JP2010057388A - 転移因子の転移が抑制された納豆菌、該納豆菌を用いて製造された納豆 - Google Patents
転移因子の転移が抑制された納豆菌、該納豆菌を用いて製造された納豆 Download PDFInfo
- Publication number
- JP2010057388A JP2010057388A JP2008223990A JP2008223990A JP2010057388A JP 2010057388 A JP2010057388 A JP 2010057388A JP 2008223990 A JP2008223990 A JP 2008223990A JP 2008223990 A JP2008223990 A JP 2008223990A JP 2010057388 A JP2010057388 A JP 2010057388A
- Authority
- JP
- Japan
- Prior art keywords
- natto
- gene
- transfer
- strain
- bacillus natto
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Granted
Links
Images




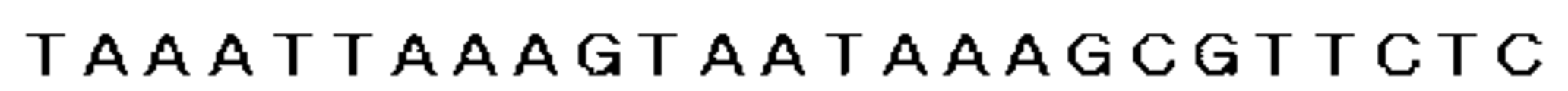



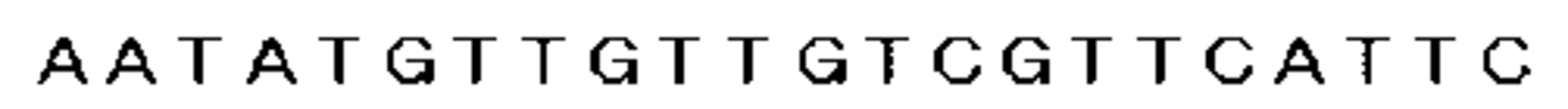






Landscapes
- Beans For Foods Or Fodder (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
【解決手段】recA遺伝子を欠損させて、comP遺伝子への転移因子の転移を抑制することにより、植え継ぎを繰り返した場合でもγ−PGA非生産性株の出現頻度が低い納豆菌を得る。また、該納豆菌を用いて納豆を製造する。
【選択図】なし
Description
このうちISは比較的単純な遺伝子構造であり、IS自身を染色体の別の位置に転移させる酵素(トランスポザーゼ)をコードするトランスポザーゼ遺伝子と、それが逆方向反復塩基配列によって挟まれた構造を有しており、染色体上の別の位置に転移して、転移先に元々存在していた遺伝子を破壊したりする特性を持っている。
ジャーナル・オブ・バクテリオロジー(J. Bacteriol.)、182巻、p.2387−2392、2000年 バイオサイエンス・バイオテクノロジー・アンド・バイオケミストリー(Biosci. Biotechnol. Biochem.)、71巻、p.2458−2464、2007年
(1)recA遺伝子を欠損させることにより、転移因子の転移が抑制されたことを特徴とする納豆菌。
(2)comP遺伝子への転移因子の転移が抑制されたことを特徴とする(1)に記載の納豆菌。
(3)12世代培養後もγ−PGA非生産株が出現しないことを特徴とする(1)又は(2)に記載の納豆菌。
(4)バシラス・サチリス REC7(Bacillus subtilis REC7)株(FERM ABP−10994)であることを特徴とする(1)〜(3)のいずれか1つに記載の納豆菌。
(5)(1)〜(4)のいずれか1つに記載の納豆菌を用いて製造されたことを特徴とする納豆。
例えば、pUC19などの多コピー数プラスミドベクターに対して当該recA遺伝子中に薬剤耐性遺伝子を挿入した遺伝子破壊用プラスミドを作製し、該プラスミドを適当な制限酵素により直線化した後、既存のコンピテンス形質転換法(例えば、フェムス・マイクロバイオロジカル・レター(FEMS Microbiol. Lett.)、236巻、p.13−20、2004年参照)を利用して納豆菌に導入して相同組換えを行わせ、当該recA遺伝子を欠損させる方法が採用される。
(1)使用菌株等
使用した納豆菌r22株は、市販納豆から分離された親株O−2株の形質転換能を高めた変異株であり、親株O−2株をニトロソグアニジン(NTG)を用いて化学変異処理することにより取得された株である(例えば、特開2000−224982号公報参照)。
以下に記載のごとく、まずrecA遺伝子を欠損させるために必要なDNA断片(recA欠損用断片)を、PCRを組み合わせた方法(例えば、ヌクレイック・アシッズ・リサーチ(Nucleic Acids Research)30巻、p.1−5、2002年参照)で構築し、納豆菌にコンピテンス法により導入して、相同組換えを起こさせることによってrecA欠損株を作製した。
染色体上のrecA遺伝子をスペクチノマイシン耐性遺伝子と置換してrecA遺伝子を欠損させるために、recA遺伝子欠損用DNA断片を、図1に示すごとく、PCRを組み合わせた以下の方法により作製した。
まず、置換に用いたスペクチノマイシン耐性遺伝子を含むDNA断片は、以下のように調製した。すなわち、GenBankに登録されているスペクチノマイシン耐性遺伝子の塩基配列(GenBank登録番号X02588)を基に、プライマー1(5´−GTATAATAAAGAATAATTATTAATC−3´、図4及び配列表の配列番号1参照)及びプライマー2(5´−TAAATTAAAGTAATAAAGCGTTCTC−3´、図5及び配列表の配列番号2参照)の2種のオリゴDNAを調製した。
上記のようにして調製したrecA遺伝子欠損用DNA断片を用いて、コンピテンス法によりr22株を形質転換した。形質転換株の選択は、スペクチノマイシン耐性を指標に行った。
納豆菌r22株およびREC7株を、5mlのLB培地を入れた試験管中で、37℃で12時間振とう培養し、これを第1世代とした。第1世代の培養液中の納豆菌を、濁度が0.05となるように新たなLB培地に植菌し、再び37℃で12時間振とう培養し、これを第2世代とした。以下、同様にして、第12世代まで培養を継続した。
このようにして培養を継続する中で、24時間ごと、すなわち2世代毎に、培養液を3枚のGSP寒天培地に100μl塗抹し、全コロニー数と、γ−PGA非生産コロニー数をそれぞれ測定した。
得られた全コロニー数とγ−PGA非生産コロニー数とから、全コロニー数に占めるγ−PGA非生産コロニーの割合を算出し、これを非γ−PGA生産株の出現頻度(%)とした。表1に、結果を示した。

これに対して、REC7株では第12世代まで植え継いでも、γ−PGA非生産株の出現頻度は常に0%である事が確認され、recA遺伝子を欠損させることにより、γ−PGA非生産性株の出現を抑制させることが出来ることが確認できた。
i)comP遺伝子へのIS転移の確認
実施例2における第3世代〜第12世代の各世代において、納豆菌r22株およびREC7株について、IS4Bsu1及びIS256Bsu1が高頻度に転移することが報告されているcomP遺伝子への転移をPCRを用いて以下のように確認した。
このようにして増幅された各世代ごとのDNA断片を電気泳動して、分子量を測定した。結果を、図2に示す。
これに対して、REC7株では第8世代以降も正常なcomP遺伝子を含むDNA断片(約2.3kb)のみが増幅したことが確認され、REC7株ではcomP遺伝子へのISの転移は確認されなかった。
上記i)における第12世代のr22株で検出されたcomP遺伝子にISが転移したと予想される約3.7kbのDNA断片、及び、PCR操作の対照としてr22株のゲノムDNAを鋳型にして、IS4Bsu1とIS256Bsu1が有する各トランスポザーゼ遺伝子をPCRにより増幅した。
12世代培養後のr22株及びREC7株から、それぞれ、常法により調製した胞子を用いて納豆を製造した。
製造された納豆について、糸引性を官能的に評価する一方、納豆1g当たりのγ−PGA量を測定した。
なお、γ−PGAは、納豆10gからトリクロロ酢酸により抽出後、エタノールにより回収し、菅野らの方法(例えば、日本食品工学会誌、42巻、p.878−886、1995年参照)に従い、γ−PGAが臭化セチルトリメチルアンモニウムに結合する性質を利用して分光学的に定量した。
表2に結果を示した。
これに対して、REC7株を用いて製造した場合は、納豆のγ−PGA量は十分であり、良好な糸引きが見られ、品質のよい納豆が安定して製造できることが確認できた。
Claims (5)
- recA遺伝子を欠損させることにより、転移因子の転移が抑制されたことを特徴とする納豆菌。
- comP遺伝子への転移因子の転移が抑制されたことを特徴とする請求項1に記載の納豆菌。
- 12世代培養後もγ−PGA非生産株が出現しないことを特徴とする請求項1又は2に記載の納豆菌。
- バシラス・サチリス REC7(Bacillus subtilis REC7)株(FERM ABP−10994)であることを特徴とする請求項1〜3のいずれか1項に記載の納豆菌。
- 請求項1〜4のいずれか1項に記載の納豆菌を用いて製造されたことを特徴とする納豆。
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2008223990A JP5319212B2 (ja) | 2008-09-01 | 2008-09-01 | 転移因子の転移が抑制された納豆菌、該納豆菌を用いて製造された納豆 |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2008223990A JP5319212B2 (ja) | 2008-09-01 | 2008-09-01 | 転移因子の転移が抑制された納豆菌、該納豆菌を用いて製造された納豆 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2010057388A true JP2010057388A (ja) | 2010-03-18 |
| JP5319212B2 JP5319212B2 (ja) | 2013-10-16 |
Family
ID=42184941
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2008223990A Active JP5319212B2 (ja) | 2008-09-01 | 2008-09-01 | 転移因子の転移が抑制された納豆菌、該納豆菌を用いて製造された納豆 |
Country Status (1)
| Country | Link |
|---|---|
| JP (1) | JP5319212B2 (ja) |
Citations (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2000224982A (ja) * | 1999-01-13 | 2000-08-15 | Mitsukan Group Honsha:Kk | 短鎖分岐脂肪酸非生産納豆菌 |
| JP2005245320A (ja) * | 2004-03-04 | 2005-09-15 | Aisin Seiki Co Ltd | 核酸組換え方法、宿主細胞、及び、発現ベクター |
| JP2007533306A (ja) * | 2004-03-19 | 2007-11-22 | ヘンケル・コマンディットゲゼルシャフト・アウフ・アクチエン | 生物工学的産生のために使用される、バシラス・リケニホルミス由来のRecA因子およびRecA不活性化安全な系統 |
-
2008
- 2008-09-01 JP JP2008223990A patent/JP5319212B2/ja active Active
Patent Citations (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2000224982A (ja) * | 1999-01-13 | 2000-08-15 | Mitsukan Group Honsha:Kk | 短鎖分岐脂肪酸非生産納豆菌 |
| JP2005245320A (ja) * | 2004-03-04 | 2005-09-15 | Aisin Seiki Co Ltd | 核酸組換え方法、宿主細胞、及び、発現ベクター |
| JP2007533306A (ja) * | 2004-03-19 | 2007-11-22 | ヘンケル・コマンディットゲゼルシャフト・アウフ・アクチエン | 生物工学的産生のために使用される、バシラス・リケニホルミス由来のRecA因子およびRecA不活性化安全な系統 |
Non-Patent Citations (1)
| Title |
|---|
| JPN6013018303; 化学と生物 Vol.46, No.5, 200805, p.302-303 * |
Also Published As
| Publication number | Publication date |
|---|---|
| JP5319212B2 (ja) | 2013-10-16 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP6648345B1 (ja) | 新規ポリペプチド及びこれを用いたimpの生産方法 | |
| CN113667682B (zh) | Yh66-rs11190基因突变体及其在制备l-缬氨酸中的应用 | |
| Chen et al. | Characterization of pMC11, a plasmid with dual origins of replication isolated from Lactobacillus casei MCJ and construction of shuttle vectors with each replicon | |
| Khunthongpan et al. | Providencia thailandensis sp. nov., isolated from seafood processing wastewater | |
| JP2004236637A (ja) | 多糖類生成に関与する遺伝子及びその利用 | |
| JPWO2004081216A1 (ja) | 酢酸菌のアルコール脱水素酵素遺伝子 | |
| JP5319212B2 (ja) | 転移因子の転移が抑制された納豆菌、該納豆菌を用いて製造された納豆 | |
| CN110846333B (zh) | 一种deoB基因改造的重组菌株及其构建方法与应用 | |
| CN110791487B (zh) | 水稻受体激酶基因LOC_Os11g47290及其编码蛋白与应用 | |
| JP4352877B2 (ja) | γ−グルタミルシステインの製造法 | |
| JP3746556B2 (ja) | プラスミド及びプラスミドベクター | |
| JP4326967B2 (ja) | 酢酸耐性に関与する遺伝子、該遺伝子を用いて育種された酢酸菌、及び該酢酸菌を用いた食酢の製造方法 | |
| CN114349831B (zh) | aspA基因突变体、重组菌及制备L-缬氨酸的方法 | |
| AU763912B2 (en) | Novel strains of the bacillus subtilis group for food fermentation | |
| KR20100008715A (ko) | 잔탄검 생합성 관련 돌연변이 유전자와 이를 포함하는잔탄검 생합성 균주 및 돌연변이 균주를 이용한 잔탄검의제조방법 | |
| EP3498727B1 (en) | Microorganism having enhanced cellulose synthase gene stability and method of producing cellulose by using the same | |
| CN114540399A (zh) | 一种制备l-缬氨酸的方法及其所用基因突变体和生物材料 | |
| JP2011508599A (ja) | イノシン生産能を有するコリネバクテリウム属微生物及びこれを利用したイノシンの生産方法 | |
| Vohra et al. | Enhanced production of penicillin G acylase from a recombinant Escherichia coli | |
| CN108018232B (zh) | 一种具有遗传改造潜力的香蕉穿孔线虫伴生细菌 | |
| JP4083455B2 (ja) | 酢酸菌のアコニターゼ遺伝子、該遺伝子を用いて育種された酢酸菌、及び該酢酸菌を用いた食酢の製造方法 | |
| CN114790436B (zh) | 基因afsR_4突变及其应用 | |
| CN114315998B (zh) | Cey17_rs00300基因突变体及其在制备l-缬氨酸中的应用 | |
| KR100953104B1 (ko) | 류코노스톡에서 유래된 신규 플라스미드 및 이를 포함하는셔틀 벡터 | |
| US20240034983A1 (en) | Asporogenous bacteria and uses thereof as a feed ingredient |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20110802 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20130423 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20130619 |
|
| TRDD | Decision of grant or rejection written | ||
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A821 Effective date: 20130620 |
|
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20130709 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20130711 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 5319212 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| S111 | Request for change of ownership or part of ownership |
Free format text: JAPANESE INTERMEDIATE CODE: R313111 |
|
| S533 | Written request for registration of change of name |
Free format text: JAPANESE INTERMEDIATE CODE: R313533 |
|
| R350 | Written notification of registration of transfer |
Free format text: JAPANESE INTERMEDIATE CODE: R350 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |