JP4427222B2 - 長波長をもつ改変蛍光タンパク質 - Google Patents
長波長をもつ改変蛍光タンパク質 Download PDFInfo
- Publication number
- JP4427222B2 JP4427222B2 JP2001586334A JP2001586334A JP4427222B2 JP 4427222 B2 JP4427222 B2 JP 4427222B2 JP 2001586334 A JP2001586334 A JP 2001586334A JP 2001586334 A JP2001586334 A JP 2001586334A JP 4427222 B2 JP4427222 B2 JP 4427222B2
- Authority
- JP
- Japan
- Prior art keywords
- fluorescent protein
- substitution
- protein
- amino acid
- fluorescence
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Expired - Lifetime
Links
- 108091006047 fluorescent proteins Proteins 0.000 title claims abstract description 151
- 102000034287 fluorescent proteins Human genes 0.000 title claims abstract description 150
- 238000000034 method Methods 0.000 claims abstract description 82
- 150000007523 nucleic acids Chemical class 0.000 claims abstract description 69
- 102000039446 nucleic acids Human genes 0.000 claims abstract description 61
- 108020004707 nucleic acids Proteins 0.000 claims abstract description 61
- 108010043121 Green Fluorescent Proteins Proteins 0.000 claims description 115
- 239000005090 green fluorescent protein Substances 0.000 claims description 113
- 102000004144 Green Fluorescent Proteins Human genes 0.000 claims description 112
- 238000006467 substitution reaction Methods 0.000 claims description 87
- 150000001450 anions Chemical class 0.000 claims description 79
- 125000003275 alpha amino acid group Chemical group 0.000 claims description 78
- 241000242583 Scyphozoa Species 0.000 claims description 49
- 150000004820 halides Chemical class 0.000 claims description 38
- 239000000523 sample Substances 0.000 claims description 38
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 claims description 36
- 239000002773 nucleotide Substances 0.000 claims description 33
- 125000003729 nucleotide group Chemical group 0.000 claims description 33
- 230000000694 effects Effects 0.000 claims description 19
- 150000001875 compounds Chemical class 0.000 claims description 16
- 238000012360 testing method Methods 0.000 claims description 13
- 238000005259 measurement Methods 0.000 claims description 7
- 102000004310 Ion Channels Human genes 0.000 claims description 5
- 238000012216 screening Methods 0.000 claims description 5
- 241000242764 Aequorea victoria Species 0.000 claims description 4
- 230000032258 transport Effects 0.000 claims description 3
- 108090000862 Ion Channels Proteins 0.000 claims 4
- 239000012190 activator Substances 0.000 claims 1
- 239000013068 control sample Substances 0.000 claims 1
- 108090000623 proteins and genes Proteins 0.000 description 141
- 230000027455 binding Effects 0.000 description 102
- 102000004169 proteins and genes Human genes 0.000 description 102
- 235000018102 proteins Nutrition 0.000 description 92
- 210000004027 cell Anatomy 0.000 description 90
- 235000001014 amino acid Nutrition 0.000 description 84
- 229940024606 amino acid Drugs 0.000 description 74
- 150000001413 amino acids Chemical class 0.000 description 67
- XMBWDFGMSWQBCA-UHFFFAOYSA-N hydrogen iodide Chemical compound I XMBWDFGMSWQBCA-UHFFFAOYSA-N 0.000 description 60
- 229910052739 hydrogen Inorganic materials 0.000 description 52
- 108091005957 yellow fluorescent proteins Proteins 0.000 description 51
- 230000014509 gene expression Effects 0.000 description 48
- 230000035772 mutation Effects 0.000 description 42
- 108090000765 processed proteins & peptides Proteins 0.000 description 40
- 102000004196 processed proteins & peptides Human genes 0.000 description 36
- 229920001184 polypeptide Polymers 0.000 description 34
- 239000013078 crystal Substances 0.000 description 33
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 32
- 230000003993 interaction Effects 0.000 description 31
- 239000000370 acceptor Substances 0.000 description 29
- 230000005284 excitation Effects 0.000 description 27
- 239000001257 hydrogen Substances 0.000 description 27
- 239000002904 solvent Substances 0.000 description 27
- VEXZGXHMUGYJMC-UHFFFAOYSA-M Chloride anion Chemical compound [Cl-] VEXZGXHMUGYJMC-UHFFFAOYSA-M 0.000 description 26
- 239000013604 expression vector Substances 0.000 description 22
- 102220566479 GDNF family receptor alpha-1_S72A_mutation Human genes 0.000 description 21
- 108091028043 Nucleic acid sequence Proteins 0.000 description 21
- 108020004414 DNA Proteins 0.000 description 20
- 102220566468 GDNF family receptor alpha-1_S65G_mutation Human genes 0.000 description 20
- 238000002866 fluorescence resonance energy transfer Methods 0.000 description 20
- 241000243290 Aequorea Species 0.000 description 19
- 108700019146 Transgenes Proteins 0.000 description 19
- 150000002500 ions Chemical class 0.000 description 19
- 229910052727 yttrium Inorganic materials 0.000 description 19
- 108020004705 Codon Proteins 0.000 description 17
- 230000006870 function Effects 0.000 description 17
- 229910052757 nitrogen Inorganic materials 0.000 description 17
- IJGRMHOSHXDMSA-UHFFFAOYSA-N Atomic nitrogen Chemical compound N#N IJGRMHOSHXDMSA-UHFFFAOYSA-N 0.000 description 16
- 229910052731 fluorine Inorganic materials 0.000 description 16
- 239000011780 sodium chloride Substances 0.000 description 16
- 230000009261 transgenic effect Effects 0.000 description 16
- 241000700605 Viruses Species 0.000 description 15
- 230000008859 change Effects 0.000 description 15
- 239000000126 substance Substances 0.000 description 15
- 229910052721 tungsten Inorganic materials 0.000 description 15
- 125000004429 atom Chemical group 0.000 description 14
- 229910052799 carbon Inorganic materials 0.000 description 14
- 102000040430 polynucleotide Human genes 0.000 description 14
- 108091033319 polynucleotide Proteins 0.000 description 14
- 239000002157 polynucleotide Substances 0.000 description 14
- 239000000758 substrate Substances 0.000 description 14
- 238000010521 absorption reaction Methods 0.000 description 13
- AYFVYJQAPQTCCC-GBXIJSLDSA-N L-threonine Chemical compound C[C@@H](O)[C@H](N)C(O)=O AYFVYJQAPQTCCC-GBXIJSLDSA-N 0.000 description 12
- 238000010586 diagram Methods 0.000 description 12
- 108020001507 fusion proteins Proteins 0.000 description 12
- 102000037865 fusion proteins Human genes 0.000 description 12
- RAXXELZNTBOGNW-UHFFFAOYSA-N imidazole Natural products C1=CNC=N1 RAXXELZNTBOGNW-UHFFFAOYSA-N 0.000 description 12
- ISWSIDIOOBJBQZ-UHFFFAOYSA-N phenol group Chemical group C1(=CC=CC=C1)O ISWSIDIOOBJBQZ-UHFFFAOYSA-N 0.000 description 12
- 235000013824 polyphenols Nutrition 0.000 description 12
- 108091026890 Coding region Proteins 0.000 description 11
- 125000003118 aryl group Chemical group 0.000 description 11
- 239000003446 ligand Substances 0.000 description 11
- 230000033001 locomotion Effects 0.000 description 11
- 239000013612 plasmid Substances 0.000 description 11
- 230000014616 translation Effects 0.000 description 11
- 239000013598 vector Substances 0.000 description 11
- 102220566483 GDNF family receptor alpha-1_V68L_mutation Human genes 0.000 description 10
- 108091005971 Wild-type GFP Proteins 0.000 description 10
- 238000003556 assay Methods 0.000 description 10
- QVGXLLKOCUKJST-UHFFFAOYSA-N atomic oxygen Chemical compound [O] QVGXLLKOCUKJST-UHFFFAOYSA-N 0.000 description 10
- 230000015572 biosynthetic process Effects 0.000 description 10
- 229910052760 oxygen Inorganic materials 0.000 description 10
- 239000001301 oxygen Substances 0.000 description 10
- 230000035897 transcription Effects 0.000 description 10
- 238000013518 transcription Methods 0.000 description 10
- 241000196324 Embryophyta Species 0.000 description 9
- 102000004190 Enzymes Human genes 0.000 description 9
- 108090000790 Enzymes Proteins 0.000 description 9
- XUJNEKJLAYXESH-REOHCLBHSA-N L-Cysteine Chemical compound SC[C@H](N)C(O)=O XUJNEKJLAYXESH-REOHCLBHSA-N 0.000 description 9
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 9
- 238000000295 emission spectrum Methods 0.000 description 9
- 229940088598 enzyme Drugs 0.000 description 9
- 239000012071 phase Substances 0.000 description 9
- 229910052700 potassium Inorganic materials 0.000 description 9
- 238000013519 translation Methods 0.000 description 9
- JKMHFZQWWAIEOD-UHFFFAOYSA-N 2-[4-(2-hydroxyethyl)piperazin-1-yl]ethanesulfonic acid Chemical compound OCC[NH+]1CCN(CCS([O-])(=O)=O)CC1 JKMHFZQWWAIEOD-UHFFFAOYSA-N 0.000 description 8
- WHUUTDBJXJRKMK-UHFFFAOYSA-N Glutamic acid Natural products OC(=O)C(N)CCC(O)=O WHUUTDBJXJRKMK-UHFFFAOYSA-N 0.000 description 8
- 239000007995 HEPES buffer Substances 0.000 description 8
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 8
- 241001465754 Metazoa Species 0.000 description 8
- -1 aromatic amino acid Chemical group 0.000 description 8
- 239000002299 complementary DNA Substances 0.000 description 8
- 238000000695 excitation spectrum Methods 0.000 description 8
- 230000036571 hydration Effects 0.000 description 8
- 238000006703 hydration reaction Methods 0.000 description 8
- 230000001965 increasing effect Effects 0.000 description 8
- 230000007935 neutral effect Effects 0.000 description 8
- 238000012552 review Methods 0.000 description 8
- 238000005415 bioluminescence Methods 0.000 description 7
- 230000029918 bioluminescence Effects 0.000 description 7
- 230000004807 localization Effects 0.000 description 7
- 230000008569 process Effects 0.000 description 7
- 238000006862 quantum yield reaction Methods 0.000 description 7
- 241000894007 species Species 0.000 description 7
- 230000003595 spectral effect Effects 0.000 description 7
- 238000012546 transfer Methods 0.000 description 7
- 230000003612 virological effect Effects 0.000 description 7
- QTBSBXVTEAMEQO-UHFFFAOYSA-N Acetic acid Chemical compound CC(O)=O QTBSBXVTEAMEQO-UHFFFAOYSA-N 0.000 description 6
- 241000894006 Bacteria Species 0.000 description 6
- LYCAIKOWRPUZTN-UHFFFAOYSA-N Ethylene glycol Chemical compound OCCO LYCAIKOWRPUZTN-UHFFFAOYSA-N 0.000 description 6
- 102220567280 GDNF family receptor alpha-1_T203F_mutation Human genes 0.000 description 6
- 108020004511 Recombinant DNA Proteins 0.000 description 6
- 238000002835 absorbance Methods 0.000 description 6
- 238000004458 analytical method Methods 0.000 description 6
- 210000002459 blastocyst Anatomy 0.000 description 6
- 125000002915 carbonyl group Chemical group [*:2]C([*:1])=O 0.000 description 6
- 230000000295 complement effect Effects 0.000 description 6
- 230000003247 decreasing effect Effects 0.000 description 6
- 230000005281 excited state Effects 0.000 description 6
- 239000012634 fragment Substances 0.000 description 6
- 230000004927 fusion Effects 0.000 description 6
- 238000007654 immersion Methods 0.000 description 6
- 230000001976 improved effect Effects 0.000 description 6
- 238000000338 in vitro Methods 0.000 description 6
- 238000001727 in vivo Methods 0.000 description 6
- 150000004694 iodide salts Chemical class 0.000 description 6
- TWRXJAOTZQYOKJ-UHFFFAOYSA-L magnesium chloride Substances [Mg+2].[Cl-].[Cl-] TWRXJAOTZQYOKJ-UHFFFAOYSA-L 0.000 description 6
- 108020004999 messenger RNA Proteins 0.000 description 6
- 239000012452 mother liquor Substances 0.000 description 6
- YBYRMVIVWMBXKQ-UHFFFAOYSA-N phenylmethanesulfonyl fluoride Chemical compound FS(=O)(=O)CC1=CC=CC=C1 YBYRMVIVWMBXKQ-UHFFFAOYSA-N 0.000 description 6
- NLKNQRATVPKPDG-UHFFFAOYSA-M potassium iodide Chemical compound [K+].[I-] NLKNQRATVPKPDG-UHFFFAOYSA-M 0.000 description 6
- 230000004044 response Effects 0.000 description 6
- 230000035945 sensitivity Effects 0.000 description 6
- 239000004289 sodium hydrogen sulphite Substances 0.000 description 6
- 230000008685 targeting Effects 0.000 description 6
- 230000005030 transcription termination Effects 0.000 description 6
- 239000004475 Arginine Substances 0.000 description 5
- OKTJSMMVPCPJKN-UHFFFAOYSA-N Carbon Chemical group [C] OKTJSMMVPCPJKN-UHFFFAOYSA-N 0.000 description 5
- 108090000056 Complement factor B Proteins 0.000 description 5
- 102000003712 Complement factor B Human genes 0.000 description 5
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 5
- 108020004711 Nucleic Acid Probes Proteins 0.000 description 5
- 102000001253 Protein Kinase Human genes 0.000 description 5
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 5
- 150000001408 amides Chemical class 0.000 description 5
- 125000000539 amino acid group Chemical group 0.000 description 5
- 125000000129 anionic group Chemical group 0.000 description 5
- 230000002547 anomalous effect Effects 0.000 description 5
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 5
- 239000000872 buffer Substances 0.000 description 5
- 125000003636 chemical group Chemical group 0.000 description 5
- 238000010367 cloning Methods 0.000 description 5
- 230000001939 inductive effect Effects 0.000 description 5
- 238000003780 insertion Methods 0.000 description 5
- 230000037431 insertion Effects 0.000 description 5
- 229910001629 magnesium chloride Inorganic materials 0.000 description 5
- 210000004962 mammalian cell Anatomy 0.000 description 5
- 238000004519 manufacturing process Methods 0.000 description 5
- 239000003550 marker Substances 0.000 description 5
- 239000000463 material Substances 0.000 description 5
- 238000000520 microinjection Methods 0.000 description 5
- 239000002853 nucleic acid probe Substances 0.000 description 5
- 230000036961 partial effect Effects 0.000 description 5
- 238000002360 preparation method Methods 0.000 description 5
- 108060006633 protein kinase Proteins 0.000 description 5
- 102000005962 receptors Human genes 0.000 description 5
- 108020003175 receptors Proteins 0.000 description 5
- 239000000243 solution Substances 0.000 description 5
- 238000003860 storage Methods 0.000 description 5
- 241001430294 unidentified retrovirus Species 0.000 description 5
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 5
- YBJHBAHKTGYVGT-ZKWXMUAHSA-N (+)-Biotin Chemical compound N1C(=O)N[C@@H]2[C@H](CCCCC(=O)O)SC[C@@H]21 YBJHBAHKTGYVGT-ZKWXMUAHSA-N 0.000 description 4
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 4
- 102100025698 Cytosolic carboxypeptidase 4 Human genes 0.000 description 4
- 102000053602 DNA Human genes 0.000 description 4
- 241000588724 Escherichia coli Species 0.000 description 4
- 102220566560 GDNF family receptor alpha-1_E222G_mutation Human genes 0.000 description 4
- 102220566687 GDNF family receptor alpha-1_F64L_mutation Human genes 0.000 description 4
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 description 4
- 241000124008 Mammalia Species 0.000 description 4
- NBIIXXVUZAFLBC-UHFFFAOYSA-N Phosphoric acid Chemical compound OP(O)(O)=O NBIIXXVUZAFLBC-UHFFFAOYSA-N 0.000 description 4
- WCUXLLCKKVVCTQ-UHFFFAOYSA-M Potassium chloride Chemical compound [Cl-].[K+] WCUXLLCKKVVCTQ-UHFFFAOYSA-M 0.000 description 4
- RJFAYQIBOAGBLC-BYPYZUCNSA-N Selenium-L-methionine Chemical compound C[Se]CC[C@H](N)C(O)=O RJFAYQIBOAGBLC-BYPYZUCNSA-N 0.000 description 4
- RJFAYQIBOAGBLC-UHFFFAOYSA-N Selenomethionine Natural products C[Se]CCC(N)C(O)=O RJFAYQIBOAGBLC-UHFFFAOYSA-N 0.000 description 4
- 239000000427 antigen Substances 0.000 description 4
- 108091007433 antigens Proteins 0.000 description 4
- 102000036639 antigens Human genes 0.000 description 4
- 230000031018 biological processes and functions Effects 0.000 description 4
- 238000004364 calculation method Methods 0.000 description 4
- 238000003776 cleavage reaction Methods 0.000 description 4
- 238000001514 detection method Methods 0.000 description 4
- 238000005516 engineering process Methods 0.000 description 4
- 210000003527 eukaryotic cell Anatomy 0.000 description 4
- 210000004602 germ cell Anatomy 0.000 description 4
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Natural products OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 description 4
- 125000000404 glutamine group Chemical group N[C@@H](CCC(N)=O)C(=O)* 0.000 description 4
- HNDVDQJCIGZPNO-UHFFFAOYSA-N histidine Natural products OC(=O)C(N)CC1=CN=CN1 HNDVDQJCIGZPNO-UHFFFAOYSA-N 0.000 description 4
- 238000012544 monitoring process Methods 0.000 description 4
- 229920000642 polymer Polymers 0.000 description 4
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 4
- 238000012545 processing Methods 0.000 description 4
- 230000006798 recombination Effects 0.000 description 4
- 230000001105 regulatory effect Effects 0.000 description 4
- 230000010076 replication Effects 0.000 description 4
- 230000007017 scission Effects 0.000 description 4
- 238000002741 site-directed mutagenesis Methods 0.000 description 4
- 238000002791 soaking Methods 0.000 description 4
- 238000003786 synthesis reaction Methods 0.000 description 4
- YNJBWRMUSHSURL-UHFFFAOYSA-N trichloroacetic acid Chemical compound OC(=O)C(Cl)(Cl)Cl YNJBWRMUSHSURL-UHFFFAOYSA-N 0.000 description 4
- 241000701161 unidentified adenovirus Species 0.000 description 4
- 239000013603 viral vector Substances 0.000 description 4
- DGVVWUTYPXICAM-UHFFFAOYSA-N β‐Mercaptoethanol Chemical compound OCCS DGVVWUTYPXICAM-UHFFFAOYSA-N 0.000 description 4
- IHPYMWDTONKSCO-UHFFFAOYSA-N 2,2'-piperazine-1,4-diylbisethanesulfonic acid Chemical compound OS(=O)(=O)CCN1CCN(CCS(O)(=O)=O)CC1 IHPYMWDTONKSCO-UHFFFAOYSA-N 0.000 description 3
- CURLTUGMZLYLDI-UHFFFAOYSA-N Carbon dioxide Chemical compound O=C=O CURLTUGMZLYLDI-UHFFFAOYSA-N 0.000 description 3
- RGHNJXZEOKUKBD-SQOUGZDYSA-N D-gluconic acid Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)[C@@H](O)C(O)=O RGHNJXZEOKUKBD-SQOUGZDYSA-N 0.000 description 3
- 241000206602 Eukaryota Species 0.000 description 3
- 102220566467 GDNF family receptor alpha-1_S65A_mutation Human genes 0.000 description 3
- 102220566451 GDNF family receptor alpha-1_Y66H_mutation Human genes 0.000 description 3
- 102220566455 GDNF family receptor alpha-1_Y66W_mutation Human genes 0.000 description 3
- 102220486246 Glucosidase 2 subunit beta_S65V_mutation Human genes 0.000 description 3
- 241000238631 Hexapoda Species 0.000 description 3
- 101000932590 Homo sapiens Cytosolic carboxypeptidase 4 Proteins 0.000 description 3
- 108700005091 Immunoglobulin Genes Proteins 0.000 description 3
- SIKJAQJRHWYJAI-UHFFFAOYSA-N Indole Chemical compound C1=CC=C2NC=CC2=C1 SIKJAQJRHWYJAI-UHFFFAOYSA-N 0.000 description 3
- 108091092195 Intron Proteins 0.000 description 3
- FFEARJCKVFRZRR-BYPYZUCNSA-N L-methionine Chemical compound CSCC[C@H](N)C(O)=O FFEARJCKVFRZRR-BYPYZUCNSA-N 0.000 description 3
- 101001033003 Mus musculus Granzyme F Proteins 0.000 description 3
- 241000699670 Mus sp. Species 0.000 description 3
- 239000007990 PIPES buffer Substances 0.000 description 3
- 102000035195 Peptidases Human genes 0.000 description 3
- 108091005804 Peptidases Proteins 0.000 description 3
- HLCFGWHYROZGBI-JJKGCWMISA-M Potassium gluconate Chemical compound [K+].OC[C@@H](O)[C@@H](O)[C@H](O)[C@@H](O)C([O-])=O HLCFGWHYROZGBI-JJKGCWMISA-M 0.000 description 3
- 241000723873 Tobacco mosaic virus Species 0.000 description 3
- 241000700618 Vaccinia virus Species 0.000 description 3
- KZSNJWFQEVHDMF-UHFFFAOYSA-N Valine Natural products CC(C)C(N)C(O)=O KZSNJWFQEVHDMF-UHFFFAOYSA-N 0.000 description 3
- 230000002159 abnormal effect Effects 0.000 description 3
- 238000000862 absorption spectrum Methods 0.000 description 3
- 230000002378 acidificating effect Effects 0.000 description 3
- 239000000556 agonist Substances 0.000 description 3
- 239000005557 antagonist Substances 0.000 description 3
- 210000000170 cell membrane Anatomy 0.000 description 3
- 230000001413 cellular effect Effects 0.000 description 3
- 239000003153 chemical reaction reagent Substances 0.000 description 3
- 210000000349 chromosome Anatomy 0.000 description 3
- 238000007796 conventional method Methods 0.000 description 3
- 230000001086 cytosolic effect Effects 0.000 description 3
- 238000013480 data collection Methods 0.000 description 3
- 238000006297 dehydration reaction Methods 0.000 description 3
- 238000002050 diffraction method Methods 0.000 description 3
- 238000004520 electroporation Methods 0.000 description 3
- 210000002257 embryonic structure Anatomy 0.000 description 3
- XAGAASDWSFGQEC-UHFFFAOYSA-N ethyl-(n-(4-methylphenyl)sulfonylanilino)mercury Chemical compound C=1C=C(C)C=CC=1S(=O)(=O)N([Hg]CC)C1=CC=CC=C1 XAGAASDWSFGQEC-UHFFFAOYSA-N 0.000 description 3
- 238000003209 gene knockout Methods 0.000 description 3
- 230000005283 ground state Effects 0.000 description 3
- 230000002209 hydrophobic effect Effects 0.000 description 3
- 125000001165 hydrophobic group Chemical group 0.000 description 3
- 238000003018 immunoassay Methods 0.000 description 3
- 230000006698 induction Effects 0.000 description 3
- 230000010354 integration Effects 0.000 description 3
- 125000005647 linker group Chemical group 0.000 description 3
- 238000004020 luminiscence type Methods 0.000 description 3
- 229930182817 methionine Natural products 0.000 description 3
- 239000000203 mixture Substances 0.000 description 3
- 230000003287 optical effect Effects 0.000 description 3
- 230000017854 proteolysis Effects 0.000 description 3
- 238000000746 purification Methods 0.000 description 3
- 238000010791 quenching Methods 0.000 description 3
- 238000005215 recombination Methods 0.000 description 3
- 238000007363 ring formation reaction Methods 0.000 description 3
- 230000019491 signal transduction Effects 0.000 description 3
- 238000004448 titration Methods 0.000 description 3
- OUYCCCASQSFEME-UHFFFAOYSA-N tyrosine Natural products OC(=O)C(N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-UHFFFAOYSA-N 0.000 description 3
- 229910052720 vanadium Inorganic materials 0.000 description 3
- BJEPYKJPYRNKOW-REOHCLBHSA-N (S)-malic acid Chemical compound OC(=O)[C@@H](O)CC(O)=O BJEPYKJPYRNKOW-REOHCLBHSA-N 0.000 description 2
- 108700028369 Alleles Proteins 0.000 description 2
- 108010083590 Apoproteins Proteins 0.000 description 2
- 102000006410 Apoproteins Human genes 0.000 description 2
- 102220543132 Bis(5'-adenosyl)-triphosphatase_Y145F_mutation Human genes 0.000 description 2
- 241000701822 Bovine papillomavirus Species 0.000 description 2
- RGHNJXZEOKUKBD-UHFFFAOYSA-N D-gluconic acid Natural products OCC(O)C(O)C(O)C(O)C(O)=O RGHNJXZEOKUKBD-UHFFFAOYSA-N 0.000 description 2
- 102220566480 GDNF family receptor alpha-1_K79R_mutation Human genes 0.000 description 2
- 102220566471 GDNF family receptor alpha-1_Q80R_mutation Human genes 0.000 description 2
- 102220566453 GDNF family receptor alpha-1_Y66F_mutation Human genes 0.000 description 2
- 108700028146 Genetic Enhancer Elements Proteins 0.000 description 2
- 108010004901 Haloalkane dehalogenase Proteins 0.000 description 2
- 101100321817 Human parvovirus B19 (strain HV) 7.5K gene Proteins 0.000 description 2
- 108010021625 Immunoglobulin Fragments Proteins 0.000 description 2
- AGPKZVBTJJNPAG-WHFBIAKZSA-N L-isoleucine Chemical compound CC[C@H](C)[C@H](N)C(O)=O AGPKZVBTJJNPAG-WHFBIAKZSA-N 0.000 description 2
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 2
- COLNVLDHVKWLRT-QMMMGPOBSA-N L-phenylalanine Chemical compound OC(=O)[C@@H](N)CC1=CC=CC=C1 COLNVLDHVKWLRT-QMMMGPOBSA-N 0.000 description 2
- KZSNJWFQEVHDMF-BYPYZUCNSA-N L-valine Chemical compound CC(C)[C@H](N)C(O)=O KZSNJWFQEVHDMF-BYPYZUCNSA-N 0.000 description 2
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 2
- 241000829100 Macaca mulatta polyomavirus 1 Species 0.000 description 2
- 102000003792 Metallothionein Human genes 0.000 description 2
- 108090000157 Metallothionein Proteins 0.000 description 2
- 241000699666 Mus <mouse, genus> Species 0.000 description 2
- 108091034117 Oligonucleotide Proteins 0.000 description 2
- 102000052812 Ornithine decarboxylases Human genes 0.000 description 2
- 108700005126 Ornithine decarboxylases Proteins 0.000 description 2
- 108091000080 Phosphotransferase Proteins 0.000 description 2
- 239000004365 Protease Substances 0.000 description 2
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 description 2
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 description 2
- 241000242739 Renilla Species 0.000 description 2
- 206010038997 Retroviral infections Diseases 0.000 description 2
- 108091028664 Ribonucleotide Proteins 0.000 description 2
- 241000283984 Rodentia Species 0.000 description 2
- 241000256251 Spodoptera frugiperda Species 0.000 description 2
- QAOWNCQODCNURD-UHFFFAOYSA-N Sulfuric acid Chemical compound OS(O)(=O)=O QAOWNCQODCNURD-UHFFFAOYSA-N 0.000 description 2
- 108020004440 Thymidine kinase Proteins 0.000 description 2
- 102220615016 Transcription elongation regulator 1_S65C_mutation Human genes 0.000 description 2
- DTQVDTLACAAQTR-UHFFFAOYSA-N Trifluoroacetic acid Chemical compound OC(=O)C(F)(F)F DTQVDTLACAAQTR-UHFFFAOYSA-N 0.000 description 2
- 208000034953 Twin anemia-polycythemia sequence Diseases 0.000 description 2
- 230000009471 action Effects 0.000 description 2
- BJEPYKJPYRNKOW-UHFFFAOYSA-N alpha-hydroxysuccinic acid Natural products OC(=O)C(O)CC(O)=O BJEPYKJPYRNKOW-UHFFFAOYSA-N 0.000 description 2
- 229910000147 aluminium phosphate Inorganic materials 0.000 description 2
- 239000012491 analyte Substances 0.000 description 2
- 210000004102 animal cell Anatomy 0.000 description 2
- 238000013459 approach Methods 0.000 description 2
- 230000001580 bacterial effect Effects 0.000 description 2
- 230000006399 behavior Effects 0.000 description 2
- 230000009286 beneficial effect Effects 0.000 description 2
- 230000004071 biological effect Effects 0.000 description 2
- 230000033228 biological regulation Effects 0.000 description 2
- 229960002685 biotin Drugs 0.000 description 2
- 235000020958 biotin Nutrition 0.000 description 2
- 239000011616 biotin Substances 0.000 description 2
- 108091005948 blue fluorescent proteins Proteins 0.000 description 2
- 210000004899 c-terminal region Anatomy 0.000 description 2
- 150000001732 carboxylic acid derivatives Chemical class 0.000 description 2
- 150000001805 chlorine compounds Chemical group 0.000 description 2
- 230000008878 coupling Effects 0.000 description 2
- 238000010168 coupling process Methods 0.000 description 2
- 238000005859 coupling reaction Methods 0.000 description 2
- 238000012926 crystallographic analysis Methods 0.000 description 2
- 210000000805 cytoplasm Anatomy 0.000 description 2
- 230000007423 decrease Effects 0.000 description 2
- 239000005547 deoxyribonucleotide Substances 0.000 description 2
- 125000002637 deoxyribonucleotide group Chemical group 0.000 description 2
- 230000001419 dependent effect Effects 0.000 description 2
- 230000005595 deprotonation Effects 0.000 description 2
- 238000010537 deprotonation reaction Methods 0.000 description 2
- 238000011161 development Methods 0.000 description 2
- 230000018109 developmental process Effects 0.000 description 2
- VLCYCQAOQCDTCN-UHFFFAOYSA-N eflornithine Chemical compound NCCCC(N)(C(F)F)C(O)=O VLCYCQAOQCDTCN-UHFFFAOYSA-N 0.000 description 2
- 230000005670 electromagnetic radiation Effects 0.000 description 2
- 239000003623 enhancer Substances 0.000 description 2
- 230000007613 environmental effect Effects 0.000 description 2
- 238000002474 experimental method Methods 0.000 description 2
- 239000000174 gluconic acid Substances 0.000 description 2
- 235000012208 gluconic acid Nutrition 0.000 description 2
- 229910052736 halogen Inorganic materials 0.000 description 2
- 150000002367 halogens Chemical class 0.000 description 2
- 125000000623 heterocyclic group Chemical group 0.000 description 2
- 238000009396 hybridization Methods 0.000 description 2
- 125000002887 hydroxy group Chemical group [H]O* 0.000 description 2
- YAMHXTCMCPHKLN-UHFFFAOYSA-N imidazolidin-2-one Chemical group O=C1NCCN1 YAMHXTCMCPHKLN-UHFFFAOYSA-N 0.000 description 2
- 230000001900 immune effect Effects 0.000 description 2
- 238000011065 in-situ storage Methods 0.000 description 2
- PZOUSPYUWWUPPK-UHFFFAOYSA-N indole Natural products CC1=CC=CC2=C1C=CN2 PZOUSPYUWWUPPK-UHFFFAOYSA-N 0.000 description 2
- RKJUIXBNRJVNHR-UHFFFAOYSA-N indolenine Natural products C1=CC=C2CC=NC2=C1 RKJUIXBNRJVNHR-UHFFFAOYSA-N 0.000 description 2
- 208000015181 infectious disease Diseases 0.000 description 2
- PNDPGZBMCMUPRI-UHFFFAOYSA-N iodine Chemical compound II PNDPGZBMCMUPRI-UHFFFAOYSA-N 0.000 description 2
- 229960000310 isoleucine Drugs 0.000 description 2
- AGPKZVBTJJNPAG-UHFFFAOYSA-N isoleucine Natural products CCC(C)C(N)C(O)=O AGPKZVBTJJNPAG-UHFFFAOYSA-N 0.000 description 2
- 239000001630 malic acid Substances 0.000 description 2
- 235000011090 malic acid Nutrition 0.000 description 2
- 210000001161 mammalian embryo Anatomy 0.000 description 2
- 239000012528 membrane Substances 0.000 description 2
- XMYQHJDBLRZMLW-UHFFFAOYSA-N methanolamine Chemical compound NCO XMYQHJDBLRZMLW-UHFFFAOYSA-N 0.000 description 2
- 244000005700 microbiome Species 0.000 description 2
- 102000035118 modified proteins Human genes 0.000 description 2
- 108091005573 modified proteins Proteins 0.000 description 2
- 238000002703 mutagenesis Methods 0.000 description 2
- 231100000350 mutagenesis Toxicity 0.000 description 2
- 210000003463 organelle Anatomy 0.000 description 2
- 239000004031 partial agonist Substances 0.000 description 2
- 230000026731 phosphorylation Effects 0.000 description 2
- 238000006366 phosphorylation reaction Methods 0.000 description 2
- 102000020233 phosphotransferase Human genes 0.000 description 2
- 229920002704 polyhistidine Polymers 0.000 description 2
- 239000001103 potassium chloride Substances 0.000 description 2
- 235000011164 potassium chloride Nutrition 0.000 description 2
- 239000004224 potassium gluconate Substances 0.000 description 2
- 229960003189 potassium gluconate Drugs 0.000 description 2
- 235000013926 potassium gluconate Nutrition 0.000 description 2
- 239000000047 product Substances 0.000 description 2
- 235000019419 proteases Nutrition 0.000 description 2
- 230000006916 protein interaction Effects 0.000 description 2
- 230000012743 protein tagging Effects 0.000 description 2
- 230000004850 protein–protein interaction Effects 0.000 description 2
- 230000000171 quenching effect Effects 0.000 description 2
- 230000005855 radiation Effects 0.000 description 2
- 238000002708 random mutagenesis Methods 0.000 description 2
- 230000002829 reductive effect Effects 0.000 description 2
- 238000011160 research Methods 0.000 description 2
- 230000001177 retroviral effect Effects 0.000 description 2
- 230000002441 reversible effect Effects 0.000 description 2
- 239000002336 ribonucleotide Substances 0.000 description 2
- 125000002652 ribonucleotide group Chemical group 0.000 description 2
- 102220243291 rs1555194026 Human genes 0.000 description 2
- 102200142969 rs398124653 Human genes 0.000 description 2
- 102200089551 rs5030826 Human genes 0.000 description 2
- FGDZQCVHDSGLHJ-UHFFFAOYSA-M rubidium chloride Chemical compound [Cl-].[Rb+] FGDZQCVHDSGLHJ-UHFFFAOYSA-M 0.000 description 2
- 150000003839 salts Chemical class 0.000 description 2
- 229960002718 selenomethionine Drugs 0.000 description 2
- 238000000926 separation method Methods 0.000 description 2
- 230000035939 shock Effects 0.000 description 2
- 238000001228 spectrum Methods 0.000 description 2
- 230000010473 stable expression Effects 0.000 description 2
- 125000003396 thiol group Chemical group [H]S* 0.000 description 2
- 238000001890 transfection Methods 0.000 description 2
- 241001515965 unidentified phage Species 0.000 description 2
- 239000004474 valine Substances 0.000 description 2
- 230000000007 visual effect Effects 0.000 description 2
- 238000002424 x-ray crystallography Methods 0.000 description 2
- DIGQNXIGRZPYDK-WKSCXVIASA-N (2R)-6-amino-2-[[2-[[(2S)-2-[[2-[[(2R)-2-[[(2S)-2-[[(2R,3S)-2-[[2-[[(2S)-2-[[2-[[(2S)-2-[[(2S)-2-[[(2R)-2-[[(2S,3S)-2-[[(2R)-2-[[(2S)-2-[[(2S)-2-[[(2S)-2-[[2-[[(2S)-2-[[(2R)-2-[[2-[[2-[[2-[(2-amino-1-hydroxyethylidene)amino]-3-carboxy-1-hydroxypropylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1-hydroxyethylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1,3-dihydroxypropylidene]amino]-1-hydroxyethylidene]amino]-1-hydroxypropylidene]amino]-1,3-dihydroxypropylidene]amino]-1,3-dihydroxypropylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1,3-dihydroxybutylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1-hydroxypropylidene]amino]-1,3-dihydroxypropylidene]amino]-1-hydroxyethylidene]amino]-1,5-dihydroxy-5-iminopentylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1,3-dihydroxybutylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1,3-dihydroxypropylidene]amino]-1-hydroxyethylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1-hydroxyethylidene]amino]hexanoic acid Chemical compound C[C@@H]([C@@H](C(=N[C@@H](CS)C(=N[C@@H](C)C(=N[C@@H](CO)C(=NCC(=N[C@@H](CCC(=N)O)C(=NC(CS)C(=N[C@H]([C@H](C)O)C(=N[C@H](CS)C(=N[C@H](CO)C(=NCC(=N[C@H](CS)C(=NCC(=N[C@H](CCCCN)C(=O)O)O)O)O)O)O)O)O)O)O)O)O)O)O)N=C([C@H](CS)N=C([C@H](CO)N=C([C@H](CO)N=C([C@H](C)N=C(CN=C([C@H](CO)N=C([C@H](CS)N=C(CN=C(C(CS)N=C(C(CC(=O)O)N=C(CN)O)O)O)O)O)O)O)O)O)O)O)O DIGQNXIGRZPYDK-WKSCXVIASA-N 0.000 description 1
- KXRMREPJUITWDU-UHFFFAOYSA-N 2-amino-4,6-dimethyl-3-oxophenoxazine-1,9-dicarboxylic acid Chemical compound N1=C2C(C(O)=O)=C(N)C(=O)C(C)=C2OC2=C1C(C(O)=O)=CC=C2C KXRMREPJUITWDU-UHFFFAOYSA-N 0.000 description 1
- QKRMFCXDTFLKKT-UHFFFAOYSA-N 2-hydroxyethanesulfonic acid Chemical compound OCCS(O)(=O)=O.OCCS(O)(=O)=O QKRMFCXDTFLKKT-UHFFFAOYSA-N 0.000 description 1
- KIUMMUBSPKGMOY-UHFFFAOYSA-N 3,3'-Dithiobis(6-nitrobenzoic acid) Chemical compound C1=C([N+]([O-])=O)C(C(=O)O)=CC(SSC=2C=C(C(=CC=2)[N+]([O-])=O)C(O)=O)=C1 KIUMMUBSPKGMOY-UHFFFAOYSA-N 0.000 description 1
- JZUWLOGLONEFKL-UHFFFAOYSA-N 4-[(4-hydroxyphenyl)methylidene]imidazolidin-2-one Chemical compound C1=CC(O)=CC=C1C=C1NC(=O)NC1 JZUWLOGLONEFKL-UHFFFAOYSA-N 0.000 description 1
- 229930024421 Adenine Natural products 0.000 description 1
- GFFGJBXGBJISGV-UHFFFAOYSA-N Adenine Chemical compound NC1=NC=NC2=C1N=CN2 GFFGJBXGBJISGV-UHFFFAOYSA-N 0.000 description 1
- 101000997963 Aequorea victoria Green fluorescent protein Proteins 0.000 description 1
- 229920000936 Agarose Polymers 0.000 description 1
- DCXYFEDJOCDNAF-UHFFFAOYSA-N Asparagine Natural products OC(=O)C(N)CC(N)=O DCXYFEDJOCDNAF-UHFFFAOYSA-N 0.000 description 1
- 241001203868 Autographa californica Species 0.000 description 1
- 108090001008 Avidin Proteins 0.000 description 1
- BVKZGUZCCUSVTD-UHFFFAOYSA-M Bicarbonate Chemical compound OC([O-])=O BVKZGUZCCUSVTD-UHFFFAOYSA-M 0.000 description 1
- 241000283690 Bos taurus Species 0.000 description 1
- 241000282472 Canis lupus familiaris Species 0.000 description 1
- 101710132601 Capsid protein Proteins 0.000 description 1
- 102000014914 Carrier Proteins Human genes 0.000 description 1
- 241000701489 Cauliflower mosaic virus Species 0.000 description 1
- 102000000844 Cell Surface Receptors Human genes 0.000 description 1
- 108010001857 Cell Surface Receptors Proteins 0.000 description 1
- 108090000317 Chymotrypsin Proteins 0.000 description 1
- 241000243321 Cnidaria Species 0.000 description 1
- 101710094648 Coat protein Proteins 0.000 description 1
- 108091033380 Coding strand Proteins 0.000 description 1
- 108091035707 Consensus sequence Proteins 0.000 description 1
- 241000938605 Crocodylia Species 0.000 description 1
- 101150074155 DHFR gene Proteins 0.000 description 1
- 102100026816 DNA-dependent metalloprotease SPRTN Human genes 0.000 description 1
- 101710175461 DNA-dependent metalloprotease SPRTN Proteins 0.000 description 1
- 102100025691 Dapper homolog 1 Human genes 0.000 description 1
- SHIBSTMRCDJXLN-UHFFFAOYSA-N Digoxigenin Natural products C1CC(C2C(C3(C)CCC(O)CC3CC2)CC2O)(O)C2(C)C1C1=CC(=O)OC1 SHIBSTMRCDJXLN-UHFFFAOYSA-N 0.000 description 1
- 108090000204 Dipeptidase 1 Proteins 0.000 description 1
- 238000002965 ELISA Methods 0.000 description 1
- YQYJSBFKSSDGFO-UHFFFAOYSA-N Epihygromycin Natural products OC1C(O)C(C(=O)C)OC1OC(C(=C1)O)=CC=C1C=C(C)C(=O)NC1C(O)C(O)C2OCOC2C1O YQYJSBFKSSDGFO-UHFFFAOYSA-N 0.000 description 1
- KRHYYFGTRYWZRS-UHFFFAOYSA-M Fluoride anion Chemical compound [F-] KRHYYFGTRYWZRS-UHFFFAOYSA-M 0.000 description 1
- PXGOKWXKJXAPGV-UHFFFAOYSA-N Fluorine Chemical compound FF PXGOKWXKJXAPGV-UHFFFAOYSA-N 0.000 description 1
- BDAGIHXWWSANSR-UHFFFAOYSA-M Formate Chemical compound [O-]C=O BDAGIHXWWSANSR-UHFFFAOYSA-M 0.000 description 1
- 241000233866 Fungi Species 0.000 description 1
- 102000004961 Furin Human genes 0.000 description 1
- 108090001126 Furin Proteins 0.000 description 1
- 108091006027 G proteins Proteins 0.000 description 1
- 102220566524 GDNF family receptor alpha-1_F99S_mutation Human genes 0.000 description 1
- 102220566496 GDNF family receptor alpha-1_M153T_mutation Human genes 0.000 description 1
- 102000030782 GTP binding Human genes 0.000 description 1
- 108091000058 GTP-Binding Proteins 0.000 description 1
- 239000004471 Glycine Substances 0.000 description 1
- 102100021181 Golgi phosphoprotein 3 Human genes 0.000 description 1
- 239000006173 Good's buffer Substances 0.000 description 1
- ZRALSGWEFCBTJO-UHFFFAOYSA-N Guanidine Chemical group NC(N)=N ZRALSGWEFCBTJO-UHFFFAOYSA-N 0.000 description 1
- 208000009889 Herpes Simplex Diseases 0.000 description 1
- 101000856043 Homo sapiens Dapper homolog 1 Proteins 0.000 description 1
- UFHFLCQGNIYNRP-UHFFFAOYSA-N Hydrogen Chemical compound [H][H] UFHFLCQGNIYNRP-UHFFFAOYSA-N 0.000 description 1
- 102000018251 Hypoxanthine Phosphoribosyltransferase Human genes 0.000 description 1
- 108010091358 Hypoxanthine Phosphoribosyltransferase Proteins 0.000 description 1
- 108060003951 Immunoglobulin Proteins 0.000 description 1
- 102000008394 Immunoglobulin Fragments Human genes 0.000 description 1
- 108010067060 Immunoglobulin Variable Region Proteins 0.000 description 1
- 108020005350 Initiator Codon Proteins 0.000 description 1
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 description 1
- 125000000998 L-alanino group Chemical class [H]N([*])[C@](C([H])([H])[H])([H])C(=O)O[H] 0.000 description 1
- DCXYFEDJOCDNAF-REOHCLBHSA-N L-asparagine Chemical compound OC(=O)[C@@H](N)CC(N)=O DCXYFEDJOCDNAF-REOHCLBHSA-N 0.000 description 1
- CKLJMWTZIZZHCS-REOHCLBHSA-N L-aspartic acid Chemical compound OC(=O)[C@@H](N)CC(O)=O CKLJMWTZIZZHCS-REOHCLBHSA-N 0.000 description 1
- FBOZXECLQNJBKD-ZDUSSCGKSA-N L-methotrexate Chemical compound C=1N=C2N=C(N)N=C(N)C2=NC=1CN(C)C1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 FBOZXECLQNJBKD-ZDUSSCGKSA-N 0.000 description 1
- 108090001090 Lectins Proteins 0.000 description 1
- 102000004856 Lectins Human genes 0.000 description 1
- 239000004472 Lysine Substances 0.000 description 1
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 1
- 101710125418 Major capsid protein Proteins 0.000 description 1
- 229910021380 Manganese Chloride Inorganic materials 0.000 description 1
- GLFNIEUTAYBVOC-UHFFFAOYSA-L Manganese chloride Chemical compound Cl[Mn]Cl GLFNIEUTAYBVOC-UHFFFAOYSA-L 0.000 description 1
- 101100261636 Methanothermobacter marburgensis (strain ATCC BAA-927 / DSM 2133 / JCM 14651 / NBRC 100331 / OCM 82 / Marburg) trpB2 gene Proteins 0.000 description 1
- 108010021466 Mutant Proteins Proteins 0.000 description 1
- 102000008300 Mutant Proteins Human genes 0.000 description 1
- 229910002651 NO3 Inorganic materials 0.000 description 1
- 241001045988 Neogene Species 0.000 description 1
- NHNBFGGVMKEFGY-UHFFFAOYSA-N Nitrate Chemical compound [O-][N+]([O-])=O NHNBFGGVMKEFGY-UHFFFAOYSA-N 0.000 description 1
- 101710141454 Nucleoprotein Proteins 0.000 description 1
- 102000016979 Other receptors Human genes 0.000 description 1
- 241001494479 Pecora Species 0.000 description 1
- 241001417958 Phialidium Species 0.000 description 1
- 102000015439 Phospholipases Human genes 0.000 description 1
- 108010064785 Phospholipases Proteins 0.000 description 1
- 101100124346 Photorhabdus laumondii subsp. laumondii (strain DSM 15139 / CIP 105565 / TT01) hisCD gene Proteins 0.000 description 1
- 229920001030 Polyethylene Glycol 4000 Polymers 0.000 description 1
- 101710083689 Probable capsid protein Proteins 0.000 description 1
- 108010076504 Protein Sorting Signals Proteins 0.000 description 1
- 244000184734 Pyrus japonica Species 0.000 description 1
- 238000001069 Raman spectroscopy Methods 0.000 description 1
- 241000700159 Rattus Species 0.000 description 1
- 241000242743 Renilla reniformis Species 0.000 description 1
- 108010003581 Ribulose-bisphosphate carboxylase Proteins 0.000 description 1
- 108091081021 Sense strand Proteins 0.000 description 1
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 1
- 241000700584 Simplexvirus Species 0.000 description 1
- 108020004682 Single-Stranded DNA Proteins 0.000 description 1
- VMHLLURERBWHNL-UHFFFAOYSA-M Sodium acetate Chemical compound [Na+].CC([O-])=O VMHLLURERBWHNL-UHFFFAOYSA-M 0.000 description 1
- 108091081024 Start codon Proteins 0.000 description 1
- 108010090804 Streptavidin Proteins 0.000 description 1
- 241000282887 Suidae Species 0.000 description 1
- 102000004183 Synaptosomal-Associated Protein 25 Human genes 0.000 description 1
- 108010057722 Synaptosomal-Associated Protein 25 Proteins 0.000 description 1
- ZMZDMBWJUHKJPS-UHFFFAOYSA-M Thiocyanate anion Chemical compound [S-]C#N ZMZDMBWJUHKJPS-UHFFFAOYSA-M 0.000 description 1
- AYFVYJQAPQTCCC-UHFFFAOYSA-N Threonine Natural products CC(O)C(N)C(O)=O AYFVYJQAPQTCCC-UHFFFAOYSA-N 0.000 description 1
- 239000004473 Threonine Substances 0.000 description 1
- 102000006601 Thymidine Kinase Human genes 0.000 description 1
- 239000007983 Tris buffer Substances 0.000 description 1
- 241000251539 Vertebrata <Metazoa> Species 0.000 description 1
- 238000002441 X-ray diffraction Methods 0.000 description 1
- 230000004913 activation Effects 0.000 description 1
- 239000013543 active substance Substances 0.000 description 1
- 125000002015 acyclic group Chemical group 0.000 description 1
- 229960000643 adenine Drugs 0.000 description 1
- 230000002776 aggregation Effects 0.000 description 1
- 238000004220 aggregation Methods 0.000 description 1
- 235000004279 alanine Nutrition 0.000 description 1
- 229940126575 aminoglycoside Drugs 0.000 description 1
- AVKUERGKIZMTKX-NJBDSQKTSA-N ampicillin Chemical compound C1([C@@H](N)C(=O)N[C@H]2[C@H]3SC([C@@H](N3C2=O)C(O)=O)(C)C)=CC=CC=C1 AVKUERGKIZMTKX-NJBDSQKTSA-N 0.000 description 1
- 229960000723 ampicillin Drugs 0.000 description 1
- 230000003321 amplification Effects 0.000 description 1
- 230000003698 anagen phase Effects 0.000 description 1
- 230000000340 anti-metabolite Effects 0.000 description 1
- 230000000692 anti-sense effect Effects 0.000 description 1
- 230000000259 anti-tumor effect Effects 0.000 description 1
- 229940100197 antimetabolite Drugs 0.000 description 1
- 239000002256 antimetabolite Substances 0.000 description 1
- 239000008346 aqueous phase Substances 0.000 description 1
- 239000003125 aqueous solvent Substances 0.000 description 1
- 125000000637 arginyl group Chemical group N[C@@H](CCCNC(N)=N)C(=O)* 0.000 description 1
- 238000000149 argon plasma sintering Methods 0.000 description 1
- 235000009582 asparagine Nutrition 0.000 description 1
- 229960001230 asparagine Drugs 0.000 description 1
- 235000003704 aspartic acid Nutrition 0.000 description 1
- 238000000958 atom scattering Methods 0.000 description 1
- 238000006701 autoxidation reaction Methods 0.000 description 1
- 230000004888 barrier function Effects 0.000 description 1
- 230000008901 benefit Effects 0.000 description 1
- 125000001797 benzyl group Chemical group [H]C1=C([H])C([H])=C(C([H])=C1[H])C([H])([H])* 0.000 description 1
- OQFSQFPPLPISGP-UHFFFAOYSA-N beta-carboxyaspartic acid Natural products OC(=O)C(N)C(C(O)=O)C(O)=O OQFSQFPPLPISGP-UHFFFAOYSA-N 0.000 description 1
- 102000006635 beta-lactamase Human genes 0.000 description 1
- 108091008324 binding proteins Proteins 0.000 description 1
- 230000008033 biological extinction Effects 0.000 description 1
- 239000012620 biological material Substances 0.000 description 1
- 150000001649 bromium compounds Chemical class 0.000 description 1
- 102220345324 c.437A>T Human genes 0.000 description 1
- 239000001506 calcium phosphate Substances 0.000 description 1
- 229910000389 calcium phosphate Inorganic materials 0.000 description 1
- 235000011010 calcium phosphates Nutrition 0.000 description 1
- 125000004432 carbon atom Chemical group C* 0.000 description 1
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 description 1
- 231100000504 carcinogenesis Toxicity 0.000 description 1
- 238000006555 catalytic reaction Methods 0.000 description 1
- 125000002091 cationic group Chemical group 0.000 description 1
- 230000011712 cell development Effects 0.000 description 1
- 230000010261 cell growth Effects 0.000 description 1
- 230000033077 cellular process Effects 0.000 description 1
- 239000013522 chelant Substances 0.000 description 1
- 238000009614 chemical analysis method Methods 0.000 description 1
- 238000001311 chemical methods and process Methods 0.000 description 1
- 238000007385 chemical modification Methods 0.000 description 1
- 238000013375 chromatographic separation Methods 0.000 description 1
- 238000004587 chromatography analysis Methods 0.000 description 1
- 229960002376 chymotrypsin Drugs 0.000 description 1
- 238000000975 co-precipitation Methods 0.000 description 1
- 239000003086 colorant Substances 0.000 description 1
- 238000004891 communication Methods 0.000 description 1
- 238000004883 computer application Methods 0.000 description 1
- 230000001143 conditioned effect Effects 0.000 description 1
- 238000010276 construction Methods 0.000 description 1
- 210000004748 cultured cell Anatomy 0.000 description 1
- 238000012258 culturing Methods 0.000 description 1
- 125000004122 cyclic group Chemical group 0.000 description 1
- 235000018417 cysteine Nutrition 0.000 description 1
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 1
- 231100000433 cytotoxic Toxicity 0.000 description 1
- 230000001472 cytotoxic effect Effects 0.000 description 1
- 230000002950 deficient Effects 0.000 description 1
- 230000018044 dehydration Effects 0.000 description 1
- 238000012217 deletion Methods 0.000 description 1
- 230000037430 deletion Effects 0.000 description 1
- 238000004925 denaturation Methods 0.000 description 1
- 230000036425 denaturation Effects 0.000 description 1
- 238000013461 design Methods 0.000 description 1
- 229910003460 diamond Inorganic materials 0.000 description 1
- 239000010432 diamond Substances 0.000 description 1
- VILAVOFMIJHSJA-UHFFFAOYSA-N dicarbon monoxide Chemical compound [C]=C=O VILAVOFMIJHSJA-UHFFFAOYSA-N 0.000 description 1
- 230000029087 digestion Effects 0.000 description 1
- QONQRTHLHBTMGP-UHFFFAOYSA-N digitoxigenin Natural products CC12CCC(C3(CCC(O)CC3CC3)C)C3C11OC1CC2C1=CC(=O)OC1 QONQRTHLHBTMGP-UHFFFAOYSA-N 0.000 description 1
- SHIBSTMRCDJXLN-KCZCNTNESA-N digoxigenin Chemical compound C1([C@@H]2[C@@]3([C@@](CC2)(O)[C@H]2[C@@H]([C@@]4(C)CC[C@H](O)C[C@H]4CC2)C[C@H]3O)C)=CC(=O)OC1 SHIBSTMRCDJXLN-KCZCNTNESA-N 0.000 description 1
- 201000010099 disease Diseases 0.000 description 1
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 1
- 238000006073 displacement reaction Methods 0.000 description 1
- 238000010494 dissociation reaction Methods 0.000 description 1
- 230000005593 dissociations Effects 0.000 description 1
- 238000009826 distribution Methods 0.000 description 1
- 235000013601 eggs Nutrition 0.000 description 1
- 238000002330 electrospray ionisation mass spectrometry Methods 0.000 description 1
- 210000002472 endoplasmic reticulum Anatomy 0.000 description 1
- 230000030583 endoplasmic reticulum localization Effects 0.000 description 1
- 238000001839 endoscopy Methods 0.000 description 1
- 230000002255 enzymatic effect Effects 0.000 description 1
- 238000000198 fluorescence anisotropy Methods 0.000 description 1
- 238000000799 fluorescence microscopy Methods 0.000 description 1
- 238000001506 fluorescence spectroscopy Methods 0.000 description 1
- 239000007850 fluorescent dye Substances 0.000 description 1
- 239000003269 fluorescent indicator Substances 0.000 description 1
- 239000011737 fluorine Substances 0.000 description 1
- 230000002068 genetic effect Effects 0.000 description 1
- 238000010353 genetic engineering Methods 0.000 description 1
- 235000013922 glutamic acid Nutrition 0.000 description 1
- 239000004220 glutamic acid Substances 0.000 description 1
- 125000000267 glycino group Chemical class [H]N([*])C([H])([H])C(=O)O[H] 0.000 description 1
- 230000013595 glycosylation Effects 0.000 description 1
- 238000006206 glycosylation reaction Methods 0.000 description 1
- 230000012010 growth Effects 0.000 description 1
- 239000003966 growth inhibitor Substances 0.000 description 1
- 238000004128 high performance liquid chromatography Methods 0.000 description 1
- 101150113423 hisD gene Proteins 0.000 description 1
- 238000002744 homologous recombination Methods 0.000 description 1
- 230000006801 homologous recombination Effects 0.000 description 1
- ZMZDMBWJUHKJPS-UHFFFAOYSA-N hydrogen thiocyanate Natural products SC#N ZMZDMBWJUHKJPS-UHFFFAOYSA-N 0.000 description 1
- 238000005286 illumination Methods 0.000 description 1
- 238000003384 imaging method Methods 0.000 description 1
- 230000014726 immortalization of host cell Effects 0.000 description 1
- 102000018358 immunoglobulin Human genes 0.000 description 1
- 229940072221 immunoglobulins Drugs 0.000 description 1
- 238000000099 in vitro assay Methods 0.000 description 1
- 239000000411 inducer Substances 0.000 description 1
- 239000003112 inhibitor Substances 0.000 description 1
- 230000005764 inhibitory process Effects 0.000 description 1
- 238000002347 injection Methods 0.000 description 1
- 239000007924 injection Substances 0.000 description 1
- 230000002452 interceptive effect Effects 0.000 description 1
- 229940125425 inverse agonist Drugs 0.000 description 1
- 229910052740 iodine Inorganic materials 0.000 description 1
- BPHPUYQFMNQIOC-NXRLNHOXSA-N isopropyl beta-D-thiogalactopyranoside Chemical compound CC(C)S[C@@H]1O[C@H](CO)[C@H](O)[C@H](O)[C@H]1O BPHPUYQFMNQIOC-NXRLNHOXSA-N 0.000 description 1
- 239000002523 lectin Substances 0.000 description 1
- 230000000670 limiting effect Effects 0.000 description 1
- 239000002502 liposome Substances 0.000 description 1
- 238000012423 maintenance Methods 0.000 description 1
- FDZZZRQASAIRJF-UHFFFAOYSA-M malachite green Chemical compound [Cl-].C1=CC(N(C)C)=CC=C1C(C=1C=CC=CC=1)=C1C=CC(=[N+](C)C)C=C1 FDZZZRQASAIRJF-UHFFFAOYSA-M 0.000 description 1
- 239000011565 manganese chloride Substances 0.000 description 1
- 230000013011 mating Effects 0.000 description 1
- 239000011159 matrix material Substances 0.000 description 1
- 230000035800 maturation Effects 0.000 description 1
- 238000010297 mechanical methods and process Methods 0.000 description 1
- 239000002609 medium Substances 0.000 description 1
- 238000002844 melting Methods 0.000 description 1
- 230000008018 melting Effects 0.000 description 1
- TWXDDNPPQUTEOV-FVGYRXGTSA-N methamphetamine hydrochloride Chemical compound Cl.CN[C@@H](C)CC1=CC=CC=C1 TWXDDNPPQUTEOV-FVGYRXGTSA-N 0.000 description 1
- 229960000485 methotrexate Drugs 0.000 description 1
- 238000000386 microscopy Methods 0.000 description 1
- 210000003470 mitochondria Anatomy 0.000 description 1
- 230000025608 mitochondrion localization Effects 0.000 description 1
- 230000004048 modification Effects 0.000 description 1
- 238000012986 modification Methods 0.000 description 1
- 238000010369 molecular cloning Methods 0.000 description 1
- 238000004776 molecular orbital Methods 0.000 description 1
- 230000000869 mutational effect Effects 0.000 description 1
- 101150091879 neo gene Proteins 0.000 description 1
- 230000010309 neoplastic transformation Effects 0.000 description 1
- 238000003199 nucleic acid amplification method Methods 0.000 description 1
- 238000007899 nucleic acid hybridization Methods 0.000 description 1
- 210000004940 nucleus Anatomy 0.000 description 1
- 230000030648 nucleus localization Effects 0.000 description 1
- 235000015097 nutrients Nutrition 0.000 description 1
- 210000000056 organ Anatomy 0.000 description 1
- 239000007800 oxidant agent Substances 0.000 description 1
- 230000003647 oxidation Effects 0.000 description 1
- 238000007254 oxidation reaction Methods 0.000 description 1
- 238000012856 packing Methods 0.000 description 1
- 230000003071 parasitic effect Effects 0.000 description 1
- VLTRZXGMWDSKGL-UHFFFAOYSA-M perchlorate Inorganic materials [O-]Cl(=O)(=O)=O VLTRZXGMWDSKGL-UHFFFAOYSA-M 0.000 description 1
- VLTRZXGMWDSKGL-UHFFFAOYSA-N perchloric acid Chemical compound OCl(=O)(=O)=O VLTRZXGMWDSKGL-UHFFFAOYSA-N 0.000 description 1
- 230000008823 permeabilization Effects 0.000 description 1
- 210000002824 peroxisome Anatomy 0.000 description 1
- 230000014318 peroxisome localization Effects 0.000 description 1
- ISWSIDIOOBJBQZ-UHFFFAOYSA-M phenolate Chemical compound [O-]C1=CC=CC=C1 ISWSIDIOOBJBQZ-UHFFFAOYSA-M 0.000 description 1
- 229940031826 phenolate Drugs 0.000 description 1
- 150000007965 phenolic acids Chemical class 0.000 description 1
- COLNVLDHVKWLRT-UHFFFAOYSA-N phenylalanine Natural products OC(=O)C(N)CC1=CC=CC=C1 COLNVLDHVKWLRT-UHFFFAOYSA-N 0.000 description 1
- 108091008695 photoreceptors Proteins 0.000 description 1
- 230000002319 phototactic effect Effects 0.000 description 1
- 230000001766 physiological effect Effects 0.000 description 1
- 238000002264 polyacrylamide gel electrophoresis Methods 0.000 description 1
- 230000008488 polyadenylation Effects 0.000 description 1
- 229920001223 polyethylene glycol Polymers 0.000 description 1
- 238000003752 polymerase chain reaction Methods 0.000 description 1
- 230000004481 post-translational protein modification Effects 0.000 description 1
- 230000001323 posttranslational effect Effects 0.000 description 1
- 230000003334 potential effect Effects 0.000 description 1
- 230000013823 prenylation Effects 0.000 description 1
- 210000001236 prokaryotic cell Anatomy 0.000 description 1
- 230000001737 promoting effect Effects 0.000 description 1
- 230000000644 propagated effect Effects 0.000 description 1
- 230000001902 propagating effect Effects 0.000 description 1
- 235000019833 protease Nutrition 0.000 description 1
- 230000013777 protein digestion Effects 0.000 description 1
- 239000011546 protein dye Substances 0.000 description 1
- 230000026447 protein localization Effects 0.000 description 1
- 230000018883 protein targeting Effects 0.000 description 1
- 230000006337 proteolytic cleavage Effects 0.000 description 1
- 230000005588 protonation Effects 0.000 description 1
- 210000001938 protoplast Anatomy 0.000 description 1
- 238000005086 pumping Methods 0.000 description 1
- 238000011002 quantification Methods 0.000 description 1
- 230000008707 rearrangement Effects 0.000 description 1
- 238000011084 recovery Methods 0.000 description 1
- 108010054624 red fluorescent protein Proteins 0.000 description 1
- 238000007670 refining Methods 0.000 description 1
- 230000008929 regeneration Effects 0.000 description 1
- 238000011069 regeneration method Methods 0.000 description 1
- 238000012857 repacking Methods 0.000 description 1
- 238000002165 resonance energy transfer Methods 0.000 description 1
- 238000006798 ring closing metathesis reaction Methods 0.000 description 1
- 102200005981 rs104894561 Human genes 0.000 description 1
- 238000007423 screening assay Methods 0.000 description 1
- 239000013606 secretion vector Substances 0.000 description 1
- 239000006152 selective media Substances 0.000 description 1
- 238000012163 sequencing technique Methods 0.000 description 1
- 230000037432 silent mutation Effects 0.000 description 1
- 239000002356 single layer Substances 0.000 description 1
- 150000003384 small molecules Chemical class 0.000 description 1
- 239000001632 sodium acetate Substances 0.000 description 1
- 235000017281 sodium acetate Nutrition 0.000 description 1
- 239000007787 solid Substances 0.000 description 1
- 238000007614 solvation Methods 0.000 description 1
- 230000009870 specific binding Effects 0.000 description 1
- 230000002269 spontaneous effect Effects 0.000 description 1
- 238000010561 standard procedure Methods 0.000 description 1
- 239000007858 starting material Substances 0.000 description 1
- 238000007619 statistical method Methods 0.000 description 1
- 238000005728 strengthening Methods 0.000 description 1
- 229910052717 sulfur Inorganic materials 0.000 description 1
- 239000006228 supernatant Substances 0.000 description 1
- 230000001629 suppression Effects 0.000 description 1
- 238000010189 synthetic method Methods 0.000 description 1
- WROMPOXWARCANT-UHFFFAOYSA-N tfa trifluoroacetic acid Chemical compound OC(=O)C(F)(F)F.OC(=O)C(F)(F)F WROMPOXWARCANT-UHFFFAOYSA-N 0.000 description 1
- 230000001225 therapeutic effect Effects 0.000 description 1
- 210000001519 tissue Anatomy 0.000 description 1
- 231100000419 toxicity Toxicity 0.000 description 1
- 230000001988 toxicity Effects 0.000 description 1
- 230000002103 transcriptional effect Effects 0.000 description 1
- 238000010361 transduction Methods 0.000 description 1
- 230000026683 transduction Effects 0.000 description 1
- 238000003151 transfection method Methods 0.000 description 1
- 230000009466 transformation Effects 0.000 description 1
- 238000011426 transformation method Methods 0.000 description 1
- QORWJWZARLRLPR-UHFFFAOYSA-H tricalcium bis(phosphate) Chemical compound [Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O QORWJWZARLRLPR-UHFFFAOYSA-H 0.000 description 1
- 101150081616 trpB gene Proteins 0.000 description 1
- 101150111232 trpB-1 gene Proteins 0.000 description 1
- 241000701447 unidentified baculovirus Species 0.000 description 1
- 210000004291 uterus Anatomy 0.000 description 1
- 210000004340 zona pellucida Anatomy 0.000 description 1
Images





























































Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/43504—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from invertebrates
- C07K14/43595—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from invertebrates from coelenteratae, e.g. medusae
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Life Sciences & Earth Sciences (AREA)
- Genetics & Genomics (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Zoology (AREA)
- Biochemistry (AREA)
- Biophysics (AREA)
- General Health & Medical Sciences (AREA)
- Toxicology (AREA)
- Medicinal Chemistry (AREA)
- Molecular Biology (AREA)
- Gastroenterology & Hepatology (AREA)
- Tropical Medicine & Parasitology (AREA)
- Peptides Or Proteins (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Investigating Or Analysing Biological Materials (AREA)
- Investigating Or Analysing Materials By The Use Of Chemical Reactions (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
- Cosmetics (AREA)
- Developing Agents For Electrophotography (AREA)
Description
発明の背景
この1997年11月19日に出願され、許可された一部継続出願第08/974,737号、1997年8月15日に出願され、米国特許第6,054,321号として発行された継続出願第08/911,825号、1996年8月30日に出願され、許可された一部継続出願第08/706,408号、これらは、1996年8月16日に出願された米国仮出願第60/024,050号を先行する出願日の恩典として主張し、各出願は参照として本明細書に組み入れられる。
【0002】
本発明は、国立科学財団(NSF)より供与された助成金番号MCB 9418479の下で政府の支援の一部としてなされた。政府は本発明の権利を有するものとする。
【0003】
蛍光分子は、感度が高くて定量が容易なため、多くのアッセイ系におけるレポーター分子として魅力ある分子である。最近、蛍光タンパク質には、生物系でインビボで作製可能で、かつ細胞内で起こる過程の追跡にマイクロインジェクションまたは透過化処理による細胞への導入を必要とすることなく使用可能であることから多くの注目を集めつつある。オワンクラゲ(Aequorea victoria)の緑色蛍光タンパク質は、蛍光タンパク質として特に関心が寄せられている。このタンパク質のcDNAは既にクローン化されている(D.C. Prasherら、「オワンクラゲの緑色蛍光タンパク質の一次構造(Primary structure of the Aequorea victoria green-fluorescent protein)」、Gene (1992) 111:229〜33)。同タンパク質の一次アミノ酸配列がcDNAから発現されているだけでなく、発現されたタンパク質は蛍光を発することもできる。これは、同タンパク質が、蛍光発生に必要と考えられる環状化および酸化を受ける可能性があることを示している。オワンクラゲの緑色蛍光タンパク質(「GFP」)は安定であり、タンパク質による分解に耐性をもつ238残基の1本鎖からなり、二つの吸収極大を395 nmと475 nm付近にもつ。2本のピークの相対的な振幅は、おそらく二つまたはそれ以上の基底状態を反映して、環境因子(W.W. Ward.、「生物発光および化学発光(Bioluminescence and Chemiluminescence)」(M.A. DeLucaおよびW.D. McElroy編)、Academic Press、pp. 235〜242 (1981);W.W. WardおよびS.H. Bokman、Biochemistry 21:4535〜4540 (1982);W.W. Wardら、Photochem. Photobiol. 35:803〜808 (1982))、および照度履歴(illumination hitrory)(A.B. Cubittら、Trends Biochem. Sci. 20:448〜455 (1995))に高感度で反応する。395 nmの一次吸収ピークで励起すると、508 nmにおける、量子収率が0.72〜0.85の発光極大が得られる(O. ShimomuraおよびF.H. Johnson、J. Cell. Comp. Physiol. 9:223 (1962);J.G. MorinおよびJ.W. Hastings、J. Cell. Physiol. 77:313 (1971);H. Moriseら、Biochemistry 13:2656 (1974);W.W. Ward、「Photochem. Photobiol. 総説」(Smith、K.C.編) 4:1 (1979);A.B. Cubittら、Trends Biochem. Sci. 20:448〜455 (1995);D.C. Prasher、Trends Genet. 11:320〜323 (1995);M. Chalfie、Photochem. Photobiol. 62:651〜656 (1995);W.W. Ward、「生物発光および化学発光(Bioluminescence and Chemiluminescence)」(M.A. DeLucaおよびW.D. McElroy編)、Academic Press、pp. 235〜242 (1981);W.W. WardおよびS.H. Bokman、Biochemistry 21:4535〜4540 (1982);W.W. Wardら、Photochem. Photobiol. 35:803〜808 (1982))。フルオロフォアは、Ser65残基とGly75残基の間にあるポリペプチド骨格の自己触媒的な環状化、およびTyr66のβ結合の酸化によって生じる(A.B. Cubittら、Trends Biochem. Sci. 20:448〜455 (1995);C.W. Codyら、Biochemistry 32:1212〜1218 (1993);R. Heimら、Proc. Natl. Acad. Sci. USA 91:12501〜12504 (1994))。Ser65がThrに変異すると(S65T)、励起スペクトルが振幅の大きい488 nmにおける1本のピークだけになり(R. Heimら、Nature 373:664〜665 (1995))、構造異性体の性質を示さなくなる(A.B. Cubittら、Trends Biochem. Sci. 20:448〜455 (1995))。
【0004】
蛍光タンパク質は、遺伝子発現のマーカー、細胞系列のトレーサー、および生細胞内におけるタンパク質の局在をモニタリングするための融合タグとして用いられている(M. Chalfieら、「遺伝子発現のマーカーとしての緑色蛍光タンパク質(Green fluorescent protein as a marker for gene expression)」、Science 263:802〜805;A.B. Cubittら、「緑色蛍光タンパク質の理解、改善、および用途(Understanding、improving and using green fluorescent proteins)」、TIBS 20、1995年11月、pp. 448〜455、M. ChalfieおよびD. Prasherによる米国特許第5,491,084号。また人為的に作製されたオワンクラゲの緑色蛍光タンパク質は、励起極大および発光極大の変化、ならびに異なる形状の励起および発光スペクトルを含む、蛍光特性の変化を示すことが明らかにされている(R. Heimら、「緑色蛍光タンパク質の波長の変異および翻訳後自己酸化(Wavelength mutations and posttranslational autoxidation of green fluorescent ptotein)」、Proc. Natl. Acad. Sci. USA、 (1994) 91:12501〜04;R. Heimら、「緑色蛍光の改善(Improved green fluorescent)」、Nature、(1995) 373:663〜665)。
【0005】
第2のクラスの応用は、いくつかの細胞特性の特異的インジケーターとしてのGFPに基づいており、したがって、使用される異型の特定のスペクトル特性に依存している。GFPの異型およびその応用に関する最近の総説は、(PalmおよびWlodawer、1999;Tsien、1998)を、また特殊な応用の総説は、(SullivanおよびKay、1999)を参照されたい。バイオセンサーへの応用には、タンパク質間相互作用(Heim、1999)、またはCa2+濃度(Miyawakiら、1999)をモニタリングするための、およびリガンド結合をモニタリングするための、GFP表面ループ内における受容体の挿入(Bairdら、1999;DoiおよびYanagawa、1999)、蛍光共鳴エネルギー移動(FRET)に対する異なる色を発するGFPの使用が含まれる。
【0006】
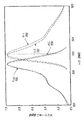
いくつかの異型の蛍光放出は、環境の酸性度に対して感度が高い(Elsligerら、1999;Wachterら、1998)。したがって、生細胞内における視覚的レポーターとして緑色蛍光タンパク質(GFP)の特に成功している応用は、オルガネラまたは細胞質のpHの決定である(Kneenら、1998;Llopisら、1998;Miesenbockら、1998;Robeyら、1998)。発色団の二種の荷電状態は、完全なタンパク質のpHに対する感度と関連することがわかっており、フェノール性末端付近の構造変化に関しては結晶学的に(Elsligerら、1999)、またラマン分光法で分光学的に(Bellら、2000)特性が解析されている。中性状態の発色団であるバンドAは、ほとんどの異型で400 nm付近で吸光を示すが、脱プロトンされたフェノール性末端をもつ発色団の陰イオン(バンドB)は、発色団付近の特定の変異では青色〜緑色を吸収する。野生型GFPは、バンドAとバンドBの組み合わせ、すなわちpH6〜10で比較的不変の組み合わせを特徴とする二種の基底状態に一致するスペクトル特性を示す(PalmおよびWlodawer、1999;Wardら、1982)。広範囲のpHで発色団のフェノレートとGlu222のカルボン酸とでプロトンが共有される内部平衡が存在することが示唆されている(Brejcら、1997;Palmら、1997)。最近実施された静電気力に関する計算では、同モデルが支持されており(Scharnaglら、1999)、完全な発色団の脱プロトンの理論的なpKaが約13(pH 11〜12における発光強度が倍加するという観察と一致する)と推定されている(BokmanおよびWard、1981;PalmおよびWlodawer、1999)。
【0007】
野生型GFPとは対照的に、大部分の異型の発色団は一つのpKaをもつとされている。色の放出および発色団のpKaは、周囲のタンパク質によって強く調節される(Llopisら、1998)。Glu222はGFP相同物で完全に保存されており(Matzら、1999)、この残基がグルタミンで置換されると、発色団形成の効率が大きく低下することが報告されている(Elsligerら、1999)。S65T、およびT203Y変異を含むGFP(YFP)のGlu222のプロトン化は、GFP S65Tでは野生型のpKaから約5.9へ(Elsligerら、1999;Kneenら、1998)、またYFP (GFP S65G/V68L/S72A/T203Y)では5.2〜5.4(Ormoら、1996;WachterおよびRemington、1999)への発色団のpKaの低下にかかわると一般に考えられている。YFPでは、発色団とTyr203との結晶学的に同一なスタッキング相互作用がスペクトルの赤色シフトに大きな役割を果たすと考えられている(Wachterら、1998)。
【0008】
他の異型とは異なり、発明者らは、YFPの発色団のpKaが、塩化物イオンなどのある種の小さな陰イオンの濃度に強く依存すること(WachterおよびRemington、1999)、ならびにpKaが140 mM NaClの存在下で約5.2から7.0へ上昇することを見出した(Elsligerら、1999)。このような感度を利用することで、生細胞内の細胞質または細胞内区画の両方に存在するイオンを測定するバイオセンサーとして新しいGFPを作製することが可能となる(WachterおよびRemington、1999)。本発明は、塩化物やヨウ化物などのハロゲン化物を含むさまざまなイオンの蛍光測定を可能とする新しいGFP異型の作製および用途を含む。このような特性は、生物学的な蛍光インジケーターの利用に多様性および利便性を加える。蛍光特性の変化を介して、さまざまな蛍光特性、およびイオン濃度に反応する能力をもつ改変蛍光タンパク質が求められている。
【0009】
発明の概要
本発明は、現行の緑色蛍光タンパク質および青色蛍光タンパク質と容易に区別可能な、さまざまな蛍光特性をもつ、機能性の改変蛍光タンパク質を提供する。このような改変蛍光タンパク質では、細胞内における二種またはそれ以上の過程を同時に測定することが可能となり、また蛍光エネルギーのドナーまたはアクセプター、ならびに陰イオンを検出するバイオセンサーとして使用することができる。長波長の改変蛍光タンパク質は、光力学性の毒性および細胞の自己蛍光が、長波長では有意に低いことから特に有用である。特に、置換T203X (Xは芳香族アミノ酸)を導入すると、オワンクラゲ関連蛍光タンパク質の励起波長および発光波長の極大が上昇する。
【0010】
一つの局面では、本発明は、アミノ酸配列がオワンクラゲの緑色蛍光タンパク質のアミノ酸配列(配列番号:2)と実質的に同一で、改変蛍光タンパク質の発色団から約0.5 nm以内に位置する少なくとも1個のアミノ酸の、発色団の電子的環境を変化させる置換が配列番号:2と異なる、オワンクラゲの緑色蛍光タンパク質とは異なった蛍光特性を有する機能性の改変蛍光タンパク質をコードするヌクレオチド配列を含む核酸分子を提供する。
【0011】
一つの局面では、本発明は、アミノ酸配列がオワンクラゲの緑色蛍光タンパク質のアミノ酸配列(配列番号:2)と実質的に同一で、少なくともT203における置換(特にT203X;XはH、Y、W、またはFからなる群より選択される芳香族アミノ酸)が配列番号:2と異なる、オワンクラゲの緑色蛍光タンパク質とは異なった蛍光特性を有する機能性の改変蛍光タンパク質をコードするヌクレオチド配列を含む核酸分子を提供する。一つの態様では、アミノ酸配列は、S65G、S65T、S65A、S65L、S65C、S65V、およびS65Iからなる群より選択される、S65における置換をさらに含む。別の態様では、アミノ酸配列は、置換S65T/T203H;S65T/T203Y;S72A/F64L/S65G/T203Y;S65G/V68L/Q69K/S72A/T203Y;S72A/S65G/V68L/T203Y;S65G/S72A/T203Y;またはS65G/S72A/T203Wのみが異なる。別の態様では、アミノ酸配列は、Y66H、Y66F、およびY66Wからなる群より選択される、Y66における置換をさらに含む。別の態様では、アミノ酸配列は、表Aに記載された変異をさらに含む。別の態様では、アミノ酸配列は、折りたたみ変異をさらに含む。別の態様では、タンパク質をコードするヌクレオチド配列は、配列番号:1のヌクレオチド配列と少なくとも一つのコドンが、哺乳類で好ましいコドンと異なる。別の態様では、核酸分子は、対象ポリペプチド、および機能性の改変蛍光タンパク質を含む融合タンパク質をコードする。
【0012】
別の局面では、本発明は、アミノ酸配列がオワンクラゲの緑色蛍光タンパク質のアミノ酸配列(配列番号:2)と実質的に同一で、L42、V61、T62、V68、Q69、Q94、N121、Y145、H148、V150、F165、I167、Q183、N185、L220、E222 (E222Gは除く)、またはV224における少なくとも一つのアミノ酸置換が配列番号:2と異なる、オワンクラゲの緑色蛍光タンパク質とは異なった蛍光特性を有する機能性の改変蛍光タンパク質をコードするヌクレオチド配列を含む核酸分子を提供する。一つの態様では、アミノ酸の置換には以下のようなものがある:
L42X (XはC、F、H、W、およびYからなる群より選択される)、
V61X (XはF、Y、H、およびCからなる群より選択される)、
T62X (XはA、V、F、S、D、N、Q、Y、H、およびCからなる群より選択される)、
V68X (XはF、Y、およびHからなる群より選択される)、
Q69X (XはK、R、E、およびGからなる群より選択される)、
Q94X (XはD、E、H、K、およびNからなる群より選択される)、
N121X (XはF、H、W、およびYからなる群より選択される)、
Y145X (XはW、C、F、L、E、H、K、およびQからなる群より選択される)、
H148X (XはF、Y、N、K、Q、およびRからなる群より選択される)、
V150X (XはF、Y、およびHからなる群より選択される)、
F165X (XはH、Q、W、およびYからなる群より選択される)、
I167X (XはF、Y、およびHからなる群より選択される)、
Q183X (XはH、Y、E、およびKからなる群より選択される)、
N185X (XはD、E、H、K、およびQからなる群より選択される)、
L220X (XはH、N、Q、およびTからなる群より選択される)、
E222X (XはNおよびQからなる群より選択される)、または
V224X (XはH、N、Q、T、F、W、およびYからなる群より選択される)。
【0013】
別の局面では、本発明は、前述の任意の核酸分子に動作可能に連結させた発現制御配列を含む発現ベクターを提供する。別の局面では、本発明は、前述の発現ベクターを含む組換え宿主細胞を提供する。
【0014】
別の局面では、本発明は、アミノ酸配列がオワンクラゲの緑色蛍光タンパク質のアミノ酸配列(配列番号:2)と実質的に同一で、改変蛍光タンパク質の発色団から約0.5 nm以内に位置する少なくとも1個のアミノ酸の、発色団の電子的環境を変化させる置換が配列番号:2と異なる、オワンクラゲの緑色蛍光タンパク質とは異なった蛍光特性を有する機能性の改変蛍光タンパク質を提供する。
【0015】
別の局面では、本発明は、アミノ酸配列がオワンクラゲの緑色蛍光タンパク質のアミノ酸配列(配列番号:2)と実質的に同一で、少なくともT203における置換(特にT203X;XはH、Y、W、またはFからなる群より選択される芳香族アミノ酸)が配列番号:2と異なる、オワンクラゲの緑色蛍光タンパク質とは異なった蛍光特性を有する機能性の改変蛍光タンパク質を提供する。一つの態様では、アミノ酸配列は、S65G、S65T、S65A、S65L、S65C、S65V、およびS65Iからなる群より選択される、S65における置換を含む。別の態様では、アミノ酸配列は、置換S65T/T203H;S65T/T203Y;S72A/F64L/S65G/T203Y;S72A/S65G/V68L/T203Y;S65G/V68L/Q69K/S72A/T203Y;S65G/S72A/T203Y;またはS65G/S72A/T203Wのみが異なる。別の態様では、アミノ酸配列は、Y66H、Y66F、およびY66Wからなる群より選択される、Y66における置換をさらに含む。別の態様では、アミノ酸配列は、折りたたみ変異をさらに含む。別の態様では、改変蛍光タンパク質は、対象ポリペプチドおよび機能性の改変蛍光タンパク質を含む融合タンパク質の一部である。
【0016】
別の局面では、本発明は、アミノ酸配列がオワンクラゲの緑色蛍光タンパク質のアミノ酸配列(配列番号:2)と実質的に同一で、L42、V61、T62、V68、Q69、Q94、N121、Y145、H148、V150、F165、I167、Q183、N185、L220、E222、またはV224における少なくとも一つのアミノ酸置換が配列番号:2と異なる、オワンクラゲの緑色蛍光タンパク質とは異なった蛍光特性を有する機能性の改変蛍光タンパク質を提供する。
【0017】
別の局面では、本発明は、前述の任意の機能性の改変蛍光タンパク質に結合させた抗体を含む、蛍光標識された抗体を提供する。一つの態様では、蛍光標識された抗体は、機能性の改変蛍光タンパク質に融合させた抗体を含む融合タンパク質である。
【0018】
別の局面では、本発明は、本発明の改変蛍光タンパク質をコードするヌクレオチド配列に融合させた抗体をコードするヌクレオチド配列を含む核酸分子を提供する。
【0019】
別の局面では、本発明は、本発明のアミノ酸配列を有する機能性の改変蛍光タンパク質に結合させた核酸プローブを含む蛍光標識された核酸プローブを提供する。このような融合は、リンカーペプチドを介する場合がある。
【0020】
別の局面では、本発明は、任意のプローブを含む蛍光標識されたプローブ、および本発明の機能性の改変蛍光タンパク質に任意の混合物を接触させる段階;および任意の標的がプローブに結合しているか否かを判定する段階を含む、混合物が標的を含むか否かを判定する方法を提供する。一つの態様では、標的分子は固体マトリックス上に捕捉される。
【0021】
別の局面では、本発明は、オワンクラゲ関連緑色蛍光タンパク質の発色団の任意の原子から0.5 nm以内の位置にあるアミノ酸を別のアミノ酸と置換する、タンパク質の蛍光特性を変化させる段階を含む、蛍光特性がオワンクラゲの緑色蛍光タンパク質とは異なる機能性の改変蛍光タンパク質を人工的に作製する方法を提供する。一つの態様では、このようなアミノ酸の置換は、発色団の電子的環境を変化させる。
【0022】
別の局面では、本発明は、オワンクラゲ関連緑色蛍光タンパク質のループドメインのアミノ酸を、リン酸化またはタンパク質分解のコンセンサス配列が作られるようなアミノ酸と置換する段階を含む、蛍光特性がオワンクラゲの緑色蛍光タンパク質とは異なる機能性の改変蛍光タンパク質を作製する方法を提供する。
【0023】
別の局面では、本発明は、本発明の機能性の改変蛍光タンパク質を含むドナー分子を提供する段階;蛍光タンパク質に適切なアクセプター分子を提供する段階;および、ドナー分子とアクセプター分子とを蛍光共鳴エネルギー移動が生じるように十分近接させる段階を含む、蛍光共鳴エネルギー移動を生じさせる方法を提供する。
【0024】
別の局面では、本発明は、本発明の機能性の改変蛍光タンパク質を含むアクセプター分子を提供する段階;蛍光タンパク質に適切なドナー分子を提供する段階;および、ドナー分子とアクセプター分子とを蛍光共鳴エネルギー移動が生じるように十分近接させる段階を含む、蛍光共鳴エネルギー移動を生じさせる方法を提供する。一つの態様では、ドナー分子は、アミノ酸配列が置換T203Iを含む改変蛍光タンパクで、またアクセプター分子が、置換T203X (XはH、Y、W、またはFからなる群より選択される芳香族アミノ酸)を含むアミノ酸配列を含む、蛍光特性がオワンクラゲの緑色蛍光タンパク質とは異なる改変蛍光タンパク質である。
【0025】
別の局面では、本発明は、アミノ酸配列が配列番号:2と実質的に同一の蛍光タンパク質を含む、少なくとも2.0〜3.0オングストロームの分解能で回折するタンパク質の結晶を提供する。
【0026】
別の態様では、本発明は、リガンドの少なくとも1個の第1の化学基と相互作用する蛍光タンパク質の少なくとも1個の相互作用性アミノ酸を、リガンドが結合した蛍光タンパク質を含む結晶化した蛍光タンパク質の三次元モデルから決定する段階、および第1の化学基の少なくとも一つの化学的修飾を選択して、相互作用性アミノ酸と第1の化学基との間の相互作用に比べて、相互作用性アミノ酸と第2の化学基との間の相互作用が低下または上昇する構造を有する第2の化学基を作製する段階を含む、蛍光タンパク質を設計するコンピュータを利用する方法を提供する。
【0027】
別の態様では、本発明は、図5-1〜5-28の原子座標に挙げた少なくとも2個の原子間の三次元関係を決定する段階を含む、蛍光タンパク質の三次元構造をモデル化するコンピュータを利用する方法を提供する。
【0028】
別の態様では、本発明は、記憶装置、および同装置において図5-1〜5-28に挙げた原子座標から選択される少なくとも10の原子座標が記憶されている、装置を提供する。一つの態様では、記憶装置は、原子座標を入力として受け取るコードを記憶するコンピュータ可読装置である。別の態様では、コンピュータ可読装置は、フロッピーディスクまたはハードドライブである。
【0029】
別の態様では、本発明は、アミノ酸配列がオワンクラゲの緑色蛍光タンパク質のアミノ酸配列(配列番号:2)と実質的に同一で、T203位における、H、Y、W、またはFからなる群より選択される少なくとも一つの第1の置換、およびH148位における少なくとも一つの第2の置換が配列番号:2と異なる機能性の改変蛍光タンパク質をコードするヌクレオチド配列を含む核酸分子を提供する。
【0030】
別の局面では、本発明は、アミノ酸配列がオワンクラゲの緑色蛍光タンパク質のアミノ酸配列(配列番号:2)と実質的に同一で、T203位における、H、Y、W、またはFからなる群より選択される少なくとも一つの第1の置換が配列番号:2と異なるアミノ酸配列を含む改変緑色蛍光タンパク質を任意の試料に導入する段階、および試料中の改変緑色蛍光タンパク質の蛍光を決定する段階を含む、試料中の対象陰イオンの存在を判定する方法を含む。
【0031】
別の局面では、本発明は、アミノ酸配列がオワンクラゲの緑色蛍光タンパク質のアミノ酸配列(配列番号:2)と実質的に同一で、T203位における、H、Y、W、またはFからなる群より選択される少なくとも一つの第1の置換、およびH148位における少なくとも一つの第2の置換が配列番号:2と異なる、オワンクラゲの緑色蛍光タンパク質とは異なった蛍光特性を有する機能性の改変蛍光タンパク質を含む。
【0032】
別の態様では、本発明は、アミノ酸配列がオワンクラゲの緑色蛍光タンパク質のアミノ酸配列(配列番号:2)と実質的に同一で、T203位における、H、Y、W、またはFからなる群より選択される少なくとも一つ第1の置換、およびH148位における少なくとも一つの第2の置換が配列番号:2と異なる、オワンクラゲの緑色蛍光タンパク質とは異なった蛍光特性を有する機能性の改変蛍光タンパク質を含む宿主細胞を含む。
【0033】
発明の詳細な説明
I.定義
文中で特に断らない限り、本明細書で用いるすべての科学技術用語は、本発明が属する技術分野の当業者によって一般に理解される用語と同じ意味をもつ。本明細書に記載した方法および材料と類似しているかまたは同一である任意の方法および材料は、本発明を実施する際または検討する際に使用されるが、好ましい方法および材料について説明する。本発明の目的上、以下の用語を定義する。
【0034】
「結合ペア」とは、相互に親和性をもつ二つの部分(例えば、化学的部分または生化学的部分)を意味する。結合ペアの例には、抗原/抗体、レクチン/アビジン、標的ポリヌクレオチド/プローブのオリゴヌクレオチド、抗体/抗-抗体、受容体/リガンド、酵素/リガンドなどがある。「結合ペアの一つの成分」とはペアの一つの部分(例えば抗原やリガンド)を意味する。
【0035】
「核酸」とは、1本鎖または2本鎖の形状をとるデオキシリボヌクレオチドまたはリボヌクレオチドの重合体を意味し、文中で特に限定されない限り、天然のヌクレオチドと同様に機能可能な天然のヌクレオチドの既知の類似体を含む。核酸分子がDNA配列である場合、これは、「U」が「T」で置換されている対応RNA配列を有するRNA分子も含むように理解される。
【0036】
「組換え核酸分子」とは、天然ではない核酸分子、および天然の状態で結合した状態にない二種のヌクレオチド配列を含む核酸分子を意味する。組換え核酸分子は、人工的な組換え法、例えば遺伝子工学的手法や化学的合成法で作製される。
【0037】
あるポリペプチドを「コードする」ヌクレオチド配列という言及は、その配列が、転写およびmRNAが翻訳されることでポリペプチドを生じることを意味する。これは、ヌクレオチド配列がmRNAと同一で同配列が通常配列表で提供されるコード鎖、ならびに、転写の鋳型として用いられる相補鎖の両方を含む。当業者であれば、この表現が、同じアミノ酸配列をコードするすべての縮重ヌクレオチド配列も含むことを理解すると思われる。任意のポリペプチドをコードするヌクレオチド配列はイントロンを含む配列を含む。
【0038】
「発現制御配列」とは、動作可能に連結されるヌクレオチド配列の発現を調節するヌクレオチド配列を意味する。発現制御配列は、同発現制御配列がヌクレオチド配列の転写、および必要に応じて翻訳を制御および調節する場合に、ヌクレオチド配列に「動作可能に連結される」。したがって発現制御配列は、適切なプロモーター、エンハンサー、転写終結配列、タンパク質コード遺伝子の前にある開始コドン(ATG)、イントロンのスプライシングシグナル、mRNAの翻訳を可能とする遺伝子の正確な読み枠の維持、および終止コドンを含む場合がある。
【0039】
本明細書で用いる、任意の物体について適用される「天然の」とは、ある物体が天然に見出されるという事実を意味する。例えば、天然の供給源から単離可能で、実験室で人によって意図して修飾されない生物体(ウイルスを含む)に存在するポリペプチドまたはポリヌクレオチドの配列は天然である。
【0040】
「使用可能に連結された」とは、このように記載された成分を、意図した様式で機能可能に関連づけるように並置することを意味する。コード配列に「使用可能に連結された」制御配列は、コード配列の発現が、適切な分子(例えば、誘導物質およびポリメラーゼ)が制御配列または調節配列に結合したときに、制御配列に適合する条件で達成されるように連結されている。
【0041】
「制御配列」とは、連結したコード配列および非コード配列の発現に作用するのに必要なポリヌクレオチド配列を意味する。制御配列の性質は宿主生物によって異なる。原核生物では、制御配列は一般に、プロモーター、リボソーム結合部位、および転写終結配列を含み、真核生物では、制御配列は一般に、プロモーター、および転写終結配列を含む。「制御配列」という用語は、存在が発現に影響を及ぼすことがあって存在が有利に作用する他の成分(例えば、リーダー配列および融合パートナー配列)を少なくとも含むことを意図する。
【0042】
「単離されたポリヌクレオチド」とは、ゲノム、cDNA、もしくは合成起源のポリヌクレオチド、またはこれらの組み合わせのポリヌクレオチドを意味し、その起源によって「単離されたポリヌクレオチド」は、(1)「単離されたポリヌクレオチド」が天然に見出される細胞と結びついていないか、または(2)天然では連結されていないポリヌクレオチドに使用可能に連結される。
【0043】
「ポリヌクレオチド」とは、少なくとも10塩基の長さの重合体状態のリボヌクレオチドもしくはデオキシリボヌクレオチド、またはいずれかのヌクレオチドの修飾型のいずれかのヌクレオチドを意味する。この用語は、1本鎖状および2本鎖状のDNAを含む。
【0044】
「プローブ」という用語は、別の物質(「標的」)に特異的に結合する物質を意味する。プローブは例えば、抗体、核酸、受容体、およびそのリガンドを含む。
【0045】
「調節」とは、生物学的な活性または過程(例えば、酵素活性または受容体結合)の機能特性を促進する能力、または抑制する能力を意味し、このような促進または抑制は、情報伝達経路の活性化などの特定の事象の発生を条件とする場合があり、および/または特定の細胞種でのみ明らかになる場合がある。
【0046】
「調節因子」という用語は、合成分子(例えば、核酸、タンパク質、非ペプチド、または有機分子)などの化学物質(天然または非天然の化学物質)、または細菌、植物、真菌、動物(特に哺乳類)の細胞もしくは組織などの生物材料から作られた抽出物を意味する。調節因子は、本明細書に記載されたスクリーニングアッセイ法に含めることで、一つの生物学的過程または複数の生物学的過程の阻害物質または活性物質としての潜在的な活性(例えば、作動物質、部分拮抗物質、部分作動物質、逆作動物質、拮抗物質、抗腫瘍物質、細胞毒性物質、腫瘍性形質転換または細胞増殖の阻害物質、細胞増殖促進性物質など)が(直接的または間接的に)評価されうる。調節因子の活性は、既知のものも、未知のものも、または部分的に既知のものもある。
【0047】
「被験化合物」という用語は、推定調節因子として本発明の一つまたは複数のスクリーニング法でによって検討される化学物質を意味する。被験化合物は通常、対象標的に結合することが不明である。「対照被験化合物」という用語は、標的に結合することが知られている化学物質(例えば、既知の作動物質、拮抗物質、部分作動物質、または逆作動物質)を意味する。通常、さまざまな所定の濃度(0.01 μM、0.1 μM、1.0 μM、および10.0 μMなど)の被験化合物をスクリーニングに用いる。
【0048】
「標的」という用語は、生物的過程にかかわる生化学的存在を意味する。標的は典型的には、ある生物の生理学的性質または生物学的性質に有用な役割を果たすタンパク質である。治療用化合物は標的に結合して、その機能を変化させたり調節したりする。本明細書で用いる標的は、細胞表面にある受容体、G-タンパク質、キナーゼ、イオンチャネル、ホスホリパーゼ、および本明細書に記載された他のタンパク質を含む場合がある。
【0049】
「標識」という用語は、分光光度的、光化学的、生化学的、免疫学的、または化学的な手法で検出可能な組成物を意味する。例えば有用な標識には、32P、蛍光色素、蛍光タンパク質、高電子密度試薬、酵素(例えば、ELISAで通常使用される酵素)、ビオチン、ジゴキシゲニン、またはハプテン、および抗血清またはモノクローナル抗体に利用可能なタンパク質などがある。例えば本発明のポリペプチドは、例えば任意のポリペプチドに組み入れることにより検出用標識として作製して、対象ポリペプチドに特異的に反応する抗体を標識する際に使用することができる。標識は、放射能、蛍光、または酵素活性などの測定可能なシグナルを生じることがあり、これを用いて結合状態の標識の量を測定することができる。
【0050】
「核酸プローブ」という用語は、他の核酸分子の特定の配列またはサブ配列に結合する核酸分子を意味する。プローブは、好ましくは標的核酸の完全配列またはサブ配列に対して相補的塩基対合を介して結合する核酸分子である。プローブは、ハイブリダイゼーション条件のストリンジェンシーに応じて、プローブ配列と完全な相補性を欠く標的配列に結合可能であると理解される。プローブは好ましくは、放射性同位元素、発色団、発光団、色素源、蛍光タンパク質として直接標識されるか、またはストレプトアビジン複合体が後に結合する可能性のあるビオチンなどで間接標識される。プローブの有無を調べることで特定の配列またはサブ配列の有無を検出することができる。
【0051】
「標識された核酸プローブ」とは、リンカーを介して共有結合的に、またはイオン結合、ファンデルワールス結合、または水素結合を介して標識に結合して、プローブに結合した標識の存在の検出によってプローブの存在を検出することができる核酸プローブである。
【0052】
「ポリペプチド」および「タンパク質」という用語は、アミノ酸残基の重合体を意味する。これらの用語は、1個または複数のアミノ酸残基が、対応する天然のアミノ酸、人工的な化学的類似体であるアミノ酸重合体、ならびに天然のアミノ酸重合体に適用される。「組換えタンパク質」という用語は、組換えDNA分子に由来するタンパク質のアミノ酸配列をコードするヌクレオチド配列の発現によって産生されるタンパク質を意味する。
【0053】
「組換え宿主細胞」という用語は、組換え核酸分子を含む細胞を意味する。したがって例えば、組換え宿主細胞は、対象細胞に由来する、天然では見出されない(非組換え)遺伝子群を発現する場合がある。
【0054】
「単離された」、「精製された」、または「生物学的に純粋な」という表現は、天然の状態では通常伴うことがわかっている成分を実質的または本質的に含まない材料を意味する。純度および均一性は、典型的には、ポリアクリルアミドゲル電気泳動、または高速液体クロマトグラフィーなどの分析化学的手法で決定される。調製物中に存在する優勢なタンパク質または核酸種であるタンパク質または核酸の分子は実質的に純粋である。一般に、単離されたタンパク質または核酸の分子は、調製物中に存在するすべての巨大分子種の80%を上回る量を含む。好ましくは、このようなタンパク質は、存在する全巨大分子種の90%を上回るように精製される。より好ましくは、このようなタンパク質は95%を上回るように精製され、また最も好ましくは、このようなタンパク質は、他の巨大分子種が従来の手法で検出されない実質的な均一状態に精製される。
【0055】
任意の物質について適用される「天然の」という表現は、物質が天然に見出されるという事実を意味する。例えば、天然の供給源から単離可能で、実験室で人によって意図して修飾されない生物体(ウイルスを含む)に存在するポリペプチドまたはポリヌクレオチドの配列は天然である。
【0056】
「抗体」という用語は、一つの免疫グロブリン遺伝子、もしくは複数の免疫グロブリン遺伝子、またはそれらの断片に実質的にコードされた、分析物(抗原)に特異的に結合して認識するポリペプチドを意味する。認識される免疫グロブリン遺伝子には、κ、λ、α、γ、δ、ε、およびμの定常領域の遺伝子群、ならびに多数の免疫グロブリン可変領域遺伝子がある。抗体は例えば、完全な免疫グロブリンとして、または、さまざまなペプチダーゼによる消化によって生じる特性がよく解析された複数の断片として存在する。これには例えば、Fab'断片、およびF(ab)'2断片が含まれる。本明細書で用いる「抗体」という用語は、組換えDNA技術を用いる抗体全体の修飾、またはデノボ合成によって作製される抗体断片も含む。
【0057】
「イムノアッセイ法」という用語は、抗体を用いて分析物に特異的に結合させるアッセイ法を意味する。イムノアッセイ法は、特定の抗体の特定の結合特性を利用して分析物を単離したり、標的としたり、および/または定量したりすることを特徴とする。
【0058】
二つの核酸配列またはポリペプチド配列に関して「同一である」という表現は、二つの配列上の残基が、最大の一致をみるように並べたときに同じであることを意味する。配列同一性のパーセンテージをタンパク質またはペプチドについて用いる場合、同一でない残基の位置は、アミノ酸残基が類似の化学的特性(例えば電荷や疎水性)を有する他のアミノ酸残基と置換されることで、したがって対象分子の機能特性を変化しないような、保存的なアミノ酸置換が異なる場合があると認識される。保存的置換の配列が異なる場合、配列同一性のパーセントは、置換の保存的性質を正すために上方に合わせられる場合がある。このような調節を行う手法は当技術分野で周知である。典型的には、これは、完全なミスマッチではなく部分的なミスマッチとして保存的置換を評点化することで配列同一性のパーセンテージを上昇させる段階を含む。したがって例えば、同一なアミノ酸の評点が1で、非保存的置換の評点が0の場合、保存的置換は0と1の間の評点となる。保存的置換の評点化は例えば、既知のアルゴリズムで計算することができる。これについては例えば、マイヤーズ(Meyers)およびミラー(Miller)、Computer Applic. Biol. Sci.、4:11〜17 (1988);スミス(Smith)およびウォーターマン(Waterman)(1981)、Adv. Appl. Math.、2:482;ニードルマン(Needleman)およびワンチ(Wunsch)(1970)、J. MoL Biol. 48:443;ピアソン(Pearson)およびリップマン(Lipman)(1988)、Proc. Natl. Acad. Sci. USA 85:2444;ヒギンス(Higgins)およびシャープ(Sharp)(1988)、Gene、73:237〜244、ならびにヒギンスおよびシャープ(1989)、CABIOS 5:151〜153;コルペット(Corpet)ら(1988)、Nucleic Acids Research 16、10881〜90;フアン(Huang)ら(1992)、Computer Applications in the Biosciences 8、155〜65、ならびにピアソンら(1994)、Methods in Molecular Biology 24、307〜31を参照されたい。アライメントは精査およびマニュアルによるアラインメントで行われることもある。
【0059】
特定の核酸配列の「保存的に修飾された変異」とは、同一なアミノ酸配列、または実質的に同一なアミノ酸配列をコードする核酸を意味し、核酸がアミノ酸配列をコードしない場合は実質的に同一な配列を意味する。遺伝暗号には縮重性があるので、機能が同一の多数の核酸は、任意のポリペプチドをコードする。例えば、コドンCGU、CGC、CGA、CGG、AGA、およびAGGはいずれもアミノ酸のアルギニンをコードする。したがって、一つのコドンによってアルギニンが指定されるあらゆる位置において、同コドンは、コードされたポリペプチドを変化させることなく、記載された任意の対応コドンに変えることができる。このような核酸の変異は、「保存的に修飾された変異」の一種である「サイレント変異」である。ポリペプチドをコードする、本明細書に記載されたあらゆる核酸配列は、可能なあらゆるサイレント変異も意味する。当業者であれば、核酸中の各コドン(通常メチオニンのみを表すコドンであるAUGを除く)が修飾されて、標準的な手法で機能的に同一な分子を生じる可能性があることを認識すると思われる。したがって、ポリペプチドをコードする核酸の個々の「サイレント変異」は、個々に記載された配列では表面化しない。また当業者であれば、コードされた配列において1個のアミノ酸またはパーセンテージの小さいアミノ酸(典型的には5%未満、より典型的には1%未満)を変化、付加、欠失させる個々の置換、欠失、または付加が、変化が化学的に類似のアミノ酸へのアミノ酸置換を生じる「保存的に修飾された変異」であることを認識すると思われる。機能が類似したアミノ酸をもたらす保存的アミノ酸の置換は当技術分野で周知である。以下の6群は、相互に保存的置換の関係にあるアミノ酸をそれぞれ含む:
1)アラニン(A)、セリン(S)、スレオニン(T);
2)アスパラギン酸(D)、グルタミン酸(E);
3)アスパラギン(N)、グルタミン(Q);
4)アルギニン(R)、リシン(K);
5)イソロイシン(I)、ロイシン(L)、メチオニン(M)、バリン(V);および
6)フェニルアラニン(F)、チロシン(Y)、トリプトファン(W)。
【0060】
「相補的」という表現は、一つの核酸分子が、他の核酸分子の結合パートナーの配列を有することを意味する。したがって、配列5'-ATGC-3'は、配列5'-GCAT-3'と相補的である。
【0061】
アミノ酸配列またはヌクレオチド配列は、アミノ酸配列またはヌクレオチド配列が、任意の比較ウィンドウにおいて標準配列に対して少なくとも80%の配列同一性を有する場合、標準配列に対して「実質的に同一」または「実質的に類似」である。したがって実質的に類似した配列は、例えば少なくとも85%の配列同一性、少なくとも90%の配列同一性、少なくとも95%の配列同一性、または少なくとも99%の配列同一性を有する配列を含む。相互に同一な二つの配列が実質的に同一であることは言うまでもない。
【0062】
あるヌクレオチド配列は、本発明のヌクレオチド配列の相補物が標準ヌクレオチド配列と実質的に同一である場合、標準ヌクレオチド配列に対して「実質的に相補的」である。
【0063】
「ストリンジェントな条件」という表現は、核酸ハイブリダイゼーションに用いられる温度およびイオン条件を意味する。ストリンジェントな条件は、配列に依存し、また異なる環境パラメータの下では異なる。一般にストリンジェントな条件は、特定のイオン強度およびpHにおいて特定配列の融点(Tm)より約5〜20℃低い温度が選択される。Tmは、標的配列の50%が完全にマッチしたプローブとハイブリッドを形成する(特定のイオン強度およびpHにおける)温度である。
【0064】
「対立遺伝子異型」とは、特定の遺伝子座にある遺伝子、ならび遺伝子のmRNA転写物に由来するcDNA、およびそれがコードするポリペプチドの多型を意味する。
【0065】
「哺乳類で好ましいコドン」という表現は、以下のリストから選択される、哺乳類細胞で発現されるタンパク質で極めて高頻度で使用される、アミノ酸をコードする一連のコドンにおけるコドンのサブセットを意味する。
【0066】

蛍光分子は、蛍光共鳴エネルギー移動(「FRET」)に有用である。FRETには、ドナー分子とアクセプター分子がかかわる。ドナー分子とアクセプター分子間におけるFRETの効率および検出率を最適なものとするためには、複数の因子の均衡を図る必要がある。ドナーの発光スペクトルは、重なり積分を最大とするためにアクセプターの励起スペクトルと可能な限り大きく重複すべきである。また、ドナー部分の量子収率、およびアクセプターの吸光係数は、エネルギー移動効率が50%となる距離であるR0を最大とするために同様にできるだけ高くすべきである。しかし、ドナーおよびアクセプターの励起スペクトルは、アクセプターを直接励起させることなくドナーが効率よく励起可能な時点で波長領域が見出されるようにするために重複を可能な限り小さくすべきである。アクセプターの直接的な励起から生じる蛍光は、FRETで生じる蛍光と区別しにくい。同様に、ドナーおよびアクセプターの発光スペクトルは、二つの発光を明らかに区別できるように、可能なかぎりわずかに重複させるべきである。アクセプター部分の高蛍光量子収率は、アクセプターに由来する発光が、唯一の読み値として、または発光比の一部として測定される場合に望ましい。ドナーとアクセプターのペアの選択時に考慮すべき一つの因子は、両者間における蛍光共鳴エネルギー移動の効率である。好ましくは、ドナーとアクセプター間のFRETの効率は少なくとも10%、より好ましくは少なくとも50%、またさらにより好ましくは少なくとも80%である。
【0068】
「蛍光特性」という用語は、適切な励起波長におけるモル吸光係数、蛍光量子効率、励起スペクトルもしくは発光スペクトルの形状、励起波長極大および発光波長極大、二種の異なる波長における励起振幅の比、二種の異なる波長における発光振幅の比、励起状態の寿命、または蛍光異方性を意味する。オワンクラゲの野生型GFPと変異型との間における上記特性の任意の一つに測定可能な差があることは有用である。測定可能な差は、任意の量的蛍光特性の量、例えば特定波長における蛍光量、または発光スペクトル全体における蛍光の積分値を決定することで決定することができる。異なる二種の波長における励起振幅または発光振幅の比を決定すること(それぞれ、「励起振幅の比の決定」および「発光振幅の比の決定」)は特に有利である。というのは、比の決定過程で内部標準が得られ、励起源の絶対明度、検出装置の感度、および光散乱または試料による消光の変動が相殺されるからである。
【0069】
II.長波長の改変蛍光タンパク質
A.蛍光タンパク質
本明細書で用いる「蛍光タンパク質」という用語は、適切な電磁放射で励起されると蛍光を発することができる任意のタンパク質を意味する。これには、アミノ酸配列が天然の配列または人工的な配列(すなわち類似体または変異体)の蛍光タンパク質が含まれる。多くの刺胞動物は、緑色蛍光タンパク質(「GFP」)を生物発光におけるエネルギー移動のアクセプターとして用いている。本明細書で用いる「緑色蛍光タンパク質」は、緑色の蛍光を発するタンパク質である。同様に「青色蛍光タンパク質」は青色の蛍光を発し、「赤色蛍光タンパク質」は赤色の蛍光を発する。GFPは、太平洋の北西部に生息するクラゲの一種、オワンクラゲ(Aequorea victoria)、ウミシイタケ(Renilla reniformis)、およびコザラクラゲ(Phialidium gregarium)から単離されている。ワード(W.W. Ward)ら、Photochem、 Photobiol.、35:803〜808 (1982);リバイン(L.D. Levine)ら、Comp. Biochem. Physiol.、72B:77〜85 (1982)を参照されたい。
【0070】
有用な励起スペクトルおよび発光スペクトルを有するさまざまなオワンクラゲ関連蛍光タンパク質が、オワンクラゲに由来する天然GFPのアミノ酸配列を修飾することで作製されている(D.C. Prasherら、Gene、111:229〜233 (1992);R. Heimら、Proc. Natl. Acad. Sci.、USA、 91:12501〜04(1994);1994年11月10日に出願された米国特許出願第08/337,915号;1995年11月10日に出願された国際出願番号PCT/US95/14692)。
【0071】
本明細書で用いるように、蛍光タンパク質の150アミノ酸の任意の隣接配列が、図3の238アミノ酸の野生型のオワンクラゲの緑色蛍光タンパク質に由来する(隣接した、または隣接していない)アミノ酸配列(配列番号:2)と少なくとも85%の配列同一性をもつ場合、その蛍光タンパク質は「オワンクラゲ関連蛍光タンパク質」である。より好ましくは、蛍光タンパク質の200アミノ酸の任意の隣接配列が、図3の野生型のオワンクラゲ緑色蛍光タンパク質に由来する(隣接した、または隣接していない)アミノ酸配列(配列番号:2)と95%の配列同一性を有する場合は、蛍光タンパク質はオワンクラゲ関連蛍光タンパク質である。同様に蛍光タンパク質は、同じ基準を用いてウミシイタケまたはコザラクラゲの野生型の蛍光タンパク質に関連する場合がある。
【0072】
オワンクラゲ関連蛍光タンパク質は例えば、野生型(天然)のオワンクラゲのGFP (D.C. Prasherら、「オワンクラゲの緑色蛍光タンパク質の一次構造(Primary Structure of the Aequorea victoria green fruolescence protein)」、Gene (1992) 111:229〜33)(このヌクレオチド配列(配列番号:1)および推定アミノ酸配列(配列番号:2)は図3に示す);同配列の対立遺伝子変異体(例えば、80位のグルタミン残基がアルギニンに置換されたQ80R)(M. Chalfieら、Science (1994) 263:802〜805);本明細書に記載された人工的に作製されたオワンクラゲ関連蛍光タンパク質、例えば表Aまたは表Fにある、アミノ末端の2個のアミノ酸が除去されたオワンクラゲの緑色蛍光タンパク質などの蛍光性の一種または複数の折りたたみ変異およびそのようなタンパク質の断片を含む異型を含むが、これらに限定されない。このようなもののいくつかは、中心となる発色団の内部に異なる芳香族アミノ酸を含み、野生型の種より明確に短い波長で蛍光を発する。例えば、改変タンパク質P4およびP4-3は(他の変異に加えて)置換Y66Hを含み、W2およびW7は(他の変異に加えて)置換Y66Wを含む。いずれもタンパク質の発色団領域に近く、一次配列では離れている他の変異がGFPのスペクトル特性に影響を及ぼす可能性があり、以下の表の最初の部分に挙げてある。
【0073】
【表A】
「折りたたみ変異体」と呼ばれる、オワンクラゲ関連蛍光タンパク質の他の変異では、蛍光タンパク質のもつ、高温における折りたたみ能力、および哺乳類細胞での発現時における、より強い蛍光発生能力が改善されているが、励起および発光のピーク波長にはほとんど影響しないか全く影響しない。これらが、スペクトル特性および折りたたみ特性が変化したタンパク質を生じるようにGFPのスペクトル特性に影響する変異と結びつく場合があることは注目すべき点である。折りたたみ変異には、F64L、V68L、S72Aのほか、T44A、F99S、Y145F、N146I、M153TまたはA、V163A、I167T、S175G、S205T、およびN212Kなどがある。
【0075】
本明細書で用いる「ループドメイン」という用語は、βバレルまたは中央部のαへリックス(56〜72残基)の11本の鎖の二次構造にかかわるアミノ酸を連結するオワンクラゲ関連蛍光タンパク質のアミノ酸配列を意味する(図1Aおよび1B参照).
【0076】
本明細書で用いる、蛍光タンパク質の「蛍光タンパク質部分」とは、蛍光タンパク質基質のアミノ酸配列が天然の蛍光タンパク質のアミノ酸配列と最適に並置され、天然の蛍光タンパク質のアミノ酸配列のアミノ末端とカルボキシ末端のアミノ酸の間に存在する(末端を含む)、蛍光タンパク質のアミノ酸配列の部分のことである。
【0077】
蛍光タンパク質は、遺伝的に他の標的タンパク質と融合可能であり、また、産生された標的タンパク質の位置および量を同定するためのマーカーとして使用可能であることがわかっている。したがって本発明は、蛍光タンパク質部分と付加的なアミノ酸配列を含む融合タンパク質を提供する。このような配列の長さは例えば、最長約15アミノ酸、最長約50アミノ酸、最長約150アミノ酸、または最長約1000アミノ酸である。融合タンパク質は、電磁放射で励起されたときに蛍光発生能力をもつ。一つの態様では、融合タンパク質は、対象タンパク質の精製を容易にするためのポリヒスチジンタグを含む。
【0078】
B.蛍光特性が変化した変異体を設計するための緑色蛍光タンパク質の結晶構造の用途
X線結晶解析およびコンピュータ処理により、発明者らは、分子内における原子の相対的な位置を示すオワンクラゲの緑色蛍光タンパク質の結晶構造モデルを構築した。このような情報は、置換がタンパク質の蛍光特性を変化させるアミノ酸を同定する際に有用である。
【0079】
オワンクラゲ関連蛍光タンパク質の蛍光特性は、発色団の電子環境に部分的に依存する。一般に、発色団の約0.5 nm以内にあるアミノ酸は発色団の電子環境に影響を及ぼす。したがって、このようなアミノ酸を置換させると、蛍光特性が変化した蛍光タンパク質が得られる可能性がある。励起状態では、電子密度は発色団のフェノール側からカルボニル末端方向へシフトする傾向がある。したがって、発色団のカルボニル末端付近に強い正電荷を配置すると、励起状態のエネルギーを低下させ、またタンパク質の吸光度および発光波長極大の赤色シフトを引き起す傾向がある。発色団のカルボニル末端付近にある正電荷を減らすと、タンパク質の波長の青色シフトを引き起す反対の作用をもたらす傾向がある。
【0080】
荷電性アミノ酸(イオン化したD、E、K、およびR)、双極性アミノ酸(H、N、Q、S、T、および無電荷のD、EおよびK)、ならびに分極性側鎖(例えば、C、F、H、M、WおよびY)は、特にアミノ酸が無電荷、非極性、または非分極性の側鎖と置換する場合において、発色団の電子環境を変化させる際に有用である。一般に、分極性側鎖をもつアミノ酸は、電子環境をほんのわずか変化させ、最終的には、蛍光特性に比較的小さな変化を引き起すと考えられている。荷電性側鎖をもつアミノ酸は、環境を最も大きく変化させ、最終的には、蛍光特性に比較的大きな変化を引き起すと考えられている。しかし、荷電性側鎖をもつアミノ酸は、発色団付近に埋没した状態の場合に、何ら追加的な溶媒和または塩橋形成なしに、タンパク質の構造を破壊する可能性、および適切な折りたたみを妨げる可能性が高い。したがって荷電性アミノ酸は、既に溶媒和されているか、または塩橋形成がかかわる他の荷電性アミノ酸または極性の高いアミノ酸と置換する際に、寛容性が大きく、有用な作用をもたらす可能性が最も高い。分極性アミノ酸との置換が選択されるような場合は、タンパク質の構造は、大きなアミノ酸(例えばW)の選択を不適当なものとする場合がある。あるいは、配向が望ましくない荷電性側鎖または極性側鎖をもつアミノ酸で占められる位置を、荷電性が弱いか、または極性が弱い側鎖をもつアミノ酸と置換することができる。別の方法では、側鎖がタンパク質の一方向に双極性配置されたアミノ酸は、異なる方向に双極性配置されたアミノ酸と置換することができる。
【0081】
より具体的には、表Bに、置換が蛍光特性の変化を生じうる発色団の約0.5 nm以内に位置する複数のアミノ酸を挙げる。この表では、タンパク質の蛍光特性を変化させる、記載位置における好ましいアミノ酸置換に下線を引いて示してある。このような置換を導入する目的で、この表は、増幅過程がかかわる部位特異的変異導入で使用されるプライマー用のコドンも示している。このようなプライマーは、経済的に好ましいアミノ酸をコードするように選択されているが、図に示すように他のアミノ酸、さらには終止コドン(Zで表す)もコードしている。このような縮重プライマーを用いて置換を導入する際に最も効率のよい方法は、集団をスクリーニングして、所望の特性を有する変異体を同定した後に、そのDNA配列を決定して可能な置換がどれかを見極めることである。コドンを、センス鎖を上にアンチセンス鎖を下にして2本鎖で示す。核酸配列では、R=(Aまたはg);Y=(CまたはT);M=(AまたはC);K=(gまたはT);S=(gまたはC);W=(AまたはT);H=(A、T、またはC);B=(g、T、またはC);V=(g、A、またはC);D=(g、A、またはT);N=(A、C、g、またはT)を示す。
【0082】
【表B】
分極性側鎖と置換可能な極性側鎖をもつアミノ酸の例には例えば、表Cに記載されたアミノ酸などがある。
【0084】
【表C】
別の態様では、第2のアミノ酸が発色団の約0.5 nm以内に近接するアミノ酸は、置換されると第2のアミノ酸の電子特性を変化させて、最終的に発色団の電子環境が変化する。表Dは、このような二種のアミノ酸を示す。アミノ酸L220およびV224はE222に近接しており、βプレート状シート内で同じ方向を向いている。
【0086】
【表D】
本発明の一つの態様は、アミノ酸配列がオワンクラゲの緑色蛍光タンパク質のアミノ酸配列(配列番号:2)と実質的に同一で、少なくともQ69における置換が配列番号:2と異なる、オワンクラゲの緑色蛍光タンパク質とは異なった蛍光特性を有する機能性の改変蛍光タンパク質をコードするヌクレオチド配列を含む核酸分子を含む。好ましくは、Q69における置換は、K、R、E、およびGからなる群より選択される。Q69における置換を他の変異(S65における機能性変異など)と組み合わせることで、タンパク質の特性を改善することができる。
【0088】
本発明の一つの態様は、アミノ酸配列がオワンクラゲの緑色蛍光タンパク質のアミノ酸配列(配列番号:2)と実質的に同一で、少なくともE222における置換(E222Gを除く)が配列番号:2と異なる、オワンクラゲの緑色蛍光タンパク質とは異なった蛍光特性を有する機能性の改変蛍光タンパク質をコードするヌクレオチド配列を含む核酸分子を含む。好ましくは、E222における置換は、NおよびQからなる群より選択される。E222における置換を他の変異(F64における機能性変異など)と組み合わせることで、タンパク質の特性を改善することができる。
【0089】
本発明の一つの態様は、アミノ酸配列がオワンクラゲの緑色蛍光タンパク質のアミノ酸配列(配列番号:2)と実質的に同一で、少なくともY145における置換が配列番号:2と異なる、オワンクラゲの緑色蛍光タンパク質とは異なった蛍光特性を有する機能性の改変蛍光タンパク質をコードするヌクレオチド配列を含む核酸分子を含む。好ましくは、Y145における置換は、W、C、F、L、E、H、K、およびQからなる群より選択される。Y145における置換を他の変異(Y66における変異など)と組み合わせることで、タンパク質の特性を改善することができる。
【0090】
本発明はまた、新しい蛍光タンパク質の変異を設計用の結晶座標を用いるコンピュータ、および座標を含む結晶データを記憶する装置を利用した方法を含むコンピュータに関連した態様も含む。例えば本発明は記憶装置を含む装置、および図5-1〜5-28に挙げた原子座標から選択される少なくとも10の原子座標に関して同装置に記憶されたデータを提供する。多くの座標を、座標を用いる計算の複雑性、または目標に応じて記憶させることができる(例えば、約100個、1,000個、またはそれより多くの座標)。例えば大量の座標が、蛍光タンパク質の構造をより詳しく表現する際に望ましい。典型的には、このような記憶装置は、原子座標を入力として受け取るコードを記憶するコンピュータ可読装置である。一方、当技術分野で周知の他の記憶手段も対象となる。コンピュータ可読装置は、フロッピーディスクまたはハードドライブである。
【0091】
C.陰イオン結合特性が変化した変異体を設計するための YFP の結晶構造の用途
別の局面では、本発明は、相対位置、および結合状態のイオンと相互作用するアミノ酸を示すYFPの結晶構造モデルを構築するためのX線結晶解析およびコンピュータ処理の使用を含む。このような情報は、置換がさまざまな陰イオンに対する結合部位の特異性および親和性を変化させるアミノ酸を同定する際に有用である。陰イオンの結合は、YFPの発色団付近で生じるので、結合によって、陰イオンの結合さらには陰イオン濃度のモニタリングに使用可能なYFPの蛍光特性が調節される。
【0092】
YFP-H148Qにある陰イオン結合部位は、他のタンパク質のハロゲン化物結合部位に一般に見出される多くの特性を示す。YFP-H148Qの陰イオンを含む空洞の場合、結合部位は天然の状態で両親媒性であり、片側に極性基および荷電性の基(Tyr203、発色団、Arg96、Gln69、およびGln183)が並んでおり、別の側に疎水性の基(Ile152、Leu201、Val163、Val150、およびPhe165)が並んでいる。
【0093】
陰イオン結合の特異性が変化した改変蛍光タンパク質の設計では、いくつかの因子を考慮する必要がある。例えば、陰イオンの親和性および選択性に寄与する最も有意な因子の一つは、結合ポケットの静電気的な配置および構成である。YFPでは、これらは表Eに挙げた基を含む。
【0094】
【表E】
原位置および推定される役割
S65、Y66、G67 発色団を形成、芳香環の端はイオンと相互作用する
Q69 イオンとの水素結合
R96 イオンとの電荷相互作用
Q183 イオンとの電荷相互作用
Y203 イオン(Y)との水素結合
【0095】
一般に陰イオン結合は、より多くの、または、より強い結合相互作用を、結合ポケット内における対象陰イオンと極性基との間に作ることで改善することができる。例えば上記の極性残基を、より極性の大きい残基と直接置換するか、または、より効率的に相互作用する可能性のある異なる大きさの残基を陰イオンと置換することでイオン結合を改善することができる。例えば、発色団の大きさや位置は、S65のG、A、C、V、L、I、またはTに置換して変えることが可能であり、Y66はH、F、またはWに置換することが可能であり、Q69はNまたはKに、R96はKに、Q183はNまたはKに置換することができる。
【0096】
水和エネルギー
また、発色団付近の埋没状態の空洞における陰イオンの結合は、イオン-溶媒相互作用とイオン-タンパク質相互作用との置換を必要とする。水和エネルギーに関連する、YFP(表J)およびYFP-H148Q(Jayaramanら、2000)に対する一価の陰イオンの相対的な結合エネルギーは、水和力が、結合に対する重要な寄与因子であることを意味している。一価の陰イオンの水和エネルギーは以下のように弱いものから強いものへ並ぶ:CO4 -<I-<NO3 -<SCN-<Br-<Cl-<F- (WrightおよびDiamond、1977)。多原子の一価の陰イオンおよびヨウ化物は、比較的弱い水和エネルギーをもつが、他のハロゲン化物は水とより強固に相互作用する。球対称のハロゲン化物の場合は、水和エネルギーは、原子体積の低下に伴って上昇し(Born、1920)、これは、なぜ大きなハロゲン化物が、タンパク質内部のより疎水性の大きな環境に容易に埋め込まれるかということの理由となる。YFPに対する陰イオンの結合に関してみられる傾向は上記の順序にほぼ従う(表J)。タンパク質の相互作用は一般に水和エネルギーの低下に伴って上昇するが、大きさが小さいことからタンパク質結合時に完全に脱水しない場合があるフッ素は例外である。
【0097】
したがって、高親和性の陰イオン結合部位の開発には、例えば、イオン結合ポケットに並んだ、極性のより大きい疎水性残基と、水素結合能力が大きい極性残基との置換による、十分なイオン-タンパク質相互作用の生成が必要である。より大きな、または小さな陰イオンのイオン結合を改善する、このようなタイプの置換の例を表Fに示す。
【0098】
【表F】
i)ヨウ化物以外のより小さな陰イオンの結合親和性を高めるイオン結合ポケットの周囲のアミノ酸の変異。
結合ポケットの大きさ
結合のサイズおよび形状も、大きな陰イオンに対して結合部位が埋没した性質をもつことから特に重要である。幾何学的直径の平均が6.2 ÅであるTCA(Halm & Frizzell、1992)は見かけ上大きすぎるのでYFPと測定可能な程度では相互作用できないが、いくぶん小さいTFAでは弱い結合を示す(表J)。したがって、より大きな陰イオンの結合親和性の改善は、表Fに概要が示されているように、結合ポケットに並ぶアミノ酸と小さな残基との置換によって、ならびに後述する溶媒接近性を上昇させることで達成される場合がある。
【0100】
陰イオン結合に伴う構造変化
YFPの結合ポケットに並んださまざまな側鎖の一連の構造変化はハロゲン化物の結合に必要である。最も大きい動きはGln69およびGln183で観察されるが、Leu201、Ile152、Val150、およびVal163の無極性側鎖(図11)はすべて、結合状態のハロゲン化物の存在下で空洞の大きさを増大させる動きに関与する。したがって、陰イオン結合を強くする別のアプローチは、結合時に極めて大きな構造変化を受ける残基を、より小さな残基と置換することである。このような変化は、結合時の構造再編成の必要性を小さくすることで、陰イオン結合をエネルギー的により望ましい状態にする可能性がある。このような変化には、上の表Fに挙げた変化、ならびにQ69のNへの置換が含まれる。
【0101】
溶媒の接触性
His148におけるさまざまな変異の構造決定で得られた結果から、同位置における特定の変異が、溶媒の接触のしやすさと結合ポケットの容積との両方に直接影響を及ぼすことがあるβバレルにおける全体的な構造調節を生じうることが示唆されている。したがって、His148における、例えばQ、N、G、A、L、V、およびIなどの、より小さなアミノ酸への置換は、発色団に対する溶媒の接触性を高めることで、より大きな陰イオンの結合を改善すると予測される。同様にHis148と、より大きなアミノ酸(FまたはWなど)との置換は、発色団に対する陰イオンの接触性を低める可能性が高い。また同様に、より小さな変化が、147位および149位における、より小さなアミノ酸、または、より大きなアミノ酸との置換によって達成される可能性がある。
【0102】
以上の変異は典型的には、オリゴを用いた部位特異的変異導入によってYFP鋳型のタンパク質に導入され、典型的には野生型アミノ酸残基を10%の確率で有する、また、さまざまな変異型残基の一つを90%の確率で有する、変異型タンパク質のライブラリーが作られる。このような方法を用いて、さまざまな組み合わせの変異体を含むライブラリーのスクリーニングを速やかに行って、特定の対象陰イオンの最良の組み合わせを同定することが可能である。典型的には、この段階を繰り返すことで、結合ポケット周囲の配列空間を、任意の特定の対象陰イオンについて完全に探索することを確実なものとすることができる。
【0103】
D.改変蛍光タンパク質の作製
蛍光タンパク質の組換え体の作製には、対象タンパク質をコードする配列を有する核酸分子を発現させる段階が含まれる。
【0104】
一つの態様では、このような核酸は、1本のポリペプチドがより長いポリペプチド中に蛍光タンパク質部分を含む融合タンパク質をコードする。より長いポリペプチドには、FRETパートナーや、第2の機能を有するタンパク質などの第2の機能性タンパク質を含めることができる(例えば、酵素、抗体、または他の結合タンパク質)。蛍光タンパク質をコードする核酸は出発材料として有用である。
【0105】
蛍光タンパク質は、組換えDNA手法で融合タンパク質として作製することができる。蛍光タンパク質の組換え体の作製では、タンパク質をコードする配列を有する核酸を発現させる。蛍光タンパク質をコードする核酸は当技術分野で周知の方法で得られる。蛍光タンパク質は、蛍光タンパク質をコードする他の核酸を対象とした部位特異的変異導入法で、または0.1 mM MnCl2 およびバランスを欠くヌクレオチド濃度で元のポリヌクレオチドのPCRのエラー発生率を上昇させるランダム変異導入法で作製することができる。これについては例えば、1994年11月10日に出願された米国特許出願第08/337,915号、または1994年11月10日に出願された国際出願番号PCT/US95/14692を参照されたい。緑色蛍光タンパク質をコードする核酸は、図3に示すオワンクラゲの緑色蛍光タンパク質DNA配列に対応するプライマー用いて、オワンクラゲに由来するcDNAを対象としたポリメラーゼ連鎖反応で単離することができる。PCR法は例えば米国特許第4,683,195号;ムリス(Mullis)ら、(1987) Cold Spring Harbor Symp. Quant. Biol. 51:263;およびエールリッヒ(Erlich)編、「PCR技術(PCR Technology)」、(Stockton Press、ニューヨーク、1989)で説明されている。
【0106】
発現ベクターの構築、およびトランスフェクトされた細胞内における遺伝子の発現には、当技術分野で周知の分子クローニング手法を用いる。これについては、サンブルック(Sambrook)ら、「分子クローニング-実験マニュアル(Molecular Cloning-A Laboratory Manual)」、Cold Spring Harbor Laboratory、Cold Spring Harbor、ニューヨーク、(1989)および「分子生物学の最新プロトコール(Current Protocols in Molecular Biology)」、F.M. Ausubelら編、(「最新プロトコール(Current Protocols)」、Greene Publishing Associates、Inc.およびJohn Wiley & Sons、Inc.の合弁事業)を参照されたい。発現ベクターは、適切なプロモーター、複製配列、マーカーなどを含めることで、原核生物または真核生物における機能に適合させることができる。
【0107】
対象ポリペプチドの発現をコードする配列を細胞にトランスフェクトする際に用いられる核酸は一般に、ポリペプチドの発現をコードするヌクレオチド配列に動作可能に連結させた発現制御配列を含む発現ベクターの形状をしている。本明細書で用いる、「(あるポリペプチド)の発現をコードするヌクレオチド配列」という表現は、転写およびmRNAの翻訳によってポリペプチドを生じる配列を意味する。これは例えばイントロンなどの配列を含む場合がある。発現制御配列は、発現制御配列が核酸配列の転写および、必要に応じて翻訳を制御および調節する際に、核酸配列に動作可能に連結される。したがって発現制御配列は、適切なプロモーター、エンハンサー、転写終結配列、タンパク質コード遺伝子の前方にある開始コドン(ATG)、イントロン用のスプライシングシグナル、mRNAの適切な翻訳を可能とする遺伝子の正確な読み枠の維持、および終止コドンを含む場合がある。
【0108】
当技術分野で周知の方法を用いて、蛍光タンパク質のコード配列、および適切な転写/翻訳制御シグナルを含む発現ベクターを構築することができる。このような方法には、インビトロ組換えDNA手法、合成法、およびインビボ組換え/遺伝子組換え法などがある(例えば、Maniatisら、「分子クローニング-実験マニュアル(Molecular Cloning-A Laboratory Manual)」、Cold Spring Harbor Laboratory、ニューヨーク、1989に記載された手法を参照)。
【0109】
組換えDNAによる宿主細胞の形質転換法は、当技術分野で周知の従来の手法で実施することができる。宿主が大腸菌などの原核生物の場合、DNAの取り込みが可能なコンピテント細胞を、指数増殖期の後に回収し、当技術分野で周知の手順によりCaCl2法で処理した細胞から調製することができる。あるいはMgCl2またはRbClを使用することもできる。形質転換は、宿主細胞のプロトプラストを形成させた後に実施できるほか、エレクトロポレーション法で実施することもできる。
【0110】
宿主が真核生物の場合、カルシウムリン酸共沈殿法、マイクロインジェクション、エレクトロポレーションなどの従来の機械的手法、リポソームに包んだプラスミドの挿入、またはウイルスベクターを用いてDNAをトランスフェクトする方法が用いられる場合がある。真核細胞にはまた、本発明の融合ポリペプチドをコードするDNA配列と、選択可能な表現型(単純ヘルペスチミジンキナーゼ遺伝子など)をコードする第2の外来DNA分子を同時にトランスフェクトすることができる。別の方法には、シミアンウイルス40(SV40)、またはウシパピローマウイルスなどの真核ウイルスベクターを用いて、真核細胞に一過的に感染させ、形質転換してタンパク質を発現させる方法がある(「真核性ウイルスベクター(Eukaryotic Viral Vectors)」、Cold Spring Harbor Laboratory、Gluzman編、1982)。好ましくは、真核宿主が本明細書に記載された宿主細胞として用いられる。
【0111】
微生物を用いるか、または真核生物で発現させた本発明のポリペプチドを単離および精製する手法は、例えば、予備的なクロマトグラフィー分離法、および免疫学的分離法(モノクローナル抗体もしくはポリクローナル抗体、または抗原を使用する方法など)などの従来の任意の手法とすることができる。一つの態様では、組換え蛍光タンパク質は、タンパク質をコードする核酸を大腸菌で発現させて作製することができる。オワンクラゲ関連蛍光タンパク質は、約15〜30℃で培養された細胞で最も良好に発現されるが、それより高い温度(例えば37℃)でも発現される。合成後、このような酵素は高温(例えば37℃)で安定であり、このような温度でアッセイ法に使用することができる。
【0112】
さまざまな宿主-発現ベクター系を用いて蛍光タンパク質のコード配列を発現させることができる。これには、蛍光タンパク質のコード配列を含む組換えバクテリオファージのDNA、プラスミドDNA、またはコスミドDNAの発現ベクターで形質転換された細菌などの微生物;蛍光タンパク質のコード配列を含む組換え酵母発現ベクターで形質転換された酵母;蛍光タンパク質コード配列を含む、組換え体ウイルス発現ベクター(例えば、カリフラワーモザイクウイルス、CaMV;タバコモザイクウイルス、TMV)を感染させた植物細胞系、もしくは組換えプラスミド発現ベクター(例えばTiプラスミド)で形質転換した植物細胞系;蛍光タンパク質のコード配列を含む組換えウイルス発現ベクター(例えばバキュロウイルス)を感染させた昆虫細胞系;または蛍光タンパク質のコード配列を含む組換えウイルス発現ベクター(例えば、レトロウイルス、アデノウイルス、ワクシニアウイルス)を感染させた動物細胞系、もしくは安定に発現するように作製された形質転換された動物細胞系が含まれるがこれらに限定されない。
【0113】
使用される宿主/ベクター系に応じて、構成的プロモーターおよび誘導性プロモーター、転写エンハンサー配列、転写終結配列などを含む任意の数の適切な転写配列および翻訳配列を発現ベクター中に用いることができる(例えば、Bitterら、Methods in Enzymology 153:516〜544、1987を参照)。例えば細菌系でクローン化を行う場合は、バクテリオファージΣのpL、plac、ptrp、ptac (ptrp-lacハイブリッドプロモーター)などの誘導性プロモーターを使用することができる。哺乳類細胞系でクローン化を行う場合は、哺乳類細胞のゲノムに由来するプロモーター(例えばメタロチオネインプロモーター)、または哺乳類ウイルスに由来するプロモーター(例えばレトロウイルスの末端反復配列(LTR);アデノウイルスの後期プロモーター;ワクシニアウイルスの7.5Kプロモーター)を使用することができる。組換えDNA手法または合成手法で作られるプロモーターを用いて、挿入された蛍光タンパク質のコード配列の転写を誘導することもできる。
【0114】
細菌系では、数種の発現ベクターを、発現させる蛍光タンパク質について意図した用途に応じて有利に選択することができる。例えば大量の蛍光タンパク質を産生させる場合、精製が容易な高レベルの融合タンパク質産物の発現を誘導するベクターが望ましい場合がある。蛍光タンパク質の回収を容易にするための切断部分を含むように改変されたベクターが好ましい。
【0115】
酵母では、構成的プロモーターまたは誘導性プロモーターを含むいくつかのベクターを用いることができる。総説は「分子生物学の最新プロトコール(Current Protocols in Molecular Biology)」、第2巻、アウシュベル(Ausubel)ら編、Greene Publish. Assoc. & Wiley Interscience、第13章、1988;グラント(Grant)ら、「酵母用発現および選択ベクター(Expression and Secretion Vectors for Yeast)」、(Methods in Enzymology、WuおよびGrossman編、31987、Acad. Press、ニューヨーク、第153巻、pp. 516〜544、1987;グローバー(Glover)、「DNAクローニング(DNA Cloning)」、第2巻、IRL Press、ワシントンDC、第3章、1986;およびビター(Bitter)、「酵母における異種遺伝子発現(Heterologous Gene Expression in Yeast)」、Methods in Enzymology、BergerおよびKimmel編、Acad. Press、ニューヨーク、第152巻、pp. 673〜684、1987;ならびに「出芽酵母の分子生物学(The Molecular Biology of the Yeast Saccharomyces)」、ストラサム(Strathem)ら編、Cold Spring Harbor Press、第I巻および第II巻、1982を参照されたい。ADHもしくはLEU2などの酵母の構成的プロモーター、またはGALなどの誘導性プロモーターを用いることができる(「酵母におけるクローニング(Cloning in Yeast)」、第3章、R. Rothstein:「DNAクローニング(DNA Cloning)」第11巻、A Practical Approach、DM Glover編、IRL Press、ワシントンDC、1986)。あるいは、外来DNA配列の酵母染色体への統合を促すベクターを用いることができる。
【0116】
植物の発現ベクターを用いる場合は、蛍光タンパク質のコード配列の発現は、任意のいくつかのプロモーターで誘導することができる。例えばCaMVの35S RNAおよび19S RNAプロモーターなどのウイルス性プロモーター(Brissonら、Nature 310:511〜514、1984)、またはTMVのコートタンパク質のプロモーター(Takamatsuら、EMBO J. 6:307〜311、1987)を用いることができるほか、RUBISCOの小サブユニットなどの植物性プロモーター(Coruzziら、1984、 EMBO J. 3:1671〜1680;Broglieら、Science 224:838〜843、1984);または例えば、ダイズのhsp17.5-Eまたはhsp17.3-Bなどの熱ショックプロモーター(Gurleyら、Mol. Cell. Biol. 6:559〜565、1986)を用いることができる。このような構築物は、Tiプラスミド、Riプラスミド、植物ウイルスベクターを用いて、また直接的な形質転換法、マイクロインジェクション法、エレクトロポレーション法などで植物細胞に導入することができる。以上の手法の総説は例えば、ワイスバハ(Weissbach)およびワイスバハ(Weissbach)、「植物分子生物学の方法(Methods for Plant Molecular Biology)」、Academic Press、ニューヨーク、第VIII章、pp. 421〜463、1988;およびグリアソン(Grierson)およびコーリー(Corey)、「植物分子生物学(Plant Molecular Biology)」、第2版、Blackie、ロンドン、第7〜9章、1988などを参照されたい。
【0117】
蛍光タンパク質を発現させる際に使用可能な他の発現系が昆虫の系である。一つの昆虫系では、キンウワバの一種(Autographa californica)の核多核体病ウイルス(AcNPV)を外来遺伝子を発現させるベクターとして用いる。このウイルスはヨトウガ(Spodoptera frugiperda)の細胞で増殖する。蛍光タンパク質のコード配列は、ウイルスの非必須領域(例えば多面体遺伝子)にクローン化して、AcNPVプロモーター(例えば多面体プロモーター)の制御下に配置することができる。蛍光タンパク質のコード配列が良好に挿入されると、多面体遺伝子が不活性化されて、非包埋状の組換えウイルス(多面体遺伝子にコードされたタンパク質性コートを欠くウイルス)が産生される。この組換えウイルスを次にヨトウガ(Spodoptera frugiperda)の細胞に感染させて、そこで挿入遺伝子を発現させる(Smithら、J. Viol. 46:584、1983;Smith、米国特許第4,215,051号を参照)。
【0118】
真核生物系、好ましくは哺乳類発現系では、発現される哺乳類タンパク質の適切な翻訳後修飾が可能である。一次転写物の適切なプロセシング、グリコシル化、リン酸化、および遺伝子産物を有利に分泌させるための細胞機構を有する真核細胞を、蛍光タンパク質を発現させる宿主細胞として使用することができる。このような宿主細胞系列には、CHO、VERO、BHK、HeLa、COS、MDCK、Jurkat、HEK-293、およびWI38などがあるがこれらに限定されない。
【0119】
組換えウイルスまたはウイルスの配列を用いて発現を誘導する哺乳類細胞系を作製することができる。例えばアデノウイルスの発現ベクターを用いる場合は、蛍光タンパク質のコード配列を、アデノウイルスの転写/翻訳の制御複合体(例えば、後期プロモーターおよび三成分からなるリーダー配列)に連結することができる。このようなキメラ遺伝子を次に、インビトロ組換え法またはインビボ組換え法でアデノウイルスのゲノムに挿入することができる。ウイルスゲノムの非必須領域(例えば領域E1またはE3)の挿入により、組換えウイルスは、感染宿主で増殖可能となり、また蛍光タンパク質を発現可能となる(例えば、LoganおよびShenk、Proc. Natl. Acad. Sci. USA、81:3655〜3659、1984を参照)。あるいは、ワクシニアウイルスの7.5Kプロモーターを用いることができる(例えば、Mackettら、Proc. Natl. Acad. Sci. USA、79:7415〜7419、1982;Mackettら、J. Virol. 49:857〜864、1984;Panicaliら、Proc. Natl. Acad. Sci. USA 79:4927〜4931、1982を参照)。染色体外因子としての複製能力をもつウシパピローマウイルスを元にしたベクターは特に重要である(Sarverら、Mol. Cell. Biol. 1:486、1981)。DNAをマウス細胞に導入して短時間後に、プラスミドは細胞1個あたり約100〜200コピーに複製される。挿入されたcDNAの転写は、プラスミドの宿主染色体への組込みを必要としないので、高レベルの発現が達成される。以上のようなベクターを用いて、プラスミド中にneo遺伝子などの選択マーカーを含めることで安定な発現が可能となる。あるいは、レトロウイルスのゲノムを修飾して、宿主細胞に導入して蛍光タンパク質遺伝子の発現を誘導可能なベクターとして使用することができる(ConeおよびMulligan、Proc. Natl. Acad. Sci. USA、81:6349〜6353、1984)。高レベルの発現は、メタロチオネインIIAプロモーター、および熱ショックプロモーターを含むがこれらに限定されない誘導性プロモーターを用いることで達成できる場合もある。
【0120】
本発明は、核局在化配列、小胞体局在化配列、ペルオキシソーム局在化配列、ミトコンドリア局在化配列などの局在化配列、または局在化タンパク質を含む場合もある。局在化配列は、例えばスタイヤー(Stryer、L.)による「タンパク質ターゲティング(Protein Targeting)」、第35章、Biochemistry、第4版、W.H. Freeman、1995に記載されているような標的配列となりうる。局在化配列は局在化タンパク質である場合もある。いくつかの重要な局在化配列には、核を標的とする配列(KKKRK)、ミトコンドリアを標的とする配列(アミノ末端MLRTSSLFTRRVQPSLFRNILRLQST-)、小胞体を標的とする配列(C末端においてKDEL、N末端に存在すると予想されるシグナル配列)、ペルオキシソームを標的とする配列(C末端においてSKF)、プレニル化または形質膜への挿入を標的とする配列(C末端においてCaaX、CC、CXC、またはCCXX)、形質膜の細胞質側を標的とする配列(SNAP-25との融合)、またはゴルジ器官を標的とする配列(フリン(furin)との融合)が含まれる。
【0121】
組換えタンパク質を長期間、高収量で産生させるためには安定した発現が好ましい。ウイルスの複製起点を含む発現ベクターを用いるより、適切な発現制御配列(例えば、プロモーター、エンハンサー配列、転写終結配列、ポリアデニル化部位など)で調節された蛍光タンパク質および選択マーカーのcDNAで宿主細胞を形質転換することができる。組換えプラスミドの選択マーカーは選択に対する耐性を付与し、細胞が、プラスミドをその染色体に安定に組み込ませ、成長させてフォーカスを形成させ、最終的にクローン化して細胞系列内で増殖させることができる。例えば外来DNAを導入することで、改変された細胞は栄養強化培地で1〜2日間増殖させた後に選択培地に切り換えることができる。単純ヘルペスウイルスのチミジンキナーゼ(Wiglerら、Cell、11:223、1977)、ヒポキサンチン-グアニンホスホリボシルトランスフェラーゼ(SzybalskaおよびSzybalski、Proc. Natl. Acad. Sci. USA、 48:2026、1962)、およびアデニンホスホリボシルトランスフェラーゼ(Lowyら、Cell、22:817、1980)の各遺伝子を含むがこれらに限定されない、いくつかの選択系を用いることが可能であり、それぞれtk-、hgprt-、またはaprt-細胞で用いることができる。また、メトトレキセート耐性を付与するdhfr(Wiglerら、Proc. Natl. Acad. Sci. USA、77:3567、1980;O'Hareら、Proc. Natl. Acad Sci. USA、8:1527、1981);ミコフェノール酸耐性を付与するgpt (MulliganおよびBerg、Proc. Natl. Acad. Sci. USA、78:2072、1981;アミノグリコシドG-418耐性を付与するneo (Colberre-Garapinら、J. Mol. Biol.、150:1、1981);およびハイグロマイシン耐性を付与するhygro(Santerreら、Gene、30:147、1984)の各遺伝子の選択の基礎として代謝拮抗剤耐性を用いることができる。最近、細胞がトリプトファンの代わりにインドールを用いることを可能とする他の選択用の遺伝子(例えばtrpB);細胞がヒスチジンの代わりにヒスチノールを利用できるようになるhisD (HartmanおよびMulligan、Proc. Natl. Acad. Sci. USA、85:8047、1988);およびオルニチンデカルボキシラーゼ阻害物質である2-(ジフルオロメチル)-DL-オルニチン(DFMO)に対する耐性を付与するODC (オルニチンデカルボキシラーゼ)(McConlogue L.、「分子生物学の最新情報(Current Communications in Molecular Biology)」、Cold Spring Harbor Laboratory編、1987)についての報告がある。
【0122】
本発明の蛍光タンパク質ポリペプチドをコードするDNA配列は、適切な宿主細胞にDNAを輸送することでインビトロで発現させることができる。「宿主細胞」は、ベクターが増殖可能で、かつそのDNAが発現される細胞である。この用語は、対象の宿主細胞の任意の子孫も含む。すべての子孫は、親の細胞と同一であるわけではないと理解されている。これは複製過程で変異が生じるためである。しかし、このような子孫は、「宿主細胞」という用語が用いられる場合に含まれる。安定な輸送法、言い換えると、外来DNAを宿主で連続して維持する方法は当技術分野で周知である。
【0123】
発現ベクターを宿主細胞にトランスフェクトさせて、組換え核酸を発現させることができる。宿主細胞は、蛍光タンパク質の融合タンパク質を精製する目的で、高レベルの発現を指標に選択することができる。大腸菌はこの目的に有用である。あるいは宿主細胞は、細胞が産生する酵素の活性を検討するために選択される原核細胞または真核細胞の場合がある。この場合、リンカーペプチドは、プロテアーゼによって認識されるアミノ酸配列を含むように選択される。このような細胞は例えば、培養細胞またはインビボの細胞である。
【0124】
蛍光タンパク質の融合タンパク質の第1の利点は、これらが通常のタンパク質生合成で調製されるために、有機合成および特注の非天然のアミノ酸類似体の要求性を完全に回避できる点である。このような構築物は、インビトロアッセイ法用に大腸菌で大規模に発現させることができる。細菌からの精製は、配列が、ニッケル-キレートクロマトグラフィーによる一段階精製用のポリヒスチジンタグを含むと単純なものとなる。あるいは、インサイチューアッセイ用に所望の宿主細胞でこのような基質を直接発現させることができる。
【0125】
別の態様では、本発明は、蛍光タンパク質をコードする核酸配列を発現するトランスジェニック非ヒト動物を提供する。
【0126】
本発明の「非ヒト動物」は、蛍光タンパク質をコードする核酸配列を有する任意の非ヒト動物を含む。非ヒト動物は、げっ歯類、非ヒト霊長類、ヒツジ、イヌ、ウシ、ブタ、両生類、および爬虫類などの脊椎動物を含む。好ましい非ヒト動物は、ラットおよびマウスを含むげっ歯類ファミリーから選択され、最も好ましくはマウスである。本発明の「トランスジェニック非ヒト動物」は、「導入遺伝子」を非ヒト動物の生殖系列に導入することで作製される。さまざまな発生段階における胚性標的細胞を用いて導入遺伝子を導入することができる。胚性標的細胞の発生段階に応じて他の方法を用いることができる。接合体はマイクロインジェクションに最も適した標的である。マウスの雄性前核は、1〜2 plのDNA溶液の再現性ある注入が可能となる直径約20 μmの大きさになる。遺伝子輸送の標的としての接合体の使用は、初回分裂前において注入されたDNAが宿主遺伝子に組み込まれる状況の多くで最も利益が大きい(Brinsterら、 Proc. Natl. Acad. Sci. USA 82:4438〜4442、1985)。したがってトランスジェニック非ヒト動物のすべての細胞は、組み込まれた導入遺伝子をもつことになる。これは一般に、創始個体の子孫への効率のよい導入遺伝子伝達を反映してもいる。というのは生殖細胞の50%が導入遺伝子をもつからである。接合体のマイクロインジェクションは、本発明を実施する上で導入遺伝子を組み込ませる好ましい方法である。
【0127】
「トランスジェニック」という用語は、外因性遺伝物質をすべての細胞に含む動物を示すものとして用いる。「トランスジェニック」動物は、再生に用いられる細胞内に外因性遺伝物質を含む二匹のキメラ動物を交配することで作製することができる。結果として得られる子孫の25%がトランスジェニック個体、すなわち外因性遺伝物質をすべての細胞の両対立遺伝子上に含む個体である。結果として得られる個体の50%が外因性遺伝物質を、1本の対立遺伝子上に含み、25%は外因性遺伝物質を含まない。
【0128】
レトロウイルスの感染も、非ヒト動物への導入遺伝子の導入に用いられる場合がある。発生段階にある非ヒト胚は胚盤胞期になるまでインビトロで培養することができる。この期間中、胚盤胞はレトロウイルス感染の標的となる(Jaenich、R.、Proc. Natl. Acad. Sci. USA 73:1260〜1264、1976)。酵素処理によって透明帯を除くと、胚盤胞への感染の効率が高まる(Hoganら (1986)、「マウス胚操作(Manipulating the Mouse Embryo)」、Cold Spring Harbor Laboratory Press、Cold Spring Harbor、ニューヨーク)。導入遺伝子の導入に用いられるウイルスベクター系は典型的には、導入遺伝子を有する複製欠損型のレトロウイルスである(Jahnerら、Proc. Natl. Acad. Sci. USA 82:6927〜6931、1985;Van der Puttenら、Proc. Natl. Acad. Sci USA 82:6148〜6152、1985)。トランスフェクションは、胚盤胞をウイルス産生細胞の単層上で培養することで容易かつ効率的に行われる(Van der Putten、前掲;Stewartら、EMBO J. 6:383〜388、1987)。あるいは感染を後期の段階で実施することができる。ウイルスまたはウイルス産生細胞は胞胚腔に注入することができる(D. Jahnerら、Nature 298:623〜628、1982)。創始個体の大部分は、導入遺伝子の組込みが、トランスジェニック非ヒト動物を形成する細胞の一部でのみ生じることから、導入遺伝子についてモザイクとなる。また創始個体では、レトロウイルスにより、子孫では通常分離してしまうゲノム上のさまざまな位置へ導入遺伝子が挿入されることがある。また、妊娠中期の胚の子宮内にレトロウイルスを感染させることで、導入遺伝子を生殖系列に導入することが低効率ながら可能である(D. Jahnerら、前掲)。
【0129】
導入遺伝子を導入する際に用いられる第3のタイプの標的細胞は胚幹(ES)細胞である。ES細胞は、インビトロで培養した着床前の胚から得られ、胚と融合させる(M.J. Evansら、Nature 292:154〜156、1981;M.O. Bradleyら、Nature 309:255〜258、1984;Gosslerら、Proc、 Natl. Acad. Sci. USA 83:9065〜9069、1986;およびRobertsonら、Nature 322:445〜448、1986)。導入遺伝子は、DNAトランスフェクション法やレトロウイルスを用いたトランスダクション法によってES細胞に効率よく導入することができる。このような形質転換されたES細胞を後に、非ヒト動物の胚盤胞と結合させることができる。このようなES細胞はその後、胚のコロニーを形成して、結果として生じる動物個体の生殖系列に寄与する(Jaenisch、R.、Science 240:1468〜1474、1988の総説を参照)。
【0130】
「形質転換された」とは、組換え核酸手法によって異種核酸分子が細胞(またはその祖先)に導入されることを意味する。「異種」とは、他の種に起源を有するか、もしくは元の状態、または細胞内で主に発現された状態から修飾される核酸配列を意味する。
【0131】
「導入遺伝子」とは、細胞に人工的に挿入されて、細胞から発生する生物体のゲノムの一部となる(すなわち、安定に組み込まれるか、または安定な染色体外因子として存在する)任意のDNA断片を意味する。このような導入遺伝子は、トランスジェニック生物に対して部分的、または全体的に異種の(すなわち外来の)遺伝子を含む場合があるか、生物体の内因性遺伝子に相同な遺伝子である場合がある。DNAに転写されてからゲノムに組み込まれるRNA配列を提供することで作製される導入遺伝子は上記の定義に含まれる。本発明の導入遺伝子は、トランスジェニック非ヒト動物で発現される可能性のある蛍光タンパク質をコードするDNA配列を含む。本明細書で用いる「トランスジェニック」という表現は、特定の遺伝子ノックアウトを誘導する、初期胚または受精卵のインビトロ操作により、またはトランスジェニック手法によりゲノムが変化した任意の生物体を追加的に含む。本明細書で用いる「遺伝子ノックアウト」という表現は、当技術分野で一般的な任意のトランスジェニック手法で達成される、完全に機能を喪失させるインビボにおける遺伝子の標的破壊を意味する。一つの態様では、遺伝子ノックアウトを有するトランスジェニック動物は、相同組換えによって非機能性となる遺伝子への標的挿入によって標的遺伝子が非機能状態となった動物である。本明細書で用いる「トランスジェニック」という用語には、導入された導入遺伝子をもつ生物、または非機能状態にされた(または「ノックアウトされた」)内因性遺伝子をもつ生物を作製可能な、当業者に一般的な任意のトランスジェニック手法が含まれる。
【0132】
III.改変蛍光タンパク質の用途
本発明のタンパク質は、蛍光タンパク質を用いる任意の方法に有用である。
【0133】
本発明の改変蛍光タンパク質は、蛍光マーカーが既に用いられている多くの方法で蛍光マーカーとして有用である。これには例えば、イムノアッセイ法またはハイブリダイゼーションアッセイ法などの検出アッセイ法に用いる、改変蛍光タンパク質と抗体、核酸、または他の受容体とのカップリングなどが含まれる。
【0134】
本発明の改変蛍光タンパク質は、細胞内におけるタンパク質の動きを追跡する際に有用である。この態様では、蛍光タンパク質をコードする核酸分子を、対象タンパク質をコードする核酸分子に発現ベクター上で融合させる。細胞内で対象タンパク質が発現すると、蛍光を手がかりにその局在を決定することができる。別のバージョンでは、二種の対象タンパク質を、蛍光特性が異なる二種の改変蛍光タンパク質と融合させる。
【0135】
本発明の改変蛍光タンパク質は、転写の誘導を検出する系で有用である。特定の態様では、改変蛍光タンパク質をコードするヌクレオチド配列を、対象の発現制御配列に融合させ、発現ベクターを細胞にトランスフェクトする。プロモーターの誘導は、蛍光の発現および/または量を検出することで測定することができる。このような構築物を用いることで、受容体からプロモーターに至る情報伝達経路を追跡することができる。
【0136】
本発明の改変蛍光タンパク質は、FRETがかかわる応用に有用である。このような応用は、相互に引き合ったり離れたりする蛍光ドナーおよび蛍光アクセプター運動に応じた事象を検出することができる。ドナー/アクセプターペアの一方または両方を蛍光タンパク質とすることができる。FRETをベースとしたアッセイ法に好ましいドナーと受容体のペアは、T203I変異をもつドナーとT203X変異をもつアクセプターである(Xは芳香族アミノ酸-39、特にT203Y、T203W、またはT203Hである)。特に有用なペアでは、ドナーは以下の変異を含む:S72A、K79R、Y145F、M153A、およびT203I (励起ピークが395 nmで発光ピークが511 nm)。またアクセプターは以下の変異を含む:S65G、S72A、K79R、およびT203Y。このような特定のペアは、ドナーの励起ピークと発光ピークの広い分離をもたらし、またドナーの発光スペクトルとアクセプターの励起スペクトルとの良好な重なりをもたらす。本明細書に記載されたような他の赤色シフト変異体も、このようなペアにおけるアクセプターとして用いることができる。
【0137】
一つの局面では、FRETを用いて、切断部位の反対側に基質に結合したドナーおよびアクセプターを有する基質の切断を検出する。基質が切断されると、ドナー/アクセプターのペアが物理的に分離してFRETを放出する。アッセイ法には、基質を試料に接触させる段階、およびFRETの量的または質的な変化を決定する段階が含まれる。一つの態様では、改変蛍光タンパク質はβ-ラクタマーゼの基質に用いられる。このような基質の例は、1995年3月20日に出願された米国特許出願第08/407,544号、および1996年3月20日に出願された国際出願番号PCT/US96/04059に記載されている。別の態様では、改変蛍光タンパク質のドナー/アクセプターペアは、タンパク質分解性の切断部位を有するペプチドが結合した融合タンパク質の一部である。このようなタンデム型の蛍光タンパク質は、1996年1月31日に出願された米国特許第08/594,575号に記載されている。
【0138】
別の局面では、FRETを用いて膜内外の電位変化を検出する。ドナーおよびアクセプターを膜の反対側に配置させて、一方が電位変化に応じて翻訳されて膜を越えるようにする。これにより測定可能なFRETが生じる。このような方法は、1995年6月7日に出願された米国特許出願第08/481,977号、および1996年6月6日に出願された国際出願番号PCT/US96/09652に記載されている。
【0139】
本発明の改変蛍光タンパク質は、試料中および生細胞内ならびにトランスジェニック生物に含まれるイオンの濃度決定用のバイオセンサーの作製に有用である。イオンが蛍光タンパク質に結合すると、改変蛍光タンパク質の少なくとも一つの測定可能な蛍光特性の変化が生じ、これが対象イオンの存在を判定する際の基礎となる。
【0140】
本発明の改変タンパク質は、タンパク質キナーゼの蛍光基質の作製に有用である。このような基質は、タンパク質キナーゼによって認識されるアミノ酸配列をもつ。リン酸化されると、改変蛍光タンパク質の蛍光特性が変化する。このような基質は、トランスフェクションおよび発現する際に、細胞試料のタンパク質キナーゼ活性を検出および測定する際に有用である。好ましくは、キナーゼ認識部位は、改変蛍光タンパク質の末端の約20アミノ酸以内に配置される。キナーゼ認識部位は、タンパク質のループドメインに配置することもできる(例えば図1B参照)。タンパク質キナーゼの蛍光性基質を作製する方法は、1996年6月16日に出願された米国特許出願第08/680,877号に記載されている。
【0141】
プロテアーゼ認識部位もループドメインに導入することができる。切断されると蛍光基質は測定可能な状態に変化する。
【0142】
本発明は、被験化合物を同定する方法も含む。典型的には、このような方法は、機能性の改変蛍光タンパク質、または機能性の改変蛍光タンパク質をコードするポリペプチドで標識された生物学的存在を含む試料に被験化合物を接触させる段階を含む。機能性の改変蛍光タンパク質を含む試料の蛍光(蛍光特性)をモニタリングすることで、被験化合物が活性であるか否かを判定することができる。シグナルの特異性を保証するために対照を含めることができる。このような対照は、被験化合物の非存在下における蛍光特性の測定、予想される活性を有する化合物(例えば既知の調節因子)、または改変された対照(例えば、改変蛍光タンパク質の非存在、使用可能に連結された蛍光タンパク質ポリヌクレオチドの非存在、または改変蛍光タンパク質の使用可能な連結の非存在)の存在下における蛍光特性の測定を含む。
【0143】
被験化合物の存在下における蛍光は、同じ被験化合物の非存在下における場合と比べて大きい場合もあれば小さい場合もある。例えば、改変蛍光タンパク質を遺伝子発現のレポーターとして用いる場合、被験化合物は遺伝子発現を上方または下方に制御する可能性がある。このようなタイプのスクリーニングでは、機能性の改変蛍光タンパク質をコードするポリヌクレオチドを、ゲノムのポリヌクレオチドに動作可能に連結させる。あるいは、機能性の改変蛍光タンパク質を第2の機能性タンパク質に融合させる。この態様を用いて、エネルギー移動を指標に第2のタンパク質の局在を追跡したり、タンパク質間相互作用を追跡したりすることができる。
【0144】
IV.手順
試料の蛍光は蛍光定量計で測定される。一般的には、第1の波長を有する励起源からの励起放射が、励起光学系を通過する。励起光学系は励起放射を生じて試料を励起させる。これに応じて、試料中の蛍光タンパク質は、波長が励起波長とは異なる光線を放出する。次に収集光学系が試料から発光を集める。この装置は、スキャン中に試料を特定の温度に維持するための温度制御装置を含む場合がある。一つの態様では、多軸トランスレーションステージが、曝露されるさまざまなウェルを配置するために複数の試料を保持するマイクロタイタープレートを動かす。多軸トランスレーションステージ、温度制御装置、オートフォーカス特性、ならびに画像化およびデータ収集に関連した電子装置を適切にプログラムされたデジタルコンピュータで管理することができる。このようなコンピュータは、アッセイ中に収集されたデータの形式を、別の形式に変えて表示させることもできる。この過程を小型化および自動化することで、数千種の化合物のスクリーニングを実施することができる。
【0145】
蛍光材料のアッセイ法を実施する方法は当技術分野で周知であり、例えば、ラコビッツ(Lakowicz、J.R.)、「蛍光分光学の原理(Principles of Fluorescence Spectroscopy)」、ニューヨーク:Plenum Press (1983);ハーマン(Herman、B.)、「培養生細胞の蛍光顕微鏡観察(Fluorescence Microscopy of Living Cells in Culture)」のパートBの「共鳴エネルギー移動顕微鏡観察(Resonance energy transfer microscopy)」、Methods in Cell Biology、第30巻、テイラー(Taylor、D.L.およびワン(Wang、Y.-L.)編、サンディエゴ:Academic Press (1989)、pp. 219〜243;チュロ(Turro、N.J.)、「最新分子光化学(Modern Molecular Photochemistry)」、メンロパーク:Benjamin/Cummings Publishing Col、Inc. (1978)、pp. 296〜361に記載されている。
【0146】
変異導入およびタンパク質の調製
YFPの異型および復帰体は、PCR法をベースとした「QuikChange(登録商標) Site-Directed Mutagenesis Kit」(Stratagene、カリフォルニア州ラホーヤ)を用いて、製造業者の指示書にしたがって、またYFPクローン10cを鋳型として用いて調製された(Ormoら、1996)。変異は遺伝子全体の配列を決定して検証し、すべてのGFP異型は文献に記載された手順で発現させて精製した(Ormoら、1996)。
【0147】
蛍光の測定
濃縮されたタンパク質の一部を、イオン強度が一定の一連の緩衝液(20 mM MES pH 6.0、MES pH 6.5、PIPES 7.0、HEPES 7.5、およびTAPS 8.0)で25倍に希釈した。緩衝液は、さまざまな濃度の塩化カリウムまたはヨウ化カリウムを含むものを使用し、イオン強度はD-グルコン酸カリウムで150 mMに調節した。pHおよびハロゲン化物濃度の関数としての蛍光の測定は、Hitachi F4500蛍光分光光度計を室温(λex=514 nm)に設定して実施し、520〜550 nmの発光を60 nm/分の速度で3回スキャンした。528 nmにおける発光極大を平均化し、ハロゲン化物が非存在下における蛍光に対して補正した。
【0148】
結晶成長およびデータ収集
YFP-H148Qを20 mM TRIS pH 7.9で15 mg/mlに濃縮し、5 μlのタンパク質および5 μlの母液を含むハンギングドロップ中で結晶を成長させた。母液は、22% PEG 1550(pH 5.5、溶媒は100 mM 酢酸ナトリウムおよび90 mM MgCl2)を含むものを用いた。棒状の結晶は、幅が約0.04 nm、長さが1.0 mmで、1.5〜2年間4℃で成長させたものを用いた。1個の結晶を、上述の成分(MgCl2は含まないが100 mM ヨウ化カリウム、および20% エチレングリコールを含む(凍結防止用))を含む合成母液に浸漬した(ヨウ化物浸漬と呼ぶ)。また別の結晶を、100 mM MgCl2、および20% エチレングリコールを含む上記の母液に浸漬した(塩化物浸漬と呼ぶ)。いずれの浸漬ともpH 4.6で4時間室温で実施し、その後直ちにデータの収集を行った。この結晶を瞬時に凍結し、X線回折データを、ミラーを備えたRigaku RUH3回転陰極型X線発生装置にマウントしたRAXIS-IIcイメージプレートを用いて100 Kで収集した。
【0149】
YFP-H148Qの構造決定、およびヨウ化物結合部位の同定
二つのデータセットをDenzo vl.9で処理し、ScalePackを用いて比較した(OtwinowskiおよびMinor、1997)。空間群P212121であり、単位格子パラメータは、ヨウ化物浸漬では、a=51.2 Å、b=62.8 Å、およびc=68.7 Å、また塩化物浸漬では、a=51.7 Å、b=62.6 Å、およびc=66.2 Åである。結晶は、YFP-H148G (Wachterら、1998)、およびGFP S65T (Ormoら、1996)の結晶とほぼ同形であり、YFP-H148Gの座標ファイル2yfp (Wachterら、1998)を位相決定のモデルとして使用した。陰イオン性発色団のモデルは、プログラム「SPARTAN」バージョン4.1に含まれる「AM1」を用いた半経験的な分子軌道計算で得た(Wavefimction Inc.、カリフォルニア州アーバイン)。
【0150】
ヨウ化物浸漬で得られたデータセットから異常差図の計算を行った(異常データの完全性は65%)。これはヨウ素が、インハウスのCuKα波長(1.54 Å)で有意に異常なシグナルを発生するためである。重原子の位相は「CCP4 program suite」のプログラム「scaleit」を用いて、タンパク質の位相計算値から90°を差引くことで概算した(Collaborative Computational Project N. 4、1994)。異常差図によって二か所のヨウ化物の位置が同定された。一か所はタンパク質内部に埋没しており、一か所はタンパク質の表面であった。
【0151】
ヨウ化物結合状態または非結合状態におけるYFP-H148Qの精密化
ヨウ化物浸漬および塩化物浸漬に由来する二つのデータセットを同様の手順で精密化した。4 Åまでの最初の剛体精密化の後に、3.0 Åまでのデータ、さらに分解能限界(表1)までのデータを対象に、プログラムTNT (Tronrudら、1987)を用いて位置精密化を行った。精密化の初期サイクルでは、結合状態のハロゲン化物のモデル化は行わず、148位のグルタミンをグリシンとしてモデル化を行った。電子密度図(2Fo-FcおよびFo-Fc)を、Oを用いて間欠的に精査した(Jonesら、1991)。Fo-Fc図から、それぞれ11 rmsおよび5.5 rmsの偏位における埋没状態のヨウ化物および表面のヨウ化物の位置が、二つの異常差電子密度ピークの中心にあるが、埋没状態の塩化物の結合と一致する正の差電子密度は認められないことがわかった。Gln148の側鎖の密度は初期に明瞭に描出されたので、元のヒスチジンとは異なる回転異性体としてのグルタミンのモデリングが可能であった。
【0152】
B因子は、デフォルトのTNT B因子相関ライブラリーを用いて精密化を行った。HisおよびPheに由来するB因子相関値を用いて発色団原子のモデル化を行った。結合状態の溶媒分子は、水素結合パートナーの差密度および近接性から適切であると判断した場合にモデルに加えた。結合状態のハロゲン化物の占有率を精密化する前に、これらのハロゲン化物のB因子を固定した。埋没状態のヨウ化物の温度因子は、それに最も近い12個の原子のB因子の平均値(30 Å2)に設定し、表面のヨウ化物の温度因子は、タンパク質表面に結合した6個の最も近い溶媒分子のB因子の平均値(39 Å2)に設定した。精密化の最終段階では、2個の埋没状態のハロゲン化物の占有率の精密化を行った。
【0153】
吸光度による発色団のpKaおよびヨウ化物の結合定数の決定
発色団のpKaを、さまざまな陰イオン濃度における吸光度スキャンから決定した。吸光度スキャンは、0.05 mg/mlのYFPを対象に、一連の緩衝液(20 mM リンゴ酸、pH 5.8、リンゴ酸、pH 6.1、MES pH 6.4、HEPES pH 7.1)から選択された特定の陰イオンに適した二つの異なるpH条件で、250〜600 nm間で室温で収集した(Shimadzu 2101分光光度計)。緩衝液のpHにおける発色団の陰イオンの光学密度(YFPおよびYFP-H148Qについて514〜515 nm)、ならびに相互作用性陰イオンの非存在下におけるpH 9における光学密度を、ヘンダーソン−ハッセルベルヒ(Henderson-Hasselbalch)式を用いて、検討した各条件における発色団のpKaを推定した。陰イオンとタンパク質の結合の微視的な結合定数は、陰イオン濃度に対する発色団のpKaのカーブフィッティングにより、二種の異なるリガンドが関与する結合平衡の式を用いて求めた。
【0154】
以下に挙げる例は、説明目的で提示するものであり、制限する意図はない。
【0155】
実施例
GFPの特性を理解する段階、また特性が変化したGFPの調整を容易にする段階として、発明者らは、オワンクラゲGFPのS65T変異体(R. Heimら、Nature 373:664〜665 (1995))の三次元構造を1.9 Åの分解能で決定した。この変異体は遍在性のQ80R置換も含む。同置換はGFP cDNAの初期の流通中に偶然生じたものであるが、タンパク質の特性に何らかの作用を及ぼしているかは不明である(M. Chalfieら、Science 263:802〜805 (1994))。
【0156】
ヒスチジンタグを付けたS65T GFP (R. Heimら、Nature 373:664〜665 (1995))を、4 lのYTブロス+アンピシリン中で37℃、450 rpmで、5 l/分のエアフローを送りながらJM109/pRSETB中で過剰に発現させた。A595が0.3となった時点で温度を25℃に下げ、次に1 mMのイソプロピルチオガラクトシドで5時間かけて誘導を行った。細胞ペーストを -80℃で一晩保存した後に、50 mM HEPES pH 7.9、0.3 M NaCl、5 mM 2-メルカプトエタノール、0.1 mM フェニルメチル-スルフォニルフルオリド(PMSF)中に再懸濁し、フレンチプレスに10,000 psiで一回通し、20K rpmで45分間遠心した。上清をNi-NTA-アガロースカラム(Qiagen)にアプライした後に、20 mM イミダゾールで洗浄し、次に100 mM イミダゾールで溶出した。緑色のフラクションをプールし、キモトリプシン(Sigma)でタンパク質の消化(1:50、重量比)を室温で22時間かけて行った。0.5 mM PMSFを添加した後に、消化物をNiカラムに再びアプライした。N末端の配列を決定して、本来のメチオニンが存在することを確認した。20 mM HEPES、pH 7.5で透析を行い、A490が20となるまで濃縮された時点で、5 μl タンパク質および5 μl ウェル溶液、22〜26% PEG 4000 (Serva)、50 mM HEPES pH 8.0〜8.5、50 mM MgCl2、および10 mM 2-メルカプトエタノールを含むハンギングドロップ中の棒状結晶を室温で5日以内に回収した。結晶は幅が0.05 mmで、長さが最大1.0 mmであった。空間群は、a=51.8 Å、b=62.8 Å、c=70.7 Å、Z=4のP21P21P21である。この状態とは無関係の野生型GFPの二種類の結晶の形状は文献に記載されている(M.A. Perrozo、K.B. Ward、R.B. Thompson、およびW.W. Ward.、J. Biol. Chem、203、7713〜7716 (1988))。
【0157】
GFPの構造を、複数の同形置換および異常散乱(表E)、溶媒平滑化(solvent flattening)、位相組み合わせ(phase combination)、ならびに結晶学的精密化により決定した。GFPの折りたたみの最も顕著な特性は、中心的な1本のらせんの周囲を包む11本鎖(各鎖は約9〜13残基を含む)のβバレル(図1Aおよび1B)である。バレルは長さが42 Åで直径が24 Åのほぼ完全な円柱である。ポリペプチドのN末端側の半分は、3本の逆平行鎖、中心のらせん、およびさらに3本の逆平行鎖を含む(残基118〜123の後者は、残基11〜23のN末端側の鎖と平行)。ポリペプチド骨格は、分子の「ボトム」と交差して、5本の鎖を含むグリークキー(Greek Key)モチーフ中におけるバレルのもう半分を形成する。円柱の先端は、3本の短いゆがんだらせん状のセグメントでキャップされており、このうち1本の短くて大きくゆがんだらせん状のセグメントが円柱のボトムをキャップしている。円柱表面に存在する主鎖の水素結合は、タンパク質が変性およびタンパク質分解に対して異常に安定であることを説明する可能性が高い。発色団周囲のシェルの完全性を保存しながら励起されうるポリペプチドの大きなセグメントはない。したがって、大きなパーセントで分子量を低下させるようにGFPを再び作り込むことは困難であると思われる(J. DopfおよびT.M. Horiagon、Gene 173:39〜43 (1996))。
【0158】
p-ヒドロキシベンジリデンイミダゾリジノンの発色団(C.W. Codyら、Biochemistry 32:1212〜1218 (1993))は、バルク溶媒から完全に保護されており、分子の中心に位置する。全体的、かつおそらく強固に封入されていることが、小さなストークスシフト(励起極大と発光極大の間の波長差)、蛍光の高量子収率、O2による励起状態の消光不能(B.D. Nageswara Raoら、Biophys. J. 32:630〜632 (1980))、および発色団の外部pH滴定に対する耐性の要因であろう(W.W. Ward、「生物発光および化学発光(Bioluminescence and Chemiluminescence)」、M.A. DeLucaおよびW.D. McElroy編、Academic Press、pp. 235〜242 (1981);W.W. WardおよびS.H. Bokman、Biochemistry 21:4535〜4540 (1982);W.W. Wardら、Photochem. Photobiol. 35:803〜808 (1982))。また、酵素が基質に接近可能となるしくみを想像することが困難であることから、フルオロフォアの形成が自発的な分子内過程であるはずである(R. Heimら、Proc. Natl. Acad. Sci. USA 91:12501〜12504 (1994))という理由を合理的に解釈することもできる。発色団のこの面は、周囲バレルの対称軸に対してほぼ垂直(60°)である。発色団の一方の側は驚くべきことに、約135 Å3の容積を占める大きな空洞に面している(B. LeeおよびF.M. Richards、J. Mol. Biol. 55:379〜400 (1971))。原子半径は、プローブ半径を1.4 ÅとしてプログラムMSで計算されたリー(Lee)およびリチャーズ(Richards)の値とした(M.L. Connolly、Science 221:709〜713 (1983))。上記の空洞はバルク溶媒に対して開いていない。4個の水分子は空洞内に位置しており、Glu222およびGln69の埋没型の側鎖を連結する水素結合の鎖を形成している。占有されていない場合には、このような大きな空洞は、数kcal/molでタンパク質を不安定化すると推定されている(S.J. Hubbardら、Protein Engineering 7:613〜626 (1994);A.E. Erikssonら、Science 255:178〜183 (1992))。空洞容積の一部は、環状化および脱水反応で生じる集密化の結果である可能性がある。また空洞は、Tyr66のα-β結合から水素を除くオキシダント(最も好ましくはO2)を一時的に収容する可能性もある(A.B. Cubittら、Trends Biochem. Sci. 20:448〜455 (1995);R. Heimら、Proc. Natl. Acad. Sci. USA 91:12501〜12504 (1994);S. InouyeおよびF.L Tsuji、FEBS Lett. 351:211〜214 (1994))。発色団、空洞、および発色団に接触する側鎖を図2Aに示さし、付近の最終的な電子密度図の一部を2Bに示す。
【0159】
発色団の反対側は、複数の芳香性側鎖および極性側鎖にもたれるように触れている。発色団との極性相互作用の複雑なネットワーク(図2C)は特に重要である。His148、Thr203、およびSer205はフェノール性ヒドロキシルと水素結合を形成しており、Arg96およびGln94はイミダゾリジノン環のカルボニルと相互作用しており、またGlu222はThr65の側鎖と水素結合を形成している。Thr62のカルボニルのArg96、およびGln183の側鎖のカルボニルとの水素結合などの別の極性相互作用は、埋没状態のArg96をプロトン化された状態で安定化させると考えられる。このような埋没状態の電荷は、過去の報告と同様に、脱プロトンされたフルオロフォアのイミダゾリジノン環のカルボニル酸素の部分的に負の電荷をもつ残基が存在することを示唆している(W.W. Ward、「生物発光および化学発光(Bioluminescence and Chemihiminescence)」(M.A. DeLucaおよびW.D. McElroy編)、Academic Press、pp. 235〜242 (1981);W.W. WardおよびS.H. Bokman. Biochemistry 21:4535〜4540 (1982);W.W. Wardら、Photochem. Photobiol. 35:803〜808 (1982))。Arg96はフルオロフォアの形成に極めて重要な役割を果たす可能性が高く、初期の環閉鎖の触媒を促進している可能性がある。最後にTyr145は、ベンジル環との典型的な安定性の端-面(edge-face)相互作用を示している。GFPの唯一のトリプトファンであるTrp57は、発色団から13〜15 Åに位置し、2個の環系の長軸はほぼ平行である。これは後者へのエネルギー移動の効率がよいことを意味しており、別個のトリプトファン発光が観察可能である理由を説明している(D.C. Prasherら、Gene 111:229〜233 (1992)。GFP中の2個のシステインCys48とCys70は24 Å離れており、距離が大きすぎてジスルフィド結合を形成することはできない。Cys70は埋没しているが、Cys48は、スルフヒドリル特異的な試薬が比較的接近可能なはずである。このような試薬5,5'-ジチオビス (2-ニトロ安息香酸)は、GFPを標識し、また蛍光を消光することが報告されている(S. InouyeおよびF.I. Tsuji、FEBS Lett 351:211〜214 (1994))。この作用は、遊離のスルフヒドリルの必要性に起因していたが、Cys48に結合する可能性のある5-チオ-2-ニトロベンゾエート部分による特異的な消光を反映している可能性もある。
【0160】
電子密度図は、シス[Z-]配置では、その大部分が発色団について提案されている構造と一致する(D.C. Prasherら、Gene 111:229〜233 (1992);C.W. Codyら、Biochemistry 32:1212〜1218 (1993))が、発色団の二重結合周辺における反対の異性体の何らかの実質的なフラクションについて証拠は得られておらず、差の特性が、完全な非環状ポリペプチドか、またはカルビノールアミン(図2Cの左上の図)のいずれかを示していると解釈可能な、最終的な(Fo-Fc)電子密度図で>4σで見出されている。これは、かなりのフラクション、おそらくは結晶中の30%ほどの分子が最終的な脱水反応を受けていないことを示唆している。脱水が不完全であることの確認はエレクトロスプレー質量分析でなされており、野生型GFPとS65T GFPの両方の平均質量(それぞれ31,086±4 Daと31,099.5±4 Da)が、完全成熟型タンパク質についての推定値(それぞれ31,079 Daと31,093 Da)より6〜7 Da高いことが一貫して示されている。このような相違は、アポタンパク質のフラクションが30〜35%モルであること、またはカルビノールアミンが18もしくは20 Da高い分子量をもつことから説明可能と考えられる。13Cおよび2Hが天然には豊富に存在し、このような測定を行う際に使用されるヒューレット-パッカード社製の5989Bエレクトロスプレー質量解析装置の分解能には限界があることから、個々のピークを分解することはできないが、その代わりに、最大値の半分における全体の幅が約15 Daのである平均質量ピークが得られる。分子量は、配列が
【0161】
過去に行われたランダム変位導入の結果は、複数のアミノ酸の側鎖が、スペクトルに実質的な作用を及ぼすことを意味しており、原子モデルでは、このような残基が発色団付近に存在することが確認されている。変異T203IおよびE222Gは重要であるが、吸収スペクトルについては反対の結果をもたらす(T. Ehrigら、FEBS Letters 367:163〜166 (1995))。T203I (野生型のSer65をもつ)は、陰イオン性発色団に通常結びつけられる475 nmの吸収ピークをもたず、中性の発色団を反映すると考えられる395 nmのピークのみを示す(R. Heimら、Proc. Natl. Acad. Sci. USA 91:12501〜12504 (1994);T. Ehrigら、FEES Letters 367:163〜166 (1995))。実際にThr203は、発色団のフェノール性酸素と水素結合を形成しており、Ileとの置換はフェノール性酸素のイオン化を妨げるはずである。Glu222のGlyへの変異(T. Ehrigら、FEBS Letters 367:163〜166 (1995))は、例えば395 nmのピークを抑えて470〜490 nmのピークに傾くように、Ser65のGly、Ala、Cys、Val、またはThrとの置換によるものとほぼ同じ分光光度的作用をもつ(R. Heimら、Nature 373:664〜665 (1995);S. Delagraveら、Biotechnology 13:151〜154 (1995))。実際にGlu222およびThr65の残りは、このような構造では相互に水素結合しており、おそらく、Thr65の側鎖の酸素に対するドナーとして作用するGlu222の無電荷のカルボキシルをもつ。変異E222G、S65G、S65A、およびS65Vは、いずれもこのような水素結合を抑制すると考えられる。なぜ野生型タンパク質のみが両方の励起ピークをもつのかを理由を説明するためには、Ser65 (Thr65ではない)を、ヒドロキシルがGlu222への水素結合のドナーとなり、陰イオンとしてGlu222を安定化する構造に選定することができる(この電荷は後に発色団のイオン化を阻害する)。このような構造から、一部の変異が中性と考えられる理由も説明される。例えばGln80は、発色団からかなり離れた表面の残基であり、これは、そのArgに対する偶然的かつ遍在性の変異が、明瞭な分子内における分光光度的作用を一見もたない理由を説明する (M. Chalfieら、Science 263:802〜805 (1994))。
【0162】
赤色シフトした励起極大および発光極大をもつGFP変異体の開発は、タンパク質工学における一つの重要な到達点である(A.B. Cubittら、Trends Biochem. Sci. 20:448〜455 (1995);R. Heimら、Nature 373:664〜665 (1995);S. Delagraveら、Bio/Technology 13:151〜154 (1995))。このような変異体は、二つまたはそれより多い細胞過程の活性を同時に多色的に明らかにする上で、またタンパク質環相互作用のシグナルとしての蛍光共鳴エネルギー移動を用いる際に、細胞による短波長における自己蛍光発生の回避にも有用であると思われる(R. HeimおよびR.Y. Tsien、Current Biol. 6:178〜182 (1996))。ランダム変異導入を用いた多くの試みで、発光極大は最大6 nm長波長側(514 nm)へシフトしており(R. HeimおよびR.Y. Tsien、Current Biol. 6:178〜182 (1996))、過去には「赤色シフト」変異体が、505 nmの発光の何らの有意な赤色化を招くことなく475 nmのピークに有利になるように395 nmの励起ピークを単に抑制したという報告がある(S. Delagraveら、Bio/Technology 13:151〜154 (1995))。Thr203は発色団のフェノール性末端に隣接することがわかっているので、発明者らは、系の分極性を増すことが隣接する発色団の励起状態のエネルギーを低める可能性のあることを期待して、His、Tyr、およびTrpなどの極性芳香族残基になるように変異を導入した。三通りの置換はすべて実際に発光ピークを520 nmを上回るようにシフトさせた(表F)。特に魅力的な変異はT203Y/S65G/V68L/S72Aで、励起ピークと発光ピークはそれぞれ513 nmと527 nmであった。これらの波長は既存のGFP変異体とは十分異なり、蛍光顕微鏡に適切なフィルターセットを使用することで容易に識別することができる。吸光係数は36,500 M-1cm-1で、量子収率は0.63であり、S65Tにおける値と同程度に高い(R. Heimら、Nature 373:664〜665 (1995))。
【0163】
オワンクラゲのGFPと他のタンパク質色素との比較は有益である。しかし残念ながら特性が最も近い相同物であるウミシイタケ(Renilla)のGFP(O. ShimomuraおよびF.H. Johnson、J. Cell. Comp. Physiol. 59:223 (1962);J.G. MorinおよびJ.W. Hastings、J. Cell. Physiol. 77:313 (1971);H. Moriseら、Biochemistry 13:2656 (1974);W.W. Ward、「Photochem. Photobiol. 総説」(Smith、K.C.編) 4:1 (1979);W.W. Ward、「生物発光および化学発光(Bioluminescence and Chemiluminescence)」(M.A. DeLucaおよびW.D. McElroy編) Academic Press pp. 235〜242 (1981);W.W. WardおよびS.H. Bokman Biochemistry 21:4535〜4540 (1982);W.W. Wardら、Photochem. Photobiol. 35:803〜808 (1982))は配列が決定されておらずクローン化もなされていないが、その発色団は、オワンクラゲの野生型GFPと同じFSYG配列に由来する(R.M. San Pietroら、Photochem. Photobiol. 57:63S (1993))。三次元構造が明らかにされている最も近い類似体が、好塩菌に由来する14 kDaの光受容体であり、光反応性をもつ黄色タンパク質(PYP、G.E.O. Borgstahlら、Biochemistry 34:6278〜6287 (1995))である。天然の暗状態でPYPは446 nmに吸収極大があり0.64の量子収率で光を伝達し、475 nm付近の野生型GFPの長波長の吸収極大、および0.72〜0.85の量子収率にマッチする。両タンパク質の基礎となる発色団は、PYPのチオエステル結合、およびGFPのヘテロ環イミノラクタム(iminolactam)を介してタンパク質に共有結合している陰イオン性のp-ヒドロキシシンナミル基である。いずれのタンパク質とも、埋没型の陽イオン性アルギニンおよび中性グルタミン基である、PYP中のArg52およびGlu46、ならびにGFP中のArg96およびGlu222の助けを借りて発色団の負電荷を安定化させるが、PYP中では、これらの残基は、GFPでは発色団のカルボニル末端の方により近いオキシフェニル環に近い。しかしPYPには、細胞の走光性反応の仲介を可能とする適切な柔軟性およびシグナル伝達ドメインを有する重複した全体的なα/βの折りたたみがある、一方GFPは、温度または構造を担う部分としての励起状態エネルギーの寄生的な散逸を最小化する、より正常かつ剛直なβバレルに近い。GFPは、視覚的訴求性および極めて有用な機能、効率のよい蛍光発生が、凝集性および経済的なタンパク質構造から自発的に生じうる優れた例の一つである。
【0164】
A.GFP 構造の決定の概要
データは、Molecular Structure社のR-axis IIまたはSan Diego Multiwire Systems (SDMS)製の検出器(CuKα)のいずれかを用いて、また後にBrookhaven National LaboratoryのビームラインX4Aにより、イメージプレートを用いたセレン吸収端(λ=0.979 Å)で、インハウスで室温において収集した。データの評価はHKLパッケージを用いて行った(Z. Otwinowski、「CCP4 Study Weekend 会報:データの収集と処理(Proceedings of the CCP4 Study Weekend:Data Collection and Processing)」、L. Sawyer、N. Issacs、S. Bailey編、(Science and Engineering Research Council (SERC)、Daresbury Laboratory、Wamngton、英国、(1991))、pp 56〜62;W. Minor、XDISPLAYF (Purdue University、インディアナ州ウェストラフィエット、1993))またはSDMSソフトウェア(A.J. Howardら Meth. Enzymol. 114:452〜471 (1985))。各データセットは1個の結晶から収集した。重原子浸漬は2日間かけて母液中で2 mMとした。最初の電子密度図は、インハウスのデータを用いた三種の重原子誘導体を元にしたものであり、後にシンクロトロンで得られたデータと置き換えた。EMTSの差パターソン図を観察して解読した後に、他の誘導体の差フーリエ図の計算に用いた。重原子パラメータの位相誤差(lack of closure)の精密化は、「Protein」パッケージを用いて行った(W. Steigemann、博士論文(Technical University、Munich、1974))。MIR図は、全体のフィギュアオブメリット(figure of merit)が示唆するよりかなり劣り、またEMTSの同形差が位相決定で優勢であることは明らかであった。シンクロトロンデータに対する異常占有率の上昇は、問題に対する部分的な解答をもたらした。位相決定パワーが、シンクロトロンデータでは低いが、フィギュアオブメリットは変化しないことに注意されたい。すべての実験的電子密度図は、溶媒接触率を38%と仮定する、CCP4(CCP4:「A Suite of Programs for Protein Crystallography」(SERC Daresbury Laboratory、Wamngton WA4 4AD UK、1979))パッケージのプログラムDMを用いた溶媒平滑化(solvent flattening)によって改善された。位相組み合わせは、原子モデルで重量1.0を用いてProteinパッケージのPHASCO2で行った。重原子パラメータは後に、組み合わされた位相に対する精密化によって改善された。モデル構築をFRODOおよびO(T.A. Jonesら、Acta. Crystallogr. Sect. A 47:110 (1991);T.A. Jones、「コンピュータ結晶学(Computational Crystallography)」、D. Sayre編、(Oxford University Press、オックスフォード、1982)、pp. 303〜317)で進め、また結晶学的精密化はTNTパッケージ(D.E. Tronrudら、Acta Cryst. A 43:489〜503 (1987))で行った。発色団の結合の長さと角度は、CHEM3D (Cambridge Scientific Computing)を用いて推定した。最終的な精密化およびモデル構築は、(2Fo-Fc)電子密度図を用いてX4Aセレノメチオニンデータセットに対して行った。1.9 Åの分解能を越えるデータは、この段階で使用しなかった。最終モデルは末端残基として残基2〜229を含み、これは電子密度図では描出されず、また複数のゆがんだ表面残基の側鎖は省略した。密度は残基156〜158については弱く、またこれらの残基の座標は信頼性に欠ける。このような無秩序化は、残基1および233〜238が不必要であるが、さらに切断を進めると蛍光発生が損なわれることを示す過去の分析結果(J. DopfおよびT.M. Horiagon、Gene 173:39〜43 (1996))と一致している。この原子モデルは、タンパク質データバンク(Protein Data Bank)に登録されている(アクセスコード=1EMA)。
【0165】
【表G】
(a)完全性(completeness)は、観察された反射の理論可能値に対する比をパーセントで表した値である。
(b)シェル(Shell)は、最高分解能のシェル(通常は幅が0.1〜0.4 Å)を意味する。
(c)
(d)
(f) 位相決定パワー=<FH>/<E>(<FH>=重原子散乱のr.m.s.、<E>=位相誤差(lack of closure)。
(g) FOM=フィギュアオブメリット(figure of merit)の平均
(h)標準的な結晶学的R因子、
B.S65T と比較したときの Thr 203 (「 T203 」)変異体の S65T スペクトル特性
変異F64L、V68L、およびS72AはGFPの折りたたみを37°改善する(B.P. Cormackら、Gene 173:33 (1996))が、発光スペクトルを有意にシフトしない。
【0167】
【表H】
C.酸性 pH および中性 pH におけるハロゲン化物センサーとしての YFP および YFP-H148Q
YFPの吸収スペクトルはNaCl濃度の関数であり(図7)、発色団の陰イオンであるバンドB(λmax 514 nm)が、塩化物の付加に伴って中性型(λmax 392 nm)に変換される。YFPでは陰イオンのみが蛍光を発するので、蛍光の抑制は[NaCl]の上昇と同時に起こる。YFPでは明瞭な等吸収点が認められる(図7)が、YFP-H148Qでは等吸収点ははっきり決まらない(Jayaramanら、2000)。この作用は完全に可逆的である。YFP-H148QおよびYFP-H148Gでは、バンドAの吸収極大は、塩の添加に伴い20 nm青色側へシフトする(415 nmから395 nmへ)が、バンドBの吸収極大は影響を受けない。詳細な結合平衡および陰イオンの特異性については後述する。
【0169】
ハロゲン化物センサーとしてのYFPの有用性を、特にCFTRのポンプ作用がアッセイされた(Jayaramanら、2000)細胞質(pH 7.4)より酸性であるオルガネラについてさらに明らかにするため、YFPおよびYFP-H148Qの発光強度を一定のイオン強度の条件でpH 6.0〜8.0(図8A、B)において0〜150 mMのNaClで測定した。その結果、酸性条件下でYFPが優れたプローブであることを発明者らは見出している。pH 6.0では、蛍光は0 mM〜20 mMのNaClで39%低下したが、細胞質のpH 7.5では、この低下は同一条件でわずか3.2%に留まった。NaClで滴定したYFP-H148Qでは、蛍光の喪失はpH 6.0でも大きく(48%)、pH 7.5ではかなり有意なまま(11%)であった。中性pHに近い低ミリモル範囲における塩化物濃度の測定では、YFP-H148Qは好ましいプローブであると思われた。ヨウ化物を塩化物に置換すると、YFP-H148Qの蛍光損失は、50%損失が認められるpH 7.5でもかなり大きい(0〜20 mM NaI)(図8B)。以上の結果は、Cl-I-を形質膜CFTRチャネルによって交換する最近の研究で、インビボで利用されている(Jayaramanら、2000)。上記の異型とは対照的に、オリジナルのYFPでは、ヨウ化物の作用規模は塩化物による作用にほぼ匹敵する(下記の結合データを参照)。
【0170】
D.ハロゲン化物結合部位の結晶学的同定および記述
発明者らは、YFP-H148Qの二つの結晶構造を決定した。一つは二か所の結合状態のヨウ化物を含むもの(100 mM ヨウ化物浸漬)で、もう一つは結合状態のハロゲン化物を全く含まないものである (200 mM 塩化物浸漬)。個々のR因子は分解能2.1 Åで18.8%と20.4%であり、幾何的性質は妥当に良好である。関連する結晶学的統計値を表Iにまとめて示す。ヨウ化物は、インハウスのCuKα波長で異常なシグナルを示すので、異常な差図を重原子の位置を同定する目的でヨウ化物浸漬について計算した。その結果、発明者らは、2本の明瞭な電子密度ピークを見出した(それぞれ7.7と5.5 rmsの偏位)。1本は発色団付近に位置してタンパク質の内部に埋没しており、もう1本はバレルのキャップにおけるTrp57付近のタンパク質表面の小さなへこみの内部にある(データは示していない)。
【0171】
100 mMのヨウ化物における結晶内における結合を示す、30 Å2に固定された温度因子で0.60の占有率に精密化された埋没状態のヨウ化物は、結合定数が2.7 mMである溶媒中におけるほど密接していない(下記参照)。ヨウ化物は発色団の複素環のカルボニル酸素から4.3 Å離れて位置し、Arg96との電荷相互作用に関与し、グアニジウム基のNE2までの距離は4.1 Åであり(図9および10)、埋没状態の正電荷は、陰イオン結合エネルギーの大きなフラクションをもたらす可能性が高い。またヨウ化物は、Trp203のフェノール性ヒドロキシルと、Gln69の側鎖のアミド窒素の両方と水素結合を形成しており、水素結合距離はそれぞれ3.3 Åと3.2 Åである(図9および10)。このような距離は、酸素または窒素と相互作用するヨウ化物について予測される水素結合距離の範囲内にある。小分子結晶構造の統計データベース分析から、ヨウ化物とフェノール性ヒドロキシル間の平均距離が3.47 Åであり、ヨウ化物とsp2混成窒素との平均距離が3.66 Åであることがわかっている(Steiner、1998)。ヨウ化物が結合したハロアルカンデハロゲナーゼの結晶構造では、ヨウ化物が2個のトリプトファンのインドール窒素から3.4 Åおよび3.6 Å離れており、溶媒分子から3.3 Å離れており、チロシンのフェノール性酸素から4.2 Å離れていることがわかっている(Verschuerenら、1993)。
【0172】
埋没状態のハロゲン化物も、発色団およびTyr203の芳香環と相互作用する(図9)。陰イオンは、芳香環面またはその付近に優先的に位置することがある。これは、芳香環の水素が部分的な正電荷をもつためである(BurleyおよびPetsko、1988)。YFP-H148Qでは、ヨウ化物は二つのπ系のいずれの平面にも存在しないが、二平面からからほぼ等距離に位置し、スタッキング(stacking)相互作用の中心から相殺され、Tyr203の芳香性CE1から4.1 Åの位置にあり、また発色団の芳香性CD2から4.5 Åの位置にある(図9)。結合部位の反対側では、一連の疎水性残基が、Ile152、Leu201、Val163、Val150、およびPhe165(いずれもヨウ化物とのファンデルワールス接触に近い)を含むハロゲン化物結合部位に並んでいる。芳香環の端における、チロシンおよびトリプトファンとの相互作用、ならびに疎水性側鎖との無極性相互作用は、ハロアルカンデハロゲナーゼでみられるように、タンパク質中の他のハロゲン化物結合部位に一般に見出されている(Pikkemaatら、1999)。
【0173】
第2の表面結合型ヨウ化物は、Trp57のアミド窒素、および規則正しく並んだ複数の溶媒分子と水素結合を形成している。このような曝露状態の陰イオンは、発色団のフェノール性酸素から16 Åの位置にあり、発色団の荷電状態に及ぼす影響は無視できることがわかる。このようなハロゲン化物の占有率は、B因子を39 Å2に固定された状態で0.41で精密化されており、発色団に隣接する一次ヨウ化物と比較して一貫して弱い結合を示す。
【0174】
E .埋没状態のヨウ化物付近における構造変化
発色団付近の陰イオン結合ポケットは、YFP-H148Qのアポ構造では空のようであり、結晶を180 mMの塩化物存在下で成長させた後に200 mM塩化物に浸漬したという事実とは無関係である。塩化物に対する溶液の結合定数が28 mMであることは、結晶母液のpHが4.6ではポケットの大部分が塩化物で占有されていることを予測させる(下記参照)。ヨウ化物浸漬から明らかなように、結晶中の分子は、おそらく結晶の充填力のために、溶液中におけるほど強く陰イオンと結合しないようである。YFP-H148Qの内部空洞の容積は、半径1.2 Åのプローブを用いて55 Å3と算出されている(Connolly、1985)。
【0175】
YFP-H148Qの結合状態の構造中におけるヨウ化物を含む空洞は大きく(91 Å3)、ファンデルワールス容積が42 Å3であるかなり大きなヨウ化物を収容する。ポケットに並ぶさまざまな側鎖の一連の構造変化が認められている(図11)。最も大きな動きはGln69で認められ、側鎖のアミドが空洞の中心から外部に振れており、ハロゲン化物と水素結合を形成するNE2の2.6 Åの動きを生じている(図11および12)。Gln183 NE2は1.0 Å動いているが、これがヨウ化物またはGln94に対する水素結合のドナーか否かは明らかではない(NE2およびOEは反対の位置に割り振ることができる)。Leu201、Ile152、Val150、およびVal163の無極性側鎖(図11)はすべて動いて、ヨウ化物の存在下で空洞の大きさを拡大させており、末端の炭素(ロイシンおよびイソロイシンではCD1、バリンではCG1)は、それぞれ2.4 Å、1.9 Å、1.6 Å、および1.2 Åシフトしている。Phe165の芳香環面は約25°回転している。
【0176】
Tyr203のフェノール性ヒドロキシルは、ヨウ化物を含む空洞方向に0.6 Åシフトしており、ハロゲン化物との水素結合による的相互作用を改善する可能性が高い(図12)。発色団に隣接するTyr203の側鎖の配置には、ある程度の柔軟性があるようである。これはおそらく、GFP S65Tの構造中で当初同定された水で満たされた大きな空洞内に突出しているためである(Ormoら、1996)。YFPおよびその異型の五つの構造のCα-炭素の重ね合わせ(Wachterら、1998)から、Tyr203のCβは良好に重なるが、フェノール性酸素は最大1.4 Å変動することがわかっている。Tyr203とハロゲン化物間の水素結合は、妥当に密接した親和性でハロゲン結合部位の生成に極めて重要な役割を果たしているようである(後述する変異分析を参照)。ハロゲン化物に対する発色団シフトは、芳香環の端と陰イオンとの相互作用の改善にかかわる可能性もある。したがって、Glu222のカルボン酸は発色団の環の窒素から回転しながら離れ(距離は3.3から3.6 Åへ上昇)、Ser205に対する密接な水素結合に関与するようになる(図12)。
【0177】
F .陰イオン結合と空洞サイズとの関係
YFP-H148Qの埋没型ヨウ化物の部位は小さな空洞であり、過去に検討されたいくつかの構造中に存在して、大きさがあまり変わらない(図13)。プローブ半径1.2 Åの球体を用いたファンデルワールス容積の計算(Connolly、1985)では、空洞の容積は野生型GFPで21 Å3であり(Brejcら、1997)、GFP S65Tで19 Å3であり(Ormoら、1996)、YFPおよびYFP-H148Gで16Å3であり(Wachterら、1998)、また500 mM KBr中に浸漬されたYFP-H148G で21Å3であり、結晶学的分析でも結合部位は空であることがわかっている(未発表データ)。これらの空洞の位置は、上述のGFPにおける位置と本質的に同じであり、Val150、Val163、Leu201、Ile152、Gln183、およびGln69は中心に近いが、発色団のメチレンブリッジから約6.6 Å、Arg96から6.1 Å離れている。野生型(下記参照)およびS65T GFP(WachterおよびRemington、1999)はNaClと相互作用しないようである。一方、検討されたすべてのYFPは、陰イオンとの相互作用を示し、YFPでは最も強い結合はCl-で認められた(下記参照)。空洞の大きさと位置が、Cl-結合と直接相関しないことは明らかである。
【0178】
上述の空洞は極めて狭いので、ファンデルワールス容積が24.8〜54 Å3の範囲にある塩化物、臭化物、またはヨウ化物と結合することができない。任意の陰イオンとの相互作用を可能とするためには構造変化が明らかに必要である。YFP-H148Qのアポ構造では、結合状態の陰イオンの非存在下でも空洞はいくぶん広く、その容積は55 Å3である。この異型では、空洞は発色団方向に広がっており、その容積は、結合部位(Gln69、Tyr203、Val150、Val163、Phe165、Arg96、His181)および発色団そのものに並んだ側鎖の原子の小さな動き(0.4 Åおよび1.2 Å)とともに拡大する。これらの残基の多くは、上述したように、また図6に示すように、ヨウ化物が結合するとさらにシフトする。Ile152の末端側鎖の炭素の相殺的な動き(2.1 Å)およびLeu201の末端側鎖の炭素の相殺的な動き(2.3 Å)により、隣接する空洞表面をそれほど変化させることなく、疎水性コアのある程度の再パッキングが生じる。YFP-H148Qの空洞の大きさが広いことは、部分的には、塩化物と比較してヨウ化物の意外なほど強い結合の原因である可能性がある。
【0179】
G . H148Q 置換に応じたβバレルの弛緩とヨウ化物の結合
ハロゲン化物非存在下におけるYFPバックグラウンドでのH148Qの導入と、ヨウ化物とYFP-H148Qとの結合はいずれも、β鎖7(残基143〜154)およびβ鎖8(残基160〜171)の構造的な調節を導く。このような調節は、YFPとYFP-H148Q のCαの重ね合わせから明らかであり、rms偏差は0.42 Åである。またI-を含む場合も含まない場合もYFP-H148Qではrms偏差は0.47 Å(図14)である。バレルの一方のキャップでは、鎖7と8が、残基158の中心にある1本のターンを介して連結されているが、バレルのもう一方のキャップの近傍では、鎖7は、残基148の周囲にβバルジを形成し、主鎖のβシートの相互作用は破壊されている(Ormoら、1996;Wachterら、1998)。それに代わって、整然と並んだ複数の溶媒分子および側鎖の接触(YFPのHis148、YFP-H148QのGln148)が、鎖間に水素結合のネットワークを形成している(図14)。His148をGlnに置換すると、残基166のα炭素はバレルの中心方向に0.94 Å引かれ、残基148のα炭素は0.94 Å押される。このような動きは、隣接するループ領域内における構造的調節によって相殺される(残基172のα炭素の1.4 Åのシフト、および残基157のα炭素の0.94 Åのシフト)。これらの残基は、両構造のいずれにおいても結晶との接触に関与しない。
【0180】
ヨウ化物がYFP-H148Qに結合すると、Lys166がバレルの中心から1.0 Å押し戻され、YFP内の原位置近くに位置するようになる(図14)。同様に、このような位置のシフトは、埋没状態の空洞の拡大に応じて生じる。Lys166は、両構造のいずれにおいても結晶との接触に関与しない。この場合、近傍のArg168は、ヨウ化物の結合に応じてArg149と分子間の塩橋を多少形成する。Gln148の側鎖とLys166の骨格の酸素の間の水素結合は、上記の動きで破壊されていない。相殺性のシフトは鎖の末端でも認められており、骨格のループの残基172と173が最大1.7 Å引かれるが、同領域における密度は詳細に決められていない。ハロゲン化物とYFPの結合が、YFP-H148Qでみられるようにβバルジ領域に類似の作用を及ぼすか否かは不明である。148位で骨格が動くことはYFP-H148Gで既に認められており(Wachterら、1998)、バレルの同部分における柔軟性が上昇することと矛盾がない。
【0181】
H . YFP-H148Q 中の発色団の溶媒接触性
YFP-H148Qの構造から、発色団のヒドロキシルと水素結合が形成されている変異導入前のヒスチジンイミダゾールとは異なり(Wachterら、1998)、Gln148の側鎖がタンパク質の外面に向かって振れていて(図9)、バルク溶媒に対する障壁となっていることがわかる。構造が明らかになる以前から、発明者らは、Gln148が溶媒中に反転する可能性があると予想していた(Elsligerら、1999)。これは、外部溶媒に対する発色団の部分的な曝露が、YFP-H148QのpKaがYFPと比べて高いことを説明する可能性があるからである(表K参照)。アポ構造およびヨウ化物結合型構造のいずれにおいても、Gln148の側鎖のアミド窒素NE2は、Lys166の骨格のカルボニル酸素、およびAsn149の骨格の窒素に対するアミド酸素OE1と水素結合を形成しており(図12および14)、発色団から十分離れている。MidasPlus(登録商標)で実行されるように、プローブ球半径1.4 Åを用いた溶媒接触面の計算(Connolly、1983)から、His148の野生型のイミダゾールが位置する、タンパク質の表面に浅いくぼみが形成されることがわかる(図13)。このような溶媒ポケットは、発色団のファンデルワールス表面とほぼ接触している。タンパク質のゆらぎ運動を考慮する際は、結晶構造中で観察されないある程度の溶媒接触が生じる可能性が高い。溶媒チャネルが発色団の空洞と直接接触するYFP-H148G (Wachterら、1998)と比べて、YFP-H148Qのチャネルは欠けており、発色団は水相消光剤に曝露されていないことを示唆した三重項状態の光退色実験(Jayaramanら、2000)と一致している。
【0182】
I .陰イオンとプロトン結合間のエネルギー分析
発色団のpKaが特定の陰イオン結合に強く依存することは、二つの異なるリガンド間における相互作用の存在を伺わせる関連した結合平衡、陰イオンがArg96と隣接して結合すること、およびプロトンが発色団のフェノール性末端と結合することから説明可能である。陰イオンの結合がプロトンの結合を促して発色団のpKaを上昇させるという事実から、正の協同作用が存在することがわかる。したがって陰イオン結合に対する結合定数はプロトン結合の量の影響を受ける。これは逆もあてはまる。したがって、一つの結合部位が二種の異なるリガンドに対応するという単純な系では、二つの微視的な結合接触を決定することができる。一つは、プロトンが存在する状態における陰イオン結合のk1であり、もう一つは、プロトンが存在しない状態における陰イオン結合のk2である。発明者らの実施した、YFP-H148Qを対象とした結晶学的解析の結果は、陰イオンに対する一つの重要な結合部位が存在することと矛盾がなく、またS65Tを対象に実施された結晶学的解析の結果は、発色団上に一か所のプロトン結合部位があることと矛盾しない(Elsligerら、1999)。認められた陰イオン結合度はpHの関数であるので、巨視的な結合定数はk1とk2の制限値の間にある。
【0183】
数学的な記述はワイマン(J. Wyman)(1964)によってなされており、カンター(Cantor)およびシメール(Schimmel)による「生物物理学化学(Biophysical Chemistry)、第III部」、(CantorおよびSchimmel、1980)に記載されている。本明細書では、発明者らは、各リガンドに対して一つの結合部位を有する特別の場合に一般的な式15-79を用いる(pKa °は、結合性陰イオンの非存在下における発色団のpKaを示す):
pKa=log{(k1+[塩化物])/k1}−log{(k2+[塩化物])/k2}+pKa °
式(1)
【0184】
pH 6.5およびpH 7.0における吸光度測定により、YFPのpKaを、多数の異なるイオンに対する陰イオン濃度の関数として決定した。対象となる特定の陰イオンの濃度は0 mMから少なくとも150 mM(ハロゲン化物では最高で400 mM、図15)の間を変動し、イオン強度はYFP群と相互作用しないグルコン酸カリウムを添加して制御した(WachterおよびRemington、1999)。相互作用性陰イオンに関する結果は式(1)に合致し、可能な微視的結合定数k1およびk2をカーブフィッティングから求めた(図15および表J)。一般に、小さな一価の陰イオンはYFPとある程度の相互作用を示すようである。結合はフッ化物で最も強くなり、k1は0.214 mMである。他のハロゲン化物を含む、相互作用することがわかっている他の陰イオンは、低ミリモル範囲の微視的結合定数を有し、中でもトリフルオロ酢酸(TFA)が最も弱い相互作用を示す(k1=21.2 mM)。
【0185】
このような相互作用に対する何ら特定の分子形状依存性は存在しないようである。というのは、三原子の直鎖状(例えばチオシアナート)、方形平面型分子(例えば過塩素酸塩)、三方晶(例えば硝酸塩)、および球状(例えばハロゲン化物)の各分子も結合することがわかっている。ギ酸塩も発色団のpKaを調節した(k1=7.47 mM)が、過去に実施された、蛍光によるいくぶん予備的な実験では、相互作用が存在しないことが報告されている(WachterおよびRemington、1999)。陰イオン性発色団と陰イオンの結合は予想通り望ましくなく、高ミリモルまたはモル範囲におけるk2は測定範囲外のことが多い(表J)。
【0186】
【表J】
YFP発色団に対する陰イオンの結合(相互作用の強度が低下する順序に記載)a
b 括弧内の数字は、Kaleidagraph(商標)で決定された標準偏差の低い方の推定値。
c 測定範囲外(おそらくモル範囲)にあったために決定できなかった結合定数。
【0187】
リン酸や硫酸などの二価の陰イオン、およびグルコン酸、グッド緩衝液(Good buffer)(例えば、HEPES、PIPES)、イセチオン酸(2-ヒドロキシエタンスルホン酸)、およびTCA (トリクロロ酢酸)などの大きな一価の陰イオンは相互作用しない(表J)。これはYFPの定数pKaが約5.4であり、測定が塩を添加することなく低イオン強度の緩衝液中で行われた場合と本質的に同じであることからわかる(WachterおよびRemington、1999)。相互作用しない、ある程度小さな一価の陰イオンには、リン酸、重炭酸、および酢酸などがある。水和エネルギーは、陰イオンを識別する際に重要な役割を果たす可能性がある。これは酢酸は強く溶媒和されるが、TCAは水性溶媒中で弱くしか水和されないことに基づく(March、1992)。表JでYFPについて示した一連の結果は、pH 7.5における蛍光を元にYFP-H148Qについて決定された結果(Jayaramanら、2000)と極めて似ており、順序にわずかな差があるだけである。例えば、YFP-H148QはCl-よりBr-に強く結合するが、YFPではこの順序は逆となる。これはYFP-H148Qの結合部位が大きいためである可能性が高い。
【0188】
J .陰イオン結合に重要な役割を果たす残基の変異分析による同定
YFPのどの置換(S65G/V68L/S72A/T203Y)が、発色団付近における特定の陰イオン結合に対する寄与因子であるかを同定するために、発明者らは変異分析を行って四種の置換を一つずつ野生型に戻した。次に発明者らは、これら復帰体のpKaを、相互作用性陰イオンの非存在下で決定し、ハロゲン化物濃度の関数としてのpKaを決定して塩化物およびヨウ化物に対する親和性を測定した後に、式(1)に対してカーブフィッティングを行った。復帰体1 (S65G/S72A/T203Y)および復帰体2 (S65G/V68L/S72A)は、YFPで認められるような良好に挙動するpHおよびハロゲン滴定挙動を示し、また塩化物およびヨウ化物の結合に関するpKaおよびk1を、表Kに記載されたYFPについて得られた値と比較した。残基68または残基203の復帰は、発色団のpKaをそれぞれ5.8と6.4に上昇させる。残基68の復帰では、塩化物に対する親和性がわずかに低下する(k1=13.2 mM、YFPでは4.69 mM)が、残基203の復帰では、相互作用が極めて大きく減じる(k1=153 mM)。表Kから明らかなように、塩化物に対する親和性は発色団のpKaと強く結びついており、pKaが上昇すると陰イオンとの相互作用は弱くなる。
【0189】
【表K】
YFPおよびその復帰体における塩化物およびヨウ化物との結合の微視的解離定数
b 塩化物 (HEPESまたはPEPES、150 mM グルコン酸のいずれかで緩衝)など任意の干渉性陰イオンの非存在下における吸光度から決定された発色団のpKa。
【0190】
上記規則の唯一の例外は復帰体2なので、T203Yが存在する場合にのみ相関は完全であると思われる。このような置換は、強い陰イオン性相互作用に不可欠であると思われる。
【0191】
ヨウ化物の結合は、検討されたすべての異型について塩化物の結合よりかなり強いようである(表K)。発色団のpKaとの相関は最高でも弱い。塩化物に対するヨウ化物の相対的な選択性はYFP-H148Qで最も強く、次にYFP-H148Gで強い。これは、ヨウ化物が塩化物より大きくて柔らかいイオンであり、特定の異型でβバレル構造の弛緩を可能としない限り、小さな空洞に適合させることが難しいという事実を反映している可能性がある(上記参照)。
【0192】
復帰体3 (S72A/T203Y)と復帰体4 (T203Y)は分析が困難であった。これは、滴定挙動が野生型GFPと似ているためである。これらの吸収スペクトルはバンドAおよびBの混合型の基底状態を示しており、pH 6.5以上ではpHにほとんど依存しない。バンドAまたはバンドBのいずれかが励起すると、上記復帰体で緑色の蛍光が発する。これは、野生型GFPについて報告されている励起状態の脱プロトンと似ている(Chattorajら、1996)。pH 6.5で、250 mMのNaClを復帰体3および復帰体4に添加すると二つの吸収バンドの比がわずかながら変化し、バンドBはバンドAに対して約20%低下する。pH 6.5における野生型GFPでは、一定のイオン強度の条件下で250 mMのNaClを添加してもスペクトルに変化は認められない。これは、イオン強度に対する感度(Wardら、1982)と矛盾しないが、特定の陰イオン結合とは一致しない。
【0193】
本発明は、新しい長波長の改変蛍光タンパク質を提供する。特定の実施例を提供したが、上記の記述は例示的なものであり、限定されるものではない。本発明の多くの変形例は、本明細書を検討することで当業者に明らかになる。したがって本発明の範囲は、上記の記述に関して決定されるのではなく、むしろ同等物の範囲とともに添付の特許請求の範囲に関して決定されるべきである。
【0194】
出版物
【図面の簡単な説明】
【図1】 (A)モルスクリプト(Molscript)(J.P. Kraulis、J. Appl. Cryst.、24:946 (1991))で作製されたGFP骨格の略図。発色団は球棒モデルで示されている。(B)GFP全体の折りたたみの略図。二次構造配列の開始端と終端の位置を示す残基番号を示す。
【図2】 (A)発色団と、すぐ近傍の残基の立体図。炭素原子を白い円で示し、酸素を黒い円で示し、窒素を斜線を付けた円で示す。溶媒分子は別に黒い円で示す。(B)発色団周囲の電子密度を示す、1.0σで描かれた最終的な2Fo-Fcの電子密度図の一部。(C)発色団の配位の第1および第2の球を示す略図。水素結合を点線で示し、長さをオングストローム(Å)で示す。挿入図は、発色団形成時に形成されると考えられているカルビノールアミン(carbinolamine)中間体について提案されている構造を示す。
【図3】 オワンクラゲの緑色蛍光タンパク質のヌクレオチド配列(配列番号:1)、および推定アミノ酸配列(配列番号:2)を示す。
【図4】 哺乳類で好ましいコドン、および最適なコザック(Kozak)配列を用いた、改変オワンクラゲ関連蛍光タンパク質S65G/S72A/T203Yのヌクレオチド配列(配列番号:3)、および推定アミノ酸配列(配列番号:4)を示す。
【図5】 オワンクラゲ関連緑色蛍光タンパク質S65T結晶構造の座標を示す。
【図6】 改変蛍光タンパク質20Aおよび10C(表F)の蛍光の励起スペクトルおよび発光スペクトルを示す。528 nmのところにある縦線は10Cの発光極大(線の左側)と20Aの発光極大(線の右側)を対比させている。
【図7】 20 mM MESで緩衝作用をもたせた、さまざまなNaCl濃度(pHは6.4で固定)におけるYFPの吸収スキャンの結果を示す(-○- 0 mM NaCl、-▽- 15 mM NaCl、-□- 50 mM NaCl、-◇- 100 mM NaCl、および-△- 400 mM NaCl)。バンドAは発色団の中性状態に対応し(λmax=392 nm)、バンドBは発色団の陰イオン(λmax=514 nm)に対応する。
【図8】 一定のイオン強度(150 mM)におけるpHおよび[Cl-]の関数としての、(a)YFP、および(b)YFP-H148Qの規格化された蛍光発光を示す。pHは、20 mM TAPS pH 8.0(○)、20 mM HEPES pH 7.5(△)、20 mM PIPES pH 7.0(◇)、ならびに20 mM MES pH 6.5(▽)およびpH 6.0(□)で調節した。図8(b)は、pH 7.5(*)における[I-]の関数としてのYFP-H148Qの蛍光発光も含む。塩化カリウム(またはヨウ化物)を記載濃度で添加し、イオン強度はグルコン酸カリウムで150 mMに調節した。約0.01 mg/mlのタンパク質を含む試料は、514 nmで励起が認められ、発光強度は528 nmで決定された。
【図9】 YFP-H148Q発色団のTyr203、Arg96、Gln69、および埋没状態のヨウ化物の、精密化後における2Fo-Fc電子密度図の立体図。2.1 Å分解能マップは+1の標準偏差で描いた。この図はプログラムBOBSCRIPTで作製した。
【図10】 YFP-H148Q (ヨウ化物浸漬)の結晶構造において、埋没状態のヨウ化物の5 Å以内にある原子を含む全ての残基を示す略図。
【図11】 ヨウ化物のある場合とない場合における、YFP-H148Qの陰イオン結合空洞(binding cavity)に並んだ一群の残基を重ね合わせた立体図(ヨウ化物が結合したときの構造=灰、アポ構造=黒)。ヨウ化物を中心にある球で示す。この図はプログラムMOLSCRIPTで描いた(Kraulis、1991)。
【図12】 (a)アポ構造、および(b)ヨウ化物が結合した状態の構造における、YFP-H148Qの発色団のすぐ近傍の環境を示す略図。
【図13】 ヨウ化物が結合した状態のYFP-H148Q構造の溶媒接触可能表面の立体図(1.4 Å プローブ半径で計算)。表面は、すべての水分子およびヨウ化物を除いた後に計算した。発色団、および発色団に接触する全ての表面セグメントも示す。タンパク質の外側表面は図の左側に沿っている。この図はプログラムMidasPlus(商標)(UCSF、1994)で作製した。
【図14】 YFPのβ鎖7および8の骨格原子トレース、YFP-148Qのアポ構造、およびヨウ化物が結合した状態のYFP-H148Qの構造を示す。His148 (YFP)およびGln148 (YFP-H148Q)の側鎖、および数個の水分子も示す。点線は形成可能な水素結合を示す。
【図15】 ハロゲン化物濃度の関数としての、YFP発色団のpKaを示す(-◇- フッ化物、-▽- ヨウ化物、-△- 塩化物、-○- 臭化物)。発色団のpKaは、さまざまなハロゲン化物濃度における吸収スキャンから推定した(「材料および方法」を参照)。このデータは方程式1に従ってカーブフィッティングを行った(本文参照)。
Claims (19)
- アミノ酸配列がオワンクラゲの緑色蛍光タンパク質のアミノ酸配列(配列番号:2)と実質的に同一で、
i) T203位における少なくとも一つの第1の置換であって、T203Yである置換、
ii)H148位における少なくとも一つの第2の置換、および
iii)次から選択される少なくとも一つの第3の置換:V150T若しくはA、V163S、T、若しくはA、I152L、またはF165Y
において配列番号:2と異なる、オワンクラゲの緑色蛍光タンパク質とは異なった蛍光特性を有し、ハロゲン化物の蛍光測定を可能にする、機能性の改変蛍光タンパク質をコードするヌクレオチド配列を含む核酸分子。 - H148位における少なくとも一つの第2の置換がH148R、H148G、H148Q、H148A、H148N、およびH148Kからなる群より選択される、請求項1記載の核酸分子。
- V150位における少なくとも一つの第3の置換がV150Tである、請求項1記載の核酸分子。
- V150位における少なくとも一つの第3の置換がV150Aである、請求項1記載の核酸分子。
- V163位における少なくとも一つの第3の置換がV163Sである、請求項1記載の核酸分子。
- V163位における少なくとも一つの第3の置換がV163Tである、請求項1記載の核酸分子。
- V163位における少なくとも一つの第3の置換がV163Aである、請求項1記載の核酸分子。
- 以下の段階を含む、試料中の対象陰イオンの存在を判定する方法:
1) オワンクラゲの緑色蛍光タンパク質のアミノ酸配列(配列番号:2)と実質的に同一で、
少なくとも一つの第1の置換T203Y、
H148位における少なくとも一つの第2の置換、および、
次から選択される少なくとも一つの第3の置換:V150T若しくはA、V163S、T、若しくはA、I152L、または、F165Y、
において配列番号:2と異なるアミノ酸配列を含む、改変緑色蛍光タンパク質であって、オワンクラゲの緑色蛍光タンパク質とは異なった蛍光特性を有し、ハロゲン化物の蛍光測定を可能にする改変緑色蛍光タンパク質を試料に導入する段階;および、
2) 試料中の改変緑色蛍光タンパク質の蛍光を判定する段階。 - 試料中の改変緑色蛍光タンパク質の蛍光を、対象陰イオンを既知濃度で含む対照試料に導入された対照となる改変緑色蛍光タンパク質の蛍光と比較する段階をさらに含む、請求項8記載の方法。
- 試料が少なくとも1個の生細胞を含む、請求項8記載の方法。
- 対象陰イオンがハロゲン化物である、請求項8記載の方法。
- 改変緑色蛍光タンパク質が請求項1ないし3のいずれか一項に記載の核酸分子によりコードされる、請求項8記載の方法。
- 請求項1ないし7のいずれか一項に記載の核酸分子によりコードされる、機能性の改変蛍光タンパク質。
- 請求項13記載の機能性の改変蛍光タンパク質を含む宿主細胞。
- 以下の段階を含む、被験化合物がイオンチャネル活性に及ぼす作用をスクリーニングする方法:
i) a)オワンクラゲの緑色蛍光タンパク質のアミノ酸配列(配列番号:2)と実質的に同一で、
少なくとも一つの第1の置換T203Y、
H148位における少なくとも一つの第2の置換、および、
以下より選択される少なくとも一つの第3の置換:V150T若しくはA、V163S、T、若しくはA、I152L、または、F165Y、
において配列番号:2と異なるアミノ酸配列を含む、改変緑色蛍光タンパク質であって、オワンクラゲの緑色蛍光タンパク質とは異なった蛍光特性を有し、ハロゲン化物の蛍光測定を可能にする改変緑色蛍光タンパク質、および、
b)対象イオンチャネル
を含む細胞を提供する段階;
ii) 細胞に被験化合物を接触させる段階;
iii)改変緑色蛍光タンパク質に由来する蛍光を決定する段階。 - 対象イオンチャネルの既知の活性化因子を細胞に接触させる段階をさらに含む、請求項15記載の方法。
- 細胞中の改変緑色蛍光タンパク質の蛍光を、対照細胞に導入された対照となる改変緑色蛍光タンパク質の蛍光と比較する段階をさらに含む、請求項15記載の方法。
- 対照イオンチャネルがハロゲン化物を輸送する、請求項15記載の方法。
- 改変緑色蛍光タンパク質が、請求項13記載の改変蛍光タンパク質である、請求項15記載の方法。
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US09/575,847 US6593135B2 (en) | 1996-08-16 | 2000-05-19 | Long wavelength engineered fluorescent proteins |
| PCT/US2001/016149 WO2001090147A2 (en) | 2000-05-19 | 2001-05-17 | Long wavelength engineered fluorescent proteins |
Publications (3)
| Publication Number | Publication Date |
|---|---|
| JP2004502410A JP2004502410A (ja) | 2004-01-29 |
| JP2004502410A5 JP2004502410A5 (ja) | 2009-08-13 |
| JP4427222B2 true JP4427222B2 (ja) | 2010-03-03 |
Family
ID=24301941
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2001586334A Expired - Lifetime JP4427222B2 (ja) | 2000-05-19 | 2001-05-17 | 長波長をもつ改変蛍光タンパク質 |
Country Status (10)
| Country | Link |
|---|---|
| US (4) | US6593135B2 (ja) |
| EP (1) | EP1285065B1 (ja) |
| JP (1) | JP4427222B2 (ja) |
| AT (1) | ATE309356T1 (ja) |
| AU (1) | AU2001263269A1 (ja) |
| CA (1) | CA2408302C (ja) |
| DE (1) | DE60114804T2 (ja) |
| DK (1) | DK1285065T3 (ja) |
| ES (1) | ES2253384T3 (ja) |
| WO (1) | WO2001090147A2 (ja) |
Families Citing this family (15)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US6803188B1 (en) * | 1996-01-31 | 2004-10-12 | The Regents Of The University Of California | Tandem fluorescent protein constructs |
| US6124128A (en) | 1996-08-16 | 2000-09-26 | The Regents Of The University Of California | Long wavelength engineered fluorescent proteins |
| US6593135B2 (en) * | 1996-08-16 | 2003-07-15 | The State Of Oregon, Acting By And Through The State Board Of Higher Education On Behalf Of The University Of Oregon | Long wavelength engineered fluorescent proteins |
| JP3829252B2 (ja) * | 2001-06-08 | 2006-10-04 | 独立行政法人理化学研究所 | 蛍光蛋白質 |
| DE03779067T1 (de) * | 2002-11-12 | 2006-06-22 | Zakrytoe Aktsionernoe Obschestvo "Evrogen" | Fluoreszenzproteine und chromoproteine aus nicht-aequorea-hydrozoa-spezies sowie verfahren zur verwendung davon |
| RU2412250C2 (ru) | 2005-11-04 | 2011-02-20 | Закрытое акционерное общество "ЕВРОГЕН" | Модифицированные зеленые флуоресцентные белки и способы их использования |
| WO2007086473A1 (ja) * | 2006-01-26 | 2007-08-02 | Nec Soft, Ltd. | 蛍光タンパク質の蛍光波長を変える方法 |
| KR100742217B1 (ko) | 2006-03-13 | 2007-07-24 | 학교법인 포항공과대학교 | 피에이치에 의한 에프알이티 효율조절을 통한항생펩타이드의 활성도 측정방법 |
| US20120122115A1 (en) * | 2006-04-25 | 2012-05-17 | Sayre Richard T | Bacterial quorum sensing biosensor |
| WO2009131957A2 (en) | 2008-04-21 | 2009-10-29 | Institute For Oneworld Health | Compounds, compositions and methods comprising oxadiazole derivatives |
| US8236838B2 (en) | 2008-04-21 | 2012-08-07 | Institute For Oneworld Health | Compounds, compositions and methods comprising isoxazole derivatives |
| SG172321A1 (en) | 2009-01-29 | 2011-07-28 | Commw Scient Ind Res Org | Measuring g protein coupled receptor activation |
| US8343976B2 (en) | 2009-04-20 | 2013-01-01 | Institute For Oneworld Health | Compounds, compositions and methods comprising pyrazole derivatives |
| WO2014020933A1 (ja) * | 2012-07-31 | 2014-02-06 | アルバイオ株式会社 | 新規蛍光物質 |
| US11341384B2 (en) * | 2014-08-28 | 2022-05-24 | Banctec, Incorporated | Document processing system and method for associating metadata with a physical document while maintaining the integrity of its content |
Family Cites Families (34)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US640374A (en) * | 1899-03-02 | 1900-01-02 | John C Fleming | Reflector for electric or other lights. |
| US5085985A (en) * | 1987-11-12 | 1992-02-04 | Becton Dickinson & Co. | Monoclonal antibodies and their use in a method for monitoring subsets of activated T cells |
| GB8916806D0 (en) | 1989-07-22 | 1989-09-06 | Univ Wales Medicine | Modified proteins |
| US5491084A (en) | 1993-09-10 | 1996-02-13 | The Trustees Of Columbia University In The City Of New York | Uses of green-fluorescent protein |
| ATE400651T1 (de) | 1993-09-10 | 2008-07-15 | Univ Columbia | Verwendung von grünem fluoreszenzprotein |
| WO1995021191A1 (en) | 1994-02-04 | 1995-08-10 | William Ward | Bioluminescent indicator based upon the expression of a gene for a modified green-fluorescent protein |
| US5795737A (en) | 1994-09-19 | 1998-08-18 | The General Hospital Corporation | High level expression of proteins |
| US5625048A (en) | 1994-11-10 | 1997-04-29 | The Regents Of The University Of California | Modified green fluorescent proteins |
| US5777079A (en) | 1994-11-10 | 1998-07-07 | The Regents Of The University Of California | Modified green fluorescent proteins |
| ATE211506T1 (de) | 1995-01-31 | 2002-01-15 | Bioimage As | Ein verfahren zum nachweis biologisch aktiver substanzen |
| WO1996027027A1 (en) | 1995-03-02 | 1996-09-06 | Rutgers The State University Of New Jersey | Improved method for purifying green fluorescent protein |
| GB9504446D0 (en) | 1995-03-06 | 1995-04-26 | Medical Res Council | Improvements in or relating to gene expression |
| US5741657A (en) * | 1995-03-20 | 1998-04-21 | The Regents Of The University Of California | Fluorogenic substrates for β-lactamase and methods of use |
| US6342379B1 (en) | 1995-06-07 | 2002-01-29 | The Regents Of The University Of California | Detection of transmembrane potentials by optical methods |
| US5661035A (en) * | 1995-06-07 | 1997-08-26 | The Regents Of The University Of California | Voltage sensing by fluorescence resonance energy transfer |
| DE69604298T2 (de) | 1995-09-22 | 2000-05-18 | Bioimage A/S, Soeborg | Varianten des grünen fluoreszenzproteins, gfp |
| US5874304A (en) | 1996-01-18 | 1999-02-23 | University Of Florida Research Foundation, Inc. | Humanized green fluorescent protein genes and methods |
| US6803188B1 (en) | 1996-01-31 | 2004-10-12 | The Regents Of The University Of California | Tandem fluorescent protein constructs |
| US5804387A (en) | 1996-02-01 | 1998-09-08 | The Board Of Trustees Of The Leland Stanford Junior University | FACS-optimized mutants of the green fluorescent protein (GFP) |
| US6027881A (en) | 1996-05-08 | 2000-02-22 | The United States Of America As Represented By The Secretary Of The Department Of Health And Human Services | Mutant Aequorea victoria fluorescent proteins having increased cellular fluorescence |
| US5912137A (en) * | 1996-07-16 | 1999-06-15 | The Regents Of The University Of California | Assays for protein kinases using fluorescent |
| US5925558A (en) * | 1996-07-16 | 1999-07-20 | The Regents Of The University Of California | Assays for protein kinases using fluorescent protein substrates |
| US6124128A (en) | 1996-08-16 | 2000-09-26 | The Regents Of The University Of California | Long wavelength engineered fluorescent proteins |
| US6593135B2 (en) * | 1996-08-16 | 2003-07-15 | The State Of Oregon, Acting By And Through The State Board Of Higher Education On Behalf Of The University Of Oregon | Long wavelength engineered fluorescent proteins |
| US5998204A (en) | 1997-03-14 | 1999-12-07 | The Regents Of The University Of California | Fluorescent protein sensors for detection of analytes |
| US6197928B1 (en) | 1997-03-14 | 2001-03-06 | The Regents Of The University Of California | Fluorescent protein sensors for detection of analytes |
| US6046925A (en) | 1997-04-14 | 2000-04-04 | The Regents Of The University Of California | Photochromic fluorescent proteins and optical memory storage devices based on fluorescent proteins |
| US6008378A (en) | 1997-10-21 | 1999-12-28 | The Regents Of The University Of California | Synthetic molecules that specifically react with target sequences |
| US6150176A (en) * | 1998-06-09 | 2000-11-21 | The Regents Of The University Of California | Fluorescent protein sensors for measuring the pH of a biological sample |
| US6140132A (en) | 1998-06-09 | 2000-10-31 | The Regents Of The University Of California | Fluorescent protein sensors for measuring the pH of a biological sample |
| US6469154B1 (en) | 1999-05-21 | 2002-10-22 | The Regents Of The University Of California | Fluorescent protein indicators |
| US6699687B1 (en) | 1999-05-21 | 2004-03-02 | The Regents Of The University Of California | Circularly permuted fluorescent protein indicators |
| US7060793B2 (en) | 1999-05-21 | 2006-06-13 | The Regents Of The University Of California | Circularly permuted fluorescent protein indicators |
| US7015310B2 (en) * | 2001-03-12 | 2006-03-21 | The State Of Oregon Acting By And Through The State Board Of Higher Education On Behalf Of The University Of Oregon | Oxidation reduction sensitive green fluorescent protein variants |
-
2000
- 2000-05-19 US US09/575,847 patent/US6593135B2/en not_active Expired - Lifetime
-
2001
- 2001-05-17 DE DE60114804T patent/DE60114804T2/de not_active Expired - Lifetime
- 2001-05-17 WO PCT/US2001/016149 patent/WO2001090147A2/en active IP Right Grant
- 2001-05-17 DK DK01937550T patent/DK1285065T3/da active
- 2001-05-17 ES ES01937550T patent/ES2253384T3/es not_active Expired - Lifetime
- 2001-05-17 EP EP01937550A patent/EP1285065B1/en not_active Expired - Lifetime
- 2001-05-17 CA CA2408302A patent/CA2408302C/en not_active Expired - Lifetime
- 2001-05-17 JP JP2001586334A patent/JP4427222B2/ja not_active Expired - Lifetime
- 2001-05-17 AT AT01937550T patent/ATE309356T1/de not_active IP Right Cessation
- 2001-05-17 AU AU2001263269A patent/AU2001263269A1/en not_active Abandoned
-
2003
- 2003-07-14 US US10/620,099 patent/US7544776B2/en not_active Expired - Fee Related
-
2009
- 2009-04-29 US US12/432,576 patent/US20100145024A1/en not_active Abandoned
-
2011
- 2011-01-21 US US13/011,432 patent/US8263412B2/en not_active Expired - Fee Related
Also Published As
| Publication number | Publication date |
|---|---|
| CA2408302A1 (en) | 2001-11-29 |
| US20110275107A1 (en) | 2011-11-10 |
| ATE309356T1 (de) | 2005-11-15 |
| US7544776B2 (en) | 2009-06-09 |
| US20040014128A1 (en) | 2004-01-22 |
| CA2408302C (en) | 2013-07-02 |
| AU2001263269A1 (en) | 2001-12-03 |
| DE60114804T2 (de) | 2006-08-10 |
| JP2004502410A (ja) | 2004-01-29 |
| US6593135B2 (en) | 2003-07-15 |
| ES2253384T3 (es) | 2006-06-01 |
| DK1285065T3 (da) | 2006-03-27 |
| EP1285065B1 (en) | 2005-11-09 |
| DE60114804D1 (de) | 2005-12-15 |
| US20030013149A1 (en) | 2003-01-16 |
| EP1285065A2 (en) | 2003-02-26 |
| US8263412B2 (en) | 2012-09-11 |
| US20100145024A1 (en) | 2010-06-10 |
| WO2001090147A3 (en) | 2002-05-02 |
| WO2001090147A2 (en) | 2001-11-29 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US8263412B2 (en) | Long wavelength engineered fluorescent proteins | |
| JP4232992B2 (ja) | 長波長の工学的に作製された蛍光タンパク質 | |
| US6608189B1 (en) | Fluorescent protein sensors for measuring the pH of a biological sample | |
| US6469154B1 (en) | Fluorescent protein indicators | |
| US20030212265A1 (en) | Fluorescent protein sensors for measuring the pH of a biological sample | |
| Zimmer | Green fluorescent protein (GFP): applications, structure, and related photophysical behavior | |
| JP4215512B2 (ja) | 非オリゴマー化タンデム蛍光性タンパク質 | |
| WO2000071565A9 (en) | Fluorescent protein indicators | |
| US7060793B2 (en) | Circularly permuted fluorescent protein indicators | |
| US6699687B1 (en) | Circularly permuted fluorescent protein indicators | |
| AU767375B2 (en) | Long wavelength engineered fluorescent proteins | |
| MXPA98002972A (en) | Long wavelength engineered fluorescent proteins | |
| AU2004200425A8 (en) | Long wavelength engineered fluorescent proteins |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20060308 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20081218 |
|
| RD04 | Notification of resignation of power of attorney |
Free format text: JAPANESE INTERMEDIATE CODE: A7424 Effective date: 20090122 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20090310 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20090317 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20090616 |
|
| A524 | Written submission of copy of amendment under article 19 pct |
Free format text: JAPANESE INTERMEDIATE CODE: A524 Effective date: 20090616 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20091118 |
|
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20091214 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 4427222 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20121218 Year of fee payment: 3 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20121218 Year of fee payment: 3 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20131218 Year of fee payment: 4 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20131218 Year of fee payment: 4 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20131218 Year of fee payment: 4 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20131218 Year of fee payment: 4 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| EXPY | Cancellation because of completion of term |