変異オリゴヌクレオチド(New England Biolabs,Beverly,MA)は、以下の配列を有していた。
突然変異誘発したプラスミドpBL−sCR1N(9.8kb)をNru IおよびBgl IIで消化し、7.8kb断片をアガロースから単離し、以下の相補鎖からなる二本鎖合成オリゴヌクレオチドに連結した。
Gaithersburg,MD)で脱リン酸化した発現ベクターpTCSgpt[Carson et al.,(1991)J.Biol.Chem.266,7883−7887]に連結した。連結ミックスのイー・コリ(E.coli)DH1受容可能な細胞への形質転換により、プラスミドpT−CR1c7が得られた。
安定な細胞系のトランスフェクションおよび選択
pT−CR1c7をFsp I消化によって線状化し、フェノール抽出し、エタノール沈殿させ、滅菌水に再懸濁した。960μFおよび230VにてのGene Pulser(Bio−Rad)でのエレクトロポレーションを用い、組換えプラスミド30μgを、3μgのpTCSdhfr[Carson et al.,(1991)J.Biol.Chem.266,7883−7887]と共に、ジヒドロ葉酸レダクターゼを欠くCHO DUKX−B11細胞[Urlaub and Chasin(1980)Proc.Natl.Acad.Sci.U.S.A.77,4216−4220]に同時トランスフェクトした。トランスフェクトした細胞を、10%加熱不活化子ウシ血清、1%ペニシリン−ストレプトマイシン、50μg/mlゲンタマイシン、4mMグルタミン(Gibco BRL)、各10μg/mlのチミジン、アデノシンおよびデオキシグアノシン(Sigma,St.Louis.MO)を加えた非選択的α−最小培地(α−MEM)に移した。2日後、細胞を、10%透析子ウシ血清、1%ペニシリン−ストレプトマイシン、50μg/mlゲンタマイシン、4mMグルタミン、20mM HEPES pH7.0、6μg/mlミコフェノール酸、250μg/mlキサンチン、および15μg/mlヒポキサンチン(Sigma)を加えたα−MEM中で選択した。sCR1[desD]LHR A、BおよびCを分泌するクローンを酵素免疫法(CellfreeR CD35;T Cell Sciences,Inc.)によって同定し、蛋白質の補体阻害性活性を溶血アッセイを用いて確認した[Yeh et al.,(1991)J.Immunol.146,250−256]。高発現クローンをメトトレキセート(Lederle,Pearl River,NY)を含有する増殖培地中で選択した。
9.実施例4
9.1 SLE X 炭水化物部位を含有するCR1の可溶性構築体の 作製
本明細書中で定義したCR1または他の補体部位の前記可溶性欠失突然変異体のいずれかを操作して、本発明の範囲内にある有用な炭水化物部位を含有させることができる。以下の例は、LHR−Aを欠きsLeX炭水化物部位を含有するCR1の可溶性欠失突然変異体sCR1[des−A]sLeXの作製を記載する。
9.1.1.sCR1[des−A]構築体のLEC−11細胞へのトラン スフェクション
Fsp I制御消化による線状化によって、pT−CR1c6A(実施例1、セクション6.2、前掲)を、メトトレキセートに対して異常に低いアフィニティーを示す改変マウス・ジヒドロ葉酸レダクターゼcDNAを含有するFsp I−線状化pTCSdhfr*(Simonsen,C.C.and Levinson,A.D.(1983)Proc.Natl.Acad.Sci.U.S.A.80:2495−2499)と共にLEC11細胞(Campbell,C.,and Stanley,P.,1984,J.Biol.Chem.259:11208−11214)に同時トランスフェクトした。sCR1[desA]sLeXを分泌するクローンを、酵素免疫法(Cellfree CD35;T Cell Diagnostics,Inc.)によって同定し、溶血アッセイ(Yeh et al.,(1991)J.Immunol.146:250−256)を用いて、蛋白質の補体阻害性活性を確認した。高発現クローンをメトトレキセート(Lederle,Pearl River,NY)を含有する増殖培地で選択した。
9.1.2.sCR1[des−A]sLe X の細胞培養産物
sCR1[desLHR−A]を分泌するCHO DUKX−B11細胞またはsCR1[desLHR−A]sLeXを分泌するCHO LEC−11細胞を、2.5%加熱不活化子ウシ血清(Hyclone,Logan,UT)を含み、高グルコース/Ham栄養素混合物F12を含むがヒポキサンチンおよびチミジン(JRH Biosciences,Lenexa,KS)を含まない1:1ダルベッコの修飾イーグル培地にて、T−225フラスコ中で増殖させた。炭酸水素ナトリウムを用いて培地のpHを7.8に調整して、順化培地に存在するシアリダーゼ活性を最小化した。これらの培養からの順化培地をデカントすることによって1週間につき3回収穫し、濾過し、精製まで−70℃で凍結した。生産はELISA法によってモニターした。
9.1.3.sCR1[desA]sLe X の精製
sCR1[desA]sLeXまたはsCR1[des−A]を含有する濾過した細胞培養上清を緩衝液交換し、交差流動限外濾過(30,000分子量カット−オフ)によって濃縮し、再度濾過し、S−セファロース・ファースト・フローカオチン交換カラムに適用し、高塩濃度(0.5M塩化ナトリウム)で溶出させた。カチオン交換溶出物を硫酸アンモニウムで沈殿させ、遠心によって分離し、PBS中に再懸濁し、濾過した。濾液を0.8M硫酸アンモニウムに調整し、ブチル−トヨパール 650Mカラムに負荷し、0.09M硫酸アンモニウムまでのステップで溶出させた。溶出物を、Centriprep−30濃縮器(Amicon)を用いて濃縮し、トヨパール HW55Fカラムのサイズ排除クロマトグラフィーに付し、Centriprep−30濃縮器を用いて再度濃縮し、滅菌濾過し、−70℃で凍結保存した。精製プロセスは280nmにおける吸光度およびELISA法によってモニターした。蛋白質純度はクーマシーブルーでまたは銀染色によるSDS−PAGEおよび走査型デンシトメトリー(光学密度計測)によって精査した。エンドトキシンのレベルは、Limulus Amebocyte Lysateアッセイ(Associates of Cape Cod,Inc,Woods Hole,MA)を用いて測定した。
10.実施例5
10.1 sCR1[des−A]sLe X およびsCR1[des−A]のin vitro補体調節活性
sCR1[des−A]sLeXのin vitro補体調節活性を、sLeX糖鎖付加を欠く以外は同一の蛋白質であって、in vitroで代替補体活性化を選択的に阻害することが示されているsCR1[des−A]のin vitro補体調節活性と比較した。sCR1[des−A]sLeXを構築し、LEC11細胞で発現させ、前記実施例で記載した細胞培養上清から精製した。pT−CR1c6aをDUKX−B11にFsp I−線状化pTCSdhfrと共に同時トランスフェクトした以外はsCR1[des−A]sLeXについて前記したごとくに、sCR1[des−A]を前記したように構築し、CHO DUKX−B11細胞で発現させた。sCR1[des−A]はsCR1[des−A]sLeXについて前記したごとくに細胞培養上清から精製した。
sCR1[des−A]sLeXおよびsCR1[des−A]は、赤血球CR1に対するダイマーC3bの結合について同等に競合した。sCR1[des−A]sLeXおよびsCR1[des−A]は、C3b α−鎖のI因子媒介分解における助因子として働くその能力において同等であった。。sCR1[des−A]sLeXおよびsCR1[des−A]は、C4−欠損モルモット血清を補体源として用いる代替補体媒介赤血球溶解を阻害するその能力において同等であった。sCR1[des−A]sLeXおよびsCR1[des−A]は、古典的経路活性化を可能とする条件下で補体媒介赤血球溶解の同等な阻害剤であった。しかしながら、共に、CR1の全細胞外配列を含有する可溶性組換え蛋白質sCR1よりも、古典的経路媒介溶血の有意に劣った阻害剤であった。かくして、sCR1[des−A]と同様に、sCR1[des−A]sLeXはin vitroにて代替補体経路の選択的阻害剤である。
10.1.1 補体蛋白質および抗体
ヒトC4、C3、C3bおよび化学的に架橋させたダイマーC3b(C3b2)は前記したごとくに調製した(Makrides et al.,1992,Scesney et al.,前掲)。C4をメチルアミンで処理して、蛋白質C4b−様形態であるC4maを得た(Makrides et al.,1992;Law and Levine,1980)。C3b、C3b2、およびC4maを、製造業者の推奨に従い、Iodo−beads(Pierce Chemical Co.)を用いて125Iで放射性同位体標識した。C4欠損モルモット血清は市販のものを得た(Sigma)。
10.1.2 C3b 2 結合実験
sCR1[des−A]sLeXおよびsCR1[des−A]の125I−C3b2への結合は、ヒト赤血球上の天然CR1との競合によってアッセイした(Weisman et al.,1990;Mekrides et al.,1992)。ヒト赤血球を同等容量のAlsever溶液(113mMデキストロース、27mMクエン酸ナトリウム、2.6mMクエン酸、72mM塩化ナトリウム、pH7)で希釈し、使用するまで4℃で保存した。新たに採血した赤血球およびAlsever溶液中で保存した赤血球へのC3b2結合で差異はなかった。使用の直前に、赤血球をPBS、0.1%BSA、および0.01%アジ化ナトリウムで3回洗浄した。125I−C3b2(0.55nM)を、濃度を変えたsCR1、sCR1[des−A]、sCR1[des−A]sLeXC3b2、またはC3bを用い、氷(0℃)上で、60分間、赤血球(4×109細胞/ml)と共にインキュベートした。結合したおよび遊離した125I−C3b2をジブチルフタレートにより遠心によって分離した。非特異的結合は、0.71mg/mlの精製したウサギ抗−sCR1抗体の存在下で測定した。
結果
C3b 2 結合実験
sCR1[des−A]、sCR1[des−A]sLeXおよびsCR1と125I−C3b2との赤血球CR1への結合の競合をアッセイした。125I−C3b2結合を最大50%阻害するのに必要な競合剤の濃度から、sCR1、C3b2およびC3bについての見掛けの解離定数(Kd,app)は、各々、2×10-9M、3×10-8M、および6×10-7Mであると見積もられ、この値は以前の研究(Weisman et al.,1990;Wong and Farrel,1991;Makrides et al.,1992)で得られた結果と同様であった。sCR1[des−A]sLeXまたはsCR1[des−A]は赤血球CR1に対する125I−C3b2結合について同等に競合した。
10.1.3 I因子による液相C3bまたはC4maの蛋白質分解 についての助因子活性
C3bまたはC4ma α−鎖の特異的蛋白質分解を促進するsCR1[desA]sLeXまたはsCR1[des−A]の能力をSDS−PAGEでアッセイした(Wong et al.,1985;Weisman et al.,1990;Scesney et al.,前掲)。125I−C3b(6.8×10-9M)または125I−C4ma(5.6×10-8M)を、I因子(0.25μM)および変化させた濃度のsCR1[des−A]sLeXもしくはsCR1[des−A]と共に、PBS中で、37℃で20分間、続いて氷上(0℃)で5分間インキュベートした。これらの条件下、C4ma C3b α−鎖の蛋白質分解は助因子の濃度に依存していた。残りの無傷C3b α−鎖を還元SDS−PAGEで分離し、バンドを切り出し、ガンマカウンターで測定した。
結果
C3b α−鎖のおよびC4ma α−鎖のI因子蛋白質分解 についての助因子活性
I因子による125I−C3bまたは125I−C4maの特異的蛋白質分解は、α−鎖切断の程度が助因子sCR1[des−A]sLeXまたはsCR1[des−A]の濃度に依存した濃度下で、SDS−PAGEでモニターした。無傷C3b α−鎖を表すバンドの喪失は、同様の濃度のsCR1[des−A]sLeXまたはsCR1[des−A]を必要とした。無傷C4ma α−鎖を表すバンドの喪失は、同様の濃度のsCR1[des−A]sLeXまたはsCR1[des−A]を必要とした。sCR1[des−A]sLeXおよびsCR1[des−A]は、C3b α−鎖のI因子媒介分解における助因子として働くその能力において同等であった。
10.1.4.古典的および代替補体活性化の阻害についての 溶血アッセイ
補体活性化の阻害は前記したごとくにアッセイした(Weisman et al.,1990;Yeh et al.,1991)。100mM HEPES、150mM塩化ナトリウム、0.1%BSA、pH7.4中に補体源としてヒト血清を用い、ウサギ抗−ヒツジ赤血球抗体(Diamedix,Miami,FL)で感作したヒツジ赤血球細胞を溶解させた。感作ヒツジ赤血球(107細胞/ml)、正常ヒト血清(400希釈中の1)、および変化させた濃度のsCR1[des−A]またはsCR1[des−A]sLeXをV底マイクロタイタープレート中、37℃にて60分間インキュベートし、細胞を遠心によってペレット化し、上清を平底マイクロタイタープレートに移し、405nmにおける吸光度を測定して、放出されたヘモグロビンを定量した。試料は、ヒト血清を欠く同一の対照(補体−非依存性溶解)と対にした。試料および対照は共に三連にて泳動させた。対照値を試料値から差し引き、阻害分率は、非阻害(sCR1[des−A]sLeXsまたはCR1[des−A]を添加せず)試料に対して求めた。
代替経路溶血の阻害は、Platts−Mills and Ishizaka(1974)の修飾方法を用いてアッセイした。EGTAおよびMg2+を各々8mMおよび5mM添加した、100mM HEPES、0.15N塩化ナトリウム、0.1%ウシ血清アルブミン、pH7.4中の補体としてC4欠損モルモット血清を用いて、ウサギ赤血球を溶解させた。ウサギ赤血球(1.2×107細胞/ml)、C4欠損モルモット血清(8希釈中の1)、およびsCR1[des−A]sLeXまたはsCR1[des−A]をV底マイクロタイタープレート中、37℃で60分間インキュベートし、放出されたヘモグロビンを前記したごとくに測定した。
結果
C4−欠損モルモット血清を用いる代替補体経路による溶 血の阻害
予め存在するまたは新しく生成したC4bからの干渉を除外するために、C4−欠損モルモット血清を補体源として用いて、ウキギ赤血球の代替経路溶解を調べた。同等濃度のsCR1、sCR1[des−A]、またはsCR1[desA]sLeXが代替補体−媒介赤血球溶解を阻害するのに必要であった。
古典的補体経路によって開始された溶血の阻害
抗体−感作ヒツジ赤血球の補体溶解の阻害は、ほぼ同等濃度のsCR1[des−A]sLeXまたはsCR1[desLHR−A]を必要とした。しかしながら、これらの濃度は、LHR A、B、CおよびDを含有するsCR1による阻害に必要なもののほぼ50倍高かった。
11.実施例6
11.1 sCR1[des−A]sLe X の分析
本実施例においては、精製した蛋白質sCR1[des−A]およびsCR1[desA]sLeXをウェスタンブロットで比較した。
11.1.1 抗体
sCR1は前記したごとくに調製した(Weisman et al.,1990;Yeh et al.,1991)。ポリクローナルウキザ抗−sCR1抗体を調製し、前記したごとくに精製した(Makrides et al.,1992)。CSLEX−1(抗−シアリル ルイスX)はBecton Dickinsonから得た。FH6(抗−シアリル ジ−LeX)はS.Hakomori博士(Biomembrane Institute,Seattle,WA)から得た。DREG−56(抗−E−セレクチン)はEndogen,Cambridge,MAから得たものであり;抗−CD15(抗−LeX)はAMAC(Westbrook,ME)から得た。
11.1.2 sCR1[desA]sLe X のウェスタンブロット分析
ウェスタンブロット分析は以下の方法に従って行った:
a)前記したLEC−11細胞のsCR1[des−A]構築体でのトランスフェクションから得られた糖蛋白質を、適当な対照および標準と共に、還元性および非還元性条件下で、SDSポリアクリルアミドゲル電気泳動に付した。
b)半乾燥電気泳動移行(Integrated Separation Systems)を介して、該糖蛋白質のバンドを固体支持膜(ImmobilonR)に移した。
c)移行した糖蛋白質を含有する膜を、1%脱脂粉乳蛋白質で2時間、(あるいは一晩)またはウシ血清アルブミン(約2.0%にて)およびゼラチン(約0.3%にて)でブロックした。後者のブロッキング剤が、(IgAのごとき)乳溶液中に存在する可能なSleX糖鎖付加蛋白質による複雑化を回避するのに好ましい。
d)次いで、複製膜を、CR1および/またはsCR1[des−A]に対する抗体、LeX(DAKO、AMAC、CD15)に対する抗体、SLeX(CSLEX−1、ATCCから入手可能、Becton Dickinsonから入手可能)に対する抗体、およびイソタイプにマッチした対照抗体と、2時間反応させた、
e)膜を、各々、緩衝液(PBS−ツィーン)中で4回約10〜15分間洗浄した。
f)次いで、膜を、HRP−標識抗−マウス抗体または抗−ウサギ抗体(すべて、Bio−Rad,Southern Biotech,Tagoのごとき種々の販売業者から入手可能)と2時間反応させた。
g)膜を(e)におけるごとくに洗浄した。
h)膜をHRP基質(Bio−Rad,Sigma)で展開して、各一次抗体と反応性の糖蛋白質バンドを可視化した(または、「ECL」という化学ルミネッセンス法(Enhanced Chemiluminescence,Amersham))によった。
結果
11.2 ウェスタンブロット分析
AmershamからのECLウェスタンブロット手法ならびにSLeXに対する抗体(CSLEX1)およびCR1に対する抗体(ウサギ・ポリクローナル抗体)を用いる前記技術は、前記した方法により得られたsCR1[des−A]と呼ばれる補体部位でトランスフェクトしたLEC−11細胞に由来する物質を用いてsCR1[desA]sLeXを得ることによって行った。このウェスタンブロット分析の結果では、LEC−11細胞(Pamela Stanley,Albert Einstein College of Medicine)に由来するsCR1[desA]sLeXはCSLEX1抗体(ATCC HB 8580、米国特許第4,752,569号参照)で染色することによって測定して、sLeX部位を有するが、sCR1[des−A]構築体のDULX.B11 CHOへのトランスフェクションに由来する物質はそうではなかった。
図1Bはこの分析の結果を示し、その中に物質を含む第1レーンは分子量標準を含有する。第2のレーンは、HL−60細胞に由来する溶解物(CSLEX1 mAbについての陽性対照)を含有する。第3のレーンは、DUKX.B11 CHO細胞に由来するcSR1[des−A]物質を含有し、第4のレーンは、LEC−11細胞に由来するsCR1[des−A]sLeX物質を含有する。sCR1[des−A]の2つの糖鎖付加形態に合致する2つの明瞭なバンドによって示されるごとく、sCR1[des−A]物質を含有する2つのレーンのうち、LEC−11細胞に由来するそのレーン(レーン4)のみがCSLEX1 mAbによって同定された。sCR1[des−A]を含有する両レーン(DUX.B11に由来するレーン3、LEC−11に由来するレーン4)は、予測されるごとくsCR1[des−A]に対するポリクローナル抗体と反応した(図1C)。図1Aは同一物質においてクーマシーブルー染色したSDS−PAGEゲルである。
11.3 第2のウェスタンブロット分析
別の実験において、sCR1[desLHR−A]およびsCR1[desA]sLeXを、4〜20%ゲル(ISS)および非還元条件を用いるSDS−PAGEに付した。半乾燥トランスブロッティング装置(ISS)を用いて該ゲルを膜(Immobilon−P)にブロットした。該膜を、2%BSA、1%正常ヤギ血清、0.05%アジ化ナトリウムを含有するトリス緩衝生理食塩水(TBS)溶液中で、室温にて一晩ブロックした。該ブロットを、室温にて2時間、FH6(抗−シアリル ジ−LeX,Hakomori上清、TBSブロッキング緩衝液中1:1希釈)でプローブした。0.05%ツィーン20を含むPBS中で十分洗浄した後、室温にて1時間、ホースラディッシュペルオキシダーゼ(HRP)結合ヤギ抗−マウスIgM(Tago,ブロッキング緩衝液中1μg/ml)で、該ブロットをプローブした。0.05%ツィーン20を含むPBS中で十分洗浄した後、該ブロットを化学ルミネッセンス基質(ECLキット、Amersham)と共に1分間インキュベートし、X−線フィルムに30〜120秒間暴露し、該フィルムを現像した。次いで、該ブロットを細片化し、ウサギ・ポリクローナル抗−sCR1(ブロッキング緩衝液中1:2000)で1時間再度プローブし、十分に洗浄し、HRP結合抗−ウサギIg(Amersham)でプローブし、洗浄し、前記したごとくに検出した。
第2ウェスタンブロット分析の結果
第2ウェスタンブロット分析において、CSLEX−1(sLeXオリゴ糖と反応するモノクローナル抗体)およびFH60(シアリル ジ−LeXと反応するモノクローナル抗体)は共にsCR1[desA]sLeXに結合したが、sCR1[desA]には結合しなかった。両オリゴ糖構造はセレクチンについてのリガンドであることが示されている(Goelz,S.et al.,J.Biol.Chem.(1994)269:1033−1040)。Parekh et al.,(1992)J.Biochem.(Tokyo)16d,137は、E−セレクチンアフィニティーカラムへの結合を担う炭水化物構造は、シアリル ジ−LeXであることを同定した。用いられたN−結合糖鎖付加部位の数、またはそれらのうちいくつがsLeX中で終わるかについての知見は、本実験では得ることができない。
図2Aないし2Cは、第2のウェスタンブロット実験の結果を詳細に示す。図2Aないし2Cは、同一ポリアクリルアミドゲルの分析である。各図のレーン1には分子量標準が入っている。各図のレーン2は、DUKX−B11細胞から得られた精製sCR1[des−A]物質である。各レーン3は、無関係な対照物質である。各ゲルのレーン4、5および6は、精製方法の間の種々の段階におけるsCR1[des−A]sLeXである。
図2Aは、クーマシーブル染色したポリアクリルアミドゲルのパターンである。レーン2、および4〜6におけるほぼ187kdの支配的バンドはsCR1[des−A]蛋白質であり、レーン2はDUKX−B11細胞から得られ、レーン4〜6はLEC−11細胞から得られた。図2Bは、ウェスタンブロットし、抗−sCR1[des−A]ポリクローナル血清でプローブした図2と同一のゲルである。予測されるごとく、DUKX−B11細胞またはLEC−11細胞に由来するかを問わず、sCR1[des−A]を含有するすべてのレーンはsCR1[des−A]について陽性である。図2Cは、細片化し、短縮表示NeuNacα2−3Galβ1−4(Fucα1−3)GlcNacβ1−3Galβ1−4(Fucα1−3)GlcNAcによって表されるシアリル ジ−ルイスX抗原について特異的な抗体で再プローブした図2Bと同一のブロットである。予測されるごとく、LEC−11細胞から得られたsCR1[des−A]sLeXを含有するレーン4−6のみが、適当な炭水化物構造について陽性である。
12.実施例7
12.1 sCR1[des−A]の単糖組成はsLeX糖鎖付加と合 致する。
気相−液相クロマトグラフィー(GLC)、続いてのReinhold,V.(1972),Methods in Enzymology,25:244−249の手法を用い、sCR1[des−A]sLeXの炭水化物部位よりなるグルカンの単糖組成を分析した。切断、誘導体化およびGLCの条件は、糖蛋白質sCR1[des−A]およびsCR1[des−A]sLeXよりなる単糖の定量的測定結果を供した。
sCR1およびsCR1[des−A]、CHO DUKX−B11の発現に用いたCHO系は、α(1,3)フコシルトランスフェラーゼ活性を欠き、かくして、sLeX糖鎖付加できない。Stanleyおよびその同僚は、突然変異体CHO細胞系LEC11を作製しており、それは内因性アルファ(1−3)フコシルトランスフェラーゼを転写し、sLeXを含めたフコシル化された末端構造を持つ炭水化物を合成できる。
本実施例において、sLeX糖鎖付加できるLEC11細胞を、sCR1[desLHR−A]をコードするプラスミドでトランスフェクトして、sCR1[des−A]sLeXを得た。該sCR1[des−A]sLeXをDUKX−B11細胞で産生されたsCR1[des−A]と比較した。2つの糖蛋白質を単糖組成について分析すると、以下の表Iに示す結果が得られた。
表Iより、sCR1[des−A]sLeXのフコースおよびシアル酸含量は、LEC11細胞系がシアリル化ルイスX抗原ならびにシアリル化ジ−ルイスX抗原に必要な適当な炭水化物を付加するものであるという予測と合致すると結論される。
sLeXはP−セレクチンおよびE−セレクチン双方についての炭水化物リガンドであり、それにより、炎症の血管部位において白血球接着を媒介する。かくして、sCR1[desA]sLeXは、補体制御蛋白質および接着分子双方の抗−炎症能力を組み合わせる。
13.実施例8
13.1 sCR1[des−A]sLeXからのオリゴ糖の質量分析 はsLeX糖鎖付加と合致する。
質量分析により、sLeX糖鎖付加と合致するフコースおよびシアル酸含有炭水化物の存在が確認された。電子顕微鏡(Fenn et al.,(1989)Science,246:64−71)、続いての質量分析により、N−アセチルノイラミン酸基(シアル酸)の評価、フコシル化、アンテナ延長および分岐の考察を行った。本実施例では、sCR1[des−A]sLeXをエンド−H脱糖し、ES−MS「指紋」が得られ、エンド−H脱糖したsCR1[des−A]糖蛋白質から得られた同様の「指紋」と比較した。
炭水化物を、各々が典型的なトリマンノースコア構造を有するビ−、トリ−およびテトラアンテナ構造に分類した。
得られたイオンプロフィールにおいて、各イオンは、各単糖について編集された組成−質量表を参照して計算された。例えば、sCR1[des−A]sLeX「指紋」で見い出されたイオンm/z1062.8は、好ましいsLeX炭水化物部位と合致するビアンテナ構造を表し、以下の構造のフコシル化シアリル化ルイスX抗原として説明できる。
表IIに示すデータは、sCR1[des−A]およびsCR1[des−A]sLeX組成に存在する特定の各炭水化物構造の各々のモルパーセント比を表す。
14.実施例9
14.1 sCR1[des−A]sLeXのin vitro機能的活性
本実施例では、精製した蛋白質sCR1[des−A]およびsCR1[des−A]sLeXの機能的活性をin vitroで比較する。sCR1[des−A]sLeXは、in vitroにて濃度依存的に、U937細胞の活性化ヒト動脈内皮細胞へのE−セレクチン媒介結合を阻害した。この静的接着アッセイにおいては、sCR1[desA]sLeXは、最終濃度250μg/mlにおいて、U937細胞の結合を50%阻害した。
14.1.1.静的接着ブロッキングアッセイ
96−ウェルマイクロタイタープレート中で密集させた動脈内皮細胞(Clonetics)を37℃で4時間、TNF(100U/ml)で刺激した。次いで、細胞を1%FBSを加えたDMEMで2回洗浄した。sCR1[des−A]sLeXおよびsCR1[des−A]の系統希釈を行って、最終濃度500、250、125、62.5、および0μg/mlとした。各ウェルに、20μlDMEM中の(ATCCから得られた)5×105U937細胞を添加し、37℃で20分間インキュベートした。ウェルを培地で満たし、シールし、低速(150×g)にて5分間逆遠心した。シールを除き、プレートをブロットし、3つの顕微鏡視野における結合細胞の数を測定した。
14.2 結果
このin vitro静的接着アッセイを用いると、sCR1[desA]sLeXは濃度依存的にE−セレクチン媒介接着を阻害した。ヒト動脈内皮細胞をTNFで刺激して、E−セレクチンの細胞表面発現を誘導した。E−セレクチンの表面発現は、免疫細胞化学染色プロトコルにてDREG−56(E−セレクチンについて特異的なモノクローナル抗体)を用いて測定した。CSLEX−1でのフローサイトメトリー分析よって表面sLeXを有することが示されたU937細胞は、活性化内皮細胞に接着することが示された。活性化動脈内皮細胞とU937細胞との間の接着現象は、セレクチン−媒介接着のホールマークであるカルシウムの存在を要することが示された。活性化内皮細胞へのU937細胞のE−セレクチン依存性接着は、濃度依存的に、sCR1[desA]sLeXによって阻害された。
図3は、この実験の結果を詳細に示す。黒い棒線はDUKX−B11細胞から得られたsCR1[des−A]物質を表す。水平線を付した棒線は、LEC−11細胞から得られたsCR1[des−A]sLeX物質を表す。該sCR1[des−A]sLeX物質は、濃度依存的に、活性化動脈内皮細胞へのU937細胞の結合を阻害した。
15.実施例10
15.1 sCR1[des−A]sLe X のin vivo機能的活性
それに対してシアリルルイスXおよびシアリル ジ−Lexが結合するセレクチンの内皮細胞上昇調節(upregulation)は、好中球についての重要な接着促進分子である。優れた補体の阻害剤である可溶性補体受容体1(sCR1)は、SLeXで修飾したあるいは修飾なしのトランケートされた形態で(各々、sCR1[desA]sLeXおよびsCR1[desA])発現された。両化合物は前記にて示したように、in vitroで実質的な補体−ブロッキング活性を有する。P−セレクチン依存性急性肺傷害のラットモデルにおいて、これらの阻害剤についての保護活性のランク順は、sCR1[desA]sLeX>sCR1≧sCR1[desA]である。セレクチン発現の部位において活性化内皮細胞への結合を引き起こすのにsCR1[desA]のオリゴ糖修飾を利用することによって、補体阻害剤を炎症部位に対して「標的化」することができる。
阻害剤製剤をラット肺傷害のCVFモデルで、in vivoにて使用した。ラットの4つの別々の群(各々n=5)を、0.4ml滅菌生理食塩水、sCR1、sCR1[desA]またはsCR1[desA]sLeX(各々、15mg/kg体重にて)で予め静脈内投与により処理し、CVFの静脈内注入の5分前に静脈内注射した。また、(CVFの不存在下で滅菌生理食塩水を注入した)陰性対照群を使用した(n=5)。CVFまたは滅菌生理食塩水の注入30分後に、ケタミンの過剰投与によって動物を殺し、下大静脈(a)から1.0ml血液を得た。
図4のデータによって示されるごとく、sCR1[desA]、sCR1[desA]sLeXまたはsCR1で動物を処理すると、各々、(パーセントとして)40±3、64±3および55±4だけ肺中MPO含量を減少させた(図4C)。図4Cは、ミエロペルオキシダーゼ活性(MPO)の測定によって見積もった肺における好中球の蓄積の測定結果である。静的に比較すると、sCR1[desA]sLeXおよびsCR1はMPO含量を減少させるにおいてsCR1[desA]よりも効果的であった。
また、図4Bは、CFVによって誘導された出血性肺傷害からのCR1、sCR1[desA]およびsCR1[desA]sLeXの保護効果を記載する。図4Bは、放射性同位体標識赤血球の血管から肺への漏れによって測定した、出血の対照よりも優れた減少の測定値である。sCR1[desA]sLeXは、対照よりも大きくほぼ65パーセント出血を阻止した。浸透性は、肺の血管からの放射性同位体標識蛋白質の漏れの測定である。sCR1[des−A]は本実験において対照よりも大きくほぼ60%浸透性を低下させた。かくして、sCR1[des−A]sLeXは、この炎症モデルで傷害を減少させるにおいて、3種の補体阻害剤のうちで最も効果的なようである。
犠牲にした時点(CVFの静脈内注入後30分)において、血漿を得て、ELISAサンドイッチ技術を用いて、sCR1[desA]抗原の濃度を評価した。その他の場合は未処理であるCVF注入動物からの血漿は、<50ng/mlの測定可能なsCR1[desA]抗原を明らかにした。一方、sCR1[desA]およびsCR1[desA]sLeX処理動物(CVF注射)におけるsCR1[desA]の血漿中抗原レベルは、各々、267±28.2および154±33.9μg/mlであった。これらのデータは、血管区画からのsCR1[desA]sLeXの加速されたセレクチン−依存性除去と合致するであろう。
これらのデータは、補体のCVF−誘導全身活性化後に起こる急性肺傷害のP−セレクチン−依存性モデルにおいて、補体阻害剤、sLeX基で修飾したsCR1[desA]が、好中球−依存性傷害のこのモデルにおいて、(sCR1[desA]またはsCR1と比較して)最も効果的な保護を提供することを示す。肺におけるMPO含量の減少は、sCR1[desA]sLeXが、P−セレクチンにつき上昇調節されることが知られている活性化内皮細胞への好中球のP−セレクチン−依存性接着をより効果的ブロックしたことを示唆する。3種の補体阻害剤の各々は、肺MPOの減少した生成に関連した保護効果を有していた。
内皮細胞活性化の量を減少させ(P−セレクチンの上昇調節)、(その結果、毒性酸素生成物を生じる)好中球活性化を減少させることによって、補体のブロックは、好中球と内皮細胞との間の傷害−促進性相互作用に干渉する。肺傷害のこのモデルにおいて、好中球および毒性酸素生成物は共に傷害の進展に必要であることが知られている。好中球と内皮細胞との間の緊密な密接性は、内皮細胞での(好中球からの)毒性酸素生成物が最も効果的に作用するために必要である。これらの接着相互作用は、P−セレクチンに対する抗体または白血球β2インテグリンで、あるいはsLeXの注入によって阻止できる。すべての場合において、これらの介入による保護効果は、組織MPOレベルの減少に関係する。sCR1[desA]sLeXの阻害活性の増強は、CVF−誘導補体活性化が起こり(かくして、P−セレクチンの内皮細胞上昇調節を引き起こす)につれて、sCR1[desA]sLeXは内皮細胞P−セレクチンへ選択的に結合でき、さらなる補体活性化に対する局所的保護を供するという解釈に合致する。sCR1[desA]sLeXの活性化内皮細胞領域への局所化は、免疫染色データによって支持され、30分におけるsCR1[desA]抗原の残存血漿中レベルが、sCR1[desA]で処理した動物よりもsCR1[des]sLeX処理動物において50%近く低いのは何故であるかも説明するであろう。
P−セレクチン(またはE−セレクチン)へのsCR1[desA]sLeXの結合を引き起こすsLeXの能力に基づき、補体阻害剤を内皮細胞に「標的化する」能力は、これらの阻害剤の保護効果を最大化するユニークな戦略を提供する。心筋に対する虚血−再潅流傷害はP−セレクチン−依存性であるようなので、ヒトにおいて、虚血−再潅流傷害の治療は、セレクチンおよび補体活性化分子が傷害に導く結果に参画する他の状況にて、かかる阻害剤の使用により益するところがあるであろう。
16.実施例11
16.1 セレクチン結合活性を持つ可溶性補体受容体1型 の作製
本発明者らは、セレクチン結合活性を持つ補体受容体1型のもう1つの可溶性形態をここに記載する。この二官能性分子は炎症反応を変調する価値ある手段である。
16.1.1.細胞系
細胞系K562はLloyd Klickstein博士(Center for Blood Research,200 Longwood Avenue,Boston,MA 02115)から供給され、一般に、American Type Culture Collection(Rockville,MD)から入手可能である。HL−60細胞はATCCから得た。
16.1.2.モノクローナル抗体
CR1について特異的なウサギ・ポリクローナル抗血清は、ウサギをヒト補体受容体1型で免疫することによって、当該分野で公知の標準的な技術によって得ることができる。CD15に対するモノクローナル抗体は商業的に入手可能であり、Dako,Californiaから得ることができ、例えば、クローン28はAMAC,Inc.,Maineから得ることができる。マウス・モノクローナル抗体3C6.D11は、元来Kohler and Milstein(1975,Nature 256:495−497)によって記載されている方法を用いて、標準的な融合から得られた。Balb/cをフロイントアジュバント中の精製組換え補体受容体1型で3〜4週間間隔で腹腔内免疫した。第3免疫接種の4週間後、マウスに10μg CR1を静脈内追加免疫注射をし、脾臓を4日後に取り出した。50%PEG−1500(Boehringer Mannheim,Indianapolis IN)1mlを添加することによって、脾臓細胞をNS0骨髄腫細胞と融合させ、次いで、OPTI−MEM培地(GIBCO)20mlで希釈した。融合後、細胞を、96ウェルの平底プレートのウェルに平板培養し、HAT(GIBCO)を含有する培地で選択した。増殖について陽性であるウェルを、CR1捕獲抗体を用いて、抗CR1 mAbsの産生についてスクリーニングした。対照抗体は、Becton Dickinson,Franklin Lakes,NJ,およびTagoから商業的に入手可能なマウスIgMおよびマウスIGg1であった。
16.1.3.トランスフェクション
補体受容体1型(CR1)を発現するK562細胞は、構築体piABCD(Klickstein,et al.,1988,168:1699)から得られた全長CR1で、エレクトロポレーションによって、宿主細胞をトランスフェクトすることにより得ることができる。0.8mlの培地に懸濁したほぼ500万のK562細胞をほぼ20μgのプラスミドDNAと混合し、SpeJで線状化し、genepulserエレクトロポレーション装置(BioRad)を用いて、200ボルト、960μFで行った。培地中数日後、T Cell Diagnostics,Inc.,Cambridge,MAから得たCELLFREERCD35ビードアッセイキットを用い、可溶性CR1遺伝子産物を発現する細胞を、可溶性CR1の発現について選択することができる。
別法として、Graham and van der Erb(1973)の方法(Virology 52:456−467)を用いて、K562細胞のリン酸カルシウム媒介トランスフェクションを行うこともできる。
16.1.4.細胞溶解物
適当な分子を発現するようにトランスフェクトされた細胞系または適当な分子を内因的に発現する細胞系を、10mMトリス pH8.0、1%ノニデットP−40(NP−40)、10mMヨードアセトアミド(I AA)、1mMフェニルメチルスルホニルフルオライド(FMSF)、0.04%アプロチニンおよび0.3mM N−トシル−L−フェニルアラニンクロロメチルケトン(TPCK)を含有する溶解緩衝液中、5×107細胞/mlにて可溶化した。
16.1.5.ウェスタンブロット分析
K562上清からのアフィニティークロマトグラフィーによって精製したCR1の反応性を、ウェスタンブロット分析によってテストした。全長CR1でトランスフェクトしたK562からの上清を4〜20%SDS−PAGEによって分画し、次いで、ニトロセルロースシートに移した。該シートをまずブロッキング緩衝液(PBS中のリン酸緩衝生理食塩水中1%ウシ血清アルブミン)でブロックした。ブロッキングの後、抗体3C6.D11(約2〜3μg/ml(抗−CR1))、抗−CD15(約20μg/ml)、または無関係なイソタイプ合致対照抗体C305(IgM、約20μg/ml)、またはW112(IgG、約2〜5μg/ml)のいずれかと共に該シートをインキュベートした。一次抗体の存在下におけるインキュベーション1〜2時間後、該シートをPBSおよび0.05%ツィーン20の溶液で洗浄した。洗浄の後、ホースラディッシュペルオキシダーゼ(HRP)結合ヤギ抗−マウス抗体と共に該シートをインキュベートした。洗浄の後、HRD基質で発色させた。
16.2 結果
予測されるごとく、K562細胞培養上清から回収した物質は、ルイスX抗原に対する抗体ならびにモノクローナル抗−CR1抗体を用い、ウェスタンブロットによって検出することができる。
16.3 KCR1の物理的特徴付け
前記の両者から回収したKCR1の特異的炭水化物構造をはっきりとさせるため、アフィニティ−精製したKCR1およびノイラミニダーゼ処理したKCR1を、96ウェルプレートのウェルに固定化した抗−CR1に結合するその能力についてテストした。検出は、LeXSLeXリガンド構造に反応性の抗−CD15抗体で行った。ノイラミニダーゼでのKCR1の処理により、末端シアル酸残基がSLeXオリゴ糖構造から除去され、ルイスX構造が得られた。この分析の結果を表IIIに示す。
1 CR1は、CD15抗体に結合するための適当な炭水化物構造を含有しないチャイニーズハムスター卵巣細胞から得られたCR1の試料を表す。
2 KCR1は、K562細胞で産生されたアフィニティ−精製CR1の試料を表す。試料はアッセイに先立ってCentriconR濃縮器によって濃縮した。
3 nKCR1は、ノイラミニダーゼで処理したK562細胞で産生されたCR1の試料を表す。ノイラミニダーゼ処理は、アッセイに先立ってノイラミニダーゼの存在下で試料をインキュベートすることにより行った。
4 KCR1は、アッセイに先立って、K562細胞からCR1アフィニティ−精製細胞で、未処理の試料を表す。
17.機能的活性を検出するための一般化されたアッセイ フォーマット
また、本発明の組成物は、ある種の細胞、例えば活性化内皮細胞に対する細胞間接着をブロックし、それにより、炎症反応における一次的事象を阻害するその能力について評価することができる。この評価は、多数の方法によって達成でき;以下の方法が、この点につき用いられ、あるいはそれに加えて有用であり得る特別の手法として記載される:
A.HL−60のEおよびPセレクチンへの結合の競合的阻害
a)EまたはPセレクチンを発現する細胞(活性化血小板またはその表面のセレクチンでトランスフェクトされるか、あるいはそのセレクチンを発現する細胞、Larsen,et al.参照)を96−ウェルのマイクロタイタープレート中で密集するまで増殖させる。
b)CR1またはCR1類似体の存在または不存在の下でHL−60細胞を4℃で加え、30分間放置し、結合させる。
c)プレートを裏返しにし、150×gで5分間遠心することによって非接着性細胞を除去する。
d)プレートを、顕微鏡視野当たりの結合HL−60の数についてスコア取りする。
B.セレクチン結合についてのin vivoアッセイ
a)Mulligan et al.,1992,J.Clin.Invest.90:1600−1607の方法に準じて、CVFにてラットでP−セレクチン上昇調節を誘導する。
b)放射性同位体標識sLeX−CR1キメラまたは類似体vsTP10を注射し、放射性同位体の分布を測定する。
C.組成物効率のin vivoアッセイ
Milligan et al.(「Role of Leukocyte Adhesion Molecules in Complement−Induced Lung Injury」,J.Immunol.Vol.150,2401−24061,No.6,March 15,1993)は、補体のコブラ毒因子(CVF)活性化後のラットにおける肺血管内皮細胞傷害でのPセレクチンの役割を記載している。補体は、CVFによって誘導された肺の急性微小血管傷害を防止するにおいて保護効果を有することが既に示されているので、セレクチンリガンドを介して傷害の部位にCR1を止めるのが望ましい。補体の局所化を評価するために、体重1kg当たりCVF20単位を300〜350グラムの雄Long Evansラットに静脈内注射した。あるいは対照動物において、肺125I−CR1−sLeX(ほぼ500μCi)に対するCR1の局所化を評価するために、125I−CR1を例えば15mg/kg体重にて注射する。CVF誘導肺傷害は瞬間的であるので、局所化は、注射の約30分後に標準的な技術によって放射性同位体標識CR1−sLeXの組織取込みを評価することによって評価することができる。
また、好中球隔離および続いてのローリングおよび強い付着のごとき炎症における一次事象を妨げるsLeXの能力を評価するために、CVFモデルを用いることができる。また、Mulligan et al.,Role Endothelial−Leukocyte Adhesion Molecule 1(ELAM−1)in Neutrophil−Mediated Lung Injury in Rats,J.Clin.Invest.,Vol.88,October 1991,1396−1406,およびMulligan et al.,「Neutrophil−dependent Acute Lung Injury」,J.Clin.Invest.,Vol.90,October 1992,1600−1607を参照されたし。
18.組成物の調製のための他の技術
18.1 突然変異誘発
sCR1を発現するCHO(チャイニーズハムスター卵巣)細胞を用いた。懸濁液中の細胞(約2×105細胞/ml)をEMSと共に18時間インキュベートし、洗浄し、相対的平板培養効率を測定することができる。突然変異誘発した細胞を7日間培養して、獲得した突然変異誘発を発現させることができる。約10%の子ウシ血清および適当な濃度の一次選択レクチン(類)を含有する培地中、約106細胞/100mm組織培養皿に、細胞を分ける。6日後、プレートをアルファ培地で2回洗浄し、10%子ウシ血清を含有するアルファ培地に二次選択レクチン(類)を添加する。さらに、インキュベーションを約4日した後、10%子ウシ血清を含有するアルファ培地に最大コロニーを拾い上げ、プレートを50%メタノール中の2%メチレンブルーで染色する。レクチンを含有しないか、あるいは一次選択レクチン(類)のみを含有する対照プレートを8日後に染色し、相対的平板培養効率を測定する。
18.2 細胞融合
α1,3−フコシルトランスフェラーゼを発現する約1×108細胞および既にDMEMに懸濁した所望の補体蛋白質を発現する5×107細胞を200×gで5分間遠心するこによってペレット化し、37℃まで加温した。37℃で穏やかに混合しつつ、5%DMSOを加えた、約1g/mlのポリエチレングリコール(PEG)4000を含有する培地1mlに該ペレットを再懸濁した。次いで、細胞を100×gで2分間回転させ、4.5mlの補足培地を次の3分間にわたって添加し、続いて、5mlの補足培地を次の2分間にわたって添加した。次いで、試験管を補足培地で満たした。当該分野でよく知られているように、これらの工程のタイミングは重要である。
室温にて100×gで5分間遠心することによって細胞をペレット化し、次いで、上清を吸引した。細胞ペレットを培地に再懸濁したが、小さな細胞の集塊を無理に分配しないように注意した。細胞を限界希釈にて96−ウェルプレート中で平板培養した(増殖を確実とするためには、プレートのウェルにフィーダー細胞を含有させることができる)。ミコフェノール酸を含有する培地を添加し、次いで、細胞選択を確実とするのに必要と思われるくらいの頻度で置き換える。sLeXまたはシアリルジ−LeXを発現する細胞を次いで選択する。
本発明は、本明細書中に記載した特別の具体例によってその範囲が限定されるものではない。事実、これまでの記載および添付した図面より、本明細書に記載したものに加えて、本発明の種々の修飾は当業者に明らかであろう。かかる修飾も添付した請求の範囲内のものである。
種々の文献を本明細書中で引用したが、その開示を出典明示して本明細書の一部をみなす。
配列表
(1)一般的情報
(i)出願人:Rittershaus,Charles W.
Roth,Carol A.
(ii)発明の名称:補体関連蛋白質および炭水化物よりなる組成物、ならびに該組成物の製法および使用方法
(iii)配列の数:4
(iv)連絡住所:
(A)宛先:T Cell Sciences,Inc.
(B)通り:38 Sidney Street
(C)都市:Cambridge
(D)州:マサチューセッツ
(E)国:米国
(F)郵便番号:02139
(v)コンピューター可読形態
(A)媒体のタイプ:ディスケット、3.5インチ、1.44Mb
(B)コンピューター:IBM PC Compatible
(C)ソフトウェア:ワードパーフェクト 5.1
(vi)現在の出願のデータ:
(A)出願番号:08/061,982
(B)出願日:1993年5月17日
(C)分類:1806
(viii)代理人の情報
(A)氏名:Matthews,Gale F.
(B)登録番号:32,269
(C)書類番号:TCS−101−P
(ix)通信情報:
(A)電話:617−621−1400
(B)ファックス:617−621−0627
(2)配列番号:1についての情報
(i)配列の特徴
(A)長さ:68塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:線形
(ii)分子の型:
(A)記載:DNA
(xi)配列の記載:配列番号:1:
(i)配列の特徴
(A)長さ:72塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:線形
(ii)分子の型:
(A)記載:DNA
(xi)配列の記載:配列番号:2:
(i)配列の特徴
(A)長さ:66塩基対
(B)型:核酸
(C)鎖の数;一本鎖
(D)トポロジー:線形
(ii)分子の型:
(A)記載:DNA
(xi)配列の記載:配列番号:3:
(i)配列の特徴
(A)長さ:56塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:線形
(ii)分子の型:
(A)記載:DNA
(xi)配列の記載:配列番号:4:
(i)配列の特徴
(A)長さ:31塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:線形
(ii)分子の型:
(A)記載:DNA
(xi)配列の記載:配列番号:5:
(i)配列の特徴
(A)長さ:56塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:線形
(ii)分子の型:
(A)記載:DNA
(xi)配列の記載:配列番号:6:
(i)配列の特徴
(A)長さ:19塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:線形
(ii)分子の型:
(A)記載:DNA
(xi)配列の記載:配列番号:7:
(i)配列の特徴
(A)長さ:19塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:線形
(ii)分子の型:
(A)記載:DNA
(xi)配列の記載:配列番号:8:
(i)配列の特徴
(A)長さ:40塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:線形
(ii)分子の型:
(A)記載:DNA
(xi)配列の記載:配列番号:9:
(i)配列の特徴
(A)長さ:13塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:線形
(ii)分子の型:
(A)記載:DNA
(xi)配列の記載:配列番号:10:
(i)配列の特徴
(A)長さ:17塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:線形
(ii)分子の型:
(A)記載:DNA
(xi)配列の記載:配列番号:11:
Claims (28)
- (a)短鎖コンセンサス反復構造モチーフを有してかつ補体成分に結合する、CR1、CR2、H因子、 C4結合蛋白質、崩壊促進因子(DAF)および膜助因子蛋 白質(MCP)からなる群から選択される可溶性補体調節蛋白質部位と、
(b)P−セレクチン、E−セレクチンおよびL−セレクチンからなる群から選択されるセレクチンと結合しうる、sLe X 、Le X 、sLe a およびLe a からなる群から選択され る炭水化物部位構造を含むセレクチンリガンドである炭水化物部位と、
がリンクしてなる補体阻害性蛋白質。 - 前記セレクチンリガンドがルイスX抗原である請求項1記載の補体阻害性蛋白質。
- 炎症反応における主要な徴候を阻害する請求項1記載の補体阻害性蛋白質。
- 炭水化物部位が活性化された内皮細胞に結合する請求項1記載の補体阻害性蛋白質。
- 補体に関連する少なくとも1つの活性化を阻害できる請求項1記載の補体阻害性蛋白質。
- 前記セレクチンリガンドがセレクチン内にあるレクチンドメインに結合し、かつ少なくとも104M-1の結合アフィニティー定数Kaを有する請求項1記載の補体阻害性蛋白質。
- 前記炭水化物部位が少なくとも1つのN−結合炭水化物を含み、その炭水化物がフコシル化されている請求項1記載の補体阻害性蛋白質。
- 前記炭水化物部位が少なくとも1つのO−結合炭水化物部位を含み、その炭水化物部位がフコシル化されている請求項1記載の補体阻害性蛋白質。
- 前記炭水化物部位が1もしくはそれ以上の以下の構造を含んでいる請求項1記載の補体阻害性蛋白質:
- 前記可溶性補体調節蛋白質部位が、補体 受容体1型の膜貫通領域を欠くかつ補体成分に結合でき る可溶性補体受容体1型(sCR1)のフラグメントである請求項1記載の補体阻害性蛋白質。
- 前記可溶性補体受容体1型のフラグメン トが、補体受容体1型の膜貫通領域の最初のアラニン残基を含み、該残基までの少なくともLHR B、LHR CおよびLHR Dを含む請求項10記載の補体阻害性蛋白質。
- 多発性硬化症、卒中、ギラン・バレー症 候群、外傷性脳傷害、パーキンソン病、
血液透析合併症、激症同種移植片拒絶、異種移植拒絶、 IL−2治療中のインターロイキン−2誘導毒性、自己免 疫疾患の炎症、クローン氏病(限局性回腸炎)、成人呼 吸困難症候群、火傷および凍傷を含めた熱傷害、心筋梗 塞、バルーン血管形成、心肺バイパスまたは心臓バイパ スでポストポンプ(post−pump)症候群、血液透析、腎 臓虚血、動脈再構成後の腸間膜動脈再灌流、移植器官再 灌流、感染症または敗血症、慢性関節リウマチ、全身性 紅斑狼瘡(SLE)、SLE腎炎、増殖性腎炎、糸球体腎炎、 溶血性貧血、重症筋無力症、からなるグループから選ば れ、補体関与の全身性病気または障害の治療に用い、医 薬上許容される担体中に請求項1記載の補体阻害性蛋白 質を分散させた医薬組成物。 - セレクチンリガンドがルイスX抗原である請求項12記載の医薬組成物。
- ルイスX原がシアリル化ルイスX抗原である請求項12記載の医薬組成物。
- 可溶性補体調節蛋白質部位が、補体受容 体1型の膜貫通領域を欠くかつ補体成分に結合できる可溶性補体受容体1型(sCR1)のフラグメントである請求項12記載の医薬組成物。
- (a)ルイスX抗原で補体調節蛋白質をグリコシル化する細胞中で、CR1、CR2、H因子、C4結合 蛋白質、崩壊促進因子(DAF)および膜助因子蛋白質(M CP)からなる群から選択される補体調節蛋白質を発現させるステップと、
(b)工程(a)で発現された補体調節蛋白質を回収するステップと、
を含む請求項1記載の補体阻害性蛋白質の製造方法。 - 前記補体調節蛋白質がシアリル化されたルイスX抗原でグリコシル化されている請求項16記載の製造方法。
- 補体調節蛋白質が補体受容体1型(CR1)である請求項17記載の製造方法。
- セレクチンリガンドと可溶性補体調節性蛋白質とを化学的もしくは酵素的に連結させることを含んでなる、請求項1に記載の補体阻害性蛋白質の製造方法。
- 補体阻害性蛋白質が補体受容体1型である請求項19記載の製造方法。
- 前記セレクチンリガンドがルイスX抗原である請求項19記載の補体阻害性蛋白質の製造方法。
- 前記ルイスX抗原がシアリル化されている請求項21記載の製造方法。
- 請求項1に記載の補体阻害性蛋白質を含む、望ましくないあるいは不適切な補体活性化に起因する疾病または疾患を有する患者を治療するために用いる薬剤を調製するためのキット。
- 前記補体阻害性蛋白質がシアリル化されたルイスX抗原を含む請求項23記載のキット。
- (1)請求項1記載の補体阻害性蛋白質と、炎症症状に関連する前記補体阻害性蛋白質に対する接着性分子リガンドを含むことが疑われる試料とを加えインキュベートする手段と、
(2)前記補体阻害性蛋白質と前記炎症症状に関連する接着性分子リガンドと間の結合量を検出する手段とを、
含む炎症症状の診断用キット。 - 請求項1に記載の補体阻害性蛋白質を含む炎症症状を有する患者を治療するための薬剤を調製するためのキット。
- 前記補体阻害性蛋白質の炭水化物部位が、P−セレクチン、E−セレクチンまたはL−セレクチンのいずれかに結合する請求項26記載のキット。
- 請求項1に記載された補体阻害性蛋白質の提供を含んでなる、炎症症状または補体活性化に使用する医薬を製造する方法。
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US6198293A | 1993-05-17 | 1993-05-17 | |
| US08/061,982 | 1993-05-17 | ||
| PCT/US1994/005285 WO1994026786A1 (en) | 1993-05-17 | 1994-05-12 | Compositions comprising complement related proteins and carbohydrates, and methods for producing and using said compositions |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JPH08510257A JPH08510257A (ja) | 1996-10-29 |
| JP3662021B2 true JP3662021B2 (ja) | 2005-06-22 |
Family
ID=22039478
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP52569594A Expired - Fee Related JP3662021B2 (ja) | 1993-05-17 | 1994-05-12 | 補体関連蛋白質および炭水化物よりなる組成物、ならびに該組成物の製法および使用方法 |
Country Status (11)
| Country | Link |
|---|---|
| US (1) | US6193979B1 (ja) |
| EP (1) | EP0730608B1 (ja) |
| JP (1) | JP3662021B2 (ja) |
| AT (1) | ATE215094T1 (ja) |
| CA (1) | CA2162600C (ja) |
| DE (1) | DE69430253T2 (ja) |
| DK (1) | DK0730608T3 (ja) |
| ES (1) | ES2174876T3 (ja) |
| PT (1) | PT730608E (ja) |
| SG (1) | SG52383A1 (ja) |
| WO (1) | WO1994026786A1 (ja) |
Families Citing this family (44)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP3662021B2 (ja) | 1993-05-17 | 2005-06-22 | アーヴァント イミュノセラピューティクス インコーポレイテッド | 補体関連蛋白質および炭水化物よりなる組成物、ならびに該組成物の製法および使用方法 |
| ES2252746T3 (es) | 1994-09-23 | 2006-05-16 | Alexion Pharmaceuticals, Inc. | Procedimientos para el tratamiento de la enfermedad inflamatoria articular. |
| GB9614871D0 (en) * | 1996-07-15 | 1996-09-04 | Smithkline Beecham Plc | Compounds |
| WO1999037149A1 (en) | 1998-01-27 | 1999-07-29 | Brigham & Women's Hospital | Methods of treating cytotoxic damage |
| GB9905503D0 (en) * | 1999-03-10 | 1999-05-05 | Adprotech Plc | Novel compound formulations and methods of delivery |
| CA2486106C (en) * | 2002-05-16 | 2013-07-16 | Glycomimetics, Inc. | Compounds and methods for inhibiting selectin-mediated function |
| CA2490703A1 (en) * | 2002-07-03 | 2004-01-15 | Glycomimetics, Inc. | Compositions and methods for diagnosis and therapy of medical conditions involving angiogenesis |
| US20040219158A1 (en) * | 2003-05-02 | 2004-11-04 | Glycomimetics, Inc. | Compositions and methods for diagnosis and therapy of medical conditions involving infection with pseudomonas bacteria |
| FR2861729B1 (fr) * | 2003-10-31 | 2006-09-08 | Fabre Pierre Dermo Cosmetique | Monomere d'alkyl-rhamnose ou d'alkyl-fucose, medicament comprenant un monomere d'alkyl-sucre reducteur |
| CA2546784A1 (en) * | 2003-11-19 | 2005-06-16 | Glycomimetics, Inc. | Glycomimetic antagonists for both e- and p-selectins |
| EP1763533B1 (en) * | 2003-11-19 | 2008-01-09 | GlycoMimetics, Inc. | Specific antagonist for both e- and p-selectins |
| US8201377B2 (en) * | 2004-11-05 | 2012-06-19 | Faus Group, Inc. | Flooring system having multiple alignment points |
| WO2006127906A1 (en) * | 2005-05-25 | 2006-11-30 | Glycomimetics, Inc. | Heterobifunctional compounds for selectin inhibition |
| CA2654440C (en) * | 2005-06-06 | 2013-09-03 | Girish J. Kotwal | Methods for treatment or prophylaxis of atherosclerosis and reperfusion injury |
| EP1738763A1 (en) | 2005-06-30 | 2007-01-03 | AVANT Immunotherapeutics, Inc. | Use of complement inhibitory proteins to treat spinal cord injury |
| US20070004625A1 (en) * | 2005-06-30 | 2007-01-04 | Li Liang-Man | Use of complement inhibitory proteins to treat spinal cord injury |
| CN101291946B (zh) * | 2005-08-09 | 2011-06-29 | 糖模拟物有限公司 | 对来自假单胞菌的pa-il凝集素、pa-iil凝集素或其两者的糖模拟物抑制剂 |
| LT2264043T (lt) | 2005-09-02 | 2017-12-11 | Glycomimetics, Inc. | Heterobifunkciniai pan-selektino inhibitoriai |
| US20090176717A1 (en) * | 2006-06-01 | 2009-07-09 | Glycomimetics, Inc. | Galactosides and thiodigalactosides as inhibitors of pa-il lectin from pseudomonas |
| EP2074132B1 (en) | 2006-10-12 | 2013-05-15 | GlycoMimetics, Inc. | Glycomimetic replacements for hexoses and n-acetyl hexosamines |
| CA2666843C (en) * | 2006-10-20 | 2015-06-16 | Celldex Therapeutics, Inc. | Treatment of age-related macular degeneration and other diseases of the eye |
| NZ598863A (en) * | 2007-02-09 | 2013-11-29 | Glycomimetics Inc | Methods of use of glycomimetics with replacements for hexoses and n-acetyl hexosamines |
| US8039442B2 (en) * | 2007-07-18 | 2011-10-18 | Glycomimetics, Inc. | Compounds and methods for treatment of sickle cell disease or complications associated therewith |
| WO2009126556A1 (en) | 2008-04-08 | 2009-10-15 | Glycomimetics, Inc. | Pan-selectin inhibitor with enhanced pharmacokinetic activity |
| ES2426784T3 (es) * | 2008-06-13 | 2013-10-25 | Glycomimetics, Inc. | Tratamiento de cánceres de la sangre usando compuestos glicomiméticos seleccionados |
| WO2010126888A1 (en) | 2009-05-01 | 2010-11-04 | Glycomimetics, Inc. | Heterobifunctional inhibitors of e-selectins and cxcr4 chemokine receptors |
| WO2012037034A1 (en) | 2010-09-14 | 2012-03-22 | Glycomimetics, Inc. | E-selectin antagonists |
| CN107661491A (zh) * | 2010-09-15 | 2018-02-06 | 赛尔戴克斯治疗公司 | 用可溶性I型补体受体(sCR1)治疗慢性肾病 |
| US9649356B2 (en) | 2010-10-27 | 2017-05-16 | Celldex Therapeutics, Inc. | Method of improving transplant function using soluble complement receptor type I (sCR1) |
| CA2858099C (en) | 2011-12-22 | 2020-12-15 | Glycomimetics, Inc. | E-selectin antagonist compounds, compositions, and methods of use |
| DK2928476T3 (en) | 2012-12-07 | 2018-05-22 | Glycomimetics Inc | RELATIONSHIPS, COMPOSITIONS AND METHODS OF USING E-SELECTIN ANTAGONISTS FOR MOBILIZATION OF HEMATOPOIETIC CELLS |
| US10238700B2 (en) | 2014-01-02 | 2019-03-26 | Genelux Corporation | Oncolytic virus adjunct therapy with agents that increase virus infectivity |
| DK3227310T3 (da) | 2014-12-03 | 2019-09-30 | Glycomimetics Inc | Heterobifunktionelle inhibitorer af e-selektiner og cxcr4-kemokin-receptorer |
| WO2017127422A1 (en) | 2016-01-22 | 2017-07-27 | Glycomimetics, Inc. | Glycomimetic inhibitors of pa-il and pa-iil lectins |
| US11291678B2 (en) | 2016-03-02 | 2022-04-05 | Glycomimetics, Inc | Methods for the treatment and/or prevention of cardiovascular disease by inhibition of E-selectin |
| US11433086B2 (en) | 2016-08-08 | 2022-09-06 | Glycomimetics, Inc. | Combination of T-cell checkpoint inhibitors with inhibitors of e-selectin or CXCR4, or with heterobifunctional inhibitors of both E-selectin and CXCR4 |
| WO2018068010A1 (en) | 2016-10-07 | 2018-04-12 | Glycomimetics, Inc. | Highly potent multimeric e-selectin antagonists |
| EP3596096A1 (en) | 2017-03-15 | 2020-01-22 | GlycoMimetics, Inc. | Galactopyranosyl-cyclohexyl derivatives as e-selectin antagonists |
| WO2019108750A1 (en) | 2017-11-30 | 2019-06-06 | Glycomimetics, Inc. | Methods of mobilizing marrow infiltrating lymphocytes and uses thereof |
| AU2018395417B2 (en) | 2017-12-29 | 2023-07-13 | Glycomimetics, Inc. | Heterobifunctional inhibitors of E-selectin and galectin-3 |
| CA3091454A1 (en) | 2018-03-05 | 2019-09-12 | Glycomimetics, Inc. | Methods for treating acute myeloid leukemia and related conditions |
| CA3100071A1 (en) | 2018-05-16 | 2019-11-21 | Csl Limited | Soluble complement receptor type 1 variants and uses thereof |
| US11845771B2 (en) | 2018-12-27 | 2023-12-19 | Glycomimetics, Inc. | Heterobifunctional inhibitors of E-selectin and galectin-3 |
| WO2021247487A1 (en) * | 2020-06-01 | 2021-12-09 | Medical University Of South Carolina | Recombinant fusion proteins targeting p-selectin, and methods of use thereof for treating diseases and disorders |
Family Cites Families (17)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US5212071A (en) | 1988-04-01 | 1993-05-18 | The Johns Hopkins University | Nucleic acids encoding a human C3b/C4b receptor (CR1) |
| US4752569A (en) | 1984-06-21 | 1988-06-21 | The Regents Of The University Of California | Sialylated Lewisx epitope, antibodies and diagnosis |
| IL82390A0 (en) | 1986-05-02 | 1987-10-30 | Genentech Inc | Nucleic acid and methods for the synthesis of novel daf compositions |
| US5079353A (en) | 1987-12-02 | 1992-01-07 | Chembiomed, Ltd. | Sialic acid glycosides, antigens, immunoadsorbents, and methods for their preparation |
| US5081034A (en) | 1988-11-14 | 1992-01-14 | Brigham & Women's Hospital | Cloned genes which encode elam-1 |
| WO1991016900A1 (en) | 1990-04-27 | 1991-11-14 | Biogen, Inc. | Fucosyl transferases involved in adhesion molecule expression |
| AU8007791A (en) | 1990-06-15 | 1992-01-07 | Cytel Corporation | Intercellular adhesion mediators |
| WO1992001718A2 (en) | 1990-07-17 | 1992-02-06 | Board Of Regents Of The University Of Oklahoma | Functionally active selectin-derived peptides and ligand for gmp-140 |
| US5211937A (en) | 1990-07-30 | 1993-05-18 | Glycomed Incorporated | Method of determining a site of inflammation utilizing elam-1 ligands |
| US5164374A (en) | 1990-12-17 | 1992-11-17 | Monsanto Company | Use of oligosaccharides for treatment of arthritis |
| US5166133A (en) | 1991-04-17 | 1992-11-24 | Cetus Corporation | Method for inhibing adhesion of white blood cells to endothelial cells |
| EP0512733A2 (en) | 1991-05-03 | 1992-11-11 | Washington University | Modified complement system regulator |
| CA2108029A1 (en) | 1991-05-06 | 1992-11-07 | Laurence A. Lasky | Glycam-1(sgp 50), a selectin ligand |
| US5227369A (en) | 1991-07-11 | 1993-07-13 | The Regents Of The University Of California | Compositions and methods for inhibiting leukocyte adhesion to cns myelin |
| US5456909A (en) * | 1992-08-07 | 1995-10-10 | T Cell Sciences, Inc. | Glycoform fractions of recombinant soluble complement receptor 1 (sCR1) having extended half-lives in vivo |
| JP3662021B2 (ja) | 1993-05-17 | 2005-06-22 | アーヴァント イミュノセラピューティクス インコーポレイテッド | 補体関連蛋白質および炭水化物よりなる組成物、ならびに該組成物の製法および使用方法 |
| US5856300A (en) * | 1994-05-12 | 1999-01-05 | T Cell Sciences, Inc. | Compositions comprising complement related proteins and carbohydrates, and methods for producing and using said compositions |
-
1994
- 1994-05-12 JP JP52569594A patent/JP3662021B2/ja not_active Expired - Fee Related
- 1994-05-12 WO PCT/US1994/005285 patent/WO1994026786A1/en active IP Right Grant
- 1994-05-12 AT AT94917961T patent/ATE215094T1/de not_active IP Right Cessation
- 1994-05-12 DK DK94917961T patent/DK0730608T3/da active
- 1994-05-12 DE DE69430253T patent/DE69430253T2/de not_active Expired - Fee Related
- 1994-05-12 EP EP94917961A patent/EP0730608B1/en not_active Expired - Lifetime
- 1994-05-12 SG SG1996003777A patent/SG52383A1/en unknown
- 1994-05-12 ES ES94917961T patent/ES2174876T3/es not_active Expired - Lifetime
- 1994-05-12 CA CA002162600A patent/CA2162600C/en not_active Expired - Fee Related
- 1994-05-12 PT PT94917961T patent/PT730608E/pt unknown
-
1995
- 1995-05-25 US US08/450,274 patent/US6193979B1/en not_active Expired - Fee Related
Also Published As
| Publication number | Publication date |
|---|---|
| WO1994026786A1 (en) | 1994-11-24 |
| EP0730608A4 (en) | 1998-12-02 |
| CA2162600A1 (en) | 1994-11-24 |
| SG52383A1 (en) | 1998-09-28 |
| AU6947594A (en) | 1994-12-12 |
| AU678486B2 (en) | 1997-05-29 |
| US6193979B1 (en) | 2001-02-27 |
| PT730608E (pt) | 2002-09-30 |
| DK0730608T3 (da) | 2002-07-22 |
| ATE215094T1 (de) | 2002-04-15 |
| EP0730608B1 (en) | 2002-03-27 |
| DE69430253D1 (de) | 2002-05-02 |
| EP0730608A1 (en) | 1996-09-11 |
| ES2174876T3 (es) | 2002-11-16 |
| DE69430253T2 (de) | 2002-11-14 |
| JPH08510257A (ja) | 1996-10-29 |
| CA2162600C (en) | 2000-07-11 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP3662021B2 (ja) | 補体関連蛋白質および炭水化物よりなる組成物、ならびに該組成物の製法および使用方法 | |
| US5976540A (en) | Compositions comprising complement related proteins and carbohydrates, and methods for producing and using said compositions | |
| US5856300A (en) | Compositions comprising complement related proteins and carbohydrates, and methods for producing and using said compositions | |
| Suzuki et al. | Molecular cloning and expression of cDNA encoding human macrophage C-type lectin. Its unique carbohydrate binding specificity for Tn antigen. | |
| Sako et al. | Expression cloning of a functional glycoprotein ligand for P-selectin | |
| JP4145356B2 (ja) | P―セレクチンリガンド蛋白 | |
| US6124267A (en) | O-glycan inhibitors of selectin mediated inflammation derived from PSGL-1 | |
| DE69630307T2 (de) | O-glycaninhibitoren von selectinvermittelten entzüngen | |
| JP3749529B2 (ja) | セレクチン変異体 | |
| JP2000325092A (ja) | 細胞接着蛋白質と糖質リガンドとの間の相互作用阻害に使用し得る抗体 | |
| Seya et al. | Purification and functional properties of soluble forms of membrane cofactor protein (CD46) of complement: identification of forms increased in cancer patients' sera | |
| JPH08502886A (ja) | 新規p−セレクチンリガンド蛋白質 | |
| CA1338878C (en) | 14-beta-gal mammalian lectins | |
| US5648465A (en) | Cloning and expression of neurocan, a chondroitin sulfate proteoglycan | |
| Lublin | Cromer and DAF: role in health and disease | |
| US20030166565A1 (en) | Compositions and methods to inhibit formation of the C5b-9 complex of complement | |
| US20020025923A1 (en) | Novel selection ligands | |
| US7399847B1 (en) | Nucleic acids encoding artificial P-selectin ligands | |
| Zhou et al. | Animal lectins: a distinct group of carbohydrate-binding proteins involved in cell adhesion, molecular recognition, and development | |
| AU678486C (en) | Compositions comprising complement related proteins and carbohydrates, and methods for producing and using said compositions | |
| KR100463567B1 (ko) | 셀렉틴매개염증의펩티드및0-글리칸억제제 | |
| CZ401497A3 (cs) | P-selektinové ligandy a příbuzné molekuly a způsoby | |
| Irimura | Molecular Cloning and Expression of cDNA Encoding Human Macrophage C-Type Lectin | |
| EP1428832A2 (en) | Peptide and O-glycan inhibitors of selectin mediated inflammation |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20031224 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20040216 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20040324 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20040831 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20041228 |
|
| A911 | Transfer to examiner for re-examination before appeal (zenchi) |
Free format text: JAPANESE INTERMEDIATE CODE: A911 Effective date: 20050120 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20050308 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20050322 |
|
| R150 | Certificate of patent or registration of utility model |
Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20090401 Year of fee payment: 4 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20100401 Year of fee payment: 5 |
|
| S533 | Written request for registration of change of name |
Free format text: JAPANESE INTERMEDIATE CODE: R313533 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20100401 Year of fee payment: 5 |
|
| R350 | Written notification of registration of transfer |
Free format text: JAPANESE INTERMEDIATE CODE: R350 |
|
| LAPS | Cancellation because of no payment of annual fees |