JP2009171981A - 中胚葉および成体型内胚葉細胞集団 - Google Patents
中胚葉および成体型内胚葉細胞集団 Download PDFInfo
- Publication number
- JP2009171981A JP2009171981A JP2009071364A JP2009071364A JP2009171981A JP 2009171981 A JP2009171981 A JP 2009171981A JP 2009071364 A JP2009071364 A JP 2009071364A JP 2009071364 A JP2009071364 A JP 2009071364A JP 2009171981 A JP2009171981 A JP 2009171981A
- Authority
- JP
- Japan
- Prior art keywords
- cells
- gfp
- cell
- endoderm
- serum
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
- 210000003716 mesoderm Anatomy 0.000 title claims abstract description 71
- 210000004039 endoderm cell Anatomy 0.000 title claims abstract description 22
- 210000004027 cell Anatomy 0.000 claims abstract description 405
- 210000002966 serum Anatomy 0.000 claims abstract description 75
- 108090000623 proteins and genes Proteins 0.000 claims abstract description 69
- 238000000034 method Methods 0.000 claims abstract description 42
- 238000012258 culturing Methods 0.000 claims abstract description 23
- 210000001671 embryonic stem cell Anatomy 0.000 claims abstract description 16
- 230000014509 gene expression Effects 0.000 claims description 118
- 230000004069 differentiation Effects 0.000 claims description 78
- 239000000488 activin Substances 0.000 claims description 73
- 108010059616 Activins Proteins 0.000 claims description 72
- 210000001900 endoderm Anatomy 0.000 claims description 53
- 239000003550 marker Substances 0.000 claims description 24
- 210000003494 hepatocyte Anatomy 0.000 claims description 18
- 239000003795 chemical substances by application Substances 0.000 claims description 11
- 210000001704 mesoblast Anatomy 0.000 claims description 11
- 241001465754 Metazoa Species 0.000 claims description 9
- 238000004519 manufacturing process Methods 0.000 claims description 8
- 108010048367 enhanced green fluorescent protein Proteins 0.000 claims description 7
- 230000012010 growth Effects 0.000 claims description 7
- 230000004083 survival effect Effects 0.000 claims description 7
- 201000010099 disease Diseases 0.000 claims description 5
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 claims description 5
- 230000009261 transgenic effect Effects 0.000 claims description 5
- 230000006378 damage Effects 0.000 claims description 4
- 230000002950 deficient Effects 0.000 claims description 4
- 230000004064 dysfunction Effects 0.000 claims description 4
- 230000006870 function Effects 0.000 claims description 4
- 230000035800 maturation Effects 0.000 claims description 4
- 108700019146 Transgenes Proteins 0.000 claims description 3
- 230000003908 liver function Effects 0.000 claims description 3
- 230000035755 proliferation Effects 0.000 claims description 3
- GACSIVHAIFQKTC-OWOJBTEDSA-N 4-fumarylacetoacetic acid Chemical compound OC(=O)CC(=O)CC(=O)\C=C\C(O)=O GACSIVHAIFQKTC-OWOJBTEDSA-N 0.000 claims description 2
- 210000004962 mammalian cell Anatomy 0.000 claims description 2
- 102000005606 Activins Human genes 0.000 claims 2
- 230000001747 exhibiting effect Effects 0.000 claims 1
- 230000006872 improvement Effects 0.000 claims 1
- 238000011161 development Methods 0.000 abstract description 55
- 238000009256 replacement therapy Methods 0.000 abstract description 7
- 239000003814 drug Substances 0.000 abstract description 4
- 230000002062 proliferating effect Effects 0.000 abstract 1
- 108010043121 Green Fluorescent Proteins Proteins 0.000 description 219
- 102000004144 Green Fluorescent Proteins Human genes 0.000 description 219
- 239000005090 green fluorescent protein Substances 0.000 description 219
- 108010053099 Vascular Endothelial Growth Factor Receptor-2 Proteins 0.000 description 137
- 102000016549 Vascular Endothelial Growth Factor Receptor-2 Human genes 0.000 description 132
- 210000002242 embryoid body Anatomy 0.000 description 92
- 102100026818 Inhibin beta E chain Human genes 0.000 description 70
- 230000018109 developmental process Effects 0.000 description 52
- 230000003394 haemopoietic effect Effects 0.000 description 33
- 239000002243 precursor Substances 0.000 description 25
- 230000003511 endothelial effect Effects 0.000 description 18
- 238000010195 expression analysis Methods 0.000 description 18
- 210000001519 tissue Anatomy 0.000 description 15
- 102100023635 Alpha-fetoprotein Human genes 0.000 description 13
- 108010014608 Proto-Oncogene Proteins c-kit Proteins 0.000 description 13
- 102000016971 Proto-Oncogene Proteins c-kit Human genes 0.000 description 13
- 239000002609 medium Substances 0.000 description 13
- 241000699666 Mus <mouse, genus> Species 0.000 description 12
- 241000699670 Mus sp. Species 0.000 description 12
- 238000004458 analytical method Methods 0.000 description 12
- 210000004413 cardiac myocyte Anatomy 0.000 description 12
- 238000002955 isolation Methods 0.000 description 12
- 210000002027 skeletal muscle Anatomy 0.000 description 12
- 101710123134 Ice-binding protein Proteins 0.000 description 11
- 101710082837 Ice-structuring protein Proteins 0.000 description 11
- 101710107540 Type-2 ice-structuring protein Proteins 0.000 description 11
- 230000020619 endoderm development Effects 0.000 description 11
- 239000012634 fragment Substances 0.000 description 11
- 108010088751 Albumins Proteins 0.000 description 10
- 102000009027 Albumins Human genes 0.000 description 10
- 230000000694 effects Effects 0.000 description 10
- 210000003566 hemangioblast Anatomy 0.000 description 10
- 210000004185 liver Anatomy 0.000 description 10
- 102100024505 Bone morphogenetic protein 4 Human genes 0.000 description 9
- 108091003079 Bovine Serum Albumin Proteins 0.000 description 9
- 238000010240 RT-PCR analysis Methods 0.000 description 9
- 238000003556 assay Methods 0.000 description 9
- 230000000747 cardiac effect Effects 0.000 description 9
- 210000003743 erythrocyte Anatomy 0.000 description 9
- 239000012894 fetal calf serum Substances 0.000 description 9
- 230000003847 mesoderm development Effects 0.000 description 9
- 230000001105 regulatory effect Effects 0.000 description 9
- 239000013598 vector Substances 0.000 description 9
- 101000762379 Homo sapiens Bone morphogenetic protein 4 Proteins 0.000 description 8
- 230000024245 cell differentiation Effects 0.000 description 8
- 210000003981 ectoderm Anatomy 0.000 description 8
- 230000011132 hemopoiesis Effects 0.000 description 8
- 230000001965 increasing effect Effects 0.000 description 8
- 230000006698 induction Effects 0.000 description 8
- 210000004165 myocardium Anatomy 0.000 description 8
- 239000013612 plasmid Substances 0.000 description 8
- 230000008685 targeting Effects 0.000 description 8
- 102100031690 Erythroid transcription factor Human genes 0.000 description 7
- 102100024785 Fibroblast growth factor 2 Human genes 0.000 description 7
- 108090000379 Fibroblast growth factor 2 Proteins 0.000 description 7
- 102000007354 PAX6 Transcription Factor Human genes 0.000 description 7
- 108010032788 PAX6 Transcription Factor Proteins 0.000 description 7
- 238000012512 characterization method Methods 0.000 description 7
- 238000000338 in vitro Methods 0.000 description 7
- 230000002107 myocardial effect Effects 0.000 description 7
- UZOVYGYOLBIAJR-UHFFFAOYSA-N 4-isocyanato-4'-methyldiphenylmethane Chemical compound C1=CC(C)=CC=C1CC1=CC=C(N=C=O)C=C1 UZOVYGYOLBIAJR-UHFFFAOYSA-N 0.000 description 6
- 108700028369 Alleles Proteins 0.000 description 6
- 101710100588 Erythroid transcription factor Proteins 0.000 description 6
- 108091034117 Oligonucleotide Proteins 0.000 description 6
- 102000016540 Tyrosine aminotransferases Human genes 0.000 description 6
- 108010042606 Tyrosine transaminase Proteins 0.000 description 6
- 210000002889 endothelial cell Anatomy 0.000 description 6
- 239000000203 mixture Substances 0.000 description 6
- 230000025366 tissue development Effects 0.000 description 6
- 238000002054 transplantation Methods 0.000 description 6
- 101150026303 HEX1 gene Proteins 0.000 description 5
- 101710150336 Protein Rex Proteins 0.000 description 5
- 206010043276 Teratoma Diseases 0.000 description 5
- 238000010009 beating Methods 0.000 description 5
- 230000011712 cell development Effects 0.000 description 5
- 210000002257 embryonic structure Anatomy 0.000 description 5
- 238000002474 experimental method Methods 0.000 description 5
- 210000004602 germ cell Anatomy 0.000 description 5
- 210000003958 hematopoietic stem cell Anatomy 0.000 description 5
- 239000000523 sample Substances 0.000 description 5
- 102000007469 Actins Human genes 0.000 description 4
- 108010085238 Actins Proteins 0.000 description 4
- CIWBSHSKHKDKBQ-JLAZNSOCSA-N Ascorbic acid Chemical compound OC[C@H](O)[C@H]1OC(=O)C(O)=C1O CIWBSHSKHKDKBQ-JLAZNSOCSA-N 0.000 description 4
- 108020004705 Codon Proteins 0.000 description 4
- 108020004414 DNA Proteins 0.000 description 4
- 108091081024 Start codon Proteins 0.000 description 4
- 102000005789 Vascular Endothelial Growth Factors Human genes 0.000 description 4
- 108010019530 Vascular Endothelial Growth Factors Proteins 0.000 description 4
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 4
- 239000000654 additive Substances 0.000 description 4
- 230000015572 biosynthetic process Effects 0.000 description 4
- 230000004663 cell proliferation Effects 0.000 description 4
- 239000002299 complementary DNA Substances 0.000 description 4
- 150000001875 compounds Chemical class 0.000 description 4
- 239000003636 conditioned culture medium Substances 0.000 description 4
- 210000003038 endothelium Anatomy 0.000 description 4
- 238000000684 flow cytometry Methods 0.000 description 4
- 210000003734 kidney Anatomy 0.000 description 4
- 210000005229 liver cell Anatomy 0.000 description 4
- 230000001537 neural effect Effects 0.000 description 4
- 230000008569 process Effects 0.000 description 4
- 230000000541 pulsatile effect Effects 0.000 description 4
- 108010010803 Gelatin Proteins 0.000 description 3
- 108010002386 Interleukin-3 Proteins 0.000 description 3
- 108090001005 Interleukin-6 Proteins 0.000 description 3
- 102100038380 Myogenic factor 5 Human genes 0.000 description 3
- 101710099061 Myogenic factor 5 Proteins 0.000 description 3
- 101100519293 Neurospora crassa (strain ATCC 24698 / 74-OR23-1A / CBS 708.71 / DSM 1257 / FGSC 987) pdx-1 gene Proteins 0.000 description 3
- 108020005067 RNA Splice Sites Proteins 0.000 description 3
- 108020004440 Thymidine kinase Proteins 0.000 description 3
- 102000040945 Transcription factor Human genes 0.000 description 3
- 108091023040 Transcription factor Proteins 0.000 description 3
- 210000000648 angioblast Anatomy 0.000 description 3
- 238000000137 annealing Methods 0.000 description 3
- 238000006243 chemical reaction Methods 0.000 description 3
- 230000001143 conditioned effect Effects 0.000 description 3
- 230000007423 decrease Effects 0.000 description 3
- 238000010586 diagram Methods 0.000 description 3
- 229940079593 drug Drugs 0.000 description 3
- 230000008913 ectoderm development Effects 0.000 description 3
- 238000011156 evaluation Methods 0.000 description 3
- 239000013604 expression vector Substances 0.000 description 3
- 239000008273 gelatin Substances 0.000 description 3
- 229920000159 gelatin Polymers 0.000 description 3
- 235000019322 gelatine Nutrition 0.000 description 3
- 235000011852 gelatine desserts Nutrition 0.000 description 3
- 239000003102 growth factor Substances 0.000 description 3
- 230000001939 inductive effect Effects 0.000 description 3
- 210000004072 lung Anatomy 0.000 description 3
- 210000002540 macrophage Anatomy 0.000 description 3
- MYWUZJCMWCOHBA-VIFPVBQESA-N methamphetamine Chemical compound CN[C@@H](C)CC1=CC=CC=C1 MYWUZJCMWCOHBA-VIFPVBQESA-N 0.000 description 3
- 229920000609 methyl cellulose Polymers 0.000 description 3
- 239000001923 methylcellulose Substances 0.000 description 3
- 238000007479 molecular analysis Methods 0.000 description 3
- 210000002241 neurite Anatomy 0.000 description 3
- 238000000059 patterning Methods 0.000 description 3
- 210000000130 stem cell Anatomy 0.000 description 3
- 230000002123 temporal effect Effects 0.000 description 3
- 238000012360 testing method Methods 0.000 description 3
- 238000011144 upstream manufacturing Methods 0.000 description 3
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 2
- -1 2-nitro-4-trifluoromethylbenzoyl Chemical group 0.000 description 2
- BPYKTIZUTYGOLE-IFADSCNNSA-N Bilirubin Chemical compound N1C(=O)C(C)=C(C=C)\C1=C\C1=C(C)C(CCC(O)=O)=C(CC2=C(C(C)=C(\C=C/3C(=C(C=C)C(=O)N\3)C)N2)CCC(O)=O)N1 BPYKTIZUTYGOLE-IFADSCNNSA-N 0.000 description 2
- 102100024506 Bone morphogenetic protein 2 Human genes 0.000 description 2
- 102000002664 Core Binding Factor Alpha 2 Subunit Human genes 0.000 description 2
- 108010043471 Core Binding Factor Alpha 2 Subunit Proteins 0.000 description 2
- RTZKZFJDLAIYFH-UHFFFAOYSA-N Diethyl ether Chemical compound CCOCC RTZKZFJDLAIYFH-UHFFFAOYSA-N 0.000 description 2
- 239000006144 Dulbecco’s modified Eagle's medium Substances 0.000 description 2
- 108700024394 Exon Proteins 0.000 description 2
- 238000012413 Fluorescence activated cell sorting analysis Methods 0.000 description 2
- 108010017213 Granulocyte-Macrophage Colony-Stimulating Factor Proteins 0.000 description 2
- 102100039620 Granulocyte-macrophage colony-stimulating factor Human genes 0.000 description 2
- 206010019663 Hepatic failure Diseases 0.000 description 2
- 102100022054 Hepatocyte nuclear factor 4-alpha Human genes 0.000 description 2
- 101000762366 Homo sapiens Bone morphogenetic protein 2 Proteins 0.000 description 2
- 101001045740 Homo sapiens Hepatocyte nuclear factor 4-alpha Proteins 0.000 description 2
- 101000612671 Homo sapiens Pulmonary surfactant-associated protein C Proteins 0.000 description 2
- 102000003815 Interleukin-11 Human genes 0.000 description 2
- 108090000177 Interleukin-11 Proteins 0.000 description 2
- 102100020880 Kit ligand Human genes 0.000 description 2
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 2
- 229930182816 L-glutamine Natural products 0.000 description 2
- 108010046938 Macrophage Colony-Stimulating Factor Proteins 0.000 description 2
- 102000007651 Macrophage Colony-Stimulating Factor Human genes 0.000 description 2
- 102000008763 Neurofilament Proteins Human genes 0.000 description 2
- 108010088373 Neurofilament Proteins Proteins 0.000 description 2
- 238000010222 PCR analysis Methods 0.000 description 2
- 108010071690 Prealbumin Proteins 0.000 description 2
- 102100040971 Pulmonary surfactant-associated protein C Human genes 0.000 description 2
- 108010039445 Stem Cell Factor Proteins 0.000 description 2
- 102000006601 Thymidine Kinase Human genes 0.000 description 2
- 102000009190 Transthyretin Human genes 0.000 description 2
- 102000004987 Troponin T Human genes 0.000 description 2
- 108090001108 Troponin T Proteins 0.000 description 2
- 241000269370 Xenopus <genus> Species 0.000 description 2
- 230000000996 additive effect Effects 0.000 description 2
- 210000001789 adipocyte Anatomy 0.000 description 2
- 102000015395 alpha 1-Antitrypsin Human genes 0.000 description 2
- 108010050122 alpha 1-Antitrypsin Proteins 0.000 description 2
- 229940024142 alpha 1-antitrypsin Drugs 0.000 description 2
- 108010026331 alpha-Fetoproteins Proteins 0.000 description 2
- 230000003321 amplification Effects 0.000 description 2
- 238000010171 animal model Methods 0.000 description 2
- 229960005070 ascorbic acid Drugs 0.000 description 2
- 235000010323 ascorbic acid Nutrition 0.000 description 2
- 239000011668 ascorbic acid Substances 0.000 description 2
- 210000002459 blastocyst Anatomy 0.000 description 2
- 210000004369 blood Anatomy 0.000 description 2
- 239000008280 blood Substances 0.000 description 2
- 239000002775 capsule Substances 0.000 description 2
- 230000001332 colony forming effect Effects 0.000 description 2
- 230000003247 decreasing effect Effects 0.000 description 2
- 230000007547 defect Effects 0.000 description 2
- 230000013020 embryo development Effects 0.000 description 2
- 230000000925 erythroid effect Effects 0.000 description 2
- 210000000777 hematopoietic system Anatomy 0.000 description 2
- 230000006801 homologous recombination Effects 0.000 description 2
- 238000002744 homologous recombination Methods 0.000 description 2
- 238000001727 in vivo Methods 0.000 description 2
- 230000000977 initiatory effect Effects 0.000 description 2
- NOESYZHRGYRDHS-UHFFFAOYSA-N insulin Chemical compound N1C(=O)C(NC(=O)C(CCC(N)=O)NC(=O)C(CCC(O)=O)NC(=O)C(C(C)C)NC(=O)C(NC(=O)CN)C(C)CC)CSSCC(C(NC(CO)C(=O)NC(CC(C)C)C(=O)NC(CC=2C=CC(O)=CC=2)C(=O)NC(CCC(N)=O)C(=O)NC(CC(C)C)C(=O)NC(CCC(O)=O)C(=O)NC(CC(N)=O)C(=O)NC(CC=2C=CC(O)=CC=2)C(=O)NC(CSSCC(NC(=O)C(C(C)C)NC(=O)C(CC(C)C)NC(=O)C(CC=2C=CC(O)=CC=2)NC(=O)C(CC(C)C)NC(=O)C(C)NC(=O)C(CCC(O)=O)NC(=O)C(C(C)C)NC(=O)C(CC(C)C)NC(=O)C(CC=2NC=NC=2)NC(=O)C(CO)NC(=O)CNC2=O)C(=O)NCC(=O)NC(CCC(O)=O)C(=O)NC(CCCNC(N)=N)C(=O)NCC(=O)NC(CC=3C=CC=CC=3)C(=O)NC(CC=3C=CC=CC=3)C(=O)NC(CC=3C=CC(O)=CC=3)C(=O)NC(C(C)O)C(=O)N3C(CCC3)C(=O)NC(CCCCN)C(=O)NC(C)C(O)=O)C(=O)NC(CC(N)=O)C(O)=O)=O)NC(=O)C(C(C)CC)NC(=O)C(CO)NC(=O)C(C(C)O)NC(=O)C1CSSCC2NC(=O)C(CC(C)C)NC(=O)C(NC(=O)C(CCC(N)=O)NC(=O)C(CC(N)=O)NC(=O)C(NC(=O)C(N)CC=1C=CC=CC=1)C(C)C)CC1=CN=CN1 NOESYZHRGYRDHS-UHFFFAOYSA-N 0.000 description 2
- 230000010354 integration Effects 0.000 description 2
- 238000012933 kinetic analysis Methods 0.000 description 2
- 208000007903 liver failure Diseases 0.000 description 2
- 231100000835 liver failure Toxicity 0.000 description 2
- 108010082117 matrigel Proteins 0.000 description 2
- 210000004379 membrane Anatomy 0.000 description 2
- 239000012528 membrane Substances 0.000 description 2
- 238000012544 monitoring process Methods 0.000 description 2
- 230000007472 neurodevelopment Effects 0.000 description 2
- 210000005044 neurofilament Anatomy 0.000 description 2
- 230000004031 neuronal differentiation Effects 0.000 description 2
- OUBCNLGXQFSTLU-UHFFFAOYSA-N nitisinone Chemical compound [O-][N+](=O)C1=CC(C(F)(F)F)=CC=C1C(=O)C1C(=O)CCCC1=O OUBCNLGXQFSTLU-UHFFFAOYSA-N 0.000 description 2
- 229960001721 nitisinone Drugs 0.000 description 2
- 210000003458 notochord Anatomy 0.000 description 2
- 238000003199 nucleic acid amplification method Methods 0.000 description 2
- 230000008488 polyadenylation Effects 0.000 description 2
- 102000027426 receptor tyrosine kinases Human genes 0.000 description 2
- 108091008598 receptor tyrosine kinases Proteins 0.000 description 2
- 230000002829 reductive effect Effects 0.000 description 2
- 230000002441 reversible effect Effects 0.000 description 2
- 238000000926 separation method Methods 0.000 description 2
- 239000012679 serum free medium Substances 0.000 description 2
- 210000002363 skeletal muscle cell Anatomy 0.000 description 2
- 230000004096 skeletal muscle tissue growth Effects 0.000 description 2
- 210000003491 skin Anatomy 0.000 description 2
- 241000894007 species Species 0.000 description 2
- 238000010186 staining Methods 0.000 description 2
- UCSJYZPVAKXKNQ-HZYVHMACSA-N streptomycin Chemical compound CN[C@H]1[C@H](O)[C@@H](O)[C@H](CO)O[C@H]1O[C@@H]1[C@](C=O)(O)[C@H](C)O[C@H]1O[C@@H]1[C@@H](NC(N)=N)[C@H](O)[C@@H](NC(N)=N)[C@H](O)[C@H]1O UCSJYZPVAKXKNQ-HZYVHMACSA-N 0.000 description 2
- 238000013518 transcription Methods 0.000 description 2
- 230000035897 transcription Effects 0.000 description 2
- 238000011830 transgenic mouse model Methods 0.000 description 2
- 238000013519 translation Methods 0.000 description 2
- 230000003827 upregulation Effects 0.000 description 2
- 108020005029 5' Flanking Region Proteins 0.000 description 1
- 102000010825 Actinin Human genes 0.000 description 1
- 108010063503 Actinin Proteins 0.000 description 1
- 101000642536 Apis mellifera Venom serine protease 34 Proteins 0.000 description 1
- 108010003415 Aspartate Aminotransferases Proteins 0.000 description 1
- 102000004625 Aspartate Aminotransferases Human genes 0.000 description 1
- 108010049955 Bone Morphogenetic Protein 4 Proteins 0.000 description 1
- 208000020084 Bone disease Diseases 0.000 description 1
- 101710113083 Carbamoyl-phosphate synthase Proteins 0.000 description 1
- 208000024172 Cardiovascular disease Diseases 0.000 description 1
- 108010051219 Cre recombinase Proteins 0.000 description 1
- 102000004127 Cytokines Human genes 0.000 description 1
- 108090000695 Cytokines Proteins 0.000 description 1
- 241000252212 Danio rerio Species 0.000 description 1
- 101100179523 Danio rerio ihhb gene Proteins 0.000 description 1
- 108010053770 Deoxyribonucleases Proteins 0.000 description 1
- 102000016911 Deoxyribonucleases Human genes 0.000 description 1
- 208000013558 Developmental Bone disease Diseases 0.000 description 1
- 206010013801 Duchenne Muscular Dystrophy Diseases 0.000 description 1
- 102000003951 Erythropoietin Human genes 0.000 description 1
- 108090000394 Erythropoietin Proteins 0.000 description 1
- 102100037680 Fibroblast growth factor 8 Human genes 0.000 description 1
- 108010028165 GATA1 Transcription Factor Proteins 0.000 description 1
- 102000016669 GATA1 Transcription Factor Human genes 0.000 description 1
- 108010017080 Granulocyte Colony-Stimulating Factor Proteins 0.000 description 1
- 102000004269 Granulocyte Colony-Stimulating Factor Human genes 0.000 description 1
- 206010019280 Heart failures Diseases 0.000 description 1
- 102100035961 Hematopoietically-expressed homeobox protein HHEX Human genes 0.000 description 1
- 102000009094 Hepatocyte Nuclear Factor 3-beta Human genes 0.000 description 1
- 108010087745 Hepatocyte Nuclear Factor 3-beta Proteins 0.000 description 1
- 241000282412 Homo Species 0.000 description 1
- 101001066268 Homo sapiens Erythroid transcription factor Proteins 0.000 description 1
- 101001027382 Homo sapiens Fibroblast growth factor 8 Proteins 0.000 description 1
- 101001021503 Homo sapiens Hematopoietically-expressed homeobox protein HHEX Proteins 0.000 description 1
- 101000851007 Homo sapiens Vascular endothelial growth factor receptor 2 Proteins 0.000 description 1
- 102100023915 Insulin Human genes 0.000 description 1
- 108090001061 Insulin Proteins 0.000 description 1
- 102000004889 Interleukin-6 Human genes 0.000 description 1
- 101150088608 Kdr gene Proteins 0.000 description 1
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 1
- 108010058398 Macrophage Colony-Stimulating Factor Receptor Proteins 0.000 description 1
- 241000124008 Mammalia Species 0.000 description 1
- 241000699660 Mus musculus Species 0.000 description 1
- 101100310648 Mus musculus Sox17 gene Proteins 0.000 description 1
- 102000003505 Myosin Human genes 0.000 description 1
- 108060008487 Myosin Proteins 0.000 description 1
- 101150079937 NEUROD1 gene Proteins 0.000 description 1
- 241001045988 Neogene Species 0.000 description 1
- 229930193140 Neomycin Natural products 0.000 description 1
- 206010028980 Neoplasm Diseases 0.000 description 1
- 108091028043 Nucleic acid sequence Proteins 0.000 description 1
- 229930182555 Penicillin Natural products 0.000 description 1
- JGSARLDLIJGVTE-MBNYWOFBSA-N Penicillin G Chemical compound N([C@H]1[C@H]2SC([C@@H](N2C1=O)C(O)=O)(C)C)C(=O)CC1=CC=CC=C1 JGSARLDLIJGVTE-MBNYWOFBSA-N 0.000 description 1
- 206010035226 Plasma cell myeloma Diseases 0.000 description 1
- 108010029485 Protein Isoforms Proteins 0.000 description 1
- 102000001708 Protein Isoforms Human genes 0.000 description 1
- 108010076504 Protein Sorting Signals Proteins 0.000 description 1
- 108091034057 RNA (poly(A)) Proteins 0.000 description 1
- 102000004278 Receptor Protein-Tyrosine Kinases Human genes 0.000 description 1
- 108090000873 Receptor Protein-Tyrosine Kinases Proteins 0.000 description 1
- 206010072610 Skeletal dysplasia Diseases 0.000 description 1
- 238000002105 Southern blotting Methods 0.000 description 1
- 108010006785 Taq Polymerase Proteins 0.000 description 1
- 102000036693 Thrombopoietin Human genes 0.000 description 1
- 102100034195 Thrombopoietin Human genes 0.000 description 1
- 108010041111 Thrombopoietin Proteins 0.000 description 1
- 102000004338 Transferrin Human genes 0.000 description 1
- 108090000901 Transferrin Proteins 0.000 description 1
- 108010073929 Vascular Endothelial Growth Factor A Proteins 0.000 description 1
- 230000001919 adrenal effect Effects 0.000 description 1
- 239000011543 agarose gel Substances 0.000 description 1
- 208000007502 anemia Diseases 0.000 description 1
- 230000008901 benefit Effects 0.000 description 1
- 210000002960 bfu-e Anatomy 0.000 description 1
- 210000001172 blastoderm Anatomy 0.000 description 1
- 210000000601 blood cell Anatomy 0.000 description 1
- 210000000988 bone and bone Anatomy 0.000 description 1
- 238000010322 bone marrow transplantation Methods 0.000 description 1
- 238000004113 cell culture Methods 0.000 description 1
- 239000002771 cell marker Substances 0.000 description 1
- 239000002458 cell surface marker Substances 0.000 description 1
- 230000008859 change Effects 0.000 description 1
- 210000004978 chinese hamster ovary cell Anatomy 0.000 description 1
- 238000010367 cloning Methods 0.000 description 1
- 238000012790 confirmation Methods 0.000 description 1
- 238000010276 construction Methods 0.000 description 1
- 230000001276 controlling effect Effects 0.000 description 1
- 239000006059 cover glass Substances 0.000 description 1
- 230000001351 cycling effect Effects 0.000 description 1
- 230000037416 cystogenesis Effects 0.000 description 1
- 230000003111 delayed effect Effects 0.000 description 1
- 238000004925 denaturation Methods 0.000 description 1
- 230000036425 denaturation Effects 0.000 description 1
- 230000001419 dependent effect Effects 0.000 description 1
- 230000001627 detrimental effect Effects 0.000 description 1
- 206010012601 diabetes mellitus Diseases 0.000 description 1
- 230000029087 digestion Effects 0.000 description 1
- 238000009510 drug design Methods 0.000 description 1
- 108010030074 endodeoxyribonuclease MluI Proteins 0.000 description 1
- 229940105423 erythropoietin Drugs 0.000 description 1
- 230000001605 fetal effect Effects 0.000 description 1
- 238000001943 fluorescence-activated cell sorting Methods 0.000 description 1
- 238000002825 functional assay Methods 0.000 description 1
- IRSCQMHQWWYFCW-UHFFFAOYSA-N ganciclovir Chemical compound O=C1NC(N)=NC2=C1N=CN2COC(CO)CO IRSCQMHQWWYFCW-UHFFFAOYSA-N 0.000 description 1
- 229960002963 ganciclovir Drugs 0.000 description 1
- 230000007045 gastrulation Effects 0.000 description 1
- 210000001654 germ layer Anatomy 0.000 description 1
- 108060003196 globin Proteins 0.000 description 1
- 150000004676 glycans Chemical class 0.000 description 1
- 238000009499 grossing Methods 0.000 description 1
- 230000036541 health Effects 0.000 description 1
- 210000002064 heart cell Anatomy 0.000 description 1
- 210000005003 heart tissue Anatomy 0.000 description 1
- 102000055590 human KDR Human genes 0.000 description 1
- 210000005260 human cell Anatomy 0.000 description 1
- 238000009396 hybridization Methods 0.000 description 1
- 210000004408 hybridoma Anatomy 0.000 description 1
- 101150010866 ihh gene Proteins 0.000 description 1
- 230000003053 immunization Effects 0.000 description 1
- 230000016784 immunoglobulin production Effects 0.000 description 1
- 238000012744 immunostaining Methods 0.000 description 1
- 238000010874 in vitro model Methods 0.000 description 1
- 230000002779 inactivation Effects 0.000 description 1
- 238000011534 incubation Methods 0.000 description 1
- 230000002401 inhibitory effect Effects 0.000 description 1
- 229940125396 insulin Drugs 0.000 description 1
- 210000004347 intestinal mucosa Anatomy 0.000 description 1
- 210000004153 islets of langerhan Anatomy 0.000 description 1
- 238000002372 labelling Methods 0.000 description 1
- 230000002045 lasting effect Effects 0.000 description 1
- 208000032839 leukemia Diseases 0.000 description 1
- 239000003446 ligand Substances 0.000 description 1
- 230000031142 liver development Effects 0.000 description 1
- 210000005265 lung cell Anatomy 0.000 description 1
- 210000001161 mammalian embryo Anatomy 0.000 description 1
- 239000000463 material Substances 0.000 description 1
- 238000010208 microarray analysis Methods 0.000 description 1
- 230000005012 migration Effects 0.000 description 1
- 238000013508 migration Methods 0.000 description 1
- 230000009456 molecular mechanism Effects 0.000 description 1
- PJUIMOJAAPLTRJ-UHFFFAOYSA-N monothioglycerol Chemical compound OCC(O)CS PJUIMOJAAPLTRJ-UHFFFAOYSA-N 0.000 description 1
- 230000000921 morphogenic effect Effects 0.000 description 1
- 230000035772 mutation Effects 0.000 description 1
- 201000000050 myeloid neoplasm Diseases 0.000 description 1
- 101150091879 neo gene Proteins 0.000 description 1
- 229960004927 neomycin Drugs 0.000 description 1
- 210000000653 nervous system Anatomy 0.000 description 1
- 210000001020 neural plate Anatomy 0.000 description 1
- 230000014511 neuron projection development Effects 0.000 description 1
- 210000004789 organ system Anatomy 0.000 description 1
- 210000000496 pancreas Anatomy 0.000 description 1
- 230000037361 pathway Effects 0.000 description 1
- 229940049954 penicillin Drugs 0.000 description 1
- 229920000642 polymer Polymers 0.000 description 1
- 102000040430 polynucleotide Human genes 0.000 description 1
- 108091033319 polynucleotide Proteins 0.000 description 1
- 239000002157 polynucleotide Substances 0.000 description 1
- 229920001184 polypeptide Polymers 0.000 description 1
- 229920001282 polysaccharide Polymers 0.000 description 1
- 239000005017 polysaccharide Substances 0.000 description 1
- OXCMYAYHXIHQOA-UHFFFAOYSA-N potassium;[2-butyl-5-chloro-3-[[4-[2-(1,2,4-triaza-3-azanidacyclopenta-1,4-dien-5-yl)phenyl]phenyl]methyl]imidazol-4-yl]methanol Chemical compound [K+].CCCCC1=NC(Cl)=C(CO)N1CC1=CC=C(C=2C(=CC=CC=2)C2=N[N-]N=N2)C=C1 OXCMYAYHXIHQOA-UHFFFAOYSA-N 0.000 description 1
- 210000002996 primitive erythroblast Anatomy 0.000 description 1
- 210000001811 primitive streak Anatomy 0.000 description 1
- 102000004196 processed proteins & peptides Human genes 0.000 description 1
- 108090000765 processed proteins & peptides Proteins 0.000 description 1
- 239000000047 product Substances 0.000 description 1
- 230000000750 progressive effect Effects 0.000 description 1
- 102000004169 proteins and genes Human genes 0.000 description 1
- 102000005962 receptors Human genes 0.000 description 1
- 108020003175 receptors Proteins 0.000 description 1
- 230000006798 recombination Effects 0.000 description 1
- 238000005215 recombination Methods 0.000 description 1
- 230000009467 reduction Effects 0.000 description 1
- 230000001172 regenerating effect Effects 0.000 description 1
- 238000011160 research Methods 0.000 description 1
- 230000004044 response Effects 0.000 description 1
- 238000010839 reverse transcription Methods 0.000 description 1
- 238000012216 screening Methods 0.000 description 1
- 239000004017 serum-free culture medium Substances 0.000 description 1
- 150000003384 small molecules Chemical class 0.000 description 1
- 210000002460 smooth muscle Anatomy 0.000 description 1
- 210000000329 smooth muscle myocyte Anatomy 0.000 description 1
- 239000007787 solid Substances 0.000 description 1
- 238000009331 sowing Methods 0.000 description 1
- 229960005322 streptomycin Drugs 0.000 description 1
- 239000000758 substrate Substances 0.000 description 1
- 239000013589 supplement Substances 0.000 description 1
- 239000000725 suspension Substances 0.000 description 1
- 230000002459 sustained effect Effects 0.000 description 1
- 239000012581 transferrin Substances 0.000 description 1
- 230000001052 transient effect Effects 0.000 description 1
- 230000007704 transition Effects 0.000 description 1
- 230000010024 tubular injury Effects 0.000 description 1
- 208000037978 tubular injury Diseases 0.000 description 1
- OUYCCCASQSFEME-UHFFFAOYSA-N tyrosine Natural products OC(=O)C(N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-UHFFFAOYSA-N 0.000 description 1
- 230000002792 vascular Effects 0.000 description 1
- 230000006459 vascular development Effects 0.000 description 1
- 208000019553 vascular disease Diseases 0.000 description 1
- 210000004509 vascular smooth muscle cell Anatomy 0.000 description 1
Images




































Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/06—Animal cells or tissues; Human cells or tissues
- C12N5/0602—Vertebrate cells
- C12N5/0603—Embryonic cells ; Embryoid bodies
- C12N5/0606—Pluripotent embryonic cells, e.g. embryonic stem cells [ES]
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P1/00—Drugs for disorders of the alimentary tract or the digestive system
- A61P1/16—Drugs for disorders of the alimentary tract or the digestive system for liver or gallbladder disorders, e.g. hepatoprotective agents, cholagogues, litholytics
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P17/00—Drugs for dermatological disorders
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P19/00—Drugs for skeletal disorders
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P21/00—Drugs for disorders of the muscular or neuromuscular system
- A61P21/04—Drugs for disorders of the muscular or neuromuscular system for myasthenia gravis
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P3/00—Drugs for disorders of the metabolism
- A61P3/08—Drugs for disorders of the metabolism for glucose homeostasis
- A61P3/10—Drugs for disorders of the metabolism for glucose homeostasis for hyperglycaemia, e.g. antidiabetics
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P35/00—Antineoplastic agents
- A61P35/02—Antineoplastic agents specific for leukemia
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P43/00—Drugs for specific purposes, not provided for in groups A61P1/00-A61P41/00
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P7/00—Drugs for disorders of the blood or the extracellular fluid
- A61P7/06—Antianaemics
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P9/00—Drugs for disorders of the cardiovascular system
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P9/00—Drugs for disorders of the cardiovascular system
- A61P9/04—Inotropic agents, i.e. stimulants of cardiac contraction; Drugs for heart failure
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/06—Animal cells or tissues; Human cells or tissues
- C12N5/0602—Vertebrate cells
- C12N5/0603—Embryonic cells ; Embryoid bodies
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/5005—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells
- G01N33/5008—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells for testing or evaluating the effect of chemical or biological compounds, e.g. drugs, cosmetics
- G01N33/5044—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells for testing or evaluating the effect of chemical or biological compounds, e.g. drugs, cosmetics involving specific cell types
- G01N33/5073—Stem cells
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2500/00—Specific components of cell culture medium
- C12N2500/90—Serum-free medium, which may still contain naturally-sourced components
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/10—Growth factors
- C12N2501/16—Activin; Inhibin; Mullerian inhibiting substance
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/40—Regulators of development
- C12N2501/415—Wnt; Frizzeled
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2506/00—Differentiation of animal cells from one lineage to another; Differentiation of pluripotent cells
- C12N2506/02—Differentiation of animal cells from one lineage to another; Differentiation of pluripotent cells from embryonic cells
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Chemical & Material Sciences (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Organic Chemistry (AREA)
- General Health & Medical Sciences (AREA)
- Biomedical Technology (AREA)
- Medicinal Chemistry (AREA)
- Veterinary Medicine (AREA)
- Pharmacology & Pharmacy (AREA)
- General Chemical & Material Sciences (AREA)
- Animal Behavior & Ethology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Public Health (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Biotechnology (AREA)
- Genetics & Genomics (AREA)
- Developmental Biology & Embryology (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Cell Biology (AREA)
- Reproductive Health (AREA)
- Gynecology & Obstetrics (AREA)
- Hematology (AREA)
- Microbiology (AREA)
- Biochemistry (AREA)
- Diabetes (AREA)
- General Engineering & Computer Science (AREA)
- Immunology (AREA)
- Cardiology (AREA)
- Molecular Biology (AREA)
- Urology & Nephrology (AREA)
- Physical Education & Sports Medicine (AREA)
- Heart & Thoracic Surgery (AREA)
- Toxicology (AREA)
- Tropical Medicine & Parasitology (AREA)
- Pathology (AREA)
- General Physics & Mathematics (AREA)
- Analytical Chemistry (AREA)
Abstract
【解決手段】血清の存在下で約2から約10日間の間ヒト胚性幹細胞を培養すること、短尾を発現する細胞を単離すること、および血清の非存在下で約1から約15日間の間、短尾を発現する前記細胞を培養することにより得られる、内胚葉細胞が濃縮された細胞集団。該細胞集団は、細胞の分化増殖に影響を及ぼす薬剤の同定、組織発達に関与する遺伝子の同定、および細胞置換療法のための分化細胞や組織の作製等に有用である。
【選択図】なし
Description
ES細胞増殖および分化。ES細胞を、15%ウシ胎児血清(FCS)、ペニシリン、ストレプトマイシン、LIF(1%ならし培地)、および1.5×10-4Mモノチオグリセロール(MTG;シグマ)を補充したダルベッコ改変イーグル培地(DMEM)中、照射胚支持細胞上に維持した。分化開始の2日前に、細胞を同じ培地中のゼラチン化プレートに移した。EBの産生のために、ES細胞をトリプシン処理し、異なる培養物中に種々の密度で播種した。EBの分化は、15%FCS、2mM L-グルタミン(ギブコ/BRL)、トランスフェリン(200μg/ml)、0.5mMアスコルビン酸(シグマ)、および4.5×10-4M MTGを補充したIMDM中、60mmペトリ等級皿中で実施した。培養物は、加湿チャンバー中で、5%CO2/空気混合物中で、37℃で維持した。
2つの異なる血清を含まない培地を、以下の実施例の異なる態様において使用した:ノックアウトSR(ギブコBRL)およびStemPro34(ギブコBRL)を補充したIIMD。
A)芽球コロニー:芽細胞コロニーを作製するために(BL-CFCアッセイ)、EB由来細胞を、10%FCS(ハイクローン)、血管内皮増殖因子(VEGF;5ng/ml)、c-kitリガンド(KL;1%ならし培地)、IL-6(5ng/ml)および25%D4T内皮細胞ならし培地(Kennedyら(1997)Nature 386:488-93)を補充した1%メチルセルロース中、0.5×1.5×105細胞/mlで播種した。移行コロニーをVEGFの非存在下で作製した。コロニーは4日間の培養後に評価した。B)造血コロニー:原始的および成体型造血コロニーの増殖のために、細胞を、10%血漿由来血清(PDS;Antech)、5%タンパク質非含有ハイブリドーマ培地(PFHM-II;ギブコ-BRL)と以下のサイトカインを含む1%メチルセルロース中に播種した:c-kitリガンド(KL;1%ならし培地)、エリスロポエチン(2U/ml)、IL-11(25ng/ml)、IL-3(1%ならし培地)、GM-CSF(3ng/ml)、G-CSF(30ng/ml)、M-CSF(5ng/ml)、IL-6(5ng/ml)およびトロンボポエチン(TPO;5ng/ml)。培養物を37℃、5%CO2中に維持した。原始赤血球コロニーを培養の5〜6日目に評価し、一方、成体型赤血球(BFU-E)、マクロファージ、および多系統コロニーは、培養の7〜10日目に計測した。c-kitリガンドは、KL発現ベクター(親切にもジェネティクス・インスチチュートから提供された)でトランスフェクトしたCHO細胞により調整された培地から得られた。IL-3は、IL-3を発現しているベクターでトランスフェクトしたX63AG8-653ミエローマ細胞により調整された培地から得られた。VEGF、GM-CSF、M-CSF、IL-6、IL-11、アクチビン、BMP2、BMP4、bFGF、FGF8、およびIhhは、R&Dシステムズから購入した。
細胞を、24ウェルのペトリ等級プレート中、15%FCS(またはノックアウトSR)、2mM L-グルタミン(ギブコ/BRL)、0.5mMアスコルビン酸(シグマ)、および4.5×10-4M MTGを補充したIMDM1mlあたり、2×105で培養した。これらを使用して、細胞がウェルの底に付着するのを防いだ。
GFP+細胞を、15%血清代替添加物で補充したIMDM中で再凝集した。20時間後、凝集物を、10%血清代替添加物(血清非含有)を有するIMDM中、24または96ウェルプレートのウェル中で培養した。ウェルはゼラチンで予め処理しておいた。培養物を、拍動細胞の発達について毎日モニタリングした。拍動細胞は、通常、培養の2から6日目に検出された。
標準的な条件を使用して、細胞を染色した。染色した懸濁液はFACScan(Becton Dickinson、CA)で解析した。
ポリA+RT-PCR解析のために、Bradyら((1990)Meth.in Mol.and Cell Bio.2:17-25)の方法を使用した。逆転写、ポリAテーリング、およびPCR手順を記載の通りに実施し、ただし、X-dtオリゴヌクレオチドは5'-GTTAACTCGAGAATTC(T)24-3'まで短くした。PCR反応からの増幅産物はアガロースゲル上で分離し、Zeta-プローブGTメンブラン(Biorad)に転写するか、または、スロットブロット装置(Schleicher&Schuell)を用いてメンブランに転写した。得られたブロットは、遺伝子の3‘領域に対応する32PランダムプライムcDNA断片(Ready-to-Goラベリング、ファルマシア)を用いてハイブリダイズした(β−H1を除いて全て)。β−H1-特異的プローブは、その3’末端において8塩基の相同性を共有する2つのオリゴヌクレオチド、(5'-TGGAGTCAAAGAGGGCATCATAGACACATGGG-3'、5'-CAGTACACTGGCAATCCCATGTG-3'をアニーリングすることにより調製した。このβ-H1特異的オリゴヌクレオチドは、クレノウ平滑化反応を使用して、32Pで標識した。遺伝子特異的PCRのために、全RNAを、RNイージーミニキットを用いて各試料から抽出し、RNase非含有DNase(キアゲン)で処理した。2μgの全RNAを、OmniscriptRTキット(キアゲン)を使用してランダムヘキサマーを用いてcDNAに逆転写した。PCRを、適切なオリゴヌクレオチドを使用して実施した。PCR反応を、2.5UのTaqポリメラーゼ(プロメガ)、PCR緩衝液、2.5mM MgCl2、0.2μMの各プライマーおよび0.2mM dNTPを用いて実施した。サイクリング条件は以下の通りであった;94℃で5分間、その後、35サイクルの増幅(94℃で1分間変性、60℃で1分間アニーリング、72℃で1分間伸張)、最後に72℃で7分間インキュベート。
適切な培養条件下で、胚幹(ES)細胞は分化し、広域な系統からの発達中の細胞集団を含む、胚様体(EB)として知られる3次元のコロニーを形成する。Smith(2001)Annu.Rev.Cell Dev.Biol.17:435-62。これらのEB由来集団の中で、造血、内皮、心筋および骨格筋系統のものを含む、中胚葉誘導体を検出できる。
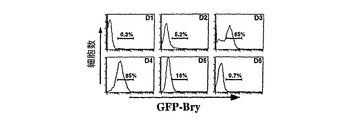
短尾+細胞をGFP発現に基づいて単離できるかどうかを決定するために、3.5日目のEBのGFP+個体を選別し、適切な遺伝子の発現について解析した。図4Aは、GFP陽性(2)および陰性(1)集団の単離に使用したゲートを示す。RT-PCR解析により、短尾発現はGFP+画分に限られることが判明し、GFP発現に基づいた細胞選別を短尾+細胞の単離に使用できることが示された。造血および内皮発達の最も古いマーカーの1つであるFlk-1は、GFP-画分よりもGFP+画分により高いレベルで存在し、少なくとも短尾+細胞の亜集団において発現されていることが示される。短尾およびFlk-1とは対照的に、初期神経発達に関与している遺伝子であるPax-6は[79、80]、GFP-画分においてより高いレベルで発現されており、この系統の前駆体は短尾陰性であることに一致している。これらの細胞選別研究により、短尾遺伝子座の制御下でのGFPの発現により、EBからの短尾+細胞の単離、特徴づけ、および操作の新規マーカーが提供されることが示される。
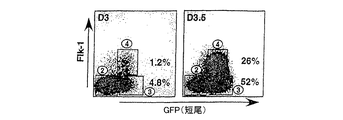
Flk-1は、初期胚において造血および内皮系統の確立に必須であることが示され、BL-CFCを含むこれらの系統の最も初期の前駆体で発現されている[Faloonら(2000)Development 127:1931-41]。血液および血管発達における中心的な役割から、GFP+集団内でのその発現は、これらの系統への特異化を受けている中胚葉の亜集団を規定すると仮定された。この可能性をさらに調査するために、EBを、いくつかの発達段階において、GFPおよびFlk-1陽性細胞の存在について解析した。図5Aに示した実験において、3.0日目のEB集団の4.8%がGFPを発現したが、Flk-1は発現せず、一方、細胞の1.2%が両方のマーカーを発現した。両方の画分のサイズは次の12時間かけて劇的に増加し、GFP+/Flk-1-およびGFP+/Flk-1+細胞は、それぞれ、全EB集団の52%および26%を示す。GFPおよびFlk-1発現により規定される3つの集団の発達能を評価するために、GFP-/Flk-1(画分2)、GFP+/Flk-1(画分3)、およびGFP+/Flk-1+(画分4)細胞を、両時点で単離し、BL-CFCおよび2゜EB含量および遺伝子発現パターンについて解析した。画分の能力を、予め選別した集団(画分1)の能力と比較した。BL-CFCの大半が、3.0および3.5日目の分化の両日にGFP+/Flk-1+画分に見出された(図5B)。以前の研究により、全てのBL-CFCがFlk-1を発現することが示されていたのでこれは驚くべきことではない。2゜EBは、GFP-/Flk-1-画分に限定され、この知見は、それらが、初期EBにおける残留の非分化ES細胞から得られるという事実と一致する。GFP+/Flk-1-画分は、これらの培養で使用した条件下では殆どコロニーを生じなかった。遺伝子発現解析により、2つの時点で単離した集団の間で興味深い差異が判明した(図5C)。ES細胞マーカーであるRex-1は、GFP-画分よりもGFP+画分においてより低いレベルで発現され、このことは、これらの集団は分化を受けていることを示す。短尾は、両時点でGFP+画分において発現されていた。レベルは、3.5日目のEBから単離したGFP+/Flk-1+画分よりもGFP+/Flk-1-の方が高いようであり、このことは、この発現は、これらの細胞が造血および内皮系統へと成熟するにつれてダウンレギュレートされ得ることを示唆する。期待したように、Flk-1は、両時点でGFP+/Flk-1+細胞において主に発現されていた。原始的および成体型の両方の造血発達に必須であるヘリックス・ループ・ヘリックス転写因子であるSclは(Shivdasaniら(1995)Nature 373:432-4)、GFP+/Flk-1+画分に限定されているようである。同様に、成体型造血系の確立に必要とされる転写因子のRunx1は(Wangら(1996)Proc.Natl.Acad.Sci.93:3444-9)、GFP+/Flk-1+画分に最も容易に検出された。3.0日目のEBから単離したGFP+/Flk-1-画分には幾分Runx1が発現されていた。分化3日目には3つの全ての画分において結節が発現されている。3.5日目には、GFP+/Flk-1+画分における発現レベルは、他の画分よりも有意に低いようである。Wnt3aおよびWnt8aは顕著に制限された発現パターンを示し、両時点においてGFP+/Flk-1-画分にのみ見出され、これは、系統制限マーカーの発現前の初期中胚葉機能に一致している。BMP2は両方のGFP+画分に発現され、一方、BMP4は、主に、GFP+Flk-1+細胞に見出され、これは、これらの分子は、この系における別個の発達段階において役割を果たしていることを示す。BL-CFC能およびGFP+/Flk-1+細胞の遺伝子発現パターンにより、それらはマウス胚に見られる胚外中胚葉を代表することが示される。
図5に観察された発現パターンは、3つの画分が、GFP-/Flk-1-細胞が、GFP+/Flk-1-細胞を生じ、これは次いでGFP+/Flk-1+細胞を生じるという発達の連続的系列であるという解釈と一致する。これらの画分が共通の発達経路内の特定の段階を示すかどうかを決定するために、各々を3日目のEBから単離し、20時間培養し、その後、GFPおよびFlk-1発現について再度解析した。BL-CFCおよびEryp-CFC能は、培養前および培養後に、各集団について決定した。単離した細胞を、EB分化に使用したのと同じ培地中、ペトリ等級24ウェルプレート中、1×105細胞またはそれ以上の密度で培養した。これらの条件下で、細胞は急速に再凝集し、EB様構造を形成し、細胞数の増殖または減少は殆ど示さずに正常な発達プログラムを追随しているようである。20時間の再凝集培養後に、GFP-/Flk-1-細胞は、有意なGFP+/Flk-1-細胞集団ならびに少数のGFP+/Flk-1+を生じた。GFP+/Flk-1-細胞は、同じ培養期間中にかなりのGFP+/Flk-1+細胞の集団を生じた。GFP+/Flk-1+集団は、再凝集培養後に幾分のGFPおよびFlk-1発現を失うようであった。結果を図6に示す。前駆体能の変化は、表面マーカーの変化と一致していた。3つの中で最も未熟であるGFP-/Flk-1-画分は、培養前または培養後に、検出不可能な数のBL-CFCおよびEryp-CFCを含んでいた。GFP+/Flk-1-画分はまた、培養前にBL-CFCおよびEryp-CFCを殆ど含んでいなかった。しかし、培養後、BL-CFC能は、劇的に、105細胞あたり74から1564まで増加し、これはFlk-1発現の増加と一致する。Eryp-CFCの頻度は、培養期間中には変化しなかった。GFP+/Flk-1+画分は、培養前に、BL-CFCを含んでいたが、Eryp-CFCは殆ど含んでいなかった。BL-CFCは培養後に検出されなかったが、しかし、集団は今ではEryp-CFCを含んでいた。表面マーカー解析と一緒に考えると、これらの前駆体データは、プレ中胚葉(GFP-/Flk-1-)から中胚葉/プレ血管芽細胞(BL-CFC)集団(GFP+/Flk-1-)への、血管芽細胞/プレ赤血球集団(GFP+/Flk-1+)への、後血管芽細胞/赤血球集団(おそらくGFP10/Flk-110)への発達進行を支持する。特定の集団における全ての細胞が20時間の培養期間後に分化するわけではなく、出発表現型を有する細胞が、GFP-/Flk-1-およびGFP+/Flk-1-培養物中には残留していた。
前の実施例により、3.0日目のEBから単離されたGFP+/Flk-1-細胞は、一晩培養した後に、GFP+/Flk-1+細胞およびBL-CFCを効率的に産生したことが実証された。このプレ-BL-CFC能は、この発達段階においてGFP+/Flk-1-画分に特異的であるかどうかを決定するために、異なる年齢のEBからのGFP+/Flk-1-細胞を、BL-CFCを生じる能力についてアッセイした。図7Aに示したように、BL-CFCを産生する能力は、3日目のGFP+/Flk-1-細胞において最も強かった。この発達能は3.5日目の分化までに劇的に減少し、4.0日目にはほぼ存在しなかった。これらの同じEB由来の新しく単離したGFP+/Flk-1+画分のBL-CFC含量はこの期間かけて増加し、このことは、分化が正常に進行していたことを示す。異なる段階のEB細胞の再凝集培養物中のFlk-1発現パターンは、BL-CFCデータと一致していた。再凝集した3.0日目のGFP+/Flk-1-細胞の培養物には、全集団の40%超過を示した別個のFlk-1画分が含まれていた(図7B)。3.5および4.0日目の培養物中のFlk-1発現は有意に低く、別個のピークよりも全集団の肩から構成されていた。
造血および内皮中胚葉の発達の後に心臓および頭蓋中胚葉が発達するマウス胚における事象の順序を考えて、単離集団の心筋能を決定した。この解析のために、異なる集団の培養物からの凝集物を、マイクロタイターまたは24ウェルプレート中の血清非含有培地中に移し、心筋細胞を示す、拍動細胞塊の発達についてモニタリングした。これらの条件は、再凝集細胞からの心筋細胞の効率的な発達を支持することが知られている。これらの塊内の細胞の心筋細胞性質の独立的な確認として、代表群を顕微鏡スライドに写し、固定し、トロポニンTの心臓特異的イソ型の存在について染色した。解析した全ての拍動細胞塊が、トロポニンT陽性細胞を含むことが判明し、これは、それらが心筋細胞であることを示す。このアッセイを使用して、異なる段階のEBからの再凝集したGFP+/Flk-1-およびGFP+/Flk-1+画分の心筋細胞能を決定した。比較のために、新しく単離したGFP+/Flk-1+細胞および培養したGFP+/Flk-1-細胞のBL-CFC能を解析した。

短尾+細胞の発達における血清の役割を評価するために、EBを血清の非存在下で分化した。EBはこれらの条件下で発達したが、正常条件で見られたものより幾分小さかった。血清の非存在下において、これらのEB内にはGFP+細胞は検出されず、このことから、中胚葉はこれらの条件下で誘導されないことが示される(図8)。有意な数のGFP+/Flk-1-およびGFP+/Flk-1+細胞が、血清を培養物に添加した場合に発達した。これらの知見により、血清内に見られる成分は、短尾+細胞の発達および分化を誘導できることが明らかに実証される。この過程に役割を果たし得る因子の同定における最初のステップとして、BMP4(20ng/ml)を、血清非含有培養物中の発達中のEBに加えた。この濃度では、BMP4は、分化3日間以内に有意な短尾+細胞の集団を誘導した。しかし、血清とは対照的に、BMP4は、この期間中に、GFP+/Flk-1+集団の発達は支持しなかった。BMP4が、血清の存在下で誘導されるGFP+/Flk-1-細胞からGFP+/Flk-1+細胞を誘導できるかどうかを決定するために、GFP+/Flk-1-細胞を、血清の存在下で3日間分化したEBから単離した。これらの細胞を、培地のみの中、血清を含む培地中、またはBMP4を含む培地中で再凝集した。図8の下の列に示したように、GFP+/Flk-1-細胞は、血清の非存在下で再凝集させた場合には実質的に分化しなかった。期待したように、同じ集団が、血清を培養物に添加した場合には、大きなGFP+/Flk-1+集団を生じた。一次分化培養物中での知見に一致して、BMP4は、培養したGFP+/Flk-1-細胞から有意な数のGFP+/Flk-1+細胞の発達を誘導できなかった。
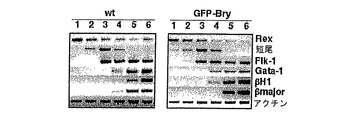
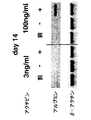
アフリカツメガエルおよびゼブラフィシュなどのモデル系を使用しての研究により、中胚葉および内胚葉は、中内胚葉として知られる共通の前駆集団から発達することが示唆された。中内胚葉発達段階がEBに存在するかどうかを決定するために、内胚葉系統の発達条件を確立した。この系統の発達のための培養条件を確立する最初のステップとして、分化培養物中の血清の量を変化させた。EBを、血清の存在下および非存在下で産生し(SP34およびその後IMDMプラスSR)、外胚葉、内胚葉および中胚葉発達に関連した遺伝子の発現について異なる段階でアッセイした。外胚葉系統では、神経系統の発達を、PAX6、Wntl、neruoD、および神経フィラメント(NFL)の発現を解析して評価した。これらの遺伝子は、異なる神経発達段階で発現されることが知られている。内胚葉発達の初期段階を、HNF3βの発現によりモニタリングした。内胚葉発達および特異化の後期段階を評価するために、肝臓発達に関与する遺伝子を解析した。これらには、Hex、α−フェトプロテイン(AFP)、HNF4、アルブミン(Alb)、α1−アンチトリプシン(AAT)およびチロシンアミノトランスフェラーゼ(TAT)が含まれた。中胚葉発達は、短尾およびGATA-1の発現によりモニタリングした。遺伝子発現パターンに加えて、神経発達を、EBからの神経突起の成長をモニタリングすることによりアッセイした。これらの神経突起の神経性質は、βIII-チューブリン発現により実証された。中胚葉発達はまた、EB中の造血前駆体の列挙により評価した。図10は、10日間の分化期間におよぶEBの発達能に対する血清の影響を示す。血清の存在下では(血清)、この系統の発達に関連した遺伝子の発現の欠如から実証されるように、神経-外胚葉の分化は殆どない。HNF3βは、分化の初期段階(2〜3日目)で発現され、その後、ダウンレギュレートされる。期待したように、GATA-1は、これらの条件下で産生されたEBで発現される。これらの遺伝子の発現パターンは、血清の非存在下で(血清-)増殖したEB中で基本的に逆転していた。これらのEBは、神経外胚葉に関連した全ての遺伝子を発現したが、中胚葉/造血マーカーGATA-1は発現しなかった。HNF3βは、これらの条件下で増殖した後期EB(10日目)で発現された。GFP発現によりモニタリングされる短尾発現パターンは、これらの知見と一致していた。血清の存在下で産生されるEBは、分化2日目から5日目に存在するかなりの短尾+集団を産生した(図11、-B-系)。短尾は、血清の非存在下で増殖させたEBには検出されなかった(図11、-H-系)。造血前駆体アッセイにより、これらの知見が確認された。血清中で産生したEBは前駆体を含んでいたが(図12、斑点の棒)、血清の非存在下で増殖させたものは含んでいなかった(図12、実線の棒、眼に見えない)。最後に、EBの神経突起能の評価は、これらの種々の解析と一致していた。血清中で増殖したEBはどれも神経突起を生じなかった。これに対し、血清の非存在下で産生されたものの85%がこの活性を示した(図13)。一緒に考えると、これらの知見により、特異的系統の産生における培養条件(血清)の重要性が実証される。それらはまた、血清完全条件も血清非含有条件も外胚葉発達に最適ではないことを実証する。
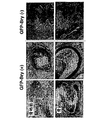
実施例6に記載した予備的な動態解析により、別個の発達運命を有する中胚葉の亜集団が、一定の時間的様式で産生されることが実証された。より詳細な動態解析により、分化の2.5日目から4.0日目の間のGFP+Flk1-(本明細書では以後GFP+集団と称される)の動的発達が示された(図18)。単離し再凝集した場合、2.5および3.0日目のGFP+画分は、最初期段階の造血および内皮関与を示す芽細胞コロニー(図19において血管芽細胞能として示す)を産生した。このGFP+細胞の集団は、殆ど心筋細胞(心臓)能を示さなかった。初期GFP+細胞とは対照的に、3.5日目のEBから単離したものは、有意に減少したBL-CFC能を示したが、心筋細胞への分化において効率的であった。4日目のEBから単離されたGFP+細胞は心筋細胞は産生せず、BL-CFCを産生する能力も殆どなく、このことは、それらはいくつかの他の中胚葉系統に運命づけられ得ることを示唆する。遺伝子発現解析により、これらの機能的アッセイが支持され、4つのGFP+画分間の分子差異が実証された(図20)。いくつかの遺伝子が全ての集団で発現されたが、他のものは興味深い異なるパターンを示した。造血発達に重要であり心臓分化に阻害的であると考えられている遺伝子であるWnt3aは、2.5および3.0日目の集団において発現され、3.5および4.0日目の細胞においてダウンレギュレートされていた。このパターンは、これらの集団の造血/血管から心筋への発達能における変化に一致している。興味深い第二のパターンは、遺伝子Mix1-1のパターンである。中胚葉および内胚葉の発達に役割を果たすこの遺伝子は、2.5および3.0日目のGFP集団には発現されているが、3.5および4.0日目の画分は発現されなかった。一緒に考えると、本実施例の知見により、異なる発達能を有する中胚葉集団は、EB内では一定の時間的パターンで産生されることが明らかに実証される。さらに、中胚葉/内胚葉遺伝子Mixl-1の発現解析により、内胚葉能を有する細胞もまた、特定の時期に、すなわち分化の2.5日目から3.0日目の間に産生されることが示される。
GFP+集団の内胚葉能をさらに評価するために、GFP+およびGFP-の2.5日目のEB細胞を、肝細胞分化を促進することが知られる条件下で14日間培養し、その後、レシピエントSCIDベージュマウスの腎臓被膜下に移植した。数匹のマウスを移植直後に屠殺し、移植片を有する腎臓を切片化し、切片を、Hep1およびAFPに対する抗体で染色した。Hep1は肝細胞の特異的マーカーであり、一方、AFPは、成体型内胚葉および肝臓系統の未熟細胞に発現される。切片内の細胞のいくつかは、Hep1に陽性で染色されたが、Hep1+細胞の近傍にある他の細胞は、AFPを発現することが判明した。Hep1+またはAFP+細胞は、GFP-陰性細胞の移植片には全く見られなかった。これらの知見によりPCRデータが支持され、培養条件が、Hep1の発現により規定される未熟肝細胞の特徴を有する細胞の発達を支持することが実証される。
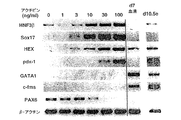
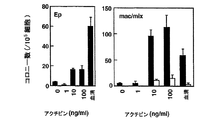
内胚葉能を有する細胞をさらに濃縮するために、他のモデルシステムでこの細胞集団を誘導することが知られる増殖因子の効果を試験した。アフリカツメガエルの研究により、アクチビンは、培養物中で、外胚葉から中胚葉および内胚葉の両方を誘導することが示された。特に興味深いのは、アクチビンが、このモデルにおいて、使用した異なる濃度で異なる細胞型を誘導するというモルフォゲンとして挙動するという観察であった。アクチビンがES/EB系で類似の能力を示すかどうかを決定するために、それを以下のプロトコルを使用してEB培養物に加えた。ES細胞は、血清を含まないStem Pro34培地中で2日間分化させた。この段階で、発達中のEBを収集し、血清代替添加物(血清非含有)および100ng/mlの濃度のアクチビンを補充したIMDM中で再度培養した。EBを異なる日に収集し、GFP+発現、ならびに、内胚葉、中胚葉および外胚葉発達を示す遺伝子の発現についてアッセイした。図22A、Bで示したように、この量のアクチビンは、GFPにより測定したところ短尾を誘導した。GFP誘導の動態は、血清中で分化したEBに比べて遅延していたが、この濃度のアクチビンは、分化6日目までにかなりの数の短尾陽性細胞(60%)を誘導した。分子解析により、アクチビン誘導細胞は、HNF3β、Mixl-1、Soxl7、Hex-1、およびpdx-1を含む内胚葉発達に関連した幅広い遺伝子を発現することが示された(図23C)。GATA-1などの造血発達に関連した遺伝子およびPAX6などの神経外胚葉分化を示すものは、アクチビンにより誘導されなかった。
前の実施例により、肝細胞系統は、中胚葉能および内胚葉能の両方を有するbry+細胞集団から発達することが明らかに示される。bry+画分の解析により、これらの細胞の亜集団は、受容体チロシンキナーゼc-kitを発現すること(図27A)、この集団は、Flk-1を発現する細胞とは別個であることが判明した。c-kit発現が、内胚葉能を有する細胞の分離に有用なマーカーであり得ることを決定するために、3日目の血清刺激EBからのbry+/c-kit-(+/-)およびbry+/c-kit+(+/+)細胞を肝細胞能についてアッセイした。図27Bに示したように、AFPおよびALB発現細胞は全てbry+/c-kit+集団から得られた。この画分の内胚葉能を推定するために、選別した細胞をカバーガラスに置き、HNF3βに対する抗体で染色した。80%を超えるbry+/c-kit+細胞がHNF3βタンパク質を発現し、10%未満のbry+/c-kit-細胞が陽性であった。これらの知見により、内胚葉前駆体は、短尾およびc-kitの両方を発現し、この集団には、内胚葉能を有する細胞が高度に濃縮されていることが示される。短尾およびc-kit発現に基づいて細胞を単離することにより、内胚葉前駆体の単離における新規な戦略が提供される。
Claims (18)
- 内胚葉細胞が濃縮された細胞集団。
- 少なくとも約50%の内胚葉細胞を含む、請求項1に記載の細胞集団。
- 少なくとも約75%の内胚葉細胞を含む、請求項1に記載の細胞集団。
- 少なくとも約90%の内胚葉細胞を含む、請求項1に記載の細胞集団。
- 血清の存在下で約2から約10日間の間ヒト胚性幹細胞を培養すること、短尾を発現する細胞を単離すること、および血清の非存在下で約1から約15日間の間、短尾を発現する前記細胞を培養することにより得られる、請求項1に記載の細胞集団。
- 血清の存在下で約2から約10日間の間ヒト胚性幹細胞を培養すること、短尾を発現する細胞を単離すること、および血清の非存在下で約1から約15日間の間、短尾を発現する前記細胞を培養することを含む、請求項1の細胞集団を単離する方法。
- 選択マーカー遺伝子を、前記胚性幹細胞の短尾遺伝子座に挿入し、選択マーカーを発現する細胞を選択することにより短尾を発現する細胞を単離する、請求項6に記載の方法。
- 前記選択マーカーは強化緑色蛍光タンパク質である、請求項7に記載の方法。
- 血清の非存在下、胚性幹細胞から内胚葉への分化を誘導するのに十分な量のアクチビンの存在下で胚性幹細胞を培養すること、および短尾を発現する細胞を単離することを含む、請求項1に記載の細胞集団を単離する方法。
- 血清の非存在下、胚性幹細胞から中胚葉への分化を誘導するには十分であるが、胚性幹細胞から内胚葉への分化を誘導するには不十分である量のアクチビンの存在下で胚性幹細胞を培養すること、および短尾を発現する細胞を単離することを含む、中胚葉細胞の濃縮された細胞集団を単離する方法。
- 請求項1に記載の細胞集団を単離すること、前記集団の亜集団を、肝細胞破壊または機能不全に関連した疾患を示す動物に移植すること、および肝機能についてアッセイすることを含む、成熟肝細胞として機能できる内胚葉細胞を同定する方法であって、前記動物の肝機能の改善は、成熟肝細胞として機能できる内胚葉細胞の同定を示す、方法。
- 前記動物はフマリルアセト酢酸の欠損したマウスである、請求項11に記載の方法。
- 試験する薬剤の存在下で請求項1に記載の細胞集団を培養すること、および前記薬剤の存在下および非存在下で前記細胞の増殖、分化、または生存を比較することを含む、内胚葉細胞の増殖、分化、または生存に影響を及ぼす薬剤を同定する方法であって、前記薬剤の存在下における差異は、前記細胞の増殖、分化、または生存に影響を及ぼす薬剤の同定を示す、方法。
- 短尾を発現する細胞が得られる条件下で胚性幹細胞をインキュベートすること、2つの異なる培養時間の後に短尾を発現する2つの細胞集団を単離すること、前記2つの集団の遺伝子発現プロファイルを比較すること、集団において独特に発現されている遺伝子を同定することを含む、細胞分化に関与する遺伝子を同定する方法であって、集団において独特に発現されている遺伝子の同定は、細胞分化に関与する遺伝子の同定を示す、方法。
- 選択マーカーを胚性幹細胞の短尾遺伝子座に挿入すること、前記細胞を培養すること、選択マーカーを発現する細胞を選択することにより短尾を発現する細胞を単離して、短尾を発現する単離細胞集団を得ること、および前記単離集団に対する抗体を産生することを含む、短尾を発現する細胞は認識するが、短尾を発現しない細胞は認識しない抗体を作製する方法。
- 抗体が、モノクローナルまたはポリクローナルである請求項15に記載の方法。
- 内胚葉から肝細胞または膵細胞への分化に有効な条件下で請求項1に記載の細胞集団を培養することを含む、哺乳動物細胞を作製する方法。
- 選択マーカーをコードするDNAが短尾遺伝子座に存在し、前記選択マーカーは短尾を発現する細胞において発現されている導入遺伝子を含むゲノムを有する、トランスジェニック非ヒト哺乳動物。
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US38161702P | 2002-05-17 | 2002-05-17 | |
| US44485103P | 2003-02-04 | 2003-02-04 |
Related Parent Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2005510829A Division JP2006513727A (ja) | 2002-05-17 | 2003-05-19 | 中胚葉および成体型内胚葉細胞集団 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| JP2009171981A true JP2009171981A (ja) | 2009-08-06 |
Family
ID=33436666
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2005510829A Pending JP2006513727A (ja) | 2002-05-17 | 2003-05-19 | 中胚葉および成体型内胚葉細胞集団 |
| JP2009071364A Pending JP2009171981A (ja) | 2002-05-17 | 2009-03-24 | 中胚葉および成体型内胚葉細胞集団 |
Family Applications Before (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2005510829A Pending JP2006513727A (ja) | 2002-05-17 | 2003-05-19 | 中胚葉および成体型内胚葉細胞集団 |
Country Status (7)
| Country | Link |
|---|---|
| US (8) | US7763466B2 (ja) |
| EP (2) | EP1543110A4 (ja) |
| JP (2) | JP2006513727A (ja) |
| AU (2) | AU2003304106C1 (ja) |
| CA (2) | CA2684022C (ja) |
| SG (1) | SG141296A1 (ja) |
| WO (1) | WO2004098490A2 (ja) |
Cited By (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2013531497A (ja) * | 2010-06-13 | 2013-08-08 | インスティチュート・オブ・バイオフィジックス,チャイニーズ・アカデミー・オブ・サイエンシズ | 幹細胞から心筋細胞を調製するための方法および組成物ならびにその使用 |
| JP2015186471A (ja) * | 2014-03-26 | 2015-10-29 | ジーイー・ヘルスケア・ユーケイ・リミテッド | 細胞分化方法 |
| JP2016521571A (ja) * | 2013-06-11 | 2016-07-25 | プルーリオミクス・ベー・フェー | 多能性哺乳動物幹細胞に由来する心筋細胞を成熟させるための培地組成物 |
| US11339371B2 (en) | 2012-07-23 | 2022-05-24 | Institute Of Biophysics, Chinese Academy Of Sciences | Method for inducing pluripotent stem cells to differentiate into ventricular myocytes in vitro |
Families Citing this family (54)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| GB2415432B (en) * | 2001-12-07 | 2006-09-06 | Geron Corp | Islet cells from human embryonic stem cells |
| AU2003304106C1 (en) * | 2002-05-17 | 2010-10-28 | Mount Sinai School Of Medicine Of New York University | Mesoderm and definitive endoderm cell populations |
| US20060003446A1 (en) * | 2002-05-17 | 2006-01-05 | Gordon Keller | Mesoderm and definitive endoderm cell populations |
| US20050266554A1 (en) * | 2004-04-27 | 2005-12-01 | D Amour Kevin A | PDX1 expressing endoderm |
| AU2013200203B2 (en) * | 2003-12-23 | 2014-11-20 | Viacyte, Inc. | Definitive endoderm |
| US7625753B2 (en) * | 2003-12-23 | 2009-12-01 | Cythera, Inc. | Expansion of definitive endoderm cells |
| US8647873B2 (en) | 2004-04-27 | 2014-02-11 | Viacyte, Inc. | PDX1 expressing endoderm |
| US8586357B2 (en) * | 2003-12-23 | 2013-11-19 | Viacyte, Inc. | Markers of definitive endoderm |
| US7541185B2 (en) | 2003-12-23 | 2009-06-02 | Cythera, Inc. | Methods for identifying factors for differentiating definitive endoderm |
| US7985585B2 (en) | 2004-07-09 | 2011-07-26 | Viacyte, Inc. | Preprimitive streak and mesendoderm cells |
| DK1709159T3 (da) | 2003-12-23 | 2019-07-29 | Viacyte Inc | Definitiv endoderm |
| EP1786896B1 (en) * | 2004-07-09 | 2018-01-10 | Viacyte, Inc. | Methods for identifying factors for differentiating definitive endoderm |
| ES2716827T3 (es) * | 2004-07-09 | 2019-06-17 | Viacyte Inc | Células mesendodérmicas y células de línea pre-primitiva |
| AU2016235001B2 (en) * | 2004-07-09 | 2018-12-13 | Viacyte, Inc. | Methods for identifying factors for differentiating definitive endoderm |
| AU2015203517B2 (en) * | 2004-07-09 | 2018-01-04 | Viacyte, Inc. | Preprimitive streak and mesendoderm cells |
| US8187878B2 (en) | 2004-08-13 | 2012-05-29 | University Of Georgia Research Foundation, Inc. | Methods for increasing definitive endoderm differentiation of pluripotent human embryonic stem cells with PI-3 kinase inhibitors |
| EP1797171A1 (en) * | 2004-09-29 | 2007-06-20 | Cellartis AB | Methods for the generation of hepatocyte-like cells from human blastocyst-derived stem (hbs) |
| EP1838843B1 (en) * | 2004-12-23 | 2019-07-10 | Viacyte, Inc. | Expansion of definitive endoderm cells |
| DK1838843T3 (da) * | 2004-12-23 | 2019-08-26 | Viacyte Inc | Ekspansion af definitive endodermceller |
| DK2674485T3 (da) * | 2005-10-27 | 2019-08-26 | Viacyte Inc | Pdx-1 udtrykkende dorsal og ventral fortarm endoderm |
| US7695965B2 (en) | 2006-03-02 | 2010-04-13 | Cythera, Inc. | Methods of producing pancreatic hormones |
| US11254916B2 (en) | 2006-03-02 | 2022-02-22 | Viacyte, Inc. | Methods of making and using PDX1-positive pancreatic endoderm cells |
| EP2650360B1 (en) * | 2006-03-02 | 2019-07-24 | Viacyte, Inc. | Endocrine precursor cells, pancreatic hormone-expressing cells and methods of production |
| US8415153B2 (en) | 2006-06-19 | 2013-04-09 | Geron Corporation | Differentiation and enrichment of islet-like cells from human pluripotent stem cells |
| US8874380B2 (en) | 2010-12-09 | 2014-10-28 | Rutgers, The State University Of New Jersey | Method of overcoming therapeutic limitations of nonuniform distribution of radiopharmaceuticals and chemotherapy drugs |
| WO2008048647A1 (en) * | 2006-10-17 | 2008-04-24 | Cythera, Inc. | Modulation of the phosphatidylinositol-3-kinase pathway in the differentiation of human embryonic stem cells |
| EP2126045A4 (en) * | 2007-01-30 | 2010-05-26 | Univ Georgia | EARLY MESODERM CELLS, STABLE POPULATION OF MESENDODERM CELLS WITH THE ABILITY TO GENERATE ENDODERM AND MESODERM CELL LINES AND MULTIPOTENTIAL MIGRATION CELLS |
| RU2467066C2 (ru) * | 2007-07-31 | 2012-11-20 | Дайити Санкио Комани, Лимитед | Способ конструирования массы миокардиальных клеток и применение массы миокардиальных клеток |
| US7695963B2 (en) | 2007-09-24 | 2010-04-13 | Cythera, Inc. | Methods for increasing definitive endoderm production |
| US8338170B2 (en) | 2008-04-21 | 2012-12-25 | Viacyte, Inc. | Methods for purifying endoderm and pancreatic endoderm cells derived from human embryonic stem cells |
| CA2723820A1 (en) * | 2008-05-09 | 2009-11-12 | Vistagen Therapeutics, Inc. | Pancreatic endocrine progenitor cells derived from pluripotent stem cells |
| JP2010075631A (ja) * | 2008-09-29 | 2010-04-08 | Japan Science & Technology Agency | 未分化細胞から肝臓を分化誘導する方法 |
| EP2356227B1 (en) | 2008-11-14 | 2018-03-28 | Viacyte, Inc. | Encapsulation of pancreatic cells derived from human pluripotent stem cells |
| CN102459575A (zh) | 2009-06-05 | 2012-05-16 | 细胞动力国际有限公司 | 重编程t细胞和造血细胞的方法 |
| US9506114B2 (en) | 2009-08-28 | 2016-11-29 | Cellular Dynamics International, Inc. | Identifying genetic variation in affected tissues |
| EP2490701B9 (en) | 2009-10-19 | 2017-11-15 | Cellular Dynamics International, Inc. | Cardiomyocyte production |
| CN103003416B (zh) | 2010-06-15 | 2016-11-16 | 细胞动力学国际有限公司 | 从小体积的外周血产生诱导性多潜能干细胞 |
| AU2011268056B2 (en) | 2010-06-18 | 2014-04-24 | Cellular Dynamics International, Inc. | Cardiomyocyte medium with dialyzed serum |
| US8765470B2 (en) | 2010-08-04 | 2014-07-01 | Cellular Dynamics International, Inc. | Reprogramming immortalized B-cells to induced pluripotent stem cells |
| US9181529B2 (en) | 2010-10-19 | 2015-11-10 | Cellular Dynamics International, Inc. | Titration of differentiation medium components |
| CA2831609C (en) | 2011-03-30 | 2019-06-11 | Cellular Dynamics International, Inc. | Priming of pluripotent stem cells for neural differentiation |
| WO2014022423A2 (en) | 2012-07-31 | 2014-02-06 | Hantash Basil M | Hla g-modified cells and methods |
| WO2014053709A1 (en) | 2012-10-05 | 2014-04-10 | Glykos Finland Oy | A method for culturing stem cells |
| USD933584S1 (en) | 2012-11-08 | 2021-10-19 | Sunpower Corporation | Solar panel |
| USD1009775S1 (en) | 2014-10-15 | 2024-01-02 | Maxeon Solar Pte. Ltd. | Solar panel |
| JP6933898B2 (ja) | 2014-04-24 | 2021-09-08 | ボード オブ リージェンツ, ザ ユニバーシティ オブ テキサス システムBoard Of Regents, The University Of Texas System | 養子細胞治療製品を製造するための誘導多能性幹細胞の適用 |
| FR3024462A1 (fr) * | 2014-07-29 | 2016-02-05 | Univ Pierre Et Marie Curie Paris 6 | Procede de production in vitro de progeniteurs adipocytaires et d'adipocytes |
| USD896747S1 (en) | 2014-10-15 | 2020-09-22 | Sunpower Corporation | Solar panel |
| USD999723S1 (en) | 2014-10-15 | 2023-09-26 | Sunpower Corporation | Solar panel |
| USD933585S1 (en) | 2014-10-15 | 2021-10-19 | Sunpower Corporation | Solar panel |
| USD913210S1 (en) | 2014-10-15 | 2021-03-16 | Sunpower Corporation | Solar panel |
| US10590389B2 (en) | 2014-12-24 | 2020-03-17 | Kyoto University | Endodermal cell production method, liver cell production method, pancreatic cell production method, endodermal cell induction promoter, liver cell induction promoting kit, pancreatic cell induction promoting kit, and microfluidic device |
| MX2018005274A (es) | 2015-10-30 | 2019-09-19 | The Regents Of The Universtiy Of California | Metodos para la generacion de celulas-t a partir de celulas madre y metodos inmunoterapeuticos que utilizan las celulas-t. |
| US10767164B2 (en) | 2017-03-30 | 2020-09-08 | The Research Foundation For The State University Of New York | Microenvironments for self-assembly of islet organoids from stem cells differentiation |
Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2001053465A1 (en) * | 2000-01-21 | 2001-07-26 | The Johns Hopkins University School Of Medicine | Human embryoid body-derived cells |
| JP2001333770A (ja) * | 2000-05-31 | 2001-12-04 | Japan Science & Technology Corp | 脊椎動物の膵臓のインビトロ形成方法 |
Family Cites Families (29)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US4997815A (en) * | 1988-11-01 | 1991-03-05 | Children's Hospital Medical Center Of Northern California | Method for augmenting fetal hemoglobin by treatment with activin and/or inhibin |
| US5563059A (en) * | 1993-02-23 | 1996-10-08 | Genentech, Inc. | Use of human inhibin and human activin to increase the number of mature primate oocytes |
| US5629194A (en) * | 1994-10-21 | 1997-05-13 | Diacrin, Inc. | Isolated porcine pancreatic cells for use in treatment of diseases characterized by insufficient insulin activity |
| US5874301A (en) | 1994-11-21 | 1999-02-23 | National Jewish Center For Immunology And Respiratory Medicine | Embryonic cell populations and methods to isolate such populations |
| US5843780A (en) | 1995-01-20 | 1998-12-01 | Wisconsin Alumni Research Foundation | Primate embryonic stem cells |
| WO1997025413A1 (en) * | 1996-01-09 | 1997-07-17 | The Regents Of The University Of California | Ungulate embryonic stem-like cells, making and using the cells to produce a transgenic ungulate |
| US5851984A (en) * | 1996-08-16 | 1998-12-22 | Genentech, Inc. | Method of enhancing proliferation or differentiation of hematopoietic stem cells using Wnt polypeptides |
| DE19837438A1 (de) * | 1998-08-18 | 2000-02-24 | Biotechnolog Forschung Gmbh | Rekombinante Humanzelle für Knochen- und Knorpelzellbildung |
| US6667176B1 (en) | 2000-01-11 | 2003-12-23 | Geron Corporation | cDNA libraries reflecting gene expression during growth and differentiation of human pluripotent stem cells |
| US6432673B1 (en) | 1998-12-07 | 2002-08-13 | Zymogenetics, Inc. | Growth factor homolog ZVEGF3 |
| GB9828383D0 (en) | 1998-12-22 | 1999-02-17 | Medical Res Council | Cell lineage markers |
| IL144654A0 (en) * | 1999-02-10 | 2002-05-23 | Curis Inc | Pancreatic progenitor cells, methods and uses related thereto |
| EP1179093A1 (en) | 1999-05-14 | 2002-02-13 | Chiron Corporation | Expression of ets-domain proteins in cancer |
| US7282366B2 (en) | 2000-04-27 | 2007-10-16 | Geron Corporation | Hepatocytes for therapy and drug screening made from embryonic stem cells |
| US6632620B1 (en) * | 2000-06-22 | 2003-10-14 | Andrew N. Makarovskiy | Compositions for identification and isolation of stem cells |
| IL154159A0 (en) | 2000-08-01 | 2003-07-31 | Yissum Res Dev Co | Directed differentiation of ebryonic cells |
| AU7753500A (en) | 2000-10-03 | 2002-04-15 | Univ North Carolina | Processes for clonal growth of hepatic progenitor cells |
| US6586251B2 (en) | 2000-10-31 | 2003-07-01 | Regeneron Pharmaceuticals, Inc. | Methods of modifying eukaryotic cells |
| US20030027331A1 (en) | 2000-11-30 | 2003-02-06 | Yan Wen Liang | Isolated homozygous stem cells, differentiated cells derived therefrom, and materials and methods for making and using same |
| JP2004527249A (ja) * | 2001-04-19 | 2004-09-09 | デヴェロゲン アクチエンゲゼルシャフト フュア エントヴィックルングスビオローギッシェ フォルシュング | 幹細胞をインスリン産生細胞に分化する方法 |
| GB2415432B (en) | 2001-12-07 | 2006-09-06 | Geron Corp | Islet cells from human embryonic stem cells |
| GB0207299D0 (en) * | 2002-03-28 | 2002-05-08 | Franks Christopher R | Stem cell therapy |
| US20060003446A1 (en) * | 2002-05-17 | 2006-01-05 | Gordon Keller | Mesoderm and definitive endoderm cell populations |
| AU2003304106C1 (en) * | 2002-05-17 | 2010-10-28 | Mount Sinai School Of Medicine Of New York University | Mesoderm and definitive endoderm cell populations |
| US20040121464A1 (en) | 2002-09-30 | 2004-06-24 | Rathjen Peter David | Method for the preparation of cells of mesodermal lineage |
| DK1709159T3 (da) | 2003-12-23 | 2019-07-29 | Viacyte Inc | Definitiv endoderm |
| WO2005097980A2 (en) | 2004-03-26 | 2005-10-20 | Geron Corporation | New protocols for making hepatocytes from embryonic stem cells |
| EP1584680A1 (en) | 2004-04-08 | 2005-10-12 | Boehringer Ingelheim Austria GmbH | Fed-batch fermentation process for the production of plasmid DNA |
| WO2009013254A1 (en) * | 2007-07-20 | 2009-01-29 | Cellartis Ab | A novel population of hepatocytes derived via definitive endoderm (de-hep) from human blastocysts stem cells |
-
2003
- 2003-05-19 AU AU2003304106A patent/AU2003304106C1/en not_active Expired
- 2003-05-19 WO PCT/US2003/015658 patent/WO2004098490A2/en active Application Filing
- 2003-05-19 SG SG200701719-7A patent/SG141296A1/en unknown
- 2003-05-19 US US10/514,759 patent/US7763466B2/en active Active
- 2003-05-19 EP EP03816210A patent/EP1543110A4/en not_active Ceased
- 2003-05-19 JP JP2005510829A patent/JP2006513727A/ja active Pending
- 2003-05-19 CA CA2684022A patent/CA2684022C/en not_active Expired - Lifetime
- 2003-05-19 CA CA2487058A patent/CA2487058C/en not_active Expired - Lifetime
- 2003-05-19 EP EP10011700A patent/EP2361966A1/en not_active Withdrawn
-
2008
- 2008-03-24 US US12/077,976 patent/US7955849B2/en not_active Expired - Lifetime
-
2009
- 2009-03-24 JP JP2009071364A patent/JP2009171981A/ja active Pending
- 2009-06-02 AU AU2009202175A patent/AU2009202175B2/en not_active Expired
-
2010
- 2010-07-14 US US12/836,275 patent/US8748171B2/en not_active Expired - Lifetime
- 2010-07-14 US US12/836,277 patent/US20110070203A1/en not_active Abandoned
-
2013
- 2013-12-17 US US14/109,675 patent/US20140287494A1/en not_active Abandoned
-
2015
- 2015-02-26 US US14/632,600 patent/US20150218512A1/en not_active Abandoned
- 2015-06-26 US US14/751,295 patent/US20150291931A1/en not_active Abandoned
-
2016
- 2016-08-15 US US15/237,079 patent/US10392600B2/en not_active Expired - Lifetime
Patent Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2001053465A1 (en) * | 2000-01-21 | 2001-07-26 | The Johns Hopkins University School Of Medicine | Human embryoid body-derived cells |
| JP2001333770A (ja) * | 2000-05-31 | 2001-12-04 | Japan Science & Technology Corp | 脊椎動物の膵臓のインビトロ形成方法 |
Non-Patent Citations (15)
| Title |
|---|
| JPN6010008437; Cytotechnology Vol.30, 1999, p.211-226 * |
| JPN6010008440; 蛋白質核酸酵素 Vol.43, No.10, 1998, p.1347-1354 * |
| JPN6010008443; Genome Res. Vol.6, 1996, p.226-233 * |
| JPN6010008445; Current Biol. Vol.11, 2001, p.514-518 * |
| JPN6010008447; 今日の移植 Vol.13, No.4, 2000, p.330-336 * |
| JPN6010008449; Exp.Cell Res. Vol.247, 1999, p.241-248 * |
| JPN6010008451; FEBS Lett. Vol.497, 2001, p.15-19 * |
| JPN6010008453; Cell Vol.105, 2001, p.169-172 * |
| JPN6010008455; Proc.Natl.Acad.Sci.USA Vol.97, No.21, 2000, p.11307-11312 * |
| JPN6010008458; Dev.Growth Differ. Vol.43, 2001, p.469-502 * |
| JPN6011018019; GI Res. Vol.7, No.4, 1999, p.3-11 * |
| JPN6011018023; GI Res. Vol.9, No.3, 2001, p.29-37 * |
| JPN6011018025; 実験医学 Vol.18, No.9, 2000, p.164-170 * |
| JPN6011064724; Science Vol.292, 2001, p.1389-1394 |
| JPN6011064729; Dev.Dynamics Vol.206, 1996, p.379-390 |
Cited By (6)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2013531497A (ja) * | 2010-06-13 | 2013-08-08 | インスティチュート・オブ・バイオフィジックス,チャイニーズ・アカデミー・オブ・サイエンシズ | 幹細胞から心筋細胞を調製するための方法および組成物ならびにその使用 |
| US9273286B2 (en) | 2010-06-13 | 2016-03-01 | Institute Of Biophysics, Chinese Academy Of Sciences | Methods and compositions for preparing cardiomyocytes from stem cells and uses thereof |
| US10590386B2 (en) | 2010-06-13 | 2020-03-17 | Insitute Of Biophysics, Chinese Academy Of Sciences | Methods and compositions for preparing cardiomyocytes from stem cells and uses thereof |
| US11339371B2 (en) | 2012-07-23 | 2022-05-24 | Institute Of Biophysics, Chinese Academy Of Sciences | Method for inducing pluripotent stem cells to differentiate into ventricular myocytes in vitro |
| JP2016521571A (ja) * | 2013-06-11 | 2016-07-25 | プルーリオミクス・ベー・フェー | 多能性哺乳動物幹細胞に由来する心筋細胞を成熟させるための培地組成物 |
| JP2015186471A (ja) * | 2014-03-26 | 2015-10-29 | ジーイー・ヘルスケア・ユーケイ・リミテッド | 細胞分化方法 |
Also Published As
| Publication number | Publication date |
|---|---|
| WO2004098490A3 (en) | 2005-04-21 |
| US20060276420A1 (en) | 2006-12-07 |
| CA2684022A1 (en) | 2004-11-18 |
| US20140287494A1 (en) | 2014-09-25 |
| US7955849B2 (en) | 2011-06-07 |
| EP2361966A1 (en) | 2011-08-31 |
| US7763466B2 (en) | 2010-07-27 |
| AU2003304106B2 (en) | 2009-06-25 |
| AU2003304106A1 (en) | 2004-11-26 |
| US10392600B2 (en) | 2019-08-27 |
| WO2004098490A2 (en) | 2004-11-18 |
| SG141296A1 (en) | 2008-04-28 |
| CA2487058A1 (en) | 2004-11-18 |
| CA2487058C (en) | 2015-04-14 |
| JP2006513727A (ja) | 2006-04-27 |
| AU2009202175B2 (en) | 2013-08-01 |
| US20150218512A1 (en) | 2015-08-06 |
| US8748171B2 (en) | 2014-06-10 |
| EP1543110A4 (en) | 2006-09-06 |
| US20120114613A1 (en) | 2012-05-10 |
| US20150291931A1 (en) | 2015-10-15 |
| US20110070203A1 (en) | 2011-03-24 |
| AU2009202175A1 (en) | 2009-06-18 |
| US20160348069A1 (en) | 2016-12-01 |
| US20080226558A1 (en) | 2008-09-18 |
| EP1543110A2 (en) | 2005-06-22 |
| AU2003304106C1 (en) | 2010-10-28 |
| CA2684022C (en) | 2014-09-23 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US10392600B2 (en) | Method of generating human pancreatic cells | |
| US10428308B2 (en) | Mesoderm and definitive endoderm cell populations | |
| US20170218332A1 (en) | Regionalised Endoderm Cells and Uses Thereof | |
| AU2012203441B2 (en) | Mesoderm and definitive endoderm cell populations |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20100218 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20100513 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20100518 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20100618 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20100623 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20100716 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20100722 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20100818 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20110414 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20110811 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20110817 |
|
| A911 | Transfer to examiner for re-examination before appeal (zenchi) |
Free format text: JAPANESE INTERMEDIATE CODE: A911 Effective date: 20110915 |
|
| A912 | Re-examination (zenchi) completed and case transferred to appeal board |
Free format text: JAPANESE INTERMEDIATE CODE: A912 Effective date: 20111209 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20140123 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20140128 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20140224 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20140227 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20140324 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20140402 |