WO2015137352A1 - Gene related to full-dark heterotrophic ability in cyanobacteria, and use of same - Google Patents
Gene related to full-dark heterotrophic ability in cyanobacteria, and use of same Download PDFInfo
- Publication number
- WO2015137352A1 WO2015137352A1 PCT/JP2015/057046 JP2015057046W WO2015137352A1 WO 2015137352 A1 WO2015137352 A1 WO 2015137352A1 JP 2015057046 W JP2015057046 W JP 2015057046W WO 2015137352 A1 WO2015137352 A1 WO 2015137352A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- protein
- gene
- cyanobacteria
- cytm
- amino acid
- Prior art date
Links
Images








Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/195—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria
Definitions
- the present specification relates to genes associated with complete dark heterotrophic ability in cyanobacteria and uses thereof.
- This application is a related application of Japanese Patent Application No. 2014-046585, which is a Japanese patent application filed on March 10, 2014, and claims the priority based on this Japanese application. Is used.
- Cyanobacteria are also called cyanobacteria (cyanobacterium) and are photoautotrophic bacteria. Utilizing the high photosynthetic ability of cyanobacteria, it is expected to be applied to the production of organic compounds such as biofuels and useful chemicals by genetic modification.
- Cyanobacteria have high photosynthetic ability, but heterotrophic ability in the dark is not necessarily high.
- natural light sources such as daylight are used to allow cyanobacteria to carry out photosynthesis
- the growth rate decreases when there is not enough sunlight, etc. at night or under cloudy weather.
- the production capacity of the substance may decrease.
- the ability to produce useful substances may increase in the dark.
- Synechocystissp. PCC 6803 is known as a standard strain. This cyanobacteria cannot grow when kept in a completely dark place and requires light irradiation for about 10 minutes a day (this phenomenon is also referred to as “Light-ActivatedActivHeterotrophic Growth” (LAHG)).
- LAHG Light-ActivatedActivHeterotrophic Growth
- Leptolyngbya boryana (L. boryana) (also called Plectonema boryanum) as a cyanobacteria.
- L. boryana belongs to the section III group of cyanobacteria, and is a type of cyanobacteria that does not differentiate into cells that specialize in nitrogen fixation called heterocysts.
- Non-patent Document 1 Unlike Synechocystis sp. PCC 6803, L. boryana is known to grow heterotrophically in the presence of sugar such as glucose even in a completely dark place. Moreover, a mutant strain having higher complete dark heterotrophic ability, that is, a faster growth in the dark has been isolated (Non-patent Document 1).
- cyanobacteria do not have or are completely dark heterotrophic like the above-mentioned standard strains, and it is generally difficult to achieve both photoautotrophic and complete dark heterotrophic capacity in cyanobacteria. there were.
- This specification provides a gene related to complete dark heterotrophic ability that can be used to impart complete dark heterotropy in cyanobacteria or to improve the nutrition, and uses thereof.
- the present inventors have identified a mutation in the gene responsible for the mutation from a mutant strain of L. boryana excellent in complete dark heterosexuality. According to this specification, the following means are provided based on such knowledge.
- a cyanobacterial transformant comprising inactivation of a cytM gene encoding cytochrome c M protein.
- the transformant according to (1), wherein the inactivation is the introduction of a mutation encoding the protein in which the cytM gene is disrupted or decreased in activity.
- the transformant according to (1) or (2), wherein the inactivation is a mutation that encodes a protein having the amino acid sequence represented by SEQ ID NO: 14 in the cytM gene.
- the cytM gene is any of the following proteins: (A) a protein having the amino acid sequence represented by SEQ ID NO: 2 (b) an amino acid sequence having 90% or more identity with the amino acid sequence represented by SEQ ID NO: 2 and functionally associated with a cytochrome c M protein Equivalent protein (c) A protein functionally equivalent to cytochrome c M protein, having an amino acid sequence having one or more amino acid substitutions, deletions, insertions or additions in the amino acid sequence represented by SEQ ID NO: 2 (D) a protein having an amino acid sequence encoded by the nucleotide sequence represented by SEQ ID NO: 1 (e) a protein encoded by a nucleotide sequence having 90% or more identity with the nucleotide sequence represented by SEQ ID NO: 1
- the transformant according to any one of (1) to (3), wherein the transformant encodes a protein functionally equivalent to the cytochrome c M protein.
- the transformant according to any one of (1) to (4) which has photosynthetic autotrophic ability and complete dark heterotrophic ability.
- the transformant according to any one of (1) to (5) further comprising a foreign gene for producing a useful substance.
- the cyanobacteria according to (7) which is a cyanobacteria other than the L. boryana dg5 strain.
- the cyanobacteria according to (7) or (8) further comprising a foreign gene for producing a useful substance.
- a transformation vector for imparting complete dark heterotrophic ability to cyanobacteria comprising a polynucleotide that imparts inactivation of the cytM gene encoding cytochrome c M protein.
- cytM gene shows a phylogenetic tree and alignment results with other cytochrome c class I protein cytochrome c M protein closely related encoding. It is a diagram showing a mutation in the gene encoding cytochrome c M protein in L. boryana IAM-M101 dg5 strain. It is a figure which shows the DNA construct for the homologous recombination with respect to the gene which may have a causal variation of complete dark heterotrophic ability. It is a figure which shows the evaluation result (agar medium) of the complete dark place heterotrophic ability after transformation. It is a figure which shows the evaluation result (liquid culture medium) of the complete dark place heterotrophic ability after transformation.
- the disclosure herein relates to genes associated with complete dark heterotrophic capacity in cyanobacteria and uses thereof. According to the disclosure herein, modification of cyanobacteria to provide for inactivation of the cytM gene encoding cytochrome c M protein provides complete darkness to cyanobacteria with photoautotrophic properties against cyanobacteria. It can give a place dependent nutrition or enhance its complete dark place nutrition.
- cyanobacteria use natural light energy such as solar energy and become mixed nutrients with complete dark heterotropy due to organic nutrition. Even in a suppressed state, it can continuously grow and produce useful substances efficiently. In addition, since the modified cyanobacteria can grow and produce useful substances even in the dark, it is possible to efficiently produce useful substances that are produced specifically in the dark.
- cyanobacteria transformant The cyanobacterial transformants disclosed herein can comprise inactivation of the cytM gene encoding cytochrome c M protein (also referred to as Cytochrome c M ).
- Cyanobacteria are gram-negative true bacteria and are prokaryotic photosynthetic microorganisms that perform oxygen-generating photosynthesis. Cyanobacteria herein refers to the ability of heterotrophic ability to grow or proliferate as a carbon source by organic compounds such as sugars, fatty acids and amino acids.
- the word “dark place” includes a complete dark place and a limited dark place.
- the complete dark place means a state in which light is blocked during the growth or multiplication period, except that it is placed under light irradiation for a time and a necessary amount necessary for artificial manipulation for growth or multiplication.
- Light irradiation conditions that are unavoidable for artificial manipulation for growth or proliferation are, for example, conditions in which light irradiation is performed within 10 to 30 minutes, preferably 20 minutes or less once every 10 days to 2 weeks. is there.
- the limited dark place means a state in which light is blocked except that it is placed under light irradiation for about 10 to 20 minutes, preferably about 10 minutes per day.
- Cyanobacteria cannot grow in complete darkness, but may exhibit dependent nutrition in limited darkness. This phenomenon is also called “Light-ActivatedActivHeterotrophic Growth” (LAHG). On the other hand, the case where it is possible to grow heterotrophically in a completely dark place is called complete dark place heterotrophic ability. The cyanobacteria may or may not have such dark heterotrophic ability.
- LAHG Light-ActivatedActivHeterotrophic Growth
- Cyanobacteria are prokaryotes that carry out oxygen-generating photosynthesis, and are not particularly limited, and include, but are not limited to, the order of the order of the order Chroococcus, Pleurocapsa, Euremo, Nemoptera, Stigonema, Groeobacter.
- Synechococcus genus such as Synechococcus elongatus PCC 7942, Prochlorococcus genus such as Prochlorococcus marinus MED4, Anabaena genus such as Anabaena sp. PCC 7120, Synechocystis genus such as Synchocystis sp.
- Gloeobacter genus such as Gloeobacter violaceus PCC 7421
- Arthrospira genus such as Arthrospira platensis
- Trichodesmium genus such as Trichodesmium erythraeum IMS101
- Acaryochloris genus such as Acaryochloris marina MBIC11017.
- CytM gene The cytochrome c M protein encoded by the cytM gene is thought to function as a Cu domain electron donor of cytochrome c oxidase, although its function is not necessarily clear (Bernroitner et al. 2009, Biochim Biophys Acta 1787 ( 3): 135-43).
- L. borissus IAM-M101 nucleotide sequence encoding a cytochrome c M protein cytM gene is represented by SEQ ID NO: 1, The amino acid sequence of the protein shown in SEQ ID NO: 2.
- L. boryana IAM-M101 and its mutant L. boryana IAM-M101 dg5 are a part of the graduate School of Bioagricultural Sciences, Nagoya University (No. 1 Fufu-cho, Chikusa-ku, Nagoya, Aichi, Japan). Is possible.
- L. boryana is a filamentous cyanobacteria. Cyanobacteria are classified into sections I to V because of their morphological characteristics, whereas L. boryana is classified into section III, which has the morphological characteristics of filamentous properties that do not form heterocysts. Section III is further subdivided according to the morphological characteristics of the filamentous body and the presence or absence of gas vesicles, but forms linear filaments (not branched) from cells of equal diameter, without gas vesicles, and between cells L. boryana, which has the characteristic of no constriction, belongs to LPP group B. L. boryana is a type that does not differentiate into cells specialized in nitrogen fixation called heterocysts.
- L. boryana can grow heterotrophically in the presence of sugar such as glucose even in a completely dark place (complete dark place heterotrophic ability).
- L. ⁇ boryana can introduce DNA into cells by electroporation and obtain mutants in which a specific gene is destroyed by homologous recombination with the genome (Fujita et al. 1992, Plant Cell Physiology). , 33: 81-92).
- a gene encoding a protein having the following amino acid sequence is known.
- SEQ ID NOs: 3 to 12 are cytochrome cM proteins.
- SEQ ID NO: 13 is a cytochrome c class I protein, but shows only about 30% identity (%) with the amino acid sequence represented by SEQ ID NO: 2, and is functionally considered as a cytochrome c6 protein.
- the cytM gene to be inactivated in the present specification has an amino acid sequence having a certain identity with the amino acid sequence of a known cytochrome c M protein in addition to those already disclosed in this way, and the cytochrome c M protein It may encode proteins that are functionally equivalent.
- Such a gene to be inactivated preferably has, for example, 60% or more identity with the amino acid sequence represented by SEQ ID NO: 2, more preferably 65% or more identity, 70% or more identity, preferably 75% or more, more preferably 80% or more, still more preferably 85% or more, more preferably 90% or more, still more preferably 95% or more, still more preferably 97% or more, More preferably, it has 98% or more, and most preferably 99% or more identity.
- the cytM gene may be a protein encoding a protein that, when aligned, comprises a -NCXXCH-motif and a -TPPMP-motif.
- These two motifs are commonly observed in cytochrome c M protein in this specification (Bernroitner et al Biochem Biophys Acta, 2009, 1787:... 135-143).
- these motifs correspond to positions 36 to 41 and positions 74 to 78 in the amino acid sequence represented by SEQ ID NO: 2, respectively.
- CytM gene for example, when the amino acid sequence alignment of SEQ ID NO: 2, found these motifs at positions corresponding to these, moreover, cytochrome c M protein functionally equivalent to a protein that is a May be encoded.
- FIG. 1 shows the phylogenetic tree and alignment results for cytochrome c M protein encoded by the cytM gene and other related cytochrome c class I proteins.
- identity in an amino acid sequence or a base sequence is used to mean the degree of coincidence of amino acid residues or bases constituting each sequence between sequences to be compared.
- amino acid sequence the existence of a gap and the nature of the amino acid are considered (Wilbur, Proc. Natl. Acad. Sci. U.S.A. 80: 726-730 (1983)).
- commercially available softwares such as BLAST (Altschul: J. Mol. Biol. 215: 403-410 (1990)), FASTA (Peasron: Methods in Enzymology 183: 63-69 (1990)), etc. Can be used.
- identity may be a numerical value calculated using a homology search program known to those skilled in the art.
- BLAST BasicBlocal alignment of the National Center for Biotechnology Information (NCBI) search) http://www.ncbi.nlm.nih.gov/BLAST/, using default (initial setting) parameters.
- Functionally equivalent to cytochrome c M protein means maintaining substantially the same level of function to express or enhance complete dark heterotrophic capacity.
- the functional equivalence is determined by comparing the gene-disrupted cyanobacteria that have disrupted the gene in cyanobacteria with the non-disrupted cyanobacteria, where the complete dark heterotrophic ability is expressed or enhanced, or limited darkness. This can be determined by the fact that the place's heterotrophic capacity is enhanced to become complete dark place heterotrophic. That is, in gene-disrupted cyanobacteria, when the complete dark heterotrophic ability is expressed and enhanced or is completely dark heterotrophic, the protein encoded by the gene is functionally equivalent to the cytochrome c M protein. Can be determined. In addition, the gene can be determined to be functionally equivalent to the cytM gene.
- CytM gene the amino acid sequences of known cytochrome c M protein, 1 or insertion of a plurality of amino acids, substitutions, cytochrome c M protein functionally equivalent have deletions and additions in the amino acid sequence It may encode a protein.
- Such proteins have, for example, 1 to 30, preferably 1 to 20, more preferably 1 to 10, more preferably 1 to 5, more preferably 1 to 2 amino acid insertions, substitutions and deletions. And additions can be provided.
- Such a protein has an amino acid sequence having a conservative substitution of one or several, preferably 1 to 10, more preferably 1 to 5, more preferably about 1 to 2 amino acids in the known cytochrome c M protein. It is preferable that
- “conservative substitution” means substitution of one or more amino acid residues with another chemically similar amino acid residue so as not to substantially alter the function of the protein. For example, when a certain hydrophobic residue is substituted by another hydrophobic residue, a certain polar residue is substituted by another polar residue having the same charge, and the like. Functionally similar amino acids that can make such substitutions are known in the art for each amino acid. Specific examples include non-polar (hydrophobic) amino acids such as alanine, valine, isoleucine, leucine, proline, tryptophan, phenylalanine, and methionine.
- Examples of polar (neutral) amino acids include glycine, serine, threonine, tyrosine, glutamine, asparagine, and cysteine.
- Examples of positively charged (basic) amino acids include arginine, histidine, and lysine.
- Examples of negatively charged (acidic) amino acids include aspartic acid and glutamic acid.
- the cytM gene is a complementary DNA that hybridizes under stringent conditions with a DNA encoding a known cytochrome c M protein, and encodes a protein functionally equivalent to the cytochrome c M protein. It may be a thing.
- “Stringent conditions” means that the reaction is carried out at a temperature of 40 ° C. to 70 ° C., preferably 60 ° C. to 65 ° C. in a hybridization buffer that can be usually used by those skilled in the art. It can be carried out according to a method of washing in a washing solution of 300 mmol / L, preferably 15 to 60 mmol / L. The temperature and salt concentration can be appropriately adjusted according to the length of the probe used. Furthermore, the conditions for washing the hybridized material can be 0.2 or 2 ⁇ SSC, 0.1% SDS, and a temperature of 20 ° C. to 68 ° C.
- a difference can be made depending on the salt concentration or temperature at the time of washing.
- 0.2 ⁇ SSC as a stringent wash buffer high ⁇ ⁇ ⁇ ⁇ ⁇ stringency wash buffer
- 0.1% SDS 0.1% SDS
- 2 ⁇ SSC as a mild washbuffer (low stringency wash buffer)
- 0.1% SDS 0.1% SDS.
- 68 ° C for stringent, 42 ° C for moderate-stringency, and room temperature (20 ° C to 25 ° C) for mild May be performed with 0.2 ⁇ SSC and 0.1% SDS.
- the cytM gene may have a base sequence having a certain identity with a base sequence encoding a known cytochrome c M protein, and may encode a protein that is functionally equivalent to the cytochrome c M protein.
- Such a gene to be inactivated preferably has, for example, 60% or more identity with the nucleotide sequence represented by SEQ ID NO: 1 encoding a known cytochrome c M protein, more preferably 65% or more. More preferably 70% or more, more preferably 75% or more, more preferably 80% or more, still more preferably 85% or more, more preferably 90% or more, and still more preferably 95%. More preferably, it has an identity of 97% or more, more preferably 98% or more, and most preferably 99% or more.
- the insertion, substitution or deletion of one or a plurality of amino acids, or addition to one or both ends thereof in the amino acid sequence is a well-known site-directed mutagenesis method or the like. Or by substitution of a plurality of amino acids to the extent that can occur naturally.
- inactivation of cytM gene It has inactivation of the cytM gene in cyanobacteria.
- the inactivation of the gene may be anything as long as the activity of the protein encoded by the gene is reduced or lost as a result. Therefore, gene inactivation includes gene disruption, transcription inhibition, transcript degradation, translation inhibition, translation of inactive proteins, and the like.
- Such gene inactivation is generally performed in the form of gene disruption, for example, insertion of foreign sequences into the cytM gene, replacement of all or part of the cytM gene by foreign sequences, cytM gene Can be done by deletion of all or part of The number of bases in the foreign sequence and the positions of substitutions, deletions and insertions are not particularly limited as long as the expression and activity of cytochrome c M protein is substantially lost.
- the foreign sequence is preferably a selection marker gene.
- the selectable marker gene can be appropriately selected from known ones and is preferably a drug resistance gene.
- the cytM gene can be disrupted preferably by replacing all or part of the cytM gene with a foreign gene.
- the part of cytM gene to be replaced for example, the form to replace the C-terminal side of the cytochrome c M protein.
- a protein with reduced activity may be produced by introducing a mutation into the protein encoded by the cytM gene.
- a mutation may be a frameshift mutation by inserting or deleting a base in the coding sequence, or a part of the protein may be substituted with a different amino acid, or a combination thereof. It may be a thing.
- Gene disruption techniques are known to those skilled in the art, and those skilled in the art can perform gene disruption according to known methods.
- Gene disruption generally involves introducing into a cell a polynucleotide (preferably a selectable marker gene) in which the nucleotide sequence of the target gene has been modified, and causing homologous recombination between the introduced polynucleotide and the target gene. By selecting cells in which homologous recombination has occurred, mutations can be introduced into the target gene.
- a polynucleotide preferably a selectable marker gene
- cytM gene in order to inactivate the cytM gene, for example, it may be a variant which encodes a protein consisting of the amino acid represented by the cytochrome c M protein in SEQ ID NO: 14.
- the transformant disclosed in this specification is a cyanobacteria equipped with inactivation of the cytM gene.
- inactivation of the cytM gene can be imparted, and dark heterotrophic ability can be imparted or enhanced to the cyanobacteria.
- a transformation vector comprising a polynucleotide for imparting inactivation of various forms of the cytM gene already described can be used.
- Polynucleotides are generally DNA and RNA, but are typically DNA.
- This transformation vector can be used as a vector for imparting dark heterotrophic ability to cyanobacteria.
- the above-described polynucleotide (for example, a polynucleotide cassette having at least a homology region for destroying and replacing the target cytM gene) is itself a dark place for imparting dark heterotrophic ability to cyanobacteria. It can be used as a heterotrophic agent.
- the polynucleotide on the transformation vector has, for example, an appropriate homologous recombination region capable of disrupting one or more cytM genes on the chromosome of cyanobacteria or introducing mutations, etc. for example, with or substituted region encoding a foreign gene or the like, a replacement area) for the mutation for the production of drug resistance marker region and inactive cytochrome c M protein.
- the transformation vector may further contain one or more enhancers or silencers, an operator, a terminator, a polyadenylation signal, one or more drug resistance genes, a cloning site for inserting a foreign gene, and the like.
- Drug resistance genes include, but are not limited to, chloramphenicol resistance gene, neomycin resistance gene, hygromycin resistance gene, kanamycin resistance gene, ampicillin resistance gene, spectinomycin resistance gene and the like.
- the transformation vector can take various forms without particularly specifying the form. Although it may be a DNA fragment, it is typically a plasmid vector, but for example, non-plasmid vectors such as viral vectors can also be used. Also preferably, the transformation vector disclosed herein is a shuttle vector for adapting to multiple host cells. For example, vectors that can be introduced into any cell of E. coli and cyanobacteria and can express foreign genes in these host cells can be mentioned. Such shuttle vectors can be replicated in large quantities using E. coli.
- the origin of the parent vector of the transformation vector disclosed in the present specification is not particularly limited as long as the gene can be expressed efficiently with cyanobacteria.
- pVZ321 Zinchenko VV, Piven IV, Melnik VA, Shestakov SV (1999) Genetika 35: 291-296
- pMB1 Kreps S, Ferino F, Mosrin C, Mergeay C, Mergeay
- vectors used in cyanobacteria Known vectors such as M, Thuriaux1990P (1990) Mol. Gen. Genet.133221: 129-133), RSF1010 (Meyer R (2009) Plasmid 62: 57-70) can be used.
- a ligase reaction or the like can be used as appropriate.
- the transformation vector may further comprise a foreign gene to be expressed in cyanobacteria.
- the foreign gene may be a gene introduced from the outside by a transformation vector or the like, and may be a gene endogenous to cyanobacteria.
- the foreign gene may encode various useful substances or one or more enzymes for producing the useful substances.
- proteins useful as pharmaceutical ingredients eg, interferon, insulin, erythropoietin, human growth hormone, various cytokines
- proteins useful as reagents eg, DNA / RNA polymerase, protease, restriction enzyme
- fluorescent proteins eg, GFP, DsRed, Phycocyanin
- bioplastic production related proteins eg polyhydroxyalkanoic acid (PHA), polyhydroxybutyric acid (PHB) or polyhydroxyvaleric acid (PHV) synthase
- amino acid synthases eg polyhydroxyalkanoic acid (PHA), polyhydroxybutyric acid (PHB) or polyhydroxyvaleric acid (PHV) synthase
- biofuel related synthases eg triglycerides, fatty acids
- hydrocarbon alkane / alkene
- thioesterase gene for fatty acid production structural gene of nitrogenase, nifE, nifN, nifH, nifU, nifS, nifB, nifP, nifM nifV, nifZ, nifX, nifW, hesA, hesB, fdxN, fdxH
- Examples include genes involved in metal-center biosynthesis of nitrogenase, hydrogenase structural genes, and genes involved in metal-center biosynthesis of hydrogenases such as hydE and hydG.
- a gene encoding such a useful substance is a known method such as a PCR method or a hybridization method using a primer or probe designed from information on the base sequence or amino acid sequence registered in an existing database such as GenBank. It can be obtained by.
- a cyanobacterial transformant can be obtained by using a known transformation method for cyanobacteria using the transformation vector disclosed herein for cyanobacteria.
- a transformation method of cyanobacteria for example, natural transformation method and electroporation method can be used. Further, a junction transmission method can be used.
- the natural transformation method is a method in which gene introduction is possible only by mixing cyanobacteria and a transformant vector.
- the electroporation method is a method for introducing a gene by applying an electric pulse to a mixed solution of cyanobacteria and a transformant vector.
- the conjugation transfer method is a method in which a plasmid into which a gene to be introduced is inserted is transformed into E. coli, and then the E. coli and cyanobacteria are mixed to introduce the gene. Those skilled in the art can appropriately implement these methods. Moreover, in order to select a transformant, the drug resistance gene etc. which were hold
- the thus obtained transformant, cyanobacteria can have the inherent photosynthetic autotrophic ability and the dark heterotrophic ability imparted for the first time or the enhanced dark heterotrophic ability.
- the presence or degree of heterotrophic ability in the dark can be determined by observing growth or proliferation when transformants are cultured under heterotrophic conditions using a medium containing organic nutrients such as glucose in the dark. Judgment can be made.
- the transformant of the present disclosure Since the transformant of the present disclosure has imparted or enhanced dark heterotrophic ability, it uses natural light energy such as sunlight and is in a depleted state (dark place) of the light energy. Because it can grow and proliferate in a heterotrophic manner, continuous growth and material production are possible throughout the day and night. That is, efficient and practical growth and material production are possible. In addition, the substances to be produced can be made different by growing them only in the dark dependent conditions or only in the presence of light energy as necessary.
- the transformant of the present disclosure can further include a foreign gene for producing a useful substance. According to such a transformant, a useful substance can be efficiently and / or selectively produced using light energy and an organic compound.
- a cyanobacteria that is not a transformant but has inactivation of the cytM gene and has dark heterotrophic ability is also provided. This is because screening for cyanobacteria based on the inactivation or absence of the cytM gene makes it possible to easily select cyanobacteria having excellent dark heterotrophic ability from natural cyanobacteria. As natural cyanobacteria, it is preferable to target cyanobacteria other than L. boryana. Moreover, a practical cyanobacteria can be obtained by introducing and transforming a foreign gene for producing a useful substance into the thus screened cyanobacteria according to the above method.
- a method for producing a culture product of cyanobacteria is also provided.
- This production method may comprise a step of culturing a cyanobacteria or cyanobacterial transformant provided with inactivation of the cytM gene disclosed in the present specification.
- cyanobacteria since the dark heterotrophic ability is imparted or enhanced, cyanobacteria can be cultured continuously, efficiently, or selectively, and the culture product can be obtained.
- This production method preferably includes a culture step under heterotrophic conditions in the dark particularly because the complete dark heterotrophic ability is imparted / enhanced or modified in the complete dark heterotrophic capacity.
- the culture product can include cultured cells and culture supernatant, but may be a part thereof.
- the useful substance is also included in the culture product.
- culture process natural light energy conditions such as solar energy, heterotrophic conditions and the like can be appropriately combined.
- culture conditions known culture conditions for cyanobacteria can be employed.
- a method for culturing cyanobacteria is also provided.
- This culture method can comprise a step of culturing cyanobacteria or cyanobacteria transformants having inactivation of the cytM gene in the dark.
- cyanobacteria can be grown and propagated without light energy.
- cyanobacteria can grow and proliferate even under autotrophic conditions under light energy, enabling efficient culture.
- the genome sequence of the wild strain was determined as described below, and compared with a known dg5 genome sequence.
- Genomic DNA was extracted using the Wizard Genomic DNA Purification kit (Promega) as shown below and partially modified from the manual. About 20 mg of WTo cells were suspended in 450 ⁇ l of a buffer solution (50 mM Tris-HCl pH 8.0, 50 mM NaCl, 5 mM NaCl, EDTA), and 50 ⁇ L of lysozyme (2.5 mg / 50 ⁇ l) was added. Then, it left still at 37 degreeC for 1 hour. The sample was centrifuged (15,000 rpm, 2 minutes, room temperature), and the supernatant was removed. From the procedure of adding NucleileLysis Solution 0.6 ml to the subsequent precipitate, the procedure was followed according to the Wizard Genomic DNA Purification kit (Promega) manual.
- a buffer solution 50 mM Tris-HCl pH 8.0, 50 mM NaCl, 5 mM NaCl, EDTA
- 50 ⁇ L of lysozyme 2.5 mg / 50 ⁇ l
- SOLiD system 5500 As a genome sequencing method, one of the high-speed sequencers, SOLiD system 5500, was used. A paired-end genomic library was created according to the manual and sequenced. As a result, sequence data from both sides of 4,802,686 library DNA fragments were obtained. At this time, as a control, sequence data was obtained using SOLiD system 5500 in the same manner as the genomic sequence of dg5. The data was mapped using the analysis software BioScope® (Applied® Biosystems) with the dg5 genomic sequence as a reference, and then single nucleotide polymorphism (SNP) and insertion deletion (Indel) analysis was performed. The initial setting was used for the setting at that time.
- SNP single nucleotide polymorphism
- Indel insertion deletion
- FIG. 2 shows an overview of mutations at dg5 in the cytM gene. It was found that truncation of the protein was caused by the frameshift mutation.
- the kanamycin resistance cassette was amplified using primer-f2 and primer-r2 using pUCK121 as a template. Each fragment has the same sequence at the end of 30 bp between the fragments to be ligated by base addition to the primer used for amplification. By performing secondary PCR using primer-f1 and primer-r3 using these three fragments as templates, the target DNA fragment in which the three fragments were ligated was obtained. Primer-f1 and primer-r3 are designed so that this fragment has a restriction enzyme site. The restriction enzyme was treated with BamHI or BamHI and KpnI, and cloned into the corresponding cloning site of pUC19.
- a plasmid in which only the drug resistance marker not having each dg5 mutation was inserted was also prepared.
- PCR was performed using the WTo genome instead of the dg5 genome as a template at the stage of PCR amplification of only the three DNA fragments that retained the dg5 mutation, and the others followed the procedure for constructing the plasmid described above. .
- Double-homologous recombination into WTo was performed by electroporation (Fujita et al. 1992, Plant Cell Physiology, 33: 81-92).
- PCR fragments obtained using primer-f1 and primer-r3 using the prepared plasmid as a template were used.
- an agar medium containing BG-11BG (20 M HEPES-KOH; containing pH 7.5, hereinafter referred to as BG-11H) and agar (BactoAgar, Difco) 15% (w / v) was used.
- a white fluorescent lamp FLR40SW / M / 36-B, Sakai Hitachi
- WTo cells that have been photosynthesised on agar at 30 ° C are collected in water, aspirated on 150 mL Bottle Top Vacuum Filter, 0.22 ⁇ m Pore (Corning), resuspended in sterile cold water, and then aspirated. The water was removed. The salt was removed from the cell suspension by performing this operation three times in total. The cells were resuspended in 0.5-2.0 ml of chilled sterilized water and used as a competent cell for electroporation.
- a 50 ⁇ l competent cell and 10 ⁇ l of each PCR fragment were mixed and subjected to an attenuation pulse treatment at a voltage of 1410 V, a resistance of 250 ⁇ , a capacitance of 25 ⁇ F, and a distance between electrodes of 1 mm.
- the pulsed cells 350 [mu] l of BG-11 (20 mM HEPES- KOH; pH8.2 containing) was recovered by the addition of a nylon membrane (Amersham Hybond TM -N +, GE Healthcare) on an agar medium BG-11H bearing the Spread out.
- the individual dg5 mutant strains selected as kanamycin resistant strains were dm1 (having Chr; 1305262 mutation), dm2 (having Chr; 3151287), dm3 (having Chr; 5175165), dm3n (Chr; 5175165 And a gene (ORF No. LBDG_45050) completely deficient strain).
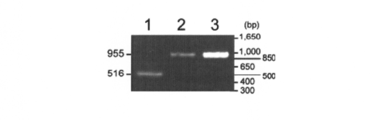
- DNA fragments were directly amplified from the cells by PCR using primer-f1 and primer-r3 shown in Table 3. At the same time, it was confirmed that the kanamycin resistance cassette was inserted into the genome by double recombination.
- the DNA fragment was purified from an agarose gel and used as a template, Chr; 1305262 was a combination of Pb1305262-f and primer-r2, Chr; 3151287 was a combination of Pb3151287-f and primer-r2, Chr: 5175165 In contrast, secondary PCR was performed with a combination of primer-f2 and Pb5175165-r, and the resulting DNA fragment was sequenced by the Sanger method to confirm the introduction of the dg5 mutation.
- the primers used for sequencing by the Sanger method are Pb1305262-f, Pb3151287-f, and Pb5175165-r shown in Table 1.
- 15 ml BG-11 medium with a final concentration of 30 mM glucose was added to 25 cm 2 Tissue Culture Flask (IWAKI) shielded from aluminum foil, and cultured with shaking at a rotational speed of 140 rpm.
- the photosynthetic growth in the light place is not significantly different in all strains regardless of the presence or absence of glucose, but in the dark place, only dm3 and dm3n are remarkably good like dg5. Showed growth. This is presumed that the mutation (single nucleotide insertion) of Chr; 5175165 to dg5 caused a frame shift in LBDG_45050 and caused gene deletion. LBDG_45050 the result of homology search, it was found that encodes a cytochrome c M. Cytochrome c M deficiency is a causal mutation that promotes dg5's complete dark heterotrophic viability.
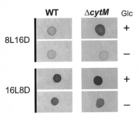
- a cytochrome cM-deficient strain of cyanobacteria (Synechocystis sp. PCC 6803) was produced and evaluated for improvement in heterotrophic growth. That is, as shown in FIG. 6, the ⁇ cytM strain was isolated by replacing the entire coding region of cytM (sll1245) with a gentamicin resistance cassette. In FIG. 6, the black bar indicates the genomic region used for the plasmid constructed for isolation of ⁇ cytM.
- ⁇ cytM was not different from WT under the photosynthetic condition (lane 1), but growth was significantly promoted under WT in the mixed nutrient condition with glucose (lane 2). Furthermore, the difference was most noticeable under LAHG conditions (lane 3).
- the WT (“YF”) used in this study is a strain that does not grow well under LAHG conditions. Furthermore, under dark heterotrophic conditions (lane 4), WT did not grow at all, but ⁇ cytM showed clear growth.
Landscapes
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Health & Medical Sciences (AREA)
- General Health & Medical Sciences (AREA)
- Biochemistry (AREA)
- Biophysics (AREA)
- Life Sciences & Earth Sciences (AREA)
- Genetics & Genomics (AREA)
- Medicinal Chemistry (AREA)
- Molecular Biology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Gastroenterology & Hepatology (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
Abstract
[Problem] To provide a gene related to full-dark heterotrophic ability that can be used to impart full-dark heterotrophic ability and to improve this trophic function in Cyanobacteria, and the use of this gene. [Solution] Cyanobacteria are transformed so as to inactivate the cytM gene that encodes cytochrome cM protein. Inactivation of this gene imparts or potentiates full-dark heterotrophic ability in Cyanobacteria.
Description
本明細書は、シアノバクテリアにおける完全暗所従属栄養能に関連する遺伝子及びその利用に関する。
本出願は、2014年3月10日に出願された日本国特許出願である特願2014-046585の関連出願であり、この日本出願に基づく優先権を主張するものであり、その全ての記載内容を援用するものである。 The present specification relates to genes associated with complete dark heterotrophic ability in cyanobacteria and uses thereof.
This application is a related application of Japanese Patent Application No. 2014-046585, which is a Japanese patent application filed on March 10, 2014, and claims the priority based on this Japanese application. Is used.
本出願は、2014年3月10日に出願された日本国特許出願である特願2014-046585の関連出願であり、この日本出願に基づく優先権を主張するものであり、その全ての記載内容を援用するものである。 The present specification relates to genes associated with complete dark heterotrophic ability in cyanobacteria and uses thereof.
This application is a related application of Japanese Patent Application No. 2014-046585, which is a Japanese patent application filed on March 10, 2014, and claims the priority based on this Japanese application. Is used.
シアノバクテリアは、藍藻(ラン藻)類とも称され、光合成独立栄養細菌である。シアノバクテリアの高い光合成能力を利用し、遺伝子改変によりバイオ燃料や有用化学物質などの有機化合物生産への適用が期待されている。
Cyanobacteria are also called cyanobacteria (cyanobacterium) and are photoautotrophic bacteria. Utilizing the high photosynthetic ability of cyanobacteria, it is expected to be applied to the production of organic compounds such as biofuels and useful chemicals by genetic modification.
シアノバクテリアは、高い光合成能力を有する一方、暗所での従属栄養能が必ずしも高くない。日中の太陽光などの天然光源を利用してシアノバクテリアに光合成を行わせようとするとき、夜間や曇天下で十分な太陽光等がないときには増殖速度が低下し、その結果、全体として有用物質の生産能が低下する可能性がある。その反面、むしろ、有用物質生産能が暗所において上昇する場合もある。
Cyanobacteria have high photosynthetic ability, but heterotrophic ability in the dark is not necessarily high. When natural light sources such as daylight are used to allow cyanobacteria to carry out photosynthesis, the growth rate decreases when there is not enough sunlight, etc. at night or under cloudy weather. There is a possibility that the production capacity of the substance may decrease. On the other hand, rather, the ability to produce useful substances may increase in the dark.
シアノバクテリアとしては、Synechocystissp. PCC 6803が標準株として知られている。このシアノバクテリアは、完全に暗所に保つと生育できず、1日10分程度の光照射を要求する(この現象を、"Light-Activated Heterotrophic Growth"(LAHG)ともいう。)。一方、シアノバクテリアとしてLeptolyngbya boryana(L. boryana)(Plectonema boryanumともいう。)がある。L. boryanaは、シアノバクテリアの分類ではセクションIIIのグループに属しており、形態的には分岐していない糸状性で、ヘテロシストと呼ばれる窒素固定専門の細胞を分化しないタイプのシアノバクテリアである。L. boryanaは、Synechocystis sp. PCC 6803とは異なり、完全な暗所であってもグルコースなどの糖があれば従属栄養的に生育することが知られている。また、より完全暗所従属栄養能のより高い、すなわち、暗所でより早く増殖する変異株も単離されている(非特許文献1)。
As a cyanobacteria, Synechocystissp. PCC 6803 is known as a standard strain. This cyanobacteria cannot grow when kept in a completely dark place and requires light irradiation for about 10 minutes a day (this phenomenon is also referred to as “Light-ActivatedActivHeterotrophic Growth” (LAHG)). On the other hand, there is Leptolyngbya boryana (L. boryana) (also called Plectonema boryanum) as a cyanobacteria. L. boryana belongs to the section III group of cyanobacteria, and is a type of cyanobacteria that does not differentiate into cells that specialize in nitrogen fixation called heterocysts. Unlike Synechocystis sp. PCC 6803, L. boryana is known to grow heterotrophically in the presence of sugar such as glucose even in a completely dark place. Moreover, a mutant strain having higher complete dark heterotrophic ability, that is, a faster growth in the dark has been isolated (Non-patent Document 1).
多くのシアノバクテリアは、上記標準株と同様に完全暗所従属栄養能を有していないか低く、シアノバクテリアにおいて光合成独立栄養能と完全暗所従属栄養能との両立は一般的には困難であった。
Many cyanobacteria do not have or are completely dark heterotrophic like the above-mentioned standard strains, and it is generally difficult to achieve both photoautotrophic and complete dark heterotrophic capacity in cyanobacteria. there were.
本明細書は、シアノバクテリアにおいて完全暗所従属栄養能を付与したり当該栄養能を向上させることができるのに用いることができる完全暗所従属栄養能に関連する遺伝子及びその利用を提供する。
This specification provides a gene related to complete dark heterotrophic ability that can be used to impart complete dark heterotropy in cyanobacteria or to improve the nutrition, and uses thereof.
本発明者らは、完全暗所従属栄養能に優れるL. boryanaの変異株から当該変異の原因となる遺伝子における変異を特定した。本明細書によれば、こうした知見に基づき以下の手段が提供される。
The present inventors have identified a mutation in the gene responsible for the mutation from a mutant strain of L. boryana excellent in complete dark heterosexuality. According to this specification, the following means are provided based on such knowledge.
(1)シトクロムcMタンパク質をコードするcytM遺伝子の不活性化を備える、シアノバクテリア形質転換体。
(2)前記不活性化は、前記cytM遺伝子の破壊又は活性が低下した前記タンパク質をコードする変異の導入である、(1)に記載の形質転換体。
(3)前記不活性化は、前記cytM遺伝子が配列番号14で表されるアミノ酸配列からなるタンパク質をコードする変異である、(1)又は(2)に記載の形質転換体。
(4)前記cytM遺伝子は、以下のいずれかに記載のタンパク質;
(a)配列番号2で表されるアミノ酸配列を有するタンパク質
(b)配列番号2で表されるアミノ酸配列と90%以上の同一性を有するアミノ酸配列を有し、シトクロムcMタンパク質と機能的に同等なタンパク質
(c)配列番号2で表されるアミノ酸配列において1又は複数個のアミノ酸の置換、欠失、挿入又は付加を有するアミノ酸配列を有し、シトクロムcMタンパク質と機能的に同等なタンパク質
(d)配列番号1で表される塩基配列でコードされるアミノ酸配列を有するタンパク質
(e)配列番号1で表される塩基配列と90%以上の同一性を有する塩基配列によってコードされるタンパク質であって、シトクロムcMタンパク質と機能的に同等なタンパク質をコードする、(1)~(3)のいずれかに記載の形質転換体。
(5)光合成独立栄養能と完全暗所従属栄養能とを備える、(1)~(4)のいずれかに記載の形質転換体。
(6)さらに、有用物質を生産するための外来遺伝子を備える、(1)~(5)のいずれかに記載の形質転換体。
(7)シトクロムcMタンパク質をコードするcytM遺伝子の不活性化を備え、完全暗所従属栄養能を有する、シアノバクテリア。
(8)L. boryana dg5株以外のシアノバクテリアである、(7)に記載のシアノバクテリア。
(9)さらに、有用物質を生産するための外来遺伝子を備える、(7)又は(8)に記載のシアノバクテリア。
(10)シトクロムcMタンパク質をコードするcytM遺伝子の不活性化を付与するポリヌクレオチドを含む、シアノバクテリアに完全暗所従属栄養能を付与するための形質転換ベクター。
(11)前記不活性化は、前記cytM遺伝子の破壊又は活性が低下した前記タンパク質をコードする変異の導入である、(10)に記載の形質転換ベクター。
(12)(1)~(6)のいずれかに記載の形質転換体の製造方法であって、
シアノバクテリアに、シトクロムcMタンパク質をコードするcytM遺伝子の不活性化を付与するポリヌクレオチドで形質転換する工程、
を備える、製造方法。
(13)前記不活性化は、前記cytM遺伝子の破壊又は活性が低下した前記タンパク質をコードする変異の導入である、(12)に記載の製造方法。
(14)前記形質転換工程は、前記ポリヌクレオチドをエレクトロポレーションで前記シアノバクテリアに導入する、(12)又は(13)に記載の製造方法。
(15)(1)~(6)のいずれかに記載の形質転換体を培養する工程、
を備える、シアノバクテリアの培養産物の生産方法。
(16)前記培養工程は、暗所での培養を含む、(15)に記載の生産方法。
(17)(1)~(6)のいずれかに記載の形質転換体を暗所で培養する工程、
を備える、シアノバクテリアの培養方法。 (1) A cyanobacterial transformant comprising inactivation of a cytM gene encoding cytochrome c M protein.
(2) The transformant according to (1), wherein the inactivation is the introduction of a mutation encoding the protein in which the cytM gene is disrupted or decreased in activity.
(3) The transformant according to (1) or (2), wherein the inactivation is a mutation that encodes a protein having the amino acid sequence represented by SEQ ID NO: 14 in the cytM gene.
(4) The cytM gene is any of the following proteins:
(A) a protein having the amino acid sequence represented by SEQ ID NO: 2 (b) an amino acid sequence having 90% or more identity with the amino acid sequence represented by SEQ ID NO: 2 and functionally associated with a cytochrome c M protein Equivalent protein (c) A protein functionally equivalent to cytochrome c M protein, having an amino acid sequence having one or more amino acid substitutions, deletions, insertions or additions in the amino acid sequence represented by SEQ ID NO: 2 (D) a protein having an amino acid sequence encoded by the nucleotide sequence represented by SEQ ID NO: 1 (e) a protein encoded by a nucleotide sequence having 90% or more identity with the nucleotide sequence represented by SEQ ID NO: 1 The transformant according to any one of (1) to (3), wherein the transformant encodes a protein functionally equivalent to the cytochrome c M protein.
(5) The transformant according to any one of (1) to (4), which has photosynthetic autotrophic ability and complete dark heterotrophic ability.
(6) The transformant according to any one of (1) to (5), further comprising a foreign gene for producing a useful substance.
(7) A cyanobacteria having inactivation of the cytM gene encoding cytochrome c M protein and having complete dark heterotrophic ability.
(8) The cyanobacteria according to (7), which is a cyanobacteria other than the L. boryana dg5 strain.
(9) The cyanobacteria according to (7) or (8), further comprising a foreign gene for producing a useful substance.
(10) A transformation vector for imparting complete dark heterotrophic ability to cyanobacteria, comprising a polynucleotide that imparts inactivation of the cytM gene encoding cytochrome c M protein.
(11) The transformation vector according to (10), wherein the inactivation is introduction of a mutation encoding the protein in which the cytM gene is disrupted or decreased in activity.
(12) A method for producing the transformant according to any one of (1) to (6),
Transforming cyanobacteria with a polynucleotide conferring inactivation of the cytM gene encoding cytochrome c M protein;
A manufacturing method comprising:
(13) The production method according to (12), wherein the inactivation is introduction of a mutation encoding the protein in which the cytM gene is disrupted or decreased in activity.
(14) The production method according to (12) or (13), wherein the transformation step introduces the polynucleotide into the cyanobacteria by electroporation.
(15) culturing the transformant according to any one of (1) to (6),
A method for producing a cyanobacterial culture product.
(16) The production method according to (15), wherein the culturing step includes culturing in a dark place.
(17) culturing the transformant according to any one of (1) to (6) in the dark,
A method for culturing cyanobacteria, comprising:
(2)前記不活性化は、前記cytM遺伝子の破壊又は活性が低下した前記タンパク質をコードする変異の導入である、(1)に記載の形質転換体。
(3)前記不活性化は、前記cytM遺伝子が配列番号14で表されるアミノ酸配列からなるタンパク質をコードする変異である、(1)又は(2)に記載の形質転換体。
(4)前記cytM遺伝子は、以下のいずれかに記載のタンパク質;
(a)配列番号2で表されるアミノ酸配列を有するタンパク質
(b)配列番号2で表されるアミノ酸配列と90%以上の同一性を有するアミノ酸配列を有し、シトクロムcMタンパク質と機能的に同等なタンパク質
(c)配列番号2で表されるアミノ酸配列において1又は複数個のアミノ酸の置換、欠失、挿入又は付加を有するアミノ酸配列を有し、シトクロムcMタンパク質と機能的に同等なタンパク質
(d)配列番号1で表される塩基配列でコードされるアミノ酸配列を有するタンパク質
(e)配列番号1で表される塩基配列と90%以上の同一性を有する塩基配列によってコードされるタンパク質であって、シトクロムcMタンパク質と機能的に同等なタンパク質をコードする、(1)~(3)のいずれかに記載の形質転換体。
(5)光合成独立栄養能と完全暗所従属栄養能とを備える、(1)~(4)のいずれかに記載の形質転換体。
(6)さらに、有用物質を生産するための外来遺伝子を備える、(1)~(5)のいずれかに記載の形質転換体。
(7)シトクロムcMタンパク質をコードするcytM遺伝子の不活性化を備え、完全暗所従属栄養能を有する、シアノバクテリア。
(8)L. boryana dg5株以外のシアノバクテリアである、(7)に記載のシアノバクテリア。
(9)さらに、有用物質を生産するための外来遺伝子を備える、(7)又は(8)に記載のシアノバクテリア。
(10)シトクロムcMタンパク質をコードするcytM遺伝子の不活性化を付与するポリヌクレオチドを含む、シアノバクテリアに完全暗所従属栄養能を付与するための形質転換ベクター。
(11)前記不活性化は、前記cytM遺伝子の破壊又は活性が低下した前記タンパク質をコードする変異の導入である、(10)に記載の形質転換ベクター。
(12)(1)~(6)のいずれかに記載の形質転換体の製造方法であって、
シアノバクテリアに、シトクロムcMタンパク質をコードするcytM遺伝子の不活性化を付与するポリヌクレオチドで形質転換する工程、
を備える、製造方法。
(13)前記不活性化は、前記cytM遺伝子の破壊又は活性が低下した前記タンパク質をコードする変異の導入である、(12)に記載の製造方法。
(14)前記形質転換工程は、前記ポリヌクレオチドをエレクトロポレーションで前記シアノバクテリアに導入する、(12)又は(13)に記載の製造方法。
(15)(1)~(6)のいずれかに記載の形質転換体を培養する工程、
を備える、シアノバクテリアの培養産物の生産方法。
(16)前記培養工程は、暗所での培養を含む、(15)に記載の生産方法。
(17)(1)~(6)のいずれかに記載の形質転換体を暗所で培養する工程、
を備える、シアノバクテリアの培養方法。 (1) A cyanobacterial transformant comprising inactivation of a cytM gene encoding cytochrome c M protein.
(2) The transformant according to (1), wherein the inactivation is the introduction of a mutation encoding the protein in which the cytM gene is disrupted or decreased in activity.
(3) The transformant according to (1) or (2), wherein the inactivation is a mutation that encodes a protein having the amino acid sequence represented by SEQ ID NO: 14 in the cytM gene.
(4) The cytM gene is any of the following proteins:
(A) a protein having the amino acid sequence represented by SEQ ID NO: 2 (b) an amino acid sequence having 90% or more identity with the amino acid sequence represented by SEQ ID NO: 2 and functionally associated with a cytochrome c M protein Equivalent protein (c) A protein functionally equivalent to cytochrome c M protein, having an amino acid sequence having one or more amino acid substitutions, deletions, insertions or additions in the amino acid sequence represented by SEQ ID NO: 2 (D) a protein having an amino acid sequence encoded by the nucleotide sequence represented by SEQ ID NO: 1 (e) a protein encoded by a nucleotide sequence having 90% or more identity with the nucleotide sequence represented by SEQ ID NO: 1 The transformant according to any one of (1) to (3), wherein the transformant encodes a protein functionally equivalent to the cytochrome c M protein.
(5) The transformant according to any one of (1) to (4), which has photosynthetic autotrophic ability and complete dark heterotrophic ability.
(6) The transformant according to any one of (1) to (5), further comprising a foreign gene for producing a useful substance.
(7) A cyanobacteria having inactivation of the cytM gene encoding cytochrome c M protein and having complete dark heterotrophic ability.
(8) The cyanobacteria according to (7), which is a cyanobacteria other than the L. boryana dg5 strain.
(9) The cyanobacteria according to (7) or (8), further comprising a foreign gene for producing a useful substance.
(10) A transformation vector for imparting complete dark heterotrophic ability to cyanobacteria, comprising a polynucleotide that imparts inactivation of the cytM gene encoding cytochrome c M protein.
(11) The transformation vector according to (10), wherein the inactivation is introduction of a mutation encoding the protein in which the cytM gene is disrupted or decreased in activity.
(12) A method for producing the transformant according to any one of (1) to (6),
Transforming cyanobacteria with a polynucleotide conferring inactivation of the cytM gene encoding cytochrome c M protein;
A manufacturing method comprising:
(13) The production method according to (12), wherein the inactivation is introduction of a mutation encoding the protein in which the cytM gene is disrupted or decreased in activity.
(14) The production method according to (12) or (13), wherein the transformation step introduces the polynucleotide into the cyanobacteria by electroporation.
(15) culturing the transformant according to any one of (1) to (6),
A method for producing a cyanobacterial culture product.
(16) The production method according to (15), wherein the culturing step includes culturing in a dark place.
(17) culturing the transformant according to any one of (1) to (6) in the dark,
A method for culturing cyanobacteria, comprising:
本明細書の開示は、シアノバクテリアにおける完全暗所従属栄養能に関連する遺伝子及びその利用に関する。本明細書の開示によれば、シトクロムcMタンパク質をコードするcytM遺伝子の不活性化を備えるようにシアノバクテリアを改変することで、シアノバクテリアに対して光合成独立栄養性を有するシアノバクテリアに完全暗所従属栄養能を付与しあるいはその完全暗所従属栄養能を増強したりできる。
The disclosure herein relates to genes associated with complete dark heterotrophic capacity in cyanobacteria and uses thereof. According to the disclosure herein, modification of cyanobacteria to provide for inactivation of the cytM gene encoding cytochrome c M protein provides complete darkness to cyanobacteria with photoautotrophic properties against cyanobacteria. It can give a place dependent nutrition or enhance its complete dark place nutrition.
これにより、シアノバクテリアは、太陽光エネルギーなどの天然の光エネルギーを利用するとともに有機栄養による完全暗所従属栄養能を備える混合栄養性となるため、天然の光エネルギーを利用しつつこうした光エネルギーが抑制された状態であっても連続的に増殖し効率的に有用物質を生産することができる。また、改変されたシアノバクテリアは、暗所においても増殖し有用物質を生産できるため、暗所特異的に生産する有用物質も効率的に生産できる。
As a result, cyanobacteria use natural light energy such as solar energy and become mixed nutrients with complete dark heterotropy due to organic nutrition. Even in a suppressed state, it can continuously grow and produce useful substances efficiently. In addition, since the modified cyanobacteria can grow and produce useful substances even in the dark, it is possible to efficiently produce useful substances that are produced specifically in the dark.
以下、本開示の代表的かつ非限定的な具体例について、適宜図面を参照して詳細に説明する。この詳細な説明は、本発明の好ましい例を実施するための詳細を当業者に示すことを単純に意図しており、本開示の範囲を限定することを意図したものではない。また、以下に開示される追加的な特徴ならびに発明は、さらに改善されたシアノバクテリア等を提供するために、他の特徴や発明とは別に、又は共に用いることができる。
Hereinafter, representative and non-limiting specific examples of the present disclosure will be described in detail with reference to the drawings as appropriate. This detailed description is intended merely to provide those skilled in the art with details for practicing the preferred embodiments of the present invention and is not intended to limit the scope of the present disclosure. Further, the additional features and inventions disclosed below can be used separately from or together with other features and inventions to provide further improved cyanobacteria and the like.
また、以下の詳細な説明で開示される特徴や工程の組み合わせは、最も広い意味において本開示を実施する際に必須のものではなく、特に本開示の代表的な具体例を説明するためにのみ記載されるものである。さらに、上記及び下記の代表的な具体例の様々な特徴、ならびに、独立及び従属クレームに記載されるものの様々な特徴は、本開示の追加的かつ有用な実施形態を提供するにあたって、ここに記載される具体例のとおりに、あるいは列挙された順番のとおりに組合せなければならないものではない。
Further, the combinations of features and steps disclosed in the following detailed description are not essential in carrying out the present disclosure in the broadest sense, and are particularly only for explaining representative specific examples of the present disclosure. It is described. Moreover, various features of the representative embodiments described above and below, as well as those described in the independent and dependent claims, are described herein in providing additional and useful embodiments of the present disclosure. They do not have to be combined in the specific examples given or in the order listed.
本明細書及び/又はクレームに記載された全ての特徴は、実施例及び/又はクレームに記載された特徴の構成とは別に、出願当初の開示ならびにクレームされた特定事項に対する限定として、個別に、かつ互いに独立して開示されることを意図するものである。さらに、全ての数値範囲及びグループ又は集団に関する記載は、出願当初の開示ならびにクレームされた特定事項に対する限定として、それらの中間の構成を開示する意図を持ってなされている。
All features described in this specification and / or claims, apart from the configuration of the features described in the examples and / or claims, are individually disclosed as limitations on the original disclosure and claimed specific matters. And are intended to be disclosed independently of each other. Further, all numerical ranges and group or group descriptions are intended to disclose intermediate configurations thereof as a limitation to the original disclosure and claimed subject matter.
以下、本明細書の開示について詳細に説明する。
Hereinafter, the disclosure of this specification will be described in detail.
(シアノバクテリア形質転換体)
本明細書に開示されるシアノバクテリア形質転換体は、シトクロムcMタンパク質(Cytochrome cMともいう。)をコードするcytM遺伝子の不活性化を備えることができる。 (Cyanobacteria transformant)
The cyanobacterial transformants disclosed herein can comprise inactivation of the cytM gene encoding cytochrome c M protein (also referred to as Cytochrome c M ).
本明細書に開示されるシアノバクテリア形質転換体は、シトクロムcMタンパク質(Cytochrome cMともいう。)をコードするcytM遺伝子の不活性化を備えることができる。 (Cyanobacteria transformant)
The cyanobacterial transformants disclosed herein can comprise inactivation of the cytM gene encoding cytochrome c M protein (also referred to as Cytochrome c M ).
(シアノバクテリア)
本明細書において、シアノバクテリアとは、グラム陰性の真性細菌であり、酸素発生型の光合成を行う原核光合成微生物である。本明細書におけるシアノバクテリアは、従属栄養能とは、炭素源として、糖、脂肪酸、アミノ酸などの有機化合物によって生育又は増殖できる能力をいう。 (Cyanobacteria)
In the present specification, cyanobacteria are gram-negative true bacteria and are prokaryotic photosynthetic microorganisms that perform oxygen-generating photosynthesis. Cyanobacteria herein refers to the ability of heterotrophic ability to grow or proliferate as a carbon source by organic compounds such as sugars, fatty acids and amino acids.
本明細書において、シアノバクテリアとは、グラム陰性の真性細菌であり、酸素発生型の光合成を行う原核光合成微生物である。本明細書におけるシアノバクテリアは、従属栄養能とは、炭素源として、糖、脂肪酸、アミノ酸などの有機化合物によって生育又は増殖できる能力をいう。 (Cyanobacteria)
In the present specification, cyanobacteria are gram-negative true bacteria and are prokaryotic photosynthetic microorganisms that perform oxygen-generating photosynthesis. Cyanobacteria herein refers to the ability of heterotrophic ability to grow or proliferate as a carbon source by organic compounds such as sugars, fatty acids and amino acids.
また、本明細書において、暗所の語は、完全暗所と限定的暗所とを含むものとする。ここで完全暗所とは、生育又は増殖期間において、生育又は増殖のための人為的な操作に必要な時間及び必要量の光照射下に置く以外は、光を遮断した状態を意味している。生育又は増殖のための人為的な操作に不可避である光照射条件とは、例えば、10日~2週間に一度、10分から30分以内程度、好ましくは20分以下の時間に光照射する条件である。一方、限定的暗所とは、1日10分~20分程度、好ましくは10分程度の光照射下に置く以外は、光を遮断した状態を意味している。
Also, in this specification, the word “dark place” includes a complete dark place and a limited dark place. Here, the complete dark place means a state in which light is blocked during the growth or multiplication period, except that it is placed under light irradiation for a time and a necessary amount necessary for artificial manipulation for growth or multiplication. . Light irradiation conditions that are unavoidable for artificial manipulation for growth or proliferation are, for example, conditions in which light irradiation is performed within 10 to 30 minutes, preferably 20 minutes or less once every 10 days to 2 weeks. is there. On the other hand, the limited dark place means a state in which light is blocked except that it is placed under light irradiation for about 10 to 20 minutes, preferably about 10 minutes per day.
シアノバクテリアは、完全暗所で生育できないが限定的暗所で従属的栄養能を発揮する場合がある。この現象を、"Light-Activated Heterotrophic Growth"(LAHG)ともいう。これに対して完全暗所においても従属栄養的に生育等可能な場合を完全暗所従属栄養能という。シアノバクテリアは、こうした暗所従属栄養能を有していてもよいし、有していなくてもよい。
Cyanobacteria cannot grow in complete darkness, but may exhibit dependent nutrition in limited darkness. This phenomenon is also called “Light-ActivatedActivHeterotrophic Growth” (LAHG). On the other hand, the case where it is possible to grow heterotrophically in a completely dark place is called complete dark place heterotrophic ability. The cyanobacteria may or may not have such dark heterotrophic ability.
シアノバクテリアとしては、酸素発生型光合成を行う原核生物であり、特に限定しないで、クロオコッカス目、プレウロカプサ目、ユレモ目、ネンジュモ目、スティゴネマ目、グロエオバクター目等が挙げられる。さらには、Synechococcus elongatus PCC 7942などのSynechococcus 属、Prochlorococcus marinus MED4などのProchlorococcus属、Anabaena sp. PCC 7120などのAnabaena属、Synechocystis sp. PCC 6803などのSynechocystis属、Thermosynechococcus elongats BP-1などのThermosynechococcus属、 Gloeobacter violaceus PCC 7421などGloeobacter属、Arthrospira platensisなどのArthrospira属、Trichodesmium erythraeum IMS101などのTrichodesmium属、Acaryochloris marina MBIC11017などのAcaryochloris属が挙げられる。
Cyanobacteria are prokaryotes that carry out oxygen-generating photosynthesis, and are not particularly limited, and include, but are not limited to, the order of the order of the order Chroococcus, Pleurocapsa, Euremo, Nemoptera, Stigonema, Groeobacter. Furthermore, Synechococcus genus such as Synechococcus elongatus PCC 7942, Prochlorococcus genus such as Prochlorococcus marinus MED4, Anabaena genus such as Anabaena sp. PCC 7120, Synechocystis genus such as Synchocystis sp. Examples include the Gloeobacter genus such as Gloeobacter violaceus PCC 7421, the Arthrospira genus such as Arthrospira platensis, the Trichodesmium genus such as Trichodesmium erythraeum IMS101, and the Acaryochloris genus such as Acaryochloris marina MBIC11017.
(cytM遺伝子)
cytM遺伝子がコードするシトクロムcMタンパク質は、その機能は必ずしも明らかではないが、シトクロムcオキシダーゼのCuドメイン電子供与体として機能するものと考えられている(Bernroitner et al. 2009, Biochim Biophys Acta 1787(3):135-43)。 (CytM gene)
The cytochrome c M protein encoded by the cytM gene is thought to function as a Cu domain electron donor of cytochrome c oxidase, although its function is not necessarily clear (Bernroitner et al. 2009, Biochim Biophys Acta 1787 ( 3): 135-43).
cytM遺伝子がコードするシトクロムcMタンパク質は、その機能は必ずしも明らかではないが、シトクロムcオキシダーゼのCuドメイン電子供与体として機能するものと考えられている(Bernroitner et al. 2009, Biochim Biophys Acta 1787(3):135-43)。 (CytM gene)
The cytochrome c M protein encoded by the cytM gene is thought to function as a Cu domain electron donor of cytochrome c oxidase, although its function is not necessarily clear (Bernroitner et al. 2009, Biochim Biophys Acta 1787 ( 3): 135-43).
シアノバクテリアにおけるcytM遺伝子の塩基配列は概して既に知られている。L. boryana IAM-M101のcytM遺伝子のシトクロムcMタンパク質をコードする塩基配列は配列番号1で表され、また、そのタンパク質のアミノ酸配列は配列番号2で表される。なお、L. boryana IAM-M101及びその変異株であるL. boryana IAM-M101 dg5は、国立大学法人名古屋大学大学院生命農学研究科(日本国愛知県名古屋市千種区不老町1番)であり分譲可能である。
In general, the base sequence of the cytM gene in cyanobacteria is already known. L. boryana IAM-M101 nucleotide sequence encoding a cytochrome c M protein cytM gene is represented by SEQ ID NO: 1, The amino acid sequence of the protein shown in SEQ ID NO: 2. L. boryana IAM-M101 and its mutant L. boryana IAM-M101 dg5 are a part of the Graduate School of Bioagricultural Sciences, Nagoya University (No. 1 Fufu-cho, Chikusa-ku, Nagoya, Aichi, Japan). Is possible.
L. boryanaは、糸状性のシアノバクテリアである。シアノバクテリアは、その形態的特徴からセクションI~Vに分類されているが、L. boryanaは、ヘテロシストを形成しない糸状性という形態的特徴をもつセクションIIIに分類されている。セクションIIIは、さらに糸状体の形態的特徴とガス胞の有無などにより細分されるが、等径の細胞から直線的な糸状体(分岐していない)を形成し、ガス胞がなく、細胞間にくびれがない、という特徴を示すL. boryanaはLPP group Bに属する。また、L. boryanaは、ヘテロシストと呼ばれる窒素固定専門の細胞を分化しないタイプである。
L. boryana is a filamentous cyanobacteria. Cyanobacteria are classified into sections I to V because of their morphological characteristics, whereas L. boryana is classified into section III, which has the morphological characteristics of filamentous properties that do not form heterocysts. Section III is further subdivided according to the morphological characteristics of the filamentous body and the presence or absence of gas vesicles, but forms linear filaments (not branched) from cells of equal diameter, without gas vesicles, and between cells L. boryana, which has the characteristic of no constriction, belongs to LPP group B. L. boryana is a type that does not differentiate into cells specialized in nitrogen fixation called heterocysts.
L. boryanaは、完全暗所であってもグルコースなどの糖があれば従属栄養的に生育することができる(完全暗所従属栄養能)。また、L. boryanaは、エレクトロポレーションによってDNAを細胞へ導入し、ゲノムとの相同的組換えによって特定の遺伝子が破壊された変異株を得ることができる(Fujita et al. 1992, Plant Cell Physiology, 33: 81-92)。
L. boryana can grow heterotrophically in the presence of sugar such as glucose even in a completely dark place (complete dark place heterotrophic ability). In addition, L. 、 boryana can introduce DNA into cells by electroporation and obtain mutants in which a specific gene is destroyed by homologous recombination with the genome (Fujita et al. 1992, Plant Cell Physiology). , 33: 81-92).
そのほか、シアノバクテリアのcytM遺伝子としては、例えば、以下のアミノ酸配列からなるタンパク質をコードする遺伝子が知られている。なお、配列番号3~12は、シトクロムcMタンパク質である。一方、配列番号13は、シトクロムcクラスIタンパク質であるが、配列番号2で表されるアミノ酸配列と30%程度の同一性(%)しか示さず、機能的にはシトクロムc6タンパク質と考えられる。
In addition, as a cyanobacterial cytM gene, for example, a gene encoding a protein having the following amino acid sequence is known. SEQ ID NOs: 3 to 12 are cytochrome cM proteins. On the other hand, SEQ ID NO: 13 is a cytochrome c class I protein, but shows only about 30% identity (%) with the amino acid sequence represented by SEQ ID NO: 2, and is functionally considered as a cytochrome c6 protein.

本明細書で不活性化対象となるcytM遺伝子は、こうして既に開示されているもののほか、公知のシトクロムcMタンパク質のアミノ酸配列と一定の同一性を有するアミノ酸配列を有し、シトクロムcMタンパク質と機能的に同等であるタンパク質をコードするものであってもよい。こうした不活性化対象遺伝子は、例えば、配列番号2で表されるアミノ酸配列と60%以上の同一性を有していることが好ましく、より好ましくは65%以上の同一性を有しており、70%以上の同一性、好ましくは75%以上、より好ましくは80%以上、さらに好ましくは85%以上、一層好ましくは90%以上、より一層好ましくは95%以上、さらに一層好ましくは97%以上、さらに好ましくは98%以上、最も好ましくは99%以上の同一性を有していることが好ましい。
The cytM gene to be inactivated in the present specification has an amino acid sequence having a certain identity with the amino acid sequence of a known cytochrome c M protein in addition to those already disclosed in this way, and the cytochrome c M protein It may encode proteins that are functionally equivalent. Such a gene to be inactivated preferably has, for example, 60% or more identity with the amino acid sequence represented by SEQ ID NO: 2, more preferably 65% or more identity, 70% or more identity, preferably 75% or more, more preferably 80% or more, still more preferably 85% or more, more preferably 90% or more, still more preferably 95% or more, still more preferably 97% or more, More preferably, it has 98% or more, and most preferably 99% or more identity.
また、例えば、cytM遺伝子は、アラインメントしたときに、-NCXXCH-モチーフと-TPPMP-モチーフとを備えることがタンパク質をコードするものであってもよい。これら2つのモチーフは、本明細書におけるシトクロムcMタンパク質に共通して観察される(Bernroitner et al. Biochem. Biophys. Acta, 2009, 1787: 135-143)。例えば、これらのモチーフは、配列番号2で表されるアミノ酸配列において第36位~41位及び第74位~78位にそれぞれ対応している。したがって、cytM遺伝子は、例えば、配列番号2で表されるアミノ酸配列とアライメントしたとき、これらに対応する位置にこれらのモチーフが見出され、しかも、シトクロムcMタンパク質と機能的に同等であるタンパク質をコードするものであってもよい。cytM遺伝子がコードするシトクロムcMタンパク質及び近縁の他のシトクロムcクラスIタンパク質についての系統樹とアライメント結果を図1に示す。
Further, for example, the cytM gene may be a protein encoding a protein that, when aligned, comprises a -NCXXCH-motif and a -TPPMP-motif. These two motifs are commonly observed in cytochrome c M protein in this specification (Bernroitner et al Biochem Biophys Acta, 2009, 1787:... 135-143). For example, these motifs correspond to positions 36 to 41 and positions 74 to 78 in the amino acid sequence represented by SEQ ID NO: 2, respectively. Therefore, CytM gene, for example, when the amino acid sequence alignment of SEQ ID NO: 2, found these motifs at positions corresponding to these, moreover, cytochrome c M protein functionally equivalent to a protein that is a May be encoded. FIG. 1 shows the phylogenetic tree and alignment results for cytochrome c M protein encoded by the cytM gene and other related cytochrome c class I proteins.
本明細書において、アミノ酸配列又は塩基配列において「同一性」とは、比較される配列間において、各々の配列を構成するアミノ酸残基又は塩基の一致の程度の意味で用いられる。このとき、アミノ酸配列については、ギャップの存在及びアミノ酸の性質が考慮される(Wilbur, Proc. Natl. Acad. Sci. U.S.A. 80:726-730 (1983))。同一性の計算には、市販のソフトであるBLAST(Altschul: J. Mol. Biol. 215:403-410 (1990))、FASTA(Peasron: Methods in Enzymology 183:63-69 (1990))等を用いることができる。「同一性」の数値はいずれも、当業者に公知の相同性検索プログラムを用いて算出される数値であればよく、例えば、全米バイオテクノロジー情報センター(NCBI)の相同性アルゴリズムBLAST(Basic local alignment search tool)http://www.ncbi.nlm.nih.gov/BLAST/においてデフォルト(初期設定)のパラメーターを用いることにより、算出することができる。
In the present specification, “identity” in an amino acid sequence or a base sequence is used to mean the degree of coincidence of amino acid residues or bases constituting each sequence between sequences to be compared. At this time, regarding the amino acid sequence, the existence of a gap and the nature of the amino acid are considered (Wilbur, Proc. Natl. Acad. Sci. U.S.A. 80: 726-730 (1983)). For the calculation of identity, commercially available softwares such as BLAST (Altschul: J. Mol. Biol. 215: 403-410 (1990)), FASTA (Peasron: Methods in Enzymology 183: 63-69 (1990)), etc. Can be used. Any numerical value of “identity” may be a numerical value calculated using a homology search program known to those skilled in the art. For example, the homology algorithm BLAST (BasicBlocal alignment) of the National Center for Biotechnology Information (NCBI) search)) http://www.ncbi.nlm.nih.gov/BLAST/, using default (initial setting) parameters.
シトクロムcMタンパク質と機能的に同等とは、完全暗所従属栄養能を発現ないし増強させる機能を実質的に同程度維持することを意味している。機能的に同等か否かは、シアノバクテリアにおいて当該遺伝子を破壊した遺伝子破壊シアノバクテリアと非破壊のシアノバクテリアとを比較して、完全暗所従属栄養能が発現若しくは増強されているか又は限定的暗所従属栄養能が増強されて完全暗所従属栄養的になっていることかで決定することができる。すなわち、遺伝子破壊シアノバクテリアにおいて、完全暗所従属栄養能が発現・増強されるか完全暗所従属栄養的になっているとき、その遺伝子がコードするタンパク質は、シトクロムcMタンパク質と機能的に同等であると決定できる。また、当該遺伝子は、cytM遺伝子と機能的に同等であると決定できる。
Functionally equivalent to cytochrome c M protein means maintaining substantially the same level of function to express or enhance complete dark heterotrophic capacity. The functional equivalence is determined by comparing the gene-disrupted cyanobacteria that have disrupted the gene in cyanobacteria with the non-disrupted cyanobacteria, where the complete dark heterotrophic ability is expressed or enhanced, or limited darkness. This can be determined by the fact that the place's heterotrophic capacity is enhanced to become complete dark place heterotrophic. That is, in gene-disrupted cyanobacteria, when the complete dark heterotrophic ability is expressed and enhanced or is completely dark heterotrophic, the protein encoded by the gene is functionally equivalent to the cytochrome c M protein. Can be determined. In addition, the gene can be determined to be functionally equivalent to the cytM gene.
また、cytM遺伝子は、公知のシトクロムcMタンパク質のアミノ酸配列において、1又は複数個のアミノ酸の挿入、置換、欠失及び付加されたアミノ酸配列を有してシトクロムcMタンパク質と機能的に同等なタンパク質をコードするものであってもよい。こうしたタンパク質は、例えば、1~30個、好ましくは1~20個、より好ましくは1~10個、さらに好ましくは1~5個、一層好ましくは1~2個のアミノ酸の挿入、置換、欠失及び付加を備えることができる。
Further, CytM gene, the amino acid sequences of known cytochrome c M protein, 1 or insertion of a plurality of amino acids, substitutions, cytochrome c M protein functionally equivalent have deletions and additions in the amino acid sequence It may encode a protein. Such proteins have, for example, 1 to 30, preferably 1 to 20, more preferably 1 to 10, more preferably 1 to 5, more preferably 1 to 2 amino acid insertions, substitutions and deletions. And additions can be provided.
こうしたタンパク質は、公知のシトクロムcMタンパク質のアミノ酸配列において1又は数個、好ましくは1~10個、より好ましくは1~5個、さらに好ましくは1~2個程度の保存的置換を有するアミノ酸配列であることが好ましい。
Such a protein has an amino acid sequence having a conservative substitution of one or several, preferably 1 to 10, more preferably 1 to 5, more preferably about 1 to 2 amino acids in the known cytochrome c M protein. It is preferable that
ここで「保存的置換」とは、タンパク質の機能を実質的に改変しないように、1または複数個のアミノ酸残基を、別の化学的に類似したアミノ酸残基で置換えることを意味する。例えば、ある疎水性残基を別の疎水性残基によって置換する場合、ある極性残基を同じ電荷を有する別の極性残基によって置換する場合などが挙げられる。このような置換を行うことができる機能的に類似のアミノ酸は、アミノ酸毎に当該技術分野において公知である。具体例を挙げると、非極性(疎水性)アミノ酸としては、アラニン、バリン、イソロイシン、ロイシン、プロリン、トリプトファン、フェニルアラニン、メチオニンなどが挙げられる。極性(中性)アミノ酸としては、グリシン、セリン、スレオニン、チロシン、グルタミン、アスパラギン、システインなどが挙げられる。陽電荷をもつ(塩基性)アミノ酸としては、アルギニン、ヒスチジン、リジンなどが挙げられる。また、負電荷をもつ(酸性)アミノ酸としては、アスパラギン酸、グルタミン酸などが挙げられる。
Here, “conservative substitution” means substitution of one or more amino acid residues with another chemically similar amino acid residue so as not to substantially alter the function of the protein. For example, when a certain hydrophobic residue is substituted by another hydrophobic residue, a certain polar residue is substituted by another polar residue having the same charge, and the like. Functionally similar amino acids that can make such substitutions are known in the art for each amino acid. Specific examples include non-polar (hydrophobic) amino acids such as alanine, valine, isoleucine, leucine, proline, tryptophan, phenylalanine, and methionine. Examples of polar (neutral) amino acids include glycine, serine, threonine, tyrosine, glutamine, asparagine, and cysteine. Examples of positively charged (basic) amino acids include arginine, histidine, and lysine. Examples of negatively charged (acidic) amino acids include aspartic acid and glutamic acid.
さらに、cytM遺伝子としては、公知のシトクロムcMタンパク質をコードするDNAとストリンジェントな条件下でハイブリダイズする相補性を有するDNAであって、シトクロムcMタンパク質と機能的に同等なタンパク質をコードするものであってもよい。
Furthermore, the cytM gene is a complementary DNA that hybridizes under stringent conditions with a DNA encoding a known cytochrome c M protein, and encodes a protein functionally equivalent to the cytochrome c M protein. It may be a thing.
「ストリンジェントな条件下」とは、当業者が通常使用し得るハイブリダイゼーション緩衝液中で、温度が40℃~70℃、好ましくは60℃~65℃などで反応を行い、塩濃度が15~300mmol/L、好ましくは15~60mmol/Lなどの洗浄液中で洗浄する方法に従って行なうことができる。温度、塩濃度は使用するプローブの長さに応じて適宜調整することが可能である。さらに、ハイブリダイズしたものを洗浄するときの条件は、0.2または2×SSC、0.1%SDS、温度20℃~68℃で行うことができる。ストリンジェント(high stringency)な条件にするかマイルド(low stringency)な条件にするかは、洗浄時の塩濃度または温度で差を設けることができる。塩濃度でハイブリダイズの差を設ける場合には、ストリンジェント洗浄バッファー(high stringency wash buffer)として0.2×SSC、0.1%SDS、マイルド洗浄バッファー(low stringency wash buffer)として2×SSC、0.1%SDSで行うことができる。また、温度でハイブリダイズの差を設ける場合には、ストリンジェントの場合は68℃、中等度(moderate stringency)の場合は42℃、マイルドの場合は室温(20℃~25℃)でいずれの場合も0.2×SSC、0.1%SDSで行えばよい。
“Stringent conditions” means that the reaction is carried out at a temperature of 40 ° C. to 70 ° C., preferably 60 ° C. to 65 ° C. in a hybridization buffer that can be usually used by those skilled in the art. It can be carried out according to a method of washing in a washing solution of 300 mmol / L, preferably 15 to 60 mmol / L. The temperature and salt concentration can be appropriately adjusted according to the length of the probe used. Furthermore, the conditions for washing the hybridized material can be 0.2 or 2 × SSC, 0.1% SDS, and a temperature of 20 ° C. to 68 ° C. Whether the conditions are stringent (high stringency) or mild (low stringency), a difference can be made depending on the salt concentration or temperature at the time of washing. When providing a difference in hybridization at the salt concentration, 0.2 × SSC as a stringent wash buffer (high バ ッ フ ァ ー stringency wash buffer), 0.1% SDS, 2 × SSC as a mild washbuffer (low stringency wash buffer), This can be done with 0.1% SDS. In addition, in the case of providing a difference in hybridization by temperature, 68 ° C for stringent, 42 ° C for moderate-stringency, and room temperature (20 ° C to 25 ° C) for mild May be performed with 0.2 × SSC and 0.1% SDS.
cytM遺伝子は、公知のシトクロムcMタンパク質をコードする塩基配列と一定の同一性を有する塩基配列を有し、シトクロムcMタンパク質と機能的に同等であるタンパク質をコードするものであってもよい。こうした不活性化対象遺伝子は、例えば、公知のシトクロムcMタンパク質をコードする配列番号1で表される塩基配列と60%以上の同一性を有していることが好ましく、より好ましくは65%以上の同一性を有しており、さらに好ましくは70%以上、より好ましくは75%以上、より好ましくは80%以上、さらに好ましくは85%以上、一層好ましくは90%以上、より一層好ましくは95%以上、さらに一層好ましくは97%以上、さらに好ましくは98%以上、最も好ましくは99%以上の同一性を有していることが好ましい。
The cytM gene may have a base sequence having a certain identity with a base sequence encoding a known cytochrome c M protein, and may encode a protein that is functionally equivalent to the cytochrome c M protein. Such a gene to be inactivated preferably has, for example, 60% or more identity with the nucleotide sequence represented by SEQ ID NO: 1 encoding a known cytochrome c M protein, more preferably 65% or more. More preferably 70% or more, more preferably 75% or more, more preferably 80% or more, still more preferably 85% or more, more preferably 90% or more, and still more preferably 95%. More preferably, it has an identity of 97% or more, more preferably 98% or more, and most preferably 99% or more.
本願明細書において、「アミノ酸配列において、1または複数個のアミノ酸の挿入、置換または欠失、あるいはその一方または両末端への付加がなされた」とは、部位特異的突然変異誘発法等の周知の技術的方法により、または天然に生じ得る程度の複数個の数のアミノ酸の置換等によりなされたことを意味する。
In the present specification, “the insertion, substitution or deletion of one or a plurality of amino acids, or addition to one or both ends thereof in the amino acid sequence” is a well-known site-directed mutagenesis method or the like. Or by substitution of a plurality of amino acids to the extent that can occur naturally.
(cytM遺伝子の不活性化)
シアノバクテリアにおいてcytM遺伝子の不活性化を備えている。ここで遺伝子の不活性化とは、結果的にその遺伝子がコードするタンパク質の活性が低下ないし喪失するものであればよい。したがって、遺伝子の不活性化とは、遺伝子破壊、転写阻害、転写産物の分解、翻訳阻害、不活性なタンパク質の翻訳等が挙げられる。 (Inactivation of cytM gene)
It has inactivation of the cytM gene in cyanobacteria. Here, the inactivation of the gene may be anything as long as the activity of the protein encoded by the gene is reduced or lost as a result. Therefore, gene inactivation includes gene disruption, transcription inhibition, transcript degradation, translation inhibition, translation of inactive proteins, and the like.
シアノバクテリアにおいてcytM遺伝子の不活性化を備えている。ここで遺伝子の不活性化とは、結果的にその遺伝子がコードするタンパク質の活性が低下ないし喪失するものであればよい。したがって、遺伝子の不活性化とは、遺伝子破壊、転写阻害、転写産物の分解、翻訳阻害、不活性なタンパク質の翻訳等が挙げられる。 (Inactivation of cytM gene)
It has inactivation of the cytM gene in cyanobacteria. Here, the inactivation of the gene may be anything as long as the activity of the protein encoded by the gene is reduced or lost as a result. Therefore, gene inactivation includes gene disruption, transcription inhibition, transcript degradation, translation inhibition, translation of inactive proteins, and the like.
こうした遺伝子の不活性化は、概して、遺伝子の破壊の形態で行われる.遺伝子破壊は、例えば、cytM遺伝子への外来配列の挿入、cytM遺伝子遺伝子全体またはその一部の外来配列による置換、cytM遺伝子の全部または一部の欠失によって行うことができる。外来配列の塩基数並びに置換、欠失、および挿入の位置は、シトクロムcMタンパク質の発現や活性が実質的に喪失する限り、特に限定されない。なお、遺伝子組換え体の選別の観点から、外来配列は選択マーカー遺伝子であることが好ましい。選択マーカー遺伝子は公知のものから適宜選択して使用でき、好ましくは薬剤に対する耐性遺伝子等が挙げられる。
Such gene inactivation is generally performed in the form of gene disruption, for example, insertion of foreign sequences into the cytM gene, replacement of all or part of the cytM gene by foreign sequences, cytM gene Can be done by deletion of all or part of The number of bases in the foreign sequence and the positions of substitutions, deletions and insertions are not particularly limited as long as the expression and activity of cytochrome c M protein is substantially lost. In addition, from the viewpoint of selecting a gene recombinant, the foreign sequence is preferably a selection marker gene. The selectable marker gene can be appropriately selected from known ones and is preferably a drug resistance gene.
cytM遺伝子の破壊にあたっては、好ましくは、cytM遺伝子の全部または一部を外来遺伝子に置換することにより行うことができる。置換されるcytM遺伝子の一部としては、例えば、シトクロムcMタンパク質のC末端側を置換する形態が挙げられる。
The cytM gene can be disrupted preferably by replacing all or part of the cytM gene with a foreign gene. The part of cytM gene to be replaced, for example, the form to replace the C-terminal side of the cytochrome c M protein.
また、cytM遺伝子の破壊にあたっては、cytM遺伝子がコードするタンパク質に変異を導入して活性が低下したタンパク質を産生させるようにしてもよい。例えば、コード配列において塩基を挿入したり、欠損させたりすることにより、フレームシフト変異であってもよいし、タンパク質の一部を異なるアミノ酸に置換するものであってもよいし、これらを組み合わせたものであってもよい。
Further, when the cytM gene is disrupted, a protein with reduced activity may be produced by introducing a mutation into the protein encoded by the cytM gene. For example, it may be a frameshift mutation by inserting or deleting a base in the coding sequence, or a part of the protein may be substituted with a different amino acid, or a combination thereof. It may be a thing.
遺伝子破壊の技術は当業者に公知であり、当業者であれば公知の方法に従って遺伝子の破壊を実施することができる。遺伝子破壊は、概して、標的となる遺伝子の塩基配列に改変を施したポリヌクレオチド(好ましくは選択マーカー遺伝子)を細胞に導入し、導入したポリヌクレオチドと標的遺伝子との間で相同組換えを生じさせ、相同組換えが生じた細胞を選択することにより、標的遺伝子に変異を導入することができる。
Gene disruption techniques are known to those skilled in the art, and those skilled in the art can perform gene disruption according to known methods. Gene disruption generally involves introducing into a cell a polynucleotide (preferably a selectable marker gene) in which the nucleotide sequence of the target gene has been modified, and causing homologous recombination between the introduced polynucleotide and the target gene. By selecting cells in which homologous recombination has occurred, mutations can be introduced into the target gene.
また、cytM遺伝子を不活性化するには、例えば、シトクロムcMタンパク質を配列番号14で表されるアミノ酸からなるタンパク質をコードする変異であってもよい。
In addition, in order to inactivate the cytM gene, for example, it may be a variant which encodes a protein consisting of the amino acid represented by the cytochrome c M protein in SEQ ID NO: 14.
本明細書に開示される形質転換体は、cytM遺伝子の不活性化を備えているシアノバクテリアである。内在性のcytM遺伝子を備える場合において、そのcytM遺伝子の不活性化を付与することで、そのシアノバクテリアに暗所従属栄養能を付与しあるいは増強することができる。
The transformant disclosed in this specification is a cyanobacteria equipped with inactivation of the cytM gene. When an endogenous cytM gene is provided, inactivation of the cytM gene can be imparted, and dark heterotrophic ability can be imparted or enhanced to the cyanobacteria.
(形質転換ベクター及び形質転換体の作製)
シアノバクテリアにcytM遺伝子の不活性化を付与するには、既に説明したcytM遺伝子の各種形態の不活性化を付与するためのポリヌクレオチドを備える形質転換ベクターを用いることができる。ポリヌクレオチドは、概してDNA及びRNAであるが、典型的にはDNAである。この形質転換ベクターは、シアノバクテリアに暗所従属栄養能を付与するためのベクターとして用いることができる。また、上記ポリヌクレオチド(例えば、少なくとも標的となるcytM遺伝子を破壊し置換するための相同性領域を備えるポリヌクレオチドカセット)は、それ自体、シアノバクテリアに暗所従属栄養能を付与するための暗所従属栄養能付与剤として利用することができる。 (Production of transformation vector and transformant)
In order to impart inactivation of the cytM gene to cyanobacteria, a transformation vector comprising a polynucleotide for imparting inactivation of various forms of the cytM gene already described can be used. Polynucleotides are generally DNA and RNA, but are typically DNA. This transformation vector can be used as a vector for imparting dark heterotrophic ability to cyanobacteria. In addition, the above-described polynucleotide (for example, a polynucleotide cassette having at least a homology region for destroying and replacing the target cytM gene) is itself a dark place for imparting dark heterotrophic ability to cyanobacteria. It can be used as a heterotrophic agent.
シアノバクテリアにcytM遺伝子の不活性化を付与するには、既に説明したcytM遺伝子の各種形態の不活性化を付与するためのポリヌクレオチドを備える形質転換ベクターを用いることができる。ポリヌクレオチドは、概してDNA及びRNAであるが、典型的にはDNAである。この形質転換ベクターは、シアノバクテリアに暗所従属栄養能を付与するためのベクターとして用いることができる。また、上記ポリヌクレオチド(例えば、少なくとも標的となるcytM遺伝子を破壊し置換するための相同性領域を備えるポリヌクレオチドカセット)は、それ自体、シアノバクテリアに暗所従属栄養能を付与するための暗所従属栄養能付与剤として利用することができる。 (Production of transformation vector and transformant)
In order to impart inactivation of the cytM gene to cyanobacteria, a transformation vector comprising a polynucleotide for imparting inactivation of various forms of the cytM gene already described can be used. Polynucleotides are generally DNA and RNA, but are typically DNA. This transformation vector can be used as a vector for imparting dark heterotrophic ability to cyanobacteria. In addition, the above-described polynucleotide (for example, a polynucleotide cassette having at least a homology region for destroying and replacing the target cytM gene) is itself a dark place for imparting dark heterotrophic ability to cyanobacteria. It can be used as a heterotrophic agent.
形質転換ベクター上のポリヌクレオチドは、例えば、シアノバクテリアの染色体上の1又は2以上のcytM遺伝子を破壊しあるいは変異等を導入可能にそれぞれ適切な相同組換え領域を備えるとともに、必要に応じて、例えば外来遺伝子等をコードする置換領域や、薬剤耐性マーカー領域や不活性シトクロムcMタンパク質を産生させるための変異のための置換領域)を備えている。
The polynucleotide on the transformation vector has, for example, an appropriate homologous recombination region capable of disrupting one or more cytM genes on the chromosome of cyanobacteria or introducing mutations, etc. for example, with or substituted region encoding a foreign gene or the like, a replacement area) for the mutation for the production of drug resistance marker region and inactive cytochrome c M protein.
形質転換ベクターは、さらに、1つ以上のエンハンサー若しくはサイレンサー、オペレーター、ターミネーター、ポリアデニル化シグナル、1つ以上の薬剤耐性遺伝子、外来遺伝子を挿入するためのクローニング部位などを含んでいてもよい。
The transformation vector may further contain one or more enhancers or silencers, an operator, a terminator, a polyadenylation signal, one or more drug resistance genes, a cloning site for inserting a foreign gene, and the like.
薬剤耐性遺伝子としては、以下に限定されるものではないが、クロラムフェニコール耐性遺伝子、ネオマイシン耐性遺伝子、ハイグロマイシン耐性遺伝子、カナマイシン耐性遺伝子、アンピシリン耐性遺伝子、スペクチノマイシン耐性遺伝子などが挙げられる。
Drug resistance genes include, but are not limited to, chloramphenicol resistance gene, neomycin resistance gene, hygromycin resistance gene, kanamycin resistance gene, ampicillin resistance gene, spectinomycin resistance gene and the like.
形質転換ベクターは、特にその形態を特定しないで、各種形態を採ることができる。DNA断片であってもよいが、典型的にはプラスミドベクターであるが、例えば、ウイルスベクターなどの非プラスミドベクターも使用できる。また好ましくは、本明細書に開示される形質転換ベクターは、複数の宿主細胞に適合させるためのシャトルベクターである。例えば、大腸菌及びシアノバクテリアのいずれの細胞にも導入可能であり、これらの宿主細胞中で外来遺伝子を発現し得るベクターが挙げられる。このようなシャトルベクターは大腸菌を用いて大量に複製することが可能である。
The transformation vector can take various forms without particularly specifying the form. Although it may be a DNA fragment, it is typically a plasmid vector, but for example, non-plasmid vectors such as viral vectors can also be used. Also preferably, the transformation vector disclosed herein is a shuttle vector for adapting to multiple host cells. For example, vectors that can be introduced into any cell of E. coli and cyanobacteria and can express foreign genes in these host cells can be mentioned. Such shuttle vectors can be replicated in large quantities using E. coli.
本明細書の開示の形質転換ベクターは、シアノバクテリアで高効率に遺伝子を発現させ得る限り、母体となるベクターの由来は特に限定されない。例えば、シアノバクテリアで使用されるベクターであるpVZ321(Zinchenko VV, Piven IV, Melnik VA, Shestakov SV (1999) Genetika 35:291-296)、pMB1(Kreps S, Ferino F, Mosrin C, Gerits J, Mergeay M, Thuriaux P (1990) Mol. Gen. Genet. 221:129-133)、RSF1010(Meyer R (2009) Plasmid 62:57-70)などの公知のベクターを利用することができる。ベクターに各領域を挿入するには、適宜リガーゼ反応等を利用することができる。
The origin of the parent vector of the transformation vector disclosed in the present specification is not particularly limited as long as the gene can be expressed efficiently with cyanobacteria. For example, pVZ321 (Zinchenko VV, Piven IV, Melnik VA, Shestakov SV (1999) Genetika 35: 291-296), pMB1 (Kreps S, Ferino F, Mosrin C, Mergeay C, Mergeay), vectors used in cyanobacteria Known vectors such as M, Thuriaux1990P (1990) Mol. Gen. Genet.133221: 129-133), RSF1010 (Meyer R (2009) Plasmid 62: 57-70) can be used. To insert each region into the vector, a ligase reaction or the like can be used as appropriate.
形質転換ベクターは、さらに、シアノバクテリアで発現させようとする外来遺伝子を備えていてもよい。外来遺伝子としては、形質転換ベクター等により外部から導入される遺伝子であればよく、シアノバクテリアに内在する遺伝子であってもよい。外来遺伝子としては、各種の有用物質あるいは当該有用物質を生産するための1又は2以上の酵素をコードするものであってもよい。例えば、医薬成分として有用なタンパク質(例えばインターフェロン、インスリン、エリスロポエチン、ヒト成長ホルモン、各種サイトカイン)、試薬として有用なタンパク質(例えばDNA/RNAポリメラーゼ、プロテアーゼ、制限酵素)、蛍光タンパク質(例えばGFP、DsRed、フィコシアニン)、バイオプラスチック製造関連タンパク質(例えばポリヒドロキシアルカン酸(PHA)、ポリヒドロキシ酪酸(PHB)又はポリヒドロキシ吉草酸(PHV)合成酵素)、アミノ酸合成酵素、バイオ燃料関連合成酵素(例えばトリグリセリド、脂肪酸(ステアリン酸若しくはオレイン酸など)又は炭化水素(アルカン/アルケン)合成酵素、水素生産酵素)、抗菌活性物質合成酵素、機能性物質合成酵素(機能性物質としては、例えばγ-リノレン酸、ドコサヘキサエン酸(DHA)、エイコサペンタエン酸、β-カロテン、アスタキサンチンがある)などをコードする遺伝子が挙げられ、特に、バイオ燃料関連合成酵素遺伝子が好ましい。また、脂肪酸生産のためのチオエステラーゼ遺伝子、ニトロゲナーゼの構造遺伝子、nifE, nifN, nifH, nifU, nifS, nifB, nifP, nifM nifV, nifZ, nifX, nifW, hesA, hesB, fdxB, fdxN, fdxHなどのニトロゲナーゼの金属中心生合成に関わる遺伝子、ヒドロゲナーゼ構造遺伝子、hydE, hydGなどのヒドロゲナーゼの金属中心生合成に関わる遺伝子が挙げられる。こうした有用物質等をコードする遺伝子は、GenBankなどの既存のデータベースに登録されている塩基配列又はアミノ酸配列の情報から設計したプライマー又はプローブを利用して、PCR法又はハイブリダイゼーション法などの公知の方法により取得することができる。
The transformation vector may further comprise a foreign gene to be expressed in cyanobacteria. The foreign gene may be a gene introduced from the outside by a transformation vector or the like, and may be a gene endogenous to cyanobacteria. The foreign gene may encode various useful substances or one or more enzymes for producing the useful substances. For example, proteins useful as pharmaceutical ingredients (eg, interferon, insulin, erythropoietin, human growth hormone, various cytokines), proteins useful as reagents (eg, DNA / RNA polymerase, protease, restriction enzyme), fluorescent proteins (eg, GFP, DsRed, Phycocyanin), bioplastic production related proteins (eg polyhydroxyalkanoic acid (PHA), polyhydroxybutyric acid (PHB) or polyhydroxyvaleric acid (PHV) synthase), amino acid synthases, biofuel related synthases (eg triglycerides, fatty acids) (Such as stearic acid or oleic acid) or hydrocarbon (alkane / alkene) synthase, hydrogen producing enzyme), antibacterial active substance synthase, functional substance synthase (for example, γ-linolenic acid, docosahexaene Acid (DHA), eicosapentaenoic acid, β-carotene, and astaxanthin) and the like, and biofuel-related synthase genes are particularly preferable. Also, thioesterase gene for fatty acid production, structural gene of nitrogenase, nifE, nifN, nifH, nifU, nifS, nifB, nifP, nifM nifV, nifZ, nifX, nifW, hesA, hesB, fdxN, fdxH, Examples include genes involved in metal-center biosynthesis of nitrogenase, hydrogenase structural genes, and genes involved in metal-center biosynthesis of hydrogenases such as hydE and hydG. A gene encoding such a useful substance is a known method such as a PCR method or a hybridization method using a primer or probe designed from information on the base sequence or amino acid sequence registered in an existing database such as GenBank. It can be obtained by.
シアノバクテリアに対して本明細書開示の形質転換ベクターを用いてシアノバクテリアについての公知の形質転換法を用いることで、シアノバクテリアの形質転換体を得ることができる。シアノバクテリアの形質転換法としては、例えば、自然形質転換法及びエレクトロポレーション法を使用することができる。また、接合伝達法を用いることができる。自然形質転換法とは、シアノバクテリアと形質転換体ベクターとを混合するだけで遺伝子導入が可能な方法である。エレクトロポレーション法とは、シアノバクテリアと形質転換体ベクターの混合溶液に電気パルスをかけることで、遺伝子導入を行う方法である。接合伝達法とは、導入したい遺伝子を挿入したプラスミドを大腸菌に形質転換した後、その大腸菌とシアノバクテリアを混合し遺伝子導入を行う方法である。これらの方法は、当業者であれば適宜実施することができる。また、形質転換体を選択するには、形質転換ベクターに保持させた薬剤耐性遺伝子等を用いることができる。
A cyanobacterial transformant can be obtained by using a known transformation method for cyanobacteria using the transformation vector disclosed herein for cyanobacteria. As a transformation method of cyanobacteria, for example, natural transformation method and electroporation method can be used. Further, a junction transmission method can be used. The natural transformation method is a method in which gene introduction is possible only by mixing cyanobacteria and a transformant vector. The electroporation method is a method for introducing a gene by applying an electric pulse to a mixed solution of cyanobacteria and a transformant vector. The conjugation transfer method is a method in which a plasmid into which a gene to be introduced is inserted is transformed into E. coli, and then the E. coli and cyanobacteria are mixed to introduce the gene. Those skilled in the art can appropriately implement these methods. Moreover, in order to select a transformant, the drug resistance gene etc. which were hold | maintained at the transformation vector can be used.
こうして得られた形質転換体であるシアノバクテリアは、本来備えている光合成独立栄養能と、初めて付与された暗所従属栄養能又は増強された暗所従属栄養能を備えることができる。暗所従属栄養能の有無や程度は、暗所において、グルコースなどの有機栄養成分を含む培地を用いた従属栄養条件下にて形質転換体を培養した際に、生育又は増殖を観察することで判断することができる。
The thus obtained transformant, cyanobacteria, can have the inherent photosynthetic autotrophic ability and the dark heterotrophic ability imparted for the first time or the enhanced dark heterotrophic ability. The presence or degree of heterotrophic ability in the dark can be determined by observing growth or proliferation when transformants are cultured under heterotrophic conditions using a medium containing organic nutrients such as glucose in the dark. Judgment can be made.
本開示の形質転換体は、暗所従属栄養能が付与ないし増強されているために、太陽光などの天然の光エネルギーを利用しつつかつ当該光エネルギーの枯渇状態(暗所)であっても、従属栄養的に生育・増殖できるため、昼夜を通して連続的な生育等と物質生産が可能となる。すなわち、効率的で実用的な生育等と物質生産が可能となっている。また、必要に応じて暗所従属条件下においてのみ、あるいは光エネルギーの存在下でのみ生育等させることで、生産させる物質対象を異ならせることもできる。
Since the transformant of the present disclosure has imparted or enhanced dark heterotrophic ability, it uses natural light energy such as sunlight and is in a depleted state (dark place) of the light energy. Because it can grow and proliferate in a heterotrophic manner, continuous growth and material production are possible throughout the day and night. That is, efficient and practical growth and material production are possible. In addition, the substances to be produced can be made different by growing them only in the dark dependent conditions or only in the presence of light energy as necessary.
本開示の形質転換体は、さらに、有用物質を生産するための外来遺伝子を備えることもできる。こうした形質転換体によれば、光エネルギーと有機化合物とを利用して効率的に及び/又は選択的に有用物質を生産することができる。
The transformant of the present disclosure can further include a foreign gene for producing a useful substance. According to such a transformant, a useful substance can be efficiently and / or selectively produced using light energy and an organic compound.
なお、以上の開示によれば、本明細書に開示される形質転換体の製造方法も提供される。また、以上の開示によれば、形質転換体でないが、cytM遺伝子の不活性化を備え、暗所従属栄養能を有する、シアノバクテリアも提供される。シアノバクテリアに対して、cytM遺伝子の不活性化や不存在に基づいてスクリーニングすることで天然のシアノバクテリアから、暗所従属栄養能に優れるシアノバクテリアを容易に選択することができるからである。天然のシアノバクテリアとしては、L. boryana以外のシアノバクテリアを対象とすることが好ましい。また、こうしてスクリーニングされたシアノバクテリアに対して有用物質を生産するための外来遺伝子を上記方法に準じて導入して形質転換することで、実用的なシアノバクテリアを得ることができる。
In addition, according to the above indication, the manufacturing method of the transformant disclosed by this specification is also provided. Moreover, according to the above disclosure, a cyanobacteria that is not a transformant but has inactivation of the cytM gene and has dark heterotrophic ability is also provided. This is because screening for cyanobacteria based on the inactivation or absence of the cytM gene makes it possible to easily select cyanobacteria having excellent dark heterotrophic ability from natural cyanobacteria. As natural cyanobacteria, it is preferable to target cyanobacteria other than L. boryana. Moreover, a practical cyanobacteria can be obtained by introducing and transforming a foreign gene for producing a useful substance into the thus screened cyanobacteria according to the above method.
(シアノバクテリアの培養産物の生産方法)
本明細書の開示によれば、シアノバクテリアの培養産物の生産方法も提供される。この生産方法は、本明細書に開示されるcytM遺伝子の不活性化を備えるシアノバクテリア又はシアノバクテリア形質転換体を培養する工程を備えることができる。この生産方法によれば、暗所従属栄養能が付与されあるいは増強されているために、連続的、効率的、あるいは選択的にシアノバクテリアを培養し、その培養産物を得ることができる。 (Production method of cyanobacterial culture product)
According to the disclosure of the present specification, a method for producing a culture product of cyanobacteria is also provided. This production method may comprise a step of culturing a cyanobacteria or cyanobacterial transformant provided with inactivation of the cytM gene disclosed in the present specification. According to this production method, since the dark heterotrophic ability is imparted or enhanced, cyanobacteria can be cultured continuously, efficiently, or selectively, and the culture product can be obtained.
本明細書の開示によれば、シアノバクテリアの培養産物の生産方法も提供される。この生産方法は、本明細書に開示されるcytM遺伝子の不活性化を備えるシアノバクテリア又はシアノバクテリア形質転換体を培養する工程を備えることができる。この生産方法によれば、暗所従属栄養能が付与されあるいは増強されているために、連続的、効率的、あるいは選択的にシアノバクテリアを培養し、その培養産物を得ることができる。 (Production method of cyanobacterial culture product)
According to the disclosure of the present specification, a method for producing a culture product of cyanobacteria is also provided. This production method may comprise a step of culturing a cyanobacteria or cyanobacterial transformant provided with inactivation of the cytM gene disclosed in the present specification. According to this production method, since the dark heterotrophic ability is imparted or enhanced, cyanobacteria can be cultured continuously, efficiently, or selectively, and the culture product can be obtained.
本生産方法としては、特に、完全暗所従属栄養能が付与・増強又は完全暗所従属栄養能的に改変されているために暗所での従属栄養条件での培養工程を備えることが好ましい。
This production method preferably includes a culture step under heterotrophic conditions in the dark particularly because the complete dark heterotrophic ability is imparted / enhanced or modified in the complete dark heterotrophic capacity.
培養産物としては、培養菌体及び培養上清を含むことができるが、その一部であってもよい。また、シアノバクテリアにおいて外来遺伝子が導入されて外来遺伝子の発現に基づく有用物質を生産している場合には、当該有用物質も培養産物に含まれる。
The culture product can include cultured cells and culture supernatant, but may be a part thereof. When a foreign gene is introduced in cyanobacteria and a useful substance based on the expression of the foreign gene is produced, the useful substance is also included in the culture product.
培養工程としては、太陽エネルギーなどの天然の光エネルギー条件、従属栄養条件等を適宜組み合わせることができる。その他の培養条件としては、公知のシアノバクテリアの培養条件を採用することができる。
As the culture process, natural light energy conditions such as solar energy, heterotrophic conditions and the like can be appropriately combined. As other culture conditions, known culture conditions for cyanobacteria can be employed.
以上の開示によれば、シアノバクテリアの培養方法も提供される。本培養方法は、cytM遺伝子の不活性化を備えるシアノバクテリア又はシアノバクテリア形質転換体を暗所で培養する工程を備えることができる。この培養方法によれば、光エネルギーがなくても、シアノバクテリアを生育・増殖させることができる。また、光エネルギー下での独立栄養条件下でもシアノバクテリアを生育・増殖させることができ、効率的な培養が可能となる。
According to the above disclosure, a method for culturing cyanobacteria is also provided. This culture method can comprise a step of culturing cyanobacteria or cyanobacteria transformants having inactivation of the cytM gene in the dark. According to this culture method, cyanobacteria can be grown and propagated without light energy. In addition, cyanobacteria can grow and proliferate even under autotrophic conditions under light energy, enabling efficient culture.
(dg5の単離)
Leptolyngbya boryana(旧名Plectonema boryanum)IAM-M101の完全暗所従属栄養能が向上した自然変異株dg5は、1991年藤田によって野生株から単離された(Fujita et al. 1996, Plant and Cell Physiology 37:313-323)。 (Isolation of dg5)
The natural mutant dg5 of Leptolyngbya boryana (formerly Plectonema boryanum) IAM-M101 with improved complete dark heterotrophic ability was isolated from a wild strain by Fujita in 1991 (Fujita et al. 1996, Plant and Cell Physiology 37: 313-323).
Leptolyngbya boryana(旧名Plectonema boryanum)IAM-M101の完全暗所従属栄養能が向上した自然変異株dg5は、1991年藤田によって野生株から単離された(Fujita et al. 1996, Plant and Cell Physiology 37:313-323)。 (Isolation of dg5)
The natural mutant dg5 of Leptolyngbya boryana (formerly Plectonema boryanum) IAM-M101 with improved complete dark heterotrophic ability was isolated from a wild strain by Fujita in 1991 (Fujita et al. 1996, Plant and Cell Physiology 37: 313-323).
(野生株のゲノム配列の再決定)
野生株(WTo)のゲノム配列を後述のようにして決定し、既知のdg5ゲノム配列との比較を行った。 (Redetermining the genome sequence of the wild strain)
The genome sequence of the wild strain (WTo) was determined as described below, and compared with a known dg5 genome sequence.
野生株(WTo)のゲノム配列を後述のようにして決定し、既知のdg5ゲノム配列との比較を行った。 (Redetermining the genome sequence of the wild strain)
The genome sequence of the wild strain (WTo) was determined as described below, and compared with a known dg5 genome sequence.
ゲノムDNAの抽出には以下のようにWizard Genomic DNA Purification kit (Promega)を用い、マニュアルを一部改変して行った。WToの細胞約20 mgを450 μlの緩衝溶液(50 mM Tris-HCl pH 8.0, 50 mM NaCl, 5 mM EDTA)に懸濁し、50 μLのリゾチーム (2.5 mg/50 μl)を加えた。その後、37 ℃で1時間静置した。サンプルを遠心し (15,000 rpm, 2分, 室温)、上清を除いた。その後の沈殿にNuclei Lysis Solution 0.6 mlを加える手順からは、Wizard Genomic DNA Purification kit (Promega)のマニュアルにそって行った。
Genomic DNA was extracted using the Wizard Genomic DNA Purification kit (Promega) as shown below and partially modified from the manual. About 20 mg of WTo cells were suspended in 450 μl of a buffer solution (50 mM Tris-HCl pH 8.0, 50 mM NaCl, 5 mM NaCl, EDTA), and 50 μL of lysozyme (2.5 mg / 50 μl) was added. Then, it left still at 37 degreeC for 1 hour. The sample was centrifuged (15,000 rpm, 2 minutes, room temperature), and the supernatant was removed. From the procedure of adding NucleileLysis Solution 0.6 ml to the subsequent precipitate, the procedure was followed according to the Wizard Genomic DNA Purification kit (Promega) manual.
ゲノム配列決定の方法として高速シークエンサーの一つSOLiD system 5500を用いた。Paired-endでのゲノムライブラリーをマニュアルに沿って作成し、シークエンスを行った。結果、ライブラリーDNA断片4,802,686本の両側からのシークエンンスデータを得た。なお、この際対照としてdg5のゲノム配列の同様にSOLiD system 5500を用いてシークエンスデータを得た。このデータを解析ソフトBioScope (Applied Biosystems)を用い、dg5ゲノム配列をレファレンスとしてデータのマッピングを行った後、一塩基多型(SNP)と挿入欠失(Indel)解析を行った。その際の設定は初期設定を用いた。
As a genome sequencing method, one of the high-speed sequencers, SOLiD system 5500, was used. A paired-end genomic library was created according to the manual and sequenced. As a result, sequence data from both sides of 4,802,686 library DNA fragments were obtained. At this time, as a control, sequence data was obtained using SOLiD system 5500 in the same manner as the genomic sequence of dg5. The data was mapped using the analysis software BioScope® (Applied® Biosystems) with the dg5 genomic sequence as a reference, and then single nucleotide polymorphism (SNP) and insertion deletion (Indel) analysis was performed. The initial setting was used for the setting at that time.
ゲノム配列の再決定を行った結果、dg5とWToのゲノムには4か所の差異が存在していた(表2)。ただし、dg5由来の他の変異株のゲノム配列も同様に再決定しており、4か所のうち1か所についてはこのdg5由来の変異株はWToと同じ配列であったため、この変異はdg5が単離された後、最近になって固定した変異だと考え、候補から除外した。その結果、残った3か所について該当部位を含む領域を表3に示したプライマーで、WToの細胞を用いたコロニーダイレクトPCRで増幅し、その断片についてサンガー法を用いて塩基配列を決定し、WToの配列がdg5の配列と異なっていることを確認した。なお、サンガー法によるシークエンスの際に用いたプライマーはPb1305262-r、Pb3151287-f、Pb5175165-fである。
As a result of re-determining the genome sequence, there were four differences between dg5 and WTo genomes (Table 2). However, the genome sequences of other mutants derived from dg5 were also redetermined in the same way, and for one of the four sites, the mutant derived from dg5 had the same sequence as WTo, so this mutation was dg5 After being isolated, it was considered a recently fixed mutation and was excluded from the candidates. As a result, the region containing the corresponding site for the remaining 3 sites was amplified by colony direct PCR using WTo cells with the primers shown in Table 3, and the nucleotide sequence of the fragment was determined using the Sanger method. It was confirmed that the sequence of WTo is different from the sequence of dg5. The primers used in the sequencing by the Sanger method are Pb1305262-r, Pb3151287-f, and Pb5175165-f.


図2に、cytM遺伝子におけるdg5における変異の概要を示す。フレームシフト変異によりタンパク質のトランケートが生じていることがわかった。
FIG. 2 shows an overview of mutations at dg5 in the cytM gene. It was found that truncation of the protein was caused by the frameshift mutation.
(形質転換によるWToへの dg5変異の導入に用いたプラスミドの構築)
実施例2で同定したdg5とWToとの3つの遺伝的差異のうちどれが完全暗所従属栄養生育能を促進するのかを突き止めるため、それぞれのdg5型の塩基配列を個別にWToに導入した。 (Construction of plasmid used to introduce dg5 mutation into WTo by transformation)
In order to determine which of the three genetic differences between dg5 and WTo identified in Example 2 promotes complete dark heterotrophic viability, each dg5-type nucleotide sequence was individually introduced into WTo.
実施例2で同定したdg5とWToとの3つの遺伝的差異のうちどれが完全暗所従属栄養生育能を促進するのかを突き止めるため、それぞれのdg5型の塩基配列を個別にWToに導入した。 (Construction of plasmid used to introduce dg5 mutation into WTo by transformation)
In order to determine which of the three genetic differences between dg5 and WTo identified in Example 2 promotes complete dark heterotrophic viability, each dg5-type nucleotide sequence was individually introduced into WTo.
方法としてはオーバーラップPCRを用いてdg5変異をもつ3つの断片を連結し、これを高コピープラスミドであるpUC19のBamHIまたはBamHIとKpnIサイトにクローニングした。この際、大腸菌JM105株をプラスミド構築に用いた。使用したprimerの塩基配列を表4に示し、目的DNA断片におけるアニーリング部位を図3に示した。相同二重組換えの足場となる二つの断片はdg5ゲノムを鋳型としてprimer-f1とprimer-r1の組み合わせ、およびprimer-f3とprimer-r3の組み合わせでPCRを行った。またカナマイシン耐性カセットはprimer-f2とprimer-r2を用いてpUCK121を鋳型にして増幅した。各断片は増幅に用いたプライマーへの塩基付加によって、連結させる断片同士の末端30 bpの同一配列をもつ。これらの3つの断片を鋳型としてprimer-f1およびprimer-r3を用いて二次PCRを行うことで、3つの断片が連結した目的のDNA断片を得た。この断片は制限酵素サイトを持つようにprimer-f1およびprimer-r3が設計されている。BamHI、またはBamHIとKpnIで制限酵素処理を行い、pUC19の対応するクローニングサイトにクローニングした。
As a method, three fragments having the dg5 mutation were ligated using overlap PCR, and this was cloned into the BamHI or BamHI and KpnI sites of pUC19, which is a high copy plasmid. At this time, Escherichia coli JM105 strain was used for plasmid construction. The base sequences of the primers used are shown in Table 4, and the annealing sites in the target DNA fragment are shown in FIG. Two fragments serving as scaffolds for homologous double recombination were subjected to PCR using the combination of primer-f1 and primer-r1 and the combination of primer-f3 and primer-r3 using the dg5 genome as a template. The kanamycin resistance cassette was amplified using primer-f2 and primer-r2 using pUCK121 as a template. Each fragment has the same sequence at the end of 30 bp between the fragments to be ligated by base addition to the primer used for amplification. By performing secondary PCR using primer-f1 and primer-r3 using these three fragments as templates, the target DNA fragment in which the three fragments were ligated was obtained. Primer-f1 and primer-r3 are designed so that this fragment has a restriction enzyme site. The restriction enzyme was treated with BamHI or BamHI and KpnI, and cloned into the corresponding cloning site of pUC19.
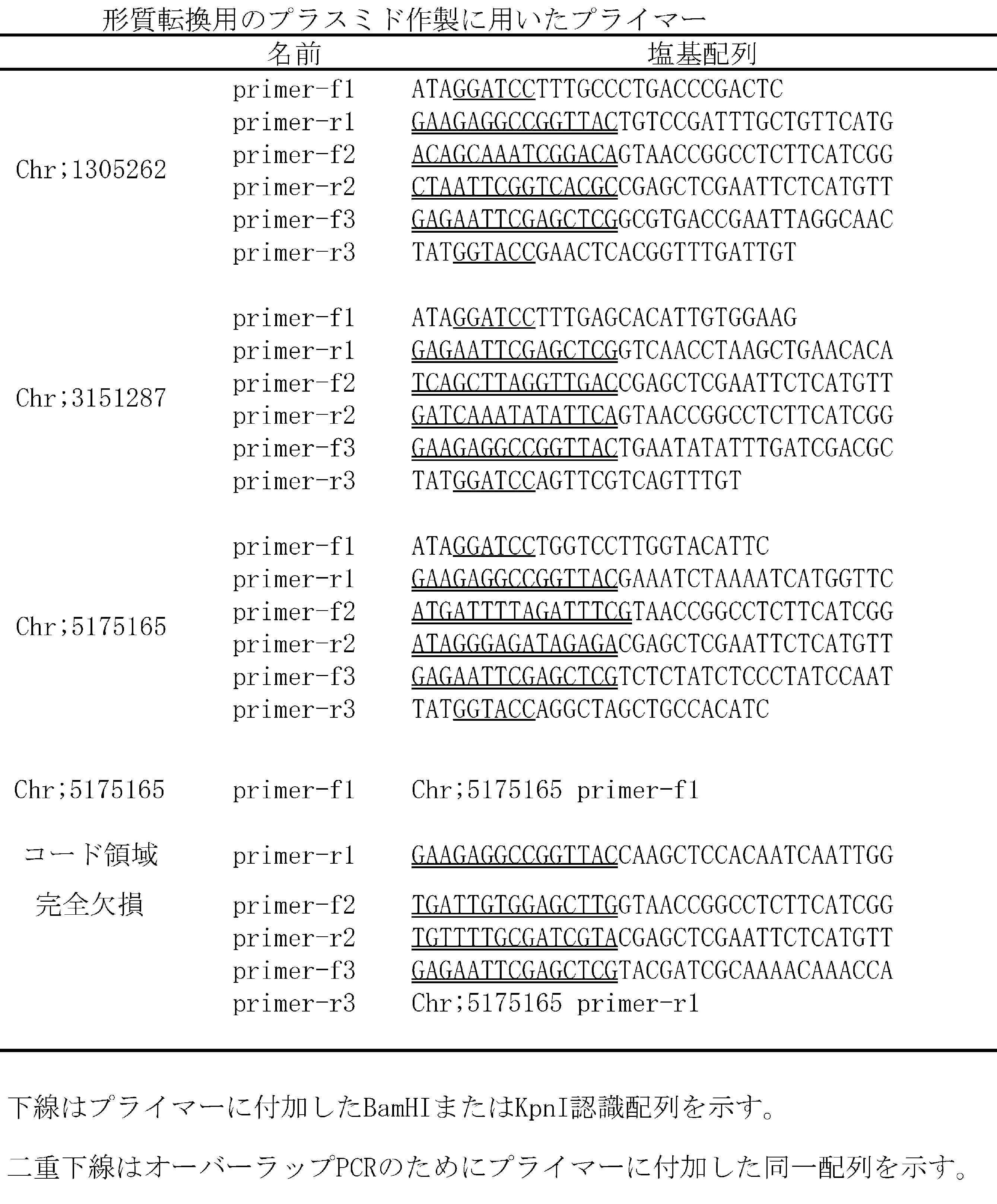
またChr;5175165の変異に関しては変異によってフレームシフトが生じ遺伝子の機能欠損を引き起こすと想定できたため、遺伝子のコード領域全てを除いた完全欠損株を作製するためのプラスミドを同様に作製した。
In addition, regarding the mutation of Chr; 5175165, it was assumed that the mutation caused a frame shift and caused a loss of function of the gene. Therefore, a plasmid for preparing a completely defective strain excluding the entire coding region of the gene was similarly prepared.
また対照株の作製のため、各々のdg5変異を有さない薬剤耐性マーカーのみを挿入されたプラスミドも同時に作成した。そのために連結する3つのDNA断片のうちdg5変異を保持した断片のみPCRによって増幅する段階でdg5ゲノムではなくWToのゲノムを鋳型としPCRを行い、その他は上記に記したプラスミド構築の手順に従った。
For the preparation of the control strain, a plasmid in which only the drug resistance marker not having each dg5 mutation was inserted was also prepared. For this purpose, PCR was performed using the WTo genome instead of the dg5 genome as a template at the stage of PCR amplification of only the three DNA fragments that retained the dg5 mutation, and the others followed the procedure for constructing the plasmid described above. .
WToへの二重相同組換えはエレクトロポレーション法を用いた(Fujita et al. 1992, Plant Cell Physiology, 33: 81-92)。形質転換には、作製したプラスミドを鋳型としてprimer-f1およびprimer-r3を用いて得たPCR断片を使用した。WToの培養はBG-11 (20 mM HEPES-KOH; pH 7.5含有、以下BG-11Hとする)、寒天(BactoAgar, Difco)15 % (w/v)の寒天培地を用いた。また、培養の光源として白色蛍光灯(FLR40SW/M/36-B, 日立)を用いた。30℃、寒天培地で光合成的に生育させたWToの細胞を水で回収し、150mL Bottle Top Vacuum Filter, 0.22 μm Pore (Corning)上において吸引した後、滅菌した冷水に再懸濁し、その後吸引して水を除いた。この操作を計3回行うことで細胞懸濁液から塩を除いた。この細胞を0.5-2.0 mlの冷却した滅菌水に再懸濁し、これをエレクトロポレーションに供するコンピテントセルとした。
Double-homologous recombination into WTo was performed by electroporation (Fujita et al. 1992, Plant Cell Physiology, 33: 81-92). For the transformation, PCR fragments obtained using primer-f1 and primer-r3 using the prepared plasmid as a template were used. For the WTo culture, an agar medium containing BG-11BG (20 M HEPES-KOH; containing pH 7.5, hereinafter referred to as BG-11H) and agar (BactoAgar, Difco) 15% (w / v) was used. In addition, a white fluorescent lamp (FLR40SW / M / 36-B, Sakai Hitachi) was used as a culture light source. WTo cells that have been photosynthesised on agar at 30 ° C are collected in water, aspirated on 150 mL Bottle Top Vacuum Filter, 0.22 μm Pore (Corning), resuspended in sterile cold water, and then aspirated. The water was removed. The salt was removed from the cell suspension by performing this operation three times in total. The cells were resuspended in 0.5-2.0 ml of chilled sterilized water and used as a competent cell for electroporation.
コンピテントセル50 μlと各PCR断片10 μlを混合し、電圧1410 V、抵抗250Ω、静電容量25 μF、電極間距離1 mmの設定で減衰パルス処理を行った。パルス処理した細胞を350 μlのBG-11 (20 mM HEPES-KOH; pH8.2含有)を加えて回収し、ナイロン膜(Amersham HybondTM-N+, GE Healthcare)を載せた寒天培地BG-11Hに広げた。30℃、光強度2 μmol photon m-2s-1で2日間生育させた後、ナイロン膜とともにカナマイシン(終濃度15 mg/L)を添加した寒天培地BG-11Hに移し、光強度50 μmol photon m-2s-1照射下で薬剤耐性株の選抜を行った。
A 50 μl competent cell and 10 μl of each PCR fragment were mixed and subjected to an attenuation pulse treatment at a voltage of 1410 V, a resistance of 250Ω, a capacitance of 25 μF, and a distance between electrodes of 1 mm. The pulsed cells 350 [mu] l of BG-11 (20 mM HEPES- KOH; pH8.2 containing) was recovered by the addition of a nylon membrane (Amersham Hybond TM -N +, GE Healthcare) on an agar medium BG-11H bearing the Spread out. After growing for 2 days at 30 ° C, light intensity 2 μmol photon m -2 s -1 , transferred to agar medium BG-11H supplemented with kanamycin (final concentration 15 mg / L) with nylon membrane, light intensity 50 μmol photon Drug-resistant strains were selected under irradiation with m −2 s −1 .
2週間後、カナマイシン耐性株として選抜されたdg5変異個別導入株をdm1 (Chr;1305262変異を有する)、dm2 (Chr;3151287を有する)、dm3 (Chr;5175165を有する)、dm3n (Chr;5175165を含む遺伝子(ORF番号LBDG_45050)の完全欠損株)と称する。これらの変異株のdg5変異導入の確認のため、表3に示したprimer-f1とprimer-r3を用いてPCRにより、細胞から直接DNA断片を増幅した。同時に、カナマイシン耐性カセットが二重組換えによってゲノムに挿入されていることを確認した。
Two weeks later, the individual dg5 mutant strains selected as kanamycin resistant strains were dm1 (having Chr; 1305262 mutation), dm2 (having Chr; 3151287), dm3 (having Chr; 5175165), dm3n (Chr; 5175165 And a gene (ORF No. LBDG_45050) completely deficient strain). In order to confirm the introduction of dg5 mutation in these mutant strains, DNA fragments were directly amplified from the cells by PCR using primer-f1 and primer-r3 shown in Table 3. At the same time, it was confirmed that the kanamycin resistance cassette was inserted into the genome by double recombination.
さらにそのDNA断片をアガロースゲルから精製後、鋳型として用い、Chr;1305262はPb1305262-fとprimer-r2の組み合わせ、Chr;3151287に対してはPb3151287-fとprimer-r2の組み合わせ、Chr;5175165に対してはprimer-f2とPb5175165-rの組み合わせで二次PCRを行い、得られたDNA断片をサンガー法によってシークエンスし、dg5変異の導入を確認した。なお、サンガー法によるシークエンスの際に用いたプライマーは表1に示したPb1305262-f、Pb3151287-f、Pb5175165-rである。
Further, the DNA fragment was purified from an agarose gel and used as a template, Chr; 1305262 was a combination of Pb1305262-f and primer-r2, Chr; 3151287 was a combination of Pb3151287-f and primer-r2, Chr: 5175165 In contrast, secondary PCR was performed with a combination of primer-f2 and Pb5175165-r, and the resulting DNA fragment was sequenced by the Sanger method to confirm the introduction of the dg5 mutation. The primers used for sequencing by the Sanger method are Pb1305262-f, Pb3151287-f, and Pb5175165-r shown in Table 1.
(形質転換体の完全暗所従属栄養生育の比較)
実施例3で得られた4種の変異株およびその対照株(dm1c, dm2c, dm3c;カナマイシン耐性を示すが該当する変異が導入されていない株)を用いて完全暗所従属栄養条件での生育比較を行った。 (Comparison of complete dark heterotrophic growth of transformants)
Growth under complete heterotrophic conditions using the four mutant strains obtained in Example 3 and their control strains (dm1c, dm2c, dm3c; strains that are resistant to kanamycin but have no corresponding mutation introduced) A comparison was made.
実施例3で得られた4種の変異株およびその対照株(dm1c, dm2c, dm3c;カナマイシン耐性を示すが該当する変異が導入されていない株)を用いて完全暗所従属栄養条件での生育比較を行った。 (Comparison of complete dark heterotrophic growth of transformants)
Growth under complete heterotrophic conditions using the four mutant strains obtained in Example 3 and their control strains (dm1c, dm2c, dm3c; strains that are resistant to kanamycin but have no corresponding mutation introduced) A comparison was made.
生育条件として30℃、暗所または明所(光強度50 μmol photon m-2s-1)における30 mMグルコース添加および無添加の寒天培地BG-11H上でのdg5、WToおよび上記の変異株の生育を確認した。完全暗所従属栄養条件では14日、明所光合成条件では3日間生育させた。なお、培養の光源として白色蛍光灯(FLR40SW/M/36-B, 日立)を用いた。結果を図4に示す。
Growth conditions of dg5, WTo and the above mutants on agar medium BG-11H with and without 30 mM glucose in the dark or light (light intensity 50 μmol photon m -2 s -1 ) Growth was confirmed. The plants were grown for 14 days under completely dark heterotrophic conditions and for 3 days under photosynthesis conditions. A white fluorescent lamp (FLR40SW / M / 36-B, Hitachi) was used as a culture light source. The results are shown in FIG.
また、WT、dg5、dm3およびdm3cの4株について、暗所従属栄養生育および光合成独立生育を液体培養によって評価した。前培養として、30 ℃、光強度60 μmol photon s-1 m-2で光合成的に3日間生育させた。この際、培養容器として25 cm2 Tissue Culture Flask (IWAKI)を用い、振盪はROTARY SHAKER NR-20 (TAITEC)で行った(回転速度140 rpm)。濁度(OD730)がおよそ0.4に達した前培養を種菌とし、初期濁度OD730=0.05となるように新しい液体培地に接種し、各条件で生育をモニターした。暗所従属栄養生育の評価では、アルミ箔で遮光した25 cm2 Tissue Culture Flask (IWAKI)に、最終濃度30 mMグルコース添加の15 ml BG-11培地を入れ,回転速度140 rpmで震盪培養した。また、光合成独立栄養生育の評価にはグルコースを含まない15 ml BG-11培地を用い、遮光せずに60 μmol photon s-1 m-2の光強度で、他の条件は同様にして培養した。なお、培養の光源として白色蛍光灯(FLR40SW/M/36-B, 日立)を用いた。結果を図5に示す。
In addition, for the four strains WT, dg5, dm3, and dm3c, dark heterotrophic growth and photosynthetic independent growth were evaluated by liquid culture. As pre-culture, the cells were grown for 3 days photosynthesis at 30 ° C. and light intensity of 60 μmol photon s −1 m −2 . At this time, 25 cm 2 Tissue Culture Flask (IWAKI) was used as a culture vessel, and shaking was performed with ROTARY SHAKER NR-20 (TAITEC) (rotation speed 140 rpm). A preculture with a turbidity (OD 730 ) of approximately 0.4 was used as an inoculum, inoculated into a new liquid medium so that the initial turbidity OD 730 = 0.05, and growth was monitored under each condition. For evaluation of dark heterotrophic growth, 15 ml BG-11 medium with a final concentration of 30 mM glucose was added to 25 cm 2 Tissue Culture Flask (IWAKI) shielded from aluminum foil, and cultured with shaking at a rotational speed of 140 rpm. In addition, for the evaluation of photosynthetic autotrophic growth, 15 ml BG-11 medium without glucose was used, and the light intensity was 60 μmol photon s -1 m -2 without shading, and the other conditions were cultured in the same manner . A white fluorescent lamp (FLR40SW / M / 36-B, Hitachi) was used as a culture light source. The results are shown in FIG.
図4に示すように、明所での光合成的生育はグルコースの有無に関わらず全ての株で有意な差は認められないが、暗所ではdm3およびdm3nのみがdg5と同様に顕著に良好な生育を示した。これはChr;5175165のdg5への変異(一塩基挿入)がLBDG_45050にフレームシフトを生じ遺伝子欠損を引き起こすことで生じたと推察される。LBDG_45050は相同性検索の結果、シトクロムcMをコードすることが分かった。シトクロム cMの欠損がdg5の完全暗所従属栄養生育能を促進した原因変異であることを示している。
As shown in FIG. 4, the photosynthetic growth in the light place is not significantly different in all strains regardless of the presence or absence of glucose, but in the dark place, only dm3 and dm3n are remarkably good like dg5. Showed growth. This is presumed that the mutation (single nucleotide insertion) of Chr; 5175165 to dg5 caused a frame shift in LBDG_45050 and caused gene deletion. LBDG_45050 the result of homology search, it was found that encodes a cytochrome c M. Cytochrome c M deficiency is a causal mutation that promotes dg5's complete dark heterotrophic viability.
また、図5に示すように、液体培地においても、明所では全ての株で有意な差は認められないが、暗所ではdm3およびdm3nのみがdg5と同様に顕著に良好な生育を示した。dg5、dm3は光独立栄養生育条件では、WT、dm3cと同様に生育するが、暗所従属栄養生育条件では、増殖速度および最終到達濁度が有意に高いことが確認された。
In addition, as shown in FIG. 5, even in the liquid medium, no significant difference was observed in all the strains in the light place, but only dm3 and dm3n showed significantly good growth like dg5 in the dark place. . dg5 and dm3 grew under the light autotrophic growth conditions in the same way as WT and dm3c, but under dark heterotrophic growth conditions, the growth rate and final turbidity were significantly higher.
本実施例では、シアノバクテリア(Synechocystis sp. PCC 6803)のチトクロムcM欠損株を作製して、従属栄養的生育の向上について評価した。すなわち、図6に示すように、cytM (sll1245) のコード領域全体をジェンタマイシン耐性カセットと置換することでΔcytM株を単離した。図6中、黒いバーはΔcytM単離のために構築したプラスミドにもちいたゲノム領域を示す。
In this example, a cytochrome cM-deficient strain of cyanobacteria (Synechocystis sp. PCC 6803) was produced and evaluated for improvement in heterotrophic growth. That is, as shown in FIG. 6, the ΔcytM strain was isolated by replacing the entire coding region of cytM (sll1245) with a gentamicin resistance cassette. In FIG. 6, the black bar indicates the genomic region used for the plasmid constructed for isolation of ΔcytM.
ΔcytM株においてcytM遺伝子がジェンタマイシン耐性カセット(gen) と完全に置換していることを、このカセットに特異的なプライマーf4, r4でWT(レーン1)とΔcytM株(レーン2)のコロニーをもちいたPCRにより確認した。結果を図7に示す。レーン3は遺伝子破壊にもちいたプラスミドを鋳型としたPCRの結果である。これらの結果により、ΔcytM株において確実にcytM遺伝子が破壊されていることがわかった。
In the ΔcytM strain, the cytM gene was completely replaced with the gentamicin resistance cassette (gen) 、, and colonies of WT (lane 1) and ΔcytM strain (lane 2) were obtained with primers f4 and r4 specific to this cassette. It was confirmed by PCR. The results are shown in FIG. Lane 3 shows the results of PCR using a plasmid used for gene disruption as a template. From these results, it was found that the cytM gene was surely disrupted in the ΔcytM strain.
次に、WTとΔcytM株について、光合成独立栄養条件(レーン1)、光合成混合栄養条件(レーン2)、LAHG条件(レーン3)および暗所従属栄養条件(レーン4)での生育を比較した。OD730が0.1の細胞懸濁液を5.0μlスポットした。図8中、レーン1と2ではBG-11H培地で3日間、レーン3と4ではグルコース含有BG-11H培地で10日間培養を行った。なお、LAHG (Light-activated heterotrophic growth光活性化従属栄養) 条件とは、24時間ごとに15分の光照射を行い、それ以外は暗所で培養を行ったことを示す。結果を図8に示す。
Next, WT and ΔcytM strains were compared in terms of growth under photosynthetic autotrophic conditions (lane 1), photosynthetic mixed nutrient conditions (lane 2), LAHG conditions (lane 3), and dark heterotrophic conditions (lane 4). 5.0 μl of a cell suspension with an OD 730 of 0.1 was spotted. In FIG. 8, lanes 1 and 2 were cultured in a BG-11H medium for 3 days, and lanes 3 and 4 were cultured in a glucose-containing BG-11H medium for 10 days. In addition, LAHG (Light-activated heterotrophic growth photoactivated heterotrophic) conditions indicate that light irradiation was performed every 24 hours for 15 minutes, and otherwise, the culture was performed in the dark. The results are shown in FIG.
図8に示すように、光合成条件(レーン1)ではΔcytMはWTと変わりがないが、グルコースによる混合栄養条件(レーン2)ではWTよりも有意に生育が促進された。さらに、LAHG条件(レーン3)ではその差はもっとも顕著となった。なお、本研究で用いたWT(“YF”)は、LAHG条件での生育がよくない株である。さらに、暗所従属栄養条件(レーン4)ではWTはまったく生育しないが、ΔcytMは明らかな生育が見られた。
As shown in FIG. 8, ΔcytM was not different from WT under the photosynthetic condition (lane 1), but growth was significantly promoted under WT in the mixed nutrient condition with glucose (lane 2). Furthermore, the difference was most noticeable under LAHG conditions (lane 3). The WT ("YF") used in this study is a strain that does not grow well under LAHG conditions. Furthermore, under dark heterotrophic conditions (lane 4), WT did not grow at all, but ΔcytM showed clear growth.
次に、グルコース含有BG-11H液体培地(15ml) にてWTとΔcytM株の培養ボトルをアルミホイルで覆い完全に光を遮蔽し140rpmの旋回培養(シェーカーNR-20、タイテック)を行い、生育はOD730(分光光度計UV-1700、島津)により評価した。結果を図9に示す。
Next, cover the culture bottle of WT and ΔcytM with aluminum foil in a glucose-containing BG-11H liquid medium (15 ml), completely shield the light, and perform a rotating culture at 140 rpm (Shaker NR-20, Tytech). Evaluation was performed by OD 730 (spectrophotometer UV-1700, Shimadzu). The results are shown in FIG.
図9に示すように、液体培地では、WTの生育は24時間以降ほとんど停止してしまうが、ΔcytM株は世代時間約33時間(24~92時間にかけての対数増殖期)で生育し、寒天培地の結果よりも明瞭に増殖が促進していることが確認された。
As shown in FIG. 9, in the liquid medium, the growth of WT almost ceases after 24 hours, but the ΔcytM strain grows at a generation time of about 33 hours (logarithmic growth phase from 24 to 92 hours), and an agar medium. It was confirmed that the proliferation was clearly promoted from the results of (1).
さらに、寒天培地BG-11Hにて光合成的に6日間生育させたSynechocystis sp. PCC 6803のWTとΔcytM株を滅菌水に懸濁し、濁度(OD730)を0.1に調製し、5.0μlを寒天培地BG-11H(光合成独立栄養)とグルコース(5mM)含有BG-11H(光合成混合栄養)寒天培地にスポットした。短日条件 (8L16D) では8時間明条件-16時間暗条件、長日条件(16L8D)では16時間明条件-8時間暗条件でいずれも4日間培養した。結果を図10に示す。
Further, WT and ΔcytM strains of Synechocystis sp. PCC 6803, photosynthesised on agar medium BG-11H for 6 days, were suspended in sterilized water, and turbidity (OD 730 ) was adjusted to 0.1. 0 μl was spotted on an agar medium BG-11H (photosynthesis autotroph) and glucose (5 mM) -containing BG-11H (photosynthesis mixed nutrition) agar medium. Under short-day conditions (8L16D), the cells were cultured for 4 days under 8 hours light conditions-16 hours dark conditions, and under long days conditions (16L8D) under 16 hours light conditions-8 hours dark conditions. The results are shown in FIG.
図10に示すように、光合成条件では短日、長日いずれもWTとΔcytM株の生育に差が見られないが、グルコース存在下の混合栄養条件では、特に短日条件においてΔcytMの生育がWTよりも促進していることがわかる。また、長日条件でもΔcytMの生育は有意にWTよりも良好に見える。この結果から、明暗周期を伴う環境においてもグルコース存在下であればΔcytM株がよりよい生育を示すことがわかった。
As shown in FIG. 10, there is no difference in the growth of the WT and ΔcytM strains on both the short and long days under the photosynthetic conditions. However, under the mixed nutrient conditions in the presence of glucose, the growth of ΔcytM is observed under the short day conditions. You can see that it promotes more. Also, ΔcytM growth appears significantly better than WT even under long day conditions. From this result, it was found that the ΔcytM strain exhibited better growth in the presence of glucose even in an environment with a light-dark cycle.
配列番号15~42:プライマー
SEQ ID NOs: 15-42: Primers
Claims (14)
- シトクロムcMタンパク質をコードするcytM遺伝子の不活性化を備える、シアノバクテリア形質転換体。 A cyanobacterial transformant comprising inactivation of the cytM gene encoding cytochrome c M protein.
- 前記不活性化は、前記cytM遺伝子の破壊又は活性が低下した前記タンパク質をコードする変異の導入である、請求項1に記載の形質転換体。 The transformant according to claim 1, wherein the inactivation is introduction of a mutation encoding the protein in which the cytM gene is disrupted or decreased in activity.
- 前記不活性化は、前記cytM遺伝子が配列番号3で表されるアミノ酸配列からなるタンパク質をコードする変異である、請求項1又は2に記載の形質転換体。 The transformant according to claim 1 or 2, wherein the inactivation is a mutation that encodes a protein having the amino acid sequence represented by SEQ ID NO: 3 in the cytM gene.
- 前記cytM遺伝子は、以下のいずれかに記載のタンパク質;
(a)配列番号2で表されるアミノ酸配列を有するタンパク質
(b)配列番号2で表されるアミノ酸配列と90%以上の同一性を有するアミノ酸配列を有し、シトクロムcMタンパク質と機能的に同等なタンパク質
(c)配列番号2で表されるアミノ酸配列において1又は複数個のアミノ酸の置換、欠失、挿入又は付加を有するアミノ酸配列を有し、シトクロムcMタンパク質と機能的に同等なタンパク質
(d)配列番号1で表される塩基配列でコードされるアミノ酸配列を有するタンパク質
(e)配列番号1で表される塩基配列と90%以上の同一性を有する塩基配列によってコードされるタンパク質であって、シトクロムcMタンパク質と機能的に同等なタンパク質をコードする、請求項1~3のいずれかに記載の形質転換体。 The cytM gene is a protein according to any of the following:
(A) a protein having the amino acid sequence represented by SEQ ID NO: 2 (b) an amino acid sequence having 90% or more identity with the amino acid sequence represented by SEQ ID NO: 2 and functionally associated with a cytochrome c M protein Equivalent protein (c) A protein functionally equivalent to cytochrome c M protein, having an amino acid sequence having one or more amino acid substitutions, deletions, insertions or additions in the amino acid sequence represented by SEQ ID NO: 2 (D) a protein having an amino acid sequence encoded by the nucleotide sequence represented by SEQ ID NO: 1 (e) a protein encoded by a nucleotide sequence having 90% or more identity with the nucleotide sequence represented by SEQ ID NO: 1 The transformant according to any one of claims 1 to 3, which encodes a protein functionally equivalent to the cytochrome c M protein. - 光合成独立栄養能と完全暗所従属栄養能とを備える、請求項1~4のいずれかに記載の形質転換体。 The transformant according to any one of claims 1 to 4, which has photosynthetic autotrophic ability and complete dark place heterotrophic ability.
- さらに、有用物質を生産するための外来遺伝子を備える、請求項1~5のいずれかに記載の形質転換体。 The transformant according to any one of claims 1 to 5, further comprising a foreign gene for producing a useful substance.
- シトクロムcMタンパク質をコードするcytM遺伝子の不活性化を付与するポリヌクレオチドを含む、シアノバクテリアに従属栄養能を付与するための形質転換ベクター。 A transformation vector for imparting heterotrophic ability to cyanobacteria, comprising a polynucleotide that imparts inactivation of a cytM gene encoding cytochrome c M protein.
- 前記不活性化は、前記cytM遺伝子の破壊又は活性が低下した前記タンパク質をコードする変異の導入である、請求項7に記載の形質転換ベクター。 The transformation vector according to claim 7, wherein the inactivation is introduction of a mutation encoding the protein in which the cytM gene is disrupted or decreased in activity.
- 請求項1~6のいずれかに記載の形質転換体の製造方法であって、
シアノバクテリアに、シトクロムcMタンパク質をコードするcytM遺伝子の不活性化を付与するポリヌクレオチドで形質転換する工程、
を備える、製造方法。 A method for producing the transformant according to any one of claims 1 to 6,
Transforming cyanobacteria with a polynucleotide conferring inactivation of the cytM gene encoding cytochrome c M protein;
A manufacturing method comprising: - 前記不活性化は、前記cytM遺伝子の破壊又は活性が低下した前記タンパク質をコードする変異の導入である、請求項9に記載の製造方法。 The production method according to claim 9, wherein the inactivation is the introduction of a mutation encoding the protein whose cytM gene has been disrupted or reduced in activity.
- 前記形質転換工程は、前記ポリヌクレオチドをエレクトロポレーションで前記シアノバクテリアに導入する、請求項9又は10に記載の製造方法。 The method according to claim 9 or 10, wherein in the transformation step, the polynucleotide is introduced into the cyanobacteria by electroporation.
- 請求項1~6のいずれかに記載の形質転換体を培養する工程、
を備える、シアノバクテリアの培養産物の生産方法。 Culturing the transformant according to any one of claims 1 to 6,
A method for producing a cyanobacterial culture product. - 前記培養工程は、暗所での培養を含む、請求項12に記載の生産方法。 The production method according to claim 12, wherein the culturing step includes culturing in a dark place.
- 請求項1~6のいずれかに記載の形質転換体を暗所で培養する工程、
を備える、シアノバクテリアの培養方法。 A step of culturing the transformant according to any one of claims 1 to 6 in a dark place;
A method for culturing cyanobacteria, comprising:
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2016507766A JPWO2015137352A1 (en) | 2014-03-10 | 2015-03-10 | Genes related to complete dark heterotrophic capacity and their utilization in cyanobacteria |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2014-046585 | 2014-03-10 | ||
| JP2014046585 | 2014-03-10 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO2015137352A1 true WO2015137352A1 (en) | 2015-09-17 |
Family
ID=54071795
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/JP2015/057046 WO2015137352A1 (en) | 2014-03-10 | 2015-03-10 | Gene related to full-dark heterotrophic ability in cyanobacteria, and use of same |
Country Status (2)
| Country | Link |
|---|---|
| JP (1) | JPWO2015137352A1 (en) |
| WO (1) | WO2015137352A1 (en) |
-
2015
- 2015-03-10 JP JP2016507766A patent/JPWO2015137352A1/en active Pending
- 2015-03-10 WO PCT/JP2015/057046 patent/WO2015137352A1/en active Application Filing
Non-Patent Citations (8)
| Title |
|---|
| DATABASE GenBank 11 December 2013 (2013-12-11), LI, T. ET AL.: "Definition: Synechococcus sp. PCC 7002, complete genome", XP055224522, retrieved from ncbi Database accession no. CP000951 * |
| FUJITA,Y. ET AL.: "Identification of the chlB gene and the gene product essential for the light-independent chlorophyll biosynthesis in the cyanobacterium Plectonema boryanum", PLANT CELL PHYSIOL., vol. 37, no. 3, 3 April 1996 (1996-04-03), pages 313 - 323, XP055224515, ISSN: 0032-0781 * |
| HIRAIDE,Y. ET AL.: "Loss of cytochrome cM stimulates cyanobacterial heterotrophic growth in the dark", PLANT CELL PHYSIOL., vol. 56, no. 2, 5 February 2015 (2015-02-05), pages 334 - 345, XP055224517, ISSN: 0032-0781 * |
| LINDBERG,P. ET AL.: "Engineering a platform for photosynthetic isoprene production in cyanobacteria, using Synechocystis as the model organism", METAB.ENG., vol. 12, no. 1, 12 January 2010 (2010-01-12), pages 70 - 9, XP026756973, ISSN: 1096-7176 * |
| MALAKHOV,M.P. ET AL.: "A New Type of Cytochrome c from Synechocystis PCC6803", JOURNAL OF PLANT PHYSIOLOGY, vol. 4, no. 3, 14 September 1994 (1994-09-14), pages 259 - 64 * |
| TAKEYAMA,H. ET AL.: "Expression of the eicosapentaenoic acid synthesis gene cluster from Shewanella sp. in a transgenic marine cyanobacterium", SYNECHOCOCCUS SP., MICROBIOLOGY, vol. 3, no. 8, 14 August 1997 (1997-08-14), pages 2725 - 2731, XP000952940 * |
| YUTO HIRAIDE ET AL.: "Cyanobacteria Leptolyngbya boryana no Genome Re-sequence ni yoru Ansho deno Juzoku Eiyo Seiiku o Sokushin suru Hen'i no Dotei", DAI 55 KAI PROCEEDINGS OF THE ANNUAL MEETING OF THE JAPANESE SOCIETY OF PLANT PHYSIOLOGISTS, 11 March 2014 (2014-03-11), pages 255 * |
| YUTO HIRAIDE ET AL.: "Cytochrome CM no Kesson wa Cyanobacteria no Juzoku Eiyo Seiiku o Sokushin suru", DAI 56 KAI PROCEEDINGS OF THE ANNUAL MEETING OF THE JAPANESE SOCIETY OF PLANT PHYSIOLOGISTS, 9 March 2015 (2015-03-09), pages 130 * |
Also Published As
| Publication number | Publication date |
|---|---|
| JPWO2015137352A1 (en) | 2017-04-06 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| Chien et al. | Solar-to-bioH2 production enhanced by homologous overexpression of hydrogenase in green alga Chlorella sp. DT | |
| CN110331146A (en) | It is a kind of regulation sgRNA transcription promoter, expression vector and its genome editing system and application | |
| WO2019094447A2 (en) | Production of mycosporine-like amino acids in cyanobacteria | |
| CN101948871B (en) | Marine microalgae chloroplast expression vector and application thereof | |
| CN106995817B (en) | Construction of high concentration CO resistance by using gene of coding chloroplast carbonic anhydrase2Application of industrial engineering microalgae capable of rapidly growing | |
| US9309541B2 (en) | Biological production of organic compounds | |
| US9902963B2 (en) | Modified microorganism having enhanced biomass synthesis capacity and a method thereof | |
| US10280428B2 (en) | Molecular biology tools for algal engineering | |
| WO2015137352A1 (en) | Gene related to full-dark heterotrophic ability in cyanobacteria, and use of same | |
| US11168329B2 (en) | Genetic control of cell size | |
| US20130302812A1 (en) | Method of microalgal chloroplast transformation using functional selection in the absence of antibiotics | |
| WO2015115583A1 (en) | Modified cyanobacteria | |
| JP6893820B2 (en) | Methods to improve resistance of nitric acid to substrate analogs in microalgae | |
| CN102718849A (en) | Protein related to chlorophyll synthesis and coding gene and application thereof | |
| RU2813283C2 (en) | Recombinant strain based on escherichia coli, a method of its construction and use | |
| EP2563927B1 (en) | L-lactate production in cyanobacteria | |
| WO2022220263A1 (en) | Ergothioneine production method | |
| US20240287443A1 (en) | Recombinant algae and production of spider silk protein from the recombinant algae | |
| CA2983636A1 (en) | Microorganisms with increased photosynthetic capacity | |
| CN105861406A (en) | Transgenic agrobacterium strain for expressing recombined human albumin genes in plants | |
| WO2024097353A1 (en) | Recombinant algae having high biomass and lipid productivity | |
| Sedelnikova et al. | Crystallization of E. coli RuvA gives insights into the symmetry of a Holliday junction/protein complex | |
| CN116323646A (en) | Recombinant algae with high lipid productivity | |
| CN116848228A (en) | Recombinant algae with high lipid productivity | |
| Class et al. | Patent application title: SUPPRESSION OF TLA2-CpFTSY GENE EXPRESSION FOR IMPROVED SOLAR ENERGY CONVERSION EFFICIENCY AND PHOTOSYNTHETIC PRODUCTIVITY IN ALGAE Inventors: Anastasios Melis (El Cerrito, CA, US) Anastasios Melis (El Cerrito, CA, US) Henning Kirst (Burgwedel, DE) Assignees: THE REGENTS OF THE UNIVERSITY OF CALIFORNIA |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 15762211 Country of ref document: EP Kind code of ref document: A1 |
|
| DPE2 | Request for preliminary examination filed before expiration of 19th month from priority date (pct application filed from 20040101) | ||
| ENP | Entry into the national phase |
Ref document number: 2016507766 Country of ref document: JP Kind code of ref document: A |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |
|
| 122 | Ep: pct application non-entry in european phase |
Ref document number: 15762211 Country of ref document: EP Kind code of ref document: A1 |