WO2022158554A1 - 二量体化した環状ペプチドをスクリーニングする方法 - Google Patents
二量体化した環状ペプチドをスクリーニングする方法 Download PDFInfo
- Publication number
- WO2022158554A1 WO2022158554A1 PCT/JP2022/002117 JP2022002117W WO2022158554A1 WO 2022158554 A1 WO2022158554 A1 WO 2022158554A1 JP 2022002117 W JP2022002117 W JP 2022002117W WO 2022158554 A1 WO2022158554 A1 WO 2022158554A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- polypeptides
- peptide
- cyclic
- dimerization domain
- leucine zipper
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Ceased
Links
Images










Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/10—Processes for the isolation, preparation or purification of DNA or RNA
- C12N15/1034—Isolating an individual clone by screening libraries
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/10—Processes for the isolation, preparation or purification of DNA or RNA
- C12N15/1034—Isolating an individual clone by screening libraries
- C12N15/1041—Ribosome/Polysome display, e.g. SPERT, ARM
-
- C—CHEMISTRY; METALLURGY
- C40—COMBINATORIAL TECHNOLOGY
- C40B—COMBINATORIAL CHEMISTRY; LIBRARIES, e.g. CHEMICAL LIBRARIES
- C40B30/00—Methods of screening libraries
- C40B30/04—Methods of screening libraries by measuring the ability to specifically bind a target molecule, e.g. antibody-antigen binding, receptor-ligand binding
-
- C—CHEMISTRY; METALLURGY
- C40—COMBINATORIAL TECHNOLOGY
- C40B—COMBINATORIAL CHEMISTRY; LIBRARIES, e.g. CHEMICAL LIBRARIES
- C40B40/00—Libraries per se, e.g. arrays, mixtures
- C40B40/04—Libraries containing only organic compounds
- C40B40/10—Libraries containing peptides or polypeptides, or derivatives thereof
-
- C—CHEMISTRY; METALLURGY
- C40—COMBINATORIAL TECHNOLOGY
- C40B—COMBINATORIAL CHEMISTRY; LIBRARIES, e.g. CHEMICAL LIBRARIES
- C40B50/00—Methods of creating libraries, e.g. combinatorial synthesis
- C40B50/06—Biochemical methods, e.g. using enzymes or whole viable microorganisms
Definitions
- the present invention relates to a method for screening dimerized cyclic peptides, a library therefor, and molecules containing cyclic peptides that can be used for these.
- growth factors which are biomolecules important for cell differentiation
- growth factors which are biomolecules important for cell differentiation
- the development of alternative molecules that improve the production cost and storage stability of molecules that exhibit biological activity as such dimers has been carried out, and the use of cyclic peptides is also being studied. Therefore, a rapid and convenient method for screening dimerized cyclic peptides is highly desired.
- a peptide dimer was obtained by expressing a molecule in which a peptide and a leucine zipper region were linked using Escherichia coli and obtaining a peptide dimer through non-covalent interactions in the leucine zipper (International Publication No. 2000/018921). issue).
- WO 2000/018921 is intended to provide a substitute molecule for bFGF, is not intended for screening peptide dimers, and does not describe peptide dimer screening.
- An object of one aspect of the present invention is to provide a rapid and convenient method for screening a dimerized cyclic peptide, or a library therefor.
- the object is to provide a rapid and convenient method for screening dimerized cyclic peptides, or a molecule containing a cyclic peptide that can be used in a library therefor.
- the present invention may include, for example, the following aspects.
- a method for screening dimerized cyclic peptides (1) in a cell-free expression system, cyclic peptides, synthesizing at least one polypeptide comprising a dimerization domain comprising a cysteine residue and a leucine zipper region, and a peptide linker present between the cyclic peptide and the dimerization domain; and (2) measuring activity based on binding of the dimer to a target molecule.
- a method for screening dimerized cyclic peptides (1) in a cell-free expression system, cyclic peptides, obtained by synthesizing at least one polypeptide comprising a dimerization domain comprising a cysteine residue and a leucine zipper region, and a peptide linker present between said cyclic peptide and said dimerization domain and (2) measuring activity based on binding of the dimer to a target molecule.
- step (1) the at least one polypeptide comprises a plurality of different polypeptides expected to have binding affinity with the target molecule; and in step (2), the plurality of different The method of [1] or [2] above, which comprises measuring the activity of the polypeptide dimer based on binding to the target molecule.
- step (1) comprises preparing a library containing a plurality of said at least one polypeptide or said dimer, or step (1) comprises said at least one polypeptide or said dimer including preparing a library containing multiple bodies, The method according to any one of [1] to [3] above, wherein the plurality of polypeptides contain different cyclic peptides.
- a method for preparing a library containing the plurality of polypeptides or dimers of the plurality of polypeptides comprising synthesizing a plurality of polypeptides in a cell-free expression system, wherein the plurality of polypeptides but each cyclic peptides, A dimerization domain comprising a cysteine residue and a leucine zipper region, and a peptide linker present between the cyclic peptide and the dimerization domain, wherein the cyclic peptides differ among the plurality of polypeptides. the method described above.
- a library for screening dimerized cyclic peptides comprising a plurality of polypeptides or dimers of the plurality of polypeptides, or a plurality of nucleic acids encoding the plurality of polypeptides, each of the plurality of polypeptides, cyclic peptides, A dimerization domain comprising a cysteine residue and a leucine zipper region, and a peptide linker present between the cyclic peptide and the dimerization domain, wherein the cyclic peptides differ among the plurality of polypeptides. said library.
- [7] The library according to [6] above, comprising 8 or more polypeptides or dimers of said 8 or more polypeptides, or 8 or more nucleic acids encoding said 8 or more polypeptides.
- a method for screening dimerized cyclic peptides (1) obtaining one or more polypeptides having binding affinity to a target molecule from a library comprising a plurality of displayed polypeptides, each of said plurality of displayed polypeptides , cyclic peptides, a dimerization domain comprising a cysteine residue and a leucine zipper region, and a peptide linker present between said cyclic peptide and said dimerization domain, wherein said cyclic peptide differs among said plurality of displayed polypeptides.
- step (2) recovering the nucleic acid associated with the polypeptide obtained in step (1); (3) amplifying the nucleic acid recovered in step (2); (4) using the nucleic acid amplified in step (3), in a cell-free expression system, cyclic peptides, Synthesizing a polypeptide comprising a dimerization domain comprising a cysteine residue and a leucine zipper region, and a peptide linker present between the cyclic peptide and the dimerization domain, and forming a dimer of the polypeptide and (5) measuring activity based on binding of the dimer to a target molecule.
- step (1) is to generate a further library comprising a plurality of displayed polypeptides based on the obtained plurality of polypeptides;
- a library for screening dimerized cyclic peptides comprising a plurality of displayed polypeptides or a plurality of nucleic acids for expressing the plurality of displayed polypeptides, wherein the plurality of each of the displayed polypeptides is cyclic peptides, a dimerization domain comprising a cysteine residue and a leucine zipper region, and a peptide linker present between said cyclic peptide and said dimerization domain, wherein said cyclic peptide differs among said plurality of displayed polypeptides.
- Said library which contains .
- the library of [11] above comprising 10 5 or more displayed polypeptides or 10 5 or more nucleic acids for expressing the 10 5 or more displayed polypeptides.
- the activity based on binding to the target molecule is measured by the method according to any one of [1] to [4] and [8] to [10] above, and the activity based on binding to the target molecule is determined.
- a method for obtaining a dimerized cyclic peptide having [14] A method for preparing a dimerized cyclic peptide, In a cell-free expression system, cyclic peptides, Synthesizing a polypeptide comprising a dimerization domain comprising a cysteine residue and a leucine zipper region, and a peptide linker present between the cyclic peptide and the dimerization domain, and forming a dimer of the polypeptide
- the above method comprising preparing.
- dimerized cyclic peptides having activity based on binding to a target molecule can be screened quickly and easily compared to conventional techniques.
- to provide highly efficient screening (high-throughput screening) for dimerized cyclic peptides having activity based on binding to a target molecule compared to conventional techniques. can be done.
- low-cost screening for dimerized cyclic peptides having activity based on binding to a target molecule can be provided compared to conventional techniques.
- dimerized cyclic peptides having activity based on binding to target molecules can be seamlessly evaluated from the acquisition of displayed candidate polypeptides, which is not available in conventional methods. Screening can be provided.
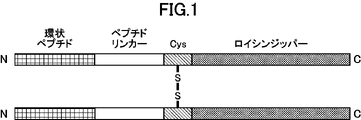
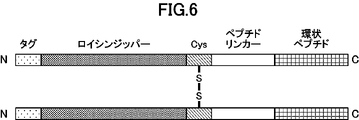
- FIG. 2 is a schematic diagram representing dimerized cyclic peptides.
- the structure “cyclic peptide” - “peptide linker” - “cysteine residue” - “leucine zipper” is shown.
- FIG. 2 is a schematic diagram representing dimerized cyclic peptides.
- the structure “cyclic peptide” - “peptide linker” - “leucine zipper” - “cysteine residue” is shown.
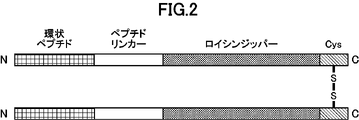
- FIG. 2 is a schematic diagram representing dimerized cyclic peptides.
- the structure “cysteine residue” - “leucine zipper” - "peptide linker” - “cyclic peptide” is shown.
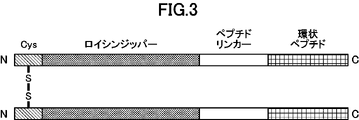
- FIG. 2 is a schematic diagram representing dimerized cyclic peptides.
- FIG. 2 is a schematic diagram representing dimerized cyclic peptides.
- the structure “tag sequence” - “cysteine residue” - “leucine zipper” - “peptide linker” - “cyclic peptide” is shown.
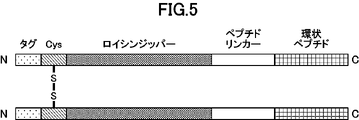
- FIG. 2 is a schematic diagram representing dimerized cyclic peptides.
- the structure “tag sequence” - “leucine zipper” - “cysteine residue” - "peptide linker” - “cyclic peptide” is shown.
- Lane 3 shows the results of SDS-polyacrylamide electrophoresis showing the synthesis of dimerized cyclic peptides by a cell-free expression system.
- Ladder is a marker
- lane 1 is a negative control (cell-free translation solution without addition of cDNA for expression)
- lane 2 is purified from the negative control using Dynabeads His-Tag Isolation & Pulldown (Thermo Fisher Scientific)
- Lane 3 is the cell-free translation solution of N11-Cys-Jun-PA8-aMD4dY
- Lane 4 is the Dynabeads His-Tag from the cell-free translation solution of N11-Cys-Jun-PA8-aMD4dY Fractions purified using Isolation & Pulldown
- lane 5 is the residual fraction
- lane 6 is N11-Cys-GCN4-PA8-aMD4dY
- lane 7 is N11-Cys-GCN4-PA8 This is a fraction pur
- Fig. 3 is a graph showing activity evaluation of dimerized cyclic peptides (aMD4dY-PA8-cys-Jun and aMD4dY-PA8-Jun) synthesized with HGF.
- the vertical axis represents the relative SRE receptor activity
- the horizontal axis represents the concentration of HGF
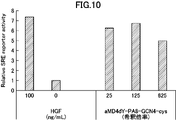
- FIG. 10 is a graph showing activity evaluation of a dimerized cyclic peptide (aMD4dY-PA8-cys-Jun ⁇ hep) synthesized with HGF.
- the vertical axis represents the relative SRE receptor activity
- the horizontal axis represents the concentration of HGF
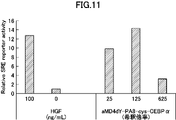
- Fig. 10 is a graph showing activity evaluation of a dimerized cyclic peptide (aMD4dY-PA8-GCN4-cys) synthesized with HGF.
- the vertical axis represents the relative SRE receptor activity
- the horizontal axis represents the concentration of HGF
- Fig. 10 is a graph showing activity evaluation of a dimerized cyclic peptide (aMD4dY-PA8-cys-CEBP ⁇ ) synthesized with HGF.
- the vertical axis represents the relative SRE receptor activity
- the horizontal axis represents the concentration of HGF
- Fig. 3 is a graph showing activity evaluation of dimerized cyclic peptides (cys-Jun-PA8-aMD4dY and Jun-cys-PA8-aMD4dY) synthesized with HGF.
- the vertical axis represents the relative SRE receptor activity
- the horizontal axis represents the concentration of HGF
- Dimerized cyclic peptides synthesized with HGF (cys-Jun-PA8-aMD4dY, N11-Jun-cys-PA8-aMD4dY, N11-cys-Jun-PA8-aMD4dY, N11-GCN4-cys-PA8-aMD4dY, and N11-cys-GCN4-PA8-aMD4dY).
- the vertical axis represents the relative SRE receptor activity
- the horizontal axis represents the concentration of HGF
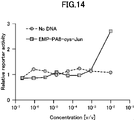
- FIG. 3 is a graph showing activity evaluation of synthesized dimerized cyclic peptide (EMP-cys-PA8-Jun).
- the vertical axis represents the relative receptor activity, and the horizontal axis represents the concentration of the reaction solution of the cell-free expression system with respect to the medium subjected to activity evaluation.
- As a negative control a cell-free expression system to which the EMP-cys-PA8-Jun synthesis gene was not added was used (No DNA).
- Lane 1 is template DNA for aMD4dY-Jun-cys-N11-SecM-GS3
- lane 2 is template DNA for TPO-Jun-cys-N11-SecM-GS3
- lane 3 is EMP'-Jun-cys- N11-SecM-GS3 template DNA
- lane 4 is DNA mix
- lane 5 is DNA mix2.
- Fig. 3 is a graph showing activity evaluation of cell-free translated EMP'-Jun-cys-N11-SecM-GS3.
- the vertical axis represents the relative receptor activity
- the horizontal axis represents the concentration of HGF
- Lane 1 is template DNA for EMP-Jun-cys-N11-SecM-GS3
- lane 2 is template DNA for aMD4dY-Jun-cys-N11-SecM-GS3
- lane 3 is TPO-Jun-cys-N11.
- lane 4 is DNA mix
- lane 5 is DNA mix2.
- DNA mix 1 is a graph showing activity evaluations of cell-free translation products of E. coli, cell-free translation products of DNA mix2, DNA-free group, and recombinant human EPO (rhEPO).
- the concentration of the cell-free expression system reaction solution in the medium subjected to activity evaluation is shown by five serial dilutions (1/110, 1/330, 1/990, 1/2970 and 1/8910).
- rhEPO 50 ⁇ g/mL of rhEPO was added to the medium used for activity evaluation at a concentration of 1/110.
- the vertical axis represents the chemiluminescence intensity (LU).
- One embodiment of the invention relates to molecules comprising cyclic peptides.
- Molecules comprising such cyclic peptides specifically include a cyclic peptide, a dimerization domain comprising a cysteine residue and a leucine zipper region, and between said cyclic peptide and said dimerization domain.
- a polypeptide comprising a peptide linker This polypeptide can be used in the methods of screening dimerized cyclic peptides and libraries therefor described herein.
- each amino acid may be either an L-amino acid or a D-amino acid, and may be either natural or non-natural.
- Each amino acid is preferably ⁇ - , ⁇ - or ⁇ -amino acids, more preferably ⁇ -amino acids.
- a polypeptide is, for example, a structure comprising "cyclic peptide” - “peptide linker” - “cysteine residue” - "leucine zipper region” from the N-terminal side, "cyclic peptide ”-“Peptide linker”-“Leucine zipper region”-“Cysteine residue” structure, from the N-terminal side “Cysteine residue”-“Leucine zipper region”-“Peptide linker”-“Cyclic peptide” structure Alternatively, it may have a structure comprising “leucine zipper region”-“cysteine residue”-“peptide linker”-“cyclic peptide” from the N-terminal side.
- polypeptide may be at the N-terminal side or the C-terminal side, or between a "cyclic peptide” and a “peptide linker", between a “peptide linker” and a “cysteine residue”, or between a "peptide linker” and a "leucine zipper”.
- Any amino acid sequence consisting of at least one amino acid residue between the "region" or between the "cysteine residue” and the "leucine zipper region” may be included.
- Any such amino acid sequence may include, but is not limited to, a tag sequence, an enzyme recognition sequence, or a sequence useful for enhancing expression of a polypeptide. Examples of tag sequences include N11 tag, FLAG tag, PA tag, histidine tag, etc.
- the tag sequence is preferably present on the N-terminal side of the above polypeptide.
- Enzyme recognition sequences include, for example, protease cleavage sequences, sugar chain modification sequences and the like. Between “cyclic peptide” and “peptide linker”, between “peptide linker” and “cysteine residue”, between “peptide linker” and “leucine zipper region”, or between “cysteine residue” and “leucine zipper region”
- the amino acid sequence that can be included between is not particularly limited, but includes, for example, one consisting of one or more glycines, preferably consisting of 1 to 5 glycines.
- the cyclic peptide is particularly limited
- the molecular weight may be 250 or greater, 500 or greater, 800 or greater, or 1000 or greater, and may be 10,000 or less, 9,000 or less, 8,000 or less, or 7,000 or less.
- the cyclic peptide is not particularly limited, but may consist of, for example, 3 or more, 6 or more, 10 or more, or 12 or more amino acid residues, 50 or less, 45 or less, 40 1 or less, or 35 or less amino acid residues.
- the cyclic peptide may form a cyclic structure by disulfide bond between two cysteine residues, for example, although there is no particular limitation on how the cyclic peptide forms a cyclic structure.
- the cyclic peptide include FGFR-binding peptide, c-Met (HGFR)-binding peptide, erythropoietin receptor-binding peptide, thrombopoietin receptor-binding peptide, albumin-binding peptide, EGFR-binding peptide, VEGFR-binding peptide, PDGFR-binding peptide, Axl binding peptide, PDGFR binding peptide, SCFR binding peptide, Flt-3 binding peptide, c-Ret binding peptide, ROR binding peptide, Tie binding peptide, NGFR binding peptide, Insulin receptor binding peptide, EphR binding peptide, Alk binding peptide, DDR binding peptid
- cyclic peptides include, but are not limited to, cyclic peptides that bind to c-Met (also simply referred to as Met).
- a cyclic peptide comprising or consisting of an amino acid sequence selected from the following (a) to (i), and forming a cyclic structure with a disulfide bond.
- (b) CRQFNRRTHEVWNLDC (SEQ ID NO: 2) also referred to herein as aMD4dY).
- amino acid sequence homology can be calculated using known analysis tools, for example, by using the homology algorithm BLAST (Basic local alignment search tool) of the National Center for Biotechnology Information (NCBI) can be calculated. For calculation of amino acid sequence homology, default (initial setting) parameters in the analysis tool may be used.
- BLAST Basic local alignment search tool
- NCBI National Center for Biotechnology Information
- amino acid modification is phosphorylation, methylation, acetylation, adenylation, ADP-ribosylation, or glycosylation.
- Met is an HGF receptor and a tyrosine kinase receptor, and such cyclic peptides that bind to Met (HGFR) are known to function as Met (HGFR) agonists by dimerization. .
- substitution, deletion or addition means a substitution with a known amino acid, preferably a conservative amino acid substitution.
- a “conservative amino acid substitution” is one in which an amino acid residue is replaced by another amino acid residue having a side chain R group with similar chemical properties (eg, charge or hydrophobicity).
- side chain R group with similar chemical properties
- amino acid groups having side chains with similar chemical properties include glycine, alanine, valine, leucine, and isoleucine with aliphatic side chains, serine and threonine with aliphatic hydroxyl side chains, amide-containing side chains.
- Each amino acid to be introduced by substitution may be an L-amino acid or a D-amino acid, and may be a natural amino acid or a non-natural amino acid.
- substitution, deletion or addition means that a known amino acid is added to the end of an amino acid sequence, or a known amino acid is inserted between amino acid residues of an amino acid sequence.
- Each amino acid to be introduced by addition may be an L-amino acid or a D-amino acid, and may be a natural amino acid or a non-natural amino acid. is an amino acid that constitutes
- cyclic peptides that bind to erythropoietin receptors are listed below, but are not limited thereto.
- a cyclic peptide comprising or consisting of an amino acid sequence selected from the following (j) to (n), and forming a cyclic structure with a disulfide bond.
- sequence identity may be 92% or greater, 95% or greater, or 98% or greater.
- amino acid modification is phosphorylation, methylation, acetylation, adenylation, ADP-ribosylation, or glycosylation.
- the peptide linker is particularly limited However, it can have a molecular weight of 10,000 or less and consist of 50 or less amino acid residues. In one embodiment, the peptide linker has a molecular weight of 10,000 or less and can consist of 50 or less amino acid residues, and at least 90%, at least 95%, or 100% of its amino acid sequence is alanine (A ), proline (P) and serine (S).
- proline residues may constitute more than 4% and less than 40% of the amino acid sequence of the peptide linker.
- proline residues are greater than about 4%, greater than about 5%, greater than about 6%, greater than about 8%, greater than about 10%, greater than about 15%, or greater than about 20% in the amino acid sequence of the peptide linker. may comprise less than about 40%, or less than about 35%.
- alanine and serine residues may each independently constitute greater than about 4%, greater than about 10%, or greater than about 20%, or less than about 50%.
- the term "about X%" is not limited to the number X, but also includes additional values of 10% to 20% more or 10% to 20% less.
- amino acid residues different from alanine, serine and proline are Arg, Asn, Asp, Cys, Gln, Glu, Gly, His, Ile, Leu, Lys, Met, Phe, Thr, Trp , Tyr and Val.
- amino acid residues different from alanine, serine and proline include those without hydrophobic side chains such as Val, Ile, Leu, Met, Phe, Tyr or Trp, and/or Lys, Arg, Those without charged side chains, such as Asp or Glu, are preferred.
- the peptide linker has a molecular weight of 10,000 or less and can consist of 50 or less amino acid residues, wherein at least 90%, at least 95%, or 100% of its amino acid sequence is alanine (A) and It may consist of amino acid residues selected from proline (P).
- proline residues may constitute more than 10% and less than 75% of the amino acid sequence of the peptide linker.
- proline residues are more than about 10%, more than about 12%, more than about 14%, more than about 18%, more than about 20%, more than about 22%, more than about 23%, may comprise greater than about 24%, or greater than about 25%, less than about 75%, less than about 70%, less than about 65%, less than about 60%, less than about 55%, less than about 50%, about 48% less than about 46%, less than about 44%, less than about 42%, less than about 41%, less than about 40%, less than about 39%, less than about 38%, less than about 37%, less than about 36%, or about 35 % may be configured.
- alanine residues are greater than about 25%, greater than about 30%, greater than about 35%, greater than about 40%, greater than about 45%, greater than about 50%, greater than about 52%, greater than about 54%, about 56% about 58%, about 59%, about 60%, about 61%, about 62%, about 63%, about 64%, or about 65%, about less than 90%, less than about 88%, less than about 86%, less than about 84%, less than about 82%, less than about 80%, less than about 79%, less than about 78%, less than about 77%, less than about 76%, or It may constitute less than about 75%.
- amino acid residues different from alanine, serine and proline are Arg, Asn, Asp, Cys, Gln, Glu, Gly, His, Ile, Leu, Lys, Met, Phe, Thr, Trp , Tyr and Val.
- amino acid residues different from alanine, serine and proline include those without hydrophobic side chains such as Val, Ile, Leu, Met, Phe, Tyr or Trp, and/or Lys, Arg, Those without charged side chains, such as Asp or Glu, are preferred.
- the peptide linker comprises a plurality of amino acid repeats, the amino acid repeats consist of alanine (A), proline (P) and serine (S), and the same amino acid residues are consecutive in the amino acid repeats.
- the number can be 6 residues or less, 5 residues or less, 4 residues or less, or 3 residues or less.
- the peptide linker comprises a plurality of amino acid repeats, the amino acid repeats consist of alanine (A) and proline (P), and the number of consecutive identical amino acid residues in the amino acid repeats is 6 residues. No more than 5 residues, 4 residues or less, or 3 residues or less.
- the peptide linker may contain two or more repeat sequences represented by AP or AAP.
- the peptide linker may comprise a sequence composed of glycine (G) and serine (S).
- the peptide linker may repeatedly comprise an amino acid sequence selected from the group consisting of GGGGS, GGGS, GGS and GS.
- the peptide linker may comprise consecutive glycines or may consist of glycines only.
- the peptide linker may repeatedly comprise the amino acid sequence designated EAAAK.
- the amino acid sequence of the peptide linker is preferably one consisting of A, P and S, one consisting of A and P, or one consisting of repeats of the sequence represented by GGGGS, but is limited to these. not a thing
- the length of the peptide linker may consist of 50 amino acid residues or less; It can consist of one or fewer amino acid residues.
- the length of the peptide linker can consist of 1 or more amino acid residues, but for example, the peptide linker can be 4 or more, 8 or more, 12 or more, or 16 or more amino acid residues. It can consist of residues.
- peptide linker examples include, but are not limited to, those containing the following amino acid sequences and those consisting of the following amino acid sequences.
- AAPAAPAPAAPAAPAPAAPAAPA SEQ ID NO: 9
- AAPAAPAPAAPAAPAPAAPAAP SEQ ID NO: 10
- AAPAAPAPAAPAAP SEQ ID NO: 11
- AAPAAAPAPAAPAAPAPAAP SEQ ID NO: 12
- AAAPAAPAAPAAAPAAPAAP SEQ ID NO: 13
- AAPAAPAAPAAPAAPAAPAAPAAP SEQ ID NO: 14
- APAAAAPAPAAAPAAPAAAPAPAAAP SEQ ID NO: 15
- AAAPAAPAAPPAAAAPAAPAAPPA SEQ ID NO: 16
- APAPAPAPAPAPAPAPAPAP SEQ ID NO: 17
- ASPAAPAPASPAAPAPSAPA SEQ ID NO: 18
- AAPASPAPAAPSAPAPAAPS SEQ ID NO: 19
- APSSPSPSAPSSPSPASPSS SAPSSPSPSAPS
- Circularly permuted sequences can be obtained, for example, by removing the first alanine of the above sequence and adding another alanine to the end of the above sequence, for example by removing the first alanine of the above sequence It can be easily made.
- Such a circularly permuted sequence of SEQ ID NO:9 becomes APAAPAPAAPAAPAPAAPAA (SEQ ID NO:28).
- non-limiting examples of circularly permuted amino acid residue sequences of SEQ ID NO:7 include the following.
- PAAPAPAAPAAPAPAAPAAA (SEQ ID NO: 29), AAPAPAAPAAPAPAAPAAPAAA (SEQ ID NO: 29), AAPAPAAPAAPAAPAAPAAAP (SEQ ID NO: 30), APAPAAPAAPAPAAPAAAPA (SEQ ID NO: 31), PAPAAPAAPAPAAPAAAPAA (SEQ ID NO: 32), APAAPAAPAPAAPAAAPAAP (SEQ ID NO: 33), PAAPAAPAPAAPAAAPAAPA (SEQ ID NO: 34), AAPAAPAPAAPAAAPAAPAP ( SEQ ID NO: 35), APAAPAPAAPAAAPAAPAPA (SEQ ID NO: 36), and PAAPAPAAPAAAPAAPAPAA (SEQ ID NO: 37).
- specific examples of the peptide linker include, but are not limited to, those containing the following amino acid sequences and those consisting of the following amino acid sequences.
- a polypeptide comprising a cyclic peptide, a dimerization domain comprising a cysteine residue and a leucine zipper region, and a peptide linker present between the cyclic peptide and the dimerization domain
- in the dimerization domain may have only one cysteine residue, or may have two or more cysteine residues.
- the leucine zipper region in the dimerization domain is a region that forms an ⁇ -helix and has a leucine residue every seven residues.
- the leucine zipper region is not particularly limited, but can consist of, for example, 60 or less, 45 or less, 35 or less, or 25 or less amino acid residues.
- the leucine zipper region in the dimerization domain is not particularly limited as long as it forms a dimer. Jun ⁇ hep (M. D. Ballinger et al., Nat. Biotechnol. 17, 1199-1204 (1999), incorporated herein by reference), deficient in heparin-binding ability by replacing the positively charged amino acids in the domain with glutamine.
- the content of the document is cited as a reference.) and the like.
- Specific examples of the leucine zipper region include, but are not limited to, those containing the following amino acid sequences and those consisting of the following amino acid sequences.
- the leucine zipper region in the dimerization domain may have its heparin binding ability removed.
- a polypeptide as described herein comprising a cyclic peptide, a dimerization domain comprising a cysteine residue and a leucine zipper region, and a peptide linker present between said cyclic peptide and said dimerization domain,
- the molecular weight may be, for example, 3,000 or more, 4,000 or more, 5,000 or more, or 6,000 or more, and may be 20,000 or less, 18,000 or less, 16,000 or less, or 14,000 or less.
- polypeptide is not particularly limited, but may consist of, for example, 30 or more, 40 or more, 50 or more, or 60 or more amino acid residues, and may consist of 200 or less, 180 or less, 160 1 or less, or 140 or less amino acid residues.
- a polypeptide as described herein comprising a cyclic peptide, a dimerization domain comprising a cysteine residue and a leucine zipper region, and a peptide linker present between said cyclic peptide and said dimerization domain, It can be prepared using well-known techniques based on the nucleic acid encoding the polypeptide.
- the polypeptides can also dimerize through non-covalent interactions at the leucine zipper region in the dimerization domain and disulfide bonds between cysteine residues in the dimerization domain. Disulfide bonds between cysteine residues contribute to tight dimerization and can greatly affect the activity of dimerized cyclic peptides.
- a nucleic acid encoding the polypeptide can be either DNA or RNA (mRNA). Synthesis of such nucleic acids can be appropriately carried out by those skilled in the art using conventional synthesis techniques based on the amino acid sequence of the desired polypeptide.
- the nucleic acid encoding the polypeptide may have a promoter sequence necessary for cell-free expression (eg, T7 promoter sequence) and a ribosome binding sequence on the 5' side.
- the nucleic acid may have a termination codon and an untranslated region on the 3' side.
- One embodiment of the invention relates to a method of preparing dimerized cyclic peptides.
- Methods for preparing such dimerized cyclic peptides include, for example, a cyclic peptide, a dimerization domain comprising a cysteine residue and a leucine zipper region, and a dimerization domain between the cyclic peptide and the dimerization domain. in a cell-free expression system, and preparing a dimer of said polypeptide, said dimer may be a homodimer or a heterodimer.
- cyclic peptides, dimerization domains (cysteine residues and leucine zipper regions), and peptide linkers are as described herein above.
- Protein synthesis in a cell-free expression system is a technique well known to those skilled in the art, and a cell extract may be used, or a commercially available cell-free expression reagent may be used.
- the components contained in the cell-free expression system include a nucleic acid (DNA or mRNA) encoding a desired template polypeptide, proteins necessary for transcription, translation, energy regeneration, and post-translational modification (RNA polymerase, ribosome, aminoacyl-tRNA, synthase, translation initiation factor, translation elongation factor, translation termination factor, ribosome regeneration factor, nucleoside diphosphate kinase, adenosine kinase, creatine kinase, prolyl isomerase, disulfide bond isomerase, chaperone, etc.), tRNA, aminoacyl-tRNA, natural amino acids , unnatural amino acids, NTPs, buffers, and the like.
- Commercially available cell-free expression reagents include, for
- One embodiment of the invention relates to a method of screening dimerized cyclic peptides.
- Methods for screening such dimerized cyclic peptides include, for example, the following steps (1A) or (1B) and step (2).
- (1A) in a cell-free expression system cyclic peptides, synthesizing at least one polypeptide comprising a dimerization domain comprising a cysteine residue and a leucine zipper region, and a peptide linker present between the cyclic peptide and the dimerization domain; preparing a dimer, or (1B) in a cell-free expression system, cyclic peptides, obtained by synthesizing at least one polypeptide comprising a dimerization domain comprising a cysteine residue and a leucine zipper region, and a peptide linker present between said cyclic peptide and said dimerization domain providing a dimer of said polypeptide; and (2) measuring activity based on binding of said dimer to a
- the method of screening dimerized cyclic peptides may further comprise the step of screening or selecting the dimers (dimerized cyclic peptides) based on the activity measured in step (2).
- one embodiment of the present invention measures the activity based on binding to the target molecule by a method of screening dimerized cyclic peptides, dimerized having activity based on binding to the target molecule It relates to a method for obtaining cyclic peptides.
- step (1A) is as described herein above with respect to the method for preparing a dimerized cyclic peptide
- step (1B) is The idea is to provide dimerized cyclic peptides (polypeptide dimers) obtained by a method as described herein above.
- a cyclic peptide, a dimerization domain comprising a cysteine residue and a leucine zipper region, and a peptide linker present between the cyclic peptide and the dimerization domain For "at least one polypeptide”, "a cyclic peptide, a dimerization domain comprising a cysteine residue and a leucine zipper region, and a dimerization domain comprising said cyclic peptide and said dimerization domain" as described herein above.
- the contents of "polypeptides with intervening peptide linkers” can be applied as such.
- polypeptide dimer is synonymous with “dimerized cyclic peptide” as described herein above.
- target molecule means a molecule to which the cyclic peptide or dimerized cyclic peptide described herein binds or is expected to bind.
- the target molecule is preferably a receptor, more preferably a receptor that is activated by dimerization, such as growth factor receptors, growth factor receptors, cytokine receptors, and the like.
- target molecules include FGFR, c-Met (HGFR), erythropoietin receptor, thrombopoietin receptor, albumin, EGFR, VEGFR, PDGFR, Axl, PDGFR, SCFR, Flt-3, c-Ret, ROR , Tie, NGFR, Insulin receptor, EphR, Alk, DDR, TGFBR, Activin receptor, BMP receptor, Interleukin receptor, T cell receptor, Transferrin receptor, Lipoprotein receptor, Ubiquitin ligase, Antibody, Complement body, GPCR, ion channel, virus, etc., but not limited to these.
- step (2) "activity based on the binding of the dimer to the target molecule” means the action and/or its strength brought about by the binding of the dimer to the target molecule, agonistic activity and Any meaning of antagonistic activity can be included.
- activity can include receptor activity, signaling activity, apoptotic activity, and the like.
- the measurement in step (2) can be performed using well-known techniques, by bioluminescence assays such as the luciferase reporter assay, by receptor dimerization assays such as the PathHunter® Receptor Dimerization assay, or It can be carried out by measuring fluorescence resonance energy transfer (FRET), or by cross-linking with a chemical linker and dimerization analysis by SDS-PAGE, or the like.
- FRET fluorescence resonance energy transfer
- the cell-free expression reaction solution obtained in step (1) may be used as it is, or may be used after being diluted as appropriate.
- the method for screening dimerized cyclic peptides comprises, in step (1), screening a plurality of different polypeptides, at least one of which is expected to have binding affinity with a target molecule. and in step (2), measuring activity based on binding to said target molecule for said plurality of dimers of different polypeptides.
- the method for screening a dimerized cyclic peptide wherein step (1) comprises preparing a library containing a plurality of at least one polypeptide or dimers thereof, wherein a plurality of said polypeptides It contains cyclic peptides that vary between peptides.
- the library containing at least one polypeptide or a plurality of dimers thereof includes the polypeptides or dimers thereof, for example, 2 or more, 8 or more, 16 or more, 32 or more, 48 Above, 96 or more, 200 or more, 1000 or more, 10000 or more, 10 5 or more, or 10 6 or more.
- a library containing a plurality of at least one polypeptide or dimers thereof may contain, for example, 10 species or less, 10 species or less, or 10 species or less of the polypeptides or dimers thereof.
- a "library” shall mean a collection containing a plurality of different substances (eg, different polypeptides or different nucleic acids).
- a method for producing such a library includes, for example, synthesizing a plurality of polypeptides in a cell-free expression system, a method for producing a library containing the plurality of polypeptides or dimers of the plurality of polypeptides wherein each of the plurality of polypeptides comprises a cyclic peptide, a dimerization domain comprising a cysteine residue and a leucine zipper region, and a peptide linker present between the cyclic peptide and the dimerization domain and wherein different cyclic peptides are included among the plurality of polypeptides.
- the step of "synthesizing a plurality of polypeptides in a cell-free expression system” is as described herein above for the method of preparing dimerized cyclic peptides.
- the plurality of polypeptides each comprise a cyclic peptide, a dimerization domain containing a cysteine residue and a leucine zipper region, and a peptide linker present between the cyclic peptide and the dimerization domain.
- references to a polypeptide "comprising” refer to "a cyclic peptide, a dimerization domain comprising a cysteine residue and a leucine zipper region, and between said cyclic peptide and said dimerization domain" described herein above. can be applied as is.
- One embodiment of the invention relates to libraries for screening dimerized cyclic peptides.
- a library comprises, for example, a plurality of polypeptides or dimers of said plurality of polypeptides, or a plurality of nucleic acids encoding said plurality of polypeptides, said plurality of polypeptides each comprising a cyclic peptide, A dimerization domain comprising a cysteine residue and a leucine zipper region, and a peptide linker present between the cyclic peptide and the dimerization domain, wherein the cyclic peptide differs between the plurality of polypeptides It is included.
- the plurality of polypeptides are each a cyclic peptide, a dimerization domain containing a cysteine residue and a leucine zipper region, and a peptide present between the cyclic peptide and the dimerization domain
- the content for a polypeptide "comprising a linker” is defined herein above as "a dimerization domain comprising a cyclic peptide, a cysteine residue and a leucine zipper region, and said cyclic peptide and said dimer can be applied as is.
- Such a library contains a plurality of polypeptides or dimers of the plurality of polypeptides, for example, 2 or more, 8 or more, 16 or more, 32 or more, 48 or more, 96 or more, 200 Above, 1000 or more, 10000 or more, 10 5 or more, or 10 6 or more. Also, such a library can comprise a plurality of polypeptides or dimers of said plurality of polypeptides, eg, 10 15 or less, 10 14 or less, or 10 13 or less.
- such a library may contain a plurality of nucleic acids encoding a plurality of polypeptides, for example, 2 or more, 8 or more, 16 or more, 32 or more, 48 or more, 96 or more, 200 or more, It may contain 1000 or more, 10000 or more, 105 or more, or 106 or more. Also, such a library can contain a plurality of nucleic acids encoding a plurality of polypeptides, eg, 10 15 or less, 10 14 or less, or 10 13 or less.
- the "nucleic acid encoding a polypeptide" is as described herein above.
- One embodiment of the invention relates to a method of screening dimerized cyclic peptides.
- Methods for screening such dimerized cyclic peptides include, for example, the following steps (1') to (5') as methods different from those described above.
- (1′) obtaining one or more polypeptides having binding affinity to a target molecule from a library comprising a plurality of displayed polypeptides, wherein the plurality of displayed polypeptides are Each, cyclic peptides, a dimerization domain comprising a cysteine residue and a leucine zipper region; and a peptide linker present between said cyclic peptide and said dimerization domain, wherein said cyclic peptide differs among said plurality of displayed polypeptides.
- step (1′) recovering the nucleic acid associated with the polypeptide obtained in step (1′), (3') amplifying the nucleic acid recovered in step (2'); (4') using the nucleic acid amplified in step (3'), in a cell-free expression system, cyclic peptides, Synthesizing a polypeptide comprising a dimerization domain comprising a cysteine residue and a leucine zipper region, and a peptide linker present between the cyclic peptide and the dimerization domain, and forming a dimer of the polypeptide (5′) measuring the activity based on the binding of said dimer to a target molecule.
- a method for screening a dimerized cyclic peptide comprising such steps (1') to (5') for a dimerized cyclic peptide having activity based on binding to a target molecule, display Evaluable screening can be provided seamlessly from the acquisition of the proposed candidate polypeptides.
- a method for screening a dimerized cyclic peptide comprising such steps (1′) to (5′) further comprises the dimer (dimeric embodied cyclic peptides).
- one embodiment of the present invention measures the activity based on binding to the target molecule by a method of screening a dimerized cyclic peptide comprising such steps (1') to (5'), It relates to a method for obtaining dimerized cyclic peptides with activity based on binding to a target molecule.
- Step (1′) is obtaining one or more polypeptides having binding affinity with the target molecule from a library containing a plurality of displayed polypeptides.
- a "library comprising a plurality of displayed polypeptides” can be produced using well-known display technology.
- Such display techniques include, for example, ribosome display, mRNA display, phage display, bacterial display, yeast display, and the like.
- the introduction of the SecM sequence into the template DNA sequence facilitates the preparation of ribosome-polypeptide (fusion protein)-mRNA complexes, allowing convenient preparation of libraries containing multiple displayed polypeptides.
- the arrays and display methods used are not limited to this.
- the library comprises a plurality of displayed polypeptides, for example, 2 or more, 8 or more, 16 or more, 32 or more, 48 or more, 96 or more, 200 or more, 1000 10,000 or more species, 10 5 or more species, 10 6 or more species, 10 7 or more species, 10 8 or more species, 10 9 or more species, or 10 10 or more species.
- a library can comprise a plurality of polypeptides or dimers of said plurality of polypeptides, eg, 10 15 or less, 10 14 or less, or 10 13 or less.
- step (1′) the “target molecule” and the “cyclic peptide” of the “plurality of displayed polypeptides”, the “dimerization domain comprising a cysteine residue and a leucine zipper region”, and “the A peptide linker present between the cyclic peptide and said dimerization domain” is as described herein above.
- step (1′) obtaining one or more polypeptides having binding affinity with the target molecule can be performed by a well-known technique, for example, expressing a recombinant target molecule or a target molecule This can be done by affinity selection on cells.
- Step (2') is a step of recovering the nucleic acid associated with the polypeptide obtained in step (1'). This can be done by well-known techniques, such as magnetic bead recovery, ELISA plate recovery, cell pellet recovery, nanodroplet recovery, microflow It can be performed by a method using a road or the like.
- Step (3') is a step of amplifying the nucleic acid recovered in step (2'). This can be done by well-known techniques, eg by PCR. In PCR, if the recovered nucleic acid is DNA, it can be used as it is, and if the recovered nucleic acid is mRNA, cDNA can be prepared by RT-PCR and the nucleic acid amplified.
- Step (4 ') is a cell-free expression system using the nucleic acid amplified in step (3 '), a dimerization domain containing a cyclic peptide, a cysteine residue and a leucine zipper region, and the cyclic peptide and the A step of synthesizing a polypeptide containing a peptide linker present between it and a dimerization domain to prepare a dimer of said polypeptide. This is as described herein above for methods of preparing dimerized cyclic peptides.
- a polypeptide comprising a cyclic peptide, a dimerization domain comprising a cysteine residue and a leucine zipper region, and a peptide linker present between the cyclic peptide and the dimerization domain is described herein. As described above.
- Step (5′) is a step of measuring the activity based on the binding of the dimer to the target molecule.
- the activity based on the binding of the dimer to the target molecule means the action and/or its strength brought about by the binding of the dimer to the target molecule, and agonist activity and antagonist activity.
- activity can include receptor activity, signaling activity, apoptotic activity, and the like.
- the "target molecule” in step (5') is as described herein above.
- Measurements in step (5′) can be performed using well-known techniques, by bioluminescence assays such as the luciferase reporter assay, by receptor dimerization assays such as the PathHunter® Receptor Dimerization assay, Alternatively, it can be performed by measurement of fluorescence resonance energy transfer (FRET), or by cross-linking with a chemical linker and dimerization analysis by SDS-PAGE, or the like.
- bioluminescence assays such as the luciferase reporter assay
- receptor dimerization assays such as the PathHunter® Receptor Dimerization assay
- FRET fluorescence resonance energy transfer
- the method for screening dimerized cyclic peptides is characterized in that step (1′) is based on the obtained plurality of polypeptides to generate a further library comprising a plurality of displayed polypeptides. and obtaining from said additional library one or more polypeptides having binding affinity with the target molecule.
- step (1′) is based on the obtained plurality of polypeptides to generate a further library comprising a plurality of displayed polypeptides. and obtaining from said additional library one or more polypeptides having binding affinity with the target molecule.
- further libraries containing the plurality of displayed polypeptides can be produced by the following steps.
- Step (1′-1) A step of recovering the nucleic acid associated with the polypeptide obtained in step (1′)
- Step (1′-2) Amplifying the nucleic acid recovered in step (1′-1) process to do.
- Step (1'-1) can be carried out in the same manner as described for step (2').
- Step (1'-2) can be carried out in the same manner as described for step (3').
- Retrieving one or more polypeptides having binding affinity to the target molecule from the additional library thus generated can be performed in the same manner as step (1').
- the method for screening dimerized cyclic peptides comprises, in step (1′), generating a further library comprising a plurality of displayed polypeptides based on the obtained plurality of polypeptides. and obtaining from said additional library one or more polypeptides having binding affinity to the target molecule, repeated 2, 3, 4, 5, 6, or 7 or more times may
- One embodiment of the invention relates to libraries for screening dimerized cyclic peptides.
- a library apart from the aforementioned library, comprises, for example, a plurality of displayed polypeptides or a plurality of nucleic acids for expressing said plurality of displayed polypeptides, and said plurality of displayed polypeptides.
- each of said polypeptides comprises a cyclic peptide, a dimerization domain comprising a cysteine residue and a leucine zipper region, and a peptide linker present between said cyclic peptide and said dimerization domain; Cyclic peptides that differ among the plurality of displayed polypeptides are included.
- a "cyclic peptide” of "a plurality of displayed polypeptides”, a “dimerization domain comprising a cysteine residue and a leucine zipper region”, and a “dimerization domain between said cyclic peptide and said dimerization domain The "intervening peptide linker” is as described herein above.
- Such libraries may contain a plurality of displayed polypeptides, e.g. species or more, 10 5 or more, 10 6 or more, 10 7 or more, 10 8 or more, 10 9 or more, or 10 10 or more.
- a library can comprise a plurality of polypeptides or dimers of said plurality of polypeptides, eg, 10 15 or less, 10 14 or less, or 10 13 or less.
- such libraries may contain a plurality of nucleic acids for expressing a plurality of displayed polypeptides, e.g., 2 or more, 8 or more, 16 or more, 32 or more, 48 or more, 96 or more.
- such a library can comprise a plurality of polypeptides or dimers of said plurality of polypeptides, eg, 10 15 or less, 10 14 or less, or 10 13 or less.
- HGFs used in Examples are recombinant human HGFs (rhHGF, R&D Systems), and in the present specification and drawings, recombinant human HGFs are simply referred to as HGFs.
- Example 1 Design of cyclic peptide-leucine zipper fusion and expression by cell-free expression system (1-1) Overview of design of aMD4dY-leucine zipper fusion Cyclic peptide aMD4dY (SEQ ID NO: 2) is dimerized It has been reported that it exhibits HGF-like activity, that is, acts as a Met agonist. Therefore, we designed a peptide sequence to express aMD4dY as a fusion with a leucine zipper region that self-dimerizes. Also, the binding is reversible, as the leucine zipper region leads to non-covalent dimerization.
- a construct with an N11 tag, which is an affinity tag, added to the N-terminus was also designed in the same manner. Specifically, the following peptide sequences were designed (Figs. 5 and 6). ⁇ "Tag sequence"-"Leucine zipper region"-"Cysteine residue"-"Peptide linker"-"aMD4dY” ⁇ "Tag sequence"-"Cysteine residue"-"Leucine zipper region"-"Peptide linker"-"aMD4dY”
- the leucine zipper region the leucine zipper domains of c-Jun, GCN4 and CEBP ⁇ proteins, which are known to form homodimers, and the positively charged amino acids of the c-Jun leucine zipper domain are replaced with glutamine.
- Jun ⁇ hep lacking heparin-binding ability was used. These leucine zipper domains are hereinafter referred to as Jun, GCN4, CEBP ⁇ and Jun ⁇ hep, respectively.
- base sequences encoding the above amino acid sequences (a) to (k) are respectively used. designed.
- a T7 promoter sequence and a ribosome binding sequence necessary for cell-free expression were added to the 5' side of these, and a termination codon and an untranslated region were added to the 3' side.
- Nucleotide sequences for cell-free expression corresponding to the above (a)-(k) are shown in (l)-(v), respectively.
- EMP-leucine zipper fusion EMP which is a cyclic peptide described in Johnson, D. L. et al. Chem. Biol. have been reported to exhibit erythropoietin (EPO)-like activity. Therefore, we designed a peptide sequence to express EMP as a fusion with a leucine zipper domain that self-dimerizes. In addition, a cysteine residue was added in the same manner as in (1-1) above, and the peptide sequence was designed to be "EMP" - "peptide linker” - "cysteine residue” - "leucine zipper region".
- EMP-PA8-cys-Jun MGGLYACHMGPMTWVCQPLRGAAPAAPAPAAGCGGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMN SEQ ID NO: 67
- EMP is as described in (1-5) above, PA8 represents a peptide linker, cys represents a cysteine residue, and Jun represents the leucine zipper domain of the c-Jun protein.
- Example 2 Activity Evaluation of aMD4dY-Leucine Zipper Fusions Dimers of each aMD4dY-leucine zipper fusion were evaluated by luciferase assay.
- 0.6 ⁇ L of Attractene Transfection Reagent (QIAGEN) was added to 25 ⁇ L of Opti-MEM medium (Thermo Fisher Scientific) and incubated at room temperature for 5 minutes.
- a mixed solution of 25 ⁇ L of Opti-MEM and 1 ⁇ L of SRE reporter vector (QIAGEN) was added to the mixed solution, and incubated at room temperature for 20 minutes.
- This mixed solution was added to a 96-well plate, HEK293E cells were seeded thereon at 40,000 cells/well, and incubated overnight at 37°C in a 5% CO 2 incubator. After transfection, all the culture supernatant was removed and Opti-MEM medium containing 0.5% FBS (Thermo Fisher Scientific), 1% non-essential amino acid solution (Thermo Fisher Scientific) and penicillin-streptomycin (Nacalai Tesque) was added. The cells were starved by adding 100 ⁇ L of evaluation basal medium) and culturing at 37° C. for 4 hours.
- each dimer when aMD4dY-PA8-GCN4-cys and aMD4dY-PA8-cys-CEBP ⁇ were expressed, each dimer was found to have signal activation ability. From this, it was understood that it is possible to widely use the leucine zipper region that forms homodimers in the dimerization domain. As shown in FIG. 12, when cys-Jun-PA8-aMD4dY and Jun-cys-PA8-aMD4dY were expressed, each dimer was found to have signal activation ability.
- the activity of the cyclic peptide (growth factor-like activity) can be obtained even if the order of the cyclic peptide, peptide linker, cysteine residue, and leucine zipper region is changed.
- FIG. 13 it was revealed that even when the construct with the N11 tag added to the N-terminus was used, the ability to activate signal was exhibited. From this, it was understood that it is also possible to add a tag sequence to the N-terminus of the expressed polypeptide.
- Example 3 Activity evaluation of EMP-leucine zipper fusion
- PathHunter registered trademark
- eXpress EpoR-JAK2 Functional Assay Kit DiscoveRx
- EPO activity evaluation cells contained in the PathHunter (registered trademark) eXpress EpoR-JAK2 Functional Assay Kit were seeded in a 96-well plate and cultured for 24 hours at 37°C under 5% CO 2 .
- a cell-free expression reaction mixture containing the EMP-leucine zipper fusion was diluted and added to the cells.
- the cell-free expression system was diluted at the same magnification as this reaction solution, and the cell-free expression system was prepared as described in (1-9) above, except that the gene with the base sequence indicated by (x) was not included.
- the treated unreacted solution was added.
- the prepared Substrate Reagent was added to the cells stimulated at room temperature for 3 hours and incubated for 60 minutes. Chemiluminescence intensity was quantified with a Nivo plate reader (Perkin Elmer). The results are shown in FIG. These results revealed that the EMP-PA8-cys-Jun dimer activates EpoR.
- Example 4 Screening to Enrich HGF-Like Active Molecules from Cyclic Peptide-Leucine Zipper Fusion Libraries Based on Binding to c-Met Molecules Cell-Free from Multiple DNAs Each Encoding a Cyclic Peptide-Leucine Zipper Fusion Molecule We expressed the fusion by translation, enriched the fusion molecule that binds to c-Met and the DNA encoding the molecule, and confirmed that the enriched fusion molecule has HGF-like activity.
- the SecM sequence (referred to as SecM below) is a sequence represented by the amino acid sequence of FSTPVWISQAQGIRAG (SEQ ID NO: 86), and inhibits the elongation reaction of the ribosome to facilitate ribosome-cyclic peptide-leucine zipper fusion. Its purpose is to form a body-mRNA complex.
- the sequence denoted as GS3 below is a sequence consisting of three repeats of "glycine"-"serine" and is a spacer sequence.
- T7 RNA polymerase T7 RNA polymerase (Takara Bio, 2540A) and rNTPs (Promega, E6000) were used for transcription.
- the reaction was carried out in the tank for 6 hours.
- a 50 mM EDTA solution was then added. It was purified with RNA Clean & Concentrator 5 (ZYMO RESEARCH, R1015) and made into a 2 ⁇ M mRNA mix.
- Target molecules were immobilized on magnetic beads.
- 5 ⁇ L of 2 pM Recombinant Human HGFR/c-MET Fc Chimera Protein (R&D systems, 8614-MT-100), 5 ⁇ L slurry of Dynabeads (invitrogen, DB10003) and 10 ⁇ L of 1x TBS-T buffer (50 mM Tris- HCl, 150 mM NaCl, 0.05% Tween 20) were mixed and mixed by inversion for 30 minutes at 4°C.
- the magnetic beads were washed three times with 100 ⁇ L of 1x TBS-T buffer, then resuspended in 10 ⁇ L of 1x TBS-T buffer and left standing on ice.
- Reporter assays were performed as follows. 0.6 ⁇ L of Attractene Transfection Reagent (QIAGEN) was added to 25 ⁇ L of Opti-MEM medium (Thermo Fisher Scientific) and incubated at room temperature for 5 minutes. A mixed solution of 25 ⁇ L of Opti-MEM and 1 ⁇ L of SRE reporter vector (QIAGEN) was added to the mixed solution, and incubated at room temperature for 20 minutes. This mixed solution was added to a 96-well plate, HEK293E cells were seeded thereon at 40,000 cells/well, and incubated overnight at 37°C in a 5% CO 2 incubator.
- QIAGEN Attractene Transfection Reagent
- Opti-MEM medium containing 0.5% FBS (Thermo Fisher Scientific), 1% non-essential amino acid solution (Thermo Fisher Scientific) and penicillin-streptomycin (Nacalai Tesque) was added.
- the cells were starved by adding 100 ⁇ L of evaluation basal medium) and culturing at 37° C. for 4 hours. Add 0 to 100 ng/mL HGF to the evaluation basal medium, or add the above cell-free expression reaction solution to the medium so that it is diluted 25 to 625 times, and add 100 ⁇ L of the evaluation basal medium to the cells. did. Cells were stimulated by culturing overnight in a 37°C, 5% CO 2 incubator.
- a Dual-Luciferase Reporter Assay System (Promega) was used for signal intensity detection. 100 ⁇ L of the medium supernatant was removed, 50 ⁇ L of Glo reagent was added, and the cells were lysed at room temperature for 10 minutes. Activation of the HGF-Erk-SRE pathway was quantified by detecting Firefly luciferase luminescence from cell lysates with a plate reader. Subsequently, 50 ⁇ L of Glo & Stop reagent solution was added, and after 10 minutes, luminescence of Renilla luciferase, an internal standard, was detected to quantify the number of cells.
- SRE activity (luminescence intensity of Firefly luciferase)/(luminescence intensity of Renilla luciferase).
- a signal activity value relative to the sample to which the evaluation compound was not added was obtained and used as the relative receptor activity (Relative SRE reporter activity).
- the results are shown in FIG. DNA mix showed no relative receptor activity, whereas DNA mix2 showed similar activity to HGF.
- aMD4dY-Jun Since only -cys-N11-SecM-GS3 showed activity, it was found that DNA mix2 was enriched with aMD4dY-Jun-cys-N11-SecM-GS3.
- Example 5 Screening Method for Enriching Active Molecules from Cyclic Peptide-Leucine Zipper Fusion Library Based on Binding to EPO Receptor Molecule The screening method performed in Example 4 is possible regardless of the target molecule To demonstrate, we enriched peptides that bind to the EPO receptor from multiple cyclic peptide-leucine zipper fusion molecules and confirmed the presence or absence of activity based on binding of the enriched DNA to the EPO receptor.
- the ribosome display method and the cyclic peptide-leucine zipper fusion were combined to recover the complex of the cyclic peptide-leucine zipper fusion that binds to the EPO receptor immobilized on the magnetic beads, the ribosome, and the mRNA complex. did. Through a series of manipulations, it was confirmed that template DNAs for cyclic peptide-leucine zipper fusions that bind to the EPO receptor were enriched from the library.
- EMP-Jun-cys-N11-SecM-GS3 MAETGGLYACHMGPMTWVCQPLRGAAPAAPAPAAGGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMNGCGGKDHLIHNHHKHEHAHAEHLFSTPVWISQAQGIRAGGSGSGS (SEQ ID NO: 87)
- Construction of template DNA for EMP-leucine zipper fusion As template DNA for expressing EMP-Jun-cys-N11-SecM-GS3, nucleotide sequences encoding the above (i) amino acid sequences were designed.
- DNA sequence of EMP-Jun-cys-N11-SecM-GS3 was prepared by PCR. Two types of DNA sequences, aMD4dY-Jun-cys-N11-SecM-GS3 and TPO-Jun-cys-N11-SecM-GS3, prepared in Example 4 were used. DNA mix of EMP-Jun-cys-N11-SecM-GS3, aMD4dY-Jun-cys-N11-SecM-GS3, and TPO-Jun-cys-N11-SecM-GS3 at a ratio of 1:50:50 was made.
- T7 RNA polymerase was used to transcribe DNA.
- T7 RNA polymerase (Takara Bio, 2540A) and rNTPs (Promega, E6000) were also used for transcription.
- the reaction was carried out in the tank for 6 hours.
- a 50 mM EDTA solution was then added. It was purified with RNA Clean & Concentrator 5 (ZYMO RESEARCH, R1015) and made into a 2 ⁇ M mRNA mix.
- Target molecules were immobilized on magnetic beads.
- 5 ⁇ L of 2 pM Recombinant Human Erythropoietin R Fc Chimera Protein, CF (R&D systems, 963-ER-50), 5 ⁇ L slurry of Dynabeads (invitrogen, DB10003) and 10 ⁇ L of 1x TBS-T buffer (50 mM Tris-HCl) , 150 mM NaCl, 0.05% Tween 20) were mixed and mixed by inversion at 4°C for 30 minutes.
- the magnetic beads were washed three times with 100 ⁇ L of 1x TBS-T buffer, then resuspended in 10 ⁇ L of 1x TBS-T buffer and left standing on ice.
- PCR Amplification of cDNA The recovered cDNA was amplified by PCR.
- KOD-Plus- Ver.2 (Toyobo, KOD-211) was used for PCR. 10 ⁇ L of 10x PCR buffer, 4 ⁇ L of 250 mM MgSO 4 , 10 ⁇ L of 2 mM dNTPs, 1 ⁇ L of 100 ⁇ M selec_PCR_F1.F39, 1 ⁇ L of 100 ⁇ M selec_PCR_R2.R24, 63 ⁇ L of ultrapure water to 4 ⁇ L of completed reverse transcription reaction. , KOD-Plus- 2 ⁇ L was mixed, and a cycle was created under the conditions of 94° C. 30 seconds ⁇ 55° C. 40 seconds ⁇ 68° C. 60 seconds, and the cycles were repeated 20 to 25 times. The collected DNA was used as DNA mix2.
- Lanes 1-5 in order, EMP-Jun-cys-N11-SecM-GS3, aMD4dY-Jun-cys-N11-SecM-GS3, TPO-Jun-cys-N11-SecM-GS3, DNA mix, DNA mix2 corresponds to When the DNA mix2 band obtained after the ribosome display method was compared with the DNA mix band, a shift toward longer DNA length was confirmed, and EMP-Jun-cys-N11-SecM The band showing the same DNA length as the -GS3-encoding DNA showed the highest intensity.
- the prepared Substrate Reagent was added to the cells stimulated at room temperature for 3 hours and incubated for 60 minutes. Chemiluminescence intensity was quantified with a Nivo plate reader (Perkin Elmer). The results are shown in FIG. Comparing the relative receptor activity calculated from the chemiluminescence intensity between DNA mix and DNA mix2, it was clarified that only DNA mix2 had activity.
Landscapes
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Organic Chemistry (AREA)
- Genetics & Genomics (AREA)
- Engineering & Computer Science (AREA)
- Biochemistry (AREA)
- Molecular Biology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biomedical Technology (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- General Engineering & Computer Science (AREA)
- Biotechnology (AREA)
- General Health & Medical Sciences (AREA)
- Microbiology (AREA)
- General Chemical & Material Sciences (AREA)
- Medicinal Chemistry (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Physics & Mathematics (AREA)
- Crystallography & Structural Chemistry (AREA)
- Plant Pathology (AREA)
- Biophysics (AREA)
- Immunology (AREA)
- Bioinformatics & Computational Biology (AREA)
- Peptides Or Proteins (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
- Investigating Or Analysing Biological Materials (AREA)
Abstract
迅速で簡便な、二量体化した環状ペプチドをスクリーニングする方法、及びそのためのライブラリ、並びにこれらに利用できる環状ペプチドを含む分子等を提供する。例えば、二量体化した環状ペプチドをスクリーニングする方法は、 (1)無細胞発現系で、 環状ペプチド、 システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに 前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含む少なくとも1種のポリペプチドを合成し、前記ポリペプチドの二量体を調製する工程、及び (2)前記二量体の標的分子との結合に基づく活性を測定する工程
を含む。
Description
本発明は、二量体化した環状ペプチドをスクリーニングする方法、及びそのためのライブラリ、並びにこれらに利用できる環状ペプチドを含む分子等に関する。
近年、医薬のモダリティとして、抗体医薬等のバイオ医薬品、再生医療、核酸医薬等が研究され、実際に用いられるようになってきた。また、新しい医薬のモダリティとして、中分子、特にペプチドが着目されている。中でも、環状ペプチド等の特殊な形状のペプチド(特殊ペプチド)が複数開発されており、医薬への応用が期待されるところである。
環状ペプチドは、直鎖ペプチドと比べて優れた結合親和性、標的選択性、分解耐性等を有し得る。そのため、医薬等の分野での環状ペプチドの利用が有望視されている。
ペプチドの中には、二量体となって生物活性を発揮するもの、又は生物活性が増強されるものが存在する。例えば、細胞分化に重要な生体分子である成長因子はその一例である。このような二量体となって生物活性を発揮する分子の生産コストや保存安定性等を向上させた代替分子の開発が近年行われており、環状ペプチドの利用も検討されているところである。そのため、二量体化した環状ペプチドのスクリーニングのための迅速で簡便な方法が切望されている。
環状ペプチドは、直鎖ペプチドと比べて優れた結合親和性、標的選択性、分解耐性等を有し得る。そのため、医薬等の分野での環状ペプチドの利用が有望視されている。
ペプチドの中には、二量体となって生物活性を発揮するもの、又は生物活性が増強されるものが存在する。例えば、細胞分化に重要な生体分子である成長因子はその一例である。このような二量体となって生物活性を発揮する分子の生産コストや保存安定性等を向上させた代替分子の開発が近年行われており、環状ペプチドの利用も検討されているところである。そのため、二量体化した環状ペプチドのスクリーニングのための迅速で簡便な方法が切望されている。
従来、環状ペプチドを二量体化したペプチドダイマーを得る手法としては、2つの環状ペプチドをリンカーで結合させた分子を合成することが行われていた。しかしながら、このような分子フォーマットの合成を生合成系で実施する場合、その鋳型となる遺伝子の、環状ペプチドに対応する2箇所の配列を変える必要があり、複数箇所の遺伝子組み換えのような煩雑な操作や遺伝子全合成が必要となる。そのため、迅速で簡便な合成は達成できず、迅速で簡便な、二量体化した環状ペプチドのスクリーニングにも応用できない。
一方で、大腸菌を用いて、ペプチドとロイシンジッパー領域を連結させた分子を発現させ、ロイシンジッパーにおける非共有結合性相互作用によりペプチドダイマーを得た例が知られている(国際公開第2000/018921号)。しかしながら、国際公開第2000/018921号は、bFGFの代替分子を提供することを目的としたものであり、ペプチドダイマーのスクリーニングを目的としたものではなく、ペプチドダイマーのスクリーニングについては何ら記載がない。
上述のとおり、二量体化した環状ペプチドのスクリーニングのための迅速で簡便な方法が望まれている。
本発明の一態様では、迅速で簡便な、二量体化した環状ペプチドをスクリーニングする方法、又はそのためのライブラリを提供することを目的とする。
また、本発明の別の態様では、迅速で簡便な、二量体化した環状ペプチドをスクリーニングする方法、又はそのためのライブラリに利用できる、環状ペプチドを含む分子を提供することを目的とする。
本発明の一態様では、迅速で簡便な、二量体化した環状ペプチドをスクリーニングする方法、又はそのためのライブラリを提供することを目的とする。
また、本発明の別の態様では、迅速で簡便な、二量体化した環状ペプチドをスクリーニングする方法、又はそのためのライブラリに利用できる、環状ペプチドを含む分子を提供することを目的とする。
本発明は、例えば、以下の態様を含み得る。
〔1〕二量体化した環状ペプチドをスクリーニングする方法であって、
(1)無細胞発現系で、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含む少なくとも1種のポリペプチドを合成し、前記ポリペプチドの二量体を調製する工程、及び
(2)前記二量体の標的分子との結合に基づく活性を測定する工程
を含む、前記方法。
〔2〕二量体化した環状ペプチドをスクリーニングする方法であって、
(1)無細胞発現系で、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含む少なくとも1種のポリペプチドを合成することにより得られた前記ポリペプチドの二量体を用意する工程、及び
(2)前記二量体の標的分子との結合に基づく活性を測定する工程
を含む、前記方法。
〔3〕工程(1)において、前記少なくとも1種のポリペプチドが、前記標的分子との結合親和性を有すると見込まれる複数の異なるポリペプチドを含み、かつ工程(2)において、前記複数の異なるポリペプチドの二量体について、前記標的分子との結合に基づく活性を測定することを含む、前記〔1〕又は〔2〕に記載の方法。
〔4〕工程(1)が、前記少なくとも1種のポリペプチド又は前記二量体を複数含むライブラリを作製することを含み、又は工程(1)が、前記少なくとも1種のポリペプチド又は前記二量体を複数含むライブラリを用意することを含み、
複数の前記ポリペプチド間で異なる環状ペプチドが含まれている、前記〔1〕~〔3〕のいずれか1項に記載の方法。
〔5〕無細胞発現系で複数のポリペプチドを合成することを含む、前記複数のポリペプチド又は前記複数のポリペプチドの二量体を含むライブラリを作製する方法であって、前記複数のポリペプチドが、各々、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含み、前記複数のポリペプチド間で異なる環状ペプチドが含まれている、前記方法。
〔6〕二量体化した環状ペプチドをスクリーニングするためのライブラリであって、複数のポリペプチド若しくは前記複数のポリペプチドの二量体、又は前記複数のポリペプチドをコードする複数の核酸を含み、前記複数のポリペプチドが、各々、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含み、前記複数のポリペプチド間で異なる環状ペプチドが含まれている、前記ライブラリ。
〔7〕8種以上のポリペプチド若しくは前記8種以上のポリペプチドの二量体、又は前記8種以上のポリペプチドをコードする8種以上の核酸を含む、前記〔6〕に記載のライブラリ。
〔8〕複数のポリペプチドの二量体の標的分子との結合に基づく活性を測定する工程を含む、前記〔6〕又は〔7〕に記載のライブラリを用いて二量体化した環状ペプチドをスクリーニングする方法。
〔9〕二量体化した環状ペプチドをスクリーニングする方法であって、
(1)複数のディスプレイされたポリペプチドを含むライブラリから、標的分子との結合親和性を有する1つ又は複数のポリペプチドを取得する工程であって、前記複数のディスプレイされたポリペプチドが、各々、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含み、前記複数のディスプレイされたポリペプチド間で異なる環状ペプチドが含まれている、該取得工程、及び
(2)工程(1)において取得した前記ポリペプチドと関連付けられた核酸を回収する工程、
(3)工程(2)において回収した前記核酸を増幅する工程、
(4)工程(3)において増幅した前記核酸を用いて、無細胞発現系で、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含むポリペプチドを合成し、前記ポリペプチドの二量体を調製する工程、及び
(5)前記二量体の標的分子との結合に基づく活性を測定する工程
を含む、前記方法。
〔10〕工程(1)が、取得した複数の前記ポリペプチドに基づき、複数のディスプレイされたポリペプチドを含む更なるライブラリを作製すること、及び前記更なるライブラリから、標的分子との結合親和性を有する1つ又は複数のポリペプチドを取得することを繰り返すことを含む、前記〔9〕に記載の方法。
〔11〕二量体化した環状ペプチドをスクリーニングするためのライブラリであって、複数のディスプレイされたポリペプチド又は前記複数のディスプレイされたポリペプチドを発現させるための複数の核酸を含み、前記複数のディスプレイされたポリペプチドが、各々、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含み、前記複数のディスプレイされたポリペプチド間で異なる環状ペプチドが含まれている、前記ライブラリ。
〔12〕105種以上のディスプレイされたポリペプチド又は前記105種以上のディスプレイされたポリペプチドを発現させるための105種以上の核酸を含む、前記〔11〕に記載のライブラリ。
〔13〕前記〔1〕~〔4〕及び〔8〕~〔10〕のいずれか1項に記載の方法によって標的分子との結合に基づく活性を測定して、標的分子との結合に基づく活性を有する二量体化した環状ペプチドを得る方法。
〔14〕二量体化した環状ペプチドを調製する方法であって、
無細胞発現系で、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含むポリペプチドを合成し、前記ポリペプチドの二量体を調製することを含む、前記方法。
〔15〕環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含むポリペプチド。
〔16〕前記二量体化ドメインにおいて、システイン残基の数が1つである、前記〔15〕に記載のポリペプチド。
〔17〕前記ペプチドリンカーが、アラニン(A)、プロリン(P)及びセリン(S)から選択される1つ又は複数のアミノ酸残基からなる、前記〔15〕又は〔16〕に記載のポリペプチド。
〔18〕前記〔15〕~〔17〕のいずれか1項に記載のポリペプチドをコードする核酸。
〔1〕二量体化した環状ペプチドをスクリーニングする方法であって、
(1)無細胞発現系で、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含む少なくとも1種のポリペプチドを合成し、前記ポリペプチドの二量体を調製する工程、及び
(2)前記二量体の標的分子との結合に基づく活性を測定する工程
を含む、前記方法。
〔2〕二量体化した環状ペプチドをスクリーニングする方法であって、
(1)無細胞発現系で、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含む少なくとも1種のポリペプチドを合成することにより得られた前記ポリペプチドの二量体を用意する工程、及び
(2)前記二量体の標的分子との結合に基づく活性を測定する工程
を含む、前記方法。
〔3〕工程(1)において、前記少なくとも1種のポリペプチドが、前記標的分子との結合親和性を有すると見込まれる複数の異なるポリペプチドを含み、かつ工程(2)において、前記複数の異なるポリペプチドの二量体について、前記標的分子との結合に基づく活性を測定することを含む、前記〔1〕又は〔2〕に記載の方法。
〔4〕工程(1)が、前記少なくとも1種のポリペプチド又は前記二量体を複数含むライブラリを作製することを含み、又は工程(1)が、前記少なくとも1種のポリペプチド又は前記二量体を複数含むライブラリを用意することを含み、
複数の前記ポリペプチド間で異なる環状ペプチドが含まれている、前記〔1〕~〔3〕のいずれか1項に記載の方法。
〔5〕無細胞発現系で複数のポリペプチドを合成することを含む、前記複数のポリペプチド又は前記複数のポリペプチドの二量体を含むライブラリを作製する方法であって、前記複数のポリペプチドが、各々、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含み、前記複数のポリペプチド間で異なる環状ペプチドが含まれている、前記方法。
〔6〕二量体化した環状ペプチドをスクリーニングするためのライブラリであって、複数のポリペプチド若しくは前記複数のポリペプチドの二量体、又は前記複数のポリペプチドをコードする複数の核酸を含み、前記複数のポリペプチドが、各々、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含み、前記複数のポリペプチド間で異なる環状ペプチドが含まれている、前記ライブラリ。
〔7〕8種以上のポリペプチド若しくは前記8種以上のポリペプチドの二量体、又は前記8種以上のポリペプチドをコードする8種以上の核酸を含む、前記〔6〕に記載のライブラリ。
〔8〕複数のポリペプチドの二量体の標的分子との結合に基づく活性を測定する工程を含む、前記〔6〕又は〔7〕に記載のライブラリを用いて二量体化した環状ペプチドをスクリーニングする方法。
〔9〕二量体化した環状ペプチドをスクリーニングする方法であって、
(1)複数のディスプレイされたポリペプチドを含むライブラリから、標的分子との結合親和性を有する1つ又は複数のポリペプチドを取得する工程であって、前記複数のディスプレイされたポリペプチドが、各々、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含み、前記複数のディスプレイされたポリペプチド間で異なる環状ペプチドが含まれている、該取得工程、及び
(2)工程(1)において取得した前記ポリペプチドと関連付けられた核酸を回収する工程、
(3)工程(2)において回収した前記核酸を増幅する工程、
(4)工程(3)において増幅した前記核酸を用いて、無細胞発現系で、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含むポリペプチドを合成し、前記ポリペプチドの二量体を調製する工程、及び
(5)前記二量体の標的分子との結合に基づく活性を測定する工程
を含む、前記方法。
〔10〕工程(1)が、取得した複数の前記ポリペプチドに基づき、複数のディスプレイされたポリペプチドを含む更なるライブラリを作製すること、及び前記更なるライブラリから、標的分子との結合親和性を有する1つ又は複数のポリペプチドを取得することを繰り返すことを含む、前記〔9〕に記載の方法。
〔11〕二量体化した環状ペプチドをスクリーニングするためのライブラリであって、複数のディスプレイされたポリペプチド又は前記複数のディスプレイされたポリペプチドを発現させるための複数の核酸を含み、前記複数のディスプレイされたポリペプチドが、各々、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含み、前記複数のディスプレイされたポリペプチド間で異なる環状ペプチドが含まれている、前記ライブラリ。
〔12〕105種以上のディスプレイされたポリペプチド又は前記105種以上のディスプレイされたポリペプチドを発現させるための105種以上の核酸を含む、前記〔11〕に記載のライブラリ。
〔13〕前記〔1〕~〔4〕及び〔8〕~〔10〕のいずれか1項に記載の方法によって標的分子との結合に基づく活性を測定して、標的分子との結合に基づく活性を有する二量体化した環状ペプチドを得る方法。
〔14〕二量体化した環状ペプチドを調製する方法であって、
無細胞発現系で、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含むポリペプチドを合成し、前記ポリペプチドの二量体を調製することを含む、前記方法。
〔15〕環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含むポリペプチド。
〔16〕前記二量体化ドメインにおいて、システイン残基の数が1つである、前記〔15〕に記載のポリペプチド。
〔17〕前記ペプチドリンカーが、アラニン(A)、プロリン(P)及びセリン(S)から選択される1つ又は複数のアミノ酸残基からなる、前記〔15〕又は〔16〕に記載のポリペプチド。
〔18〕前記〔15〕~〔17〕のいずれか1項に記載のポリペプチドをコードする核酸。
本発明の一実施態様によれば、従来の手法と比較して、迅速で簡便に、標的分子との結合に基づく活性を有する二量体化した環状ペプチドをスクリーニングすることができる。
本発明の一実施態様によれば、従来の手法と比較して、標的分子との結合に基づく活性を有する二量体化した環状ペプチドについて、高効率なスクリーニング(ハイスループットスクリーニング)を提供することができる。
本発明の一実施態様によれば、従来の手法と比較して、標的分子との結合に基づく活性を有する二量体化した環状ペプチドについて、低コストなスクリーニングを提供することができる。
本発明の一実施態様によれば、従来の手法にはない、標的分子との結合に基づく活性を有する二量体化した環状ペプチドについて、ディスプレイされた候補ポリペプチドの取得からシームレスに評価可能なスクリーニングを提供することができる。
本発明の一実施態様によれば、従来の手法と比較して、標的分子との結合に基づく活性を有する二量体化した環状ペプチドについて、高効率なスクリーニング(ハイスループットスクリーニング)を提供することができる。
本発明の一実施態様によれば、従来の手法と比較して、標的分子との結合に基づく活性を有する二量体化した環状ペプチドについて、低コストなスクリーニングを提供することができる。
本発明の一実施態様によれば、従来の手法にはない、標的分子との結合に基づく活性を有する二量体化した環状ペプチドについて、ディスプレイされた候補ポリペプチドの取得からシームレスに評価可能なスクリーニングを提供することができる。
本発明の一実施態様は、環状ペプチドを含む分子に関する。このような環状ペプチドを含む分子は、具体的には、環状ペプチド、システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカーを含むポリペプチドである。このポリペプチドは、本明細書に記載の、二量体化した環状ペプチドをスクリーニングする方法や、そのためのライブラリにおいて利用することができる。なお、ポリペプチドにおいて、各アミノ酸は、L-アミノ酸であってもD-アミノ酸であってもよく、天然型であっても非天然型であってもよいし、各アミノ酸は、好ましくはα-、β-又はγ-アミノ酸であり、より好ましくはα-アミノ酸である。
一実施態様において、このようなポリペプチドは、例えば、N末端側から「環状ペプチド」-「ペプチドリンカー」-「システイン残基」-「ロイシンジッパー領域」を含む構造、N末端側から「環状ペプチド」-「ペプチドリンカー」-「ロイシンジッパー領域」-「システイン残基」を含む構造、N末端側から「システイン残基」-「ロイシンジッパー領域」-「ペプチドリンカー」-「環状ペプチド」を含む構造、又はN末端側から「ロイシンジッパー領域」-「システイン残基」-「ペプチドリンカー」-「環状ペプチド」を含む構造を有し得る。更に、当該ポリペプチドは、N末端側もしくはC末端側において、又は「環状ペプチド」と「ペプチドリンカー」の間、「ペプチドリンカー」と「システイン残基」の間、「ペプチドリンカー」と「ロイシンジッパー領域」の間、もしくは「システイン残基」と「ロイシンジッパー領域」の間において、少なくとも1つのアミノ酸残基からなる任意のアミノ酸配列を含んでもよい。このような任意のアミノ酸配列は、特に限定されないが、タグ配列、酵素認識配列、又はポリペプチドの発現を高めるのに有用な配列を含むものであってもよい。タグ配列としては、例えば、N11タグ、FLAGタグ、PAタグ、ヒスチジンタグ等が挙げられ、タグ配列は、上記ポリペプチドのN末端側に存在することが好ましい。酵素認識配列としては、例えば、プロテアーゼ切断配列、糖鎖修飾配列等が挙げられる。「環状ペプチド」と「ペプチドリンカー」の間、「ペプチドリンカー」と「システイン残基」の間、「ペプチドリンカー」と「ロイシンジッパー領域」の間、もしくは「システイン残基」と「ロイシンジッパー領域」の間に含まれ得るアミノ酸配列は特に限定されないが、例えば、1つ以上のグリシンからなるものが挙げられ、好ましくは1~5個のグリシンからなる。
一実施態様において、このようなポリペプチドは、例えば、N末端側から「環状ペプチド」-「ペプチドリンカー」-「システイン残基」-「ロイシンジッパー領域」を含む構造、N末端側から「環状ペプチド」-「ペプチドリンカー」-「ロイシンジッパー領域」-「システイン残基」を含む構造、N末端側から「システイン残基」-「ロイシンジッパー領域」-「ペプチドリンカー」-「環状ペプチド」を含む構造、又はN末端側から「ロイシンジッパー領域」-「システイン残基」-「ペプチドリンカー」-「環状ペプチド」を含む構造を有し得る。更に、当該ポリペプチドは、N末端側もしくはC末端側において、又は「環状ペプチド」と「ペプチドリンカー」の間、「ペプチドリンカー」と「システイン残基」の間、「ペプチドリンカー」と「ロイシンジッパー領域」の間、もしくは「システイン残基」と「ロイシンジッパー領域」の間において、少なくとも1つのアミノ酸残基からなる任意のアミノ酸配列を含んでもよい。このような任意のアミノ酸配列は、特に限定されないが、タグ配列、酵素認識配列、又はポリペプチドの発現を高めるのに有用な配列を含むものであってもよい。タグ配列としては、例えば、N11タグ、FLAGタグ、PAタグ、ヒスチジンタグ等が挙げられ、タグ配列は、上記ポリペプチドのN末端側に存在することが好ましい。酵素認識配列としては、例えば、プロテアーゼ切断配列、糖鎖修飾配列等が挙げられる。「環状ペプチド」と「ペプチドリンカー」の間、「ペプチドリンカー」と「システイン残基」の間、「ペプチドリンカー」と「ロイシンジッパー領域」の間、もしくは「システイン残基」と「ロイシンジッパー領域」の間に含まれ得るアミノ酸配列は特に限定されないが、例えば、1つ以上のグリシンからなるものが挙げられ、好ましくは1~5個のグリシンからなる。
環状ペプチド、システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカーを含むポリペプチドにおいて、当該環状ペプチドは、特に限定されないが、例えば、分子量250以上、500以上、800以上、又は1000以上であり得、10000以下、9000以下、8000以下、又は7000以下であり得る。また、当該環状ペプチドは、特に限定されないが、例えば、3個以上、6個以上、10個以上、又は12個以上のアミノ酸残基からなるものであり得、50個以下、45個以下、40個以下、又は35個以下のアミノ酸残基からなるものであり得る。
当該環状ペプチドは、どのような結合様式で環状構造となっているかは特に限定されないが、例えば2つのシステイン残基間でジスルフィド結合により環状構造を形成し得る。
当該環状ペプチドとしては、例えば、FGFR結合ペプチド、c-Met(HGFR)結合ペプチド、エリスロポエチン受容体結合ペプチド、トロンボポエチン受容体結合ペプチド、アルブミン結合ペプチド、EGFR結合ペプチド、VEGFR結合ペプチド、PDGFR結合ペプチド、Axl結合ペプチド、PDGFR結合ペプチド、SCFR結合ペプチド、Flt-3結合ペプチド、c-Ret結合ペプチド、ROR結合ペプチド、Tie結合ペプチド、NGFR結合ペプチド、Insulin受容体結合ペプチド、EphR結合ペプチド、Alk結合ペプチド、DDR結合ペプチド、TGFBR結合ペプチド、Activin受容体結合ペプチド、BMP受容体結合ペプチド、インターロイキン受容体結合ペプチド、T細胞受容体結合ペプチド、トランスフェリン受容体結合ペプチド、リポタンパク質受容体結合ペプチド、ユビキチンリガーゼ結合ペプチド、抗体結合ペプチド、補体結合ペプチド、GPCR結合ペプチド、イオンチャネル結合ペプチド、ウイルス結合ペプチド、或いはこれらのペプチドとして作用すると見込まれるペプチド等が挙げられるが、これらに限定されるものではない。
当該環状ペプチドは、どのような結合様式で環状構造となっているかは特に限定されないが、例えば2つのシステイン残基間でジスルフィド結合により環状構造を形成し得る。
当該環状ペプチドとしては、例えば、FGFR結合ペプチド、c-Met(HGFR)結合ペプチド、エリスロポエチン受容体結合ペプチド、トロンボポエチン受容体結合ペプチド、アルブミン結合ペプチド、EGFR結合ペプチド、VEGFR結合ペプチド、PDGFR結合ペプチド、Axl結合ペプチド、PDGFR結合ペプチド、SCFR結合ペプチド、Flt-3結合ペプチド、c-Ret結合ペプチド、ROR結合ペプチド、Tie結合ペプチド、NGFR結合ペプチド、Insulin受容体結合ペプチド、EphR結合ペプチド、Alk結合ペプチド、DDR結合ペプチド、TGFBR結合ペプチド、Activin受容体結合ペプチド、BMP受容体結合ペプチド、インターロイキン受容体結合ペプチド、T細胞受容体結合ペプチド、トランスフェリン受容体結合ペプチド、リポタンパク質受容体結合ペプチド、ユビキチンリガーゼ結合ペプチド、抗体結合ペプチド、補体結合ペプチド、GPCR結合ペプチド、イオンチャネル結合ペプチド、ウイルス結合ペプチド、或いはこれらのペプチドとして作用すると見込まれるペプチド等が挙げられるが、これらに限定されるものではない。
環状ペプチドの具体例として、c-Met(単にMetとも言う。)に結合する環状ペプチドを以下に挙げるが、これに限定されるものではない。
以下の(a)~(i)から選択されるアミノ酸配列を含む又はからなる、ジスルフィド結合により環状構造を形成している環状ペプチド。
(a)CYRQFNRRTHEVWNLDC(配列番号:1)
(b)CRQFNRRTHEVWNLDC(配列番号:2)(本明細書において、aMD4dYとも称される。)
(c)CYWYYAWDQTYKAFPC(配列番号:3)
(d)CWYYAWDQTYKAFPC(配列番号:4)
(e)CYISWNEFNSPNWRFITC(配列番号:5)
(f)CISWNEFNSPNWRFITC(配列番号:6)
(g)(a)~(f)のいずれかのアミノ酸配列において、システイン残基以外の1又は2個のアミノ酸残基が置換、欠失又は付加されており、かつc-Metタンパク質に結合するペプチド。
(h)(a)~(f)のいずれかのアミノ酸配列と90%以上の配列同一性を有し、但し両端のシステイン残基を有しているアミノ酸配列からなり、かつc-Metタンパク質に結合するペプチド。例えば、配列同一性は、92%以上、95%以上、又は98%以上であってもよい。本明細書において、アミノ酸配列相同性は、公知の解析ツールを用いて算出することができ、例えば、全米バイオテクノロジー情報センター(NCBI)の相同性アルゴリズムBLAST(Basic local alignment search tool)を用いることにより算出することができる。また、アミノ酸配列相同性の算出には、解析ツールにおけるデフォルト(初期設定)のパラメータを用いればよい。
(i)(a)~(h)のいずれかのアミノ酸配列において、両端のシステイン残基以外の少なくとも1つのアミノ酸が修飾されているペプチド。ここで、アミノ酸の修飾は、リン酸化、メチル化、アセチル化、アデニリル化、ADPリボシル化又は糖鎖付加である。
なお、MetはHGF受容体で、チロシンキナーゼ型受容体であり、このようなMet(HGFR)に結合する環状ペプチドは、ダイマー化することでMet(HGFR)アゴニストとして機能することが知られている。
以下の(a)~(i)から選択されるアミノ酸配列を含む又はからなる、ジスルフィド結合により環状構造を形成している環状ペプチド。
(a)CYRQFNRRTHEVWNLDC(配列番号:1)
(b)CRQFNRRTHEVWNLDC(配列番号:2)(本明細書において、aMD4dYとも称される。)
(c)CYWYYAWDQTYKAFPC(配列番号:3)
(d)CWYYAWDQTYKAFPC(配列番号:4)
(e)CYISWNEFNSPNWRFITC(配列番号:5)
(f)CISWNEFNSPNWRFITC(配列番号:6)
(g)(a)~(f)のいずれかのアミノ酸配列において、システイン残基以外の1又は2個のアミノ酸残基が置換、欠失又は付加されており、かつc-Metタンパク質に結合するペプチド。
(h)(a)~(f)のいずれかのアミノ酸配列と90%以上の配列同一性を有し、但し両端のシステイン残基を有しているアミノ酸配列からなり、かつc-Metタンパク質に結合するペプチド。例えば、配列同一性は、92%以上、95%以上、又は98%以上であってもよい。本明細書において、アミノ酸配列相同性は、公知の解析ツールを用いて算出することができ、例えば、全米バイオテクノロジー情報センター(NCBI)の相同性アルゴリズムBLAST(Basic local alignment search tool)を用いることにより算出することができる。また、アミノ酸配列相同性の算出には、解析ツールにおけるデフォルト(初期設定)のパラメータを用いればよい。
(i)(a)~(h)のいずれかのアミノ酸配列において、両端のシステイン残基以外の少なくとも1つのアミノ酸が修飾されているペプチド。ここで、アミノ酸の修飾は、リン酸化、メチル化、アセチル化、アデニリル化、ADPリボシル化又は糖鎖付加である。
なお、MetはHGF受容体で、チロシンキナーゼ型受容体であり、このようなMet(HGFR)に結合する環状ペプチドは、ダイマー化することでMet(HGFR)アゴニストとして機能することが知られている。
本明細書において、「置換、欠失又は付加」との表現において、置換とは、公知のアミノ酸への置換を意味し、好ましくは保存的アミノ酸置換である。「保存的アミノ酸置換」は、類似した化学的性質(例えば、電荷又は疎水性)を伴う側鎖R基を有する別のアミノ酸残基によって、アミノ酸残基が置換された置換である。類似した化学的性質を伴う側鎖を有するアミノ酸基の例には、脂肪族側鎖を有するグリシン、アラニン、バリン、ロイシン、及びイソロイシン、脂肪族ヒドロキシル側鎖を有するセリン及びスレオニン、アミド含有側鎖を有するアスパラギン及びグルタミン、芳香族側鎖を有するフェニルアラニン、チロシン、及びトリプトファン、塩基性側鎖を有するリシン、アルギニン、及びヒスチジン、並びに酸性側鎖を有するアスパラギン酸及びグルタミン酸が含まれる。置換により導入される各アミノ酸は、L-アミノ酸であってもD-アミノ酸であってもよく、天然型アミノ酸であっても非天然型であってもよいが、好ましくは、ヒトや動物においてタンパク質を構成するアミノ酸である。
本明細書において、「置換、欠失又は付加」との表現において、付加とは、公知のアミノ酸がアミノ酸配列の末端に付加されること、又は公知のアミノ酸がアミノ酸配列のアミノ酸残基間に挿入されることを意味する。付加により導入される各アミノ酸は、L-アミノ酸であってもD-アミノ酸であってもよく、天然型アミノ酸であっても非天然型であってもよいが、好ましくは、ヒトや動物においてタンパク質を構成するアミノ酸である。
本明細書において、「置換、欠失又は付加」との表現において、付加とは、公知のアミノ酸がアミノ酸配列の末端に付加されること、又は公知のアミノ酸がアミノ酸配列のアミノ酸残基間に挿入されることを意味する。付加により導入される各アミノ酸は、L-アミノ酸であってもD-アミノ酸であってもよく、天然型アミノ酸であっても非天然型であってもよいが、好ましくは、ヒトや動物においてタンパク質を構成するアミノ酸である。
また、環状ペプチドの具体例として、エリスロポエチン受容体に結合する環状ペプチドを以下に挙げるが、これに限定されるものではない。
以下の(j)~(n)から選択されるアミノ酸配列を含む又はからなる、ジスルフィド結合により環状構造を形成している環状ペプチド。
(j)GGLYACHMGPMTWVCQPLRG(配列番号:7)
(k)CISWNEFNSPNWRFITC(配列番号:8)
(l)(j)又は(k)のアミノ酸配列において、システイン残基以外の1又は2個のアミノ酸残基が置換、欠失又は付加されており、かつエリスロポエチン受容体に結合するペプチド。
(m)(j)又は(k)のいずれかのアミノ酸配列と90%以上の配列同一性を有し、但しシステイン残基を(j)又は(k)におけるそのままの位置で有しているアミノ酸配列からなり、かつエリスロポエチン受容体に結合するペプチド。例えば、配列同一性は、92%以上、95%以上、又は98%以上であってもよい。
(n)(j)~(m)のいずれかのアミノ酸配列において、システイン残基以外の少なくとも1つのアミノ酸が修飾されているペプチド。ここで、アミノ酸の修飾は、リン酸化、メチル化、アセチル化、アデニリル化、ADPリボシル化又は糖鎖付加である。
以下の(j)~(n)から選択されるアミノ酸配列を含む又はからなる、ジスルフィド結合により環状構造を形成している環状ペプチド。
(j)GGLYACHMGPMTWVCQPLRG(配列番号:7)
(k)CISWNEFNSPNWRFITC(配列番号:8)
(l)(j)又は(k)のアミノ酸配列において、システイン残基以外の1又は2個のアミノ酸残基が置換、欠失又は付加されており、かつエリスロポエチン受容体に結合するペプチド。
(m)(j)又は(k)のいずれかのアミノ酸配列と90%以上の配列同一性を有し、但しシステイン残基を(j)又は(k)におけるそのままの位置で有しているアミノ酸配列からなり、かつエリスロポエチン受容体に結合するペプチド。例えば、配列同一性は、92%以上、95%以上、又は98%以上であってもよい。
(n)(j)~(m)のいずれかのアミノ酸配列において、システイン残基以外の少なくとも1つのアミノ酸が修飾されているペプチド。ここで、アミノ酸の修飾は、リン酸化、メチル化、アセチル化、アデニリル化、ADPリボシル化又は糖鎖付加である。
環状ペプチド、システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカーを含むポリペプチドにおいて、当該ペプチドリンカーは、特に限定されないが、分子量10,000以下であり、かつ50個以下のアミノ酸残基からなり得る。
また、一実施態様において、当該ペプチドリンカーは、分子量10,000以下であり、かつ50個以下のアミノ酸残基からなり得て、そのアミノ酸配列の少なくとも90%、少なくとも95%、又は100%がアラニン(A)、プロリン(P)及びセリン(S)から選択されるアミノ酸残基からなり得る。この場合、プロリン残基は、当該ペプチドリンカーのアミノ酸配列において、4%超かつ40%未満を構成し得る。例えば、プロリン残基は、当該ペプチドリンカーのアミノ酸配列において、約4%超、約5%超、約6%超、約8%超、約10%超、約15%超、又は約20%超を構成してもよく、約40%未満、又は約35%未満を構成してもよい。また、アラニン残基及びセリン残基は、それぞれ独立に、約4%超、約10%超、又は約20%超を構成してもよく、約50%未満を構成してもよい。
本明細書において、用語「約X%」とは、そのXという数に制限されるのではなく、さらなる10%~20%多い値、又は10%~20%少ない値も含む。例えば、用語「約10%」はまた、それぞれ、11%又は12%、及び9%又は8%にも関する。
当該ペプチドリンカーのアミノ酸配列において、アラニン、セリンおよびプロリンとは異なるアミノ酸残基は、Arg、Asn、Asp、Cys、Gln、Glu、Gly、His、Ile、Leu、Lys、Met、Phe、Thr、Trp、Tyr及びValからなる群から選択することができる。このようなアラニン、セリンおよびプロリンとは異なるアミノ酸残基としては、Val、Ile、Leu、Met、Phe、TyrもしくはTrpのような疎水性側鎖を有さないもの、及び/又はLys、Arg、AspもしくはGluのような荷電を有する側鎖を有さないものが好ましい。
また、一実施態様において、当該ペプチドリンカーは、分子量10,000以下であり、かつ50個以下のアミノ酸残基からなり得て、そのアミノ酸配列の少なくとも90%、少なくとも95%、又は100%がアラニン(A)、プロリン(P)及びセリン(S)から選択されるアミノ酸残基からなり得る。この場合、プロリン残基は、当該ペプチドリンカーのアミノ酸配列において、4%超かつ40%未満を構成し得る。例えば、プロリン残基は、当該ペプチドリンカーのアミノ酸配列において、約4%超、約5%超、約6%超、約8%超、約10%超、約15%超、又は約20%超を構成してもよく、約40%未満、又は約35%未満を構成してもよい。また、アラニン残基及びセリン残基は、それぞれ独立に、約4%超、約10%超、又は約20%超を構成してもよく、約50%未満を構成してもよい。
本明細書において、用語「約X%」とは、そのXという数に制限されるのではなく、さらなる10%~20%多い値、又は10%~20%少ない値も含む。例えば、用語「約10%」はまた、それぞれ、11%又は12%、及び9%又は8%にも関する。
当該ペプチドリンカーのアミノ酸配列において、アラニン、セリンおよびプロリンとは異なるアミノ酸残基は、Arg、Asn、Asp、Cys、Gln、Glu、Gly、His、Ile、Leu、Lys、Met、Phe、Thr、Trp、Tyr及びValからなる群から選択することができる。このようなアラニン、セリンおよびプロリンとは異なるアミノ酸残基としては、Val、Ile、Leu、Met、Phe、TyrもしくはTrpのような疎水性側鎖を有さないもの、及び/又はLys、Arg、AspもしくはGluのような荷電を有する側鎖を有さないものが好ましい。
一実施態様において、当該ペプチドリンカーは、分子量10,000以下であり、かつ50個以下のアミノ酸残基からなり得て、そのアミノ酸配列の少なくとも90%、少なくとも95%、又は100%がアラニン(A)及びプロリン(P)から選択されるアミノ酸残基からなり得る。この場合、プロリン残基は、当該ペプチドリンカーのアミノ酸配列において、10%超かつ75%未満を構成し得る。例えば、プロリン残基は、当該ペプチドリンカーのアミノ酸配列において、約10%超、約12%超、約14%超、約18%超、約20%超、約22%超、約23%超、約24%超、又は約25%超を構成してもよく、約75%未満、約70%未満、約65%未満、約60%未満、約55%未満、約50%未満、約48%未満、約46%未満、約44%未満、約42%未満、約41%未満、約40%未満、約39%未満、約38%未満、約37%未満、約36%未満、又は約35%未満を構成してもよい。また、アラニン残基は、約25%超、約30%超、約35%超、約40%超、約45%超、約50%超、約52%超、約54%超、約56%超、約58%超、約59%超、約60%超、約61%超、約62%超、約63%超、約64%超、又は約65%超を構成してもよく、約90%未満、約88%未満、約86%未満、約84%未満、約82%未満、約80%未満、約79%未満、約78%未満、約77%未満、約76%未満、又は約75%未満を構成してもよい。
当該ペプチドリンカーのアミノ酸配列において、アラニン、セリンおよびプロリンとは異なるアミノ酸残基は、Arg、Asn、Asp、Cys、Gln、Glu、Gly、His、Ile、Leu、Lys、Met、Phe、Thr、Trp、Tyr及びValからなる群から選択することができる。このようなアラニン、セリンおよびプロリンとは異なるアミノ酸残基としては、Val、Ile、Leu、Met、Phe、TyrもしくはTrpのような疎水性側鎖を有さないもの、及び/又はLys、Arg、AspもしくはGluのような荷電を有する側鎖を有さないものが好ましい。
当該ペプチドリンカーのアミノ酸配列において、アラニン、セリンおよびプロリンとは異なるアミノ酸残基は、Arg、Asn、Asp、Cys、Gln、Glu、Gly、His、Ile、Leu、Lys、Met、Phe、Thr、Trp、Tyr及びValからなる群から選択することができる。このようなアラニン、セリンおよびプロリンとは異なるアミノ酸残基としては、Val、Ile、Leu、Met、Phe、TyrもしくはTrpのような疎水性側鎖を有さないもの、及び/又はLys、Arg、AspもしくはGluのような荷電を有する側鎖を有さないものが好ましい。
一実施態様において、当該ペプチドリンカーは、複数のアミノ酸リピートを含み、前記アミノ酸リピートがアラニン(A)、プロリン(P)及びセリン(S)からなり、前記アミノ酸リピートにおいて同一のアミノ酸残基が連続する数が6残基以下、5残基以下、4残基以下、又は3残基以下であり得る。
一実施態様において、当該ペプチドリンカーは、複数のアミノ酸リピートを含み、前記アミノ酸リピートがアラニン(A)及びプロリン(P)からなり、前記アミノ酸リピートにおいて同一のアミノ酸残基が連続する数が6残基以下、5残基以下、4残基以下、又は3残基以下であり得る。
一実施態様において、当該ペプチドリンカーは、AP又はAAPで表される配列を2つ以上繰り返して含むものであり得る。
一実施態様において、当該ペプチドリンカーは、グリシン(G)とセリン(S)から構成される配列を含むものであり得る。この場合、当該ペプチドリンカーは、GGGGS、GGGS、GGS、及びGSからなる群から選択されるアミノ酸配列を繰り返し含み得る。
一実施態様において、当該ペプチドリンカーは、グリシンを連続して含むものであり得、また、グリシンのみからなるものであり得る。
一実施態様において、当該ペプチドリンカーは、EAAAKで示されるアミノ酸配列を繰り返し含み得る。
一実施態様において、当該ペプチドリンカーのアミノ酸配列としては、A、P及びSからなるもの、A及びPからなるもの、又はGGGGSで示される配列の繰り返しからなるものが好ましいが、これらに限定されるものではない。
一実施態様において、当該ペプチドリンカーの長さは、50個以下のアミノ酸残基からなり得るが、例えば、当該ペプチドリンカーは、49個以下、40個以下、35個以下、30個以下、又は25個以下のアミノ酸残基からなることができる。
一実施態様において、当該ペプチドリンカーの長さは、1個以上のアミノ酸残基からなり得るが、例えば、当該ペプチドリンカーは、4個以上、8個以上、12個以上、又は16個以上のアミノ酸残基からなることができる。
一実施態様において、当該ペプチドリンカーは、複数のアミノ酸リピートを含み、前記アミノ酸リピートがアラニン(A)及びプロリン(P)からなり、前記アミノ酸リピートにおいて同一のアミノ酸残基が連続する数が6残基以下、5残基以下、4残基以下、又は3残基以下であり得る。
一実施態様において、当該ペプチドリンカーは、AP又はAAPで表される配列を2つ以上繰り返して含むものであり得る。
一実施態様において、当該ペプチドリンカーは、グリシン(G)とセリン(S)から構成される配列を含むものであり得る。この場合、当該ペプチドリンカーは、GGGGS、GGGS、GGS、及びGSからなる群から選択されるアミノ酸配列を繰り返し含み得る。
一実施態様において、当該ペプチドリンカーは、グリシンを連続して含むものであり得、また、グリシンのみからなるものであり得る。
一実施態様において、当該ペプチドリンカーは、EAAAKで示されるアミノ酸配列を繰り返し含み得る。
一実施態様において、当該ペプチドリンカーのアミノ酸配列としては、A、P及びSからなるもの、A及びPからなるもの、又はGGGGSで示される配列の繰り返しからなるものが好ましいが、これらに限定されるものではない。
一実施態様において、当該ペプチドリンカーの長さは、50個以下のアミノ酸残基からなり得るが、例えば、当該ペプチドリンカーは、49個以下、40個以下、35個以下、30個以下、又は25個以下のアミノ酸残基からなることができる。
一実施態様において、当該ペプチドリンカーの長さは、1個以上のアミノ酸残基からなり得るが、例えば、当該ペプチドリンカーは、4個以上、8個以上、12個以上、又は16個以上のアミノ酸残基からなることができる。
一実施態様において、当該ペプチドリンカーの具体例としては、以下のアミノ酸配列を含むものや、以下のアミノ酸配列からなるものが挙げられるが、これらに限定されるものではない。AAPAAPAPAAPAAPAPAAPA(配列番号:9)、AAPAAPAPAAPAAPAPAAPAAP(配列番号:10)、AAPAAPAPAAPAAP(配列番号:11)、AAPAAAPAPAAPAAPAPAAP(配列番号12)、AAAPAAAPAAAPAAAPAAAP(配列番号13)、AAPAAPAAPAAPAAPAAPAAPAAP(配列番号14)、APAAAPAPAAAPAPAAAPAPAAAP(配列番号15)、AAAPAAPAAPPAAAAPAAPAAPPA(配列番号16)、及びAPAPAPAPAPAPAPAPAPAP(配列番号17)、ASPAAPAPASPAAPAPSAPA(配列番号18)、AAPASPAPAAPSAPAPAAPS(配列番号19)、APSSPSPSAPSSPSPASPSS(配列番号20)、SAPSSPSPSAPSSPSPASPS(配列番号21)、SSPSAPSPSSPASPSPSSPA(配列番号22)、AASPAAPSAPPAAASPAAPSAPPA(配列番号23)、ASAAAPAAASAAASAPSAAA(配列番号24)、AAPAAPAPAAPAAPAP(配列番号:25)、AAPAAPAPAAPAAPAPAAPAAPAPAAPAAPAPAAPAAPAPAAPAAPAPA(配列番号:26)、及びSAPSPSSAPASASAPASPASASASASPASSPASASSASPAASSASPASS(配列番号:27)、並びにこれらの円順列変異配列から選択されるアミノ酸配列。
円順列変異配列は、AAPAAPAPAAPAAPAPAAPA(配列番号:9)を例にして説明すれば、例えば、上記の配列の第1のアラニンを除去すること及び上記の配列の末端に別のアラニンを付加することによって容易に作製され得る。配列番号:9のこのような円順列変異配列は、APAAPAPAAPAAPAPAAPAA(配列番号:28)となる。さらに、配列番号7の環状にアミノ酸残基の順序を変えられた円順列変異配列の限定されない例として、以下のものが挙げられる。PAAPAPAAPAAPAPAAPAAA(配列番号:29)、AAPAPAAPAAPAPAAPAAAP(配列番号:30)、APAPAAPAAPAPAAPAAAPA(配列番号:31)、PAPAAPAAPAPAAPAAAPAA(配列番号:32)、APAAPAAPAPAAPAAAPAAP(配列番号:33)、PAAPAAPAPAAPAAAPAAPA(配列番号:34)、AAPAAPAPAAPAAAPAAPAP(配列番号:35)、APAAPAPAAPAAAPAAPAPA(配列番号:36)、及びPAAPAPAAPAAAPAAPAPAA(配列番号:37)。
一実施態様において、当該ペプチドリンカーの具体例としては、以下のアミノ酸配列を含むものや、以下のアミノ酸配列からなるものが挙げられるが、これらに限定されるものではない。AAPAAPAPAA(配列番号:38)、AAPAAPAP(配列番号:39)、及びAAPAAPAPA(配列番号:40)、並びにこれらの円順列変異配列から選択されるアミノ酸配列。
円順列変異配列は、AAPAAPAPAAPAAPAPAAPA(配列番号:9)を例にして説明すれば、例えば、上記の配列の第1のアラニンを除去すること及び上記の配列の末端に別のアラニンを付加することによって容易に作製され得る。配列番号:9のこのような円順列変異配列は、APAAPAPAAPAAPAPAAPAA(配列番号:28)となる。さらに、配列番号7の環状にアミノ酸残基の順序を変えられた円順列変異配列の限定されない例として、以下のものが挙げられる。PAAPAPAAPAAPAPAAPAAA(配列番号:29)、AAPAPAAPAAPAPAAPAAAP(配列番号:30)、APAPAAPAAPAPAAPAAAPA(配列番号:31)、PAPAAPAAPAPAAPAAAPAA(配列番号:32)、APAAPAAPAPAAPAAAPAAP(配列番号:33)、PAAPAAPAPAAPAAAPAAPA(配列番号:34)、AAPAAPAPAAPAAAPAAPAP(配列番号:35)、APAAPAPAAPAAAPAAPAPA(配列番号:36)、及びPAAPAPAAPAAAPAAPAPAA(配列番号:37)。
一実施態様において、当該ペプチドリンカーの具体例としては、以下のアミノ酸配列を含むものや、以下のアミノ酸配列からなるものが挙げられるが、これらに限定されるものではない。AAPAAPAPAA(配列番号:38)、AAPAAPAP(配列番号:39)、及びAAPAAPAPA(配列番号:40)、並びにこれらの円順列変異配列から選択されるアミノ酸配列。
環状ペプチド、システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカーを含むポリペプチドにおいて、当該二量体化ドメイン中のシステイン残基は、1つのみであってもよいし、2個以上であってもよい。好ましくは、当該二量体化ドメイン中のシステイン残基は、1つのみである。
当該二量体化ドメイン中のロイシンジッパー領域とは、αヘリックスを形成し、7残基ごとにロイシン残基を有する領域のことである。当該ロイシンジッパー領域は、特に限定されないが、例えば、60個以下、45個以下、35個以下、又は25個以下のアミノ酸残基からなることができる。
当該二量体化ドメイン中のロイシンジッパー領域としては、ダイマーを形成するものであれば特に限定されないが、例えば、c-Junタンパク質、GCN4タンパク質又はCEBPαタンパク質のロイシンジッパードメイン、及びc-Junロイシンジッパードメインの正電荷アミノ酸をグルタミンに置換することでヘパリン結合能を欠損させたJunΔhep(M. D. Ballinger et al., Nat. Biotechnol. 17, 1199-1204 (1999)、ここに本明細書の一部を構成するものとして当該文献の内容を参照により引用する。)等が挙げられる。当該ロイシンジッパー領域の具体例としては、以下のアミノ酸配列を含むものや、以下のアミノ酸配列からなるものが挙げられるが、これらに限定されるものではない。RIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMN(配列番号:41)、QIAQLEEKVQTLQAQNSELASTANMLREQVAQLKQKVMN(配列番号:42)、RMKQLEDKVEELLSKNYHLENEVARLKKLVGER(配列番号:43)、RNVETQQKVLELTSDNDRLRKRVEQLSRELDTLRGIFRQ(配列番号:44)。
一実施態様において、当該二量体化ドメイン中のロイシンジッパー領域は、ヘパリン結合能を除去したものであってもよい。
当該二量体化ドメイン中のロイシンジッパー領域とは、αヘリックスを形成し、7残基ごとにロイシン残基を有する領域のことである。当該ロイシンジッパー領域は、特に限定されないが、例えば、60個以下、45個以下、35個以下、又は25個以下のアミノ酸残基からなることができる。
当該二量体化ドメイン中のロイシンジッパー領域としては、ダイマーを形成するものであれば特に限定されないが、例えば、c-Junタンパク質、GCN4タンパク質又はCEBPαタンパク質のロイシンジッパードメイン、及びc-Junロイシンジッパードメインの正電荷アミノ酸をグルタミンに置換することでヘパリン結合能を欠損させたJunΔhep(M. D. Ballinger et al., Nat. Biotechnol. 17, 1199-1204 (1999)、ここに本明細書の一部を構成するものとして当該文献の内容を参照により引用する。)等が挙げられる。当該ロイシンジッパー領域の具体例としては、以下のアミノ酸配列を含むものや、以下のアミノ酸配列からなるものが挙げられるが、これらに限定されるものではない。RIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMN(配列番号:41)、QIAQLEEKVQTLQAQNSELASTANMLREQVAQLKQKVMN(配列番号:42)、RMKQLEDKVEELLSKNYHLENEVARLKKLVGER(配列番号:43)、RNVETQQKVLELTSDNDRLRKRVEQLSRELDTLRGIFRQ(配列番号:44)。
一実施態様において、当該二量体化ドメイン中のロイシンジッパー領域は、ヘパリン結合能を除去したものであってもよい。
環状ペプチド、システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカーを含む、本明細書に記載のポリペプチドは、特に限定されないが、例えば、分子量3000以上、4000以上、5000以上、又は6000以上であり得、20000以下、18000以下、16000以下、又は14000以下であり得る。また、当該ポリペプチドは、特に限定されないが、例えば、30個以上、40個以上、50個以上、又は60個以上のアミノ酸残基からなるものであり得、200個以下、180個以下、160個以下、又は140個以下のアミノ酸残基からなるものであり得る。
環状ペプチド、システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカーを含む、本明細書に記載のポリペプチドは、当該ポリペプチドをコードする核酸に基づき、周知技術を用いて調製し得る。また、当該ポリペプチドは、二量体化ドメイン中のロイシンジッパー領域における非共有結合性相互作用と、二量体化ドメイン中のシステイン残基間でのジスルフィド結合により、二量体化し得る。システイン残基間でのジスルフィド結合は、強固な二量体化に寄与し、二量体化した環状ペプチドの活性に大きな影響を与え得る。
当該ポリペプチドをコードする核酸は、DNAでもRNA(mRNA)でもあり得る。このような核酸の合成は、目的とするポリペプチドのアミノ酸配列に基づき、従来の合成技術を用いて当業者が適宜行い得る。
一実施態様において、当該ポリペプチドをコードする核酸は、5’側に無細胞発現に必要なプロモーター配列(例えばT7プロモーター配列)及びリボソーム結合配列を有し得る。また、当該核酸は、3’側に終止コドン及び非翻訳領域を有し得る。
従来、環状ペプチドを二量体化したペプチドダイマーを生合成系を用いてワンポットに得る手法としては、2つの環状ペプチドをリンカーで結合させた分子フォーマットについて遺伝子から発現する方法が用いられていたが、このような合成における鋳型遺伝子の調製には、2箇所の配列を環状ペプチドをコードするものへと変える必要があり、遺伝子全合成が必要となる。一方、本明細書に記載のポリペプチドは、ポリペプチドの合成後に二量化するものであり、そのようなポリペプチドの合成に用いられる核酸は、環状ペプチドをコードする配列を1ヶ所のみ有する。そのため、本明細書に記載の発明においては、核酸の合成、及びポリペプチドの合成を迅速で簡便に行うことができる。
当該ポリペプチドをコードする核酸は、DNAでもRNA(mRNA)でもあり得る。このような核酸の合成は、目的とするポリペプチドのアミノ酸配列に基づき、従来の合成技術を用いて当業者が適宜行い得る。
一実施態様において、当該ポリペプチドをコードする核酸は、5’側に無細胞発現に必要なプロモーター配列(例えばT7プロモーター配列)及びリボソーム結合配列を有し得る。また、当該核酸は、3’側に終止コドン及び非翻訳領域を有し得る。
従来、環状ペプチドを二量体化したペプチドダイマーを生合成系を用いてワンポットに得る手法としては、2つの環状ペプチドをリンカーで結合させた分子フォーマットについて遺伝子から発現する方法が用いられていたが、このような合成における鋳型遺伝子の調製には、2箇所の配列を環状ペプチドをコードするものへと変える必要があり、遺伝子全合成が必要となる。一方、本明細書に記載のポリペプチドは、ポリペプチドの合成後に二量化するものであり、そのようなポリペプチドの合成に用いられる核酸は、環状ペプチドをコードする配列を1ヶ所のみ有する。そのため、本明細書に記載の発明においては、核酸の合成、及びポリペプチドの合成を迅速で簡便に行うことができる。
本発明の一実施態様は、二量体化した環状ペプチドを調製する方法に関する。このような二量体化した環状ペプチドを調製する方法は、例えば、環状ペプチド、システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカーを含むポリペプチドを無細胞発現系で合成し、前記ポリペプチドの二量体を調製することを含み、前記二量体はホモダイマーであってもヘテロダイマーであってもよい。ここで、環状ペプチド、二量体化ドメイン(システイン残基及びロイシンジッパー領域)、ペプチドリンカーについては本明細書に上述されているとおりである。
無細胞発現系でのタンパク質の合成は、当業者に周知の技術であり、細胞の抽出液を用いてもよいし、市販の無細胞発現用試薬を用いてもよい。無細胞発現系に含まれる成分としては、テンプレートである所望のポリペプチドをコードする核酸(DNA又はmRNA)、転写・翻訳・エネルギー再生・翻訳後修飾に必要なタンパク質(RNAポリメラーゼ、リボソーム、アミノアシルtRNA合成酵素、翻訳開始因子、翻訳伸長因子、翻訳終結因子、リボソーム再生因子、ヌクレオシド二リン酸キナーゼ、アデノシンキナーゼ、クレアチンキナーゼ、プロリルイソメラーゼ、ジスルフィド結合イソメラーゼ、シャペロンなど)、tRNA、アミノアシルtRNA、天然アミノ酸、非天然アミノ酸、NTP、バッファー等が挙げられる。
市販の無細胞発現用試薬としては、例えば、PUREfrex2.0(ジーンフロンティア社)、PURExpress In Vitro Protein Synthesis Kit(New England BioLabs社)等が挙げられる。
無細胞発現系でのタンパク質の合成は、当業者に周知の技術であり、細胞の抽出液を用いてもよいし、市販の無細胞発現用試薬を用いてもよい。無細胞発現系に含まれる成分としては、テンプレートである所望のポリペプチドをコードする核酸(DNA又はmRNA)、転写・翻訳・エネルギー再生・翻訳後修飾に必要なタンパク質(RNAポリメラーゼ、リボソーム、アミノアシルtRNA合成酵素、翻訳開始因子、翻訳伸長因子、翻訳終結因子、リボソーム再生因子、ヌクレオシド二リン酸キナーゼ、アデノシンキナーゼ、クレアチンキナーゼ、プロリルイソメラーゼ、ジスルフィド結合イソメラーゼ、シャペロンなど)、tRNA、アミノアシルtRNA、天然アミノ酸、非天然アミノ酸、NTP、バッファー等が挙げられる。
市販の無細胞発現用試薬としては、例えば、PUREfrex2.0(ジーンフロンティア社)、PURExpress In Vitro Protein Synthesis Kit(New England BioLabs社)等が挙げられる。
本発明の一実施態様は、二量体化した環状ペプチドをスクリーニングする方法に関する。このような二量体化した環状ペプチドをスクリーニングする方法は、例えば、以下の工程(1A)又は(1B)、及び工程(2)を含む。
(1A)無細胞発現系で、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含む少なくとも1種のポリペプチドを合成し、前記ポリペプチドの二量体を調製する工程、又は
(1B)無細胞発現系で、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含む少なくとも1種のポリペプチドを合成することにより得られた前記ポリペプチドの二量体を用意する工程、
及び
(2)前記二量体の標的分子との結合に基づく活性を測定する工程。
二量体化した環状ペプチドをスクリーニングする方法は、更に、工程(2)で測定された活性に基づいて前記二量体(二量体化した環状ペプチド)をスクリーニングする又は選別する工程を含み得る。
また、本発明の一実施態様は、二量体化した環状ペプチドをスクリーニングする方法によって標的分子との結合に基づく活性を測定して、標的分子との結合に基づく活性を有する二量体化した環状ペプチドを得る方法に関する。
(1A)無細胞発現系で、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含む少なくとも1種のポリペプチドを合成し、前記ポリペプチドの二量体を調製する工程、又は
(1B)無細胞発現系で、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含む少なくとも1種のポリペプチドを合成することにより得られた前記ポリペプチドの二量体を用意する工程、
及び
(2)前記二量体の標的分子との結合に基づく活性を測定する工程。
二量体化した環状ペプチドをスクリーニングする方法は、更に、工程(2)で測定された活性に基づいて前記二量体(二量体化した環状ペプチド)をスクリーニングする又は選別する工程を含み得る。
また、本発明の一実施態様は、二量体化した環状ペプチドをスクリーニングする方法によって標的分子との結合に基づく活性を測定して、標的分子との結合に基づく活性を有する二量体化した環状ペプチドを得る方法に関する。
二量体化した環状ペプチドをスクリーニングする方法において、工程(1A)は、二量体化した環状ペプチドを調製する方法に関して本明細書に上述されているとおりであり、工程(1B)は、本明細書に上述されているとおりの方法で得られた二量体化した環状ペプチド(ポリペプチドの二量体)を用意するというものである。工程(1A)及び(1B)における、「環状ペプチド、システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカーを含む少なくとも1種のポリペプチド」については、本明細書に上述されている「環状ペプチド、システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカーを含むポリペプチド」の内容をそのまま適用し得る。なお、工程(1A)及び(1B)における少なくとも1種のポリペプチドは、標的分子との結合親和性を有すると見込まれる複数の異なるポリペプチドを含み得る。
また、本明細書において、「ポリペプチドの二量体」は、本明細書に上述されている「二量体化した環状ペプチド」と同義である。
また、本明細書において、「ポリペプチドの二量体」は、本明細書に上述されている「二量体化した環状ペプチド」と同義である。
工程(2)において、「標的分子」とは、本明細書に記載の環状ペプチドもしくは二量体化した環状ペプチドが結合する又は結合することが見込まれる分子を意味する。標的分子は、好ましくは受容体であり、より好ましくは二量化により活性化する受容体、例えば、増殖因子受容体、成長因子受容体、サイトカイン受容体等である。標的分子の具体例としては、例えば、FGFR、c-Met(HGFR)、エリスロポエチン受容体、トロンボポエチン受容体、アルブミン、EGFR、VEGFR、PDGFR、Axl、PDGFR、SCFR、Flt-3、c-Ret、ROR、Tie、NGFR、Insulin受容体、EphR、Alk、DDR、TGFBR、Activin受容体、BMP受容体、インターロイキン受容体、T細胞受容体、トランスフェリン受容体、リポタンパク質受容体、ユビキチンリガーゼ、抗体、補体、GPCR、イオンチャネル、ウイルス等が挙げられるが、これらに限定されるものではない。
工程(2)において、「二量体の標的分子との結合に基づく活性」とは、二量体が標的分子と結合することによりもたらされる作用及び/又はその強さを意味し、アゴニスト活性及びアンタゴニスト活性のいずれの意味をも含み得る。例えば、活性には、受容体活性、シグナル伝達活性、アポトーシス活性等が含まれ得る。
工程(2)における測定は、周知の手法を用いて行うことができ、ルシフェラーゼレポーターアッセイ等のバイオルミネセンスアッセイにより、PathHunter(登録商標) Receptor Dimerization アッセイ等の受容体二量体化アッセイにより、又は蛍光共鳴エネルギー移動(FRET)の測定により、またはケミカルリンカーによるクロスリンクおよびSDS-PAGEによる二量化分析等により行うことができる。
工程(2)における測定には、工程(1)で得られた無細胞発現の反応液をそのまま用いてもよいし、適宜希釈して用いてもよい。
工程(2)において、「二量体の標的分子との結合に基づく活性」とは、二量体が標的分子と結合することによりもたらされる作用及び/又はその強さを意味し、アゴニスト活性及びアンタゴニスト活性のいずれの意味をも含み得る。例えば、活性には、受容体活性、シグナル伝達活性、アポトーシス活性等が含まれ得る。
工程(2)における測定は、周知の手法を用いて行うことができ、ルシフェラーゼレポーターアッセイ等のバイオルミネセンスアッセイにより、PathHunter(登録商標) Receptor Dimerization アッセイ等の受容体二量体化アッセイにより、又は蛍光共鳴エネルギー移動(FRET)の測定により、またはケミカルリンカーによるクロスリンクおよびSDS-PAGEによる二量化分析等により行うことができる。
工程(2)における測定には、工程(1)で得られた無細胞発現の反応液をそのまま用いてもよいし、適宜希釈して用いてもよい。
一実施態様において、二量体化した環状ペプチドをスクリーニングする方法は、工程(1)で、少なくとも1種のポリペプチドが、標的分子との結合親和性を有すると見込まれる複数の異なるポリペプチドを含み、かつ工程(2)で、前記複数の異なるポリペプチドの二量体について、前記標的分子との結合に基づく活性を測定することを含む。
一実施態様において、二量体化した環状ペプチドをスクリーニングする方法は、工程(1)が、少なくとも1種のポリペプチド又はその二量体を複数含むライブラリを作製することを含み、複数の前記ポリペプチド間で異なる環状ペプチドが含まれている。ここで、少なくとも1種のポリペプチド又はその二量体を複数含むライブラリは、当該ポリペプチド又はその二量体を、例えば、2種以上、8種以上、16種以上、32種以上、48種以上、96種以上、200種以上、1000種以上、10000種以上、105種以上、又は106種以上含み得る。また、少なくとも1種のポリペプチド又はその二量体を複数含むライブラリは、当該ポリペプチド又はその二量体を、例えば、1015種以下、1014種以下、又は1013種以下含み得る
本明細書において、「ライブラリ」とは、複数の異なる物質(例えば、異なるポリペプチド又は異なる核酸)を含む集合体を意味するものである。
一実施態様において、二量体化した環状ペプチドをスクリーニングする方法は、工程(1)が、少なくとも1種のポリペプチド又はその二量体を複数含むライブラリを作製することを含み、複数の前記ポリペプチド間で異なる環状ペプチドが含まれている。ここで、少なくとも1種のポリペプチド又はその二量体を複数含むライブラリは、当該ポリペプチド又はその二量体を、例えば、2種以上、8種以上、16種以上、32種以上、48種以上、96種以上、200種以上、1000種以上、10000種以上、105種以上、又は106種以上含み得る。また、少なくとも1種のポリペプチド又はその二量体を複数含むライブラリは、当該ポリペプチド又はその二量体を、例えば、1015種以下、1014種以下、又は1013種以下含み得る
本明細書において、「ライブラリ」とは、複数の異なる物質(例えば、異なるポリペプチド又は異なる核酸)を含む集合体を意味するものである。
本発明の一実施態様は、ライブラリを作製する方法に関する。このようなライブラリを作製する方法は、例えば、無細胞発現系で複数のポリペプチドを合成することを含む、前記複数のポリペプチド又は前記複数のポリペプチドの二量体を含むライブラリを作製する方法であって、前記複数のポリペプチドが、各々、環状ペプチド、システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカーを含み、前記複数のポリペプチド間で異なる環状ペプチドが含まれている方法であり得る。
このライブラリを作製する方法において、「無細胞発現系で複数のポリペプチドを合成する」という工程は、二量体化した環状ペプチドを調製する方法に関して本明細書に上述されているとおりである。また、「前記複数のポリペプチドが、各々、環状ペプチド、システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカーを含み」というポリペプチドに関する内容は、本明細書に上述されている「環状ペプチド、システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカーを含むポリペプチド」の内容をそのまま適用し得る。
このライブラリを作製する方法において、「無細胞発現系で複数のポリペプチドを合成する」という工程は、二量体化した環状ペプチドを調製する方法に関して本明細書に上述されているとおりである。また、「前記複数のポリペプチドが、各々、環状ペプチド、システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカーを含み」というポリペプチドに関する内容は、本明細書に上述されている「環状ペプチド、システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカーを含むポリペプチド」の内容をそのまま適用し得る。
本発明の一実施態様は、二量体化した環状ペプチドをスクリーニングするためのライブラリに関する。このようなライブラリは、例えば、複数のポリペプチド若しくは前記複数のポリペプチドの二量体、又は前記複数のポリペプチドをコードする複数の核酸を含み、前記複数のポリペプチドが、各々、環状ペプチド、システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカーを含むものであり、前記複数のポリペプチド間で異なる環状ペプチドが含まれている。
このライブラリにおいて、「前記複数のポリペプチドが、各々、環状ペプチド、システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカーを含むものであり」というポリペプチドに関する内容は、本明細書に上述されている「環状ペプチド、システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカーを含むポリペプチド」の内容をそのまま適用し得る。
このライブラリにおいて、「前記複数のポリペプチドが、各々、環状ペプチド、システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカーを含むものであり」というポリペプチドに関する内容は、本明細書に上述されている「環状ペプチド、システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカーを含むポリペプチド」の内容をそのまま適用し得る。
このようなライブラリは、複数のポリペプチド若しくは前記複数のポリペプチドの二量体を、例えば、2種以上、8種以上、16種以上、32種以上、48種以上、96種以上、200種以上、1000種以上、10000種以上、105種以上、又は106種以上含み得る。また、このようなライブラリは、複数のポリペプチド若しくは前記複数のポリペプチドの二量体を、例えば、1015種以下、1014種以下、又は1013種以下含み得る。
或いは、このようなライブラリは、複数のポリペプチドをコードする複数の核酸を、例えば、2種以上、8種以上、16種以上、32種以上、48種以上、96種以上、200種以上、1000種以上、10000種以上、105種以上、又は106種以上含み得る。また、このようなライブラリは、複数のポリペプチドをコードする複数の核酸を、例えば、1015種以下、1014種以下、又は1013種以下含み得る。なお、「ポリペプチドをコードする核酸」は、本明細書に上述されているとおりである。
或いは、このようなライブラリは、複数のポリペプチドをコードする複数の核酸を、例えば、2種以上、8種以上、16種以上、32種以上、48種以上、96種以上、200種以上、1000種以上、10000種以上、105種以上、又は106種以上含み得る。また、このようなライブラリは、複数のポリペプチドをコードする複数の核酸を、例えば、1015種以下、1014種以下、又は1013種以下含み得る。なお、「ポリペプチドをコードする核酸」は、本明細書に上述されているとおりである。
本発明の一実施態様は、二量体化した環状ペプチドをスクリーニングする方法に関する。このような二量体化した環状ペプチドをスクリーニングする方法は、前述の方法とは別の方法として、例えば、以下の工程(1’)~(5’)を含む。
(1’)複数のディスプレイされたポリペプチドを含むライブラリから、標的分子との結合親和性を有する1つ又は複数のポリペプチドを取得する工程であって、前記複数のディスプレイされたポリペプチドが、各々、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含み、前記複数のディスプレイされたポリペプチド間で異なる環状ペプチドが含まれている、該取得工程、及び
(2’)工程(1’)において取得した前記ポリペプチドと関連付けられた核酸を回収する工程、
(3’)工程(2’)において回収した前記核酸を増幅する工程、
(4’)工程(3’)において増幅した前記核酸を用いて、無細胞発現系で、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含むポリペプチドを合成し、前記ポリペプチドの二量体を調製する工程、及び
(5’)前記二量体の標的分子との結合に基づく活性を測定する工程。
(1’)複数のディスプレイされたポリペプチドを含むライブラリから、標的分子との結合親和性を有する1つ又は複数のポリペプチドを取得する工程であって、前記複数のディスプレイされたポリペプチドが、各々、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含み、前記複数のディスプレイされたポリペプチド間で異なる環状ペプチドが含まれている、該取得工程、及び
(2’)工程(1’)において取得した前記ポリペプチドと関連付けられた核酸を回収する工程、
(3’)工程(2’)において回収した前記核酸を増幅する工程、
(4’)工程(3’)において増幅した前記核酸を用いて、無細胞発現系で、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含むポリペプチドを合成し、前記ポリペプチドの二量体を調製する工程、及び
(5’)前記二量体の標的分子との結合に基づく活性を測定する工程。
このような工程(1’)~(5’)を含む二量体化した環状ペプチドをスクリーニングする方法によれば、標的分子との結合に基づく活性を有する二量体化した環状ペプチドについて、ディスプレイされた候補ポリペプチドの取得からシームレスに評価可能なスクリーニングを提供することができる。
このような工程(1’)~(5’)を含む二量体化した環状ペプチドをスクリーニングする方法は、更に、工程(5’)で測定された活性に基づいて前記二量体(二量体化した環状ペプチド)をスクリーニングする又は選別する工程を含み得る。
また、本発明の一実施態様は、このような工程(1’)~(5’)を含む二量体化した環状ペプチドをスクリーニングする方法によって標的分子との結合に基づく活性を測定して、標的分子との結合に基づく活性を有する二量体化した環状ペプチドを得る方法に関する。
このような工程(1’)~(5’)を含む二量体化した環状ペプチドをスクリーニングする方法は、更に、工程(5’)で測定された活性に基づいて前記二量体(二量体化した環状ペプチド)をスクリーニングする又は選別する工程を含み得る。
また、本発明の一実施態様は、このような工程(1’)~(5’)を含む二量体化した環状ペプチドをスクリーニングする方法によって標的分子との結合に基づく活性を測定して、標的分子との結合に基づく活性を有する二量体化した環状ペプチドを得る方法に関する。
工程(1’)は、複数のディスプレイされたポリペプチドを含むライブラリから、標的分子との結合親和性を有する1つ又は複数のポリペプチドを取得する工程である。ここで、「複数のディスプレイされたポリペプチドを含むライブラリ」は、周知のディスプレイ技術を用いて作製し得るものである。このようなディスプレイ技術としては、例えば、リボソームディスプレイ、mRNAディスプレイ、ファージディスプレイ、細菌ディスプレイ、酵母ディスプレイ等が挙げられる。具体的には、鋳型DNA配列にSecM配列を導入することで、リボソーム-ポリペプチド(融合タンパク質)-mRNA複合体の作製が容易になり、簡便に複数のディスプレイされたポリペプチドを含むライブラリを作製可能であるが、用いる配列およびディスプレイ方法はこれに限定されるものではない。
工程(1’)において、ライブラリは、複数のディスプレイされたポリペプチドを、例えば、2種以上、8種以上、16種以上、32種以上、48種以上、96種以上、200種以上、1000種以上、10000種以上、105種以上、106種以上、107種以上、108種以上、109種以上、又は1010種以上含み得る。また、このようなライブラリは、複数のポリペプチド若しくは前記複数のポリペプチドの二量体を、例えば、1015種以下、1014種以下、又は1013種以下含み得る。
また、工程(1’)における、「標的分子」、並びに「複数のディスプレイされたポリペプチド」の「環状ペプチド」、「システイン残基及びロイシンジッパー領域を含む二量体化ドメイン」、及び「前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー」は、本明細書に上述されているとおりである。
工程(1’)において、標的分子との結合親和性を有する1つ又は複数のポリペプチドを取得することは、周知の手法で行われ得るが、例えば、リコンビナントの標的分子や標的分子を発現する細胞に対するアフィニティセレクションにより行われ得る。
工程(1’)において、ライブラリは、複数のディスプレイされたポリペプチドを、例えば、2種以上、8種以上、16種以上、32種以上、48種以上、96種以上、200種以上、1000種以上、10000種以上、105種以上、106種以上、107種以上、108種以上、109種以上、又は1010種以上含み得る。また、このようなライブラリは、複数のポリペプチド若しくは前記複数のポリペプチドの二量体を、例えば、1015種以下、1014種以下、又は1013種以下含み得る。
また、工程(1’)における、「標的分子」、並びに「複数のディスプレイされたポリペプチド」の「環状ペプチド」、「システイン残基及びロイシンジッパー領域を含む二量体化ドメイン」、及び「前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー」は、本明細書に上述されているとおりである。
工程(1’)において、標的分子との結合親和性を有する1つ又は複数のポリペプチドを取得することは、周知の手法で行われ得るが、例えば、リコンビナントの標的分子や標的分子を発現する細胞に対するアフィニティセレクションにより行われ得る。
工程(2’)は、工程(1’)において取得したポリペプチドと関連付けられた核酸を回収する工程である。これは、周知の手法で行われ得るが、例えば、磁気ビーズを用いた回収法、ELISAプレートを用いた回収法、細胞のペレットを用いた回収法、ナノドロップレットを用いた回収法、マイクロ流路を用いた手法等により行われ得る。
工程(3’)は、工程(2’)において回収した核酸を増幅する工程である。これは、周知の手法で行われ得るが、例えば、PCRにより行われ得る。PCRにおいて、回収した核酸がDNAであればそのまま使用でき、回収した核酸がmRNAであれば、RT-PCRによりcDNAを作製し、核酸を増幅すればよい。
工程(4’)は、工程(3’)において増幅した核酸を用いて、無細胞発現系で、環状ペプチド、システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカーを含むポリペプチドを合成し、前記ポリペプチドの二量体を調製する工程である。これは、二量体化した環状ペプチドを調製する方法に関して本明細書に上述されているとおりである。「環状ペプチド、システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカーを含むポリペプチド」については、本明細書に上述されているとおりである。
工程(3’)は、工程(2’)において回収した核酸を増幅する工程である。これは、周知の手法で行われ得るが、例えば、PCRにより行われ得る。PCRにおいて、回収した核酸がDNAであればそのまま使用でき、回収した核酸がmRNAであれば、RT-PCRによりcDNAを作製し、核酸を増幅すればよい。
工程(4’)は、工程(3’)において増幅した核酸を用いて、無細胞発現系で、環状ペプチド、システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカーを含むポリペプチドを合成し、前記ポリペプチドの二量体を調製する工程である。これは、二量体化した環状ペプチドを調製する方法に関して本明細書に上述されているとおりである。「環状ペプチド、システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカーを含むポリペプチド」については、本明細書に上述されているとおりである。
工程(5’)は、二量体の標的分子との結合に基づく活性を測定する工程である。
工程(5’)において、「二量体の標的分子との結合に基づく活性」とは、二量体が標的分子と結合することによりもたらされる作用及び/又はその強さを意味し、アゴニスト活性及びアンタゴニスト活性のいずれの意味をも含み得る。例えば、活性には、受容体活性、シグナル伝達活性、アポトーシス活性等が含まれ得る。なお、工程(5’)における「標的分子」は、本明細書に上述されているとおりである。
工程(5’)における測定は、周知の手法を用いて行うことができ、ルシフェラーゼレポーターアッセイ等のバイオルミネセンスアッセイにより、PathHunter(登録商標) Receptor Dimerization アッセイ等の受容体二量体化アッセイにより、又は蛍光共鳴エネルギー移動(FRET)の測定により、またはケミカルリンカーによるクロスリンクおよびSDS-PAGEによる二量化分析等により行うことができる。
工程(5’)において、「二量体の標的分子との結合に基づく活性」とは、二量体が標的分子と結合することによりもたらされる作用及び/又はその強さを意味し、アゴニスト活性及びアンタゴニスト活性のいずれの意味をも含み得る。例えば、活性には、受容体活性、シグナル伝達活性、アポトーシス活性等が含まれ得る。なお、工程(5’)における「標的分子」は、本明細書に上述されているとおりである。
工程(5’)における測定は、周知の手法を用いて行うことができ、ルシフェラーゼレポーターアッセイ等のバイオルミネセンスアッセイにより、PathHunter(登録商標) Receptor Dimerization アッセイ等の受容体二量体化アッセイにより、又は蛍光共鳴エネルギー移動(FRET)の測定により、またはケミカルリンカーによるクロスリンクおよびSDS-PAGEによる二量化分析等により行うことができる。
一実施態様において、二量体化した環状ペプチドをスクリーニングする方法は、工程(1’)が、取得した複数のポリペプチドに基づき、複数のディスプレイされたポリペプチドを含む更なるライブラリを作製すること、及び前記更なるライブラリから、標的分子との結合親和性を有する1つ又は複数のポリペプチドを取得することを繰り返すことを含む。
ここで、取得した複数のポリペプチドに基づき、複数のディスプレイされたポリペプチドを含む更なるライブラリの作製は、以下のような工程により行われ得る。
工程(1’-1):工程(1’)において取得したポリペプチドと関連付けられた核酸を回収する工程、及び
工程(1’-2):工程(1’-1)において回収した核酸を増幅する工程。
工程(1’-1)は、工程(2’)として記載の方法と同じように実施することができる。工程(1’-2)は、工程(3’)として記載の方法と同じように実施することができる。このように作製された更なるライブラリから、標的分子との結合親和性を有する1つ又は複数のポリペプチドを取得することは、工程(1’)と同じように実施することができる。
一実施態様において、二量体化した環状ペプチドをスクリーニングする方法は、工程(1’)において、取得した複数のポリペプチドに基づき、複数のディスプレイされたポリペプチドを含む更なるライブラリを作製すること、及び前記更なるライブラリから、標的分子との結合親和性を有する1つ又は複数のポリペプチドを取得することを、2回、3回、4回、5回、6回、又は7回以上繰り返してもよい。
ここで、取得した複数のポリペプチドに基づき、複数のディスプレイされたポリペプチドを含む更なるライブラリの作製は、以下のような工程により行われ得る。
工程(1’-1):工程(1’)において取得したポリペプチドと関連付けられた核酸を回収する工程、及び
工程(1’-2):工程(1’-1)において回収した核酸を増幅する工程。
工程(1’-1)は、工程(2’)として記載の方法と同じように実施することができる。工程(1’-2)は、工程(3’)として記載の方法と同じように実施することができる。このように作製された更なるライブラリから、標的分子との結合親和性を有する1つ又は複数のポリペプチドを取得することは、工程(1’)と同じように実施することができる。
一実施態様において、二量体化した環状ペプチドをスクリーニングする方法は、工程(1’)において、取得した複数のポリペプチドに基づき、複数のディスプレイされたポリペプチドを含む更なるライブラリを作製すること、及び前記更なるライブラリから、標的分子との結合親和性を有する1つ又は複数のポリペプチドを取得することを、2回、3回、4回、5回、6回、又は7回以上繰り返してもよい。
本発明の一実施態様は、二量体化した環状ペプチドをスクリーニングするためのライブラリに関する。このようなライブラリは、前述のライブラリとは別のものとして、例えば、複数のディスプレイされたポリペプチド又は前記複数のディスプレイされたポリペプチドを発現させるための複数の核酸を含み、前記複数のディスプレイされたポリペプチドが、各々、環状ペプチド、システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカーを含むものであり、前記複数のディスプレイされたポリペプチド間で異なる環状ペプチドが含まれている。
このライブラリにおいて、「複数のディスプレイされたポリペプチド」の「環状ペプチド」、「システイン残基及びロイシンジッパー領域を含む二量体化ドメイン」、及び「前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー」は、本明細書に上述されているとおりである。
このライブラリにおいて、「複数のディスプレイされたポリペプチド」の「環状ペプチド」、「システイン残基及びロイシンジッパー領域を含む二量体化ドメイン」、及び「前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー」は、本明細書に上述されているとおりである。
このようなライブラリは、複数のディスプレイされたポリペプチドを、例えば、2種以上、8種以上、16種以上、32種以上、48種以上、96種以上、200種以上、1000種以上、10000種以上、105種以上、106種以上、107種以上、108種以上、109種以上、又は1010種以上含み得る。また、このようなライブラリは、複数のポリペプチド若しくは前記複数のポリペプチドの二量体を、例えば、1015種以下、1014種以下、又は1013種以下含み得る。
或いは、このようなライブラリは、複数のディスプレイされたポリペプチドを発現させるための複数の核酸を、例えば、2種以上、8種以上、16種以上、32種以上、48種以上、96種以上、200種以上、1000種以上、10000種以上、105種以上、106種以上、107種以上、108種以上、109種以上、又は1010種以上含み得る。また、このようなライブラリは、複数のポリペプチド若しくは前記複数のポリペプチドの二量体を、例えば、1015種以下、1014種以下、又は1013種以下含み得る。
或いは、このようなライブラリは、複数のディスプレイされたポリペプチドを発現させるための複数の核酸を、例えば、2種以上、8種以上、16種以上、32種以上、48種以上、96種以上、200種以上、1000種以上、10000種以上、105種以上、106種以上、107種以上、108種以上、109種以上、又は1010種以上含み得る。また、このようなライブラリは、複数のポリペプチド若しくは前記複数のポリペプチドの二量体を、例えば、1015種以下、1014種以下、又は1013種以下含み得る。
以下、実施例により本発明を更に詳細に説明するが、本発明はこれに限定されるものではない。
なお、実施例において用いたHGFは、全てリコンビナントヒトHGF(rhHGF, R&D Systems)であり、本明細書及び図面において、リコンビナントヒトHGFを単にHGFと表記する。
なお、実施例において用いたHGFは、全てリコンビナントヒトHGF(rhHGF, R&D Systems)であり、本明細書及び図面において、リコンビナントヒトHGFを単にHGFと表記する。
実施例1:環状ペプチド-ロイシンジッパー融合体の設計及び無細胞発現系による発現
(1-1)aMD4dY-ロイシンジッパー融合体の設計の概要
環状ペプチドであるaMD4dY(配列番号:2)はダイマーにすることでHGF様の活性を発揮する、すなわちMetのアゴニストとして作用することが報告されている。そこで、aMD4dYを自己二量化するロイシンジッパー領域との融合体として発現するためにペプチド配列の設計を行った。
また、ロイシンジッパー領域は非共有結合により二量化をもたらすため、結合は可逆的である。そのため、システイン残基を付加し、分子間でジスルフィド結合を形成することでより強固に二量化をするようなペプチド配列を設計した。
具体的には、以下のようなペプチド配列を設計した(図1~4)。
・「aMD4dY」-「ペプチドリンカー」-「システイン残基」-「ロイシンジッパー領域」
・「aMD4dY」-「ペプチドリンカー」-「ロイシンジッパー領域」-「システイン残基」
・「ロイシンジッパー領域」-「システイン残基」-「ペプチドリンカー」-「aMD4dY」
・「システイン残基」-「ロイシンジッパー領域」-「ペプチドリンカー」-「aMD4dY」
このように、ロイシンジッパー領域のN末端側もしくはC末端側でジスルフィド結合を形成し、共有結合で二量体化することが可能である。
(1-1)aMD4dY-ロイシンジッパー融合体の設計の概要
環状ペプチドであるaMD4dY(配列番号:2)はダイマーにすることでHGF様の活性を発揮する、すなわちMetのアゴニストとして作用することが報告されている。そこで、aMD4dYを自己二量化するロイシンジッパー領域との融合体として発現するためにペプチド配列の設計を行った。
また、ロイシンジッパー領域は非共有結合により二量化をもたらすため、結合は可逆的である。そのため、システイン残基を付加し、分子間でジスルフィド結合を形成することでより強固に二量化をするようなペプチド配列を設計した。
具体的には、以下のようなペプチド配列を設計した(図1~4)。
・「aMD4dY」-「ペプチドリンカー」-「システイン残基」-「ロイシンジッパー領域」
・「aMD4dY」-「ペプチドリンカー」-「ロイシンジッパー領域」-「システイン残基」
・「ロイシンジッパー領域」-「システイン残基」-「ペプチドリンカー」-「aMD4dY」
・「システイン残基」-「ロイシンジッパー領域」-「ペプチドリンカー」-「aMD4dY」
このように、ロイシンジッパー領域のN末端側もしくはC末端側でジスルフィド結合を形成し、共有結合で二量体化することが可能である。
また、アフィニティータグであるN11タグをN末端に付加したコンストラクトについても同様に設計を行った。具体的には、以下のようなペプチド配列を設計した(図5及び6)。
・「タグ配列」-「ロイシンジッパー領域」-「システイン残基」-「ペプチドリンカー」-「aMD4dY」
・「タグ配列」-「システイン残基」-「ロイシンジッパー領域」-「ペプチドリンカー」-「aMD4dY」
ロイシンジッパー領域としては、ホモ二量体を形成することが知られているc-Jun、GCN4及びCEBPαタンパク質のロイシンジッパードメイン、並びにc-Junロイシンジッパードメインの正電荷アミノ酸をグルタミンに置換することでヘパリン結合能を欠損させたJunΔhep(M. D. Ballinger et al., Nat. Biotechnol. 17, 1199-1204 (1999))を用いた。なお、これらのロイシンジッパードメインを、以下、それぞれJun、GCN4、CEBPα及びJunΔhepと表す。
・「タグ配列」-「ロイシンジッパー領域」-「システイン残基」-「ペプチドリンカー」-「aMD4dY」
・「タグ配列」-「システイン残基」-「ロイシンジッパー領域」-「ペプチドリンカー」-「aMD4dY」
ロイシンジッパー領域としては、ホモ二量体を形成することが知られているc-Jun、GCN4及びCEBPαタンパク質のロイシンジッパードメイン、並びにc-Junロイシンジッパードメインの正電荷アミノ酸をグルタミンに置換することでヘパリン結合能を欠損させたJunΔhep(M. D. Ballinger et al., Nat. Biotechnol. 17, 1199-1204 (1999))を用いた。なお、これらのロイシンジッパードメインを、以下、それぞれJun、GCN4、CEBPα及びJunΔhepと表す。
(1-2)aMD4dY-ロイシンジッパー融合体の設計
設計したaMD4dY-ロイシンジッパー融合体の配列を以下に示す。aMD4dY、Jun、GCN4、CEBPα及びJunΔhepは上記(1-1)のとおりであり、PA8はペプチドリンカーを表し、cysはシステイン残基を表す。
(a)aMD4dY-PA8-cys-Jun
MAETCRQFNRRTHEVWNLDCGAAPAAPAPAAGCGGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMN(配列番号:45)
(b)aMD4dY-PA8-Jun
MAETCRQFNRRTHEVWNLDCGAAPAAPAPAAGGGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMN(配列番号:46)
(c)aMD4dY-PA8-cys-JunΔhep
MAETCRQFNRRTHEVWNLDCGAAPAAPAPAAGCGGQIAQLEEKVQTLQAQNSELASTANMLREQVAQLKQKVMN(配列番号:47)
(d)aMD4dY-PA8-GCN4-cys
MAETCRQFNRRTHEVWNLDCGAAPAAPAPGRMKQLEDKVEELLSKNYHLENEVARLKKLVGERGGCGG(配列番号:48)
(e)aMD4dY-PA8-cys-CEBPα
MAETCRQFNRRTHEVWNLDCGAAPAAPAPAAGCGGRNVETQQKVLELTSDNDRLRKRVEQLSRELDTLRGIFRQ(配列番号:49)
(f)cys-Jun-PA8-aMD4dY
MGGCGGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMNAAPAAPAPACRQFNRRTHEVWNLDC(配列番号:50)
(g)Jun-cys-PA8-aMD4dY
MGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMNGGCGGAAPAAPAPACRQFNRRTHEVWNLDC(配列番号:51)
(h)N11-Jun-cys-PA8-aMD4dY
MKDHLIHNHHKHEHAHAEHLGGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMNGGCGGAAPAAPAPACRQFNRRTHEVWNLDC(配列番号:52)
(i)N11-cys-Jun-PA8-aMD4dY
MKDHLIHNHHKHEHAHAEHLGGCGGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMNAAPAAPAPACRQFNRRTHEVWNLDC(配列番号:53)
(j)N11-GCN4-cys-PA8-aMD4dY
MKDHLIHNHHKHEHAHAEHLGGRMKQLEDKVEELLSKNYHLENEVARLKKLVGERGGCGGAAPAAPAPACRQFNRRTHEVWNLDC(配列番号:54)
(k)N11-cys-GCN4-PA8-aMD4dY
MKDHLIHNHHKHEHAHAEHLGGCGGRMKQLEDKVEELLSKNYHLENEVARLKKLVGERAAPAAPAPACRQFNRRTHEVWNLDC(配列番号:55)
設計したaMD4dY-ロイシンジッパー融合体の配列を以下に示す。aMD4dY、Jun、GCN4、CEBPα及びJunΔhepは上記(1-1)のとおりであり、PA8はペプチドリンカーを表し、cysはシステイン残基を表す。
(a)aMD4dY-PA8-cys-Jun
MAETCRQFNRRTHEVWNLDCGAAPAAPAPAAGCGGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMN(配列番号:45)
(b)aMD4dY-PA8-Jun
MAETCRQFNRRTHEVWNLDCGAAPAAPAPAAGGGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMN(配列番号:46)
(c)aMD4dY-PA8-cys-JunΔhep
MAETCRQFNRRTHEVWNLDCGAAPAAPAPAAGCGGQIAQLEEKVQTLQAQNSELASTANMLREQVAQLKQKVMN(配列番号:47)
(d)aMD4dY-PA8-GCN4-cys
MAETCRQFNRRTHEVWNLDCGAAPAAPAPGRMKQLEDKVEELLSKNYHLENEVARLKKLVGERGGCGG(配列番号:48)
(e)aMD4dY-PA8-cys-CEBPα
MAETCRQFNRRTHEVWNLDCGAAPAAPAPAAGCGGRNVETQQKVLELTSDNDRLRKRVEQLSRELDTLRGIFRQ(配列番号:49)
(f)cys-Jun-PA8-aMD4dY
MGGCGGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMNAAPAAPAPACRQFNRRTHEVWNLDC(配列番号:50)
(g)Jun-cys-PA8-aMD4dY
MGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMNGGCGGAAPAAPAPACRQFNRRTHEVWNLDC(配列番号:51)
(h)N11-Jun-cys-PA8-aMD4dY
MKDHLIHNHHKHEHAHAEHLGGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMNGGCGGAAPAAPAPACRQFNRRTHEVWNLDC(配列番号:52)
(i)N11-cys-Jun-PA8-aMD4dY
MKDHLIHNHHKHEHAHAEHLGGCGGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMNAAPAAPAPACRQFNRRTHEVWNLDC(配列番号:53)
(j)N11-GCN4-cys-PA8-aMD4dY
MKDHLIHNHHKHEHAHAEHLGGRMKQLEDKVEELLSKNYHLENEVARLKKLVGERGGCGGAAPAAPAPACRQFNRRTHEVWNLDC(配列番号:54)
(k)N11-cys-GCN4-PA8-aMD4dY
MKDHLIHNHHKHEHAHAEHLGGCGGRMKQLEDKVEELLSKNYHLENEVARLKKLVGERAAPAAPAPACRQFNRRTHEVWNLDC(配列番号:55)
(1-3)aMD4dY-ロイシンジッパー融合体の発現
無細胞発現系を用いてaMD4dY-ロイシンジッパー融合体の発現検討を行った。以下、発現検討の実施例を記載する。
無細胞発現系を用いてaMD4dY-ロイシンジッパー融合体の発現検討を行った。以下、発現検討の実施例を記載する。
(1-4)無細胞発現用aMD4dY-ロイシンジッパー融合体の遺伝子コンストラクトの構築
aMD4dY-ロイシンジッパー融合体を発現させる核酸として、上記(a)-(k)のアミノ酸配列をコードする塩基配列をそれぞれ設計した。さらに、これらの5’側には無細胞発現に必要なT7プロモーター配列及びリボソーム結合配列を付加し、3’側には終止コドンと非翻訳領域を付加した。上記(a)-(k)に対応する無細胞発現用塩基配列をそれぞれ(l)-(v)に示す。
(l)aMD4dY-PA8-cys-Junを発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGCAGAGACCTGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTGGCGCAGCCCCCGCAGCACCAGCTCCAGCAGCAGGCTGCGGTGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAACTAGTGAATAACTAATCC(配列番号:56)
(m)aMD4dY-PA8-Junを発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGCAGAGACCTGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTGGCGCAGCCCCCGCAGCACCAGCTCCAGCAGCAGGCGGTGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAACTAGTGAATAACTAATCC(配列番号:57)
(n)aMD4dY-PA8-cys-JunΔhepを発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGCAGAGACCTGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTGGCGCAGCCCCCGCAGCACCAGCTCCAGCAGCAGGCTGCGGTGGTCAGATTGCCCAACTGGAAGAGAAAGTGCAGACGTTACAAGCGCAAAACAGCGAACTGGCTTCGACCGCGAACATGTTGCGCGAACAGGTTGCACAGCTGAAACAGAAGGTCATGAATTAGTGAATAACTAATCC(配列番号:58)
(o)aMD4dY-PA8-GCN4-cysを発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGCAGAGACCTGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTGGCGCAGCACCCGCAGCTCCAGCACCAGGCCGCATGAAACAGTTGGAGGATAAAGTCGAAGAGCTTCTGAGCAAGAACTATCACCTCGAAAACGAAGTTGCTCGTCTGAAGAAACTCGTTGGAGAACGTGGTGGTTGTGGCGGGTAGTGAATAACTAATCC(配列番号:59)
(p)aMD4dY-PA8-cys-CEBPαを発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGCAGAGACCTGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTGGCGCAGCCCCCGCAGCACCAGCTCCAGCAGCAGGCTGCGGTGGTCGCAATGTGGAGACTCAGCAGAAGGTCCTGGAACTGACGTCTGACAACGATCGGTTACGCAAACGGGTTGAACAGCTGTCTCGTGAACTCGACACTCTTCGTGGCATCTTCCGCCAATAGTGAATAACTAATCC(配列番号:60)
(q)cys-Jun-PA8-aMD4dYを発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGGTGGTTGCGGTGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAACGCAGCCCCCGCAGCACCAGCTCCAGCATGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTTAGTGAATAACTAATCC(配列番号:61)
(r)Jun-cys-PA8-aMD4dYを発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAACGGTGGTTGCGGTGGTGCAGCCCCCGCAGCACCAGCTCCAGCATGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTTAGTGAATAACTAATCC(配列番号:62)
(s)N11-Jun-cys-PA8-aMD4dYを発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGAAAGATCATCTCATCCACAATCATCACAAACATGAGCACGCTCATGCCGAACACCTGGGTGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAACGGTGGTTGCGGTGGTGCAGCACCAGCAGCTCCGGCCCCTGCTTGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTTAGTGAATAACTAATCC(配列番号:63)
(t)N11-cys-Jun-PA8-aMD4dYを発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGAAAGATCATCTCATCCACAATCATCACAAACATGAGCACGCTCATGCCGAACACCTGGGTGGTTGCGGTGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAACGCAGCACCAGCAGCTCCGGCCCCTGCTTGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTTAGTGAATAACTAATCC(配列番号:64)
(u)N11-GCN4-cys-PA8-aMD4dYを発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGAAAGATCATCTCATCCACAATCATCACAAACATGAGCACGCTCATGCCGAACACCTGGGTGGTCGCATGAAACAGTTGGAGGATAAAGTCGAAGAGCTTCTGAGCAAGAACTATCACCTCGAAAACGAAGTTGCTCGTCTGAAGAAACTCGTTGGAGAACGTGGTGGTTGCGGTGGTGCAGCACCAGCAGCTCCGGCCCCTGCTTGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTTAGTGAATAACTAATCC(配列番号:65)
(v)N11-cys-GCN4-PA8-aMD4dYを発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGAAAGATCATCTCATCCACAATCATCACAAACATGAGCACGCTCATGCCGAACACCTGGGTGGTTGCGGTGGTCGCATGAAACAGTTGGAGGATAAAGTCGAAGAGCTTCTGAGCAAGAACTATCACCTCGAAAACGAAGTTGCTCGTCTGAAGAAACTCGTTGGAGAACGTGCAGCACCAGCAGCTCCGGCCCCTGCTTGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTTAGTGAATAACTAATCC(配列番号:66)
(l)-(v)に示すDNA配列は、ユーロフィンジェノミクス株式会社にて全合成およびpEX-K4J2プラスミドベクターへのクローニングを行った。
aMD4dY-ロイシンジッパー融合体を発現させる核酸として、上記(a)-(k)のアミノ酸配列をコードする塩基配列をそれぞれ設計した。さらに、これらの5’側には無細胞発現に必要なT7プロモーター配列及びリボソーム結合配列を付加し、3’側には終止コドンと非翻訳領域を付加した。上記(a)-(k)に対応する無細胞発現用塩基配列をそれぞれ(l)-(v)に示す。
(l)aMD4dY-PA8-cys-Junを発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGCAGAGACCTGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTGGCGCAGCCCCCGCAGCACCAGCTCCAGCAGCAGGCTGCGGTGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAACTAGTGAATAACTAATCC(配列番号:56)
(m)aMD4dY-PA8-Junを発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGCAGAGACCTGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTGGCGCAGCCCCCGCAGCACCAGCTCCAGCAGCAGGCGGTGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAACTAGTGAATAACTAATCC(配列番号:57)
(n)aMD4dY-PA8-cys-JunΔhepを発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGCAGAGACCTGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTGGCGCAGCCCCCGCAGCACCAGCTCCAGCAGCAGGCTGCGGTGGTCAGATTGCCCAACTGGAAGAGAAAGTGCAGACGTTACAAGCGCAAAACAGCGAACTGGCTTCGACCGCGAACATGTTGCGCGAACAGGTTGCACAGCTGAAACAGAAGGTCATGAATTAGTGAATAACTAATCC(配列番号:58)
(o)aMD4dY-PA8-GCN4-cysを発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGCAGAGACCTGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTGGCGCAGCACCCGCAGCTCCAGCACCAGGCCGCATGAAACAGTTGGAGGATAAAGTCGAAGAGCTTCTGAGCAAGAACTATCACCTCGAAAACGAAGTTGCTCGTCTGAAGAAACTCGTTGGAGAACGTGGTGGTTGTGGCGGGTAGTGAATAACTAATCC(配列番号:59)
(p)aMD4dY-PA8-cys-CEBPαを発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGCAGAGACCTGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTGGCGCAGCCCCCGCAGCACCAGCTCCAGCAGCAGGCTGCGGTGGTCGCAATGTGGAGACTCAGCAGAAGGTCCTGGAACTGACGTCTGACAACGATCGGTTACGCAAACGGGTTGAACAGCTGTCTCGTGAACTCGACACTCTTCGTGGCATCTTCCGCCAATAGTGAATAACTAATCC(配列番号:60)
(q)cys-Jun-PA8-aMD4dYを発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGGTGGTTGCGGTGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAACGCAGCCCCCGCAGCACCAGCTCCAGCATGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTTAGTGAATAACTAATCC(配列番号:61)
(r)Jun-cys-PA8-aMD4dYを発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAACGGTGGTTGCGGTGGTGCAGCCCCCGCAGCACCAGCTCCAGCATGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTTAGTGAATAACTAATCC(配列番号:62)
(s)N11-Jun-cys-PA8-aMD4dYを発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGAAAGATCATCTCATCCACAATCATCACAAACATGAGCACGCTCATGCCGAACACCTGGGTGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAACGGTGGTTGCGGTGGTGCAGCACCAGCAGCTCCGGCCCCTGCTTGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTTAGTGAATAACTAATCC(配列番号:63)
(t)N11-cys-Jun-PA8-aMD4dYを発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGAAAGATCATCTCATCCACAATCATCACAAACATGAGCACGCTCATGCCGAACACCTGGGTGGTTGCGGTGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAACGCAGCACCAGCAGCTCCGGCCCCTGCTTGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTTAGTGAATAACTAATCC(配列番号:64)
(u)N11-GCN4-cys-PA8-aMD4dYを発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGAAAGATCATCTCATCCACAATCATCACAAACATGAGCACGCTCATGCCGAACACCTGGGTGGTCGCATGAAACAGTTGGAGGATAAAGTCGAAGAGCTTCTGAGCAAGAACTATCACCTCGAAAACGAAGTTGCTCGTCTGAAGAAACTCGTTGGAGAACGTGGTGGTTGCGGTGGTGCAGCACCAGCAGCTCCGGCCCCTGCTTGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTTAGTGAATAACTAATCC(配列番号:65)
(v)N11-cys-GCN4-PA8-aMD4dYを発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGAAAGATCATCTCATCCACAATCATCACAAACATGAGCACGCTCATGCCGAACACCTGGGTGGTTGCGGTGGTCGCATGAAACAGTTGGAGGATAAAGTCGAAGAGCTTCTGAGCAAGAACTATCACCTCGAAAACGAAGTTGCTCGTCTGAAGAAACTCGTTGGAGAACGTGCAGCACCAGCAGCTCCGGCCCCTGCTTGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTTAGTGAATAACTAATCC(配列番号:66)
(l)-(v)に示すDNA配列は、ユーロフィンジェノミクス株式会社にて全合成およびpEX-K4J2プラスミドベクターへのクローニングを行った。
(1-5)EMP-ロイシンジッパー融合体の設計の概要
Johnson, D. L. et al. Chem. Biol.1997, 4, 939-950に記載の環状ペプチドであるEMPは、二量体化することで強力なエリスロポエチン(EPO)様の活性を発揮することが報告されている。そこで、EMPを自己二量化するロイシンジッパー領域との融合体として発現するためにペプチド配列の設計を行った。また、上記(1-1)と同様にシステイン残基を付加し、「EMP」-「ペプチドリンカー」-「システイン残基」-「ロイシンジッパー領域」となるようペプチド配列を設計した。
Johnson, D. L. et al. Chem. Biol.1997, 4, 939-950に記載の環状ペプチドであるEMPは、二量体化することで強力なエリスロポエチン(EPO)様の活性を発揮することが報告されている。そこで、EMPを自己二量化するロイシンジッパー領域との融合体として発現するためにペプチド配列の設計を行った。また、上記(1-1)と同様にシステイン残基を付加し、「EMP」-「ペプチドリンカー」-「システイン残基」-「ロイシンジッパー領域」となるようペプチド配列を設計した。
(1-6)EMP-ロイシンジッパー融合体の設計
設計したaMD4dY-ロイシンジッパー融合体の配列を以下に示す。
(w)EMP-PA8-cys-Jun
MGGLYACHMGPMTWVCQPLRGAAPAAPAPAAGCGGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMN(配列番号:67)
EMPは上記(1-5)のとおりであり、PA8はペプチドリンカーを表し、cysはシステイン残基を表し、Junはc-Junタンパク質のロイシンジッパードメインを表す。
設計したaMD4dY-ロイシンジッパー融合体の配列を以下に示す。
(w)EMP-PA8-cys-Jun
MGGLYACHMGPMTWVCQPLRGAAPAAPAPAAGCGGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMN(配列番号:67)
EMPは上記(1-5)のとおりであり、PA8はペプチドリンカーを表し、cysはシステイン残基を表し、Junはc-Junタンパク質のロイシンジッパードメインを表す。
(1-7)EMP-ロイシンジッパー融合体の発現
無細胞発現系を用いてEMP-ロイシンジッパー融合体の発現検討を行った。以下、発現検討の実施例を記載する。
無細胞発現系を用いてEMP-ロイシンジッパー融合体の発現検討を行った。以下、発現検討の実施例を記載する。
(1-8)無細胞発現用EMP-ロイシンジッパー融合体の遺伝子コンストラクトの構築
EMP-ロイシンジッパー融合体を発現させる核酸として、上記(w)のアミノ酸配列をコードする塩基配列を設計した。さらに、その5’側には無細胞発現に必要なT7プロモーター配列及びリボソーム結合配列を付加し、3’側には終止コドンと非翻訳領域を付加した。上記(w)に対応する無細胞発現用塩基配列を(x)に示す。
(x)EMP-PA8-cys-Junを発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGGTGGCCTGTATGCGTGCCATATGGGGCCAATGACCTGGGTGTGTCAGCCGTTACGCGGCGCAGCCCCCGCAGCACCAGCTCCAGCAGCAGGCTGCGGTGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAACTAGTGAATAACTAATCC(配列番号:68)
(x)に示すDNA配列は、ユーロフィンジェノミクス株式会社にて全合成およびpEX-K4J2プラスミドベクターへのクローニングを行った。
EMP-ロイシンジッパー融合体を発現させる核酸として、上記(w)のアミノ酸配列をコードする塩基配列を設計した。さらに、その5’側には無細胞発現に必要なT7プロモーター配列及びリボソーム結合配列を付加し、3’側には終止コドンと非翻訳領域を付加した。上記(w)に対応する無細胞発現用塩基配列を(x)に示す。
(x)EMP-PA8-cys-Junを発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGGTGGCCTGTATGCGTGCCATATGGGGCCAATGACCTGGGTGTGTCAGCCGTTACGCGGCGCAGCCCCCGCAGCACCAGCTCCAGCAGCAGGCTGCGGTGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAACTAGTGAATAACTAATCC(配列番号:68)
(x)に示すDNA配列は、ユーロフィンジェノミクス株式会社にて全合成およびpEX-K4J2プラスミドベクターへのクローニングを行った。
(1-9)環状ペプチド-ロイシンジッパー融合体の無細胞発現
上記で構築した環状ペプチド-ロイシンジッパー融合体の遺伝子をPCRにより増幅した。PCRは以下の(y)及び(z)に示す配列のプライマーと、PCR酵素としてKOD -Plus- Ver.2(東洋紡)を用いて行った。
(y)Cell-free primer Fwd:
GAAATTAATACGACTCACTATAGGG(配列番号:69)
(z)Cell-free primer Rev:
CACTGACTGGATTAGTTATTCAC(配列番号:70)
次に、PCRにより増幅した遺伝子断片を基に、環状ペプチド-ロイシンジッパー融合体を無細胞発現系によって発現した。無細胞発現にはPUREfrex2.0(ジーンフロンティア)又はPURExpress In Vitro Protein Synthesis Kit(New England BioLabs)を用いた。
PUREfrex2.0で無細胞発現を行う場合、(l)-(v)のうちのいずれかまたは(x)のDNA配列断片4 ng、もしくは(l)-(v)のうちのいずれかまたは(x)のDNA配列を含む各プラスミドベクター90 ngに対し、10 μLのSolution I、1 μLのSolution II、2 μLのSolution III、7 μLの水を添加し、37 ℃で6時間インキュベーションすることで翻訳反応を行った。
PURExpress In Vitro Protein Synthesis Kitで無細胞発現を行う場合、(l)-(v)のうちのいずれかまたは(x)のDNA配列を含む各プラスミドベクター90 ngに対し、8 μLのSolution A、6 μLのSolution B、6 μLの水を添加し、37 ℃で6時間インキュベーションすることで翻訳反応を行った。
上記で構築した環状ペプチド-ロイシンジッパー融合体の遺伝子をPCRにより増幅した。PCRは以下の(y)及び(z)に示す配列のプライマーと、PCR酵素としてKOD -Plus- Ver.2(東洋紡)を用いて行った。
(y)Cell-free primer Fwd:
GAAATTAATACGACTCACTATAGGG(配列番号:69)
(z)Cell-free primer Rev:
CACTGACTGGATTAGTTATTCAC(配列番号:70)
次に、PCRにより増幅した遺伝子断片を基に、環状ペプチド-ロイシンジッパー融合体を無細胞発現系によって発現した。無細胞発現にはPUREfrex2.0(ジーンフロンティア)又はPURExpress In Vitro Protein Synthesis Kit(New England BioLabs)を用いた。
PUREfrex2.0で無細胞発現を行う場合、(l)-(v)のうちのいずれかまたは(x)のDNA配列断片4 ng、もしくは(l)-(v)のうちのいずれかまたは(x)のDNA配列を含む各プラスミドベクター90 ngに対し、10 μLのSolution I、1 μLのSolution II、2 μLのSolution III、7 μLの水を添加し、37 ℃で6時間インキュベーションすることで翻訳反応を行った。
PURExpress In Vitro Protein Synthesis Kitで無細胞発現を行う場合、(l)-(v)のうちのいずれかまたは(x)のDNA配列を含む各プラスミドベクター90 ngに対し、8 μLのSolution A、6 μLのSolution B、6 μLの水を添加し、37 ℃で6時間インキュベーションすることで翻訳反応を行った。
(1-10)SDS-PAGEによる環状ペプチド-ロイシンジッパー融合体の発現確認
上記(1-9)で無細胞発現した各環状ペプチド-ロイシンジッパー融合体を含む反応液3μLをNuPAGE(登録商標)12% Bis-Tirs Gel(Thermo Fisher Scientific)を用いてSDS-PAGEに供してから、Quick-CBB(Wako)にて染色を行った。PUREfrex2.0を用いて無細胞発現した、(i)N11-cys-Jun-PA8-aMD4dY、(k)N11-cys-GCN4-PA8-aMD4dYの各反応液を用いた場合、また各反応液をDynabeads His-Tag Isolation & Pulldownで精製実施後PAGEに供した場合について、結果を図7に示す。N11タグ精製を実施したレーンについて、それぞれのaMD4dY-ロイシンジッパー融合体(Monomer)、またそのジスルフィド二量体(Dimer)と推定されるタンパク質バンドが検出された。
上記(1-9)で無細胞発現した各環状ペプチド-ロイシンジッパー融合体を含む反応液3μLをNuPAGE(登録商標)12% Bis-Tirs Gel(Thermo Fisher Scientific)を用いてSDS-PAGEに供してから、Quick-CBB(Wako)にて染色を行った。PUREfrex2.0を用いて無細胞発現した、(i)N11-cys-Jun-PA8-aMD4dY、(k)N11-cys-GCN4-PA8-aMD4dYの各反応液を用いた場合、また各反応液をDynabeads His-Tag Isolation & Pulldownで精製実施後PAGEに供した場合について、結果を図7に示す。N11タグ精製を実施したレーンについて、それぞれのaMD4dY-ロイシンジッパー融合体(Monomer)、またそのジスルフィド二量体(Dimer)と推定されるタンパク質バンドが検出された。
実施例2:aMD4dY-ロイシンジッパー融合体の活性評価
各aMD4dY-ロイシンジッパー融合体の二量体をルシフェラーゼアッセイによって評価した。Opti-MEM培地(Thermo Fisher Scientific社)25 μLに対し、0.6 μLのAttractene Transfection Reagent(QIAGEN社)を添加し、室温で5分インキュベートを行った。上記混合溶液に対し、Opti-MEM 25 μLと1 μLのSRE reporter vector(QIAGEN社)の混合溶液を加え、室温で20分インキュベートを行った。この混合溶液を96-well plateに添加し、その上から40,000 cell/wellのHEK293E細胞を播種し、37 ℃、5% CO2インキュベータ内で終夜インキュベートを行った。トランスフェクション後、培養上清を全て除去し、0.5% FBS(Thermo Fisher Scientific社)、1% 非必須アミノ酸溶液(Thermo Fisher Scientific社)及びペニシリン-ストレプトマイシン(ナカライテスク社)を含むOpti-MEM培地(評価基礎培地)を100 μL添加し、37 ℃で4時間培養することで、細胞を飢餓状態にした。
評価基礎培地に、0~100 ng/mLのHGFを加えるか、又は無細胞発現の反応液を当該培地に対して25~625倍希釈されるように加え、その評価基礎培地を細胞に100 μL添加した。37 ℃、5% CO2インキュベータ内で終夜培養することで細胞への刺激を行った。
シグナル強度の検出にはDual-Luciferase Reporter Assay System(Promega社)を使用した。100 μLの培地上清を除去し、Glo reagentを50 μL添加し、室温で10分細胞を溶解した。細胞の溶解液から発せられるFirefly luciferaseの発光をプレートリーダーで検出することで、HGF-Erk-SRE経路の活性化を定量した。続いて、50 μLのGlo & Stop reagent溶液を添加し、10分後に内部標準であるRenilla luciferaseの発光を検出し、細胞数の定量を行った。SRE activity = (Firefly luciferaseの発光強度)/(Renilla luciferaseの発光強度)として、各サンプルのシグナル活性を定量した。評価化合物を加えていないサンプルに対する相対的なシグナル活性値を求め、相対的受容体活性(Relative reporter activity)とした。
各aMD4dY-ロイシンジッパー融合体の二量体をルシフェラーゼアッセイによって評価した。Opti-MEM培地(Thermo Fisher Scientific社)25 μLに対し、0.6 μLのAttractene Transfection Reagent(QIAGEN社)を添加し、室温で5分インキュベートを行った。上記混合溶液に対し、Opti-MEM 25 μLと1 μLのSRE reporter vector(QIAGEN社)の混合溶液を加え、室温で20分インキュベートを行った。この混合溶液を96-well plateに添加し、その上から40,000 cell/wellのHEK293E細胞を播種し、37 ℃、5% CO2インキュベータ内で終夜インキュベートを行った。トランスフェクション後、培養上清を全て除去し、0.5% FBS(Thermo Fisher Scientific社)、1% 非必須アミノ酸溶液(Thermo Fisher Scientific社)及びペニシリン-ストレプトマイシン(ナカライテスク社)を含むOpti-MEM培地(評価基礎培地)を100 μL添加し、37 ℃で4時間培養することで、細胞を飢餓状態にした。
評価基礎培地に、0~100 ng/mLのHGFを加えるか、又は無細胞発現の反応液を当該培地に対して25~625倍希釈されるように加え、その評価基礎培地を細胞に100 μL添加した。37 ℃、5% CO2インキュベータ内で終夜培養することで細胞への刺激を行った。
シグナル強度の検出にはDual-Luciferase Reporter Assay System(Promega社)を使用した。100 μLの培地上清を除去し、Glo reagentを50 μL添加し、室温で10分細胞を溶解した。細胞の溶解液から発せられるFirefly luciferaseの発光をプレートリーダーで検出することで、HGF-Erk-SRE経路の活性化を定量した。続いて、50 μLのGlo & Stop reagent溶液を添加し、10分後に内部標準であるRenilla luciferaseの発光を検出し、細胞数の定量を行った。SRE activity = (Firefly luciferaseの発光強度)/(Renilla luciferaseの発光強度)として、各サンプルのシグナル活性を定量した。評価化合物を加えていないサンプルに対する相対的なシグナル活性値を求め、相対的受容体活性(Relative reporter activity)とした。
図8に示されるとおり、aMD4dY-PA8-cys-Junを発現させた場合にはHGFと同程度の活性を示した一方で、aMD4dY-PA8-Junを発現させた場合には活性を示さなかった。このことから、システインが形成するジスルフィド結合による強固な二量体化が環状ペプチドの活性(成長因子様の活性)を発揮するのに必要であることが明らかになった。
図9に示されるとおり、ヘパリン結合能を除去したaMD4dY-PA8-cys-JunΔhepを発現させた場合も活性を示した。このことから、環状ペプチドの活性(成長因子様の活性)にロイシンジッパー領域のヘパリン結合能は必要ないことが明らかになった。
図10及び11に示されるとおり、aMD4dY-PA8-GCN4-cys及びaMD4dY-PA8-cys-CEBPαを発現させた場合、それぞれの二量体はシグナル活性化能を有することが明らかになった。このことから、二量体化ドメインにおいては、ホモダイマーを形成するロイシンジッパー領域を広く用いることが可能であると理解できた。
図12に示されるとおり、cys-Jun-PA8-aMD4dY及びJun-cys-PA8-aMD4dYを発現させた場合、それぞれの二量体はシグナル活性化能を有することが明らかになった。このことから、環状ペプチド、ペプチドリンカー、システイン残基、ロイシンジッパー領域の順序を入れ替えても環状ペプチドの活性(成長因子様の活性)が得られることが理解できた。
図13に示されるとおり、N11タグをN末端に付加したコンストラクトを用いた場合でもシグナル活性化能を示すことが明らかになった。このことから、発現ポリぺプチドのN末端にタグ配列を付加することも可能であると理解できた。
図9に示されるとおり、ヘパリン結合能を除去したaMD4dY-PA8-cys-JunΔhepを発現させた場合も活性を示した。このことから、環状ペプチドの活性(成長因子様の活性)にロイシンジッパー領域のヘパリン結合能は必要ないことが明らかになった。
図10及び11に示されるとおり、aMD4dY-PA8-GCN4-cys及びaMD4dY-PA8-cys-CEBPαを発現させた場合、それぞれの二量体はシグナル活性化能を有することが明らかになった。このことから、二量体化ドメインにおいては、ホモダイマーを形成するロイシンジッパー領域を広く用いることが可能であると理解できた。
図12に示されるとおり、cys-Jun-PA8-aMD4dY及びJun-cys-PA8-aMD4dYを発現させた場合、それぞれの二量体はシグナル活性化能を有することが明らかになった。このことから、環状ペプチド、ペプチドリンカー、システイン残基、ロイシンジッパー領域の順序を入れ替えても環状ペプチドの活性(成長因子様の活性)が得られることが理解できた。
図13に示されるとおり、N11タグをN末端に付加したコンストラクトを用いた場合でもシグナル活性化能を示すことが明らかになった。このことから、発現ポリぺプチドのN末端にタグ配列を付加することも可能であると理解できた。
実施例3:EMP-ロイシンジッパー融合体の活性評価
EMP-ロイシンジッパー融合体の二量体のEpoR活性化能をPathHunter(登録商標) eXpress EpoR-JAK2 Functional Assay Kit(DiscoveRx社)を用いて評価した。
96-wellプレートにPathHunter(登録商標) eXpress EpoR-JAK2 Functional Assay Kitに含まれるEPO活性評価用細胞を播種し、24時間、37 ℃、5% CO2下で培養を行った。細胞に対し、EMP-ロイシンジッパー融合体を含む無細胞発現の反応液を希釈して添加した。陰対照としては、この反応液と同じ倍率で希釈した、無細胞発現系に(x)で示される塩基配列の遺伝子を含まないこと以外は上記(1-9)に記載のとおり無細胞発現の処理を行った未反応液を添加した。3時間室温で刺激した細胞に対し、調製したSubstrate Reagentを添加し、60分インキュベートした。化学発光強度をNivoプレートリーダー(Perkin Elmer社)で定量した。
結果を図14に示す。この結果より、EMP-PA8-cys-Junの二量体はEpoRを活性化することが明らかになった。
EMP-ロイシンジッパー融合体の二量体のEpoR活性化能をPathHunter(登録商標) eXpress EpoR-JAK2 Functional Assay Kit(DiscoveRx社)を用いて評価した。
96-wellプレートにPathHunter(登録商標) eXpress EpoR-JAK2 Functional Assay Kitに含まれるEPO活性評価用細胞を播種し、24時間、37 ℃、5% CO2下で培養を行った。細胞に対し、EMP-ロイシンジッパー融合体を含む無細胞発現の反応液を希釈して添加した。陰対照としては、この反応液と同じ倍率で希釈した、無細胞発現系に(x)で示される塩基配列の遺伝子を含まないこと以外は上記(1-9)に記載のとおり無細胞発現の処理を行った未反応液を添加した。3時間室温で刺激した細胞に対し、調製したSubstrate Reagentを添加し、60分インキュベートした。化学発光強度をNivoプレートリーダー(Perkin Elmer社)で定量した。
結果を図14に示す。この結果より、EMP-PA8-cys-Junの二量体はEpoRを活性化することが明らかになった。
実施例4:c-Met分子との結合に基づき、環状ペプチド-ロイシンジッパー融合体ライブラリからHGF様活性分子を濃縮するスクリーニング
各々が環状ペプチド-ロイシンジッパー融合体分子をコードする複数のDNAから無細胞翻訳により当該融合体を発現し、c-Metに結合する融合体分子および当該分子をコードするDNAを濃縮し、濃縮した融合体分子がHGF様活性を有することを確認した。当該実施例ではリボソームディスプレイ法と環状ペプチド-ロイシンジッパー融合体を組み合わせることにより、磁気ビーズに固定化したc-Metに結合する環状ペプチド-ロイシンジッパー融合体とリボソーム、mRNA複合体との複合体を単離した。一連の操作により、c-Metに結合する環状ペプチド-ロイシンジッパー融合体の鋳型DNAがライブラリより濃縮されること、および濃縮されたDNAの無細胞翻訳産物がc-Metを活性化するHGF様活性を有することを確認した。
各々が環状ペプチド-ロイシンジッパー融合体分子をコードする複数のDNAから無細胞翻訳により当該融合体を発現し、c-Metに結合する融合体分子および当該分子をコードするDNAを濃縮し、濃縮した融合体分子がHGF様活性を有することを確認した。当該実施例ではリボソームディスプレイ法と環状ペプチド-ロイシンジッパー融合体を組み合わせることにより、磁気ビーズに固定化したc-Metに結合する環状ペプチド-ロイシンジッパー融合体とリボソーム、mRNA複合体との複合体を単離した。一連の操作により、c-Metに結合する環状ペプチド-ロイシンジッパー融合体の鋳型DNAがライブラリより濃縮されること、および濃縮されたDNAの無細胞翻訳産物がc-Metを活性化するHGF様活性を有することを確認した。
(4-1)リボソームディスプレイ法に供する環状ペプチド-ロイシンジッパー融合体の設計
設計した環状ペプチド-ロイシンジッパー融合体の配列を以下に示す。各融合体は「標的受容体結合環状ペプチド」―「ペプチドリンカー」-「ロイシンジッパー」-「システイン」-「N11タグ」-「SecM配列」となるよう設計した。標的受容体結合環状ペプチドはc-Met結合環状ペプチド(aMD4dY)またはトロンボポエチン受容体結合環状ペプチド(TPO)またはエリスロポエチン受容体結合環状ペプチド(EMP’)を選択した。SecM配列(下記においてSecMと表記される。)は、FSTPVWISQAQGIRAG(配列番号:86)のアミノ酸配列で表される配列であり、リボソームの伸長反応を阻害することで、リボソーム-環状ペプチド-ロイシンジッパー融合体-mRNA複合体を形成させることを目的とするものである。下記においてGS3と表記される配列は、「グリシン」-「セリン」の3回繰り返しからなる配列で、スペーサー配列である。
(A)aMD4dY-Jun-cys-N11-SecM-GS3
MAETCRQFNRRTHEVWNLDCGAAPAAPAPAAGGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMNGCGGKDHLIHNHHKHEHAHAEHLFSTPVWISQAQGIRAGGSGSGS(配列番号:71)
(B)TPO-Jun-cys-N11-SecM-GS3
MAETCADGPTLREWISFCGAAPAAPAPAAGGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMNGCGGKDHLIHNHHKHEHAHAEHLFSTPVWISQAQGIRAGGSGSGS(配列番号:72)
(C)EMP’-Jun-cys-N11-SecM-GS3
MAETCHMGPMTWVCGAAPAAPAPAAGGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMNGCGGKDHLIHNHHKHEHAHAEHLFSTPVWISQAQGIRAGGSGSGS(配列番号:73)
設計した環状ペプチド-ロイシンジッパー融合体の配列を以下に示す。各融合体は「標的受容体結合環状ペプチド」―「ペプチドリンカー」-「ロイシンジッパー」-「システイン」-「N11タグ」-「SecM配列」となるよう設計した。標的受容体結合環状ペプチドはc-Met結合環状ペプチド(aMD4dY)またはトロンボポエチン受容体結合環状ペプチド(TPO)またはエリスロポエチン受容体結合環状ペプチド(EMP’)を選択した。SecM配列(下記においてSecMと表記される。)は、FSTPVWISQAQGIRAG(配列番号:86)のアミノ酸配列で表される配列であり、リボソームの伸長反応を阻害することで、リボソーム-環状ペプチド-ロイシンジッパー融合体-mRNA複合体を形成させることを目的とするものである。下記においてGS3と表記される配列は、「グリシン」-「セリン」の3回繰り返しからなる配列で、スペーサー配列である。
(A)aMD4dY-Jun-cys-N11-SecM-GS3
MAETCRQFNRRTHEVWNLDCGAAPAAPAPAAGGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMNGCGGKDHLIHNHHKHEHAHAEHLFSTPVWISQAQGIRAGGSGSGS(配列番号:71)
(B)TPO-Jun-cys-N11-SecM-GS3
MAETCADGPTLREWISFCGAAPAAPAPAAGGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMNGCGGKDHLIHNHHKHEHAHAEHLFSTPVWISQAQGIRAGGSGSGS(配列番号:72)
(C)EMP’-Jun-cys-N11-SecM-GS3
MAETCHMGPMTWVCGAAPAAPAPAAGGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMNGCGGKDHLIHNHHKHEHAHAEHLFSTPVWISQAQGIRAGGSGSGS(配列番号:73)
(4-2)環状ペプチド-ロイシンジッパー融合体の鋳型DNAの構築
環状ペプチド-ロイシンジッパー融合体を発現させる鋳型DNAとして、上記(A)-(C)のアミノ酸配列をコードする塩基配列をそれぞれ設計した。さらに、これらの5’側には無細胞発現に必要なT7プロモーター配列及びリボソーム結合配列を付加し、3’側には終止コドンと非翻訳領域を付加した(D)-(F)を設計した。また、これらの鋳型DNAをPCRによって作成すべく、一重鎖DNA(G)-(O)を設計した。
(G)-(O)に示す一重鎖DNA配列は、ユーロフィンジェノミクス株式会社にて全合成を行った。
(D)aMD4dY-Jun-cys-N11-SecM-GS3を発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGCAGAGACCTGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTGGCGCAGCTCCCGCAGCACCAGCTCCAGCAGCAGGCGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAACGGCTGCGGTGGCAAAGATCATCTCATCCACAATCATCACAAACATGAGCACGCTCATGCCGAACACCTGTTTTCTACTCCTGTTTGGATTTCTCAAGCTCAAGGTATTCGTGCTGGTGGTAGCGGTAGCGGTAGCTAGTGAATAACTAATCC(配列番号:74)
(E)TPO-Jun-cys-N11-SecM-GS3を発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGCAGAGACCTGTGCGGACGGTCCGACGTTACGCGAATGGATCAGCTTTTGTGGCGCAGCTCCCGCAGCACCAGCTCCAGCAGCAGGCGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAACGGCTGCGGTGGCAAAGATCATCTCATCCACAATCATCACAAACATGAGCACGCTCATGCCGAACACCTGTTTTCTACTCCTGTTTGGATTTCTCAAGCTCAAGGTATTCGTGCTGGTGGTAGCGGTAGCGGTAGCTAGTGAATAACTAATCC(配列番号:75)
(F)EMP’-Jun-cys-N11-SecM-GS3を発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGCAGAGACCTGTCATATGGGGCCAATGACCTGGGTGTGTGGCGCAGCTCCCGCAGCACCAGCTCCAGCAGCAGGCGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAACGGCTGCGGTGGCAAAGATCATCTCATCCACAATCATCACAAACATGAGCACGCTCATGCCGAACACCTGTTTTCTACTCCTGTTTGGATTTCTCAAGCTCAAGGTATTCGTGCTGGTGGTAGCGGTAGCGGTAGCTAGTGAATAACTAATCC(配列番号:76)
(G)(D)-(F)を作製するための一重鎖DNAの塩基配列1
CAATGGCAGAGACCTGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTGGCGCAGCTCCCGC(配列番号:77)
(H)(D)-(F)を作製するための一重鎖DNAの塩基配列2
CAATGGCAGAGACCTGTGCGGACGGTCCGACGTTACGCGAATGGATCAGCTTTTGTGGCGCAGCTCCCGC(配列番号:78)
(I)(D)-(F)を作製するための一重鎖DNAの塩基配列3
CAATGGCAGAGACCTGTCATATGGGGCCAATGACCTGGGTGTGTGGCGCAGCTCCCGC(配列番号:79)
(J)(D)-(F)を作製するための一重鎖DNAの塩基配列4
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGCAGAGACCTGT(配列番号:80)
(K)(D)-(F)を作製するための一重鎖DNAの塩基配列5
TGTGGCGCAGCTCCCGCAGCACCAGCTCCAGCAGCAGGCGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAAC(配列番号:81)
(L)(D)-(F)を作製するための一重鎖DNAの塩基配列6
ACCAGCACGAATACCTTGAGCTTGAGAAATCCAAACAGGAGTAGAAAACAGGTGTTCGGCATGAGCGTGCTCATGTTTGTGATGATTGTGGATGAGATGATCTTTGCCACCGCAGCCGTTCATAACTTTCTGTTTCAGC(配列番号:82)
(M)(D)-(F)を作製するための一重鎖DNAの塩基配列7
GGATTAGTTATTCACTAGCTACCGCTACCGCTACCACCAGCACGAATACCTTG(配列番号:83)
(N)(D)-(F)を作製するための一重鎖DNAの塩基配列8(selec_PCR_F1.F39)
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTT(配列番号:84)
(O)(D)-(F)を作製するための一重鎖DNAの塩基配列9(selec_PCR_R2.R24)
GGATTAGTTATTCACTAGCTACCG(配列番号:85)
環状ペプチド-ロイシンジッパー融合体を発現させる鋳型DNAとして、上記(A)-(C)のアミノ酸配列をコードする塩基配列をそれぞれ設計した。さらに、これらの5’側には無細胞発現に必要なT7プロモーター配列及びリボソーム結合配列を付加し、3’側には終止コドンと非翻訳領域を付加した(D)-(F)を設計した。また、これらの鋳型DNAをPCRによって作成すべく、一重鎖DNA(G)-(O)を設計した。
(G)-(O)に示す一重鎖DNA配列は、ユーロフィンジェノミクス株式会社にて全合成を行った。
(D)aMD4dY-Jun-cys-N11-SecM-GS3を発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGCAGAGACCTGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTGGCGCAGCTCCCGCAGCACCAGCTCCAGCAGCAGGCGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAACGGCTGCGGTGGCAAAGATCATCTCATCCACAATCATCACAAACATGAGCACGCTCATGCCGAACACCTGTTTTCTACTCCTGTTTGGATTTCTCAAGCTCAAGGTATTCGTGCTGGTGGTAGCGGTAGCGGTAGCTAGTGAATAACTAATCC(配列番号:74)
(E)TPO-Jun-cys-N11-SecM-GS3を発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGCAGAGACCTGTGCGGACGGTCCGACGTTACGCGAATGGATCAGCTTTTGTGGCGCAGCTCCCGCAGCACCAGCTCCAGCAGCAGGCGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAACGGCTGCGGTGGCAAAGATCATCTCATCCACAATCATCACAAACATGAGCACGCTCATGCCGAACACCTGTTTTCTACTCCTGTTTGGATTTCTCAAGCTCAAGGTATTCGTGCTGGTGGTAGCGGTAGCGGTAGCTAGTGAATAACTAATCC(配列番号:75)
(F)EMP’-Jun-cys-N11-SecM-GS3を発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGCAGAGACCTGTCATATGGGGCCAATGACCTGGGTGTGTGGCGCAGCTCCCGCAGCACCAGCTCCAGCAGCAGGCGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAACGGCTGCGGTGGCAAAGATCATCTCATCCACAATCATCACAAACATGAGCACGCTCATGCCGAACACCTGTTTTCTACTCCTGTTTGGATTTCTCAAGCTCAAGGTATTCGTGCTGGTGGTAGCGGTAGCGGTAGCTAGTGAATAACTAATCC(配列番号:76)
(G)(D)-(F)を作製するための一重鎖DNAの塩基配列1
CAATGGCAGAGACCTGTCGTCAGTTTAACCGTCGTACGCACGAAGTTTGGAATCTGGACTGTGGCGCAGCTCCCGC(配列番号:77)
(H)(D)-(F)を作製するための一重鎖DNAの塩基配列2
CAATGGCAGAGACCTGTGCGGACGGTCCGACGTTACGCGAATGGATCAGCTTTTGTGGCGCAGCTCCCGC(配列番号:78)
(I)(D)-(F)を作製するための一重鎖DNAの塩基配列3
CAATGGCAGAGACCTGTCATATGGGGCCAATGACCTGGGTGTGTGGCGCAGCTCCCGC(配列番号:79)
(J)(D)-(F)を作製するための一重鎖DNAの塩基配列4
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGCAGAGACCTGT(配列番号:80)
(K)(D)-(F)を作製するための一重鎖DNAの塩基配列5
TGTGGCGCAGCTCCCGCAGCACCAGCTCCAGCAGCAGGCGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAAC(配列番号:81)
(L)(D)-(F)を作製するための一重鎖DNAの塩基配列6
ACCAGCACGAATACCTTGAGCTTGAGAAATCCAAACAGGAGTAGAAAACAGGTGTTCGGCATGAGCGTGCTCATGTTTGTGATGATTGTGGATGAGATGATCTTTGCCACCGCAGCCGTTCATAACTTTCTGTTTCAGC(配列番号:82)
(M)(D)-(F)を作製するための一重鎖DNAの塩基配列7
GGATTAGTTATTCACTAGCTACCGCTACCGCTACCACCAGCACGAATACCTTG(配列番号:83)
(N)(D)-(F)を作製するための一重鎖DNAの塩基配列8(selec_PCR_F1.F39)
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTT(配列番号:84)
(O)(D)-(F)を作製するための一重鎖DNAの塩基配列9(selec_PCR_R2.R24)
GGATTAGTTATTCACTAGCTACCG(配列番号:85)
(4-3)ライブラリの作製
PCRによりaMD4dY-Jun-cys-N11-SecM-GS3、TPO-Jun-cys-N11-SecM-GS3、EMP’-Jun-cys-N11-SecM-GS3の3種類のDNA配列を作製した。aMD4dY-Jun-cys-N11-SecM-GS3、TPO-Jun-cys-N11-SecM-GS3、EMP’-Jun-cys-N11-SecM-GS3のDNAを1:50:50の割合で混合したDNA mixを作製した。
PCRによりaMD4dY-Jun-cys-N11-SecM-GS3、TPO-Jun-cys-N11-SecM-GS3、EMP’-Jun-cys-N11-SecM-GS3の3種類のDNA配列を作製した。aMD4dY-Jun-cys-N11-SecM-GS3、TPO-Jun-cys-N11-SecM-GS3、EMP’-Jun-cys-N11-SecM-GS3のDNAを1:50:50の割合で混合したDNA mixを作製した。
(4-4)転写反応によるmRNAライブラリの作製
T7 RNA polymeraseを使用してDNAの転写を実施した。転写には、T7 RNA polymerase(タカラバイオ、2540A)とrNTPs(プロメガ、E6000)を使用した。11 μLの超純水、2 μLの10x T7 RNA polymerase buffer、2 μLの50 mM DTT、2 μLの25 mM rNTPs、1 μLのT7 RNA polymeraseと1 μLのDNA mixを混ぜて、37℃の恒温槽で6時間反応させた。その後、50 mM EDTA溶液を加えた。RNA Clean & Concentrator 5(ZYMO RESEARCH、R1015)で精製し、2μMのmRNA mixとした。
T7 RNA polymeraseを使用してDNAの転写を実施した。転写には、T7 RNA polymerase(タカラバイオ、2540A)とrNTPs(プロメガ、E6000)を使用した。11 μLの超純水、2 μLの10x T7 RNA polymerase buffer、2 μLの50 mM DTT、2 μLの25 mM rNTPs、1 μLのT7 RNA polymeraseと1 μLのDNA mixを混ぜて、37℃の恒温槽で6時間反応させた。その後、50 mM EDTA溶液を加えた。RNA Clean & Concentrator 5(ZYMO RESEARCH、R1015)で精製し、2μMのmRNA mixとした。
(4-5)mRNAライブラリの翻訳およびリボソーム-環状ペプチド-ロイシンジッパー融合体-mRNA複合体の形成
調製したライブラリの翻訳発現にはPURExpress ΔRF1,2,3 kitを用いた。2 μLのsolution A、1.5 μLのsolution B、1 μLのmRNA mixと0.5 μLの超純水を混ぜて、37 ℃恒温槽で2時間培養した。反応後、反応容器を氷上に静置し、形成したリボソーム-環状ペプチド-ロイシンジッパー融合体-mRNA複合体に氷冷した1x TBSM-T buffer(50 mM Tris-HCl、150 mM NaCl、50 mM MgCl2、0.05% Tween 20)を20 μL加えた。この複合体反応溶液を下記(4-7)で用いた。
調製したライブラリの翻訳発現にはPURExpress ΔRF1,2,3 kitを用いた。2 μLのsolution A、1.5 μLのsolution B、1 μLのmRNA mixと0.5 μLの超純水を混ぜて、37 ℃恒温槽で2時間培養した。反応後、反応容器を氷上に静置し、形成したリボソーム-環状ペプチド-ロイシンジッパー融合体-mRNA複合体に氷冷した1x TBSM-T buffer(50 mM Tris-HCl、150 mM NaCl、50 mM MgCl2、0.05% Tween 20)を20 μL加えた。この複合体反応溶液を下記(4-7)で用いた。
(4-6)標的分子の固定化
磁気ビーズへの標的分子の固定化を実施した。5 μLの2 pM Recombinant Human HGFR/c-MET Fc Chimera Protein (R&D systems、8614-MT-100)、5 μL slurryのDynabeads(invitrogen、DB10003)と10 μL の1x TBS-T buffer(50 mM Tris-HCl、150 mM NaCl、0.05% Tween 20)を混ぜ合わせ、4℃で30分転倒混和した。磁気ビーズを1x TBS-T buffer 100 μLで3回洗浄し、その後10 μLの1x TBS-T bufferで再懸濁して氷上に静置した。
磁気ビーズへの標的分子の固定化を実施した。5 μLの2 pM Recombinant Human HGFR/c-MET Fc Chimera Protein (R&D systems、8614-MT-100)、5 μL slurryのDynabeads(invitrogen、DB10003)と10 μL の1x TBS-T buffer(50 mM Tris-HCl、150 mM NaCl、0.05% Tween 20)を混ぜ合わせ、4℃で30分転倒混和した。磁気ビーズを1x TBS-T buffer 100 μLで3回洗浄し、その後10 μLの1x TBS-T bufferで再懸濁して氷上に静置した。
(4-7)リボソーム-環状ペプチド-ロイシンジッパー融合体-mRNA複合体と標的分子との混合、単離
磁気ビーズ懸濁液から1x TBS-T bufferを除去し、そこへ上記(4-5)の複合体反応溶液を全量入れた。ゆっくりと再懸濁後、4℃で30分転倒混和した。磁気ビーズ懸濁液を氷冷した1x TBSM-T buffer 200 μLで3回洗浄した。ネオジム磁石を用いて単離した磁気ビーズに20 μL の1x TBS-T buffer, 5 μL 500 mM EDTA水溶液を加え、よく混ぜた後室温に5分静置し、リボソーム-環状ペプチド-ロイシンジッパー融合体-mRNA複合体からmRNAを遊離させた。
磁気ビーズ懸濁液から1x TBS-T bufferを除去し、そこへ上記(4-5)の複合体反応溶液を全量入れた。ゆっくりと再懸濁後、4℃で30分転倒混和した。磁気ビーズ懸濁液を氷冷した1x TBSM-T buffer 200 μLで3回洗浄した。ネオジム磁石を用いて単離した磁気ビーズに20 μL の1x TBS-T buffer, 5 μL 500 mM EDTA水溶液を加え、よく混ぜた後室温に5分静置し、リボソーム-環状ペプチド-ロイシンジッパー融合体-mRNA複合体からmRNAを遊離させた。
(4-8)mRNAの精製
回収した溶液からRNA Clean & Concentrator 5を用いてmRNAを精製した。
回収した溶液からRNA Clean & Concentrator 5を用いてmRNAを精製した。
(4-9)精製したmRNAの逆転写
精製したmRNAをSuperScriptTM II Reverse Transcriptase(Invitrogen、18064022)によって逆転写した。精製したmRNA溶液 1 μLと、100 μMのselec_PCR_R2.R24 0.5 μL、dNTPs 2.5 μL、超純水 2 μLとを混ぜ合わせ、70℃で 5分加熱した。室温でゆっくりと冷却し、卓上遠心装置で遠心を行った。5x First-strand buffer 2 μL、0.1 M DTT溶液 1 μL、超純水0.5 μLとSuperScript II Reverse Transcriptase 0.5 μLを順番に添加し、42℃の条件で50分反応させてmRNA-cDNA複合体を得た。
精製したmRNAをSuperScriptTM II Reverse Transcriptase(Invitrogen、18064022)によって逆転写した。精製したmRNA溶液 1 μLと、100 μMのselec_PCR_R2.R24 0.5 μL、dNTPs 2.5 μL、超純水 2 μLとを混ぜ合わせ、70℃で 5分加熱した。室温でゆっくりと冷却し、卓上遠心装置で遠心を行った。5x First-strand buffer 2 μL、0.1 M DTT溶液 1 μL、超純水0.5 μLとSuperScript II Reverse Transcriptase 0.5 μLを順番に添加し、42℃の条件で50分反応させてmRNA-cDNA複合体を得た。
(4-10)cDNAのPCR増幅
回収したcDNAをPCRによって増幅した。PCRには、KOD -Plus- Ver.2(東洋紡、KOD-211)を使用した。逆転写反応の完了した溶液4 μLに、10x PCR buffer 10 μL、250 mM MgSO4 4 μL、2mM dNTPs 10 μL、100 μM selec_PCR_F1.F39 1 μL、100 μM selec_PCR_R2.R24 1 μL、超純水 63 μL、KOD -Plus- 2 μLを混ぜ、94℃ 30秒→55℃ 40秒→68℃ 60 秒の条件でサイクルを作成し、20-25サイクル繰り返した。回収したDNAをDNA mix2とした。
回収したcDNAをPCRによって増幅した。PCRには、KOD -Plus- Ver.2(東洋紡、KOD-211)を使用した。逆転写反応の完了した溶液4 μLに、10x PCR buffer 10 μL、250 mM MgSO4 4 μL、2mM dNTPs 10 μL、100 μM selec_PCR_F1.F39 1 μL、100 μM selec_PCR_R2.R24 1 μL、超純水 63 μL、KOD -Plus- 2 μLを混ぜ、94℃ 30秒→55℃ 40秒→68℃ 60 秒の条件でサイクルを作成し、20-25サイクル繰り返した。回収したDNAをDNA mix2とした。
(4-11)cDNAのDNA長分析
aMD4dY-Jun-cys-N11-SecM-GS3、TPO-Jun-cys-N11-SecM-GS3、EMP’-Jun-cys-N11-SecM-GS3の3種類のDNAとDNA mix、DNA mix2を2100 Bioanalyzer(Agilent technologies)で評価した。結果を図15に示す。レーン1-5はそれぞれ順番に、aMD4dY-Jun-cys-N11-SecM-GS3、TPO-Jun-cys-N11-SecM-GS3、EMP’-Jun-cys-N11-SecM-GS3、DNA mix、DNA mix2に対応する。リボソームディスプレイ法を実施した後に得られたDNA mix2のバンドをDNA mixのバンドと比較したところ、DNA長が長くなる方向へのシフトが確認され、DNA mix2において、aMD4dY-Jun-cys-N11-SecM-GS3をコードするDNAと同一のDNA長を示すバンドが最も高い強度を示した。
aMD4dY-Jun-cys-N11-SecM-GS3、TPO-Jun-cys-N11-SecM-GS3、EMP’-Jun-cys-N11-SecM-GS3の3種類のDNAとDNA mix、DNA mix2を2100 Bioanalyzer(Agilent technologies)で評価した。結果を図15に示す。レーン1-5はそれぞれ順番に、aMD4dY-Jun-cys-N11-SecM-GS3、TPO-Jun-cys-N11-SecM-GS3、EMP’-Jun-cys-N11-SecM-GS3、DNA mix、DNA mix2に対応する。リボソームディスプレイ法を実施した後に得られたDNA mix2のバンドをDNA mixのバンドと比較したところ、DNA長が長くなる方向へのシフトが確認され、DNA mix2において、aMD4dY-Jun-cys-N11-SecM-GS3をコードするDNAと同一のDNA長を示すバンドが最も高い強度を示した。
(4-12)増幅したcDNAから無細胞翻訳した産物の活性評価
aMD4dY-Jun-cys-N11-SecM-GS3、TPO-Jun-cys-N11-SecM-GS3、EMP’-Jun-cys-N11-SecM-GS3の3種類のDNAとDNA mix、DNA mix2を用い、PUREfrex2.0によって無細胞翻訳を行い、それぞれの無細胞翻訳産物をルシフェラーゼアッセイ活性により評価した。無細胞翻訳は各DNA20 ngに対し、2.5 μLのSolution I、0.25 μLのSolution II、0.5μLのSolution III、1.75μLの水を添加し、37 ℃で6時間インキュベーションすることで翻訳反応を行った。
レポーターアッセイは以下の通り実施した。Opti-MEM培地(Thermo Fisher Scientific社)25 μLに対し、0.6 μLのAttractene Transfection Reagent(QIAGEN社)を添加し、室温で5分インキュベートを行った。上記混合溶液に対し、Opti-MEM 25 μLと1 μLのSRE reporter vector(QIAGEN社)の混合溶液を加え、室温で20分インキュベートを行った。この混合溶液を96-well plateに添加し、その上から40,000 cell/wellのHEK293E細胞を播種し、37 ℃、5% CO2インキュベータ内で終夜インキュベートを行った。トランスフェクション後、培養上清を全て除去し、0.5% FBS(Thermo Fisher Scientific社)、1% 非必須アミノ酸溶液(Thermo Fisher Scientific社)及びペニシリン-ストレプトマイシン(ナカライテスク社)を含むOpti-MEM培地(評価基礎培地)を100 μL添加し、37 ℃で4時間培養することで、細胞を飢餓状態にした。
評価基礎培地に、0~100 ng/mLHGFを加えるか、又は上記無細胞発現の反応液を当該培地に対して25~625倍希釈されるように加え、その評価基礎培地を細胞に100 μL添加した。37 ℃、5% CO2インキュベータ内で終夜培養することで細胞への刺激を行った。
シグナル強度の検出にはDual-Luciferase Reporter Assay System(Promega社)を使用した。100 μLの培地上清を除去し、Glo reagentを50 μL添加し、室温で10分細胞を溶解した。細胞の溶解液から発せられるFirefly luciferaseの発光をプレートリーダーで検出することで、HGF-Erk-SRE経路の活性化を定量した。続いて、50 μLのGlo & Stop reagent溶液を添加し、10分後に内部標準であるRenilla luciferaseの発光を検出し、細胞数の定量を行った。SRE activity = (Firefly luciferaseの発光強度)/(Renilla luciferaseの発光強度)として、各サンプルのシグナル活性を定量した。評価化合物を加えていないサンプルに対する相対的なシグナル活性値を求め、相対的受容体活性(Relative SRE reporter activity)とした。
結果を図16に示す。DNA mixは相対的受容体活性を示さないのに対し、DNA mix2ではHGFと同様の活性を持つことが明らかとなった。また、aMD4dY-Jun-cys-N11-SecM-GS3、TPO-Jun-cys-N11-SecM-GS3、EMP’-Jun-cys-N11-SecM-GS3それぞれの無細胞翻訳産物の中ではaMD4dY-Jun-cys-N11-SecM-GS3しか活性を示さないことから、DNA mix2はaMD4dY-Jun-cys-N11-SecM-GS3が濃縮されたものであることがわかった。以上の結果より、当該手法により複数のディスプレイされた環状ペプチド-ロイシンジッパー融合体を含むライブラリから、標的分子との結合親和性を持ちかつ結合に起因する活性を有する環状ペプチド-ロイシンジッパー融合体を取得できることが明らかとなった。
aMD4dY-Jun-cys-N11-SecM-GS3、TPO-Jun-cys-N11-SecM-GS3、EMP’-Jun-cys-N11-SecM-GS3の3種類のDNAとDNA mix、DNA mix2を用い、PUREfrex2.0によって無細胞翻訳を行い、それぞれの無細胞翻訳産物をルシフェラーゼアッセイ活性により評価した。無細胞翻訳は各DNA20 ngに対し、2.5 μLのSolution I、0.25 μLのSolution II、0.5μLのSolution III、1.75μLの水を添加し、37 ℃で6時間インキュベーションすることで翻訳反応を行った。
レポーターアッセイは以下の通り実施した。Opti-MEM培地(Thermo Fisher Scientific社)25 μLに対し、0.6 μLのAttractene Transfection Reagent(QIAGEN社)を添加し、室温で5分インキュベートを行った。上記混合溶液に対し、Opti-MEM 25 μLと1 μLのSRE reporter vector(QIAGEN社)の混合溶液を加え、室温で20分インキュベートを行った。この混合溶液を96-well plateに添加し、その上から40,000 cell/wellのHEK293E細胞を播種し、37 ℃、5% CO2インキュベータ内で終夜インキュベートを行った。トランスフェクション後、培養上清を全て除去し、0.5% FBS(Thermo Fisher Scientific社)、1% 非必須アミノ酸溶液(Thermo Fisher Scientific社)及びペニシリン-ストレプトマイシン(ナカライテスク社)を含むOpti-MEM培地(評価基礎培地)を100 μL添加し、37 ℃で4時間培養することで、細胞を飢餓状態にした。
評価基礎培地に、0~100 ng/mLHGFを加えるか、又は上記無細胞発現の反応液を当該培地に対して25~625倍希釈されるように加え、その評価基礎培地を細胞に100 μL添加した。37 ℃、5% CO2インキュベータ内で終夜培養することで細胞への刺激を行った。
シグナル強度の検出にはDual-Luciferase Reporter Assay System(Promega社)を使用した。100 μLの培地上清を除去し、Glo reagentを50 μL添加し、室温で10分細胞を溶解した。細胞の溶解液から発せられるFirefly luciferaseの発光をプレートリーダーで検出することで、HGF-Erk-SRE経路の活性化を定量した。続いて、50 μLのGlo & Stop reagent溶液を添加し、10分後に内部標準であるRenilla luciferaseの発光を検出し、細胞数の定量を行った。SRE activity = (Firefly luciferaseの発光強度)/(Renilla luciferaseの発光強度)として、各サンプルのシグナル活性を定量した。評価化合物を加えていないサンプルに対する相対的なシグナル活性値を求め、相対的受容体活性(Relative SRE reporter activity)とした。
結果を図16に示す。DNA mixは相対的受容体活性を示さないのに対し、DNA mix2ではHGFと同様の活性を持つことが明らかとなった。また、aMD4dY-Jun-cys-N11-SecM-GS3、TPO-Jun-cys-N11-SecM-GS3、EMP’-Jun-cys-N11-SecM-GS3それぞれの無細胞翻訳産物の中ではaMD4dY-Jun-cys-N11-SecM-GS3しか活性を示さないことから、DNA mix2はaMD4dY-Jun-cys-N11-SecM-GS3が濃縮されたものであることがわかった。以上の結果より、当該手法により複数のディスプレイされた環状ペプチド-ロイシンジッパー融合体を含むライブラリから、標的分子との結合親和性を持ちかつ結合に起因する活性を有する環状ペプチド-ロイシンジッパー融合体を取得できることが明らかとなった。
実施例5:EPO受容体分子との結合に基づいた環状ペプチド-ロイシンジッパー融合体ライブラリから活性分子の濃縮をするスクリーニング方法
実施例4にて実施したスクリーニング方法が標的分子に関わらず可能であることを示すため、複数の環状ペプチド-ロイシンジッパー融合体分子からEPO受容体に結合するペプチドを濃縮し、濃縮したDNAのEPO受容体に対する結合に基づいた活性の有無を確認した。当該実施例ではリボソームディスプレイ法と環状ペプチド-ロイシンジッパー融合体を組み合わせて、磁気ビーズに固定化したEPO受容体に結合する環状ペプチド-ロイシンジッパー融合体とリボソーム、mRNA複合体との複合体を回収した。一連の操作により、EPO受容体に結合する環状ペプチド-ロイシンジッパー融合体の鋳型DNAがライブラリより濃縮されることを確認した。
実施例4にて実施したスクリーニング方法が標的分子に関わらず可能であることを示すため、複数の環状ペプチド-ロイシンジッパー融合体分子からEPO受容体に結合するペプチドを濃縮し、濃縮したDNAのEPO受容体に対する結合に基づいた活性の有無を確認した。当該実施例ではリボソームディスプレイ法と環状ペプチド-ロイシンジッパー融合体を組み合わせて、磁気ビーズに固定化したEPO受容体に結合する環状ペプチド-ロイシンジッパー融合体とリボソーム、mRNA複合体との複合体を回収した。一連の操作により、EPO受容体に結合する環状ペプチド-ロイシンジッパー融合体の鋳型DNAがライブラリより濃縮されることを確認した。
(5-1)リボソームディスプレイ法に供する環状ペプチド-ロイシンジッパー融合体の設計
実施例4に加えて新たに設計した環状ペプチド-ロイシンジッパー融合体の配列を以下に示す。
(i)EMP-Jun-cys-N11-SecM-GS3
MAETGGLYACHMGPMTWVCQPLRGAAPAAPAPAAGGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMNGCGGKDHLIHNHHKHEHAHAEHLFSTPVWISQAQGIRAGGSGSGS(配列番号:87)
(5-2)EMP-ロイシンジッパー融合体の鋳型DNAの構築
EMP-Jun-cys-N11-SecM-GS3を発現させる鋳型DNAとして、上記(i)のアミノ酸配列をコードする塩基配列をそれぞれ設計した。さらに、これらの5’側には無細胞発現に必要なT7プロモーター配列及びリボソーム結合配列を付加し、3’側には終止コドンと非翻訳領域を付加した(ii)を設計した。また、これらの鋳型DNAをPCRによって作成すべく、一重鎖DNA(iii)-(ix)を設計した。
(iii)-(ix)に示す一重鎖DNA配列は、ユーロフィンジェノミクス株式会社にて全合成を行った。
(ii)EMP-Jun-cys-N11-SecM-GS3を発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGCAGAGACCGGTGGCCTGTATGCGTGCCATATGGGGCCAATGACCTGGGTGTGTCAGCCGTTACGCGGCGCAGCTCCCGCAGCACCAGCTCCAGCAGCAGGCGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAACGGCTGCGGTGGCAAAGATCATCTCATCCACAATCATCACAAACATGAGCACGCTCATGCCGAACACCTGTTTTCTACTCCTGTTTGGATTTCTCAAGCTCAAGGTATTCGTGCTGGTGGTAGCGGTAGCGGTAGCTAGTGAATAACTAATCC(配列番号:88)
(iii)(ii)を作製するための一重鎖DNAの塩基配列1(配列番号:77で示される)
(iv)(ii)を作製するための一重鎖DNAの塩基配列2
ATACCAATGGCAGAGACCGGTGGCCTGTATGCGTGCCATATGGGGCCAATGACCTGGGTGTGTCAGCCGTTACGCGGCGCAGCTCCCGCAG(配列番号:89)
(v)(ii)を作製するための一重鎖DNAの塩基配列3
GGCGCAGCTCCCGCAGCACCAGCTCCAGCAGCAGGCGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAAC(配列番号:90)
(vi)(ii)を作製するための一重鎖DNAの塩基配列4(配列番号:82で示される)
(vii)(ii)を作製するための一重鎖DNAの塩基配列5(配列番号:83で示される)
(viii)(ii)を作製するための一重鎖DNAの塩基配列6(selec_PCR_F1.F39、配列番号:84で示される)
(ix)(ii)を作製するための一重鎖DNAの塩基配列7(selec_PCR_R2.R24、配列番号:85で示される)
実施例4に加えて新たに設計した環状ペプチド-ロイシンジッパー融合体の配列を以下に示す。
(i)EMP-Jun-cys-N11-SecM-GS3
MAETGGLYACHMGPMTWVCQPLRGAAPAAPAPAAGGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMNGCGGKDHLIHNHHKHEHAHAEHLFSTPVWISQAQGIRAGGSGSGS(配列番号:87)
(5-2)EMP-ロイシンジッパー融合体の鋳型DNAの構築
EMP-Jun-cys-N11-SecM-GS3を発現させる鋳型DNAとして、上記(i)のアミノ酸配列をコードする塩基配列をそれぞれ設計した。さらに、これらの5’側には無細胞発現に必要なT7プロモーター配列及びリボソーム結合配列を付加し、3’側には終止コドンと非翻訳領域を付加した(ii)を設計した。また、これらの鋳型DNAをPCRによって作成すべく、一重鎖DNA(iii)-(ix)を設計した。
(iii)-(ix)に示す一重鎖DNA配列は、ユーロフィンジェノミクス株式会社にて全合成を行った。
(ii)EMP-Jun-cys-N11-SecM-GS3を発現するための塩基配列
GAAATTAATACGACTCACTATAGGGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACCAATGGCAGAGACCGGTGGCCTGTATGCGTGCCATATGGGGCCAATGACCTGGGTGTGTCAGCCGTTACGCGGCGCAGCTCCCGCAGCACCAGCTCCAGCAGCAGGCGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAACGGCTGCGGTGGCAAAGATCATCTCATCCACAATCATCACAAACATGAGCACGCTCATGCCGAACACCTGTTTTCTACTCCTGTTTGGATTTCTCAAGCTCAAGGTATTCGTGCTGGTGGTAGCGGTAGCGGTAGCTAGTGAATAACTAATCC(配列番号:88)
(iii)(ii)を作製するための一重鎖DNAの塩基配列1(配列番号:77で示される)
(iv)(ii)を作製するための一重鎖DNAの塩基配列2
ATACCAATGGCAGAGACCGGTGGCCTGTATGCGTGCCATATGGGGCCAATGACCTGGGTGTGTCAGCCGTTACGCGGCGCAGCTCCCGCAG(配列番号:89)
(v)(ii)を作製するための一重鎖DNAの塩基配列3
GGCGCAGCTCCCGCAGCACCAGCTCCAGCAGCAGGCGGTCGTATCGCTCGTCTGGAAGAAAAAGTTAAAACCCTGAAAGCTCAGAACTCCGAACTGGCTTCCACCGCTAACATGCTGCGTGAACAGGTTGCTCAGCTGAAACAGAAAGTTATGAAC(配列番号:90)
(vi)(ii)を作製するための一重鎖DNAの塩基配列4(配列番号:82で示される)
(vii)(ii)を作製するための一重鎖DNAの塩基配列5(配列番号:83で示される)
(viii)(ii)を作製するための一重鎖DNAの塩基配列6(selec_PCR_F1.F39、配列番号:84で示される)
(ix)(ii)を作製するための一重鎖DNAの塩基配列7(selec_PCR_R2.R24、配列番号:85で示される)
(5-3)ライブラリの作製
PCRによりEMP-Jun-cys-N11-SecM-GS3のDNA配列を作製した。aMD4dY-Jun-cys-N11-SecM-GS3及びTPO-Jun-cys-N11-SecM-GS3の2種類のDNA配列は実施例4にて作製したものを使用した。EMP-Jun-cys-N11-SecM-GS3、aMD4dY-Jun-cys-N11-SecM-GS3、TPO-Jun-cys-N11-SecM-GS3のDNAを1:50:50の割合で混合したDNA mixを作製した。
PCRによりEMP-Jun-cys-N11-SecM-GS3のDNA配列を作製した。aMD4dY-Jun-cys-N11-SecM-GS3及びTPO-Jun-cys-N11-SecM-GS3の2種類のDNA配列は実施例4にて作製したものを使用した。EMP-Jun-cys-N11-SecM-GS3、aMD4dY-Jun-cys-N11-SecM-GS3、TPO-Jun-cys-N11-SecM-GS3のDNAを1:50:50の割合で混合したDNA mixを作製した。
(5-4)転写反応によるmRNAライブラリの作製
T7 RNA polymeraseを使用してDNAの転写を実施した。転写には、T7 RNA polymerase(タカラバイオ、2540A)とrNTPs(プロメガ、E6000)も使用した。11 μLの超純水、2 μLの10x T7 RNA polymerase buffer、2 μLの50 mM DTT、2 μLの25 mM rNTPs、1 μLのT7 RNA polymeraseと1 μLのDNA mixを混ぜて、37℃の恒温槽で6時間反応させた。その後、50 mM EDTA溶液を加えた。RNA Clean & Concentrator 5(ZYMO RESEARCH、R1015)で精製し、2μMのmRNA mixとした。
T7 RNA polymeraseを使用してDNAの転写を実施した。転写には、T7 RNA polymerase(タカラバイオ、2540A)とrNTPs(プロメガ、E6000)も使用した。11 μLの超純水、2 μLの10x T7 RNA polymerase buffer、2 μLの50 mM DTT、2 μLの25 mM rNTPs、1 μLのT7 RNA polymeraseと1 μLのDNA mixを混ぜて、37℃の恒温槽で6時間反応させた。その後、50 mM EDTA溶液を加えた。RNA Clean & Concentrator 5(ZYMO RESEARCH、R1015)で精製し、2μMのmRNA mixとした。
(5-5)mRNAライブラリの翻訳およびリボソーム-環状ペプチド-ロイシンジッパー融合体-mRNA複合体の形成
調製したライブラリの翻訳発現にはPURExpress ΔRF1,2,3 kitを用いた。2 μLのsolution A、1.5 μLのsolution B、1 μLのmRNA mixと0.5 μLの超純水を混ぜて、37 ℃恒温槽で2時間培養した。反応後、反応容器を氷上に静置し、形成したリボソーム-環状ペプチド-ロイシンジッパー融合体-mRNA複合体に氷冷した1x TBSM-T buffer(50 mM Tris-HCl、150 mM NaCl、50 mM MgCl2、0.05% Tween 20)を20 μL加えた。
調製したライブラリの翻訳発現にはPURExpress ΔRF1,2,3 kitを用いた。2 μLのsolution A、1.5 μLのsolution B、1 μLのmRNA mixと0.5 μLの超純水を混ぜて、37 ℃恒温槽で2時間培養した。反応後、反応容器を氷上に静置し、形成したリボソーム-環状ペプチド-ロイシンジッパー融合体-mRNA複合体に氷冷した1x TBSM-T buffer(50 mM Tris-HCl、150 mM NaCl、50 mM MgCl2、0.05% Tween 20)を20 μL加えた。
(5-6)標的分子の固定化
磁気ビーズへの標的分子の固定化を実施した。5 μLの2 pM Recombinant Human Erythropoietin R Fc Chimera Protein, CF(R&D systems、963-ER-50)、5 μL slurryのDynabeads(invitrogen、DB10003)と10 μL の1x TBS-T buffer(50 mM Tris-HCl、150 mM NaCl、0.05% Tween 20)を混ぜ合わせ、4℃で30分転倒混和した。磁気ビーズを1x TBS-T buffer 100 μLで3回洗浄し、その後10 μLの1x TBS-T bufferで再懸濁して氷上に静置した。
磁気ビーズへの標的分子の固定化を実施した。5 μLの2 pM Recombinant Human Erythropoietin R Fc Chimera Protein, CF(R&D systems、963-ER-50)、5 μL slurryのDynabeads(invitrogen、DB10003)と10 μL の1x TBS-T buffer(50 mM Tris-HCl、150 mM NaCl、0.05% Tween 20)を混ぜ合わせ、4℃で30分転倒混和した。磁気ビーズを1x TBS-T buffer 100 μLで3回洗浄し、その後10 μLの1x TBS-T bufferで再懸濁して氷上に静置した。
(5-7)リボソーム-環状ペプチド-ロイシンジッパー融合体-mRNA複合体と標的分子との混合、回収
磁気ビーズ懸濁液から1x TBS-T bufferを除去し、そこへ上記(5-5)の複合体反応溶液を全量入れた。ゆっくりと再懸濁後、4℃で30分転倒混和した。磁気ビーズ懸濁液を氷冷した1x TBSM-T buffer 200 μLで3回洗浄した。ネオジム磁石を用いて単離した磁気ビーズに20 μL の1x TBS-T buffer, 5 μL 500 mM EDTA水溶液を加え、よく混ぜた後室温に5分静置し、リボソーム-環状ペプチド-ロイシンジッパー融合体-mRNA複合体からmRNAを遊離させた。
磁気ビーズ懸濁液から1x TBS-T bufferを除去し、そこへ上記(5-5)の複合体反応溶液を全量入れた。ゆっくりと再懸濁後、4℃で30分転倒混和した。磁気ビーズ懸濁液を氷冷した1x TBSM-T buffer 200 μLで3回洗浄した。ネオジム磁石を用いて単離した磁気ビーズに20 μL の1x TBS-T buffer, 5 μL 500 mM EDTA水溶液を加え、よく混ぜた後室温に5分静置し、リボソーム-環状ペプチド-ロイシンジッパー融合体-mRNA複合体からmRNAを遊離させた。
(5-8)mRNAの精製
回収した溶液からRNA Clean & Concentrator 5を用いてmRNAを精製した。
回収した溶液からRNA Clean & Concentrator 5を用いてmRNAを精製した。
(5-9)精製したmRNAの逆転写
精製したmRNAをSuperScriptTM II Reverse Transcriptase(Invitrogen、18064022)によって逆転写した。精製したmRNA溶液 1 μLと、100 μMのselec_PCR_R2.R24 0.5 μL、dNTPs 2.5 μL、超純水 2 μLとを混ぜ合わせ、70℃で 5分加熱した。室温でゆっくりと冷却し、卓上遠心装置にかけた。5x First-strand buffer 2 μL、0.1 M DTT溶液 1 μL、超純水0.5 μLとSuperScript II Reverse Transcriptase 0.5 μLを順番に添加し、42℃の条件で50分反応させてmRNA-cDNA複合体を得た。
精製したmRNAをSuperScriptTM II Reverse Transcriptase(Invitrogen、18064022)によって逆転写した。精製したmRNA溶液 1 μLと、100 μMのselec_PCR_R2.R24 0.5 μL、dNTPs 2.5 μL、超純水 2 μLとを混ぜ合わせ、70℃で 5分加熱した。室温でゆっくりと冷却し、卓上遠心装置にかけた。5x First-strand buffer 2 μL、0.1 M DTT溶液 1 μL、超純水0.5 μLとSuperScript II Reverse Transcriptase 0.5 μLを順番に添加し、42℃の条件で50分反応させてmRNA-cDNA複合体を得た。
(5-10)cDNAのPCR増幅
回収したcDNAをPCRによって増幅した。PCRには、KOD -Plus- Ver.2(東洋紡、KOD-211)を使用した。逆転写反応の完了した溶液4 μLに、10x PCR buffer 10 μL、250 mM MgSO4 4 μL、2mM dNTPs 10 μL、100 μM selec_PCR_F1.F39 1 μL、100 μM selec_PCR_R2.R24 1 μL、超純水 63 μL、KOD -Plus- 2 μLを混ぜ、94℃ 30秒→55℃ 40秒→68℃ 60 秒の条件でサイクルを作成し、20-25サイクル繰り返した。回収したDNAをDNA mix2とした。
回収したcDNAをPCRによって増幅した。PCRには、KOD -Plus- Ver.2(東洋紡、KOD-211)を使用した。逆転写反応の完了した溶液4 μLに、10x PCR buffer 10 μL、250 mM MgSO4 4 μL、2mM dNTPs 10 μL、100 μM selec_PCR_F1.F39 1 μL、100 μM selec_PCR_R2.R24 1 μL、超純水 63 μL、KOD -Plus- 2 μLを混ぜ、94℃ 30秒→55℃ 40秒→68℃ 60 秒の条件でサイクルを作成し、20-25サイクル繰り返した。回収したDNAをDNA mix2とした。
(5-11)cDNAのDNA長分析
EMP-Jun-cys-N11-SecM-GS3、aMD4dY-Jun-cys-N11-SecM-GS3、TPO-Jun-cys-N11-SecM-GS3の3種類のDNAとDNA mix、DNA mix2を2100 Bioanalyzer(Agilent technologies)で評価した。結果を図17に示す。レーン1-5はそれぞれ順番に、EMP-Jun-cys-N11-SecM-GS3、aMD4dY-Jun-cys-N11-SecM-GS3、TPO-Jun-cys-N11-SecM-GS3、DNA mix、DNA mix2に対応する。リボソームディスプレイ法を実施した後に得られたDNA mix2のバンドをDNA mixのバンドと比較したところ、DNA長が長くなる方向へのシフトが確認され、DNA mix2において、EMP-Jun-cys-N11-SecM-GS3をコードするDNAと同一のDNA長を示すバンドが最も高い強度を示した。
EMP-Jun-cys-N11-SecM-GS3、aMD4dY-Jun-cys-N11-SecM-GS3、TPO-Jun-cys-N11-SecM-GS3の3種類のDNAとDNA mix、DNA mix2を2100 Bioanalyzer(Agilent technologies)で評価した。結果を図17に示す。レーン1-5はそれぞれ順番に、EMP-Jun-cys-N11-SecM-GS3、aMD4dY-Jun-cys-N11-SecM-GS3、TPO-Jun-cys-N11-SecM-GS3、DNA mix、DNA mix2に対応する。リボソームディスプレイ法を実施した後に得られたDNA mix2のバンドをDNA mixのバンドと比較したところ、DNA長が長くなる方向へのシフトが確認され、DNA mix2において、EMP-Jun-cys-N11-SecM-GS3をコードするDNAと同一のDNA長を示すバンドが最も高い強度を示した。
(5-12)増幅したcDNAから無細胞翻訳した産物の活性評価
EMP-Jun-cys-N11-SecM-GS3、aMD4dY-Jun-cys-N11-SecM-GS3、TPO-Jun-cys-N11-SecM-GS3の3種類のDNAとDNA mix、DNA mix2、更には無添加群に相当するDNA不含の蒸留水を用い、それぞれPUREfrex2.0(ジーンフロンティア)によって無細胞翻訳を行った。各DNA20 ngに対し、2.5 μLのSolution I、0.25 μLのSolution II、0.5μLのSolution III、1.75μLの水を添加し、37 ℃で6時間インキュベーションすることで翻訳反応を行った。
これらの無細胞翻訳産物をPathHunter(登録商標) eXpress EpoR-JAK2 Functional Assay Kit(DiscoveRx社)により評価した。96-wellプレートにPathHunter(登録商標) eXpress EpoR-JAK2 Functional Assay Kitに含まれるEPO活性評価用細胞を播種し、24時間、37 ℃、5% CO2下で培養を行った。細胞に対し、培地中に上記無細胞発現系の反応液の濃度が5段階希釈(1/110、1/330、1/990、1/2970及び1/8910)となるように、またはrecombinant human EPO 50 μg/mLが培地中に1/110の濃度となるように添加した。3時間室温で刺激した細胞に対し、調製したSubstrate Reagentを添加し、60分インキュベートした。化学発光強度をNivoプレートリーダー(Perkin Elmer社)で定量した。
結果を図18に示す。化学発光強度から算出される相対的受容体活性についてDNA mixとDNA mix2とを比較して、DNA mix2のみ活性を持つことが明らかとなった。また、EMP-Jun-cys-N11-SecM-GS3、aMD4dY-Jun-cys-N11-SecM-GS3、TPO-Jun-cys-N11-SecM-GS3それぞれの無細胞翻訳産物の活性から、DNA mix2はEMP-Jun-cys-N11-SecM-GS3が濃縮されたものであることがわかった。
以上の結果より、当該手法により複数のディスプレイされたポリペプチド(環状ペプチド-ロイシンジッパー融合体)を含むライブラリから、標的分子との結合親和性を持ちかつ結合に基づいた活性を有する環状ペプチド-ロイシンジッパー融合体を取得できることが明らかとなった。また、実施例4の結果と合わせて、標的分子も限定されないことが示された。
EMP-Jun-cys-N11-SecM-GS3、aMD4dY-Jun-cys-N11-SecM-GS3、TPO-Jun-cys-N11-SecM-GS3の3種類のDNAとDNA mix、DNA mix2、更には無添加群に相当するDNA不含の蒸留水を用い、それぞれPUREfrex2.0(ジーンフロンティア)によって無細胞翻訳を行った。各DNA20 ngに対し、2.5 μLのSolution I、0.25 μLのSolution II、0.5μLのSolution III、1.75μLの水を添加し、37 ℃で6時間インキュベーションすることで翻訳反応を行った。
これらの無細胞翻訳産物をPathHunter(登録商標) eXpress EpoR-JAK2 Functional Assay Kit(DiscoveRx社)により評価した。96-wellプレートにPathHunter(登録商標) eXpress EpoR-JAK2 Functional Assay Kitに含まれるEPO活性評価用細胞を播種し、24時間、37 ℃、5% CO2下で培養を行った。細胞に対し、培地中に上記無細胞発現系の反応液の濃度が5段階希釈(1/110、1/330、1/990、1/2970及び1/8910)となるように、またはrecombinant human EPO 50 μg/mLが培地中に1/110の濃度となるように添加した。3時間室温で刺激した細胞に対し、調製したSubstrate Reagentを添加し、60分インキュベートした。化学発光強度をNivoプレートリーダー(Perkin Elmer社)で定量した。
結果を図18に示す。化学発光強度から算出される相対的受容体活性についてDNA mixとDNA mix2とを比較して、DNA mix2のみ活性を持つことが明らかとなった。また、EMP-Jun-cys-N11-SecM-GS3、aMD4dY-Jun-cys-N11-SecM-GS3、TPO-Jun-cys-N11-SecM-GS3それぞれの無細胞翻訳産物の活性から、DNA mix2はEMP-Jun-cys-N11-SecM-GS3が濃縮されたものであることがわかった。
以上の結果より、当該手法により複数のディスプレイされたポリペプチド(環状ペプチド-ロイシンジッパー融合体)を含むライブラリから、標的分子との結合親和性を持ちかつ結合に基づいた活性を有する環状ペプチド-ロイシンジッパー融合体を取得できることが明らかとなった。また、実施例4の結果と合わせて、標的分子も限定されないことが示された。
Claims (18)
- 二量体化した環状ペプチドをスクリーニングする方法であって、
(1)無細胞発現系で、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含む少なくとも1種のポリペプチドを合成し、前記ポリペプチドの二量体を調製する工程、及び
(2)前記二量体の標的分子との結合に基づく活性を測定する工程
を含む、前記方法。 - 二量体化した環状ペプチドをスクリーニングする方法であって、
(1)無細胞発現系で、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含む少なくとも1種のポリペプチドを合成することにより得られた前記ポリペプチドの二量体を用意する工程、及び
(2)前記二量体の標的分子との結合に基づく活性を測定する工程
を含む、前記方法。 - 工程(1)において、前記少なくとも1種のポリペプチドが、前記標的分子との結合親和性を有すると見込まれる複数の異なるポリペプチドを含み、かつ工程(2)において、前記複数の異なるポリペプチドの二量体について、前記標的分子との結合に基づく活性を測定することを含む、請求項1又は2に記載の方法。
- 工程(1)が、前記少なくとも1種のポリペプチド又は前記二量体を複数含むライブラリを作製することを含み、又は工程(1)が、前記少なくとも1種のポリペプチド又は前記二量体を複数含むライブラリを用意することを含み、
複数の前記ポリペプチド間で異なる環状ペプチドが含まれている、請求項1~3のいずれか1項に記載の方法。 - 無細胞発現系で複数のポリペプチドを合成することを含む、前記複数のポリペプチド又は前記複数のポリペプチドの二量体を含むライブラリを作製する方法であって、前記複数のポリペプチドが、各々、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含み、前記複数のポリペプチド間で異なる環状ペプチドが含まれている、前記方法。 - 二量体化した環状ペプチドをスクリーニングするためのライブラリであって、複数のポリペプチド若しくは前記複数のポリペプチドの二量体、又は前記複数のポリペプチドをコードする複数の核酸を含み、前記複数のポリペプチドが、各々、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含み、前記複数のポリペプチド間で異なる環状ペプチドが含まれている、前記ライブラリ。 - 8種以上のポリペプチド若しくは前記8種以上のポリペプチドの二量体、又は前記8種以上のポリペプチドをコードする8種以上の核酸を含む、請求項6に記載のライブラリ。
- 複数のポリペプチドの二量体の標的分子との結合に基づく活性を測定する工程を含む、請求項6又は7に記載のライブラリを用いて二量体化した環状ペプチドをスクリーニングする方法。
- 二量体化した環状ペプチドをスクリーニングする方法であって、
(1)複数のディスプレイされたポリペプチドを含むライブラリから、標的分子との結合親和性を有する1つ又は複数のポリペプチドを取得する工程であって、前記複数のディスプレイされたポリペプチドが、各々、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含み、前記複数のディスプレイされたポリペプチド間で異なる環状ペプチドが含まれている、該取得工程、及び
(2)工程(1)において取得した前記ポリペプチドと関連付けられた核酸を回収する工程、
(3)工程(2)において回収した前記核酸を増幅する工程、
(4)工程(3)において増幅した前記核酸を用いて、無細胞発現系で、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含むポリペプチドを合成し、前記ポリペプチドの二量体を調製する工程、及び
(5)前記二量体の標的分子との結合に基づく活性を測定する工程
を含む、前記方法。 - 工程(1)が、取得した複数の前記ポリペプチドに基づき、複数のディスプレイされたポリペプチドを含む更なるライブラリを作製すること、及び前記更なるライブラリから、標的分子との結合親和性を有する1つ又は複数のポリペプチドを取得することを繰り返すことを含む、請求項9に記載の方法。
- 二量体化した環状ペプチドをスクリーニングするためのライブラリであって、複数のディスプレイされたポリペプチド又は前記複数のディスプレイされたポリペプチドを発現させるための複数の核酸を含み、前記複数のディスプレイされたポリペプチドが、各々、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含み、前記複数のディスプレイされたポリペプチド間で異なる環状ペプチドが含まれている、前記ライブラリ。 - 105種以上のディスプレイされたポリペプチド又は前記105種以上のディスプレイされたポリペプチドを発現させるための105種以上の核酸を含む、請求項11に記載のライブラリ。
- 請求項1~4及び8~10のいずれか1項に記載の方法によって標的分子との結合に基づく活性を測定して、標的分子との結合に基づく活性を有する二量体化した環状ペプチドを得る方法。
- 二量体化した環状ペプチドを調製する方法であって、
無細胞発現系で、
環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含むポリペプチドを合成し、前記ポリペプチドの二量体を調製することを含む、前記方法。 - 環状ペプチド、
システイン残基及びロイシンジッパー領域を含む二量体化ドメイン、並びに
前記環状ペプチドと前記二量体化ドメインとの間に存在するペプチドリンカー
を含むポリペプチド。 - 前記二量体化ドメインにおいて、システイン残基の数が1つである、請求項15に記載のポリペプチド。
- 前記ペプチドリンカーが、アラニン(A)、プロリン(P)及びセリン(S)から選択される1つ又は複数のアミノ酸残基からなる、請求項15又は16に記載のポリペプチド。
- 請求項15~17のいずれか1項に記載のポリペプチドをコードする核酸。
Priority Applications (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| EP22742673.1A EP4282873A4 (en) | 2021-01-22 | 2022-01-21 | METHOD FOR SCREENING DIMERIZED CYCLIC PEPTIDES |
| JP2022576754A JPWO2022158554A1 (ja) | 2021-01-22 | 2022-01-21 | |
| US18/356,516 US20240026343A1 (en) | 2021-01-22 | 2023-07-21 | Method for screening dimerized cyclic peptide |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2021008420 | 2021-01-22 | ||
| JP2021-008420 | 2021-01-22 |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| US18/356,516 Continuation US20240026343A1 (en) | 2021-01-22 | 2023-07-21 | Method for screening dimerized cyclic peptide |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO2022158554A1 true WO2022158554A1 (ja) | 2022-07-28 |
Family
ID=82549756
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/JP2022/002117 Ceased WO2022158554A1 (ja) | 2021-01-22 | 2022-01-21 | 二量体化した環状ペプチドをスクリーニングする方法 |
Country Status (4)
| Country | Link |
|---|---|
| US (1) | US20240026343A1 (ja) |
| EP (1) | EP4282873A4 (ja) |
| JP (1) | JPWO2022158554A1 (ja) |
| WO (1) | WO2022158554A1 (ja) |
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2024214800A1 (ja) | 2023-04-11 | 2024-10-17 | 富士フイルム株式会社 | 生理活性評価用のポリペプチド溶液を調製する方法、及びポリペプチドの生理活性を評価する方法 |
Citations (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2000018921A2 (en) | 1998-09-30 | 2000-04-06 | Chiron Corporation | Synthetic peptides having fgf receptor affinity |
| JP2014199261A (ja) * | 2008-10-29 | 2014-10-23 | ビージー メディシン, インコーポレイテッド | ガレクチン−3の免疫アッセイ |
| JP2018172415A (ja) * | 2012-06-07 | 2018-11-08 | プレジデント・アンド・フェロウズ・オブ・ハーバード・カレッジ | 薬物標的指向化のためのナノ療法 |
| JP2019509054A (ja) * | 2016-01-08 | 2019-04-04 | メディトープ バイオサイセンシーズ インク.Meditope Biosciences, Inc. | 自己架橋性抗体 |
| JP2020040886A (ja) * | 2018-09-06 | 2020-03-19 | 国立大学法人 東京大学 | 環状ペプチド |
Family Cites Families (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US5773569A (en) * | 1993-11-19 | 1998-06-30 | Affymax Technologies N.V. | Compounds and peptides that bind to the erythropoietin receptor |
| CA2490411A1 (en) * | 2002-06-28 | 2004-01-08 | Centocor, Inc. | Mammalian epo mimetic ch1 deleted mimetibodies, compositions, methods and uses |
| EP3911662A4 (en) * | 2019-01-14 | 2022-10-26 | New York University | CYCLIC PEPTIDE DIMERS |
| CN110101880A (zh) * | 2019-05-08 | 2019-08-09 | 北京大学 | 一种基于2PisoDGR2多肽的放射性药物及其制备方法 |
-
2022
- 2022-01-21 EP EP22742673.1A patent/EP4282873A4/en active Pending
- 2022-01-21 WO PCT/JP2022/002117 patent/WO2022158554A1/ja not_active Ceased
- 2022-01-21 JP JP2022576754A patent/JPWO2022158554A1/ja active Pending
-
2023
- 2023-07-21 US US18/356,516 patent/US20240026343A1/en active Pending
Patent Citations (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2000018921A2 (en) | 1998-09-30 | 2000-04-06 | Chiron Corporation | Synthetic peptides having fgf receptor affinity |
| JP2014199261A (ja) * | 2008-10-29 | 2014-10-23 | ビージー メディシン, インコーポレイテッド | ガレクチン−3の免疫アッセイ |
| JP2018172415A (ja) * | 2012-06-07 | 2018-11-08 | プレジデント・アンド・フェロウズ・オブ・ハーバード・カレッジ | 薬物標的指向化のためのナノ療法 |
| JP2019509054A (ja) * | 2016-01-08 | 2019-04-04 | メディトープ バイオサイセンシーズ インク.Meditope Biosciences, Inc. | 自己架橋性抗体 |
| JP2020040886A (ja) * | 2018-09-06 | 2020-03-19 | 国立大学法人 東京大学 | 環状ペプチド |
Non-Patent Citations (3)
| Title |
|---|
| JOHNSON, D. L. ET AL., CHEM. BIOL., vol. 4, 1997, pages 939 - 950 |
| M. D. BALLINGER ET AL., NAT. BIOTECHNOL., vol. 17, 1999, pages 1199 - 1204 |
| WU X.-Q., XU L., HECHT N. B.: "Dimerization of the testis brain RNA-binding protein (translin) is mediated through its C-terminus and is required for DNA- and RNA-binding", NUCLEIC ACIDS RESEARCH, OXFORD UNIVERSITY PRESS, GB, vol. 26, no. 7, 1 April 1998 (1998-04-01), GB , pages 1675 - 1680, XP055953108, ISSN: 0305-1048, DOI: 10.1093/nar/26.7.1675 * |
Cited By (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2024214800A1 (ja) | 2023-04-11 | 2024-10-17 | 富士フイルム株式会社 | 生理活性評価用のポリペプチド溶液を調製する方法、及びポリペプチドの生理活性を評価する方法 |
| EP4678650A1 (en) | 2023-04-11 | 2026-01-14 | FUJIFILM Corporation | Method for preparing polypeptide solution for evaluating physiological activity, and method for evaluating physiological activity of polypeptide |
Also Published As
| Publication number | Publication date |
|---|---|
| US20240026343A1 (en) | 2024-01-25 |
| JPWO2022158554A1 (ja) | 2022-07-28 |
| EP4282873A4 (en) | 2024-11-27 |
| EP4282873A1 (en) | 2023-11-29 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| KR102481388B1 (ko) | 내열성의 역전사 효소 돌연변이체 | |
| US5723584A (en) | Biotinylation of proteins | |
| JP2025113367A (ja) | 生理活性を有するペプチドの製造方法及び短鎖リンカーを含むペプチド | |
| AU2013300549B2 (en) | Peptide library and use thereof | |
| TW201835328A (zh) | 用於定序反應之聚合酶 | |
| US11136574B2 (en) | Methods for in vitro ribosome synthesis and evolution | |
| KR20240042497A (ko) | 폴리펩타이드의 제작 방법, 태그, 발현 벡터, 폴리펩타이드의 평가 방법, 핵산 디스플레이 라이브러리의 제작 방법 및 스크리닝 방법 | |
| US20170305988A1 (en) | Method of using a peptide library | |
| CN109689681B (zh) | 源自鲎属的重组蛋白及编码该重组蛋白的dna | |
| JP2019163270A (ja) | 親和性タンパク質及びその使用 | |
| US20240026343A1 (en) | Method for screening dimerized cyclic peptide | |
| CN115850507B (zh) | 一种正交、双组份蛋白质自组装偶联体系及其构建方法 | |
| CN109748970B (zh) | α-酮戊二酸光学探针及其制备方法和应用 | |
| CN110938610B (zh) | 一种转座酶突变体、融合蛋白、其制备方法和应用 | |
| US20220073908A1 (en) | Methods of producing high diversity peptide libraries and promoting protein folding | |
| KR20220023985A (ko) | 효율적인 피피알 단백질의 작제방법 및 그의 이용 | |
| CN121046336A (zh) | 海萤荧光素酶突变体及其应用 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 22742673 Country of ref document: EP Kind code of ref document: A1 |
|
| ENP | Entry into the national phase |
Ref document number: 2022576754 Country of ref document: JP Kind code of ref document: A |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |
|
| ENP | Entry into the national phase |
Ref document number: 2022742673 Country of ref document: EP Effective date: 20230822 |