JP2021528059A - グリコシル化されたComPピリンバリアント、製造方法及びその使用 - Google Patents
グリコシル化されたComPピリンバリアント、製造方法及びその使用 Download PDFInfo
- Publication number
- JP2021528059A JP2021528059A JP2020569986A JP2020569986A JP2021528059A JP 2021528059 A JP2021528059 A JP 2021528059A JP 2020569986 A JP2020569986 A JP 2020569986A JP 2020569986 A JP2020569986 A JP 2020569986A JP 2021528059 A JP2021528059 A JP 2021528059A
- Authority
- JP
- Japan
- Prior art keywords
- comp
- seq
- protein
- glycosylation
- adp1
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
- 238000004519 manufacturing process Methods 0.000 title claims abstract description 26
- 108010000916 Fimbriae Proteins Proteins 0.000 title description 27
- 239000012634 fragment Substances 0.000 claims abstract description 134
- 229960005486 vaccine Drugs 0.000 claims abstract description 103
- 108020001507 fusion proteins Proteins 0.000 claims abstract description 90
- 102000037865 fusion proteins Human genes 0.000 claims abstract description 90
- 238000000034 method Methods 0.000 claims abstract description 66
- 201000010099 disease Diseases 0.000 claims abstract description 9
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 claims abstract description 9
- 208000035143 Bacterial infection Diseases 0.000 claims abstract description 5
- 108090000623 proteins and genes Proteins 0.000 claims description 218
- 230000013595 glycosylation Effects 0.000 claims description 217
- 238000006206 glycosylation reaction Methods 0.000 claims description 203
- 102000004169 proteins and genes Human genes 0.000 claims description 194
- 150000004676 glycans Chemical class 0.000 claims description 183
- 229920001282 polysaccharide Polymers 0.000 claims description 167
- 239000005017 polysaccharide Substances 0.000 claims description 167
- 101100388296 Arabidopsis thaliana DTX51 gene Proteins 0.000 claims description 162
- 101100215626 Saccharomyces cerevisiae (strain ATCC 204508 / S288c) ADP1 gene Proteins 0.000 claims description 162
- 125000003607 serino group Chemical group [H]N([H])[C@]([H])(C(=O)[*])C(O[H])([H])[H] 0.000 claims description 141
- 108090000765 processed proteins & peptides Proteins 0.000 claims description 118
- 210000004027 cell Anatomy 0.000 claims description 102
- 102000004196 processed proteins & peptides Human genes 0.000 claims description 90
- 229920001184 polypeptide Polymers 0.000 claims description 81
- 125000003275 alpha amino acid group Chemical group 0.000 claims description 69
- 150000001413 amino acids Chemical class 0.000 claims description 67
- 229920001542 oligosaccharide Polymers 0.000 claims description 58
- 150000002482 oligosaccharides Chemical class 0.000 claims description 58
- 108010089072 Dolichyl-diphosphooligosaccharide-protein glycotransferase Proteins 0.000 claims description 56
- 150000007523 nucleic acids Chemical class 0.000 claims description 41
- 102000014914 Carrier Proteins Human genes 0.000 claims description 40
- 108010078791 Carrier Proteins Proteins 0.000 claims description 40
- 101100434240 Schizosaccharomyces pombe (strain 972 / ATCC 24843) act1 gene Proteins 0.000 claims description 40
- 102000039446 nucleic acids Human genes 0.000 claims description 39
- 108020004707 nucleic acids Proteins 0.000 claims description 39
- 230000004044 response Effects 0.000 claims description 37
- 238000006467 substitution reaction Methods 0.000 claims description 34
- 230000028993 immune response Effects 0.000 claims description 29
- 239000000203 mixture Substances 0.000 claims description 27
- 241000894006 Bacteria Species 0.000 claims description 26
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 claims description 26
- 125000000539 amino acid group Chemical group 0.000 claims description 26
- 230000001580 bacterial effect Effects 0.000 claims description 23
- 230000005875 antibody response Effects 0.000 claims description 22
- 239000008103 glucose Substances 0.000 claims description 22
- 108010060123 Conjugate Vaccines Proteins 0.000 claims description 21
- 241000588748 Klebsiella Species 0.000 claims description 21
- 241000193998 Streptococcus pneumoniae Species 0.000 claims description 21
- 238000007792 addition Methods 0.000 claims description 21
- 229940031670 conjugate vaccine Drugs 0.000 claims description 21
- 238000001727 in vivo Methods 0.000 claims description 21
- 229940031000 streptococcus pneumoniae Drugs 0.000 claims description 21
- 241000588747 Klebsiella pneumoniae Species 0.000 claims description 20
- 239000002671 adjuvant Substances 0.000 claims description 20
- 238000012217 deletion Methods 0.000 claims description 20
- 230000037430 deletion Effects 0.000 claims description 20
- 208000015181 infectious disease Diseases 0.000 claims description 18
- 230000021615 conjugation Effects 0.000 claims description 16
- 239000012528 membrane Substances 0.000 claims description 16
- 239000013598 vector Substances 0.000 claims description 16
- 108091035707 Consensus sequence Proteins 0.000 claims description 14
- 229940031999 pneumococcal conjugate vaccine Drugs 0.000 claims description 13
- 108010071134 CRM197 (non-toxic variant of diphtheria toxin) Proteins 0.000 claims description 12
- 241000588724 Escherichia coli Species 0.000 claims description 12
- 241000606768 Haemophilus influenzae Species 0.000 claims description 12
- 108091006106 transcriptional activators Proteins 0.000 claims description 12
- 210000004369 blood Anatomy 0.000 claims description 11
- 239000008280 blood Substances 0.000 claims description 11
- 229960000814 tetanus toxoid Drugs 0.000 claims description 11
- 102000009016 Cholera Toxin Human genes 0.000 claims description 10
- 108010049048 Cholera Toxin Proteins 0.000 claims description 10
- 102100037840 Dehydrogenase/reductase SDR family member 2, mitochondrial Human genes 0.000 claims description 10
- 101710188053 Protein D Proteins 0.000 claims description 10
- 101710132893 Resolvase Proteins 0.000 claims description 10
- 230000027455 binding Effects 0.000 claims description 10
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 claims description 9
- XFTWUNOVBCHBJR-UHFFFAOYSA-N Aspergillomarasmine A Chemical group OC(=O)C(N)CNC(C(O)=O)CNC(C(O)=O)CC(O)=O XFTWUNOVBCHBJR-UHFFFAOYSA-N 0.000 claims description 9
- 208000035473 Communicable disease Diseases 0.000 claims description 9
- 101900161471 Pseudomonas aeruginosa Exotoxin A Proteins 0.000 claims description 9
- 108010055044 Tetanus Toxin Proteins 0.000 claims description 9
- 108091008053 gene clusters Proteins 0.000 claims description 9
- 229940118376 tetanus toxin Drugs 0.000 claims description 9
- -1 tetanus toxoid Proteins 0.000 claims description 9
- 241000589517 Pseudomonas aeruginosa Species 0.000 claims description 8
- 241000226935 Klebsiella pneumoniae subsp. pneumoniae NTUH-K2044 Species 0.000 claims description 7
- 206010010356 Congenital anomaly Diseases 0.000 claims description 6
- 241000588749 Klebsiella oxytoca Species 0.000 claims description 6
- 206010035664 Pneumonia Diseases 0.000 claims description 6
- 230000005867 T cell response Effects 0.000 claims description 6
- 230000008348 humoral response Effects 0.000 claims description 6
- 230000009885 systemic effect Effects 0.000 claims description 6
- 208000035109 Pneumococcal Infections Diseases 0.000 claims description 5
- 241000194017 Streptococcus Species 0.000 claims description 5
- 244000052616 bacterial pathogen Species 0.000 claims description 5
- 229960003983 diphtheria toxoid Drugs 0.000 claims description 5
- 210000003719 b-lymphocyte Anatomy 0.000 claims description 4
- 238000012258 culturing Methods 0.000 claims description 4
- 230000005745 host immune response Effects 0.000 claims description 4
- 230000006054 immunological memory Effects 0.000 claims description 4
- 230000001939 inductive effect Effects 0.000 claims description 4
- 238000002347 injection Methods 0.000 claims description 4
- 239000007924 injection Substances 0.000 claims description 4
- 230000001404 mediated effect Effects 0.000 claims description 4
- 102000004127 Cytokines Human genes 0.000 claims description 3
- 108090000695 Cytokines Proteins 0.000 claims description 3
- 230000008649 adaptation response Effects 0.000 claims description 3
- 230000001363 autoimmune Effects 0.000 claims description 3
- 230000036755 cellular response Effects 0.000 claims description 3
- 230000003828 downregulation Effects 0.000 claims description 3
- 210000000987 immune system Anatomy 0.000 claims description 3
- 230000007787 long-term memory Effects 0.000 claims description 3
- 210000001806 memory b lymphocyte Anatomy 0.000 claims description 3
- 210000000056 organ Anatomy 0.000 claims description 3
- 210000003491 skin Anatomy 0.000 claims description 3
- 210000004872 soft tissue Anatomy 0.000 claims description 3
- 230000003827 upregulation Effects 0.000 claims description 3
- 208000022362 bacterial infectious disease Diseases 0.000 claims description 2
- 238000010255 intramuscular injection Methods 0.000 claims description 2
- 239000007927 intramuscular injection Substances 0.000 claims description 2
- 239000007928 intraperitoneal injection Substances 0.000 claims description 2
- 238000010253 intravenous injection Methods 0.000 claims description 2
- 238000010254 subcutaneous injection Methods 0.000 claims description 2
- 239000007929 subcutaneous injection Substances 0.000 claims description 2
- 206010043376 Tetanus Diseases 0.000 claims 2
- 210000004072 lung Anatomy 0.000 claims 1
- 210000003071 memory t lymphocyte Anatomy 0.000 claims 1
- 101150065984 Comp gene Proteins 0.000 abstract description 12
- 235000018102 proteins Nutrition 0.000 description 165
- 101150051775 PGLS gene Proteins 0.000 description 124
- 101100321796 Dictyostelium discoideum pgl gene Proteins 0.000 description 123
- 235000001014 amino acid Nutrition 0.000 description 85
- 229940024606 amino acid Drugs 0.000 description 59
- 241000699670 Mus sp. Species 0.000 description 56
- 210000002966 serum Anatomy 0.000 description 37
- 238000001262 western blot Methods 0.000 description 29
- 235000004400 serine Nutrition 0.000 description 27
- 229940124950 Prevnar 13 Drugs 0.000 description 25
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 25
- 108091033319 polynucleotide Proteins 0.000 description 23
- 102000040430 polynucleotide Human genes 0.000 description 23
- 239000002157 polynucleotide Substances 0.000 description 23
- 230000004927 fusion Effects 0.000 description 21
- RAXXELZNTBOGNW-UHFFFAOYSA-N imidazole Natural products C1=CNC=N1 RAXXELZNTBOGNW-UHFFFAOYSA-N 0.000 description 18
- 235000000346 sugar Nutrition 0.000 description 17
- 230000001965 increasing effect Effects 0.000 description 16
- LFQSCWFLJHTTHZ-UHFFFAOYSA-N Ethanol Chemical compound CCO LFQSCWFLJHTTHZ-UHFFFAOYSA-N 0.000 description 15
- 241000699666 Mus <mouse, genus> Species 0.000 description 15
- 239000000872 buffer Substances 0.000 description 15
- 230000014509 gene expression Effects 0.000 description 15
- 108091006146 Channels Proteins 0.000 description 14
- 108010015899 Glycopeptides Proteins 0.000 description 14
- 102000002068 Glycopeptides Human genes 0.000 description 14
- 238000004458 analytical method Methods 0.000 description 14
- 238000012546 transfer Methods 0.000 description 13
- 239000000047 product Substances 0.000 description 12
- 238000002255 vaccination Methods 0.000 description 12
- 108090000288 Glycoproteins Proteins 0.000 description 11
- 102000003886 Glycoproteins Human genes 0.000 description 11
- WEVYAHXRMPXWCK-UHFFFAOYSA-N Acetonitrile Chemical compound CC#N WEVYAHXRMPXWCK-UHFFFAOYSA-N 0.000 description 10
- 238000002965 ELISA Methods 0.000 description 10
- 230000015572 biosynthetic process Effects 0.000 description 10
- LOKCTEFSRHRXRJ-UHFFFAOYSA-I dipotassium trisodium dihydrogen phosphate hydrogen phosphate dichloride Chemical compound P(=O)(O)(O)[O-].[K+].P(=O)(O)([O-])[O-].[Na+].[Na+].[Cl-].[K+].[Cl-].[Na+] LOKCTEFSRHRXRJ-UHFFFAOYSA-I 0.000 description 10
- 238000013467 fragmentation Methods 0.000 description 10
- 238000006062 fragmentation reaction Methods 0.000 description 10
- 238000004949 mass spectrometry Methods 0.000 description 10
- 239000002953 phosphate buffered saline Substances 0.000 description 10
- 230000001105 regulatory effect Effects 0.000 description 10
- 241000589291 Acinetobacter Species 0.000 description 9
- 108020004414 DNA Proteins 0.000 description 9
- 230000003053 immunization Effects 0.000 description 9
- 230000005847 immunogenicity Effects 0.000 description 9
- 230000002147 killing effect Effects 0.000 description 9
- 108020004999 messenger RNA Proteins 0.000 description 9
- 239000013612 plasmid Substances 0.000 description 9
- 238000013518 transcription Methods 0.000 description 9
- 230000035897 transcription Effects 0.000 description 9
- 108091026890 Coding region Proteins 0.000 description 8
- 102000004163 DNA-directed RNA polymerases Human genes 0.000 description 8
- 108090000626 DNA-directed RNA polymerases Proteins 0.000 description 8
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 description 8
- 235000004279 alanine Nutrition 0.000 description 8
- 238000003556 assay Methods 0.000 description 8
- 238000011161 development Methods 0.000 description 8
- 230000018109 developmental process Effects 0.000 description 8
- 238000002649 immunization Methods 0.000 description 8
- 239000000758 substrate Substances 0.000 description 8
- 238000003786 synthesis reaction Methods 0.000 description 8
- 239000012130 whole-cell lysate Substances 0.000 description 8
- 230000000844 anti-bacterial effect Effects 0.000 description 7
- 238000012512 characterization method Methods 0.000 description 7
- 230000001419 dependent effect Effects 0.000 description 7
- 238000009826 distribution Methods 0.000 description 7
- 239000000902 placebo Substances 0.000 description 7
- 229940068196 placebo Drugs 0.000 description 7
- 150000003355 serines Chemical class 0.000 description 7
- 206010057249 Phagocytosis Diseases 0.000 description 6
- 238000000502 dialysis Methods 0.000 description 6
- 238000002474 experimental method Methods 0.000 description 6
- 230000004048 modification Effects 0.000 description 6
- 238000012986 modification Methods 0.000 description 6
- 229910052757 nitrogen Inorganic materials 0.000 description 6
- 230000008782 phagocytosis Effects 0.000 description 6
- 239000013641 positive control Substances 0.000 description 6
- 230000008569 process Effects 0.000 description 6
- 238000001228 spectrum Methods 0.000 description 6
- 150000008163 sugars Chemical class 0.000 description 6
- 238000012360 testing method Methods 0.000 description 6
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 5
- SBISABLNPMVJHR-SQGDDOFFSA-N 2-aminoacetic acid (2S)-2-amino-3-hydroxypropanoic acid Chemical compound NCC(O)=O.NCC(O)=O.NCC(O)=O.OC[C@H](N)C(O)=O SBISABLNPMVJHR-SQGDDOFFSA-N 0.000 description 5
- 241000589875 Campylobacter jejuni Species 0.000 description 5
- 108010067770 Endopeptidase K Proteins 0.000 description 5
- 108010093488 His-His-His-His-His-His Proteins 0.000 description 5
- 230000004988 N-glycosylation Effects 0.000 description 5
- 241000283973 Oryctolagus cuniculus Species 0.000 description 5
- 210000001744 T-lymphocyte Anatomy 0.000 description 5
- 239000000427 antigen Substances 0.000 description 5
- 108091007433 antigens Proteins 0.000 description 5
- 102000036639 antigens Human genes 0.000 description 5
- 238000006243 chemical reaction Methods 0.000 description 5
- 238000012790 confirmation Methods 0.000 description 5
- 238000010494 dissociation reaction Methods 0.000 description 5
- 230000005593 dissociations Effects 0.000 description 5
- 244000052637 human pathogen Species 0.000 description 5
- 230000036039 immunity Effects 0.000 description 5
- 238000011068 loading method Methods 0.000 description 5
- 238000001819 mass spectrum Methods 0.000 description 5
- 244000052769 pathogen Species 0.000 description 5
- 230000001717 pathogenic effect Effects 0.000 description 5
- 238000000746 purification Methods 0.000 description 5
- 239000000126 substance Substances 0.000 description 5
- 239000000725 suspension Substances 0.000 description 5
- 230000002103 transcriptional effect Effects 0.000 description 5
- DQJCDTNMLBYVAY-ZXXIYAEKSA-N (2S,5R,10R,13R)-16-{[(2R,3S,4R,5R)-3-{[(2S,3R,4R,5S,6R)-3-acetamido-4,5-dihydroxy-6-(hydroxymethyl)oxan-2-yl]oxy}-5-(ethylamino)-6-hydroxy-2-(hydroxymethyl)oxan-4-yl]oxy}-5-(4-aminobutyl)-10-carbamoyl-2,13-dimethyl-4,7,12,15-tetraoxo-3,6,11,14-tetraazaheptadecan-1-oic acid Chemical compound NCCCC[C@H](C(=O)N[C@@H](C)C(O)=O)NC(=O)CC[C@H](C(N)=O)NC(=O)[C@@H](C)NC(=O)C(C)O[C@@H]1[C@@H](NCC)C(O)O[C@H](CO)[C@H]1O[C@H]1[C@H](NC(C)=O)[C@@H](O)[C@H](O)[C@@H](CO)O1 DQJCDTNMLBYVAY-ZXXIYAEKSA-N 0.000 description 4
- PZUPAGRIHCRVKN-UHFFFAOYSA-N 5-[5-[3,4-dihydroxy-6-[(3,4,5-trihydroxyoxan-2-yl)oxymethyl]-5-[3,4,5-trihydroxy-6-[(3,4,5-trihydroxyoxan-2-yl)oxymethyl]oxan-2-yl]oxyoxan-2-yl]oxy-3,4-dihydroxy-6-[(3,4,5-trihydroxyoxan-2-yl)oxymethyl]oxan-2-yl]oxy-6-(hydroxymethyl)oxane-2,3,4-triol Chemical compound OCC1OC(O)C(O)C(O)C1OC1C(O)C(O)C(OC2C(C(O)C(OC3C(C(O)C(O)C(COC4C(C(O)C(O)CO4)O)O3)O)C(COC3C(C(O)C(O)CO3)O)O2)O)C(COC2C(C(O)C(O)CO2)O)O1 PZUPAGRIHCRVKN-UHFFFAOYSA-N 0.000 description 4
- 101150013062 CHRNA1 gene Proteins 0.000 description 4
- 101710175625 Maltose/maltodextrin-binding periplasmic protein Proteins 0.000 description 4
- 108091028043 Nucleic acid sequence Proteins 0.000 description 4
- 108700026244 Open Reading Frames Proteins 0.000 description 4
- ONIBWKKTOPOVIA-UHFFFAOYSA-N Proline Natural products OC(=O)C1CCCN1 ONIBWKKTOPOVIA-UHFFFAOYSA-N 0.000 description 4
- 108010076504 Protein Sorting Signals Proteins 0.000 description 4
- 229940037003 alum Drugs 0.000 description 4
- 210000004899 c-terminal region Anatomy 0.000 description 4
- 239000000969 carrier Substances 0.000 description 4
- 229940028617 conventional vaccine Drugs 0.000 description 4
- 239000000284 extract Substances 0.000 description 4
- 238000009472 formulation Methods 0.000 description 4
- 230000006870 function Effects 0.000 description 4
- 239000000499 gel Substances 0.000 description 4
- 230000002163 immunogen Effects 0.000 description 4
- 238000011534 incubation Methods 0.000 description 4
- BDAGIHXWWSANSR-UHFFFAOYSA-N methanoic acid Natural products OC=O BDAGIHXWWSANSR-UHFFFAOYSA-N 0.000 description 4
- 239000013642 negative control Substances 0.000 description 4
- 235000013930 proline Nutrition 0.000 description 4
- 230000001681 protective effect Effects 0.000 description 4
- 101150110106 rmpA gene Proteins 0.000 description 4
- 238000002741 site-directed mutagenesis Methods 0.000 description 4
- 235000020183 skimmed milk Nutrition 0.000 description 4
- 230000001225 therapeutic effect Effects 0.000 description 4
- 230000009466 transformation Effects 0.000 description 4
- MTCFGRXMJLQNBG-REOHCLBHSA-N (2S)-2-Amino-3-hydroxypropansäure Chemical compound OC[C@H](N)C(O)=O MTCFGRXMJLQNBG-REOHCLBHSA-N 0.000 description 3
- NHBKXEKEPDILRR-UHFFFAOYSA-N 2,3-bis(butanoylsulfanyl)propyl butanoate Chemical compound CCCC(=O)OCC(SC(=O)CCC)CSC(=O)CCC NHBKXEKEPDILRR-UHFFFAOYSA-N 0.000 description 3
- DOBIZWYVJFIYOV-UHFFFAOYSA-N 7-hydroxynaphthalene-1,3-disulfonic acid Chemical compound C1=C(S(O)(=O)=O)C=C(S(O)(=O)=O)C2=CC(O)=CC=C21 DOBIZWYVJFIYOV-UHFFFAOYSA-N 0.000 description 3
- DLFVBJFMPXGRIB-UHFFFAOYSA-N Acetamide Chemical compound CC(N)=O DLFVBJFMPXGRIB-UHFFFAOYSA-N 0.000 description 3
- QTBSBXVTEAMEQO-UHFFFAOYSA-N Acetic acid Chemical compound CC(O)=O QTBSBXVTEAMEQO-UHFFFAOYSA-N 0.000 description 3
- 241001165345 Acinetobacter baylyi Species 0.000 description 3
- 241000588625 Acinetobacter sp. Species 0.000 description 3
- 108010077805 Bacterial Proteins Proteins 0.000 description 3
- 102000004190 Enzymes Human genes 0.000 description 3
- 108090000790 Enzymes Proteins 0.000 description 3
- 101710082714 Exotoxin A Proteins 0.000 description 3
- HNDVDQJCIGZPNO-YFKPBYRVSA-N L-histidine Chemical compound OC(=O)[C@@H](N)CC1=CN=CN1 HNDVDQJCIGZPNO-YFKPBYRVSA-N 0.000 description 3
- COLNVLDHVKWLRT-QMMMGPOBSA-N L-phenylalanine Chemical compound OC(=O)[C@@H](N)CC1=CC=CC=C1 COLNVLDHVKWLRT-QMMMGPOBSA-N 0.000 description 3
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 description 3
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 3
- 239000004472 Lysine Substances 0.000 description 3
- 241000124008 Mammalia Species 0.000 description 3
- OKKJLVBELUTLKV-UHFFFAOYSA-N Methanol Chemical compound OC OKKJLVBELUTLKV-UHFFFAOYSA-N 0.000 description 3
- OVRNDRQMDRJTHS-UHFFFAOYSA-N N-acetylhexosamine Chemical compound CC(=O)NC1C(O)OC(CO)C(O)C1O OVRNDRQMDRJTHS-UHFFFAOYSA-N 0.000 description 3
- 230000004989 O-glycosylation Effects 0.000 description 3
- 241001494479 Pecora Species 0.000 description 3
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 3
- 241000251539 Vertebrata <Metazoa> Species 0.000 description 3
- PYMYPHUHKUWMLA-WDCZJNDASA-N arabinose Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)C=O PYMYPHUHKUWMLA-WDCZJNDASA-N 0.000 description 3
- PYMYPHUHKUWMLA-UHFFFAOYSA-N arabinose Natural products OCC(O)C(O)C(O)C=O PYMYPHUHKUWMLA-UHFFFAOYSA-N 0.000 description 3
- SRBFZHDQGSBBOR-UHFFFAOYSA-N beta-D-Pyranose-Lyxose Natural products OC1COC(O)C(O)C1O SRBFZHDQGSBBOR-UHFFFAOYSA-N 0.000 description 3
- 210000000349 chromosome Anatomy 0.000 description 3
- 230000004186 co-expression Effects 0.000 description 3
- 238000003745 diagnosis Methods 0.000 description 3
- NLEBIOOXCVAHBD-QKMCSOCLSA-N dodecyl beta-D-maltoside Chemical compound O[C@@H]1[C@@H](O)[C@H](OCCCCCCCCCCCC)O[C@H](CO)[C@H]1O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 NLEBIOOXCVAHBD-QKMCSOCLSA-N 0.000 description 3
- 239000003623 enhancer Substances 0.000 description 3
- 150000002402 hexoses Chemical class 0.000 description 3
- HNDVDQJCIGZPNO-UHFFFAOYSA-N histidine Natural products OC(=O)C(N)CC1=CN=CN1 HNDVDQJCIGZPNO-UHFFFAOYSA-N 0.000 description 3
- 230000006698 induction Effects 0.000 description 3
- 150000002500 ions Chemical class 0.000 description 3
- 210000000265 leukocyte Anatomy 0.000 description 3
- 230000004576 lipid-binding Effects 0.000 description 3
- 150000002632 lipids Chemical class 0.000 description 3
- 210000004962 mammalian cell Anatomy 0.000 description 3
- 239000002773 nucleotide Substances 0.000 description 3
- 125000003729 nucleotide group Chemical group 0.000 description 3
- 230000014207 opsonization Effects 0.000 description 3
- 210000001322 periplasm Anatomy 0.000 description 3
- COLNVLDHVKWLRT-UHFFFAOYSA-N phenylalanine Natural products OC(=O)C(N)CC1=CC=CC=C1 COLNVLDHVKWLRT-UHFFFAOYSA-N 0.000 description 3
- 239000002243 precursor Substances 0.000 description 3
- 238000002360 preparation method Methods 0.000 description 3
- 230000002829 reductive effect Effects 0.000 description 3
- 210000003786 sclera Anatomy 0.000 description 3
- 238000000926 separation method Methods 0.000 description 3
- 238000004885 tandem mass spectrometry Methods 0.000 description 3
- OUYCCCASQSFEME-UHFFFAOYSA-N tyrosine Natural products OC(=O)C(N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-UHFFFAOYSA-N 0.000 description 3
- 235000002374 tyrosine Nutrition 0.000 description 3
- OSWFIVFLDKOXQC-UHFFFAOYSA-N 4-(3-methoxyphenyl)aniline Chemical compound COC1=CC=CC(C=2C=CC(N)=CC=2)=C1 OSWFIVFLDKOXQC-UHFFFAOYSA-N 0.000 description 2
- LRDIEHDJWYRVPT-UHFFFAOYSA-N 4-amino-5-hydroxynaphthalene-1-sulfonic acid Chemical compound C1=CC(O)=C2C(N)=CC=C(S(O)(=O)=O)C2=C1 LRDIEHDJWYRVPT-UHFFFAOYSA-N 0.000 description 2
- 101100463771 Acinetobacter baylyi (strain ATCC 33305 / BD413 / ADP1) pglL1 gene Proteins 0.000 description 2
- 101100463772 Acinetobacter baylyi (strain ATCC 33305 / BD413 / ADP1) pglL2 gene Proteins 0.000 description 2
- 241000115154 Aerococcus suis Species 0.000 description 2
- DCXYFEDJOCDNAF-UHFFFAOYSA-N Asparagine Natural products OC(=O)C(N)CC(N)=O DCXYFEDJOCDNAF-UHFFFAOYSA-N 0.000 description 2
- 241000271566 Aves Species 0.000 description 2
- 108020004705 Codon Proteins 0.000 description 2
- 241000195493 Cryptophyta Species 0.000 description 2
- BWGNESOTFCXPMA-UHFFFAOYSA-N Dihydrogen disulfide Chemical compound SS BWGNESOTFCXPMA-UHFFFAOYSA-N 0.000 description 2
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 2
- 241000238631 Hexapoda Species 0.000 description 2
- 241000347881 Kadua laxiflora Species 0.000 description 2
- 238000001282 Kruskal–Wallis one-way analysis of variance Methods 0.000 description 2
- ONIBWKKTOPOVIA-BYPYZUCNSA-N L-Proline Chemical compound OC(=O)[C@@H]1CCCN1 ONIBWKKTOPOVIA-BYPYZUCNSA-N 0.000 description 2
- DCXYFEDJOCDNAF-REOHCLBHSA-N L-asparagine Chemical compound OC(=O)[C@@H](N)CC(N)=O DCXYFEDJOCDNAF-REOHCLBHSA-N 0.000 description 2
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 2
- AGPKZVBTJJNPAG-WHFBIAKZSA-N L-isoleucine Chemical compound CC[C@H](C)[C@H](N)C(O)=O AGPKZVBTJJNPAG-WHFBIAKZSA-N 0.000 description 2
- FFEARJCKVFRZRR-BYPYZUCNSA-N L-methionine Chemical compound CSCC[C@H](N)C(O)=O FFEARJCKVFRZRR-BYPYZUCNSA-N 0.000 description 2
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 2
- KZSNJWFQEVHDMF-BYPYZUCNSA-N L-valine Chemical compound CC(C)[C@H](N)C(O)=O KZSNJWFQEVHDMF-BYPYZUCNSA-N 0.000 description 2
- 108010052285 Membrane Proteins Proteins 0.000 description 2
- 241000588653 Neisseria Species 0.000 description 2
- 241000588650 Neisseria meningitidis Species 0.000 description 2
- 239000000020 Nitrocellulose Substances 0.000 description 2
- 102000015636 Oligopeptides Human genes 0.000 description 2
- 108010038807 Oligopeptides Proteins 0.000 description 2
- 108091093037 Peptide nucleic acid Proteins 0.000 description 2
- 241001377010 Pila Species 0.000 description 2
- 108010026552 Proteome Proteins 0.000 description 2
- 102220473708 Ras-related protein Rab-5A_S84A_mutation Human genes 0.000 description 2
- 241001522306 Serinus serinus Species 0.000 description 2
- 238000000692 Student's t-test Methods 0.000 description 2
- KZSNJWFQEVHDMF-UHFFFAOYSA-N Valine Natural products CC(C)C(N)C(O)=O KZSNJWFQEVHDMF-UHFFFAOYSA-N 0.000 description 2
- 229960000723 ampicillin Drugs 0.000 description 2
- AVKUERGKIZMTKX-NJBDSQKTSA-N ampicillin Chemical compound C1([C@@H](N)C(=O)N[C@H]2[C@H]3SC([C@@H](N3C2=O)C(O)=O)(C)C)=CC=CC=C1 AVKUERGKIZMTKX-NJBDSQKTSA-N 0.000 description 2
- 244000037640 animal pathogen Species 0.000 description 2
- 235000009582 asparagine Nutrition 0.000 description 2
- 229960001230 asparagine Drugs 0.000 description 2
- 230000001851 biosynthetic effect Effects 0.000 description 2
- 239000006161 blood agar Substances 0.000 description 2
- 150000001720 carbohydrates Chemical class 0.000 description 2
- 235000014633 carbohydrates Nutrition 0.000 description 2
- 210000004671 cell-free system Anatomy 0.000 description 2
- 230000005754 cellular signaling Effects 0.000 description 2
- 239000003153 chemical reaction reagent Substances 0.000 description 2
- 239000003795 chemical substances by application Substances 0.000 description 2
- 230000008045 co-localization Effects 0.000 description 2
- 238000010835 comparative analysis Methods 0.000 description 2
- 230000000295 complement effect Effects 0.000 description 2
- 230000029087 digestion Effects 0.000 description 2
- 230000000694 effects Effects 0.000 description 2
- 238000010828 elution Methods 0.000 description 2
- 238000005516 engineering process Methods 0.000 description 2
- 210000003527 eukaryotic cell Anatomy 0.000 description 2
- 230000007717 exclusion Effects 0.000 description 2
- 235000019253 formic acid Nutrition 0.000 description 2
- 238000005194 fractionation Methods 0.000 description 2
- 230000002538 fungal effect Effects 0.000 description 2
- 229930182830 galactose Natural products 0.000 description 2
- 230000002068 genetic effect Effects 0.000 description 2
- 230000002209 hydrophobic effect Effects 0.000 description 2
- 230000001900 immune effect Effects 0.000 description 2
- 238000003780 insertion Methods 0.000 description 2
- 230000037431 insertion Effects 0.000 description 2
- AGPKZVBTJJNPAG-UHFFFAOYSA-N isoleucine Natural products CCC(C)C(N)C(O)=O AGPKZVBTJJNPAG-UHFFFAOYSA-N 0.000 description 2
- 229960000310 isoleucine Drugs 0.000 description 2
- BPHPUYQFMNQIOC-NXRLNHOXSA-N isopropyl beta-D-thiogalactopyranoside Chemical compound CC(C)S[C@@H]1O[C@H](CO)[C@H](O)[C@H](O)[C@H]1O BPHPUYQFMNQIOC-NXRLNHOXSA-N 0.000 description 2
- 238000004895 liquid chromatography mass spectrometry Methods 0.000 description 2
- 239000003550 marker Substances 0.000 description 2
- 230000007246 mechanism Effects 0.000 description 2
- 229930182817 methionine Natural products 0.000 description 2
- 150000002772 monosaccharides Chemical class 0.000 description 2
- 238000002703 mutagenesis Methods 0.000 description 2
- 231100000350 mutagenesis Toxicity 0.000 description 2
- 229920001220 nitrocellulos Polymers 0.000 description 2
- 230000003287 optical effect Effects 0.000 description 2
- 230000037361 pathway Effects 0.000 description 2
- 239000008188 pellet Substances 0.000 description 2
- 239000008194 pharmaceutical composition Substances 0.000 description 2
- 229910052698 phosphorus Inorganic materials 0.000 description 2
- 239000013600 plasmid vector Substances 0.000 description 2
- 229940124733 pneumococcal vaccine Drugs 0.000 description 2
- 229940031937 polysaccharide vaccine Drugs 0.000 description 2
- 150000004804 polysaccharides Polymers 0.000 description 2
- 230000004481 post-translational protein modification Effects 0.000 description 2
- 230000003389 potentiating effect Effects 0.000 description 2
- 230000002265 prevention Effects 0.000 description 2
- 230000002797 proteolythic effect Effects 0.000 description 2
- 230000002685 pulmonary effect Effects 0.000 description 2
- 239000013014 purified material Substances 0.000 description 2
- 238000010188 recombinant method Methods 0.000 description 2
- 230000009467 reduction Effects 0.000 description 2
- 230000010076 replication Effects 0.000 description 2
- 230000002441 reversible effect Effects 0.000 description 2
- 230000003248 secreting effect Effects 0.000 description 2
- 230000028327 secretion Effects 0.000 description 2
- 239000000243 solution Substances 0.000 description 2
- 241000894007 species Species 0.000 description 2
- 230000001954 sterilising effect Effects 0.000 description 2
- 238000004659 sterilization and disinfection Methods 0.000 description 2
- 238000007920 subcutaneous administration Methods 0.000 description 2
- 238000012353 t test Methods 0.000 description 2
- 150000004044 tetrasaccharides Chemical group 0.000 description 2
- 241001430294 unidentified retrovirus Species 0.000 description 2
- 229940125575 vaccine candidate Drugs 0.000 description 2
- 239000004474 valine Substances 0.000 description 2
- 239000003643 water by type Substances 0.000 description 2
- 210000005253 yeast cell Anatomy 0.000 description 2
- DGVVWUTYPXICAM-UHFFFAOYSA-N β‐Mercaptoethanol Chemical compound OCCS DGVVWUTYPXICAM-UHFFFAOYSA-N 0.000 description 2
- XSYUPRQVAHJETO-WPMUBMLPSA-N (2s)-2-[[(2s)-2-[[(2s)-2-[[(2s)-2-[[(2s)-2-[[(2s)-2-amino-3-(1h-imidazol-5-yl)propanoyl]amino]-3-(1h-imidazol-5-yl)propanoyl]amino]-3-(1h-imidazol-5-yl)propanoyl]amino]-3-(1h-imidazol-5-yl)propanoyl]amino]-3-(1h-imidazol-5-yl)propanoyl]amino]-3-(1h-imidaz Chemical compound C([C@H](N)C(=O)N[C@@H](CC=1NC=NC=1)C(=O)N[C@@H](CC=1NC=NC=1)C(=O)N[C@@H](CC=1NC=NC=1)C(=O)N[C@@H](CC=1NC=NC=1)C(=O)N[C@@H](CC=1NC=NC=1)C(O)=O)C1=CN=CN1 XSYUPRQVAHJETO-WPMUBMLPSA-N 0.000 description 1
- UKAUYVFTDYCKQA-UHFFFAOYSA-N -2-Amino-4-hydroxybutanoic acid Natural products OC(=O)C(N)CCO UKAUYVFTDYCKQA-UHFFFAOYSA-N 0.000 description 1
- OWEGMIWEEQEYGQ-UHFFFAOYSA-N 100676-05-9 Natural products OC1C(O)C(O)C(CO)OC1OCC1C(O)C(O)C(O)C(OC2C(OC(O)C(O)C2O)CO)O1 OWEGMIWEEQEYGQ-UHFFFAOYSA-N 0.000 description 1
- BRMWTNUJHUMWMS-UHFFFAOYSA-N 3-Methylhistidine Natural products CN1C=NC(CC(N)C(O)=O)=C1 BRMWTNUJHUMWMS-UHFFFAOYSA-N 0.000 description 1
- 229940117976 5-hydroxylysine Drugs 0.000 description 1
- GGZZISOUXJHYOY-UHFFFAOYSA-N 8-amino-4-hydroxynaphthalene-2-sulfonic acid Chemical compound C1=C(S(O)(=O)=O)C=C2C(N)=CC=CC2=C1O GGZZISOUXJHYOY-UHFFFAOYSA-N 0.000 description 1
- UBDHSURDYAETAL-UHFFFAOYSA-N 8-aminonaphthalene-1,3,6-trisulfonic acid Chemical compound OS(=O)(=O)C1=CC(S(O)(=O)=O)=C2C(N)=CC(S(O)(=O)=O)=CC2=C1 UBDHSURDYAETAL-UHFFFAOYSA-N 0.000 description 1
- 241001527178 Acinetobacter puyangensis Species 0.000 description 1
- 241000271152 Acinetobacter soli Species 0.000 description 1
- 241001372176 Acinetobacter soli CIP 110264 Species 0.000 description 1
- 241000897241 Acinetobacter sp. ADP1 Species 0.000 description 1
- 102000007469 Actins Human genes 0.000 description 1
- 108010085238 Actins Proteins 0.000 description 1
- 239000004475 Arginine Substances 0.000 description 1
- 241000283690 Bos taurus Species 0.000 description 1
- 108091003079 Bovine Serum Albumin Proteins 0.000 description 1
- 241000510930 Brachyspira pilosicoli Species 0.000 description 1
- 206010006500 Brucellosis Diseases 0.000 description 1
- 241001136175 Burkholderia pseudomallei Species 0.000 description 1
- 241000283707 Capra Species 0.000 description 1
- 241000700199 Cavia porcellus Species 0.000 description 1
- 241000282693 Cercopithecidae Species 0.000 description 1
- 206010008631 Cholera Diseases 0.000 description 1
- 241000701022 Cytomegalovirus Species 0.000 description 1
- 108010016626 Dipeptides Proteins 0.000 description 1
- 241000283073 Equus caballus Species 0.000 description 1
- 241001131785 Escherichia coli HB101 Species 0.000 description 1
- 241000282326 Felis catus Species 0.000 description 1
- 230000005526 G1 to G0 transition Effects 0.000 description 1
- 239000004471 Glycine Substances 0.000 description 1
- 102000002812 Heat-Shock Proteins Human genes 0.000 description 1
- 108010004889 Heat-Shock Proteins Proteins 0.000 description 1
- 102100021519 Hemoglobin subunit beta Human genes 0.000 description 1
- 108091005904 Hemoglobin subunit beta Proteins 0.000 description 1
- PMMYEEVYMWASQN-DMTCNVIQSA-N Hydroxyproline Chemical compound O[C@H]1CN[C@H](C(O)=O)C1 PMMYEEVYMWASQN-DMTCNVIQSA-N 0.000 description 1
- 108091092195 Intron Proteins 0.000 description 1
- 208000037836 K. pneumoniae liver abscess Diseases 0.000 description 1
- XUJNEKJLAYXESH-REOHCLBHSA-N L-Cysteine Chemical compound SC[C@H](N)C(O)=O XUJNEKJLAYXESH-REOHCLBHSA-N 0.000 description 1
- AHLPHDHHMVZTML-BYPYZUCNSA-N L-Ornithine Chemical compound NCCC[C@H](N)C(O)=O AHLPHDHHMVZTML-BYPYZUCNSA-N 0.000 description 1
- ODKSFYDXXFIFQN-BYPYZUCNSA-P L-argininium(2+) Chemical compound NC(=[NH2+])NCCC[C@H]([NH3+])C(O)=O ODKSFYDXXFIFQN-BYPYZUCNSA-P 0.000 description 1
- CKLJMWTZIZZHCS-REOHCLBHSA-N L-aspartic acid Chemical compound OC(=O)[C@@H](N)CC(O)=O CKLJMWTZIZZHCS-REOHCLBHSA-N 0.000 description 1
- WHUUTDBJXJRKMK-VKHMYHEASA-N L-glutamic acid Chemical compound OC(=O)[C@@H](N)CCC(O)=O WHUUTDBJXJRKMK-VKHMYHEASA-N 0.000 description 1
- 229930182816 L-glutamine Natural products 0.000 description 1
- UKAUYVFTDYCKQA-VKHMYHEASA-N L-homoserine Chemical compound OC(=O)[C@@H](N)CCO UKAUYVFTDYCKQA-VKHMYHEASA-N 0.000 description 1
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 1
- KDXKERNSBIXSRK-YFKPBYRVSA-N L-lysine Chemical compound NCCCC[C@H](N)C(O)=O KDXKERNSBIXSRK-YFKPBYRVSA-N 0.000 description 1
- AYFVYJQAPQTCCC-GBXIJSLDSA-N L-threonine Chemical compound C[C@@H](O)[C@H](N)C(O)=O AYFVYJQAPQTCCC-GBXIJSLDSA-N 0.000 description 1
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 1
- 206010024652 Liver abscess Diseases 0.000 description 1
- 102220471249 M-phase inducer phosphatase 1_S82A_mutation Human genes 0.000 description 1
- 241000829100 Macaca mulatta polyomavirus 1 Species 0.000 description 1
- GUBGYTABKSRVRQ-PICCSMPSSA-N Maltose Natural products O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@@H](CO)OC(O)[C@H](O)[C@H]1O GUBGYTABKSRVRQ-PICCSMPSSA-N 0.000 description 1
- 102000018697 Membrane Proteins Human genes 0.000 description 1
- 201000009906 Meningitis Diseases 0.000 description 1
- 206010027476 Metastases Diseases 0.000 description 1
- 241000069157 Miconia aeruginosa Species 0.000 description 1
- 239000012901 Milli-Q water Substances 0.000 description 1
- 108010021466 Mutant Proteins Proteins 0.000 description 1
- 102000008300 Mutant Proteins Human genes 0.000 description 1
- JDHILDINMRGULE-LURJTMIESA-N N(pros)-methyl-L-histidine Chemical compound CN1C=NC=C1C[C@H](N)C(O)=O JDHILDINMRGULE-LURJTMIESA-N 0.000 description 1
- OVRNDRQMDRJTHS-FMDGEEDCSA-N N-acetyl-beta-D-glucosamine Chemical compound CC(=O)N[C@H]1[C@H](O)O[C@H](CO)[C@@H](O)[C@@H]1O OVRNDRQMDRJTHS-FMDGEEDCSA-N 0.000 description 1
- MBLBDJOUHNCFQT-LXGUWJNJSA-N N-acetylglucosamine Natural products CC(=O)N[C@@H](C=O)[C@@H](O)[C@H](O)[C@H](O)CO MBLBDJOUHNCFQT-LXGUWJNJSA-N 0.000 description 1
- 125000001429 N-terminal alpha-amino-acid group Chemical group 0.000 description 1
- 108091061960 Naked DNA Proteins 0.000 description 1
- 241000588652 Neisseria gonorrhoeae Species 0.000 description 1
- AHLPHDHHMVZTML-UHFFFAOYSA-N Orn-delta-NH2 Natural products NCCCC(N)C(O)=O AHLPHDHHMVZTML-UHFFFAOYSA-N 0.000 description 1
- UTJLXEIPEHZYQJ-UHFFFAOYSA-N Ornithine Natural products OC(=O)C(C)CCCN UTJLXEIPEHZYQJ-UHFFFAOYSA-N 0.000 description 1
- 101710116435 Outer membrane protein Proteins 0.000 description 1
- 229910019142 PO4 Inorganic materials 0.000 description 1
- 108010090127 Periplasmic Proteins Proteins 0.000 description 1
- 241000009328 Perro Species 0.000 description 1
- 108010002747 Pfu DNA polymerase Proteins 0.000 description 1
- 241000709664 Picornaviridae Species 0.000 description 1
- 229940124158 Protease/peptidase inhibitor Drugs 0.000 description 1
- 108010001267 Protein Subunits Proteins 0.000 description 1
- 102000002067 Protein Subunits Human genes 0.000 description 1
- 101100464097 Pseudomonas aeruginosa (strain ATCC 15692 / DSM 22644 / CIP 104116 / JCM 14847 / LMG 12228 / 1C / PRS 101 / PAO1) pilO gene Proteins 0.000 description 1
- 239000012980 RPMI-1640 medium Substances 0.000 description 1
- 241000700159 Rattus Species 0.000 description 1
- 101100510671 Rattus norvegicus Lnpep gene Proteins 0.000 description 1
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 description 1
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 description 1
- 108091081062 Repeated sequence (DNA) Proteins 0.000 description 1
- 241000714474 Rous sarcoma virus Species 0.000 description 1
- 241000607768 Shigella Species 0.000 description 1
- 241000147000 Shigella flexneri 2a Species 0.000 description 1
- QAOWNCQODCNURD-UHFFFAOYSA-L Sulfate Chemical compound [O-]S([O-])(=O)=O QAOWNCQODCNURD-UHFFFAOYSA-L 0.000 description 1
- 241000282898 Sus scrofa Species 0.000 description 1
- 108010008038 Synthetic Vaccines Proteins 0.000 description 1
- 108020005038 Terminator Codon Proteins 0.000 description 1
- 239000004098 Tetracycline Substances 0.000 description 1
- AYFVYJQAPQTCCC-UHFFFAOYSA-N Threonine Natural products CC(O)C(N)C(O)=O AYFVYJQAPQTCCC-UHFFFAOYSA-N 0.000 description 1
- 239000004473 Threonine Substances 0.000 description 1
- 108700029229 Transcriptional Regulatory Elements Proteins 0.000 description 1
- 241000700605 Viruses Species 0.000 description 1
- 238000002835 absorbance Methods 0.000 description 1
- 230000001133 acceleration Effects 0.000 description 1
- 230000021736 acetylation Effects 0.000 description 1
- 238000006640 acetylation reaction Methods 0.000 description 1
- 230000002378 acidificating effect Effects 0.000 description 1
- 230000009471 action Effects 0.000 description 1
- 230000004913 activation Effects 0.000 description 1
- 239000004480 active ingredient Substances 0.000 description 1
- 230000003044 adaptive effect Effects 0.000 description 1
- 210000005006 adaptive immune system Anatomy 0.000 description 1
- 230000007815 allergy Effects 0.000 description 1
- WNROFYMDJYEPJX-UHFFFAOYSA-K aluminium hydroxide Chemical compound [OH-].[OH-].[OH-].[Al+3] WNROFYMDJYEPJX-UHFFFAOYSA-K 0.000 description 1
- 230000009435 amidation Effects 0.000 description 1
- 238000007112 amidation reaction Methods 0.000 description 1
- 150000003862 amino acid derivatives Chemical class 0.000 description 1
- 230000003698 anagen phase Effects 0.000 description 1
- RJGDLRCDCYRQOQ-UHFFFAOYSA-N anthrone Chemical compound C1=CC=C2C(=O)C3=CC=CC=C3CC2=C1 RJGDLRCDCYRQOQ-UHFFFAOYSA-N 0.000 description 1
- 239000003242 anti bacterial agent Substances 0.000 description 1
- 229940088710 antibiotic agent Drugs 0.000 description 1
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 1
- 125000003118 aryl group Chemical group 0.000 description 1
- 235000003704 aspartic acid Nutrition 0.000 description 1
- 229960001212 bacterial vaccine Drugs 0.000 description 1
- OQFSQFPPLPISGP-UHFFFAOYSA-N beta-carboxyaspartic acid Natural products OC(=O)C(N)C(C(O)=O)C(O)=O OQFSQFPPLPISGP-UHFFFAOYSA-N 0.000 description 1
- 230000003115 biocidal effect Effects 0.000 description 1
- 238000003766 bioinformatics method Methods 0.000 description 1
- 230000004071 biological effect Effects 0.000 description 1
- 229960000074 biopharmaceutical Drugs 0.000 description 1
- 230000000903 blocking effect Effects 0.000 description 1
- 238000010241 blood sampling Methods 0.000 description 1
- 108010006025 bovine growth hormone Proteins 0.000 description 1
- 210000004556 brain Anatomy 0.000 description 1
- 239000002775 capsule Substances 0.000 description 1
- 239000006143 cell culture medium Substances 0.000 description 1
- 229960005091 chloramphenicol Drugs 0.000 description 1
- WIIZWVCIJKGZOK-RKDXNWHRSA-N chloramphenicol Chemical compound ClC(Cl)C(=O)N[C@H](CO)[C@H](O)C1=CC=C([N+]([O-])=O)C=C1 WIIZWVCIJKGZOK-RKDXNWHRSA-N 0.000 description 1
- 230000002759 chromosomal effect Effects 0.000 description 1
- 238000003776 cleavage reaction Methods 0.000 description 1
- 238000010367 cloning Methods 0.000 description 1
- 239000013599 cloning vector Substances 0.000 description 1
- 239000011248 coating agent Substances 0.000 description 1
- 238000000576 coating method Methods 0.000 description 1
- 230000001332 colony forming effect Effects 0.000 description 1
- 238000010959 commercial synthesis reaction Methods 0.000 description 1
- 235000021310 complex sugar Nutrition 0.000 description 1
- 230000001276 controlling effect Effects 0.000 description 1
- 239000012050 conventional carrier Substances 0.000 description 1
- 101150019098 cps gene Proteins 0.000 description 1
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 1
- 235000018417 cysteine Nutrition 0.000 description 1
- 230000001086 cytosolic effect Effects 0.000 description 1
- 230000034994 death Effects 0.000 description 1
- 231100000517 death Toxicity 0.000 description 1
- 230000002950 deficient Effects 0.000 description 1
- YSMODUONRAFBET-UHFFFAOYSA-N delta-DL-hydroxylysine Natural products NCC(O)CCC(N)C(O)=O YSMODUONRAFBET-UHFFFAOYSA-N 0.000 description 1
- 238000004925 denaturation Methods 0.000 description 1
- 230000036425 denaturation Effects 0.000 description 1
- 238000001212 derivatisation Methods 0.000 description 1
- 238000013461 design Methods 0.000 description 1
- 238000001514 detection method Methods 0.000 description 1
- 239000003599 detergent Substances 0.000 description 1
- 239000000032 diagnostic agent Substances 0.000 description 1
- 229940039227 diagnostic agent Drugs 0.000 description 1
- 238000010790 dilution Methods 0.000 description 1
- 239000012895 dilution Substances 0.000 description 1
- XPPKVPWEQAFLFU-UHFFFAOYSA-J diphosphate(4-) Chemical compound [O-]P([O-])(=O)OP([O-])([O-])=O XPPKVPWEQAFLFU-UHFFFAOYSA-J 0.000 description 1
- 235000011180 diphosphates Nutrition 0.000 description 1
- PMMYEEVYMWASQN-UHFFFAOYSA-N dl-hydroxyproline Natural products OC1C[NH2+]C(C([O-])=O)C1 PMMYEEVYMWASQN-UHFFFAOYSA-N 0.000 description 1
- 239000012636 effector Substances 0.000 description 1
- 238000004520 electroporation Methods 0.000 description 1
- 210000004696 endometrium Anatomy 0.000 description 1
- 239000002158 endotoxin Substances 0.000 description 1
- YSMODUONRAFBET-UHNVWZDZSA-N erythro-5-hydroxy-L-lysine Chemical compound NC[C@H](O)CC[C@H](N)C(O)=O YSMODUONRAFBET-UHNVWZDZSA-N 0.000 description 1
- 239000013613 expression plasmid Substances 0.000 description 1
- 239000013604 expression vector Substances 0.000 description 1
- 238000000605 extraction Methods 0.000 description 1
- 239000012091 fetal bovine serum Substances 0.000 description 1
- 239000007789 gas Substances 0.000 description 1
- 238000003197 gene knockdown Methods 0.000 description 1
- 238000012252 genetic analysis Methods 0.000 description 1
- 238000012268 genome sequencing Methods 0.000 description 1
- 229930195712 glutamate Natural products 0.000 description 1
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Natural products OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 description 1
- 235000004554 glutamine Nutrition 0.000 description 1
- 125000003147 glycosyl group Chemical group 0.000 description 1
- 102000035122 glycosylated proteins Human genes 0.000 description 1
- 108091005608 glycosylated proteins Proteins 0.000 description 1
- 230000012010 growth Effects 0.000 description 1
- 229940047650 haemophilus influenzae Drugs 0.000 description 1
- 230000036541 health Effects 0.000 description 1
- 229920000669 heparin Polymers 0.000 description 1
- ZFGMDIBRIDKWMY-PASTXAENSA-N heparin Chemical compound CC(O)=N[C@@H]1[C@@H](O)[C@H](O)[C@@H](COS(O)(=O)=O)O[C@@H]1O[C@@H]1[C@@H](C(O)=O)O[C@@H](O[C@H]2[C@@H]([C@@H](OS(O)(=O)=O)[C@@H](O[C@@H]3[C@@H](OC(O)[C@H](OS(O)(=O)=O)[C@H]3O)C(O)=O)O[C@@H]2O)CS(O)(=O)=O)[C@H](O)[C@H]1O ZFGMDIBRIDKWMY-PASTXAENSA-N 0.000 description 1
- 229960001008 heparin sodium Drugs 0.000 description 1
- 229910052739 hydrogen Inorganic materials 0.000 description 1
- 239000001257 hydrogen Substances 0.000 description 1
- 238000002013 hydrophilic interaction chromatography Methods 0.000 description 1
- 229960002591 hydroxyproline Drugs 0.000 description 1
- 238000000338 in vitro Methods 0.000 description 1
- 238000003331 infrared imaging Methods 0.000 description 1
- 238000001802 infusion Methods 0.000 description 1
- 239000004615 ingredient Substances 0.000 description 1
- 230000010354 integration Effects 0.000 description 1
- 238000007918 intramuscular administration Methods 0.000 description 1
- 238000007912 intraperitoneal administration Methods 0.000 description 1
- 238000001990 intravenous administration Methods 0.000 description 1
- 229940065638 intron a Drugs 0.000 description 1
- PGLTVOMIXTUURA-UHFFFAOYSA-N iodoacetamide Chemical compound NC(=O)CI PGLTVOMIXTUURA-UHFFFAOYSA-N 0.000 description 1
- 229960000318 kanamycin Drugs 0.000 description 1
- 229930027917 kanamycin Natural products 0.000 description 1
- SBUJHOSQTJFQJX-NOAMYHISSA-N kanamycin Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CN)O[C@@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](N)[C@H](O)[C@@H](CO)O2)O)[C@H](N)C[C@@H]1N SBUJHOSQTJFQJX-NOAMYHISSA-N 0.000 description 1
- 229930182823 kanamycin A Natural products 0.000 description 1
- 230000001665 lethal effect Effects 0.000 description 1
- 238000001638 lipofection Methods 0.000 description 1
- 229920006008 lipopolysaccharide Polymers 0.000 description 1
- 230000004807 localization Effects 0.000 description 1
- 239000006166 lysate Substances 0.000 description 1
- VTHJTEIRLNZDEV-UHFFFAOYSA-L magnesium dihydroxide Chemical compound [OH-].[OH-].[Mg+2] VTHJTEIRLNZDEV-UHFFFAOYSA-L 0.000 description 1
- 239000000347 magnesium hydroxide Substances 0.000 description 1
- 229910001862 magnesium hydroxide Inorganic materials 0.000 description 1
- 239000000463 material Substances 0.000 description 1
- 238000005259 measurement Methods 0.000 description 1
- 230000015654 memory Effects 0.000 description 1
- 230000031864 metaphase Effects 0.000 description 1
- 230000009401 metastasis Effects 0.000 description 1
- 230000011987 methylation Effects 0.000 description 1
- 238000007069 methylation reaction Methods 0.000 description 1
- 238000001823 molecular biology technique Methods 0.000 description 1
- 239000000178 monomer Substances 0.000 description 1
- 238000010172 mouse model Methods 0.000 description 1
- 229950006780 n-acetylglucosamine Drugs 0.000 description 1
- 238000010899 nucleation Methods 0.000 description 1
- 229960003104 ornithine Drugs 0.000 description 1
- XLYOFNOQVPJJNP-UHFFFAOYSA-O oxonium Chemical compound [OH3+] XLYOFNOQVPJJNP-UHFFFAOYSA-O 0.000 description 1
- 230000036961 partial effect Effects 0.000 description 1
- 239000002245 particle Substances 0.000 description 1
- 230000007918 pathogenicity Effects 0.000 description 1
- 239000000137 peptide hydrolase inhibitor Substances 0.000 description 1
- NBIIXXVUZAFLBC-UHFFFAOYSA-K phosphate Chemical compound [O-]P([O-])([O-])=O NBIIXXVUZAFLBC-UHFFFAOYSA-K 0.000 description 1
- 239000010452 phosphate Substances 0.000 description 1
- 230000026731 phosphorylation Effects 0.000 description 1
- 238000006366 phosphorylation reaction Methods 0.000 description 1
- 229940033515 pneumovax 23 Drugs 0.000 description 1
- 229920002401 polyacrylamide Polymers 0.000 description 1
- 230000008488 polyadenylation Effects 0.000 description 1
- 229920000136 polysorbate Polymers 0.000 description 1
- 230000001124 posttranscriptional effect Effects 0.000 description 1
- 238000004321 preservation Methods 0.000 description 1
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 1
- 210000001236 prokaryotic cell Anatomy 0.000 description 1
- 230000000069 prophylactic effect Effects 0.000 description 1
- 238000011321 prophylaxis Methods 0.000 description 1
- 230000018570 protein O-linked glycosylation Effects 0.000 description 1
- 230000004952 protein activity Effects 0.000 description 1
- 230000012743 protein tagging Effects 0.000 description 1
- 230000017854 proteolysis Effects 0.000 description 1
- 230000006337 proteolytic cleavage Effects 0.000 description 1
- 238000003259 recombinant expression Methods 0.000 description 1
- 229940124551 recombinant vaccine Drugs 0.000 description 1
- 238000011160 research Methods 0.000 description 1
- 230000000717 retained effect Effects 0.000 description 1
- 230000007017 scission Effects 0.000 description 1
- 238000013207 serial dilution Methods 0.000 description 1
- 230000001568 sexual effect Effects 0.000 description 1
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 1
- 230000010473 stable expression Effects 0.000 description 1
- 238000005728 strengthening Methods 0.000 description 1
- 125000001424 substituent group Chemical group 0.000 description 1
- 239000006228 supernatant Substances 0.000 description 1
- 239000006188 syrup Substances 0.000 description 1
- 235000020357 syrup Nutrition 0.000 description 1
- 150000003505 terpenes Chemical group 0.000 description 1
- 229960002180 tetracycline Drugs 0.000 description 1
- 229930101283 tetracycline Natural products 0.000 description 1
- 235000019364 tetracycline Nutrition 0.000 description 1
- 150000003522 tetracyclines Chemical class 0.000 description 1
- 235000008521 threonine Nutrition 0.000 description 1
- 125000000341 threoninyl group Chemical group [H]OC([H])(C([H])([H])[H])C([H])(N([H])[H])C(*)=O 0.000 description 1
- 210000001519 tissue Anatomy 0.000 description 1
- 231100000331 toxic Toxicity 0.000 description 1
- 230000002588 toxic effect Effects 0.000 description 1
- 230000005030 transcription termination Effects 0.000 description 1
- 238000001890 transfection Methods 0.000 description 1
- 230000010474 transient expression Effects 0.000 description 1
- 230000007704 transition Effects 0.000 description 1
- 238000013519 translation Methods 0.000 description 1
- 230000014621 translational initiation Effects 0.000 description 1
- 230000005945 translocation Effects 0.000 description 1
- 238000011282 treatment Methods 0.000 description 1
- 125000001493 tyrosinyl group Chemical group [H]OC1=C([H])C([H])=C(C([H])=C1[H])C([H])([H])C([H])(N([H])[H])C(*)=O 0.000 description 1
- 238000004704 ultra performance liquid chromatography Methods 0.000 description 1
- 208000019206 urinary tract infection Diseases 0.000 description 1
- 239000013603 viral vector Substances 0.000 description 1
- 230000001018 virulence Effects 0.000 description 1
- 239000000304 virulence factor Substances 0.000 description 1
- 230000007923 virulence factor Effects 0.000 description 1
- 238000005406 washing Methods 0.000 description 1
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 1
Images











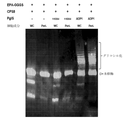


























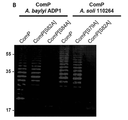




Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/195—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/1048—Glycosyltransferases (2.4)
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/02—Bacterial antigens
- A61K39/025—Enterobacteriales, e.g. Enterobacter
- A61K39/0266—Klebsiella
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/02—Bacterial antigens
- A61K39/05—Actinobacteria, e.g. Actinomyces, Streptomyces, Nocardia, Bifidobacterium, Gardnerella, Corynebacterium; Propionibacterium
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/02—Bacterial antigens
- A61K39/09—Lactobacillales, e.g. aerococcus, enterococcus, lactobacillus, lactococcus, streptococcus
- A61K39/092—Streptococcus
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/04—Antibacterial agents
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/195—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria
- C07K14/21—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria from Pseudomonadaceae (F)
- C07K14/212—Moraxellaceae, e.g. Acinetobacter, Moraxella, Oligella, Psychrobacter
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/57—Medicinal preparations containing antigens or antibodies characterised by the type of response, e.g. Th1, Th2
- A61K2039/575—Medicinal preparations containing antigens or antibodies characterised by the type of response, e.g. Th1, Th2 humoral response
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/60—Medicinal preparations containing antigens or antibodies characteristics by the carrier linked to the antigen
- A61K2039/6031—Proteins
- A61K2039/6037—Bacterial toxins, e.g. diphteria toxoid [DT], tetanus toxoid [TT]
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/55—Fusion polypeptide containing a fusion with a toxin, e.g. diphteria toxin
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/90—Fusion polypeptide containing a motif for post-translational modification
- C07K2319/91—Fusion polypeptide containing a motif for post-translational modification containing a motif for glycosylation
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Medicinal Chemistry (AREA)
- General Health & Medical Sciences (AREA)
- Organic Chemistry (AREA)
- Mycology (AREA)
- Veterinary Medicine (AREA)
- Public Health (AREA)
- Animal Behavior & Ethology (AREA)
- Pharmacology & Pharmacy (AREA)
- Microbiology (AREA)
- Epidemiology (AREA)
- Immunology (AREA)
- Genetics & Genomics (AREA)
- Biochemistry (AREA)
- Molecular Biology (AREA)
- Biophysics (AREA)
- Gastroenterology & Hepatology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Engineering & Computer Science (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Wood Science & Technology (AREA)
- Zoology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Communicable Diseases (AREA)
- Oncology (AREA)
- General Chemical & Material Sciences (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Biomedical Technology (AREA)
- General Engineering & Computer Science (AREA)
- Biotechnology (AREA)
- Peptides Or Proteins (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Medicinal Preparation (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
Abstract
Description
この国際出願は、2018年6月16日に出願された米国仮出願第62/685,970号及び2018年12月21日に出願された米国仮出願第62/783,971号の権益を主張しており、これらは両方ともその全体が本明細書に組み込まれている。
本発明は、国立アレルギー感染症研究所(NIAID)によって授与されたR41 AI131742助成金の下で政府の支援を受けて行われた。政府は、本発明において一定の権利を有する。
本明細書で提供されるのは、バイオコンジュゲートであり、例えば、その特定の態様及び特徴がこの段落に記載されており、融合タンパク質に共有結合したオリゴ糖または多糖を含み、ここで融合タンパク質は、ComPタンパク質(ComP)またはそのグリコシル化タグフラグメントを含む。特定の態様では、融合タンパク質は、配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基において、ComPタンパク質上のオリゴ糖もしくは多糖、またはそのグリコシル化タグフラグメントでグリコシル化される。特定の態様では、ComPタンパク質は、配列番号7(ComPΔ28ADP1)に対して、少なくとも50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%、または100%同一であるアミノ酸配列を含み、配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基を含む。特定の態様では、ComPタンパク質は、配列番号7(ComPΔ28ADP1)、配列番号8(ComPΔ28110264)、配列番号9(ComPΔ28GFJ−2)、配列番号10(ComPΔ28P50v1)、配列番号11(ComPΔ284466)、または配列番号12(ComPΔ28SFC)を含む。特定の態様では、ComPタンパク質は、配列番号1(ComPADP1:AAC45886.1)に対して、少なくとも50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%、または100%同一であるアミノ酸配列を含み、配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基を含む。特定の態様では、ComPタンパク質は、配列番号1(ComPADP1:AAC45886.1)、配列番号2(ComP110264:ENV58402.1)、配列番号3(ComPGFJ−2:APV36638.1)、配列番号4(ComP50v1:PKD82822.1)、配列番号5(ComP4466:SNX44537.1)、または配列番号6(ComPSFC:OAL75955.1)を含む。特定の態様では、ComPタンパク質のグリコシル化タグフラグメントは、配列番号1(ComPADP1:AAC45886.1)のアミノ酸残基1から28に対応するアミノ酸残基を欠くComPΔ28ポリペプチドである。特定の実施形態では、ComPΔ28ポリペプチドは、配列番号7、配列番号8、配列番号9、配列番号10、配列番号11、及び配列番号12からなる群から選択される。特定の態様では、ComPタンパク質のグリコシル化タグフラグメントは、アルファベータループをベータストランド領域に接続するジスルフィド結合に隣接する、配列番号2(ComP110264:ENV58402.1)の82位のセリン残基を含む、配列番号2(ComP110264:ENV58402.1)の領域に対応する領域を含む。特定の態様では、ComPタンパク質のグリコシル化タグフラグメントは、VGVQEISASNATTNVATAT(配列番号39)、TGVTQIASGASAATTNVASAQ(配列番号40)、VGVQEINASSSTSNVATAT(配列番号41)、AGVETIGASNKTKNVESAA(配列番号42)、VGVQTIAASNATKNVATAT(配列番号43)、及びNGVISASATTNVASSA(配列番号44)からなる群から選択されるアミノ酸配列に対して、少なくとも70%、75%、80%、85%、90%、95%、96%、97%、98%、99%、または100%同一であるアミノ酸配列を含み、上記グリコシル化タグフラグメントは、配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基を含む。特定の態様では、ComPタンパク質のグリコシル化タグフラグメントは、VGVQEISASNATTNVATAT(配列番号39)、TGVTQIASGASAATTNVASAQ(配列番号40)、VGVQEINASSSTSNVATAT(配列番号41)、AGVETIGASNKTKNVESAA(配列番号42)、VGVQTIAASNATKNVATAT(配列番号43)、及びNGVISASATTNVASSA(配列番号44)、または1、2、3、4、5、6、または7個のアミノ酸置換、付加、及び/または欠失を有するそれらのバリアントからなる群から選択されるアミノ酸配列を含み、ここでバリアントは配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基を維持する。特定の態様では、ComPタンパク質のグリコシル化タグフラグメントは、配列番号37のアミノ酸コンセンサス配列、または少なくとも5、10、15、20、30、35、または40個の連続的なアミノ酸のそのフラグメントを含み、上記グリコシル化タグフラグメントは、配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基を含み、またはComPタンパク質のグリコシル化タグフラグメントは、1、2、3、4、5、6、または7個のアミノ酸置換、付加、及び/または欠失を有するそれらのバリアントを含み、ここでバリアントは配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基を維持する。特定の態様では、ComPタンパク質のグリコシル化タグフラグメントは、配列番号38または45のアミノ酸コンセンサス配列、または少なくとも5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、または20個の連続的なアミノ酸のそのフラグメントを含み、ここで上記グリコシル化タグフラグメントは、配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基を含み、またはComPタンパク質のグリコシル化タグフラグメントは、1、2、3、4、5、6、または7個のアミノ酸置換、付加、及び/または欠失を有するそれらのバリアントを含み、ここでバリアントは配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基を維持する。特定の態様では、オリゴ糖または多糖は、ストレプトコッカス属からの細菌によって産生される。特定の態様では、多糖は、S. pneumoniae、S. agalactiae、またはS. suisのきょう膜多糖である。特定の態様では、きょう膜多糖は、CPS14、CPS8、CPS9V、またはCPS15bである。特定の態様では、きょう膜多糖は、CPS8である。特定の態様では、オリゴ糖または多糖は、クレブシエラ属からの細菌によって産生される。特定の態様では、オリゴ糖または多糖は、Klebsiella pneumoniae、Klebsiella varricola、Klebsiella michinganenis、またはKlebsiella oxytocaのきょう膜多糖である。特定の態様では、多糖は、Klebsiella pneumoniaeのきょう膜多糖である。特定の態様では、多糖は、Klebsiella pneumoniaeの血清型K1または血清型K2のきょう膜多糖である。特定の態様では、オリゴ糖または多糖は、その還元末端にグルコースを含む。特定の態様では、バイオコンジュゲートはin vivoで産生される。特定の態様では、バイオコンジュゲートは、細菌細胞において産生される。特定の態様では、融合タンパク質は、ジフテリアトキソイドCRM197、破傷風トキソイド、Pseudomonas aeruginosa外毒素A(EPA)、破傷風毒素Cフラグメント、コレラ毒素Bサブユニット、及びHaemophilus influenzaタンパク質D、またはそれらのフラグメントからなる群から選択される担体タンパク質を含む。特定の態様では、ComPタンパク質またはそのグリコシル化タグフラグメントは、融合タンパク質のN末端、融合タンパク質のC末端、及び/または融合タンパク質の内部に位置する。特定の態様では、担体タンパク質またはそのフラグメントは、アミノ酸リンカーを介してComPタンパク質またはそのグリコシル化タグフラグメントに連結されている。特定の態様では、融合タンパク質は、2以上、3以上、4以上、5以上、6以上、8以上、10以上、15以上、または20以上のComPタンパク質のグリコシル化タグフラグメントを含む。特定の態様では、融合タンパク質は、2、3、4、5、6、7、8、9、10、11、12、13、14、15、または20のうちのいずれかから3、4、5、6、7、8、9、10、11、12、13、14、15、20、または25のうちのいずれかまでのComPタンパク質のグリコシル化タグフラグメントを含む。特定の態様では、バイオコンジュゲートはコンジュゲートワクチンである。特定の態様では、コンジュゲートワクチンは、Streptococcus pneumoniae血清型8に対するワクチンである。特定の態様では、コンジュゲートワクチンは対象に投与され、それは免疫応答を誘導する。特定の態様では、免疫応答は長期記憶(記憶B及びT細胞)を誘発し、抗体応答である。特定の態様では、抗体反応は、血清型特異的抗体応答である。特定の態様では、抗体応答はIgGまたはIgM応答である。特定の態様では、抗体応答はIgG応答である。特定の態様では、IgG応答はIgG1応答である。特定の態様では、コンジュゲートワクチンは、ワクチンを投与された対象において免疫学的記憶を生成する。

X1は、V、A、またはアミノ酸不在であり;
X2は、A、G、T、またはアミノ酸不在であり;
X5は、P、S、またはQであり;
X6は、S、M、またはIであり;
X7は、T、P、またはVであり;
X8は、A、S、またはTであり;
X9は、G、N、S、またはTであり;
X10は、N、またはアミノ酸不在であり;
X11は、S、G、またはAであり;
X12は、SまたはNであり;
X14は、V、T、またはAであり;
X17は、Q、T、またはEであり;
X18は、E、Q、またはTであり;
X20は、S、N、A、またはGであり;
X21は、S、またはアミノ酸不在であり;
X22は、G、またはアミノ酸不在であり;
X25は、N、S、またはAであり;
X26は、A、S、またはKであり;
X28は、T、S、またはKであり;
X31は、AまたはEであり;
X32は、TまたはSであり;
X34は、T、Q、またはAであり;
X36は、G、S、またはTであり;
X37は、A、G、またはDであり;
X38は、S、L、またはAであり;
X39は、S、G、D、またはTであり;
X40は、A、V、またはGであり;
X41は、G、I、またはVであり;
X42は、Q、T、またはIであり;
X43は、I、V、T、またはLであり;
X44は、I、T、またはVであり;
X45は、M、またはアミノ酸不在であり;
X46は、D、またはアミノ酸不在である。
X14は、V、T、またはA、任意選択でVであり;
X17は、Q、T、またはE、任意選択でQであり;
X18は、E、Q、またはTであり;
X20は、S、N、A、またはGであり;
X21は、S、またはアミノ酸不在であり;
X22は、G、またはアミノ酸不在であり;
X25は、N、S、またはA、任意選択でNであり;
X26は、A、S、またはK、任意選択でAであり;
X28は、T、S、またはKであり;
X31は、AまたはE、任意選択でAであり;
X32は、TまたはS、任意選択でTであり;あるいは
X34は、T、Q、またはA、任意選択でTである。
1. O’Brien, K. L. et al. Burden of disease caused by Streptococcus pneumoniae in children younger than 5 years: global estimates. Lancet 374, 893−902, doi:10.1016/S0140−6736(09)61204−6 (2009).
2. Pneumococcal conjugate vaccine for childhood immunization−−WHO position paper. Wkly Epidemiol Rec 82, 93−104 (2007).
3. Prevention, C. f. D. C. a. Pneumococcal Vaccination, <https://www.cdc.gov/vaccines/vpd/pneumo/index.html>
4. Pace, D. Glycoconjugate vaccines. Expert Opin Biol Ther 13, 11−33, doi:10.1517/14712598.2012.725718 (2013).
5. Vella, M. & Pace, D. Glycoconjugate vaccines: an update. Expert Opin Biol Ther 15, 529−546, doi:10.1517/14712598.2015.993375 (2015).
6. Pollard, A. J., Perrett, K. P. & Beverley, P. C. Maintaining protection against invasive bacteria with protein−polysaccharide conjugate vaccines. Nat Rev Immunol 9, 213−220, doi:10.1038/nri2494 (2009).
7. Avci, F. Y., Li, X., Tsuji, M. & Kasper, D. L. A mechanism for glycoconjugate vaccine activation of the adaptive immune system and its implications for vaccine design. Nat Med 17, 1602−1609, doi:10.1038/nm.2535 (2011).
8. Package Insert − Prevnar 13 − FDA, <https://www.fda.gov/downloads/BiologicsBloodVaccines/Vaccines/ApprovedProducts/UCM201669.pdf>
9. Prevention, C. f. D. C. a. Vaccines for Children Program (VFC), <https://www.cdc.gov/vaccines/programs/vfc/awardees/vaccine−management/price−list/index.html> (2018).
10. Pfizer Inc. 2017 Financial Report, <https://www.sec.gov/Archives/edgar/data/78003/000007800318000027/pfe−exhibit13x12312017x10k.htm> (2018).
11. Frasch, C. E. Preparation of bacterial polysaccharide−protein conjugates: analytical and manufacturing challenges. Vaccine 27, 6468−6470, doi:10.1016/j.vaccine.2009.06.013 (2009).
12. Huttner, A. & Gambillara, V. The development and early clinical testing of the ExPEC4V conjugate vaccine against uropathogenic Escherichia coli. Clin Microbiol Infect, doi:10.1016/j.cmi.2018.05.009 (2018).
13. Huttner, A. et al. Safety, immunogenicity, and preliminary clinical efficacy of a vaccine against extraintestinal pathogenic Escherichia coli in women with a history of recurrent urinary tract infection: a randomised, single−blind, placebo−controlled phase 1b trial. Lancet Infect Dis 17, 528−537, doi:10.1016/S1473−3099(17)30108−1 (2017).
14. Riddle, M. S. et al. Safety and Immunogenicity of a Candidate Bioconjugate Vaccine against Shigella flexneri 2a Administered to Healthy Adults: a Single−Blind, Randomized Phase I Study. Clin Vaccine Immunol 23, 908−917, doi:10.1128/CVI.00224−16 (2016).
15. Apweiler, R., Hermjakob, H. & Sharon, N. On the frequency of protein glycosylation, as deduced from analysis of the SWISS−PROT database. Biochim Biophys Acta 1473, 4−8 (1999).
16. Nothaft, H. & Szymanski, C. M. Protein glycosylation in bacteria: sweeter than ever. Nat Rev Microbiol 8, 765−778, doi:10.1038/nrmicro2383 (2010).
17. Iwashkiw, J. A., Vozza, N. F., Kinsella, R. L. & Feldman, M. F. Pour some sugar on it: the expanding world of bacterial protein O−linked glycosylation. Mol Microbiol 89, 14−28, doi:10.1111/mmi.12265 (2013).
18. Ciocchini, A. E. et al. A bacterial engineered glycoprotein as a novel antigen for diagnosis of bovine brucellosis. Vet Microbiol 172, 455−465, doi:10.1016/j.vetmic.2014.04.014 (2014).
19. Garcia−Quintanilla, F., Iwashkiw, J. A., Price, N. L., Stratilo, C. & Feldman, M. F. Production of a recombinant vaccine candidate against Burkholderia pseudomallei exploiting the bacterial N−glycosylation machinery. Front Microbiol 5, 381, doi:10.3389/fmicb.2014.00381 (2014).
20. Iwashkiw, J. A. et al. Exploiting the Campylobacter jejuni protein glycosylation system for glycoengineering vaccines and diagnostic tools directed against brucellosis. Microb Cell Fact 11, 13, doi:10.1186/1475−2859−11−13 (2012).
21. Wacker, M. et al. Substrate specificity of bacterial oligosaccharyltransferase suggests a common transfer mechanism for the bacterial and eukaryotic systems. Proc Natl Acad Sci U S A 103, 7088−7093, doi:10.1073/pnas.0509207103 (2006).
22. Feldman, M. F. et al. Engineering N−linked protein glycosylation with diverse O antigen lipopolysaccharide structures in Escherichia coli. Proc Natl Acad Sci U S A 102, 3016−3021, doi:10.1073/pnas.0500044102 (2005).
23. Faridmoayer, A., Fentabil, M. A., Mills, D. C., Klassen, J. S. & Feldman, M. F. Functional characterization of bacterial oligosaccharyltransferases involved in O−linked protein glycosylation. J Bacteriol 189, 8088−8098, doi:10.1128/JB.01318−07 (2007).
24. Geno, K. A. et al. Pneumococcal Capsules and Their Types: Past, Present, and Future. Clin Microbiol Rev 28, 871−899, doi:10.1128/CMR.00024−15 (2015).
25. Ihssen, J. et al. Increased efficiency of Campylobacter jejuni N−oligosaccharyltransferase PglB by structure−guided engineering. Open Biol 5, 140227, doi:10.1098/rsob.140227 (2015).
26. Pan, C. et al. Biosynthesis of Conjugate Vaccines Using an O−Linked Glycosylation System. MBio 7, e00443−00416, doi:10.1128/mBio.00443−16 (2016).
27. Wacker, M. et al. N−linked glycosylation in Campylobacter jejuni and its functional transfer into E. coli. Science 298, 1790−1793, doi:10.1126/science.298.5599.1790 (2002).
28. Vik, A. et al. Broad spectrum O−linked protein glycosylation in the human pathogen Neisseria gonorrhoeae. Proc Natl Acad Sci U S A 106, 4447−4452, doi:10.1073/pnas.0809504106 (2009).
29. Harding, C. M. et al. Acinetobacter strains carry two functional oligosaccharyltransferases, one devoted exclusively to type IV pilin, and the other one dedicated to O−glycosylation of multiple proteins. Mol Microbiol 96, 1023−1041, doi:10.1111/mmi.12986 (2015).
30. Pan, Y. J. et al. Genetic analysis of capsular polysaccharide synthesis gene clusters in 79 capsular types of Klebsiella spp. Sci Rep 5, 15573, doi:10.1038/srep15573 (2015).
31. Kovach, M. E. et al. Four new derivatives of the broad−host−range cloning vector pBBR1MCS, carrying different antibiotic−resistance cassettes. Gene 166, 175−176 (1995).
32. Wu, K. M. et al. Genome sequencing and comparative analysis of Klebsiella pneumoniae NTUH−K2044, a strain causing liver abscess and meningitis. J Bacteriol 191, 4492−4501, doi:10.1128/JB.00315−09 (2009).
33. Lery, L. M. et al. Comparative analysis of Klebsiella pneumoniae genomes identifies a phospholipase D family protein as a novel virulence factor. BMC Biol 12, 41, doi:10.1186/1741−7007−12−41 (2014).
34. Dykxhoorn, D. M., St Pierre, R. & Linn, T. A set of compatible tac promoter expression vectors. Gene 177, 133−136 (1996).
35. Arakawa, Y. et al. Biosynthesis of Klebsiella K2 capsular polysaccharide in Escherichia coli HB101 requires the functions of rmpA and the chromosomal cps gene cluster of the virulent strain Klebsiella pneumoniae Chedid (O1:K2). Infect Immun 59, 2043−2050 (1991).
36. Yeh, K. M. et al. Capsular serotype K1 or K2, rather than magA and rmpA, is a major virulence determinant for Klebsiella pneumoniae liver abscess in Singapore and Taiwan. J Clin Microbiol 45, 466−471, doi:10.1128/JCM.01150−06 (2007).
37. Kowarik, M. et al. Definition of the bacterial N−glycosylation site consensus sequence. EMBO J 25, 1957−1966, doi:10.1038/sj.emboj.7601087 (2006).
38. Comer, J. E., Marshall, M. A., Blanch, V. J., Deal, C. D. & Castric, P. Identification of the Pseudomonas aeruginosa 1244 pilin glycosylation site. Infect Immun 70, 2837−2845 (2002).
39. Scott, N. E. et al. Diversity within the O−linked protein glycosylation systems of acinetobacter species. Mol Cell Proteomics 13, 2354−2370, doi:10.1074/mcp.M114.038315 (2014).
40. Schwarz, F. et al. A combined method for producing homogeneous glycoproteins with eukaryotic N−glycosylation. Nat Chem Biol 6, 264−266, doi:10.1038/nchembio.314 (2010).
41. Porstendorfer, D., Drotschmann, U. & Averhoff, B. A novel competence gene, comP, is essential for natural transformation of Acinetobacter sp. strain BD413. Appl Environ Microbiol 63, 4150−4157 (1997).
42. Giltner, C. L., Nguyen, Y. & Burrows, L. L. Type IV pilin proteins: versatile molecular modules. Microbiol Mol Biol Rev 76, 740−772, doi:10.1128/MMBR.00035−12 (2012).
43. Pelicic, V. Type IV pili: e pluribus unum? Mol Microbiol 68, 827−837, doi:10.1111/j.1365−2958.2008.06197.x (2008).
44. Malik, A. Protein fusion tags for efficient expression and purification of recombinant proteins in the periplasmic space of E. coli. 3 Biotech 6, 44, doi:10.1007/s13205−016−0397−7 (2016).
45. Ravenscroft, N. et al. Purification and characterization of a Shigella conjugate vaccine, produced by glycoengineering Escherichia coli. Glycobiology 26, 51−62, doi:10.1093/glycob/cwv077 (2016).
46. Schulz, B. L. et al. Identification of bacterial protein O−oligosaccharyltransferases and their glycoprotein substrates. PLoS One 8, e62768, doi:10.1371/journal.pone.0062768 (2013).
47. Castric, P. pilO, a gene required for glycosylation of Pseudomonas aeruginosa 1244 pilin. Microbiology 141 ( Pt 5), 1247−1254, doi:10.1099/13500872−141−5−1247 (1995).
48. Power, P. M. et al. Genetic characterization of pilin glycosylation and phase variation in Neisseria meningitidis. Mol Microbiol 49, 833−847 (2003).
49. Stimson, E. et al. Meningococcal pilin: a glycoprotein substituted with digalactosyl 2,4−diacetamido−2,4,6−trideoxyhexose. Mol Microbiol 17, 1201−1214 (1995).
50. Ishihama, Y., Rappsilber, J. & Mann, M. Modular stop and go extraction tips with stacked disks for parallel and multidimensional Peptide fractionation in proteomics. J Proteome Res 5, 988−994, doi:10.1021/pr050385q (2006).
51. Rappsilber, J., Mann, M. & Ishihama, Y. Protocol for micro−purification, enrichment, pre−fractionation and storage of peptides for proteomics using StageTips. Nature protocols 2, 1896−1906, doi:10.1038/nprot.2007.261 (2007).
52. Roepstorff, P. & Fohlman, J. Proposal for a common nomenclature for sequence ions in mass spectra of peptides. Biomed Mass Spectrom 11, 601, doi:10.1002/bms.1200111109 (1984).
53. Haurat, M. F. et al. Selective sorting of cargo proteins into bacterial membrane vesicles. J Biol Chem 286, 1269−1276, doi:10.1074/jbc.M110.185744 (2011).
54. Price, N. L. et al. Glycoengineered Outer Membrane Vesicles: A Novel Platform for Bacterial Vaccines. Sci Rep 6, 24931, doi:10.1038/srep24931 (2016).
55. Kay, E. J., Yates, L. E., Terra, V. S., Cuccui, J. & Wren, B. W. Recombinant expression of Streptococcus pneumoniae capsular polysaccharides in Escherichia coli. Open Biol 6, 150243, doi:10.1098/rsob.150243 (2016).
Claims (80)
- 融合タンパク質に共有結合したオリゴ糖または多糖を含み、前記融合タンパク質が、ComPタンパク質(ComP)またはそのグリコシル化タグフラグメントを含む、バイオコンジュゲート。
- 前記融合タンパク質が、配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基において、前記ComPタンパク質上のオリゴ糖もしくは多糖、またはそのグリコシル化タグフラグメントでグリコシル化される、請求項1に記載のバイオコンジュゲート。
- 前記ComPタンパク質が、配列番号7(ComPΔ28ADP1)に対して、少なくとも50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%、または100%同一であるアミノ酸配列を含み、配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基を含み;任意選択で、
前記ComPタンパク質が、配列番号7(ComPΔ28ADP1)、配列番号8(ComPΔ28110264)、配列番号9(ComPΔ28GFJ−2)、配列番号10(ComPΔ28P50v1)、配列番号11(ComPΔ284466)、または配列番号12(ComPΔ28SFC)を含む、請求項1または2に記載のバイオコンジュゲート。 - 前記ComPタンパク質が、配列番号1(ComPADP1:AAC45886.1)に対して、少なくとも50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%、または100%同一であるアミノ酸配列を含み、配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基を含む、請求項1〜3のいずれか1項に記載のバイオコンジュゲート。
- 前記ComPタンパク質が、配列番号1(ComPADP1:AAC45886.1)、配列番号2(ComP110264:ENV58402.1)、配列番号3(ComPGFJ−2:APV36638.1)、配列番号4(ComP50v1:PKD82822.1)、配列番号5(ComP4466:SNX44537.1)、または配列番号6(ComPSFC:OAL75955.1)を含む、請求項1〜4のいずれか1項に記載のバイオコンジュゲート。
- 前記ComPタンパク質のグリコシル化タグフラグメントが、配列番号1(ComPADP1:AAC45886.1)のアミノ酸残基1から28に対応するアミノ酸残基を欠くComPΔ28ポリペプチドである、請求項1〜5のいずれか1項に記載のバイオコンジュゲート。
- 前記ComPΔ28ポリペプチドが、配列番号7、配列番号8、配列番号9、配列番号10、配列番号11、及び配列番号12からなる群から選択される、請求項6に記載のバイオコンジュゲート。
- 前記ComPタンパク質のグリコシル化タグフラグメントが、アルファベータループをベータストランド領域に接続するジスルフィド結合に隣接する、配列番号2(ComP110264:ENV58402.1)の82位のセリン残基を含む、配列番号2(ComP110264:ENV58402.1)の領域に対応する領域を含む、請求項1〜7のいずれか1項に記載のバイオコンジュゲート。
- 前記ComPタンパク質のグリコシル化タグフラグメントが、VGVQEISASNATTNVATAT(配列番号39)、TGVTQIASGASAATTNVASAQ(配列番号40)、VGVQEINASSSTSNVATAT(配列番号41)、AGVETIGASNKTKNVESAA(配列番号42)、VGVQTIAASNATKNVATAT(配列番号43)、及びNGVISASATTNVASSA(配列番号44)からなる群から選択されるアミノ酸配列に対して、少なくとも70%、75%、80%、85%、90%、95%、96%、97%、98%、99%、または100%同一であるアミノ酸配列を含み、前記グリコシル化タグフラグメントが、配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基を含み、任意選択で、
前記ComPタンパク質のグリコシル化タグフラグメントが、VGVQEISASNATTNVATAT(配列番号39)、TGVTQIASGASAATTNVASAQ(配列番号40)、VGVQEINASSSTSNVATAT(配列番号41)、AGVETIGASNKTKNVESAA(配列番号42)、VGVQTIAASNATKNVATAT(配列番号43)、及びNGVISASATTNVASSA(配列番号44)、または1、2、3、4、5、6、または7個のアミノ酸置換、付加、及び/または欠失を有するそれらのバリアントからなる群から選択されるアミノ酸配列を含み、前記バリアントが配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基を維持する、請求項1〜8のいずれか1項に記載のバイオコンジュゲート。 - 前記ComPタンパク質のグリコシル化タグフラグメントが、配列番号37のアミノ酸コンセンサス配列、または少なくとも5、10、15、20、30、35、または40個の連続的なアミノ酸のそのフラグメントを含み、前記グリコシル化タグフラグメントが、配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基を含み、または前記グリコシル化タグが、1、2、3、4、5、6、または7個のアミノ酸置換、付加、及び/または欠失を有するそれらのバリアントを含み、前記バリアントが配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基を維持する、請求項1〜8のいずれか1項に記載のバイオコンジュゲート。
- 前記ComPタンパク質のグリコシル化タグフラグメントが、配列番号38、もしくは配列番号45のアミノ酸コンセンサス配列、または少なくとも5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、または20個の連続的なアミノ酸のそのフラグメントを含み、前記グリコシル化タグフラグメントが、配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基を含み、または前記グリコシル化タグが、1、2、3、4、5、6、または7個のアミノ酸置換、付加、及び/または欠失を有するそれらのバリアントを含み、前記バリアントが配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基を維持する、請求項1〜8のいずれか1項に記載のバイオコンジュゲート。
- 前記オリゴ糖または多糖が、ストレプトコッカス属の細菌によって産生され、任意選択で、
前記多糖が、S.pneumoniae、S.agalactiae、またはS.suisのきょう膜多糖である、請求項1〜11のいずれか1項に記載のバイオコンジュゲート。 - 前記きょう膜多糖が、CPS14、CPS8、CPS9V、またはCPS15bである、請求項12に記載のバイオコンジュゲート。
- 前記オリゴ糖または多糖が、クレブシエラ属の細菌によって産生され、任意選択で、
前記オリゴ糖または多糖が、Klebsiella pneumoniae、Klebsiella varricola、Klebsiella michinganenis、またはKlebsiella oxytocaのきょう膜多糖である、請求項1〜11のいずれか1項に記載のバイオコンジュゲート。 - 前記多糖が、Klebsiella pneumoniaeのきょう膜多糖である、請求項1〜11のいずれか1項に記載のバイオコンジュゲート。
- 前記多糖が、Klebsiella pneumoniaeの血清型K1または血清型K2のきょう膜多糖である、請求項15に記載のバイオコンジュゲート。
- 前記オリゴ糖または多糖が、その還元末端にグルコースを含む、請求項1〜16のいずれか1項に記載のバイオコンジュゲート。
- 前記バイオコンジュゲートが、in vivoで産生され、任意選択で、
細菌細胞で産生される、請求項1〜17のいずれか1項に記載のバイオコンジュゲート。 - 前記融合タンパク質が、ジフテリアトキソイドCRM197、破傷風トキソイド、Pseudomonas aeruginosa外毒素A(EPA)、破傷風毒素Cフラグメント、コレラ毒素Bサブユニット、及びHaemophilus influenzaタンパク質D、またはそれらのフラグメントからなる群から選択される担体タンパク質を含む、請求項1〜18のいずれか1項に記載のバイオコンジュゲート。
- 前記ComPタンパク質またはそのグリコシル化タグフラグメントが前記融合タンパク質のN末端、前記融合タンパク質のC末端、及び/または前記融合タンパク質の内部に位置し、任意選択で、
前記担体タンパク質またはそのフラグメントが、アミノ酸リンカーを介して、前記ComPタンパク質またはそのグリコシル化タグフラグメントに連結する、請求項19に記載のバイオコンジュゲート。 - 前記融合タンパク質が、2以上、3以上、4以上、5以上、6以上、8以上、10以上、15以上、または20以上のComPタンパク質のグリコシル化タグフラグメントを含む、請求項1〜20のいずれか1項に記載のバイオコンジュゲート。
- 前記融合タンパク質が、2、3、4、5、6、7、8、9、10、11、12、13、14、15、または20のうちのいずれかから3、4、5、6、7、8、9、10、11、12、13、14、15、20、または25のうちのいずれかまでのComPタンパク質のグリコシル化タグフラグメントを含む、請求項1〜20のいずれか1項に記載のバイオコンジュゲート。
- 前記バイオコンジュゲートが、コンジュゲートワクチンであり、任意選択で、
前記コンジュゲートワクチンが、Streptococcus pneumoniae血清型8に対するワクチンである、請求項1〜22のいずれか1項に記載のバイオコンジュゲート。 - 前記コンジュゲートワクチンが対象に投与され、それは免疫応答を誘導する、請求項23に記載のバイオコンジュゲート。
- 前記免疫応答が、長期記憶(メモリーB細胞及びメモリーT細胞)を誘発し、抗体応答であり、任意選択で、血清型特異的免疫応答である、請求項23または24に記載のバイオコンジュゲート。
- 前記抗体応答がIgGまたはIgM応答である、請求項25に記載のバイオコンジュゲート。
- 前記抗体応答がIgG応答であり、任意選択でIgG1応答である、請求項26に記載のバイオコンジュゲート。
- 前記コンジュゲートワクチンが、ワクチンを投与された対象において免疫学的記憶を生成する、請求項23〜27のいずれか1項に記載のバイオコンジュゲート。
- ComPグリコシル化タグであって、ComPタンパク質の単離されたフラグメントを含み、前記フラグメントが、配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基を含む、前記グリコシル化タグ。
- 前記フラグメントが、ComPタンパク質の少なくとも5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27を含む、28、29、30、35、または40のアミノ酸を含む、請求項29に記載のグリコシル化タグ。
- 前記ComPタンパク質が、請求項3〜5のいずれか1項に記載のComPタンパク質である、請求項29または30に記載のグリコシル化タグ。
- 異種担体タンパク質に結合している請求項29〜31のいずれか1項に記載のグリコシル化タグ。
- 前記異種担体タンパク質が、ジフテリアトキソイドCRM197、破傷風トキソイド、Pseudomonas aeruginosa外毒素A(EPA)、破傷風毒素Cフラグメント、コレラ毒素Bサブユニット、及びHaemophilus influenzaタンパク質D、またはそれらのフラグメントからなる群から選択される、請求項32に記載のグリコシル化タグ。
- 前記グリコシル化タグが、配列番号1(ComPADP1:AAC45886.1)のアミノ酸残基1から28に対応するアミノ酸残基を欠くComPΔ28ポリペプチドである、請求項29〜33のいずれか1項に記載のグリコシル化タグ。
- 前記ComPΔ28ポリペプチドが、配列番号7(ComPΔ28ADP1)、配列番号8(ComPΔ28110264)、配列番号9(ComPΔ28GFJ−2)、配列番号10(ComPΔ28P50v1)、配列番号11(ComPΔ284466)、及び配列番号12(ComPΔ28SFC)からなる群から選択される、請求項34に記載のグリコシル化タグ。
- 前記グリコシル化タグが、アルファベータループをベータストランド領域に接続するジスルフィド結合に隣接する、配列番号2(ComP110264:ENV58402.1)の82位のセリン残基を含む、配列番号2(ComP110264:ENV58402.1)の領域に対応する領域を含む、請求項29〜35のいずれか1項に記載のグリコシル化タグ。
- 前記グリコシル化タグが、VGVQEISASNATTNVATAT(配列番号39)、TGVTQIASGASAATTNVASAQ(配列番号40)、VGVQEINASSSTSNVATAT(配列番号41)、AGVETIGASNKTKNVESAA(配列番号42)、VGVQTIAASNATKNVATAT(配列番号43)、及びNGVISASATTNVASSA(配列番号44)からなる群から選択されるアミノ酸配列に対して、少なくとも80%、85%、90%、95%、96%、97%、98%、99%、または100%同一であるアミノ酸配列を含み、前記グリコシル化タグが、配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基を含み、任意選択で、
前記グリコシル化タグが、VGVQEISASNATTNVATAT(配列番号39)、TGVTQIASGASAATTNVASAQ(配列番号40)、VGVQEINASSSTSNVATAT(配列番号41)、AGVETIGASNKTKNVESAA(配列番号42)、VGVQTIAASNATKNVATAT(配列番号43)、及びNGVISASATTNVASSA(配列番号44)、または1、2、3、4、5、6、または7個のアミノ酸置換、付加、及び/または欠失を有するそれらのバリアントからなる群から選択されるアミノ酸配列を含み、前記バリアントが配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基を維持する、請求項29〜36のいずれか1項に記載のバイオコンジュゲート。 - 前記グリコシル化タグが、配列番号37のアミノ酸コンセンサス配列、または少なくとも5、10、15、20、30、35、または40個の連続的なアミノ酸のそのフラグメントを含み、前記グリコシル化タグフラグメントが、配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基を含み、または前記グリコシル化タグが、1、2、3、4、5、6、または7個のアミノ酸置換、付加、及び/または欠失を有するそれらのバリアントを含み、前記バリアントが配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基を維持する、請求項29〜36のいずれか1項に記載のグリコシル化タグ。
- 前記グリコシル化タグが、配列番号38、もしくは配列番号45のアミノ酸コンセンサス配列、または少なくとも5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、または20個の連続的なアミノ酸のそのフラグメントを含み、前記グリコシル化タグフラグメントが、配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基を含み、または前記グリコシル化タグが、1、2、3、4、5、6、または7個のアミノ酸置換、付加、及び/または欠失を有するそれらのバリアントを含み、前記バリアントが配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基を維持する、請求項29〜36のいずれか1項に記載のグリコシル化タグ。
- 前記グリコシル化タグが、配列番号1(ComPADP1:AAC45886.1)のアミノ酸残基24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、または82のうちのいずれかから、配列番号1(ComPADP1:AAC45886.1)のアミノ酸残基86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、または147のうちのいずれかまでに対応するComPタンパク質アミノ酸配列を含み、任意選択で、
前記グリコシル化タグが、配列番号1(ComPADP1:AAC45886.1)のアミノ酸残基24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、または82のうちのいずれかから、配列番号1(ComPADP1:AAC45886.1)のアミノ酸残基86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、または147のうちのいずれかまでを含むアミノ酸配列を含む、請求項29に記載のグリコシル化タグ。 - 前記グリコシル化タグが、124、120、115、110、100、90、80、75、70、60、50、40、30、25、20、15、10、または5アミノ酸以下の長さである、請求項28〜40のいずれか1項に記載のグリコシル化タグ。
- 前記グリコシル化タグが、配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するセリン残基でオリゴ糖または多糖に共有結合している、請求項28〜41のいずれか1項に記載のグリコシル化タグ。
- 前記オリゴ糖または多糖が、ストレプトコッカス属の細菌によって産生され、任意選択で、
前記多糖が、S.pneumoniae、S.agalactiae、またはS.suisのきょう膜多糖である、請求項42に記載のグリコシル化タグ。 - 前記きょう膜多糖が、CPS14、CPS8、CPS9V、またはCPS15bである、請求項43に記載のグリコシル化タグ。
- 前記オリゴ糖または多糖が、クレブシエラ属の細菌によって産生され、任意選択で、
前記オリゴ糖または多糖が、Klebsiella pneumoniae、Klebsiella varricola、Klebsiella michinganenis、またはKlebsiella oxytocaのきょう膜多糖である、請求項42に記載のグリコシル化タグ。 - 前記多糖が、Klebsiella pneumoniaeのきょう膜多糖である、請求項45に記載のグリコシル化タグ。
- 前記多糖が、Klebsiella pneumoniaeの血清型K1または血清型K2のきょう膜多糖である、請求項46に記載のグリコシル化タグ。
- 前記オリゴ糖または多糖が、その還元末端にグルコースを含む、請求項42〜47のいずれか1項に記載のグリコシル化タグ。
- 請求項29〜48のいずれか1項に記載のComPグリコシル化タグを含む融合タンパク質であって、任意選択で、
配列番号1(ComPADP1:AAC45886.1)の84位の保存されたセリン残基に対応するグリコシル化タグのセリン残基でグリコシル化されている、前記融合タンパク質。 - ジフテリアトキソイドCRM197、破傷風トキソイド、Pseudomonas aeruginosa外毒素A(EPA)、破傷風毒素Cフラグメント、コレラ毒素Bサブユニット、及びHaemophilus influenzaタンパク質D、またはそれらのフラグメントからなる群から選択される担体タンパク質を含む、請求項49に記載の融合タンパク質。
- アミノ酸リンカー配列を含む、請求項49または50に記載の融合タンパク質。
- オリゴ糖または多糖をアクセプターポリペプチドにin vivoで結合する方法であって、PglSオリゴサッカリルトランスフェラーゼ(OTase)によって、前記オリゴ糖または多糖を前記アクセプターポリペプチドに共有結合させることを含み、前記アクセプターポリペプチドがComPタンパク質またはそのグリコシル化タグフラグメントを含み、任意選択で、
前記ComPタンパク質またはそのグリコシル化タグフラグメントが、異種担体タンパク質に連結されている、前記方法。 - 前記PglS OTaseが、PglS110264、PglSADP1、PglSGFJ−2、PglS50v1、PglS4466、またはPglSSFCであり、任意選択で、
前記ComPタンパク質が、ComP110264、ComPADP1、ComPGFJ−2、ComP50v1、ComP4466、またはComPSFCであり、任意選択で、
前記PglS OTaseが、PglSADP1である、請求項52に記載の方法。 - 前記PglS OTaseがPglSADP1であるが、前記ComPタンパク質がComPADP1ではなく、任意選択で、
前記ComPタンパク質がComP110264である、請求項53に記載の方法。 - 前記オリゴ糖または多糖が、配列番号1(ComPADP1:AAC45886.1)の84位のセリン残基に対応するセリン残基において、前記ComPタンパク質またはそのグリコシル化タグフラグメントに連結している、請求項54に記載の方法。
- 前記in vivoのコンジュゲーションが宿主細胞内で起こる、請求項52〜54のいずれか1項に記載の方法。
- 前記細胞が細菌細胞である、請求項56に記載の方法。
- 前記細菌宿主細胞がE.coliである、請求項57に記載の方法。
- 以下を含む宿主細胞を培養することを含む、請求項54〜56のいずれか1項に記載の方法:(a)前記オリゴ糖または多糖を合成するために必要なタンパク質をコードする遺伝子クラスター;(b)PglS OTase;及び(3)前記アクセプターポリペプチド。
- 前記オリゴ糖または多糖の産生が、K. pneumoniaeの転写活性化因子rmpA(K.pneumoniae NTUH K−2044)、またはK.pneumoniaeの転写活性化因子rmpA(K.pneumoniae NTUH K−2044)の相同体によって増強される、請求項52〜59のいずれか1項に記載の方法。
- 前記担体タンパク質が、ジフテリアトキソイドCRM197、破傷風トキソイド、Pseudomonas aeruginosa外毒素A(EPA)、破傷風毒素Cフラグメント、コレラ毒素Bサブユニット、及びHaemophilus influenzaタンパク質D、またはそれらのフラグメントからなる群から選択される、請求項52〜60のいずれか1項に記載の方法。
- 前記方法がコンジュゲートワクチンを産生する、請求項52〜61のいずれか1項に記載の方法。
- 宿主細胞であって、(a)オリゴ糖または多糖を合成するために必要なタンパク質をコードする遺伝子クラスター;(b)PglS OTase;及び(3)ComPタンパク質またはそのグリコシル化タグフラグメントを含むアクセプターポリペプチドを含む、前記宿主細胞。
- 前記アクセプターポリペプチドが融合タンパク質である、請求項63に記載の宿主細胞。
- 前記宿主細胞がPglS OTaseをコードする核酸を含む、請求項63または64に記載の宿主細胞。
- 前記宿主細胞が、前記アクセプターポリペプチドをコードする核酸を含む、請求項63〜65のいずれか1項に記載の宿主細胞。
- 請求項29〜48のいずれか1項に記載のComPグリコシル化タグ、及び/または請求項49〜51のいずれか1項に記載の融合タンパク質をコードし、任意選択で
ベクターである、単離された核酸。 - 請求項67に記載の単離された核酸を含む、宿主細胞。
- 請求項23〜28のいずれか1項に記載のコンジュゲートワクチンまたは請求項48〜51のいずれか1項に記載の融合タンパク質、及びアジュバントを含む組成物。
- 細菌性病原体に対する宿主免疫応答を誘導する方法であって、免疫応答を必要とする対象に、請求項23〜28のいずれか1項に記載のコンジュゲートワクチン、請求項49〜51のいずれか1項に記載の融合タンパク質、または請求項69に記載の組成物の有効量を投与することを含む、前記方法。
- 前記免疫応答が抗体応答である、請求項70に記載の方法。
- 前記免疫応答が、先天性応答、適応応答、体液性応答、抗体応答、細胞媒介性応答、B細胞応答、T細胞応答、サイトカインの上方制御または下方制御、免疫系クロストーク、及び前記免疫応答の2つ以上の組み合わせからなる群から選択される、請求項71に記載の方法。
- 前記免疫応答が、先天性応答、体液性応答、抗体応答、T細胞応答、及び前記免疫応答の2つ以上の組み合わせからなる群から選択される、請求項72に記載の方法。
- 対象における細菌性疾患及び/または感染症を予防または治療する方法であって、それを必要とする対象に、請求項23〜28のいずれか1項に記載のコンジュゲートワクチン、請求項49〜51のいずれか1項に記載の融合タンパク質、または請求項69に記載の組成物を投与することを含む、前記方法。
- 前記感染症が、皮膚、軟部組織、血液、または臓器の局所的または全身的な感染症であるか、または本質的に自己免疫性である、請求項73に記載の方法。
- 前記疾患が肺炎である、請求項74に記載の方法。
- 前記感染症が、全身感染症及び/または血液の感染症である、請求項75に記載の方法。
- 前記対象がヒトである、請求項70〜77のいずれか1項に記載の方法。
- 前記組成物が、筋肉内注射、皮内注射、腹腔内注射、皮下注射、静脈内注射、経口投与、粘膜投与、鼻腔内投与、または肺投与を介して投与される、請求項70〜78のいずれか1項に記載の方法。
- 肺炎球菌感染症に対する肺炎球菌コンジュゲートワクチンを製造する方法であって、
(a)請求項23〜28のいずれか1項に記載のバイオコンジュゲートまたは請求項49〜51のいずれか1項に記載のグリコシル化融合タンパク質を単離することと、
(b)前記単離されたコンジュゲートワクチンまたは単離されたグリコシル化融合タンパク質をアジュバントと組み合わせることと、を含む、前記方法。
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2024061674A JP2024079845A (ja) | 2018-06-16 | 2024-04-05 | グリコシル化されたComPピリンバリアント、製造方法及びその使用 |
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201862685970P | 2018-06-16 | 2018-06-16 | |
| US62/685,970 | 2018-06-16 | ||
| US201862783971P | 2018-12-21 | 2018-12-21 | |
| US62/783,971 | 2018-12-21 | ||
| PCT/US2019/037251 WO2019241672A2 (en) | 2018-06-16 | 2019-06-14 | Glycosylated comp pilin variants, methods of making and uses thereof |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2024061674A Division JP2024079845A (ja) | 2018-06-16 | 2024-04-05 | グリコシル化されたComPピリンバリアント、製造方法及びその使用 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2021528059A true JP2021528059A (ja) | 2021-10-21 |
| JPWO2019241672A5 JPWO2019241672A5 (ja) | 2022-06-22 |
Family
ID=68843212
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2020569986A Pending JP2021528059A (ja) | 2018-06-16 | 2019-06-14 | グリコシル化されたComPピリンバリアント、製造方法及びその使用 |
| JP2024061674A Pending JP2024079845A (ja) | 2018-06-16 | 2024-04-05 | グリコシル化されたComPピリンバリアント、製造方法及びその使用 |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2024061674A Pending JP2024079845A (ja) | 2018-06-16 | 2024-04-05 | グリコシル化されたComPピリンバリアント、製造方法及びその使用 |
Country Status (6)
| Country | Link |
|---|---|
| US (2) | US11932670B2 (ja) |
| EP (1) | EP3806881A4 (ja) |
| JP (2) | JP2021528059A (ja) |
| AU (1) | AU2019287659A1 (ja) |
| CA (1) | CA3103474A1 (ja) |
| WO (1) | WO2019241672A2 (ja) |
Families Citing this family (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2016134485A1 (en) | 2015-02-26 | 2016-09-01 | Vaxalta Inc. | Acinetobacter o-oligosaccharyltransferases and uses thereof |
| US10729763B2 (en) | 2017-06-10 | 2020-08-04 | Inventprise, Llc | Mixtures of polysaccharide-protein pegylated compounds |
| US11932670B2 (en) | 2018-06-16 | 2024-03-19 | Vaxnewmo Llc | Glycosylated ComP pilin variants, methods of making and uses thereof |
| US20220088211A1 (en) * | 2018-12-21 | 2022-03-24 | Vaxnewmo Llc | O-linked glycosylation recognition motifs |
| WO2022232256A1 (en) * | 2021-04-28 | 2022-11-03 | Vaxnewmo Llc | Minimal sequons sufficient for o-linking glycosylation |
Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2015533511A (ja) * | 2012-11-07 | 2015-11-26 | グリコヴァキシン アーゲー | 酵素的コンジュゲート化による大腸菌での組み換えワクチンの製造 |
| JP2018507693A (ja) * | 2015-02-26 | 2018-03-22 | ヴァックスニューモ エルエルシーVaxnewmo Llc | アシネトバクター(Acinetobacter)O−オリゴサッカリルトランスフェラーゼおよびその使用 |
Family Cites Families (9)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| ES2652416T3 (es) | 2006-12-13 | 2018-02-02 | The Governors Of The University Of Alberta | Métodos y sistemas para la O-glicosilación de proteínas |
| WO2011109600A1 (en) | 2010-03-05 | 2011-09-09 | Alnylam Pharmaceuticals, Inc. | Compositions and methods for modifying the glycosylation pattern of a polypeptide |
| CN112980907A (zh) | 2011-11-04 | 2021-06-18 | 康奈尔大学 | 一种用于糖蛋白合成的基于原核生物的无细胞系统 |
| CA2887133C (en) | 2012-10-12 | 2022-05-03 | Glycovaxyn Ag | Methods of host cell modification |
| US9499614B2 (en) | 2013-03-14 | 2016-11-22 | Abbvie Inc. | Methods for modulating protein glycosylation profiles of recombinant protein therapeutics using monosaccharides and oligosaccharides |
| WO2016044773A1 (en) | 2014-09-18 | 2016-03-24 | University Of Maryland, Baltimore | Broad spectrum conjugate vaccine to prevent klebsiella pneumoniae and pseudomonas aeruginosa infections |
| MX2017008817A (es) | 2014-12-30 | 2017-10-24 | Glaxosmithkline Biologicals Sa | Composiciones y metodos para la glicosilacion de proteinas. |
| US11932670B2 (en) | 2018-06-16 | 2024-03-19 | Vaxnewmo Llc | Glycosylated ComP pilin variants, methods of making and uses thereof |
| US20220088211A1 (en) | 2018-12-21 | 2022-03-24 | Vaxnewmo Llc | O-linked glycosylation recognition motifs |
-
2019
- 2019-06-14 US US17/251,994 patent/US11932670B2/en active Active
- 2019-06-14 AU AU2019287659A patent/AU2019287659A1/en active Pending
- 2019-06-14 JP JP2020569986A patent/JP2021528059A/ja active Pending
- 2019-06-14 CA CA3103474A patent/CA3103474A1/en active Pending
- 2019-06-14 EP EP19820087.5A patent/EP3806881A4/en active Pending
- 2019-06-14 WO PCT/US2019/037251 patent/WO2019241672A2/en unknown
-
2024
- 2024-03-14 US US18/605,436 patent/US20240262870A1/en active Pending
- 2024-04-05 JP JP2024061674A patent/JP2024079845A/ja active Pending
Patent Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2015533511A (ja) * | 2012-11-07 | 2015-11-26 | グリコヴァキシン アーゲー | 酵素的コンジュゲート化による大腸菌での組み換えワクチンの製造 |
| JP2018507693A (ja) * | 2015-02-26 | 2018-03-22 | ヴァックスニューモ エルエルシーVaxnewmo Llc | アシネトバクター(Acinetobacter)O−オリゴサッカリルトランスフェラーゼおよびその使用 |
Non-Patent Citations (1)
| Title |
|---|
| HARDING, C. M. ET AL.: "Acinetobacter strains carry two functional oligosaccharyltransferases, one devoted exclusively to ty", MOLECULAR MICROBIOLOGY, vol. 96, no. 5, JPN6023021512, 2015, pages 1023 - 1041, XP009506444, ISSN: 0005070930, DOI: 10.1111/mmi.12986 * |
Also Published As
| Publication number | Publication date |
|---|---|
| EP3806881A2 (en) | 2021-04-21 |
| WO2019241672A3 (en) | 2020-02-06 |
| CA3103474A1 (en) | 2019-12-19 |
| JP2024079845A (ja) | 2024-06-11 |
| EP3806881A4 (en) | 2022-07-27 |
| US20210324017A1 (en) | 2021-10-21 |
| US11932670B2 (en) | 2024-03-19 |
| US20240262870A1 (en) | 2024-08-08 |
| WO2019241672A2 (en) | 2019-12-19 |
| AU2019287659A1 (en) | 2021-01-07 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US11932670B2 (en) | Glycosylated ComP pilin variants, methods of making and uses thereof | |
| JP6339366B2 (ja) | 莢膜グラム陽性菌のバイオコンジュゲートワクチン | |
| US20190248841A1 (en) | Immunogenic Composition | |
| WO2017067964A1 (en) | P. aeruginosa pcrv-linked antigen vaccines | |
| US20220088211A1 (en) | O-linked glycosylation recognition motifs | |
| US20220242920A1 (en) | Immunogenic composition | |
| US11819544B2 (en) | Immunogenic composition | |
| JP2023531059A (ja) | 肺炎桿菌o抗原ワクチン | |
| CN112153980A (zh) | 包含葡萄球菌抗原的免疫原性组合物 | |
| JP7551618B2 (ja) | O-結合型グリコシル化のための修飾キャリアタンパク質 | |
| US12048743B2 (en) | Glyconjugate vaccines | |
| JP2024517754A (ja) | O-結合型グリコシル化に十分な最小シークオン | |
| WO2023118033A1 (en) | Vaccine | |
| Mayer et al. | A platform for glycoengineering a polyvalent pneumococcal bioconjugate vaccine using E. coli as a host | |
| WO2024175620A1 (en) | Immunogenic composition | |
| WO2024182291A2 (en) | Compositions and methods for producing glycoconjugate polypeptides having isopeptide bonds with a second polypeptide partner and uses thereof |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20220614 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20220614 |
|
| A977 | Report on retrieval |
Free format text: JAPANESE INTERMEDIATE CODE: A971007 Effective date: 20230419 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20230530 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20230829 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20231205 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20240405 |
|
| A911 | Transfer to examiner for re-examination before appeal (zenchi) |
Free format text: JAPANESE INTERMEDIATE CODE: A911 Effective date: 20240419 |
|
| A912 | Re-examination (zenchi) completed and case transferred to appeal board |
Free format text: JAPANESE INTERMEDIATE CODE: A912 Effective date: 20240517 |