JP6280300B2 - 脳疾患治療剤 - Google Patents
脳疾患治療剤 Download PDFInfo
- Publication number
- JP6280300B2 JP6280300B2 JP2012185867A JP2012185867A JP6280300B2 JP 6280300 B2 JP6280300 B2 JP 6280300B2 JP 2012185867 A JP2012185867 A JP 2012185867A JP 2012185867 A JP2012185867 A JP 2012185867A JP 6280300 B2 JP6280300 B2 JP 6280300B2
- Authority
- JP
- Japan
- Prior art keywords
- amino acid
- antigen
- binding
- region
- ala
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
- 208000014644 Brain disease Diseases 0.000 title claims description 35
- 230000027455 binding Effects 0.000 claims description 704
- 102000036639 antigens Human genes 0.000 claims description 648
- 108091007433 antigens Proteins 0.000 claims description 648
- 239000000427 antigen Substances 0.000 claims description 646
- 150000001413 amino acids Chemical class 0.000 claims description 193
- 125000003275 alpha amino acid group Chemical group 0.000 claims description 108
- 102100026120 IgG receptor FcRn large subunit p51 Human genes 0.000 claims description 81
- 101710177940 IgG receptor FcRn large subunit p51 Proteins 0.000 claims description 81
- 229910001424 calcium ion Inorganic materials 0.000 claims description 79
- BHPQYMZQTOCNFJ-UHFFFAOYSA-N Calcium cation Chemical compound [Ca+2] BHPQYMZQTOCNFJ-UHFFFAOYSA-N 0.000 claims description 78
- 150000002500 ions Chemical class 0.000 claims description 45
- 239000003814 drug Substances 0.000 claims description 40
- 229940124597 therapeutic agent Drugs 0.000 claims description 34
- 230000000694 effects Effects 0.000 claims description 31
- 239000013598 vector Substances 0.000 claims description 21
- 230000000295 complement effect Effects 0.000 claims description 19
- 210000005013 brain tissue Anatomy 0.000 claims description 14
- 238000004519 manufacturing process Methods 0.000 claims description 14
- 108010073807 IgG Receptors Proteins 0.000 claims description 7
- 239000001963 growth medium Substances 0.000 claims description 7
- 108091033319 polynucleotide Proteins 0.000 claims description 7
- 102000040430 polynucleotide Human genes 0.000 claims description 7
- 239000002157 polynucleotide Substances 0.000 claims description 7
- 102000009490 IgG Receptors Human genes 0.000 claims description 5
- 239000003795 chemical substances by application Substances 0.000 claims description 2
- 230000002829 reductive effect Effects 0.000 claims description 2
- 235000001014 amino acid Nutrition 0.000 description 173
- 229940024606 amino acid Drugs 0.000 description 166
- 210000004027 cell Anatomy 0.000 description 158
- 238000000034 method Methods 0.000 description 99
- 108090000623 proteins and genes Proteins 0.000 description 95
- 125000000539 amino acid group Chemical group 0.000 description 63
- -1 His Chemical group 0.000 description 61
- 108090000765 processed proteins & peptides Proteins 0.000 description 48
- 238000012360 testing method Methods 0.000 description 48
- 230000007935 neutral effect Effects 0.000 description 42
- 102100035360 Cerebellar degeneration-related antigen 1 Human genes 0.000 description 41
- 108010021472 Fc gamma receptor IIB Proteins 0.000 description 39
- 102100029205 Low affinity immunoglobulin gamma Fc region receptor II-b Human genes 0.000 description 39
- 206010028980 Neoplasm Diseases 0.000 description 38
- 230000002378 acidificating effect Effects 0.000 description 38
- 235000018102 proteins Nutrition 0.000 description 38
- 102000004169 proteins and genes Human genes 0.000 description 38
- GPRLSGONYQIRFK-UHFFFAOYSA-N hydron Chemical compound [H+] GPRLSGONYQIRFK-UHFFFAOYSA-N 0.000 description 35
- 230000000670 limiting effect Effects 0.000 description 32
- 210000004408 hybridoma Anatomy 0.000 description 31
- 125000003295 alanine group Chemical group N[C@@H](C)C(=O)* 0.000 description 29
- MTCFGRXMJLQNBG-REOHCLBHSA-N (2S)-2-Amino-3-hydroxypropansäure Chemical compound OC[C@H](N)C(O)=O MTCFGRXMJLQNBG-REOHCLBHSA-N 0.000 description 28
- 108020004414 DNA Proteins 0.000 description 27
- AYFVYJQAPQTCCC-GBXIJSLDSA-N L-threonine Chemical compound C[C@@H](O)[C@H](N)C(O)=O AYFVYJQAPQTCCC-GBXIJSLDSA-N 0.000 description 27
- 238000010494 dissociation reaction Methods 0.000 description 26
- 230000005593 dissociations Effects 0.000 description 26
- 239000013604 expression vector Substances 0.000 description 26
- 102000005962 receptors Human genes 0.000 description 26
- 108020003175 receptors Proteins 0.000 description 26
- DCXYFEDJOCDNAF-REOHCLBHSA-N L-asparagine Chemical compound OC(=O)[C@@H](N)CC(N)=O DCXYFEDJOCDNAF-REOHCLBHSA-N 0.000 description 24
- 125000000174 L-prolyl group Chemical group [H]N1C([H])([H])C([H])([H])C([H])([H])[C@@]1([H])C(*)=O 0.000 description 24
- 239000011575 calcium Substances 0.000 description 24
- 241000699666 Mus <mouse, genus> Species 0.000 description 23
- 239000013078 crystal Substances 0.000 description 23
- CKLJMWTZIZZHCS-REOHCLBHSA-N L-aspartic acid Chemical compound OC(=O)[C@@H](N)CC(O)=O CKLJMWTZIZZHCS-REOHCLBHSA-N 0.000 description 21
- 210000004556 brain Anatomy 0.000 description 21
- 201000011510 cancer Diseases 0.000 description 20
- 229920001184 polypeptide Polymers 0.000 description 20
- 102000004196 processed proteins & peptides Human genes 0.000 description 20
- 210000004602 germ cell Anatomy 0.000 description 19
- 101100112922 Candida albicans CDR3 gene Proteins 0.000 description 18
- 239000002299 complementary DNA Substances 0.000 description 18
- 230000004048 modification Effects 0.000 description 18
- 238000012986 modification Methods 0.000 description 18
- WHUUTDBJXJRKMK-UHFFFAOYSA-N Glutamic acid Natural products OC(=O)C(N)CCC(O)=O WHUUTDBJXJRKMK-UHFFFAOYSA-N 0.000 description 17
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 17
- 230000001235 sensitizing effect Effects 0.000 description 17
- OYPRJOBELJOOCE-UHFFFAOYSA-N Calcium Chemical compound [Ca] OYPRJOBELJOOCE-UHFFFAOYSA-N 0.000 description 16
- WHUUTDBJXJRKMK-VKHMYHEASA-N L-glutamic acid Chemical compound OC(=O)[C@@H](N)CCC(O)=O WHUUTDBJXJRKMK-VKHMYHEASA-N 0.000 description 16
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 16
- 229910052791 calcium Inorganic materials 0.000 description 16
- 230000014509 gene expression Effects 0.000 description 16
- 239000003446 ligand Substances 0.000 description 16
- 241001465754 Metazoa Species 0.000 description 15
- 125000002707 L-tryptophyl group Chemical group [H]C1=C([H])C([H])=C2C(C([C@](N([H])[H])(C(=O)[*])[H])([H])[H])=C([H])N([H])C2=C1[H] 0.000 description 14
- 206010035226 Plasma cell myeloma Diseases 0.000 description 14
- 238000002649 immunization Methods 0.000 description 14
- 230000003053 immunization Effects 0.000 description 14
- 230000036961 partial effect Effects 0.000 description 14
- 238000012216 screening Methods 0.000 description 14
- NFGXHKASABOEEW-UHFFFAOYSA-N 1-methylethyl 11-methoxy-3,7,11-trimethyl-2,4-dodecadienoate Chemical compound COC(C)(C)CCCC(C)CC=CC(C)=CC(=O)OC(C)C NFGXHKASABOEEW-UHFFFAOYSA-N 0.000 description 13
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 13
- 210000002865 immune cell Anatomy 0.000 description 13
- 239000000243 solution Substances 0.000 description 13
- 210000004369 blood Anatomy 0.000 description 12
- 239000008280 blood Substances 0.000 description 12
- 230000007910 cell fusion Effects 0.000 description 12
- 230000004927 fusion Effects 0.000 description 12
- 201000000050 myeloid neoplasm Diseases 0.000 description 12
- 230000003472 neutralizing effect Effects 0.000 description 12
- 238000001727 in vivo Methods 0.000 description 11
- 108060003951 Immunoglobulin Proteins 0.000 description 10
- 241000124008 Mammalia Species 0.000 description 10
- 210000001218 blood-brain barrier Anatomy 0.000 description 10
- 102000018358 immunoglobulin Human genes 0.000 description 10
- 230000035772 mutation Effects 0.000 description 10
- 102000005701 Calcium-Binding Proteins Human genes 0.000 description 9
- 108010045403 Calcium-Binding Proteins Proteins 0.000 description 9
- 102000001301 EGF receptor Human genes 0.000 description 9
- 108060006698 EGF receptor Proteins 0.000 description 9
- COLNVLDHVKWLRT-QMMMGPOBSA-N L-phenylalanine Chemical compound OC(=O)[C@@H](N)CC1=CC=CC=C1 COLNVLDHVKWLRT-QMMMGPOBSA-N 0.000 description 9
- 102100024598 Tumor necrosis factor ligand superfamily member 10 Human genes 0.000 description 9
- 238000004458 analytical method Methods 0.000 description 9
- 239000002585 base Substances 0.000 description 9
- 210000001175 cerebrospinal fluid Anatomy 0.000 description 9
- 229910021645 metal ion Inorganic materials 0.000 description 9
- 108010068617 neonatal Fc receptor Proteins 0.000 description 9
- 108091008146 restriction endonucleases Proteins 0.000 description 9
- 108010059616 Activins Proteins 0.000 description 8
- 102100025475 Carcinoembryonic antigen-related cell adhesion molecule 5 Human genes 0.000 description 8
- 238000002965 ELISA Methods 0.000 description 8
- 102100026818 Inhibin beta E chain Human genes 0.000 description 8
- 125000000393 L-methionino group Chemical group [H]OC(=O)[C@@]([H])(N([H])[*])C([H])([H])C(SC([H])([H])[H])([H])[H] 0.000 description 8
- 102100040247 Tumor necrosis factor Human genes 0.000 description 8
- 102100036922 Tumor necrosis factor ligand superfamily member 13B Human genes 0.000 description 8
- 102100022203 Tumor necrosis factor receptor superfamily member 25 Human genes 0.000 description 8
- 239000000488 activin Substances 0.000 description 8
- 230000008499 blood brain barrier function Effects 0.000 description 8
- 230000002860 competitive effect Effects 0.000 description 8
- 238000001943 fluorescence-activated cell sorting Methods 0.000 description 8
- 229910052739 hydrogen Inorganic materials 0.000 description 8
- 239000001257 hydrogen Substances 0.000 description 8
- 239000000047 product Substances 0.000 description 8
- 239000000126 substance Substances 0.000 description 8
- 241000283707 Capra Species 0.000 description 7
- 101000935587 Homo sapiens Flavin reductase (NADPH) Proteins 0.000 description 7
- 125000000570 L-alpha-aspartyl group Chemical group [H]OC(=O)C([H])([H])[C@]([H])(N([H])[H])C(*)=O 0.000 description 7
- KDXKERNSBIXSRK-YFKPBYRVSA-N L-lysine Chemical compound NCCCC[C@H](N)C(O)=O KDXKERNSBIXSRK-YFKPBYRVSA-N 0.000 description 7
- 108700012411 TNFSF10 Proteins 0.000 description 7
- 238000010586 diagram Methods 0.000 description 7
- 210000003722 extracellular fluid Anatomy 0.000 description 7
- 230000006870 function Effects 0.000 description 7
- 108020004999 messenger RNA Proteins 0.000 description 7
- 238000012546 transfer Methods 0.000 description 7
- 108010032595 Antibody Binding Sites Proteins 0.000 description 6
- 102100032937 CD40 ligand Human genes 0.000 description 6
- 108010022366 Carcinoembryonic Antigen Proteins 0.000 description 6
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 6
- 101000679903 Homo sapiens Tumor necrosis factor receptor superfamily member 25 Proteins 0.000 description 6
- 102100029204 Low affinity immunoglobulin gamma Fc region receptor II-a Human genes 0.000 description 6
- IQFYYKKMVGJFEH-XLPZGREQSA-N Thymidine Chemical compound O=C1NC(=O)C(C)=CN1[C@@H]1O[C@H](CO)[C@@H](O)C1 IQFYYKKMVGJFEH-XLPZGREQSA-N 0.000 description 6
- KZSNJWFQEVHDMF-UHFFFAOYSA-N Valine Chemical compound CC(C)C(N)C(O)=O KZSNJWFQEVHDMF-UHFFFAOYSA-N 0.000 description 6
- 241000700605 Viruses Species 0.000 description 6
- 230000004913 activation Effects 0.000 description 6
- 230000001747 exhibiting effect Effects 0.000 description 6
- 108020001507 fusion proteins Proteins 0.000 description 6
- 102000037865 fusion proteins Human genes 0.000 description 6
- 208000005017 glioblastoma Diseases 0.000 description 6
- 230000001225 therapeutic effect Effects 0.000 description 6
- 101710181478 Envelope glycoprotein GP350 Proteins 0.000 description 5
- 108010021468 Fc gamma receptor IIA Proteins 0.000 description 5
- 102000003886 Glycoproteins Human genes 0.000 description 5
- 108090000288 Glycoproteins Proteins 0.000 description 5
- 101000868215 Homo sapiens CD40 ligand Proteins 0.000 description 5
- 101000611183 Homo sapiens Tumor necrosis factor Proteins 0.000 description 5
- 101000851370 Homo sapiens Tumor necrosis factor receptor superfamily member 9 Proteins 0.000 description 5
- 241000701024 Human betaherpesvirus 5 Species 0.000 description 5
- 108090001005 Interleukin-6 Proteins 0.000 description 5
- 102000004889 Interleukin-6 Human genes 0.000 description 5
- AGPKZVBTJJNPAG-WHFBIAKZSA-N L-isoleucine Chemical compound CC[C@H](C)[C@H](N)C(O)=O AGPKZVBTJJNPAG-WHFBIAKZSA-N 0.000 description 5
- 102100026890 Tumor necrosis factor ligand superfamily member 4 Human genes 0.000 description 5
- 102100028785 Tumor necrosis factor receptor superfamily member 14 Human genes 0.000 description 5
- 102100040403 Tumor necrosis factor receptor superfamily member 6 Human genes 0.000 description 5
- 102100036856 Tumor necrosis factor receptor superfamily member 9 Human genes 0.000 description 5
- 230000009471 action Effects 0.000 description 5
- 238000003556 assay Methods 0.000 description 5
- 125000004429 atom Chemical group 0.000 description 5
- 239000000872 buffer Substances 0.000 description 5
- 230000008859 change Effects 0.000 description 5
- 239000012228 culture supernatant Substances 0.000 description 5
- 238000013461 design Methods 0.000 description 5
- 210000001163 endosome Anatomy 0.000 description 5
- 239000012530 fluid Substances 0.000 description 5
- 239000012634 fragment Substances 0.000 description 5
- 210000004698 lymphocyte Anatomy 0.000 description 5
- 238000005259 measurement Methods 0.000 description 5
- 229910052751 metal Inorganic materials 0.000 description 5
- 230000031998 transcytosis Effects 0.000 description 5
- 230000009261 transgenic effect Effects 0.000 description 5
- 230000032258 transport Effects 0.000 description 5
- 102000013455 Amyloid beta-Peptides Human genes 0.000 description 4
- 108010090849 Amyloid beta-Peptides Proteins 0.000 description 4
- 102000000584 Calmodulin Human genes 0.000 description 4
- 108010041952 Calmodulin Proteins 0.000 description 4
- 102100023126 Cell surface glycoprotein MUC18 Human genes 0.000 description 4
- 108090000368 Fibroblast growth factor 8 Proteins 0.000 description 4
- 102000053171 Glial Fibrillary Acidic Human genes 0.000 description 4
- 101710193519 Glial fibrillary acidic protein Proteins 0.000 description 4
- 101000623903 Homo sapiens Cell surface glycoprotein MUC18 Proteins 0.000 description 4
- 101000798130 Homo sapiens Tumor necrosis factor receptor superfamily member 11B Proteins 0.000 description 4
- 101000801234 Homo sapiens Tumor necrosis factor receptor superfamily member 18 Proteins 0.000 description 4
- 101000597785 Homo sapiens Tumor necrosis factor receptor superfamily member 6B Proteins 0.000 description 4
- 102100034349 Integrase Human genes 0.000 description 4
- 102000001399 Kallikrein Human genes 0.000 description 4
- 108060005987 Kallikrein Proteins 0.000 description 4
- ODKSFYDXXFIFQN-BYPYZUCNSA-N L-arginine Chemical compound OC(=O)[C@@H](N)CCCN=C(N)N ODKSFYDXXFIFQN-BYPYZUCNSA-N 0.000 description 4
- 102100026894 Lymphotoxin-beta Human genes 0.000 description 4
- 108010063954 Mucins Proteins 0.000 description 4
- 102000015728 Mucins Human genes 0.000 description 4
- 102000014128 RANK Ligand Human genes 0.000 description 4
- 108010025832 RANK Ligand Proteins 0.000 description 4
- 201000000582 Retinoblastoma Diseases 0.000 description 4
- 108060008682 Tumor Necrosis Factor Proteins 0.000 description 4
- 102100024584 Tumor necrosis factor ligand superfamily member 12 Human genes 0.000 description 4
- 102100024587 Tumor necrosis factor ligand superfamily member 15 Human genes 0.000 description 4
- 102100032236 Tumor necrosis factor receptor superfamily member 11B Human genes 0.000 description 4
- 102100033728 Tumor necrosis factor receptor superfamily member 18 Human genes 0.000 description 4
- 102100035284 Tumor necrosis factor receptor superfamily member 6B Human genes 0.000 description 4
- 239000012491 analyte Substances 0.000 description 4
- 230000010056 antibody-dependent cellular cytotoxicity Effects 0.000 description 4
- 239000007864 aqueous solution Substances 0.000 description 4
- 235000003704 aspartic acid Nutrition 0.000 description 4
- OQFSQFPPLPISGP-UHFFFAOYSA-N beta-carboxyaspartic acid Natural products OC(=O)C(N)C(C(O)=O)C(O)=O OQFSQFPPLPISGP-UHFFFAOYSA-N 0.000 description 4
- 230000004071 biological effect Effects 0.000 description 4
- 230000015572 biosynthetic process Effects 0.000 description 4
- 238000010367 cloning Methods 0.000 description 4
- 208000029742 colonic neoplasm Diseases 0.000 description 4
- 230000000875 corresponding effect Effects 0.000 description 4
- 238000012258 culturing Methods 0.000 description 4
- 230000007812 deficiency Effects 0.000 description 4
- 230000002950 deficient Effects 0.000 description 4
- 229940079593 drug Drugs 0.000 description 4
- 230000001605 fetal effect Effects 0.000 description 4
- 210000005046 glial fibrillary acidic protein Anatomy 0.000 description 4
- 235000013922 glutamic acid Nutrition 0.000 description 4
- 239000004220 glutamic acid Substances 0.000 description 4
- FDGQSTZJBFJUBT-UHFFFAOYSA-N hypoxanthine Chemical compound O=C1NC=NC2=C1NC=N2 FDGQSTZJBFJUBT-UHFFFAOYSA-N 0.000 description 4
- 230000001900 immune effect Effects 0.000 description 4
- 239000003550 marker Substances 0.000 description 4
- 239000002184 metal Substances 0.000 description 4
- 235000013336 milk Nutrition 0.000 description 4
- 210000004080 milk Anatomy 0.000 description 4
- 239000008267 milk Substances 0.000 description 4
- 208000027831 neuroepithelial neoplasm Diseases 0.000 description 4
- 210000002569 neuron Anatomy 0.000 description 4
- 239000008194 pharmaceutical composition Substances 0.000 description 4
- 230000035755 proliferation Effects 0.000 description 4
- 210000002966 serum Anatomy 0.000 description 4
- 238000006467 substitution reaction Methods 0.000 description 4
- 238000003786 synthesis reaction Methods 0.000 description 4
- 229940104230 thymidine Drugs 0.000 description 4
- 230000007704 transition Effects 0.000 description 4
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 3
- 208000024827 Alzheimer disease Diseases 0.000 description 3
- 206010003571 Astrocytoma Diseases 0.000 description 3
- 102100022718 Atypical chemokine receptor 2 Human genes 0.000 description 3
- 208000023275 Autoimmune disease Diseases 0.000 description 3
- 108010028006 B-Cell Activating Factor Proteins 0.000 description 3
- 102100021663 Baculoviral IAP repeat-containing protein 5 Human genes 0.000 description 3
- DWRXFEITVBNRMK-UHFFFAOYSA-N Beta-D-1-Arabinofuranosylthymine Natural products O=C1NC(=O)C(C)=CN1C1C(O)C(O)C(CO)O1 DWRXFEITVBNRMK-UHFFFAOYSA-N 0.000 description 3
- 102100032367 C-C motif chemokine 5 Human genes 0.000 description 3
- 108090000342 C-Type Lectins Proteins 0.000 description 3
- 102000003930 C-Type Lectins Human genes 0.000 description 3
- 102100025248 C-X-C motif chemokine 10 Human genes 0.000 description 3
- 102100025277 C-X-C motif chemokine 13 Human genes 0.000 description 3
- 102100025221 CD70 antigen Human genes 0.000 description 3
- 102000000905 Cadherin Human genes 0.000 description 3
- 108050007957 Cadherin Proteins 0.000 description 3
- 206010009944 Colon cancer Diseases 0.000 description 3
- 102100031111 Disintegrin and metalloproteinase domain-containing protein 17 Human genes 0.000 description 3
- 102100037354 Ectodysplasin-A Human genes 0.000 description 3
- 241000196324 Embryophyta Species 0.000 description 3
- 102000004190 Enzymes Human genes 0.000 description 3
- 108090000790 Enzymes Proteins 0.000 description 3
- 241000588724 Escherichia coli Species 0.000 description 3
- 102100020997 Fractalkine Human genes 0.000 description 3
- 102100041003 Glutamate carboxypeptidase 2 Human genes 0.000 description 3
- 102100035379 Growth/differentiation factor 5 Human genes 0.000 description 3
- 102400001369 Heparin-binding EGF-like growth factor Human genes 0.000 description 3
- 101800001649 Heparin-binding EGF-like growth factor Proteins 0.000 description 3
- 101000678892 Homo sapiens Atypical chemokine receptor 2 Proteins 0.000 description 3
- 101000858064 Homo sapiens C-X-C motif chemokine 13 Proteins 0.000 description 3
- 101000880080 Homo sapiens Ectodysplasin-A Proteins 0.000 description 3
- 101000892862 Homo sapiens Glutamate carboxypeptidase 2 Proteins 0.000 description 3
- 101000917826 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor II-a Proteins 0.000 description 3
- 101000764294 Homo sapiens Lymphotoxin-beta Proteins 0.000 description 3
- 101000830596 Homo sapiens Tumor necrosis factor ligand superfamily member 15 Proteins 0.000 description 3
- 101000764263 Homo sapiens Tumor necrosis factor ligand superfamily member 4 Proteins 0.000 description 3
- 101000610604 Homo sapiens Tumor necrosis factor receptor superfamily member 10B Proteins 0.000 description 3
- 101000610602 Homo sapiens Tumor necrosis factor receptor superfamily member 10C Proteins 0.000 description 3
- 101000610609 Homo sapiens Tumor necrosis factor receptor superfamily member 10D Proteins 0.000 description 3
- 101000795167 Homo sapiens Tumor necrosis factor receptor superfamily member 13B Proteins 0.000 description 3
- 101000611023 Homo sapiens Tumor necrosis factor receptor superfamily member 6 Proteins 0.000 description 3
- 101000851376 Homo sapiens Tumor necrosis factor receptor superfamily member 8 Proteins 0.000 description 3
- 108010091358 Hypoxanthine Phosphoribosyltransferase Proteins 0.000 description 3
- 102100029098 Hypoxanthine-guanine phosphoribosyltransferase Human genes 0.000 description 3
- 102100032818 Integrin alpha-4 Human genes 0.000 description 3
- 108010041012 Integrin alpha4 Proteins 0.000 description 3
- 102100025390 Integrin beta-2 Human genes 0.000 description 3
- 108010002616 Interleukin-5 Proteins 0.000 description 3
- 102100039897 Interleukin-5 Human genes 0.000 description 3
- 108010038501 Interleukin-6 Receptors Proteins 0.000 description 3
- 102000010781 Interleukin-6 Receptors Human genes 0.000 description 3
- 125000003338 L-glutaminyl group Chemical group O=C([*])[C@](N([H])[H])([H])C([H])([H])C([H])([H])C(=O)N([H])[H] 0.000 description 3
- 102100029185 Low affinity immunoglobulin gamma Fc region receptor III-B Human genes 0.000 description 3
- 108010061593 Member 14 Tumor Necrosis Factor Receptors Proteins 0.000 description 3
- 108010052285 Membrane Proteins Proteins 0.000 description 3
- 241000699670 Mus sp. Species 0.000 description 3
- 108010025020 Nerve Growth Factor Proteins 0.000 description 3
- 102100027347 Neural cell adhesion molecule 1 Human genes 0.000 description 3
- 108091028043 Nucleic acid sequence Proteins 0.000 description 3
- 102100040557 Osteopontin Human genes 0.000 description 3
- 108010081689 Osteopontin Proteins 0.000 description 3
- 206010033128 Ovarian cancer Diseases 0.000 description 3
- 206010061535 Ovarian neoplasm Diseases 0.000 description 3
- 239000002202 Polyethylene glycol Substances 0.000 description 3
- 102100033237 Pro-epidermal growth factor Human genes 0.000 description 3
- 206010060862 Prostate cancer Diseases 0.000 description 3
- 208000000236 Prostatic Neoplasms Diseases 0.000 description 3
- 108010076504 Protein Sorting Signals Proteins 0.000 description 3
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 3
- 241000700584 Simplexvirus Species 0.000 description 3
- 102100021669 Stromal cell-derived factor 1 Human genes 0.000 description 3
- 108010002687 Survivin Proteins 0.000 description 3
- 101710097155 Tumor necrosis factor ligand superfamily member 12 Proteins 0.000 description 3
- 102100032100 Tumor necrosis factor ligand superfamily member 8 Human genes 0.000 description 3
- 102100040113 Tumor necrosis factor receptor superfamily member 10A Human genes 0.000 description 3
- 102100040112 Tumor necrosis factor receptor superfamily member 10B Human genes 0.000 description 3
- 102100040115 Tumor necrosis factor receptor superfamily member 10C Human genes 0.000 description 3
- 102100040110 Tumor necrosis factor receptor superfamily member 10D Human genes 0.000 description 3
- 102100029675 Tumor necrosis factor receptor superfamily member 13B Human genes 0.000 description 3
- 102100033725 Tumor necrosis factor receptor superfamily member 16 Human genes 0.000 description 3
- 102100033726 Tumor necrosis factor receptor superfamily member 17 Human genes 0.000 description 3
- 102100033760 Tumor necrosis factor receptor superfamily member 19 Human genes 0.000 description 3
- 102100033732 Tumor necrosis factor receptor superfamily member 1A Human genes 0.000 description 3
- 102100033733 Tumor necrosis factor receptor superfamily member 1B Human genes 0.000 description 3
- 102100040245 Tumor necrosis factor receptor superfamily member 5 Human genes 0.000 description 3
- 102100036857 Tumor necrosis factor receptor superfamily member 8 Human genes 0.000 description 3
- 108010053100 Vascular Endothelial Growth Factor Receptor-3 Proteins 0.000 description 3
- 102100033177 Vascular endothelial growth factor receptor 2 Human genes 0.000 description 3
- IQFYYKKMVGJFEH-UHFFFAOYSA-N beta-L-thymidine Natural products O=C1NC(=O)C(C)=CN1C1OC(CO)C(O)C1 IQFYYKKMVGJFEH-UHFFFAOYSA-N 0.000 description 3
- 210000004204 blood vessel Anatomy 0.000 description 3
- 239000013522 chelant Substances 0.000 description 3
- 230000001085 cytostatic effect Effects 0.000 description 3
- 238000003113 dilution method Methods 0.000 description 3
- LOKCTEFSRHRXRJ-UHFFFAOYSA-I dipotassium trisodium dihydrogen phosphate hydrogen phosphate dichloride Chemical compound P(=O)(O)(O)[O-].[K+].P(=O)(O)([O-])[O-].[Na+].[Na+].[Cl-].[K+].[Cl-].[Na+] LOKCTEFSRHRXRJ-UHFFFAOYSA-I 0.000 description 3
- 230000002708 enhancing effect Effects 0.000 description 3
- 229940088598 enzyme Drugs 0.000 description 3
- 230000002163 immunogen Effects 0.000 description 3
- 230000001965 increasing effect Effects 0.000 description 3
- 230000002401 inhibitory effect Effects 0.000 description 3
- 102000006495 integrins Human genes 0.000 description 3
- 108010044426 integrins Proteins 0.000 description 3
- 230000003834 intracellular effect Effects 0.000 description 3
- 210000003292 kidney cell Anatomy 0.000 description 3
- 238000002372 labelling Methods 0.000 description 3
- 239000007788 liquid Substances 0.000 description 3
- 210000004962 mammalian cell Anatomy 0.000 description 3
- 210000001161 mammalian embryo Anatomy 0.000 description 3
- 238000000691 measurement method Methods 0.000 description 3
- 238000007857 nested PCR Methods 0.000 description 3
- 239000002773 nucleotide Substances 0.000 description 3
- 125000003729 nucleotide group Chemical group 0.000 description 3
- 238000004091 panning Methods 0.000 description 3
- 230000002093 peripheral effect Effects 0.000 description 3
- 102000013415 peroxidase activity proteins Human genes 0.000 description 3
- 108040007629 peroxidase activity proteins Proteins 0.000 description 3
- 239000002953 phosphate buffered saline Substances 0.000 description 3
- 102000005162 pleiotrophin Human genes 0.000 description 3
- 229920001223 polyethylene glycol Polymers 0.000 description 3
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 3
- 208000029340 primitive neuroectodermal tumor Diseases 0.000 description 3
- 230000008569 process Effects 0.000 description 3
- 238000004064 recycling Methods 0.000 description 3
- 235000004400 serine Nutrition 0.000 description 3
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Chemical compound O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 3
- YBJHBAHKTGYVGT-ZKWXMUAHSA-N (+)-Biotin Chemical compound N1C(=O)N[C@@H]2[C@H](CCCCC(=O)O)SC[C@@H]21 YBJHBAHKTGYVGT-ZKWXMUAHSA-N 0.000 description 2
- MZOFCQQQCNRIBI-VMXHOPILSA-N (3s)-4-[[(2s)-1-[[(2s)-1-[[(1s)-1-carboxy-2-hydroxyethyl]amino]-4-methyl-1-oxopentan-2-yl]amino]-5-(diaminomethylideneamino)-1-oxopentan-2-yl]amino]-3-[[2-[[(2s)-2,6-diaminohexanoyl]amino]acetyl]amino]-4-oxobutanoic acid Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CC(O)=O)NC(=O)CNC(=O)[C@@H](N)CCCCN MZOFCQQQCNRIBI-VMXHOPILSA-N 0.000 description 2
- PJOHVEQSYPOERL-SHEAVXILSA-N (e)-n-[(4r,4as,7ar,12br)-3-(cyclopropylmethyl)-9-hydroxy-7-oxo-2,4,5,6,7a,13-hexahydro-1h-4,12-methanobenzofuro[3,2-e]isoquinoline-4a-yl]-3-(4-methylphenyl)prop-2-enamide Chemical compound C1=CC(C)=CC=C1\C=C\C(=O)N[C@]1(CCC(=O)[C@@H]2O3)[C@H]4CC5=CC=C(O)C3=C5[C@]12CCN4CC1CC1 PJOHVEQSYPOERL-SHEAVXILSA-N 0.000 description 2
- JKMHFZQWWAIEOD-UHFFFAOYSA-N 2-[4-(2-hydroxyethyl)piperazin-1-yl]ethanesulfonic acid Chemical compound OCC[NH+]1CCN(CCS([O-])(=O)=O)CC1 JKMHFZQWWAIEOD-UHFFFAOYSA-N 0.000 description 2
- BGFTWECWAICPDG-UHFFFAOYSA-N 2-[bis(4-chlorophenyl)methyl]-4-n-[3-[bis(4-chlorophenyl)methyl]-4-(dimethylamino)phenyl]-1-n,1-n-dimethylbenzene-1,4-diamine Chemical compound C1=C(C(C=2C=CC(Cl)=CC=2)C=2C=CC(Cl)=CC=2)C(N(C)C)=CC=C1NC(C=1)=CC=C(N(C)C)C=1C(C=1C=CC(Cl)=CC=1)C1=CC=C(Cl)C=C1 BGFTWECWAICPDG-UHFFFAOYSA-N 0.000 description 2
- TVZGACDUOSZQKY-LBPRGKRZSA-N 4-aminofolic acid Chemical compound C1=NC2=NC(N)=NC(N)=C2N=C1CNC1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 TVZGACDUOSZQKY-LBPRGKRZSA-N 0.000 description 2
- 102100033400 4F2 cell-surface antigen heavy chain Human genes 0.000 description 2
- 102100033350 ATP-dependent translocase ABCB1 Human genes 0.000 description 2
- 102100031934 Adhesion G-protein coupled receptor G1 Human genes 0.000 description 2
- 101710096372 Adhesion G-protein coupled receptor G1 Proteins 0.000 description 2
- 102100024321 Alkaline phosphatase, placental type Human genes 0.000 description 2
- 102100022749 Aminopeptidase N Human genes 0.000 description 2
- 102100025672 Angiopoietin-related protein 2 Human genes 0.000 description 2
- 241000252073 Anguilliformes Species 0.000 description 2
- 102000000412 Annexin Human genes 0.000 description 2
- 108050008874 Annexin Proteins 0.000 description 2
- 108050009514 Antigen peptide transporter 1 Proteins 0.000 description 2
- CJLHTKGWEUGORV-UHFFFAOYSA-N Artemin Chemical compound C1CC2(C)C(O)CCC(=C)C2(O)C2C1C(C)C(=O)O2 CJLHTKGWEUGORV-UHFFFAOYSA-N 0.000 description 2
- 102000005427 Asialoglycoprotein Receptor Human genes 0.000 description 2
- DCXYFEDJOCDNAF-UHFFFAOYSA-N Asparagine Natural products OC(=O)C(N)CC(N)=O DCXYFEDJOCDNAF-UHFFFAOYSA-N 0.000 description 2
- 241000228245 Aspergillus niger Species 0.000 description 2
- IJGRMHOSHXDMSA-UHFFFAOYSA-N Atomic nitrogen Chemical compound N#N IJGRMHOSHXDMSA-UHFFFAOYSA-N 0.000 description 2
- 108010008014 B-Cell Maturation Antigen Proteins 0.000 description 2
- 102100024222 B-lymphocyte antigen CD19 Human genes 0.000 description 2
- 102100023995 Beta-nerve growth factor Human genes 0.000 description 2
- 102100021257 Beta-secretase 1 Human genes 0.000 description 2
- 108010049870 Bone Morphogenetic Protein 7 Proteins 0.000 description 2
- 102100022544 Bone morphogenetic protein 7 Human genes 0.000 description 2
- 206010006187 Breast cancer Diseases 0.000 description 2
- 208000026310 Breast neoplasm Diseases 0.000 description 2
- 102100023701 C-C motif chemokine 18 Human genes 0.000 description 2
- 102100036850 C-C motif chemokine 23 Human genes 0.000 description 2
- 102100021933 C-C motif chemokine 25 Human genes 0.000 description 2
- 102100021936 C-C motif chemokine 27 Human genes 0.000 description 2
- 102100032366 C-C motif chemokine 7 Human genes 0.000 description 2
- 102100031658 C-X-C chemokine receptor type 5 Human genes 0.000 description 2
- 102100025279 C-X-C motif chemokine 11 Human genes 0.000 description 2
- 102100036153 C-X-C motif chemokine 6 Human genes 0.000 description 2
- 102100036170 C-X-C motif chemokine 9 Human genes 0.000 description 2
- 102000002110 C2 domains Human genes 0.000 description 2
- 108050009459 C2 domains Proteins 0.000 description 2
- 102100028743 CAP-Gly domain-containing linker protein 2 Human genes 0.000 description 2
- 102100032912 CD44 antigen Human genes 0.000 description 2
- 108010078791 Carrier Proteins Proteins 0.000 description 2
- 102100024940 Cathepsin K Human genes 0.000 description 2
- 241000282693 Cercopithecidae Species 0.000 description 2
- 108010055166 Chemokine CCL5 Proteins 0.000 description 2
- 102100035294 Chemokine XC receptor 1 Human genes 0.000 description 2
- 108010066813 Chitinase-3-Like Protein 1 Proteins 0.000 description 2
- 102000018704 Chitinase-3-Like Protein 1 Human genes 0.000 description 2
- 108010059480 Chondroitin Sulfate Proteoglycans Proteins 0.000 description 2
- 102000005598 Chondroitin Sulfate Proteoglycans Human genes 0.000 description 2
- 102100022641 Coagulation factor IX Human genes 0.000 description 2
- 102100023804 Coagulation factor VII Human genes 0.000 description 2
- 108020004705 Codon Proteins 0.000 description 2
- 208000001333 Colorectal Neoplasms Diseases 0.000 description 2
- 241000699800 Cricetinae Species 0.000 description 2
- 102100039498 Cytotoxic T-lymphocyte protein 4 Human genes 0.000 description 2
- WQZGKKKJIJFFOK-QTVWNMPRSA-N D-mannopyranose Chemical compound OC[C@H]1OC(O)[C@@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-QTVWNMPRSA-N 0.000 description 2
- 108010037897 DC-specific ICAM-3 grabbing nonintegrin Proteins 0.000 description 2
- 108700022150 Designed Ankyrin Repeat Proteins Proteins 0.000 description 2
- IAZDPXIOMUYVGZ-UHFFFAOYSA-N Dimethylsulphoxide Chemical compound CS(C)=O IAZDPXIOMUYVGZ-UHFFFAOYSA-N 0.000 description 2
- 102100025012 Dipeptidyl peptidase 4 Human genes 0.000 description 2
- 102100024074 Dystrobrevin alpha Human genes 0.000 description 2
- 102000012545 EGF-like domains Human genes 0.000 description 2
- 108050002150 EGF-like domains Proteins 0.000 description 2
- 238000012286 ELISA Assay Methods 0.000 description 2
- 101150029707 ERBB2 gene Proteins 0.000 description 2
- 108010009900 Endothelial Protein C Receptor Proteins 0.000 description 2
- 102000009839 Endothelial Protein C Receptor Human genes 0.000 description 2
- 101710121417 Envelope glycoprotein Proteins 0.000 description 2
- 102100023688 Eotaxin Human genes 0.000 description 2
- 206010014967 Ependymoma Diseases 0.000 description 2
- 108010076282 Factor IX Proteins 0.000 description 2
- 108010023321 Factor VII Proteins 0.000 description 2
- 102000030914 Fatty Acid-Binding Human genes 0.000 description 2
- 102000009123 Fibrin Human genes 0.000 description 2
- 108010073385 Fibrin Proteins 0.000 description 2
- BWGVNKXGVNDBDI-UHFFFAOYSA-N Fibrin monomer Chemical compound CNC(=O)CNC(=O)CN BWGVNKXGVNDBDI-UHFFFAOYSA-N 0.000 description 2
- 108090000379 Fibroblast growth factor 2 Proteins 0.000 description 2
- 108090000385 Fibroblast growth factor 7 Proteins 0.000 description 2
- 102000003972 Fibroblast growth factor 7 Human genes 0.000 description 2
- 102100037362 Fibronectin Human genes 0.000 description 2
- 102100028461 Frizzled-9 Human genes 0.000 description 2
- 102100035184 General transcription and DNA repair factor IIH helicase subunit XPD Human genes 0.000 description 2
- 102000034615 Glial cell line-derived neurotrophic factor Human genes 0.000 description 2
- 108091010837 Glial cell line-derived neurotrophic factor Proteins 0.000 description 2
- 201000010915 Glioblastoma multiforme Diseases 0.000 description 2
- 206010018338 Glioma Diseases 0.000 description 2
- 108010090254 Growth Differentiation Factor 5 Proteins 0.000 description 2
- 102100034221 Growth-regulated alpha protein Human genes 0.000 description 2
- 102100040896 Growth/differentiation factor 15 Human genes 0.000 description 2
- 102100035368 Growth/differentiation factor 6 Human genes 0.000 description 2
- 102100039939 Growth/differentiation factor 8 Human genes 0.000 description 2
- 239000007995 HEPES buffer Substances 0.000 description 2
- 102000006354 HLA-DR Antigens Human genes 0.000 description 2
- 108010058597 HLA-DR Antigens Proteins 0.000 description 2
- 102100022623 Hepatocyte growth factor receptor Human genes 0.000 description 2
- NTYJJOPFIAHURM-UHFFFAOYSA-N Histamine Chemical compound NCCC1=CN=CN1 NTYJJOPFIAHURM-UHFFFAOYSA-N 0.000 description 2
- 101000800023 Homo sapiens 4F2 cell-surface antigen heavy chain Proteins 0.000 description 2
- 101000980825 Homo sapiens B-lymphocyte antigen CD19 Proteins 0.000 description 2
- 101000894895 Homo sapiens Beta-secretase 1 Proteins 0.000 description 2
- 101000777558 Homo sapiens C-C chemokine receptor type 10 Proteins 0.000 description 2
- 101000978371 Homo sapiens C-C motif chemokine 18 Proteins 0.000 description 2
- 101000713081 Homo sapiens C-C motif chemokine 23 Proteins 0.000 description 2
- 101000897486 Homo sapiens C-C motif chemokine 25 Proteins 0.000 description 2
- 101000797758 Homo sapiens C-C motif chemokine 7 Proteins 0.000 description 2
- 101000922405 Homo sapiens C-X-C chemokine receptor type 5 Proteins 0.000 description 2
- 101000858088 Homo sapiens C-X-C motif chemokine 10 Proteins 0.000 description 2
- 101000868273 Homo sapiens CD44 antigen Proteins 0.000 description 2
- 101000934356 Homo sapiens CD70 antigen Proteins 0.000 description 2
- 101000914324 Homo sapiens Carcinoembryonic antigen-related cell adhesion molecule 5 Proteins 0.000 description 2
- 101000804783 Homo sapiens Chemokine XC receptor 1 Proteins 0.000 description 2
- 101000777461 Homo sapiens Disintegrin and metalloproteinase domain-containing protein 17 Proteins 0.000 description 2
- 101000854520 Homo sapiens Fractalkine Proteins 0.000 description 2
- 101001061405 Homo sapiens Frizzled-9 Proteins 0.000 description 2
- 101000876511 Homo sapiens General transcription and DNA repair factor IIH helicase subunit XPD Proteins 0.000 description 2
- 101000599951 Homo sapiens Insulin-like growth factor I Proteins 0.000 description 2
- 101000878605 Homo sapiens Low affinity immunoglobulin epsilon Fc receptor Proteins 0.000 description 2
- 101000917858 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor III-A Proteins 0.000 description 2
- 101000573526 Homo sapiens Membrane protein MLC1 Proteins 0.000 description 2
- 101000934338 Homo sapiens Myeloid cell surface antigen CD33 Proteins 0.000 description 2
- 101000582950 Homo sapiens Platelet factor 4 Proteins 0.000 description 2
- 101000650117 Homo sapiens Protein Wnt-9a Proteins 0.000 description 2
- 101000836286 Homo sapiens Solute carrier organic anion transporter family member 1C1 Proteins 0.000 description 2
- 101100369992 Homo sapiens TNFSF10 gene Proteins 0.000 description 2
- 101000830565 Homo sapiens Tumor necrosis factor ligand superfamily member 10 Proteins 0.000 description 2
- 101000851434 Homo sapiens Tumor necrosis factor ligand superfamily member 13B Proteins 0.000 description 2
- 101000597779 Homo sapiens Tumor necrosis factor ligand superfamily member 18 Proteins 0.000 description 2
- 101000638255 Homo sapiens Tumor necrosis factor ligand superfamily member 8 Proteins 0.000 description 2
- 101000610605 Homo sapiens Tumor necrosis factor receptor superfamily member 10A Proteins 0.000 description 2
- 101000648507 Homo sapiens Tumor necrosis factor receptor superfamily member 14 Proteins 0.000 description 2
- 101000801254 Homo sapiens Tumor necrosis factor receptor superfamily member 16 Proteins 0.000 description 2
- 101000801227 Homo sapiens Tumor necrosis factor receptor superfamily member 19 Proteins 0.000 description 2
- 101000801228 Homo sapiens Tumor necrosis factor receptor superfamily member 1A Proteins 0.000 description 2
- 101000801232 Homo sapiens Tumor necrosis factor receptor superfamily member 1B Proteins 0.000 description 2
- 101000679921 Homo sapiens Tumor necrosis factor receptor superfamily member 21 Proteins 0.000 description 2
- 101000679851 Homo sapiens Tumor necrosis factor receptor superfamily member 4 Proteins 0.000 description 2
- 102000002265 Human Growth Hormone Human genes 0.000 description 2
- 108010000521 Human Growth Hormone Proteins 0.000 description 2
- 239000000854 Human Growth Hormone Substances 0.000 description 2
- 241001135569 Human adenovirus 5 Species 0.000 description 2
- UFHFLCQGNIYNRP-UHFFFAOYSA-N Hydrogen Chemical compound [H][H] UFHFLCQGNIYNRP-UHFFFAOYSA-N 0.000 description 2
- UGQMRVRMYYASKQ-UHFFFAOYSA-N Hypoxanthine nucleoside Natural products OC1C(O)C(CO)OC1N1C(NC=NC2=O)=C2N=C1 UGQMRVRMYYASKQ-UHFFFAOYSA-N 0.000 description 2
- 102000004877 Insulin Human genes 0.000 description 2
- 108090001061 Insulin Proteins 0.000 description 2
- 108090000723 Insulin-Like Growth Factor I Proteins 0.000 description 2
- 102000004218 Insulin-Like Growth Factor I Human genes 0.000 description 2
- 102100037920 Insulin-like growth factor 2 mRNA-binding protein 3 Human genes 0.000 description 2
- 101710126182 Insulin-like growth factor 2 mRNA-binding protein 3 Proteins 0.000 description 2
- 102100037852 Insulin-like growth factor I Human genes 0.000 description 2
- 102100025323 Integrin alpha-1 Human genes 0.000 description 2
- 102100032817 Integrin alpha-5 Human genes 0.000 description 2
- 108010041014 Integrin alpha5 Proteins 0.000 description 2
- 102100037877 Intercellular adhesion molecule 1 Human genes 0.000 description 2
- 108010050904 Interferons Proteins 0.000 description 2
- 102000014150 Interferons Human genes 0.000 description 2
- 102100039064 Interleukin-3 Human genes 0.000 description 2
- 108010002386 Interleukin-3 Proteins 0.000 description 2
- 102000004388 Interleukin-4 Human genes 0.000 description 2
- 108090000978 Interleukin-4 Proteins 0.000 description 2
- 125000003798 L-tyrosyl group Chemical group [H]N([H])[C@]([H])(C(=O)[*])C([H])([H])C1=C([H])C([H])=C(O[H])C([H])=C1[H] 0.000 description 2
- 108010001831 LDL receptors Proteins 0.000 description 2
- 208000009625 Lesch-Nyhan syndrome Diseases 0.000 description 2
- 108010051335 Lipocalin-2 Proteins 0.000 description 2
- 102000013519 Lipocalin-2 Human genes 0.000 description 2
- 102100038007 Low affinity immunoglobulin epsilon Fc receptor Human genes 0.000 description 2
- 108060001084 Luciferase Proteins 0.000 description 2
- 239000005089 Luciferase Substances 0.000 description 2
- 206010058467 Lung neoplasm malignant Diseases 0.000 description 2
- 108090000362 Lymphotoxin-beta Proteins 0.000 description 2
- 108010000684 Matrix Metalloproteinases Proteins 0.000 description 2
- 102000002274 Matrix Metalloproteinases Human genes 0.000 description 2
- 208000000172 Medulloblastoma Diseases 0.000 description 2
- 108010047230 Member 1 Subfamily B ATP Binding Cassette Transporter Proteins 0.000 description 2
- 102000018697 Membrane Proteins Human genes 0.000 description 2
- 102100039373 Membrane cofactor protein Human genes 0.000 description 2
- 102100026290 Membrane protein MLC1 Human genes 0.000 description 2
- 101100366137 Mesembryanthemum crystallinum SODCC.1 gene Proteins 0.000 description 2
- 102100039364 Metalloproteinase inhibitor 1 Human genes 0.000 description 2
- 102100023174 Methionine aminopeptidase 2 Human genes 0.000 description 2
- 208000034578 Multiple myelomas Diseases 0.000 description 2
- 101100066431 Mus musculus Fcgr2 gene Proteins 0.000 description 2
- 101000913072 Mus musculus Low affinity immunoglobulin gamma Fc region receptor III Proteins 0.000 description 2
- 102100025243 Myeloid cell surface antigen CD33 Human genes 0.000 description 2
- 108010067385 Myosin Light Chains Proteins 0.000 description 2
- 102000016349 Myosin Light Chains Human genes 0.000 description 2
- 108010056852 Myostatin Proteins 0.000 description 2
- 108090000028 Neprilysin Proteins 0.000 description 2
- 102000003729 Neprilysin Human genes 0.000 description 2
- 102000008730 Nestin Human genes 0.000 description 2
- 108010088225 Nestin Proteins 0.000 description 2
- 108010069196 Neural Cell Adhesion Molecules Proteins 0.000 description 2
- 206010029260 Neuroblastoma Diseases 0.000 description 2
- 102100021852 Neuronal cell adhesion molecule Human genes 0.000 description 2
- 101710130688 Neuronal cell adhesion molecule Proteins 0.000 description 2
- 108010011536 PTEN Phosphohydrolase Proteins 0.000 description 2
- 101100096142 Panax ginseng SODCC gene Proteins 0.000 description 2
- 108060005874 Parvalbumin Proteins 0.000 description 2
- 102000001675 Parvalbumin Human genes 0.000 description 2
- 206010057249 Phagocytosis Diseases 0.000 description 2
- 102100032543 Phosphatidylinositol 3,4,5-trisphosphate 3-phosphatase and dual-specificity protein phosphatase PTEN Human genes 0.000 description 2
- 102100035194 Placenta growth factor Human genes 0.000 description 2
- 102100036154 Platelet basic protein Human genes 0.000 description 2
- 102100030304 Platelet factor 4 Human genes 0.000 description 2
- 102100037265 Podoplanin Human genes 0.000 description 2
- 101710118150 Podoplanin Proteins 0.000 description 2
- 101710098940 Pro-epidermal growth factor Proteins 0.000 description 2
- 102100022661 Pro-neuregulin-1, membrane-bound isoform Human genes 0.000 description 2
- 102000003923 Protein Kinase C Human genes 0.000 description 2
- 108090000315 Protein Kinase C Proteins 0.000 description 2
- 102100027503 Protein Wnt-9a Human genes 0.000 description 2
- 102100031269 Putative peripheral benzodiazepine receptor-related protein Human genes 0.000 description 2
- 108010092799 RNA-directed DNA polymerase Proteins 0.000 description 2
- 108010038036 Receptor Activator of Nuclear Factor-kappa B Proteins 0.000 description 2
- 102000010498 Receptor Activator of Nuclear Factor-kappa B Human genes 0.000 description 2
- 101150099493 STAT3 gene Proteins 0.000 description 2
- 241000235070 Saccharomyces Species 0.000 description 2
- 102100027229 Solute carrier organic anion transporter family member 1C1 Human genes 0.000 description 2
- 208000005718 Stomach Neoplasms Diseases 0.000 description 2
- 101710088580 Stromal cell-derived factor 1 Proteins 0.000 description 2
- 102100035721 Syndecan-1 Human genes 0.000 description 2
- 108091008874 T cell receptors Proteins 0.000 description 2
- 102000016266 T-Cell Antigen Receptors Human genes 0.000 description 2
- 102100038126 Tenascin Human genes 0.000 description 2
- 108010008125 Tenascin Proteins 0.000 description 2
- 108020005038 Terminator Codon Proteins 0.000 description 2
- 102100040874 Tetraspanin-3 Human genes 0.000 description 2
- AYFVYJQAPQTCCC-UHFFFAOYSA-N Threonine Natural products CC(O)C(N)C(O)=O AYFVYJQAPQTCCC-UHFFFAOYSA-N 0.000 description 2
- 239000004473 Threonine Substances 0.000 description 2
- 108090000190 Thrombin Proteins 0.000 description 2
- 108010000499 Thromboplastin Proteins 0.000 description 2
- 102000002938 Thrombospondin Human genes 0.000 description 2
- 108060008245 Thrombospondin Proteins 0.000 description 2
- 102100030859 Tissue factor Human genes 0.000 description 2
- 108010033576 Transferrin Receptors Proteins 0.000 description 2
- 102100026144 Transferrin receptor protein 1 Human genes 0.000 description 2
- 102000004887 Transforming Growth Factor beta Human genes 0.000 description 2
- 108090001012 Transforming Growth Factor beta Proteins 0.000 description 2
- 102000013534 Troponin C Human genes 0.000 description 2
- 101710090322 Truncated surface protein Proteins 0.000 description 2
- 102100024568 Tumor necrosis factor ligand superfamily member 11 Human genes 0.000 description 2
- 102100024585 Tumor necrosis factor ligand superfamily member 13 Human genes 0.000 description 2
- 101710181056 Tumor necrosis factor ligand superfamily member 13B Proteins 0.000 description 2
- 102100024586 Tumor necrosis factor ligand superfamily member 14 Human genes 0.000 description 2
- 102100035283 Tumor necrosis factor ligand superfamily member 18 Human genes 0.000 description 2
- 102100031988 Tumor necrosis factor ligand superfamily member 6 Human genes 0.000 description 2
- 102100032101 Tumor necrosis factor ligand superfamily member 9 Human genes 0.000 description 2
- 102100029690 Tumor necrosis factor receptor superfamily member 13C Human genes 0.000 description 2
- 102100022205 Tumor necrosis factor receptor superfamily member 21 Human genes 0.000 description 2
- 102100022156 Tumor necrosis factor receptor superfamily member 3 Human genes 0.000 description 2
- 102100022153 Tumor necrosis factor receptor superfamily member 4 Human genes 0.000 description 2
- 101150117115 V gene Proteins 0.000 description 2
- 108091008605 VEGF receptors Proteins 0.000 description 2
- 102000016663 Vascular Endothelial Growth Factor Receptor-3 Human genes 0.000 description 2
- 108010019530 Vascular Endothelial Growth Factors Proteins 0.000 description 2
- 102000005789 Vascular Endothelial Growth Factors Human genes 0.000 description 2
- HCHKCACWOHOZIP-UHFFFAOYSA-N Zinc Chemical compound [Zn] HCHKCACWOHOZIP-UHFFFAOYSA-N 0.000 description 2
- 239000002253 acid Substances 0.000 description 2
- 208000009956 adenocarcinoma Diseases 0.000 description 2
- 239000002671 adjuvant Substances 0.000 description 2
- 229960003896 aminopterin Drugs 0.000 description 2
- 238000012197 amplification kit Methods 0.000 description 2
- 210000004102 animal cell Anatomy 0.000 description 2
- 229940125644 antibody drug Drugs 0.000 description 2
- 210000000628 antibody-producing cell Anatomy 0.000 description 2
- 108010006523 asialoglycoprotein receptor Proteins 0.000 description 2
- 235000009582 asparagine Nutrition 0.000 description 2
- 229960001230 asparagine Drugs 0.000 description 2
- 210000003719 b-lymphocyte Anatomy 0.000 description 2
- 230000001580 bacterial effect Effects 0.000 description 2
- 230000023555 blood coagulation Effects 0.000 description 2
- 210000002798 bone marrow cell Anatomy 0.000 description 2
- 238000010804 cDNA synthesis Methods 0.000 description 2
- 238000004422 calculation algorithm Methods 0.000 description 2
- 238000004364 calculation method Methods 0.000 description 2
- 239000006285 cell suspension Substances 0.000 description 2
- 238000006243 chemical reaction Methods 0.000 description 2
- BFPSDSIWYFKGBC-UHFFFAOYSA-N chlorotrianisene Chemical compound C1=CC(OC)=CC=C1C(Cl)=C(C=1C=CC(OC)=CC=1)C1=CC=C(OC)C=C1 BFPSDSIWYFKGBC-UHFFFAOYSA-N 0.000 description 2
- 210000001072 colon Anatomy 0.000 description 2
- 201000010897 colon adenocarcinoma Diseases 0.000 description 2
- 108010036922 cytoplasmic linker protein 115 Proteins 0.000 description 2
- 230000001472 cytotoxic effect Effects 0.000 description 2
- 230000001419 dependent effect Effects 0.000 description 2
- 230000004069 differentiation Effects 0.000 description 2
- 238000007865 diluting Methods 0.000 description 2
- 238000010790 dilution Methods 0.000 description 2
- 239000012895 dilution Substances 0.000 description 2
- 210000002889 endothelial cell Anatomy 0.000 description 2
- 230000002327 eosinophilic effect Effects 0.000 description 2
- 229960004222 factor ix Drugs 0.000 description 2
- 229940012413 factor vii Drugs 0.000 description 2
- 108091022862 fatty acid binding Proteins 0.000 description 2
- 229950003499 fibrin Drugs 0.000 description 2
- MHMNJMPURVTYEJ-UHFFFAOYSA-N fluorescein-5-isothiocyanate Chemical compound O1C(=O)C2=CC(N=C=S)=CC=C2C21C1=CC=C(O)C=C1OC1=CC(O)=CC=C21 MHMNJMPURVTYEJ-UHFFFAOYSA-N 0.000 description 2
- 230000002538 fungal effect Effects 0.000 description 2
- 201000005649 gangliocytoma Diseases 0.000 description 2
- 201000008361 ganglioneuroma Diseases 0.000 description 2
- GIVLTTJNORAZON-HDBOBKCLSA-N ganglioside GM2 (18:0) Chemical compound O[C@@H]1[C@@H](O)[C@H](OC[C@H](NC(=O)CCCCCCCCCCCCCCCCC)[C@H](O)\C=C\CCCCCCCCCCCCC)O[C@H](CO)[C@H]1O[C@H]1[C@H](O)[C@@H](O[C@]2(O[C@H]([C@H](NC(C)=O)[C@@H](O)C2)[C@H](O)[C@H](O)CO)C(O)=O)[C@@H](O[C@H]2[C@@H]([C@@H](O)[C@@H](O)[C@@H](CO)O2)NC(C)=O)[C@@H](CO)O1 GIVLTTJNORAZON-HDBOBKCLSA-N 0.000 description 2
- 201000006585 gastric adenocarcinoma Diseases 0.000 description 2
- 206010017758 gastric cancer Diseases 0.000 description 2
- 238000010353 genetic engineering Methods 0.000 description 2
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Natural products OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 description 2
- 235000004554 glutamine Nutrition 0.000 description 2
- 125000003630 glycyl group Chemical group [H]N([H])C([H])([H])C(*)=O 0.000 description 2
- 230000012010 growth Effects 0.000 description 2
- 239000003102 growth factor Substances 0.000 description 2
- 230000036541 health Effects 0.000 description 2
- 229940088597 hormone Drugs 0.000 description 2
- 239000005556 hormone Substances 0.000 description 2
- 210000001822 immobilized cell Anatomy 0.000 description 2
- 230000028993 immune response Effects 0.000 description 2
- 230000006058 immune tolerance Effects 0.000 description 2
- 229940072221 immunoglobulins Drugs 0.000 description 2
- 102000028416 insulin-like growth factor binding Human genes 0.000 description 2
- 108091022911 insulin-like growth factor binding Proteins 0.000 description 2
- 229940079322 interferon Drugs 0.000 description 2
- 201000005202 lung cancer Diseases 0.000 description 2
- 208000020816 lung neoplasm Diseases 0.000 description 2
- 239000002609 medium Substances 0.000 description 2
- 201000001441 melanoma Diseases 0.000 description 2
- 210000000274 microglia Anatomy 0.000 description 2
- 230000005012 migration Effects 0.000 description 2
- 238000013508 migration Methods 0.000 description 2
- 230000009149 molecular binding Effects 0.000 description 2
- 238000010369 molecular cloning Methods 0.000 description 2
- 229940053128 nerve growth factor Drugs 0.000 description 2
- 210000005055 nestin Anatomy 0.000 description 2
- 230000001537 neural effect Effects 0.000 description 2
- 102000039446 nucleic acids Human genes 0.000 description 2
- 108020004707 nucleic acids Proteins 0.000 description 2
- 150000007523 nucleic acids Chemical class 0.000 description 2
- 239000002245 particle Substances 0.000 description 2
- 230000037361 pathway Effects 0.000 description 2
- 238000002823 phage display Methods 0.000 description 2
- 230000008782 phagocytosis Effects 0.000 description 2
- 230000026731 phosphorylation Effects 0.000 description 2
- 238000006366 phosphorylation reaction Methods 0.000 description 2
- 108010031345 placental alkaline phosphatase Proteins 0.000 description 2
- 230000036470 plasma concentration Effects 0.000 description 2
- 230000001737 promoting effect Effects 0.000 description 2
- 238000000746 purification Methods 0.000 description 2
- 150000003230 pyrimidines Chemical class 0.000 description 2
- 238000003127 radioimmunoassay Methods 0.000 description 2
- 230000006798 recombination Effects 0.000 description 2
- BOLDJAUMGUJJKM-LSDHHAIUSA-N renifolin D Natural products CC(=C)[C@@H]1Cc2c(O)c(O)ccc2[C@H]1CC(=O)c3ccc(O)cc3O BOLDJAUMGUJJKM-LSDHHAIUSA-N 0.000 description 2
- 230000028327 secretion Effects 0.000 description 2
- 239000006152 selective media Substances 0.000 description 2
- 230000035945 sensitivity Effects 0.000 description 2
- 238000002741 site-directed mutagenesis Methods 0.000 description 2
- 229940126586 small molecule drug Drugs 0.000 description 2
- 101150017120 sod gene Proteins 0.000 description 2
- 230000000638 stimulation Effects 0.000 description 2
- 201000011549 stomach cancer Diseases 0.000 description 2
- 235000000346 sugar Nutrition 0.000 description 2
- ZRKFYGHZFMAOKI-QMGMOQQFSA-N tgfbeta Chemical compound C([C@H](NC(=O)[C@H](C(C)C)NC(=O)CNC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@H](CC(C)C)NC(=O)CNC(=O)[C@H](C)NC(=O)[C@H](CO)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@@H](NC(=O)[C@H](C)NC(=O)[C@H](C)NC(=O)[C@@H](NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CCSC)C(C)C)[C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)N[C@@H](C)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(=O)N1[C@@H](CCC1)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O)C1=CC=C(O)C=C1 ZRKFYGHZFMAOKI-QMGMOQQFSA-N 0.000 description 2
- 235000008521 threonine Nutrition 0.000 description 2
- 229960004072 thrombin Drugs 0.000 description 2
- 239000003053 toxin Substances 0.000 description 2
- 231100000765 toxin Toxicity 0.000 description 2
- 238000013518 transcription Methods 0.000 description 2
- 230000035897 transcription Effects 0.000 description 2
- 230000005758 transcription activity Effects 0.000 description 2
- 230000009466 transformation Effects 0.000 description 2
- 238000002054 transplantation Methods 0.000 description 2
- ARLKVQYMFRECLV-JSGCOSHPSA-N (2s)-2-[[(2s)-2-amino-3-(1h-indol-3-yl)propanoyl]amino]-4-methylsulfanylbutanamide Chemical compound C1=CC=C2C(C[C@H](N)C(=O)N[C@@H](CCSC)C(N)=O)=CNC2=C1 ARLKVQYMFRECLV-JSGCOSHPSA-N 0.000 description 1
- LAQPKDLYOBZWBT-NYLDSJSYSA-N (2s,4s,5r,6r)-5-acetamido-2-{[(2s,3r,4s,5s,6r)-2-{[(2r,3r,4r,5r)-5-acetamido-1,2-dihydroxy-6-oxo-4-{[(2s,3s,4r,5s,6s)-3,4,5-trihydroxy-6-methyloxan-2-yl]oxy}hexan-3-yl]oxy}-3,5-dihydroxy-6-(hydroxymethyl)oxan-4-yl]oxy}-4-hydroxy-6-[(1r,2r)-1,2,3-trihydrox Chemical compound O[C@H]1[C@H](O)[C@H](O)[C@H](C)O[C@H]1O[C@H]([C@@H](NC(C)=O)C=O)[C@@H]([C@H](O)CO)O[C@H]1[C@H](O)[C@@H](O[C@]2(O[C@H]([C@H](NC(C)=O)[C@@H](O)C2)[C@H](O)[C@H](O)CO)C(O)=O)[C@@H](O)[C@@H](CO)O1 LAQPKDLYOBZWBT-NYLDSJSYSA-N 0.000 description 1
- PXGPLTODNUVGFL-BRIYLRKRSA-N (E,Z)-(1R,2R,3R,5S)-7-(3,5-Dihydroxy-2-((3S)-(3-hydroxy-1-octenyl))cyclopentyl)-5-heptenoic acid Chemical compound CCCCC[C@H](O)C=C[C@H]1[C@H](O)C[C@H](O)[C@@H]1CC=CCCCC(O)=O PXGPLTODNUVGFL-BRIYLRKRSA-N 0.000 description 1
- NDJNDUULNXNRQD-XKBRQERYSA-N 1-[(2r,4s,5s)-5-[bromo(hydroxy)methyl]-4-hydroxyoxolan-2-yl]pyrimidine-2,4-dione Chemical compound C1[C@H](O)[C@@H](C(Br)O)O[C@H]1N1C(=O)NC(=O)C=C1 NDJNDUULNXNRQD-XKBRQERYSA-N 0.000 description 1
- NVKAWKQGWWIWPM-ABEVXSGRSA-N 17-β-hydroxy-5-α-Androstan-3-one Chemical compound C1C(=O)CC[C@]2(C)[C@H]3CC[C@](C)([C@H](CC4)O)[C@@H]4[C@@H]3CC[C@H]21 NVKAWKQGWWIWPM-ABEVXSGRSA-N 0.000 description 1
- VKUYLANQOAKALN-UHFFFAOYSA-N 2-[benzyl-(4-methoxyphenyl)sulfonylamino]-n-hydroxy-4-methylpentanamide Chemical compound C1=CC(OC)=CC=C1S(=O)(=O)N(C(CC(C)C)C(=O)NO)CC1=CC=CC=C1 VKUYLANQOAKALN-UHFFFAOYSA-N 0.000 description 1
- DTFAJAKTSMLKAT-JDCCYXBGSA-N 2-deoxystreptamine Chemical compound N[C@H]1C[C@@H](N)[C@H](O)[C@@H](O)[C@@H]1O DTFAJAKTSMLKAT-JDCCYXBGSA-N 0.000 description 1
- 238000005084 2D-nuclear magnetic resonance Methods 0.000 description 1
- ZOOGRGPOEVQQDX-UUOKFMHZSA-N 3',5'-cyclic GMP Chemical compound C([C@H]1O2)OP(O)(=O)O[C@H]1[C@@H](O)[C@@H]2N1C(N=C(NC2=O)N)=C2N=C1 ZOOGRGPOEVQQDX-UUOKFMHZSA-N 0.000 description 1
- AZKSAVLVSZKNRD-UHFFFAOYSA-M 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide Chemical compound [Br-].S1C(C)=C(C)N=C1[N+]1=NC(C=2C=CC=CC=2)=NN1C1=CC=CC=C1 AZKSAVLVSZKNRD-UHFFFAOYSA-M 0.000 description 1
- 102100021834 3-hydroxyacyl-CoA dehydrogenase Human genes 0.000 description 1
- QXZBMSIDSOZZHK-DOPDSADYSA-N 31362-50-2 Chemical compound C([C@@H](C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCSC)C(N)=O)NC(=O)CNC(=O)[C@@H](NC(=O)[C@H](C)NC(=O)[C@H](CC=1C2=CC=CC=C2NC=1)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@H](CC(N)=O)NC(=O)CNC(=O)[C@H](CC(C)C)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@H]1NC(=O)CC1)C(C)C)C1=CNC=N1 QXZBMSIDSOZZHK-DOPDSADYSA-N 0.000 description 1
- 108010082808 4-1BB Ligand Proteins 0.000 description 1
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 description 1
- AVPYQKSLYISFPO-UHFFFAOYSA-N 4-chlorobenzaldehyde Chemical compound ClC1=CC=C(C=O)C=C1 AVPYQKSLYISFPO-UHFFFAOYSA-N 0.000 description 1
- 102100030310 5,6-dihydroxyindole-2-carboxylic acid oxidase Human genes 0.000 description 1
- KFGOFTHODYBSGM-IJCBKZNRSA-N 6-Keto-prostaglandin F1a Chemical compound CCCCC[C@H](O)C=C[C@H]1[C@H](O)C[C@H](O)[C@@H]1CC(=O)CCCCC(O)=O KFGOFTHODYBSGM-IJCBKZNRSA-N 0.000 description 1
- WYWHKKSPHMUBEB-UHFFFAOYSA-N 6-Mercaptoguanine Natural products N1C(N)=NC(=S)C2=C1N=CN2 WYWHKKSPHMUBEB-UHFFFAOYSA-N 0.000 description 1
- 102100026802 72 kDa type IV collagenase Human genes 0.000 description 1
- 101710151806 72 kDa type IV collagenase Proteins 0.000 description 1
- HCAJQHYUCKICQH-VPENINKCSA-N 8-Oxo-7,8-dihydro-2'-deoxyguanosine Chemical compound C1=2NC(N)=NC(=O)C=2NC(=O)N1[C@H]1C[C@H](O)[C@@H](CO)O1 HCAJQHYUCKICQH-VPENINKCSA-N 0.000 description 1
- LPXQRXLUHJKZIE-UHFFFAOYSA-N 8-azaguanine Chemical compound NC1=NC(O)=C2NN=NC2=N1 LPXQRXLUHJKZIE-UHFFFAOYSA-N 0.000 description 1
- 229960005508 8-azaguanine Drugs 0.000 description 1
- ZKRFOXLVOKTUTA-KQYNXXCUSA-N 9-(5-phosphoribofuranosyl)-6-mercaptopurine Chemical compound O[C@@H]1[C@H](O)[C@@H](COP(O)(O)=O)O[C@H]1N1C(NC=NC2=S)=C2N=C1 ZKRFOXLVOKTUTA-KQYNXXCUSA-N 0.000 description 1
- 102100027400 A disintegrin and metalloproteinase with thrombospondin motifs 4 Human genes 0.000 description 1
- 102000029791 ADAM Human genes 0.000 description 1
- 108091022885 ADAM Proteins 0.000 description 1
- 108091007504 ADAM10 Proteins 0.000 description 1
- 108091007507 ADAM12 Proteins 0.000 description 1
- 108091007505 ADAM17 Proteins 0.000 description 1
- 108091022879 ADAMTS Proteins 0.000 description 1
- 102000029750 ADAMTS Human genes 0.000 description 1
- 108091005664 ADAMTS4 Proteins 0.000 description 1
- 102000051389 ADAMTS5 Human genes 0.000 description 1
- 108091005663 ADAMTS5 Proteins 0.000 description 1
- 101150007969 ADORA1 gene Proteins 0.000 description 1
- 102100031585 ADP-ribosyl cyclase/cyclic ADP-ribose hydrolase 1 Human genes 0.000 description 1
- 102100033793 ALK tyrosine kinase receptor Human genes 0.000 description 1
- 101100192359 Acinetobacter johnsonii ptk gene Proteins 0.000 description 1
- 206010069754 Acquired gene mutation Diseases 0.000 description 1
- 108010075348 Activated-Leukocyte Cell Adhesion Molecule Proteins 0.000 description 1
- 102100034111 Activin receptor type-1 Human genes 0.000 description 1
- 102100034134 Activin receptor type-1B Human genes 0.000 description 1
- 101710173011 Activin receptor type-1B Proteins 0.000 description 1
- 102100034135 Activin receptor type-1C Human genes 0.000 description 1
- 101710173005 Activin receptor type-1C Proteins 0.000 description 1
- ORILYTVJVMAKLC-UHFFFAOYSA-N Adamantane Natural products C1C(C2)CC3CC1CC2C3 ORILYTVJVMAKLC-UHFFFAOYSA-N 0.000 description 1
- 208000010507 Adenocarcinoma of Lung Diseases 0.000 description 1
- 108010054404 Adenylyl-sulfate kinase Proteins 0.000 description 1
- 102100024439 Adhesion G protein-coupled receptor A2 Human genes 0.000 description 1
- 102100026423 Adhesion G protein-coupled receptor E5 Human genes 0.000 description 1
- 108010088751 Albumins Proteins 0.000 description 1
- 102000009027 Albumins Human genes 0.000 description 1
- 102100025677 Alkaline phosphatase, germ cell type Human genes 0.000 description 1
- 102100035248 Alpha-(1,3)-fucosyltransferase 4 Human genes 0.000 description 1
- 102100026882 Alpha-synuclein Human genes 0.000 description 1
- 102100034452 Alternative prion protein Human genes 0.000 description 1
- 101710085851 Angiopoietin-related protein 2 Proteins 0.000 description 1
- 108010049777 Ankyrins Proteins 0.000 description 1
- 102000008102 Ankyrins Human genes 0.000 description 1
- 102100036523 Anoctamin-6 Human genes 0.000 description 1
- 102100031323 Anthrax toxin receptor 1 Human genes 0.000 description 1
- 102100021253 Antileukoproteinase Human genes 0.000 description 1
- 102000004411 Antithrombin III Human genes 0.000 description 1
- 108090000935 Antithrombin III Proteins 0.000 description 1
- 108010059886 Apolipoprotein A-I Proteins 0.000 description 1
- 102000005666 Apolipoprotein A-I Human genes 0.000 description 1
- 108010087614 Apolipoprotein A-II Proteins 0.000 description 1
- 102000009081 Apolipoprotein A-II Human genes 0.000 description 1
- 108010062544 Apoptotic Protease-Activating Factor 1 Proteins 0.000 description 1
- 102100034524 Apoptotic protease-activating factor 1 Human genes 0.000 description 1
- 101100339431 Arabidopsis thaliana HMGB2 gene Proteins 0.000 description 1
- 102100026376 Artemin Human genes 0.000 description 1
- 101710205806 Artemin Proteins 0.000 description 1
- 206010003445 Ascites Diseases 0.000 description 1
- 101150075175 Asgr1 gene Proteins 0.000 description 1
- 241000228212 Aspergillus Species 0.000 description 1
- 101000605172 Aspergillus niger (strain CBS 513.88 / FGSC A1513) Probable endopolygalacturonase E Proteins 0.000 description 1
- 101000605171 Aspergillus niger Endopolygalacturonase E Proteins 0.000 description 1
- 101800001288 Atrial natriuretic factor Proteins 0.000 description 1
- 102400001282 Atrial natriuretic peptide Human genes 0.000 description 1
- 101800001890 Atrial natriuretic peptide Proteins 0.000 description 1
- 241000271566 Aves Species 0.000 description 1
- 108090001008 Avidin Proteins 0.000 description 1
- 102100038080 B-cell receptor CD22 Human genes 0.000 description 1
- 102100022005 B-lymphocyte antigen CD20 Human genes 0.000 description 1
- 101150033765 BAG1 gene Proteins 0.000 description 1
- PCLCDPVEEFVAAQ-UHFFFAOYSA-N BCA 1 Chemical compound CC(CO)CCCC(C)C1=CCC(C)(O)C1CC2=C(O)C(O)CCC2=O PCLCDPVEEFVAAQ-UHFFFAOYSA-N 0.000 description 1
- 108091007065 BIRCs Proteins 0.000 description 1
- 244000063299 Bacillus subtilis Species 0.000 description 1
- 235000014469 Bacillus subtilis Nutrition 0.000 description 1
- 102100021676 Baculoviral IAP repeat-containing protein 1 Human genes 0.000 description 1
- 102100028239 Basal cell adhesion molecule Human genes 0.000 description 1
- 108010064528 Basigin Proteins 0.000 description 1
- 102000015279 Basigin Human genes 0.000 description 1
- 102100032412 Basigin Human genes 0.000 description 1
- 102100032305 Bcl-2 homologous antagonist/killer Human genes 0.000 description 1
- 102100031109 Beta-catenin-like protein 1 Human genes 0.000 description 1
- 101710164563 Beta-catenin-like protein 1 Proteins 0.000 description 1
- 102000013585 Bombesin Human genes 0.000 description 1
- 108010051479 Bombesin Proteins 0.000 description 1
- 108010049931 Bone Morphogenetic Protein 2 Proteins 0.000 description 1
- 108010049951 Bone Morphogenetic Protein 3 Proteins 0.000 description 1
- 108010049955 Bone Morphogenetic Protein 4 Proteins 0.000 description 1
- 108010049976 Bone Morphogenetic Protein 5 Proteins 0.000 description 1
- 108010049974 Bone Morphogenetic Protein 6 Proteins 0.000 description 1
- 102100037086 Bone marrow stromal antigen 2 Human genes 0.000 description 1
- 102100024506 Bone morphogenetic protein 2 Human genes 0.000 description 1
- 102100024504 Bone morphogenetic protein 3 Human genes 0.000 description 1
- 102100024505 Bone morphogenetic protein 4 Human genes 0.000 description 1
- 102100022526 Bone morphogenetic protein 5 Human genes 0.000 description 1
- 102100022525 Bone morphogenetic protein 6 Human genes 0.000 description 1
- 102100025423 Bone morphogenetic protein receptor type-1A Human genes 0.000 description 1
- 101710120270 Bone morphogenetic protein receptor type-1A Proteins 0.000 description 1
- 101710120271 Bone morphogenetic protein receptor type-1B Proteins 0.000 description 1
- 102100025422 Bone morphogenetic protein receptor type-2 Human genes 0.000 description 1
- 108050008407 Bone morphogenetic protein receptor type-2 Proteins 0.000 description 1
- 108030001720 Bontoxilysin Proteins 0.000 description 1
- ZOXJGFHDIHLPTG-UHFFFAOYSA-N Boron Chemical compound [B] ZOXJGFHDIHLPTG-UHFFFAOYSA-N 0.000 description 1
- 101000941281 Bos taurus Gastric triacylglycerol lipase Proteins 0.000 description 1
- 208000003174 Brain Neoplasms Diseases 0.000 description 1
- 108090000715 Brain-derived neurotrophic factor Proteins 0.000 description 1
- 102000004219 Brain-derived neurotrophic factor Human genes 0.000 description 1
- 208000011691 Burkitt lymphomas Diseases 0.000 description 1
- 102100031172 C-C chemokine receptor type 1 Human genes 0.000 description 1
- 101710149814 C-C chemokine receptor type 1 Proteins 0.000 description 1
- 102100031151 C-C chemokine receptor type 2 Human genes 0.000 description 1
- 101710149815 C-C chemokine receptor type 2 Proteins 0.000 description 1
- 102100024167 C-C chemokine receptor type 3 Human genes 0.000 description 1
- 101710149862 C-C chemokine receptor type 3 Proteins 0.000 description 1
- 101710149863 C-C chemokine receptor type 4 Proteins 0.000 description 1
- 102100035875 C-C chemokine receptor type 5 Human genes 0.000 description 1
- 101710149870 C-C chemokine receptor type 5 Proteins 0.000 description 1
- 102100036305 C-C chemokine receptor type 8 Human genes 0.000 description 1
- 102100025074 C-C chemokine receptor-like 2 Human genes 0.000 description 1
- 102100036841 C-C motif chemokine 1 Human genes 0.000 description 1
- 102100023702 C-C motif chemokine 13 Human genes 0.000 description 1
- 102100023705 C-C motif chemokine 14 Human genes 0.000 description 1
- 102100023703 C-C motif chemokine 15 Human genes 0.000 description 1
- 102100023700 C-C motif chemokine 16 Human genes 0.000 description 1
- 102100023698 C-C motif chemokine 17 Human genes 0.000 description 1
- 102100036842 C-C motif chemokine 19 Human genes 0.000 description 1
- 102100021943 C-C motif chemokine 2 Human genes 0.000 description 1
- 102100036848 C-C motif chemokine 20 Human genes 0.000 description 1
- 102100036846 C-C motif chemokine 21 Human genes 0.000 description 1
- 102100036849 C-C motif chemokine 24 Human genes 0.000 description 1
- 102100021935 C-C motif chemokine 26 Human genes 0.000 description 1
- 101710112538 C-C motif chemokine 27 Proteins 0.000 description 1
- 102100021942 C-C motif chemokine 28 Human genes 0.000 description 1
- 102100031092 C-C motif chemokine 3 Human genes 0.000 description 1
- 101710155856 C-C motif chemokine 3 Proteins 0.000 description 1
- 102100034871 C-C motif chemokine 8 Human genes 0.000 description 1
- 102100036166 C-X-C chemokine receptor type 1 Human genes 0.000 description 1
- 102100028989 C-X-C chemokine receptor type 2 Human genes 0.000 description 1
- 102100028990 C-X-C chemokine receptor type 3 Human genes 0.000 description 1
- 102100031650 C-X-C chemokine receptor type 4 Human genes 0.000 description 1
- 101710098275 C-X-C motif chemokine 10 Proteins 0.000 description 1
- 101710098272 C-X-C motif chemokine 11 Proteins 0.000 description 1
- 102100025250 C-X-C motif chemokine 14 Human genes 0.000 description 1
- 102100039396 C-X-C motif chemokine 16 Human genes 0.000 description 1
- 102100039398 C-X-C motif chemokine 2 Human genes 0.000 description 1
- 102100036189 C-X-C motif chemokine 3 Human genes 0.000 description 1
- 102100036150 C-X-C motif chemokine 5 Human genes 0.000 description 1
- 101710085504 C-X-C motif chemokine 6 Proteins 0.000 description 1
- 101710085500 C-X-C motif chemokine 9 Proteins 0.000 description 1
- 102000007269 CA-125 Antigen Human genes 0.000 description 1
- 108010008629 CA-125 Antigen Proteins 0.000 description 1
- 102100024217 CAMPATH-1 antigen Human genes 0.000 description 1
- 108091005932 CCKBR Proteins 0.000 description 1
- 101150049756 CCL6 gene Proteins 0.000 description 1
- 101150011672 CCL9 gene Proteins 0.000 description 1
- 102100031168 CCN family member 2 Human genes 0.000 description 1
- 102100032976 CCR4-NOT transcription complex subunit 6 Human genes 0.000 description 1
- 102100024210 CD166 antigen Human genes 0.000 description 1
- 108010059108 CD18 Antigens Proteins 0.000 description 1
- 102100038077 CD226 antigen Human genes 0.000 description 1
- 108010046080 CD27 Ligand Proteins 0.000 description 1
- 102100027207 CD27 antigen Human genes 0.000 description 1
- 102000017420 CD3 protein, epsilon/gamma/delta subunit Human genes 0.000 description 1
- 108050005493 CD3 protein, epsilon/gamma/delta subunit Proteins 0.000 description 1
- 108010029697 CD40 Ligand Proteins 0.000 description 1
- 101150013553 CD40 gene Proteins 0.000 description 1
- 108010065524 CD52 Antigen Proteins 0.000 description 1
- 108010009575 CD55 Antigens Proteins 0.000 description 1
- 108010084313 CD58 Antigens Proteins 0.000 description 1
- 102100027221 CD81 antigen Human genes 0.000 description 1
- 101150107439 CDR3 gene Proteins 0.000 description 1
- 108010021064 CTLA-4 Antigen Proteins 0.000 description 1
- 229940045513 CTLA4 antagonist Drugs 0.000 description 1
- 108090000835 CX3C Chemokine Receptor 1 Proteins 0.000 description 1
- 102100039196 CX3C chemokine receptor 1 Human genes 0.000 description 1
- 102100036364 Cadherin-2 Human genes 0.000 description 1
- 102100036360 Cadherin-3 Human genes 0.000 description 1
- 101100381481 Caenorhabditis elegans baz-2 gene Proteins 0.000 description 1
- 101100289995 Caenorhabditis elegans mac-1 gene Proteins 0.000 description 1
- 102400000113 Calcitonin Human genes 0.000 description 1
- 108060001064 Calcitonin Proteins 0.000 description 1
- OKTJSMMVPCPJKN-UHFFFAOYSA-N Carbon Chemical compound [C] OKTJSMMVPCPJKN-UHFFFAOYSA-N 0.000 description 1
- 102100024423 Carbonic anhydrase 9 Human genes 0.000 description 1
- 201000009030 Carcinoma Diseases 0.000 description 1
- 102100028892 Cardiotrophin-1 Human genes 0.000 description 1
- 102000014914 Carrier Proteins Human genes 0.000 description 1
- 108010076119 Caseins Proteins 0.000 description 1
- 102000011632 Caseins Human genes 0.000 description 1
- 108010059081 Cathepsin A Proteins 0.000 description 1
- 102000005572 Cathepsin A Human genes 0.000 description 1
- 108090000712 Cathepsin B Proteins 0.000 description 1
- 102000004225 Cathepsin B Human genes 0.000 description 1
- 102000003902 Cathepsin C Human genes 0.000 description 1
- 108090000267 Cathepsin C Proteins 0.000 description 1
- 102000003908 Cathepsin D Human genes 0.000 description 1
- 108090000258 Cathepsin D Proteins 0.000 description 1
- 102000004178 Cathepsin E Human genes 0.000 description 1
- 108090000611 Cathepsin E Proteins 0.000 description 1
- 108090000619 Cathepsin H Proteins 0.000 description 1
- 102400001321 Cathepsin L Human genes 0.000 description 1
- 108090000624 Cathepsin L Proteins 0.000 description 1
- 102100026540 Cathepsin L2 Human genes 0.000 description 1
- 101710177066 Cathepsin O Proteins 0.000 description 1
- 108090000613 Cathepsin S Proteins 0.000 description 1
- 102100035654 Cathepsin S Human genes 0.000 description 1
- 102100026657 Cathepsin Z Human genes 0.000 description 1
- 108010061117 Cathepsin Z Proteins 0.000 description 1
- 101150075117 Ccl12 gene Proteins 0.000 description 1
- 108010082548 Chemokine CCL11 Proteins 0.000 description 1
- 108010078239 Chemokine CX3CL1 Proteins 0.000 description 1
- 108010009685 Cholinergic Receptors Proteins 0.000 description 1
- 208000004139 Choroid Plexus Neoplasms Diseases 0.000 description 1
- 108010038447 Chromogranin A Proteins 0.000 description 1
- 102000010792 Chromogranin A Human genes 0.000 description 1
- 108010038439 Chromogranin B Proteins 0.000 description 1
- 102000010791 Chromogranin B Human genes 0.000 description 1
- 108010005939 Ciliary Neurotrophic Factor Proteins 0.000 description 1
- 102100031614 Ciliary neurotrophic factor Human genes 0.000 description 1
- 101000862089 Clarkia lewisii Glucose-6-phosphate isomerase, cytosolic 1A Proteins 0.000 description 1
- 241000193155 Clostridium botulinum Species 0.000 description 1
- 241000193468 Clostridium perfringens Species 0.000 description 1
- 108091026890 Coding region Proteins 0.000 description 1
- 108010048623 Collagen Receptors Proteins 0.000 description 1
- 102100027995 Collagenase 3 Human genes 0.000 description 1
- 108050005238 Collagenase 3 Proteins 0.000 description 1
- 208000035473 Communicable disease Diseases 0.000 description 1
- 108010053085 Complement Factor H Proteins 0.000 description 1
- 102000016550 Complement Factor H Human genes 0.000 description 1
- 102100025680 Complement decay-accelerating factor Human genes 0.000 description 1
- 108090000056 Complement factor B Proteins 0.000 description 1
- 102000003712 Complement factor B Human genes 0.000 description 1
- 102100032768 Complement receptor type 2 Human genes 0.000 description 1
- 108010047041 Complementarity Determining Regions Proteins 0.000 description 1
- 108010002947 Connectin Proteins 0.000 description 1
- RYGMFSIKBFXOCR-UHFFFAOYSA-N Copper Chemical group [Cu] RYGMFSIKBFXOCR-UHFFFAOYSA-N 0.000 description 1
- 108050006400 Cyclin Proteins 0.000 description 1
- 206010011732 Cyst Diseases 0.000 description 1
- 102000015833 Cystatin Human genes 0.000 description 1
- 108010079245 Cystic Fibrosis Transmembrane Conductance Regulator Proteins 0.000 description 1
- 102000012605 Cystic Fibrosis Transmembrane Conductance Regulator Human genes 0.000 description 1
- 102100035298 Cytokine SCM-1 beta Human genes 0.000 description 1
- 101150082208 DIABLO gene Proteins 0.000 description 1
- 230000004544 DNA amplification Effects 0.000 description 1
- 102100033215 DNA nucleotidylexotransferase Human genes 0.000 description 1
- 230000004543 DNA replication Effects 0.000 description 1
- 102100035619 DNA-(apurinic or apyrimidinic site) lyase Human genes 0.000 description 1
- 102100027700 DNA-directed RNA polymerase I subunit RPA2 Human genes 0.000 description 1
- 101100203200 Danio rerio shha gene Proteins 0.000 description 1
- 101100317380 Danio rerio wnt4a gene Proteins 0.000 description 1
- 208000031124 Dementia Alzheimer type Diseases 0.000 description 1
- 102100033189 Diablo IAP-binding mitochondrial protein Human genes 0.000 description 1
- 102100030074 Dickkopf-related protein 1 Human genes 0.000 description 1
- LTMHDMANZUZIPE-AMTYYWEZSA-N Digoxin Natural products O([C@H]1[C@H](C)O[C@H](O[C@@H]2C[C@@H]3[C@@](C)([C@@H]4[C@H]([C@]5(O)[C@](C)([C@H](O)C4)[C@H](C4=CC(=O)OC4)CC5)CC3)CC2)C[C@@H]1O)[C@H]1O[C@H](C)[C@@H](O[C@H]2O[C@@H](C)[C@H](O)[C@@H](O)C2)[C@@H](O)C1 LTMHDMANZUZIPE-AMTYYWEZSA-N 0.000 description 1
- 102100029921 Dipeptidyl peptidase 1 Human genes 0.000 description 1
- 101710087078 Dipeptidyl peptidase 1 Proteins 0.000 description 1
- 108010043648 Discoidin Domain Receptors Proteins 0.000 description 1
- 102000002706 Discoidin Domain Receptors Human genes 0.000 description 1
- 102100039673 Disintegrin and metalloproteinase domain-containing protein 10 Human genes 0.000 description 1
- 102100031107 Disintegrin and metalloproteinase domain-containing protein 11 Human genes 0.000 description 1
- 101710121366 Disintegrin and metalloproteinase domain-containing protein 11 Proteins 0.000 description 1
- 102100031112 Disintegrin and metalloproteinase domain-containing protein 12 Human genes 0.000 description 1
- 102100031113 Disintegrin and metalloproteinase domain-containing protein 15 Human genes 0.000 description 1
- 102100024364 Disintegrin and metalloproteinase domain-containing protein 8 Human genes 0.000 description 1
- 102100024361 Disintegrin and metalloproteinase domain-containing protein 9 Human genes 0.000 description 1
- 101000968267 Drosophila melanogaster Protein dachsous Proteins 0.000 description 1
- 101000708615 Drosophila melanogaster Protein smoothened Proteins 0.000 description 1
- 101100261976 Drosophila melanogaster trk gene Proteins 0.000 description 1
- 101710192866 Dystrobrevin alpha Proteins 0.000 description 1
- 108010024212 E-Selectin Proteins 0.000 description 1
- 102100023471 E-selectin Human genes 0.000 description 1
- 102100037024 E3 ubiquitin-protein ligase XIAP Human genes 0.000 description 1
- 108010059397 ENA-VASP proteins Proteins 0.000 description 1
- 101150039808 Egfr gene Proteins 0.000 description 1
- 102100040897 Embryonic growth/differentiation factor 1 Human genes 0.000 description 1
- 102100038083 Endosialin Human genes 0.000 description 1
- 102000010180 Endothelin receptor Human genes 0.000 description 1
- 108050001739 Endothelin receptor Proteins 0.000 description 1
- 101800004490 Endothelin-1 Proteins 0.000 description 1
- 102100031983 Ephrin type-B receptor 4 Human genes 0.000 description 1
- 102100023721 Ephrin-B2 Human genes 0.000 description 1
- 108010044090 Ephrin-B2 Proteins 0.000 description 1
- 101800003838 Epidermal growth factor Proteins 0.000 description 1
- 102000018651 Epithelial Cell Adhesion Molecule Human genes 0.000 description 1
- 108010066687 Epithelial Cell Adhesion Molecule Proteins 0.000 description 1
- 241000402754 Erythranthe moschata Species 0.000 description 1
- 101100172469 Escherichia coli (strain K12) envZ gene Proteins 0.000 description 1
- 101000759376 Escherichia phage Mu Tail sheath protein Proteins 0.000 description 1
- 101150021185 FGF gene Proteins 0.000 description 1
- 108091008794 FGF receptors Proteins 0.000 description 1
- 108010029144 Factor IIa Proteins 0.000 description 1
- 108010048049 Factor IXa Proteins 0.000 description 1
- 108010014172 Factor V Proteins 0.000 description 1
- 108010054218 Factor VIII Proteins 0.000 description 1
- 102000001690 Factor VIII Human genes 0.000 description 1
- 108010061932 Factor VIIIa Proteins 0.000 description 1
- 108010054265 Factor VIIa Proteins 0.000 description 1
- 108010074105 Factor Va Proteins 0.000 description 1
- 108010014173 Factor X Proteins 0.000 description 1
- 108010074864 Factor XI Proteins 0.000 description 1
- 108010080865 Factor XII Proteins 0.000 description 1
- 102000000429 Factor XII Human genes 0.000 description 1
- 108010071289 Factor XIII Proteins 0.000 description 1
- 108010000196 Factor XIIIa Proteins 0.000 description 1
- 108010071241 Factor XIIa Proteins 0.000 description 1
- 108010080805 Factor XIa Proteins 0.000 description 1
- 108010074860 Factor Xa Proteins 0.000 description 1
- 102000008857 Ferritin Human genes 0.000 description 1
- 108050000784 Ferritin Proteins 0.000 description 1
- 238000008416 Ferritin Methods 0.000 description 1
- 108010049003 Fibrinogen Proteins 0.000 description 1
- 102000008946 Fibrinogen Human genes 0.000 description 1
- 108010088842 Fibrinolysin Proteins 0.000 description 1
- 108090000386 Fibroblast Growth Factor 1 Proteins 0.000 description 1
- 102100031706 Fibroblast growth factor 1 Human genes 0.000 description 1
- 102100031734 Fibroblast growth factor 19 Human genes 0.000 description 1
- 102100024785 Fibroblast growth factor 2 Human genes 0.000 description 1
- 102000003974 Fibroblast growth factor 2 Human genes 0.000 description 1
- 102100028043 Fibroblast growth factor 3 Human genes 0.000 description 1
- 102100037680 Fibroblast growth factor 8 Human genes 0.000 description 1
- 108010067306 Fibronectins Proteins 0.000 description 1
- 201000008808 Fibrosarcoma Diseases 0.000 description 1
- 108090000652 Flap endonucleases Proteins 0.000 description 1
- 102000004150 Flap endonucleases Human genes 0.000 description 1
- 102000012673 Follicle Stimulating Hormone Human genes 0.000 description 1
- 108010079345 Follicle Stimulating Hormone Proteins 0.000 description 1
- 102100021259 Frizzled-1 Human genes 0.000 description 1
- 102100021261 Frizzled-10 Human genes 0.000 description 1
- 102100021265 Frizzled-2 Human genes 0.000 description 1
- 102100039820 Frizzled-4 Human genes 0.000 description 1
- 102100039818 Frizzled-5 Human genes 0.000 description 1
- 102100039799 Frizzled-6 Human genes 0.000 description 1
- 102100039676 Frizzled-7 Human genes 0.000 description 1
- 102100028466 Frizzled-8 Human genes 0.000 description 1
- 241000233866 Fungi Species 0.000 description 1
- 101150022345 GAS6 gene Proteins 0.000 description 1
- 101000934641 Gallus gallus Bone morphogenetic protein receptor type-1B Proteins 0.000 description 1
- 101100181195 Gallus gallus RPS6KA gene Proteins 0.000 description 1
- 101710115997 Gamma-tubulin complex component 2 Proteins 0.000 description 1
- 208000003098 Ganglion Cysts Diseases 0.000 description 1
- 102000004878 Gelsolin Human genes 0.000 description 1
- 108090001064 Gelsolin Proteins 0.000 description 1
- 108700039691 Genetic Promoter Regions Proteins 0.000 description 1
- 206010071602 Genetic polymorphism Diseases 0.000 description 1
- CEAZRRDELHUEMR-URQXQFDESA-N Gentamicin Chemical class O1[C@H](C(C)NC)CC[C@@H](N)[C@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](NC)[C@@](C)(O)CO2)O)[C@H](N)C[C@@H]1N CEAZRRDELHUEMR-URQXQFDESA-N 0.000 description 1
- 229930182566 Gentamicin Natural products 0.000 description 1
- 101000757797 Geobacillus stearothermophilus Aminopeptidase 2 Proteins 0.000 description 1
- 208000000527 Germinoma Diseases 0.000 description 1
- 208000032612 Glial tumor Diseases 0.000 description 1
- 201000005409 Gliomatosis cerebri Diseases 0.000 description 1
- 102400000321 Glucagon Human genes 0.000 description 1
- 108060003199 Glucagon Proteins 0.000 description 1
- 102000018899 Glutamate Receptors Human genes 0.000 description 1
- 108010027915 Glutamate Receptors Proteins 0.000 description 1
- 102000005720 Glutathione transferase Human genes 0.000 description 1
- 108010070675 Glutathione transferase Proteins 0.000 description 1
- PEDCQBHIVMGVHV-UHFFFAOYSA-N Glycerine Chemical compound OCC(O)CO PEDCQBHIVMGVHV-UHFFFAOYSA-N 0.000 description 1
- 102100032530 Glypican-3 Human genes 0.000 description 1
- 102100039619 Granulocyte colony-stimulating factor Human genes 0.000 description 1
- 108010017213 Granulocyte-Macrophage Colony-Stimulating Factor Proteins 0.000 description 1
- 102100039620 Granulocyte-macrophage colony-stimulating factor Human genes 0.000 description 1
- 108010090296 Growth Differentiation Factor 1 Proteins 0.000 description 1
- 108010041834 Growth Differentiation Factor 15 Proteins 0.000 description 1
- 108010090293 Growth Differentiation Factor 3 Proteins 0.000 description 1
- 108010090250 Growth Differentiation Factor 6 Proteins 0.000 description 1
- 239000000095 Growth Hormone-Releasing Hormone Substances 0.000 description 1
- 102000004858 Growth differentiation factor-9 Human genes 0.000 description 1
- 108090001086 Growth differentiation factor-9 Proteins 0.000 description 1
- 101710194460 Growth/differentiation factor 15 Proteins 0.000 description 1
- 102100035364 Growth/differentiation factor 3 Human genes 0.000 description 1
- 101710204282 Growth/differentiation factor 5 Proteins 0.000 description 1
- 101710204281 Growth/differentiation factor 6 Proteins 0.000 description 1
- 102100030595 HLA class II histocompatibility antigen gamma chain Human genes 0.000 description 1
- 108700010013 HMGB1 Proteins 0.000 description 1
- 101150021904 HMGB1 gene Proteins 0.000 description 1
- 101000691214 Haloarcula marismortui (strain ATCC 43049 / DSM 3752 / JCM 8966 / VKM B-1809) 50S ribosomal protein L44e Proteins 0.000 description 1
- 102100034051 Heat shock protein HSP 90-alpha Human genes 0.000 description 1
- 102100031573 Hematopoietic progenitor cell antigen CD34 Human genes 0.000 description 1
- 102100024025 Heparanase Human genes 0.000 description 1
- 102100031465 Hepatocyte growth factor activator Human genes 0.000 description 1
- 101710085796 Hepatocyte growth factor activator Proteins 0.000 description 1
- 241000238631 Hexapoda Species 0.000 description 1
- 101710121996 Hexon protein p72 Proteins 0.000 description 1
- 102100026122 High affinity immunoglobulin gamma Fc receptor I Human genes 0.000 description 1
- 102100037907 High mobility group protein B1 Human genes 0.000 description 1
- 108010000487 High-Molecular-Weight Kininogen Proteins 0.000 description 1
- 101000773083 Homo sapiens 5,6-dihydroxyindole-2-carboxylic acid oxidase Proteins 0.000 description 1
- 101000777636 Homo sapiens ADP-ribosyl cyclase/cyclic ADP-ribose hydrolase 1 Proteins 0.000 description 1
- 101000779641 Homo sapiens ALK tyrosine kinase receptor Proteins 0.000 description 1
- 101000799140 Homo sapiens Activin receptor type-1 Proteins 0.000 description 1
- 101000833358 Homo sapiens Adhesion G protein-coupled receptor A2 Proteins 0.000 description 1
- 101000718243 Homo sapiens Adhesion G protein-coupled receptor E5 Proteins 0.000 description 1
- 101000574440 Homo sapiens Alkaline phosphatase, germ cell type Proteins 0.000 description 1
- 101001022185 Homo sapiens Alpha-(1,3)-fucosyltransferase 4 Proteins 0.000 description 1
- 101000757160 Homo sapiens Aminopeptidase N Proteins 0.000 description 1
- 101000693081 Homo sapiens Angiopoietin-related protein 2 Proteins 0.000 description 1
- 101000928362 Homo sapiens Anoctamin-6 Proteins 0.000 description 1
- 101000796095 Homo sapiens Anthrax toxin receptor 1 Proteins 0.000 description 1
- 101000615334 Homo sapiens Antileukoproteinase Proteins 0.000 description 1
- 101000884305 Homo sapiens B-cell receptor CD22 Proteins 0.000 description 1
- 101000897405 Homo sapiens B-lymphocyte antigen CD20 Proteins 0.000 description 1
- 101100325746 Homo sapiens BAK1 gene Proteins 0.000 description 1
- 101000935638 Homo sapiens Basal cell adhesion molecule Proteins 0.000 description 1
- 101000798441 Homo sapiens Basigin Proteins 0.000 description 1
- 101000740785 Homo sapiens Bone marrow stromal antigen 2 Proteins 0.000 description 1
- 101000766294 Homo sapiens Branched-chain-amino-acid aminotransferase, mitochondrial Proteins 0.000 description 1
- 101000716068 Homo sapiens C-C chemokine receptor type 6 Proteins 0.000 description 1
- 101000716063 Homo sapiens C-C chemokine receptor type 8 Proteins 0.000 description 1
- 101000716070 Homo sapiens C-C chemokine receptor type 9 Proteins 0.000 description 1
- 101000713104 Homo sapiens C-C motif chemokine 1 Proteins 0.000 description 1
- 101000978379 Homo sapiens C-C motif chemokine 13 Proteins 0.000 description 1
- 101000978381 Homo sapiens C-C motif chemokine 14 Proteins 0.000 description 1
- 101000978376 Homo sapiens C-C motif chemokine 15 Proteins 0.000 description 1
- 101000978375 Homo sapiens C-C motif chemokine 16 Proteins 0.000 description 1
- 101000978362 Homo sapiens C-C motif chemokine 17 Proteins 0.000 description 1
- 101000713106 Homo sapiens C-C motif chemokine 19 Proteins 0.000 description 1
- 101000897480 Homo sapiens C-C motif chemokine 2 Proteins 0.000 description 1
- 101000713099 Homo sapiens C-C motif chemokine 20 Proteins 0.000 description 1
- 101000713085 Homo sapiens C-C motif chemokine 21 Proteins 0.000 description 1
- 101000713078 Homo sapiens C-C motif chemokine 24 Proteins 0.000 description 1
- 101000897493 Homo sapiens C-C motif chemokine 26 Proteins 0.000 description 1
- 101000897494 Homo sapiens C-C motif chemokine 27 Proteins 0.000 description 1
- 101000897477 Homo sapiens C-C motif chemokine 28 Proteins 0.000 description 1
- 101000797762 Homo sapiens C-C motif chemokine 5 Proteins 0.000 description 1
- 101000946794 Homo sapiens C-C motif chemokine 8 Proteins 0.000 description 1
- 101000947174 Homo sapiens C-X-C chemokine receptor type 1 Proteins 0.000 description 1
- 101000916050 Homo sapiens C-X-C chemokine receptor type 3 Proteins 0.000 description 1
- 101000922348 Homo sapiens C-X-C chemokine receptor type 4 Proteins 0.000 description 1
- 101000858060 Homo sapiens C-X-C motif chemokine 11 Proteins 0.000 description 1
- 101000858068 Homo sapiens C-X-C motif chemokine 14 Proteins 0.000 description 1
- 101000889133 Homo sapiens C-X-C motif chemokine 16 Proteins 0.000 description 1
- 101000889128 Homo sapiens C-X-C motif chemokine 2 Proteins 0.000 description 1
- 101000947193 Homo sapiens C-X-C motif chemokine 3 Proteins 0.000 description 1
- 101000947186 Homo sapiens C-X-C motif chemokine 5 Proteins 0.000 description 1
- 101000947177 Homo sapiens C-X-C motif chemokine 6 Proteins 0.000 description 1
- 101000947172 Homo sapiens C-X-C motif chemokine 9 Proteins 0.000 description 1
- 101100165850 Homo sapiens CA9 gene Proteins 0.000 description 1
- 101000914211 Homo sapiens CASP8 and FADD-like apoptosis regulator Proteins 0.000 description 1
- 101000777550 Homo sapiens CCN family member 2 Proteins 0.000 description 1
- 101000884298 Homo sapiens CD226 antigen Proteins 0.000 description 1
- 101000914511 Homo sapiens CD27 antigen Proteins 0.000 description 1
- 101000914479 Homo sapiens CD81 antigen Proteins 0.000 description 1
- 101000714537 Homo sapiens Cadherin-2 Proteins 0.000 description 1
- 101000714553 Homo sapiens Cadherin-3 Proteins 0.000 description 1
- 101000761509 Homo sapiens Cathepsin K Proteins 0.000 description 1
- 101000983577 Homo sapiens Cathepsin L2 Proteins 0.000 description 1
- 101000910979 Homo sapiens Cathepsin Z Proteins 0.000 description 1
- 101000856022 Homo sapiens Complement decay-accelerating factor Proteins 0.000 description 1
- 101000941929 Homo sapiens Complement receptor type 2 Proteins 0.000 description 1
- 101000856395 Homo sapiens Cullin-9 Proteins 0.000 description 1
- 101000804771 Homo sapiens Cytokine SCM-1 beta Proteins 0.000 description 1
- 101000889276 Homo sapiens Cytotoxic T-lymphocyte protein 4 Proteins 0.000 description 1
- 101001137256 Homo sapiens DNA-(apurinic or apyrimidinic site) lyase Proteins 0.000 description 1
- 101000650600 Homo sapiens DNA-directed RNA polymerase I subunit RPA2 Proteins 0.000 description 1
- 101000864646 Homo sapiens Dickkopf-related protein 1 Proteins 0.000 description 1
- 101000908391 Homo sapiens Dipeptidyl peptidase 4 Proteins 0.000 description 1
- 101000777455 Homo sapiens Disintegrin and metalloproteinase domain-containing protein 15 Proteins 0.000 description 1
- 101000832767 Homo sapiens Disintegrin and metalloproteinase domain-containing protein 8 Proteins 0.000 description 1
- 101000832769 Homo sapiens Disintegrin and metalloproteinase domain-containing protein 9 Proteins 0.000 description 1
- 101001053689 Homo sapiens Dystrobrevin alpha Proteins 0.000 description 1
- 101001010541 Homo sapiens Electron transfer flavoprotein subunit alpha, mitochondrial Proteins 0.000 description 1
- 101000884275 Homo sapiens Endosialin Proteins 0.000 description 1
- 101000978392 Homo sapiens Eotaxin Proteins 0.000 description 1
- 101000851181 Homo sapiens Epidermal growth factor receptor Proteins 0.000 description 1
- 101000846394 Homo sapiens Fibroblast growth factor 19 Proteins 0.000 description 1
- 101001060280 Homo sapiens Fibroblast growth factor 3 Proteins 0.000 description 1
- 101000819438 Homo sapiens Frizzled-1 Proteins 0.000 description 1
- 101000819451 Homo sapiens Frizzled-10 Proteins 0.000 description 1
- 101000819477 Homo sapiens Frizzled-2 Proteins 0.000 description 1
- 101000819458 Homo sapiens Frizzled-3 Proteins 0.000 description 1
- 101000885581 Homo sapiens Frizzled-4 Proteins 0.000 description 1
- 101000885585 Homo sapiens Frizzled-5 Proteins 0.000 description 1
- 101000885673 Homo sapiens Frizzled-6 Proteins 0.000 description 1
- 101000885797 Homo sapiens Frizzled-7 Proteins 0.000 description 1
- 101001061408 Homo sapiens Frizzled-8 Proteins 0.000 description 1
- 101001014668 Homo sapiens Glypican-3 Proteins 0.000 description 1
- 101000746367 Homo sapiens Granulocyte colony-stimulating factor Proteins 0.000 description 1
- 101001069921 Homo sapiens Growth-regulated alpha protein Proteins 0.000 description 1
- 101001082627 Homo sapiens HLA class II histocompatibility antigen gamma chain Proteins 0.000 description 1
- 101001016865 Homo sapiens Heat shock protein HSP 90-alpha Proteins 0.000 description 1
- 101000777663 Homo sapiens Hematopoietic progenitor cell antigen CD34 Proteins 0.000 description 1
- 101000913074 Homo sapiens High affinity immunoglobulin gamma Fc receptor I Proteins 0.000 description 1
- 101000878602 Homo sapiens Immunoglobulin alpha Fc receptor Proteins 0.000 description 1
- 101001078158 Homo sapiens Integrin alpha-1 Proteins 0.000 description 1
- 101001046686 Homo sapiens Integrin alpha-M Proteins 0.000 description 1
- 101000935043 Homo sapiens Integrin beta-1 Proteins 0.000 description 1
- 101000935040 Homo sapiens Integrin beta-2 Proteins 0.000 description 1
- 101001015004 Homo sapiens Integrin beta-3 Proteins 0.000 description 1
- 101000599852 Homo sapiens Intercellular adhesion molecule 1 Proteins 0.000 description 1
- 101001057504 Homo sapiens Interferon-stimulated gene 20 kDa protein Proteins 0.000 description 1
- 101001055144 Homo sapiens Interleukin-2 receptor subunit alpha Proteins 0.000 description 1
- 101000998120 Homo sapiens Interleukin-3 receptor subunit alpha Proteins 0.000 description 1
- 101000599048 Homo sapiens Interleukin-6 receptor subunit alpha Proteins 0.000 description 1
- 101001055222 Homo sapiens Interleukin-8 Proteins 0.000 description 1
- 101001034314 Homo sapiens Lactadherin Proteins 0.000 description 1
- 101000605020 Homo sapiens Large neutral amino acids transporter small subunit 1 Proteins 0.000 description 1
- 101000945751 Homo sapiens Leukocyte cell-derived chemotaxin-2 Proteins 0.000 description 1
- 101000917824 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor II-b Proteins 0.000 description 1
- 101000917839 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor III-B Proteins 0.000 description 1
- 101000804764 Homo sapiens Lymphotactin Proteins 0.000 description 1
- 101000764535 Homo sapiens Lymphotoxin-alpha Proteins 0.000 description 1
- 101001106413 Homo sapiens Macrophage-stimulating protein receptor Proteins 0.000 description 1
- 101000798109 Homo sapiens Melanotransferrin Proteins 0.000 description 1
- 101000961414 Homo sapiens Membrane cofactor protein Proteins 0.000 description 1
- 101000694615 Homo sapiens Membrane primary amine oxidase Proteins 0.000 description 1
- 101000669513 Homo sapiens Metalloproteinase inhibitor 1 Proteins 0.000 description 1
- 101000628547 Homo sapiens Metalloreductase STEAP1 Proteins 0.000 description 1
- 101000979001 Homo sapiens Methionine aminopeptidase 2 Proteins 0.000 description 1
- 101000969087 Homo sapiens Microtubule-associated protein 2 Proteins 0.000 description 1
- 101000946889 Homo sapiens Monocyte differentiation antigen CD14 Proteins 0.000 description 1
- 101001133056 Homo sapiens Mucin-1 Proteins 0.000 description 1
- 101000616778 Homo sapiens Myelin-associated glycoprotein Proteins 0.000 description 1
- 101000635885 Homo sapiens Myosin light chain 1/3, skeletal muscle isoform Proteins 0.000 description 1
- 101100405240 Homo sapiens NRG1 gene Proteins 0.000 description 1
- 101000581981 Homo sapiens Neural cell adhesion molecule 1 Proteins 0.000 description 1
- 101000996111 Homo sapiens Neuroligin-4, X-linked Proteins 0.000 description 1
- 101000595923 Homo sapiens Placenta growth factor Proteins 0.000 description 1
- 101000947178 Homo sapiens Platelet basic protein Proteins 0.000 description 1
- 101001126417 Homo sapiens Platelet-derived growth factor receptor alpha Proteins 0.000 description 1
- 101001096065 Homo sapiens Plexin domain-containing protein 1 Proteins 0.000 description 1
- 101001056707 Homo sapiens Proepiregulin Proteins 0.000 description 1
- 101000684208 Homo sapiens Prolyl endopeptidase FAP Proteins 0.000 description 1
- 101001098868 Homo sapiens Proprotein convertase subtilisin/kexin type 9 Proteins 0.000 description 1
- 101001136592 Homo sapiens Prostate stem cell antigen Proteins 0.000 description 1
- 101000814371 Homo sapiens Protein Wnt-10a Proteins 0.000 description 1
- 101000770799 Homo sapiens Protein Wnt-10b Proteins 0.000 description 1
- 101000804728 Homo sapiens Protein Wnt-2b Proteins 0.000 description 1
- 101000804792 Homo sapiens Protein Wnt-5a Proteins 0.000 description 1
- 101000804804 Homo sapiens Protein Wnt-5b Proteins 0.000 description 1
- 101000855002 Homo sapiens Protein Wnt-6 Proteins 0.000 description 1
- 101000855004 Homo sapiens Protein Wnt-7a Proteins 0.000 description 1
- 101000814380 Homo sapiens Protein Wnt-7b Proteins 0.000 description 1
- 101000814350 Homo sapiens Protein Wnt-8a Proteins 0.000 description 1
- 101000650149 Homo sapiens Protein Wnt-8b Proteins 0.000 description 1
- 101000650119 Homo sapiens Protein Wnt-9b Proteins 0.000 description 1
- 101000781955 Homo sapiens Proto-oncogene Wnt-1 Proteins 0.000 description 1
- 101000954762 Homo sapiens Proto-oncogene Wnt-3 Proteins 0.000 description 1
- 101000845206 Homo sapiens Putative peripheral benzodiazepine receptor-related protein Proteins 0.000 description 1
- 101001099199 Homo sapiens RalA-binding protein 1 Proteins 0.000 description 1
- 101001012157 Homo sapiens Receptor tyrosine-protein kinase erbB-2 Proteins 0.000 description 1
- 101000738771 Homo sapiens Receptor-type tyrosine-protein phosphatase C Proteins 0.000 description 1
- 101000606506 Homo sapiens Receptor-type tyrosine-protein phosphatase eta Proteins 0.000 description 1
- 101001092206 Homo sapiens Replication protein A 32 kDa subunit Proteins 0.000 description 1
- 101001133085 Homo sapiens Sialomucin core protein 24 Proteins 0.000 description 1
- 101000652846 Homo sapiens Single Ig IL-1-related receptor Proteins 0.000 description 1
- 101000708614 Homo sapiens Smoothened homolog Proteins 0.000 description 1
- 101000631760 Homo sapiens Sodium channel protein type 1 subunit alpha Proteins 0.000 description 1
- 101000654356 Homo sapiens Sodium channel protein type 10 subunit alpha Proteins 0.000 description 1
- 101000640020 Homo sapiens Sodium channel protein type 11 subunit alpha Proteins 0.000 description 1
- 101000684826 Homo sapiens Sodium channel protein type 2 subunit alpha Proteins 0.000 description 1
- 101000684820 Homo sapiens Sodium channel protein type 3 subunit alpha Proteins 0.000 description 1
- 101000693993 Homo sapiens Sodium channel protein type 4 subunit alpha Proteins 0.000 description 1
- 101000694017 Homo sapiens Sodium channel protein type 5 subunit alpha Proteins 0.000 description 1
- 101000654381 Homo sapiens Sodium channel protein type 8 subunit alpha Proteins 0.000 description 1
- 101000654386 Homo sapiens Sodium channel protein type 9 subunit alpha Proteins 0.000 description 1
- 101000617130 Homo sapiens Stromal cell-derived factor 1 Proteins 0.000 description 1
- 101000874179 Homo sapiens Syndecan-1 Proteins 0.000 description 1
- 101000914496 Homo sapiens T-cell antigen CD7 Proteins 0.000 description 1
- 101000934376 Homo sapiens T-cell differentiation antigen CD6 Proteins 0.000 description 1
- 101000934346 Homo sapiens T-cell surface antigen CD2 Proteins 0.000 description 1
- 101000980827 Homo sapiens T-cell surface glycoprotein CD1a Proteins 0.000 description 1
- 101000716149 Homo sapiens T-cell surface glycoprotein CD1b Proteins 0.000 description 1
- 101000716124 Homo sapiens T-cell surface glycoprotein CD1c Proteins 0.000 description 1
- 101000946860 Homo sapiens T-cell surface glycoprotein CD3 epsilon chain Proteins 0.000 description 1
- 101000716102 Homo sapiens T-cell surface glycoprotein CD4 Proteins 0.000 description 1
- 101000934341 Homo sapiens T-cell surface glycoprotein CD5 Proteins 0.000 description 1
- 101000946843 Homo sapiens T-cell surface glycoprotein CD8 alpha chain Proteins 0.000 description 1
- 101000914514 Homo sapiens T-cell-specific surface glycoprotein CD28 Proteins 0.000 description 1
- 101000914484 Homo sapiens T-lymphocyte activation antigen CD80 Proteins 0.000 description 1
- 101000712674 Homo sapiens TGF-beta receptor type-1 Proteins 0.000 description 1
- 101100369999 Homo sapiens TNFSF13 gene Proteins 0.000 description 1
- 101100207070 Homo sapiens TNFSF8 gene Proteins 0.000 description 1
- 101000612990 Homo sapiens Tetraspanin-3 Proteins 0.000 description 1
- 101000845170 Homo sapiens Thymic stromal lymphopoietin Proteins 0.000 description 1
- 101000834948 Homo sapiens Tomoregulin-2 Proteins 0.000 description 1
- 101000845233 Homo sapiens Translocator protein Proteins 0.000 description 1
- 101000638154 Homo sapiens Transmembrane protease serine 2 Proteins 0.000 description 1
- 101000830603 Homo sapiens Tumor necrosis factor ligand superfamily member 11 Proteins 0.000 description 1
- 101000830598 Homo sapiens Tumor necrosis factor ligand superfamily member 12 Proteins 0.000 description 1
- 101000830600 Homo sapiens Tumor necrosis factor ligand superfamily member 13 Proteins 0.000 description 1
- 101000638161 Homo sapiens Tumor necrosis factor ligand superfamily member 6 Proteins 0.000 description 1
- 101000638251 Homo sapiens Tumor necrosis factor ligand superfamily member 9 Proteins 0.000 description 1
- 101000795169 Homo sapiens Tumor necrosis factor receptor superfamily member 13C Proteins 0.000 description 1
- 101000801255 Homo sapiens Tumor necrosis factor receptor superfamily member 17 Proteins 0.000 description 1
- 101000762805 Homo sapiens Tumor necrosis factor receptor superfamily member 19L Proteins 0.000 description 1
- 101000679907 Homo sapiens Tumor necrosis factor receptor superfamily member 27 Proteins 0.000 description 1
- 101000679857 Homo sapiens Tumor necrosis factor receptor superfamily member 3 Proteins 0.000 description 1
- 101000611185 Homo sapiens Tumor necrosis factor receptor superfamily member 5 Proteins 0.000 description 1
- 101000920026 Homo sapiens Tumor necrosis factor receptor superfamily member EDAR Proteins 0.000 description 1
- 101000956263 Homo sapiens Uncharacterized protein C19orf48 Proteins 0.000 description 1
- 206010020649 Hyperkeratosis Diseases 0.000 description 1
- 241000235789 Hyperoartia Species 0.000 description 1
- 208000018121 Hypoxanthine-guanine phosphoribosyltransferase deficiency Diseases 0.000 description 1
- 101150106931 IFNG gene Proteins 0.000 description 1
- 108010042653 IgA receptor Proteins 0.000 description 1
- 102000009438 IgE Receptors Human genes 0.000 description 1
- 102000018071 Immunoglobulin Fc Fragments Human genes 0.000 description 1
- 108010091135 Immunoglobulin Fc Fragments Proteins 0.000 description 1
- 102100038005 Immunoglobulin alpha Fc receptor Human genes 0.000 description 1
- 206010061218 Inflammation Diseases 0.000 description 1
- 102000002746 Inhibins Human genes 0.000 description 1
- 108010004250 Inhibins Proteins 0.000 description 1
- 108010001127 Insulin Receptor Proteins 0.000 description 1
- 102000003746 Insulin Receptor Human genes 0.000 description 1
- 102000048143 Insulin-Like Growth Factor II Human genes 0.000 description 1
- 108090001117 Insulin-Like Growth Factor II Proteins 0.000 description 1
- 102100025306 Integrin alpha-IIb Human genes 0.000 description 1
- 101710149643 Integrin alpha-IIb Proteins 0.000 description 1
- 102100022338 Integrin alpha-M Human genes 0.000 description 1
- 102100022297 Integrin alpha-X Human genes 0.000 description 1
- 108010041341 Integrin alpha1 Proteins 0.000 description 1
- 108010055795 Integrin alpha1beta1 Proteins 0.000 description 1
- 102000000507 Integrin alpha2 Human genes 0.000 description 1
- 102000000510 Integrin alpha3 Human genes 0.000 description 1
- 108010041357 Integrin alpha3 Proteins 0.000 description 1
- 108010008212 Integrin alpha4beta1 Proteins 0.000 description 1
- 108010042918 Integrin alpha5beta1 Proteins 0.000 description 1
- 102000000426 Integrin alpha6 Human genes 0.000 description 1
- 108010041100 Integrin alpha6 Proteins 0.000 description 1
- 102100025304 Integrin beta-1 Human genes 0.000 description 1
- 102100032999 Integrin beta-3 Human genes 0.000 description 1
- 102100033016 Integrin beta-7 Human genes 0.000 description 1
- 102000012355 Integrin beta1 Human genes 0.000 description 1
- 108010022222 Integrin beta1 Proteins 0.000 description 1
- 102000008607 Integrin beta3 Human genes 0.000 description 1
- 108010020950 Integrin beta3 Proteins 0.000 description 1
- 108010064593 Intercellular Adhesion Molecule-1 Proteins 0.000 description 1
- 108010064600 Intercellular Adhesion Molecule-3 Proteins 0.000 description 1
- 102100037871 Intercellular adhesion molecule 3 Human genes 0.000 description 1
- 102000008070 Interferon-gamma Human genes 0.000 description 1
- 108010074328 Interferon-gamma Proteins 0.000 description 1
- 102000000589 Interleukin-1 Human genes 0.000 description 1
- 108010002352 Interleukin-1 Proteins 0.000 description 1
- 102000003777 Interleukin-1 beta Human genes 0.000 description 1
- 108090000193 Interleukin-1 beta Proteins 0.000 description 1
- 102000003814 Interleukin-10 Human genes 0.000 description 1
- 108090000174 Interleukin-10 Proteins 0.000 description 1
- 102000013462 Interleukin-12 Human genes 0.000 description 1
- 108010065805 Interleukin-12 Proteins 0.000 description 1
- 102000003816 Interleukin-13 Human genes 0.000 description 1
- 108090000176 Interleukin-13 Proteins 0.000 description 1
- 102000003812 Interleukin-15 Human genes 0.000 description 1
- 108090000172 Interleukin-15 Proteins 0.000 description 1
- 108050003558 Interleukin-17 Proteins 0.000 description 1
- 102000013691 Interleukin-17 Human genes 0.000 description 1
- 102000003810 Interleukin-18 Human genes 0.000 description 1
- 108090000171 Interleukin-18 Proteins 0.000 description 1
- 108010002350 Interleukin-2 Proteins 0.000 description 1
- 102000000588 Interleukin-2 Human genes 0.000 description 1
- 102100026878 Interleukin-2 receptor subunit alpha Human genes 0.000 description 1
- 102000013264 Interleukin-23 Human genes 0.000 description 1
- 108010065637 Interleukin-23 Proteins 0.000 description 1
- 102100033493 Interleukin-3 receptor subunit alpha Human genes 0.000 description 1
- 102100021596 Interleukin-31 Human genes 0.000 description 1
- 101710181613 Interleukin-31 Proteins 0.000 description 1
- 102000004890 Interleukin-8 Human genes 0.000 description 1
- 108090001007 Interleukin-8 Proteins 0.000 description 1
- 102100026236 Interleukin-8 Human genes 0.000 description 1
- 108010018951 Interleukin-8B Receptors Proteins 0.000 description 1
- 108010002335 Interleukin-9 Proteins 0.000 description 1
- 102100026871 Interleukin-9 Human genes 0.000 description 1
- 102100024319 Intestinal-type alkaline phosphatase Human genes 0.000 description 1
- 102000004310 Ion Channels Human genes 0.000 description 1
- 108090000862 Ion Channels Proteins 0.000 description 1
- 102100027612 Kallikrein-11 Human genes 0.000 description 1
- 101710115807 Kallikrein-11 Proteins 0.000 description 1
- 102100038318 Kallikrein-12 Human genes 0.000 description 1
- 101710115809 Kallikrein-12 Proteins 0.000 description 1
- 102100038298 Kallikrein-14 Human genes 0.000 description 1
- 101710115806 Kallikrein-14 Proteins 0.000 description 1
- 102100038301 Kallikrein-15 Human genes 0.000 description 1
- 101710115873 Kallikrein-15 Proteins 0.000 description 1
- 102100038356 Kallikrein-2 Human genes 0.000 description 1
- 101710176220 Kallikrein-2 Proteins 0.000 description 1
- 102100034868 Kallikrein-5 Human genes 0.000 description 1
- 101710176223 Kallikrein-5 Proteins 0.000 description 1
- 102100034866 Kallikrein-6 Human genes 0.000 description 1
- 101710176224 Kallikrein-6 Proteins 0.000 description 1
- 102100035792 Kininogen-1 Human genes 0.000 description 1
- 241000235058 Komagataella pastoris Species 0.000 description 1
- 108010092694 L-Selectin Proteins 0.000 description 1
- 125000003412 L-alanyl group Chemical group [H]N([H])[C@@](C([H])([H])[H])(C(=O)[*])[H] 0.000 description 1
- 125000002059 L-arginyl group Chemical group O=C([*])[C@](N([H])[H])([H])C([H])([H])C([H])([H])C([H])([H])N([H])C(=N[H])N([H])[H] 0.000 description 1
- 125000000010 L-asparaginyl group Chemical group O=C([*])[C@](N([H])[H])([H])C([H])([H])C(=O)N([H])[H] 0.000 description 1
- 125000000415 L-cysteinyl group Chemical group O=C([*])[C@@](N([H])[H])([H])C([H])([H])S[H] 0.000 description 1
- 125000002066 L-histidyl group Chemical group [H]N1C([H])=NC(C([H])([H])[C@](C(=O)[*])([H])N([H])[H])=C1[H] 0.000 description 1
- 125000002061 L-isoleucyl group Chemical group [H]N([H])[C@]([H])(C(=O)[*])[C@](C([H])([H])[H])([H])C(C([H])([H])[H])([H])[H] 0.000 description 1
- 125000003440 L-leucyl group Chemical group O=C([*])[C@](N([H])[H])([H])C([H])([H])C(C([H])([H])[H])([H])C([H])([H])[H] 0.000 description 1
- 125000001176 L-lysyl group Chemical group [H]N([H])[C@]([H])(C(=O)[*])C([H])([H])C([H])([H])C([H])([H])C(N([H])[H])([H])[H] 0.000 description 1
- 125000002435 L-phenylalanyl group Chemical group O=C([*])[C@](N([H])[H])([H])C([H])([H])C1=C([H])C([H])=C([H])C([H])=C1[H] 0.000 description 1
- 102100033467 L-selectin Human genes 0.000 description 1
- 125000002842 L-seryl group Chemical group O=C([*])[C@](N([H])[H])([H])C([H])([H])O[H] 0.000 description 1
- 125000000769 L-threonyl group Chemical group [H]N([H])[C@]([H])(C(=O)[*])[C@](O[H])(C([H])([H])[H])[H] 0.000 description 1
- 125000003580 L-valyl group Chemical group [H]N([H])[C@]([H])(C(=O)[*])C(C([H])([H])[H])(C([H])([H])[H])[H] 0.000 description 1
- 102000000853 LDL receptors Human genes 0.000 description 1
- 102100039648 Lactadherin Human genes 0.000 description 1
- 102100027000 Latent-transforming growth factor beta-binding protein 1 Human genes 0.000 description 1
- 101710178954 Latent-transforming growth factor beta-binding protein 1 Proteins 0.000 description 1
- 102100023487 Lens fiber major intrinsic protein Human genes 0.000 description 1
- 108010006444 Leucine-Rich Repeat Proteins Proteins 0.000 description 1
- 102100034762 Leukocyte cell-derived chemotaxin-2 Human genes 0.000 description 1
- 102000019298 Lipocalin Human genes 0.000 description 1
- 108050006654 Lipocalin Proteins 0.000 description 1
- 101710089435 Lipopolysaccharide-binding protein Proteins 0.000 description 1
- 102000057248 Lipoprotein(a) Human genes 0.000 description 1
- 108010033266 Lipoprotein(a) Proteins 0.000 description 1
- 102000004895 Lipoproteins Human genes 0.000 description 1
- 108090001030 Lipoproteins Proteins 0.000 description 1
- 102100029193 Low affinity immunoglobulin gamma Fc region receptor III-A Human genes 0.000 description 1
- 101710099301 Low affinity immunoglobulin gamma Fc region receptor III-A Proteins 0.000 description 1
- 102100024640 Low-density lipoprotein receptor Human genes 0.000 description 1
- 102000009151 Luteinizing Hormone Human genes 0.000 description 1
- 108010073521 Luteinizing Hormone Proteins 0.000 description 1
- 108010064548 Lymphocyte Function-Associated Antigen-1 Proteins 0.000 description 1
- 206010025323 Lymphomas Diseases 0.000 description 1
- 102100035304 Lymphotactin Human genes 0.000 description 1
- 108010091221 Lymphotoxin beta Receptor Proteins 0.000 description 1
- 102000004083 Lymphotoxin-alpha Human genes 0.000 description 1
- 108090000542 Lymphotoxin-alpha Proteins 0.000 description 1
- 102100026238 Lymphotoxin-alpha Human genes 0.000 description 1
- 102000043136 MAP kinase family Human genes 0.000 description 1
- 108091054455 MAP kinase family Proteins 0.000 description 1
- 102000034655 MIF Human genes 0.000 description 1
- 108060004872 MIF Proteins 0.000 description 1
- 108010046938 Macrophage Colony-Stimulating Factor Proteins 0.000 description 1
- 102100028123 Macrophage colony-stimulating factor 1 Human genes 0.000 description 1
- 102100021435 Macrophage-stimulating protein receptor Human genes 0.000 description 1
- 108700018351 Major Histocompatibility Complex Proteins 0.000 description 1
- 101710125418 Major capsid protein Proteins 0.000 description 1
- 102100030417 Matrilysin Human genes 0.000 description 1
- 108090000855 Matrilysin Proteins 0.000 description 1
- 102000000380 Matrix Metalloproteinase 1 Human genes 0.000 description 1
- 108010016113 Matrix Metalloproteinase 1 Proteins 0.000 description 1
- 108010076557 Matrix Metalloproteinase 14 Proteins 0.000 description 1
- 102100030216 Matrix metalloproteinase-14 Human genes 0.000 description 1
- 102100030201 Matrix metalloproteinase-15 Human genes 0.000 description 1
- 108090000560 Matrix metalloproteinase-15 Proteins 0.000 description 1
- 102100024129 Matrix metalloproteinase-24 Human genes 0.000 description 1
- 108050005214 Matrix metalloproteinase-24 Proteins 0.000 description 1
- 102100030412 Matrix metalloproteinase-9 Human genes 0.000 description 1
- 108010015302 Matrix metalloproteinase-9 Proteins 0.000 description 1
- 108010071463 Melanoma-Specific Antigens Proteins 0.000 description 1
- 102000007557 Melanoma-Specific Antigens Human genes 0.000 description 1
- 102000000440 Melanoma-associated antigen Human genes 0.000 description 1
- 108050008953 Melanoma-associated antigen Proteins 0.000 description 1
- 101710151321 Melanostatin Proteins 0.000 description 1
- 102100032239 Melanotransferrin Human genes 0.000 description 1
- 108010060408 Member 25 Tumor Necrosis Factor Receptors Proteins 0.000 description 1
- 102100027159 Membrane primary amine oxidase Human genes 0.000 description 1
- 206010027406 Mesothelioma Diseases 0.000 description 1
- 102000005741 Metalloproteases Human genes 0.000 description 1
- 108010006035 Metalloproteases Proteins 0.000 description 1
- 102100026712 Metalloreductase STEAP1 Human genes 0.000 description 1
- 108090000192 Methionyl aminopeptidases Proteins 0.000 description 1
- 102100025825 Methylated-DNA-protein-cysteine methyltransferase Human genes 0.000 description 1
- 108010011756 Milk Proteins Proteins 0.000 description 1
- 102000014171 Milk Proteins Human genes 0.000 description 1
- 108010047660 Mitochondrial intermediate peptidase Proteins 0.000 description 1
- 102100035877 Monocyte differentiation antigen CD14 Human genes 0.000 description 1
- 241000711408 Murine respirovirus Species 0.000 description 1
- 101100165563 Mus musculus Bmp8a gene Proteins 0.000 description 1
- 101100437777 Mus musculus Bmpr1a gene Proteins 0.000 description 1
- 101000713102 Mus musculus C-C motif chemokine 1 Proteins 0.000 description 1
- 101100222387 Mus musculus Cxcl15 gene Proteins 0.000 description 1
- 101000935589 Mus musculus Flavin reductase (NADPH) Proteins 0.000 description 1
- 101100335081 Mus musculus Flt3 gene Proteins 0.000 description 1
- 101100175313 Mus musculus Gdf3 gene Proteins 0.000 description 1
- 101100175325 Mus musculus Gdf7 gene Proteins 0.000 description 1
- 101000746363 Mus musculus Granulocyte colony-stimulating factor receptor Proteins 0.000 description 1
- 101000913073 Mus musculus High affinity immunoglobulin gamma Fc receptor I Proteins 0.000 description 1
- 101100011750 Mus musculus Hsp90b1 gene Proteins 0.000 description 1
- 101100288960 Mus musculus Lefty1 gene Proteins 0.000 description 1
- 101100153533 Mus musculus Ltbr gene Proteins 0.000 description 1
- 101100346932 Mus musculus Muc1 gene Proteins 0.000 description 1
- 101100239613 Mus musculus Myadm gene Proteins 0.000 description 1
- 101100153523 Mus musculus Tnfrsf22 gene Proteins 0.000 description 1
- 101100153524 Mus musculus Tnfrsf23 gene Proteins 0.000 description 1
- 101100153526 Mus musculus Tnfrsf26 gene Proteins 0.000 description 1
- 101100207071 Mus musculus Tnfsf8 gene Proteins 0.000 description 1
- 101000866339 Mus musculus Transcription factor E2F6 Proteins 0.000 description 1
- 101000597780 Mus musculus Tumor necrosis factor ligand superfamily member 18 Proteins 0.000 description 1
- 101100264116 Mus musculus Xcl1 gene Proteins 0.000 description 1
- 101710190051 Muscle, skeletal receptor tyrosine protein kinase Proteins 0.000 description 1
- 102100038168 Muscle, skeletal receptor tyrosine-protein kinase Human genes 0.000 description 1
- 108060008487 Myosin Proteins 0.000 description 1
- 102000003505 Myosin Human genes 0.000 description 1
- 101001055320 Myxine glutinosa Insulin-like growth factor Proteins 0.000 description 1
- 102000038100 NR2 subfamily Human genes 0.000 description 1
- 108020002076 NR2 subfamily Proteins 0.000 description 1
- 229930193140 Neomycin Natural products 0.000 description 1
- 108010032605 Nerve Growth Factor Receptors Proteins 0.000 description 1
- 102000007072 Nerve Growth Factors Human genes 0.000 description 1
- 102400000054 Neuregulin-3 Human genes 0.000 description 1
- 101800000673 Neuregulin-3 Proteins 0.000 description 1
- 208000009277 Neuroectodermal Tumors Diseases 0.000 description 1
- 208000033383 Neuroendocrine tumor of pancreas Diseases 0.000 description 1
- 102100034441 Neuroligin-4, X-linked Human genes 0.000 description 1
- 108010006696 Neuronal Apoptosis-Inhibitory Protein Proteins 0.000 description 1
- 102400000064 Neuropeptide Y Human genes 0.000 description 1
- 102000004230 Neurotrophin 3 Human genes 0.000 description 1
- 108090000742 Neurotrophin 3 Proteins 0.000 description 1
- 102000003683 Neurotrophin-4 Human genes 0.000 description 1
- 108090000099 Neurotrophin-4 Proteins 0.000 description 1
- 108090000095 Neurotrophin-6 Proteins 0.000 description 1
- 102100021584 Neurturin Human genes 0.000 description 1
- 108010015406 Neurturin Proteins 0.000 description 1
- 102100030411 Neutrophil collagenase Human genes 0.000 description 1
- 101710118230 Neutrophil collagenase Proteins 0.000 description 1
- 241000208125 Nicotiana Species 0.000 description 1
- 244000061176 Nicotiana tabacum Species 0.000 description 1
- 235000002637 Nicotiana tabacum Nutrition 0.000 description 1
- 102100022397 Nitric oxide synthase, brain Human genes 0.000 description 1
- 101710111444 Nitric oxide synthase, brain Proteins 0.000 description 1
- 102100028452 Nitric oxide synthase, endothelial Human genes 0.000 description 1
- 101710090055 Nitric oxide synthase, endothelial Proteins 0.000 description 1
- 102100029438 Nitric oxide synthase, inducible Human genes 0.000 description 1
- 101710089543 Nitric oxide synthase, inducible Proteins 0.000 description 1
- 108091060545 Nonsense suppressor Proteins 0.000 description 1
- 108010042215 OX40 Ligand Proteins 0.000 description 1
- 201000010133 Oligodendroglioma Diseases 0.000 description 1
- 108091034117 Oligonucleotide Proteins 0.000 description 1
- 108700020796 Oncogene Proteins 0.000 description 1
- 108090000630 Oncostatin M Proteins 0.000 description 1
- 102100031942 Oncostatin-M Human genes 0.000 description 1
- 241000283973 Oryctolagus cuniculus Species 0.000 description 1
- 101150044441 PECAM1 gene Proteins 0.000 description 1
- 229910019142 PO4 Inorganic materials 0.000 description 1
- 101150071808 PTHLH gene Proteins 0.000 description 1
- 206010061902 Pancreatic neoplasm Diseases 0.000 description 1
- 108090000445 Parathyroid hormone Proteins 0.000 description 1
- 102000003982 Parathyroid hormone Human genes 0.000 description 1
- 102000035195 Peptidases Human genes 0.000 description 1
- 108091005804 Peptidases Proteins 0.000 description 1
- 102100026918 Phospholipase A2 Human genes 0.000 description 1
- 101710096328 Phospholipase A2 Proteins 0.000 description 1
- 102100021768 Phosphoserine aminotransferase Human genes 0.000 description 1
- 241000235648 Pichia Species 0.000 description 1
- 201000007286 Pilocytic astrocytoma Diseases 0.000 description 1
- 102000014743 Pituitary Adenylate Cyclase-Activating Polypeptide Receptors Human genes 0.000 description 1
- 108010064032 Pituitary Adenylate Cyclase-Activating Polypeptide Receptors Proteins 0.000 description 1
- 108010082093 Placenta Growth Factor Proteins 0.000 description 1
- 108090000113 Plasma Kallikrein Proteins 0.000 description 1
- 102100038124 Plasminogen Human genes 0.000 description 1
- 108010051456 Plasminogen Proteins 0.000 description 1
- 108010022233 Plasminogen Activator Inhibitor 1 Proteins 0.000 description 1
- 102000004179 Plasminogen Activator Inhibitor 2 Human genes 0.000 description 1
- 108090000614 Plasminogen Activator Inhibitor 2 Proteins 0.000 description 1
- 102100039418 Plasminogen activator inhibitor 1 Human genes 0.000 description 1
- 102100030485 Platelet-derived growth factor receptor alpha Human genes 0.000 description 1
- 102100037891 Plexin domain-containing protein 1 Human genes 0.000 description 1
- 101710179684 Poly [ADP-ribose] polymerase Proteins 0.000 description 1
- 102100023712 Poly [ADP-ribose] polymerase 1 Human genes 0.000 description 1
- 229920000776 Poly(Adenosine diphosphate-ribose) polymerase Polymers 0.000 description 1
- 108010071690 Prealbumin Proteins 0.000 description 1
- 206010057846 Primitive neuroectodermal tumour Diseases 0.000 description 1
- 108091000054 Prion Proteins 0.000 description 1
- 102100025974 Pro-cathepsin H Human genes 0.000 description 1
- 101710093543 Probable non-specific lipid-transfer protein Proteins 0.000 description 1
- 102100025498 Proepiregulin Human genes 0.000 description 1
- 108010076181 Proinsulin Proteins 0.000 description 1
- 102100036691 Proliferating cell nuclear antigen Human genes 0.000 description 1
- 102100038567 Properdin Human genes 0.000 description 1
- 108010005642 Properdin Proteins 0.000 description 1
- 102100038955 Proprotein convertase subtilisin/kexin type 9 Human genes 0.000 description 1
- 102100036735 Prostate stem cell antigen Human genes 0.000 description 1
- 108010072866 Prostate-Specific Antigen Proteins 0.000 description 1
- 101800004937 Protein C Proteins 0.000 description 1
- 102000002727 Protein Tyrosine Phosphatase Human genes 0.000 description 1
- 102100039461 Protein Wnt-10a Human genes 0.000 description 1
- 102100029062 Protein Wnt-10b Human genes 0.000 description 1
- 102100035289 Protein Wnt-2b Human genes 0.000 description 1
- 102100035331 Protein Wnt-5b Human genes 0.000 description 1
- 102100020732 Protein Wnt-6 Human genes 0.000 description 1
- 102100020729 Protein Wnt-7a Human genes 0.000 description 1
- 102100039470 Protein Wnt-7b Human genes 0.000 description 1
- 102100039453 Protein Wnt-8a Human genes 0.000 description 1
- 102100027542 Protein Wnt-8b Human genes 0.000 description 1
- 102100027502 Protein Wnt-9b Human genes 0.000 description 1
- 102100027378 Prothrombin Human genes 0.000 description 1
- 108010094028 Prothrombin Proteins 0.000 description 1
- 108010089836 Proto-Oncogene Proteins c-met Proteins 0.000 description 1
- 102100036385 Protocadherin-12 Human genes 0.000 description 1
- 101710158929 Protocadherin-12 Proteins 0.000 description 1
- 108010052562 RELT Proteins 0.000 description 1
- 102000018795 RELT Human genes 0.000 description 1
- 102000003890 RNA-binding protein FUS Human genes 0.000 description 1
- 108090000292 RNA-binding protein FUS Proteins 0.000 description 1
- 239000012980 RPMI-1640 medium Substances 0.000 description 1
- 102100038914 RalA-binding protein 1 Human genes 0.000 description 1
- 241000700159 Rattus Species 0.000 description 1
- 101100372762 Rattus norvegicus Flt1 gene Proteins 0.000 description 1
- 101100517381 Rattus norvegicus Ntrk1 gene Proteins 0.000 description 1
- 102100030086 Receptor tyrosine-protein kinase erbB-2 Human genes 0.000 description 1
- 101710100969 Receptor tyrosine-protein kinase erbB-3 Proteins 0.000 description 1
- 101710100963 Receptor tyrosine-protein kinase erbB-4 Proteins 0.000 description 1
- 102100037422 Receptor-type tyrosine-protein phosphatase C Human genes 0.000 description 1
- 102100039808 Receptor-type tyrosine-protein phosphatase eta Human genes 0.000 description 1
- 102400000834 Relaxin A chain Human genes 0.000 description 1
- 101800000074 Relaxin A chain Proteins 0.000 description 1
- 102400000610 Relaxin B chain Human genes 0.000 description 1
- 101710109558 Relaxin B chain Proteins 0.000 description 1
- 102100028255 Renin Human genes 0.000 description 1
- 108090000783 Renin Proteins 0.000 description 1
- 108700008625 Reporter Genes Proteins 0.000 description 1
- 241000283984 Rodentia Species 0.000 description 1
- 101150110386 SLC2A4 gene Proteins 0.000 description 1
- 108091007602 SLC58A1 Proteins 0.000 description 1
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 1
- 101000733766 Saccharomyces cerevisiae (strain ATCC 204508 / S288c) Aminopeptidase 2, mitochondrial Proteins 0.000 description 1
- 101001117144 Saccharomyces cerevisiae (strain ATCC 204508 / S288c) [Pyruvate dehydrogenase (acetyl-transferring)] kinase 1, mitochondrial Proteins 0.000 description 1
- 102100036546 Salivary acidic proline-rich phosphoprotein 1/2 Human genes 0.000 description 1
- 101800001700 Saposin-D Proteins 0.000 description 1
- 101100537955 Schizosaccharomyces pombe (strain 972 / ATCC 24843) trk1 gene Proteins 0.000 description 1
- 102100034201 Sclerostin Human genes 0.000 description 1
- 108050006698 Sclerostin Proteins 0.000 description 1
- 102100030053 Secreted frizzled-related protein 3 Human genes 0.000 description 1
- 108050007990 Secreted frizzled-related protein 3 Proteins 0.000 description 1
- 102100034136 Serine/threonine-protein kinase receptor R3 Human genes 0.000 description 1
- 101710082813 Serine/threonine-protein kinase receptor R3 Proteins 0.000 description 1
- 102000008847 Serpin Human genes 0.000 description 1
- 108050000761 Serpin Proteins 0.000 description 1
- 102000007562 Serum Albumin Human genes 0.000 description 1
- 108010071390 Serum Albumin Proteins 0.000 description 1
- 102100034258 Sialomucin core protein 24 Human genes 0.000 description 1
- 102000008115 Signaling Lymphocytic Activation Molecule Family Member 1 Human genes 0.000 description 1
- 108010074687 Signaling Lymphocytic Activation Molecule Family Member 1 Proteins 0.000 description 1
- 102100030929 Single Ig IL-1-related receptor Human genes 0.000 description 1
- 102100032799 Smoothened homolog Human genes 0.000 description 1
- 102100028910 Sodium channel protein type 1 subunit alpha Human genes 0.000 description 1
- 102100031374 Sodium channel protein type 10 subunit alpha Human genes 0.000 description 1
- 102100033974 Sodium channel protein type 11 subunit alpha Human genes 0.000 description 1
- 102100023150 Sodium channel protein type 2 subunit alpha Human genes 0.000 description 1
- 102100023720 Sodium channel protein type 3 subunit alpha Human genes 0.000 description 1
- 102100027195 Sodium channel protein type 4 subunit alpha Human genes 0.000 description 1
- 102100027198 Sodium channel protein type 5 subunit alpha Human genes 0.000 description 1
- 102100031371 Sodium channel protein type 8 subunit alpha Human genes 0.000 description 1
- 102100031367 Sodium channel protein type 9 subunit alpha Human genes 0.000 description 1
- 102100022831 Somatoliberin Human genes 0.000 description 1
- 101710142969 Somatoliberin Proteins 0.000 description 1
- 102100039024 Sphingosine kinase 1 Human genes 0.000 description 1
- 101000879712 Streptomyces lividans Protease inhibitor Proteins 0.000 description 1
- 102100030416 Stromelysin-1 Human genes 0.000 description 1
- 101710108790 Stromelysin-1 Proteins 0.000 description 1
- 102100028848 Stromelysin-2 Human genes 0.000 description 1
- 101710108792 Stromelysin-2 Proteins 0.000 description 1
- 102100028847 Stromelysin-3 Human genes 0.000 description 1
- 108050005271 Stromelysin-3 Proteins 0.000 description 1
- 102000019259 Succinate Dehydrogenase Human genes 0.000 description 1
- 108010012901 Succinate Dehydrogenase Proteins 0.000 description 1
- 108010021188 Superoxide Dismutase-1 Proteins 0.000 description 1
- 102100038836 Superoxide dismutase [Cu-Zn] Human genes 0.000 description 1
- 108090000058 Syndecan-1 Proteins 0.000 description 1
- 108090000054 Syndecan-2 Proteins 0.000 description 1
- 102100026087 Syndecan-2 Human genes 0.000 description 1
- 108090000068 Syndecan-3 Proteins 0.000 description 1
- 102100026084 Syndecan-3 Human genes 0.000 description 1
- 108010055215 Syndecan-4 Proteins 0.000 description 1
- 102100037220 Syndecan-4 Human genes 0.000 description 1
- 101100342402 Synechocystis sp. (strain PCC 6803 / Kazusa) prk gene Proteins 0.000 description 1
- 208000005400 Synovial Cyst Diseases 0.000 description 1
- 102100027208 T-cell antigen CD7 Human genes 0.000 description 1
- 102100025131 T-cell differentiation antigen CD6 Human genes 0.000 description 1
- 102100025237 T-cell surface antigen CD2 Human genes 0.000 description 1
- 102100024219 T-cell surface glycoprotein CD1a Human genes 0.000 description 1
- 102100035794 T-cell surface glycoprotein CD3 epsilon chain Human genes 0.000 description 1
- 102100036011 T-cell surface glycoprotein CD4 Human genes 0.000 description 1
- 102100025244 T-cell surface glycoprotein CD5 Human genes 0.000 description 1
- 102100034922 T-cell surface glycoprotein CD8 alpha chain Human genes 0.000 description 1
- 102100027213 T-cell-specific surface glycoprotein CD28 Human genes 0.000 description 1
- 210000001744 T-lymphocyte Anatomy 0.000 description 1
- 102100027222 T-lymphocyte activation antigen CD80 Human genes 0.000 description 1
- 102100040347 TAR DNA-binding protein 43 Human genes 0.000 description 1
- 101150014554 TARDBP gene Proteins 0.000 description 1
- 102100033456 TGF-beta receptor type-1 Human genes 0.000 description 1
- 101150081494 TMPO gene Proteins 0.000 description 1
- 108091007178 TNFRSF10A Proteins 0.000 description 1
- 101150077103 TPO gene Proteins 0.000 description 1
- 206010043276 Teratoma Diseases 0.000 description 1
- 208000024313 Testicular Neoplasms Diseases 0.000 description 1
- 206010057644 Testis cancer Diseases 0.000 description 1
- 101710151650 Tetraspanin-3 Proteins 0.000 description 1
- 108010041111 Thrombopoietin Proteins 0.000 description 1
- 102100034195 Thrombopoietin Human genes 0.000 description 1
- 102100031294 Thymic stromal lymphopoietin Human genes 0.000 description 1
- 102000006601 Thymidine Kinase Human genes 0.000 description 1
- 108020004440 Thymidine kinase Proteins 0.000 description 1
- 102000011923 Thyrotropin Human genes 0.000 description 1
- 108010061174 Thyrotropin Proteins 0.000 description 1
- 102100030951 Tissue factor pathway inhibitor Human genes 0.000 description 1
- 108050006955 Tissue-type plasminogen activator Proteins 0.000 description 1
- 102100026260 Titin Human genes 0.000 description 1
- 102100026160 Tomoregulin-2 Human genes 0.000 description 1
- 102000040945 Transcription factor Human genes 0.000 description 1
- 108091023040 Transcription factor Proteins 0.000 description 1
- 102000007238 Transferrin Receptors Human genes 0.000 description 1
- 108050003222 Transferrin receptor protein 1 Proteins 0.000 description 1
- 102000046299 Transforming Growth Factor beta1 Human genes 0.000 description 1
- 102000011117 Transforming Growth Factor beta2 Human genes 0.000 description 1
- 108010009583 Transforming Growth Factors Proteins 0.000 description 1
- 102000009618 Transforming Growth Factors Human genes 0.000 description 1
- 102400001320 Transforming growth factor alpha Human genes 0.000 description 1
- 101800004564 Transforming growth factor alpha Proteins 0.000 description 1
- 101800002279 Transforming growth factor beta-1 Proteins 0.000 description 1
- 101800000304 Transforming growth factor beta-2 Proteins 0.000 description 1
- 108090000097 Transforming growth factor beta-3 Proteins 0.000 description 1
- 102000056172 Transforming growth factor beta-3 Human genes 0.000 description 1
- 108050005134 Translocation protein Sec62 Proteins 0.000 description 1
- 101710166801 Translocator protein Proteins 0.000 description 1
- 102100031989 Transmembrane protease serine 2 Human genes 0.000 description 1
- 102000009190 Transthyretin Human genes 0.000 description 1
- 241000219793 Trifolium Species 0.000 description 1
- GLNADSQYFUSGOU-GPTZEZBUSA-J Trypan blue Chemical compound [Na+].[Na+].[Na+].[Na+].C1=C(S([O-])(=O)=O)C=C2C=C(S([O-])(=O)=O)C(/N=N/C3=CC=C(C=C3C)C=3C=C(C(=CC=3)\N=N\C=3C(=CC4=CC(=CC(N)=C4C=3O)S([O-])(=O)=O)S([O-])(=O)=O)C)=C(O)C2=C1N GLNADSQYFUSGOU-GPTZEZBUSA-J 0.000 description 1
- 108010065158 Tumor Necrosis Factor Ligand Superfamily Member 14 Proteins 0.000 description 1
- 102000000852 Tumor Necrosis Factor-alpha Human genes 0.000 description 1
- 101710097161 Tumor necrosis factor ligand superfamily member 11 Proteins 0.000 description 1
- 108090000138 Tumor necrosis factor ligand superfamily member 15 Proteins 0.000 description 1
- 108050002568 Tumor necrosis factor ligand superfamily member 6 Proteins 0.000 description 1
- 101710178300 Tumor necrosis factor receptor superfamily member 13C Proteins 0.000 description 1
- 101710187887 Tumor necrosis factor receptor superfamily member 19 Proteins 0.000 description 1
- 102100026716 Tumor necrosis factor receptor superfamily member 19L Human genes 0.000 description 1
- 101710187743 Tumor necrosis factor receptor superfamily member 1A Proteins 0.000 description 1
- 101710187830 Tumor necrosis factor receptor superfamily member 1B Proteins 0.000 description 1
- 102100022202 Tumor necrosis factor receptor superfamily member 27 Human genes 0.000 description 1
- 102100030810 Tumor necrosis factor receptor superfamily member EDAR Human genes 0.000 description 1
- 208000035896 Twin-reversed arterial perfusion sequence Diseases 0.000 description 1
- 208000006391 Type 1 Hyper-IgM Immunodeficiency Syndrome Diseases 0.000 description 1
- 102100038573 Uncharacterized protein C19orf48 Human genes 0.000 description 1
- 208000007097 Urinary Bladder Neoplasms Diseases 0.000 description 1
- 102000003990 Urokinase-type plasminogen activator Human genes 0.000 description 1
- 108090000435 Urokinase-type plasminogen activator Proteins 0.000 description 1
- 208000002495 Uterine Neoplasms Diseases 0.000 description 1
- 102000008790 VE-cadherin Human genes 0.000 description 1
- 238000005411 Van der Waals force Methods 0.000 description 1
- 108010000134 Vascular Cell Adhesion Molecule-1 Proteins 0.000 description 1
- 102100023543 Vascular cell adhesion protein 1 Human genes 0.000 description 1
- 102100033179 Vascular endothelial growth factor receptor 3 Human genes 0.000 description 1
- 108010003533 Viral Envelope Proteins Proteins 0.000 description 1
- 101150010310 WNT-4 gene Proteins 0.000 description 1
- 101150019524 WNT2 gene Proteins 0.000 description 1
- 102000013814 Wnt Human genes 0.000 description 1
- 108050003627 Wnt Proteins 0.000 description 1
- 101710194167 Wnt inhibitory factor 1 Proteins 0.000 description 1
- 102100038258 Wnt inhibitory factor 1 Human genes 0.000 description 1
- 102000052547 Wnt-1 Human genes 0.000 description 1
- 102000052556 Wnt-2 Human genes 0.000 description 1
- 108700020986 Wnt-2 Proteins 0.000 description 1
- 102000052549 Wnt-3 Human genes 0.000 description 1
- 102000052548 Wnt-4 Human genes 0.000 description 1
- 108700020984 Wnt-4 Proteins 0.000 description 1
- 102000043366 Wnt-5a Human genes 0.000 description 1
- 102000044880 Wnt3A Human genes 0.000 description 1
- 108700013515 Wnt3A Proteins 0.000 description 1
- 108700031544 X-Linked Inhibitor of Apoptosis Proteins 0.000 description 1
- 201000001696 X-linked hyper IgM syndrome Diseases 0.000 description 1
- 241000269370 Xenopus <genus> Species 0.000 description 1
- 101100485099 Xenopus laevis wnt2b-b gene Proteins 0.000 description 1
- 238000002835 absorbance Methods 0.000 description 1
- 102000034337 acetylcholine receptors Human genes 0.000 description 1
- 150000007513 acids Chemical class 0.000 description 1
- 230000003213 activating effect Effects 0.000 description 1
- 108010023082 activin A Proteins 0.000 description 1
- 108010023079 activin B Proteins 0.000 description 1
- 208000021841 acute erythroid leukemia Diseases 0.000 description 1
- 230000002411 adverse Effects 0.000 description 1
- 230000009824 affinity maturation Effects 0.000 description 1
- 229910052783 alkali metal Inorganic materials 0.000 description 1
- 150000001340 alkali metals Chemical class 0.000 description 1
- 229910052784 alkaline earth metal Inorganic materials 0.000 description 1
- 150000001342 alkaline earth metals Chemical class 0.000 description 1
- 108010050122 alpha 1-Antitrypsin Proteins 0.000 description 1
- 102000015395 alpha 1-Antitrypsin Human genes 0.000 description 1
- 229940024142 alpha 1-antitrypsin Drugs 0.000 description 1
- ANBQYFIVLNNZCU-CQCLMDPOSA-N alpha-L-Fucp-(1->2)-[alpha-D-GalpNAc-(1->3)]-beta-D-Galp-(1->3)-[alpha-L-Fucp-(1->4)]-beta-D-GlcpNAc-(1->3)-beta-D-Galp Chemical compound O[C@H]1[C@H](O)[C@H](O)[C@H](C)O[C@H]1O[C@H]1[C@H](O[C@H]2[C@@H]([C@@H](O[C@@H]3[C@@H]([C@@H](O)[C@@H](O)[C@@H](CO)O3)NC(C)=O)[C@@H](O)[C@@H](CO)O2)O[C@H]2[C@H]([C@H](O)[C@H](O)[C@H](C)O2)O)[C@@H](NC(C)=O)[C@H](O[C@H]2[C@H]([C@@H](CO)O[C@@H](O)[C@@H]2O)O)O[C@@H]1CO ANBQYFIVLNNZCU-CQCLMDPOSA-N 0.000 description 1
- SRHNADOZAAWYLV-XLMUYGLTSA-N alpha-L-Fucp-(1->2)-beta-D-Galp-(1->4)-[alpha-L-Fucp-(1->3)]-beta-D-GlcpNAc Chemical compound O[C@H]1[C@H](O)[C@H](O)[C@H](C)O[C@H]1O[C@H]1[C@H](O[C@H]2[C@@H]([C@@H](NC(C)=O)[C@H](O)O[C@@H]2CO)O[C@H]2[C@H]([C@H](O)[C@H](O)[C@H](C)O2)O)O[C@H](CO)[C@H](O)[C@@H]1O SRHNADOZAAWYLV-XLMUYGLTSA-N 0.000 description 1
- MXKCYTKUIDTFLY-ZNNSSXPHSA-N alpha-L-Fucp-(1->2)-beta-D-Galp-(1->4)-[alpha-L-Fucp-(1->3)]-beta-D-GlcpNAc-(1->3)-D-Galp Chemical compound O[C@H]1[C@H](O)[C@H](O)[C@H](C)O[C@H]1O[C@H]1[C@H](O[C@H]2[C@@H]([C@@H](NC(C)=O)[C@H](O[C@H]3[C@H]([C@@H](CO)OC(O)[C@@H]3O)O)O[C@@H]2CO)O[C@H]2[C@H]([C@H](O)[C@H](O)[C@H](C)O2)O)O[C@H](CO)[C@H](O)[C@@H]1O MXKCYTKUIDTFLY-ZNNSSXPHSA-N 0.000 description 1
- 108090000185 alpha-Synuclein Proteins 0.000 description 1
- 230000003321 amplification Effects 0.000 description 1
- 239000005557 antagonist Substances 0.000 description 1
- 239000003242 anti bacterial agent Substances 0.000 description 1
- 229940088710 antibiotic agent Drugs 0.000 description 1
- 230000000890 antigenic effect Effects 0.000 description 1
- 229910052787 antimony Inorganic materials 0.000 description 1
- WATWJIUSRGPENY-UHFFFAOYSA-N antimony atom Chemical compound [Sb] WATWJIUSRGPENY-UHFFFAOYSA-N 0.000 description 1
- 229960005348 antithrombin iii Drugs 0.000 description 1
- 108010073614 apolipoprotein A-IV Proteins 0.000 description 1
- 210000001130 astrocyte Anatomy 0.000 description 1
- 230000035578 autophosphorylation Effects 0.000 description 1
- 239000012752 auxiliary agent Substances 0.000 description 1
- 210000003050 axon Anatomy 0.000 description 1
- 230000004888 barrier function Effects 0.000 description 1
- 102000055102 bcl-2-Associated X Human genes 0.000 description 1
- 108700000707 bcl-2-Associated X Proteins 0.000 description 1
- 230000008901 benefit Effects 0.000 description 1
- DQEPMTIXHXSFOR-UHFFFAOYSA-N benzo[a]pyrene diol epoxide I Chemical compound C1=C2C(C3OC3C(C3O)O)=C3C=C(C=C3)C2=C2C3=CC=CC2=C1 DQEPMTIXHXSFOR-UHFFFAOYSA-N 0.000 description 1
- CXQCLLQQYTUUKJ-ALWAHNIESA-N beta-D-GalpNAc-(1->4)-[alpha-Neup5Ac-(2->8)-alpha-Neup5Ac-(2->3)]-beta-D-Galp-(1->4)-beta-D-Glcp-(1<->1')-Cer(d18:1/18:0) Chemical compound O[C@@H]1[C@@H](O)[C@H](OC[C@H](NC(=O)CCCCCCCCCCCCCCCCC)[C@H](O)\C=C\CCCCCCCCCCCCC)O[C@H](CO)[C@H]1O[C@H]1[C@H](O)[C@@H](O[C@]2(O[C@H]([C@H](NC(C)=O)[C@@H](O)C2)[C@H](O)[C@@H](CO)O[C@]2(O[C@H]([C@H](NC(C)=O)[C@@H](O)C2)[C@H](O)[C@H](O)CO)C(O)=O)C(O)=O)[C@@H](O[C@H]2[C@@H]([C@@H](O)[C@@H](O)[C@@H](CO)O2)NC(C)=O)[C@@H](CO)O1 CXQCLLQQYTUUKJ-ALWAHNIESA-N 0.000 description 1
- 230000002457 bidirectional effect Effects 0.000 description 1
- 230000033228 biological regulation Effects 0.000 description 1
- 229960000074 biopharmaceutical Drugs 0.000 description 1
- 229920001222 biopolymer Polymers 0.000 description 1
- 229960002685 biotin Drugs 0.000 description 1
- 235000020958 biotin Nutrition 0.000 description 1
- 239000011616 biotin Substances 0.000 description 1
- 229910052797 bismuth Inorganic materials 0.000 description 1
- JCXGWMGPZLAOME-UHFFFAOYSA-N bismuth atom Chemical compound [Bi] JCXGWMGPZLAOME-UHFFFAOYSA-N 0.000 description 1
- 230000017531 blood circulation Effects 0.000 description 1
- 210000000988 bone and bone Anatomy 0.000 description 1
- 229910052796 boron Inorganic materials 0.000 description 1
- 229940053031 botulinum toxin Drugs 0.000 description 1
- 210000000481 breast Anatomy 0.000 description 1
- 238000010805 cDNA synthesis kit Methods 0.000 description 1
- 108010018828 cadherin 5 Proteins 0.000 description 1
- BBBFJLBPOGFECG-VJVYQDLKSA-N calcitonin Chemical compound N([C@H](C(=O)N[C@@H](CC(C)C)C(=O)NCC(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC=1NC=NC=1)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)NCC(=O)N[C@@H](CO)C(=O)NCC(=O)N[C@@H]([C@@H](C)O)C(=O)N1[C@@H](CCC1)C(N)=O)C(C)C)C(=O)[C@@H]1CSSC[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)O)C(=O)N1 BBBFJLBPOGFECG-VJVYQDLKSA-N 0.000 description 1
- 229960004015 calcitonin Drugs 0.000 description 1
- 150000001720 carbohydrates Chemical class 0.000 description 1
- 235000014633 carbohydrates Nutrition 0.000 description 1
- 229910052799 carbon Inorganic materials 0.000 description 1
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 description 1
- 108010041776 cardiotrophin 1 Proteins 0.000 description 1
- NSQLIUXCMFBZME-MPVJKSABSA-N carperitide Chemical compound C([C@H]1C(=O)NCC(=O)NCC(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@H](C(NCC(=O)N[C@@H](C)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CO)C(=O)NCC(=O)N[C@@H](CC(C)C)C(=O)NCC(=O)N[C@@H](CSSC[C@@H](C(=O)N1)NC(=O)[C@H](CO)NC(=O)[C@H](CO)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CO)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(O)=O)=O)[C@@H](C)CC)C1=CC=CC=C1 NSQLIUXCMFBZME-MPVJKSABSA-N 0.000 description 1
- 108010027904 cartilage-derived-morphogenetic protein-2 Proteins 0.000 description 1
- 150000003943 catecholamines Chemical class 0.000 description 1
- 125000002091 cationic group Chemical group 0.000 description 1
- 150000001768 cations Chemical class 0.000 description 1
- 230000020411 cell activation Effects 0.000 description 1
- 238000010370 cell cloning Methods 0.000 description 1
- 238000004113 cell culture Methods 0.000 description 1
- 238000003163 cell fusion method Methods 0.000 description 1
- 230000010261 cell growth Effects 0.000 description 1
- 210000000170 cell membrane Anatomy 0.000 description 1
- 230000004663 cell proliferation Effects 0.000 description 1
- 230000036755 cellular response Effects 0.000 description 1
- 210000003169 central nervous system Anatomy 0.000 description 1
- 238000005119 centrifugation Methods 0.000 description 1
- 239000003153 chemical reaction reagent Substances 0.000 description 1
- 210000004978 chinese hamster ovary cell Anatomy 0.000 description 1
- 102000006834 complement receptors Human genes 0.000 description 1
- 108010047295 complement receptors Proteins 0.000 description 1
- 150000001875 compounds Chemical class 0.000 description 1
- 238000004590 computer program Methods 0.000 description 1
- 238000012790 confirmation Methods 0.000 description 1
- 239000000470 constituent Substances 0.000 description 1
- LBJNMUFDOHXDFG-UHFFFAOYSA-N copper;hydrate Chemical compound O.[Cu].[Cu] LBJNMUFDOHXDFG-UHFFFAOYSA-N 0.000 description 1
- 230000002596 correlated effect Effects 0.000 description 1
- 210000003792 cranial nerve Anatomy 0.000 description 1
- 108040004564 crotonyl-CoA reductase activity proteins Proteins 0.000 description 1
- 210000004748 cultured cell Anatomy 0.000 description 1
- 208000031513 cyst Diseases 0.000 description 1
- 108050004038 cystatin Proteins 0.000 description 1
- 235000018417 cysteine Nutrition 0.000 description 1
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 1
- 230000007423 decrease Effects 0.000 description 1
- 230000000593 degrading effect Effects 0.000 description 1
- 230000008021 deposition Effects 0.000 description 1
- 230000002074 deregulated effect Effects 0.000 description 1
- 238000011161 development Methods 0.000 description 1
- 230000018109 developmental process Effects 0.000 description 1
- 239000005546 dideoxynucleotide Substances 0.000 description 1
- 229960005156 digoxin Drugs 0.000 description 1
- MOAVUYWYFFCBNM-PUGKRICDSA-N digoxin(1-) Chemical compound C[C@H]([C@H]([C@H](C1)O)O)O[C@H]1O[C@H]([C@@H](C)O[C@H](C1)O[C@H]([C@@H](C)O[C@H](C2)O[C@@H](CC3)C[C@@H](CC4)[C@@]3(C)[C@@H](C[C@H]([C@]3(C)[C@H](CC5)C([CH-]O6)=CC6=O)O)[C@@H]4[C@]35O)[C@H]2O)[C@H]1O MOAVUYWYFFCBNM-PUGKRICDSA-N 0.000 description 1
- LTMHDMANZUZIPE-UHFFFAOYSA-N digoxine Natural products C1C(O)C(O)C(C)OC1OC1C(C)OC(OC2C(OC(OC3CC4C(C5C(C6(CCC(C6(C)C(O)C5)C=5COC(=O)C=5)O)CC4)(C)CC3)CC2O)C)CC1O LTMHDMANZUZIPE-UHFFFAOYSA-N 0.000 description 1
- 239000000539 dimer Substances 0.000 description 1
- 201000010099 disease Diseases 0.000 description 1
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 1
- ADDTYLVLXWBJNH-UHFFFAOYSA-M disodium 4-[2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)tetrazol-2-ium-5-yl]benzene-1,3-disulfonate Chemical compound [Na]OS(=O)(=O)C1=C(C=CC(=C1)S(=O)(=O)O[Na])C=1N=[N+](N(N=1)C1=CC=C(C=C1)[N+](=O)[O-])C1=C(C=C(C=C1)[N+](=O)[O-])OC ADDTYLVLXWBJNH-UHFFFAOYSA-M 0.000 description 1
- 238000009826 distribution Methods 0.000 description 1
- 101150007302 dntt gene Proteins 0.000 description 1
- 238000003134 dye exclusion method Methods 0.000 description 1
- 230000009881 electrostatic interaction Effects 0.000 description 1
- 210000002257 embryonic structure Anatomy 0.000 description 1
- 210000001900 endoderm Anatomy 0.000 description 1
- 238000005516 engineering process Methods 0.000 description 1
- 239000003623 enhancer Substances 0.000 description 1
- 238000006911 enzymatic reaction Methods 0.000 description 1
- 229940116977 epidermal growth factor Drugs 0.000 description 1
- 208000010932 epithelial neoplasm Diseases 0.000 description 1
- 210000000981 epithelium Anatomy 0.000 description 1
- 210000003743 erythrocyte Anatomy 0.000 description 1
- 210000003527 eukaryotic cell Anatomy 0.000 description 1
- 230000028023 exocytosis Effects 0.000 description 1
- 210000000887 face Anatomy 0.000 description 1
- 229940012414 factor viia Drugs 0.000 description 1
- 229960000301 factor viii Drugs 0.000 description 1
- 229940012444 factor xiii Drugs 0.000 description 1
- 230000002349 favourable effect Effects 0.000 description 1
- 210000003754 fetus Anatomy 0.000 description 1
- 229940012952 fibrinogen Drugs 0.000 description 1
- 210000002950 fibroblast Anatomy 0.000 description 1
- 102000052178 fibroblast growth factor receptor activity proteins Human genes 0.000 description 1
- 238000000684 flow cytometry Methods 0.000 description 1
- 238000001917 fluorescence detection Methods 0.000 description 1
- 229940028334 follicle stimulating hormone Drugs 0.000 description 1
- QPJBWNIQKHGLAU-IQZHVAEDSA-N ganglioside GM1 Chemical compound O[C@@H]1[C@@H](O)[C@H](OC[C@H](NC(=O)CCCCCCCCCCCCCCCCC)[C@H](O)\C=C\CCCCCCCCCCCCC)O[C@H](CO)[C@H]1O[C@H]1[C@H](O)[C@@H](O[C@]2(O[C@H]([C@H](NC(C)=O)[C@@H](O)C2)[C@H](O)[C@H](O)CO)C(O)=O)[C@@H](O[C@H]2[C@@H]([C@@H](O[C@H]3[C@@H]([C@@H](O)[C@@H](O)[C@@H](CO)O3)O)[C@@H](O)[C@@H](CO)O2)NC(C)=O)[C@@H](CO)O1 QPJBWNIQKHGLAU-IQZHVAEDSA-N 0.000 description 1
- PFJKOHUKELZMLE-VEUXDRLPSA-N ganglioside GM3 Chemical compound O[C@@H]1[C@@H](O)[C@H](OC[C@@H]([C@H](O)/C=C/CCCCCCCCCCCCC)NC(=O)CCCCCCCCCCCCC\C=C/CCCCCCCC)O[C@H](CO)[C@H]1O[C@H]1[C@H](O)[C@@H](O[C@]2(O[C@H]([C@H](NC(C)=O)[C@@H](O)C2)[C@H](O)[C@H](O)CO)C(O)=O)[C@@H](O)[C@@H](CO)O1 PFJKOHUKELZMLE-VEUXDRLPSA-N 0.000 description 1
- 150000002270 gangliosides Chemical class 0.000 description 1
- 238000001415 gene therapy Methods 0.000 description 1
- 238000007429 general method Methods 0.000 description 1
- 229960002518 gentamicin Drugs 0.000 description 1
- 201000003115 germ cell cancer Diseases 0.000 description 1
- MASNOZXLGMXCHN-ZLPAWPGGSA-N glucagon Chemical compound C([C@@H](C(=O)N[C@H](C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O)C(C)C)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@H](C)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CO)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CO)NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CO)NC(=O)[C@@H](NC(=O)[C@H](CC=1C=CC=CC=1)NC(=O)[C@@H](NC(=O)CNC(=O)[C@H](CCC(N)=O)NC(=O)[C@H](CO)NC(=O)[C@@H](N)CC=1NC=NC=1)[C@@H](C)O)[C@@H](C)O)C1=CC=CC=C1 MASNOZXLGMXCHN-ZLPAWPGGSA-N 0.000 description 1
- 229960004666 glucagon Drugs 0.000 description 1
- PCHJSUWPFVWCPO-UHFFFAOYSA-N gold Chemical compound [Au] PCHJSUWPFVWCPO-UHFFFAOYSA-N 0.000 description 1
- 229910052737 gold Inorganic materials 0.000 description 1
- 239000010931 gold Substances 0.000 description 1
- 201000005787 hematologic cancer Diseases 0.000 description 1
- 208000024200 hematopoietic and lymphoid system neoplasm Diseases 0.000 description 1
- 108010037536 heparanase Proteins 0.000 description 1
- 229960001340 histamine Drugs 0.000 description 1
- 210000003630 histaminocyte Anatomy 0.000 description 1
- 102000045108 human EGFR Human genes 0.000 description 1
- 102000052623 human IL6R Human genes 0.000 description 1
- 235000020256 human milk Nutrition 0.000 description 1
- 210000004251 human milk Anatomy 0.000 description 1
- 150000002431 hydrogen Chemical class 0.000 description 1
- 125000004435 hydrogen atom Chemical group [H]* 0.000 description 1
- 230000002209 hydrophobic effect Effects 0.000 description 1
- 208000026095 hyper-IgM syndrome type 1 Diseases 0.000 description 1
- 101150026046 iga gene Proteins 0.000 description 1
- 230000014726 immortalization of host cell Effects 0.000 description 1
- 210000000987 immune system Anatomy 0.000 description 1
- 230000009851 immunogenic response Effects 0.000 description 1
- 230000001976 improved effect Effects 0.000 description 1
- 238000000338 in vitro Methods 0.000 description 1
- 230000001939 inductive effect Effects 0.000 description 1
- 208000015181 infectious disease Diseases 0.000 description 1
- 208000027866 inflammatory disease Diseases 0.000 description 1
- 230000004054 inflammatory process Effects 0.000 description 1
- 239000000893 inhibin Substances 0.000 description 1
- 239000003112 inhibitor Substances 0.000 description 1
- 230000005764 inhibitory process Effects 0.000 description 1
- ZPNFWUPYTFPOJU-LPYSRVMUSA-N iniprol Chemical compound C([C@H]1C(=O)NCC(=O)NCC(=O)N[C@H]2CSSC[C@H]3C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@H](C(N[C@H](C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC=4C=CC(O)=CC=4)C(=O)N[C@@H](CC=4C=CC=CC=4)C(=O)N[C@@H](CC=4C=CC(O)=CC=4)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C)C(=O)NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CSSC[C@H](NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](C)NC(=O)[C@H](CO)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CC=4C=CC=CC=4)NC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CCCCN)NC(=O)[C@H](C)NC(=O)[C@H](CCCNC(N)=N)NC2=O)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CSSC[C@H](NC(=O)[C@H](CC=2C=CC=CC=2)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H]2N(CCC2)C(=O)[C@@H](N)CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(=O)N2[C@@H](CCC2)C(=O)N2[C@@H](CCC2)C(=O)N[C@@H](CC=2C=CC(O)=CC=2)C(=O)N[C@@H]([C@@H](C)O)C(=O)NCC(=O)N2[C@@H](CCC2)C(=O)N3)C(=O)NCC(=O)NCC(=O)N[C@@H](C)C(O)=O)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@H](C(=O)N[C@@H](CC=2C=CC=CC=2)C(=O)N[C@H](C(=O)N1)C(C)C)[C@@H](C)O)[C@@H](C)CC)=O)[C@@H](C)CC)C1=CC=C(O)C=C1 ZPNFWUPYTFPOJU-LPYSRVMUSA-N 0.000 description 1
- 230000010354 integration Effects 0.000 description 1
- 108010021315 integrin beta7 Proteins 0.000 description 1
- 229960003130 interferon gamma Drugs 0.000 description 1
- 102000004114 interleukin 20 Human genes 0.000 description 1
- 108040006732 interleukin-1 receptor activity proteins Proteins 0.000 description 1
- 102000014909 interleukin-1 receptor activity proteins Human genes 0.000 description 1
- 108040001304 interleukin-17 receptor activity proteins Proteins 0.000 description 1
- 102000053460 interleukin-17 receptor activity proteins Human genes 0.000 description 1
- 108040002014 interleukin-18 receptor activity proteins Proteins 0.000 description 1
- 102000008625 interleukin-18 receptor activity proteins Human genes 0.000 description 1
- 108040006849 interleukin-2 receptor activity proteins Proteins 0.000 description 1
- 108040001834 interleukin-20 receptor activity proteins Proteins 0.000 description 1
- 108040006852 interleukin-4 receptor activity proteins Proteins 0.000 description 1
- 108040006859 interleukin-5 receptor activity proteins Proteins 0.000 description 1
- 210000002490 intestinal epithelial cell Anatomy 0.000 description 1
- 210000004347 intestinal mucosa Anatomy 0.000 description 1
- 230000010189 intracellular transport Effects 0.000 description 1
- 238000011835 investigation Methods 0.000 description 1
- 238000002955 isolation Methods 0.000 description 1
- 108010028309 kalinin Proteins 0.000 description 1
- 108010045069 keyhole-limpet hemocyanin Proteins 0.000 description 1
- 108010012808 leiomyoma-derived growth factor Proteins 0.000 description 1
- 210000004901 leucine-rich repeat Anatomy 0.000 description 1
- 208000032839 leukemia Diseases 0.000 description 1
- 210000000265 leukocyte Anatomy 0.000 description 1
- VNYSSYRCGWBHLG-AMOLWHMGSA-N leukotriene B4 Chemical compound CCCCC\C=C/C[C@@H](O)\C=C\C=C\C=C/[C@@H](O)CCCC(O)=O VNYSSYRCGWBHLG-AMOLWHMGSA-N 0.000 description 1
- 108010013555 lipoprotein-associated coagulation inhibitor Proteins 0.000 description 1
- 230000033001 locomotion Effects 0.000 description 1
- 210000004072 lung Anatomy 0.000 description 1
- 201000005249 lung adenocarcinoma Diseases 0.000 description 1
- 229940040129 luteinizing hormone Drugs 0.000 description 1
- 230000003211 malignant effect Effects 0.000 description 1
- 208000015486 malignant pancreatic neoplasm Diseases 0.000 description 1
- 238000013507 mapping Methods 0.000 description 1
- 230000007246 mechanism Effects 0.000 description 1
- 230000001404 mediated effect Effects 0.000 description 1
- 210000004379 membrane Anatomy 0.000 description 1
- 239000012528 membrane Substances 0.000 description 1
- QSHDDOUJBYECFT-UHFFFAOYSA-N mercury Chemical compound [Hg] QSHDDOUJBYECFT-UHFFFAOYSA-N 0.000 description 1
- 229910052753 mercury Inorganic materials 0.000 description 1
- CWWARWOPSKGELM-SARDKLJWSA-N methyl (2s)-2-[[(2s)-2-[[2-[[(2s)-2-[[(2s)-2-[[(2s)-5-amino-2-[[(2s)-5-amino-2-[[(2s)-1-[(2s)-6-amino-2-[[(2s)-1-[(2s)-2-amino-5-(diaminomethylideneamino)pentanoyl]pyrrolidine-2-carbonyl]amino]hexanoyl]pyrrolidine-2-carbonyl]amino]-5-oxopentanoyl]amino]-5 Chemical compound C([C@@H](C(=O)NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCSC)C(=O)OC)NC(=O)[C@H](CC=1C=CC=CC=1)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@H]1N(CCC1)C(=O)[C@H](CCCCN)NC(=O)[C@H]1N(CCC1)C(=O)[C@@H](N)CCCN=C(N)N)C1=CC=CC=C1 CWWARWOPSKGELM-SARDKLJWSA-N 0.000 description 1
- 108040008770 methylated-DNA-[protein]-cysteine S-methyltransferase activity proteins Proteins 0.000 description 1
- 108010071421 milk fat globule Proteins 0.000 description 1
- 235000021239 milk protein Nutrition 0.000 description 1
- 210000003470 mitochondria Anatomy 0.000 description 1
- 201000004058 mixed glioma Diseases 0.000 description 1
- 238000002156 mixing Methods 0.000 description 1
- 239000000203 mixture Substances 0.000 description 1
- 238000010172 mouse model Methods 0.000 description 1
- 229940126619 mouse monoclonal antibody Drugs 0.000 description 1
- 210000004877 mucosa Anatomy 0.000 description 1
- 101150069922 mug gene Proteins 0.000 description 1
- 230000004118 muscle contraction Effects 0.000 description 1
- 210000004165 myocardium Anatomy 0.000 description 1
- VMGAPWLDMVPYIA-HIDZBRGKSA-N n'-amino-n-iminomethanimidamide Chemical compound N\N=C\N=N VMGAPWLDMVPYIA-HIDZBRGKSA-N 0.000 description 1
- 229960004927 neomycin Drugs 0.000 description 1
- 239000003900 neurotrophic factor Substances 0.000 description 1
- 229940032018 neurotrophin 3 Drugs 0.000 description 1
- 229940097998 neurotrophin 4 Drugs 0.000 description 1
- 238000006386 neutralization reaction Methods 0.000 description 1
- 210000000440 neutrophil Anatomy 0.000 description 1
- 229910052757 nitrogen Inorganic materials 0.000 description 1
- 238000003199 nucleic acid amplification method Methods 0.000 description 1
- 235000015097 nutrients Nutrition 0.000 description 1
- 210000000287 oocyte Anatomy 0.000 description 1
- 108010071584 oxidized low density lipoprotein Proteins 0.000 description 1
- 210000000496 pancreas Anatomy 0.000 description 1
- 201000002528 pancreatic cancer Diseases 0.000 description 1
- 208000008443 pancreatic carcinoma Diseases 0.000 description 1
- 230000005298 paramagnetic effect Effects 0.000 description 1
- 239000000199 parathyroid hormone Substances 0.000 description 1
- 229960001319 parathyroid hormone Drugs 0.000 description 1
- 230000001575 pathological effect Effects 0.000 description 1
- 230000007170 pathology Effects 0.000 description 1
- SVHOVVJFOWGYJO-UHFFFAOYSA-N pentabromophenol Chemical compound OC1=C(Br)C(Br)=C(Br)C(Br)=C1Br SVHOVVJFOWGYJO-UHFFFAOYSA-N 0.000 description 1
- LUYQYZLEHLTPBH-UHFFFAOYSA-N perfluorobutanesulfonyl fluoride Chemical compound FC(F)(F)C(F)(F)C(F)(F)C(F)(F)S(F)(=O)=O LUYQYZLEHLTPBH-UHFFFAOYSA-N 0.000 description 1
- 210000003668 pericyte Anatomy 0.000 description 1
- 239000000825 pharmaceutical preparation Substances 0.000 description 1
- 229940127557 pharmaceutical product Drugs 0.000 description 1
- 230000000144 pharmacologic effect Effects 0.000 description 1
- NBIIXXVUZAFLBC-UHFFFAOYSA-K phosphate Chemical compound [O-]P([O-])([O-])=O NBIIXXVUZAFLBC-UHFFFAOYSA-K 0.000 description 1
- 239000010452 phosphate Substances 0.000 description 1
- 239000002504 physiological saline solution Substances 0.000 description 1
- 230000003114 pinocytic effect Effects 0.000 description 1
- 210000002826 placenta Anatomy 0.000 description 1
- 230000003169 placental effect Effects 0.000 description 1
- 229940012957 plasmin Drugs 0.000 description 1
- BASFCYQUMIYNBI-UHFFFAOYSA-N platinum Chemical group [Pt] BASFCYQUMIYNBI-UHFFFAOYSA-N 0.000 description 1
- 229910052699 polonium Inorganic materials 0.000 description 1
- HZEBHPIOVYHPMT-UHFFFAOYSA-N polonium atom Chemical compound [Po] HZEBHPIOVYHPMT-UHFFFAOYSA-N 0.000 description 1
- 229920000155 polyglutamine Polymers 0.000 description 1
- 102000054765 polymorphisms of proteins Human genes 0.000 description 1
- 239000011148 porous material Substances 0.000 description 1
- 230000035935 pregnancy Effects 0.000 description 1
- 238000002360 preparation method Methods 0.000 description 1
- 210000001236 prokaryotic cell Anatomy 0.000 description 1
- 108010087851 prorelaxin Proteins 0.000 description 1
- KAQKFAOMNZTLHT-OZUDYXHBSA-N prostaglandin I2 Chemical compound O1\C(=C/CCCC(O)=O)C[C@@H]2[C@@H](/C=C/[C@@H](O)CCCCC)[C@H](O)C[C@@H]21 KAQKFAOMNZTLHT-OZUDYXHBSA-N 0.000 description 1
- UQOQENZZLBSFKO-POPPZSFYSA-N prostaglandin J2 Chemical compound CCCCC[C@H](O)\C=C\[C@@H]1[C@@H](C\C=C/CCCC(O)=O)C=CC1=O UQOQENZZLBSFKO-POPPZSFYSA-N 0.000 description 1
- 210000002307 prostate Anatomy 0.000 description 1
- 229960000856 protein c Drugs 0.000 description 1
- 108020000494 protein-tyrosine phosphatase Proteins 0.000 description 1
- 229940039716 prothrombin Drugs 0.000 description 1
- GPTFURBXHJWNHR-UHFFFAOYSA-N protopine Chemical compound C1=C2C(=O)CC3=CC=C4OCOC4=C3CN(C)CCC2=CC2=C1OCO2 GPTFURBXHJWNHR-UHFFFAOYSA-N 0.000 description 1
- 230000002685 pulmonary effect Effects 0.000 description 1
- NGVDGCNFYWLIFO-UHFFFAOYSA-N pyridoxal 5'-phosphate Chemical compound CC1=NC=C(COP(O)(O)=O)C(C=O)=C1O NGVDGCNFYWLIFO-UHFFFAOYSA-N 0.000 description 1
- 230000001850 reproductive effect Effects 0.000 description 1
- 230000000241 respiratory effect Effects 0.000 description 1
- 230000002207 retinal effect Effects 0.000 description 1
- 238000002702 ribosome display Methods 0.000 description 1
- 239000000523 sample Substances 0.000 description 1
- 238000000926 separation method Methods 0.000 description 1
- 239000003001 serine protease inhibitor Substances 0.000 description 1
- 101150088976 shh gene Proteins 0.000 description 1
- 230000019491 signal transduction Effects 0.000 description 1
- 229910052710 silicon Inorganic materials 0.000 description 1
- 239000010703 silicon Substances 0.000 description 1
- 210000002027 skeletal muscle Anatomy 0.000 description 1
- 210000000813 small intestine Anatomy 0.000 description 1
- 150000003384 small molecules Chemical class 0.000 description 1
- 210000002460 smooth muscle Anatomy 0.000 description 1
- JUJBNYBVVQSIOU-UHFFFAOYSA-M sodium;4-[2-(4-iodophenyl)-3-(4-nitrophenyl)tetrazol-2-ium-5-yl]benzene-1,3-disulfonate Chemical compound [Na+].C1=CC([N+](=O)[O-])=CC=C1N1[N+](C=2C=CC(I)=CC=2)=NC(C=2C(=CC(=CC=2)S([O-])(=O)=O)S([O-])(=O)=O)=N1 JUJBNYBVVQSIOU-UHFFFAOYSA-M 0.000 description 1
- JACYMBNQPPWQML-UHFFFAOYSA-N sodium;4-methoxy-5-[3-(2-methoxy-4-nitro-5-sulfophenyl)-5-(phenylcarbamoyl)tetrazol-3-ium-2-yl]-2-nitrobenzenesulfonate Chemical compound [Na+].COC1=CC([N+]([O-])=O)=C(S([O-])(=O)=O)C=C1N1[N+](C=2C(=CC(=C(C=2)S(O)(=O)=O)[N+]([O-])=O)OC)=NC(C(=O)NC=2C=CC=CC=2)=N1 JACYMBNQPPWQML-UHFFFAOYSA-N 0.000 description 1
- 229950007874 solanezumab Drugs 0.000 description 1
- 230000037439 somatic mutation Effects 0.000 description 1
- 238000004611 spectroscopical analysis Methods 0.000 description 1
- 150000003408 sphingolipids Chemical class 0.000 description 1
- 210000004989 spleen cell Anatomy 0.000 description 1
- 238000010186 staining Methods 0.000 description 1
- 230000002739 subcortical effect Effects 0.000 description 1
- 239000000758 substrate Substances 0.000 description 1
- 239000006228 supernatant Substances 0.000 description 1
- 230000020382 suppression by virus of host antigen processing and presentation of peptide antigen via MHC class I Effects 0.000 description 1
- 230000002194 synthesizing effect Effects 0.000 description 1
- 102000013498 tau Proteins Human genes 0.000 description 1
- 108010026424 tau Proteins Proteins 0.000 description 1
- 230000002381 testicular Effects 0.000 description 1
- 201000003120 testicular cancer Diseases 0.000 description 1
- 125000003831 tetrazolyl group Chemical group 0.000 description 1
- 229940126585 therapeutic drug Drugs 0.000 description 1
- XNRNNGPBEPRNAR-JQBLCGNGSA-N thromboxane B2 Chemical compound CCCCC[C@H](O)\C=C\[C@H]1OC(O)C[C@H](O)[C@@H]1C\C=C/CCCC(O)=O XNRNNGPBEPRNAR-JQBLCGNGSA-N 0.000 description 1
- 210000001541 thymus gland Anatomy 0.000 description 1
- 210000001578 tight junction Anatomy 0.000 description 1
- 229960003087 tioguanine Drugs 0.000 description 1
- MNRILEROXIRVNJ-UHFFFAOYSA-N tioguanine Chemical compound N1C(N)=NC(=S)C2=NC=N[C]21 MNRILEROXIRVNJ-UHFFFAOYSA-N 0.000 description 1
- 101150117196 tra-1 gene Proteins 0.000 description 1
- 108010042974 transforming growth factor beta4 Proteins 0.000 description 1
- 238000013519 translation Methods 0.000 description 1
- 210000004881 tumor cell Anatomy 0.000 description 1
- 238000005199 ultracentrifugation Methods 0.000 description 1
- 241000701161 unidentified adenovirus Species 0.000 description 1
- VBEQCZHXXJYVRD-GACYYNSASA-N uroanthelone Chemical compound C([C@@H](C(=O)N[C@H](C(=O)N[C@@H](CS)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CS)C(=O)N[C@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)NCC(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(=O)N[C@@H](CO)C(=O)NCC(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CS)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O)C(C)C)[C@@H](C)O)NC(=O)[C@H](CO)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CO)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](NC(=O)[C@H](CC=1NC=NC=1)NC(=O)[C@H](CCSC)NC(=O)[C@H](CS)NC(=O)[C@@H](NC(=O)CNC(=O)CNC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CS)NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)CNC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)[C@H](CO)NC(=O)[C@H](CO)NC(=O)[C@H]1N(CCC1)C(=O)[C@H](CS)NC(=O)CNC(=O)[C@H]1N(CCC1)C(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)[C@H](CO)NC(=O)[C@@H](N)CC(N)=O)C(C)C)[C@@H](C)CC)C1=CC=C(O)C=C1 VBEQCZHXXJYVRD-GACYYNSASA-N 0.000 description 1
- 229960005356 urokinase Drugs 0.000 description 1
- 102000009816 urokinase plasminogen activator receptor activity proteins Human genes 0.000 description 1
- 108040001269 urokinase plasminogen activator receptor activity proteins Proteins 0.000 description 1
- 206010046766 uterine cancer Diseases 0.000 description 1
- 210000003556 vascular endothelial cell Anatomy 0.000 description 1
- 230000002861 ventricular Effects 0.000 description 1
- 108700026220 vif Genes Proteins 0.000 description 1
- 230000003612 virological effect Effects 0.000 description 1
- 101150068520 wnt3a gene Proteins 0.000 description 1
- 238000002424 x-ray crystallography Methods 0.000 description 1
- 235000021247 β-casein Nutrition 0.000 description 1
Images



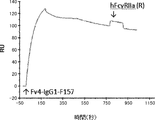


































Description
〔1〕脳疾患に固有に脳組織内に存在する抗原に対する結合活性がイオン濃度の条件によって変化する抗原結合ドメイン、ならびにpH中性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインおよびpH酸性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインを含む抗原結合分子の、脳疾患を治療するための使用、
〔2〕前記抗原結合ドメインが、前記抗原に対する結合活性がカルシウムイオン濃度の条件によって変化する抗原結合ドメインである、〔1〕に記載の使用、
〔3〕前記抗原結合ドメインが、前記抗原に対する低カルシウムイオン濃度の条件下での結合活性が前記抗原に対する高カルシウムイオン濃度の条件下での結合活性よりも低いように結合活性が変化する抗原結合ドメインである、〔2〕に記載の使用、
〔4〕前記抗原結合ドメインが、前記抗原に対する結合活性がpHの条件によって変化する抗原結合ドメインである、〔1〕に記載の使用、
〔5〕前記抗原結合ドメインが、前記抗原に対するpH酸性域の条件下での結合活性がpH中性域の条件下での結合活性よりも低いように結合活性が変化する抗原結合ドメインである、〔4〕に記載の使用、
〔6〕前記抗原結合ドメインが抗体の可変領域である、〔1〕から〔5〕のいずれかに記載の使用、
〔7〕pH酸性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインが、抗体のFc領域である、〔1〕から〔6〕のいずれかに記載の使用、
〔8〕前記Fc領域が、配列番号:14、15、16または17のいずれかに含まれるFc領域である、〔7〕に記載の使用、
〔9〕前記Fc領域が、pH酸性域の条件下でのFcRnに対する結合活性が、配列番号:14、15、16または17のいずれかに含まれるFc領域のFcRnに対する結合活性より増強されているFc領域である、〔7〕に記載の使用、
〔10〕前記Fc領域が、配列番号:14、15、16、または17のいずれかに含まれるFc領域のアミノ酸配列のうち、EUナンバリングで表される238位、244位、245位、249位、250位、251位、252位、253位、254位、255位、256位、257位、258位、260位、262位、265位、270位、272位、279位、283位、285位、286位、288位、293位、303位、305位、307位、308位、309位、311位、312位、314位、316位、317位、318位、332位、339位、340位、341位、343位、356位、360位、362位、375位、376位、377位、378位、380位、382位、385位、386位、387位、388位、389位、400位、413位、415位、423位、424位、427位、428位、430位、431位、433位、434位、435位、436位、438位、439位、440位、442位または447位の群から選択される少なくとも一つ以上のアミノ酸が置換されているFc領域である、〔9〕に記載の使用、
〔11〕前記Fc領域が、配列番号:14、15、16、または17に含まれるFc領域のアミノ酸配列のうち、EUナンバリングで表される;
238位のアミノ酸がLeu、
244位のアミノ酸がLeu、
245位のアミノ酸がArg、
249位のアミノ酸がPro、
250位のアミノ酸がGlnまたはGluのいずれか、もしくは
251位のアミノ酸がArg、Asp、Glu、またはLeuのいずれか、
252位のアミノ酸がPhe、Ser、Thr、またはTyrのいずれか、
254位のアミノ酸がSerまたはThrのいずれか、
255位のアミノ酸がArg、Gly、Ile、またはLeuのいずれか、
256位のアミノ酸がAla、Arg、Asn、Asp、Gln、Glu、Pro、またはThrのいずれか、
257位のアミノ酸がAla、Ile、Met、Asn、Ser、またはValのいずれか、
258位のアミノ酸がAsp、
260位のアミノ酸がSer、
262位のアミノ酸がLeu、
270位のアミノ酸がLys、
272位のアミノ酸がLeu、またはArgのいずれか、
279位のアミノ酸がAla、Asp、Gly、His、Met、Asn、Gln、Arg、Ser、Thr、Trp、またはTyrのいずれか、
283位のアミノ酸がAla、Asp、Phe、Gly、His、Ile、Lys、Leu、Asn、Pro、Gln、Arg、Ser、Thr、Trp、またはTyrのいずれか、
285位のアミノ酸がAsn、
286位のアミノ酸がPhe、
288位のアミノ酸がAsn、またはProのいずれか、
293位のアミノ酸がVal、
307位のアミノ酸がAla、Glu、Gln、またはMetのいずれか、
311位のアミノ酸がAla、Glu、Ile、Lys、Leu、Met、Ser 、Val、またはTrpのいずれか、
309位のアミノ酸がPro、
312位のアミノ酸がAla、Asp、またはProのいずれか、
314位のアミノ酸がAlaまたはLeuのいずれか、
316位のアミノ酸がLys、
317位のアミノ酸がPro、
318位のアミノ酸がAsn、またはThrのいずれか、
332位のアミノ酸がPhe、His、Lys、Leu、Met、Arg、Ser、またはTrpのいずれか、
339位のアミノ酸がAsn、Thr、またはTrpのいずれか、
341位のアミノ酸がPro、
343位のアミノ酸がGlu、His、Lys、Gln、Arg、Thr、またはTyrのいずれか、
375位のアミノ酸がArg、
376位のアミノ酸がGly、Ile、Met、Pro、Thr、またはValのいずれか、
377位のアミノ酸がLys、
378位のアミノ酸がAsp、Asn、またはValのいずれか、
380位のアミノ酸がAla、Asn、Ser、またはThrのいずれか、
382位のアミノ酸がPhe、His、Ile、Lys、Leu、Met、Asn、Gln、Arg、Ser、Thr、Val、Trp、またはTyrのいずれか、
385位のアミノ酸がAla、Arg、Asp、Gly、His、Lys、Ser、またはThrのいずれか、
386位のアミノ酸がArg、Asp、Ile、Lys、Met、Pro、Ser、またはThrのいずれか、
387位のアミノ酸がAla、Arg、His、Pro、Ser、またはThrのいずれか、
389位のアミノ酸がAsn、Pro、またはSerのいずれか、
423位のアミノ酸がAsn、
427位のアミノ酸がAsn、
428位のアミノ酸がLeu、Met、Phe、Ser、またはThrのいずれか、
430位のアミノ酸がAla、Phe、Gly、His、Ile、Lys、Leu、Met、Asn、Gln、Arg、Ser、Thr、Val、またはTyrのいずれか、
431位のアミノ酸がHis、またはAsnのいずれか、
433位のアミノ酸がArg、Gln、His、Ile、Lys、Pro、またはSerのいずれか、
434位のアミノ酸がAla、Gly、His、Phe、Ser、Trp、またはTyrのいずれか、
436位のアミノ酸がArg、Asn、His、Ile、Leu、Lys、Met、またはThrのいずれか、
438位のアミノ酸がLys、Leu、Thr、またはTrpのいずれか、
440位のアミノ酸がLys、もしくは、
442位のアミノ酸がLys、308位のアミノ酸がIle、Pro、またはThrのいずれか、
の群から選択される少なくとも一つ以上のアミノ酸を含む、〔10〕に記載の使用、
〔12〕前記pH中性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインが、抗体のFc領域である、〔1〕から〔11〕のいずれかに記載の使用、
〔13〕前記Fc領域が、配列番号:14、15、16、または17に含まれるFc領域のアミノ酸配列のうち、EUナンバリングで表される237位、248位、250位、252位、254位、255位、256位、257位、258位、265位、286位、289位、297位、298位、303位、305位、307位、308位、309位、311位、312位、314位、315位、317位、332位、334位、360位、376位、380位、382位、384位、385位、386位、387位、389位、424位、428位、433位、434位または436位の群から選択される少なくとも一つ以上のアミノ酸が置換されているFc領域である、〔12〕に記載の使用、
〔14〕前記Fc領域が、配列番号:14、15、16、または17に含まれるFc領域のアミノ酸配列のうち、EUナンバリングで表される;
237位のアミノ酸がMet、
248位のアミノ酸がIle、
250位のアミノ酸がAla、Phe、Ile、Met、Gln、Ser、Val、Trp、またはTyrのいずれか、
252位のアミノ酸がPhe、Trp、またはTyrのいずれか、
254位のアミノ酸がThr、
255位のアミノ酸がGlu、
256位のアミノ酸がAsp、Asn、Glu、またはGlnのいずれか、
257位のアミノ酸がAla、Gly、Ile、Leu、Met、Asn、Ser、Thr、またはValのいずれか、
258位のアミノ酸がHis、
265位のアミノ酸がAla、
286位のアミノ酸がAlaまたはGluのいずれか、
289位のアミノ酸がHis、
297位のアミノ酸がAla、
303位のアミノ酸がAla、
305位のアミノ酸がAla、
307位のアミノ酸がAla、Asp、Phe、Gly、His、Ile、Lys、Leu、Met、Asn、Pro、Gln、Arg、Ser、Val、Trp、またはTyrのいずれか、
308位のアミノ酸がAla、Phe、Ile、Leu、Met、Pro、Gln、またはThrのいずれか、
309位のアミノ酸がAla、Asp、Glu、Pro、またはArgのいずれか、
311位のアミノ酸がAla、His、またはIleのいずれか、
312位のアミノ酸がAlaまたはHisのいずれか、
314位のアミノ酸がLysまたはArgのいずれか、
315位のアミノ酸がAla、AspまたはHisのいずれか、
317位のアミノ酸がAla、
332位のアミノ酸がVal、
334位のアミノ酸がLeu、
360位のアミノ酸がHis、
376位のアミノ酸がAla、
380位のアミノ酸がAla、
382位のアミノ酸がAla、
384位のアミノ酸がAla、
385位のアミノ酸がAspまたはHisのいずれか、
386位のアミノ酸がPro、
387位のアミノ酸がGlu、
389位のアミノ酸がAlaまたはSerのいずれか、
424位のアミノ酸がAla、
428位のアミノ酸がAla、Asp、Phe、Gly、His、Ile、Lys、Leu、Asn、Pro、Gln、Ser、Thr、Val、Trp、またはTyrのいずれか、
433位のアミノ酸がLys、
434位のアミノ酸がAla、Phe、His、Ser、Trp、またはTyrのいずれか、もしくは
436位のアミノ酸がHis 、Ile、Leu、Phe、Thr、またはVal、
の群から選択される少なくとも一つ以上のアミノ酸を含む、〔13〕に記載の使用、
〔15〕前記Fc領域が、FcγR結合活性が低いまたは低減されたFc領域である、〔12〕から〔14〕に記載の使用、
〔16〕前記pH中性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインが、抗体の可変領域である、〔1〕から〔11〕のいずれかに記載の使用、
〔17〕前記抗原結合分子がさらにFcγR結合ドメインを含む、〔1〕から〔16〕のいずれかに記載の使用、
〔18〕前記FcγR結合ドメインが抗体のFc領域である、〔17〕に記載の使用、
〔19〕前記Fc領域が、天然型ヒトIgGのFc領域のFcγレセプターに対する結合活性よりもFcγレセプターに対する結合活性が高いFcγR結合改変Fc領域を含む、〔18〕に記載の使用、
〔20〕前記抗原結合分子がさらに補体結合ドメインを含む、〔1〕から〔17〕のいずれかに記載の使用、
〔21〕前記補体結合ドメインが抗体のFc領域である、〔20〕に記載の使用、
〔22〕前記Fc領域が、天然型ヒトIgGのFc領域の補体に対する結合活性よりも補体に対する結合活性が高い補体結合改変Fc領域を含む、〔21〕に記載の使用、
〔23〕前記FcγR結合ドメインが抗体の可変領域である、〔17〕に記載の使用、
〔24〕前記補体結合ドメインが抗体の可変領域である、〔20〕に記載の使用、
〔25〕pH中性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインおよびpH酸性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインが、抗体の可変領域である、〔1〕から〔6〕のいずれかに記載の使用、
〔26〕前記抗原結合分子が抗体である、〔1〕から〔24〕のいずれかに記載の使用、
〔27〕〔1〕から〔26〕のいずれかに記載の抗原結合分子をコードするポリヌクレオチドが作用可能に連結されたベクターが導入された細胞の培養液から抗原結合分子を回収する工程を含む、脳疾患治療剤の製造方法。
アミノ酸
本明細書において、たとえば、Ala/A、Leu/L、Arg/R、Lys/K、Asn/N、Met/M、Asp/D、Phe/F、Cys/C、Pro/P、Gln/Q、Ser/S、Glu/E、Thr/T、Gly/G、Trp/W、His/H、Tyr/Y、Ile/I、Val/Vと表されるように、アミノ酸は1文字コードまたは3文字コード、またはその両方で表記されている。
抗原結合分子のアミノ酸配列中のアミノ酸の改変のためには、部位特異的変異誘発法(Kunkelら(Proc. Natl. Acad. Sci. USA (1985) 82, 488-492))やOverlap extension PCR等の公知の方法が適宜採用され得る。また、天然のアミノ酸以外のアミノ酸に置換するアミノ酸の改変方法として、複数の公知の方法もまた採用され得る(Annu. Rev. Biophys. Biomol. Struct. (2006) 35, 225-249、Proc. Natl. Acad. Sci. U.S.A. (2003) 100 (11), 6353-6357)。例えば、終止コドンの1つであるUAGコドン(アンバーコドン)の相補的アンバーサプレッサーtRNAに非天然アミノ酸が結合されたtRNAが含まれる無細胞翻訳系システム(Clover Direct(Protein Express))等も好適に用いられる。また、アミノ酸の改変を表す表現として、特定の位置を表す数字の前後に改変前と改変後のアミノ酸の1文字コードを用いた表現が適宜使用され得る。例えば、抗体定常領域に含まれるFc領域にアミノ酸の置換を加える際に用いられるP238Dという改変は、EUナンバリングで表される238位のProのAspへの置換を表す。すなわち、数字はEUナンバリングで表されるアミノ酸の位置を表し、その前に記載されるアミノ酸の一文字コードは置換前のアミノ酸、そのあとに記載されるアミノ酸の1文字コードは置換後のアミノ酸を表す。
(a) 33位、(b) 55位、(c) 96位、(d) 33位および55位、(e) 33位および96位、(f) 55位および96位、(g) 33位および55位および96位。
本明細書において「抗原」は抗原結合ドメインが結合するエピトープを含む限りその構造は特定の構造に限定されない。別の意味では、抗原は無機物でもあり得るし有機物でもあり得る。抗原としては下記のような分子;17-IA、4-1BB、4Dc、6-ケト-PGF1a、8-イソ-PGF2a、8-オキソ-dG、A1 アデノシン受容体、A33、ACE、ACE-2、アクチビン、アクチビンA、アクチビンAB、アクチビンB、アクチビンC、アクチビンRIA、アクチビンRIA ALK-2、アクチビンRIB ALK-4、アクチビンRIIA、アクチビンRIIB、ADAM、ADAM10、ADAM12、ADAM15、ADAM17/TACE、ADAM8、ADAM9、ADAMTS、ADAMTS4、ADAMTS5、アドレシン、aFGF、ALCAM、ALK、ALK-1、ALK-7、アルファ-1-アンチトリプシン、アルファ−V/ベータ-1アンタゴニスト、ANG、Ang、APAF-1、APE、APJ、APP、APRIL、AR、ARC、ART、アルテミン、抗Id、ASPARTIC、心房性ナトリウム利尿因子、av/b3インテグリン、Axl、b2M、B7-1、B7-2、B7-H、B-リンパ球刺激因子(BlyS)、BACE、BACE-1、Bad、BAFF、BAFF-R、Bag-1、BAK、Bax、BCA-1、BCAM、Bcl、BCMA、BDNF、b-ECGF、bFGF、BID、Bik、BIM、BLC、BL-CAM、BLK、BMP、BMP-2 BMP-2a、BMP-3 オステオゲニン(Osteogenin)、BMP-4 BMP-2b、BMP-5、BMP-6 Vgr-1、BMP-7(OP-1)、BMP-8(BMP-8a、OP-2)、BMPR、BMPR-IA(ALK-3)、BMPR-IB(ALK-6)、BRK-2、RPK-1、BMPR-II(BRK-3)、BMP、b-NGF、BOK、ボンベシン、骨由来神経栄養因子、BPDE、BPDE-DNA、BTC、補体因子3(C3)、C3a、C4、C5、C5a、C10、CA125、CAD-8、カルシトニン、cAMP、癌胎児性抗原(CEA)、癌関連抗原、カテプシンA、カテプシンB、カテプシンC/DPPI、カテプシンD、カテプシンE、カテプシンH、カテプシンL、カテプシンO、カテプシンS、カテプシンV、カテプシンX/Z/P、CBL、CCI、CCK2、CCL、CCL1、CCL11、CCL12、CCL13、CCL14、CCL15、CCL16、CCL17、CCL18、CCL19、CCL2、CCL20、CCL21、CCL22、CCL23、CCL24、CCL25、CCL26、CCL27、CCL28、CCL3、CCL4、CCL5、CCL6、CCL7、CCL8、CCL9/10、CCR、CCR1、CCR10、CCR10、CCR2、CCR3、CCR4、CCR5、CCR6、CCR7、CCR8、CCR9、CD1、CD2、CD3、CD3E、CD4、CD5、CD6、CD7、CD8、CD10、CD11a、CD11b、CD11c、CD13、CD14、CD15、CD16、CD18、CD19、CD20、CD21、CD22、CD23、CD25、CD27L、CD28、CD29、CD30、CD30L、CD32、CD33(p67タンパク質)、CD34、CD38、CD40、CD40L、CD44、CD45、CD46、CD49a、CD52、CD54、CD55、CD56、CD61、CD64、CD66e、CD74、CD80(B7-1)、CD89、CD95、CD123、CD137、CD138、CD140a、CD146、CD147、CD148、CD152、CD164、CEACAM5、CFTR、cGMP、CINC、ボツリヌス菌毒素、ウェルシュ菌毒素、CKb8-1、CLC、CMV、CMV UL、CNTF、CNTN-1、COX、C-Ret、CRG-2、CT-1、CTACK、CTGF、CTLA-4、CX3CL1、CX3CR1、CXCL、CXCL1、CXCL2、CXCL3、CXCL4、CXCL5、CXCL6、CXCL7、CXCL8、CXCL9、CXCL10、CXCL11、CXCL12、CXCL13、CXCL14、CXCL15、CXCL16、CXCR、CXCR1、CXCR2、CXCR3、CXCR4、CXCR5、CXCR6、サイトケラチン腫瘍関連抗原、DAN、DCC、DcR3、DC-SIGN、補体制御因子(Decay accelerating factor)、des(1-3)-IGF-I(脳IGF-1)、Dhh、ジゴキシン、DNAM-1、Dnase、Dpp、DPPIV/CD26、Dtk、ECAD、EDA、EDA-A1、EDA-A2、EDAR、EGF、EGFR(ErbB-1)、EMA、EMMPRIN、ENA、エンドセリン受容体、エンケファリナーゼ、eNOS、Eot、エオタキシン1、EpCAM、エフリンB2/EphB4、EPO、ERCC、E-セレクチン、ET-1、ファクターIIa、ファクターVII、ファクターVIIIc、ファクターIX、線維芽細胞活性化タンパク質(FAP)、Fas、FcR1、FEN-1、フェリチン、FGF、FGF-19、FGF-2、FGF3、FGF-8、FGFR、FGFR-3、フィブリン、FL、FLIP、Flt-3、Flt-4、卵胞刺激ホルモン、フラクタルカイン、FZD1、FZD2、FZD3、FZD4、FZD5、FZD6、FZD7、FZD8、FZD9、FZD10、G250、Gas6、GCP-2、GCSF、GD2、GD3、GDF、GDF-1、GDF-3(Vgr-2)、GDF-5(BMP-14、CDMP-1)、GDF-6(BMP-13、CDMP-2)、GDF-7(BMP-12、CDMP-3)、GDF-8(ミオスタチン)、GDF-9、GDF-15(MIC-1)、GDNF、GDNF、GFAP、GFRa-1、GFR-アルファ1、GFR-アルファ2、GFR-アルファ3、GITR、グルカゴン、Glut4、糖タンパク質IIb/IIIa(GPIIb/IIIa)、GM-CSF、gp130、gp72、GRO、成長ホルモン放出因子、ハプテン(NP-capまたはNIP-cap)、HB-EGF、HCC、HCMV gBエンベロープ糖タンパク質、HCMV gHエンベロープ糖タンパク質、HCMV UL、造血成長因子(HGF)、Hep B gp120、ヘパラナーゼ、Her2、Her2/neu(ErbB-2)、Her3(ErbB-3)、Her4(ErbB-4)、単純ヘルペスウイルス(HSV) gB糖タンパク質、HSV gD糖タンパク質、HGFA、高分子量黒色腫関連抗原(HMW-MAA)、HIV gp120、HIV IIIB gp 120 V3ループ、HLA、HLA-DR、HM1.24、HMFG PEM、HRG、Hrk、ヒト心臓ミオシン、ヒトサイトメガロウイルス(HCMV)、ヒト成長ホルモン(HGH)、HVEM、I-309、IAP、ICAM、ICAM-1、ICAM-3、ICE、ICOS、IFNg、Ig、IgA受容体、IgE、IGF、IGF結合タンパク質、IGF-1R、IGFBP、IGF-I、IGF-II、IL、IL-1、IL-1R、IL-2、IL-2R、IL-4、IL-4R、IL-5、IL-5R、IL-6、IL-6R、IL-8、IL-9、IL-10、IL-12、IL-13、IL-15、IL-18、IL-18R、IL-23、インターフェロン(INF)-アルファ、INF-ベータ、INF-ガンマ、インヒビン、iNOS、インスリンA鎖、インスリンB鎖、インスリン様増殖因子1、インテグリンアルファ2、インテグリンアルファ3、インテグリンアルファ4、インテグリンアルファ4/ベータ1、インテグリンアルファ4/ベータ7、インテグリンアルファ5(アルファV)、インテグリンアルファ5/ベータ1、インテグリンアルファ5/ベータ3、インテグリンアルファ6、インテグリンベータ1、インテグリンベータ2、インターフェロンガンマ、IP-10、I-TAC、JE、カリクレイン2、カリクレイン5、カリクレイン6、カリクレイン11、カリクレイン12、カリクレイン14、カリクレイン15、カリクレインL1、カリクレインL2、カリクレインL3、カリクレインL4、KC、KDR、ケラチノサイト増殖因子(KGF)、ラミニン5、LAMP、LAP、LAP(TGF-1)、潜在的TGF-1、潜在的TGF-1 bp1、LBP、LDGF、LECT2、レフティ、ルイス−Y抗原、ルイス−Y関連抗原、LFA-1、LFA-3、Lfo、LIF、LIGHT、リポタンパク質、LIX、LKN、Lptn、L-セレクチン、LT-a、LT-b、LTB4、LTBP-1、肺表面、黄体形成ホルモン、リンホトキシンベータ受容体、Mac-1、MAdCAM、MAG、MAP2、MARC、MCAM、MCAM、MCK-2、MCP、M-CSF、MDC、Mer、METALLOPROTEASES、MGDF受容体、MGMT、MHC(HLA-DR)、MIF、MIG、MIP、MIP-1-アルファ、MK、MMAC1、MMP、MMP-1、MMP-10、MMP-11、MMP-12、MMP-13、MMP-14、MMP-15、MMP-2、MMP-24、MMP-3、MMP-7、MMP-8、MMP-9、MPIF、Mpo、MSK、MSP、ムチン(Muc1)、MUC18、ミュラー管抑制物質、Mug、MuSK、NAIP、NAP、NCAD、N-Cアドヘリン、NCA 90、NCAM、NCAM、ネプリライシン、ニューロトロフィン-3、-4、または-6、ニュールツリン、神経成長因子(NGF)、NGFR、NGF−ベータ、nNOS、NO、NOS、Npn、NRG-3、NT、NTN、OB、OGG1、OPG、OPN、OSM、OX40L、OX40R、p150、p95、PADPr、副甲状腺ホルモン、PARC、PARP、PBR、PBSF、PCAD、P-カドヘリン、PCNA、PDGF、PDGF、PDK-1、PECAM、PEM、PF4、PGE、PGF、PGI2、PGJ2、PIN、PLA2、胎盤性アルカリホスファターゼ(PLAP)、PlGF、PLP、PP14、プロインスリン、プロレラキシン、プロテインC、PS、PSA、PSCA、前立腺特異的膜抗原(PSMA)、PTEN、PTHrp、Ptk、PTN、R51、RANK、RANKL、RANTES、RANTES、レラキシンA鎖、レラキシンB鎖、レニン、呼吸器多核体ウイルス(RSV)F、RSV Fgp、Ret、リウマイド因子、RLIP76、RPA2、RSK、S100、SCF/KL、SDF-1、SERINE、血清アルブミン、sFRP-3、Shh、SIGIRR、SK-1、SLAM、SLPI、SMAC、SMDF、SMOH、SOD、SPARC、Stat、STEAP、STEAP-II、TACE、TACI、TAG-72(腫瘍関連糖タンパク質−72)、TARC、TCA-3、T細胞受容体(例えば、T細胞受容体アルファ/ベータ)、TdT、TECK、TEM1、TEM5、TEM7、TEM8、TERT、睾丸PLAP様アルカリホスファターゼ、TfR、TGF、TGF-アルファ、TGF-ベータ、TGF-ベータ Pan Specific、TGF-ベータRI(ALK-5)、TGF-ベータRII、TGF-ベータRIIb、TGF-ベータRIII、TGF-ベータ1、TGF-ベータ2、TGF-ベータ3、TGF-ベータ4、TGF-ベータ5、トロンビン、胸腺Ck-1、甲状腺刺激ホルモン、Tie、TIMP、TIQ、組織因子、TMEFF2、Tmpo、TMPRSS2、TNF、TNF-アルファ、TNF-アルファベータ、TNF-ベータ2、TNFc、TNF-RI、TNF-RII、TNFRSF10A(TRAIL R1 Apo-2、DR4)、TNFRSF10B(TRAIL R2 DR5、KILLER、TRICK-2A、TRICK-B)、TNFRSF10C(TRAIL R3 DcR1、LIT、TRID)、TNFRSF10D(TRAIL R4 DcR2、TRUNDD)、TNFRSF11A(RANK ODF R、TRANCE R)、TNFRSF11B(OPG OCIF、TR1)、TNFRSF12(TWEAK R FN14)、TNFRSF13B(TACI)、TNFRSF13C(BAFF R)、TNFRSF14(HVEM ATAR、HveA、LIGHT R、TR2)、TNFRSF16(NGFR p75NTR)、TNFRSF17(BCMA)、TNFRSF18(GITR AITR)、TNFRSF19(TROY TAJ、TRADE)、TNFRSF19L(RELT)、TNFRSF1A(TNF RI CD120a、p55-60)、TNFRSF1B(TNF RII CD120b、p75-80)、TNFRSF26(TNFRH3)、TNFRSF3(LTbR TNF RIII、TNFC R)、TNFRSF4(OX40 ACT35、TXGP1 R)、TNFRSF5(CD40 p50)、TNFRSF6(Fas Apo-1、APT1、CD95)、TNFRSF6B(DcR3 M68、TR6)、TNFRSF7(CD27)、TNFRSF8(CD30)、TNFRSF9(4-1BB CD137、ILA)、TNFRSF21(DR6)、TNFRSF22(DcTRAIL R2 TNFRH2)、TNFRST23(DcTRAIL R1 TNFRH1)、TNFRSF25(DR3 Apo-3、LARD、TR-3、TRAMP、WSL-1)、TNFSF10(TRAIL Apo-2リガンド、TL2)、TNFSF11(TRANCE/RANKリガンド ODF、OPGリガンド)、TNFSF12(TWEAK Apo-3リガンド、DR3リガンド)、TNFSF13(APRIL TALL2)、TNFSF13B(BAFF BLYS、TALL1、THANK、TNFSF20)、TNFSF14(LIGHT HVEMリガンド、LTg)、TNFSF15(TL1A/VEGI)、TNFSF18(GITRリガンド AITRリガンド、TL6)、TNFSF1A(TNF-a コネクチン(Conectin)、DIF、TNFSF2)、TNFSF1B(TNF-b LTa、TNFSF1)、TNFSF3(LTb TNFC、p33)、TNFSF4(OX40リガンド gp34、TXGP1)、TNFSF5(CD40リガンド CD154、gp39、HIGM1、IMD3、TRAP)、TNFSF6(Fasリガンド Apo-1リガンド、APT1リガンド)、TNFSF7(CD27リガンド CD70)、TNFSF8(CD30リガンド CD153)、TNFSF9(4-1BBリガンド CD137リガンド)、TP-1、t-PA、Tpo、TRAIL、TRAIL R、TRAIL-R1、TRAIL-R2、TRANCE、トランスフェリン受容体、TRF、Trk、TROP-2、TSG、TSLP、腫瘍関連抗原CA125、腫瘍関連抗原発現ルイスY関連炭水化物、TWEAK、TXB2、Ung、uPAR、uPAR-1、ウロキナーゼ、VCAM、VCAM-1、VECAD、VE-Cadherin、VE-cadherin-2、VEFGR-1(flt-1)、VEGF、VEGFR、VEGFR-3(flt-4)、VEGI、VIM、ウイルス抗原、VLA、VLA-1、VLA-4、VNRインテグリン、フォン・ヴィレブランド因子、WIF-1、WNT1、WNT2、WNT2B/13、WNT3、WNT3A、WNT4、WNT5A、WNT5B、WNT6、WNT7A、WNT7B、WNT8A、WNT8B、WNT9A、WNT9A、WNT9B、WNT10A、WNT10B、WNT11、WNT16、XCL1、XCL2、XCR1、XCR1、XEDAR、XIAP、XPD、HMGB1、IgA、Aβ、CD81, CD97, CD98, DDR1, DKK1, EREG、Hsp90, IL-17/IL-17R、IL-20/IL-20R、酸化LDL, PCSK9, prekallikrein , RON, TMEM16F、SOD1, Chromogranin A, Chromogranin B、tau, VAP1、高分子キニノーゲン、IL-31、IL-31R、Nav1.1、Nav1.2、Nav1.3、Nav1.4、Nav1.5、Nav1.6、Nav1.7、Nav1.8、Nav1.9、EPCR、C1, C1q, C1r, C1s, C2, C2a, C2b, C3, C3a, C3b, C4, C4a, C4b, C5, C5a, C5b, C6, C7, C8, C9, factor B, factor D, factor H, properdin、sclerostin、fibrinogen, fibrin, prothrombin, thrombin, 組織因子, factor V, factor Va, factor VII, factor VIIa, factor VIII, factor VIIIa, factor IX, factor IXa, factor X, factor Xa, factor XI, factor XIa, factor XII, factor XIIa, factor XIII, factor XIIIa, TFPI, antithrombin III, EPCR, トロンボモデュリン、TAPI, tPA, plasminogen, plasmin, PAI-1, PAI-2、GPC3、Syndecan-1、Syndecan-2、Syndecan-3、Syndecan-4、LPA、S1Pならびにホルモンおよび成長因子のための受容体が例示され得る。
抗原中に存在する抗原決定基を意味するエピトープは、本明細書において開示される抗原結合分子中の抗原結合ドメインが結合する抗原上の部位を意味する。よって、例えば、エピトープは、その構造によって定義され得る。また、当該エピトープを認識する抗原結合分子中の抗原に対する結合活性によっても当該エピトープが定義され得る。抗原がペプチド又はポリペプチドである場合には、エピトープを構成するアミノ酸残基によってエピトープを特定することも可能である。また、エピトープが糖鎖である場合には、特定の糖鎖構造によってエピトープを特定することも可能である。
下記にIL-6Rに対する抗原結合ドメインを含む被験抗原結合分子によるエピトープへの結合の確認方法が例示されるが、IL-6R以外の抗原に対する抗原結合ドメインを含む被験抗原結合分子によるエピトープへの結合の確認方法も下記の例示に準じて適宜実施され得る。
FACSCantoTM II
FACSAriaTM
FACSArrayTM
FACSVantageTM SE
FACSCaliburTM (いずれもBD Biosciences社の商品名)
EPICS ALTRA HyPerSort
Cytomics FC 500
EPICS XL-MCL ADC EPICS XL ADC
Cell Lab Quanta / Cell Lab Quanta SC(いずれもBeckman Coulter社の商品名)
本明細書において、「抗原結合ドメイン」は目的とする抗原に結合するかぎりどのような構造のドメインも使用され得る。そのようなドメインの例として、例えば、抗体の重鎖および軽鎖の可変領域、生体内に存在する細胞膜タンパクであるAvimerに含まれる35アミノ酸程度のAドメインと呼ばれるモジュール(国際公開WO2004/044011、WO2005/040229)、細胞膜に発現する糖たんぱく質であるfibronectin中のタンパク質に結合するドメインである10Fn3ドメインを含むAdnectin(国際公開WO2002/032925)、ProteinAの58アミノ酸からなる3つのヘリックスの束(bundle)を構成するIgG結合ドメインをscaffoldとするAffibody(国際公開WO1995/001937)、33アミノ酸残基を含むターンと2つの逆並行ヘリックスおよびループのサブユニットが繰り返し積み重なった構造を有するアンキリン反復(ankyrin repeat:AR)の分子表面に露出する領域であるDARPins(Designed Ankyrin Repeat proteins)(国際公開WO2002/020565)、好中球ゲラチナーゼ結合リポカリン(neutrophil gelatinase-associated lipocalin(NGAL))等のリポカリン分子において高度に保存された8つの逆並行ストランドが中央方向にねじれたバレル構造の片側を支える4つのループ領域であるAnticalin等(国際公開WO2003/029462)、ヤツメウナギ、ヌタウナギなど無顎類の獲得免疫システムとしてイムノグロブリンの構造を有さない可変性リンパ球受容体(variable lymphocyte receptor(VLR))のロイシン残基に富んだリピート(leucine-rich-repeat(LRR))モジュールが繰り返し積み重なった馬てい形の構造の内部の並行型シート構造のくぼんだ領域(国際公開WO2008/016854)が好適に挙げられる。本発明の抗原結合ドメインの好適な例として、抗体の重鎖および軽鎖の可変領域を含む抗原結合ドメインが挙げられる。こうした抗原結合ドメインの例としては、「scFv(single chain Fv)」、「単鎖抗体(single chain antibody)」、「Fv」、「scFv2(single chain Fv 2)」、「Fab」または「F(ab')2」等が好適に挙げられる。
本発明によって提供される抗原結合分子の抗原に対する結合に関して「特異的」とは、特異的に結合する分子の一方の分子がその一または複数の結合する相手方の分子以外の分子に対しては実質的に結合しない状態をいう。ここで、実質的に結合しないとは、前記の結合活性の項で記載されるように、当該相手方の分子に対する結合活性の80%以下、通常50%以下、好ましくは30%以下、特に好ましくは15%以下の結合活性を当該相手方の分子以外の分子に対して示すことをいう。また、抗原結合ドメインが、ある抗原中に含まれる複数のエピトープのうち特定のエピトープに対して特異的である場合にも用いられる。また、抗原結合ドメインが結合するエピトープが複数の異なる抗原に含まれる場合には、当該抗原結合ドメインを有する抗原結合分子は当該エピトープを含む様々な抗原と結合することができる。
本発明の非限定な一態様では、脳疾患に固有に脳組織内に存在する抗原に対する結合活性がイオン濃度の条件によって変化する抗原結合ドメインを含み、当該抗原に対する中和活性を有する抗原結合分子の当該脳疾患を治療するための使用が提供される。一般的に、中和活性とは、ウイルスや毒素など、細胞に対して生物学的活性を有するリガンドの当該生物学的活性を阻害する活性をいう。即ち、中和活性を有する物質とは、当該リガンド又は当該リガンドが結合するレセプターに結合し、当該リガンドとレセプターの結合を阻害する物質をさす。中和活性によりリガンドとの結合を阻止されたレセプターは、当該レセプターを通じた生物学的活性を発揮することができなくなる。抗原結合分子が抗体である場合、このような中和活性を有する抗体は一般に中和抗体と呼ばれる。ある被検物質の中和活性は、リガンドの存在下における生物学的活性をその被検物質の存在又は非存在下の条件の間で比較することにより測定され得る。
本発明の非限定な一態様では、脳疾患に固有に脳組織内に存在する抗原を発現する細胞に対する細胞傷害活性を有する抗原結合分子の当該脳疾患を治療するための使用が提供される。本発明において細胞傷害活性とは、例えば、抗体のFc領域に含まれるFcγレセプター(FcγR)結合ドメインに、免疫細胞に発現するFcγRを介して結合した免疫細胞が、当該抗原を発現する標的細胞に対して傷害を与える抗体依存性細胞介在性細胞傷害(antibody-dependent cell-mediated cytotoxicity:ADCC)活性、および抗体のFc領域に含まれる補体結合ドメインに結合した補体と免疫細胞に発現する補体受容体を介して結合した免疫細胞が、当該抗原または当該抗原を発現する標的細胞を貪食する補体依存性貪食(complement-dependent phagocytosis:CDP)活性等が挙げられる。脳組織において、こうした機能を発揮し得る免疫細胞として、インテグリン、FcγRIおよびFcγRIII等のFcγR、またはCR3等の補体受容体を発現するミクログリア細胞が挙げられる。本発明の抗原結合分子がミクログリアによるADCC活性、またはCDP活性を誘導するか否かは公知の方法により測定され得る(例えば、Pathobiology (1991) 59 (4), 254-258、またはJ. Neuroscience (2004) 24 (44), 9838-9846)。
本明細書において、抗体とは、天然のものであるかまたは部分的もしくは完全合成により製造された免疫グロブリンをいう。抗体はそれが天然に存在する血漿や血清等の天然資源や抗体を産生するハイブリドーマ細胞の培養上清から単離され得るし、または遺伝子組換え等の手法を用いることによって部分的にもしくは完全に合成され得る。抗体の例としては免疫グロブリンのアイソタイプおよびそれらのアイソタイプのサブクラスが好適に挙げられる。ヒトの免疫グロブリンとして、IgG1、IgG2、IgG3、IgG4、IgA1、IgA2、IgD、IgE、IgMの9種類のクラス(アイソタイプ)が知られている。本発明の抗体には、これらのアイソタイプのうちIgG1、IgG2、IgG3、IgG4が含まれ得る。ヒトIgG1、ヒトIgG2、ヒトIgG3、ヒトIgG4定常領域としては、遺伝子多型による複数のアロタイプ配列がSequences of proteins of immunological interest, NIH Publication No.91-3242 に記載されているが、本発明においてはそのいずれであっても良い。特にヒトIgG1の配列としては、EUナンバリングで表される356-358位のアミノ酸配列がDELであってもEEMであってもよい。また、ヒトIgκ(Kappa)定常領域とヒトIgλ (Lambda)定常領域としては、遺伝子多型による複数のアロタイプ配列がSequences of proteins of immunological interest, NIH Publication No.91-3242に記載されているが、本発明においてはそのいずれであっても良い。
−IL-6Rのような膜蛋白質の構造を維持して免疫刺激が与えられ得る
−免疫抗原を精製する必要が無い
−グアニジン超遠心法(Biochemistry (1979) 18 (24), 5294-5299)
−AGPC法(Anal. Biochem. (1987) 162 (1), 156-159)
(1)ハイブリドーマから得られたcDNAによってコードされるV領域を含む抗体をIL-6R発現細胞に接触させる工程、
(2)IL-6R発現細胞と抗体との結合を検出する工程、および
(3)IL-6R発現細胞に結合する抗体を選択する工程。
(1)哺乳類細胞、:CHO(Chinese hamster ovary cell line)、COS(Monkey kidney cell line)、ミエローマ(Sp2/O、NS0等)、BHK (baby hamster kidney cell line)、Hela、Vero、HEK293(human embryonic kidney cell line with sheared adenovirus (Ad)5 DNA)、Freestyle293、PER.C6 cell (human embryonic retinal cell line transformed with the Adenovirus Type 5 (Ad5) E1A and E1B genes)など(Current Protocols in Protein Science (May, 2001, Unit 5.9, Table 5.9.1))
(2)両生類細胞:アフリカツメガエル卵母細胞など
(3)昆虫細胞:sf9、sf21、Tn5など
−酵母:サッカロミセス・セレビシエ(Saccharomyces serevisiae)などのサッカロミセス(Saccharomyces )属、メタノール資化酵母(Pichia pastoris)などのPichia属
−糸状菌:アスペスギルス・ニガー(Aspergillus niger)などのアスペルギルス(Aspergillus )属
本発明で使用されている方法によると、抗体のCDRとFRに割り当てられるアミノ酸位置はKabatにしたがって規定される(Sequences of Proteins of Immunological Interest(National Institute of Health, Bethesda, Md., 1987年および1991年)。本明細書において、抗原結合分子が抗体または抗原結合断片である場合、可変領域のアミノ酸はKabatナンバリングにしたがい、定常領域のアミノ酸はKabatのアミノ酸位置に準じたEUナンバリングにしたがって表される。
金属イオン濃度の条件
本発明の一つの態様では、イオン濃度とは金属イオン濃度のことをいう。「金属イオン」とは、水素を除くアルカリ金属および銅族等の第I族、アルカリ土類金属および亜鉛族等の第II族、ホウ素を除く第III族、炭素とケイ素を除く第IV族、鉄族および白金族等の第VIII族、V、VIおよびVII族の各A亜族に属する元素と、アンチモン、ビスマス、ポロニウム等の金属元素のイオンをいう。金属原子は原子価電子を放出して陽イオンになる性質を有しており、これをイオン化傾向という。イオン化傾向の大きい金属は、化学的に活性に富むとされる。
ある一態様によれば、本発明の抗原結合ドメイン又は抗原結合分子は、イオン濃度の条件によって抗原に対する抗原結合分子の結合活性を変化させる少なくとも一つのアミノ酸残基が抗原結合ドメインに含まれている互いに配列の異なる複数の抗原結合分子から主としてなるライブラリから取得され得る。イオン濃度の例としては金属イオン濃度や水素イオン濃度が好適に挙げられる。
前記のスクリーニング方法によってスクリーニングされる本発明の抗原結合ドメインまたは抗原結合分子はどのように調製されてもよく、例えば、金属イオンがカルシウムイオン濃度である場合には、あらかじめ存在している抗原結合ドメインまたは抗原結合分子、あらかじめ存在しているライブラリ(ファージライブラリ等)、動物への免疫から得られたハイブリドーマや免疫動物からのB細胞から作製された抗体またはライブラリ、これらの抗体やライブラリにカルシウムをキレート可能なアミノ酸(例えばアスパラギン酸やグルタミン酸)や非天然アミノ酸変異を導入した抗体またはライブラリ(カルシウムをキレート可能なアミノ酸(例えばアスパラギン酸やグルタミン酸)または非天然アミノ酸の含有率を高くしたライブラリや特定箇所にカルシウムをキレート可能なアミノ酸(例えばアスパラギン酸やグルタミン酸)または非天然アミノ酸変異を導入したライブラリ等)などを用いることが可能である。

また、本発明の一つの態様では、イオン濃度の条件とは水素イオン濃度の条件またはpHの条件をいう。本発明で、プロトンすなわち水素原子の原子核の濃度の条件は、水素指数(pH)の条件とも同義に取り扱われる。水溶液中の水素イオンの活動量をaH+で表すと、pHは-log10aH+と定義される。水溶液中のイオン強度が(例えば10-3より)低ければ、aH+は水素イオン強度にほぼ等しい。例えば25℃、1気圧における水のイオン積はKw=aH+aOH=10-14であるため、純水ではaH+=aOH=10-7である。この場合のpH=7が中性であり、pHが7より小さい水溶液は酸性、pHが7より大きい水溶液はアルカリ性である。
(a) pH酸性域の条件における抗原結合ドメインまたは抗原結合分子の抗原結合活性を得る工程、
(b) pH中性域の条件における抗原結合ドメインまたは抗原結合分子の抗原結合活性を得る工程、
(c) pH酸性域の条件における抗原結合活性が、pH中性域の条件における抗原結合活性より低い抗原結合ドメインまたは抗原結合分子を選択する工程。
(a) pH中性域の条件における抗原結合ドメインまたは抗原結合分子もしくはそれらのライブラリを抗原に接触させる工程、
(b) 前記工程(a)で抗原に結合した抗原結合ドメインまたは抗原結合分子をpH酸性域の条件に置く工程、
(c) 前記工程(b)で解離した抗原結合ドメインまたは抗原結合分子を単離する工程。
(a) pH酸性域の条件で抗原結合ドメイン又は抗原結合分子のライブラリを抗原に接触させる工程、
(b) 前記工程(a)で抗原に結合しない抗原結合ドメイン又は抗原結合分子を選択する工程、
(c) 前記工程(b)で選択された抗原結合ドメイン又は抗原結合分子をpH中性域の条件で抗原に結合させる工程、
(d) 前記工程(c)で抗原に結合した抗原結合ドメイン又は抗原結合分子を単離する工程。
(a) 抗原を固定したカラムにpH中性域の条件で抗原結合ドメイン又は抗原結合分子のライブラリを接触させる工程、
(b) 前記工程(a)でカラムに結合した抗原結合ドメイン又は抗原結合分子をpH酸性域の条件でカラムから溶出する工程、
(c) 前記工程(b)で溶出された抗原結合ドメイン又は抗原結合分子を単離する工程。
(a) 抗原を固定したカラムにpH酸性域の条件で抗原結合ドメイン又は抗原結合分子のライブラリを通過させる工程、
(b) 前記工程(a)でカラムに結合せずに溶出した抗原結合ドメイン又は抗原結合分子を回収する工程、
(c) 前記工程(b)で回収された抗原結合ドメイン又は抗原結合分子をpH中性域の条件で抗原に結合させる工程、
(d) 前記工程(c)で抗原に結合した抗原結合ドメイン又は抗原結合分子を単離する工程。
(a) pH中性域の条件で抗原結合ドメイン又は抗原結合分子のライブラリを抗原に接触させる工程、
(b) 前記工程(a)で抗原に結合した抗原結合ドメイン又は抗原結合分子を取得する工程、
(c) 前記工程(b)で取得した抗原結合ドメイン又は抗原結合分子をpH酸性域の条件に置く工程、
(d) 前記工程(c)で抗原結合活性が、前記工程(b)で選択した基準より弱い抗原結合ドメイン又は抗原結合分子を単離する工程。
前記のスクリーニング方法によってスクリーニングされる本発明の抗原結合ドメイン又は抗原結合分子はどのように調製されてもよく、例えば、あらかじめ存在している抗原結合分子、あらかじめ存在しているライブラリ(ファージライブラリ等)、動物への免疫から得られたハイブリドーマや免疫動物からのB細胞から作製された抗体又はライブラリ、これらの抗体やライブラリに側鎖のpKaが4.0-8.0であるアミノ酸(例えばヒスチジンやグルタミン酸)や非天然アミノ酸変異を導入した抗体又はライブラリ(側鎖のpKaが4.0-8.0であるアミノ酸(例えばヒスチジンやグルタミン酸)又は非天然アミノ酸の含有率を高くしたライブラリや特定箇所に側鎖のpKaが4.0-8.0であるアミノ酸(例えばヒスチジンやグルタミン酸)又は非天然アミノ酸変異を導入したライブラリ等)などを用いることが可能である。
前記のスクリーニング方法によってスクリーニングされる本発明の抗原結合ドメインまたは抗原結合分子はどのように調製されてもよく、例えば、イオン濃度の条件が水素イオン濃度の条件もしくはpHの条件である場合には、あらかじめ存在している抗原結合ドメインまたは抗原結合分子、あらかじめ存在しているライブラリ(ファージライブラリ等)、動物への免疫から得られたハイブリドーマや免疫動物からのB細胞から作製された抗体またはライブラリ、これらの抗体やライブラリに側鎖のpKaが4.0-8.0であるアミノ酸(例えばヒスチジンやグルタミン酸)や非天然アミノ酸の変異を導入した抗体またはライブラリ(側鎖のpKaが4.0-8.0であるアミノ酸(例えばヒスチジンやグルタミン酸)や非天然アミノ酸の含有率を高くしたライブラリや特定箇所に側鎖のpKaが4.0-8.0であるアミノ酸(例えばヒスチジンやグルタミン酸)や非天然アミノ酸の変異を導入したライブラリ等)などを用いることが可能である 。
免疫グロブリンスーパーファミリーに属するFcγレセプターと異なり、FcRn特にヒトFcRnは構造的には主要組織不適合性複合体(MHC)クラスIのポリペプチドに構造的に類似しクラスIのMHC分子と22から29%の配列同一性を有する(Ghetieら,Immunol. Today (1997) 18 (12), 592-598)。FcRnは、可溶性βまたは軽鎖(β2マイクログロブリン)と複合体化された膜貫通αまたは重鎖よりなるヘテロダイマーとして発現される。MHCのように、FcRnのα鎖は3つの細胞外ドメイン(α1、α2、α3)よりなり、短い細胞質ドメインはタンパク質を細胞表面に繋留する。α1およびα2ドメインが抗体のFc領域中のFcRn結合ドメインと相互作用する(Raghavanら(Immunity (1994) 1, 303-315)。
本発明により提供される抗原結合分子に含まれるFcRn結合ドメインのFcRn、特にヒトFcRnに対する結合活性は、前記結合活性の項で述べられているように、当業者に公知の方法により測定することが可能であり、pH以外の条件については当業者が適宜決定することが可能である。抗原結合ドメインまたは当該ドメイン含む抗原結合分子とヒトFcRnとの結合活性は、KD(Dissociation constant:解離定数)、見かけのKD(Apparent dissociation constant:見かけの解離定数)、解離速度であるkd(Dissociation rate:解離速度)、又は見かけのkd(Apparent dissociation:見かけの解離速度)等として評価され得る。これらは当業者公知の方法で測定され得る。例えばBiacore(GE healthcare)、スキャッチャードプロット、フローサイトメーター等が使用され得る。
本発明の抗原結合分子は、pH酸性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインを含む。FcRn結合ドメインは、抗原結合分子がpH酸性域においてFcRnに対する結合活性を有していれば特に限定されず、また、直接または間接的にFcRnに対して結合活性を有するドメインであってもよい。そのようなドメインとしては、例えば、直接的にFcRnに対する結合活性を有するFcRn結合ドメインとして、例えばIgG型免疫グロブリンのFc領域、アルブミン、アルブミンドメイン3、Christiansonら(mAbs (2012) 4 (2), 208-216)等で記載される抗FcRn抗体、WO2007/098420等で記載される抗FcRnペプチド、抗FcRn足場(Scaffold)分子等、あるいは間接的にFcRnに対する結合活性を有するIgGやアルブミンに結合する分子等が好適に挙げられる。当該ドメインは、あらかじめpH酸性域においてFcRnに対する結合活性を有しているドメインであればそのまま好適に使用され得る。当該ドメインがpH酸性域においてFcRnに対する結合活性がないまたは弱い場合には、抗原結合分子中のFcRn結合ドメインを形成するアミノ酸を改変してFcRnに対する結合活性を付与することが可能である。また、あらかじめpH酸性域においてFcRnに対する結合活性を有しているドメイン中のアミノ酸を改変して、pH酸性域におけるFcRnに対する結合活性を高めてもよい。FcRn結合ドメインのアミノ酸の改変は、アミノ酸改変前と改変後のpH酸性域におけるFcRnに対する結合活性を比較することによって目的の改変を見出すことが可能である。
Fc領域は、抗体重鎖の定常領域に由来するアミノ酸配列を含む。Fc領域は、EUナンバリングで表されるおよそ216位のアミノ酸における、パパイン切断部位のヒンジ領域のN末端から、当該ヒンジ、CH2およびCH3ドメインを含める抗体の重鎖定常領域の部分である。Fc領域は、ヒトIgG1から取得され得るが、IgGの特定のサブクラスに限定されるものでもない。そのようなFc領域の非限定な一態様として、ヒトIgG1(配列番号:14)、IgG2(配列番号:15)、IgG3(配列番号:16)、またはIgG4(配列番号:17)で表されるFc領域が例示される。
前記のように、本発明の非限定な一態様においては、pH酸性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインとしてヒトIgG型免疫グロブリンのFc領域が使用される。当該ドメインとして、あらかじめpH酸性域の条件下でFcRnに対する結合活性を有しているFc領域であればそのまま用いられ得るが、そのようなFc領域として、例えばヒトIgG(IgG1、IgG2、IgG3、またはIgG4、およびそれらの改変体)のFc領域が挙げられる。また、Fc領域のpH酸性域の条件下におけるFcRnに対する結合活性がない若しくは弱い場合には、当該Fc領域中のアミノ酸を改変することによって所望のFcRnに対する結合活性を有するFc領域が取得され得る。また、Fc領域中のアミノ酸を改変することによってpH酸性域の条件下でFcRnに対する所望の結合活性を有する、または増強されたFc領域も好適に取得され得る。そのような所望の結合活性をもたらすFc領域のアミノ酸改変は、アミノ酸改変前と改変後のpH酸性域の条件下でFcRnに対する結合活性を比較することによって見出され得る。例えば、前記「FcRn、特にヒトFcRnに対するFcRn結合ドメインまたは当該ドメインを含む抗原結合分子の結合活性」の項で記載された方法によって、そのようなアミノ酸改変が見出され得る。
251位のアミノ酸がArgまたはLeuのいずれか、
252位のアミノ酸がPhe、Ser、Thr、またはTyrのいずれか、
254位のアミノ酸がSerまたはThrのいずれか、
255位のアミノ酸がArg、Gly、Ile、またはLeuのいずれか、
256位のアミノ酸がAla、Arg、Asn、Asp、Gln、Glu、またはThrのいずれか、
308位のアミノ酸がIleまたはThrのいずれか、
309位のアミノ酸がPro、
311位のアミノ酸がGlu、Leu、またはSerのいずれか、
312位のアミノ酸がAlaまたはAspのいずれか、
314位のアミノ酸がAlaまたはLeuのいずれか、
385位のアミノ酸がAla、Arg、Asp、Gly、His、Lys、Ser、またはThrのいずれか、
386位のアミノ酸がArg、Asp、Ile、Lys、Met、Pro、Ser、またはThrのいずれか、
387位のアミノ酸がAla、Arg、His、Pro、Ser、またはThrのいずれか、
389位のアミノ酸がAsn、Pro、またはSerのいずれか、
428位のアミノ酸がLeu、Met、Phe、Ser、またはThrのいずれか
433位のアミノ酸がArg、Gln、His、Ile、Lys、Pro、またはSerのいずれか、
434位のアミノ酸がHis、Phe、またはTyrのいずれか、もしくは
436位のアミノ酸がArg、Asn、His、Lys、Met、またはThrのいずれか、
の群から選択される少なくとも一つ以上のアミノ酸の改変が挙げられる。また、改変されるアミノ酸の数は特に限定されず、一箇所のみのアミノ酸が改変され得るし、二箇所以上のアミノ酸が改変され得る。
250位のアミノ酸がGlnまたはGluのいずれか、もしくは
428位のアミノ酸がLeuまたはPheのいずれか、
の群から選択される少なくとも一つ以上のアミノ酸の改変が挙げられる。また、改変されるアミノ酸の数は特に限定されず、一箇所のみのアミノ酸が改変され得るし、二箇所のアミノ酸が改変され得る。
251位のアミノ酸がAspまたはGluのいずれか、
252位のアミノ酸がTyr、
307位のアミノ酸がGln、
308位のアミノ酸がPro、
378位のアミノ酸がVal、
380位のアミノ酸がAla、
428位のアミノ酸がLeu、
430位のアミノ酸がAla、またはLysのいずれか、
434位のアミノ酸がAla、His、Ser、またはTyrのいずれか、もしくは
436位のアミノ酸がIle、
の群から選択される少なくとも二つ以上のアミノ酸の改変が挙げられる。また、改変されるアミノ酸の数は特に限定されず、二箇所のみのアミノ酸が改変され得るし、三箇所以上のアミノ酸が改変され得る。
238位のアミノ酸がLeu、
244位のアミノ酸がLeu、
245位のアミノ酸がArg、
249位のアミノ酸がPro、
252位のアミノ酸がTyr、
256位のアミノ酸がPro、
257位のアミノ酸がAla、Ile、Met、Asn、Ser、またはValのいずれか、
258位のアミノ酸がAsp、
260位のアミノ酸がSer、
262位のアミノ酸がLeu、
270位のアミノ酸がLys、
272位のアミノ酸がLeu、またはArgのいずれか、
279位のアミノ酸がAla、Asp、Gly、His、Met、Asn、Gln、Arg、Ser、Thr、Trp、またはTyrのいずれか、
283位のアミノ酸がAla、Asp、Phe、Gly、His、Ile、Lys、Leu、Asn、Pro、Gln、Arg、Ser、Thr、Trp、またはTyrのいずれか、
285位のアミノ酸がAsn、
286位のアミノ酸がPhe、
288位のアミノ酸がAsn、またはProのいずれか、
293位のアミノ酸がVal、
307位のアミノ酸がAla、Glu、またはMetのいずれか、
311位のアミノ酸がAla、Ile、Lys、Leu、Met、Val、またはTrpのいずれか、
312位のアミノ酸がPro、
316位のアミノ酸がLys、
317位のアミノ酸がPro、
318位のアミノ酸がAsn、またはThrのいずれか、
332位のアミノ酸がPhe、His、Lys、Leu、Met、Arg、Ser、またはTrpのいずれか、
339位のアミノ酸がAsn、Thr、またはTrpのいずれか、
341位のアミノ酸がPro、
343位のアミノ酸がGlu、His、Lys、Gln、Arg、Thr、またはTyrのいずれか、
375位のアミノ酸がArg、
376位のアミノ酸がGly、Ile、Met、Pro、Thr、またはValのいずれか、
377位のアミノ酸がLys、
378位のアミノ酸がAsp、またはAsnのいずれか、
380位のアミノ酸がAsn、Ser、またはThrのいずれか、
382位のアミノ酸がPhe、His、Ile、Lys、Leu、Met、Asn、Gln、Arg、Ser、Thr、Val、Trp、またはTyrのいずれか、
423位のアミノ酸がAsn、
427位のアミノ酸がAsn、
430位のアミノ酸がAla、Phe、Gly、His、Ile、Lys、Leu、Met、Asn、Gln、Arg、Ser、Thr、Val、またはTyrのいずれか、
431位のアミノ酸がHis、またはAsnのいずれか、
434位のアミノ酸がPhe、Gly、His、Trp、またはTyrのいずれか、
436位のアミノ酸がIle、Leu、、またはThrのいずれか、
438位のアミノ酸がLys、Leu、Thr、またはTrpのいずれか、
440位のアミノ酸がLys、もしくは、
442位のアミノ酸がLys、
の群から選択される少なくとも二つ以上のアミノ酸の改変が挙げられる。また、改変されるアミノ酸の数は特に限定されず、二箇所のみのアミノ酸が改変され得るし、三箇所以上のアミノ酸が改変され得る。
本発明の抗原結合分子は、pH中性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインを含む。FcRn結合ドメインは、抗原結合分子がpH中性域においてFcRnに対する結合活性を有していれば特に限定されず、また、直接または間接的にFcRnに対して結合活性を有するドメインであってもよい。そのようなドメインとしては、例えば、直接的にFcRnに対する結合活性を有するFcRn結合ドメインとして、例えばWO2011/122011等で記載されるIgG型免疫グロブリンのFc領域、Christiansonら(mAbs (2012) 4 (2), 208-216)等で記載される抗FcRn抗体、WO2007/098420等で記載される抗FcRnペプチド、抗FcRn足場(Scaffold)分子等が好適に挙げられる。当該ドメインは、あらかじめpH中性域においてFcRnに対する結合活性を有しているドメインであればそのまま好適に使用され得る。当該ドメインがpH中性域においてFcRnに対する結合活性がないまたは弱い場合には、抗原結合分子中のFcRn結合ドメインを形成するアミノ酸を改変してFcRnに対する結合活性を付与することが可能である。また、あらかじめpH中性域においてFcRnに対する結合活性を有しているドメイン中のアミノ酸を改変して、pH中性域におけるFcRnに対する結合活性を高めてもよい。FcRn結合ドメインのアミノ酸の改変は、アミノ酸改変前と改変後のpH中性域におけるFcRnに対する結合活性を比較することによって目的の改変を見出すことが可能である。
237位のアミノ酸がMet、
248位のアミノ酸がIle、
250位のアミノ酸がAla、Phe、Ile、Met、Gln、Ser、Val、Trp、またはTyrのいずれか、
252位のアミノ酸がPhe、Trp、またはTyrのいずれか、
254位のアミノ酸がThr、
255位のアミノ酸がGlu、
256位のアミノ酸がAsp、Asn、Glu、またはGlnのいずれか、
257位のアミノ酸がAla、Gly、Ile、Leu、Met、Asn、Ser、Thr、またはValのいずれか、
258位のアミノ酸がHis、
265位のアミノ酸がAla、
286位のアミノ酸がAlaまたはGluのいずれか、
289位のアミノ酸がHis、
297位のアミノ酸がAla、
303位のアミノ酸がAla、
305位のアミノ酸がAla、
307位のアミノ酸がAla、Asp、Phe、Gly、His、Ile、Lys、Leu、Met、Asn、Pro、Gln、Arg、Ser、Val、Trp、またはTyrのいずれか、
308位のアミノ酸がAla、Phe、Ile、Leu、Met、Pro、Gln、またはThrのいずれか、
309位のアミノ酸がAla、Asp、Glu、Pro、またはArgのいずれか、
311位のアミノ酸がAla、His、またはIleのいずれか、
312位のアミノ酸がAlaまたはHisのいずれか、
314位のアミノ酸がLysまたはArgのいずれか、
315位のアミノ酸がAla、AspまたはHisのいずれか、
317位のアミノ酸がAla、
332位のアミノ酸がVal、
334位のアミノ酸がLeu、
360位のアミノ酸がHis、
376位のアミノ酸がAla、
380位のアミノ酸がAla、
382位のアミノ酸がAla、
384位のアミノ酸がAla、
385位のアミノ酸がAspまたはHisのいずれか、
386位のアミノ酸がPro、
387位のアミノ酸がGlu、
389位のアミノ酸がAlaまたはSerのいずれか、
424位のアミノ酸がAla、
428位のアミノ酸がAla、Asp、Phe、Gly、His、Ile、Lys、Leu、Asn、Pro、Gln、Ser、Thr、Val、Trp、またはTyrのいずれか、
433位のアミノ酸がLys、
434位のアミノ酸がAla、Phe、His、Ser、Trp、またはTyrのいずれか、もしくは
436位のアミノ酸がHis 、Ile、Leu、Phe、Thr、またはVal、
が挙げられる。また、改変されるアミノ酸の数は特に限定されず、一箇所のみのアミノ酸が改変され得るし、二箇所以上のアミノ酸が改変され得る。これらのアミノ酸の改変の組合せとしては、例えば表5−1〜5−33に示すアミノ酸の改変が挙げられる。
Fcγレセプターとは、IgG1、IgG2、IgG3、IgG4モノクローナル抗体のFc領域に結合し得るレセプターをいい、実質的にFcγレセプター遺伝子にコードされるタンパク質のファミリーのいかなるメンバーをも意味する。ヒトでは、このファミリーには、アイソフォームFcγRIa、FcγRIbおよびFcγRIcを含むFcγRI(CD64);アイソフォームFcγRIIa(アロタイプH131およびR131を含む)、FcγRIIb(FcγRIIb-1およびFcγRIIb-2を含む)およびFcγRIIcを含むFcγRII(CD32);およびアイソフォームFcγRIIIa(アロタイプV158およびF158を含む)およびFcγRIIIb(アロタイプFcγRIIIb-NA1およびFcγRIIIb-NA2を含む)を含むFcγRIII(CD16)、並びにいかなる未発見のヒトFcγR類またはFcγRアイソフォームまたはアロタイプも含まれるが、これらに限定されるものではない。FcγRは、ヒト、マウス、ラット、ウサギおよびサルを含むが、これらに限定されるものではない、いかなる生物由来でもよい。マウスFcγR類には、FcγRI(CD64)、FcγRII(CD32)、FcγRIII(CD16)およびFcγRIII-2(FcγRIV、CD16-2)、並びにいかなる未発見のマウスFcγR類またはFcγRアイソフォームまたはアロタイプも含まれるが、これらに限定されない。こうしたFcγレセプターの好適な例としてはヒトFcγRI(CD64)、FcγRIIa(CD32)、FcγRIIb(CD32)、FcγRIIIa(CD16)及び/又はFcγRIIIb(CD16)が挙げられる。FcγRIのポリヌクレオチド配列及びアミノ酸配列はそれぞれ配列番号:18(NM_000566.3)及び19(NP_000557.1)に、FcγRIIaのポリヌクレオチド配列及びアミノ酸配列はそれぞれ配列番号:20(BC020823.1)及び21(AAH20823.1)に、FcγRIIbのポリヌクレオチド配列及びアミノ酸配列はそれぞれ配列番号:22(BC146678.1)及び23(AAI46679.1)に、FcγRIIIaのポリヌクレオチド配列及びアミノ酸配列はそれぞれ配列番号:24(BC033678.1)及び25(AAH33678.1)に、及びFcγRIIIbのポリヌクレオチド配列及びアミノ酸配列は、それぞれ配列番号:26(BC128562.1)及び27(AAI28563.1)に記載されている(カッコ内はRefSeq登録番号を示す)。Fcγ受容体が、IgG1、IgG2、IgG3、IgG4モノクローナル抗体のFc領域に結合活性を有するか否かは、上記に記載されるFACSやELISAフォーマットのほか、ALPHAスクリーン(Amplified Luminescent Proximity Homogeneous Assay)や表面プラズモン共鳴(SPR)現象を利用したBIACORE法等によって確認され得る(Proc.Natl.Acad.Sci.USA (2006) 103 (11), 4005-4010)。
本発明の抗原結合分子に含まれるFcγR結合ドメインのFcγRI、FcγRIIa、FcγRIIb、FcγRIIIa及び/又はFcγRIIIbのいずれかのヒトFcγレセプターに対する結合活性は、上記に記載されるFACSやELISAフォーマットのほか、ALPHAスクリーン(Amplified Luminescent Proximity Homogeneous Assay)や表面プラズモン共鳴(SPR)現象を利用したBIACORE法等によって確認することができる(Proc.Natl.Acad.Sci.USA (2006) 103 (11), 4005-4010)。
本発明の抗原結合分子は、脳疾患に固有に脳組織内に存在する抗原に対する結合活性がイオン濃度の条件によって変化する抗原結合ドメイン、ならびにpH中性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインおよびpH酸性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインに加えてFcγR結合ドメインを含むことも可能である。FcγR結合ドメインとしては、例えば、IgG型免疫グロブリンのFc領域、抗FcγR抗体、抗FcγR足場(Scaffold)分子等が好適に挙げられる。当該ドメインは、あらかじめFcγRに対する結合活性を有しているドメインであればそのまま好適に使用され得る。当該ドメインがFcγRに対する結合活性がないまたは弱い場合には、抗原結合分子中のFcγR結合ドメインを形成するアミノ酸を改変してFcγRに対する結合活性を付与することが可能である。また、あらかじめFcγRに対する結合活性を有しているドメイン中のアミノ酸を改変して、FcγRに対する結合活性を高めてもよい。FcγR結合ドメインのアミノ酸の改変は、アミノ酸改変前と改変後のFcγRに対する結合活性を比較することによって目的の改変を見出すことが可能である。例えば、pH中性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインおよびpH酸性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインとしてIgG型抗体のFc領域が使用される場合には、FcγR結合ドメインとして当該Fc領域に含まれるFcγR結合ドメインが使用され得る。
221位のアミノ酸がLysまたはTyrのいずれか、
222位のアミノ酸がPhe、Trp、GluまたはTyrのいずれか、
223位のアミノ酸がPhe、Trp、GluまたはLysのいずれか、
224位のアミノ酸がPhe、Trp、GluまたはTyrのいずれか、
225位のアミノ酸がGlu、LysまたはTrpのいずれか、
227位のアミノ酸がGlu、Gly、LysまたはTyrのいずれか、
228位のアミノ酸がGlu、Gly、LysまたはTyrのいずれか、
230位のアミノ酸がAla、Glu、GlyまたはTyrのいずれか、
231位のアミノ酸がGlu、Gly、Lys、ProまたはTyrのいずれか、
232位のアミノ酸がGlu、Gly、LysまたはTyrのいずれか、
233位のアミノ酸がAla、Asp、Phe、Gly、His、Ile、Lys、Leu、Met、Asn、Gln、Arg、Ser、Thr、Val、TrpまたはTyrのいずれか、
234位のアミノ酸がAla、Asp、Glu、Phe、Gly、His、Ile、Lys、Met、Asn、Pro、Gln、Arg、Ser、Thr、Val、TrpまたはTyrのいずれか、
235位のアミノ酸がAla、Asp、Glu、Phe、Gly、His、Ile、Lys、Met、Asn、Pro、Gln、Arg、Ser、Thr、Val、TrpまたはTyrのいずれか、
236位のアミノ酸がAla、Asp、Glu、Phe、His、Ile、Lys、Leu、Met、Asn、Pro、Gln、Arg、Ser、Thr、Val、TrpまたはTyrのいずれか、
237位のアミノ酸がAsp、Glu、Phe、His、Ile、Lys、Leu、Met、Asn、Pro、Gln、Arg、Ser、Thr、Val、TrpまたはTyrのいずれか、
238位のアミノ酸がAsp、Glu、Phe、Gly、His、Ile、Lys、Leu、Met、Asn、Gln、Arg、Ser、Thr、Val、TrpまたはTyrのいずれか、
239位のアミノ酸がAsp、Glu、Phe、Gly、His、Ile、Lys、Leu、Met、Asn、Pro、Gln、Arg、Thr、Val、TrpまたはTyrのいずれか、
240位のアミノ酸がAla、Ile、MetまたはThrのいずれか、
241位のアミノ酸がAsp、Glu、Leu、Arg、TrpまたはTyrのいずれか、
243位のアミノ酸がLeu、Glu、Leu、Gln、Arg、TrpまたはTyrのいずれか、
244位のアミノ酸がHis、
245位のアミノ酸がAla、
246位のアミノ酸がAsp、Glu、HisまたはTyrのいずれか、
247位のアミノ酸がAla、Phe、Gly、His、Ile、Leu、Met、Thr、ValまたはTyrのいずれか、
249位のアミノ酸がGlu、His、GlnまたはTyrのいずれか、
250位のアミノ酸がGluまたはGlnのいずれか、
251位のアミノ酸がPhe、
254位のアミノ酸がPhe、MetまたはTyrのいずれか、
255位のアミノ酸がGlu、LeuまたはTyrのいずれか、
256位のアミノ酸がAla、MetまたはProのいずれか、
258位のアミノ酸がAsp、Glu、His、SerまたはTyrのいずれか、
260位のアミノ酸がAsp、Glu、HisまたはTyrのいずれか、
262位のアミノ酸がAla、Glu、Phe、IleまたはThrのいずれか、
263位のアミノ酸がAla、Ile、MetまたはThrのいずれか、
264位のアミノ酸がAsp、Glu、Phe、Gly、His、Ile、Lys、Leu、Met、Asn、Pro、Gln、Arg、Ser、Thr、TrpまたはTyrのいずれか、
265位のアミノ酸がAla、Leu、Phe、Gly、His、Ile、Lys、Leu、Met、Asn、Pro、Gln、Arg、Ser、Thr、Val、Val、TrpまたはTyrのいずれか、
266位のアミノ酸がAla、Ile、MetまたはThrのいずれか、
267位のアミノ酸がAsp、Glu、Phe、His、Ile、Lys、Leu、Met、Asn、Pro、Gln、Arg、Thr、Val、TrpまたはTyrのいずれか、
268位のアミノ酸がAsp、Glu、Phe、Gly、Ile、Lys、Leu、Met、Pro、Gln、Arg、Thr、ValまたはTrpのいずれか、
269位のアミノ酸がPhe、Gly、His、Ile、Lys、Leu、Met、Asn、Pro、Arg、Ser、Thr、Val、TrpまたはTyrのいずれか、
270位のアミノ酸がGlu、Phe、Gly、His、Ile、Leu、Met、Pro、Gln、Arg、Ser、Thr、TrpまたはTyrのいずれか、
271位のアミノ酸がAla、Asp、Glu、Phe、Gly、His、Ile、Lys、Leu、Met、Asn、Gln、Arg、Ser、Thr、Val、TrpまたはTyrのいずれか、
272位のアミノ酸がAsp、Phe、Gly、His、Ile、Lys、Leu、Met、Pro、Arg、Ser、Thr、Val、TrpまたはTyrのいずれか、
273位のアミノ酸がPheまたはIleのいずれか、
274位のアミノ酸がAsp、Glu、Phe、Gly、His、Ile、Leu、Met、Asn、Pro、Arg、Ser、Thr、Val、TrpまたはTyrのいずれか、
275位のアミノ酸がLeuまたはTrpのいずれか、
276位のアミノ酸が、Asp、Glu、Phe、Gly、His、Ile、Leu、Met、Pro、Arg、Ser、Thr、Val、TrpまたはTyrのいずれか、
278位のアミノ酸がAsp、Glu、Gly、His、Ile、Lys、Leu、Met、Asn、Pro、Gln、Arg、Ser、Thr、ValまたはTrpのいずれか、
279位のアミノ酸がAla、
280位のアミノ酸がAla、Gly、His、Lys、Leu、Pro、Gln、TrpまたはTyrのいずれか、
281位のアミノ酸がAsp、Lys、ProまたはTyrのいずれか、
282位のアミノ酸がGlu、Gly、Lys、ProまたはTyrのいずれか、
283位のアミノ酸がAla、Gly、His、Ile、Lys、Leu、Met、Pro、ArgまたはTyrのいずれか、
284位のアミノ酸がAsp、Glu、Leu、Asn、ThrまたはTyrのいずれか、
285位のアミノ酸がAsp、Glu、Lys、Gln、TrpまたはTyrのいずれか、
286位のアミノ酸がGlu、Gly、ProまたはTyrのいずれか、
288位のアミノ酸がAsn、Asp、GluまたはTyrのいずれか、
290位のアミノ酸がAsp、Gly、His、Leu、Asn、Ser、Thr、TrpまたはTyrのいずれか、291位のアミノ酸がAsp、Glu、Gly、His、Ile、GlnまたはThrのいずれか、
292位のアミノ酸がAla、Asp、Glu、Pro、ThrまたはTyrのいずれか、
293位のアミノ酸がPhe、Gly、His、Ile、Leu、Met、Asn、Pro、Arg、Ser、Thr、Val、TrpまたはTyrのいずれか、
294位のアミノ酸がPhe、Gly、His、Ile、Lys、Leu、Met、Asn、Pro、Arg、Ser、Thr、Val、TrpまたはTyrのいずれか、
295位のアミノ酸がAsp、Glu、Phe、Gly、His、Ile、Lys、Met、Asn、Pro、Arg、Ser、Thr、Val、TrpまたはTyrのいずれか、
296位のアミノ酸がAla、Asp、Glu、Gly、His、Ile、Lys、Leu、Met、Asn、Gln、Arg、Ser、ThrまたはValのいずれか、
297位のアミノ酸がAsp、Glu、Phe、Gly、His、Ile、Lys、Leu、Met、Pro、Gln、Arg、Ser、Thr、Val、TrpまたはTyrのいずれか、
298位のアミノ酸がAla、Asp、Glu、Phe、His、Ile、Lys、Met、Asn、Gln、Arg、Thr、Val、TrpまたはTyrのいずれか、
299位のアミノ酸がAla、Asp、Glu、Phe、Gly、His、Ile、Lys、Leu、Met、Asn、Pro、Gln、Arg、Ser、Val、TrpまたはTyrのいずれか、
300位のアミノ酸がAla、Asp、Glu、Gly、His、Ile、Lys、Leu、Met、Asn、Pro、Gln、Arg、Ser、Thr、ValまたはTrpのいずれか、
301位のアミノ酸がAsp、Glu、HisまたはTyrのいずれか、
302位のアミノ酸がIle、
303位のアミノ酸がAsp、GlyまたはTyrのいずれか、
304位のアミノ酸がAsp、His、Leu、AsnまたはThrのいずれか、
305位のアミノ酸がGlu、Ile、ThrまたはTyrのいずれか、
311位のアミノ酸がAla、Asp、Asn、Thr、ValまたはTyrのいずれか、
313位のアミノ酸がPhe、
315位のアミノ酸がLeu、
317位のアミノ酸がGluまたはGln、
318位のアミノ酸がHis、Leu、Asn、Pro、Gln、Arg、Thr、ValまたはTyrのいずれか、
320位のアミノ酸がAsp、Phe、Gly、His、Ile、Leu、Asn、Pro、Ser、Thr、Val、TrpまたはTyrのいずれか、
322位のアミノ酸がAla、Asp、Phe、Gly、His、Ile、Pro、Ser、Thr、Val、TrpまたはTyrのいずれか、
323位のアミノ酸がIle、
324位のアミノ酸がAsp、Phe、Gly、His、Ile、Leu、Met、Pro、Arg、Thr、Val、TrpまたはTyrのいずれか、
325位のアミノ酸がAla、Asp、Glu、Phe、Gly、His、Ile、Lys、Leu、Met、Pro、Gln、Arg、Ser、Thr、Val、TrpまたはTyrのいずれか、
326位のアミノ酸がAla、Asp、Glu、Gly、Ile、Leu、Met、Asn、Pro、Gln、Ser、Thr、Val、TrpまたはTyrのいずれか、
327位のアミノ酸がAla、Asp、Glu、Phe、Gly、His、Ile、Lys、Leu、Met、Asn、Pro、Arg、Thr、Val、TrpまたはTyrのいずれか、
328位のアミノ酸がAla、Asp、Glu、Phe、Gly、His、Ile、Lys、Met、Asn、Pro、Gln、Arg、Ser、Thr、Val、TrpまたはTyrのいずれか、
329位のアミノ酸がAsp、Glu、Phe、Gly、His、Ile、Lys、Leu、Met、Asn、Gln、Arg、Ser、Thr、Val、TrpまたはTyrのいずれか、
330位のアミノ酸がCys、Glu、Phe、Gly、His、Ile、Lys、Leu、Met、Asn、Pro、Arg、Ser、Thr、Val、TrpまたはTyrのいずれか、
331位のアミノ酸がAsp、Phe、His、Ile、Leu、Met、Gln、Arg、Thr、Val、TrpまたはTyrのいずれか、
332位のアミノ酸がAla、Asp、Glu、Phe、Gly、His、Lys、Leu、Met、Asn、Pro、Gln、Arg、Ser、Thr、Val、TrpまたはTyrのいずれか、
333位のアミノ酸がAla、Asp、Glu、Phe、Gly、His、Ile、Leu、Met、Pro、Ser、Thr、ValまたはTyrのいずれか、
334位のアミノ酸がAla、Glu、Phe、Ile、Leu、ProまたはThrのいずれか、
335位のアミノ酸がAsp、Phe、Gly、His、Ile、Leu、Met、Asn、Pro、Arg、Ser、Val、TrpまたはTyrのいずれか、
336位のアミノ酸がGlu、LysまたはTyrのいずれか、
337位のアミノ酸がGlu、HisまたはAsnのいずれか、
339位のアミノ酸がAsp、Phe、Gly、Ile、Lys、Met、Asn、Gln、Arg、SerまたはThrのいずれか、
376位のアミノ酸がAlaまたはValのいずれか、
377位のアミノ酸がGlyまたはLysのいずれか、
378位のアミノ酸がAsp、
379位のアミノ酸がAsn、
380位のアミノ酸がAla、AsnまたはSerのいずれか、
382位のアミノ酸がAlaまたはIleのいずれか、
385位のアミノ酸がGlu、
392位のアミノ酸がThr、
396位のアミノ酸がLeu、
421位のアミノ酸がLys、
427位のアミノ酸がAsn、
428位のアミノ酸がPheまたはLeuのいずれか、
429位のアミノ酸がMet、
434位のアミノ酸がTrp、
436位のアミノ酸がIle、
440位のアミノ酸がGly、His、Ile、LeuまたはTyrのいずれか、
が挙げられる。また、改変されるアミノ酸の数は特に限定されず、一箇所のみのアミノ酸が改変され得るし、二箇所以上のアミノ酸が改変され得る。二箇所以上のアミノ酸の改変の組合せとしては、例えば表6(表6−1〜表6−3)に記載されるような組合せが挙げられる。
本発明が提供する抗原結合分子に含まれる「Fc領域のFcγレセプターに対する結合活性がFcγレセプターに対する天然型Fc領域の結合活性よりも高いFc領域」として、Fc領域に結合した糖鎖の組成がフコース欠損糖鎖を結合したFc領域の割合が高くなるように、またはバイセクティングN-アセチルグルコサミンが付加したFc領域の割合が高くなるように修飾されたFc領域も使用され得る。抗体Fc領域に結合するN -グリコシド結合複合型糖鎖還元末端のN -アセチルグルコサミンからフコース残基を除去すると、FcγRIIIaに対する親和性が増強されることが知られている(Satohら(Expert Opin. Biol. Ther. (2006) 6 (11), 1161-1173))。よって、当該Fc領域を含む抗原結合分子は、本発明の医薬組成物に含まれる抗原結合分子としても有用である。抗体Fc領域に結合するN -グリコシド結合複合型糖鎖還元末端のN -アセチルグルコサミンからフコース残基が除去された抗体としては、例えば、次のような抗体;
グリコシル化が修飾された抗体(国際公開WO1999/054342等)、
糖鎖に付加するフコースが欠損した抗体(国際公開WO2000/061739、WO2002/031140、WO2006/067913等)、
補体レセプターとは、単核球、好中級、好酸球、好塩基球、B細胞、T細胞、NK細胞等の赤血球、および血小板等の免疫細胞に発現する補体系を形成する補体タンパク質リガンドのいずれかに結合することが可能なレセプターをいい、リガンドとレセプターとの結合によって当該レセプターを発現する免疫細胞の化学誘因(chemoattraction)、脱顆粒、貪食の促進、免疫複合体の除去、免疫複合体に結合したC3bのプロセッシング、B細胞の増殖、代替経路の活性化、NOの生成、活性酸素代謝物の生成、貪食、白血球遊走(leucocyte migration)、血小板凝集等の機能が発揮される(Leslieら(eLS (2009) 1-8))。補体系は、約30のタンパク質の集まりから構成され、免疫系の主要なエフェクター機構の一つである。補体カスケードは、基本的に古典経路(classical pathway)または代替経路(alterenative pathway)のいずれか一方を介して活性化される。いずれの経路を介した活性化であっても、C3転換酵素の生成をもたらす。この酵素は、上記カスケードの中心的な酵素複合体である。C3転換酵素は、血清C3を切断してC3aとC3bとに分け、後者は活性部位に共有結合してC3転移酵素(増幅ループ)をさらに生成する。活性化産物であるC3b(および古典経路を介してのみ生成されるC4bも同様)ならびにその分解産物は、重要なオプソニンであり、免疫結合体の輸送および可溶化を促進させることに関与する。C3/C4活性化産物および免疫系の種々の細胞上にあるそのレセプターもまた、細胞性免疫反応の調節にとって重要である。ミクログリア細胞の細胞膜には補体レセプターであるC5aR、CR3、CR4、およびC1qRpが、星状膠細胞(astrocyte)にはC5aR、およびCR2が発現することが知られている(Leslieら(eLS (2009) 1-8))。
本発明の抗原結合分子は、脳疾患に固有に脳組織内に存在する抗原に対する結合活性がイオン濃度の条件によって変化する抗原結合ドメイン、ならびにpH中性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインおよびpH酸性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインに加えて補体結合ドメインを含むことも可能である。当該ドメインが結合する補体としては、補体カスケードを形成するポリペプチドであればいずれの補体成分も使用され得るが、好ましい補体としては、オプソニンの結合に関与するC1q、C1rまたはC1sの補体成分が好適に挙げられる。補体結合ドメインとしては、例えば、IgG型免疫グロブリンのFc領域、抗補体抗体、、抗補体足場(Scaffold)分子等が好適に挙げられる。当該ドメインは、あらかじめ補体に対する結合活性を有しているドメインであればそのまま好適に使用され得る。当該ドメインが補体に対する結合活性がないまたは弱い場合には、抗原結合分子中の補体結合ドメインを形成するアミノ酸を改変して補体に対する結合活性を付与することが可能である。また、あらかじめ補体に対する結合活性を有しているドメイン中のアミノ酸を改変して、補体に対する結合活性を高めてもよい。補体結合ドメインのアミノ酸の改変は、アミノ酸改変前と改変後の補体に対する結合活性を比較することによって目的の改変を見出すことが可能である。例えば、pH中性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインおよびpH酸性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインとしてIgG型抗体のFc領域が使用される場合には、FcγR結合ドメインとして当該Fc領域に含まれる補体結合ドメインが使用され得る。
267位のアミノ酸がGlu、
268位のアミノ酸がPheまたはTyrのいずれか、
276位のアミノ酸がArg、
324位のアミノ酸がThr、
327位のアミノ酸がGly、
331位のアミノ酸がPro、もしくは
333位のアミノ酸がAla、Asp、Gly、SerまたはValのいずれか、
が挙げられる。また、改変されるアミノ酸の数は特に限定されず、一箇所のみのアミノ酸が改変され得るし、これらが任意に組み合わせられた二箇所以上のアミノ酸が改変され得る。
本発明では、神経細胞をとりまく間質液(組織間液、細胞外液)もしくは脳脊髄液または髄液の中に可溶型で存在する抗原に対する結合活性がイオン濃度の条件によって変化する抗原結合ドメイン、ならびにpH中性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインおよびpH酸性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインを含む抗原結合分子の脳疾患を治療するための使用が提供される。前記のようなFcRn欠損マウスを用いた検討によって、BBBを形成する内皮細胞に発現するFcRnが、血中から脳中へのIgG抗体の移行に関与していないことが示されていた従来技術(非特許文献10)に反して、pH中性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインを含む本発明の抗原結合分子が、当該結合活性を有しないFcRn結合ドメインを含む抗原結合分子と比較して、血中から脳中への移行が増強されていることが見出された。さらに、驚くべきことに、前記ドメインに加えて、脳疾患に固有に脳組織内に存在する抗原に対する結合活性がイオン濃度の条件によって変化する抗原結合ドメイン、およびpH酸性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインを含む抗原結合分子が、神経細胞をとりまく間質液(組織間液、細胞外液)もしくは脳脊髄液または髄液中の当該抗原濃度を低減する。
FcRnとIgG抗体との結晶学的研究によって、FcRn-IgG複合体は、二分子のFcRnに対して一分子のIgGから構成され、IgGのFc領域の両側に位置するCH2およびCH3ドメインの接触面付近において、二分子の結合が起こると考えられている(Burmeisterら(Nature (1994) 372, 336-343)。一方、後述する参考実施例1において確認されたように、抗体のFc領域が二分子のFcRnおよび一分子の活性型Fcγレセプターの四者を含む複合体を形成できることが明らかとなった(図14)。このヘテロ複合体の形成は、pH中性域の条件下でFcRnに対する結合活性を有するFc領域を含む抗原結合分子の性質について解析を進めた結果明らかとなった現象である。
前記Fc領域のアミノ酸のうちEUナンバリングで表される234、235、236、237、238、239、270、297、298、325、328、および329のいずれかひとつ以上のアミノ酸が天然型Fc領域と異なるアミノ酸に改変されているFc領域が好適に挙げられるが、Fc領域の改変は上記改変に限定されず、例えばCurrent Opinion in Biotechnology (2009) 20 (6), 685-691に記載されている脱糖鎖 (N297A, N297Q)、IgG1-L234A/L235A、IgG1-A325A/A330S/P331S、IgG1-C226S/C229S、IgG1-C226S/C229S/E233P/L234V/L235A、IgG1-L234F/L235E/P331S、IgG1-S267E/L328F、IgG2-V234A/G237A、IgG2-H268Q/V309L/A330S/A331S、IgG4-L235A/G237A/E318A、IgG4-L236E等の改変、および、WO 2008/092117に記載されているG236R/L328R、L235G/G236R、N325A/L328R、N325LL328R等の改変、および、EUナンバリング233位、234位、235位、237位におけるアミノ酸の挿入、WO 2000/042072に記載されている個所の改変であってもよい。
234位のアミノ酸をAla、Arg、Asn、Asp、Gln、Glu、Gly、His、Lys、Met、Phe、Pro、Ser、ThrまたはTrpのいずれか、
235位のアミノ酸をAla、Asn、Asp、Gln、Glu、Gly、His、Ile、Lys、Met、Pro、Ser、Thr、ValまたはArgのいずれか、
236位のアミノ酸をArg、Asn、Gln、His、Leu、Lys、Met、Phe、ProまたはTyrのいずれか、
237位のアミノ酸をAla、Asn、Asp、Gln、Glu、His、Ile、Leu、Lys、Met、Pro、Ser、Thr、Val、TyrまたはArgのいずれか、
238位のアミノ酸をAla、Asn、Gln、Glu、Gly、His、Ile、Lys、Thr、TrpまたはArgのいずれか、
239位のアミノ酸をGln、His、Lys、Phe、Pro、Trp、TyrまたはArgのいずれか、
265位のアミノ酸をAla、Arg、Asn、Gln、Gly、His、Ile、Leu、Lys、Met、Phe、Ser、Thr、Trp、TyrまたはValのいずれか、
266位のアミノ酸をAla、Arg、Asn、Asp、Gln、Glu、Gly、His、Lys、Phe、Pro、Ser、Thr、TrpまたはTyrのいずれか、
267位のアミノ酸をArg、His、Lys、Phe、Pro、TrpまたはTyrのいずれか、
269位のアミノ酸をAla、Arg、Asn、Gln、Gly、His、Ile、Leu、Lys、Met、Phe、Pro、Ser、Thr、Trp、TyrまたはValのいずれか、
270位のアミノ酸をAla、Arg、Asn、Gln、Gly、His、Ile、Leu、Lys、Met、Phe、Pro、Ser、Thr、Trp、TyrまたはValのいずれか、
271位のアミノ酸をArg、His、Phe、Ser、Thr、TrpまたはTyrのいずれか、
295位のアミノ酸をArg、Asn、Asp、Gly、His、Phe、Ser、TrpまたはTyrのいずれか、
296位のアミノ酸をArg、Gly、LysまたはProのいずれか、
297位のアミノ酸をAla、
298位のアミノ酸をArg、Gly、Lys、Pro、TrpまたはTyrのいずれか、
300位のアミノ酸をArg、LysまたはProのいずれか、
324位のアミノ酸をLysまたはProのいずれか、
325位のアミノ酸をAla、Arg、Gly、His、Ile、Lys、Phe、Pro、Thr、TrpTyr、もしくはValのいずれか、
327位のアミノ酸をArg、Gln、His、Ile、Leu、Lys、Met、Phe、Pro、Ser、Thr、Trp、TyrまたはValのいずれか、
328位のアミノ酸をArg、Asn、Gly、His、LysまたはProのいずれか、
329位のアミノ酸をAsn、Asp、Gln、Glu、Gly、His、Ile、Leu、Lys、Met、Phe、Ser、Thr、Trp、Tyr、ValまたはArgのいずれか、
330位のアミノ酸をProまたはSerのいずれか、
331位のアミノ酸をArg、GlyまたはLysのいずれか、もしくは
332位のアミノ酸をArg、LysまたはProのいずれか、
のいずれかひとつ以上に改変されているFc領域が好適に挙げられる。
233位のアミノ酸がAsp、
234位のアミノ酸がTyr、
237位のアミノ酸がAsp、
264位のアミノ酸がIle、
265位のアミノ酸がGlu、
266位のアミノ酸がPhe、Met、またはLeuのいずれか、
267位のアミノ酸がAla、Glu、Gly、またはGlnのいずれか、
268位のアミノ酸がAsp、またはGluのいずれか、
269位のアミノ酸がAsp、
272位のアミノ酸がAsp、Phe、Ile、Met、Asn、またはGlnのいずれか
296位のアミノ酸がAsp、
326位のアミノ酸がAla、またはAspのいずれか、
327位のアミノ酸がGly、
330位のアミノ酸がLys、またはArgのいずれか、
331位のアミノ酸がSer、
332位のアミノ酸がThr、
333位のアミノ酸がThr、Lys、またはArgのいずれか、
396位のアミノ酸がAsp、Glu、Phe、Ile、Lys、Leu、Met、Gln、Arg、またはTyrのいずれか、
のいずれかひとつ以上である改変Fc領域が例示される。
本発明では、神経細胞をとりまく間質液(組織間液、細胞外液)もしくは脳脊髄液または髄液の中に存在する疾患をもたらす細胞表面に存在する受容体型抗原に対する結合活性がイオン濃度の条件によって変化する抗原結合ドメイン、ならびにpH中性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインおよびpH酸性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインを含む抗原結合分子の脳疾患を治療するための使用が提供される。前記のようなFcRn欠損マウスを用いた検討によって、BBBを形成する内皮細胞に発現するFcRnが、血中から脳中へのIgG抗体の移行に関与していないことが示されていた従来技術(非特許文献10)に反して、pH中性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインを含む本発明の抗原結合分子が、当該結合活性を有しないFcRn結合ドメインを含む抗原結合分子と比較して、血中から脳中への移行が増強されていることが見出された。さらに、前記ドメインに加えて、脳疾患に固有に脳組織内に存在する抗原に対する結合活性がイオン濃度の条件によって変化する抗原結合ドメイン、およびpH酸性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインに加えて、FcγR結合ドメインまたは補体レセプター結合ドメインを含む抗原結合分子が、神経細胞をとりまく間質液(組織間液、細胞外液)もしくは脳脊髄液または髄液中に存在する疾患をもたらす細胞表面に存在する受容体型抗原を低減する機能を奏する。
本発明において、抗原結合分子は、脳疾患に固有に脳組織内に存在する抗原に対する結合活性がイオン濃度の条件によって変化する抗原結合ドメイン、ならびにpH中性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインおよびpH酸性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインを含むを含む分子を表す最も広義な意味として使用されており、具体的には、それらが抗原に対する結合活性を示す限り、様々な分子型が含まれる。例えば、当該抗原結合ドメインがpH中性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインおよびpH酸性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインと結合した分子の例として、抗体が挙げられる。抗体には、単一のモノクローナル抗体(アゴニストおよびアンタゴニスト抗体を含む)、ヒト抗体、ヒト化抗体、キメラ抗体等が含まれ得る。また抗体の断片として使用される場合としては、抗原結合ドメインおよび抗原結合断片(例えば、Fab、F(ab')2、scFvおよびFv)が好適に挙げられ得る。既存の安定なα/βバレルタンパク質構造等の立体構造が scaffold(土台)として用いられ、その一部分の構造のみが抗原結合ドメインの構築のためにライブラリ化されたスキャフォールド分子も、本発明の抗原結合分子に含まれ得る 。
Ser
Gly・Ser
Gly・Gly・Ser
Ser・Gly・Gly
Gly・Gly・Gly・Ser(配列番号:28)
Ser・Gly・Gly・Gly(配列番号:29)
Gly・Gly・Gly・Gly・Ser(配列番号:30)
Ser・Gly・Gly・Gly・Gly(配列番号:31)
Gly・Gly・Gly・Gly・Gly・Ser(配列番号:32)
Ser・Gly・Gly・Gly・Gly・Gly(配列番号:33)
Gly・Gly・Gly・Gly・Gly・Gly・Ser(配列番号:34)
Ser・Gly・Gly・Gly・Gly・Gly・Gly(配列番号:35)
(Gly・Gly・Gly・Gly・Ser(配列番号:30))n
(Ser・Gly・Gly・Gly・Gly(配列番号:31))n
[nは1以上の整数である]等が好適に挙げられる。但し、ペプチドリンカーの長さや配列は目的に応じて当業者が適宜選択することができる。
(1)脳疾患に特異的に脳組織内において可溶型で存在する抗原に対する結合活性がイオン濃度の条件によって変化する抗原結合ドメイン、ならびに、pH中性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインおよびpH酸性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインを含むFc領域を含む抗体
(1−1)可溶型抗原に対する結合活性がイオン濃度の条件によって変化する二つの可変領域を含むF(ab')2とpH中性域および酸性域の条件下でFcRnに対する結合活性を有するFc領域を含む抗体
(1−2)F(ab')2を形成する可変領域のうち一方の可変領域が、可溶型抗原に対する結合活性がイオン濃度の条件によって変化する可変領域であり、もう一方の可変領域がFcRnに対する結合活性がイオン濃度の条件によって変化する可変領域であるF(ab')2とpH酸性域の条件下でFcRnに対する結合活性を有するFc領域を含む抗体
(1−3)可変領域を少なくとも二つ含む抗体断片の一方の可変領域が、可溶型抗原に対する結合活性がイオン濃度の条件によって変化する可変領域であり、もう一方の可変領域がpH中性域および酸性域でのFcRnに対する結合活性を有する可変領域を含む抗体断片
(a)L234F、L235E、P331S、
(b)C226S、C229S、P238S、
(c)C226S、C229S、
(d)C226S、C229S、E233P、L234V、L235A
の改変を有するFc領域、又は、231位から238位のアミノ酸配列が欠失したFc領域を有する抗原結合分子も適宜使用され得る。
(e)H268Q、V309L、A330S、P331S
(f)V234A
(g)G237A
(h)V234A、G237A
(i)A235E、G237A
(j)V234A、A235E、G237A
の改変を有するFc領域を有するポリペプチド会合体も適宜使用され得る。
(k)F241A
(l)D265A
(m)V264A
の改変を有するFc領域を有する抗原結合分子も適宜使用され得る。
(n)L235A、G237A、E318A
(o)L235E
(p)F234A、L235A
の改変を有するFc領域を有する抗原結合分子も適宜使用され得る。
(1−4)可溶型抗原に対する結合活性がイオン濃度の条件によって変化する二つの可変領域を含むF(ab')2とFcγRに対する結合活性ならびにpH中性域および酸性域の条件下でFcRnに対する結合活性を有するFc領域を含む抗体
(1−5)F(ab')2を形成する可変領域のうち一方の可変領域が、可溶型抗原に対する結合活性がイオン濃度の条件によって変化する可変領域であり、もう一方の可変領域がFcRnに対する結合活性がイオン濃度の条件によって変化する可変領域であるF(ab')2と FcγRに対する結合活性ならびにpH酸性域の条件下でFcRnに対する結合活性を有するFc領域を含む抗体
(1−6)F(ab')2を形成する可変領域のうち一方の可変領域が、可溶型抗原に対する結合活性がイオン濃度の条件によって変化する可変領域であり、もう一方の可変領域がFcγRに対する結合活性を有する可変領域であるF(ab')2とpH中性域および酸性域の条件下でFcRnに対する結合活性を有するFc領域を含む抗体
(1−7)可溶型抗原に対する結合活性がイオン濃度の条件によって変化する二つの可変領域を含むF(ab')2とC1q等の補体に対する結合活性ならびにpH中性域および酸性域の条件下でFcRnに対する結合活性を有するFc領域を含む抗体
(1−8)F(ab')2を形成する可変領域のうち一方の可変領域が、可溶型抗原に対する結合活性がイオン濃度の条件によって変化する可変領域であり、もう一方の可変領域がFcRnに対する結合活性がイオン濃度の条件によって変化する可変領域であるF(ab')2とC1q等の補体に対する結合活性ならびにpH酸性域の条件下でFcRnに対する結合活性を有するFc領域を含む抗体
(1−9)F(ab')2を形成する可変領域のうち一方の可変領域が、可溶型抗原に対する結合活性がイオン濃度の条件によって変化する可変領域であり、もう一方の可変領域がC1q等の補体に対する結合活性を有する可変領域であるF(ab')2とpH中性域および酸性域の条件下でFcRnに対する結合活性を有するFc領域を含む抗体
(2−1)受容体型抗原に対する結合活性がイオン濃度の条件によって変化する二つの可変領域を含むF(ab')2とFcγRに対する結合活性ならびにpH中性域および酸性域の条件下でFcRnに対する結合活性を有するFc領域を含む抗体
(2−2)F(ab')2を形成する可変領域のうち一方の可変領域が、受容体型抗原に対する結合活性がイオン濃度の条件によって変化する可変領域であり、もう一方の可変領域がFcRnに対する結合活性がイオン濃度の条件によって変化する可変領域であるF(ab')2と FcγRに対する結合活性ならびにpH酸性域の条件下でFcRnに対する結合活性を有するFc領域を含む抗体
(2−3)F(ab')2を形成する可変領域のうち一方の可変領域が、受容体型抗原に対する結合活性がイオン濃度の条件によって変化する可変領域であり、もう一方の可変領域がFcγRに対する結合活性を有する可変領域であるF(ab')2とpH中性域および酸性域の条件下でFcRnに対する結合活性を有するFc領域を含む抗体
(2−4)受容体型抗原に対する結合活性がイオン濃度の条件によって変化する二つの可変領域を含むF(ab')2とC1q等の補体に対する結合活性ならびにpH中性域および酸性域の条件下でFcRnに対する結合活性を有するFc領域を含む抗体
(2−5)F(ab')2を形成する可変領域のうち一方の可変領域が、受容体型抗原に対する結合活性がイオン濃度の条件によって変化する可変領域であり、もう一方の可変領域がFcRnに対する結合活性がイオン濃度の条件によって変化する可変領域であるF(ab')2とC1q等の補体に対する結合活性ならびにpH酸性域の条件下でFcRnに対する結合活性を有するFc領域を含む抗体
(2−6)F(ab')2を形成する可変領域のうち一方の可変領域が、受容体型抗原に対する結合活性がイオン濃度の条件によって変化する可変領域であり、もう一方の可変領域がC1q等の補体に対する結合活性を有する可変領域であるF(ab')2とpH中性域および酸性域の条件下でFcRnに対する結合活性を有するFc領域を含む抗体
前記の(1−2)、(1−5)、(1−8)、(2−2)、(2−5)の構造を含む抗体の調製方法の一態様として、二重特異性抗体の作製方法が適用され得る。二重特異性抗体とは、異なるエピトープに対して特異的に結合する二種類の可変領域を含む抗体である。IgG型の二重特異性抗体はIgG抗体を産生するハイブリドーマ二種を融合することによって生じるhybrid hybridoma(quadroma)によって分泌させることが可能である(Milsteinら(Nature (1983) 305, 537-540)。
(i) Fc領域を形成する二つのポリペプチドであって、その一方のポリペプチドのアミノ酸配列のうちEUナンバリングで表される409のアミノ酸がAsp、370のアミノ酸がGluであり、他方のポリペプチドのアミノ酸配列のうちEUナンバリングで表される399のアミノ酸がLys、357のアミノ酸がLysであることを特徴とする二つのポリペプチド(本態様では、EUナンバリングで表される370のアミノ酸のGluに代えてAspであってもよく、EUナンバリングで表される370のアミノ酸のGluに代えて392のアミノ酸のAspであってもよい)、
(ii) Fc領域を形成する二つのポリペプチドであって、その一方のポリペプチドのアミノ酸配列のうちEUナンバリングで表される409のアミノ酸がAsp、439のアミノ酸がGluであり、他方のポリペプチドのアミノ酸配列のうちEUナンバリングで表される399のアミノ酸がLys、356のアミノ酸がLysであることを特徴とする二つのポリペプチド(本態様では、EUナンバリングで表される439のアミノ酸のGluに代えて360のアミノ酸のAsp、EUナンバリングで表される392のアミノ酸のAsp又は439のアミノ酸のAspであってもよい)、
(iii) Fc領域を形成する二つのポリペプチドであって、その一方のポリペプチドのアミノ酸配列のうちEUナンバリングで表される370のアミノ酸がGlu、439のアミノ酸がGluであり、他方のポリペプチドのアミノ酸配列のうちEUナンバリングで表される357のアミノ酸がLys、356のアミノ酸がLysであることを特徴とする二つのポリペプチド、または、
Fc領域を形成する二つのポリペプチドであって、その一方のポリペプチドのアミノ酸配列のうちEUナンバリングで表される409のアミノ酸がAsp、370のアミノ酸がGlu、439のアミノ酸がGluであり、他方のポリペプチドのアミノ酸配列のうちEUナンバリングで表される399のアミノ酸がLys、357のアミノ酸がLys、356のアミノ酸がLysであることを特徴とする二つのポリペプチド(本態様では、EUナンバリングで表される370のアミノ酸をGluに置換しなくてもよく、更に、370のアミノ酸をGluに置換しない上で、439のアミノ酸のGluに代えてAsp又は439のアミノ酸のGluに代えて392のアミノ酸のAspであってもよい)、
が好適に用いられる。
本発明において、本発明の抗原結合分子の「脳疾患を治療するための使用」とは、本発明の抗原結合分子に含まれる抗原結合ドメインが結合する抗原が脳疾患に特異的に脳組織内において可溶型で存在する抗原である場合、「当該抗原を脳組織液から消失させるための使用」と表現することも可能である。または、本発明の抗原結合分子を投与することを含む「当該抗原を脳組織液から消失させる方法」と表現することも可能である。また、本発明の抗原結合分子に含まれる抗原結合ドメインが結合する抗原が脳疾患をもたらす細胞表面に存在する受容体型で存在する抗原である場合も、「当該抗原を脳組織液から消失させるための使用」と表現することも可能である。また、本発明の抗原結合分子を投与することを含む「当該抗原を脳組織液から消失させる方法」と表現することも可能である。
A値=各時点での抗原のモル濃度
B値=各時点での抗原結合分子のモル濃度
C値=各時点での抗原結合分子のモル濃度あたりの抗原のモル濃度(抗原/抗原結合分子モル比)
C=A/B。
本発明は、また以下の工程(a)〜(i)を含む、脳疾患に固有に脳組織内に存在する抗原を消失する機能を有する抗原結合分子の製造方法;
(a) 脳疾患に固有に脳組織内に存在する抗原に対する結合活性がイオン濃度の条件によって変化する抗原結合ドメインを得る工程、
(b) 前記工程(a)で選択された抗原結合ドメインをコードする遺伝子を得る工程、
(c) 前記工程(b)で得られた遺伝子を、pH中性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインおよびpH酸性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインをコードする遺伝子と作動可能に連結する工程、
(d) 前記工程(c)で作動可能に連結された遺伝子を含む宿主細胞を培養する工程、
(e) 前記工程(d)で得られた培養液から抗原結合分子を単離する工程、
(f) 前記工程(e)で得られた抗原結合分子による当該抗原の脳組織液からの消失を評価する工程、
(g) 前記工程(f)で当該抗原の脳組織液からの消失が確認された抗原結合ドメインをコードする遺伝子を含むベクターを含む宿主細胞を培養する工程、
(h) 前記工程(g)で得られた培養液から抗原結合分子を単離する工程、
を提供する。
(a) 脳疾患に固有に脳組織内に存在する抗原に対する結合活性がイオン濃度の条件によって変化する抗原結合ドメインを得る工程、
(b) 前記工程(a)で選択された抗原結合ドメインをコードする遺伝子を得る工程、
(c) 前記工程(b)で得られた遺伝子を、pH中性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインおよびpH酸性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインならびにFcγR結合ドメインをコードする遺伝子と作動可能に連結する工程、
(d) 前記工程(c)で作動可能に連結された遺伝子を含む宿主細胞を培養する工程、
(e) 前記工程(d)で得られた培養液から抗原結合分子を単離する工程、
(f) 前記工程(e)で得られた抗原結合分子による当該抗原の脳組織液からの消失を評価する工程、
(g) 前記工程(f)で当該抗原の脳組織液からの消失が確認された抗原結合ドメインをコードする遺伝子を含むベクターを含む宿主細胞を培養する工程、
(h) 前記工程(g)で得られた培養液から抗原結合分子を単離する工程、
を提供する。
(a) 脳疾患に固有に脳組織内に存在する抗原に対する結合活性がイオン濃度の条件によって変化する抗原結合ドメインを得る工程、
(b) 前記工程(a)で選択された抗原結合ドメインをコードする遺伝子を得る工程、
(c) 前記工程(b)で得られた遺伝子を、pH中性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインおよびpH酸性域の条件下でFcRnに対する結合活性を有するFcRn結合ドメインならびに補体結合ドメインをコードする遺伝子と作動可能に連結する工程、
(d) 前記工程(c)で作動可能に連結された遺伝子を含む宿主細胞を培養する工程、
(e) 前記工程(d)で得られた培養液から抗原結合分子を単離する工程、
(f) 前記工程(e)で得られた抗原結合分子による当該抗原の脳組織液からの消失を評価する工程、
(g) 前記工程(f)で当該抗原の脳組織液からの消失が確認された抗原結合ドメインをコードする遺伝子を含むベクターを含む宿主細胞を培養する工程、
(h) 前記工程(g)で得られた培養液から抗原結合分子を単離する工程、
を提供する。
本発明は、本発明の抗原結合分子、または本発明の製造方法により製造された抗原結合分子を含む医薬組成物も提供する。本発明の抗原結合分子または本発明の製造方法により製造された抗原結合分子はその投与により通常の抗原結合分子と比較して脳組織液中の抗原濃度を低下させる作用が高い上に、投与された生体による免疫応答や当該生体中の薬物動態等が改変されていることから医薬組成物として有用である。本発明の医薬組成物には医薬的に許容される担体が含まれ得る。
なお本明細書において引用されたすべての先行技術文献は、参照として本明細書に組み入れられる。
(1−1)中性域FcRn結合増強IgG抗体の調製
Fv4-IgG1(重鎖配列番号:36、軽鎖配列番号:37)に対して、表8に示すFc領域に中性域におけるFcRn結合を増強するアミノ酸置換が導入されたFv4-hF22(重鎖配列番号:38、軽鎖配列番号:39)およびFv4-hF1(重鎖配列番号:40、軽鎖配列番号:41)をコードするDNA配列が動物発現用プラスミドに当該業者公知の方法で組み込まれた。抗体は以下の方法を用いて発現された。FreeStyle 293 Expression Medium培地(Invitrogen)に1.33 x 106/mLの細胞密度で懸濁され6well plateの各ウェルへ3 mLずつ播種されたヒト胎児腎細胞由来FreeStyle 293-F株(Invitrogen)に、リポフェクション法により調製されたプラスミドが導入された。CO2インキュベーター(37℃、8%CO2, 90 rpm)で4日間培養された培養上清から、rProtein A SepharoseTM Fast Flow(Amersham Biosciences)を用いて当業者公知の方法で抗体が精製された。精製された抗体の濃度は、分光光度計を用いた280 nmでの吸光度の測定値からPACE法により算出された吸光係数を用いて算出された(Protein Science (1995) 4, 2411-2423)。
調製された抗体のマウスFcRnに対するアフィニティが以下に示す方法で測定された。Biacore T100(GE Healthcare)を用いて、マウスFcRnと抗体との結合が速度論的に解析された。Sensor chip CM4(GE Healthcare)上にアミンカップリング法でprotein L(ACTIGEN)を適当量固定化し、そこへ目的の抗体をキャプチャーさせた。次に、FcRn希釈液とブランクであるランニングバッファーをインジェクトし、センサーチップ上にキャプチャーさせた抗体にヒトFcRnを相互作用させた。ランニングバッファーには50 mmol/L sodium phosphate、150 mmol/L NaCl、0.05% (w/v) Tween20、pH7.0を用い、FcRnの希釈にもそれぞれのバッファーが使用された。センサーチップの再生には10 mmol/L Glycine-HCl, pH1.5が用いられた。測定は全て25℃で実施された。測定で得られたセンサーグラムから算出された、カイネティクスパラメーターである結合速度定数 ka(1/Ms)、および解離速度定数 kd(1/s)をもとに各抗体のヒトFcRnに対する KD(M)(アフィニティ) が算出された。各パラメーターの算出にはBiacore T100 Evaluation Software(GE Healthcare)が用いられた。Fv4-IgG1、Fv4-hF22およびFv4-hF1のマウスFcRnに対する中性条件下(pH7.0)におけるアフィニティを表8に示した。hF22およびhF1のマウスFcRnへの中性条件下におけるアフィニティはIgG1と比較してそれぞれ約4.5倍、24.7倍、向上していることが確認された。
ノーマルマウス(C57BL/6J mouse、Charles River Japan)にFv4-IgG1、Fv4-hF22およびFv4-hF1が5mg/kgの用量で尾静脈に10 mL/kgで単回投与された。
血液サンプルは以下のとおり採取された。検体が尾静脈内に投与された後28日目に脳が採材される個体から、抗体の投与後5分、7時間、4日、7日、14日、28日の時点で採血された。
マウス血漿中の検体濃度はELISA法にて測定された。まず、Anti-Human IgG(γ-chain specific) F(ab')2 Fragment of Antibody(SIGMA)が各ウェルに分注されたNunc-Immuno Plate, MaxiSoup(Nalge nunc International)を4℃で1晩静置することによってAnti-Human IgG固相化プレートが作成された。血漿中濃度として0.08、0.04、 0.02、0.01、0.005、0.0025、0.00125μg/mLの検体を含む検量線試料と100倍以上希釈されたマウス血漿測定試料が調製された。これらの検量線試料および血漿測定試料100μLが分注されたAnti-Human IgG固相化プレートが25℃で1時間保温(incubation)された。その後Biotinylated Anti-human IL-6 R Antibody(R&D)と25℃で1時間反応させ、さらにStreptavidin-PolyHRP80(Stereospecific Detection Technologies)と25℃で1時間反応させた。その後、TMB One Component HRP Microwell Substrate(BioFX Laboratories)を基質として用いた発色反応が1N-Sulfuric acid(Showa Chemical)で停止された後、マイクロプレートリーダーにて反応液の450 nmの吸光度が測定された。マウス血漿中濃度は検量線の吸光度から解析ソフトウェアSOFTmax PRO(Molecular Devices)を用いて算出された。この方法で測定された静脈内投与後のノーマルマウスにおける血漿中抗体濃度推移を図1に示した。
脳は以下のとおり採取された。ペントバルビタールナトリウム70 mg/kgが腹腔内に投与され、その全身麻酔の導入が確認されたC57BL/6J雄性マウスが、開胸された。心突部から穿刺されたダルベッコリン酸緩衝生理食塩水(PBS) が充填されたシリンジに接続された静脈留置針(外套針)が大動脈に留置された。直ちに、切開された右心耳から、シリンジポンプにて PBS 60 mLを流速15 mL/分で送液することによって灌流された。右心耳から流れ出る液体が透明になったのが確認された後に、速やかに当該マウスから脳が採取された。小脳を除去した後に大脳が半球に分割された。このうちカミソリを用いて軽く刻まれた後に2mLチューブ中で液体窒素にて急速凍結された半球が、ホモジネート(homogenate)が調製されるまで-80℃の冷凍庫で保管された。
脳組織のホモジネートは以下のとおり作製された。予め調製されたNo-detergent buffer(25 mM Tris-HCl (pH 7.6)(Wako))、150 mM NaCl)と、2xRIPA buffer(25 mM Tris-HCl (pH 7.6)(Wako))、150 mM NaCl、2% NP-40、2% sodium deoxycholate、0.2% SDS)の各々に、実験直前に1 vol%のprotease inhibitor(Thermo scientific, Halt protease inhibitor cocktail)が加えられ氷冷された。以下の操作は特に断りがない限り、氷冷して行われた。その重量が測定された刻んだ脳組織に、333 mg/mLとなるようにNo-detergent bufferが添加された各チューブに一つずつアルミナボール(直径5 mm)が投入された。4℃の低温にてミキサーミル(MM300・Retsch)を用いて、20 Hzで5分間振動させた。4℃、3000 gで2分間遠心分離された内容物の混合物が、150μLずつ新しいチューブに添加された。更に350μLずつNo-detergent bufferが添加された各チューブに一つずつアルミナボール(直径5 mm)が投入された。4℃の低温にてミキサーミル(MM300・Retsch)を用いて、20 Hzで5分間振動させた。2xRIPA bufferが500μLずつ添加された各チューブが、4℃で1時間転倒混和された。4℃、14000 gで20分間遠心分離された上清が移された新しいチューブは、ホモジネート中の抗体濃度の測定に用いるまで-80℃の冷凍庫で保存された。
マウス脳ホモジネート中の検体濃度はELISA法にて測定された。まず、Anti-Human IgG(γ-chain specific) F(ab')2 Fragment of Antibody (SIGMA) をNunc-Immuno Plate, MaxiSorp(Nalge nunc International)に分注し、4℃で1晩静置することによってAnti-Human IgG固相化プレートが作成された。脳ホモジネート中濃度として16、8、 4、2、1、0.5、0.25 ng/mLの検体を含む検量線試料と20倍以上希釈されたマウス脳ホモジナイズサンプル測定試料が調製された。これらの検量線試料および脳ホモジネート測定試料100μLが各ウェルに分注されたAnti-Human IgG固相化プレートが25℃で1時間保温された。その後Biotinylated Anti-human IL-6 R Antibody(R&D)と25℃で1時間反応させ、さらにStreptavidin-PolyHRP80(Stereospecific Detection Technologies)と25℃で1時間反応させた。その後、TMB One Component HRP Microwell Substrate(BioFX Laboratories)を基質として用いた発色反応が、1N-Sulfuric acid(Showa Chemical)で停止された後、マイクロプレートリーダーにて反応液の450 nmの吸光度が測定された。マウス血漿中濃度は検量線の吸光度から解析ソフトウェアSOFTmax PRO(Molecular Devices)を用いて算出された。この方法で測定された静脈内投与後のノーマルマウスにおける脳中抗体濃度推移を図2に示した。
図1に各抗体の血漿中抗体濃度推移を示した。また、図2に各抗体の脳中抗体濃度推移を示した。また、これらの濃度推移から、解析ソフトウェアWinNonlin versions 5.1を用いて、脳中AUC(血中濃度−時間曲線下面積(area under the concentration-time curve))および血漿中AUCが算出された。表9に、各抗体の脳移行率の指標となる脳中AUC/血漿中AUC×100(%)の値を示した。これらの結果から、中性(pH7.0)条件下におけるマウスFcRnへの結合を24.7倍増強し、KD 150 nMを有するFv4-hF1は、Fv4-hIgG1と比較して、脳中AUC/血漿中AUC×100が12.7倍向上したことから、中性(pH7.0)条件下におけるFcRnの結合を増強することによって、IgG抗体の脳移行性を向上できることが示された。
(2−1)中性条件下におけるFcRnへの結合を増強したpH依存的結合抗体を用いた血漿中からの抗原の除去
抗体一分子あたりが中和できる抗原量はアフィニティに依存し、アフィニティを強くすることで少ない抗体量で抗原を中和することが可能であり、さらに抗原に共有結合的に結合し、アフィニティを無限大にすることができれば一分子の抗体で一分子の抗原(二価の場合は二抗原)を中和することが可能である。しかしながら、これまでの方法では一分子の抗体は、一分子の抗原(二価の場合は二抗原)に結合することが限界であったが、最近、抗原に対してpH依存的に結合する抗体を用いることで、一分子の抗体が複数分子の抗原に結合することが可能であることが報告された(WO 2009/125825)。pH依存的抗原結合抗体は、抗原に対して血漿中の中性条件下においては強く結合し、エンドソーム内の酸性条件下において抗原を解離する。さらに抗原を解離した後に抗体がFcRnによって血漿中にリサイクルされると再び抗原に結合することが可能であるため、一つのpH依存的抗原結合抗体で複数の抗原に繰り返し結合することが可能となる。
FcRn欠損マウスにおいては抗体の血漿中滞留性が大幅に低下することから、FcRnが抗体の血漿中滞留性に重要な役割を果たしていることが明らかであり、このFcRnに対する中性条件下における結合を増強したpH依存的結合抗体を用いることで血漿中から抗原を除去することが可能であることはすでに報告されている(WO 2011/122011)。しかしながら、FcRn欠損マウスにおいてはIgG抗体の脳移行性は、正常マウスと差が無いことから(非特許文献10)、脳脊髄液中におけるIgG抗体に対するFcRnの役割は不明であった。そのため、中性条件下におけるFcRn結合を増強したpH依存的結合抗体を用いることで、脳脊髄液中の抗原を除去できるかは不明であった。実施例1においてFcRnへの中性における結合を増強することで、脳内への移行(トランスサイトーシス)を増大出来ることが示されたことから、脳内にFcRnがトランスサイトーシス機能を発揮していることが確認された。そのため、このFcRnのトランスサイトーシス機能を用いることで、脳脊髄液中の抗原を血漿中と同様に除去することが出来ることが示唆された。
(2−3−1)被検物質及び抗原の投与
脳組織における抗原の除去を観察するために、被検物質(抗体)及び抗原が脳室内または静脈内に投与される。マウスの脳室内への被験物質および抗原を含む薬液の投与の方法として、(1)インジェクションカニューレを刺入して当該薬液を注入する、(2)あらかじめ留置しておいたガイドカニューレに、インジェクションカニューレを挿入して当該薬液を注入する、(3)当該薬液を充填した浸透圧ポンプを接続した脳室内投与用カニューレを埋め込む、の上記のいずれかの方法が用いられる。静脈内投与の場合は尾静脈から当該薬液が投与される。
麻酔下で、後頭部正中切開が行われたマウスの後頭骨一環椎間の硬膜が露出される。硬膜表面および周囲組織を充分止血した後、当該硬膜に加えられた小切開部にガラスチューブを接触させる。毛細管現象を利用して血液の混入のない髄液が採取される。
マウス脳脊髄液中の検体濃度がELISA法にて測定される。まず、Anti-Human IgG(γ-chain specific) F(ab')2 Fragment of Antibody(SIGMA)が各ウェルに分注されたNunc-Immuno Plate, MaxiSorp(Nalge nunc International)を4℃で1晩静置することによってAnti-Human IgG固相化プレートが作成される。脳脊髄液中濃度として0.08、0.04、 0.02、0.01、0.005、0.0025、0.00125μg/mLの検体を含む検量線試料と100倍以上希釈されたマウス脳脊髄液測定試料が調製される。これらの検量線試料および脳脊髄液測定試料100μLが分注されたAnti-Human IgG固相化プレートが25℃で1時間保温される。その後Biotinylated Anti-human IL-6 R Antibody(R&D)と25℃で1時間反応させ、さらにStreptavidin-PolyHRP80(Stereospecific Detection Technologies)と25℃で1時間反応させた。TMB One Component HRP Microwell Substrate(BioFX Laboratories)を基質として用いた発色反応が1N-Sulfuric acid(Showa Chemical)で反応停止された後、マイクロプレートリーダーにて反応液の450 nmの吸光度が測定される。マウス血漿中濃度は検量線の吸光度から解析ソフトウェアSOFTmax PRO(Molecular Devices)を用いて算出される。
マウスの脳脊髄液中のhsIL-6R濃度は電気化学発光法にて測定される。脳髄液中の濃度として12.5、6.25、3.125、1.561、0.781、0.390、0.195 ng/mLに調製されるhsIL-6R検量線試料および50倍以上希釈したマウス脳髄液測定試料を調製される。これらの試料にSULFO-TAG NHS Ester(Meso Scale Discovery)でルテニウム化したMonoclonal Anti-human IL-6R Antibody(R&D)およびBiotinylated Anti-human IL-6 R Antibody (R&D)を混合し4℃で1晩反応させる。その際のAssay bufferには可溶型ヒトIL-6レセプターに対して大過剰のトシリズマブが含まれており、サンプル中のほぼ全てのhsIL-6Rが検体から乖離し、トシリズマブとの複合体として存在する状態とすることを目的とした。その後、反応液はMA400 PR Streptavidin Plate(Meso Scale Discovery)に分注される。さらに反応液を25℃で1時間反応させた後、洗浄後の各ウェルに、Read Buffer T(×4)(Meso Scale Discovery)が分注される。ただちに反応液の吸光度がSECTOR PR 400 reader(Meso Scale Discovery)で測定される。hSIL-6R濃度は検量線のレスポンスから解析ソフトウェアSOFTmax PRO(Molecular Devices)を用いて算出される。
(1−1)Biacoreを用いたFcRnとFcγRに対する同時結合の評価
Biacore T100又はT200(GE Healthcare)を用いて、ヒト又はマウスFcRnとヒト又はマウスFcγRsとが抗原結合分子に同時に結合するかが評価された。Sensor chip CM4 (GE Healthcare) 上にアミンカップリング法によって固定化されたヒト又はマウスFcRnに、被験対象の抗原結合分子をキャプチャーさせた。次に、ヒト又はマウスFcγRsの希釈液とブランクとして使用されたランニングバッファーが注入され、センサーチップ上のFcRnに結合した抗原結合分子にヒト又はマウスFcγRsを相互作用させた。ランニングバッファーとして50 mmol/L sodium phosphate、150 mmol/L NaCl、0.05% (w/v) Tween20、pH7.4が用いられ、FcγRsの希釈にもこのバッファーが使用された。センサーチップの再生には10 mmol/L Trsi-HCl、pH9.5が用いられた。結合の測定は全て25℃で実施された。
pH中性域における条件下でヒトFcRnに対する結合能を有する抗体である、VH3-IgG1-F157(配列番号:55)とVL3-CK(配列番号:42) を含むFv4-IgG1-F157が、ヒトFcRnに結合するのと同時に、各種ヒトFcγRまたは各種マウスFcγRに結合するか否かが評価された。その結果、Fv4-IgG1-F157が、 ヒトFcRnに結合するのと同時に、ヒトFcγRIa、FcγRIIa(R)、FcγRIIa(H)、FcγRIIb、FcγRIIIa(F)に対して結合できることが示された(図3、4、5、6、7)。また、Fv4-IgG1-F157は同様に、ヒトFcRnに結合するのと同時に、マウスFcγRI、FcγRIIb、FcγRIII、FcγRIVに対しても結合できることが示された。(図8、9、10、11)
以上のことから、pH中性域の条件下におけるヒトFcRnに対する結合活性を有するヒト抗体が、 ヒトFcRnに結合するのと同時に、ヒトFcγRIa、FcγRIIa(R)、FcγRIIa(H)、FcγRIIb、FcγRIIIa(F)やマウスFcγRI、FcγRIIb、FcγRIII、FcγRIV等の各種ヒトFcγRおよび各種マウスFcγRに対しても結合できることが示された。
pH中性域における条件下でマウスFcRnに対する結合活性を有する抗体であるVH3-IgG1-F20(配列番号:43)とVL3-CK(配列番号:42)を含むFv4-IgG1-F20が、マウスFcRnに結合するのと同時に、各種マウスFcγRに結合するか否かが評価された。その結果、Fv4-IgG1-F20が、マウスFcRnに結合するのと同時に、マウスFcγRI、FcγRIIb、FcγRIII、FcγRIVに対して結合できることが示された(図12)。
pH中性域における条件下でマウスFcRnに対する結合能を有する抗体であるmPM1H-mIgG1-mF3(配列番号:44)とmPM1L-mk1(配列番号:45)を含むmPM1-mIgG1-mF3が、マウスFcRnに結合するのと同時に、各種マウスFcγRに結合するか否かが評価された。その結果、mPM1-mIgG1-mF3は、マウスFcRnに結合するのと同時に、マウスFcγRIIbおよびFcγRIIIに対して結合できることが示された(図13)。マウスFcγRIおよびIVに対して結合が確認されなかった結果は、マウスIgG1抗体はマウスFcγRIおよびIVに対して結合能を持たないとの報告(J. Immunol. (2011) 187 (4), 1754-1763))から判断すると、妥当な結果であると考えられる。
pH中性域におけるヒトFcRnに対する結合アフィニティを増大させることを目的として、様々な変異がVH3-IgG1(配列番号:46)に導入され、評価がなされた。作製された重鎖と軽鎖であるL (WT)-CK(配列番号:47)とを各々含む改変体(IgG1-F1からIgG1-F1379)が、実施例(1−1)に記載された方法に準じて発現および精製された。これらの抗体とヒトFcRnとの結合が、参考実施例1に記載される方法に準じて解析された。すなわち、Biacoreを用いた中性条件下(pH7.0)におけるヒトFcRnに対する改変体の結合活性を表5−1〜5−33に示した。
ヒト天然型IgG1に対してEUナンバリングで表される238位のProがAspに置換されたFcに対し、さらにFcγRIIbへの結合を上げると天然型抗体の解析から予測される他の改変を組み合わせても、期待される組合せ効果は得られなかった。そこで、EUナンバリングで表される238位のProがAspに置換された改変Fcに対して網羅的改変を導入することによって、さらにFcγRIIbへの結合を増強する改変体を見出すことが試みられた。抗体H鎖として用いられたIL6R-G1d(配列番号:48)のEUナンバリングで表される252位のMetをTyrに置換する改変、EUナンバリングで表される434位のAsnをTyrに置換する改変が導入されたIL6R-F11(配列番号:49)が作製された。さらに、IL6R-F11に対してEUナンバリングで表される238位のProをAspに置換する改変が導入されたIL6R-F652(配列番号:50)が作製された。L6R-F652に対し、EUナンバリングで表される238位の残基の近傍の領域(EUナンバリングで表される234位から237位、239位)が元のアミノ酸とシステインを除く18種類のアミノ酸にそれぞれ置換された抗体H鎖配列を含む発現プラスミドがそれぞれ調製された。抗体L鎖としてはIL6R-L(配列番号:51)が共通して用いられた。これらの改変体が実施例(1−1)と同様の方法により発現、精製された。これらのFc変異体はPD variantと呼ばれる。参考実施例1と同様の方法により各PD variantのFcγRIIa R型およびFcγRIIbに対する相互作用が網羅的に評価された。
〔式5〕
KD =C x Rmax / (Req - RI) - C
の式を利用して算出した値である。
先の参考実施例3に示した通り、P238Dを含むFcに対して、FcγRIIbとの結合活性を向上する、あるいはFcγRIIbへの選択性を向上させると天然型IgG1抗体の解析から予測された改変を導入しても、FcγRIIbに対する結合活性が減弱してしまうことが明らかとなり、この原因としてFcとFcγRIIbとの相互作用界面の構造がP238Dを導入することで変化していることが考えられた。そこで、この現象の原因を追及するためP238Dの変異をもつIgG1のFc(以下、Fc(P238D)と呼ぶ)とFcγRIIb細胞外領域との複合体の立体構造をX線結晶構造解析により明らかにし、天然型 IgG1のFc(以下、Fc (WT)と呼ぶ)とFcγRIIb細胞外領域との複合体との立体構造を対比することによって、これらの結合様式が比較された。なお、FcとFcγR細胞外領域との複合体の立体構造に関する複数の報告がすでにあり、Fc (WT) / FcγRIIIb細胞外領域複合体(Nature (2000) 400, 267-273、J. Biol. Chem. (2011) 276, 16469-16477)、Fc (WT) / FcγRIIIa細胞外領域複合体(Proc. Natl. Acad. Sci. USA (2011) 108, 12669-126674)、およびFc (WT) / FcγRIIa細胞外領域複合体(J. Immunol. (2011) 187, 3208-3217)の立体構造が解析されている。これまでにFc (WT) / FcγRIIb細胞外領域複合体の立体構造は解析されていないが、Fc(WT)との複合体の立体構造が既知であるFcγRIIaとFcγRIIbでは細胞外領域においてアミノ酸配列の93%が一致し、非常に高い相同性を有していることから、Fc (WT) / FcγRIIb細胞外領域複合体の立体構造はFc (WT) / FcγRIIa細胞外領域複合体の結晶構造からモデリングにより推定された。
P238D改変を含むFcの調製は以下のように行われた。まず、hIL6R-IgG1-v1(配列番号:53)のEUナンバリングで表される220位のCysをSerに置換し、EUナンバリングで表される236位のGluからそのC末端をPCRによってクローニングした遺伝子配列Fc (P238D)を参考実施例(1−1)に記載された方法と同様な方法で発現ベクターの作製、発現、精製が行われた。なお、EUナンバリングで表される220位のCysは通常のIgG1においては、L鎖のCysとdisulfide bondを形成しているが、Fcのみを調製する場合にはL鎖を共発現させないことから、不要なdisulfide bond形成を回避するために当該Cys残基はSerに置換された。
FcγRIIb細胞外領域は、公知のタンパク質の調製方法に準じて調製された。
結晶化に使用するため得られたFcγRIIb細胞外領域サンプル 2 mgに対し、glutathione S-transferaseとの融合蛋白として大腸菌により発現精製したEndo F1 (Protein Science (1996) 5, 2617-2622) 0.29 mgを加え、0.1M Bis-Tris pH6.5のBuffer条件で、室温にて3日間静置することにより、FcγRIIb細胞外領域のAsnに直接結合したN-acetylglucosamine以外のN型糖鎖が切断された。次に5000MWCOの限外ろ過膜により濃縮された糖鎖切断処理が施されたFcγRIIb細胞外領域サンプルが、20mM HEPS pH7.5, 0.05M NaClで平衡化したゲルろ過カラムクロマトグラフィー(Superdex200 10/300)により精製された。さらに得られた糖鎖切断FcγRIIb細胞外領域画分にFc (P238D)をモル比でFcγRIIb細胞外領域のほうが若干過剰となるよう混合された。10000MWCOの限外ろ過膜により濃縮された前記混合液を20 mM HEPS pH7.5、0.05M NaClで平衡化したゲルろ過カラムクロマトグラフィー(Superdex200 10/300)を用いて精製することによって、Fc (P238D) / FcγRIIb細胞外領域複合体のサンプルが得られた。
10000MWCOの限外ろ過膜 により約10 mg/mlまで濃縮された前記のFc (P238D) / FcγRIIb細胞外領域複合体の試料を用いて、シッティングドロップ蒸気拡散法により当該複合体が結晶化された。結晶化にはHydra II Plus One (MATRIX)を用い、100 mM Bis-Tris pH6.5、17% PEG3350、0.2 M Ammonium acetate、および2.7%(w/v) D-Galactoseのリザーバー溶液に対し、リザーバー溶液:結晶化サンプルを0.2μl:0.2μlで混合して結晶化ドロップを作成した。シールされた当該結晶化ドロップを20℃に静置することによって、薄い板状の結晶が得られた。
得られたFc (P238D) / FcγRIIb細胞外領域複合体の単結晶一つが100 mM Bis-Tris pH6.5、20% PEG3350、Ammonium acetate、2.7% (w/v) D-Galactose、Ethylene glycol 22.5%(v/v) の溶液に浸漬された。微小なナイロンループ付きのピンを用いて溶液ごとすくいとられた単結晶を液体窒素中で凍結させた。高エネルギー加速器研究機構の放射光施設フォトンファクトリーBL-1Aにて当該結晶のX線回折データが測定された。なお、測定中は常に-178℃の窒素気流中に置くことによって凍結状態を維持し、ビームラインに備え付けられたCCDディテクタQuantum 270(ADSC)によって、結晶を0.8°ずつ回転させながらトータル225枚のX線回折画像が収集された。得られた回折画像からの格子定数の決定、回折斑点の指数付け、ならびに回折データの処理には、プログラムXia2(CCP4 Software Suite)、XDS Package(Walfgang Kabsch)ならびにScala(CCP4 Software Suite)を用い、最終的に分解能2.46Åまでの当該結晶の回折強度データが得られた。本結晶は、空間群P21に属し、格子定数a=48.85Å、b=76.01Å、c=115.09Å、α=90°、β=100.70°、γ=90°であった。
Fc (P238D) / FcγRIIb細胞外領域複合体の結晶構造決定は、プログラムPhaser(CCP4 Software Suite)を用いた分子置換法によりおこなわれた。得られた結晶格子の大きさとFc (P238D) / FcγRIIb細胞外領域複合体の分子量から非対称単位中の複合体の数は一個と予想された。Fc (WT) / FcγRIIIa細胞外領域複合体の結晶構造であるPDB code:3SGJの構造座標から、A鎖239-340番ならびにB鎖239-340番のアミノ酸残基部分を別座標として取り出し、それぞれFc CH2ドメインの探索用モデルと設定した。同じくPDB code:3SGJの構造座標から、A鎖341-444番とB鎖341-443番のアミノ酸残基部分を一つの座標として取り出し、Fc CH3ドメインの探索用モデルと設定した。最後にFcγRIIb細胞外領域の結晶構造であるPDB code:2FCBの構造座標からA鎖6-178番のアミノ酸残基部分を取り出しFcγRIIb細胞外領域の探索用モデルと設定した。Fc CH3ドメイン、FcγRIIb細胞外領域、Fc CH2ドメインの順番に各探索用モデルの結晶格子内での向きと位置を、回転関数および並進関数から決定し、Fc (P238D) / FcγRIIb細胞外領域複合体結晶構造の初期モデルが得られた。得られた初期モデルに対し2つのFc CH2ドメイン、2つのFc CH3ドメインならびにFcγRIIb細胞外領域を動かす剛体精密化をおこなったところ、この時点で25-3.0Åの回折強度データに対し、結晶学的信頼度因子R値は40.4%、Free R値は41.9%となった。さらにプログラムRefmac5(CCP4 Software Suite)を用いた構造精密化と、実験的に決定された構造因子Foとモデルから計算された構造因子Fcならびにモデルから計算された位相をもとに算出された2Fo-Fc、Fo-Fcを係数とする電子密度マップを見ながらのモデル修正をプログラムCoot(Paul Emsley)でおこなった。これらの作業を繰り返すことによってモデルの精密化がおこなわれた。最後に2Fo-Fc、Fo-Fcを係数とする電子密度マップをもとに水分子をモデルに組み込み、精密化をおこなうことによって、最終的に分解能25-2.6Åの24291個の回折強度データを用い、4846個の非水素原子を含むモデルに対し、結晶学的信頼度因子R値は23.7%、Free R値は27.6%となった。
Fc (WT) / FcγRIIa細胞外領域複合体の結晶構造であるPDB code:3RY6の構造座標をベースに、プログラムDisovery Studio 3.1(Accelrys)のBuild Mutants機能を使い、FcγRIIbのアミノ酸配列と一致するように構造座標中のFcγRIIaに変異が導入された。その際、Optimization LevelをHigh、Cut Radiusを4.5とし、5つのモデルを発生させ、その中から最もエネルギースコアが良いものを採用し、Fc (WT) / FcγRIIb細胞外領域複合体のモデル構造と設定した。
参考実施例4で得られたFc (P238D)とFcγRIIb細胞外領域との複合体のX線結晶構造解析の結果に基づき、EUナンバリングで表される238位のProがAspに置換された改変FcにおいてFcγRIIbとの相互作用に影響を与えることが予測される部位(EUナンバリングで表される233位、240位、241位、263位、265位、266位、267位、268位、271位、273位、295位、296位、298位、300位、323位、325位、326位、327位、328位、330位、332位、334位の残基)に対して網羅的な改変が導入された改変体を構築することによって、P238D 改変に加えてさらにFcγRIIbとの結合を増強する改変の組合せを得ることが可能であるか検討された。
〔式5〕
KD =C x Rmax / (Req - RI) - C
の式を利用して算出した値である。
参考実施例3および5において得られた改変の中で、FcγRIIbへの結合を増強する効果もしくはFcγRIIbへの結合を維持し、他のFcγRへの結合を抑制する効果がみられた改変同士を組み合わせることによる効果が検証された。
〔式5〕
KD =C x Rmax / (Req - RI) - C
の式を利用して算出した値である。
FcγRIIbへの結合を増強する際に、他の活性型FcγRに対する結合を可能な限り抑制したうえで、FcγRIIbへの結合を増強することが好ましい。そこで、FcγRIIbへの結合を増強する、または選択性を向上する効果がある改変同士を組み合わせ、さらにFcγRIIbへの結合が増強されまたは選択性が向上した改変体が作製された。具体的には、FcγRIIbへの結合の増強、選択性の向上において優れた効果を示すP238Dの改変を基礎とし、参考実施例3、参考実施例5、参考実施例6においてP238Dの改変と組み合わせることで効果が見られた改変同士がさらに組み合わされた。
〔式5〕
KD =C x Rmax / (Req - RI) - C
の式を利用して算出した値である。
参考実施例7において最もFcγRIIbへの結合が増強された改変体IL6R-BP230/IL6R-LのFcγRIIbへの結合は、改変導入前のIL6R-B3/IL6R-Lと比較して約150倍に増強され、FcγRIIaRへの結合も1.9倍程度に抑えられている。従ってIL6R-BP230/IL6R-LはFcγRIIbへの結合、選択性共に優れた改変体であるが、可能な限りFcγRIIaRへの結合を抑制したうえでFcγRIIbへの結合がさらに増強されたより好ましい改変体を作製する可能性が模索された。
[Fc (P208) の発現精製]
Fc (P208)は以下のように調製された。まず、IL6R-BP208の、EUナンバリングで表される439位のGluを天然型ヒトIgG1の配列であるLysにしたIL6R-P208が作製された。次にEUナンバリングで表される220位のCysがSerに置換された改変体をコードするDNAを鋳型として、EUナンバリングで表される216位のGluからそのC末端がPCRによってクローニングされた遺伝子配列Fc (P208)がクローニングされた。参考実施例14に記載の方法にしたがって発現ベクターの作製、発現、精製を行った。なお、通常のIgG1のEUナンバリングで表される220位のCysは、L鎖に存在するCysとジスルフィド結合を形成しているが、Fc領域のみを調製する場合、L鎖を共発現させないため、不要なdisulfide bond形成を回避するために当該220位のCysはSerに置換された。
FcγRIIb細胞外領域は、参考実施例15の方法にしたがって調製された。
glutathione S-transferaseとの融合蛋白として大腸菌内で発現されたEndo F1(Protein Science (1996) 5, 2617-2622)の精製産物0.15 mgが加えられた、結晶化のために得られた1.5 mgのFcγRIIb細胞外領域サンプルを、0.1M Bis-Tris pH6.5のバッファー中で、室温にて3日間静置することにより、当該FcγRIIb細胞外領域サンプル中のN型糖鎖がAsnに直接結合したN-acetylglucosamineを残して切断された。次に5000MWCOの限外ろ過膜により濃縮された前記の糖鎖切断処理が施されたFcγRIIb細胞外領域サンプルは、20mM HEPES pH7.5, 0.1M NaClで平衡化されたゲルろ過カラムクロマトグラフィー(Superdex200 10/300)によって精製された。さらに、Fc (P208) がモル比でFcγRIIb細胞外領域のほうが若干過剰となるよう加えられた、前記の糖鎖切断FcγRIIb細胞外領域の精製画分は、10000MWCOの限外ろ過膜により濃縮され、その後、25mM HEPES pH7.5, 0.1M NaClで平衡化されたゲルろ過カラムクロマトグラフィー(Superdex200 10/300)により精製された。前記のように得られた精製画分が、Fc (P208) / FcγRIIb細胞外領域複合体のサンプルとして以後の検討に使用された。
10000MWCOの限外ろ過膜 により約10 mg/mlまで濃縮されたFc (P208) / FcγRIIb細胞外領域複合体のサンプルは、ハンギングドロップ蒸気拡散法によりSeeding法を併用して結晶化された。結晶化にはVDXmプレート(Hampton Research)を用い、0.1M Bis-Tris pH6.5、19%(w/v) PEG3350、0.2 M Potassium Phosphate dibasicのリザーバー溶液に対し、リザーバー溶液:結晶化サンプル=0.85μl:0.85μlで混合して作成された結晶化ドロップに、同様な条件で得られた同複合体の結晶をSeed Bead(Hampton Research)を用いて砕いた種結晶溶液から作成した希釈溶液0.15μlを添加し、リザーバーの入ったウェルに密閉、20℃に静置することにより、板状の結晶が得られた。
前記のように得られたFc (P208) / FcγRIIb細胞外領域複合体の単結晶の一つを0.1M Bis-Tris pH6.5、24% (w/v) PEG3350、0.2 M Potassium Phosphate dibasic、20% (v/v) Ethylene glycolの溶液に浸した後、微小なナイロンループ付きのピンで溶液ごとすくいとり、液体窒素中で凍結させた当該単結晶からのX線回折データは、Spring-8のBL32XUにて測定された。なお、測定中、常に-178℃の窒素気流中に置くことで凍結状態が維持され、ビームライン備え付けのCCDディテクタMX-225HE(RAYONIX)により、当該単結晶を0.6°ずつ回転させながら計300枚の当該単結晶のX線回折画像が収集された。得られた回折画像からの格子定数の決定、回折斑点の指数付け、および回折データの処理には、プログラムXia2(J. Appl. Cryst. (2010) 43, 186-190)、XDS Package(Acta Cryst. (2010) D66, 125-132)およびScala(Acta Cryst. (2006) D62, 72-82)を用い、最終的に分解能2.81Åまでの当該単結晶の回折強度データが得られた。本結晶は、空間群C2221に属し、格子定数a=156.69Å、b=260.17Å、c=56.85Å、α=90°、β=90°、γ=90°であった。
Fc (P208) / FcγRIIb細胞外領域複合体の構造は、プログラムPhaser(J. Appl. Cryst. (2007) 40, 658-674)を用いた分子置換法により決定された。得られた結晶格子の大きさとFc (P208) / FcγRIIb細胞外領域複合体の分子量から非対称単位中の複合体の数は一個と予想された。Fc (WT) / FcγRIIIa細胞外領域複合体の結晶構造であるPDB code:3SGJの構造座標から別座標として取り出された、A鎖239-340番ならびにB鎖239-340番のアミノ酸残基部分が、Fc領域のCH2ドメインの探索用モデルとして使用された。同じくPDB code:3SGJの構造座標から一つの座標として取り出された、A鎖341-444番とB鎖341-443番のアミノ酸残基部分がFc領域のCH3ドメインの探索用モデルとして使用された。最後にFcγRIIb細胞外領域の結晶構造であるPDB code:2FCB の構造座標から取り出されたA鎖6-178番のアミノ酸残基部分がFc (P208) の探索用モデルとして使用された。Fc領域のCH3ドメイン、FcγRIIb細胞外領域、Fc領域のCH2ドメインの各探索用モデルの結晶格子内での向きと位置を、回転関数および並進関数から決定しようとしたところ、CH2ドメインの一つの位置がうまく決定されなかった。そこで残りの三つの部分から計算された位相をもとに計算された電子密度マップから、Fc (WT) / FcγRIIIa細胞外領域複合体の結晶構造を参考にしながら最後のCH2ドメインAの位置が決定され、Fc (P208) / FcγRIIb細胞外領域複合体結晶構造の初期モデルが得られた。得られた初期モデルに対し二つのFc領域のCH2ドメイン、二つのFc領域のCH3ドメインおよびFcγRIIb細胞外領域を動かす剛体精密化をおこなったところ、剛体精密化が行われた時点で25-3.0Åの回折強度データを用いた構造モデルの結晶学的信頼度因子R値は42.6%、Free R値は43.7%となった。さらにプログラムREFMAC5(Acta Cryst. (2011) D67, 355-367)を用いた構造精密化と、実験的に決定された構造因子Foと構造モデルから計算された構造因子Fc領域および構造モデルから計算された位相をもとに算出された2Fo-Fc、Fo-Fcを係数とする電子密度マップをみながら、構造モデルをプログラムCoot(Acta Cryst. (2010) D66, 486-501)を用いて修正する作業を繰り返すことで構造モデルの精密化が行われた。さらに2Fo-Fc、Fo-Fcを係数とする電子密度マップをもとに水分子を構造モデルに組み込み、精密化を行うことで、最終的に分解能25-2.81Åの27259個の回折強度データを用いた、4786個の非水素原子を含む構造モデルの、結晶学的信頼度因子R値は24.5%、Free R値は28.2%となった。
[FcγRIIaR細胞外領域の発現精製]
FcγRIIaR細胞外領域は、参考実施例15の方法にしたがって調製された。
S-transferaseとの融合蛋白として大腸菌内で発現されたEndo F1(Protein Science (1996) 5, 2617-2622)の精製産物0.15 mg、20μlの5 U/ml 濃度のEndo F2(QA-bio)および20μlの5 U/ml 濃度のEndo F3(QA-bio)が加えられた、1.5 mgの精製されたFcγRIIa Rの細胞外領域サンプルを、0.1M Na Acetate pH4.5のバッファー中で、室温にて9日間静置の後、さらに、0.07 mgの前記Endo F1、7.5μl の前記Endo F2および7.5μl の前記Endo F3の追加後3日間静置することにより、当該FcγRIIa R細胞外領域サンプル中のN型糖鎖がAsnに直接結合したN-acetylglucosamineを残して切断された。次に10000MWCOの限外ろ過膜により濃縮された前記の糖鎖切断処理が施されたFcγRIIaR細胞外領域サンプルは、25 mM HEPES pH7、0.1M NaClで平衡化されたゲルろ過カラムクロマトグラフィー(Superdex200 10/300)によって精製された。さらに、Fc (P208) をモル比でFcγRIIaR細胞外領域のほうが若干過剰となるよう加えられた、前記の糖鎖切断FcγRIIaR細胞外領域の精製画分は、10000MWCOの限外ろ過膜による濃縮後、25mM HEPES pH7、0.1M NaClで平衡化されたゲルろ過カラムクロマトグラフィー(Superdex200 10/300)により精製された。前記のように得られた精製画分が、Fc (P208)/FcγRIIaR型細胞外領域複合体のサンプルとして以後の検討に使用された。
10000MWCOの限外ろ過膜 により約10 mg/mlまで濃縮されたFc (P208)/FcγRIIa R型細胞外領域複合体のサンプルは、シッテイングドロップ蒸気拡散法にて結晶化された。0.1 M Bis-Tris pH7.5、26% (w/v) PEG3350、0.2 M Ammonium Sulfaeのリザーバー溶液に対し、リザーバー溶液:結晶化サンプル=0.8μl:1.0μlで混合して作成された結晶化ドロップを、シールで密閉して20℃に静置することにより、板状の結晶が得られた。
前記のように得られたFc (P208)/FcγRIIaR細胞外領域複合体の単結晶の一つを0.1 M Bis-Tris pH7.5、27.5% (w/v) PEG3350、0.2 M Ammonium Sulfate、20% (v/v) Glycerolの溶液に浸した後、微小なナイロンループ付きのピンで溶液ごとすくいとり、液体窒素中で凍結させた当該単結晶中からのX線回折データは、高エネルギー加速器研究機構の放射光施設フォトンファクトリーBL-17Aにて測定された。なお、測定中、常に-178℃の窒素気流中に置くことで凍結状態が維持され、ビームライン備え付けのCCDディテクタQuantum 315r(ADSC)により、当該単結晶を0.6°ずつ回転させながら計225枚の当該単結晶のX線回折画像が収集された。得られた回折画像からの格子定数の決定、回折斑点の指数付け、および回折データの処理には、プログラムXia2(J. Appl. Cryst. (2010) 43, 186-190)、XDS Package(Acta Cryst. (2010) D66, 125-132)およびScala(Acta Cryst. (2006) D62, 72-82)を用い、最終的に分解能2.87Åまでの回折強度データが得られた。本結晶は、空間群C2221に属し、格子定数a=154.31Å、b=257.61Å、c=56.19Å、α=90°、β=90°、γ=90°であった。
Fc (P208)/FcγRIIaR型細胞外領域複合体の構造は、プログラムPhaser(J. Appl. Cryst. (2007) 40, 658-674)を用いた分子置換法により決定された。得られた結晶格子の大きさとFc (P208)/FcγRIIaR細胞外領域複合体の分子量から非対称単位中の複合体の数は一個と予想された。実施例(8−1)で得られたFc (P208)/FcγRIIb細胞外領域複合体の結晶構造を探索用モデルとして用いて、Fc (P208)/FcγRIIaR細胞外領域複合体の結晶格子内での向きと位置が、回転関数および並進関数から決定された。さらに得られた初期構造モデルに対し、二つのFc領域のCH2ドメイン、二つのFc領域のCH3ドメインおよびFcγRIIaR細胞外領域を独立に動かす剛体精密化をおこなったところ、剛体精密化が終了した時点で25-3.0Åの回折強度データを用いた構造モデルの結晶学的信頼度因子R値は38.4%、Free R値は30.0%となった。さらにプログラムREFMAC5(Acta Cryst. (2011) D67, 355-367)を用いた構造精密化と、実験的に決定された構造因子Foとモデルから計算された構造因子Fcならびにモデルから計算された位相をもとに算出された2Fo-Fc、Fo-Fcを係数とする電子密度マップをみながら当該構造モデルをプログラムCoot(Acta Cryst. (2010) D66, 486-501)を用いて修正する作業を繰り返すことで構造モデルの精密化が行われた。最後に2Fo-Fc、Fo-Fcを係数とする電子密度マップをもとに水分子をモデルに組み込み、精密化をおこなうことで、最終的に分解能25-2.87Åの24838個の回折強度データを用いた、4758個の非水素原子を含む構造モデルの、結晶学的信頼度因子R値は26.3%、Free R値は38.0%となった。
参考実施例8においてFcγRIIbへの結合が増強された改変体(P208)のドメインB内で、P271G改変の導入にともなう周囲の構造変化の結果として、EUナンバリングで表される268位のAspが、EUナンバリングで表される292位のArgと静電相互作用していることが示唆された(図33)。この相互作用形成がEUナンバリングで表される266位から271位のループ構造の安定化に働き、結果としてFcγRIIbへの結合増強に寄与した可能性が考えられた。そこで、当該改変体のEUナンバリングで表される268位のAspをGluに置換することによって、前記の静電相互作用が強化され、当該改変体のループ構造をさらに安定化させることで、FcγRIIbへの結合増強が可能か否かが検討された。また図32に示したとおり、FcγRIIbのEUナンバリングで表される160位のTyrは、P208のドメインAのEUナンバリングで表される237位のAspの主鎖と相互作用している。一方、EUナンバリングで表される237位のAspの側鎖は、分子内のEUナンバリングで表される332位のIle、333位のGlu、334位のLysの近傍に位置しているが特に目立った相互作用は形成していない。そこで、これらの部位を親水性アミノ酸残基に置換することでEUナンバリングで表される237位のAspの側鎖との相互作用を強めて、EUナンバリングで表される266位から271位のループ構造を安定化することで、FcγRIIbの160番目のTyrとの相互作用が増強されるか否かが、併せて検証された。
〔式5〕
KD =C x Rmax / (Req - RI) - C
の式を利用して算出した値である。
Fc (P208)とFcγRIIbの構造とFc (P238D)/FcγRIIbの構造の比較において最も違いがみられるのは、Fc領域のCH2ドメインBのEUナンバリングで表される271位の近傍の構造である(図32)。参考実施例4に示したようにFc (P238D)ではEUナンバリングで表される270位のAspがFcγRIIbの131番目のArgと強固な静電相互作用を形成する際に、EUナンバリングで表される271位のPro部分に立体化学的なストレスがかかっている可能性が示唆された。Fc (P208)/FcγRIIbの構造においては、EUナンバリングで表される271位のProのGlyへの置換により、この構造的なひずみを解消するよう主鎖レベルでの位置変化がおきており、その結果EUナンバリングで表される271位の近傍のループ構造が大きく変化したと考えられる。この変化した271位の近傍の構造をさらに安定化させることができれば、FcγRIIbの131番目のArgとの静電相互作用を形成結合にともなうエントロピー的なエネルギー損失をさらに軽減できる可能性も考えられた。そこで、EUナンバリングで表される271位の周辺のアミノ酸残基に対する改変を網羅的に導入することにより、Fc領域のFcγRIIbに対する結合の増強あるいは選択性の向上を示す改変が探索された。
〔式5〕
KD =C x Rmax / (Req - RI) - C
の式を利用して算出した値である。
EUナンバリングで表される396位のProをLeuに置換する改変はFcγRIIbへの結合を増強することが報告されている(Cancer Res. (2007) 67, 8882-8890)。EUナンバリングで表される396位のアミノ酸はFcγRとの相互作用には直接関与しない部位であるが、抗体の構造を変化させることでFcγRとの相互作用に影響を与えるとも考えられる。そこで、Fc領域のEUナンバリングで表される396位のアミノ酸に改変を網羅的に導入することにより、Fc慮域のFcγRIIbへの結合あるいは選択性が向上するか否かが検証された。
〔式5〕
KD =C x Rmax / (Req - RI) - C
の式を利用して算出した値である。
ヒトIgGに存在するサブクラスによって、FcγRへの結合プロファイルが異なる。IgG1とIgG4の各FcγRへの結合活性の違いを、FcγRIIbへの結合活性の増強、および/または選択性の向上に利用できるか否かが検証された。はじめに、IgG1とIgG4の各FcγRへの結合活性が解析された。抗体H鎖としては、ヒトIgG4 のEUナンバリングで表される228位のSerがProに置換され、C末端のGlyおよびLysが除去されたFc領域であるG4dを含むIL6R-G4d(配列番号:56)が抗体H鎖として作製された。抗体L鎖としてはIL6R-L(配列番号:51)が用いられた。参考実施例14の方法に従い発現した、IL6R-Lの軽鎖とIL6R-G1d/IL6R-LまたはIL6R-G4d/IL6R-Lを含む抗体が精製された。精製された抗体の各FcγR(FcγRIa、FcγRIIaH、FcγRIIaR、FcγRIIb、FcγRIIIaV)に対する結合が、参考実施例15の方法によって評価された。得られた改変体の各FcgRへの結合を表21にまとめた。
〔式5〕
KD =C x Rmax / (Req - RI) - C
の式を利用して算出した値である。
参考実施例12までの実施例においてFcγRIIbへの結合の増強あるいは選択性の向上の面で、効果がみられた改変のさらなる組合せが検討された。具体的には、IL6R-B3(配列番号:54)に対してこれまでの検討の中でFcγRIIbへの結合の増強、および/または選択性の向上をもたらした改変の組合せが導入された。また比較対照として既存のFcγRIIbへの結合を増強する改変(Seungら(Mol. Immunol. (2008) 45, 3926-3933))である、S267E、およびL328Fの改変がIL6R-B3に導入されたIL6R-BP253が作製された。抗体L鎖としてはIL6R-L(配列番号:51)が用いられた。参考実施例14の方法に従い発現した、前記の重鎖の改変体とIL6R-Lの軽鎖を含む抗体が精製された。精製された抗体の各FcγR(FcγRIa、FcγRIIaH、FcγRIIaR、FcγRIIb、FcγRIIIaV)に対する結合が、参考実施例15の方法によって評価された。
〔式5〕
KD =C x Rmax / (Req - RI) - C
の式を利用して算出した値である。
抗体の可変領域のH鎖およびL鎖の塩基配列をコードする全長の遺伝子の合成は、Assemble PCR等を用いて、当業者公知の方法で作製した。アミノ酸置換の導入はPCR等を用いて当業者公知の方法で行った。得られたプラスミド断片を動物細胞発現ベクターに挿入し、H鎖発現ベクターおよびL鎖発現ベクターを作製した。得られた発現ベクターの塩基配列は当業者公知の方法で決定した。作製したプラスミドをヒト胎児腎癌細胞由来HEK293H株(Invitrogen社)、またはFreeStyle293細胞(Invitrogen社)に、一過性に導入し、抗体の発現を行った。得られた培養上清を回収した後、0.22μmフィルターMILLEX(R)-GV(Millipore)、または0.45μmフィルターMILLEX(R)-GV(Millipore)を通して培養上清を得た。得られた培養上清から、rProtein A Sepharose Fast Flow(GEヘルスケア)またはProtein G Sepharose 4 Fast Flow(GEヘルスケア)を用いて当業者公知の方法で、抗体を精製した。精製抗体濃度は、分光光度計を用いて280 nmでの吸光度を測定し、得られた値からPACE等の方法により算出された吸光係数を用いて抗体濃度を算出した(Protein Science (1995); 4, 2411-2423)。
FcγRの細胞外ドメインを以下の方法で調製した。まずFcγRの細胞外ドメインの遺伝子の合成を当業者公知の方法で実施した。その際、各FcγRの配列はNCBIに登録されている情報に基づき作製した。具体的には、FcγRIについてはNCBIのアクセッション番号NM_000566.3(バージョン番号NM_000566.3)の配列、FcγRIIaについてはNCBIのアクセッション番号NM_001136219.1(バージョン番号NM_001136219.1)の配列、FcγRIIbについてはNCBIのアクセッション番号NM_004001.3(バージョン番号NM_004001.3)の配列、FcγRIIIaについてはNCBIのアクセッション番号NM_001127593.1(バージョン番号NM_001127593.1)の配列、FcγRIIIbについてはNCBIのアクセッション番号NM_000570.3(バージョン番号NM_000570.3)の配列に基づいて作製し、C末端にHisタグを付加した。またFcγRIIa、FcγRIIIa、FcγRIIIbは多型が存在することが知られているが、FcγRIIaの多型部位はWarmerdamら(J. Exp. Med. (1990) 172, 19-25)、FcγRIIIaの多型部位はWuら(J. Clin. Invest. (1997) 100 (5), 1059-1070)、FcγRIIIbの多型部位はOryら(J. Clin. Invest. (1989) 84, 1688-1691)を参考にして作製した。
〔式6〕
Req =C x Rmax / (KD + C) + RI
Req: a plot of steady state binding levels against analyte concentration
C: concentration
RI: bulk refractive index contribution in the sample
Rmax: analyte binding capacity of the surface
〔式5〕
KD =C x Rmax / (Req - RI) - C
Claims (22)
- 脳組織内に存在し、脳疾患に固有な抗原に対する結合活性がイオン濃度の条件によって変化するヒトIgG可変領域の抗原結合ドメイン、ならびにpH7.0の条件下で天然型のヒトIgGのFc領域より高いFcRnに対する結合活性を有するヒトIgG由来のFc領域およびpH5.8の条件下でFcRnに対する結合活性を有するヒトIgG由来のFc領域を含む抗原結合分子を含む、脳疾患治療剤。
- 前記抗原結合ドメインが、前記抗原に対する結合活性がカルシウムイオン濃度の条件によって変化する抗原結合ドメインである、請求項1に記載の治療剤。
- 前記抗原結合ドメインが、前記抗原に対する低カルシウムイオン濃度の条件下での結合活性が前記抗原に対する高カルシウムイオン濃度の条件下での結合活性よりも低いように結合活性が変化する抗原結合ドメインである、請求項2に記載の治療剤。
- 前記抗原結合ドメインが、前記抗原に対する結合活性がpHの条件によって変化する抗原結合ドメインである、請求項1に記載の治療剤。
- 前記抗原結合ドメインが、前記抗原に対するpH5.8の条件下での結合活性がpH7.0の条件下での結合活性よりも低いように結合活性が変化する抗原結合ドメインである、請求項4に記載の治療剤。
- 前記pH5.8の条件下でFcRnに対する結合活性を有するヒトIgG由来のFc領域が、配列番号:14、15、16または17のいずれかに含まれるFc領域である、請求項1に記載の治療剤。
- 前記pH5.8の条件下でFcRnに対する結合活性を有するヒトIgG由来のFc領域が、pH5.8の条件下でのFcRnに対する結合活性が、配列番号:14、15、16または17のいずれかに含まれるFc領域のFcRnに対する結合活性より増強されているFc領域である、請求項1に記載の治療剤。
- 前記Fc領域が、配列番号:14、15、16、または17のいずれかに含まれるFc領域のアミノ酸配列のうち、EUナンバリングで表される238位、244位、245位、249位、250位、251位、252位、253位、254位、255位、256位、257位、258位、260位、262位、265位、270位、272位、279位、283位、285位、286位、288位、293位、303位、305位、307位、308位、309位、311位、312位、314位、316位、317位、318位、332位、339位、340位、341位、343位、356位、360位、362位、375位、376位、377位、378位、380位、382位、385位、386位、387位、388位、389位、400位、413位、415位、423位、424位、427位、428位、430位、431位、433位、434位、435位、436位、438位、439位、440位、442位または447位の群から選択される少なくとも一つ以上のアミノ酸が置換されているFc領域である、請求項7に記載の治療剤。
- 前記Fc領域が、配列番号:14、15、16、または17に含まれるFc領域のアミノ酸配列のうち、EUナンバリングで表される;
238位のアミノ酸がLeu、
244位のアミノ酸がLeu、
245位のアミノ酸がArg、
249位のアミノ酸がPro、
250位のアミノ酸がGlnまたはGluのいずれか、もしくは
251位のアミノ酸がArg、Asp、Glu、またはLeuのいずれか、
252位のアミノ酸がPhe、Ser、Thr、またはTyrのいずれか、
254位のアミノ酸がSerまたはThrのいずれか、
255位のアミノ酸がArg、Gly、Ile、またはLeuのいずれか、
256位のアミノ酸がAla、Arg、Asn、Asp、Gln、Glu、Pro、またはThrのいずれか、
257位のアミノ酸がAla、Ile、Met、Asn、Ser、またはValのいずれか、
258位のアミノ酸がAsp、
260位のアミノ酸がSer、
262位のアミノ酸がLeu、
270位のアミノ酸がLys、
272位のアミノ酸がLeu、またはArgのいずれか、
279位のアミノ酸がAla、Asp、Gly、His、Met、Asn、Gln、Arg、Ser、Thr、Trp、またはTyrのいずれか、
283位のアミノ酸がAla、Asp、Phe、Gly、His、Ile、Lys、Leu、Asn、Pro、Gln、Arg、Ser、Thr、Trp、またはTyrのいずれか、
285位のアミノ酸がAsn、
286位のアミノ酸がPhe、
288位のアミノ酸がAsn、またはProのいずれか、
293位のアミノ酸がVal、
307位のアミノ酸がAla、Glu、Gln、またはMetのいずれか、
311位のアミノ酸がAla、Glu、Ile、Lys、Leu、Met、Ser 、Val、またはTrpのいずれか、
309位のアミノ酸がPro、
312位のアミノ酸がAla、Asp、またはProのいずれか、
314位のアミノ酸がAlaまたはLeuのいずれか、
316位のアミノ酸がLys、
317位のアミノ酸がPro、
318位のアミノ酸がAsn、またはThrのいずれか、
332位のアミノ酸がPhe、His、Lys、Leu、Met、Arg、Ser、またはTrpのいずれか、
339位のアミノ酸がAsn、Thr、またはTrpのいずれか、
341位のアミノ酸がPro、
343位のアミノ酸がGlu、His、Lys、Gln、Arg、Thr、またはTyrのいずれか、
375位のアミノ酸がArg、
376位のアミノ酸がGly、Ile、Met、Pro、Thr、またはValのいずれか、
377位のアミノ酸がLys、
378位のアミノ酸がAsp、Asn、またはValのいずれか、
380位のアミノ酸がAla、Asn、Ser、またはThrのいずれか、
382位のアミノ酸がPhe、His、Ile、Lys、Leu、Met、Asn、Gln、Arg、Ser、Thr、Val、Trp、またはTyrのいずれか、
385位のアミノ酸がAla、Arg、Asp、Gly、His、Lys、Ser、またはThrのいずれか、
386位のアミノ酸がArg、Asp、Ile、Lys、Met、Pro、Ser、またはThrのいずれか、
387位のアミノ酸がAla、Arg、His、Pro、Ser、またはThrのいずれか、
389位のアミノ酸がAsn、Pro、またはSerのいずれか、
423位のアミノ酸がAsn、
427位のアミノ酸がAsn、
428位のアミノ酸がLeu、Met、Phe、Ser、またはThrのいずれか、
430位のアミノ酸がAla、Phe、Gly、His、Ile、Lys、Leu、Met、Asn、Gln、Arg、Ser、Thr、Val、またはTyrのいずれか、
431位のアミノ酸がHis、またはAsnのいずれか、
433位のアミノ酸がArg、Gln、His、Ile、Lys、Pro、またはSerのいずれか、
434位のアミノ酸がAla、Gly、His、Phe、Ser、Trp、またはTyrのいずれか、
436位のアミノ酸がArg、Asn、His、Ile、Leu、Lys、Met、またはThrのいずれか、
438位のアミノ酸がLys、Leu、Thr、またはTrpのいずれか、
440位のアミノ酸がLys、もしくは、
442位のアミノ酸がLys、308位のアミノ酸がIle、Pro、またはThrのいずれか、
の群から選択される少なくとも一つ以上のアミノ酸を含む、請求項8に記載の治療剤。 - 前記pH7.0の条件下で天然型のヒトIgGのFc領域より高いFcRnに対する結合活性を有するヒトIgG由来のFc領域が、配列番号:14、15、16、または17に含まれるFc領域のアミノ酸配列のうち、EUナンバリングで表される237位、248位、250位、252位、254位、255位、256位、257位、258位、265位、286位、289位、297位、298位、303位、305位、307位、308位、309位、311位、312位、314位、315位、317位、332位、334位、360位、376位、380位、382位、384位、385位、386位、387位、389位、424位、428位、433位、434位または436位の群から選択される少なくとも一つ以上のアミノ酸が置換されているFc領域である、請求項1に記載の治療剤。
- 前記Fc領域が、配列番号:14、15、16、または17に含まれるFc領域のアミノ酸配列のうち、EUナンバリングで表される;
237位のアミノ酸がMet、
248位のアミノ酸がIle、
250位のアミノ酸がAla、Phe、Ile、Met、Gln、Ser、Val、Trp、またはTyrのいずれか、
252位のアミノ酸がPhe、Trp、またはTyrのいずれか、
254位のアミノ酸がThr、
255位のアミノ酸がGlu、
256位のアミノ酸がAsp、Asn、Glu、またはGlnのいずれか、
257位のアミノ酸がAla、Gly、Ile、Leu、Met、Asn、Ser、Thr、またはValのいずれか、
258位のアミノ酸がHis、
265位のアミノ酸がAla、
286位のアミノ酸がAlaまたはGluのいずれか、
289位のアミノ酸がHis、
297位のアミノ酸がAla、
303位のアミノ酸がAla、
305位のアミノ酸がAla、
307位のアミノ酸がAla、Asp、Phe、Gly、His、Ile、Lys、Leu、Met、Asn、Pro、Gln、Arg、Ser、Val、Trp、またはTyrのいずれか、
308位のアミノ酸がAla、Phe、Ile、Leu、Met、Pro、Gln、またはThrのいずれか、
309位のアミノ酸がAla、Asp、Glu、Pro、またはArgのいずれか、
311位のアミノ酸がAla、His、またはIleのいずれか、
312位のアミノ酸がAlaまたはHisのいずれか、
314位のアミノ酸がLysまたはArgのいずれか、
315位のアミノ酸がAla、AspまたはHisのいずれか、
317位のアミノ酸がAla、
332位のアミノ酸がVal、
334位のアミノ酸がLeu、
360位のアミノ酸がHis、
376位のアミノ酸がAla、
380位のアミノ酸がAla、
382位のアミノ酸がAla、
384位のアミノ酸がAla、
385位のアミノ酸がAspまたはHisのいずれか、
386位のアミノ酸がPro、
387位のアミノ酸がGlu、
389位のアミノ酸がAlaまたはSerのいずれか、
424位のアミノ酸がAla、
428位のアミノ酸がAla、Asp、Phe、Gly、His、Ile、Lys、Leu、Asn、Pro、Gln、Ser、Thr、Val、Trp、またはTyrのいずれか、
433位のアミノ酸がLys、
434位のアミノ酸がAla、Phe、His、Ser、Trp、またはTyrのいずれか、もしくは
436位のアミノ酸がHis 、Ile、Leu、Phe、Thr、またはVal、
の群から選択される少なくとも一つ以上のアミノ酸を含む、請求項10に記載の治療剤。 - 前記Fc領域が、FcγR結合活性が低いまたは低減されたFc領域である、請求項10または11に記載の治療剤。
- 前記抗原結合分子がさらにFcγR結合ドメインを含む、請求項1から12のいずれか一項に記載の治療剤。
- 前記FcγR結合ドメインが抗体のFc領域である、請求項13に記載の治療剤。
- 前記Fc領域が、天然型ヒトIgGのFc領域のFcγレセプターに対する結合活性よりもFcγレセプターに対する結合活性が高いFcγR結合改変Fc領域を含む、請求項14に記載の治療剤。
- 前記抗原結合分子がさらに補体結合ドメインを含む、請求項1から15のいずれか一項に記載の治療剤。
- 前記補体結合ドメインが抗体のFc領域である、請求項16に記載の治療剤。
- 前記Fc領域が、天然型ヒトIgGのFc領域の補体に対する結合活性よりも補体に対する結合活性が高い補体結合改変Fc領域を含む、請求項17に記載の治療剤。
- 前記FcγR結合ドメインが抗体の可変領域である、請求項13に記載の治療剤。
- 前記補体結合ドメインが抗体の可変領域である、請求項16に記載の治療剤。
- 前記抗原結合分子が抗体である、請求項1から20のいずれか一項に記載の治療剤。
- 請求項1から21のいずれか一項に記載の抗原結合分子をコードするポリヌクレオチドが作用可能に連結されたベクターが導入された細胞の培養液から抗原結合分子を回収する工程を含む、脳疾患治療剤の製造方法。
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2012185867A JP6280300B2 (ja) | 2012-08-24 | 2012-08-24 | 脳疾患治療剤 |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2012185867A JP6280300B2 (ja) | 2012-08-24 | 2012-08-24 | 脳疾患治療剤 |
Publications (3)
| Publication Number | Publication Date |
|---|---|
| JP2014043405A JP2014043405A (ja) | 2014-03-13 |
| JP2014043405A5 JP2014043405A5 (ja) | 2015-10-01 |
| JP6280300B2 true JP6280300B2 (ja) | 2018-02-14 |
Family
ID=50394958
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2012185867A Active JP6280300B2 (ja) | 2012-08-24 | 2012-08-24 | 脳疾患治療剤 |
Country Status (1)
| Country | Link |
|---|---|
| JP (1) | JP6280300B2 (ja) |
Families Citing this family (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US9840553B2 (en) | 2014-06-28 | 2017-12-12 | Kodiak Sciences Inc. | Dual PDGF/VEGF antagonists |
| RU2744860C2 (ru) | 2015-12-30 | 2021-03-16 | Кодиак Сайенсиз Инк. | Антитела и их конъюгаты |
| US11858980B2 (en) | 2016-08-02 | 2024-01-02 | Visterra, Inc. | Engineered polypeptides and uses thereof |
| EP3898667A2 (en) * | 2018-12-20 | 2021-10-27 | F. Hoffmann-La Roche AG | Modified antibody fcs and methods of use |
| CA3157509A1 (en) | 2019-10-10 | 2021-04-15 | Kodiak Sciences Inc. | Methods of treating an eye disorder |
Family Cites Families (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| AU2011337704B2 (en) * | 2010-11-30 | 2017-06-15 | Chugai Seiyaku Kabushiki Kaisha | Antigen-binding molecule capable of binding to plurality of antigen molecules repeatedly |
-
2012
- 2012-08-24 JP JP2012185867A patent/JP6280300B2/ja active Active
Also Published As
| Publication number | Publication date |
|---|---|
| JP2014043405A (ja) | 2014-03-13 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP7411698B2 (ja) | FcγRIIBを介して抗原の消失を促進する抗原結合分子 | |
| JP7441284B2 (ja) | 免疫複合体を形成する細胞内への運搬体(キャリア)を含む医薬 | |
| JP6826620B2 (ja) | 抗原結合分子の血漿中滞留性と免疫原性を改変する方法 | |
| JP6261785B2 (ja) | 抗原の消失を促進する抗原結合分子 | |
| JP2021074002A (ja) | 抗原結合分子の血漿中滞留性と免疫原性を改変する方法 | |
| TWI681970B (zh) | 包含依離子濃度之條件對抗原之結合活性會改變之抗原結合分域、及於pH中性之條件下對FcRn有結合活性之FcRn結合分域、且誘導對標的抗原的免疫反應之抗原結合分子 | |
| WO2012133782A1 (ja) | 抗原結合分子の血漿中滞留性と免疫原性を改変する方法 | |
| JP6280300B2 (ja) | 脳疾患治療剤 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| RD04 | Notification of resignation of power of attorney |
Free format text: JAPANESE INTERMEDIATE CODE: A7424 Effective date: 20150326 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20150814 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20150814 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20160606 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20160803 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20170302 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20170428 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A821 Effective date: 20170630 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20171204 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20171221 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20180109 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20180119 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 6280300 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |