JP2023543803A - プライム編集ガイドrna、その組成物、及びその使用方法 - Google Patents
プライム編集ガイドrna、その組成物、及びその使用方法 Download PDFInfo
- Publication number
- JP2023543803A JP2023543803A JP2023519084A JP2023519084A JP2023543803A JP 2023543803 A JP2023543803 A JP 2023543803A JP 2023519084 A JP2023519084 A JP 2023519084A JP 2023519084 A JP2023519084 A JP 2023519084A JP 2023543803 A JP2023543803 A JP 2023543803A
- Authority
- JP
- Japan
- Prior art keywords
- pegrna
- nucleotides
- napdnabp
- editing
- nucleic acid
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
- 108020005004 Guide RNA Proteins 0.000 title claims description 215
- 239000000203 mixture Substances 0.000 title claims description 19
- 125000003729 nucleotide group Chemical group 0.000 claims abstract description 384
- 239000002773 nucleotide Substances 0.000 claims abstract description 381
- 238000000034 method Methods 0.000 claims abstract description 172
- 108091028043 Nucleic acid sequence Proteins 0.000 claims abstract description 58
- 108091023037 Aptamer Proteins 0.000 claims abstract description 42
- 230000004048 modification Effects 0.000 claims abstract description 34
- 238000012986 modification Methods 0.000 claims abstract description 34
- 108020004566 Transfer RNA Proteins 0.000 claims abstract description 22
- 239000013604 expression vector Substances 0.000 claims abstract description 18
- 108020004422 Riboswitch Proteins 0.000 claims abstract description 12
- 239000008194 pharmaceutical composition Substances 0.000 claims abstract 3
- 108020004414 DNA Proteins 0.000 claims description 425
- 108091033409 CRISPR Proteins 0.000 claims description 278
- 108010092799 RNA-directed DNA polymerase Proteins 0.000 claims description 267
- 102100034343 Integrase Human genes 0.000 claims description 238
- 102000053602 DNA Human genes 0.000 claims description 156
- 230000027455 binding Effects 0.000 claims description 149
- 150000007523 nucleic acids Chemical class 0.000 claims description 137
- 102000039446 nucleic acids Human genes 0.000 claims description 126
- 108020004707 nucleic acids Proteins 0.000 claims description 126
- 230000006820 DNA synthesis Effects 0.000 claims description 118
- 108091032973 (ribonucleotides)n+m Proteins 0.000 claims description 115
- 230000000694 effects Effects 0.000 claims description 97
- 102000007260 Deoxyribonuclease I Human genes 0.000 claims description 95
- 108010008532 Deoxyribonuclease I Proteins 0.000 claims description 95
- 230000035772 mutation Effects 0.000 claims description 77
- 125000006850 spacer group Chemical group 0.000 claims description 76
- 230000000295 complement effect Effects 0.000 claims description 75
- 108010014303 DNA-directed DNA polymerase Proteins 0.000 claims description 59
- 102000016928 DNA-directed DNA polymerase Human genes 0.000 claims description 59
- 102000037865 fusion proteins Human genes 0.000 claims description 43
- 108020001507 fusion proteins Proteins 0.000 claims description 43
- 230000003993 interaction Effects 0.000 claims description 42
- 125000003275 alpha amino acid group Chemical group 0.000 claims description 34
- 239000013598 vector Substances 0.000 claims description 26
- 230000001965 increasing effect Effects 0.000 claims description 25
- 102000040430 polynucleotide Human genes 0.000 claims description 23
- 108091033319 polynucleotide Proteins 0.000 claims description 23
- 239000002157 polynucleotide Substances 0.000 claims description 23
- 102100034746 Cyclin-dependent kinase-like 5 Human genes 0.000 claims description 19
- 101000945692 Homo sapiens Cyclin-dependent kinase-like 5 Proteins 0.000 claims description 19
- 239000012634 fragment Substances 0.000 claims description 19
- 102000052510 DNA-Binding Proteins Human genes 0.000 claims description 17
- 101710096438 DNA-binding protein Proteins 0.000 claims description 14
- 101710125418 Major capsid protein Proteins 0.000 claims description 13
- 150000001413 amino acids Chemical class 0.000 claims description 13
- 101150032457 CDKL5 gene Proteins 0.000 claims description 11
- 108700004991 Cas12a Proteins 0.000 claims description 11
- 101100439048 Homo sapiens CDKL5 gene Proteins 0.000 claims description 11
- 108010088141 Argonaute Proteins Proteins 0.000 claims description 10
- 102000008682 Argonaute Proteins Human genes 0.000 claims description 10
- 238000006467 substitution reaction Methods 0.000 claims description 10
- 108091081406 G-quadruplex Proteins 0.000 claims description 9
- 230000008878 coupling Effects 0.000 claims description 9
- 238000010168 coupling process Methods 0.000 claims description 9
- 238000005859 coupling reaction Methods 0.000 claims description 9
- 230000037452 priming Effects 0.000 claims description 8
- 102220100740 rs878854050 Human genes 0.000 claims description 8
- 101710132601 Capsid protein Proteins 0.000 claims description 7
- 101710094648 Coat protein Proteins 0.000 claims description 7
- 102100021181 Golgi phosphoprotein 3 Human genes 0.000 claims description 7
- 101710141454 Nucleoprotein Proteins 0.000 claims description 7
- 101710083689 Probable capsid protein Proteins 0.000 claims description 7
- 241001515965 unidentified phage Species 0.000 claims description 7
- 108090000994 Catalytic RNA Proteins 0.000 claims description 5
- 102000053642 Catalytic RNA Human genes 0.000 claims description 5
- -1 G-quadruplexes Proteins 0.000 claims description 5
- 239000000546 pharmaceutical excipient Substances 0.000 claims description 5
- 108091092562 ribozyme Proteins 0.000 claims description 5
- 230000007812 deficiency Effects 0.000 claims description 3
- 241001430294 unidentified retrovirus Species 0.000 claims description 3
- 101100206286 Caenorhabditis elegans tns-1 gene Proteins 0.000 claims description 2
- 101150016028 apc2 gene Proteins 0.000 claims description 2
- 230000009467 reduction Effects 0.000 claims description 2
- 238000000205 computational method Methods 0.000 claims 1
- 238000005094 computer simulation Methods 0.000 claims 1
- 102200119069 rs137852869 Human genes 0.000 claims 1
- 241000713869 Moloney murine leukemia virus Species 0.000 abstract description 39
- 238000010839 reverse transcription Methods 0.000 abstract description 25
- 230000001976 improved effect Effects 0.000 abstract description 23
- 238000001727 in vivo Methods 0.000 abstract description 5
- 230000003612 virological effect Effects 0.000 abstract description 4
- 108090000623 proteins and genes Proteins 0.000 description 188
- 102000004169 proteins and genes Human genes 0.000 description 159
- 235000018102 proteins Nutrition 0.000 description 157
- 210000004027 cell Anatomy 0.000 description 129
- 239000013615 primer Substances 0.000 description 126
- 101150104383 ALOX5AP gene Proteins 0.000 description 96
- 101100236114 Mus musculus Lrrfip1 gene Proteins 0.000 description 96
- 238000003780 insertion Methods 0.000 description 91
- 230000037431 insertion Effects 0.000 description 91
- 238000012217 deletion Methods 0.000 description 66
- 230000037430 deletion Effects 0.000 description 66
- 230000017730 intein-mediated protein splicing Effects 0.000 description 60
- 108020004682 Single-Stranded DNA Proteins 0.000 description 58
- 101000898676 Homo sapiens Ephrin type-A receptor 8 Proteins 0.000 description 55
- 230000008569 process Effects 0.000 description 55
- 102100021601 Ephrin type-A receptor 8 Human genes 0.000 description 53
- 108090000765 processed proteins & peptides Proteins 0.000 description 52
- 101710163270 Nuclease Proteins 0.000 description 51
- 230000015572 biosynthetic process Effects 0.000 description 50
- 238000010586 diagram Methods 0.000 description 42
- 239000000047 product Substances 0.000 description 41
- 108700028369 Alleles Proteins 0.000 description 38
- 230000008859 change Effects 0.000 description 36
- 230000001404 mediated effect Effects 0.000 description 36
- 239000013612 plasmid Substances 0.000 description 32
- 239000005090 green fluorescent protein Substances 0.000 description 30
- 238000010348 incorporation Methods 0.000 description 30
- 239000000758 substrate Substances 0.000 description 30
- 238000012163 sequencing technique Methods 0.000 description 29
- 108010043121 Green Fluorescent Proteins Proteins 0.000 description 28
- 102000004144 Green Fluorescent Proteins Human genes 0.000 description 28
- 230000006870 function Effects 0.000 description 28
- 230000000875 corresponding effect Effects 0.000 description 26
- 238000000338 in vitro Methods 0.000 description 26
- 230000007115 recruitment Effects 0.000 description 25
- 230000008685 targeting Effects 0.000 description 25
- 230000004927 fusion Effects 0.000 description 24
- 102000004196 processed proteins & peptides Human genes 0.000 description 23
- 238000011144 upstream manufacturing Methods 0.000 description 23
- 238000006243 chemical reaction Methods 0.000 description 22
- 238000013461 design Methods 0.000 description 22
- 230000002068 genetic effect Effects 0.000 description 22
- 229920001184 polypeptide Polymers 0.000 description 22
- 108091079001 CRISPR RNA Proteins 0.000 description 21
- 230000001419 dependent effect Effects 0.000 description 21
- 238000003786 synthesis reaction Methods 0.000 description 21
- 102100023823 Homeobox protein EMX1 Human genes 0.000 description 20
- 101001048956 Homo sapiens Homeobox protein EMX1 Proteins 0.000 description 20
- 235000001014 amino acid Nutrition 0.000 description 20
- 238000010362 genome editing Methods 0.000 description 20
- 238000006116 polymerization reaction Methods 0.000 description 20
- 238000012937 correction Methods 0.000 description 19
- 230000033616 DNA repair Effects 0.000 description 18
- 102000004150 Flap endonucleases Human genes 0.000 description 18
- 108090000652 Flap endonucleases Proteins 0.000 description 18
- 108091028113 Trans-activating crRNA Proteins 0.000 description 18
- 230000000692 anti-sense effect Effects 0.000 description 18
- 230000007704 transition Effects 0.000 description 18
- 102000004190 Enzymes Human genes 0.000 description 17
- 108090000790 Enzymes Proteins 0.000 description 17
- 229940088598 enzyme Drugs 0.000 description 17
- 238000002474 experimental method Methods 0.000 description 17
- 239000003446 ligand Substances 0.000 description 17
- 230000016434 protein splicing Effects 0.000 description 17
- 102100037964 E3 ubiquitin-protein ligase RING2 Human genes 0.000 description 16
- 101001095815 Homo sapiens E3 ubiquitin-protein ligase RING2 Proteins 0.000 description 16
- 229940024606 amino acid Drugs 0.000 description 16
- 230000010076 replication Effects 0.000 description 16
- 108091093088 Amplicon Proteins 0.000 description 15
- 102100022548 Beta-hexosaminidase subunit alpha Human genes 0.000 description 15
- 241000282414 Homo sapiens Species 0.000 description 15
- 101001045440 Homo sapiens Beta-hexosaminidase subunit alpha Proteins 0.000 description 15
- 102100027281 Fanconi anemia group F protein Human genes 0.000 description 14
- 210000004899 c-terminal region Anatomy 0.000 description 14
- 125000002887 hydroxy group Chemical group [H]O* 0.000 description 14
- 230000001717 pathogenic effect Effects 0.000 description 14
- 108010022012 Fanconi Anemia Complementation Group F protein Proteins 0.000 description 13
- 239000003795 chemical substances by application Substances 0.000 description 13
- 238000006731 degradation reaction Methods 0.000 description 13
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 13
- 230000007246 mechanism Effects 0.000 description 13
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 12
- 235000014680 Saccharomyces cerevisiae Nutrition 0.000 description 12
- 230000015556 catabolic process Effects 0.000 description 12
- 238000003776 cleavage reaction Methods 0.000 description 12
- 230000014509 gene expression Effects 0.000 description 12
- 230000001939 inductive effect Effects 0.000 description 12
- 108020004999 messenger RNA Proteins 0.000 description 12
- 230000007017 scission Effects 0.000 description 12
- 239000000126 substance Substances 0.000 description 12
- 230000035897 transcription Effects 0.000 description 12
- 238000013518 transcription Methods 0.000 description 12
- 108020004705 Codon Proteins 0.000 description 11
- 108010009540 DNA (Cytosine-5-)-Methyltransferase 1 Proteins 0.000 description 11
- 102100036279 DNA (cytosine-5)-methyltransferase 1 Human genes 0.000 description 11
- 102100021519 Hemoglobin subunit beta Human genes 0.000 description 11
- 102000029797 Prion Human genes 0.000 description 11
- 108091000054 Prion Proteins 0.000 description 11
- 102100031780 Endonuclease Human genes 0.000 description 10
- 108010042407 Endonucleases Proteins 0.000 description 10
- 102100026121 Flap endonuclease 1 Human genes 0.000 description 10
- 108050002219 Flap endonuclease 1 Proteins 0.000 description 10
- 238000007792 addition Methods 0.000 description 10
- 230000037433 frameshift Effects 0.000 description 10
- 238000012545 processing Methods 0.000 description 10
- 108091027305 Heteroduplex Proteins 0.000 description 9
- 108010052160 Site-specific recombinase Proteins 0.000 description 9
- 238000004458 analytical method Methods 0.000 description 9
- 238000005516 engineering process Methods 0.000 description 9
- 239000000463 material Substances 0.000 description 9
- 241000894007 species Species 0.000 description 9
- 238000011282 treatment Methods 0.000 description 9
- 108020004635 Complementary DNA Proteins 0.000 description 8
- 241000191967 Staphylococcus aureus Species 0.000 description 8
- 241000193996 Streptococcus pyogenes Species 0.000 description 8
- 238000010804 cDNA synthesis Methods 0.000 description 8
- 239000002299 complementary DNA Substances 0.000 description 8
- 201000010099 disease Diseases 0.000 description 8
- 210000005260 human cell Anatomy 0.000 description 8
- 238000002264 polyacrylamide gel electrophoresis Methods 0.000 description 8
- 230000008439 repair process Effects 0.000 description 8
- 208000024891 symptom Diseases 0.000 description 8
- 108010043471 Core Binding Factor Alpha 2 Subunit Proteins 0.000 description 7
- 230000007018 DNA scission Effects 0.000 description 7
- 238000011529 RT qPCR Methods 0.000 description 7
- 102100025373 Runt-related transcription factor 1 Human genes 0.000 description 7
- 108091081021 Sense strand Proteins 0.000 description 7
- 238000003556 assay Methods 0.000 description 7
- 230000008901 benefit Effects 0.000 description 7
- UYTPUPDQBNUYGX-UHFFFAOYSA-N guanine Chemical compound O=C1NC(N)=NC2=C1N=CN2 UYTPUPDQBNUYGX-UHFFFAOYSA-N 0.000 description 7
- 230000006872 improvement Effects 0.000 description 7
- 238000001890 transfection Methods 0.000 description 7
- 230000004568 DNA-binding Effects 0.000 description 6
- XSQUKJJJFZCRTK-UHFFFAOYSA-N Urea Chemical compound NC(N)=O XSQUKJJJFZCRTK-UHFFFAOYSA-N 0.000 description 6
- 230000003197 catalytic effect Effects 0.000 description 6
- 231100000219 mutagenic Toxicity 0.000 description 6
- 230000003505 mutagenic effect Effects 0.000 description 6
- 229920000642 polymer Polymers 0.000 description 6
- 238000000746 purification Methods 0.000 description 6
- 238000003762 quantitative reverse transcription PCR Methods 0.000 description 6
- 230000002829 reductive effect Effects 0.000 description 6
- 238000011160 research Methods 0.000 description 6
- 150000003384 small molecules Chemical class 0.000 description 6
- 101000808011 Homo sapiens Vascular endothelial growth factor A Proteins 0.000 description 5
- 238000000636 Northern blotting Methods 0.000 description 5
- 108091034117 Oligonucleotide Proteins 0.000 description 5
- 239000004698 Polyethylene Substances 0.000 description 5
- 102000044126 RNA-Binding Proteins Human genes 0.000 description 5
- 108091027544 Subgenomic mRNA Proteins 0.000 description 5
- 102100039037 Vascular endothelial growth factor A Human genes 0.000 description 5
- 239000006227 byproduct Substances 0.000 description 5
- 229910052799 carbon Inorganic materials 0.000 description 5
- 230000001413 cellular effect Effects 0.000 description 5
- 208000035475 disorder Diseases 0.000 description 5
- 239000012636 effector Substances 0.000 description 5
- 238000012165 high-throughput sequencing Methods 0.000 description 5
- 239000000543 intermediate Substances 0.000 description 5
- 230000030648 nucleus localization Effects 0.000 description 5
- 238000005457 optimization Methods 0.000 description 5
- 230000002441 reversible effect Effects 0.000 description 5
- 238000003860 storage Methods 0.000 description 5
- 101150075675 tatC gene Proteins 0.000 description 5
- 101710159080 Aconitate hydratase A Proteins 0.000 description 4
- 101710159078 Aconitate hydratase B Proteins 0.000 description 4
- 108091032955 Bacterial small RNA Proteins 0.000 description 4
- 101710203526 Integrase Proteins 0.000 description 4
- 241001465754 Metazoa Species 0.000 description 4
- 101710105008 RNA-binding protein Proteins 0.000 description 4
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 description 4
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 description 4
- 102000006382 Ribonucleases Human genes 0.000 description 4
- 108010083644 Ribonucleases Proteins 0.000 description 4
- IQFYYKKMVGJFEH-XLPZGREQSA-N Thymidine Chemical compound O=C1NC(=O)C(C)=CN1[C@@H]1O[C@H](CO)[C@@H](O)C1 IQFYYKKMVGJFEH-XLPZGREQSA-N 0.000 description 4
- 241000700605 Viruses Species 0.000 description 4
- OIRDTQYFTABQOQ-KQYNXXCUSA-N adenosine Chemical compound C1=NC=2C(N)=NC=NC=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O OIRDTQYFTABQOQ-KQYNXXCUSA-N 0.000 description 4
- 238000000137 annealing Methods 0.000 description 4
- 230000001580 bacterial effect Effects 0.000 description 4
- 239000013043 chemical agent Substances 0.000 description 4
- 238000010367 cloning Methods 0.000 description 4
- 239000000499 gel Substances 0.000 description 4
- 238000009396 hybridization Methods 0.000 description 4
- 230000000670 limiting effect Effects 0.000 description 4
- 210000004940 nucleus Anatomy 0.000 description 4
- 229920002401 polyacrylamide Polymers 0.000 description 4
- 238000012216 screening Methods 0.000 description 4
- 238000013515 script Methods 0.000 description 4
- 238000000926 separation method Methods 0.000 description 4
- 238000012384 transportation and delivery Methods 0.000 description 4
- 238000012800 visualization Methods 0.000 description 4
- 102000040650 (ribonucleotides)n+m Human genes 0.000 description 3
- 241000894006 Bacteria Species 0.000 description 3
- 108091033380 Coding strand Proteins 0.000 description 3
- 102220605874 Cytosolic arginine sensor for mTORC1 subunit 2_D10A_mutation Human genes 0.000 description 3
- 108020001019 DNA Primers Proteins 0.000 description 3
- 239000003155 DNA primer Substances 0.000 description 3
- 108700020911 DNA-Binding Proteins Proteins 0.000 description 3
- 101710191360 Eosinophil cationic protein Proteins 0.000 description 3
- 108060002716 Exonuclease Proteins 0.000 description 3
- 108091092195 Intron Proteins 0.000 description 3
- 102000018697 Membrane Proteins Human genes 0.000 description 3
- 108010052285 Membrane Proteins Proteins 0.000 description 3
- 241000714177 Murine leukemia virus Species 0.000 description 3
- 241000699670 Mus sp. Species 0.000 description 3
- 102100036007 Ribonuclease 3 Human genes 0.000 description 3
- 101710192197 Ribonuclease 3 Proteins 0.000 description 3
- 241000194020 Streptococcus thermophilus Species 0.000 description 3
- 238000010459 TALEN Methods 0.000 description 3
- 108010017070 Zinc Finger Nucleases Proteins 0.000 description 3
- 230000009471 action Effects 0.000 description 3
- 239000011543 agarose gel Substances 0.000 description 3
- 230000003321 amplification Effects 0.000 description 3
- 238000013459 approach Methods 0.000 description 3
- 238000003491 array Methods 0.000 description 3
- 239000004202 carbamide Substances 0.000 description 3
- 238000007385 chemical modification Methods 0.000 description 3
- 239000007795 chemical reaction product Substances 0.000 description 3
- 230000009977 dual effect Effects 0.000 description 3
- 230000010856 establishment of protein localization Effects 0.000 description 3
- 102000013165 exonuclease Human genes 0.000 description 3
- 238000001943 fluorescence-activated cell sorting Methods 0.000 description 3
- 231100000221 frame shift mutation induction Toxicity 0.000 description 3
- 238000012239 gene modification Methods 0.000 description 3
- 230000005017 genetic modification Effects 0.000 description 3
- 235000013617 genetically modified food Nutrition 0.000 description 3
- 230000002779 inactivation Effects 0.000 description 3
- 230000002401 inhibitory effect Effects 0.000 description 3
- 239000006166 lysate Substances 0.000 description 3
- 238000001768 microscale thermophoresis Methods 0.000 description 3
- 238000010369 molecular cloning Methods 0.000 description 3
- 231100000350 mutagenesis Toxicity 0.000 description 3
- 238000002703 mutagenesis Methods 0.000 description 3
- 239000013642 negative control Substances 0.000 description 3
- 238000003199 nucleic acid amplification method Methods 0.000 description 3
- 230000037048 polymerization activity Effects 0.000 description 3
- 239000002243 precursor Substances 0.000 description 3
- 230000001681 protective effect Effects 0.000 description 3
- 230000008707 rearrangement Effects 0.000 description 3
- 230000003362 replicative effect Effects 0.000 description 3
- 108020004418 ribosomal RNA Proteins 0.000 description 3
- 239000000523 sample Substances 0.000 description 3
- 230000037432 silent mutation Effects 0.000 description 3
- 230000000087 stabilizing effect Effects 0.000 description 3
- 230000005030 transcription termination Effects 0.000 description 3
- 230000002103 transcriptional effect Effects 0.000 description 3
- 230000001052 transient effect Effects 0.000 description 3
- 230000014616 translation Effects 0.000 description 3
- 238000013519 translation Methods 0.000 description 3
- 238000010200 validation analysis Methods 0.000 description 3
- RHFUOMFWUGWKKO-XVFCMESISA-N 2-thiocytidine Chemical compound S=C1N=C(N)C=CN1[C@H]1[C@H](O)[C@H](O)[C@@H](CO)O1 RHFUOMFWUGWKKO-XVFCMESISA-N 0.000 description 2
- ZAYHVCMSTBRABG-JXOAFFINSA-N 5-methylcytidine Chemical compound O=C1N=C(N)C(C)=CN1[C@H]1[C@H](O)[C@H](O)[C@@H](CO)O1 ZAYHVCMSTBRABG-JXOAFFINSA-N 0.000 description 2
- 238000010356 CRISPR-Cas9 genome editing Methods 0.000 description 2
- 108091035707 Consensus sequence Proteins 0.000 description 2
- 238000010442 DNA editing Methods 0.000 description 2
- 102100033215 DNA nucleotidylexotransferase Human genes 0.000 description 2
- 230000004543 DNA replication Effects 0.000 description 2
- 108090000626 DNA-directed RNA polymerases Proteins 0.000 description 2
- 102000004163 DNA-directed RNA polymerases Human genes 0.000 description 2
- 108010007577 Exodeoxyribonuclease I Proteins 0.000 description 2
- 102000008779 Exonuclease 1 Human genes 0.000 description 2
- 101150058673 FANCF gene Proteins 0.000 description 2
- 208000001914 Fragile X syndrome Diseases 0.000 description 2
- 208000024412 Friedreich ataxia Diseases 0.000 description 2
- 108700007698 Genetic Terminator Regions Proteins 0.000 description 2
- NYHBQMYGNKIUIF-UUOKFMHZSA-N Guanosine Chemical compound C1=NC=2C(=O)NC(N)=NC=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O NYHBQMYGNKIUIF-UUOKFMHZSA-N 0.000 description 2
- HVLSXIKZNLPZJJ-TXZCQADKSA-N HA peptide Chemical compound C([C@@H](C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](C(C)C)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(=O)N[C@@H](C)C(O)=O)NC(=O)[C@H]1N(CCC1)C(=O)[C@@H](N)CC=1C=CC(O)=CC=1)C1=CC=C(O)C=C1 HVLSXIKZNLPZJJ-TXZCQADKSA-N 0.000 description 2
- 102000029812 HNH nuclease Human genes 0.000 description 2
- 108060003760 HNH nuclease Proteins 0.000 description 2
- 208000026350 Inborn Genetic disease Diseases 0.000 description 2
- 108700011259 MicroRNAs Proteins 0.000 description 2
- 241000699666 Mus <mouse, genus> Species 0.000 description 2
- 102220626214 Periaxin_R1335Q_mutation Human genes 0.000 description 2
- CZPWVGJYEJSRLH-UHFFFAOYSA-N Pyrimidine Chemical compound C1=CN=CN=C1 CZPWVGJYEJSRLH-UHFFFAOYSA-N 0.000 description 2
- 108091008103 RNA aptamers Proteins 0.000 description 2
- 108010091086 Recombinases Proteins 0.000 description 2
- 102000018120 Recombinases Human genes 0.000 description 2
- 102000004389 Ribonucleoproteins Human genes 0.000 description 2
- 108010081734 Ribonucleoproteins Proteins 0.000 description 2
- 102000039471 Small Nuclear RNA Human genes 0.000 description 2
- 108020004459 Small interfering RNA Proteins 0.000 description 2
- 101000910035 Streptococcus pyogenes serotype M1 CRISPR-associated endonuclease Cas9/Csn1 Proteins 0.000 description 2
- 238000000692 Student's t-test Methods 0.000 description 2
- 102220644232 Tenascin-N_D1135E_mutation Human genes 0.000 description 2
- DRTQHJPVMGBUCF-XVFCMESISA-N Uridine Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C=C1 DRTQHJPVMGBUCF-XVFCMESISA-N 0.000 description 2
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 2
- 230000004913 activation Effects 0.000 description 2
- 108010013829 alpha subunit DNA polymerase III Proteins 0.000 description 2
- 125000000539 amino acid group Chemical group 0.000 description 2
- 229910052785 arsenic Inorganic materials 0.000 description 2
- 239000011230 binding agent Substances 0.000 description 2
- 230000008512 biological response Effects 0.000 description 2
- 230000000981 bystander Effects 0.000 description 2
- 230000003833 cell viability Effects 0.000 description 2
- 108091092259 cell-free RNA Proteins 0.000 description 2
- 108091092356 cellular DNA Proteins 0.000 description 2
- 230000002860 competitive effect Effects 0.000 description 2
- 230000008602 contraction Effects 0.000 description 2
- 210000003618 cortical neuron Anatomy 0.000 description 2
- 230000001186 cumulative effect Effects 0.000 description 2
- 238000005520 cutting process Methods 0.000 description 2
- 238000000326 densiometry Methods 0.000 description 2
- 238000001514 detection method Methods 0.000 description 2
- 230000005782 double-strand break Effects 0.000 description 2
- 239000003814 drug Substances 0.000 description 2
- 238000000605 extraction Methods 0.000 description 2
- 230000005714 functional activity Effects 0.000 description 2
- 208000016361 genetic disease Diseases 0.000 description 2
- 230000036039 immunity Effects 0.000 description 2
- 230000002163 immunogen Effects 0.000 description 2
- 230000000415 inactivating effect Effects 0.000 description 2
- 210000004263 induced pluripotent stem cell Anatomy 0.000 description 2
- 230000000977 initiatory effect Effects 0.000 description 2
- 239000007924 injection Substances 0.000 description 2
- 238000002347 injection Methods 0.000 description 2
- 230000010354 integration Effects 0.000 description 2
- 230000003834 intracellular effect Effects 0.000 description 2
- 230000009545 invasion Effects 0.000 description 2
- 238000005304 joining Methods 0.000 description 2
- 108020001756 ligand binding domains Proteins 0.000 description 2
- FDZZZRQASAIRJF-UHFFFAOYSA-M malachite green Chemical compound [Cl-].C1=CC(N(C)C)=CC=C1C(C=1C=CC=CC=1)=C1C=CC(=[N+](C)C)C=C1 FDZZZRQASAIRJF-UHFFFAOYSA-M 0.000 description 2
- 229940107698 malachite green Drugs 0.000 description 2
- 238000004519 manufacturing process Methods 0.000 description 2
- 239000003550 marker Substances 0.000 description 2
- 239000002609 medium Substances 0.000 description 2
- 238000002844 melting Methods 0.000 description 2
- 230000008018 melting Effects 0.000 description 2
- 239000002679 microRNA Substances 0.000 description 2
- 230000000394 mitotic effect Effects 0.000 description 2
- 239000003607 modifier Substances 0.000 description 2
- 239000002777 nucleoside Substances 0.000 description 2
- 230000036961 partial effect Effects 0.000 description 2
- 125000002467 phosphate group Chemical group [H]OP(=O)(O[H])O[*] 0.000 description 2
- 239000013641 positive control Substances 0.000 description 2
- 230000001737 promoting effect Effects 0.000 description 2
- 239000013636 protein dimer Substances 0.000 description 2
- 125000004219 purine nucleobase group Chemical group 0.000 description 2
- 238000011002 quantification Methods 0.000 description 2
- 230000001105 regulatory effect Effects 0.000 description 2
- 238000009877 rendering Methods 0.000 description 2
- 102200008469 rs267606980 Human genes 0.000 description 2
- RHFUOMFWUGWKKO-UHFFFAOYSA-N s2C Natural products S=C1N=C(N)C=CN1C1C(O)C(O)C(CO)O1 RHFUOMFWUGWKKO-UHFFFAOYSA-N 0.000 description 2
- 238000007480 sanger sequencing Methods 0.000 description 2
- 230000003007 single stranded DNA break Effects 0.000 description 2
- 239000004055 small Interfering RNA Substances 0.000 description 2
- 108091029842 small nuclear ribonucleic acid Proteins 0.000 description 2
- 238000005063 solubilization Methods 0.000 description 2
- 230000007928 solubilization Effects 0.000 description 2
- 230000002123 temporal effect Effects 0.000 description 2
- 230000001225 therapeutic effect Effects 0.000 description 2
- 210000001519 tissue Anatomy 0.000 description 2
- 238000010361 transduction Methods 0.000 description 2
- 238000012546 transfer Methods 0.000 description 2
- 230000035899 viability Effects 0.000 description 2
- 239000002023 wood Substances 0.000 description 2
- RIFDKYBNWNPCQK-IOSLPCCCSA-N (2r,3s,4r,5r)-2-(hydroxymethyl)-5-(6-imino-3-methylpurin-9-yl)oxolane-3,4-diol Chemical compound C1=2N(C)C=NC(=N)C=2N=CN1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O RIFDKYBNWNPCQK-IOSLPCCCSA-N 0.000 description 1
- RKSLVDIXBGWPIS-UAKXSSHOSA-N 1-[(2r,3r,4s,5r)-3,4-dihydroxy-5-(hydroxymethyl)oxolan-2-yl]-5-iodopyrimidine-2,4-dione Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C(I)=C1 RKSLVDIXBGWPIS-UAKXSSHOSA-N 0.000 description 1
- QLOCVMVCRJOTTM-TURQNECASA-N 1-[(2r,3r,4s,5r)-3,4-dihydroxy-5-(hydroxymethyl)oxolan-2-yl]-5-prop-1-ynylpyrimidine-2,4-dione Chemical compound O=C1NC(=O)C(C#CC)=CN1[C@H]1[C@H](O)[C@H](O)[C@@H](CO)O1 QLOCVMVCRJOTTM-TURQNECASA-N 0.000 description 1
- PISWNSOQFZRVJK-XLPZGREQSA-N 1-[(2r,4s,5r)-4-hydroxy-5-(hydroxymethyl)oxolan-2-yl]-5-methyl-2-sulfanylidenepyrimidin-4-one Chemical compound S=C1NC(=O)C(C)=CN1[C@@H]1O[C@H](CO)[C@@H](O)C1 PISWNSOQFZRVJK-XLPZGREQSA-N 0.000 description 1
- UHDGCWIWMRVCDJ-UHFFFAOYSA-N 1-beta-D-Xylofuranosyl-NH-Cytosine Natural products O=C1N=C(N)C=CN1C1C(O)C(O)C(CO)O1 UHDGCWIWMRVCDJ-UHFFFAOYSA-N 0.000 description 1
- GFYLSDSUCHVORB-IOSLPCCCSA-N 1-methyladenosine Chemical compound C1=NC=2C(=N)N(C)C=NC=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O GFYLSDSUCHVORB-IOSLPCCCSA-N 0.000 description 1
- UTAIYTHAJQNQDW-KQYNXXCUSA-N 1-methylguanosine Chemical compound C1=NC=2C(=O)N(C)C(N)=NC=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O UTAIYTHAJQNQDW-KQYNXXCUSA-N 0.000 description 1
- RFCQJGFZUQFYRF-UHFFFAOYSA-N 2'-O-Methylcytidine Natural products COC1C(O)C(CO)OC1N1C(=O)N=C(N)C=C1 RFCQJGFZUQFYRF-UHFFFAOYSA-N 0.000 description 1
- SXUXMRMBWZCMEN-UHFFFAOYSA-N 2'-O-methyl uridine Natural products COC1C(O)C(CO)OC1N1C(=O)NC(=O)C=C1 SXUXMRMBWZCMEN-UHFFFAOYSA-N 0.000 description 1
- RFCQJGFZUQFYRF-ZOQUXTDFSA-N 2'-O-methylcytidine Chemical compound CO[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)N=C(N)C=C1 RFCQJGFZUQFYRF-ZOQUXTDFSA-N 0.000 description 1
- YKBGVTZYEHREMT-KVQBGUIXSA-N 2'-deoxyguanosine Chemical compound C1=NC=2C(=O)NC(N)=NC=2N1[C@H]1C[C@H](O)[C@@H](CO)O1 YKBGVTZYEHREMT-KVQBGUIXSA-N 0.000 description 1
- CKTSBUTUHBMZGZ-SHYZEUOFSA-N 2'‐deoxycytidine Chemical compound O=C1N=C(N)C=CN1[C@@H]1O[C@H](CO)[C@@H](O)C1 CKTSBUTUHBMZGZ-SHYZEUOFSA-N 0.000 description 1
- ZDTFMPXQUSBYRL-UUOKFMHZSA-N 2-Aminoadenosine Chemical compound C12=NC(N)=NC(N)=C2N=CN1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O ZDTFMPXQUSBYRL-UUOKFMHZSA-N 0.000 description 1
- JRYMOPZHXMVHTA-DAGMQNCNSA-N 2-amino-7-[(2r,3r,4s,5r)-3,4-dihydroxy-5-(hydroxymethyl)oxolan-2-yl]-1h-pyrrolo[2,3-d]pyrimidin-4-one Chemical compound C1=CC=2C(=O)NC(N)=NC=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O JRYMOPZHXMVHTA-DAGMQNCNSA-N 0.000 description 1
- FWBHETKCLVMNFS-UHFFFAOYSA-N 4',6-Diamino-2-phenylindol Chemical compound C1=CC(C(=N)N)=CC=C1C1=CC2=CC=C(C(N)=N)C=C2N1 FWBHETKCLVMNFS-UHFFFAOYSA-N 0.000 description 1
- YBRVSVVVWCFQMG-UHFFFAOYSA-N 4,4'-diaminodiphenylmethane Chemical compound C1=CC(N)=CC=C1CC1=CC=C(N)C=C1 YBRVSVVVWCFQMG-UHFFFAOYSA-N 0.000 description 1
- BCZUPRDAAVVBSO-MJXNYTJMSA-N 4-acetylcytidine Chemical compound C1=CC(C(=O)C)(N)NC(=O)N1[C@H]1[C@H](O)[C@H](O)[C@@H](CO)O1 BCZUPRDAAVVBSO-MJXNYTJMSA-N 0.000 description 1
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 description 1
- XXSIICQLPUAUDF-TURQNECASA-N 4-amino-1-[(2r,3r,4s,5r)-3,4-dihydroxy-5-(hydroxymethyl)oxolan-2-yl]-5-prop-1-ynylpyrimidin-2-one Chemical compound O=C1N=C(N)C(C#CC)=CN1[C@H]1[C@H](O)[C@H](O)[C@@H](CO)O1 XXSIICQLPUAUDF-TURQNECASA-N 0.000 description 1
- UVGCZRPOXXYZKH-QADQDURISA-N 5-(carboxyhydroxymethyl)uridine Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C(C(O)C(O)=O)=C1 UVGCZRPOXXYZKH-QADQDURISA-N 0.000 description 1
- ZAYHVCMSTBRABG-UHFFFAOYSA-N 5-Methylcytidine Natural products O=C1N=C(N)C(C)=CN1C1C(O)C(O)C(CO)O1 ZAYHVCMSTBRABG-UHFFFAOYSA-N 0.000 description 1
- MMUBPEFMCTVKTR-IBNKKVAHSA-N 5-[(2s,3r,4s,5r)-3,4-dihydroxy-5-(hydroxymethyl)-2-methyloxolan-2-yl]-1h-pyrimidine-2,4-dione Chemical compound C=1NC(=O)NC(=O)C=1[C@]1(C)O[C@H](CO)[C@@H](O)[C@H]1O MMUBPEFMCTVKTR-IBNKKVAHSA-N 0.000 description 1
- AGFIRQJZCNVMCW-UAKXSSHOSA-N 5-bromouridine Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C(Br)=C1 AGFIRQJZCNVMCW-UAKXSSHOSA-N 0.000 description 1
- FHIDNBAQOFJWCA-UAKXSSHOSA-N 5-fluorouridine Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C(F)=C1 FHIDNBAQOFJWCA-UAKXSSHOSA-N 0.000 description 1
- KDOPAZIWBAHVJB-UHFFFAOYSA-N 5h-pyrrolo[3,2-d]pyrimidine Chemical compound C1=NC=C2NC=CC2=N1 KDOPAZIWBAHVJB-UHFFFAOYSA-N 0.000 description 1
- BXJHWYVXLGLDMZ-UHFFFAOYSA-N 6-O-methylguanine Chemical compound COC1=NC(N)=NC2=C1NC=N2 BXJHWYVXLGLDMZ-UHFFFAOYSA-N 0.000 description 1
- UEHOMUNTZPIBIL-UUOKFMHZSA-N 6-amino-9-[(2r,3r,4s,5r)-3,4-dihydroxy-5-(hydroxymethyl)oxolan-2-yl]-7h-purin-8-one Chemical compound O=C1NC=2C(N)=NC=NC=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O UEHOMUNTZPIBIL-UUOKFMHZSA-N 0.000 description 1
- HCAJQHYUCKICQH-VPENINKCSA-N 8-Oxo-7,8-dihydro-2'-deoxyguanosine Chemical compound C1=2NC(N)=NC(=O)C=2NC(=O)N1[C@H]1C[C@H](O)[C@@H](CO)O1 HCAJQHYUCKICQH-VPENINKCSA-N 0.000 description 1
- HDZZVAMISRMYHH-UHFFFAOYSA-N 9beta-Ribofuranosyl-7-deazaadenin Natural products C1=CC=2C(N)=NC=NC=2N1C1OC(CO)C(O)C1O HDZZVAMISRMYHH-UHFFFAOYSA-N 0.000 description 1
- 108020005037 A-Form DNA Proteins 0.000 description 1
- 108010066676 Abrin Proteins 0.000 description 1
- 241000251468 Actinopterygii Species 0.000 description 1
- 241000380131 Ammophila arenaria Species 0.000 description 1
- 108020005544 Antisense RNA Proteins 0.000 description 1
- 101100108891 Arabidopsis thaliana PRMT11 gene Proteins 0.000 description 1
- 241000203069 Archaea Species 0.000 description 1
- 241000271566 Aves Species 0.000 description 1
- 108020003591 B-Form DNA Proteins 0.000 description 1
- DWRXFEITVBNRMK-UHFFFAOYSA-N Beta-D-1-Arabinofuranosylthymine Natural products O=C1NC(=O)C(C)=CN1C1C(O)C(O)C(CO)O1 DWRXFEITVBNRMK-UHFFFAOYSA-N 0.000 description 1
- 102100028728 Bone morphogenetic protein 1 Human genes 0.000 description 1
- 241000283690 Bos taurus Species 0.000 description 1
- 239000002126 C01EB10 - Adenosine Substances 0.000 description 1
- 108010040467 CRISPR-Associated Proteins Proteins 0.000 description 1
- 238000010453 CRISPR/Cas method Methods 0.000 description 1
- 241000283707 Capra Species 0.000 description 1
- 102000014914 Carrier Proteins Human genes 0.000 description 1
- 108010078791 Carrier Proteins Proteins 0.000 description 1
- 206010008342 Cervix carcinoma Diseases 0.000 description 1
- 108091006146 Channels Proteins 0.000 description 1
- 108010062580 Concanavalin A Proteins 0.000 description 1
- VPAXJOUATWLOPR-UHFFFAOYSA-N Conferone Chemical compound C1=CC(=O)OC2=CC(OCC3C4(C)CCC(=O)C(C)(C)C4CC=C3C)=CC=C21 VPAXJOUATWLOPR-UHFFFAOYSA-N 0.000 description 1
- 241000186227 Corynebacterium diphtheriae Species 0.000 description 1
- 229920000742 Cotton Polymers 0.000 description 1
- MIKUYHXYGGJMLM-GIMIYPNGSA-N Crotonoside Natural products C1=NC2=C(N)NC(=O)N=C2N1[C@H]1O[C@@H](CO)[C@H](O)[C@@H]1O MIKUYHXYGGJMLM-GIMIYPNGSA-N 0.000 description 1
- PMATZTZNYRCHOR-CGLBZJNRSA-N Cyclosporin A Chemical class CC[C@@H]1NC(=O)[C@H]([C@H](O)[C@H](C)C\C=C\C)N(C)C(=O)[C@H](C(C)C)N(C)C(=O)[C@H](CC(C)C)N(C)C(=O)[C@H](CC(C)C)N(C)C(=O)[C@@H](C)NC(=O)[C@H](C)NC(=O)[C@H](CC(C)C)N(C)C(=O)[C@H](C(C)C)NC(=O)[C@H](CC(C)C)N(C)C(=O)CN(C)C1=O PMATZTZNYRCHOR-CGLBZJNRSA-N 0.000 description 1
- UHDGCWIWMRVCDJ-PSQAKQOGSA-N Cytidine Natural products O=C1N=C(N)C=CN1[C@@H]1[C@@H](O)[C@@H](O)[C@H](CO)O1 UHDGCWIWMRVCDJ-PSQAKQOGSA-N 0.000 description 1
- NYHBQMYGNKIUIF-UHFFFAOYSA-N D-guanosine Natural products C1=2NC(N)=NC(=O)C=2N=CN1C1OC(CO)C(O)C1O NYHBQMYGNKIUIF-UHFFFAOYSA-N 0.000 description 1
- SHZGCJCMOBCMKK-UHFFFAOYSA-N D-mannomethylose Natural products CC1OC(O)C(O)C(O)C1O SHZGCJCMOBCMKK-UHFFFAOYSA-N 0.000 description 1
- HMFHBZSHGGEWLO-SOOFDHNKSA-N D-ribofuranose Chemical compound OC[C@H]1OC(O)[C@H](O)[C@@H]1O HMFHBZSHGGEWLO-SOOFDHNKSA-N 0.000 description 1
- 102000011724 DNA Repair Enzymes Human genes 0.000 description 1
- 108010076525 DNA Repair Enzymes Proteins 0.000 description 1
- 108091008102 DNA aptamers Proteins 0.000 description 1
- 108010008286 DNA nucleotidylexotransferase Proteins 0.000 description 1
- 238000001712 DNA sequencing Methods 0.000 description 1
- CKTSBUTUHBMZGZ-UHFFFAOYSA-N Deoxycytidine Natural products O=C1N=C(N)C=CN1C1OC(CO)C(O)C1 CKTSBUTUHBMZGZ-UHFFFAOYSA-N 0.000 description 1
- 241000702421 Dependoparvovirus Species 0.000 description 1
- 241000588724 Escherichia coli Species 0.000 description 1
- 241000206602 Eukaryota Species 0.000 description 1
- 108700024394 Exon Proteins 0.000 description 1
- 108050001049 Extracellular proteins Proteins 0.000 description 1
- 101150054617 FCPF gene Proteins 0.000 description 1
- XZWYTXMRWQJBGX-VXBMVYAYSA-N FLAG peptide Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CCCCN)NC(=O)[C@@H](NC(=O)[C@@H](N)CC(O)=O)CC1=CC=C(O)C=C1 XZWYTXMRWQJBGX-VXBMVYAYSA-N 0.000 description 1
- 241000282326 Felis catus Species 0.000 description 1
- 241000724791 Filamentous phage Species 0.000 description 1
- 208000034951 Genetic Translocation Diseases 0.000 description 1
- 102100041003 Glutamate carboxypeptidase 2 Human genes 0.000 description 1
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 1
- 108091027874 Group I catalytic intron Proteins 0.000 description 1
- 108091005904 Hemoglobin subunit beta Proteins 0.000 description 1
- 241000238631 Hexapoda Species 0.000 description 1
- 101000892862 Homo sapiens Glutamate carboxypeptidase 2 Proteins 0.000 description 1
- 241000713772 Human immunodeficiency virus 1 Species 0.000 description 1
- 208000023105 Huntington disease Diseases 0.000 description 1
- UGQMRVRMYYASKQ-KQYNXXCUSA-N Inosine Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C2=NC=NC(O)=C2N=C1 UGQMRVRMYYASKQ-KQYNXXCUSA-N 0.000 description 1
- 229930010555 Inosine Natural products 0.000 description 1
- 108010001127 Insulin Receptor Proteins 0.000 description 1
- 102100036721 Insulin receptor Human genes 0.000 description 1
- 108010015268 Integration Host Factors Proteins 0.000 description 1
- CKLJMWTZIZZHCS-REOHCLBHSA-N L-aspartic acid Chemical compound OC(=O)[C@@H](N)CC(O)=O CKLJMWTZIZZHCS-REOHCLBHSA-N 0.000 description 1
- PNNNRSAQSRJVSB-UHFFFAOYSA-N L-rhamnose Natural products CC(O)C(O)C(O)C(O)C=O PNNNRSAQSRJVSB-UHFFFAOYSA-N 0.000 description 1
- 101710173438 Late L2 mu core protein Proteins 0.000 description 1
- 108090001090 Lectins Proteins 0.000 description 1
- 102000004856 Lectins Human genes 0.000 description 1
- 241000270322 Lepidosauria Species 0.000 description 1
- 241000124008 Mammalia Species 0.000 description 1
- 102000003939 Membrane transport proteins Human genes 0.000 description 1
- 108090000301 Membrane transport proteins Proteins 0.000 description 1
- 201000009906 Meningitis Diseases 0.000 description 1
- 101710145242 Minor capsid protein P3-RTD Proteins 0.000 description 1
- VQAYFKKCNSOZKM-IOSLPCCCSA-N N(6)-methyladenosine Chemical compound C1=NC=2C(NC)=NC=NC=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O VQAYFKKCNSOZKM-IOSLPCCCSA-N 0.000 description 1
- 125000000729 N-terminal amino-acid group Chemical group 0.000 description 1
- VQAYFKKCNSOZKM-UHFFFAOYSA-N NSC 29409 Natural products C1=NC=2C(NC)=NC=NC=2N1C1OC(CO)C(O)C1O VQAYFKKCNSOZKM-UHFFFAOYSA-N 0.000 description 1
- 241000588653 Neisseria Species 0.000 description 1
- 241000588654 Neisseria cinerea Species 0.000 description 1
- 241000244206 Nematoda Species 0.000 description 1
- 241000424623 Nostoc punctiforme Species 0.000 description 1
- 108010077850 Nuclear Localization Signals Proteins 0.000 description 1
- 102000007999 Nuclear Proteins Human genes 0.000 description 1
- 108010089610 Nuclear Proteins Proteins 0.000 description 1
- 108091007494 Nucleic acid- binding domains Proteins 0.000 description 1
- 108091005461 Nucleic proteins Proteins 0.000 description 1
- 108700026244 Open Reading Frames Proteins 0.000 description 1
- 241001494479 Pecora Species 0.000 description 1
- 241000009328 Perro Species 0.000 description 1
- 101100484946 Petunia hybrida VPY gene Proteins 0.000 description 1
- 108010010677 Phosphodiesterase I Proteins 0.000 description 1
- ZLMJMSJWJFRBEC-UHFFFAOYSA-N Potassium Chemical compound [K] ZLMJMSJWJFRBEC-UHFFFAOYSA-N 0.000 description 1
- 208000024777 Prion disease Diseases 0.000 description 1
- 101800001494 Protease 2A Proteins 0.000 description 1
- 101800001066 Protein 2A Proteins 0.000 description 1
- 101710188315 Protein X Proteins 0.000 description 1
- 108020004518 RNA Probes Proteins 0.000 description 1
- 239000003391 RNA probe Substances 0.000 description 1
- 108700020471 RNA-Binding Proteins Proteins 0.000 description 1
- 230000004570 RNA-binding Effects 0.000 description 1
- 108091081062 Repeated sequence (DNA) Proteins 0.000 description 1
- 102000003661 Ribonuclease III Human genes 0.000 description 1
- 108010057163 Ribonuclease III Proteins 0.000 description 1
- PYMYPHUHKUWMLA-LMVFSUKVSA-N Ribose Natural products OC[C@@H](O)[C@@H](O)[C@@H](O)C=O PYMYPHUHKUWMLA-LMVFSUKVSA-N 0.000 description 1
- 101150062997 Rnf2 gene Proteins 0.000 description 1
- 241000283984 Rodentia Species 0.000 description 1
- 238000012300 Sequence Analysis Methods 0.000 description 1
- 101800004236 Ssp dnaE intein Proteins 0.000 description 1
- 101150020213 Stard3 gene Proteins 0.000 description 1
- 241000194017 Streptococcus Species 0.000 description 1
- 102000001435 Synapsin Human genes 0.000 description 1
- 108050009621 Synapsin Proteins 0.000 description 1
- 208000022292 Tay-Sachs disease Diseases 0.000 description 1
- 102000007000 Tenascin Human genes 0.000 description 1
- 108010008125 Tenascin Proteins 0.000 description 1
- 241000248384 Tetrahymena thermophila Species 0.000 description 1
- 108090000190 Thrombin Proteins 0.000 description 1
- 102000040945 Transcription factor Human genes 0.000 description 1
- 108091023040 Transcription factor Proteins 0.000 description 1
- 241000589892 Treponema denticola Species 0.000 description 1
- 208000006105 Uterine Cervical Neoplasms Diseases 0.000 description 1
- 241000251539 Vertebrata <Metazoa> Species 0.000 description 1
- 108020005202 Viral DNA Proteins 0.000 description 1
- 108020005172 Z-Form DNA Proteins 0.000 description 1
- 239000002253 acid Substances 0.000 description 1
- 150000007513 acids Chemical class 0.000 description 1
- 230000003044 adaptive effect Effects 0.000 description 1
- 210000005006 adaptive immune system Anatomy 0.000 description 1
- 229960005305 adenosine Drugs 0.000 description 1
- 235000004279 alanine Nutrition 0.000 description 1
- 125000003295 alanine group Chemical group N[C@@H](C)C(=O)* 0.000 description 1
- PNNNRSAQSRJVSB-BXKVDMCESA-N aldehydo-L-rhamnose Chemical compound C[C@H](O)[C@H](O)[C@@H](O)[C@@H](O)C=O PNNNRSAQSRJVSB-BXKVDMCESA-N 0.000 description 1
- HMFHBZSHGGEWLO-UHFFFAOYSA-N alpha-D-Furanose-Ribose Natural products OCC1OC(O)C(O)C1O HMFHBZSHGGEWLO-UHFFFAOYSA-N 0.000 description 1
- 230000004075 alteration Effects 0.000 description 1
- 150000001408 amides Chemical group 0.000 description 1
- 238000010171 animal model Methods 0.000 description 1
- PYMYPHUHKUWMLA-WDCZJNDASA-N arabinose Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)C=O PYMYPHUHKUWMLA-WDCZJNDASA-N 0.000 description 1
- PYMYPHUHKUWMLA-UHFFFAOYSA-N arabinose Natural products OCC(O)C(O)C(O)C=O PYMYPHUHKUWMLA-UHFFFAOYSA-N 0.000 description 1
- RQNWIZPPADIBDY-UHFFFAOYSA-N arsenic atom Chemical compound [As] RQNWIZPPADIBDY-UHFFFAOYSA-N 0.000 description 1
- 229940009098 aspartate Drugs 0.000 description 1
- 238000000429 assembly Methods 0.000 description 1
- 230000000712 assembly Effects 0.000 description 1
- 230000008970 bacterial immunity Effects 0.000 description 1
- 230000037429 base substitution Effects 0.000 description 1
- SRBFZHDQGSBBOR-UHFFFAOYSA-N beta-D-Pyranose-Lyxose Natural products OC1COC(O)C(O)C1O SRBFZHDQGSBBOR-UHFFFAOYSA-N 0.000 description 1
- IQFYYKKMVGJFEH-UHFFFAOYSA-N beta-L-thymidine Natural products O=C1NC(=O)C(C)=CN1C1OC(CO)C(O)C1 IQFYYKKMVGJFEH-UHFFFAOYSA-N 0.000 description 1
- DRTQHJPVMGBUCF-PSQAKQOGSA-N beta-L-uridine Natural products O[C@H]1[C@@H](O)[C@H](CO)O[C@@H]1N1C(=O)NC(=O)C=C1 DRTQHJPVMGBUCF-PSQAKQOGSA-N 0.000 description 1
- 230000008275 binding mechanism Effects 0.000 description 1
- 239000012867 bioactive agent Substances 0.000 description 1
- 238000010256 biochemical assay Methods 0.000 description 1
- 230000033228 biological regulation Effects 0.000 description 1
- 210000003995 blood forming stem cell Anatomy 0.000 description 1
- 210000002798 bone marrow cell Anatomy 0.000 description 1
- 102220354910 c.4C>G Human genes 0.000 description 1
- 238000004422 calculation algorithm Methods 0.000 description 1
- 125000000837 carbohydrate group Chemical group 0.000 description 1
- 150000001768 cations Chemical class 0.000 description 1
- 210000003855 cell nucleus Anatomy 0.000 description 1
- 238000012054 celltiter-glo Methods 0.000 description 1
- 108091092328 cellular RNA Proteins 0.000 description 1
- 230000030570 cellular localization Effects 0.000 description 1
- 230000033077 cellular process Effects 0.000 description 1
- 201000010881 cervical cancer Diseases 0.000 description 1
- 238000012512 characterization method Methods 0.000 description 1
- 150000005829 chemical entities Chemical class 0.000 description 1
- 230000002759 chromosomal effect Effects 0.000 description 1
- 230000008045 co-localization Effects 0.000 description 1
- 239000003184 complementary RNA Substances 0.000 description 1
- 230000001010 compromised effect Effects 0.000 description 1
- JECGPMYZUFFYJW-UHFFFAOYSA-N conferone Natural products CC1=CCC2C(C)(C)C(=O)CCC2(C)C1COc3cccc4C=CC(=O)Oc34 JECGPMYZUFFYJW-UHFFFAOYSA-N 0.000 description 1
- 230000021615 conjugation Effects 0.000 description 1
- 239000000356 contaminant Substances 0.000 description 1
- UHDGCWIWMRVCDJ-ZAKLUEHWSA-N cytidine Chemical compound O=C1N=C(N)C=CN1[C@H]1[C@H](O)[C@@H](O)[C@H](CO)O1 UHDGCWIWMRVCDJ-ZAKLUEHWSA-N 0.000 description 1
- SUYVUBYJARFZHO-RRKCRQDMSA-N dATP Chemical compound C1=NC=2C(N)=NC=NC=2N1[C@H]1C[C@H](O)[C@@H](COP(O)(=O)OP(O)(=O)OP(O)(O)=O)O1 SUYVUBYJARFZHO-RRKCRQDMSA-N 0.000 description 1
- SUYVUBYJARFZHO-UHFFFAOYSA-N dATP Natural products C1=NC=2C(N)=NC=NC=2N1C1CC(O)C(COP(O)(=O)OP(O)(=O)OP(O)(O)=O)O1 SUYVUBYJARFZHO-UHFFFAOYSA-N 0.000 description 1
- HAAZLUGHYHWQIW-KVQBGUIXSA-N dGTP Chemical compound C1=NC=2C(=O)NC(N)=NC=2N1[C@H]1C[C@H](O)[C@@H](COP(O)(=O)OP(O)(=O)OP(O)(O)=O)O1 HAAZLUGHYHWQIW-KVQBGUIXSA-N 0.000 description 1
- 238000012350 deep sequencing Methods 0.000 description 1
- 230000007547 defect Effects 0.000 description 1
- ZPTBLXKRQACLCR-XVFCMESISA-N dihydrouridine Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)CC1 ZPTBLXKRQACLCR-XVFCMESISA-N 0.000 description 1
- 239000000539 dimer Substances 0.000 description 1
- 230000005750 disease progression Effects 0.000 description 1
- 238000009826 distribution Methods 0.000 description 1
- 231100000673 dose–response relationship Toxicity 0.000 description 1
- 229940079593 drug Drugs 0.000 description 1
- 238000002337 electrophoretic mobility shift assay Methods 0.000 description 1
- 230000009881 electrostatic interaction Effects 0.000 description 1
- 108010048367 enhanced green fluorescent protein Proteins 0.000 description 1
- 238000011067 equilibration Methods 0.000 description 1
- 238000011156 evaluation Methods 0.000 description 1
- 230000001747 exhibiting effect Effects 0.000 description 1
- 125000004030 farnesyl group Chemical group [H]C([*])([H])C([H])=C(C([H])([H])[H])C([H])([H])C([H])([H])C([H])=C(C([H])([H])[H])C([H])([H])C([H])([H])C([H])=C(C([H])([H])[H])C([H])([H])[H] 0.000 description 1
- 125000005313 fatty acid group Chemical group 0.000 description 1
- 210000002950 fibroblast Anatomy 0.000 description 1
- 238000000684 flow cytometry Methods 0.000 description 1
- 238000000799 fluorescence microscopy Methods 0.000 description 1
- 239000007850 fluorescent dye Substances 0.000 description 1
- 102000034287 fluorescent proteins Human genes 0.000 description 1
- 108091006047 fluorescent proteins Proteins 0.000 description 1
- 125000000524 functional group Chemical group 0.000 description 1
- 238000007306 functionalization reaction Methods 0.000 description 1
- 229940029575 guanosine Drugs 0.000 description 1
- 230000036541 health Effects 0.000 description 1
- 150000002402 hexoses Chemical class 0.000 description 1
- 229920001519 homopolymer Polymers 0.000 description 1
- 229940098197 human immunoglobulin g Drugs 0.000 description 1
- 208000010544 human prion disease Diseases 0.000 description 1
- 230000002209 hydrophobic effect Effects 0.000 description 1
- 230000007124 immune defense Effects 0.000 description 1
- 230000001900 immune effect Effects 0.000 description 1
- 238000003018 immunoassay Methods 0.000 description 1
- 230000008676 import Effects 0.000 description 1
- 238000011534 incubation Methods 0.000 description 1
- 239000000411 inducer Substances 0.000 description 1
- 208000015181 infectious disease Diseases 0.000 description 1
- 229960003786 inosine Drugs 0.000 description 1
- 230000004155 insulin signaling pathway Effects 0.000 description 1
- 210000005061 intracellular organelle Anatomy 0.000 description 1
- 210000003734 kidney Anatomy 0.000 description 1
- 239000002523 lectin Substances 0.000 description 1
- 230000003902 lesion Effects 0.000 description 1
- 208000032839 leukemia Diseases 0.000 description 1
- 230000007774 longterm Effects 0.000 description 1
- 238000004020 luminiscence type Methods 0.000 description 1
- 210000004962 mammalian cell Anatomy 0.000 description 1
- 230000035800 maturation Effects 0.000 description 1
- 238000005259 measurement Methods 0.000 description 1
- 230000009061 membrane transport Effects 0.000 description 1
- 230000000813 microbial effect Effects 0.000 description 1
- 230000033607 mismatch repair Effects 0.000 description 1
- 239000000178 monomer Substances 0.000 description 1
- 230000000877 morphologic effect Effects 0.000 description 1
- 230000001537 neural effect Effects 0.000 description 1
- 210000002569 neuron Anatomy 0.000 description 1
- 230000037434 nonsense mutation Effects 0.000 description 1
- 210000000633 nuclear envelope Anatomy 0.000 description 1
- 230000025308 nuclear transport Effects 0.000 description 1
- 108091008104 nucleic acid aptamers Proteins 0.000 description 1
- 150000003833 nucleoside derivatives Chemical class 0.000 description 1
- 125000003835 nucleoside group Chemical group 0.000 description 1
- 230000009437 off-target effect Effects 0.000 description 1
- 230000008520 organization Effects 0.000 description 1
- 201000008968 osteosarcoma Diseases 0.000 description 1
- 101150091418 pam1 gene Proteins 0.000 description 1
- 239000002245 particle Substances 0.000 description 1
- 150000004713 phosphodiesters Chemical class 0.000 description 1
- 150000008300 phosphoramidites Chemical class 0.000 description 1
- 230000026731 phosphorylation Effects 0.000 description 1
- 238000006366 phosphorylation reaction Methods 0.000 description 1
- 230000007505 plaque formation Effects 0.000 description 1
- 229920001481 poly(stearyl methacrylate) Polymers 0.000 description 1
- 230000000379 polymerizing effect Effects 0.000 description 1
- 230000029279 positive regulation of transcription, DNA-dependent Effects 0.000 description 1
- 230000004481 post-translational protein modification Effects 0.000 description 1
- 230000001124 posttranscriptional effect Effects 0.000 description 1
- 230000001323 posttranslational effect Effects 0.000 description 1
- 239000011591 potassium Substances 0.000 description 1
- 229910052700 potassium Inorganic materials 0.000 description 1
- 210000001236 prokaryotic cell Anatomy 0.000 description 1
- 230000001915 proofreading effect Effects 0.000 description 1
- 230000004952 protein activity Effects 0.000 description 1
- 235000004252 protein component Nutrition 0.000 description 1
- 108020001580 protein domains Proteins 0.000 description 1
- 238000001742 protein purification Methods 0.000 description 1
- 230000017854 proteolysis Effects 0.000 description 1
- 102000005962 receptors Human genes 0.000 description 1
- 108020003175 receptors Proteins 0.000 description 1
- 238000010188 recombinant method Methods 0.000 description 1
- 108010054624 red fluorescent protein Proteins 0.000 description 1
- 230000004044 response Effects 0.000 description 1
- 230000001177 retroviral effect Effects 0.000 description 1
- 238000012552 review Methods 0.000 description 1
- 102220044145 rs144094037 Human genes 0.000 description 1
- 102220094365 rs776407427 Human genes 0.000 description 1
- 102220169418 rs886047594 Human genes 0.000 description 1
- 239000010979 ruby Substances 0.000 description 1
- 229910001750 ruby Inorganic materials 0.000 description 1
- 230000019491 signal transduction Effects 0.000 description 1
- 230000002269 spontaneous effect Effects 0.000 description 1
- 235000000346 sugar Nutrition 0.000 description 1
- 150000008163 sugars Chemical class 0.000 description 1
- CCEKAJIANROZEO-UHFFFAOYSA-N sulfluramid Chemical group CCNS(=O)(=O)C(F)(F)C(F)(F)C(F)(F)C(F)(F)C(F)(F)C(F)(F)C(F)(F)C(F)(F)F CCEKAJIANROZEO-UHFFFAOYSA-N 0.000 description 1
- 238000012360 testing method Methods 0.000 description 1
- 238000002560 therapeutic procedure Methods 0.000 description 1
- RYYWUUFWQRZTIU-UHFFFAOYSA-K thiophosphate Chemical compound [O-]P([O-])([O-])=S RYYWUUFWQRZTIU-UHFFFAOYSA-K 0.000 description 1
- 229960004072 thrombin Drugs 0.000 description 1
- 229940104230 thymidine Drugs 0.000 description 1
- 238000013334 tissue model Methods 0.000 description 1
- 239000003053 toxin Substances 0.000 description 1
- 231100000765 toxin Toxicity 0.000 description 1
- 108700012359 toxins Proteins 0.000 description 1
- 230000005026 transcription initiation Effects 0.000 description 1
- 108091006106 transcriptional activators Proteins 0.000 description 1
- 230000026683 transduction Effects 0.000 description 1
- 230000009466 transformation Effects 0.000 description 1
- 230000001960 triggered effect Effects 0.000 description 1
- HDZZVAMISRMYHH-KCGFPETGSA-N tubercidin Chemical compound C1=CC=2C(N)=NC=NC=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O HDZZVAMISRMYHH-KCGFPETGSA-N 0.000 description 1
- 238000012762 unpaired Student’s t-test Methods 0.000 description 1
- DRTQHJPVMGBUCF-UHFFFAOYSA-N uracil arabinoside Natural products OC1C(O)C(CO)OC1N1C(=O)NC(=O)C=C1 DRTQHJPVMGBUCF-UHFFFAOYSA-N 0.000 description 1
- 229940045145 uridine Drugs 0.000 description 1
- 108700026220 vif Genes Proteins 0.000 description 1
- 239000013603 viral vector Substances 0.000 description 1
- 210000005253 yeast cell Anatomy 0.000 description 1
Images














































































































































































































































































































































































































Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/113—Non-coding nucleic acids modulating the expression of genes, e.g. antisense oligonucleotides; Antisense DNA or RNA; Triplex- forming oligonucleotides; Catalytic nucleic acids, e.g. ribozymes; Nucleic acids used in co-suppression or gene silencing
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/16—Hydrolases (3) acting on ester bonds (3.1)
- C12N9/22—Ribonucleases RNAses, DNAses
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/10—Type of nucleic acid
- C12N2310/20—Type of nucleic acid involving clustered regularly interspaced short palindromic repeats [CRISPRs]
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/30—Chemical structure
- C12N2310/35—Nature of the modification
- C12N2310/351—Conjugate
- C12N2310/3519—Fusion with another nucleic acid
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/50—Physical structure
- C12N2310/53—Physical structure partially self-complementary or closed
- C12N2310/531—Stem-loop; Hairpin
Landscapes
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Engineering & Computer Science (AREA)
- Genetics & Genomics (AREA)
- Chemical & Material Sciences (AREA)
- Wood Science & Technology (AREA)
- Molecular Biology (AREA)
- Organic Chemistry (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biomedical Technology (AREA)
- Zoology (AREA)
- General Engineering & Computer Science (AREA)
- Biotechnology (AREA)
- Microbiology (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Medicinal Chemistry (AREA)
- Physics & Mathematics (AREA)
- Biophysics (AREA)
- Plant Pathology (AREA)
- Pharmaceuticals Containing Other Organic And Inorganic Compounds (AREA)
- Enzymes And Modification Thereof (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Medicinal Preparation (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
Abstract
本開示は、プライム編集中の編集効率を増加させ、in vivo半減期を増加させ、細胞の寿命を増加させる、1つ又は複数の付加されたヌクレオチド構造モチーフを含む修飾pegRNAを提供する。修飾は、アプタマー(例として、プレキノシン1-1リボスイッチアプタマー又は「evopreQ1-1」)又はそのバリアント、シュードノット(MMLVウイルスゲノムシュードノット又は「Mpknot-1」)又はそのバリアント、tRNA(例として、逆転写のためのプライマーとしてMMLVによって使用される修飾tRNA)又はそのバリアント、あるいはG-四重鎖又はそのバリアントを包含するが、これらに限定されない。本開示は更に、修飾されたpegRNAを含み、安定性、改善された細胞寿命、及び改善された編集効率を包含する改善された特性及び/又は性能を有するプライム編集因子複合体を提供する。本開示はまた、修飾されたpegRNAとのプライム編集因子複合体を使用してゲノムを編集する方法、並びに該プライム編集因子及び修飾されたpegRNAをコードするヌクレオチド配列及び発現ベクター、並びに改善されたプライム編集因子複合体を含む細胞、キット及び医薬組成物を提供する。
Description
関連出願
本出願は、2021年8月9日に出願された米国仮特許出願第63/231,231号、2021年4月30日に出願された米国仮特許出願第63/182,633号、及び2020年9月24日に出願された米国仮特許出願第63/083,067号の利益を主張する。更に、本出願は、2020年10月13日に出願された米国仮特許出願第63/091,272号の利益を主張し、その各々の内容は参照により本明細書に組み込まれる。
本出願は、2021年8月9日に出願された米国仮特許出願第63/231,231号、2021年4月30日に出願された米国仮特許出願第63/182,633号、及び2020年9月24日に出願された米国仮特許出願第63/083,067号の利益を主張する。更に、本出願は、2020年10月13日に出願された米国仮特許出願第63/091,272号の利益を主張し、その各々の内容は参照により本明細書に組み込まれる。
政府支援
本発明は、国立衛生研究所によって授与された助成金番号AI142756、HG009490、EB022376及び、GM118062の下で政府の支援を受けてなされた。政府は、本発明に一定の権利を有する。
本発明は、国立衛生研究所によって授与された助成金番号AI142756、HG009490、EB022376及び、GM118062の下で政府の支援を受けてなされた。政府は、本発明に一定の権利を有する。
プライム編集(PE)は、所望の遺伝子座におけるヌクレオチド配列における定義された変化の標的化されたプログラム型組み入れを可能にする核酸編集プラットフォームである。それは、プライム編集因子をゲノム内の標的部位に標的化することを伴い、プライム編集因子は、プライム編集ガイドRNA(pegRNA)に関連するポリメラーゼ(例として、逆転写酵素(RT))に融合された核酸プログラム型DNA結合タンパク質(napDNAbp)を含む。pegRNAは、骨格(napDNAbpに結合する)、スペーサ配列(ゲノム部位に相補的である)、及びpegRNAの3'又は5'末端の伸長アームを含む。伸長アームは、所望の編集の配列を包含するDNA合成鋳型を包含する。プライム編集中、pegRNAと複合体を形成したプライム編集因子がゲノム部位に局在化すると、ポリメラーゼ(例として、逆転写酵素)は、DNA合成鋳型を使用して所望の編集を含有するDNAの新しい鎖を合成する。次いで、DNAの新しい鎖は、ゲノム部位で対応する内生DNA鎖を置き換え、それによって所望の編集されたヌクレオチド配列を編集部位でゲノムに組み入れする。
所望の編集を指定するためのDNA合成鋳型のプログラミングの容易さ等、ゲノム編集の他の様式を超えるプライム編集の多くの利点にもかかわらず、例えば、所望の編集を組み入れする効率及び/又はインデル形成を減少させる効率を包含する、プライム編集の特徴及び性能を更に高めることが依然として望ましい。
編集効率の増加及び/又は特異性の増加をもたらすプライム編集及び/又はその構成要素の修飾は、ゲノム編集の分野を著しく進歩させるであろう。
本開示は、増加した安定性、増加したin vivo半減期、並びに/あるいはnapDNAbp及び/又は標的DNA配列に対する改善された結合親和性を包含するが、これらに限定されない改善された特性を有する次世代pegRNAを提供する。これらの改善された特性は、pegRNAの安定性を増加させるためのステムループ等の三次元RNA構造をpegRNAに付加すること、又はpegRNAのスペーサ配列に対するpegRNA伸長アームのプライマー結合部位(PBS)の結合親和性を低下させるための修飾(例として、本明細書に更に記載されるように、napDNAbp結合時に解離する足がかりでPBSを閉塞することによって、トランスで3'伸長アームを提供することによって、あるいはpegRNAに化学修飾及び/又は遺伝子修飾を導入することによって)を包含するが、それらに限定されない様々な方法で達成されてもよい。これらの修飾されたpegRNAは、napDNAbpドメイン(例として、Cas9ドメイン)及びポリメラーゼドメイン(例として、逆転写酵素ドメイン)を含む融合タンパク質等のプライム編集因子と組み合わせて使用されると、プライム編集の活性及び/又は効率の改善をもたらす。特に、本発明者らは、pegRNAが、核酸プログラム型DNA結合タンパク質(例として、Cas9ニッカーゼ)に対する親和性の低下、標準的なシングルガイドRNA(sgRNA)と比較して分解に対する感受性の増加(特に、伸長アームの劣化)、及び伸長アーム(具体的には、伸長アームのプライマー結合部位)とpegRNA中のスペーサ配列との間の望ましくない二重鎖形成による不活性化の傾向を包含する様々な欠陥を被り、それによって標的DNAへのpegRNAの結合と競合することもあることを発見した。いかなる特定の理論にも束縛されることを望むものではないが、これらの問題は、典型的なsgRNAには存在しないpegRNAの不可欠な部分である伸長アームの存在のために生じる。これらの欠点を克服するために、本発明者らは、pegRNAが、プライム編集におけるそれらの全体的な安定性及び/又は性能を改善するために1つ又はそれを超える方法で修飾されてもよいことを発見した。
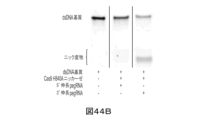
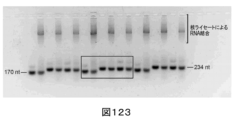
最初に、本発明者らは、1つ以上のRNA構造モチーフをpegRNAに付加することが、pegRNAの分解から保護し得ることを発見した。そのようなRNA構造モチーフは、プレケオシン1-1リボスイッチアプタマー(evopreQ1)及びその変異体、以下「mpknot」と呼ばれるモロニーマウス白血病ウイルス(MMLV)22由来のフレームシフトシュードノット及びその変異体、G-四重鎖、ヘアピン構造(例として、15bpヘアピン)、並びにグループIイントロンのP4-P6ドメインを包含し得るが、これらに限定されない。
第2に、本発明者らは、伸長アームのプライマー結合部位(PBS)とpegRNAのスペーサ配列との間の二重鎖の形成を低減させる(すなわち、PBS/スペーサ結合相互作用の低減させる)様々な方式を発見した。一態様では、PBS/スペーサバインダー相互作用は、(i)napDNAbp(例として、Cas9ニッカーゼ)結合時に解離する足がかりでPBSを閉塞すること、(ii)3'伸長アームをトランスで提供すること、すなわち、3'伸長アーム又はその部分(例として、PBS並びに/あるいはPBS及びDNA鋳型部分)をpegRNAから別の分子、例として、ニッキングgRNAに移動させること、及び(iii)RNA/DNA二重鎖形成を促進するがRNA/RNA二重鎖形成を阻害するpegRNAへの化学修飾及び/又は遺伝子修飾を導入し、それにより、pegRNAのPBSと標的DNAとの間の所望の相互作用を促進すること、を包含するがそれらに限定されない、3'伸長アームを安定化することによって回避される。
まとめると、これらの戦略の実施から生じる本明細書に開示される修飾されたpegRNAは、本明細書では「操作された」pegRNA又は「epegRNA」と呼ばれる。
本開示の別の側面では、本発明者らは、アプタマー(例として、プレキノシン1-1リボスイッチアプタマー又は「evopreQ1-1」)又はその変異体、シュードノット(MMLVウイルスゲノムシュードノット又は「Mpknot-1」)又はその変異体、tRNA(例として、逆転写のためのプライマーとしてMMLVによって使用される修飾tRNA)又はその変異体、あるいはG-四重鎖又はその変異体等であるが、これらに限定されない核酸部分にプライム編集ガイドRNA(pegRNA)をカップリングして、操作されたpegRNAを形成又はもたらすための1つ又は複数のヌクレオチドリンカーを同定するためのソフトウェアで具体化されてもよい新規な計算アルゴリズムを開発した。本明細書においてpegRNAリンカー同定ツール(「pegLIT」)と呼ばれることもある計算技術は、核酸リンカー候補を効率的に評価して、pegRNAの他の領域(例として、プライマー結合部位、スペーサ、DNA合成鋳型、及び/又はgRNAコアを含む領域)に対する塩基対形成の傾向がより低いものを同定することを伴う。
更に、本開示は、epegRNAをコード及び/又は発現する核酸分子、並びに本明細書に記載のepegRNAを発現するための発現ベクター及び構築物、当該核酸分子及び発現ベクターを含む宿主細胞、並びに本明細書に記載のプライム編集システムと併せてepegRNAを送達及び/又は投与するための組成物を提供する。更に、本開示は、単離されたepegRNA、並びに本明細書に記載の当該epegRNAを含む組成物を提供する。なお更に、本開示は、(a)プライム編集因子(例として、napDNAbp(例として、Cas9ニッカーゼ)及び逆転写酵素又は他のRNA依存性DNAポリメラーゼを含む複合体又は融合タンパク質)及び(b)本明細書に開示されるepegRNAを含むプライム編集因子システムを提供する。なお更に、本開示は、本明細書中に開示されるepegRNAを作製する方法、並びに1つ又はそれを超える変化を標的核酸分子(例として、ゲノム)に導入するためのプライム編集の方法において、プライム編集因子と比較して改善された効率でepegRNAを使用する方法を提供し、pegRNAを使用する。本明細書はまた、本明細書に記載のプライム編集システム(例として、本明細書中に記載されるようなプライム編集因子又はそれをコードするベクター若しくは構築物及び本明細書中に記載されるepegRNAの形態で)又は前述の任意のプライム編集システムを用いて、標的核酸分子、例として、ゲノムの単一核酸塩基を効率的に編集する方法を提供する。更に、本明細書は、標的核酸分子、例として、ゲノムを、本明細書に記載されるか、又は本明細書に記載されるepegRNAを利用する以前に記載されたプライム編集システムと接触させることによって、遺伝的疾患を処置するため、及び/又は遺伝的形質若しくは状態を修飾若しくは変化させるための処置方法を提供する。
特定の態様では、驚くべきことに、アプタマー(例として、プレキノシン1-1リボスイッチアプタマー又は「evopreQ1-1」)又はその変異体、シュードノット(MMLVウイルスゲノムシュードノット又は「Mpknot-1」)又はその変異体、tRNA(例として、逆転写のためのプライマーとしてMMLVによって使用される修飾tRNA)又はその変異体、あるいはG四重鎖又はその変異体を包含するがこれらに限定されないヌクレオチド構造モチーフを、pegRNAの伸長アームの末端に付加することによって、編集効率の一貫した増加が達成されたことが見出された。したがって、本開示は、プライム編集因子と複合体を形成したときに編集効率を改善する1つ以上の付加ヌクレオチド構造モチーフを含む修飾pegRNAを提供する。更に、本開示は、本明細書に開示される操作されたpegRNAと複合体化されたプライム編集因子を含むプライム編集複合体、並びに当該修飾されたpegRNAをコードするヌクレオチド配列及び発現ベクターを提供し、任意選択で、同じ又は異なるベクター分子上のプライム編集因子もコードしてもよい。なお更に、本開示は、pegRNA(すなわち、本明細書に記載の様式で修飾されていないそれらのpegRNA)を使用するプライム編集よりも高い編集効率を特徴とする核酸内の所望の部位に所望のヌクレオチド配列変化を組み入れするための、本明細書に開示される修飾pegRNAに関連するプライム編集因子の使用を伴う、プライム編集に基づくゲノム編集方法を提供する。本開示はまた、開示された修飾pegRNA、又は当該修飾pegRNAを含むプライム編集複合体を含む細胞及びキットを提供する。本開示はまた、任意選択でヌクレオチドリンカーを介して、1つ又は複数の構造ヌクレオチドモチーフ(例として、アプタマー、G-四重鎖、tRNA、又はシュードノット)をpegRNAの伸長アームの末端にカップリングさせることを含む、開示された修飾pegRNAを作製する方法を提供する。本開示は更に、所望の標的部位でゲノム編集を行うために修飾pegRNA及び任意選択でプライム編集因子を標的細胞に送達する方法、並びに開示される修飾pegRNAと組み合わせてプライム編集を使用して遺伝性障害を処置する方法を提供する。
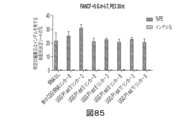
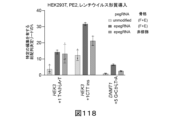
プライム編集のプロセスは、以下の遺伝子変化:トランスバーション、移行、欠失、及び挿入の少なくとも1つ又は複数を核酸(例として、ゲノム)に導入してもよい。更に、プライム編集は、特定の用途のために実施されてもよい。例えば、プライム編集は、(a)ヌクレオチド配列に対する変異補正変化を組み入れするため、(b)タンパク質及びRNAタグを組み入れするため、(c)目的のタンパク質に免疫エピトープを組み入れするため、(d)タンパク質中の二量体化ドメインを組み入れするため、(e)生体分子の活性を変化させる配列を組み入れ又は除去するため、(f)特定の遺伝子変化を指示するリコンビナーゼ標的部位を組み入れするため、(g)エラープローンRTを使用することによって標的配列を変異誘発するため、並びに他の目的のために使用され得る。また、本明細書中に記載される修飾されたpegRNAを用いて、プライム編集のこれらの適用が、インデルの高効率及び/又は低減した発生で行われてもよい。
第1の側面では、本開示は、ガイドRNAと、DNA合成鋳型及びプライマー結合部位を含む少なくとも1つの核酸伸長アームとを含むプライム編集のためのpegRNAを提供し、伸長アームは、トゥーループ、ヘアピン、ステムループ、シュードノット、アプタマー、G-四重鎖、tRNA、リボスイッチ、又はリボザイムからなる群から選択される、それに取り付けられた核酸部分を含む。特定の態様では、核酸部分は、pegRNAの伸長アームの3'末端に取り付けられている。他の態様では、核酸部分は、pegRNAの伸長アームの5'末端に取り付けられる。
様々な態様では、核酸部分は、配列番号195(Mpknot1)、配列番号196(3'トリミングしたMpknot1)、配列番号197(5'extraを有するMpknot1)、配列番号198(Mpknot1 U38A)、配列番号199(Mpknot1 U38A A29C)、配列番号200(MMLC A29C)、配列番号201(5'extra及びU38Aを有するMpknot1)、配列番号202(5'extra及びU38 A29Cを有するMpknot1)及び配列番号203(5'extra及びA29Cを有するMpknot1)からなる群から選択されるヌクレオチド配列であるか、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有するMpknot1部分である。
他の態様では、核酸部分は、配列番号204(tns1)、配列番号205(stk40)、配列番号206(apc2)、配列番号207(ceacam4)、配列番号208(pitpnm3)、配列番号209(rlf)、配列番号210(erc1)、配列番号211(ube3c)、配列番号212(taf15)、配列番号213(stard3)、及び配列番号214(g2)からなる群から選択されるヌクレオチド配列、又はそれらと少なくとも80%の配列同一性を有するヌクレオチド配列を有するG-四重鎖である。
更に他の態様では、pegRNAを修飾する核酸部分は、配列番号215(evopreq1)、配列番号216(evopreq1motif1)、配列番号217(evopreq1motif2)、配列番号218(evopreq1motif3)、配列番号219(より短いpreq 1-1)、配列番号220(preq 1-1 G5C(mut1))、及び配列番号221(preq1-1 G15C(mut2))からなる群から選択されるヌクレオチド配列、又はそれらと少なくとも80%の配列同一性を有するヌクレオチド配列を有するevopreq 1アプタマーである。
更に他の態様では、核酸部分は、配列番号222のヌクレオチド配列、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有するtRNA部分である。
更に他の態様では、核酸部分は、配列番号223(xrn1)のヌクレオチド配列、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有する。
他の態様では、核酸部分は、配列番号224(grp1イントロンP4P6)のヌクレオチド配列、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有する。
本明細書に記載される核酸部分のいずれも、リンカー、例としてヌクレオチドリンカーによって、pegRNAに、例として、pegRNAの3'末端に取り付けられ得る。リンカーは、配列番号225~236からなる群から選択されるヌクレオチド配列を有し得る。リンカーは、任意の適切な配列であり得る。任意選択で、リンカー配列は、各pegRNAについて経験的に決定され得る。
リンカーは、任意の適切な長さであり得る。特定の態様では、リンカーは、少なくとも3ヌクレオチド、少なくとも4ヌクレオチド、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、少なくとも15ヌクレオチド、少なくとも16ヌクレオチド、少なくとも17ヌクレオチド、少なくとも18ヌクレオチド、少なくとも19ヌクレオチド、少なくとも20ヌクレオチド、少なくとも21ヌクレオチド、少なくとも22ヌクレオチド、少なくとも23ヌクレオチド、少なくとも24ヌクレオチド、少なくとも25ヌクレオチド、少なくとも26ヌクレオチド、少なくとも27ヌクレオチド、少なくとも28ヌクレオチド、少なくとも29ヌクレオチド、又は少なくとも30ヌクレオチド長である。
好ましい態様では、リンカーは少なくとも8ヌクレオチド長である。
様々な態様では、pegRNAの伸長アームは、ガイドRNAの3'若しくは5'末端又はガイドRNAの分子内位置に配置され、核酸伸長アームはDNA又はRNAである。
様々な態様では、pegRNAは、napDNAbpに結合し、napDNAbpを標的DNA配列に向けることができる。標的DNA配列は、標的鎖及び相補的な非標的鎖を含み得る。ガイドRNAは、標的鎖にハイブリダイズして、RNA-DNAハイブリッド及びRループを形成し得る。
様々な態様において、伸長アームの長さは様々であり得、DNA合成鋳型の長さに依存する。特定の態様では、核酸伸長アームは、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、少なくとも15ヌクレオチド、少なくとも16ヌクレオチド、少なくとも17ヌクレオチド、少なくとも18ヌクレオチド、少なくとも19ヌクレオチド、少なくとも20ヌクレオチド、少なくとも21ヌクレオチド、少なくとも22ヌクレオチド、少なくとも23ヌクレオチド、少なくとも24ヌクレオチド、少なくとも25ヌクレオチド、少なくとも26ヌクレオチド、少なくとも27ヌクレオチド、少なくとも28ヌクレオチド、少なくとも29ヌクレオチド、少なくとも30ヌクレオチド、少なくとも31ヌクレオチド、少なくとも32ヌクレオチド、少なくとも33ヌクレオチド、少なくとも34ヌクレオチド、少なくとも35ヌクレオチド、少なくとも36ヌクレオチド、少なくとも37ヌクレオチド、少なくとも38ヌクレオチド、少なくとも39ヌクレオチド、少なくとも40ヌクレオチド、少なくとも41ヌクレオチド、少なくとも42ヌクレオチド、少なくとも43ヌクレオチド、少なくとも44ヌクレオチド、少なくとも45ヌクレオチド、少なくとも46ヌクレオチド、少なくとも47ヌクレオチド、少なくとも48ヌクレオチド、少なくとも49ヌクレオチド、又は少なくとも50ヌクレオチドである。
DNA合成鋳型はまた、所望の編集に応じて異なり得、少なくとも3ヌクレオチド、少なくとも4ヌクレオチド、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、又は少なくとも15ヌクレオチド長であり得る。
様々な態様において、所望の編集は、単一ヌクレオチド置換、又は単一ヌクレオチド欠失、又は挿入である。所望の編集はまた、プライム編集によって組み入れされることができる任意の長さであり得、削除、挿入、又は反転を包含し得る。
プライマー結合部位はまた、長さが異なり得、例えば、少なくとも3ヌクレオチド、少なくとも4ヌクレオチド、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、又は少なくとも15ヌクレオチド長であり得る。
別の側面において、本開示は、(i)スペーサを含むガイドRNA、及び(ii)DNA合成鋳型、プライマー結合部位、足がかりモチーフ、及び更なる核酸部分を含む少なくとも1つの核酸伸長アームを含む、プライム編集のためのpegRNAを提供し、ここで、その足がかりモチーフは、PEgRNAがプライム編集因子によって結合されていないときはプライマー結合部位とスペーサとの相互作用を遮り、PEgRNAがプライム編集因子によって結合されているときはプライマー結合部位と標的DNA分子上のプロトスペーサ配列との相互作用を遮らない。いくつかの態様では、足がかりモチーフ及び追加の核酸部分は、伸長アームの3'末端に取り付けられる。いくつかの態様では、足がかりモチーフは、伸長アームの3'末端に取り付けられ、追加の核酸部分は、足がかりモチーフの3'末端に取り付けられる。いくつかの態様では、足がかりモチーフは、リンカーによってPEgRNAに取り付けられる。
別の側面において、本開示は、(i)ガイドRNAを含む第1のPEgRNAであって、ガイドRNAがスペーサを含む、第1のPEgRNAと、(ii)第2の鎖ニッキングガイドRNAを含む第2のPEgRNAであって、DNA合成鋳型及びプライマー結合部位を含む少なくとも1つの核酸伸長アームを含む、第2の鎖ニッキングガイドRNAと、を含む、プライム編集のためのPEgRNA対を提供する。いくつかの態様では、第1のPEgRNA及び第2のPEgRNAはそれぞれ、プライム編集因子の核酸プログラム型DNA結合タンパク質(napDNAbp)に結合し、napDNAbpを標的DNA配列に向けることができる。
別の側面では、本開示は、(i)スペーサを含むガイドRNAと、(ii)DNA合成鋳型及びプライマー結合部位を含む少なくとも1つの核酸伸長アームとを含むPEgRNAを提供し、プライマー結合部位は1つ又は複数の修飾ヌクレオチドを含み、1つ又は複数の修飾ヌクレオチドは、標的DNA分子上のプロトスペーサ配列に対するプライマー結合部位よりもスペーサに対するプライマー結合部位の結合親和性のより大きな低減をもたらす。いくつかの態様において、1つ又は複数の修飾ヌクレオチドは遺伝子変異を含む。いくつかの態様において、1つ又は複数の修飾ヌクレオチドは化学修飾ヌクレオチドを含む。
別の側面では、本開示は、プライム編集のための複合体であって、
(a)核酸プログラム型DNA結合タンパク質(napDNAbp)及びRNA依存性DNAポリメラーゼ活性を含むドメインを含む融合タンパク質;及び
(b)伸長アームの末端に付加された核酸部分を含む上記の任意のpegRNA、を含む、プライム編集のための複合体を提供する。
(a)核酸プログラム型DNA結合タンパク質(napDNAbp)及びRNA依存性DNAポリメラーゼ活性を含むドメインを含む融合タンパク質;及び
(b)伸長アームの末端に付加された核酸部分を含む上記の任意のpegRNA、を含む、プライム編集のための複合体を提供する。
いくつかの態様では、プライム編集複合体のnapDNAbpは、核酸プログラム型DNA結合能力を有するエンドヌクレアーゼを含む。いくつかの態様では、napDNAbpは、二本鎖標的DNAの両方の鎖を切断することができる活性エンドヌクレアーゼを含む。いくつかの態様では、napDNAbpは、各鎖にニックを生成することによって二本鎖標的DNAの両方の鎖を切断し得る、ヌクレアーゼ活性エンドヌクレアーゼ、例としてヌクレアーゼ活性Casタンパク質である。例えば、ヌクレアーゼ活性Casタンパク質は、二本鎖標的DNAの各鎖に切断(ニック)を生成し得る。いくつかの態様では、両方の鎖上の2つのニックは、例えばCas12a又はCas12b1を含むnapDNAbpによって生成される互い違いのニックである。いくつかの態様では、両方の鎖上の2つのニックは、同じゲノム位置にあり、例えば、ヌクレアーゼ活性Cas9を含むnapDNAbpによって生成される。いくつかの態様では、napDNAbpは、ニッカーゼであるエンドヌクレアーゼを含む。例えば、いくつかの態様では、napDNAbpは、エンドヌクレアーゼのヌクレアーゼ活性を低下させ、それをニッカーゼにする1つ以上の突然変異を含むエンドヌクレアーゼを含む。いくつかの態様では、napDNAbpは不活性エンドヌクレアーゼを含み、例えば、いくつかの態様では、napDNAbpは、ヌクレアーゼ活性を無効にする1つ以上の突然変異を含むエンドヌクレアーゼを含む。様々な態様では、napDNAbpはCas9タンパク質又はその変異体である。napDNAbpはまた、ヌクレアーゼ活性Cas9、ヌクレアーゼ不活性Cas9(dCas9)、又はCas9ニッカーゼ(nCas9)であり得る。好ましい態様では、napDNAbpは、一本鎖のみにニックを入れるCas9ニッカーゼ(nCas9)である。他の態様では、napDNAbpは、Cas9、Cas12e、Cas12d、Cas12a、Cas12b1、Cas12b2、Cas13a、Cas12c、Cas12d、Cas12e、Cas12h、Cas12i、Cas12g、Cas12f(Cas14)、Cas12f1、Cas12j(CasΦ)、及びアルゴノートからなる群から選択され得、任意選択で、1本の鎖のみが切断されるようにニッカーゼ活性を有しする。いくつかの態様では、napDNAbpは、Cas9、Cas12e、Cas12d、Cas12a、Cas12b1、Cas12b2、Cas13a、Cas12c、Cas12d、Cas12e、Cas12h、Cas12i、Cas12g、Cas12f(Cas14)、Cas12f1、Cas12j(CasΦ)、及びアルゴノートから選択され、任意選択で、一方のDNA鎖が他方のDNA鎖に対して優先的に切断されるようにニッカーゼ活性を有する。様々な態様では、RNA依存性DNAポリメラーゼ活性を含むドメインは、配列番号32、34、36、102~128及び132のアミノ酸配列のいずれか1つを含む逆転写酵素である。
RNA依存性DNAポリメラーゼ活性を含むドメインは、いくつかの態様では、配列番号32、34、36、102~128及び132のいずれか1つのアミノ酸配列と少なくとも80%、85%、90%、95%、98%、又は99%の配列同一性を有するアミノ酸配列を含む逆転写酵素である。他の態様では、RNA依存性DNAポリメラーゼ活性を含むドメインは、レトロウイルス又はレトロトランスポゾン由来の天然に存在する逆転写酵素である。
別の側面では、本開示は、上記及び本開示で提供される修飾pegRNAをコードする核酸分子を提供する。
更に別の側面では、本開示は、上記核酸分子を含む発現ベクターを提供する。核酸分子は、プロモータの制御下にあり得る。プロモータは、polIIIプロモータであり得る。プロモータはまた、配列番号3915~3918のいずれかのヌクレオチド配列を有するプロモータを包含する、U6、U6v4、U6v7、若しくはU6v9プロモータ又はそのフラグメントであり得る。
更に別の側面では、本開示は、上記の修飾pegRNAを含む細胞(例として、形質トランスバーション細胞株)を提供する。細胞はまた、上記のプライム編集複合体を含み得る(例として、細胞は、修飾されたpegRNA及びプライム編集因子の両方を含む)。細胞はまた、修飾されたpegRNAを発現し、任意選択でプライム編集因子を発現する上記の核酸分子のいずれかを含み得る。更に、細胞は、修飾されたpegRNAを発現し、任意選択でプライム編集因子を発現する上記の発現ベクターのいずれかを含み得る。
別の側面では、本開示は、(i)上記の修飾されたpegRNA、又は上記のプライム編集複合体、上記の核酸分子、又は上記の発現ベクター、又は上記の細胞のいずれかと、(ii)薬学的に許容される賦形剤と、を含む医薬組成物を提供する。
更に別の側面では、本開示は、(i)上記の修飾されたpegRNA、又は上記のプライム編集複合体、上記の核酸分子、又は上記の発現ベクター、又は上記の細胞のいずれかと、(ii)プライム編集を行うための説明書のセットと、を含むキットを提供する。
別の側面では、本開示は、(i)本明細書に開示される任意のpegRNA又はepegRNAと、(ii)napDNAbp及びDNAポリメラーゼを含む少なくとも1つのプライム編集因子と、を含むシステムを提供する。
別の側面では、本開示は、標的DNA配列を上記の修飾されたpegRNAと接触させることを含むプライム編集の方法、及びnapDNAbpとRNA依存性DNAポリメラーゼ活性を有するドメインとを含むプライム編集因子を提供し、ここで編集効率は、修飾を含まないpegRNAを使用する同じ方法と比較して増加される。特定の態様では、編集効率は、少なくとも1.5倍増加する。他の態様では、編集効率は少なくとも2.0倍増加する。更に他の態様では、編集効率は少なくとも3.0倍増加する。更に他の態様では、編集効率は、少なくとも4、5、6、7、8、9、又は10倍増加する。
別の側面では、本開示は、ガイドRNA(pegRNA)と組み合わせてプライム編集因子(例として、PE1、PE2、又はPE3)を使用してプライム編集を実行して、CDKL5欠損症を引き起こすCDKL5遺伝子の突然変異を直接組み入れするか、又は修正する。様々な態様では、本開示は、プライム編集因子(例として、PE1、PE2、又はPE3)と、複数の対象においてCDKL5遺伝子に2つ以上の突然変異を直接組み入れするか、又は修正することができるpegRNAと、を含む複合体を提供する。
本明細書に開示されるプライム編集の方法では、napDNAbpはニッカーゼ活性を有し得る。napDNAbpは、Cas9タンパク質又はその変異体であり得る。napDNAbpはまた、ヌクレアーゼ活性Cas9、ヌクレアーゼ不活性Cas9(dCas9)、又はCas9ニッカーゼ(nCas9)であり得る。napDNAbpはまた、Cas9、Cas12e、Cas12d、Cas12a、Cas12b1、Cas12b2、Cas13a、Cas12c、Cas12d、Cas12e、Cas12h、Cas12i、Cas12g、Cas12f(Cas14)、Cas12f1、Cas12j(CasΦ)、及びアルゴノートであり得、任意選択でニッカーゼ活性を有し得る。
プライム編集の方法において、RNA依存性DNAポリメラーゼ活性は、配列番号32、34、36、102~128及び132のアミノ酸配列のいずれか1つを含む逆転写酵素であり得る。他の態様では、RNA依存性DNAポリメラーゼ活性は、配列番号32、34、36、102~128、及び132のいずれか1つのアミノ酸配列と少なくとも80%、85%、90%、95%、98%、又は99%の配列同一性を有するアミノ酸配列を含む逆転写酵素であり得る。
本明細書は更に、プライム編集に関する以下の参照の出願、すなわち、2019年3月19日に出願された米国仮出願第62/820,813号(代理人整理番号B1195.70074 US 00)、2019年6月7日に出願された米国仮出願第62/858,958号(代理人整理番号B1195.70074 US 01)、2019年8月21日に出願された米国仮出願第62/889,996号(代理人整理番号B1195.70074 US 02)、2019年8月21日に出願された米国仮出願第62/922,654号(代理人整理番号B1195.70083 US 00)、2019年10月10日に出願された米国仮出願第62/913,553号(代理人整理番号B1195.70074 US 03)、2019年10月10日に出願された米国仮出願第62/973,558号(代理人整理番号B1195.70083 US 01)、2019年11月5日に出願された米国仮出願第62/931,195号(代理人整理番号B1195.70074 US 04)、2019年12月5日に出願された米国仮出願第62/944,231号(代理人整理番号B1195.70074 US 05)、2019年12月5日に出願された米国仮出願第62/974,537号(代理人整理番号B1195.70083 US 02)、2020年3月17日に出願された米国仮出願第62/991,069号(代理人整理番号B1195.70074 US 06)、及び2020年3月17日に出願された米国仮出願第(63/100,548)(代理人整理番号B1195.70083 US 03)を参照し、組み込む。更に、この米国仮出願は、それぞれ2020年3月19日に出願された、国際PCT出願番号PCT/US 20/23721;PCT/US 20/23730;PCT/US 20/23713;PCT/US 20/23712;PCT/US 20/23727;PCT/US 20/23724;PCT/US 20/23725;PCT/US 20/23728;PCT/US 20/23732;PCT/US 20/23723;PCT/US 20/23553;及びPCT/US 20/23583に記載されている。
前述の概念、及び後述する追加の概念は、本開示がこの点に関して限定されないため、任意の適切な組み合わせで配置されてもよいことを了解されるべきである。更に、本開示の他の利点及び新規な特徴は、添付の図面と併せて考慮すると、様々な非限定的な態様の以下の詳細な説明から明らかになるであろう。
以下の図面は、本明細書の一部を形成し、本明細書に提示される特定の側面の詳細な説明と組み合わせてこれらの図面の1つ又は複数を参照することによってよりよく理解され得る本開示の特定の態様を更に実証するために包含される。
図3Eの態様は、本明細書で企図される別のpegRNAの構造を提供する。pegRNAは、5'から3'方向に並べられた3つの主要な構成要素、すなわち、スペーサ、gRNAコア、及び3'末端の伸長アームを含む。伸長アームは、5'から3'方向の以下の構造要素、すなわち、任意選択の相同アーム、DNA合成鋳型、及びプライマー結合部位(PBS)に更に分割されてもよい。更に、pegRNAは、任意選択の3'末端修飾領域(e1)及び任意選択の5'末端修飾領域(e2)を含んでもよい。更に、pegRNAは、pegRNAの3'末端上に転写終結シグナルを含んでもよい(図示せず)。これらの構造要素は、本明細書で更に定義される。pegRNAの構造の描写は、限定することを意味されるものではなく、要素の配置のバリエーションを包含する。例えば、任意選択の配列修飾因子(e1)及び(e2)は、3'末端及び5'末端に位置することに限定されず、示されている他の領域のいずれかの中又は間に配置され得る。pegRNAは、特定の態様では、ヘアピン、ステム/ループ、トゥーループ、RNA結合タンパク質動員ドメイン(例として、MS2cpタンパク質を動員し、MS2cpタンパク質に結合するMS2アプタマー)等であるがこれらに限定されない二次RNA構造を含み得る。これらの二次構造は、pegRNA分子内のどこにでも配置され得る。例えば、そのような二次構造は、スペーサ、gRNAコア、又は伸長アーム内、特にe1及び/又はe2修飾領域内の位置であり得る。二次RNA構造に加えて、pegRNAは、化学リンカー又はポリ(N)リンカー若しくはテールを含むことができ(例として、e1及び/又はe2修飾領域内)、「N」は任意の核酸塩基であり得る。いくつかの態様(例として、図72(c)に示すように)では、化学リンカーは、sgRNA骨格又はコアの逆転写を防止するように機能してもよい。更に、特定の態様(例として、図72(c)を参照されたい)では、伸長アーム(3)は、RNA又はDNAで構成され得、及び/又は1つ以上の核酸塩基類似体(例として、温度回復力等の機能を追加する可能性がある)を包含し得る。更に、伸長アーム(3)の配向は、天然の5'から3'方向であり得、又は(全体的なpegRNA分子の配向に対して)3'から5'方向の反対の配向で合成され得る。当業者は、所望の編集を包含する所望の鋳型コード3'一本鎖DNAフラップを合成するために、napDNAbpとの融合として実施されてもよいか、又は別個の部分としてトランスで提供されるプライム編集での使用のために、伸長アーム(すなわち、DNA又はRNA)の核酸材料の性質に応じて、適切なDNAポリメラーゼを選択することができることにも留意されたい。例えば、伸長アームがRNAである場合、DNAポリメラーゼは逆転写酵素又は任意の他の適切なRNA依存性DNAポリメラーゼであり得る。しかしながら、伸長アームがDNAである場合、DNAポリメラーゼはDNA依存性DNAポリメラーゼであり得る。様々な態様において、DNAポリメラーゼの提供は、例として、RNA-タンパク質動員ドメイン(例として、pegRNAに組み入れされたMS2ヘアピン(例として、e1若しくはe2領域、又は他の場所で、及びDNAポリメラーゼに融合したMS2cpタンパク質、それによってDNAポリメラーゼをpegRNAに共局在化させる)の使用を介してトランスであり得る。プライマー結合部位は、一般に、所望の編集を包含する得られた3'一本鎖DNAフラップをコードするためにDNAポリメラーゼ(例として、逆転写酵素)によって使用される鋳型の一部を形成しないことにも留意されたい。したがって、「DNA合成鋳型」の名称は、編集を含有する所望の3'一本鎖DNAフラップと、プライム編集DNA合成の3'一本鎖DNA鎖産物によって置き換えられる5'内生一本鎖DNAフラップと相同性の領域と、をコードするためにDNAポリメラーゼによって鋳型として使用される伸長アーム(3)の領域又は部分を指す。いくつかの態様では、DNA合成鋳型は、「編集鋳型」及び「相同アーム」、又は例として編集鋳型の前後の1つ若しくは複数の相同アームを包含する。編集鋳型は、単一ヌクレオチド置換ほど小さくあり得、又はDNAの挿入若しくは逆位であってもよい。更に、編集鋳型はまた、所望の欠失を含有する相同アームをコードすることによって操作され得る欠失を包含してもよい。他の態様では、DNA合成鋳型はまた、e2領域又はその一部を包含してもよい。例えば、e2領域がDNAポリメラーゼ活性の終結を引き起こす二次構造を含む場合、e2領域の任意の部分が実際にDNAにコードされる前にDNAポリメラーゼ機能が終結される可能性がある。e2領域の一部又は更には全部がDNAにコードされることも可能である。e2のどの程度が実際に鋳型として使用されるかは、その構成及びその構成がDNAポリメラーゼ機能を中断させるかどうかに依存する。
図3Fの概略図は、典型的なpegRNAと二本鎖DNAの標的部位との相互作用、及び目的の遺伝的変化を含有する3'一本鎖DNAフラップの同時生成を示す。二本鎖DNAは、3'から5'方向の上部鎖(すなわち、標的鎖)及び5'から3'方向の下部鎖(すなわち、PAM鎖又は非標的鎖)で示されている。上部鎖は、「プロトスペーサ」の相補体とPAM配列の相補体とを含み、それは、それがpegRNAのスペーサによる標的であり、それにアニールする鎖であるので、「標的鎖」と呼ばれる。相補的な下部鎖は、PAM配列(例として、NGG)及びプロトスペーサを含有するため、「非標的鎖」又は「PAM鎖」又は「プロトスペーサ鎖」と呼ばれる。示されていないが、図示されたpegRNAは、プライム編集因子のCas9又は同等のドメインと複合体化されるであろう。概略図(図3F)に示すように、pegRNAのスペーサ配列は、標的鎖上のプロトスペーサの相補領域にアニールする。この相互作用は、スペーサRNAとプロトスペーサDNAの相補体との間でDNA/RNAハイブリッドとして形成し、プロトスペーサにおけるRループの形成を誘導する。本明細書の他の箇所で教示されるように、Cas9タンパク質(図示せず)は、次いで、示されるように、非標的鎖にニックを誘導する。これにより、ニック部位のすぐ上流に3'ssDNAフラップ領域の形成を導き、これは*z*に従って、プライマー結合部位でpegRNAの3'末端と相互作用する。ssDNAフラップ(すなわち、逆転写酵素プライマー配列)の3'末端は、pegRNA上のプライマー結合部位(A)にアニーリングし、それによって逆転写酵素をプライミングする。次に、逆転写酵素(例として、トランスで提供されるか、又はCas9構築物に取り付けられた融合タンパク質としてシスで提供される)は、DNA合成鋳型(編集鋳型(B)及び相同アーム(C)を包含する)によってコードされるDNAの一本鎖を重合する。重合は、伸張アームの5'末端に向かって継続する。ssDNAの重合鎖は、ssDNA 3'末端フラップを形成し、これは、他の箇所に記載されているように(例として、図1Gに示すように)、内生DNAに侵入し、対応する内生鎖を置き換え(内生DNAの5'末端DNAフラップとして除去される)、DNA修復/複製ラウンドを通して所望のヌクレオチド編集(単一ヌクレオチド塩基対の変化、欠失、挿入(全遺伝子を包含する))を組み入れする。
図3Gは、本明細書で企図されるプライム編集の更に別の態様を示す。特に、上の概略図は、リンカーによって一緒にされたnapDNAbp(例として、SpCas9)とポリメラーゼ(例として、逆転写酵素)との融合タンパク質を含む、プライム編集因子(PE)の一態様を示す。PEは、pegRNAのgRNAコアに結合することによって、pegRNAと複合体を形成する。示されている態様では、pegRNAには、3'末端から開始してプライマー結合部位(PBS)とそれに続くDNA合成鋳型とを含む3'伸長アームが装備されている。下の概略図は、「トランスプライム編集因子(tPE)」と呼ばれるプライム編集因子の変形を示す。この態様では、DNA合成鋳型及びPBSは、pegRNAから切り離され、RNA-タンパク質動員ドメイン(例として、MS2ヘアピン)を含むトランスプライム編集因子RNA鋳型(「tPERT」)と呼ばれる別個の分子上に提示される。PE自体は、RNA-タンパク質動員ドメインを特異的に認識し、それに結合するタンパク質であるrPERT動員タンパク質(「RP」)に対する融合物を含むように更に修飾される。RNA-タンパク質動員ドメインがMS2ヘアピンである例では、対応するrPERT動員タンパク質は、MS2タグ付けシステムのMS2cpであり得る。MS2タグ付けシステムは、MS2バクテリオファージコートタンパク質(「MCP」又は「MS2cp」)と、ファージのゲノムに存在するステムループ又はヘアピン構造、すなわち「MS2ヘアピン」又は「MS2アプタマー」との天然の相互作用に基づく。トランスプライム編集の場合、RP-PE:gRNA複合体は、適切なRNA-タンパク質動員ドメインを有するtPERTを「動員」してPE:gRNA複合体と共局在化し、それにより、図3Hに表される例に示されるように、プライム編集に使用するためのPBS及びDNA合成鋳型をトランスで提供する。
図3Hは、トランスプライム編集のプロセスを示す。この態様では、トランスプライム編集因子は、MS2cpタンパク質(すなわち、MS2アプタマーを認識し、MS2アプタマーに結合する動員タンパク質のタイプ)に融合し、sgRNA(すなわち、pegRNAとは対照的に標準的なガイドRNA)と複合体化した「PE2」プライム編集因子(すなわち、Cas9(H840A)と変異体MMLV RTとの融合物)を含む。トランスプライム編集因子は標的DNAに結合し、非標的鎖にニックを入れる。MS2cpタンパク質は、tPERT分子上のRNA-タンパク質動員ドメインとの特異的相互作用を介してtPERTをトランスで動員する。tPERTは、トランスプライム編集因子と共局在化し、それによって、逆転写酵素ポリメラーゼによる使用のためにPBS及びDNA合成鋳型機能をトランスで提供して、3'末端を有し、DNA合成鋳型によってコードされる所望の遺伝情報を含有する一本鎖DNAフラップを合成する。
9のアミノ酸741-743(C末端エクステインの最初の残基)の配列であり、これらはインテイントランススプライシング反応にとって重要である。「SMP」はネイティブな残基である。我々はこれらを「CFN」コンセンサススプライシング配列へもまた変異させた。プライム編集パーセンテージによって測定される通り、コンセンサス配列は最も高い再構成を生むことが示されている。
図84Bは、スペーサ、gRNAコア、及び伸長アーム(RT鋳型+プライマー結合部位)を有する例示のpegRNAを図示する。これは、UCUリンカーを介してカップリングされたtRNA分子によってpegRNAの3'端を修飾されている。tRNAは種々の転写後修飾を包含する。しかしながら、該修飾は要求されない。例2を見よ。
定義
他に定義されない限り、本明細書で使用される全ての技術用語及び科学用語は、本発明が属する技術分野の当業者によって一般的に理解される意味を有する。以下の参考文献は、当業者に、本発明で使用される多くの用語の一般的な定義を提供する:Singleton et al.,Dictionary of Microbiology and Molecular Biology(2nd ed.1994);The Cambridge Dictionary of Science and Technology(Walker ed.,1988);The Glossary of Genetics,5th Ed.,R.Rieger et al.(eds.),Springer Verlag(1991);and Hale&Marham,The Harper Collins Dictionary of Biology(1991)。本明細書で使用される場合、以下の用語は、特に指定されない限り、それらに帰する意味を有する。
他に定義されない限り、本明細書で使用される全ての技術用語及び科学用語は、本発明が属する技術分野の当業者によって一般的に理解される意味を有する。以下の参考文献は、当業者に、本発明で使用される多くの用語の一般的な定義を提供する:Singleton et al.,Dictionary of Microbiology and Molecular Biology(2nd ed.1994);The Cambridge Dictionary of Science and Technology(Walker ed.,1988);The Glossary of Genetics,5th Ed.,R.Rieger et al.(eds.),Springer Verlag(1991);and Hale&Marham,The Harper Collins Dictionary of Biology(1991)。本明細書で使用される場合、以下の用語は、特に指定されない限り、それらに帰する意味を有する。
アンチセンス鎖
遺伝学において、二本鎖のDNA内のセグメントの「アンチセンス」鎖は、鋳型鎖であって、3'→5'配向に伸びる(runs)と考えられる。これに反して、「センス」鎖は、5'から3'へ伸びる二本鎖のDNA内のセグメントであって、3'から5'へ伸びるDNAのアンチセンス鎖又は鋳型鎖に相補的である。タンパク質をコードするDNAセグメントのケースにおいて、センス鎖は、転写の最中にその鋳型としてアンチセンス鎖を取り結局(典型的には、常にではない)翻訳を経てタンパク質になるmRNAと同じ配列を有するDNAの鎖である。よって、アンチセンス鎖は、後にタンパク質へ翻訳されるRNAを担い、一方センス鎖は、mRNAとほぼ同一の組成(makeup)を保有する。dsDNAの各セグメントについて、一方がどの方向から読まれるかに応じて、おそらく(センス及びアンチセンスは視点に相対するため)センス及びアンチセンスが2組存在するであろう点に留意する。遺伝子産物又はmRNAは結局、dsDNAの一方のセグメントのどちらかの鎖がセンス又はアンチセンスと呼ばれることを示す。
遺伝学において、二本鎖のDNA内のセグメントの「アンチセンス」鎖は、鋳型鎖であって、3'→5'配向に伸びる(runs)と考えられる。これに反して、「センス」鎖は、5'から3'へ伸びる二本鎖のDNA内のセグメントであって、3'から5'へ伸びるDNAのアンチセンス鎖又は鋳型鎖に相補的である。タンパク質をコードするDNAセグメントのケースにおいて、センス鎖は、転写の最中にその鋳型としてアンチセンス鎖を取り結局(典型的には、常にではない)翻訳を経てタンパク質になるmRNAと同じ配列を有するDNAの鎖である。よって、アンチセンス鎖は、後にタンパク質へ翻訳されるRNAを担い、一方センス鎖は、mRNAとほぼ同一の組成(makeup)を保有する。dsDNAの各セグメントについて、一方がどの方向から読まれるかに応じて、おそらく(センス及びアンチセンスは視点に相対するため)センス及びアンチセンスが2組存在するであろう点に留意する。遺伝子産物又はmRNAは結局、dsDNAの一方のセグメントのどちらかの鎖がセンス又はアンチセンスと呼ばれることを示す。
アプタマー
「アプタマー」は、特定の標的分子に結合するオリゴヌクレオチド又はペプチド分子を指す。アプタマーには、それらの固有の三次元構造に折り畳まれたときに、低分子リガンド又はタンパク質標的に高い親和性及び特異性で選択的に結合し得る短い一本鎖DNA又はRNAベースのオリゴヌクレオチドであるDNA又はRNAアプタマーを包含する。分子レベルでは、アプタマーは、様々な非共有結合相互作用、静電相互作用、疎水性相互作用、及び誘導された適合を介してその同族標的に結合する。更なる参照が、Ku et al.,「Nucleic Acid Aptamers:An Emerging Tool for Biotechnology and Biomedical Sensing,」Sensors,2015,15(7):16281-16313に対してなされ得る。本開示は、商業的供給源から得られるものを包含する、任意のアプタマーの使用を企図する。例えば、多数のアプタマーがAPTAGEN(www.aptagen.com)から得られてもよく、これらは、トロンビン(15量体)、HIV-1 TAR RNAヘアピンループ(B22-19)、ヒト免疫グロブリンG(IgG)(Apt 8)、反応性緑色19(GR-30)、アブリン毒素(TA6)、マラカイトグリーン(MG-4)、PSMAアプタマー(A10-3)、テネイシン-C(GBI-10)及びメチレンジアニリン(M1)を包含するが、これらに限定されない。別の例は、最小の天然の三次RNA構造(evopreQ1-1としても知られる)の1つであるプレケオシン1-1リボスイッチアプタマーである。
「アプタマー」は、特定の標的分子に結合するオリゴヌクレオチド又はペプチド分子を指す。アプタマーには、それらの固有の三次元構造に折り畳まれたときに、低分子リガンド又はタンパク質標的に高い親和性及び特異性で選択的に結合し得る短い一本鎖DNA又はRNAベースのオリゴヌクレオチドであるDNA又はRNAアプタマーを包含する。分子レベルでは、アプタマーは、様々な非共有結合相互作用、静電相互作用、疎水性相互作用、及び誘導された適合を介してその同族標的に結合する。更なる参照が、Ku et al.,「Nucleic Acid Aptamers:An Emerging Tool for Biotechnology and Biomedical Sensing,」Sensors,2015,15(7):16281-16313に対してなされ得る。本開示は、商業的供給源から得られるものを包含する、任意のアプタマーの使用を企図する。例えば、多数のアプタマーがAPTAGEN(www.aptagen.com)から得られてもよく、これらは、トロンビン(15量体)、HIV-1 TAR RNAヘアピンループ(B22-19)、ヒト免疫グロブリンG(IgG)(Apt 8)、反応性緑色19(GR-30)、アブリン毒素(TA6)、マラカイトグリーン(MG-4)、PSMAアプタマー(A10-3)、テネイシン-C(GBI-10)及びメチレンジアニリン(M1)を包含するが、これらに限定されない。別の例は、最小の天然の三次RNA構造(evopreQ1-1としても知られる)の1つであるプレケオシン1-1リボスイッチアプタマーである。
Cas9
用語「Cas9」又は「Cas9ヌクレアーゼ」は、Cas9ドメイン又はそのフラグメントを含む、RNAにガイドされるヌクレアーゼ(例として、Cas9の活性若しくは不活性なDNA切断ドメイン及び/又はCas9のgRNA結合ドメインを含むタンパク質)を指す。「Cas9ドメイン」は、本明細書に使用されるとき、Cas9の活性若しくは不活性な切断ドメイン及び/又はCas9のgRNA結合ドメインを含むタンパク質フラグメントである。「Cas9タンパク質」は、全長Cas9タンパク質である。Cas9ヌクレアーゼはまた、ときにcasn1ヌクレアーゼ又はCRISPR(クラスター化して規則的な配置の短い回文配列反復(Clustered Regularly Interspaced Short Palindromic Repeat))関連ヌクレアーゼとしても言及される。CRISPRは、可動遺伝因子(ウイルス、転移因子、及び接合性プラスミド)に対して保護を提供する適応免疫系である。CRISPRクラスターは、スペーサ、先行する可動因子に相補的な配列、及び標的化侵入核酸を含有する。CRISPRクラスターは転写され、プロセスを受けて(processed)CRISPR RNA(crRNA)になる。II型CRISPR系において、pre-crRNAの修正プロセシングは、transでコードされた低分子(small)RNA(tracrRNA)、内生リボヌクレアーゼ3(rnc)、及びCas9ドメインを要する。tracrRNAは、pre-crRNAの、リボヌクレアーゼ3に補助された(-aided)プロセシングのためのガイドとして働く。続いて、Cas9/crRNA/tracrRNAは、スペーサに相補的な線状又は環状のdsDNA標的をエンドヌクレアーゼ的に切断する。crRNAに相補的ではない標的鎖は、最初にエンドヌクレアーゼ的に切られ、次いで3'-5'エキソヌクレアーゼ的に切り取られる。本来、DNAの結合及び切断は、典型的には、タンパク質及び両RNAを要する。しかしながら、単一ガイドRNA(「sgRNA」、又は単に「gRNA」)は、crRNAとtracrRNAとの両方の側面を単一RNA種中へ組み込むために、操作され得る。例として、Jinek M.,Chylinski K.,Fonfara I.,Hauer M.,Doudna J.A.,Charpentier E.Science 337:816-821(2012)を参照(この内容全体は参照により本明細書に組み込まれる)。Cas9は、CRISPR反復配列(PAM又はプロトスペーサ隣接モチーフ)中の短いモチーフを認識することで、自己・対・非自己を区別するのに役立つ。Cas9ヌクレアーゼ配列及び構造は当業者に周知である(例として、「Complete genome sequence of an M1 strain of Streptococcus pyogenes.」Ferretti et al.,J.J.,McShan W.M.,Ajdic D.J.,Savic D.J.,Savic G.,Lyon K.,Primeaux C.,Sezate S.,Suvorov A.N.,Kenton S.,Lai H.S.,Lin S.P.,Qian Y.,Jia H.G.,Najar F.Z.,Ren Q.,Zhu H.,Song L.,White J.,Yuan X.,Clifton S.W.,Roe B.A.,McLaughlin R.E.,Proc.Natl.Acad.Sci.U.S.A.98:4658-4663(2001);「CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III.」Deltcheva E.,Chylinski K.,Sharma C.M.,Gonzales K.,Chao Y.,Pirzada Z.A.,Eckert M.R.,Vogel J.,Charpentier E.,Nature 471:602-607(2011);及び「A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity.」Jinek M.,Chylinski K.,Fonfara I.,Hauer M.,Doudna J.A.,Charpentier E.Science 337:816-821(2012)を参照(これら各々の内容全体は参照により本明細書に組み込まれる))。Cas9オルソログは、これらに限定されないが、S.pyogenes及びS.thermophilusを包含する様々な種において記載されている。追加の好適なCas9ヌクレアーゼ及び配列は、本開示に基づき当業者に明らかであろう。またかかるCas9ヌクレアーゼ及び配列は、Chylinski,Rhun,and Charpentier,「The tracrRNA and Cas9 families of type II CRISPR-Cas immunity systems」(2013)RNA Biology 10:5,726-737(この内容全体は参照により本明細書に組み込まれる)に開示された生物及び遺伝子座からのCas9配列を包含する。いくつかの態様において、Cas9ヌクレアーゼは、DNA切断ドメインを部分的に損なうか又は不活性化させる1以上の突然変異を含む。
用語「Cas9」又は「Cas9ヌクレアーゼ」は、Cas9ドメイン又はそのフラグメントを含む、RNAにガイドされるヌクレアーゼ(例として、Cas9の活性若しくは不活性なDNA切断ドメイン及び/又はCas9のgRNA結合ドメインを含むタンパク質)を指す。「Cas9ドメイン」は、本明細書に使用されるとき、Cas9の活性若しくは不活性な切断ドメイン及び/又はCas9のgRNA結合ドメインを含むタンパク質フラグメントである。「Cas9タンパク質」は、全長Cas9タンパク質である。Cas9ヌクレアーゼはまた、ときにcasn1ヌクレアーゼ又はCRISPR(クラスター化して規則的な配置の短い回文配列反復(Clustered Regularly Interspaced Short Palindromic Repeat))関連ヌクレアーゼとしても言及される。CRISPRは、可動遺伝因子(ウイルス、転移因子、及び接合性プラスミド)に対して保護を提供する適応免疫系である。CRISPRクラスターは、スペーサ、先行する可動因子に相補的な配列、及び標的化侵入核酸を含有する。CRISPRクラスターは転写され、プロセスを受けて(processed)CRISPR RNA(crRNA)になる。II型CRISPR系において、pre-crRNAの修正プロセシングは、transでコードされた低分子(small)RNA(tracrRNA)、内生リボヌクレアーゼ3(rnc)、及びCas9ドメインを要する。tracrRNAは、pre-crRNAの、リボヌクレアーゼ3に補助された(-aided)プロセシングのためのガイドとして働く。続いて、Cas9/crRNA/tracrRNAは、スペーサに相補的な線状又は環状のdsDNA標的をエンドヌクレアーゼ的に切断する。crRNAに相補的ではない標的鎖は、最初にエンドヌクレアーゼ的に切られ、次いで3'-5'エキソヌクレアーゼ的に切り取られる。本来、DNAの結合及び切断は、典型的には、タンパク質及び両RNAを要する。しかしながら、単一ガイドRNA(「sgRNA」、又は単に「gRNA」)は、crRNAとtracrRNAとの両方の側面を単一RNA種中へ組み込むために、操作され得る。例として、Jinek M.,Chylinski K.,Fonfara I.,Hauer M.,Doudna J.A.,Charpentier E.Science 337:816-821(2012)を参照(この内容全体は参照により本明細書に組み込まれる)。Cas9は、CRISPR反復配列(PAM又はプロトスペーサ隣接モチーフ)中の短いモチーフを認識することで、自己・対・非自己を区別するのに役立つ。Cas9ヌクレアーゼ配列及び構造は当業者に周知である(例として、「Complete genome sequence of an M1 strain of Streptococcus pyogenes.」Ferretti et al.,J.J.,McShan W.M.,Ajdic D.J.,Savic D.J.,Savic G.,Lyon K.,Primeaux C.,Sezate S.,Suvorov A.N.,Kenton S.,Lai H.S.,Lin S.P.,Qian Y.,Jia H.G.,Najar F.Z.,Ren Q.,Zhu H.,Song L.,White J.,Yuan X.,Clifton S.W.,Roe B.A.,McLaughlin R.E.,Proc.Natl.Acad.Sci.U.S.A.98:4658-4663(2001);「CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III.」Deltcheva E.,Chylinski K.,Sharma C.M.,Gonzales K.,Chao Y.,Pirzada Z.A.,Eckert M.R.,Vogel J.,Charpentier E.,Nature 471:602-607(2011);及び「A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity.」Jinek M.,Chylinski K.,Fonfara I.,Hauer M.,Doudna J.A.,Charpentier E.Science 337:816-821(2012)を参照(これら各々の内容全体は参照により本明細書に組み込まれる))。Cas9オルソログは、これらに限定されないが、S.pyogenes及びS.thermophilusを包含する様々な種において記載されている。追加の好適なCas9ヌクレアーゼ及び配列は、本開示に基づき当業者に明らかであろう。またかかるCas9ヌクレアーゼ及び配列は、Chylinski,Rhun,and Charpentier,「The tracrRNA and Cas9 families of type II CRISPR-Cas immunity systems」(2013)RNA Biology 10:5,726-737(この内容全体は参照により本明細書に組み込まれる)に開示された生物及び遺伝子座からのCas9配列を包含する。いくつかの態様において、Cas9ヌクレアーゼは、DNA切断ドメインを部分的に損なうか又は不活性化させる1以上の突然変異を含む。
ヌクレアーゼが不活性化されたCas9ドメインは、互換的に「dCas9」タンパク質(ヌクレアーゼが「不活性型」のCas9を表す)と呼ばれてもよい。不活性なDNA切断ドメインを有するCas9ドメイン(又はそのフラグメント)を生成するための方法は知られている(例として、Jinek et al.,Science.337:816-821(2012);Qi et al.,「Repurposing CRISPR as an RNA-Guided Platform for Sequence-Specific Control of Gene Expression」(2013)Cell.28;152(5):1173-83を参照(これら各々の内容全体は参照により本明細書に組み込まれる))。例えば、Cas9のDNA切断ドメインは、2つのサブドメイン、HNHヌクレアーゼサブドメイン及びRuvC1サブドメインを包含することが知られている。HNHサブドメインはgRNAに相補的な鎖を切断するのに対し、RuvC1サブドメインは非相補鎖を切断する。これらのサブドメイン内の突然変異は、Cas9のヌクレアーゼ活性をサイレンシングさせ(silence)得る。例えば、突然変異D10A及びH840Aは、S.pyogenes Cas9のヌクレアーゼ活性を完全に不活性化する(Jinek et al.,Science.337:816-821(2012);Qi et al.,Cell.28;152(5):1173-83(2013))。いくつかの態様において、Cas9のフラグメントを含むタンパク質が提供される。例えば、いくつかの態様において、タンパク質は、2つのCas9ドメイン:(1)Cas9のgRNA結合ドメイン;又は(2)Cas9のDNA切断ドメインのうち一方を含む。いくつかの態様において、Cas9を含むタンパク質又はそのフラグメントは、「Cas9バリアント」と呼ばれる。Cas9バリアントは、Cas9又はそのフラグメントとの相同性を共有する。例えば、Cas9バリアントは、野生型Cas9(例として、配列番号37のSpCas9)と、少なくとも約70%同一、少なくとも約80%同一、少なくとも約90%同一、少なくとも約95%同一、少なくとも約96%同一、少なくとも約97%同一、少なくとも約98%同一、少なくとも約99%同一、少なくとも約99.5%同一、少なくとも約99.8%同一、又は少なくとも約99.9%同一である。いくつかの態様において、Cas9バリアントは、野生型Cas9(例として、配列番号37のSpCas9)と比較して、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、21、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、又はそれ以上のアミノ酸変化を有していてもよい。いくつかの態様において、Cas9バリアントは、そのフラグメントが、野生型Cas9(例として、配列番号37のSpCas9)の対応するフラグメントと、少なくとも約70%同一、少なくとも約80%同一、少なくとも約90%同一、少なくとも約95%同一、少なくとも約96%同一、少なくとも約97%同一、少なくとも約98%同一、少なくとも約99%同一、少なくとも約99.5%同一、又は少なくとも約99.9%同一となるように、配列番号37 Cas9のフラグメント(例として、gRNA結合ドメイン又はDNA切断ドメイン)を含む。いくつかの態様において、フラグメントは、対応する野生型Cas9(例として、配列番号37のSpCas9)のアミノ酸の長さの、少なくとも30%、少なくとも35%、少なくとも40%、少なくとも45%、少なくとも50%、少なくとも55%、少なくとも60%、少なくとも65%、少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%同一の、少なくとも96%、少なくとも97%、少なくとも98%、少なくとも99%、又は少なくとも99.5%である。
cDNA
用語「cDNA」は、RNA鋳型からコピーされたDNAの鎖を指す。cDNAは、RNA鋳型に相補的である。
用語「cDNA」は、RNA鋳型からコピーされたDNAの鎖を指す。cDNAは、RNA鋳型に相補的である。
循環置換体
本明細書に使用されるとき、用語「循環置換体(circular permutant)」は、タンパク質のアミノ酸配列中に現れるアミノ酸の順序の変化を伴うタンパク質の構造上の立体配置における変化である循環置換(circular permutation)を含むタンパク質又はポリペプチド(例として、Cas9)を指す。換言すれば、循環置換体は、野生型対応物と比較してN末端及びC末端が変更されたタンパク質であり、例として、野生型タンパク質のC末端半分が新しいN末端半分になっている。循環置換(又はCP)は本質的に、そのN末端及びC末端が接続され、しばしばペプチドリンカーをもつ、タンパク質1次配列の形態上の再配置であるが、同時にその配列を異なる位置にて分裂させることで、隣接していた新しいN末端及びC末端を作成する。その結果は、接続が異なるが、しばしば同じか全体的には同様の3次元の(3D)形状を有し得るタンパク質構造であって、おそらく、改善又は変更された特徴(タンパク分解への低減された感受性、改善された触媒活性、変更された基質結合若しくはリガンド結合、及び/又は改善された熱安定性を包含する)を包含する。循環置換タンパク質は天然に存在し得る(例として、コンカナバリンA及びレクチン)。加えて、循環置換は、翻訳後修飾の結果として生じ得るか、又は組換え技法を使用して操作されてもよい。
本明細書に使用されるとき、用語「循環置換体(circular permutant)」は、タンパク質のアミノ酸配列中に現れるアミノ酸の順序の変化を伴うタンパク質の構造上の立体配置における変化である循環置換(circular permutation)を含むタンパク質又はポリペプチド(例として、Cas9)を指す。換言すれば、循環置換体は、野生型対応物と比較してN末端及びC末端が変更されたタンパク質であり、例として、野生型タンパク質のC末端半分が新しいN末端半分になっている。循環置換(又はCP)は本質的に、そのN末端及びC末端が接続され、しばしばペプチドリンカーをもつ、タンパク質1次配列の形態上の再配置であるが、同時にその配列を異なる位置にて分裂させることで、隣接していた新しいN末端及びC末端を作成する。その結果は、接続が異なるが、しばしば同じか全体的には同様の3次元の(3D)形状を有し得るタンパク質構造であって、おそらく、改善又は変更された特徴(タンパク分解への低減された感受性、改善された触媒活性、変更された基質結合若しくはリガンド結合、及び/又は改善された熱安定性を包含する)を包含する。循環置換タンパク質は天然に存在し得る(例として、コンカナバリンA及びレクチン)。加えて、循環置換は、翻訳後修飾の結果として生じ得るか、又は組換え技法を使用して操作されてもよい。
循環置換されたCas9
用語「循環置換されたCas9」は、循環置換体として生じるものであり、これによってそのN末端及びC末端が局所的に再配置された、いずれのCas9タンパク質又はそのバリアントも指す。循環置換されたかかるCas9タンパク質(「CP-Cas9」)、又はそれらのバリアントは、ガイドRNA(gRNA)と複合体化されたときのDNAへの結合能を保持している。Oakes et al.,「Protein Engineering of Cas9 for enhanced function,」Methods Enzymol,2014,546:491-511及びOakes et al.,「CRISPR-Cas9 Circular Permutants as Programmable Scaffolds for Genome Modification,」Cell,January10,2019,176:254-267を参照(これらの各々は参照により本明細書に組み込まれる)。本開示は、これまでに知られているいずれのCP-Cas9も企図するか、又は新しいCP-Cas9を、その結果得られた循環置換タンパク質がガイドRNA(gRNA)と複合体化されたときのDNAへの結合能を保持している限り、使用する。例示のCP-Cas9タンパク質は、配列番号88~97である。
用語「循環置換されたCas9」は、循環置換体として生じるものであり、これによってそのN末端及びC末端が局所的に再配置された、いずれのCas9タンパク質又はそのバリアントも指す。循環置換されたかかるCas9タンパク質(「CP-Cas9」)、又はそれらのバリアントは、ガイドRNA(gRNA)と複合体化されたときのDNAへの結合能を保持している。Oakes et al.,「Protein Engineering of Cas9 for enhanced function,」Methods Enzymol,2014,546:491-511及びOakes et al.,「CRISPR-Cas9 Circular Permutants as Programmable Scaffolds for Genome Modification,」Cell,January10,2019,176:254-267を参照(これらの各々は参照により本明細書に組み込まれる)。本開示は、これまでに知られているいずれのCP-Cas9も企図するか、又は新しいCP-Cas9を、その結果得られた循環置換タンパク質がガイドRNA(gRNA)と複合体化されたときのDNAへの結合能を保持している限り、使用する。例示のCP-Cas9タンパク質は、配列番号88~97である。
CRISPR
CRISPRは、原核生物に侵入したウイルスによる先行する感染のスニペットを表す細菌及び古細菌におけるDNA配列(すなわちCRISPRクラスター)のファミリーである。DNAのスニペットは、類似のウイルスによる続く攻撃からのDNAを検出及び破壊するために原核細胞によって用いられ、CRISPR関連タンパク質(Cas9及びそのホモログを包含する)及びCRISPR関連RNAのアレイと併せて、有効に原核生物免疫防御システムを成す。天然では、CRISPRクラスターはCRISPR RNA(crRNA)へと転写及びプロセシングされる。ある種の型のCRISPRシステム(例として、II型CRISPRシステム)では、プレcrRNAの正しいプロセシングは、トランスにコードされる低分子RNA(tracrRNA)、内生リボヌクレアーゼ3(rnc)、及びCas9タンパク質を要求する。tracrRNAは、pre-crRNAの、リボヌクレアーゼ3に補助された(-aided)プロセシングのためのガイドとして働く。続いてCas9/crRNA/tracrRNAはRNAに対して相補的な直線状又は環状dsDNA標的をエンド的に切断する。具体的には、crRNAに対して相補的でない標的鎖が第1にエンド的にカットされ、次いで3'-5'エキソ的にトリミングされる。本来、DNAの結合及び切断は、典型的には、タンパク質及び両RNAを要する。しかしながら、シングルガイドRNA(「sgRNA」又は単純に「gNRA」)が、crRNA及びtracrRNA両方の側面を単一のRNA種のガイドRNAに組み込むようにして操作され得る。例として、Jinek M.,Chylinski K.,Fonfara I.,Hauer M.,Doudna J.A.,Charpentier E.Science 337:816-821(2012)を見よ。これの内容全体は参照によってここに組み込まれる。Cas9は、CRISPR反復配列(PAM又はプロトスペーサ隣接モチーフ)中の短いモチーフを認識することで、自己・対・非自己を区別するのに役立つ。CRISPR生物学並びにCas9ヌクレアーゼ配列及び構造は当業者には周知である(例として、「Complete genome sequence of an M1 strain of Streptococcus pyogenes.」Ferretti et al.,J.J.,McShan W.M.,Ajdic D.J.,Savic D.J.,Savic G.,Lyon K.,Primeaux C.,Sezate S.,Suvorov A.N.,Kenton S.,Lai H.S.,Lin S.P.,Qian Y.,Jia H.G.,Najar F.Z.,Ren Q.,Zhu H.,Song L.,White J.,Yuan X.,Clifton S.W.,Roe B.A.,McLaughlin R.E.,Proc.Natl.Acad.Sci.U.S.A.98:4658-4663(2001);「CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III.」Deltcheva E.,Chylinski K.,Sharma C.M.,Gonzales K.,Chao Y.,Pirzada Z.A.,Eckert M.R.,Vogel J.,Charpentier E.,Nature 471:602-607(2011);及び「A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity.」Jinek M.,Chylinski K.,Fonfara I.,Hauer M.,Doudna J.A.,Charpentier E.Science 337:816-821(2012)を見よ。これらの夫々の内容全体は参照によって本明細書に組み込まれる)。Cas9オルソログは、これらに限定されないが、S.pyogenes及びS.thermophilusを包含する様々な種において記載されている。追加の好適なCas9ヌクレアーゼ及び配列は、本開示に基づき当業者に明らかであろう。またかかるCas9ヌクレアーゼ及び配列は、Chylinski,Rhun,and Charpentier,「The tracrRNA and Cas9 families of type II CRISPR-Cas immunity systems」(2013)RNA Biology 10:5,726-737(この内容全体は参照により本明細書に組み込まれる)に開示された生物及び遺伝子座からのCas9配列を包含する。
CRISPRは、原核生物に侵入したウイルスによる先行する感染のスニペットを表す細菌及び古細菌におけるDNA配列(すなわちCRISPRクラスター)のファミリーである。DNAのスニペットは、類似のウイルスによる続く攻撃からのDNAを検出及び破壊するために原核細胞によって用いられ、CRISPR関連タンパク質(Cas9及びそのホモログを包含する)及びCRISPR関連RNAのアレイと併せて、有効に原核生物免疫防御システムを成す。天然では、CRISPRクラスターはCRISPR RNA(crRNA)へと転写及びプロセシングされる。ある種の型のCRISPRシステム(例として、II型CRISPRシステム)では、プレcrRNAの正しいプロセシングは、トランスにコードされる低分子RNA(tracrRNA)、内生リボヌクレアーゼ3(rnc)、及びCas9タンパク質を要求する。tracrRNAは、pre-crRNAの、リボヌクレアーゼ3に補助された(-aided)プロセシングのためのガイドとして働く。続いてCas9/crRNA/tracrRNAはRNAに対して相補的な直線状又は環状dsDNA標的をエンド的に切断する。具体的には、crRNAに対して相補的でない標的鎖が第1にエンド的にカットされ、次いで3'-5'エキソ的にトリミングされる。本来、DNAの結合及び切断は、典型的には、タンパク質及び両RNAを要する。しかしながら、シングルガイドRNA(「sgRNA」又は単純に「gNRA」)が、crRNA及びtracrRNA両方の側面を単一のRNA種のガイドRNAに組み込むようにして操作され得る。例として、Jinek M.,Chylinski K.,Fonfara I.,Hauer M.,Doudna J.A.,Charpentier E.Science 337:816-821(2012)を見よ。これの内容全体は参照によってここに組み込まれる。Cas9は、CRISPR反復配列(PAM又はプロトスペーサ隣接モチーフ)中の短いモチーフを認識することで、自己・対・非自己を区別するのに役立つ。CRISPR生物学並びにCas9ヌクレアーゼ配列及び構造は当業者には周知である(例として、「Complete genome sequence of an M1 strain of Streptococcus pyogenes.」Ferretti et al.,J.J.,McShan W.M.,Ajdic D.J.,Savic D.J.,Savic G.,Lyon K.,Primeaux C.,Sezate S.,Suvorov A.N.,Kenton S.,Lai H.S.,Lin S.P.,Qian Y.,Jia H.G.,Najar F.Z.,Ren Q.,Zhu H.,Song L.,White J.,Yuan X.,Clifton S.W.,Roe B.A.,McLaughlin R.E.,Proc.Natl.Acad.Sci.U.S.A.98:4658-4663(2001);「CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III.」Deltcheva E.,Chylinski K.,Sharma C.M.,Gonzales K.,Chao Y.,Pirzada Z.A.,Eckert M.R.,Vogel J.,Charpentier E.,Nature 471:602-607(2011);及び「A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity.」Jinek M.,Chylinski K.,Fonfara I.,Hauer M.,Doudna J.A.,Charpentier E.Science 337:816-821(2012)を見よ。これらの夫々の内容全体は参照によって本明細書に組み込まれる)。Cas9オルソログは、これらに限定されないが、S.pyogenes及びS.thermophilusを包含する様々な種において記載されている。追加の好適なCas9ヌクレアーゼ及び配列は、本開示に基づき当業者に明らかであろう。またかかるCas9ヌクレアーゼ及び配列は、Chylinski,Rhun,and Charpentier,「The tracrRNA and Cas9 families of type II CRISPR-Cas immunity systems」(2013)RNA Biology 10:5,726-737(この内容全体は参照により本明細書に組み込まれる)に開示された生物及び遺伝子座からのCas9配列を包含する。
ある種の型のCRISPRシステム(例として、II型CRISPRシステム)では、プレcrRNAの正しいプロセシングは、トランスにコードされる低分子RNA(tracrRNA)、内生リボヌクレアーゼ3(rnc)、及びCas9タンパク質を要求する。tracrRNAは、pre-crRNAの、リボヌクレアーゼ3に補助された(-aided)プロセシングのためのガイドとして働く。続いてCas9/crRNA/tracrRNAはRNAに対して相補的な直線状又は環状核酸標的をエンド的に切断する。具体的には、crRNAに対して相補的でない標的鎖が第1にエンド的にカットされ、次いで3'-5'エキソ的にトリミングされる。本来、DNAの結合及び切断は、典型的には、タンパク質及び両RNAを要する。しかしながら、シングルガイドRNA(「sgRNA」又は単純に「gRNA」)が、crRNA及びtracrRNA両方の態様を単一のRNA種のガイドRNAに組み込むようにして操作され得る。
一般的に、「CRISPRシステム」は、集合的に、CRISPR関連(「Cas」)遺伝子の発現を伴うか、又はそれの活性を導く転写物及び他のエレメントを指し、Cas遺伝子、tracr(トランス活性化CRISPR)配列(例として、tracrRNA又は活性な部分的なtracrRNA)、tracr mate配列(内生CRISPRシステムという文脈において、「ダイレクト反復」及びtracrRNAによってプロセシングされる部分的なダイレクト反復を包摂する)、ガイド配列(内生CRISPRシステムという文脈において「スペーサ」ともまた呼ばれる)、又はCRISPR遺伝子座からの他の配列及び転写物をコードする配列を包含する。システムのtracrRNAはガイドRNA上に存在するtracr mate配列に対して(完全に又は部分的に)相補的である。
DNA合成鋳型
本明細書に使用されるとき、用語「DNA合成鋳型」は、プライム編集因子のポリメラーゼによって鋳型鎖(所望の編集を含有しており、次いでプライム編集の機序を通して、対応する内生DNAの鎖を標的部位にて置き換える、3'単鎖DNAフラップをコードする)として利用される、pegRNAの伸長アームの領域又は部分を指す。様々な態様において、DNA合成鋳型は、図3A(5'伸長アームを含むpegRNAの文脈(context)において)、図3B(3'伸長アームを含むpegRNAの文脈において)、図3C(内部伸長アームの文脈において)、図3D(3'伸長アームの文脈において)、及び図3E(5'伸長アームの文脈において)に示される。伸長アームは、DNA合成鋳型を包含しており、DNA又はRNAから構成されていてもよい。RNAのケースにおいて、プライム編集因子のポリメラーゼは、RNA依存性DNAポリメラーゼ(例として、逆転写酵素)であり得る。DNAのケースにおいて、プライム編集因子のポリメラーゼは、DNA依存性DNAポリメラーゼであり得る。様々な(例として、図3D~図3Eに描かれる通りの)態様において、DNA合成鋳型は、「編集鋳型」及び「相同アーム」を含む。様々な(例として、図3D~図3Eに描かれる通りの)態様において、DNA合成鋳型(4)は、「編集鋳型」及び「相同アーム」、並びに任意選択の5'末修飾領域e2の全部又は一部を含んでいてもよい。つまり、e2領域の性質に応じて(例として、これが、ヘアピン、トゥーループ、又はステム/ループの2次構造を包含するかどうかにかかわらず)、ポリメラーゼは、e2領域を何もコードしていなくても、又はその一部若しくは全部をコードしていてもよい。別の言い方をすれば、3'伸長アームのケースにおいて、DNA合成鋳型(3)は、ポリメラーゼ(例として、逆転写酵素)によるDNAの単鎖の合成のための鋳型として動作してもよい、プライマー結合部位(PBS)の5'末からgRNAコアの3'末までに及ぶ伸長アーム(3)の部分を包含し得る。5'伸長アームのケースにおいて、DNA合成鋳型(3)は、pegRNA分子の5'末から編集鋳型の3'末までに及ぶ伸長アームの部分を包含し得る。いくつかの態様では、DNA合成鋳型は、3'伸長アーム又は5'伸長アームのいずれかを有するpegRNAのプライマー結合部位(PBS)を排除する。本明細書に記載のある態様(例として、図71A)は、編集鋳型及び相同アーム、すなわち、DNA合成の最中に鋳型として実際に使用されるpegRNA伸長アームの配列を包含する「RT鋳型」を指す。「RT鋳型」という用語は、「DNA合成鋳型」という用語と同等である。ある特定の態様において、RT鋳型は、逆転写のための鋳型ポリヌクレオチドを、例として、逆転写酵素であるポリメラーゼを有するプライム編集因子を使用するプライム編集システム、複合体又は方法において指すために使用されてもよい。いくつかの態様では、DNA合成鋳型は、例として、RNA依存性DNAポリメラーゼ又はDNA依存性DNAポリメラーゼであるポリメラーゼを有するプライム編集因子を使用するプライム編集システム、複合体又は方法における、DNA重合、例としてRNA依存性DNA重合又はDNA依存性重合のための鋳型ポリヌクレオチドを指すために使用されてもよい。
本明細書に使用されるとき、用語「DNA合成鋳型」は、プライム編集因子のポリメラーゼによって鋳型鎖(所望の編集を含有しており、次いでプライム編集の機序を通して、対応する内生DNAの鎖を標的部位にて置き換える、3'単鎖DNAフラップをコードする)として利用される、pegRNAの伸長アームの領域又は部分を指す。様々な態様において、DNA合成鋳型は、図3A(5'伸長アームを含むpegRNAの文脈(context)において)、図3B(3'伸長アームを含むpegRNAの文脈において)、図3C(内部伸長アームの文脈において)、図3D(3'伸長アームの文脈において)、及び図3E(5'伸長アームの文脈において)に示される。伸長アームは、DNA合成鋳型を包含しており、DNA又はRNAから構成されていてもよい。RNAのケースにおいて、プライム編集因子のポリメラーゼは、RNA依存性DNAポリメラーゼ(例として、逆転写酵素)であり得る。DNAのケースにおいて、プライム編集因子のポリメラーゼは、DNA依存性DNAポリメラーゼであり得る。様々な(例として、図3D~図3Eに描かれる通りの)態様において、DNA合成鋳型は、「編集鋳型」及び「相同アーム」を含む。様々な(例として、図3D~図3Eに描かれる通りの)態様において、DNA合成鋳型(4)は、「編集鋳型」及び「相同アーム」、並びに任意選択の5'末修飾領域e2の全部又は一部を含んでいてもよい。つまり、e2領域の性質に応じて(例として、これが、ヘアピン、トゥーループ、又はステム/ループの2次構造を包含するかどうかにかかわらず)、ポリメラーゼは、e2領域を何もコードしていなくても、又はその一部若しくは全部をコードしていてもよい。別の言い方をすれば、3'伸長アームのケースにおいて、DNA合成鋳型(3)は、ポリメラーゼ(例として、逆転写酵素)によるDNAの単鎖の合成のための鋳型として動作してもよい、プライマー結合部位(PBS)の5'末からgRNAコアの3'末までに及ぶ伸長アーム(3)の部分を包含し得る。5'伸長アームのケースにおいて、DNA合成鋳型(3)は、pegRNA分子の5'末から編集鋳型の3'末までに及ぶ伸長アームの部分を包含し得る。いくつかの態様では、DNA合成鋳型は、3'伸長アーム又は5'伸長アームのいずれかを有するpegRNAのプライマー結合部位(PBS)を排除する。本明細書に記載のある態様(例として、図71A)は、編集鋳型及び相同アーム、すなわち、DNA合成の最中に鋳型として実際に使用されるpegRNA伸長アームの配列を包含する「RT鋳型」を指す。「RT鋳型」という用語は、「DNA合成鋳型」という用語と同等である。ある特定の態様において、RT鋳型は、逆転写のための鋳型ポリヌクレオチドを、例として、逆転写酵素であるポリメラーゼを有するプライム編集因子を使用するプライム編集システム、複合体又は方法において指すために使用されてもよい。いくつかの態様では、DNA合成鋳型は、例として、RNA依存性DNAポリメラーゼ又はDNA依存性DNAポリメラーゼであるポリメラーゼを有するプライム編集因子を使用するプライム編集システム、複合体又は方法における、DNA重合、例としてRNA依存性DNA重合又はDNA依存性重合のための鋳型ポリヌクレオチドを指すために使用されてもよい。
トランスプライム編集のケースでは(例として、図3G及び図3H)、プライマー結合部位(PBS)及びDNA合成鋳型は、トランスプライム編集因子RNA鋳型(tPERT)と呼ばれる別個の分子へと操作され得る。
いくつかの態様では、DNA合成鋳型は、PBSの5'であり、PAM鎖に対する相補性領域(すなわち、非標的鎖又は編集鎖)を含み、二本鎖標的DNAの内生配列と比較して1つ又は複数のヌクレオチド編集を含む、PEgRNAの一本鎖部分である。いくつかの態様において、DNA合成鋳型は、意図されたヌクレオチド編集位置における1つ又は複数の非相補的ヌクレオチドを除いて、ニック部位の下流にある非標的鎖上の配列に相補的又は実質的に相補的である。いくつかの態様において、DNA合成鋳型は、意図されたヌクレオチド編集位置における1つ又は複数の非相補的ヌクレオチドを除いて、ニック部位のすぐ下流(すなわち、直接下流に)にある非標的鎖上の配列に相補的又は実質的に相補的である。いくつかの態様において、意図されたヌクレオチド編集位置における非相補的ヌクレオチドの1つ又は複数は、ニック部位のすぐ下流にある。いくつかの態様では、DNA合成鋳型は、二本鎖標的DNA配列に対して1つ以上のヌクレオチド編集を含む。いくつかの態様では、DNA合成鋳型は、二本鎖標的DNA配列の非標的鎖に対して1つ又は複数のヌクレオチド編集を含む。本明細書に記載の各PEgRNAについて、ニック部位は、PEgRNAのgRNAコアが会合する特定のnapDNAbpに特徴的であり、napDNAbpの認識及び機能に必要な特定のPAMに特徴的である。例えば、SpCas9と会合するgRNAコアを含むPEgRNAの場合、3塩基(PAM配列の1位に対して「-3」位置)と4塩基(PAM配列の1位に対して「-4」位置)との間のホスホジエステル結合におけるニック部位。いくつかの態様において、DNA合成鋳型及びプライマー結合部位は、互いに直接隣接している。「ヌクレオチド編集」、「ヌクレオチド変化」、「所望のヌクレオチド変化」、及び「所望のヌクレオチド編集」という用語は、特定のヌクレオチド編集、例として、標的DNA配列に組み込まれるべきPEgRNAのDNA合成鋳型の特定の位置における1つ又は複数のヌクレオチドの特定の欠失、1つ又は複数のヌクレオチドの特定の挿入、1つ又は複数のヌクレオチドの特定の置換、又はそれらの組み合わせを指すために互換的に使用される。いくつかの態様では、DNA合成鋳型は、二本鎖標的DNA配列に対して2つ以上のヌクレオチド編集を含む。そのような態様では、各ヌクレオチド編集は、DNA合成鋳型内の特定の位置における特定のヌクレオチド編集であり、各ヌクレオチド編集は、DNA合成鋳型内の他のヌクレオチド編集のいずれかと比較して異なる特定の位置にあり、各ヌクレオチド編集は、1つ以上のヌクレオチドの特定の欠失、1つ以上のヌクレオチドの特定の挿入、1つ以上のヌクレオチドの特定の置換、又はそれらの組み合わせから独立して選択される。ヌクレオチド編集は、標的遺伝子の標的鎖上の配列と比較してDNA合成鋳型上の編集を指してもよく、又は非標的鎖上の内生標的DNA配列を置き換える新たに合成された一本鎖DNA上のDNA合成鋳型によってコードされる編集を指してもよく、いずれの場合も、標的DNA配列と比較されるヌクレオチド編集を指してもよい。
下流
本明細書に使用されるとき、用語「上流」及び「下流」は、5'→3'方向に配向された核酸分子(単鎖又は二本鎖であるかにかかわらず)中に位置付けられた少なくとも2つの要素の線状の位置を定義する相対的な用語である。特に、第1要素が第2要素に対して5'のどこかに位置付けられている場合、第1要素は、核酸分子中、第2要素の上流にある。例えば、SNPがニック部位の5'側にある場合、SNPは、Cas9に誘導されたニック部位の上流にある。逆に言うと、第1要素が第2要素に対して3'のどこかに位置付けられている場合、第1要素は、核酸分子中、第2要素の下流にある。例えば、SNPがニック部位の3'側にある場合、SNPは、Cas9に誘導されたニック部位の下流にある。核酸分子は、DNA(二本鎖若しくは単鎖)、RNA(二本鎖若しくは単鎖)、又はDNAとRNAとのハイブリッドであり得る。二本鎖分子のどちらの鎖を考慮するのか選択する必要がある場合を除き、上流及び下流という用語が核酸分子の単鎖にのみ言及するため、分析は単鎖核酸分子及び二本鎖分子について同じである。しばしば、少なくとも2つの要素の相対的な位置を決定するのに使用され得る二本鎖のDNAの鎖は、「センス」鎖又は「コード」鎖である。遺伝学において、「センス」鎖は、5'から3'へ伸びる二本鎖DNA内セグメントであって、3'から5'へ伸びるDNAのアンチセンス鎖又は鋳型鎖に相補的である。よって、例として、SNP核酸塩基がセンス鎖又はコード鎖上のプロモータの3'側にある場合、SNP核酸塩基はゲノムDNA(二本鎖である)中のプロモータ配列の「下流」にある。
本明細書に使用されるとき、用語「上流」及び「下流」は、5'→3'方向に配向された核酸分子(単鎖又は二本鎖であるかにかかわらず)中に位置付けられた少なくとも2つの要素の線状の位置を定義する相対的な用語である。特に、第1要素が第2要素に対して5'のどこかに位置付けられている場合、第1要素は、核酸分子中、第2要素の上流にある。例えば、SNPがニック部位の5'側にある場合、SNPは、Cas9に誘導されたニック部位の上流にある。逆に言うと、第1要素が第2要素に対して3'のどこかに位置付けられている場合、第1要素は、核酸分子中、第2要素の下流にある。例えば、SNPがニック部位の3'側にある場合、SNPは、Cas9に誘導されたニック部位の下流にある。核酸分子は、DNA(二本鎖若しくは単鎖)、RNA(二本鎖若しくは単鎖)、又はDNAとRNAとのハイブリッドであり得る。二本鎖分子のどちらの鎖を考慮するのか選択する必要がある場合を除き、上流及び下流という用語が核酸分子の単鎖にのみ言及するため、分析は単鎖核酸分子及び二本鎖分子について同じである。しばしば、少なくとも2つの要素の相対的な位置を決定するのに使用され得る二本鎖のDNAの鎖は、「センス」鎖又は「コード」鎖である。遺伝学において、「センス」鎖は、5'から3'へ伸びる二本鎖DNA内セグメントであって、3'から5'へ伸びるDNAのアンチセンス鎖又は鋳型鎖に相補的である。よって、例として、SNP核酸塩基がセンス鎖又はコード鎖上のプロモータの3'側にある場合、SNP核酸塩基はゲノムDNA(二本鎖である)中のプロモータ配列の「下流」にある。
編集鋳型
用語「編集鋳型」は、ポリメラーゼ、例として、DNA依存性DNAポリメラーゼ、RNA依存性DNAポリメラーゼ(例として、逆転写酵素)によって合成される一本鎖3'DNAフラップ上に所望の編集をコードする伸長アームの部分を指す。本明細書に記載のある態様(例として、図71A)は、編集鋳型と相同アームとの両方一緒、すなわち、DNA合成の最中に鋳型として実際に使用されるpegRNA伸長アームの配列を指す「RT鋳型」を指す。用語「RT編集鋳型」はまた、用語「DNA合成鋳型」とも等価であるが、ここでRT編集鋳型は、逆転写酵素であるポリメラーゼを有するプライム編集因子の使用を反映し、DNA合成鋳型は、いずれのポリメラーゼも有するプライム編集因子の使用をより広く反映する。
用語「編集鋳型」は、ポリメラーゼ、例として、DNA依存性DNAポリメラーゼ、RNA依存性DNAポリメラーゼ(例として、逆転写酵素)によって合成される一本鎖3'DNAフラップ上に所望の編集をコードする伸長アームの部分を指す。本明細書に記載のある態様(例として、図71A)は、編集鋳型と相同アームとの両方一緒、すなわち、DNA合成の最中に鋳型として実際に使用されるpegRNA伸長アームの配列を指す「RT鋳型」を指す。用語「RT編集鋳型」はまた、用語「DNA合成鋳型」とも等価であるが、ここでRT編集鋳型は、逆転写酵素であるポリメラーゼを有するプライム編集因子の使用を反映し、DNA合成鋳型は、いずれのポリメラーゼも有するプライム編集因子の使用をより広く反映する。
有効量
本明細書において用いられる用語「有効量」は、所望の生物学的応答を誘起するために十分である生物活性薬剤の量を指す。例えば、いくつかの態様では、プライム編集因子(PE)の有効量は、標的部位ヌクレオチド配列、例としてゲノムを編集するために十分である編集因子の量を指してもよい。いくつかの態様では、本明細書において提供されるプライム編集因子(PE)の、例としてニッカーゼCas9ドメイン及び逆転写酵素を含む融合タンパク質の有効量は、融合タンパク質によって特異的に結合及び編集される標的部位の編集を誘導するために十分である融合タンパク質の量を指してもよい。当業者には了解されるであろう通り、薬剤、例として融合タンパク質、ヌクレアーゼ、ハイブリッドタンパク質、タンパク質二量体、タンパク質(又はタンパク質二量体)及びポリヌクレオチドの複合体、又はポリヌクレオチドの有効量は、例えば、所望の生物学的反応、例として編集されるべき特定のアレル、ゲノム、又は標的部位、標的化されようとする細胞又は組織、及び用いられようとする薬剤のような種々の因子に依存して変わってもよい。
本明細書において用いられる用語「有効量」は、所望の生物学的応答を誘起するために十分である生物活性薬剤の量を指す。例えば、いくつかの態様では、プライム編集因子(PE)の有効量は、標的部位ヌクレオチド配列、例としてゲノムを編集するために十分である編集因子の量を指してもよい。いくつかの態様では、本明細書において提供されるプライム編集因子(PE)の、例としてニッカーゼCas9ドメイン及び逆転写酵素を含む融合タンパク質の有効量は、融合タンパク質によって特異的に結合及び編集される標的部位の編集を誘導するために十分である融合タンパク質の量を指してもよい。当業者には了解されるであろう通り、薬剤、例として融合タンパク質、ヌクレアーゼ、ハイブリッドタンパク質、タンパク質二量体、タンパク質(又はタンパク質二量体)及びポリヌクレオチドの複合体、又はポリヌクレオチドの有効量は、例えば、所望の生物学的反応、例として編集されるべき特定のアレル、ゲノム、又は標的部位、標的化されようとする細胞又は組織、及び用いられようとする薬剤のような種々の因子に依存して変わってもよい。
エラーを起こしやすい逆転写酵素
本明細書に使用されるとき、用語「エラーを起こしやすい」逆転写酵素(又はより広くは、いずれのポリメラーゼ)は、天然に存在するか、又は野生型M-MLV逆転写酵素の誤差率未満の誤差率を有する別の逆転写酵素(例として、野生型M-MLV逆転写酵素)に由来される逆転写酵素(又はより広くは、いずれのポリメラーゼ)を指す。野生型M-MLV逆転写酵素の誤差率は、1誤差の範囲が15,000(より高い)~27,000(より低い)にあると報告されている。15,000中の1の誤差率は6.7×10-5の誤差率に相当する。27,000中の1の誤差率は3.7×10-5の誤差率に相当する。Boutabout et al.(2001)「DNA synthesis fidelity by the reverse transcriptase of the yeast retrotransposon Ty1,」Nucleic Acids Res 29(11):2217-2222を参照(これは参照により本明細書に組み込まれる)。よって、本出願の目的において、用語「エラーを起こしやすい」は、15,000核酸塩基の組み込み中1より大きい誤差(6.7×10-5又はこれより高い)、例として、14,000核酸塩基中1誤差(7.14×10-5若しくはこれより高い)、13,000核酸塩基又はこれより少ない中で1誤差(7.7×10-5若しくはこれより高い)、12,000核酸塩基又はこれより少ない中で1誤差(7.7×10-5若しくはこれより高い)、11,000核酸塩基又はこれより少ない中で1誤差(9.1×10-5若しくはこれより高い)、10,000核酸塩基又はこれより少ない中で1誤差(1×10-4又は0.0001若しくはこれより高い)、9,000核酸塩基又はこれより少ない中で1誤差(0.00011若しくはこれより高い)、8,000核酸塩基又はこれより少ない中で1誤差(0.00013若しくはこれより高い)、7,000核酸塩基又はこれより少ない中で1誤差(0.00014若しくはこれより高い)、6,000核酸塩基又はこれより少ない中で1誤差(0.00016若しくはこれより高い)、5,000核酸塩基又はこれより少ない中で1誤差(0.0002若しくはこれより高い)、4,000核酸塩基又はこれより少ない中で1誤差(0.00025若しくはこれより高い)、3,000核酸塩基又はこれより少ない中で1誤差(0.00033若しくはこれより高い)、2,000核酸塩基又はこれより少ない中で1誤差(0.00050若しくはこれより高い)、あるいは1,000核酸塩基又はこれより少ない中で1誤差(0.001若しくはこれより高い)、あるいは500核酸塩基又はこれより少ない中で1誤差(0.002若しくはこれより高い)、あるいは250核酸塩基又はこれより少ない中で1誤差(0.004若しくはこれより高い)である誤差率を有する、それらRTを指す。
本明細書に使用されるとき、用語「エラーを起こしやすい」逆転写酵素(又はより広くは、いずれのポリメラーゼ)は、天然に存在するか、又は野生型M-MLV逆転写酵素の誤差率未満の誤差率を有する別の逆転写酵素(例として、野生型M-MLV逆転写酵素)に由来される逆転写酵素(又はより広くは、いずれのポリメラーゼ)を指す。野生型M-MLV逆転写酵素の誤差率は、1誤差の範囲が15,000(より高い)~27,000(より低い)にあると報告されている。15,000中の1の誤差率は6.7×10-5の誤差率に相当する。27,000中の1の誤差率は3.7×10-5の誤差率に相当する。Boutabout et al.(2001)「DNA synthesis fidelity by the reverse transcriptase of the yeast retrotransposon Ty1,」Nucleic Acids Res 29(11):2217-2222を参照(これは参照により本明細書に組み込まれる)。よって、本出願の目的において、用語「エラーを起こしやすい」は、15,000核酸塩基の組み込み中1より大きい誤差(6.7×10-5又はこれより高い)、例として、14,000核酸塩基中1誤差(7.14×10-5若しくはこれより高い)、13,000核酸塩基又はこれより少ない中で1誤差(7.7×10-5若しくはこれより高い)、12,000核酸塩基又はこれより少ない中で1誤差(7.7×10-5若しくはこれより高い)、11,000核酸塩基又はこれより少ない中で1誤差(9.1×10-5若しくはこれより高い)、10,000核酸塩基又はこれより少ない中で1誤差(1×10-4又は0.0001若しくはこれより高い)、9,000核酸塩基又はこれより少ない中で1誤差(0.00011若しくはこれより高い)、8,000核酸塩基又はこれより少ない中で1誤差(0.00013若しくはこれより高い)、7,000核酸塩基又はこれより少ない中で1誤差(0.00014若しくはこれより高い)、6,000核酸塩基又はこれより少ない中で1誤差(0.00016若しくはこれより高い)、5,000核酸塩基又はこれより少ない中で1誤差(0.0002若しくはこれより高い)、4,000核酸塩基又はこれより少ない中で1誤差(0.00025若しくはこれより高い)、3,000核酸塩基又はこれより少ない中で1誤差(0.00033若しくはこれより高い)、2,000核酸塩基又はこれより少ない中で1誤差(0.00050若しくはこれより高い)、あるいは1,000核酸塩基又はこれより少ない中で1誤差(0.001若しくはこれより高い)、あるいは500核酸塩基又はこれより少ない中で1誤差(0.002若しくはこれより高い)、あるいは250核酸塩基又はこれより少ない中で1誤差(0.004若しくはこれより高い)である誤差率を有する、それらRTを指す。
エクテイン
本明細書において用いられる用語「エクステイン」は、インテインによってフランキングされ、タンパク質スプライシングのプロセスの間に別のエクステインにライゲーションされて成熟したスプライシングされたタンパク質を形成するポリペプチド配列を指す。典型的には、インテインは2つのエクステイン配列によってフランキングされ、これらはインテインがそれ自体の切除を触媒するときに一緒にライゲーションされる。エクステインは、従って、mRNA上に見出されるエキソンに対するタンパク質アナログである。例えば、インテインを含むポリペプチドは構造エクステイン(N)-インテイン-エクステイン(C)であってもよい。インテインの切除及び2つのエクステインのスプライシング後に、もたらされる構造はエクステイン(N)-エクステイン(C)及び遊離のインテインである。種々の構成では、エクステインは、夫々が分裂インテインに融合された別個のタンパク質(例として、Cas9又はプライム編集因子の半分)であってもよく、分裂インテインの切除はエクステイン配列のスプライシングを一緒に引き起こす。
本明細書において用いられる用語「エクステイン」は、インテインによってフランキングされ、タンパク質スプライシングのプロセスの間に別のエクステインにライゲーションされて成熟したスプライシングされたタンパク質を形成するポリペプチド配列を指す。典型的には、インテインは2つのエクステイン配列によってフランキングされ、これらはインテインがそれ自体の切除を触媒するときに一緒にライゲーションされる。エクステインは、従って、mRNA上に見出されるエキソンに対するタンパク質アナログである。例えば、インテインを含むポリペプチドは構造エクステイン(N)-インテイン-エクステイン(C)であってもよい。インテインの切除及び2つのエクステインのスプライシング後に、もたらされる構造はエクステイン(N)-エクステイン(C)及び遊離のインテインである。種々の構成では、エクステインは、夫々が分裂インテインに融合された別個のタンパク質(例として、Cas9又はプライム編集因子の半分)であってもよく、分裂インテインの切除はエクステイン配列のスプライシングを一緒に引き起こす。
伸長アーム
用語「伸長アーム」は、プライマー結合部位及びポリメラーゼ(例として、逆転写酵素)のためのDNA合成鋳型(例として、編集鋳型及び相同アーム)を含むpegRNAのヌクレオチド配列構成要素を指す。いくつかの態様、例として、図3Dにおいて、伸長アームは、ガイドRNAの3'末にて位置付けられる。他の態様、例として、図3Eにおいて、伸長アームは、ガイドRNAの5'末にて位置付けられる。いくつかの態様において、伸長アームは、DNA合成鋳型及びプライマー結合部位を含む。いくつかの態様では、伸長アームは、5'から3'方向に以下の構成要素:DNA合成鋳型及びプライマー結合部位を含む。いくつかの態様において、伸長アームはまた、相同アームも包含する。様々な態様において、伸長アームは、5'→3'方向に以下の構成要素:相同アーム、編集鋳型、及びプライマー結合部位を含む。様々な態様において、伸長アームは、5'→3'方向に以下の構成要素:相同アーム、編集鋳型、及びプライマー結合部位を含む。逆転写酵素の重合活性が5'→3'方向であるから、相同アーム、編集鋳型、及びプライマー結合部位の好ましい配置は、逆転写酵素が、アニールされたプライマー配列によって一旦プライムされると、編集鋳型を相補的な鋳型鎖として使用して単鎖DNAを重合するように、5'→3'方向にある。
用語「伸長アーム」は、プライマー結合部位及びポリメラーゼ(例として、逆転写酵素)のためのDNA合成鋳型(例として、編集鋳型及び相同アーム)を含むpegRNAのヌクレオチド配列構成要素を指す。いくつかの態様、例として、図3Dにおいて、伸長アームは、ガイドRNAの3'末にて位置付けられる。他の態様、例として、図3Eにおいて、伸長アームは、ガイドRNAの5'末にて位置付けられる。いくつかの態様において、伸長アームは、DNA合成鋳型及びプライマー結合部位を含む。いくつかの態様では、伸長アームは、5'から3'方向に以下の構成要素:DNA合成鋳型及びプライマー結合部位を含む。いくつかの態様において、伸長アームはまた、相同アームも包含する。様々な態様において、伸長アームは、5'→3'方向に以下の構成要素:相同アーム、編集鋳型、及びプライマー結合部位を含む。様々な態様において、伸長アームは、5'→3'方向に以下の構成要素:相同アーム、編集鋳型、及びプライマー結合部位を含む。逆転写酵素の重合活性が5'→3'方向であるから、相同アーム、編集鋳型、及びプライマー結合部位の好ましい配置は、逆転写酵素が、アニールされたプライマー配列によって一旦プライムされると、編集鋳型を相補的な鋳型鎖として使用して単鎖DNAを重合するように、5'→3'方向にある。
伸長アームの長さ等の更なる詳細は、本明細書に記載の他のどこかに記載されている。
伸長アームはまた、図3G(最上列)に実例として示される通り、一般に2つの領域:プライマー結合部位(PBS)及びDNA合成鋳型を含むものとしても記載されていてもよい。プライマー結合部位は、プライム編集因子複合体によってニックが入れられるようになり、これによってニックが入った内生鎖上に3'末が晒されたとき、標的部位の内生DNA鎖から形成されるプライマー配列へ結合する。本明細書に説明される通り、プライマー配列の、pegRNAの伸長アーム上のプライマー結合部位への結合は、晒された3'末(すなわち、プライマー配列の3')をもつ二重鎖領域を作成し、これは次いで、晒された3'末からDNA合成鋳型の長さに沿ってDNAの単鎖を重合し始めるポリメラーゼのための基質を提供する。単鎖DNA産物の配列は、DNA合成鋳型の相補体である。重合は、重合が終止するまでDNA合成鋳型(又は伸長アーム)の5'へ続く。よって、DNA合成鋳型は、プライム編集因子複合体のポリメラーゼによって単鎖DNA産物(すなわち、所望の遺伝子編集情報を含有する3'単鎖DNAフラップ)中へコードされる伸長アームの部分を表し、単鎖DNA産物は、PEに誘導されたニック部位のすぐ下流にある標的部位の対応する内生DNA鎖に置き換える。理論に拘泥されないが、DNA合成鋳型の重合は、終止という事象まで、伸長アームの5'末へ続く。重合は、これらに限定されないが、(a)pegRNAの5'末端に達すること(例として、5'伸長アームのケースにおいて、DNAポリメラーゼが単に鋳型を使い果たす)、(b)通り抜けられないRNA2次構造(例として、ヘアピン若しくはステム/ループ)に達すること、あるいは(c)複製終止シグナル、例として、ポリメラーゼを遮断若しくは阻害する特定のヌクレオチド配列、又はスーパーコイル状のDNA若しくはRNA等の核酸形態上のシグナルに達すること、を包含する様々な形で終止してもよい。
フラップエンドヌクレアーゼ(例として、FEN1)
本明細書に使用されるとき、用語「フラップエンドヌクレアーゼ」は、5'単鎖DNAフラップの除去を触媒する酵素を指す。これらは、細胞プロセス(DNA複製を包含する)の最中に形成された5'フラップの除去を処理する酵素である。本明細書に記載のプライム編集方法は、内生で供給されるフラップエンドヌクレアーゼ、又はプライム編集の最中に標的部位にて形成される内生DNAの5'フラップを除去するのにtransで提供されるものを利用してもよい。フラップエンドヌクレアーゼは、当該技術分野において知られており、Patel et al.,「Flap endonucleases pass 5'-flaps through a flexible arch using a disorder-thread-order mechanism to confer specificity for free 5'-ends,」Nucleic Acids Research,2012,40(10):4507-4519及びTsutakawa et al.,「Human flap endonuclease structures,DNA double-base flipping,and a unified understanding of the FEN1 superfamily,」Cell,2011,145(2):198-211、及びBalakrishnan et al., 「Flap Endonuclease 1,」Annu Rev Biochem,2013,Vol 82:119-138(これらの各々は参照により本明細書に組み込まれる)の記載から見出され得る。例示のフラップエンドヌクレアーゼは、以下のアミノ酸配列によって表され得るFEN1である:

本明細書に使用されるとき、用語「フラップエンドヌクレアーゼ」は、5'単鎖DNAフラップの除去を触媒する酵素を指す。これらは、細胞プロセス(DNA複製を包含する)の最中に形成された5'フラップの除去を処理する酵素である。本明細書に記載のプライム編集方法は、内生で供給されるフラップエンドヌクレアーゼ、又はプライム編集の最中に標的部位にて形成される内生DNAの5'フラップを除去するのにtransで提供されるものを利用してもよい。フラップエンドヌクレアーゼは、当該技術分野において知られており、Patel et al.,「Flap endonucleases pass 5'-flaps through a flexible arch using a disorder-thread-order mechanism to confer specificity for free 5'-ends,」Nucleic Acids Research,2012,40(10):4507-4519及びTsutakawa et al.,「Human flap endonuclease structures,DNA double-base flipping,and a unified understanding of the FEN1 superfamily,」Cell,2011,145(2):198-211、及びBalakrishnan et al., 「Flap Endonuclease 1,」Annu Rev Biochem,2013,Vol 82:119-138(これらの各々は参照により本明細書に組み込まれる)の記載から見出され得る。例示のフラップエンドヌクレアーゼは、以下のアミノ酸配列によって表され得るFEN1である:
機能的等価物
用語「機能的等価物」は、機能は等価であるが構造は第1生体分子と必ずしも等価ではない第2生体分子を指す。例えば、「Cas9等価物」は、Cas9と同じか又は実質的に同じ機能を有するが、必ずしも同じアミノ酸配列は有さないタンパク質を指す。本開示の文脈において、本明細書は通してずっと「タンパク質X、又はその機能的等価物」を指す。これに関連して、タンパク質Xの「機能的等価物」は、タンパク質Xの等価機能を担持する、いずれのホモログ、パラログ、フラグメント、天然に存在するバージョン、操作バージョン、突然変異バージョン、又は合成バージョンを受け入れる。
用語「機能的等価物」は、機能は等価であるが構造は第1生体分子と必ずしも等価ではない第2生体分子を指す。例えば、「Cas9等価物」は、Cas9と同じか又は実質的に同じ機能を有するが、必ずしも同じアミノ酸配列は有さないタンパク質を指す。本開示の文脈において、本明細書は通してずっと「タンパク質X、又はその機能的等価物」を指す。これに関連して、タンパク質Xの「機能的等価物」は、タンパク質Xの等価機能を担持する、いずれのホモログ、パラログ、フラグメント、天然に存在するバージョン、操作バージョン、突然変異バージョン、又は合成バージョンを受け入れる。
融合タンパク質
用語「融合タンパク質」は、本明細書に使用されるとき、少なくとも2つの異なるタンパク質からタンパク質ドメインを含むハイブリッドポリペプチドを指す。あるタンパク質は、融合タンパク質のアミノ末端(N末端)部分にて、又はカルボキシ末端(C末端)タンパク質にて位置付けられ、よって「アミノ末端の融合タンパク質」又は「カルボキシ末端の融合タンパク質」の夫々を形成してもよい。タンパク質は、異なるドメイン、例えば、核酸結合ドメイン(例として、タンパク質を標的部位への結合へ向かわせる、Cas9のgRNA結合ドメイン)と、核酸編集タンパク質の核酸切断ドメイン又は触媒ドメインとを含んでいてもよい。別の例は、逆転写酵素と融合したCas9又はその等価物を包含する。本明細書に提供されるタンパク質のいずれも、当該技術分野において知られているいずれの方法によって産生されてもよい。例えば、本明細書に提供されるタンパク質は、ペプチドリンカーを含む融合タンパク質に特に適した、組換えタンパク質の発現及び精製を介して産生されてもよい。組換えタンパク質の発現及び精製のための方法は周知であり、Green and Sambrook,Molecular Cloning:A Laboratory Manual(4th ed.,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,N.Y.(2012)(これらの内容全体は参照により本明細書に組み込まれる))によって記載されるものを包含する。
用語「融合タンパク質」は、本明細書に使用されるとき、少なくとも2つの異なるタンパク質からタンパク質ドメインを含むハイブリッドポリペプチドを指す。あるタンパク質は、融合タンパク質のアミノ末端(N末端)部分にて、又はカルボキシ末端(C末端)タンパク質にて位置付けられ、よって「アミノ末端の融合タンパク質」又は「カルボキシ末端の融合タンパク質」の夫々を形成してもよい。タンパク質は、異なるドメイン、例えば、核酸結合ドメイン(例として、タンパク質を標的部位への結合へ向かわせる、Cas9のgRNA結合ドメイン)と、核酸編集タンパク質の核酸切断ドメイン又は触媒ドメインとを含んでいてもよい。別の例は、逆転写酵素と融合したCas9又はその等価物を包含する。本明細書に提供されるタンパク質のいずれも、当該技術分野において知られているいずれの方法によって産生されてもよい。例えば、本明細書に提供されるタンパク質は、ペプチドリンカーを含む融合タンパク質に特に適した、組換えタンパク質の発現及び精製を介して産生されてもよい。組換えタンパク質の発現及び精製のための方法は周知であり、Green and Sambrook,Molecular Cloning:A Laboratory Manual(4th ed.,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,N.Y.(2012)(これらの内容全体は参照により本明細書に組み込まれる))によって記載されるものを包含する。
着目した遺伝子(GOI)
用語「着目した遺伝子」又は「GOI」は、着目した生体分子(例として、タンパク質又はRNA分子)をコードする遺伝子を指す。着目したタンパク質は、いずれの細胞内タンパク質、膜タンパク質、又は細胞外タンパク質、例として、核内タンパク質、転写因子、核膜輸送体、細胞内オルガネラ関連タンパク質、膜受容体、触媒タンパク質、及び酵素、処置用タンパク質、膜タンパク質、膜輸送タンパク質、シグナル伝達タンパク質、又は免疫学的タンパク質(例として、IgG又は他の抗体タンパク質)、等々を包含し得る。着目した遺伝子はまた、これらに限定されないが、メッセンジャーRNA(mRNA)、トランスファーRNA(tRNA)、リボソームRNA(rRNA)、低分子核内RNA(snRNA)、アンチセンスRNA、ガイドRNA、マイクロRNA(miRNA)、低分子干渉RNA(siRNA)、及び無細胞RNA(cfRNA)を包含するRNA分子もコードしていてもよい。
用語「着目した遺伝子」又は「GOI」は、着目した生体分子(例として、タンパク質又はRNA分子)をコードする遺伝子を指す。着目したタンパク質は、いずれの細胞内タンパク質、膜タンパク質、又は細胞外タンパク質、例として、核内タンパク質、転写因子、核膜輸送体、細胞内オルガネラ関連タンパク質、膜受容体、触媒タンパク質、及び酵素、処置用タンパク質、膜タンパク質、膜輸送タンパク質、シグナル伝達タンパク質、又は免疫学的タンパク質(例として、IgG又は他の抗体タンパク質)、等々を包含し得る。着目した遺伝子はまた、これらに限定されないが、メッセンジャーRNA(mRNA)、トランスファーRNA(tRNA)、リボソームRNA(rRNA)、低分子核内RNA(snRNA)、アンチセンスRNA、ガイドRNA、マイクロRNA(miRNA)、低分子干渉RNA(siRNA)、及び無細胞RNA(cfRNA)を包含するRNA分子もコードしていてもよい。
ガイドRNA(「gRNA」)
本明細書に使用されるとき、用語「ガイドRNA」は、大部分が一般的にCRISPR-Cas9のCasタンパク質に関連する具体的な型のガイド核酸であって、Cas9に結び付いて、Cas9タンパク質をDNA分子中の特定の配列(ガイドRNAのプロトスペース配列との相補性を包含する)へ向かわせる。しかしながら、この用語はまた、天然に存在するか若しくは天然には存在しない(例として、操作又は組換え)かにかかわらず、Cas9等価物、ホモログ、オルソログ、又はパラログに結び付き、またCas9等価物を特定の標的ヌクレオチド配列へ局在化するよう別にプログラムする、等価なガイド核酸分子も受け入れる。Cas9等価物は、Cpf1(V型CRISPR-Cas系)、C2c1(V型CRISPR-Cas系)、C2c2(VI型CRISPR-Cas系)及びC2c3(V型CRISPR-Cas系)を包含する、いずれの型のCRISPR系(例として、II型、V型、VI型)からの他のnapDNAbpも包含していてもよい。更なるCas等価物は、Makarova et al.,「C2c2 is a single-component programmable RNA-guided RNA-targeting CRISPR effector,」Science 2016;353(6299)(この内容は参照により本明細書に組み込まれる)に記載されている。ガイドRNAの例示の配列及び構造は本明細書に提供されている。加えて、適切なガイドRNA配列をデザインするための方法も、本明細書に提供されている。本明細書に使用されるとき、「ガイドRNA」はまた、本明細書に開示のプライム編集方法及び組成物のために発明された「プライム編集ガイドRNA」(又は「pegRNA」)と呼ばれる修飾形態のガイドRNAと対比させるために「既存のガイドRNA」とも呼ばれてもよい。
本明細書に使用されるとき、用語「ガイドRNA」は、大部分が一般的にCRISPR-Cas9のCasタンパク質に関連する具体的な型のガイド核酸であって、Cas9に結び付いて、Cas9タンパク質をDNA分子中の特定の配列(ガイドRNAのプロトスペース配列との相補性を包含する)へ向かわせる。しかしながら、この用語はまた、天然に存在するか若しくは天然には存在しない(例として、操作又は組換え)かにかかわらず、Cas9等価物、ホモログ、オルソログ、又はパラログに結び付き、またCas9等価物を特定の標的ヌクレオチド配列へ局在化するよう別にプログラムする、等価なガイド核酸分子も受け入れる。Cas9等価物は、Cpf1(V型CRISPR-Cas系)、C2c1(V型CRISPR-Cas系)、C2c2(VI型CRISPR-Cas系)及びC2c3(V型CRISPR-Cas系)を包含する、いずれの型のCRISPR系(例として、II型、V型、VI型)からの他のnapDNAbpも包含していてもよい。更なるCas等価物は、Makarova et al.,「C2c2 is a single-component programmable RNA-guided RNA-targeting CRISPR effector,」Science 2016;353(6299)(この内容は参照により本明細書に組み込まれる)に記載されている。ガイドRNAの例示の配列及び構造は本明細書に提供されている。加えて、適切なガイドRNA配列をデザインするための方法も、本明細書に提供されている。本明細書に使用されるとき、「ガイドRNA」はまた、本明細書に開示のプライム編集方法及び組成物のために発明された「プライム編集ガイドRNA」(又は「pegRNA」)と呼ばれる修飾形態のガイドRNAと対比させるために「既存のガイドRNA」とも呼ばれてもよい。
ガイドRNA又はpegRNAは、これらに限定されないが、以下を包含する様々な構造要素を含んでいてもよい:
スペーサ配列-標的DNA中のプロトスペーサへ結合するガイドRNA又はpegRNA(長さが約20ntを有する)中の配列。
gRNAコア(又はgRNA骨格若しくはバックボーン配列)-は、Cas9結合を担うgRNA内の配列を指し、これは、Cas9を標的DNAへガイドするのに使用される20bpのスペーサ/標的化配列を包含していない。
伸長アーム-pegRNAの3'端又は5'端の一本鎖伸長。これは、プライマー結合部位とポリメラーゼ(例として、逆転写酵素)によって目当ての遺伝子変化を含有する一本鎖DNAフラップをコードするDNA合成鋳型配列とを含む。これが、次いで、対応する内生鎖を置き換えることによって内生DNA上に取り入れし、それによって所望の遺伝子変化を組み入れる。
転写ターミネータ-ガイドRNA又はpegRNAは分子の3'に転写終結配列を含んでもよい。
G四重鎖
「G四重鎖」という用語は、その通常及び通例の意味を指す。G四重鎖は、グアニン(G)が豊富な核酸配列に形成された複雑な三次元核酸部分である。それらはらせん形状であり、グアニンテトラッド(又は「G-テトラッド」)の相互接続スタックから形成され、個々には4つのグアニンから形成された平坦なリング状構造であり、G-テトラッドの対の間の中央チャネルに位置するカチオン(例として、カリウム)の存在によって安定化され得る。G-四重極は、構造の多様な集合であり、単一の構造ではない。G-四重鎖への更なる参照は、(1)Kwok et al.,「G-Quadruplexes:Prediction,Characterization,and Biological Application,」Trends in Biotechnology,2017,Vol.35(10;pp.997-1013;(2)Hansel-Hertsch R.et al.,「DNA G-quadruplexes in the human genome:detection,functions and therapeutic potential,」Nat.Rev.Mol.Cell Biol.,2017;18:279-284;and(3)Millevoi S.et al.,「G-quadruplexes in RNA biology,」Wiley InterdiscipRev.RNA.,2012;3:495-507に見出され得、それぞれが参照により本明細書に組み込まれる。
「G四重鎖」という用語は、その通常及び通例の意味を指す。G四重鎖は、グアニン(G)が豊富な核酸配列に形成された複雑な三次元核酸部分である。それらはらせん形状であり、グアニンテトラッド(又は「G-テトラッド」)の相互接続スタックから形成され、個々には4つのグアニンから形成された平坦なリング状構造であり、G-テトラッドの対の間の中央チャネルに位置するカチオン(例として、カリウム)の存在によって安定化され得る。G-四重極は、構造の多様な集合であり、単一の構造ではない。G-四重鎖への更なる参照は、(1)Kwok et al.,「G-Quadruplexes:Prediction,Characterization,and Biological Application,」Trends in Biotechnology,2017,Vol.35(10;pp.997-1013;(2)Hansel-Hertsch R.et al.,「DNA G-quadruplexes in the human genome:detection,functions and therapeutic potential,」Nat.Rev.Mol.Cell Biol.,2017;18:279-284;and(3)Millevoi S.et al.,「G-quadruplexes in RNA biology,」Wiley InterdiscipRev.RNA.,2012;3:495-507に見出され得、それぞれが参照により本明細書に組み込まれる。
相同アーム
用語「相同アーム」は、結果として得られる、逆転写酵素にコードされた単鎖DNAフラップ(内生鎖を置き換えることによって標的DNA部位中へ取り入れられる)の部分をコードする伸長アームの部分を指す。相同アームによってコードされる単鎖DNAフラップの部分は、標的DNA配列の非編集済鎖に相補的であって、内生鎖に取って代わること及びその場所において単鎖DNAフラップとアニールすることを容易にし、これによって編集が組み入れられる。この構成要素は他のどこかで更に定義される。定義上、それは本明細書に記載のプライム編集因子のポリメラーゼによってコードされるので、相同アームはDNA合成鋳型の一部である。
用語「相同アーム」は、結果として得られる、逆転写酵素にコードされた単鎖DNAフラップ(内生鎖を置き換えることによって標的DNA部位中へ取り入れられる)の部分をコードする伸長アームの部分を指す。相同アームによってコードされる単鎖DNAフラップの部分は、標的DNA配列の非編集済鎖に相補的であって、内生鎖に取って代わること及びその場所において単鎖DNAフラップとアニールすることを容易にし、これによって編集が組み入れられる。この構成要素は他のどこかで更に定義される。定義上、それは本明細書に記載のプライム編集因子のポリメラーゼによってコードされるので、相同アームはDNA合成鋳型の一部である。
宿主細胞
用語「宿主細胞」は、本明細書に使用されるとき、本明細書に記載のベクター、例として、Cas9若しくはCas9等価物と逆転写酵素とを含む融合タンパク質をコードする核酸分子を含むベクターの宿主となり(host)複製してこれを発現し得る細胞を指す。
用語「宿主細胞」は、本明細書に使用されるとき、本明細書に記載のベクター、例として、Cas9若しくはCas9等価物と逆転写酵素とを含む融合タンパク質をコードする核酸分子を含むベクターの宿主となり(host)複製してこれを発現し得る細胞を指す。
インテイン
本明細書において用いられる用語「インテイン」は、生命の全てのドメインからの生物に見出される自己プロセシングポリペプチドドメインを指す。インテイン(介在タンパク質)は、タンパク質スプライシングとして公知の固有の自己プロセシングイベントを実行する。これにおいては、それは、2つのペプチド結合の切断によってより大きい前駆体ポリペプチドからそれ自体を切除、プロセスにおいて、新たなペプチド結合の形成によってフランキングするエクステイン(外部タンパク質)配列同士をライゲーションする。インテイン遺伝子は他のタンパク質コード遺伝子とインフレームで埋め込まれて見出されるので、この再構成は翻訳後に(又は共翻訳的に)生起する。更にその上、インテインによって媒介されるタンパク質スプライシングは自発的である;それは、外部の因子又はエネルギー源ではなくインテインドメインのフォールディングのみを要求する。このプロセスは、「分裂インテイン」によるトランスタンパク質スプライシングの天然のプロセスと対比して、シスタンパク質スプライシングとしてもまた公知である。インテインは自己スプライシングRNAイントロンのタンパク質等価物であり(Perler et al.,Nucleic Acids Res.22:1125-1127(1994)を見よ)、これらは前駆体タンパク質からのそれら自体の切除を触媒し、エクステインとして公知のフランキングするタンパク質配列の付随的融合を有する(Perler et al.,Curr.Opin.Chem.Biol.1:292-299(1997);Perler,F.B.Cell 92(1):1-4(1998);Xu et al.,EMBO J.15(19):5146-5153(1996)において総説されている)。
本明細書において用いられる用語「インテイン」は、生命の全てのドメインからの生物に見出される自己プロセシングポリペプチドドメインを指す。インテイン(介在タンパク質)は、タンパク質スプライシングとして公知の固有の自己プロセシングイベントを実行する。これにおいては、それは、2つのペプチド結合の切断によってより大きい前駆体ポリペプチドからそれ自体を切除、プロセスにおいて、新たなペプチド結合の形成によってフランキングするエクステイン(外部タンパク質)配列同士をライゲーションする。インテイン遺伝子は他のタンパク質コード遺伝子とインフレームで埋め込まれて見出されるので、この再構成は翻訳後に(又は共翻訳的に)生起する。更にその上、インテインによって媒介されるタンパク質スプライシングは自発的である;それは、外部の因子又はエネルギー源ではなくインテインドメインのフォールディングのみを要求する。このプロセスは、「分裂インテイン」によるトランスタンパク質スプライシングの天然のプロセスと対比して、シスタンパク質スプライシングとしてもまた公知である。インテインは自己スプライシングRNAイントロンのタンパク質等価物であり(Perler et al.,Nucleic Acids Res.22:1125-1127(1994)を見よ)、これらは前駆体タンパク質からのそれら自体の切除を触媒し、エクステインとして公知のフランキングするタンパク質配列の付随的融合を有する(Perler et al.,Curr.Opin.Chem.Biol.1:292-299(1997);Perler,F.B.Cell 92(1):1-4(1998);Xu et al.,EMBO J.15(19):5146-5153(1996)において総説されている)。
本明細書において用いられる用語「タンパク質スプライシング」は、前駆体タンパク質の内側領域(インテイン)が切り出され、タンパク質のフランキング領域(エクステイン)同士がライゲーションされて、成熟したタンパク質を形成するプロセスを指す。この天然のプロセスは、原核生物及び真核生物両方からの数々のタンパク質で観察されている(Perler,F.B.,Xu,M.Q.,Paulus,H.Current Opinion in Chemical Biology 1997,1,292-299;Perler,F.B.Nucleic Acids Research 1999,27,346-347)。インテイン単位は、タンパク質スプライシングを触媒するために必要とされる必要な構成要素を含有し、多くの場合には、インテイン可動性に関与するエンドヌクレアーゼドメインを含有する(Perler,F.B.,Davis,E.O.,Dean,G.E.,Gimble,F.S.,Jack,W.E.,Neff,N.,Noren,C.J.,Thomer,J.,Belfort,M.Nucleic Acids Research 1994,22,1127-1127)。しかしながら、もたらされるタンパク質は連結されており、別個のタンパク質としては発現されない。タンパク質スプライシングは、別個のポリペプチド上で発現される分裂インテインによって、トランスでもまた行われてもよい。これらは自発的に組み合わさって単一のインテインを形成し、これが次いでタンパク質スプライシングプロセスを経過して別個のタンパク質に連結する。
タンパク質スプライシングの機序の解明は、インテインに基づく数多の用途へ繋がっている(Comb,et al.,米国特許第5,496,714号;Comb,et al.,米国特許第5,834,247号;Camarero and Muir,J.Amer.Chem.Soc.,121:5597-5598(1999);Chong,et al.,Gene,192:271-281(1997),Chong,et al.,Nucleic Acids Res.,26:5109-5115(1998);Chong,et al.,J.Biol.Chem.,273:10567-10577(1998);Cotton,et al.J.Am.Chem.Soc.,121:1100-1101(1999);Evans,et al.,J.Biol.Chem.,274:18359-18363(1999);Evans,et al.,J.Biol.Chem.,274:3923-3926(1999);Evans,et al.,Protein Sci.,7:2256-2264(1998);Evans,et al.,J.Biol.Chem.,275:9091-9094(2000);Iwai and Pluckthun,FEBS Lett.459:166-172(1999);Mathys,et al.,Gene,231:1-13(1999);Mills,et al.,Proc.Natl.Acad.Sci.USA 95:3543-3548(1998);Muir,et al.,Proc.Natl.Acad.Sci.USA 95:6705-6710(1998);Otomo,et al.,Biochemistry 38:16040-16044(1999);Otomo,et al.,J.Biolmol.NMR 14:105-114(1999);Scott,et al.,Proc.Natl.Acad.Sci.USA 96:13638-13643(1999);Severinov and Muir,J.Biol.Chem.,273:16205-16209(1998);Shingledecker,et al.,Gene,207:187-195(1998);Southworth,et al.,EMBO J.17:918-926(1998);Southworth,et al.,Biotechniques,27:110-120(1999);Wood,et al.,Nat.Biotechnol.,17:889-892(1999);Wu,et al.,Proc.Natl.Acad.Sci.USA 95:9226-9231(1998a);Wu,et al.,Biochim Biophys Acta 1387:422-432(1998b);Xu,et al.,Proc.Natl.Acad.Sci.USA 96:388-393(1999);Yamazaki,et al.,J.Am.Chem.Soc.,120:5591-5592(1998))。各参照文献は参照により本明細書に組み込まれる。
リガンド依存性インテイン
本明細書において用いられる用語「リガンド依存性インテイン」は、リガンド結合ドメインを含むインテインを指す。典型的には、リガンド結合ドメインはインテインのアミノ酸配列上に挿入され、構造インテイン(N)-リガンド結合ドメイン-インテイン(C)をもたらす。典型的には、リガンド依存性インテインは、適当なリガンドの不在下においてはタンパク質スプライシング活性を発揮しないか又は最小限のみ発揮し、リガンドの存在下においてはタンパク質スプライシング活性の顕著な増大を発揮する。いくつかの態様では、リガンド依存性インテインはリガンドの不在下においては観察可能なスプライシング活性を発揮しないが、リガンドの存在下においてはスプライシング活性を発揮する。いくつかの態様では、リガンド依存性インテインはリガンドの不在下においては観察可能なタンパク質スプライシング活性を、適当なリガンド存在下においては、リガンドの不在下において観察される活性よりも少なくとも5倍、少なくとも10倍、少なくとも50倍、少なくとも100倍、少なくとも150倍、少なくとも200倍、少なくとも250倍、少なくとも500倍、少なくとも1000倍、少なくとも1500倍、少なくとも2000倍、少なくとも2500倍、少なくとも5000倍、少なくとも10000倍、少なくとも20000倍、少なくとも25000倍、少なくとも50000倍、少なくとも100000倍、少なくとも500000倍、又は少なくとも1000000倍多大であるタンパク質スプライシング活性を発揮する。いくつかの態様では、活性の増大は少なくとも1桁、少なくとも2桁、少なくとも3桁、少なくとも4桁、又は少なくとも5桁にわたって用量依存的であり、リガンドの濃度を調整することによってインテイン活性の微調整を許す。好適なリガンド依存性インテインは当該技術分野において知られており、下で提供されるもの及び公開米国特許出願第U.S.2014/0065711A1号;Mootz et al.,「Protein splicng triggered by a small molecule.」J.Am.Chem.Soc.2002;124,9044-9045;Mootz et al.,「Conditional protein splicng:a new tool to control protein structure and function in vitro and in vivo.」J.Am.Chem.Soc.2003;125,10561-10569;Buskirk et al.,Proc.Natl.Acad.Sci.USA.2004;101,10505-10510);Skretas&Wood,「Regulation of protein activity with small-molecule-controlled inteins.」Protein Sci.2005;14,523-532;Schwartz,et al.,「Post-translational enzyme activation in an animal via optimized conditional protein splicing.」Nat.Chem.Biol.2007;3,50-54;Peck et al.,Chem.Biol.2011;18(5),619-630に記載されているものを包含する;夫々の内容全体は参照によってここに組み込まれる。例示の配列は次の通りである:

本明細書において用いられる用語「リガンド依存性インテイン」は、リガンド結合ドメインを含むインテインを指す。典型的には、リガンド結合ドメインはインテインのアミノ酸配列上に挿入され、構造インテイン(N)-リガンド結合ドメイン-インテイン(C)をもたらす。典型的には、リガンド依存性インテインは、適当なリガンドの不在下においてはタンパク質スプライシング活性を発揮しないか又は最小限のみ発揮し、リガンドの存在下においてはタンパク質スプライシング活性の顕著な増大を発揮する。いくつかの態様では、リガンド依存性インテインはリガンドの不在下においては観察可能なスプライシング活性を発揮しないが、リガンドの存在下においてはスプライシング活性を発揮する。いくつかの態様では、リガンド依存性インテインはリガンドの不在下においては観察可能なタンパク質スプライシング活性を、適当なリガンド存在下においては、リガンドの不在下において観察される活性よりも少なくとも5倍、少なくとも10倍、少なくとも50倍、少なくとも100倍、少なくとも150倍、少なくとも200倍、少なくとも250倍、少なくとも500倍、少なくとも1000倍、少なくとも1500倍、少なくとも2000倍、少なくとも2500倍、少なくとも5000倍、少なくとも10000倍、少なくとも20000倍、少なくとも25000倍、少なくとも50000倍、少なくとも100000倍、少なくとも500000倍、又は少なくとも1000000倍多大であるタンパク質スプライシング活性を発揮する。いくつかの態様では、活性の増大は少なくとも1桁、少なくとも2桁、少なくとも3桁、少なくとも4桁、又は少なくとも5桁にわたって用量依存的であり、リガンドの濃度を調整することによってインテイン活性の微調整を許す。好適なリガンド依存性インテインは当該技術分野において知られており、下で提供されるもの及び公開米国特許出願第U.S.2014/0065711A1号;Mootz et al.,「Protein splicng triggered by a small molecule.」J.Am.Chem.Soc.2002;124,9044-9045;Mootz et al.,「Conditional protein splicng:a new tool to control protein structure and function in vitro and in vivo.」J.Am.Chem.Soc.2003;125,10561-10569;Buskirk et al.,Proc.Natl.Acad.Sci.USA.2004;101,10505-10510);Skretas&Wood,「Regulation of protein activity with small-molecule-controlled inteins.」Protein Sci.2005;14,523-532;Schwartz,et al.,「Post-translational enzyme activation in an animal via optimized conditional protein splicing.」Nat.Chem.Biol.2007;3,50-54;Peck et al.,Chem.Biol.2011;18(5),619-630に記載されているものを包含する;夫々の内容全体は参照によってここに組み込まれる。例示の配列は次の通りである:
リンカー
本明細書において用いられる用語「リンカー」は、2つの他の分子又は部分を連結するある分子を指す。リンカーは、2つの融合タンパク質を連結するリンカーのケースではアミノ酸配列であり得る。例えば、Cas9はアミノ酸リンカー配列によってポリメラーゼ(例として、逆転写酵素)に融合され得る。リンカーは、2つのヌクレオチド配列を一緒に連結するケースではヌクレオチド配列でもまたあり得る。例えば、本ケースでは、従来のガイドRNAは、スペーサ又はリンカーヌクレオチド配列を介して、RT鋳型配列とRTプライマー結合部位とを含んでもよいプライム編集ガイドRNAのRNA伸長に連結される。他の態様において、リンカーは有機分子、基、ポリマー、又は化学的部分である。いくつかの態様では、リンカーは、長さが5-100アミノ酸、例えば長さが5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、30~35、35~40、40~45、45~50、50~60、60~70、70~80、80~90、90~100、100~150、又は150~200アミノ酸である。より長い又はより短いリンカーもまた企図される。
本明細書において用いられる用語「リンカー」は、2つの他の分子又は部分を連結するある分子を指す。リンカーは、2つの融合タンパク質を連結するリンカーのケースではアミノ酸配列であり得る。例えば、Cas9はアミノ酸リンカー配列によってポリメラーゼ(例として、逆転写酵素)に融合され得る。リンカーは、2つのヌクレオチド配列を一緒に連結するケースではヌクレオチド配列でもまたあり得る。例えば、本ケースでは、従来のガイドRNAは、スペーサ又はリンカーヌクレオチド配列を介して、RT鋳型配列とRTプライマー結合部位とを含んでもよいプライム編集ガイドRNAのRNA伸長に連結される。他の態様において、リンカーは有機分子、基、ポリマー、又は化学的部分である。いくつかの態様では、リンカーは、長さが5-100アミノ酸、例えば長さが5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、30~35、35~40、40~45、45~50、50~60、60~70、70~80、80~90、90~100、100~150、又は150~200アミノ酸である。より長い又はより短いリンカーもまた企図される。
単離された
「単離された」は、自然状態から変更又は除去されることを意味する。例えば、生きている動物中に天然に存在する核20酸又はペプチドは「単離され」ていないが、共存材料のその自然状態から部分的又は完全に分離された同じ核酸又はペプチドは、「単離され」ている。単離された核酸又はタンパク質は、実質的に精製された形態で存在し得るか、又は非ネイティブ(non-native)的環境(例えば、宿主細胞等)で存在し得る。
「単離された」は、自然状態から変更又は除去されることを意味する。例えば、生きている動物中に天然に存在する核20酸又はペプチドは「単離され」ていないが、共存材料のその自然状態から部分的又は完全に分離された同じ核酸又はペプチドは、「単離され」ている。単離された核酸又はタンパク質は、実質的に精製された形態で存在し得るか、又は非ネイティブ(non-native)的環境(例えば、宿主細胞等)で存在し得る。
いくつかの態様において、着目する遺伝子は、単離された核酸によってコードされる。本明細書に使用されるとき、用語「単離された」は、その元の又はネイティブ(native)の環境(例として、天然に存在する場合は自然環境)から除去されている、本明細書に提供される通りの材料の特徴を指す。したがって、生きている動物中に存在する、天然に存在するポリヌクレオチド又はタンパク質若しくはポリペプチドは単離されていないが、ヒトの介入によって自然系中の共存材料のいくつか又は全てから分離されている同じポリヌクレオチド又はポリペプチドは、単離されている。人工材料又は操作された材料、例えば、本明細書に記載の発現コンストラクト及びベクター等の、天然には存在しない核酸構築物はまた、結果的に、単離されたとも呼ばれる。材料は、単離されるために精製される必要はない。結果的に、材料はベクターの一部及び/又は組成物の一部であってもよく、かかるベクター又は組成物が、材料が自然界から見出される環境の一部にない点で、依然単離されている。
MS2タグ付け技術
種々の態様では(例として、図72~図73及び例19の態様に図示されている通り)、用語「MS2タグ付け技術」は、RNA-タンパク質相互作用ドメイン、例として特定のヘアピン構造を特異的に認識及び結合するRNA結合タンパク質とペアリングされた「RNA-タンパク質相互作用ドメイン」(「RNA-タンパク質動員ドメイン又はタンパク質」としてもまた公知)の組み合わせを指す。これらの型のシステムは、標的部位に結合されているプライム編集因子複合体に種々の機能性を動員するように活用され得る。MS2タグ付け技術は、ファージのゲノム上に存在するステムループ又はヘアピン構造、すなわち「MS2ヘアピン」とのMS2バクテリオファージコートタンパク質(「MCP」又は「MS2cp」)の天然の相互作用に基づく。プライム編集のケースでは、MS2タグ付け技術は、プライム編集を伴う所望のRNA分子(例として、pegRNA又はtPERT)上にMS2ヘアピンを導入することを含む。これは、次いで、その構造を認識及び結合するRNA結合タンパク質にとっての特異的な相互作用可能な結合標的を構成する。MS2ヘアピンのケースでは、それはMS2バクテリオファージコートタンパク質(MCP)によって認識及び結合される。そして、MCPが別のタンパク質(例として、逆転写酵素又は他のDNAポリメラーゼ)に融合される場合には、MS2ヘアピンが、プライム編集複合体によって占有される標的部位にその他のタンパク質をトランスで「動員」するために使用されてもよい。
種々の態様では(例として、図72~図73及び例19の態様に図示されている通り)、用語「MS2タグ付け技術」は、RNA-タンパク質相互作用ドメイン、例として特定のヘアピン構造を特異的に認識及び結合するRNA結合タンパク質とペアリングされた「RNA-タンパク質相互作用ドメイン」(「RNA-タンパク質動員ドメイン又はタンパク質」としてもまた公知)の組み合わせを指す。これらの型のシステムは、標的部位に結合されているプライム編集因子複合体に種々の機能性を動員するように活用され得る。MS2タグ付け技術は、ファージのゲノム上に存在するステムループ又はヘアピン構造、すなわち「MS2ヘアピン」とのMS2バクテリオファージコートタンパク質(「MCP」又は「MS2cp」)の天然の相互作用に基づく。プライム編集のケースでは、MS2タグ付け技術は、プライム編集を伴う所望のRNA分子(例として、pegRNA又はtPERT)上にMS2ヘアピンを導入することを含む。これは、次いで、その構造を認識及び結合するRNA結合タンパク質にとっての特異的な相互作用可能な結合標的を構成する。MS2ヘアピンのケースでは、それはMS2バクテリオファージコートタンパク質(MCP)によって認識及び結合される。そして、MCPが別のタンパク質(例として、逆転写酵素又は他のDNAポリメラーゼ)に融合される場合には、MS2ヘアピンが、プライム編集複合体によって占有される標的部位にその他のタンパク質をトランスで「動員」するために使用されてもよい。
本明細書に記載のプライム編集因子は、側面として、目当ての特定の機能をプライム編集因子複合体に動員又は「共局在」させるためにいずれかの公知のRNA-タンパク質相互作用ドメインを組み込んでもよい。RNA-タンパク質相互作用の他のモジュールドメインの総説は、当該技術分野において、例えば、Johansson et al.,「RNA recognition by the MS2 phage coat protein,」Sem Virol.,1997,Vol.8(3):176-185;Delebecque et al.,「Organization of intracellular reactions with rationally designed RNA assemblies,」Science,2011,Vol.333:470-474;Mali et al.,「Cas9 transcriptional activators for target specificity screening and paired nickases for cooperative genome engineering,」Nat.Biotechnol.,2013,Vol.31:833-838;及びZalatan et al.,「Engineering complex synthetic transcriptional programs with CRISPR RNA scaffolds,」Cell,2015,Vol.160:339-350(これら各々はその全体が参照されることによって本明細書に組み込まれる)において記載されている。他の系も、PCPタンパク質を特異的に動員するPP7ヘアピン、及びComタンパク質を特異的に動員する「com」ヘアピンを包含する。Zalatan et al.を参照。
MS2ヘアピン(すなわち、「MS2アプタマー」と呼ばれる)のヌクレオチド配列は以下:GCCAACATGAGGATCACCCATGTCTGCAGGGCC(配列番号24)である。
MCP又はMS2cpのアミノ酸配列は以下:GSASNFTQFVLVDNGGTGDVTVAPSNFANGVAEWISSNSRSQAYKVTCSVRQSSAQNRKYTIKVEVPKVATQTVGGEELPVAGWRSYLNMELTIPIFATNSDCELIVKAMQGLLKDGNPIPSAIAANSGIY(配列番号25)である。
MS2ヘアピン(又は「MS2アプタマー」)は、ある型の「RNAエフェクター動員ドメイン」(又は同等に「RNA結合タンパク質動員ドメイン」若しくは単純に「動員ドメイン」)ともまた呼ばれてもよい。なぜなら、それは、pegRNA又はtPERT上に組み入れされる物理的構造(例としてヘアピン)であり、これはそのように修飾されたpegRNA又はrPERTに他のエフェクター機能(例として、種々の機能を有するRNA結合タンパク質、例えばDNAポリメラーゼ、又は他のDNA修飾酵素)を有効に動員し、それゆえにプライム編集機構にエフェクター機能をトランスで共局在させるからである。この適用は、いずれかの具体的なRNAエフェクター動員ドメインに限定されることを決して意図されず、MS2ヘアピンを包含するいずれかの利用可能なかかるドメインを包含してもよい。例19及び図72(b)は、標的DNA部位に結合したプライム編集因子複合体(MS2cp-PE2:sgRNA複合体)とtPERT分子のDNA合成ドメインとの共局在が成就することを引き起こすための、DNA合成ドメイン(すなわちtPERT分子)に連結されたMS2アプタマーとPE2に融合されたMS2cpタンパク質を含むプライム編集因子との使用を図示する。
napDNAbp
本明細書に使用されるとき、用語「核酸プログラム型DNA結合タンパク質」又は「napDNAbp」(そのCas9が一例である)は、DNA分子中の特定の配列を標的にし結合するためにRNA:DNAハイブリダイゼーションを使用するタンパク質を指す。各napDNAbpは、ガイド核酸又はこの一部(例として、ガイドRNAのプロトスペーサ)に相補的なDNA鎖(すなわち、標的鎖)を含むDNA配列へnapDNAbpを局在化する、少なくとも1つのガイド核酸(例として、ガイドRNA)に関連されている。換言すれば、ガイド核酸は、相補的な配列へ局在化し結合するようnapDNAbp(例として、Cas9又は等価物)を「プログラム」する。
本明細書に使用されるとき、用語「核酸プログラム型DNA結合タンパク質」又は「napDNAbp」(そのCas9が一例である)は、DNA分子中の特定の配列を標的にし結合するためにRNA:DNAハイブリダイゼーションを使用するタンパク質を指す。各napDNAbpは、ガイド核酸又はこの一部(例として、ガイドRNAのプロトスペーサ)に相補的なDNA鎖(すなわち、標的鎖)を含むDNA配列へnapDNAbpを局在化する、少なくとも1つのガイド核酸(例として、ガイドRNA)に関連されている。換言すれば、ガイド核酸は、相補的な配列へ局在化し結合するようnapDNAbp(例として、Cas9又は等価物)を「プログラム」する。
理論に拘泥されないが、napDNAbp-ガイドRNA複合体の結合機序は、一般に、napDNAbpが二本鎖DNA標的の巻き戻しを誘導するRループを形成する(これによってnapDNAbpによって結合された領域中の鎖が分離される)工程を包含する。ガイドRNAプロトスペーサは次いで「標的鎖」とハイブリダイズする。これは、標的鎖に相補的な「非標的鎖」に取って代わり、これはRループの単鎖領域を形成する。いくつかの態様において、napDNAbpは1以上のヌクレアーゼ活性を包含しており、次いでDNAを切ることで、様々な型の病変がなくなる。例えば、napDNAbpは、非標的鎖を第1の場所にて切るか、及び/又は標的鎖を第2の場所にて切る、ヌクレアーゼ活性を含んでいてもよい。ヌクレアーゼ活性に応じて標的DNAが切られることで、両鎖とも切られた「二本鎖切断」を形成し得る。他の態様において、標的DNAは単一の部位でしか切られ得ない、すなわち、DNAは一方の鎖上に「ニックが入っている(nicked)」。種々のヌクレアーゼ活性をもつ例示のnapDNAbpは、「Cas9ニッカーゼ」(「nCas9」)、及びヌクレアーゼ活性を一切有さない不活性化Cas9(「不活性型Cas9」又は「dCas9」)を包含する。これらの及び他のnapDNAbpへの例示の配列は、本明細書に提供されている。
ニッカーゼ
本明細書で使用される場合、「ニッカーゼ」は、二本鎖標的DNA配列の2つの相補鎖の一方のみを切断し、それによってその鎖にニックを生成することができるnapDNAbp(例として、Casタンパク質)を指す。いくつかの態様では、ニッカーゼは、二本鎖標的DNA配列の非標的鎖を切断する。いくつかの態様では、ニッカーゼは、標準的なnapDNAbp(例として、Casタンパク質)の触媒ドメインに1つ以上の変異を有するアミノ酸配列を含み、1つ以上の変異は、触媒ドメインのヌクレアーゼ活性を低減又は消失させる。いくつかの態様において、ニッカーゼは、野生型Cas9配列に対する、又は他のCas9バリアント若しくはCas9等価物における等価なアミノ酸位置に対するRuvC様ドメインにおける1つ又は複数の変異を含むCas9である。いくつかの態様において、ニッカーゼは、野生型Cas9配列に対する、又は他のCas9バリアント若しくはCas9等価物における等価なアミノ酸位置に対するHNH様ドメインにおける1つ又は複数の変異を含むCas9である。いくつかの態様において、ニッカーゼは、標準的なCas9配列に対する、又は他のCas9バリアント若しくはCas9等価物における等価なアミノ酸位置に対する、Cas9のRuvC I触媒ドメインにおけるアスパラギン酸からアラニンへの置換(D10A)を含むCas9である。いくつかの態様において、ニッカーゼは、標準的なCas9配列に対する、又は他のCas9バリアント若しくはCas9等価物における等価なアミノ酸位置に対するH840A、N854A及び/又はN863A変異を含むCas9である。いくつかの態様において、「Cas9ニッカーゼ」という用語は、2つのヌクレアーゼドメインのうちの1つが不活性化されたCas9を指す。この酵素は、標的DNAの一方の鎖のみを切断することができる。いくつかの態様において、ニッカーゼはCas9ニッカーゼではないCasタンパク質である。核局在化配列(NLS)
本明細書で使用される場合、「ニッカーゼ」は、二本鎖標的DNA配列の2つの相補鎖の一方のみを切断し、それによってその鎖にニックを生成することができるnapDNAbp(例として、Casタンパク質)を指す。いくつかの態様では、ニッカーゼは、二本鎖標的DNA配列の非標的鎖を切断する。いくつかの態様では、ニッカーゼは、標準的なnapDNAbp(例として、Casタンパク質)の触媒ドメインに1つ以上の変異を有するアミノ酸配列を含み、1つ以上の変異は、触媒ドメインのヌクレアーゼ活性を低減又は消失させる。いくつかの態様において、ニッカーゼは、野生型Cas9配列に対する、又は他のCas9バリアント若しくはCas9等価物における等価なアミノ酸位置に対するRuvC様ドメインにおける1つ又は複数の変異を含むCas9である。いくつかの態様において、ニッカーゼは、野生型Cas9配列に対する、又は他のCas9バリアント若しくはCas9等価物における等価なアミノ酸位置に対するHNH様ドメインにおける1つ又は複数の変異を含むCas9である。いくつかの態様において、ニッカーゼは、標準的なCas9配列に対する、又は他のCas9バリアント若しくはCas9等価物における等価なアミノ酸位置に対する、Cas9のRuvC I触媒ドメインにおけるアスパラギン酸からアラニンへの置換(D10A)を含むCas9である。いくつかの態様において、ニッカーゼは、標準的なCas9配列に対する、又は他のCas9バリアント若しくはCas9等価物における等価なアミノ酸位置に対するH840A、N854A及び/又はN863A変異を含むCas9である。いくつかの態様において、「Cas9ニッカーゼ」という用語は、2つのヌクレアーゼドメインのうちの1つが不活性化されたCas9を指す。この酵素は、標的DNAの一方の鎖のみを切断することができる。いくつかの態様において、ニッカーゼはCas9ニッカーゼではないCasタンパク質である。核局在化配列(NLS)
用語「核局在化配列」又は「NLS」は、タンパク質の細胞核中への、例えば核輸送による、インポート(import)を促進するアミノ酸配列を指す。核局在化配列は当該技術分野において知られており、当業者に明らかであろう。例えば、NLS配列は、2000年11月23日に出願され、2001年5月31日にWO/2001/038547として公開された、Plankらの国際PCT出願PCT/EP2000/011690に記載されている(この内容は、例示の核局在化配列のその開示について参照により本明細書に組み込まれる)。いくつかの態様において、NLSは、アミノ酸配列PKKKRKV(配列番号26)又はMDSLLMNRRKFLYQFKNVRWAKGRRETYLC(配列番号27)を含む。
核酸分子
用語「核酸」は、本明細書に使用されるとき、ヌクレオチドのポリマーを指す。ポリマーは、天然のヌクレオシド(すなわち、アデノシン、チミジン、グアノシン、シチジン、ウリジン、デオキシアデノシン、デオキシチミジン、デオキシグアノシン、及びデオキシシチジン)、ヌクレオシド類似体(例として、2-アミノアデノシン、2-チオチミジン、イノシン、ピロロ-ピリミジン、3-メチルアデノシン、5-メチルシチジン、C5ブロモウリジン、C5フルオロウリジン、C5ヨードウリジン、C5プロピニルウリジン、C5プロピニルシチジン、C5メチルシチジン、7デアザアデノシン、7デアザグアノシン、8オキソアデノシン、8オキソグアノシン、O(6)メチルグアニン、4-アセチルシチジン、5-(カルボキシヒドロキシメチル)ウリジン、ジヒドロウリジン、メチルシュードウリジン、1-メチルアデノシン、1-メチルグアノシン、N6-メチルアデノシン、及び2-チオシチジン)、化学修飾された塩基、生物学的に修飾された塩基(例として、メチル化された塩基)、インターカレートされた(intercalated)塩基、修飾された糖(例として、2'-フルオロリボース、リボース、2'-デオキシリボース、2'-O-メチルシチジン、アラビノース、及びヘキソース)、又は修飾されたホスファート基(例として、ホスホロチオアート及び5'-Nホスホラミダイト連結)を包含していてもよい。
用語「核酸」は、本明細書に使用されるとき、ヌクレオチドのポリマーを指す。ポリマーは、天然のヌクレオシド(すなわち、アデノシン、チミジン、グアノシン、シチジン、ウリジン、デオキシアデノシン、デオキシチミジン、デオキシグアノシン、及びデオキシシチジン)、ヌクレオシド類似体(例として、2-アミノアデノシン、2-チオチミジン、イノシン、ピロロ-ピリミジン、3-メチルアデノシン、5-メチルシチジン、C5ブロモウリジン、C5フルオロウリジン、C5ヨードウリジン、C5プロピニルウリジン、C5プロピニルシチジン、C5メチルシチジン、7デアザアデノシン、7デアザグアノシン、8オキソアデノシン、8オキソグアノシン、O(6)メチルグアニン、4-アセチルシチジン、5-(カルボキシヒドロキシメチル)ウリジン、ジヒドロウリジン、メチルシュードウリジン、1-メチルアデノシン、1-メチルグアノシン、N6-メチルアデノシン、及び2-チオシチジン)、化学修飾された塩基、生物学的に修飾された塩基(例として、メチル化された塩基)、インターカレートされた(intercalated)塩基、修飾された糖(例として、2'-フルオロリボース、リボース、2'-デオキシリボース、2'-O-メチルシチジン、アラビノース、及びヘキソース)、又は修飾されたホスファート基(例として、ホスホロチオアート及び5'-Nホスホラミダイト連結)を包含していてもよい。
ヌクレオチド構造モチーフ(又は核酸部分)
本明細書で使用される場合、「ヌクレオチド構造モチーフ」又は同等に「核酸部分」という用語は、単一の核酸ポリマー内又は2つ以上の核酸ポリマー間の塩基対形成相互作用により二次又は三次構造を形成する核酸分子又はその一部を指す。そのようなヌクレオチド構造モチーフは、DNA、RNA、又はDNAとRNAとのハイブリッドから形成され得る。この用語は、標準的なDNA二重らせんを指すことを意味しない。核酸部分の例としては、限定されないが、トゥーループ、ヘアピン、ステムループ、シュードノット、アプタマー、G四重鎖、tRNA、リボザイム、リボスイッチ、A型DNA、B型DNA又はZ型DNAを包含する。
本明細書で使用される場合、「ヌクレオチド構造モチーフ」又は同等に「核酸部分」という用語は、単一の核酸ポリマー内又は2つ以上の核酸ポリマー間の塩基対形成相互作用により二次又は三次構造を形成する核酸分子又はその一部を指す。そのようなヌクレオチド構造モチーフは、DNA、RNA、又はDNAとRNAとのハイブリッドから形成され得る。この用語は、標準的なDNA二重らせんを指すことを意味しない。核酸部分の例としては、限定されないが、トゥーループ、ヘアピン、ステムループ、シュードノット、アプタマー、G四重鎖、tRNA、リボザイム、リボスイッチ、A型DNA、B型DNA又はZ型DNAを包含する。
pegRNA
本明細書に使用されるとき、用語「プライム編集ガイドRNA」又は「pegRNA」又は「伸長されたガイドRNA」は、本明細書に記載のプライム編集方法及び組成物を実践するための1以上の追加の配列を包含するよう修飾されたガイドRNAの特化された形態を指す。本明細書に記載の通り、プライム編集ガイドRNAは、核酸配列の1以上の「伸長された領域」を含む。伸長された領域は、これらに限定されないが、単鎖のRNA又はDNAを含んでいてもよい。更に、伸長された領域は、既存のガイドRNAの3'末にて生じていてもよい。他の配置において、伸長された領域は、既存のガイドRNAの5'末にて生じていてもよい。また他の配置において、伸長された領域は、既存のガイドRNAの末端の分子内のある領域にて(例えば、napDNAbpへ結び付き及び/又は結合するgRNAコア領域中)生じていてもよい。伸長された領域は、単鎖DNAを(プライム編集因子のポリメラーゼによって)コードする「DNA合成鋳型」を含むが、分子は次に(a)編集されるべき内生標的DNAと相同であるようデザインされており、及び(b)内生標的DNA中へ誘導又は取り入れられるべき少なくとも1の所望のヌクレオチド変化(例として、トランジション、トランスバーション、欠失、若しくは挿入)を含んでいる。伸長された領域はまた、他の機能的配列要素(これらに限定されないが、「プライマー結合部位」及び「スペーサ若しくはリンカー」配列等)、又は他の構造的要素(これらに限定されないが、アプタマー、ステムループ、ヘアピン、トゥーループ(例として、3'トゥーループ)、若しくはRNA-タンパク質動員ドメイン(例として、MS2ヘアピン))も含んでいてもよい。本明細書に使用されるとき「プライマー結合部位」は、Rループのニックが入ったDNAから生成された3'末を有する単鎖DNA配列とハイブリダイズする配列を含む。
本明細書に使用されるとき、用語「プライム編集ガイドRNA」又は「pegRNA」又は「伸長されたガイドRNA」は、本明細書に記載のプライム編集方法及び組成物を実践するための1以上の追加の配列を包含するよう修飾されたガイドRNAの特化された形態を指す。本明細書に記載の通り、プライム編集ガイドRNAは、核酸配列の1以上の「伸長された領域」を含む。伸長された領域は、これらに限定されないが、単鎖のRNA又はDNAを含んでいてもよい。更に、伸長された領域は、既存のガイドRNAの3'末にて生じていてもよい。他の配置において、伸長された領域は、既存のガイドRNAの5'末にて生じていてもよい。また他の配置において、伸長された領域は、既存のガイドRNAの末端の分子内のある領域にて(例えば、napDNAbpへ結び付き及び/又は結合するgRNAコア領域中)生じていてもよい。伸長された領域は、単鎖DNAを(プライム編集因子のポリメラーゼによって)コードする「DNA合成鋳型」を含むが、分子は次に(a)編集されるべき内生標的DNAと相同であるようデザインされており、及び(b)内生標的DNA中へ誘導又は取り入れられるべき少なくとも1の所望のヌクレオチド変化(例として、トランジション、トランスバーション、欠失、若しくは挿入)を含んでいる。伸長された領域はまた、他の機能的配列要素(これらに限定されないが、「プライマー結合部位」及び「スペーサ若しくはリンカー」配列等)、又は他の構造的要素(これらに限定されないが、アプタマー、ステムループ、ヘアピン、トゥーループ(例として、3'トゥーループ)、若しくはRNA-タンパク質動員ドメイン(例として、MS2ヘアピン))も含んでいてもよい。本明細書に使用されるとき「プライマー結合部位」は、Rループのニックが入ったDNAから生成された3'末を有する単鎖DNA配列とハイブリダイズする配列を含む。
ある態様において、pegRNAは、5'伸長アーム、スペーサ、及びgRNAコアを有するpegRNAを示す図3Aによって表されている。5'伸長は、5'→3'方向に逆転写酵素鋳型、プライマー結合部位、及びリンカーを更に含む。示されている通り、逆転写酵素鋳型はより幅広く「DNA合成鋳型」ともまた呼ばれてもよく、ここで、本明細書に記載のプライム編集因子のポリメラーゼはRTではなく別の型のポリメラーゼである。
ある他の態様において、pegRNAは、5'伸長アーム、スペーサ、及びgRNAコアを有するpegRNAを示す図3Bによって表されている。5'伸長は、5'→3'方向に逆転写酵素鋳型、プライマー結合部位、及びリンカーを更に含む。示されている通り、逆転写酵素鋳型はより幅広く「DNA合成鋳型」ともまた呼ばれてもよく、ここで、本明細書に記載のプライム編集因子のポリメラーゼはRTではなく別の型のポリメラーゼである。
なお他の態様において、PegRNAは図3Dによって表され、これは5'→3'方向にスペーサ(1)、gRNAコア(2)、及び伸長アーム(3)を有するpegRNAを示す。伸長アーム(3)はPegRNAの3'端にある。伸長アーム(3)は更に3'→5'方向に「プライマー結合部位」(A)、「編集鋳型」(B)、及び「相同アーム」(C)を含む。伸長アーム(3)は任意選択の修飾領域をもまた3'及び5'端に含んでもよく、これらは同じ配列又は異なる配列であってもよい。pegRNAの3'末は、転写ターミネータ配列を含んでいてもよい。pegRNAのこれらの配列要素は更に、本明細書に記載、定義される。
なお他の態様において、pegRNAは図3Eによって表され、これは5'→3'方向に伸長アーム(3)、スペーサ(1)、及びgRNAコア(2)を有するpegRNAを示す。伸長アーム(3)はpegRNAの5'端にある。伸長アーム(3)は更に3'→5'方向に「プライマー結合部位」(A)、「編集鋳型」(B)、及び「相同アーム」(C)を含む。伸長アーム(3)は任意選択の修飾領域をもまた3'及び5'端に含んでもよく、これらは同じ配列又は異なる配列であってもよい。pegRNAの3'末は、転写ターミネータ配列を含んでいてもよい。pegRNAのこれらの配列要素は更に、本明細書に記載、定義される。
PE1
本明細書において用いられる「PE1」は、次の構造を有するCas9(H840A)と野生型MMLV RTとを含む融合タンパク質を含むPE複合体を指す:[NLS]-[Cas9(H840A)]-[リンカー]-[MMLV_RT(wt)]+所望のpegRNA。PE融合体は配列番号28のアミノ酸配列を有する。
本明細書において用いられる「PE1」は、次の構造を有するCas9(H840A)と野生型MMLV RTとを含む融合タンパク質を含むPE複合体を指す:[NLS]-[Cas9(H840A)]-[リンカー]-[MMLV_RT(wt)]+所望のpegRNA。PE融合体は配列番号28のアミノ酸配列を有する。
PE2
本明細書において用いられる「PE2」は、次の構造を有するCas9(H840A)とバリアントMMLV RTとを含む融合タンパク質を含むPE複合体を指す:[NLS]-[Cas9(H840A)]-[リンカー]-[MMLV_RT(D200N)(T330P)(L603W)(T306K)(W313F)]+所望のpegRNA。PE融合体は配列番号33のアミノ酸配列を有する。
本明細書において用いられる「PE2」は、次の構造を有するCas9(H840A)とバリアントMMLV RTとを含む融合タンパク質を含むPE複合体を指す:[NLS]-[Cas9(H840A)]-[リンカー]-[MMLV_RT(D200N)(T330P)(L603W)(T306K)(W313F)]+所望のpegRNA。PE融合体は配列番号33のアミノ酸配列を有する。
PE3
本明細書において用いられる「PE3」は、PE2、プラスPE2と複合体化する第2鎖ニッキングガイドRNAを指し、編集される鎖の選好的置換えを誘導するために非編集DNA鎖上にニックを導入する。
本明細書において用いられる「PE3」は、PE2、プラスPE2と複合体化する第2鎖ニッキングガイドRNAを指し、編集される鎖の選好的置換えを誘導するために非編集DNA鎖上にニックを導入する。
PE3b
本明細書において用いられる「PE3b」はPE3を指すが、第2鎖ニッキングガイドRNAは、所望の編集の組み入れ後までは第2鎖ニックが組み入れされないようにして時間的コントロールのためにデザインされる。これは、元のアレルではなく編集された鎖のみにマッチするスペーサ配列を有するgRNAをデザインすることによって達成される。以降ではPE3bと呼ばれるこの戦略を用いると、プロトスペーサと編集されないアレルとの間のミスマッチは、PAM鎖上の編集イベントが生起する後までは、sgRNAによるニッキングを好まないはずである。
本明細書において用いられる「PE3b」はPE3を指すが、第2鎖ニッキングガイドRNAは、所望の編集の組み入れ後までは第2鎖ニックが組み入れされないようにして時間的コントロールのためにデザインされる。これは、元のアレルではなく編集された鎖のみにマッチするスペーサ配列を有するgRNAをデザインすることによって達成される。以降ではPE3bと呼ばれるこの戦略を用いると、プロトスペーサと編集されないアレルとの間のミスマッチは、PAM鎖上の編集イベントが生起する後までは、sgRNAによるニッキングを好まないはずである。
PE4
本明細書で使用される場合、「PE4」は、PE2と、トランスで発現されるMLH1ドミナントネガティブタンパク質(すなわち、本明細書で「MLH1 Δ754-756」又は「MLH1dn」と呼ばれてもよいアミノ酸754~756が短縮された野生型MLH1)とを含むシステムを指す。いくつかの態様では、PE4は、任意選択のリンカーを介して連結されたPE2及びMLH1ドミナントネガティブタンパク質を含む融合タンパク質を指す。
本明細書で使用される場合、「PE4」は、PE2と、トランスで発現されるMLH1ドミナントネガティブタンパク質(すなわち、本明細書で「MLH1 Δ754-756」又は「MLH1dn」と呼ばれてもよいアミノ酸754~756が短縮された野生型MLH1)とを含むシステムを指す。いくつかの態様では、PE4は、任意選択のリンカーを介して連結されたPE2及びMLH1ドミナントネガティブタンパク質を含む融合タンパク質を指す。
PE5
本明細書で使用される場合、「PE5」は、PE3と、トランスで発現されるMLH1ドミナントネガティブタンパク質(すなわち、「MLH1 Δ754-756」又は「MLH1dn」と呼ばれてもよい、本明細書で更に記載されるようにアミノ酸754~756が短縮された野生型MLH1)とを含むシステムを指す。いくつかの態様では、PE5は、任意選択のリンカーを介して連結されたPE3及びMLH1ドミナントネガティブタンパク質を含む融合タンパク質を指す。
本明細書で使用される場合、「PE5」は、PE3と、トランスで発現されるMLH1ドミナントネガティブタンパク質(すなわち、「MLH1 Δ754-756」又は「MLH1dn」と呼ばれてもよい、本明細書で更に記載されるようにアミノ酸754~756が短縮された野生型MLH1)とを含むシステムを指す。いくつかの態様では、PE5は、任意選択のリンカーを介して連結されたPE3及びMLH1ドミナントネガティブタンパク質を含む融合タンパク質を指す。
PE-短
本明細書で使用される場合、「PE-短」は、C末端切断逆転写酵素に融合され、以下のアミノ酸配列を有するPE構築物を指す:
鍵:
核局在化配列(NLS)上部:(配列番号29)、下部:(配列番号30)
CAS9(H840A)(配列番号31)
33アミノ酸リンカー1(配列番号11)
M-MLV TRUNCATED逆転写酵素
(配列番号36)
本明細書で使用される場合、「PE-短」は、C末端切断逆転写酵素に融合され、以下のアミノ酸配列を有するPE構築物を指す:
核局在化配列(NLS)上部:(配列番号29)、下部:(配列番号30)
CAS9(H840A)(配列番号31)
33アミノ酸リンカー1(配列番号11)
M-MLV TRUNCATED逆転写酵素
(配列番号36)
ペプチドタグ
用語「ペプチドタグ」は、遺伝子学的にタンパク質配列と融合してタンパク質上へ1以上の機能を付与するペプチドアミノ酸配列を指し、これによって可視化、精製、可溶化、分離等々等の様々な目的でのタンパク質の操作が容易になる。ペプチドタグは、目的又は機能によって分類される様々な型のタグを包含し得、これは、「親和性タグ」(タンパク質精製を容易にするための)、「可溶化タグ」(タンパク質の正しい折り畳みを補助するための)、「クロマトグラフィータグ」(タンパク質のクロマトグラフ特性を変更するための)、「エピトープタグ」(高親和性抗体へ結合するための)、「蛍光タグ」(細胞中又はin vitroでのタンパク質の可視化を容易にするための)を包含していてもよい。
用語「ペプチドタグ」は、遺伝子学的にタンパク質配列と融合してタンパク質上へ1以上の機能を付与するペプチドアミノ酸配列を指し、これによって可視化、精製、可溶化、分離等々等の様々な目的でのタンパク質の操作が容易になる。ペプチドタグは、目的又は機能によって分類される様々な型のタグを包含し得、これは、「親和性タグ」(タンパク質精製を容易にするための)、「可溶化タグ」(タンパク質の正しい折り畳みを補助するための)、「クロマトグラフィータグ」(タンパク質のクロマトグラフ特性を変更するための)、「エピトープタグ」(高親和性抗体へ結合するための)、「蛍光タグ」(細胞中又はin vitroでのタンパク質の可視化を容易にするための)を包含していてもよい。
ポリメラーゼ
本明細書において用いられる用語「ポリメラーゼ」は、本明細書に記載のプライム編集因子システムに関して用いられてもよいヌクレオチド鎖を合成する酵素を指す。ポリメラーゼは「鋳型依存性」ポリメラーゼであり得る(すなわち、鋳型鎖のヌクレオチド塩基の順序に基づいてヌクレオチド鎖を合成するポリメラーゼ)。ポリメラーゼは「鋳型非依存性」ポリメラーゼでもまたあり得る(すなわち、鋳型鎖の要件なしにヌクレオチド鎖を合成するポリメラーゼ)。ポリメラーゼは更に「DNAポリメラーゼ」又は「RNAポリメラーゼ」としてもまた類別されてもよい。種々の態様において、プライム編集因子システムはDNAポリメラーゼを含む。種々の態様において、DNAポリメラーゼは「DNA依存性DNAポリメラーゼ」であり得る(すなわち、それによると、鋳型分子はDNAの鎖である)。かかるケースでは、DNA鋳型分子はpegRNAであり得、伸長アームはDNAの鎖を含む。かかるケースでは、pegRNAはキメラ又はハイブリッドpegRNAと呼ばれてもよく、これはRNA部分(すなわち、スペーサ及びgRNAコアを包含するガイドRNA構成要素)とDNA部分(すなわち伸長アーム)とを含む。種々の他の態様において、DNAポリメラーゼは「RNA依存性DNAポリメラーゼ」であり得る(すなわち、それによると、鋳型分子はRNAの鎖である)。かかるケースでは、pegRNAはRNAであり、すなわちRNA伸長を包含する。用語「ポリメラーゼ」はまた、ヌクレオチドの重合を触媒する酵素を指してもよい(すなわちポリメラーゼ活性)。一般的に、酵素は、ポリヌクレオチド鋳型配列にアニーリングしたプライマー(例として、pegRNAのプライマー結合部位にアニーリングしたプライマー配列等)の3'端において合成を開始し、鋳型鎖の5'端の方へ進むであろう。「DNAポリメラーゼ」はデオキシヌクレオチドの重合を触媒する。DNAポリメラーゼを参照して本明細書において用いられる用語DNAポリメラーゼは、「その機能的フラグメント」を包含する。「その機能的フラグメント」は、少なくとも1つのセットの条件下においてはポリヌクレオチドの重合を触媒する能力を保持する、ポリメラーゼのアミノ酸配列全体未満を包摂する野生型又は変異体DNAポリメラーゼのいずれかの部分を指す。かかる機能的フラグメントは別個の実態として存在し得てもよいか、又はそれは融合タンパク質等のより大きいポリペプチドの構成要素であってもよい。
本明細書において用いられる用語「ポリメラーゼ」は、本明細書に記載のプライム編集因子システムに関して用いられてもよいヌクレオチド鎖を合成する酵素を指す。ポリメラーゼは「鋳型依存性」ポリメラーゼであり得る(すなわち、鋳型鎖のヌクレオチド塩基の順序に基づいてヌクレオチド鎖を合成するポリメラーゼ)。ポリメラーゼは「鋳型非依存性」ポリメラーゼでもまたあり得る(すなわち、鋳型鎖の要件なしにヌクレオチド鎖を合成するポリメラーゼ)。ポリメラーゼは更に「DNAポリメラーゼ」又は「RNAポリメラーゼ」としてもまた類別されてもよい。種々の態様において、プライム編集因子システムはDNAポリメラーゼを含む。種々の態様において、DNAポリメラーゼは「DNA依存性DNAポリメラーゼ」であり得る(すなわち、それによると、鋳型分子はDNAの鎖である)。かかるケースでは、DNA鋳型分子はpegRNAであり得、伸長アームはDNAの鎖を含む。かかるケースでは、pegRNAはキメラ又はハイブリッドpegRNAと呼ばれてもよく、これはRNA部分(すなわち、スペーサ及びgRNAコアを包含するガイドRNA構成要素)とDNA部分(すなわち伸長アーム)とを含む。種々の他の態様において、DNAポリメラーゼは「RNA依存性DNAポリメラーゼ」であり得る(すなわち、それによると、鋳型分子はRNAの鎖である)。かかるケースでは、pegRNAはRNAであり、すなわちRNA伸長を包含する。用語「ポリメラーゼ」はまた、ヌクレオチドの重合を触媒する酵素を指してもよい(すなわちポリメラーゼ活性)。一般的に、酵素は、ポリヌクレオチド鋳型配列にアニーリングしたプライマー(例として、pegRNAのプライマー結合部位にアニーリングしたプライマー配列等)の3'端において合成を開始し、鋳型鎖の5'端の方へ進むであろう。「DNAポリメラーゼ」はデオキシヌクレオチドの重合を触媒する。DNAポリメラーゼを参照して本明細書において用いられる用語DNAポリメラーゼは、「その機能的フラグメント」を包含する。「その機能的フラグメント」は、少なくとも1つのセットの条件下においてはポリヌクレオチドの重合を触媒する能力を保持する、ポリメラーゼのアミノ酸配列全体未満を包摂する野生型又は変異体DNAポリメラーゼのいずれかの部分を指す。かかる機能的フラグメントは別個の実態として存在し得てもよいか、又はそれは融合タンパク質等のより大きいポリペプチドの構成要素であってもよい。
プライム編集
本明細書において用いられる用語「プライム編集」は、遺伝子編集のための新規のアプローチを指し、napDNAbp、ポリメラーゼ(例として、逆転写酵素)、及び次いで標的DNA配列上に組み込まれる所望の新たな遺伝情報をコードする(又は遺伝情報を欠失する)ためのDNA合成鋳型を包含する特殊化したガイドRNAを用いる。プライム編集のある種の態様は他の図の中でも図1A~図1H及び図72(a)~図72(c)の態様に記載される。
本明細書において用いられる用語「プライム編集」は、遺伝子編集のための新規のアプローチを指し、napDNAbp、ポリメラーゼ(例として、逆転写酵素)、及び次いで標的DNA配列上に組み込まれる所望の新たな遺伝情報をコードする(又は遺伝情報を欠失する)ためのDNA合成鋳型を包含する特殊化したガイドRNAを用いる。プライム編集のある種の態様は他の図の中でも図1A~図1H及び図72(a)~図72(c)の態様に記載される。
プライム編集は、規定されたDNA部位に新たな遺伝情報を直接的に書き込む汎用的なかつ精密なゲノム編集法であるゲノム編集のための完全に新しいプラットフォームを表し、ポリメラーゼ(すなわち、融合タンパク質の形態で、又はさもなければnapDNAbpとトランスで提供されて)と共同で働く核酸プログラム型DNA結合タンパク質(「napDNAbp」)を用いる。プライム編集システムはプライム編集(PE)ガイドRNA(「pegRNA」)によってプログラムされ、これは、標的部位を規定すると共に、ガイドRNA上に(例として、ガイドRNAの5'若しくは3'端に又は内部の部分に)操作された伸長(DNA又はRNAどちらか)によって置換えDNA鎖の形態での所望の編集の合成の鋳型となる。所望の編集(例として、1核酸塩基置換)を含有する置換え鎖は、編集されるべき標的部位の(ニック部位の直ちに下流の)内生鎖と同じ配列を共有する(又はそれに対して相同的である)(それが所望の編集を包含するということを例外とする)。DNA修復及び/又は複製機構によって、ニック部位の下流の内生鎖は、所望の編集を含有する新たに合成された置換え鎖によって置き換えられる。いくつかのケースでは、プライム編集は「検索置換」ゲノム編集テクノロジーと考えられてもよい。なぜなら、本明細書に記載のプライム編集因子は、編集されるべき所望の標的部位を検索し位置付けるのみならず、同時に、対応する標的部位の内生DNA鎖の代わりに組み入れされる所望の編集を含有する置換え鎖をコードするからである。本開示のプライム編集因子は、部分的には、プライム標的にプライムされた逆転写(TPRT)又は「プライム編集」の機序が、高い効率及び遺伝子フレキシビリティーを有する精密なCRISPR/Casに基づくゲノム編集を行うために活用又は適合され得るという発見に関する(例として、図1A-図1Fの種々の態様に図示されている通り)。天然には、TPRTは、可動DNAエレメント、例えば哺乳動物非LTRレトロトランスポゾン及び細菌グループIIイントロンによって用いられる28,29。本発明者らは、本明細書において、Casタンパク質-逆転写酵素融合体又は関係するシステムを用いて、ガイドRNAによって特定のDNA配列を標的化し、標的部位に一本鎖ニックを生成し、ガイドRNAに取り入れされた操作された逆転写酵素鋳型の逆転写のためのプライマーとして、ニックが入ったDNAを用いた。しかしながら、概念はDNAポリメラーゼ構成要素として逆転写酵素を用いるプライム編集因子から始まるが、本明細書に記載のプライム編集因子は逆転写酵素に限定されず、実質的にいずれかのDNAポリメラーゼの使用を包含してもよい。実に、本明細書は、至る所で、「逆転写酵素」を有するプライム編集因子を指してもよいが、逆転写酵素は、プライム編集で働いてもよいDNAポリメラーゼの1つの型であるのみであるということがここで明示される。それゆえに、本明細書が「逆転写酵素」に言及するところではいつも、当業者は、いずれかの好適なDNAポリメラーゼが逆転写酵素の代わりに用いられてもよいということを了解するべきである。それゆえに、一側面において、プライム編集因子はCas9(又は同等のnapDNAbp)を含んでもよく、これは、標的DNA上の相補的なプロトスペーサにアニールするスペーサ配列を含有する特殊化したガイドRNA(すなわちpegRNA)とそれを会合させることによって、DNA配列を標的化するようにプログラムされる。特殊化したガイドRNAは、所望の遺伝子変改を含有するDNAの置換え鎖をコードする伸長の形態の新たな遺伝情報をもまた含有し、これは、標的部位における対応する内生DNA鎖を置き換えるために用いられる。pegRNAからの情報を標的DNAに移入するために、プライム編集の機序には、DNAの1つの鎖上の標的部位へニックを入れて3'ヒドロキシル基を暴露することを伴う。次いで、暴露された3'ヒドロキシル基は、直接的に標的部位へのpegRNA上の編集をコードする伸長のDNA重合をプライムするために用いられ得る。種々の態様において、編集を含有する置換え鎖の重合のための鋳型を提供する伸長は、RNA又はDNAから形成され得る。RNA伸長のケースでは、プライム編集因子のポリメラーゼはRNA依存性DNAポリメラーゼ(例えば逆転写酵素)であり得る。DNA伸長のケースでは、プライム編集因子のポリメラーゼはDNA依存性DNAポリメラーゼであってもよい。本明細書に開示のプライム編集因子によって形成される新たに合成された鎖(すなわち、所望の編集を含有する置換えDNA鎖)は、所望のヌクレオチド変化(例として、1ヌクレオチド変化、欠失、若しくは挿入、又はそれらの組み合わせ)の包含を例外として、ゲノム標的配列に対して相同的である(すなわち、それと同じ配列を有する)であろう。DNAの新たに合成された(又は置き換え)鎖は一本鎖DNAフラップともまた呼ばれてもよい。これは相補的な相同的な内生DNA鎖とのハイブリダイゼーションについて競合し、それによって対応する内生鎖に取って代わるであろう。ある態様において、システムは、エラーを起こしやすい逆転写酵素酵素の使用と組み合わせられ得る(例として、Cas9ドメインとの融合タンパク質として提供されるか、又はCas9ドメインに対してトランスで提供される)。エラーを起こしやすい逆転写酵素酵素は一本鎖DNAフラップの合成の間に変改を導入し得る。それゆえに、ある態様において、エラーを起こしやすい逆転写酵素は、標的DNAへのヌクレオチド変化を導入するために利用され得る。システムに用いられるエラーを起こしやすい逆転写酵素に依存して、変化はランダム又は非ランダムであり得る。ハイブリダイズした中間体(内生DNA鎖にハイブリダイズした逆転写酵素によって合成される一本鎖DNAフラップを含む)の分解は、内生DNAのもたらされる取って代わられたフラップの除去(例として、5'端DNAフラップエンドヌクレアーゼFEN1による)、標的DNAに対する合成された一本鎖DNAフラップのライゲーション、並びに細胞性のDNA修復及び/又は複製プロセスの結果としての所望のヌクレオチド変化の同化を包含し得る。鋳型によるDNA合成は挿入及び欠失を包含するいずれかのヌクレオチドの修飾について1ヌクレオチド精度をオファーするので、このアプローチの範囲は非常に幅広く、予見可能には、基礎研究及び処置学において無数の適用に用いられ得る。
種々の態様において、プライム編集は、プライム編集ガイドRNA(PEgRNA)と複合体化した核酸プログラム型DNA結合タンパク質(napDNAbp)と標的DNA分子(これについて、ヌクレオチド配列の変化が導入されることが望まれる)を接触させることによって作動する。図1Gを参照すると、プライム編集ガイドRNA(pegRNA)は、ガイドRNAの3'若しくは5'端に又はガイドRNAの分子内の位置付けに伸長を含み、所望のヌクレオチド変化(例として、1ヌクレオチド変化、挿入、又は欠失)をコードする。工程(a)では、napDNAbp/pegRNA複合体がDNA分子に接触し、伸長されたpegRNAが、標的遺伝子座に結合するようにnapDNAbpをガイドする。工程(b)では、標的遺伝子座のDNAの鎖の1つにニックが導入され(例として、ヌクレアーゼ又は化学的薬剤による)、それによって利用可能な3'端を標的遺伝子座の鎖の1つに作成する。ある態様において、ニックは、Rループ鎖に対応するDNAの鎖、すなわちガイドRNA配列にハイブリダイズされない鎖、すなわち「非標的鎖」上に作成される。しかしながら、ニックは鎖のどちらかに導入され得る。つまり、ニックはRループ「標的鎖」(すなわち、pegRNAにハイブリダイズされる鎖)又は「非標的鎖」(すなわち、標的鎖に対して相補的であるRループの一本鎖部分を形成する鎖)に導入され得る。工程(c)では、DNA鎖の3'端(ニックによって形成される)は、逆転写をプライムするためにガイドRNAの伸長された部分と相互作用する(すなわち、「プライム標的にプライムされたRT」)。ある態様において、3'端DNA鎖は、ガイドRNAの伸長された部分の特異的なRTプライム配列、すなわちpegRNA上の「逆転写酵素プライム配列」又は「プライマー結合部位」にハイブリダイズする。工程(d)では、逆転写酵素(又は他の好適なDNAポリメラーゼ)が導入される。これは、プライムされた部位の3'端からプライム編集ガイドRNAの5'端の方へDNAの一本鎖を合成する。DNAポリメラーゼ(例として逆転写酵素)はnapDNAbpに融合され得るか、又は代替的にはnapDNAbpに対してトランスで提供され得る。これは、ニック部位の又はそれに隣接する内生DNAに対してさもなければ相同的である所望のヌクレオチド変化(例として、1塩基変化、挿入、若しくは欠失、又はそれらの組み合わせ)を含む一本鎖DNAフラップを形成する。工程(e)では、napDNAbp及びガイドRNAがリリースされる。工程(f)及び(g)は一本鎖DNAフラップの分解に関し、その結果、所望のヌクレオチド変化が標的遺伝子座に組み込まれるようになる。このプロセスは、ひとたび3'一本鎖DNAフラップが内生DNA配列に侵入及びハイブリダイズすると形成する対応する5'内生DNAフラップを除去することによって、所望の産物形成の方へ駆動され得る。理論によって拘束されることなしに、細胞の内生DNA修復及び複製プロセスは、ミスマッチのDNAを分解して、ヌクレオチド変化(単数又は複数)を組み込んで、所望の変改された産物を形成する。プロセスは、図1Fに例示されている通り「第2鎖ニッキング」によってもまた産物形成の方へ駆動され得る。このプロセスは次の遺伝子変化の少なくとも1以上を導入してもよい:トランスバーション、トランジション、欠失、及び挿入。
用語「プライム編集因子(PE)システム」又は「プライム編集因子(PE)」又は「PEシステム」又は「PE編集システム」は、本明細書に記載のプライム編集を用いるゲノム編集の方法を伴う組成物を指し、napDNAbp、逆転写酵素、融合タンパク質(例として、napDNAbp及び逆転写酵素を含む)、プライム編集ガイドRNA、並びに融合タンパク質及びプライム編集ガイドRNAを含む複合体、並びに補助的な要素、例えば、プライム編集プロセスを編集された産物形成の方へ駆動することを助けるための第2鎖ニッキング構成要素(例として、第2鎖sgRNA)及び5'内生DNAフラップ除去エンドヌクレアーゼ(例としてFEN1)を包含するが、これらに限定されない。
ここまで記載された態様では、pegRNAは、ガイドRNA(これは、それ自体が、スペーサ配列及びgRNAコア又は骨格を含む)とプライマー結合部位及びDNA合成鋳型を含む5'又は3'伸長アームとを含む単一分子を構成するが(例として図3Dを見よ)、pegRNAは2つの個々の分子の形態をもまたとってもよく、ガイドRNAと、トランスプライム編集因子RNA鋳型(tPERT)とからなる。これは、本質的に、伸長アーム(特に、プライマー結合部位及びDNA合成ドメインを包含する)及びRNA-タンパク質動員ドメイン(例として、MS2アプタマー又はヘアピン)を同じ分子上に収容する。これは、tPERT動員タンパク質(例として、MS2アプタマーに結合するMS2cpタンパク質)を含む修飾されたプライム編集因子複合体へと共局在又は動員されるようになる。プライム編集に用いられてもよいtPERTの例としては図3G及び図3Hを見よ。
プライム編集因子
用語「プライム編集因子」は、napDNAbp(例として、Cas9ニッカーゼ)及び逆転写酵素を含む本明細書に記載の融合構築物を指し、pegRNA(又は「伸長されたガイドRNA」)の存在下で標的ヌクレオチド配列上にプライム編集を遂行することが可能である。用語「プライム編集因子」は、融合タンパク質、又はpegRNAと複合体化された融合タンパク質、及び/又は第2鎖へニックを入れるsgRNAと更に複合体化された融合タンパク質を指してもよい。いくつかの態様において、プライム編集因子はまた、本明細書に記載の通りの非編集済鎖の第2部位へのニック入れ工程へ向かうことが可能な、融合タンパク質(napDNAbpと融合した逆転写酵素)、pegRNA、及び定例の(regular)ガイドRNAを含む複合体も指してもよい。他の態様において、「プライマー編集因子」の逆転写酵素構成要素は、transで提供されてもよい。
用語「プライム編集因子」は、napDNAbp(例として、Cas9ニッカーゼ)及び逆転写酵素を含む本明細書に記載の融合構築物を指し、pegRNA(又は「伸長されたガイドRNA」)の存在下で標的ヌクレオチド配列上にプライム編集を遂行することが可能である。用語「プライム編集因子」は、融合タンパク質、又はpegRNAと複合体化された融合タンパク質、及び/又は第2鎖へニックを入れるsgRNAと更に複合体化された融合タンパク質を指してもよい。いくつかの態様において、プライム編集因子はまた、本明細書に記載の通りの非編集済鎖の第2部位へのニック入れ工程へ向かうことが可能な、融合タンパク質(napDNAbpと融合した逆転写酵素)、pegRNA、及び定例の(regular)ガイドRNAを含む複合体も指してもよい。他の態様において、「プライマー編集因子」の逆転写酵素構成要素は、transで提供されてもよい。
プライマー結合部位
「プライマー結合部位」又は「PBS」という用語は、伸長アーム(典型的には、例えば、伸張アームの3'端部において)の構成要素としてのpegRNA上に位置するヌクレオチド配列の部分を指す。「プライマー結合部位」という用語は、非標的鎖上の配列と相補性の領域を含む伸長アームの構成要素としてのPEgRNAの一本鎖部分を指す。いくつかの態様において、プライマー結合部位は、非標的鎖におけるニック部位の上流の領域に相補的である。いくつかの態様において、プライマー結合部位は、非標的鎖におけるニック部位のすぐ上流の領域に相補的である。いくつかの態様において、プライマー結合部位は、プライム編集因子による標的配列のニッキング後に形成されるプライマー配列に結合することができる。プライム編集因子が標的DNA配列の1つの鎖を切断すると(例として、プライム編集因子のCasニッカーゼ構成要素)、3'末端ssDNAフラップが形成され、これは、逆転写をプライミングするためにpegRNA上のプライマー結合部位にアニーリングするプライマー配列に役立つ。図27及び図28は、夫々3'及び5'伸長アーム上に位置するプライマー結合部位の態様を示す。いくつかの態様では、PBSは、ニック部位の二本鎖標的DNAの非標的鎖上の遊離3'末端に相補的又は実質的に相補的であり、アニールし得る。いくつかの態様では、非標的鎖上の遊離3'末端にアニーリングしたPBSは、標的プライミングDNA合成を開始し得る。
「プライマー結合部位」又は「PBS」という用語は、伸長アーム(典型的には、例えば、伸張アームの3'端部において)の構成要素としてのpegRNA上に位置するヌクレオチド配列の部分を指す。「プライマー結合部位」という用語は、非標的鎖上の配列と相補性の領域を含む伸長アームの構成要素としてのPEgRNAの一本鎖部分を指す。いくつかの態様において、プライマー結合部位は、非標的鎖におけるニック部位の上流の領域に相補的である。いくつかの態様において、プライマー結合部位は、非標的鎖におけるニック部位のすぐ上流の領域に相補的である。いくつかの態様において、プライマー結合部位は、プライム編集因子による標的配列のニッキング後に形成されるプライマー配列に結合することができる。プライム編集因子が標的DNA配列の1つの鎖を切断すると(例として、プライム編集因子のCasニッカーゼ構成要素)、3'末端ssDNAフラップが形成され、これは、逆転写をプライミングするためにpegRNA上のプライマー結合部位にアニーリングするプライマー配列に役立つ。図27及び図28は、夫々3'及び5'伸長アーム上に位置するプライマー結合部位の態様を示す。いくつかの態様では、PBSは、ニック部位の二本鎖標的DNAの非標的鎖上の遊離3'末端に相補的又は実質的に相補的であり、アニールし得る。いくつかの態様では、非標的鎖上の遊離3'末端にアニーリングしたPBSは、標的プライミングDNA合成を開始し得る。
プロモータ
用語「プロモータ」は当該技術分野において認識されており、細胞性の転写機構によって認識され、下流遺伝子の転写を開始する能力がある配列を有する核酸分子を指す。プロモータは、プロモータが所与の細胞性という文脈において常に活性であるということを意味する恒常的に活性、又はプロモータが特定の条件の存在下においてのみ活性であるということを意味する条件付きで活性であり得る。例えば、条件付きプロモータは、プロモータ上の制御エレメントと会合したタンパク質を基本的な転写機構に接続する特定のタンパク質の存在下においてのみ、又は阻害分子の不在下においてのみ活性であってもよい。条件付きで活性なプロモータのサブクラスは、活性のために低分子「誘導因子」の存在を要求する誘導性プロモータである。誘導性プロモータの例は、アラビノース誘導性プロモータ、Tet-onプロモータ、及びタモキシフェン誘導性プロモータを包含するが、これらに限定されない。種々の恒常的、条件付き、及び誘導性プロモータが当業者に周知であり、当業者は本発明を実行することに有用な種々のかかるプロモータを確かめる能力があるであろう。これはこの点について限定されない。
用語「プロモータ」は当該技術分野において認識されており、細胞性の転写機構によって認識され、下流遺伝子の転写を開始する能力がある配列を有する核酸分子を指す。プロモータは、プロモータが所与の細胞性という文脈において常に活性であるということを意味する恒常的に活性、又はプロモータが特定の条件の存在下においてのみ活性であるということを意味する条件付きで活性であり得る。例えば、条件付きプロモータは、プロモータ上の制御エレメントと会合したタンパク質を基本的な転写機構に接続する特定のタンパク質の存在下においてのみ、又は阻害分子の不在下においてのみ活性であってもよい。条件付きで活性なプロモータのサブクラスは、活性のために低分子「誘導因子」の存在を要求する誘導性プロモータである。誘導性プロモータの例は、アラビノース誘導性プロモータ、Tet-onプロモータ、及びタモキシフェン誘導性プロモータを包含するが、これらに限定されない。種々の恒常的、条件付き、及び誘導性プロモータが当業者に周知であり、当業者は本発明を実行することに有用な種々のかかるプロモータを確かめる能力があるであろう。これはこの点について限定されない。
プロトスペーサ
本明細書において用いられる用語「プロトスペーサ」はPAM(プロトスペーサ隣接モチーフ)配列に隣接するDNA上の配列(~20bp)を指す。プロトスペーサはガイドRNAのスペーサ配列と同じ配列を共有する。ガイドRNAは標的DNA上のプロトスペーサ配列の相補体(具体的には、それの1つの鎖、すなわち標的DNA配列の「非標的鎖」に対する「標的鎖」)にアニールする。いくつかの態様では、プライム編集因子のCasニッカーゼ構成要素が機能するためには、特定のプロトスペーサ隣接モチーフ(PAM)も必要であり、これはCasタンパク質構成要素自体、例としてCasタンパク質の種類及びそれが由来される細菌種に応じて異なる。例えば、S.pyogenesに由来する最も普通に用いられるCas9ヌクレアーゼは、ゲノムDNA上の標的配列の直接的に下流である非標的鎖上のNGGというPAM配列を認識する。当業者は、現況技術の文献が、場合によっては、それを「スペーサ」と指すよりもむしろ、ガイドRNAそれ自体の上の~20nt標的特異的ガイド配列として「プロトスペーサ」を指すということを了解するであろう。それゆえに、いくつかのケースでは、本明細書において用いられる用語「プロトスペーサ」は用語「スペーサ」と交換可能に用いられてもよい。「プロトスペーサ」又は「スペーサ」どちらかの出現の周囲の記載という文脈は、用語がgRNA又はDNA標的を参照しているかどうかについて読み手に情報提供することを助けるであろう。
本明細書において用いられる用語「プロトスペーサ」はPAM(プロトスペーサ隣接モチーフ)配列に隣接するDNA上の配列(~20bp)を指す。プロトスペーサはガイドRNAのスペーサ配列と同じ配列を共有する。ガイドRNAは標的DNA上のプロトスペーサ配列の相補体(具体的には、それの1つの鎖、すなわち標的DNA配列の「非標的鎖」に対する「標的鎖」)にアニールする。いくつかの態様では、プライム編集因子のCasニッカーゼ構成要素が機能するためには、特定のプロトスペーサ隣接モチーフ(PAM)も必要であり、これはCasタンパク質構成要素自体、例としてCasタンパク質の種類及びそれが由来される細菌種に応じて異なる。例えば、S.pyogenesに由来する最も普通に用いられるCas9ヌクレアーゼは、ゲノムDNA上の標的配列の直接的に下流である非標的鎖上のNGGというPAM配列を認識する。当業者は、現況技術の文献が、場合によっては、それを「スペーサ」と指すよりもむしろ、ガイドRNAそれ自体の上の~20nt標的特異的ガイド配列として「プロトスペーサ」を指すということを了解するであろう。それゆえに、いくつかのケースでは、本明細書において用いられる用語「プロトスペーサ」は用語「スペーサ」と交換可能に用いられてもよい。「プロトスペーサ」又は「スペーサ」どちらかの出現の周囲の記載という文脈は、用語がgRNA又はDNA標的を参照しているかどうかについて読み手に情報提供することを助けるであろう。
プロトスペーサ隣接モチーフ(PAM)
本明細書において用いられる用語「プロトスペーサ隣接配列」又は「PAM」は、Cas9ヌクレアーゼの重要な標的化構成要素であるおよそ2-6塩基対のDNA配列を指す。典型的には、PAM配列はどちらかの鎖上にあり、Cas9によってカットされる部位の5'→3'方向においては下流である。標準的なPAM配列(すなわち、Streptococcus pyogenesのCas9ヌクレアーゼ又はSpCas9に関連するPAM配列)は5'-NGG-3'であり、「N」は2つのグアニン(「G」)核酸塩基によって後続されるいずれかの核酸塩基である。異なるPAM配列が異なる生物からの異なるCas9ヌクレアーゼ又は同等タンパク質と関連され得る。加えて、いずれかの所与のCas9ヌクレアーゼ、例としてSpCas9は、ヌクレアーゼが代替的なPAM配列を認識するようにしてヌクレアーゼのPAM特異性を変改するように修飾されてもよい。
本明細書において用いられる用語「プロトスペーサ隣接配列」又は「PAM」は、Cas9ヌクレアーゼの重要な標的化構成要素であるおよそ2-6塩基対のDNA配列を指す。典型的には、PAM配列はどちらかの鎖上にあり、Cas9によってカットされる部位の5'→3'方向においては下流である。標準的なPAM配列(すなわち、Streptococcus pyogenesのCas9ヌクレアーゼ又はSpCas9に関連するPAM配列)は5'-NGG-3'であり、「N」は2つのグアニン(「G」)核酸塩基によって後続されるいずれかの核酸塩基である。異なるPAM配列が異なる生物からの異なるCas9ヌクレアーゼ又は同等タンパク質と関連され得る。加えて、いずれかの所与のCas9ヌクレアーゼ、例としてSpCas9は、ヌクレアーゼが代替的なPAM配列を認識するようにしてヌクレアーゼのPAM特異性を変改するように修飾されてもよい。
例えば、配列番号37の(is)標準的なSpCas9アミノ酸配列を参照して、PAM配列は1以上の変異を導入することによって修飾され得る。(a)PAM特異性をNGAN又はNGNGへと変改するD1135V、R1335Q、及びT1337R「VQRバリアント」、(b)PAM特異性をNGAGへと変改するD1135E、R1335Q、及びT1337R「EQRバリアント」、並びに(c)PAM特異性をNGCGへと変改するD1135V、G1218R、R1335E、及びT1337R「VRERバリアント」を包含する。加えて、標準的なSpCas9のD1135EバリアントはなおNGGを認識するが、それは野生型SpCas9タンパク質と比較して選択的である。
異なる細菌種からのCas9酵素(すなわち、Cas9オーソログ)は様々なPAM特異性を有し得るということもまた了解されるであろう。例えば、Staphylococcus aureusからのCas9(SaCas9)はNGRRT又はNGRRNを認識する。加えて、Neisseria meningitisからのCas9(NmCas)はNNNNGATTを認識する。別の例では、Streptococcus thermophilisからのCas9(StCas9)はNNAGAAWを認識する。なお別の例では、Treponema denticolaからのCas9(TdCas)はNAAAACを認識する。これらは例であって、限定することを意味されない。更に、非SpCas9が種々のPAM配列に結合するということは了解されるであろう。これは、好適なSpCas9 PAM配列が所望の標的のカットされる部位に存在しないときにそれらを有用にする。更にその上、非SpCas9は、それらをSpCas9よりも有用にする他の特徴を有してもよい。例えば、Staphylococcus aureusからのCas9(SaCas9)はSpCas9よりも約1キロベース小さい。そのため、それはアデノ随伴ウイルス(AAV)にパッケージングされ得る。Shah et al.,「Protospacer recognition motifs:mixed identities and functional diversity,」RNA Biology,10(5):891-899の更なる参照がなされてもよい(これは参照によって本明細書に組み込まれる)。
逆転写酵素
用語「逆転写酵素」はRNA依存性DNAポリメラーゼとして特徴付けられるポリメラーゼのあるクラスを記述する。全ての公知の逆転写酵素はRNA鋳型からDNA転写物を合成するためにはプライマーを要求する。歴史的には、逆転写酵素は主としてmRNAをcDNAに転写するために用いられている。これは次いで更なる操作のためにベクター上にクローニングされ得る。トリ骨髄芽球症(myoblastosis)ウイルス(AMV)逆転写酵素は最初の広く用いられるRNA依存性DNAポリメラーゼであった(Verma,Biochim.Biophys.Acta 473:1(1977))。酵素は、5'-3'RNA指令型DNAポリメラーゼ活性、5'-3'DNA指令型DNAポリメラーゼ活性、及びRNase H活性を有する。RNase HはRNA-DNAハイブリッドのRNA鎖に特異的なプロセッシブな5'及び3'リボヌクレアーゼである(Perbal,A Practical Guide to Molecular Cloning,New York:Wiley&Sons(1984))。公知のウイルス逆転写酵素はプルーフリーディングにとって必要な3'-5'エキソヌクレアーゼ活性を欠くので、転写のエラーは逆転写酵素によって修正され得ない(Saunders and Saunders,Microbial Genetics Applied to Biotechnology,London:Croom Helm(1987))。AMV逆転写酵素の活性及びその関連するRNase H活性の詳細な研究は、Berger et al.,Biochemistry 22:2365-2372(1983)によって提示されている。分子生物学に鋭意用いられる別の逆転写酵素はモロニーマウス白血病ウイルス(M-MLV)を起源とする逆転写酵素である。例として、Gerard,G.R.,DNA 5:271-279(1986)及びKotewicz,M.L.,et al.,Gene 35:249-258(1985)を見よ。RNase H活性を実質的に欠くM-MLV逆転写酵素もまた記載されている。例としてU.S.Pat.No.5,244,797を見よ。本発明はいずれかのかかる逆転写酵素又はそれらのバリアント若しくは変異体の使用を企図する。
用語「逆転写酵素」はRNA依存性DNAポリメラーゼとして特徴付けられるポリメラーゼのあるクラスを記述する。全ての公知の逆転写酵素はRNA鋳型からDNA転写物を合成するためにはプライマーを要求する。歴史的には、逆転写酵素は主としてmRNAをcDNAに転写するために用いられている。これは次いで更なる操作のためにベクター上にクローニングされ得る。トリ骨髄芽球症(myoblastosis)ウイルス(AMV)逆転写酵素は最初の広く用いられるRNA依存性DNAポリメラーゼであった(Verma,Biochim.Biophys.Acta 473:1(1977))。酵素は、5'-3'RNA指令型DNAポリメラーゼ活性、5'-3'DNA指令型DNAポリメラーゼ活性、及びRNase H活性を有する。RNase HはRNA-DNAハイブリッドのRNA鎖に特異的なプロセッシブな5'及び3'リボヌクレアーゼである(Perbal,A Practical Guide to Molecular Cloning,New York:Wiley&Sons(1984))。公知のウイルス逆転写酵素はプルーフリーディングにとって必要な3'-5'エキソヌクレアーゼ活性を欠くので、転写のエラーは逆転写酵素によって修正され得ない(Saunders and Saunders,Microbial Genetics Applied to Biotechnology,London:Croom Helm(1987))。AMV逆転写酵素の活性及びその関連するRNase H活性の詳細な研究は、Berger et al.,Biochemistry 22:2365-2372(1983)によって提示されている。分子生物学に鋭意用いられる別の逆転写酵素はモロニーマウス白血病ウイルス(M-MLV)を起源とする逆転写酵素である。例として、Gerard,G.R.,DNA 5:271-279(1986)及びKotewicz,M.L.,et al.,Gene 35:249-258(1985)を見よ。RNase H活性を実質的に欠くM-MLV逆転写酵素もまた記載されている。例としてU.S.Pat.No.5,244,797を見よ。本発明はいずれかのかかる逆転写酵素又はそれらのバリアント若しくは変異体の使用を企図する。
加えて、本発明は、エラーを起こしやすい、すなわちエラーを起こしやすい逆転写酵素と呼ばれてもよい逆転写酵素、又は重合の間にヌクレオチドの高忠実性組み込みを支持しない逆転写酵素の使用を企図する。ガイドRNAと取り入れされたRT鋳型に基づく一本鎖DNAフラップの合成の間に、エラーを起こしやすい逆転写酵素は、RT鋳型配列とミスマッチである1以上のヌクレオチドを導入し得、それによって、一本鎖DNAフラップの誤りがちな重合によってヌクレオチド配列に変化を導入する。次いで、一本鎖DNAフラップの合成の間に導入されるこれらのエラーは、対応する内生標的鎖に対するハイブリダイゼーション、取って代わられた内生鎖の除去、ライゲーション、並びに次いで内生DNA修復及び/又は配列決定プロセスのもう1つのラウンドによって、二本鎖分子上に取り入れられるようになる。
逆転写
本明細書において用いられる用語「逆転写」は、酵素がRNAを鋳型として用いてDNA鎖(つまり、相補的なDNA又はcDNA)を合成する能力を指示する。いくつかの態様では、逆転写は「エラーを起こしやすい逆転写」であり得る。これは、それらのDNA重合活性がエラーを起こしやすい、ある種の逆転写酵素の特性を指す。
本明細書において用いられる用語「逆転写」は、酵素がRNAを鋳型として用いてDNA鎖(つまり、相補的なDNA又はcDNA)を合成する能力を指示する。いくつかの態様では、逆転写は「エラーを起こしやすい逆転写」であり得る。これは、それらのDNA重合活性がエラーを起こしやすい、ある種の逆転写酵素の特性を指す。
タンパク質、ペプチド、及びポリペプチド
用語「タンパク質」、「ペプチド」、及び「ポリペプチド」は本明細書中、互換的に使用され、ペプチド(アミド)結合によって一緒に連結されたアミノ酸残基の重合体を指す。用語は、いずれのサイズ、構造、若しくは機能の、タンパク質、ペプチド、又はポリペプチドを指す。典型的には、タンパク質、ペプチド、又はポリペプチドは、少なくとも3アミノ酸長であろう。タンパク質、ペプチド、又はポリペプチドは、個々のタンパク質又はタンパク質の一群を指してもよい。タンパク質、ペプチド、又はポリペプチド中のアミノ酸の1以上は、例えば、炭水化物基、ヒドロキシル基、ホスファート基、ファルネシル基、イソファルネシル基、脂肪酸基、抱合のためのリンカー、官能基化、又は他の修飾等々等の化学的実体(chemical entity)の付加によって修飾されていてもよい。タンパク質、ペプチド、又はポリペプチドはまた、単一分子であってもよく、又は多分子複合体であってもよい。タンパク質、ペプチド、又はポリペプチドは、天然に存在するタンパク質又はペプチドのただのフラグメントであってもよい。タンパク質、ペプチド、又はポリペプチドは、天然に存在するもの、組換え体、又は合成物、又はいずれのそれらの組み合わせであってもよい。本明細書に提供されるタンパク質のいずれも、当該技術分野において知られているいずれの方法によって産生されてもよい。例えば、本明細書に提供されるタンパク質は、ペプチドリンカーを含む融合タンパク質に特に適した、組換えタンパク質の発現及び精製を介して産生されてもよい。組換えタンパク質の発現及び精製のための方法は周知であり、Green and Sambrook,Molecular Cloning:A Laboratory Manual(4th ed.,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,N.Y.(2012)(これらの内容全体は参照により本明細書に組み込まれる))によって記載されるものを包含する。
用語「タンパク質」、「ペプチド」、及び「ポリペプチド」は本明細書中、互換的に使用され、ペプチド(アミド)結合によって一緒に連結されたアミノ酸残基の重合体を指す。用語は、いずれのサイズ、構造、若しくは機能の、タンパク質、ペプチド、又はポリペプチドを指す。典型的には、タンパク質、ペプチド、又はポリペプチドは、少なくとも3アミノ酸長であろう。タンパク質、ペプチド、又はポリペプチドは、個々のタンパク質又はタンパク質の一群を指してもよい。タンパク質、ペプチド、又はポリペプチド中のアミノ酸の1以上は、例えば、炭水化物基、ヒドロキシル基、ホスファート基、ファルネシル基、イソファルネシル基、脂肪酸基、抱合のためのリンカー、官能基化、又は他の修飾等々等の化学的実体(chemical entity)の付加によって修飾されていてもよい。タンパク質、ペプチド、又はポリペプチドはまた、単一分子であってもよく、又は多分子複合体であってもよい。タンパク質、ペプチド、又はポリペプチドは、天然に存在するタンパク質又はペプチドのただのフラグメントであってもよい。タンパク質、ペプチド、又はポリペプチドは、天然に存在するもの、組換え体、又は合成物、又はいずれのそれらの組み合わせであってもよい。本明細書に提供されるタンパク質のいずれも、当該技術分野において知られているいずれの方法によって産生されてもよい。例えば、本明細書に提供されるタンパク質は、ペプチドリンカーを含む融合タンパク質に特に適した、組換えタンパク質の発現及び精製を介して産生されてもよい。組換えタンパク質の発現及び精製のための方法は周知であり、Green and Sambrook,Molecular Cloning:A Laboratory Manual(4th ed.,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,N.Y.(2012)(これらの内容全体は参照により本明細書に組み込まれる))によって記載されるものを包含する。
タンパク質スプライシング
本明細書において用いられる用語「タンパク質スプライシング」は、インテイン(又は場合に応じて分裂インテイン)の配列がアミノ酸配列内から切り出され、アミノ酸配列の残りのフラグメントのエクステインがアミド結合によってライゲーションされて連続的なアミノ酸配列を形成するプロセスを指す。用語「トランス」タンパク質スプライシングは、インテインが分裂インテインであり、それらが異なるタンパク質上に位置付けられる特定のケースを指す。
本明細書において用いられる用語「タンパク質スプライシング」は、インテイン(又は場合に応じて分裂インテイン)の配列がアミノ酸配列内から切り出され、アミノ酸配列の残りのフラグメントのエクステインがアミド結合によってライゲーションされて連続的なアミノ酸配列を形成するプロセスを指す。用語「トランス」タンパク質スプライシングは、インテインが分裂インテインであり、それらが異なるタンパク質上に位置付けられる特定のケースを指す。
第2鎖ニッキング
プライム編集の結果として形成されるヘテロ二重鎖DNA(すなわち、1つの編集された及び1つの非編集の鎖を含有する)の分解は、長期的な編集アウトカムを決定する。言葉にすると、プライム編集のゴールは、相補的な内生鎖上に編集された鎖を永久的に取り入れすることによって、PEの中間体として形成されるヘテロ二重鎖DNA(内生非編集鎖と対形成した編集された鎖)を分解することである。DNA分子上への編集された鎖の永久的な取り入れに有利にヘテロ二重鎖DNAの分解を駆動することを助けるために、「第2鎖ニッキング」のアプローチが本明細書において用いられ得る。本明細書において用いられる「第2鎖ニッキング」の概念は、第1のニック(すなわち、ガイドRNAの伸長された部分上の逆転写酵素のプライムへの使用のための遊離の3'端を提供する当初のニック部位)の下流の位置付けにおける、好ましくは編集されない鎖上の第2のニックの導入を指す。ある態様において、第1のニック及び第2のニックは対向する鎖上にある。他の態様において、第1のニック及び第2のニックは対向する鎖上にある。更に別の態様では、第1のニックは非標的鎖(すなわち、Rループの一本鎖部分を形成する鎖)上にあり、第2のニックは標的鎖上にある。なお他の態様において、第1のニックは編集される鎖上にあり、第2のニックは編集されない鎖上にある。第2のニックは、第1のニックの少なくとも5ヌクレオチド下流に、あるいは第1のニックの少なくとも6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、40、50、60、70、80、90、100、110、120、130、140、若しくは150ヌクレオチド、又はより多く下流に配置され得る。第2のニックは、ある態様において、編集されない鎖上において、pegRNAによって誘導されるニックの部位から約5~150ヌクレオチドの間、又は約5~140の間、若しくは約5~130の間、若しくは約5~120の間、若しくは約5~110の間、若しくは約5~100の間、若しくは約5~90の間、若しくは約5~80の間、若しくは約5~70の間、若しくは約5~60の間、若しくは約5~50の間、若しくは約5~40の間、若しくは約5~30の間、若しくは約5~20の間、若しくは約5~10の間離れて導入され得る。1つの態様では、第2のニックは、pegRNAによって誘導されるニックから14~116ヌクレオチドの間離れて導入される。理論によって拘束されることなしに、第2のニックは細胞の内生DNA修復及び複製プロセスを編集されない鎖の置換え又は編集の方へ誘導し、それによって、両鎖上に編集された配列を永久的に組み入れし、PEの結果として形成されるヘテロ二重鎖を分解する。いくつかの態様では、編集される鎖は非標的鎖であり、編集されない鎖は標的鎖である。他の態様において、編集される鎖は標的鎖であり、編集されない鎖は非標的鎖である。
プライム編集の結果として形成されるヘテロ二重鎖DNA(すなわち、1つの編集された及び1つの非編集の鎖を含有する)の分解は、長期的な編集アウトカムを決定する。言葉にすると、プライム編集のゴールは、相補的な内生鎖上に編集された鎖を永久的に取り入れすることによって、PEの中間体として形成されるヘテロ二重鎖DNA(内生非編集鎖と対形成した編集された鎖)を分解することである。DNA分子上への編集された鎖の永久的な取り入れに有利にヘテロ二重鎖DNAの分解を駆動することを助けるために、「第2鎖ニッキング」のアプローチが本明細書において用いられ得る。本明細書において用いられる「第2鎖ニッキング」の概念は、第1のニック(すなわち、ガイドRNAの伸長された部分上の逆転写酵素のプライムへの使用のための遊離の3'端を提供する当初のニック部位)の下流の位置付けにおける、好ましくは編集されない鎖上の第2のニックの導入を指す。ある態様において、第1のニック及び第2のニックは対向する鎖上にある。他の態様において、第1のニック及び第2のニックは対向する鎖上にある。更に別の態様では、第1のニックは非標的鎖(すなわち、Rループの一本鎖部分を形成する鎖)上にあり、第2のニックは標的鎖上にある。なお他の態様において、第1のニックは編集される鎖上にあり、第2のニックは編集されない鎖上にある。第2のニックは、第1のニックの少なくとも5ヌクレオチド下流に、あるいは第1のニックの少なくとも6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、40、50、60、70、80、90、100、110、120、130、140、若しくは150ヌクレオチド、又はより多く下流に配置され得る。第2のニックは、ある態様において、編集されない鎖上において、pegRNAによって誘導されるニックの部位から約5~150ヌクレオチドの間、又は約5~140の間、若しくは約5~130の間、若しくは約5~120の間、若しくは約5~110の間、若しくは約5~100の間、若しくは約5~90の間、若しくは約5~80の間、若しくは約5~70の間、若しくは約5~60の間、若しくは約5~50の間、若しくは約5~40の間、若しくは約5~30の間、若しくは約5~20の間、若しくは約5~10の間離れて導入され得る。1つの態様では、第2のニックは、pegRNAによって誘導されるニックから14~116ヌクレオチドの間離れて導入される。理論によって拘束されることなしに、第2のニックは細胞の内生DNA修復及び複製プロセスを編集されない鎖の置換え又は編集の方へ誘導し、それによって、両鎖上に編集された配列を永久的に組み入れし、PEの結果として形成されるヘテロ二重鎖を分解する。いくつかの態様では、編集される鎖は非標的鎖であり、編集されない鎖は標的鎖である。他の態様において、編集される鎖は標的鎖であり、編集されない鎖は非標的鎖である。
センス鎖
遺伝学において、「センス」鎖は、5'から3'へ伸びる二本鎖DNA内セグメントであって、3'から5'へ伸びるDNAのアンチセンス鎖又は鋳型鎖に相補的である。タンパク質をコードするDNAセグメントのケースにおいて、センス鎖は、転写の最中にその鋳型としてアンチセンス鎖を取り結局(典型的には、常にではない)翻訳を経てタンパク質になるmRNAと同じ配列を有するDNAの鎖である。よって、アンチセンス鎖は、後にタンパク質へ翻訳されるRNAを担い、一方センス鎖は、mRNAとほぼ同一の組成(makeup)を保有する。dsDNAの各セグメントについて、一方がどの方向から読まれるかに応じて、おそらく(センス及びアンチセンスは視点に相対するため)センス及びアンチセンスが2組存在するであろう点に留意する。遺伝子産物又はmRNAは結局、dsDNAの一方のセグメントのどちらかの鎖がセンス又はアンチセンスと呼ばれることを示す。
遺伝学において、「センス」鎖は、5'から3'へ伸びる二本鎖DNA内セグメントであって、3'から5'へ伸びるDNAのアンチセンス鎖又は鋳型鎖に相補的である。タンパク質をコードするDNAセグメントのケースにおいて、センス鎖は、転写の最中にその鋳型としてアンチセンス鎖を取り結局(典型的には、常にではない)翻訳を経てタンパク質になるmRNAと同じ配列を有するDNAの鎖である。よって、アンチセンス鎖は、後にタンパク質へ翻訳されるRNAを担い、一方センス鎖は、mRNAとほぼ同一の組成(makeup)を保有する。dsDNAの各セグメントについて、一方がどの方向から読まれるかに応じて、おそらく(センス及びアンチセンスは視点に相対するため)センス及びアンチセンスが2組存在するであろう点に留意する。遺伝子産物又はmRNAは結局、dsDNAの一方のセグメントのどちらかの鎖がセンス又はアンチセンスと呼ばれることを示す。
pegRNAの文脈において、第1工程は、5'→3'方向に配向される単鎖相補DNA(すなわち、組み込まれることになる3'ssDNAフラップ)の合成であるが、合成鎖はpegRNA伸長アームの鋳型から外れる。3'ssDNAフラップがセンス鎖と見なされるべきか又はアンチセンス鎖と見なされるべきかについては、DNAの両鎖が転写のための鋳型として働いてもよい(が同時ではない)ことが十分に許容されているため、転写の方向に依存する。よって、いくつかの態様において、3'ssDNAフラップ(全体的に5'→3'方向に伸びる)は、コード鎖であることから、センス鎖として働くであろう。他の態様において、3'ssDNAフラップ(全体的に5'→3'方向に伸びる)はアンチセンス鎖として働き、よって転写の鋳型であろう。
スペーサ配列
本明細書で使用される場合、ガイドRNA又はpegRNAに関連する「スペーサ配列」という用語は、標的鎖に相補的なヌクレオチド配列を含有する約20ヌクレオチド(例として、16、17、18、19、20、21、22、23又は24ヌクレオチド)のガイドRNA又はpegRNAの部分を指す。いくつかの態様では、スペーサ配列は、非標的鎖上のプロトスペーサに相補的な標的鎖上の領域にハイブリダイズして、標的部位にssRNA/ssDNAハイブリッド構造を形成し、非標的鎖上に相補的な内生DNA鎖の対応するRループssDNA構造を形成する。
本明細書で使用される場合、ガイドRNA又はpegRNAに関連する「スペーサ配列」という用語は、標的鎖に相補的なヌクレオチド配列を含有する約20ヌクレオチド(例として、16、17、18、19、20、21、22、23又は24ヌクレオチド)のガイドRNA又はpegRNAの部分を指す。いくつかの態様では、スペーサ配列は、非標的鎖上のプロトスペーサに相補的な標的鎖上の領域にハイブリダイズして、標的部位にssRNA/ssDNAハイブリッド構造を形成し、非標的鎖上に相補的な内生DNA鎖の対応するRループssDNA構造を形成する。
対象
本明細書で使用される「対象」という用語は、個々の生物、例えば個々の哺乳動物を指す。いくつかの態様において、対象はヒトである。いくつかの態様では、対象は、非ヒト哺乳動物である。いくつかの態様では、対象は、非ヒト霊長類である。いくつかの態様では、対象はげっ歯類である。いくつかの態様では、対象は、ヒツジ、ヤギ、ウシ、ネコ、又はイヌである。いくつかの態様では、対象は、脊椎動物、両生類、爬虫類、魚類、昆虫、ハエ又は線形動物である。いくつかの態様では、対象は研究動物である。いくつかの態様において、対象は遺伝子操作されており、例として、遺伝子操作された非ヒト対象である。対象は、いずれかの性別及び任意の発達段階であってもよい。
本明細書で使用される「対象」という用語は、個々の生物、例えば個々の哺乳動物を指す。いくつかの態様において、対象はヒトである。いくつかの態様では、対象は、非ヒト哺乳動物である。いくつかの態様では、対象は、非ヒト霊長類である。いくつかの態様では、対象はげっ歯類である。いくつかの態様では、対象は、ヒツジ、ヤギ、ウシ、ネコ、又はイヌである。いくつかの態様では、対象は、脊椎動物、両生類、爬虫類、魚類、昆虫、ハエ又は線形動物である。いくつかの態様では、対象は研究動物である。いくつかの態様において、対象は遺伝子操作されており、例として、遺伝子操作された非ヒト対象である。対象は、いずれかの性別及び任意の発達段階であってもよい。
分裂型インテイン
インテインは連続ドメインとして最も頻繁に見出されるが、いくつかは自然に分裂された形態で存在する。この場合、2つのフラグメントは別個のポリペプチドとして発現され、スプライシングが起こる前に会合しなければならない、いわゆるタンパク質トランススプライシングである。
インテインは連続ドメインとして最も頻繁に見出されるが、いくつかは自然に分裂された形態で存在する。この場合、2つのフラグメントは別個のポリペプチドとして発現され、スプライシングが起こる前に会合しなければならない、いわゆるタンパク質トランススプライシングである。
例示的な分裂インテインは、2つのサブユニット、すなわちDnaE-N及びDnaE-Cを含むSsp DnaEインテインである。2つの異なるサブユニットは、DnaE-N及びDnaE-Cサブユニットを夫々コードする別々の遺伝子、すなわちdnaE-n及びdnaE-cによってコードされる。DnaEは、Synechocytis sp.PCC6803における天然に存在する分裂インテインであり、それぞれがDnaE-N又はDnaE-Cのいずれかとの融合物を含む2つの別個のタンパク質のトランススプライシングを指示することができる。
更なる天然に存在するか又は操作されたスプリットインテイン配列は、本明細書に記載の全インテイン配列又は当技術分野で利用可能なものから公知であるか、又は作製され得る。スプリットインテイン配列の例は、Stevens et al.,「A promiscuous split intein with expanded protein engineering applications,」 PNAS,2017,Vol.114:8538-8543;Iwai et al.,「Highly efficient protein trans-splicing by a naturally split DnaE intein from Nostoc punctiforme,FEBS Lett,580:1853-1858に見出され得、これらの各々は参照により本明細書に組み込まれる)。更なる分裂インテイン配列は、例えば、国際公開第2013/045632号、国際公開第2014/055782号、国際公開第2016/069774号、及び欧州特許第2877490号に見出され得、これらの各々の内容は参照により本明細書に組み込まれる。
更に、トランスでのタンパク質スプライシングがin vivo及びin vitroで記載されている(Shingledecker,et al.,Gene 207:187(1998),Southworth,et al.,EMBO J.17:918(1998);Mills,et al.,Proc.Natl.Acad.Sci.USA,95:3543-3548(1998);Lew,et al.,J.Biol.Chem.,273:15887-15890(1998);Wu,et al.,Biochim.Biophys.Acta 35732:1(1998b),Yamazaki,et al.,J.Am.Chem.Soc.120:5591(1998),Evans,et al.,J.Biol.Chem.275:9091(2000);Otomo,et al.,Biochemistry 38:16040-16044(1999);Otomo,et al.,J.Biolmol.NMR 14:105-114(1999);Scott,et al.,Proc.Natl.Acad.Sci.USA 96:13638-13643(1999))、例として、別々に発現される二つの半分からの完全なプライム編集因子の形成に関する図66及び図67に示されるように、機能的産物を形成するためにその後ライゲーションを受ける2つの不活性フラグメントに関するタンパク質を発現する機会を提供する。
標的部位
用語「標的部位」は、本明細書に開示のプライム編集因子(PE)によって編集される核酸分子上の配列を指す。更に、標的部位は、プライム編集因子(PE)及びgRNAの複合体が結合する核酸分子上の配列を指す。
用語「標的部位」は、本明細書に開示のプライム編集因子(PE)によって編集される核酸分子上の配列を指す。更に、標的部位は、プライム編集因子(PE)及びgRNAの複合体が結合する核酸分子上の配列を指す。
tPERT
「transプライム編集因子「RNA型(tPERT)」に係る定義を参照。
「transプライム編集因子「RNA型(tPERT)」に係る定義を参照。
一時的な第2鎖へのニック入れ
本明細書に使用されるとき、用語「一時的な第2鎖へのニック入れ」は、未編集鎖中の第2ニックの組み入れが、所望の編集が編集済鎖に組み入れられた後でしか生じない、第2鎖へのニック入れのバリアントを指す。これによって、二本鎖DNA切断へ繋がり得る両鎖上の同時ニックが回避される。第2鎖にニックを入れるガイドRNAは、第2鎖ニックが所望の編集の組み入れ後まで組み入れされないように、一時的制御のためにデザインされている。これは、元のアレルではなく編集された鎖のみにマッチするスペーサ配列を有するgRNAをデザインすることによって達成される。このストラテジーを使用すると、プロトスペーサと未編集アレルとの間のミスマッチによって、PAM鎖上の編集事象が起きた後まで、sgRNAによるニック入れが疎まれるはずである。
本明細書に使用されるとき、用語「一時的な第2鎖へのニック入れ」は、未編集鎖中の第2ニックの組み入れが、所望の編集が編集済鎖に組み入れられた後でしか生じない、第2鎖へのニック入れのバリアントを指す。これによって、二本鎖DNA切断へ繋がり得る両鎖上の同時ニックが回避される。第2鎖にニックを入れるガイドRNAは、第2鎖ニックが所望の編集の組み入れ後まで組み入れされないように、一時的制御のためにデザインされている。これは、元のアレルではなく編集された鎖のみにマッチするスペーサ配列を有するgRNAをデザインすることによって達成される。このストラテジーを使用すると、プロトスペーサと未編集アレルとの間のミスマッチによって、PAM鎖上の編集事象が起きた後まで、sgRNAによるニック入れが疎まれるはずである。
Transプライム編集
本明細書に使用されるとき、用語「transプライム編集」は、分裂pegRNAを利用するプライム編集の修飾形態を指すが、すなわち、ここでpegRNAは、2つ別々の分子:sgRNA及びtransプライム編集RNA鋳型(tPERT)に分離される。sgRNAが、所望のゲノムの標的部位へ、プライム編集因子を標的化させる(又はより一般に、プライム編集因子のnapDNAbp構成要素を標的化させる)よう働く一方で、一旦tPERTが、プライム編集因子上及びtPERT上に位置付けられた結合ドメインの相互作用によって、プライム編集因子へtransに動員されると、tPERTはポリメラーゼ(例として、逆転写酵素)によって使用されて新しいDNA配列を標的遺伝子座中へ書き込む。一態様において、結合ドメインは、tPERT上に位置付けられたMS2アプタマー及びプライム編集因子と融合したMS2cpタンパク質等の、RNA-タンパク質動員部分を包含し得る。transプライム編集の利点は、DNA合成鋳型をガイドRNAから分離させることによって、潜在的により長い長さの鋳型を使用し得る点である。
本明細書に使用されるとき、用語「transプライム編集」は、分裂pegRNAを利用するプライム編集の修飾形態を指すが、すなわち、ここでpegRNAは、2つ別々の分子:sgRNA及びtransプライム編集RNA鋳型(tPERT)に分離される。sgRNAが、所望のゲノムの標的部位へ、プライム編集因子を標的化させる(又はより一般に、プライム編集因子のnapDNAbp構成要素を標的化させる)よう働く一方で、一旦tPERTが、プライム編集因子上及びtPERT上に位置付けられた結合ドメインの相互作用によって、プライム編集因子へtransに動員されると、tPERTはポリメラーゼ(例として、逆転写酵素)によって使用されて新しいDNA配列を標的遺伝子座中へ書き込む。一態様において、結合ドメインは、tPERT上に位置付けられたMS2アプタマー及びプライム編集因子と融合したMS2cpタンパク質等の、RNA-タンパク質動員部分を包含し得る。transプライム編集の利点は、DNA合成鋳型をガイドRNAから分離させることによって、潜在的により長い長さの鋳型を使用し得る点である。
transプライム編集の態様は図3G及び図3Hに示される。図3Gは左に、transプライム編集因子複合体の組成物(「RP-PE:gRNA複合体」)を示すが、これは、ポリメラーゼ(例として、逆転写酵素)及びrPERT動員タンパク質(例として、MS2sc)の各々と融合したnapDNAbpを含み、ガイドRNAと複合体化されている。図3Gは更に、DNA合成鋳型及びプライマー結合配列を包含するpegRNAの伸長アーム特色を含む、別々のtPERT分子を示す。tPERT分子はまた、RNA-タンパク質動員ドメイン(これは、このケースにおいて、ステムループ構造であり、例えば、MS2アプタマーであり得る)も包含する。図3Hに記載のプロセスに描かれる通り、RP-PE:gRNA複合体は、標的DNA配列へ結合してニックを入れる。次いで、動員タンパク質(RP)は、tPERTを動員して、DNA標的部位へ結合したプライム編集因子複合体へ共局在化させ、それによってプライマー結合部位が、ニックが入った鎖上のプライマー配列へ結合できるようになり、続いて、ポリメラーゼ(例として、RT)が、DNA合成鋳型に対し、tPERTの5'に至るまでDNAの単鎖を合成できるようになる。
tPERTは、RNA-タンパク質動員ドメインの5'末上にPBS及びDNA合成鋳型を含むように、図3G及び図3Hに示されているが、他の立体配置のtPERTも、RNA-タンパク質動員ドメインの3'末上に位置付けられるPBS及びDNA合成鋳型をもつようデザインされてもよい。しかしながら、5'伸長をもつtPERTは、DNAの単鎖の合成がtPERTの5'末にて自然と終始するであろうという利点を有し、よって、プライム編集のDNA合成段階の最中に、RNA-タンパク質動員ドメインのいずれの部分も鋳型として使用するリスクがない。
トランジション
本明細書に使用されるとき、「トランジション」は、プリン核酸塩基(A⇔G)の相互変換(interchange)又はピリミジン核酸塩基(C⇔T)の相互変換を指す。このクラスの相互変換は、同様の形状の核酸塩基を伴う。本明細書に開示の組成物及び方法は、標的DNA分子において1以上のトランジションを誘導することが可能である。本明細書に開示の組成物及び方法はまた、同じ標的DNA分子においてトランジションとトランスバーションとの両方を誘導することも可能である。これらの変化は、A⇔G、G⇔A、C⇔T、又はT⇔Cを伴う。ワトソン・クリック対の核酸塩基をもつ二本鎖DNAの文脈において、トランスバーションは、以下の塩基対交換:A:T⇔G:C、G:G⇔A:T、C:G⇔T:A、又はT:A⇔C:Gを指す。本明細書に開示の組成物及び方法は、標的DNA分子において1以上のトランジションを誘導することが可能である。本明細書に開示の組成物及び方法はまた、同じ標的DNA分子においてトランジションとトランスバーションとの両方、並びに欠失及び挿入を包含する他のヌクレオチド変化を誘導することも可能である。
本明細書に使用されるとき、「トランジション」は、プリン核酸塩基(A⇔G)の相互変換(interchange)又はピリミジン核酸塩基(C⇔T)の相互変換を指す。このクラスの相互変換は、同様の形状の核酸塩基を伴う。本明細書に開示の組成物及び方法は、標的DNA分子において1以上のトランジションを誘導することが可能である。本明細書に開示の組成物及び方法はまた、同じ標的DNA分子においてトランジションとトランスバーションとの両方を誘導することも可能である。これらの変化は、A⇔G、G⇔A、C⇔T、又はT⇔Cを伴う。ワトソン・クリック対の核酸塩基をもつ二本鎖DNAの文脈において、トランスバーションは、以下の塩基対交換:A:T⇔G:C、G:G⇔A:T、C:G⇔T:A、又はT:A⇔C:Gを指す。本明細書に開示の組成物及び方法は、標的DNA分子において1以上のトランジションを誘導することが可能である。本明細書に開示の組成物及び方法はまた、同じ標的DNA分子においてトランジションとトランスバーションとの両方、並びに欠失及び挿入を包含する他のヌクレオチド変化を誘導することも可能である。
トランスバーション
本明細書に使用されるとき、「トランスバーション」は、プリン核酸塩基のピリミジン核酸塩基への相互変換、又はその逆を指し、よって、形状が似ていない核酸塩基の相互変換を伴う。これらの変化は、T⇔A、T⇔G、C⇔G、C⇔A、A⇔T、A⇔C、G⇔C、及びG⇔Tを伴う。ワトソン・クリック対の核酸塩基をもつ二本鎖DNAの文脈において、トランスバーションは、以下の塩基対交換:T:A⇔A:T、T:A⇔G:C、C:G⇔G:C、C:G⇔A:T、A:T⇔T:A、A:T⇔C:G、G:C⇔C:G、及びG:C⇔T:Aを指す。本明細書に開示の組成物及び方法は、標的DNA分子において1以上のトランスバーションを誘導することが可能である。本明細書に開示の組成物及び方法はまた、同じ標的DNA分子においてトランジションとトランスバーションとの両方、並びに欠失及び挿入を包含する他のヌクレオチド変化を誘導することも可能である。
本明細書に使用されるとき、「トランスバーション」は、プリン核酸塩基のピリミジン核酸塩基への相互変換、又はその逆を指し、よって、形状が似ていない核酸塩基の相互変換を伴う。これらの変化は、T⇔A、T⇔G、C⇔G、C⇔A、A⇔T、A⇔C、G⇔C、及びG⇔Tを伴う。ワトソン・クリック対の核酸塩基をもつ二本鎖DNAの文脈において、トランスバーションは、以下の塩基対交換:T:A⇔A:T、T:A⇔G:C、C:G⇔G:C、C:G⇔A:T、A:T⇔T:A、A:T⇔C:G、G:C⇔C:G、及びG:C⇔T:Aを指す。本明細書に開示の組成物及び方法は、標的DNA分子において1以上のトランスバーションを誘導することが可能である。本明細書に開示の組成物及び方法はまた、同じ標的DNA分子においてトランジションとトランスバーションとの両方、並びに欠失及び挿入を包含する他のヌクレオチド変化を誘導することも可能である。
処置
用語「処置」、「処置する」、及び「処置すること」は、本明細書に記載の通りの疾患若しくは障害、又は1以上のその症状の発病を、食い止め、緩和し、遅延させるか、あるいはその進行を阻害することを目的とした臨床的介入を指す。本明細書に使用されるとき、用語「処置」、「処置する」、及び「処置すること」は、本明細書に記載の通りの疾患若しくは障害、又は1以上のその症状の発病を、食い止め、緩和し、遅延させるか、あるいはその進行を阻害することを目的とした臨床的介入を指す。いくつかの態様において、処置は、1以上の症状が発症した後、及び/又は疾患と診断された後、施されてもよい。他の態様において、処置は、症状がない中、例として、症状の発病を予防若しくは遅延させるか、又は疾患の発病若しくは進行を阻害するために、施されてもよい。例えば、処置は、症状の発現に先立ち(例として、症状の経歴に照らして、及び/又は遺伝因子若しくは他の感受性因子に照らして)、感受性のある個体へ施されてもよい。処置はまた、症状が消散した後も、例えば、それらの再発を予防又は遅延するため、続けられてもよい。
用語「処置」、「処置する」、及び「処置すること」は、本明細書に記載の通りの疾患若しくは障害、又は1以上のその症状の発病を、食い止め、緩和し、遅延させるか、あるいはその進行を阻害することを目的とした臨床的介入を指す。本明細書に使用されるとき、用語「処置」、「処置する」、及び「処置すること」は、本明細書に記載の通りの疾患若しくは障害、又は1以上のその症状の発病を、食い止め、緩和し、遅延させるか、あるいはその進行を阻害することを目的とした臨床的介入を指す。いくつかの態様において、処置は、1以上の症状が発症した後、及び/又は疾患と診断された後、施されてもよい。他の態様において、処置は、症状がない中、例として、症状の発病を予防若しくは遅延させるか、又は疾患の発病若しくは進行を阻害するために、施されてもよい。例えば、処置は、症状の発現に先立ち(例として、症状の経歴に照らして、及び/又は遺伝因子若しくは他の感受性因子に照らして)、感受性のある個体へ施されてもよい。処置はまた、症状が消散した後も、例えば、それらの再発を予防又は遅延するため、続けられてもよい。
上流
本明細書に使用されるとき、用語「上流」及び「下流」は、5'→3'方向に配向された核酸分子(単鎖又は二本鎖であるかにかかわらず)中に位置付けられた少なくとも2つの要素の線状の位置を定義する相対的な用語である。特に、第1要素が第2要素に対して5'のどこかに位置付けられている場合、第1要素は、核酸分子中、第2要素の上流にある。例えば、SNPがニック部位の5'側にある場合、SNPは、Cas9に誘導されたニック部位の上流にある。逆に言うと、第1要素が第2要素に対して3'のどこかに位置付けられている場合、第1要素は、核酸分子中、第2要素の下流にある。例えば、SNPがニック部位の3'側にある場合、SNPは、Cas9に誘導されたニック部位の下流にある。核酸分子は、DNA(二本鎖若しくは単鎖)、RNA(二本鎖若しくは単鎖)、又はDNAとRNAとのハイブリッドであり得る。二本鎖分子のどちらの鎖を考慮するのか選択する必要がある場合を除き、上流及び下流という用語が核酸分子の単鎖にのみ言及するため、分析は単鎖核酸分子及び二本鎖分子について同じである。しばしば、少なくとも2つの要素の相対的な位置を決定するのに使用され得る二本鎖のDNAの鎖は、「センス」鎖又は「コード」鎖である。遺伝学において、「センス」鎖は、5'から3'へ伸びる二本鎖DNA内セグメントであって、3'から5'へ伸びるDNAのアンチセンス鎖又は鋳型鎖に相補的である。よって、例として、SNP核酸塩基がセンス鎖又はコード鎖上のプロモータの3'側にある場合、SNP核酸塩基はゲノムDNA(二本鎖である)中のプロモータ配列の「下流」にある。
本明細書に使用されるとき、用語「上流」及び「下流」は、5'→3'方向に配向された核酸分子(単鎖又は二本鎖であるかにかかわらず)中に位置付けられた少なくとも2つの要素の線状の位置を定義する相対的な用語である。特に、第1要素が第2要素に対して5'のどこかに位置付けられている場合、第1要素は、核酸分子中、第2要素の上流にある。例えば、SNPがニック部位の5'側にある場合、SNPは、Cas9に誘導されたニック部位の上流にある。逆に言うと、第1要素が第2要素に対して3'のどこかに位置付けられている場合、第1要素は、核酸分子中、第2要素の下流にある。例えば、SNPがニック部位の3'側にある場合、SNPは、Cas9に誘導されたニック部位の下流にある。核酸分子は、DNA(二本鎖若しくは単鎖)、RNA(二本鎖若しくは単鎖)、又はDNAとRNAとのハイブリッドであり得る。二本鎖分子のどちらの鎖を考慮するのか選択する必要がある場合を除き、上流及び下流という用語が核酸分子の単鎖にのみ言及するため、分析は単鎖核酸分子及び二本鎖分子について同じである。しばしば、少なくとも2つの要素の相対的な位置を決定するのに使用され得る二本鎖のDNAの鎖は、「センス」鎖又は「コード」鎖である。遺伝学において、「センス」鎖は、5'から3'へ伸びる二本鎖DNA内セグメントであって、3'から5'へ伸びるDNAのアンチセンス鎖又は鋳型鎖に相補的である。よって、例として、SNP核酸塩基がセンス鎖又はコード鎖上のプロモータの3'側にある場合、SNP核酸塩基はゲノムDNA(二本鎖である)中のプロモータ配列の「下流」にある。
バリアント
本明細書に使用されるとき、用語「バリアント」は、天然に存在するものから逸脱したパターンを有する属性(qualities)を呈するものを意味するとして解釈されるはずであり、例として、バリアントCas9は、野生型Cas9アミノ酸配列と比較してアミノ酸残基中に1以上の変化を含むCas9である。用語「バリアント」は、参照配列と少なくとも75%、又は少なくとも80%、又は少なくとも85%、又は少なくとも90%、又は少なくとも95%、又は少なくとも99%のパーセント同一性を有し、参照配列と同じか又は実質的に同じ機能活性(単数若しくは複数)を有する、相同のタンパク質を包括する(encompasses)。用語はまた、参照配列の突然変異体、トランケーション体(truncations)、又はドメインであって、参照配列と同じか又は実質的に同じ機能活性(単数若しくは複数)を呈示するものも包括する。
本明細書に使用されるとき、用語「バリアント」は、天然に存在するものから逸脱したパターンを有する属性(qualities)を呈するものを意味するとして解釈されるはずであり、例として、バリアントCas9は、野生型Cas9アミノ酸配列と比較してアミノ酸残基中に1以上の変化を含むCas9である。用語「バリアント」は、参照配列と少なくとも75%、又は少なくとも80%、又は少なくとも85%、又は少なくとも90%、又は少なくとも95%、又は少なくとも99%のパーセント同一性を有し、参照配列と同じか又は実質的に同じ機能活性(単数若しくは複数)を有する、相同のタンパク質を包括する(encompasses)。用語はまた、参照配列の突然変異体、トランケーション体(truncations)、又はドメインであって、参照配列と同じか又は実質的に同じ機能活性(単数若しくは複数)を呈示するものも包括する。
ベクター
用語「ベクター」は、本明細書に使用されるとき、着目する遺伝子をコードするよう修飾され得る核酸であって、宿主細胞中へ入って宿主細胞内で突然変異又は複製する(次いでベクターの複製形態を別の宿主細胞中へ移す)ことができる核酸を指す。例示の好適なベクターは、レトロウイルスベクター又はバクテリオファージ及び繊維状ファージ、及び接合性プラスミド等の、ウイルスベクターを包含する。追加の好適なベクターは、本開示に基づき当業者に明らかであろう。
用語「ベクター」は、本明細書に使用されるとき、着目する遺伝子をコードするよう修飾され得る核酸であって、宿主細胞中へ入って宿主細胞内で突然変異又は複製する(次いでベクターの複製形態を別の宿主細胞中へ移す)ことができる核酸を指す。例示の好適なベクターは、レトロウイルスベクター又はバクテリオファージ及び繊維状ファージ、及び接合性プラスミド等の、ウイルスベクターを包含する。追加の好適なベクターは、本開示に基づき当業者に明らかであろう。
野生型
本明細書に使用されるとき、用語「野生型」は、当業者によって理解される専門用語であって、突然変異体若しくはバリアントの形態とは区別されるような、天然に存在する通りの生物、株、遺伝子、又は特徴の典型的な形態を意味する。
本明細書に使用されるとき、用語「野生型」は、当業者によって理解される専門用語であって、突然変異体若しくはバリアントの形態とは区別されるような、天然に存在する通りの生物、株、遺伝子、又は特徴の典型的な形態を意味する。
5'内生DNAフラップ
本明細書において用いられる用語「内生5'DNAフラップ」は、標的DNA上のPEによって誘導されるニック部位の直ちに下流に所在するDNAの鎖を指す。PEによる標的DNA鎖のニッキングは、ニック部位の上流側の3'ヒドロキシル基及びニック部位の下流側の5'ヒドロキシル基を暴露する。3'ヒドロキシル基で終わる内生鎖は、プライム編集因子のDNAポリメラーゼをプライムするために用いられる(例として、DNAポリメラーゼは逆転写酵素である)。ニック部位の下流側のかつ暴露された5'ヒドロキシル基で始まる内生鎖は、「5'内生DNAフラップ」と呼ばれ、究極的には除去され、pegRNAの伸長によってコードされる新たに合成された置換え鎖(すなわち、「3'置換えDNAフラップ」)によって置き換えられる。
本明細書において用いられる用語「内生5'DNAフラップ」は、標的DNA上のPEによって誘導されるニック部位の直ちに下流に所在するDNAの鎖を指す。PEによる標的DNA鎖のニッキングは、ニック部位の上流側の3'ヒドロキシル基及びニック部位の下流側の5'ヒドロキシル基を暴露する。3'ヒドロキシル基で終わる内生鎖は、プライム編集因子のDNAポリメラーゼをプライムするために用いられる(例として、DNAポリメラーゼは逆転写酵素である)。ニック部位の下流側のかつ暴露された5'ヒドロキシル基で始まる内生鎖は、「5'内生DNAフラップ」と呼ばれ、究極的には除去され、pegRNAの伸長によってコードされる新たに合成された置換え鎖(すなわち、「3'置換えDNAフラップ」)によって置き換えられる。
5'内生DNAフラップ除去
本明細書に使用されるとき、用語「5'内生DNAフラップ除去」又は「5'フラップ除去」は、RTによって合成された単鎖DNAフラップが競合的に侵入して内生DNAとハイブリダイズしたときに形成される5'内生DNAフラップの除去を指し、そのプロセスにおいて内生鎖に取って代わる。取って代わられたこの内生鎖を除去することは、所望のヌクレオチド変化を含む所望の産物を形成する反応を推進し得る。細胞自身のDNA修復酵素は、5'内生フラップの除去又は切除を触媒してもよい(例として、EXO1又はFEN1等の、フラップエンドヌクレアーゼ)。また、宿主細胞も、該5'内生フラップの除去を触媒する1以上の酵素を発現するよう形質トランスバーションされていてもよく、これによって産物を形成するプロセスを推進する(例として、フラップエンドヌクレアーゼ)。フラップエンドヌクレアーゼは当該技術分野において知られており、Patel et al.,「Flap endonucleases pass 5'-flaps through a flexible arch using a disorder-thread-order mechanism to confer specificity for free 5'-ends,」Nucleic Acids Research,2012,40(10):4507-4519及びTsutakawa et al.,「Human flap endonuclease structures,DNA double-base flipping,and a unified understanding of the FEN1 superfamily,」Cell,2011,145(2):198-211(これらの各々は参照により本明細書に組み込まれる)に記載のものから見出され得る。
本明細書に使用されるとき、用語「5'内生DNAフラップ除去」又は「5'フラップ除去」は、RTによって合成された単鎖DNAフラップが競合的に侵入して内生DNAとハイブリダイズしたときに形成される5'内生DNAフラップの除去を指し、そのプロセスにおいて内生鎖に取って代わる。取って代わられたこの内生鎖を除去することは、所望のヌクレオチド変化を含む所望の産物を形成する反応を推進し得る。細胞自身のDNA修復酵素は、5'内生フラップの除去又は切除を触媒してもよい(例として、EXO1又はFEN1等の、フラップエンドヌクレアーゼ)。また、宿主細胞も、該5'内生フラップの除去を触媒する1以上の酵素を発現するよう形質トランスバーションされていてもよく、これによって産物を形成するプロセスを推進する(例として、フラップエンドヌクレアーゼ)。フラップエンドヌクレアーゼは当該技術分野において知られており、Patel et al.,「Flap endonucleases pass 5'-flaps through a flexible arch using a disorder-thread-order mechanism to confer specificity for free 5'-ends,」Nucleic Acids Research,2012,40(10):4507-4519及びTsutakawa et al.,「Human flap endonuclease structures,DNA double-base flipping,and a unified understanding of the FEN1 superfamily,」Cell,2011,145(2):198-211(これらの各々は参照により本明細書に組み込まれる)に記載のものから見出され得る。
3'置換えDNAフラップ
本明細書に使用されるとき、用語「3'置換えDNAフラップ」、又は単に「置換えDNAフラップ」は、プライム編集因子によって合成され、プライム編集因子pegRNAの伸長アームによってコードされるDNAの鎖を指す。より具体的には、3'置換えDNAフラップは、pegRNAのポリメラーゼ鋳型によってコードされる。3'置換えDNAフラップは、編集済配列(例として、単一ヌクレオチドの変化)も含有することを除くと、5'内生DNAフラップと同じ配列を含む。3'置換えDNAフラップは標的DNAとアニールして、5'内生DNAフラップ(これは、例えば、FEN1若しくはEXO1等の5'フラップエンドヌクレアーゼによって、切除され得る)に取って代わるか又はこれを置き換え、次いで、3'置換えDNAフラップの3'末を、内生DNAの晒された(5'内生DNAフラップの切除後に晒される)5'ヒドロキシル末と結び合わせるようにライゲートされ、これによってホスホジエステル結合を再形成して3'置換えDNAフラップが組み入れし、1編集済鎖と1未編集鎖とを含有するヘテロ二重鎖DNAを形成する。DNA修復プロセスはヘテロ二重鎖を分解し、編集済鎖の情報を相補鎖へコピーすることによってDNA中へ編集を永続的に組み入れする。この分解プロセスは、完了まで(to completion)、未編集鎖へニックを入れることによって、すなわち、本明細書に記載の通りの「第2鎖へのニック入れ」を経由して、更に推進され得る。
本明細書に使用されるとき、用語「3'置換えDNAフラップ」、又は単に「置換えDNAフラップ」は、プライム編集因子によって合成され、プライム編集因子pegRNAの伸長アームによってコードされるDNAの鎖を指す。より具体的には、3'置換えDNAフラップは、pegRNAのポリメラーゼ鋳型によってコードされる。3'置換えDNAフラップは、編集済配列(例として、単一ヌクレオチドの変化)も含有することを除くと、5'内生DNAフラップと同じ配列を含む。3'置換えDNAフラップは標的DNAとアニールして、5'内生DNAフラップ(これは、例えば、FEN1若しくはEXO1等の5'フラップエンドヌクレアーゼによって、切除され得る)に取って代わるか又はこれを置き換え、次いで、3'置換えDNAフラップの3'末を、内生DNAの晒された(5'内生DNAフラップの切除後に晒される)5'ヒドロキシル末と結び合わせるようにライゲートされ、これによってホスホジエステル結合を再形成して3'置換えDNAフラップが組み入れし、1編集済鎖と1未編集鎖とを含有するヘテロ二重鎖DNAを形成する。DNA修復プロセスはヘテロ二重鎖を分解し、編集済鎖の情報を相補鎖へコピーすることによってDNA中へ編集を永続的に組み入れする。この分解プロセスは、完了まで(to completion)、未編集鎖へニックを入れることによって、すなわち、本明細書に記載の通りの「第2鎖へのニック入れ」を経由して、更に推進され得る。
プライム編集の文脈において本明細書で互換的に使用される「切断部位」、「ニック部位」及び「切断部位」という用語は、二本鎖標的DNA配列中の2つのヌクレオチド又は2つの塩基対の間の特定の位置を指す。いくつかの態様において、ニック部位の位置は、特定のPAM配列の位置に対して決定される。いくつかの態様では、ニック部位は、特定のPAM配列を認識する二本鎖標的DNAが、napDNAbp、例としてCasニッカーゼ等のニッカーゼと接触されたときにニックが発生する特定の位置である。本明細書に記載の各PEgRNAについて、ニック部位は、PEgRNAのgRNAコアが会合する特定のnapDNAbpに特徴的であり、napDNAbpの認識及び機能に必要な特定のPAMに特徴的である。例えば、SpCas9と会合するgRNAコアを含むPEgRNAの場合、3塩基(PAM配列の1位に対して「-3」位置)と4塩基(PAM配列の1位に対して「-4」位置)との間のホスホジエステル結合におけるニック部位。
いくつかの態様では、ニック部位は、二本鎖標的DNA配列の標的鎖にある。いくつかの態様では、ニック部位は、二本鎖標的DNA配列の非標的鎖にある。いくつかの態様では、ニック部位はプロトスペーサ配列にある。いくつかの態様では、ニック部位はプロトスペーサ配列に隣接している。いくつかの態様では、ニック部位は、PEgRNAのプライマー結合部位に相補的な、例として、非標的鎖上の領域の下流にある。いくつかの態様では、ニック部位は、PEgRNAのプライマー結合部位に結合する領域の下流、例として、非標的鎖上にある。いくつかの態様では、ニック部位は、PEgRNAのプライマー結合部位に相補的な、例として、非標的鎖上の領域のすぐ下流にある。いくつかの態様では、ニック部位は、二本鎖標的DNAの非標的鎖上の特定のPAM配列の上流にあり、PAM配列は、PEgRNAのgRNAコアと会合するnapDNAbpによる認識に特異的である。いくつかの態様では、ニック部位は、二本鎖標的DNAの非標的鎖上の特定のPAM配列の下流にあり、PAM配列は、PEgRNAのgRNAコアと会合するnapDNAbpによる認識に特異的である。いくつかの態様では、ニック部位はPAM配列の3ヌクレオチド上流であり、PAM配列は、Streptococcus pyogene Cas9ニッカーゼ、.lavamentivorans Cas9ニッカーゼ、C.diphtheriae Cas9ニッカーゼ、N.cinerea Cas9、S.aureus Cas9、又はN.lari Cas9ニッカーゼによって認識される。いくつかの態様において、ニック部位はPAM配列の3ヌクレオチド上流にあり、PAM配列はCas9ニッカーゼによって認識され、Cas9ニッカーゼはヌクレアーゼ活性HNHドメイン及びヌクレアーゼ不活性RuvCドメインを含む。いくつかの態様において、ニック部位はPAM配列の2塩基対上流にあり、PAM配列はS.thermophilus Cas9ニッカーゼによって認識される。
特定の態様の詳細な説明
本開示は、限定するものではないが、安定性の増加、in vivoでの寿命の増加、及び/又はnapDNAbpに対する結合親和性の改善を包含する、改善された特性を有する次世代修飾pegRNAを提供する。これらの修飾されたpegRNAは、Cas9ニッカーゼドメイン及び逆転写酵素ドメインを含む融合タンパク質等のプライム編集因子と組み合わせて使用される場合、プライム編集の活性及び/又は効率の改善をもたらす。特に、本発明者らは、pegRNAが、核酸プログラム型DNA結合タンパク質(例として、Cas9ニッカーゼ)に対する親和性の低下、標準的なシングルガイドRNA(sgRNA)と比較して分解に対する感受性の増加(特に、伸長アームの劣化)、及び伸長アーム(具体的には、伸長アームのプライマー結合部位)とpegRNA中のスペーサ配列との間の望ましくない二重鎖形成による不活性化の傾向を包含する様々な欠陥を被り、それによって標的DNAの鎖に対するpegRNAのスペーサ及びプライマー結合部位の結合と競合してもよいことを発見した。理論に束縛されるものではないが、これらの問題は、標準的なsgRNA中に存在しないpegRNAの不可欠な部分である伸長アーム特徴の存在のために生じる。これらの欠点を克服するために、本発明者らは、pegRNAが、プライム編集におけるそれらの全体的な安定性及び/又は性能を改善するために1つ又はそれを超える方法で修飾されてもよいことを発見した。最初に、本発明者らは、1つ以上のRNA構造モチーフをpegRNAに付加することが、pegRNAの分解から保護し得ることを発見した。そのようなRNA構造モチーフは、限定されないが、(i)プレケオシン1-1リボスイッチアプタマー(evopreQ1)及びそのバリアント、(ii)モロニーマウス白血病ウイルス(MMLV)22由来のフレームシフトシュードノット、以下「mpknot」と呼ばれる、及びそのバリアント、(iii)G-四重鎖、(iv)ヘアピン構造(例として、15bpヘアピン)、(v)xrRNA、及び(vi)グループIイントロンのP 4-P 6ドメインを包含し得る。第2に、本発明者らは、伸長アームのプライマー結合部位(PBS)とpegRNAのスペーサ配列との間の二重鎖の形成を低減させる(すなわち、PBS/スペーサ結合相互作用の低減させる)様々な方式を発見した。一態様では、PBS/スペーサバインダー相互作用は、(i)napDNAbp(例として、Cas9ニッカーゼ)結合時に解離する足がかりでPBSを閉塞すること、(ii)3'伸長アームをトランスで提供すること、すなわち、3'伸長アーム又はその部分(例として、PBS並びに/あるいはPBS及びDNA鋳型部分)をpegRNAから別の分子、例として、ニッキングgRNAに移動させること、及び(iii)RNA/DNA二重鎖形成を促進するがRNA/RNA二重鎖形成を阻害するpegRNAへの化学修飾を導入し、それにより、pegRNAのPBSと標的DNAとの間の所望の相互作用を促進すること、を包含するがそれらに限定されない、3'伸長アームを安定化することによって回避される。まとめると、これらの戦略の実施から生じる本明細書に開示される修飾pegRNAは、本明細書では「操作された」pegRNA若しくは「epegRNA」又は同等に「修飾pegRNA」と呼ばれる。
本開示は、限定するものではないが、安定性の増加、in vivoでの寿命の増加、及び/又はnapDNAbpに対する結合親和性の改善を包含する、改善された特性を有する次世代修飾pegRNAを提供する。これらの修飾されたpegRNAは、Cas9ニッカーゼドメイン及び逆転写酵素ドメインを含む融合タンパク質等のプライム編集因子と組み合わせて使用される場合、プライム編集の活性及び/又は効率の改善をもたらす。特に、本発明者らは、pegRNAが、核酸プログラム型DNA結合タンパク質(例として、Cas9ニッカーゼ)に対する親和性の低下、標準的なシングルガイドRNA(sgRNA)と比較して分解に対する感受性の増加(特に、伸長アームの劣化)、及び伸長アーム(具体的には、伸長アームのプライマー結合部位)とpegRNA中のスペーサ配列との間の望ましくない二重鎖形成による不活性化の傾向を包含する様々な欠陥を被り、それによって標的DNAの鎖に対するpegRNAのスペーサ及びプライマー結合部位の結合と競合してもよいことを発見した。理論に束縛されるものではないが、これらの問題は、標準的なsgRNA中に存在しないpegRNAの不可欠な部分である伸長アーム特徴の存在のために生じる。これらの欠点を克服するために、本発明者らは、pegRNAが、プライム編集におけるそれらの全体的な安定性及び/又は性能を改善するために1つ又はそれを超える方法で修飾されてもよいことを発見した。最初に、本発明者らは、1つ以上のRNA構造モチーフをpegRNAに付加することが、pegRNAの分解から保護し得ることを発見した。そのようなRNA構造モチーフは、限定されないが、(i)プレケオシン1-1リボスイッチアプタマー(evopreQ1)及びそのバリアント、(ii)モロニーマウス白血病ウイルス(MMLV)22由来のフレームシフトシュードノット、以下「mpknot」と呼ばれる、及びそのバリアント、(iii)G-四重鎖、(iv)ヘアピン構造(例として、15bpヘアピン)、(v)xrRNA、及び(vi)グループIイントロンのP 4-P 6ドメインを包含し得る。第2に、本発明者らは、伸長アームのプライマー結合部位(PBS)とpegRNAのスペーサ配列との間の二重鎖の形成を低減させる(すなわち、PBS/スペーサ結合相互作用の低減させる)様々な方式を発見した。一態様では、PBS/スペーサバインダー相互作用は、(i)napDNAbp(例として、Cas9ニッカーゼ)結合時に解離する足がかりでPBSを閉塞すること、(ii)3'伸長アームをトランスで提供すること、すなわち、3'伸長アーム又はその部分(例として、PBS並びに/あるいはPBS及びDNA鋳型部分)をpegRNAから別の分子、例として、ニッキングgRNAに移動させること、及び(iii)RNA/DNA二重鎖形成を促進するがRNA/RNA二重鎖形成を阻害するpegRNAへの化学修飾を導入し、それにより、pegRNAのPBSと標的DNAとの間の所望の相互作用を促進すること、を包含するがそれらに限定されない、3'伸長アームを安定化することによって回避される。まとめると、これらの戦略の実施から生じる本明細書に開示される修飾pegRNAは、本明細書では「操作された」pegRNA若しくは「epegRNA」又は同等に「修飾pegRNA」と呼ばれる。
更に、本開示は、本明細書に開示される操作されたpegRNAと複合体化したプライム編集因子を含むプライム編集複合体、並びに該操作されたpegRNAをコードするヌクレオチド配列及び発現ベクター、並びに操作されたpegRNAを含むプライム編集複合体を提供する。更に、本開示は、標準的なpegRNA(すなわち、本明細書に記載の様式で修飾されていないそれらのpegRNA)を使用するプライム編集よりも高い編集効率を特徴とするゲノムの所望の部位に所望のヌクレオチド配列変化を組み入れするために、操作されたpegRNAと複合体化された本明細書に開示されるプライム編集融合タンパク質の使用を伴う、プライム編集に基づくゲノム編集方法を提供する。本開示はまた、本明細書に開示された修飾pegRNA、又は当該修飾pegRNAを含むプライム編集複合体を含む細胞及びキットを提供する。本開示はまた、任意選択でヌクレオチドリンカーを介して、1つ又は複数の構造ヌクレオチドモチーフ(例として、evopreQ1-1、evopreQ1-1又は修飾MMLV tRNA)をpegRNAの伸長アームの末端にカップリングさせることを含む、開示された修飾pegRNAを作製する方法を提供する。本開示は更に、所望の編集部位でゲノム編集を行うための標的細胞への修飾pegRNA及びプライム編集因子構成要素の送達方法、並びに本明細書に開示される修飾pegRNAと組み合わせてプライム編集を使用して遺伝性障害を処置する方法を提供する。
[1]プライム編集
本発明は、プライム編集の安定性、細胞寿命、Cas9(又はより広義には、napDNAbp)に対する親和性、又は標的DNAとの相互作用(例として、プライマー結合部位と標的DNAとの間の相互作用の改善)を包含する1つ又は複数の特性を改善する1つ又は複数の構造的修飾を含むように操作された、修飾又は同等の操作されたpegRNAを利用し、それによってプライム編集の編集効率を高める「プライム編集」の改良版に関する。本発明者らは、「検索及び置換え」ゲノム編集ツールとしてプライム編集を開発し、これはAnzalone et al.,「Search-and-replace genome editing without double-strand breaks or donor DNA,」Nature,October 21,2019,576,pp.149-157に更に記載されており、その内容は参照によりその全体が本明細書に組み込まれる。
本発明は、プライム編集の安定性、細胞寿命、Cas9(又はより広義には、napDNAbp)に対する親和性、又は標的DNAとの相互作用(例として、プライマー結合部位と標的DNAとの間の相互作用の改善)を包含する1つ又は複数の特性を改善する1つ又は複数の構造的修飾を含むように操作された、修飾又は同等の操作されたpegRNAを利用し、それによってプライム編集の編集効率を高める「プライム編集」の改良版に関する。本発明者らは、「検索及び置換え」ゲノム編集ツールとしてプライム編集を開発し、これはAnzalone et al.,「Search-and-replace genome editing without double-strand breaks or donor DNA,」Nature,October 21,2019,576,pp.149-157に更に記載されており、その内容は参照によりその全体が本明細書に組み込まれる。
プライム編集は、規定されたDNA部位に新たな遺伝情報を直接的に書き込む汎用的なかつ精密なゲノム編集法であり、ポリメラーゼ(すなわち、融合タンパク質の形態で、又はさもなければnapDNAbpとトランスで提供されて)と共同で働く核酸プログラム型DNA結合タンパク質(「napDNAbp」)を用いる。プライム編集システムはプライム編集(PE)ガイドRNA(「PegRNA」)(又は本開示のように、操作されたpegRNAでプログラムされた)によってプログラムされ、これは、標的部位を規定すると共に、ガイドRNA上に(例として、ガイドRNAの5'若しくは3'端に又は内部の部分に)操作された伸長(DNA又はRNAどちらか)によって置換えDNA鎖の形態での所望の編集の合成の鋳型となる。所望の編集(例として、単一の核酸塩基の置換、欠失又は挿入)を含有する置換鎖は、(所望の編集を包含することを除いて)編集される標的部位の内生鎖と同じ配列を共有する。DNA修復及び/又は複製機構を介して、標的部位の内生鎖は、所望の編集を含有する新たに合成された置換鎖によって置き換えられる。いくつかのケースでは、プライム編集は「検索置換」ゲノム編集テクノロジーと考えられてもよい。なぜなら、本明細書に記載のプライム編集因子は、編集されるべき所望の標的部位を検索し位置付けるのみならず、同時に、対応する標的部位の内生DNA鎖の代わりに組み入れされる所望の編集を含有する置換え鎖をコードするからである。
様々な態様では、プライム編集は、標的DNA分子(ヌクレオチド配列の変化が導入されることが望ましい)を、pegRNA(又は本開示のような操作されたepegRNA)と複合体化した核酸プログラム型DNA結合タンパク質(napDNAbp)と接触させることによって作動する。図1Gを参照すると、pegRNA(又はepegRNA)は、ガイドRNAの3'若しくは5'端に又はガイドRNAの分子内の位置付けに伸長を含み、所望のヌクレオチド変化(例として、1ヌクレオチド変化、挿入、又は欠失)をコードする。工程(a)において、napDNAbp/pegRNA複合体(又は本開示のようなnapDNAbp/epegRNA複合体)はDNA分子と接触し、e/pegRNAはnapDNAbpを誘導して標的遺伝子座に結合させる。工程(b)では、標的遺伝子座のDNAの鎖の1つにニックが導入され(例として、ヌクレアーゼ又は化学的薬剤による)、それによって利用可能な3'端を標的遺伝子座の鎖の1つに作成する。ある態様において、ニックは、Rループ鎖に対応するDNAの鎖、すなわちガイドRNA配列にハイブリダイズされない鎖、すなわち「非標的鎖」上に作成される。しかしながら、ニックは鎖のどちらかに導入され得る。すなわち、ニックは、Rループの「標的鎖」(すなわち、pegRNAのスペーサ配列にハイブリダイズした鎖)又は「非標的鎖」(すなわち、Rループの一本鎖部分を形成し、標的鎖に相補的な鎖)に導入され得た。工程(c)では、DNA鎖の3'端(ニックによって形成される)は、逆転写をプライムするためにガイドRNAの伸長された部分と相互作用する(すなわち、「プライム標的にプライムされたRT」)。特定の態様では、3'末端DNA鎖は、ガイドRNAの伸長部分上の特定のRTプライミング配列、すなわち「逆転写酵素プライミング配列」にハイブリダイズする。工程(d)において、プライミングされた部位の3'末端からe/pegRNAの5'末端に向かってDNAの一本鎖を合成する逆転写酵素が(napDNAbpとの融合タンパク質として又はトランスで)導入される。これは、ニック部位の又はそれに隣接する内生DNAに対してさもなければ相同的である所望のヌクレオチド変化(例として、1塩基変化、挿入、若しくは欠失、又はそれらの組み合わせ)を含む一本鎖DNAフラップを形成する。工程(e)において、napDNAbp及びe/pegRNAが放出される。工程(f)及び(g)は一本鎖DNAフラップの分解に関し、その結果、所望のヌクレオチド変化が標的遺伝子座に組み込まれるようになる。このプロセスは、3'一本鎖DNAフラップが侵入し、内生DNA配列にハイブリダイズすると形成する、対応する5'内生DNAフラップ(例として、FEN1又はトランスで提供される類似の酵素によって、プライム編集因子との融合物として、又は内因的に提供される)を除去することによって所望の産物形成に向けて推進され得る。理論によって拘束されることなしに、細胞の内生DNA修復及び複製プロセスは、ミスマッチのDNAを分解して、ヌクレオチド変化(単数又は複数)を組み込んで、所望の変改された産物を形成する。プロセスはまた、図1Gに例示されるような「第2の鎖ニッキング」、又は図1Iに例示され本明細書で論じられるような「一時的な第2の鎖ニッキング」による産物形成に向けて推進され得る。
プライム編集の別の態様では、図3Fは、典型的なpegRNA(本明細書に開示されるepegRNAで置換されてもよい)と二本鎖DNAの標的部位との相互作用、及び目的の遺伝子変化を含有する3'一本鎖DNAフラップの同時生成を示す。二本鎖DNAは、上部鎖が3'から5'方向に、下部鎖が5'から3'方向に示されている。上部鎖は、「プロトスペーサ」及びPAM配列を含み、「標的鎖」と呼ばれる。相補的な下部鎖は、「非標的鎖」と呼ばれる。示されていないが、図示されたpegRNAは、Cas9又は同等物と複合体化されるであろう。概略図に示すように、pegRNAのスペーサ配列は、PAM配列のすぐ下流に位置し、約20ヌクレオチド長であるプロトスペーサと呼ばれる標的鎖上の相補的領域にアニールする。この相互作用は、スペーサRNAとプロトスペーサDNAとの間にDNA/RNAハイブリッドを形成し、プロトスペーサとは反対側の領域にRループの形成を誘導する。本明細書の他の箇所で教示されるように、Cas9タンパク質(図示せず)は、次いで、示されるように、非標的鎖にニックを誘導する。次いで、これは、*z*に従って、プライマー結合部位でpegRNAの3'末端と相互作用する3'ssDNAフラップ領域の形成をもたらす。ssDNAフラップ(すなわち、逆転写酵素プライマー配列)の3'末端は、pegRNA上のプライマー結合部位(A)にアニーリングし、それによって逆転写酵素をプライミングする。次に、逆転写酵素(例として、トランスで提供されるか、又はCas9構築物に取り付けられた融合タンパク質として提供されるシス)は、編集鋳型(B)及び相同アーム(C)によってコードされるDNAの一本鎖を重合する(これらは一緒になってDNA合成鋳型を構成する)。重合は、伸張アームの5'末端に向かって継続する。ssDNAの重合鎖は、ssDNA 3'末端フラップを形成し、これは、他の箇所に記載されているように(例として、図1Gに示すように)、内生DNAに侵入し、対応する内生鎖を置き換え(内生DNAの5'DNAフラップとして除去される)、DNA修復/複製ラウンドを通して所望のヌクレオチド編集(単一ヌクレオチド塩基対の変化、欠失、挿入(全遺伝子を包含する))を組み入れする。
様々な態様において、プライム編集因子は、プライム編集の機構に依存する(例として、図1A~図1Fの様々な態様に示されるように)。様々な態様では、プライム編集因子は、Casタンパク質-逆転写酵素融合体又は関係するシステムを含んで、ガイドRNAによって特定のDNA配列を標的化し、標的部位に一本鎖ニックを生成し、ガイドRNAに取り入れされた操作された逆転写酵素鋳型の逆転写のためのプライマーとして、ニックが入ったDNAを用いた。本明細書に記載されるプライム編集因子は、逆転写酵素に限定されず、実質的に任意のDNAポリメラーゼの使用を包含してもよい。実に、本明細書は、至る所で、「逆転写酵素」を有するプライム編集因子を指してもよいが、逆転写酵素は、プライム編集で働いてもよいDNAポリメラーゼの1つの型であるのみであるということがここで明示される。それゆえに、本明細書が「逆転写酵素」に言及する場合、当業者は、いずれかの好適なDNAポリメラーゼが逆転写酵素の代わりに用いられてもよいということを了解するべきである。それゆえに、一側面において、プライム編集因子はCas9(又は同等のnapDNAbp)を含んでもよく、これは、標的DNA上の相補的なプロトスペーサにアニールするスペーサ配列を含有する特殊化したガイドRNA(すなわちpegRNA)とそれを会合させることによって、DNA配列を標的化するようにプログラムされる。特殊化したガイドRNAは、所望の遺伝子変改を含有するDNAの置換え鎖をコードする伸長の形態の新たな遺伝情報をもまた含有し、これは、標的部位における対応する内生DNA鎖を置き換えるために用いられる。pegRNAからの情報を標的DNAに移入するために、プライム編集の機序には、DNAの1つの鎖上の標的部位へニックを入れて3'ヒドロキシル基を暴露することを伴う。次いで、暴露された3'ヒドロキシル基は、直接的に標的部位へのpegRNA上の編集をコードする伸長のDNA重合をプライムするために用いられ得る。種々の態様において、編集を含有する置換え鎖の重合のための鋳型を提供する伸長は、RNA又はDNAから形成され得る。RNA伸長のケースでは、プライム編集因子のポリメラーゼはRNA依存性DNAポリメラーゼ(例えば逆転写酵素)であり得る。DNA伸長のケースでは、プライム編集因子のポリメラーゼはDNA依存性DNAポリメラーゼであってもよい。
本明細書に開示のプライム編集因子によって形成される新たに合成された鎖(すなわち、所望の編集を含有する置換えDNA鎖)は、所望のヌクレオチド変化(例として、1ヌクレオチド変化、欠失、若しくは挿入、又はそれらの組み合わせ)の包含を例外として、ゲノム標的配列に対して相同的である(すなわち、それと同じ配列を有する)であろう。DNAの新たに合成された(又は置き換え)鎖は一本鎖DNAフラップともまた呼ばれてもよい。これは相補的な相同的な内生DNA鎖とのハイブリダイゼーションについて競合し、それによって対応する内生鎖に取って代わるであろう。ある態様において、システムは、エラーを起こしやすい逆転写酵素酵素の使用と組み合わせられ得る(例として、Cas9ドメインとの融合タンパク質として提供されるか、又はCas9ドメインに対してトランスで提供される)。エラーを起こしやすい逆転写酵素酵素は一本鎖DNAフラップの合成の間に変改を導入し得る。それゆえに、ある態様において、エラーを起こしやすい逆転写酵素は、標的DNAへのヌクレオチド変化を導入するために利用され得る。システムに用いられるエラーを起こしやすい逆転写酵素に依存して、変化はランダム又は非ランダムであり得る。
ハイブリダイズした中間体(内生DNA鎖にハイブリダイズした逆転写酵素によって合成される一本鎖DNAフラップを含む)の分解は、内生DNAのもたらされる取って代わられたフラップの除去(例として、5'端DNAフラップエンドヌクレアーゼFEN1による)、標的DNAに対する合成された一本鎖DNAフラップのライゲーション、並びに細胞性のDNA修復及び/又は複製プロセスの結果としての所望のヌクレオチド変化の同化を包含し得る。鋳型によるDNA合成は挿入及び欠失を包含するいずれかのヌクレオチドの修飾について1ヌクレオチド精度をオファーするので、このアプローチの範囲は非常に幅広く、予見可能には、基礎研究及び処置学において無数の適用に用いられ得る。
プライム編集のこれらの態様のそれぞれにおいて、本明細書中に記載される修飾又は操作されたpegRNAは、プライム編集の編集効率を高めるために標準的なpegRNAの代わりに使用され得る。理論に束縛されるものではないが、編集効率の増加は、改善されたpegRNA安定性、pegRNAの改善された細胞寿命、pegRNAに対するCas9の結合親和性の増加、又はプライマー結合部位とepegRNAのスペーサとの間の結合相互作用の減少(したがって、プライマー結合部位と標的DNAとの間のより良好な相互作用)のうちのいずれか1つ以上に由来されると考えられる。
ここで、詳細な説明は、本明細書中で企図され、プライム編集の編集効率を高めるために本明細書中に記載される修飾又は操作されたpegRNAと共に使用されてもよいプライム編集因子の様々な構成要素を記載する。
[2]napDNAbp
本明細書に記載のプライム編集因子は、核酸プログラム型DNA結合タンパク質(napDNAbp)を含んでもよい。
本明細書に記載のプライム編集因子は、核酸プログラム型DNA結合タンパク質(napDNAbp)を含んでもよい。
一側面では、napDNAbpは、ガイド核酸又はその一部(例として、DNA標的のプロトスペーサにアニーリングするガイドRNAのスペーサ)に相補的なDNA鎖(すなわち、標的鎖)を含むDNA配列にnapDNAbpを局在化する少なくとも1つのガイド核酸(例として、ガイドRNA又はpegRNA)と会合又は複合体化され得る。換言すれば、ガイド核酸は、DNA中のプロトスペーサの相補的配列に局在して結合するようにnapDNAbp(例として、Cas9又は同等物)を「プログラム」する。
任意の適切なnapDNAbpは、本明細書に記載のプライム編集因子で使用されてもよい。様々な態様では、napDNAbpは、任意のII型、V型、又はVI型CRISPR-Cas酵素を包含する、任意のクラス2 CRISPR-Casシステムであってもよい。ゲノム編集のためのツールとしてのCRISPR-Casの急速な発展を考えると、Cas9及びCas9オルソログ等のCRISPR-Cas酵素を記載及び/又は同定するために使用される命名法が絶えず開発されてきた。本出願は、古い及び/又は新しくてもよい命名法を有するCRISPR-Cas酵素を参照する。当業者は、使用されている命名法に基づいて、それが古い(すなわち、「レガシー」)命名法であるか新しい命名法であるかにかかわらず、本出願で言及されている特定のCRISPR-Cas酵素を同定することができる。CRISPR-Cas命名法は、Makarova et al.,「Classification and Nomenclature of CRISPR-Cas Systems:Where from Here?,」The CRISPR Journal,Vol.1.No.5,2018で広く論じられており、これらの内容は参照によって本明細書に組み込まれる。本出願の任意の所与の例で使用される特定のCRISPR-Cas命名法は、決して限定するものではなく、当業者は、どのCRISPR-Cas酵素が参照されているかを識別することができる。
例えば、以下のタイプII、タイプV及びタイプVIクラス2 CRISPR-Cas酵素は、当技術分野で認識されている以下の古い(すなわち、レガシー)及び新しい名称を有する。これらの酵素及び/又はそのバリアントのそれぞれは、本明細書に記載のプライム編集因子と使用されてもよい。

理論に束縛されるものではないが、本明細書で企図される特定のnapDNAbpの作用機序は、Rループを形成し、それによってnapDNAbpが二本鎖DNA標的の巻き戻しを誘導し、それによってnapDNAbpによって結合された領域内の鎖を分離する工程を包含する。次いで、ガイドRNAスペーサは、プロトスペーサ配列に相補的な領域で「標的鎖」にハイブリダイズする。これは、標的鎖に相補的な「非標的鎖」に取って代わり、これはRループの単鎖領域を形成する。いくつかの態様において、napDNAbpは1以上のヌクレアーゼ活性を包含しており、次いでDNAを切ることで、様々な型の病変がなくなる。例えば、napDNAbpは、非標的鎖を第1の場所にて切るか、及び/又は標的鎖を第2の場所にて切る、ヌクレアーゼ活性を含んでいてもよい。ヌクレアーゼ活性に応じて標的DNAが切られることで、両鎖とも切られた「二本鎖切断」を形成し得る。他の態様において、標的DNAは単一の部位でしか切られ得ない、すなわち、DNAは一方の鎖上に「ニックが入っている(nicked)」。種々のヌクレアーゼ活性をもつ例示のnapDNAbpは、「Cas9ニッカーゼ」(「nCas9」)、及びヌクレアーゼ活性を一切有さない不活性化Cas9(「不活性型Cas9」又は「dCas9」)を包含する。
本開示のプライム編集因子に関して用いられ得る種々のnapDNAbpの下の記載は、決して限定することを意味されない。プライム編集因子は、公知であるか又は指向性進化若しくは別様の変異プロセスによって作られ得るか若しくは進化させられ得る標準的なSpCas9又はいずれかのオーソログCas9タンパク質又はいずれかのバリアントCas9タンパク質を含み得、Cas9のいずれかの天然に存在するバリアント、変異体、又は別様に操作されたバージョンを包含する。種々の態様において、Cas9又はCas9バリアントはニッカーゼ活性を有し、すなわち標的DNA配列の一方の鎖のみを切断する。他の態様において、Cas9又はCas9バリアントは不活性なヌクレアーゼを有し、すなわち「不活性型」Cas9タンパク質である。用いられてもよい他のバリアントCas9タンパク質は、(例として、より容易な送達のために)標準的なSpCas9よりも小さい分子量を有するか又は修飾若しくは再配置された一次アミノ酸構造(例として循環置換体フォーマット)を有するものである。
本明細書に記載のプライム編集因子は、収斂進化の結果であるCas12a(Cpf1)及びCas12b1タンパク質を包含するCas9等価物をもまた含んでもよい。本明細書において用いられるnapDNAbp(例として、SpCas9、Cas9バリアント、又はCas9等価物)は、それらのPAM特異性を変改/増強する種々の修飾をもまた含有してもよい。最後に、本明細書は、少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも91%、少なくとも92%、少なくとも93%、少なくとも94%、少なくとも95%、少なくとも96%、少なくとも97%、少なくとも98%、少なくとも99%、又は少なくとも99.9%配列同一性を参照Cas9配列、例えば参照SpCas9標準的な配列又は参照Cas9等価物(例として、Cas12a(Cpf1))に対して有するいずれかのCas9、Cas9バリアント、又はCas9等価物を企図する。
napDNAbpは、CRISPR(clustered regularly interspaced short palindromic repeat)関連ヌクレアーゼであり得る。上に概説したように、CRISPRは、可動遺伝因子(ウイルス、転移因子、及び接合性プラスミド)に対して保護を提供する適応免疫系である。CRISPRクラスターは、スペーサ、先行する可動因子に相補的な配列、及び標的化侵入核酸を含有する。CRISPRクラスターは転写され、プロセスを受けて(processed)CRISPR RNA(crRNA)になる。II型CRISPR系において、pre-crRNAの修正プロセシングは、transでコードされた低分子(small)RNA(tracrRNA)、内生リボヌクレアーゼ3(rnc)、及びCas9タンパク質を要する。tracrRNAは、pre-crRNAの、リボヌクレアーゼ3に補助された(-aided)プロセシングのためのガイドとして働く。続いて、Cas9/crRNA/tracrRNAは、スペーサに相補的な線状又は環状のdsDNA標的をエンドヌクレアーゼ的に切断する。crRNAに相補的ではない標的鎖は、最初にエンドヌクレアーゼ的に切られ、次いで3'-5'エキソヌクレアーゼ的に切り取られる。本来、DNAの結合及び切断は、典型的には、タンパク質及び両RNAを要する。しかしながら、単一ガイドRNA(「sgRNA」、又は単に「gRNA」)は、crRNAとtracrRNAとの両方の側面を単一RNA種中へ組み込むために、操作され得る。例として、その全内容が参照により本明細書に組み込まれるJinek M.et al.,Science 337:816-821(2012)を参照されたい。
いくつかの態様では、napDNAbpは、標的配列上において及び/又は標的配列の相補体上等、標的配列の位置付けにおける一方又は両方の鎖の切断を導く。いくつかの態様では、napDNAbpは、標的配列の最初の又は最後のヌクレオチドから約1、2、3、4、5、6、7、8、9、10、15、20、25、50、100、200、500、又はより多くの塩基対以内において、一方又は両方の鎖の切断を導く。いくつかの態様では、ベクターは、変異したnapDNAbpが標的配列を含有する標的ポリヌクレオチドの一方又は両方の鎖を切断する能力を欠くようにして対応する野生型酵素に対して変異しているnapDNAbpをコードする。例えば、S.pyogenesからのCas9のRuvC I触媒ドメイン上のアスパラギン酸からアラニンの置換(D10A)は、両鎖を切断するヌクレアーゼからニッカーゼ(一本鎖を切断する)へとCas9を変換する。Cas9をニッカーゼにする変異の他の例は、限定なしに、標準的なSpCas9配列を又は他のCas9バリアント若しくはCas9等価物の同等のアミノ酸位置を参照して、H840A、N854A、及びN863Aを包含する。
本明細書において用いられる用語「Casタンパク質」は、天然から得られる全長Casタンパク質、天然に存在するCasタンパク質とは異なる配列を有する組換えCasタンパク質、又は本開示の方法に必要とされる全て若しくは有意な量の不可欠の基本機能、すなわち(i)標的DNAに対するCasタンパク質の核酸プログラム型結合の所有及び(ii)1つの鎖上で標的DNA配列へニックを入れる能力をそれでもなお保持するCasタンパク質のいずれかのフラグメントを指す。本明細書において企図されるCasタンパク質は、CRISPR Cas9タンパク質、及びCas9等価物、バリアント(例として、Cas9ニッカーゼ(nCas9)又はヌクレアーゼ不活性型Cas9(dCas9))ホモログ、オーソログ、又はパラログを、天然に存在するか又は天然に存在しないか(例として、操作されたか又は組み換えか)にかかわらず包摂し、いずれかのクラス2 CRISPRシステム(例としてII、V、VI型)からのCas9等価物を包含してもよく、Cas12a(Cpf1)、Cas12e(CasX)、Cas12b1(C2c1)、Cas12b2、Cas12c(C2c3)、C2c4、C2c8、C2c5、C2c10、C2c9 Cas13a(C2c2)、Cas13d、Cas13c(C2c7)、Cas13b(C2c6)、及びCas13bを包含する。更なるCas等価物はMakarova et al.,「C2c2 is a single-component programmable RNA-guided RNA-targeting CRISPR effector,」Science 2016;353(6299)及びMakarova et al.,「Classification and Nomenclature of CRISPR-Cas Systems:Where from Here?,」The CRISPR Journal,Vol.1.No.5,2018に記載されている。これらの内容は参照によって本明細書に組み込まれる。
用語「Cas9」又は「Cas9ヌクレアーゼ」又は「Cas9部分」又は「Cas9ドメイン」は、いずれかの生物からのいずれかの天然に存在するCas9、いずれかの天然に存在するCas9等価物又はその機能的フラグメント、いずれかの生物からのいずれかのCas9ホモログ、オーソログ、又はパラログ、及び天然に存在するか又は操作されたCas9のいずれかの変異体又はバリアントを包摂する。用語Cas9は特に限定することを意味されず、「Cas9又は等価物」と呼ばれてもよい。例示のCas9タンパク質は更に本明細書に記載及び/又は当該技術分野において記載されており、参照によって本明細書に組み込まれる。本開示は、本発明のプライム編集因子(PE)に使用される具体的なCas9について限定されない。
本明細書に記されるCas9ヌクレアーゼ配列及び構造は当業者には周知である(例として、「Complete genome sequence of an M1 strain of Streptococcus pyogenes.」Ferretti et al.,J.J.,McShan W.M.,Ajdic D.J.,Savic D.J.,Savic G.,Lyon K.,Primeaux C.,Sezate S.,Suvorov A.N.,Kenton S.,Lai H.S.,Lin S.P.,Qian Y.,Jia H.G.,Najar F.Z.,Ren Q.,Zhu H.,Song L.,White J.,Yuan X.,Clifton S.W.,Roe B.A.,McLaughlin R.E.,Proc.Natl.Acad.Sci.U.S.A.98:4658-4663(2001);「CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III.」Deltcheva E.,Chylinski K.,Sharma C.M.,Gonzales K.,Chao Y.,Pirzada Z.A.,Eckert M.R.,Vogel J.,Charpentier E.,Nature 471:602-607(2011);及び「A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity.」Jinek M.,Chylinski K.,Fonfara I.,Hauer M.,Doudna J.A.,Charpentier E.Science 337:816-821(2012)(これら各々の内容全体は参照により本明細書に組み込まれる)を参照)。
Cas9及びCas9等価物の例が次の通り提供される;しかしながら、これらの特定の例は限定することを意味されない。本開示のプライム編集因子は、いずれかの好適なCas9又はCas9等価物を包含するいずれかの好適なnapDNAbpを使用してもよい。
A.標準的な野生型SpCas9
1つの態様では、本明細書に記載のプライム編集因子構築物は、S.pyogenesからの「標準的なSpCas9」ヌクレアーゼを含んでもよい。これはゲノム工学のツールとして広く用いられており、クラス2 CRISPR-Casシステムの酵素のII型サブグループとして類別される。このCas9タンパク質は、2つの別物のヌクレアーゼドメインを含有する大きいマルチドメインタンパク質である。1つの又は両方のヌクレアーゼ活性を廃止するための点変異がCas9上に導入され得、夫々ニッカーゼCas9(nCas9)又は不活性型Cas9(dCas9)をもたらす。これは、sgRNAによってプログラムされる様式でDNAに結合するその能力をなお保持する。原理的には、別のタンパク質又はドメインに融合されるときに、Cas9又はそのバリアント(例としてnCas9)は、単純に適当なsgRNAとの共発現によって、そのタンパク質を実質的にいずれかのDNA配列へと標的化し得る。本明細書において用いられる標準的なSpCas9タンパク質は、次のアミノ酸配列を有するStreptococcus pyogenesからの野生型タンパク質を指す:


1つの態様では、本明細書に記載のプライム編集因子構築物は、S.pyogenesからの「標準的なSpCas9」ヌクレアーゼを含んでもよい。これはゲノム工学のツールとして広く用いられており、クラス2 CRISPR-Casシステムの酵素のII型サブグループとして類別される。このCas9タンパク質は、2つの別物のヌクレアーゼドメインを含有する大きいマルチドメインタンパク質である。1つの又は両方のヌクレアーゼ活性を廃止するための点変異がCas9上に導入され得、夫々ニッカーゼCas9(nCas9)又は不活性型Cas9(dCas9)をもたらす。これは、sgRNAによってプログラムされる様式でDNAに結合するその能力をなお保持する。原理的には、別のタンパク質又はドメインに融合されるときに、Cas9又はそのバリアント(例としてnCas9)は、単純に適当なsgRNAとの共発現によって、そのタンパク質を実質的にいずれかのDNA配列へと標的化し得る。本明細書において用いられる標準的なSpCas9タンパク質は、次のアミノ酸配列を有するStreptococcus pyogenesからの野生型タンパク質を指す:
本明細書に記載のプライム編集因子は、標準的なSpCas9、又は上で提供されている野生型Cas9配列との少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、又は少なくとも99%配列同一性を有するそのいずれかのバリアントを包含してもよい。これらのバリアントは、1以上の変異を含有するSpCas9バリアントを包含し得、SwissProtアクセッションNo.Q99ZW2(配列番号37)エントリーで報告されているいずれかの公知の変異を包含してもよい。これらは以下を包含する。

本開示に用いられてもよい他の野生型SpCas9配列は以下を包含する。




本明細書に記載のプライム編集因子は、上のSpCas9配列のいずれか、又は少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、又は少なくとも99%配列同一性をそれに対して有するそのいずれかのバリアントを包含してもよい。
B.野生型Cas9オーソログ
他の態様において、Cas9タンパク質は、S.pyogenesからの標準的なCas9とは異なる別の細菌種からの野生型Cas9オーソログであり得る。例えば、次のCas9オーソログが本明細書に記載されるプライム編集因子構築物に関して用いられ得る。加えて、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、又は少なくとも99%配列同一性を下のオーソログのいずれかに対して有するいずれかのバリアントCas9オーソログもまた本プライム編集因子に用いられてもよい。




他の態様において、Cas9タンパク質は、S.pyogenesからの標準的なCas9とは異なる別の細菌種からの野生型Cas9オーソログであり得る。例えば、次のCas9オーソログが本明細書に記載されるプライム編集因子構築物に関して用いられ得る。加えて、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、又は少なくとも99%配列同一性を下のオーソログのいずれかに対して有するいずれかのバリアントCas9オーソログもまた本プライム編集因子に用いられてもよい。
本明細書に記載のプライム編集因子は、上のCas9オーソログ配列のいずれか、又は少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、又は少なくとも99%配列同一性をそれに対して有するそのいずれかのバリアントを包含してもよい。
napDNAbpは、Cas9等の天然に存在する酵素の(or)いずれかの好適なホモログ及び/又はオーソログを包含してもよい。Cas9ホモログ及び/又はオーソログはS.pyogenes及びS.thermophilusを包含するがこれらに限定されない種々の種において記載されている。好ましくは、Cas部分はニッカーゼとして構成される(例として、変異導入されるか、組換え操作されるか、又は別様に天然から得られる)。すなわち、標的二本鎖DNAの一本鎖のみを切断することができる。追加の好適なCas9ヌクレアーゼ及び配列は、本開示に基づき当業者に明らかであろう。またかかるCas9ヌクレアーゼ及び配列は、Chylinski,Rhun,and Charpentier,「The tracrRNA and Cas9 families of type II CRISPR-Cas immunity systems」(2013)RNA Biology 10:5,726-737(この内容全体は参照により本明細書に組み込まれる)に開示された生物及び遺伝子座からのCas9配列を包含する。いくつかの態様では、Cas9ヌクレアーゼは不活性な(例として不活性化された)DNA切断ドメインを有する。つまり、Cas9はニッカーゼである。いくつかの態様では、Cas9タンパク質は、表3のバリアントのいずれか1つによって提供されるCas9タンパク質のアミノ酸配列と少なくとも80%同一であるアミノ酸配列を含む。いくつかの態様では、Cas9タンパク質は、上の表のCas9オーソログのいずれか1つによって提供されるCas9タンパク質のアミノ酸配列と少なくとも85%、少なくとも90%、少なくとも92%、少なくとも95%、少なくとも96%、少なくとも97%、少なくとも98%、少なくとも99%、又は少なくとも99.5%同一であるアミノ酸配列を含む。
C.不活性型napDNAbpバリアント
いくつかの態様では、開示されるプライム編集因子は、触媒的に不活性な又は「不活性型」napDNAbpドメインを含んでもよい。ある態様において、本明細書に記載の多重フラッププライム編集因子は、不活性型Cas9、例として不活性型SpCas9を包含してもよい。これは、Cas9の両方のヌクレアーゼドメイン、つまりRuvCドメイン(これは、非プロトスペーサDNA鎖を切断する)及びHNHドメイン(これはプロトスペーサDNA鎖を切断する)を不活性化する1以上の変異を原因として、ヌクレアーゼ活性を有さない。ヌクレアーゼ不活性化は、コードされるタンパク質、又は少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、若しくは少なくとも99%配列同一性をそれに対して有するそのいずれかのバリアントのアミノ酸配列に1以上の置換及び/又は欠失をもたらす1以上の変異を原因としてもよい。
いくつかの態様では、開示されるプライム編集因子は、触媒的に不活性な又は「不活性型」napDNAbpドメインを含んでもよい。ある態様において、本明細書に記載の多重フラッププライム編集因子は、不活性型Cas9、例として不活性型SpCas9を包含してもよい。これは、Cas9の両方のヌクレアーゼドメイン、つまりRuvCドメイン(これは、非プロトスペーサDNA鎖を切断する)及びHNHドメイン(これはプロトスペーサDNA鎖を切断する)を不活性化する1以上の変異を原因として、ヌクレアーゼ活性を有さない。ヌクレアーゼ不活性化は、コードされるタンパク質、又は少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、若しくは少なくとも99%配列同一性をそれに対して有するそのいずれかのバリアントのアミノ酸配列に1以上の置換及び/又は欠失をもたらす1以上の変異を原因としてもよい。
本明細書において用いられる用語「dCas9」は、ヌクレアーゼ不活性(inactive)Cas9若しくはヌクレアーゼ不活性型(dead)Cas9又はその機能的フラグメントを指し、いずれかの生物からのいずれかの天然に存在するdCas9、いずれかの天然に存在するdCas9等価物又はその機能的フラグメント、いずれかの生物からのいずれかのdCas9ホモログ、オーソログ、又はパラログ、及び天然に存在するか又は操作されたdCas9のいずれかの変異体又はバリアントを包摂する。用語dCas9は特に限定することを意味されず、「dCas9又は等価物」と呼ばれてもよい。例示のdCas9タンパク質とdCas9タンパク質を作るための方法とは、更に本明細書に記載及び/又は当該技術分野において記載されており、参照によって本明細書に組み込まれる。
他の態様において、dCas9は、部分的に又は丸ごとで、Cas9ヌクレアーゼ活性を不活性化する1以上の変異を有するCas9アミノ酸配列に対応するか又はそれを含む。他の態様において、D10A及びH840A以外の変異を有するCas9バリアントが提供され、これらは内生Cas9ヌクレアーゼ活性の完全な又は部分的な不活性化物をもたらしてもよい(例として、夫々nCas9又はdCas9)。かかる変異は、例として、D10及びH820における他のアミノ酸置換、又はCas9のヌクレアーゼドメイン上の他の置換(例えば、HNHヌクレアーゼサブドメイン及び/又はRuvC1サブドメイン上の置換)を、Streptococcus pyogenesからのCas9(NCBI参照配列:NC_017053.1)等の野生型配列を参照して包含する。いくつかの態様では、Cas9のバリアント又はホモログ(例として、Streptococcus pyogenesからのCas9(NCBI参照配列:NC_017053.1(配列番号39))のバリアント)が提供され、これらは、NCBI参照配列:NC_017053.1と少なくとも約70%同一、少なくとも約80%同一、少なくとも約90%同一、少なくとも約95%同一、少なくとも約98%同一、少なくとも約99%同一、少なくとも約99.5%同一、又は少なくとも約99.9%同一である。いくつかの態様では、dCas9のバリアント(例として、NCBI参照配列:NC_017053.1(配列番号39)のバリアント)が提供され、NC_017053.1(配列番号39)よりも約5アミノ酸、約10アミノ酸、約15アミノ酸、約20アミノ酸、約25アミノ酸、約30アミノ酸、約40アミノ酸、約50アミノ酸、約75アミノ酸、約100アミノ酸、又はより多くだけ短いか又は長いアミノ酸配列を有する。
1つの態様では、不活性型Cas9はQ99ZW2の標準的なSpCas9配列に基づいてもよく、D10X及びH810X置換(下線付きかつ太字)を含む次の配列を有してもよく、Xはいずれかのアミノ酸であってもよく、又はバリアントは少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、若しくは少なくとも99%配列同一性をそれに対して有する配列番号57のバリアントであり得る。
1つの態様では、不活性型Cas9はQ99ZW2の標準的なSpCas9配列に基づいてもよく、D10A及びH810A置換(下線付きかつ太字)を含む次の配列を有し得、又は少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、若しくは少なくとも99%配列同一性をそれに対して有する配列番号58のバリアントであってもよい。

D.napDNAbpニッカーゼバリアント
いくつかの態様では、開示される塩基編集因子は、ニッカーゼを含むnapDNAbpドメインを含んでもよい。1つの態様において、本明細書に記載のプライム編集因子はCas9ニッカーゼを含む。用語「Cas9ニッカーゼ」又は「nCas9」は、二本鎖DNA分子標的上に一本鎖切断を導入することができるCas9のバリアントを指す。いくつかの態様では、Cas9ニッカーゼは単一の機能するヌクレアーゼドメインのみを含む。野生型Cas9(例として標準的なSpCas9)は、2つの別個のヌクレアーゼドメイン、つまりRuvCドメイン(これは非プロトスペーサDNA鎖を切断する)及びHNHドメイン(これはプロトスペーサDNA鎖を切断する)を含む。1つの態様では、Cas9ニッカーゼはRuvCヌクレアーゼ活性を不活性化するRuvCドメインの変異を含む。例えば、アスパラギン酸(D)10、ヒスチジン(H)983、アスパラギン酸(D)986、又はグルタミン酸(E)762の変異は、RuvCヌクレアーゼドメインの機能喪失変異及び機能的なCas9ニッカーゼの作成として報告されている(例として、Nishimasu et al.,「Crystal structure of Cas9 in complex with guide RNA and target DNA,」Cell 156(5),935-949。これは参照によって本明細書に組み込まれる)。それゆえに、RuvCドメインのニッカーゼ変異はD10X、H983X、D986X、又はE762Xを包含し得、Xは野生型アミノ酸以外のいずれかのアミノ酸である。ある態様において、ニッカーゼはD10A、若しくは(of)H983A、若しくはD986A、若しくはE762A、又はそれらの組み合わせであり得る。
いくつかの態様では、開示される塩基編集因子は、ニッカーゼを含むnapDNAbpドメインを含んでもよい。1つの態様において、本明細書に記載のプライム編集因子はCas9ニッカーゼを含む。用語「Cas9ニッカーゼ」又は「nCas9」は、二本鎖DNA分子標的上に一本鎖切断を導入することができるCas9のバリアントを指す。いくつかの態様では、Cas9ニッカーゼは単一の機能するヌクレアーゼドメインのみを含む。野生型Cas9(例として標準的なSpCas9)は、2つの別個のヌクレアーゼドメイン、つまりRuvCドメイン(これは非プロトスペーサDNA鎖を切断する)及びHNHドメイン(これはプロトスペーサDNA鎖を切断する)を含む。1つの態様では、Cas9ニッカーゼはRuvCヌクレアーゼ活性を不活性化するRuvCドメインの変異を含む。例えば、アスパラギン酸(D)10、ヒスチジン(H)983、アスパラギン酸(D)986、又はグルタミン酸(E)762の変異は、RuvCヌクレアーゼドメインの機能喪失変異及び機能的なCas9ニッカーゼの作成として報告されている(例として、Nishimasu et al.,「Crystal structure of Cas9 in complex with guide RNA and target DNA,」Cell 156(5),935-949。これは参照によって本明細書に組み込まれる)。それゆえに、RuvCドメインのニッカーゼ変異はD10X、H983X、D986X、又はE762Xを包含し得、Xは野生型アミノ酸以外のいずれかのアミノ酸である。ある態様において、ニッカーゼはD10A、若しくは(of)H983A、若しくはD986A、若しくはE762A、又はそれらの組み合わせであり得る。
種々の態様において、Cas9ニッカーゼはRuvCヌクレアーゼドメインに変異を有し得、次のアミノ酸配列の1つ、又は少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、若しくは少なくとも99%配列同一性をそれに対して有するアミノ酸配列を有するそのバリアントを有し得る。



別の態様では、Cas9ニッカーゼはHNHヌクレアーゼ活性を不活性化するHNHドメインの変異を含む。例えば、ヒスチジン(H)840又はアスパラギン(R)863の変異は、HNHヌクレアーゼドメインの機能喪失変異及び機能的なCas9ニッカーゼの作成として報告されている(例として、Nishimasu et al.,「Crystal structure of Cas9 in complex with guide RNA and target DNA,」Cell 156(5),935-949。これは参照によって本明細書に組み込まれる)。それゆえに、HNHドメインのニッカーゼ変異はH840X及びR863Xを包含し得、Xは野生型アミノ酸以外のいずれかのアミノ酸である。ある態様において、ニッカーゼはH840A若しくはR863A又はそれらの組み合わせであり得る。
種々の態様において、Cas9ニッカーゼはHNHヌクレアーゼドメインの変異を有し得、次のアミノ酸配列の1つ、又は少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、又は少なくとも99%配列同一性をそれに対して有するアミノ酸配列を有するそのバリアントを有し得る。


いくつかの態様では、N末端メチオニンが、Cas9ニッカーゼから、又は本明細書において開示若しくは企図されるいずれかのCas9バリアント、オーソログ、若しくは等価物から除去される。例えば、メチオニンマイナスCas9ニッカーゼは、次の配列、又は少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、若しくは少なくとも99%配列同一性をそれらに対して有するアミノ酸配列を有するそれらのバリアントを包含する。


E.他のCas9バリアント
不活性型Cas9及びCas9ニッカーゼバリアントの他に、本明細書において用いられるCas9タンパク質は、本明細書に開示のか又は当該技術分野において公知のいずれかの野生型Cas9又は変異体Cas9(例として、不活性型Cas9又はCas9ニッカーゼ)、又はフラグメントCas9、又は循環置換体Cas9、又はCas9の他のバリアントを包含するいずれかの参照Cas9タンパク質と少なくとも約70%同一、少なくとも約80%同一、少なくとも約90%同一、少なくとも約95%同一、少なくとも約96%同一、少なくとも約97%同一、少なくとも約98%同一、少なくとも約99%同一、少なくとも約99.5%同一、又は少なくとも約99.9%同一である他の「Cas9バリアント」をもまた包含してもよい。いくつかの態様では、Cas9バリアントは、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、21、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、又はより多くのアミノ酸変化を、参照Cas9と比較して有してもよい。いくつかの態様では、Cas9バリアントは参照Cas9のフラグメント(例として、gRNA結合ドメイン又はDNA切断ドメイン)を含み、その結果、フラグメントは、野生型Cas9の対応するフラグメントと少なくとも約70%同一、少なくとも約80%同一、少なくとも約90%同一、少なくとも約95%同一、少なくとも約96%同一、少なくとも約97%同一、少なくとも約98%同一、少なくとも約99%同一、少なくとも約99.5%同一、又は少なくとも約99.9%同一である。いくつかの態様では、フラグメントは、対応する野生型Cas9(例として配列番号37)のアミノ酸長さの少なくとも30%、少なくとも35%、少なくとも40%、少なくとも45%、少なくとも50%、少なくとも55%、少なくとも60%、少なくとも65%、少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%同一、少なくとも96%、少なくとも97%、少なくとも98%、少なくとも99%、又は少なくとも99.5%である。
不活性型Cas9及びCas9ニッカーゼバリアントの他に、本明細書において用いられるCas9タンパク質は、本明細書に開示のか又は当該技術分野において公知のいずれかの野生型Cas9又は変異体Cas9(例として、不活性型Cas9又はCas9ニッカーゼ)、又はフラグメントCas9、又は循環置換体Cas9、又はCas9の他のバリアントを包含するいずれかの参照Cas9タンパク質と少なくとも約70%同一、少なくとも約80%同一、少なくとも約90%同一、少なくとも約95%同一、少なくとも約96%同一、少なくとも約97%同一、少なくとも約98%同一、少なくとも約99%同一、少なくとも約99.5%同一、又は少なくとも約99.9%同一である他の「Cas9バリアント」をもまた包含してもよい。いくつかの態様では、Cas9バリアントは、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、21、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、又はより多くのアミノ酸変化を、参照Cas9と比較して有してもよい。いくつかの態様では、Cas9バリアントは参照Cas9のフラグメント(例として、gRNA結合ドメイン又はDNA切断ドメイン)を含み、その結果、フラグメントは、野生型Cas9の対応するフラグメントと少なくとも約70%同一、少なくとも約80%同一、少なくとも約90%同一、少なくとも約95%同一、少なくとも約96%同一、少なくとも約97%同一、少なくとも約98%同一、少なくとも約99%同一、少なくとも約99.5%同一、又は少なくとも約99.9%同一である。いくつかの態様では、フラグメントは、対応する野生型Cas9(例として配列番号37)のアミノ酸長さの少なくとも30%、少なくとも35%、少なくとも40%、少なくとも45%、少なくとも50%、少なくとも55%、少なくとも60%、少なくとも65%、少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%同一、少なくとも96%、少なくとも97%、少なくとも98%、少なくとも99%、又は少なくとも99.5%である。
いくつかの態様では、本開示は、本明細書に開示のいずれかのCas9タンパク質のフラグメントであるそれらの機能性を保持するCas9フラグメントをもまた利用してもよい。いくつかの態様では、Cas9フラグメントは長さが少なくとも100アミノ酸である。いくつかの態様では、フラグメントは長さが少なくとも100、150、200、250、300、350、400、450、500、550、600、650、700、750、800、850、900、950、1000、1050、1100、1150、1200、1250、又は少なくとも1300アミノ酸である。
種々の態様において、本明細書に開示のプライム編集因子は、次の通り記載されるCas9バリアントの1つ、又はいずれかの参照Cas9バリアントと少なくとも約70%同一、少なくとも約80%同一、少なくとも約90%同一、少なくとも約95%同一、少なくとも約96%同一、少なくとも約97%同一、少なくとも約98%同一、少なくとも約99%同一、少なくとも約99.5%同一、若しくは少なくとも約99.9%同一であるそのCas9バリアントを含んでもよい。
F.小さいサイズのCas9バリアント
いくつかの態様では、本明細書において企図される多重フラッププライム編集因子は、標準的なSpCas9配列よりも小さい分子量であるCas9タンパク質を包含し得る。いくつかの態様では、より小さいサイズのCas9バリアントは、例として発現ベクター、ナノ粒子、又は送達の他の手段による細胞への送達を容易化してもよい。ある態様において、より小さいサイズのCas9バリアントは、クラス2 CRISPR-CasシステムのII型酵素として類別される酵素を包含し得る。いくつかの態様では、より小さいサイズのCas9バリアントは、クラス2 CRISPR-CasシステムのV型酵素として類別される酵素を包含し得る。他の態様において、より小さいサイズのCas9バリアントは、クラス2 CRISPR-CasシステムのVI型酵素として類別される酵素を包含し得る。
いくつかの態様では、本明細書において企図される多重フラッププライム編集因子は、標準的なSpCas9配列よりも小さい分子量であるCas9タンパク質を包含し得る。いくつかの態様では、より小さいサイズのCas9バリアントは、例として発現ベクター、ナノ粒子、又は送達の他の手段による細胞への送達を容易化してもよい。ある態様において、より小さいサイズのCas9バリアントは、クラス2 CRISPR-CasシステムのII型酵素として類別される酵素を包含し得る。いくつかの態様では、より小さいサイズのCas9バリアントは、クラス2 CRISPR-CasシステムのV型酵素として類別される酵素を包含し得る。他の態様において、より小さいサイズのCas9バリアントは、クラス2 CRISPR-CasシステムのVI型酵素として類別される酵素を包含し得る。
標準的なSpCas9タンパク質は、1368アミノ酸長であり、158キロダルトンの予測分子量を有する。本明細書で使用される「小型Cas9変異体」という用語は、少なくとも1300アミノ酸未満、又は少なくとも1290アミノ酸未満、又は1280アミノ酸未満、又は1270アミノ酸未満、又は1260アミノ酸未満、又は1250アミノ酸未満、又は1240アミノ酸未満、又は1230アミノ酸未満、又は1220アミノ酸未満、又は1210アミノ酸未満、又は1200アミノ酸未満、又は1190アミノ酸未満、又は1180アミノ酸未満、又は1170アミノ酸未満、又は1160アミノ酸未満、又は1150アミノ酸未満、又は1140アミノ酸未満、又は1130アミノ酸未満、又は1120アミノ酸未満、又は1110アミノ酸未満、又は1100アミノ酸未満、又は1050アミノ酸未満、又は1000アミノ酸未満、又は950アミノ酸未満、又は900アミノ酸未満、又は850アミノ酸未満、又は800アミノ酸未満、又は750アミノ酸未満、又は700アミノ酸未満、又は650アミノ酸未満、又は600アミノ酸未満、又は550アミノ酸未満、又は500アミノ酸未満であるが、少なくとも400アミノ酸超であり、Cas9タンパク質の必要な機能を保持する、天然に存在する、操作された、又はその他の任意のCas9変異体を指す。Cas9変異体は、クラス2 CRISPR-Cas系のII型、V型又はVI型酵素として分類されるものを包含し得る。
様々な態様において、本明細書に開示されるプライム編集因子は、以下のように記載される小型Cas9変異体、又は任意の参照小型Cas9タンパク質と少なくとも約70%同一、少なくとも約80%同一、少なくとも約90%同一、少なくとも約95%同一、少なくとも約96%同一、少なくとも約97%同一、少なくとも約98%同一、少なくとも約99%同一、少なくとも約99.5%同一、又は少なくとも約99.9%同一であるそのCas9変異体の1つを含んでもよい。


G.Cas9等価物
いくつかの態様では、本明細書に記載のプライム編集因子はいずれかのCas9等価物を包含し得る。本明細書において用いられる用語「Cas9等価物」は、幅広い用語であり、これは、そのアミノ酸一次配列及び/又はその三次元構造が異なってもよく及び/又は進化的立場からは無関係であってもよいにもかかわらず、本プライム編集因子においてCas9と同じ機能を供するいずれかのnapDNAbpタンパク質を包摂する。それゆえに、Cas9等価物は進化的に関係する本明細書に記載又は包摂されるいずれかのCas9オーソログ、ホモログ、変異体、又はバリアントを包含するが、Cas9等価物は、Cas9と同じか又は類似の機能を有するように収斂進化プロセスによって進化してもよいが、アミノ酸配列及び/又は三次元構造についていずれかの類似性を必ずしも有さないタンパク質をもまた包摂する。Cas9等価物が収斂進化によって発生したタンパク質に基づいてもよいにもかかわらず、ここで記載されるプライム編集因子は、Cas9と同じか又は類似の機能を提供するであろういずれかのCas9等価物を包摂する。例えば、Cas9がCRISPR-CasシステムのII型酵素を指す場合には、Cas9等価物はCRISPR-CasシステムのV型又はVI型酵素を指し得る。
いくつかの態様では、本明細書に記載のプライム編集因子はいずれかのCas9等価物を包含し得る。本明細書において用いられる用語「Cas9等価物」は、幅広い用語であり、これは、そのアミノ酸一次配列及び/又はその三次元構造が異なってもよく及び/又は進化的立場からは無関係であってもよいにもかかわらず、本プライム編集因子においてCas9と同じ機能を供するいずれかのnapDNAbpタンパク質を包摂する。それゆえに、Cas9等価物は進化的に関係する本明細書に記載又は包摂されるいずれかのCas9オーソログ、ホモログ、変異体、又はバリアントを包含するが、Cas9等価物は、Cas9と同じか又は類似の機能を有するように収斂進化プロセスによって進化してもよいが、アミノ酸配列及び/又は三次元構造についていずれかの類似性を必ずしも有さないタンパク質をもまた包摂する。Cas9等価物が収斂進化によって発生したタンパク質に基づいてもよいにもかかわらず、ここで記載されるプライム編集因子は、Cas9と同じか又は類似の機能を提供するであろういずれかのCas9等価物を包摂する。例えば、Cas9がCRISPR-CasシステムのII型酵素を指す場合には、Cas9等価物はCRISPR-CasシステムのV型又はVI型酵素を指し得る。
例えば、Cas12e(CasX)は、報告によるとCas9と同じ機能を有するが、収斂進化によって進化したCas9等価物である。それゆえに、Liu et al.,「CasX enzymes comprises a distinct family of RNA-guided genome editors,」Nature,2019,Vol.566:218-223に記載されているCas12e(CasX)タンパク質は、本明細書に記載のプライム編集因子に用いられることを企図される。加えて、Cas12e(CasX)のいずれかのバリアント又は修飾が想像され、本開示の範囲内である。
Cas9は広い種々の種において進化した細菌酵素である。しかしながら、本明細書において企図されるCas9等価物は古細菌からもまた得られてもよい。これらは細菌とは異なる単細胞原核微生物のドメイン及び界を構成する。
いくつかの態様では、Cas9等価物はCas12e(CasX)又はCas12d(CasY)を指してもよい。これらは例えばBurstein et al.,「New CRISPR-Cas systems from uncultivated microbes.」Cell Res.2017 Feb 21.doi:10.1038/cr.2017.21に記載されている。これの内容全体は参照によってここに組み込まれる。ゲノム分解能メタゲノミクスを用いて、生命の古細菌ドメインにおける最初に報告されたCas9を包含するいくつものCRISPR-Casシステムが同定された。この分岐したCas9タンパク質は、わずかにしか研究されていないナノ古細菌から活性なCRISPR-Casシステムの一部として見出された。細菌では、2つの先には未知のシステムCRISPR-Cas12e及びCRISPR-Cas12dが発見された。これらはこれまでに発見された最もコンパクトなシステムのうちである。いくつかの態様では、Cas9はCas12e又はCas12eのバリアントを指す。いくつかの態様では、Cas9はCas12d又はCas12dのバリアントを指す。他のRNAによってガイドされるDNA結合タンパク質が核酸プログラム型DNA結合タンパク質(napDNAbp)として用いられてもよく、本開示の範囲内であるということは了解されるはずである。Liu et al.,「CasX enzymes comprises a distinct family of RNA-guided genome editors,」Nature,2019,Vol.566:218-223をもまた見よ。これらのCas9等価物のいずれかが企図される。
いくつかの態様では、Cas9等価物は、天然に存在するCas12e(CasX)又はCas12d(CasY)タンパク質と少なくとも85%、少なくとも90%、少なくとも91%、少なくとも92%、少なくとも93%、少なくとも94%、少なくとも95%、少なくとも96%、少なくとも97%、少なくとも98%、少なくとも99%、又は少なくとも99.5%同一であるアミノ酸配列を含む。いくつかの態様では、napDNAbpは天然に存在するCas12e(CasX)又はCas12d(CasY)タンパク質である。いくつかの態様では、napDNAbpは、本明細書において提供される野生型Cas部分又はいずれかのCas部分と少なくとも85%、少なくとも90%、少なくとも91%、少なくとも92%、少なくとも93%、少なくとも94%、少なくとも95%、少なくとも96%、少なくとも97%、少なくとも98%、少なくとも99%、又は少なくとも99.5%同一であるアミノ酸配列を含む。
種々の態様において、核酸プログラム型DNA結合タンパク質は、限定なしに、Cas9(例として、dCas9及びnCas9)、Cas12e(CasX)、Cas12d(CasY)、Cas12a(Cpf1)、Cas12b1(C2c1)、Cas13a(C2c2)、Cas12c(C2c3)、アルゴノート、及びCas12b1を包含する。Cas9とは異なるPAM特異性を有する核酸プログラム型DNA結合タンパク質の1つの例は、Prevotella及びFrancisellaからのクラスター化規則的間隔短回文反復1である(すなわちCas12a(Cpf1))。Cas9と類似に、Cas12a(Cpf1)もまたクラス2 CRISPRエフェクターであるが、それはII型サブグループよりもむしろ酵素のV型サブグループである。Cas12a(Cpf1)はCas9とは別物の特色を有するロバストなDNA干渉を媒介するということが示されている。Cas12a(Cpf1)は、tracrRNAを欠く単一のRNAによってガイドされるエンドヌクレアーゼであり、それは、Tリッチなプロトスペーサ隣接モチーフ(TTN、TTTN、又はYTN)を利用する。その上、Cpf1は互い違いのDNA二本鎖切断によってDNAを切断する。16のCpf1ファミリータンパク質のうち、Acidaminococcus及びLachnospiraceaeからの2つの酵素はヒト細胞における効率的なゲノム編集活性を有することが示される。Cpf1タンパク質は当該技術分野で知られており、先に記載されている。例えばYamano et al.,「Crystal structure of Cpf1 in complex with guide RNA and target DNA.」Cell(165)2016,p.949-962;これの内容全体は参照によってここに組み込まれる。
なお他の態様において、Casタンパク質はいずれかのCRISPR関連タンパク質を包含してもよく、Cas12a、Cas12b1、Cas1、Cas1B、Cas2、Cas3、Cas4、Cas5、Cas6、Cas7、Cas8、Cas9(Csn1及びCsx12としてもまた公知)、Cas10、Csy1、Csy2、Csy3、Cse1、Cse2、Csc1、Csc2、Csa5、Csn2、Csm2、Csm3、Csm4、Csm5、Csm6、Cmr1、Cmr3、Cmr4、Cmr5、Cmr6、Csb1、Csb2、Csb3、Csx17、Csx14、Csx10、Csx16、CsaX、Csx3、Csx1、Csx15、Csf1、Csf2、Csf3、Csf4、それらのホモログ、又はそれらの修飾されたバージョンを包含するが、これらに限定されず、好ましくはニッカーゼ変異を含む(例として、配列番号37の野生型Cas9ポリペプチドのD10A変異に対応する変異)。
種々の他の態様において、napDNAbpは次のタンパク質のいずれかであり得る:Cas9、Cas12a(Cpf1)、Cas12e(CasX)、Cas12d(CasY)、Cas12b1(C2c1)、Cas13a(C2c2)、Cas12c(C2c3)、GeoCas9、CjCas9、Cas12g、Cas12h、Cas12i、Cas13b、Cas13c、Cas13d、Cas14、Csn2、xCas9、SpCas9-NG、循環置換Cas9、若しくはアルゴノート(Ago)ドメイン、又はそのバリアント。
例示のCas9等価物タンパク質配列は次を包含し得る:



本明細書に記載のプライム編集因子は、ガイドヌクレオチド配列によってプログラム型DNA結合タンパク質ドメインとして用いられてもよいCas12a(Cpf1)(dCpf1)バリアントをもまた含んでもよい。Cas12a(Cpf1)タンパク質は、Cas9のRuvCドメインに類似であるRuvC様エンドヌクレアーゼドメインを有するが、HNHエンドヌクレアーゼドメインは有さず、Cas12a(Cpf1)のN末端はCas9のアルファヘリックス系の認識ローブを有さない。Zetsche et al.,Cell,163,759-771,2015(これは参照によって本明細書に組み込まれる)においては、Cas12a(Cpf1)のRuvC様ドメインは両方のDNA鎖を切断することを担い、RuvC様ドメインの不活性化がCas12a(Cpf1)ヌクレアーゼ活性を不活性化するということが示された。
いくつかの態様では、napDNAbpは微生物CRISPR-Casシステムの単一のエフェクターである。微生物CRISPR-Casシステムの単一のエフェクターは、限定なしに、Cas9、Cas12a(Cpf1)、Cas12b1(C2c1)、Cas13a(C2c2)、及びCas12c(C2c3)を包含する。典型的には、微生物CRISPR-Casシステムはクラス1及びクラス2システムに分けられる。クラス1システムはマルチサブユニットエフェクター複合体を有するが、クラス2システムは単一のタンパク質エフェクターを有する。例えば、Cas9及びCas12a(Cpf1)はクラス2エフェクターである。Cas9及びCas12a(Cpf1)に加えて、3つの別物のクラス2 CRISPR-Casシステム(Cas12b1、Cas13a、及びCas12c)がShmakov et al.,「Discovery and Functional Characterization of Diverse Class 2 CRISPR Cas Systems」,Mol.Cell,2015 Nov 5;60(3):385-397によって記載されている。これの内容全体は参照によってここに組み込まれる。
システムの2つCas12b1及びCas12cのエフェクターは、Cas12aに関係するRuvC様エンドヌクレアーゼドメインを含有する。第3のシステムのCas13aは、2つの予測される(predicted)HEPN RNaseドメインを有するエフェクターを含有する。Cas12b1によるCRISPR RNAの生産とは違って、成熟CRISPR RNAの生産はtracrRNA非依存的である。Cas12b1はDNA切断についてはCRISPR RNA及びtracrRNA両方に依存する。細菌Cas13aは、そのRNAによって活性化される一本鎖RNA分解活性とは別物のCRISPR RNA成熟のための固有のRNase活性を所有することが示されている。これらのRNase機能は、互いとは、及びCas12aのCRISPR RNAプロセシング挙動とは異なる。例として、East-Seletsky,et al.,「Two distinct RNase activities of CRISPR-Cas13a enable guide-RNA processing and RNA detection」,Nature,2016 Oct 13;538(7624):270-273を見よ。これの内容全体は参照によってここに組み込まれる。Leptotrichia shahiiのCas13aのin vitroの生化学分析は、Cas13aが単一のCRISPR RNAによってガイドされ、相補的なプロトスペーサを持つssRNA標的を切断するようにプログラムされ得るということを示している。2つの保存されたHEPNドメイン上の触媒残基が切断を媒介する。触媒残基の変異は触媒的に不活性なRNA結合タンパク質を生成する。例として、Abudayyeh et al.,「C2c2 is a single-component programmable RNA-guided RNA-targeting CRISPR effector」,Science,2016 Aug 5;353(6299)を見よ。これの内容全体は参照によってここに組み込まれる。
Alicyclobaccillus acidoterrastris Cas12b1(AacC2c1)の結晶構造がキメラ単分子ガイドRNA(sgRNA)との複合体として報告されている。例として、Liu et al.,「C2c1-sgRNA Complex Structure Reveals RNA-Guided DNA Cleavage Mechanism」,Mol.Cell,2017 Jan 19;65(2):310-322を見よ。これの内容全体は参照によってここに組み込まれる。結晶構造は三者複合体として標的DNAに結合したAlicyclobacillus acidoterrestris C2c1としてもまた報告されている。例として、Yang et al.,「PAM-dependent Target DNA Recognition and Cleavage by C2C1 CRISPR-Cas endonuclease」,Cell,2016 Dec 15;167(7):1814-1828を見よ。これの内容全体は参照によってここに組み込まれる。AacC2c1の触媒的に有能な立体配座は、独立して単一のRuvC触媒ポケット内に配置された標的及び非標的DNA鎖両方によって、キャプチャされている。C2c1によって媒介される切断は、標的DNAの互い違いの7ヌクレオチド切断をもたらす。C2c1三者複合体と先に同定されたCas9及びCpf1カウンターパートとの間の構造比較は、CRISPR-Cas9システムによって用いられる機序の多様性を実証する。
いくつかの態様では、napDNAbpはC2c1、C2c2、又はC2c3タンパク質であってもよい。いくつかの態様では、napDNAbpはC2c1タンパク質である。いくつかの態様では、napDNAbpはCas13aタンパク質である。いくつかの態様では、napDNAbpはCas12cタンパク質である。いくつかの態様では、napDNAbpは、天然に存在するCas12b1(C2c1)、Cas13a(C2c2)、又はCas12c(C2c3)タンパク質と少なくとも85%、少なくとも90%、少なくとも91%、少なくとも92%、少なくとも93%、少なくとも94%、少なくとも95%、少なくとも96%、少なくとも97%、少なくとも98%、少なくとも99%、又は少なくとも99.5%同一であるアミノ酸配列を含む。いくつかの態様では、napDNAbpは天然に存在するCas12b1(C2c1)、Cas13a(C2c2)、又はCas12c(C2c3)タンパク質である。
H.Cas9循環置換体
種々の態様において、本明細書に開示のプライム編集因子はCas9の循環置換体を含んでもよい。
種々の態様において、本明細書に開示のプライム編集因子はCas9の循環置換体を含んでもよい。
用語「循環置換Cas9」又はCas9の「循環置換体」又は「CP-Cas9」は、循環置換体バリアントとして生起するか又は操作されるように修飾されているいずれかのCas9タンパク質又はそのバリアントを指す。これは、Cas9タンパク質(例として、野生型Cas9タンパク質)のN末端及びC末端が局所的に再配置されているということを意味する。かかる循環置換Cas9タンパク質又はそれらのバリアントは、ガイドRNA(gRNA)と複合体化したときにDNAに結合する能力を保持する。Oakes et al.,「Protein Engineering of Cas9 for enhanced function,」Methods Enzymol,2014,546:491-511及びOakes et al.,「CRISPR-Cas9 Circular Permutants as Programmable Scaffolds for Genome Modification,」Cell,January 10,2019,176:254-267を見よ。夫々は参照によって本明細書に組み込まれる。もたらされる循環置換タンパク質がガイドRNA(gRNA)と複合体化したときにDNAに結合する能力を保持する限り、本開示は、いずれかの先に公知のCP-Cas9を企図するか、又は新たなCP-Cas9を用いる。
いずれかのバリアント、オーソログ、若しくは天然に存在するCas9を包含する本明細書に記載のCas9タンパク質のいずれか、又はその等価物が、循環置換体バリアントとして再構成されてもよい。
種々の態様において、Cas9の循環置換体は次の構造を有してもよい:
N末端-[元のC末端]-[任意選択のリンカー]-[元のN末端]-C末端。
N末端-[元のC末端]-[任意選択のリンカー]-[元のN末端]-C末端。
例として、本開示は標準的なS.pyogenes Cas9の次の循環置換体を企図する(UniProtKB-Q99ZW2(CAS9_STRP1)の1368アミノ酸(付番は配列番号37のアミノ酸位置に基づく)):
N末端-[1268~1368]-[任意選択のリンカー]-[1~1267]-C末端;
N末端-[1168~1368]-[任意選択のリンカー]-[1~1167]-C末端;
N末端-[1068~1368]-[任意選択のリンカー]-[1~1067]-C末端;
N末端-[968~1368]-[任意選択のリンカー]-[1~967]-C末端;
N末端-[868~1368]-[任意選択のリンカー]-[1~867]-C末端;
N末端-[768~1368]-[任意選択のリンカー]-[1~767]-C末端;
N末端-[668~1368]-[任意選択のリンカー]-[1~667]-C末端;
N末端-[568~1368]-[任意選択のリンカー]-[1~567]-C末端;
N末端-[468~1368]-[任意選択のリンカー]-[1~467]-C末端;
N末端-[368~1368]-[任意選択のリンカー]-[1~367]-C末端;
N末端-[268~1368]-[任意選択のリンカー]-[1~267]-C末端;
N末端-[168~1368]-[任意選択のリンカー]-[1~167]-C末端;
N末端-[68~1368]-[任意選択のリンカー]-[1~67]-C末端;若しくは
N末端-[10~1368]-[任意選択のリンカー]-[1~9]-C末端、又は
他のCas9タンパク質(他のCas9オーソログ、バリアント等を包含する)の対応する循環置換体。
N末端-[1268~1368]-[任意選択のリンカー]-[1~1267]-C末端;
N末端-[1168~1368]-[任意選択のリンカー]-[1~1167]-C末端;
N末端-[1068~1368]-[任意選択のリンカー]-[1~1067]-C末端;
N末端-[968~1368]-[任意選択のリンカー]-[1~967]-C末端;
N末端-[868~1368]-[任意選択のリンカー]-[1~867]-C末端;
N末端-[768~1368]-[任意選択のリンカー]-[1~767]-C末端;
N末端-[668~1368]-[任意選択のリンカー]-[1~667]-C末端;
N末端-[568~1368]-[任意選択のリンカー]-[1~567]-C末端;
N末端-[468~1368]-[任意選択のリンカー]-[1~467]-C末端;
N末端-[368~1368]-[任意選択のリンカー]-[1~367]-C末端;
N末端-[268~1368]-[任意選択のリンカー]-[1~267]-C末端;
N末端-[168~1368]-[任意選択のリンカー]-[1~167]-C末端;
N末端-[68~1368]-[任意選択のリンカー]-[1~67]-C末端;若しくは
N末端-[10~1368]-[任意選択のリンカー]-[1~9]-C末端、又は
他のCas9タンパク質(他のCas9オーソログ、バリアント等を包含する)の対応する循環置換体。
具体的な態様では、循環置換体(circular permutant)Cas9は次の構造を有する(S.pyogenes Cas9(UniProtKB-Q99ZW2(CAS9_STRP1)の1368アミノ酸(付番は配列番号37のアミノ酸位置に基づく)に基づく:
N末端-[102~1368]-[任意選択のリンカー]-[1~101]-C末端;
N末端-[1028~1368]-[任意選択のリンカー]-[1~1027]-C末端;
N末端-[1041~1368]-[任意選択のリンカー]-[1~1043]-C末端;
N末端-[1249~1368]-[任意選択のリンカー]-[1~1248]-C末端;若しくは
N末端-[1300~1368]-[任意選択のリンカー]-[1~1299]-C末端、又は
他のCas9タンパク質(他のCas9オーソログ、バリアント等を包含する)の対応する循環置換体。
N末端-[102~1368]-[任意選択のリンカー]-[1~101]-C末端;
N末端-[1028~1368]-[任意選択のリンカー]-[1~1027]-C末端;
N末端-[1041~1368]-[任意選択のリンカー]-[1~1043]-C末端;
N末端-[1249~1368]-[任意選択のリンカー]-[1~1248]-C末端;若しくは
N末端-[1300~1368]-[任意選択のリンカー]-[1~1299]-C末端、又は
他のCas9タンパク質(他のCas9オーソログ、バリアント等を包含する)の対応する循環置換体。
なお他の態様において、循環置換体(circular permutant)Cas9は次の構造を有する(S.pyogenes Cas9(UniProtKB-Q99ZW2(CAS9_STRP1)の1368アミノ酸(付番は配列番号37のアミノ酸位置に基づく)に基づく:
N末端-[103~1368]-[任意選択のリンカー]-[1~102]-C末端;
N末端-[1029~1368]-[任意選択のリンカー]-[1~1028]-C末端;
N末端-[1042~1368]-[任意選択のリンカー]-[1~1041]-C末端;
N末端-[1250~1368]-[任意選択のリンカー]-[1~1249]-C末端;若しくは
N末端-[1301~1368]-[任意選択のリンカー]-[1~1300]-C末端、又は
他のCas9タンパク質(他のCas9オーソログ、バリアント等を包含する)の対応する循環置換体。
N末端-[103~1368]-[任意選択のリンカー]-[1~102]-C末端;
N末端-[1029~1368]-[任意選択のリンカー]-[1~1028]-C末端;
N末端-[1042~1368]-[任意選択のリンカー]-[1~1041]-C末端;
N末端-[1250~1368]-[任意選択のリンカー]-[1~1249]-C末端;若しくは
N末端-[1301~1368]-[任意選択のリンカー]-[1~1300]-C末端、又は
他のCas9タンパク質(他のCas9オーソログ、バリアント等を包含する)の対応する循環置換体。
いくつかの態様では、循環置換体は、直接的に又はアミノ酸リンカー等のリンカーを用いることによってどちらかで、Cas9のC末端フラグメントをCas9のN末端フラグメントに連結することによって形成され得る。いくつかの態様では、C末端フラグメントは、Cas9のアミノ酸(例として、アミノ酸約1300-1368)のC末端95%以上、あるいはCas9(例として、配列番号88-97のいずれか1つ)のC末端90%、85%、80%、75%、70%、65%、60%、55%、50%、45%、40%、35%、30%、25%、20%、15%、10%、若しくは5%、又はより多くに対応してもよい。N末端部分は、Cas9のアミノ酸(例として、アミノ酸約1-1300)のN末端95%以上、あるいはCas9の(例として配列番号37の)N末端90%、85%、80%、75%、70%、65%、60%、55%、50%、45%、40%、35%、30%、25%、20%、15%、10%、若しくは5%、又はより多くに対応してもよい。
いくつかの態様では、循環置換体は、直接的に又はアミノ酸リンカー等のリンカーを用いることによってどちらかで、Cas9のC末端フラグメントをCas9のN末端フラグメントに連結することによって形成され得る。いくつかの態様では、N末端へと再配置されるC末端フラグメントは、Cas9のアミノ酸のC末端30%以下(例として、配列番号37のアミノ酸1012~1368)を包含するか又はそれらに対応する。いくつかの態様では、N末端へと再配置されるC末端フラグメントは、Cas9のアミノ酸(例として、配列番号37のCas9)のC末端30%、29%、28%、27%、26%、25%、24%、23%、22%、21%、20%、19%、18%、17%、16%、15%、14%、13%、12%、11%、10%、9%、8%、7%、6%、5%、4%、3%、2%、又は1%を包含するか又はそれらに対応する。いくつかの態様では、N末端へと再配置されるC末端フラグメントは、Cas9(例として、配列番号37のCas9)のC末端410残基以下を包含するか又はそれらに対応する。いくつかの態様では、N末端へと再配置されるC末端部分は、Cas9(例として、配列番号37のCas9)のC末端410、400、390、380、370、360、350、340、330、320、310、300、290、280、270、260、250、240、230、220、210、200、190、180、170、160、150、140、130、120、110、100、90、80、70、60、50、40、30、20、又は10残基を包含するか又はそれらに対応する。いくつかの態様では、N末端へと再配置されるC末端フラグメントは、Cas9(例として、配列番号37のCas9)のC末端357、341、328、120、又は69残基を包含するか又はそれらに対応する。
他の態様において、循環置換体Cas9バリアントは、次の方法に基づくCas9一次構造のトポロジー的再配置として定められてもよく、これは配列番号37のS.pyogenes Cas9に基づく:(a)Cas9一次構造の内部のアミノ酸残基に対応する循環置換体(CP)部位を選択し、これが元のタンパク質を2つの半分:N末端領域及びC末端領域へと切り分けること;(b)元のN末端領域に先立つ(precede)ように元のC末端領域(CP部位アミノ酸を含む)を動かすことによって、Cas9タンパク質配列を修飾し(例として、遺伝子工学技術による)、それによって、今やCP部位アミノ酸残基で始まるCas9タンパク質の新たなN末端を形成すること。CP部位は、例えばヘリカルIIドメイン、RuvCIIIドメイン、又はCTDドメインを包含するCas9タンパク質のいずれかのドメイン上に位置付られ得る。例えば、CP部位は、(配列番号37のS.pyogenes Cas9に対して相対的に)元のアミノ酸残基181、199、230、270、310、1010、1016、1023、1029、1041、1247、1249、又は1282に位置付けられてもよい。それゆえに、ひとたびN末端に位置替えされると、元のアミノ酸181、199、230、270、310、1010、1016、1023、1029、1041、1247、1249、又は1282は新たなN末端アミノ酸になるであろう。これらのCP-Cas9タンパク質の命名体系は、夫々Cas9-CP181、Cas9-CP199、Cas9-CP230、Cas9-CP270、Cas9-CP310、Cas9-CP1010、Cas9-CP1016、Cas9-CP1023、Cas9-CP1029、Cas9-CP1041、Cas9-CP1247、Cas9-CP1249、及びCas9-CP1282と呼ばれてもよい。この記載は配列番号37からのCPバリアントを作ることに限定されることを意味されず、いずれかのCas9配列において、これらの位置に対応するCP部位において又は全く(entirely)他のCP部位においてどちらかでCPバリアントを作るために実装されてもよい。この記載は決して特定のCP部位を限定することを意味されない。実質的にいずれかのCP部位がCP-Cas9バリアントを形成するために用いられてもよい。
配列番号37のCas9に基づく例示のCP-Cas9アミノ酸配列が下で提供される。これにおいては、リンカー配列は下線付きによって指示され、任意選択のメチオニン(M)残基は太字で指示されている。本開示はリンカー配列を包含しないか又は異なるリンカー配列を包含するCP-Cas9配列を提供するということは了解されるはずである。CP-Cas9配列が配列番号37以外のCas9配列に基づいてもよいということは了解されるはずであり、本明細書において提供されるいずれかの例は限定することを意味されない。例示のCP-Cas9配列は次の通りである:


Cas9循環置換体は本明細書に記載のプライム編集構築物に有用であってもよい。Cas9のN末端へと再配置されてもよい配列番号37のCas9に基づくCas9の例示のC末端フラグメントが、下で提供される。Cas9のかかるC末端フラグメントは例示的であり、限定することを意味されないということは了解されるはずである。これらの例示のCP-Cas9フラグメントは次の配列を有する:

I.修飾されたPAM特異性を有するCas9バリアント
本開示のプライム編集因子はまた、修飾されたPAM特異性を有するCas9変異体を含んでもよい。本開示のいくつかの側面は、その3'末端に標準的なPAM(5'-NGG-3'(式中、NはA、C、G又はTである))を含まない標的配列に対して活性を示すCas9タンパク質を提供する。いくつかの態様において、Cas9タンパク質は、その3'末端に5'-NGG-3'PAM配列を含む標的配列に対して活性を示す。いくつかの態様において、Cas9タンパク質は、その3'末端に5'-NNG-3'PAM配列を含む標的配列に対して活性を示す。いくつかの態様において、Cas9タンパク質は、その3'末端に5'-NNA-3'PAM配列を含む標的配列に対して活性を示す。いくつかの態様において、Cas9タンパク質は、その3'末端に5'-NNC-3'PAM配列を含む標的配列に対して活性を示す。いくつかの態様において、Cas9タンパク質は、その3'末端に5'-NNT-3'PAM配列を含む標的配列に対して活性を示す。いくつかの態様において、Cas9タンパク質は、その3'末端に5'-NGT-3'PAM配列を含む標的配列に対して活性を示す。いくつかの態様において、Cas9タンパク質は、その3'末端に5'-NGA-3'PAM配列を含む標的配列に対して活性を示す。いくつかの態様において、Cas9タンパク質は、その3'末端に5'-NGC-3'PAM配列を含む標的配列に対して活性を示す。いくつかの態様において、Cas9タンパク質は、その3'末端に5'-NAA-3'PAM配列を含む標的配列に対して活性を示す。いくつかの態様において、Cas9タンパク質は、その3'末端に5'-NAC-3'PAM配列を含む標的配列に対して活性を示す。いくつかの態様において、Cas9タンパク質は、その3'末端に5'-NAT-3'PAM配列を含む標的配列に対して活性を示す。更に他の態様では、Cas9タンパク質は、その3'末端に5'-NAG-3'PAM配列を含む標的配列に対して活性を示す。
本開示のプライム編集因子はまた、修飾されたPAM特異性を有するCas9変異体を含んでもよい。本開示のいくつかの側面は、その3'末端に標準的なPAM(5'-NGG-3'(式中、NはA、C、G又はTである))を含まない標的配列に対して活性を示すCas9タンパク質を提供する。いくつかの態様において、Cas9タンパク質は、その3'末端に5'-NGG-3'PAM配列を含む標的配列に対して活性を示す。いくつかの態様において、Cas9タンパク質は、その3'末端に5'-NNG-3'PAM配列を含む標的配列に対して活性を示す。いくつかの態様において、Cas9タンパク質は、その3'末端に5'-NNA-3'PAM配列を含む標的配列に対して活性を示す。いくつかの態様において、Cas9タンパク質は、その3'末端に5'-NNC-3'PAM配列を含む標的配列に対して活性を示す。いくつかの態様において、Cas9タンパク質は、その3'末端に5'-NNT-3'PAM配列を含む標的配列に対して活性を示す。いくつかの態様において、Cas9タンパク質は、その3'末端に5'-NGT-3'PAM配列を含む標的配列に対して活性を示す。いくつかの態様において、Cas9タンパク質は、その3'末端に5'-NGA-3'PAM配列を含む標的配列に対して活性を示す。いくつかの態様において、Cas9タンパク質は、その3'末端に5'-NGC-3'PAM配列を含む標的配列に対して活性を示す。いくつかの態様において、Cas9タンパク質は、その3'末端に5'-NAA-3'PAM配列を含む標的配列に対して活性を示す。いくつかの態様において、Cas9タンパク質は、その3'末端に5'-NAC-3'PAM配列を含む標的配列に対して活性を示す。いくつかの態様において、Cas9タンパク質は、その3'末端に5'-NAT-3'PAM配列を含む標的配列に対して活性を示す。更に他の態様では、Cas9タンパク質は、その3'末端に5'-NAG-3'PAM配列を含む標的配列に対して活性を示す。
第1のアミノ酸残基(例としてA)から第2のアミノ酸残基(例としてT)への本明細書に記載のアミノ酸変異のいずれか(例として、A262T)は、第1のアミノ酸残基から(例として、保存された)第2のアミノ酸残基に類似であるアミノ酸残基への変異をもまた包含してもよいということは了解されるはずである。例えば、疎水性側鎖を有するアミノ酸(例として、アラニン、バリン、イソロイシン、ロイシン、メチオニン、フェニルアラニン、チロシン、又はトリプトファン)の変異は、異なる疎水性側鎖を有する第2のアミノ酸(例として、アラニン、バリン、イソロイシン、ロイシン、メチオニン、フェニルアラニン、チロシン、又はトリプトファン)への変異であってもよい。例えば、アラニンからトレオニンへの変異(例としてA262T変異)はまた、アラニンからサイズ及び化学的特性がトレオニンに類似であるアミノ酸、例えばセリンへの変異であってもよい。別の例として、正荷電側鎖を有するアミノ酸(例として、アルギニン、ヒスチジン、又はリジン)の変異は、異なる正荷電側鎖を有する第2のアミノ酸(例として、アルギニン、ヒスチジン、又はリジン)への変異であってもよい。別の例として、極性側鎖を有するアミノ酸(例として、セリン、トレオニン、アスパラギン、又はグルタミン)の変異は、異なる極性側鎖を有する第2のアミノ酸(例として、セリン、トレオニン、アスパラギン、又はグルタミン)への変異であってもよい。追加の類似のアミノ酸ペアは次を包含するが、これらに限定されない:フェニルアラニン及びチロシン;アスパラギン及びグルタミン;メチオニン及びシステイン;アスパラギン酸及びグルタミン酸;並びにアルギニン及びリジン。当業者は、かかる保存的アミノ酸置換が蓋然的にはタンパク質構造に対して軽度の効果を有し、機能を毀損することなしに良く忍容されることが蓋然的であるということを認識するであろう。いくつかの態様では、1つのアミノ酸からトレオニンへの本明細書で提供されるアミノ酸変異の任意のアミノは、セリンへのアミノ酸変異であってもよい。いくつかの態様では、1つのアミノ酸からアルギニンへの本明細書で提供されるアミノ酸変異の任意のアミノは、リジンへのアミノ酸変異であってもよい。いくつかの態様では、1つのアミノ酸からイソロイシンへの本明細書において提供されるアミノ酸変異の任意のアミノは、アラニン、バリン、メチオニン、又はロイシンへのアミノ酸変異であってもよい。いくつかの態様では、1つのアミノ酸からリジンへの本明細書で提供されるアミノ酸変異の任意のアミノは、アルギニンへのアミノ酸変異であってもよい。いくつかの態様では、1つのアミノ酸からアスパラギン酸への本明細書で提供されるアミノ酸変異の任意のアミノは、グルタミン酸又はアスパラギンへのアミノ酸変異であってもよい。いくつかの態様では、1つのアミノ酸からバリンへの本明細書において提供されるアミノ酸変異の任意のアミノは、アラニン、イソロイシン、メチオニン、又はロイシンへのアミノ酸変異であってもよい。いくつかの態様では、1つのアミノ酸からグリシンへの本明細書で提供されるアミノ酸変異の任意のアミノは、アラニンへのアミノ酸変異であってもよい。しかしながら、追加の保存されたアミノ酸残基は当業者によって認識されるであろうということは了解されるはずであり、他の保存されたアミノ酸残基へのアミノ酸変異のいずれかもまた本開示の範囲内である。
いくつかの態様では、Cas9タンパク質は、5'-NAA-3'PAM配列をその3'端に含む標的配列に対して活性を発揮する変異の組み合わせを含む。いくつかの態様では、変異の組み合わせは、表1の列記されているクローンのいずれか1つに存在する。いくつかの態様では、変異の組み合わせは、表1の列記されているクローンの保存的変異である。いくつかの態様では、Cas9タンパク質は、表1に列記されるCas9クローンのいずれか1つの変異の組み合わせを含む。

いくつかの態様では、Cas9タンパク質は、表1のバリアントのいずれか1つによって提供されるCas9タンパク質のアミノ酸配列と少なくとも80%同一であるアミノ酸配列を含む。いくつかの態様では、Cas9タンパク質は、表1のバリアントのいずれか1つによって提供されるCas9タンパク質のアミノ酸配列と少なくとも85%、少なくとも90%、少なくとも92%、少なくとも95%、少なくとも96%、少なくとも97%、少なくとも98%、少なくとも99%、又は少なくとも99.5%同一であるアミノ酸配列を含む。
いくつかの態様では、Cas9タンパク質は、配列番号37によって提供されるStreptococcus pyogenes Cas9と比較して、標準的なPAM(5'-NGG-3')をその3'端に含まない標的配列に対して増大した活性を発揮する。いくつかの態様では、Cas9タンパク質は、標準的なPAM配列(5'-NGG-3')に直接的に隣接しない3'端を有する標的配列に対して、同じ標的配列に対する配列番号37によって提供されるStreptococcus pyogenes Cas9の活性と比較して、少なくとも5倍増大した活性を発揮する。いくつかの態様では、Cas9タンパク質は、標準的なPAM配列(5'-NGG-3')に直接的に隣接しない標的配列に対して、同じ標的配列に対する配列番号37によって提供されるStreptococcus pyogenesの活性と比較して、少なくとも10倍、少なくとも50倍、少なくとも100倍、少なくとも500倍、少なくとも1,000倍、少なくとも5,000倍、少なくとも10,000倍、少なくとも50,000倍、少なくとも100,000倍、少なくとも500,000倍、又は少なくとも1,000,000倍増大した活性を発揮する。いくつかの態様では、標的配列の3'端はAAA、GAA、CAA、又はTAA配列に直接的に隣接する。いくつかの態様では、Cas9タンパク質は、5'-NAC-3'PAM配列をその3'端に含む標的配列に対して活性を発揮する変異の組み合わせを含む。いくつかの態様では、変異の組み合わせは、表2の列記されているクローンのいずれか1つに存在する。いくつかの態様では、変異の組み合わせは、表2の列記されているクローンの保存的変異である。いくつかの態様では、Cas9タンパク質は、表2の列記されているCas9クローンのいずれか1つの変異の組み合わせを含む。

いくつかの態様では、Cas9タンパク質は、表2のバリアントのいずれか1つによって提供されるCas9タンパク質のアミノ酸配列と少なくとも80%同一であるアミノ酸配列を含む。いくつかの態様では、Cas9タンパク質は、表2のバリアントのいずれか1つによって提供されるCas9タンパク質のアミノ酸配列と少なくとも85%、少なくとも90%、少なくとも92%、少なくとも95%、少なくとも96%、少なくとも97%、少なくとも98%、少なくとも99%、又は少なくとも99.5%同一であるアミノ酸配列を含む。
いくつかの態様では、Cas9タンパク質は、配列番号37によって提供されるStreptococcus pyogenes Cas9と比較して、標準的なPAM(5'-NGG-3')をその3'端に含まない標的配列に対して増大した活性を発揮する。いくつかの態様では、Cas9タンパク質は、標準的なPAM配列(5'-NGG-3')に直接的に隣接しない3'端を有する標的配列に対して、同じ標的配列に対する配列番号37によって提供されるStreptococcus pyogenes Cas9の活性と比較して、少なくとも5倍増大した活性を発揮する。いくつかの態様では、Cas9タンパク質は、標準的なPAM配列(5'-NGG-3')に直接的に隣接しない標的配列に対して、同じ標的配列に対する配列番号37によって提供されるStreptococcus pyogenesの活性と比較して、少なくとも10倍、少なくとも50倍、少なくとも100倍、少なくとも500倍、少なくとも1,000倍、少なくとも5,000倍、少なくとも10,000倍、少なくとも50,000倍、少なくとも100,000倍、少なくとも500,000倍、又は少なくとも1,000,000倍増大した活性を発揮する。いくつかの態様では、標的配列の3'端はAAC、GAC、CAC、又はTAC配列に直接的に隣接する。
いくつかの態様では、Cas9タンパク質は、5'-NAT-3'PAM配列をその3'端に含む標的配列に対して活性を発揮する変異の組み合わせを含む。いくつかの態様では、変異の組み合わせは表3の列記されているクローンのいずれか1つに存在する。いくつかの態様では、変異の組み合わせは、表3の列記されているクローンの保存的変異である。いくつかの態様では、Cas9タンパク質は、表3に列記されるCas9クローンのいずれか1つの変異の組み合わせを含む。

本開示のプライム編集因子に関して用いられ得る種々のnapDNAbpの上の記載は、決して限定することを意味されない。プライム編集因子は、公知であるか又は指向性進化若しくは別様の変異プロセスによって作られ得るか若しくは進化させられ得る標準的なSpCas9又はいずれかのオーソログCas9タンパク質又はいずれかのバリアントCas9タンパク質を含み得、Cas9のいずれかの天然に存在するバリアント、変異体、又は別様に操作されたバージョンを包含する。種々の態様において、Cas9又はCas9バリアント(variant)はニッカーゼ活性を有し、すなわち標的DNA配列の1つの(of)鎖のみを切断する。他の態様において、Cas9又はCas9バリアントは不活性なヌクレアーゼを有し、すなわち「不活性型」Cas9タンパク質である。用いられてもよい他のバリアントCas9タンパク質は、(例として、より容易な送達のために)標準的なSpCas9よりも小さい分子量を有するか又は修飾若しくは再配置された一次アミノ酸構造(例として循環置換体フォーマット)を有するものである。本明細書に記載のプライム編集因子は、収斂進化の結果であるCas12a/Cpf1及びCas12bタンパク質を包含するCas9等価物をもまた含んでもよい。本明細書において用いられるnapDNAbp(例として、SpCas9、Cas9バリアント、又はCas9等価物)は、それらのPAM特異性を変改/増強する種々の修飾をもまた含有してもよい。最後に、本明細書は、少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも91%、少なくとも92%、少なくとも93%、少なくとも94%、少なくとも95%、少なくとも96%、少なくとも97%、少なくとも98%、少なくとも99%、又は少なくとも99.9%配列同一性を参照Cas9配列、例えば参照SpCas9標準的な配列又は参照Cas9等価物(例として、Cas12a/Cpf1)に対して有するいずれかのCas9、Cas9バリアント、又はCas9等価物を企図する。
具体的な態様では、拡張されたPAM能力を有するCas9バリアントはSpCas9(H840A)VRQR(配列番号98)であり、これは次のアミノ酸配列を有する(配列番号68のSpCas9(H840A)に対して相対的なV、R、Q、R置換が太字の下線で示されている。加えて、SpCas9(H840)のメチオニン残基はSpCas9(H840A)VRQRでは除去された):

別の具体的な態様では、拡張されたPAM能力を有するCas9バリアントはSpCas9(H840A)VRERであり、これは次のアミノ酸配列を有する(配列番号68のSpCas9(H840A)に対して相対的なV、R、E、R置換が太字の下線で示されている。加えて、SpCas9(H840)のメチオニン残基はSpCas9(H840A)VRERでは除去された):

いくつかの態様では、非標準的なPAM配列について機能するnapDNAbpはアルゴノートタンパク質である。かかる核酸プログラム型DNA結合タンパク質の1つの例は、Natronobacterium gregoryiからのアルゴノートタンパク質(NgAgo)である。NgAgoはssDNAによってガイドされるエンドヌクレアーゼである。NgAgoは~24ヌクレオチドの5'リン酸化されたssDNA(gDNA)に結合してそれをその標的部位へとガイドし、gDNA部位においてDNA二本鎖切断を作製するであろう。Cas9とは対照的に、NgAgo-gDNAシステムはプロトスペーサ隣接モチーフ(PAM)を要求しない。ヌクレアーゼ不活性なNgAgo(dNgAgo)を用いることは、標的化され得る塩基を多大に拡張してもよい。NgAgoの特徴付け及び使用はGao et al.,Nat Biotechnol.,2016 Jul;34(7):768-73.PubMed PMID:27136078;Swarts et al.,Nature.507(7491)(2014):258-61;及びSwarts et al.,Nucleic Acids Res.43(10)(2015):5120-9に記載されている。これらの夫々は参照によって本明細書に組み込まれる。
いくつかの態様では、napDNAbpはアルゴノートタンパク質の原核生物ホモログである。アルゴノートタンパク質の原核生物ホモログは知られており、例えばMakarova K.,et al.,「Prokaryotic homologs of Argonaute proteins are predicted to function as key components of a novel system of defense against mobile genetic elements」,Biol Direct.2009 Aug 25;4:29.doi:10.1186/1745-6150-4-29に記載されている。これの内容全体は参照によってここに組み込まれる。いくつかの態様では、napDNAbpはMarinitoga piezophila アルゴノート(MpAgo)タンパク質である。CRISPR関連Marinitoga piezophila アルゴノート(MpAgo)タンパク質は、5'-リン酸化されたガイドを用いて一本鎖標的配列を切断する。5'ガイドは全ての公知のアルゴノートによって用いられる。MpAgo-RNA複合体の結晶構造は、5'リン酸相互作用をブロックする残基を含むガイド鎖結合部位を示す。このデータは、5'ヒドロキシル化されたガイドに対する非標準的な特異性を有するアルゴノートサブクラスの進化を示唆する。例として、Kaya et al.,「A bacterial Argonaute with noncanonical guide RNA specificity」,Proc Natl Acad Sci U S A.2016 Apr 12;113(15):4057-62を見よ。これの内容全体は参照によってここに組み込まれる)。他のアルゴノートタンパク質が用いられてもよく、本開示の範囲内であるということは了解されるはずである。
本開示のいくつかの側面は、異なるPAM特異性を有するCas9ドメインを提供する。典型的には、S.pyogenesからのCas9(spCas9)等のCas9タンパク質は、具体的な核酸領域に結合するためには標準的なNGG PAM配列を要求する。これはゲノム上の所望の塩基を編集する能力を限定してもよい。いくつかの態様では、本明細書において提供される塩基編集融合タンパク質は、精密な位置付けに置かれることを必要としてもよい。例えば、ここでは、標的塩基が4塩基領域(例として、「編集窓」)上に置かれ、これがPAMのおよそ15塩基上流にある。Komor,A.C.,et al.,「Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage」Nature 533,420-424(2016)を見よ。これの内容全体は参照によってここに組み込まれる。従って、いくつかの態様では、本明細書において提供される融合タンパク質のいずれかは、標準的な(例として、NGG)PAM配列を含有しないヌクレオチド配列に結合することができるCas9ドメインを含有してもよい。非標準的なPAM配列に結合するCas9ドメインは当該技術分野において記載されており、当業者には明らかであろう。例えば、非標準的なPAM配列に結合するCas9ドメインは、Kleinstiver,B.P.,et al.,「Engineered CRISPR-Cas9 nucleases with altered PAM specificities」Nature 523,481-485(2015);及びKleinstiver,B.P.,et al.,「Broadening the targeting range of Staphylococcus aureus CRISPR-Cas9 by modifying PAM recognition」 Nature Biotechnology 33,1293-1298(2015)に記載されている;夫々の内容全体は参照によってここに組み込まれる。
例えば、次のアミノ酸配列を有する野生型Francisella novicida Cpf1(D917、E1006、及びD1255)(配列番号100)と少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、又は少なくとも99%配列同一性を有するドメイン等の変改されたPAM特異性を有するnapDNAbpドメイン:

野生型Geobacillus thermodenitrificans Cas9(配列番号55)と少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、又は少なくとも99%配列同一性を有するドメイン等の変改されたPAM特異性を有する追加のnapDNAbpドメイン。いくつかの態様では、核酸プログラム型DNA結合タンパク質(napDNAbp)は、標準的な(NGG)PAM配列を要求しない核酸プログラム型DNA結合タンパク質である。いくつかの態様では、napDNAbpはアルゴノートタンパク質である。かかる核酸プログラム型DNA結合タンパク質の1つの例は、Natronobacterium gregoryiからのアルゴノートタンパク質(NgAgo)である。NgAgoはssDNAによってガイドされるエンドヌクレアーゼである。NgAgoは~24ヌクレオチドの5'リン酸化されたssDNA(gDNA)に結合してそれをその標的部位へとガイドし、gDNA部位においてDNA二本鎖切断を作製するであろう。Cas9とは対照的に、NgAgo-gDNAシステムはプロトスペーサ隣接モチーフ(PAM)を要求しない。ヌクレアーゼ不活性なNgAgo(dNgAgo)を用いることは、標的化され得る塩基を多大に拡張してもよい。NgAgoの特徴付け及び使用はGao et al.,Nat Biotechnol.,34(7):768-73(2016),PubMed PMID:27136078;Swarts et al.,Nature,507(7491):258-61(2014);及びSwarts et al.,Nucleic Acids Res.43(10)(2015):5120-9に記載されている。これらの夫々は参照によって本明細書に組み込まれる。Natronobacterium gregoryi Argonauteの配列は配列番号101によって提供される。
開示される融合タンパク質は、次のアミノ酸配列を有する野生型Natronobacterium gregoryi Argonaute(配列番号101)と少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、又は少なくとも99%配列同一性を有するnapDNAbpドメインを含んでもよい:

加えて、いずれかの利用可能な方法が、バリアント又は変異体Cas9タンパク質を得るか又は構築するために利用されてもよい。本明細書において用いられる用語「変異」は、別の残基によるある配列、例として核酸若しくはアミノ酸配列上の残基の置換、又はある配列上の1以上の残基の欠失若しくは挿入を指す。変異は、典型的には、元の残基、次に配列上の残基の位置及び新たに置換された残基の同一性を同定することによって本明細書に記載される。本明細書において提供されるアミノ酸置換(変異)を作るための種々の方法が当該技術分野において周知であり、例えばGreen and Sambrook,Molecular Cloning:A Laboratory Manual(4th ed.,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,N.Y.(2012))によって提供される。変異は、種々のカテゴリー、例えば1塩基多型、微小重複領域、インデル、及び逆位を包含し得、決して限定することを意味されない。変異は、タンパク質活性を縮減又は廃止する変異の正常の結果である「機能喪失」変異を包含し得る。ほとんどの機能喪失変異は劣性である。なぜなら、ヘテロ接合体では、第2の染色体コピーが完全に機能的なタンパク質をコードする遺伝子の未変異のバージョンを持ち、この存在が変異の効果を補償するからである。変異は、「機能獲得」変異をもまた包摂する。これは、正常な条件下ではさもなければ存在しない異常な活性をタンパク質又は細胞に付与するものである。多くの機能獲得変異はコード領域よりもむしろ制御配列にあり、よって、いくつもの帰結を有し得る。例えば、変異は、1以上の遺伝子が間違った組織において発現されることに至り得、これらの組織は、それらが正常では欠く機能を獲得する。それらの性質ゆえに、機能獲得変異は通常は優性である。
変異は部位特異的変異導入を用いて参照Cas9タンパク質上に導入され得る。当該技術分野で公知の部位特異的変異導入のより古い方法は、一本鎖DNA鋳型の単離を許すM13バクテリオファージベクター等のベクター上への変異させられるべき配列のサブクローニングに依拠する。これらの方法では、変異誘発プライマー(すなわち、変異させられるべき部位にアニールすることができるが、変異させられるべき部位に1以上のミスマッチのヌクレオチドを持つプライマー)を一本鎖鋳型にアニーリングさせ、次いで、変異誘発プライマーの3'端からスタートして鋳型の相補体を重合する。次いで、もたらされた二重鎖はホスト細菌に形質トランスバーションされ、プラークが所望の変異についてスクリーニングされる。より最近では、部位特異的変異導入はPCR方法論を使用している。これらは一本鎖鋳型を要求しないという利点を有する。加えて、サブクローニングを要求しない方法が開発されている。PCRに基づく部位特異的変異導入が行われるときには、いくつかのイシューが考慮されなければならない。第1に、これらの方法では、ポリメラーゼによって導入される望まれない変異の拡張を防止するために、PCRサイクル数を縮減することが望ましい。第2に、反応中に残存する変異していない親の分子の数を縮減するために、セレクションが使用されなければならない。第3に、単一のPCRプライマーセットの使用を許すために、伸長された長さのPCR法が好ましい。そして第4に、いくつかの熱安定性ポリメラーゼの非鋳型依存性末端伸長活性ゆえに、多くの場合には、PCRによって生成された変異体産物の平滑末端ライゲーションに先行して、端を磨く工程を手続きに組み込むことが必要である。
変異は、指向性進化プロセス、例えばファージによって補助される連続的進化(PACE)又はファージによって補助される非連続的進化(PANCE)によってもまた導入されてもよい。本明細書において用いられる用語「ファージによって補助される連続的進化(PACE)」は、ファージをウイルスベクターとして使用する連続的進化を指す。PACEテクノロジーの一般概念は、例えば、2010年3月11日にWO2010/028347として公開された2009年9月8日出願の国際PCT出願PCT/US2009/056194;2012年6月28日にWO2012/088381として公開された2011年12月22日出願の国際PCT出願PCT/US2011/066747;2015年5月5日登録のU.S.出願U.S.特許No.9,023,594、2015年9月11日にWO2015/134121として公開された2015年1月20日出願の国際PCT出願PCT/US2015/012022、及び2016年10月20日にWO2016/168631として公開された2016年4月15日出願の国際PCT出願PCT/US2016/027795に記載されている。これらの夫々の内容全体は参照によって本明細書に組み込まれる。バリアントCas9はファージによって補助される非連続的進化(PANCE)によってもまた得られてもよい。本明細書において用いられるこれは、ファージをウイルスベクターとして使用する非連続的進化を指す。PANCEは急速なin vivoの指向性進化のための単純化された技術であり、新しいE.coliホスト細胞による進化させられるべき目当ての遺伝子を含有する進化する「セレクションファージ」(SP)の連続フラスコ植え継ぎを用い、それによって、ホストE.coli内の遺伝子が一定に保たれることを許しながら、SPに含有される遺伝子は連続的に進化する。連続フラスコ植え継ぎは微生物の実験室進化のための広くアクセス可能なアプローチとして長く用をなしており、より最近では、バクテリオファージ進化のための類縁のアプローチが開発されている。PANCEシステムはPACEシステムよりも低いストリンジェンシーを特色とする。
Cas9又はCas9等価物に関する上に記されている参照のいずれかは、すでにそのように申し立てられていない場合には、それらの全体が参照によってここに組み込まれる。
J.分裂されたPE送達のための割られたnapDNAbpドメイン
種々の態様において、本明細書に記載のプライム編集因子は2以上のフラグメントとして細胞に送達されてもよく、これらは、細胞内において、再構成されたプライム編集因子へとアセンブリされるようになる(受動的アセンブリによるか、又は分裂インテイン配列を用いること等の能動的アセンブリによる)。いくつかのケースでは、自己アセンブリは受動的であってもよく、これによって、2以上のプライム編集因子フラグメントは細胞内で共有結合的に又は非共有結合的に会合してプライム編集因子を再構成する。他のケースでは、自己アセンブリはフラグメントの夫々に組み入れされた二量体化ドメインによって触媒されてもよい。二量体化ドメインの例は本明細書に記載される。なお他のケースでは、自己アセンブリは、プライム編集因子フラグメントの夫々に組み入れされた分裂インテイン配列によって触媒されてもよい。
種々の態様において、本明細書に記載のプライム編集因子は2以上のフラグメントとして細胞に送達されてもよく、これらは、細胞内において、再構成されたプライム編集因子へとアセンブリされるようになる(受動的アセンブリによるか、又は分裂インテイン配列を用いること等の能動的アセンブリによる)。いくつかのケースでは、自己アセンブリは受動的であってもよく、これによって、2以上のプライム編集因子フラグメントは細胞内で共有結合的に又は非共有結合的に会合してプライム編集因子を再構成する。他のケースでは、自己アセンブリはフラグメントの夫々に組み入れされた二量体化ドメインによって触媒されてもよい。二量体化ドメインの例は本明細書に記載される。なお他のケースでは、自己アセンブリは、プライム編集因子フラグメントの夫々に組み入れされた分裂インテイン配列によって触媒されてもよい。
分裂されたPE送達は、異なる送達アプローチの種々のサイズ制約に対処するために有利であってもよい。例えば、送達アプローチは、ウイルスに基づく送達方法、メッセンジャーRNAに基づく送達方法、又はRNPに基づく送達(リボヌクレオタンパク質に基づく送達)を包含してもよい。そして、送達のこれらの方法の夫々は、プライム編集因子をより小さいピースへと割ることによって、より効率的及び/又は有効であってもよい。ひとたび細胞内においては、より小さいピースは機能的なプライム編集因子へとアセンブリし得る。分裂の手段に依存して、割られたプライム編集因子フラグメントは非共有結合的様式又は共有結合的様式で再アセンブリされてプライム編集因子を再形成し得る。1つの態様では、プライム編集因子は1以上の分裂部位において2以上のフラグメントへと分裂され得る。フラグメントは(分裂される以外は)未修飾であり得る。ひとたびフラグメントが細胞に送達されると(例として、リボヌクレオタンパク質複合体の直接的送達によるか、又は核酸送達、例としてmRNA送達若しくはウイルスベクターに基づく送達による)、フラグメントは共有結合的に又は非共有結合的に再会合してプライム編集因子を再構成し得る。別の態様では、プライム編集因子は1以上の分裂部位において2以上のフラグメントへと分裂され得る。フラグメントの夫々は二量体化ドメインを含むように修飾され得、これによって、形成された各フラグメントは二量体化ドメインにカップリングされる。ひとたび細胞内に送達又は発現されると、異なるフラグメントの二量体化ドメイン同士は会合し、互いに結合し、異なるプライム編集因子フラグメントを一緒に持って来て、機能的なプライム編集因子を再形成する。更に別の態様では、プライム編集因子フラグメントは分裂インテインを含むように修飾されてもよい。ひとたび細胞内に送達又は発現されると、異なるフラグメントの分裂インテインドメイン同士は会合し、互いに結合し、次いでトランススプライシングを経過する。これが、フラグメントの夫々からの分裂インテインドメインの切除と、フラグメント間のペプチド結合の付随的形成とをもたらし、それによってプライム編集因子を復元する。
1つの態様では、プライム編集因子は分裂インテインアプローチを用いて送達され得る。
分裂部位の位置付けは、プライム編集因子上の残基のいずれか1以上のペアの間に、並びにnapDNAbpドメイン、ポリメラーゼドメイン(例として、RTドメイン)、napDNAbpドメイン及びポリメラーゼドメインを連結するリンカードメイン内を包含するその中のいずれかのドメイン上に、配置され得る。
図66に図示される1つの態様では、プライム編集因子(PE)はnapDNAbp上の分裂部位において割られる。
ある態様において、napDNAbpは配列番号37の標準的なSpCas9ポリペプチドである。ある態様において、SpCas9は、配列番号37の標準的なSpCas9の残基1及び2、若しくは2及び3、若しくは3及び4、若しくは4及び5、若しくは5及び6、若しくは6及び7、若しくは7及び8、若しくは8及び9、若しくは9及び10の間に位置付けられた分裂部位において、又は残基1~10、10~20、20~30、30~40、40~50、50~60、60~70、70~80、80~90、90~100、100~200、200~300、300~400、400~500、500~600、600~700、700~800、800~900、1000~1100、1100~1200、1200~1300、若しくは1300~1368の間のどこかに位置付けられた残基のいずれかの2つのペアの間において、2つのフラグメントへと分裂される。
ある態様において、napDNAbpは、配列番号37の標準的なSpCas9の位置1~10、10~20、20~30、30~40、40~50、50~60、60~70、70~80、80~90、90~100、100~200、200~300、300~400、400~500、500~600、600~700、700~800、800~900、1000~1100、1100~1200、1200~1300、又は1300~1368の間のどこかに位置付けられた残基のいずれか2つのペアに対応する残基のペアに位置付けられる分裂部位において、2つのフラグメントへと分裂される。
ある態様において、SpCas9は、配列番号37の標準的なSpCas9の残基1及び2、若しくは2及び3、若しくは3及び4、若しくは4及び5、若しくは5及び6、若しくは6及び7、若しくは7及び8、若しくは8及び9、若しくは9及び10の間に位置付けられた分裂部位において、又は残基1~10、10~20、20~30、30~40、40~50、50~60、60~70、70~80、80~90、90~100、100~200、200~300、300~400、400~500、500~600、600~700、700~800、800~900、1000~1100、1100~1200、1200~1300、若しくは1300~1368の間のどこかに位置付けられた残基のいずれか2つのペアの間において、2つのフラグメントへと分裂される。ある態様において、分裂部位は、1以上のポリペプチド結合部位(すなわち、「分裂部位又は分裂インテイン分裂部位」)に位置付けられ、分裂インテインに融合され、次いで、別個にコードされる融合タンパク質として細胞に送達される。ひとたび分裂インテイン融合タンパク質同士(すなわち、タンパク質の半分同士)が細胞内で発現されると、タンパク質はトランススプライシングを経過して、連結された分裂インテイン配列の付随的除去によって完全な又は丸ごとのPEを形成する。
例えば、図66に示されている通り、N末端エクステインは第1の分裂インテイン(例として、Nインテイン)に融合され得、C末端エクステインは第2の分裂インテイン(例として、Cインテイン)に融合され得る。細胞内におけるNインテイン及びCインテインの自己会合、次にそれらの自己切除と丸ごとのプライム編集因子(PE)のN末端エクステイン及びC末端エクステイン部分の間のペプチド結合の付随的形成とによって、N末端エクステインはC末端エクステインに融合されるようになって、napDNAbpドメイン及びポリメラーゼドメイン(例として、RTドメイン)を含む丸ごとのプライム編集因子を再形成する。
分裂インテインを用いる分裂されたPE送達戦略を利用するためには、プライム編集因子は、1以上の分裂部位において割られて、プライム編集因子の少なくとも2つの別個の半分を作成することを必要とする。これらの夫々は、各半分が分裂インテイン配列に融合される場合には、細胞内で再連結されてもよい。
ある態様において、プライム編集因子は単一の分裂部位において分裂される。ある種の他の態様において、プライム編集因子は2つの分裂部位、若しくは3つの分裂部位、若しくは4つの分裂部位、又はより多くにおいて分裂される。
好ましい態様では、プライム編集因子は単一の分裂部位において分裂されて、プライム編集因子の2つの別個の半分を作成する。これらの夫々は分裂インテイン配列に融合され得る。
例示的な分裂インテインは、2つのサブユニット、すなわちDnaE-N及びDnaE-Cを含むSsp DnaEインテインである。2つの異なるサブユニットは、DnaE-N及びDnaE-Cサブユニットを夫々コードする別々の遺伝子、すなわちdnaE-n及びdnaE-cによってコードされる。DnaEは、Synechocytis sp.PCC6803における天然に存在する分裂インテインであり、それぞれがDnaE-N又はDnaE-Cのいずれかとの融合物を含む2つの別個のタンパク質のトランススプライシングを指示することができる。
更なる天然に存在するか又は操作されたスプリットインテイン配列は、本明細書に記載の全インテイン配列又は当技術分野で利用可能なものから公知であるか、又は作製され得る。スプリットインテイン配列の例は、Stevens et al.,「A promiscuous split intein with expanded protein engineering applications,」 PNAS,2017,Vol.114:8538-8543;Iwai et al.,「Highly efficient protein trans-splicing by a naturally split DnaE intein from Nostoc punctiforme,FEBS Lett,580:1853-1858に見出され得、これらの各々は参照により本明細書に組み込まれる)。更なる分裂インテイン配列は、例えば、国際公開第2013/045632号、国際公開第2014/055782号、国際公開第2016/069774号、及び欧州特許第2877490号に見出され得、これらの各々の内容は参照により本明細書に組み込まれる。
更に、トランスでのタンパク質スプライシングがin vivo及びin vitroで記載されている(Shingledecker,et al.,Gene 207:187(1998),Southworth,et al.,EMBO J.17:918(1998);Mills,et al.,Proc.Natl.Acad.Sci.USA,95:3543-3548(1998);Lew,et al.,J.Biol.Chem.,273:15887-15890(1998);Wu,et al.,Biochim.Biophys.Acta 35732:1(1998b),Yamazaki,et al.,J.Am.Chem.Soc.120:5591(1998),Evans,et al.,J.Biol.Chem.275:9091(2000);Otomo,et al.,Biochemistry 38:16040-16044(1999);Otomo,et al.,J.Biolmol.NMR 14:105-114(1999);Scott,et al.,Proc.Natl.Acad.Sci.USA 96:13638-13643(1999))、例として、別々に発現される二つの半分からの完全なプライム編集因子の形成に関する図66及び図67に示されるように、機能的産物を形成するためにその後ライゲーションを受ける2つの不活性フラグメントに関するタンパク質を発現する機会を提供する。
本明細書に記載の様々な態様では、連続進化法(例として、PACE)が使用されて、ベースエディタの第1の部分を進化させてもよい。第1の部分は、単一の構成要素又はドメイン、例としてCas9ドメイン、デアミナーゼドメイン、又はUGIドメインを包含し得た。次いで、別々に進化した構成要素又はドメインが、分裂インテインポリペプチドドメインを有する進化した部分及び残りの未進化の部分の両方を別々に発現させることによって、細胞内のベースエディタの残りの部分に融合され得る。第1の部分は、本明細書に記載の連続進化法を使用して進化されることが望まれるベースエディタの任意の第1のアミノ酸部分をより広く包含し得た。第2の部分は、この態様では、本明細書の方法を使用して進化されていないベースエディタの残りのアミノ酸部分を指す。進化したベースエディタの第1の部分及び第2の部分はそれぞれ、細胞内の分裂インテインポリペプチドドメインで発現され得た。細胞の天然のタンパク質スプライシング機構は、進化した第1の部分及び未進化の第2の部分を再構築して、単一の融合タンパク質進化型塩基編集因子を形成するであろう。進化した第1の部分は、単一の融合タンパク質のN末端部分又はC末端部分のいずれかを含んでもよい。同様に、第2の直交トランススプライシングインテイン対の使用は、進化した第1の部分が単一の融合タンパク質の内部部分を含むことを可能にし得た。
したがって、本明細書に記載のベースエディタの進化したコンポーネント及び進化していないコンポーネントのいずれも、細胞内の進化したコンポーネント及び進化していないコンポーネントを含む完全なベースエディタの形成を容易にするために、分裂インテインタグで表現されてもよい。
タンパク質スプライシングプロセスの機序は多大に詳細に研究されており(Chong,et al.,J.Biol.Chem.1996,271,22159-22168;Xu,M-Q&Perler,F.B.EMBO Journal,1996,15,5146-5153)、保存されたアミノ酸がインテイン及びエクステインスプライシング点に見出されている(Xu,et al.,EMBO Journal,1994,13 5517-522)。本明細書に記載の構築物は、第1の遺伝子(例として、塩基編集因子の進化した部分)の5'末端に融合されたインテイン配列を含有する。好適なインテイン配列は、タンパク質スプライシングエレメントを含有することが公知のタンパク質のいずれかから選択され得る。全ての公知のインテインを含有するデータベースはワールドワイドウェブ上に見出され得る(Perler,F.B.Nucleic Acids Research,1999,27,346-347)。インテイン配列は3'端において第2の遺伝子の5'端に融合される。ある種のオルガネラへのこの遺伝子の標的化のためには、ペプチドシグナルが遺伝子のコード配列に融合され得る。第2の遺伝子の後に、インテイン遺伝子配列は、同じ細胞における複数のタンパク質の発現のために所望の回数だけ繰り返され得る。マルチインテイン含有構築物では、異なるソースからのインテイン要素を用いることが有用であってもよい。発現されるべき最後の遺伝子の配列の後には、転写終結配列が挿入されなければならない。1つの態様では、それがインテインからのエクステインの切除を触媒すると共にエクステインのライゲーションを防止し得るようにして、修飾されたインテインスプライシング単位がデザインされる。Pyrococcus種GB-D DNAポリメラーゼ上のC末端エクステインジャンクションの変異導入は、変改されたスプライシングエレメントを生ずることが見出された。これは、エクステイン及びインテインの切断を誘導するが、エクステインの続くライゲーションを防止する(Xu,M-Q&Perler,F.B.EMBO Journal,1996,15,5146-5153)。セリン538からアラニン又はグリシンどちらかへの変異は、切断を誘導したが、ライゲーションを防止した。インテインへのC末端エクステインジャンクションにおけるアミノ酸の保存を原因として、他のインテインスプライシング単位における同等残基の変異もまたエクステインライゲーションを防止するはずである。エンドヌクレアーゼドメインを含有しない好ましいインテインは、Mycobacterium xenopi GyrAタンパク質である(Telenti,et al.J.Bacteriol.1997,179,6378-6382)。他は天然に見出されているか、又はインテインを含有するエンドヌクレアーゼからエンドヌクレアーゼドメインを除去することによって人工的に作成されている(Chong,et al.J.Biol.Chem.1997,272,15587-15590)。好ましい態様では、インテインは、それがスプライシング機能を果たすために必要とされる最小限の数のアミノ酸からなるようにして選択され、例えばMycobacterium xenopi GyrAタンパク質からのインテインである(Telenti,A.,et al.,J.Bacteriol.1997,179,6378-6382)。代替的な態様では、エンドヌクレアーゼ活性なしのインテインが選択され、例えばMycobacterium xenopi GyrAタンパク質からのインテイン、又はエンドヌクレアーゼドメインを除去するように修飾されたSaccharomyces cerevisiae VMAインテインである(Chong,1997)。インテインスプライシング単位の更なる修飾は、切断反応の反応速度が変改されることを許してもよく、スプライシング単位の遺伝子配列を単純に修飾することによって、タンパク質用量がコントロールされることを許す。
インテインは2つの別個に転写及び翻訳される遺伝子によってコードされる2つのフラグメントとしてもまた存在し得る。これらのいわゆる分裂インテインは自己会合し、タンパク質スプライシング活性をトランスで触媒する。分裂インテインは多様なシアノバクテリア及び古細菌において同定されているが(Caspi et al,Mol Microbiol.50:1569-1577(2003);Choi J.et al,J Mol Biol.556:1093-1106(2006.);Dassa B.et al,Biochemistry.46:322-330(2007.);Liu X.and Yang J.,J Biol Chem.275:26315-26318(2003);Wu H.et al.
Proc Natl Acad Sci USA.£5:9226-9231(1998.);及びZettler J.et al,FEBS Letters.553:909-914(2009))、真核生物ではここまで見出されていない。最近、環境メタゲノムデータのバイオインフォマティクス分析は、新規のゲノム配置を有する26の異なる遺伝子座を明らかにした。各遺伝子座においては、保存された酵素コード領域が分裂インテインによって分断され、自立したエンドヌクレアーゼ遺伝子が、インテインサブドメインをコードするセクション間に挿入されている。それらのうち、5つの遺伝子座が完全にアセンブリされた:DNAヘリカーゼ(gp41-1、gp41-8);イノシン-5'-一リン酸デヒドロゲナーゼ(IMPDH-1);及びリボヌクレオチドレダクターゼ触媒サブユニット(NrdA-2及びNrdJ-1)である。このばらばらになった遺伝子編成は主にファージに存在するように見える(Dassa et al,Nucleic Acids Research.57:2560-2573(2009))。
分裂インテインNpu DnaEは、タンパク質トランススプライシング反応について報告された最も高い速度を有するとして特徴付けられた。加えて、Npu DnaEタンパク質スプライシング反応は、異なるエクステイン配列、6~37℃の温度、及び最高で6Mの尿素の存在についてロバストかつ高収量と考慮される(Zettler J.et al,FEBS Letters.553:909-914(2009);Iwai I.et al,FEBS Letters 550:1853-1858(2006))。予想された通り、これらのインテインのNドメインにおけるCysl Ala変異が導入されたときには、当初のN→Sのアシルシフト、よってタンパク質スプライシングがブロックされた。残念ながら、C末端切断反応もまたほとんど完全に阻害された。N末端の切れやすいペプチド結合におけるアシルシフトに対するC末端スプライスジャンクションにおけるアスパラギン環化の依存性は、天然に分裂されたDnaEインテインアレルに普通の固有の特性であるように見える(Zettler J.et al.FEBS Letters.555:909-914(2009))。
タンパク質スプライシングの機序は、典型的には4つの工程を有する[29-30]:1)インテインN末端におけるN-S又はN-Oアシルシフト。これは上流のペプチド結合を切断し、Nエクステインとインテインの第1のアミノ酸(Cys又はSer)の側鎖との間にエステル結合を形成する;2)NエクステインをインテインC末端へと位置替えするトランスエステル化。新たなエステル結合を形成し、NエクステインをCエクステインの第1のアミノ酸(Cys、Ser、又はThr)の側鎖に連結する;3)インテイン及びCエクステインの間のペプチド結合を切断するAsn環化;並びに4)Nエクステイン及びCエクステインの間においてペプチド結合によってエステル結合を置き換えるS-N又はO-Nアシルシフト。
分裂インテインによって触媒されるタンパク質トランススプライシングは、タンパク質ライゲーションのための全く酵素的な方法を提供する[31]。分裂インテインは本質的には一続きのインテイン(例としてミニインテイン)であり、夫々Nインテイン及びCインテインという名称の2つのピースへと分裂されている。分裂インテインのNインテイン及びCインテインは、非共有結合的に会合して活性なインテインを形成し、一続きのインテインがするのと本質的に同じやり方でスプライシング反応を触媒し得る。分裂インテインは天然に見出され、実験室でもまた操作されている[31-35]。本明細書において用いられる用語「分裂インテイン」は、1以上のペプチド結合切断がN末端及びC末端アミノ酸配列の間に存在するいずれかのインテインを指し、その結果、N末端及びC末端配列が別個の分子になり、これらは、トランススプライシング反応について機能的であるインテインへと非共有結合的に再会合又は再構成し得る。いずれかの触媒的に活性なインテイン又はそのフラグメントが、本発明の方法への使用のための分裂インテインを導出するために用いられてもよい。例えば、一側面において、分裂インテインは真核生物インテインに由来されてもよい。別の側面において、分裂インテインは細菌インテインに由来されてもよい。別の側面において、分裂インテインは古細菌インテインに由来されてもよい。好ましくは、そのように導出された分裂インテインは、トランススプライシング反応を触媒することにとって必須のアミノ酸配列のみを所有するであろう。
本明細書において用いられる「N末端分裂インテイン(In)」は、トランススプライシング反応について機能的であるN末端アミノ酸配列を含むいずれかのインテイン配列を指す。Inは、それゆえに、トランススプライシングが生起するときにスプライスアウトされる配列をもまた含む。Inは、天然に存在するインテイン配列のN末端部分の修飾である配列を含み得る。例えば、かかる追加の及び/又は変異した残基の包含がInをトランススプライシングにおいて非機能的にしない限り、Inは追加のアミノ酸残基及び/又は変異した残基を含み得る。好ましくは、追加の及び/又は変異した残基の包含は、Inのトランススプライシング活性を改善又は増強する。
本明細書において用いられる「C末端分裂インテイン(Ic)」は、トランススプライシング反応について機能的であるC末端アミノ酸配列を含むいずれかのインテイン配列を指す。一側面において、Icは一続きの4~7アミノ酸残基を含み、これらの少なくとも4アミノ酸はそれが由来されたインテイン最後のβ鎖からである。Icは、それゆえに、トランススプライシングが生起するときにスプライスアウトされる配列をもまた含む。Icは、天然に存在するインテイン配列のC末端部分の修飾である配列を含み得る。例えば、かかる追加の及び/又は変異した残基の包含がInをトランススプライシングにおいて非機能的にしない限り、Icは追加のアミノ酸残基及び/又は変異した残基を含み得る。好ましくは、追加の及び/又は変異した残基の包含は、Icのトランススプライシング活性を改善又は増強する。
本発明のいくつかの態様では、Ic又はInに連結されたペプチドは、とりわけ蛍光基、ビオチン、ポリエチレングリコール(PEG)、アミノ酸アナログ、非天然のアミノ酸、リン酸基、グリコシル基、放射性同位体標識、及び医薬分子を包含する追加の化学的部分を含み得る。他の態様において、Icに連結されたペプチドは、とりわけケトン、アルデヒド、Cys残基、及びLys残基を包含する1以上の化学反応性基を含み得る。「インテインスプライシングポリペプチド(ISP)」が存在するときには、分裂インテインのNインテイン及びCインテインは非共有結合的に会合して活性なインテインを形成し、スプライシング反応を触媒し得る。本明細書において用いられる「インテインスプライシングポリペプチド(ISP)」は、Ic、In、又は両方が分裂インテインから除去されるときに留まる分裂インテインのアミノ酸配列の部分を指す。ある態様において、InはISPを含む。別の態様では、IcはISPを含む。更に別の態様では、ISPはInにもIcにも共有結合的に連結されていない別個のペプチドである。
分裂インテインは、ミニインテインの構造上に見出される-12の保存されたベータ鎖間の構造化されていないループ又は介在アミノ酸配列中に1以上の分裂部位を操作することによって、一続きのインテインから作成されてもよい[25-28]。分裂の作成が、タンパク質スプライシング活性が失われる十分な度合まではインテインの構造、特に構造化されたベータ鎖を遮断しないであろうという条件で、ベータ鎖間の領域内の分裂部位の位置のいくらかのフレキシビリティーが存在してもよい。
タンパク質トランススプライシングでは、1つの前駆体タンパク質はNインテインによって後続されるNエクステインパーツからなり、別の前駆体タンパク質はCエクステインパーツによって後続されるCインテインからなり、トランススプライシング反応(一緒にN及びCインテインによって触媒される)が2つのインテイン配列を切除、2つのエクステイン配列をペプチド結合によって連結する。酵素反応であるタンパク質トランススプライシングは、非常に低い(例としてマイクロモル)濃度のタンパク質で働き得、生理条件下において実行され得る。
K.他のプログラム型ヌクレアーゼ
本明細書に記載の種々の態様において、プライム編集因子はCas9タンパク質等のnapDNAbpを含む。これらのタンパク質は、それらがガイドRNA(又は場合に応じてpegRNA)と複合体化するようになることによって「プログラム型」である。これは、gRNA(又はpegRNA)のスペーサ部分に対して相補的である配列を所有し、かつ要求されるPAM配列をもまた所有するDNA上の標的部位へとCas9タンパク質をガイドする。しかしながら、ここで想定されるある態様において、napDNAbpは、異なる型のプログラム型タンパク質、例えばジンクフィンガーヌクレアーゼ又は転写活性化因子様エフェクターヌクレアーゼ(TALEN)によって置換されてもよい。
本明細書に記載の種々の態様において、プライム編集因子はCas9タンパク質等のnapDNAbpを含む。これらのタンパク質は、それらがガイドRNA(又は場合に応じてpegRNA)と複合体化するようになることによって「プログラム型」である。これは、gRNA(又はpegRNA)のスペーサ部分に対して相補的である配列を所有し、かつ要求されるPAM配列をもまた所有するDNA上の標的部位へとCas9タンパク質をガイドする。しかしながら、ここで想定されるある態様において、napDNAbpは、異なる型のプログラム型タンパク質、例えばジンクフィンガーヌクレアーゼ又は転写活性化因子様エフェクターヌクレアーゼ(TALEN)によって置換されてもよい。
図1Jは、napDNAbp(例としてSpCas9ニッカーゼ)をいずれかのプログラム型ヌクレアーゼドメイン、例えばジンクフィンガーヌクレアーゼ(ZFN)又は転写活性化因子様エフェクターヌクレアーゼ(TALEN)によって置き換える本明細書において企図されるプライム編集のかかるバリエーションを図示する。そのため、好適なヌクレアーゼは必ずしも核酸標的化分子(ガイドRNA等)によって「プログラム」されることを必要とせず、むしろ、DNA結合ドメイン、例えば特にヌクレアーゼの特異性を定めることによってプログラムされてもよいということが企図される。ちょうどnapDNAbp部分によるプライム編集のように、かかる代替的なプログラム型ヌクレアーゼは、標的DNAの1つの鎖のみがカットされるようにして修飾されるということが好ましい。換言すると、プログラム型ヌクレアーゼは好ましくはニッカーゼとして機能するべきである。ひとたびプログラム型ヌクレアーゼが選択されると(例として、ZFN又はTALEN)、次いで、それがプライム編集様機序に従って作動することを許すように、追加の機能性がシステム上に操作されてもよい。例えば、プログラム型ヌクレアーゼは(例として、化学的リンカーを介して)RNA又はDNA伸長アームをそれにカップリングすることによって修飾されてもよく、伸長アームはプライマー結合部位(PBS)及びDNA合成鋳型を含む。プログラム型ヌクレアーゼは(例として、化学的又はアミノ酸リンカーを介して)ポリメラーゼにもまたカップリングされてもよい。これの性質は、伸長アームがDNA又はRNAであるかどうかに依存するであろう。RNA伸長アームのケースでは、ポリメラーゼはRNA依存性DNAポリメラーゼであり得る(例として逆転写酵素)。DNA伸長アームのケースでは、ポリメラーゼはDNA依存性DNAポリメラーゼであり得る(例として、Pol I、Pol II、若しくはPol IIIを包含する原核生物ポリメラーゼ、又はPol a、Pol b、Pol g、Pol d、Pol e、若しくはPol zを包含する真核生物ポリメラーゼ)。システムは、丸ごととしての反応を容易化するために、プログラム型ヌクレアーゼへの融合体として追加又はトランスで追加される他の機能性をもまた包含し得る(例として、(a)カットされた部位においてDNAをほどいて、プライマーとして利用可能な3'端を有するカットされた鎖を作るためのヘリカーゼ、(b)カットされた鎖上の内生鎖を除去することを助けて、合成された鎖による内生鎖の置換えの方に反応を駆動するためのFEN1、又は(c)非編集鎖の有利な細胞性修復によって合成修復の取り入れを駆動することを助けてもよい、対向する鎖上の第2部位ニックを作成するためのnCas9:gRNA複合体)。napDNAbpによるプライム編集と類縁の様式で、別様にプログラム型ヌクレアーゼによるかかる複合体は、目当ての編集を持つDNAの新たに合成された置換え鎖を合成し、次いで永久的にDNAの標的部位上に組み入れるために用いられ得る。
好適な代替的なプログラム型ヌクレアーゼは当該技術分野で周知であり、これらは、DNAの標的部位に選択的に結合するようにプログラムされ得る代替的なプライム編集因子システムを構築するために、napDNAbp:gRNA複合体の代わりに用いられてもよい。かつ、これらは、ポリメラーゼとプライマー結合部位及びDNA合成鋳型を含むRNA又はDNA伸長アームとを特定のニック部位へと共局在させるために、上に記載されている様式で更に修飾され得る。例えば、図1Jによって表される通り、転写活性化因子様エフェクターヌクレアーゼ(TALEN)が、プログラム型ヌクレアーゼとして、本明細書に記載のプライム編集法及び組成物に用いられてもよい。TALENは、TALエフェクターDNA結合ドメインをDNA切断ドメインに融合することによって生成される人工の制限酵素である。これらの試薬は効率的な、プログラム型、かつ特異的なDNA切断を可能化し、in situのゲノム編集のための強力なツールを表す。転写活性化因子様エフェクター(TALE)は、事実上いずれかのDNA配列に結合するように迅速に操作され得る。本明細書において用いられる用語TALENは幅広く、別のTALENからの補助なしに二本鎖DNAを切断し得るモノマーTALENを包含する。用語TALENは、同じ部位においてDNAを切断するために一緒に働くように操作されているTALENのペアの1つ又は両方のメンバーを指すためにもまた用いられる。一緒に働くTALENは左TALEN及び右TALENと呼ばれてもよい。これはDNAの掌性を参照している。米国第12/965,590号;米国第13/426,991号(米国特許第8,450,471号);米国第13/427,040号(米国特許第8,440,431号);米国第13/427,137号(米国特許第8,440,432号);及び米国第13/738,381号を見よ。これらの全てはそれらの全体が参照によって本明細書に組み込まれる。加えて、TALENはWO2015/027134、US 9,181,535、Boch et al.,「Breaking the Code of DNA Binding Specificity of TAL-Type III Effectors」,Science,vol.326,pp.1509-1512(2009),Bogdanove et al.,TAL Effectors:Customizable Proteins for DNA Targeting,Science,vol.333,pp.1843-1846(2011),Cade et al.,「Highly efficient generation of heritable zebrafish gene mutations using homo-and heterodimeric TALENs」,Nucleic Acids Research,vol.40,pp.8001-8010(2012),及びCermak et al.,「Efficient design and assembly of custom TALEN and other TAL effector-based constructs for DNA targeting」,Nucleic Acids Research,vol.39,No.17,e82(2011)に記載されている。これらの夫々は参照によって本明細書に組み込まれる。
図1Jに表されている通り、ジンクフィンガーヌクレアーゼもまた、プライム編集への使用のための代替的なプログラム型ヌクレアーゼとして、Cas9ニッカーゼ等のnapDNAbpの代わりに用いられてもよい。TALENのように、ZFNタンパク質は、それらがニッカーゼとして機能するようにして修飾されてもよい。すなわち、本明細書に記載のプライム編集因子に用いられるnapDNAbpと類似の様式で、それが標的DNAの1つの鎖のみを切断するようにして、ZFNを操作する。ZFNタンパク質は、当該技術分野において、例えば、Carroll et al.,「Genome Engineering with Zinc-Finger Nucleases,」Genetics,Aug 2011,Vol.188:773-782;Durai et al.,「Zinc finger nucleases:custom-designed molecular scissors for genome engineering of plant and mammalian cells,」Nucleic Acids Res,2005,Vol.33:5978-90;及びGaj et al.,「ZFN,TALEN,and CRISPR/Cas-based methods for genome engineering,」Trends Biotechnol.2013,Vol.31:397-405(これらの各々はその全体が参照されることによって本明細書に組み込まれる)に広範囲に記載されている。
[3]ポリメラーゼ(例として逆転写酵素)
種々の態様において、本明細書に開示のプライム編集因子(PE)システムはポリメラーゼ(例として、DNA依存性DNAポリメラーゼ又はRNA依存性DNAポリメラーゼ、例えば逆転写酵素)又はそのバリアントを包含し、これは、napDNAbp若しくは他のプログラム型ヌクレアーゼとの融合タンパク質として提供又はトランスで提供され得る。
種々の態様において、本明細書に開示のプライム編集因子(PE)システムはポリメラーゼ(例として、DNA依存性DNAポリメラーゼ又はRNA依存性DNAポリメラーゼ、例えば逆転写酵素)又はそのバリアントを包含し、これは、napDNAbp若しくは他のプログラム型ヌクレアーゼとの融合タンパク質として提供又はトランスで提供され得る。
いずれかのポリメラーゼが、本明細書に開示のプライム編集因子に用いられてもよい。ポリメラーゼは野生型ポリメラーゼ、機能的フラグメント、変異体、バリアント、又は短縮されたバリアント、及び同類であってもよい。ポリメラーゼは、真核、原核、古細菌、若しくはウイルス生物からの野生型ポリメラーゼを包含してもよく、及び/又はポリメラーゼは遺伝子工学、変異導入、指向性進化に基づくプロセスによって修飾されてもよい。ポリメラーゼはT7 DNAポリメラーゼ、T5 DNAポリメラーゼ、T4 DNAポリメラーゼ、KlenowフラグメントDNAポリメラーゼ、DNAポリメラーゼIII、及び同類を包含してもよい。ポリメラーゼはまた、熱安定性であってもよく、Taq、Tne、Tma、Pfu、Tfl、Tth、Stoffelフラグメント、VENT(登録商標)及びDEEPVENT(登録商標)DNAポリメラーゼ、KOD、Tgo、JDF3、並びにそれらの変異体、バリアント、及び誘導体を包含してもよい(米国特許第5,436,149号;米国特許第4,889,818号;米国特許第4,965,185号;米国特許第5,079,352号;米国特許第5,614,365号;米国特許第5,374,553号;米国特許第5,270,179号;米国特許第5,047,342号;米国特許第5,512,462号;WO 92/06188;WO 92/06200;WO 96/10640;Barnes,W.M.,Gene 112:29-35(1992);Lawyer,F.C.,et al.,PCR Meth.Appl.2:275-287(1993);Flaman,J.-M,et al.,Nuc.Acids Res.22(15):3259-3260(1994)(これらの各々は参照により本明細書に組み込まれる)を参照)。より長い核酸分子(例として、長さが約3-5Kbよりも長い核酸分子)の合成には、少なくとも2つのDNAポリメラーゼが使用され得る。ある態様において、ポリメラーゼの1つは3'エキソヌクレアーゼ活性を実質的に欠き得、他は3'エキソヌクレアーゼ活性を有してもよい。かかるペアリングは同じか又は異なるポリメラーゼを包含してもよい。3'エキソヌクレアーゼ活性を実質的に欠くDNAポリメラーゼの例は、Taq、Tne(exo-)、Tma(exo-)、Pfu(exo-)、Pwo(exo-)、exo-KOD、及びTth DNAポリメラーゼ、並びにそれらの変異体、バリアント、及び誘導体を包含するが、これらに限定されない。
好ましくは、本明細書に開示のプライム編集因子に使用可能なポリメラーゼは「鋳型依存性」ポリメラーゼである(なぜなら、ポリメラーゼは、プライム編集の間に合成中のDNA鎖の配列を規定するためにはDNA合成鋳型に依拠することを意図されるからである。本明細書において用いられる用語「鋳型DNA分子」は、相補的な核酸鎖がDNAポリメラーゼによって例えばpegRNAのDNA合成鋳型のプライマー伸長反応によって次いで合成される核酸の鎖を指す。
本明細書において用いられる用語「鋳型依存的様式」は、プライマー分子の鋳型依存的伸長(例として、DNAポリメラーゼによるDNA合成)を伴うプロセスを指すことを意図される。用語「鋳型依存的様式」は、ポリヌクレオチドの新たに合成される鎖の配列が相補的な塩基対形成の周知のルールによって記述されるRNA又はDNAのポリヌクレオチド合成を指す(例えば、Watson,J.D.et al.,In:Molecular Biology of the Gene,4th Ed.,W.A.Benjamin,Inc.,Menlo Park,Calif.(1987)を見よ)。用語「相補的」は、塩基対形成による2つのポリヌクレオチド鎖の領域間の又は2つのヌクレオチド間の配列相補性の幅広い概念を指す。アデニンヌクレオチドは、チミン又はウラシルであるヌクレオチドと特異的な水素結合を形成する(「塩基対形成する」)ことができるということが公知である。類似に、シトシンヌクレオチドは、グアニンヌクレオチドと塩基対形成することができるということが公知である。そのため、プライム編集のケースでは、DNA合成鋳型に対してプライム編集因子のポリメラーゼによって合成されるDNAの一本鎖は、DNA合成鋳型の配列に対して「相補的」であると言われるということが言われ得る。
A.例示のポリメラーゼ
種々の態様において、本明細書に記載のプライム編集因子はポリメラーゼを含む。本開示は、いずれかの天然に存在する生物若しくはウイルスから得られるか、又は市販の若しくは非市販のソースから得られるいずれかの野生型ポリメラーゼを企図する。加えて、本開示のプライム編集因子に使用可能なポリメラーゼは、いずれかの天然に存在する変異体ポリメラーゼ、操作された変異体ポリメラーゼ、又は他のバリアントポリメラーゼを包含し得、機能を保持する短縮されたバリアントを包含する。本明細書において使用可能なポリメラーゼは、特定のアミノ酸置換、例えば具体的に本明細書に開示のものを含有するようにもまた操作されてもよい。ある種の好ましい態様では、本開示のプライム編集因子に使用可能なポリメラーゼは、鋳型に基づくポリメラーゼである。すなわち、それらは鋳型依存的な様式でヌクレオチド配列を合成する。
種々の態様において、本明細書に記載のプライム編集因子はポリメラーゼを含む。本開示は、いずれかの天然に存在する生物若しくはウイルスから得られるか、又は市販の若しくは非市販のソースから得られるいずれかの野生型ポリメラーゼを企図する。加えて、本開示のプライム編集因子に使用可能なポリメラーゼは、いずれかの天然に存在する変異体ポリメラーゼ、操作された変異体ポリメラーゼ、又は他のバリアントポリメラーゼを包含し得、機能を保持する短縮されたバリアントを包含する。本明細書において使用可能なポリメラーゼは、特定のアミノ酸置換、例えば具体的に本明細書に開示のものを含有するようにもまた操作されてもよい。ある種の好ましい態様では、本開示のプライム編集因子に使用可能なポリメラーゼは、鋳型に基づくポリメラーゼである。すなわち、それらは鋳型依存的な様式でヌクレオチド配列を合成する。
ポリメラーゼは、本明細書に記載のプライム編集因子システムに関して用いられてもよい、ヌクレオチド鎖を合成する酵素である。ポリメラーゼは好ましくは「鋳型依存性」ポリメラーゼである(すなわち、鋳型鎖のヌクレオチド塩基の順序に基づいてヌクレオチド鎖を合成するポリメラーゼ)。ある種の構成では、ポリメラーゼは「鋳型非依存性」でもまたあり得る(すなわち、鋳型鎖の要件なしにヌクレオチド鎖を合成するポリメラーゼ)。ポリメラーゼは更に「DNAポリメラーゼ」又は「RNAポリメラーゼ」としてもまた類別されてもよい。種々の態様において、プライム編集因子システムはDNAポリメラーゼを含む。種々の態様において、DNAポリメラーゼは「DNA依存性DNAポリメラーゼ」であり得る(すなわち、それによると、鋳型分子はDNAの鎖である)。かかるケースでは、DNA鋳型分子はpegRNAであり得、伸長アームはDNAの鎖を含む。かかるケースでは、pegRNAはキメラ又はハイブリッドpegRNAと呼ばれてもよく、これはRNA部分(すなわち、スペーサ及びgRNAコアを包含するガイドRNA構成要素)とDNA部分(すなわち伸長アーム)とを含む。種々の他の態様において、DNAポリメラーゼは「RNA依存性DNAポリメラーゼ」であり得る(すなわち、それによると、鋳型分子はRNAの鎖である)。かかるケースでは、pegRNAはRNAであり、すなわちRNA伸長を包含する。用語「ポリメラーゼ」はまた、ヌクレオチドの重合を触媒する酵素を指してもよい(すなわちポリメラーゼ活性)。一般的に、酵素は、ポリヌクレオチド鋳型配列にアニーリングしたプライマー(例として、pegRNAのプライマー結合部位にアニーリングしたプライマー配列等)の3'端において合成を開始し、鋳型鎖の5'端の方へ進むであろう。「DNAポリメラーゼ」はデオキシヌクレオチドの重合を触媒する。DNAポリメラーゼを参照して本明細書において用いられる用語DNAポリメラーゼは、「その機能的フラグメント」を包含する。「その機能的フラグメント」は、少なくとも1つのセットの条件下においてはポリヌクレオチドの重合を触媒する能力を保持する、ポリメラーゼのアミノ酸配列全体未満を包摂する野生型又は変異体DNAポリメラーゼのいずれかの部分を指す。かかる機能的フラグメントは別個の実態として存在し得てもよいか、又はそれは融合タンパク質等のより大きいポリペプチドの構成要素であってもよい。
いくつかの態様では、ポリメラーゼはバクテリオファージからであり得る。バクテリオファージDNAポリメラーゼは5'→3'のエキソヌクレアーゼ活性を一般的に欠いている。なぜなら、この活性は別個のポリペプチドによってコードされるからである。好適なDNAポリメラーゼの例はT4、T7、及びファイ29 DNAポリメラーゼである。市販で利用可能な酵素は:T4(多くのソース、例としてEpicentreから利用可能)及びT7(多くのソース、例として未修飾についてはEpicentre及び3'→5'のエキソT7「Sequenase」DNAポリメラーゼについてはUSBから利用可能)である。
他の態様、ポリメラーゼは古細菌ポリメラーゼである。古細菌において同定されているDNAポリメラーゼの2つの異なるクラスがある:1.ファミリーB/pol I型(Pyrococcus furiosusからのPfuのホモログ)及び2.pol II型(P.furiosus DP1/DP2 2サブユニットポリメラーゼのホモログ)である。両クラスからのDNAポリメラーゼは、関連する5'→3'のエキソヌクレアーゼ活性を天然に欠くことと、3'→5'のエキソヌクレアーゼ(プルーフリーディング)活性を所有することとが示されている。好適なDNAポリメラーゼ(pol I又はpol II)は、所望のアッセイ温度に類似である最適な成長温度を有する古細菌に由来され得る。
熱安定性古細菌DNAポリメラーゼはPyrococcus種(furiosus、sp.GB-D、woesii、abysii、horikoshii)、Thermococcus種(kodakaraensis KOD1、litoralis、sp.9 degrees North-7、sp.JDF-3、gorgonarius)、Pyrodictium occultum、及びArchaeoglobus fulgidusから単離される。
ポリメラーゼはまた、ユーバクテリウム種からであってもよいユーバクテリウムDNAポリメラーゼの3つのクラス、pol I、II、及びIIIがある。Pol I DNAポリメラーゼファミリーの酵素は5'→3'のエキソヌクレアーゼ活性を所有し、ある種のメンバーは3'→5'のエキソヌクレアーゼ活性をもまた発揮する。Pol II DNAポリメラーゼは5'→3'のエキソヌクレアーゼ活性を天然に欠くが、3'→5'のエキソヌクレアーゼ活性を発揮する。Pol III DNAポリメラーゼは細胞の主要な複製DNAポリメラーゼを表し、複数のサブユニットから成る。pol III触媒サブユニットは5'→3'のエキソヌクレアーゼ活性を欠くが、いくつかのケースでは、3'→5'のエキソヌクレアーゼ活性が同じポリペプチド上に位置付けられる。
種々の市販で利用可能なPol I DNAポリメラーゼがあり、これらのいくつかは5'→3'のエキソヌクレアーゼ活性を縮減又は廃止するように修飾されている。
好適な熱安定性pol I DNAポリメラーゼは、Thermus種及びThermotoga maritima、例えばThermus aquaticus(Taq)、Thermus thermophilus(Tth)、及びThermotoga maritima(Tma UlTma)を包含する種々の好熱ユーバクテリウムから単離され得る。
上に列記されているものに関係する追加のユーバクテリウムはThermophilic Bacteria(Kristjansson,J.K.,ed.)CRC Press,Inc.,Boca Raton,Fla.,1992に記載されている。
本発明は、更に、米国特許第5,677,152号、第6,479,264号、及び第6,183,998号に開示されている方法に従って化学的に修飾されているキメラ又は非キメラDNAポリメラーゼを可能にする。これらの内容はそれらの全体が参照によってここに組み込まれる。
上に列記されているものに関係する追加の古細菌DNAポリメラーゼは次の参照に記載されている:Archaea:A Laboratory Manual(Robb,F.T.and Place,A.R.,eds.),Cold Spring Harbor Laboratory Press,Cold Spring Harbor,N.Y.,1995及びThermophilic Bacteria(Kristjansson,J.K.,ed.)CRC Press,Inc.,Boca Raton,Fla.,1992に記載されている。
B.例示の逆転写酵素
種々の態様において、本明細書に記載のプライム編集因子はポリメラーゼとして逆転写酵素を含む。本開示は、いずれかの天然に存在する生物若しくはウイルスから得られるか又は市販の若しくは非市販のソースから得られるいずれかの野生型逆転写酵素を企図する。加えて、本開示のプライム編集因子に使用可能な逆転写酵素は、いずれかの天然に存在する変異体RT、操作された変異体RT、又は他のバリアントRTを包含し得、機能を保持する短縮されたバリアントを包含する。RTは、特定のアミノ酸置換、例えば具体的に本明細書に開示のものを含有するようにもまた操作されてもよい。
種々の態様において、本明細書に記載のプライム編集因子はポリメラーゼとして逆転写酵素を含む。本開示は、いずれかの天然に存在する生物若しくはウイルスから得られるか又は市販の若しくは非市販のソースから得られるいずれかの野生型逆転写酵素を企図する。加えて、本開示のプライム編集因子に使用可能な逆転写酵素は、いずれかの天然に存在する変異体RT、操作された変異体RT、又は他のバリアントRTを包含し得、機能を保持する短縮されたバリアントを包含する。RTは、特定のアミノ酸置換、例えば具体的に本明細書に開示のものを含有するようにもまた操作されてもよい。
逆転写酵素は多機能酵素であり、典型的には、RNA及びDNA依存性DNA重合活性、並びにRNA-DNAハイブリッド中のRNAの切断を触媒するRNaseH活性を包含する3つの酵素活性を有する。逆転写酵素のいくつかの変異体は、mRNAの意図されない損傷を防止するためにRNaseH部分を不能化している。mRNAを鋳型として用いて相補的なDNA(cDNA)を合成するこれらの酵素は、RNAウイルスにおいて最初に同定された。続いて逆転写酵素は直接的にウイルス粒子、細胞、又は組織から単離及び精製された(例として、Kacian et al.,1971,Biochim.Biophys.Acta 46:365-83;Yang et al.,1972,Biochem.Biophys.Res.Comm.47:505-11;Gerard et al.,1975,J.Virol.15:785-97;Liu et al.,1977,Arch.Virol.55 187-200;Kato et al.,1984,J.Virol.Methods 9:325-39;Luke et al.,1990,Biochem.29:1764-69及びLe Grice et al.,1991,J.Virol.65:7004-07(これらの各々は参照により組み込まれる)を参照)。より最近では、改善された特性、例えば熱安定性、忠実性、及び活性を求めて、変異体及び融合タンパク質が作成されている。当該技術分野で公知であるか又は当該技術分野で公知の方法を用いて作られ得る逆転写酵素の野生型、バリアント、及び/又は変異体形態のいずれかは、本明細書において企図される。
逆転写酵素(RT)遺伝子(又はそれに含有される遺伝情報)はいくつもの異なるソースから得られ得る。例えば、遺伝子は、レトロウイルスに感染している真核細胞から、又はレトロウイルスゲノム全体若しくはそれの部分どちらかを含有するいくつものプラスミドから得られてもよい。加えて、RT遺伝子を含有するメッセンジャーRNA様のRNAがレトロウイルスから得られ得る。RTのソースの例は、モロニーマウス白血病ウイルス(M-MLV又はMLVRT);ヒトT細胞白血病ウイルス1型(HTLV-1);ウシ白血病ウイルス(BLV);ラウス肉腫ウイルス(RSV);ヒト免疫不全ウイルス(HIV);Saccharomycesを包含する酵母、Neurospora、Drosophila;霊長類;及び齧歯類を包含するが、これらに限定されない。例えば、Weiss,et al.,米国特許第4,663,290号(1987);Gerard,G.R.,DNA:271-79(1986);Kotewicz,M.L.,et al.,Gene 35:249-58(1985);Tanese,N.,et al.,Proc.Natl.Acad.Sci.(USA):4944-48(1985);Roth,M.J.,at al.,J.Biol.Chem.260:9326-35(1985);Michel,F.,et al.,Nature 316:641-43(1985);Akins,R.A.,et al.,Cell 47:505-16(1986),EMBO J.4:1267-75(1985);及びFawcett,D.F.,Cell 47:1007-15(1986)(これらの各々はその全体が参照されることによって本明細書に組み込まれる)を参照。
野生型RT
本明細書に開示のプライム編集因子による使用のための例示の酵素は、M-MLV逆転写酵素及びRSV逆転写酵素を包含し得るが、これらに限定されない。逆転写酵素活性を有する酵素は市販で利用可能である。ある態様において、逆転写酵素は、プライム編集因子(PE)システムの他の構成要素に対してトランスで提供される。つまり、逆転写酵素は、個々の構成要素として、すなわちnapDNAbpとの融合タンパク質としてではなく、発現又は別様に提供される。
本明細書に開示のプライム編集因子による使用のための例示の酵素は、M-MLV逆転写酵素及びRSV逆転写酵素を包含し得るが、これらに限定されない。逆転写酵素活性を有する酵素は市販で利用可能である。ある態様において、逆転写酵素は、プライム編集因子(PE)システムの他の構成要素に対してトランスで提供される。つまり、逆転写酵素は、個々の構成要素として、すなわちnapDNAbpとの融合タンパク質としてではなく、発現又は別様に提供される。
当業者は、野生型逆転写酵素が(モロニーマウス白血病ウイルス(M-MLV);ヒト免疫不全ウイルス(HIV)逆転写酵素、及びトリ白血病・肉腫ウイルス(ASLV)逆転写酵素を包含するが、これらに限定されず、これはラウス肉腫ウイルス(RSV)逆転写酵素、トリ骨髄芽球症ウイルス(AMV)逆転写酵素、トリ赤芽球症ウイルス(AEV)ヘルパーウイルスMCAV逆転写酵素、トリ骨髄球腫症ウイルスMC29ヘルパーウイルスMCAV逆転写酵素、トリ細網内皮症ウイルス(REV-T)ヘルパーウイルスREV-A逆転写酵素、トリ肉腫ウイルスUR2ヘルパーウイルスUR2AV逆転写酵素、トリ肉腫ウイルスY73ヘルパーウイルスYAV逆転写酵素、ラウス随伴ウイルス(RAV)逆転写酵素、及び骨髄芽球症ウイルス随伴ウイルス(MAV)逆転写酵素を包含するが、これらに限定されない)、本明細書に記載の対象の方法及び組成物に好適に用いられてもよいということを認識するであろう。
例示の野生型RT酵素は次の通りである:


バリアント及びエラーを起こしやすいRT
逆転写酵素は、相補的なDNA(cDNA)鎖をRNA鋳型から合成することにとって必須である。逆転写酵素は、異なる生化学的活性を発揮する別物のドメインから成る酵素である。酵素は次の通りRNA鋳型からのDNAの合成を触媒する:アニーリングしたプライマーの存在下において、逆転写酵素はRNA鋳型に結合し、重合反応を開始する。RNA依存性DNAポリメラーゼ活性が相補的なDNA(cDNA)鎖を合成し、dNTPを組み込む。RNase H活性がDNA:RNA複合体のRNA鋳型を分解する。それゆえに、逆転写酵素は、(a)RNA/DNAハイブリッドを認識及び結合する結合活性、(b)RNA依存性DNAポリメラーゼ活性、及び(c)RNase H活性を含む。加えて、逆転写酵素は、一般的に、それらの熱安定性、処理能力(dNTP組み込みの速度)、及び忠実性(又はエラー率)を包含する種々の属性を有すると見なされる。本明細書において企図される逆転写酵素バリアントは、これらの酵素活性(例として、RNA依存性DNAポリメラーゼ活性、RNase H活性、又はDNA/RNAハイブリッド結合活性)又は酵素特性(例として、熱安定性、処理能力、又は忠実性)のいずれか1以上にインパクトを及ぼすか又は変化させる、逆転写酵素のいずれかの変異を包含してもよい。かかるバリアントは、当該技術分野においてパブリックドメインにおいて利用可能、市販で利用可能であってもよいか、又は指向性進化プロセス(例として、PACE又はPANCE)を包含する変異導入の公知の方法を用いて作製されてもよい。
逆転写酵素は、相補的なDNA(cDNA)鎖をRNA鋳型から合成することにとって必須である。逆転写酵素は、異なる生化学的活性を発揮する別物のドメインから成る酵素である。酵素は次の通りRNA鋳型からのDNAの合成を触媒する:アニーリングしたプライマーの存在下において、逆転写酵素はRNA鋳型に結合し、重合反応を開始する。RNA依存性DNAポリメラーゼ活性が相補的なDNA(cDNA)鎖を合成し、dNTPを組み込む。RNase H活性がDNA:RNA複合体のRNA鋳型を分解する。それゆえに、逆転写酵素は、(a)RNA/DNAハイブリッドを認識及び結合する結合活性、(b)RNA依存性DNAポリメラーゼ活性、及び(c)RNase H活性を含む。加えて、逆転写酵素は、一般的に、それらの熱安定性、処理能力(dNTP組み込みの速度)、及び忠実性(又はエラー率)を包含する種々の属性を有すると見なされる。本明細書において企図される逆転写酵素バリアントは、これらの酵素活性(例として、RNA依存性DNAポリメラーゼ活性、RNase H活性、又はDNA/RNAハイブリッド結合活性)又は酵素特性(例として、熱安定性、処理能力、又は忠実性)のいずれか1以上にインパクトを及ぼすか又は変化させる、逆転写酵素のいずれかの変異を包含してもよい。かかるバリアントは、当該技術分野においてパブリックドメインにおいて利用可能、市販で利用可能であってもよいか、又は指向性進化プロセス(例として、PACE又はPANCE)を包含する変異導入の公知の方法を用いて作製されてもよい。
種々の態様において、逆転写酵素はバリアント逆転写酵素であってもよい。本明細書において用いられる「バリアント逆転写酵素」は、参照配列(例として、参照野生型配列)に対して相対的に1以上の変異(単数の変異、逆位、欠失、挿入、及び再構成を包含する)を含む、いずれかの天然に存在するか又は遺伝子操作されたバリアントを包含する。RTは、RNA依存性DNAポリメラーゼ活性、リボヌクレアーゼH活性、及びDNA依存性DNAポリメラーゼ活性を包含するいくつかの活性を天然に有する。集合的に、これらの活性は酵素が一本鎖RNAを二本鎖cDNAへと変換することを可能化する。レトロウイルス及びレトロトランスポゾンでは、このcDNAは次いでホストゲノム上に取り入れし得、これから新たなRNAのコピーがホスト細胞転写によって作製され得る。バリアントRTは、これらの活性の1以上にインパクトを及ぼす変異を含んでもよい(これはこれらの活性を縮減若しくは増大させるか、又はこれはこれらの活性をすっかり消去するかどちらかである)。加えて、バリアントRTは、RTを多かれ少なかれ安定に、凝集をよりしにくくし、精製及び/若しくは検出を容易化する1以上の変異、並びに/又は特性若しくは特徴の他の(other the)修飾を含んでもよい。
当業者は、他の逆転写酵素(モロニーマウス白血病ウイルス(M-MLV);ヒト免疫不全ウイルス(HIV)逆転写酵素、及びトリ白血病・肉腫ウイルス(ASLV)逆転写酵素を包含するが、これらに限定されず、これはラウス肉腫ウイルス(RSV)逆転写酵素、トリ骨髄芽球症ウイルス(AMV)逆転写酵素、トリ赤芽球症ウイルス(AEV)ヘルパーウイルスMCAV逆転写酵素、トリ骨髄球腫症ウイルスMC29ヘルパーウイルスMCAV逆転写酵素、トリ細網内皮症ウイルス(REV-T)ヘルパーウイルスREV-A逆転写酵素、トリ肉腫ウイルスUR2ヘルパーウイルスUR2AV逆転写酵素、トリ肉腫ウイルスY73ヘルパーウイルスYAV逆転写酵素、ラウス随伴ウイルス(RAV)逆転写酵素、及び骨髄芽球症関連ウイルス(MAV)逆転写酵素を包含するが、これらに限定されない)に由来するバリアント逆転写酵素が、本明細書に記載の対象の方法及び組成物に好適に用いられてもよいということを認識するであろう。
バリアントRTを調製する1つの方法は、遺伝子修飾によって(例として、野生型逆転写酵素のDNA配列を修飾することによって)である。DNA配列のランダムな及び標的化された変異を許すいくつもの方法が当該技術分野において公知である(例えば、Ausubel et.al.Short Protocols in Molecular Biology(1995)3.sup.rd Ed.John Wiley&Sons,Inc.を見よ)。加えて、従来の及びPCRに基づく方法両方を包含する部位特異的変異導入のためのいくつもの市販で利用可能なキットがある。例は、QuikChange部位特異的変異導入キット(AGILENT(登録商標))、Q5(登録商標)部位特異的変異導入キット(NEW ENGLAND BIOLABS(登録商標))、及びGeneArt(商標)部位特異的変異導入システム(THERMOFISHER SCIENTIFIC(登録商標))を包含する。
加えて、変異体逆転写酵素が、挿入変異又は短縮(N末端の、内部の、又はC末端の挿入又は短縮)によって当業者に公知の方法論に従って生成されてもよい。本明細書において用いられる用語「変異」は、別の残基によるある配列、例として核酸若しくはアミノ酸配列上の残基の置換、又はある配列上の1以上の残基の欠失若しくは挿入を指す。変異は、典型的には、元の残基、次に配列上の残基の位置及び新たに置換された残基の同一性を同定することによって本明細書に記載される。本明細書において提供されるアミノ酸置換(変異)を作るための種々の方法が当該技術分野において周知であり、例えばGreen and Sambrook,Molecular Cloning:A Laboratory Manual(4th ed.,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,N.Y.(2012))によって提供される。変異は、種々のカテゴリー、例えば1塩基多型、微小重複領域、インデル、及び逆位を包含し得、決して限定することを意味されない。変異は、タンパク質活性を縮減又は廃止する変異の正常の結果である「機能喪失」変異を包含し得る。ほとんどの機能喪失変異は劣性である。なぜなら、ヘテロ接合体では、第2の染色体コピーが完全に機能的なタンパク質をコードする遺伝子の未変異のバージョンを持ち、この存在が変異の効果を補償するからである。変異は、「機能獲得」変異をもまた包摂する。これは、正常な条件下ではさもなければ存在しない異常な活性をタンパク質又は細胞に付与するものである。多くの機能獲得変異はコード領域よりもむしろ制御配列にあり、よって、いくつもの帰結を有し得る。例えば、変異は、1以上の遺伝子が間違った組織において発現されることに至り得、これらの組織は、それらが正常では欠く機能を獲得する。それらの性質ゆえに、機能獲得変異は通常は優性である。
当該技術分野で公知の部位特異的変異導入のより古い方法は、一本鎖DNA鋳型の単離を許すM13バクテリオファージベクター等のベクター上への変異させられるべき配列のサブクローニングに依拠する。これらの方法では、変異誘発プライマー(すなわち、変異させられるべき部位にアニールすることができるが、変異させられるべき部位に1以上のミスマッチのヌクレオチドを持つプライマー)を一本鎖鋳型にアニーリングさせ、次いで、変異誘発プライマーの3'端からスタートして鋳型の相補体を重合する。次いで、もたらされた二重鎖はホスト細菌に形質トランスバーションされ、プラークが所望の変異についてスクリーニングされる。
より最近では、部位特異的変異導入はPCR方法論を使用している。これらは一本鎖鋳型を要求しないという利点を有する。加えて、サブクローニングを要求しない方法が開発されている。PCRに基づく部位特異的変異導入が行われるときには、いくつかのイシューが考慮されなければならない。第1に、これらの方法では、ポリメラーゼによって導入される望まれない変異の拡張を防止するために、PCRサイクル数を縮減することが望ましい。第2に、反応中に残存する変異していない親の分子の数を縮減するために、セレクションが使用されなければならない。第3に、単一のPCRプライマーセットの使用を許すために、伸長された長さのPCR法が好ましい。そして第4に、いくつかの熱安定性ポリメラーゼの非鋳型依存性末端伸長活性ゆえに、多くの場合には、PCRによって生成された変異体産物の平滑末端ライゲーションに先行して、端を磨く工程を手続きに組み込むことが必要である。
ランダム変異導入の方法が当該技術分野に存在し、これらは1以上のランダムに所在する変異を持つ変異体のパネルをもたらすであろう。次いで、変異体のかかるパネルは、所望の特性、例えば野生型逆転写酵素に対して相対的に増大した安定性を発揮するものについてスクリーニングされてもよい。
ランダムな変異導入のための方法の例はいわゆる「エラーを起こしやすいPCR法」である。名称が含意する通り、方法は、DNAポリメラーゼが高忠実性組み込みを支持しない条件下において所与の配列を増幅する。エラーを起こしやすい組み込みを促す条件は異なるDNAポリメラーゼについて変わるが、当業者は所与の酵素についてかかる条件を決定してもよい。増幅の忠実性においての多くのDNAポリメラーゼの鍵の変数は、例えば緩衝液中の二価金属イオンの型及び濃度である。よって、マンガンイオンの使用及び/又はマグネシウム若しくはマンガンイオン濃度のバリエーションが、ポリメラーゼのエラー率に影響するように適用されてもよい。
種々の側面では、プライム編集因子のRTは「エラーを起こしやすい」逆転写酵素バリアントであってもよい。当該技術分野において公知及び/又は利用可能であるエラーを起こしやすい逆転写酵素が用いられてもよい。逆転写酵素は天然にはいずれかのプルーフリーディング機能を有さず;それゆえに、逆転写酵素のエラー率はプルーフリーディング活性を含むDNAポリメラーゼよりも一般的に高いということは了解されるであろう。いずれかの具体的な逆転写酵素のエラー率は酵素の「忠実性」の特性であり、これはそのRNA鋳型に対してDNAの鋳型によって導かれる重合の正確度を表す。高忠実性を有するRTは低いエラー率を有する。反対に、低い忠実性を有するRTは高いエラー率を有する。M-MLVに基づく逆転写酵素の忠実性は、合成される15,000~27,000ヌクレオチドに1エラーの範囲のエラー率を有することが報告される。Boutabout et al.,「DNA synthesis fidelity by the reverse transcriptase of the yeast retrotransposon Ty1,」Nucleic Acids Res,2001,29:2217-2222を見よ。これは参照によって組み込まれる。それゆえに、この適用の目的には、「エラーを起こしやすい」であると考慮されるか又は「エラーを起こしやすい忠実性」を有すると考慮される逆転写酵素は、合成される15,000ヌクレオチドに1つのエラー未満であるエラー率を有するものである。
エラーを起こしやすい逆転写酵素は、出発RT酵素(例として、野生型M-MLV RT)の変異導入によってもまた作成されてもよい。変異導入の方法は限定されず、指向性進化プロセス、例えばファージによって補助される連続的進化(PACE)又はファージによって補助される非連続的進化(PANCE)を包含してもよい。本明細書において用いられる用語「ファージによって補助される連続的進化(PACE)」は、ファージをウイルスベクターとして使用する連続的進化を指す。PACEテクノロジーの一般概念は、例えば、2010年3月11日にWO2010/028347として公開された2009年9月8日出願の国際PCT出願PCT/US2009/056194;2012年6月28日にWO2012/088381として公開された2011年12月22日出願の国際PCT出願PCT/US2011/066747;2015年5月5日登録のU.S.出願U.S.特許No.9,023,594、2015年9月11日にWO2015/134121として公開された2015年1月20日出願の国際PCT出願PCT/US2015/012022、及び2016年10月20日にWO2016/168631として公開された2016年4月15日出願の国際PCT出願PCT/US2016/027795に記載されている。これらの夫々の内容全体は参照によって本明細書に組み込まれる。
エラーを起こしやすい逆転写酵素は、「ファージによって補助される非連続的進化(PANCE)」によってもまた得られてもよく、これは本明細書で使用される場合、ファージをウイルスベクターとして使用する非連続進化を指す。PANCEは急速なin vivoの指向性進化のための単純化された技術であり、新しいE.coliホスト細胞による進化させられるべき目当ての遺伝子を含有する進化する「セレクションファージ」(SP)の連続フラスコ植え継ぎを用い、それによって、ホストE.coli内の遺伝子が一定に保たれることを許しながら、SPに含有される遺伝子は連続的に進化する。連続フラスコ植え継ぎは微生物の実験室進化のための広くアクセス可能なアプローチとして長く用をなしており、より最近では、バクテリオファージ進化のための類縁のアプローチが開発されている。PANCEシステムはPACEシステムよりも低いストリンジェンシーを特色とする。
他のエラーを起こしやすい逆転写酵素が文献に記載されている。これらの夫々は本明細書の方法及び組成物への使用を企図される。例えば、エラーを起こしやすい逆転写酵素は、Bebenek et al.,「Error-prone Polymerization by HIV-1 Reverse Transcriptase,」J Biol Chem,1993,Vol.268:10324-10334及びSebastian-Martin et al.,「Transcriptional inaccuracy threshold attenuates differences in RNA-dependent DNA synthesis fidelity between retroviral reverse transcriptases,」Scientific Reports,2018,Vol.8:627に記載されている。これらの夫々は参照によって組み込まれる。なお、更に、エラーを起こしやすい逆転写酵素を包含する逆転写酵素は市販のサプライヤーから得られ得、全てNEW ENGLAND BIOLABS(登録商標)からのProtoScript(登録商標)(II)逆転写酵素、AMV逆転写酵素、WarmStart(登録商標)逆転写酵素、及びM-MuLV逆転写酵素、又は全てTAKARA BIO USA,INC.(以前はCLONTECH)からのAMV逆転写酵素XL、SMARTScribe逆転写酵素、GPRウルトラピュアMMLV逆転写酵素を包含する。
本明細書の開示はRNaseHドメインに変異を有する逆転写酵素をもまた企図する。上で言及されている通り、逆転写酵素の本来的な特性の1つはRNase H活性であり、これはRNA:cDNAハイブリッドのRNA鋳型を重合と同時的に切断する。RNA鋳型が全長の逆転写の完了前に分解されてもよいため、RNase H活性は長いcDNAの合成にとっては望ましくなくあり得る。おそらくは酵素のポリメラーゼ活性とのその競合を原因として、RNase H活性は逆転写効率をもまた低下させてもよい。それゆえに、本開示は修飾されたRNaseH活性を含むいずれかの逆転写酵素バリアントを企図する。
本明細書の開示は、RNA依存性DNAポリメラーゼドメインに変異を有する逆転写酵素をもまた企図する。上で言及されている通り、逆転写酵素の本来的な特性の1つはRNA依存性DNAポリメラーゼ活性である。これは、RNA:cDNAハイブリッドの鋳型RNA鎖によってコードされる新生cDNA鎖に核酸塩基を組み込む。RNA依存性DNAポリメラーゼ活性は、酵素の処理能力を増大させるか又は減少させるかどちらかのために(すなわち、その組み込み速度の観点から)増大又は減少させられ得る。それゆえに、本開示は、酵素の処理能力が未修飾のバージョンに対して相対的に増大させられるか又は減少させられるかどちらかのようにして、修飾されたRNA依存性DNAポリメラーゼ活性を含むいずれかの逆転写酵素バリアントを企図する。
変改された熱安定性特徴を有する逆転写酵素バリアントもまた本明細書において企図される。逆転写酵素が高い温度に耐える能力はcDNA合成の重要な側面である。上昇した反応温度は、強い二次構造及び/又は高いGC含量を有するRNAを変性することを助け、逆転写酵素が配列を読み通すことを許す。結果として、より高い温度における逆転写は全長cDNA合成及びより高い収量を可能化し、これはプライム編集プロセスの結果としての3'フラップssDNAの改善された生成に至り得る。野生型M-MLV逆転写酵素は典型的には37~48℃の範囲に至適温度を有する;しかしながら、49℃、50℃、51℃、52℃、53℃、54℃、55℃、56℃、57℃、58℃、59℃、60℃、61℃、62℃、63℃、64℃、65℃、66℃、及びより高くを包含する48℃超のより高い温度において逆転写活性を許す変異が導入されてもよい。
エラーを起こしやすいRT、熱安定性RT、処理能力増大RTを包含する本明細書において企図されるバリアント逆転写酵素は、変異導入又は進化プロセスを包含する種々のルーチン戦略によって操作され得る。いくつかのケースでは、バリアントは単一の変異を導入することによって産生され得る。他のケースでは、バリアントは1より多くの変異を要求してもよい。1より多くの変異を含む変異体では、所与の変異の効果は、具体的な変異体によって持たれる他の変異からは単離して、部位特異的変異導入による野生型遺伝子への同定された変異の導入によって評価されてもよい。次いで、それゆえに産生された一重変異体のスクリーニングアッセイは、その変異単独の効果の決定を許すであろう。
本明細書において用いられるバリアントRT酵素は、本明細書において開示若しくは企図されるか又は当該技術分野で公知のいずれかの野生型RT又は変異体RT又はフラグメントRT又はRTの他のバリアントを包含するいずれかの参照RTタンパク質と少なくとも約70%同一、少なくとも約80%同一、少なくとも約90%同一、少なくとも約95%同一、少なくとも約96%同一、少なくとも約97%同一、少なくとも約98%同一、少なくとも約99%同一、少なくとも約99.5%同一、又は少なくとも約99.9%同一である他の「RTバリアント」をもまた包含してもよい。
いくつかの態様では、RTバリアントは、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、21、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、又は最高で100、又は最高で200、又は最高で300、又は最高で400、又は最高で500、又はより多くのアミノ酸変化を、参照RTと比較して有してもよい。いくつかの態様では、フラグメントが参照RTの対応するフラグメントと少なくとも約70%同一、少なくとも約80%同一、少なくとも約90%同一、少なくとも約95%同一、少なくとも約96%同一、少なくとも約97%同一、少なくとも約98%同一、少なくとも約99%同一、少なくとも約99.5%同一、又は少なくとも約99.9%同一であるようにして、RTバリアントは参照RTのフラグメントを含む。いくつかの態様では、フラグメントは、対応する野生型RT(M-MLV逆転写酵素)(例として、配列番号32)又は配列番号102-112の逆転写酵素のいずれかのアミノ酸長さの少なくとも30%、少なくとも35%、少なくとも40%、少なくとも45%、少なくとも50%、少なくとも55%、少なくとも60%、少なくとも65%、少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%同一、少なくとも96%、少なくとも97%、少なくとも98%、少なくとも99%、又は少なくとも99.5%である。
いくつかの態様では、本開示は、それらの機能性を保持し、かついずれかの本明細書に開示のRTタンパク質のフラグメントであるRTフラグメントをもまた利用してもよい。いくつかの態様では、RTフラグメントは長さが少なくとも100アミノ酸である。いくつかの態様では、フラグメントは長さが少なくとも100、150、200、250、300、350、400、450、500、550、若しくは最高で600、又はより多くのアミノ酸である。
なお他の態様において、本開示は、N末端若しくはC末端又は両方がある種のアミノ酸数だけ短縮されているRTバリアントをもまた利用してもよく、これは、なお十分なポリメラーゼ機能を保持する短縮バリアントをもたらす。いくつかの態様では、RT短縮バリアントは、少なくとも1、少なくとも2、少なくとも3、少なくとも4、少なくとも5、少なくとも6、少なくとも7、少なくとも8、少なくとも9、少なくとも10、少なくとも11、少なくとも12、少なくとも13、少なくとも14、少なくとも15、少なくとも16、少なくとも17、少なくとも18、少なくとも19、少なくとも20、少なくとも21、少なくとも22、少なくとも23、少なくとも24、少なくとも25、少なくとも30、40、50、60、70、80、90、100、110、120、130、140、150、160、170、180、190、200、210、220、230、240、又は250アミノ酸の短縮を、タンパク質のN末端の端に有する。他の態様において、RT短縮バリアントは、少なくとも1、少なくとも2、少なくとも3、少なくとも4、少なくとも5、少なくとも6、少なくとも7、少なくとも8、少なくとも9、少なくとも10、少なくとも11、少なくとも12、少なくとも13、少なくとも14、少なくとも15、少なくとも16、少なくとも17、少なくとも18、少なくとも19、少なくとも20、少なくとも21、少なくとも22、少なくとも23、少なくとも24、少なくとも25、少なくとも30、40、50、60、70、80、90、100、110、120、130、140、150、160、170、180、190、200、210、220、230、240、又は250アミノ酸の短縮を、タンパク質のC末端の端に有する。なお他の態様において、RT短縮バリアントは同じか又は異なる長さである短縮をN末端及びC末端の端に有する。
例えば、本明細書に開示のプライム編集因子は、M-MLV逆転写酵素の短縮されたバージョンを包含してもよい。この態様では、逆転写酵素は4つの変異を含有する(D200N、T306K、W313F、T330P;PE2に存在するL603W変異は短縮を原因としてもはや存在しないということに注意)。この短縮された編集因子をコードするDNA配列は、PE2よりも522bp小さく、よって、DNA配列の送達がそのサイズを原因として難しい適用(すなわち、アデノ随伴ウイルス及びレンチウイルス送達)にとってそれを可能性として有用にする。この態様はMMLV-RT(trunc)と呼ばれ、次のアミノ酸配列を有する:

種々の態様において、本明細書に開示のプライム編集因子は、本明細書に記載のRTバリアントの1つ、又はいずれかの参照Cas9バリアントと少なくとも約70%同一、少なくとも約80%同一、少なくとも約90%同一、少なくとも約95%同一、少なくとも約96%同一、少なくとも約97%同一、少なくとも約98%同一、少なくとも約99%同一、少なくとも約99.5%同一、又は少なくとも約99.9%同一であるそのRTバリアントを含んでもよい。
なお他の態様において、本方法及び組成物は、Effefson et al.,「Synthetic evolutionary origin of a proofreading reverse transcriptase,」Science,June 24,2016,Vol.352:1590-1593に記載されている通り、逆転写酵素へと進化させられたDNAポリメラーゼを利用してもよい。これの内容は参照によって本明細書に組み込まれる。
ある種の他の態様において、逆転写酵素は、napDNAbpをもまた含む融合タンパク質の構成要素として提供される。換言すると、いくつかの態様では、逆転写酵素は融合タンパク質としてnapDNAbpに融合される。
種々の態様において、バリアント逆転写酵素は配列番号32によって表される野生型M-MLV逆転写酵素から操作され得る。
種々の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、次の変異の1以上を含むバリアントRTを包含し得る:配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるP51L、S67K、E69K、L139P、T197A、D200N、H204R、F209N、E302K、E302R、T306K、F309N、W313F、T330P、L345G、L435G、N454K、D524G、E562Q、D583N、H594Q、L603W、E607K、又はD653N。
本開示の種々の態様に従ってnapDNAbpタンパク質に融合又は個々のタンパク質として提供され得るいくつかの例示の逆転写酵素が、下で提供される。例示の逆転写酵素は、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、又は少なくとも99%配列同一性を次の野生型酵素又は部分的な酵素に対して有するバリアントを包含する:



種々の他の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、次の変異の1以上を含むバリアントRTを包含し得る:配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるP51X、S67X、E69X、L139X、T197X、D200X、H204X、F209X、E302X、T306X、F309X、W313X、T330X、L345X、L435X、N454X、D524X、E562X、D583X、H594X、L603X、E607X、又はD653Xであって、「X」はいずれかのアミノ酸であり得る。
種々の他の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるP51X変異を含むバリアントRTを包含し得、「X」はいずれかのアミノ酸であり得る。ある態様において、XはLである。
種々の他の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるS67X変異を含むバリアントRTを包含し得、「X」はいずれかのアミノ酸であり得る。ある態様において、XはKである。
種々の他の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるE69X変異を含むバリアントRTを包含し得、「X」はいずれかのアミノ酸であり得る。ある態様において、XはKである。
種々の他の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるL139X変異を含むバリアントRTを包含し得、「X」はいずれかのアミノ酸であり得る。ある態様において、XはPである。
種々の他の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるT197X変異を含むバリアントRTを包含し得、「X」はいずれかのアミノ酸であり得る。ある態様において、XはAである。
種々の他の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるD200X変異を含むバリアントRTを包含し得、「X」はいずれかのアミノ酸であり得る。ある態様において、XはNである。
種々の他の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるH204X変異を含むバリアントRTを包含し得、「X」はいずれかのアミノ酸であり得る。ある態様において、XはRである。
種々の他の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるF209X変異を含むバリアントRTを包含し得、「X」はいずれかのアミノ酸であり得る。ある態様において、XはNである。
種々の他の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるE302X変異を含むバリアントRTを包含し得、「X」はいずれかのアミノ酸であり得る。ある態様において、XはKである。
種々の他の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるE302X変異を含むバリアントRTを包含し得、「X」はいずれかのアミノ酸であり得る。ある態様において、XはRである。
種々の他の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるT306X変異を含むバリアントRTを包含し得、「X」はいずれかのアミノ酸であり得る。ある態様において、XはKである。
種々の他の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるF309X変異を含むバリアントRTを包含し得、「X」はいずれかのアミノ酸であり得る。ある態様において、XはNである。
種々の他の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるW313X変異を含むバリアントRTを包含し得、「X」はいずれかのアミノ酸であり得る。ある態様において、XはFである。
種々の他の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるT330X変異を含むバリアントRTを包含し得、「X」はいずれかのアミノ酸であり得る。ある態様において、XはPである。
種々の他の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるL345X変異を含むバリアントRTを包含し得、「X」はいずれかのアミノ酸であり得る。ある態様において、XはGである。
種々の他の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるL435X変異を含むバリアントRTを包含し得、「X」はいずれかのアミノ酸であり得る。ある態様において、XはGである。
種々の他の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるN454X変異を含むバリアントRTを包含し得、「X」はいずれかのアミノ酸であり得る。ある態様において、XはKである。
種々の他の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるD524X変異を含むバリアントRTを包含し得、「X」はいずれかのアミノ酸であり得る。ある態様において、XはGである。
種々の他の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるE562X変異を含むバリアントRTを包含し得、「X」はいずれかのアミノ酸であり得る。ある態様において、XはQである。
種々の他の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるD583X変異を含むバリアントRTを包含し得、「X」はいずれかのアミノ酸であり得る。ある態様において、XはNである。
種々の他の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるH594X変異を含むバリアントRTを包含し得、「X」はいずれかのアミノ酸であり得る。ある態様において、XはQである。
種々の他の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるL603X変異を含むバリアントRTを包含し得、「X」はいずれかのアミノ酸であり得る。ある態様において、XはWである。
種々の他の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるE607X変異を含むバリアントRTを包含し得、「X」はいずれかのアミノ酸であり得る。ある態様において、XはKである。
種々の他の態様において、本明細書に記載のプライム編集因子(RTが融合パートナーとして又はトランスでどちらかで提供される)は、配列番号32の野生型M-MLV RTにおける又は別の野生型RTポリペプチド配列上の対応するアミノ酸位置におけるD653X変異を含むバリアントRTを包含し得、「X」はいずれかのアミノ酸であり得る。ある態様において、XはNである。
本開示の種々の態様に従ってnapDNAbpタンパク質に融合又は個々のタンパク質として提供され得るいくつかの例示の逆転写酵素が、下で提供される。例示の逆転写酵素は、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、又は少なくとも99%配列同一性を、配列番号32、34、113~128によって表される野生型酵素又は部分的な酵素に対して有するバリアントを包含する。ここに記載されるプライム編集因子(PE)システムは、次の米国特許(これらの夫々はそれらの全体が参照によって組み込まれる):米国特許第10,202,658号;第10,189,831号;第10,150,955号;第9,932,567号;第9,783,791号;第9,580,698号;第9,534,201号;及び第9,458,484号のいずれかに記載又は開示されているいずれかの公に利用可能な逆転写酵素と、変異を組み入れるための公知の方法又はタンパク質を進化させるための公知の方法を用いて作られ得るそのいずれかのバリアントとを企図する。次の参照は当該技術分野の逆転写酵素を記載する。それらの開示の夫々はそれらの全体が参照によって本明細書に組み込まれる。
Herzig,E.,Voronin,N.,Kucherenko,N.&Hizi,A.A Novel Leu92 Mutant of HIV-1 Reverse Transcriptase with a Selective Deficiency in Strand Transfer Causes a Loss of Viral Replication.J.Virol.89,8119-8129(2015).
Mohr,G.et al.A Reverse Transcriptase-Cas1 Fusion Protein Contains a Cas6 Domain Required for Both CRISPR RNA Biogenesis and RNA Spacer Acquisition.Mol.Cell 72,700-714.e8(2018).
Zhao,C.,Liu,F.&Pyle,A.M.An ultraprocessive,accurate reverse transcriptase encoded by a metazoan group II intron.RNA 24,183-195(2018).
Zimmerly,S.&Wu,L.An Unexplored Diversity of Reverse Transcriptases in Bacteria.Microbiol Spectr 3,MDNA3-0058-2014(2015).
Ostertag,E.M.&Kazazian Jr,H.H.Biology of Mammalian L1 Retrotransposons.Annual Review of Genetics 35,501-538(2001).
Perach,M.&Hizi,A.Catalytic Features of the Recombinant Reverse Transcriptase of Bovine Leukemia Virus Expressed in Bacteria.Virology 259,176-189(1999).
Lim,D.et al.Crystal structure of the moloney murine leukemia virus RNase H domain.J.Virol.80,8379-8389(2006).
Zhao,C.&Pyle,A.M.Crystal structures of a group II intron maturase reveal a missing link in spliceosome evolution.Nature Structural&Molecular Biology 23,558-565(2016).
Griffiths,D.J.Endogenous retroviruses in the human genome sequence.Genome Biol.2,REVIEWS1017(2001).
Baranauskas,A.et al.Generation and characterization of new highly thermostable and processive M-MuLV reverse transcriptase variants.Protein Eng Des Sel 25,657-668(2012).
Zimmerly,S.,Guo,H.,Perlman,P.S.&Lambowltz,A.M.Group II intron mobility occurs by target DNA-primed reverse transcription.Cell 82,545-554(1995).
Feng,Q.,Moran,J.V.,Kazazian,H.H.&Boeke,J.D.Human L1 retrotransposon encodes a conserved endonuclease required for retrotransposition.Cell 87,905-916(1996).
Berkhout,B.,Jebbink,M.&Zsiros,J.Identification of an Active Reverse Transcriptase Enzyme Encoded by a Human Endogenous HERV-K Retrovirus.Journal of Virology 73,2365-2375(1999).
Kotewicz,M.L.,Sampson,C.M.,D'Alessio,J.M.&Gerard,G.F.Isolation of cloned Moloney murine leukemia virus reverse transcriptase lacking ribonuclease H activity.Nucleic Acids Res 16,265-277(1988).
Arezi,B.&Hogrefe,H.Novel mutations in Moloney Murine Leukemia Virus reverse transcriptase increase thermostability through tighter binding to template-primer.Nucleic Acids Res 37,473-481(2009).
Blain,S.W.&Goff,S.P.Nuclease activities of Moloney murine leukemia virus reverse transcriptase.Mutants with altered substrate specificities.J.Biol.Chem.268,23585-23592(1993).
Xiong,Y.&Eickbush,T.H.Origin and evolution of retroelements based upon their reverse transcriptase sequences.EMBO J 9,3353-3362(1990).
Herschhorn,A.&Hizi,A.Retroviral reverse transcriptases.Cell.Mol.Life Sci.67,2717-2747(2010).
Taube,R.,Loya,S.,Avidan,O.,Perach,M.&Hizi,A.Reverse transcriptase of mouse mammary tumour virus:expression in bacteria,purification and biochemical characterization.Biochem.J.329(Pt 3),579-587(1998).
Liu,M.et al.Reverse Transcriptase-Mediated Tropism Switching in Bordetella Bacteriophage.Science 295,2091-2094(2002).
Luan,D.D.,Korman,M.H.,Jakubczak,J.L.&Eickbush,T.H.Reverse transcription of R2Bm RNA is primed by a nick at the chromosomal target site:a mechanism for non-LTR retrotransposition.Cell 72,595-605(1993).
Nottingham,R.M.et al.RNA-seq of human reference RNA samples using a thermostable group II intron reverse transcriptase.RNA 22,597-613(2016).
Telesnitsky,A.&Goff,S.P.RNase H domain mutations affect the interaction between Moloney murine leukemia virus reverse transcriptase and its primer-template.Proc.Natl.Acad.Sci.U.S.A.90,1276-1280(1993).
Halvas,E.K.,Svarovskaia,E.S.&Pathak,V.K.Role of Murine Leukemia Virus Reverse Transcriptase Deoxyribonucleoside Triphosphate-Binding Site in Retroviral Replication and In Vivo Fidelity.Journal of Virology 74,10349-10358(2000).
Nowak,E.et al.Structural analysis of monomeric retroviral reverse transcriptase in complex with an RNA/DNA hybrid.Nucleic Acids Res 41,3874-3887(2013).
Stamos,J.L.,Lentzsch,A.M.&Lambowitz,A.M.Structure of a Thermostable Group II Intron Reverse Transcriptase with Template-Primer and Its Functional and Evolutionary Implications.Molecular Cell 68,926-939.e4(2017).
Das,D.&Georgiadis,M.M.The Crystal Structure of the Monomeric Reverse Transcriptase from Moloney Murine Leukemia Virus.Structure 12,819-829(2004).
Avidan,O.,Meer,M.E.,Oz,I.&Hizi,A.The processivity and fidelity of DNA synthesis exhibited by the reverse transcriptase of bovine leukemia virus.European Journal of Biochemistry 269,859-867(2002).
Gerard,G.F.et al.The role of template-primer in protection of reverse transcriptase from thermal inactivation.Nucleic Acids Res 30,3118-3129(2002).
Monot,C.et al.The Specificity and Flexibility of L1 Reverse Transcription Priming at Imperfect T-Tracts.PLOS Genetics 9,e1003499(2013).
Mohr,S.et al.Thermostable group II intron reverse transcriptase fusion proteins and their use in cDNA synthesis and next-generation RNA sequencing.RNA 19,958-970(2013).
逆転写酵素に関する上に記されている参照のいずれかは、すでにそのように申し立てられていない場合には、それらの全体が参照によってここに組み込まれる。
[4]プライム編集因子
本明細書に記載のプライム編集因子(PE)システムは、(A)少なくとも2つのタンパク質:(1)napDNAbp(例として、Cas9ニッカーゼ)及び(2)ポリメラーゼ(例として、逆転写酵素等の、DNA依存性DNAポリメラーゼ又はRNA依存性DNAポリメラーゼ)並びに(B)標準的なpegRNAと比較して少なくとも1つの性能向上修飾を含む操作されたpegRNAを含むシステムを指す。napDNAbp及びポリメラーゼ構成要素は、別々に、すなわち互いにトランスで提供されてもよく、又はnapDNAbp及びポリメラーゼ構成要素が、例としてポリペプチドリンカーを介してカップリングされる融合タンパク質として提供されてもよい。
本明細書に記載のプライム編集因子(PE)システムは、(A)少なくとも2つのタンパク質:(1)napDNAbp(例として、Cas9ニッカーゼ)及び(2)ポリメラーゼ(例として、逆転写酵素等の、DNA依存性DNAポリメラーゼ又はRNA依存性DNAポリメラーゼ)並びに(B)標準的なpegRNAと比較して少なくとも1つの性能向上修飾を含む操作されたpegRNAを含むシステムを指す。napDNAbp及びポリメラーゼ構成要素は、別々に、すなわち互いにトランスで提供されてもよく、又はnapDNAbp及びポリメラーゼ構成要素が、例としてポリペプチドリンカーを介してカップリングされる融合タンパク質として提供されてもよい。
本出願は、本明細書に開示される操作されたpegRNAと共に使用するための単一の融合タンパク質に組み合わされる任意の適切なnapDNAbp及びポリメラーゼ(例として、逆転写酵素等の、DNA依存性DNAポリメラーゼ又はRNA依存性DNAポリメラーゼ)を企図する。napDNAbps及びポリメラーゼ(例として、逆転写酵素等の、DNA依存性DNAポリメラーゼ又はRNA依存性DNAポリメラーゼ)の例はそれぞれ本明細書で定義される。ポリメラーゼは当技術分野で周知であり、アミノ酸配列は容易に入手可能であるため、本開示は、本明細書で同定された特定のポリメラーゼに限定されることを決して意味されない。
種々の態様において、融合タンパク質はいずれかの好適な構造的な構成を含んでもよい。例えば、融合タンパク質は、N末端→C末端方向に、napDNAbpをポリメラーゼ(例として、DNA依存性DNAポリメラーゼ又はRNA依存性DNAポリメラーゼ、例えば逆転写酵素)に融合されて含んでもよい。他の態様において、融合タンパク質は、N末端→C末端方向に、ポリメラーゼ(例として、逆転写酵素)をnapDNAbpに融合されて含んでもよい。融合したドメインは、任意選択で、リンカー、例としてアミノ酸配列によって連結されてもよい。他の態様において、融合タンパク質は構造NH2-[napDNAbp]-[ポリメラーゼ]-COOH;又はNH2-[ポリメラーゼ]-[napDNAbp]-COOHを含んでもよく、「]-[」の夫々のものは任意選択のリンカー配列の存在を指示する。ポリメラーゼが逆転写酵素である態様では、融合タンパク質は構造NH2-[napDNAbp]-[RT]-COOH;又はNH2-[RT]-[napDNAbp]-COOHを含んでもよく、「]-[」の夫々のものは任意選択のリンカー配列の存在を指示する。
例示の融合タンパク質が図14に図示されている。これはリンカー配列を介してニッカーゼCas9(「Cas9(H840A)」)に融合されたMLV逆転写酵素(「MLV-RT」)を含む融合タンパク質を示す。この例は、本明細書に記載のプライム編集因子(PE)システムに利用されてもよい融合タンパク質の範囲を限定することを意図されない。
種々の態様において、プライム編集因子は次のアミノ酸配列(本明細書においては「PE1」と呼ばれる)を有してもよく、これは、H840A変異を含むCas9バリアント(すなわち、Cas9ニッカーゼ)及びM-MLV RT野生型と、N末端NLS配列(19アミノ酸)及びCas9ニッカーゼドメインのC末端をRTドメインのN末端に連結するアミノ酸リンカー(32アミノ酸)とを包含する。PE1融合タンパク質は次の構造を有する:[NLS]-[Cas9(H840A)]-[リンカー]-[MMLV_RT(wt)]。PE1及びその個々の構成要素のアミノ酸配列は次の通りである:


別の態様では、プライム編集因子は次のアミノ酸配列(本明細書においては「PE2」と呼ばれる)を有してもよく、これは、H840A変異を含むCas9バリアント(すなわち、Cas9ニッカーゼ)並びに変異D200N、T330P、L603W、T306K、及びW313Fを含むM-MLV RTと、N末端NLS配列(19アミノ酸)並びにCas9ニッカーゼドメインのC末端をRTドメインのN末端に連結するアミノ酸リンカー(33アミノ酸)とを包含する。PE2融合タンパク質は次の構造を有する:[NLS]-[Cas9(H840A)]-[リンカー]-[MMLV_RT(D200N)(T330P)(L603W)(T306K)(W313F)]。PE2のアミノ酸配列は次の通りである:

更なる他の態様において、プライム編集因子は、以下のアミノ酸配列を有していてもよい:


他の態様において、プライム編集因子は、次の例示の配列等のSaCas9に又は変改されたPAM特異性を有するSpCas9ニッカーゼに基づき得る:


更に他の態様において、本明細書において企図されるプライム編集因子は、M-MLV逆転写酵素の短縮されたバージョンに融合されたCas9ニッカーゼ(例として、Cas9(H840A))を包含してもよい。この態様では、逆転写酵素は4つの変異をもまた含有する(D200N、T306K、W313F、T330P;PE2に存在するL603W変異は短縮を原因としてもはや存在しないということに注意)。この短縮された編集因子をコードするDNA配列はPE2よりも522bp小さく、よって、DNA配列の送達がそのサイズを原因として難しい適用(すなわち、アデノ随伴ウイルス及びレンチウイルス送達)にとってそれを可能性として有用にする。この態様はCas9(H840A)-MMLV-RT(trunc)又は「短いPE2(PE2-short)」又は「PE2-trunc」と呼ばれ、次のアミノ酸配列を有する:

図75を見よ。これは、PE2、PE2-trunc、PE3、及びPE3-truncの効率(すなわち、「規定された編集又はインデルを有する配列決定総読取の%」)を種々の細胞株において異なる標的部位について比較する棒グラフを提供する。データは、短縮されたRTバリアントを含むプライム編集因子が、短縮されていないRTタンパク質を含むプライム編集因子と約同じくらい効率的であったということを示す。
種々の態様において、本明細書において企図されるプライム編集因子は、PE1、PE2、又は上で指示されているプライム編集因子融合体配列のいずれかと少なくとも約70%同一、少なくとも約80%同一、少なくとも約90%同一、少なくとも約95%同一、少なくとも約96%同一、少なくとも約97%同一、少なくとも約98%同一、少なくとも約99%同一、少なくとも約99.5%同一、又は少なくとも約99.9%同一であるアミノ酸配列を有する上で開示されている配列のいずれかのバリアントをもまた包含してもよい。
ある態様において、リンカーが、本発明のペプチド又はペプチドドメイン若しくは部分のいずれかを連結するために用いられてもよい(例として、逆転写酵素等のポリメラーゼに連結又は融合されたnapDNAbp)。
[5]リンカー及び他のドメイン
プライム編集因子は、napDNAbp(例として、Cas9ドメイン)及びポリメラーゼドメイン(例として、RTドメイン)以外に種々の他のドメインを含んでもよい。例えば、napDNAbpがCas9でありかつポリメラーゼがRTであるケースでは、プライム編集因子は、Cas9ドメインをRTドメインと連結する1以上のリンカーを含んでもよい。リンカーは、他の機能ドメイン、例えば核局在配列(NLS)又はFEN1(又は他のフラップエンドヌクレアーゼ)をもまたプライム編集因子又はそのドメインに連結してもよい。
プライム編集因子は、napDNAbp(例として、Cas9ドメイン)及びポリメラーゼドメイン(例として、RTドメイン)以外に種々の他のドメインを含んでもよい。例えば、napDNAbpがCas9でありかつポリメラーゼがRTであるケースでは、プライム編集因子は、Cas9ドメインをRTドメインと連結する1以上のリンカーを含んでもよい。リンカーは、他の機能ドメイン、例えば核局在配列(NLS)又はFEN1(又は他のフラップエンドヌクレアーゼ)をもまたプライム編集因子又はそのドメインに連結してもよい。
加えて、トランスプライム編集を伴う態様では、リンカーが、tPERT動員タンパク質をプライム編集因子に例としてtPERt動員タンパク質及びnapDNAbpの間において連結するために用いられてもよい。例として、ポリメラーゼドメイン及び動員タンパク質ドメインをnapDNAbpに別個に融合するためのリンカーを包含するトランスプライム編集因子(tPE)の例示の模式図については、図3Gを見よ。
A.リンカー
上で定められている通り、本明細書において用いられる用語「リンカー」は、2つの分子又は部分、例として、ヌクレアーゼの結合ドメイン及び切断ドメインを連結する化学基又は分子を指す。いくつかの態様では、リンカーはRNAによってプログラム型ヌクレアーゼのgRNA結合ドメイン及びポリメラーゼ(例として、逆転写酵素)の触媒ドメインを連結する。いくつかの態様では、リンカーはdCas9及び逆転写酵素を連結する。典型的には、リンカーは2つの基、分子、又は他の部分の間に配置されるか又はそれらによってフランキングされ、共有結合を介して夫々の1つに接続され、それゆえに2つを接続する。いくつかの態様では、リンカーはアミノ酸又は複数のアミノ酸(例として、ペプチド又はタンパク質)である。いくつかの態様では、リンカーは有機分子、基、ポリマー、又は化学的部分である。いくつかの態様では、リンカーは、長さが5-100アミノ酸、例えば長さが5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、30~35、35~40、40~45、45~50、50~60、60~70、70~80、80~90、90~100、100~150、又は150~200アミノ酸である。より長い又はより短いリンカーもまた企図される。
上で定められている通り、本明細書において用いられる用語「リンカー」は、2つの分子又は部分、例として、ヌクレアーゼの結合ドメイン及び切断ドメインを連結する化学基又は分子を指す。いくつかの態様では、リンカーはRNAによってプログラム型ヌクレアーゼのgRNA結合ドメイン及びポリメラーゼ(例として、逆転写酵素)の触媒ドメインを連結する。いくつかの態様では、リンカーはdCas9及び逆転写酵素を連結する。典型的には、リンカーは2つの基、分子、又は他の部分の間に配置されるか又はそれらによってフランキングされ、共有結合を介して夫々の1つに接続され、それゆえに2つを接続する。いくつかの態様では、リンカーはアミノ酸又は複数のアミノ酸(例として、ペプチド又はタンパク質)である。いくつかの態様では、リンカーは有機分子、基、ポリマー、又は化学的部分である。いくつかの態様では、リンカーは、長さが5-100アミノ酸、例えば長さが5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、30~35、35~40、40~45、45~50、50~60、60~70、70~80、80~90、90~100、100~150、又は150~200アミノ酸である。より長い又はより短いリンカーもまた企図される。
特定の態様では、リンカーは、ヌクレオチドリンカーであり、限定するものではないが、アプタマー(例として、プレキノシン1-1リボスイッチアプタマー又は「evopreQ1-1」)若しくはそのバリアント、シュードノット(MMLVウイルスゲノムシュードノット又は「Mpknot-1」)若しくはそのバリアント、tRNA(例として、逆転写のためのプライマーとしてMMLVによって使用される修飾tRNA)若しくはそのバリアント、又はG-四重鎖若しくはそのバリアント等の本明細書に記載されるように、pegRNAを追加のヌクレオチド部分に結合するリンカーを指し得る。そのようなリンカーの例示的なヌクレオチド配列は、本明細書全体にわたって提供され、それだけに限らないが、配列番号225~236を包含する。
リンカーは共有結合ほども単純であってもよいか、又はそれは長さが多くの原子であるポリマー性リンカーであってもよい。ある態様において、リンカーはポリ(pol)ペプチドであるか又はアミノ酸に基づく。他の態様において、リンカーはペプチド様ではない。ある態様において、リンカーは共有結合である(例として、炭素-炭素結合、ジスルフィド結合、炭素-ヘテロ原子結合等)。ある態様において、リンカーはアミド連結部の炭素-窒素結合である。ある態様において、リンカーは、環状又は非環状の、置換又は無置換の、分枝又は非分枝の脂肪族又はヘテロ脂肪族リンカーである。ある態様において、リンカーはポリマー性である(例として、ポリエチレン、ポリエチレングリコール、ポリアミド、ポリエステル等)。ある態様において、リンカーはアミノアルカン酸のモノマー、二量体、又はポリマーを含む。ある態様において、リンカーはアミノアルカン酸を含む(例として、グリシン、エタン酸、アラニン、ベータアラニン、3-アミノプロパン酸、4-アミノ酪酸、5-ペンタン酸等)。ある態様において、リンカーはアミノヘキサン酸(Ahx)のモノマー、二量体、又はポリマーを含む。ある態様において、リンカーは炭素環部分(例として、シクロペンタン、シクロヘキサン)に基づく。他の態様において、リンカーはポリエチレングリコール部分(PEG)を含む。他の態様において、リンカーはアミノ酸を含む。ある態様において、リンカーはペプチドを含む。ある態様において、リンカーはアリール又はヘテロアリール部分を含む。ある態様において、リンカーはフェニル環に基づく。リンカーは、ペプチドからリンカーへの求核剤の取り付けを容易化するための官能化された部分を包含してもよい(例として、チオール、アミノ)。いずれかの求電子剤がリンカーの一部として用いられてもよい。例示の求電子剤は、活性化エステル、活性化アミド、マイケルアクセプター、ハロゲン化アルキル、ハロゲン化アリール、ハロゲン化アシル、及びイソチオシアネートを包含するが、これらに限定されない。
いくつかの他の態様において、リンカーはアミノ酸配列(GGGGS)n(配列番号138)、(G)n(配列番号139)、(EAAAK)n(配列番号12)、(GGS)n(配列番号140)、(SGGS)n(配列番号8)、(XP)n(配列番号141)、又はそれらのいずれかの組み合わせを含み、nは独立して1及び30の間の整数であり、Xはいずれかのアミノ酸である。いくつかの態様では、リンカーはアミノ酸配列(GGS)N(配列番号140)を含み、nは1、3、又は7である。いくつかの態様では、リンカーはアミノ酸配列SGSETPGTSESATPES(配列番号142)を含む。いくつかの態様では、リンカーはアミノ酸配列SGGSSGGSSGSETPGTSESATPESSGGSSGGS(配列番号143)を含む。いくつかの態様では、リンカーはアミノ酸配列SGGSGGSGGS(配列番号144)を含む。いくつかの態様では、リンカーはアミノ酸配列SGGS(配列番号8)を含む。他の態様において、リンカーはアミノ酸配列SGGSSGGSSGSETPGTSESATPESAGSYPYDVPDYAGSAAPAAKKKKLDGSGSGGSSGGS(配列番号131)を含む。
ある態様において、リンカーが、本発明のペプチド又はペプチドドメイン若しくは部分のいずれかを連結するために用いられてもよい(例として、逆転写酵素等のポリメラーゼに連結又は融合されたnapDNAbp)。
上で定められている通り、本明細書において用いられる用語「リンカー」は、2つの分子又は部分、例として、ヌクレアーゼの結合ドメイン及び切断ドメインを連結する化学基又は分子を指す。いくつかの態様では、リンカーはRNAによってプログラム型ヌクレアーゼのgRNA結合ドメイン及びリコンビナーゼの触媒ドメインを連結する。いくつかの態様では、リンカーはdCas9及び逆転写酵素を連結する。典型的には、リンカーは2つの基、分子、又は他の部分の間に配置されるか又はそれらによってフランキングされ、共有結合を介して夫々の1つに接続され、それゆえに2つを接続する。いくつかの態様では、リンカーはアミノ酸又は複数のアミノ酸(例として、ペプチド又はタンパク質)である。いくつかの態様では、リンカーは有機分子、基、ポリマー、又は化学的部分である。いくつかの態様では、リンカーは、長さが5-100アミノ酸、例えば長さが5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、30~35、35~40、40~45、45~50、50~60、60~70、70~80、80~90、90~100、100~150、又は150~200アミノ酸である。より長い又はより短いリンカーもまた企図される。
リンカーは共有結合ほども単純であってもよいか、又はそれは長さが多くの原子であるポリマー性リンカーであってもよい。ある態様において、リンカーはポリ(pol)ペプチドであるか又はアミノ酸に基づく。他の態様において、リンカーはペプチド様ではない。ある態様において、リンカーは共有結合である(例として、炭素-炭素結合、ジスルフィド結合、炭素-ヘテロ原子結合等)。ある態様において、リンカーはアミド連結部の炭素-窒素結合である。ある態様において、リンカーは、環状又は非環状の、置換又は無置換の、分枝又は非分枝の脂肪族又はヘテロ脂肪族リンカーである。ある態様において、リンカーはポリマー性である(例として、ポリエチレン、ポリエチレングリコール、ポリアミド、ポリエステル等)。ある態様において、リンカーはアミノアルカン酸のモノマー、二量体、又はポリマーを含む。ある態様において、リンカーはアミノアルカン酸を含む(例として、グリシン、エタン酸、アラニン、ベータアラニン、3-アミノプロパン酸、4-アミノ酪酸、5-ペンタン酸等)。ある態様において、リンカーはアミノヘキサン酸(Ahx)のモノマー、二量体、又はポリマーを含む。ある態様において、リンカーは炭素環部分(例として、シクロペンタン、シクロヘキサン)に基づく。他の態様において、リンカーはポリエチレングリコール部分(PEG)を含む。他の態様において、リンカーはアミノ酸を含む。ある態様において、リンカーはペプチドを含む。ある態様において、リンカーはアリール又はヘテロアリール部分を含む。ある態様において、リンカーはフェニル環に基づく。リンカーは、ペプチドからリンカーへの求核剤の取り付けを容易化するための官能化された部分を包含してもよい(例として、チオール、アミノ)。いずれかの求電子剤がリンカーの一部として用いられてもよい。例示の求電子剤は、活性化エステル、活性化アミド、マイケルアクセプター、ハロゲン化アルキル、ハロゲン化アリール、ハロゲン化アシル、及びイソチオシアネートを包含するが、これらに限定されない。
いくつかの他の態様において、リンカーはアミノ酸配列(GGGGS)n(配列番号138)、(G)n(配列番号139)、(EAAAK)n(配列番号12)、(GGS)n(配列番号140)、(SGGS)n(配列番号8)、(XP)n(配列番号141)、又はそれらのいずれかの組み合わせを含み、nは独立して1及び30の間の整数であり、Xはいずれかのアミノ酸である。いくつかの態様では、リンカーはアミノ酸配列(GGS)N(配列番号140)を含み、nは1、3、又は7である。いくつかの態様では、リンカーはアミノ酸配列SGSETPGTSESATPES(配列番号142)を含む。いくつかの態様では、リンカーはアミノ酸配列SGGSSGGSSGSETPGTSESATPESSGGSSGGS(配列番号143)を含む。いくつかの態様では、リンカーはアミノ酸配列SGGSGGSGGS(配列番号144)を含む。いくつかの態様では、リンカーはアミノ酸配列SGGS(配列番号8)を含む。
特に、次のリンカーが、プライム編集因子ドメインを互いと連結するために種々の態様に用いられ得る:
GGS(配列番号140);
GGSGGS(配列番号145);
GGSGGSGGS(配列番号146);
SGGSSGGSSGSETPGTSESATPESSGGSSGGSS(配列番号11);
SGSETPGTSESATPES(配列番号142);
SGGSSGGSSGSETPGTSESATPESAGSYPYDVPDYAGSAAPAAKKKKLDGSGSGGSSGGS(配列番号131).
GGS(配列番号140);
GGSGGS(配列番号145);
GGSGGSGGS(配列番号146);
SGGSSGGSSGSETPGTSESATPESSGGSSGGSS(配列番号11);
SGSETPGTSESATPES(配列番号142);
SGGSSGGSSGSETPGTSESATPESAGSYPYDVPDYAGSAAPAAKKKKLDGSGSGGSSGGS(配列番号131).
B.核局在配列(NLS)
種々の態様において、プライム編集因子は1以上の核局在配列(NLS)を含んでもよく、これらは細胞核内へのタンパク質の移行を促進することを助ける。かかる配列は当該技術分野で周知であり、次の例を包含し得る:

種々の態様において、プライム編集因子は1以上の核局在配列(NLS)を含んでもよく、これらは細胞核内へのタンパク質の移行を促進することを助ける。かかる配列は当該技術分野で周知であり、次の例を包含し得る:
上のNLS例は限定しない。プライム編集因子はいずれかの公知のNLS配列を含んでもよく、Cokol et al.,「Finding nuclear localization signals,」EMBO Rep.,2000,1(5):411-415及びFreitas et al.,「Mechanisms and Signals for the Nuclear Import of Proteins,」Current Genomics,2009,10(8):550-7に記載されているもののいずれかを包含する。これらの夫々は参照によって本明細書に組み込まれる。
種々の態様において、本明細書に開示のプライム編集因子及びプライム編集因子をコードする構築物は、更に、1以上の、好ましくは少なくとも2つの核局在シグナルを含む。ある態様において、プライム編集因子は少なくとも2つのNLSを含む。少なくとも2つのNLSによる態様では、NLSは同じNLSであり得るか、又はそれらは異なるNLSであり得る。加えて、NLSはプライム編集因子の残りの部分との融合タンパク質の一部として発現されてもよい。いくつかの態様では、NLSの1以上は双節型NLS(「bpNLS」)である。ある態様において、本開示の融合タンパク質は2つの双節型NLSを含む。いくつかの態様では、本開示の融合タンパク質は2つよりも多くの双節型NLSを含む。
NLS融合の位置付けはプライム編集因子のN末端、C末端、又は配列内であり得る(例としてコードされるnapDNAbp構成要素(例としてCas9)及びポリメラーゼドメイン(例として逆転写酵素ドメイン)の間に挿入される。
NLSは当該技術分野のいずれかの公知のNLS配列であってもよい。NLSはまた、核局在のためのいずれかの将来に発見されるNLSであってもよい。NLSはまた、いずれかの天然に存在するNLS又はいずれかの天然に存在しないNLS(例として、1以上の所望の変異を有するNLS)であってもよい。
用語「核局在化配列」又は「NLS」は、タンパク質の細胞核中への、例えば核輸送による、インポート(import)を促進するアミノ酸配列を指す。核局在化配列は当該技術分野において知られており、当業者に明らかであろう。例えば、NLS配列は、2001年5月31日にWO/2001/038547として公開された2000年11月23日出願のPlank et al.の国際PCT出願PCT/EP2000/011690に記載されている。これの内容は参照によって本明細書に組み込まれる。いくつかの態様では、NLSはアミノ酸配列PKKKRKV(配列番号26)、MDSLLMNRRKFLYQFKNVRWAKGRRETYLC(配列番号27)、KRTADGSEFESPKKKRKV(配列番号154)、又はKRTADGSEFEPKKKRKV(配列番号155)を含む。他の態様において、NLSはアミノ酸配列NLSKRPAAIKKAGQAKKKK(配列番号156)、PAAKRVKLD(配列番号149)、RQRRNELKRSF(配列番号157)、NQSSNFGPMKGGNFGGRSSGPYGGGGQYFAKPRNQGGY(配列番号158)を含む。
本開示の一側面において、プライム編集因子は、1以上の核局在シグナル(NLS)、好ましくは少なくとも2つのNLSによって修飾されてもよい。ある態様において、プライム編集因子は2以上のNLSによって修飾される。本開示は、本開示のときに当該技術分野で公知のいずれかの核局在シグナル又は本出願時の後の現況技術によって同定されるか若しくはさもなければ利用可能にされるいずれかの核局在シグナルの使用を企図する。代表的な核局在シグナルは、配列が発現される細胞の核にタンパク質を導くペプチド配列である。核局在シグナルは支配的には塩基性であり、タンパク質のアミノ酸配列のほとんどどこにでも配置され得、一般的には、4アミノ酸(Autieri&Agrawal,(1998)J.Biol.Chem.273:14731-37、参照によって本明細書に組み込まれる)~8アミノ酸の短い配列を含み、典型的にはリジン及びアルギニン残基がリッチである(Magin et al.,(2000)Virology 274:11-16、参照によって本明細書に組み込まれる)。核局在シグナルは多くの場合にはプロリン残基を含む。種々の核局在シグナルが同定されており、細胞質から細胞の核への生物学的分子の輸送を成し遂げるために用いられている。例として、Tinland et al.,(1992)Proc.Natl.Acad.Sci.U.S.A.89:7442-46;Moede et al.,(1999)FEBS Lett.461:229-34を見よ。これは参照によって組み込まれる。移行は現行では核膜孔タンパク質を伴うと考えられる。
ほとんどのNLSは3つの一般的な群に分類され得る:(i)SV40ラージT抗原NLS(PKKKRKV(配列番号26))によって例示される単節型NLS;(ii)XenopusヌクレオプラスミンNLS(KRXXXXXXXXXXKKKL(配列番号159))によって例示される、可変の数のスペーサアミノ酸によって分離された2つの塩基性ドメインからなる双節型モチーフ;並びに(iii)非標準的な配列、例えばhnRNP A1タンパク質のM9、インフルエンザウイルス核タンパク質NLS、及び酵母Gal4タンパク質NLS(Dingwall and Laskey 1991)。
核局在シグナルはタンパク質のアミノ酸配列上の種々の点に出現する。NLSはタンパク質のN末端、C末端、及び中心領域から同定されている。それゆえに、本開示は、プライム編集因子のC末端、N末端、及び内部の領域において1以上のNLSによって修飾されてもよいプライム編集因子を提供する。核局在シグナルそれ自体と例えば強直的に又は立体的に干渉しないようにして、構成要素NLS残基として機能しないより長い配列の残基が選択されるべきである。よって、NLSを含む配列の組成に厳格な限定はないが、事実上、かかる配列は長さ及び組成が機能的に限定され得る。
本開示は、1以上のNLSを包含するようにプライム編集因子を修飾するためのいずれかの好適な手段を企図する。一側面において、プライム編集因子は、そのN末端又はそのC末端(又は両方)において1以上のNLSに翻訳的に融合されているプライム編集因子タンパク質を発現するように、すなわちプライム編集因子-NLS融合体構築物を形成するように、操作されてもよい。他の態様において、プライム編集因子をコードするヌクレオチド配列は、コードされるプライム編集因子の内部の領域に1以上のNLSをコードする読み枠を組み込むように遺伝子修飾されてもよい。加えて、NLSは、プライム編集因子とN末端に、C末端に、又は内部に、例としてそしてタンパク質の中心領域に取り付けられたNLSアミノ酸配列との間にコードされる種々のアミノ酸リンカー又はスペーサ領域を包含してもよい。それゆえに、本開示は、プライム編集因子及び1以上のNLSを含む融合タンパク質を発現するためのヌクレオチド構築物、ベクター、及びホスト細胞をもまた可能にする。
本明細書に記載のプライム編集因子は核局在シグナルをもまた含んでもよく、これらは、1以上のリンカー、例としてそしてポリマー性、アミノ酸、核酸、多糖、化学的、又は核酸リンカー要素を介してプライム編集因子に連結される。本開示の企図される範囲内のリンカーはいずれかの限定を有することを意図されず、いずれかの好適な型の分子(例として、ポリマー、アミノ酸、多糖、核酸、脂質、又はいずれかの合成化学的リンカードメイン)であり得、プライム編集因子及び1以上のNLSの間に結合(例として、共有結合的連結、水素結合)を形成することを成就するいずれかの好適な戦略によって、プライム編集因子に連結され得る。
C.フラップエンドヌクレアーゼ(例としてFEN1)
種々の態様において、プライム編集因子は1以上のフラップエンドヌクレアーゼ(例としてFEN1)を含んでもよく、これは5'一本鎖DNAフラップの除去を触媒する酵素を指す。これらは、細胞プロセス(DNA複製を包含する)の最中に形成された5'フラップの除去を処理する酵素である。本明細書に記載のプライム編集方法は、内生で供給されるフラップエンドヌクレアーゼ、又はプライム編集の最中に標的部位にて形成される内生DNAの5'フラップを除去するのにtransで提供されるものを利用してもよい。フラップエンドヌクレアーゼは当該技術分野において知られており、Patel et al.,「Flap endonucleases pass 5'-flaps through a flexible arch using a disorder-thread-order mechanism to confer specificity for free 5'-ends,」Nucleic Acids Research,2012,40(10):4507-4519及びTsutakawa et al.,「Human flap endonuclease structures,DNA double-base flipping,and a unified understanding of the FEN1 superfamily,」Cell,2011,145(2):198-211(これらの各々は参照により本明細書に組み込まれる)に記載のものから見出され得る。例示のフラップエンドヌクレアーゼは、配列番号15のアミノ酸配列によって表され得るFEN1である:
種々の態様において、プライム編集因子は1以上のフラップエンドヌクレアーゼ(例としてFEN1)を含んでもよく、これは5'一本鎖DNAフラップの除去を触媒する酵素を指す。これらは、細胞プロセス(DNA複製を包含する)の最中に形成された5'フラップの除去を処理する酵素である。本明細書に記載のプライム編集方法は、内生で供給されるフラップエンドヌクレアーゼ、又はプライム編集の最中に標的部位にて形成される内生DNAの5'フラップを除去するのにtransで提供されるものを利用してもよい。フラップエンドヌクレアーゼは当該技術分野において知られており、Patel et al.,「Flap endonucleases pass 5'-flaps through a flexible arch using a disorder-thread-order mechanism to confer specificity for free 5'-ends,」Nucleic Acids Research,2012,40(10):4507-4519及びTsutakawa et al.,「Human flap endonuclease structures,DNA double-base flipping,and a unified understanding of the FEN1 superfamily,」Cell,2011,145(2):198-211(これらの各々は参照により本明細書に組み込まれる)に記載のものから見出され得る。例示のフラップエンドヌクレアーゼは、配列番号15のアミノ酸配列によって表され得るFEN1である:
フラップエンドヌクレアーゼは、いずれかのFEN1バリアント、変異体、又は他のフラップエンドヌクレアーゼオーソログ、ホモログ、若しくはバリアントをもまた包含してもよい。非限定的なFEN1バリアントの例は以下の通りである。

種々の態様において、本明細書において企図されるプライム編集因子は、上の配列のいずれかと少なくとも約70%同一、少なくとも約80%同一、少なくとも約90%同一、少なくとも約95%同一、少なくとも約96%同一、少なくとも約97%同一、少なくとも約98%同一、少なくとも約99%同一、少なくとも約99.5%同一、又は少なくとも約99.9%同一であるアミノ酸配列を有する上に開示されている配列のいずれかのフラップエンドヌクレアーゼバリアントを包含してもよい。
5'端一本鎖DNAフラップの除去を容易化するために本方法によって利用されてもよい他のエンドヌクレアーゼは、(1)trex2、(2)exo1エンドヌクレアーゼを包含するが、これらに限定されない(例として、Keijzers et al.,Biosci Rep.2015,35(3):e00206)。
Trex2
3'threeプライム修復エキソヌクレアーゼ2(TREX 2)-ヒト
3'threeプライム修復エキソヌクレアーゼ2(TREX 2)-ヒト
受託番号NM_080701
MSEAPRAETFVFLDLEATGLPSVEPEIAELSLFAVHRSSLENPEHDESGALVLPRVLDKLTLCMCPERPFTAKASEITGLSSEGLARCRKAGFDGAVVRTLQAFLSRQAGPICLVAHNGFDYDFPLLCAELRRLGARLPRDTVCLDTLPALRGLDRAHSHGTRARGRQGYSLGSLFHRYFRAEPSAAHSAEGDVHTLLLIFLHRAAELLAWADEQARGWAHIEPMYLPPDDPSLEA(配列番号165).
MSEAPRAETFVFLDLEATGLPSVEPEIAELSLFAVHRSSLENPEHDESGALVLPRVLDKLTLCMCPERPFTAKASEITGLSSEGLARCRKAGFDGAVVRTLQAFLSRQAGPICLVAHNGFDYDFPLLCAELRRLGARLPRDTVCLDTLPALRGLDRAHSHGTRARGRQGYSLGSLFHRYFRAEPSAAHSAEGDVHTLLLIFLHRAAELLAWADEQARGWAHIEPMYLPPDDPSLEA(配列番号165).
3'three修復エキソヌクレアーゼ2(TREX 2)-マウス
受託番号NM_011907
MSEPPRAETFVFLDLEATGLPNMDPEIAEISLFAVHRSSLENPERDDSGSLVLPRVLDKLTLCMCPERPFTAKASEITGLSSESLMHCGKAGFNGAVVRTLQGFLSRQEGPICLVAHNGFDYDFPLLCTELQRLGAHLPQDTVCLDTLPALRGLDRAHSHGTRAQGRKSYSLASLFHRYFQAEPSAAHSAEGDVHTLLLIFLHRAPELLAWADEQARSWAHIEPMYVPPDGPSLEA(配列番号166).
3'threeプライム修復エキソヌクレアーゼ2(TREX 2)-ラット
受託番号NM_001107580
MSEPLRAETFVFLDLEATGLPNMDPEIAEISLFAVHRSSLENPERDDSGSLVLPRVLDKLTLCMCPERPFTAKASEITGLSSEGLMNCRKAAFNDAVVRTLQGFLSRQEGPICLVAHNGFDYDFPLLCTELQRLGAHLPRDTVCLDTLPALRGLDRVHSHGTRAQGRKSYSLASLFHRYFQAEPSAAHSAEGDVNTLLLIFLHRAPELLAWADEQARSWAHIEPMYVPPDGPSLEA(配列番号167).
ExoI
ヒトエキソヌクレアーゼ1(EXO1)は、DNAミスマッチ修復(MMR)、マイクロ媒介末端結合(micro-mediated end-joining、相同組換え(HR)、及び複製を包含する、多くの種々のDNA代謝プロセスに関係する。ヒトEXO1は、真核生物ヌクレアーゼRad2/XPGのファミリーに属し、これはまたFEN1及びGEN1も包含する。Rad2/XPGファミリーは、ファージからヒトまでの種を通してヌクレアーゼドメインが保存されている。EXO1遺伝子産物は、5'エキソヌクレアーゼと5'フラップ活性との両方を呈する。加えて、EXO1は、固有の5'RNase H活性も含有する。ヒトEXO1は、二本鎖DNA(dsDNA)、ニック、ギャップ、シュードY構造のプロセシングに対し高親和性を有し、遺伝で受け継いだそのフラップ活性を使用してホリデイ接点を分解し得る。ヒトEXO1はMMRに関係し、MLH1及びMSH2と直接相互作用する保存された結合ドメインを含有する。EXO1核酸分解活性は、PCNA、MutSα(MSH2/MSH6複合体)、14-3-3、MRN、及び9-1-1複合体によって正に刺激される。
ヒトエキソヌクレアーゼ1(EXO1)は、DNAミスマッチ修復(MMR)、マイクロ媒介末端結合(micro-mediated end-joining、相同組換え(HR)、及び複製を包含する、多くの種々のDNA代謝プロセスに関係する。ヒトEXO1は、真核生物ヌクレアーゼRad2/XPGのファミリーに属し、これはまたFEN1及びGEN1も包含する。Rad2/XPGファミリーは、ファージからヒトまでの種を通してヌクレアーゼドメインが保存されている。EXO1遺伝子産物は、5'エキソヌクレアーゼと5'フラップ活性との両方を呈する。加えて、EXO1は、固有の5'RNase H活性も含有する。ヒトEXO1は、二本鎖DNA(dsDNA)、ニック、ギャップ、シュードY構造のプロセシングに対し高親和性を有し、遺伝で受け継いだそのフラップ活性を使用してホリデイ接点を分解し得る。ヒトEXO1はMMRに関係し、MLH1及びMSH2と直接相互作用する保存された結合ドメインを含有する。EXO1核酸分解活性は、PCNA、MutSα(MSH2/MSH6複合体)、14-3-3、MRN、及び9-1-1複合体によって正に刺激される。
エキソヌクレアーゼ1(EXO1)受託番号NM_003686(Homo sapiensエキソヌクレアーゼ1(EXO1)、転写産物バリアント3)-アイソフォームA
MGIQGLLQFIKEASEPIHVRKYKGQVVAVDTYCWLHKGAIACAEKLAKGEPTDRYVGFCMKFVNMLLSHGIKPILVFDGCTLPSKKEVERSRRERRQANLLKGKQLLREGKVSEARECFTRSINITHAMAHKVIKAARSQGVDCLVAPYEADAQLAYLNKAGIVQAIITEDSDLLAFGCKKVILKMDQFGNGLEIDQARLGMCRQLGDVFTEEKFRYMCILSGCDYLSSLRGIGLAKACKVLRLANNPDIVKVIKKIGHYLKMNITVPEDYINGFIRANNTFLYQLVFDPIKRKLIPLNAYEDDVDPETLSYAGQYVDDSIALQIALGNKDINTFEQIDDYNPDTAMPAHSRSHSWDDKTCQKSANVSSIWHRNYSPRPESGTVSDAPQLKENPSTVGVERVISTKGLNLPRKSSIVKRPRSAELSEDDLLSQYSLSFTKKTKKNSSEGNKSLSFSEVFVPDLVNGPTNKKSVSTPPRTRNKFATFLQRKNEESGAVVVPGTRSRFFCSSDSTDCVSNKVSIQPLDETAVTDKENNLHESEYGDQEGKRLVDTDVARNSSDDIPNNHIPGDHIPDKATVFTDEESYSFESSKFTRTISPPTLGTLRSCFSWSGGLGDFSRTPSPSPSTALQQFRRKSDSPTSLPENNMSDVSQLKSEESSDDESHPLREEACSSQSQESGEFSLQSSNASKLSQCSSKDSDSEESDCNIKLLDSQSDQTSKLRLSHFSKKDTPLRNKVPGLYKSSSADSLSTTKIKPLGPARASGLSKKPASIQKRKHHNAENKPGLQIKLNELWKNFGFKKF(配列番号168).
MGIQGLLQFIKEASEPIHVRKYKGQVVAVDTYCWLHKGAIACAEKLAKGEPTDRYVGFCMKFVNMLLSHGIKPILVFDGCTLPSKKEVERSRRERRQANLLKGKQLLREGKVSEARECFTRSINITHAMAHKVIKAARSQGVDCLVAPYEADAQLAYLNKAGIVQAIITEDSDLLAFGCKKVILKMDQFGNGLEIDQARLGMCRQLGDVFTEEKFRYMCILSGCDYLSSLRGIGLAKACKVLRLANNPDIVKVIKKIGHYLKMNITVPEDYINGFIRANNTFLYQLVFDPIKRKLIPLNAYEDDVDPETLSYAGQYVDDSIALQIALGNKDINTFEQIDDYNPDTAMPAHSRSHSWDDKTCQKSANVSSIWHRNYSPRPESGTVSDAPQLKENPSTVGVERVISTKGLNLPRKSSIVKRPRSAELSEDDLLSQYSLSFTKKTKKNSSEGNKSLSFSEVFVPDLVNGPTNKKSVSTPPRTRNKFATFLQRKNEESGAVVVPGTRSRFFCSSDSTDCVSNKVSIQPLDETAVTDKENNLHESEYGDQEGKRLVDTDVARNSSDDIPNNHIPGDHIPDKATVFTDEESYSFESSKFTRTISPPTLGTLRSCFSWSGGLGDFSRTPSPSPSTALQQFRRKSDSPTSLPENNMSDVSQLKSEESSDDESHPLREEACSSQSQESGEFSLQSSNASKLSQCSSKDSDSEESDCNIKLLDSQSDQTSKLRLSHFSKKDTPLRNKVPGLYKSSSADSLSTTKIKPLGPARASGLSKKPASIQKRKHHNAENKPGLQIKLNELWKNFGFKKF(配列番号168).
エキソヌクレアーゼ1(EXO1)受託番号NM_006027(Homo sapiensエキソヌクレアーゼ1(EXO1)、転写産物バリアント3)-アイソフォームB
MGIQGLLQFIKEASEPIHVRKYKGQVVAVDTYCWLHKGAIACAEKLAKGEPTDRYVGFCMKFVNMLLSHGIKPILVFDGCTLPSKKEVERSRRERRQANLLKGKQLLREGKVSEARECFTRSINITHAMAHKVIKAARSQGVDCLVAPYEADAQLAYLNKAGIVQAIITEDSDLLAFGCKKVILKMDQFGNGLEIDQARLGMCRQLGDVFTEEKFRYMCILSGCDYLSSLRGIGLAKACKVLRLANNPDIVKVIKKIGHYLKMNITVPEDYINGFIRANNTFLYQLVFDPIKRKLIPLNAYEDDVDPETLSYAGQYVDDSIALQIALGNKDINTFEQIDDYNPDTAMPAHSRSHSWDDKTCQKSANVSSIWHRNYSPRPESGTVSDAPQLKENPSTVGVERVISTKGLNLPRKSSIVKRPRSAELSEDDLLSQYSLSFTKKTKKNSSEGNKSLSFSEVFVPDLVNGPTNKKSVSTPPRTRNKFATFLQRKNEESGAVVVPGTRSRFFCSSDSTDCVSNKVSIQPLDETAVTDKENNLHESEYGDQEGKRLVDTDVARNSSDDIPNNHIPGDHIPDKATVFTDEESYSFESSKFTRTISPPTLGTLRSCFSWSGGLGDFSRTPSPSPSTALQQFRRKSDSPTSLPENNMSDVSQLKSEESSDDESHPLREEACSSQSQESGEFSLQSSNASKLSQCSSKDSDSEESDCNIKLLDSQSDQTSKLRLSHFSKKDTPLRNKVPGLYKSSSADSLSTTKIKPLGPARASGLSKKPASIQKRKHHNAENKPGLQIKLNELWKNFGFKKDSEKLPPCKKPLSPVRDNIQLTPEAEEDIFNKPECGRVQRAIFQ(配列番号169).
エキソヌクレアーゼ1(EXO1)受託番号NM_001319224(Homo sapiensエキソヌクレアーゼ1(EXO1)、転写産物バリアント4)-アイソフォームC
MGIQGLLQFIKEASEPIHVRKYKGQVVAVDTYCWLHKGAIACAEKLAKGEPTDRYVGFCMKFVNMLLSHGIKPILVFDGCTLPSKKEVERSRRERRQANLLKGKQLLREGKVSEARECFTRSINITHAMAHKVIKAARSQGVDCLVAPYEADAQLAYLNKAGIVQAIITEDSDLLAFGCKKVILKMDQFGNGLEIDQARLGMCRQLGDVFTEEKFRYMCILSGCDYLSSLRGIGLAKACKVLRLANNPDIVKVIKKIGHYLKMNITVPEDYINGFIRANNTFLYQLVFDPIKRKLIPLNAYEDDVDPETLSYAGQYVDDSIALQIALGNKDINTFEQIDDYNPDTAMPAHSRSHSWDDKTCQKSANVSSIWHRNYSPRPESGTVSDAPQLKENPSTVGVERVISTKGLNLPRKSSIVKRPRSELSEDDLLSQYSLSFTKKTKKNSSEGNKSLSFSEVFVPDLVNGPTNKKSVSTPPRTRNKFATFLQRKNEESGAVVVPGTRSRFFCSSDSTDCVSNKVSIQPLDETAVTDKENNLHESEYGDQEGKRLVDTDVARNSSDDIPNNHIPGDHIPDKATVFTDEESYSFESSKFTRTISPPTLGTLRSCFSWSGGLGDFSRTPSPSPSTALQQFRRKSDSPTSLPENNMSDVSQLKSEESSDDESHPLREEACSSQSQESGEFSLQSSNASKLSQCSSKDSDSEESDCNIKLLDSQSDQTSKLRLSHFSKKDTPLRNKVPGLYKSSSADSLSTTKIKPLGPARASGLSKKPASIQKRKHHNAENKPGLQIKLNELWKNFGFKKDSEKLPPCKKPLSPVRDNIQLTPEAEEDIFNKPECGRVQRAIFQ(配列番号170).
D.インテイン及び分裂インテイン
いくつかの態様では(例として、AAV粒子を用いるin vivoのプライム編集因子の送達)、ポリペプチド(例として、デアミナーゼ又はnapDNAbp)又は融合タンパク質(例としてプライム編集因子)をN末端の半分及びC末端の半分へと分裂し、それらを別個に送達し、次いでそれらの共局在が完全なタンパク質(又は場合に応じて融合タンパク質)を細胞内で再形成することを許すことが有利であってもよいということは理解されるであろう。タンパク質又は融合タンパク質の別個の半分同士は、夫々が、タンパク質トランススプライシングの機序による完全なタンパク質又は融合タンパク質の再形成を容易化するための分裂インテインタグを含んでもよい。
いくつかの態様では(例として、AAV粒子を用いるin vivoのプライム編集因子の送達)、ポリペプチド(例として、デアミナーゼ又はnapDNAbp)又は融合タンパク質(例としてプライム編集因子)をN末端の半分及びC末端の半分へと分裂し、それらを別個に送達し、次いでそれらの共局在が完全なタンパク質(又は場合に応じて融合タンパク質)を細胞内で再形成することを許すことが有利であってもよいということは理解されるであろう。タンパク質又は融合タンパク質の別個の半分同士は、夫々が、タンパク質トランススプライシングの機序による完全なタンパク質又は融合タンパク質の再形成を容易化するための分裂インテインタグを含んでもよい。
分裂インテインによって触媒されるタンパク質トランススプライシングは、タンパク質ライゲーションのための全く酵素的な方法を提供する。分裂インテインは本質的には一続きのインテイン(例としてミニインテイン)であり、夫々Nインテイン及びCインテインという名称の2つのピースへと分裂されている。分裂インテインのNインテイン及びCインテインは、非共有結合的に会合して活性なインテインを形成し、一続きのインテインがするのと本質的に同じやり方でスプライシング反応を触媒し得る。分裂インテインは天然に見出され、実験室でもまた操作されている。本明細書において用いられる用語「分裂インテイン」は、1以上のペプチド結合切断がN末端及びC末端アミノ酸配列の間に存在するいずれかのインテインを指し、その結果、N末端及びC末端配列が別個の分子になり、これらは、トランススプライシング反応について機能的であるインテインへと非共有結合的に再会合又は再構成し得る。いずれかの触媒的に活性なインテイン又はそのフラグメントが、本発明の方法への使用のための分裂インテインを導出するために用いられてもよい。例えば、一側面において、分裂インテインは真核生物インテインに由来されてもよい。別の側面において、分裂インテインは細菌インテインに由来されてもよい。別の側面において、分裂インテインは古細菌インテインに由来されてもよい。好ましくは、そのように導出された分裂インテインは、トランススプライシング反応を触媒することにとって必須のアミノ酸配列のみを所有するであろう。
本明細書において用いられる「N末端分裂インテイン(In)」は、トランススプライシング反応について機能的であるN末端アミノ酸配列を含むいずれかのインテイン配列を指す。Inは、それゆえに、トランススプライシングが生起するときにスプライスアウトされる配列をもまた含む。Inは、天然に存在するインテイン配列のN末端部分の修飾である配列を含み得る。例えば、かかる追加の及び/又は変異した残基の包含がInをトランススプライシングにおいて非機能的にしない限り、Inは追加のアミノ酸残基及び/又は変異した残基を含み得る。好ましくは、追加の及び/又は変異した残基の包含は、Inのトランススプライシング活性を改善又は増強する。
本明細書において用いられる「C末端分裂インテイン(Ic)」は、トランススプライシング反応について機能的であるC末端アミノ酸配列を含むいずれかのインテイン配列を指す。一側面において、Icは一続きの4~7アミノ酸残基を含み、これらの少なくとも4アミノ酸はそれが由来されたインテイン最後のβ鎖からである。Icは、それゆえに、トランススプライシングが生起するときにスプライスアウトされる配列をもまた含む。Icは、天然に存在するインテイン配列のC末端部分の修飾である配列を含み得る。例えば、かかる追加の及び/又は変異した残基の包含がInをトランススプライシングにおいて非機能的にしない限り、Icは追加のアミノ酸残基及び/又は変異した残基を含み得る。好ましくは、追加の及び/又は変異した残基の包含は、Icのトランススプライシング活性を改善又は増強する。
本発明のいくつかの態様では、Ic又はInに連結されたペプチドは、とりわけ蛍光基、ビオチン、ポリエチレングリコール(PEG)、アミノ酸アナログ、非天然のアミノ酸、リン酸基、グリコシル基、放射性同位体標識、及び医薬分子を包含する追加の化学的部分を含み得る。他の態様において、Icに連結されたペプチドは、とりわけケトン、アルデヒド、Cys残基、及びLys残基を包含する1以上の化学反応性基を含み得る。「インテインスプライシングポリペプチド(ISP)」が存在するときには、分裂インテインのNインテイン及びCインテインは非共有結合的に会合して活性なインテインを形成し、スプライシング反応を触媒し得る。本明細書において用いられる「インテインスプライシングポリペプチド(ISP)」は、Ic、In、又は両方が分裂インテインから除去されるときに留まる分裂インテインのアミノ酸配列の部分を指す。ある態様において、InはISPを含む。別の態様では、IcはISPを含む。更に別の態様では、ISPはInにもIcにも共有結合的に連結されていない別個のペプチドである。
分裂インテインは、ミニインテインの構造上に見出される-12の保存されたベータ鎖間の構造化されていないループ又は介在アミノ酸配列中に1以上の分裂部位を操作することによって、一続きのインテインから作成されてもよい。分裂の作成が、タンパク質スプライシング活性が失われる十分な度合まではインテインの構造、特に構造化されたベータ鎖を遮断しないであろうという条件で、ベータ鎖間の領域内の分裂部位の位置のいくらかのフレキシビリティーが存在してもよい。
タンパク質トランススプライシングでは、1つの前駆体タンパク質はNインテインによって後続されるNエクステインパーツからなり、別の前駆体タンパク質はCエクステインパーツによって後続されるCインテインからなり、トランススプライシング反応(一緒にN及びCインテインによって触媒される)が2つのインテイン配列を切除、2つのエクステイン配列をペプチド結合によって連結する。酵素反応であるタンパク質トランススプライシングは、非常に低い(例としてマイクロモル)濃度のタンパク質で働き得、生理条件下において実行され得る。
例示的な配列は、配列番号16~配列番号23で表される。
インテインは連続ドメインとして最も頻繁に見出されるが、いくつかは自然に分裂された形態で存在する。この場合、2つのフラグメントは別個のポリペプチドとして発現され、スプライシングが起こる前に会合しなければならない、いわゆるタンパク質トランススプライシングである。
例示的な分裂インテインは、2つのサブユニット、すなわちDnaE-N及びDnaE-Cを含むSsp DnaEインテインである。2つの異なるサブユニットは、DnaE-N及びDnaE-Cサブユニットを夫々コードする別々の遺伝子、すなわちdnaE-n及びdnaE-cによってコードされる。DnaEは、Synechocytis sp.PCC6803における天然に存在する分裂インテインであり、それぞれがDnaE-N又はDnaE-Cのいずれかとの融合物を含む2つの別個のタンパク質のトランススプライシングを指示することができる。
更なる天然に存在するか又は操作されたスプリットインテイン配列は、本明細書に記載の全インテイン配列又は当技術分野で利用可能なものから公知であるか、又は作製され得る。スプリットインテイン配列の例は、Stevens et al.,「A promiscuous split intein with expanded protein engineering applications,」 PNAS,2017,Vol.114:8538-8543;Iwai et al.,「Highly efficient protein trans-splicing by a naturally split DnaE intein from Nostoc punctiforme,FEBS Lett,580:1853-1858に見出され得、これらの各々は参照により本明細書に組み込まれる)。更なる分裂インテイン配列は、例えば、国際公開第2013/045632号、国際公開第2014/055782号、国際公開第2016/069774号、及び欧州特許第2877490号に見出され得、これらの各々の内容は参照により本明細書に組み込まれる。
更に、トランスでのタンパク質スプライシングがin vivo及びin vitroで記載されている(Shingledecker,et al.,Gene 207:187(1998),Southworth,et al.,EMBO J.17:918(1998);Mills,et al.,Proc.Natl.Acad.Sci.USA,95:3543-3548(1998);Lew,et al.,J.Biol.Chem.,273:15887-15890(1998);Wu,et al.,Biochim.Biophys.Acta 35732:1(1998b),Yamazaki,et al.,J.Am.Chem.Soc.120:5591(1998),Evans,et al.,J.Biol.Chem.275:9091(2000);Otomo,et al.,Biochemistry 38:16040-16044(1999);Otomo,et al.,J.Biolmol.NMR 14:105-114(1999);Scott,et al.,Proc.Natl.Acad.Sci.USA 96:13638-13643(1999))、例として、別々に発現される二つの半分からの完全なプライム編集因子の形成に関する図66及び図67に示されるように、機能的産物を形成するためにその後ライゲーションを受ける2つの不活性フラグメントに関するタンパク質を発現する機会を提供する。
E.RNA-タンパク質相互作用ドメイン
種々の態様において、2つの別個のタンパク質ドメイン(例として、Cas9ドメイン及びポリメラーゼドメイン)は、「RNA-タンパク質動員システム」、例えば「MS2タグ付け技術」を用いることによって、機能的な複合体(2つの別個のタンパク質ドメインを含む融合タンパク質の機能と同様)を形成するように互いに共局在させられてもよい。かかるシステムは、一般的には、1つのタンパク質ドメインを「RNA-タンパク質相互作用ドメイン」(「RNA-タンパク質動員ドメイン」としてもまた公知)によって、他をRNA-タンパク質相互作用ドメイン、例として特定のヘアピン構造を特異的に認識及び結合する「RNA結合タンパク質」によってタグ付けする。これらの型のシステムは、プライム編集因子のドメイン同士を共局在させるために及びUGIドメイン等の追加の機能性を多重フラッププライム編集因子に動員するために活用され得る。1つの例では、MS2タグ付け技術は、ファージのゲノム上に存在するステムループ又はヘアピン構造、すなわち「MS2ヘアピン」とのMS2バクテリオファージコートタンパク質(「MCP」又は「MS2cp」)の天然の相互作用に基づく。MS2ヘアピンのケースでは、それはMS2バクテリオファージコートタンパク質(MCP)によって認識及び結合される。それゆえに、1つの例示的シナリオでは、デアミナーゼ-MS2融合体はCas9-MCP融合体を動員し得る。
種々の態様において、2つの別個のタンパク質ドメイン(例として、Cas9ドメイン及びポリメラーゼドメイン)は、「RNA-タンパク質動員システム」、例えば「MS2タグ付け技術」を用いることによって、機能的な複合体(2つの別個のタンパク質ドメインを含む融合タンパク質の機能と同様)を形成するように互いに共局在させられてもよい。かかるシステムは、一般的には、1つのタンパク質ドメインを「RNA-タンパク質相互作用ドメイン」(「RNA-タンパク質動員ドメイン」としてもまた公知)によって、他をRNA-タンパク質相互作用ドメイン、例として特定のヘアピン構造を特異的に認識及び結合する「RNA結合タンパク質」によってタグ付けする。これらの型のシステムは、プライム編集因子のドメイン同士を共局在させるために及びUGIドメイン等の追加の機能性を多重フラッププライム編集因子に動員するために活用され得る。1つの例では、MS2タグ付け技術は、ファージのゲノム上に存在するステムループ又はヘアピン構造、すなわち「MS2ヘアピン」とのMS2バクテリオファージコートタンパク質(「MCP」又は「MS2cp」)の天然の相互作用に基づく。MS2ヘアピンのケースでは、それはMS2バクテリオファージコートタンパク質(MCP)によって認識及び結合される。それゆえに、1つの例示的シナリオでは、デアミナーゼ-MS2融合体はCas9-MCP融合体を動員し得る。
RNA-タンパク質相互作用の他のモジュールドメインの総説は、当該技術分野において、例えば、Johansson et al.,「RNA recognition by the MS2 phage coat protein,」Sem Virol.,1997,Vol.8(3):176-185;Delebecque et al.,「Organization of intracellular reactions with rationally designed RNA assemblies,」Science,2011,Vol.333:470-474;Mali et al.,「Cas9 transcriptional activators for target specificity screening and paired nickases for cooperative genome engineering,」Nat.Biotechnol.,2013,Vol.31:833-838;及びZalatan et al.,「Engineering complex synthetic transcriptional programs with CRISPR RNA scaffolds,」Cell,2015,Vol.160:339-350(これら各々はその全体が参照されることによって本明細書に組み込まれる)において記載されている。他の系も、PCPタンパク質を特異的に動員するPP7ヘアピン、及びComタンパク質を特異的に動員する「com」ヘアピンを包含する。Zalatan et al.を参照。
MS2ヘアピンの塩基配列は、配列番号24で表される。
MCP又はMS2cpのアミノ酸配列は、配列番号25で表される。
F.UGIドメイン
他の態様において、本明細書に記載のプライム編集因子は1以上のウラシルグリコシラーゼ阻害剤ドメインを含んでもよい。本明細書において用いられる用語「ウラシルグリコシラーゼ阻害剤(UGI)」又は「UGIドメイン」は、ウラシルDNAグリコシラーゼ塩基除去修復酵素を阻害することができるタンパク質を指す。いくつかの態様では、UGIドメインは、野生型UGI又は配列番号171で明示されるUGIを含む。いくつかの態様では、本明細書において提供されるUGIタンパク質は、UGIのフラグメント、及びUGI又はUGIフラグメントに対して相同的なタンパク質を包含する。例えば、いくつかの態様では、UGIドメインは、配列番号171で明示されるアミノ酸配列のフラグメントを含む。いくつかの態様では、UGIフラグメントは、配列番号171で明示されるアミノ酸配列の少なくとも60%、少なくとも65%、少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、少なくとも96%、少なくとも97%、少なくとも98%、少なくとも99%、又は少なくとも99.5%を含むアミノ酸配列を含む。いくつかの態様では、UGIは、配列番号171で明示されるアミノ酸配列に対して相同的なアミノ酸配列、又は配列番号171で明示されるアミノ酸配列のフラグメントに対して相同的なアミノ酸配列を含む。いくつかの態様では、UGI若しくはUGIのフラグメント又はUGI若しくはUGIフラグメントのホモログを含むタンパク質は、「UGIバリアント」と呼ばれる。UGIバリアントはUGI又はそのフラグメントに対する相同性を共有する。例えば、UGIバリアントは、野生型UGI又は配列番号171で明示されるUGIと少なくとも70%同一、少なくとも75%同一、少なくとも80%同一、少なくとも85%同一、少なくとも90%同一、少なくとも95%同一、少なくとも96%同一、少なくとも97%同一、少なくとも98%同一、少なくとも99%同一、少なくとも99.5%同一、又は少なくとも99.9%同一である。いくつかの態様では、UGIバリアントは、フラグメントが野生型UGI又は配列番号171で明示されるUGIの対応するフラグメントと少なくとも70%同一、少なくとも80%同一、少なくとも90%同一、少なくとも95%同一、少なくとも96%同一、少なくとも97%同一、少なくとも98%同一、少なくとも99%同一、少なくとも99.5%同一、又は少なくとも99.9%であるようにして、UGIのフラグメントを含む。いくつかの態様では、UGIは次のアミノ酸配列を含む:
他の態様において、本明細書に記載のプライム編集因子は1以上のウラシルグリコシラーゼ阻害剤ドメインを含んでもよい。本明細書において用いられる用語「ウラシルグリコシラーゼ阻害剤(UGI)」又は「UGIドメイン」は、ウラシルDNAグリコシラーゼ塩基除去修復酵素を阻害することができるタンパク質を指す。いくつかの態様では、UGIドメインは、野生型UGI又は配列番号171で明示されるUGIを含む。いくつかの態様では、本明細書において提供されるUGIタンパク質は、UGIのフラグメント、及びUGI又はUGIフラグメントに対して相同的なタンパク質を包含する。例えば、いくつかの態様では、UGIドメインは、配列番号171で明示されるアミノ酸配列のフラグメントを含む。いくつかの態様では、UGIフラグメントは、配列番号171で明示されるアミノ酸配列の少なくとも60%、少なくとも65%、少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、少なくとも96%、少なくとも97%、少なくとも98%、少なくとも99%、又は少なくとも99.5%を含むアミノ酸配列を含む。いくつかの態様では、UGIは、配列番号171で明示されるアミノ酸配列に対して相同的なアミノ酸配列、又は配列番号171で明示されるアミノ酸配列のフラグメントに対して相同的なアミノ酸配列を含む。いくつかの態様では、UGI若しくはUGIのフラグメント又はUGI若しくはUGIフラグメントのホモログを含むタンパク質は、「UGIバリアント」と呼ばれる。UGIバリアントはUGI又はそのフラグメントに対する相同性を共有する。例えば、UGIバリアントは、野生型UGI又は配列番号171で明示されるUGIと少なくとも70%同一、少なくとも75%同一、少なくとも80%同一、少なくとも85%同一、少なくとも90%同一、少なくとも95%同一、少なくとも96%同一、少なくとも97%同一、少なくとも98%同一、少なくとも99%同一、少なくとも99.5%同一、又は少なくとも99.9%同一である。いくつかの態様では、UGIバリアントは、フラグメントが野生型UGI又は配列番号171で明示されるUGIの対応するフラグメントと少なくとも70%同一、少なくとも80%同一、少なくとも90%同一、少なくとも95%同一、少なくとも96%同一、少なくとも97%同一、少なくとも98%同一、少なくとも99%同一、少なくとも99.5%同一、又は少なくとも99.9%であるようにして、UGIのフラグメントを含む。いくつかの態様では、UGIは次のアミノ酸配列を含む:
ウラシル-DNAグリコシラーゼ阻害剤:
>sp|P14739|UNGI_BPPB2 MTNLSDIIEKETGKQLVIQESILMLPEEVEEVIGNKPESDILVHTAYDESTDENVMLLTSDAPEYKPWALVIQDSNGENKIKML(配列番号171).
本明細書に記載のプライム編集因子は1より多くのUGIドメインを含んでもよく、これらは本明細書に記載の1以上のリンカーによって分離されてもよい。
G.追加のPEエレメント
ある態様において、本明細書に記載のプライム編集因子は塩基修復の阻害剤を含んでもよい。用語「塩基修復の阻害剤」又は「IBR」は、核酸修復酵素、例えば塩基除去修復酵素の活性を阻害することができるタンパク質を指す。いくつかの態様では、IBRはOGG塩基除去修復の阻害剤である。いくつかの態様では、IBRは塩基除去修復の阻害剤(「iBER」)である。塩基除去修復の例示の阻害剤は、APE1、Endo III、Endo IV、Endo V、Endo VIII、Fpg、hOGG1、hNEIL1、T7 EndoI、T4PDG、UDG、hSMUG1、及びhAAGの阻害剤を包含する。いくつかの態様では、IBRはEndo V又はhAAGの阻害剤である。いくつかの態様では、IBRは、触媒的に不活性なグリコシラーゼ又は触媒的に不活性なジオキシゲナーゼ、又はオキシダーゼの低分子若しくはペプチド阻害剤、あるいはそれらのバリアントであってもよいiBERである。いくつかの態様では、IBRは、TDG阻害剤、MBD4阻害剤、又はAlkBH酵素の阻害剤であってもよいiBERである。いくつかの態様では、IBRは、触媒的に不活性なTDG又は触媒的に不活性なMBD4を含むiBERである。例示の触媒的に不活性なTDGは配列番号175(ヒトTDG)のN140A変異体である。
ある態様において、本明細書に記載のプライム編集因子は塩基修復の阻害剤を含んでもよい。用語「塩基修復の阻害剤」又は「IBR」は、核酸修復酵素、例えば塩基除去修復酵素の活性を阻害することができるタンパク質を指す。いくつかの態様では、IBRはOGG塩基除去修復の阻害剤である。いくつかの態様では、IBRは塩基除去修復の阻害剤(「iBER」)である。塩基除去修復の例示の阻害剤は、APE1、Endo III、Endo IV、Endo V、Endo VIII、Fpg、hOGG1、hNEIL1、T7 EndoI、T4PDG、UDG、hSMUG1、及びhAAGの阻害剤を包含する。いくつかの態様では、IBRはEndo V又はhAAGの阻害剤である。いくつかの態様では、IBRは、触媒的に不活性なグリコシラーゼ又は触媒的に不活性なジオキシゲナーゼ、又はオキシダーゼの低分子若しくはペプチド阻害剤、あるいはそれらのバリアントであってもよいiBERである。いくつかの態様では、IBRは、TDG阻害剤、MBD4阻害剤、又はAlkBH酵素の阻害剤であってもよいiBERである。いくつかの態様では、IBRは、触媒的に不活性なTDG又は触媒的に不活性なMBD4を含むiBERである。例示の触媒的に不活性なTDGは配列番号175(ヒトTDG)のN140A変異体である。
いくつかの例示のグリコシラーゼは下で提供される。これらのグリコシラーゼドメインのいずれかの触媒的に不活性化されたバリアントは、本開示で提供されるプライム編集因子のnapDNAbp又はポリメラーゼドメインに融合されてもよいiBERである。
OGG(ヒト)
MPARALLPRRMGHRTLASTPALWASIPCPRSELRLDLVLPSGQSFRWREQSPAHWSGVLADQVWTLTQTEEQLHCTVYRGDKSQASRPTPDELEAVRKYFQLDVTLAQLYHHWGSVDSHFQEVAQKFQGVRLLRQDPIECLFSFICSSNNNIARITGMVERLCQAFGPRLIQLDDVTYHGFPSLQALAGPEVEAHLRKLGLGYRARYVSASARAILEEQGGLAWLQQLRESSYEEAHKALCILPGVGTKVADCICLMALDKPQAVPVDVHMWHIAQRDYSWHPTTSQAKGPSPQTNKELGNFFRSLWGPYAGWAQAVLFSADLRQSRHAQEPPAKRRKGSKGPEG(配列番号172)
MPG(ヒト)
MVTPALQMKKPKQFCRRMGQKKQRPARAGQPHSSSDAAQAPAEQPHSSSDAAQAPCPRERCLGPPTTPGPYRSIYFSSPKGHLTRLGLEFFDQPAVPLARAFLGQVLVRRLPNGTELRGRIVETEAYLGPEDEAAHSRGGRQTPRNRGMFMKPGTLYVYIIYGMYFCMNISSQGDGACVLLRALEPLEGLETMRQLRSTLRKGTASRVLKDRELCSGPSKLCQALAINKSFDQRDLAQDEAVWLERGPLEPSEPAVVAAARVGVGHAGEWARKPLRFYVRGSPWVSVVDRVAEQDTQA(配列番号173)
MBD4(ヒト)
MGTTGLESLSLGDRGAAPTVTSSERLVPDPPNDLRKEDVAMELERVGEDEEQMMIKRSSECNPLLQEPIASAQFGATAGTECRKSVPCGWERVVKQRLFGKTAGRFDVYFISPQGLKFRSKSSLANYLHKNGETSLKPEDFDFTVLSKRGIKSRYKDCSMAALTSHLQNQSNNSNWNLRTRSKCKKDVFMPPSSSSELQESRGLSNFTSTHLLLKEDEGVDDVNFRKVRKPKGKVTILKGIPIKKTKKGCRKSCSGFVQSDSKRESVCNKADAESEPVAQKSQLDRTVCISDAGACGETLSVTSEENSLVKKKERSLSSGSNFCSEQKTSGIINKFCSAKDSEHNEKYEDTFLESEEIGTKVEVVERKEHLHTDILKRGSEMDNNCSPTRKDFTGEKIFQEDTIPRTQIERRKTSLYFSSKYNKEALSPPRRKAFKKWTPPRSPFNLVQETLFHDPWKLLIATIFLNRTSGKMAIPVLWKFLEKYPSAEVARTADWRDVSELLKPLGLYDLRAKTIVKFSDEYLTKQWKYPIELHGIGKYGNDSYRIFCVNEWKQVHPEDHKLNKYHDWLWENHEKLSLS(配列番号174)
TDG(ヒト)
MEAENAGSYSLQQAQAFYTFPFQQLMAEAPNMAVVNEQQMPEEVPAPAPAQEPVQEAPKGRKRKPRTTEPKQPVEPKKPVESKKSGKSAKSKEKQEKITDTFKVKRKVDRFNGVSEAELLTKTLPDILTFNLDIVIIGINPGLMAAYKGHHYPGPGNHFWKCLFMSGLSEVQLNHMDDHTLPGKYGIGFTNMVERTTPGSKDLSSKEFREGGRILVQKLQKYQPRIAVFNGKCIYEIFSKEVFGVKVKNLEFGLQPHKIPDTETLCYVMPSSSARCAQFPRAQDKVHYYIKLKDLRDQLKGIERNMDVQEVQYTFDLQLAQEDAKKMAVKEEKYDPGYEAAYGGAYGENPCSSEPCGFSSNGLIESVELRGESAFSGIPNGQWMTQSFTDQIPSFSNHCGTQEQEEESHA(配列番号175)
いくつかの態様では、本明細書に記載の融合タンパク質は1以上の異種タンパク質ドメインを含んでもよい(例として、プライム編集因子構成要素に加えて、約又は約1、2、3、4、5、6、7、8、9、10以上の又はそれよりも多くのドメイン)。融合タンパク質は、いずれかの追加のタンパク質配列と、任意選択でいずれかの2つのドメイン間のリンカー配列とを含んでもよい。存在してもよい他の例示の特色は、局在配列、例えば細胞質局在配列、搬出配列、例えば核搬出配列、又は他の局在配列、及び融合タンパク質の可溶化、精製、又は検出に有用である配列タグである。
プライム編集因子又はその構成要素(例として、napDNAbpドメイン、ポリメラーゼドメイン、又はNLSドメイン)に融合されてもよいタンパク質ドメインの例は、限定なしに、エピトープタグ、及びレポーター遺伝子配列を包含する。エピトープタグの限定しない例はヒスチジン(His)タグ、V5タグ、FLAGタグ、インフルエンザヘマグルチニン(HA)タグ、Mycタグ、VSV-Gタグ、及びチオレドキシン(Trx)タグを包含する。レポーター遺伝子の例は、グルタチオン-5-トランスフェラーゼ(GST)、西洋ワサビペルオキシダーゼ(HRP)、クロラムフェニコールアセチルトランスフェラーゼ(CAT)、ベータガラクトシダーゼ、ベータグルクロニダーゼ、ルシフェラーゼ、緑色蛍光タンパク質(GFP)、HcRed、DsRed、シアン蛍光タンパク質(CFP)、黄色蛍光タンパク質(YFP)、及び青色蛍光タンパク質(BFP)を包含する自家蛍光タンパク質を包含するが、これらに限定されない。プライム編集因子は、DNA分子に結合又は他の細胞分子に結合するタンパク質又はタンパク質のフラグメントをコードする遺伝子配列に融合されてもよく、マルトース結合タンパク質(MBP)、Sタグ、Lex A DNA結合ドメイン(DBD)融合体、GAL4 DNA結合ドメイン融合体、及び単純ヘルペスウイルス(HSV)BP16タンパク質融合体を包含するがこれらに限定されない。プライム編集因子の一部を形成してもよい追加のドメインは2011年3月10日公開のUS特許公開No.2011/0059502に記載され、その全体が参照によって本明細書に組み込まれる。
本開示のある側面では、グルタチオン-5-トランスフェラーゼ(GST)、西洋ワサビペルオキシダーゼ(HRP)、クロラムフェニコールアセチルトランスフェラーゼ(CAT)ベータガラクトシダーゼ、ベータグルクロニダーゼ、ルシフェラーゼ、緑色蛍光タンパク質(GFP)、HcRed、DsRed、シアン蛍光タンパク質(CFP)、黄色蛍光タンパク質(YFP)、及び青色蛍光タンパク質(BFP)を包含する自家蛍光タンパク質を包含するがこれらに限定されないレポーター遺伝子が、遺伝子産物の発現の変改又は修飾を測定するためのマーカーとしての用をなす遺伝子産物をコードするように、細胞に導入されてもよい。本開示のある態様において、遺伝子産物はルシフェラーゼである。本開示の更なる態様では、遺伝子産物の発現は減少させられる。
本明細書において提供される好適なタンパク質タグは、ビオチンカルボキシラーゼキャリアタンパク質(BCCP)タグ、mycタグ、カルモジュリンタグ、FLAGタグ、ヘマグルチニン(HA)タグ、ヒスチジンタグ又はHisタグともまた呼ばれるポリヒスチジンタグ、マルトース結合タンパク質(MBP)タグ、nusタグ、グルタチオン-S-トランスフェラーゼ(GST)タグ、緑色蛍光タンパク質(GFP)タグ、チオレドキシンタグ、Sタグ、Softag(例として、Softag 1、Softag 3)、strepタグ、ビオチンリガーゼタグ、FlAsHタグ、V5タグ、及びSBPタグを包含するが、これらに限定されない。追加の好適な配列は当業者には明らかであろう。いくつかの態様では、融合タンパク質は1以上のHisタグを含む。
本開示のいくつかの態様では、PEシステムの発現される構成要素の滞留時間、量、及び/又は活性を調整することによって、プライム編集システムの活性は時間的に制御されてもよい。例えば、本明細書に記載のPEは、PEの細胞内半減期を修飾することができるタンパク質ドメインと融合されてもよい。2以上のベクターを伴うある種の態様(例として、本明細書に記載の構成要素が2以上の別個のベクター上にコードされるベクターシステム)では、PEシステムの活性は、ベクターが送達されるタイミングをコントロールすることによって時間的に制御されてもよい。例えば、いくつかの態様では、鋳型をコードするベクターに先行して、ヌクレアーゼシステムをコードするベクターがPEを送達してもよい。他の態様において、PEシステムをコードするベクターに先行して、pegRNAをコードするベクターはガイドを送達してもよい。いくつかの態様では、PEシステム及びpegRNAをコードするベクターが同時に送達される。ある態様において、同時に送達されるベクターは、例としてPE、pegRNA、及び/又は第2の鎖ガイドRNA構成要素を時間的に送達する。更なる態様では、ベクター上のコード配列から転写されるRNA(例として、例えばヌクレアーゼ転写物)は、更に、RNAの細胞内半減期を修飾及び/又は翻訳コントロールを調節することができる少なくとも1つのエレメントを含んでもよい。いくつかの態様では、RNAの半減期は増大させられてもよい。いくつかの態様では、RNAの半減期は減少させられてもよい。いくつかの態様では、エレメントはRNAの安定性を増大させることができてもよい。いくつかの態様では、エレメントはRNAの安定性を減少させることができてもよい。いくつかの態様では、エレメントはRNAの3'UTR内にあってもよい。いくつかの態様では、エレメントはポリアデニル化シグナル(PA)を包含してもよい。いくつかの態様では、エレメントは、キャップ、例として上流のmRNA又はpegRNA端を包含してもよい。いくつかの態様では、RNAはPAを含まなくてもよく、その結果、それは転写後に細胞内でより迅速な分解を受ける。いくつかの態様では、エレメントは少なくとも1つのAUリッチエレメント(ARE)を包含してもよい。組織型、細胞型、タイミング、細胞局在、及び環境に依存的である様式で、AREはARE結合タンパク質(ARE-BP)によって結合されてもよい。いくつかの態様では、不安定化エレメントがRNA崩壊を促進、RNA安定性に影響、又は翻訳を活性化してもよい。いくつかの態様では、AREは長さが50~150ヌクレオチドを含んでもよい。いくつかの態様では、AREは少なくとも1コピーの配列AUUUAを含んでもよい。いくつかの態様では、少なくとも1つのAREがRNAの3'UTRに追加されてもよい。いくつかの態様では、エレメントはウッドチャック肝炎ウイルス(WHP)であってもよい。
転写物からの発現を増強するための三次構造を作成する転写後制御エレメント(WPRE)。更なる態様では、エレメントは、転写物からの発現を増強することができる修飾及び/又は短縮されたWPRE配列であり、例えばZufferey et al.,J Virol,73(4):2886-92(1999)及びFlajolet et al.,J Virol,72(7):6175-80(1998)に記載されている通りである。いくつかの態様では、WPRE又は等価物はRNAの3'UTRに追加されてもよい。いくつかの態様では、エレメントは、高速で又は低速で崩壊する転写物どちらかに濃縮された他のRNA配列モチーフから選択されてもよい。
いくつかの態様では、PE又はpegRNAをコードするベクターは、PEシステムによるベクター上に存在する標的配列の切断によって自己破壊されてもよい。切断はベクターからのPE又はpegRNAの連続した転写を防止してもよい。転写は直線化されたベクター上において何らかの時間量にわたって生起してもよいが、細胞内分解を受ける発現された転写物又はタンパク質は、コードするベクターの発現からの連続した供給なしには、オフターゲット効果を生ずるためのより少ない時間を有するであろう。
[6]修飾pegRNA
本明細書中に記載されるプライム編集システムは、改善された特徴(例えば、Cas9に対する増大した安定性及び/又は増大した親和性等)を付与する本明細書中に開示される構造モチーフの1つ又は複数を包含するように修飾される任意の適切なpegRNA、特に、pegRNAの使用を企図する。本発明者らは、驚くべきことに、限定されるものだが、プレキノシン1-1リボスイッチアプタマー(「evopreQ1-1」)、MMLVウイルスゲノム由来のシュードノット(「evopreQ1-1」)、及び逆転写のためのプライマーとしてMMLV RTによって使用される修飾tRNAを包含する、特定のヌクレオチド構造モチーフをpegRNA、例として、pegRNAの伸長アームの末端に付加することによって、編集活性の一貫した増加が達成されたことを見出した。
本明細書中に記載されるプライム編集システムは、改善された特徴(例えば、Cas9に対する増大した安定性及び/又は増大した親和性等)を付与する本明細書中に開示される構造モチーフの1つ又は複数を包含するように修飾される任意の適切なpegRNA、特に、pegRNAの使用を企図する。本発明者らは、驚くべきことに、限定されるものだが、プレキノシン1-1リボスイッチアプタマー(「evopreQ1-1」)、MMLVウイルスゲノム由来のシュードノット(「evopreQ1-1」)、及び逆転写のためのプライマーとしてMMLV RTによって使用される修飾tRNAを包含する、特定のヌクレオチド構造モチーフをpegRNA、例として、pegRNAの伸長アームの末端に付加することによって、編集活性の一貫した増加が達成されたことを見出した。
標準的なpegRNAアーキテクチャ
図3Aは、修飾され、次いで本明細書に開示されてもよいプライム編集システムで使用され得る標準的なpegRNAの一態様を示す。標準的なpegRNA(すなわち、本明細書に記載される修飾のいずれも包含しないpegRNA)は、約20ntのスペーサ配列及びgRNAコア領域を包含し、napDNAbpと結合する従来のガイドRNA(緑色部分)を含む。標準的なpegRNAはまた、5'末端、すなわち5'伸長、又は3'末端、すなわち3'伸長に伸長RNAセグメントを包含する。5'伸長は、逆転写鋳型配列、逆転写プライマー結合部位、及び任意選択の5~20ヌクレオチドリンカー配列を包含する。図1A~図1Bに示される通り、RTプライマー結合部位の、ニックの後に形成される遊離3'末とのハイブリッドは、Rループの非標的鎖に形成され、これによって5'-3'方向におけるDNA重合のために逆転写酵素をプライムする。
図3Aは、修飾され、次いで本明細書に開示されてもよいプライム編集システムで使用され得る標準的なpegRNAの一態様を示す。標準的なpegRNA(すなわち、本明細書に記載される修飾のいずれも包含しないpegRNA)は、約20ntのスペーサ配列及びgRNAコア領域を包含し、napDNAbpと結合する従来のガイドRNA(緑色部分)を含む。標準的なpegRNAはまた、5'末端、すなわち5'伸長、又は3'末端、すなわち3'伸長に伸長RNAセグメントを包含する。5'伸長は、逆転写鋳型配列、逆転写プライマー結合部位、及び任意選択の5~20ヌクレオチドリンカー配列を包含する。図1A~図1Bに示される通り、RTプライマー結合部位の、ニックの後に形成される遊離3'末とのハイブリッドは、Rループの非標的鎖に形成され、これによって5'-3'方向におけるDNA重合のために逆転写酵素をプライムする。
図3Bは、本明細書に開示のプライム編集システムに使用可能なpegRNAの別の態様を示すが、既存のガイドRNA(緑色部分)は、~20ntプロトスペーサ配列とgRNAコアとを包含し、napDNAbpとともに結合する。この態様において、ガイドRNAは、3'末にて伸長されたRNAセグメント、すなわち、3'伸長を包含する。この態様において、3'伸長は、逆転写鋳型配列及び逆転写プライマー結合部位を包含する。図1C~図1Dに示される通り、RTプライマー結合部位の、ニックの後に形成される遊離3'末とのハイブリッドは、Rループの非標的鎖に形成され、これによって5'-3'方向におけるDNA重合のために逆転写酵素をプライムする。
図3Cは、本明細書に開示のプライム編集システムに使用可能な、pegRNAの別の態様を示すが、既存のガイドRNA(緑色部分)は、~20ntプロトスペーサ配列とgRNAコアとを包含し、napDNAbpとともに結合する。この態様において、ガイドRNAは、gRNAコア内の分子間のある位置にて伸長されたRNAセグメント、すなわち、分子内伸長を包含する。この態様において、分子内伸長は、逆転写鋳型配列及び逆転写プライマー結合部位を包含する。RTプライマー結合部位はニックの後に形成される遊離3'末とハイブリダイズし、Rループの非標的鎖に形成され、これによって5'-3'方向におけるDNA重合のために逆転写酵素をプライムする。
これらの標準的なpegRNAのいずれかは、プライム編集の効率を高めるために、本明細書中に記載される修飾の1つ又は複数を包含するように更に修飾され得る。
一態様において、分子内RNA伸長の位置は、ガイドRNAのプロトスペーサ配列中にはない。別の態様において、分子内RNA伸長の位置は、gRNAコア中にある。また別の態様において、分子内RNA伸長の位置は、プロトスペーサ配列内を除きガイドRNA分子内のどこにでも、又はプロトスペーサ配列を妨害しない位置にある。
一態様では、分子間RNA伸長は、プロトスペーサ配列の3'末端の下流に挿入される。別の態様では、分子間RNA伸長は、プロトスペーサ配列の3'末端の、少なくとも1ヌクレオチド、少なくとも2ヌクレオチド、少なくとも3ヌクレオチド、少なくとも4ヌクレオチド、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、少なくとも15ヌクレオチド、少なくとも16ヌクレオチド、少なくとも17ヌクレオチド、少なくとも18ヌクレオチド、少なくとも19ヌクレオチド、少なくとも20ヌクレオチド、少なくとも21ヌクレオチド、少なくとも22ヌクレオチド、少なくとも23ヌクレオチド、少なくとも24ヌクレオチド、少なくとも25ヌクレオチド下流に挿入される。
他の態様では、分子間RNA伸長は、gRNAに挿入され、cas 9タンパク質又はその等価物(すなわち、異なるnapDNAbp)と結合及び/又は相互作用する、tracrRNAに対応する又はtracrRNAを含むガイドRNAの部分を指す。好ましくは、分子間RNA伸長の挿入は、tracrRNA部分とnapDNAbpとの間の相互作用を破壊しないか、又は最小限に破壊する。
RNA伸長の長さ(少なくともRT鋳型及びプライマー結合部位を包含する)は、任意の有用な長さであり得る。様々な態様において、RNA伸長は、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、少なくとも15ヌクレオチド、少なくとも16ヌクレオチド、少なくとも17ヌクレオチド、少なくとも18ヌクレオチド、少なくとも19ヌクレオチド、少なくとも20ヌクレオチド、少なくとも21ヌクレオチド、少なくとも22ヌクレオチド、少なくとも23ヌクレオチド、少なくとも24ヌクレオチド、少なくとも25ヌクレオチド、少なくとも30ヌクレオチド、少なくとも40ヌクレオチド、少なくとも50ヌクレオチド、少なくとも60ヌクレオチド、少なくとも70ヌクレオチド、少なくとも80ヌクレオチド、少なくとも90ヌクレオチド、少なくとも100ヌクレオチド、少なくとも200ヌクレオチド、少なくとも300ヌクレオチド、少なくとも400ヌクレオチド、又は少なくとも500ヌクレオチド長である。
RT鋳型配列はまた、任意の適切な長さであり得る。例えば、RT鋳型配列は、少なくとも3ヌクレオチド、少なくとも4ヌクレオチド、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、少なくとも15ヌクレオチド、少なくとも16ヌクレオチド、少なくとも17ヌクレオチド、少なくとも18ヌクレオチド、少なくとも19ヌクレオチド、少なくとも20ヌクレオチド、少なくとも30ヌクレオチド、少なくとも40ヌクレオチド、少なくとも50ヌクレオチド、少なくとも60ヌクレオチド、少なくとも70ヌクレオチド、少なくとも80ヌクレオチド、少なくとも90ヌクレオチド、少なくとも100ヌクレオチド、少なくとも200ヌクレオチド、少なくとも300ヌクレオチド、少なくとも400ヌクレオチド、or 少なくとも500ヌクレオチド長であり得る。
更に他の態様において、逆転写プライマー結合部位配列は、少なくとも3ヌクレオチド、少なくとも4ヌクレオチド、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、少なくとも15ヌクレオチド、少なくとも16ヌクレオチド、少なくとも17ヌクレオチド、少なくとも18ヌクレオチド、少なくとも19ヌクレオチド、少なくとも20ヌクレオチド、少なくとも30ヌクレオチド、少なくとも40ヌクレオチド、少なくとも50ヌクレオチド、少なくとも60ヌクレオチド、少なくとも70ヌクレオチド、少なくとも80ヌクレオチド、少なくとも90ヌクレオチド、少なくとも100ヌクレオチド、少なくとも200ヌクレオチド、少なくとも300ヌクレオチド、少なくとも400ヌクレオチド、or 少なくとも500ヌクレオチド長である。
他の態様では、任意選択のリンカー又はスペーサ配列は、少なくとも3ヌクレオチド、少なくとも4ヌクレオチド、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、少なくとも15ヌクレオチド、少なくとも16ヌクレオチド、少なくとも17ヌクレオチド、少なくとも18ヌクレオチド、少なくとも19ヌクレオチド、少なくとも20ヌクレオチド、少なくとも30ヌクレオチド、少なくとも40ヌクレオチド、少なくとも50ヌクレオチド、少なくとも60ヌクレオチド、少なくとも70ヌクレオチド、少なくとも80ヌクレオチド、少なくとも90ヌクレオチド、少なくとも100ヌクレオチド、少なくとも200ヌクレオチド、少なくとも300ヌクレオチド、少なくとも400ヌクレオチド、or 少なくとも500ヌクレオチド長である。
RT鋳型配列は、ある態様において、単鎖DNA分子をコードするが、これは、非標的鎖と相同である(よって、標的鎖の対応する部位に相補的である)が1以上のヌクレオチド変化を包含する。少なくとも1ヌクレオチド変化は、1以上の単一塩基ヌクレオチド変化、1以上の欠失、及び1以上の挿入を包含していてもよい。
図1Gに描かれる通り、RT鋳型配列の合成された単鎖DNA産物は、非標的鎖と相同であって、1以上のヌクレオチド変化を含有する。RT鋳型配列の単鎖DNA産物は、平衡状態で、相補的な標的鎖配列とハイブリダイズし、これによって相同の内生標的鎖配列に取って代わる。取って代わられた内生鎖は、いくつかの態様において、5'内生DNAフラップ種として言及されてもよい(例として、図1Eを参照)。この5'内生DNAフラップ種は、5'フラップエンドヌクレアーゼ(例として、FEN1)によって除去され得、単鎖DNA産物は、今や内生標的鎖とハイブリダイズされており、ライゲートされて内生配列と新しく合成された鎖との間にミスマッチを作成してもよい。ミスマッチは、細胞固有のDNA修復及び/又は複製プロセスによって分解されてもよい。
様々な態様において、RT鋳型配列のヌクレオチド配列は、5'フラップ種と取って代わられ、かつ編集されるべき部位と重複する、非標的鎖のヌクレオチド配列に対応する。
pegRNAの様々な態様において、逆転写鋳型配列は、ニック部位に隣接する内生DNA配列に相補的な単鎖DNAフラップをコードしていてもよく、ここで単鎖DNAフラップは、所望のヌクレオチド変化を含む。単鎖DNAフラップは、ニック部位にて内生単鎖DNAに取って代わってもよい。ニック部位にて取って代わられた内生単鎖DNAは、5'末を有し、内生フラップを形成し得るが、これは細胞によって切除され得る。様々な態様において、5'末内生フラップを除去することが、単鎖3'DNAフラップの、対応する相補的なDNA鎖とのハイブリダイゼーション、及び単鎖3'DNAフラップが持つ所望のヌクレオチド変化の標的DNA中への組み込み又は同化を促すため、5'末内生フラップの切除は、産物形成を推進するのに役立ち得る。
pegRNAの様々な態様において、単鎖DNAフラップの細胞修復は、所望のヌクレオチド変化の組み入れをもたらし、これによって所望の産物を形成する。
なお他の態様において、所望のヌクレオチド変化は、ニック部位の約-5~+5の間、又はニック部位の約-10~+10の間、又はニック部位の約-20~+20の間、又はニック部位の約-30~+30の間、又はニック部位の約-40~+40の間、又はニック部位の約-50~+50の間、又はニック部位の約-60~+60の間、又はニック部位の約-70~+70の間、又はニック部位の約-80~+80の間、又はニック部位の約-90~+90の間、又はニック部位の約-100~+100の間、又はニック部位の約-200~+200の間である編集窓上に組み入れされる。
他の態様では、所望のヌクレオチド変化は、ニック部位から約+1~+2、又はニック部位から約+1~+3、+1~+4、+1~+5、+1~+6、+1~+7、+1~+8、+1~+9、+1~+10、+1~+11、+1~+12、+1~+13、+1~+14、+1~+15、+1~+16、+1~+17、+1~+18、+1~+19、+1~+20、+1~+21、+1~+22、+1~+23、+1~+24、+1~+25、+1~+26、+1~+27、+1~+28、+1~+29、+1~+30、+1~+31、+1~+32、+1~+33、+1~+34、+1~+35、+1~+36、+1~+37、+1~+38、+1~+39、+1~+40、+1~+41、+1~+42、+1~+43、+1~+44、+1~+45、+1~+46、+1~+47、+1~+48、+1~+49、+1~+50、+1~+51、+1~+52、+1~+53、+1~+54、+1~+55、+1~+56、+1~+57、+1~+58、+1~+59、+1~+60、+1~+61、+1~+62、+1~+63、+1~+64、+1~+65、+1~+66、+1~+67、+1~+68、+1~+69、+1~+70、+1~+71、+1~+72、+1~+73、+1~+74、+1~+75、+1~+76、+1~+77、+1~+78、+1~+79、+1~+80、+1~+81、+1~+82、+1~+83、+1~+84、+1~+85、+1~+86、+1~+87、+1~+88、+1~+89、+1~+90、+1~+90、+1~+91、+1~+92、+1~+93、+1~+94、+1~+95、+1~+96、+1~+97、+1~+98、+1~+99、+1~+100、+1~+101、+1~+102、+1~+103、+1~+104、+1~+105、+1~+106、+1~+107、+1~+108、+1~+109、+1~+110、+1~+111、+1~+112、+1~+113、+1~+114、+1~+115、+1~+116、+1~+117、+1~+118、+1~+119、+1~+120、+1~+121、+1~+122、+1~+123、+1~+124、又は+1~+125の編集ウインドウに組み入れされる。
他の態様では、所望のヌクレオチド変化は、ニック部位から約+1~+2、又はニック部位から約+1~+3、+1~+4、+1~+5、+1~+6、+1~+7、+1~+8、+1~+9、+1~+10、+1~+11、+1~+12、+1~+13、+1~+14、+1~+15、+1~+16、+1~+17、+1~+18、+1~+19、+1~+20、+1~+21、+1~+22、+1~+23、+1~+24、+1~+25、+1~+26、+1~+27、+1~+28、+1~+29、+1~+30、+1~+31、+1~+32、+1~+33、+1~+34、+1~+35、+1~+36、+1~+37、+1~+38、+1~+39、+1~+40、+1~+41、+1~+42、+1~+43、+1~+44、+1~+45、+1~+46、+1~+47、+1~+48、+1~+49、+1~+50、+1~+51、+1~+52、+1~+53、+1~+54、+1~+55、+1~+56、+1~+57、+1~+58、+1~+59、+1~+60、+1~+61、+1~+62、+1~+63、+1~+64、+1~+65、+1~+66、+1~+67、+1~+68、+1~+69、+1~+70、+1~+71、+1~+72、+1~+73、+1~+74、+1~+75、+1~+76、+1~+77、+1~+78、+1~+79、+1~+80、+1~+81、+1~+82、+1~+83、+1~+84、+1~+85、+1~+86、+1~+87、+1~+88、+1~+89、+1~+90、+1~+90、+1~+91、+1~+92、+1~+93、+1~+94、+1~+95、+1~+96、+1~+97、+1~+98、+1~+99、+1~+100、+1~+101、+1~+102、+1~+103、+1~+104、+1~+105、+1~+106、+1~+107、+1~+108、+1~+109、+1~+110、+1~+111、+1~+112、+1~+113、+1~+114、+1~+115、+1~+116、+1~+117、+1~+118、+1~+119、+1~+120、+1~+121、+1~+122、+1~+123、+1~+124、又は+1~+125の編集ウインドウに組み入れされる。
更に他の態様では、所望のヌクレオチド変化は、ニック部位から約+1~+2、又はニック部位から約+1~+5、+1~+10、+1~+15、+1~+20、+1~+25、+1~+30、+1~+35、+1~+40、+1~+45、+1~+50、+1~+55、+1~+100、+1~+105、+1~+110、+1~+115、+1~+120、+1~+125、+1~+130、+1~+135、+1~+140、+1~+145、+1~+150、+1~+155、+1~+160、+1~+165、+1~+170、+1~+175、+1~+180、+1~+185、+1~+190、+1~+195、又は+1~+200の編集ウインドウに組み入れされる。
様々な側面において、pegRNAは、ガイドRNAの修飾バージョンである。ガイドRNAは、コードする核酸から発現されても、又は化学的に合成されてもよい。着目するゲノムの標的部位の標的鎖と相互作用してハイブリダイズするプロトスペーサ配列を包含する、ガイドRNAを入手又はそうでなければ合成するための方法、及びガイドRNAの適切な配列を決定するための方法は、当該技術分野において周知である。
様々な態様において、ガイドRNA配列の具体的なデザイン側面は、PAM配列の場所、標的配列中のパーセントG/C含量、マイクロ相同領域の程度、2次構造等々等の他の因子の中でも、着目するゲノム標的部位のヌクレオチド配列(すなわち、編集されるべき所望の部位)と、本明細書に記載のプライム編集システムに存在するnapDNAbpの型(例として、Cas9タンパク質)とに依存するであろう。
一般に、ガイド配列は、標的ポリヌクレオチド配列との十分な相補性を有するいずれのポリヌクレオチド配列であって、標的配列とハイブリダイズし、かつnapDNAbp(例として、Cas9、Cas9ホモログ、又はCas9バリアント)の標的配列への配列特異的結合に向けるものである。いくつかの態様において、ガイド配列とその対応する標的配列との間の相補性の程度は、好適な整列(alignment)アルゴリズムを使用して最適に整列されたとき、約50%、60%、75%、80%、85%、90%、95%、97.5%、99%以上、又はこれらより高い。最適な整列は、配列を整列させるためのいずれか好適なアルゴリズムの使用で決定されてもよく、アルゴリズムの非限定例は、Smith-Watermanアルゴリズム、Needleman-Wunschアルゴリズム、Burrows-Wheeler Transform(例として、Burrows Wheeler Aligner)、ClustalW、Clustal X、BLAT、Novoalign(Novocraft Technologies)、ELAND(Illumina,San Diego,Calif.)、SOAP(soap.genomics.org.cnにて利用可能)、及びMaq(maq.sourceforge.netにて利用可能)に基づくアルゴリズムを包含する。いくつかの態様において、ガイド配列は、長さが約5、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、35、40、45、50、75ヌクレオチド以上であるか、又はこれらより長い。
いくつかの態様において、ガイド配列は、長さが約75、50、45、40、35、30、25、20、15、12ヌクレオチド未満であるか、又はこれらより短い。ガイド配列の、プライム編集因子(PE)の標的配列への配列特異的結合に向ける能力は、いずれの好適なアッセイによっても査定されてもよい。例えば、プライム編集因子(PE)の構成要素(試験されるべきガイド配列を包含する)は、本明細書に開示のプライム編集因子(PE)の構成要素をコードするベクターでのトランスフェクション、これに続く、本明細書に記載の通りのSurveyorアッセイ等による標的配列内の優先的な切断の査定等によって、対応する標的配列を有する宿主細胞へ提供されてもよい。同様に、標的ポリヌクレオチド配列の切断は試験管中、標的配列、プライム編集因子(PE)の構成要素(試験されるべきガイド配列を包含する)、及び試験ガイド配列とは異なる制御ガイド配列を提供すること、並びに試験ガイド配列と及び制御ガイド配列との間の反応の、標的配列での結合又は切断率を比較することによって、評価されてもよい。他のアッセイもなし得、当業者に思い当たるものであろう。
ガイド配列は、いずれの標的配列をも標的にするのに選択されてもよい。いくつかの態様において、標的配列は、細胞のゲノム内の配列である。例示の標的配列は、標的ゲノム中にユニークな配列を包含する。例えば、S.pyogenes Cas9について、ゲノム中のユニークな標的配列は、形態MMMMMMMMNNNNNNNNNNNNXGG(配列番号176)のCas9標的部位を包含していてもよく、ここでNNNNNNNNNNNNXGG(配列番号177)(Nは、A、G、T、又はCである;及びXは、何でもあり得る)。ゲノム中のユニークな標的配列は、形態MMMMMMMMMNNNNNNNNNNNXGG(配列番号178)のS.pyogenes Cas9標的部位を包含していてもよく、ここでNNNNNNNNNNNXGG(配列番号179)(Nは、A、G、T、又はCである;及びXは、何でもあり得る)。S.thermophilus CRISPR1Cas9について、ゲノム中のユニークな標的配列は、形態MMMMMMMMNNNNNNNNNNNNXXAGAAW(配列番号180)のCas9標的部位を包含していてもよく、ここでNNNNNNNNNNNNXXAGAAW(配列番号181)(Nは、A、G、T、又はCである;Xは、何でもあり得る;及びWは、A又はTである)。ゲノム中のユニークな標的配列は、形態MMMMMMMMMNNNNNNNNNNNXXAGAAW(配列番号182)のS.thermophilus CRISPR1 Cas9標的部位を包含していてもよく、ここでNNNNNNNNNNNXXAGAAW(配列番号183)(Nは、A、G、T、又はCである;Xは、何でもあり得る;及びWは、A又はTである)。S.pyogenes Cas9について、ゲノム中のユニークな標的配列は、形態MMMMMMMMNNNNNNNNNNNNXGGXG(配列番号184)のCas9標的部位を包含していてもよく、ここでNNNNNNNNNNNNXGGXG(配列番号185)(Nは、A、G、T、又はCである;及びXは、何でもあり得る)。ゲノム中のユニークな標的配列は、形態MMMMMMMMMNNNNNNNNNNNXGGXG(配列番号186)のS.pyogenes Cas9標的部位を包含していてもよく、ここでNNNNNNNNNNNXGGXG(配列番号187)(Nは、A、G、T、又はC;及びXは、何でもあり得る)。これらの配列の各々において、「M」は、A、G、T、又はCであってもよく、配列をユニークなものとして同定する際に考慮される必要はない。
いくつかの態様において、ガイド配列は、ガイド配列内の2次構造の程度を低減するよう選択される。2次構造は、いずれの好適なポリヌクレオチド折り畳みアルゴリズムによっても決定されてもよい。いくつかのプログラムは、最小限のギブス(Gibbs)自由エネルギーを算出することに基づく。かかるアルゴリズムの1つの例は、Zuker及びStiegler(Nucleic Acids Res.9(1981),133-148)によって記載される通りの、mFoldである。別の例の折り畳みアルゴリズムは、重心構造予測アルゴリズムを使用する、ウィーン大学(University of Vienna)の理論化学研究所(Institute for Theoretical Chemistry)にて開発された、オンラインウェブサーバRNA foldである(例として、A.R.Gruber et al.,2008,Cell 106(1):23-24;及びPA Carr and GM Church,2009,Nature Biotechnology 27(12):1151-62を参照)。更なるアルゴリズムは、米国出願第61/836,080号;Broad参照BI-2013/004A)に見出されるものであってもよい(参照により本明細書に組み込まれる)。
一般的に、tracrメイト配列は:(1)対応するtracr配列を含有する細胞におけるtracrメイト配列によってフランキングされるガイド配列の切除;及び(2)標的配列における複合体の形成の1以上を促進するために、tracr配列との十分な相補性を有するいずれかの配列を包含し、複合体は、tracr配列にハイブリダイズしたtracrメイト配列を含む。一般的に、相補性の度合は、2つの配列の短い方の長さに沿って、tracrメイト配列及びtracr配列の最適な整列の参照によってである。最適な整列は、いずれの好適な整列アルゴリズムによっても決定されてもよく、更にtracr配列又はtracrメイト配列のいずれかの配列内に自己相補性等の2次構造を構成していてもよい。いくつかの態様において、tracr配列とtracrメイト配列との間の相補性の程度は、その2つのより短い長さに沿って最適に整列されたとき、約25%、30%、40%、50%、60%、70%、80%、90%、95%、97.5%、99%以上、又はこれらより高い。いくつかの態様において、tracr配列は、長さが約5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、25、30、40、50ヌクレオチド以上、又はこれらより長い。いくつかの態様において、tracr配列及びtracrメイト配列は、その2つの間のハイブリダイゼーションがヘアピン等の2次構造を有する転写産物を産生するように、単一転写産物内に含有される。ヘアピン構造における使用のための好ましいループ形成配列は、長さが4ヌクレオチドであり、最も好ましくは配列GAAAを有する。しかしながら、より長い又はより短いループ配列も、代替の配列になり得るよう(as may alternative sequences)使用されてもよい。配列は、好ましくは、ヌクレオチドトリプレット(例えば、AAA)、及び追加のヌクレオチド(例えばC又はG)を包含する。ループ形成配列の例は、CAAA及びAAAGを包含する。本発明の態様において、転写産物又は転写されたポリヌクレオチド配列は、少なくとも2つの、又はそれより多くのヘアピンを有する。好ましい一態様において、転写産物は、2、3、4又は5つのヘアピンを有する。本発明の更なる態様において、転写産物は、最大でも5つのヘアピンを有する。いくつかの態様において、単一の転写産物は更に、転写終止配列を包含する;好ましくは、これは、ポリT配列、例えば6つのTヌクレオチドを包含する。ガイド配列、tracrメイト配列、及びtracr配列を含む単一ポリヌクレオチドの更なる非限定例は、以下の通りであるが(5'→3'に列挙)、ここで「N」は、ガイド配列の塩基を表し、第1ブロックの小文字は、tracrブロック配列を表し、及び第2ブロックの小文字は、tracr配列を表し、及び最後のポリT配列は、転写ターミネータを表す:(1)NNNNNNNNGTTTTTGTACTCTCAAGATTTAGAAATAAATCTTGCAGAAGCTACAAAGATAAGGCTTCATGCCGAAATCAACACCCTGTCATTTTATGGCAGGGTGTTTTCGTTATTTAATTTTTT(配列番号188);
(2)
NNNNNNNNNNNNNNNNNNGTTTTTGTACTCTCAGAAATGCAGAAGCTACAAAGATAAGGCTTCATGCCGAAATCAACACCCTGTCATTTTATGGCAGGGTGTTTTCGTTATTTAATTTTTT(配列番号189);
NNNNNNNNNNNNNNNNNNGTTTTTGTACTCTCAGAAATGCAGAAGCTACAAAGATAAGGCTTCATGCCGAAATCAACACCCTGTCATTTTATGGCAGGGTGTTTTCGTTATTTAATTTTTT(配列番号189);
(3)
NNNNNNNNNNNNNNNNNNNNGTTTTTGTACTCTCAGAAATGCAGAAGCTACAAAGATAAGGCTTCATGCCGAAATCAACACCCTGTCATTTTATGGCAGGGTGTTTTTT(配列番号190);
NNNNNNNNNNNNNNNNNNNNGTTTTTGTACTCTCAGAAATGCAGAAGCTACAAAGATAAGGCTTCATGCCGAAATCAACACCCTGTCATTTTATGGCAGGGTGTTTTTT(配列番号190);
(4)
NNNNNNNNNNNNNNNNNNNNGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTTTTT(配列番号191);
NNNNNNNNNNNNNNNNNNNNGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTTTTT(配列番号191);
(5)
NNNNNNNNNNNNNNNNNNNNGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGTTTTTTT(配列番号192);及び
NNNNNNNNNNNNNNNNNNNNGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGTTTTTTT(配列番号192);及び
(6)
NNNNNNNNNNNNNNNNNNNNGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCATTTTTTTT(配列番号193).
NNNNNNNNNNNNNNNNNNNNGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCATTTTTTTT(配列番号193).
いくつかの態様において、配列(1)~(3)は、S.thermophilus CRISPR1からのCas9と組み合わせて使用される。いくつかの態様において、配列(4)~(6)は、S.pyogenesからのCas9と組み合わせて使用される。いくつかの態様において、tracr配列は、tracrメイト配列を含む転写産物とは別々の転写産物である。
Cas9ドメインと単鎖DNA結合タンパク質とを含む本明細書に開示の通りの融合タンパク質のいずれも、標的部位(例として、編集されるべき点突然変異を含む部位)に対して、標的にさせるために、典型的には、融合タンパク質をガイドRNA(例として、sgRNA)と一緒に共発現させることが必要であることは、当業者には明らかであろう。本明細書中、他のどこかでより詳細に説明される通り、ガイドRNAは、典型的には、Cas9結合を可能にさせるtracrRNAフレームワークと、配列特異性をCas9:核酸編集酵素/ドメイン融合タンパク質へ付与するガイド配列とを含む。
いくつかの態様において、ガイドRNAは、構造5'-[ガイド配列]-GUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGCUUUUU-3'(配列番号194)を含み、ここでガイド配列は、標的配列に相補的な配列を含む。ガイド配列は、典型的には20ヌクレオチド長である。Cas9:核酸編集酵素/ドメイン融合タンパク質を特定のゲノム標的部位に対して標的にさせるための好適なガイドRNAの配列は、本開示に基づき当業者に明らかであろう。かかる好適なガイドRNA配列は、典型的には、編集されるべき標的ヌクレオチドから50ヌクレオチド以内上流又は下流の核酸配列に相補的なガイド配列を含む。提供される融合タンパク質に、特定の標的配列を標的にさせるのに好適ないくつかの例示のガイドRNA配列は、本明細書に提供される。追加のガイド配列は、当該技術分野において周知であり、本明細書に記載のプライム編集因子(PE)とともに使用され得る。
他の態様において、pegRNAは、図3Dに描かれるものを包含する。
更なる他の態様において、pegRNAは、図3Eに描かれるものを包含していてもよい。
図3Dは、本明細書に企図され、例2に定義される方法論に従ってデザインされてもよい、ある態様のpegRNAの構造を提供する。pegRNAは、5'から3'方向に並べられた3つの主要な構成要素、すなわち、スペーサ、gRNAコア、及び3'末端の伸長アームを含む。伸長アームは、5'から3'方向の以下の構造要素、すなわち、任意選択の相同アーム、DNA合成鋳型、及びプライマー結合部位(PBS)に更に分割されてもよい。更に、pegRNAは、任意選択の3'末端修飾領域(e1)及び任意選択の5'末端修飾領域(e2)を含んでもよい。更に、pegRNAは、pegRNAの3'末端に転写終結シグナルを含んでもよい(図示せず)。これらの構造要素は、本明細書で更に定義される。pegRNAの構造の描写は、限定することを意味されるものではなく、要素の配置のバリエーションを包含する。例えば、任意選択の配列修飾因子(e1)及び(e2)は、3'末端及び5'末端に位置することに限定されず、示されている他の領域のいずれかの中又は間に配置され得る。
図3Eは、本明細書に企図される別の態様のpegRNAの構造を提供し、これは例2に定義される方法論に従ってデザインされてもよい。pegRNAは、5'から3'方向に並べられた3つの主要な構成要素、すなわち、スペーサ、gRNAコア、及び3'末端の伸長アームを含む。伸長アームは、5'から3'方向の以下の構造要素、すなわち、任意選択の相同アーム、DNA合成鋳型、及びプライマー結合部位(PBS)に更に分割されてもよい。更に、pegRNAは、任意選択の3'末端修飾領域(e1)及び任意選択の5'末端修飾領域(e2)を含んでもよい。更に、pegRNAは、pegRNAの3'末端上に転写終結シグナルを含んでもよい(図示せず)。これらの構造要素は、本明細書で更に定義される。pegRNAの構造の描写は、限定することを意味されるものではなく、要素の配置のバリエーションを包含する。例えば、任意選択の配列修飾因子(e1)及び(e2)は、3'末端及び5'末端に位置することに限定されず、示されている他の領域のいずれかの中又は間に配置され得る。
いくつかの態様では、本明細書に記載のPEgRNA又はニッキングガイドRNAは、化学修飾核酸塩基又は核酸塩基類似体を含む。いくつかの態様では、PEgRNA又はニッキングガイドRNAは、修飾塩基(例として、メチル化塩基)、インターカレート塩基、修飾糖(例として、2'-フルオロリボース、リボース、2'-デオキシリボース、2'-O-メチルシチジン、アラビノース及びヘキソース)又は修飾リン酸基(例として、ホスホロチオアート及び5'Nホスホロアミダイト結合)を含む。いくつかの態様では、PEgRNAは、2'-O-メチル修飾を含む。いくつかの態様において、PEgRNAは、RNAの最初の3ヌクレオチドと最後の3ヌクレオチドとの間にホスホロチオアート結合を含む。
いくつかの態様では、本明細書に記載のPEgRNA又はニッキングガイドRNAは、ネブラリン又はデオキシネブラリンを含む化学修飾を含む。いくつかの態様において、PEgRNA又はニッキングガイドRNAは、ホスホロチオアート連結を含む化学修飾を含む。いくつかの態様では、PEgRNA又はニッキングガイドRNAは、5'末端又は3'末端にホスホロチオアート連結を含む。いくつかの態様では、PEgRNA又はニッキングガイドRNAは、5'末端又は3'末端に2個及び2個以下の連続するホスホロチオアート連結を含む。いくつかの態様において、PEgRNA又はニッキングガイドRNAは、5'末端又は3'末端に3つの連続するホスホロチオアート連結を含む。いくつかの態様では、PEgRNA又はニッキングガイドRNAは、3'末端又は5'末端に配列5'-UsusU-3'を含み、Uはウリジンを示し、sはホスホロチオアート連結を示す。いくつかの態様では、核酸塩基は化学修飾されていてもよい。核酸塩基に対する化学修飾の例は、2-チオウリジン、4-チオウリジン、N 6-メチルアデノシン、シュードウリジン、2,6-ジアミノプリン、イノシン、チミジン、5-メチルシトシン、5-置換ピリミジン、イソグアニン、イソシトシン又はハロゲン化芳香族基を包含するが、これらに限定されない。
修飾の非限定的な例は、2'-O-メチル(2'-O-Me)、2'-O-(2-メトキシエチル)(2'-O-MOE)、2'-フルオロ(2'-F)、ヌクレオチド間のホスホロチオアート(PS)結合、G-C置換、及びヌクレオチドとその等価物との間の逆無塩基結合を包含してもよい。いくつかの態様では、PEgRNAは、ジヒドロウリジン、イノシン、7-メチルグアノシン、5-メチルシチジン(5 mC)、5'リン酸リボチミジン、2'-O-メチルリボチミジン、2'-O-エチルリボチミジン、2'-フルオロリボチミジン、C-5プロピニル-デオキシシチジン(pdC)、C-5プロピニル-デオキシウリジン(pdU)、C-5プロピニル-ウリジン(pU)、5-メチルシチジン、5-メチルウリジン、5-メチルデオキシシチジン、5-メチルデオキシウリジンメトキシ、2,6-ジアミノプリン、5'-ジメトキシトリチル-N4-エチル-2'-デオキシシチジン、C-5プロピニル-f-ウリジン(pfU)、5-メチルf-シチジン、5-メチルf-ウリジン、C-5プロピニル-m-シチジン(pmC)、5-メチルm-シチジン、5-メチルm-ウリジン、MGB(副溝結合剤)シュードウリジン(Ψ)、1-N-メチルシュードウリジン(1-Me-Ψ)、5-メトキシウリジン(5-MO-U)2'-O-メチル修飾、2'-O-(2-メトキシエチル)修飾、2'-フルオロ修飾、ホスホロチオアート修飾、逆位塩基修飾、デオキシリボヌクレオチド、二環リボース類似体(例として、ロックド核酸(LNA)、C-エチレン架橋核酸(ENA)、架橋核酸(BNA)、ロック解除核酸(UNA))、塩基又は核酸塩基修飾、ヌクレオシド間結合修飾、リボネブラリン、2'-O-メチルネブラリン、又は2'-デオキシネブラリンを含む。修飾の他の例は、限定されないが、5'アデニレート、5'グアノシン-トリホスファートキャップ、5'N7-メチルグアノシン-トリホスファートキャップ、5'トリホスファートキャップ、3'ホスファート、3'チオホスファート、5'ホスファート、5'チオホスファート、シス-シンチミジン二量体、三量体、無塩基性、アクリジン、アゾベンゼン、ビオチン、ビオチンBB、ビオチンTEG、コレステリルTEG、デスチオビオチン、TEG、DNP-TEG、DNP-X、DOTA、dT-ビオチン、二重ビオチン、PCビオチン、ソラレンC2、ソラレンC6、TINA、3'DABCYL、ブラックホールクエンチャ1、ブラックホールクエンチャ2、DABCYL SE、dT-DABCYL、IRDye QC-1、QSY-21、QSY-35、QSY-7、QSY-9、カルボキシルリンカー、チオールリンカー、2'デオキシリボヌクレオシド類似体プリン、2'デオキシリボヌクレオシド類似体ピリミジン、リボヌクレオシド類似体、2'-0-メチルリボヌクレオシド類似体、糖修飾類似体、ゆらぎ/ユニバーサル塩基、蛍光色素標識、2'フルオロRNA、2'O-メチルRNA、メチルホスホネート、ホスホジエステルDNA、ホスホジエステルRNA、ホスホロチオアートDNA、ホスホロチオアートRNA、UNA、シュードウリジン-5'-トリホスファート、5-メチルシチジン-5'-トリホスファート、2-O-メチル-3-ホスホロチオアート又はそれらの任意の組み合わせを包含する。
いくつかの態様では、本開示で提供されるPEgRNA及び/又はニッキングガイドRNAは、修飾、例として、化学修飾又は生物学的修飾を受けていてもよい。修飾は、PEgRNA又はニッキングガイドRNA内の任意の位置で行われてもよく、核酸塩基、リボース構成要素、リン酸バックボーン、又はそれらの任意の組み合わせに対する1つ以上の修飾を包含してもよい。いくつかの態様では、修飾は、構造ガイド修飾であり得る。いくつかの態様では、修飾は、PEgRNAの5'末端及び/又は3'末端にある。いくつかの態様では、化学修飾は、ニッキングガイドRNAの5'末端及び/又は3'末端にある。いくつかの態様では、修飾は、スペーサ配列、伸長アーム、DNA合成鋳型、及び/又はPEgRNAのプライマー結合部位内にあってもよい。いくつかの態様では、修飾は、PEgRNA又はニッキングガイドRNAのスペーサ配列又はgRNAコア内にあってもよい。いくつかの態様では、修飾は、PEgRNA又はニッキングガイドRNAの3'最末端内にあってもよい。いくつかの態様では、修飾は、PEgRNA又はニッキングガイドRNAの5'最末端内にあってもよい。いくつかの態様では、PEgRNA又はニッキングガイドRNAは、3'末端に1、2、3、4、5、6、7、8、9、10個又はそれ以上の修飾ヌクレオチドを含む。いくつかの態様では、PEgRNA又はニッキングガイドRNAは、5'末端に1、2、3、4、5、6、7、8、9、10個又はそれ以上の修飾ヌクレオチドを含む。いくつかの態様では、PEgRNA又はニッキングガイドRNAは、3'末端に1、2、3、4、5個又はそれ以上の修飾ヌクレオチドを含む。いくつかの態様では、PEgRNA又はニッキングガイドRNAは、5'末端に1、2、3、4、又は5個以上の修飾ヌクレオチドを含む。いくつかの態様では、PEgRNA又はニッキングガイドRNAは、3'末端に1、2、3以上の修飾ヌクレオチドを含む。いくつかの態様では、PEgRNA又はニッキングガイドRNAは、5'末端に1、2、3以上の修飾ヌクレオチドを含む。いくつかの態様では、PEgRNA又はニッキングガイドRNAは、3'末端に1、2、3、4、5、6、7、8、9、10個又はそれ以上の連続する修飾ヌクレオチドを含む。いくつかの態様では、PEgRNA又はニッキングガイドRNAは、5'末端に1、2、3、4、5、6、7、8、9、10個又はそれ以上の連続する修飾ヌクレオチドを含む。いくつかの態様では、PEgRNA又はニッキングガイドRNAは、3'末端に1、2、3、4、又は5個の連続する修飾ヌクレオチドを含む。いくつかの態様では、PEgRNA又はニッキングガイドRNAは、5'末端に1、2、3、4、又は5個の連続する修飾ヌクレオチドを含む。いくつかの態様では、PEgRNA又はニッキングガイドRNAは、3'末端に1、2、又は3個の連続する修飾ヌクレオチドを含む。いくつかの態様では、PEgRNA又はニッキングガイドRNAは、5'末端に1、2、又は3個の連続する修飾ヌクレオチドを含む。いくつかの態様では、PEgRNA又はニッキングガイドRNAは、3'末端に3つの連続する修飾ヌクレオチドを含む。いくつかの態様では、PEgRNA又はニッキングガイドRNAは、3'末端付近に1、2、3、4、5個、又はそれ以上の修飾ヌクレオチドを含む。いくつかの態様では、PEgRNA又はニッキングガイドRNAは、3'末端に3つの連続する修飾ヌクレオチドを含む。いくつかの態様では、PEgRNA又はニッキングガイドRNAは、5'末端に3つの連続する修飾ヌクレオチドを含む。いくつかの態様では、PEgRNA又はニッキングガイドRNAは、3'末端付近に1、2、3、4、5個、又はそれ以上の修飾ヌクレオチドを含む。いくつかの態様では、PEgRNA又はニッキングガイドRNAは、3'末端付近に1、2、3、4、5又はそれ以上の連続した修飾ヌクレオチドを含む。
pegRNAデザイン法
本開示はまた、pegRNAをデザインするための方法に関する。
デザインの一側面において、デザインアプローチは、プライム編集が用いられようとする具体的な適用を考慮に入れ得る。例えば、本明細書において例示され論じられる通り、プライム編集は、限定なしに、(a)変異を修正する変化をヌクレオチド配列に組み入れ、(b)タンパク質及びRNAタグを組み入れ、(c)免疫エピトープを目当てのタンパク質に組み入れ、(d)誘導可能な二量体化ドメインをタンパク質に組み入れ、(e)バイオ分子のその活性を変改するために配列を組み入れ又は除去、(f)特定の遺伝子変化を導くためにリコンビナーゼ標的部位を組み入れ、及び(g)エラーを起こしやすいRTを用いることによる標的配列の変異導入をするために用いられ得る。目当ての標的部位におけるヌクレオチド配列を一般的に挿入、変化、又は欠失させるこれらの方法に加えて、プライム編集因子は、高度にプログラム型ライブラリーを構築するために、並びに細胞データ記録及び系統トレース研究を行うためにもまた用いられ得る。これらの種々の使用では、本明細書に記載の通り、これらの適用のいずれかの所与のものにとって特に有用であるpegRNAの調製に当てはまる具体的なデザイン側面があり得る。
プライム編集のいずれかの具体的な適用又は使用のためのpegRNAをデザインするときには、いくつもの考慮事項が考慮に入れられてもよい。これらは:
(a)1以上の核酸塩基修飾がプライム編集因子によって組み入れされることを望まれる標的配列、すなわちヌクレオチド配列;
(b)標的配列上のカットされる部位の位置付け、すなわち、プライム編集因子が一本鎖ニックを誘導して、ニックの1つの側に3'端RTプライマー配列及びニックの他の側に5'端の内生フラップ(これは究極的にはFEN1又はその等価物によって除去され、3'ssDNAフラップによって置き換えられる)を作成するであろう特定の核酸塩基位置。カットされる部位は「編集の位置付け」に類縁である。なぜなら、これは3'端RTプライマー配列を作成し、これはRNA依存的なDNA重合の間にRTによって伸長されるようになって、所望の編集を含有する3'ssDNAフラップを作成し、これが次いで標的配列上の5'内生DNAフラップを置き換えるからである;
(c)利用可能なPAM配列(標準的なSpCas9 PAM部位、及び拡張された又は異なるPAM特異性を有するCas9バリアント及び等価物によって認識される非標準的なPAM部位を包含する);
(d)標的配列上の利用可能なPAM配列とカットされる部位の位置付けとの間の間隔;
(e)用いられようとするプライム編集因子の具体的なCas9、Cas9バリアント、又はCas9等価物;
(f)プライマー結合部位の配列及び長さ;
(g)編集鋳型の配列及び長さ;
(h)相同アームの配列及び長さ;
(i)スペーサ配列及び長さ;並びに
(j)コア配列、
を包含するが、これらに限定されない。
(a)1以上の核酸塩基修飾がプライム編集因子によって組み入れされることを望まれる標的配列、すなわちヌクレオチド配列;
(b)標的配列上のカットされる部位の位置付け、すなわち、プライム編集因子が一本鎖ニックを誘導して、ニックの1つの側に3'端RTプライマー配列及びニックの他の側に5'端の内生フラップ(これは究極的にはFEN1又はその等価物によって除去され、3'ssDNAフラップによって置き換えられる)を作成するであろう特定の核酸塩基位置。カットされる部位は「編集の位置付け」に類縁である。なぜなら、これは3'端RTプライマー配列を作成し、これはRNA依存的なDNA重合の間にRTによって伸長されるようになって、所望の編集を含有する3'ssDNAフラップを作成し、これが次いで標的配列上の5'内生DNAフラップを置き換えるからである;
(c)利用可能なPAM配列(標準的なSpCas9 PAM部位、及び拡張された又は異なるPAM特異性を有するCas9バリアント及び等価物によって認識される非標準的なPAM部位を包含する);
(d)標的配列上の利用可能なPAM配列とカットされる部位の位置付けとの間の間隔;
(e)用いられようとするプライム編集因子の具体的なCas9、Cas9バリアント、又はCas9等価物;
(f)プライマー結合部位の配列及び長さ;
(g)編集鋳型の配列及び長さ;
(h)相同アームの配列及び長さ;
(i)スペーサ配列及び長さ;並びに
(j)コア配列、
を包含するが、これらに限定されない。
本開示は上のこれらの側面を論ずる。
1つの態様では、好適なpegRNAと任意選択で第2部位ニッキングのためのニッキングsgRNAデザインガイドとをデザインするアプローチが、ここに提供される。この態様は、プライム編集のためのpegRNA及びニッキングsgRNAをデザインするための説明書の工程バイ工程のセットを提供し、これは上の考慮事項の1以上を考慮に入れる。工程は図70A-図70Iに示されている例を参照する。
1.標的配列及び編集を定める。所望の編集(点変異、挿入、欠失、又はそれらの組み合わせ)の位置付けを中心とした標的DNA領域(~200bp)の配列をリトリーブする。図70Aを見よ。
2.標的PAMを位置付ける。所望の編集位置付けに対して近位のPAMを同定する。PAMは所望の編集位置付けに対して近位のDNAのどちらかの鎖上に同定され得る。編集位置に近接するPAMが好ましいが(すなわち、ニック部位は編集位置から30nt未満である。又は編集位置からニック部位まで29nt、28nt、27nt、26nt、25nt、24nt、23nt、22nt、21nt、20nt、19nt、18nt、17nt、16nt、15nt、14nt、13nt、12nt、11nt、10nt、9nt、8nt、7nt、6nt、5nt、4nt、3nt、若しくは2nt未満)、編集位置から≧30ntのニックを置くプロトスペーサ及びPAMを用いて編集を組み入れることが可能である。図70Bを見よ。
3.ニック部位を位置付ける。考慮されている各PAMについて、対応するニック部位とどの鎖上かとを同定する。Sp Cas9 H840Aニッカーゼでは、切断はNGG PAMの5'の第3及び第4の塩基の間においてPAM含有鎖に生起する。全ての編集されるヌクレオチドはニック部位の3'に存在しなければならない。そのため、適当なPAMはPAM含有鎖上の標的の編集の5'にニックを置かなければならない。下で示されている例では、2つの可能なPAMがある。単純のために、残りの工程はPAM1のみを用いるpegRNAのデザインを実証する。図70Cを見よ。
4.スペーサ配列をデザインする。Sp Cas9のプロトスペーサは、PAM含有鎖上のNGG PAMの5'の20ヌクレオチドに対応する。効率的なPol III転写開始はGが第1の転写されるヌクレオチドであることを要求する。プロトスペーサの第1のヌクレオチドがGである場合には、pegRNAのスペーサ配列は単純にプロトスペーサ配列である。プロトスペーサの第1のヌクレオチドがGではない場合には、pegRNAのスペーサ配列は、G、次にプロトスペーサ配列である。図70Dを見よ。
5.プライマー結合部位(PBS)をデザインする。出発アレル配列を用いて、PAM含有鎖上のDNAプライマーを同定する。DNAプライマーの3'端はニック部位のちょうど上流のヌクレオチドである(すなわち、Sp Cas9ではNGG PAMの5'の第4の塩基)。PE2及びPE3への使用のための一般的なデザイン原理としては、DNAプライマーに対する相補性の12~13ヌクレオチドを含有するpegRNAプライマー結合部位(PBS)が、~40-60%のGC含量を含有する配列に用いられ得る。低いGC含量を有する配列では、より長い(14~15nt)PBSが試験されるべきである。より高いGC含量を有する配列では、より短い(8~11nt)PBSが試験されるべきである。GC含量にかかわらず、最適なPBS配列は経験的に決定されるはずである。長さpのPBS配列をデザインするためには、出発アレル配列を用いて、PAM含有鎖上のニック部位の5'の最初のpヌクレオチドの逆相補体を取る。図70Eを見よ。
6.RT鋳型(又はDNA合成鋳型)をデザインする。RT鋳型(又は、ポリメラーゼが逆転写酵素ではないところでは、DNA合成鋳型)は、デザインされた編集と編集に隣接する配列に対する相同性とをコードする。1つの態様では、これらの領域は図3D及び図3EのDNA合成鋳型に対応し、DNA合成鋳型は「編集鋳型」及び「相同アーム」を含む。最適なRT鋳型長さは標的部位に基づいて変わる。ショートレンジ編集(位置+1~+6)では、短い(9~12nt)、中程度の(13~16nt)、及び長い(17~20nt)RT鋳型を試験することが推奨される。ロングレンジ編集(位置+7以降)では、十分な3'DNAフラップ相同性を許すように、編集の位置から少なくとも5nt(好ましくは10nt以上)伸長するRT鋳型を用いることが推奨される。ロングレンジ編集では、機能的なデザインを同定するためには、いくつかのRT鋳型がスクリーニングされるべきである。より大きい挿入及び欠失(≧5nt)では、RT鋳型上へのより多大な3'相同性(~20nt以上)の組み込みが推奨される。RT鋳型が、逆転写されるDNA産物上の最後のヌクレオチドとしてG(pegRNAのRT鋳型ではCに対応する)の合成をコードするときには、編集効率は典型的には損なわれる。多くのRT鋳型が効率的なプライム編集を支持するので、RT鋳型をデザインするときには、最後の合成されるヌクレオチドとしてのGの回避が推奨される。長さrのRT鋳型配列をデザインするためには、所望のアレル配列を用い、元々はPAMを含有した鎖上のニック部位の3'の最初のrヌクレオチドの逆相補体を取る。SNP編集と比較して、同じ長さのRT鋳型を用いる挿入又は欠失編集は同一の相同性を含有しないであろうということに注意。図70Fを見よ。
7.完全なpegRNA配列をアセンブリする。pegRNA構成要素を次の順序でコンカテネーションする(5'→3'):スペーサ、骨格、RT鋳型、及びPBS。図70Gを見よ。
8.PE3のためのニッキングsgRNAをデザインする。編集の上流及び下流の非編集鎖上のPAMを同定する。最適なニッキング位置は高度に遺伝子座依存性であり、経験的に決定されるはずである。一般的に、pegRNAによって誘導されるニックの向かいの位置の40~90ヌクレオチド5'に置かれたニックは、より高い編集収量及びより少数のインデルに至る。ニッキングsgRNAは、出発アレル上の20ntプロトスペーサにマッチするスペーサ配列を有する。プロトスペーサがGで始まらない場合には5'Gの追加を有する。図70Hを見よ。
9.PE3bニッキングsgRNAをデザインする。PAMが相補鎖上に存在し、かつその対応するプロトスペーサが編集のために標的化される配列とオーバーラップする場合には、この編集はPE3bシステムの候補であり得る。PE3bシステムでは、ニッキングsgRNAのスペーサ配列は出発アレルではなく所望の編集されるアレルの配列にマッチする。編集されるヌクレオチド(単数又は複数)がニッキングsgRNAプロトスペーサのシード領域(PAMに隣接する~10nt)内に収まるときには、PE3bシステムは効率的に作動する。これは、編集された鎖の組み入れ後まで相補鎖のニッキングを防止し、標的DNAに結合することについてのpegRNA及びsgRNAの間の競合を防止する。PE3bは、両鎖上の同時のニックの生成をもまた回避し、それゆえに、高い編集効率を維持しながらインデル形成を有意に縮減する。PE3b sgRNAは、所望のアレルの20ntプロトスペーサにマッチするスペーサ配列を有するはずである。必要とされる場合には5'Gの追加を有する。図70Iを見よ。
1.標的配列及び編集を定める。所望の編集(点変異、挿入、欠失、又はそれらの組み合わせ)の位置付けを中心とした標的DNA領域(~200bp)の配列をリトリーブする。図70Aを見よ。
2.標的PAMを位置付ける。所望の編集位置付けに対して近位のPAMを同定する。PAMは所望の編集位置付けに対して近位のDNAのどちらかの鎖上に同定され得る。編集位置に近接するPAMが好ましいが(すなわち、ニック部位は編集位置から30nt未満である。又は編集位置からニック部位まで29nt、28nt、27nt、26nt、25nt、24nt、23nt、22nt、21nt、20nt、19nt、18nt、17nt、16nt、15nt、14nt、13nt、12nt、11nt、10nt、9nt、8nt、7nt、6nt、5nt、4nt、3nt、若しくは2nt未満)、編集位置から≧30ntのニックを置くプロトスペーサ及びPAMを用いて編集を組み入れることが可能である。図70Bを見よ。
3.ニック部位を位置付ける。考慮されている各PAMについて、対応するニック部位とどの鎖上かとを同定する。Sp Cas9 H840Aニッカーゼでは、切断はNGG PAMの5'の第3及び第4の塩基の間においてPAM含有鎖に生起する。全ての編集されるヌクレオチドはニック部位の3'に存在しなければならない。そのため、適当なPAMはPAM含有鎖上の標的の編集の5'にニックを置かなければならない。下で示されている例では、2つの可能なPAMがある。単純のために、残りの工程はPAM1のみを用いるpegRNAのデザインを実証する。図70Cを見よ。
4.スペーサ配列をデザインする。Sp Cas9のプロトスペーサは、PAM含有鎖上のNGG PAMの5'の20ヌクレオチドに対応する。効率的なPol III転写開始はGが第1の転写されるヌクレオチドであることを要求する。プロトスペーサの第1のヌクレオチドがGである場合には、pegRNAのスペーサ配列は単純にプロトスペーサ配列である。プロトスペーサの第1のヌクレオチドがGではない場合には、pegRNAのスペーサ配列は、G、次にプロトスペーサ配列である。図70Dを見よ。
5.プライマー結合部位(PBS)をデザインする。出発アレル配列を用いて、PAM含有鎖上のDNAプライマーを同定する。DNAプライマーの3'端はニック部位のちょうど上流のヌクレオチドである(すなわち、Sp Cas9ではNGG PAMの5'の第4の塩基)。PE2及びPE3への使用のための一般的なデザイン原理としては、DNAプライマーに対する相補性の12~13ヌクレオチドを含有するpegRNAプライマー結合部位(PBS)が、~40-60%のGC含量を含有する配列に用いられ得る。低いGC含量を有する配列では、より長い(14~15nt)PBSが試験されるべきである。より高いGC含量を有する配列では、より短い(8~11nt)PBSが試験されるべきである。GC含量にかかわらず、最適なPBS配列は経験的に決定されるはずである。長さpのPBS配列をデザインするためには、出発アレル配列を用いて、PAM含有鎖上のニック部位の5'の最初のpヌクレオチドの逆相補体を取る。図70Eを見よ。
6.RT鋳型(又はDNA合成鋳型)をデザインする。RT鋳型(又は、ポリメラーゼが逆転写酵素ではないところでは、DNA合成鋳型)は、デザインされた編集と編集に隣接する配列に対する相同性とをコードする。1つの態様では、これらの領域は図3D及び図3EのDNA合成鋳型に対応し、DNA合成鋳型は「編集鋳型」及び「相同アーム」を含む。最適なRT鋳型長さは標的部位に基づいて変わる。ショートレンジ編集(位置+1~+6)では、短い(9~12nt)、中程度の(13~16nt)、及び長い(17~20nt)RT鋳型を試験することが推奨される。ロングレンジ編集(位置+7以降)では、十分な3'DNAフラップ相同性を許すように、編集の位置から少なくとも5nt(好ましくは10nt以上)伸長するRT鋳型を用いることが推奨される。ロングレンジ編集では、機能的なデザインを同定するためには、いくつかのRT鋳型がスクリーニングされるべきである。より大きい挿入及び欠失(≧5nt)では、RT鋳型上へのより多大な3'相同性(~20nt以上)の組み込みが推奨される。RT鋳型が、逆転写されるDNA産物上の最後のヌクレオチドとしてG(pegRNAのRT鋳型ではCに対応する)の合成をコードするときには、編集効率は典型的には損なわれる。多くのRT鋳型が効率的なプライム編集を支持するので、RT鋳型をデザインするときには、最後の合成されるヌクレオチドとしてのGの回避が推奨される。長さrのRT鋳型配列をデザインするためには、所望のアレル配列を用い、元々はPAMを含有した鎖上のニック部位の3'の最初のrヌクレオチドの逆相補体を取る。SNP編集と比較して、同じ長さのRT鋳型を用いる挿入又は欠失編集は同一の相同性を含有しないであろうということに注意。図70Fを見よ。
7.完全なpegRNA配列をアセンブリする。pegRNA構成要素を次の順序でコンカテネーションする(5'→3'):スペーサ、骨格、RT鋳型、及びPBS。図70Gを見よ。
8.PE3のためのニッキングsgRNAをデザインする。編集の上流及び下流の非編集鎖上のPAMを同定する。最適なニッキング位置は高度に遺伝子座依存性であり、経験的に決定されるはずである。一般的に、pegRNAによって誘導されるニックの向かいの位置の40~90ヌクレオチド5'に置かれたニックは、より高い編集収量及びより少数のインデルに至る。ニッキングsgRNAは、出発アレル上の20ntプロトスペーサにマッチするスペーサ配列を有する。プロトスペーサがGで始まらない場合には5'Gの追加を有する。図70Hを見よ。
9.PE3bニッキングsgRNAをデザインする。PAMが相補鎖上に存在し、かつその対応するプロトスペーサが編集のために標的化される配列とオーバーラップする場合には、この編集はPE3bシステムの候補であり得る。PE3bシステムでは、ニッキングsgRNAのスペーサ配列は出発アレルではなく所望の編集されるアレルの配列にマッチする。編集されるヌクレオチド(単数又は複数)がニッキングsgRNAプロトスペーサのシード領域(PAMに隣接する~10nt)内に収まるときには、PE3bシステムは効率的に作動する。これは、編集された鎖の組み入れ後まで相補鎖のニッキングを防止し、標的DNAに結合することについてのpegRNA及びsgRNAの間の競合を防止する。PE3bは、両鎖上の同時のニックの生成をもまた回避し、それゆえに、高い編集効率を維持しながらインデル形成を有意に縮減する。PE3b sgRNAは、所望のアレルの20ntプロトスペーサにマッチするスペーサ配列を有するはずである。必要とされる場合には5'Gの追加を有する。図70Iを見よ。
好適なpegRNA及び第2部位ニッキングsgRNAをデザインするための上の工程バイ工程のプロセスは、決して限定することを意味されない。本開示は上に記載されている工程バイ工程のプロセスのバリエーションを企図し、これらは当業者によってそこから導出可能であろう。pegRNA修飾
本開示は、限定するものではないが、安定性及び細胞寿命の増加、並びにnapDNAbpに対する結合親和性の改善を包含する、改善された特性を有する次世代修飾pegRNAを提供する。これらの修飾されたpegRNAは、多種多様なゲノム部位での編集効率の増加によって実証されるように、改良されたゲノム編集をもたらす。本発明者らは、驚くべきことに、限定されるものだが、プレキノシン1-1リボスイッチアプタマー(「evopreQ1-1」)又はそのバリアント、MMLVウイルスゲノム由来のシュードノット(「evopreQ1-1」)又はそのバリアント、逆転写のためのプライマーとしてMMLV RTによって使用される修飾tRNA又はそのバリアント、及びG四重鎖又はそのバリアントを包含する特定の核酸構造モチーフをpegRNAの伸長アームの末端に付加することによって、編集活性の一貫した増加が達成されることを見出した。
一態様では、修飾pegRNAは、図98に従って、pegRNAの3'末端に核酸部分を包含する。任意選択で、pegRNAの3'末端は、ヌクレオチドリンカーを介して核酸部分に融合される。様々な態様では、多種多様なヌクレオチド配列が各ゲノム標的部位に対して合理的に良好に機能することが了解されよう。リンカーの長さも可変であり得る。いくつかの場合、3~18ヌクレオチドの長さの範囲のリンカーが機能する。他のケースでは、リンカーは、少なくとも3ヌクレオチド、少なくとも4ヌクレオチド、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、少なくとも15ヌクレオチド、少なくとも16ヌクレオチド、少なくとも17ヌクレオチド、少なくとも18ヌクレオチド、少なくとも19ヌクレオチド、少なくとも20ヌクレオチド、少なくとも21ヌクレオチド、少なくとも22ヌクレオチド、少なくとも23ヌクレオチド、少なくとも24ヌクレオチド、少なくとも25ヌクレオチド、少なくとも26ヌクレオチド、少なくとも27ヌクレオチド、少なくとも28ヌクレオチド、少なくとも29ヌクレオチド、又は少なくとも30ヌクレオチドであってもよい。
一般に、例えば、それをpegRNAの3'末端に取り付けることによって、pegRNAを修飾するために使用されてもよい核酸部分は、例えば、二重らせん部分、トゥーループ部分、ヘアピン部分、ステムループ部分、シュードノット部分、アプタマー部分、G四重鎖部分、tRNA部分又はリボザイム部分を含むか又はそれらを形成する核酸分子を包含する任意の核酸部分を包含してもよい。核酸部分は、二次核酸構造、三次核酸構造、又は四倍核酸構造を形成することを特徴としてもよい。換言すれば、核酸部分は、そのような構造によって形成されることが知られている任意の二次元又は三次元構造を形成してもよい。核酸部分は、DNA又はRNAであってもよい。
限定されないが、以下は、pegRNAの伸長アームの末端に付加されてもよいヌクレオチドモチーフの具体例である。したがって、3'伸長アームの場合、ヌクレオチドモチーフは、任意選択的にリンカーを介して、pegRNAの3'にカップリング、取り付け、又は他の方法で連結されるであろう。5'伸長アームの場合、ヌクレオチドモチーフは、任意選択的にリンカーを介して、pegRNAの5'末端にカップリング、取り付け、又は他の方法で連結されるであろう。





上記のように、これらのモチーフは、標準的なpegRNAにリンカーを介してカップリング、取り付け、又は他の方法で連結されてもよい。例示的なリンカーは、それだけに限らないが、以下を包含する。

いくつかの態様において、リンカーは、プライム編集によって標的とされるゲノム部位及び修飾されたpegRNAに基づいてデザイン及び/又は選択されるであろう。
様々な態様では、多種多様なヌクレオチド配列が各ゲノム標的部位に対して合理的に良好に機能することが了解されよう。リンカーの長さも可変である可能性が高い。いくつかの場合、3~18ヌクレオチドの長さの範囲のリンカーが機能する。他のケースでは、リンカーは、少なくとも3ヌクレオチド、少なくとも4ヌクレオチド、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、少なくとも15ヌクレオチド、少なくとも16ヌクレオチド、少なくとも17ヌクレオチド、少なくとも18ヌクレオチド、少なくとも19ヌクレオチド、少なくとも20ヌクレオチド、少なくとも21ヌクレオチド、少なくとも22ヌクレオチド、少なくとも23ヌクレオチド、少なくとも24ヌクレオチド、少なくとも25ヌクレオチド、少なくとも26ヌクレオチド、少なくとも27ヌクレオチド、少なくとも28ヌクレオチド、少なくとも29ヌクレオチド、又は少なくとも30ヌクレオチドであってもよい。
一態様では、リンカーは8ヌクレオチド長である。
本開示はまた、上記のモチーフ及びリンカー配列のいずれかと少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも91%、少なくとも92%、少なくとも93%、少なくとも94%、少なくとも95%、少なくとも96%、少なくとも97%、少なくとも98%、少なくとも99%、又は少なくとも99.9%の配列同一性を有する上記のヌクレオチドモチーフ及びリンカーのバリアントを企図する。
pegRNAはまた、pegRNAの特性及び/又は特徴を修飾してもよく、それによってプライム編集の有効性を改善する追加のデザイン改善を包含してもよい。様々な態様において、これらの改善は、(1)面倒な配列要件なしにより長いpegRNAの発現を可能にする、非ポリメラーゼIII(pol III)プロモータからの機能性pegRNAの効率的な発現を可能にするデザイン;(2)有効性を改善し得るコア、Cas9結合pegRNA骨格に対する改善;(3)RTプロセス性を改善し、標的ゲノム遺伝子座でのより長い配列の挿入を可能にするためのpegRNAへの修飾;(4)pegRNAの安定性を改善し、RTプロセス性を増強し、pegRNAのミスフォールディングを防止し、又はゲノム編集に重要な更なる因子を動員するRNAモチーフの、pegRNAの5'又は3'末端への付加を包含するがこれらに限定されない、多数の異なるカテゴリーの1つ又は複数に属してもよい。
一態様では、より大きな伸長アームを有するより長い長さのpegRNAの発現を改善するために、polIIIプロモータを用いてpegRNAがデザインされ得た。sgRNAは、典型的には、U6 snRNAプロモータから発現される。このプロモータは、pol IIIを動員して関連するRNAを発現させ、核内に保持される短いRNAの発現に有用である。しかし、pol IIIは高度にプロセッシブではなく、効率的なゲノム編集に必要なレベルで数百ヌクレオチド長よりも長いRNAを発現することができない。更に、pol IIIは、Uのストレッチで失速又は終結し得、pegRNAを使用して挿入され得た配列多様性を潜在的に制限する。ポリメラーゼII(pCMV等)又はポリメラーゼI(U1 snRNAプロモータ等)を動員する他のプロモータは、より長いsgRNAを発現する能力について調べられている。しかしながら、これらのプロモータは、典型的には部分的に転写され、発現されたpegRNA中のスペーサの余分な配列5'をもたらし、これにより、部位依存的にCas9:sgRNA活性が著しく低下することが示されている。更に、pol III転写されたpegRNAは、6~7 Uの連続で単純に終結し得るが、pol II又はpol Iから転写されたpegRNAは、異なる終結シグナルを必要とするであろう。多くの場合、そのようなシグナルはまた、核からのpegRNAの望ましくない輸送をもたらすポリアデニル化をもたらす。同様に、pCMV等のpol IIプロモータから発現されるRNAは、典型的には5'キャップされ、その核外輸送ももたらす。
例示的なU6プロモータは、それだけに限らないが、以下を包含する。
U6プロモータ:
GAGGGCCTATTTCCCATGATTCCTTCATATTTGCATATACGATACAAGGCTGTTAGAGAGATAATTAGAATTAATTTGACTGTAAACACAAAGATATTAGTACAAAATACGTGACGTAGAAAGTAATAATTTCTTGGGTAGTTTGCAGTTTTAAAATTATGTTTTAAAATGGACTATCATATGCTTACCGTAACTTGAAAGTATTTCGATTTCTTGGCTTTATATATCTTGTGGAAAGGACGAAACACCG(配列番号237)
GAGGGCCTATTTCCCATGATTCCTTCATATTTGCATATACGATACAAGGCTGTTAGAGAGATAATTAGAATTAATTTGACTGTAAACACAAAGATATTAGTACAAAATACGTGACGTAGAAAGTAATAATTTCTTGGGTAGTTTGCAGTTTTAAAATTATGTTTTAAAATGGACTATCATATGCTTACCGTAACTTGAAAGTATTTCGATTTCTTGGCTTTATATATCTTGTGGAAAGGACGAAACACCG(配列番号237)
U6v9プロモータ:
GCCTGAGGCGTGGGGCCGCCTCCCAAAGACTTCTGGGAGGGCGGTGCGGCTCAGGCTCTGCCCCGCCTCCGGGGCTATTTGCATACGACCATTTCCAGTAATTCCCAGCAGCCACCGTAGCTATATTTGGTAGAACAACGAGCACTTTCTCAACTCCAGTCAATAACTACGTTAGTTGCATTACACATTGGGCTAATATAAATAGAGGTTAAATCTCTAGGTCATTTAAGAGAAGTCGGCCTATGTGTACAGACATTTGTTCCAGGGGCTTTAAATAGCTGGTGGTGGAACTCAATATTCG(配列番号238)
GCCTGAGGCGTGGGGCCGCCTCCCAAAGACTTCTGGGAGGGCGGTGCGGCTCAGGCTCTGCCCCGCCTCCGGGGCTATTTGCATACGACCATTTCCAGTAATTCCCAGCAGCCACCGTAGCTATATTTGGTAGAACAACGAGCACTTTCTCAACTCCAGTCAATAACTACGTTAGTTGCATTACACATTGGGCTAATATAAATAGAGGTTAAATCTCTAGGTCATTTAAGAGAAGTCGGCCTATGTGTACAGACATTTGTTCCAGGGGCTTTAAATAGCTGGTGGTGGAACTCAATATTCG(配列番号238)
U6v7プロモータ:
AAGTCCGCGGCACGAGAAATCAAAGCCCCGGGGCCTGGGTCCCACGCGGGGTCCCTTACCCAGGGTGCCCCGGGCGCTCATTTGCATGTCCCACCCAACAGGTAAACCTGACAGATCGGTCGCGGCCAGGTACGGCCTGGCGGTCAGAGCACCAAACTTACGAGCCTTGTGATGAGTTCCGTTACATGAAATTCTCCTAAAGGCTCCAAGATGGACAGGAAAGCGCTCGATTAGGTTACCGTAAGGAAAACAAATGAGAAACTCCCGTGCCTTATAAGACCTGGGGACGGACTTATTTGCG(配列番号239)
AAGTCCGCGGCACGAGAAATCAAAGCCCCGGGGCCTGGGTCCCACGCGGGGTCCCTTACCCAGGGTGCCCCGGGCGCTCATTTGCATGTCCCACCCAACAGGTAAACCTGACAGATCGGTCGCGGCCAGGTACGGCCTGGCGGTCAGAGCACCAAACTTACGAGCCTTGTGATGAGTTCCGTTACATGAAATTCTCCTAAAGGCTCCAAGATGGACAGGAAAGCGCTCGATTAGGTTACCGTAAGGAAAACAAATGAGAAACTCCCGTGCCTTATAAGACCTGGGGACGGACTTATTTGCG(配列番号239)
U6v4プロモータ:
AAATTGAGTCATCTGACAGAAATTATCTTTGGCAAGGTTTTAGTCCTAGGGTTACCAGATGGAATACAGGACATCCATTTAAATTTGAATTTCAGATAAACAGTTAACACTTCTCAAGGATAAATATGCCTCAAATATTGCACGGGACATATTTATACTAAAAAAAAAGTGTTTTTTTTTTTCCTGCGATTCAAACTTAACTGGTGTCCTGCATTTGTATTTGTTAAATCTGTCAATCCTATCTCAGTTTCCTTTGATGGAATGTACCTCTGTGCTAATATTTAAAAATAGGTTACATTTG(配列番号240)
AAATTGAGTCATCTGACAGAAATTATCTTTGGCAAGGTTTTAGTCCTAGGGTTACCAGATGGAATACAGGACATCCATTTAAATTTGAATTTCAGATAAACAGTTAACACTTCTCAAGGATAAATATGCCTCAAATATTGCACGGGACATATTTATACTAAAAAAAAAGTGTTTTTTTTTTTCCTGCGATTCAAACTTAACTGGTGTCCTGCATTTGTATTTGTTAAATCTGTCAATCCTATCTCAGTTTCCTTTGATGGAATGTACCTCTGTGCTAATATTTAAAAATAGGTTACATTTG(配列番号240)
当業者は、これらのプロモータ配列が5'でトリミングされ、依然として同じ又はほぼ同じレベルで機能し得ることを理解するであろう。例えば、U6プロモータのいずれかは、5'末端から最大1、10、20、30、40、50、60、70、80、90又は100ヌクレオチド、すなわちプロモータ長の約30%を除去することによって5'末端でトリミングされ得た。他の態様では、5'末端からプロモータの長さの最大1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、11%、12%、13%、14%、15%、16%、17%、18%、19%、19%、20%、21%、22%、23%、24%、25%、26%、27%、28%、29%又は最大30%である。
当業者はまた、より大きな伸長アームを有するより長い長さのpegRNAの発現を改善するために他のプロモータが使用され得たことを理解するであろう。例えば、異なる細胞型では、他のプロモータが好ましくてもよく、より長いpegRNAのより大きな発現をもたらす。
以前に、Rinn及び共同研究者らは、長鎖非コードRNA-(lncRNA)タグ付きsgRNAの産生のために様々な発現プラットフォームをスクリーニングした183。これらのプラットフォームは、pCMVから発現され、ヒト由来のMALAT1 ncRNA由来のENEエレメント184、KSHV由来のPAN ENEエレメント185、又はU1 snRNA由来の3'ボックス186で終結するRNAを包含する。注意すべきことに、MALAT1 ncRNA及びPAN ENEは、ポリAテールを保護する三重らせんを形成する184,187。これらの構築物はまた、RNA安定性を高め得た。これらの発現系は、より長いpegRNAの発現も可能にすることが企図される。
更に、pegRNAの一部として転写されることになるpol IIプロモータの部分を切断するための一連の方法がデザインされており、ハンマーヘッド188、ピストル189、ハチェット189、ヘアピン190、VS191、ツイスター192、若しくはツイスターシスター192リボザイム等の自己切断リボザイム、又は転写されたガイドを処理するための他の自己切断要素、又はCsy4193によって認識され、ガイドの処理にもつながるヘアピンのいずれかを付加する。また、複数のENEモチーフの組み込みは、改善されたpegRNA発現及び安定性をもたらし得る。KSHV PAN RNA及び要素について以前に実証されたように、循環させる185。環状イントロンRNA(ciRNA)の形態のpegRNAを環状化すると、RNA発現及び安定性の増強、並びに核局在化をもたらし得ることも予想される。
様々な態様では、pegRNAは、以下の配列によって例示されるように、様々な上記要素を包含してもよい。
非限定的実施例1-pCMV、Csy4ヘアピン、pegRNA及びMALAT1 ENEからなるpegRNA発現プラットフォーム
TAGTTATTAATAGTAATCAATTACGGGGTCATTAGTTCATAGCCCATATATGGAGTTCCGCGTTACATAACTTACGGTAAATGGCCCGCCTGGCTGACCGCCCAACGACCCCCGCCCATTGACGTCAATAATGACGTATGTTCCCATAGTAACGCCAATAGGGACTTTCCATTGACGTCAATGGGTGGAGTATTTACGGTAAACTGCCCACTTGGCAGTACATCAAGTGTATCATATGCCAAGTACGCCCCCTATTGACGTCAATGACGGTAAATGGCCCGCCTGGCATTATGCCCAGTACATGACCTTATGGGACTTTCCTACTTGGCAGTACATCTACGTATTAGTCATCGCTATTACCATGGTGATGCGGTTTTGGCAGTACATCAATGGGCGTGGATAGCGGTTTGACTCACGGGGATTTCCAAGTCTCCACCCCATTGACGTCAATGGGAGTTTGTTTTGGCACCAAAATCAACGGGACTTTCCAAAATGTCGTAACAACTCCGCCCCATTGACGCAAATGGGCGGTAGGCGTGTACGGTGGGAGGTCTATATAAGCAGAGCTGGTTTAGTGAACCGTCAGATCGTTCACTGCCGTATAGGCAGGGCCCAGACTGAGCACGTGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCGTGCTCAGTCTGTTTTAGGGTCATGAAGGTTTTTCTTTTCCTGAGAAAACAACACGTATTGTTTTCTCAGGTTTTGCTTTTTGGCCTTTTTCTAGCTTAAAAAAAAAAAAAGCAAAAGATGCTGGTGGTTGGCACTCCTGGTTTCCAGGACGGGGTTCAAATCCCTGCGGCGTCTTTGCTTTGACT(配列番号241)
TAGTTATTAATAGTAATCAATTACGGGGTCATTAGTTCATAGCCCATATATGGAGTTCCGCGTTACATAACTTACGGTAAATGGCCCGCCTGGCTGACCGCCCAACGACCCCCGCCCATTGACGTCAATAATGACGTATGTTCCCATAGTAACGCCAATAGGGACTTTCCATTGACGTCAATGGGTGGAGTATTTACGGTAAACTGCCCACTTGGCAGTACATCAAGTGTATCATATGCCAAGTACGCCCCCTATTGACGTCAATGACGGTAAATGGCCCGCCTGGCATTATGCCCAGTACATGACCTTATGGGACTTTCCTACTTGGCAGTACATCTACGTATTAGTCATCGCTATTACCATGGTGATGCGGTTTTGGCAGTACATCAATGGGCGTGGATAGCGGTTTGACTCACGGGGATTTCCAAGTCTCCACCCCATTGACGTCAATGGGAGTTTGTTTTGGCACCAAAATCAACGGGACTTTCCAAAATGTCGTAACAACTCCGCCCCATTGACGCAAATGGGCGGTAGGCGTGTACGGTGGGAGGTCTATATAAGCAGAGCTGGTTTAGTGAACCGTCAGATCGTTCACTGCCGTATAGGCAGGGCCCAGACTGAGCACGTGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCGTGCTCAGTCTGTTTTAGGGTCATGAAGGTTTTTCTTTTCCTGAGAAAACAACACGTATTGTTTTCTCAGGTTTTGCTTTTTGGCCTTTTTCTAGCTTAAAAAAAAAAAAAGCAAAAGATGCTGGTGGTTGGCACTCCTGGTTTCCAGGACGGGGTTCAAATCCCTGCGGCGTCTTTGCTTTGACT(配列番号241)
非限定的実施例2-pCMV、Csy4ヘアピン、pegRNA及びPAN ENEからなるpegRNA発現プラットフォーム
TAGTTATTAATAGTAATCAATTACGGGGTCATTAGTTCATAGCCCATATATGGAGTTCCGCGTTACATAACTTACGGTAAATGGCCCGCCTGGCTGACCGCCCAACGACCCCCGCCCATTGACGTCAATAATGACGTATGTTCCCATAGTAACGCCAATAGGGACTTTCCATTGACGTCAATGGGTGGAGTATTTACGGTAAACTGCCCACTTGGCAGTACATCAAGTGTATCATATGCCAAGTACGCCCCCTATTGACGTCAATGACGGTAAATGGCCCGCCTGGCATTATGCCCAGTACATGACCTTATGGGACTTTCCTACTTGGCAGTACATCTACGTATTAGTCATCGCTATTACCATGGTGATGCGGTTTTGGCAGTACATCAATGGGCGTGGATAGCGGTTTGACTCACGGGGATTTCCAAGTCTCCACCCCATTGACGTCAATGGGAGTTTGTTTTGGCACCAAAATCAACGGGACTTTCCAAAATGTCGTAACAACTCCGCCCCATTGACGCAAATGGGCGGTAGGCGTGTACGGTGGGAGGTCTATATAAGCAGAGCTGGTTTAGTGAACCGTCAGATCGTTCACTGCCGTATAGGCAGGGCCCAGACTGAGCACGTGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCGTGCTCAGTCTGTTTTGTTTTGGCTGGGTTTTTCCTTGTTCGCACCGGACACCTCCAGTGACCAGACGGCAAGGTTTTTATCCCAGTGTATATTGGAAAAACATGTTATACTTTTGACAATTTAACGTGCCTAGAGCTCAAATTAAACTAATACCATAACGTAATGCAACTTACAACATAAATAAAGGTCAATGTTTAATCCATAAAAAAAAAAAAAAAAAAA(配列番号242)
TAGTTATTAATAGTAATCAATTACGGGGTCATTAGTTCATAGCCCATATATGGAGTTCCGCGTTACATAACTTACGGTAAATGGCCCGCCTGGCTGACCGCCCAACGACCCCCGCCCATTGACGTCAATAATGACGTATGTTCCCATAGTAACGCCAATAGGGACTTTCCATTGACGTCAATGGGTGGAGTATTTACGGTAAACTGCCCACTTGGCAGTACATCAAGTGTATCATATGCCAAGTACGCCCCCTATTGACGTCAATGACGGTAAATGGCCCGCCTGGCATTATGCCCAGTACATGACCTTATGGGACTTTCCTACTTGGCAGTACATCTACGTATTAGTCATCGCTATTACCATGGTGATGCGGTTTTGGCAGTACATCAATGGGCGTGGATAGCGGTTTGACTCACGGGGATTTCCAAGTCTCCACCCCATTGACGTCAATGGGAGTTTGTTTTGGCACCAAAATCAACGGGACTTTCCAAAATGTCGTAACAACTCCGCCCCATTGACGCAAATGGGCGGTAGGCGTGTACGGTGGGAGGTCTATATAAGCAGAGCTGGTTTAGTGAACCGTCAGATCGTTCACTGCCGTATAGGCAGGGCCCAGACTGAGCACGTGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCGTGCTCAGTCTGTTTTGTTTTGGCTGGGTTTTTCCTTGTTCGCACCGGACACCTCCAGTGACCAGACGGCAAGGTTTTTATCCCAGTGTATATTGGAAAAACATGTTATACTTTTGACAATTTAACGTGCCTAGAGCTCAAATTAAACTAATACCATAACGTAATGCAACTTACAACATAAATAAAGGTCAATGTTTAATCCATAAAAAAAAAAAAAAAAAAA(配列番号242)
非限定的実施例3-pCMV、Csy4ヘアピン、pegRNA及び3xPAN ENEからなるpegRNA発現プラットフォーム
TAGTTATTAATAGTAATCAATTACGGGGTCATTAGTTCATAGCCCATATATGGAGTTCCGCGTTACATAACTTACGGTAAATGGCCCGCCTGGCTGACCGCCCAACGACCCCCGCCCATTGACGTCAATAATGACGTATGTTCCCATAGTAACGCCAATAGGGACTTTCCATTGACGTCAATGGGTGGAGTATTTACGGTAAACTGCCCACTTGGCAGTACATCAAGTGTATCATATGCCAAGTACGCCCCCTATTGACGTCAATGACGGTAAATGGCCCGCCTGGCATTATGCCCAGTACATGACCTTATGGGACTTTCCTACTTGGCAGTACATCTACGTATTAGTCATCGCTATTACCATGGTGATGCGGTTTTGGCAGTACATCAATGGGCGTGGATAGCGGTTTGACTCACGGGGATTTCCAAGTCTCCACCCCATTGACGTCAATGGGAGTTTGTTTTGGCACCAAAATCAACGGGACTTTCCAAAATGTCGTAACAACTCCGCCCCATTGACGCAAATGGGCGGTAGGCGTGTACGGTGGGAGGTCTATATAAGCAGAGCTGGTTTAGTGAACCGTCAGATCGTTCACTGCCGTATAGGCAGGGCCCAGACTGAGCACGTGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCGTGCTCAGTCTGTTTTGTTTTGGCTGGGTTTTTCCTTGTTCGCACCGGACACCTCCAGTGACCAGACGGCAAGGTTTTTATCCCAGTGTATATTGGAAAAACATGTTATACTTTTGACAATTTAACGTGCCTAGAGCTCAAATTAAACTAATACCATAACGTAATGCAACTTACAACATAAATAAAGGTCAATGTTTAATCCATAAAAAAAAAAAAAAAAAAAACACACTGTTTTGGCTGGGTTTTTCCTTGTTCGCACCGGACACCTCCAGTGACCAGACGGCAAGGTTTTTATCCCAGTGTATATTGGAAAAACATGTTATACTTTTGACAATTTAACGTGCCTAGAGCTCAAATTAAACTAATACCATAACGTAATGCAACTTACAACATAAATAAAGGTCAATGTTTAATCCATAAAAAAAAAAAAAAAAAAATCTCTCTGTTTTGGCTGGGTTTTTCCTTGTTCGCACCGGACACCTCCAGTGACCAGACGGCAAGGTTTTTATCCCAGTGTATATTGGAAAAACATGTTATACTTTTGACAATTTAACGTGCCTAGAGCTCAAATTAAACTAATACCATAACGTAATGCAACTTACAACATAAATAAAGGTCAATGTTTAATCCATAAAAAAAAAAAAAAAAAAA(配列番号243)
TAGTTATTAATAGTAATCAATTACGGGGTCATTAGTTCATAGCCCATATATGGAGTTCCGCGTTACATAACTTACGGTAAATGGCCCGCCTGGCTGACCGCCCAACGACCCCCGCCCATTGACGTCAATAATGACGTATGTTCCCATAGTAACGCCAATAGGGACTTTCCATTGACGTCAATGGGTGGAGTATTTACGGTAAACTGCCCACTTGGCAGTACATCAAGTGTATCATATGCCAAGTACGCCCCCTATTGACGTCAATGACGGTAAATGGCCCGCCTGGCATTATGCCCAGTACATGACCTTATGGGACTTTCCTACTTGGCAGTACATCTACGTATTAGTCATCGCTATTACCATGGTGATGCGGTTTTGGCAGTACATCAATGGGCGTGGATAGCGGTTTGACTCACGGGGATTTCCAAGTCTCCACCCCATTGACGTCAATGGGAGTTTGTTTTGGCACCAAAATCAACGGGACTTTCCAAAATGTCGTAACAACTCCGCCCCATTGACGCAAATGGGCGGTAGGCGTGTACGGTGGGAGGTCTATATAAGCAGAGCTGGTTTAGTGAACCGTCAGATCGTTCACTGCCGTATAGGCAGGGCCCAGACTGAGCACGTGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCGTGCTCAGTCTGTTTTGTTTTGGCTGGGTTTTTCCTTGTTCGCACCGGACACCTCCAGTGACCAGACGGCAAGGTTTTTATCCCAGTGTATATTGGAAAAACATGTTATACTTTTGACAATTTAACGTGCCTAGAGCTCAAATTAAACTAATACCATAACGTAATGCAACTTACAACATAAATAAAGGTCAATGTTTAATCCATAAAAAAAAAAAAAAAAAAAACACACTGTTTTGGCTGGGTTTTTCCTTGTTCGCACCGGACACCTCCAGTGACCAGACGGCAAGGTTTTTATCCCAGTGTATATTGGAAAAACATGTTATACTTTTGACAATTTAACGTGCCTAGAGCTCAAATTAAACTAATACCATAACGTAATGCAACTTACAACATAAATAAAGGTCAATGTTTAATCCATAAAAAAAAAAAAAAAAAAATCTCTCTGTTTTGGCTGGGTTTTTCCTTGTTCGCACCGGACACCTCCAGTGACCAGACGGCAAGGTTTTTATCCCAGTGTATATTGGAAAAACATGTTATACTTTTGACAATTTAACGTGCCTAGAGCTCAAATTAAACTAATACCATAACGTAATGCAACTTACAACATAAATAAAGGTCAATGTTTAATCCATAAAAAAAAAAAAAAAAAAA(配列番号243)
非限定的実施例4-pCMV、Csy4ヘアピン、pegRNA及び3'boxからなるpegRNA発現プラットフォーム
TAGTTATTAATAGTAATCAATTACGGGGTCATTAGTTCATAGCCCATATATGGAGTTCCGCGTTACATAACTTACGGTAAATGGCCCGCCTGGCTGACCGCCCAACGACCCCCGCCCATTGACGTCAATAATGACGTATGTTCCCATAGTAACGCCAATAGGGACTTTCCATTGACGTCAATGGGTGGAGTATTTACGGTAAACTGCCCACTTGGCAGTACATCAAGTGTATCATATGCCAAGTACGCCCCCTATTGACGTCAATGACGGTAAATGGCCCGCCTGGCATTATGCCCAGTACATGACCTTATGGGACTTTCCTACTTGGCAGTACATCTACGTATTAGTCATCGCTATTACCATGGTGATGCGGTTTTGGCAGTACATCAATGGGCGTGGATAGCGGTTTGACTCACGGGGATTTCCAAGTCTCCACCCCATTGACGTCAATGGGAGTTTGTTTTGGCACCAAAATCAACGGGACTTTCCAAAATGTCGTAACAACTCCGCCCCATTGACGCAAATGGGCGGTAGGCGTGTACGGTGGGAGGTCTATATAAGCAGAGCTGGTTTAGTGAACCGTCAGATCGTTCACTGCCGTATAGGCAGGGCCCAGACTGAGCACGTGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCGTGCTCAGTCTGTTTGTTTCAAAAGTAGACTGTACGCTAAGGGTCATATCTTTTTTTGTTTGGTTTGTGTCTTGGTTGGCGTCTTAAA(配列番号244)
TAGTTATTAATAGTAATCAATTACGGGGTCATTAGTTCATAGCCCATATATGGAGTTCCGCGTTACATAACTTACGGTAAATGGCCCGCCTGGCTGACCGCCCAACGACCCCCGCCCATTGACGTCAATAATGACGTATGTTCCCATAGTAACGCCAATAGGGACTTTCCATTGACGTCAATGGGTGGAGTATTTACGGTAAACTGCCCACTTGGCAGTACATCAAGTGTATCATATGCCAAGTACGCCCCCTATTGACGTCAATGACGGTAAATGGCCCGCCTGGCATTATGCCCAGTACATGACCTTATGGGACTTTCCTACTTGGCAGTACATCTACGTATTAGTCATCGCTATTACCATGGTGATGCGGTTTTGGCAGTACATCAATGGGCGTGGATAGCGGTTTGACTCACGGGGATTTCCAAGTCTCCACCCCATTGACGTCAATGGGAGTTTGTTTTGGCACCAAAATCAACGGGACTTTCCAAAATGTCGTAACAACTCCGCCCCATTGACGCAAATGGGCGGTAGGCGTGTACGGTGGGAGGTCTATATAAGCAGAGCTGGTTTAGTGAACCGTCAGATCGTTCACTGCCGTATAGGCAGGGCCCAGACTGAGCACGTGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCGTGCTCAGTCTGTTTGTTTCAAAAGTAGACTGTACGCTAAGGGTCATATCTTTTTTTGTTTGGTTTGTGTCTTGGTTGGCGTCTTAAA(配列番号244)
非限定的実施例5-pU1、Csy4ヘアピン、pegRNA及び3'boxからなるpegRNA発現プラットフォーム
CTAAGGACCAGCTTCTTTGGGAGAGAACAGACGCAGGGGCGGGAGGGAAAAAGGGAGAGGCAGACGTCACTTCCCCTTGGCGGCTCTGGCAGCAGATTGGTCGGTTGAGTGGCAGAAAGGCAGACGGGGACTGGGCAAGGCACTGTCGGTGACATCACGGACAGGGCGACTTCTATGTAGATGAGGCAGCGCAGAGGCTGCTGCTTCGCCACTTGCTGCTTCACCACGAAGGAGTTCCCGTGCCCTGGGAGCGGGTTCAGGACCGCTGATCGGAAGTGAGAATCCCAGCTGTGTGTCAGGGCTGGAAAGGGCTCGGGAGTGCGCGGGGCAAGTGACCGTGTGTGTAAAGAGTGAGGCGTATGAGGCTGTGTCGGGGCAGAGGCCCAAGATCTCAGTTCACTGCCGTATAGGCAGGGCCCAGACTGAGCACGTGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCGTGCTCAGTCTGTTTCAGCAAGTTCAGAGAAATCTGAACTTGCTGGATTTTTGGAGCAGGGAGATGGAATAGGAGCTTGCTCCGTCCACTCCACGCATCGACCTGGTATTGCAGTACCTCCAGGAACGGTGCACCCACTTTCTGGAGTTTCAAAAGTAGACTGTACGCTAAGGGTCATATCTTTTTTTGTTTGGTTTGTGTCTTGGTTGGCGTCTTAAA(配列番号245).
CTAAGGACCAGCTTCTTTGGGAGAGAACAGACGCAGGGGCGGGAGGGAAAAAGGGAGAGGCAGACGTCACTTCCCCTTGGCGGCTCTGGCAGCAGATTGGTCGGTTGAGTGGCAGAAAGGCAGACGGGGACTGGGCAAGGCACTGTCGGTGACATCACGGACAGGGCGACTTCTATGTAGATGAGGCAGCGCAGAGGCTGCTGCTTCGCCACTTGCTGCTTCACCACGAAGGAGTTCCCGTGCCCTGGGAGCGGGTTCAGGACCGCTGATCGGAAGTGAGAATCCCAGCTGTGTGTCAGGGCTGGAAAGGGCTCGGGAGTGCGCGGGGCAAGTGACCGTGTGTGTAAAGAGTGAGGCGTATGAGGCTGTGTCGGGGCAGAGGCCCAAGATCTCAGTTCACTGCCGTATAGGCAGGGCCCAGACTGAGCACGTGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCGTGCTCAGTCTGTTTCAGCAAGTTCAGAGAAATCTGAACTTGCTGGATTTTTGGAGCAGGGAGATGGAATAGGAGCTTGCTCCGTCCACTCCACGCATCGACCTGGTATTGCAGTACCTCCAGGAACGGTGCACCCACTTTCTGGAGTTTCAAAAGTAGACTGTACGCTAAGGGTCATATCTTTTTTTGTTTGGTTTGTGTCTTGGTTGGCGTCTTAAA(配列番号245).
様々な他の態様では、骨格又はコア配列に改善を導入することによって、pegRNAが改善されてもよい。
コアであるCas9結合pegRNA骨格は、PE活性を増強するために改善され得る。例示的なアプローチでは、骨格(P1)の第1の対合要素は、GTTTT-AAAAC(配列番号246)対合要素を含有する。そのようなTsの実行は、pol IIIの休止及びRNA転写物の早期終結をもたらし得る。P1のこの部分におけるT-A対の1つのG-C対への合理的突然変異は、sgRNA活性を増強し得る。このアプローチは、pegRNAを改善するために使用され得る。更に、P1の長さを増加させることにより、sgRNAフォールディングを増強し、改善されたpegRNA活性をもたらし得る。コアの改良例としては、以下を包含し得る。
P1に対する6nt伸長を含有するpegRNA
GGCCCAGACTGAGCACGTGAGTTTTAGAGCTAGCTCATGAAAATGAGCTAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCGTGCTCAGTCTGTTTTTTT(配列番号247)
GGCCCAGACTGAGCACGTGAGTTTTAGAGCTAGCTCATGAAAATGAGCTAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCGTGCTCAGTCTGTTTTTTT(配列番号247)
P1内にT-AからG-Cへの変異を含有するpegRNA
GGCCCAGACTGAGCACGTGAGTTTGAGAGCTAGAAATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCGTGCTCAGTCTGTTTTTTT(配列番号248)
GGCCCAGACTGAGCACGTGAGTTTGAGAGCTAGAAATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCGTGCTCAGTCTGTTTTTTT(配列番号248)
pegRNAはCRISPR構成要素及びtracrRNA構成要素を分裂する。
GGCCCAGACTGAGCACGTGAGTTTTAGAGCTAGA(配列番号249)
AATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCGTGCTCAGTCTG(配列番号250)
GGCCCAGACTGAGCACGTGAGTTTTAGAGCTAGA(配列番号249)
AATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCGTGCTCAGTCTG(配列番号250)
様々な他の態様において、pegRNAは、編集鋳型領域に修飾を導入することによって改善されてもよい。pegRNAによって鋳型される挿入のサイズが増加するにつれて、それはエンドヌクレアーゼによって分解され、自発的加水分解を受け、又はRTによって逆転写され得ないか若しくはpegRNA骨格の折り畳み及びその後のCas9-RT結合を破壊する二次構造に折り畳まれる可能性がより高い。したがって、遺伝子全体の挿入等の大きな挿入に影響を及ぼすために、pegRNAの鋳型の修飾が必要である可能性がある。そうするためのいくつかの戦略は、RNAを分解又は加水分解に対してより耐性にするか、又は阻害性二次構造を採用する可能性を低くする、合成又は半合成pegRNA内への修飾ヌクレオチドの組み込みを包含する196。そのような修飾は、Gリッチ配列中のRNA二次構造を減少させる8-アザ-7-デアザグアノシン;分解を低減し、ある種のRNA二次構造を増強するロックド核酸(LNA);RNA安定性を高める2'-O-メチル、2'-フルオロ又は2'-O-メトキシエトキシ修飾を包含し得た。このような修飾はまた、安定性及び活性を高めるために、pegRNAの他の場所に包含され得た。代替的又は追加的に、pegRNAの鋳型は、それが所望のタンパク質産物をコードし、RTによって展開されることができる単純な二次構造を採用する可能性も高くなるようにデザインされ得る。そのような単純な構造は熱力学的シンクとして作用し、逆転写を妨げるより複雑な構造が生じる可能性を低くする。最後に、鋳型を2つの別個のpegRNAに分裂し得た。そのようなデザインでは、PEが使用されて転写を開始し、Cas9に融合したRNA結合タンパク質又はMS2アプタマー等のpegRNA自体のRNA認識エレメントを介して別個の鋳型RNAを標的部位に動員する。RTは、この別個の鋳型RNAに直接結合するか、又は第2の鋳型に交換する前に元のpegRNA上で逆転写を開始し得た。そのようなアプローチは、長い鋳型の添加時のpegRNAのミスフォールディングを防止すること、及び、おそらくPEに基づく長い挿入を阻害し得る長い挿入が起こるためにゲノムからのCas9の解離を必要としないことの両方によって、長い挿入を可能にし得た。
更に他の態様では、pegRNAは、pegRNAの5'及び3'末端に、又は更にはその間の位置(例として、gRNAコア領域、又はスペーサ)に更なるRNAモチーフを導入することによって改善されてもよい。KSHV由来のPAN ENE及びMALAT1由来のENE等のいくつかのこのようなモチーフは、非pol IIIプロモータ由来のより長いpegRNAの発現を終結させる可能な手段として上に論じられた。これらの要素は、ポリAテールを包み込むRNA三重らせんを形成し、その結果、それらは核内に保持される184,187。しかしながら、末端ヌクレオチドを閉塞するpegRNAの3'末端に複雑な構造を形成することによって、これらの構造は、おそらく、pegRNAのエキソヌクレアーゼ媒介性分解の防止にも役立つだろう。
非pol IIIプロモータからの終結を可能にすることなく、3'末端に挿入された他の構造要素もRNA安定性を高め得た。そのようなモチーフは、3'末端を閉塞するヘアピン若しくはRNA四重鎖197、又は3'末端に2'-3'-環状リン酸の形成をもたらし、またpegRNAをエキソヌクレアーゼによって分解されにくくする可能性があるHDV等の自己切断リボザイムを包含し得た。198不完全なスプライシングを介して環状化するように(ciRNAを形成するように)pegRNAを誘導することはまた、pegRNAの安定性を高め、pegRNAが核内に保持されるようにすることもできる194。
更なるRNAモチーフはまた、RTプロセス性を改善するか、又はDNA-RNA二重鎖へのRT結合を増強することによってpegRNA活性を増強し得た。RTによって結合された天然配列をその同族レトロウイルスゲノムに追加すると、RT活性を増強し得た199。これには、天然プライマー結合部位(PBS)、ポリプリントラクト(PPT)、又はレトロウイルスゲノム二量体化及び転写の開始を伴うキスループを包含し得た199。
pegRNAの5'及び3'末端に、キスループ又はGNRAテトラループ/テトラループ受容体ペア200等の二量体化モチーフを付加することも、pegRNAの効果的な環状化をもたらし、安定性を改善し得た。更に、これらのモチーフの付加は、pegRNAスペーサ及びプライマーの物理的分離、PE活性を妨げる可能性があるスペーサの閉塞の防止を可能にし得ることが想定される。スペーサ領域又はプライマー結合部位に沿って小さな足がかりヘアピンを形成する、pegRNAに対する短い5'伸長又は3'伸長もまた、pegRNAの長さに沿った、例としてスペーサとプライマー結合部位との間の起こり得る相互作用に沿った、相補内領域のアニーリングに対して有利に競合し得た。最後に、また、キスループが使用されて、他の鋳型RNAをゲノム部位に動員し、一方のRNAから他方のRNAへのRT活性の交換を可能にし得た。様々な二次構造の例示的な態様として、図3D及び図3Eに示されるpegRNAは、示されるように、伸長アーム(すなわち、e1及びe2)の末端部分を包含する、pegRNAの任意の領域に操作されてもよいいくつかの二次RNA構造を列挙する。
例示的な改善は、それだけに限らないが、以下を包含する。
pegRNA-HDV融合
GGCCCAGACTGAGCACGTGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCGTGCTCAGTCTGGGCCGGCATGGTCCCAGCCTCCTCGCTGGCGCCGGCTGGGCAACATGCTTCGGCATGGCGAATGGGACTTTTTTT(配列番号251)
GGCCCAGACTGAGCACGTGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCGTGCTCAGTCTGGGCCGGCATGGTCCCAGCCTCCTCGCTGGCGCCGGCTGGGCAACATGCTTCGGCATGGCGAATGGGACTTTTTTT(配列番号251)
pegRNA-MMLVキスループ
GGTGGGAGACGTCCCACCGGCCCAGACTGAGCACGTGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCTTCGACCGTGCTCAGTCTGGTGGGAGACGTCCCACCTTTTTTT(配列番号252)
GGTGGGAGACGTCCCACCGGCCCAGACTGAGCACGTGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCTTCGACCGTGCTCAGTCTGGTGGGAGACGTCCCACCTTTTTTT(配列番号252)
pegRNA-VSリボザイムのキスループ
GAGCAGCATGGCGTCGCTGCTCACGGCCCAGACTGAGCACGTGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCTTCGACCGTGCTCAGTCTCCATCAGTTGACACCCTGAGGTTTTTTT(配列番号253)
GAGCAGCATGGCGTCGCTGCTCACGGCCCAGACTGAGCACGTGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCTTCGACCGTGCTCAGTCTCCATCAGTTGACACCCTGAGGTTTTTTT(配列番号253)
pegRNA-GNRAテトラループ/テトラループ受容体
GCAGACCTAAGTGGUGACATATGGTCTGGGCCCAGACTGAGCACGTGAGTTTTAGAGCTAUACGTAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTUACGAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCTTCGACCGTGCTCAGTCTGCATGCGATTAGAAATAATCGCATGTTTTTTT(配列番号254)
GCAGACCTAAGTGGUGACATATGGTCTGGGCCCAGACTGAGCACGTGAGTTTTAGAGCTAUACGTAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTUACGAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCTTCGACCGTGCTCAGTCTGCATGCGATTAGAAATAATCGCATGTTTTTTT(配列番号254)
二次RNA-HDV融合を切り替えるpegRNA鋳型
TCTGCCATCAAAGCTGCGACCGTGCTCAGTCTGGTGGGAGACGTCCCACCGGCCGGCATGGTCCCAGCCTCCTCGCTGGCGCCGGCTGGGCAACATGCTTCGGCATGGCGAATGGGACTTTTTTT(配列番号255)
TCTGCCATCAAAGCTGCGACCGTGCTCAGTCTGGTGGGAGACGTCCCACCGGCCGGCATGGTCCCAGCCTCCTCGCTGGCGCCGGCTGGGCAACATGCTTCGGCATGGCGAATGGGACTTTTTTT(配列番号255)
SpCas9及びプライム編集因子(PE)が改善された方法と類似の様式で、指向性進化によってpegRNA骨格が更に改善され得た。指向性進化は、Cas9又は進化したCas9バリアントによるpegRNA認識を増強し得た。更に、異なるpegRNA骨格配列が異なるゲノム遺伝子座で最適であり、問題の部位でPE活性を増強するか、オフターゲット活性を減少させるか、又はその両方である可能性が高い。最後に、他のRNAモチーフが付加されたpegRNA骨格の進化は、未進化の融合RNAと比較して、融合pegRNAの活性をほぼ確実に改善するであろう。例えば、c-ジ-GMP-Iアプタマー及びハンマーヘッド型リボザイムから構成されるアロステリックリボザイムの進化は、劇的に改善された活性をもたらし202、進化がハンマーヘッド-pegRNA融合の活性も改善することを示唆した。加えて、Cas9は現在、sgRNAの5'伸長を一般に許容しないが、指向性進化は、この不耐性を緩和し、更なるRNAモチーフが利用されることを可能にする可能性のある突然変異を生成する可能性が高い。
様々な態様では、標準的なsgRNA骨格と比較して活性を改善することが示されている他の骨格が、本明細書に記載のpegRNA及びepegRNAに使用されてもよい。そのような改善は、例えば、Chen,B.et al.Dynamic Imaging of Genomic Loci in Living Human Cells by an Optimized CRISPR/Cas System.Cell.2013,155(7),1479-1471 and Jost,M.et al.Titrating expression using libraries of systematically attenuated CRISPR guide RNAs.Nat.Biotechnol.2020,38,355-364に開示されているものを包含してもよく、それらの全体が参照により本明細書に組み込まれる。これらの改善は、プライム編集因子への結合の改善及び/又は発現の改善を通して、epegRNA活性を増強してもよい。sgRNA骨格の安定化はまた、pegRNA及びepegRNA活性を阻害するPBS/スペーサ相互作用を減少させ得た。
改善されたsgRNA骨格を組み込んだ例示的なepegRNAとしては、限定されないが、以下を包含する。
HEK3 1-15del standard scaffold evopreQ1
GGCCCAGACTGAGCACGTGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCGCCCTCTGGAGGAAGCAGGGCTTCCCGTGCTCAGTCTGTCTCTCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号256)
GGCCCAGACTGAGCACGTGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCGCCCTCTGGAGGAAGCAGGGCTTCCCGTGCTCAGTCTGTCTCTCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号256)
HEK3 1-15del cr748 evopreQ1
GGCCCAGACTGAGCACGTGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTGCCCTCTGGAGGAAGCAGGGCTTCCCGTGCTCAGTCTGTCTCTCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号257)
GGCCCAGACTGAGCACGTGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTGCCCTCTGGAGGAAGCAGGGCTTCCCGTGCTCAGTCTGTCTCTCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号257)
HEK3 1-15del cr289 evopreQ1
GGCCCAGACTGAGCACGTGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTGCCCTCTGGAGGAAGCAGGGCTTCCCGTGCTCAGTCTGTCTCTCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号258)
GGCCCAGACTGAGCACGTGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTGCCCTCTGGAGGAAGCAGGGCTTCCCGTGCTCAGTCTGTCTCTCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号258)
HEK3 1-15del cr622 evopreQ1
GGCCCAGACTGAGCACGTGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTGCCCTCTGGAGGAAGCAGGGCTTCCCGTGCTCAGTCTGTCTCTCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号259)
GGCCCAGACTGAGCACGTGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTGCCCTCTGGAGGAAGCAGGGCTTCCCGTGCTCAGTCTGTCTCTCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号259)
HEK3 1-15del cr772 evopreQ1
GGCCCAGACTGAGCACGTGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTGCCCTCTGGAGGAAGCAGGGCTTCCCGTGCTCAGTCTGTCTCTCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号260)
GGCCCAGACTGAGCACGTGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTGCCCTCTGGAGGAAGCAGGGCTTCCCGTGCTCAGTCTGTCTCTCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号260)
HEK3 1-15del cr532 evopreQ1
GGCCCAGACTGAGCACGTGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTGCCCTCTGGAGGAAGCAGGGCTTCCCGTGCTCAGTCTGTCTCTCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号261)
GGCCCAGACTGAGCACGTGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTGCCCTCTGGAGGAAGCAGGGCTTCCCGTGCTCAGTCTGTCTCTCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号261)
HEK3 1-15del cr961 evopreQ1
GGCCCAGACTGAGCACGTGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTGCCCTCTGGAGGAAGCAGGGCTTCCCGTGCTCAGTCTGTCTCTCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号262)
GGCCCAGACTGAGCACGTGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTGCCCTCTGGAGGAAGCAGGGCTTCCCGTGCTCAGTCTGTCTCTCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号262)
HEK3 1-15delフリップ及び骨格evopreQ1
GGCCCAGACTGAGCACGTGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTGCCCTCTGGAGGAAGCAGGGCTTCCCGTGCTCAGTCTGTCTCTCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号263)
GGCCCAGACTGAGCACGTGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTGCCCTCTGGAGGAAGCAGGGCTTCCCGTGCTCAGTCTGTCTCTCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号263)
RNF2 1-15del cr748 evopreQ1
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTATGGGAACTCAGTTTATATGAGTTAGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号264)
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTATGGGAACTCAGTTTATATGAGTTAGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号264)
RNF2 1-15del cr289 evopreQ1
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTATGGGAACTCAGTTTATATGAGTTAGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号265)
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTATGGGAACTCAGTTTATATGAGTTAGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号265)
RNF2 1-15del cr622 evopreQ1
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTATGGGAACTCAGTTTATATGAGTTAGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号266)
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTATGGGAACTCAGTTTATATGAGTTAGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号266)
RNF2 1-15del cr772 evopreQ1
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTATGGGAACTCAGTTTATATGAGTTAGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号267)
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTATGGGAACTCAGTTTATATGAGTTAGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号267)
RNF2 1-15del cr532 evopreQ1
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTATGGGAACTCAGTTTATATGAGTTAGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号268)
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTATGGGAACTCAGTTTATATGAGTTAGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号268)
RNF2 1-15del cr961 evopreQ1
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTATGGGAACTCAGTTTATATGAGTTAGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号269)
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTATGGGAACTCAGTTTATATGAGTTAGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号269)
RNF2 1-15delフリップ及び伸長骨格evopreQ1
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTATGGGAACTCAGTTTATATGAGTTAGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号270)
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTATGGGAACTCAGTTTATATGAGTTAGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号270)
RUNX1 1-15del standard scaffold evopreQ1
GCATTTTCAGGAGGAAGCGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTACGAAGGAAATGACTCAAATATGCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号271)
GCATTTTCAGGAGGAAGCGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTACGAAGGAAATGACTCAAATATGCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号271)
RUNX1 1-15del cr748 evopreQ1
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTTACGAAGGAAATGACTCAAATATGCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号272)
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTTACGAAGGAAATGACTCAAATATGCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号272)
RUNX1 1-15del cr289 evopreQ1
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTTACGAAGGAAATGACTCAAATATGCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号273)
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTTACGAAGGAAATGACTCAAATATGCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号273)
RUNX1 1-15del cr622 evopreQ1
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTTACGAAGGAAATGACTCAAATATGCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号274)
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTTACGAAGGAAATGACTCAAATATGCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号274)
RUNX1 1-15del cr772 evopreQ1
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTTACGAAGGAAATGACTCAAATATGCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号275)
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTTACGAAGGAAATGACTCAAATATGCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号275)
RUNX1 1-15del cr532 evopreQ1
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTTACGAAGGAAATGACTCAAATATGCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号276)
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTTACGAAGGAAATGACTCAAATATGCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号276)
RUNX1 1-15del cr961 evopreQ1
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTTACGAAGGAAATGACTCAAATATGCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号277)
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTTACGAAGGAAATGACTCAAATATGCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号277)
RUNX1 1-15del flip and extension scaffold evopreQ1
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTACGAAGGAAATGACTCAAATATGCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号278)
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTACGAAGGAAATGACTCAAATATGCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号278)
RUNX1+5G-T standard scaffold evopreQ1
GCATTTTCAGGAGGAAGCGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTGTCTGAAGCAATCGCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号279)
GCATTTTCAGGAGGAAGCGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTGTCTGAAGCAATCGCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号279)
RUNX1+5G-T cr748 evopreQ1
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTTGTCTGAAGCAATCGCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号280)
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTTGTCTGAAGCAATCGCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号280)
RUNX1+5G-T cr289 evopreQ1
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTTGTCTGAAGCAATCGCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号281)
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTTGTCTGAAGCAATCGCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号281)
RUNX1+5G-T cr622 evopreQ1
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTTGTCTGAAGCAATCGCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号282)
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTTGTCTGAAGCAATCGCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号282)
RUNX1+5G-T cr772 evopreQ1
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTTGTCTGAAGCAATCGCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号283)
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTTGTCTGAAGCAATCGCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号283)
RUNX1+5G-T cr532 evopreQ1
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTTGTCTGAAGCAATCGCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号284)
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTTGTCTGAAGCAATCGCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号284)
RUNX1+5G-T cr961 evopreQ1
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTTGTCTGAAGCAATCGCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号285)
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTTGTCTGAAGCAATCGCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号285)
RUNX1+5G-T flip and extension scaffold evopreQ1
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTGTCTGAAGCAATCGCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号286)
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTGTCTGAAGCAATCGCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号286)
DNMT1 1-15del standard scaffold evopreQ1
GATTCCTGGTGCCAGAAACAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTGCTAAGGACTAGTTCTGCCCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号287)
GATTCCTGGTGCCAGAAACAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTGCTAAGGACTAGTTCTGCCCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号287)
DNMT1 1-15del cr748 evopreQ1
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTTGCTAAGGACTAGTTCTGCCCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号288)
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTTGCTAAGGACTAGTTCTGCCCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号288)
DNMT1 1-15del cr289 evopreQ1
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTTGCTAAGGACTAGTTCTGCCCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号289)
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTTGCTAAGGACTAGTTCTGCCCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号289)
DNMT1 1-15del cr622 evopreQ1
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTTGCTAAGGACTAGTTCTGCCCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号290)
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTTGCTAAGGACTAGTTCTGCCCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号290)
DNMT1 1-15del cr772 evopreQ1
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTTGCTAAGGACTAGTTCTGCCCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号291)
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTTGCTAAGGACTAGTTCTGCCCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号291)
DNMT1 1-15del cr532 evopreQ1
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTTGCTAAGGACTAGTTCTGCCCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号292)
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTTGCTAAGGACTAGTTCTGCCCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号292)
DNMT1 1-15del cr961 evopreQ1
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTTGCTAAGGACTAGTTCTGCCCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号293)
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTTGCTAAGGACTAGTTCTGCCCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号293)
DNMT1 1-15del flip and extension scaffold evopreQ1
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTGCTAAGGACTAGTTCTGCCCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号294)
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTGCTAAGGACTAGTTCTGCCCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号294)
DNMT1+5 G--T standard scaffold evopreQ1
GATTCCTGGTGCCAGAAACAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCGTCACCACTGTTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号295)
GATTCCTGGTGCCAGAAACAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCGTCACCACTGTTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号295)
DNMT1+5 G--T cr748 evopreQ1
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTGTCACCACTGTTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号296)
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTGTCACCACTGTTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号296)
DNMT1+5 G--T cr289 evopreQ1
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTGTCACCACTGTTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号297)
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTGTCACCACTGTTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号297)
DNMT1+5 G--T cr622 evopreQ1
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTGTCACCACTGTTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号298)
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTGTCACCACTGTTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号298)
DNMT1+5 G--T cr772 evopreQ1
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTGTCACCACTGTTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号299)
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTGTCACCACTGTTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号299)
DNMT1+5 G--T cr532 evopreQ1
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTGTCACCACTGTTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号300)
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTGTCACCACTGTTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号300)
DNMT1+5 G--T cr961 evopreQ1
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTGTCACCACTGTTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号301)
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTGTCACCACTGTTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号301)
DNMT1+5 G--T flip and extension scaffold evopreQ1
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTGTCACCACTGTTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号302)
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTGTCACCACTGTTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号302)
FANCF 1-15del standard scaffold evopreQ1
GGAATCCCTTCTGCAGCACCGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTAGTGCTTGAGACCGCCAGAAGCTCGGGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号303)
GGAATCCCTTCTGCAGCACCGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTAGTGCTTGAGACCGCCAGAAGCTCGGGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号303)
FANCF 1-15del cr748 evopreQ1
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTTAGTGCTTGAGACCGCCAGAAGCTCGGGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号304)
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTTAGTGCTTGAGACCGCCAGAAGCTCGGGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号304)
FANCF 1-15del cr289 evopreQ1
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTTAGTGCTTGAGACCGCCAGAAGCTCGGGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号305)
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTTAGTGCTTGAGACCGCCAGAAGCTCGGGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号305)
FANCF 1-15del cr622 evopreQ1
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTTAGTGCTTGAGACCGCCAGAAGCTCGGGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号306)
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTTAGTGCTTGAGACCGCCAGAAGCTCGGGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号306)
FANCF 1-15del cr772 evopreQ1
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTTAGTGCTTGAGACCGCCAGAAGCTCGGGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号307)
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTTAGTGCTTGAGACCGCCAGAAGCTCGGGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号307)
FANCF 1-15del cr532 evopreQ1
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTTAGTGCTTGAGACCGCCAGAAGCTCGGGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号308)
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTTAGTGCTTGAGACCGCCAGAAGCTCGGGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号308)
FANCF 1-15del cr961 evopreQ1
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTTAGTGCTTGAGACCGCCAGAAGCTCGGGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号309)
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTTAGTGCTTGAGACCGCCAGAAGCTCGGGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号309)
FANCF 1-15del flip and extension scaffold evopreQ1
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTAGTGCTTGAGACCGCCAGAAGCTCGGGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号310)
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTAGTGCTTGAGACCGCCAGAAGCTCGGGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号310)
FANCF+5 G--T cr748 evopreQ1
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTGGAAAAGCGATCAAGGTGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号311)
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTGGAAAAGCGATCAAGGTGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号311)
FANCF+5 G--T cr289 evopreQ1
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTGGAAAAGCGATCAAGGTGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号312)
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTGGAAAAGCGATCAAGGTGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号312)
FANCF+5 G--T cr622 evopreQ1
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTGGAAAAGCGATCAAGGTGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号313)
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTGGAAAAGCGATCAAGGTGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号313)
FANCF+5 G--T cr772 evopreQ1
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTGGAAAAGCGATCAAGGTGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号314)
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTGGAAAAGCGATCAAGGTGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号314)
FANCF+5 G--T cr532 evopreQ1
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTGGAAAAGCGATCAAGGTGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号315)
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTGGAAAAGCGATCAAGGTGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号315)
FANCF+5 G--T cr961 evopreQ1
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTGGAAAAGCGATCAAGGTGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号316)
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTGGAAAAGCGATCAAGGTGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号316)
FANCF+5 G--T flip and extension scaffold evopreQ1
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTGGAAAAGCGATCAAGGTGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号317)
GGAATCCCTTCTGCAGCACCGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTGGAAAAGCGATCAAGGTGCTGCAGAAGGGACAATCACTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号317)
EMX1 1-15del standard scaffold evopreQ1
GAGTCCGAGCAGAAGAAGAAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTCGTGGCAATGCGCCACCGGTTGATGTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号318)
GAGTCCGAGCAGAAGAAGAAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTCGTGGCAATGCGCCACCGGTTGATGTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号318)
EMX1 1-15del cr748 evopreQ1
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTTCGTGGCAATGCGCCACCGGTTGATGTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号319)
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTTCGTGGCAATGCGCCACCGGTTGATGTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号319)
EMX1 1-15del cr289 evopreQ1
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTTCGTGGCAATGCGCCACCGGTTGATGTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号320)
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTTCGTGGCAATGCGCCACCGGTTGATGTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号320)
EMX1 1-15del cr622 evopreQ1
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTTCGTGGCAATGCGCCACCGGTTGATGTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号321)
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTTCGTGGCAATGCGCCACCGGTTGATGTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号321)
EMX1 1-15del cr772 evopreQ1
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTTCGTGGCAATGCGCCACCGGTTGATGTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号322)
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTTCGTGGCAATGCGCCACCGGTTGATGTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号322)
EMX1 1-15del cr532 evopreQ1
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTTCGTGGCAATGCGCCACCGGTTGATGTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号323)
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTTCGTGGCAATGCGCCACCGGTTGATGTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号323)
EMX1 1-15del cr961 evopreQ1
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTTCGTGGCAATGCGCCACCGGTTGATGTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号324)
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTTCGTGGCAATGCGCCACCGGTTGATGTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号324)
EMX1 1-15del flip and extension scaffold evopreQ1
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTCGTGGCAATGCGCCACCGGTTGATGTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号325)
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTCGTGGCAATGCGCCACCGGTTGATGTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号325)
EMX1+5 G--T standard scaffold evopreQ1
GAGTCCGAGCAGAAGAAGAAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCGTGATGGGAGCACTTCTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号326)
GAGTCCGAGCAGAAGAAGAAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCGTGATGGGAGCACTTCTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号326)
EMX1+5 G--T cr748 evopreQ1
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTGTGATGGGAGCACTTCTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号327)
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTGTGATGGGAGCACTTCTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号327)
EMX1+5 G--T cr289 evopreQ1
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTGTGATGGGAGCACTTCTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号328)
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTGTGATGGGAGCACTTCTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号328)
EMX1+5 G--T cr622 evopreQ1
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTGTGATGGGAGCACTTCTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号329)
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTGTGATGGGAGCACTTCTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号329)
EMX1+5 G--T cr772 evopreQ1
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTGTGATGGGAGCACTTCTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号330)
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTGTGATGGGAGCACTTCTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号330)
EMX1+5 G--T cr532 evopreQ1
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTGTGATGGGAGCACTTCTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号331)
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTGTGATGGGAGCACTTCTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号331)
EMX1+5 G--T cr961 evopreQ1
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTGTGATGGGAGCACTTCTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号332)
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTGTGATGGGAGCACTTCTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号332)
EMX1+5 G--T flip and extension scaffold evopreQ1
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTGTGATGGGAGCACTTCTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号333)
GAGTCCGAGCAGAAGAAGAAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTGTGATGGGAGCACTTCTTCTTCTGCTCGGAAACAATCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号333)
RNF2+1FLAG standard scaffold evopreQ1
GTCATCTTAGTCATTACCTGGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTGAGTTACAACGAACACCTCAGCTTATCGTCGTCATCCTTGTAATCGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号334)
GTCATCTTAGTCATTACCTGGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTGAGTTACAACGAACACCTCAGCTTATCGTCGTCATCCTTGTAATCGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号334)
RNF2+1FLAG cr748 evopreQ1
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTTGAGTTACAACGAACACCTCAGCTTATCGTCGTCATCCTTGTAATCGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号335)
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTTGAGTTACAACGAACACCTCAGCTTATCGTCGTCATCCTTGTAATCGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号335)
RNF2+1FLAG cr289 evopreQ1
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTTGAGTTACAACGAACACCTCAGCTTATCGTCGTCATCCTTGTAATCGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号336)
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTTGAGTTACAACGAACACCTCAGCTTATCGTCGTCATCCTTGTAATCGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号336)
RNF2+1FLAG cr622 evopreQ1
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTTGAGTTACAACGAACACCTCAGCTTATCGTCGTCATCCTTGTAATCGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号337)
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTTGAGTTACAACGAACACCTCAGCTTATCGTCGTCATCCTTGTAATCGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号337)
RNF2+1FLAG cr772 evopreQ1
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTTGAGTTACAACGAACACCTCAGCTTATCGTCGTCATCCTTGTAATCGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号338)
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTTGAGTTACAACGAACACCTCAGCTTATCGTCGTCATCCTTGTAATCGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号338)
RNF2+1FLAG cr532 evopreQ1
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTTGAGTTACAACGAACACCTCAGCTTATCGTCGTCATCCTTGTAATCGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号339)
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTTGAGTTACAACGAACACCTCAGCTTATCGTCGTCATCCTTGTAATCGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号339)
RNF2+1FLAG cr961 evopreQ1
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTTGAGTTACAACGAACACCTCAGCTTATCGTCGTCATCCTTGTAATCGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号340)
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTTGAGTTACAACGAACACCTCAGCTTATCGTCGTCATCCTTGTAATCGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号340)
RNF2+1FLAG flip and extension scaffold evopreQ1
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTGAGTTACAACGAACACCTCAGCTTATCGTCGTCATCCTTGTAATCGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号341)
GTCATCTTAGTCATTACCTGGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTGAGTTACAACGAACACCTCAGCTTATCGTCGTCATCCTTGTAATCGTAATGACTAAGATGTCATCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号341)
VEGFA+5 G--T cr748 evopreQ1
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTAATGTGCCATCTGGAGCACTCATCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号342)
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTAATGTGCCATCTGGAGCACTCATCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号342)
VEGFA+5 G--T cr289 evopreQ1
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTAATGTGCCATCTGGAGCACTCATCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号343)
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTAATGTGCCATCTGGAGCACTCATCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号343)
VEGFA+5 G--T cr622 evopreQ1
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTAATGTGCCATCTGGAGCACTCATCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号344)
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTAATGTGCCATCTGGAGCACTCATCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号344)
VEGFA+5 G--T cr772 evopreQ1
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTAATGTGCCATCTGGAGCACTCATCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号345)
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTAATGTGCCATCTGGAGCACTCATCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号345)
VEGFA+5 G--T cr532 evopreQ1
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTAATGTGCCATCTGGAGCACTCATCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号346)
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTAATGTGCCATCTGGAGCACTCATCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号346)
VEGFA+5 G--T cr961 evopreQ1
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTAATGTGCCATCTGGAGCACTCATCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号347)
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTAATGTGCCATCTGGAGCACTCATCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号347)
VEGFA+5 G--T flip and extension scaffold evopreQ1
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTAATGTGCCATCTGGAGCACTCATCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号348)
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTAATGTGCCATCTGGAGCACTCATCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号348)
VEGFA+1FLAG standard scaffold evopreQ1
GATGTCTGCAGGCCAGATGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCAATGTGCCATCTGGAGCACTCACTTATCGTCGTCATCCTTGTAATCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号349)
GATGTCTGCAGGCCAGATGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCAATGTGCCATCTGGAGCACTCACTTATCGTCGTCATCCTTGTAATCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号349)
VEGFA+1FLAG cr748 evopreQ1
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTAATGTGCCATCTGGAGCACTCACTTATCGTCGTCATCCTTGTAATCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号350)
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTAATGTGCCATCTGGAGCACTCACTTATCGTCGTCATCCTTGTAATCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号350)
VEGFA+1FLAG cr289 evopreQ1
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTAATGTGCCATCTGGAGCACTCACTTATCGTCGTCATCCTTGTAATCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号351)
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTAATGTGCCATCTGGAGCACTCACTTATCGTCGTCATCCTTGTAATCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号351)
VEGFA+1FLAG cr622 evopreQ1
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTAATGTGCCATCTGGAGCACTCACTTATCGTCGTCATCCTTGTAATCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号352)
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTAATGTGCCATCTGGAGCACTCACTTATCGTCGTCATCCTTGTAATCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号352)
VEGFA+1FLAG cr772 evopreQ1
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTAATGTGCCATCTGGAGCACTCACTTATCGTCGTCATCCTTGTAATCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号353)
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTAATGTGCCATCTGGAGCACTCACTTATCGTCGTCATCCTTGTAATCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号353)
VEGFA+1FLAG cr532 evopreQ1
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTAATGTGCCATCTGGAGCACTCACTTATCGTCGTCATCCTTGTAATCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号354)
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTAATGTGCCATCTGGAGCACTCACTTATCGTCGTCATCCTTGTAATCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号354)
VEGFA+1FLAG cr961 evopreQ1
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTAATGTGCCATCTGGAGCACTCACTTATCGTCGTCATCCTTGTAATCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号355)
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTAATGTGCCATCTGGAGCACTCACTTATCGTCGTCATCCTTGTAATCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号355)
VEGFA+1FLAG flip and extension scaffold evopreQ1
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTAATGTGCCATCTGGAGCACTCACTTATCGTCGTCATCCTTGTAATCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号356)
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTAATGTGCCATCTGGAGCACTCACTTATCGTCGTCATCCTTGTAATCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号356)
VEGFA 1-15del standard scaffold evopreQ1
GATGTCTGCAGGCCAGATGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCGTGTGTCCCTCTGACAATGTGCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号357)
GATGTCTGCAGGCCAGATGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCGTGTGTCCCTCTGACAATGTGCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号357)
VEGFA 1-15del cr748 evopreQ1
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTGTGTGTCCCTCTGACAATGTGCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号358)
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTGTGTGTCCCTCTGACAATGTGCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号358)
VEGFA 1-15del cr289 evopreQ1
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTGTGTGTCCCTCTGACAATGTGCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号359)
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTGTGTGTCCCTCTGACAATGTGCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号359)
VEGFA 1-15del cr622 evopreQ1
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTGTGTGTCCCTCTGACAATGTGCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号360)
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTGTGTGTCCCTCTGACAATGTGCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号360)
VEGFA 1-15del cr772 evopreQ1
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTGTGTGTCCCTCTGACAATGTGCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号361)
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTGTGTGTCCCTCTGACAATGTGCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号361)
VEGFA 1-15del cr532 evopreQ1
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTGTGTGTCCCTCTGACAATGTGCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号362)
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTGTGTGTCCCTCTGACAATGTGCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号362)
VEGFA 1-15del cr961 evopreQ1
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTGTGTGTCCCTCTGACAATGTGCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号363)
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTGTGTGTCCCTCTGACAATGTGCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号363)
VEGFA 1-15del flip and extension scaffold evopreQ1
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTGTGTGTCCCTCTGACAATGTGCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号364)
GATGTCTGCAGGCCAGATGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTGTGTGTCCCTCTGACAATGTGCTCTGGCCTGCAGAACAATCTCTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号364)
RUNX1+1FLAG standard scaffold evopreQ1
GCATTTTCAGGAGGAAGCGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTGTCTGAAGCCATCCCTTATCGTCGTCATCCTTGTAATCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号365)
GCATTTTCAGGAGGAAGCGAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTGTCTGAAGCCATCCCTTATCGTCGTCATCCTTGTAATCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号365)
RUNX1+1FLAG cr748 evopreQ1
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTTGTCTGAAGCCATCCCTTATCGTCGTCATCCTTGTAATCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号366)
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTTGTCTGAAGCCATCCCTTATCGTCGTCATCCTTGTAATCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号366)
RUNX1+1FLAG cr289 evopreQ1
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTTGTCTGAAGCCATCCCTTATCGTCGTCATCCTTGTAATCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号367)
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTTGTCTGAAGCCATCCCTTATCGTCGTCATCCTTGTAATCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号367)
RUNX1+1FLAG cr622 evopreQ1
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTTGTCTGAAGCCATCCCTTATCGTCGTCATCCTTGTAATCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号368)
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTTGTCTGAAGCCATCCCTTATCGTCGTCATCCTTGTAATCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号368)
RUNX1+1FLAG cr772 evopreQ1
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTTGTCTGAAGCCATCCCTTATCGTCGTCATCCTTGTAATCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号369)
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTTGTCTGAAGCCATCCCTTATCGTCGTCATCCTTGTAATCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号369)
RUNX1+1FLAG cr532 evopreQ1
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTTGTCTGAAGCCATCCCTTATCGTCGTCATCCTTGTAATCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号370)
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTTGTCTGAAGCCATCCCTTATCGTCGTCATCCTTGTAATCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号370)
RUNX1+1FLAG cr961 evopreQ1
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTTGTCTGAAGCCATCCCTTATCGTCGTCATCCTTGTAATCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号371)
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTTGTCTGAAGCCATCCCTTATCGTCGTCATCCTTGTAATCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号371)
RUNX1+1FLAG flip and extension scaffold evopreQ1
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTGTCTGAAGCCATCCCTTATCGTCGTCATCCTTGTAATCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号372)
GCATTTTCAGGAGGAAGCGAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTGTCTGAAGCCATCCCTTATCGTCGTCATCCTTGTAATCCTTCCTCCTGAAAATAACTCTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号372)
DNMT1+1FLAG standard scaffold evopreQ1
GATTCCTGGTGCCAGAAACAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTCTGCCCTCCCGTCACCCCTGTCTTATCGTCGTCATCCTTGTAATCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号373)
GATTCCTGGTGCCAGAAACAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTCTGCCCTCCCGTCACCCCTGTCTTATCGTCGTCATCCTTGTAATCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号373)
DNMT1+1FLAG cr748 evopreQ1
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTTCTGCCCTCCCGTCACCCCTGTCTTATCGTCGTCATCCTTGTAATCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号374)
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAGAGAGTGGCACCGAGTCGGTGCTTCTGCCCTCCCGTCACCCCTGTCTTATCGTCGTCATCCTTGTAATCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号374)
DNMT1+1FLAG cr289 evopreQ1
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTTCTGCCCTCCCGTCACCCCTGTCTTATCGTCGTCATCCTTGTAATCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号375)
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTGGAAACAGTGGCACCGAGTCGGTGCTTCTGCCCTCCCGTCACCCCTGTCTTATCGTCGTCATCCTTGTAATCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号375)
DNMT1+1FLAG cr622 evopreQ1
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTTCTGCCCTCCCGTCACCCCTGTCTTATCGTCGTCATCCTTGTAATCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号376)
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAGCTGGAAACAGCGGCACCGAGTCGGTGCTTCTGCCCTCCCGTCACCCCTGTCTTATCGTCGTCATCCTTGTAATCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号376)
DNMT1+1FLAG cr772 evopreQ1
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTTCTGCCCTCCCGTCACCCCTGTCTTATCGTCGTCATCCTTGTAATCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号377)
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGGCACCGAGTCGGTGCTTCTGCCCTCCCGTCACCCCTGTCTTATCGTCGTCATCCTTGTAATCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号377)
DNMT1+1FLAG cr532 evopreQ1
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTTCTGCCCTCCCGTCACCCCTGTCTTATCGTCGTCATCCTTGTAATCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号378)
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACGTGAAAACGTGACTCCGAGTCGGAGTTTCTGCCCTCCCGTCACCCCTGTCTTATCGTCGTCATCCTTGTAATCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号378)
DNMT1+1FLAG cr961 evopreQ1
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTTCTGCCCTCCCGTCACCCCTGTCTTATCGTCGTCATCCTTGTAATCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号379)
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTCGAAAGAGTGCAACCGAGTCGGTTGTTCTGCCCTCCCGTCACCCCTGTCTTATCGTCGTCATCCTTGTAATCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号379)
DNMT1+1FLAG flip and extension scaffold evopreQ1
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTCTGCCCTCCCGTCACCCCTGTCTTATCGTCGTCATCCTTGTAATCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号380)
GATTCCTGGTGCCAGAAACAGTTTAAGAGCTAAGCTGGAAACAGCATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTCTGCCCTCCCGTCACCCCTGTCTTATCGTCGTCATCCTTGTAATCTTCTGGCACCAGGACCTCTTCTTTGACGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAAATTTTTT(配列番号380)
本開示は、本明細書に開示されるプライム編集システムの有効性を更に改善するための任意のそのような方法を企図する。
様々な態様において、Tの連続系列は転写されるべきpegRNAの能力を制限することもあるため、伸長アームからのTsの連続配列の出現を制限することが有利であることもある。例えば、少なくとも連続した3つのT、少なくとも連続した4つのT、少なくとも連続した5つのT、少なくとも連続した6つのT、少なくとも連続した7つのT、少なくとも連続した8つのT、少なくとも連続した9つのT、少なくとも連続した10 T、少なくとも連続した11のT、少なくとも連続した12のT、少なくとも連続した13のT、少なくとも連続した14のT、又は少なくとも連続した15のTのストリングは、pegRNAをデザインするときに回避されるべきであり、又は最終デザイン配列から少なくとも除去されるべきである。一態様では、連続するA:T核酸塩基対が豊富な標的部位を回避しながら、連続するTの望ましくないストリングがpegRNA伸長アームに包含されることを回避し得る。
他の態様では、プライム編集システムは、プライム編集効率を改善し得るpegRNAデザイン及び戦略の使用を包含してもよい。これらの戦略は、プライム編集に必要な多段階プロセスのために存在するいくつかの問題を克服しようとするものである。例えば、pegRNA内に形成され得る好ましくないRNA構造は、DNA編集の阻害をもたらし得、DNA編集は、pegRNAからゲノム遺伝子座にコピーされる。これらの制限は、pegRNA構成要素の再デザイン及び操作によって克服され得た。これらの再デザインは、プライム編集因子効率を改善し、ゲノムへのより長い挿入配列の組み入れを可能にし得た。
したがって、様々な態様において、pegRNAデザインは、非ポリメラーゼIII(pol III)プロモータからの機能的pegRNAの効率的な発現を可能にすることによって、より長いpegRNAをもたらし得、これは面倒な配列要件の必要性を回避するであろう。他の態様では、システムの有効性を改善するために、コアCas9結合pegRNA骨格が改善され得る。更に他の態様では、標的ゲノム遺伝子座でのより長い配列の挿入を可能にする逆転写酵素(RT)プロセス性を改善するために、pegRNAに修飾がなされ得る。他の態様では、RNAモチーフは、安定性を改善し、RTプロセス性を増強し、pegRNAのミスフォールディングを防止し、及び/又はゲノム編集に重要な更なる因子を動員するために、pegRNAの5'及び/又は3'末端に付加され得る。更に別の態様では、pegRNA骨格を改善し、プライム編集因子効率を高め得た所与の配列標的に対するpegRNAの進化のためのプラットフォームが提供される。これらのデザインは、任意のCas9又はその進化したバリアントによって認識される任意のpegRNAを改善するために使用され得た。
プライム編集のこの適用は、実施例2において更に記載され得る。
pegRNAは、pegRNAの特性及び/又は特徴を修飾してもよく、それによってプライム編集の有効性を改善する追加のデザイン改善を包含してもよい。様々な態様において、これらの改善は、(1)面倒な配列要件なしにより長いpegRNAの発現を可能にする、非ポリメラーゼIII(pol III)プロモータからの機能性pegRNAの効率的な発現を可能にするデザイン;(2)有効性を改善し得るコア、Cas9結合pegRNA骨格に対する改善;(3)RTプロセス性を改善し、標的ゲノム遺伝子座でのより長い配列の挿入を可能にするためのpegRNAへの修飾;(4)pegRNAの安定性を改善し、RTプロセス性を増強し、pegRNAのミスフォールディングを防止し、又はゲノム編集に重要な更なる因子を動員するRNAモチーフの、pegRNAの5'又は3'末端への付加を包含するがこれらに限定されない、多数の異なるカテゴリーの1つ又は複数に属してもよい。
一態様では、より大きな伸長アームを有するより長い長さのpegRNAの発現を改善するために、polIIIプロモータを用いてpegRNAがデザインされ得た。sgRNAは、典型的には、U6 snRNAプロモータから発現される。このプロモータは、pol IIIを動員して関連するRNAを発現させ、核内に保持される短いRNAの発現に有用である。しかし、pol IIIは高度にプロセッシブではなく、効率的なゲノム編集に必要なレベルで数百ヌクレオチド長よりも長いRNAを発現することができない。更に、pol IIIは、Uのストレッチで失速又は終結し得、pegRNAを使用して挿入され得た配列多様性を潜在的に制限する。ポリメラーゼII(pCMV等)又はポリメラーゼI(U1 snRNAプロモータ等)を動員する他のプロモータは、より長いsgRNAを発現する能力について調べられている。しかしながら、これらのプロモータは、典型的には部分的に転写され、発現されたpegRNA中のスペーサの余分な配列5'をもたらし、これにより、部位依存的にCas9:sgRNA活性が著しく低下することが示されている。更に、pol III転写されたpegRNAは、6~7 Uの連続で単純に終結し得るが、pol II又はpol Iから転写されたpegRNAは、異なる終結シグナルを必要とするであろう。多くの場合、そのようなシグナルはまた、核からのpegRNAの望ましくない輸送をもたらすポリアデニル化をもたらす。同様に、pCMV等のpol IIプロモータから発現されるRNAは、典型的には5'キャップされ、その核外輸送ももたらす。
以前に、Rinn及び共同研究者らは、長鎖非コードRNA-(lncRNA)タグ付きsgRNAの産生のために様々な発現プラットフォームをスクリーニングした183。これらのプラットフォームは、pCMVから発現され、ヒト由来のMALAT1 ncRNA由来のENEエレメント184、KSHV由来のPAN ENEエレメント185、又はU1 snRNA由来の3'ボックス186で終結するRNAを包含する。注意すべきことに、MALAT1 ncRNA及びPAN ENEは、ポリAテールを保護する三重らせんを形成する184,187。これらの構築物はまた、RNA安定性を高め得た。これらの発現系は、より長いpegRNAの発現も可能にすることが企図される。
更に、pegRNAの一部として転写されることになるpol IIプロモータの部分を切断するための一連の方法がデザインされており、ハンマーヘッド188、ピストル189、ハチェット189、ヘアピン190、VS191、ツイスター192、若しくはツイスターシスター192リボザイム等の自己切断リボザイム、又は転写されたガイドを処理するための他の自己切断要素、又はCsy4193によって認識され、ガイドの処理にもつながるヘアピンのいずれかを付加する。また、複数のENEモチーフの組み込みは、改善されたpegRNA発現及び安定性をもたらし得る。環状イントロンRNA(ciRNA)の形態でpegRNAを循環させると、RNA発現及び安定性の増強、並びに核局在化をもたらし得る。
様々な態様では、pegRNAは、配列番号241~245によって例示されるように、様々な上記要素を包含してもよい。
様々な他の態様では、骨格又はコア配列に改善を導入することによって、pegRNAが改善されてもよい。これは、既知の導入によって行われ得る。
コアであるCas9結合pegRNA骨格は、PE活性を増強するために改善され得る。例示的なアプローチでは、骨格(P1)の第1の対合要素は、GTTTT-AAAAC(配列番号246)対合要素を含有する。そのようなTsの実行は、pol IIIの休止及びRNA転写物の早期終結をもたらし得る。P1のこの部分におけるT-A対の1つのG-C対への合理的突然変異は、sgRNA活性を増強し得る。このアプローチは、pegRNAを改善するために使用され得る。更に、P1の長さを増加させることにより、sgRNAフォールディングを増強し、pegRNA活性を改善し得る。コアの改良例としては、以下を包含し得る。
P1に対する6nt伸長を含有するpegRNA
GGCCCAGACTGAGCACGTGAGTTTTAGAGCTAGCTCATGAAAATGAGCTAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCGTGCTCAGTCTGTTTTTTT(配列番号228)
GGCCCAGACTGAGCACGTGAGTTTTAGAGCTAGCTCATGAAAATGAGCTAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCGTGCTCAGTCTGTTTTTTT(配列番号228)
P1内にT-AからG-Cへの変異を含有するpegRNA
GGCCCAGACTGAGCACGTGAGTTTGAGAGCTAGAAATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCGTGCTCAGTCTGTTTTTTT(SEQ ID NOS:247及び248.
GGCCCAGACTGAGCACGTGAGTTTGAGAGCTAGAAATAGCAAGTTTAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGGACCGAGTCGGTCCTCTGCCATCAAAGCGTGCTCAGTCTGTTTTTTT(SEQ ID NOS:247及び248.
様々な他の態様において、pegRNAは、編集鋳型領域に修飾を導入することによって改善されてもよい。pegRNAによって鋳型される挿入のサイズが増加するにつれて、それはエンドヌクレアーゼによって分解され、自発的加水分解を受け、又はRTによって逆転写され得ないか若しくはpegRNA骨格の折り畳み及びその後のCas9-RT結合を破壊する二次構造に折り畳まれる可能性がより高い。したがって、遺伝子全体の挿入等の大きな挿入に影響を及ぼすために、pegRNAの鋳型の修飾が必要である可能性がある。そうするためのいくつかの戦略は、RNAを分解又は加水分解に対してより耐性にするか、又は阻害性二次構造を採用する可能性を低くする、合成又は半合成pegRNA内への修飾ヌクレオチドの組み込みを包含する196。そのような修飾は、Gリッチ配列中のRNA二次構造を減少させる8-アザ-7-デアザグアノシン;分解を低減し、ある種のRNA二次構造を増強するロックド核酸(LNA);RNA安定性を高める2'-O-メチル、2'-フルオロ又は2'-O-メトキシエトキシ修飾を包含し得た。このような修飾はまた、安定性及び活性を高めるために、pegRNAの他の場所に包含され得た。代替的又は追加的に、pegRNAの鋳型は、それが所望のタンパク質産物をコードし、RTによって展開されることができる単純な二次構造を採用する可能性も高くなるようにデザインされ得る。そのような単純な構造は熱力学的シンクとして作用し、逆転写を妨げるより複雑な構造が生じる可能性を低くする。最後に、鋳型を2つの別個のpegRNAに分裂し得た。そのようなデザインでは、PEが使用されて転写を開始し、Cas9に融合したRNA結合タンパク質又はMS2アプタマー等のpegRNA自体のRNA認識エレメントを介して別個の鋳型RNAを標的部位に動員する。RTは、この別個の鋳型RNAに直接結合するか、又は第2の鋳型に交換する前に元のpegRNA上で逆転写を開始し得た。そのようなアプローチは、長い鋳型の添加時のpegRNAのミスフォールディングを防止すること、及び、おそらくPEに基づく長い挿入を阻害し得る長い挿入が起こるためにゲノムからのCas9の解離を必要としないことの両方によって、長い挿入を可能にし得た。
更に他の態様では、pegRNAの5'及び3'末端に追加のRNAモチーフを導入することによって、pegRNAが改善されてもよい。KSHV由来のPAN ENE及びMALAT1由来のENE等のいくつかのこのようなモチーフは、非pol IIIプロモータ由来のより長いpegRNAの発現を終結させる可能な手段として上に論じられた。これらの要素は、ポリAテールを包み込むRNA三重らせんを形成し、その結果、それらは核内に保持される184,187。しかしながら、末端ヌクレオチドを閉塞するpegRNAの3'末端に複雑な構造を形成することによって、これらの構造は、おそらく、pegRNAのエキソヌクレアーゼ媒介性分解の防止にも役立つだろう。
非pol IIIプロモータからの終結を可能にすることなく、3'末端に挿入された他の構造要素もRNA安定性を高め得た。そのようなモチーフは、3'末端を閉塞するヘアピン若しくはRNA四重鎖197、又は3'末端に2'-3'-環状リン酸の形成をもたらし、またpegRNAをエキソヌクレアーゼによって分解されにくくする可能性があるHDV等の自己切断リボザイムを包含し得た。198不完全なスプライシングを介して環状化するように(ciRNAを形成するように)pegRNAを誘導することはまた、pegRNAの安定性を高め、pegRNAが核内に保持されるようにすることもできる194。
更なるRNAモチーフはまた、RTプロセス性を改善するか、又はDNA-RNA二重鎖へのRT結合を増強することによってpegRNA活性を増強し得た。RTによって結合された天然配列をその同族レトロウイルスゲノムに追加すると、RT活性を増強し得た199。これには、天然プライマー結合部位(PBS)、ポリプリントラクト(PPT)、又はレトロウイルスゲノム二量体化及び転写の開始を伴うキスループを包含し得た199。
pegRNAの5'及び3'末端に、キスループ又はGNRAテトラループ/テトラループ受容体ペア200等の二量体化モチーフを付加することも、pegRNAの効果的な環状化をもたらし、安定性を改善し得た。更に、これらのモチーフの付加は、pegRNAスペーサ及びプライマーの物理的分離、PE活性を妨げる可能性があるスペーサの閉塞の防止を可能にし得ることが想定される。スペーサ領域に小さな足がかりヘアピンを形成する、pegRNAに対する短い5'又は3'伸長もまた、スペーサに結合するpegRNAのアニーリング領域と有利に競合し得た。最後に、また、キスループが使用されて、他の鋳型RNAをゲノム部位に動員し、一方のRNAから他方のRNAへのRT活性の交換を可能にし得た。改善例は、配列番号251~255を包含するが、これらに限定されない。
SpCas9及びプライム編集因子(PE)が改善された方法と類似の様式で、指向性進化によってpegRNA骨格が更に改善され得た。指向性進化は、Cas9又は進化したCas9バリアントによるpegRNA認識を増強し得た。更に、異なるpegRNA骨格配列が異なるゲノム遺伝子座で最適であり、問題の部位でPE活性を増強するか、オフターゲット活性を減少させるか、又はその両方である可能性が高い。最後に、他のRNAモチーフが付加されたpegRNA骨格の進化は、未進化の融合RNAと比較して、融合pegRNAの活性をほぼ確実に改善するであろう。例えば、c-ジ-GMP-Iアプタマー及びハンマーヘッド型リボザイムから構成されるアロステリックリボザイムの進化は、劇的に改善された活性をもたらし202、進化がハンマーヘッド-pegRNA融合の活性も改善することを示唆した。加えて、Cas9は現在、sgRNAの5'伸長を一般に許容しないが、指向性進化は、この不耐性を緩和し、更なるRNAモチーフを利用することを可能にする可能性のある突然変異を生成する可能性が高い。
本開示は、本明細書に開示されるプライム編集システムの有効性を更に改善するための任意のそのような方法を企図する。
[7]ヌクレオチドリンカーデザインのための計算方法
本開示の一側面では、本発明者らは、アプタマー(例として、プレキノシン1-1リボスイッチアプタマー又は「evopreQ1-1」)又はその変異体、シュードノット(MMLVウイルスゲノムシュードノット又は「Mpknot-1」)又はその変異体、tRNA(例として、逆転写のためのプライマーとしてMMLVによって使用される修飾tRNA)又はその変異体、あるいはG-四重鎖又はその変異体等であるが、これらに限定されない核酸部分にプライム編集ガイドRNAをカップリングするための、1つ又は複数のヌクレオチドリンカーを同定するためのソフトウェアで具体化されてもよい新規な計算技術を開発した。そのようなリンカーの例示的なヌクレオチド配列は、本明細書全体にわたって提供され、それだけに限らないが、配列番号225~236を包含する。
本開示の一側面では、本発明者らは、アプタマー(例として、プレキノシン1-1リボスイッチアプタマー又は「evopreQ1-1」)又はその変異体、シュードノット(MMLVウイルスゲノムシュードノット又は「Mpknot-1」)又はその変異体、tRNA(例として、逆転写のためのプライマーとしてMMLVによって使用される修飾tRNA)又はその変異体、あるいはG-四重鎖又はその変異体等であるが、これらに限定されない核酸部分にプライム編集ガイドRNAをカップリングするための、1つ又は複数のヌクレオチドリンカーを同定するためのソフトウェアで具体化されてもよい新規な計算技術を開発した。そのようなリンカーの例示的なヌクレオチド配列は、本明細書全体にわたって提供され、それだけに限らないが、配列番号225~236を包含する。
本明細書においてpegRNAリンカー同定ツール(「pegLIT」)と呼ばれることもある計算技術は、核酸リンカー候補を効率的に評価して、pegRNAの他の領域(例として、プライマー結合部位、スペーサ、DNA合成鋳型、及び/又はgRNAコアを含む領域)に対する塩基対形成の傾向がより低いものを同定することを伴う。いくつかの態様では、特定のリンカー候補がpegRNAの1つ又は複数の領域と塩基対を形成する傾向は、RNAの二次構造を考慮しながらRNA-RNA相互作用をモデル化するための計算ツールを使用して決定されてもよい。そのような計算ツールの例示的な一例はViennaRNAであり、その側面はLorenz,R.et al.ViennaRNA package 2.0.Algorithms Mol Biol 6,26(2011)に記載されており、その全体が参照により本明細書に組み込まれる。
本発明者らは、可能な各核酸リンカー候補の適合性を評価することは、(1)単一のリンカー候補の適合性の評価が計算的に高価であるため(例として、RNA二次構造の物理学ベースのモデリングを伴うため)、及び(2)考慮されるリンカー候補の数は、それらの長さと共に指数関数的に増加するため、計算上非現実的であることを認識した。更に、スクリーニングとの関連において、全てのリンカー候補は、(例として、PBS、スペーサ、鋳型及び/又はコア領域において)pegRNAの何らかの変化について再評価されなければならないであろう。
したがって、本発明者らは、核酸リンカー候補の空間を効率的に探索して、pegRNAを核酸部分にカップリングさせるのに適したリンカーを同定するための最適化技術を開発した。いくつかの態様において、最適化技術は、反復最適化アプローチ(例として、模擬アニーリング)を使用して、多数のリンカー候補を同定することを伴う。いくつかの態様において、最適化技術を用いて同定されたリンカー候補は、リンカークラスターを得るためにクラスター化されてもよく、各クラスター中の1つ又はそれを超える代表的リンカーが戻されてもよく、これは、同定されたリンカー間の多様性を促進するのに役立ってもよい。
いくつかの態様において、最適化技術は、考慮される各リンカー候補について計算された複数のスコアを伴う。複数のスコアのそれぞれは、リンカー候補がpegRNAの領域と相互作用し得る程度を示してもよい。このようにして、単一のリンカー候補に対する複数のスコアは、リンカー候補がpegRNAの複数の領域と相互作用する程度を表す。領域ごとにリンカー-pegRNA相互作用を考慮することは、他の方法によって可能であるよりも正確に各リンカー候補の適合性を決定するのに役立つ。
実際、本発明者らによって開発された計算技術は、総当たり探索アプローチと比較して計算的改善(例として、プロセッサ及びメモリコンピュータリソースの利用の低減)をもたらすだけでなく、プライマー結合部位と相互作用するように計算ツールによって予測されたヒトデザインリンカー又はリンカーと比較して全体的なPE編集効率を改善するリンカーも同定する。編集効率の改善は、図113A~図113Eを参照して示され、説明される。
したがって、いくつかの態様は、プライム編集ガイドRNA(pegRNA)を核酸部分にカップリングさせるための少なくとも1つの核酸リンカーを同定するための方法を提供し、方法は、(1)第1の核酸リンカー候補を包含する複数の核酸リンカー候補を生成する工程と、(2)(a)複数の核酸リンカー候補のうちの少なくともいくつかのそれぞれについて複数のスコアを計算することであって、計算することが、第1の核酸リンカー候補について第1のスコアセットを計算することを含み、第1のスコアセットが、第1の核酸リンカー候補とpegRNAの第1の領域との間の相互作用の程度を示す第1のスコア;第1の核酸リンカー候補とpegRNAの第2の領域との間の相互作用の程度を示す第2のスコア(例として、第1及び第2の領域が異なる領域である場合)を含む、計算することと、(b)計算された複数のスコアを使用して、複数の核酸リンカー候補の少なくともいくつかの中から少なくとも1つの核酸リンカーを同定することと、によって複数の核酸リンカー候補の中から少なくとも1つの核酸リンカーを少なくとも部分的に同定する工程と、(3)少なくとも1つの核酸リンカーを示す情報を出力する工程と、を含む。
いくつかの態様では、第1のスコアは、第1の核酸リンカー候補がpegRNAの第1の領域との相互作用を回避すると予測される程度を示し、第2のスコアは、第1の核酸リンカー候補がpegRNAの第2の領域との相互作用を回避すると予測される程度を示す。例えば、第1のスコアが0.8の値を有する場合、それは、平均して、リンカー候補の任意のヌクレオチドと第1の領域との間の塩基対形成を欠くpegRNA折り畳み状態の予測確率が80%であることを示してもよい。別の例として、第2のスコアが0.9の値を有する場合、これは、平均して、リンカー候補の任意のヌクレオチドと第2の領域との間の塩基対形成を欠くpegRNA折り畳み状態の予測確率が90%であることを示してもよい。
いくつかの態様では、第1の領域は、pegRNAのプライマー結合部位、pegRNAのスペーサ、pegRNAのDNA合成鋳型、又はpegRNAのgRNAコアを含んでもよい。第2の領域はまた、pegRNAのプライマー結合部位、pegRNAのスペーサ、pegRNAのDNA合成鋳型、又はpegRNAのgRNAコアを含んでもよい。
いくつかの態様では、スコアの第1のセットは、第1の核酸リンカー候補がpegRNAの第3の領域との相互作用を回避すると予測される程度を示す第3のスコアと、第1の核酸リンカー候補がpegRNAの第4の領域との相互作用を回避すると予測される程度を示す第4のスコアと、を更に含む。いくつかの態様では、第1、第2、第3及び第4の領域は、夫々、pegRNAのPBS、pegRNAのスペーサ、pegRNAのDNA合成鋳型、及びpegRNAのgRNAコアを含んでもよい。
いくつかの態様では、pegRNAは、二本鎖標的DNA配列にヌクレオチド編集を組み入れするためのものである。pegRNAは、二本鎖標的DNA配列の第1の鎖にハイブリダイズするスペーサ、二本鎖標的DNA配列の第2の鎖にハイブリダイズする伸長アームを含んでもよく、伸長アームは、プライマー結合部位(PBS)及びヌクレオチド編集を含むDNA合成鋳型、並びに核酸プログラム型DNA結合タンパク質napDNAbpと相互作用するgRNAコアを含む。いくつかのこのような態様では、第1の領域はPBSを含み、第2の領域はスペーサを含み、第3の領域はDNA合成鋳型を含み、第4の領域はgRNAコアを含む。
いくつかの態様において、様々なリンカー候補の適合性は、それらのスコアに基づいて互いに対して評価されてもよい。いくつかの態様では、複数の核酸リンカー候補は、第2の核酸リンカー候補を含み、計算された複数のスコアを用いて複数の核酸リンカー候補の少なくともいくつかの中から少なくとも1つの核酸リンカーを同定することは、第1の核酸リンカー候補に対するスコアの第1のセットを第2の核酸リンカー候補に対するスコアの第2のセットと比較することを含む。
2つの異なるリンカー候補のスコアセットが比較されてもよい複数の方法がある。例えば、いくつかの態様では、各スコアセットは、特定の領域(例として、PBSを含む領域、スペーサを含む領域)の構成スコアを有してもよく、候補は、特定の領域(例として、PBSを含む領域)のそれぞれのスコアに基づいて最初に比較されてもよい。特定の領域のスコアが等しいか又は閾値内にある場合(例として、それらが互いに近接していると見なされてもよいように)、別の領域のスコアが比較されてもよい(例として、スペーサを含む領域)。他の領域のスコアが等しいか又は閾値内にある場合、第3の領域(例として、DNA合成鋳型を含む領域)のスコアが比較されてもよい。第3の領域のスコアが等しいか又は閾値内にある場合、第4の領域(例として、gRNAコアを含む領域)のスコアが比較されてもよい。以下同様である。
したがって、いくつかの態様では、第1の領域はプライマー結合部位(PBS)を含み、第1のスコアセットにおける第1のスコアは、第1の核酸リンカー候補がpegRNAの第1の領域との相互作用を回避すると予測される程度を示し、第2のスコアセットにおける第3のスコアは、第2の核酸リンカー候補がpegRNAの第1の領域との相互作用を回避すると予測される程度を示し、第1のスコアセットを第2のスコアセットと比較することは、第1のスコアと第3のスコアとを比較することを含む。いくつかの態様では、第1のスコアが第3のスコアの閾値距離に等しいか、又は閾値距離内にある場合、第1のスコアセットと第2のスコアセットとを比較する工程は、第1のスコアのセットにおける第1のスコア以外のスコアと、第2のスコアのセットにおける第3のスコア以外の別のスコアとを比較することを含む。
いくつかの態様において、候補リンカーを同定するための技術は、反復的に実施されてもよい。いくつかの態様において、複数の核酸リンカー候補を生成製し、複数の核酸リンカー候補の中から少なくとも1つの核酸リンカーを決定することは、反復最適化アルゴリズム(例として、アニーリングをシミュレートすることを伴うもの)に従って行われてもよい。
いくつかの態様では、複数の核酸リンカー候補は第2の核酸リンカー候補を含み、反復最適化アルゴリズムに従って生成及び決定を行うことは、第1の核酸リンカー候補を生成することと、第1の核酸リンカー候補についての第1のスコアを決定することであって、特定することが、第1のスコアに基づいて、少なくとも1つの核酸リンカーに第1の核酸リンカー候補を包含するかどうかを決定することを含む、ことと、第1の核酸リンカー候補に対する第1のスコアを決定した後、第2の酸リンカー候補を生成することと、第2の核酸リンカー候補の第2のスコアを決定することであって、同定することが、第2のスコアに基づいて、少なくとも1つの核酸リンカーに第2の核酸リンカー候補を包含するかどうかを決定することを含む、ことと、を含む。そのような反復的な態様の側面は、図119の参照を包含する本明細書で説明される。
いくつかの態様において、スコアの第1のセットにおけるスコアの計算は、RNA-RNA相互作用をモデル化するためのソフトウェア(例として、ViennaRNA)を使用して行われる。
いくつかの態様において、本技術は、1つ又は複数のフィルタリングルールを用いて複数の核酸リンカー候補をフィルタリングすることを更に包含する。例えば、連続して現れる同じヌクレオチド(例として、一列に並んだ4つのウリジン)の閾値数を少なくとも有するリンカー候補は、フィルタリングルールに従って更なる検討から除外されてもよい。別の例として、閾値パーセンテージ未満(例として、50%未満)のAC含有量を有するリンカー候補は、フィルタリングルールに従って更なる検討から除外されてもよい。本明細書に記載の技術の側面はこの点に関して限定されないので、上記の2つの例示的なフィルタリングルールに加えて、又はその代わりに、1つ又は複数の他のフィルタリングルールが使用されてもよい。
いくつかの態様では、本技術は、多様なリンカー候補集団を得るために、同定されたリンカー候補をクラスタリングすること、及び異なるクラスターを表すリンカーを決定することを更に包含する。したがって、いくつかの態様では、少なくとも1つの核酸リンカーを同定することは、それぞれのスコアに基づいて複数の核酸リンカー候補のサブセットを同定することと、複数のクラスターを得るために核酸リンカー候補のサブセットをクラスター化する(例として、階層的凝集クラスタリング又は任意の他の適切なクラスタリング技術を使用する)ことと、少なくとも1つの核酸リンカー中に、複数のクラスターの各々の少なくとも1つの代表的なメンバーを包含することと、を含む。
いくつかの態様では、同定された少なくとも1つの核酸リンカーの1つ又は複数が製造されてもよく、本明細書に記載の様々な用途に使用されてもよい。
図118は、本明細書中に記載される技術のいくつかの態様に従って、プライム編集ガイドRNAを核酸部分にカップリングさせるための1つ又はそれを超える核酸リンカーを同定するための例示的なプロセス11800のフローチャートである。プロセス11800は、本明細書に記載の技術の側面がこの点に関して限定されないので、任意の適切なコンピューティングデバイス(複数可)を使用して実施されてもよい。
プロセス11800は、1又は複数の核酸リンカー候補が生成されてもよい行為11802から開始する。任意の適切な数のリンカー候補が、任意の適切な方法で行為11802において生成されてもよい。いくつかの態様において、複数のリンカー候補が、行為11802において生成されてもよく、これらの候補のいくつか又は全てが、行為11810及びそのサブ行為において更に評価されてもよい。他の態様において、1つ又は少数のリンカー候補が、行為11802において生成されてもよく、更なるリンカー候補が必要であると決定された場合(例として、行為11818において)、プロセス11800は、追加のリンカー候補が生成されるように、行為11802に戻ってもよい。
生成されるリンカーの各々は、任意の適切な長さを有してもよい。例えば、リンカー候補は、4ヌクレオチド、8ヌクレオチド、15ヌクレオチド、又は4~32ヌクレオチド、8~16ヌクレオチドの範囲内の任意の適切な数のヌクレオチド、又はこれらの範囲内の任意の他の適切な範囲からなってもよい。
いくつかの態様において、リンカー候補は、ヌクレオチドの1つ又はそれを超える(又は全ての)それぞれをランダムに選択することによって生成されてもよい。各ヌクレオチドは、ランダムに又は指定された分布(例として、均一又は任意の他の離散分布)に従って均一に選択されてもよい。いくつかの態様において、ヌクレオチドの各々は、他のリンカーヌクレオチドとは独立して選択されてもよい。いくつかの態様において、2つ又はそれを超えるヌクレオチドは、例えば、2つ又はそれを超えるヌクレオチドの配列(連続しているかどうかにかかわらず)に対して定義された共同分布からのサンプリングによって、相関のある様式で選択されてもよい。
次に、プロセス11800は行為11810に進み、行為11802中に生成された核酸リンカー候補の中から少なくとも1つの核酸リンカーが同定される。同定は、(1)行為11812において、行為11802の間に生成されたリンカー候補の少なくともいくつかの各々について複数のスコアを計算することと、(2)行為11818において、行為11812において計算された複数のスコアを用いて少なくとも1つの核酸リンカー候補を同定することと、を伴う。これらの各行為について順に説明される。いくつかの態様において、リンカー候補は、そうでなければ不適切であるリンカー候補についてのスコアを計算するために計算リソースを費やす必要がないように、1つ又はそれを超えるフィルタリング規則(その例が本明細書に記載される)を使用してフィルタリングされてもよい。
本明細書に記載されるように、特定のリンカー候補について複数のスコアを計算することは、それぞれの複数のpegRNA領域について複数のスコアを決定することを伴う。いくつかの態様において、複数のスコアのそれぞれは、リンカー候補と複数の領域のそれぞれ1つとの間の相互作用の程度を示してもよい。例えば、特定のスコアは、リンカーが特定の領域と相互作用するか、又は特定の領域との相互作用を回避すると予測される程度を示してもよい。例示的な例において、行為11812は、リンカー候補の少なくともいくつかの各々について少なくとも2つのスコアを計算することを伴う。特に、行為11814において、第1のリンカー候補とpegRNAの第1の領域(例として、PBSを含む領域又はその例が本明細書に提供される任意の他の適切な領域)との間の相互作用の程度を示す第1のスコアが計算されてもよく、行為11816において、第1のリンカー候補とpegRNAの第2の領域(例として、スペーサを含む領域、又はその例が本明細書に提供される任意の他の適切な領域)との間の相互作用の程度を示す第2のスコアが計算されてもよい。本明細書に記載の技術の側面はこの点に関して限定されないので、スコアの各々は、RNA-RNA相互作用モデリングソフトウェア(例として、ViennaRNA)又は任意の他の適切なソフトウェアを使用して計算されてもよい。
例示的な実施例では、行為11812は2つのスコアを計算することを伴うが、いくつかの態様では、3、4、5、6、又は任意の他の適切な数のスコアが各リンカー候補について計算されて、3、4、5、6、又は任意の他の適切な数のpegRNA領域との相互作用の程度の尺度を得てもよいが、本明細書に記載の技術の側面はこの点に関して限定されない。例えば、1つの例示的な態様では、4つのスコアがリンカー候補について計算されてもよく、リンカー候補とPBS、スペーサ、DNA合成鋳型、及びpegRNAのgRNAコア領域との間の相互作用の程度が示されてもよい。
行為11818において、計算されたスコアが使用されて、「最良の」核酸リンカー候補を同定してもよい。例えば、スコアが使用されて、pegRNAの1つ以上の領域との相互作用が最も少ないと予測されるリンカー候補のサブセットを同定してもよい。pegRNAのいくつかの領域との相互作用は、pegRNAの他の領域との相互作用よりも悪いと考えられてもよい。したがって、いくつかの態様において、スコアは、その後の使用のためのリンカー候補を同定するために領域ごとに検査されてもよい。例えば、いくつかの態様では、リンカー候補は、それらのPBSスコア-リンカーがPBSを含むpegRNAの領域との相互作用を回避すると予測される程度を示すスコアに基づいて比較されてもよい。そのような領域と最も相互作用しない候補リンカーの閾値数が保持されてもよい(例として、PBSとの相互作用が最も少ないと予測される100個のリンカー候補が保持されてもよい)。複数のリンカー候補が同じ領域に対して同じスコアを有する場合、これらの候補は、本明細書中に記載されるように、他の領域に対するそれらのスコアを使用して比較/ランク付けされてもよい。
いくつかの態様では、行為11812及び11818は、反復最適化アルゴリズムに従って(図118の11818から11812までの矢印で示すように)実行されてもよい。例えば、行為11812及び11818は、模擬アニーリング技術に従って実行されてもよい。この側面は、図119を参照することを包含する本明細書に記載される。
核酸候補のサブセットがそれらの対応するスコア(識別された任意の適切な数のそのような候補が存在してもよく、例えば、これは、所望の候補数を示すパラメータ設定によって制御されてもよい)に基づいて同定された後、いくつかの態様では、同定された核酸リンカー候補が更にふるい分けされて、配列構成が多様なリンカー候補のサブセットを同定してもよい。この目的のために、いくつかの態様では、同定された核酸リンカー候補はクラスター化されて、複数のクラスターを得てもよく、各クラスター内の1つ又は複数の代表的リンカーが行為11818で出力されてもよく、これにより、出力リンカー候補間の配列多様性を促進する。本明細書に記載の技術の側面はこの点に関して限定されないので、任意の適切なクラスタリング技術(例として、凝集型階層的クラスタリング)がこれに使用されてもよい。
行為11810中に特定されたリンカー候補に関する情報は、行為11820において出力されてもよい。情報は、リンカー候補の配列、それらのスコア、及び/又は任意の他の関連情報を包含してもよい。情報は、通信ネットワークを介して1つ又は複数の他のコンピューティングデバイスに送信されてもよく、後続のアクセスのために少なくとも1つの非一時的コンピュータ可読記憶媒体(例として、メモリ、ハードドライブ、ファイル等)に記憶され、グラフィカルユーザインターフェースに提示され、及び/又は任意の他の適切な方法で出力されてもよい。
上述したように、プロセス18000の側面は、反復的に実行されてもよい。この一例が図119に示される。図119は、本明細書中に記載される技術のいくつかの態様に従って、プライム編集ガイドRNAを核酸部分にカップリングさせるための1つ又はそれを超える核酸リンカーを反復的に同定するための例示的なプロセス11900のフローチャートである。プロセス11900は、本明細書に記載の技術の側面がこの点に関して限定されないので、任意の適切なコンピューティングデバイス(複数可)を使用して実施されてもよい。
プロセス11900は、核酸リンカー候補が生成される行為11902から開始する。リンカー候補は、任意の適切な長さを有していてもよく、長さの例が本明細書中に提供される。リンカー候補は、行為11802を参照して説明した方法のいずれかを包含する任意の適切な方法で生成されてもよい。
次に、プロセス11900は決定ブロック11904に進み、そこで、行為11902において生成されたリンカー候補が1つ又は複数のフィルタリングルールに合格するかどうかが決定される。望ましくないリンカー候補を排除するために、任意の適切なフィルタリングルールが使用されてもよい。例えば、連続して現れる同じヌクレオチド(例として、一列に並んだ4つのウリジン)の閾値数を少なくとも有する候補リンカーは、(このフィルタリングルールを通さないことによって)更なる検討から除外されてもよい。別の例として、閾値パーセンテージ未満(例として、50%未満)のAC含有量を有する候補リンカーは、(このフィルタリングルールを通過させないことによって)更なる検討から除外されてもよい。
決定ブロック11904において、リンカー候補が1つ又は複数のフィルタルールを通過しないと決定されると、プロセス11900は行為11902に戻り、別のリンカー候補が生成される。一方、決定ブロック11904において、リンカー候補がフィルタルール(複数可)に通過すると決定された場合、プロセス11900は行為11906に進み、リンカー候補に対する複数のスコアが計算される。本明細書に記載されるように、複数のスコアは、それぞれの複数のpegRNA領域について、候補リンカーと領域との間の相互作用の程度を示す。リンカー候補についての複数のスコアをどのように計算するかの側面が、本明細書中に記載される。
次に、プロセス11900は行為11908に進み、行為11906で決定されたリンカー候補についてのスコアが、以前に保持されていたリンカー候補についてのスコアと比較される。例えば、リンカー候補のセット(例として、100個の候補)が、これまでに調べられた候補リンカーの中からすでに同定されていてもよく、新しいリンカー候補(行為11902で生成された候補)についてのスコアが、保持された候補について以前に決定されたスコアと比較されてもよい。リンカー候補のスコアを他のリンカー候補のそれぞれのスコアと比較する方法の側面が本明細書中に記載される。
この比較に基づいて、決定ブロック11910において、新しいリンカー候補が保持されるべきかどうかが決定される。行為11908の比較が、新たなリンカー候補が保持された候補の少なくとも一方よりも良好であることを示す場合、行為11912において、新たなリンカー候補が保持されてもよい。任意選択で、以前に保持されたリンカー候補のうちの1つがリストから削除されてもよい(例として、保持されてもよい固定数のリンカー候補が存在し、新たなリンカーをリストに追加すると、保持されたリンカー候補の総数が固定数を超えることになる場合)。
一方、行為11908の比較が、新たなリンカー候補が保持された候補のいずれよりも優れていないことを示す場合、行為11912において、新たなリンカー候補は、ある確率でのみ保持され、そうでなければ、それは削除され、プロセス11900は、新たなリンカー候補が生成されてもよい行為11902に戻る。その確率は、いくつかの態様では、模擬アニーリング・スケジュールに従って選択されてもよい。その意味で、プロセス11900の反復最適化スキームは、模擬アニーリングを伴うと考えられてもよい。
リンカー候補が行為11912において保持された後、プロセス11900は決定ブロック11914に進み、そこで更なるリンカー候補が生成されるべきかどうかが決定される。この決定は、任意の適切な方法で、例えば、行われた反復回数/時間、保持された候補リンカーの数、保持された候補間の品質及び/又は多様性の推定値、並びに/あるいは任意の適切なメトリックに基づいて行われてもよい。追加のリンカー候補が生成されるべきであると決定されたとき、プロセス11900は行為11902に戻り、そこで新しいリンカー候補が生成されてもよい。そうでない場合、プロセス11900は、保持されたリンカー候補の少なくともいくつか(例として、全ての、全てのクラスターのうちの少なくとも1つの代表的なメンバー)を示す情報が出力される行為11916に進む。保持された候補リンカーに関する情報を出力する例が本明細書で説明される。
図120は、本明細書に記載の技術の態様が実装されてもよいコンピュータシステム12000の例示的な実装形態を示す。例えば、本明細書に記載のコンピューティングデバイスのいずれかは、コンピューティングシステム12000として実装されてもよい。コンピューティングシステム12000は、1つ又は複数のコンピュータハードウェアプロセッサ12002と、非一時的コンピュータ可読記憶媒体(例として、メモリ12004及び1つ以上の不揮発性記憶装置12006)を備える1つ又は複数の製品とを包含してもよい。プロセッサ12002(複数可)は、任意の適切な方法で、メモリ12004及び不揮発性記憶装置(複数可)12006へのデータの書き込み及びメモリ及びそれらからのデータの読み出しを制御してもよい。本明細書で説明する機能のいずれかを実行するために、プロセッサ(複数可)12002は、プロセッサ(複数可)12002による実行のためのプロセッサ実行可能命令を格納する非一時的コンピュータ可読記憶媒体として機能してもよい、1つ又は複数の非一時的コンピュータ可読記憶媒体(例として、メモリ12004)に格納された1つ又は複数のプロセッサ実行可能命令を実行してもよい。
「プログラム」又は「ソフトウェア」という用語は、本明細書では一般的な意味で使用されて、上述のような態様の様々な側面を実施するためにコンピュータ又は他のプロセッサをプログラムするために使用されてもよい任意のタイプのコンピュータコード又はプロセッサ実行可能命令のセットを指す。更に、一側面によれば、実行されると本明細書で提供される本開示の方法を実行する1つ又は複数のコンピュータプログラムは、単一のコンピュータ又はプロセッサ上に存在する必要はなく、本明細書で提供される本開示の様々な側面を実施するために異なるコンピュータ又はプロセッサ間でモジュール方式で分散されてもよい。
プロセッサ実行可能命令は、1つ又は複数のコンピュータ又は他のデバイスによって実行されるプログラムモジュール等の多くの形態であってもよい。一般に、プログラムモジュールは、特定のタスクを実行するか、又は特定の抽象データ型を実装するルーチン、プログラム、オブジェクト、構成要素、データ構造等を包含する。典型的には、プログラムモジュールの機能は組み合わされてもよく、又は分散されてもよい。
また、データ構造は、任意の適切な形態で1つ以上の非一時的コンピュータ可読記憶媒体に記憶されてもよい。説明を簡単にするために、データ構造は、データ構造内の位置を介して関連するフィールドを有するように示されてもよい。そのような関係は、同様に、フィールド間の関係を伝達する非一時的コンピュータ可読媒体内の位置を有するフィールドに記憶を割り当てることによって達成されてもよい。しかしながら、ポインタ、タグ、又はデータ要素間の関係を確立する他のメカニズムの使用を包含する、任意の適切なメカニズムが使用されて、データ構造のフィールド内の情報間の関係を確立してもよい。
本明細書及び特許請求の範囲で使用される場合、1つ又は複数の要素のリストに関連して「少なくとも1つ」という語句は、要素のリスト内の要素のいずれか1つ又は複数から選択される少なくとも1つの要素を意味するが、要素のリスト内に具体的に列挙されたありとあらゆる要素の少なくとも1つを必ずしも包含せず、要素のリスト内の要素の任意の組み合わせを除外しないことを理解されたい。この定義はまた、具体的に特定された要素に関連するか否かにかかわらず、「少なくとも1つ」という語句が指す要素のリスト内で具体的に特定された要素以外の要素が任意選択的に存在してもよいことを可能にする。したがって、例えば、「A及びBのうちの少なくとも1つ」(又は、同等に、「A又はBの少なくとも1つ」、又は同等に、「A及び/又はBの少なくとも1つ」)は、一態様では、Bが存在しない(及び任意選択的にB以外の要素を包含する)少なくとも1つの、任意選択的に2つ以上のAを指し得、別の態様では、Aが存在しない(及び任意選択的にA以外の元素を包含する)少なくとも1つの、任意選択的に2つ以上のB;更に別の態様では、少なくとも1つの、任意選択的に2つ以上を含むA、及び少なくとも1つの、任意選択的に2つ以上を含むB(及び任意選択的に他の要素を含む)等である。
本明細書及び特許請求の範囲で使用される「及び/又は」という語句は、そのように結合された要素、すなわち、場合によっては結合的に存在し、他の場合には選言的に存在する要素の「いずれか又は両方」を意味すると理解されるべきである。「及び/又は」で列挙された複数の要素は、同じように、すなわちそのように結合された要素の「1つ又は複数」と解釈されるべきである。「及び/又は」節によって具体的に特定される要素以外に、具体的に特定される要素に関連するか否かにかかわらず、他の要素が任意選択的に存在してもよい。したがって、非限定的な例として、「A及び/又はB」への言及は、「含む」等のオープンエンド言語と組み合わせて使用される場合、一態様では、Aのみ(B以外の要素を任意選択的に包含する)、別の態様では、Bのみ(任意選択的にA以外の要素を包含する)、更に別の態様では、A及びBの両方に(任意選択的に他の要素を包含する)等を指し得る。
クレーム要素を修飾するための特許請求の範囲における「第1」、「第2」、「第3」等の序数の用語の使用は、それ自体では、あるクレーム要素の別のクレーム要素に対する優先順位、過程、若しくは順序、又は方法の動作が実行される時間的順序を意味しない。そのような用語は、特定の名前を有する1つのクレーム要素を同じ名前を有する別の要素と区別するためのラベルとしてのみ使用される(ただし、序数の用語を使用するため)。本明細書で使用される表現及び用語は、説明のためのものであり、限定と見なされるべきではない。「包含する(including)」、「含む(comprising)」、「有する(having)」、「含有する(containing)」、「伴う(involving)」、及びそれらの変形の使用は、その後に列挙される項目及び追加の項目を包含することが意味される。
本明細書に記載された技術のいくつかの態様を詳細に説明してきたが、様々な修正及び改善が当業者には容易に想起されるであろう。そのような修正及び改善は、本開示の趣旨及び範囲内にあることが意図されている。したがって、前述の説明は単なる例であり、限定を意図されるものではない。これらの技術は、添付の特許請求の範囲及びその均等物によって定義されるようにのみ限定される。[9]トランスプライム編集のためのスプリットpegRNAデザイン
本開示はまた、pegRNAを2つの異なる分子:ガイドRNA及びtPERT分子に分離することによって作用するプライム編集の修飾版を指すトランスプライム編集も企図する。tPERT分子は、標的DNA部位でプライム編集因子複合体と共局在するようにプログラムされ、プライマー結合部位及びDNA合成鋳型をトランスでプライム編集因子にもたらす。例えば、(1)動員タンパク質(RP)-PE:gRNA複合体、並びに(2)プライマー結合部位及びRNA-タンパク質動員ドメイン(例として、ステムループ又はヘアピン)に連結されたDNA合成鋳型を包含するtPERTを含む2構成要素系を示すトランスプライム編集因子(tPE)の態様については図3Gを参照されたく、RP-PE:gRNA複合体の動員タンパク質構成要素は、tPERTを編集される標的部位に動員し、それによってPBS及びDNA合成鋳型をトランスプライム編集因子と会合する。指し換えれば、tPERTは、プライマー結合部位及びDNA合成鋳型を包含するpegRNAの伸長アーム(の全部又は一部)を含有するように操作される。このアプローチの1つの利点は、ガイドRNAからpegRNAの伸長アームを分離し、それによって伸長アームのPBSとガイドRNAのスペーサ配列との間に生じる傾向があるアニーリング相互作用を最小限に抑えることである。
トランスプライム編集は、本明細書に記載の任意のpegRNA(PE編集効率の改善をもたらす本明細書に記載の修飾pegRNAを包含する)を用いて行われてもよい。
トランスプライム編集の重要な特徴は、トランスプライム編集因子がtPERTをDNA編集部位に動員し、それにより、プライム編集部位におけるpegRNAの機能の全てを効果的に共局在化させる能力である。動員は、MS2アプタマー等のRNA-タンパク質動員ドメインをtPERTに組み入れし、RNA-タンパク質動員ドメインに特異的に結合することができる対応する動員タンパク質をプライム編集因子に(例として、リンカーを介してnapDNAbpに、又はリンカーを介してポリメラーゼに)融合し、それによってtPERT分子をプライム編集因子複合体に動員することによって達成され得る。図3Hに記載のプロセスに描かれる通り、RP-PE:gRNA複合体は、標的DNA配列へ結合してニックを入れる。次いで、動員タンパク質(RP)は、tPERTを動員して、DNA標的部位に結合したプライム編集因子複合体に共局在化し、それによって、tPERT上に位置するプライマー結合部位がニック鎖上のプライマー配列に結合することを可能にし、続いて、ポリメラーゼ(例として、RT)が、tPERT上に位置するDNA合成鋳型に対して、tPERTの5'末端までDNAの一本鎖を合成することを可能にする。
tPERTは、RNA-タンパク質動員ドメインの5'末上にPBS及びDNA合成鋳型を含むように、図3G及び図3Hに示されているが、他の立体配置のtPERTも、RNA-タンパク質動員ドメインの3'末上に位置付けられるPBS及びDNA合成鋳型をもつようデザインされてもよい。しかしながら、5'伸長をもつtPERTは、DNAの単鎖の合成がtPERTの5'末にて自然と終始するであろうという利点を有し、よって、プライム編集のDNA合成段階の最中に、RNA-タンパク質動員ドメインのいずれの部分も鋳型として使用するリスクがない。
[8]プライム編集因子の送達
別の側面において、本開示は、種々の戦略を用いてin vitro及びin vivoのプライム編集因子の送達を可能にし、分裂インテインを用いる別個のベクター、並びにエレクトロポレーション等の技術を用いるリボヌクレオタンパク質複合体(すなわち、pegRNA及び/又は第2部位gRNAと複合体化したプライム編集因子)、カチオン性脂質によって媒介される製剤の使用、及びリボヌクレオタンパク質複合体に融合された受容体リガンドを用いる誘導性エンドサイトーシス法という直接的送達戦略を包含する。いずれかのかかる方法が本明細書において企図される。
別の側面において、本開示は、種々の戦略を用いてin vitro及びin vivoのプライム編集因子の送達を可能にし、分裂インテインを用いる別個のベクター、並びにエレクトロポレーション等の技術を用いるリボヌクレオタンパク質複合体(すなわち、pegRNA及び/又は第2部位gRNAと複合体化したプライム編集因子)、カチオン性脂質によって媒介される製剤の使用、及びリボヌクレオタンパク質複合体に融合された受容体リガンドを用いる誘導性エンドサイトーシス法という直接的送達戦略を包含する。いずれかのかかる方法が本明細書において企図される。
送達オプションの概観
いくつかの側面では、本発明は、1以上のプライム編集因子をコードするポリヌクレオチド、例えば本明細書に記載のプライム編集システムの1以上の構成要素をコードする本明細書に記載の1以上のベクター、その1以上の転写物、及び/又は次いで転写される1以上のタンパク質を、ホスト細胞に送達することを含む方法を提供する。いくつかの側面では、本発明は、更に、かかる方法によって産生される細胞、及びかかる細胞を含むか又は次いで産生される生物(例えば動物、植物、又は真菌)を提供する。いくつかの態様では、本明細書に記載のプライム編集因子は、ガイド配列との組み合わせで(任意選択で、それと複合体化されて)、細胞に送達される。従来のウイルス及び非ウイルスに基づく遺伝子移入方法が、哺乳動物細胞又は標的組織において核酸を導入するために用いられ得る。かかる方法は、プライム編集因子の構成要素をコードする核酸を培養中の又はホスト生物中の細胞に投与するために用いられ得る。非ウイルスベクター送達システムは、DNAプラスミド、RNA(例として、本明細書に記載のベクターの転写物)、裸の核酸、及びリポソーム等の送達基剤と複合体化した核酸を包含する。ウイルスベクター送達システムはDNA及びRNAウイルスを包含し、これらは細胞への送達後にエピソーム性の又は取り入れされたゲノムどちらかを有する。遺伝子処置手順の総説については、Anderson,Science 256:808-813(1992);Nabel&Felgner,TIBTECH 11:211-217(1993);Mitani&Caskey,TIBTECH 11:162-166(1993);Dillon,TIBTECH 11:167-175(1993);Miller,Nature 357:455-460(1992);Van Brunt,Biotechnology 6(10):1149-1154(1988);Vigne,Restorative Neurology and Neuroscience 8:35-36(1995);Kremer&Perricaudet,British Medical Bulletin 51(1):31-44(1995);Haddada et al.,in Current Topics in Microbiology and Immunology Doerfler and Bihm(eds)(1995);及びYu et al.,Gene Therapy 1:13-26(1994)を参照されたい。
いくつかの側面では、本発明は、1以上のプライム編集因子をコードするポリヌクレオチド、例えば本明細書に記載のプライム編集システムの1以上の構成要素をコードする本明細書に記載の1以上のベクター、その1以上の転写物、及び/又は次いで転写される1以上のタンパク質を、ホスト細胞に送達することを含む方法を提供する。いくつかの側面では、本発明は、更に、かかる方法によって産生される細胞、及びかかる細胞を含むか又は次いで産生される生物(例えば動物、植物、又は真菌)を提供する。いくつかの態様では、本明細書に記載のプライム編集因子は、ガイド配列との組み合わせで(任意選択で、それと複合体化されて)、細胞に送達される。従来のウイルス及び非ウイルスに基づく遺伝子移入方法が、哺乳動物細胞又は標的組織において核酸を導入するために用いられ得る。かかる方法は、プライム編集因子の構成要素をコードする核酸を培養中の又はホスト生物中の細胞に投与するために用いられ得る。非ウイルスベクター送達システムは、DNAプラスミド、RNA(例として、本明細書に記載のベクターの転写物)、裸の核酸、及びリポソーム等の送達基剤と複合体化した核酸を包含する。ウイルスベクター送達システムはDNA及びRNAウイルスを包含し、これらは細胞への送達後にエピソーム性の又は取り入れされたゲノムどちらかを有する。遺伝子処置手順の総説については、Anderson,Science 256:808-813(1992);Nabel&Felgner,TIBTECH 11:211-217(1993);Mitani&Caskey,TIBTECH 11:162-166(1993);Dillon,TIBTECH 11:167-175(1993);Miller,Nature 357:455-460(1992);Van Brunt,Biotechnology 6(10):1149-1154(1988);Vigne,Restorative Neurology and Neuroscience 8:35-36(1995);Kremer&Perricaudet,British Medical Bulletin 51(1):31-44(1995);Haddada et al.,in Current Topics in Microbiology and Immunology Doerfler and Bihm(eds)(1995);及びYu et al.,Gene Therapy 1:13-26(1994)を参照されたい。
核酸の非ウイルス的送達の方法は、リポフェクション、ヌクレオフェクション、マイクロインジェクション、バイオリスティック、ビロソーム、リポソーム、イムノリポソーム、ポリカチオン又は脂質:核酸コンジュゲート、裸のDNA、人工ビリオン、及びDNAの薬剤によって増強された取り込みを包含する。リポフェクションは例として米国特許第5,049,386号、第4,946,787号;及び第4,897,355号に記載され、リポフェクション試薬は市販で販売されている(例として、Transfectam(商標)及びLipofectin(商標))。ポリヌクレオチドの効率的な受容体認識リポフェクションにとって好適であるカチオン性及び中性脂質は、FeignerのWO91/17424;WO91/16024のものを包含する。送達は細胞(例として、in vitroの又はex vivoの投与)又は標的組織(例としてin vivoの投与)であり得る。
イムノ脂質複合体等の標的化リポソームを包含する脂質:核酸複合体の調製は当業者に周知である(例として、Crystal,Science 270:404-410(1995);Blaese et al.,Cancer Gene Ther.2:291-297(1995);Behr et al.,Bioconjugate Chem.5:382-389(1994);Remy et al.,Bioconjugate Chem.5:647-654(1994);Gao et al.,Gene Therapy 2:710-722(1995);Ahmad et al.,Cancer Res.52:4817-4820(1992);米国特許第4,186,183号、第4,217,344号、第4,235,871号、第4,261,975号、第4,485,054号、第4,501,728号、第4,774,085号、第4,837,028号、及び第4,946,787を参照)。
核酸の送達のためのRNA又はDNAウイルスに基づくシステムの使用は、体の特定の細胞へとウイルスを標的化及びウイルスペイロードを核に輸送するための高度に進化したプロセスを利用する。ウイルスベクターは、直接的に患者に投与され得るか(in vivo)、又はそれらはin vitroの細胞を処置するために用いられ得、修飾された細胞が任意選択で患者に投与されてもよい(ex vivo)。従来のウイルスに基づくシステムは、遺伝子移入のためのレトロウイルス、レンチウイルス、アデノウイルス、アデノ随伴、及び単純ヘルペスウイルスベクターを包含し得る。ホストゲノムへの取り入れはレトロウイルス、レンチウイルス、及びアデノ随伴ウイルス遺伝子移入法で可能であり、多くの場合には、挿入されたトランスジーンの長期的発現をもたらす。加えて、高い形質導入効率が多くの異なる細胞型及び標的組織で観察されている。
ウイルスのトロピズムは外生のエンベロープタンパク質を組み込むことによって変改され得、標的細胞の可能性ある標的集団を拡張する。レンチウイルスベクターは、非分裂細胞に形質導入又は感染する能力があるレトロウイルスベクターであり、典型的には高いウイルス力価を生ずる。よって、レトロウイルス遺伝子移入システムの選択は標的組織に依存するであろう。レトロウイルスベクターはシスに作用する長い末端反復からなり、最高で6-10kbの外生の配列のパッケージング容量を有する。最小のシスに作用するLTRはベクターの複製及びパッケージングにとって十分であり、これらは次いで処置学的な遺伝子を標的細胞に取り入れするために用いられて、永久的なトランスジーン発現を提供する。広く使用されているレトロウイルスベクターは、マウス白血病ウイルス(MuLV)、テナガザル白血病ウイルス(GaLV)、サル免疫不全ウイルス(SIV)、ヒト免疫不全ウイルス(HIV)、及びそれらの組み合わせに基づくものを包含する(例として、Buchscher et al.,J.Virol.66:2731-2739(1992);Johann et al.,J.Virol.66:1635-1640(1992);Sommnerfelt et al.,Virol.176:58-59(1990);Wilson et al.,J.Virol.63:2374-2378(1989);Miller et al.,J.Virol.65:2220-2224(1991);PCT/US94/05700)。一過性の発現が好ましい適用では、アデノウイルスに基づくシステムが用いられてもよい。アデノウイルスに基づくベクターは、多くの細胞型において非常に高い形質導入効率ができ、細胞分裂を要求しない。かかるベクターによって、高い力価及び発現レベルが得られている。このベクターは相対的に単純なシステムによって大きい数量で産生され得る。アデノ随伴ウイルス(「AAV」)ベクターはまた、例として、核酸及びペプチドのin vitro産生において、並びにin vivo及びex vivo遺伝子処置手順のために、細胞に標的核酸を形質導入するために使用されてもよい(例として、West et al.,Virology 160:38-47(1987);U.S.Pat.No.4,797,368;WO 93/24641;Kotin,Human Gene Therapy 5:793-801(1994);Muzyczka,J.Clin.Invest.94:1351(1994)を参照されたい)。組換えAAVベクターの構築は、米国特許第5,173,414号;Tratschin et al.,Mol.Cell.Biol.5:3251-3260(1985);Tratschin,et al.,Mol.Cell.Biol.4:2072-2081(1984);Hermonat&Muzyczka,PNAS 81:6466-6470(1984);and Samulski et al.,J.Virol.63:03822-3828(1989)を包含する多数の刊行物に記載されている。
パッケージング細胞は、ホスト細胞に感染することができるウイルス粒子を形成するために典型的に用いられる。かかる細胞は、アデノウイルスをパッケージングする293細胞及びレトロウイルスをパッケージングするψ2細胞又はPA317細胞を包含する。遺伝子処置に用いられるウイルスベクターは、通常は、核酸ベクターをウイルス粒子にパッケージングする細胞株を生ずることによって生成される。ベクターは、典型的には、パッケージング及びホストへの続く取り入れに要求される最小限のウイルス配列を含有し、他のウイルス配列は発現されるべきポリヌクレオチド(単数又は複数)の発現カセットによって置き換えられる。欠けているウイルス機能は典型的にはパッケージング細胞株によってトランスで供給される。例えば、遺伝子処置に用いられるAAVベクターは、典型的には、パッケージング及びホストゲノムへの取り入れに要求されるAAVゲノムからのITR配列のみを所有する。ウイルスDNAは細胞株によってパッケージングされ、これは、他のAAV遺伝子つまりrep及びcapをコードするがITR配列を欠くヘルパープラスミドを含有する。細胞株はヘルパーとしてのアデノウイルスにもまた感染されてもよい。ヘルパーウイルスはAAVベクターの複製及びヘルパープラスミドからのAAV遺伝子の発現を促進する。ヘルパープラスミドはITR配列の欠如を原因として有意な量ではパッケージングされない。アデノウイルスによるコンタミネーションは、例として、アデノウイルスがAAVよりも敏感である熱処置によって縮減され得る。細胞への核酸の送達のための追加の方法は当業者に公知である。例えば、参照によって本明細書に組み込まれるUS20030087817を見よ。
種々の態様において、PE構築物(分裂構築物を包含する)は1以上のrAAVベクターによる送達のために操作されてもよい。本明細書において提供される方法及び組成物のいずれかに関するrAAVは、いずれかの誘導体又はシュードタイプを包含するいずれかの血清型であってもよい(例として、1、2、3、4、5、6、7、8、9、10、11、12、13、2/1、2/5、2/8、2/9、3/1、3/5、3/8、又は3/9)。rAAVは細胞に送達されるべきである遺伝子積荷(すなわち、rAAVによって細胞内に持ち込まれる丸ごとの又は分裂のプライム編集因子等の目当ての遺伝子を発現する組換え核酸ベクター)を含んでもよい。rAAVはキメラであってもよい。
rAAVの本明細書において用いられる血清型は、組換えウイルスのカプシドタンパク質の血清型を指す。誘導体及びシュードタイプの限定しない例は、rAAV2/1、rAAV2/5、rAAV2/8、rAAV2/9、AAV2-AAV3ハイブリッド、AAVrh.10、AAVhu.14、AAV3a/3b、AAVrh32.33、AAV-HSC15、AAV-HSC17、AAVhu.37、AAVrh.8、CHt-P6、AAV2.5、AAV6.2、AAV2i8、AAV-HSC15/17、AAVM41、AAV9.45、AAV6(Y445F/Y731F)、AAV2.5T、AAV-HAE1/2、AAVクローン32/83、AAVShH10、AAV2(Y->F)、AAV8(Y733F)、AAV2.15、AAV2.4、AAVM41、及びAAVr3.45を包含する。キメラVP1タンパク質を有する誘導体及びシュードタイプの限定しない例はrAAV2/5-1VP1uであり、これはAAV2のゲノム、AAV5のカプシドバックボーン、及びAAV1のVP1uを有する。キメラVP1タンパク質を有する誘導体及びシュードタイプの他の限定しない例はrAAV2/5-8VP1u、rAAV2/9-1VP1u、及びrAAV2/9-8VP1uである。
AAV誘導体/シュードタイプ、及びかかる誘導体/シュードタイプを生ずるための方法は、当該技術分野で公知である(例として、Mol Ther.2012 Apr;20(4):699-708.doi:10.1038/mt.2011.287.Epub 2012 Jan 24.The AAV vector toolkit:poised at the clinical crossroads.Asokan A1,Schaffer DV,Samulski RJ.を見よ)。シュードタイプのrAAVベクターを生ずる及び用いるための方法は当該技術分野で公知である(例として、Duan et al.,J.Virol.,75:7662-7671,2001;Halbert et al.,J.Virol.,74:1524-1532,2000;Zolotukhin et al.,Methods,28:158-167,2002;及びAuricchio et al.,Hum.Molec.Genet.,10:3075-3081,2001を見よ)。
rAAV粒子を作る又はパッケージングする方法は当該技術分野で知られており、試薬が市販で利用可能である(例として、Zolotukhin et al.Production and purification of serotype 1,2,and 5 recombinant adeno-associated viral vectors.Methods 28(2002)158-167;及び米国特許公開番号US20070015238及びUS20120322861を見よ。これらは参照によって本明細書に組み込まれる;プラスミド及びキットはATCC及びCell Biolabs,Inc.から利用可能)。例えば、目当ての遺伝子を含むプラスミドが、例としてrep遺伝子(例として、Rep78、Rep68、Rep52、及びRep40をコードする)及びcap遺伝子(本明細書に記載の修飾されたVP2領域を包含するVP1、VP2、及びVP3をコードする)を含有する1以上のヘルパープラスミドと組み合わせられてもよく、組換え細胞にトランスフェクションされ得る。その結果、rAAV粒子がパッケージングされ、続いて精製され得る。
組換えAAVは核酸ベクターを含んでもよく、これは、最小で:(a)目当てのタンパク質若しくはポリペプチド又は目当てのRNA(例として、siRNA若しくはmicroRNA)をコードする配列を含む1以上の異種核酸領域と、(b)1以上の核酸領域(例として、異種核酸領域)をフランキングする逆位末端反復(ITR)配列(例として、野生型ITR配列又は操作されたITR配列)を含む1以上の領域とを含んでもよい。本明細書において、目当てのタンパク質又は目当てのRNAをコードする配列を含む異種核酸領域は目当ての遺伝子と呼ばれる。
本明細書において提供されるrAAV粒子のいずれか1つは、VP1u領域の外に異なる血清型のアミノ酸を有するカプシドタンパク質を有してもよい。いくつかの態様では、VP1タンパク質のバックボーンの血清型はITR及び/又はRep遺伝子の血清型とは異なる。いくつかの態様では、粒子のVP1カプシドタンパク質のバックボーンの血清型はITRの血清型と同じである。いくつかの態様では、粒子のVP1カプシドタンパク質のバックボーンの血清型はRep遺伝子の血清型と同じである。いくつかの態様では、rAAV粒子のカプシドタンパク質は改善された形質導入効率をもたらすアミノ酸変異を含む。
いくつかの態様では、核酸ベクターは、核酸(例として、異種核酸)の発現を容易化する配列、例として核酸に作動可能に連結された発現コントロール配列を含む1以上の領域を含む。数々のかかる配列が当該技術分野で公知である。発現コントロール配列の限定しない例は、プロモータ、インシュレーター、サイレンサー、応答エレメント、イントロン、エンハンサー、開始部位、終結シグナル、及びポリ(A)テールを包含する。かかるコントロール配列のいずれかの組み合わせは本明細書において企図される(例として、プロモータ及びエンハンサー)。
最終的なAAV構築物はpegRNAをコードする配列を組み込んでもよい。他の態様において、AAV構築物は、第2部位ニッキングガイドRNAをコードする配列を組み込んでもよい。なお他の態様において、AAV構築物は、第2部位ニッキングガイドRNAをコードする配列とpegRNAをコードする配列とを組み込んでもよい。
種々の態様において、pegRNA及び第2部位ニッキングガイドRNAは、適当なプロモータ、例えばヒトU6(hU6)プロモータ、マウスU6(mU6)プロモータ、又は他の適当なプロモータから発現され得る。pegRNA及び第2部位ニッキングガイドRNAは同じプロモータ又は異なるプロモータから駆動され得る。
いくつかの態様では、rAAV構築物又は本明細書の組成物は対象に経腸的に投与される。いくつかの態様では、rAAV構築物又は本明細書の組成物は対象に非経口的に投与される。いくつかの態様では、rAAV粒子又は本明細書の組成物は、対象に皮下で、眼内で、硝子体内で、網膜下で、静脈内で(IV)、脳室内で、筋肉内で、髄腔内で(IT)、脳槽内で、腹腔内で、吸入によって、局所的に、又は1以上の細胞、組織、若しくは臓器への直接的注射によって投与される。いくつかの態様では、rAAV粒子又は本明細書の組成物は、対象に肝動脈又は門脈への注射によって投与される。
分裂PEベクターに基づく戦略
この側面では、プライム編集因子は分裂部位において割られ得、丸ごとの/完全なプライム編集因子の2つの半分として提供され得る。2つの半分は細胞に送達され得(例として、発現されたタンパク質として、又は別個の発現ベクター上において)、ひとたび細胞内で接触をすると、2つの半分は各プライム編集因子半分のインテインの自己スプライシング作用によって完全なプライム編集因子を形成する。細胞内におけるそれらのトランススプライシングと完全な機能するPEの付随的復元とを容易化するために、分裂インテイン配列が、コードされるプライム編集因子の半分の夫々に操作され得る。
この側面では、プライム編集因子は分裂部位において割られ得、丸ごとの/完全なプライム編集因子の2つの半分として提供され得る。2つの半分は細胞に送達され得(例として、発現されたタンパク質として、又は別個の発現ベクター上において)、ひとたび細胞内で接触をすると、2つの半分は各プライム編集因子半分のインテインの自己スプライシング作用によって完全なプライム編集因子を形成する。細胞内におけるそれらのトランススプライシングと完全な機能するPEの付随的復元とを容易化するために、分裂インテイン配列が、コードされるプライム編集因子の半分の夫々に操作され得る。
これらの分裂インテインに基づく方法はin vivo送達のいくつかの関門を克服する。例えば、プライム編集因子をコードするDNAはrAAVパッケージング限界よりも大きく、そのため、特殊な解決を要求する。1つのかかる解決は、分裂インテインペアに融合された編集因子を製剤することであり、これらが2つの別個のrAAV粒子にパッケージングされ、これらは細胞に同時送達されるときに機能的な編集因子タンパク質を再構成する。プライム編集の固有の特色を説明するいくつかの他の特殊な考慮事項が記載され、第2部位ニッキング標的の最適化と、レンチウイルス及びrAAVを包含するウイルスベクターにプライム編集因子を適切にパッケージングすることとを包含する。
この側面では、プライム編集因子は分裂部位において割られ得、丸ごとの/完全なプライム編集因子の2つの半分として提供され得る。2つの半分は細胞に送達され得(例として、発現されたタンパク質として、又は別個の発現ベクター上において)、ひとたび細胞内で接触をすると、2つの半分は各プライム編集因子半分のインテインの自己スプライシング作用によって完全なプライム編集因子を形成する。細胞内におけるそれらのトランススプライシングと完全な機能するPEの付随的復元とを容易化するために、分裂インテイン配列が、コードされるプライム編集因子の半分の夫々に操作され得る。
図66は、2つのPEの半分のタンパク質として提供されているプライム編集因子の1つの態様を図示する。これらは、プライム編集因子の半分のタンパク質の夫々の終わり又は始まりに位置付けられた分裂インテインの半分同士の自己スプライシング作用によって、丸ごとのプライム編集因子を再生する。本明細書において用いられる用語「PE N末端半分」は、完全なプライム編集因子のPE N末端半分のC末端の端に「Nインテイン」を含む、完全なプライム編集因子のN末端半分を指す(すなわち、N末端エクステイン)。「Nインテイン」は、完全な十分に形成された分裂インテイン部分のN末端半分を指す。本明細書において用いられる用語「PE C末端半分」は、完全なプライム編集因子のC末端半分のN末端の端に「Cインテイン」を含む完全なプライム編集因子のC末端半分を指す(すなわち、C末端エクステイン)。2つの半分タンパク質、すなわちPE N末端半分及びPE C末端半分が例として細胞内において互いと接触をするときには、Nインテイン及びCインテインは、自己切除とPE N末端半分のC末端の端及びPE C末端半分のN末端の端の間のペプチド結合の形成との同時プロセスを経過して、完全なnapDNAbpドメイン(例としてCas9ニッカーゼ)及びRTドメインを含む完全なプライム編集因子タンパク質を再形成する。図面には示されていないが、プライム編集因子は、N末端及び/又はC末端のNLSと各ドメインを連結するアミノ酸リンカー配列とを包含する追加の配列をもまた含んでもよい。
種々の態様において、プライム編集因子は、丸ごとのプライム編集因子を「分裂部位」において(as)「分裂」することによって、2つの半分タンパク質(すなわち、PE N末端半分及びPE C末端半分)として操作されてもよい。「分裂部位」は、プライム編集因子上の2つの隣接するアミノ酸残基の間における分裂インテイン配列(すなわち、Nインテイン及びCインテイン)の挿入の位置付けを指す。より具体的には、「分裂部位」は丸ごとのプライム編集因子を2つの別個の半分へと割る位置付けを指し、各半分は分裂部位においてNインテイン又はCインテインモチーフどちらかに融合される。分裂部位はプライム編集因子上のいずれかの好適な位置付けであり得るが、好ましくは、分裂部位は送達(例として発現ベクターによる)にとって適当なサイズである2つの半分タンパク質の形成を許す位置に位置付けられ、分裂部位末端において各半分タンパク質に融合されているインテインは、1つの半分タンパク質が細胞内において他の半分タンパク質に接触するときに互いと十分に相互作用するように利用可能である。
いくつかの態様では、分裂部位はnapDNAbpドメイン上に位置付けられる。他の態様において、分裂部位はRTドメイン上に位置付けられる。他の態様において、分裂部位はnapDNAbpドメイン及びRTドメインを連結するリンカー上に位置付けられる。
種々の態様において、分裂部位デザインは、2つの半分のプライム編集因子ドメインを2つの異なるAAVゲノムにパッケージングする目的を両方が構造的に許容するN及びC末端インテインを分裂及び挿入するための部位を見出すことを要求する。加えて、トランススプライシングに必要なインテイン残基は、C末端エクステインのN末端の残基を変異させること又はインテインの「傷跡」を残すであろう残基を挿入することによって組み込まれ得る。
SpCas9ニッカーゼ又はSaCas9ニッカーゼどちらかを含む分裂されたプライム編集因子の例示の分裂構成は次の通りである。


種々の態様では、例としてSpCas9ニッカーゼ(配列番号37、1368アミノ酸)を用いて、分裂は、1及び1368の間のいずれかの2つのアミノ酸の間においてであり得る。しかしながら、好ましい分裂は、タンパク質の中心領域の間に、例として、配列番号37のアミノ酸50~1250、又は100~1200、又は150~1150、又は200~1100、又は250~1050、又は300~1000、又は350~950、又は400~900、又は450~850、又は500~800、又は550~750、又は600~700に位置付けられるであろう。特定の例示の態様では、分裂部位は740/741、又は801/802、又は1010/1011、又は1041/1042の間であってもよい。他の態様では、分裂部位は、配列番号37のSpCas9に対して、少なくとも80%、85%、90%、95%、98%、99%、or 99.9%の配列同一性を有するアミノ酸配列中の任意の2つの対応する残基の間、又は配列番号31、37~38、40、42、44~99のいずれかのアミノ酸配列のSpCas9のバリアント若しくは等価物中の任意の2つの対応する残基の間、又は配列番号31、37~38、40、42、44~99のいずれかと少なくとも80%、85%、90%、95%、98%、99%、若しくは99.9%の配列同一性を有するアミノ酸配列において、1/2、2/3、3/4、4/5、5/6、6/7、7/8、8/9、9/10、10/11、12/13、14/15、15/16、17/18、19/20、20/21、21/22、22/23、23/24、24/25、25/26、26/27、27/28、28/29、29/30、30/31、31/32、32/33、33/34、34/35、35/36、36/37、38/39、39/40、41/42、42/43、43/44、44/45、45/46、46/47、47/48、48/49、49/50、51/52、52/53、53/54、54/55、55/56、56/57、57/58、58/59、59/60、61/62、62/63、63/64、64/65、65/66、66/67、67/68、68/69、69/70、71/72、72/73、73/74、74/75、75/76、76/77、77/78、78/79、79/80、81/82、82/83、83/84、84/85、85/86、86/87、87/88、88/89、89/90、又は90~100、100~150、150~200、200~250、250~300、300~350、350~400、450~500、500~550、550~600、600~650、650~700、700~750、750~800、800~850、850~900、900~950、950~1000、1000~1050、1050~1100、1100~1150、1150~1200、1200~1250、1250~1300、1300~1350、及び1350-1368の間の隣接残基の任意の2対の間であってもよい。
様々な態様において、分裂インテイン配列は、配列番号16~23によって表されるインテイン配列から操作され得る。
様々な他の態様において、分裂インテイン配列は、以下のように使用され得る:

種々の態様において、分裂インテインは完全なプライム編集因子の別個の部分を細胞に別個に送達するために用いられ得、これらは、細胞内での発現によって、トランススプライシングによって完全なプライム編集因子として再構成されるようになる。
いくつかの態様では、本開示は、プライム編集因子を細胞に送達する方法を提供し:
(a)第1の分裂インテイン配列に融合されたプライム編集因子のN末端フラグメントをコードする第1の発現ベクターを構築すること;
(b)第2の分裂インテイン配列に融合されたプライム編集因子のC末端フラグメントをコードする第2の発現ベクターを構築すること;
(c)第1及び第2の発現ベクターを細胞に送達すること、
を含み、
第1及び第2の分裂インテイン配列の自己切除を引き起こすトランススプライシング活性の結果として、N末端及びC末端フラグメントは、細胞内においてプライム編集因子として再構成される。
(a)第1の分裂インテイン配列に融合されたプライム編集因子のN末端フラグメントをコードする第1の発現ベクターを構築すること;
(b)第2の分裂インテイン配列に融合されたプライム編集因子のC末端フラグメントをコードする第2の発現ベクターを構築すること;
(c)第1及び第2の発現ベクターを細胞に送達すること、
を含み、
第1及び第2の分裂インテイン配列の自己切除を引き起こすトランススプライシング活性の結果として、N末端及びC末端フラグメントは、細胞内においてプライム編集因子として再構成される。
分裂部位は、いくつかの態様では、napDNAbpドメイン、リンカー、又は逆転写酵素ドメインを包含するプライム編集因子融合体のどこかであり得る。
他の態様において、分裂部位はnapDNAbpドメイン上である。
なお他の態様において、分裂部位は逆転写酵素又はポリメラーゼドメイン上である。
更に他の態様において、分裂部位はリンカー上である。
種々の態様において、本開示は、napDNAbp(例としてCas9ドメイン)と逆転写酵素とを含む多重フラッププライム編集因子を提供し、napDNAbp及び/又は逆転写酵素の1つ又は両方は、インテイン、例えばリガンド依存性インテインを含む。典型的には、インテインはリガンド依存性インテインであり、これはリガンド(例として、4-ヒドロキシタモキシフェン等の低分子、ペプチド、タンパク質、ポリヌクレオチド、アミノ酸、及びヌクレオチド)の不在下ではタンパク質スプライシング活性を発揮しないか、又は最小限発揮する。リガンド依存性インテインは知られており、米国2014/0065711A1として公開された米国特許出願第(U.S.S.N.)14/004,280号に記載されているものを包含する。これの内容全体は参照によって本明細書に組み込まれる。加えて、分裂Cas9アーキテクチャの使用、いくつかの態様では、インテインは配列番号16~23、382、385、388からなる群から選択されるアミノ酸配列を含む。
種々の態様において、napDNAbpドメインは配列番号37の標準的なSpCas9ドメインと比較して小さいサイズのnapDNAbpドメインである。
標準的なSpCas9タンパク質は、1368アミノ酸長であり、158キロダルトンの予測分子量を有する。本明細書で使用される「小型Cas9変異体」という用語は、少なくとも1300アミノ酸未満、又は少なくとも1290アミノ酸未満、又は1280アミノ酸未満、又は1270アミノ酸未満、又は1260アミノ酸未満、又は1250アミノ酸未満、又は1240アミノ酸未満、又は1230アミノ酸未満、又は1220アミノ酸未満、又は1210アミノ酸未満、又は1200アミノ酸未満、又は1190アミノ酸未満、又は1180アミノ酸未満、又は1170アミノ酸未満、又は1160アミノ酸未満、又は1150アミノ酸未満、又は1140アミノ酸未満、又は1130アミノ酸未満、又は1120アミノ酸未満、又は1110アミノ酸未満、又は1100アミノ酸未満、又は1050アミノ酸未満、又は1000アミノ酸未満、又は950アミノ酸未満、又は900アミノ酸未満、又は850アミノ酸未満、又は800アミノ酸未満、又は750アミノ酸未満、又は700アミノ酸未満、又は650アミノ酸未満、又は600アミノ酸未満、又は550アミノ酸未満、又は500アミノ酸未満であるが、少なくとも400アミノ酸超であり、Cas9タンパク質の必要な機能を保持する、天然に存在する、操作された、又はその他の任意のCas9変異体を指す。
1つの態様では、例20に図示される通り、本明細書は次の分裂インテインPE構築物を包摂する。これらは標準的なSpCas9(配列番号37)の残基1024及び1025の間で分裂される(又は、これらはMetマイナスの配列番号37に対して相対的に夫々残基1023及び1024と呼ばれてもよい)。
第1に、配列番号37のアミノ酸配列は次の通り示される。1024(「K」)及び1025(「S」)残基の間の分裂部位の位置付けを指示している:

この構成において、N末端半分のアミノ酸配列(アミノ酸1~1024)は以下の通りである:
MDKKYSIGLDIGTNSVGWAVITDEYKVPSKKFKVLGNTDRHSIKKNLIGALLFDSGETAEATRLKRTARRRYTRRKNRICYLQEIFSNEMAKVDDSFFHRLEESFLVEEDKKHERHPIFGNIVDEVAYHEKYPTIYHLRKKLVDSTDKADLRLIYLALAHMIKFRGHFLIEGDLNPDNSDVDKLFIQLVQTYNQLFEENPINASGVDAKAILSARLSKSRRLENLIAQLPGEKKNGLFGNLIALSLGLTPNFKSNFDLAEDAKLQLSKDTYDDDLDNLLAQIGDQYADLFLAAKNLSDAILLSDILRVNTEITKAPLSASMIKRYDEHHQDLTLLKALVRQQLPEKYKEIFFDQSKNGYAGYIDGGASQEEFYKFIKPILEKMDGTEELLVKLNREDLLRKQRTFDNGSIPHQIHLGELHAILRRQEDFYPFLKDNREKIEKILTFRIPYYVGPLARGNSRFAWMTRKSEETITPWNFEEVVDKGASAQSFIERMTNFDKNLPNEKVLPKHSLLYEYFTVYNELTKVKYVTEGMRKPAFLSGEQKKAIVDLLFKTNRKVTVKQLKEDYFKKIECFDSVEISGVEDRFNASLGTYHDLLKIIKDKDFLDNEENEDILEDIVLTLTLFEDREMIEERLKTYAHLFDDKVMKQLKRRRYTGWGRLSRKLINGIRDKQSGKTILDFLKSDGFANRNFMQLIHDDSLTFKEDIQKAQVSGQGDSLHEHIANLAGSPAIKKGILQTVKVVDELVKVMGRHKPENIVIEMARENQTTQKGQKNSRERMKRIEEGIKELGSQILKEHPVENTQLQNEKLYLYYLQNGRDMYVDQELDINRLSDYDVDAIVPQSFLKDDSIDNKVLTRSDKNRGKSDNVPSEEVVKKMKNYWRQLLNAKLITQRKFDNLTKAERGGLSELDKAGFIKRQLVETRQITKHVAQILDSRMNTKYDENDKLIREVKVITLKSKLVSDFRKDFQFYKVREINNYHHAHDAYLNAVVGTALIKKYPKLESEFVYGDYKVYDVRKMIAK(配列番号391).
この構成において、N末端半分のアミノ酸配列(アミノ酸1~1023)(ここでタンパク質は位置1にてMetマイナスである)は以下の通りである:
DKKYSIGLDIGTNSVGWAVITDEYKVPSKKFKVLGNTDRHSIKKNLIGALLFDSGETAEATRLKRTARRRYTRRKNRICYLQEIFSNEMAKVDDSFFHRLEESFLVEEDKKHERHPIFGNIVDEVAYHEKYPTIYHLRKKLVDSTDKADLRLIYLALAHMIKFRGHFLIEGDLNPDNSDVDKLFIQLVQTYNQLFEENPINASGVDAKAILSARLSKSRRLENLIAQLPGEKKNGLFGNLIALSLGLTPNFKSNFDLAEDAKLQLSKDTYDDDLDNLLAQIGDQYADLFLAAKNLSDAILLSDILRVNTEITKAPLSASMIKRYDEHHQDLTLLKALVRQQLPEKYKEIFFDQSKNGYAGYIDGGASQEEFYKFIKPILEKMDGTEELLVKLNREDLLRKQRTFDNGSIPHQIHLGELHAILRRQEDFYPFLKDNREKIEKILTFRIPYYVGPLARGNSRFAWMTRKSEETITPWNFEEVVDKGASAQSFIERMTNFDKNLPNEKVLPKHSLLYEYFTVYNELTKVKYVTEGMRKPAFLSGEQKKAIVDLLFKTNRKVTVKQLKEDYFKKIECFDSVEISGVEDRFNASLGTYHDLLKIIKDKDFLDNEENEDILEDIVLTLTLFEDREMIEERLKTYAHLFDDKVMKQLKRRRYTGWGRLSRKLINGIRDKQSGKTILDFLKSDGFANRNFMQLIHDDSLTFKEDIQKAQVSGQGDSLHEHIANLAGSPAIKKGILQTVKVVDELVKVMGRHKPENIVIEMARENQTTQKGQKNSRERMKRIEEGIKELGSQILKEHPVENTQLQNEKLYLYYLQNGRDMYVDQELDINRLSDYDVDAIVPQSFLKDDSIDNKVLTRSDKNRGKSDNVPSEEVVKKMKNYWRQLLNAKLITQRKFDNLTKAERGGLSELDKAGFIKRQLVETRQITKHVAQILDSRMNTKYDENDKLIREVKVITLKSKLVSDFRKDFQFYKVREINNYHHAHDAYLNAVVGTALIKKYPKLESEFVYGDYKVYDVRKMIAK(配列番号392).
この構成において、C末端半分のアミノ酸配列(アミノ酸1024~1368)(又はMetマイナスCas9中アミノ酸1023~1367として数えた)は以下の通りである:
SEQEIGKATAKYFFYSNIMNFFKTEITLANGEIRKRPLIETNGETGEIVWDKGRDFATVRKVLSMPQVNIVKKTEVQTGGFSKESILPKRNSDKLIARKKDWDPKKYGGFDSPTVAYSVLVVAKVEKGKSKKLKSVKELLGITIMERSSFEKNPIDFLEAKGYKEVKKDLIIKLPKYSLFELENGRKRMLASAGELQKGNELALPSKYVNFLYLASHYEKLKGSPEDNEQKQLFVEQHKHYLDEIIEQISEFSKRVILADANLDKVLSAYNKHRDKPIREQAENIIHLFTLTNLGAPAAFKYFDTTIDRKRYTSTKEVLDATLIHQSITGLYETRIDLSQLGGD(配列番号393).
例20に示されている通り、PE2(これは配列番号37のSpCas9に基づく)構築物を、次の通り、(Metマイナス配列番号37に対して相対的に)位置1023/1024で2つの別個の構築物へと分裂した:
1023/1024のN末端半分でのSpPE2分裂
SpPE2は1023/1024 Cの末端半分で分裂
本開示は、分裂インテイン多重フラッププライム編集因子を細胞に送達及び/又は分裂インテインプライム編集因子によって細胞を処置する方法をもまた企図する。
いくつかの態様では、本開示は、プライム編集因子を細胞に送達する方法を提供し:
(a)第1の分裂インテイン配列に融合されたプライム編集因子のN末端フラグメントをコードする第1の発現ベクターを構築すること;
(b)第2の分裂インテイン配列に融合されたプライム編集因子のC末端フラグメントをコードする第2の発現ベクターを構築すること;
(c)第1及び第2の発現ベクターを細胞に送達すること、
を含み、
第1及び第2の分裂インテイン配列の自己切除を引き起こすトランススプライシング活性の結果として、N末端及びC末端フラグメントは、細胞内においてプライム編集因子として再構成される。
(a)第1の分裂インテイン配列に融合されたプライム編集因子のN末端フラグメントをコードする第1の発現ベクターを構築すること;
(b)第2の分裂インテイン配列に融合されたプライム編集因子のC末端フラグメントをコードする第2の発現ベクターを構築すること;
(c)第1及び第2の発現ベクターを細胞に送達すること、
を含み、
第1及び第2の分裂インテイン配列の自己切除を引き起こすトランススプライシング活性の結果として、N末端及びC末端フラグメントは、細胞内においてプライム編集因子として再構成される。
ある態様において、第1の分裂インテイン配列に融合されたプライム編集因子のN末端フラグメントは、配列番号394、又は配列番号394との少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、少なくとも98%、又は少なくとも99.9%配列同一性を有するアミノ酸配列である。
他の態様において、第1の分裂インテイン配列に融合されたプライム編集因子のC端フラグメントは、配列番号395、又は配列番号395の少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、少なくとも98%、又は少なくとも99.9%配列同一性を有するアミノ酸配列である。
他の態様では、本開示は、細胞内の標的DNA配列を編集する方法を提供し、
(a)第1の分裂インテイン配列に融合されたプライム編集因子のN末端フラグメントをコードする第1の発現ベクターを構築すること;
(b)第2の分裂インテイン配列に融合されたプライム編集因子のC末端フラグメントをコードする第2の発現ベクターを構築すること;
(c)第1及び第2の発現ベクターを細胞に送達すること、
を含み、
第1及び第2の分裂インテイン配列の自己切除を引き起こすトランススプライシング活性の結果として、N末端及びC末端フラグメントは、細胞内においてプライム編集因子として再構成される。
(a)第1の分裂インテイン配列に融合されたプライム編集因子のN末端フラグメントをコードする第1の発現ベクターを構築すること;
(b)第2の分裂インテイン配列に融合されたプライム編集因子のC末端フラグメントをコードする第2の発現ベクターを構築すること;
(c)第1及び第2の発現ベクターを細胞に送達すること、
を含み、
第1及び第2の分裂インテイン配列の自己切除を引き起こすトランススプライシング活性の結果として、N末端及びC末端フラグメントは、細胞内においてプライム編集因子として再構成される。
ある態様において、第1の分裂インテイン配列に融合されたプライム編集因子のN末端フラグメントは、配列番号394、又は配列番号394との少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、少なくとも98%、又は少なくとも99.9%配列同一性を有するアミノ酸配列である。
他の態様において、第1の分裂インテイン配列に融合されたプライム編集因子のC端フラグメントは、配列番号395、又は配列番号395の少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、少なくとも98%、又は少なくとも99.9%配列同一性を有するアミノ酸配列である。
PEリボヌクレオタンパク質複合体の送達
この側面では、多重フラッププライム編集因子は、エレクトロポレーション及び脂質ナノ粒子を包含する種々の方法によって、pegRNAと複合体化した多重フラッププライム編集因子(すなわち、PEリボヌクレオタンパク質複合体)の送達を伴う非ウイルス的送達戦略によって送達されてもよい。核酸の非ウイルス的送達の方法は、リポフェクション、ヌクレオフェクション、マイクロインジェクション、バイオリスティック、ビロソーム、リポソーム、イムノリポソーム、ポリカチオン又は脂質:核酸コンジュゲート、裸のDNA、人工ビリオン、及びDNAの薬剤によって増強された取り込みを包含する。リポフェクションは例として米国特許第5,049,386号、第4,946,787号;及び第4,897,355号に記載され、リポフェクション試薬は市販で販売されている(例として、Transfectam(商標)及びLipofectin(商標))。ポリヌクレオチドの効率的な受容体認識リポフェクションにとって好適であるカチオン性及び中性脂質は、FeignerのWO91/17424;WO91/16024のものを包含する。送達は細胞(例として、in vitroの又はex vivoの投与)又は標的組織(例としてin vivoの投与)であり得る。
この側面では、多重フラッププライム編集因子は、エレクトロポレーション及び脂質ナノ粒子を包含する種々の方法によって、pegRNAと複合体化した多重フラッププライム編集因子(すなわち、PEリボヌクレオタンパク質複合体)の送達を伴う非ウイルス的送達戦略によって送達されてもよい。核酸の非ウイルス的送達の方法は、リポフェクション、ヌクレオフェクション、マイクロインジェクション、バイオリスティック、ビロソーム、リポソーム、イムノリポソーム、ポリカチオン又は脂質:核酸コンジュゲート、裸のDNA、人工ビリオン、及びDNAの薬剤によって増強された取り込みを包含する。リポフェクションは例として米国特許第5,049,386号、第4,946,787号;及び第4,897,355号に記載され、リポフェクション試薬は市販で販売されている(例として、Transfectam(商標)及びLipofectin(商標))。ポリヌクレオチドの効率的な受容体認識リポフェクションにとって好適であるカチオン性及び中性脂質は、FeignerのWO91/17424;WO91/16024のものを包含する。送達は細胞(例として、in vitroの又はex vivoの投与)又は標的組織(例としてin vivoの投与)であり得る。
イムノ脂質複合体等の標的化リポソームを包含する脂質:核酸複合体の調製は当業者に周知である(例として、Crystal,Science 270:404-410(1995);Blaese et al.,Cancer Gene Ther.2:291-297(1995);Behr et al.,Bioconjugate Chem.5:382-389(1994);Remy et al.,Bioconjugate Chem.5:647-654(1994);Gao et al.,Gene Therapy 2:710-722(1995);Ahmad et al.,Cancer Res.52:4817-4820(1992);米国特許第4,186,183号、第4,217,344号、第4,235,871号、第4,261,975号、第4,485,054号、第4,501,728号、第4,774,085号、第4,837,028号、及び第4,946,787を参照)。
リボヌクレオタンパク質複合体の非ウイルス的送達のためのアプローチを論ずる次の参照の追加の参照がなされてもよい。これらの夫々は参照によって本明細書に組み込まれる。
Chen,Sean,et al.「Highly efficient mouse genome editing by CRISPR ribonucleoprotein electroporation of zygotes.」Journal of Biological Chemistry(2016):jbc-M116.PubMed
Zuris,John A.,et al.「Cationic lipid-mediated delivery of proteins enables efficient protein-based genome editing in vitro and in vivo.」 Nature biotechnology 33.1(2015):73.PubMed
Rouet,Romain,et al.「Receptor-Mediated Delivery of CRISPR-Cas9 Endonuclease for Cell-Type-Specific Gene Editing.」Journal of the American Chemical Society 140.21(2018):6596-6603.PubMed.
図68Cは、種々の開示されるPEリボヌクレオタンパク質複合体(高濃度のPE2、高濃度のPE3、及び低濃度のPE3)がこの様式で送達され得るということを示すデータを提供する。
mRNAによるPEの送達
プライム編集に基づくゲノム編集が望まれる細胞に、プライム編集因子及び/又はpegRNAを送達するために使用されてもよい別の方法は、メッセンジャーRNA(mRNA)送達方法及びテクノロジーの使用を使用することによってである。本開示に利用されてもよいmRNA送達方法及び組成物の例は、例えばPCT/US2014/028330、US8822663B2、NZ700688A、ES2740248T3、EP2755693A4、EP2755986A4、WO2014152940A1、EP3450553B1、BR112016030852A2、及びEP3362461A1を包含する。これらの夫々はそれらの全体が参照によって本明細書に組み込まれる。参照によってここに組み込まれる追加の開示はKowalski et al.,「Delivering the Messenger:Advances in Technologies for Therapeutic mRNA Delivery,」Mol Therap.,2019;27(4):710-728に見出され得る。
プライム編集に基づくゲノム編集が望まれる細胞に、プライム編集因子及び/又はpegRNAを送達するために使用されてもよい別の方法は、メッセンジャーRNA(mRNA)送達方法及びテクノロジーの使用を使用することによってである。本開示に利用されてもよいmRNA送達方法及び組成物の例は、例えばPCT/US2014/028330、US8822663B2、NZ700688A、ES2740248T3、EP2755693A4、EP2755986A4、WO2014152940A1、EP3450553B1、BR112016030852A2、及びEP3362461A1を包含する。これらの夫々はそれらの全体が参照によって本明細書に組み込まれる。参照によってここに組み込まれる追加の開示はKowalski et al.,「Delivering the Messenger:Advances in Technologies for Therapeutic mRNA Delivery,」Mol Therap.,2019;27(4):710-728に見出され得る。
プライム編集因子をコードするDNAベクターとは対照的に、プライム編集因子の送達物質としてのRNAの使用は、遺伝物質がその機能を果たすために核に入らなくてもよいという利点を有する。送達されたmRNAは細胞質において直接的に所望のタンパク質(例として、プライム編集因子)及び核酸産物(例としてpegRNA)へと翻訳されてもよい。しかしながら、より安定である(例として、細胞質のRNA分解酵素に抵抗する)ためには、いくつかの態様では、送達効率を改善するためにmRNAを安定化することが必要である。ある種の送達担体、例えばカチオン性脂質又はポリマー性の送達担体は、トランスフェクションされたmRNAを内生RNase酵素から防護することをもまた助け得る。これらは、さもなければ、所望のプライム編集因子をコードする処置学的なmRNAを分解し得る。加えて、修飾されたmRNAの増大した安定性にもかかわらず、処置学的レベルのタンパク質生産を許す様式でのin vivoの細胞へのmRNA、特に全長タンパク質をコードするmRNAの送達は、課題のままに留まっている。
いくつかの例外はあるが、mRNAの細胞内送達は小さいオリゴヌクレオチドのものよりも一般的に難しく、それは送達ナノ粒子へのカプセル化を要求する。部分的には、他の型のRNA(低分子干渉RNA[siRNA]、~14kDa;アンチセンスオリゴヌクレオチド[ASO]、4~10kDa)と比較して、mRNA分子の有意に大きいサイズ(300~5,000kDa、~1~15kb)を原因とする。
mRNAは細胞質に達するためには細胞膜を横断しなければならない。細胞膜は細胞内送達への動的なかつ手強い関門である。それは主として双性イオン性の及び負荷電のリン脂質の脂質二重層から作り上げられており、ここで、リン脂質の極性頭部は水性環境の方を向き、疎水性テールは疎水性コアを形成する。
いくつかの態様では、本開示のmRNA組成物は、mRNA(プライム編集因子及び/又はpegRNAをコードする)、輸送基剤、及び任意選択で標的細胞との接触と続くトランスフェクションとを容易化する薬剤を含む。
いくつかの態様では、mRNAは、(例として、mRNAの野生型の又はネイティブなバージョンと比較して)mRNAに安定性を付与し、タンパク質の関連する異常な発現を伴う1以上の修飾を包含し得る。欠陥を修正する野生型の1以上の修飾もまた包含されてもよい。例えば、本発明の核酸は5'非翻訳領域又は3'非翻訳領域の1つ又は両方の修飾を包含し得る。かかる修飾は、サイトメガロウイルス(CMV)最初期1(IE1)遺伝子、ポリAテール、キャップ1構造、又はヒト成長ホルモン(hGH)の部分配列をコードする配列の包含を包含してもよい。いくつかの態様では、mRNAはmRNA免疫原性を縮減するように修飾される。
1つの態様では、本発明の組成物中の「プライム編集因子」mRNAは、標的細胞への送達を容易化するためにリポソーム移入基剤によって製剤され得る。企図される移入基剤は、1以上のカチオン性脂質、非カチオン性脂質、及び/又はPEG修飾脂質を包含し得る。例えば、移入基剤は次のカチオン性脂質の少なくとも1つを包含し得る:C12-200、DLin-KC2-DMA、DODAP、HGT4003、ICE、HGT5000、又はHGT5001。態様では、移入基剤はコレステロール(chol)及び/又はPEG修飾脂質を含む。いくつかの態様では、移入基剤はDMG-PEG2Kを含む。ある態様において、移入基剤は次の脂質製剤を有する:C12-200、DOPE、chol、DMG-PEG2K;DODAP、DOPE、コレステロール、DMG-PEG2K;HGT5000、DOPE、chol、DMG-PEG2K、HGT5001、DOPE、chol、DMG-PEG2Kの1つ。
本開示は、1以上のPEをコードするmRNA分子による標的細胞のトランスフェクションを容易化することにとって有用な組成物及び方法をもまた提供する。例えば、本発明の組成物及び方法は、1以上の標的細胞に対する組成物の親和性を増大させ得る標的化リガンドの使用を企図する。1つの態様では、標的化リガンドはアポリポタンパク質B又はアポリポタンパク質Eであり、対応する標的細胞は低密度リポタンパク質受容体を発現し、それゆえに標的化リガンドの認識を促進する。膨大な数の標的細胞が本開示の方法及び組成物を用いて選好的に標的化され得る。例えば、企図される標的細胞は、肝細胞、上皮細胞、造血細胞、上皮細胞、内皮細胞、肺細胞、骨細胞、幹細胞、間葉系細胞、神経細胞、心臓細胞、脂肪細胞、血管平滑筋包含(Includes)細胞、心筋細胞、骨格筋細胞、ベータ細胞、下垂体細胞、滑膜表層細胞、卵巣細胞、精巣細胞、線維芽細胞、B細胞、T細胞、網赤血球、白血球、顆粒球、及び腫瘍細胞を包含する。しかしながら、それはこれらに限定されない。
いくつかの態様では、PEをコードするmRNAは、任意選択で、例えばかかるmRNAの安定性及び/若しくは半減期を改善するか又はタンパク質生産を改善若しくは別様に容易化する化学的又は生物学的修飾を有してもよい。トランスフェクションによって、本発明の組成物中の天然のmRNAは30分及び何日かの間の半減期で崩壊してもよい。本開示の組成物中のmRNAは翻訳される少なくともいくらかの能力を保持してもよく、それによって機能的なタンパク質又は酵素を生ずる。従って、本発明は、安定化されたmRNAを含む組成物及びそれを投与する方法を提供する。いくつかの態様では、mRNAの活性は伸長された期間にわたって遷延される。例えば、本開示の組成物が半週毎若しくは隔週毎の基準で、又はより好ましくは毎月、隔月毎、四半期毎、若しくは毎年の基準で対象に投与されるようにして、mRNAの活性は遷延されてもよい。本発明のmRNAの伸長又は遷延した活性は、かかるmRNAから産生されるタンパク質又は酵素の数量に直接的に関係する。類似に、本開示の組成物の活性は、mRNAの翻訳を改善又は増強するためになされる修飾によって更に伸長又は遷延されてもよい。更にその上、標的細胞によって産生される機能的なタンパク質又は酵素の数量は、標的細胞に送達されるmRNAの数量及びかかるmRNAの安定性の関数である。本発明のmRNAの安定性が改善又は増強されてもよい程度に合わせて、半減期、産生されるタンパク質又は酵素の活性、及び組成物の投薬頻度は更に伸長されてもよい。
従って、いくつかの態様では、本開示の組成物中のmRNAは、例えばin vivoのヌクレアーゼ消化に対する改善された耐性を包含する増大又は増強された安定性を核酸に付与する少なくとも1つの修飾を含む。本明細書において用いられる用語「修飾」及び「修飾される」は、かかる用語が本明細書において提供される核酸に関すると、好ましくは安定性を増強し、mRNAの野生型又は天然に存在するバージョンよりもmRNAを安定に(例として、ヌクレアーゼ消化に対して耐性に)する少なくとも1つの変改を包含する。本明細書において用いられる用語「安定」及び「安定性」は、かかる用語が特にmRNAについて本発明の核酸に関すると、例えば正常ではかかるmRNAを分解することができるヌクレアーゼ(すなわち、エンドヌクレアーゼ又はエキソヌクレアーゼ)による分解に対する増大又は増強した耐性を指す。増大した安定性は、例えば、標的細胞又は組織内の内生酵素(例として、エンドヌクレアーゼ若しくはエキソヌクレアーゼ)又は条件による加水分解又は他の破壊に対するより少ない感度を包含し得、それによって、標的細胞、組織、対象、及び/又は細胞質におけるかかるmRNAの滞留を増大又は増強する。本明細書において提供される安定化されたmRNA分子は、それらの天然に存在する未修飾のカウンターパート(例として、mRNAの野生型バージョン)に対して相対的に、長い半減期を実証する。また、かかる用語が本発明のmRNAに関係すると、用語「修飾」及び「修飾される」によって、mRNA核酸の翻訳を改善又は増強する変改が企図される。例えば、タンパク質翻訳の開始に機能する配列(例として、Kozakコンセンサス配列)の包含を包含する。(Kozak,M.,Nucleic Acids Res 15(20):8125-48(1987))。
いくつかの態様では、本開示の組成物に用いられるmRNAは、それらをより安定にするための化学的又は生物学的修飾を経過している。mRNAの例示の修飾は、塩基の枯渇(例として、1つのヌクレオチドの欠失若しくは別のものへの置換による)又は塩基の修飾、例えば塩基の化学修飾を包含する。本明細書において用いられる指し回し「化学修飾」は、天然に存在するmRNAに見られるものとは異なるケミストリーを導入する修飾、例えば共有結合的修飾、例えば修飾ヌクレオチドの導入(例として、ヌクレオチドアナログ、又はかかるmRNA分子に天然には見出されないペンダント基の包含)を包含する。
本開示の組成物に用いられるPEをコードするmRNAに組み込まれてもよい他の好適なポリヌクレオチド修飾は、4'-チオ修飾塩基:4'-チオアデノシン、4'-チオグアノシン、4'-チオシチジン、4'-チオウリジン、4'-チオ-5-メチル-シチジン、4'-チオシュードウリジン、及び4'-チオ2-チオウリジン、ピリジン-4-オンリボヌクレオシド、5-アザ-ウリジン、2-チオ-5-アザ-ウリジン、2-チオウリジン、4-チオシュードウリジン、2-チオシュードウリジン、5-ヒドロキシウリジン、3-メチルウリジン、5-カルボキシメチル-ウリジン、1-カルボキシメチル-シュードウリジン、5-プロピニルウリジン、1-プロピニルシュードウリジン、5-タウリノメチルウリジン、1-タウリノメチル-シュードウリジン、5-タウリノメチル-2-チオウリジン、1-タウリノメチル-4-チオウリジン、5-メチル-ウリジン、1-メチル-シュードウリジン、4-チオ-1-メチル-シュードウリジン、2-チオ-1-メチル-シュードウリジン、1-メチル-1-デアザ-シュードウリジン、2-チオ-1-メチル-1-デアザ-シュードウリジン、ジヒドロウリジン、ジヒドロシュードウリジン、2-チオジヒドロウリジン、2-チオジヒドロシュードウリジン、2-メトキシウリジン、2-メトキシ-4-チオウリジン、4-メトキシ-シュードウリジン、4-メトキシ-2-チオシュードウリジン、5-アザ-シチジン、シュードイソシチジン、3-メチル-シチジン、N4-アセチルシチジン、5-ホルミルシチジン、N4-メチルシチジン、5-ヒドロキシメチルシチジン、1-メチル-シュードイソシチジン、ピロロ-シチジン、ピロロ-シュードイソシチジン、2-チオシチジン、2-チオ-5-メチル-シチジン、4-チオシュードイソシチジン、4-チオ-1-メチル-シュードイソシチジン、4-チオ-1-メチル-1-デアザ-シュードイソシチジン、1-メチル-1-デアザ-シュードイソシチジン、ゼブラリン、5-アザ-ゼブラリン、5-メチルゼブラリン、5-アザ-2-チオゼブラリン、2-チオゼブラリン、2-メトキシ-シチジン、2-メトキシ-5-メチル-シチジン、4-メトキシ-シュードイソシチジン、4-メトキシ-1-メチル-シュードイソシチジン、2-アミノプリン、2,6-ジアミノプリン、7-デアザ-アデニン、7-デアザ-8-アザ-アデニン、7-デアザ-2-アミノプリン、7-デアザ-8-アザ-2-アミノプリン、7-デアザ-2,6-ジアミノプリン、7-デアザ-8-アザ-2,6-ジアミノプリン、1-メチルアデノシン、N6-メチルアデノシン、N6-イソペンテニルアデノシン、N6-(シス-ヒドロキシイソペンテニル)アデノシン、2-メチルチオ-N6-(シス-ヒドロキシイソペンテニル)アデノシン、N6-グリシニルカルバモイルアデノシン、N6-スレオニルカルバモイルアデノシン、2-メチルチオ-N6-スレオニルカルバモイルアデノシン、N6,N6-ジメチルアデノシン、7-メチルアデニン、2-メチルチオアデニン、及び2-メトキシ-アデニン、イノシン、1-メチル-イノシン、ワイオシン、ワイブトシン、7-デアザ-グアノシン、7-デアザ-8-アザ-グアノシン、6-チオグアノシン、6-チオ-7-デアザ-グアノシン、6-チオ-7-デアザ-8-アザ-グアノシン、7-メチル-グアノシン、6-チオ-7-メチル-グアノシン、7-メチルイノシン、6-メトキシ-グアノシン、1-メチルグアノシン、N2-メチルグアノシン、N2,N2-ジメチルグアノシン、8-オキソ-グアノシン、7-メチル-8-オキソ-グアノシン、1-メチル-6-チオグアノシン、N2-メチル-6-チオグアノシン、及びN2,N2-ジメチル-6-チオグアノシン、並びにそれらの組み合わせを包含するが、これらに限定されない。用語修飾は、例えば、本発明のmRNA配列上への非ヌクレオチド連結部又は修飾ヌクレオチドの組み込みをもまた包含する(例として、機能的なタンパク質又は酵素をコードするmRNA分子の3'及び5'端の1つ又は両方の修飾)。かかる修飾は、mRNA配列への塩基の追加(例として、ポリAテール又はより長いポリAテールの包含)、3'UTR又は5'UTRの変改、mRNAを薬剤(例として、タンパク質又は相補的な核酸分子)と複合体化すること、mRNA分子の構造を変化させるエレメント(例として、これは二次構造を形成する)の包含を包含する。
いくつかの態様では、PEをコードするmRNAは5'キャップ構造を包含する。5'キャップは典型的には次の通り追加される:第1に、RNA末端ホスファターゼが末端リン酸基の1つを5'ヌクレオチドから除去し、2つの末端リン酸を残す;次いで、グアノシン三リン酸(GTP)がグアニリルトランスフェラーゼによって末端リン酸に追加され、5'5'5三リン酸連結部を生ずる;次いで、グアニンの7-窒素がメチルトランスフェラーゼによってメチル化される。キャップ構造の例は、m7G(5')ppp(5'(A、G(5')ppp(5')A、及びG(5')ppp(5')Gを包含するが、これらに限定されない。天然に存在するキャップ構造は、三リン酸ブリッジを介して最初に転写されるヌクレオチドの5'端に連結されている7-メチルグアノシンを含み、m7G(5')ppp(5')Nのジヌクレオチドキャップをもたらし、Nはいずれかのヌクレオシドである。in vivoでは、キャップは酵素的に追加される。キャップは核内で追加され、酵素グアニリルトランスフェラーゼによって触媒される。RNAの5'末端の端へのキャップの追加は転写の開始後に直ちに生起する。末端ヌクレオシドは典型的にはグアノシンであり、全ての他のヌクレオチドとは逆向き、すなわちG(5')ppp(5')GpNpNpである。
追加のキャップアナログは、m7GpppG、m7GpppA、m7GpppC;非メチル化キャップアナログ(例として、GpppG);ジメチル化キャップアナログ(例として、m2,7GpppG)、トリメチル化キャップアナログ(例として、m2,2,7GpppG)、ジメチル化対称キャップアナログ(例として、m7Gpppm7G)、又はアンチリバースキャップアナログ(例として、ARCA;m7,2'OmeGpppG、m72'dGpppG、m7,3'OmeGpppG、m7,3'dGpppG、及びそれらの四リン酸誘導体)からなる群から選択される化学構造を包含するが、これらに限定されない(例として、Jemielity,J.et al.,「Novel'anti-reverse'cap analogs with superior translational properties」,RNA,9:1108-1122(2003)を見よ)。
典型的には、「テール」の存在はエキソヌクレアーゼ分解からmRNAを防護するための用をなす。ポリA又はポリUテールは天然のメッセンジャー及び合成センスRNAを安定化すると考えられる。よって、ある態様において、長いポリA又はポリUテールがmRNA分子に追加され得、それゆえにRNAをより安定にする。ポリA又はポリUテールは種々の当該技術分野で認識される技術を用いて追加され得る。例えば、長いポリAテールは、ポリAポリメラーゼを用いて合成の又はin vitro転写されたRNAに追加され得る(Yokoe,et al.Nature Biotechnology.1996;14:1252-1256)。転写ベクターもまた長いポリAテールをコードし得る。加えて、ポリAテールは直接的にPCR産物からの転写によって追加され得る。ポリAはRNAリガーゼによってもまたセンスRNAの3'端にライゲーションされてもよい(例として、Sambrook、Fritsch及びManiatisによるMolecular Cloning A Laboratory Manual,2nd Ed.,ed.(Cold Spring Harbor Laboratory Press:1991 edition)を見よ)。
典型的には、ポリA又はポリUテールの長さは少なくとも約10、50、100、200、300、400、少なくとも500ヌクレオチドであり得る。いくつかの態様では、mRNAの3'末端のポリAテールは典型的には約10~300アデノシンヌクレオチドを包含する(例として、約10~200アデノシンヌクレオチド、約10~150アデノシンヌクレオチド、約10~100アデノシンヌクレオチド、約20~70アデノシンヌクレオチド、又は約20~60アデノシンヌクレオチド)。いくつかの態様では、mRNAは3'ポリ(C)テール構造を包含する。mRNAの3'末端の好適なポリCテールは典型的には約10~200シトシンヌクレオチドを包含する(例として、約10~150シトシンヌクレオチド、約10~100シトシンヌクレオチド、約20~70シトシンヌクレオチド、約20~60シトシンヌクレオチド、又は約10~40シトシンヌクレオチド)。ポリCテールはポリA又はポリUテールに追加されてもよいか、あるいはポリA又はポリUテールを置換してもよい。
本開示に従うPEをコードするmRNAは種々の公知の方法のいずれかに従って合成されてもよい。例えば、本発明に従うmRNAはin vitro転写(IVT)によって合成されてもよい。手短かには、IVTは、典型的には、プロモータを含有する直線状又は環状DNA鋳型、リボヌクレオチド三リン酸のプール、DTT及びマグネシウムイオンを包含してもよい緩衝液系、並びに適当なRNAポリメラーゼ(例として、T3、T7、又はSP6 RNAポリメラーゼ)、DNAse I、ピロホスファターゼ、及び/又はRNAse阻害剤によって行われる。厳密な条件は特定の適用に従って変わるであろう。
mRNA送達を伴う態様では、プライム編集因子をコードするmRNA対pegRNAの比は効率的な編集にとって重要であってもよい。ある態様において、mRNA(プライム編集因子をコードする)対pegRNAの重量比は1:1である。ある種の他の態様において、mRNA(プライム編集因子をコードする)対pegRNAの重量比は2:1である。なお他の態様において、mRNA(プライム編集因子をコードする)対pegRNAの重量比は1:2である。なお更なる態様では、mRNA(プライム編集因子をコードする)対pegRNAの重量比は、約1:1000、1:900;1:800;1:700;1:600;1:500;1:400;1:300;1:200;1:100;1:90;1:80;1:70;1:60;1:50;1:40;1:30;1:20;1:10;及び1:1からなる群から選択される。他の態様において、mRNA(プライム編集因子をコードする)対pegRNAの重量比は、約1:1000、1:900;800:1;700:1;600:1;500:1;400:1;300:1;200:1;100:1;90:1;80:1;70:1;60:1;50:1;40:1;30:1;20:1;10:1;及び1:1からなる群から選択される。
[9]処置方法
本開示は、例示される通り本明細書において提供されるプライム編集システムによって修正され得る点変異又は他の変異(例として、欠失、挿入、逆位、重複等)に関連するか又はそれによって引き起こされるが、プリオン病、トリヌクレオチドリピート伸長病、又はCDKL5欠損症(CDD)(例として、本明細書の例6)に限定されない疾患と診断された対象の処置のための方法を提供する。
本開示は、例示される通り本明細書において提供されるプライム編集システムによって修正され得る点変異又は他の変異(例として、欠失、挿入、逆位、重複等)に関連するか又はそれによって引き起こされるが、プリオン病、トリヌクレオチドリピート伸長病、又はCDKL5欠損症(CDD)(例として、本明細書の例6)に限定されない疾患と診断された対象の処置のための方法を提供する。
実質的にいずれかの疾患を引き起こす遺伝子欠陥は、プライム編集を用いることによって修復されてもよい。これは、適当なプライム編集因子(napDNAbp及びポリメラーゼ(例として逆転写酵素を包含する)の選択と適当なpegRNAのデザインとを包含し、(a)編集部位を含有する適当な標的DNAを標的化及び(b)ニック部位の直ちに下流の内生鎖を押し除けかつ置き換える所望の編集を包含するニック部位の3'端からのDNAの一本鎖の合成のための鋳型を提供するようにデザインされる。プライム編集は、限定なしに、(a)変異を修正する変化をヌクレオチド配列に組み入れる、(b)タンパク質及びRNAタグを組み入れる、(c)免疫エピトープを目当てのタンパク質に組み入れる、(d)誘導性の二量体化ドメインをタンパク質に組み入れる、(e)バイオ分子の活性を変改するための配列を組み入れ又は除去する、(f)特定の遺伝学的変化を導くためのリコンビナーゼ標的部位を組み入れる、及び(g)エラーを起こしやすいRTを用いることによって標的配列に変異導入するために用いられ得る。
障害を処置する方法には、早期の工程として、本明細書に記載の方法に従う適当なpegRNA及びプライム編集因子のデザインを伴い得、これらは、考慮に入れられてもよいいくつもの考慮事項を包含する。例えば:
(a)1以上の核酸塩基修飾がプライム編集因子によって組み入れされることを望まれる標的配列、すなわちヌクレオチド配列;
(b)標的配列上のカットされる部位の位置付け、すなわち、プライム編集因子が一本鎖ニックを誘導して、ニックの1つの側に3'端RTプライマー配列及びニックの他の側に5'端の内生フラップ(これは究極的にはFEN1又はその等価物によって除去され、3'ssDNAフラップによって置き換えられる)を作成するであろう特定の核酸塩基位置。カットされた部位は3'端プライマー配列を作成し、これはRNA依存的なDNA重合の間にプライム編集因子(例としてRT酵素)のポリメラーゼによって伸長されるようになって、所望の編集を含有する3'ssDNAフラップを作成し、これが次いで標的配列上の5'内生DNAフラップを置き換える;
(c)利用可能なPAM配列(標準的なSpCas9 PAM部位、及び拡張された又は異なるPAM特異性を有するCas9バリアント及び等価物によって認識される非標準的なPAM部位を包含する);
(d)PAM鎖上の利用可能なPAM配列とカットされる部位の位置付けとの間の間隔;
(e)用いられるべき利用可能なプライム編集因子の具体的なCas9、Cas9バリアント、又はCas9等価物(これは部分的には利用可能なPAMによって記述される);
(f)プライマー結合部位の配列及び長さ;
(g)編集鋳型の配列及び長さ;
(h)相同アームの配列及び長さ;
(i)スペーサ配列及び長さ;並びに
(j)gRNAコア配列。
(a)1以上の核酸塩基修飾がプライム編集因子によって組み入れされることを望まれる標的配列、すなわちヌクレオチド配列;
(b)標的配列上のカットされる部位の位置付け、すなわち、プライム編集因子が一本鎖ニックを誘導して、ニックの1つの側に3'端RTプライマー配列及びニックの他の側に5'端の内生フラップ(これは究極的にはFEN1又はその等価物によって除去され、3'ssDNAフラップによって置き換えられる)を作成するであろう特定の核酸塩基位置。カットされた部位は3'端プライマー配列を作成し、これはRNA依存的なDNA重合の間にプライム編集因子(例としてRT酵素)のポリメラーゼによって伸長されるようになって、所望の編集を含有する3'ssDNAフラップを作成し、これが次いで標的配列上の5'内生DNAフラップを置き換える;
(c)利用可能なPAM配列(標準的なSpCas9 PAM部位、及び拡張された又は異なるPAM特異性を有するCas9バリアント及び等価物によって認識される非標準的なPAM部位を包含する);
(d)PAM鎖上の利用可能なPAM配列とカットされる部位の位置付けとの間の間隔;
(e)用いられるべき利用可能なプライム編集因子の具体的なCas9、Cas9バリアント、又はCas9等価物(これは部分的には利用可能なPAMによって記述される);
(f)プライマー結合部位の配列及び長さ;
(g)編集鋳型の配列及び長さ;
(h)相同アームの配列及び長さ;
(i)スペーサ配列及び長さ;並びに
(j)gRNAコア配列。
好適なpegRNAと任意選択で第2部位ニッキングのためのニッキングsgRNAデザインガイドとが、説明書の次の例示の工程バイ工程のセットによってデザインされ得る。これは上の考慮事項の1以上を考慮に入れる。工程は図70A-図70Iに示されている例を参照する。
1.標的配列及び編集を定める。所望の編集(点変異、挿入、欠失、又はそれらの組み合わせ)の位置付けを中心とした標的DNA領域(~200bp)の配列をリトリーブする。図70Aを見よ。
2.標的PAMを位置付ける。所望の編集位置付けに対して近位のPAMを同定する。PAMは所望の編集位置付けに対して近位のDNAのどちらかの鎖上に同定され得る。編集位置に近接するPAMが好ましいが(すなわち、ニック部位は編集位置から30nt未満である。又は編集位置からニック部位まで29nt、28nt、27nt、26nt、25nt、24nt、23nt、22nt、21nt、20nt、19nt、18nt、17nt、16nt、15nt、14nt、13nt、12nt、11nt、10nt、9nt、8nt、7nt、6nt、5nt、4nt、3nt、若しくは2nt未満)、編集位置から≧30ntのニックを置くプロトスペーサ及びPAMを用いて編集を組み入れることが可能である。図70Bを見よ。
3.ニック部位を位置付ける。考慮されている各PAMについて、対応するニック部位とどの鎖上かとを同定する。Sp Cas9 H840Aニッカーゼでは、切断はNGG PAMの5'の第3及び第4の塩基の間においてPAM含有鎖に生起する。全ての編集されるヌクレオチドはニック部位の3'に存在しなければならない。そのため、適当なPAMはPAM含有鎖上の標的の編集の5'にニックを置かなければならない。下で示されている例では、2つの可能なPAMがある。単純のために、残りの工程はPAM1のみを用いるpegRNAのデザインを実証する。図70Cを見よ。
4.スペーサ配列をデザインする。SpCas9のプロトスペーサは、PAM含有鎖上のNGG PAMの5'の20ヌクレオチドに対応する。効率的なPol III転写開始はGが第1の転写されるヌクレオチドであることを要求する。プロトスペーサの第1のヌクレオチドがGである場合には、pegRNAのスペーサ配列は単純にプロトスペーサ配列である。プロトスペーサの第1のヌクレオチドがGではない場合には、pegRNAのスペーサ配列は、G、次にプロトスペーサ配列である。図70Dを見よ。
5.プライマー結合部位(PBS)をデザインする。出発アレル配列を用いて、PAM含有鎖上のDNAプライマーを同定する。DNAプライマーの3'端はニック部位のちょうど上流のヌクレオチドである(すなわち、Sp Cas9ではNGG PAMの5'の第4の塩基)。PE2及びPE3への使用のための一般的なデザイン原理としては、DNAプライマーに対する相補性の12~13ヌクレオチドを含有するpegRNAプライマー結合部位(PBS)が、~40-60%のGC含量を含有する配列に用いられ得る。低いGC含量を有する配列では、より長い(14~15nt)PBSが試験されるべきである。より高いGC含量を有する配列では、より短い(8~11nt)PBSが試験されるべきである。GC含量にかかわらず、最適なPBS配列は経験的に決定されるはずである。長さpのPBS配列をデザインするためには、出発アレル配列を用いて、PAM含有鎖上のニック部位の5'の最初のpヌクレオチドの逆相補体を取る。図70Eを見よ。
6.RT鋳型(又はDNA合成鋳型)をデザインする。RT鋳型(又は、ポリメラーゼが逆転写酵素ではないところでは、DNA合成鋳型)は、デザインされた編集と編集に隣接する配列に対する相同性とをコードする。1つの態様では、これらの領域は図3D及び図3EのDNA合成鋳型に対応し、DNA合成鋳型は「編集鋳型」及び「相同アーム」を含む。最適なRT鋳型長さは標的部位に基づいて変わる。ショートレンジ編集(位置+1~+6)では、短い(9~12nt)、中程度の(13~16nt)、及び長い(17~20nt)RT鋳型を試験することが推奨される。ロングレンジ編集(位置+7以降)では、十分な3'DNAフラップ相同性を許すように、編集の位置から少なくとも5nt(好ましくは10nt以上)伸長するRT鋳型を用いることが推奨される。ロングレンジ編集では、機能的なデザインを同定するためには、いくつかのRT鋳型がスクリーニングされるべきである。より大きい挿入及び欠失(≧5nt)では、RT鋳型上へのより多大な3'相同性(~20nt以上)の組み込みが推奨される。RT鋳型が、逆転写されるDNA産物上の最後のヌクレオチドとしてG(pegRNAのRT鋳型ではCに対応する)の合成をコードするときには、編集効率は典型的には損なわれる。多くのRT鋳型が効率的なプライム編集を支持するので、RT鋳型をデザインするときには、最後の合成されるヌクレオチドとしてのGの回避が推奨される。長さrのRT鋳型配列をデザインするためには、所望のアレル配列を用い、元々はPAMを含有した鎖上のニック部位の3'の最初のrヌクレオチドの逆相補体を取る。SNP編集と比較して、同じ長さのRT鋳型を用いる挿入又は欠失編集は同一の相同性を含有しないであろうということに注意。図70Fを見よ。
7.完全なpegRNA配列をアセンブリする。pegRNA構成要素を次の順序でコンカテネーションする(5'→3'):スペーサ、骨格、RT鋳型、及びPBS。図70Gを見よ。
8.PE3のためのニッキングsgRNAをデザインする。編集の上流及び下流の非編集鎖上のPAMを同定する。最適なニッキング位置は高度に遺伝子座依存性であり、経験的に決定されるはずである。一般的に、pegRNAによって誘導されるニックの向かいの位置の40~90ヌクレオチド5'に置かれたニックは、より高い編集収量及びより少数のインデルに至る。ニッキングsgRNAは、出発アレル上の20ntプロトスペーサにマッチするスペーサ配列を有する。プロトスペーサがGで始まらない場合には5'Gの追加を有する。図70Hを見よ。
9.PE3bニッキングsgRNAをデザインする。PAMが相補鎖上に存在し、かつその対応するプロトスペーサが編集のために標的化される配列とオーバーラップする場合には、この編集はPE3bシステムの候補であり得る。PE3bシステムでは、ニッキングsgRNAのスペーサ配列は出発アレルではなく所望の編集されるアレルの配列にマッチする。編集されるヌクレオチド(単数又は複数)がニッキングsgRNAプロトスペーサのシード領域(PAMに隣接する~10nt)内に収まるときには、PE3bシステムは効率的に作動する。これは、編集された鎖の組み入れ後まで相補鎖のニッキングを防止し、標的DNAに結合することについてのpegRNA及びsgRNAの間の競合を防止する。PE3bは、両鎖上の同時のニックの生成をもまた回避し、それゆえに、高い編集効率を維持しながらインデル形成を有意に縮減する。PE3b sgRNAは、所望のアレルの20ntプロトスペーサにマッチするスペーサ配列を有するはずである。必要とされる場合には5'Gの追加を有する。図70Iを見よ。
1.標的配列及び編集を定める。所望の編集(点変異、挿入、欠失、又はそれらの組み合わせ)の位置付けを中心とした標的DNA領域(~200bp)の配列をリトリーブする。図70Aを見よ。
2.標的PAMを位置付ける。所望の編集位置付けに対して近位のPAMを同定する。PAMは所望の編集位置付けに対して近位のDNAのどちらかの鎖上に同定され得る。編集位置に近接するPAMが好ましいが(すなわち、ニック部位は編集位置から30nt未満である。又は編集位置からニック部位まで29nt、28nt、27nt、26nt、25nt、24nt、23nt、22nt、21nt、20nt、19nt、18nt、17nt、16nt、15nt、14nt、13nt、12nt、11nt、10nt、9nt、8nt、7nt、6nt、5nt、4nt、3nt、若しくは2nt未満)、編集位置から≧30ntのニックを置くプロトスペーサ及びPAMを用いて編集を組み入れることが可能である。図70Bを見よ。
3.ニック部位を位置付ける。考慮されている各PAMについて、対応するニック部位とどの鎖上かとを同定する。Sp Cas9 H840Aニッカーゼでは、切断はNGG PAMの5'の第3及び第4の塩基の間においてPAM含有鎖に生起する。全ての編集されるヌクレオチドはニック部位の3'に存在しなければならない。そのため、適当なPAMはPAM含有鎖上の標的の編集の5'にニックを置かなければならない。下で示されている例では、2つの可能なPAMがある。単純のために、残りの工程はPAM1のみを用いるpegRNAのデザインを実証する。図70Cを見よ。
4.スペーサ配列をデザインする。SpCas9のプロトスペーサは、PAM含有鎖上のNGG PAMの5'の20ヌクレオチドに対応する。効率的なPol III転写開始はGが第1の転写されるヌクレオチドであることを要求する。プロトスペーサの第1のヌクレオチドがGである場合には、pegRNAのスペーサ配列は単純にプロトスペーサ配列である。プロトスペーサの第1のヌクレオチドがGではない場合には、pegRNAのスペーサ配列は、G、次にプロトスペーサ配列である。図70Dを見よ。
5.プライマー結合部位(PBS)をデザインする。出発アレル配列を用いて、PAM含有鎖上のDNAプライマーを同定する。DNAプライマーの3'端はニック部位のちょうど上流のヌクレオチドである(すなわち、Sp Cas9ではNGG PAMの5'の第4の塩基)。PE2及びPE3への使用のための一般的なデザイン原理としては、DNAプライマーに対する相補性の12~13ヌクレオチドを含有するpegRNAプライマー結合部位(PBS)が、~40-60%のGC含量を含有する配列に用いられ得る。低いGC含量を有する配列では、より長い(14~15nt)PBSが試験されるべきである。より高いGC含量を有する配列では、より短い(8~11nt)PBSが試験されるべきである。GC含量にかかわらず、最適なPBS配列は経験的に決定されるはずである。長さpのPBS配列をデザインするためには、出発アレル配列を用いて、PAM含有鎖上のニック部位の5'の最初のpヌクレオチドの逆相補体を取る。図70Eを見よ。
6.RT鋳型(又はDNA合成鋳型)をデザインする。RT鋳型(又は、ポリメラーゼが逆転写酵素ではないところでは、DNA合成鋳型)は、デザインされた編集と編集に隣接する配列に対する相同性とをコードする。1つの態様では、これらの領域は図3D及び図3EのDNA合成鋳型に対応し、DNA合成鋳型は「編集鋳型」及び「相同アーム」を含む。最適なRT鋳型長さは標的部位に基づいて変わる。ショートレンジ編集(位置+1~+6)では、短い(9~12nt)、中程度の(13~16nt)、及び長い(17~20nt)RT鋳型を試験することが推奨される。ロングレンジ編集(位置+7以降)では、十分な3'DNAフラップ相同性を許すように、編集の位置から少なくとも5nt(好ましくは10nt以上)伸長するRT鋳型を用いることが推奨される。ロングレンジ編集では、機能的なデザインを同定するためには、いくつかのRT鋳型がスクリーニングされるべきである。より大きい挿入及び欠失(≧5nt)では、RT鋳型上へのより多大な3'相同性(~20nt以上)の組み込みが推奨される。RT鋳型が、逆転写されるDNA産物上の最後のヌクレオチドとしてG(pegRNAのRT鋳型ではCに対応する)の合成をコードするときには、編集効率は典型的には損なわれる。多くのRT鋳型が効率的なプライム編集を支持するので、RT鋳型をデザインするときには、最後の合成されるヌクレオチドとしてのGの回避が推奨される。長さrのRT鋳型配列をデザインするためには、所望のアレル配列を用い、元々はPAMを含有した鎖上のニック部位の3'の最初のrヌクレオチドの逆相補体を取る。SNP編集と比較して、同じ長さのRT鋳型を用いる挿入又は欠失編集は同一の相同性を含有しないであろうということに注意。図70Fを見よ。
7.完全なpegRNA配列をアセンブリする。pegRNA構成要素を次の順序でコンカテネーションする(5'→3'):スペーサ、骨格、RT鋳型、及びPBS。図70Gを見よ。
8.PE3のためのニッキングsgRNAをデザインする。編集の上流及び下流の非編集鎖上のPAMを同定する。最適なニッキング位置は高度に遺伝子座依存性であり、経験的に決定されるはずである。一般的に、pegRNAによって誘導されるニックの向かいの位置の40~90ヌクレオチド5'に置かれたニックは、より高い編集収量及びより少数のインデルに至る。ニッキングsgRNAは、出発アレル上の20ntプロトスペーサにマッチするスペーサ配列を有する。プロトスペーサがGで始まらない場合には5'Gの追加を有する。図70Hを見よ。
9.PE3bニッキングsgRNAをデザインする。PAMが相補鎖上に存在し、かつその対応するプロトスペーサが編集のために標的化される配列とオーバーラップする場合には、この編集はPE3bシステムの候補であり得る。PE3bシステムでは、ニッキングsgRNAのスペーサ配列は出発アレルではなく所望の編集されるアレルの配列にマッチする。編集されるヌクレオチド(単数又は複数)がニッキングsgRNAプロトスペーサのシード領域(PAMに隣接する~10nt)内に収まるときには、PE3bシステムは効率的に作動する。これは、編集された鎖の組み入れ後まで相補鎖のニッキングを防止し、標的DNAに結合することについてのpegRNA及びsgRNAの間の競合を防止する。PE3bは、両鎖上の同時のニックの生成をもまた回避し、それゆえに、高い編集効率を維持しながらインデル形成を有意に縮減する。PE3b sgRNAは、所望のアレルの20ntプロトスペーサにマッチするスペーサ配列を有するはずである。必要とされる場合には5'Gの追加を有する。図70Iを見よ。
好適なpegRNA及び第2部位ニッキングsgRNAをデザインするための上の工程バイ工程のプロセスは、決して限定することを意味されない。本開示は上に記載されている工程バイ工程のプロセスのバリエーションを企図し、これらは当業者によって次いで導出可能であろう。
ひとたび好適なpegRNA及びプライム編集因子が選択/デザインされると、それらは、好適な方法論によって、例えば、ベクターに基づくトランスフェクション(これにおいては、ベクターによるトランスフェクションによって細胞内で発現される、pegRNA及びプライム編集因子をコードするDNAを含む1以上のベクター)、送達フォーマット(例として、脂質粒子、ナノ粒子)でpegRNAと複合体化したプライム編集因子の直接的送達(例としてRNP送達)によって、又はmRNAに基づく送達システムによって、投与されてもよい。かかる方法は本明細書において本開示によって記載され、いずれかの公知の方法が利用されてもよい。
目当ての標的DNAに接触することによって、所望の編集がそれに組み入れされるようになるようにして、pegRNA及びプライム編集因子(又は一緒にPE複合体と呼ばれる)は処置学的有効量で細胞に送達され得る。
適当な細胞への送達が実現可能である限りは、いずれかの疾患は、着想上はかかる方法によって処置可能である。当業者は、意図される目的及び意図される標的細胞に適するようにPE送達方法論を選ぶ及び/又はセレクションする能力があるであろう。
例えば、いくつかの態様において、かかる疾患(例として、上に記載の通りの点突然変異に関連するがん)を有する対象へ、有効量の、本明細書に記載のプライム編集システム(所望の遺伝子変化を含むドナーDNA分子の存在下での相同特異的(homology-directed)修復によって媒介される通りに、点突然変異を修正するか又は疾患関連遺伝子中へ不活性化突然変異を導入する)を投与することを含む方法が提供される。いくつかの態様において、かかる疾患(例として、上に記載の通りの点突然変異に関連するがん)を有する対象へ、有効量の、本明細書に記載のプライム編集システム(点突然変異を修正するか又は疾患関連遺伝子中へ不活性化突然変異を導入する)を投与することを含む方法が提供される。いくつかの態様において、疾患は、増殖性疾患である。いくつかの態様において、疾患は、遺伝的疾患である。いくつかの態様において、疾患は、新生物疾患である。いくつかの態様において、疾患は、代謝性疾患である。いくつかの態様において、疾患は、リソソーム蓄積症である。点突然変異を修正するか又は疾患関連遺伝子中へ不活性化突然変異を導入することによって処置され得る他の疾患は当業者に知られており、本開示はこの点において限定されない。
別の側面では、ガイドRNA(pegRNA)と組み合わせてプライム編集因子(例として、PE1、PE2、又はPE3)を使用してプライム編集を実行して、CDKL5欠損症を引き起こすCDKL5遺伝子の突然変異を直接組み入れするか、又は修正する方法が提供される。様々な態様では、本開示は、プライム編集因子(例として、PE1、PE2、又はPE3)と、複数の対象においてCDKL5遺伝子に2つ以上の突然変異を直接組み入れするか、又は修正することができるpegRNAと、を含む複合体を提供する。
本開示は、追加の疾患又は障害(例として、プライム編集によって修正され得る点突然変異に関連されるか若しくはこれによって引き起こされる疾患又は障害)の処置のための方法を提供する。いくつかのかかる疾患は本明細書に記載されており、本明細書に提供されるストラテジー及び融合タンパク質で処置され得る追加の好適な疾患は本開示に基づき当業者に明らかであろう。例示の好適な疾患及び障害は下に列挙される。夫々の配列における特定の位置又は残基のナンバリングが、使用される具体的なタンパク質及びナンバリングスキームに依存することは、理解されるであろう。ナンバリングは、例として、成熟したタンパク質の前駆体及び成熟したタンパク質それ自体において異なることもあり、種ごと(from species to species)の配列の差異はナンバリングに影響を及ぼしてもよい。当業者は、当該技術分野において周知である方法によって(例として、配列の整列及び相同の残基の決定によって)、いずれの相同のタンパク質及び夫々のコード核酸における夫々の残基を同定することができるであろう。例示の好適な疾患及び障害は、限定せずに以下を包含する:2-メチル-3-ヒドロキシ酪酸尿症;3ベータ-ヒドロキシステロイドデヒドロゲナーゼ欠損症;3-メチルグルタコン酸尿症;3-オキソ-5アルファ-ステロイドデルタ4-デヒドロゲナーゼ欠損症;46,XY性トランスバーション、1型,3型及び5型;5-オキソプロリナーゼ欠損症;6-ピルボイル-テトラヒドロプテリンシンターゼ欠損症;アースコグ症候群;アーセ症候群;2型軟骨無発生症;色覚異常2及び7;後天性QT延長症候群;シンゲル(Schinzel)型先端脳梁症候群;先端大腿骨頭異形成症;ホルモン耐性の有無に関わらない先端骨形成不全症2;先端紅斑角皮症(Acroerythrokeratoderma);先端短肢異形成症;Acth-非依存性の副腎皮質大結節性過形成2;活性化PI3K-delta症候群;急性間欠性ポルフィリン症;アシル-CoAデヒドロゲナーゼファミリー、メンバー9の欠損症;アダムス-オリバー症候群5及び6;アデニンホスホリボシルトランスフェラーゼ欠損症;アデニル酸キナーゼ欠損症;アデニロコハク酸リアーゼ欠損症に起因する溶血性貧血;青年期のネフロン癆;腎・肝・膵異形成症;7型メッケル症候群;副腎白質ジストロフィー;成人接合部型表皮水疱症;表皮水疱症、接合部型、限局性バリアント;成人神経セロイドリポフスチン症;成人神経セロイドリポフスチン症;成人発症の、眼球運動失効を伴う運動失調症;ADULT症候群;無フィブリノーゲン血症及び先天性無フィブリノーゲン血症;常染色体潜性無ガンマグロブリン血症2;加齢性黄斑変性症3,6,11及び12;アイカルディ・グティエール症候群1,4及び5;凍傷状狼瘡1;アラジール症候群1及び2;アレキサンダー病;アルカプトン尿症;アラン・ハーンドン・ダドリー症候群;先天性の汎発性脱毛症;アルパース脳症;アルファ-1-抗トリプシン欠損症;常染色体優性の、常染色体劣性の、及びX連鎖劣性のアルポート症候群;アルツハイマー病、家族性、3、痙性対麻痺及び運動失行あり;アルツハイマー病、1型,3型及び4型;低石灰化型及び低成熟型、IIA1遺伝性エナメル質形成不全症;アミノアシラーゼ1欠損症;アーミッシュ小児てんかん症候群;アミロイド形成性トランスサイレチンアミロイドーシス;トランスサイレチンに関する、アミロイド心筋症;心筋症;筋萎縮性側索硬化症1型,6型,15型(前頭側頭型認知症の有無に関わらない),22型(前頭側頭型認知症の有無に関わらない),及び10型;TARDBPに関する、TDP43封入体を伴う前頭側頭型認知症;アンダーマン(Andermann)症候群;アンダーセン・タウィル症候群;先天性QT延長症候群;G6PD欠損症に起因する非球形溶血性の貧血症;アンジェルマン(Angelman)症候群;小頭症を伴う、新生児発症の重症脳症;自閉症、X連鎖性3に対する感受性;腎症、動脈瘤、及び筋痙攣を伴う遺伝性の血管症;アンジオテンシンi変換酵素、穏やかな血清増加;無虹彩症、小脳性運動失調症、及び精神遅滞;無爪症;アンチトロンビンIII欠損症;性器奇形及びステロイド産生異常を伴うアントレー・ビクスラー症候群;家族性胸部大動脈瘤4,6及び9;胸部大動脈瘤及び大動脈解離;多臓器平滑筋機能障害症候群(multisystemic smooth muscle dysfunction syndrome);もやもや病5;再生不良性貧血;見かけの鉱質コルチコイド過剰;アルギナーゼ欠損症;アルギニノコハク酸リアーゼ欠損症;アロマターゼ欠損症;5型,8型及び10型不整脈原性右室心筋症;原発性家族性肥大型心筋症;先天性多発性関節拘縮症、末梢の、X連鎖性;関節拘縮-腎機能障害-胆汁うっ滞症候群;関節拘縮、腎機能障害、及び胆汁うっ滞2;アスパラギン合成酵素欠損症;神経細胞移動の異常性;ビタミンE欠損症を伴う運動失調症;感覚性の、常染色体優性の、運動失調症;運動失調症-毛細血管拡張症候群;遺伝性がん素因症候群(hereditary cancer-predisposing syndrome);無トランスフェリン血症;家族性の心房細動11,12,13及び16;心房中隔欠損症2,4及び7(心室伝導欠陥の有無に関わらない);心房停止2;房室中隔欠損症4;遺伝性眼球萎縮;ATR-X症候群;耳介下顎骨症候群2;自己免疫疾患、多臓器、小児発症;1a型自己免疫リンパ増殖性症候群;常染色体優性の無汗性外胚葉形成不全症;ミトコンドリアDNA欠失1及び3を伴う、常染色体優性の進行性外眼筋麻痺;常染色体優性の捻転ジストニア4;常染色体劣性の中心核ミオパチー;常染色体劣性の先天性魚鱗癬1,2,3,4A及び4B;常染色体劣性のIA型及び1B型皮膚弛緩症;常染色体劣性の無汗性外胚葉形成不全症候群;外胚葉異形成症11b;発汗低下/毛髪/歯型、常染色体劣性;常染色体劣性低リン酸血症性の骨疾患;3型アクセンフェルト・リーガー症候群;ベインブリッジ・ロパーズ症候群;バナヤン・ライリー・ルバルカ症候群;PTEN過誤腫症候群;バライスター(Baraitser)・ウインター症候群1及び2;バラカート症候群;バルデ・ビードル症候群1,11,16及び19;2型裸リンパ球症候群、相補群E;出生前2型バーター症候群;バーター症候群の3型,低カルシウム尿を伴う3型,及び4型;特発性基底核石灰化症4;連珠毛;良性家族性血尿;良性家族性新生児発作1及び2;発作、良性家族性新生児1、及び/又はミオキミア;発作、早期小児てんかん性脳症7;良性家族性新生児-小児発作;良性遺伝性舞踏病;心筋症を伴う良性肩甲骨骨膜筋ジストロフィー;A1型及びA2型(常染色体優性)ベルナール・スーリエ症候群;ベストロフィノパチー(常染色体劣性);ベータ サラセミア;ベスレムミオパチー及びベスレムミオパチー2;ビエッティ結晶性角膜網膜ジストロフィー;胆汁酸合成欠損症、先天性2;ビオチニダーゼ欠損症;バーク・バレル(Birk Barel)精神遅滞異形症候群;眼裂縮小、眼瞼下垂症、及び眼瞼内反症;ブルーム症候群;ベルエソン・フォルスマン・レーマン症候群;バウチャー・ノイハウザー(Boucher Neuhauser)症候群;A1型及びA2型乳頭症;高血圧症を伴う乳頭症;出血を伴う脳小血管疾患;分岐鎖ケト酸デヒドロゲナーゼキナーゼ欠損症;分岐鎖症候群2及び3;乳がん、早期発症;乳房卵巣がん、家族性1,2及び4;角膜脆性症候群2;ブロディミオパチー;汗の塩化物上昇の有無に関わらない気管支拡張症3;ブラウン・ビアレット・ヴァン レアレ症候群及びブラウン・ビアレット・ヴァン レアレ症候群2;ブルガダ症候群;ブルガダ症候群1;心室細動;発作性家族性心室細動;ブルガダ症候群及びブルガダ症候群4;QT延長症候群;心臓性突然死;標的(Bull eye)黄斑ジストロフィー;スターガート病4;錐体桿体ジストロフィー12;水疱性魚鱗癬様紅皮症;バーン・マッキューン(Burn-Mckeown)症候群;家族性カンジダ症2,5,6及び8;I型及びII型糖タンパク質糖鎖不全症候群;高アンモニア血症に起因する炭酸脱水酵素VA欠損症;結腸癌;心筋不整脈;QT延長症候群、LQT1亜型;チトクロームcオキシダーゼ欠損症に起因する致死性乳児心臓脳筋症;心筋症;ダノン病;肥大型心筋症;左心室非圧縮心筋症;カルネヴァーレ症候群;1型カーニー複合体;カルニチンアシルカルニチントランスロカーゼ欠損症;カルニチンパルミトイルトランスフェラーゼI,II,II(遅発性)及びII(小児性)欠損症;白内障1,4、常染色体優性、常染色体優性の多型、微小角膜を伴う、コポック様、若年性、微小角膜と糖尿を伴う、及び核びまん性非進行性;カテコラミン作動性多形心室頻拍;尾状回帰症候群;家族性Cd8欠損症;中心核疾患;1,9及び16番染色体の中心核不安定性並びに免疫不全;進行性外眼筋を伴う小児小脳性運動失調症及び小脳性運動失調症、精神遅滞、並びに平衡障害症候群2;APPに関する脳アミロイド血管症;大脳皮質下梗塞及び白質脳症を伴う、脳の常染色体優性及び劣性の動脈症;脳海綿状奇形2;脳・眼・顔・骨格症候群2;脳-眼-顔-骨格症候群;石灰化及び嚢胞を伴う脳網膜微小血管症;セロイドリポフスチン症神経細胞2,6,7及び10;Ch\xc3\xa9diak・東症候群;成人型チェディアック・東症候群;シャルコー・マリー・トゥース病の1B型,2B2型,2C型,2F型,2I型,2U型(軸索性),1C型(脱髄性),優性中間C型,劣性中間A型,2A2型,4C型,4D型,4H型,IF型,IVF型及びX型;肩甲骨腹側脊髄筋萎縮症;遠位型脊髄性筋萎縮症、先天性非進行性;脊髄筋萎縮症、遠位型、常染色体劣性、5;CHARGE関連;小児低リン酸塩症;成人低リン酸塩症;胆嚢炎;進行性家族性肝内胆汁うっ滞3;妊娠性肝内胆汁うっ滞3;コレスタノール蓄積症;コレステロールモノオキシゲナーゼ(側鎖切断)欠損症;ブロムストランド型軟骨異形成症;穿刺性軟骨異形成症1、X連鎖劣性及び2 X連鎖優性;CHOPS症候群;慢性肉芽腫性疾患、常染色体劣性チトクロームb陽性、1型及び2型;チュドリー・マッカラー症候群;原発性毛様体ジスキネジア、7,11,15,20及び22;I型シトルリン血症;I型及びII型シトルリン血症;頭蓋骨裂傷症;C様症候群;A型コケイン症候群;原発性補酵素Q10欠乏症、1,4及び7;コフィン・シリス/知的障害;コフィン・ローリー症候群;コーエン症候群;寒冷時発汗症候群1;コール・カーペンター症候群2;
肉芽腫を伴う、細胞性及び体液性免疫複合欠損症;d-2-及びl-2-ヒドロキシグルタル酸複合尿症;マロン酸及びメチルマロン酸複合尿症;酸化的リン酸化複合欠損症1,3,4,12,15及び25;一部及び全部の17-アルファ-ヒドロキシラーゼ/17,20-リアーゼ複合欠損症;共通可変性免疫不全症9;c1インヒビターの機能不全に起因する補体構成要素4の部分欠損;補体因子B欠損症;錐体単色症;錐体桿体ジストロフィー2及び6;錐体桿体ジストロフィー遺伝性エナメル質形成不全症;先天性副腎過形成及び先天性副腎低形成、X連鎖性;先天性無巨核球性血小板減少症;先天性無虹彩症;先天性中枢性低換気;ヒルシュスプルング病3;先天性収縮性腕十二指腸症;四肢及び顔面の先天性拘縮、低緊張、及び発育遅延;グリコシル化の先天性障害1B型,1D型,1G型,1H型,1J型,1K型,1N型,1P型,2C型,2J型,2K型,IIm型;I型及びII型先天性赤血球異形成貧血;顔面の先天性外胚葉異形成症;先天性赤血球生成性ポルフィリン症;2型先天性全身性リポジストロフィー;先天性心疾患、多発型2;先天性心疾患;大動脈弓離断症;先天性脂肪腫性過成長、血管奇形、及び表皮母斑;非小細胞肺がん;卵巣の新生物;心伝導系障害、非特異性;先天性微絨毛萎縮症;先天性筋ジストロフィー;LAMA2の部分的欠損に起因する先天性筋ジストロフィー;先天性筋ジストロフィー-脳及び眼の異常を伴うジストログリカノパチー、A2型,A7型,A8型,A11型及びA14型;先天性筋ジストロフィー-精神遅滞を伴うジストログリカノパチー、B2型,B3型,B5型及びB15型;先天性筋ジストロフィー-精神遅滞を伴わないジストログリカノパチー、B5型;先天性筋肥大-大脳症候群;先天性筋無力症候群、アセタゾールアミド反応性;繊維型不均衡を伴う先天性ミオパチー;先天性眼球コロボマ;先天性静止型夜盲症、1A型,1B型,1C型,1E型,1F型及び2A型;コプロポルフィリン症;角膜平板症2;角膜ジストロフィー、フックス内皮性4;2型角膜内皮性ジストロフィー;角膜脆弱性ケラトグロブ、青色強膜、及び関節過度可動性;コーネリア ド ランゲ症候群1及び5;常染色体優性の冠動脈疾患2;冠状動脈疾患;高αリポタンパク質血症2;皮質異形成症(他の脳奇形との複合)5及び6;後頭部の皮質奇形;コルチコステロイド結合グロブリン欠損症;2型コルチコステロンメチルオキシダーゼ欠損症;コステロ症候群;カウデン症候群1;扁平股;常染色体優性の頭蓋骨異形成症;頭蓋縫合早期癒合症1及び4;頭蓋縫合早期癒合症及び歯異常;クレアチン欠損症、X連鎖;クルーゾン症候群;潜在眼球症候群;停留精巣、片側又は両側;クッシング指節癒合症;皮膚悪性黒色腫1;骨異栄養症と、肺、胃腸、及び泌尿器の重度の異常とを伴う皮膚弛緩症;チアノーゼ、一過的な新生児の及び非定型の腎症;嚢胞性線維症;シスチン尿症;シトクロムcオキシダーゼi欠損症;シトクロムcオキシダーゼ欠損症;D-2-ヒドロキシグルタル酸尿症2;分節性ダリエー病;内耳形成不全、小耳症、及び小歯症(LAMM)を伴う難聴;難聴、常染色体優性3a,4,12,13,15、常染色体優性の非症候性感音性17,20及び65;難聴、常染色体劣性1A,2,3,6,8,9,12,15,16,18b,22,28,31,44,49,63,77,86及び89;難聴、蝸牛の、近視及び知的障害あり、前庭障害なし、常染色体優性、X連鎖2;2-メチルブチリル-CoA脱水素酵素の欠損症;3-ヒドロキシアシル-CoA脱水素酵素の欠損症;アルファ-マンノシダーゼの欠損症;芳香族-L-アミノ酸脱炭酸酵素の欠損症;ビスホスホグリセリン酸ムターゼの欠損症;ブチリル-CoA脱水素酵素の欠損症;フェロキシダーゼの欠損症;ガラクトキナーゼの欠損症;グアニジノ酢酸メチルトランスフェラーゼの欠損症;ヒアルロノグルコサミニダーゼの欠損症;リボース-5-リン酸イソメラーゼの欠損症;ステロイド11-ベータ-モノオキシゲナーゼの欠損症;UDPグルコース-ヘキソース-1-リン酸ウリジリルトランスフェラーゼの欠損症;キサンチンオキシダーゼの欠損症;デジェリン・ソッタス病;シャルコー・マリー・トゥース病、ID型及びIVF型;デジェリン・ソッタス症候群、常染色体優性;樹状細胞、単球、Bリンパ球、及びナチュラルキラーリンパ球の欠損症;デビュクワ(Desbuquois)異形成症2;デビュクワ症候群;DFNA 2非症候性の聴力低下;視神経萎縮及び難聴を伴う糖尿病及び尿崩症;2型糖尿病、及びインスリン依存性20;ダイアモンド・ブラックファン貧血1,5,8及び10;下痢3(分泌性ナトリウム、先天性、症候性)、及び5(タフティング腸症(tufting enteropathy)を伴う、先天性);ジカルボキシルアミノ酸尿症;びまん性掌蹠角化症、ボスニア型;デジトレノ脳症候群;ジヒドロプテリジン還元酵素欠損症;拡張型心筋症1A,1AA,1C,1G,1BB,1DD,1FF,1HH,1I,1KK,1N,1S,1Y及び3B;左心室非圧縮3;シトクロムp450酸化還元酵素欠損症に起因するステロイド産生異常;遠位型関節拘縮2B型;遠位型遺伝性運動ニューロン症2B型;遠位型ミオパチーMarkesbery-Griggs型;遠位型脊髄性筋萎縮症、X連鎖3;二重睫毛・リンパ浮腫症候群;皮膚の欠如を伴う、優性栄養傷害性表皮水疱症;優性遺伝性視神経萎縮症;ドンナイ・バロウ症候群;ドーパミン ベータ水酸化酵素欠損症;ドーパミン受容体d2、脳内密度低下;ダウリング・デゴス病4;ドイン蜂巣状網膜ジストロフィー;マラティア・レベンチーヌ(Malattia leventinese);2型デュアン症候群;デュビン・ジョンソン症候群;デュシェンヌ型筋ジストロフィー;ベッカー型筋ジストロフィー;異常フィブリノーゲン血症;常染色体優性の先天性角化異常症及び常染色体優性、3;常染色体劣性の先天性角化異常症1,3,4及び5;X連鎖性の先天性角化異常症;顔面ミオキミアを伴う、家族性ジスキネジア;プラスミノーゲン分子異常症;ジストニア2(捻転、常染色体劣性),3(捻転、X連鎖),5(ドーパ反応型),10,12,16,25,26(ミオクロニー);良性家族性乳児の発作2;早期乳児てんかん性脳症2,4,7,9,10,11,13及び14;非定型レット症候群;初期T細胞前駆体急性リンパ性白血病;外胚葉性異形成表皮水疱症候群;外胚葉性異形成-合指症候群1;偏位水晶体、単独で(isolated)常染色体劣性及び優性;裂手裂足(Ectrodactyly)、外胚葉性異形成、及び口唇口蓋裂症候群3;エーラス・ダンロス症候群7型(常染色体劣性)、古典型、2型(早老性)、ヒドロキシリジン欠損型、4型、4型バリアント、及びテナシンX欠損に起因するもの;アイヒスフェルト型先天性筋ジストロフィー;内分泌-大脳骨異形成症(cerebroosteodysplasia);S錐体増強症候群;拡大前庭水管症候群;エンテロキナーゼ欠損症;疣贅状表皮発育異常症;単純型表皮水疱症及び肢帯筋ジストロフィー、斑状色素沈着(mottled pigmentation)を伴う単純型、幽門閉鎖を伴う単純型、単純型、常染色体劣性、及び幽門閉鎖を伴う;表皮剥離性掌蹠角化症;家族性熱性痙攣8;てんかん、小児欠神2,12(特発性全身、感受性),5(夜間前頭葉型)、夜間前頭葉型1、部分性、可変の病巣があるもの(with variable foci)、進行性ミオクロニー3、及びX連鎖性、可変の学習障害及び行動障害があるもの;てんかん性脳症、小児期発症、乳児期早期、1,19,23,25,30及び32;骨端異形成症、多発性、近視及び伝音難聴を伴う;一過性運動失調症2型;一過性疼痛症候群、家族性、3;エプスタイン症候群;フェクトナー症候群;赤芽球増殖性プロトポルフィリン症;エストロゲン抵抗性;滲出性硝子体網膜症6;ファブリー病、及びファブリー病、心臓変異型;第H因子、第VII因子、第X因子、第v因子、及び第viii因子の、2つの複合欠損症、第xiii因子、サブユニット、欠損症;家族性腺腫性ポリポーシス1及び3;蕁麻疹及び難聴を伴う家族性アミロイド腎症;家族性寒冷蕁麻疹;家族性の小脳虫部無発生;家族性良性天疱瘡;家族性乳がん;乳がん感受性(Breast cancer,susceptibility to);骨肉腫;膵臓がん3;家族性心筋症;家族性寒冷自己炎症性症候群2;家族性大腸がん;家族性滲出性硝子体網膜症、X連鎖性;家族性片麻痺性片頭痛1型及び2型;家族性高コレステロール血症;家族性肥大型心筋症1,2,3,4,7,10,23及び24;家族性低カリウム血症-低マグネシウム血症;家族性低形成の、糸球体嚢胞腎;家族性乳児筋無力症;家族性若年性痛風;家族性地中海熱、及び家族性地中海熱、常染色体優性;家族性多弁症;家族性晩発性皮膚ポルフィリン症;家族性肺毛細血管腫症;家族性腎性糖尿;家族性腎性低尿酸血症;家族性拘束性心筋症1;家族性1型及び3型高リポ蛋白血症;ファンコニー貧血,補欠群E,I,N及びO;ファンコニー・ビッケル症候群;ファビズム感受性(Favism,susceptibility to);熱性痙攣、家族性、11;フェインゴールド症候群1;胎児ヘモグロビン量的形質遺伝子座1;FG症候群及びFG症候群4;眼球外筋の線維症、先天性、1,2,3a(眼球外病変の有無に関わらない),3b;魚眼病;フレック角膜ジストロフィー;フローティング・ハーバー症候群;精神遅滞の有無に関わらない、言語障害を伴う焦点てんかん;巣状分節性糸球体硬化症5;前脳欠損症;フランク・テル・ハール症候群;Borrone Di Rocco Crovato症候群;フレイジャー症候群;ウィルムス腫瘍1;フリーマン・シェルドン症候群;前頭骨幹端異形成症1及び3;前頭側頭型認知症;前頭側頭型認知症及び/又は筋萎縮性側索硬化症3及び4;前頭側頭型認知症第3染色体連鎖性及び前頭側頭型認知症ユビキチン陽性;フルクトース-ビホスファターゼ欠損症;フールマン(Fuhrmann)症候群;ガンマ-アミノ酪酸トランスアミナーゼ欠損症;ガムストープ・ウォールファールト(Gamstorp-Wohlfart)症候群;ゴーシェ病1型及び亜急性神経障害性;進行性の脊椎側弯症を伴う、家族性水平注視麻痺;全般性優性ジストロフィー型表皮水疱症;全般性てんかん、熱性痙攣プラス3,1型,2型を伴う;
レノックス・ガストー型てんかん脳症;巨大軸索ニューロパチー;グランツマン血栓症;緑内障1、開放角、e,F及びG;緑内障3、原発性先天性、d;先天性緑内障、及び先天性緑内障、コロボマ;緑内障、原発性開放角、若年性発症;神経膠腫感受性1;グルコーストランスポーター1型欠損症候群;ルコース-6-リン酸輸送欠損症;GLUT1欠損症候群2;特発性全般性てんかん感受性、12;グルタミン酸ホルミノトランスフェラーゼ欠損症;グルタル酸血症IIA及びIIB;グルタル酸尿症1型;グルタチオン合成酵素欠損症;糖原病0(筋肉),II(成人形),IXa2,IXc,1A型;II型,IV型,IV(肝とミオパチーとの複合),V型及びVI型;ゴールドマン・ファーブル症候群;ゴードン症候群;ゴーリン症候群;全前脳胞症シークエンス;全前脳胞症7;顆粒腫性疾患、慢性、X連鎖性、バリアント;卵巣の顆粒膜細胞腫瘍;灰色血小板症候群;グリセリ症候群3型;グレーノー角膜ジストロフィーI型;成長及び精神遅滞、下顎顔面異形症、小頭症、及び口蓋裂;下垂体異常を伴う成長ホルモン欠損症;免疫不全を伴う成長ホルモン不感受性;GTPシクロハイドロラーゼI欠損症;ハジュ・チーニー症候群;手足子宮症候群;聴覚障害;小児毛細血管腫;血液腫瘍;ヘモクロマトーシス1型,2B型及び3型;糖尿病の微小血管合併症7;トランスフェリン血清レベル量的形質座2;ヘモグロビンH病、無欠損;グルコースリン酸イソメラーゼ欠損症に起因する、溶血性貧血、非球状赤血球性;血球貪食性リンパ組織球症、家族性、2;血球貪食性リンパ組織球症、家族性、3;ヘパリン補因子II欠損症;遺伝性腸性先端皮膚炎;遺伝性乳がん・卵巣がん症候群;運動失調・毛細血管拡張症様障害;遺伝性びまん性胃がん;スフェロイドを伴う遺伝性びまん性白質脳症;遺伝性第II,第IX,第VIII因子欠損症;遺伝性出血性毛細血管拡張症2型;無汗症を伴う遺伝性無痛症;遺伝性リンパ浮腫I型;視神経萎縮を伴う遺伝性運動・感覚神経症;早期呼吸不全を伴う遺伝性ミオパチー;遺伝性神経痛性筋萎縮症;遺伝性非ポリポーシス大腸新生物;リンチ症候群I及びII;遺伝性膵炎;慢性膵炎感受性;遺伝性感覚・自律神経障害IIB型及びIIA型;遺伝性鉄芽球性貧血;ヘルマンスキー・ポドラック症候群1,3,4及び6;内臓錯位、2,4及び6、常染色体;内臓錯位、X連鎖性;組織球性髄様細網症;異所形成;組織球症-リンパ節腫大プラス症候群;ホロカルボキシラーゼ合成酵素欠損症;全前脳胞症2,3,7及び9;ホルト・オラム症候群;MTHFR欠損症、CBS欠損症、及びホモシステイン尿症に起因するホモシステイン血症、ピリドキシン応答性;コバラミン代謝異常に起因する、ホモシステイン尿症-巨赤芽球性貧血、cblE相補型;ハウエル・エバンス(Howel-Evans)症候群;ハーラー症候群;ハッチンソン・ギルフォード症候群;水頭症;高アンモニア血症、III型;高コレステロール血症、及び高コレステロール血症、常染色体劣性;過剰驚愕症2及び遺伝性の過剰驚愕症;高フェリチン血症白内障症候群;高グリシン尿症;周期性発熱を伴う高免疫グロブリンD;メバロン酸尿症;高免疫グロブリンE症候群;家族性の高インスリン血症性低血糖症3,4及び5;高インスリン血症-高アンモニア血症症候群;高リシン血症;ジストニア、赤血球増加症、及び肝硬変を伴う高マンガン血症;高オルニチン血症-高アンモニア血症-ホモシトルリン尿症症候群;副甲状腺機能亢進症1及び2;副甲状腺機能亢進症、新生児重度;部分的pts欠損症、BH4欠損症、D、非pkuに起因する、高フェニルアラニン血症、bh4欠損症、a;精神遅滞症候群2,3及び4を伴う高リン酸血症;多毛性骨軟骨異形成症;apob32に関連する、低ベータリポ蛋白血症、家族性;低カルシウム血症、常染色体優性1;低カルシウム尿性高カルシウム血症、家族性、1型及び3型;低軟骨形成症;鉄過剰症を伴う低クロム小球性貧血;肝臓中グリコーゲン合成酵素の欠損症を伴う低血糖症;臭覚障害の有無に関わらない低ゴナドトロピン性性腺機能低下症11;免疫不全を伴う無汗性外胚葉形成不全症;発汗低下のX連鎖性外胚葉異形成;低カリウム性周期性四肢麻痺1及び2;低マグネシウム血症1、腸の;低マグネシウム血症、発作、及び精神遅滞;低髄性白質ジストロフィー7;左心低形成症候群;房室中隔欠損症及び共通房室接合部;尿道下裂1及び2、X連鎖性;甲状腺機能低下症、先天性非甲状腺腫、1;減毛症8及び12;減毛症-リンパ浮腫-血管拡張症候群;I血液型系;シーメンス型水疱性魚鱗癬;剥脱性魚鱗癬;未熟児魚鱗癬症候群;特発性基底核石灰化症5;特発性線維性肺胞炎、慢性型;先天性角化不全症、常染色体優性、2及び5;乳児期の特発性高カルシウム血症;カルシウム流入障害2に起因するT細胞不活性化を伴う免疫機能障害;1型及び2型高IgMを伴う、cd3-ゼータの欠陥に起因する、免疫不全15,16,19,30,31C,38,40,8、並びにマグネシウム欠陥を伴う、X連鎖性、エプスタイン・バーウイルス感染、及び新形成;免疫不全-動原体不安定性-顔面奇形症候群2;封入体ミオパチー2及び3;野中ミオパチー;乳児痙攣及び発作性舞踏症、家族性;乳児皮質過骨症;乳児GM1ガングリオシドーシス;乳児低ホスファタシアーゼ血症;乳児ネフロン癆;乳児眼振、X連鎖性;乳児パーキンソン病-ジストニア;多尾性精子及び過剰なDNAに関連する不妊症;インスリン抵抗性;インスリン抵抗性糖尿病及び黒色表皮腫;インスリン依存性糖尿病分泌性下痢症候群;間質性腎炎、核巨大化(karyomegalic);子宮内発育遅延、骨幹端異形成症、先天性副腎低形成、及び性器奇形;ヨードチロシン結合欠損症;IRAK4欠損症;虹彩隅角異発生優性型及び1型;脳内鉄蓄積;坐骨膝蓋骨異形成;膵島細胞過形成;17,20-リアーゼ単独欠損症;ルトロピン単独欠損症;イソバレリル-CoAデヒドロゲナーゼ欠損症;ヤンコヴィッチ・リベラ症候群;ジャーベル及びランゲ・ニールセン症候群2;ジュベール症候群1,6,7,9/15(2遺伝子性),14,16及び17、並びに口顔面指症候群xiv;ヘルリッツの接合部表皮水疱症;若年性GM>1<ガングリオシドーシス;若年性ポリポーシス症候群;若年性ポリポーシス/遺伝性出血性毛細血管拡張症症候群;若年性網膜炎;歌舞伎メーキャップ症候群;カルマン症候群1,2及び6;思春期遅延症;神崎病;カラック症候群;カルタゲナー症候群;ケニー・ケイフィ症候群2型;ケッペン・ルビンスキー(Keppen-Lubinsky)症候群;円錐角膜1;毛包性角化症;掌蹠膿疱症1;キンドラー症候群;L-2-ヒドロキシグルタル酸尿症;ラーセン症候群、優性型;格子状角膜ジストロフィーIII型;レーバー黒内障;ツェルウェガー症候群;ペルオキシソーム生合成異常症;ツェルウェガー症候群スペクトラム;レーバー先天性黒内障11,12,13,16,4,7及び9;レーバー視神経萎縮症;アミノグリコシド誘発性難聴;難聴、非症候性感音性、ミトコンドリア;左心室非圧縮症5;左右軸奇形;リー病;ミトコンドリア短鎖エノイル-CoAヒドラターゼ1欠損症;ミトコンドリア複合体I欠損症に起因するリー症候群;ライナー病;レリー・ワイル軟骨骨異形成症;致死性先天性拘縮症候群6;白血球接着不全症I型及びIII型;白骨ジストロフィー、ミエリン形成不全性、11及び6;白質脳症、運動失調を伴う、脳幹・脊髄障害と乳酸値上昇とを伴う、白質の消失を伴う、及び進行性の、卵巣不全を伴う;全白爪;レビー小体型認知症;リヒテンシュタイン・クノール症候群;リー・フラウメニ症候群1;Lig4症候群;肢帯筋ジストロフィー1B型,2A型,2B型,2D型,C1型,C5型,C9型,C14型;脳及び眼の異常を伴う先天性筋ジストロフィー-ジストログリカノパチー、A14型及びB14型;複合型リパーゼ欠損症;脂肪蛋白症;リポジストロフィー、家族性部分的、2型及び3型;脳回欠損1,2(X連鎖性),3,6(小頭症を伴う),X連鎖性;皮質下帯状ヘテロトピア、X連鎖性;急性乳児肝不全;ロイス・ディーツ症候群1,2,3;QT延長症候群1,2,2/9,2/5,(2遺伝子性),3,5及び5、後天性、感受性;肺がん;リンパ浮腫、遺伝性、id;リンパ浮腫、原発性、骨髄異形成を伴う;リンパ増殖症候群1,1(X連鎖性)及び2;リソソーム酸リパーゼ欠損症;大頭症、巨人症、顔面異形症候群;黄斑変性症、卵黄様、成人期に発症;悪性高熱感受性1型;悪性リンパ腫、非ホジキン;悪性黒色腫;前立腺の悪性腫瘍;下顎骨異形成症;A型又はB型リポジストロフィーを伴う下顎骨異形成症、非定型;下顎顔面異形成症、トレーチャー・コリンズ型、常染色体劣性;マンノース結合タンパク質欠乏症;メープルシロップ尿症1A型及び3型;マルデン・ウォーカー様症候群;マルファン症候群;マリネスコ・Sj\xc3\xb6gren症候群;マートソフル(Martsolf)症候群;成熟期に発症する若年性糖尿病、1型,2型,11型,3型及び9型;メイ・ヘグリン異常症;MYH9関連疾患;セバスチャン症候群;マキュン・オルブライト症候群;体細胞腺腫;性索-間質性腫瘍;クッシング症候群;
マクシック・カウフマン症候群;マクラウド神経赤血球症症候群;メッケル・グルーバー症候群;中鎖アシル-補酵素A脱水素酵素欠損症;髄芽腫;皮質下嚢胞1及び2aを伴う大脳白質脳症;大頭-先天性大理石様皮膚;PIK3CA関連過成長スペクトラム;巨脳-多小脳回-多指-水頭症候群2;巨大芽球性貧血、チアミン反応性、糖尿病及び感音性難聴を伴う;マイヤー・ゴーリン症候群1及び4;メルニック・ニードルズ症候群;髄膜腫;精神遅滞、X連鎖性、3,21,30及び72;精神遅滞及び小頭症、脳橋及び小脳の低形成を伴う;X連鎖精神遅滞症候群5;精神遅滞、上顎前突、及び斜視;精神遅滞、常染色体優性12,13,15,24,3,30,4,5,6及び9;精神遅滞、常染色体劣性15,44,46及び5;精神遅滞、定型動作、てんかん、及び/又は脳奇形;精神遅滞、症候群性、クラース・ジェンセン(Claes-Jensen)型、X連鎖性;精神遅滞、X連鎖、非特異的、症候性、ヘデラ(Hedera)型、及び症候性、ウー(wu)型;メロシン欠損型先天性筋ジストロフィー;異染性白質ジストロフィー若年型、乳児期後期型、及び成人型;異染性白質ジストロフィー;変容性骨異形成症;メトヘモグロビン血症I型及び2型;メチオニンアデノシルトランスフェラーゼ欠損症、常染色体優性;ホモシスチン尿症を伴うメチルマロン酸血症;メチルマロン酸尿症cblB型;メチルマロニル-CoAムターゼ欠損症に起因するメチルマロン酸尿症;メチルマロン酸尿症mut(0)型;小頭症性骨異形成原基性小人症2型;絨毛網膜症、リンパ浮腫、又は精神遅滞の有無に関わらない小頭症;小頭症、裂孔ヘルニア、及びネフローゼ症候群;小頭症;脳梁の低形成症;痙性対麻痺50、常染色体劣性;全身性発育遅延;CNSミエリン形成不全症;脳萎縮;小頭症、正常な知能及び免疫不全;小頭症-毛細血管奇形症候群;小球性貧血;症候群性小眼球症5,7及び9;小眼球症、単独3,5,6,8、及びコロボマ6を伴う;微小球症;片頭痛、家族性片麻痺性;ミラー症候群;外眼筋麻痺を伴うミニコアミオパチー;コアを伴う先天性ミオパチー;ミッチェル・ライリー症候群;ミトコンドリア3-ヒドロキシ-3-メチルグルタリル-CoA合成酵素欠損症;ミトコンドリア複合体I,II,III,III(核型2,4又は8)欠損症;ミトコンドリアDNA枯渇症候群11,12(心筋症型),2,4B(MNGIE型),8B(MNGIE型);ミトコンドリアDNA枯渇症候群3及び7,肝脳型及び13(脳筋症型);ミトコンドリアリン酸担体及びピルビン酸担体欠損症;ミトコンドリア三機能性タンパク質欠損症;長鎖3-ヒドロキシアシル-CoAデヒドロゲナーゼ欠損症;三好型筋ジストロフィー1;遠位型ミオパチー、前脛骨発症を伴う;モーア・トラネジャーグ(Mohr-Tranebjaerg)症候群;モリブデン補因子欠損症、相補群A;モワット・ウィルソン症候群;ムコリピドーシスIIIガンマ;ムコ多糖症VI型,VI型(重症)及びVII型;ムコ多糖症、MPS-I-H/S,MPS-II,MPS-III-A,MPS-III-B,MPS-III-C、MPS-IV-A,MPS-IV-B;網膜色素変性症73;ガングリオシドーシスGM1型1(心臓障害を伴う)3;多中心性骨溶解腎症;多中心性骨溶解、結節症、及び関節症;多発性先天異常;心房中隔欠損症2;多発性先天異常-筋緊張低下-発作症候群3;多発性の皮膚及び粘膜静脈奇形;多発性内分泌腺新形成1型及び4型;多発性骨端異形成症5型又は優性;多発性腸管閉鎖症;多発性翼状片症候群エスコバール型;多発性スルファターゼ欠損症;多発性骨癒合症症候群3;筋AMPグアニンオキシダーゼ欠損症;筋眼脳疾患;筋ジストロフィー、先天性、巨円錐体型;筋無力症、家族性小児、1;アセチルコリン受容体欠損症に関連する、先天性筋無力症症候群11;先天性筋無力症症候群17,2A(スローチャネル),4B(ファストチャネル)及び管状集合体を伴わない;ミエロペルオキシダーゼ欠損症;MYH関連ポリポーシス;子宮内膜癌;心筋梗塞1;ミオクローヌス性ジストニア;ミオクローヌス性-無緊張性てんかん;赤色ボロ繊維・ミオクローヌスてんかん;筋原線維ミオパチー1及びZASP関連;ミオグロビン尿、急性再発性、常染色体劣性;筋神経胃腸管性脳症症候群;進行性外眼筋麻痺を伴う小児小脳性運動失調症;ミトコンドリアDNA枯渇症候群4B、MNGIE型;ミオパチー、中心核、1、先天性、過剰の筋紡錘を伴う、遠位の、1、乳酸アシドーシス、及び鉄芽球性貧血1、先天性白内障を伴うミトコンドリア進行性の、聴覚消失、及び発育遅延、及び管状集合体、2;近視6;筋硬化症、常染色体劣性;先天性筋強直症;先天性筋強直症、常染色体優性型及び劣性型;爪膝蓋骨症候群;ナンス・ホラン症候群;小眼球症2;ナバホ神経肝障害;ネマリンミオパチー3及び9;新生児筋緊張低下;知的障害;痙攣;言語発達遅延;精神遅滞、常染色体優性31;シトリン欠損症によって引き起こされる新生児肝内胆汁うっ滞;腎性尿崩症、腎性尿崩症、X連鎖性;腎結石症/骨粗鬆症、低リン酸血症性、2;ネフロン癆13,15及び4;不妊症;脳-眼-腎症候群(ネフロン癆、眼球運動失効、及び小脳異常);ネフローゼ症候群、3型,5型、眼球異常の有無に関わらない、7型及び9型;ネスター・グレルモプロジェリア症候群;ノイ・ラクソバ(Neu-Laxova)症候群1;脳鉄蓄積を伴う神経変性症4及び6;神経フェリチン症;神経線維腫症、1型及び2型;神経線維肉腫;神経下垂体性尿崩症;ニューロパチー、遺伝性感覚性、IC型;中性1アミノ酸輸送障害;ミオパチーを伴う中性脂質蓄積症;好中球免疫不全症候群;ニコライデス・バライスター症候群;ニーマン・ピック病C1型,C2型,A型及びC1、成人型;非ケトーシス性高グリシン血症;ヌーナン症候群1及び4、LEOPARD症候群1;若年性骨髄単球性白血病の有無に関わらないヌーナン症候群様障害;正常カリウム血性周期性四肢麻痺、カリウム感受性;ノルム病;てんかん、聴覚消失、及び精神遅滞症候群;精神遅滞、X連鎖性102及び症候性13;肥満;眼白子症、I型;眼皮膚白子症1B型,3型及び4型;眼歯指異形成症;歯限局型低ホスファターゼ症;歯-毛髪-四肢症候群(Odontotrichomelic syndrome);大口病;減歯症-大腸直腸癌症候群;オピッツG/BBB症候群;視神経萎縮症9;口腔顔面指趾症候群;オルニチンアミノトランスフェラーゼ欠損症;口腔顔面裂11及び7、口唇/口蓋外胚葉異形成症候群;オルスタビック・リンデマン・ソルベルグ症候群;軽度の軟骨異形成症を伴う骨関節炎;離断性骨軟骨炎;骨形成不全症12型,5型,7型,8型,I型,III型、正常な強膜を伴う、優性型、劣性周産期致死性;頭蓋骨硬化症を伴う線条骨症;常染色体優性の大理石骨症1型及び2型,劣性4型,劣性1型,劣性6型;偽神経膠腫を伴う骨粗鬆症;耳口蓋指症候群I型及びII型;卵巣形成不全1;卵巣白質脳症;先天性爪肥厚症4及び2型;骨パジェット病、家族性;パリスター・ホール症候群;掌蹠角化症、非表皮剥離性、局所性又はびまん性;膵臓無形成及び先天性心疾患;パピロン・Lef\xc3\xa8vre症候群;傍神経膠節腫3;フォン・オレインブルグ(von Eulenburg)の先天性異常筋強直症;副甲状腺癌;パーキンソン病14,15,19(若年性発症),2,20(早期発症),6,(常染色体劣性の早期発症)及び9;限局性白皮証;ヒポキサンチン-グアニンホスホリボシルトランスフェラーゼ部分欠損症;網膜色素上皮の模様ジストロフィー;PC-K6a;ペリザウス・メルツバッハー病;ペンドレッド症候群;末梢性脱髄性神経障害、中枢性脱髄性;ヒルシュスプルング病;永続型新生児糖尿病;永続型新生児糖尿病、神経学的特色を伴う;新生児インスリン依存性糖尿病;若年発症成人型糖尿病、2型;ペルオキシソーム生合成障害14B,2A,4A,5B,6A,7A及び7B;ペロー症候群4;ペリー症候群;新生児の持続性高インスリン性低血糖症;家族性高インスリン症;表現型;フェニルケトン尿症;褐色細胞腫;遺伝性パラガングリオーマ-褐色細胞腫症候群;パラガングリオーマ1;腸のカルチノイド腫瘍;カウデン症候群3;ホスホグリセリン酸脱水素酵素欠損症;ホスホグリセリン酸キナーゼ1欠損症;光感受性硫黄欠乏性毛髪発育異常症;フィタン酸蓄積病;ピック病;ピアソン症候群;色素性網膜ジストロフィー;色素性結節状副腎皮質病変、原発性、1;石灰化上皮腫;ピット・ホプキンス症候群;下垂体依存性高コルチゾール症;下垂体ホルモン欠損症、複合型1,2,3及び4;プラスミノーゲン活性化因子インヒビター1型欠損症;プラスミノーゲン欠損症、I型;血小板型出血障害15及び8;多形皮膚萎縮症、遺伝性線維化、腱性拘縮、ミオパチー、及び肺線維症を伴う;多発性嚢胞腎疾患2、成人型及び乳児型;硬化性白質脳症を伴う多嚢胞性脂肪膜性骨異形成症;免疫不全の有無に関わらないポリグルコサン体ミオパチー1;多小脳回、非対称性、両側性前頭頭頂部;多発性神経炎、聴覚消失、運動失調、網膜色素変性、及び白内障;4型橋小脳形成不全症;膝窩部贅皮症候群;孔脳症2;汗孔角化症8、伝染性表層化学活性型;
ポルフォビリノーゲン合成酵素欠損症;晩発性皮膚ポルフィリン症;網膜色素変性症を伴う脊髄後索性失調症;後極白内障2型;プラダー・ウィリー様症候群;早発卵巣不全4,5,7及び9;原発性常染色体劣性小頭症10,2,3及び5;原発性線毛機能不全24;原発性拡張型心筋症;左心緻密化障害6;4、左心緻密化障害10;発作性心房細動;原発性高シュウ酸尿症、I型及びIII型;原発性肥大性骨関節症、常染色体劣性2;原発性低マグネシウム血症;原発性開放隅角緑内障若年発症1;原発性肺高血圧症;プリムローズ症候群;進行性家族性心ブロック1B型;進行性家族性肝内胆汁うっ滞2及び3;進行性肝内胆汁うっ滞;運動失調症を伴う進行性ミオクローヌスてんかん;進行性偽リウマチ性異形成症;進行性硬化性灰白異栄養症;プロリダーゼ欠損症;プロリン脱水素酵素欠損症;統合失調症4;プロパジン欠損症、X連鎖性;プロピオン酸血症;プロタンパク質変換酵素1/3欠損症;前立腺がん、遺伝性、2;プロタン欠損症;蛋白尿;フィンランド型先天性ネフローゼ症候群;プロテウス症候群;乳腺癌;偽軟骨発育不全脊椎骨端異形成症;偽性低アルドステロン症1型常染色体優性及び劣性及び2型;偽性副甲状腺機能低下症1A型;偽性偽性副甲状腺機能低下症;仮性新生児副腎白質ジストロフィー;偽原発性高アルドステロン症;弾性線維性仮性黄色腫;乳児の全身性動脈石灰化症2;多発性凝固因子欠損症を伴う弾性線維性仮性黄色腫様障害;乾癬感受性2;PTEN過誤腫症候群;遺伝性出血性毛細血管拡張症に関連する肺動脈性高血圧症;肺線維症及び/又は骨髄不全、テロメアに関連する、1及び3;遺伝性出血性毛細血管拡張症を伴う、肺高血圧症、原発性、1;プリン-ヌクレオシドホスホリラーゼ欠損症;ピルビン酸カルボキシラーゼ欠損症;ピルビン酸脱水素酵素E1-アルファ欠損症;赤血球のピルビン酸キナーゼ欠損症;レイン症候群;ラスオパチー;劣性栄養障害性表皮水疱症;爪障害、非症候性先天性、8;ライフェンスタイン症候群;腎欠損症;腎カルニチン輸送障害;腎コロボマ症候群;腎異形成症;腎異形成症、網膜色素性ジストロフィー、小脳性運動失調症、及び骨格異形成症;遅発性感音性聴覚消失を伴うか、又は溶血性貧血を伴う、腎尿細管性アシドーシス、遠位型、常染色体劣性;眼球異常及び精神遅滞を伴う、腎尿細管性アシドーシス、近位型;網膜錐体ジストロフィー3B;網膜色素変性症;網膜色素変性症10,11,12,14,15,17及び19;網膜色素変性症2,20,25,35,36,38,39,4,40,43,45,48,66,7,70,72;網膜芽細胞腫;レット障害;ラブドイド腫瘍素因症候群2;裂孔原生網膜剥離、常染色体優性;肢根型点状軟骨異形成症2型及び3型;ロバーツ-SCあざらし肢奇形症候群;ロビノー・ソラウフ(Robinow Sorauf)症候群;ロビノー症候群、常染色体劣性、短合多指症を伴う常染色体劣性;ロスムンド・トムソン症候群;ラパデリノ症候群;RRM2B関連ミトコンドリア病;ルービンシュタイン・タイビ症候群;サラ病;サンドホフ病、成人型及び乳児型;サルコイドーシス、早期発症;ブラウ症候群;シンドラー病、1型;裂脳症;統合失調症15;蝸牛様骨盤異形成症;神経鞘腫症2;シュワルツ・ヤンペル症候群1型;角膜硬化、常染色体劣性;硬結性骨化症;二次性甲状腺機能低下症;瀬川症候群、常染色体劣性;セニオール・ローケン症候群4及び5;感覚性運動失調型ニューロパチー、構音障害、及び眼麻痺;セピアプテリン還元酵素欠損症;SeSAME症候群;小頭症、成長遅延、電離放射線感受性を伴う、ADA欠損症に起因する重症複合免疫不全症、非定型、常染色体劣性、T細胞陰性、B細胞陽性、NK細胞陽性のNK細胞陰性;重症先天性好中球減少症;重症先天性好中球減少症3、常染色体劣性又は優性;重症先天性好中球減少症及び6、常染色体劣性;乳児期の重篤なミオクロニーてんかん;熱性痙攣プラスの全般てんかん、1型及び2型;重篤なX連鎖性筋細管ミオパチー;QT短縮症候群3型;非特異的な骨格異常を伴う低身長;低身長、耳道狭窄症、下顎形成不全、骨格異常;低身長、爪甲形成不全、顔異形、及び貧毛症;原基性小人症;多指症の有無に関わらない短肋胸郭異形成症11又は3;シアリドーシスI型及びII型;シルバー痙性対麻痺症候群;神経伝導速度遅延、常染色体優性;スミス・レムリ・オピッツ症候群;スナイダー・ロビンソン症候群;成長ホルモン分泌腺腫;プロラクチノーマ;家族性、下垂体腺腫素因;ソトス症候群1又は2;痙性失調症5、常染色体劣性、シャルルボワ・サグネ型、1,10又は11、常染色体劣性;筋萎縮性側索硬化症5型;痙性対麻痺15,2,3,35,39,4、常染色体優性、55、常染色体劣性、及び5A;胆汁酸合成障害、先天性、3;精子形成不全11,3及び8;球状赤血球症4型及び5型;球状体ミオパチー;脊髄性筋萎縮症、下肢優位2、常染色体優性;脊髄性筋萎縮症、II型;脊髄小脳失調症14,21,35,40及び6;脊髄小脳失調症、常染色体劣性1及び16;脾臓低形成;脊椎手根骨足根骨癒合症候群;脊椎手掌異形成、エーラス・ダンロス症候群様、免疫調節異常を伴う、アグリカン型、先天性関節脱臼を伴う、短肢-手型、セダガッテン(Sedaghatian)型、錐体桿体ジストロフィーを伴う、及びコズロウスキー型;捩れ小人症;シュタルガルト病1;錐体桿体ジストロフィー3;スティックラー症候群1型;クニースト(Kniest)異形成症;スティックラー症候群、1型(非症候性眼性)及び4型;スティング関連脈管障害、小児期発症;ストームオルケン症候群;スタージ・ウェーバー症候群、毛細血管奇形、先天性、1;サクシニル-CoAアセト酢酸転移酵素欠損症;スクラーゼ-イソマルターゼ欠損症;乳児突然死症候群;亜硫酸酸化酵素欠損症、独立型;大動脈弁上狭窄;肺サーファクタント代謝機能障害、2及び3;指節癒合症、近位、1b;セナニー・レンツ型合指症;3型合指症;症候性X連鎖性精神遅滞16;内反尖足;タンジール病;TARP症候群;テイ・サックス病、B1バリアント、Gm2ガングリオシドーシス(成人)、Gm2ガングリオシドーシス(成人発症);テムタリー(Temtamy)症候群;テノリオ症候群;肢端骨異形成;テストステロン17-ベータ-デヒドロゲナーゼ欠損症;無四肢症、常染色体劣性;ファロー四徴症;左心低形成症候群2;動脈瘤;心臓及び大血管の奇形;心室中隔欠損症1;ティール・ベーンケ(Thiel-Behnke)角膜ジストロフィー;胸部大動脈瘤及び大動脈解離;マルファン様体型;3M症候群2;血小板減少症、血小板機能障害、溶血、及びグロビン合成不均衡;血小板減少症、X連鎖性;血栓症、遺伝性、プロテインC欠損症に起因、常染色体優性及び劣性;甲状腺無形成;甲状腺がん、濾胞性;甲状腺ホルモン代謝異常;甲状腺ホルモン抵抗性、汎発性、常染色体優性;甲状腺中毒性周期性四肢麻痺及び甲状腺中毒性周期性四肢麻痺2;サイトロピン放出ホルモン抵抗性、汎発性;チモシー症候群;TNF受容体関連周期熱症候群(TRAPS);歯無発生、選択的、3及び4;多形性心室頻拍(Torsades de pointes);タウンズ・ブロックス鰓弓耳腎様症候群;新生児の一過性表皮水疱症;トリーチャー・コリンズ症候群1;精神遅滞、小人症、及び網膜の色素変性を伴う長睫毛症;三叉神経節異形成症I型;三叉神経節症候群3型;トリメチルアミン尿症;結節性硬化症症候群;リンパ管筋腫症;結節性硬化症1及び2;チロシナーゼ陰性眼皮膚白皮症;チロシナーゼ陽性眼皮膚白皮症;チロシン血症I型;UDPグルコース-4-エピメラーゼ欠損症;ウルリッヒ型先天性筋ジストロフィー;重度の四肢欠損を伴う尺骨・腓骨欠損症;アップショー・シュールマン症候群;ウロカニン酸加水酵素欠損症;アッシャー症候群、1型,1B型,1D型,1G型,2A型,2C型及び2D型;網膜色素変性症39;紫外線感受性症候群;ファン・デル・ウーデ(Van der Woude)症候群;ヴァン・マルダーゲム(Van Maldergem)症候群2;ヘンカムリンパ管拡張症-リンパ浮腫症候群2;異型ポルフィリン症;嚢胞性腎疾患を伴う脳室拡大;ヴェルヘイ(Verheij)症候群;超長鎖アシル-CoA脱水素酵素欠損症;膀胱尿管逆流症8;内臓逆位5、常染色体;腹部ミオパチー;ビタミンD依存性くる病、1型及び2型;卵黄型ジストロフィー;フォン・ヴィルブランド病2M型及び3型;ワールデンブルグ症候群1型,4C型及び2E型(神経学的病変を伴う);クライン・ワールデンブルグ症候群;ウォーカー・ワールブルグ先天性筋ジストロフィー;ワールブルグ・マイクロ症候群2及び4;イボ、低ガンマグロブリン血症、感染症、及び骨髄性細胞貯留;ウィーバー症候群;ヴァイユ・マルケザーニ症候群1及び3;ヴァイユ・マルケザーニ様症候群;ワイセンバッハー・ツウェイミュラー(Weissenbacher-Zweymuller)症候群;ウェルドニッヒ・ホフマン病;シャルコー・マリー・トゥース病;ウェルナー症候群;WFS1関連障害;ウィデマン・スタイナー症候群;ウィルソン病;ウォルフラム様症候群、常染色体優性;ワース(Worth)病;ファン・ブッヘム病2型;色素性乾皮症、相補群b,群D,群E及び群G;X連鎖無ガンマグロブリン血症;X連鎖性 遺伝性運動感覚性ニューロパチー;ステリルスルファターゼ欠損症を伴うX連鎖性魚鱗癬;X連鎖性脳室周囲異所性灰白質;耳口蓋指症候群、I型;X連鎖性重症複合免疫不全症;チンメルマン・レーバンド(Zimmermann-Laband)症候群及びチンメルマン・レーバンド症候群2;並びに小帯(Zonular)粉状白内障3。
肉芽腫を伴う、細胞性及び体液性免疫複合欠損症;d-2-及びl-2-ヒドロキシグルタル酸複合尿症;マロン酸及びメチルマロン酸複合尿症;酸化的リン酸化複合欠損症1,3,4,12,15及び25;一部及び全部の17-アルファ-ヒドロキシラーゼ/17,20-リアーゼ複合欠損症;共通可変性免疫不全症9;c1インヒビターの機能不全に起因する補体構成要素4の部分欠損;補体因子B欠損症;錐体単色症;錐体桿体ジストロフィー2及び6;錐体桿体ジストロフィー遺伝性エナメル質形成不全症;先天性副腎過形成及び先天性副腎低形成、X連鎖性;先天性無巨核球性血小板減少症;先天性無虹彩症;先天性中枢性低換気;ヒルシュスプルング病3;先天性収縮性腕十二指腸症;四肢及び顔面の先天性拘縮、低緊張、及び発育遅延;グリコシル化の先天性障害1B型,1D型,1G型,1H型,1J型,1K型,1N型,1P型,2C型,2J型,2K型,IIm型;I型及びII型先天性赤血球異形成貧血;顔面の先天性外胚葉異形成症;先天性赤血球生成性ポルフィリン症;2型先天性全身性リポジストロフィー;先天性心疾患、多発型2;先天性心疾患;大動脈弓離断症;先天性脂肪腫性過成長、血管奇形、及び表皮母斑;非小細胞肺がん;卵巣の新生物;心伝導系障害、非特異性;先天性微絨毛萎縮症;先天性筋ジストロフィー;LAMA2の部分的欠損に起因する先天性筋ジストロフィー;先天性筋ジストロフィー-脳及び眼の異常を伴うジストログリカノパチー、A2型,A7型,A8型,A11型及びA14型;先天性筋ジストロフィー-精神遅滞を伴うジストログリカノパチー、B2型,B3型,B5型及びB15型;先天性筋ジストロフィー-精神遅滞を伴わないジストログリカノパチー、B5型;先天性筋肥大-大脳症候群;先天性筋無力症候群、アセタゾールアミド反応性;繊維型不均衡を伴う先天性ミオパチー;先天性眼球コロボマ;先天性静止型夜盲症、1A型,1B型,1C型,1E型,1F型及び2A型;コプロポルフィリン症;角膜平板症2;角膜ジストロフィー、フックス内皮性4;2型角膜内皮性ジストロフィー;角膜脆弱性ケラトグロブ、青色強膜、及び関節過度可動性;コーネリア ド ランゲ症候群1及び5;常染色体優性の冠動脈疾患2;冠状動脈疾患;高αリポタンパク質血症2;皮質異形成症(他の脳奇形との複合)5及び6;後頭部の皮質奇形;コルチコステロイド結合グロブリン欠損症;2型コルチコステロンメチルオキシダーゼ欠損症;コステロ症候群;カウデン症候群1;扁平股;常染色体優性の頭蓋骨異形成症;頭蓋縫合早期癒合症1及び4;頭蓋縫合早期癒合症及び歯異常;クレアチン欠損症、X連鎖;クルーゾン症候群;潜在眼球症候群;停留精巣、片側又は両側;クッシング指節癒合症;皮膚悪性黒色腫1;骨異栄養症と、肺、胃腸、及び泌尿器の重度の異常とを伴う皮膚弛緩症;チアノーゼ、一過的な新生児の及び非定型の腎症;嚢胞性線維症;シスチン尿症;シトクロムcオキシダーゼi欠損症;シトクロムcオキシダーゼ欠損症;D-2-ヒドロキシグルタル酸尿症2;分節性ダリエー病;内耳形成不全、小耳症、及び小歯症(LAMM)を伴う難聴;難聴、常染色体優性3a,4,12,13,15、常染色体優性の非症候性感音性17,20及び65;難聴、常染色体劣性1A,2,3,6,8,9,12,15,16,18b,22,28,31,44,49,63,77,86及び89;難聴、蝸牛の、近視及び知的障害あり、前庭障害なし、常染色体優性、X連鎖2;2-メチルブチリル-CoA脱水素酵素の欠損症;3-ヒドロキシアシル-CoA脱水素酵素の欠損症;アルファ-マンノシダーゼの欠損症;芳香族-L-アミノ酸脱炭酸酵素の欠損症;ビスホスホグリセリン酸ムターゼの欠損症;ブチリル-CoA脱水素酵素の欠損症;フェロキシダーゼの欠損症;ガラクトキナーゼの欠損症;グアニジノ酢酸メチルトランスフェラーゼの欠損症;ヒアルロノグルコサミニダーゼの欠損症;リボース-5-リン酸イソメラーゼの欠損症;ステロイド11-ベータ-モノオキシゲナーゼの欠損症;UDPグルコース-ヘキソース-1-リン酸ウリジリルトランスフェラーゼの欠損症;キサンチンオキシダーゼの欠損症;デジェリン・ソッタス病;シャルコー・マリー・トゥース病、ID型及びIVF型;デジェリン・ソッタス症候群、常染色体優性;樹状細胞、単球、Bリンパ球、及びナチュラルキラーリンパ球の欠損症;デビュクワ(Desbuquois)異形成症2;デビュクワ症候群;DFNA 2非症候性の聴力低下;視神経萎縮及び難聴を伴う糖尿病及び尿崩症;2型糖尿病、及びインスリン依存性20;ダイアモンド・ブラックファン貧血1,5,8及び10;下痢3(分泌性ナトリウム、先天性、症候性)、及び5(タフティング腸症(tufting enteropathy)を伴う、先天性);ジカルボキシルアミノ酸尿症;びまん性掌蹠角化症、ボスニア型;デジトレノ脳症候群;ジヒドロプテリジン還元酵素欠損症;拡張型心筋症1A,1AA,1C,1G,1BB,1DD,1FF,1HH,1I,1KK,1N,1S,1Y及び3B;左心室非圧縮3;シトクロムp450酸化還元酵素欠損症に起因するステロイド産生異常;遠位型関節拘縮2B型;遠位型遺伝性運動ニューロン症2B型;遠位型ミオパチーMarkesbery-Griggs型;遠位型脊髄性筋萎縮症、X連鎖3;二重睫毛・リンパ浮腫症候群;皮膚の欠如を伴う、優性栄養傷害性表皮水疱症;優性遺伝性視神経萎縮症;ドンナイ・バロウ症候群;ドーパミン ベータ水酸化酵素欠損症;ドーパミン受容体d2、脳内密度低下;ダウリング・デゴス病4;ドイン蜂巣状網膜ジストロフィー;マラティア・レベンチーヌ(Malattia leventinese);2型デュアン症候群;デュビン・ジョンソン症候群;デュシェンヌ型筋ジストロフィー;ベッカー型筋ジストロフィー;異常フィブリノーゲン血症;常染色体優性の先天性角化異常症及び常染色体優性、3;常染色体劣性の先天性角化異常症1,3,4及び5;X連鎖性の先天性角化異常症;顔面ミオキミアを伴う、家族性ジスキネジア;プラスミノーゲン分子異常症;ジストニア2(捻転、常染色体劣性),3(捻転、X連鎖),5(ドーパ反応型),10,12,16,25,26(ミオクロニー);良性家族性乳児の発作2;早期乳児てんかん性脳症2,4,7,9,10,11,13及び14;非定型レット症候群;初期T細胞前駆体急性リンパ性白血病;外胚葉性異形成表皮水疱症候群;外胚葉性異形成-合指症候群1;偏位水晶体、単独で(isolated)常染色体劣性及び優性;裂手裂足(Ectrodactyly)、外胚葉性異形成、及び口唇口蓋裂症候群3;エーラス・ダンロス症候群7型(常染色体劣性)、古典型、2型(早老性)、ヒドロキシリジン欠損型、4型、4型バリアント、及びテナシンX欠損に起因するもの;アイヒスフェルト型先天性筋ジストロフィー;内分泌-大脳骨異形成症(cerebroosteodysplasia);S錐体増強症候群;拡大前庭水管症候群;エンテロキナーゼ欠損症;疣贅状表皮発育異常症;単純型表皮水疱症及び肢帯筋ジストロフィー、斑状色素沈着(mottled pigmentation)を伴う単純型、幽門閉鎖を伴う単純型、単純型、常染色体劣性、及び幽門閉鎖を伴う;表皮剥離性掌蹠角化症;家族性熱性痙攣8;てんかん、小児欠神2,12(特発性全身、感受性),5(夜間前頭葉型)、夜間前頭葉型1、部分性、可変の病巣があるもの(with variable foci)、進行性ミオクロニー3、及びX連鎖性、可変の学習障害及び行動障害があるもの;てんかん性脳症、小児期発症、乳児期早期、1,19,23,25,30及び32;骨端異形成症、多発性、近視及び伝音難聴を伴う;一過性運動失調症2型;一過性疼痛症候群、家族性、3;エプスタイン症候群;フェクトナー症候群;赤芽球増殖性プロトポルフィリン症;エストロゲン抵抗性;滲出性硝子体網膜症6;ファブリー病、及びファブリー病、心臓変異型;第H因子、第VII因子、第X因子、第v因子、及び第viii因子の、2つの複合欠損症、第xiii因子、サブユニット、欠損症;家族性腺腫性ポリポーシス1及び3;蕁麻疹及び難聴を伴う家族性アミロイド腎症;家族性寒冷蕁麻疹;家族性の小脳虫部無発生;家族性良性天疱瘡;家族性乳がん;乳がん感受性(Breast cancer,susceptibility to);骨肉腫;膵臓がん3;家族性心筋症;家族性寒冷自己炎症性症候群2;家族性大腸がん;家族性滲出性硝子体網膜症、X連鎖性;家族性片麻痺性片頭痛1型及び2型;家族性高コレステロール血症;家族性肥大型心筋症1,2,3,4,7,10,23及び24;家族性低カリウム血症-低マグネシウム血症;家族性低形成の、糸球体嚢胞腎;家族性乳児筋無力症;家族性若年性痛風;家族性地中海熱、及び家族性地中海熱、常染色体優性;家族性多弁症;家族性晩発性皮膚ポルフィリン症;家族性肺毛細血管腫症;家族性腎性糖尿;家族性腎性低尿酸血症;家族性拘束性心筋症1;家族性1型及び3型高リポ蛋白血症;ファンコニー貧血,補欠群E,I,N及びO;ファンコニー・ビッケル症候群;ファビズム感受性(Favism,susceptibility to);熱性痙攣、家族性、11;フェインゴールド症候群1;胎児ヘモグロビン量的形質遺伝子座1;FG症候群及びFG症候群4;眼球外筋の線維症、先天性、1,2,3a(眼球外病変の有無に関わらない),3b;魚眼病;フレック角膜ジストロフィー;フローティング・ハーバー症候群;精神遅滞の有無に関わらない、言語障害を伴う焦点てんかん;巣状分節性糸球体硬化症5;前脳欠損症;フランク・テル・ハール症候群;Borrone Di Rocco Crovato症候群;フレイジャー症候群;ウィルムス腫瘍1;フリーマン・シェルドン症候群;前頭骨幹端異形成症1及び3;前頭側頭型認知症;前頭側頭型認知症及び/又は筋萎縮性側索硬化症3及び4;前頭側頭型認知症第3染色体連鎖性及び前頭側頭型認知症ユビキチン陽性;フルクトース-ビホスファターゼ欠損症;フールマン(Fuhrmann)症候群;ガンマ-アミノ酪酸トランスアミナーゼ欠損症;ガムストープ・ウォールファールト(Gamstorp-Wohlfart)症候群;ゴーシェ病1型及び亜急性神経障害性;進行性の脊椎側弯症を伴う、家族性水平注視麻痺;全般性優性ジストロフィー型表皮水疱症;全般性てんかん、熱性痙攣プラス3,1型,2型を伴う;
レノックス・ガストー型てんかん脳症;巨大軸索ニューロパチー;グランツマン血栓症;緑内障1、開放角、e,F及びG;緑内障3、原発性先天性、d;先天性緑内障、及び先天性緑内障、コロボマ;緑内障、原発性開放角、若年性発症;神経膠腫感受性1;グルコーストランスポーター1型欠損症候群;ルコース-6-リン酸輸送欠損症;GLUT1欠損症候群2;特発性全般性てんかん感受性、12;グルタミン酸ホルミノトランスフェラーゼ欠損症;グルタル酸血症IIA及びIIB;グルタル酸尿症1型;グルタチオン合成酵素欠損症;糖原病0(筋肉),II(成人形),IXa2,IXc,1A型;II型,IV型,IV(肝とミオパチーとの複合),V型及びVI型;ゴールドマン・ファーブル症候群;ゴードン症候群;ゴーリン症候群;全前脳胞症シークエンス;全前脳胞症7;顆粒腫性疾患、慢性、X連鎖性、バリアント;卵巣の顆粒膜細胞腫瘍;灰色血小板症候群;グリセリ症候群3型;グレーノー角膜ジストロフィーI型;成長及び精神遅滞、下顎顔面異形症、小頭症、及び口蓋裂;下垂体異常を伴う成長ホルモン欠損症;免疫不全を伴う成長ホルモン不感受性;GTPシクロハイドロラーゼI欠損症;ハジュ・チーニー症候群;手足子宮症候群;聴覚障害;小児毛細血管腫;血液腫瘍;ヘモクロマトーシス1型,2B型及び3型;糖尿病の微小血管合併症7;トランスフェリン血清レベル量的形質座2;ヘモグロビンH病、無欠損;グルコースリン酸イソメラーゼ欠損症に起因する、溶血性貧血、非球状赤血球性;血球貪食性リンパ組織球症、家族性、2;血球貪食性リンパ組織球症、家族性、3;ヘパリン補因子II欠損症;遺伝性腸性先端皮膚炎;遺伝性乳がん・卵巣がん症候群;運動失調・毛細血管拡張症様障害;遺伝性びまん性胃がん;スフェロイドを伴う遺伝性びまん性白質脳症;遺伝性第II,第IX,第VIII因子欠損症;遺伝性出血性毛細血管拡張症2型;無汗症を伴う遺伝性無痛症;遺伝性リンパ浮腫I型;視神経萎縮を伴う遺伝性運動・感覚神経症;早期呼吸不全を伴う遺伝性ミオパチー;遺伝性神経痛性筋萎縮症;遺伝性非ポリポーシス大腸新生物;リンチ症候群I及びII;遺伝性膵炎;慢性膵炎感受性;遺伝性感覚・自律神経障害IIB型及びIIA型;遺伝性鉄芽球性貧血;ヘルマンスキー・ポドラック症候群1,3,4及び6;内臓錯位、2,4及び6、常染色体;内臓錯位、X連鎖性;組織球性髄様細網症;異所形成;組織球症-リンパ節腫大プラス症候群;ホロカルボキシラーゼ合成酵素欠損症;全前脳胞症2,3,7及び9;ホルト・オラム症候群;MTHFR欠損症、CBS欠損症、及びホモシステイン尿症に起因するホモシステイン血症、ピリドキシン応答性;コバラミン代謝異常に起因する、ホモシステイン尿症-巨赤芽球性貧血、cblE相補型;ハウエル・エバンス(Howel-Evans)症候群;ハーラー症候群;ハッチンソン・ギルフォード症候群;水頭症;高アンモニア血症、III型;高コレステロール血症、及び高コレステロール血症、常染色体劣性;過剰驚愕症2及び遺伝性の過剰驚愕症;高フェリチン血症白内障症候群;高グリシン尿症;周期性発熱を伴う高免疫グロブリンD;メバロン酸尿症;高免疫グロブリンE症候群;家族性の高インスリン血症性低血糖症3,4及び5;高インスリン血症-高アンモニア血症症候群;高リシン血症;ジストニア、赤血球増加症、及び肝硬変を伴う高マンガン血症;高オルニチン血症-高アンモニア血症-ホモシトルリン尿症症候群;副甲状腺機能亢進症1及び2;副甲状腺機能亢進症、新生児重度;部分的pts欠損症、BH4欠損症、D、非pkuに起因する、高フェニルアラニン血症、bh4欠損症、a;精神遅滞症候群2,3及び4を伴う高リン酸血症;多毛性骨軟骨異形成症;apob32に関連する、低ベータリポ蛋白血症、家族性;低カルシウム血症、常染色体優性1;低カルシウム尿性高カルシウム血症、家族性、1型及び3型;低軟骨形成症;鉄過剰症を伴う低クロム小球性貧血;肝臓中グリコーゲン合成酵素の欠損症を伴う低血糖症;臭覚障害の有無に関わらない低ゴナドトロピン性性腺機能低下症11;免疫不全を伴う無汗性外胚葉形成不全症;発汗低下のX連鎖性外胚葉異形成;低カリウム性周期性四肢麻痺1及び2;低マグネシウム血症1、腸の;低マグネシウム血症、発作、及び精神遅滞;低髄性白質ジストロフィー7;左心低形成症候群;房室中隔欠損症及び共通房室接合部;尿道下裂1及び2、X連鎖性;甲状腺機能低下症、先天性非甲状腺腫、1;減毛症8及び12;減毛症-リンパ浮腫-血管拡張症候群;I血液型系;シーメンス型水疱性魚鱗癬;剥脱性魚鱗癬;未熟児魚鱗癬症候群;特発性基底核石灰化症5;特発性線維性肺胞炎、慢性型;先天性角化不全症、常染色体優性、2及び5;乳児期の特発性高カルシウム血症;カルシウム流入障害2に起因するT細胞不活性化を伴う免疫機能障害;1型及び2型高IgMを伴う、cd3-ゼータの欠陥に起因する、免疫不全15,16,19,30,31C,38,40,8、並びにマグネシウム欠陥を伴う、X連鎖性、エプスタイン・バーウイルス感染、及び新形成;免疫不全-動原体不安定性-顔面奇形症候群2;封入体ミオパチー2及び3;野中ミオパチー;乳児痙攣及び発作性舞踏症、家族性;乳児皮質過骨症;乳児GM1ガングリオシドーシス;乳児低ホスファタシアーゼ血症;乳児ネフロン癆;乳児眼振、X連鎖性;乳児パーキンソン病-ジストニア;多尾性精子及び過剰なDNAに関連する不妊症;インスリン抵抗性;インスリン抵抗性糖尿病及び黒色表皮腫;インスリン依存性糖尿病分泌性下痢症候群;間質性腎炎、核巨大化(karyomegalic);子宮内発育遅延、骨幹端異形成症、先天性副腎低形成、及び性器奇形;ヨードチロシン結合欠損症;IRAK4欠損症;虹彩隅角異発生優性型及び1型;脳内鉄蓄積;坐骨膝蓋骨異形成;膵島細胞過形成;17,20-リアーゼ単独欠損症;ルトロピン単独欠損症;イソバレリル-CoAデヒドロゲナーゼ欠損症;ヤンコヴィッチ・リベラ症候群;ジャーベル及びランゲ・ニールセン症候群2;ジュベール症候群1,6,7,9/15(2遺伝子性),14,16及び17、並びに口顔面指症候群xiv;ヘルリッツの接合部表皮水疱症;若年性GM>1<ガングリオシドーシス;若年性ポリポーシス症候群;若年性ポリポーシス/遺伝性出血性毛細血管拡張症症候群;若年性網膜炎;歌舞伎メーキャップ症候群;カルマン症候群1,2及び6;思春期遅延症;神崎病;カラック症候群;カルタゲナー症候群;ケニー・ケイフィ症候群2型;ケッペン・ルビンスキー(Keppen-Lubinsky)症候群;円錐角膜1;毛包性角化症;掌蹠膿疱症1;キンドラー症候群;L-2-ヒドロキシグルタル酸尿症;ラーセン症候群、優性型;格子状角膜ジストロフィーIII型;レーバー黒内障;ツェルウェガー症候群;ペルオキシソーム生合成異常症;ツェルウェガー症候群スペクトラム;レーバー先天性黒内障11,12,13,16,4,7及び9;レーバー視神経萎縮症;アミノグリコシド誘発性難聴;難聴、非症候性感音性、ミトコンドリア;左心室非圧縮症5;左右軸奇形;リー病;ミトコンドリア短鎖エノイル-CoAヒドラターゼ1欠損症;ミトコンドリア複合体I欠損症に起因するリー症候群;ライナー病;レリー・ワイル軟骨骨異形成症;致死性先天性拘縮症候群6;白血球接着不全症I型及びIII型;白骨ジストロフィー、ミエリン形成不全性、11及び6;白質脳症、運動失調を伴う、脳幹・脊髄障害と乳酸値上昇とを伴う、白質の消失を伴う、及び進行性の、卵巣不全を伴う;全白爪;レビー小体型認知症;リヒテンシュタイン・クノール症候群;リー・フラウメニ症候群1;Lig4症候群;肢帯筋ジストロフィー1B型,2A型,2B型,2D型,C1型,C5型,C9型,C14型;脳及び眼の異常を伴う先天性筋ジストロフィー-ジストログリカノパチー、A14型及びB14型;複合型リパーゼ欠損症;脂肪蛋白症;リポジストロフィー、家族性部分的、2型及び3型;脳回欠損1,2(X連鎖性),3,6(小頭症を伴う),X連鎖性;皮質下帯状ヘテロトピア、X連鎖性;急性乳児肝不全;ロイス・ディーツ症候群1,2,3;QT延長症候群1,2,2/9,2/5,(2遺伝子性),3,5及び5、後天性、感受性;肺がん;リンパ浮腫、遺伝性、id;リンパ浮腫、原発性、骨髄異形成を伴う;リンパ増殖症候群1,1(X連鎖性)及び2;リソソーム酸リパーゼ欠損症;大頭症、巨人症、顔面異形症候群;黄斑変性症、卵黄様、成人期に発症;悪性高熱感受性1型;悪性リンパ腫、非ホジキン;悪性黒色腫;前立腺の悪性腫瘍;下顎骨異形成症;A型又はB型リポジストロフィーを伴う下顎骨異形成症、非定型;下顎顔面異形成症、トレーチャー・コリンズ型、常染色体劣性;マンノース結合タンパク質欠乏症;メープルシロップ尿症1A型及び3型;マルデン・ウォーカー様症候群;マルファン症候群;マリネスコ・Sj\xc3\xb6gren症候群;マートソフル(Martsolf)症候群;成熟期に発症する若年性糖尿病、1型,2型,11型,3型及び9型;メイ・ヘグリン異常症;MYH9関連疾患;セバスチャン症候群;マキュン・オルブライト症候群;体細胞腺腫;性索-間質性腫瘍;クッシング症候群;
マクシック・カウフマン症候群;マクラウド神経赤血球症症候群;メッケル・グルーバー症候群;中鎖アシル-補酵素A脱水素酵素欠損症;髄芽腫;皮質下嚢胞1及び2aを伴う大脳白質脳症;大頭-先天性大理石様皮膚;PIK3CA関連過成長スペクトラム;巨脳-多小脳回-多指-水頭症候群2;巨大芽球性貧血、チアミン反応性、糖尿病及び感音性難聴を伴う;マイヤー・ゴーリン症候群1及び4;メルニック・ニードルズ症候群;髄膜腫;精神遅滞、X連鎖性、3,21,30及び72;精神遅滞及び小頭症、脳橋及び小脳の低形成を伴う;X連鎖精神遅滞症候群5;精神遅滞、上顎前突、及び斜視;精神遅滞、常染色体優性12,13,15,24,3,30,4,5,6及び9;精神遅滞、常染色体劣性15,44,46及び5;精神遅滞、定型動作、てんかん、及び/又は脳奇形;精神遅滞、症候群性、クラース・ジェンセン(Claes-Jensen)型、X連鎖性;精神遅滞、X連鎖、非特異的、症候性、ヘデラ(Hedera)型、及び症候性、ウー(wu)型;メロシン欠損型先天性筋ジストロフィー;異染性白質ジストロフィー若年型、乳児期後期型、及び成人型;異染性白質ジストロフィー;変容性骨異形成症;メトヘモグロビン血症I型及び2型;メチオニンアデノシルトランスフェラーゼ欠損症、常染色体優性;ホモシスチン尿症を伴うメチルマロン酸血症;メチルマロン酸尿症cblB型;メチルマロニル-CoAムターゼ欠損症に起因するメチルマロン酸尿症;メチルマロン酸尿症mut(0)型;小頭症性骨異形成原基性小人症2型;絨毛網膜症、リンパ浮腫、又は精神遅滞の有無に関わらない小頭症;小頭症、裂孔ヘルニア、及びネフローゼ症候群;小頭症;脳梁の低形成症;痙性対麻痺50、常染色体劣性;全身性発育遅延;CNSミエリン形成不全症;脳萎縮;小頭症、正常な知能及び免疫不全;小頭症-毛細血管奇形症候群;小球性貧血;症候群性小眼球症5,7及び9;小眼球症、単独3,5,6,8、及びコロボマ6を伴う;微小球症;片頭痛、家族性片麻痺性;ミラー症候群;外眼筋麻痺を伴うミニコアミオパチー;コアを伴う先天性ミオパチー;ミッチェル・ライリー症候群;ミトコンドリア3-ヒドロキシ-3-メチルグルタリル-CoA合成酵素欠損症;ミトコンドリア複合体I,II,III,III(核型2,4又は8)欠損症;ミトコンドリアDNA枯渇症候群11,12(心筋症型),2,4B(MNGIE型),8B(MNGIE型);ミトコンドリアDNA枯渇症候群3及び7,肝脳型及び13(脳筋症型);ミトコンドリアリン酸担体及びピルビン酸担体欠損症;ミトコンドリア三機能性タンパク質欠損症;長鎖3-ヒドロキシアシル-CoAデヒドロゲナーゼ欠損症;三好型筋ジストロフィー1;遠位型ミオパチー、前脛骨発症を伴う;モーア・トラネジャーグ(Mohr-Tranebjaerg)症候群;モリブデン補因子欠損症、相補群A;モワット・ウィルソン症候群;ムコリピドーシスIIIガンマ;ムコ多糖症VI型,VI型(重症)及びVII型;ムコ多糖症、MPS-I-H/S,MPS-II,MPS-III-A,MPS-III-B,MPS-III-C、MPS-IV-A,MPS-IV-B;網膜色素変性症73;ガングリオシドーシスGM1型1(心臓障害を伴う)3;多中心性骨溶解腎症;多中心性骨溶解、結節症、及び関節症;多発性先天異常;心房中隔欠損症2;多発性先天異常-筋緊張低下-発作症候群3;多発性の皮膚及び粘膜静脈奇形;多発性内分泌腺新形成1型及び4型;多発性骨端異形成症5型又は優性;多発性腸管閉鎖症;多発性翼状片症候群エスコバール型;多発性スルファターゼ欠損症;多発性骨癒合症症候群3;筋AMPグアニンオキシダーゼ欠損症;筋眼脳疾患;筋ジストロフィー、先天性、巨円錐体型;筋無力症、家族性小児、1;アセチルコリン受容体欠損症に関連する、先天性筋無力症症候群11;先天性筋無力症症候群17,2A(スローチャネル),4B(ファストチャネル)及び管状集合体を伴わない;ミエロペルオキシダーゼ欠損症;MYH関連ポリポーシス;子宮内膜癌;心筋梗塞1;ミオクローヌス性ジストニア;ミオクローヌス性-無緊張性てんかん;赤色ボロ繊維・ミオクローヌスてんかん;筋原線維ミオパチー1及びZASP関連;ミオグロビン尿、急性再発性、常染色体劣性;筋神経胃腸管性脳症症候群;進行性外眼筋麻痺を伴う小児小脳性運動失調症;ミトコンドリアDNA枯渇症候群4B、MNGIE型;ミオパチー、中心核、1、先天性、過剰の筋紡錘を伴う、遠位の、1、乳酸アシドーシス、及び鉄芽球性貧血1、先天性白内障を伴うミトコンドリア進行性の、聴覚消失、及び発育遅延、及び管状集合体、2;近視6;筋硬化症、常染色体劣性;先天性筋強直症;先天性筋強直症、常染色体優性型及び劣性型;爪膝蓋骨症候群;ナンス・ホラン症候群;小眼球症2;ナバホ神経肝障害;ネマリンミオパチー3及び9;新生児筋緊張低下;知的障害;痙攣;言語発達遅延;精神遅滞、常染色体優性31;シトリン欠損症によって引き起こされる新生児肝内胆汁うっ滞;腎性尿崩症、腎性尿崩症、X連鎖性;腎結石症/骨粗鬆症、低リン酸血症性、2;ネフロン癆13,15及び4;不妊症;脳-眼-腎症候群(ネフロン癆、眼球運動失効、及び小脳異常);ネフローゼ症候群、3型,5型、眼球異常の有無に関わらない、7型及び9型;ネスター・グレルモプロジェリア症候群;ノイ・ラクソバ(Neu-Laxova)症候群1;脳鉄蓄積を伴う神経変性症4及び6;神経フェリチン症;神経線維腫症、1型及び2型;神経線維肉腫;神経下垂体性尿崩症;ニューロパチー、遺伝性感覚性、IC型;中性1アミノ酸輸送障害;ミオパチーを伴う中性脂質蓄積症;好中球免疫不全症候群;ニコライデス・バライスター症候群;ニーマン・ピック病C1型,C2型,A型及びC1、成人型;非ケトーシス性高グリシン血症;ヌーナン症候群1及び4、LEOPARD症候群1;若年性骨髄単球性白血病の有無に関わらないヌーナン症候群様障害;正常カリウム血性周期性四肢麻痺、カリウム感受性;ノルム病;てんかん、聴覚消失、及び精神遅滞症候群;精神遅滞、X連鎖性102及び症候性13;肥満;眼白子症、I型;眼皮膚白子症1B型,3型及び4型;眼歯指異形成症;歯限局型低ホスファターゼ症;歯-毛髪-四肢症候群(Odontotrichomelic syndrome);大口病;減歯症-大腸直腸癌症候群;オピッツG/BBB症候群;視神経萎縮症9;口腔顔面指趾症候群;オルニチンアミノトランスフェラーゼ欠損症;口腔顔面裂11及び7、口唇/口蓋外胚葉異形成症候群;オルスタビック・リンデマン・ソルベルグ症候群;軽度の軟骨異形成症を伴う骨関節炎;離断性骨軟骨炎;骨形成不全症12型,5型,7型,8型,I型,III型、正常な強膜を伴う、優性型、劣性周産期致死性;頭蓋骨硬化症を伴う線条骨症;常染色体優性の大理石骨症1型及び2型,劣性4型,劣性1型,劣性6型;偽神経膠腫を伴う骨粗鬆症;耳口蓋指症候群I型及びII型;卵巣形成不全1;卵巣白質脳症;先天性爪肥厚症4及び2型;骨パジェット病、家族性;パリスター・ホール症候群;掌蹠角化症、非表皮剥離性、局所性又はびまん性;膵臓無形成及び先天性心疾患;パピロン・Lef\xc3\xa8vre症候群;傍神経膠節腫3;フォン・オレインブルグ(von Eulenburg)の先天性異常筋強直症;副甲状腺癌;パーキンソン病14,15,19(若年性発症),2,20(早期発症),6,(常染色体劣性の早期発症)及び9;限局性白皮証;ヒポキサンチン-グアニンホスホリボシルトランスフェラーゼ部分欠損症;網膜色素上皮の模様ジストロフィー;PC-K6a;ペリザウス・メルツバッハー病;ペンドレッド症候群;末梢性脱髄性神経障害、中枢性脱髄性;ヒルシュスプルング病;永続型新生児糖尿病;永続型新生児糖尿病、神経学的特色を伴う;新生児インスリン依存性糖尿病;若年発症成人型糖尿病、2型;ペルオキシソーム生合成障害14B,2A,4A,5B,6A,7A及び7B;ペロー症候群4;ペリー症候群;新生児の持続性高インスリン性低血糖症;家族性高インスリン症;表現型;フェニルケトン尿症;褐色細胞腫;遺伝性パラガングリオーマ-褐色細胞腫症候群;パラガングリオーマ1;腸のカルチノイド腫瘍;カウデン症候群3;ホスホグリセリン酸脱水素酵素欠損症;ホスホグリセリン酸キナーゼ1欠損症;光感受性硫黄欠乏性毛髪発育異常症;フィタン酸蓄積病;ピック病;ピアソン症候群;色素性網膜ジストロフィー;色素性結節状副腎皮質病変、原発性、1;石灰化上皮腫;ピット・ホプキンス症候群;下垂体依存性高コルチゾール症;下垂体ホルモン欠損症、複合型1,2,3及び4;プラスミノーゲン活性化因子インヒビター1型欠損症;プラスミノーゲン欠損症、I型;血小板型出血障害15及び8;多形皮膚萎縮症、遺伝性線維化、腱性拘縮、ミオパチー、及び肺線維症を伴う;多発性嚢胞腎疾患2、成人型及び乳児型;硬化性白質脳症を伴う多嚢胞性脂肪膜性骨異形成症;免疫不全の有無に関わらないポリグルコサン体ミオパチー1;多小脳回、非対称性、両側性前頭頭頂部;多発性神経炎、聴覚消失、運動失調、網膜色素変性、及び白内障;4型橋小脳形成不全症;膝窩部贅皮症候群;孔脳症2;汗孔角化症8、伝染性表層化学活性型;
ポルフォビリノーゲン合成酵素欠損症;晩発性皮膚ポルフィリン症;網膜色素変性症を伴う脊髄後索性失調症;後極白内障2型;プラダー・ウィリー様症候群;早発卵巣不全4,5,7及び9;原発性常染色体劣性小頭症10,2,3及び5;原発性線毛機能不全24;原発性拡張型心筋症;左心緻密化障害6;4、左心緻密化障害10;発作性心房細動;原発性高シュウ酸尿症、I型及びIII型;原発性肥大性骨関節症、常染色体劣性2;原発性低マグネシウム血症;原発性開放隅角緑内障若年発症1;原発性肺高血圧症;プリムローズ症候群;進行性家族性心ブロック1B型;進行性家族性肝内胆汁うっ滞2及び3;進行性肝内胆汁うっ滞;運動失調症を伴う進行性ミオクローヌスてんかん;進行性偽リウマチ性異形成症;進行性硬化性灰白異栄養症;プロリダーゼ欠損症;プロリン脱水素酵素欠損症;統合失調症4;プロパジン欠損症、X連鎖性;プロピオン酸血症;プロタンパク質変換酵素1/3欠損症;前立腺がん、遺伝性、2;プロタン欠損症;蛋白尿;フィンランド型先天性ネフローゼ症候群;プロテウス症候群;乳腺癌;偽軟骨発育不全脊椎骨端異形成症;偽性低アルドステロン症1型常染色体優性及び劣性及び2型;偽性副甲状腺機能低下症1A型;偽性偽性副甲状腺機能低下症;仮性新生児副腎白質ジストロフィー;偽原発性高アルドステロン症;弾性線維性仮性黄色腫;乳児の全身性動脈石灰化症2;多発性凝固因子欠損症を伴う弾性線維性仮性黄色腫様障害;乾癬感受性2;PTEN過誤腫症候群;遺伝性出血性毛細血管拡張症に関連する肺動脈性高血圧症;肺線維症及び/又は骨髄不全、テロメアに関連する、1及び3;遺伝性出血性毛細血管拡張症を伴う、肺高血圧症、原発性、1;プリン-ヌクレオシドホスホリラーゼ欠損症;ピルビン酸カルボキシラーゼ欠損症;ピルビン酸脱水素酵素E1-アルファ欠損症;赤血球のピルビン酸キナーゼ欠損症;レイン症候群;ラスオパチー;劣性栄養障害性表皮水疱症;爪障害、非症候性先天性、8;ライフェンスタイン症候群;腎欠損症;腎カルニチン輸送障害;腎コロボマ症候群;腎異形成症;腎異形成症、網膜色素性ジストロフィー、小脳性運動失調症、及び骨格異形成症;遅発性感音性聴覚消失を伴うか、又は溶血性貧血を伴う、腎尿細管性アシドーシス、遠位型、常染色体劣性;眼球異常及び精神遅滞を伴う、腎尿細管性アシドーシス、近位型;網膜錐体ジストロフィー3B;網膜色素変性症;網膜色素変性症10,11,12,14,15,17及び19;網膜色素変性症2,20,25,35,36,38,39,4,40,43,45,48,66,7,70,72;網膜芽細胞腫;レット障害;ラブドイド腫瘍素因症候群2;裂孔原生網膜剥離、常染色体優性;肢根型点状軟骨異形成症2型及び3型;ロバーツ-SCあざらし肢奇形症候群;ロビノー・ソラウフ(Robinow Sorauf)症候群;ロビノー症候群、常染色体劣性、短合多指症を伴う常染色体劣性;ロスムンド・トムソン症候群;ラパデリノ症候群;RRM2B関連ミトコンドリア病;ルービンシュタイン・タイビ症候群;サラ病;サンドホフ病、成人型及び乳児型;サルコイドーシス、早期発症;ブラウ症候群;シンドラー病、1型;裂脳症;統合失調症15;蝸牛様骨盤異形成症;神経鞘腫症2;シュワルツ・ヤンペル症候群1型;角膜硬化、常染色体劣性;硬結性骨化症;二次性甲状腺機能低下症;瀬川症候群、常染色体劣性;セニオール・ローケン症候群4及び5;感覚性運動失調型ニューロパチー、構音障害、及び眼麻痺;セピアプテリン還元酵素欠損症;SeSAME症候群;小頭症、成長遅延、電離放射線感受性を伴う、ADA欠損症に起因する重症複合免疫不全症、非定型、常染色体劣性、T細胞陰性、B細胞陽性、NK細胞陽性のNK細胞陰性;重症先天性好中球減少症;重症先天性好中球減少症3、常染色体劣性又は優性;重症先天性好中球減少症及び6、常染色体劣性;乳児期の重篤なミオクロニーてんかん;熱性痙攣プラスの全般てんかん、1型及び2型;重篤なX連鎖性筋細管ミオパチー;QT短縮症候群3型;非特異的な骨格異常を伴う低身長;低身長、耳道狭窄症、下顎形成不全、骨格異常;低身長、爪甲形成不全、顔異形、及び貧毛症;原基性小人症;多指症の有無に関わらない短肋胸郭異形成症11又は3;シアリドーシスI型及びII型;シルバー痙性対麻痺症候群;神経伝導速度遅延、常染色体優性;スミス・レムリ・オピッツ症候群;スナイダー・ロビンソン症候群;成長ホルモン分泌腺腫;プロラクチノーマ;家族性、下垂体腺腫素因;ソトス症候群1又は2;痙性失調症5、常染色体劣性、シャルルボワ・サグネ型、1,10又は11、常染色体劣性;筋萎縮性側索硬化症5型;痙性対麻痺15,2,3,35,39,4、常染色体優性、55、常染色体劣性、及び5A;胆汁酸合成障害、先天性、3;精子形成不全11,3及び8;球状赤血球症4型及び5型;球状体ミオパチー;脊髄性筋萎縮症、下肢優位2、常染色体優性;脊髄性筋萎縮症、II型;脊髄小脳失調症14,21,35,40及び6;脊髄小脳失調症、常染色体劣性1及び16;脾臓低形成;脊椎手根骨足根骨癒合症候群;脊椎手掌異形成、エーラス・ダンロス症候群様、免疫調節異常を伴う、アグリカン型、先天性関節脱臼を伴う、短肢-手型、セダガッテン(Sedaghatian)型、錐体桿体ジストロフィーを伴う、及びコズロウスキー型;捩れ小人症;シュタルガルト病1;錐体桿体ジストロフィー3;スティックラー症候群1型;クニースト(Kniest)異形成症;スティックラー症候群、1型(非症候性眼性)及び4型;スティング関連脈管障害、小児期発症;ストームオルケン症候群;スタージ・ウェーバー症候群、毛細血管奇形、先天性、1;サクシニル-CoAアセト酢酸転移酵素欠損症;スクラーゼ-イソマルターゼ欠損症;乳児突然死症候群;亜硫酸酸化酵素欠損症、独立型;大動脈弁上狭窄;肺サーファクタント代謝機能障害、2及び3;指節癒合症、近位、1b;セナニー・レンツ型合指症;3型合指症;症候性X連鎖性精神遅滞16;内反尖足;タンジール病;TARP症候群;テイ・サックス病、B1バリアント、Gm2ガングリオシドーシス(成人)、Gm2ガングリオシドーシス(成人発症);テムタリー(Temtamy)症候群;テノリオ症候群;肢端骨異形成;テストステロン17-ベータ-デヒドロゲナーゼ欠損症;無四肢症、常染色体劣性;ファロー四徴症;左心低形成症候群2;動脈瘤;心臓及び大血管の奇形;心室中隔欠損症1;ティール・ベーンケ(Thiel-Behnke)角膜ジストロフィー;胸部大動脈瘤及び大動脈解離;マルファン様体型;3M症候群2;血小板減少症、血小板機能障害、溶血、及びグロビン合成不均衡;血小板減少症、X連鎖性;血栓症、遺伝性、プロテインC欠損症に起因、常染色体優性及び劣性;甲状腺無形成;甲状腺がん、濾胞性;甲状腺ホルモン代謝異常;甲状腺ホルモン抵抗性、汎発性、常染色体優性;甲状腺中毒性周期性四肢麻痺及び甲状腺中毒性周期性四肢麻痺2;サイトロピン放出ホルモン抵抗性、汎発性;チモシー症候群;TNF受容体関連周期熱症候群(TRAPS);歯無発生、選択的、3及び4;多形性心室頻拍(Torsades de pointes);タウンズ・ブロックス鰓弓耳腎様症候群;新生児の一過性表皮水疱症;トリーチャー・コリンズ症候群1;精神遅滞、小人症、及び網膜の色素変性を伴う長睫毛症;三叉神経節異形成症I型;三叉神経節症候群3型;トリメチルアミン尿症;結節性硬化症症候群;リンパ管筋腫症;結節性硬化症1及び2;チロシナーゼ陰性眼皮膚白皮症;チロシナーゼ陽性眼皮膚白皮症;チロシン血症I型;UDPグルコース-4-エピメラーゼ欠損症;ウルリッヒ型先天性筋ジストロフィー;重度の四肢欠損を伴う尺骨・腓骨欠損症;アップショー・シュールマン症候群;ウロカニン酸加水酵素欠損症;アッシャー症候群、1型,1B型,1D型,1G型,2A型,2C型及び2D型;網膜色素変性症39;紫外線感受性症候群;ファン・デル・ウーデ(Van der Woude)症候群;ヴァン・マルダーゲム(Van Maldergem)症候群2;ヘンカムリンパ管拡張症-リンパ浮腫症候群2;異型ポルフィリン症;嚢胞性腎疾患を伴う脳室拡大;ヴェルヘイ(Verheij)症候群;超長鎖アシル-CoA脱水素酵素欠損症;膀胱尿管逆流症8;内臓逆位5、常染色体;腹部ミオパチー;ビタミンD依存性くる病、1型及び2型;卵黄型ジストロフィー;フォン・ヴィルブランド病2M型及び3型;ワールデンブルグ症候群1型,4C型及び2E型(神経学的病変を伴う);クライン・ワールデンブルグ症候群;ウォーカー・ワールブルグ先天性筋ジストロフィー;ワールブルグ・マイクロ症候群2及び4;イボ、低ガンマグロブリン血症、感染症、及び骨髄性細胞貯留;ウィーバー症候群;ヴァイユ・マルケザーニ症候群1及び3;ヴァイユ・マルケザーニ様症候群;ワイセンバッハー・ツウェイミュラー(Weissenbacher-Zweymuller)症候群;ウェルドニッヒ・ホフマン病;シャルコー・マリー・トゥース病;ウェルナー症候群;WFS1関連障害;ウィデマン・スタイナー症候群;ウィルソン病;ウォルフラム様症候群、常染色体優性;ワース(Worth)病;ファン・ブッヘム病2型;色素性乾皮症、相補群b,群D,群E及び群G;X連鎖無ガンマグロブリン血症;X連鎖性 遺伝性運動感覚性ニューロパチー;ステリルスルファターゼ欠損症を伴うX連鎖性魚鱗癬;X連鎖性脳室周囲異所性灰白質;耳口蓋指症候群、I型;X連鎖性重症複合免疫不全症;チンメルマン・レーバンド(Zimmermann-Laband)症候群及びチンメルマン・レーバンド症候群2;並びに小帯(Zonular)粉状白内障3。
標的ヌクレオチド配列は、疾患、障害、又は疾病に関連する標的配列(例として点変異)を含んでもよい。標的配列は疾患、障害、又は疾病に関連するT→C(又はA→G)の点変異を含んでもよく、変異体C塩基の脱アミノ化は、疾患、障害、又は疾病に関連しない配列へのミスマッチ修復によって媒介される修正をもたらす。標的配列は疾患、障害、又は疾病に関連するG→A(又はC→T)の点変異を含んでもよく、変異体A塩基の脱アミノ化は、疾患、障害、又は疾病に関連しない配列へのミスマッチ修復によって媒介される修正をもたらす。標的配列はタンパク質をコードしてもよく、点変異がコドンにあるところでは、野生型コドンと比較して変異体コドンによってコードされるアミノ酸の変化をもたらす。標的配列はスプライス部位にもあってもよく、点変異は野生型転写物と比較してmRNA転写物のスプライシングの変化をもたらす。加えて、標的は遺伝子の非コード配列、例えばプロモータにもあってもよく、点変異は遺伝子の増大又は減少した発現をもたらす。
それゆえに、いくつかの側面では、変異体Cの脱アミノ化は変異体コドンによってコードされるアミノ酸の変化をもたらし、これはいくつかのケースでは野生型アミノ酸の発現をもたらし得る。他の側面では、変異体Aの脱アミノ化は変異体コドンによってコードされるアミノ酸の変化をもたらし、これはいくつかのケースでは野生型アミノ酸の発現をもたらし得る。
細胞を組成物又はrAAV粒子と接触させることを伴う本明細書に記載の方法は、in vitroで、ex vivoで、又はin vivoで生起し得る。ある態様において、接触させる工程は対象に生起する。ある態様において、対象は疾患、障害、又は疾病と診断されている。
いくつかの態様では、本明細書に開示の方法には、哺乳動物細胞を組成物又はrAAV粒子と接触させることを伴う。具体的な態様では、方法には、網膜細胞、皮質細胞、又は小脳細胞に接触することを伴う。
本明細書に記載の方法を用いて送達される分裂されたCas9タンパク質又は分裂されたプライム編集因子は、好ましくは、元のCas9タンパク質又はプライム編集因子(すなわち、丸ごと細胞に送達又は細胞によって発現される未分裂のタンパク質)と比較して遜色ない活性を有する。例えば、分裂されたCas9タンパク質又は分裂されたプライム編集因子は、元のCas9タンパク質又はプライム編集因子の活性の少なくとも50%(例として、少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%、又は100%)を保持する。いくつかの態様では、分裂されたCas9タンパク質又は分裂されたプライム編集因子は、元のCas9タンパク質又はプライム編集因子のものよりも(例として、2倍、5倍、10倍、100倍、1000倍、又はより多く)活性である。
本明細書に記載の組成物は、対象が患っている疾患又は障害を処置及び/又は防止するための処置学的有効量でその必要がある対象に投与されてもよい。CRISPR/Cas9に基づくゲノム編集テクノロジーを用いて処置及び/又は防止されてもよいいずれかの疾患又は障害は、本明細書に記載の分裂されたCas9タンパク質又は分裂されたプライム編集因子によって処置されてもよい。分裂されたCas9タンパク質又はプライム編集因子をコードするヌクレオチド配列が更にgRNAをコードしない場合には、gRNAをコードする別個の核酸ベクターが本明細書に記載の組成物と一緒に投与されてもよいということは理解されるはずである。
例示の好適な疾患、障害、又は疾病は、限定なしに:嚢胞性線維症、フェニルケトン尿症、表皮剥離性角質増殖症(EHK)、慢性閉塞性肺疾患(COPD)、シャルコー・マリー・トゥース病4J型、神経芽腫(NB)、フォン・ヴィレブランド病(vWD)、先天性ミオトニー、遺伝性腎アミロイドーシス、拡張型心筋症、遺伝性リンパ浮腫、家族性アルツハイマー病、プリオン病、慢性乳児神経皮膚関節症候群(CINCA)、先天性難聴、ニーマン・ピック病C型(NPC)疾患、及びデスミン関連ミオパチー(DRM)からなる群から選択される疾患又は障害を包含する。具体的な態様では、疾患又は疾病はニーマン・ピック病C型(NPC)疾患である。
いくつかの態様では、疾患、障害、又は状態はNPC遺伝子、DNMT1遺伝子、PCSK9遺伝子、又はTMC1遺伝子の点変異に関連される。ある態様において、点変異はNPCのT3182C変異であり、これはI1061Tアミノ酸置換をもたらす。
ある態様において、点変異はTMC1のA545G変異であり、これはY182Cアミノ酸置換をもたらす。TMC1は内耳の感覚有毛細胞の機械受容イオンチャネルを形成するタンパク質をコードし、正常な聴覚機能に要求される。Y182Cアミノ酸置換は先天性難聴に関連される。
いくつかの態様では、疾患、障害、又は疾病は、停止コドン、例えば遺伝子のコード領域内の未成熟停止コドンを生成する点変異に関連される。
追加の例示の疾患、障害、又は疾病は、嚢胞性線維症(例として、Schwank et al.,Functional repair of CFTR by CRISPR/Cas9 in intestinal stem cell organoids of cystic fibrosis patients.Cell stem cell.2013;13:653-658;及びWu et.al.,Correction of a genetic disease in mouse via use of CRISPR-Cas9.Cell stem cell.2013;13:659-662を見よ。これらのいずれも、遺伝子欠陥を修正するためにデアミナーゼ融合タンパク質を用いていない);フェニルケトン尿症-例として、フェニルアラニンヒドロキシラーゼ遺伝子の位置835(マウス)若しくは240(ヒト)又は相同残基におけるフェニルアラニン→セリンの変異(T>C変異)-例としてMcDonald et al.,Genomics.1997;39:402-405を見よ;ベルナール・スーリエ症候群(BSS)-例として、血小板膜糖タンパク質IXの位置55若しくは相同残基におけるフェニルアラニン→セリンの変異又は残基24若しくは相同残基におけるシステイン→アルギニン(T>C変異)-例としてNoris et al.,British Journal of Haematology.1997;97:312-320及びAli et al.,Hematol.2014;93:381-384を見よ;表皮剥離性角質増殖症(EHK)-例として、ケラチン1の位置160若しくは161(開始メチオニンをカウントする場合)又は相同残基におけるロイシン→プロリンの変異(T>C変異)-例として、Chipev et al.,Cell.1992;70:821-828を見よ。www[dot]uniprot[dot]orgのUNIPROTデータベースのアクセッション番号P04264をもまた見よ;慢性閉塞性肺疾患(COPD)-例として、α1-アンチトリプシンのプロセシングされた形態の位置54若しくは55(開始メチオニンをカウントする場合)又は相同残基あるいはプロセシングされない形態の残基78又は相同残基におけるロイシン→プロリンの変異(T>C変異)-例として、Poller et al.,Genomics.1993;17:740-743を見よ。UNIPROTデータベースのアクセッション番号P01011をもまた見よ;シャルコー・マリー・トゥース病4J型-例としてFIG4の位置41又は相同残基におけるイソロイシン→トレオニンの変異(T>C変異)-例として、Lenk et al.,PLoS Genetics.2011;7:e1002104を見よ;神経芽腫(NB)-例として、カスパーゼ-9の位置197又は相同残基におけるロイシン→プロリンの変異(T>C変異)-例として、Kundu et al.,3 Biotech.2013,3:225-234を見よ;フォン・ヴィレブランド病(vWD)-例として、フォン・ヴィレブランド因子のプロセシングされた形態の位置509若しくは相同残基又はフォン・ヴィレブランド因子のプロセシングされない形態の位置1272若しくは相同残基におけるシステイン→アルギニンの変異(T>C変異)-例として、Lavergne et al.,Br.J.Haematol.1992を見よ。UNIPROTデータベースのアクセッション番号P04275をもまた見よ;82:66-72;先天性ミオトニー-例として、筋肉塩素チャネル遺伝子CLCN1の位置277又は相同残基におけるシステイン→アルギニンの変異(T>C変異)-例として、Weinberger et al.,The J.of Physiology.2012;590:3449-3464を見よ;遺伝性腎アミロイドーシス-例として、アポリポタンパク質AIIのプロセシングされた形態の位置78若しくは相同残基又はプロセシングされない形態の位置101若しくは相同残基における停止コドン→アルギニンの変異(T>C変異)-例として、Yazaki et al.,Kidney Int.2003;64:11-16を見よ;拡張型心筋症(DCM)-例として、FOXD4遺伝子の位置148又は相同残基におけるトリプトファン→アルギニンの変異(T>C変異)、例として、Minoretti et.al.,Int.J.of Mol.Med.2007;19:369-372を見よ;遺伝性リンパ浮腫-例として、VEGFR3チロシンキナーゼの位置1035又は相同残基におけるヒスチジン→アルギニンの変異(A>G変異)、例として、Irrthum et al.,Am.J.Hum.Genet.2000;67:295-301を見よ;家族性アルツハイマー病-例として、プレセニリン1の位置143又は相同残基におけるイソロイシン→バリンの変異(A>G変異)、例として、Gallo et.al.,J.Alzheimer's Disease.2011;25:425-431を見よ;プリオン病-例として、プリオンタンパク質の位置129又は相同残基におけるメチオニン→バリンの変異(A>G変異)-例として、Lewis et.al.,J.of General Virology.2006;87:2443-2449を見よ;慢性乳児神経皮膚関節症候群(CINCA)-例として、クリオピリンの位置570又は相同残基におけるチロシン→システインの変異(A>G変異)-例として、Fujisawa et.al.Blood.2007;109:2903-2911を見よ;並びにデスミン関連ミオパチー(DRM)-例として、αβクリスタリンの位置120又は相同残基におけるアルギニン→グリシンの変異(A>G変異)を包含する。例として、Kumar et al.,J.Biol.Chem.1999;274:24137-24141を見よ。全ての参照及びデータベースエントリーの内容全体は参照によって本明細書に組み込まれる。
トリヌクレオチド反復拡大疾患
トリヌクレオチド反復拡大は、ハンチントン病、脆弱X症候群、及びフリードライヒ運動失調症を包含するいくつものヒト疾患に関連される。最も一般的なトリヌクレオチドリピートはCAGトリプレットを含有するが、GAAトリプレット(フリードライヒ運動失調症)及びCGGトリプレット(脆弱X症候群)も生じる。伸長に対する素因を継承すること、又はすでに伸長した親対立遺伝子を獲得することは、疾患を獲得する可能性を増加させる。トリヌクレオチドリピートの病原性伸長は、プライム編集を使用して仮説的に修正され得た。
トリヌクレオチド反復拡大は、ハンチントン病、脆弱X症候群、及びフリードライヒ運動失調症を包含するいくつものヒト疾患に関連される。最も一般的なトリヌクレオチドリピートはCAGトリプレットを含有するが、GAAトリプレット(フリードライヒ運動失調症)及びCGGトリプレット(脆弱X症候群)も生じる。伸長に対する素因を継承すること、又はすでに伸長した親対立遺伝子を獲得することは、疾患を獲得する可能性を増加させる。トリヌクレオチドリピートの病原性伸長は、プライム編集を使用して仮説的に修正され得た。
図1G又は図22に概説されている一般的な機序に従って、反復領域の上流の領域が、RNAによってガイドされるヌクレアーゼによってニックが入れられ得、次いで、健康な反復数(これは具体的な遺伝子及び疾患に依存する)を含有する新たなDNA鎖の合成をプライムするために用いられ得る。反復配列の後には、反復の他端に隣接する配列の同一性にマッチする相同性の短いストレッチが追加される(赤色の鎖)。プライム編集因子による新たに合成された鎖の侵入、及び新たに合成されたフラップによる内生DNAの続く置換えは、短縮された反復アレルに至る。用語「短縮された」はヌクレオチド反復領域の長さの短くなることを指し、それによって、トリヌクレオチド反復領域を修復することをもたらす。
本明細書に記載のプライム編集システム又はプライム編集(PE)システムは、ハンチントン病及び他のトリヌクレオチド反復障害等の疾病を処置するために、トリヌクレオチド反復変異(又は「トリプレット拡大疾患」)を短縮するために用いられてもよい。トリヌクレオチド反復拡大障害は発達神経生物学を伴う複雑な進行性の障害であり、多くの場合には、認知及び感覚運動機能に影響する。障害は遺伝学的な表現促進現象(すなわち、各世代での増大した重症度)を示す。DNA伸長又は短縮は、通常は減数分裂的に(すなわち、配偶子形成の時の間に、又は胚発生中に早期に)起こり、多くの場合には性バイアスを有し、いくつかの遺伝子は女性から受け継がれるときにのみ、他は男性からのみ伸長するということを意味する。ヒトでは、トリヌクレオチド反復拡大障害は転写又は翻訳レベルどちらかで遺伝子サイレンシングを引き起こし得、これは本質的に遺伝子機能をノックアウトする。代替的には、トリヌクレオチド反復拡大障害は、大きい繰り返しアミノ酸配列を有して生成された変改されたタンパク質を引き起こし得る。これは、多くの場合にはドミナントネガティブな様式で(例としてポリグルタミン疾患)、タンパク質機能を無効化するか又は変化させるかどちらかである。
理論によって拘束されることなしに、トリプレット拡大はDNA複製又はDNA修復合成の間のスリップによって引き起こされる。タンデム反復は互いに同一の配列を有するので、2つのDNA鎖間の塩基対形成は配列上の複数の点において生起し得る。これはDNA複製又はDNA修復合成の間の「ループアウト」構造の形成に至ってもよい。これは繰り返し配列の繰り返しのコピーに至ってもよく、反復数を伸長させる。ハイブリッドRNA:DNA中間体を伴う追加の機序が提案されている。プライム編集は、害をなす繰り返しのコドントリプレットの1以上を欠失させることによって、これらのトリプレット拡大領域を縮減又は消去するために用いられてもよい。この使用のある態様では、図23が、プライム編集によってトリヌクレオチド反復配列を短縮又は縮減するためのpegRNAデザインの模式図を提供する。
プライム編集は、カットされる部位へと標的化される充当されたpegRNAを含むプライム編集因子によって、トリプレット反復領域の上流の領域へニックを入れることによって、トリプレット拡大領域を短縮するように実装されてもよい。次いで、プライム編集因子は、健康な数のトリプレット反復(これは具体的な遺伝子及び疾患に依存する)をコードする鋳型(すなわち、その編集鋳型)としてのpegRNAに基づく新たなDNA鎖(ssDNAフラップ)を合成する。新たに合成されたssDNA鎖は健康なトリプレット反復配列を含み、また、反復の他端に隣接する配列にマッチする相同性の短いストレッチ(すなわち相同アーム)を包含するように合成される(赤色の鎖)。新たに合成された鎖の侵入、及び新たに合成されたssDNAフラップによる内生DNAの続く置換えは、短縮された反復アレルに至る。
具体的なトリヌクレオチド伸長障害に依存して、欠陥を誘導するトリプレット拡大は「トリヌクレオチド反復拡大タンパク質」において生起してもよい。トリヌクレオチド反復拡大タンパク質は、トリヌクレオチド反復拡大障害を発生する感受性、トリヌクレオチド反復拡大障害の存在、トリヌクレオチド反復拡大障害の重症度、又はそれらのいずれかの組み合わせに関連するタンパク質の多様なセットである。トリヌクレオチド反復拡大障害は反復の型によって決定される2つのカテゴリーに分けられる。最も普通の反復はトリプレットCAGであり、これは遺伝子のコード領域に存在するときにはアミノ酸のグルタミン(Q)をコードする。よって、これらの障害はポリグルタミン(ポリQ)障害と呼ばれ、次の疾患を含む:ハンチントン病(HD);球脊髄性筋萎縮症(SBMA);脊髄小脳失調症(SCA1、2、3、6、7、及び17型);及び歯状核赤核淡蒼球ルイ体萎縮症(DRPLA)。残りのトリヌクレオチド反復拡大障害にはCAGトリプレットを伴わないか、又はCAGトリプレットが遺伝子のコード領域にはないかどちらかであり、よって、非ポリグルタミン障害と呼ばれる。非ポリグルタミン障害は、脆弱X症候群(FRAXA);脆弱XE精神遅滞(FRAXE);フリードライヒ運動失調症(FRDA);筋強直性ジストロフィー(DM);及び脊髄小脳失調症(SCA 8及び12型)を含む。
トリヌクレオチド反復拡大障害に関連するタンパク質は、トリヌクレオチド反復拡大障害に対するトリヌクレオチド反復拡大障害に関連するタンパク質の実験的関連に基づいて選択され得る。例えば、トリヌクレオチド反復拡大障害に関連するタンパク質の生産速度又は循環濃度は、トリヌクレオチド反復拡大障害を欠く集団に対して相対的に、トリヌクレオチド反復拡大障害を有する集団において上昇又は降下されてもよい。タンパク質レベルの違いは、ウエスタンブロット、免疫組織化学染色、酵素結合免疫吸着アッセイ(ELISA)、及び質量分析を包含するがこれらに限定されないプロテオミクス技術を用いて評価されてもよい。代替的には、トリヌクレオチド反復拡大障害に関連するタンパク質はタンパク質をコードする遺伝子の遺伝子発現プロファイルを得ることによって同定されてもよく、DNAマイクロアレイ分析、遺伝子発現の連続分析(SAGE)、及び定量的リアルタイムポリメラーゼ連鎖反応(Q-PCR)を包含するがこれらに限定されないゲノム技術を用いる。
プライム編集によって修正され得るトリヌクレオチド反復拡大障害に関連するタンパク質の限定しない例は、AR(アンドロゲン受容体)、FMR1(脆弱X精神遅滞1)、HTT(ハンチンチン)、DMPK(筋緊張性ジストロフィータンパク質キナーゼ)、FXN(フラタキシン)、ATXN2(アタキシン2)、ATN1(アトロフィン1)、FEN1(フラップ構造特異的エンドヌクレアーゼ1)、TNRC6A(トリヌクレオチド反復含有6A)、PABPN1(核ポリ(A)結合タンパク質1)、JPH3(ジャンクトフィリン3)、MED15(メディエーター複合体サブユニット15)、ATXN1(アタキシン1)、ATXN3(アタキシン3)、TBP(TATAボックス結合タンパク質)、CACNA1A(電位依存性カルシウムチャネルP/Q型アルファ1Aサブユニット)、ATXN80S(ATXN8対向鎖(非タンパク質コード))、PPP2R2B(タンパク質ホスファターゼ2制御サブユニットBベータ)、ATXN7(アタキシン7)、TNRC6B(トリヌクレオチド反復含有6B)、TNRC6C(トリヌクレオチド反復含有6C)、CELF3(CUGBP Elav様ファミリーメンバー3)、MAB21L1(mab-21様1(C.elegans))、MSH2(mutSホモログ2結腸がん非ポリポーシス1型(E.coli))、TMEM185A(膜貫通タンパク質185A)、SIX5(SIXホメオボックス5)、CNPY3(canopy 3ホモログ(ゼブラフィッシュ))、FRAXE(脆弱部位、葉酸型、稀、fra(X)(q28)E)、GNB2(グアニンヌクレオチド結合タンパク質(Gタンパク質)ベータポリペプチド2)、RPL14(リボソームタンパク質L14)、ATXN8(アタキシン8)、INSR(インスリン受容体)、TTR(トランスサイレチン)、EP400(E1A結合タンパク質p400)、GIGYF2(GRB10相互作用GYFタンパク質2)、OGG1(8-オキソグアニンDNAグリコシラーゼ)、STC1(スタニオカルシン1)、CNDP1(カルノシンジペプチダーゼ1(メタロペプチダーゼM20ファミリー))、C10orf2(染色体10オープンリーディングフレーム2)、MAML3 mastermind様3(Drosophila)、DKC1(先天性角化不全症1、ジスケリン)、PAXIP1(PAX相互作用(転写活性化ドメイン)タンパク質1)、CASK(カルシウム/カルモジュリン依存性セリンタンパク質キナーゼ(MAGUKファミリー))、MAPT(微小管関連タンパク質tau)、SP1(Sp1転写因子)、POLG(ポリメラーゼ(DNA依存性)ガンマ)、AFF2(AF4/FMR2ファミリーメンバー2)、THBS1(トロンボスポンジン1)、TP53(腫瘍タンパク質p53)、ESR1(エストロゲン受容体1)、CGGBP1(CGGリプレット反復結合タンパク質1)、ABT1(基本転写活性化因子1)、KLK3(カリクレイン関連ペプチダーゼ3)、PRNP(プリオンタンパク質)、JUN(junがん遺伝子)、KCNN3(カリウム中/小コンダクタンスカルシウム活性化チャネルサブファミリーNメンバー3)、BAX(BCL2関連Xタンパク質)、FRAXA(脆弱部位、葉酸型、稀、fra(X)(q27.3)A(巨睾丸、精神遅滞))、KBTBD10(kelch反復及びBTB(POZ)ドメイン含有10)、MBNL1(muscleblind様(Drosophila))、RAD51(RAD51ホモログ(RecAホモログ、E.coli)(S.cerevisiae))、NCOA3(核受容体コアクチベーター3)、ERDA1(伸長した反復ドメインCAG/CTG1)、TSC1(結節性硬化症1)、COMP(軟骨オリゴマーマトリックスタンパク質)、GCLC(グルタミン酸-システインリガーゼ触媒サブユニット)、RRAD(糖尿病に関連するRas関連)、MSH3(mutSホモログ3(E.coli))、DRD2(ドーパミン受容体D2)、CD44(CD44分子(インド人血液型))、CTCF(CCCTC結合因子(ジンクフィンガータンパク質))、CCND1(サイクリンD1)、CLSPN(クラスピンホモログ(Xenopus laevis))、MEF2A(筋細胞エンハンサー因子2A)、PTPRU(タンパク質チロシンホスファターゼ受容体型U)、GAPDH(グリセルアルデヒド-3-リン酸デヒドロゲナーゼ)、TRIM22(三節型モチーフ含有22)、WT1(ウィルムス腫瘍1)、AHR(アリール炭化水素受容体)、GPX1(グルタチオンペルオキシダーゼ1)、TPMT(チオプリンS-メチルトランスフェラーゼ)、NDP(ノリエ病(偽神経膠腫))、ARX(aristaless関連ホメオボックス)、MUS81(MUS81エンドヌクレアーゼホモログ(S.cerevisiae))、TYR(チロシナーゼ(眼皮膚白皮症IA))、EGR1(初期増殖応答1)、UNG(ウラシル-DNAグリコシラーゼ)、NUMBL(numbホモログ(Drosophila)様)、FABP2(腸脂肪酸結合タンパク質2)、EN2(engrailedホメオボックス2)、CRYGC(クリスタリンガンマC)、SRP14(シグナル認識粒子14kDa(相同的Alu RNA結合タンパク質))、CRYGBクリスタリンガンマB)、PDCD1(プログラム細胞死1)、HOXA1(ホメオボックスA1)、ATXN2L(アタキシン2様)、PMS2(PMS2減数分裂後分離増大2(S.cerevisiae))、GLA(ガラクトシダーゼアルファ)、CBL(Cas-Br-M(ネズミ)同種指向性レトロウイルス形質トランスバーション配列)、FTH1(フェリチン重鎖ポリペプチド1)、IL12RB2(インターロイキン12受容体ベータ2)、OTX2(orthodenticleホメオボックス2)、HOXA5(ホメオボックスA5)、POLG2(ポリメラーゼ(DNA依存性)ガンマ2補助的サブユニット)、DLX2(distal-lessホメオボックス2)、SIRPA(シグナル制御タンパク質アルファ)、OTX1(orthodenticleホメオボックス1)、AHRR(アリール炭化水素受容体リプレッサー)、MANF(中脳アストロサイト由来神経栄養因子)、TMEM158(膜貫通タンパク質158(遺伝子/偽遺伝子))、及びENSG00000078687を包含する。
具体的な側面において、本開示は、拡大反復障害(また反復拡大障害若しくはトリヌクレオチド反復障害としても知られている)と診断された対象の処置のためのプライム編集を提供する。拡大反復障害は、マイクロサテライト反復が長さ閾値を超えて拡大したときに生じる。目下、少なくとも30の遺伝的疾患は、反復拡大によって引き起こされるものと考えられている。この多様な群の障害への科学的理解は、1990年代初頭に、トリヌクレオチド反復が、脆弱X、球脊髄性筋萎縮症、筋強直症ジストロフィー、及びハンチントン病(Nelson et al,「The unstable repeats-three evolving faces of neurological disease,」Neuron,March 6,2013,Vol.77;825-843(これは参照により本明細書に組み込まれる))、並びにホー・リバー(Haw River)症候群、ヤコブセン症候群、歯状核赤核淡蒼球ルイ体萎縮症(DRPLA)、マシャド・ジョセフ病、合多指症(SPD II)、手足生殖器症候群(HFGS)、鎖骨頭蓋異形成症(CCD)、全前脳欠損障害(HPE)、先天性中枢性低換気症候群(CCHS)、ARX非症候性のX連鎖精神遅滞(XLMR)、及び眼咽頭型筋ジストロフィー(OPMD)(参照を包含する、数種の主要な遺伝性疾病にあるという発見で明るみに出た。マイクロサテライト反復の不安定性は、反復拡大が、後に続く各世代に生じ得、子孫におけるより重度の表現型及び発症の低年齢へ繋がるという現象が予想された通り、これらの疾病の証であることが見出された。反復拡大は、数種の異なる機序を介して疾患を引き起こすものと考えられている。つまり、拡大は、遺伝子、mRNA転写産物、及び/又はコードされたタンパク質のレベルにて機能する細胞に干渉してもよい。いくつかの疾病において、突然変異は、反復含有遺伝子をサイレンシングすることによる機能喪失型の機序を介して作用する。その他において、疾患は、mRNA転写産物又はタンパク質のいずれかが新しい異常な機能を帯びるようになる機能獲得型の機序の結果として生じる。
一態様において、トリヌクレオチド反復障害を処置する方法は、図23に描かれている。一般に、アプローチは、プライム編集プロセスの機序を通して内生の罹患トリヌクレオチド反復配列を置き換えることが意図される、所望されかつ健常な置換えトリヌクレオチド反復配列をコードする領域を含むpegRNAと組み合わせて、プライム編集を使用することを伴う。例示の、トリヌクレオチド反復配列を縮小するためのgRNAデザイン及びプライム編集でのトリヌクレオチド反復縮小の概略図が、図23に示される。
プリオン病
プライム編集は、疾患の過程においてミスフォールディングされるようになるプリオンタンパク質(PRNP)上への1以上の防護性変異の組み入れによって、プリオン病を防止又はその進行を停止させるためにもまた用いられ得る。プリオン病又は伝達性海綿状脳症(TSE)は、ヒト及び動物両方を冒す稀な進行性の神経変性障害のファミリーである。それらは、長い潜伏期間、ニューロン喪失に関連する特徴的な海綿状変化、及び炎症性応答を誘導することの失敗によって見分けられる。
プライム編集は、疾患の過程においてミスフォールディングされるようになるプリオンタンパク質(PRNP)上への1以上の防護性変異の組み入れによって、プリオン病を防止又はその進行を停止させるためにもまた用いられ得る。プリオン病又は伝達性海綿状脳症(TSE)は、ヒト及び動物両方を冒す稀な進行性の神経変性障害のファミリーである。それらは、長い潜伏期間、ニューロン喪失に関連する特徴的な海綿状変化、及び炎症性応答を誘導することの失敗によって見分けられる。
ヒトでは、プリオン病は、クロイツフェルト・ヤコブ病(CJD)、変異型クロイツフェルト・ヤコブ病(vCJD)、ゲルストマン・ストロイスラー・シャインカー症候群、致死性家族性不眠症、及びクールー病を包含する。動物では、プリオン病は牛海綿状脳症(BSE又は「狂牛病」)、慢性消耗病(CWD)、スクレイピー、伝達性ミンク脳症、ネコ海綿状脳症、及び有蹄類海綿状脳症を包含する。これらのプリオン病のいずれか1つを防止又はその進行を停止させるために、プライム編集は防護性の点変異をプリオンタンパク質上に組み入れるために用いられてもよい。
古典的CJDはヒトプリオン病である。それは、特徴的な臨床及び診断特色を有する神経変性障害である。この疾患は急速に進行性であり、常に致死的である。この疾患による感染は通常は病気の発症から1年以内に死に至る。CJDは急速に進行性の変わることなく致死的な神経変性障害であり、プリオンタンパク質として公知の細胞性の糖タンパク質の異常なアイソフォームによって引き起こされると信じられている。CJDは世界的に生起し、米国を包含する多くの国の見積もられる年間発生率は100万人の集団当たり約1ケースであることが報告されている。CJD患者の大多数は通常は病気の発症から1年以内に死ぬ。CJDは、ヒト及び動物において生起する他のプリオン病と併せて、伝達性海綿状脳症(TSE)として分類される。患者の約85%では、CJDは伝達の認識可能なパターンなしに特発性疾患として生起する。患者のより小さい割合(5~15%)は、プリオンタンパク質遺伝子の受け継がれた変異ゆえにCJDを発生する。これらの受け継がれる形態は、ゲルストマン・ストロイスラー・シャインカー症候群及び致死性家族性不眠症を包含する。CJDについて、処置は現行では公知ではない。
変異型クロイツフェルト・ヤコブ病(vCJD)は英国で1996年に最初に記載されたプリオン病である。今や、牛のプリオン病の牛海綿状脳症(BSE又は「狂牛」病)のアウトブレイクを担う物質が、ヒトのvCJDのアウトブレイクを担う同じ物質であるという強い科学的証拠がある。バリアントCJD(vCJD)は古典的CJD(多くの場合には単純にCJDと呼ばれる)と同じ疾患ではない。それは古典的CJDとは異なる臨床的及び病理学的特徴を有する。各疾患はプリオンタンパク質遺伝子の具体的な遺伝子プロファイルをもまた有する。両方の障害は、年で測定される普通でなく長い潜伏期間を有する変わることなく致死的な脳疾患であり、プリオンと呼ばれる非従来的な伝達性物質によって引き起こされる。vCJDについて、処置は現行では公知ではない。
BSE(牛海綿状脳症又は「狂牛病」)は、プリオンと呼ばれる普通でない伝達性物質による感染からもたらされる畜牛の進行性の神経学的障害である。伝達性物質の性質は良く理解されてはいない。現行では、最も受け入れられた理論は、物質が、プリオンタンパク質として公知の正常タンパク質の修飾された形態であるということである。まだ理解されていない理由から、正常なプリオンタンパク質は病原性の(有害な)形態へと変化し、これは次いで畜牛の中枢神経系を損傷する。BSEの異なる株があるという増大して行く証拠がある:英国におけるアウトブレイクを担った典型的又は古典的BSE株及び2つの非典型的な株(H及びL株)である。BSEについて、処置は現行では公知ではない。
慢性消耗病(CWD)は、鹿、ヘラジカ、トナカイ、ニホンジカ、及びムースを冒すプリオン病である。それはカナダ及び米国を包含する北米、ノルウェー、並びに韓国のいくつかのエリアにおいて見出されている。感染動物が症状を発生する前には1年超がかかってもよく、これらは激しい体重減少(消耗)、よろめき、無気力、及び他の神経症状を包含し得る。CWDは全ての年齢の動物を冒し得、いくつかの感染動物はいつか疾患を発生することなしに死に至ることもある。CWDは動物にとって致死的であり、処置又はワクチンはない。
TSEの原因物質はプリオンであると信じられている。用語「プリオン」は、伝達性であり、脳に最も豊富に見出されるプリオンタンパク質と呼ばれる特定の正常な細胞性タンパク質の異常なフォールディングを誘導する能力がある異常な病原性物質を指す。これらの正常なプリオンタンパク質の機能はなお完全には理解されていない。プリオンタンパク質の異常なフォールディングは脳損傷と疾患の特徴的な徴候及び症状とに至る。プリオン病は通常は急速に進行性であり、常に致死的である。
本明細書において用いられる用語「プリオン」は、ヒト及び動物の疾患(海綿状脳症)を引き起こすことが公知の感染性粒子を意味する。用語「プリオン」は単語「タンパク質」及び「感染」の短縮であり、粒子は、専らではないにしても、大きくは、立体配座を変化させてPRNPScになるPRNPCを発現するPRNP遺伝子によってコードされるPRNPSc分子からなる。プリオンは細菌、ウイルス、及びウイロイドとは別物である。公知のプリオンは、動物に感染して、羊及び山羊の神経系の伝達性の変性疾患スクレイピー、並びに牛海綿状脳症(BSE)又は狂牛病及び猫のネコ海綿状脳症を引き起こすものを包含する。ヒトを冒すことが公知の上で論じられている4つのプリオン病は(1)クールー病、(2)クロイツフェルト・ヤコブ病(CJD)、(3)ゲルストマン・ストロイスラー・シャインカー病(GSS)、及び(4)致死性家族性不眠症(FFI)である。本明細書において用いられるプリオンは、用いられるいずれかの動物、特にヒト及び飼育畜産動物においてこれらの疾患又は他の全て又はいずれかを引き起こすプリオンの全ての形態を包含する。
一般的に、理論によって拘束されることなしに、プリオン病はプリオンタンパク質のミスフォールディングによって引き起こされる。多くの場合に沈着性疾患と呼ばれるかかる疾患、プリオンタンパク質のミスフォールディングは次の通り説明され得る。Aが、意図される生理的役割をモノマー又はオリゴマー状態で実行する正常に合成された遺伝子産物であり、かつA*が、劇的な立体配座変化を経過するために有能であるAの立体配座的に活性化された形態である場合には、Bは多量体アセンブリを好む立体配座的に変改された状態であり(すなわち、沈着を形成するミスフォールディングされた形態)、かつBnは、病原性であり、リサイクルすることが相対的に困難である多量体材料である。プリオン病では、PRNPC及びPRNPScは状態A及びBnに対応し、ここで、Aは大きくはヘリカルかつモノマー性であり、Bnはβリッチかつ多量体性である。
プリオンタンパク質のある種の変異はプリオン(prior)病の増大したリスクに関連し得るということが公知である。反対に、プリオンタンパク質のある種の変異は天然に防護性であり得る。Bagynszky et al.,「Characterization of mutations in PRNP(prion)gene and their possible roles in neurodegenerative diseases,」Neuropsychiatr Dis Treat.,2018;14:2067-2085を見よ。これの内容は参照によって本明細書に組み込まれる。
PRNP(NCBI RefSeq No.NP_000302.1(配列番号396))ヒトプリオンタンパク質は、染色体20(4686151-4701588)上に位置付けられた16kb長の遺伝子によってコードされる。それは2つのエキソンを含有し、エキソン2はオープンリーディングフレームを持ち、これは253アミノ酸(AA)長のPrPタンパク質をコードする。エキソン1は非コードエキソンであり、これは転写開始部位としての用をなしてもよい。翻訳後修飾は最初の22AA N末端フラグメント(NTF)及び最後の23AA C末端フラグメント(CTF)の除去をもたらす。NTFは小胞体(ER)へのPrP輸送後に切断され、CTF(グリコシルホスファチジルイノシトール[GPI]シグナルペプチド[GPI-SP])はGPIアンカーによって切断される。GPIアンカーはPrPタンパク質輸送を伴い得た。それは細胞膜の外側表面へのプリオンタンパク質の取り付けの役割をもまた果たしてもよい。正常なPrPは、長いN末端ループ(これはオクタペプチド反復領域を含有する)、2つの短いβシート、3つのαヘリックス、及びC末端領域(これはGPIアンカーを含有する)から成る。PrPの切断は、細胞膜にアンカー固定される208AA長の糖タンパク質をもたらす。
PRNP(NP_000302.1)の253アミノ酸配列は以下の通りである:
MANLGCWMLVLFVATWSDLGLCKKRPKPGGWNTGGSRYPGQGSPGGNRYPPQGGGGWGQPHGGGWGQPHGGGWGQPHGGGWGQPHGGGWGQGGGTHSQWNKPSKPKTNMKHMAGAAAAGAVVGGLGGYMLGSAMSRPIIHFGSDYEDRYYRENMHRYPNQVYYRPMDEYSNQNNFVHDCVNITIKQHTVTTTTKGENFTETDVKMMERVVEQMCITQYERESQAYYQRGSSMVLFSSPPV(配列番号396)
MANLGCWMLVLFVATWSDLGLCKKRPKPGGWNTGGSRYPGQGSPGGNRYPPQGGGGWGQPHGGGWGQPHGGGWGQPHGGGWGQPHGGGWGQGGGTHSQWNKPSKPKTNMKHMAGAAAAGAVVGGLGGYMLGSAMSRPIIHFGSDYEDRYYRENMHRYPNQVYYRPMDEYSNQNNFVHDCVNITIKQHTVTTTTKGENFTETDVKMMERVVEQMCITQYERESQAYYQRGSSMVLFSSPPV(配列番号396)
PRNP(NP_000302.1)の253アミノ酸配列は、以下のヌクレオチド配列(NCBI Ref.Seq No.NM_000311.5,「homo sapiens prion protein(PRNP),transcript variant 1,mRNA)によってコードされており、以下の通りである:
GCGAACCTTGGCTGCTGGATGCTGGTTCTCTTTGTGGCCACATGGAGTGACCTGGGCCTCTGCAAGAAGCGCCCGAAGCCTGGAGGATGGAACACTGGGGGCAGCCGATACCCGGGGCAGGGCAGCCCTGGAGGCAACCGCTACCCACCTCAGGGCGGTGGTGGCTGGGGGCAGCCTCATGGTGGTGGCTGGGGGCAGCCTCATGGTGGTGGCTGGGGGCAGCCCCATGGTGGTGGCTGGGGACAGCCTCATGGTGGTGGCTGGGGTCAAGGAGGTGGCACCCACAGTCAGTGGAACAAGCCGAGTAAGCCAAAAACCAACATGAAGCACATGGCTGGTGCTGCAGCAGCTGGGGCAGTGGTGGGGGGCCTTGGCGGCTACATGCTGGGAAGTGCCATGAGCAGGCCCATCATACATTTCGGCAGTGACTATGAGGACCGTTACTATCGTGAAAACATGCACCGTTACCCCAACCAAGTGTACTACAGGCCCATGGATGAGTACAGCAACCAGAACAACTTTGTGCACGACTGCGTCAATATCACAATCAAGCAGCACACGGTCACCACAACCACCAAGGGGGAGAACTTCACCGAGACCGACGTTAAGATGATGGAGCGCGTGGTTGAGCAGATGTGTATCACCCAGTACGAGAGGGAATCTCAGGCCTATTACCAGAGAGGATCGAGCATGGTCCTCTTCTCCTCTCCACCTGTGATCCTCCTGATCTCTTTCCTCATCTTCCTGATAGTGGGATGAGGAAGGTCTTCCTGTTTTCACCATCTTTCTAATCTTTTTCCAGCTTGAGGGAGGCGGTATCCACCTGCAGCCCTTTTAGTGGTGGTGTCTCACTCTTTCTTCTCTCTTTGTCCCGGATAGGCTAATCAATACCCTTGGCACTGATGGGCACTGGAAAACATAGAGTAGACCTGAGATGCTGGTCAAGCCCCCTTTGATTGAGTTCATCATGAGCCGTTGCTAATGCCAGGCCAGTAAAAGTATAACAGCAAATAACCATTGGTTAATCTGGACTTATTTTTGGACTTAGTGCAACAGGTTGAGGCTAAAACAAATCTCAGAACAGTCTGAAATACCTTTGCCTGGATACCTCTGGCTCCTTCAGCAGCTAGAGCTCAGTATACTAATGCCCTATCTTAGTAGAGATTTCATAGCTATTTAGAGATATTTTCCATTTTAAGAAAACCCGACAACATTTCTGCCAGGTTTGTTAGGAGGCCACATGATACTTATTCAAAAAAATCCTAGAGATTCTTAGCTCTTGGGATGCAGGCTCAGCCCGCTGGAGCATGAGCTCTGTGTGTACCGAGAACTGGGGTGATGTTTTACTTTTCACAGTATGGGCTACACAGCAGCTGTTCAACAAGAGTAAATATTGTCACAACACTGAACCTCTGGCTAGAGGACATATTCACAGTGAACATAACTGTAACATATATGAAAGGCTTCTGGGACTTGAAATCAAATGTTTGGGAATGGTGCCCTTGGAGGCAACCTCCCATTTTAGATGTTTAAAGGACCCTATATGTGGCATTCCTTTCTTTAAACTATAGGTAATTAAGGCAGCTGAAAAGTAAATTGCCTTCTAGACACTGAAGGCAAATCTCCTTTGTCCATTTACCTGGAAACCAGAATGATTTTGACATACAGGAGAGCTGCAGTTGTGAAAGCACCATCATCATAGAGGATGATGTAATTAAAAAATGGTCAGTGTGCAAAGAAAAGAACTGCTTGCATTTCTTTATTTCTGTCTCATAATTGTCAAAAACCAGAATTAGGTCAAGTTCATAGTTTCTGTAATTGGCTTTTGAATCAAAGAATAGGGAGACAATCTAAAAAATATCTTAGGTTGGAGATGACAGAAATATGATTGATTTGAAGTGGAAAAAGAAATTCTGTTAATGTTAATTAAAGTAAAATTATTCCCTGAATTGTTTGATATTGTCACCTAGCAGATATGTATTACTTTTCTGCAATGTTATTATTGGCTTGCACTTTGTGAGTATTCTATGTAAAAATATATATGTATATAAAATATATATTGCATAGGACAGACTTAGGAGTTTTGTTTAGAGCAGTTAACATCTGAAGTGTCTAATGCATTAACTTTTGTAAGGTACTGAATACTTAATATGTGGGAAACCCTTTTGCGTGGTCCTTAGGCTTACAATGTGCACTGAATCGTTTCATGTAAGAATCCAAAGTGGACACCATTAACAGGTCTTTGAAATATGCATGTACTTTATATTTTCTATATTTGTAACTTTGCATGTTCTTGTTTTGTTATATAAAAAAATTGTAAATGTTTAATATCTGACTGAAATTAAACGAGCGAAGATGAGCACCA(配列番号397)
GCGAACCTTGGCTGCTGGATGCTGGTTCTCTTTGTGGCCACATGGAGTGACCTGGGCCTCTGCAAGAAGCGCCCGAAGCCTGGAGGATGGAACACTGGGGGCAGCCGATACCCGGGGCAGGGCAGCCCTGGAGGCAACCGCTACCCACCTCAGGGCGGTGGTGGCTGGGGGCAGCCTCATGGTGGTGGCTGGGGGCAGCCTCATGGTGGTGGCTGGGGGCAGCCCCATGGTGGTGGCTGGGGACAGCCTCATGGTGGTGGCTGGGGTCAAGGAGGTGGCACCCACAGTCAGTGGAACAAGCCGAGTAAGCCAAAAACCAACATGAAGCACATGGCTGGTGCTGCAGCAGCTGGGGCAGTGGTGGGGGGCCTTGGCGGCTACATGCTGGGAAGTGCCATGAGCAGGCCCATCATACATTTCGGCAGTGACTATGAGGACCGTTACTATCGTGAAAACATGCACCGTTACCCCAACCAAGTGTACTACAGGCCCATGGATGAGTACAGCAACCAGAACAACTTTGTGCACGACTGCGTCAATATCACAATCAAGCAGCACACGGTCACCACAACCACCAAGGGGGAGAACTTCACCGAGACCGACGTTAAGATGATGGAGCGCGTGGTTGAGCAGATGTGTATCACCCAGTACGAGAGGGAATCTCAGGCCTATTACCAGAGAGGATCGAGCATGGTCCTCTTCTCCTCTCCACCTGTGATCCTCCTGATCTCTTTCCTCATCTTCCTGATAGTGGGATGAGGAAGGTCTTCCTGTTTTCACCATCTTTCTAATCTTTTTCCAGCTTGAGGGAGGCGGTATCCACCTGCAGCCCTTTTAGTGGTGGTGTCTCACTCTTTCTTCTCTCTTTGTCCCGGATAGGCTAATCAATACCCTTGGCACTGATGGGCACTGGAAAACATAGAGTAGACCTGAGATGCTGGTCAAGCCCCCTTTGATTGAGTTCATCATGAGCCGTTGCTAATGCCAGGCCAGTAAAAGTATAACAGCAAATAACCATTGGTTAATCTGGACTTATTTTTGGACTTAGTGCAACAGGTTGAGGCTAAAACAAATCTCAGAACAGTCTGAAATACCTTTGCCTGGATACCTCTGGCTCCTTCAGCAGCTAGAGCTCAGTATACTAATGCCCTATCTTAGTAGAGATTTCATAGCTATTTAGAGATATTTTCCATTTTAAGAAAACCCGACAACATTTCTGCCAGGTTTGTTAGGAGGCCACATGATACTTATTCAAAAAAATCCTAGAGATTCTTAGCTCTTGGGATGCAGGCTCAGCCCGCTGGAGCATGAGCTCTGTGTGTACCGAGAACTGGGGTGATGTTTTACTTTTCACAGTATGGGCTACACAGCAGCTGTTCAACAAGAGTAAATATTGTCACAACACTGAACCTCTGGCTAGAGGACATATTCACAGTGAACATAACTGTAACATATATGAAAGGCTTCTGGGACTTGAAATCAAATGTTTGGGAATGGTGCCCTTGGAGGCAACCTCCCATTTTAGATGTTTAAAGGACCCTATATGTGGCATTCCTTTCTTTAAACTATAGGTAATTAAGGCAGCTGAAAAGTAAATTGCCTTCTAGACACTGAAGGCAAATCTCCTTTGTCCATTTACCTGGAAACCAGAATGATTTTGACATACAGGAGAGCTGCAGTTGTGAAAGCACCATCATCATAGAGGATGATGTAATTAAAAAATGGTCAGTGTGCAAAGAAAAGAACTGCTTGCATTTCTTTATTTCTGTCTCATAATTGTCAAAAACCAGAATTAGGTCAAGTTCATAGTTTCTGTAATTGGCTTTTGAATCAAAGAATAGGGAGACAATCTAAAAAATATCTTAGGTTGGAGATGACAGAAATATGATTGATTTGAAGTGGAAAAAGAAATTCTGTTAATGTTAATTAAAGTAAAATTATTCCCTGAATTGTTTGATATTGTCACCTAGCAGATATGTATTACTTTTCTGCAATGTTATTATTGGCTTGCACTTTGTGAGTATTCTATGTAAAAATATATATGTATATAAAATATATATTGCATAGGACAGACTTAGGAGTTTTGTTTAGAGCAGTTAACATCTGAAGTGTCTAATGCATTAACTTTTGTAAGGTACTGAATACTTAATATGTGGGAAACCCTTTTGCGTGGTCCTTAGGCTTACAATGTGCACTGAATCGTTTCATGTAAGAATCCAAAGTGGACACCATTAACAGGTCTTTGAAATATGCATGTACTTTATATTTTCTATATTTGTAACTTTGCATGTTCTTGTTTTGTTATATAAAAAAATTGTAAATGTTTAATATCTGACTGAAATTAAACGAGCGAAGATGAGCACCA(配列番号397)
CJD及びFFIにリンクしているPRNP(NP_000302.1)に対して相対的な変異部位は報告されており、次の通りである。これらの変異は本明細書に開示のプライム編集因子を用いて除去又は組み入れられ得る。


GSSに連結されたPRNP(NP_000302.1)(配列番号396)に対する突然変異部位が以下のように報告される。

プリオン病に対する可能な防護性の性質にリンクしているPRNP(NP_000302.1)(配列番号396)に対して相対的な変異部位は次の通り:

それゆえに、種々の態様において、プライム編集は、プリオン病にリンクしているPRNPの変異を除去するか又はプリオン病に対して防護性であると考慮されるPRNPの変異を組み入れるために用いられてもよい。例えば、プライム編集は、PRNPタンパク質のD178N、V180I、T188K、E196K、E196A、E200K、E200G、V203I、R208H、V210I、E211Q、I215V、又はM232R変異を除去又は復元するために用いられてもよい(NP_000302.1のPRNPに対して相対的)(配列番号396)。他の態様において、プライム編集は、PRNPタンパク質のP102L、P105L、A117V、G131V、V176G、H187R、F198S、D202N、Q212P、Q217R、又はM232T変異を除去又は復元するために用いられてもよい(NP_000302.1のPRNPに対して相対的)(配列番号396)。プライム編集を用いてPRNPのかかる変異の存在を除去又は修正することによって、プリオン病のリスクが縮減又は消去されてもよい。
他の態様において、プライム編集は、1以上のプリオン病に対する防護性の効果にリンクしているPRNPの防護性変異を組み入れるために用いられてもよい。例えば、プライム編集は、PRNPのG127S、G127V、M129V、D167G、D167N、N171S、E219K、又はP238S防護性変異を組み入れるために用いられてもよい(NP_000302.1のPRNPに対して相対的)(配列番号396)。なお他の態様において、防護性変異は、PRNPのG127、G127、M129、D167、D167、N171、E219、又はP238に組み入れされるいずれかの代替アミノ酸であってもよい(NP_000302.1のPRNPに対して相対的)(配列番号396)。
具体的な態様では、図27において例解され例5において論じられる通り、プライム編集はG127V防護性変異をPRNPに組み入れるために用いられてもよい。
別の態様では、プライム編集はE219K防護性変異をPRNPに組み入れるために用いられてもよい。
PRNPタンパク質及び防護性変異部位は哺乳動物において保存されている。そのため、ヒト疾患を処置することに加えて、それは、プリオン病に対して免疫がある牛及び羊を生成するか、又は更にはプリオン病を患っている動物の野生集団を治癒させることを助けるためにもまた用いられ得る。プライム編集はヒト細胞における天然に存在する防護性アレルの~25%組み入れを達成するために用いられ得、マウス実験は組み入れのこのレベルがプリオン病に対する免疫を引き起こすために十分であるということを指示している。この方法は、ほとんどの細胞型においてかかる高い効率でこのアレルを組み入れるための第1のかつ可能性としては唯一の現行のやり方である。処置のための別の可能な戦略は、プライム編集を用いて、早期停止コドンを遺伝子に組み入れることによってPRNPの発現を縮減又は消去することである。
pegRNAデザインのための本明細書に記載の原理を用いて、適当なpegRNAが、所望の防護性変異を組み入れるために、又はPRNPからプリオン病関連変異を除去するためにデザインされてもよい。例えば、pegRNAの下のリストは、G127V防護性アレル及びE219K防護性アレルをヒトPRNPに、並びにG127V防護性アレルを種々の動物のPRNPに組み入れるために用いられ得る。
[10]医薬組成物
他の本開示の側面は、本明細書に記載のプライム編集システムの様々な構成要素のいずれも(例として、これらに限定されないが、napDNAbp、逆転写酵素、融合タンパク質(例として、napDNAbpsと逆転写酵素とを含む)、pegRNA、及び融合タンパク質とpegRNAとを含む複合体、並びに第2鎖へニックを入れる構成要素及び5'内生DNAフラップ除去エンドヌクレアーゼ等の、編集済産物形成へのプライム編集プロセスを推進するのに役立つ付属要素を包含する)含む医薬組成物に関する。
他の本開示の側面は、本明細書に記載のプライム編集システムの様々な構成要素のいずれも(例として、これらに限定されないが、napDNAbp、逆転写酵素、融合タンパク質(例として、napDNAbpsと逆転写酵素とを含む)、pegRNA、及び融合タンパク質とpegRNAとを含む複合体、並びに第2鎖へニックを入れる構成要素及び5'内生DNAフラップ除去エンドヌクレアーゼ等の、編集済産物形成へのプライム編集プロセスを推進するのに役立つ付属要素を包含する)含む医薬組成物に関する。
用語「医薬組成物」は、本明細書に使用されるとき、医薬使用のために製剤化された組成物を指す。いくつかの態様において、医薬組成物は更に、薬学的に許容し得る担体を含む。いくつかの態様において、医薬組成物は、追加の剤(例として、特定の送達、半減期の増大、又は他の処置的化合物のための)を含む。
ここで使用されるとき、用語「薬学的に許容し得る担体」は、液体又は固体の充填剤、希釈剤、賦形剤、製造補助剤(例として、潤滑剤、タルクマグネシウム、カルシウム、又はステアリン酸亜鉛若しくはステアリン酸)、あるいは身体のある部位(例として、送達部位)から別の部位(例として、器官、組織、若しくは身体部分)へ化合物を運搬又は輸送することを伴う溶媒封入材料等の、薬学的に許容し得る材料、組成物、又はビヒクルを意味する。薬学的に許容し得る担体は、製剤の他の構成要素と相溶性があるが対象の組織に対しては傷害性がない(例として、生理学的に相溶性のある、滅菌された、生理学的なpH、等々)という意味で「許容し得る」ものである。薬学的に許容し得る担体として働き得る材料のいくつかの例は以下を包含する:(1)ラクトース、グルコース、及びスクロース等の、糖;(2)コーンスターチ及びジャガイモデンプン等の、デンプン;(3)ナトリウムカルボキシメチルセルロース、メチルセルロース、エチルセルロース、微結晶性セルロース、及び酢酸セルロース等の、セルロース及びその誘導体;(4)粉末状トラガカント;(5)麦芽;(6)ゼラチン;(7)ステアリン酸マグネシウム、ラウリル硫酸ナトリウム、及びタルク等の、平滑剤;(8)カカオバター及び坐薬蝋等の、賦形剤;(9)落花生油、綿実油、紅花油、ゴマ油、オリーブ油、トウモロコシ油、及び大豆油等の、油;(10)プロピレングリコール等の、グリコール;(11)グリセリン、ソルビトール、マンニトール、及びポリエチレングリコール(PEG)等の、ポリオール;(12)オレイン酸エチル及びラウリン酸エチル等の、エステル;(13)寒天;(14)水酸化マグネシウム及び水酸化アルミニウム等の、緩衝剤;(15)アルギン酸;(16)パイロジェンフリー水;(17)等浸透圧の生理食塩水;(18)リンガー溶液;(19)エチルアルコール;(20)pH緩衝溶液;(21)ポリエステル、ポリカーボネート、及び/又はポリ酸無水物;(22)ポリペプチド及びアミノ酸等の、増量剤;(23)血清アルブミン、HDL、及びLDL等の、血清構成要素;(22)エタノール等の、C2~C12アルコール;並びに(23)医薬製剤に採用される、相溶性のある他の非毒性物質。湿潤剤、着色剤、離型剤、コーティング剤、甘味剤、香味剤、芳香剤、保存料、及び抗酸化剤もまた、製剤中に存在し得る。「賦形剤」、「担体」、「薬学的に許容し得る担体」又は同種のもの等の用語は本明細書中、互換的に使用される。
いくつかの態様において、医薬組成物は、対象への送達のために、例として、遺伝子編集のために、製剤化される。本明細書に記載の医薬組成物を投与する好適なルートは、限定せずに、以下を包含する:局所、皮下、経皮、皮内、病巣内、関節内、腹腔内、膀胱内、経粘膜、歯肉、歯内(intradental)、蝸牛内、経鼓膜、器官内、硬膜外、髄腔内、筋肉内、静脈内、血管内、骨内、眼周囲、腫瘍内、脳内、及び脳室内の投与。
いくつかの態様において、本明細書に記載の医薬組成物は、罹患部位(例として、腫瘍部位)へ局部的に投与される。いくつかの態様において、本明細書に記載の医薬組成物は、注射によって、カテーテルを用いて、坐薬を用いて、あるいは移植片(多孔性材料、非多孔性材料、又はゼラチン材料(シラスティック膜等の膜若しくは繊維を包含する)の移植片)を用いて、対象へ投与される。
他の態様において、本明細書に記載の医薬組成物は、制御放出系において送達される。一態様において、ポンプが使用されてもよい(例として、Langer,1990,Science 249:1527-1533;Sefton,1989,CRC Crit.Ref.Biomed.Eng.14:201;Buchwald et al.,1980,Surgery 88:507;Saudek et al.,1989,N.Engl.J.Med.321:574を参照)。別の態様において、ポリマー材料が使用され得る。(例として、Langer and Wise eds.,CRC Press,Boca Raton,Fla.,1974);Controlled Drug Bioavailability,Drug Product Design and Performance(Smolen and Ball eds.,Wiley,New York,1984);Ranger and Peppas,1983,Macromol.Sci.Rev.Macromol.Chem.23:61を参照。またLevy et al.,1985,Science 228:190;During et al.,1989,Ann.Neurol.25:351;Howard et al.,1989,J.Neurosurg.71:105も参照)。他の制御放出系も、例えば、Langer(上記を参照)において考察されている。
いくつかの態様において、医薬組成物は、対象(例として、ヒト)への静脈内投与又は皮下投与に適した組成物として、定型的な手順に従って製剤化される。いくつかの態様において、注射による投与のための医薬組成物は、滅菌された等浸透圧の水性緩衝液中の溶液である。必要なら、医薬はまた、可溶化剤、及び注射部位での疼痛を和らげるリグノカイン等の局所麻酔薬も包含し得る。一般に、構成要素は、個別に又は一緒に混合されて、例えば、活性剤の分量を示したアンプル若しくは小袋(sachette)等の密閉容器中の凍結乾燥された(dry lyophilized)粉末又は水がない濃縮物として単位剤形で供給される。医薬が注入によって投与されることになっている場合、滅菌医薬グレードの水又は生理食塩水を含有する注入瓶で分配され得る。医薬組成物が注射によって投与される場合、注射用滅菌水又は生理食塩水のアンプルは、構成要素が投与に先立ち混合され得るように、提供され得る。
全身投与のための医薬組成物は、液体、例として、滅菌された生理食塩水、乳酸リンガー溶液又はハンクス溶液であってもよい。加えて、医薬組成物は固体形態であって、使用に先立ち即時に再溶解又は懸濁され得る。凍結乾燥された形態もまた企図される。
医薬組成物は、非経口投与にも好適なリポソーム若しくはマイクロ結晶等の脂質粒子又はベシクル内に含有され得る。粒子は、組成物がこれに含有される限り、単層(unilamellar)又は多層(plurilamellar)等の、いずれの好適な構造でもあり得る。化合物は、膜融合(fusogenic)脂質ジオレオイルホスファチジルエタノールアミン(DOPE)、低レベル(5~10mol%)のカチオン性脂質を含有し、かつポリエチレングリコール(PEG)コーティングによって安定化された「安定化プラスミド-脂質粒子」(SPLP)に封入され得る(Zhang Y.P.et al.,Gene Ther.1999,6:1438-47)。N-[1-(2,3-ジオレオイルオキシ)プロピル]-N,N,N-トリメチル-アンモニウムメチルスルファート、又は「DOTAP」等の、正に帯電した脂質は、かかる粒子及びベシクルにとって具体的に好ましい。かかる脂質粒子の調製は周知である。例として、米国特許第4,880,635号;第4,906,477号;第4,911,928号;第4,917,951号;第4,920,016号;及び第4,921,757号を参照;これらの各々は参照により本明細書に組み込まれる。
本明細書に記載の医薬組成物は、例えば、単位用量として投与又はパッケージされてもよい。用語「単位用量」は、本開示の医薬組成物に関して使用されるとき、まとまった投薬量として対象に好適な、物理的に不連続な単位を指し、予め決められた分量の活性材料を含有する各単位は、要求される希釈剤;すなわち、担体又はビヒクルを伴って所望の処置効果を産生するよう算出される。
更に、医薬組成物は、(a)本発明の化合物を凍結乾燥形態で含有する容器、及び(b)注射のための薬学的に許容し得る希釈剤(例として、滅菌水)を含有する第2容器を含む医薬のキットとして提供され得る。薬学的に許容し得る希釈剤は、凍結乾燥された本発明の化合物の再構成又は希釈のために使用され得る。任意選択で、かかる容器(単数又は複数)に関連する通知は、医薬若しくは生物学的産物の製造、使用、又は販売を規制する政府機関によって規定される形態であり得、通知は、ヒト投与のための製造、使用、又は販売の機関による承認を反映する。
別の側面において、上に記載の疾患の処置に有用な材料を含有する製造物品が、包含される。いくつかの態様において、製造物品は、容器及びラベルを含む。好適な容器は、例えば、瓶、バイアル、シリンジ、及び試験管を包含する。容器は、ガラス又はプラスチック等の様々な材料から形成されてもよい。いくつかの態様において、容器は、本明細書に記載の疾患を処置するのに有効であって滅菌アクセスポートを有していてもよい組成物をとどめておく。例えば、容器は、皮下注射針によって穿刺可能なストッパーを有する静脈内溶液バッグ又はバイアルであってもよい。組成物中の活性剤は本発明の化合物である。いくつかの態様において、容器上の又はこれに関連したラベルは、組成物が、選んだ疾患を処置するのに使用されることを示す。製造物品は更に、リン酸緩衝生理食塩水、リンガー溶液、又はデキストロース溶液等の薬学的に許容し得る緩衝液を含む第2容器を含んでもよい。これは更に、他の緩衝液、希釈剤、フィルター、針、シリンジ、及び使用のための指示がある添付文書を包含する、商業的観点及びユーザーの観点から所望される他の材料も包含していてもよい。
[11]キット、ベクター、細胞、及び送達
キット
本開示の組成物は、キット中へ集められ得る。いくつかの態様において、キットは、本明細書に記載のプライム編集因子の発現のための核酸ベクターを含む。他の態様において、キットは更に、適切なガイドヌクレオチド配列(例として、pegRNA及び第2部位gRNA)、又はかかるガイドヌクレオチド配列(Cas9タンパク質又はプライム編集因子に所望の標的配列を標的にさせる)の発現のための核酸ベクターを含む。
キット
本開示の組成物は、キット中へ集められ得る。いくつかの態様において、キットは、本明細書に記載のプライム編集因子の発現のための核酸ベクターを含む。他の態様において、キットは更に、適切なガイドヌクレオチド配列(例として、pegRNA及び第2部位gRNA)、又はかかるガイドヌクレオチド配列(Cas9タンパク質又はプライム編集因子に所望の標的配列を標的にさせる)の発現のための核酸ベクターを含む。
本明細書に記載のキットは、本明細書に記載の方法を実施するための構成要素及び任意選択で使用のための指示を収納する1以上の容器を包含していてもよい。本明細書に記載のキットのいずれも、アッセイ方法を実施するのに必要とされる構成要素を更に含んでいてもよい。キットの各構成要素は、適用可能であれば、液体形態(例として、溶液中)で、又は固体形態(例として、乾燥粉末)で提供されてもよい。あるケースにおいて、構成要素のいくつかは、例えば、好適な溶媒又は他の種(例えば、水)(これらはキットとともに提供されても又はされなくてもよい)の添加によって、再構成可能又は別様に(例として、活性形態へ)処理可能であってもよい。
いくつかの態様において、キットは任意選択で、提供される構成要素の使用のための指示及び/又は促進(promotion)を包含していてもよい。本明細書に使用されるとき、「指示」は、指示及び/又は促進の構成要素を定義し得るものであって、典型的には、本開示のパッケージング上に又はこれに関連して書面の指示を伴い得る。指示はまた、ユーザーが明確に、指示がキットに関連するものであることを認識できるように(例えば、視聴覚に訴えるもの(例として、ビデオテープ、DVD、等々)、インターネット、及び/又はウェブベースの連絡、等々)、形はどうあれ、いずれの口授又は電子的指示も包含し得る。書面の指示は、医薬若しくは生物学的産物の製造、使用、又は販売を規制する政府機関によって規定される形態であってもよく、これは動物投与のための製造、使用、又は販売の機関による承認も反映し得る。本明細書に使用されるとき、「促進された」は、本開示に関連する、教育方法、病院の及び他の臨床の指示、科学研究、薬物の発見又は開発、学術研究、医薬産業活動(医薬販売を包含する)、並びにいずれの広告活動又は他の宣伝活動(いずれの形態の書面の、口述の、及び電子的な連絡を包含する)を包含する、営業行為の全ての方法を包含する。加えて、キットは、本明細書に記載の通りの特定の用途に応じて、他の構成要素も包含していてもよい。
キットは、1以上の容器中に、本明細書に記載のいずれの構成要素の1以上を含有していてもよい。構成要素は、滅菌的に調製され、シリンジ中にパッケージされ、及び冷凍保存されて出荷されてもよい。代替的に、これは、バイアル又は保管のための他の容器に収納されてもよい。第2容器は、滅菌的に調製された他の構成要素も有していてもよい。代替的に、キットは、予め混合されて、バイアル、管、又は他の容器中で出荷される活性剤を包含していてもよい。
キットは、パウチ、1以上の管、容器、箱、又は袋内に緩くパックされた付属物とともに、ブリスターパウチ(blister pouch)、収縮包装パウチ、真空シール可能なパウチ、シール可能な熱成形トレイ、又は同様のパウチ若しくはトレイの形態等の様々な形態を有してもよい。キットは、付属物が加えられた後に滅菌されてもよく、これによって容器中の個々の付属物が別様に開封され得る。キットは、放射線滅菌、加熱滅菌、又は当該技術分野において知られている他の滅菌方法等の、いずれの適切な滅菌技法も使用して滅菌され得る。キットはまた、特定の用途に応じて、他の構成要素、例えば、容器、細胞培地、塩、緩衝剤、試薬、シリンジ、針、殺菌剤を適用又は除去するための生地(ガーゼ等)、使い捨て手袋、投与に先立ち剤を支持するためのもの等々も包含していてもよい。本開示のいくつかの側面は、本明細書に記載のプライム編集システムの様々な構成要素(例として、これらに限定されないが、napDNAbp、逆転写酵素、ポリメラーゼ、融合タンパク質(例として、napDNAbpと逆転写酵素(又はより広くは、ポリメラーゼ)とを含む)、pegRNA、及び融合タンパク質とpegRNAとを含む複合体、並びに第2鎖へニックを入れる構成要素(例として、第2鎖へニックを入れるgRNA)及び5'内生DNAフラップ除去エンドヌクレアーゼ等の付属要素(編集済産物形成への多重フラッププライム編集プロセスを推進するのに役立つ)を包含する)をコードするヌクレオチド配列を含む核酸構築物を含むキットを提供する。いくつかの態様において、ヌクレオチド配列(単数又は複数)は、プライム編集システム構成要素の発現を推進する異種プロモータ(又は単一のプロモータより多く)を含む。
本開示の他の側面は、本明細書に記載のプライム編集システムの様々な構成要素(例として、標的DNA配列を修飾することが可能なプライム編集システムの構成要素をコードするヌクレオチド配列を含む)をコードする1以上の核酸構築物を含むキットを提供する。いくつかの態様において、ヌクレオチド配列は、プライム編集システム構成要素の発現を推進する異種プロモータを含む。
本開示のいくつかの側面は、(a)逆転写酵素等のポリメラーゼと融合したnapDNAbp(例として、Cas9ドメイン)をコードするヌクレオチド配列、及び(b)(a)の配列の発現を推進する異種プロモータを含む核酸構築物を含むキットを提供する。
細胞
本明細書に記載の組成物のいずれも含有していてもよい細胞は、原核細胞及び真核細胞を包含する。本明細書に記載の方法は、Cas9タンパク質又はプライム編集因子を真核細胞(例として、ヒト細胞等の哺乳動物の細胞)中へ送達するのに使用される。いくつかの態様において、細胞は、in vitroにある(例として、培養された細胞。いくつかの態様において、細胞は、in vivoにある(例として、ヒト対象等の対象中にある)。いくつかの態様において、細胞は、ex vivoにある(例として、対象から単離され、同じ対象へ戻すか又は異なる対象へ投与されてもよい)。
本明細書に記載の組成物のいずれも含有していてもよい細胞は、原核細胞及び真核細胞を包含する。本明細書に記載の方法は、Cas9タンパク質又はプライム編集因子を真核細胞(例として、ヒト細胞等の哺乳動物の細胞)中へ送達するのに使用される。いくつかの態様において、細胞は、in vitroにある(例として、培養された細胞。いくつかの態様において、細胞は、in vivoにある(例として、ヒト対象等の対象中にある)。いくつかの態様において、細胞は、ex vivoにある(例として、対象から単離され、同じ対象へ戻すか又は異なる対象へ投与されてもよい)。
本開示の哺乳動物の細胞は、ヒト細胞、霊長目の動物細胞(例として、ベロ細胞)、ラット細胞(例として、GH3細胞、OC23細胞)、又はマウス細胞(例として、MC3T3細胞)を包含する。様々なヒト細胞株には、限定せずに、ヒト胚性腎臓(HEK)細胞、HeLa細胞、国立がん研究所の60のがん細胞株(NCI60)からのがん細胞、DU145(前立腺がん)細胞、Lncap(前立腺がん)細胞、MCF-7(乳がん)細胞、MDA-MB-438(乳がん)細胞、PC3(前立腺がん)細胞、T47D(乳がん)細胞、THP-1(急性骨髄性白血病)細胞、U87(膠芽腫)細胞、SHSY5Yヒト神経芽細胞腫細胞(骨髄腫からクローン化)、及びSaos-2(骨がん)細胞が包含される。いくつかの態様において、rAAVベクターは、ヒト胚性腎臓(HEK)細胞(例として、HEK293又はHEK 293T細胞)中へ送達される。いくつかの態様において、rAAVベクターは、例えば、多能性幹細胞(例として、ヒト人工(induced)多能性幹細胞(hiPSC)を包含するヒト多能性幹細胞)等の、幹細胞(例として、ヒト幹細胞)中へ送達される。幹細胞は培養中、無期限に分裂して、特化された細胞を生じさせる能力をもつ細胞を指す。多能性幹細胞は、分化して生物の全ての組織になることが可能であるが、完全な生物の発生を単独では支えることが可能でないある種の幹細胞を指す。ヒト人工多能性幹細胞は、胚性幹細胞を定義する特性を維持するのに重要な遺伝子及び因子の発現が強いられることによって胚性幹細胞様状態へ再プログラムされる体細胞(例として、成熟した(mature)細胞又は成熟(adult)細胞)を指す(例として、Takahashi and Yamanaka,Cell 126(4):663-76,2006を参照(参照により本明細書に組み込まれる))。ヒト人工多能性幹細胞は、幹細胞マーカーを発現しており、全3種の胚葉(外胚葉、内胚葉、中胚葉)の細胞特徴を生成するのが可能である。
本開示に従って使用されてもよい追加の細胞株の非限定例は、293-T、293-T、3T3、4T1、721、9L、A-549、A172、A20、A253、A2780、A2780ADR、A2780cis、A431、ALC、B16、B35、BCP-1、BEAS-2B、bEnd.3、BHK-21、BR 293、BxPC3、C2C12、C3H-10T1/2、C6、C6/36、Cal-27、CGR8、CHO、CML T1、CMT、COR-L23、COR-L23/5010、COR-L23/CPR、COR-L23/R23、COS-7、COV-434、CT26、D17、DH82、DU145、DuCaP、E14Tg2a、EL4、EM2、EM3、EMT6/AR1、EMT6/AR10.0、FM3、H1299、H69、HB54、HB55、HCA2、Hepa1c1c7、High Five細胞、HL-60、HMEC、HT-29、HUVEC、J558L細胞、Jurkat、JY細胞、K562細胞、KCL22、KG1、Ku812、KYO1、LNCap、Ma-Mel 1、2、3....48、MC-38、MCF-10A、MCF-7、MDA-MB-231、MDA-MB-435、MDA-MB-468、MDCK II、MG63、MONO-MAC 6、MOR/0.2R、MRC5、MTD-1A、MyEnd、NALM-1、NCI-H69/CPR、NCI-H69/LX10、NCI-H69/LX20、NCI-H69/LX4、NIH-3T3、NW-145、OPCN/OPCT Peer、PNT-1A/PNT 2、PTK2、Raji、RBL細胞、RenCa、RIN-5F、RMA/RMAS、S2、Saos-2細胞、Sf21、Sf9、SiHa、SKBR3、SKOV-3、T-47D、T2、T84、THP1、U373、U87、U937、VCaP、WM39、WT-49、X63、YAC-1、及びYAR細胞を包含する。
本開示のいくつかの側面は、本明細書に開示される構築物のいずれも含む細胞を提供する。いくつかの態様において、宿主細胞は、本明細書に記載の1以上のベクターで一過的又は非一過的にトランスフェクトされている。いくつかの態様において、細胞は、対象中に天然に存在するとき、トランスフェクトされる。いくつかの態様において、トランスフェクトされる細胞は、対象から採られる。いくつかの態様において、細胞は、細胞株等の、対象から採られた細胞に由来される。組織培養のための多種多様の細胞株が当該技術分野において知られている。細胞株の例は、これらに限定されないが、C8161、CCRF-CEM、MOLT、mIMCD-3、NHDF、HeLa-S3、Huh1、Huh4、Huh7、HUVEC、HASMC、HEKn、HEKa、MiaPaCell、Panc1、PC-3、TF1、CTLL-2、C1R、Rat6、CV1、RPTE、A10、T24、J82、A375、ARH-77、Calu1、SW480、SW620、SKOV3、SK-UT、CaCo2、P388D1、SEM-K2、WEHI-231、HB56、TIB55、Jurkat、J45.01、LRMB、Bcl-1、BC-3、IC21、DLD2、Raw264.7、NRK、NRK-52E、MRC5、MEF、Hep G2、HeLa B、HeLa T4、COS、COS-1、COS-6、COS-M6A、BS-C-1サル腎臓上皮、BALB/3T3マウス胚線維芽細胞、3T3 Swiss、3T3-L1、132-d5ヒト胎児線維芽細胞;10.1マウス線維芽細胞、293-T、3T3、721、9L、A2780、A2780ADR、A2780cis、A 172、A20、A253、A431、A-549、ALC、B16、B35、BCP-1細胞、BEAS-2B、bEnd.3、BHK-21、BR 293.BxPC3.C3H-10T1/2、C6/36、Cal-27、CHO、CHO-7、CHO-IR、CHO-K1、CHO-K2、CHO-T、CHO Dhfr-/-、COR-L23、COR-L23/CPR、COR-L23/5010、COR-L23/R23、COS-7、COV-434、CML T1、CMT、CT26、D17、DH82、DU145、DuCaP、EL4、EM2、EM3、EMT6/AR1、EMT6/AR10.0、FM3、H1299、H69、HB54、HB55、HCA2、HEK-293、HeLa、Hepa1c1c7、HL-60、HMEC、HT-29、Jurkat、JY細胞、K562細胞、Ku812、KCL22、KG1、KYO1、LNCap、Ma-Mel 1-48、MC-38、MCF-7、MCF-10A、MDA-MB-231、MDA-MB-468、MDA-MB-435、MDCK II、MDCK 11、MOR/0.2R、MONO-MAC 6、MTD-1A、MyEnd、NCI-H69/CPR、NCI-H69/LX10、NCI-H69/LX20、NCI-H69/LX4、NIH-3T3、NALM-1、NW-145、OPCN/OPCT細胞株、Peer、PNT-1A/PNT 2、RenCa、RIN-5F、RMA/RMAS、Saos-2細胞、Sf-9、SkBr3、T2、T-47D、T84、THP1細胞株、U373、U87、U937、VCaP、Vero細胞、WM39、WT-49、X63、YAC-1、YAR、及びそれらのトランスジェニック種を包含する。
細胞株は、当業者に知られている様々な供給源から利用可能である(例として、American Type Culture Collection(ATCC)(Manassus,Va.)を参照)。いくつかの態様において、本明細書に記載の1以上のベクターでトランスフェクトされた細胞は、1以上のベクター由来配列を含む新しい細胞株を確立するのに使用される。いくつかの態様において、本明細書に記載の通りのCRISPR系の構成要素で(1以上のベクターの一過的なトランスフェクション、又はRNAでのトランスフェクション等によって)一過的にトランスフェクトされて、CRISPR複合体の活性を通して修飾された細胞は、修飾は含有するが他の外来配列のいずれも欠く細胞を含む新しい細胞株を確立するのに使用される。いくつかの態様において、本明細書に記載の1以上のベクターで一過的若しくは非一過的にトランスフェクトされた細胞、又はかかる細胞に由来する細胞株は、1以上の試験化合物を査定するのに使用される。
ベクター
本開示のいくつかの側面は、本明細書に記載のプライム編集因子又はその構成要素(例として、分裂Cas9タンパク質若しくは分裂核酸塩基プライム編集因子)の細胞中への送達のための、組換えウイルスベクター(例として、アデノ随伴ウイルスベクター、アデノウイルスベクター、又は単純ヘルペスウイルスベクター)の使用に関する。分裂-PEアプローチのケースにおいて、全長Cas9タンパク質又はプライム編集因子が様々なウイルスベクター(例として、rAAV(~4.9kb))のパッケージング限界を超えることから、プライム編集因子のN末端部分及びPE融合のC末端部分は、別々の組換えウイルスベクター(例として、アデノ随伴ウイルスベクター、アデノウイルスベクター、又は単純ヘルペスウイルスベクター)によって同じ細胞中へ送達される。
本開示のいくつかの側面は、本明細書に記載のプライム編集因子又はその構成要素(例として、分裂Cas9タンパク質若しくは分裂核酸塩基プライム編集因子)の細胞中への送達のための、組換えウイルスベクター(例として、アデノ随伴ウイルスベクター、アデノウイルスベクター、又は単純ヘルペスウイルスベクター)の使用に関する。分裂-PEアプローチのケースにおいて、全長Cas9タンパク質又はプライム編集因子が様々なウイルスベクター(例として、rAAV(~4.9kb))のパッケージング限界を超えることから、プライム編集因子のN末端部分及びPE融合のC末端部分は、別々の組換えウイルスベクター(例として、アデノ随伴ウイルスベクター、アデノウイルスベクター、又は単純ヘルペスウイルスベクター)によって同じ細胞中へ送達される。
よって、一態様において、本開示は、分裂プライム編集因子又はそれらの分裂構成要素を送達するのか可能なベクターを企図する。いくつかの態様において、分裂Cas9タンパク質又は分裂プライム編集因子を細胞(例として、哺乳動物の細胞、ヒト細胞)中へ送達するための組成物が提供される。いくつかの態様において、本開示の組成物は以下を含む:(i)そのC末端にてインテイン-Nと融合したCas9タンパク質又はプライム編集因子のN末端部分をコードする第1ヌクレオチド配列を含む第1組換えアデノ随伴ウイルス(rAAV)粒子;及び(ii)Cas9のタンパク質又はプライム編集因子のC末端部分のN末端と融合したインテイン-Cをコードする第2ヌクレオチド配列を含む第2組換えアデノ随伴ウイルス(rAAV)粒子。本開示のrAAV粒子は、ウイルスのカプシドタンパク質で包まれた(encapsulated)rAAVベクター(すなわち、rAAVの組換えゲノム)を含む。
いくつかの態様において、rAAVベクターは以下を含む:(1)本明細書に記載の通りのいずれの形態の分裂Cas9タンパク質若しくは分裂プライム編集因子のN末端部分又はC末端部分をコードする第1あるいは第2ヌクレオチド配列を含む異種核酸領域、(2)異種核酸領域の発現を容易にさせる配列(例として、プロモータ)を含む1以上のヌクレオチド配列、及び(3)異種核酸領域(任意選択で、発現を容易にさせる配列を含む1以上の核酸領域をもつ)の細胞ゲノム中への組み入れを容易にさせる配列を含む1以上の核酸領域。いくつかの態様において、組み入れを容易にさせるウイルスの配列は、逆方向末端反復(ITR)配列を含む。いくつかの態様において、分裂Cas9タンパク質若しくは分裂プライム編集因子のN末端部分又はC末端部分をコードする第1あるいは第2ヌクレオチド配列は、ITR配列によって両側から挟まれている。いくつかの態様において、核酸ベクターは更に、本明細書に記載の通りのAAV Repタンパク質をコードする領域を含むが、領域は、ITRによって挟まれる領域内又はその領域外のいずれかに含有される。ITR配列は、いずれかのAAV血清型(例として、1、2、3、4、5、6、7、8、9若しくは10)に由来し得るか、又は1より多くの血清型に由来され得る。いくつかの態様において、ITR配列は、AAV2又はAAV6に由来される。
よって、いくつかの態様において、本明細書に開示のrAAV粒子は、少なくとも1のrAAV2粒子、rAAV6粒子、rAAV8粒子、rPHP.B粒子、rPHP.eB粒子、若しくはrAAV9粒子、又はそのバリアントを含む。具体的な態様において、開示されたrAAV粒子は、rPHP.B粒子、rPHP.eB粒子、rAAV9粒子である。
ITR配列及びITR配列含有プラスミドは当該技術分野において知られており、市販されている(例として、Vector Biolabs,Philadelphia,PA;Cellbiolabs,San Diego,CA;Agilent Technologies,Santa Clara,Ca;及びAddgene,Cambridge,MAから利用可能な製品並びにサービス;並びに、骨格筋への遺伝子送達は処置用タンパク質の持続発現及び全身送達をもたらす。Kessler PD,Podsakoff GM,Chen X,McQuiston SA,Colosi PC,Matelis LA,Kurtzman GJ,Byrne BJ.Proc Natl Acad Sci USA.1996 Nov 26;93(24):14082-7;及びCurtis A.Machida.Methods in Molecular Medicine(商標).Viral Vectors for Gene Therapy Methods and Protocols.10.1385/1-59259-304-6:201cHumana Press Inc.2003.Chapter 10.Targeted Integration by Adeno-Associated Virus.Matthew D.Weitzman,Samuel M.Young Jr.,Toni Cathomen and Richard Jude Samulski;米国特許第5,139,941号及び第5,962,313号を参照(これら全ては参照により本明細書に組み込まれる))。
いくつかの態様において、本開示のrAAVベクターは、異種核酸領域の発現を制御する1以上の調節要素(例として、プロモータ、転写ターミネータ、及び/又は他の調節要素)を含む。いくつかの態様において、第1及び/又は第2ヌクレオチド配列は、1以上の(例として、1、2、3、4、5、又はそれ以上の)転写ターミネータへ作動可能に(operably)連結されている。本開示に従って使用されてもよい転写ターミネータの非限定例は、ウシ成長ホルモン遺伝子(bGH)、ヒト成長ホルモン遺伝子(hGH)、SV40、CW3、Φ、又はそれらの組み合わせの転写ターミネータを包含する。数種の転写ターミネータの効率が試験され、分裂Cas9タンパク質又は分裂プライム編集因子の発現レベルにおけるそれら夫々の効果を決定した。いくつかの態様において、本開示に使用される転写ターミネータは、bGH転写ターミネータである。いくつかの態様において、rAAVベクターは更に、ウッドチャック肝炎ウイルス転写後調節要素(WPRE)を含む。ある態様において、WPREは、「W3」等のトランケートされたWPRE配列である。いくつかの態様において、WPREは、転写ターミネータの5'に挿入されている。かかる配列は、転写されたとき、発現、特にウイルスベクターからの発現を増強する3次構造を作成する。
いくつかの態様において、本明細書に使用されるベクターは、プライム編集因子、又はそれら構成要素(例として、napDNAbp、リンカー、若しくはポリメラーゼ)のいずれもコードしていてもよい。加えて、本明細書に使用されるベクターは、pegRNA、及び/又は第2鎖へニックを入れるための付属(accessory)gRNAをコードしていてもよい。ベクターは、細胞中の1以上のコード配列の発現を推進するのが可能であってもよい。いくつかの態様において、細胞は、例として細菌性細胞等の、原核細胞であってもよい。いくつかの態様において、細胞は、例として、酵母、植物、昆虫、又は哺乳動物の細胞等の、真核細胞であってもよい。いくつかの態様において、真核細胞は、哺乳動物の細胞であってもよい。いくつかの態様において、真核細胞は、齧歯類の動物の細胞であってもよい。いくつかの態様において、真核細胞は、ヒトの細胞であってもよい。種々の型の細胞中の発現を推進する好適なプロモータは当該技術分野において知られている。いくつかの態様において、プロモータは、野生型であってもよい。他の態様において、プロモータは、より効率的又はより効果的な発現のために修飾されていてもよい。もう1つの他の態様において、プロモータは、トランケートされていてもよいが、依然としてその機能を保持する。例えば、プロモータは、正常なサイズ、又はベクターのウイルス中への正しいパッケージングに好適な低減されたサイズを有していてもよい。
いくつかの態様において、プライム編集因子ベクターに使用されてもよいプロモータは、構成的、誘導的、又は組織特異的であってもよい。いくつかの態様において、プロモータは、構成的プロモータであってもよい。非限定的な例示の構成的プロモータは、サイトメガロウイルス前初期プロモータ(CMV)、サルウイルス(SV40)プロモータ、アデノウイルス主要後期(MLP)プロモータ、ラウス肉腫ウイルス(RSV)プロモータ、マウス乳腺腫瘍ウイルス(MMTV)プロモータ、ホスホグリセリン酸キナーゼ(PGK)プロモータ、伸長(elongation)因子-アルファ(EFla)プロモータ、ユビキチンプロモータ、アクチンプロモータ、チューブリンプロモータ、免疫グロブリンプロモータ、それらの機能的フラグメント、又は上記のいずれの組み合わせを包含する。いくつかの態様において、プロモータは、CMVプロモータであってもよい。いくつかの態様において、プロモータは、トランケートされたCMVプロモータであってもよい。他の態様において、プロモータは、EFlaプロモータであってもよい。いくつかの態様において、プロモータは、誘導性プロモータであってもよい。非限定的な例示の誘導性プロモータは、熱ショック、光、化学物質、ペプチド、金属、ステロイド、抗生物質、又はアルコールによって誘導可能なものを包含する。いくつかの態様において、誘導性プロモータは、基底(非誘導)発現レベルが低いもの、例として、Tet-On(登録商標)プロモータ(Clontech)等であってもよい。いくつかの態様において、プロモータは、組織特異的プロモータであってもよい。いくつかの態様において、組織特異的プロモータは、専ら又は主として、肝臓組織に発現される。非限定的な例示の組織特異的プロモータは、B29プロモータ、CD14プロモータ、CD43プロモータ、CD45プロモータ、CD68プロモータ、デスミンプロモータ、エラスターゼ-1プロモータ、エンドグリンプロモータ、フィブロネクチンプロモータ、Flt-1プロモータ、GFAPプロモータ、GPIIbプロモータ、ICAM-2プロモータ、INF-βプロモータ、Mbプロモータ、Nphslプロモータ、OG-2プロモータ、SP-Bプロモータ、SYN1プロモータ、及びWASPプロモータを包含する。
いくつかの態様において、プライム編集因子ベクター(例として、プライム編集因子及び/又はpegRNA、及び/又は第2鎖へニックを入れる付属gRNAをコードするいずれのベクターも包含する)は、標的細胞へ送達された後にしか発現を開始しない誘導性プロモータを含んでいてもよい。非限定的な例示の誘導性プロモータは、熱ショック、光、化学物質、ペプチド、金属、ステロイド、抗生物質、又はアルコールによって誘導可能なものを包含する。いくつかの態様において、誘導性プロモータは、基底(非誘導)発現レベルが低いもの、例として、Tet-On(登録商標)プロモータ(Clontech)等であってもよい。
追加の態様において、プライム編集因子ベクター(例として、プライム編集因子及び/又はpegRNA、及び/又は第2鎖へニックを入れる付属gRNAをコードするいずれのベクターも包含する)は、特定の組織へ送達された後にしか発現を開始しない組織特異的なプロモータを含んでいてもよい。非限定的な例示の組織特異的プロモータは、B29プロモータ、CD14プロモータ、CD43プロモータ、CD45プロモータ、CD68プロモータ、デスミンプロモータ、エラスターゼ-1プロモータ、エンドグリンプロモータ、フィブロネクチンプロモータ、Flt-1プロモータ、GFAPプロモータ、GPIIbプロモータ、ICAM-2プロモータ、INF-βプロモータ、Mbプロモータ、Nphslプロモータ、OG-2プロモータ、SP-Bプロモータ、SYN1プロモータ、及びWASPプロモータを包含する。
いくつかの態様において、pegRNA(又はプライム編集に関係して使用されるいずれのガイドRNA)をコードするヌクレオチド配列は、少なくとも1の転写又は翻訳制御配列へ作動可能に連結されていてもよい。いくつかの態様において、ガイドRNAをコードするヌクレオチド配列は、少なくとも1のプロモータへ作動可能に連結されていてもよい。いくつかの態様において、プロモータは、RNAポリメラーゼ III(Pol III)によって認識されてもよい。Pol IIIプロモータの非限定例は、U6、HI、及びtRNAプロモータを包含する。いくつかの態様において、ガイドRNAをコードするヌクレオチド配列は、マウス又はヒトU6プロモータへ作動可能に連結されていてもよい。他の態様において、ガイドRNAをコードするヌクレオチド配列は、マウス又はヒトHIプロモータへ作動可能に連結されていてもよい。いくつかの態様において、ガイドRNAをコードするヌクレオチド配列は、マウス又はヒトtRNAプロモータへ作動可能に連結されていてもよい。1より多くのガイドRNAを用いる態様において、発現を推進するのに使用されるプロモータは、同じであっても、又は異なっていてもよい。いくつかの態様において、ガイドRNAのcrRNAをコードするヌクレオチド、及びガイドRNAのtracr RNAをコードするヌクレオチドは、同じベクター上に提供されていてもよい。いくつかの態様において、crRNAをコードするヌクレオチド、及びtracr RNAをコードするヌクレオチドは、同じプロモータによって推進されてもよい。いくつかの態様において、crRNA及びtracr RNAは、転写されて単一の転写産物になってもよい。例えば、crRNA及びtracr RNAは、単一の転写産物からプロセシングされて二本鎖分子(double-molecule)ガイドRNAを形成してもよい。代替的に、crRNA及びtracr RNAは、転写されて単一分子ガイドRNAになってもよい。
いくつかの態様において、ガイドRNAをコードするヌクレオチド配列は、プライム編集因子をコードするヌクレオチド配列を含む同じベクター上に位置付けられていてもよい。いくつかの態様において、ガイドRNAの発現及びプライム編集因子の発現は、それらの対応するプロモータによって推進されてもよい。いくつかの態様において、ガイドRNAの発現は、プライム編集因子の発現を推進する同じプロモータによって推進されてもよい。いくつかの態様において、ガイドRNA及びプライム編集因子の転写産物は、単一の転写産物内に含有されていてもよい。例えば、ガイドRNAは、Cas9タンパク質転写産物の非翻訳領域(UTR)内にあってもよい。いくつかの態様において、ガイドRNAは、プライム編集因子転写産物の5'UTR内にあってもよい。他の態様において、ガイドRNAは、プライム編集因子転写産物の3'UTR内にあってもよい。いくつかの態様において、プライム編集因子転写産物の細胞内半減期は、ガイドRNAをその3'UTR内に含有することによって低減されてもよく、これによってその3'UTRの長さを短縮し得る。追加の態様において、ガイドRNAは、プライム編集因子転写産物のイントロン内にあってもよい。いくつかの態様において、好適なスプライシング部位は、ガイドRNAが転写産物から正しくスプライシングされるように、ガイドRNAが位置付けられるイントロンに付加されてもよい。いくつかの態様において、同じベクター上の極めて近いところにあるCas9タンパク質及びガイドRNAの発現は、CRISPR複合体のより効率的な形成を容易にさせてもよい。
プライム編集因子ベクター系は、1つのベクター、若しくは2つのベクター、若しくは3つのベクター、若しくは4つのベクター、若しくは5つのベクター、又はそれ以上を含んでいてもよい。いくつかの態様において、ベクター系は、プライム編集因子とpegRNAとの両方をコードする1つの単一ベクターを含んでいてもよい。他の態様において、ベクター系は2つのベクターを含んでいてもよいが、ここで一方のベクターはプライム編集因子をコードし、他方がpegRNAをコードする。追加の態様において、ベクター系は3つのベクターを含んでいてもよいが、ここで第3ベクターは、本明細書の方法に使用される、第2鎖へニックを入れるgRNAをコードする。
いくつかの態様において、rAAV粒子を含む組成物(本明細書に企図されるいずれの形態)は更に、薬学的に許容し得る担体を含む。いくつかの態様において、組成物は、ヒト又は動物の対象への投与に適切な医薬ビヒクル中で製剤化される。
薬学的に許容し得る担体として働き得る材料のいくつかの例は以下を包含する:(1)ラクトース、グルコース、及びスクロース等の、糖;(2)コーンスターチ及びジャガイモデンプン等の、デンプン;(3)ナトリウムカルボキシメチルセルロース、メチルセルロース、エチルセルロース、微結晶性セルロース、及び酢酸セルロース等の、セルロース及びその誘導体;(4)粉末状トラガカント;(5)麦芽;(6)ゼラチン;(7)ステアリン酸マグネシウム、ラウリル硫酸ナトリウム、及びタルク等の、平滑剤;(8)カカオバター及び坐薬蝋等の、賦形剤;(9)落花生油、綿実油、紅花油、ゴマ 油、オリーブ油、トウモロコシ油、及び大豆油等の、油;(10)プロピレングリコール等の、グリコール;(11)グリセリン、ソルビトール、マンニトール、及びポリエチレングリコール(PEG)等の、ポリオール;(12)オレイン酸エチル及びラウリン酸エチル等の、エステル;(13)寒天;(14)水酸化マグネシウム及び水酸化アルミニウム等の、緩衝剤;(15)アルギン酸;(16)パイロジェンフリー水;(17)等浸透圧の生理食塩水;(18)リンガー溶液;(19)エチルアルコール;(20)pH緩衝溶液;(21)ポリエステル、ポリカーボネート、及び/又はポリ酸無水物;(22)ポリペプチド及びアミノ酸等の、増量剤;(23)血清アルブミン、HDL、及びLDL等の、血清構成要素;(22)エタノール等の、C2~C12アルコール;並びに(23)医薬製剤に採用される、相溶性のある他の非毒性物質。湿潤剤、着色剤、離型剤、コーティング剤、甘味剤、香味剤、芳香剤、保存料、及び抗酸化剤もまた、製剤中に存在し得る。「賦形剤」、「担体」、「薬学的に許容し得る担体」又は同種のもの等の用語は本明細書中、互換的に使用される。
送達方法
いくつかの側面では、本発明は、本明細書に記載の1つ又は複数のベクター、その1つ又は複数の転写物、並びに/あるいはそれから転写された1つ又は複数のタンパク質等の1つ又は複数のポリヌクレオチドを宿主細胞に送達することを含む方法を提供する。いくつかの側面では、本発明は、更に、かかる方法によって産生される細胞、及びかかる細胞を含むか又は次いで産生される生物(例えば動物、植物、又は真菌)を提供する。いくつかの態様において、本明細書に記載の通りの塩基編集因子は、ガイド配列と組み合わせて(任意選択でこれと複合化されて)、細胞へ送達される。
いくつかの側面では、本発明は、本明細書に記載の1つ又は複数のベクター、その1つ又は複数の転写物、並びに/あるいはそれから転写された1つ又は複数のタンパク質等の1つ又は複数のポリヌクレオチドを宿主細胞に送達することを含む方法を提供する。いくつかの側面では、本発明は、更に、かかる方法によって産生される細胞、及びかかる細胞を含むか又は次いで産生される生物(例えば動物、植物、又は真菌)を提供する。いくつかの態様において、本明細書に記載の通りの塩基編集因子は、ガイド配列と組み合わせて(任意選択でこれと複合化されて)、細胞へ送達される。
例示の送達ストラテジーは、本明細書のどこかに記載されるが、ベクターベースのストラテジー、PEリボヌクレオタンパク質複合体の送達、及びmRNA方法によるPEの送達を包含する。
いくつかの態様において、提供される送達方法は、ヌクレオフェクション、マイクロ注射、微粒子銃(biolistics)、ビロソーム、リポソーム、イムノリポソーム、ポリカチオン又は脂質:核酸抱合体、裸のDNA、人工ビリオン、及びDNAの取り込みを増強する剤を包含する。
例示の核酸送達方法は、リポフェクション、ヌクレオフェクション、エレクトロポレーション、安定したゲノム組み入れ(例として、piggybac)、マイクロ注射、微粒子銃(biolistics)、ビロソーム、リポソーム、イムノリポソーム、ポリカチオン又は脂質:核酸抱合体、裸のDNA、人工ビリオン、及びDNAの取り込みを増強する剤を包含する。リポフェクションは、例として、米国特許第5,049,386号、第4,946,787号;及び第4,897,355号)に記載されており、リポフェクション試薬は商業的に販売されている(例として、Transfectam(商標)、Lipofectin(商標)、及びSF細胞株4D-Nucleofector X Kit(商標)(Lonza))。ポリヌクレオチドの効率的な受容体認識リポフェクションにとって好適であるカチオン性及び中性脂質は、FeignerのWO91/17424;WO91/16024のものを包含する。送達は、細胞(例として、in vitro投与若しくはex vivo投与)、又は標的組織(例として、in vivo投与)に対するものであってもよい。送達は、RNP複合体の使用を通して達成されてもよい。
イムノ脂質複合体等の標的化リポソームを包含する脂質:核酸複合体の調製は当業者に周知である(例として、Crystal,Science 270:404-410(1995);Blaese et al.,Cancer Gene Ther.2:291-297(1995);Behr et al.,Bioconjugate Chem.5:382-389(1994);Remy et al.,Bioconjugate Chem.5:647-654(1994);Gao et al.,Gene Therapy 2:710-722(1995);Ahmad et al.,Cancer Res.52:4817-4820(1992);米国特許第4,186,183号、第4,217,344号、第4,235,871号、第4,261,975号、第4,485,054号、第4,501,728号、第4,774,085号、第4,837,028号、及び第4,946,787を参照)。
他の態様において、本明細書に提供される送達方法及びベクターは、RNP複合体である。融合タンパク質のRNP送達は、塩基編集のDNA特異性を著しく増大させる。融合タンパク質のRNP送達は、オン-及びオフ-ターゲットDNA編集の分断(decoupling)へ繋がる。RNP送達は、プラスミド送達に匹敵するオン-ターゲット編集を維持しながら、非反復部位にてオフターゲット編集を小さくし(ablates)、高度に反復したVEGFA部位2でさえもオフ-ターゲットDNA編集を大いに低減させる。Rees,H.A.et al.,Improving the DNA specificity and applicability of base editing through protein engineering and protein delivery,Nat.Commun.8,15790(2017)、2016年12月27日に発行された米国特許第9,526,784号、及び2017年8月22日に発行された米国特許第9,737,604号を参照(これらの各々は参照により本明細書に組み込まれる)。
細胞への核酸の送達のための追加の方法は当業者に公知である。例えば、US 2003/0087817を参照(参照により本明細書に組み込まれる)。
他の本開示の側面は、プライム編集因子構築物を細胞中へ送達して、完全かつ機能的なプライム編集因子を細胞内に形成する方法を提供する。例えば、いくつかの態様において、細胞は、本明細書に記載の組成物(例として、分裂Cas9若しくは分裂プライム編集因子をコードするヌクレオチド配列、又はかかるヌクレオチド配列を含む核酸ベクターを含有するAAV粒子を含む組成物)と接触させられる。いくつかの態様において、接触させることは、かかるヌクレオチド配列の細胞中への送達をもたらすが、ここでCas9タンパク質又はプライム編集因子のN末端部分、及びCas9タンパク質又はプライム編集因子のC末端部分は、細胞中で発現され結び合わされて、完全なCas9タンパク質又は完全なプライム編集因子を形成する。
本明細書に提供されるいずれのrAAV粒子、核酸分子、又は組成物も、安定的又は一過的のいずれかのいずれか好適なやり方で、細胞中へ導入されてもよいことは了解されるはずである。いくつかの態様において、開示されたタンパク質は、細胞中へトランスフェクトされてもよい。いくつかの態様において、細胞は、核酸分子で形質導入又はトランスフェクトされてもよい。例えば、細胞は、分裂タンパク質をコードする核酸分子、若しくは1以上の核酸分子をコードするウイルスのゲノムを含有するrAAV粒子で、形質導入されても(例として、分裂タンパク質をコードするウイルスで)、又はトランスフェクトされてもよい(例として、分裂タンパク質をコードするプラスミドで)。かかる形質導入は、安定的な形質導入又は一過的な形質導入であってもよい。いくつかの態様において、分裂タンパク質を発現するか又は分裂タンパク質を含有する細胞は、例えば、分裂Cas9(例として、nCas9)タンパク質の送達において、1以上のガイドRNA配列で形質導入又はトランスフェクトされてもよい。いくつかの態様において、分裂タンパク質を発現するプラスミドは、エレクトロポレーション、一過的な(例として、リポフェクション)及び安定したゲノム取り入れ(例として、piggybac)、及びウイルスの形質導入又は当業者に知られている他の方法を通して、細胞中へ導入されてもよい。
ある態様において、本明細書に提供される組成物は、脂質及び/又はポリマーを含む。ある態様において、脂質及び/又はポリマーはカチオン性である。かかる脂質粒子の調製は周知である。例として、米国特許第4,880,635号;第4,906,477号;第4,911,928号;第4,917,951号;第4,920,016号;第4,921,757号;及び第9,737,604号を参照(これらの各々は参照により本明細書に組み込まれる)。
ガイドRNA配列は長さが15~100ヌクレオチドであってもよく、標的ヌクレオチド配列に相補的な少なくとも10、少なくとも15、又は少なくとも20の連続したヌクレオチドの配列を含む。ガイドRNAは、標的ヌクレオチド配列に相補的な15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、又は40の連続したヌクレオチドの配列を含んでいてもよい。ガイドRNAは、長さが15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、又は50ヌクレオチドであってもよい。
いくつかの態様において、標的ヌクレオチド配列は、ゲノム(例として、真核生物のゲノム)中のDNA配列である。ある態様において、標的ヌクレオチド配列は、哺乳動物の動物(例として、ヒト)ゲノム中にある。
本開示の組成物は、例えば、単位用量として投与又はパッケージされてもよい。用語「単位用量」は、本開示の医薬組成物に関して使用されるとき、まとまった投薬量として対象に好適な、物理的に不連続な単位を指し、予め決められた分量の活性材料を含有する各単位は、要求される希釈剤;すなわち、担体又はビヒクルを伴って所望の処置効果を産生するよう算出される。
疾患又は障害の処置は、疾患の発症若しくは進行を遅延させるか、又は疾患の重症度を低減させることを包含する。疾患を処置することは、根治した(curative)結果を必ずしも要さない。
そこで使用された通り、疾患の発症を「遅延すること」は、疾患の進行を、先送りする(defer)、妨げる(hinder)、遅らせる(slow)、妨害する(retard)、留める(stabilize)、及び/又は延ばす(postpone)ことを意味する。この遅延は、病歴及び/又は処置される個体に応じて、時間の長さを変動させることでもあり得る。疾患の発症を「遅延」若しくは緩和する方法、又は疾患の発病を遅延する方法は、この方法を使用しない場合と比較して、所定の時間枠での疾患の1以上の症状を発症する確率を低減するか、及び/又は所定の時間枠での症状の程度を低減する方法である。かかる比較は、典型的には、統計的に有意な結果を与えるのに十分数多の対象を使用する臨床研究に基づく。
疾患の「発症」又は「進行」は、最初の兆候及び/又はその後の疾患の進行を意味する。疾患の発症は、当該技術分野において周知の通りの標準の臨床技法を使用し検出可能であり、かつ査定され得る。しかしながら、発症はまた、検出不能であってもよい進行も指す。本開示の目的上、発症又は進行は、症状の生物学的な経過を指す。「発症」は、発生、再発、及び発病を包含する。
本明細書に使用されるとき、疾患の「発病」又は「発生」は、最初の発病及び/又は再発を包含する。医学の当業者に知られている従来の方法は、処置されるべき疾患の型又は疾患の部位に応じて、単離されたポリペプチド又は医薬組成物を対象へ投与するのに使用され得る。
更なる詳細化はしないが、当業者は、上の記載に基づき、本開示を最大限に利用し得るものと思われる。したがって、以下の特定の態様は、単なる説明として解釈されるべきであって、いずれにしても本開示の残りを何ら限定するものではない。本明細書に引用される全ての刊行物は、本明細書に言及される目的又は主題のため、参照により組み込まれる。
態様
本開示は更に、以下の非限定的な番号付き段落に関する。
1.二本鎖標的DNA配列の標的鎖に対して相補性の領域を含むスペーサ配列と、
核酸プログラム型DNA結合タンパク質(napDNAbp)と会合するDNA合成鋳型コアを含む核酸伸長アームと、
を含む、プライム編集ガイドRNA(PEgRNA)であって、
プライマー結合部位は、二本鎖標的DNA配列の非標的鎖に対して相補性の領域を含み、
DNA合成鋳型は、二本鎖標的DNA配列の非標的鎖に対して相補性の領域を含み、二本鎖標的DNA配列と比較して1つ以上のヌクレオチド編集を含み、
伸長アームは、トゥ-ループ、ヘアピン、ステムループ、シュードノット、アプタマー、G-四重鎖、tRNA、リボスイッチ、又はリボザイムからなる群から選択される核酸部分を更に含む、
プライム編集ガイドRNA(PEgRNA)。
2.核酸部分が伸長アームの3'末端にある、段落1に記載のPEgRNA。
3.核酸部分が伸長アームの5'末端にある、段落1に記載のPEgRNA。
4.核酸部分が、モロニーマウス白血病ウイルス(M-MLV)ゲノム由来のフレームシフトシュードノット(Mpknot)を含み、任意選択で、Mpknotが、配列番号3930(Mpknot1)、配列番号3931(Mpknot1 3'トリミング)、配列番号3932(5'extraを有するMpknot1)、配列番号3933(Mpknot1 U38A)、配列番号3934(Mpknot1 U38A A29C)、配列番号3935(MMLC A29C)、配列番号3936(5'extra及びU 38 Aを有するMpknot1)、配列番号3937(5'extra及びU 38 A A29Cを有するMpknot1)、及び配列番号3938(5'extra及びA29Cを有するMpknot1)からなる群から選択されるヌクレオチド配列、又はそれらと少なくとも80%の配列同一性を有するヌクレオチド配列を有するMpknot1部分である、段落1に記載のPEgRNA。
5.核酸部分がG-四重極を含み、任意選択で、G-四重極が、配列番号3939(tns1)、配列番号3940(stk40)、配列番号3941(apc2)、配列番号3942(ceacam4)、配列番号3943(pitpnm3)、配列番号3944(rlf)、配列番号3945(erc1)、配列番号3946(ube3c)、配列番号3947(taf15)、配列番号3948(stard3)、及び配列番号3949(g2)からなる群から選択されるヌクレオチド配列、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有する、段落1に記載のPEgRNA。
6.核酸部分がプレケオシン1リボスイッチアプタマーを含む、段落1に記載のPEgRNA。
7.核酸部分が、進化したプレケオシン1-1リボスイッチアプタマー(evopreQ1)を含み、任意選択で、evopreQ1が、配列番号3950(evopreq1)、配列番号3951(evopreq1motif1)、配列番号3952(evopreq1motif2)、配列番号3953(evopreq1motif3)、配列番号3954(shorter preq1-1)、配列番号3955(preq1-1 G5C(mut1))、及び配列番号3956(preq1-1 G15C(mut2))からなる群から選択されるヌクレオチド配列、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有する、段落6に記載のPEgRNA。
8.核酸部分が、配列番号3957のヌクレオチド配列、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有するtRNA部分を含む、段落1に記載のPEgRNA。
9.核酸部分が、配列番号3958(xrn1)のヌクレオチド配列、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有する、段落1に記載のPEgRNA。
10.核酸部分が、グループIイントロンのP4-P6ドメインを含み、任意選択で、P4-P6ドメインが、配列番号3959のヌクレオチド配列、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有する、段落1に記載のPEgRNA。
11.PEgRNAがリンカーを更に含む、段落1~10のいずれかに記載のPEgRNA。
12.リンカーが核酸部分とPEgRNAの別の構成要素との間にある、段落11に記載のPEgRNA。
13.リンカーが核酸部分とプライマー結合部位との間又はgRNAコアと核酸部分との間にある、段落11に記載のPEgRNA。PEgRNA
14.リンカーが、配列番号3960、配列番号3961、配列番号3962、配列番号3963、配列番号3964、配列番号3965、配列番号3966、配列番号3967、配列番号3968、配列番号3969、配列番号3970、及び配列番号3971からなる群から選択されるヌクレオチド配列を含む、段落11に記載のPEgRNA。
15.リンカーが、少なくとも3ヌクレオチド、少なくとも4ヌクレオチド、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、少なくとも15ヌクレオチド、少なくとも16ヌクレオチド、少なくとも17ヌクレオチド、少なくとも18ヌクレオチド、少なくとも19ヌクレオチド、少なくとも20ヌクレオチド、少なくとも21ヌクレオチド、少なくとも22ヌクレオチド、少なくとも23ヌクレオチド、少なくとも24ヌクレオチド、少なくとも25ヌクレオチド、少なくとも26ヌクレオチド、少なくとも27ヌクレオチド、少なくとも28ヌクレオチド、少なくとも29ヌクレオチド、又は少なくとも30ヌクレオチド長であり、リンカーが50ヌクレオチド以下である、段落11に記載のPEgRNA。
16.リンカーが、1~5ヌクレオチド、5~10ヌクレオチド、10~20ヌクレオチド、15~25ヌクレオチド、20~30ヌクレオチド、25~35ヌクレオチド、30~40ヌクレオチド、35~45ヌクレオチド、又は40~50ヌクレオチド長であるか、あるいはリンカーが、1~50、3~50、5~50、又は8~50ヌクレオチド長である、段落11に記載のPEgRNA。
17.リンカーが8ヌクレオチド長である、段落11に記載のPEgRNA。
18.伸長アームがガイドRNAの3'末端又は5'末端に配置され、核酸伸長アームがDNA又はRNAを含む、段落4~17のいずれか1つに記載のPEgRNA。
19.プライマー結合部位が、標的DNA配列の非標的鎖のニック部位の上流の領域と相補性の領域を含み、ニック部位がnapDNAbpの特徴である、段落18に記載のPEgRNA。
20.DNA合成鋳型が、標的DNA配列の非標的鎖のニック部位の下流の領域と相補性の領域を含む、段落19に記載のPEgRNA。
21.プライマー結合部位が、標的DNA配列の非標的鎖のニック部位のすぐ上流の領域と相補性の領域を含む、段落18に記載のPEgRNA。
22.核酸伸長アームが、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、少なくとも15ヌクレオチド、少なくとも16ヌクレオチド、少なくとも17ヌクレオチド、少なくとも18ヌクレオチド、少なくとも19ヌクレオチド、少なくとも20ヌクレオチド、少なくとも21ヌクレオチド、少なくとも22ヌクレオチド、少なくとも23ヌクレオチド、少なくとも24ヌクレオチド、少なくとも25ヌクレオチド、少なくとも26ヌクレオチド、少なくとも27ヌクレオチド、少なくとも28ヌクレオチド、少なくとも29ヌクレオチド、少なくとも30ヌクレオチド、少なくとも31ヌクレオチド、少なくとも32ヌクレオチド、少なくとも33ヌクレオチド、少なくとも34ヌクレオチド、少なくとも35ヌクレオチド、少なくとも36ヌクレオチド、少なくとも37ヌクレオチド、少なくとも38ヌクレオチド、少なくとも39ヌクレオチド、少なくとも40ヌクレオチド、少なくとも41ヌクレオチド、少なくとも42ヌクレオチド、少なくとも43ヌクレオチド、少なくとも44ヌクレオチド、少なくとも45ヌクレオチド、少なくとも46ヌクレオチド、少なくとも47ヌクレオチド、少なくとも48ヌクレオチド、少なくとも49ヌクレオチド、又は少なくとも50ヌクレオチドであり、あるいは核酸伸長アームが、10~20、20~30、30~40、40~50、50~60、60~70、70~80、80~90、90~100、100~110、110~120、20~120、40~120、60~120、80~120、100~120、40~100、60~100、80~100、又は60~80ヌクレオチド長であり、あるいは核酸伸長アームが、15~300、20~250、20~200、20~150、25~150、15~100、20~100又は25~100ヌクレオチド長であり、あるいは核酸伸長アームが、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、又は100ヌクレオチド長である、段落18に記載のPEgRNA。
23.DNA合成鋳型が、少なくとも3ヌクレオチド、少なくとも4ヌクレオチド、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、又は少なくとも15ヌクレオチド長であり、あるいはDNA合成鋳型が、1~10、5~15、10~20、20~30、30~40、40~50、50~60、60~70、70~80、80~90、90~100、10~30、10~40、10~50、10~60、20~40、20~60、30~100、40~100、50~100、60~100、又は70~100ヌクレオチド長であり、DNA合成鋳型が、5~300、5~250、15~200、15~150、5~100、10~100、又は15~100ヌクレオチド長であり、あるいはDNA合成鋳型が、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、又は80ヌクレオチド長である、段落18に記載のPEgRNA。
24.DNA合成鋳型が15~35ヌクレオチド長である、段落23に記載のPEgRNA。
25.プライマー結合部位が、少なくとも3ヌクレオチド、少なくとも4ヌクレオチド、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、又は少なくとも15ヌクレオチド長であり、あるいはプライマー結合部位が、1~10ヌクレオチド、5~10ヌクレオチド、10~15ヌクレオチド、10~20ヌクレオチド、8~20ヌクレオチド、15~25ヌクレオチド、20~30ヌクレオチド、又は25~30ヌクレオチド長であり、プライマー結合部位が、3~60、5~60、8~50、又は12~50ヌクレオチド長であり、あるいはプライマー結合部位が、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、又は25ヌクレオチド長である、段落18に記載のPEgRNA。
26.gRNAコアが直接反復を含み、直接反復が4つ以上の連続するA-U塩基対を含有しない、段落1~25のいずれか1つに記載のPEgRNA。
27.直接反復がヌクレオチド配列UUUAを含む、段落26に記載のPEgRNA。
28.化学的若しくは生物学的に修飾されたヌクレオチド又はヌクレオチドアナログを含む、段落1~27のいずれか1つに記載のPEgRNA。
29.PEgRNAの5'末端の3つの連続するヌクレオチドが1つ又は複数の化学修飾ヌクレオチドを含み、及び/又はPEgRNAの3'末端の3つの連続するヌクレオチドが1つ又は複数の化学修飾ヌクレオチドを含む、段落28に記載のPEgRNA。
30.
(a)核酸プログラム型DNA結合タンパク質(napDNAbp)と、
(b)DNAポリメラーゼ活性を含むドメインと、
(c)段落1~29のいずれか1つに記載のPEgRNAと、
を含む、プライム編集因子システム。
31.PEgRNA及びnapDNAbp及び/又はDNAポリメラーゼ活性を含むドメインが複合体を形成する、段落30に記載のプライム編集因子システム。
32.DNAポリメラーゼ活性を有するドメインとnapDNAbpとが融合されて融合タンパク質を形成する、段落30又は31に記載のプライム編集因子システム。
33.napDNAbpがニッカーゼ活性を有する、段落30~32のいずれか1つに記載のプライム編集因子システム。
34.napDNAbpがCas9タンパク質又はそのバリアントである、段落30~32のいずれか1つに記載のプライム編集因子システム。
35.napDNAbpが、ヌクレアーゼ活性Cas9、ヌクレアーゼ不活性Cas9(dCas9)、又はCas9ニッカーゼ(nCas9)である、段落34に記載のプライム編集因子システム。
36.napDNAbpがCas9ニッカーゼ(nCas9)である、段落35に記載のプライム編集因子システム。
37.napDNAbpが、Cas9、Cas12e、Cas12d、Cas12a、Cas12b1、Cas13a、Cas12c及びアルゴノートからなる群から選択され、任意選択でニッカーゼ活性を有する、段落30~32のいずれか1つに記載のプライム編集因子システム。
38.RNA依存性DNAポリメラーゼ活性を含むドメインが、配列番号89~100、105~122、128~129、132、139、143、149、154、159、235、454、471、516、662、700、701~716、739~741、及び766のいずれか1つのアミノ酸配列と少なくとも80%、85%、90%、95%、98%、又は99%の配列同一性を有するアミノ酸配列を含む逆転写酵素である、段落30~37のいずれか1つに記載のプライム編集因子システム。
39.RNA依存性DNAポリメラーゼ活性を含むドメインが、配列番号89~100、105~122、128~129、132、139、143、149、154、159、235、454、471、516、662、700、701~716、739~741及び766のアミノ酸配列のいずれか1つを含む逆転写酵素である、段落30~37のいずれか1つに記載のプライム編集因子システム。
40.逆転写酵素がモロニーマウス白血病ウイルス逆転写酵素(M-MLVRT)である、段落38に記載のプライム編集因子システム。
41.RNA依存性DNAポリメラーゼドメインがバリアントモロニーマウス白血病ウイルス逆転写酵素(M-MLV RT)ドメインを含み、バリアントM-MLV RTドメインが、配列番号89のアミノ酸配列に対して以下の変異:P51X、S67X、E69X、L139X、T197X、D200X、H204X、F209X、E302X、T306X、F309X、W313X、T330X、L345X、L435X、N454X、D524X、E562X、D583X、H594X、L603X、E607X、又はD653Xのうちの1つ又は複数を含み、Xが任意のアミノ酸である、段落40に記載のプライム編集因子システム。
42.変異体M-MLV RTドメインが、配列番号89のアミノ酸配列に対して以下の変異:P51L、S67K、E69K、L139P、T197A、D200N、H204R、F209N、E302K、E302R、T306K、F309N、W313F、T330P、L345G、L435G、N454K、D524G、E562Q、D583N、H594Q、L603W、E607K、又はD653Nのうちの1つ又は複数を含む、段落41に記載のプライム編集因子システム。
43.バリアントM-MLV RTドメインが、配列番号89のアミノ酸配列に対してアミノ酸置換D200N、T330P及びL603Wを含み、任意選択でM-MLV RTドメインが、配列番号89のアミノ酸配列に対してアミノ酸置換D200N、T306K、W313F、T330P及びL603Wを含む、段落41に記載のプライム編集因子システム。
44.RNA依存性DNAポリメラーゼ活性を含むドメインが、レトロウイルス又はレトロトランスポゾン由来の天然に存在する逆転写酵素である、段落30~37のいずれか1つに記載のプライム編集因子システム。
45.段落1~29のいずれか1つに記載のPEgRNAをコードする核酸分子。
46.段落30~45のいずれか1つのnapDNAbp及び/又はDNAポリメラーゼ活性を有するドメインをコードする核酸分子。
47.段落45に記載の核酸分子及び/又は段落46に記載の核酸分子を含む発現ベクターであって、任意選択で、核酸分子はプロモータの制御下にある、発現ベクター。
48.プロモータがpolIIIプロモータである、段落47に記載の発現ベクター。
49.プロモータがU6プロモータである、段落47に記載の発現ベクター。
50.プロモータが、U6、U6v4、U6v7若しくはU6v9プロモータ又はそのフラグメントである、段落47に記載の発現ベクター。
51.段落1~29のいずれか1つに記載のPEgRNAを含む細胞。
52.段落30~44のいずれか1つに記載のプライム編集因子システム、段落45若しくは46に記載の核酸分子、又は段落47~50のいずれか1つに記載の発現ベクターを含む細胞。
53.段落1~29のいずれか1つに記載のPEgRNA、段落30~44のいずれか1つに記載のプライム編集因子システム、又は段落45若しくは46に記載の核酸分子を含む脂質ナノ粒子(LNP)。
54.段落1~29のいずれか1つのPEgRNA、段落30~44のいずれか1つのプライム編集因子システム、又は段落45若しくは46の核酸分子を含む、リボ核タンパク質複合体(RNP)。
55.(i)段落1~29のいずれか1つのPEgRNA、段落30~44のいずれか1つのプライム編集因子システム、又は段落45若しくは46の核酸分子、段落47~50のいずれか1つの発現ベクター、段落51若しくは52の細胞、段落53のLNP、又は段落54のRNP、及び(ii)薬学的に許容される賦形剤を含む医薬組成物。
56.(i)段落1~29のいずれか1つに記載のPEgRNA、段落30~44のいずれか1つに記載のプライム編集因子システム、又は段落45若しくは46に記載の核酸分子、段落47~50のいずれか1つに記載の発現ベクター、段落51若しくは52に記載の細胞、段落53に記載のLNP、又は段落54に記載のRNP、PEgRNA(ii)プライム編集を行うための説明書のセットを含むキット組成物。
57.標的DNA配列を段落1~29のいずれかのPEgRNA並びにnapDNAbp及びDNAポリメラーゼ活性を有するドメインを含むプライム編集因子と接触させることを含み、接触させることが、標的DNA配列に1つ又は複数のヌクレオチド編集を組み入れる、プライム編集の方法。
58.編集効率が、標的DNAをプライム編集因子及び核酸部分PEgRNAを含有しない対照PEgRNAと接触させた場合の編集効率と比較して増加する、段落57に記載の方法。
59.編集効率が少なくとも1.5倍増加する、段落58に記載の方法。
60.編集効率が少なくとも2倍増加する、段落58に記載の方法。
61.編集効率が、少なくとも3倍、4倍、5倍、6倍、7倍、8倍、9倍又は10倍増加する、段落58の方法[1255]、[1270]。
62.napDNAbpがニッカーゼ活性を有する、段落57~61のいずれか1つに記載の方法。
63.napDNAbpがCas9タンパク質又はその変異体である、段落57~62のいずれか1つに記載の方法。
64.napDNAbpが、ヌクレアーゼ活性Cas9、ヌクレアーゼ不活性Cas9(dCas9)、又はCas9ニッカーゼ(nCas9)である、段落63に記載の方法。
65.napDNAbpがCas9ニッカーゼ(nCas9)である、段落64に記載の方法。
66.napDNAbpが、Cas9、Cas12e、Cas12d、Cas12a、Cas12b1、Cas13a、Cas12c及びアルゴノートからなる群から選択され、任意選択的にニッカーゼ活性を有する、段落57~62のいずれか1つに記載の方法。
67.RNA依存性DNAポリメラーゼ活性を含むドメインが、配列番号89~100、105~122、128~129、132、139、143、149、154、159、235、454、471、516、662、700、701~716、739~741及び766のアミノ酸配列のいずれか1つを含む逆転写酵素である、段落57~66のいずれか1つに記載の方法。
68.RNA依存性DNAポリメラーゼ活性を含むドメインが、配列番号89~100、105~122、128~129、132、139、143、149、154、159、235、454、471、516、662、700、701~716、739~741、及び766のいずれか1つのアミノ酸配列と少なくとも80%、85%、90%、95%、98%、又は99%の配列同一性を有するアミノ酸配列を含む逆転写酵素である、段落57~66のいずれか1つに記載の方法。
69.逆転写酵素がモロニーマウス白血病ウイルス逆転写酵素(M-MLVRT)である、段落68に記載の方法。
70.RNA依存性DNAポリメラーゼドメインがバリアントモロニーマウス白血病ウイルス逆転写酵素(M-MLV RT)ドメインを含み、バリアントM-MLV RTドメインが、配列番号89のアミノ酸配列に対して以下の変異:P51X、S67X、E69X、L139X、T197X、D200X、H204X、F209X、E302X、T306X、F309X、W313X、T330X、L345X、L435X、N454X、D524X、E562X、D583X、H594X、L603X、E607X、又はD653Xのうちの1つ又は複数を含み、Xが任意のアミノ酸である、段落69に記載の方法。
71.変異体M-MLV RTドメインが、配列番号89のアミノ酸配列に対して以下の変異:P51L、S67K、E69K、L139P、T197A、D200N、H204R、F209N、E302K、E302R、T306K、F309N、W313F、T330P、L345G、L435G、N454K、D524G、E562Q、D583N、H594Q、L603W、E607K、又はD653Nのうちの1つ又は複数を含む、段落70に記載の方法。
72.バリアントM-MLV RTドメインが、配列番号89のアミノ酸配列に対してアミノ酸置換D200N、T330P及びL603Wを含み、任意選択でM-MLV RTドメインが、配列番号89のアミノ酸配列に対してアミノ酸置換D200N、T306K、W313F、T330P及びL603Wを含む、段落70に記載の方法。
73.RNA依存性DNAポリメラーゼ活性を含むドメインが、レトロウイルス又はレトロトランスポゾン由来の天然に存在する逆転写酵素である、段落57~66に記載の方法。
74.二本鎖標的DNA配列にヌクレオチド編集を組み入れする方法であって、二本鎖標的DNA配列を、核酸プログラム型DNA結合タンパク質(napDNAbp)、DNAポリメラーゼ、及びプライム編集ガイドRNA(PEgRNA)を含むプライム編集因子と接触させる工程を含み、PEgRNAが、
(a)二本鎖標的DNA配列の標的鎖にハイブリダイズする相補性領域を含むスペーサ配列、
(b)DNA合成鋳型及びプライマー結合部位を含む無核酸伸長アーム、
(c)核酸プログラム型DNA結合タンパク質(napDNAbp)と会合するgRNAコア、
(d)トゥーループ、ヘアピン、ステムループ、シュードノット、アプタマー、G-四重鎖、tRNA、リボスイッチ又はリボザイムからなる群から選択される核酸部分、及び
(e)核酸部分をPEgRNAの別の構成要素に連結するリンカーと、
を含み、
プライマー結合部位は、二本鎖標的DNA配列の非標的鎖に対して相補性の領域を含み、
DNA合成鋳型は、二本鎖標的DNA配列の非標的鎖と相補性の領域を含み、二本鎖標的DNA配列と比較して1つ以上のヌクレオチド編集を含み、リンカーは計算モデルによってデザインされる、
二本鎖標的DNA配列にヌクレオチド編集を組み入れする方法。PEgRNA.
75.リンカーが、配列番号3960、配列番号3961、配列番号3962、配列番号3963、配列番号3964、配列番号3965、配列番号3966、配列番号3967、配列番号3968、配列番号3969、配列番号3970、及び配列番号3971からなる群から選択されるヌクレオチド配列を含む、段落75に記載のPEgRNA。
76.プライム編集ガイドRNA(PEgRNA)の構成要素を核酸部分に連結するための少なくとも1つの核酸リンカーを同定するための方法であって、
少なくとも1つのコンピュータ・ハードウェア・プロセッサを使用して、
第1の核酸リンカー候補を包含する複数の核酸リンカー候補を生成することと、
複数の核酸リンカー候補の中から少なくとも1つの核酸リンカーを、少なくとも部分的に、
複数の核酸リンカー候補の少なくともいくつかのそれぞれについて複数のスコアを計算することであって、計算することが、第1の核酸リンカー候補について第1のスコアセットを計算することを含み、第1のスコアセットが、
第1の核酸リンカー候補とPEgRNAの第1の領域との間の相互作用の程度を示す第1のスコアと、
第1の核酸リンカー候補とPEgRNAの第2の領域との間の相互作用の程度を示す第2のスコアと、
を含む、計算することと、
計算された複数のスコアを使用して、複数の核酸リンカー候補の少なくともいくつかの中から少なくとも1つの核酸リンカーを同定することと、
によって同定することと、
少なくとも1つの核酸リンカーを示す情報を出力することと、
を行うことを含む、プライム編集ガイドRNA(PEgRNA)の構成要素を核酸部分に連結するための少なくとも1つの核酸リンカーを同定するための方法。
77.第1のスコアが、第1の核酸リンカー候補がPEgRNAの第1の領域との相互作用を回避すると予測される程度を示し、第2のスコアが、第1の核酸リンカー候補がPEgRNAの第2の領域との相互作用を回避すると予測される程度を示す、段落77に記載の方法。
78.第1の領域がPEgRNAのプライマー結合部位(PBS)を含む、段落78に記載の方法。
79.第2の領域がPEgRNAのスペーサを含む、段落79に記載の方法。
80.スコアの第1のセットが、第1の核酸リンカー候補がPEgRNAの第3の領域との相互作用を回避すると予測される程度を示す第3のスコアと、第1の核酸リンカー候補がPEgRNAの第4の領域との相互作用を回避すると予測される程度を示す第4のスコアと、を更に含む、段落78に記載の方法。
81.第3の領域がDNA合成鋳型を含む、段落81に記載の方法。
82.第4の領域が、核酸プログラム型DNA結合タンパク質(napDNAbp)と相互作用するgRNAコアを含む、段落82に記載の方法。
83.PEgRNAが、二本鎖標的DNA配列にヌクレオチド編集を組み入れするためのものであり、
PEgRNAが、
二本鎖標的DNA配列の標的鎖にハイブリダイズする相補性領域を含むスペーサ配列、
DNA合成鋳型及びプライマー結合部位を含む核酸伸長アーム、
核酸プログラム型DNA結合タンパク質napDNAbpと相互作用するgRNAコア、
を含み、
プライマー結合部位が、二本鎖標的DNA配列の非標的鎖に対して相補性の領域を含み、
DNA合成鋳型は、二本鎖標的DNA配列の非標的鎖と相補性の領域を含み、二本鎖標的DNA配列と比較して1つ以上のヌクレオチド編集を含み、第1の領域はPBSを含み、第2の領域はスペーサを含み、第3の領域はDNA合成鋳型を含み、第4の領域はgRNAコアを含む、段落81に記載の方法。
84.複数の核酸リンカー候補が第2の核酸リンカー候補を含み、計算された複数のスコアを用いて複数の核酸リンカー候補の少なくともいくつかの中から少なくとも1つの核酸リンカーを同定することが、
第1の核酸リンカー候補に対するスコアの第1のセットを第2の核酸リンカー候補に対するスコアの第2のセットと比較することを含む、
段落77に記載の方法。
85.第1の領域がプライマー結合部位(PBS)を含み、
第1のスコアセットにおける第1のスコアが、第1の核酸リンカー候補がPEgRNAの第1の領域との相互作用を回避すると予測される程度を示し、
第2のスコアセットにおける第3のスコアは、第2の核酸リンカー候補がPEgRNAの第1の領域との相互作用を回避すると予測される程度を示し、
第1のスコアセットを第2のスコアセットと比較することが、
第1のスコアと第3のスコアとを比較することを含む、
段落85に記載の方法。
86.第1のスコアが第3のスコアの閾値距離に等しいか、又は閾値距離内にある場合、第1のスコアセットと第2のスコアセットとを比較する工程は、
第1のスコアのセットにおける第1のスコア以外のスコアと、第2のスコアのセットにおける第3のスコア以外の別のスコアとを比較することとを含む、
段落86に記載の方法。
本開示は更に、以下の非限定的な番号付き段落に関する。
1.二本鎖標的DNA配列の標的鎖に対して相補性の領域を含むスペーサ配列と、
核酸プログラム型DNA結合タンパク質(napDNAbp)と会合するDNA合成鋳型コアを含む核酸伸長アームと、
を含む、プライム編集ガイドRNA(PEgRNA)であって、
プライマー結合部位は、二本鎖標的DNA配列の非標的鎖に対して相補性の領域を含み、
DNA合成鋳型は、二本鎖標的DNA配列の非標的鎖に対して相補性の領域を含み、二本鎖標的DNA配列と比較して1つ以上のヌクレオチド編集を含み、
伸長アームは、トゥ-ループ、ヘアピン、ステムループ、シュードノット、アプタマー、G-四重鎖、tRNA、リボスイッチ、又はリボザイムからなる群から選択される核酸部分を更に含む、
プライム編集ガイドRNA(PEgRNA)。
2.核酸部分が伸長アームの3'末端にある、段落1に記載のPEgRNA。
3.核酸部分が伸長アームの5'末端にある、段落1に記載のPEgRNA。
4.核酸部分が、モロニーマウス白血病ウイルス(M-MLV)ゲノム由来のフレームシフトシュードノット(Mpknot)を含み、任意選択で、Mpknotが、配列番号3930(Mpknot1)、配列番号3931(Mpknot1 3'トリミング)、配列番号3932(5'extraを有するMpknot1)、配列番号3933(Mpknot1 U38A)、配列番号3934(Mpknot1 U38A A29C)、配列番号3935(MMLC A29C)、配列番号3936(5'extra及びU 38 Aを有するMpknot1)、配列番号3937(5'extra及びU 38 A A29Cを有するMpknot1)、及び配列番号3938(5'extra及びA29Cを有するMpknot1)からなる群から選択されるヌクレオチド配列、又はそれらと少なくとも80%の配列同一性を有するヌクレオチド配列を有するMpknot1部分である、段落1に記載のPEgRNA。
5.核酸部分がG-四重極を含み、任意選択で、G-四重極が、配列番号3939(tns1)、配列番号3940(stk40)、配列番号3941(apc2)、配列番号3942(ceacam4)、配列番号3943(pitpnm3)、配列番号3944(rlf)、配列番号3945(erc1)、配列番号3946(ube3c)、配列番号3947(taf15)、配列番号3948(stard3)、及び配列番号3949(g2)からなる群から選択されるヌクレオチド配列、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有する、段落1に記載のPEgRNA。
6.核酸部分がプレケオシン1リボスイッチアプタマーを含む、段落1に記載のPEgRNA。
7.核酸部分が、進化したプレケオシン1-1リボスイッチアプタマー(evopreQ1)を含み、任意選択で、evopreQ1が、配列番号3950(evopreq1)、配列番号3951(evopreq1motif1)、配列番号3952(evopreq1motif2)、配列番号3953(evopreq1motif3)、配列番号3954(shorter preq1-1)、配列番号3955(preq1-1 G5C(mut1))、及び配列番号3956(preq1-1 G15C(mut2))からなる群から選択されるヌクレオチド配列、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有する、段落6に記載のPEgRNA。
8.核酸部分が、配列番号3957のヌクレオチド配列、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有するtRNA部分を含む、段落1に記載のPEgRNA。
9.核酸部分が、配列番号3958(xrn1)のヌクレオチド配列、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有する、段落1に記載のPEgRNA。
10.核酸部分が、グループIイントロンのP4-P6ドメインを含み、任意選択で、P4-P6ドメインが、配列番号3959のヌクレオチド配列、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有する、段落1に記載のPEgRNA。
11.PEgRNAがリンカーを更に含む、段落1~10のいずれかに記載のPEgRNA。
12.リンカーが核酸部分とPEgRNAの別の構成要素との間にある、段落11に記載のPEgRNA。
13.リンカーが核酸部分とプライマー結合部位との間又はgRNAコアと核酸部分との間にある、段落11に記載のPEgRNA。PEgRNA
14.リンカーが、配列番号3960、配列番号3961、配列番号3962、配列番号3963、配列番号3964、配列番号3965、配列番号3966、配列番号3967、配列番号3968、配列番号3969、配列番号3970、及び配列番号3971からなる群から選択されるヌクレオチド配列を含む、段落11に記載のPEgRNA。
15.リンカーが、少なくとも3ヌクレオチド、少なくとも4ヌクレオチド、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、少なくとも15ヌクレオチド、少なくとも16ヌクレオチド、少なくとも17ヌクレオチド、少なくとも18ヌクレオチド、少なくとも19ヌクレオチド、少なくとも20ヌクレオチド、少なくとも21ヌクレオチド、少なくとも22ヌクレオチド、少なくとも23ヌクレオチド、少なくとも24ヌクレオチド、少なくとも25ヌクレオチド、少なくとも26ヌクレオチド、少なくとも27ヌクレオチド、少なくとも28ヌクレオチド、少なくとも29ヌクレオチド、又は少なくとも30ヌクレオチド長であり、リンカーが50ヌクレオチド以下である、段落11に記載のPEgRNA。
16.リンカーが、1~5ヌクレオチド、5~10ヌクレオチド、10~20ヌクレオチド、15~25ヌクレオチド、20~30ヌクレオチド、25~35ヌクレオチド、30~40ヌクレオチド、35~45ヌクレオチド、又は40~50ヌクレオチド長であるか、あるいはリンカーが、1~50、3~50、5~50、又は8~50ヌクレオチド長である、段落11に記載のPEgRNA。
17.リンカーが8ヌクレオチド長である、段落11に記載のPEgRNA。
18.伸長アームがガイドRNAの3'末端又は5'末端に配置され、核酸伸長アームがDNA又はRNAを含む、段落4~17のいずれか1つに記載のPEgRNA。
19.プライマー結合部位が、標的DNA配列の非標的鎖のニック部位の上流の領域と相補性の領域を含み、ニック部位がnapDNAbpの特徴である、段落18に記載のPEgRNA。
20.DNA合成鋳型が、標的DNA配列の非標的鎖のニック部位の下流の領域と相補性の領域を含む、段落19に記載のPEgRNA。
21.プライマー結合部位が、標的DNA配列の非標的鎖のニック部位のすぐ上流の領域と相補性の領域を含む、段落18に記載のPEgRNA。
22.核酸伸長アームが、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、少なくとも15ヌクレオチド、少なくとも16ヌクレオチド、少なくとも17ヌクレオチド、少なくとも18ヌクレオチド、少なくとも19ヌクレオチド、少なくとも20ヌクレオチド、少なくとも21ヌクレオチド、少なくとも22ヌクレオチド、少なくとも23ヌクレオチド、少なくとも24ヌクレオチド、少なくとも25ヌクレオチド、少なくとも26ヌクレオチド、少なくとも27ヌクレオチド、少なくとも28ヌクレオチド、少なくとも29ヌクレオチド、少なくとも30ヌクレオチド、少なくとも31ヌクレオチド、少なくとも32ヌクレオチド、少なくとも33ヌクレオチド、少なくとも34ヌクレオチド、少なくとも35ヌクレオチド、少なくとも36ヌクレオチド、少なくとも37ヌクレオチド、少なくとも38ヌクレオチド、少なくとも39ヌクレオチド、少なくとも40ヌクレオチド、少なくとも41ヌクレオチド、少なくとも42ヌクレオチド、少なくとも43ヌクレオチド、少なくとも44ヌクレオチド、少なくとも45ヌクレオチド、少なくとも46ヌクレオチド、少なくとも47ヌクレオチド、少なくとも48ヌクレオチド、少なくとも49ヌクレオチド、又は少なくとも50ヌクレオチドであり、あるいは核酸伸長アームが、10~20、20~30、30~40、40~50、50~60、60~70、70~80、80~90、90~100、100~110、110~120、20~120、40~120、60~120、80~120、100~120、40~100、60~100、80~100、又は60~80ヌクレオチド長であり、あるいは核酸伸長アームが、15~300、20~250、20~200、20~150、25~150、15~100、20~100又は25~100ヌクレオチド長であり、あるいは核酸伸長アームが、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、又は100ヌクレオチド長である、段落18に記載のPEgRNA。
23.DNA合成鋳型が、少なくとも3ヌクレオチド、少なくとも4ヌクレオチド、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、又は少なくとも15ヌクレオチド長であり、あるいはDNA合成鋳型が、1~10、5~15、10~20、20~30、30~40、40~50、50~60、60~70、70~80、80~90、90~100、10~30、10~40、10~50、10~60、20~40、20~60、30~100、40~100、50~100、60~100、又は70~100ヌクレオチド長であり、DNA合成鋳型が、5~300、5~250、15~200、15~150、5~100、10~100、又は15~100ヌクレオチド長であり、あるいはDNA合成鋳型が、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、又は80ヌクレオチド長である、段落18に記載のPEgRNA。
24.DNA合成鋳型が15~35ヌクレオチド長である、段落23に記載のPEgRNA。
25.プライマー結合部位が、少なくとも3ヌクレオチド、少なくとも4ヌクレオチド、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、又は少なくとも15ヌクレオチド長であり、あるいはプライマー結合部位が、1~10ヌクレオチド、5~10ヌクレオチド、10~15ヌクレオチド、10~20ヌクレオチド、8~20ヌクレオチド、15~25ヌクレオチド、20~30ヌクレオチド、又は25~30ヌクレオチド長であり、プライマー結合部位が、3~60、5~60、8~50、又は12~50ヌクレオチド長であり、あるいはプライマー結合部位が、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、又は25ヌクレオチド長である、段落18に記載のPEgRNA。
26.gRNAコアが直接反復を含み、直接反復が4つ以上の連続するA-U塩基対を含有しない、段落1~25のいずれか1つに記載のPEgRNA。
27.直接反復がヌクレオチド配列UUUAを含む、段落26に記載のPEgRNA。
28.化学的若しくは生物学的に修飾されたヌクレオチド又はヌクレオチドアナログを含む、段落1~27のいずれか1つに記載のPEgRNA。
29.PEgRNAの5'末端の3つの連続するヌクレオチドが1つ又は複数の化学修飾ヌクレオチドを含み、及び/又はPEgRNAの3'末端の3つの連続するヌクレオチドが1つ又は複数の化学修飾ヌクレオチドを含む、段落28に記載のPEgRNA。
30.
(a)核酸プログラム型DNA結合タンパク質(napDNAbp)と、
(b)DNAポリメラーゼ活性を含むドメインと、
(c)段落1~29のいずれか1つに記載のPEgRNAと、
を含む、プライム編集因子システム。
31.PEgRNA及びnapDNAbp及び/又はDNAポリメラーゼ活性を含むドメインが複合体を形成する、段落30に記載のプライム編集因子システム。
32.DNAポリメラーゼ活性を有するドメインとnapDNAbpとが融合されて融合タンパク質を形成する、段落30又は31に記載のプライム編集因子システム。
33.napDNAbpがニッカーゼ活性を有する、段落30~32のいずれか1つに記載のプライム編集因子システム。
34.napDNAbpがCas9タンパク質又はそのバリアントである、段落30~32のいずれか1つに記載のプライム編集因子システム。
35.napDNAbpが、ヌクレアーゼ活性Cas9、ヌクレアーゼ不活性Cas9(dCas9)、又はCas9ニッカーゼ(nCas9)である、段落34に記載のプライム編集因子システム。
36.napDNAbpがCas9ニッカーゼ(nCas9)である、段落35に記載のプライム編集因子システム。
37.napDNAbpが、Cas9、Cas12e、Cas12d、Cas12a、Cas12b1、Cas13a、Cas12c及びアルゴノートからなる群から選択され、任意選択でニッカーゼ活性を有する、段落30~32のいずれか1つに記載のプライム編集因子システム。
38.RNA依存性DNAポリメラーゼ活性を含むドメインが、配列番号89~100、105~122、128~129、132、139、143、149、154、159、235、454、471、516、662、700、701~716、739~741、及び766のいずれか1つのアミノ酸配列と少なくとも80%、85%、90%、95%、98%、又は99%の配列同一性を有するアミノ酸配列を含む逆転写酵素である、段落30~37のいずれか1つに記載のプライム編集因子システム。
39.RNA依存性DNAポリメラーゼ活性を含むドメインが、配列番号89~100、105~122、128~129、132、139、143、149、154、159、235、454、471、516、662、700、701~716、739~741及び766のアミノ酸配列のいずれか1つを含む逆転写酵素である、段落30~37のいずれか1つに記載のプライム編集因子システム。
40.逆転写酵素がモロニーマウス白血病ウイルス逆転写酵素(M-MLVRT)である、段落38に記載のプライム編集因子システム。
41.RNA依存性DNAポリメラーゼドメインがバリアントモロニーマウス白血病ウイルス逆転写酵素(M-MLV RT)ドメインを含み、バリアントM-MLV RTドメインが、配列番号89のアミノ酸配列に対して以下の変異:P51X、S67X、E69X、L139X、T197X、D200X、H204X、F209X、E302X、T306X、F309X、W313X、T330X、L345X、L435X、N454X、D524X、E562X、D583X、H594X、L603X、E607X、又はD653Xのうちの1つ又は複数を含み、Xが任意のアミノ酸である、段落40に記載のプライム編集因子システム。
42.変異体M-MLV RTドメインが、配列番号89のアミノ酸配列に対して以下の変異:P51L、S67K、E69K、L139P、T197A、D200N、H204R、F209N、E302K、E302R、T306K、F309N、W313F、T330P、L345G、L435G、N454K、D524G、E562Q、D583N、H594Q、L603W、E607K、又はD653Nのうちの1つ又は複数を含む、段落41に記載のプライム編集因子システム。
43.バリアントM-MLV RTドメインが、配列番号89のアミノ酸配列に対してアミノ酸置換D200N、T330P及びL603Wを含み、任意選択でM-MLV RTドメインが、配列番号89のアミノ酸配列に対してアミノ酸置換D200N、T306K、W313F、T330P及びL603Wを含む、段落41に記載のプライム編集因子システム。
44.RNA依存性DNAポリメラーゼ活性を含むドメインが、レトロウイルス又はレトロトランスポゾン由来の天然に存在する逆転写酵素である、段落30~37のいずれか1つに記載のプライム編集因子システム。
45.段落1~29のいずれか1つに記載のPEgRNAをコードする核酸分子。
46.段落30~45のいずれか1つのnapDNAbp及び/又はDNAポリメラーゼ活性を有するドメインをコードする核酸分子。
47.段落45に記載の核酸分子及び/又は段落46に記載の核酸分子を含む発現ベクターであって、任意選択で、核酸分子はプロモータの制御下にある、発現ベクター。
48.プロモータがpolIIIプロモータである、段落47に記載の発現ベクター。
49.プロモータがU6プロモータである、段落47に記載の発現ベクター。
50.プロモータが、U6、U6v4、U6v7若しくはU6v9プロモータ又はそのフラグメントである、段落47に記載の発現ベクター。
51.段落1~29のいずれか1つに記載のPEgRNAを含む細胞。
52.段落30~44のいずれか1つに記載のプライム編集因子システム、段落45若しくは46に記載の核酸分子、又は段落47~50のいずれか1つに記載の発現ベクターを含む細胞。
53.段落1~29のいずれか1つに記載のPEgRNA、段落30~44のいずれか1つに記載のプライム編集因子システム、又は段落45若しくは46に記載の核酸分子を含む脂質ナノ粒子(LNP)。
54.段落1~29のいずれか1つのPEgRNA、段落30~44のいずれか1つのプライム編集因子システム、又は段落45若しくは46の核酸分子を含む、リボ核タンパク質複合体(RNP)。
55.(i)段落1~29のいずれか1つのPEgRNA、段落30~44のいずれか1つのプライム編集因子システム、又は段落45若しくは46の核酸分子、段落47~50のいずれか1つの発現ベクター、段落51若しくは52の細胞、段落53のLNP、又は段落54のRNP、及び(ii)薬学的に許容される賦形剤を含む医薬組成物。
56.(i)段落1~29のいずれか1つに記載のPEgRNA、段落30~44のいずれか1つに記載のプライム編集因子システム、又は段落45若しくは46に記載の核酸分子、段落47~50のいずれか1つに記載の発現ベクター、段落51若しくは52に記載の細胞、段落53に記載のLNP、又は段落54に記載のRNP、PEgRNA(ii)プライム編集を行うための説明書のセットを含むキット組成物。
57.標的DNA配列を段落1~29のいずれかのPEgRNA並びにnapDNAbp及びDNAポリメラーゼ活性を有するドメインを含むプライム編集因子と接触させることを含み、接触させることが、標的DNA配列に1つ又は複数のヌクレオチド編集を組み入れる、プライム編集の方法。
58.編集効率が、標的DNAをプライム編集因子及び核酸部分PEgRNAを含有しない対照PEgRNAと接触させた場合の編集効率と比較して増加する、段落57に記載の方法。
59.編集効率が少なくとも1.5倍増加する、段落58に記載の方法。
60.編集効率が少なくとも2倍増加する、段落58に記載の方法。
61.編集効率が、少なくとも3倍、4倍、5倍、6倍、7倍、8倍、9倍又は10倍増加する、段落58の方法[1255]、[1270]。
62.napDNAbpがニッカーゼ活性を有する、段落57~61のいずれか1つに記載の方法。
63.napDNAbpがCas9タンパク質又はその変異体である、段落57~62のいずれか1つに記載の方法。
64.napDNAbpが、ヌクレアーゼ活性Cas9、ヌクレアーゼ不活性Cas9(dCas9)、又はCas9ニッカーゼ(nCas9)である、段落63に記載の方法。
65.napDNAbpがCas9ニッカーゼ(nCas9)である、段落64に記載の方法。
66.napDNAbpが、Cas9、Cas12e、Cas12d、Cas12a、Cas12b1、Cas13a、Cas12c及びアルゴノートからなる群から選択され、任意選択的にニッカーゼ活性を有する、段落57~62のいずれか1つに記載の方法。
67.RNA依存性DNAポリメラーゼ活性を含むドメインが、配列番号89~100、105~122、128~129、132、139、143、149、154、159、235、454、471、516、662、700、701~716、739~741及び766のアミノ酸配列のいずれか1つを含む逆転写酵素である、段落57~66のいずれか1つに記載の方法。
68.RNA依存性DNAポリメラーゼ活性を含むドメインが、配列番号89~100、105~122、128~129、132、139、143、149、154、159、235、454、471、516、662、700、701~716、739~741、及び766のいずれか1つのアミノ酸配列と少なくとも80%、85%、90%、95%、98%、又は99%の配列同一性を有するアミノ酸配列を含む逆転写酵素である、段落57~66のいずれか1つに記載の方法。
69.逆転写酵素がモロニーマウス白血病ウイルス逆転写酵素(M-MLVRT)である、段落68に記載の方法。
70.RNA依存性DNAポリメラーゼドメインがバリアントモロニーマウス白血病ウイルス逆転写酵素(M-MLV RT)ドメインを含み、バリアントM-MLV RTドメインが、配列番号89のアミノ酸配列に対して以下の変異:P51X、S67X、E69X、L139X、T197X、D200X、H204X、F209X、E302X、T306X、F309X、W313X、T330X、L345X、L435X、N454X、D524X、E562X、D583X、H594X、L603X、E607X、又はD653Xのうちの1つ又は複数を含み、Xが任意のアミノ酸である、段落69に記載の方法。
71.変異体M-MLV RTドメインが、配列番号89のアミノ酸配列に対して以下の変異:P51L、S67K、E69K、L139P、T197A、D200N、H204R、F209N、E302K、E302R、T306K、F309N、W313F、T330P、L345G、L435G、N454K、D524G、E562Q、D583N、H594Q、L603W、E607K、又はD653Nのうちの1つ又は複数を含む、段落70に記載の方法。
72.バリアントM-MLV RTドメインが、配列番号89のアミノ酸配列に対してアミノ酸置換D200N、T330P及びL603Wを含み、任意選択でM-MLV RTドメインが、配列番号89のアミノ酸配列に対してアミノ酸置換D200N、T306K、W313F、T330P及びL603Wを含む、段落70に記載の方法。
73.RNA依存性DNAポリメラーゼ活性を含むドメインが、レトロウイルス又はレトロトランスポゾン由来の天然に存在する逆転写酵素である、段落57~66に記載の方法。
74.二本鎖標的DNA配列にヌクレオチド編集を組み入れする方法であって、二本鎖標的DNA配列を、核酸プログラム型DNA結合タンパク質(napDNAbp)、DNAポリメラーゼ、及びプライム編集ガイドRNA(PEgRNA)を含むプライム編集因子と接触させる工程を含み、PEgRNAが、
(a)二本鎖標的DNA配列の標的鎖にハイブリダイズする相補性領域を含むスペーサ配列、
(b)DNA合成鋳型及びプライマー結合部位を含む無核酸伸長アーム、
(c)核酸プログラム型DNA結合タンパク質(napDNAbp)と会合するgRNAコア、
(d)トゥーループ、ヘアピン、ステムループ、シュードノット、アプタマー、G-四重鎖、tRNA、リボスイッチ又はリボザイムからなる群から選択される核酸部分、及び
(e)核酸部分をPEgRNAの別の構成要素に連結するリンカーと、
を含み、
プライマー結合部位は、二本鎖標的DNA配列の非標的鎖に対して相補性の領域を含み、
DNA合成鋳型は、二本鎖標的DNA配列の非標的鎖と相補性の領域を含み、二本鎖標的DNA配列と比較して1つ以上のヌクレオチド編集を含み、リンカーは計算モデルによってデザインされる、
二本鎖標的DNA配列にヌクレオチド編集を組み入れする方法。PEgRNA.
75.リンカーが、配列番号3960、配列番号3961、配列番号3962、配列番号3963、配列番号3964、配列番号3965、配列番号3966、配列番号3967、配列番号3968、配列番号3969、配列番号3970、及び配列番号3971からなる群から選択されるヌクレオチド配列を含む、段落75に記載のPEgRNA。
76.プライム編集ガイドRNA(PEgRNA)の構成要素を核酸部分に連結するための少なくとも1つの核酸リンカーを同定するための方法であって、
少なくとも1つのコンピュータ・ハードウェア・プロセッサを使用して、
第1の核酸リンカー候補を包含する複数の核酸リンカー候補を生成することと、
複数の核酸リンカー候補の中から少なくとも1つの核酸リンカーを、少なくとも部分的に、
複数の核酸リンカー候補の少なくともいくつかのそれぞれについて複数のスコアを計算することであって、計算することが、第1の核酸リンカー候補について第1のスコアセットを計算することを含み、第1のスコアセットが、
第1の核酸リンカー候補とPEgRNAの第1の領域との間の相互作用の程度を示す第1のスコアと、
第1の核酸リンカー候補とPEgRNAの第2の領域との間の相互作用の程度を示す第2のスコアと、
を含む、計算することと、
計算された複数のスコアを使用して、複数の核酸リンカー候補の少なくともいくつかの中から少なくとも1つの核酸リンカーを同定することと、
によって同定することと、
少なくとも1つの核酸リンカーを示す情報を出力することと、
を行うことを含む、プライム編集ガイドRNA(PEgRNA)の構成要素を核酸部分に連結するための少なくとも1つの核酸リンカーを同定するための方法。
77.第1のスコアが、第1の核酸リンカー候補がPEgRNAの第1の領域との相互作用を回避すると予測される程度を示し、第2のスコアが、第1の核酸リンカー候補がPEgRNAの第2の領域との相互作用を回避すると予測される程度を示す、段落77に記載の方法。
78.第1の領域がPEgRNAのプライマー結合部位(PBS)を含む、段落78に記載の方法。
79.第2の領域がPEgRNAのスペーサを含む、段落79に記載の方法。
80.スコアの第1のセットが、第1の核酸リンカー候補がPEgRNAの第3の領域との相互作用を回避すると予測される程度を示す第3のスコアと、第1の核酸リンカー候補がPEgRNAの第4の領域との相互作用を回避すると予測される程度を示す第4のスコアと、を更に含む、段落78に記載の方法。
81.第3の領域がDNA合成鋳型を含む、段落81に記載の方法。
82.第4の領域が、核酸プログラム型DNA結合タンパク質(napDNAbp)と相互作用するgRNAコアを含む、段落82に記載の方法。
83.PEgRNAが、二本鎖標的DNA配列にヌクレオチド編集を組み入れするためのものであり、
PEgRNAが、
二本鎖標的DNA配列の標的鎖にハイブリダイズする相補性領域を含むスペーサ配列、
DNA合成鋳型及びプライマー結合部位を含む核酸伸長アーム、
核酸プログラム型DNA結合タンパク質napDNAbpと相互作用するgRNAコア、
を含み、
プライマー結合部位が、二本鎖標的DNA配列の非標的鎖に対して相補性の領域を含み、
DNA合成鋳型は、二本鎖標的DNA配列の非標的鎖と相補性の領域を含み、二本鎖標的DNA配列と比較して1つ以上のヌクレオチド編集を含み、第1の領域はPBSを含み、第2の領域はスペーサを含み、第3の領域はDNA合成鋳型を含み、第4の領域はgRNAコアを含む、段落81に記載の方法。
84.複数の核酸リンカー候補が第2の核酸リンカー候補を含み、計算された複数のスコアを用いて複数の核酸リンカー候補の少なくともいくつかの中から少なくとも1つの核酸リンカーを同定することが、
第1の核酸リンカー候補に対するスコアの第1のセットを第2の核酸リンカー候補に対するスコアの第2のセットと比較することを含む、
段落77に記載の方法。
85.第1の領域がプライマー結合部位(PBS)を含み、
第1のスコアセットにおける第1のスコアが、第1の核酸リンカー候補がPEgRNAの第1の領域との相互作用を回避すると予測される程度を示し、
第2のスコアセットにおける第3のスコアは、第2の核酸リンカー候補がPEgRNAの第1の領域との相互作用を回避すると予測される程度を示し、
第1のスコアセットを第2のスコアセットと比較することが、
第1のスコアと第3のスコアとを比較することを含む、
段落85に記載の方法。
86.第1のスコアが第3のスコアの閾値距離に等しいか、又は閾値距離内にある場合、第1のスコアセットと第2のスコアセットとを比較する工程は、
第1のスコアのセットにおける第1のスコア以外のスコアと、第2のスコアのセットにおける第3のスコア以外の別のスコアとを比較することとを含む、
段落86に記載の方法。
本開示はまた、以下の番号が付けられた態様を提供する。
1.疾患関連遺伝子の2つ以上のコピーを編集するための方法であって、疾患関連遺伝子の各コピーが二本鎖標的DNA配列を含み、疾患関連遺伝子の2つ以上のコピーの各々を、
(a)核酸プログラム型DNA結合タンパク質(napDNAbp)ドメイン又はnapDNAbpドメインをコードするポリヌクレオチド;
(b)ポリメラーゼドメイン又はポリメラーゼドメインをコードするポリヌクレオチド;及び
(c)プライム編集ガイドRNA(PEgRNA)、
を含む、プライム編集因子システムと接触させることを含み、
PEgRNAは、
二本鎖DNA配列の標的鎖と相補性の領域を含むスペーサ;
napDNAbpドメインと会合するgRNAコア;並びに
プライマー結合部位及びDNA合成鋳型を含む核酸伸長アーム
を含み、プライマー結合部位は、二本鎖標的DNA配列の非標的鎖に対して相補性の領域を含み、DNA合成鋳型は、二本鎖標的DNA配列の非標的鎖に対して相補性の領域を含み、二本鎖標的DNAと比較して1つ又は複数のヌクレオチド編集を含み、非標的鎖は標的鎖に相補的であり、
疾患関連遺伝子の各コピーは、病原性バリアントを含み、疾患関連遺伝子の2つ以上のコピーが2つ以上の異なる病原性バリアントを含み、接触させることが、疾患関連遺伝子の2つ以上のコピーのそれぞれに1つ以上のヌクレオチド編集を組み入れし、組み入れが、疾患関連遺伝子のそれぞれの病原性バリアントを修正し、それにより、疾患関連遺伝子の2つ以上のコピーのそれぞれを編集する、
疾患関連遺伝子の2つ以上のコピーを編集するための方法。
2.疾患関連遺伝子の2つ又はそれを超えるコピーが1人の対象に存在する、態様1に記載の方法。
3.疾患関連遺伝子の2つ以上のコピーが、2人以上の異なる対象に存在する、態様1に記載の方法。
4.疾患関連遺伝子をそれぞれ含む2人以上の対象において疾患を処置する方法であって、2人以上の対象における疾患関連遺伝子が二本鎖標的DNA配列を含み、方法が、2人以上の対象に、
(a)核酸プログラム型DNA結合タンパク質(napDNAbp)ドメイン又はnapDNAbpドメインをコードするポリヌクレオチド;
(b)ポリメラーゼドメイン又はポリメラーゼドメインをコードするポリヌクレオチド;及び
(c)プライム編集ガイドRNA(PEgRNA)
を含むプライム編集因子システムを投与することを含み、
PEgRNAは、
(i)二本鎖DNA配列の標的鎖と相補性の領域を含むスペーサ、
(ii)napDNAbpドメインと会合するgRNAコア;及び
(iii)プライマー結合部位及びDNA合成鋳型を含む核酸伸長アーム
を含み、
プライマー結合部位は、二本鎖標的DNA配列の非標的鎖に対して相補性の領域を含み、DNA合成鋳型は、二本鎖標的DNA配列の非標的鎖に対して相補性の領域を含み、二本鎖標的DNAと比較して1つ又は複数のヌクレオチド編集を含み、非標的鎖は標的鎖に相補的であり
2人以上の対象が、疾患関連遺伝子に2つ以上の異なる病原性バリアントを含み、投与が、2人以上の対象の各々の疾患関連遺伝子に1つ以上のヌクレオチド編集を組み入れし、組み入れが、2人以上の対象の各々の疾患関連遺伝子の病原性変バリアントを修正し、それにより、2人以上の対象の疾患を処置する、
疾患関連遺伝子をそれぞれ含む2人以上の対象において疾患を処置する方法。
5.napDNAbpドメインをコードするポリヌクレオチド及び/又はポリメラーゼドメインをコードするポリヌクレオチドがRNAを含み、任意選択でnapDNAbpドメインをコードするポリヌクレオチド及び/又はポリメラーゼドメインをコードするポリヌクレオチドがmRNAである、態様1~4のいずれか一項に記載の方法。
6.ポリメラーゼドメインがRNA依存性DNAポリメラーゼドメインである、態様1~5のいずれか1つに記載の方法。
7.ポリメラーゼドメインが逆転写酵素であり、任意選択で逆転写酵素がレトロウイルス又はレトロトランスポゾン由来の逆転写酵素である、態様6に記載の方法。
8.逆転写酵素が、89~100、105~122、128~129、132、139、143、149、154、159、235、454、471、516、662、700、701~716、739~741及び766のいずれか1つのアミノ酸配列と少なくとも80%、85%、90%、95%、98%又は99%の配列同一性を有する、態様6に記載の方法。
9.逆転写酵素がモロニーマウス白血病ウイルス逆転写酵素(M-MLVRT)である、態様6に記載の方法。
10.RNA依存性DNAポリメラーゼドメインがバリアントモロニーマウス白血病ウイルス逆転写酵素(M-MLV RT)ドメインを含み、バリアントM-MLV RTドメインが、配列番号89のアミノ酸配列に対して以下の変異:P51X、S67X、E69X、L139X、T197X、D200X、H204X、F209X、E302X、T306X、F309X、W313X、T330X、L345X、L435X、N454X、D524X、E562X、D583X、H594X、L603X、E607X、又はD653Xのうちの1つ又は複数を含み、Xが任意のアミノ酸である、態様9に記載の方法。
11.変異体M-MLV RTドメインが、配列番号89のアミノ酸配列に対して以下の変異:P51L、S67K、E69K、L139P、T197A、D200N、H204R、F209N、E302K、E302R、T306K、F309N、W313F、T330P、L345G、L435G、N454K、D524G、E562Q、D583N、H594Q、L603W、E607K、又はD653Nのうちの1つ又は複数を含む、態様9に記載の方法。
12.バリアントM-MLV RTドメインが、配列番号89のアミノ酸配列に対してアミノ酸置換D200N、T330P及びL603Wを含む、態様11に記載の方法。
13.M-MLV RTドメインが、配列番号89のアミノ酸配列に対してアミノ酸置換D200N、T306K、W313F、T330P及びL603Wを含む、態様11に記載の方法。
14.バリアントM-MLV RTドメインが、配列番号106~122、143、701~716又は740~741のアミノ酸配列のいずれか1つを含む、態様11に記載の方法。
15.M-MLV RTドメインが、配列番号741の配列を有する、態様11に記載の方法。
16.バリアントM-MLV RTドメインが、D200N、T306K、W313F及びT330P変異を含有するM-MLV RTの短縮バリアントである、態様9に記載の方法。
17.バリアントM-MLV RTドメインが配列番号766の配列を有する、態様16に記載の方法。
18.napDNAbpドメインがニッカーゼ活性を有する、態様1~4のいずれか1つに記載の方法。
19.napDNAbpドメインが、Cas9、Cas12e、Cas12d、Cas12a、Cas12b1、Cas13a、Cas12c、及びアルゴノートからなる群から選択され、任意選択でニッカーゼ活性を有する、態様1~4のいずれか1つに記載の方法。
20.napDNAbpドメインがCas9タンパク質又はそのバリアントである、態様1~4のいずれか1つに記載の方法。
21.napDNAbpドメインが、ヌクレアーゼ活性Cas9、ヌクレアーゼ不活性Cas9(dCas9)、又はCas9ニッカーゼ(nCas9)である、態様20に記載の方法。
22.napDNAbpドメインがCas9ニッカーゼ(nCas9)である、態様21に記載の方法。
23.napDNAbpドメインが、配列番号18、19、21、25、26、126、137、141、147、153、157、445、460、467、及び482~487のいずれか1つのアミノ酸配列と少なくとも80%、85%、90%、95%、98%、又は99%の配列同一性を有するアミノ酸配列を含む、態様1から4のいずれか1つに記載のプライム編集因子。
24.napDNAbpドメインが、配列番号18のアミノ酸配列と少なくとも80%、85%、90%、95%、98%、又は99%の配列同一性を有するアミノ酸配列を含む、態様23に記載の方法。
25.napDNAbpドメインとRNA依存性DNAポリメラーゼドメインとが連結されて融合タンパク質を形成する、態様1~24のいずれか1つに記載の方法。
26.napDNAbpドメインとRNA依存性DNAポリメラーゼドメインとがペプチドリンカーを介してつなげられて、融合タンパク質を形成する、態様25に記載の方法。
27.融合タンパク質が、構造NH 2-[napDNAbpドメイン]-[RNA依存性DNAポリメラーゼドメイン]-COOH又はNH2-[RNA依存性DNAポリメラーゼドメイン]-[napDNAbpドメイン]-COOHを含み、「]-[」の各実例が、任意選択のリンカー配列の存在を示す、態様25又は26に記載の方法。
28.ペプチドリンカーが、SGGS、(2×SGGS)、(3×SGGS)、XTEN、EAAAK、(2×EAAAK)及び(3×EAAAK)から選択されるアミノ酸配列を含む、態様26又は27に記載の方法。
29.ペプチドリンカーが、1×XTENのアミノ酸配列からなる、態様28に記載の方法。
30.融合タンパク質が、配列番号134のアミノ酸配列、又は配列番号134のアミノ酸配列と少なくとも80%の配列同一性を有するアミノ酸配列を含む、態様25に記載の方法。
31.ニック部位が、二本鎖標的DNAの非標的鎖上のプロトスペーサ内にあり、プロトスペーサがプロトスペーサ隣接モチーフ(PAM)に直接隣接している、態様1~30のいずれか1つの方法。
32.スペーサ、核酸伸長アーム及びgRNAコアが単一分子内にある、態様1~31のいずれか1つに記載の方法。
33.核酸伸長アームがgRNAコアの3'若しくは5'末端又はgRNAコアの分子内位置に配置され、任意選択で核酸伸長アームがDNA又はRNAを含む、態様32に記載の方法。
34.核酸伸長アームが、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、少なくとも15ヌクレオチド、少なくとも16ヌクレオチド、少なくとも17ヌクレオチド、少なくとも18ヌクレオチド、少なくとも19ヌクレオチド、少なくとも20ヌクレオチド、少なくとも21ヌクレオチド、少なくとも22ヌクレオチド、少なくとも23ヌクレオチド、少なくとも24ヌクレオチド、少なくとも25ヌクレオチド、少なくとも26ヌクレオチド、少なくとも27ヌクレオチド、少なくとも28ヌクレオチド、少なくとも29ヌクレオチド、少なくとも30ヌクレオチド、少なくとも31ヌクレオチド、少なくとも32ヌクレオチド、少なくとも33ヌクレオチド、少なくとも34ヌクレオチド、少なくとも35ヌクレオチド、少なくとも36ヌクレオチド、少なくとも37ヌクレオチド、少なくとも38ヌクレオチド、少なくとも39ヌクレオチド、少なくとも40ヌクレオチド、少なくとも41ヌクレオチド、少なくとも42ヌクレオチド、少なくとも43ヌクレオチド、少なくとも44ヌクレオチド、少なくとも45ヌクレオチド、少なくとも46ヌクレオチド、少なくとも47ヌクレオチド、少なくとも48ヌクレオチド、少なくとも49ヌクレオチド、又は少なくとも50ヌクレオチドであり、任意選択で、核酸伸長アームが、10 10~20、20~30、30~40、40~50、50~60、60~70、70~80、80~90、90~100、100~110、110~120、20~120、40~120、60~120、80~120、100~120、40~100、60~100、80~100、又は60~80ヌクレオチド長である、態様32に記載の方法。
35.プライマー結合部位が、少なくとも3ヌクレオチド、少なくとも4ヌクレオチド、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、又は少なくとも15ヌクレオチド長であり、任意選択で、プライマー結合部位が、1~10ヌクレオチド、5~10ヌクレオチド、10~15ヌクレオチド、10~20ヌクレオチド、8~20ヌクレオチド、15~25ヌクレオチド、20~30ヌクレオチド、又は25~30ヌクレオチド長である、態様32に記載の方法。
36.プライマー結合部位が8ヌクレオチド~15ヌクレオチド長である、態様35に記載の方法。
37.プライマー結合部位が、(a)8ヌクレオチド~11ヌクレオチド長であり、約60%を超えるGC含有量を含有し、(b)12ヌクレオチド~13ヌクレオチド長であり、約40~60%のGC含有量を含み、又は(c)14ヌクレオチド~15ヌクレオチド長であり、約40%未満のGC含有量を含有する、態様32に記載の方法。
38.DNA合成鋳型が逆転写鋳型配列である、態様32に記載の方法。
39.DNA合成鋳型が疾患関連遺伝子の野生型配列を有する、態様1~38のいずれか1つの方法。
40.DNA合成鋳型が、少なくとも3ヌクレオチド、少なくとも4ヌクレオチド、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、又は少なくとも15ヌクレオチド長である、態様1~39のいずれか1つに記載の方法。
41.DNA合成鋳型が5~10、5~15、10~20、20~30、30~40、40~50、50~60、60~70、70~80、80~90、90~100、10~30、10~40、10~50、10~60、20~40、20~60、30~100、40~100、50~100、60~100又は70~100ヌクレオチド長であり、任意選択でDNA合成鋳型が10~35ヌクレオチド長である、態様1~39のいずれか1つに記載の方法。
42.DNA合成鋳型が少なくとも3~58ヌクレオチド長である、態様1~39のいずれか1つに記載の方法。
43.DNA合成鋳型が8ヌクレオチド~31ヌクレオチド長である、態様1~39のいずれか1つに記載の方法。
44.DNA合成鋳型が、(a)10ヌクレオチド~16ヌクレオチド長又は(b)12ヌクレオチド~17ヌクレオチド長である、態様1~39のいずれか一項に記載の方法。
45.DNA合成鋳型が、二本鎖標的DNA配列と80%、又は85%、又は90%、又は95%、又は99%同一であるヌクレオチド配列を含む、態様1~39のいずれか1つに記載の方法。
46.PEgRNAが、トゥ-ループ、ヘアピン、ステムループ、シュードノット、アプタマー、G-四重鎖、tRNA、リボスイッチ又はリボザイムからなる群から選択される少なくとも1つの核酸部分を更に含む、態様1~45のいずれか1つに記載の方法。
47.核酸部分がPEgRNAの3'又は5'末端に位置する、態様46に記載の方法。
48.伸長アームが核酸部分を含む、態様46に記載の方法。
49.核酸部分が、伸長アームの3'末端又は5'末端に位置する、態様48に記載の方法。
50.核酸部分が、モロニーマウス白血病ウイルス(M-MLV)ゲノム由来のフレームシフトシュードノット(Mpknot)を含み、任意選択で、Mpknotが、配列番号3930(Mpknot1)、配列番号3931(Mpknot1 3'トリミング)、配列番号3932(5'extraを有するMpknot1)、配列番号3933(Mpknot1 U38A)、配列番号3934(Mpknot1 U38A A29C)、配列番号3935(MMLC A29C)、配列番号3936(5'extra及びU 38 Aを有するMpknot1)、配列番号3937(5'extra及びU 38 A A29Cを有するMpknot1)、及び配列番号3938(5'extra及びA29Cを有するMpknot1)からなる群から選択されるヌクレオチド配列、又はそれらと少なくとも80%の配列同一性を有するヌクレオチド配列を有するMpknot1部分である、態様46に記載の方法。
51.核酸部分がG-四重極を含み、任意選択で、G-四重極が、配列番号3939(tns1)、配列番号3940(stk40)、配列番号3941(apc2)、配列番号3942(ceacam4)、配列番号3943(pitpnm3)、配列番号3944(rlf)、配列番号3945(erc1)、配列番号3946(ube3c)、配列番号3947(taf15)、配列番号3948(stard3)、及び配列番号3949(g2)からなる群から選択されるヌクレオチド配列、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有する、態様46に記載の方法。
52.核酸部分がプレケオシン1リボスイッチアプタマーを含み、任意選択で核酸部分が進化型プレケオシン1-1リボスイッチアプタマー(配列番号3950(evopreq 1)、配列番号3951(evopreq 1モチーフ1)、配列番号3952(evopreq 1モチーフ2)、配列番号3953(evopreq 1モチーフ3)、配列番号3954(より短いpreq 1-1)、配列番号3955(preq 1-1 G5C(mut1))、及び配列番号3956(preq 1-1 G15 C(mut2))からなる群から選択されるヌクレオチド配列を含むevopreQ1)、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を含む、態様46に記載の方法。
53.核酸部分が、配列番号3957のヌクレオチド配列、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有するtRNA部分を含む、態様46に記載の方法。
54.核酸部分が、配列番号3958(xrn1)のヌクレオチド配列、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有する、態様46に記載の方法。
55.核酸部分が、グループIイントロンのP4-P6ドメインを含み、任意選択で、P 4-P 6ドメインが、配列番号3959のヌクレオチド配列、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有する、態様46に記載の方法。
56.PEgRNAがリンカーを更に含む、態様46~55のいずれか1つに記載の方法。
57.リンカーが核酸部分とPEgRNAの別の構成要素との間にある、態様56に記載の方法。
58.リンカーが核酸部分とプライマー結合部位との間又はgRNAコアと核酸部分との間にある、態様57に記載の方法。
59.リンカーが、配列番号3960、配列番号3961、配列番号3962、配列番号3963、配列番号3964、配列番号3965、配列番号3966、配列番号3967、配列番号3968、配列番号3969、配列番号3970、及び配列番号3971からなる群から選択されるヌクレオチド配列を含む、態様58に記載の方法。
60.1つ又は複数のヌクレオチド編集が、二本鎖DNA配列と比較して1つ又は複数のヌクレオチドの挿入を含む、態様1~59のいずれか1つに記載の方法。
61.1つ又は複数のヌクレオチド編集が、二本鎖DNA配列と比較して1つ又は複数のヌクレオチドの欠失を含む、態様1~59のいずれか1つに記載の方法。
62.1つ又は複数のヌクレオチド編集が、二本鎖DNA配列と比較してヌクレオチド置換を含む、態様1~59のいずれか1つに記載の方法。
63.1つ又は複数のヌクレオチド編集が、二本鎖標的DNA配列と比較して、1つ又は複数のヌクレオチドの1つ又は複数の挿入、ヌクレオチド置換、1つ又は複数のヌクレオチドの欠失、又は任意のそのようなヌクレオチド編集の組み合わせを含む、態様1~59のいずれか1つに記載の方法。
64.1つ又は複数のヌクレオチド置換が一塩基ヌクレオチド置換である、態様62~63のいずれか1つに記載の方法。
65.投与が、2人以上の対象の疾患関連遺伝子における2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19又は20個の病原性バリアントを補正するか、又は投与が、2人以上の対象の疾患関連遺伝子における2~5、2~7、3~10、3~12、4~15又は4~20の病原性バリアントを補正する、態様2~65のいずれか1つに記載の方法。
66.PEgRNAが、修飾核酸塩基、修飾糖、修飾リン酸基又はヌクレオシド類似体を含む、態様1~63のいずれか1つに記載の方法。
67.方法及び薬学的に許容される賦形剤を含む医薬組成物を2人以上の対象に投与することを含む、態様4~64に記載の方法。
68.疾患関連遺伝子がCDKL5である、態様1~65のいずれか1つに記載の方法。
69.異なる病原性バリアントのそれぞれが、野生型CDKL5タンパク質と比較して、V172I、A173D、R175S、W176G、W176R、Y177C、R178P、P180L、E181A、及びL182Pからなる群から選択される変異をコードする、態様66に記載の方法。
70.PEgRNAが図2のPEgRNA配列の配列を含む、態様66に記載の方法。
71.PEgRNAが図4のPEgRNA配列の配列を含む、態様66に記載の方法。
1.疾患関連遺伝子の2つ以上のコピーを編集するための方法であって、疾患関連遺伝子の各コピーが二本鎖標的DNA配列を含み、疾患関連遺伝子の2つ以上のコピーの各々を、
(a)核酸プログラム型DNA結合タンパク質(napDNAbp)ドメイン又はnapDNAbpドメインをコードするポリヌクレオチド;
(b)ポリメラーゼドメイン又はポリメラーゼドメインをコードするポリヌクレオチド;及び
(c)プライム編集ガイドRNA(PEgRNA)、
を含む、プライム編集因子システムと接触させることを含み、
PEgRNAは、
二本鎖DNA配列の標的鎖と相補性の領域を含むスペーサ;
napDNAbpドメインと会合するgRNAコア;並びに
プライマー結合部位及びDNA合成鋳型を含む核酸伸長アーム
を含み、プライマー結合部位は、二本鎖標的DNA配列の非標的鎖に対して相補性の領域を含み、DNA合成鋳型は、二本鎖標的DNA配列の非標的鎖に対して相補性の領域を含み、二本鎖標的DNAと比較して1つ又は複数のヌクレオチド編集を含み、非標的鎖は標的鎖に相補的であり、
疾患関連遺伝子の各コピーは、病原性バリアントを含み、疾患関連遺伝子の2つ以上のコピーが2つ以上の異なる病原性バリアントを含み、接触させることが、疾患関連遺伝子の2つ以上のコピーのそれぞれに1つ以上のヌクレオチド編集を組み入れし、組み入れが、疾患関連遺伝子のそれぞれの病原性バリアントを修正し、それにより、疾患関連遺伝子の2つ以上のコピーのそれぞれを編集する、
疾患関連遺伝子の2つ以上のコピーを編集するための方法。
2.疾患関連遺伝子の2つ又はそれを超えるコピーが1人の対象に存在する、態様1に記載の方法。
3.疾患関連遺伝子の2つ以上のコピーが、2人以上の異なる対象に存在する、態様1に記載の方法。
4.疾患関連遺伝子をそれぞれ含む2人以上の対象において疾患を処置する方法であって、2人以上の対象における疾患関連遺伝子が二本鎖標的DNA配列を含み、方法が、2人以上の対象に、
(a)核酸プログラム型DNA結合タンパク質(napDNAbp)ドメイン又はnapDNAbpドメインをコードするポリヌクレオチド;
(b)ポリメラーゼドメイン又はポリメラーゼドメインをコードするポリヌクレオチド;及び
(c)プライム編集ガイドRNA(PEgRNA)
を含むプライム編集因子システムを投与することを含み、
PEgRNAは、
(i)二本鎖DNA配列の標的鎖と相補性の領域を含むスペーサ、
(ii)napDNAbpドメインと会合するgRNAコア;及び
(iii)プライマー結合部位及びDNA合成鋳型を含む核酸伸長アーム
を含み、
プライマー結合部位は、二本鎖標的DNA配列の非標的鎖に対して相補性の領域を含み、DNA合成鋳型は、二本鎖標的DNA配列の非標的鎖に対して相補性の領域を含み、二本鎖標的DNAと比較して1つ又は複数のヌクレオチド編集を含み、非標的鎖は標的鎖に相補的であり
2人以上の対象が、疾患関連遺伝子に2つ以上の異なる病原性バリアントを含み、投与が、2人以上の対象の各々の疾患関連遺伝子に1つ以上のヌクレオチド編集を組み入れし、組み入れが、2人以上の対象の各々の疾患関連遺伝子の病原性変バリアントを修正し、それにより、2人以上の対象の疾患を処置する、
疾患関連遺伝子をそれぞれ含む2人以上の対象において疾患を処置する方法。
5.napDNAbpドメインをコードするポリヌクレオチド及び/又はポリメラーゼドメインをコードするポリヌクレオチドがRNAを含み、任意選択でnapDNAbpドメインをコードするポリヌクレオチド及び/又はポリメラーゼドメインをコードするポリヌクレオチドがmRNAである、態様1~4のいずれか一項に記載の方法。
6.ポリメラーゼドメインがRNA依存性DNAポリメラーゼドメインである、態様1~5のいずれか1つに記載の方法。
7.ポリメラーゼドメインが逆転写酵素であり、任意選択で逆転写酵素がレトロウイルス又はレトロトランスポゾン由来の逆転写酵素である、態様6に記載の方法。
8.逆転写酵素が、89~100、105~122、128~129、132、139、143、149、154、159、235、454、471、516、662、700、701~716、739~741及び766のいずれか1つのアミノ酸配列と少なくとも80%、85%、90%、95%、98%又は99%の配列同一性を有する、態様6に記載の方法。
9.逆転写酵素がモロニーマウス白血病ウイルス逆転写酵素(M-MLVRT)である、態様6に記載の方法。
10.RNA依存性DNAポリメラーゼドメインがバリアントモロニーマウス白血病ウイルス逆転写酵素(M-MLV RT)ドメインを含み、バリアントM-MLV RTドメインが、配列番号89のアミノ酸配列に対して以下の変異:P51X、S67X、E69X、L139X、T197X、D200X、H204X、F209X、E302X、T306X、F309X、W313X、T330X、L345X、L435X、N454X、D524X、E562X、D583X、H594X、L603X、E607X、又はD653Xのうちの1つ又は複数を含み、Xが任意のアミノ酸である、態様9に記載の方法。
11.変異体M-MLV RTドメインが、配列番号89のアミノ酸配列に対して以下の変異:P51L、S67K、E69K、L139P、T197A、D200N、H204R、F209N、E302K、E302R、T306K、F309N、W313F、T330P、L345G、L435G、N454K、D524G、E562Q、D583N、H594Q、L603W、E607K、又はD653Nのうちの1つ又は複数を含む、態様9に記載の方法。
12.バリアントM-MLV RTドメインが、配列番号89のアミノ酸配列に対してアミノ酸置換D200N、T330P及びL603Wを含む、態様11に記載の方法。
13.M-MLV RTドメインが、配列番号89のアミノ酸配列に対してアミノ酸置換D200N、T306K、W313F、T330P及びL603Wを含む、態様11に記載の方法。
14.バリアントM-MLV RTドメインが、配列番号106~122、143、701~716又は740~741のアミノ酸配列のいずれか1つを含む、態様11に記載の方法。
15.M-MLV RTドメインが、配列番号741の配列を有する、態様11に記載の方法。
16.バリアントM-MLV RTドメインが、D200N、T306K、W313F及びT330P変異を含有するM-MLV RTの短縮バリアントである、態様9に記載の方法。
17.バリアントM-MLV RTドメインが配列番号766の配列を有する、態様16に記載の方法。
18.napDNAbpドメインがニッカーゼ活性を有する、態様1~4のいずれか1つに記載の方法。
19.napDNAbpドメインが、Cas9、Cas12e、Cas12d、Cas12a、Cas12b1、Cas13a、Cas12c、及びアルゴノートからなる群から選択され、任意選択でニッカーゼ活性を有する、態様1~4のいずれか1つに記載の方法。
20.napDNAbpドメインがCas9タンパク質又はそのバリアントである、態様1~4のいずれか1つに記載の方法。
21.napDNAbpドメインが、ヌクレアーゼ活性Cas9、ヌクレアーゼ不活性Cas9(dCas9)、又はCas9ニッカーゼ(nCas9)である、態様20に記載の方法。
22.napDNAbpドメインがCas9ニッカーゼ(nCas9)である、態様21に記載の方法。
23.napDNAbpドメインが、配列番号18、19、21、25、26、126、137、141、147、153、157、445、460、467、及び482~487のいずれか1つのアミノ酸配列と少なくとも80%、85%、90%、95%、98%、又は99%の配列同一性を有するアミノ酸配列を含む、態様1から4のいずれか1つに記載のプライム編集因子。
24.napDNAbpドメインが、配列番号18のアミノ酸配列と少なくとも80%、85%、90%、95%、98%、又は99%の配列同一性を有するアミノ酸配列を含む、態様23に記載の方法。
25.napDNAbpドメインとRNA依存性DNAポリメラーゼドメインとが連結されて融合タンパク質を形成する、態様1~24のいずれか1つに記載の方法。
26.napDNAbpドメインとRNA依存性DNAポリメラーゼドメインとがペプチドリンカーを介してつなげられて、融合タンパク質を形成する、態様25に記載の方法。
27.融合タンパク質が、構造NH 2-[napDNAbpドメイン]-[RNA依存性DNAポリメラーゼドメイン]-COOH又はNH2-[RNA依存性DNAポリメラーゼドメイン]-[napDNAbpドメイン]-COOHを含み、「]-[」の各実例が、任意選択のリンカー配列の存在を示す、態様25又は26に記載の方法。
28.ペプチドリンカーが、SGGS、(2×SGGS)、(3×SGGS)、XTEN、EAAAK、(2×EAAAK)及び(3×EAAAK)から選択されるアミノ酸配列を含む、態様26又は27に記載の方法。
29.ペプチドリンカーが、1×XTENのアミノ酸配列からなる、態様28に記載の方法。
30.融合タンパク質が、配列番号134のアミノ酸配列、又は配列番号134のアミノ酸配列と少なくとも80%の配列同一性を有するアミノ酸配列を含む、態様25に記載の方法。
31.ニック部位が、二本鎖標的DNAの非標的鎖上のプロトスペーサ内にあり、プロトスペーサがプロトスペーサ隣接モチーフ(PAM)に直接隣接している、態様1~30のいずれか1つの方法。
32.スペーサ、核酸伸長アーム及びgRNAコアが単一分子内にある、態様1~31のいずれか1つに記載の方法。
33.核酸伸長アームがgRNAコアの3'若しくは5'末端又はgRNAコアの分子内位置に配置され、任意選択で核酸伸長アームがDNA又はRNAを含む、態様32に記載の方法。
34.核酸伸長アームが、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、少なくとも15ヌクレオチド、少なくとも16ヌクレオチド、少なくとも17ヌクレオチド、少なくとも18ヌクレオチド、少なくとも19ヌクレオチド、少なくとも20ヌクレオチド、少なくとも21ヌクレオチド、少なくとも22ヌクレオチド、少なくとも23ヌクレオチド、少なくとも24ヌクレオチド、少なくとも25ヌクレオチド、少なくとも26ヌクレオチド、少なくとも27ヌクレオチド、少なくとも28ヌクレオチド、少なくとも29ヌクレオチド、少なくとも30ヌクレオチド、少なくとも31ヌクレオチド、少なくとも32ヌクレオチド、少なくとも33ヌクレオチド、少なくとも34ヌクレオチド、少なくとも35ヌクレオチド、少なくとも36ヌクレオチド、少なくとも37ヌクレオチド、少なくとも38ヌクレオチド、少なくとも39ヌクレオチド、少なくとも40ヌクレオチド、少なくとも41ヌクレオチド、少なくとも42ヌクレオチド、少なくとも43ヌクレオチド、少なくとも44ヌクレオチド、少なくとも45ヌクレオチド、少なくとも46ヌクレオチド、少なくとも47ヌクレオチド、少なくとも48ヌクレオチド、少なくとも49ヌクレオチド、又は少なくとも50ヌクレオチドであり、任意選択で、核酸伸長アームが、10 10~20、20~30、30~40、40~50、50~60、60~70、70~80、80~90、90~100、100~110、110~120、20~120、40~120、60~120、80~120、100~120、40~100、60~100、80~100、又は60~80ヌクレオチド長である、態様32に記載の方法。
35.プライマー結合部位が、少なくとも3ヌクレオチド、少なくとも4ヌクレオチド、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、又は少なくとも15ヌクレオチド長であり、任意選択で、プライマー結合部位が、1~10ヌクレオチド、5~10ヌクレオチド、10~15ヌクレオチド、10~20ヌクレオチド、8~20ヌクレオチド、15~25ヌクレオチド、20~30ヌクレオチド、又は25~30ヌクレオチド長である、態様32に記載の方法。
36.プライマー結合部位が8ヌクレオチド~15ヌクレオチド長である、態様35に記載の方法。
37.プライマー結合部位が、(a)8ヌクレオチド~11ヌクレオチド長であり、約60%を超えるGC含有量を含有し、(b)12ヌクレオチド~13ヌクレオチド長であり、約40~60%のGC含有量を含み、又は(c)14ヌクレオチド~15ヌクレオチド長であり、約40%未満のGC含有量を含有する、態様32に記載の方法。
38.DNA合成鋳型が逆転写鋳型配列である、態様32に記載の方法。
39.DNA合成鋳型が疾患関連遺伝子の野生型配列を有する、態様1~38のいずれか1つの方法。
40.DNA合成鋳型が、少なくとも3ヌクレオチド、少なくとも4ヌクレオチド、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、又は少なくとも15ヌクレオチド長である、態様1~39のいずれか1つに記載の方法。
41.DNA合成鋳型が5~10、5~15、10~20、20~30、30~40、40~50、50~60、60~70、70~80、80~90、90~100、10~30、10~40、10~50、10~60、20~40、20~60、30~100、40~100、50~100、60~100又は70~100ヌクレオチド長であり、任意選択でDNA合成鋳型が10~35ヌクレオチド長である、態様1~39のいずれか1つに記載の方法。
42.DNA合成鋳型が少なくとも3~58ヌクレオチド長である、態様1~39のいずれか1つに記載の方法。
43.DNA合成鋳型が8ヌクレオチド~31ヌクレオチド長である、態様1~39のいずれか1つに記載の方法。
44.DNA合成鋳型が、(a)10ヌクレオチド~16ヌクレオチド長又は(b)12ヌクレオチド~17ヌクレオチド長である、態様1~39のいずれか一項に記載の方法。
45.DNA合成鋳型が、二本鎖標的DNA配列と80%、又は85%、又は90%、又は95%、又は99%同一であるヌクレオチド配列を含む、態様1~39のいずれか1つに記載の方法。
46.PEgRNAが、トゥ-ループ、ヘアピン、ステムループ、シュードノット、アプタマー、G-四重鎖、tRNA、リボスイッチ又はリボザイムからなる群から選択される少なくとも1つの核酸部分を更に含む、態様1~45のいずれか1つに記載の方法。
47.核酸部分がPEgRNAの3'又は5'末端に位置する、態様46に記載の方法。
48.伸長アームが核酸部分を含む、態様46に記載の方法。
49.核酸部分が、伸長アームの3'末端又は5'末端に位置する、態様48に記載の方法。
50.核酸部分が、モロニーマウス白血病ウイルス(M-MLV)ゲノム由来のフレームシフトシュードノット(Mpknot)を含み、任意選択で、Mpknotが、配列番号3930(Mpknot1)、配列番号3931(Mpknot1 3'トリミング)、配列番号3932(5'extraを有するMpknot1)、配列番号3933(Mpknot1 U38A)、配列番号3934(Mpknot1 U38A A29C)、配列番号3935(MMLC A29C)、配列番号3936(5'extra及びU 38 Aを有するMpknot1)、配列番号3937(5'extra及びU 38 A A29Cを有するMpknot1)、及び配列番号3938(5'extra及びA29Cを有するMpknot1)からなる群から選択されるヌクレオチド配列、又はそれらと少なくとも80%の配列同一性を有するヌクレオチド配列を有するMpknot1部分である、態様46に記載の方法。
51.核酸部分がG-四重極を含み、任意選択で、G-四重極が、配列番号3939(tns1)、配列番号3940(stk40)、配列番号3941(apc2)、配列番号3942(ceacam4)、配列番号3943(pitpnm3)、配列番号3944(rlf)、配列番号3945(erc1)、配列番号3946(ube3c)、配列番号3947(taf15)、配列番号3948(stard3)、及び配列番号3949(g2)からなる群から選択されるヌクレオチド配列、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有する、態様46に記載の方法。
52.核酸部分がプレケオシン1リボスイッチアプタマーを含み、任意選択で核酸部分が進化型プレケオシン1-1リボスイッチアプタマー(配列番号3950(evopreq 1)、配列番号3951(evopreq 1モチーフ1)、配列番号3952(evopreq 1モチーフ2)、配列番号3953(evopreq 1モチーフ3)、配列番号3954(より短いpreq 1-1)、配列番号3955(preq 1-1 G5C(mut1))、及び配列番号3956(preq 1-1 G15 C(mut2))からなる群から選択されるヌクレオチド配列を含むevopreQ1)、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を含む、態様46に記載の方法。
53.核酸部分が、配列番号3957のヌクレオチド配列、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有するtRNA部分を含む、態様46に記載の方法。
54.核酸部分が、配列番号3958(xrn1)のヌクレオチド配列、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有する、態様46に記載の方法。
55.核酸部分が、グループIイントロンのP4-P6ドメインを含み、任意選択で、P 4-P 6ドメインが、配列番号3959のヌクレオチド配列、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有する、態様46に記載の方法。
56.PEgRNAがリンカーを更に含む、態様46~55のいずれか1つに記載の方法。
57.リンカーが核酸部分とPEgRNAの別の構成要素との間にある、態様56に記載の方法。
58.リンカーが核酸部分とプライマー結合部位との間又はgRNAコアと核酸部分との間にある、態様57に記載の方法。
59.リンカーが、配列番号3960、配列番号3961、配列番号3962、配列番号3963、配列番号3964、配列番号3965、配列番号3966、配列番号3967、配列番号3968、配列番号3969、配列番号3970、及び配列番号3971からなる群から選択されるヌクレオチド配列を含む、態様58に記載の方法。
60.1つ又は複数のヌクレオチド編集が、二本鎖DNA配列と比較して1つ又は複数のヌクレオチドの挿入を含む、態様1~59のいずれか1つに記載の方法。
61.1つ又は複数のヌクレオチド編集が、二本鎖DNA配列と比較して1つ又は複数のヌクレオチドの欠失を含む、態様1~59のいずれか1つに記載の方法。
62.1つ又は複数のヌクレオチド編集が、二本鎖DNA配列と比較してヌクレオチド置換を含む、態様1~59のいずれか1つに記載の方法。
63.1つ又は複数のヌクレオチド編集が、二本鎖標的DNA配列と比較して、1つ又は複数のヌクレオチドの1つ又は複数の挿入、ヌクレオチド置換、1つ又は複数のヌクレオチドの欠失、又は任意のそのようなヌクレオチド編集の組み合わせを含む、態様1~59のいずれか1つに記載の方法。
64.1つ又は複数のヌクレオチド置換が一塩基ヌクレオチド置換である、態様62~63のいずれか1つに記載の方法。
65.投与が、2人以上の対象の疾患関連遺伝子における2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19又は20個の病原性バリアントを補正するか、又は投与が、2人以上の対象の疾患関連遺伝子における2~5、2~7、3~10、3~12、4~15又は4~20の病原性バリアントを補正する、態様2~65のいずれか1つに記載の方法。
66.PEgRNAが、修飾核酸塩基、修飾糖、修飾リン酸基又はヌクレオシド類似体を含む、態様1~63のいずれか1つに記載の方法。
67.方法及び薬学的に許容される賦形剤を含む医薬組成物を2人以上の対象に投与することを含む、態様4~64に記載の方法。
68.疾患関連遺伝子がCDKL5である、態様1~65のいずれか1つに記載の方法。
69.異なる病原性バリアントのそれぞれが、野生型CDKL5タンパク質と比較して、V172I、A173D、R175S、W176G、W176R、Y177C、R178P、P180L、E181A、及びL182Pからなる群から選択される変異をコードする、態様66に記載の方法。
70.PEgRNAが図2のPEgRNA配列の配列を含む、態様66に記載の方法。
71.PEgRNAが図4のPEgRNA配列の配列を含む、態様66に記載の方法。
実施例1:プライム編集:二本鎖DNA切断なしのヒト細胞における高度に汎用的かつ精密な検索置換ゲノム編集
背景
現行のゲノム編集法は、プログラム型ヌクレアーゼを用いて二本鎖DNA切断の随伴する副産物と共に標的遺伝子を遮断、欠失、又は挿入し得、塩基編集因子を用いて標的遺伝子座に4つのトランジション点変異を組み入れ得る。しかしながら、小さい挿入、小さい欠失、及び8つのトランスバーション点変異は、集合的にはほとんどの病原性の遺伝子バリアントを表すが、ほとんどの細胞型において、効率的にかつ余分な副産物なしに修正はされ得ない。標的部位を規定すること及び所望の編集をコードすることの両方をする操作されたプライム編集ガイドRNA(PEgRNA)によってプログラムされる操作された逆転写酵素に融合された触媒的に損なわれたCas9を用いて、新たな遺伝情報を規定されたDNA部位に直接的に書き込む高度に汎用的なかつ精密なゲノム編集法、プライム編集が本明細書に記載される。ヒト細胞の175よりも多大な別物の編集が行われて、プライム編集が、標的化された挿入、欠失、全ての12の可能な型の点変異、及びそれらの組み合わせを、効率的に(未ソーティングの細胞において典型的には20-60%、最高で77%)かつ低い副産物(典型的には1-10%)によって、二本鎖切断又はドナーDNA鋳型を要求することなしになし得るということを確立した。プライム編集がヒト細胞において適用されて、鎌状赤血球症(HBBのA・T→T・Aのトランスバーションを要求する)及びテイ・サックス病(HEXAの4塩基欠失を要求する)の一次的な遺伝学的原因を修正し、両方のケースにおいて、最小限の副産物によって病原性のゲノムアレルを野生型に効率的に復帰させた。これらの病原性のHBBトランスバーション及びHEXA挿入変異を有するヒト細胞株を作成するために、プリオン病に対する耐性を付与するG127V変異をPRNPに組み入れるために(G・C→T・Aのトランスバーションを要求する)、並びにHis6タグ、FLAGエピトープタグ、及び伸長されたLoxP部位をヒト細胞の標的遺伝子座に効率的に挿入するためにもまた、プライム編集が用いられた。プライム編集は、HDRと比べて効率及び産物純度の利点、並びに塩基編集と比較して補完的な強さ及び弱さをオファーする。3つの別物の塩基対形成イベントを要求するその検索置換機序と整合して、プライム編集は公知のCas9オフターゲット部位においてCas9よりもかなりオフターゲットDNA修飾をしにくい。プライム編集はゲノム編集の範囲及び能力を実質的に拡張し、原理的には、公知の病原性のヒト遺伝子バリアントの~89%を修正し得る。
背景
現行のゲノム編集法は、プログラム型ヌクレアーゼを用いて二本鎖DNA切断の随伴する副産物と共に標的遺伝子を遮断、欠失、又は挿入し得、塩基編集因子を用いて標的遺伝子座に4つのトランジション点変異を組み入れ得る。しかしながら、小さい挿入、小さい欠失、及び8つのトランスバーション点変異は、集合的にはほとんどの病原性の遺伝子バリアントを表すが、ほとんどの細胞型において、効率的にかつ余分な副産物なしに修正はされ得ない。標的部位を規定すること及び所望の編集をコードすることの両方をする操作されたプライム編集ガイドRNA(PEgRNA)によってプログラムされる操作された逆転写酵素に融合された触媒的に損なわれたCas9を用いて、新たな遺伝情報を規定されたDNA部位に直接的に書き込む高度に汎用的なかつ精密なゲノム編集法、プライム編集が本明細書に記載される。ヒト細胞の175よりも多大な別物の編集が行われて、プライム編集が、標的化された挿入、欠失、全ての12の可能な型の点変異、及びそれらの組み合わせを、効率的に(未ソーティングの細胞において典型的には20-60%、最高で77%)かつ低い副産物(典型的には1-10%)によって、二本鎖切断又はドナーDNA鋳型を要求することなしになし得るということを確立した。プライム編集がヒト細胞において適用されて、鎌状赤血球症(HBBのA・T→T・Aのトランスバーションを要求する)及びテイ・サックス病(HEXAの4塩基欠失を要求する)の一次的な遺伝学的原因を修正し、両方のケースにおいて、最小限の副産物によって病原性のゲノムアレルを野生型に効率的に復帰させた。これらの病原性のHBBトランスバーション及びHEXA挿入変異を有するヒト細胞株を作成するために、プリオン病に対する耐性を付与するG127V変異をPRNPに組み入れるために(G・C→T・Aのトランスバーションを要求する)、並びにHis6タグ、FLAGエピトープタグ、及び伸長されたLoxP部位をヒト細胞の標的遺伝子座に効率的に挿入するためにもまた、プライム編集が用いられた。プライム編集は、HDRと比べて効率及び産物純度の利点、並びに塩基編集と比較して補完的な強さ及び弱さをオファーする。3つの別物の塩基対形成イベントを要求するその検索置換機序と整合して、プライム編集は公知のCas9オフターゲット部位においてCas9よりもかなりオフターゲットDNA修飾をしにくい。プライム編集はゲノム編集の範囲及び能力を実質的に拡張し、原理的には、公知の病原性のヒト遺伝子バリアントの~89%を修正し得る。
実質的にいずれかの標的化された変化をいずれかの生細胞又は生物のゲノムになす能力は、生命科学の長年の熱望である。ゲノム編集テクノロジーの急速な進歩にもかかわらず、疾患に関連する>75,000の公知のヒト遺伝子バリアントの多数は111、ほとんどの処置学的に該当する細胞においては修正又は組み入れされ得ない(図38A)。CRISPR-Cas9等のプログラム型ヌクレアーゼは二本鎖DNA切断(DSB)を作り、これは挿入及び欠失の混合物(インデル)を標的部位において誘導することによって遺伝子を遮断し得る112-114。ヌクレアーゼは、相同性非依存的プロセスによって、標的遺伝子を欠失115,116又は外因的な遺伝子を挿入117-119するためにもまた用いられ得る。しかしながら、二本鎖DNA切断は、産物の複雑な混合物、転座120、及びp53活性化121,122を包含する望まれないアウトカムにもまた関連される。その上、病原性のアレルの大多数は、それらの非病原性のカウンターパートとは小さい挿入、欠失、又は塩基置換が異なり、これらは修正するためにはかなりより精密な編集テクノロジーを要求する(図38A)。ヌクレアーゼによって誘導されるDSBによって刺激される相同組換え修復(HDR)123は、種々の精密なDNA変化を組み入れるために広く用いられている。しかしながら、HDRは外因的なドナーDNA修復鋳型に依拠し、典型的にはDSBの末端結合修復からの余分なインデル副産物を生成し、ほとんどの処置学的に該当する細胞型では非効率的である(T細胞及びいくつかの幹細胞は重要な例外である)124,125。DSBによって媒介されるゲノム編集の効率及び精度を増強することは有望な作業のフォーカスのままに留まっており126-130、これらの課題は代替的な精密ゲノム編集戦略の調査を要する。
塩基編集は、哺乳動物を包含する広い種々の細胞型及び生物において、DSBを要求することなしに、4つの型のトランジション変異(C→T、G→A、A→G、及びT→C)を効率的に組み入れ又は修正し得るが128-131、現行では、8つのトランスバーション変異(C→A、C→G、G→C、G→T、A→C、A→T、T→A、及びT→G)のいずれか、例えば鎌状赤血球症の最も普通の原因(HBB E6V)を直接的に修正するために必要とされるT・A→A・Tの変異を達成し得ない132。加えて、標的の欠失、例えばテイ・サックス病を引き起こす4塩基重複(HEXA 1278+TATC)の除去133、又は標的化された挿入、例えば嚢胞性線維症の最も普通の原因(CFTR ΔF508)を直接的に修正するために要求される精密な3塩基挿入134を行うためのDSBフリーの方法は報告されていない。それゆえに、それらは集合的にほとんどの公知の病原性アレルを説明するにもかかわらず、標的化されたトランスバーション点変異、挿入、及び欠失は、ほとんどの細胞型においては効率的にかつ余分な副産物なしに組み入れ又は修正することが困難である(図38A)。
二本鎖DNA切断又はドナーDNA鋳型を要求することなしに、ヒト細胞の標的遺伝子座における標的化された挿入、欠失、及び12全ての可能な塩基から塩基の変換を媒介する新たな「検索置換」ゲノム編集テクノロジー、プライム編集の開発が本明細書に記載される。当初にPE1によって例示されたプライム編集因子は、プログラム型ニッカーゼに融合された逆転写酵素とプライム編集pegRNA(pegRNA)とを用いて、遺伝情報をpegRNA上の伸長から標的ゲノム遺伝子座に直接的にコピーする。第2世代プライム編集因子(PE2)は、操作された逆転写酵素を用いて最小限の(典型的には<2%)インデル形成によって編集効率を実質的に増大させ、第3世代PE3システムは第2のガイドRNAを追加して非編集鎖にニッキングし、それによって非編集鎖の置換えを好み、約1~10%インデル形成によってヒト細胞において典型的には約20~50%まで編集効率を更に増大させる。PE3は最適化されたCas9ヌクレアーゼによって開始されるHDRと比較してずっと少数の副産物及び高いか又は類似の効率をオファーし、現行世代の塩基編集因子と比較して補完的な強さ及び弱さをオファーする。
PE3がヒトHEK293T細胞のゲノム遺伝子座において適用されて、HBB E6Vから野生型HBBへの効率的変換、HEXA 1278+TATCを野生型HEXAに復元するための挿入されたTATCの欠失、プリオン病に対する耐性を付与するG127V変異のPRNPへの組み入れ135(G・C→T・Aのトランスバーションを要求する)、並びにHis6タグ(18bp)、FLAGエピトープタグ(24bp)、及びCreによって媒介される組換えのための伸長されたLoxP部位(44bp)の標的化された挿入を達成した。プライム編集は3つの他のヒト細胞株及び有糸分裂後初代マウス皮質ニューロンにおいてもまた様々な効率で首尾良かった。当初のニック及び編集の位置付けの間の距離の高い度合のフレキシビリティーを原因として、プライム編集はCas9のPAM要件によって実質的に制約されず、原理的にはゲノム遺伝子座の大多数を標的化し得る。蓋然的には、生産的なプライム編集が生起するための3つの別物のDNA塩基対形成イベントという要件を原因として、オフターゲットプライム編集は公知のCas9オフターゲット遺伝子座におけるオフターゲットCas9編集よりもかなり稀である。精密な標的化された挿入、欠失、及び全ての12の可能なクラスの点変異を広い種々のゲノム遺伝子座においてDSB又はドナーDNA鋳型の必要なしに可能化することによって、プライム編集は多くの遺伝子バリアントの研究及び修正を進歩させるポテンシャルを有する。
結果
pegRNAからの情報を標的DNA遺伝子座上に移入するための戦略
Cas9は、標的DNA部位にハイブリダイズするスペーサ配列を含有するガイドRNAを用いてDNAを標的化する112~114,136,137。狙いは、天然のCRISPRシステムのようにDNA標的を規定するように138,139、また、標的遺伝子座の対応するDNAヌクレオチドを置換える新たな遺伝情報を含有するように両方でガイドRNAを操作することであった。pegRNAから規定されたDNA部位への遺伝情報の直接的移入、次に元の編集されないDNAの置換えは、原理的には、DSB又はドナーDNA鋳型への依存性なしに、標的化されたDNA配列変化を生細胞に組み入れる一般的手段を提供し得る。この直接的情報移入を達成するために、狙いは、3'-ヒドロキシル基を暴露するように標的部位においてニックが入ったゲノムDNAを用いて、操作されたガイドRNA(これ以降では、プライム編集ガイドRNA又はPEgRNAと呼ばれる)の伸長から直接的に標的部位への遺伝情報の逆転写をプライムすることであった(図38A)。
pegRNAからの情報を標的DNA遺伝子座上に移入するための戦略
Cas9は、標的DNA部位にハイブリダイズするスペーサ配列を含有するガイドRNAを用いてDNAを標的化する112~114,136,137。狙いは、天然のCRISPRシステムのようにDNA標的を規定するように138,139、また、標的遺伝子座の対応するDNAヌクレオチドを置換える新たな遺伝情報を含有するように両方でガイドRNAを操作することであった。pegRNAから規定されたDNA部位への遺伝情報の直接的移入、次に元の編集されないDNAの置換えは、原理的には、DSB又はドナーDNA鋳型への依存性なしに、標的化されたDNA配列変化を生細胞に組み入れる一般的手段を提供し得る。この直接的情報移入を達成するために、狙いは、3'-ヒドロキシル基を暴露するように標的部位においてニックが入ったゲノムDNAを用いて、操作されたガイドRNA(これ以降では、プライム編集ガイドRNA又はPEgRNAと呼ばれる)の伸長から直接的に標的部位への遺伝情報の逆転写をプライムすることであった(図38A)。
いくつかの天然の可動遺伝因子によって用いられる機序に似るニッキング及び逆転写のこれらの当初の工程は140、2つの冗長性の一本鎖DNAフラップを1つの鎖上に有する分岐した中間体をもたらす:編集されないDNA配列を含有する5'フラップ及びpegRNAからコピーされた編集された配列を含有する3'フラップである(図38B)。首尾良い編集を達成するためには、編集された3'フラップが編集されない5'フラップを置き換えるようにして、この分岐した中間体は分解されなければならない。編集された3'フラップは編集されない鎖とより少数の塩基対を作り得るので、編集されない鎖との5'フラップのハイブリダイゼーションは熱力学的に有利であることが蓋然的であるが、5'フラップはFEN1等の構造特異的エンドヌクレアーゼの好ましい基質である141。これは、ラギング鎖DNA合成及びロングパッチ塩基除去修復の間に生成された5'フラップを切り出す。選好的5'フラップ切除及び3'フラップライゲーションは、編集されたDNA鎖の組み込みを駆動し得、1つの編集された鎖及び1つの編集されない鎖を含有するヘテロ二重鎖DNAを作成するということが推論された(図38B)。
編集の永久的な組み入れは、編集された鎖から相補的なDNA鎖に情報をコピーする様式で、2つのDNA鎖間のミスマッチを分解する続くDNA修復から発生し得る(図38C)。DNA塩基編集の効率を最大化するために開発された類似の戦略に基づいて131-133、二本鎖切断形成を最小化するために当初のニックの部位から十分遠くにおける非編集DNA鎖へニックを入れることは、非編集鎖を選好的に置き換えるようにDNA修復を偏らせ得るということが想定された。
in vitro及び酵母細胞におけるプライム編集工程の検証
Cas9のRuvCヌクレアーゼドメインによるPAM含有DNA鎖の切断後に、この鎖のPAM遠位フラグメントは、さもなければ安定なCas9:sgRNA:DNA複合体から解離し得る143。この解放された鎖の3'端は、DNA重合をプライムするために十分にアクセス可能であり得る。ガイドRNA操作作業144-146及びCas9:sgRNA:DNA複合体の結晶構造147-149は、sgRNAの5'及び3'末端がCas9:sgRNA活性を廃止することなしに伸長され得るということを示唆する。2つの重大な構成要素を包含するようにsgRNAを伸長することによって、PEgRNAがデザインされた:ニックが入ったDNA鎖の3'端がPEgRNAにハイブリダイズすることを許すプライマー結合部位(PBS)、及びニックが入ったDNA鎖の3'端がポリメラーゼによってRNA鋳型上を伸長されるとゲノムDNA部位に直接的にコピーされるであろう所望の編集を含有する逆転写酵素(RT)鋳型である(図38C)。
Cas9のRuvCヌクレアーゼドメインによるPAM含有DNA鎖の切断後に、この鎖のPAM遠位フラグメントは、さもなければ安定なCas9:sgRNA:DNA複合体から解離し得る143。この解放された鎖の3'端は、DNA重合をプライムするために十分にアクセス可能であり得る。ガイドRNA操作作業144-146及びCas9:sgRNA:DNA複合体の結晶構造147-149は、sgRNAの5'及び3'末端がCas9:sgRNA活性を廃止することなしに伸長され得るということを示唆する。2つの重大な構成要素を包含するようにsgRNAを伸長することによって、PEgRNAがデザインされた:ニックが入ったDNA鎖の3'端がPEgRNAにハイブリダイズすることを許すプライマー結合部位(PBS)、及びニックが入ったDNA鎖の3'端がポリメラーゼによってRNA鋳型上を伸長されるとゲノムDNA部位に直接的にコピーされるであろう所望の編集を含有する逆転写酵素(RT)鋳型である(図38C)。
これらの仮説は、精製されたS.pyogenes Cas9タンパク質を用いてin vitroで試験された。sgRNAをどちらかの末端においてPBS配列(5~6ヌクレオチド、nt)及びRT鋳型(7~22nt)によって伸長することによって、一連のpegRNA候補が構築された。5'伸長されたPEgRNAは標的DNAに対するCas9結合を導くということ、並びに5'伸長されたPEgRNA及び3'伸長されたPEgRNA両方がin vitroのCas9によって媒介される標的ニッキングと哺乳動物細胞におけるDNA切断活性とを支持するということが確認された(図44A~図44C)。これらの候補pegRNAデザインは、予めニックが入った5'-Cy5標識dsDNA基質、触媒的に不活性型Cas9(dCas9)、及びモロニーマウス白血病ウイルス(M-MLV)逆転写酵素の市販のバリアントを用いて試験された(図44D)。全ての構成要素が存在するときには、ゲル移動度によって、蛍光標識されたDNA鎖からより長いDNA産物への効率的な変換が観察され、RT鋳型に沿った逆転写と整合した(図38D、図44D~図44E)。所望の長さの産物が、5'伸長又は3'伸長されたPEgRNAどちらかで形成された(図38D~図38E)。dCas9の省略は、pegRNA情報移入なしに、DNA鋳型上の逆転写酵素によって媒介されるDNA重合に由来するニックトランスレーション産物に至った(図38D)。pegRNAが従来のsgRNAによって置き換えられたときには、DNA重合産物は観察されず、pegRNAのPBS及びRT鋳型構成要素の必要性を確認した(図38D)。これらの結果は、Cas9によって媒介されるDNA融解が一本鎖Rループを暴露し、これは、ニッキングされる場合には、5'伸長又は3'伸長されたPEgRNAどちらかからの逆転写をプライムすることに有能であるということを実証している。
次に、ニッキングされないdsDNA基質が、専らPAM含有鎖へニックを入れるCas9ニッカーゼ(H840A変異体)によって試験された112。これらの反応では、ことによると損なわれたCas9ニッカーゼ活性を原因として、5'伸長されたPEgRNAは逆転写産物を非効率的に生成した(図44F)。しかしながら、3'伸長されたPEgRNAはロバストなCas9ニッキング及び効率的な逆転写を可能化した(図38E)。原理的には逆転写がPEgRNAの残りのどこかで終結するポテンシャルにもかかわらず、3'伸長されたPEgRNAの使用は単一の明らかな産物のみを生成した。Cas9ニッカーゼ、RT、及び3'伸長されたPEgRNAによる反応の産物のDNA配列決定は、完全なRT鋳型配列がDNA基質へと逆転写されるということを明らかにした(図44G)。これらの実験は、3'伸長されたPEgRNAが、Cas9ニッカーゼ活性を導く能力を保持しながら、新たなDNA鎖の逆転写の鋳型となり得るということを確立した。
in vitroのpegRNAによってプログラムされる逆転写によって産生される3'フラップの真核細胞DNA修復アウトカムを評価するために、レポータープラスミド基質に対するin vitroのpegRNA、Cas9ニッカーゼ、及びRTを用いるDNAニッキング及び逆転写が行われ、次いで、反応産物が酵母(S.cerevisiae)細胞に形質トランスバーションされた(図45A)。心強いことに、プラスミドが、未成熟停止コドンを修正するT・A→A・Tのトランスバーションをコードする3'伸長されたPEgRNAによってin vitroで編集されたときには、酵母形質トランスバーション体の37%がGFP及びmCherryタンパク質両方を発現した(図38F、図45C)。図38E及び図44Fの結果と整合して、5'伸長されたPEgRNAによってin vitroで実行された編集反応は、3'伸長されたPEgRNAによるものよりも少数のGFP及びmCherry二重陽性コロニー(9%)を生んだ(図38F及び図45D)。生産的な編集は、フレームシフト変異を修正するために1ヌクレオチドを挿入するか(15%の二重陽性形質トランスバーション体)又は1ヌクレオチドを欠失する(29%の二重陽性形質トランスバーション体)3'伸長されたPEgRNAを用いてもまた観察された(図38F及び図45E~図45F)。二重陽性の酵母コロニーから回復した編集されたプラスミドのDNA配列決定は、コードされるトランスバーション編集が所望の配列位置において生起するということを確認した(図45G)。これらの結果は、真核細胞のDNA修復がプライム編集から発生する3'DNAフラップを分解して、トランスバーション、挿入、及び欠失を包含する精密なDNA編集を組み込み得るということを実証している。
プライム編集因子1(PE1)のデザイン
in vitroの及び酵母の結果によって促されて、哺乳動物細胞のゲノムDNAを編集することができる最小の数の構成要素によるプライム編集システムが、開発のために探求された。3'伸長されたPEgRNA(これ以降では単純にPEgRNAと呼ばれる、図39A)とフレキシブルなリンカーを介した逆転写酵素へのCas9 H840Aの直接的融合体は、機能的な2構成要素プライム編集システムを構成し得る。HEK293T(不死化ヒト胎児腎臓)細胞は、Cas9 H840Aニッカーゼのどちらかの末端への野生型M-MLV逆転写酵素の融合体をコードする1つのプラスミドと、PEgRNAをコードする第2のプラスミドとによってトランスフェクションされた。当初の試みはHEK3標的遺伝子座における検出可能なT・A→A・Tの変換には至らなかった。
in vitroの及び酵母の結果によって促されて、哺乳動物細胞のゲノムDNAを編集することができる最小の数の構成要素によるプライム編集システムが、開発のために探求された。3'伸長されたPEgRNA(これ以降では単純にPEgRNAと呼ばれる、図39A)とフレキシブルなリンカーを介した逆転写酵素へのCas9 H840Aの直接的融合体は、機能的な2構成要素プライム編集システムを構成し得る。HEK293T(不死化ヒト胎児腎臓)細胞は、Cas9 H840Aニッカーゼのどちらかの末端への野生型M-MLV逆転写酵素の融合体をコードする1つのプラスミドと、PEgRNAをコードする第2のプラスミドとによってトランスフェクションされた。当初の試みはHEK3標的遺伝子座における検出可能なT・A→A・Tの変換には至らなかった。
しかしながら、8-15塩基へのpegRNA上のPBSの伸長は(図39A)、HEK3標的部位における検出可能なT・A→A・Tの編集に至った(図39B)。RTがCas9ニッカーゼのC末端に融合されたプライム編集因子構築物(8~15ntの範囲であるPBS長さによって、3.7%の最大のT・A→A・Tの変換)では、N末端RT-Cas9ニッカーゼ融合体(1.3%の最大のT・A→A・Tの変換)と比較して高い効率であった(図39B;別様に規定されない限り、本明細書の全ての哺乳動物細胞データは、セレクション又はソーティングなしの処置された細胞集団全体の値を報告している)。これらの結果は、トランスで供給されるM-MLV RTの市販のバリアントを用いてin vitroで要求されるものと比較して、Cas9に融合された野生型M-MLV RTがヒト細胞におけるゲノム編集のための長いPBS配列を要求するということを示唆する。Cas9 H840AニッカーゼのC末端に融合されたこの第1世代野生型M-MLV逆転写酵素はPE1として指定された。
pegRNAによって規定される4つの追加のゲノム標的部位におけるトランスバーション点変異を精密に導入するPE1の能力(図39C)が試験された。HEK3遺伝子座における編集と類似に、これらのゲノム部位における効率はPBS長さに依存的であり、最大の編集効率は0.7-5.5%の範囲であった(図39C)。PE1からのインデルは低く、各部位の編集効率を最大化する条件下において、5つの部位について平均で0.2±0.1%であった(図46A)。PE1は、HEK3遺伝子座において、1ヌクレオチド欠失(4.0%効率)、1ヌクレオチド挿入(9.7%)、及び3ヌクレオチド挿入(17%)によって例示される標的化された挿入及び欠失を組み入れる能力があった(図39C)。これらの結果は、二本鎖DNA切断又はDNA鋳型を要求することなしに、標的化されたトランスバーション、挿入、及び欠失を直接的に組み入れるPE1の能力を確立する。
プライム編集因子2(PE2)のデザイン
PE1は種々の編集をHEK293T細胞のいくつかの遺伝子座において組み入れ得るが、編集効率は一般的に低かった(典型的には≦5%)(図39C)。PE1の逆転写酵素を操作することは、プライム編集複合体の固有の立体配座制約内でDNA合成の効率を改善し得、より高いゲノム編集収量をもたらす。酵素熱安定性150,151、処理能力150、及びDNA:RNAヘテロ二重鎖基質親和性152を増大させる、並びにRNaseH活性を不活性化する153M-MLV RT変異が先に報告されている。種々の逆転写酵素変異を含有する19のPE1バリアントが構築されて、それらのプライム編集効率をヒト細胞において評価した。
PE1は種々の編集をHEK293T細胞のいくつかの遺伝子座において組み入れ得るが、編集効率は一般的に低かった(典型的には≦5%)(図39C)。PE1の逆転写酵素を操作することは、プライム編集複合体の固有の立体配座制約内でDNA合成の効率を改善し得、より高いゲノム編集収量をもたらす。酵素熱安定性150,151、処理能力150、及びDNA:RNAヘテロ二重鎖基質親和性152を増大させる、並びにRNaseH活性を不活性化する153M-MLV RT変異が先に報告されている。種々の逆転写酵素変異を含有する19のPE1バリアントが構築されて、それらのプライム編集効率をヒト細胞において評価した。
第1に、上昇した温度における逆転写を支持するそれらの能力について先に実験室進化から新興した一連のM-MLV RTバリアント150が取り調べられた。これ以降ではM3と呼ばれるM-MLV RT上へのこれらのアミノ酸置換の3つ(D200N、L603W、及びT330P)の順次の導入は、PE1のものと比較して、HEK293T細胞の5つのゲノム遺伝子座におけるトランスバーション及び挿入編集効率の6.8倍の平均の増大に至った(図47A-図47S)。
次に、M3との組み合わせで、鋳型:PBS複合体に対する結合、酵素処理能力、及び熱安定性を増強することが先に示された追加の逆転写酵素変異152が試験された。分析された14の追加の変異体のうち、M3変異に加えてT306K及びW313F置換を有するバリアントは、ヒト細胞の5つのゲノム部位における6つのトランスバーション又は挿入編集について、M3と比較して編集効率を追加の1.3倍~3.0倍改善された(図47A~図47S)。PE1アーキテクチャに組み込まれたM-MLV逆転写酵素のこの五重変異体(Cas9 H840A-M-MLV RT(D200N L603W T330P T306K W313F))はこれ以降ではPE2と呼ばれる。
PE2は、1ヌクレオチドトランスバーション、挿入、及び欠失変異をPE1よりも実質的に高い効率で組み入れし(図39C)、より短いPBS pegRNA配列と適合性であり(図39C)、一過性のゲノムDNA:PBS複合体に生産的に係合する増強された能力と整合する。平均で、PE2はPE1と比べてプライム編集点変異効率の1.6~5.1倍の改善に至り(図39C)、いくつかのケースでは、編集収量を最高で46倍まで劇的に改善された(図47F及び図47I)。PE2は標的化された挿入及び欠失をPE1よりも効率的に成し遂げ、PE1によってこの挿入を組み入れる効率と比べて15倍の改善の4.5%の効率でHEK3遺伝子座における24bp FLAGエピトープタグの標的化された挿入を達成し(図47D)、PE1のものよりも2.1倍高い8.6%の効率でHEK3の1bp欠失を媒介した(図39C)。これらの結果はPE2をPE1よりも効率的なプライム編集因子として確立する。
pegRNA特色の最適化
pegRNAアーキテクチャ及びプライム編集効率の間の関係性がHEK293T細胞の5つのゲノム遺伝子座においてPE2によって体系的に探査された(図39C)。一般的に、より低いGC含量を有するプライム部位はより長いPBS配列を要求したが(EMX1及びRNF2。夫々40%及び30%GC含量をニックの上流の最初の10ntに含有する)、より多大なGC含量を有するものはより短いPBS配列によるプライム編集を支持し(HEK4及びFANCF。夫々80%及び60%GC含量をニックの上流の最初の10ntに含有する)(図39C)、pegRNA PBSに対するニックが入ったDNA鎖のハイブリダイゼーションのためのエネルギー的要件と整合した。PBS長さ又はGC含量レベルはプライム編集効率を厳格には予測せず、DNAプライマー又はpegRNA伸長の二次構造等の他の因子もまた編集活性に影響を及ぼしてもよい。典型的な標的配列では~13ntのPBS長さから出発し、配列が、~40~60%GC含量から逸脱する場合には異なるPBS長さを調査することが推奨される。必要なときには、最適なPBS配列は経験的に決定されるはずである。
pegRNAアーキテクチャ及びプライム編集効率の間の関係性がHEK293T細胞の5つのゲノム遺伝子座においてPE2によって体系的に探査された(図39C)。一般的に、より低いGC含量を有するプライム部位はより長いPBS配列を要求したが(EMX1及びRNF2。夫々40%及び30%GC含量をニックの上流の最初の10ntに含有する)、より多大なGC含量を有するものはより短いPBS配列によるプライム編集を支持し(HEK4及びFANCF。夫々80%及び60%GC含量をニックの上流の最初の10ntに含有する)(図39C)、pegRNA PBSに対するニックが入ったDNA鎖のハイブリダイゼーションのためのエネルギー的要件と整合した。PBS長さ又はGC含量レベルはプライム編集効率を厳格には予測せず、DNAプライマー又はpegRNA伸長の二次構造等の他の因子もまた編集活性に影響を及ぼしてもよい。典型的な標的配列では~13ntのPBS長さから出発し、配列が、~40~60%GC含量から逸脱する場合には異なるPBS長さを調査することが推奨される。必要なときには、最適なPBS配列は経験的に決定されるはずである。
次に、pegRNAのRT鋳型部分の性能決定要因が研究された。長さが10~20ntの範囲であるRT鋳型を有するpegRNAを、5つのゲノム標的部位においてPE2を用いて(図39D)及び31ntほども長いより長いRT鋳型によって3つのゲノム部位において(図48A~図48C)体系的に評価された。PBS長さのように、RT鋳型長さもまたプライム編集効率を最大化するために変えられ得るが、一般的には、多くのRT鋳型長さ≧10nt長はより効率的なプライム編集を支持する(図39D)。いくつかの標的部位はより高い編集効率(FANCF、EMX1)を達成するためにはより長いRT鋳型(>15nt)を好み、他の遺伝子座は短いRT鋳型(HEK3、HEK4)を好んだので(図39D)、~10~16ntから出発して、PEgRNAを最適化するときには、短い及び長いRT鋳型両方が試験されるということが推奨される。
重要なことに、sgRNA骨格の末端ヘアピンに隣接するヌクレオチドとしてCを置くRT鋳型は、一般的に、類似の長さのRT鋳型を有する他のPEgRNAと比較して低い編集効率をもたらした(図48A~図48C)。Cas9に結合したsgRNAの構造に基づいて148,149、標準的なsgRNAの3'伸長の最初のヌクレオチドとしてのCの存在は、Cas9のTyr 1356とのパイスタック及びsgRNA A68との非標準的な塩基対をネイティブに形成するヌクレオチドG81との対形成によって、sgRNA骨格折り畳みを遮断し得るということが考えられた。多くのRT鋳型長さはプライム編集を支持するので、3'伸長の最初の塩基(RT鋳型の最後の逆転写される塩基)がCではないPEgRNAを選ぶことが推奨される。
プライム編集因子3システム(PE3及びPE3b)のデザイン
PE2は遺伝情報をpegRNAから標的遺伝子座にPE1よりも効率的に移入し得るが、細胞が1つの編集された鎖及び1つの編集されない鎖によって作成されるもたらされるヘテロ二重鎖DNAを分解する様式は、編集が耐久性であるかどうかを決定する。シトシン又はアデニン脱アミノ化の当初の産物は1つの編集された及び1つの非編集の鎖を含有するヘテロ二重鎖DNAであるので、塩基編集の先の開発は類似の課題に当面した。塩基編集の効率を増大させるために、Cas9 D10Aニッカーゼが用いられてニックを非編集鎖に導入し、編集された鎖を鋳型として用いるDNA修復をその鎖へと導いた129,130,142。この原理を有効利用してプライム編集効率を増強するために、細胞による非編集鎖の選好的置換えを誘導するために、すでにPE2に存在するCas9 H840Aニッカーゼと単純なsgRNAとを用いて非編集鎖へニックを入れる類似の戦略(図40A)が試験された。編集されたDNA鎖もまたプライム編集を開始するためにニックを入れられたので、種々のsgRNAによってプログラムされるニックの位置付けが非編集鎖について試験されて、インデルに至る二本鎖DNA切断の生産を最小化した。
PE2は遺伝情報をpegRNAから標的遺伝子座にPE1よりも効率的に移入し得るが、細胞が1つの編集された鎖及び1つの編集されない鎖によって作成されるもたらされるヘテロ二重鎖DNAを分解する様式は、編集が耐久性であるかどうかを決定する。シトシン又はアデニン脱アミノ化の当初の産物は1つの編集された及び1つの非編集の鎖を含有するヘテロ二重鎖DNAであるので、塩基編集の先の開発は類似の課題に当面した。塩基編集の効率を増大させるために、Cas9 D10Aニッカーゼが用いられてニックを非編集鎖に導入し、編集された鎖を鋳型として用いるDNA修復をその鎖へと導いた129,130,142。この原理を有効利用してプライム編集効率を増強するために、細胞による非編集鎖の選好的置換えを誘導するために、すでにPE2に存在するCas9 H840Aニッカーゼと単純なsgRNAとを用いて非編集鎖へニックを入れる類似の戦略(図40A)が試験された。編集されたDNA鎖もまたプライム編集を開始するためにニックを入れられたので、種々のsgRNAによってプログラムされるニックの位置付けが非編集鎖について試験されて、インデルに至る二本鎖DNA切断の生産を最小化した。
PAMの5'又は3'どちらかのPEgRNAによって誘導されるニックの部位から14~116塩基に位置付けられるニックを誘導するsgRNAをスクリーニングすることによって、このPE3戦略は最初にHEK293T細胞の5つのゲノム部位において試験された。試験された5つの部位のうち4つでは、非編集鎖へニックを入れることは、インデルフリーなプライム編集産物の量を、PE2システムと比較して1.5~4.2倍、55%ほども高くまで増大させた(図40B)。最適なニッキング位置はゲノム部位に依存して変わったが、pegRNAによって誘導されるニックからおよそ40~90bpのPAMの3'に配置されるニックは(図40Bの正の距離)、一般的には、プライム編集効率の好ましい増大(平均で41%である)を余分なインデル形成なしに産生した(試験された5つの部位の夫々において、最も高い編集効率をもたらすsgRNAでは6.8%の平均インデル)(図40B)。予想される通り、いくつかの部位では、PEgRNAによって誘導されるニックの40bp以内に非編集鎖ニックを置くことは、最高で22%のインデル形成の大きい増大に至った(図40B)。おそらくは、近接する両鎖に一緒へニックを入れることからの二本鎖切断の形成を原因とする。しかしながら、他の部位では、pegRNAによって誘導されるニックから14bpほども近接するニッキングは5%インデルのみを産生し(図40B)、遺伝子座依存性因子が近位の二元的なニックから二本鎖DNA切断への変換をコントロールするということを示唆した。1つの試験された部位(HEK4)では、PEgRNAによって誘導されるニックから距離>70bpに置かれるときでさえも、相補鎖ニックは利益を提供しないか又は編集効率を越えるインデルレベル(最高で26%)に至るかどちらかであり、その部位の編集された鎖が細胞によってニッキングされるか又は非効率的にライゲーションされる普通でない傾向と整合した。PEgRNAによって媒介されるニックからおよそ50bpの非編集鎖ニックから出発することと、インデル度数が許容されるレベルを超過する場合には代替的なニック位置付けを試験することとが推奨される。
どのようにして相補鎖ニッキングがプライム編集効率を改善するのかについてのこのモデルは(図40A)、編集された鎖のフラップ分解後にのみ非編集鎖へニックを入れることが、同時的なニックの存在を最小化し得、引き続きインデルを形成する二本鎖切断の度数を減少させるということを予測した。非編集鎖ニッキングの時間的コントロールを達成するために、元のアレルではなく編集された鎖にマッチするスペーサ配列を有するsgRNAがデザインされた。これ以降ではPE3bと呼ばれるこの戦略を用いて、スペーサ及び編集されないアレルの間のミスマッチは、PAM鎖上の編集イベントが生起する後までは、sgRNAによるニッキングを好まないはずである。このPE3bアプローチは、HEK293T細胞の3つのゲノム部位における5つの異なる編集によって試験され、アウトカムをPE2及びPE3システムによって達成されるものと比較された。全てのケースで、PE3bは、PE3と比較して総体的な編集効率のいずれかの明白な減少なしに、PE3と比較して実質的に低いレベルのインデルに関連された(3.5~30倍。平均で12倍低いインデル又は0.85%である)(図40C)。よって、編集が第2のプロトスペーサ上に在ったときには、PE3bシステムは、PE2と比較して編集効率を多くの場合にはPE3のものと類似のレベルまでなお改善しながら、インデルを減少させ得た(図40C)。
一緒になって、これらの知見は、PE3システム(Cas9ニッカーゼ-最適化逆転写酵素+pegRNA+sgRNA)が編集効率をPE2と比較して~3倍改善するということを確立した(図40B~40C)。PE3の追加のニッキング活性から、予想される通り、PE3はPE2よりも広い範囲のインデルによって随伴された。PE3の使用はプライム編集効率を優先するときに推奨される。インデルの最小化が重大であるときには、PE2は~10倍低いインデル度数をオファーする。組み入れされた編集を認識して非編集鎖へニックを入れるsgRNAを用いることが可能であるときには、PE3bシステムはインデル形成を多大に縮減しながらPE3様の編集レベルを達成し得る。
PE3によるプライム編集の標的化範囲及び汎用性を実証するために、HEK3標的部位の+1~+8位置(PEgRNAによって誘導されるニックの3'の最初の塩基を位置+1としてカウントする)における全ての可能な1ヌクレオチド置換の組み入れが、PE3と10ヌクレオチドRT鋳型を有するPEgRNAとを用いて(図41A)調査された。集合的に、これらの24の別物の編集は全ての4つのトランジション変異及び全ての8つのトランスバーション変異をカバーし、平均で33±7.9%である(14%及び48%の間の範囲である)編集効率(インデルを含有しない)で、平均で7.5±1.8%のインデルによって進む。
重要なことに、長距離RT鋳型もまたPE3による効率的なプライム編集を生じさせ得る。例えば、PE3を34nt RT鋳型と共に用いて、点変異をHEK3遺伝子座の位置+12、+14、+17、+20、+23、+24、+26、+30、及び+33(PEgRNAによって誘導されるニックから12~33塩基)に組み入れされ、平均で36±8.7%の効率及び8.6±2.0%のインデルを有した(図41B)。他の遺伝子座における+10位置以降の編集は試みられなかったが、3つの代替的な部位における他のRT鋳型≧30ntもまた効率的編集を支持する(図48A~図48C)。長いRT鋳型の実行可能性は、当初のニック部位から何ダースものヌクレオチドについて効率的なプライム編集を可能化した。どちらかのDNA鎖上のNGG PAMは、効率的なプライム編集を支持する編集及びPAMの間の最大距離よりもずっと少ない平均で~8bp毎に生起するので、他の精密ゲノム編集法とは対照的に、プライム編集は近傍のPAM配列の利用可能性によって実質的に制約されない125,142,154。RNA二次構造とプライム編集効率との間の推定される関係性から、ロングレンジ編集のためのPEgRNAをデザインするときには、種々の長さ及び必要な場合には配列組成(例として、同義コドン)のRT鋳型を試験して編集効率を最適化することが賢明である。
トランジション及びトランスバーション点変異を導入するためのPE3システムの範囲及び限定を更に試験するために、全ての12の可能な方の点変異をカバーする72の追加の編集を6つの追加のゲノム標的部位(図41C~図41H)について試験された。総体的に、インデルフリーな編集効率は平均で25±14%であり、インデル形成は平均で8.3±7.5%であった。pegRNA RT鋳型はPAM配列を包含したので、プライム編集はPAM配列の変化を誘導し得た。これらのケースでは、より高い編集効率(平均で39±9.7%である)及びより低いインデル生成(平均で5.0±2.9%である)が観察された(図41A~図41K。位置+5又は+6における点変異)。PAM編集の効率のこの増大及びインデル形成の減少は、Cas9ニッカーゼが相補鎖の修復に先行して編集された鎖に再結合及びニッキングすることの不能から発生してもよい。プライム編集は編集効率の明らかな損失なしに組み合わせ編集を支持するので、可能なときには、他の所望の変化に加えてPAMを編集することが推奨される。
次に、14の標的化された小さい挿入及び14の標的化された小さい欠失を、7つのゲノム部位において、PE3を用いて(図41I)行われた。標的化された1bp挿入は32±9.8%の平均効率で進み、3bp挿入は39±16%の平均効率で組み入れされた。標的化された1bp及び3bp欠失もまた効率的であり、夫々29±14%及び32±11%の平均収量で進んだ。インデル生成(標的化された挿入又は欠失以外)は平均で6.8±5.4%であった。位置+1及び+6の間に導入された挿入及び欠失はPAMの位置又は構造を変改するので、PAMを編集する点変異と類似に、Cas9ニッカーゼが相補鎖の修復に先行して編集された鎖に再結合及びニッキングすることの不能を原因として、この範囲の挿入及び欠失編集は典型的にはより効率的であるということが考えられた。
PE3は、HEK3部位における5bp~80bpのより大きい精密な欠失を媒介するその能力についてもまた試験された(図41J)。13nt PBSと夫々29、24、又は19bpの相同性を標的遺伝子座に対して含有するRT鋳型とを用いたときには、非常に高い編集効率(52~78%)が5、10、及び15bp欠失について観察された。26nt RT鋳型を用いることは、72±4.2%の効率で25bpというより大きい欠失を支持し、20nt RT鋳型は52±3.8%の効率で80bp欠失を可能化した。これらの標的化された欠失は平均で11±4.8%であるインデル度数によって随伴された(図41J)。
最後に、3つのゲノム部位における挿入及び欠失、挿入及び点変異、欠失及び点変異、又は2つの点変異からなる同じ標的遺伝子座における複数の編集の12の組み合わせを媒介するPE3の能力が試験された。これらの組み合わせ編集は非常に効率的であり、6.4%のインデルによって平均で55%の標的編集であり(図41K)、精密な挿入、欠失、及び点変異の組み合わせを個々の標的部位において高い効率及び低いインデル度数でなすプライム編集の能力を実証した。
一緒になって、図41A-図41Kの例は7つのヒトゲノム遺伝子座における156の別物のトランジション、トランスバーション、挿入、欠失、及び組み合わせ編集を表す。これらの知見はプライム編集の汎用性、精度、及び標的化フレキシビリティーを確立する。
塩基編集と比較したプライム編集
現行世代のシチジン塩基編集因子(CBE)及びアデニン塩基編集因子(ABE)は、C・G→T・Aのトランジション変異及びA・T→G・Cのトランジション変異を高い効率及び低いインデルで組み入れ得る129,130,142。塩基編集の適用は、欲されないバイスタンダー編集を生じさせる塩基編集活性窓(典型的には~5bpの幅)上の複数のシチジン又はアデニン塩基の存在によって129,130,142,155、又は標的ヌクレオチドからおよそ15±2ntに配置されるPAMの不在によって142,156限定され得る。プライム編集は、バイスタンダー編集なしの、又は好適に配置されたPAMの欠如が標的ヌクレオチドをCBE又はABE活性窓上に好ましく位置取らせることを不可能にするときの、トランジション変異の精密な組み入れにとって特に有用であり得る。
現行世代のシチジン塩基編集因子(CBE)及びアデニン塩基編集因子(ABE)は、C・G→T・Aのトランジション変異及びA・T→G・Cのトランジション変異を高い効率及び低いインデルで組み入れ得る129,130,142。塩基編集の適用は、欲されないバイスタンダー編集を生じさせる塩基編集活性窓(典型的には~5bpの幅)上の複数のシチジン又はアデニン塩基の存在によって129,130,142,155、又は標的ヌクレオチドからおよそ15±2ntに配置されるPAMの不在によって142,156限定され得る。プライム編集は、バイスタンダー編集なしの、又は好適に配置されたPAMの欠如が標的ヌクレオチドをCBE又はABE活性窓上に好ましく位置取らせることを不可能にするときの、トランジション変異の精密な組み入れにとって特に有用であり得る。
プライム編集及びシトシン塩基編集が、複数の標的シチジンを標準的な塩基編集窓(プロトスペーサ位置4~8。PAMを位置21~23としてカウントする)上に含有する3つのゲノム遺伝子座を編集することによって比較された。ニッカーゼ活性なしの(BE2max)若しくはニッカーゼ活性ありの(BE4max)最適化されたCBE157を用いるか、又は類縁のPE2及びPE3プライム編集システムを用いた。3つの部位の塩基編集窓上の合計9つの標的シトシンのうち、BE4maxは、PE3よりも2.2倍高い平均のC・G→T・Aの総変換を塩基編集窓の中心の塩基(プロトスペーサ位置5~7。図42A)について生んだ。同様に、非ニッキングBE2maxはこれらの良く配置された塩基においてPE2に平均で1.4倍優った(図42A)。しかしながら、塩基編集窓の中心以外のシトシンについて、PE3はBE4maxに2.7倍優り、PE2はBE2maxに2.0倍優った(PE3の40±17%対BE4maxの15±18%、及びPE2の22±11%対BE2maxの11±13%の平均編集)。総体的に、PE2のインデル度数は非常に低く(平均で0.86±0.47%であった)、PE3では、BE4maxのものに類似か又はそれよりも控えめに高かった(BE4max範囲:2.5%~14%;PE3範囲:2.5%~21%)(図42B)。
精密なC・G→T・Aの編集の組み入れ(いずれかのバイスタンダー編集なし)について塩基編集の効率をプライム編集と比較したときには、プライム編集の効率は上の部位において塩基編集のものを多大に超過した。これらは、ほとんどのゲノムDNA部位のように、複数のシトシンを~5bp塩基編集窓上に含有する(図42C)。シトシンをプロトスペーサ位置C5、C6、及びC7に含有するEMX1等のこれらの部位では、BE4maxは、バイスタンダー編集なしの単一の標的塩基対変換のみを含有する少数の産物を生成した。対照的に、この部位におけるプライム編集は、C・G→T・Aの編集をいずれかの位置又は位置の組み合わせ(C5、C6、C7、C5+C6、C6+C7、C5+C7、又はC5+C6+C7)において選択的に組み入れるために用いられ得た(図42C)。全ての精密な1塩基又は2塩基編集(つまり、いずれかの他の近傍塩基を修飾しない編集)は、PE3又はPE2では夫々BE4max又はBE2よりもかなり効率的であったが、3塩基のC・G→T・Aの編集はBE4maxでより効率的であり(図42C)、活性窓上の全ての標的塩基を編集する塩基編集因子の傾向を反映した。一緒にすると、これらの結果は、シトシン塩基編集因子は最適に配置された標的塩基においてPE2又はPE3よりも高いレベルの編集をもたらし得るが、プライム編集は非最適に配置された標的塩基において塩基編集に優り得、複数の編集可能な塩基ではかなり高い精度で編集し得るということを実証している。
最適化された非ニッキングABE(Cas9ニッカーゼの代わりにdCas9によるABEmax152。これ以降ではABEdmaxと呼ばれる)対PE2による及び最適化されたニッキングアデニン塩基編集因子ABEmax対PE3によるA・T→G・Cの編集が2つのゲノム遺伝子座において比較された。2つの標的アデニンを塩基編集窓上に含有する部位(HEK3)では、ABEはA5の変換についてPE2又はPE3よりも効率的であったが、PE3はABEmax編集窓の縁部に在るA8の変換についてより効率的であった(図42D)。単一のアデニンのみが変換される精密編集の効率を比較するときには、PE3はA5及びA8両方においてABEmaxに優った(図42E)。総体的には、ABEはプライム編集因子よりもずっと少数のインデルをHEK3において産生した(ABEdmaxの0.19±0.02%対PE2の1.5±0.46%、及びABEmaxの0.53±0.16%対PE3の11±2.3%、図42F)。単一のAのみが塩基編集窓上に存在するFANCFにおいては、ABE2及びABEmaxは全標的塩基対変換がそれらのプライム編集カウンターパートに1.8~2.9倍優り、塩基編集及びプライム編集両方からの実質的に全ての編集された産物は精密な編集のみを含有した(図42D~図42E)。HEK3部位のように、ABEはFANCF部位においてずっと少数のインデルを産生した(図42F)。
集合的に、これらの結果は、標的化されたトランジション変異を作ることについて、塩基編集及びプライム編集が補完的な強さ及び弱さをオファーするということを指示する。単一の標的ヌクレオチドが塩基編集窓上に存在するケースでは、又はバイスタンダー編集が許容されるときには、現行の塩基編集因子は典型的にはプライム編集因子よりも効率的であり、少数のインデルをもたらす。複数のシトシン又はアデニンが存在しかつバイスタンダー編集が望ましくないときには、又は標的塩基が利用可能なPAMに対して相対的に塩基編集にとって不良に配置されるときには、プライム編集因子は実質的な利点をオファーする。
オフターゲットプライム編集
生産的な編集をもたらすためには、プライム編集は、Cas9ドメインが結合するための相補的な標的遺伝子座:pegRNAスペーサ、pegRNAによってプライムされる逆転写が開始するための標的遺伝子座:pegRNA PBS相補性、及びフラップ分解のための標的遺伝子座:逆転写酵素産物相補性を要求する。これらの3つの別物のDNAハイブリダイゼーション要件は、他のゲノム編集法のものと比較してオフターゲットプライム編集を最小化し得る。これを実証するために、HEK293T細胞は、PE3又はPE2と4つのオンターゲットゲノム遺伝子座を標的化するようにデザインされた合計16のPEgRNAとによって、Cas9と同じプロトスペーサを標的化する4つの対応するsgRNAとによって、又はCas9と同じ16のPEgRNAとによって処置された。夫々が、Cas9及び対応するオンターゲットsgRNAがHEK293T細胞において実質的なオフターゲットDNA修飾を引き起こすことが公知である少なくとも4つの良く特徴付けられたオフターゲット部位を有するので118,159、これらの4つの標的遺伝子座が選ばれた。処置後に、各オンターゲットスペーサにとっての4つのオンターゲット遺伝子座及び上位4つの公知のCas9オフターゲット部位が合計16のオフターゲット部位について配列決定された(表1)。
生産的な編集をもたらすためには、プライム編集は、Cas9ドメインが結合するための相補的な標的遺伝子座:pegRNAスペーサ、pegRNAによってプライムされる逆転写が開始するための標的遺伝子座:pegRNA PBS相補性、及びフラップ分解のための標的遺伝子座:逆転写酵素産物相補性を要求する。これらの3つの別物のDNAハイブリダイゼーション要件は、他のゲノム編集法のものと比較してオフターゲットプライム編集を最小化し得る。これを実証するために、HEK293T細胞は、PE3又はPE2と4つのオンターゲットゲノム遺伝子座を標的化するようにデザインされた合計16のPEgRNAとによって、Cas9と同じプロトスペーサを標的化する4つの対応するsgRNAとによって、又はCas9と同じ16のPEgRNAとによって処置された。夫々が、Cas9及び対応するオンターゲットsgRNAがHEK293T細胞において実質的なオフターゲットDNA修飾を引き起こすことが公知である少なくとも4つの良く特徴付けられたオフターゲット部位を有するので118,159、これらの4つの標的遺伝子座が選ばれた。処置後に、各オンターゲットスペーサにとっての4つのオンターゲット遺伝子座及び上位4つの公知のCas9オフターゲット部位が合計16のオフターゲット部位について配列決定された(表1)。
先の研究と整合して118、Cas9及び4つの標的sgRNAは、先に報告されたオフターゲット遺伝子座の全ての16を修飾した(図42G)。HEK3標的遺伝子座について、4つのオフターゲット部位におけるCas9オフターゲット修飾効率は平均で16%であった。Cas9及びHEK4を標的化するsgRNAは、4つの試験された公知のオフターゲット部位の平均で60%の修飾をもたらした。同様に、EMX1及びFANCFのオフターゲット部位は、Cas9:sgRNAによって夫々48%及び4.3%の平均度数で修飾された(図42G)。sgRNAと比較して、pegRNAはCas9ヌクレアーゼと共にオンターゲット部位を平均して類似の(1~1.5倍低い)効率で修飾されるが、pegRNAはCas9ヌクレアーゼと共にオフターゲット部位をsgRNAよりも~4倍低い平均効率で修飾されるということが留意された。
著しくは、これらの4つの標的スペーサを含有する同じ16の試験されたpegRNAによるPE3又はPE2はかなり低いオフターゲット編集をもたらした(図42H)。Cas9+sgRNAによるオフターゲット編集を経過することが公知の16の部位のうち、PE3+pegRNA又はPE2+pegRNAは16のオフターゲット部位のうち3つのみにおいて検出可能なオフターゲットプライム編集をもたらし、16のうち1つのみがオフターゲット編集効率≧1%を示した(図42H)。これらの16の公知のCas9オフターゲット部位におけるHEK3、HEK4、EMX1、及びFANCFを標的化するpegRNAの平均オフターゲットプライム編集は、夫々<0.1%、<2.2±5.2%、<0.1%、及び<0.13±0.11%であった(図42H)。注意すべきことに、Cas9+pegRNA1が97%効率で編集するHEK4オフターゲット3部位では、PE2+pegRNA1は同じスペーサ配列を共有するにもかかわらず0.7%オフターゲット編集のみをもたらし、Cas9編集と比較してプライム編集に要求される2つの追加のDNAハイブリダイゼーションイベントが、どのようにしてオフターゲット編集を多大に縮減し得るのかということを実証する。一緒にすると、これらの結果は、PE3及びpegRNAが、Cas9及び同じプロトスペーサを標的化するsgRNAよりもヒト細胞におけるかなり低いオフターゲットDNA編集を誘導するということを示唆する。
3'伸長されたpegRNAの逆転写は原理的にはガイドRNA骨格上へと進み得る。もたらされる3'フラップが、編集されないDNA鎖とのその3'端における相補性の欠如にもかかわらず、標的遺伝子座に組み込まれる場合には、アウトカムはpegRNA骨格ヌクレオチドの挿入であり、これはインデル度数に寄与する。我々は、HEK293T細胞の4つの遺伝子座における66のPE3によって媒介される編集実験からの配列決定データを分析し、いずれかの数のpegRNA骨格ヌクレオチドの平均で1.7±1.5%の全挿入である低い度数で、pegRNA骨格挿入を観察した(図56A~図56D)。Cas9ドメイン結合を原因とする逆転写酵素にとってのガイドRNA骨格のアクセス不可能性と、pegRNA骨格逆転写からもたらされる3'フラップのミスマッチの3'端のフラップ分解の間の細胞性の切除とは、pegRNA骨格ヌクレオチドを組み込む産物を最小化し得る。かかるイベントは稀であるが、pegRNA骨格組み込みを最小化するpegRNA又はプライム編集因子タンパク質を操作するための将来の作業は、インデル度数を更に減少させてもよい。
デアミナーゼはいくつかの塩基編集因子ではCas9非依存的様式で作用し得、低レベルだが広範囲のオフターゲットDNA編集を(ABEではなく)第1世代CBEにおいて160~162、並びにオフターゲットRNA編集を第1世代CBE及びABEにおいてもたらすが163~165、操作されたデアミナーゼによるより新たなCBE及びABEバリアントはCas9非依存的なオフターゲットDNA及びRNA編集を多大に縮減する163~165。プライム編集因子はデアミナーゼ等の塩基修飾酵素を欠き、よって、Cas9非依存的様式でDNA又はRNA塩基を修飾する内在的能力を有さない。
原理的には、プライム編集因子の逆転写酵素ドメインは適切にプライムされたRNA又はDNA鋳型を細胞内でプロセシングし得るが、レトロトランスポゾン、例えばLINE-1ファミリー166、内生レトロウイルス167,168、及びヒトテロメラーゼのものは全て、活性な内生ヒト逆転写酵素を提供するということが留意された。ヒト細胞におけるそれらの天然の存在は、逆転写酵素活性それ自体は実質的に毒性ではないということを示唆する。実に、dCas9、Cas9 H840Aニッカーゼ、又は逆転写酵素を不活性化するR110S+K103L変異を有するPE2(PE2-dRT)を発現する対照のものと比較して、HEK293T細胞生存性のPE3依存的な違いは観察されなかった169(図49A~図49B)。
上のデータ及び分析にもかかわらず、オフターゲットプライム編集をバイアスがないゲノムワイドな様式で評価するために、及びプライム編集因子又はプライム編集中間体の逆転写酵素バリアントが細胞に影響してもよい程度を特徴付けるために、追加の研究が必要とされる。
ヒト細胞の病原性のトランスバーション、挿入、及び欠失変異をプライム編集する
遺伝子疾患を引き起こすトランスバーション、小さい挿入、及び小さい欠失変異をヒト細胞に直接的に組み入れ又は修正するPE3の能力が試験された。鎌状赤血球症は最も普通にはHBBのA・T→T・Aのトランスバーション変異によって引き起こされ、ベータグロビンのGlu6→Valの変異をもたらす。Cas9ヌクレアーゼとHDRのためのドナーDNA鋳型とによるex vivoの造血幹細胞の処置、次に編集された細胞の濃縮、移植、及び定着は、鎌状赤血球症の処置のための有望な可能性ある戦略である170。しかしながら、このアプローチは、正しく編集されたHBBアレルに加えてなお多くのインデル含有副産物を生成する170~171。塩基編集因子は一般的にはずっと少数のインデルを生ずるが、それらはHBBの正常配列を直接的に復元するために必要とされるT・A→A・Tのトランスバーション変異を現行ではなし得ない。
遺伝子疾患を引き起こすトランスバーション、小さい挿入、及び小さい欠失変異をヒト細胞に直接的に組み入れ又は修正するPE3の能力が試験された。鎌状赤血球症は最も普通にはHBBのA・T→T・Aのトランスバーション変異によって引き起こされ、ベータグロビンのGlu6→Valの変異をもたらす。Cas9ヌクレアーゼとHDRのためのドナーDNA鋳型とによるex vivoの造血幹細胞の処置、次に編集された細胞の濃縮、移植、及び定着は、鎌状赤血球症の処置のための有望な可能性ある戦略である170。しかしながら、このアプローチは、正しく編集されたHBBアレルに加えてなお多くのインデル含有副産物を生成する170~171。塩基編集因子は一般的にはずっと少数のインデルを生ずるが、それらはHBBの正常配列を直接的に復元するために必要とされるT・A→A・Tのトランスバーション変異を現行ではなし得ない。
PE3が用いられて、44%の効率及び4.8%インデルでHEK293T細胞にHBB E6V変異を組み入れした(図43A)。PE3処置された細胞の混合物から、我々はHBB E6Vアレルについてホモ接合(三倍体)である6つのHEK293T細胞株を単離し(図53A~図53D)、プライム編集が病原性の変異を有するヒト細胞株を生成する能力を実証した。HBB E6Vアレルを野生型HBBに修正するために、我々は、HBB E6V変異を野生型HBBに直接的に復帰させるようにプログラムされたPE3及びpegRNAによってホモ接合HBB E6V HEK293T細胞を処置した。合計14のpegRNAデザインが試験された。3日後に、DNA配列決定は、全ての14のpegRNAがPE3と組み合わせられたときに、HBB E6Vから野生型HBBへの効率的な修正と(インデルなしの≧26%野生型HBB)平均で2.8±0.70%であるインデルレベルとを与えるということを明らかにした(図50A)。最も良好なpegRNAはHBB E6Vから野生型への52%修正を2.4%インデルでもたらした(図43A)。pegRNAによって認識されるPAMを修飾するサイレントな変異の導入は、編集効率及び産物純度を1.4%インデルによる58%修正まで控えめに改善した(図43A)。これらの結果は、プライム編集がヒト細胞株の病原性のトランスバーション点変異を高い効率及び最小限の副産物によって組み入れ及び修正し得るということを確立する。
テイ・サックス病は最も多くの場合にはHEXA遺伝子上への4bp挿入(HEXA 1278+TATC)によって引き起こされる136。PE3を用いて、この4bp挿入をHEK293T細胞に31%の効率及び0.8%のインデルで組み入れし(図43B)、HEXA 1278+TATCアレルについてホモ接合である2つのHEK293T細胞株を単離した(図53A-図53D)。これらの細胞が用いられて、43のpegRNA及び3つのニッキングsgRNAを、PE3又はPE3bシステムによって、HEXAの病原性の挿入の修正について試験した(図50B)。野生型アレルへの完璧な復帰によるか、又はPAMを遮断しかつサイレントな変異を組み入れるシフトした4bp欠失によるかどちらかである。試験された43のpegRNAのうち19は≧20%編集をもたらした。PE3又はPE3b及び最も良好なpegRNAによる野生型HEXAへの完璧な修正は類似の平均効率で進んだが(PE3の30%対PE3bの33%)、PE3bシステムは5.3倍少数のインデル産物によって随伴された(PE3の1.7%対PE3bの0.32%)(図43B及び図50B)。これらの知見は、哺乳動物細胞の病原性のアレルを効率的にかつ最小の副産物によって組み入れ又は修正する精密な小さい挿入及び欠失をなすプライム編集の能力を実証する。
最後に、ヒトプリオンタンパク質(PrP)をコードする遺伝子PRNP上への防護性のSNPの組み入れが試験された。PrPミスフォールディングは進行性のかつ致死的な神経変性プリオン病を引き起こし、これはPRNP遺伝子上の受け継がれた優性変異によって又はミスフォールディングしたPrPに対する暴露によって自発的に発生し得る172。PRNP G127V変異体アレルはヒト138及びマウス173においてプリオン病に対する耐性を付与する。PE3が用いられてG127VをHEK293T細胞のヒトPRNPアレルに組み入れした。これはG・C→T・Aのトランスバーションを要求する。4つのpegRNA及び3つのニッキングsgRNAがPE3システムによって評価された。最も有効なPE3及びpegRNAに対する3日の暴露後に、DNA配列決定は、G127V変異を組み入れる53±11%の効率と1.7±0.7%のインデルレベルとを明らかにした(図43C)。一緒にすると、これらの結果は、プライム編集がヒト細胞においてトランスバーション、挿入、又は欠失変異を組み入れ又は修正する能力を確立する。これらは、疾患に対する耐性を効率的にかつ最小の副産物によって引き起こすか又は付与する。
種々のヒト細胞株及び初代マウスニューロンのプライム編集
次に、プライム編集が、3つの追加のヒト細胞株の内生部位を編集するその能力について試験された。K562(白血病骨髄)細胞では、PE3が用いられて、HEK3、EMX1、及びFANCF部位のトランスバーション編集、並びにHEK3における6xHisタグの18bp挿入を行った。15-30%の平均編集効率がこれらの4つのPE3によって媒介される編集の夫々について観察され、インデルは平均で0.85~2.2%であった(図43A)。U2OS(骨肉腫)細胞では、HEK3及びFANCFのトランスバーション変異、並びにHEK3への3bp挿入及び6xHisタグ挿入を、インデル形成が10~76倍超過する7.9~22%の編集効率によって組み入れされた(図43A)。最後に、HeLa(子宮頸がん)細胞では、HEK3への3bp挿入が12%平均効率及び1.3%インデルによって行われた(図43A)。集合的に、これらのデータはHEK293T細胞以外の複数の細胞株がプライム編集を支持するということを指示しているが、編集効率は細胞型によって変わり、一般的にはHEK293T細胞よりも効率的ではない。編集:インデル比は、全ての試験されたヒト細胞株において高いままに留まった。
次に、プライム編集が、3つの追加のヒト細胞株の内生部位を編集するその能力について試験された。K562(白血病骨髄)細胞では、PE3が用いられて、HEK3、EMX1、及びFANCF部位のトランスバーション編集、並びにHEK3における6xHisタグの18bp挿入を行った。15-30%の平均編集効率がこれらの4つのPE3によって媒介される編集の夫々について観察され、インデルは平均で0.85~2.2%であった(図43A)。U2OS(骨肉腫)細胞では、HEK3及びFANCFのトランスバーション変異、並びにHEK3への3bp挿入及び6xHisタグ挿入を、インデル形成が10~76倍超過する7.9~22%の編集効率によって組み入れされた(図43A)。最後に、HeLa(子宮頸がん)細胞では、HEK3への3bp挿入が12%平均効率及び1.3%インデルによって行われた(図43A)。集合的に、これらのデータはHEK293T細胞以外の複数の細胞株がプライム編集を支持するということを指示しているが、編集効率は細胞型によって変わり、一般的にはHEK293T細胞よりも効率的ではない。編集:インデル比は、全ての試験されたヒト細胞株において高いままに留まった。
プライム編集が有糸分裂後の終末分化した初代細胞において可能であるかどうかを決定するために、E18.5マウスから収穫された初代皮質ニューロンが二元的な分裂PE3レンチウイルス送達システムによって形質導入された。これにおいては、分裂インテインスプライシング203が、夫々が別個のウイルスから送達されるN末端及びC末端半分からPE2タンパク質を再構成する。編集を有糸分裂後ニューロンに制限するために、成熟したニューロンに高度に特異的であるヒトシナプシンプロモータ204が用いられて両方のPE2タンパク質構成要素の発現を駆動した。GFPが自己切断P2Aペプチド205を介してPE2のN末端半分に融合された。ニューロンからの核が二元的なウイルス形質導入の2週後に単離され、直接的に配列決定されるか又は配列決定前にGFP発現についてソーティングされた。ソーティングされた核において0.58±0.14%の平均インデルによってDNMT1遺伝子座におけるトランスバーションを組み入れるための7.1±1.2%の平均プライム編集(図43Dが観察された。同じ分裂インテインの二元的レンチウイルスシステムにおけるCas9ヌクレアーゼは、ソーティングされた皮質ニューロン核において31±5.5%のインデルをもたらした(図43D。これらのデータは、有糸分裂後の終末分化した初代細胞がプライム編集を支持し得るということを指示し、それゆえにプライム編集が細胞複製を要求しないということを確立する。
Cas9によって開始されるHDRと比較したプライム編集
PE3の性能が、HDRを支持する有糸分裂細胞株において128、最適化されたCas9によって開始されるHDR128,125のものと比較された。HEK293T、HeLa、K562、及びU2OS細胞が、種々のトランスバーション及び挿入編集を組み入れるようにデザインされたCas9ヌクレアーゼ、sgRNA、及びssDNAドナーオリゴヌクレオチド鋳型によって処置された(図43E-図43G及び図51A-図51F)。Cas9によって開始されるHDRは、全てのケースにおいて、二本鎖切断を引き起こす処置から予想される通りずっと高いレベルの副産物(支配的にはインデル)によって、所望の編集を首尾良く組み入れした。PE3をHEK293T細胞に用いて、HBB E6V組み入れ及び修正は夫々42%及び58%の平均編集効率で2.6%及び1.4%の平均インデルによって進んだ(図43E及び図43G)。対照的に、Cas9ヌクレアーゼ及びHDR鋳型による同じ編集は、5.2%及び6.7%の平均編集効率を79%及び51%の平均インデル度数によってもたらした(図43E及び図43G)。類似に、PE3はPRNP G127Vを53%の効率及び1.7%インデルで組み入れしたが、Cas9によって開始されるHDRはこの変異を6.9%の効率及び53%インデルで組み入れした(図43E及び図43G)。それゆえに、HBB E6V組み入れ、HBB E6V修正、及びPRNP G127V組み入れの編集:インデルの比は、Cas9によって開始されるHDRよりもPE3では平均で270倍高かった。
PE3の性能が、HDRを支持する有糸分裂細胞株において128、最適化されたCas9によって開始されるHDR128,125のものと比較された。HEK293T、HeLa、K562、及びU2OS細胞が、種々のトランスバーション及び挿入編集を組み入れるようにデザインされたCas9ヌクレアーゼ、sgRNA、及びssDNAドナーオリゴヌクレオチド鋳型によって処置された(図43E-図43G及び図51A-図51F)。Cas9によって開始されるHDRは、全てのケースにおいて、二本鎖切断を引き起こす処置から予想される通りずっと高いレベルの副産物(支配的にはインデル)によって、所望の編集を首尾良く組み入れした。PE3をHEK293T細胞に用いて、HBB E6V組み入れ及び修正は夫々42%及び58%の平均編集効率で2.6%及び1.4%の平均インデルによって進んだ(図43E及び図43G)。対照的に、Cas9ヌクレアーゼ及びHDR鋳型による同じ編集は、5.2%及び6.7%の平均編集効率を79%及び51%の平均インデル度数によってもたらした(図43E及び図43G)。類似に、PE3はPRNP G127Vを53%の効率及び1.7%インデルで組み入れしたが、Cas9によって開始されるHDRはこの変異を6.9%の効率及び53%インデルで組み入れした(図43E及び図43G)。それゆえに、HBB E6V組み入れ、HBB E6V修正、及びPRNP G127V組み入れの編集:インデルの比は、Cas9によって開始されるHDRよりもPE3では平均で270倍高かった。
HEK293T以外のヒト細胞株におけるPE3及びHDRの間の比較は、より低いPE3編集効率でだが、類似の結果を示した。例えば、K562細胞では、HEK3上へのPE3によって媒介される3bp挿入は、Cas9によって開始されるHDRの17%編集及び72%インデルと比較して25%の効率及び2.8%のインデル、PE3を好む40倍の編集:インデル率という利点によって進んだ(図43F~図43G)。U2OS細胞では、PE3はこの3bp挿入を22%効率及び2.2%インデルで行い、Cas9によって開始されるHDRは、74%のインデルによる15%の編集、49倍低い編集:インデル比をもたらした(図43F-図43G)。HeLa細胞では、Cas9によって開始されるHDRの3.0%の編集及び69%のインデルに対して、PE3はこの挿入を12%の効率及び1.3%のインデル、210倍の編集:インデル比の違いによってなした(図43F~図43G)。集合的に、これらのデータは、HDRが、典型的には、試験された4つの細胞株においてPE3よりも低いか又は類似の編集効率及びずっと高いインデルをもたらすということを指示した(図51A~図51F)。
考察及び将来の方向
1ヌクレオチド精度でDNA配列を挿入する能力は、特に有能なプライム編集能力である。例えば、PE3が用いられて、HEK293T細胞のHEK3遺伝子座にHis6タグ(18bp、65%平均効率)、FLAGエピトープタグ(24bp、18%平均効率)、及びCreリコンビナーゼのネイティブな基質である伸長されたLoxP部位(44bp、23%平均効率)を精密に挿入した。平均インデルはこれらの例では3.0%及び5.9%の間の範囲であった(図43H)。多くのバイオテクノロジー、合成生物学、及び処置学的適用が、新たなDNA配列を生細胞の目当ての標的部位に効率的にかつ精密に導入する能力から発生することが想定される。
1ヌクレオチド精度でDNA配列を挿入する能力は、特に有能なプライム編集能力である。例えば、PE3が用いられて、HEK293T細胞のHEK3遺伝子座にHis6タグ(18bp、65%平均効率)、FLAGエピトープタグ(24bp、18%平均効率)、及びCreリコンビナーゼのネイティブな基質である伸長されたLoxP部位(44bp、23%平均効率)を精密に挿入した。平均インデルはこれらの例では3.0%及び5.9%の間の範囲であった(図43H)。多くのバイオテクノロジー、合成生物学、及び処置学的適用が、新たなDNA配列を生細胞の目当ての標的部位に効率的にかつ精密に導入する能力から発生することが想定される。
集合的に、本明細書に記載のプライム編集実験は、最高で44bpの18の挿入、最高で80bpの22の欠失、77のトランスバーションを包含する113の点変異、及び18の組み合わせ編集を、ヒト及びマウスゲノムの12の内生遺伝子座において、PAMのスタートの3bp上流から29bp下流の範囲である位置付けにおいて、あからさまな二本鎖DNA切断を作ることなしに組み入れした。これらの結果はプライム編集を著しく汎用的なゲノム編集法として確立する。ClinVarの挿入、欠失、インデル、及び重複の圧倒的多数(85-99%)は≦30bpであるので(図52A~図52D)、原理的には、プライム編集はClinVarの75,122の現行で公知の病原性のヒト遺伝子バリアントの最高で~89%を修正し得(図38Aのトランジション、トランスバーション、挿入、欠失、インデル、及び重複)、コピー数獲得又は損失によって引き起こされる疾患を改善する追加のポテンシャルを有する。
重要なことに、いずれかの所望の編集について、プライム編集のフレキシビリティーは、pegRNAによって誘導されるニックの位置付け、sgRNAによって誘導される第2のニックの位置付け、PBS長さ、RT鋳型長さ、及びどの鎖が最初に編集されるかの多くの可能な選択肢を、本明細書において鋭意実証される通りオファーする。他の精密ゲノム編集法で典型的に利用可能なより限定されたオプション125,142,154とは対照をなすこのフレキシビリティーは、編集効率、産物純度、DNA特異性、又は他のパラメータが、所与の適用の必要に適するように最適化されることを許す。図50A~図50Bに示されている通りであり、これらにおいては、ある範囲のプライム編集戦略をカバーする14及び43のpegRNAを試験することが、夫々病原性のHBB及びHEXAアレルの修正を最適化した。
しかしながら、二本鎖切断又はHDRを要求することなしに、哺乳動物細胞のゲノム上の高度に精密な標的化されたトランジション、トランスバーション、小さい挿入、及び小さい欠失を可能化することによって、プライム編集は、ゲノム編集の範囲を実質的に拡張する新たな「検索置換」能力を提供する。
実施例2:pegRNA修飾
プライム編集(PE)効率を改善し得る一連のpegRNAデザイン及び戦略が本明細書に記載される。
プライム編集(PE)効率を改善し得る一連のpegRNAデザイン及び戦略が本明細書に記載される。
プライム編集(PE)は、プライム編集ガイドRNA(pegRNA)内にコードされた情報を使用して、標的遺伝子座内の定義されたDNA配列を置換、挿入、又は除去し得るゲノム編集技術である。プライム編集因子(PE)は、逆転写酵素(RT)酵素等のポリメラーゼに融合されたヌクレアーゼ活性を有する配列プログラム型DNA結合タンパク質(Cas9)からなる。PEは、スペーサ配列内の特定のDNA遺伝子座を標的化するための情報、並びに標準的なsgRNA骨格に組み込まれた操作された伸長における所望の編集を指定する情報を含有する、pegRNAと複合体を形成する。PE:pegRNA複合体は、プログラムされた標的DNA遺伝子座に結合してニックを入れ、ニックが入ったDNA鎖とpegRNAの操作されたプライマー結合配列(PBS)とのハイブリダイゼーションを可能にする。次いで、逆転写酵素ドメインは、ニックが入ったゲノムDNAをDNA重合のためのプライマーとして使用して、pegRNAのRT鋳型部分内で編集コード情報をコピーする。その後のDNA修復プロセスは、新しく合成された編集されたDNA鎖をゲノム遺伝子座に組み込む。これらのpegRNAのデザインの改善は、PE効率の改善をもたらし得、ゲノムへのより長い挿入配列の組み入れを可能にする。
PEの有効性を改善するために想定される一連のpegRNAデザインが本明細書に記載される。これらのデザインは、sgRNAの有効性及び/又は安定性を改善するための多くの以前に公開されたアプローチを利用するとともに、多くの新規な戦略を利用する。これらの改善は、多数の異なるカテゴリーのうちの1つ又は複数に属し得る。
(1)より長いpegRNA。このカテゴリーは、非ポリメラーゼIII(pol III)プロモータからの機能的なpegRNAの効率的な発現を可能にする改良されたデザインに関し、これは、厄介な配列要件なしに、より長いpegRNAの発現を可能にするであろう。
(2)コアの改良。このカテゴリーは、有効性を改善し得るコア、Cas9結合pegRNA骨格の改善に関する。
(3)RTプロセス性。このカテゴリーは、RTプロセス性を改善し、標的ゲノム遺伝子座でのより長い配列の挿入を可能にする、pegRNAに対する修飾に関する。
(4)末端モチーフ。このカテゴリーは、pegRNAの安定性を改善し、RTプロセス性を増強し、pegRNAのミスフォールディングを防止し、又はゲノム編集に重要な更なる因子を動員する、pegRNAの5'及び/又は3'末端へのRNAモチーフの付加に関する。
本明細書には、各カテゴリーにおける多数の潜在的なそのようなpegRNAデザインが記載される。これらのデザインのいくつかは、Cas9によるsgRNA活性を改善するために以前に記載されており、そのようなものとして示されている。本明細書には、pegRNA骨格の研磨を可能にし、PE活性を増強する、所与の配列標的に対するpegRNAの進化のためのプラットフォームも記載される(5)。注意すべきことに、これらのデザインはまた、任意のCas9又はその進化したバリアントによって認識されるpegRNAを改善するために容易に適用されてもよい。
(1)より長いpegRNA。
sgRNAは、典型的には、U6 snRNAプロモータから発現される。このプロモータは、pol IIIを動員して関連するRNAを発現させ、核内に保持される短いRNAの発現に有用である。しかし、pol IIIは高度にプロセッシブではなく、効率的なゲノム編集に必要なレベルで数百ヌクレオチド長よりも長いRNAを発現することができない183。更に、pol IIIは、Uのストレッチで失速又は終結し得、pegRNAを使用して挿入され得た配列多様性を潜在的に制限する。ポリメラーゼII(pCMV等)又はポリメラーゼI(U1 snRNAプロモータ等)を動員する他のプロモータは、より長いsgRNAを発現する能力について調べられている183。しかしながら、これらのプロモータは、典型的には部分的に転写され、発現されたpegRNA中のスペーサの余分な配列5'をもたらし、これにより、部位依存的にCas9:sgRNA活性が著しく低下することが示されている。更に、pol III転写されたpegRNAは、6~7 Uの連続で単純に終結し得るが、pol II又はpol Iから転写されたpegRNAは、異なる終結シグナルを必要とするであろう。多くの場合、そのようなシグナルはまた、核からのpegRNAの望ましくない輸送をもたらすポリアデニル化をもたらす。同様に、pCMV等のpol IIプロモータから発現されるRNAは、典型的には5'キャップされ、その核外輸送ももたらす。
sgRNAは、典型的には、U6 snRNAプロモータから発現される。このプロモータは、pol IIIを動員して関連するRNAを発現させ、核内に保持される短いRNAの発現に有用である。しかし、pol IIIは高度にプロセッシブではなく、効率的なゲノム編集に必要なレベルで数百ヌクレオチド長よりも長いRNAを発現することができない183。更に、pol IIIは、Uのストレッチで失速又は終結し得、pegRNAを使用して挿入され得た配列多様性を潜在的に制限する。ポリメラーゼII(pCMV等)又はポリメラーゼI(U1 snRNAプロモータ等)を動員する他のプロモータは、より長いsgRNAを発現する能力について調べられている183。しかしながら、これらのプロモータは、典型的には部分的に転写され、発現されたpegRNA中のスペーサの余分な配列5'をもたらし、これにより、部位依存的にCas9:sgRNA活性が著しく低下することが示されている。更に、pol III転写されたpegRNAは、6~7 Uの連続で単純に終結し得るが、pol II又はpol Iから転写されたpegRNAは、異なる終結シグナルを必要とするであろう。多くの場合、そのようなシグナルはまた、核からのpegRNAの望ましくない輸送をもたらすポリアデニル化をもたらす。同様に、pCMV等のpol IIプロモータから発現されるRNAは、典型的には5'キャップされ、その核外輸送ももたらす。
以前に、Rinn及び共同研究者らは、長鎖非コードRNA-(lncRNA)タグ付きsgRNAの産生のために様々な発現プラットフォームをスクリーニングした183。これらのプラットフォームは、pCMVから発現され、ヒト由来のMALAT1 ncRNA由来のENEエレメント184、KSHV由来のPAN ENEエレメント185、又はU1 snRNA由来の3'ボックス186で終結するRNAを包含する。注意すべきことに、MALAT1 ncRNA及びPAN ENEは、ポリAテールを保護する三重らせんを形成する184,187。RNAの発現を可能にすることに加えて、これらの構築物はRNA安定性も高め得た(セクションiv参照)。これらのより長いsgRNA183の発現を可能にするためにU 1 snRNAからのプロモータを使用することも検討された。これらの発現系はまた、より長いpegRNAの発現を可能にする。更に、pegRNAの一部として転写されることになるpol IIプロモータの部分を切断するための一連の方法がデザインされており、ハンマーヘッド188、ピストル189、ハチェット189、ヘアピン190、VS191、ツイスター192、若しくはツイスターシスター192リボザイム等の自己切断リボザイム、又は転写されたガイドを処理するための他の自己切断要素、又はCsy4193によって認識され、ガイドの処理にもつながるヘアピンのいずれかを付加する。また、複数のENEモチーフの組み込みは、改善されたpegRNA発現及び安定性をもたらし得る。環状イントロンRNA(ciRNA)の形態でpegRNAを循環させると、RNA発現及び安定性の増強、並びに核局在化をもたらし得る194。例示的なpegRNA発現プラットフォームは、配列番号241~245によって表される。
(2)コア/骨格の改善。
コアであるCas9結合pegRNA骨格は、PE活性を増強するために改善され得る可能性が高い。例示的なアプローチでは、骨格(P1)の第1の対合要素は、GTTTT-AAAAC(配列番号246)対合要素を含有する。そのようなTsの実行は、pol IIIの休止及びRNA転写物の早期終結をもたらし得る。P1のこの部分におけるT-A対の1つのG-C対への合理的突然変異は、sgRNA活性を増強し得る。このアプローチは、pegRNAを改善するために使用され得る。更に、P1の長さを増加させることにより、sgRNAフォールディングを増強し、改善された活性をもたらし得る。最後に、所与のDNA標的上でのpegRNAの指向性進化によるpegRNA骨格の研磨もまた、改善された活性をもたらす可能性が高い。これはセクション(v)で説明される。例示的な修飾されたpegRNAは、配列番号247及び248によって表される。
コアであるCas9結合pegRNA骨格は、PE活性を増強するために改善され得る可能性が高い。例示的なアプローチでは、骨格(P1)の第1の対合要素は、GTTTT-AAAAC(配列番号246)対合要素を含有する。そのようなTsの実行は、pol IIIの休止及びRNA転写物の早期終結をもたらし得る。P1のこの部分におけるT-A対の1つのG-C対への合理的突然変異は、sgRNA活性を増強し得る。このアプローチは、pegRNAを改善するために使用され得る。更に、P1の長さを増加させることにより、sgRNAフォールディングを増強し、改善された活性をもたらし得る。最後に、所与のDNA標的上でのpegRNAの指向性進化によるpegRNA骨格の研磨もまた、改善された活性をもたらす可能性が高い。これはセクション(v)で説明される。例示的な修飾されたpegRNAは、配列番号247及び248によって表される。
gRNA骨格に対する多数の構造修飾も試験されたが、いずれも編集活性の有意な増加を示さなかった(3.30と比較して、X軸における3.30.13~3.30.19の図82を参照されたい。)。しかしながら、このデータには、注目に値する2つの注意点を有する。第1に、このガイドはすでに非常にうまく機能しており、あまり効果的でないガイドの方が試験には優れていた。第2に、HEK細胞では、トランスフェクションは非常に効率的であり、トランスフェクトされたガイドRNAの量は必要とされる量と比較して非常に過剰であることが認められた(量を約1/8~1/4に減少させることは編集に影響を及ぼさない)。これらの改善は、トランスフェクション効率がより低い、又はあまり効果的でないガイドを有する他の細胞型においてのみ見られ得る。これらの変化の多くは、他の細胞株におけるsgRNA活性を改善するために先行される。
図82の構築物の配列は以下の通りである。


末端モチーフもUの連続で終端しない末端モチーフも存在しない場合、転写物は、以下のHDVリボザイム
GGCCGGCAUGGUCCCAGCCUCCUCGCUGGCGCCGGCUGGGCAACAUGCUUCGGCAUGGCGAAUGGGAC:(配列番号457)を使用して終端されたことに留意されたい。
GGCCGGCAUGGUCCCAGCCUCCUCGCUGGCGCCGGCUGGGCAACAUGCUUCGGCAUGGCGAAUGGGAC:(配列番号457)を使用して終端されたことに留意されたい。
(3)pegRNAの鋳型領域への修飾によるRT加工性の改善
pegRNAによって鋳型される挿入のサイズが増加するにつれて、それはエンドヌクレアーゼによって分解され、自発的加水分解を受け、又はRTによって逆転写され得ないか若しくはpegRNA骨格の折り畳み及びその後のCas9-RT結合を破壊する二次構造に折り畳まれる可能性がより高い。したがって、遺伝子全体の挿入等の大きな挿入に影響を及ぼすために、pegRNAの鋳型の修飾が必要である可能性がある。そうするためのいくつかの戦略は、RNAを分解又は加水分解に対してより耐性にするか、又は阻害性二次構造を採用する可能性を低くする、合成又は半合成pegRNA内への修飾ヌクレオチドの組み込みを包含する196。そのような修飾は、Gリッチ配列中のRNA二次構造を減少させる8-アザ-7-デアザグアノシン;分解を低減し、ある種のRNA二次構造を増強するロックド核酸(LNA);RNA安定性を高める2'-O-メチル、2'-フルオロ又は2'-O-メトキシエトキシ修飾を包含し得た。このような修飾はまた、安定性及び活性を高めるために、pegRNAの他の場所に包含され得た。代替的又は追加的に、pegRNAの鋳型は、それが所望のタンパク質産物をコードし、RTによって展開されることができる単純な二次構造を採用する可能性も高くなるようにデザインされ得る。そのような単純な構造は熱力学的シンクとして作用し、逆転写を妨げるより複雑な構造が生じる可能性を低くする。最後に、鋳型を2つの別個のpegRNAに分裂することも考えられ得た。そのようなデザインでは、PEが使用されて転写を開始し、Cas9に融合したRNA結合タンパク質又はMS2アプタマー等のpegRNA自体のRNA認識エレメントを介して別個の鋳型RNAを標的部位に動員する。RTは、この別個の鋳型RNAに直接結合するか、又は第2の鋳型に交換する前に元のpegRNA上で逆転写を開始し得た。そのようなアプローチは、長い鋳型の添加時のpegRNAのミスフォールディングを防止すること、及び、おそらくPEに基づく長い挿入を阻害し得る長い挿入が起こるためにゲノムからのCas9の解離を必要としないことの両方によって、長い挿入を可能にし得た。
pegRNAによって鋳型される挿入のサイズが増加するにつれて、それはエンドヌクレアーゼによって分解され、自発的加水分解を受け、又はRTによって逆転写され得ないか若しくはpegRNA骨格の折り畳み及びその後のCas9-RT結合を破壊する二次構造に折り畳まれる可能性がより高い。したがって、遺伝子全体の挿入等の大きな挿入に影響を及ぼすために、pegRNAの鋳型の修飾が必要である可能性がある。そうするためのいくつかの戦略は、RNAを分解又は加水分解に対してより耐性にするか、又は阻害性二次構造を採用する可能性を低くする、合成又は半合成pegRNA内への修飾ヌクレオチドの組み込みを包含する196。そのような修飾は、Gリッチ配列中のRNA二次構造を減少させる8-アザ-7-デアザグアノシン;分解を低減し、ある種のRNA二次構造を増強するロックド核酸(LNA);RNA安定性を高める2'-O-メチル、2'-フルオロ又は2'-O-メトキシエトキシ修飾を包含し得た。このような修飾はまた、安定性及び活性を高めるために、pegRNAの他の場所に包含され得た。代替的又は追加的に、pegRNAの鋳型は、それが所望のタンパク質産物をコードし、RTによって展開されることができる単純な二次構造を採用する可能性も高くなるようにデザインされ得る。そのような単純な構造は熱力学的シンクとして作用し、逆転写を妨げるより複雑な構造が生じる可能性を低くする。最後に、鋳型を2つの別個のpegRNAに分裂することも考えられ得た。そのようなデザインでは、PEが使用されて転写を開始し、Cas9に融合したRNA結合タンパク質又はMS2アプタマー等のpegRNA自体のRNA認識エレメントを介して別個の鋳型RNAを標的部位に動員する。RTは、この別個の鋳型RNAに直接結合するか、又は第2の鋳型に交換する前に元のpegRNA上で逆転写を開始し得た。そのようなアプローチは、長い鋳型の添加時のpegRNAのミスフォールディングを防止すること、及び、おそらくPEに基づく長い挿入を阻害し得る長い挿入が起こるためにゲノムからのCas9の解離を必要としないことの両方によって、長い挿入を可能にし得た。
(4)5'末端又は3'末端における更なるRNAモチーフの組み入れ
pegRNAデザインは、RNAの末端のいずれかの末端に追加のモチーフを組み入れすることによっても改善され得た。KSHV由来のPAN ENE及びMALAT1由来のENE等のいくつかのこのようなモチーフは、非pol IIIプロモータ由来のより長いpegRNAの発現を終結させる可能性のある手段として、部分(i)184,185で前述された。これらの要素は、ポリAテールを包み込むRNA三重らせんを形成し、その結果、それらは核内に保持される184,187。しかしながら、末端ヌクレオチドを閉塞するpegRNAの3'末端に複雑な構造を形成することによって、これらの構造は、おそらく、pegRNAのエキソヌクレアーゼ媒介性分解の防止にも役立つだろう。非pol IIIプロモータからの終結を可能にすることなく、3'末端に挿入された他の構造要素もRNA安定性を高め得た。そのようなモチーフは、3'末端を閉塞するヘアピン若しくはRNA四重鎖197、又は3'末端に2'-3'-環状リン酸の形成をもたらし、またpegRNAをエキソヌクレアーゼによって分解されにくくする可能性があるHDV等の自己切断リボザイムを包含し得た。198不完全なスプライシングを介して環状化するように(ciRNAを形成するように)pegRNAを誘導することはまた、pegRNAの安定性を高め、pegRNAが核内に保持されるようにすることもできる194。
pegRNAデザインは、RNAの末端のいずれかの末端に追加のモチーフを組み入れすることによっても改善され得た。KSHV由来のPAN ENE及びMALAT1由来のENE等のいくつかのこのようなモチーフは、非pol IIIプロモータ由来のより長いpegRNAの発現を終結させる可能性のある手段として、部分(i)184,185で前述された。これらの要素は、ポリAテールを包み込むRNA三重らせんを形成し、その結果、それらは核内に保持される184,187。しかしながら、末端ヌクレオチドを閉塞するpegRNAの3'末端に複雑な構造を形成することによって、これらの構造は、おそらく、pegRNAのエキソヌクレアーゼ媒介性分解の防止にも役立つだろう。非pol IIIプロモータからの終結を可能にすることなく、3'末端に挿入された他の構造要素もRNA安定性を高め得た。そのようなモチーフは、3'末端を閉塞するヘアピン若しくはRNA四重鎖197、又は3'末端に2'-3'-環状リン酸の形成をもたらし、またpegRNAをエキソヌクレアーゼによって分解されにくくする可能性があるHDV等の自己切断リボザイムを包含し得た。198不完全なスプライシングを介して環状化するように(ciRNAを形成するように)pegRNAを誘導することはまた、pegRNAの安定性を高め、pegRNAが核内に保持されるようにすることもできる194。
更なるRNAモチーフはまた、RTプロセス性を改善するか、又はDNA-RNA二重鎖へのRT結合を増強することによってpegRNA活性を増強し得た。RTによって結合された天然配列をその同族レトロウイルスゲノムに追加すると、RT活性を増強し得た199。これには、天然プライマー結合部位(PBS)、ポリプリントラクト(PPT)、又はレトロウイルスゲノム二量体化及び転写の開始を伴うキスループを包含し得た199。pegRNAの5'及び3'末端に、キスループ又はGNRAテトラループ/テトラループ受容体ペア200等の二量体化モチーフを付加することも、pegRNAの効果的な環状化をもたらし、安定性を改善し得た。更に、これらのモチーフの付加は、pegRNAスペーサ及びプライマーの物理的分離、PE活性を妨げる可能性があるスペーサの閉塞の防止を可能にし得ることが想定される。スペーサ領域に小さな足がかりヘアピンを形成する、pegRNAに対する短い5'伸長もまた、スペーサに結合するpegRNAのアニーリング領域と有利に競合し得た。最後に、また、キスループが使用されて、他の鋳型RNAをゲノム部位に動員し、一方のRNAから他方のRNAへのRT活性の交換を可能にし得た(セクションiii)。例示的なpegRNA構築物は、配列番号251~255によって表される。
(5)pegRNAの進化
SpCas9及び塩基編集因子が改善された方法と類似の様式で、指向性進化によってpegRNA骨格が更に改善され得る可能性がある201。指向性進化は、Cas9又は進化したCas9バリアントによるpegRNA認識を増強し得た。更に、異なるpegRNA骨格配列が異なるゲノム遺伝子座で最適であり、問題の部位でPE活性を増強するか、オフターゲット活性を減少させるか、又はその両方である可能性が高い。最後に、他のRNAモチーフが付加されたpegRNA骨格の進化は、未進化の融合RNAと比較して、融合pegRNAの活性をほぼ確実に改善するであろう。例えば、c-ジ-GMP-Iアプタマー及びハンマーヘッド型リボザイムから構成されるアロステリックリボザイムの進化は、劇的に改善された活性をもたらし202、進化がハンマーヘッド-pegRNA融合の活性も改善することを示唆した。加えて、Cas9は現在、sgRNAの5'伸長を一般に許容しないが、指向性進化は、この不耐性を緩和し、更なるRNAモチーフが利用されることを可能にする可能性のある突然変異を生成する可能性が高い。
SpCas9及び塩基編集因子が改善された方法と類似の様式で、指向性進化によってpegRNA骨格が更に改善され得る可能性がある201。指向性進化は、Cas9又は進化したCas9バリアントによるpegRNA認識を増強し得た。更に、異なるpegRNA骨格配列が異なるゲノム遺伝子座で最適であり、問題の部位でPE活性を増強するか、オフターゲット活性を減少させるか、又はその両方である可能性が高い。最後に、他のRNAモチーフが付加されたpegRNA骨格の進化は、未進化の融合RNAと比較して、融合pegRNAの活性をほぼ確実に改善するであろう。例えば、c-ジ-GMP-Iアプタマー及びハンマーヘッド型リボザイムから構成されるアロステリックリボザイムの進化は、劇的に改善された活性をもたらし202、進化がハンマーヘッド-pegRNA融合の活性も改善することを示唆した。加えて、Cas9は現在、sgRNAの5'伸長を一般に許容しないが、指向性進化は、この不耐性を緩和し、更なるRNAモチーフが利用されることを可能にする可能性のある突然変異を生成する可能性が高い。
本明細書に記載されているように、Cas9:sgRNA複合体とともに使用するための多数のこれらのアプローチはすでに記載されているが、pegRNA活性を改善するためのデザインは報告されていない。プログラム型突然変異をゲノムに組み入れするための他の戦略は、塩基編集、相同組換え(HDR)、精密なマイクロホモロジー媒介末端結合(MMEJ)又はトランスポザーゼ媒介編集を包含する。しかしながら、これらのアプローチは全て、PEと比較した場合に重大な欠点を有する。現在の塩基編集因子は、既存のPEよりも効率的であるが、特定のクラスのゲノム変異のみを組み入れ得、目的の部位で更なる望ましくないヌクレオチド変換をもたらし得る。HDRは、ごく少数の細胞型においてのみ実現可能であり、比較的高い割合のランダムな挿入及び欠失変異(インデル)をもたらす。精密なMMEJは、二本鎖切断の予測可能な修復をもたらし得るが、主に欠失の組み入れに限定され、非常に部位依存性であり、また、比較的高い割合の望ましくないインデルを有し得る。トランスポザーゼ媒介編集は、これまで、細菌において機能することが示されているだけである。したがって、PEに対する改善は、ゲノム変異の幅広い見本の処置的補正のための最良の道を表す可能性がある。
(5)PBS足がかり
PE活性を更に改善するために、本発明者らは、3'伸長アームを有するpegRNAの3'末端にトゥーループ配列を付加することを企図した。図71Aは、3'伸長アーム(上部分子)を有する一般的なSpCas9 pegRNAの一例を提供する。次いで、3'伸長アームは、RT鋳型(所望の編集を包含する)及び分子の3'末端にプライマー結合部位(PBS)を含む。分子は、3つのU核酸塩基(すなわち、5'-UUU-3')を含むポリ(U)配列で終結する。
PE活性を更に改善するために、本発明者らは、3'伸長アームを有するpegRNAの3'末端にトゥーループ配列を付加することを企図した。図71Aは、3'伸長アーム(上部分子)を有する一般的なSpCas9 pegRNAの一例を提供する。次いで、3'伸長アームは、RT鋳型(所望の編集を包含する)及び分子の3'末端にプライマー結合部位(PBS)を含む。分子は、3つのU核酸塩基(すなわち、5'-UUU-3')を含むポリ(U)配列で終結する。
対照的に、図71Aの下部は、図71Aの上部と同じpegRNA分子を示すが、5'-GAAANNNNN-3'の9核酸塩基配列がプライマー結合部位の3'末端と末端ポリ(U)配列の5'末端との間に挿入されている。この構造は180°折り返されて「トゥーループ」RNA構造を形成し、9-核酸塩基挿入の5'-NNNNN-3'の配列がプライマー結合部位の相補的配列とアニーリングし、5'-GAAA-3'部分が180°ターンを形成する。図71Aに示すトゥーループ配列の特徴は、その場所で使用され得た可能なトゥーループの範囲を限定又は狭めることを意図されていない。更に、トゥーループの配列は、プライマー結合部位の相補的配列に依存する。しかし、本質的に、トゥーループ配列は、様々な態様において、180°を形成する第1の配列部分と、プライマー結合部位の一部に相補的な配列を有する第2の配列部分とを有してもよい。
理論に束縛されるものではないが、トゥーループ配列は、そうでなければ可能であるよりも長いプライマー結合部位を有するpegRNAの使用を可能にすると考えられる。更に、より長いPBS配列は、PE活性を改善すると考えられる。より具体的には、トゥーループの機能は、PBSがスペーサと相互作用するのを閉塞するか、又は少なくとも最小限に抑えることである。PBSとスペーサとの間の安定なヘアピン形成は、不活性なpegRNAをもたらし得る。トゥーループなしでは、この相互作用は、PBSの長さを制限することを必要としてもよい。3'末端トゥーループを使用してスペーサとPBSとの間の相互作用を遮断又は最小化することは、PE活性の改善につながってもよい。
(6)非pol IIIプロモータからのpegRNAの発現
種々のpegRNA発現系を、読み取りとしてFKBPからの102ヌクレオチド配列の挿入を使用して、pegRNAを生成する能力について試験された。
種々のpegRNA発現系を、読み取りとしてFKBPからの102ヌクレオチド配列の挿入を使用して、pegRNAを生成する能力について試験された。
pegRNAの転写は、U6プロモータ等の典型的な構成的プロモータによって指示され得る。U6プロモータは、ほとんどの場合、pegRNAの転写を指令するのに有効であるが、U6プロモータは、より長いpegRNA又はUリッチRNAの転写を指令するのにあまり有効ではない。UリッチRNAストレッチは、転写の早期終結を引き起こす。この実施例は、CMVプロモータ又はU 1プロモータから発現されたガイドの編集結果をU6プロモータと比較された。これらのプロモータは、以下に提供されるように、MASC ENE又はPAN ENE等の異なるターミネータ配列を必要とする。pCMV/MASC-ENEシステムでは編集の増加が観察されたが、これらのガイドは配列の不完全な挿入をもたらしたが、U6プロモータではより低いレベルの編集で完全な挿入が観察された。図81を見よ。データは、代替発現系が長時間の挿入に有用であってもよい可能性を示唆している。
pCMV/MASC-ENE発現系のヌクレオチド配列は以下の通りである(5'から3'方向)(参照する領域の直前に太字のモチーフの名称を有する):
[pCMVプロモータ]-pol II RNAポリメラーゼに結合する
[Csy4ループ]-Csy4タンパク質によって結合されると、ループの3'側の切断をもたらす。[CMVプロモータ]の一部が転写され、この配列がgRNAの5'に取り付けられる場合、活性を低下させる/排除するので必要である(既知である)。
pegRNAの[スペーサ配列]
[pegRNA骨格]
[DNA合成鋳型]
[挿入編集(FKBPから108nt)]
[プライマー結合部位]
[リンカー](可変性が高い)-PBSとターミネータ要素とを接続する
[MASC ENE転写ターミネータ]-このエレメントの転写は転写の終結をもたらす;ポリAテールがエンコードされ、次いでENE要素によって隔離される
[非重要配列]
[Ubcプロモータ]-Csy4タンパク質の発現に必要
[Csy4タンパク質及びNLS]-ガイドの5'末端のプロセシングに必要である。大きなタンパク質の発現(スペーサのリボザイム媒介切断等)を必要としない他の戦略も使用されることができるが、これらは異なるスペーサ配列に対してより個別の調整を必要とするので、これを使用した。
[SV40ターミネータ]-Csy4タンパク質の終結のため。
(7)追加のRNAモチーフ
pegRNAのHDVリボザイム3'又はG-四重鎖挿入、P1伸長、鋳型ヘアピン、及びテトラループcirc'd等、その性能を改善するためにpegRNAに導入されてもよい特定のモチーフの詳細については図82を参照されたい。
pegRNAのHDVリボザイム3'又はG-四重鎖挿入、P1伸長、鋳型ヘアピン、及びテトラループcirc'd等、その性能を改善するためにpegRNAに導入されてもよい特定のモチーフの詳細については図82を参照されたい。
特に、この実施例では、プライマー結合部位のtRNAモチーフ3'を組み入れする効果を試験した。この要素は、複数の潜在的な機能のために選択された。
(1)tRNAモチーフは非常に安定なRNAモチーフであるため、pegRNA分解を潜在的に減少させ得た。
(2)MMLV RTは、転写中にウイルスゲノムをDNAに変換するときにプロリル-tRNAをプライマーとして使用するので、同じキャップがRTによって結合され、PEによるpegRNAの結合、RNA安定性を改善し、PBSをゲノム部位に近接して戻すことができ、潜在的に活性も改善すると疑われた。
これらの構築物では、tRNAのP1(図84参照)が伸長された。P1は、tRNAの第1のステム/塩基対合エレメントを指す(図84参照)。これは、P1のtRNA 5'のRNAseP媒介性切断を防止するために必要であると考えられ、その結果、pegRNAからのその除去をもたらすであろう。
このデザインでは、tRNAとPBSとの間に伸長したP1及び短い3ntリンカーを有するプロリル-tRNA(コドンCGG)が使用された。様々なtRNAデザインが試験され、tRNAキャップを有さないpegRNAと比較して編集効率が試験された-図83(HEK3遺伝子の編集を標的とし、具体的にはニック部位に対して+1の位置での10nt挿入の挿入を標的とし、PE3を使用したPE実験を示す)、図85(FANCF遺伝子の編集を標的とし、具体的にはニック部位に対して+5位でのGからTへの変換を標的とし、PE3構築物を使用するPE実験を示す)及び図86(HEK3遺伝子の編集を標的とし、具体的にはニック部位に対して+1位の位置での71nt FLAGタグ挿入の挿入を標的とし、PE3構築物を使用したPE実験を示す)の比較データを参照のこと。tRNA修飾pegRNAは非修飾pegRNA対照に対して試験された。
UGG/CGGは使用されるコドンを指し、数字は付加されたP1伸長の長さを指し、longは8ntリンカーを示し、3ntリンカーの指定はない。
このデータは、tRNAの組み入れがより短いPBSの使用を可能にしてもよいことを示唆しており、これは更なる活性改善をもたらす可能性がある。RNF2の場合、使用されるリンカーは、スペーサへのPBS結合を改善し、結果として活性を低下させる見込み/可能性が高い。
使用したいくつかの配列:
HEK3+1 FLAGタグ挿入、proly-tRNA{UGG}P1 ext 5nt、リンカー3nt
GGCCCAGACUGAGCACGUGAGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGCUGGAGGAAGCAGGGCUUCCUUUCCUCUGCCAUCACUUAUCGUCGUCAUCCUUGUAAUCCGUGCUCAGUCUGUCUGGCGGGGCUCGUUGGUCUAGGGGUAUGAUUCUCGCUUCGGGUGCGAGAGGUCCCGGGUUCAAAUCCCGGACGAGCCCCGCCUUUU(配列番号459)
FANCF+5 GからT proly-tRNA{CGG}P1 ext 5nt、リンカー3nt
GGAAUCCCUUCUGCAGCACCGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGCGGAAAAGCGAUCAAGGUGCUGCAGAAGGGAUCUGGCGGGGCUCGUUGGUCUAGGGGUAUGAUUCUCGCUUCGGGUGCGAGAGGUCCCGGGUUCAAAUCCCGGACGAGCCCCGCCUUUU(配列番号460)
HEK3++1 10nt挿入、proly-tRNA{UGG}P1 ext 5nt、リンカー3nt
GGCCCAGACUGAGCACGUGAGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGGACCGAGUCGGUCCUCUGCCAUCAAAGCUUCGACCGUGCUCAGUCUUCUGCUCGAGGCGGGGCUCGUUGGUCUAGGGGUAUGAUUCUCGCUUCGGGUGCGAGAGGUCCCGGGUUCAAAUCCCGGACGAGCCCCGCCUCGAGCUUUU(配列番号461)
図85及び図86のデータで報告された配列は以下の通りである。


実施例3:プライム編集効率を改善するための次世代PEGRNA修飾
背景
プライム編集因子複合体は2つの構成要素からなる。第1に、プライム編集因子(PE)自体は、一態様では、逆転写酵素等のポリメラーゼに融合され、HNHヌクレアーゼドメインを不活性化する突然変異を有する、Streptococcus pyogenes Cas9(SpCas9)等のプログラム型ヌクレアーゼである。第2の構成要素は、編集因子をプログラムされたゲノム部位に標的化し、プログラムされた編集を組み入れするために逆転写酵素によって使用される鋳型を含有するpegRNAである。実質的に任意のプログラム型編集を作成する能力にもかかわらず、PEは一般に、同等の編集について塩基エディタ(BE)よりも低い活性を有する。pegRNAの合理的な操作はまた、編集結果の改善をもたらし、細胞のゲノム、例として非HEK293T細胞株におけるより多様なプライム編集を可能にし得ると考えられる。
背景
プライム編集因子複合体は2つの構成要素からなる。第1に、プライム編集因子(PE)自体は、一態様では、逆転写酵素等のポリメラーゼに融合され、HNHヌクレアーゼドメインを不活性化する突然変異を有する、Streptococcus pyogenes Cas9(SpCas9)等のプログラム型ヌクレアーゼである。第2の構成要素は、編集因子をプログラムされたゲノム部位に標的化し、プログラムされた編集を組み入れするために逆転写酵素によって使用される鋳型を含有するpegRNAである。実質的に任意のプログラム型編集を作成する能力にもかかわらず、PEは一般に、同等の編集について塩基エディタ(BE)よりも低い活性を有する。pegRNAの合理的な操作はまた、編集結果の改善をもたらし、細胞のゲノム、例として非HEK293T細胞株におけるより多様なプライム編集を可能にし得ると考えられる。
pegRNAは、標準的な単一ガイドRNA(sgRNA)と比較して、Cas9親和性の低下及び安定性の低下の両方を被ると考えられる。このCas9親和性の低下は、3'伸長だけでなく、Cas9結合を阻害するスペーサとプライマー結合部位(PBS)との間のRNA二重鎖の形成にも起因する可能性が高い。実際、より長いPBS長は、おそらくこの機構を介して、試験した全ての部位でPE活性を完全に排除する。更に、SpCas9ヌクレアーゼによるpegRNAのトランスフェクションは、同じ部位を標的とするsgRNAよりも少ないインデルをもたらし、3'伸長がCas9結合及び潜在的に触媒活性を低下させることを更に示唆する。Cas9によって結合及び保護されていない3'伸長は、エキソヌクレアーゼによって潜在的に分解され得るか、又はCas9若しくはRT結合と競合し得る他の細胞因子によって結合され得るか、又はさもなければプライム編集を阻害し得る可能性もあるようであった。実際、RT-qPCRによってpegRNAの細胞寿命を調べると、骨格領域と比較して3'伸長に対する安定性の有意な減少が観察された(図90A~図90C)。
既存のsgRNA改善戦略
ヒト細胞における編集を改善するsgRNAに対する多数の修飾が報告されている。最も一般的なそのような修飾の中には、sgRNA骨格に対する「フリップ」及び「伸長」突然変異がある。足場の直接反復配列(DR)中の4個のウリジン(U)ヌクレオチド伸長が可能なポリメラーゼIII(pol III)終結配列であり、A・U塩基対に対する末端ウリジン・アデノシン(A)塩基対のフリップがsgRNAの発現及び活性の増加をもたらすことは以前に指摘されている。同様に、DRの伸長は、おそらくsgRNAのCas9結合コンピテント構造の安定化を介して、sgRNA活性を改善することも示されている。そのような修飾は、sgRNA活性と同様に、pegRNA活性を増加させ得ることが見出されている(図91A~図91D)。これはおそらく、HEK293T細胞におけるpegRNAをコードするプラスミドの効率的なトランスフェクションによるものであり、これらの修飾は他の細胞型における活性を広く改善することが予想される。別の修飾は、足がかりステムの組み込みを介して、スペーサとPBSとの間の相互作用を減少させることが求められた。
ヒト細胞における編集を改善するsgRNAに対する多数の修飾が報告されている。最も一般的なそのような修飾の中には、sgRNA骨格に対する「フリップ」及び「伸長」突然変異がある。足場の直接反復配列(DR)中の4個のウリジン(U)ヌクレオチド伸長が可能なポリメラーゼIII(pol III)終結配列であり、A・U塩基対に対する末端ウリジン・アデノシン(A)塩基対のフリップがsgRNAの発現及び活性の増加をもたらすことは以前に指摘されている。同様に、DRの伸長は、おそらくsgRNAのCas9結合コンピテント構造の安定化を介して、sgRNA活性を改善することも示されている。そのような修飾は、sgRNA活性と同様に、pegRNA活性を増加させ得ることが見出されている(図91A~図91D)。これはおそらく、HEK293T細胞におけるpegRNAをコードするプラスミドの効率的なトランスフェクションによるものであり、これらの修飾は他の細胞型における活性を広く改善することが予想される。別の修飾は、足がかりステムの組み込みを介して、スペーサとPBSとの間の相互作用を減少させることが求められた。
次世代sgRNA改善戦略
上記の知見を考慮して、3'伸長の安定性を改善する戦略に焦点を当てることが決定された。PBSの分解は、いかなる分解によってもpegRNAをRTによって結合できなくするが、それでもなおCas9による結合を可能にし得るので、pegRNAにとって特に有害である。したがって、分解されたpegRNAは、Cas9及び標的部位への結合の両方について競合し得、更にその部位でのニッキングを可能にし、編集を減少させ、インデル形成を増加させる可能性がある。
上記の知見を考慮して、3'伸長の安定性を改善する戦略に焦点を当てることが決定された。PBSの分解は、いかなる分解によってもpegRNAをRTによって結合できなくするが、それでもなおCas9による結合を可能にし得るので、pegRNAにとって特に有害である。したがって、分解されたpegRNAは、Cas9及び標的部位への結合の両方について競合し得、更にその部位でのニッキングを可能にし、編集を減少させ、インデル形成を増加させる可能性がある。
したがって、PBSの構造モチーフ3'の取り込みは、sgRNAに付加されたRNA G-四重鎖について報告されているように、改善された安定性をもたらし得ることが発見された。しかし、プリンに富む配列はpegRNAのミスフォールディングを引き起こす可能性があるので、追加の構造モチーフをスクリーニングすることも決定された。したがって、短い構造化されていないヌクレオチドリンカーによってPBSに付加されたいくつかの他の構造モチーフがスクリーニングされた。
第1に、より安定になるように進化したプレケオシン1-1リボスイッチアプタマー(最小の天然の三次RNA構造の1つ)は、以後evopreQ1-1と呼ばれる。第2に、MMLV RTと相互作用する可能性があり、それによってPEに対する改善された安定性及び親和性の両方をもたらし得る2つの構造モチーフ、すなわち、MMLVウイルスゲノム由来のシュードノット(ここではMpknot-1と呼ばれる)及び逆転写のためのプライマーとしてMMLV RTによって使用される修飾tRNAが選択された。
試験アッセイは、様々なゲノム遺伝子座に組み入れされるFLAGタグ挿入配列-困難な編集-をコードするように構成されたpegRNAガイドのスクリーニングを伴った(図92A~図92C)。興味深いことに、編集活性の有意な増加が、HEK293T細胞において試験された全ての部位において、短い(G 2)四重鎖、同様にまた、evopreQ1-1及びMpknot-1の両方について観察され、このことは、これらのモチーフが様々なゲノム遺伝子座における活性を改善することを示唆した。
構造モチーフをpegRNAの3'末端に付加することは、より長い伸長を有するpegRNAに対する活性のみを改善し得ると考えられた。これが該当するかどうかを決定するために、点突然変異又は欠失のいずれかをコードし、6つの更なるゲノム部位に長さが増加する鋳型を含有するpegRNAの小さなライブラリーがスクリーニングされた。試験された実質的に全てのガイド(図93A~図93H)で大幅に改善された編集が観察され、部位、編集タイプ又は鋳型の長さに関係なく、1.5~6倍の範囲の改善があり、それらの一般的な有用性を示唆している。興味深いことに、構造モチーフの組み込みは、リンカー単独の添加と比較して改善された編集をもたらしたが、リンカーの添加は、親pegRNAと比較して改善された編集活性をしばしばもたらした(図94)。これらの構造タグ付きpegRNAが他の細胞株において改善された編集をもたらすかどうかを決定するために、K562、U2OS及びHeLa細胞におけるHEK3遺伝子座にFLAGタグを組み入れする修飾pegRNAの能力が試験された。PBSのevopreQ1-1又はMpknot-1シュードノット3'のいずれかを付加すると、これらの細胞で編集効力の大きな改善が観察された(図95 A-B)。
初期デザインを改善するために、これらのモチーフがどのように活性を改善しているかを理解することが求められた。それらは改善された細胞寿命を介して機能する可能性があるように思われたが、短い非構造化リンカーの添加は、親pegRNAと比較してpegRNA活性を改善するのに十分な場合があることが観察された(図94)。同時に、モチーフ構造を破壊すると予測される突然変異は、編集の減少(図96)evoPreQ 1(mut1)及びevoPreQ 1(mut2)をもたらし、モチーフの構造が活性にとって重要であることを示唆した。これは、これらのモチーフがPE効率を増加させた複数の可能な機構があり得ることを示唆した。
第1の工程として、修飾されたpegRNAがpegRNAの細胞寿命を改善することを確認することが求められた。これまで、RT-qPCRが使用されて、pegRNA骨格及び鋳型の相対量を測定し、構造化モチーフをpegRNAの3'テールに付加すると、鋳型の量が有意に改善されることが見出された(図97)。これらのpegRNAのPBS長を長くすることができ、編集活性を更に向上させることができると考えられる。
これらの次世代のpegRNAのデザインが更に改善され得ると考えられる。そうするために、多数の更なる3'モチーフがスクリーニングされる。これらは、更なる進化したpreq1-アプタマー、Mpknot-1に対する修飾、安定性が改善された更なる天然G-四重鎖、グループIイントロンのP 4-P 6ドメイン、及び自己切断性HDVリボザイムを包含する。このリボザイムは、それ自体のすぐ5'側でRNAプロセシングをもたらし、エキソヌクレアーゼに耐性であるRNAの3'末端に2'-3'-環状リン酸を残す。更に、HEK細胞及び他の細胞型における活性を改善するかどうかを調べるために、Cas9ヌクレアーゼ切断の編集効力を増加させることが報告されている標準的なsgRNA骨格に対する変異が試験される。
これらの研究は、8ヌクレオチド(nt)のリンカー長を伴っているが、例えば、4、5、6、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39又は40以上の使用され得るヌクレオチドを包含する、他のリンカー長が可能である。いくつかの場合、リンカーの長さ及び配列は、各部位について経験的に決定されなければならないであろう。他の場合には、pegRNA配列非依存性である単一のリンカーが使用されてもよい。このプロセスを支援するために、pegRNA構造に干渉しないリンカー配列をデザインするために、計算スクリプトが使用されてもよい。
追加のデザイン
最後の工程として、pegRNAの更なるデザインが求められた。以下を包含する、pegRNA構造のいくつかの側面が検討された:pegRNAを発現するために使用されるpol IIIプロモータ、pegRNA骨格、及び反対の鎖をニッキングすることによって編集効力を増強するためにPE3で使用されるニッキングガイド。様々なpol IIIプロモータが、ヒト細胞において低分子RNAを発現させるために使用されている。歴史的に、2つの-u6及びh1-がプレ-マイクロRNAの発現に使用されてきた。これら2つのうち、u6はsgRNA発現に関して優れていることが見出された。しかしながら、他のプロモータは、pegRNAの発現の改善をもたらし得る。これが当てはまるかどうかを決定するために、HEK293T細胞における編集有効性について、h1及びu6の他のホモログを包含する多数のpol IIIプロモータがスクリーニングされた。u6-9と呼ばれるu6の1つのホモログを包含するいくつかのプロモータは、編集効力を劇的に改善することが見出された(図100A~図100Eを参照されたい)。これらのプロモータは、以下のように同定される。
最後の工程として、pegRNAの更なるデザインが求められた。以下を包含する、pegRNA構造のいくつかの側面が検討された:pegRNAを発現するために使用されるpol IIIプロモータ、pegRNA骨格、及び反対の鎖をニッキングすることによって編集効力を増強するためにPE3で使用されるニッキングガイド。様々なpol IIIプロモータが、ヒト細胞において低分子RNAを発現させるために使用されている。歴史的に、2つの-u6及びh1-がプレ-マイクロRNAの発現に使用されてきた。これら2つのうち、u6はsgRNA発現に関して優れていることが見出された。しかしながら、他のプロモータは、pegRNAの発現の改善をもたらし得る。これが当てはまるかどうかを決定するために、HEK293T細胞における編集有効性について、h1及びu6の他のホモログを包含する多数のpol IIIプロモータがスクリーニングされた。u6-9と呼ばれるu6の1つのホモログを包含するいくつかのプロモータは、編集効力を劇的に改善することが見出された(図100A~図100Eを参照されたい)。これらのプロモータは、以下のように同定される。
U6プロモータの非限定的な例は、配列番号237~240によって表されるものを包含する。
結論
一般的にPE活性を改善することとは別に、修飾されたpegRNAは、pegRNAをデザインするプロセスを単純化し得た。現在、最適なpegRNAのデザインは、多くの場合、10秒~100秒のpegRNA構築物のスクリーニングを必要とし得る。そのような試験は、特に時間がかかり、高価であり、pegRNAのライブラリーを構築する場合には実行不可能である。そのようなライブラリーの1つの潜在的な用途は、所与のセット内の全てのタンパク質の体系的なタグ付けである。本明細書に記載される修飾されたpegRNAの重要な利点は、HEK3での編集について見られるように、それらが不十分な鋳型選択の悪影響を制限することによってpegRNAデザインを単純化することである(図93B;図93E)。更に、3'モチーフがPBSをその最大可能長さ(17)まで延長することを可能にする場合、これはpegRNAデザインを大幅に単純化するはずである。
一般的にPE活性を改善することとは別に、修飾されたpegRNAは、pegRNAをデザインするプロセスを単純化し得た。現在、最適なpegRNAのデザインは、多くの場合、10秒~100秒のpegRNA構築物のスクリーニングを必要とし得る。そのような試験は、特に時間がかかり、高価であり、pegRNAのライブラリーを構築する場合には実行不可能である。そのようなライブラリーの1つの潜在的な用途は、所与のセット内の全てのタンパク質の体系的なタグ付けである。本明細書に記載される修飾されたpegRNAの重要な利点は、HEK3での編集について見られるように、それらが不十分な鋳型選択の悪影響を制限することによってpegRNAデザインを単純化することである(図93B;図93E)。更に、3'モチーフがPBSをその最大可能長さ(17)まで延長することを可能にする場合、これはpegRNAデザインを大幅に単純化するはずである。
結論として、改善された編集活性を有する修飾されたpegRNAのデザインが検証された。これらのpegRNAは、PBSの構造化RNA 3'を含有し、それらの改善された活性は、改善された細胞寿命及びCas9結合活性に由来される。これらの修飾は、多種多様なゲノム遺伝子座、コードされた編集、及び細胞型におけるPE活性を広く改善する。
実施例4:プライム編集効率を改善する操作されたpegRNA
生命系のゲノムに標的化された変化を加える能力は、生命科学及び医学を進歩させ続けている。ZFN、TALEN、又はCRISPR-Casヌクレアーゼ等のプログラム型ヌクレアーゼを使用する二本鎖切断(DSB)媒介DNA編集戦略は、標的部位での挿入又は欠失(インデル)を誘導することによって遺伝子を効率的に破壊し得るが、DSBはまた、編集結果1、2、より大きなDNA再編成3~5、p 53活性化6~8、及びクロモトリプシン9、10の制御されていない混合物を包含する、しばしば望ましくない結果をもたらす。標的化DSBは相同指向修復を介して正確な遺伝子補正を刺激し得るが、このプロセスはほとんどの処置的に関連する細胞型では非効率的である11。対照的に、塩基エディタ12、13及びプライム編集因子14は、DSBを必要とせずに、処置的に関連する細胞に正確な変化を効率的に組み入れし得る。シトシン及びアデノシン塩基編集因子は、夫々C・GからT・A及びA・TからG・Cへの変換を可能にするが、プライム編集因子は、標的DNA部位での最大数十塩基対の置換、挿入及び/又は欠失を包含する実質的に任意の局所突然変異の組み入れを可能にする。
生命系のゲノムに標的化された変化を加える能力は、生命科学及び医学を進歩させ続けている。ZFN、TALEN、又はCRISPR-Casヌクレアーゼ等のプログラム型ヌクレアーゼを使用する二本鎖切断(DSB)媒介DNA編集戦略は、標的部位での挿入又は欠失(インデル)を誘導することによって遺伝子を効率的に破壊し得るが、DSBはまた、編集結果1、2、より大きなDNA再編成3~5、p 53活性化6~8、及びクロモトリプシン9、10の制御されていない混合物を包含する、しばしば望ましくない結果をもたらす。標的化DSBは相同指向修復を介して正確な遺伝子補正を刺激し得るが、このプロセスはほとんどの処置的に関連する細胞型では非効率的である11。対照的に、塩基エディタ12、13及びプライム編集因子14は、DSBを必要とせずに、処置的に関連する細胞に正確な変化を効率的に組み入れし得る。シトシン及びアデノシン塩基編集因子は、夫々C・GからT・A及びA・TからG・Cへの変換を可能にするが、プライム編集因子は、標的DNA部位での最大数十塩基対の置換、挿入及び/又は欠失を包含する実質的に任意の局所突然変異の組み入れを可能にする。
プライム編集(PE)系は、2つの構成要素:操作された逆転写酵素(RT)に融合したプログラム型DNAニッカーゼを含有するタンパク質、及びプライム編集ガイドRNA、又はpegRNA(図104A)14から最小限で構成される。pegRNAは、標的部位を指定するスペーサ、sgRNA骨格、及び所望の編集をコードする3'伸長を含有する。この伸長部は、DNAプロトスペーサの一部に相補的なプライマー結合部位(PBS)と、所望の編集及び下流ゲノム配列をコードするRT鋳型とを含有する。PEリボ核タンパク質(RNP)が標的部位に結合し、PAM含有DNA鎖にニックが入った後、得られたニックを入れられたDNA鎖は、pegRNA中のPBSと塩基対を形成し、RT鋳型の逆転写を標的DNA部位に直接プライミングする14。次いで、編集されたDNAの新たに合成された3'フラップは、細胞DNA修復経路によって分離され、標的部位に所望の編集が組み入れされる。
プライム編集の汎用性は、多種多様な編集された配列をコードするpegRNAの3'伸長の能力から生じる。その汎用性にもかかわらず、現在のプライム編集因子の効率は、標的部位及び細胞型の間で実質的に異なる14。この実施例では、pegRNAの3'伸長の推定上の分解がプライム編集効率を損ない得ることが記載される。得られた切断型pegRNAは標的部位の関与について競合するが、プライム編集には不適である。この脆弱性に対処するために、pegRNAの完全性を保護し、プライム編集効率を広く改善するRNAモチーフが、複数の細胞株における様々な標的部位において、複数の送達様式を介して同定された。得られた操作されたpegRNA(epegRNA)は、プライム編集の有効性及び適用範囲を実質的に進歩させた。
結果
RNA安定性はpegRNAの有効性を制限する
保護されていない核RNAは、エキソヌクレアーゼによる5'及び3'末端の両方からの分解を受けやすい15。ガイドRNA全体が関連するCas9タンパク質16によって保護されているsgRNAとは対照的に、pegRNAの3'伸長は細胞内に露出される可能性が高く、したがってエキソヌクレアーゼ分解を受けやすい。部分的に分解されたpegRNAは、Cas9に結合し、標的DNA部位に係合する能力を保持し得るが、PBSの喪失又は切断は、所望の編集を組み入れする能力を妨げ、それによってPEタンパク質及び標的部位をプライム編集を媒介することができないガイドRNAで占有する。
RNA安定性はpegRNAの有効性を制限する
保護されていない核RNAは、エキソヌクレアーゼによる5'及び3'末端の両方からの分解を受けやすい15。ガイドRNA全体が関連するCas9タンパク質16によって保護されているsgRNAとは対照的に、pegRNAの3'伸長は細胞内に露出される可能性が高く、したがってエキソヌクレアーゼ分解を受けやすい。部分的に分解されたpegRNAは、Cas9に結合し、標的DNA部位に係合する能力を保持し得るが、PBSの喪失又は切断は、所望の編集を組み入れする能力を妨げ、それによってPEタンパク質及び標的部位をプライム編集を媒介することができないガイドRNAで占有する。
この可能性を実証するために、HEK293T細胞は、T・AからA・TトランスバーションをコードするRT鋳型を含有する完全長pegRNA、又はT・AからG・Cトランスバーションをコードするが3'末端にPBSを欠くRT鋳型を含有する切断型pegRNAのいずれかを生成する様々な比の2つのプラスミドの混合物でトランスフェクトされた。2つのpegRNAは、ヒト細胞において同じ又は異なるゲノム遺伝子座のいずれかを標的とした。SpCas9 pegRNAをコードするプラスミドと転写について競合するはずであるが、プライム編集因子タンパク質とは相互作用しない、相互作用しないSaCas9 pegRNAを生成するプラスミドを添加する効果も試験された。切断型pegRNAの産生の増加は、全長及び切断型pegRNAが同じ部位に標的化された場合にPE活性を阻害した(図104B)。対照的に、異なるゲノム部位を標的とする切断型pegRNAも非標的SpCas9 sgRNAも、SaCas9 pegRNAよりもPE活性を妨げなかった(図104B)。これらのデータは、切断型3'伸長を有する分解されたpegRNAが、編集能力のないプライム編集因子リボヌクレオタンパク質(RNP)が標的ゲノム遺伝子座について競合することを可能にすることによってPE活性を阻害することを示唆する。
プライム編集効率を改善する操作されたpegRNA(epegRNA)のデザイン
切断型pegRNAをプライム編集の強力な阻害剤として同定したので、次に、pegRNA分解を最小限に抑えることが求められた。pegRNAの3'末端の構造化RNAモチーフは、5'又は3'末端のRNA構造がヒト細胞及び酵母のmRNA安定性を増強する能力と一致して、pegRNA安定性を改善し得ることが想定された17、18。例えば、長鎖非コードRNA MALAT 1は、そのポリ(A)尾部を捕捉する三重らせんによって安定化され、分解と核外輸送の両方を制限する19。
切断型pegRNAをプライム編集の強力な阻害剤として同定したので、次に、pegRNA分解を最小限に抑えることが求められた。pegRNAの3'末端の構造化RNAモチーフは、5'又は3'末端のRNA構造がヒト細胞及び酵母のmRNA安定性を増強する能力と一致して、pegRNA安定性を改善し得ることが想定された17、18。例えば、長鎖非コードRNA MALAT 1は、そのポリ(A)尾部を捕捉する三重らせんによって安定化され、分解と核外輸送の両方を制限する19。
追加のRNA構造を組み込むことによってプライム編集効率が改善されることができるかどうかが、pegRNAの3'末端にある2つの安定なシュードノットのうちの1つを使用して試験された:修飾されたプレケオシン1-1リボスピッチアプタマー20、21(evopreQ1)、又はモロニーマウス白血病ウイルス(MMLV)22由来のフレームシフトシュードノット(以下mpknotと呼ばれる)(図108)。EvopreQ 1は、定義された三次構造(長さ42ヌクレオチド、nt)を有する最小の天然由来RNA構造モチーフの1つであるので選択した20,21。より小さいモチーフは、pegRNA機能を妨害し得る二次構造の形成を最小限に抑えると推論された。更に、より短いpegRNAは化学合成によってより容易に産生され得る。Mpknotは、その三次構造のために、及びそれが、標準的なプライム編集因子におけるRTが操作されたMMLV RTの内生鋳型であるために選択され、mpknotがRTを動員するのを助ける可能性を高める。
これらのepegRNAが、HEK293T細胞において5つのゲノム遺伝子座でPE3を使用してFLAGエピトープタグ配列を挿入することができるかどうかが試験された(図105A)。プライム編集中にモチーフがpegRNA機能を妨害する可能性を低下させるために、evopreQ1又はmpknotのいずれかをepegRNA PBSの3'末端に接続するための8ntリンカーが包含された。リンカーとPBSとの間、又はリンカーとpegRNAスペーサとの間の潜在的な塩基対形成相互作用を回避するために、ViennaRNA23を使用してリンカー配列がデザインされた14。試験した5つ全てのゲノム部位で、標準的なpegRNAと比較して平均2.1倍のFLAGタグ挿入効率の増加が、epegRNAを使用した場合に観察され、編集:インデル比に明らかな変化はなく(図109A~図109C)、3'末端シュードノットモチーフがPE有効性を改善し得ることを示唆した。
編集効率におけるリンカー配列の役割が、8-ntリンカーを含む又は含まないepegRNAがトランスバーション又はFLAGタグ挿入を媒介する能力を比較することによって特性決定された。PE3編集効率の低下は、mpknotを含有するepegRNAについてのリンカーを除去すると観察されたが(p=0.022)、evopreQ1を含有するepegRNAについては有意差は観察されなかった(図110)。これはおそらく、evopreQ1がmpknotよりも小さく、RTとの立体的衝突が起こりにくいためである。evopreQ1を有するepegRNAについての全体的な平均編集効率は類似していたが(リンカーあり又はなしで)、リンカーを有さないepegRNAについては時折性能低下が認められた(図110)。したがって、その後の全てのepegRNAデザインについて特に明記しない限り、8-ntリンカーが使用された。
PE有効性のこの改善がより長い伸長を有するepegRNAに限定されないことを確実にするために、PE3を使用してHEK293T細胞において7つの異なるゲノム部位で様々なRT鋳型長を有する様々な点突然変異又は欠失をコードする148個の追加のepegRNAが試験された。いずれかのモチーフの使用は、HEK293T細胞における全ての試験部位及びpegRNAにわたって、標準的なpegRNAと比較してプライム編集効率の平均1.5倍の改善をもたらし、編集:インデル比に明らかな変化はなかった(図105B~図105C、図111A~図111K、及び図112A~図112C)。まとめると、これらの結果は、epegRNAがHEK293T細胞におけるPE有効性を広く改善することを確立する。
操作されたpegRNAは、複数の哺乳動物細胞株においてプライム編集を改善する
PE効率が哺乳動物細胞型間で実質的に異なることが以前に観察されており14、様々な細胞において改善されたPE系を試験する必要性を強調した。3'evopreQ1又はmpknotモチーフを含有するepegRNAが、HEK3に24-bpのFLAGエピトープタグを挿入し、DNMT1で15bpを欠失させ、又はK562、U2OS及びHeLa細胞においてPE3を介してRNF2にC・GからA・Tトランスバーションを組み入れする能力が試験された。これらの細胞株の各々において、epegRNAは、pegRNAと比較して編集効率の大きな改善をもたらし、編集:インデル比の減少を伴うことなく、全ての試験された編集(図105D)にわたって、K562細胞において平均2.4倍高い編集、HeLa細胞において3.1倍高い編集、及びU2OS細胞において5.6倍高い編集をもたらした(図109A~図109C)。これらの結果は、複数の哺乳動物細胞株において、epegRNAが使用されてプライム編集を増強し得ることを示している。更に、epegRNAは、HEK293T細胞よりも非HEK293T細胞において編集効率を大幅に改善し(図105Dと比較した、図105A及び図111A~図111K)、元のPE系によってあまり効率的にトランスフェクト又は編集されていない細胞株において特に有益であることを示唆した。
PE効率が哺乳動物細胞型間で実質的に異なることが以前に観察されており14、様々な細胞において改善されたPE系を試験する必要性を強調した。3'evopreQ1又はmpknotモチーフを含有するepegRNAが、HEK3に24-bpのFLAGエピトープタグを挿入し、DNMT1で15bpを欠失させ、又はK562、U2OS及びHeLa細胞においてPE3を介してRNF2にC・GからA・Tトランスバーションを組み入れする能力が試験された。これらの細胞株の各々において、epegRNAは、pegRNAと比較して編集効率の大きな改善をもたらし、編集:インデル比の減少を伴うことなく、全ての試験された編集(図105D)にわたって、K562細胞において平均2.4倍高い編集、HeLa細胞において3.1倍高い編集、及びU2OS細胞において5.6倍高い編集をもたらした(図109A~図109C)。これらの結果は、複数の哺乳動物細胞株において、epegRNAが使用されてプライム編集を増強し得ることを示している。更に、epegRNAは、HEK293T細胞よりも非HEK293T細胞において編集効率を大幅に改善し(図105Dと比較した、図105A及び図111A~図111K)、元のPE系によってあまり効率的にトランスフェクト又は編集されていない細胞株において特に有益であることを示唆した。
オフターゲットプライム編集に対する操作されたpegRNAの効果
プライム編集により、他のCRISPR遺伝子編集戦略よりもオフターゲット編集が実質的に少なくなることが以前に実証された14,24-27。evopreQ1又はmpknotの添加がオフターゲット編集の程度を変化させたかどうかを決定するために、HEK293T細胞が、トランスバーション(HEK3でのT・AからA・T又はEMX1及びFANCFでのG・CからT・A)又はPE3を使用した15bp欠失のいずれかを鋳型とする、HEK3、EMX1又はFANCFを標的とするpegRNA又はepegRNAで処理された。インデル生成の程度、並びに各標的遺伝子座について、上位4つの実験的に確認されたオフターゲット部位28でのプライム編集から合理的に生じ得る任意のヌクレオチド変化が測定され、epegRNAと非修飾pegRNAとの間のオフターゲット編集の程度はPE3での処置後に比較された。全ての場合において、epegRNA及びpegRNAは両方とも、調べた部位において≦0.1%のオフターゲットプライム編集及び/又はインデルを示し(図113)、このことは、epegRNA及びpegRNAが同様のレベルのオフターゲット編集を示すことを示唆していた。
プライム編集により、他のCRISPR遺伝子編集戦略よりもオフターゲット編集が実質的に少なくなることが以前に実証された14,24-27。evopreQ1又はmpknotの添加がオフターゲット編集の程度を変化させたかどうかを決定するために、HEK293T細胞が、トランスバーション(HEK3でのT・AからA・T又はEMX1及びFANCFでのG・CからT・A)又はPE3を使用した15bp欠失のいずれかを鋳型とする、HEK3、EMX1又はFANCFを標的とするpegRNA又はepegRNAで処理された。インデル生成の程度、並びに各標的遺伝子座について、上位4つの実験的に確認されたオフターゲット部位28でのプライム編集から合理的に生じ得る任意のヌクレオチド変化が測定され、epegRNAと非修飾pegRNAとの間のオフターゲット編集の程度はPE3での処置後に比較された。全ての場合において、epegRNA及びpegRNAは両方とも、調べた部位において≦0.1%のオフターゲットプライム編集及び/又はインデルを示し(図113)、このことは、epegRNA及びpegRNAが同様のレベルのオフターゲット編集を示すことを示唆していた。
操作されたpegRNAによる強化されたプライム編集の基礎
EpegRNAは、分解に対する耐性、より高い発現レベル、より効率的なCas9結合、及び/又はCas9と複合体化した場合の標的DNA結合を包含する様々な機構によってプライム編集結果を増強してもよい。これらの可能性のそれぞれが調査された。
EpegRNAは、分解に対する耐性、より高い発現レベル、より効率的なCas9結合、及び/又はCas9と複合体化した場合の標的DNA結合を包含する様々な機構によってプライム編集結果を増強してもよい。これらの可能性のそれぞれが調査された。
evopreQ1又はmpknotがpegRNA 3'伸長の分解を妨げるかどうかを決定するために、内在性エキソヌクレアーゼを含有するHEK293T核ライセートとのin vitroインキュベーション後に、epegRNA及びpegRNAの安定性が比較された。pegRNAは、epegRNAと比較して、この処理からより大きな程度まで分解されることが見出された(evopreQ1と比較して1.9倍及びmpknotと比較して1.8倍、p<0.005、図106A)。逆に、ガイドRNA骨格に結合し、コアsgRNAを分解から保護する可能性が高いCas9の添加は、ガイドRNA骨格のRT-qPCR定量によって決定されるように、いずれかのepegRNAと比較して、pegRNA存在量を救済した(図106B)。
3'構造モチーフが上流骨格領域(図106B)の存在量を増加させる能力は、核内のpegRNA分解が3'指向性分解によって支配されていることを示唆している。このモデルは、核におけるRNAターンオーバーの主要な供給源である核エキソソームの特徴的な挙動と一致している29。しかし、部分的に分解されたpegRNAは、プライム編集を阻害することが以前に示された編集不適格RNPを生成するであろう(図104C)。細胞中の部分的に分解されたRNAを検出するために、PE2をコードするプラスミド、及びHEK3での+1 FLAGタグ挿入又はEMX1でのヌクレオチドトランスバーションのいずれかを鋳型とするpegRNA又はepegRNAのいずれかをトランスフェクトしたHEK293T細胞の溶解物がノーザンブロットによって分析された。sgRNA骨格を含有し、sgRNAと同等のサイズのRNA種が観察され、Cas9結合が3'指向性分解から骨格を保護するという以前の知見(図106B)と一致した(図114A~図114C)。しかしながら、異なる総レベルのpegRNA又はepegRNAを有するライセートは、同様のレベルのsgRNA様切断種を有し、これはライセートのガイドRNA含有量のごく一部を表した(図114A~図114C)。核ライセートに曝露されたpegRNAの強力な分解がin vitroで観察され(図106A~図106B)、pegRNAは、HEK293T細胞においてPE2を超えるレベルで存在するので(図104B)、部分的に分解されたpegRNA種は、ノーザンブロット検出に適したレベルで蓄積しない可能性が高い。
次に、ゲノムプライム編集中間体が調べられて、どのようにしてepegRNAが改善された編集効率を媒介し得るかをよりよく理解した。現在のモデルでは、ニックを入れられた標的部位のRT伸長によって生成された3'フラップ中間体は5'フラップ中間体に変換され、元のゲノム配列を新しく合成されたもので置き換える14。次いで、この5'フラップは5'-3'エキソヌクレアーゼによって除去され、得られたゲノムニックをライゲーションしてプライム編集を組み入れする14。完全長pegRNAはニックが入ったゲノム鎖のRT伸長を効率的に鋳型化すると予想されるが、PBSを含まない切断型pegRNAはそうすることができないはずであり、その結果、標的化された鎖のニッキングが起こり、その後、DNA修復酵素による鎖の再生又は伸長が起こる(いずれの場合も鋳型編集がない)。RT伸長されたプライム編集中間体のより大きな割合が、pegRNAよりもepegRNAで観察されるならば、これは、3'RNAモチーフの付加がPBSの完全性を改善することを示唆するであろう。
これらの中間体を捕捉するために、HEK293T細胞は、PE2をコードするプラスミド、及びHEK3、DNMT1、EMX1又はRNF2でトランスバーションするevopreQ1又はmpknotを含有する非修飾pegRNA又はepegRNAのいずれかでトランスフェクトされた。次に、末端トランスフェラーゼが使用されて、まだライゲーションを受けていないプライム編集の中間体を包含するはずのゲノムDNAの3'末端をオリゴdGで標識した。いずれの場合も、epegRNAは、標的部位での編集不適格な中間体の程度を4つの部位にわたって平均2.2倍減少させた(図106C及び図115A~図115C)。主要な逆転写産物は、3'伸長によって鋳型化された完全な配列と、pegRNA骨格の最後の2つのヌクレオチドによって鋳型化された2つのヌクレオチドとを含有し、PE中間体の以前のin vitro特性評価と一致した14。骨格を鋳型としたヌクレオチドは、おそらく、PEの優性産物を表すきれいに編集された対立遺伝子を生成するために、標的遺伝子座のDNA修復中に除去される。これらのデータは、短縮型pegRNAに結合したプライム編集因子からの非生産的な標的部位ニッキングの頻度を減少させることによって、epegRNAが標的部位へのpegRNA伸長の逆転写を改善するモデルと一致している。
一本鎖3'末端は3'エキソヌクレアーゼ基質の共通の特徴であるので30、これらのモチーフによって付与される分解耐性が、シュードノットのより機械的に安定な三次構造によって説明され得るかどうかが次に試験された。注意すべきことに、3'末端に15bp(34-nt)のヘアピンを付加することは、シュードノット(図116 A~図116 D)を付加することと比較して、PE効率に対する一貫しない改善をもたらし、三次構造が実際にepegRNAの重要な特徴であることを示唆した。
PE効率のepegRNA媒介改善のために三次シュードノット構造が必要とされるかどうかを試験するために、シュードノット形成を破壊することが知られている突然変異であるevopreQ1内のG15C点突然変異を含有するepegRNAの編集効率が調べられた(図108のM1)23。epegRNAが使用されて、PE3を使用してHEK293T細胞のHEK3又はRNF2に24bpのFLAGエピトープタグ挿入、15bpの欠失、又はトランスバーションのいずれかを組み入れした。実際、G15C変異のevopreQ1への組み込みは、編集効率の増加を無効にした(図106D)。これらの結果は、モチーフの二次又は三次構造が、おそらく3'伸長を安定化することによって、epegRNA媒介PEの改善に重要であることを確立する。
次に、epegRNA中の構造化3'モチーフが試験されて、それらがpegRNAと比較してそれらの発現レベルを増加させ得るかどうかを決定した。pegRNA骨格のRT-qPCR定量により、非修飾pegRNAと比較して、epegRNA発現レベルの標的依存的差異を明らかにした(図114 A~図114C)。HEK3での+1 FLAGタグ挿入を鋳型とするpegRNAについて、evopreQ1又はmpknotの添加は、HEK3でのFLAGタグエピトープ挿入の効率の1.9倍の改善をもたらしたにもかかわらず、pegRNA発現を9.2~9.6倍減少させることが観察された(図105A)。同様に、DNMT1でトランスバーションを鋳型化するepegRNAも、発現の低下を示した(1.6~2.1倍)。しかし、RNF2又はEMX1でトランスバーションを鋳型するepegRNAは、非修飾pegRNAよりも高いレベルで発現された(夫々2.2~2.4倍及び1.4~3.7倍。図114A-図114C)。これらのデータは、3'モチーフがpegRNA発現に一貫を示すことなく影響を与え、これらのトランスフェクション条件下でのPE効率がHEK293T細胞におけるpegRNA発現によって制限されないという以前の知見(図104B)と一致することを示唆する。しかしながら、epegRNA発現がより限定的である場合、epegRNA発現の改善は編集効率を更に改善し得る。
次に、3'RNA構造モチーフの付加が、dCas9-VP64-p65-Rta(dCas9-VPR)融合物による転写活性化を支持するepegRNA及びpegRNAの能力を比較することによって標的DNA部位の関与を減少させるかどうかが試験された32、33。HEK293T細胞が、dCas9-VPR、HEK3、DNMT1、RNF2又はEMX1標的プロトスペーサのいずれかの下流のGFP、及び対応する部位を標的とするpegRNA、epegRNA又はsgRNAのいずれかをコードするプラスミドでトランスフェクトされた。転写活性化が、3日後に細胞GFP蛍光を介して測定された。PE活性を増強するそれらの能力とは対照的に(図105A)、epegRNAは、pegRNAと同様のCas9依存性転写活性化をHEK293T細胞において示した(図106 F)。epegRNAと標準的なpegRNAの両方が、同じ部位を標的とするsgRNAと比較して低い転写活性化をもたらし(4つの部位にわたるpegRNAについて3.0倍、evopreQ1 epegRNAについて2.3倍、及びmpknot epegRNAについて1.9倍)、pegRNA及びepegRNAの3'伸長が標的部位の関与をわずかに妨げることを示唆している。
標的部位結合の潜在的変化並びにpegRNA及びepegRNA発現の差をデコンボリュートするために、マイクロスケール熱泳動(MST)が実施されて、触媒的に不活性なCas9(dCas9)と、dsDNA基質に対するpegRNA又はepegRNAとのプレインキュベートしたRNP複合体の親和性を測定した。mpknot又はevopreQ1の添加は、夫々非修飾pegRNAと比較して、dsDNAに対する同等又はやや低下した結合親和性をもたらすことが見出された(KD=evopreQ1 epegRNAについては10 nM及びmpknot pegRNAについては21 nM対非修飾pegRNAについては8.1 nM、図106 E)。Cas9 H840Aニッカーゼに対するpegRNAの親和性も、いずれかのモチーフによってわずかに低下された(KD=evopreQ1 epegRNAについては18 nM、mpknot pegRNAについては11 nM、及び非修飾pegRNAについては5 nM;図106G)。これらの所見は、epegRNAからのPE効率の増加が、標的部位へのpegRNAのCas9又はPE RNP複合体の結合の改善から生じないことを示唆している。
まとめると、これらの結果は、epegRNAがpegRNAよりも細胞分解に対して耐性であり、したがって、プライム編集効率を損なう切断型pegRNA種をより少なく生成することを示唆する。epegRNAからの改善の背後にある更なる機構は排除され得ない。
操作されたpegRNA 3'モチーフの最適化
epegRNAがエキソヌクレアーゼ分解に抵抗することによって編集効率を改善することを確立して、より安定なRNAモチーフがPE活性を更に改善し得ると仮定された。25個の更なる構造化RNAモチーフが、24bp FLAGエピトープタグ挿入、15bp欠失、又はHEK3若しくはRNF2でのトランスバーションのいずれかをコードするepegRNAにわたるepegRNA編集効率を改善する能力についてスクリーニングされた(図116 A~図116D、図117A~図117C)。これらのモチーフは、追加の進化したプレケオシン1-1リボスイッチアプタマー21、改善されたシュードノット安定性を有するmpknot変異体22、安定性が増加したG四重鎖34、15bpヘアピン、xrRNA35、及びグループIイントロンのP4-P6ドメイン36を包含していた。試験した137個のepegRNAのうち123個が、対応するpegRNAと比較して改善された全体的なプライム編集を示したが、試験した編集の大部分にわたってevopreQ1又はmpknotに対する一貫した改善を実証したものはなかった(図116A~図116D、図117A~図117C)。
epegRNAがエキソヌクレアーゼ分解に抵抗することによって編集効率を改善することを確立して、より安定なRNAモチーフがPE活性を更に改善し得ると仮定された。25個の更なる構造化RNAモチーフが、24bp FLAGエピトープタグ挿入、15bp欠失、又はHEK3若しくはRNF2でのトランスバーションのいずれかをコードするepegRNAにわたるepegRNA編集効率を改善する能力についてスクリーニングされた(図116 A~図116D、図117A~図117C)。これらのモチーフは、追加の進化したプレケオシン1-1リボスイッチアプタマー21、改善されたシュードノット安定性を有するmpknot変異体22、安定性が増加したG四重鎖34、15bpヘアピン、xrRNA35、及びグループIイントロンのP4-P6ドメイン36を包含していた。試験した137個のepegRNAのうち123個が、対応するpegRNAと比較して改善された全体的なプライム編集を示したが、試験した編集の大部分にわたってevopreQ1又はmpknotに対する一貫した改善を実証したものはなかった(図116A~図116D、図117A~図117C)。
次に、付加されたevopreQ1及びmpknotモチーフから不要な配列をトリミングすると、構造化RNA内の無関係な配列を除去するとミスフォールドの傾向を減少させ得るので、epegRNAデザインを更に改善し得る37。evopreQ1又はmpknotから5 ntの過剰配列をトリミングすると、全長epegRNAと比較して平均PE3編集効率がわずかに増加することが見出された(図117A~図117C)。これらのRNAモチーフのトリミングは編集効率に悪影響を及ぼさず、より短いepegRNAは化学合成によってより容易に調製されるので、処置的に関連する突然変異を組み入れするためにepegRNAを適用する場合、トリミングされたevopreQ1(tevopreQ1)がepegRNAに使用された(下記参照)。
「フリップと伸長」(F+E)sgRNA骨格38がepegRNA編集効率を更に改善するかどうかも調べた。このガイドRNA骨格は、直接反復の第4の塩基対をU・AからA・Uに変異させて潜在的なpol IIIターミネータを除去し、直接反復を5塩基対伸長させてCas9結合を改善する38。HEK293T細胞は、HEK3若しくはDNMT1でのトランスバーション、又はHEK3での3nt挿入を鋳型とする標準的な骨格を用いて、非修飾(F+E)pegRNA、tevopreQ1を含有する(F+E)epegRNA、又はtevopreQ1 epegRNAのいずれかをコードするレンチウイルスで形質導入された。tevopreQ1の使用は、編集効率を実質的に改善した(ヌクレオチドトランスバーションについては3.8倍、HEK3での3nt挿入については2.6倍、DNMT1では6.8倍)(図118)。tevopreQ1 epegRNAにおける(F+E)骨格の使用は、編集効率を更に改善した(ヌクレオチドトランスバーションについては1.1倍、HEK3での3-nt挿入については1.5倍、及びDNMT1では2.5倍)。プラスミドの量を減少させたトランスフェクション条件下でCas9-ヌクレアーゼ活性39を増加させることが以前に示されたsgRNA骨格バリアントも特徴付けられ、より大きな変動性を有するにもかかわらず、同様の全体的な利益が観察された(図119)。これらの知見は、発現が制限されている場合、epegRNAがPE効率のより大きな改善を媒介することを更に示唆する。更に、これらのデータは、修飾された骨格がepegRNAと併せてPE効率を改善する可能性を強調している。
epegRNAリンカーをデザインするための計算ツール
タンパク質リンカーとは対照的に、RNAリンカーは配列依存性である可能性が高く、その結果、同じリンカーが1つのepegRNAに対して機能するが別のepegRNAを妨げる可能性がある。epegRNAリンカーからの干渉の可能性を最小限に抑えるために、epegRNAの残りの部分と最小塩基対合すると予測されるリンカー配列を同定する計算ツールであるpegLIT(pegRNAリンカー同定ツール)が開発された(図120A~図120F)。最初の検証のために、RNF2でのC・GからA・Tトランスバーション又はDNMT1での15bp欠失のいずれかを鋳型とする異なるリンカーを用いて、15 evopreQ1 epegRNAの2つのセットが試験された。各セット内で、5つのリンカーがpegLITによって推奨された;5個がスペーサと塩基対を形成すると予測され、5個がPBSと塩基対を形成すると予測された。pegLITデザインリンカーの使用は、手動デザインリンカーの使用よりもPE3編集効率の適度な増加をもたらした(RNF2では1.2倍高く、DNMT1では1.1倍高い)(図120A~図120F)。スペーサ相互作用は編集効率に有意に影響しなかったが、リンカー-PBS相互作用はPE3編集効率の低下と相関し、夫々RNF2及びDNMT1のpegLITリンカーと比較して1.3倍及び1.1倍低い編集効率をもたらした。最適なリンカー配列と比較してRNF2で1.9倍及び3.4倍低い効率のPE3編集をもたらした2つの最悪性能のリンカーは、PBS相互作用について不十分なスコアリングとしてpegLITによって正確に同定された(図120A~図120F)。スペーサと比較して、リンカーがPBSに近接していることは、リンカー:スペーサの対形成と比較して、リンカー:PBS相互作用にエントロピー的利点を与えてもよい。
タンパク質リンカーとは対照的に、RNAリンカーは配列依存性である可能性が高く、その結果、同じリンカーが1つのepegRNAに対して機能するが別のepegRNAを妨げる可能性がある。epegRNAリンカーからの干渉の可能性を最小限に抑えるために、epegRNAの残りの部分と最小塩基対合すると予測されるリンカー配列を同定する計算ツールであるpegLIT(pegRNAリンカー同定ツール)が開発された(図120A~図120F)。最初の検証のために、RNF2でのC・GからA・Tトランスバーション又はDNMT1での15bp欠失のいずれかを鋳型とする異なるリンカーを用いて、15 evopreQ1 epegRNAの2つのセットが試験された。各セット内で、5つのリンカーがpegLITによって推奨された;5個がスペーサと塩基対を形成すると予測され、5個がPBSと塩基対を形成すると予測された。pegLITデザインリンカーの使用は、手動デザインリンカーの使用よりもPE3編集効率の適度な増加をもたらした(RNF2では1.2倍高く、DNMT1では1.1倍高い)(図120A~図120F)。スペーサ相互作用は編集効率に有意に影響しなかったが、リンカー-PBS相互作用はPE3編集効率の低下と相関し、夫々RNF2及びDNMT1のpegLITリンカーと比較して1.3倍及び1.1倍低い編集効率をもたらした。最適なリンカー配列と比較してRNF2で1.9倍及び3.4倍低い効率のPE3編集をもたらした2つの最悪性能のリンカーは、PBS相互作用について不十分なスコアリングとしてpegLITによって正確に同定された(図120A~図120F)。スペーサと比較して、リンカーがPBSに近接していることは、リンカー:スペーサの対形成と比較して、リンカー:PBS相互作用にエントロピー的利点を与えてもよい。
PEgLITデザインリンカー配列が研究されて、それらが2つのepegRNAの有効性を改善することができるかどうかを決定し(EMX1においてG・CからT・Aのトランスバーション及びVEGFAにおいて15bpの欠失を鋳型化)、これは最初は改善された編集を示すことに失敗した(図111A~図111K)。実際、pegLITデザインリンカーを使用すると、PE3編集効率が、これらの2つの編集についてのpegRNAのものよりも夫々1.3倍及び1.4倍増加した(図120A-図120G)。まとめると、これらの知見は、pegLITが、プライム編集結果を一貫して改善するためのepegRNAの使用を促進することを実証している。
PegLITでデザインされたリンカーも研究されて、それらがリンカーを含まないepegRNAと比較して、epegRNAの活性を改善したかどうかを決定した。リンカーを含まないmpknot epegRNAと比較して、pegLITデザインリンカーを添加すると、手動デザインリンカーを使用した場合よりも編集効率がわずかに増加した(図110及び図120A~図120F)。対照的に、evopreQ1又はtevopreQ1のepegRNAを有するpegLITリンカーの使用は、リンカーを含まないepegRNAと比較して編集を有意に増加させなかった(図120A-図120G)。
化学修飾されたepegRNAによる編集効率の改善
化学的に合成されたgRNAは、細胞をmRNA又はRNPでトランスフェクトする場合に一般的に使用される40。合成gRNAは、エキソヌクレアーゼ分解に対する耐性を促進する化学修飾を組み込み得るが16、40、構造モチーフは、そのような修飾と併せて更なる改善を依然として媒介し得ると考えられた。
化学的に合成されたgRNAは、細胞をmRNA又はRNPでトランスフェクトする場合に一般的に使用される40。合成gRNAは、エキソヌクレアーゼ分解に対する耐性を促進する化学修飾を組み込み得るが16、40、構造モチーフは、そのような修飾と併せて更なる改善を依然として媒介し得ると考えられた。
この可能性を実証するために、合成tevopreQ1 epegRNAのプライム編集効率が、HEK293T細胞の5つのゲノム部位(HEK3、RNF2、DNMT1、RUNX1、及びEMX1)に点突然変異又は15bp欠失のいずれかを組み入れする合成pegRNAのものと比較された。epegRNA及びpegRNAの両方は、RNAの最初の3ヌクレオチドと最後の3ヌクレオチドとの間に2'-O-メチル修飾及びホスホロチオアート結合を含有していた。試験した7つのpegRNAのうち6つについて、対応するepegRNAは、変化しない編集:インデル比で1.1~3.1倍高い編集を示した(図121A~図121B)。これらのデータは、化学合成及び修飾されたpegRNAを使用する用途において、epegRNAが、pegRNAと比較してPE転帰も増強することを示唆している。
操作されたpegRNAは、処置関連変異のプライム編集を改善する
PE活性を広く改善するための戦略としてのepegRNAの使用を検証したので、本発明者らは次に、様々な保護的又は処置的遺伝子変異を組み入れするために、tevopreQ1を含有するepegRNAの活性をpegRNAの活性と比較した。標準的なpegRNAよりも1.4倍高い効率で、HEK293T細胞においてヒトプリオン病41、42から保護するPRNP G127V対立遺伝子を組み入れするために、epegRNAが首尾よく使用された(図107A)。更に、以前に構築されたHEXA1278+TATC HEK293T細胞株14においてプラスミドリポフェクションを介して、及びin vitro転写mRNA及び合成pegRNAのヌクレオフェクションを介して一次患者由来線維芽細胞において、epegRNAを使用されて、テイ・サックス病の最も一般的な原因(HEXA1278+TATC)を修正した(図107B~107C)。両方の場合において、pegLITデザイン8-ntリンカーを含有するtevopreQ1epegRNAについて、標準的なpegRNAよりも改善された編集効率が観察された(HEK293T細胞において2.8倍高く、患者由来線維芽細胞において2.3倍高い)。
PE活性を広く改善するための戦略としてのepegRNAの使用を検証したので、本発明者らは次に、様々な保護的又は処置的遺伝子変異を組み入れするために、tevopreQ1を含有するepegRNAの活性をpegRNAの活性と比較した。標準的なpegRNAよりも1.4倍高い効率で、HEK293T細胞においてヒトプリオン病41、42から保護するPRNP G127V対立遺伝子を組み入れするために、epegRNAが首尾よく使用された(図107A)。更に、以前に構築されたHEXA1278+TATC HEK293T細胞株14においてプラスミドリポフェクションを介して、及びin vitro転写mRNA及び合成pegRNAのヌクレオフェクションを介して一次患者由来線維芽細胞において、epegRNAを使用されて、テイ・サックス病の最も一般的な原因(HEXA1278+TATC)を修正した(図107B~107C)。両方の場合において、pegLITデザイン8-ntリンカーを含有するtevopreQ1epegRNAについて、標準的なpegRNAよりも改善された編集効率が観察された(HEK293T細胞において2.8倍高く、患者由来線維芽細胞において2.3倍高い)。
最適化されていないepegRNAを使用した処置的に関連する編集の組み入れ
異なるPBS鋳型及びRT鋳型を有する多くのpegRNAのデザイン及びスクリーニングは、プライム編集の成功した使用における重要な第1の工程である14。PBS及びRT鋳型の長さ及び組成を導くための一般的な規則が記載されているが14、43、最適なpegRNAを同定するには、しばしば、pegRNA構築物の広範なスクリーニングが必要である。epegRNAは、広範なpegRNA最適化なしでも、処置的に関連するプライム編集のより効率的な組み入れを支援し得ると考えられた。最適化されていないpegRNA及びepegRNAが、PE2を使用して9つの保護的又は病原的な点変異の組み入れを鋳型化する能力。全ての場合において、この実験において使用されたpegRNA及びepegRNAは、3'伸長がシトシン14で始まる場合を除いて、最後に編集されたヌクレオチドの後の標的部位に対して10ntの相同性を含有する13ntのPBS及びRT鋳型を含有し、その場合、それは最も近い非Cヌクレオチドに伸長された。
異なるPBS鋳型及びRT鋳型を有する多くのpegRNAのデザイン及びスクリーニングは、プライム編集の成功した使用における重要な第1の工程である14。PBS及びRT鋳型の長さ及び組成を導くための一般的な規則が記載されているが14、43、最適なpegRNAを同定するには、しばしば、pegRNA構築物の広範なスクリーニングが必要である。epegRNAは、広範なpegRNA最適化なしでも、処置的に関連するプライム編集のより効率的な組み入れを支援し得ると考えられた。最適化されていないpegRNA及びepegRNAが、PE2を使用して9つの保護的又は病原的な点変異の組み入れを鋳型化する能力。全ての場合において、この実験において使用されたpegRNA及びepegRNAは、3'伸長がシトシン14で始まる場合を除いて、最後に編集されたヌクレオチドの後の標的部位に対して10ntの相同性を含有する13ntのPBS及びRT鋳型を含有し、その場合、それは最も近い非Cヌクレオチドに伸長された。
アルツハイマー病44、冠動脈心疾患45、46、2型糖尿病47、先天性免疫48、CDKL5欠損症49、ラミンA欠損50、及びレット症候群51、52に関連する処置上関連する変異を組み入れするpegRNAが調査された。これらの9つの変異は、APP、PCSK 9、SLC30A8、CD209及びCETPにおける保護対立遺伝子、並びにCDKL5、LMNA及びMECP254における病原性変異を包含する。HEK293T細胞(図111D)におけるpegRNA及び8-nt pegLITリンカーを有する対応するtevopreQ1 epegRNAによるプライム編集の結果が比較された。単一のpegRNA又はepegRNAデザインのみは標的ごとに試験された。いずれの場合も、編集効率において、epegRNAはpegRNAよりも優れていた。試験した9つの処置的に関連する編集のうち5つについて、epegRNAは20%以上の編集効率をもたらし、これは典型的にはモデル細胞株を生成するのに十分である。比較すると、9つのpegRNAのうち3つのみがこのレベルの編集効率を達成した。epegRNAによって媒介されるより高い編集効率(平均でpegRNAより2.8倍高い)は、ホモ接合性細胞株の産生を合理化するはずであり、これは劣性変異をモデル化するための重要な考慮事項である。同様に、非最適化epegRNAは、15個の試験部位のうち5個で10%以上の効率で24bp FLAGタグの挿入を媒介し、対応するpegRNAは、試験したいずれの部位でも10%以上の効率を達成しなかった(図122A~図122B)。まとめると、これらの知見は、epegRNAがPEを有するモデル細胞株の産生を合理化することを実証している。
考察
プライム編集における重要なボトルネックに対処するための操作されたpegRNAのデザイン、特徴付け及び検証が、本明細書中に提示される。これらのepegRNAは、標的ゲノム部位へのアクセスについて競合する、pegRNA伸長の分解及びその後の編集不能なPE複合体の形成を防止するPBSの構造化RNAモチーフ3'を含有する。epegRNAは、試験した5つ全ての細胞株及び初代細胞型においてプライム編集効率を広く改善し、トランスフェクトがより困難な細胞株においてより大きな改善が観察されることが見出された。更に、epegRNAの使用は、化学修飾されたpegRNAを使用する場合、ヒト細胞に処置的に関連する編集を組み入れする場合、及び最適化されていないpegRNAデザインを使用する場合、プライム編集性能を高め得ることが観察された。最後に、逆効果の二次構造のリスクを最小限に抑えるリンカーを同定することによって、epegRNAデザインを促進する計算プログラムが記載された。全体として、これらの知見は、epegRNAが、多種多様なゲノム遺伝子座、編集型(置換、挿入及び欠失)及び細胞型においてプライム編集結果を広く改善することを立証している。
プライム編集における重要なボトルネックに対処するための操作されたpegRNAのデザイン、特徴付け及び検証が、本明細書中に提示される。これらのepegRNAは、標的ゲノム部位へのアクセスについて競合する、pegRNA伸長の分解及びその後の編集不能なPE複合体の形成を防止するPBSの構造化RNAモチーフ3'を含有する。epegRNAは、試験した5つ全ての細胞株及び初代細胞型においてプライム編集効率を広く改善し、トランスフェクトがより困難な細胞株においてより大きな改善が観察されることが見出された。更に、epegRNAの使用は、化学修飾されたpegRNAを使用する場合、ヒト細胞に処置的に関連する編集を組み入れする場合、及び最適化されていないpegRNAデザインを使用する場合、プライム編集性能を高め得ることが観察された。最後に、逆効果の二次構造のリスクを最小限に抑えるリンカーを同定することによって、epegRNAデザインを促進する計算プログラムが記載された。全体として、これらの知見は、epegRNAが、多種多様なゲノム遺伝子座、編集型(置換、挿入及び欠失)及び細胞型においてプライム編集結果を広く改善することを立証している。
epegRNAによって可能になるプライム編集の改善は、送達戦略に依存する可能性が高い。一部のウイルスベクター等の低発現送達様式は、pegRNA濃度が制限的である場合(図119)、epegRNAの使用からより強く利益を得る可能性がある。同様に、化学修飾RNAの合成の更なる改善は、pegRNA 3'分解を緩和することによって、epegRNAの利益を減少させ得る。更に、より長い長さのepegRNA(tevopreQ1を使用する場合、更に37 nt)は、より長いRNAを化学的に合成するという現在の課題を考慮すると、合成epegRNAを使用する場合の重要な考慮事項である。
epegRNAの使用は、やや長いpegRNAを支持し得るプライム編集実験のために推奨される。編集効率を最大化することが優先事項ではない場合、広範なスクリーニングは必要でなくてもよい。これらの場合、トリミングされたevopreQ1モチーフ及び13のPBS長を有する8ntのpegLITデザインリンカーと、小さな挿入、欠失及び点変異についての標的編集を超えた10ntの相同性(又はより大きな挿入若しくは欠失についての25ntの相同性)を包含する鋳型とを含有するepegRNAは、epegRNAデザインの有望な出発点を提供する。観察された編集効率が不十分である場合、PBS、RT鋳型長及びニッキングsgRNAが最適化され得る。
最適なリンカー配列を同定するためのpegLIT戦略
pegLITは、シミュレートされたアニーリングを使用して、分析されたリンカー空間を効率的にサンプリングする1。アデノシン又はシトシンに富むリンカーは、これらのヌクレオチドが柔軟なRNAリンカーとしてより良好に機能することが報告されているので、pegLITによって好ましい2。更に、pegLITは、そのような配列が時期尚早の転写終結を引き起こし得るので、4つ以上のウリジンの連続を含有するリンカーを除外する3。
pegLITは、シミュレートされたアニーリングを使用して、分析されたリンカー空間を効率的にサンプリングする1。アデノシン又はシトシンに富むリンカーは、これらのヌクレオチドが柔軟なRNAリンカーとしてより良好に機能することが報告されているので、pegLITによって好ましい2。更に、pegLITは、そのような配列が時期尚早の転写終結を引き起こし得るので、4つ以上のウリジンの連続を含有するリンカーを除外する3。
次いで、pegLITツールは、ViennaRNA4を使用してこれらの要件を満たすリンカーを分析して、リンカー配列とpegRNAスペーサ、PBS、鋳型又は骨格との間の潜在的な相互作用を予測する。これらの予測された相互作用の塩基対確率が使用されて、pegRNAの各領域のサブスコアを生成し、そのそれぞれは、リンカーが関連領域との相互作用を回避すると予測される程度を表す。例えば、PBSのサブスコア0.95は、本質的に、平均して、任意のリンカーヌクレオチドとPBSとの間の塩基対形成を欠くpegRNA折り畳み状態の予測確率が95%であることを示す。
pegLITの使用がリンカーデザインについて検証され、pegLITによって同定されたどの相互作用が編集効率に最も有害であるかを調べた。RNF2でのC・GからA・Tトランスバーション又はDNMT1での15bp欠失のいずれかを鋳型とするevopreQ1 epegRNAで試験するために、30個のリンカー配列が生成された(10はpegLITによって推奨され、10はスペーサと相互作用し、10はPBSと相互作用する)。平均スペーサ及びPBSサブコアは、最適な配列については0.94及び0.97であり、スペーサ配列については0.66及び.95であり、PBS配列については0.86及び0.21であった。推奨されるデザインと比較して、PBSと相互作用するリンカーの使用は、RNF2及びDNMT1で夫々1.3倍及び1.1倍低い編集効率に関連されていた(図120A~図120G)が、スペーサと相互作用するリンカーは、編集効率に対して無視できるほどの効果を有していた。この違いは、スペーサと比較してリンカーがPBSに近接していると、リンカー:スペーサ対形成と比較してリンカー:PBS相互作用がエントロピー的に有利になってもよいためであり得る。
HEK293T細胞における最適化されたガイドRNA足場によるプラスミドトランスフェクションを介して送達されたepegRNA
低発現条件を模倣するために、PEに対する「フリップ及び伸長」(F+E)sgRNA骨格バリアントの適用性を評価する場合、HEK293T細胞は、20ngのPE2プラスミド及び4ngのpegRNA又はepegRNAプラスミドをトランスフェクトされた。標準的なsgRNA骨格、(F+E)骨格5、又はCas9ヌクレアーゼ活性6を増加させることが以前に示された突然変異を有する6つの(F+E)骨格のうちの1つを含有したPRNP、HEK3、RUNX1及びEMX1を標的とするepegRNAの編集効率が比較された。これらの代替的な骨格は、標準的な骨格と比較して全体的にPE効率を維持又は改善し、cr772が最良の改善を示すことが見出された(図119)。効率の改善は、レンチウイルス形質導入と比較してこれらの条件下で一貫性が低かったが(図118)、これは発現の違いに起因してもよい。プラスミドトランスフェクション後のEpegRNA発現は、単一コピーのレンチウイルス形質導入後よりも数倍高い可能性があり、より効率的な転写及びCas9結合親和性の利点を部分的に不明瞭にしてもよい。特にプラスミドトランスフェクションよりも発現が低い用途では、epegRNAを用いてPE効率を更に改善するためにcr772又は元の(F+E)骨格を試験することが推奨される。
低発現条件を模倣するために、PEに対する「フリップ及び伸長」(F+E)sgRNA骨格バリアントの適用性を評価する場合、HEK293T細胞は、20ngのPE2プラスミド及び4ngのpegRNA又はepegRNAプラスミドをトランスフェクトされた。標準的なsgRNA骨格、(F+E)骨格5、又はCas9ヌクレアーゼ活性6を増加させることが以前に示された突然変異を有する6つの(F+E)骨格のうちの1つを含有したPRNP、HEK3、RUNX1及びEMX1を標的とするepegRNAの編集効率が比較された。これらの代替的な骨格は、標準的な骨格と比較して全体的にPE効率を維持又は改善し、cr772が最良の改善を示すことが見出された(図119)。効率の改善は、レンチウイルス形質導入と比較してこれらの条件下で一貫性が低かったが(図118)、これは発現の違いに起因してもよい。プラスミドトランスフェクション後のEpegRNA発現は、単一コピーのレンチウイルス形質導入後よりも数倍高い可能性があり、より効率的な転写及びCas9結合親和性の利点を部分的に不明瞭にしてもよい。特にプラスミドトランスフェクションよりも発現が低い用途では、epegRNAを用いてPE効率を更に改善するためにcr772又は元の(F+E)骨格を試験することが推奨される。
最適化されていないepegRNAを用いたFLAGタグの組み入れ
24-bp FLAGエピトープタグ(図105A)の挿入等のより困難な編集の組み入れについて、epegRNA及びpegRNAが比較された。PE2を使用したHEK293T細胞における15個の遺伝子座でのFLAGエピトープタグの組み入れを鋳型化する、2つの遺伝子座特異的pegLITデザイン8-ntリンカーのうちの1つを含有する非最適化pegRNA及びtevopreQ1 epegRNAの能力が評価された(図122A~図122B)。最適化されていないepegRNA及びpegRNAが、3'伸長がシトシン7で始まる場合を除いて、挿入されたFLAGエピトープタグの下流に25ntの相同性を含有する13ntのPBS及びRT鋳型を用いてデザインされ、その場合、最も近い非Cヌクレオチドに伸長された。epegRNAの使用により、FLAGタグを10%以上の効率でPE2と共に組み入れすることが可能になり、PBS及びRT鋳型の最適化は15の部位のうち5つで行われなかったが、10%以上の効率はいずれのpegRNAでも観察されなかった(図122A~図122B)。これらの観察結果は、epegRNAが、多くの異なる内生ヒトゲノム遺伝子座における様々な編集のためのプライム編集性能を高め得ることを更に実証している。
24-bp FLAGエピトープタグ(図105A)の挿入等のより困難な編集の組み入れについて、epegRNA及びpegRNAが比較された。PE2を使用したHEK293T細胞における15個の遺伝子座でのFLAGエピトープタグの組み入れを鋳型化する、2つの遺伝子座特異的pegLITデザイン8-ntリンカーのうちの1つを含有する非最適化pegRNA及びtevopreQ1 epegRNAの能力が評価された(図122A~図122B)。最適化されていないepegRNA及びpegRNAが、3'伸長がシトシン7で始まる場合を除いて、挿入されたFLAGエピトープタグの下流に25ntの相同性を含有する13ntのPBS及びRT鋳型を用いてデザインされ、その場合、最も近い非Cヌクレオチドに伸長された。epegRNAの使用により、FLAGタグを10%以上の効率でPE2と共に組み入れすることが可能になり、PBS及びRT鋳型の最適化は15の部位のうち5つで行われなかったが、10%以上の効率はいずれのpegRNAでも観察されなかった(図122A~図122B)。これらの観察結果は、epegRNAが、多くの異なる内生ヒトゲノム遺伝子座における様々な編集のためのプライム編集性能を高め得ることを更に実証している。
方法
一般方法pegRNA及びepegRNAを発現するプラスミドは、Gibsonアセンブリ、Golden Gateアセンブリのいずれかによって、以前に記載されたカスタムアクセプタープラスミド14又はトリミングされたevopreQ1若しくはmpknotを含有する新規にデザインされたカスタムアクセプタープラスミド(その使用は以下に記載される)のいずれかを用いてクローニングされたか、又はTwist Biosciencesによって合成及びクローニングされた。sgRNAを発現するプラスミドは、Gibson又はUSERアセンブリを介してクローニングされた。DNA増幅は、Phusion U又はHigh Fidelity Phusion Green Hot Start II(New England Biolabs)を用いたPCRによって達成された。pegRNAを発現するプラスミドは、HEK293T細胞をトランスフェクトする場合はPureYieldプラスミドミニプレップキット(Promega)又は他の細胞型をトランスフェクトする場合はPlasmid Plus Midiprepキット(Qiagen)を使用して精製したが、プライム編集因子を発現するプラスミドは、Plasmid Plus Midiprepキットのみを使用して精製された。Twist Biosciencesから注文したプラスミドをヌクレアーゼフリー水に再懸濁され、直接使用された。プライマー及びdsDNAフラグメントは、Integrated DNA Technologies(IDT)から注文された。
一般方法pegRNA及びepegRNAを発現するプラスミドは、Gibsonアセンブリ、Golden Gateアセンブリのいずれかによって、以前に記載されたカスタムアクセプタープラスミド14又はトリミングされたevopreQ1若しくはmpknotを含有する新規にデザインされたカスタムアクセプタープラスミド(その使用は以下に記載される)のいずれかを用いてクローニングされたか、又はTwist Biosciencesによって合成及びクローニングされた。sgRNAを発現するプラスミドは、Gibson又はUSERアセンブリを介してクローニングされた。DNA増幅は、Phusion U又はHigh Fidelity Phusion Green Hot Start II(New England Biolabs)を用いたPCRによって達成された。pegRNAを発現するプラスミドは、HEK293T細胞をトランスフェクトする場合はPureYieldプラスミドミニプレップキット(Promega)又は他の細胞型をトランスフェクトする場合はPlasmid Plus Midiprepキット(Qiagen)を使用して精製したが、プライム編集因子を発現するプラスミドは、Plasmid Plus Midiprepキットのみを使用して精製された。Twist Biosciencesから注文したプラスミドをヌクレアーゼフリー水に再懸濁され、直接使用された。プライマー及びdsDNAフラグメントは、Integrated DNA Technologies(IDT)から注文された。
Golden Gate DNA assemblyによるepegRNAクローニングのためのガイドライン61。Golden Gate法を用いてepegRNAをクローニングする場合、以前に記載されたプロトコル14と同じプロトコルが適切であり、重要な注記として、3'伸長オリゴとプラスミドバックボーンとの間の連結配列は、下記に示されるように、tevopreQ1及びトリミングされたmpknot(tmpknot)を用いたepegRNAでは異なる。pegRNAのデザイン及びクローニングに関する更なる詳細は、liugroup.us.で入手可能である。Golden Gateクローニングに使用されるプラスミドバックボーンはAddgeneに寄託されている。配列番号486~489(上から下):
pegRNA及びepegRNAの3'伸長のフォワードオリゴ 5'-GTGCNNNNNNNNNNNNNNNNNNNNNNNN-3'
pegRNAの3'伸長のリバースオリゴ 3'-NNNNNNNNNNNNNNNNNNNNNNNNAAAA-5'
tevoprep1-1を有するepegRNAの3'伸長のリバースオリゴ 3'-NNNNNNNNNNNNNNNNNNNNNNNNGCGC-5'
tmpknotを有するepegRNAの3'伸長のリバースオリゴ 3'-NNNNNNNNNNNNNNNNNNNNNNNNGGGAGTC-5'
pegRNA及びepegRNAの3'伸長のフォワードオリゴ 5'-GTGCNNNNNNNNNNNNNNNNNNNNNNNN-3'
pegRNAの3'伸長のリバースオリゴ 3'-NNNNNNNNNNNNNNNNNNNNNNNNAAAA-5'
tevoprep1-1を有するepegRNAの3'伸長のリバースオリゴ 3'-NNNNNNNNNNNNNNNNNNNNNNNNGCGC-5'
tmpknotを有するepegRNAの3'伸長のリバースオリゴ 3'-NNNNNNNNNNNNNNNNNNNNNNNNGGGAGTC-5'
合成pegRNA及びin vitro転写mRNA生成。合成pegRNAをIDTから注文され、最初と最後の3つのヌクレオチドに2'-O-メチル修飾を含有し、最初と最後の3つのヌクレオチド間にホスホロチオアート結合を含み、直接使用された。合成ニッキングsgRNAをSynthegoから注文し、最初と最後の3つのヌクレオチドに2'-O-メチル修飾を含有し、最初の3つのヌクレオチドと最後の2つのヌクレオチドとの間にホスホロチオアート結合を含有した。PEコードmRNAは、Gaudelli et al.(2020)のプロトコルを使用してin vitroで転写された。簡潔には、5'UTR、コザック配列、PE2 ORF及び3'UTRからなるPE2カセットは、不活性T7(dT7)プロモータを含有するプラスミドにクローニングされた。mRNA転写鋳型は、正しいT7プロモータ配列を組み入れするためのプライマー及びポリA尾部を組み入れしたリバースプライマーを使用してPCRによって生成された。mRNAは、N 1-メチルシュードウリジントリホスファート(Trilink)をウリジントリホスファートの代わりに使用し、共転写キャッピングを可能にするためにCleanCapAG(Trilink)が添加されたことを除いて、HiScribe T7 High-Yield RNA Kit(New England Biolabs)を製造者の説明書に従って使用して生成された。得られたmRNAは塩化リチウム沈殿によって精製され、TE緩衝液(10 mMトリス、1 mM EDTA、25℃でpH 8.0)中で再構成した。この実施例で使用されるpegRNA及びsgRNAの配列は、表E1に見出され得る。この実施例で調べた構造化RNAモチーフのリストは、表E2に見出され得る。
一般的な哺乳動物細胞培養条件。HEK293T細胞(ATCC CRL-3216)、U2OS細胞(ATCC HTB-96)、K562細胞(CCL-243)及びHeLa細胞(CCL-2)はATCCから購入され、それぞれ10%(v/v)ウシ胎児血清(認定Gibco)を補充した、GlutaMax(Thermo Fisher Scientific)、McCoyの5 A培地(Gibco)、RPMI培地1640+GlutaMAX(Gibco)又はEagleの最小必須培地(EMEM,ATCC)を夫々補充したダルベッコ修飾イーグル培地(DMEM)で培養し、継代した。初代テイ・サックス病患者線維芽細胞が、Coriell Institute(カタログ番号GM 00221)から入手され、更なる2 mM L-グルタミン(Thermo Fisher Scientific)を補充した低グルコースDMEM(Sigma Aldrich)及び10%(v/v)FBS中で増殖された。全ての細胞型がインキュベートされ、維持され、5% CO2を含む37℃で培養された。各細胞株はそれぞれの供給者によって認証され、マイコプラズマについて陰性と試験された。
組織培養トランスフェクション及びヌクレオフェクションプロトコル並びにゲノムDNA調製。トランスフェクションのために、10,000個のHEK293T細胞を96ウェルプレート(Corning)にウェル当たり播種した。播種の16~24時間後、製造業者のプロトコルに従って、0.5μLのリポフェクタミン2000(Thermo Fisher Scientific)、並びに200ngのPEプラスミド、40ngのpegRNAプラスミド、及び13ngのsgRNAプラスミド(PE3用)を用いて、細胞は約60%の集密度でトランスフェクトされた。
ヌクレオフェクションのために、SF細胞株キット(Lonza)を備えたLonza 4D Nucleofectorを使用して、HEK293T細胞にin vitro転写mRNA及び合成pegRNAは電気穿孔された。エレクトロポレーション当たり200,000個の細胞は120×gで8分間遠心分離され、次いで、1 mLのPBS(Thermo Fisher Scientific)で洗浄された。第2の遠心分離後、細胞を試料当たり5μLの再構成SF緩衝液に再懸濁され、マイクロキュベットに添加された。
各キュベットについて、17μLのカーゴミックス(0.5μL中1 ugのPE2 mRNA、0.9μL中90 pmolのpegRNA、及び0.6μL中60 pmolのニッキングsgRNA、及び15μLの再構成SF緩衝液)が添加され、上下に3回ピペッティングして混合された。プログラムCM-130を使用して細胞が電気穿孔され、次いで、80μLの温培地が添加され、細胞が室温で10分間インキュベートされた。次いで、混合物がピペットされて混合し、25μLが48ウェルプレートのウェルに添加され、最終培養体積をウェル当たり250μLとした。HeLa、U2OS、及びK562細胞での実験のために、800ngのPE2発現プラスミド、200ngのpegRNA発現プラスミド、及び83ngのニッキングsgRNA発現プラスミドが、16ウェルヌクレオベットストリップ(Lonza)中で20μLの最終体積でヌクレオフェクトされた。HeLa細胞が、SE Cell Line 4D-Nucleofector X Kit(Lonza)を使用して、製造元のプロトコルに従って、サンプル当たり2×105細胞(プログラムCN-114)でクレオフェクトされた。U2OS細胞を、SE Cell Line 4D-Nucleofector X Kit(Lonza)を使用して、製造元のプロトコルに従って、サンプル当たり2×105細胞(プログラムDN-100)でクレオフェクトされた。K562細胞を、SE Cell Line 4D-Nucleofector X Kit(Lonza)を使用して、製造元のプロトコルに従って、サンプル当たり2×105細胞(プログラムFF-120)でクレオフェクトされた。
患者由来線維芽細胞に、HEK293T細胞について上記のようにmRNAをコードするPE2及び合成pegRNA及びニッキングsgRNAが、SE細胞株キットを使用して電気穿孔され、100,000個の細胞が100×gで10分間遠心分離された。更に、40μLの回収した細胞が25の代わりに48ウェルプレートに加えられた。全ての場合において、トランスフェクションの3日後に細胞が培養され、その後、培地が除去され、細胞がPBSで洗浄され、続いて、新たに調製した溶解緩衝液(10 mM Tris-HCl、25℃でpH 8;0.05% SDS;25μg mL-1プロテイナーゼK(Qiagen))の96ウェルプレートについて50μL又は48ウェルプレートについて150μLを添加し、37℃で1時間以上インキュベートすることによって溶解し、その後、プロテイナーゼKが80℃で30分間にわたって不活性化された。得られたgDNAは使用されるまで-20℃で保存された。
ゲノムDNA試料のハイスループットDNA配列決定。目的のゲノム部位がゲノムDNA試料から増幅され、前述のようにIllumina MiSeqで配列決定された14。HEK3、EMX1及びFANCFのCas9オフターゲット部位は、ガイド配列29によって以前に同定された。哺乳動物細胞ゲノムDNA増幅に使用されるプライマーは表E 3に列挙され、アンプリコンは表E4に列挙される。配列決定リードは、MiSeq Reporter(Illumina)を使用して逆多重化された。アンプリコン配列の参照配列へのアラインメントは、CRISPReso259を用いて行われた。全てのプライム編集収率の定量について、編集効率が、少なくとも30の平均phredスコアを有するリードの総数からインデルなしの所望の編集を有するリードのパーセンテージとして計算された。点突然変異編集の定量化のために、CRISPResso 2が「discard_indel_reads」をオンにして標準モードで実行された。編集収率は、編集を含有する廃棄されていないリードのパーセンテージを総リードで割ったものとして計算された。挿入又は欠失編集のために、所望の対立遺伝子を予想される対立遺伝子として使用し、「discard_indel_reads」をオンにして、CRISPResso 2はHDRモードで実行された。編集収率は、HDRアラインメントされたリードのパーセンテージを全リードで割ったものとして計算された。全ての実験について、インデル頻度は、廃棄されたリードの数をリードの総数で割ったものとして計算された。PE2を伴う実験では、pegRNAニック部位の上流及び下流の10ヌクレオチド以内のインデルについてリードが分析された(両端を含む)。PE3を伴う実験では、pegRNAニック部位の上流及びsgRNAニック部位の下流の10ヌクレオチド間のインデルについてリードが分析された(両端を含む)。オフターゲット編集は、前述のように定量化された14。
in vitroエキソヌクレアーゼ感受性アッセイ。製造業者のプロトコルに従って、T7プロモータ配列を含有するPCR増幅鋳型から、HiScribeT7 Quick High Yield RNA合成キット(New England Biolabs)を使用して、mpknot又はevopreQ1のいずれかを含有するpegRNA又はepegRNAが調製された。EpiQuik核抽出キット(EpiGentek)を使用して、製造業者のプロトコルに従って70~80%のコンフルエントまで増殖させた300万個のHEK293T細胞から核抽出物が調製された。エンドヌクレアーゼ活性を阻害するために、20 mM Tris-HCl(pH 7.5)、5 mM MgCl2、50 mM NaCl、2 mM DTT、1 mM NTP及び0.8 U/μl RNaseOUT組換えリボヌクレアーゼ阻害剤(40 U/μL;ThermoFisher Scientific)を含有する10μL反応物中でアッセイが行われた。3μLの新鮮な核溶解物が使用されて、反応当たり0.5μgのRNA基質を分解した。続いて、反応混合物を37℃で20分間インキュベートし、SYBR Goldで染色した2.0%アガロースゲルで分解産物が分離された。ImageJソフトウェア(NIH)を使用して分解の程度が決定された。
全RNAのRTqPCR。ウェル当たり10,000個のHEK293T細胞が96ウェルプレートに播種された。播種の16~24時間後、製造業者のプロトコルに従って0.5μLのリポフェクタミン2000及び200ngのPE2プラスミド及び40ngのpegRNA又はepegRNAプラスミドのいずれかが細胞に約60%のコンフルエンシーでトランスフェクトされた。3日後、Power SYBR Green CellsからCTキット(Thermo Fisher Scientific)が使用されて、全RNAが抽出されて、全cDNAをランダムヘキサマーで逆転写し、sgRNA骨格を増幅するフォワードプライマー及びリバースプライマーでqPCRを実施した。プライマー配列は表E5において入手可能である。
Cas9に基づく転写活性化。96ウェル黒壁プレート(Corning)に、ウェル当たり10,000個のHEK293T細胞が播種された。播種の16~24時間後、製造業者のプロトコルに従って0.5μLのリポフェクタミン2000及び100ngのdCas9-VPRプラスミド、30ngのGFPレポータープラスミド、15ngのiRFPプラスミド及び20ngのsgRNA、pegRNA又はepegRNAプラスミドを用いて細胞が約60%のコンフルエンシーでトランスフェクトされた。3日後、Infinite M1000 Proマイクロプレートリーダー(Tecan)を使用して、GFP及びiRFP蛍光について細胞が測定された。未処理細胞からバックグラウンド蛍光シグナルを差し引いた後、GFP蛍光はiRFP蛍光に対して正規化された。
pegLITによるリンカーデザイン。epegRNAリンカー配列をデザインするために、pegRNAの残りの部分との塩基対合を最小限に抑える指定された長さのリンカー配列を検索するカスタムアルゴリズム、pegRNAリンカー同定ツール、又はpegLITが記載された。この手順は、擬似アニーリングを使用してサブスコアを最大化し、その各々は、pegRNA:スペーサ、PBS、鋳型又は骨格の部分配列に対応する。最適化の間、任意のリンカー対におけるよりスコアの高いリンカーが、以下の部分配列優先順位:スペーサ、PBS、鋳型、次いで骨格の順にそれらの離散化された部分配列を比較することによって決定された。各サブスコアは、リンカー中のヌクレオチドが検討中のpegRNA部分配列中の任意のヌクレオチドと塩基対を形成する平均確率の補数として、標準パラメータ(37℃、1M NaCl、0.05M MgCl2)下でViennaRNA 2.025によって計算された塩基対確率を使用して計算され、ここで平均はリンカー中の全塩基にわたってとられる。AC含有量が50%未満のリンカー配列、及び4つの同じヌクレオチドを連続して含有するpegRNAをもたらすリンカー配列を検討から削除する39,40。任意選択で、アルゴリズムは、100個の最高スコアリンカーに対して階層的凝集クラスタリングを実行し、最終出力における配列多様性を促進するために、クラスター当たり1つのリンカーを出力する。pegLITisのコードが以下に示される。
math import prodから
random import choice,randint,randomから
import heapq
import numpy as np
tqdm import trangeから
scipy.special import expitからsigmoidとして
sklearn.cluster import AgglomerativeClusteringからHACとして
Levenshtein import distanceからlevenshtein_distanceとして
import RNA
BASE_SYMBOLS={
”A”:(”A”,),”C”:(”C”,),”G”:(”G”,),”T”:(”T”,),”U”:(”T”,),
”W”:(”A”,”T”),”S”:(”C”,”G”),”M”:(”A”,”C”),
”K”:(”G”,”T”),”R”:(”A”,”G”),”Y”:(”C”,”T”),
”B”:(”C”,”G”,”T”),”D”:(”A”,”G”,”T”),”H”:(”A”,”C”,”T”),”V”:(”A”,”C”,”G”),
”N”:(”A”,”C”,”G”,”T”)}
def apply_filters(seq_pre,seq_linker,seq_post,ac_thresh,u_thresh,n_thresh):
”””
任意のフィルターが失敗された場合、すなわちAC含有量<ac_thresh OR consecutive Us>u_threshである場合、Falseに戻る。
OR consecutive Ns>n_thresh.そうでなければ、全てのフィルターが合格されればTrueである。全てのしきい値は、
単位nt(すなわち、ac_threshはパーセントではない)。TはUとして扱われる。
”””
#AC含有量
if seq_linker.count(”A”)+seq_linker.count(”C”)<ac_thresh:
Falseに戻る
#Consecutive U
seq_neighborhood=seq_pre[-(u_thresh):]+seq_linker+seq_post[:u_thresh]
seq_neighborhood=seq_neighborhood.replace(”T”,”U”)
seq_neighborhood内の「U」*(u_thresh+1)の場合:
Falseに戻る
#Consecutive N
seq_neighborhood=seq_pre[-(n_thresh):]+seq_linker+seq_post[:n_thresh]
seq_neighborhood=seq_neighborhood.replace(”T”,”U”)
セット(seq_linker)内のntについて、seq_neighborhood内に何らかの(nt*(n_thresh+1))がある場合:
Falseに戻る
Trueに戻る
def calc_subscores(linker_pos,*sequence_components):
”””
各ヌクレオチドについて周縁化された塩基対合プローブを計算する
”””
#ViennaRNAからbppを計算する
pegrna=RNA.fold_compound(””.join(sequence_components))
_=pegrna.pf()#need to first internally calculate partition function
basepair_probs=np.array(pegrna.bpp())[1:,1:]
#ViennaRNAの上三角bppマトリックスの下三角及び対角線を記入する
unpaired_probs=1.-(basepair_probs.sum(axis=0)+basepair_probs.sum(axis=1))
#データをコピーして対称にする
i_lower=np.tril_indices(len(basepair_probs),-1)
i_diag=np.eye(len(basepair_probs),dtype=bool)
basepair_probs[i_lower]=basepair_probs.T[i_lower]
basepair_probs[i_diag]=unpaired_probs
#部分列のトラックインデックス
idx_cur=0
seq_idx=[]
sequence_componentsの部分列について:
idx_prev=idx_cur
idx_cur+=len(subseq)
seq_idx.append(slice(idx_prev,idx_cur))
#部分列のサブスコアを抽出する
bpp_subseq=np.ma.masked_all(len(sequence_components))
iの場合、enumerate(sequence_components)の部分列:
bpp_within_subseq=basepair_probs[seq_idx[i],seq_idx[linker_pos]]
bpp_subseq[i]=np.mean(np.sum(bpp_within_subseq,axis=0))
return bpp_subseq
def apply_score(seq_spacer,seq_scaffold,seq_template,seq_pbs,seq_linker,
score_to_beat=None,epsilon=0.01):
”””
サブコアを計算し、次いでハッシュスコアを出力する。スコアがscore_to_beat未満である場合は早期に計算を終了。
PBS、スペーサ、鋳型、骨格に優先順位を付ける。
”””
Rループサブスコアにおける#Cas9複合体
bpp_subseq1=calc_subscores(2,seq_template,seq_pbs,seq_linker)
subscore_pbs=1.-bpp_subseq1[1]
subscore_template=1.-bpp_subseq1[0]
#遊離pegRNAサブスコア
もし((score_to_beatはNoneではない)
及び(epsilon*int(subscore_pbs/epsilon)<score_to_beat[0])):
subscore_spacer=0.
subscore_scaffold=0.
その他:
bpp_subseq2=calc_subscores(4,seq_spacer,seq_scaffold,
seq_template,seq_pbs,seq_linker)
subscore_spacer=1.-bpp_subseq2[0]
subscore_scaffold=1.-bpp_subseq2[1]
#サブスコアを単一のスコアに変換する
tuple(
epsilon*int(val/epsilon)に戻る
valがNoneでない場合0
(subscore_pbs,subscore_spacer,subscore_template,subscore_scaffold)のvalについて
)
def optimize(seq_spacer,seq_scaffold,seq_template,seq_pbs,seq_motif,
linker_pattern,ac_thresh,u_thresh,n_thresh,topn,epsilon,
num_repeats,num_steps,temp_init,temp_decay):
”””
リンカーの模擬アニーリング最適化
”””
##前処理入力
seq_pre=seq_spacer+seq_scaffold+seq_template+seq_pbs
seq_post=seq_motif
ac_thresh=ac_thresh*len(linker_pattern)
##リンカー配列を最適化するための模擬アニーリング
#すでに考慮されたシーケンスのハッシュマップを初期化する
linker_skip={}
len_sequence_space=prod(len(BASE_SYMBOLS[nt])fornt in linker_pattern)
#トップリンカーの最小の山を初期化する
linker_heap=[]
for_in trange(num_repeats,desc=”Repeats”,position=0,leave=False):
#擬似アニーリングの初期化
seq_linker_prev=””.join([choice(BASE_SYMBOLS[nt])fornt in linker_pattern])
score_prev=None
temp=temp_init
for_in trange(num_steps,desc=”Steps”,position=1,leave=False):
#パスフィルターまで文字を順番に置換することによって新しい配列を生成する
seq_linker=seq_linker_prev
keep_going=True
while keep_going:
char_pos=randint(0,len(linker_pattern)-1)
seq_linker=(
seq_linker[:char_pos]
+choice(BASE_SYMBOLS[linker_pattern[char_pos]])
+seq_linker[(char_pos+1):])
keep_going=(
seq_linker in linker_skip
or not apply_filters(seq_pre,seq_linker,seq_post,
ac_thresh,u_thresh,n_thresh)
or len(linker_skip)>=len_sequence_space)#already screened whole seq space
linker_skip[seq_linker]=True
#リンカー配列のスコアを計算する
score_to_beat=linker_heap[0][0]if len(linker_heap)>=topn else None
score=apply_score(seq_spacer,seq_scaffold,seq_template,seq_pbs,seq_linker,
score_to_beat=score_to_beat,epsilon=epsilon)
#min heapに加算する、すなわち、上位「トップ」の最大エントリーを維持する
score_to_beatがNoneの場合:#heapはまだ一杯ではない
heapq.heappush(linker_heap,(score,seq_linker))
elif score>score_to_beat:
heapq.heapreplace(linker_heap,(score,seq_linker))
#提案を続行するか否かを決定
(score_prev is None#initialize
又はスコア>score_prev#エクスプロイト改善
or random()<sigmoid(#explore
sum((s1-s2)*(epsilon**i)
iの場合、列挙(zip(スコア、score_prev)))の(s1,s 2))/temp
)):
seq_linker_prev=seq_linker
score_prev=score
#擬似アニーリングのパラメータを更新
temp *=temp_decay
linker_heap_scores,linker_heap=zip(*linker_heap)
return linker_heap_scores,linker_heap
def apply_bottleneck(heap_scores,heap,bottleneck):
”””
クラスター配列及びクラスターごとの最高得点配列の出力。
”””
#最良の出力を選択するだけでよい
ボトルネック==1の場合:
return heap[np.argmax(heap_scores)]
#各リンカー配列の特徴を計算する、すなわち他の全てのリンカー配列との編集距離
features=np.zeros((len(heap),len(heap)),dtype=int)
for i,seq_x in enumerate(heap):
for j,seq_y in enumerate(heap):
features[i,j]=levenshtein_distance(seq_x,seq_y)
#クラスターリンカー配列
clusters=HAC(n_clusters=bottleneck,linkage=”complete”).fit_predict(features)
#各クラスターから最高得点のリンカー配列を出力する
output=[]
heap_scores=np.array(heap_scores)
範囲内のcluster_num(ボトルネック)について:
cluster_scores=clusters==cluster_num
for subscore_num in range(heap_scores.shape[1]):
subscore_maxed=(heap_scores[:,subscore_num]
==np.max(heap_scores[cluster_scores,subscore_num]))
cluster_scores=np.logical_and(cluster_scores,subscore_maxed)
idx_maxed=np.where(cluster_scores)[0]
output.append(heap[np.random.choice(idx_maxed)])
出力に戻る
def pegLIT(seq_spacer,seq_scaffold,seq_template,seq_pbs,seq_motif,
linker_pattern=”NNNNNNNN”,ac_thresh=0.5,u_thresh=3,n_thresh=3,topn=100,
epsilon=1e-2,num_repeats=10,num_steps=250,temp_init=0.15,temp_decay=0.95,
bottleneck=1):
”””
入力されたpegRNAに対して+ボトルネックのリンカーを最適化する。リンカー推奨(複数可)を出力する。
”””
##リンカー配列を最適化するための模擬アニーリング
linker_heap_scores,linker_heap=optimize(
seq_spacer,seq_scaffold,seq_template,seq_pbs,seq_motif,
linker_pattern=linker_pattern,ac_thresh=ac_thresh,u_thresh=u_thresh,
n_thresh=n_thresh,topn=topn,epsilon=epsilon,num_repeats=num_repeats,
num_steps=num_steps,temp_init=temp_init,temp_decay=temp_decay)
#多様な配列のサンプル
linker_output=apply_bottleneck(linker_heap_scores,linker_heap,bottleneck=bottleneck)
return linker_output
#HEK3+1 FLAGインの使用例
print(pegLIT(
seq_spacer=”GGCCCAGACTGAGCACGTGA”,
seq_scaffold=”GTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTAT”
”CAACTTGAAAAAGTGGCACCGAGTCGGTGC”,
seq_template=”TGGAGGAAGCAGGGCTTCCTTTCCTCTGCCATCACTTATCG”
”TCGTCATCCTTGTAATC”,
seq_pbs=”CGTGCTCAGTCTG”,
seq_motif=”CGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAA”))
示されている配列は、seq_spacer(配列番号490);seq_scaffold(top)(配列番号491);seq_scaffold(bottom)(配列番号492);seq_template(top)(配列番号493);seq_template(bottom)(配列番号494);seq_pbs(配列番号495);seq_motif(配列番号219)である。
math import prodから
random import choice,randint,randomから
import heapq
import numpy as np
tqdm import trangeから
scipy.special import expitからsigmoidとして
sklearn.cluster import AgglomerativeClusteringからHACとして
Levenshtein import distanceからlevenshtein_distanceとして
import RNA
BASE_SYMBOLS={
”A”:(”A”,),”C”:(”C”,),”G”:(”G”,),”T”:(”T”,),”U”:(”T”,),
”W”:(”A”,”T”),”S”:(”C”,”G”),”M”:(”A”,”C”),
”K”:(”G”,”T”),”R”:(”A”,”G”),”Y”:(”C”,”T”),
”B”:(”C”,”G”,”T”),”D”:(”A”,”G”,”T”),”H”:(”A”,”C”,”T”),”V”:(”A”,”C”,”G”),
”N”:(”A”,”C”,”G”,”T”)}
def apply_filters(seq_pre,seq_linker,seq_post,ac_thresh,u_thresh,n_thresh):
”””
任意のフィルターが失敗された場合、すなわちAC含有量<ac_thresh OR consecutive Us>u_threshである場合、Falseに戻る。
OR consecutive Ns>n_thresh.そうでなければ、全てのフィルターが合格されればTrueである。全てのしきい値は、
単位nt(すなわち、ac_threshはパーセントではない)。TはUとして扱われる。
”””
#AC含有量
if seq_linker.count(”A”)+seq_linker.count(”C”)<ac_thresh:
Falseに戻る
#Consecutive U
seq_neighborhood=seq_pre[-(u_thresh):]+seq_linker+seq_post[:u_thresh]
seq_neighborhood=seq_neighborhood.replace(”T”,”U”)
seq_neighborhood内の「U」*(u_thresh+1)の場合:
Falseに戻る
#Consecutive N
seq_neighborhood=seq_pre[-(n_thresh):]+seq_linker+seq_post[:n_thresh]
seq_neighborhood=seq_neighborhood.replace(”T”,”U”)
セット(seq_linker)内のntについて、seq_neighborhood内に何らかの(nt*(n_thresh+1))がある場合:
Falseに戻る
Trueに戻る
def calc_subscores(linker_pos,*sequence_components):
”””
各ヌクレオチドについて周縁化された塩基対合プローブを計算する
”””
#ViennaRNAからbppを計算する
pegrna=RNA.fold_compound(””.join(sequence_components))
_=pegrna.pf()#need to first internally calculate partition function
basepair_probs=np.array(pegrna.bpp())[1:,1:]
#ViennaRNAの上三角bppマトリックスの下三角及び対角線を記入する
unpaired_probs=1.-(basepair_probs.sum(axis=0)+basepair_probs.sum(axis=1))
#データをコピーして対称にする
i_lower=np.tril_indices(len(basepair_probs),-1)
i_diag=np.eye(len(basepair_probs),dtype=bool)
basepair_probs[i_lower]=basepair_probs.T[i_lower]
basepair_probs[i_diag]=unpaired_probs
#部分列のトラックインデックス
idx_cur=0
seq_idx=[]
sequence_componentsの部分列について:
idx_prev=idx_cur
idx_cur+=len(subseq)
seq_idx.append(slice(idx_prev,idx_cur))
#部分列のサブスコアを抽出する
bpp_subseq=np.ma.masked_all(len(sequence_components))
iの場合、enumerate(sequence_components)の部分列:
bpp_within_subseq=basepair_probs[seq_idx[i],seq_idx[linker_pos]]
bpp_subseq[i]=np.mean(np.sum(bpp_within_subseq,axis=0))
return bpp_subseq
def apply_score(seq_spacer,seq_scaffold,seq_template,seq_pbs,seq_linker,
score_to_beat=None,epsilon=0.01):
”””
サブコアを計算し、次いでハッシュスコアを出力する。スコアがscore_to_beat未満である場合は早期に計算を終了。
PBS、スペーサ、鋳型、骨格に優先順位を付ける。
”””
Rループサブスコアにおける#Cas9複合体
bpp_subseq1=calc_subscores(2,seq_template,seq_pbs,seq_linker)
subscore_pbs=1.-bpp_subseq1[1]
subscore_template=1.-bpp_subseq1[0]
#遊離pegRNAサブスコア
もし((score_to_beatはNoneではない)
及び(epsilon*int(subscore_pbs/epsilon)<score_to_beat[0])):
subscore_spacer=0.
subscore_scaffold=0.
その他:
bpp_subseq2=calc_subscores(4,seq_spacer,seq_scaffold,
seq_template,seq_pbs,seq_linker)
subscore_spacer=1.-bpp_subseq2[0]
subscore_scaffold=1.-bpp_subseq2[1]
#サブスコアを単一のスコアに変換する
tuple(
epsilon*int(val/epsilon)に戻る
valがNoneでない場合0
(subscore_pbs,subscore_spacer,subscore_template,subscore_scaffold)のvalについて
)
def optimize(seq_spacer,seq_scaffold,seq_template,seq_pbs,seq_motif,
linker_pattern,ac_thresh,u_thresh,n_thresh,topn,epsilon,
num_repeats,num_steps,temp_init,temp_decay):
”””
リンカーの模擬アニーリング最適化
”””
##前処理入力
seq_pre=seq_spacer+seq_scaffold+seq_template+seq_pbs
seq_post=seq_motif
ac_thresh=ac_thresh*len(linker_pattern)
##リンカー配列を最適化するための模擬アニーリング
#すでに考慮されたシーケンスのハッシュマップを初期化する
linker_skip={}
len_sequence_space=prod(len(BASE_SYMBOLS[nt])fornt in linker_pattern)
#トップリンカーの最小の山を初期化する
linker_heap=[]
for_in trange(num_repeats,desc=”Repeats”,position=0,leave=False):
#擬似アニーリングの初期化
seq_linker_prev=””.join([choice(BASE_SYMBOLS[nt])fornt in linker_pattern])
score_prev=None
temp=temp_init
for_in trange(num_steps,desc=”Steps”,position=1,leave=False):
#パスフィルターまで文字を順番に置換することによって新しい配列を生成する
seq_linker=seq_linker_prev
keep_going=True
while keep_going:
char_pos=randint(0,len(linker_pattern)-1)
seq_linker=(
seq_linker[:char_pos]
+choice(BASE_SYMBOLS[linker_pattern[char_pos]])
+seq_linker[(char_pos+1):])
keep_going=(
seq_linker in linker_skip
or not apply_filters(seq_pre,seq_linker,seq_post,
ac_thresh,u_thresh,n_thresh)
or len(linker_skip)>=len_sequence_space)#already screened whole seq space
linker_skip[seq_linker]=True
#リンカー配列のスコアを計算する
score_to_beat=linker_heap[0][0]if len(linker_heap)>=topn else None
score=apply_score(seq_spacer,seq_scaffold,seq_template,seq_pbs,seq_linker,
score_to_beat=score_to_beat,epsilon=epsilon)
#min heapに加算する、すなわち、上位「トップ」の最大エントリーを維持する
score_to_beatがNoneの場合:#heapはまだ一杯ではない
heapq.heappush(linker_heap,(score,seq_linker))
elif score>score_to_beat:
heapq.heapreplace(linker_heap,(score,seq_linker))
#提案を続行するか否かを決定
(score_prev is None#initialize
又はスコア>score_prev#エクスプロイト改善
or random()<sigmoid(#explore
sum((s1-s2)*(epsilon**i)
iの場合、列挙(zip(スコア、score_prev)))の(s1,s 2))/temp
)):
seq_linker_prev=seq_linker
score_prev=score
#擬似アニーリングのパラメータを更新
temp *=temp_decay
linker_heap_scores,linker_heap=zip(*linker_heap)
return linker_heap_scores,linker_heap
def apply_bottleneck(heap_scores,heap,bottleneck):
”””
クラスター配列及びクラスターごとの最高得点配列の出力。
”””
#最良の出力を選択するだけでよい
ボトルネック==1の場合:
return heap[np.argmax(heap_scores)]
#各リンカー配列の特徴を計算する、すなわち他の全てのリンカー配列との編集距離
features=np.zeros((len(heap),len(heap)),dtype=int)
for i,seq_x in enumerate(heap):
for j,seq_y in enumerate(heap):
features[i,j]=levenshtein_distance(seq_x,seq_y)
#クラスターリンカー配列
clusters=HAC(n_clusters=bottleneck,linkage=”complete”).fit_predict(features)
#各クラスターから最高得点のリンカー配列を出力する
output=[]
heap_scores=np.array(heap_scores)
範囲内のcluster_num(ボトルネック)について:
cluster_scores=clusters==cluster_num
for subscore_num in range(heap_scores.shape[1]):
subscore_maxed=(heap_scores[:,subscore_num]
==np.max(heap_scores[cluster_scores,subscore_num]))
cluster_scores=np.logical_and(cluster_scores,subscore_maxed)
idx_maxed=np.where(cluster_scores)[0]
output.append(heap[np.random.choice(idx_maxed)])
出力に戻る
def pegLIT(seq_spacer,seq_scaffold,seq_template,seq_pbs,seq_motif,
linker_pattern=”NNNNNNNN”,ac_thresh=0.5,u_thresh=3,n_thresh=3,topn=100,
epsilon=1e-2,num_repeats=10,num_steps=250,temp_init=0.15,temp_decay=0.95,
bottleneck=1):
”””
入力されたpegRNAに対して+ボトルネックのリンカーを最適化する。リンカー推奨(複数可)を出力する。
”””
##リンカー配列を最適化するための模擬アニーリング
linker_heap_scores,linker_heap=optimize(
seq_spacer,seq_scaffold,seq_template,seq_pbs,seq_motif,
linker_pattern=linker_pattern,ac_thresh=ac_thresh,u_thresh=u_thresh,
n_thresh=n_thresh,topn=topn,epsilon=epsilon,num_repeats=num_repeats,
num_steps=num_steps,temp_init=temp_init,temp_decay=temp_decay)
#多様な配列のサンプル
linker_output=apply_bottleneck(linker_heap_scores,linker_heap,bottleneck=bottleneck)
return linker_output
#HEK3+1 FLAGインの使用例
print(pegLIT(
seq_spacer=”GGCCCAGACTGAGCACGTGA”,
seq_scaffold=”GTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTAT”
”CAACTTGAAAAAGTGGCACCGAGTCGGTGC”,
seq_template=”TGGAGGAAGCAGGGCTTCCTTTCCTCTGCCATCACTTATCG”
”TCGTCATCCTTGTAATC”,
seq_pbs=”CGTGCTCAGTCTG”,
seq_motif=”CGCGGTTCTATCTAGTTACGCGTTAAACCAACTAGAA”))
示されている配列は、seq_spacer(配列番号490);seq_scaffold(top)(配列番号491);seq_scaffold(bottom)(配列番号492);seq_template(top)(配列番号493);seq_template(bottom)(配列番号494);seq_pbs(配列番号495);seq_motif(配列番号219)である。
実施例5:プライム編集を改善するための追加戦略
プライム編集を改善するための更なる戦略も開発された。これらは、図131に示されるように、プライム編集が改善され得る3つの広い領域を包含する:1)標的核酸の認識;2)編集の組み入れ;及び3)編集されたDNAヘテロ二本鎖の分割。以下の実施例は、標的核酸の認識を改善することによって、特にPBSとpegRNA上のスペーサ配列との間の相互作用を減少させることによって、編集効率を高めることに焦点を合わせている。sgRNAと比較して、pegRNA及びepegRNAは、標的部位のエンゲージメントが低下し、Cas9への結合が低下してもよい。PBS:スペーサ相互作用は、Cas9親和性を低下させることによってプライム編集効率を制限し得る(図132)。しかしながら、そのような相互作用は、PBS:プロトスペーサ結合が起こるためにも必要である。図132に示されるように、より短いPBSは、Cas9に対する改善された結合親和性をもたらし得る。したがって、PBS:スペーサ相互作用を減少させるための戦略は、1)Cas9結合時に解離する足がかりによるPBSの閉塞、;2)ニッキングsgRNAを介したトランスでのpegRNA鋳型の送達;及び3)PBS:スペーサ及びPBS:プロトスペーサ相互作用に差次的に影響を及ぼす化学修飾及び/又は遺伝子修飾を導入することを包含して調査された。
プライム編集を改善するための更なる戦略も開発された。これらは、図131に示されるように、プライム編集が改善され得る3つの広い領域を包含する:1)標的核酸の認識;2)編集の組み入れ;及び3)編集されたDNAヘテロ二本鎖の分割。以下の実施例は、標的核酸の認識を改善することによって、特にPBSとpegRNA上のスペーサ配列との間の相互作用を減少させることによって、編集効率を高めることに焦点を合わせている。sgRNAと比較して、pegRNA及びepegRNAは、標的部位のエンゲージメントが低下し、Cas9への結合が低下してもよい。PBS:スペーサ相互作用は、Cas9親和性を低下させることによってプライム編集効率を制限し得る(図132)。しかしながら、そのような相互作用は、PBS:プロトスペーサ結合が起こるためにも必要である。図132に示されるように、より短いPBSは、Cas9に対する改善された結合親和性をもたらし得る。したがって、PBS:スペーサ相互作用を減少させるための戦略は、1)Cas9結合時に解離する足がかりによるPBSの閉塞、;2)ニッキングsgRNAを介したトランスでのpegRNA鋳型の送達;及び3)PBS:スペーサ及びPBS:プロトスペーサ相互作用に差次的に影響を及ぼす化学修飾及び/又は遺伝子修飾を導入することを包含して調査された。
最初に、Cas9結合時に解離する足がかりでPBSを閉塞する戦略が調査された。足がかりは、Cas9結合とは無関係であれば、PBS:スペーサ及びPBS:プロトスペーサ相互作用の両方を阻害し得ることが観察された(図133)。MS2ヘアピンがpegRNAの3'末端に融合され、MS2バクテリオファージコートタンパク質は、プライム編集因子の逆転写酵素に融合された。図134に示すように、足がかりは、競合的RNA-タンパク質相互作用に起因するPE2結合によって競合され得る。この戦略を使用する場合、1)Cas9-RTリンカー及びRT-MS2リンカーの両方の長さ、pegRNA伸長及びPBSリンカー、足がかりリンカー、並びにMS2アプタマーと足がかりとの間のリンカーの相互依存性;2)足がかり長さのPBS融解温度及び部位アクセスしやすさに対する依存性;3)部位毎の最適化;及び4)非相互作用17ヌクレオチドPBSに対する寛容を包含する、いくつかのデザイン上の考慮がされるべきである。MS2とPE2とのN末端及びC末端融合物の両方が試験された。C末端MS2融合物の使用は、HEK3におけるN末端融合物よりも優れた編集効率をもたらすことが発見された(図135)。PE2のMS2タグ付けは、様々なpegRNAを使用するタグなしのPE2と比較して、編集効率において利点を提供することが観察された(図136)。xten-16 aaリンカー又はxten-33 aaリンカーのいずれかを含むPE2-MS2融合物を両方とも、MS2融合物を含まないPE2-xtenに対して試験された。MS2及びトゥーループタグ付けが、長いプライマー結合部位を救済することも観察された(図137)。全体として、Cas9結合時に解離する足がかりでPBSを閉塞する戦略は、様々なゲノム部位での編集効率においていくつかの利点を示し、特に、低いPBS:プロトスペーサ安定性に起因してより低い効率で通常編集されるものを示す。また、epegRNAモチーフは二機能性であり得、pegRNA安定性を改善することが示された。
次に、ニッキングsgRNAを介してpegRNA鋳型をトランスで送達する戦略が調査された。PBS-スペーサ相互作用を完全に回避するために、pegRNA伸長をニッキングガイド上に移動させなければならないという仮説が立てられた(図138)。この戦略を使用する場合、:1)フラップ分解能に対するリンカーとしての拡張鋳型の影響;2)ニッキングスペーサの最適化;及び3)両方のPE複合体がゲノム上に同時に存在する必要性、を包含するいくつかのデザイン上の考慮事項も考慮に入れられるべきである。このストラテジーは、DMNT 1、HEK3、PRNP、RUNX1及びVEGFAにおけるプライム編集を可能にすることが観察された(図139)。
図140は、ニックに対するPBS内のミスマッチ同一性及び位置に基づくモデルを示す。
図141は、PBSに対する突然変異が許容されるか、又はいくつかの状況ではPE活性を増強し、突然変異の位置及び同一性がPE効率を決定する初期モデルに適合することを示す。
図142は、より長いPBS(RNF2、15 nt)は、潜在的にPBS:プロトスペーサ相互作用を過度に阻害するので、突然変異を許容しないことを示す。
図143は、PBSに対する変異が、より短い最適PBSを有するpegRNAについてのPE効率を改善し得ることを示す。MutPBS epegRNAは、4つの連続した突然変異(HEK3、DNMT1、PRNP)を有する17のmutPBS、又は4つの連続した突然変異(RNF2)を有する15のmutPBS、続いて8 ntリンカー及びtevopreQ1を有する。
図144は、mutPBSの改善が、epegRNAと組み合わせて使用された場合、編集効率の更なる向上を提供し得ることを示す。
図145は、ツインプライム編集の概略図を提供する。フラップは外因性であり、互いに塩基対を形成することしかできないため、ツインプライム編集は大きな編集を行うのに特に有用である。
図146は、編集効率を改善するために競合的相同性を減少させるためのツインプライム編集における介在領域のニッキングを示す。余分なニック(又は複数のニック)は、2つのフラップ間のゲノムの領域を分解し、中間体の複雑さを低減し、収量を改善する。
図147は、アクセサリニックを有するMECP2ツインプライム編集を示す。
実施例6:プライム編集によるCDKL5欠損障害の処置
CDKL5欠損症は、出生後の人生の早い時期に始まる発作を特徴とする遺伝性疾患であり、その後、発達の多くの側面において遅延する。CDKL5欠損症に関連する発作は、年齢とともに性質が変化することがしばしば見出される。罹患者が経験する最も一般的なタイプの発作は、全般性強直間代発作(別名、大発作)と呼ばれ、意識消失、筋硬直、及び身体痙攣を伴う。強直発作は、CDKL5欠損症に関連する別の主要なタイプの発作を表し、異常な筋収縮により特徴付けられ得る。別の一般的な発作タイプはてんかん性攣縮であり、これは不随意筋攣縮の短いエピソードを伴う。発作は、CDKL5欠損症のほとんどの人々において毎日起こるが、発作のない期間が経験され得る。CDKL5欠損症における発作は、典型的には、処置に対して抵抗性である。
CDKL5欠損症は、出生後の人生の早い時期に始まる発作を特徴とする遺伝性疾患であり、その後、発達の多くの側面において遅延する。CDKL5欠損症に関連する発作は、年齢とともに性質が変化することがしばしば見出される。罹患者が経験する最も一般的なタイプの発作は、全般性強直間代発作(別名、大発作)と呼ばれ、意識消失、筋硬直、及び身体痙攣を伴う。強直発作は、CDKL5欠損症に関連する別の主要なタイプの発作を表し、異常な筋収縮により特徴付けられ得る。別の一般的な発作タイプはてんかん性攣縮であり、これは不随意筋攣縮の短いエピソードを伴う。発作は、CDKL5欠損症のほとんどの人々において毎日起こるが、発作のない期間が経験され得る。CDKL5欠損症における発作は、典型的には、処置に対して抵抗性である。
CDKL5欠損症はまた、小児における発達障害に関連される。そのような小児は、著しく制限された発話を伴う重度の知的障害を有する。更に、特定の個体では、粗大な運動技能(例として、歩くこと、座ること、立つこと)の発達が遅れているか、又は完全に欠如している。実際、影響を受けた個体の約1/3しか補助なしで歩行することができない。微細な運動技能も損なわれており、罹患した個人の約半分のみが手を有意に使用している。視力もまた、CDKL5欠損症に罹患した多くの個体において損なわれている。
CDKL5欠損症は、CDKL5遺伝子の変異によって引き起こされる。この遺伝子は、正常な脳の発達及び機能に不可欠なタンパク質を作製するための指示を提供する。特に、CDKL5遺伝子の突然変異は、機能性CDKL5タンパク質の量を減少させるか、又はニューロンにおけるその活性を変化させる。CDKL5の不足(欠乏)又はその機能の障害は脳の発達を乱すが、これらの変化がCDKL5欠乏障害の特定の特徴をどのように引き起こすかは不明である。
CDKL5変異/欠損の現在の処置は、主に症状の管理に焦点を当てられている。しかしながら、現在のところ、CDKL5の突然変異又は欠損を有する対象の神経学的結果を改善する処置法、又は障害を引き起こすCDKL5遺伝子の突然変異を修正し得る処置法はない。したがって、CDKL5欠損症を処置するための遺伝子処置アプローチが必要とされている。
本開示では、図148に示すように、プライム編集因子(例として、PE2)がpegRNAと複合体化して使用されて、CDKL5遺伝子の複数の病原性変異を同時に修正した(V172I、A173D、R175S、W176G、W176R、Y177C、R178P、P180L、E181A、及びL182P突然変異の修正を包含する)。単一のpegRNAと複合体を形成した単一のプライム編集因子(例として、PE2)も、PAM配列の1位に対して+4、+8、+12、+17、+21、及び+25の位置で多数の病原性変異を修正することができることが示された(すなわち、ヌクレオチドは、5'-最も高い位置にある;図149)。
実施例7:単一のガイドRNAを使用してマウスのCDKL5における複数の変異を補正する方法
以下の実施例は、マウス細胞における病原性1412delA変異の組み入れの最適化を記載する。これらの細胞は神経芽細胞腫に由来され、CDKL5欠損症(CDD)は主に神経疾患であるので、N2A細胞がこの研究に使用された。pegRNA及びニッキングガイドを最適化し、この突然変異を組み入れするために、広範な努力が行われた。DNAプラスミドトランスフェクションを使用するこのような最適化の一例が図150に提供される。PE系、ニッキングガイド、及びpegRNAパラメータは、X軸上に詳述されている。「13_20」のpegRNAは、エレクトロポレーションのためのその後の合成pegRNAに使用された。
以下の実施例は、マウス細胞における病原性1412delA変異の組み入れの最適化を記載する。これらの細胞は神経芽細胞腫に由来され、CDKL5欠損症(CDD)は主に神経疾患であるので、N2A細胞がこの研究に使用された。pegRNA及びニッキングガイドを最適化し、この突然変異を組み入れするために、広範な努力が行われた。DNAプラスミドトランスフェクションを使用するこのような最適化の一例が図150に提供される。PE系、ニッキングガイド、及びpegRNAパラメータは、X軸上に詳述されている。「13_20」のpegRNAは、エレクトロポレーションのためのその後の合成pegRNAに使用された。
次に、N2A細胞は、in vitro転写されたPE mRNA、合成epegRNA、及び合成ガイドRNA(PE3)、又はmMLH1 neg mRNAを有する上述の基質(PE5)のいずれかと電気穿孔された(図151)。ニッキングガイド位置(NG1対NG3)も変化された。NG1を含むPE5系が最も高い組み入れ率を与え、NG3を含むPE5が最良の所望の編集対インデル比を与えると結論付けられた。
その後、同様の実験が、epegRNAによってコードされるシード編集、同様にまた、所望される1412delA変異(図152)を加えて行われた。シード編集はサイレントであり、したがって、アミノ酸配列が変化しないので病原性であると予想されない。PE5b戦略のためのニッキングガイドの試験に加えて、2つの標準的なニッキングガイドがPE5(NG1、NG3)に使用された。逆転写酵素鋳型の長さは、pegRNA 081と082とで異なる。pegRNA 081が最も効率的であり、約70%の効率で所望の突然変異を組み入れしたと結論付けられた。
CDKL5はX染色体上の優性突然変異によって引き起こされるので、女性患者は通常、1つの健康な対立遺伝子を有する。この健康な対立遺伝子におけるインデルの効果は不明であり、標準的なPE(SpCas9 PE)を標的とすることができる。SpCas9-NRCH PEを必要とする新しいpegRNAをデザインし(表E7)、1)健康な対立遺伝子を標的とせず、2)PAMが破壊されるので、最初の編集事象が起こると、その後の編集のための良好な基質にならないであろう。
最後に、1つのpegRNAが使用されて、複数の病原性対立遺伝子を補正した。突然変異はデノボであるため、任意の2人の患者が同じ突然変異を有することは極めてまれである。しかしながら、CDKL5遺伝子には、これらの病原性変異を有する可能性が他よりも高い遺伝子座が存在する。そのような遺伝子座の1つはエクソン8である。プラスミドトランスフェクション(図153)によって、複数の病原性CDKL5対立遺伝子がHEK293T細胞に組み入れされた。2つのpegRNAパラメータがX軸に記載される。
実施例8:PE4及びPE5を用いたCDKL5軌跡のプライム編集
PE4max及びPE5max(図154及び図155)が適用されて、CDKL5欠損症、重度の神経発達状態(Olson et al.,2019)の原因突然変異を含有することが知られているCDKL5部位にサイレントC・GからT・A突然変異を導入した。PE4maxは、平均プライム編集効率をPE2よりもHeLa細胞で29倍及びHEK293T細胞で2.1倍増加させることが観察された(図156)。注意すべきことに、PE4max編集効率(HeLa細胞では0.19%インデルで8.6%編集、HEK293T細胞では0.26%インデルで20%編集)は、PE3(HeLa細胞では1.5%インデルで4.5%編集、HEK293T細胞では5.4%インデルで24%編集)のものと同様又はそれを超えたが、インデルははるかに少なかった。更に、PE5maxは、疾患に関連する対立遺伝子変換をPE3よりもHeLa細胞で平均6.1倍及びHEK293T細胞で1.5倍改善し、編集:インデル純度をHeLa細胞で6.4倍及びHEK293T細胞で3.5倍向上させた(図156)。
PE4max及びPE5max(図154及び図155)が適用されて、CDKL5欠損症、重度の神経発達状態(Olson et al.,2019)の原因突然変異を含有することが知られているCDKL5部位にサイレントC・GからT・A突然変異を導入した。PE4maxは、平均プライム編集効率をPE2よりもHeLa細胞で29倍及びHEK293T細胞で2.1倍増加させることが観察された(図156)。注意すべきことに、PE4max編集効率(HeLa細胞では0.19%インデルで8.6%編集、HEK293T細胞では0.26%インデルで20%編集)は、PE3(HeLa細胞では1.5%インデルで4.5%編集、HEK293T細胞では5.4%インデルで24%編集)のものと同様又はそれを超えたが、インデルははるかに少なかった。更に、PE5maxは、疾患に関連する対立遺伝子変換をPE3よりもHeLa細胞で平均6.1倍及びHEK293T細胞で1.5倍改善し、編集:インデル純度をHeLa細胞で6.4倍及びHEK293T細胞で3.5倍向上させた(図156)。
次に、PE4及びPE5編集系を、遺伝性疾患の細胞モデル及び初代ヒト細胞において評価された。この対立遺伝子についてヘテロ接合性の患者に由来するヒト人工多能性幹細胞(iPSC)における病原性CDKL5 c.1412delA変異が修正された(Chen et al.,2021)。これらのiPSCのPE3構成要素(in vitro転写されたPE2 mRNA及び合成pegRNA並びにニッキングsgRNA)とのエレクトロポレーションにより、編集可能な病原性対立遺伝子の17%の補正及び20%の総インデル産物が得られた(図157及び図158)。PE5編集のためのMLH1dn mRNAによるこれらの構成要素の同時エレクトロポレーションは、補正効率を34%に上昇させ、インデルの頻度を6.1%に低下させた。インデルを更に最小化するために、PE4及びPE5b戦略も使用された。相補鎖ニッキングなしでは、MLH1dnは対立遺伝子補正を4.0%(PE2)から10%(PE4)に改善し、インデルはほとんどなかった(<0.34%)ことが観察された(図157及び図158)。同様に、PE3bは4.8%インデルで変異対立遺伝子の13%の編集をもたらしたが、PE5bは3.8%インデルで編集を27%に上昇させた。試験したPE4、PE5及びPE5b系にわたって、MLH1dnは、患者由来iPSCにおいて効率が2.2倍及び転帰純度が3.6倍の病原性CDKL5 c.1412delA変異の補正を増強する。
MLH1dn及びepegRNAもCDKL5編集のために組み合わせられた(図159)。HEK293T細胞におけるCDKL5 c.1412 A~G変異の編集効率は、MLH1dn(PE4及びPE5)及びepegRNAの使用によって増加された。最後に、ニッキングsgRNAは、CDKL5でのプライム編集のために最適化された(図160)。HEK293T細胞におけるCDKL5サイレント+1 CからTへの変異(c.1412delA部位)の組み入れの編集効率は、このような最適化によって増加された。この実施例で使用したガイドRNAの配列は表E8に提供される。
実施例9:CDKL5軌跡を編集するためのPAMバリアントプライム編集因子
PE4及びPE5が使用されてヘテロ接合ヒト患者由来人工多能性幹細胞のCDKL5 c.1412delA変異を修正した場合、意図したプライム編集に加えて高レベルの挿入及び欠失副産物(インデル)が観察された。これらのインデルの多くは、c.1412delA変異を含有しない野生型対立遺伝子の試みられたプライム編集によって引き起こされると仮定された。具体的には、c.1412delA変異はSpCas9-PEの標的プロトスペーサから遠く離れており、このプロトスペーサは野生型対立遺伝子であっても依然としてニックを入れられ、それによってインデル副産物を生成する(図161)。
PE4及びPE5が使用されてヘテロ接合ヒト患者由来人工多能性幹細胞のCDKL5 c.1412delA変異を修正した場合、意図したプライム編集に加えて高レベルの挿入及び欠失副産物(インデル)が観察された。これらのインデルの多くは、c.1412delA変異を含有しない野生型対立遺伝子の試みられたプライム編集によって引き起こされると仮定された。具体的には、c.1412delA変異はSpCas9-PEの標的プロトスペーサから遠く離れており、このプロトスペーサは野生型対立遺伝子であっても依然としてニックを入れられ、それによってインデル副産物を生成する(図161)。
インデルを緩和するために、c.1412delA変異の存在下でのみDNAを標的としてニックを入れるプライム編集因子が開発された。したがって、NRCH及びNRTH SpCas9変異体を使用するプライム編集因子を作製した(国際公開第2020/041751号に記載されているように)。NRCH SpCas9-PE及びNRTH SpCas9-PEは、野生型CDKL5対立遺伝子に結合し、ニックを入れられないように、c.1412delA変異を特異的に標的とし得る(図162及び図163)。
したがって、NRCH SpCas9-PE及びNRTH SpCas9-PEは、突然変異が存在する場合にのみCDKL5 c.1412delA突然変異を修正し、インデル副産物を最小限に抑えるはずである。この戦略で使用されるpegRNA及びニッキングsgRNA配列は表E9に提供される。
実施例10:CDKL5変異の組み入れのためのプライム編集因子
様々なプライマー結合部位(PBS)及び鋳型長を有するプライム編集ガイドRNA(pegRNA)がスクリーニングされて、PE2プライム編集因子を使用して、HEK293T細胞のCDKL5遺伝子におけるc.1412のトランジション点変異の最も効率的な組み入れを可能にするものが同定された(図164)次に、PE3プライム編集因子システムで使用されるニッキングガイドの選択が最適化され、c.1412で編集の効率を更に改善した(図165)。編集効率を更に改善するために、コードサイレントトランジションもまた、pegRNAによって標的化されたプロトスペーサのシード領域内に組み込まれた(図165)。この戦略で使用されるガイドRNA配列を表E10に提供する。全体として、pegRNA CDKL5h37及びepegRNA JNpeg 0953が最も高い編集効率を示した。
配列
表E1~E6の以下の配列は、実施例4及び関連する図面を通して参照される。


























































































































































































































実施例4の参考文献
1.Komor,A.C.,Badran,A.H.&Liu,D.R.CRISPR-Based Technologies for the Manipulation of Eukaryotic Genomes.Cell 168,20-36(2017).
2.Anzalone,A.V.,Koblan,L.W.&Liu,D.R.Genome editing with CRISPR-Cas nucleases,base editors,transposases and prime editors.Nat Biotechnol 38,824-844(2020).
3.Cullot,G.et al.CRISPR-Cas9 genome editing induces megabase-scale chromosomal truncations.Nat Commun 10,1136(2019).
4.Kosicki,M.,Tomberg,K.&Bradley,A.Repair of double-strand breaks induced by CRISPR-Cas9 leads to large deletions and complex rearrangements.Nat Biotechnol 36,765-771(2018).
5.Boroviak,K.,Fu,B.,Yang,F.,Doe,B.&Bradley,A.Revealing hidden complexities of genomic rearrangements generated with Cas9.Sci Rep 7,12867(2017).
6.Enache,O.M.et al.Cas9 activates the p53 pathway and selects for p53-inactivating mutations.Nat Genet 52,662-668(2020).
7.Haapaniemi,E.,Botla,S.,Persson,J.,Schmierer,B.&Taipale,J.CRISPR-Cas9 genome editing induces a p53-mediated DNA damage response.Nat Med 24,927-930(2018).
8.Ihry,R.J.et al.p53 inhibits CRISPR-Cas9 engineering in human pluripotent stem cells.Nat Med 24,939-946(2018).
9.Leibowitz,M.L.et al.Chromothripsis as an on-target consequence of CRISPR-Cas9 genome editing.Preprint at https://www.biorxiv.org/content/10.1101/2020.07.13.200998v1(2020).
10.Burgio,G.&Teboul,L.Anticipating and Identifying Collateral Damage in Genome Editing.Trends Genet 36,905-914(2020).
11.Cox,D.B.,Platt,R.J.&Zhang,F.Therapeutic genome editing:prospects and challenges.Nat Med 21,121-131(2015).
12.Komor,A.C.,Kim,Y.B.,Packer,M.S.,Zuris,J.A.&Liu,D.R.Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage.Nature 533,420-424(2016).
13.Gaudelli,N.M.et al.Programmable base editing of A*T to G*C in genomic DNA without DNA cleavage.Nature 551,464-471(2017).
14.Anzalone,A.V.et al.Search-and-replace genome editing without double-strand breaks or donor DNA.Nature 576,149-157(2019).
15.Houseley,J.&Tollervey,D.The many pathways of RNA degradation.Cell 136,763-776(2009).
16.Hendel,A.et al.Chemically modified guide RNAs enhance CRISPR-Cas genome editing in human primary cells.Nat Biotechnol 33,985-989(2015).
17.Geisberg,J.V.,Moqtaderi,Z.,Fan,X.,Ozsolak,F.&Struhl,K.Global analysis of mRNA isoform half-lives reveals stabilizing and destabilizing elements in yeast.Cell 156,812-824(2014).
18.Wu,X.&Bartel,D.P.Widespread Influence of 3'-End Structures on Mammalian mRNA Processing and Stability.Cell 169,905-917 e911(2017).
19.Brown,J.A.et al.Structural insights into the stabilization of MALAT1 noncoding RNA by a bipartite triple helix.Nat Struct Mol Biol 21,633-640(2014).
20.MacFadden,A.et al.Mechanism and structural diversity of exoribonuclease-resistant RNA structures in flaviviral RNAs.Nat Commun 9,119(2018).
21.Pijlman,G.P.et al.A highly structured,nuclease-resistant,noncoding RNA produced by flaviviruses is required for pathogenicity.Cell Host Microbe 4,579-591(2008).
22.Roth,A.et al.A riboswitch selective for the queuosine precursor preQ1 contains an unusually small aptamer domain.Nat Struct Mol Biol 14,308-317(2007).
23.Anzalone,A.V.,Lin,A.J.,Zairis,S.,Rabadan,R.&Cornish,V.W.Reprogramming eukaryotic translation with ligand-responsive synthetic RNA switches.Nat Methods 13,453-458(2016).
24.Houck-Loomis,B.et al.An equilibrium-dependent retroviral mRNA switch regulates translational recoding.Nature 480,561-564(2011).
25.Lorenz,R.et al.ViennaRNA package 2.0.Algorithms Mol Biol 6,26(2011).
26.Schene,I.F.et al.Prime editing for functional repair in patient-derived disease models.Nat Commun 11,5352(2020).
27.Kim,D.Y.,Moon,S.B.,Ko,J.H.,Kim,Y.S.&Kim,D.Unbiased investigation of specificities of prime editing systems in human cells.Nucleic Acids Res 48,10576-10589(2020).
28.Gao,P.et al.Prime editing in mice reveals the essentiality of a single base in driving tissue specific gene expression.Preprint at www.biorxiv.org/content/10.1101/2020.11.07.372748v3.full.pdf(2020).
29.Tsai,S.Q.et al.GUIDE-seq enables genome-wide profiling of off-target cleavage by CRISPR-Cas nucleases.Nat Biotechnol 33,187-197(2015).
30.Ibrahim,H.,Wilusz,J.&Wilusz,C.J.RNA recognition by 3'-to-5'exonucleases:the substrate perspective.Biochim biophys acta 1779,256-265(2008).
31.Green,L.,Kim,C.H.,Bustamante,C.&Tinoco,I.,Jr.Characterization of the mechanical unfolding of RNA pseudoknots.J Mol Biol 375,511-528(2008).
32.Chavez,A.et al.Highly efficient Cas9-mediated transcriptional programming.Nat Methods 12,326-328(2015).
33.Hu,J.H.et al.Evolved Cas9 variants with broad PAM compatibility and high DNA specificity.Nature 556,57-63(2018).
34.Nahar,S.et al.A G-quadruplex motif at the 3' end of sgRNAs improves CRISPR-Cas9 based genome editing efficiency.Chem Commun 54,2377-2380(2018).
35.Steckelberg,A.L.et al.A folded viral noncoding RNA blocks host cell exoribonucleases through a conformationally dynamic RNA structure.Proc Natl Acad Sci USA 115,6404-6409(2018).
36.Cate,J.H.et al.Crystal structure of a group I ribozyme domain:principles of RNA packing.Science 273,1678-1685(1996).
37.Fedor,M.J.&Westhof,E.Ribozymes:the first 20 years.Mol Cell 10,703-704(2002).
38.Chen,X.,Zaro,J.L.&Shen,W.C.Fusion protein linkers:property,design and functionality.Adv Drug Deliv Rev 65,1357-1369(2013).
39.Win,M.N.&Smolke,C.D.A modular and extensible RNA-based gene-regulatory platform for engineering cellular function.Proc Natl Acad Sci USA 104,14283-14288(2007).
40.Nielsen,S.,Yuzenkova,Y.&Zenkin,N.Mechanism of eukaryotic RNA polymerase III transcription termination.Science 340,1577-1580(2013).
41.Huang,T.P.,Newby,G.A.&Liu,D.R.Precision genome editing using cytosine and adenine base editors in mammalian cells.Nat Protoc(2021).
42.Basila,M.,Kelley,M.L.&Smith,A.V.B.Minimal 2'-O-methyl phosphorothioate linkage modification pattern of synthetic guide RNAs for increased stability and efficient CRISPR-Cas9 gene editing avoiding cellular toxicity.PLoS One 12,e0188593(2017).
43.Mead,S.et al.A novel protective prion protein variant that colocalizes with Kuru exposure.N Engl J Med 361,2056-2065(2009).
44.Asante,E.A.et al.A naturally occurring variant of the human prion protein completely prevents prion disease.Nature 522,478-481(2015).
45.Kim,H.K.et al.Predicting the efficiency of prime editing guide RNAs in human cells.Nat Biotechnol(2020).
46.Jonsson,T.et al.A mutation in APP protects against Alzheimer's disease and age-related cognitive decline.Nature 488,96-99(2012).
47.Abifadel,M.et al.Mutations in PCSK9 cause autosomal dominant hypercholesterolemia.Nat Genet 34,154-156(2003).
48.Bustami,J.et al.Cholesteryl ester transfer protein(CETP)I405V polymorphism and cardiovascular disease in eastern European Caucasians-a cross-sectional study.BMC Geriatr 16,144(2016).
49.Flannick,J.et al.Loss-of-function mutations in SLC30A8 protect against type 2 diabetes.Nat Genet 46,357-363(2014).
50.Sakuntabhai,A.et al.A variant in the CD209 promoter is associated with severity of dengue disease.Nat Genet 37,507-513(2005).
51.Olson,H.E.et al.Cyclin-Dependent Kinase-Like 5 Deficiency Disorder:Clinical Review.Pediatr Neurol 97,18-25(2019).
52.Al-Saaidi,R.et al.The LMNA mutation p.Arg321Ter associated with dilated cardiomyopathy leads to reduced expression and a skewed ratio of lamin A and lamin C proteins.Exp Cell Res 319,3010-3019(2013).
53.Ip,J.P.K.,Mellios,N.&Sur,M.Rett syndrome:insights into genetic,molecular and circuit mechanisms.Nat Rev Neurosci 19,368-382(2018).
54.Christodoulou,J.,Grimm,A.,Maher,T.&Bennetts,B.RettBASE:The IRSA MECP2 variation database-a new mutation database in evolution.Hum Mutat 21,466-472(2003).
55.Dwivedi,O.P.et al.Loss of ZnT8 function protects against diabetes by enhanced insulin secretion.Nat Genet 51,1596-1606(2019).
56.Thyme,S.B.,Akhmetova,L.,Montague,T.G.,Valen,E.&Schier,A.F.Internal guide RNA interactions interfere with Cas9-mediated cleavage.Nat Commun 7,11750(2016).
57.Boyle,E.A.et al.Quantification of Cas9 binding and cleavage across diverse guide sequences maps landscapes of target engagement.Science Advances,in press(2021).
58.Gaudelli,N.M.et al.Directed evolution of adenine base editors with increased activity and therapeutic application.Nat Biotechnol 38,892-900(2020).
59.Clement,K.et al.CRISPResso2 provides accurate and rapid genome editing sequence analysis.Nat Biotechnol 37,224-226(2019).
60.Pandey,S.,Agarwala,P.&Maiti,S.Effect of loops and G-quartets on the stability of RNA G-quadruplexes.J Phys Chem B 117,6896-6905(2013).
61.Engler,C.,Gruetzner,R.,Kandzia,R.&Marillonnet,S.Golden gate shuffling:a one-pot DNA shuffling method based on type IIs restriction enzymes.PLoS One 4,e5553(2009).
参考文献
以下の参考文献は全て、それらの全体が参照により本明細書にそれぞれ組み込まれる。
1.Jinek,M.et al.A Programmable Dual-RNA-Guided DNA Endonuclease in Adaptive Bacterial Immunity.Science 337,816-821(2012).
2.Cong,L.et al.Multiplex Genome Engineering Using CRISPR/Cas Systems.Science 339,819-823(2013).
3.Komor,A.C.,Badran,A.H.&Liu,D.R.CRISPR-Based Technologies for the Manipulation of Eukaryotic Genomes.Cell 168,20-36(2017).
4.Komor,A.C.,Kim,Y.B.,Packer,M.S.,Zuris,J.A.&Liu,D.R.Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage.Nature 533,420-424(2016).
5.Nishida,K.et al.Targeted nucleotide editing using hybrid prokaryotic and vertebrate adaptive immune systems.Science 353,aaf8729(2016).
6.Gaudelli,N.M.et al.Programmable base editing of A・T to G・C in genomic DNA without DNA cleavage.Nature 551,464-471(2017).
7.ClinVar,July 2019.
8.Dunbar,C.E.et al.Gene therapy comes of age.Science 359,eaan4672(2018).
9.Cox,D.B.T.,Platt,R.J.&Zhang,F.Therapeutic genome editing:prospects and challenges.Nat.Med.21,121-131(2015).
10.Adli,M.The CRISPR tool kit for genome editing and beyond.Nat.Commun.9,1911(2018).
11.Kleinstiver,B.P.et al.Engineered CRISPR-Cas9 nucleases with altered PAM specificities.Nature 523,481-485(2015).
12.Kleinstiver,B.P.et al.High-fidelity CRISPR-Cas9 nucleases with no detectable genome-wide off-target effects.Nature 529,490-495(2016).
13.Hu,J.H.et al.Evolved Cas9 variants with broad PAM compatibility and high DNA specificity.Nature 556,57-63(2018).
14.Nishimasu,H.et al.Engineered CRISPR-Cas9 nuclease with expanded targeting space.Science 361,1259-1262(2018).
15.Jasin,M.&Rothstein,R.Repair of strand breaks by homologous recombination.Cold Spring Harb.Perspect.Biol.5,a012740(2013).
16.Paquet,D.et al.Efficient introduction of specific homozygous and heterozygous mutations using CRISPR/Cas9.Nature 533,125-129(2016).
17.Kosicki,M.,Tomberg,K.&Bradley,A.Repair of double-strand breaks induced by CRISPR-Cas9 leads to large deletions and complex rearrangements.Nat.Biotechnol.36,765-771(2018).
18.Haapaniemi,E.,Botla,S.,Persson,J.,Schmierer,B.&Taipale,J.CRISPR-Cas9 genome editing induces a p53-mediated DNA damage response.Nat.Med.24,927-930(2018).
19.Ihry,R.J.et al.p53 inhibits CRISPR-Cas9 engineering in human pluripotent stem cells.Nat.Med.24,939-946(2018).
20.Richardson,C.D.,Ray,G.J.,DeWitt,M.A.,Curie,G.L.&Corn,J.E.Enhancing homology-directed genome editing by catalytically active and inactive CRISPR-Cas9 using asymmetric donor DNA.Nat.Biotechnol.34,339-344(2016).
21.Srivastava,M.et al.An Inhibitor of Nonhomologous End-Joining Abrogates Double-Strand Break Repair and Impedes Cancer Progression.Cell 151,1474-1487(2012).
22.Chu,V.T.et al.Increasing the efficiency of homology-directed repair for CRISPR-Cas9-induced precise gene editing in mammalian cells.Nat.Biotechnol.33,543-548(2015).
23.Maruyama,T.et al.Increasing the efficiency of precise genome editing with CRISPR-Cas9 by inhibition of nonhomologous end joining.Nat.Biotechnol.33,538-542(2015).
24.Kim,Y.B.et al.Increasing the genome-targeting scope and precision of base editing with engineered Cas9-cytidine deaminase fusions.Nat.Biotechnol.35,371-376(2017).
25.Li,X.et al.Base editing with a Cpf1-cytidine deaminase fusion.Nat.Biotechnol.36,324-327(2018).
26.Gehrke,J.M.et al.An APOBEC3A-Cas9 base editor with minimized bystander and off-target activities.Nat.Biotechnol.(2018).doi:10.1038/nbt.4199
27.Rees,H.A.&Liu,D.R.Base editing:precision chemistry on the genome and transcriptome of living cells.Nat.Rev.Genet.1(2018).doi:10.1038/s41576-018-0059-1.
28.Ostertag,E.M.&Kazazian Jr,H.H.Biology of Mammalian L1 Retrotransposons.Annu.Rev.Genet.35,501-538(2001).
29.Zimmerly,S.,Guo,H.,Perlman,P.S.&Lambowltz,A.M.Group II intron mobility occurs by target DNA-primed reverse transcription.Cell 82,545-554(1995).
30.Luan,D.D.,Korman,M.H.,Jakubczak,J.L.&Eickbush,T.H.Reverse transcription of R2Bm RNA is primed by a nick at the chromosomal target site:a mechanism for non-LTR retrotransposition.Cell 72,595-605(1993).
31.Feng,Q.,Moran,J.V.,Kazazian,H.H.&Boeke,J.D.Human L1 retrotransposon encodes a conserved endonuclease required for retrotransposition.Cell 87,905-916(1996).
32.Jinek,M.et al.Structures of Cas9 Endonucleases Reveal RNA-Mediated Conformational Activation.Science 343,1247997(2014).
33.Jiang,F.et al.Structures of a CRISPR-Cas9 R-loop complex primed for DNA cleavage.Science aad8282(2016).doi:10.1126/science.aad8282
34.Qi,L.S.et al.Repurposing CRISPR as an RNA-Guided Platform for Sequence-Specific Control of Gene Expression.Cell 152,1173-1183(2013).
35.Tang,W.,Hu,J.H.&Liu,D.R.Aptazyme-embedded guide RNAs enable ligand-responsive genome editing and transcriptional activation.Nat.Commun.8,15939(2017).
36.Shechner,D.M.,Hacisuleyman,E.,Younger,S.T.&Rinn,J.L.Multiplexable,locus-specific targeting of long RNAs with CRISPR-Display.Nat.Methods 12,664-670(2015).
37.Anders,C.&Jinek,M.Chapter One-In vitro Enzymology of Cas9.in Methods in Enzymology(eds.Doudna,J.A.&Sontheimer,E.J.)546,1-20(Academic Press,2014).
38.Briner,A.E.et al.Guide RNA Functional Modules Direct Cas9 Activity and Orthogonality.Mol.Cell 56,333-339(2014).
39.Nowak,C.M.,Lawson,S.,Zerez,M.&Bleris,L.Guide RNA engineering for versatile Cas9 functionality.Nucleic Acids Res.44,9555-9564(2016).
40.Sternberg,S.H.,Redding,S.,Jinek,M.,Greene,E.C.&Doudna,J.A.DNA interrogation by the CRISPR RNA-guided endonuclease Cas9.Nature 507,62-67(2014).
41.Mohr,S.et al.Thermostable group II intron reverse transcriptase fusion proteins and their use in cDNA synthesis and next-generation RNA sequencing.RNA 19,958-970(2013).
42.Stamos,J.L.,Lentzsch,A.M.&Lambowitz,A.M.Structure of a Thermostable Group II Intron Reverse Transcriptase with Template-Primer and Its Functional and Evolutionary Implications.Mol.Cell 68,926-939.e4(2017).
43.Zhao,C.&Pyle,A.M.Crystal structures of a group II intron maturase reveal a missing link in spliceosome evolution.Nat.Struct.Mol.Biol.23,558-565(2016).
44.Zhao,C.,Liu,F.&Pyle,A.M.An ultraprocessive,accurate reverse transcriptase encoded by a metazoan group II intron.RNA 24,183-195(2018).
45.Ran,F.A.et al.Genome engineering using the CRISPR-Cas9 system.Nat.Protoc.8,2281-2308(2013).
46.Liu,Y.,Kao,H.-I.&Bambara,R.A.Flap endonuclease 1:a central component of DNA metabolism.Annu.Rev.Biochem.73,589-615(2004).
47.Krokan,H.E.&Bjoras,M.Base Excision Repair.Cold Spring Harb.Perspect.Biol.5,(2013).
48.Kelman,Z.PCNA:structure,functions and interactions.Oncogene 14,629-640(1997).
49.Choe,K.N.&Moldovan,G.-L.Forging Ahead through Darkness:PCNA,Still the Principal Conductor at the Replication Fork.Mol.Cell 65,380-392(2017).
50.Li,X.,Li,J.,Harrington,J.,Lieber,M.R.&Burgers,P.M.Lagging strand DNA synthesis at the eukaryotic replication fork involves binding and stimulation of FEN-1 by proliferating cell nuclear antigen.J.Biol.Chem.270,22109-22112(1995).
51.Tom,S.,Henricksen,L.A.&Bambara,R.A.Mechanism whereby proliferating cell nuclear antigen stimulates flap endonuclease 1.J.Biol.Chem.275,10498-10505(2000).
52.Tanenbaum,M.E.,Gilbert,L.A.,Qi,L.S.,Weissman,J.S.&Vale,R.D.A protein-tagging system for signal amplification in gene expression and fluorescence imaging.Cell 159,635-646(2014).
53.Bertrand,E.et al.Localization of ASH1 mRNA particles in living yeast.Mol.Cell 2,437-445(1998).
54.Dahlman,J.E.et al.Orthogonal gene knockout and activation with a catalytically active Cas9 nuclease.Nat.Biotechnol.33,1159-1161(2015).
55.Tsai,S.Q.et al.GUIDE-seq enables genome-wide profiling of off-target cleavage by CRISPR-Cas nucleases.Nat.Biotechnol.33,187-197(2015).
56.Tsai,S.Q.et al.CIRCLE-seq:a highly sensitive in vitro screen for genome-wide CRISPR-Cas9 nuclease off-targets.Nat.Methods 14,607-614(2017).
57.Schek N,Cooke C,Alwine JC.Molecular and Cellular Biology.(1992).
58.Gil A,Proudfoot NJ.Cell.(1987).
59.Zhao,B.S.,Roundtree,I.A.,He,C.Nat Rev Mol Cell Biol.(2017).
60.Rubio,M.A.T.,Hopper,A.K.Wiley Interdiscip Rev RNA(2011).
61.Shechner,D.M.,Hacisuleyman E.,Younger,S.T.,Rinn,J.L.Nat Methods.(2015).
62.Paige,J.S.,Wu,K.Y.,Jaffrey,S.R.Science(2011).
63.Ray D.,… Hughes TR.Nature(2013).
64.Chadalavada,D.M.,Cerrone-Szakal,A.L.,Bevilacqua,P.C.RNA(2007).
65.Forster AC,Symons RH.Cell.(1987).
66.Weinberg Z,Kim PB,Chen TH,Li S,Harris KA,Lunse CE,Breaker RR.Nat.Chem.Biol.(2015).
67.Feldstein PA,Buzayan JM,Bruening G.Gene(1989).
68.Saville BJ,Collins RA.Cell.(1990).
69.Winkler WC,Nahvi A,Roth A,Collins JA,Breaker RR.Nature(2004).
70.Roth A,Weinberg Z,Chen AG,Kim PG,Ames TD,Breaker RR.Nat Chem Biol.(2013).
71.Choudhury R,Tsai YS,Dominguez D,Wang Y,Wang Z.Nat Commun.(2012).
72.MacRae IJ,Doudna JA.Curr Opin Struct Biol.(2007).
73.Bernstein E,Caudy AA,Hammond SM,Hannon GJ Nature(2001).
74.Filippov V,Solovyev V,Filippova M,Gill SS.Gene(2000).
75.Cadwell RC and Joyce GF.PCR Methods Appl.(1992).
76.McInerney P,Adams P,and Hadi MZ.Mol Biol Int.(2014).
77.Esvelt KM,Carlson JC,and Liu DR.Nature.(2011).
78.Naorem SS,Hin J,Wang S,Lee WR,Heng X,Miller JF,Guo H.Proc Natl Acad Sci USA(2017).
79.Martinez MA,Vartanian JP,Wain-Hobson S.Proc Natl Acad Sci USA(1994).
80.Meyer AJ,Ellefson JW,Ellington AD.Curr Protoc Mol Biol.(2014).
81.Wang HH,Isaacs FJ,Carr PA,Sun ZZ,Xu G,Forest CR,Church GM.Nature.(2009).
82.Nyerges A et al.Proc Natl Acad Sci USA.(2016).
83.Mascola JR,Haynes BF.Immunol Rev.(2013).
84.X.Wen,K.Wen,D.Cao,G.Li,R.W.Jones,J.Li,S.Szu,Y.Hoshino,L.Yuan,Inclusion of a universal tetanus toxoid CD4(+)T cell epitope P2 significantly enhanced the immunogenicity of recombinant rotavirusΔVP8*subunit parenteral vaccines.Vaccine 32,4420-4427(2014).
85.G.Ada,D.Isaacs,Carbohydrate-protein conjugate vaccines.Clin Microbiol Infect 9,79-85(2003).
86.E.Malito,B.Bursulaya,C.Chen,P.L.Surdo,M.Picchianti,E.Balducci,M.Biancucci,A.Brock,F.Berti,M.J.Bottomley,M.Nissum,P.Costantino,R.Rappuoli,G.Spraggon,Structural basis for lack of toxicity of the diphtheria toxin mutant CRM197.Proceedings of the National Academy of Sciences 109,5229(2012).
87.J.de Wit,M.E.Emmelot,M.C.M.Poelen,J.Lanfermeijer,W.G.H.Han,C.van Els,P.Kaaijk,The Human CD4(+)T Cell Response against Mumps Virus Targets a Broadly Recognized Nucleoprotein Epitope.J Virol 93,(2019).
88.M.May,C.A.Rieder,R.J.Rowe,Emergent lineages of mumps virus suggest the need for a polyvalent vaccine.Int J Infect Dis 66,1-4(2018).
89.M.Ramamurthy,P.Rajendiran,N.Saravanan,S.Sankar,S.Gopalan,B.Nandagopal,Identification of immunogenic B-cell epitope peptides of rubella virus E1 glycoprotein towards development of highly specific immunoassays and/or vaccine.Conference Abstract,(2019).
90.U.S.F.Tambunan,F.R.P.Sipahutar,A.A.Parikesit,D.Kerami,Vaccine Design for H5N1 Based on B-and T-cell Epitope Predictions.Bioinform Biol Insights 10,27-35(2016).
91.Asante,EA.et.al.「A naturally occurring variant of the human prion protein completely prevents prion disease」.Nature.(2015).
92.Crabtree,G.R.&Schreiber,S.L.Three-part inventions:intracellular signaling and induced proximity.Trends Biochem.Sci.21,418-22(1996).
93.Liu,J.et al.Calcineurin Is a Common Target of A and FKBP-FK506 Complexes.Cell 66,807-815(1991).
94.Keith,C.T.et al.A mammalian protein targeted by G1-arresting rapamycin-receptor complex.Nature 369,756-758(2003).
95.Spencer,D.M.,Wandless,T.J.,Schreiber,S.L.S.&Crabtree,G.R.Controlling signal transduction with synthetic ligands.Science 262,1019-24(1993).
96.Pruschy,M.N.et al.Mechanistic studies of a signaling pathway activated by the organic dimerizer FK1012.Chem.Biol.1,163-172(1994).
97.Spencer,D.M.et al.Functional analysis of Fas signaling in vivo using synthetic inducers of dimerization.Curr.Biol.6,839-847(1996).
98.Belshaw,P.J.,Spencer,D.M.,Crabtree,G.R.&Schreiber,S.L.Controlling programmed cell death with a cyclophilin-cyclosporin-based chemical inducer of dimerization.Chem.Biol.3,731-738(1996).
99.Yang,J.X.,Symes,K.,Mercola,M.&Schreiber,S.L.Small-molecule control of insulin and PDGF receptor signaling and the role of membrane attachment.Curr.Biol.8,11-18(1998).
100.Belshaw,P.J.,Ho,S.N.,Crabtree,G.R.&Schreiber,S.L.Controlling protein association and subcellular localization with a synthetic ligand that induces heterodimerization of proteins.Proc.Natl.Acad.Sci.93,4604-4607(2002).
101.Stockwell,B.R.&Schreiber,S.L.Probing the role of homomeric and heteromeric receptor interactions in TGF-βsignaling using small molecule dimerizers.Curr.Biol.8,761-773(2004).
102.Spencer,D.M.,Graef,I.,Austin,D.J.,Schreiber,S.L.&Crabtree,G.R.A general strategy for producing conditional alleles of Src-like tyrosine kinases.Proc.Natl.Acad.Sci.92,9805-9809(2006).
103.Holsinger,L.J.,Spencer,D.M.,Austin,D.J.,Schreiber,S.L.&Crabtree,G.R.Signal transduction in T lymphocytes using a conditional allele of Sos.Proc.Natl.Acad.Sci.92,9810-9814(2006).
104.Myers,M.G.Insulin Signal Transduction and the IRS Proteins.Annu.Rev.Pharmacol.Toxicol.36,615-658(1996).
105.Watowich,S.S.The erythropoietin receptor:Molecular structure and hematopoietic signaling pathways.J.Investig.Med.59,1067-1072(2011).
106.Blau,C.A.,Peterson,K.R.,Drachman,J.G.&Spencer,D.M.A proliferation switch for genetically modified cells.Proc.Natl.Acad.Sci.94,3076-3081(2002).
107.Clackson,T.et al.Redesigning an FKBP-ligand interface to generate chemical dimerizers with novel specificity.Proc.Natl.Acad.Sci.95,10437-10442(1998).
108.Diver,S.T.&Schreiber,S.L.Single-step synthesis of cell-permeable protein dimerizers that activate signal transduction and gene expression.J.Am.Chem.Soc.119,5106-5109(1997).
109.Guo,Z.F.,Zhang,R.&Liang,F.Sen.Facile functionalization of FK506 for biological studies by the thiol-ene'click'reaction.RSC Adv.4,11400-11403(2014).
110.Robinson,D.R.,Wu,Y.-M.&Lin,S.-F.The protein tyrosine kinase family of the human genome.Oncogene 19,5548-5557(2000).
111.Landrum,M.J.et al.ClinVar:public archive of interpretations of clinically relevant variants.Nucleic Acids Res.44,D862-D868(2016).
112.Jinek,M.et al.A Programmable Dual-RNA-Guided DNA Endonuclease in Adaptive Bacterial Immunity.Science 337,816-821(2012).
113.Cong,L.et al.Multiplex Genome Engineering Using CRISPR/Cas Systems.Science
339,819-823(2013).
114.Mali,P.et al.RNA-Guided Human Genome Engineering via Cas9.Science 339,823-826(2013).
115.Yang,H.et al.One-Step Generation of Mice Carrying Reporter and Conditional Alleles by CRISPR/Cas-Mediated Genome Engineering.Cell 154,1370-1379(2013).
116.Kim,S.,Kim,D.,Cho,S.W.,Kim,J.&Kim,J.-S.Highly efficient RNA-guided genome editing in human cells via delivery of purified Cas9 ribonucleoproteins.Genome Res.24,1012-1019(2014).
117.Orlando,S.J.et al.Zinc-finger nuclease-driven targeted integration into mammalian genomes using donors with limited chromosomal homology.Nucleic Acids Res.38,e152-e152(2010).
118.Tsai,S.Q.et al.GUIDE-seq enables genome-wide profiling of off-target cleavage by CRISPR-Cas nucleases.Nat.Biotechnol.33,187-197(2015).
119.Suzuki,K.et al.In vivo genome editing via CRISPR/Cas9 mediated homology-independent targeted integration.Nature 540,144-149(2016).
120.Kosicki,M.,Tomberg,K.&Bradley,A.Repair of double-strand breaks induced by CRISPR-Cas9 leads to large deletions and complex rearrangements.Nat.Biotechnol.36,765-771(2018).
121.Haapaniemi,E.,Botla,S.,Persson,J.,Schmierer,B.&Taipale,J.CRISPR-Cas9 genome editing induces a p53-mediated DNA damage response.Nat.Med.24,927-930(2018).
122.Ihry,R.J.et al.p53 inhibits CRISPR-Cas9 engineering in human pluripotent stem cells.Nat.Med.24,939-946(2018).
123.Chapman,J.R.,Taylor,M.R.G.&Boulton,S.J.Playing the end game:DNA double-strand break repair pathway choice.Mol.Cell 47,497-510(2012).
124.Cox,D.B.T.,Platt,R.J.&Zhang,F.Therapeutic genome editing:prospects and challenges.Nat.Med.21,121-131(2015).
125.Paquet,D.et al.Efficient introduction of specific homozygous and heterozygous mutations using CRISPR/Cas9.Nature 533,125-129(2016).
126.Chu,V.T.et al.Increasing the efficiency of homology-directed repair for CRISPR-Cas9-induced precise gene editing in mammalian cells.Nat.Biotechnol.33,543-548(2015).
127.Maruyama,T.et al.Increasing the efficiency of precise genome editing with CRISPR-Cas9 by inhibition of nonhomologous end joining.Nat.Biotechnol.33,538-542(2015).
128.Rees,H.A.,Yeh,W.-H.&Liu,D.R.Development of hRad51-Cas9 nickase fusions that mediate HDR without double-stranded breaks.Nat.Commun.10,1-12(2019).
129.Komor,A.C.,Kim,Y.B.,Packer,M.S.,Zuris,J.A.&Liu,D.R.Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage.Nature 533,420-424(2016).
130.Gaudelli,N.M.et al.Programmable base editing of A・T to G・C in genomic DNA without DNA cleavage.Nature 551,464-471(2017).
131.Gao,X.et al.Treatment of autosomal dominant hearing loss by in vivo delivery of genome editing agents.Nature 553,217-221(2018).
132.Ingram,V.M.A specific chemical difference between the globins of normal human and sickle-cell anaemia haemoglobin.Nature 178,792-794(1956).
133.Myerowitz,R.&Costigan,F.C.The major defect in Ashkenazi Jews with Tay-Sachs disease is an insertion in the gene for the alpha-chain of beta-hexosaminidase.J.Biol.Chem.263,18587-18589(1988).
134.Zielenski,J.Genotype and Phenotype in Cystic Fibrosis.Respiration 67,117-133(2000).
135.Mead,S.et al.A Novel Protective Prion Protein Variant that Colocalizes with Kuru Exposure.N.Engl.J.Med.361,2056-2065(2009).
136.Marraffini,L.A.&Sontheimer,E.J.CRISPR interference limits horizontal gene transfer in staphylococci by targeting DNA.Science 322,1843-1845(2008).
137.Barrangou,R.et al.CRISPR provides acquired resistance against viruses in prokaryotes.Science 315,1709-1712(2007).
138.Jiang,F.&Doudna,J.A.CRISPR-Cas9 Structures and Mechanisms.Annu.Rev.Biophys.46,505-529(2017).
139.Hille,F.et al.The Biology of CRISPR-Cas:Backward and Forward.Cell 172,1239-1259(2018).
140.Luan,D.D.,Korman,M.H.,Jakubczak,J.L.&Eickbush,T.H.Reverse transcription of R2Bm RNA is primed by a nick at the chromosomal target site:a mechanism for non-LTR retrotransposition.Cell 72,595-605(1993).
141.Liu,Y.,Kao,H.-I.&Bambara,R.A.Flap endonuclease 1:a central component of DNA metabolism.Annu.Rev.Biochem.73,589-615(2004).
142.Rees,H.A.&Liu,D.R.Base editing:precision chemistry on the genome and transcriptome of living cells.Nat.Rev.Genet.19,770(2018).
143.Richardson,C.D.,Ray,G.J.,DeWitt,M.A.,Curie,G.L.&Corn,J.E.Enhancing homology-directed genome editing by catalytically active and inactive CRISPR-Cas9 using asymmetric donor DNA.Nat.Biotechnol.34,339-344(2016).
144.Qi,L.S.et al.Repurposing CRISPR as an RNA-Guided Platform for Sequence-Specific Control of Gene Expression.Cell 152,1173-1183(2013).
145.Shechner,D.M.,Hacisuleyman,E.,Younger,S.T.&Rinn,J.L.Multiplexable,locus-specific targeting of long RNAs with CRISPR-Display.Nat.Methods 12,664-670(2015).
146.Tang,W.,Hu,J.H.&Liu,D.R.Aptazyme-embedded guide RNAs enable ligand-responsive genome editing and transcriptional activation.Nat.Commun.8,15939(2017).
147.Jinek,M.et al.Structures of Cas9 Endonucleases Reveal RNA-Mediated Conformational Activation.Science 343,1247997(2014).
148.Nishimasu,H.et al.Crystal Structure of Cas9 in Complex with Guide RNA and Target DNA.Cell 156,935-949(2014).
149.Jiang,F.,Zhou,K.,Ma,L.,Gressel,S.&Doudna,J.A.A Cas9-guide RNA complex preorganized for target DNA recognition.Science 348,1477-1481(2015).
150.Baranauskas,A.et al.Generation and characterization of new highly thermostable and processive M-MuLV reverse transcriptase variants.Protein Eng.Des.Sel.25,657-668(2012).
151.Gerard,G.F.et al.The role of template-primer in protection of reverse transcriptase from thermal inactivation.Nucleic Acids Res.30,3118-3129(2002).
152.Arezi,B.&Hogrefe,H.Novel mutations in Moloney Murine Leukemia Virus reverse transcriptase increase thermostability through tighter binding to template-primer.Nucleic Acids Res.37,473-481(2009).
153.Kotewicz,M.L.,Sampson,C.M.,D'Alessio,J.M.&Gerard,G.F.Isolation of cloned Moloney murine leukemia virus reverse transcriptase lacking ribonuclease H activity.Nucleic Acids Res.16,265-277(1988).
154.Shen,M.W.et al.Predictable and precise template-free CRISPR editing of pathogenic variants.Nature 563,646-651(2018).
155.Thuronyi,B.W.et al.Continuous evolution of base editors with expanded target compatibility and improved activity.Nat.Biotechnol.(2019).doi:10.1038/s41587-019-0193-0
156.Kim,Y.B.et al.Increasing the genome-targeting scope and precision of base editing with engineered Cas9-cytidine deaminase fusions.Nat.Biotechnol.35,371-376(2017).
157.Koblan,L.W.et al.Improving cytidine and adenine base editors by expression optimization and ancestral reconstruction.Nat.Biotechnol.(2018).doi:10.1038/nbt.4172
158.Komor,A.C.et al.Improved base excision repair inhibition and bacteriophage Mu Gam protein yields C:GからT:A base editors with higher efficiency and product purity.Sci.Adv.3,eaao4774(2017).
159.Kleinstiver,B.P.et al.High-fidelity CRISPR-Cas9 nucleases with no detectable genome-wide off-target effects.Nature 529,490-495(2016).
160.Zuo,E.et al.Cytosine base editor generates substantial off-target single-nucleotide variants in mouse embryos.Science 364,289-292(2019).
161.Jin,S.et al.Cytosine,but not adenine,base editors induce genome-wide off-target mutations in rice.Science 364,292-295(2019).
162.Kim,D.,Kim,D.,Lee,G.,Cho,S.-I.&Kim,J.-S.Genome-wide target specificity of CRISPR RNA-guided adenine base editors.Nat.Biotechnol.37,430-435(2019).
163.Grunewald,J.et al.Transcriptome-wide off-target RNA editing induced by CRISPR-guided DNA base editors.Nature 569,433-437(2019).
164.Zhou,C.et al.Off-target RNA mutation induced by DNA base editing and its elimination by mutagenesis.Nature 571,275-278(2019).
165.Rees,H.A.,Wilson,C.,Doman,J.L.&Liu,D.R.Analysis and minimization of cellular RNA editing by DNA adenine base editors.Sci.Adv.5,eaax5717(2019).
166.Ostertag,E.M.&Kazazian Jr,H.H.Biology of Mammalian L1 Retrotransposons.Annu.Rev.Genet.35,501-538(2001).
167.Griffiths,D.J.Endogenous retroviruses in the human genome sequence.Genome Biol.2,REVIEWS1017(2001).
168.Berkhout,B.,Jebbink,M.&Zsiros,J.Identification of an Active Reverse Transcriptase Enzyme Encoded by a Human Endogenous HERV-K Retrovirus.J.Virol.73,2365-2375(1999).
169.Halvas,E.K.,Svarovskaia,E.S.&Pathak,V.K.Role of Murine Leukemia Virus Reverse Transcriptase Deoxyribonucleoside Triphosphate-Binding Site in Retroviral Replication and In Vivo Fidelity.J.Virol.74,10349-10358(2000).
170.Dever,D.P.et al.CRISPR/Cas9 Beta-globin Gene Targeting in Human Hematopoietic Stem Cells.Nature 539,384-389(2016).
171.Park,S.H.et al.Highly efficient editing of the β-globin gene in patient-derived hematopoietic stem and progenitor cells to treat sickle cell disease.Nucleic Acids Res.doi:10.1093/nar/gkz475
172.Collinge,J.Prion diseases of humans and animals:their causes and molecular basis.Annu.Rev.Neurosci.24,519-550(2001).
173.Asante,E.A.et al.A naturally occurring variant of the human prion protein completely prevents prion disease.Nature 522,478-481(2015).
174.Anzalone,A.V.,Lin,A.J.,Zairis,S.,Rabadan,R.&Cornish,V.W.Reprogramming eukaryotic translation with ligand-responsive synthetic RNA switches.Nat.Methods 13,453-458(2016).
175.Badran,A.H.et al.Continuous evolution of Bacillus thuringiensis toxins overcomes insect resistance.Nature 533,58-63(2016).
176.Anders,C.&Jinek,M.Chapter One-In Vitro Enzymology of Cas9.in Methods in Enzymology(eds.Doudna,J.A.&Sontheimer,E.J.)546,1-20(Academic Press,2014).
177.Pirakitikulr,N.,Ostrov,N.,Peralta-Yahya,P.&Cornish,V.W.PCRless library mutagenesis via oligonucleotide recombination in yeast.Protein Sci.Publ.Protein Soc.19,2336-2346(2010).
178.Clement,K.et al.CRISPResso2 provides accurate and rapid genome editing sequence analysis.Nat.Biotechnol.37,224-226(2019).
179.Tsai,S.Q.et al.GUIDE-seq enables genome-wide profiling of off-target cleavage by CRISPR-Cas nucleases.Nat.Biotechnol.33,187-197(2015).
180.Kleinstiver,B.P.et al.High-fidelity CRISPR-Cas9 nucleases with no detectable genome-wide off-target effects.Nature 529,490-495(2016).
181.Koblan,L.W.et al.Improving cytidine and adenine base editors by expression optimization and ancestral reconstruction.Nat.Biotechnol.(2018).doi:10.1038/nbt.4172
182.Baranauskas,A.et al.Generation and characterization of new highly thermostable and processive M-MuLV reverse transcriptase variants.Protein Eng.Des.Sel.25,657-668(2012).
183.Schechner,DM,Hacisuleyman E.,Younger ST,Rinn JL.Nat Methods 664-70(2015).
184.Brown JA,et al.Nat Struct Mol Biol 633-40(2014).
185.Conrad NA and Steitz JA.EMBO J 1831-41(2005).
186.Bartlett JS,et al.Proc Natl Acad Sci USA 8852-7(1996).
187.Mitton-Fry RM,DeGregorio SJ,Wang J,Steitz TA,Steitz JA.Science 1244-7(2010).
188.Forster AC,Symons RH.Cell.1987.
189.Weinberg Z,Kim PB,Chen TH,Li S,Harris KA,Lunse CE,Breaker RR.Nat.Chem.Biol.2015.
190.Feldstein PA,Buzayan JM,Bruening G.Gene 1989.
191.Saville BJ,Collins RA.Cell.1990.
192.Roth A,Weinberg Z,Chen AG,Kim PG,Ames TD,Breaker RR.Nat Chem Biol.2013.
193.Borchardt EK,et al.RNA 1921-30(2015).
194.Zhang Y,et al.Mol Cell 792-806(2013).
195.Dang Y,et al.Genome Biol 280(2015).
196.Schaefer M,Kapoor U,and Jantsch MF.Open Biol 170077(2017).
197.Nahar S,et al.Chem Comm 2377-80(2018).
198.Gao Y and Zhao Y.J Integr Plant Biol 343-9(2014).
199.Dubois N,Marquet R,Paillart J,Bernacchi S.Front Microbiol 527(2018).
200.Costa M and Michel F.EMBO J 1276-85(1995).
201.Hu JH,et al.Nature 57-63(2018).
202.Furukawa K,Gu H,Breaker RR.Methods Mol Biol 209-20(2014).
203.Zettler,J.,Schutz,V.&Mootz,H.D.The naturally split Npu DnaE intein exhibits an extraordinarily high rate in the protein trans-splicing reaction.FEBS Lett.583,909-914(2009).
204.Kugler,S.,Kilic,E.&Bahr,M.Human synapsin 1 gene promoter confers highly neuron-specific long-term transgene expression from an adenoviral vector in the adult rat brain depending on the transduced area.Gene Ther.10,337-347(2003).
205.de Felipe,P.,Hughes,L.E.,Ryan,M.D.&Brown,J.D.Co-translational,intraribosomal cleavage of polypeptides by the foot-and-mouth disease virus 2A peptide.J.Biol.Chem.278,11441-11448(2003).
206.Levy,J.M.&Nicoll,R.A.Membrane-associated guanylate kinase dynamics reveal regional and developmental specificity of synapse stability.J.Physiol.595,1699-1709(2017).
207.Li,B.&Dewey,C.N.RSEM:accurate transcript quantification from RNA-Seq data with or without a reference genome.BMC Bioinformatics 12,323(2011).
208.Ritchie,M.E.et al.limma powers differential expression analyses for RNA-sequencing and microarray studies.Nucleic Acids Res.43,e47-e47(2015).
様々なプライマー結合部位(PBS)及び鋳型長を有するプライム編集ガイドRNA(pegRNA)がスクリーニングされて、PE2プライム編集因子を使用して、HEK293T細胞のCDKL5遺伝子におけるc.1412のトランジション点変異の最も効率的な組み入れを可能にするものが同定された(図164)次に、PE3プライム編集因子システムで使用されるニッキングガイドの選択が最適化され、c.1412で編集の効率を更に改善した(図165)。編集効率を更に改善するために、コードサイレントトランジションもまた、pegRNAによって標的化されたプロトスペーサのシード領域内に組み込まれた(図165)。この戦略で使用されるガイドRNA配列を表E10に提供する。全体として、pegRNA CDKL5h37及びepegRNA JNpeg 0953が最も高い編集効率を示した。
配列
表E1~E6の以下の配列は、実施例4及び関連する図面を通して参照される。
1.Komor,A.C.,Badran,A.H.&Liu,D.R.CRISPR-Based Technologies for the Manipulation of Eukaryotic Genomes.Cell 168,20-36(2017).
2.Anzalone,A.V.,Koblan,L.W.&Liu,D.R.Genome editing with CRISPR-Cas nucleases,base editors,transposases and prime editors.Nat Biotechnol 38,824-844(2020).
3.Cullot,G.et al.CRISPR-Cas9 genome editing induces megabase-scale chromosomal truncations.Nat Commun 10,1136(2019).
4.Kosicki,M.,Tomberg,K.&Bradley,A.Repair of double-strand breaks induced by CRISPR-Cas9 leads to large deletions and complex rearrangements.Nat Biotechnol 36,765-771(2018).
5.Boroviak,K.,Fu,B.,Yang,F.,Doe,B.&Bradley,A.Revealing hidden complexities of genomic rearrangements generated with Cas9.Sci Rep 7,12867(2017).
6.Enache,O.M.et al.Cas9 activates the p53 pathway and selects for p53-inactivating mutations.Nat Genet 52,662-668(2020).
7.Haapaniemi,E.,Botla,S.,Persson,J.,Schmierer,B.&Taipale,J.CRISPR-Cas9 genome editing induces a p53-mediated DNA damage response.Nat Med 24,927-930(2018).
8.Ihry,R.J.et al.p53 inhibits CRISPR-Cas9 engineering in human pluripotent stem cells.Nat Med 24,939-946(2018).
9.Leibowitz,M.L.et al.Chromothripsis as an on-target consequence of CRISPR-Cas9 genome editing.Preprint at https://www.biorxiv.org/content/10.1101/2020.07.13.200998v1(2020).
10.Burgio,G.&Teboul,L.Anticipating and Identifying Collateral Damage in Genome Editing.Trends Genet 36,905-914(2020).
11.Cox,D.B.,Platt,R.J.&Zhang,F.Therapeutic genome editing:prospects and challenges.Nat Med 21,121-131(2015).
12.Komor,A.C.,Kim,Y.B.,Packer,M.S.,Zuris,J.A.&Liu,D.R.Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage.Nature 533,420-424(2016).
13.Gaudelli,N.M.et al.Programmable base editing of A*T to G*C in genomic DNA without DNA cleavage.Nature 551,464-471(2017).
14.Anzalone,A.V.et al.Search-and-replace genome editing without double-strand breaks or donor DNA.Nature 576,149-157(2019).
15.Houseley,J.&Tollervey,D.The many pathways of RNA degradation.Cell 136,763-776(2009).
16.Hendel,A.et al.Chemically modified guide RNAs enhance CRISPR-Cas genome editing in human primary cells.Nat Biotechnol 33,985-989(2015).
17.Geisberg,J.V.,Moqtaderi,Z.,Fan,X.,Ozsolak,F.&Struhl,K.Global analysis of mRNA isoform half-lives reveals stabilizing and destabilizing elements in yeast.Cell 156,812-824(2014).
18.Wu,X.&Bartel,D.P.Widespread Influence of 3'-End Structures on Mammalian mRNA Processing and Stability.Cell 169,905-917 e911(2017).
19.Brown,J.A.et al.Structural insights into the stabilization of MALAT1 noncoding RNA by a bipartite triple helix.Nat Struct Mol Biol 21,633-640(2014).
20.MacFadden,A.et al.Mechanism and structural diversity of exoribonuclease-resistant RNA structures in flaviviral RNAs.Nat Commun 9,119(2018).
21.Pijlman,G.P.et al.A highly structured,nuclease-resistant,noncoding RNA produced by flaviviruses is required for pathogenicity.Cell Host Microbe 4,579-591(2008).
22.Roth,A.et al.A riboswitch selective for the queuosine precursor preQ1 contains an unusually small aptamer domain.Nat Struct Mol Biol 14,308-317(2007).
23.Anzalone,A.V.,Lin,A.J.,Zairis,S.,Rabadan,R.&Cornish,V.W.Reprogramming eukaryotic translation with ligand-responsive synthetic RNA switches.Nat Methods 13,453-458(2016).
24.Houck-Loomis,B.et al.An equilibrium-dependent retroviral mRNA switch regulates translational recoding.Nature 480,561-564(2011).
25.Lorenz,R.et al.ViennaRNA package 2.0.Algorithms Mol Biol 6,26(2011).
26.Schene,I.F.et al.Prime editing for functional repair in patient-derived disease models.Nat Commun 11,5352(2020).
27.Kim,D.Y.,Moon,S.B.,Ko,J.H.,Kim,Y.S.&Kim,D.Unbiased investigation of specificities of prime editing systems in human cells.Nucleic Acids Res 48,10576-10589(2020).
28.Gao,P.et al.Prime editing in mice reveals the essentiality of a single base in driving tissue specific gene expression.Preprint at www.biorxiv.org/content/10.1101/2020.11.07.372748v3.full.pdf(2020).
29.Tsai,S.Q.et al.GUIDE-seq enables genome-wide profiling of off-target cleavage by CRISPR-Cas nucleases.Nat Biotechnol 33,187-197(2015).
30.Ibrahim,H.,Wilusz,J.&Wilusz,C.J.RNA recognition by 3'-to-5'exonucleases:the substrate perspective.Biochim biophys acta 1779,256-265(2008).
31.Green,L.,Kim,C.H.,Bustamante,C.&Tinoco,I.,Jr.Characterization of the mechanical unfolding of RNA pseudoknots.J Mol Biol 375,511-528(2008).
32.Chavez,A.et al.Highly efficient Cas9-mediated transcriptional programming.Nat Methods 12,326-328(2015).
33.Hu,J.H.et al.Evolved Cas9 variants with broad PAM compatibility and high DNA specificity.Nature 556,57-63(2018).
34.Nahar,S.et al.A G-quadruplex motif at the 3' end of sgRNAs improves CRISPR-Cas9 based genome editing efficiency.Chem Commun 54,2377-2380(2018).
35.Steckelberg,A.L.et al.A folded viral noncoding RNA blocks host cell exoribonucleases through a conformationally dynamic RNA structure.Proc Natl Acad Sci USA 115,6404-6409(2018).
36.Cate,J.H.et al.Crystal structure of a group I ribozyme domain:principles of RNA packing.Science 273,1678-1685(1996).
37.Fedor,M.J.&Westhof,E.Ribozymes:the first 20 years.Mol Cell 10,703-704(2002).
38.Chen,X.,Zaro,J.L.&Shen,W.C.Fusion protein linkers:property,design and functionality.Adv Drug Deliv Rev 65,1357-1369(2013).
39.Win,M.N.&Smolke,C.D.A modular and extensible RNA-based gene-regulatory platform for engineering cellular function.Proc Natl Acad Sci USA 104,14283-14288(2007).
40.Nielsen,S.,Yuzenkova,Y.&Zenkin,N.Mechanism of eukaryotic RNA polymerase III transcription termination.Science 340,1577-1580(2013).
41.Huang,T.P.,Newby,G.A.&Liu,D.R.Precision genome editing using cytosine and adenine base editors in mammalian cells.Nat Protoc(2021).
42.Basila,M.,Kelley,M.L.&Smith,A.V.B.Minimal 2'-O-methyl phosphorothioate linkage modification pattern of synthetic guide RNAs for increased stability and efficient CRISPR-Cas9 gene editing avoiding cellular toxicity.PLoS One 12,e0188593(2017).
43.Mead,S.et al.A novel protective prion protein variant that colocalizes with Kuru exposure.N Engl J Med 361,2056-2065(2009).
44.Asante,E.A.et al.A naturally occurring variant of the human prion protein completely prevents prion disease.Nature 522,478-481(2015).
45.Kim,H.K.et al.Predicting the efficiency of prime editing guide RNAs in human cells.Nat Biotechnol(2020).
46.Jonsson,T.et al.A mutation in APP protects against Alzheimer's disease and age-related cognitive decline.Nature 488,96-99(2012).
47.Abifadel,M.et al.Mutations in PCSK9 cause autosomal dominant hypercholesterolemia.Nat Genet 34,154-156(2003).
48.Bustami,J.et al.Cholesteryl ester transfer protein(CETP)I405V polymorphism and cardiovascular disease in eastern European Caucasians-a cross-sectional study.BMC Geriatr 16,144(2016).
49.Flannick,J.et al.Loss-of-function mutations in SLC30A8 protect against type 2 diabetes.Nat Genet 46,357-363(2014).
50.Sakuntabhai,A.et al.A variant in the CD209 promoter is associated with severity of dengue disease.Nat Genet 37,507-513(2005).
51.Olson,H.E.et al.Cyclin-Dependent Kinase-Like 5 Deficiency Disorder:Clinical Review.Pediatr Neurol 97,18-25(2019).
52.Al-Saaidi,R.et al.The LMNA mutation p.Arg321Ter associated with dilated cardiomyopathy leads to reduced expression and a skewed ratio of lamin A and lamin C proteins.Exp Cell Res 319,3010-3019(2013).
53.Ip,J.P.K.,Mellios,N.&Sur,M.Rett syndrome:insights into genetic,molecular and circuit mechanisms.Nat Rev Neurosci 19,368-382(2018).
54.Christodoulou,J.,Grimm,A.,Maher,T.&Bennetts,B.RettBASE:The IRSA MECP2 variation database-a new mutation database in evolution.Hum Mutat 21,466-472(2003).
55.Dwivedi,O.P.et al.Loss of ZnT8 function protects against diabetes by enhanced insulin secretion.Nat Genet 51,1596-1606(2019).
56.Thyme,S.B.,Akhmetova,L.,Montague,T.G.,Valen,E.&Schier,A.F.Internal guide RNA interactions interfere with Cas9-mediated cleavage.Nat Commun 7,11750(2016).
57.Boyle,E.A.et al.Quantification of Cas9 binding and cleavage across diverse guide sequences maps landscapes of target engagement.Science Advances,in press(2021).
58.Gaudelli,N.M.et al.Directed evolution of adenine base editors with increased activity and therapeutic application.Nat Biotechnol 38,892-900(2020).
59.Clement,K.et al.CRISPResso2 provides accurate and rapid genome editing sequence analysis.Nat Biotechnol 37,224-226(2019).
60.Pandey,S.,Agarwala,P.&Maiti,S.Effect of loops and G-quartets on the stability of RNA G-quadruplexes.J Phys Chem B 117,6896-6905(2013).
61.Engler,C.,Gruetzner,R.,Kandzia,R.&Marillonnet,S.Golden gate shuffling:a one-pot DNA shuffling method based on type IIs restriction enzymes.PLoS One 4,e5553(2009).
参考文献
以下の参考文献は全て、それらの全体が参照により本明細書にそれぞれ組み込まれる。
1.Jinek,M.et al.A Programmable Dual-RNA-Guided DNA Endonuclease in Adaptive Bacterial Immunity.Science 337,816-821(2012).
2.Cong,L.et al.Multiplex Genome Engineering Using CRISPR/Cas Systems.Science 339,819-823(2013).
3.Komor,A.C.,Badran,A.H.&Liu,D.R.CRISPR-Based Technologies for the Manipulation of Eukaryotic Genomes.Cell 168,20-36(2017).
4.Komor,A.C.,Kim,Y.B.,Packer,M.S.,Zuris,J.A.&Liu,D.R.Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage.Nature 533,420-424(2016).
5.Nishida,K.et al.Targeted nucleotide editing using hybrid prokaryotic and vertebrate adaptive immune systems.Science 353,aaf8729(2016).
6.Gaudelli,N.M.et al.Programmable base editing of A・T to G・C in genomic DNA without DNA cleavage.Nature 551,464-471(2017).
7.ClinVar,July 2019.
8.Dunbar,C.E.et al.Gene therapy comes of age.Science 359,eaan4672(2018).
9.Cox,D.B.T.,Platt,R.J.&Zhang,F.Therapeutic genome editing:prospects and challenges.Nat.Med.21,121-131(2015).
10.Adli,M.The CRISPR tool kit for genome editing and beyond.Nat.Commun.9,1911(2018).
11.Kleinstiver,B.P.et al.Engineered CRISPR-Cas9 nucleases with altered PAM specificities.Nature 523,481-485(2015).
12.Kleinstiver,B.P.et al.High-fidelity CRISPR-Cas9 nucleases with no detectable genome-wide off-target effects.Nature 529,490-495(2016).
13.Hu,J.H.et al.Evolved Cas9 variants with broad PAM compatibility and high DNA specificity.Nature 556,57-63(2018).
14.Nishimasu,H.et al.Engineered CRISPR-Cas9 nuclease with expanded targeting space.Science 361,1259-1262(2018).
15.Jasin,M.&Rothstein,R.Repair of strand breaks by homologous recombination.Cold Spring Harb.Perspect.Biol.5,a012740(2013).
16.Paquet,D.et al.Efficient introduction of specific homozygous and heterozygous mutations using CRISPR/Cas9.Nature 533,125-129(2016).
17.Kosicki,M.,Tomberg,K.&Bradley,A.Repair of double-strand breaks induced by CRISPR-Cas9 leads to large deletions and complex rearrangements.Nat.Biotechnol.36,765-771(2018).
18.Haapaniemi,E.,Botla,S.,Persson,J.,Schmierer,B.&Taipale,J.CRISPR-Cas9 genome editing induces a p53-mediated DNA damage response.Nat.Med.24,927-930(2018).
19.Ihry,R.J.et al.p53 inhibits CRISPR-Cas9 engineering in human pluripotent stem cells.Nat.Med.24,939-946(2018).
20.Richardson,C.D.,Ray,G.J.,DeWitt,M.A.,Curie,G.L.&Corn,J.E.Enhancing homology-directed genome editing by catalytically active and inactive CRISPR-Cas9 using asymmetric donor DNA.Nat.Biotechnol.34,339-344(2016).
21.Srivastava,M.et al.An Inhibitor of Nonhomologous End-Joining Abrogates Double-Strand Break Repair and Impedes Cancer Progression.Cell 151,1474-1487(2012).
22.Chu,V.T.et al.Increasing the efficiency of homology-directed repair for CRISPR-Cas9-induced precise gene editing in mammalian cells.Nat.Biotechnol.33,543-548(2015).
23.Maruyama,T.et al.Increasing the efficiency of precise genome editing with CRISPR-Cas9 by inhibition of nonhomologous end joining.Nat.Biotechnol.33,538-542(2015).
24.Kim,Y.B.et al.Increasing the genome-targeting scope and precision of base editing with engineered Cas9-cytidine deaminase fusions.Nat.Biotechnol.35,371-376(2017).
25.Li,X.et al.Base editing with a Cpf1-cytidine deaminase fusion.Nat.Biotechnol.36,324-327(2018).
26.Gehrke,J.M.et al.An APOBEC3A-Cas9 base editor with minimized bystander and off-target activities.Nat.Biotechnol.(2018).doi:10.1038/nbt.4199
27.Rees,H.A.&Liu,D.R.Base editing:precision chemistry on the genome and transcriptome of living cells.Nat.Rev.Genet.1(2018).doi:10.1038/s41576-018-0059-1.
28.Ostertag,E.M.&Kazazian Jr,H.H.Biology of Mammalian L1 Retrotransposons.Annu.Rev.Genet.35,501-538(2001).
29.Zimmerly,S.,Guo,H.,Perlman,P.S.&Lambowltz,A.M.Group II intron mobility occurs by target DNA-primed reverse transcription.Cell 82,545-554(1995).
30.Luan,D.D.,Korman,M.H.,Jakubczak,J.L.&Eickbush,T.H.Reverse transcription of R2Bm RNA is primed by a nick at the chromosomal target site:a mechanism for non-LTR retrotransposition.Cell 72,595-605(1993).
31.Feng,Q.,Moran,J.V.,Kazazian,H.H.&Boeke,J.D.Human L1 retrotransposon encodes a conserved endonuclease required for retrotransposition.Cell 87,905-916(1996).
32.Jinek,M.et al.Structures of Cas9 Endonucleases Reveal RNA-Mediated Conformational Activation.Science 343,1247997(2014).
33.Jiang,F.et al.Structures of a CRISPR-Cas9 R-loop complex primed for DNA cleavage.Science aad8282(2016).doi:10.1126/science.aad8282
34.Qi,L.S.et al.Repurposing CRISPR as an RNA-Guided Platform for Sequence-Specific Control of Gene Expression.Cell 152,1173-1183(2013).
35.Tang,W.,Hu,J.H.&Liu,D.R.Aptazyme-embedded guide RNAs enable ligand-responsive genome editing and transcriptional activation.Nat.Commun.8,15939(2017).
36.Shechner,D.M.,Hacisuleyman,E.,Younger,S.T.&Rinn,J.L.Multiplexable,locus-specific targeting of long RNAs with CRISPR-Display.Nat.Methods 12,664-670(2015).
37.Anders,C.&Jinek,M.Chapter One-In vitro Enzymology of Cas9.in Methods in Enzymology(eds.Doudna,J.A.&Sontheimer,E.J.)546,1-20(Academic Press,2014).
38.Briner,A.E.et al.Guide RNA Functional Modules Direct Cas9 Activity and Orthogonality.Mol.Cell 56,333-339(2014).
39.Nowak,C.M.,Lawson,S.,Zerez,M.&Bleris,L.Guide RNA engineering for versatile Cas9 functionality.Nucleic Acids Res.44,9555-9564(2016).
40.Sternberg,S.H.,Redding,S.,Jinek,M.,Greene,E.C.&Doudna,J.A.DNA interrogation by the CRISPR RNA-guided endonuclease Cas9.Nature 507,62-67(2014).
41.Mohr,S.et al.Thermostable group II intron reverse transcriptase fusion proteins and their use in cDNA synthesis and next-generation RNA sequencing.RNA 19,958-970(2013).
42.Stamos,J.L.,Lentzsch,A.M.&Lambowitz,A.M.Structure of a Thermostable Group II Intron Reverse Transcriptase with Template-Primer and Its Functional and Evolutionary Implications.Mol.Cell 68,926-939.e4(2017).
43.Zhao,C.&Pyle,A.M.Crystal structures of a group II intron maturase reveal a missing link in spliceosome evolution.Nat.Struct.Mol.Biol.23,558-565(2016).
44.Zhao,C.,Liu,F.&Pyle,A.M.An ultraprocessive,accurate reverse transcriptase encoded by a metazoan group II intron.RNA 24,183-195(2018).
45.Ran,F.A.et al.Genome engineering using the CRISPR-Cas9 system.Nat.Protoc.8,2281-2308(2013).
46.Liu,Y.,Kao,H.-I.&Bambara,R.A.Flap endonuclease 1:a central component of DNA metabolism.Annu.Rev.Biochem.73,589-615(2004).
47.Krokan,H.E.&Bjoras,M.Base Excision Repair.Cold Spring Harb.Perspect.Biol.5,(2013).
48.Kelman,Z.PCNA:structure,functions and interactions.Oncogene 14,629-640(1997).
49.Choe,K.N.&Moldovan,G.-L.Forging Ahead through Darkness:PCNA,Still the Principal Conductor at the Replication Fork.Mol.Cell 65,380-392(2017).
50.Li,X.,Li,J.,Harrington,J.,Lieber,M.R.&Burgers,P.M.Lagging strand DNA synthesis at the eukaryotic replication fork involves binding and stimulation of FEN-1 by proliferating cell nuclear antigen.J.Biol.Chem.270,22109-22112(1995).
51.Tom,S.,Henricksen,L.A.&Bambara,R.A.Mechanism whereby proliferating cell nuclear antigen stimulates flap endonuclease 1.J.Biol.Chem.275,10498-10505(2000).
52.Tanenbaum,M.E.,Gilbert,L.A.,Qi,L.S.,Weissman,J.S.&Vale,R.D.A protein-tagging system for signal amplification in gene expression and fluorescence imaging.Cell 159,635-646(2014).
53.Bertrand,E.et al.Localization of ASH1 mRNA particles in living yeast.Mol.Cell 2,437-445(1998).
54.Dahlman,J.E.et al.Orthogonal gene knockout and activation with a catalytically active Cas9 nuclease.Nat.Biotechnol.33,1159-1161(2015).
55.Tsai,S.Q.et al.GUIDE-seq enables genome-wide profiling of off-target cleavage by CRISPR-Cas nucleases.Nat.Biotechnol.33,187-197(2015).
56.Tsai,S.Q.et al.CIRCLE-seq:a highly sensitive in vitro screen for genome-wide CRISPR-Cas9 nuclease off-targets.Nat.Methods 14,607-614(2017).
57.Schek N,Cooke C,Alwine JC.Molecular and Cellular Biology.(1992).
58.Gil A,Proudfoot NJ.Cell.(1987).
59.Zhao,B.S.,Roundtree,I.A.,He,C.Nat Rev Mol Cell Biol.(2017).
60.Rubio,M.A.T.,Hopper,A.K.Wiley Interdiscip Rev RNA(2011).
61.Shechner,D.M.,Hacisuleyman E.,Younger,S.T.,Rinn,J.L.Nat Methods.(2015).
62.Paige,J.S.,Wu,K.Y.,Jaffrey,S.R.Science(2011).
63.Ray D.,… Hughes TR.Nature(2013).
64.Chadalavada,D.M.,Cerrone-Szakal,A.L.,Bevilacqua,P.C.RNA(2007).
65.Forster AC,Symons RH.Cell.(1987).
66.Weinberg Z,Kim PB,Chen TH,Li S,Harris KA,Lunse CE,Breaker RR.Nat.Chem.Biol.(2015).
67.Feldstein PA,Buzayan JM,Bruening G.Gene(1989).
68.Saville BJ,Collins RA.Cell.(1990).
69.Winkler WC,Nahvi A,Roth A,Collins JA,Breaker RR.Nature(2004).
70.Roth A,Weinberg Z,Chen AG,Kim PG,Ames TD,Breaker RR.Nat Chem Biol.(2013).
71.Choudhury R,Tsai YS,Dominguez D,Wang Y,Wang Z.Nat Commun.(2012).
72.MacRae IJ,Doudna JA.Curr Opin Struct Biol.(2007).
73.Bernstein E,Caudy AA,Hammond SM,Hannon GJ Nature(2001).
74.Filippov V,Solovyev V,Filippova M,Gill SS.Gene(2000).
75.Cadwell RC and Joyce GF.PCR Methods Appl.(1992).
76.McInerney P,Adams P,and Hadi MZ.Mol Biol Int.(2014).
77.Esvelt KM,Carlson JC,and Liu DR.Nature.(2011).
78.Naorem SS,Hin J,Wang S,Lee WR,Heng X,Miller JF,Guo H.Proc Natl Acad Sci USA(2017).
79.Martinez MA,Vartanian JP,Wain-Hobson S.Proc Natl Acad Sci USA(1994).
80.Meyer AJ,Ellefson JW,Ellington AD.Curr Protoc Mol Biol.(2014).
81.Wang HH,Isaacs FJ,Carr PA,Sun ZZ,Xu G,Forest CR,Church GM.Nature.(2009).
82.Nyerges A et al.Proc Natl Acad Sci USA.(2016).
83.Mascola JR,Haynes BF.Immunol Rev.(2013).
84.X.Wen,K.Wen,D.Cao,G.Li,R.W.Jones,J.Li,S.Szu,Y.Hoshino,L.Yuan,Inclusion of a universal tetanus toxoid CD4(+)T cell epitope P2 significantly enhanced the immunogenicity of recombinant rotavirusΔVP8*subunit parenteral vaccines.Vaccine 32,4420-4427(2014).
85.G.Ada,D.Isaacs,Carbohydrate-protein conjugate vaccines.Clin Microbiol Infect 9,79-85(2003).
86.E.Malito,B.Bursulaya,C.Chen,P.L.Surdo,M.Picchianti,E.Balducci,M.Biancucci,A.Brock,F.Berti,M.J.Bottomley,M.Nissum,P.Costantino,R.Rappuoli,G.Spraggon,Structural basis for lack of toxicity of the diphtheria toxin mutant CRM197.Proceedings of the National Academy of Sciences 109,5229(2012).
87.J.de Wit,M.E.Emmelot,M.C.M.Poelen,J.Lanfermeijer,W.G.H.Han,C.van Els,P.Kaaijk,The Human CD4(+)T Cell Response against Mumps Virus Targets a Broadly Recognized Nucleoprotein Epitope.J Virol 93,(2019).
88.M.May,C.A.Rieder,R.J.Rowe,Emergent lineages of mumps virus suggest the need for a polyvalent vaccine.Int J Infect Dis 66,1-4(2018).
89.M.Ramamurthy,P.Rajendiran,N.Saravanan,S.Sankar,S.Gopalan,B.Nandagopal,Identification of immunogenic B-cell epitope peptides of rubella virus E1 glycoprotein towards development of highly specific immunoassays and/or vaccine.Conference Abstract,(2019).
90.U.S.F.Tambunan,F.R.P.Sipahutar,A.A.Parikesit,D.Kerami,Vaccine Design for H5N1 Based on B-and T-cell Epitope Predictions.Bioinform Biol Insights 10,27-35(2016).
91.Asante,EA.et.al.「A naturally occurring variant of the human prion protein completely prevents prion disease」.Nature.(2015).
92.Crabtree,G.R.&Schreiber,S.L.Three-part inventions:intracellular signaling and induced proximity.Trends Biochem.Sci.21,418-22(1996).
93.Liu,J.et al.Calcineurin Is a Common Target of A and FKBP-FK506 Complexes.Cell 66,807-815(1991).
94.Keith,C.T.et al.A mammalian protein targeted by G1-arresting rapamycin-receptor complex.Nature 369,756-758(2003).
95.Spencer,D.M.,Wandless,T.J.,Schreiber,S.L.S.&Crabtree,G.R.Controlling signal transduction with synthetic ligands.Science 262,1019-24(1993).
96.Pruschy,M.N.et al.Mechanistic studies of a signaling pathway activated by the organic dimerizer FK1012.Chem.Biol.1,163-172(1994).
97.Spencer,D.M.et al.Functional analysis of Fas signaling in vivo using synthetic inducers of dimerization.Curr.Biol.6,839-847(1996).
98.Belshaw,P.J.,Spencer,D.M.,Crabtree,G.R.&Schreiber,S.L.Controlling programmed cell death with a cyclophilin-cyclosporin-based chemical inducer of dimerization.Chem.Biol.3,731-738(1996).
99.Yang,J.X.,Symes,K.,Mercola,M.&Schreiber,S.L.Small-molecule control of insulin and PDGF receptor signaling and the role of membrane attachment.Curr.Biol.8,11-18(1998).
100.Belshaw,P.J.,Ho,S.N.,Crabtree,G.R.&Schreiber,S.L.Controlling protein association and subcellular localization with a synthetic ligand that induces heterodimerization of proteins.Proc.Natl.Acad.Sci.93,4604-4607(2002).
101.Stockwell,B.R.&Schreiber,S.L.Probing the role of homomeric and heteromeric receptor interactions in TGF-βsignaling using small molecule dimerizers.Curr.Biol.8,761-773(2004).
102.Spencer,D.M.,Graef,I.,Austin,D.J.,Schreiber,S.L.&Crabtree,G.R.A general strategy for producing conditional alleles of Src-like tyrosine kinases.Proc.Natl.Acad.Sci.92,9805-9809(2006).
103.Holsinger,L.J.,Spencer,D.M.,Austin,D.J.,Schreiber,S.L.&Crabtree,G.R.Signal transduction in T lymphocytes using a conditional allele of Sos.Proc.Natl.Acad.Sci.92,9810-9814(2006).
104.Myers,M.G.Insulin Signal Transduction and the IRS Proteins.Annu.Rev.Pharmacol.Toxicol.36,615-658(1996).
105.Watowich,S.S.The erythropoietin receptor:Molecular structure and hematopoietic signaling pathways.J.Investig.Med.59,1067-1072(2011).
106.Blau,C.A.,Peterson,K.R.,Drachman,J.G.&Spencer,D.M.A proliferation switch for genetically modified cells.Proc.Natl.Acad.Sci.94,3076-3081(2002).
107.Clackson,T.et al.Redesigning an FKBP-ligand interface to generate chemical dimerizers with novel specificity.Proc.Natl.Acad.Sci.95,10437-10442(1998).
108.Diver,S.T.&Schreiber,S.L.Single-step synthesis of cell-permeable protein dimerizers that activate signal transduction and gene expression.J.Am.Chem.Soc.119,5106-5109(1997).
109.Guo,Z.F.,Zhang,R.&Liang,F.Sen.Facile functionalization of FK506 for biological studies by the thiol-ene'click'reaction.RSC Adv.4,11400-11403(2014).
110.Robinson,D.R.,Wu,Y.-M.&Lin,S.-F.The protein tyrosine kinase family of the human genome.Oncogene 19,5548-5557(2000).
111.Landrum,M.J.et al.ClinVar:public archive of interpretations of clinically relevant variants.Nucleic Acids Res.44,D862-D868(2016).
112.Jinek,M.et al.A Programmable Dual-RNA-Guided DNA Endonuclease in Adaptive Bacterial Immunity.Science 337,816-821(2012).
113.Cong,L.et al.Multiplex Genome Engineering Using CRISPR/Cas Systems.Science
339,819-823(2013).
114.Mali,P.et al.RNA-Guided Human Genome Engineering via Cas9.Science 339,823-826(2013).
115.Yang,H.et al.One-Step Generation of Mice Carrying Reporter and Conditional Alleles by CRISPR/Cas-Mediated Genome Engineering.Cell 154,1370-1379(2013).
116.Kim,S.,Kim,D.,Cho,S.W.,Kim,J.&Kim,J.-S.Highly efficient RNA-guided genome editing in human cells via delivery of purified Cas9 ribonucleoproteins.Genome Res.24,1012-1019(2014).
117.Orlando,S.J.et al.Zinc-finger nuclease-driven targeted integration into mammalian genomes using donors with limited chromosomal homology.Nucleic Acids Res.38,e152-e152(2010).
118.Tsai,S.Q.et al.GUIDE-seq enables genome-wide profiling of off-target cleavage by CRISPR-Cas nucleases.Nat.Biotechnol.33,187-197(2015).
119.Suzuki,K.et al.In vivo genome editing via CRISPR/Cas9 mediated homology-independent targeted integration.Nature 540,144-149(2016).
120.Kosicki,M.,Tomberg,K.&Bradley,A.Repair of double-strand breaks induced by CRISPR-Cas9 leads to large deletions and complex rearrangements.Nat.Biotechnol.36,765-771(2018).
121.Haapaniemi,E.,Botla,S.,Persson,J.,Schmierer,B.&Taipale,J.CRISPR-Cas9 genome editing induces a p53-mediated DNA damage response.Nat.Med.24,927-930(2018).
122.Ihry,R.J.et al.p53 inhibits CRISPR-Cas9 engineering in human pluripotent stem cells.Nat.Med.24,939-946(2018).
123.Chapman,J.R.,Taylor,M.R.G.&Boulton,S.J.Playing the end game:DNA double-strand break repair pathway choice.Mol.Cell 47,497-510(2012).
124.Cox,D.B.T.,Platt,R.J.&Zhang,F.Therapeutic genome editing:prospects and challenges.Nat.Med.21,121-131(2015).
125.Paquet,D.et al.Efficient introduction of specific homozygous and heterozygous mutations using CRISPR/Cas9.Nature 533,125-129(2016).
126.Chu,V.T.et al.Increasing the efficiency of homology-directed repair for CRISPR-Cas9-induced precise gene editing in mammalian cells.Nat.Biotechnol.33,543-548(2015).
127.Maruyama,T.et al.Increasing the efficiency of precise genome editing with CRISPR-Cas9 by inhibition of nonhomologous end joining.Nat.Biotechnol.33,538-542(2015).
128.Rees,H.A.,Yeh,W.-H.&Liu,D.R.Development of hRad51-Cas9 nickase fusions that mediate HDR without double-stranded breaks.Nat.Commun.10,1-12(2019).
129.Komor,A.C.,Kim,Y.B.,Packer,M.S.,Zuris,J.A.&Liu,D.R.Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage.Nature 533,420-424(2016).
130.Gaudelli,N.M.et al.Programmable base editing of A・T to G・C in genomic DNA without DNA cleavage.Nature 551,464-471(2017).
131.Gao,X.et al.Treatment of autosomal dominant hearing loss by in vivo delivery of genome editing agents.Nature 553,217-221(2018).
132.Ingram,V.M.A specific chemical difference between the globins of normal human and sickle-cell anaemia haemoglobin.Nature 178,792-794(1956).
133.Myerowitz,R.&Costigan,F.C.The major defect in Ashkenazi Jews with Tay-Sachs disease is an insertion in the gene for the alpha-chain of beta-hexosaminidase.J.Biol.Chem.263,18587-18589(1988).
134.Zielenski,J.Genotype and Phenotype in Cystic Fibrosis.Respiration 67,117-133(2000).
135.Mead,S.et al.A Novel Protective Prion Protein Variant that Colocalizes with Kuru Exposure.N.Engl.J.Med.361,2056-2065(2009).
136.Marraffini,L.A.&Sontheimer,E.J.CRISPR interference limits horizontal gene transfer in staphylococci by targeting DNA.Science 322,1843-1845(2008).
137.Barrangou,R.et al.CRISPR provides acquired resistance against viruses in prokaryotes.Science 315,1709-1712(2007).
138.Jiang,F.&Doudna,J.A.CRISPR-Cas9 Structures and Mechanisms.Annu.Rev.Biophys.46,505-529(2017).
139.Hille,F.et al.The Biology of CRISPR-Cas:Backward and Forward.Cell 172,1239-1259(2018).
140.Luan,D.D.,Korman,M.H.,Jakubczak,J.L.&Eickbush,T.H.Reverse transcription of R2Bm RNA is primed by a nick at the chromosomal target site:a mechanism for non-LTR retrotransposition.Cell 72,595-605(1993).
141.Liu,Y.,Kao,H.-I.&Bambara,R.A.Flap endonuclease 1:a central component of DNA metabolism.Annu.Rev.Biochem.73,589-615(2004).
142.Rees,H.A.&Liu,D.R.Base editing:precision chemistry on the genome and transcriptome of living cells.Nat.Rev.Genet.19,770(2018).
143.Richardson,C.D.,Ray,G.J.,DeWitt,M.A.,Curie,G.L.&Corn,J.E.Enhancing homology-directed genome editing by catalytically active and inactive CRISPR-Cas9 using asymmetric donor DNA.Nat.Biotechnol.34,339-344(2016).
144.Qi,L.S.et al.Repurposing CRISPR as an RNA-Guided Platform for Sequence-Specific Control of Gene Expression.Cell 152,1173-1183(2013).
145.Shechner,D.M.,Hacisuleyman,E.,Younger,S.T.&Rinn,J.L.Multiplexable,locus-specific targeting of long RNAs with CRISPR-Display.Nat.Methods 12,664-670(2015).
146.Tang,W.,Hu,J.H.&Liu,D.R.Aptazyme-embedded guide RNAs enable ligand-responsive genome editing and transcriptional activation.Nat.Commun.8,15939(2017).
147.Jinek,M.et al.Structures of Cas9 Endonucleases Reveal RNA-Mediated Conformational Activation.Science 343,1247997(2014).
148.Nishimasu,H.et al.Crystal Structure of Cas9 in Complex with Guide RNA and Target DNA.Cell 156,935-949(2014).
149.Jiang,F.,Zhou,K.,Ma,L.,Gressel,S.&Doudna,J.A.A Cas9-guide RNA complex preorganized for target DNA recognition.Science 348,1477-1481(2015).
150.Baranauskas,A.et al.Generation and characterization of new highly thermostable and processive M-MuLV reverse transcriptase variants.Protein Eng.Des.Sel.25,657-668(2012).
151.Gerard,G.F.et al.The role of template-primer in protection of reverse transcriptase from thermal inactivation.Nucleic Acids Res.30,3118-3129(2002).
152.Arezi,B.&Hogrefe,H.Novel mutations in Moloney Murine Leukemia Virus reverse transcriptase increase thermostability through tighter binding to template-primer.Nucleic Acids Res.37,473-481(2009).
153.Kotewicz,M.L.,Sampson,C.M.,D'Alessio,J.M.&Gerard,G.F.Isolation of cloned Moloney murine leukemia virus reverse transcriptase lacking ribonuclease H activity.Nucleic Acids Res.16,265-277(1988).
154.Shen,M.W.et al.Predictable and precise template-free CRISPR editing of pathogenic variants.Nature 563,646-651(2018).
155.Thuronyi,B.W.et al.Continuous evolution of base editors with expanded target compatibility and improved activity.Nat.Biotechnol.(2019).doi:10.1038/s41587-019-0193-0
156.Kim,Y.B.et al.Increasing the genome-targeting scope and precision of base editing with engineered Cas9-cytidine deaminase fusions.Nat.Biotechnol.35,371-376(2017).
157.Koblan,L.W.et al.Improving cytidine and adenine base editors by expression optimization and ancestral reconstruction.Nat.Biotechnol.(2018).doi:10.1038/nbt.4172
158.Komor,A.C.et al.Improved base excision repair inhibition and bacteriophage Mu Gam protein yields C:GからT:A base editors with higher efficiency and product purity.Sci.Adv.3,eaao4774(2017).
159.Kleinstiver,B.P.et al.High-fidelity CRISPR-Cas9 nucleases with no detectable genome-wide off-target effects.Nature 529,490-495(2016).
160.Zuo,E.et al.Cytosine base editor generates substantial off-target single-nucleotide variants in mouse embryos.Science 364,289-292(2019).
161.Jin,S.et al.Cytosine,but not adenine,base editors induce genome-wide off-target mutations in rice.Science 364,292-295(2019).
162.Kim,D.,Kim,D.,Lee,G.,Cho,S.-I.&Kim,J.-S.Genome-wide target specificity of CRISPR RNA-guided adenine base editors.Nat.Biotechnol.37,430-435(2019).
163.Grunewald,J.et al.Transcriptome-wide off-target RNA editing induced by CRISPR-guided DNA base editors.Nature 569,433-437(2019).
164.Zhou,C.et al.Off-target RNA mutation induced by DNA base editing and its elimination by mutagenesis.Nature 571,275-278(2019).
165.Rees,H.A.,Wilson,C.,Doman,J.L.&Liu,D.R.Analysis and minimization of cellular RNA editing by DNA adenine base editors.Sci.Adv.5,eaax5717(2019).
166.Ostertag,E.M.&Kazazian Jr,H.H.Biology of Mammalian L1 Retrotransposons.Annu.Rev.Genet.35,501-538(2001).
167.Griffiths,D.J.Endogenous retroviruses in the human genome sequence.Genome Biol.2,REVIEWS1017(2001).
168.Berkhout,B.,Jebbink,M.&Zsiros,J.Identification of an Active Reverse Transcriptase Enzyme Encoded by a Human Endogenous HERV-K Retrovirus.J.Virol.73,2365-2375(1999).
169.Halvas,E.K.,Svarovskaia,E.S.&Pathak,V.K.Role of Murine Leukemia Virus Reverse Transcriptase Deoxyribonucleoside Triphosphate-Binding Site in Retroviral Replication and In Vivo Fidelity.J.Virol.74,10349-10358(2000).
170.Dever,D.P.et al.CRISPR/Cas9 Beta-globin Gene Targeting in Human Hematopoietic Stem Cells.Nature 539,384-389(2016).
171.Park,S.H.et al.Highly efficient editing of the β-globin gene in patient-derived hematopoietic stem and progenitor cells to treat sickle cell disease.Nucleic Acids Res.doi:10.1093/nar/gkz475
172.Collinge,J.Prion diseases of humans and animals:their causes and molecular basis.Annu.Rev.Neurosci.24,519-550(2001).
173.Asante,E.A.et al.A naturally occurring variant of the human prion protein completely prevents prion disease.Nature 522,478-481(2015).
174.Anzalone,A.V.,Lin,A.J.,Zairis,S.,Rabadan,R.&Cornish,V.W.Reprogramming eukaryotic translation with ligand-responsive synthetic RNA switches.Nat.Methods 13,453-458(2016).
175.Badran,A.H.et al.Continuous evolution of Bacillus thuringiensis toxins overcomes insect resistance.Nature 533,58-63(2016).
176.Anders,C.&Jinek,M.Chapter One-In Vitro Enzymology of Cas9.in Methods in Enzymology(eds.Doudna,J.A.&Sontheimer,E.J.)546,1-20(Academic Press,2014).
177.Pirakitikulr,N.,Ostrov,N.,Peralta-Yahya,P.&Cornish,V.W.PCRless library mutagenesis via oligonucleotide recombination in yeast.Protein Sci.Publ.Protein Soc.19,2336-2346(2010).
178.Clement,K.et al.CRISPResso2 provides accurate and rapid genome editing sequence analysis.Nat.Biotechnol.37,224-226(2019).
179.Tsai,S.Q.et al.GUIDE-seq enables genome-wide profiling of off-target cleavage by CRISPR-Cas nucleases.Nat.Biotechnol.33,187-197(2015).
180.Kleinstiver,B.P.et al.High-fidelity CRISPR-Cas9 nucleases with no detectable genome-wide off-target effects.Nature 529,490-495(2016).
181.Koblan,L.W.et al.Improving cytidine and adenine base editors by expression optimization and ancestral reconstruction.Nat.Biotechnol.(2018).doi:10.1038/nbt.4172
182.Baranauskas,A.et al.Generation and characterization of new highly thermostable and processive M-MuLV reverse transcriptase variants.Protein Eng.Des.Sel.25,657-668(2012).
183.Schechner,DM,Hacisuleyman E.,Younger ST,Rinn JL.Nat Methods 664-70(2015).
184.Brown JA,et al.Nat Struct Mol Biol 633-40(2014).
185.Conrad NA and Steitz JA.EMBO J 1831-41(2005).
186.Bartlett JS,et al.Proc Natl Acad Sci USA 8852-7(1996).
187.Mitton-Fry RM,DeGregorio SJ,Wang J,Steitz TA,Steitz JA.Science 1244-7(2010).
188.Forster AC,Symons RH.Cell.1987.
189.Weinberg Z,Kim PB,Chen TH,Li S,Harris KA,Lunse CE,Breaker RR.Nat.Chem.Biol.2015.
190.Feldstein PA,Buzayan JM,Bruening G.Gene 1989.
191.Saville BJ,Collins RA.Cell.1990.
192.Roth A,Weinberg Z,Chen AG,Kim PG,Ames TD,Breaker RR.Nat Chem Biol.2013.
193.Borchardt EK,et al.RNA 1921-30(2015).
194.Zhang Y,et al.Mol Cell 792-806(2013).
195.Dang Y,et al.Genome Biol 280(2015).
196.Schaefer M,Kapoor U,and Jantsch MF.Open Biol 170077(2017).
197.Nahar S,et al.Chem Comm 2377-80(2018).
198.Gao Y and Zhao Y.J Integr Plant Biol 343-9(2014).
199.Dubois N,Marquet R,Paillart J,Bernacchi S.Front Microbiol 527(2018).
200.Costa M and Michel F.EMBO J 1276-85(1995).
201.Hu JH,et al.Nature 57-63(2018).
202.Furukawa K,Gu H,Breaker RR.Methods Mol Biol 209-20(2014).
203.Zettler,J.,Schutz,V.&Mootz,H.D.The naturally split Npu DnaE intein exhibits an extraordinarily high rate in the protein trans-splicing reaction.FEBS Lett.583,909-914(2009).
204.Kugler,S.,Kilic,E.&Bahr,M.Human synapsin 1 gene promoter confers highly neuron-specific long-term transgene expression from an adenoviral vector in the adult rat brain depending on the transduced area.Gene Ther.10,337-347(2003).
205.de Felipe,P.,Hughes,L.E.,Ryan,M.D.&Brown,J.D.Co-translational,intraribosomal cleavage of polypeptides by the foot-and-mouth disease virus 2A peptide.J.Biol.Chem.278,11441-11448(2003).
206.Levy,J.M.&Nicoll,R.A.Membrane-associated guanylate kinase dynamics reveal regional and developmental specificity of synapse stability.J.Physiol.595,1699-1709(2017).
207.Li,B.&Dewey,C.N.RSEM:accurate transcript quantification from RNA-Seq data with or without a reference genome.BMC Bioinformatics 12,323(2011).
208.Ritchie,M.E.et al.limma powers differential expression analyses for RNA-sequencing and microarray studies.Nucleic Acids Res.43,e47-e47(2015).
均等物及び範囲
クレームにおいて、「a」、「an」、及び「the」等の冠詞は、1又は1より多いことを意味してもよいが、それと反する指示がないか、又はそれとは別に、文脈から明らかでない場合に限る。ある群の1以上のメンバー間に「又は」を包含する態様又は記載は、その群のメンバーのうち、1つ、1つより多いか、又は、全てが、所定の産物又はプロセスに存在するか、それに採用されるか、又はそれとは別に、それに関係があるか、を満たすと考えられるが、それと反する指示がないか、又はそれとは別に、文脈から明らかでない場合に限る。本発明は、その群のうち、正確に1つのメンバーが、所定の産物又はプロセスに存在するか、それに採用されるか、又はそれとは別に、それに関係がある態様を包含する。本発明は、その群のメンバーのうち、1つより多いか又は全てが、所定の産物又はプロセスに存在するか、それに採用されるか、又はそれとは別に、それに関係がある態様を包含する。
クレームにおいて、「a」、「an」、及び「the」等の冠詞は、1又は1より多いことを意味してもよいが、それと反する指示がないか、又はそれとは別に、文脈から明らかでない場合に限る。ある群の1以上のメンバー間に「又は」を包含する態様又は記載は、その群のメンバーのうち、1つ、1つより多いか、又は、全てが、所定の産物又はプロセスに存在するか、それに採用されるか、又はそれとは別に、それに関係があるか、を満たすと考えられるが、それと反する指示がないか、又はそれとは別に、文脈から明らかでない場合に限る。本発明は、その群のうち、正確に1つのメンバーが、所定の産物又はプロセスに存在するか、それに採用されるか、又はそれとは別に、それに関係がある態様を包含する。本発明は、その群のメンバーのうち、1つより多いか又は全てが、所定の産物又はプロセスに存在するか、それに採用されるか、又はそれとは別に、それに関係がある態様を包含する。
更には、本開示は、列挙されたクレームの1以上からの1以上の限定、要素、節、及び記述用語(descriptive terms)が、別のクレーム中へ導入される、全ての変動、組み合わせ、及び順列を包括する。例えば、別のクレームに従属するいずれのクレームも、同じ基本クレームに従属するいずれか他のクレーム中に見出される1以上の限定を包含するように修飾され得る。要素が、例として、マーカッシュ群形式において列挙されたものとして提示されている場合、要素の各下位群もまた開示されており、いずれの要素(単数又は複数)も群から除去され得る。一般に、本発明、又は本発明の側面が、特定の要素及び/又は特長を含むとして見なされる場合、本発明のある側面又は本発明のある態様は、かかる要素及び/又は特長からなるか、又は実質的に次いでなると理解されるべきである。簡潔さを目的として、それらの態様は、本明細書中、in haec verbaで具体的に表明されていない。用語「含むこと(comprising)」及び「含有すること(containing)」が、オープンであることを意図し、追加の要素又は工程の包含を容認することにもまた留意されたい。範囲が与えられるとき、エンドポイントも包含される。更には、別段の指示がないか、又はそれとは別に、文脈及び当業者の理解から明らかでない場合に限り、範囲として表現された値は、いずれか具体的な値、又は本発明の異なる態様において述べられた範囲内の部分範囲を、文脈が明確に別段の指図をしない限り範囲の下限の単位の10倍まで、想定し得る。
本出願は、種々の発行済み特許、公開特許出願、雑誌記事、及び他の公刊物を参照し、これらの全ては参照によって本明細書に組み込まれる。組み込まれる参照のいずれかと本明細書との間に矛盾がある場合、本明細書がコントロールする(control)ものとする。加えて、先行技術に属する本発明のいずれか特定の態様は、クレームのいずれかの1以上からはっきりと除外されてもよい。かかる態様は当業者に知られていると思われるので、それらは、除外が本明細書においてはっきりと表明されていない場合であっても、除外されてもよい。本発明のいずれか特定の態様は、先行技術の存在に関するか否かにかかわらず、いずれかのクレームから、いずれかの理由で除外され得る。
当業者は、本明細書に記載の具体的な態様の多くの均等物を認識するか、又はせいぜいルーチンな実験法を使用して確かめる能力があるであろう。本明細書に記載の本態様の範囲は、上の記載に限定されることを意図されず、むしろ添付のクレームに表明されている通りである。当業者は、以下の態様において定義される通り、本発明の精神又は範囲から逸脱せずに、この記載への種々の変化及び修飾がなされてもよいことを了解するであろう。
Claims (145)
- ガイドRNAと、DNA合成鋳型及びプライマー結合部位を含む少なくとも1つの核酸伸長アームとを含むプライム編集のための、pegRNAであって、伸長アームは、それに取り付けられた、トゥーループ、ヘアピン、ステムループ、シュードノット、アプタマー、G-四重鎖、tRNA、リボスイッチ、又はリボザイムからなる群から選択される核酸部分を含む、pegRNA。
- 核酸部分が伸長アームの3'末端に取り付けられている、請求項1に記載のpegRNA。
- 核酸部分が伸長アームの5'末端に取り付けられている、請求項1に記載のpegRNA。
- シュードノットが、配列番号195(Mpknot1)、配列番号196(3'トリミングしたMpknot1)、配列番号197(5'extraを有するMpknot1)、配列番号198(Mpknot1 U38A)、配列番号199(Mpknot1 U38A A29C)、配列番号200(MMLC A29C)、配列番号201(5'extra及びU38Aを有するMpknot1)、配列番号202(5'extra及びU38A A29Cを有するMpknot1)及び配列番号203(5'extra及びA29Cを有するMpknot1)からなる群から選択されるヌクレオチド配列、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有するMpknot1部分である、請求項1に記載のpegRNA。
- G-四重鎖が、配列番号204(tns1)、配列番号205(stk40)、配列番号206(apc2)、配列番号207(ceacam4)、配列番号208(pitpnm3)、配列番号209(rlf)、配列番号210(erc1)、配列番号211(ube3c)、配列番号212(taf15)、配列番号213(stard3)、及び配列番号214(g2)からなる群から選択されるヌクレオチド配列、又はそれらと少なくとも80%の配列同一性を有するヌクレオチド配列を有する、請求項1に記載のpegRNA。
- evopreq1が、配列番号215(evopreq1)、配列番号216(evopreq1motif1)、配列番号217(evopreq1motif2)、配列番号218(evopreq1motif3)、配列番号219(より短いpreq 1-1)、配列番号220(preq 1-1 G5C(mut1))、及び配列番号221(preq1-1 G15C(mut2))からなる群から選択されるヌクレオチド配列、又はそれらと少なくとも80%の配列同一性を有するヌクレオチド配列を有する、請求項1に記載のpegRNA。
- tRNA部分が、配列番号222のヌクレオチド配列、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有する、請求項1に記載のpegRNA。
- 核酸部分が、配列番号223(xrn1)のヌクレオチド配列、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有する、請求項1に記載のpegRNA。
- 核酸部分が、配列番号224(grp1イントロンP4P6)のヌクレオチド配列、又はそれと少なくとも80%の配列同一性を有するヌクレオチド配列を有する、請求項1に記載のpegRNA。
- 核酸部分が、リンカーによってpegRNAに取り付けれられている、請求項1~9のいずれか一項に記載のpegRNA。
- リンカーが、配列番号225~236からなる群より選択されるヌクレオチド配列を有する、請求項10に記載のpegRNA。
- リンカーが、少なくとも3ヌクレオチド、少なくとも4ヌクレオチド、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、少なくとも15ヌクレオチド、少なくとも16ヌクレオチド、少なくとも17ヌクレオチド、少なくとも18ヌクレオチド、少なくとも19ヌクレオチド、少なくとも20ヌクレオチド、少なくとも21ヌクレオチド、少なくとも22ヌクレオチド、少なくとも23ヌクレオチド、少なくとも24ヌクレオチド、少なくとも25ヌクレオチド、少なくとも26ヌクレオチド、少なくとも27ヌクレオチド、少なくとも28ヌクレオチド、少なくとも29ヌクレオチド、又は少なくとも30ヌクレオチド長であり、リンカーが50ヌクレオチド以下である、請求項10に記載のpegRNA。
- リンカーが8ヌクレオチド長である、請求項10に記載のpegRNA。
- 伸長アームがガイドRNAの3'又は5'末端に配置され、核酸伸長アームがDNA又はRNAである、請求項1に記載のpegRNA。
- pegRNAが、napDNAbpに結合し、napDNAbpを標的DNA配列に向けることができる、請求項1に記載のpegRNA。
- 標的DNA配列が、標的鎖及び相補的な非標的鎖を含む、請求項15に記載のpegRNA。
- ガイドRNAが標的鎖にハイブリダイズして、RNA-DNAハイブリッド及びR-ループを形成する、請求項16に記載のpegRNA。
- 核酸伸長アームが、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、少なくとも15ヌクレオチド、少なくとも16ヌクレオチド、少なくとも17ヌクレオチド、少なくとも18ヌクレオチド、少なくとも19ヌクレオチド、少なくとも20ヌクレオチド、少なくとも21ヌクレオチド、少なくとも22ヌクレオチド、少なくとも23ヌクレオチド、少なくとも24ヌクレオチド、少なくとも25ヌクレオチド、少なくとも26ヌクレオチド、少なくとも27ヌクレオチド、少なくとも28ヌクレオチド、少なくとも29ヌクレオチド、少なくとも30ヌクレオチド、少なくとも31ヌクレオチド、少なくとも32ヌクレオチド、少なくとも33ヌクレオチド、少なくとも34ヌクレオチド、少なくとも35ヌクレオチド、少なくとも36ヌクレオチド、少なくとも37ヌクレオチド、少なくとも38ヌクレオチド、少なくとも39ヌクレオチド、少なくとも40ヌクレオチド、少なくとも41ヌクレオチド、少なくとも42ヌクレオチド、少なくとも43ヌクレオチド、少なくとも44ヌクレオチド、少なくとも45ヌクレオチド、少なくとも46ヌクレオチド、少なくとも47ヌクレオチド、少なくとも48ヌクレオチド、少なくとも49ヌクレオチド、又は少なくとも50ヌクレオチドである、請求項1に記載のpegRNA。
- DNA合成鋳型が、少なくとも3ヌクレオチド、少なくとも4ヌクレオチド、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、又は少なくとも15ヌクレオチド長である、請求項1に記載のpegRNA。
- DNA合成鋳型が所望の編集をコードする、請求項1に記載のpegRNA。
- プライマー結合部位が、少なくとも3ヌクレオチド、少なくとも4ヌクレオチド、少なくとも5ヌクレオチド、少なくとも6ヌクレオチド、少なくとも7ヌクレオチド、少なくとも8ヌクレオチド、少なくとも9ヌクレオチド、少なくとも10ヌクレオチド、少なくとも11ヌクレオチド、少なくとも12ヌクレオチド、少なくとも13ヌクレオチド、少なくとも14ヌクレオチド、又は少なくとも15ヌクレオチド長である、請求項1に記載のpegRNA。
- (a)核酸プログラム型DNA結合タンパク質(napDNAbp)と、RNA依存性DNAポリメラーゼ活性を含むドメインと、を含む融合タンパク質;及び
(b)請求項1~21のいずれか一項に記載のpegRNA
を含む、プライム編集のための複合体。 - napDNAbpがニッカーゼ活性を有する、請求項22に記載の複合体。
- napDNAbpがCas9タンパク質又はそのバリアントである、請求項22に記載の複合体。
- napDNAbpが、ヌクレアーゼ活性Cas9、ヌクレアーゼ不活性Cas9(dCas9)、又はCas9ニッカーゼ(nCas9)である、請求項22に記載の複合体。
- napDNAbpがCas9ニッカーゼ(nCas9)である、請求項22に記載の複合体。
- napDNAbpが、Cas9、Cas12e、Cas12d、Cas12a、Cas12b1、Cas12b2、Cas13a、Cas12c、Cas12d、Cas12e、Cas12h、Cas12i、Cas12g、Cas12f(Cas14)、Cas12f1、Cas12j(CasΦ)、及びアルゴノートからなる群から選択され、任意選択でニッカーゼ活性を有する、請求項22に記載の複合体。
- RNA依存性DNAポリメラーゼ活性を含むドメインが、配列番号32、34、36、102~128及び132のアミノ酸配列のいずれか1つを含む逆転写酵素である、請求項22に記載の複合体。
- RNA依存性DNAポリメラーゼ活性を含むドメインが、配列番号32、34、36、102~128及び132のいずれか1つのアミノ酸配列と少なくとも80%、85%、90%、95%、98%、又は99%の配列同一性を有するアミノ酸配列を含む逆転写酵素である、請求項22に記載の複合体。
- RNA依存性DNAポリメラーゼ活性を含むドメインが、レトロウイルス又はレトロトランスポゾン由来の天然に存在する逆転写酵素である、請求項22に記載の複合体。
- 請求項1~19のいずれか一項に記載のpegRNAをコードする核酸分子。
- 核酸分子がプロモータの制御下にある、請求項31に記載の核酸分子を含む発現ベクター。
- プロモータがpolIIIプロモータである、請求項32に記載の発現ベクター。
- プロモータがU6プロモータである、請求項32に記載の発現ベクター。
- プロモータが、U6、U6v4、U6v7、若しくはU6v9プロモータ、又はそのフラグメントである、請求項32に記載の発現ベクター。
- 請求項1~21のいずれか一項に記載のpegRNAを含む細胞。
- 請求項22~30のいずれか一項に記載の複合体を含む細胞。
- 請求項31に記載の核酸分子を含む細胞。
- 請求項32~35のいずれか一項に記載の発現ベクターを含む細胞。
- (i)請求項1~21のいずれか一項に記載のpegRNA、請求項22~30のいずれか一項に記載の複合体、請求項31に記載の核酸分子、請求項32~35のいずれか一項に記載の発現ベクター、又は請求項36~39のいずれか一項に記載の細胞、及び(ii)薬学的に許容される賦形剤を含む医薬組成物。
- (i)請求項1~21のいずれか一項に記載のpegRNA、請求項22~30のいずれか一項に記載の複合体、請求項31に記載の核酸分子、請求項32~35のいずれか一項に記載の発現ベクター、又は請求項36~39のいずれか一項に記載の細胞、及び(ii)プライム編集を行うための説明書のセット、を含むキット組成物。
- 標的DNA配列を、請求項1~21のいずれか一項に記載のpegRNA及びnapDNAbpとRNA依存性DNAポリメラーゼ活性を有するドメインとを含むプライム編集因子と接触させることを含む、プライム編集の方法であって、修飾を含まないpegRNAを使用する同じ方法と比較して編集効率が増加される、プライム編集の方法。
- 編集効率が少なくとも1.5倍増加する、請求項42に記載の方法。
- 編集効率が少なくとも2倍増加する、請求項42に記載の方法。
- 編集効率が少なくとも3倍増加する、請求項42に記載の方法。
- napDNAbpがニッカーゼ活性を有する、請求項42に記載の方法。
- napDNAbpがCas9タンパク質又はそのバリアントである、請求項42に記載の方法。
- napDNAbpが、ヌクレアーゼ活性Cas9、ヌクレアーゼ不活性Cas9(dCas9)、又はCas9ニッカーゼ(nCas9)である、請求項47に記載の方法。
- napDNAbpがCas9ニッカーゼ(nCas9)である、請求項48に記載の方法。
- napDNAbpが、Cas9、Cas12e、Cas12d、Cas12a、Cas12b1、Cas12b2、Cas13a、Cas12c、Cas12d、Cas12e、Cas12h、Cas12i、Cas12g、Cas12f(Cas14)、Cas12f1、Cas12j(CasΦ)、及びアルゴノートからなる群から選択され、任意選択でニッカーゼ活性を有する、請求項42に記載の方法。
- RNA依存性DNAポリメラーゼ活性を含むドメインが、配列番号32、34、36、102~128及び132のアミノ酸配列のいずれか1つを含む逆転写酵素である、請求項42に記載の方法。
- RNA依存性DNAポリメラーゼ活性を含むドメインが、配列番号32、34、36、102~128及び132のいずれか1つのアミノ酸配列と少なくとも80%、85%、90%、95%、98%、又は99%の配列同一性を有するアミノ酸配列を含む逆転写酵素である、請求項42に記載の方法。
- RNA依存性DNAポリメラーゼ活性を含むドメインが、レトロウイルス又はレトロトランスポゾン由来の天然に存在する逆転写酵素である、請求項42に記載の方法。
- リンカーが、請求項56に記載の計算方法によってデザインされる、請求項10に記載のpegRNA。
- 二本鎖標的DNA配列にヌクレオチド編集を正確に組み入れする方法であって、方法は、二本鎖標的DNA配列を、核酸プログラム型DNA結合タンパク質(napDNAbp)、DNAポリメラーゼ、及びプライム編集ガイドRNA(PEgRNA)を含むプライム編集因子と接触させることを含み、PEgRNAは、
(a)二本鎖標的DNA配列の第1鎖にハイブリダイズするスペーサと、
(b)二本鎖標的DNA配列の第2鎖にハイブリダイズする伸長アームと、
(c)ヌクレオチド編集を含むDNA合成鋳型と、
(d)napDNAbpと相互作用するgRNAコアと、
(e)核酸部分に取り付けられた、トゥーループ、ヘアピン、ステムループ、シュードノット、アプタマー、G-四重鎖、tRNA、リボスイッチ、又はリボザイムからなる群から選択される核酸部分と、
(f)核酸部分をpegRNAにカップリングするリンカーと、
を含み、
リンカーは、計算モデルによってデザインされ、
PEgRNAは、二本鎖標的DNA配列にヌクレオチド編集を組み入れするようにプライム編集因子に指示する、
二本鎖標的DNA配列にヌクレオチド編集を正確に組み入れする方法。 - プライム編集ガイドRNA(pegRNA)を核酸部分にカップリングするための少なくとも1つの核酸リンカーを同定するための方法であって、
少なくとも1つのコンピュータ・ハードウェア・プロセッサを使用して、
第1の核酸リンカー候補を包含する複数の核酸リンカー候補を生成することと、
複数の核酸リンカー候補の中から少なくとも1つの核酸リンカーを、少なくとも部分的に、
複数の核酸リンカー候補の少なくともいくつかのそれぞれについて複数のスコアを計算することであって、計算することが、第1の核酸リンカー候補について第1のスコアセットを計算することを含み、第1のスコアセットが、
第1の核酸リンカー候補とpegRNAの第1の領域との間の相互作用の程度を示す第1のスコアと、
第1の核酸リンカー候補とpegRNAの第2の領域との間の相互作用の程度を示す第2のスコアと、
を含む、計算することと、
計算された複数のスコアを使用して、複数の核酸リンカー候補の少なくともいくつかの中から少なくとも1つの核酸リンカーを同定することと、
によって同定することと、
少なくとも1つの核酸リンカーを示す情報を出力することと、
を行うことを含む、プライム編集ガイドRNA(pegRNA)を核酸部分にカップリングするための少なくとも1つの核酸リンカーを同定するための方法。 - 第1のスコアが、第1の核酸リンカー候補がpegRNAの第1の領域との相互作用を回避すると予測される程度を示し、第2のスコアが、第1の核酸リンカー候補がpegRNAの第2の領域との相互作用を回避すると予測される程度を示す、請求項56に記載の方法。
- 第1の領域がpegRNAのプライマー結合部位(PBS)を含む、請求項57に記載の方法。
- 第2の領域がpegRNAのスペーサを含む、請求項58に記載の方法。
- スコアの第1のセットが、第1の核酸リンカー候補がpegRNAの第3の領域との相互作用を回避すると予測される程度を示す第3のスコアと、第1の核酸リンカー候補がpegRNAの第4の領域との相互作用を回避すると予測される程度を示す第4のスコアと、を更に含む、請求項57に記載の方法。
- 第3の領域がDNA合成鋳型を含む、請求項60に記載の方法。
- 第4の領域が、核酸プログラム型DNA結合タンパク質(napDNAbp)と相互作用するgRNAコアを含む、請求項61に記載の方法。
- pegRNAが、二本鎖標的DNA配列にヌクレオチド編集を組み入れするためのものであり、
pegRNAが、
二本鎖標的DNA配列の第1鎖にハイブリダイズするスペーサと、
二本鎖標的DNA配列の第2の鎖にハイブリダイズする伸長アームであって、プライマー結合部位(PBS)と、ヌクレオチド編集を含むDNA合成鋳型とを含む、伸長アームと、
核酸プログラム型DNA結合タンパク質napDNAbpと相互作用するgRNAコアと、
を含み、
第1の領域がPBSを含み、第2の領域がスペーサを含み、
第3の領域がDNA合成鋳型を含み、第4の領域がgRNAコアを含む、
請求項60に記載の方法。 - 複数の核酸リンカー候補が第2の核酸リンカー候補を含み、計算された複数のスコアを用いて複数の核酸リンカー候補の少なくともいくつかの中から少なくとも1つの核酸リンカーを同定することが、
第1の核酸リンカー候補に対するスコアの第1のセットを第2の核酸リンカー候補に対するスコアの第2のセットと比較することを含む、
請求項56に記載の方法。 - 第1の領域がプライマー結合部位(PBS)を含み、
第1のスコアセットにおける第1のスコアが、第1の核酸リンカー候補がpegRNAの第1の領域との相互作用を回避すると予測される程度を示し、
第2のスコアセットにおける第3のスコアは、第2の核酸リンカー候補がpegRNAの第1の領域との相互作用を回避すると予測される程度を示し、
第1のスコアセットを第2のスコアセットと比較することが、
第1のスコアと第3のスコアとを比較することを含む、
請求項64に記載の方法。 - 第1のスコアが第3のスコアの閾値距離に等しいか、又は閾値距離内にある場合、第1のスコアセットと第2のスコアセットとを比較する工程は、
第1のスコアのセットにおける第1のスコア以外のスコアと、第2のスコアのセットにおける第3のスコア以外の別のスコアとを比較することとを含む、
請求項65に記載の方法。 - (i)スペーサを含むガイドRNA、及び(ii)DNA合成鋳型、プライマー結合部位、足がかりモチーフ、及び更なる核酸部分を含む少なくとも1つの核酸伸長アームを含む、プライム編集のためのPEgRNA。
- 足がかりモチーフ及び更なる核酸部分が伸長アームの3'末端に取り付けられる、請求項67に記載のPEgRNA。
- 足がかりモチーフが伸長アームの3'末端に取り付けられ、更なる核酸部分が足がかりモチーフの3'末端に取り付けられる、請求項67又は68に記載のPEgRNA。
- 足がかりモチーフが、リンカーによってPEgRNAに取り付けられる、請求項67~69のいずれか一項に記載のPEgRNA。
- リンカーが、少なくとも3、少なくとも4、少なくとも5、少なくとも6、少なくとも7、少なくとも8、少なくとも9、少なくとも10、少なくとも11、少なくとも12、少なくとも13、少なくとも14、少なくとも15、少なくとも16、少なくとも17、少なくとも18、少なくとも19、少なくとも20、少なくとも21、少なくとも22、少なくとも23、少なくとも24、少なくとも25、少なくとも26、少なくとも27、少なくとも28、少なくとも29、又は少なくとも30ヌクレオチド長である、請求項70に記載のPEgRNA。
- PEgRNAが、プライム編集因子の核酸プログラム型DNA結合タンパク質(napDNAbp)に結合し、napDNAbpを標的DNA配列に向けることができる、請求項67~71のいずれか一項に記載のPEgRNA。
- (a)請求項67~72のいずれか一項に記載のPEgRNAと、(b)(i)napDNAbp、(ii)DNAポリメラーゼ、及び(iii)PEgRNAの足がかりモチーフに結合する部分を含むプライム編集因子と、を含む、部位特異的ゲノム修飾のためのプライム編集システム。
- PEgRNAの足がかりモチーフに結合するプライム編集因子の部分が、プライム編集因子のN末端に融合される、請求項73に記載のシステム。
- PEgRNAの足がかりモチーフに結合するプライム編集因子の部分が、プライム編集因子のC末端に融合される、請求項73に記載のシステム。
- PEgRNAの足がかりモチーフに結合するプライム編集因子の部分が、リンカーによってプライム編集因子に融合される、請求項73~75のいずれか一項に記載のシステム。
- リンカーが、少なくとも3、少なくとも4、少なくとも5、少なくとも6、少なくとも7、少なくとも8、少なくとも9、少なくとも10、少なくとも11、少なくとも12、少なくとも13、少なくとも14、少なくとも15、少なくとも16、少なくとも17、少なくとも18、少なくとも19、少なくとも20、少なくとも21、少なくとも22、少なくとも23、少なくとも24、少なくとも25、少なくとも26、少なくとも27、少なくとも28、少なくとも29、少なくとも30、又は30超の、アミノ酸長である、請求項76に記載のシステム。
- リンカーが、xtenリンカーを含む、請求項76又は77に記載のシステム。
- PEgRNAの足がかりモチーフに結合するプライム編集因子の部分がMS2バクテリオファージコートタンパク質を含む、請求項73~78のいずれか一項に記載のシステム。
- napDNAbpがニッカーゼ活性を有する、請求項73~79のいずれか一項に記載のシステム。
- napDNAbpがCas9タンパク質又はそのバリアントである、請求項73~80のいずれか一項に記載のシステム。
- napDNAbpが、ヌクレアーゼ活性Cas9、ヌクレアーゼ不活性Cas9(dCas9)、又はCas9ニッカーゼ(nCas9)である、請求項73~79のいずれか一項に記載のシステム。
- napDNAbpがCas9ニッカーゼ(nCas9)である、請求項73~79のいずれか一項に記載のシステム。
- napDNAbpが、Cas9、Cas12e、Cas12d、Cas12a、Cas12b1、Cas12b2、Cas13a、Cas12c、Cas12d、Cas12e、Cas12h、Cas12i、Cas12g、Cas12f(Cas14)、Cas12f1、Cas12j(CasΦ)、及びアルゴノートからなる群から選択され、任意選択でニッカーゼ活性を有する、請求項73~79のいずれか一項に記載のシステム。
- 請求項67~72のいずれか一項に記載のPEgRNAを含むポリヌクレオチド。
- 請求項85に記載のポリヌクレオチドを含むベクター。
- 請求項67~72のいずれか一項に記載のPEgRNA、請求項73~84のいずれか一項に記載のシステム、請求項85に記載のポリヌクレオチド、又は請求項86に記載のベクターを含む、細胞。
- (i)請求項67~72のいずれか一項に記載のPEgRNA、請求項73~84のいずれか一項に記載のシステム、請求項85に記載のポリヌクレオチド、又は請求項86に記載のベクターと、(ii)薬学的に許容される賦形剤と、を含む医薬組成物。
- 請求項67~72のいずれか一項に記載のPEgRNA、請求項73~84のいずれか一項に記載のシステム、請求項85に記載のポリヌクレオチド、請求項86に記載のベクター、又は請求項87に記載の細胞を含むキット。
- 標的DNA配列を請求項73~84のいずれか一項記載のシステムに提供することを含む、プライム編集の方法であって、標的DNA配列が、PEgRNA及びシステムのプライム編集因子と接触される、プライム編集の方法。
- プライム編集のための1対のPEgRNAであって、
(i)ガイドRNAと、DNA合成鋳型及びプライマー結合部位を含む少なくとも1つの核酸伸長アームとを含む第1のPEgRNAであって、伸長アームが、それに取り付けられた、トゥーループ、ヘアピン、ステムループ、シュードノット、アプタマー、G-四重鎖、tRNA、リボスイッチ又はリボザイムからなる群から選択される、核酸部分を含む、第1のPEgRNAと、
(ii)第2の鎖ニッキングガイドRNAを含む第2のPEgRNAであって、第2の鎖ニッキングガイドRNAが、DNA合成鋳型及びプライマー結合部位を含む少なくとも1つの核酸伸長アームを含む、第2のPEgRNAと、
を含む、プライム編集のための1対のPEgRNA。 - 第1のPEgRNA及び第2のPEgRNAがそれぞれ、プライム編集因子の核酸プログラム型DNA結合タンパク質(napDNAbp)に結合し、napDNAbpを標的DNA配列に向けることができる、請求項91に記載の1対のPEgRNA。
- (a)請求項91又は92に記載の1対のPEgRNAと、(b)napDNAbp及びDNAポリメラーゼを含む少なくとも1つのプライム編集因子とを含む、部位特異的ゲノム修飾のためのプライム編集システム。
- システムが、各々がnapDNAbp及びDNAポリメラーゼを含む第1のプライム編集因子及び第2のプライム編集因子を含む、請求項93に記載のシステム。
- 第1のプライム編集因子のnapDNAbpが1対のPEgRNAの第1のPEgRNAに結合し、第2のプライム編集因子のnapDNAbpが1対のPEgRNAの第2のPEgRNAに結合する、請求項94に記載のシステム。
- napDNAbpがニッカーゼ活性を有する、請求項93~95のいずれか一項に記載のシステム。
- napDNAbpがCas9タンパク質又はその変異体である、請求項93~95のいずれか一項に記載のシステム。
- napDNAbpが、ヌクレアーゼ活性Cas9、ヌクレアーゼ不活性Cas9(dCas9)、又はCas9ニッカーゼ(nCas9)である、請求項93~95のいずれか一項に記載のシステム。
- napDNAbpがCas9ニッカーゼ(nCas9)である、請求項93~95のいずれか一項に記載のシステム。
- napDNAbpが、Cas9、Cas12e、Cas12d、Cas12a、Cas12b1、Cas12b2、Cas13a、Cas12c、Cas12d、Cas12e、Cas12h、Cas12i、Cas12g、Cas12f(Cas14)、Cas12f1、Cas12j(CasΦ)、及びアルゴノートからなる群から選択され、任意選択でニッカーゼ活性を有する、請求項93~95のいずれか一項に記載のシステム。
- 請求項91又は92に記載のPEgRNAを含むポリヌクレオチド。
- 請求項101に記載のポリヌクレオチドを含むベクター。
- 請求項91又は92に記載のPEgRNA、請求項93~100のいずれか一項に記載のシステム、請求項101に記載のポリヌクレオチド、又は請求項102に記載のベクターを含む細胞。
- (i)請求項91又は92に記載のPEgRNA、請求項93~100のいずれか一項に記載のシステム、請求項101に記載のポリヌクレオチド、又は請求項102に記載のベクターと、(ii)薬学的に許容される賦形剤と、を含む医薬組成物。
- 請求項91又は92に記載のPEgRNA、請求項93~100のいずれか一項に記載のシステム、請求項101に記載のポリヌクレオチド、請求項102に記載のベクター、又は請求項103に記載の細胞を含むキット。
- 標的DNA配列を請求項93~100のいずれか一項記載のシステムに提供することを含むプライム編集の方法であって、標的DNA配列は、1対のPEgRNA及びシステムの1つ又は複数のプライム編集因子と接触される、プライム編集の方法。
- (i)スペーサを含むガイドRNAと、(ii)DNA合成鋳型及びプライマー結合部位を含む少なくとも1つの核酸伸長アームとを含む、PEgRNAであって、プライマー結合部位が1つ又は複数の修飾ヌクレオチドスイッチを含み、修飾ヌクレオチドスイッチは、標的DNA分子上のプロトスペーサ配列に対するプライマー結合部位の結合親和性よりもスペーサに対するプライマー結合部位の結合親和性のより大きな低下をもたらす、PEgRNA。
- 1つ又は複数の修飾ヌクレオチドが遺伝子変異を含む、請求項107に記載のPEgRNA。
- 1つ又は複数の修飾ヌクレオチドが化学修飾ヌクレオチドを含む、請求項107に記載のPEgRNA。
- 部位特異的ゲノム修飾のためのプライム編集システムであって、(a)請求項107~109のいずれか一項に記載の1対のPEgRNAと、(b)napDNAbp及びDNAポリメラーゼを含む少なくとも1つのプライム編集因子とを含む、プライム編集システム。
- システムが、各々がnapDNAbp及びDNAポリメラーゼを含む第1のプライム編集因子及び第2のプライム編集因子を含む、請求項110に記載のシステム。
- 第1のプライム編集因子のnapDNAbpが1対のPEgRNAの第1のPEgRNAに結合し、第2のプライム編集因子のnapDNAbpが1対のPEgRNAの第2のPEgRNAに結合する、請求項111に記載のシステム。
- napDNAbpがニッカーゼ活性を有する、請求項110~112のいずれか一項に記載のシステム。
- napDNAbpがCas9タンパク質又はそのバリアントである、請求項110~112のいずれか一項に記載のシステム。
- napDNAbpが、ヌクレアーゼ活性Cas9、ヌクレアーゼ不活性Cas9(dCas9)、又はCas9ニッカーゼ(nCas9)である、請求項110~112のいずれか一項に記載のシステム。
- napDNAbpがCas9ニッカーゼ(nCas9)である、請求項110~112のいずれか一項に記載のシステム。
- napDNAbpが、Cas9、Cas12e、Cas12d、Cas12a、Cas12b1、Cas12b2、Cas13a、Cas12c、Cas12d、Cas12e、Cas12h、Cas12i、Cas12g、Cas12f(Cas14)、Cas12f1、Cas12j(CasΦ)、及びアルゴノートからなる群から選択され、任意選択でニッカーゼ活性を有する、請求項110~112のいずれか一項に記載のシステム。
- 請求項107~109のいずれか一項に記載のPEgRNAを含むポリヌクレオチド。
- 請求項118に記載のポリヌクレオチドを含むベクター。
- 請求項107~109のいずれか一項に記載のPEgRNA、請求項110~117のいずれか一項に記載のシステム、請求項118に記載のポリヌクレオチド、又は請求項119に記載のベクターを含む、細胞。
- (i)請求項107~109のいずれか一項に記載のPEgRNA、請求項110~117のいずれか一項に記載のシステム、請求項118に記載のポリヌクレオチド、又は請求項119に記載のベクターと、(ii)薬学的に許容される賦形剤と、を含む医薬組成物。
- 請求項107~109のいずれか一項に記載のPEgRNA、請求項110~117のいずれか一項に記載のシステム、請求項118に記載のポリヌクレオチド、請求項119に記載のベクター、又は請求項120に記載の細胞を含むキット。
- 標的DNA配列を請求項110~117のいずれか一項記載のシステムに提供することを含むプライム編集の方法であって、標的DNA配列は、1対のPEgRNA及びシステムの1つ又は複数のプライム編集因子と接触される、プライム編集の方法。
- 単一のpegRNAを使用するプライム編集によってCDKL5遺伝子の1つ又は複数の突然変異を修正する方法であって、標的DNA配列を、(i)napDNAbp及び(ii)RNA依存性DNAポリメラーゼ活性を有するドメインと、pegRNAとを含むプライム編集因子と接触させることを含み、pegRNAが、プライム編集因子を、1つ又は複数の突然変異を含むCDKL5遺伝子に標的化する、方法。
- pegRNAが図146に提供される、請求項124に記載の方法。
- pegRNAが図148に提供される、請求項124に記載の方法。
- CDKL5遺伝子の変異が1412delA変異を含む、請求項124に記載の方法。
- 1つ又は複数の変異が、V172I、A173D、R175S、W176G、W176R、Y177C、R178P、P180L、E181A、又はL182P置換をコードする、請求項124に記載の方法。
- napDNAbpがニッカーゼ活性を有する、請求項124に記載の方法。
- napDNAbpがCas9タンパク質又はそのバリアントである、請求項124に記載の方法。
- napDNAbpが、ヌクレアーゼ活性Cas9、ヌクレアーゼ不活性Cas9(dCas9)、又はCas9ニッカーゼ(nCas9)である、請求項124に記載の方法。
- napDNAbpがCas9ニッカーゼ(nCas9)である、請求項124に記載の方法。
- napDNAbpが、Cas9、Cas12e、Cas12d、Cas12a、Cas12b1、Cas12b2、Cas13a、Cas12c、Cas12d、Cas12e、Cas12h、Cas12i、Cas12g、Cas12f(Cas14)、Cas12f1、Cas12j(CasΦ)、及びアルゴノートからなる群から選択され、任意選択でニッカーゼ活性を有する、請求項124に記載の方法。
- RNA依存性DNAポリメラーゼ活性を含むドメインが逆転写酵素である、請求項124に記載の方法。
- CDKL5遺伝子の異なる変異によって引き起こされるCDKL5欠損症を有する複数の対象を処置する方法であって、標的DNA配列を、(i)napDNAbpと、(ii)RNA依存性DNAポリメラーゼ活性を有するドメイン及び単一pegRNAと、を含むプライム編集因子と接触させることを含み、単一pegRNAが、変異非依存的な様式で修復されたCDKL5遺伝子をもたらすように複数の対象のいずれかにおけるCDKL5遺伝子に対してプライム編集因子を標的化することができる、方法。
- pegRNAが図148に提供される、請求項135に記載の方法。
- pegRNAが図150に提供される、請求項135に記載の方法。
- CDKL5遺伝子の変異が1412delA変異を含む、請求項135に記載の方法。
- 1つ又は複数の変異が、V172I、A173D、R175S、W176G、W176R、Y177C、R178P、P180L、E181A、又はL182P置換をコードする、請求項135に記載の方法。
- napDNAbpがニッカーゼ活性を有する、請求項135に記載の方法。
- napDNAbpがCas9タンパク質又はそのバリアントである、請求項135に記載の方法。
- napDNAbpが、ヌクレアーゼ活性Cas9、ヌクレアーゼ不活性Cas9(dCas9)、又はCas9ニッカーゼ(nCas9)である、請求項135に記載の方法。
- napDNAbpがCas9ニッカーゼ(nCas9)である、請求項135に記載の方法。
- napDNAbpが、Cas9、Cas12e、Cas12d、Cas12a、Cas12b1、Cas12b2、Cas13a、Cas12c、Cas12d、Cas12e、Cas12h、Cas12i、Cas12g、Cas12f(Cas14)、Cas12f1、Cas12j(CasΦ)、及びアルゴノートからなる群から選択され、任意選択でニッカーゼ活性を有する、請求項135に記載の方法。
- RNA依存性DNAポリメラーゼ活性を含むドメインが逆転写酵素である、請求項135に記載の方法。
Applications Claiming Priority (9)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US202063083067P | 2020-09-24 | 2020-09-24 | |
| US63/083,067 | 2020-09-24 | ||
| US202063091272P | 2020-10-13 | 2020-10-13 | |
| US63/091,272 | 2020-10-13 | ||
| US202163182633P | 2021-04-30 | 2021-04-30 | |
| US63/182,633 | 2021-04-30 | ||
| US202163231231P | 2021-08-09 | 2021-08-09 | |
| US63/231,231 | 2021-08-09 | ||
| PCT/US2021/052097 WO2022067130A2 (en) | 2020-09-24 | 2021-09-24 | Prime editing guide rnas, compositions thereof, and methods of using the same |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2023543803A true JP2023543803A (ja) | 2023-10-18 |
| JPWO2022067130A5 JPWO2022067130A5 (ja) | 2024-10-02 |
Family
ID=78302966
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2023519084A Pending JP2023543803A (ja) | 2020-09-24 | 2021-09-24 | プライム編集ガイドrna、その組成物、及びその使用方法 |
Country Status (6)
| Country | Link |
|---|---|
| US (1) | US20230357766A1 (ja) |
| EP (1) | EP4217490A2 (ja) |
| JP (1) | JP2023543803A (ja) |
| AU (1) | AU2021350835A1 (ja) |
| CA (1) | CA3193099A1 (ja) |
| WO (1) | WO2022067130A2 (ja) |
Families Citing this family (41)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US11542509B2 (en) | 2016-08-24 | 2023-01-03 | President And Fellows Of Harvard College | Incorporation of unnatural amino acids into proteins using base editing |
| EP4146804A1 (en) | 2020-05-08 | 2023-03-15 | The Broad Institute Inc. | Methods and compositions for simultaneous editing of both strands of a target double-stranded nucleotide sequence |
| US20220145293A1 (en) | 2020-10-21 | 2022-05-12 | Massachusetts Institute Of Technology | Systems, methods, and compositions for site-specific genetic engineering using programmable addition via site-specific targeting elements (paste) |
| WO2022251712A1 (en) | 2021-05-28 | 2022-12-01 | Sana Biotechnology, Inc. | Lipid particles containing a truncated baboon endogenous retrovirus (baev) envelope glycoprotein and related methods and uses |
| EP4352215A1 (en) * | 2021-06-11 | 2024-04-17 | LifeEDIT Therapeutics, Inc. | Rna polymerase iii promoters and methods of use |
| AU2022325232A1 (en) | 2021-08-11 | 2024-02-08 | Sana Biotechnology, Inc. | Genetically modified primary cells for allogeneic cell therapy |
| AU2022326565A1 (en) | 2021-08-11 | 2024-02-08 | Sana Biotechnology, Inc. | Genetically modified cells for allogeneic cell therapy |
| WO2023019227A1 (en) | 2021-08-11 | 2023-02-16 | Sana Biotechnology, Inc. | Genetically modified cells for allogeneic cell therapy to reduce complement-mediated inflammatory reactions |
| EP4384193A2 (en) | 2021-08-11 | 2024-06-19 | Sana Biotechnology, Inc. | Genetically modified cells for allogeneic cell therapy to reduce instant blood mediated inflammatory reactions |
| EP4419117A1 (en) | 2021-10-22 | 2024-08-28 | Sana Biotechnology, Inc. | Methods of engineering allogeneic t cells with a transgene in a tcr locus and associated compositions and methods |
| WO2023115039A2 (en) | 2021-12-17 | 2023-06-22 | Sana Biotechnology, Inc. | Modified paramyxoviridae fusion glycoproteins |
| TW202342757A (zh) | 2021-12-17 | 2023-11-01 | 美商薩那生物科技公司 | 經修飾副黏液病毒科附著醣蛋白 |
| WO2023133595A2 (en) | 2022-01-10 | 2023-07-13 | Sana Biotechnology, Inc. | Methods of ex vivo dosing and administration of lipid particles or viral vectors and related systems and uses |
| WO2023150518A1 (en) | 2022-02-01 | 2023-08-10 | Sana Biotechnology, Inc. | Cd3-targeted lentiviral vectors and uses thereof |
| WO2023150647A1 (en) | 2022-02-02 | 2023-08-10 | Sana Biotechnology, Inc. | Methods of repeat dosing and administration of lipid particles or viral vectors and related systems and uses |
| IL315000A (en) | 2022-02-17 | 2024-10-01 | Sana Biotechnology Inc | Transgenic CD47 proteins and their uses |
| WO2023192655A2 (en) * | 2022-04-01 | 2023-10-05 | Prime Medicine, Inc. | Methods and compositions for editing nucleotide sequences |
| WO2023205694A2 (en) * | 2022-04-20 | 2023-10-26 | Tacit Therapeutics, Inc. | Stabilization of therapeutic trans-splicing rna molecules in human cells |
| WO2023215831A1 (en) * | 2022-05-04 | 2023-11-09 | Tome Biosciences, Inc. | Guide rna compositions for programmable gene insertion |
| WO2023220672A1 (en) * | 2022-05-13 | 2023-11-16 | The Regents Of The University Of California | Compositions and methods for rna editing |
| WO2023232024A1 (en) * | 2022-05-30 | 2023-12-07 | Wuhan University | System and methods for duplicating target fragments |
| WO2023235501A1 (en) * | 2022-06-02 | 2023-12-07 | University Of Massachusetts | High fidelity nucleotide polymerase chimeric prime editor systems |
| CN114958767B (zh) * | 2022-06-02 | 2022-12-27 | 健颐生物科技发展(山东)有限公司 | 基于hiPSC细胞构建的神经干细胞制剂的制备方法 |
| WO2024026344A1 (en) * | 2022-07-27 | 2024-02-01 | Inscripta, Inc. | Modulating cellular repair mechanisms for genomic editing |
| CN116064517A (zh) * | 2022-07-29 | 2023-05-05 | 之江实验室 | 一种先导编辑gRNA的产生方式及其用途 |
| WO2024044655A1 (en) | 2022-08-24 | 2024-02-29 | Sana Biotechnology, Inc. | Delivery of heterologous proteins |
| CN115948449A (zh) * | 2022-09-20 | 2023-04-11 | 浙江大学杭州国际科创中心 | 一种适用于酵母先导编辑的双质粒系统及应用 |
| WO2024064761A2 (en) * | 2022-09-20 | 2024-03-28 | The Board Of Trustees Of The Leland Stanford Junior University | Methods and compositions to control gene using genome editing |
| WO2024064838A1 (en) | 2022-09-21 | 2024-03-28 | Sana Biotechnology, Inc. | Lipid particles comprising variant paramyxovirus attachment glycoproteins and uses thereof |
| WO2024077267A1 (en) | 2022-10-07 | 2024-04-11 | The Broad Institute, Inc. | Prime editing methods and compositions for treating triplet repeat disorders |
| WO2024081820A1 (en) | 2022-10-13 | 2024-04-18 | Sana Biotechnology, Inc. | Viral particles targeting hematopoietic stem cells |
| WO2024086586A2 (en) * | 2022-10-18 | 2024-04-25 | Flagship Pioneering Innovations Vi, Llc | Improved gene editing systems utilizing trans recruiting components |
| WO2024097314A2 (en) | 2022-11-02 | 2024-05-10 | Sana Biotechnology, Inc. | Methods and systems for determining donor cell features and formulating cell therapy products based on cell features |
| WO2024119157A1 (en) | 2022-12-02 | 2024-06-06 | Sana Biotechnology, Inc. | Lipid particles with cofusogens and methods of producing and using the same |
| WO2024151541A1 (en) | 2023-01-09 | 2024-07-18 | Sana Biotechnology, Inc. | Type-1 diabetes autoimmune mouse |
| WO2024168116A1 (en) | 2023-02-08 | 2024-08-15 | Prime Medicine, Inc. | Genome editing compositions and methods for treatment of myotonic dystrophy |
| WO2024168147A2 (en) | 2023-02-09 | 2024-08-15 | The Broad Institute, Inc. | Evolved recombinases for editing a genome in combination with prime editing |
| WO2024170866A1 (en) * | 2023-02-16 | 2024-08-22 | Genome Research Limited | Methods for modifying a cell genome |
| WO2024206125A1 (en) * | 2023-03-24 | 2024-10-03 | The Broad Institute, Inc. | Use of prime editing for treating sickle cell disease |
| CN117497092B (zh) * | 2024-01-02 | 2024-05-14 | 微观纪元(合肥)量子科技有限公司 | 基于动态规划和量子退火的rna结构预测方法及系统 |
| CN118581153A (zh) * | 2024-08-05 | 2024-09-03 | 崖州湾国家实验室 | Dna聚合酶介导的核苷酸序列编辑方法及组合物 |
Family Cites Families (45)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US4663290A (en) | 1982-01-21 | 1987-05-05 | Molecular Genetics, Inc. | Production of reverse transcriptase |
| US4889818A (en) | 1986-08-22 | 1989-12-26 | Cetus Corporation | Purified thermostable enzyme |
| US5079352A (en) | 1986-08-22 | 1992-01-07 | Cetus Corporation | Purified thermostable enzyme |
| US5374553A (en) | 1986-08-22 | 1994-12-20 | Hoffmann-La Roche Inc. | DNA encoding a thermostable nucleic acid polymerase enzyme from thermotoga maritima |
| US5244797B1 (en) | 1988-01-13 | 1998-08-25 | Life Technologies Inc | Cloned genes encoding reverse transcriptase lacking rnase h activity |
| FR2632821B1 (fr) | 1988-06-22 | 1990-11-16 | Inst Kriobiologii | Procede de conservation a basse temperature des embryons |
| US5047342A (en) | 1989-08-10 | 1991-09-10 | Life Technologies, Inc. | Cloning and expression of T5 DNA polymerase |
| US5270179A (en) | 1989-08-10 | 1993-12-14 | Life Technologies, Inc. | Cloning and expression of T5 DNA polymerase reduced in 3'- to-5' exonuclease activity |
| JP2709311B2 (ja) | 1990-09-28 | 1998-02-04 | エフ.ホフマン−ラ ロシュ アクチェンゲゼルシャフト | 熱安定性dnaポリメラーゼの5→3′のエキソヌクレアーゼ突然変異 |
| EP0553264A4 (en) | 1990-10-05 | 1994-07-13 | Wayne M Barnes | Thermostable dna polymerase |
| US5834247A (en) | 1992-12-09 | 1998-11-10 | New England Biolabs, Inc. | Modified proteins comprising controllable intervening protein sequences or their elements methods of producing same and methods for purification of a target protein comprised by a modified protein |
| US5496714A (en) | 1992-12-09 | 1996-03-05 | New England Biolabs, Inc. | Modification of protein by use of a controllable interveining protein sequence |
| US5436149A (en) | 1993-02-19 | 1995-07-25 | Barnes; Wayne M. | Thermostable DNA polymerase with enhanced thermostability and enhanced length and efficiency of primer extension |
| US5512462A (en) | 1994-02-25 | 1996-04-30 | Hoffmann-La Roche Inc. | Methods and reagents for the polymerase chain reaction amplification of long DNA sequences |
| US5912155A (en) | 1994-09-30 | 1999-06-15 | Life Technologies, Inc. | Cloned DNA polymerases from Thermotoga neapolitana |
| US5614365A (en) | 1994-10-17 | 1997-03-25 | President & Fellow Of Harvard College | DNA polymerase having modified nucleotide binding site for DNA sequencing |
| US5773258A (en) | 1995-08-25 | 1998-06-30 | Roche Molecular Systems, Inc. | Nucleic acid amplification using a reversibly inactivated thermostable enzyme |
| US6183998B1 (en) | 1998-05-29 | 2001-02-06 | Qiagen Gmbh Max-Volmer-Strasse 4 | Method for reversible modification of thermostable enzymes |
| GB9920194D0 (en) | 1999-08-27 | 1999-10-27 | Advanced Biotech Ltd | A heat-stable thermostable DNA polymerase for use in nucleic acid amplification |
| EP1235914A2 (en) | 1999-11-24 | 2002-09-04 | Joseph Rosenecker | Polypeptides comprising multimers of nuclear localization signals or of protein transduction domains and their use for transferring molecules into cells |
| WO2006089045A2 (en) | 2005-02-18 | 2006-08-24 | Monogram Biosciences, Inc. | Methods and compositions for determining hypersusceptibility of hiv-1 to non-nucleoside reverse transcriptase inhibitors |
| US9783791B2 (en) | 2005-08-10 | 2017-10-10 | Agilent Technologies, Inc. | Mutant reverse transcriptase and methods of use |
| CN101668848B (zh) | 2007-04-26 | 2014-10-22 | 雷蒙特亚特特拉维夫大学有限公司 | 来自口腔粘膜的多能自体干细胞和使用方法 |
| EP2342336B1 (en) | 2008-09-05 | 2016-12-14 | President and Fellows of Harvard College | Continuous directed evolution of proteins and nucleic acids |
| BRPI1009221A2 (pt) | 2009-03-04 | 2016-03-15 | Univ Texas | proteínas de fusão transcriptase reversa estabilizadas. |
| US8889394B2 (en) | 2009-09-07 | 2014-11-18 | Empire Technology Development Llc | Multiple domain proteins |
| CN106834320B (zh) | 2009-12-10 | 2021-05-25 | 明尼苏达大学董事会 | Tal效应子介导的dna修饰 |
| WO2012054727A1 (en) | 2010-10-22 | 2012-04-26 | Bio-Rad Laboratories, Inc. | Reverse transcriptase mixtures with improved storage stability |
| JP6088438B2 (ja) | 2010-12-22 | 2017-03-01 | プレジデント アンド フェローズ オブ ハーバード カレッジ | 連続的定向進化 |
| WO2012125445A2 (en) | 2011-03-11 | 2012-09-20 | President And Fellows Of Harvard College | Small molecule-dependent inteins and uses thereof |
| WO2013045632A1 (en) | 2011-09-28 | 2013-04-04 | Era Biotech, S.A. | Split inteins and uses thereof |
| EP3431497B1 (en) | 2012-06-27 | 2022-07-27 | The Trustees of Princeton University | Split inteins, conjugates and uses thereof |
| US9181535B2 (en) | 2012-09-24 | 2015-11-10 | The Chinese University Of Hong Kong | Transcription activator-like effector nucleases (TALENs) |
| WO2014055782A1 (en) | 2012-10-03 | 2014-04-10 | Agrivida, Inc. | Intein-modified proteases, their production and industrial applications |
| JO3470B1 (ar) | 2012-10-08 | 2020-07-05 | Merck Sharp & Dohme | مشتقات 5- فينوكسي-3h-بيريميدين-4-أون واستخدامها كمثبطات ناسخ عكسي ل hiv |
| US9359599B2 (en) | 2013-08-22 | 2016-06-07 | President And Fellows Of Harvard College | Engineered transcription activator-like effector (TALE) domains and uses thereof |
| WO2015134121A2 (en) | 2014-01-20 | 2015-09-11 | President And Fellows Of Harvard College | Negative selection and stringency modulation in continuous evolution systems |
| EP3212778B1 (en) | 2014-10-28 | 2019-08-28 | Agrivida, Inc. | Methods and compositions for stabilizing trans-splicing intein modified proteases |
| US11299729B2 (en) | 2015-04-17 | 2022-04-12 | President And Fellows Of Harvard College | Vector-based mutagenesis system |
| WO2018049168A1 (en) * | 2016-09-09 | 2018-03-15 | The Board Of Trustees Of The Leland Stanford Junior University | High-throughput precision genome editing |
| US9580698B1 (en) | 2016-09-23 | 2017-02-28 | New England Biolabs, Inc. | Mutant reverse transcriptase |
| US10011849B1 (en) * | 2017-06-23 | 2018-07-03 | Inscripta, Inc. | Nucleic acid-guided nucleases |
| US11268092B2 (en) * | 2018-01-12 | 2022-03-08 | GenEdit, Inc. | Structure-engineered guide RNA |
| CN114729365A (zh) * | 2019-03-19 | 2022-07-08 | 布罗德研究所股份有限公司 | 用于编辑核苷酸序列的方法和组合物 |
| EP4146804A1 (en) * | 2020-05-08 | 2023-03-15 | The Broad Institute Inc. | Methods and compositions for simultaneous editing of both strands of a target double-stranded nucleotide sequence |
-
2021
- 2021-09-24 WO PCT/US2021/052097 patent/WO2022067130A2/en active Application Filing
- 2021-09-24 EP EP21795122.7A patent/EP4217490A2/en active Pending
- 2021-09-24 US US18/028,183 patent/US20230357766A1/en active Pending
- 2021-09-24 JP JP2023519084A patent/JP2023543803A/ja active Pending
- 2021-09-24 CA CA3193099A patent/CA3193099A1/en active Pending
- 2021-09-24 AU AU2021350835A patent/AU2021350835A1/en active Pending
Also Published As
| Publication number | Publication date |
|---|---|
| AU2021350835A2 (en) | 2023-04-27 |
| CA3193099A1 (en) | 2022-03-31 |
| WO2022067130A2 (en) | 2022-03-31 |
| US20230357766A1 (en) | 2023-11-09 |
| WO2022067130A3 (en) | 2022-06-23 |
| EP4217490A2 (en) | 2023-08-02 |
| AU2021350835A1 (en) | 2023-04-27 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP2023543803A (ja) | プライム編集ガイドrna、その組成物、及びその使用方法 | |
| CN114127285B (zh) | 编辑核苷酸序列的方法和组合物 | |
| US11912985B2 (en) | Methods and compositions for simultaneous editing of both strands of a target double-stranded nucleotide sequence | |
| JPWO2020191233A5 (ja) | ||
| JPWO2020191234A5 (ja) | ||
| JPWO2020191243A5 (ja) | ||
| CN116685682A (zh) | 引导编辑向导rna、其组合物以及使用它们的方法 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20240924 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20240924 |