JP2010528653A - 植物における非生物的ストレスに対する抵抗性 - Google Patents
植物における非生物的ストレスに対する抵抗性 Download PDFInfo
- Publication number
- JP2010528653A JP2010528653A JP2010511137A JP2010511137A JP2010528653A JP 2010528653 A JP2010528653 A JP 2010528653A JP 2010511137 A JP2010511137 A JP 2010511137A JP 2010511137 A JP2010511137 A JP 2010511137A JP 2010528653 A JP2010528653 A JP 2010528653A
- Authority
- JP
- Japan
- Prior art keywords
- plant
- rks
- gene
- rks4
- plants
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
Images



Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8241—Phenotypically and genetically modified plants via recombinant DNA technology
- C12N15/8261—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield
- C12N15/8271—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield for stress resistance, e.g. heavy metal resistance
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8241—Phenotypically and genetically modified plants via recombinant DNA technology
- C12N15/8261—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield
- C12N15/8271—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield for stress resistance, e.g. heavy metal resistance
- C12N15/8273—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield for stress resistance, e.g. heavy metal resistance for drought, cold, salt resistance
Landscapes
- Genetics & Genomics (AREA)
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Organic Chemistry (AREA)
- Biomedical Technology (AREA)
- Chemical & Material Sciences (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- Molecular Biology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Microbiology (AREA)
- Physics & Mathematics (AREA)
- Plant Pathology (AREA)
- Biophysics (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Cell Biology (AREA)
- Breeding Of Plants And Reproduction By Means Of Culturing (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
本発明は、植物または植物細胞に非生物的ストレスに対する耐性を与えるための方法に関する。これは、RKSタンパク質をコードしている遺伝子(特にRKSサブグループIIタンパク質をコードしている遺伝子、さらに具体的にはRKS1、RKS4もしくは切断されたRKS4、またはRKSサブグループIIIの遺伝子、より好ましくはRKS12)を導入することによってなされる。RKS遺伝子の過剰発現の効果は、当該植物をブラシノステロイド化合物でさらに処理することによって促進され得る。
【選択図】なし
【選択図】なし
Description
本発明は、植物遺伝学、特に非生物的ストレスに対して自身の耐性を増加させた遺伝子工学植物の分野に関する。
植物は、自身の環境(の変化)に効率的かつ速やかに適応することを強いられる高度な柔軟性を持つ生命体である。移動ができないため、植物は、厳しい環境下であっても成長が可能な、形態的かつ生理学的な戦略を進化させた。しかし、環境的な適応は、農業従事者または消費者が望む最適な経済的特性とは必ずしも調和しない。特定の環境に完全に適応した植物は、相対的に低収量もしくは低栄養価であり、または鑑賞植物特性に欠けることが多い。逆に、農業従事者および消費者のニーズに適するように設計された大規模に繁殖された変種は、環境の状態および/または環境の変化の影響を受けることが多い。
非生物的ストレスまたは環境ストレスは、生命体を介さない他の方法で植物にもたらされるストレスである。非生物的ストレスの例として、高塩分、浸透圧ストレス、酸化的ストレス、(極度の)高温および(極度の)低温、ならびに乾燥といった環境条件が挙げられる。これらのストレスが原因となる米、とうもろこし(maize、corn)および小麦などの主要農作物の作物損失および作物収量損失は、重要な経済的および政治的要因であり、かつ多くの発展途上国の食料不足の一因となる。
植物は、一般的に、自身のライフサイクルの間、減少した環境水量の条件に曝露されている。植物の多くは、これらの乾燥状態から自身を保護するための戦略を進化させてきた。しかし、乾燥状態の過酷さおよび期間が過度である場合には、植物の発育、多くの作物の成長および収穫高への影響は深刻である。さらに、作物の多くは、土壌のより高い塩分濃度に非常に影響を受けやすい。乾燥および高塩分濃度に継続して曝露されることは、植物の代謝に大きな変化が生じる原因となる。同様の変化は、極端な高温または低温への長期にわたる曝露によっても認められ得る。代謝のこのような大きな変化は、究極的には細胞死、そしてその結果として収量損失の原因となる。
1979年、ブラシノリドと名付けられた新しい植物成長促進因子が、セイヨウアブラナ(アブラナ属セイヨウアブラナ(Brassica napus))の花粉から分離され、新タイプのステロイドラクトンとして同定された。ブラシノリド様ステロイド化合物(ブラシノステロイドと呼ばれる)は、試験された植物種全てに非常に低濃度で生じ、植物を生物的ストレスおよび非生物的ストレスの両方に抵抗するよう適応させる機能を持つことが分かった(総説として非特許文献1を参照のこと)。ブラシノリドの生理的作用に関する初期の研究は、この特定の因子は、(i)低温で植苗の発芽および成長を促進し、(ii)細胞の表面上の、樹皮の微小管およびセルロースのマイクロフィラメントを縦に配列させることにより、細胞のサイズおよび細胞伸長の増加を促進し、(iii)導管要素の増殖により、木部分化を促進し、(iv)結果として、植物およびそれらの果実の乾燥重量が著しく増加し、(v)葉の広がりおよび拡張を促進し、(vi)オーキシンに誘導された細胞成長に特徴付けられる、H+輸送および膜過分極を誘導し、(vii)クラウンゴール腫瘍細胞の分裂および茎の放射状の成長を抑制し、(viii)明所で生育する植物のアントシアニン生成を抑制し、(ix)例えば、暗所でサイトカイニンにより誘導される脱黄化を抑制し、(x)暗所で細胞組織の老化を促進するが、明所での植物の寿命を延長し、かつ(xi)多数の細菌種および真菌種(非特許文献2に記載されている)に対する植物の病原体耐性応答を誘導したことを示した。最近の研究で、さらに、幅広い非生物的ストレスに対するブラシノステロイドの保護的役割が確認された(非特許文献3)。
ブラシノリドの初期の分離および生理学的研究に続き、推定上の生合成中間体を表わす多数のブラシノステロイド化合物が、異なる植物種で同定された。これらの化合物のin vivoの濃度が極端に低いことが分かったため、これらの化合物の化学合成法を開発するための取り組みがなされてきた(総説として、非特許文献4を参照のこと)。生理学的研究の結果を確認するために、大豆、とうもろこし、米および他の農作物、さらに木を使用した野外実験で、これらの化合物が試験された。しかし、植物の表皮を通じたステロイドの吸収が十分ではなかったため、散布または施肥に必要なステロイドの量が多量になり、その結果、植物に(非)生物的ストレスへの抵抗性を与えるためのブラシノステロイドの使用が事実上不可能であることが、野外実験により明らかとなった。
ストレス耐性植物の開発は、少なくともこれらの問題のいくつかを解決する、または仲介する可能性を有する戦略である。しかし、このようなタイプのストレスに対する抵抗性(耐性)を示す新しい系統の植物を開発するための、従来の植物育種戦略は、比較的進みが遅く、望ましい系統と交配するために特定の耐性のある系統が必要とされている。ストレス耐性についての生殖質資源が限定されていること、および遠縁植物種間の交配が不適合であることが、従来の育種に見られる重要な問題を示している。さらに、乾燥、高温/低温、塩分および他の耐性につながる、モデル耐性植物の細胞内プロセスは、本来複雑で、また複数の細胞適応機構および多数の代謝経路と関係する。ストレス耐性の、この多要素からなる性質は、耐性を目的とした交配をほとんど不成功に導いただけではなく、バイオテクノロジー的手法を用いて、ストレス耐性植物を遺伝的に操作する可能性を制限した。
従って、必要なことは、ストレス耐性につながるこれらの多要素のプロセスに関わる遺伝子およびタンパク質を特定することである。ストレス耐性植物に発現する遺伝子の機能の解明により、環境的ストレスへの植物の適応および耐性についての我々の理解が進展するばかりでなく、作物の改良のための新戦略を計画するための重要な情報が提供される可能性がある。
非生物的ストレス誘導遺伝子の発現よび機能については、分子レベルでの研究が進んでいる。ストレス応答における遺伝子の発現およびシグナル伝達には、複雑な機構が関与しているように思われる。これらには、非生物的ストレスの感知機構、細胞内シグナルへのストレスシグナルの調整、核への移行、ストレスシグナル伝達に関与する第2のメッセンジャー、ストレス誘導遺伝子の転写調節、ならびにストレス誘導遺伝子の機能および共同作用が含まれる。
動物の細胞では、ホスファチジルイノシトール特異的なホスホリパーゼC(PI−PLC)が、多様なシグナル伝達経路の初期の段階で、重要な役割を果たす。ホルモンや成長因子のような細胞外刺激が、PI−PLCを活性化する。PI−PLCは、ホスファチジルイノシトール4,5−二リン酸(PIP2)を加水分解し、イノシトール 4,5−三リン酸(IP3)および1,2−ジアシルグリセロール(DG)の2個の第2メッセンジャーを生成する。IP3は、細胞内Ca<2+>の細胞質への放出を誘導し、それに引き続いて細胞質内で多様な反応を引き起こす。DGおよびPIP2は、また、第2メッセンジャーとして機能し、多様な細胞内反応を制御する。
植物の場合、非生物的ストレス応答では、類似のシステムが機能すると考えられる。切断部位次第で、ホスホリパーゼA、CまたはD(PLA、PLCまたはPLD)は、植物のストレス耐性に重要な、植物の浸透圧ストレスおよび浸透圧調節に関わる細胞容積変化を促進する初期のシグナル伝達事象で役割を果たすことが明確に示されている(非特許文献5;非特許文献6)。例えば、孔辺細胞の場合、アブシジン酸(ABA)に誘導された気孔閉鎖は、PIP2−PLCの急速な活性化により介される。これは、IP3レベルの増加および細胞質カルシウム濃度の上昇を引き起こし、それに続いてK+チャネルが抑制される。例えば、シロイヌナズナ(Arabidopsis thaliana)においては、ホスホリパーゼCの遺伝子である、AtPLCが、乾燥および塩分ストレスにより急速に誘導されることが示された(非特許文献7)。
上記のように、Ca2+イオンは、植物の多様なシグナル伝達経路において、第2メッセンジャーとして重要な役割を果たす。風、接触、非生物的ストレス(低温、乾燥および塩分濃度)または真菌性エリシターによる刺激を受けると、細胞内のCa2+濃度が著しく増加することが観察されている。典型的なEFハンドドメインを持ったCa2+結合タンパク質のいくつかの遺伝子が分離されており、これらは非生物的ストレス処理に対して、発現レベルの増加を示した(非特許文献8;非特許文献9)。
不思議な名前を持つ14−3−3タンパク質も、最近、粘菌から高等植物にわたる真核生物の様々な生理的プロセスの調節に関わっているとして、大きな注目の的となっている。植物では、14−3−3タンパク質に対する多くの生物学上の役割が示唆されている。これらの中でもっとも重要なものには、核でコードされた葉緑体タンパク質の取り込み、転写因子複合体の組立、および細胞内シグナル伝達カスケードに応答した酵素活性の調節における役割が含まれる(非特許文献10)。天然14−3−3タンパク質は、ホモダイマーまたはヘテロダイマーであり、各モノマーには結合部位が一つあるため、ダイマーは二つの標的を強力に結合することができ、それらの結合を促進する。その他に、標的タンパク質には、1超の14−3−3結合部位を持つ可能性がある。
14−3−3タンパク質について、植物ストレス耐性の関与という面から、いくつかの機能が提案されている。14−3−3タンパク質は、ストレスシグナル伝達におけるレギュレーターとして機能することができるかもしれない。例えば、RCI14A遺伝子およびRCI14B遺伝子は、シロイヌナズナ(シロイヌナズナ)を低温処理することにより誘導され、14−3−3タンパク質との類似性が非常に高い。低温処理に応答して観察されるRCI転写レベルの上昇は、ストレスシグナル伝達経路におけるRCIタンパク質の役割を示唆する(非特許文献11)。
作物に対する環境被害の商業上の重要性から、植物のストレス応答シグナル伝達機構を理解し、また環境被害への植物の応答の改善のための操作方法に対して、関心が高まっている。従って、ホスト植物および他の植物種にストレス抵抗性を与える能力を持つストレス耐性植物に発現する遺伝子を同定する必要がある。新規に生み出されるストレス耐性植物には、例えば、植物種の水分要求量を減少させることにより、作物植物を栽培できる範囲を拡大できるなどの多くの利点がある。
Mandava、Ann.Rev.Plant Physiol.Plant Mol.Biol.39(1988)、23−52
Mandava(1988)、loc.cit.
drought、cold and salt、Kagaleら、Planta 225(2007)、353−364
AdamおよびMarquardt、Phytochem.25(1986)、1787−1799
Wang X.ら、2000、Biochemical Society Transactions.28;813−816
Chapman KD、1998 Tr. Plant Sci.3:419−426
Hirayama、T.ら、1995 Proc.Natl.Acad.Sci.92:3903−3907
Frandsen G.ら、1996 J Biol.Chem.271:343−348
Takahashi S.ら、2000 Pant Cell Physiol.41:898−903
Chung H J.ら、1999 Tr.Plant Sci.4:367−371
Jarillo J Aら、1994 Plant Mol.Biol.25:693−704
本発明は、RKS遺伝子をコードするヌクレオチド配列を植物に与えることにより、その植物に非生物的ストレス耐性を与える方法を含む。恐らくそのRKS遺伝子は、切断されたRKS遺伝子で、受容体の細胞外ドメインをコードする。
RKS遺伝子は、RKSサブグループII(RKS1、RKS4、RKS5、RKS7、RKS11およびRKS14)(さらに具体的にはRKS1もしくはRKS4、および切断されたRKS4)で構成される群、またはRKSサブグループIII(RKS0、RKS8、RKS10、RKS12およびRKS13)(さらに具体的にはRKS12)で構成される群から選択されることが望ましい。
さらに好適なのは、植物がブラシノステロイドでさらに処理される方法であり、当該ブラシノステロイドは、好ましくは、ブラシノリド、エピブラシノリド、ホモブラシノリドおよび類似体で構成される群から選択されることが望ましい。
また、本発明の一部は、非生物的ストレスへの耐性を植物または植物細胞に与えるための、RKS遺伝子の使用に関する。RKS遺伝子については、さらに具体的には、RKSサブグループIIの遺伝子(さらに好ましくは、RKS1もしくはRKS4、もしくは切断されたRKS4)であり、またはRKSのサブグループIII(RKS0、RKS8、RKS10、RKS12およびRKS13)(さらに具体的にはRKS12)で構成される群の遺伝子であることが望ましい。
非生物的ストレスは多くの形態をとることができ、多くの作用を持ち得る。表1は、非生物的ストレスの主要な形態、実験でのそれらの再現方法、および植物への影響についての簡単な概説を示す。表に示されているように、ほぼ全ての非生物的ストレスの形態が、植物の成長障害の原因となっており、従ってこれは、支配的な影響を及ぼしていると考えられる。

導入部で詳説したように、ブラシノステロイドは、植物の成長に関連する特徴に有益な影響を及ぼすことが分かっている。
ブラシノステロイド受容体BRI1(ブラシノステロイド非感受性1(BRassinosteroid Insensitive 1))は、LRR(ロイシンリッチな反復を含む)膜貫通型受容体キナーゼである(Cell、1997、90、929−938)。それは、BRI1(At4g39400);BRL1(At1g55610)、BRL2(At2g01950)およびBRL3(At3g13380)で構成されるシロイヌナズナ内の小さなファミリーに属す(Development、2004、131、5341−5351)。BRI1および相同体は、ステロイド感知に直接的に関係するばかりでなく(Nature 2005、433、167−171)、病原体抵抗性反応の浸透移行性の情報伝達に関与するペプチドホルモンである、システミン(シロイヌナズナ由来のプロシステミン相同体:At2g22940)にも高い親和性を持って結合する(PNAS、2002、99、9090−9092)。植物ステロイド情報伝達の下流に存在する細胞内経路については記載がある(Bioassays、2001、23、1028−1036;Trends in Plant Science、2004、9、91−95)。
ブラシノステロイド感知に関与する他の受容体ファミリーは、RKS(受容体キナーゼ様SERK;Development、1997、124、2049−2062)遺伝子産物により定義される(国際公開第04/007712号パンフレット)。これらのRKS遺伝子産物は、また、植物のブラシノステロイド情報伝達の媒介に関与し、BRI1様受容体と複合体を形成すると思われる(The Plant Cell、2004、16、3216−3229;Cell、2002、110、213−222;Cell、2002、110、203−212)。それらは、また、14種のシロイヌナズナRKS遺伝子産物に直接結合すると仮定されている14種のシロイヌナズナGASA(Gibberelic Acid Stimulated Arabidopsis;Plant Mol Biol.、1995、27、743−752)遺伝子産物のような候補ペプチドリガンドに代表される細胞外ペプチドリガンドとの結合に関係している(国際公開第04/007712号パンフレット)。GASAタンパク質は、高い親和性でブラシノステロイド結合に関与すると仮定されるポケットを、その構造中に含む。GASAペプチドリガンドは、それにより、RKS/BRI二量体とブラシノステロイド分子との媒介物の役を務めることになる。RKSとBRI1のような他の受容体との二量体複合物は、動的細胞膜複合体であり、そこでは異なるファミリーメンバーが二量化のパートナーとして関与できる。
このようなクラスの受容体キナーゼの活性の調整は、ペプチドリガンドおよびステロイドホルモンの両方により調節される。色々な形態の植物ブラシノステロイドが手に入る(J.Exp.Botany、1999、50、275−282;The Plant Cell、2002、S97−S110;Plant Physiol.、2003、131、287−297に記載されている)。これらとは別に、これらの受容体の活性を調節するために、多数の合成アゴニストまたはアンタゴニスト(Trends in Plant Science、1999、4、348−353)を使用することができる。
上記のタンパク質受容体複合体では、ELSタンパク質(国際公開第04/007712号パンフレット)が、また、ブラシノステロイドの感知およびシグナルの伝達に関与し、従って、植物全体の抵抗性反応の媒介に関与する。シロイヌナズナELS遺伝子産物のトマト相同体であるLRPは、発症中に特異的に誘導され、驚くべきことにタンパク質分解的に処理されることもある(Mol.Gen.Genet.、1994、243、47−53;Plant J.、1996、10、315−330)。ELSタンパク質産物は明らかに病原菌抵抗性反応に関与し、ブラシノステロイドの耐性の調整において何らかの役割を果たすと考えられる。
植物が環境の脅威に応答する、いくつかの経路の収斂についての証拠が増加してきている。生物的および非生物的ストレスは、その大部分が一定のストレスに対して特異的な、多数の一連の遺伝子の調節につながる複雑な機構により感知されるが、共通する遺伝子もまた、いくつかの独立したストレスによって調節される(Fujita、Mら、2006、Curr.Opin.Plant Biol.9(4):436−442)。
本発明で用いられる用語「核酸」は、デオキシリボヌレオチドまたはリボヌクレオチドポリマー、すなわち、一本鎖型または二本鎖型のポリヌクレオチドという意味を含み、特に別に限定されていない限り、自然に生じるヌクレオチド(例えばペプチド核酸)と類似した方法で一本鎖型核酸とハイブリダイズするという点において、天然ヌクレオチドの本質的な性質を持つ公知の類似物質を含む。ポリヌクレオチドは、天然のまたは異種性の構造遺伝子もしくは調節遺伝子の全長または部分配列でありえる。他に示されていない限り、その用語には、特定の塩基配列と同時にその相補配列をも含む。従って、安定性または他の理由のために主鎖修飾を施されたDNAまたはRNAは、本発明でその用語が意図する通りの「ポリヌクレオチド」である。さらに、2例のみを挙げるとすれば、イノシンのような独特の塩基、もしくはトリチル基のような修飾塩基を含むDNAまたはRNAは、本発明でその用語が意図する通りのポリヌクレオチドである。DNAとRNAには、当業者に知られている多数の有用な目的に適う非常に多様な修飾が施されてきていることが理解されるだろう。本発明で使用する用語、ポリヌクレオチドは、ポリヌクレオチドを化学的に、酵素的にまたは代謝的に修飾を施した形態、その他、とりわけ単純細胞および複雑な細胞を含む、ウイルスや細胞に典型的なDNAやRNAの化学形態を包含する。
本発明では、「ポリペプチド」、「ペプチド」および「タンパク質」の用語は、アミノ酸残基のポリマーを指して同じ意味で使われる。それらの用語は、一つ以上のアミノ酸残基が、対応する自然に存在するアミノ酸の人工的な化学類似体であるアミノ酸ポリマーに適用されると同時に、自然に存在するアミノ酸ポリマーにも適用される。このような自然起源のアミノ酸類似体の基本的な性質は、タンパク質に組み込まれると、そのタンパク質が、そのタンパク質により誘発される抗体に特異的に反応することである。ただし、完全に自然界に存在するアミノ酸からなるものに限定される。「ポリペプチド」、「ペプチド」および「タンパク質」の用語は、また、例えば糖鎖付加、脂質付着、硫酸化、グルタミン酸残基のガンマカルボキシル化、水酸化およびADPリボシル化の修飾を含むが、これらに限定されない。
「コード」または「エンコード」配列は、タンパク質のアミノ酸配列、またはtRNAもしくはrRNAのような機能的RNAをコードする遺伝子の一部であり、核酸配列には、ある特定のタンパク質に翻訳するための情報が含まれるという事実を特に示している。タンパク質をエンコードする核酸は、核酸の翻訳領域内の非翻訳配列(例えばイントロン)を含む場合もあり、またはこのような介在する非翻訳配列がない場合もある(例えばcDNAに見られるように)。タンパク質がエンコードされる情報は、コドンの使用により特定される。典型的に、アミノ酸配列は、「普遍的」遺伝子コードを使用した核酸によりエンコードされる。しかし、そこでは、核酸が発現する際に、いくつかの植物、動物および真菌ミトコンドリア、バクテリアのマイコプラズマ・カプリコラム(Mycoplasma capricolum)または繊毛虫の大核中に存在する、普遍的コードの変異体が使用される可能性がある。核酸を合成的に調製する、または変化させる場合には、その核酸の発現が意図されるホストにとって好ましいと知られているコドンを利用することができる。例えば、本発明の核酸配列は、単子葉植物および双子葉植物の両方に発現させることができるが、単子葉植物または双子葉類の、特定のコドンの優先度およびGC含量の優先度は、異なっていることが示されているので、これらを明らかにするために配列を修飾することができる。
「発現」とは、構造RNA(rRNA、tRNA)またはメッセンジャーRNA(mRNA)への遺伝子の転写、およびそれに続くタンパク質への遺伝子の翻訳を指す。
本発明で用いられる用語「配列の同一性」は、2つ以上のポリヌクレオチド間、または2つ以上のポリペプチド間に同一性が存在することを意味する。それぞれアミノ酸配列を有するポリヌクレオチドまたはポリペプチドは、ヌクレオチドの配列が、最大一致で整列した時に同じである場合に、「同一の」配列を持つ。2つ以上のポリヌクレオチドまたはポリペプチド間の配列比較は、通常、比較窓を使って、2つの配列の一部を比較し、配列類似性の局所領域を同定し、比較することによって行われる。比較窓は通常、約20〜200の連続ヌクレオチドまたは約7〜70の連続アミノ酸である。ポリヌクレオチドまたはポリペプチドの「配列同一性のパーセンテージ」(50、60、70、80、90、95、98、99または100パーセントなどの配列同一性)は、最適に整列した2つの配列を比較窓によって比較することにより決定される。その比較窓内のポリヌクレオチドまたはポリペプチド配列の一部は、2つの配列の最適な配列のための参考配列(付加または欠損を含まない)と比較した場合、付加または欠損(すなわち、ギャップ)を含み得る。パーセンテージは、(a)一致する位置の数を得るために、同一の核酸塩基またはアミノ酸残基が両配列内に現れる位置の数を決定し、(b)その一致した位置の数を、比較窓内の総位置数で割り、かつ(c)配列相同性を得るためにその結果に100を乗ずることにより算出される。比較のための最適な配列整列は、公知のアルゴリズムのコンピューター処理または検査で実施することができる。配列の相同性または同一性を比較および計算するための、配列の整列における使用に適するアルゴリズムおよびソフトウエアーは当業者に公知であるだろう。このような手段の中でも注目される例は、PearsonおよびLipmanのサーチに基づくFASTAおよびBLASTプログラムであり、これらの詳細については、Altschulら(1997)、Nucleic Acid Res.25:3389−3402;Altschulら(1990)、J.Mol.Biol.215:403−10;PearsonおよびLipman(1988)、Proc.Natl.Acad.Sci.USA 85:2444−8;LipmanおよびPearson(1985)、Science 227:l435−41に見出され得る。他にふさわしいプログラムとしては、以下が挙げられる:ウィスコンシン大学 Genetics Computer Group(米国、ウィスコンシン州、マディソン)のGCG(登録商標)Wisconsin Package(登録商標)におけるPILEUP、LINEUP、GAP、BESTFITおよびFASTAプログラム。現在は、アクセルリス(Accelrys)社より入手できる。上記のプログラムの詳細は、次のURLを通じてインターネット上で入手可能である:http://www.ncbi.nlm.nih.gov/BLASTまたはミラーサイト、およびhttp://www.accelrys.com/products/gcg_wisconsin_package。従って、このような相同性および同一性のパーセンテージは、公にもしくは商業的に入手可能なソフトウエアーパッケージを使用して、またはインターネット上のコンピューターのサーバーを通して確かめることができる。「同一性」という用語は、アミノ酸または塩基の最高パーセンテージでの整列を可能にするために、ギャップの導入を必要とする特定の位置において、配列が欠損または付加を持ち得るという事実にも関わらず、配列が最適に整列された場合に、請求項のアミノ酸配列または核酸配列の一定のパーセンテージが、同じ相対位置の参考配列においても見出されることを意味する。配列は、10以下のギャップ(すなわち、一緒に付加されるものが10以下である場合に、2つの配列に導入されるギャップの総数)を使用することによって整列させることが望ましい。このようなギャップの長さは、特に重要ではないが、通常、アミノ酸は10未満、さらには5未満が好ましい。塩基の場合は、30未満、さらには15未満が好ましい。
用語「遺伝子コードの縮重」は、多数の機能的に同一の核酸が、いずれかの所定のタンパク質をエンコードするという事実を指す。例えば、コドンGCA、GCC、GCGおよびGCUは全てアミノ酸アラニンをエンコードする。従って、アラニンがコドンにより特定される全ての位置で、コドンは、エンコードされたポリペプチドを変化させることなく上記の対応コドンのいずれかに変えることができる。このような核酸の変異体は、「サイレントな変異体」である。本発明においては、ポリペプチドをエンコードする全ての核酸配列はまた、遺伝子コードを参照することによって、核酸の、可能性のある全てのサイレントな変異体をも表わすことになる。
用語「相補鎖」の「相補」は、核酸鎖が、ワトソン・クリック塩基対のルールに基づき、他のヌクレオチド配列と水素結合二本鎖を形成するヌクレオチド配列を持つことを意味している。例えば、5’−AAGGCT−3’の相補塩基配列は、3’−TTCCGA−5’である。
アミノ酸に関して使用される「保存置換」という表現は、同じクラス内の物理化学的特性をもつアミノ酸による一定のアミノ酸の置換に関する。従って、RKS配列内のアミノ酸は疎水基を持っており、保存置換により、疎水基を持つ別のアミノ酸によりそれが置き換えられる。他のこのようなクラスは、特性基が親水性、陽イオン性、陰イオン性であり、またはチオールもしくはチオエーテルを含有するものである。このような置換は、通常の当業者によく知られているものである(すなわち、米国特許第5,380,712号明細書を参照)。保存アミノ酸置換は、例えば、脂肪族非極性のアミノ酸群(Gly、Ala、Pro、Ile、Leu、Val)、極性無荷電アミノ酸群(Cys、Ser、Thr、Met、Asn、Gln)、極性荷電アミノ酸群(Asp、Glu、Lys、Arg)または芳香族アミノ酸の群(His、Phe、Tyr、Trp)内で行われ得る。
用語「選択マーカー」は、遺伝子組み換え体と遺伝子非組み換え体の分離を可能にする代謝特性をエンコードするポリヌクレオチド配列を指し、また多くの場合、抗生物質抵抗性の供給を指す。選択可能なマーカーは、例えば、NPTIIにエンコードされたカナマイシン抵抗性マーカーであるHPT遺伝子であり、この遺伝子はハイグロマイシン耐性をコードする。他にも選択マーカーとして、クロラムフェニコールアセチルトランスフェラーゼ、b−ガラクトシダーゼ、ルシフェラーゼおよび緑色蛍光タンパク質のようなレポーター遺伝子が例として挙げられる。レポーター遺伝子産物の同定方法は、酵素的分析法および蛍光分析法を含むが、それらに限定されない。それらの産物を検出するためのレポーター遺伝子および分析法は、当該技術分野においてよく知られており、例えば、Current Protocols in Molecular Biology、編集、Ausubelら、Greene Publishing and Wiley−Interscience:New York(1987)および定期的に発行される最新情報に記載されている。
本発明で用いられる用語「ベクター」には、ホスト細胞の形質転換あるいは形質移入に使用される核酸への参照が含まれ、その中にポリヌクレオチドを挿入することができる。ベクターは多くの場合、レプリコンである。発現ベクターは、その中に挿入された核酸の転写を可能にする。
本発明で用いられる用語「作動可能に連結した」は、機能的連結または機能的並置を指し、そこでは記載された構成要素が、それらが意図した方法で機能することを可能にする関係にある。別の制御配列および/またはコード配列に「作動可能に連結した」制御配列は、制御配列と適合する条件でコード配列の転写および/または発現が達成されるように連結される。通常、作動可能に連結したとは、連結された核酸配列が隣接していて、2つのタンパク質コード領域を結合するために必要な場合には、隣接し、同じリーディングフレームにあることを意味する。
「ホスト細胞」は、ベクターを包含し、当該ベクターの複製および/または発現を補助する一つの細胞を意味する。ホスト細胞は、大腸菌のような原核細胞か、または植物、酵母、昆虫、両生類もしくは哺乳類の細胞のような真核細胞の場合がある。ホスト細胞は細菌細胞または植物細胞であることが好ましく、植物細胞がより好ましい。
本発明で核酸に関連して用いられる「異種性の」は、外来種から生じた核酸、または同種から生じたものの場合には、意図的な人間の介在により、構成および/またはゲノム遺伝子座内で、その天然型から実質的に修飾されたものである。例えば、異種性構造遺伝子に作動可能に連結したプロモーターは、構造遺伝子が由来する種とは異なる種からのものであり、または同種起源である場合には、一方もしくは両方が、その原形から実質的に修飾されたものである。異種性タンパク質は、外来種が起源である場合でもよく、または同種起源である場合には、人間が故意に介在してその原形から実質的に修飾されたものである。
用語「調節配列」または「制御配列」は、本発明では、コード配列の発現に必要なまたは好都合ないずれの成分を含むように定義する。調節配列は、コード配列由来である場合もあれば無関係な場合もある。このような調節配列は、リーダー、ポリアデニル化配列、プロペプチド配列、プロモーター、シグナル配列および転写ターミネーターを含む、ただしそれらに限定されない。調節配列は、少なくとも、プロモーター、ならびに転写シグナルならびに翻訳開始シグナルおよび停止シグナルを含む。調節配列には、ポリペプチドをエンコードする核酸配列のコード領域を使って調節配列の連結を容易にする特定の制限部位を導入することを目的として、リンカーが供給される場合がある。
本発明で用いられる用語「プロモーター」は、当該技術分野において承認されている意味で使用され、RNAポリメラーゼの結合および転写の開始を導くDNA配列を含む遺伝子の一部を意味する。プロモーター配列は、必ずしもではないが、通常、遺伝子の5’側非コード領域に見られる。「植物プロモーター」は、その開始点が植物細胞由来であるかどうかを問わず、植物細胞内での転写を開始することができるプロモーターである。典型的な植物プロモーターには、アグロバクテリウム(Agrobacterium)またはリゾビウム(Rhizobium)のような植物細胞内で発現する遺伝子を含む、植物、植物ウイルスおよびバクテリアから得られるものが含まれる。ただし、それらに限定されない。適切なプロモーターの例としては、カリフラワーモザイクウイルスの35Sプロモーターおよびその誘導体、フェレドキシンプロモーター、ノパリンシンターゼ(nos)、マノピンシンターゼ(mas)およびオクトピンシンターゼ(ocs)プロモーター(欧州特許第0 122 791号明細書、欧州特許第0 126 546号明細書、欧州特許第0 145 338号明細書)、ユビキチンプロモーター(欧州特許第0 342 926号明細書)、キャッサバ・ベイン・モザイク・ウイルス・プロモーター、ならびにルビスコの短いサブユニットのためのキクプロモーターが挙げられる。
用語「遺伝子組み換え植物または植物細胞」は、そのゲノム内に異種性ポリヌクレオチドを含む植物または植物細胞への参照を含む。通常、異種性ポリヌクレオチドは、安定的にゲノム内に統合され、そのため、ポリヌクレオチドは次世代に受け継がれる。異種性ポリヌクレオチドは、ゲノムに、単独で、または組み換え発現カセットの一部として統合される場合がある。また、異種性ポリヌクレオチドが、形質転換植物のゲノムに統合されない、または安定に統合されない可能性もある。その場合、遺伝子を「一時的に」発現させることができ、それは、一定の時間の間、発現が起こり、その後導入されたポリヌクレオチドが当該細胞から失われることを意味する。本発明の目的のために、遺伝子組み換え植物または植物細胞は、異種性ポリペプチドを一時的に発現する植物または植物細胞をも含む。本発明で用いられる「遺伝子組み換え」は、遺伝子型が異種性核酸の存在によって変化させられたいずれかの細胞、細胞株、カルス、組織、植物の部分または植物を含み、その異種性核酸は、元々そのように変化された遺伝子組み換え体、さらにその元の組み換え体との交配または無性繁殖により生み出された遺伝子組み換え体に見られるものである。本発明で用いられる用語「遺伝子組み換え」は、従来の植物繁殖方法または、乱交雑受精、非組み換え型のウイルス感染、非組み換え型の細菌形質転換、非組み換え型の転移または自然突然変異のような自然に起こる事象によるゲノム(染色体の、または染色体外の)の変化を含まない。
用語「挿入」は、核酸の細胞への導入に関連して、「形質移入」、「形質転換」または「形質導入」を意味する。また、核酸が細胞のゲノムに取り込まれ(例えば、染色体、プラスミド、プラスチドまたはミトコンドリアのDNA)、自律的レプリコンに変換され、または一時的に発現させられる(例えば、トランスフェクトされたmRNA)、真核または原核細胞への核酸の取り込みへの参照を含む。
本発明で用いられる用語「植物」は、全ての植物、植物器官(例えば、葉、茎、根など)、種および植物細胞、ならびにそれら植物または植物細胞の子孫への参照を含む。本発明で用いられる用語の植物細胞には、制限なく、種、懸濁培養液、胚、分裂組織領域、カルス組織、葉、根、新芽、配偶体、胞子体、花粉および小胞子が含まれる。本発明の方法に使用され得る植物のクラスは、通常、単子葉植物および双子葉植物を両方含み、形質転換技術を受け入れやすい高等植物のクラスと同様に幅広い。
本発明は、植物に非生物的ストレス耐性を与えるために、RKS遺伝子(好ましくはRKSサブグループIIの遺伝子(さらに具体的にはRKS1もしくはRKS)、またはRKSサブグループIIIの遺伝子(さらに具体的にはRKS12))を有するヌクレオチドコンストラクトを導入することに関連する。本明細書全体では、耐性という用語は、植物非生物的ストレスに耐えるまたは反応しなくなる能力、という意味で使用されている。非生物的ストレスに対する耐性および抵抗性という用語は、互換的に使われている。
RKS遺伝子産物またはRKSタンパク質の異なるドメインは(国際公開第2004/007712号パンフレットをも参照のこと)、基本的に次の機能を持つ:N−端末部に予測されるタンパク質構造の最初のドメインは、細胞膜に向かうタンパク質を標的にすることに関するシグナル配列からなる。この配列が、タンパク質切断により最終的な成熟タンパク質産物から除去される(Jainら 1994、J.Biol.Chemistry 269:16306−16310)。第2のドメインは、異なる数のロイシンジッパーモチーフで構成され、タンパク質間二量化に関与する可能性がある。次のドメインは、保存された対のシステイン残基を含み、ジスルフィド架橋に関与する。次のドメインは、5つ(RKS3の場合は4つのみ)のロイシンリッチな反復(LRR)から構成され、リガンド結合に関与する可能性がある(KobeおよびDeisenhofer 1994、TIBS 19:415−420)。一方、このドメインは、ジスルフィド架橋に関与する対の保存されたシステイン残基を含むドメインと再び隣接し、しばしばセリン/プロリンリッチ領域がそれに続く。次のドメインは、ただ一つの膜貫通ドメインの特性の全てを示す。タンパク質の予想される細胞質部位では、一つのドメインが未知の機能を備えて位置し、続いてセリン/トレオニンキナーゼ活性を持つドメインが位置する(Schmidtら 1997、Development 124:2049−2062、国際公開第01/29240号パンフレット)。キナーゼドメインの次には、未知の機能を備えたドメインがあり、一方で、タンパク質のC‐末端にはロイシンリッチな反復の一部が位置し、恐らくタンパク質−タンパク質間の相互作用に関与する。
RKSファミリー(受容体キナーゼ様SERK)は、ロイシンリッチな反復(LRR)のコピー数および構造配列に基づいてShiuおよびBleeker(2001、PNAS、98:10763−10768)が定義したように、LRRII RLKサブファミリーを形成する。それは、シロイヌナズナでは14メンバーで構成され、対応する遺伝子については国際公開第01/29240号パンフレットに最初に記載されており(国際公開第2004/007712号パンフレットも参照のこと)、それらは以下の通りである。
RKS受容体は全て、このサブファミリーの3つの特徴的なドメイン、すなわち、一つの連続したブロック内のタンデムに配置される5つのLRRで構成される細胞外ドメイン、膜貫通ドメインおよび細胞内キナーゼドメインを包含している。細胞外ドメインの最初の4つのLRRは、完全長(24のアミノ酸)で、一方、LRR5は切断されて、16の残基のみで構成されている。RKS3内には、LRR4が欠けている。イントロンの位置および数は、RKS3の細胞外ドメイン、ならびにRKS2およびRKS6のキナーゼドメイン以外で保存されている。
キナーゼドメインを見てみると、そのファミリーは、アミノ酸配列に基づいて、さらに3つの群(国際公開第01/29240号パンフレトおよび国際公開第2004/007712号パンフレットを参照のこと)に細分することができる。また最近Zhangらにより記載されている(J.Mol.Evol.(2006)63:612−621)。これらのサブグループは次の通りである。グループI:RKS2、3および6、グループII:RKS1、4、5、7、11および14、ならびにグループIII:RKS0、8、10、12および13。さらに、サブグループIIIは、膜貫通ドメインの前に、他のサブグループにはない一つの共通のSPPボックスを持つ(Schmidtら(1997)Dev.124:2049−2062)。一方、サブグループIIは、例えば種を越えて保存されているLRR1内の「PSQ」モチーフ、またはLRR2内の「LQNNxI」の存在により、自身を他のグループと区別する。
また、RKS遺伝子の定義には、異なるRKS遺伝子のドメイン、または上記のRKS遺伝子のドメインに相同する、合成的に作製されたドメインからさえも構成されている核酸が含まれる。このように、例えば、サブグループIIの一つのRKS遺伝子の膜貫通ドメインをエンコードする核酸のフラグメントを、SPPボックスを含むサブグループIIIの一つの膜貫通ドメインをエンコードする核酸のフラグメントで置き換えることができる。RKS遺伝子のコドンの使用を、形質転換される植物に最適なコドンの使用に適合させることも可能である。シロイヌナズナのRKS遺伝子の植物相同体は、多様な植物のデーターベースの比較により見出し得る。数ある中から数例を以下に記す:
Y14600|SBRLK1|モロコシ
BF004020|BF004020|EST432518 KV1 タルウマゴヤシ
AW934655|AW934655|EST353547 トマト
AW617954|AW617954|EST314028 野生種トマト(L.pennellii)
AA738544|AA738544|SbRLK2 モロコシ
AA738545|AA738545|SbRLK3 モロコシ
BG595415|BG595415|EST494093 cSTS ジャガイモ
AI896277|AI896277|EST265720 トマト
BF643238|BF643238|NF002H05EC1F1045
AA738546|AA738546|SbRLK4 モロコシ
BE658174|BE658174|GM700005A20D5 Gm−r1070 ダイズ
BF520845|BF520845|EST458318 DSILタルウマゴヤシ(Medicago truncata)
AC069324|AC069324|イネ
AW761055|AW761055|sl70d06.y1 Gm−c1027 ダイズ
BE352622|BE352622|WHE0425_G11_M21ZS コムギ
BG647340|BG647340|EST508959 HOGA タルウマゴヤシ
AY028699|AY028699|セイヨウアブラナ
AW666082|AW666082|sk31h04.y1 Gm−c1028 ダイズ
AA738547|AA738547|SbRLK5 モロコシ
BG127658|BG127658|EST473220 トマト
L27821|RICPRKI|イネ
BG238468|BG238468|sab51a09.y1 Gm−c1043 ダイズ
BG441204|BG441204|GA__Ea0012C15f ワタ(Gossypium arbo)
AW667985|AW667985|GA__Ea0012C15 ワタ(Gossypium arbore)
AW233982|AW233982|sf32g05.y1 Gm−c1028 ダイズ
AP003235|AP003235|イネ
BF460294|BF460294|074A05 成熟塊茎(Mature tuber)
AY007545|AY007545|セイヨウアブラナ
AC087544|AC087544|イネ
AB041503|AB041503|ポプラ
Y14600|SBRLK1|モロコシ
BF004020|BF004020|EST432518 KV1 タルウマゴヤシ
AW934655|AW934655|EST353547 トマト
AW617954|AW617954|EST314028 野生種トマト(L.pennellii)
AA738544|AA738544|SbRLK2 モロコシ
AA738545|AA738545|SbRLK3 モロコシ
BG595415|BG595415|EST494093 cSTS ジャガイモ
AI896277|AI896277|EST265720 トマト
BF643238|BF643238|NF002H05EC1F1045
AA738546|AA738546|SbRLK4 モロコシ
BE658174|BE658174|GM700005A20D5 Gm−r1070 ダイズ
BF520845|BF520845|EST458318 DSILタルウマゴヤシ(Medicago truncata)
AC069324|AC069324|イネ
AW761055|AW761055|sl70d06.y1 Gm−c1027 ダイズ
BE352622|BE352622|WHE0425_G11_M21ZS コムギ
BG647340|BG647340|EST508959 HOGA タルウマゴヤシ
AY028699|AY028699|セイヨウアブラナ
AW666082|AW666082|sk31h04.y1 Gm−c1028 ダイズ
AA738547|AA738547|SbRLK5 モロコシ
BG127658|BG127658|EST473220 トマト
L27821|RICPRKI|イネ
BG238468|BG238468|sab51a09.y1 Gm−c1043 ダイズ
BG441204|BG441204|GA__Ea0012C15f ワタ(Gossypium arbo)
AW667985|AW667985|GA__Ea0012C15 ワタ(Gossypium arbore)
AW233982|AW233982|sf32g05.y1 Gm−c1028 ダイズ
AP003235|AP003235|イネ
BF460294|BF460294|074A05 成熟塊茎(Mature tuber)
AY007545|AY007545|セイヨウアブラナ
AC087544|AC087544|イネ
AB041503|AB041503|ポプラ
RKS4過剰発現植物内では、ブラシノステロイド誘導マーカーであって、オーキシン誘導マーカーではないAt2g14560遺伝子産物が、上方調節される(国際公開第2004/007712号パンフレットの図9および10を参照のこと)。RSK4の機能を詳しく調べるために、機能獲得型および機能喪失型のアプローチを続けた。RKS4の完全長のcDNAは、CaMV 35Sプロモーターの制御下のシロイヌナズナのWs−0植物内において異所的に発現し、発明者らはSALK収集物内にT−DNA挿入系統を探した(Alonsoら、2003、NASC ヨーロッパ シロイヌナズナ種子保存センターから入手可能)。挿入系統SALK_066568(改名されてrks4−1)が、過剰発現系統(RKS4−OX)とともに研究された。RKS4定常状態mRNAレベルの変化が、植えてから12日目の苗でRT−PCRにより立証され、それにより、RKS4遺伝子が事実RKS4−OX植物内では過剰発現しており、かつrks4−1植物内では、その完全長メッセンジャーはもはや検知できないことが明らかになった(データは示されていない)。それにもかかわらず、RKS4 mRNAの5’末端(T−DNA挿入の上流部)は依然としてrks4−1系統内で転写されていて、生産された切断されたメッセンジャーのレベルは他の全てのサンプルと比較してより高かった。このフラグメントは、RKS4受容体の細胞外ドメインの一部(コード配列の最初の531塩基対)に相当する。変異系統が、形態学的レベルでも病害抵抗性の点においても、RKS4−OX植物のそれに類似する表現型を示していたので、この状態を模倣し、その影響を詳しく調べるために、切断型RKS4受容体(RKS4m1−OX(rks4−1内のように最初の531塩基対)およびRKS4m2−OX(全てのLRRドメインを含む最初の618塩基))に相当する過剰発現コンストラクトを調製し、シロイヌナズナに形質転換した。この適用では、切断されたRKS遺伝子は、完全長RKS遺伝子の少なくとも最初の531塩基対を含むRKS遺伝子として定義される。同様に、切断されたRKSタンパク質は、完全長RKS遺伝子の少なくとも最初の531塩基対によりエンコードされたタンパク質として定義される。
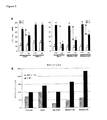
RKS4の過剰発現は、非生物ストレス(具体的に言えば、高塩分ストレス(図1A)、およびより少ない程度の浸透圧ストレス(図2)の場合)に対する耐性を与えるように思われた。一方、切断型RKS形態の過剰発現は、rks4−1植物にも観察されるように、霜害に対する保護の役割を果たした(図3)。非生物的ストレスにおけるRKS受容体の役割は、サブグループIIの別のメンバーであるRKS1、およびサブグループIIIのメンバーであるRKS12の過剰発現でさらに確認された。共に高塩分ストレスに対して増加した耐性をもたらした(図1C)。
RKS4遺伝子組み換えシロイヌナズナ植物のトランスクリプトームおよびメタボロームの分析に基づき、発明者らは、非生物的ストレスに対するRKS介在耐性が特に以下を経由して達成されると仮定する:
1)活性酸素種の生産の原因となるいくつかの非生物的ストレスにより、急速に活性化されるとして知られるGABAシャント代謝経路の調整((ROS、BoucheおよびFromm(2004)TIPS 9:110−115)。ストレス耐性における役割についてのエビデンスが増えてきており(KinnersleyおよびTurano(2000)Crit.Rev.Plant Sci.19:479−509)、γ−アミノ酪酸(GABA)のレベルが、例えばグルタミン酸、フマル酸、アラニンおよびプロリンのレベルとともにRKS4植物で増加する。これらの化合物の増加は、グルタミン酸をGABAに変換する酵素グルタミン酸デカルボキシラーゼ(GAD)を抑制する没食子酸の減少、およびTCAサイクル内でコハク酸からフマル酸への変換を抑制するギ酸の減少を伴う。興味深いことに、GADは、ストレスに曝露された場合に大腸菌内で活発に誘導され、GABAの生成を通して細胞質型pH調節に寄与すると仮定されている(BoucheおよびFromm(2004)TIPS 9:110−115)。次に、GABAはアラニンの前駆体でもあり、そのレベルはRKS4植物においてより高く、またグルタミン酸はプロリンの前駆体で、そのレベルも同じ植物において上昇している。増加したプロリンレベルはまた、通常、浸透圧および塩分ストレスに関係している(それぞれ、Roosenら(1998)Plant Phys.117:263−271およびArmengaudら(2004)Plant Phys.120:442−450)。
1)活性酸素種の生産の原因となるいくつかの非生物的ストレスにより、急速に活性化されるとして知られるGABAシャント代謝経路の調整((ROS、BoucheおよびFromm(2004)TIPS 9:110−115)。ストレス耐性における役割についてのエビデンスが増えてきており(KinnersleyおよびTurano(2000)Crit.Rev.Plant Sci.19:479−509)、γ−アミノ酪酸(GABA)のレベルが、例えばグルタミン酸、フマル酸、アラニンおよびプロリンのレベルとともにRKS4植物で増加する。これらの化合物の増加は、グルタミン酸をGABAに変換する酵素グルタミン酸デカルボキシラーゼ(GAD)を抑制する没食子酸の減少、およびTCAサイクル内でコハク酸からフマル酸への変換を抑制するギ酸の減少を伴う。興味深いことに、GADは、ストレスに曝露された場合に大腸菌内で活発に誘導され、GABAの生成を通して細胞質型pH調節に寄与すると仮定されている(BoucheおよびFromm(2004)TIPS 9:110−115)。次に、GABAはアラニンの前駆体でもあり、そのレベルはRKS4植物においてより高く、またグルタミン酸はプロリンの前駆体で、そのレベルも同じ植物において上昇している。増加したプロリンレベルはまた、通常、浸透圧および塩分ストレスに関係している(それぞれ、Roosenら(1998)Plant Phys.117:263−271およびArmengaudら(2004)Plant Phys.120:442−450)。
2)フラボノール合成経路の調整、さらに具体的に言えば、ケルセチンおよびケンフェロール生合成レベルにおける調整。癌や循環器疾患の予防への関与(Grafら(2005)J.Med.Food 8:281−290)とは別に、フラボノールを含むフラボノイド一般は、植物内で、紫外線保護、生物的および非生物的薬剤に対する防御ならびに抵抗性、ならびに植物ホルモンとの相互作用、といった機能を有すると提唱されている(Winkel−Shirley(2002)Curr.Opin.Plant Biol.5:218−223)。RKS4遺伝子組み換え植物において、様々な形態のケンフェロールレベルの上昇が、ケルセチングルコシドレベルの上昇と合わせて認められている。両方のフラボノール誘導体は、自身に解毒剤の役割を果たす能力を与える強力な抗酸化能を持ち(Torresら(2006)J.Exp.Bot.57:1933−1947)、GABAに関しては、ROS生産に対して保護する役割を果たす。さらに、GABAとストレス誘導性フェニルプロパノイド(フラボノイドを含む)生成との間の関連は、ストレスに対し、フラボノイド生成を可能にするために、GABAが別の炭素源を供給するTCAサイクルを介すると提唱されている(KinnersleyおよびTurano(2000)Crit.Rev.Plant Sci.19:479−509)。
3)RKS4植物においてレベルが上昇するコリン生合成の調整、および浸透圧保護剤とは別に、例えばグリシンベタイン(beatine)の生合成のための前駆体としてのその使用は、塩分濃度、乾燥および他の環境ストレスに対する耐性を与える(McNeilら(2001)PNAS 98:10001−10005)。興味深いことには、グリシンはまたRKS4植物においてより豊富であり、一方、ベタイン(beatine)類似体のレベルは減少している。観察された上昇コリンレベルは、また、シナピン酸(RKS4により多い)およびコリン(Strack(1981)Z.Naturforsch.36c:215−221)の生成につながるシナピンの加水分解が進むことにより、調整されたシナパートエステル生合成経路由来であり得る。
植物核内のRKS遺伝子のような遺伝子の発現のためのポリヌクレオチドコンストラクトは、異種性のヌクレオチド配列下流に作動可能に連結した3’調節配列、または転写ターミネーターのような適切な3’配列を持つものであることが望ましい。いくつかのこのようなターミネーターは、当該技術分野で既知のものであり、入手可能である(例えば、CaMVのtm1、ジャガイモのPotPI II、rbcSのE9)。本発明に関しては、植物内で機能することが知られているいずれの入手可能なターミネーターを使用できる。本発明に記載されるDNA分子の発現のために、多数の他の配列をポリヌクレオチドコンストラクトに組み込むことができる。これらには、イントロン配列(例えばAdh1およびbronzelからのもの)およびウイルス性リーダー配列(例えばTMV、MCMVおよびAMVからのもの)などの、発現を高めることが明らかになっている配列が含まれる。
ポリヌクレオチドコンストラクトは、RKS遺伝子、好ましくは、サブグループIIのRKS遺伝子、さらに好ましくはRKS4遺伝子の発現のための組み換え型ポリヌクレオチドを含む。上記遺伝子は、RKSタンパク質、相同性RKSタンパク質、またはそれらの機能的フラグメントをコードする核酸を含むことが好ましい、上記タンパク質の機能的フラグメントは、野生型RSKタンパク質と相同であり、ある植物において発現した際に機能性を維持しているタンパク質として定義され、上記機能性は、非生物的ストレス抵抗性を与えることができることを意味する。この意味でもまた、切断されたRKS4タンパク質は機能的フラグメントと見なすことができる。
上記の意味で、相同性とは、あるアミノ酸配列が上記の配列と50%超(好ましくは70%超、もっと好ましくは80%超、最も好ましくは90%超)の配列の同一性を持っていることを意味する。あるいは、相同はヌクレオチドのレベルで判断する。その際、相同性はヌクレオチドの配列が、野生型のRKS遺伝子またはそのフラグメントと、50%超(好ましくは70%超、さらに好ましくは80%超、一番好ましくは90%超)の配列の同一性があることを意味する。
本発明のポリヌクレオチドコンストラクトは、好ましくは、少なくとも、作動可能に連結した、植物内で機能する第1のプロモーター、RKS遺伝子(好ましくはRKS4)をエンコードするヌクレチオド配列、およびターミネーターを含むように構成される。必要に応じて、ポリヌクレオチドは発現のために、調節配列に作動可能に連結した、選択可能なまたはスクリーニングできるマーカーをエンコードした遺伝子配列を含む場合がある。
好ましくは、ウイルスのプロモーターであり、キャッサバ・ベイン・モザイク・ウイルス(CVMV)由来のプロモーター、またはカリフラワーモザイクウイルス(CMV)由来のプロモーターなどが使用される。しかし、構成的発現をもたらすいずれのプロモーター(35Sまたは増強された35Sプロモーターなど)も使用可能である。
組み換え遺伝子コンストラクトは市販され、植物への形質転換に適しており、また形質転換された細胞内での遺伝子産物の発現に適しているベクターへ挿入され得る。アグロバクテリウム(Agrobacterium)を使用した植物形質転換に有用なバイナリーベクター(Goddijn、O.J.M.ら、1993、Plant J、4:863−873で知られているpMOG22など)の使用が望ましい。
原則として、本発明のキメラDNAをいずれの適切な祖先細胞に導入するために、形質転換の方法を使用することができる。方法は、プロトプラストに対するカルシウム/ポリエチレングリコール法、プロトプラストのエレクトロポレーション法、植物物質へのマイクロインジェクション、種々の植物物質の(DNAまたはRNAをコーティングした)粒子銃、(非組み込み型の)ウイルスによる感染、成熟植物への侵入、または成熟花粉もしくは小胞子(欧州特許第0 301 316号明細書)の形質転換による、植物体のアグロバクテリウム・トゥメファシエンス(Agrobacterium tumefaciens)を介した遺伝子導入、などから適切に選ばれ得る。本発明による好ましい方法は、アグロバクテリウム(Agrobacterium)を介したDNAトランスファーを含む。特に好ましいのは、欧州特許第0 120 516号明細書および米国特許第4,940,838号明細書で公開されている、いわゆるバイナリーベクター技術の使用である。
本発明の遺伝子組み換えの植物または植物の部位を作り出す方法は、形質転換植物または植物の部位を選択するステップを含み得る。一般に形質転換の後、多様な酵素をエンコードするまたは本発明の機構を阻害する遺伝子を持ったDNA配列を含むポリヌクレオチドコンストラクトによって形質転換するために、植物細胞または細胞群が選択され、その後、形質転換された物質が植物全体の中で再生される、当業者に既知のステップ、およびRKSタンパク質の過剰生産に関する形質転換された植物の評価が続く。
導入された遺伝子組み換え型DNAの一部として含まれる選択可能なマーカーは、非形質転換細胞に対して形質転換細胞(組み換え型DNAを含むもの)を選択するために使用され得る。適切なマーカーの例には、抗生物質または除草剤に対する抵抗性をもたらす遺伝子が含まれる。組み換え型DNAを含む細胞は、非形質転換細胞を殺すような濃度の抗生物質または除草剤が存在したとしても生き延びることができる。選択可能なマーカー遺伝子の例として、除草剤Bastaに対する抵抗性をもたらすbar遺伝子、カナマイシンに対する耐性を与えるnptII遺伝子、ハイグロマイシンに対する耐性を与えるhpt遺伝子、およびシアナミドに対する耐性を与えるcah遺伝子が挙げられる。一つの形質転換植物細胞から、当業者に既知の細胞培養技術を通して、1本の植物全体を生成することが出来る。
本発明の遺伝子組み換え植物を得るためのプロセスは、別の態様では、本発明のベクターを祖先植物に導入し、さらにこの祖先植物から遺伝子組み換え植物を作り出すことを含む。
さらに、本発明の遺伝子組み換え植物を得るためのもう一つ別の態様には、本発明のポリヌクレオチドコンストラクトを適切なベクターへ導入し、形質転換された植物部位を作るために植物部位を形質転換し、さらにその形質転換した植物部位から当該遺伝子組み換え植物を再生することが含まれ得る。
DNAトランスファーおよび再生に続いて、例えば、サザン解析を使用して、形質転換されたと推定される植物の、本発明による組み換え型DNAの存在、コピー数および/またはゲノム構成について評価することができる。さらに、または別の方法として、新しく導入されたDNAの発現レベルを、当業者に既知の技術であるノーザン解析、および/またはウェスタン解析で解析することができる。さらに、非生物ストレス条件下における表現型分析により、あるRKS遺伝子が導入されたために耐性を持つようになった植物が明らかになるかもしれない。
RKS遺伝子(望ましくはRKS4)のコード配列、が挿入されている遺伝子組み換え植物は、ブラシノステロイド化合物、すなわち、ブラシノステロイド受容体を活性化することができる化合物がその植物に供給されることにより、非生物ストレスに対してより強い耐性を持つであろうとも言われている。そのような化合物は、ブラシノリド、エピブラシノリド、ホモブラシノリドおよび類似体を含む群から選択されるのが望ましい。非生物ストレスに対するブラシノステロイドの保護的役割、およびブラシノステロイドのシグナル伝達におけるRKS遺伝子の役割を考慮すると、調整されたRKS受容体の発現とブラシノステロイドの適用を組み合わせることで、2つの方法をそれぞれ独立して適用する場合に比べて、非生物ストレスに対する耐性がさらにいっそう強化されることが可能である、との提唱することは受け入れやすいものである。
ブラシノステロイド化合物を植物に適合させる方法は、従来の適用方法(例えば、散布または散水による)を使用することでなされる。
(実施例1 種子発芽に対する非生物的ストレスの影響(高塩分および浸透圧ストレス))
全ての実験に対して、生態型 Wassilewskija(Ws−0)、またはColumbia(Col−0)のシロイヌナズナの種(野生型、およびホモ接合性の遺伝子組み換え系統)を2%の漂白剤+0.01%のTween 20で表面殺菌して、無菌水で5回すすぎ、そして180もしくは200mMのNaCl、もしくは400もしくは450mMマンニトールを補充したもの、または補充していないMS+ビタミン(0.8%の寒天 w/v)を撒いた。暗所において24時間、4℃で層別化を実施した後、プレートを、20℃で16時間の光周期(100umol.m−2.s−1)のグロースチャンバーに移した。発芽のパーセンテージは、層別化の3日後から2日おきに、それぞれの処置ごとに、14日〜17日目まで測定した。
全ての実験に対して、生態型 Wassilewskija(Ws−0)、またはColumbia(Col−0)のシロイヌナズナの種(野生型、およびホモ接合性の遺伝子組み換え系統)を2%の漂白剤+0.01%のTween 20で表面殺菌して、無菌水で5回すすぎ、そして180もしくは200mMのNaCl、もしくは400もしくは450mMマンニトールを補充したもの、または補充していないMS+ビタミン(0.8%の寒天 w/v)を撒いた。暗所において24時間、4℃で層別化を実施した後、プレートを、20℃で16時間の光周期(100umol.m−2.s−1)のグロースチャンバーに移した。発芽のパーセンテージは、層別化の3日後から2日おきに、それぞれの処置ごとに、14日〜17日目まで測定した。
(結果)
種子発芽に対する高濃度塩分の影響を、導入遺伝子の可能性のある微妙な効果を明確にするために、期間内に観察し、異なるNaCl濃度を使用してモニターした。NaCl濃度は、実際には、異なる遺伝子組み換え系統に対応する野生型の対照群の種の系統の反応に対応して調整した。例えば、RKS1およびRKS12の過剰発現系統の野生型は、180mMのNaClにおいてでさえかなりよく発芽し、従って差異が見られたのは200mMのみであった。一方、RKS4系統とその野生型では180mMで十分であった。全ての系統ではないが、RKS4−OX系統では、より高頻度の発芽が観察された(図1A)。恐らくそれは、高濃度塩分の存在下での発芽能力とかなりよく関連する発現レベルの差異に起因すると思われる(図1Bを参照)。例えば、導入遺伝子の最も高い発現レベルも示したOX2系統は、発芽中、高度の塩分濃度に対して一貫してより耐性があることが示された。塩分に対する耐性のこの増加がRKS4に限定されるかどうかを確かめるために、同じサブグループIIの別の2つのRKS遺伝子であるRKS1およびRKS14の過剰発現系統が、RKSファミリーのサブグループIIIに属するRKS12の過剰発現系統とともに調べられた。塩分に対する耐性の増加が、また、これらの系統(図1C)で観察され、RKS4以外の他のRKS遺伝子の過剰発現が、同時に塩分に対する改善された耐性ももたらし得ることを示す。これらの結果は、両生態型の系統を並行して調べられたRKS1の場合に、一層納得できる。RKS14−OXにおける発芽は、野生型対照群に比して有意に高くなかった。しかし、たった1系統しか試験できなかったので、RKS14の発現があまりにも低いため塩分に対する耐性が改良されなかったことは排除できない。種子発芽に対する浸透圧ストレスの影響も、浸透圧調節物質マンニトールの濃度を変えて同様に期間内にモニターした。NaClの場合と同様に、高濃度(450mM)の場合がもっとも明瞭な結果を示した(図2)。塩分で判明したことに似て、450mMマンニトールで最も高いパーセンテージの種子発芽が示されているように、RKS4過剰発現系統OX2が、浸透圧調節物質に対してもっとも高いレベルの耐性を示した。一方、切断型RKS4の過剰発現系統はもっと影響を受けやすいことがあきらかになった。
種子発芽に対する高濃度塩分の影響を、導入遺伝子の可能性のある微妙な効果を明確にするために、期間内に観察し、異なるNaCl濃度を使用してモニターした。NaCl濃度は、実際には、異なる遺伝子組み換え系統に対応する野生型の対照群の種の系統の反応に対応して調整した。例えば、RKS1およびRKS12の過剰発現系統の野生型は、180mMのNaClにおいてでさえかなりよく発芽し、従って差異が見られたのは200mMのみであった。一方、RKS4系統とその野生型では180mMで十分であった。全ての系統ではないが、RKS4−OX系統では、より高頻度の発芽が観察された(図1A)。恐らくそれは、高濃度塩分の存在下での発芽能力とかなりよく関連する発現レベルの差異に起因すると思われる(図1Bを参照)。例えば、導入遺伝子の最も高い発現レベルも示したOX2系統は、発芽中、高度の塩分濃度に対して一貫してより耐性があることが示された。塩分に対する耐性のこの増加がRKS4に限定されるかどうかを確かめるために、同じサブグループIIの別の2つのRKS遺伝子であるRKS1およびRKS14の過剰発現系統が、RKSファミリーのサブグループIIIに属するRKS12の過剰発現系統とともに調べられた。塩分に対する耐性の増加が、また、これらの系統(図1C)で観察され、RKS4以外の他のRKS遺伝子の過剰発現が、同時に塩分に対する改善された耐性ももたらし得ることを示す。これらの結果は、両生態型の系統を並行して調べられたRKS1の場合に、一層納得できる。RKS14−OXにおける発芽は、野生型対照群に比して有意に高くなかった。しかし、たった1系統しか試験できなかったので、RKS14の発現があまりにも低いため塩分に対する耐性が改良されなかったことは排除できない。種子発芽に対する浸透圧ストレスの影響も、浸透圧調節物質マンニトールの濃度を変えて同様に期間内にモニターした。NaClの場合と同様に、高濃度(450mM)の場合がもっとも明瞭な結果を示した(図2)。塩分で判明したことに似て、450mMマンニトールで最も高いパーセンテージの種子発芽が示されているように、RKS4過剰発現系統OX2が、浸透圧調節物質に対してもっとも高いレベルの耐性を示した。一方、切断型RKS4の過剰発現系統はもっと影響を受けやすいことがあきらかになった。
(実施例2 植物成長に対する低温の影響)
全ての実験に対して、生態型 Wassilewskija(Ws−0)、またはColumbia(Col−0)のシロイヌナズナの種(野生型、およびホモ接合性の遺伝子組み換え系統)を2%の漂白剤+0.01%のTween 20で表面殺菌して、殺菌水で5回すすぎ、そして10g/lスクロース(=MS10)を補充したもの、または補充しないMS+ビタミン(0.8%の寒天 w/v)に撒いた。暗所において24時間、4℃で層別化を実施した後、プレートを、21℃で16時間の光周期(100umol.m−2.s−1)のグロースチャンバーに移した。10日後に、苗をサンプルごとに20本ずつ、同じ培地を含む新しいプレートに移した。苗はさらに10日間、同一の温度、同一の光の条件下で育成した。その後、全てのプレートをアルミホイルでラップして、1時間、−25℃または室温におく(対照群プレート)。次に苗を土壌に移し、次いで、位置効果を補正するためにランダム化し、各苗のロゼット直径を測定した。その後、植物を土壌で、21℃でかつ16時間の光周期(100umol.m−2.s−1)で育成し、再度、処置後7日目と14日目にロゼット直径を測定した。処置の結果としての生育低下の違いは、霜害に対する耐性の変化に関する基準として考慮された。統計的有意性は、スチューデントのt検定を使用することによって判定した(p値<0.05)。
全ての実験に対して、生態型 Wassilewskija(Ws−0)、またはColumbia(Col−0)のシロイヌナズナの種(野生型、およびホモ接合性の遺伝子組み換え系統)を2%の漂白剤+0.01%のTween 20で表面殺菌して、殺菌水で5回すすぎ、そして10g/lスクロース(=MS10)を補充したもの、または補充しないMS+ビタミン(0.8%の寒天 w/v)に撒いた。暗所において24時間、4℃で層別化を実施した後、プレートを、21℃で16時間の光周期(100umol.m−2.s−1)のグロースチャンバーに移した。10日後に、苗をサンプルごとに20本ずつ、同じ培地を含む新しいプレートに移した。苗はさらに10日間、同一の温度、同一の光の条件下で育成した。その後、全てのプレートをアルミホイルでラップして、1時間、−25℃または室温におく(対照群プレート)。次に苗を土壌に移し、次いで、位置効果を補正するためにランダム化し、各苗のロゼット直径を測定した。その後、植物を土壌で、21℃でかつ16時間の光周期(100umol.m−2.s−1)で育成し、再度、処置後7日目と14日目にロゼット直径を測定した。処置の結果としての生育低下の違いは、霜害に対する耐性の変化に関する基準として考慮された。統計的有意性は、スチューデントのt検定を使用することによって判定した(p値<0.05)。
(結果)
発明者らの実験で用いた霜処置では、成長が著しく阻害される結果となり、ロゼットの直径の観点から判定した。より耐性のある植物は処置の影響をあまり受けず、影響を受けやすい植物に比べると、処置を施していない植物に近い処置後のロゼットサイズを有するだろう。発明者らの測定結果は、処置後14日目のロゼットサイズが対照群の植物のそれと違いがない(図3A、左パネル)ため、rks4−1植物の方が野生型(Col−0)に比べ、霜害に対してより耐性があることを明確に示している。しかし、Col−0植物は、非処置対照群の植物に比べて、処置後の成長が遅い。興味深いことに、rks4−1で作られた、部分的に転写物に対応する切断型RKS4遺伝子の過剰発現系統もまた、霜害に対する耐性が改善されたことを示している(図3A、右パネル)。これらの結果はまた、使用された2つの生態型の間の処置に対する感度の違いを明らかにしている。Col−0よりもWs−0の方が影響ははるかに大きく、そのため、RKS4−m1−OXおよびRKS4m2−OXの耐性の増加は、rks4−1より重要ではないように見える。にもかかわらず、もし処置植物と非処置植物との間のロゼットの直径の違いを相対的成長度合いで言い換えると、示された(図3B)全ての遺伝子組み換え系統で耐性が実質的に増加していることが明らかである。これは、この違いを野生型の相対的成長度合いと相関させた場合に、特に明らかである。従って、霜害の影響については、RKS4−m1−OXおよびRKS4m2−OXは、rks4−1に比してそれほど重要ではないようにさえ見える。それらは、野生型に比べて、それぞれ1.6倍および2.3倍実際に成長した。一方、rks4−1の場合は1.4倍である。従って、切断型RKS4遺伝子(細胞外LRR)の過剰発現系統によって、霜害に対する耐性の改善を得ることができる。切断されたRKS4受容体の影響に基づけば、細胞外LRRの下流のコード配列の変異原性を通じて、同じ結果が達成され得る、と仮定するのは妥当である。
発明者らの実験で用いた霜処置では、成長が著しく阻害される結果となり、ロゼットの直径の観点から判定した。より耐性のある植物は処置の影響をあまり受けず、影響を受けやすい植物に比べると、処置を施していない植物に近い処置後のロゼットサイズを有するだろう。発明者らの測定結果は、処置後14日目のロゼットサイズが対照群の植物のそれと違いがない(図3A、左パネル)ため、rks4−1植物の方が野生型(Col−0)に比べ、霜害に対してより耐性があることを明確に示している。しかし、Col−0植物は、非処置対照群の植物に比べて、処置後の成長が遅い。興味深いことに、rks4−1で作られた、部分的に転写物に対応する切断型RKS4遺伝子の過剰発現系統もまた、霜害に対する耐性が改善されたことを示している(図3A、右パネル)。これらの結果はまた、使用された2つの生態型の間の処置に対する感度の違いを明らかにしている。Col−0よりもWs−0の方が影響ははるかに大きく、そのため、RKS4−m1−OXおよびRKS4m2−OXの耐性の増加は、rks4−1より重要ではないように見える。にもかかわらず、もし処置植物と非処置植物との間のロゼットの直径の違いを相対的成長度合いで言い換えると、示された(図3B)全ての遺伝子組み換え系統で耐性が実質的に増加していることが明らかである。これは、この違いを野生型の相対的成長度合いと相関させた場合に、特に明らかである。従って、霜害の影響については、RKS4−m1−OXおよびRKS4m2−OXは、rks4−1に比してそれほど重要ではないようにさえ見える。それらは、野生型に比べて、それぞれ1.6倍および2.3倍実際に成長した。一方、rks4−1の場合は1.4倍である。従って、切断型RKS4遺伝子(細胞外LRR)の過剰発現系統によって、霜害に対する耐性の改善を得ることができる。切断されたRKS4受容体の影響に基づけば、細胞外LRRの下流のコード配列の変異原性を通じて、同じ結果が達成され得る、と仮定するのは妥当である。
Claims (12)
- 植物にRKS遺伝子またはその相同体をエンコードするヌクレオチド配列を与えることによって、前記植物に非生物的ストレス耐性を与えるための方法。
- 前記RKS遺伝子は切断されたRKS遺伝子である請求項1記載の方法。
- 前記RKS遺伝子は、RKSサブグループII(RKS1、RKS4、RKS5、RKS7、RKS11、およびRKS14)、さらに具体的にはRKS1、RKS4および切断されたRKS4からなる群から選択される請求項1または2記載の方法。
- 前記RKS遺伝子は、RKSサブグループIII(RKS0、RKS8、RKS10、RKS12およびRKS13)、さらに具体的にはRKS12からなる群から選択される請求項1または2記載の方法。
- 前記ヌクレオチド配列は過剰発現している請求項1〜4いずれか1項に記載の方法。
- 前記植物は、さらにブラシノステロイドで処理される請求項1〜5いずれか1項に記載の方法。
- 前記ブラシノステロイドは、ブラシノリド、エピブラシノリド、ホモブラシノリドおよび類似体からなる群から選択される請求項6記載の方法。
- 前記非生物的ストレスは高塩分である請求項1〜7いずれか1項に記載の方法。
- 前記非生物的ストレスは浸透圧ストレスである請求項1〜7いずれか1項に記載の方法。
- 前記非生物的ストレスは霜害である請求項1〜7いずれか1項に記載の方法。
- 植物または植物細胞に非生物的ストレスに対する耐性を与えるための、RKS遺伝子(さらに具体的には、RKSサブグループIIの遺伝子、より好ましくはRKS1、RKS4または切断されたRKS4)の使用。
- 植物または植物細胞に非生物的ストレスに対する耐性を与えるための、RKS遺伝子(さらに具体的にはRKSサブグループIIIの遺伝子、より好ましくはRKS12)の使用。
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| EP07109621A EP2000539A1 (en) | 2007-06-05 | 2007-06-05 | Resistance to abiotic stress in plants |
| PCT/NL2008/050350 WO2008150165A1 (en) | 2007-06-05 | 2008-06-04 | Resistance to abiotic stress in plants |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| JP2010528653A true JP2010528653A (ja) | 2010-08-26 |
Family
ID=38650152
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2010511137A Pending JP2010528653A (ja) | 2007-06-05 | 2008-06-04 | 植物における非生物的ストレスに対する抵抗性 |
Country Status (12)
| Country | Link |
|---|---|
| US (1) | US8119855B2 (ja) |
| EP (2) | EP2000539A1 (ja) |
| JP (1) | JP2010528653A (ja) |
| CN (1) | CN101743313A (ja) |
| AR (1) | AR066883A1 (ja) |
| AU (1) | AU2008260738B2 (ja) |
| BR (1) | BRPI0813896A2 (ja) |
| CA (1) | CA2690855A1 (ja) |
| IL (1) | IL202472A0 (ja) |
| NZ (1) | NZ581766A (ja) |
| WO (1) | WO2008150165A1 (ja) |
| ZA (1) | ZA200908682B (ja) |
Families Citing this family (7)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| NZ601341A (en) | 2010-01-22 | 2014-02-28 | Bayer Ip Gmbh | Acaricide and/or insecticide active substance combinations |
| US9265252B2 (en) | 2011-08-10 | 2016-02-23 | Bayer Intellectual Property Gmbh | Active compound combinations comprising specific tetramic acid derivatives |
| RU2522519C1 (ru) * | 2013-02-06 | 2014-07-20 | Федеральное государственное бюджетное образовательное учреждение высшего профессионального образования "Национальный исследовательский Томский государственный университет" (ТГУ) | Способ повышения устойчивости растений рапса к интенсивному хлоридному засолению |
| RU2711577C1 (ru) * | 2019-08-07 | 2020-01-17 | Федеральное государственное автономное образовательное учреждение высшего образования "Национальный исследовательский Томский государственный университет" (ТГУ, НИ ТГУ) | Способ повышения продуктивности растений картофеля в оптимальных и стрессовых условиях выращивания |
| CN110592137B (zh) * | 2019-10-12 | 2021-02-23 | 中国科学院昆明植物研究所 | 拟南芥at5g10290基因及其突变体在提高植物耐旱性的应用 |
| CN113969289A (zh) * | 2021-12-08 | 2022-01-25 | 河南大学 | 芥子酸在植物气孔开闭调节中的应用 |
| CN114277014B (zh) * | 2021-12-27 | 2023-09-12 | 中国科学院昆明植物研究所 | 拟南芥at5g10290基因在调控植物生长中的应用 |
Family Cites Families (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US6063731A (en) * | 1999-03-30 | 2000-05-16 | Agritope, Inc. | Brassinosteroid analogs useful as plant growth regulators |
| DE60041006D1 (de) * | 1999-12-22 | 2009-01-15 | Basf Plant Science Gmbh | Stress-gekoppelte kalium kanal proteine sowie methoden zu deren verwendung in pflanzen |
| EP1621629A1 (en) * | 2004-07-28 | 2006-02-01 | Expressive Research B.V. | A method to increase pathogen resistance in plants |
| MX2007015417A (es) * | 2005-06-08 | 2008-02-19 | Cropdesign Nv | Plantas que tienen caracteristicas de crecimiento mejoradas y metodo para elaborar las mismas. |
-
2007
- 2007-06-05 EP EP07109621A patent/EP2000539A1/en not_active Withdrawn
-
2008
- 2008-06-04 CN CN200880019032A patent/CN101743313A/zh active Pending
- 2008-06-04 AU AU2008260738A patent/AU2008260738B2/en not_active Ceased
- 2008-06-04 WO PCT/NL2008/050350 patent/WO2008150165A1/en active Application Filing
- 2008-06-04 BR BRPI0813896-6A2A patent/BRPI0813896A2/pt not_active Application Discontinuation
- 2008-06-04 EP EP08766773A patent/EP2171067A1/en not_active Withdrawn
- 2008-06-04 CA CA2690855A patent/CA2690855A1/en not_active Abandoned
- 2008-06-04 NZ NZ581766A patent/NZ581766A/en not_active IP Right Cessation
- 2008-06-04 JP JP2010511137A patent/JP2010528653A/ja active Pending
- 2008-06-05 AR ARP080102402A patent/AR066883A1/es unknown
-
2009
- 2009-12-02 IL IL202472A patent/IL202472A0/en unknown
- 2009-12-03 US US12/630,012 patent/US8119855B2/en not_active Expired - Fee Related
- 2009-12-07 ZA ZA200908682A patent/ZA200908682B/xx unknown
Also Published As
| Publication number | Publication date |
|---|---|
| AU2008260738B2 (en) | 2014-03-27 |
| EP2000539A1 (en) | 2008-12-10 |
| ZA200908682B (en) | 2010-08-25 |
| IL202472A0 (en) | 2011-08-01 |
| BRPI0813896A2 (pt) | 2014-10-07 |
| US8119855B2 (en) | 2012-02-21 |
| WO2008150165A1 (en) | 2008-12-11 |
| AR066883A1 (es) | 2009-09-16 |
| CN101743313A (zh) | 2010-06-16 |
| US20100146664A1 (en) | 2010-06-10 |
| NZ581766A (en) | 2011-08-26 |
| AU2008260738A1 (en) | 2008-12-11 |
| EP2171067A1 (en) | 2010-04-07 |
| CA2690855A1 (en) | 2008-12-11 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US8153863B2 (en) | Transgenic plants expressing GLK1 and CCA1 having increased nitrogen assimilation capacity | |
| US9624502B2 (en) | Methods of controlling plant seed and organ size | |
| Li et al. | GsGASA1 mediated root growth inhibition in response to chronic cold stress is marked by the accumulation of DELLAs | |
| EP2935315B1 (en) | Transcriptional regulation for improved plant productivity | |
| Zhang et al. | Molecular characterization of magnesium chelatase in soybean [Glycine max (L.) Merr.] | |
| US8119855B2 (en) | Resistance to abiotic stress in plants | |
| US7432416B2 (en) | Generation of plants with improved drought tolerance | |
| CA2955191A1 (en) | Methods of increasing crop yield under abiotic stress | |
| CN106471124B (zh) | 液泡膜质子/糖逆向转运蛋白及其增加植物蔗糖贮存器官的蔗糖浓度的用途 | |
| Shui et al. | Characterization of the R2R3-MYB transcription factor CsMYB113 regulates anthocyanin biosynthesis in Tea plants (Camellia sinensis) | |
| WO2016130087A1 (en) | Controlling timing of plant flowering | |
| US7968768B2 (en) | Generation of plants with improved drought tolerance | |
| KR102026764B1 (ko) | 안토시아닌 생합성을 증진시키기 위한 재조합 벡터 및 이의 용도 | |
| CA2854069C (en) | Methods using acyl-coenzyme a-binding proteins to enhance drought tolerance in genetically modified plants | |
| US10421975B2 (en) | 14-3-3 proteins enhance WRINKLED1 transcription factor-regulated oil production | |
| AU1368801A (en) | Transgenic plants carrying neoxanthin cleavage enzyme gene | |
| KR20150045611A (ko) | 식물의 지베렐린 메커니즘에 관련된 OsGASD 유전자 및 이의 용도 | |
| Fan et al. | Overexpression of rice F-box phloem protein gene OsPP12-A13 confers salinity tolerance in Arabidopsis | |
| WO2014037735A1 (en) | Atsp1, an e3 ubiquitin ligase, and its use | |
| US6558922B1 (en) | Methods and compositions for production of floral scent compounds | |
| EP3246336B1 (en) | Nucleic acid molecule encoding gle1 protein variant having increased phytic acid sensitivity, and use thereof | |
| KR101756180B1 (ko) | AtHPY2 유전자를 이용한 식물체의 FLC 단백질의 수모화를 촉진하는 방법 및 그에 따른 식물체 | |
| KR101791584B1 (ko) | 향상된 수확량과 관련된 형질을 가지는 형질전환 식물체 및 이의 제조방법 | |
| WO2015193653A1 (en) | Oxidative resistance chimeric genes and proteins, and transgenic plants including the same |