JP2006525994A - Methods for inducing regulated pancreatic hormone production in non-islet tissue - Google Patents
Methods for inducing regulated pancreatic hormone production in non-islet tissue Download PDFInfo
- Publication number
- JP2006525994A JP2006525994A JP2006506631A JP2006506631A JP2006525994A JP 2006525994 A JP2006525994 A JP 2006525994A JP 2006506631 A JP2006506631 A JP 2006506631A JP 2006506631 A JP2006506631 A JP 2006506631A JP 2006525994 A JP2006525994 A JP 2006525994A
- Authority
- JP
- Japan
- Prior art keywords
- pdx
- cell
- cells
- expression
- pancreatic
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
Images






























































Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/85—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells
- C12N15/86—Viral vectors
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K48/00—Medicinal preparations containing genetic material which is inserted into cells of the living body to treat genetic diseases; Gene therapy
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P3/00—Drugs for disorders of the metabolism
- A61P3/08—Drugs for disorders of the metabolism for glucose homeostasis
- A61P3/10—Drugs for disorders of the metabolism for glucose homeostasis for hyperglycaemia, e.g. antidiabetics
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P43/00—Drugs for specific purposes, not provided for in groups A61P1/00-A61P41/00
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P5/00—Drugs for disorders of the endocrine system
- A61P5/48—Drugs for disorders of the endocrine system of the pancreatic hormones
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2710/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA dsDNA viruses
- C12N2710/00011—Details
- C12N2710/10011—Adenoviridae
- C12N2710/10311—Mastadenovirus, e.g. human or simian adenoviruses
- C12N2710/10341—Use of virus, viral particle or viral elements as a vector
- C12N2710/10343—Use of virus, viral particle or viral elements as a vector viral genome or elements thereof as genetic vector
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2830/00—Vector systems having a special element relevant for transcription
- C12N2830/008—Vector systems having a special element relevant for transcription cell type or tissue specific enhancer/promoter combination
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2830/00—Vector systems having a special element relevant for transcription
- C12N2830/80—Vector systems having a special element relevant for transcription from vertebrates
- C12N2830/85—Vector systems having a special element relevant for transcription from vertebrates mammalian
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2840/00—Vectors comprising a special translation-regulating system
- C12N2840/20—Vectors comprising a special translation-regulating system translation of more than one cistron
Abstract
本出願は非膵および非内分泌細胞、特に肝細胞/組織における膵ホルモン生産を含む膵内分泌表現型および機能を誘導する方法に関する。これは、最終的にはニコチンアミド、EGF、アクチビンA、HGF、エキセジンGLP−1またはベータセルリンの存在下にPDX−1ポリペプチド、ニューロDポリペプチドまたはベータセルリンペプチドをコードする核酸のようなPDX−1インデューサー化合物に該非膵および非内分泌細胞を接触させることにより達成される。This application relates to methods of inducing pancreatic endocrine phenotypes and functions, including pancreatic hormone production in non-pancreatic and non-endocrine cells, particularly hepatocytes / tissues. This ultimately results in PDX such as nucleic acids encoding PDX-1 polypeptide, neuro D polypeptide or betacellulin peptide in the presence of nicotinamide, EGF, activin A, HGF, exedin GLP-1 or betacellulin. Achieved by contacting the non-pancreatic and non-endocrine cells with the -1 inducer compound.
Description
(発明の分野)
本発明は一般的には非内分泌組織における膵ホルモン生産を含む膵内分泌表現型および機能を誘導する方法、そして特に、内分泌関連疾患を治療するための方法および医薬組成物に関する。
(Field of Invention)
The present invention relates generally to methods of inducing pancreatic endocrine phenotypes and functions, including pancreatic hormone production in non-endocrine tissues, and in particular to methods and pharmaceutical compositions for treating endocrine related diseases.
(発明の背景)
内分泌膵臓は主にペプチドホルモングルカゴン、インスリン、ソマトスタチンおよび膵ポリペプチドを合成して分泌する島細胞よりなる。インスリン遺伝子の発現は特定の転写因子により部分的には媒介されている制御機序を介して哺乳類膵の膵島β細胞に限定されている。他の細胞においては、インスリン、他の膵ホルモンおよび特定のペプチダーゼの遺伝子は転写的にサイレントである。ホメオドメイン蛋白PDX−1(膵および十二指腸ホメオボックス遺伝子−1、別称IDX−1、IPF−1、STF−1またはIUF−1)は膵島の発生および機能の調節において中枢的な役割を果たす。PDX−1は例えばインスリン、グルカゴンソマトスタチン、プロインスリン変換酵素1/3(PC1/3)、GLUT−2およびグルコキナーゼのような種々の遺伝子の島細胞特異的発現に直接または間接的に関与している。更にまた、PDX−1はグルコースに応答したインスリン遺伝子の転写を媒介する。
(Background of the Invention)
The endocrine pancreas consists mainly of islet cells that synthesize and secrete the peptide hormones glucagon, insulin, somatostatin, and pancreatic polypeptide. Insulin gene expression is restricted to islet β cells in the mammalian pancreas through a regulatory mechanism that is mediated in part by specific transcription factors. In other cells, insulin, other pancreatic hormones and certain peptidase genes are transcriptionally silent. Homeodomain protein PDX-1 (pancreatic and duodenal homeobox gene-1, aka IDX-1, IPF-1, STF-1, or IUF-1) plays a central role in the regulation of islet development and function. PDX-1 is directly or indirectly involved in islet cell-specific expression of various genes such as insulin, glucagon somatostatin,
(発明の要旨)
本発明は部分的には肝における膵および十二指腸のホメオボックス遺伝子(PDX−1)の異所性の発現がサイレント膵ホルモン遺伝子の発現およびプロホルモンを成熟した生物学的に活性なホルモンに変換するプロセシング機序を誘導するということの発見に基づいている。
(Summary of the Invention)
The present invention relates in part to the ectopic expression of the pancreatic and duodenal homeobox gene (PDX-1) in the liver, the processing of silent pancreatic hormone gene expression and conversion of the prohormone into a mature biologically active hormone. It is based on the discovery that it induces a mechanism.
本発明はPDX−1インデューサー化合物を細胞内に導入することによる細胞における膵遺伝子の発現を誘導する方法を提供する。本発明は更に、PDX−1インデューサー化合物に非膵細胞を接触させることによる膵細胞に非膵細胞を変換する方法を提供する。非膵細胞を、該非膵細胞におけるC/EBPβ、アルブミンまたはADH−1の発現を抑制するような、内因性PDX−1、胚マーカー、インスリン、グルコゴンまたはソマトスタチンの発現を誘導する量のPDX−1インデューサー化合物に接触させる。胚マーカーは例えばアルファ−1フェトプロテインまたはGata−4である。 The present invention provides a method for inducing pancreatic gene expression in a cell by introducing a PDX-1 inducer compound into the cell. The present invention further provides a method of converting non-pancreatic cells into pancreatic cells by contacting the non-pancreatic cells with a PDX-1 inducer compound. PDX-1 in an amount that induces expression of endogenous PDX-1, an embryonic marker, insulin, glucogon or somatostatin, such that the non-pancreatic cell suppresses expression of C / EBPβ, albumin or ADH-1 in the non-pancreatic cell Contact with inducer compound. The embryo marker is, for example, alpha-1 fetoprotein or Gata-4.
PDX−1インデューサー化合物は内因性のPDX−1の発現を誘導する何れかの化合物である。PDX−1インデューサー化合物は核酸、ポリペプチドまたは小型分子である。例示されるPDX−1インデューサー化合物は、膵および十二指腸ホモボックス(PDX−1)ポリペプチド、ニューロDポリペプチドまたはベータセルリンポリペプチドをコードする核酸である。 A PDX-1 inducer compound is any compound that induces endogenous PDX-1 expression. PDX-1 inducer compounds are nucleic acids, polypeptides or small molecules. Exemplary PDX-1 inducer compounds are nucleic acids encoding pancreatic and duodenal homobox (PDX-1) polypeptides, neuro D polypeptides or betacellulin polypeptides.
核酸は例えばサイトメガロウィルス(CMV)プロモーター、BOSプロモーター、トランスサイレチンプロモーター、グルコース6−ホスファターゼプロモーター、アルブミン腸脂肪酸結合蛋白プロモーター、チログロブリンプロモーター、界面活性剤Aプロモーター、界面活性剤cプロモーターまたはホスホグリセレートキナーゼ1プロモーターのようなプロモーターに作動可能に連結している。核酸の方法はプラスミドまたはベクターに存在する。ベクターはウィルスベクター、例えばアデノウィルスベクターまたはレンチウィルスベクターである。アデノウィルスベクターは例えばgutless組み換えアデノウィルスベクターである。
Examples of nucleic acids include cytomegalovirus (CMV) promoter, BOS promoter, transthyretin promoter, glucose 6-phosphatase promoter, albumin intestinal fatty acid binding protein promoter, thyroglobulin promoter, surfactant A promoter, surfactant c promoter, or phosphoglycease. It is operably linked to a promoter such as the
「発現を誘導する」とは、遺伝子の発現が化合物非存在下と比較して化合物存在下において増大することを意味する。該細胞に対し、膵または十二指腸ホモボックス1(PDX−1)ポリペプチドをコードする核酸を含む組成物は該細胞において該遺伝子を誘導するのに十分な量である。「発現を抑制する」とは、化合物非存在下と比較して化合物存在下において遺伝子の発現が減少することを意味する。 “Induces expression” means that the expression of a gene is increased in the presence of the compound as compared to in the absence of the compound. For the cell, a composition comprising a nucleic acid encoding a pancreatic or duodenal homobox 1 (PDX-1) polypeptide is in an amount sufficient to induce the gene in the cell. “Suppressing expression” means that the expression of a gene is decreased in the presence of a compound compared to the absence of the compound.
膵遺伝子は例えば、膵転写因子、例えばPDX−1、ベータ2、ISL−2、Nkx6.1、Ngn3.1またはNKx2.2、内分泌遺伝子、例えばSCG2、SGNE1、CHGN、PTPRN、AMPH、NBEA、ニューロDまたはフォリスタチン、または外分泌遺伝子、例えばセリンプロテアーゼ阻害剤、KazalI型、エラスターゼ、p48因子または再生島誘導1アルファを包含する。
Pancreatic genes include, for example, pancreatic transcription factors such as PDX-1,
細胞は哺乳類対象からインビボ、インビトロまたはエクスビボで提供される。細胞は非膵細胞である。細胞は分化した細胞である。細胞は内胚葉細胞、外胚葉細胞または中胚葉細胞である。例えば細胞は肝細胞、皮膚細胞または骨髄細胞である。或いは、細胞は更に、トランスフェクション剤またはニコチンアミド、表皮成長因子、アクチビンA、肝成長因子、エキセンジン、GLP−1またはベータセルリンを含む組成物に接触させる。 The cell is provided from a mammalian subject in vivo, in vitro or ex vivo. The cell is a non-pancreatic cell. The cell is a differentiated cell. The cell is an endoderm cell, ectoderm cell or mesoderm cell. For example, the cell is a hepatocyte, skin cell or bone marrow cell. Alternatively, the cells are further contacted with a transfection agent or a composition comprising nicotinamide, epidermal growth factor, activin A, liver growth factor, exendin, GLP-1 or betacellulin.
本発明は膵ホルモン、例えばインスリン、グルカゴンおよびソマトスタチンの対象における濃度を誘導する方法を提供する。1つの特徴において、方法は、対象において膵ホルモン生産を誘導するのに十分な量の、PDXの発現または活性を増大させる化合物を、それを必要とする対象に投与することを含む。別の特徴において、方法は膵ホルモンを発現することができる細胞を準備すること、PDXの発現または活性を増大させる化合物に細胞を接触させること、および、細胞を対象に導入することにより対象における膵ホルモン生産を誘導することを含む。 The present invention provides methods for inducing concentrations in a subject of pancreatic hormones such as insulin, glucagon and somatostatin. In one aspect, the method comprises administering to a subject in need thereof a compound that increases PDX expression or activity in an amount sufficient to induce pancreatic hormone production in the subject. In another aspect, the method provides for a cell capable of expressing pancreatic hormones, contacting the cell with a compound that increases PDX expression or activity, and introducing the cell into the subject. Including inducing hormone production.
本発明において更に提供されるものは、対象における膵関連の障害、例えば糖尿病、例えばI型またはII型の治療、症状緩解または発症遅延の方法である。方法はPDXの発現を増大させる化合物の治療有効量を対象に投与することを含む。例えば、化合物は膵および十二指腸のホモボックス1(PDX−1)ポリペプチドをコードする核酸である。糖尿病の症状は高血糖症、血中グルコース(血糖値)上昇、頻尿、飲水渇望亢進、飢餓感亢進、異常体重減少、疲労亢進、刺激感または眼のかすみを包含する。糖尿病は例えば空腹時血漿中グルコース試験またはランダム血糖値試験により診断される。 Further provided in the present invention is a method for the treatment of pancreatic-related disorders, such as diabetes, eg type I or type II, symptom relief or delayed onset in a subject. The method includes administering to the subject a therapeutically effective amount of a compound that increases PDX expression. For example, the compound is a nucleic acid encoding pancreatic and duodenal homobox 1 (PDX-1) polypeptide. Symptoms of diabetes include hyperglycemia, increased blood glucose (blood glucose level), frequent urination, increased drinking craving, increased hunger, abnormal weight loss, increased fatigue, irritation, or blurred vision. Diabetes is diagnosed, for example, by a fasting plasma glucose test or a random blood glucose test.
別の特徴において、本発明は対象における膵島遺伝子発現の特徴を誘導する方法を提供する。方法は膵島遺伝子発現を誘導するのに十分な量の、PDXの発現または活性を増大させる化合物を、それを必要とする対象に投与することを含む。 In another aspect, the present invention provides a method of inducing characteristics of islet gene expression in a subject. The method includes administering to a subject in need thereof a compound that increases PDX expression or activity in an amount sufficient to induce islet gene expression.
本発明の更に別の特徴は細胞における膵島細胞表現型の誘導または増強の方法である。方法は該細胞において膵島細胞表現型を誘導または増強するのに十分な量の、PDXの発現または活性を増大させる化合物に細胞を接触させることを含む。 Yet another feature of the invention is a method of inducing or enhancing an islet cell phenotype in a cell. The method comprises contacting the cell with an amount of a compound that increases PDX expression or activity sufficient to induce or enhance an islet cell phenotype in the cell.
更にまたPDXの発現を増大させる化合物および製薬上許容しうる担体を含む医薬組成物も包含される。 Also included are pharmaceutical compositions comprising a compound that increases PDX expression and a pharmaceutically acceptable carrier.
特段の記載が無い限り、本明細書において使用する全ての専門技術用語は本発明が属する技術分野で通常理解されている通りの意味を有する。本明細書に記載したものと同様または等価な方法および材料を本発明の実施または試験において使用できるが、適当な方法および材料は以下に記載するとおりである。本明細書に記載する全ての出版物、特許出願、特許および他の参考文献は参照によりその全体が本明細書に組み込まれる。矛盾がある場合は、定義を含む本明細書が優先する。更に、材料、方法および実施例は例示に過ぎず限定を意図しない。 Unless defined otherwise, all technical terms used herein have the same meaning as commonly understood in the technical field to which this invention belongs. Although methods and materials similar or equivalent to those described herein can be used in the practice or testing of the present invention, suitable methods and materials are as described below. All publications, patent applications, patents and other references mentioned herein are hereby incorporated by reference in their entirety. In case of conflict, the present specification, including definitions, will control. In addition, the materials, methods, and examples are illustrative only and not intended to be limiting.
本発明の他の特徴および利点は以下の詳細な説明および請求項から明らかになる。 Other features and advantages of the invention will be apparent from the following detailed description and claims.
(発明の詳細な説明)
本発明は部分的には、肝および皮膚における膵および十二指腸のホモボックス遺伝子1(PDX−1)の異所性発現が肝および皮膚細胞における膵島細胞表現型を誘導し、そして、膵ホルモンの発現、生産およびプロセシングをもたらすという発見に基づいている。PDX−1はまたIDX−1、IPF−1、STF−1およびIUF−1としても知られており、これらは全て本明細書においては「PDX」と総称する。更にまた、本発明は膵障害を治療するための方法および医薬組成物を提供する。
(Detailed description of the invention)
In part, the ectopic expression of pancreatic and duodenal homobox gene 1 (PDX-1) in the liver and skin induces an islet cell phenotype in liver and skin cells and expression of pancreatic hormones Based on the discovery that it brings production and processing. PDX-1 is also known as IDX-1, IPF-1, STF-1, and IUF-1, all of which are collectively referred to herein as “PDX”. Furthermore, the present invention provides methods and pharmaceutical compositions for treating pancreatic disorders.
糖尿病に対する臨床的膵島移植の結果における最近の進歩は血中グルコース濃度の継続的制御が膵島細胞移植により達成できることを示唆している。しかしながら、この良好な治療方法は死体ドナーからの限定された組織の供給により、そして、一生に亘る免疫抑制の必要性により多大な制約を受けている。糖尿病患者の治療としての島細胞の移植は島細胞またはβ細胞の新しい原料が発見された場合のみ広範に使用可能となる。I型自己免疫糖尿病におけるβ細胞の機能を代替するために考案された組織の最適原料は容易に単離され、広範に増殖され、そして、自己免疫の攻撃に優先して抵抗できなければならず、このような細胞は潜在的には膵臓外組織に存在すると考えられる。 Recent advances in the results of clinical islet transplantation for diabetes suggest that continued control of blood glucose levels can be achieved by islet cell transplantation. However, this good method of treatment is severely limited by the limited supply of tissue from cadaveric donors and the need for lifelong immunosuppression. Islet cell transplantation as a treatment for diabetics is only widely available when new sources of islet cells or beta cells are discovered. The optimal source of tissue designed to replace β-cell function in type I autoimmune diabetes must be easily isolated, extensively proliferated, and able to resist preferential autoimmune attacks Such cells are thought to be potentially present in extrapancreatic tissue.
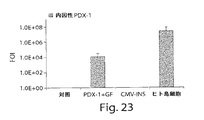
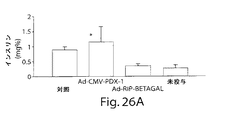
第1世代のE1欠失組み換えアデノウィルス(Ad−CMV−PDX−1)により送達されるインビボのPDX−1の異所性発現は、遺伝子発現の内分泌および外分泌の膵レパートリーおよびプロセシングされた生物学的に活性なインスリンの生産と分泌の両方を誘導した。これらの結果は、成熟し完全に分化した臓器において異所性に発現されたPDX−1、PDX−1が膵分化因子として機能することを示している。更にまた、PDX−1は第1世代の組み換えアデノウィルスを用いてインビボで送達されたが、膵ホルモンの発現および生産は投与後8ヶ月超に亘り持続している。 In vivo ectopic expression of PDX-1 delivered by the first generation E1-deleted recombinant adenovirus (Ad-CMV-PDX-1) is the endocrine and exocrine pancreatic repertoire of gene expression and processed biological Induced both active insulin production and secretion. These results indicate that PDX-1 and PDX-1 expressed ectopically in mature and fully differentiated organs function as pancreatic differentiation factors. Furthermore, although PDX-1 was delivered in vivo using a first generation recombinant adenovirus, pancreatic hormone expression and production has been sustained for over 8 months after administration.
機能的内分泌膵臓を発生させるための組織の潜在的原料として機能する肝臓のこの意外な能力は、異所性PDX−1遺伝子発現を用いたインビボのマウスにおいて、本発明者等により最初に明らかにされた。マウスにおけるPDX−1トランスジーンの短期間発現が肝細胞のサブ集団において包括的、非可逆的および機能的な転移分化過程を誘導(即ち1つの成熟細胞の特徴および機能を別の完全に分化した細胞に変換)したことがわかった。更に、インビトロ培養下の制御された条件下の新しく単離された成熟並びに胎児のヒト肝細胞は機能的インスリン生産組織に転移分化するように誘導できることがわかった。PDX−1トランスジーンを発現した肝細胞の約50%が他では負活性なインスリンプロモーターを活性化した。転移分化したヒト肝細胞はホルモンを生産し、これを分泌顆粒中に保存し、そしてグルコース調節態様においてプロセシングされたインスリンを放出した。インスリン生産ヒト肝細胞は機能的であり、糖尿病免疫不全マウスおよび非肥満糖尿病マウスにおいて誘導されたシクロホスファミド加速糖尿病において、正常血糖を回復した。 This surprising ability of the liver to function as a potential source of tissue to generate functional endocrine pancreas was first revealed by the inventors in vivo in mice using ectopic PDX-1 gene expression. It was done. Short-term expression of the PDX-1 transgene in mice induces a global, irreversible and functional metastatic differentiation process in a subpopulation of hepatocytes (ie one fully differentiated characteristic and function of one mature cell) It was found that it was converted to cells. Furthermore, it has been found that newly isolated mature as well as fetal human hepatocytes under controlled conditions in vitro culture can be induced to metastasize into functional insulin producing tissues. Approximately 50% of the hepatocytes expressing the PDX-1 transgene activated the otherwise negatively active insulin promoter. Metastatic human hepatocytes produced hormones that were stored in secretory granules and released insulin processed in a glucose-regulated manner. Insulin-producing human hepatocytes were functional and restored normoglycemia in cyclophosphamide accelerated diabetes induced in diabetic immunodeficient and non-obese diabetic mice.
その多様な特徴および実施形態において、本発明はPDXの発現または活性を増大させる化合物(本明細書においてはPDXインデューサー化合物とも称する)を対象に投与するか、これを細胞に接触させることを含む。PDX発現または活性は例えば内因性PDX発現を活性化させる化合物により増大される。化合物は例えば(i)PDX、ニューロDまたはベータセルリンポリペプチド;(ii)PDX、ニューロDまたはベータセルリンポリペプチドをコードずる核酸;(iii)PDXポリペプチドをコードする核酸の発現を増大させる核酸、および、それらの誘導体、フラグメント、類縁体および相同体であることができる。PDXポリペプチドをコードする核酸の発現を増大させる核酸は例えばプロモーター、エンハンサーを包含する。核酸は内因性または外因性のいずれかであることができる。場合により、細胞を更にニコチンアミド、表皮成長因子、アクチビンA、肝成長因子、エキセンジン、GLP−1またはベータセルリンに接触させる。 In its various features and embodiments, the invention includes administering to a subject a compound that increases PDX expression or activity (also referred to herein as a PDX inducer compound) or contacting it with a cell. . PDX expression or activity is increased, for example, by compounds that activate endogenous PDX expression. The compound may be, for example, (i) PDX, neuroD or betacellulin polypeptide; (ii) a nucleic acid encoding PDX, neuroD or betacellulin polypeptide; (iii) a nucleic acid that increases expression of a nucleic acid encoding PDX polypeptide; And their derivatives, fragments, analogs and homologues. Nucleic acids that increase the expression of a nucleic acid encoding a PDX polypeptide include, for example, promoters and enhancers. Nucleic acids can be either endogenous or exogenous. Optionally, the cells are further contacted with nicotinamide, epidermal growth factor, activin A, liver growth factor, exendin, GLP-1 or betacellulin.
本明細書においては、「核酸」という用語はDNA分子(例えばcDNAまたはゲノムDNA)、RNA分子(例えばmRNA)、ヌクレオチド類縁体を用いて作成されたDNAまたはRNAの類縁体、および、その誘導体、フラグメントおよび相同体を包含するものとする。核酸分子は1本鎖または2本鎖であることができる。好ましくは、核酸はDNAである。PDXポリペプチドをコードする核酸の発現を増大させる核酸は例えばプロモーター、エンハンサーを包含する。核酸は内因性または外因性の何れかであることができる。 As used herein, the term “nucleic acid” refers to DNA molecules (eg, cDNA or genomic DNA), RNA molecules (eg, mRNA), DNA or RNA analogs made using nucleotide analogs, and derivatives thereof, It is intended to include fragments and homologues. The nucleic acid molecule can be single-stranded or double-stranded. Preferably, the nucleic acid is DNA. Nucleic acids that increase the expression of a nucleic acid encoding a PDX polypeptide include, for example, promoters and enhancers. Nucleic acids can be either endogenous or exogenous.
PDXをコードする核酸の適当な原料は例えばそれぞれゲンバンクアクセッション番号U35632およびAAA88820として入手可能なヒトPDX核酸(およびコードされる蛋白配列)を包含する。他の原料はラットPDX核酸を包含し、そして蛋白配列はそれぞれゲンバンクアクセッション番号U35632およびAAA18355に示されており、参照によりその全体が本明細書に組み込まれる。別の原料はカサゴPDX核酸を包含し、そして蛋白配列はそれぞれゲンバンクアクセッション番号AF036325およびAAC41260に示されており、参照によりその全体が本明細書に組み込まれる。 Suitable sources of nucleic acids encoding PDX include, for example, human PDX nucleic acids (and encoded protein sequences) available as Genbank accession numbers U35632 and AAA88820, respectively. Other sources include rat PDX nucleic acid and protein sequences are shown in Genbank Accession Nos. U35632 and AAA18355, respectively, which are incorporated herein by reference in their entirety. Another source includes scorpion PDX nucleic acid and the protein sequences are shown in Genbank Accession Nos. AF036325 and AAC41260, respectively, which are incorporated herein by reference in their entirety.
化合物は対象に対して直接(即ち対象を核酸または核酸含有ベクターに直接曝露する)または間接的に(即ちまずインビトロで細胞を核酸で形質転換し、次に対象に移植する)投与できる。例えば1つの実施形態において、哺乳類細胞を対象から単離し、そしてPDX核酸をインビトロで単離された細胞に導入する。細胞を適当な哺乳類対象に再導入する。好ましくは、細胞は自己由来の対象に導入する。化合物の投与経路は例えば非経口、静脈内、皮内、皮下、経口(例えば吸入)、経皮(局所)、経粘膜、および直腸投与を包含する。1つの実施形態において、化合物は静脈内投与する。好ましくは化合物は腎被膜下に移植するか、または、門脈に注入する。 The compound can be administered directly to the subject (ie, directly exposing the subject to the nucleic acid or nucleic acid-containing vector) or indirectly (ie, first transforming the cell with the nucleic acid in vitro and then transplanting it into the subject). For example, in one embodiment, mammalian cells are isolated from the subject and PDX nucleic acid is introduced into the cells isolated in vitro. Cells are reintroduced into an appropriate mammalian subject. Preferably, the cell is introduced into an autologous subject. The routes of administration of the compounds include, for example, parenteral, intravenous, intradermal, subcutaneous, oral (eg inhalation), transdermal (topical), transmucosal, and rectal administration. In one embodiment, the compound is administered intravenously. Preferably the compound is implanted under the renal capsule or injected into the portal vein.
細胞は膵ホルモンを生産できる何れかの細胞、例えば骨髄、筋肉、脾臓、腎臓、血液、皮膚、膵臓または肝臓のものであることができる。1つの実施形態において、細胞は膵島細胞として機能できるものであり、即ち、膵ホルモン、好ましくはインスリンを、細胞外からのトリガーにより保存、プロセシングまたは分泌できるものである。別の実施形態においては、細胞は肝細胞である。別の実施形態においては、細胞は全能または多能である。別の実施形態においては細胞は造血幹細胞、胚性幹細胞または、好ましくは肝幹細胞である。 The cell can be any cell capable of producing pancreatic hormones, such as bone marrow, muscle, spleen, kidney, blood, skin, pancreas or liver. In one embodiment, the cells are capable of functioning as islet cells, i.e., capable of storing, processing, or secreting pancreatic hormones, preferably insulin, by an extracellular trigger. In another embodiment, the cell is a hepatocyte. In another embodiment, the cell is totipotent or pluripotent. In another embodiment, the cells are hematopoietic stem cells, embryonic stem cells, or preferably hepatic stem cells.
対象は好ましくは哺乳類である。哺乳類は例えばヒト、非ヒト霊長類、マウス、ラット、イヌ、ネコ、ウマまたはウシであることができる。 The subject is preferably a mammal. The mammal can be, for example, a human, non-human primate, mouse, rat, dog, cat, horse or cow.
(膵ホルモン生産を誘導する方法)
種々の特徴において、本発明は対象における膵ホルモン生産を誘導する方法を提供する。例えば、方法は対象に膵ホルモン生産を誘導するのに十分な量の、PDXの発現または活性を増大させる化合物を投与することを包含する。
(Method of inducing pancreatic hormone production)
In various aspects, the present invention provides a method of inducing pancreatic hormone production in a subject. For example, the method includes administering to the subject a compound that increases PDX expression or activity in an amount sufficient to induce pancreatic hormone production.
別の特徴において、方法は、対象由来の細胞を準備すること、膵ホルモン生産を増大させるのに十分な量のPDX発現を増大させる化合物に細胞を接触させること、および、細胞を対象に導入することを含む。1つの実施形態において、膵ホルモン生産は対象への細胞の導入により、インビトロまたはインビボで起こる。別の実施形態においては、膵ホルモン生産は対象への細胞の導入によりインビボで起こる。 In another aspect, the method provides for preparing cells from a subject, contacting the cells with a compound that increases PDX expression in an amount sufficient to increase pancreatic hormone production, and introducing the cells into the subject. Including that. In one embodiment, pancreatic hormone production occurs in vitro or in vivo by introduction of cells into the subject. In another embodiment, pancreatic hormone production occurs in vivo upon introduction of cells into the subject.
膵ホルモンは例えばインスリン、グルコゴン、ソマトスタチンまたは島アミロイドポリペプチド(IAPP)であることができる。インスリンは肝インスリンまたは血清インスリンであることができる。他の実施形態においては、膵ホルモンは肝インスリンである。別の実施形態においては膵ホルモンは血清インスリン(即ち例えばグルコース利用、炭水化物、脂肪および蛋白の代謝を促進することが可能なインスリンの完全にプロセシングされた形態)である。 The pancreatic hormone can be, for example, insulin, glucogon, somatostatin or islet amyloid polypeptide (IAPP). Insulin can be liver insulin or serum insulin. In other embodiments, the pancreatic hormone is liver insulin. In another embodiment, the pancreatic hormone is serum insulin (ie, a fully processed form of insulin capable of promoting glucose utilization, carbohydrate, fat and protein metabolism, for example).
一部の実施形態においては、膵ホルモンは「プロホルモン」型である。他の実施形態においては、膵ホルモンはホルモンの完全にプロセシングされた生物学的に活性な形態である。他の実施形態においては、膵ホルモンは調節制御下にあり、即ち、ホルモンの分泌は内因性に生産される膵ホルモンの場合と同様に栄養およびホルモンの制御下にある。例えば、本発明の1つの特徴において、ホルモンはグルコースの調節制御下にある。 In some embodiments, the pancreatic hormone is of the “prohormone” type. In other embodiments, the pancreatic hormone is a fully processed biologically active form of the hormone. In other embodiments, the pancreatic hormone is under regulatory control, that is, the secretion of the hormone is under nutritional and hormonal control, as is the endogenously produced pancreatic hormone. For example, in one aspect of the invention, the hormone is under the regulatory control of glucose.
化合物に曝露、例えば接触させる細胞集団は何れかの数量の細胞、即ち1つ以上の細胞であることができ、そして、インビトロ、インビボまたはエクスビボで提供される。 The cell population exposed to, eg, contacted with, the compound can be any quantity of cells, ie, one or more cells, and is provided in vitro, in vivo or ex vivo.
(膵関連障害を治療または防止する方法)
本発明は更に対象における膵関連障害を治療、即ち防止、または発症を遅延または症状を緩解する方法を包含する。種々の特徴において、方法はPDXの発現または活性をモジュレートする化合物を対象に投与することを包含する。「モジュレートする」とはPDXの発現または活性を増大または低減することを包含する。好ましくは、モジュレーションにより、膵障害に罹患していない対象と同様または同一の水準まで、対象におけるPDXの発現または活性の改変が起こる。別の特徴において、方法は非膵細胞に膵島細胞機能、例えばインスリン、ソマトスタチンまたはグルカゴンの発現能力を導き出す化合物を対象に投与することを包含する。1つの実施形態において、化合物はPDXの発現または活性をモジュレートする。
(Methods for treating or preventing pancreatic-related disorders)
The invention further encompasses methods for treating, ie preventing, or delaying onset or ameliorating symptoms in a subject. In various aspects, the method includes administering to the subject a compound that modulates PDX expression or activity. “Modulate” includes increasing or decreasing PDX expression or activity. Preferably, the modulation results in alteration of PDX expression or activity in the subject to the same or the same level as a subject not suffering from a pancreatic disorder. In another aspect, the method includes administering to the subject a compound that induces non-pancreatic cells to express pancreatic islet cell function, eg, insulin, somatostatin or glucagon. In one embodiment, the compound modulates PDX expression or activity.
膵障害は膵に関連する何れかの障害であることができる。例えば方法は膵ホルモン不全(例えば糖尿病(I型およびII型))、インスリノーマ、および高血糖症を治療する際に有用である。基本的に、PDX活性に病因的に関連している何れかの障害が治療に感受性であると考えられる。 The pancreatic disorder can be any disorder associated with the pancreas. For example, the methods are useful in treating pancreatic hormone dysfunction (eg, diabetes (type I and type II)), insulinoma, and hyperglycemia. Basically, any disorder that is etiologically related to PDX activity is considered sensitive to treatment.
糖尿病に罹患しているか発症の危険性を有する対象は血糖値の測定のような当該分野で知られた方法により識別される。例えば、少なくとも2回の機会において一夜絶食後に140mg/dLより高値の血糖値は糖尿病患者であることを意味する。糖尿病に罹患しても発症の危険性を有してもいない者は70〜110mg/dLの空腹時血糖値を有するものとして特徴付けられる。 Subjects with or at risk for developing diabetes are identified by methods known in the art, such as measuring blood glucose levels. For example, a blood glucose level higher than 140 mg / dL after an overnight fast on at least two occasions means that the patient is diabetic. Those who have diabetes or who are not at risk of developing are characterized as having fasting blood glucose levels of 70-110 mg / dL.
糖尿病の症状は疲労、嘔気、頻尿、飲水渇望亢進、体重減少、眼のかすみ、感染頻発および創傷や潰瘍の治癒緩徐、140/90以上の血圧持続、HDLコレステロール35mg/dL未満またはトリグリセリド250mg/dL超、高血糖症、低血糖症、インスリン不全または耐性を包含する。化合物を投与する糖尿病または前糖尿病の患者は当該分野で知られた診断方法を用いて識別される。 Symptoms of diabetes include fatigue, nausea, frequent urination, increased drinking craving, weight loss, blurred vision, frequent infection and slow healing of wounds and ulcers, sustained blood pressure above 140/90, HDL cholesterol less than 35 mg / dL or triglyceride 250 mg / Includes over dL, hyperglycemia, hypoglycemia, insulin failure or tolerance. Diabetic or pre-diabetic patients receiving the compound are identified using diagnostic methods known in the art.
本明細書に記載したPDXモジュレート化合物は治療に使用された場合、本明細書では「治療薬」と称する。治療薬の投与方法は、例えば皮内、筋肉内、腹腔内、静脈内、皮下、鼻内、硬膜外および経口の経路を包含する。本発明の治療薬は何れかの好都合な経路、例えば注入または単回注射により、上皮または粘膜表皮ライニング(例えば口腔粘膜、直腸および腸の粘膜等)を経由する吸収により投与してよく、そして、他の生物学的に活性な薬剤と共に投与してよい。投与は全身または局所であることができる。更にまた、何れかの適当な経路、例えば脳室内およびくも膜下腔内注射により中枢神経系に治療薬を投与することが好都合な場合がある。脳室内注射はリザーバ(例えばOmmayaリザーバ)に連結した脳室内カテーテルにより容易に行える。肺投与は吸入器またはネブライザーおよびエアロゾル化剤を含有する処方を用いることにより使用してもよい。治療を要する領域に治療薬を局所投与することが望ましい場合があり、それは例えば手術中の局所注入、局所適用、注射、カテーテル使用、座薬使用またはインプラントの使用により行える。種々の送達系が知られており、本発明の治療薬の投与に使用することができ、その例は(i)リポソーム、微粒子、マイクロカプセル内への封入、(ii)治療薬を発現できる組み換え細胞、(iii)受容体媒介エンドサイトーシス(例えばWu and Wu,1987,J.Biol.Chem.262:4429−4432参照)、(iv)レトロウィルス、アデノウィルスまたは他のベクターの部分としての治療薬核酸の構築等である。本発明の1つの実施形態においては、治療薬はベシクル、特にリポソーム中で送達してよい。リポソームにおいて、本発明の蛋白は他の製薬上許容しうる担体のほかに、例えば、ミセルとして凝集した形態で存在する脂質などの両親媒性の薬剤、不溶性単層、液晶またはラメラ層などと、水溶液中で組み合わせられる。リポソーム処方に適する脂質は例えばモノグリセリド、ジグリセリド、スルファチド、リソレシチン、リン脂質、サポニン、胆汁酸等を包含する。このようなリポソーム処方の調製は当該分野で知られるとおりであり、例えば米国特許4,837,028;および米国特許4,737,323に記載されており、これらは全て参照により本明細書に組み込まれる。更に別の実施形態においては、治療薬は例えば送達ポンプ(例えばSaudek et al.,1989,New Engl.J.Med.321:572参照)および半透過性重合体物質(例えばHoward et al.,1989,J.Neurosurg.71:105参照)のような制御放出系中で送達できる。更にまた、制御放出系は治療標的(例えば脳)の近接部に入れることができ、即ち、全身用量の僅か一部を要するのみとなる。例えばGoodson、Medical Applications of Controlled Release 1984(CRC Press,Boca Raton,FL)参照。 The PDX-modulating compounds described herein are referred to herein as “therapeutic agents” when used in therapy. Methods of administering therapeutic agents include, for example, intradermal, intramuscular, intraperitoneal, intravenous, subcutaneous, intranasal, epidural and oral routes. The therapeutics of the invention may be administered by any convenient route, such as by infusion or single injection, by absorption via epithelial or mucosal epidermal linings (eg, oral mucosa, rectum and intestinal mucosa, etc.) It may be administered with other biologically active agents. Administration can be systemic or local. Furthermore, it may be advantageous to administer the therapeutic agent to the central nervous system by any suitable route, such as intraventricular and intrathecal injection. Intraventricular injection can be facilitated by an intraventricular catheter connected to a reservoir (eg, Ommaya reservoir). Pulmonary administration may be used by using a formulation containing an inhaler or nebulizer and an aerosolizing agent. It may be desirable to administer the therapeutic agent locally to the area in need of treatment, for example by local injection during surgery, topical application, injection, catheterization, suppository use or the use of implants. Various delivery systems are known and can be used to administer the therapeutic agents of the present invention, examples of which are (i) encapsulated in liposomes, microparticles, microcapsules, (ii) recombinant capable of expressing therapeutic agents. A therapeutic agent as part of a cell, (iii) receptor mediated endocytosis (see eg Wu and Wu, 1987, J. Biol. Chem. 262: 4429-4432), (iv) retrovirus, adenovirus or other vector Such as the construction of nucleic acids. In one embodiment of the invention, the therapeutic agent may be delivered in vesicles, particularly liposomes. In liposomes, the protein of the present invention is in addition to other pharmaceutically acceptable carriers, for example, amphipathic drugs such as lipids present in aggregated form as micelles, insoluble monolayers, liquid crystals or lamellar layers, Combine in aqueous solution. Lipids suitable for liposome formulations include, for example, monoglycerides, diglycerides, sulfatides, lysolecithin, phospholipids, saponins, bile acids and the like. The preparation of such liposome formulations is as known in the art and is described, for example, in US Pat. No. 4,837,028; and US Pat. No. 4,737,323, all of which are incorporated herein by reference. It is. In yet another embodiment, the therapeutic agent is, for example, a delivery pump (see, eg, Saudek et al., 1989, New Engl. J. Med. 321: 572) and a semi-permeable polymeric material (eg, Howard et al., 1989). , J. Neurosurg. 71: 105). Furthermore, the controlled release system can be placed in close proximity to the therapeutic target (eg, the brain), ie, only a small portion of the systemic dose is required. See, for example, Goodson, Medical Applications of Controlled Release 1984 (CRC Press, Boca Raton, FL).
治療薬が蛋白をコードする核酸であるような本発明の特定の実施形態においては、治療薬核酸は、それが細胞内(例えばレトロウィルスベクターの使用による)となるように適当な核酸発現ベクターの部分として構築して投与することにより、直接注射により、微粒子衝突の使用により、脂質コーティングまたは細胞表面コーティングまたはトランスフェクション剤により、または、核内に進入することがわかっているホメオボックス様ペプチドに連結させて投与すること(例えばJoliot et al.,1991,Proc.Natl.Acad.Sci.USA88:1864−1868参照)等により、そのコードする蛋白の発現を促進するようにインビボで投与してよい。或いは、核酸治療薬は細胞内に導入されて相同組み換えにより発現用宿主細胞DNA内に取り込まれるか、または、エピソームとしてとどまることができる。 In certain embodiments of the invention where the therapeutic agent is a nucleic acid encoding a protein, the therapeutic nucleic acid is a suitable nucleic acid expression vector such that it is intracellular (eg, by use of a retroviral vector). Constructed and administered as a part, linked by direct injection, by use of microparticle bombardment, by lipid coating or cell surface coating or transfection agent, or linked to a homeobox-like peptide known to enter the nucleus (See, for example, Joliot et al., 1991, Proc. Natl. Acad. Sci. USA 88: 1864-1868) and the like, and may be administered in vivo so as to promote expression of the encoded protein. Alternatively, the nucleic acid therapeutic can be introduced into the cell and taken up into the host cell DNA for expression by homologous recombination, or can remain as an episome.
本明細書においては、「治療有効量」という用語は意味のある患者の利益、即ち、該当する医療状態の治療、治癒、防止または緩解、または、そのような状態の治療、治癒、防止または緩解の速度の増大を示すのに十分な医薬組成物または方法の各活性化合物の総量を意味する。単独で投与される個々の活性成分に適用する場合は、用語はその成分のみを指す。組み合わせに適用する場合は、用語は、複合、連続または同時の投与に関わらず、治療効果をもたらすような活性成分の複合量を指す。 As used herein, the term “therapeutically effective amount” means meaningful patient benefit, ie, treatment, cure, prevention or amelioration of the relevant medical condition, or treatment, cure, prevention or amelioration of such condition. Means the total amount of each active compound in a pharmaceutical composition or method sufficient to show an increase in the rate of When applied to an individual active ingredient administered alone, the term refers only to that ingredient. When applied to a combination, the term refers to a combined amount of active ingredients that provides a therapeutic effect, whether combined, sequential or simultaneous administration.
特定の障害または状態の治療において有効である本発明の治療薬の量は障害または状態の性質により変化し、そして当該分野における平均的な技術による標準的な臨床手法により決定してよい。更に、インビトロ試験を場合により使用することにより最適用量範囲を発見するのを助けることができる。製剤中に使用すべき厳密な用量は投与経路および疾患または障害の全体的重症度により異なり、そして、医師の判断および各患者の状況に従って決定すべきである。最終的には、担当医師が各個々の患者の治療に用いる本発明の蛋白の量を決定する。初期においては、担当医師は本発明の蛋白の少用量を投与し、そして、患者の応答を観察する。本発明の蛋白の漸増量を患者に対する最適治療効果が得られるまで投与し、その後は更に用量を増加させない。しかしながら、本発明の治療薬の静脈内投与のための適当な用量範囲は一般的にはキログラム(Kg)体重当たり活性化合物約20〜500マイクログラム(μg)である。鼻内投与の適当な用量範囲は一般的には約0.01pg/kg体重〜1mg/kg体重である。有効用量はインビトロまたは動物モデル試験系から誘導された用量−応答曲線から推定してよい。座剤は一般的には0.5%〜10重量%の範囲の活性成分を含有し、経口用製剤は好ましくは10〜95%活性成分を含有する。 The amount of the therapeutic agent of the invention that is effective in the treatment of a particular disorder or condition will vary with the nature of the disorder or condition and may be determined by standard clinical techniques according to average techniques in the art. In addition, in vitro testing can optionally be used to help find the optimal dose range. The exact dose to be used in the formulation will depend on the route of administration and the overall severity of the disease or disorder and should be determined according to the judgment of the physician and the circumstances of each patient. Ultimately, the attending physician will decide the amount of protein of the invention to be used in the treatment of each individual patient. Initially, the attending physician will administer small doses of the protein of the invention and observe the patient's response. Increasing doses of the protein of the invention are administered until an optimal therapeutic effect is obtained for the patient, after which the dose is not increased further. However, suitable dosage ranges for intravenous administration of the therapeutic agents of the invention are generally about 20-500 micrograms (μg) of active compound per kilogram (Kg) body weight. Suitable dosage ranges for intranasal administration are generally about 0.01 pg / kg body weight to 1 mg / kg body weight. Effective doses may be extrapolated from dose-response curves derived from in vitro or animal model test systems. Suppositories generally contain active ingredient in the range of 0.5% to 10% by weight; oral formulations preferably contain 10-95% active ingredient.
本発明の治療薬を用いた静脈内療法の期間は治療すべき疾患の重症度および状態および各個体間での潜在的な特異体質応答により変動する。本発明の蛋白の各適用の期間は連続静脈内投与の12〜24時間の範囲であると考えられる。最終的には担当医師が本発明の医薬組成物を使用する静脈内療法の適切な期間について決定する。 The duration of intravenous therapy using the therapeutic agents of the present invention will vary depending on the severity and condition of the disease to be treated and the potential idiosyncratic response between individuals. It is believed that the duration of each application of the protein of the invention ranges from 12 to 24 hours of continuous intravenous administration. Ultimately, the attending physician will decide on the appropriate duration of intravenous therapy using the pharmaceutical composition of the present invention.
細胞はまた、本発明の治療薬または蛋白の存在下にエクスビボで培養することにより、その細胞に対する所望の作用または細胞における活性を増大させる、またはもたらすことができる。 A cell can also be increased or brought to a desired effect or activity on the cell by culturing ex vivo in the presence of a therapeutic agent or protein of the invention.
(島細胞の表現型または機能を誘導する方法)
本発明はまた細胞における膵島細胞の表現型1つ以上を誘導または増強する方法を包含する。1つの実施形態において、膵細胞表現型は非島細胞型において誘導される。例えば、PDX−1インデューサー化合物に細胞を接触させることにより非膵細胞を膵細胞に変換(即ち転移分化)する。細胞は非膵細胞において内因性PDX−1、胚マーカー、インスリン、グルコゴンまたはソマトスタチンの発現を誘導するような量のPDXインデューサーと接触させる。或いは、非膵細胞におけるC/EBPβ、アルブミンまたはADH−1の発現を抑制する量のPDXインデューサーに細胞を接触させる。方法は、膵島細胞の表現型、例えばベータ、アルファおよびデルタ島細胞を誘導または増強するのに十分な量の、例えばPDX−1、ベータ2、ISL−2、Nkx6.1、Ngn3.1またはNkx2.2のような島細胞特異的転写因子をモジュレートする化合物に細胞を接触させることを包含する。好ましくは、化合物はPDXの発現(例えば内因性PDX−1の発現)、生産または活性を増大させる。好ましくは方法は膵島β細胞表現型を誘導する。
(Methods of inducing islet cell phenotype or function)
The invention also encompasses a method of inducing or enhancing one or more islet cell phenotypes in a cell. In one embodiment, the pancreatic cell phenotype is induced in a non-islet cell type. For example, non-pancreatic cells are converted to pancreatic cells (ie, metastasized) by contacting the cells with a PDX-1 inducer compound. The cells are contacted with an amount of PDX inducer that induces expression of endogenous PDX-1, embryonic marker, insulin, glucogon or somatostatin in non-pancreatic cells. Alternatively, the cells are contacted with an amount of PDX inducer that suppresses the expression of C / EBPβ, albumin or ADH-1 in non-pancreatic cells. The method involves an amount sufficient to induce or enhance an islet cell phenotype, eg, beta, alpha and delta islet cells, eg, PDX-1,
「膵島細胞表現型」とは、膵島細胞に典型的な特性の1つ以上、即ち、ホルモンの生産、プロセシング、分泌顆粒中の保存、栄養的およびホルモン的に調節される分泌または特徴的な島細胞の遺伝子発現の特性を示す細胞である。膵島細胞表現型は例えば膵ホルモンの生産、例えばインスリン、ソマトスタチンまたはグルカゴンを測定することにより調べることができる。ホルモンの生産は当該分野で知られた方法、例えばイムノアッセイ、ウエスタンブロット、受容体結合試験によるか、または、糖尿病宿主への移植後に高血糖症を緩解する能力により機能的に、調べることができる。 “Islet cell phenotype” refers to one or more of the characteristics typical of islet cells, namely hormone production, processing, preservation in secretory granules, nutritionally and hormonally regulated secretion or characteristic islands. It is a cell that exhibits the gene expression characteristics of the cell. The islet cell phenotype can be examined, for example, by measuring pancreatic hormone production, such as insulin, somatostatin or glucagon. Hormone production can be examined by methods known in the art, such as immunoassays, Western blots, receptor binding tests, or functionally by the ability to ameliorate hyperglycemia after transplantation into a diabetic host.
細胞は膵島細胞の表現型を発現することができる何れかの細胞、例えば筋肉、骨髄、脾臓、腎臓、皮膚、膵臓または肝臓のものであることができる。1つの実施形態において、細胞は膵島細胞として機能できるものであり、即ち、膵ホルモン、好ましくはインスリンを、細胞外からのトリガーにより保存、プロセシングまたは分泌できるものである。別の実施形態においては、細胞は肝細胞である。細胞は成熟細胞、即ち分化した細胞である。別の実施形態においては、全能または多能である。別の実施形態においては細胞は造血幹細胞、胚性幹細胞、または、好ましくは肝幹細胞である。 The cell can be any cell capable of expressing an islet cell phenotype, such as muscle, bone marrow, spleen, kidney, skin, pancreas or liver. In one embodiment, the cells are capable of functioning as islet cells, i.e., capable of storing, processing, or secreting pancreatic hormones, preferably insulin, by an extracellular trigger. In another embodiment, the cell is a hepatocyte. The cell is a mature cell, ie a differentiated cell. In another embodiment, it is totipotent or pluripotent. In another embodiment, the cells are hematopoietic stem cells, embryonic stem cells, or preferably hepatic stem cells.
化合物に曝露、即ち接触させられる細胞集団は何れかの数量の細胞、即ち1つ以上の細胞であることができ、そして、インビトロ、インビボまたはエクスビボで提供される。 The cell population exposed to, or contacted with, the compound can be any number of cells, ie, one or more cells, and is provided in vitro, in vivo or ex vivo.
(膵島遺伝子発現特性を誘導する方法)
本発明はまた対象または細胞において膵島遺伝子発現特性を誘導または増強する方法を包含する。「膵遺伝子発現特性」とは非内分泌組織において通常は転写的にサイレントである遺伝子、例えば膵転写因子、内分泌遺伝子または外分泌遺伝子の1つ以上を包含するものとする。例えば、PC1/3、インスリン、グルカゴン、ソマトスタチンまたは内因性PDX−1の発現である。方法は膵島または内分泌遺伝子発現特性を誘導するのに十分な量の、PDXの発現または活性を増大させる化合物を対象に投与することを包含する。1つの実施形態において、方法は対象においてPC1/3遺伝子発現を誘導する。
(Method of inducing islet gene expression characteristics)
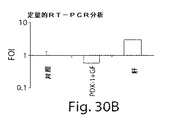
The invention also encompasses a method of inducing or enhancing islet gene expression characteristics in a subject or cell. “Pancreatic gene expression profile” is intended to encompass one or more genes that are normally transcriptionally silent in non-endocrine tissues, such as pancreatic transcription factors, endocrine genes or exocrine genes. For example, expression of PC1 / 3, insulin, glucagon, somatostatin or endogenous PDX-1. The method includes administering to the subject a compound that increases PDX expression or activity in an amount sufficient to induce islet or endocrine gene expression characteristics. In one embodiment, the method induces PC1 / 3 gene expression in the subject.
膵遺伝子発現特性の誘導は、当該分野で知られた手法を用いて検出できる。例えば、膵ホルモンRNA配列はノーザンブロットハイブリダイゼーション分析、増幅系の検出方法、例えば逆転写系のポリメラーゼ連鎖反応またはマイクロアレイチップ分析による系統的検出などにおいて検出できる。或いは、発現は蛋白レベルにおいて、即ち、遺伝子によりコードされるポリペプチドの濃度を測定することによっても測定できる。特定の実施形態においては、PC1/3遺伝子または蛋白の発現はプロホルモンをその活性成熟型にプロセシングする場合のその活性により測定できる。このような方法は当該分野で知られており、そして例えば遺伝子によりコードされる蛋白に対する抗体に基づいたイムノアッセイまたはプロセシングされたホルモンのHPLCを包含する。 Induction of pancreatic gene expression characteristics can be detected using techniques known in the art. For example, pancreatic hormone RNA sequences can be detected by Northern blot hybridization analysis, amplification system detection methods such as systematic detection by reverse transcription system polymerase chain reaction or microarray chip analysis. Alternatively, expression can also be measured at the protein level, i.e. by measuring the concentration of the polypeptide encoded by the gene. In certain embodiments, the expression of the PC1 / 3 gene or protein can be measured by its activity when processing the prohormone to its active mature form. Such methods are known in the art and include, for example, antibody-based immunoassays or processed hormone HPLC for the protein encoded by the gene.
(発現がPDXによりモジュレートされる遺伝子の識別方法)
本発明はまた発現がPDXによりモジュレートされる核酸を識別する方法を包含する。方法はPDXの活性または発現をモジュレートする化合物に曝露された被験細胞集団において核酸1つ以上の発現を測定することを包含する。次に被験細胞集団における核酸配列の発現を化合物に曝露されていない細胞集団、または、一部の実施形態においては化合物に曝露されている細胞集団である比較対照細胞集団における核酸配列の発現と比較する。比較は、同時に、または異なる時間に測定された被験および比較対照の試料に対して実施する。後者の例はコンパイルされた発現の情報、例えば種々の薬剤の投与後の既知配列の発現濃度に関する情報を集積した配列データベースの使用である。例えば、化合物の投与後の発現濃度の改変をPDX核酸のような対照薬剤の投与後の核酸配列において観察される発現の変化と比較する。
(Method for identifying genes whose expression is modulated by PDX)
The invention also includes a method of identifying nucleic acids whose expression is modulated by PDX. The method includes measuring the expression of one or more nucleic acids in a test cell population exposed to a compound that modulates PDX activity or expression. The expression of the nucleic acid sequence in the test cell population is then compared to the expression of the nucleic acid sequence in a control cell population that is a cell population that has not been exposed to the compound or, in some embodiments, a cell population that has been exposed to the compound. To do. Comparisons are performed on test and comparative samples measured at the same time or at different times. The latter example is the use of a sequence database that compiles compiled expression information, for example information on the expression concentration of known sequences after administration of various drugs. For example, modification of the expression concentration after administration of the compound is compared to the change in expression observed in the nucleic acid sequence after administration of a control agent such as PDX nucleic acid.
化合物に曝露されていない比較対照細胞集団における核酸配列の発現と比較した場合に被験細胞集団において核酸配列の発現が改変されていることは、核酸の発現がPDXによりモジュレートされたことを示している。 The altered expression of the nucleic acid sequence in the test cell population when compared to the expression of the nucleic acid sequence in a control cell population not exposed to the compound indicates that the nucleic acid expression was modulated by PDX. Yes.
被験細胞はPDXによりモジュレートされることができる何れかの組織、例えば膵臓、肝臓、脾臓または腎臓から得ることができる。1つの実施形態において、細胞は非内分泌組織に由来する。好ましくは、細胞は肝組織である。 Test cells can be obtained from any tissue that can be modulated by PDX, such as the pancreas, liver, spleen or kidney. In one embodiment, the cells are derived from non-endocrine tissue. Preferably, the cell is liver tissue.
好ましくは、比較対照細胞集団における細胞は被験細胞と可能な限り同じ組織型、例えば肝組織から得る。一部の実施形態においては、対照細胞は被験細胞と同じ対象から、例えば被験細胞の原料の領域に近接する領域から得る。別の実施形態においては、対照細胞集団は試験されたパラメーターまたは条件が既知である細胞から得られた分子情報のデータベースから得る。 Preferably, the cells in the control cell population are obtained from the same tissue type as possible as the test cells, eg liver tissue. In some embodiments, the control cells are obtained from the same subject as the test cells, eg, from an area proximate to the source area of the test cells. In another embodiment, the control cell population is obtained from a database of molecular information obtained from cells with known parameters or conditions tested.
核酸の発現は当該分野で知られた何れかの方法を用いてRNAレベルで測定できる。例えばこれらの配列の1つ以上を特異的に認識するプローブを用いたノーザンハイブリダイゼーション分析を用いて遺伝子発現を測定できる。或いは、発現は逆転写系PCR分析を用いて測定できる。発現はまた蛋白レベルで、即ち、遺伝子産物によりコードされるポリペプチドの濃度を測定することにより測定できる。このような方法は当該分野で知られており、例えば遺伝子によりコードされる蛋白に対する抗体に基づいたイムノアッセイが挙げられる。 Nucleic acid expression can be measured at the RNA level using any method known in the art. For example, gene expression can be measured using Northern hybridization analysis using probes that specifically recognize one or more of these sequences. Alternatively, expression can be measured using reverse transcription-based PCR analysis. Expression can also be measured at the protein level, ie by measuring the concentration of the polypeptide encoded by the gene product. Such methods are known in the art and include, for example, immunoassays based on antibodies to proteins encoded by genes.
遺伝子発現の改変が遺伝子の増幅または欠失に関連する場合は、被験および比較対照集団の配列の比較は被験および比較対照細胞集団における検査されたDNA配列の相対量を比較することにより行うことができる。 Where the altered gene expression is associated with gene amplification or deletion, the sequence comparison of the test and control populations may be made by comparing the relative amounts of the tested DNA sequences in the test and control cell populations. it can.
本発明はまた本スクリーニング方法に従って識別されるPDXモジュレート核酸およびそのようにして識別されたPDXモジュレート核酸を含む医薬組成物を包含する。 The present invention also encompasses PDX modulated nucleic acids identified according to the present screening methods and pharmaceutical compositions comprising PDX modulated nucleic acids so identified.
(PDX組み換え発現ベクターおよび宿主細胞)
本発明の別の特徴はPDX蛋白をコードする核酸またはその誘導体、フラグメント、類縁体または相同体を含有するベクター、好ましくは発現ベクターに関する。本明細書においては、「ベクター」という用語はそれが連結されている別の核酸を輸送することができる核酸分子を指す。ベクターの1つの型は「プラスミド」であり、これは、別のDNAセグメントをライゲーションすることができる線状または環状の2本鎖DNAループを指す。ベクターの別の型はウィルスベクターであり、この場合は別のDNAセグメントをウィルスゲノム内にライゲーションできる。特定のベクターはそれらが導入される宿主細胞内で自律複製が可能である(例えば細菌複製起点を有する細菌ベクターおよびエピソーム哺乳類ベクター)。他のベクター(例えば非エピソーム哺乳類ベクター)は宿主細胞への導入時に宿主細胞のゲノム内に組み込まれ、これにより宿主ゲノムと共に複製される。更にまた、特定のベクターはそれらが作動可能に連結している遺伝子の発現を指向することができる。このようなベクターは本明細書においては「発現ベクター」と称する。一般的に、組み換えDNA法において使用される発現ベクターはプラスミドの形態をとる場合が多い。プラスミドは最も一般的に使用されているベクターの形態であるため、本明細書においては、「プラスミド」および「ベクター」は互換的に使用するものとする。しかしながら、本発明は等しく機能するウィルスベクター(例えば複製欠損レトロウィルス、レンチウィルス、アデノウィルスおよびアデノ関連ウィルス)のような発現ベクターの他の形態も包含するものとする。更に、一部のウィルスベクターは特異的または非特異的に特定の細胞型をターゲティングすることができる。
(PDX recombinant expression vector and host cell)
Another feature of the present invention relates to a vector, preferably an expression vector, containing a nucleic acid encoding PDX protein or a derivative, fragment, analog or homologue thereof. As used herein, the term “vector” refers to a nucleic acid molecule capable of transporting another nucleic acid to which it has been linked. One type of vector is a “plasmid”, which refers to a linear or circular double stranded DNA loop into which another DNA segment can be ligated. Another type of vector is a viral vector, in which another DNA segment can be ligated into the viral genome. Certain vectors are capable of autonomous replication in a host cell into which they are introduced (eg, bacterial vectors having a bacterial origin of replication and episomal mammalian vectors). Other vectors (eg, non-episomal mammalian vectors) are integrated into the genome of the host cell upon introduction into the host cell, and thereby are replicated along with the host genome. Furthermore, certain vectors can direct the expression of genes to which they are operably linked. Such vectors are referred to herein as “expression vectors”. In general, expression vectors used in recombinant DNA methods often take the form of plasmids. In the present specification, “plasmid” and “vector” are used interchangeably as the plasmid is the most commonly used form of vector. However, the present invention is intended to encompass other forms of expression vectors such as viral vectors that function equally (eg, replication defective retroviruses, lentiviruses, adenoviruses and adeno-associated viruses). In addition, some viral vectors can target specific cell types either specifically or non-specifically.
本発明の組み換え発現ベクターは宿主細胞内の核酸の発現に適する形態の本発明の核酸を含み、このことは、組み換え発現ベクターが、発現すべき核酸配列に作動可能に連結された状態で、発現のために使用されることとなる宿主細胞に基づいて選択される調節配列1つ以上を含むことを意味する。組み換え発現ベクター内において、「作動可能に連結」とは、目的のヌクレオチド配列が、核酸配列の発現(例えばインビトロ転写/翻訳系において、または、ベクターを宿主細胞に導入する場合は宿主細胞において)を可能にするような態様で調節配列に連結していることを意味する。「調節配列」という用語はプロモーター、エンハンサーおよび他の発現制御エレメント(例えばポリアデニル化シグナル)を包含するものとする。このような調節配列は例えばGoeddel;Gene Expression Technology:Methods in Enzymology 185,Academic Press,San Diego,Calif.(1990)に記載されている。調節配列は多くの型の宿主細胞においてヌクレオチド配列の構成的発現を指向するもの、および、特定の宿主細胞においてのみヌクレオチド配列の発現を指向するもの(例えば組織特異的調節配列)を包含する。当業者の知るとおり、発現ベクターの設計は形質転換すべき宿主細胞の選択肢、所望の蛋白の発現濃度等の要因により変動する。本発明の発現ベクターは宿主細胞に導入することにより、本明細書に記載する核酸によりコードされる融合蛋白またはペプチドを含む蛋白またはペプチドを生産することができる(例えばPDX蛋白、PDXの突然変異体、融合蛋白等)。 A recombinant expression vector of the present invention comprises a nucleic acid of the present invention in a form suitable for expression of the nucleic acid in a host cell, wherein the recombinant expression vector is expressed operably linked to the nucleic acid sequence to be expressed. It is meant to include one or more regulatory sequences selected based on the host cell to be used for the purpose. Within a recombinant expression vector, “operably linked” refers to expression of a nucleic acid sequence of interest in a nucleic acid sequence (eg, in an in vitro transcription / translation system or in a host cell if the vector is introduced into a host cell). It means linked to the regulatory sequence in such a way as to allow. The term “regulatory sequence” is intended to include promoters, enhancers and other expression control elements (eg, polyadenylation signals). Such regulatory sequences are described in, for example, Goeddel; Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, Calif. (1990). Regulatory sequences include those that direct constitutive expression of nucleotide sequences in many types of host cells and those that direct expression of nucleotide sequences only in specific host cells (eg, tissue-specific regulatory sequences). As is known to those skilled in the art, the design of an expression vector varies depending on factors such as the choice of host cell to be transformed and the expression level of the desired protein. By introducing the expression vector of the present invention into a host cell, a protein or peptide containing a fusion protein or peptide encoded by the nucleic acid described herein can be produced (eg, PDX protein, PDX mutant). , Fusion proteins, etc.).
本発明の組み換え発現ベクターは原核細胞または真核細胞におけるPDXの発現のために設計できる。例えばPDXは細菌細胞、例えばE.coli、昆虫細胞(バキュロウィルス発現ベクターを使用)、コウボ細胞または哺乳類細胞中で発現できる。適当な宿主細胞はGoeddel;Gene Expression Technology:Methods in Enzymology 185,Academic Press,San Diego,Calif.(1990)にさらに記載されている。或いは、組み換え発現ベクターは例えばT7プロモーター調節配列およびT7ポリメラーゼを用いてインビトロで転写および翻訳できる。 The recombinant expression vectors of the invention can be designed for the expression of PDX in prokaryotic or eukaryotic cells. For example, PDX is a bacterial cell such as E. coli. It can be expressed in E. coli, insect cells (using baculovirus expression vectors), yeast cells or mammalian cells. Suitable host cells include Goeddel; Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, Calif. (1990). Alternatively, the recombinant expression vector can be transcribed and translated in vitro, for example using T7 promoter regulatory sequences and T7 polymerase.
原核細胞における蛋白の発現は最も頻繁には融合または非融合蛋白の発現を指向する構成または誘導プロモーターを含むベクターを用いてE.coli中で行われる。融合ベクターは、通常は組み換え蛋白のアミノ末端においてそこにコードされる蛋白に多くのアミノ酸を付加する。このような融合ベクターは典型的には3つの目的、即ち(1)組み換え蛋白の発現を増大させるため、(2)組み換え蛋白の溶解度を上昇させるため、および(3)アフィニティー精製におけるリガンドとして作用することにより組み換え蛋白の精製に寄与するために機能する。多くの場合、融合発現ベクターにおいては、蛋白分解切断部位を融合部分と組み換え蛋白の接合部に導入することにより融合蛋白の精製後の融合部分からの組み換え蛋白の分離を可能にする。このような酵素およびその同属体認識配列は第Xa因子、トロンビンおよびエンテロキナーゼを包含する。典型的な融合発現ベクターはそれぞれグルタチオンS−トランスフェラーゼ(GST)、マルトースE結合蛋白またはプロテインAを標的組み換え蛋白に融合させるpGEX(Pharmacia Biotech Inc.,Smith and Johnson(1988)Gene 67:31−40)、pMAL(New England Biolabs,Beverly,Mass.)およびpRIT5(Pharmacia,Piscataway,NJ)を包含する。 Expression of proteins in prokaryotic cells is most often performed in E. coli using vectors containing a constitutive or inducible promoter that directs the expression of fused or unfused proteins. performed in E. coli. Fusion vectors usually add many amino acids to the protein encoded therein at the amino terminus of the recombinant protein. Such fusion vectors typically act as a ligand for three purposes: (1) to increase expression of the recombinant protein, (2) to increase the solubility of the recombinant protein, and (3) to affinity purification. It functions to contribute to the purification of recombinant proteins. In many cases, in fusion expression vectors, a proteolytic cleavage site is introduced at the junction between the fusion part and the recombinant protein, thereby allowing the recombinant protein to be separated from the fusion part after purification of the fusion protein. Such enzymes and their congener recognition sequences include Factor Xa, thrombin and enterokinase. Typical fusion expression vectors are glutathione S-transferase (GST), maltose E binding protein, or pGEX (Pharmacia Biotech Inc., Smith and Johnson (1988) Gene 67: 31-40), which fuses protein A to a target recombinant protein, respectively. PMAL (New England Biolabs, Beverly, Mass.) And pRIT5 (Pharmacia, Piscataway, NJ).
適当な誘導非融合E.coli発現ベクターの例は、pTrc(Amrann et al.,(1988)Gene 69:301−315)およびpET 11d(Studier et al.,Gene Expression Technology:Methods in Enzymology 185,Academic Press,San Diego,Calif。(1990)60−89)を包含する。 Suitable induction non-fusion E. Examples of E. coli expression vectors include pTrc (Amann et al., (1988) Gene 69: 301-315) and pET 11d (Studenter et al., Gene Expression Technology: Methods in Enzymology 185, Academy. (1990) 60-89).
E.coli中の組み換え蛋白発現を最大限にするための1つの方法は組み換え蛋白を蛋白分解的に切断する能力が損なわれている宿主細菌中で蛋白を発現させることである。Gottesman,Gene Expression Technology:Methods in Enzymology 185,Academic Press,San Diego,Calif.(1990)119−128を参照できる。別の方法は各アミノ酸に対する個々のコドンがE.coliにより優先的に利用されるものであるように発現ベクター内に挿入すべき核酸の核酸配列を改変することである(Wada et al.,(1992)Nucleic Acids Res.20:2111−2118)。本発明の核酸配列のこのような改変は標準的なDNA合成手法により行うことができる。 E. One way to maximize recombinant protein expression in E. coli is to express the protein in a host bacterium that has impaired ability to proteolytically cleave the recombinant protein. Gottesman, Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, Calif. (1990) 119-128. Another method is that the individual codons for each amino acid are E. coli. modifying the nucleic acid sequence of the nucleic acid to be inserted into the expression vector so that it is preferentially used by E. coli (Wada et al., (1992) Nucleic Acids Res. 20: 2111-2118). Such modifications of the nucleic acid sequences of the present invention can be performed by standard DNA synthesis techniques.
別の実施形態においては、PDX発現ベクターはコウボ発現ベクターである。コウボS・セレビシアエ中における発現のためのベクターの例はpYepSec1(Baldari,et al.,(1987)EMBO J6:229−234)、pMFa(Kurjan and Herskowitz,(1982)Cell 30:933−943),pJRY88(Schultz et al.,(1987)Gene 54:113−123),pYES2(Invitrogen Corporation,San Diego,Calif.)およびpicZ(InVitrogen Corp,San Diego,Calif.)を包含する。 In another embodiment, the PDX expression vector is a yeast expression vector. Examples of vectors for expression in S. cerevisiae are pYepSec1 (Baldari, et al., (1987) EMBO J6: 229-234), pMFa (Kurjan and Herskowitz, (1982) Cell 30: 933-943), pJRY88 (Schultz et al., (1987) Gene 54: 113-123), pYES2 (Invitrogen Corporation, San Diego, Calif.) and picZ (InVitrogen Corp, San Diego, Calif.).
或いは、PDXはバキュロウィルス発現ベクターを用いて昆虫細胞内で発現できる。培養昆虫細胞(例えばSF9細胞)中の蛋白の発現のために使用できるバキュロウィルスベクターはpAcシリーズ(Smith et al.,(1983)Mol Cell Biol 3:2156−2165)およびpVLシリーズ(Lucklow and Summers(1989)Virology170:31−39)を包含する。 Alternatively, PDX can be expressed in insect cells using baculovirus expression vectors. Baculovirus vectors that can be used for the expression of proteins in cultured insect cells (eg SF9 cells) are the pAc series (Smith et al., (1983) Mol Cell Biol 3: 2156-2165) and the pVL series (Lucklow and Summers ( 1989) Virology 170: 31-39).
更に別の実施形態においては、本発明の核酸は哺乳類発現ベクターを用いて哺乳類細胞内で発現させる。哺乳類発現ベクターの例はpCDM8(Seed(1987)Nature 329:840)およびpMT2PC(Kaufman et al.(1987)EMBO J 6:187−195)を包含する。哺乳類細胞において使用する場合は、発現ベクターの制御機能はウィルス調節エレメントにより与えられる場合が多い。例えば一般的に使用されているプロモーターはポリオーマ、アデノウィルス2、サイトメガロウィルスおよびサルウイルス40に由来する。原核細胞と真核細胞双方についての他の適当な発現系については、例えばSambrook et al.,Molecular Cloning:A Laboratory Manual.2nd ed.,Cold Spring Harbor Laboratory,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,N.Y.,1989の第16および17章を参照できる。
In yet another embodiment, the nucleic acids of the invention are expressed in mammalian cells using mammalian expression vectors. Examples of mammalian expression vectors include pCDM8 (Seed (1987) Nature 329: 840) and pMT2PC (Kaufman et al. (1987) EMBO J 6: 187-195). When used in mammalian cells, the expression vector's control functions are often provided by viral regulatory elements. For example, commonly used promoters are derived from polyoma,
別の実施形態においては、組み換え哺乳類発現ベクターは特定の細胞型において優先的に核酸発現を指向することができる(例えば組織特異的調節エレメントを使用して核酸を発現させる)。組織特異的調節エレメントは当該分野で知られている。適当な組織特異的プロモーターの非限定的な例は、アルブミンプロモーター(肝特異的;Pinkert et al.(1987)Genes Dev 1:268−277)、リンパ特異的プロモーター(Calame and Eaton(1988)Adv Immunol 43:235−275)、特にT細胞受容体のプロモーター(Winoto and Baltimore(1989)EMBO J8:729−733)および免疫グロブリン(Banerji et al.,(1983)Cell 33:729−740;Queen and Baltimore(1983)Cell 33:741−748)、ニューロン特異的プロモーター(例えばニューロフィラメントプロモーター、Byrne and Ruddle(1989)PNAS 86:5473−5477)、膵特異的プロモーター(Edlund et al.,(1985)Science 230:912−916)および乳腺特異的プロモーター(例えば乳精プロモーター、米国特許4,873,316および欧州特許出願264,166)を包含する。発生調節されたプロモーターもまた包含され、例えばネズミhoxプロモーター(Kessel and Gruss(1990)Science 249:374−379)およびα−フェトプロテインプラスミド(Campes and Tilghman(1989)Genes Dev3:537−546)が挙げられる。 In another embodiment, the recombinant mammalian expression vector can direct nucleic acid expression preferentially in certain cell types (eg, use a tissue-specific regulatory element to express the nucleic acid). Tissue specific regulatory elements are known in the art. Non-limiting examples of suitable tissue specific promoters include albumin promoters (liver specific; Pinkert et al. (1987) Genes Dev 1: 268-277), lymphoid specific promoters (Callame and Eaton (1988) Adv Immunol). 43: 235-275), in particular the promoters of the T cell receptor (Winoto and Baltimore (1989) EMBO J8: 729-733) and immunoglobulins (Banerji et al., (1983) Cell 33: 729-740; Queen and Baltimore. (1983) Cell 33: 741-748), neuron-specific promoters (eg, neurofilament promoters, Byrne and Ruddle). 1989) PNAS 86: 5473-5477), pancreatic specific promoters (Edrund et al., (1985) Science 230: 912-916) and mammary gland specific promoters (eg, the whey promoter, US Pat. No. 4,873,316 and Europe) Patent application 264,166). Developmentally regulated promoters are also encompassed, including the murine hox promoter (Kessel and Gross (1990) Science 249: 374-379) and the alpha-fetoprotein plasmid (Campes and Tilghman (1989) Genes Dev 3: 537-546). .
本発明は更に、アンチセンス方向に発現ベクターにクローニングされた本発明のDNA分子を含む組み換え発現ベクターを提供する。即ち、DNA分子は、PDXmRNAに対してアンチセンスであるRNA分子の発現(DNA分子の転写による)を可能にする態様で調節配列に作動可能に連結している。アンチセンス方向にクローニングされた核酸に作動可能に連結した調節配列は種々の細胞型においてアンチセンスRNA分子の持続的発現を指向するように選択でき、例えばウィルスプロモーターおよび/またはエンハンサーまたは調節配列は、構成的な、組織特異的または細胞特異的なアンチセンスRNAの発現を指向するように選択できる。アンチセンス発現ベクターは、高効率の調節領域の制御下にアンチセンス核酸が生産される組み換えプラスミド、ファージミドまたは力価低減ウィルスの形態であることができ、その活性はベクターが導入される細胞型により決定される。アンチセンス遺伝子を用いた遺伝子発現の調節に関する考察は、Weintraub et al.,”Antisense RNA as a molecular tool for genetic analysis,”Reviews−Trends in Genetics,Vol.1(1)1986に記載されている。 The present invention further provides a recombinant expression vector comprising the DNA molecule of the present invention cloned in the antisense direction into the expression vector. That is, the DNA molecule is operably linked to a regulatory sequence in a manner that allows expression of an RNA molecule that is antisense to PDX mRNA (by transcription of the DNA molecule). Regulatory sequences operably linked to nucleic acids cloned in the antisense orientation can be selected to direct the sustained expression of antisense RNA molecules in various cell types, for example viral promoters and / or enhancers or regulatory sequences include One can select to direct expression of constitutive, tissue-specific or cell-specific antisense RNA. Antisense expression vectors can be in the form of recombinant plasmids, phagemids or titer-reducing viruses that produce antisense nucleic acids under the control of highly efficient regulatory regions, the activity depending on the cell type into which the vector is introduced. It is determined. A discussion on the regulation of gene expression using antisense genes can be found in Weintraub et al. "Antisense RNA as a molecular tool for genetic analysis," Reviews-Trends in Genetics, Vol. 1 (1) 1986.
本発明の別の特徴は、本発明の組み換え発現ベクターが導入されている宿主細胞に関する。「宿主細胞」および「組み換え宿主細胞」という用語は本明細書においては互換的に使用する。このような用語は特定の対象細胞のみならず、そのような細胞の子孫または潜在的子孫も指すものとする。突然変異または環境の影響により特定の修飾が継代の間に生じる場合があるため、そのような子孫は実際は親細胞とは同一ではない場合があるが、なお本明細書において使用する用語の範囲に包含されるものとする。更にまた、宿主細胞はPDXを発現した後にモジュレートされる場合があり、そして元の特性を維持または喪失する場合がある。 Another feature of the invention relates to host cells into which a recombinant expression vector of the invention has been introduced. The terms “host cell” and “recombinant host cell” are used interchangeably herein. Such terms are intended to refer not only to the particular subject cell, but also to the progeny or potential progeny of such a cell. Such progeny may not actually be identical to the parent cell because certain modifications may occur during passage due to mutations or environmental effects, but the scope of the terms used herein is still To be included. Furthermore, the host cell may be modulated after expressing PDX and may retain or lose its original properties.
宿主細胞は原核細胞または真核細胞であることができる。例えばPDX蛋白はE.coliのような細菌細胞、昆虫細胞、コウボまたは哺乳類の細胞(例えばチャイニーズハムスター卵巣細胞(CHO)またはCOS細胞)中で発現されることができる。或いは、宿主細胞は未成熟の哺乳類細胞、即ち多能幹細胞であることができる。宿主細胞はまた他のヒト組織に由来することできる。他の適当な宿主細胞は当該分野で知られている。 The host cell can be a prokaryotic cell or a eukaryotic cell. For example, PDX protein is E. coli. It can be expressed in bacterial cells such as E. coli, insect cells, yeast or mammalian cells (eg Chinese hamster ovary cells (CHO) or COS cells). Alternatively, the host cell can be an immature mammalian cell, ie a pluripotent stem cell. Host cells can also be derived from other human tissues. Other suitable host cells are known in the art.
ベクターDNAは従来の形質転換、形質導入、感染またはトランスフェクションの手法により原核細胞または真核細胞に導入できる。本明細書においては、「形質転換」、「形質導入」、「感染」および「トランスフェクション」という用語は宿主細胞に外来の核酸(例えばDNA)を導入するための当該分野でよく知られた手法の種々のもの、例えばリン酸カルシウムまたは塩化カルシウム共沈殿、DEAEデキストラン媒介トランスフェクション、リポフェクションまたはエレクトロポレーションを指すものとする。更にまた、トランスフェクションはトランスフェクション剤により媒介することができる。「トランスフェクション剤」とは宿主細胞内へのDNAの取り込みを媒介する何れかの化合物、例えばリポソームを包含するものを意味する。宿主細胞を形質転換またはトランスフェクトするための適当な方法はSambrook et al.(Molecular Cloning:A Laboratory Manual.2nd ed.,Cold Spring Harbor Laboratory,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,N.Y.,1989)および他の実験室マニュアルに記載されている。 Vector DNA can be introduced into prokaryotic or eukaryotic cells via conventional transformation, transduction, infection or transfection techniques. As used herein, the terms “transformation”, “transduction”, “infection” and “transfection” are well-known techniques in the art for introducing foreign nucleic acid (eg, DNA) into a host cell. A variety of such as calcium phosphate or calcium chloride coprecipitation, DEAE dextran mediated transfection, lipofection or electroporation. Furthermore, transfection can be mediated by a transfection agent. By “transfection agent” is meant any compound that mediates uptake of DNA into a host cell, such as those comprising liposomes. Suitable methods for transforming or transfecting host cells can be found in Sambrook et al. (Molecular Cloning: A Laboratory Manual. 2nd ed., Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, et al., 1989) and in the Manual of Lab., 1989).
トランスフェクションは「安定」(即ち宿主細胞ゲノムへの外来DNAの組み込み)または「一過性」(即ちDNAは宿主細胞内でエピソーム発現される)であってよい。 Transfection may be “stable” (ie, the integration of foreign DNA into the host cell genome) or “transient” (ie, the DNA is episomally expressed in the host cell).
哺乳類細胞の安定なトランスフェクションのためには、使用する発現ベクターおよびトランスフェクション方法に応じて、細胞の僅か小区分のみが外来DNAをそのゲノム内に組み込み、DNAの残余はエピソームとなることがわかっている。これらの組み込み体を識別して選択するためには、選択可能なマーカー(例えば抗生物質への耐性)をコードする遺伝子を一般的には目的の遺伝子と共に宿主細胞に導入する。種々の選択可能なマーカーには薬剤、例えばG418、ハイグロマイシンおよびメトトレキセートへの耐性を付与するものが挙げられる。選択可能なマーカーをコードする核酸は、PDXをコードするベクターと同様のベクター上で宿主細胞に導入するか、または、別のベクター上で導入することができる。導入された核酸で安定にトランスフェクトされた細胞は薬剤選択性により識別できる(例えば選択可能なマーカーを取り込んでいる細胞は生存し、他の細胞は死滅する)。別の実施形態においては、PDXによりモジュレートされた細胞またはトランスフェクトされた細胞は内因性のレポーター遺伝子の発現の誘導により識別される。特定の実施形態においては、プロモーターは緑色蛍光蛋白(GFP)の発現を駆動するインスリンプロモーターである。 For stable transfection of mammalian cells, depending on the expression vector and transfection method used, only a small fraction of the cell is found to incorporate foreign DNA into its genome and the remainder of the DNA is episomal. ing. In order to identify and select these integrants, a gene that encodes a selectable marker (eg, resistance to antibiotics) is generally introduced into the host cells along with the gene of interest. Various selectable markers include those that confer resistance to drugs such as G418, hygromycin and methotrexate. Nucleic acid encoding a selectable marker can be introduced into a host cell on a vector similar to the vector encoding PDX or on a separate vector. Cells stably transfected with the introduced nucleic acid can be distinguished by drug selectivity (eg, cells incorporating a selectable marker survive and other cells die). In another embodiment, cells modulated or transfected with PDX are identified by induction of expression of an endogenous reporter gene. In certain embodiments, the promoter is an insulin promoter that drives expression of green fluorescent protein (GFP).
1つの実施形態において、PDX核酸はウィルスベクター中に存在する。別の実施形態においては、PDX核酸はウィルス内に封入される。一部の実施形態においては、ウィルスは好ましくは種々の組織型の多能細胞、例えば造血幹細胞、ニューロン幹細胞、肝幹細胞または胚性幹細胞に感染し、好ましくはウィルスは肝向性である。「肝向性」とは、ウィルスが特異的または非特異的に肝臓の細胞を好ましくターゲティングする能力を有することを意味する。更に別の実施形態においては、ウィルスはモジュレートされた肝炎ウィルス、SV−40またはエプスタイン・バーウィルスである。更に別の実施形態においては、ウィルスはアデノウィルスである。 In one embodiment, the PDX nucleic acid is present in a viral vector. In another embodiment, the PDX nucleic acid is encapsulated within the virus. In some embodiments, the virus preferably infects pluripotent cells of various tissue types, such as hematopoietic stem cells, neuronal stem cells, hepatic stem cells or embryonic stem cells, preferably the virus is hepatic tropic. “Liver tropism” means that the virus has the ability to specifically or non-specifically target liver cells. In yet another embodiment, the virus is a modulated hepatitis virus, SV-40 or Epstein-Barr virus. In yet another embodiment, the virus is an adenovirus.
(遺伝子療法)
本発明の1つの特徴において、核酸またはその機能的誘導体をコードする核酸は遺伝子療法により投与される。遺伝子療法は対象に特定の核酸を投与することにより実施される治療法である。本発明のこの特徴において、核酸はそのコードされたペプチドを生産し、次にこれが上記した疾患または障害、例えば糖尿病の機能をモジュレートすることにより治療効果を発揮する。当該分野で使用できる遺伝子療法に関わる何れかの方法を本発明の実施において使用してよい。例えばGoldspiel et al.,1993,Clin Pharm 12:488−505を参照できる。
(Gene therapy)
In one aspect of the invention, the nucleic acid encoding the nucleic acid or functional derivative thereof is administered by gene therapy. Gene therapy is a treatment performed by administering a specific nucleic acid to a subject. In this aspect of the invention, the nucleic acid produces its encoded peptide, which then exerts a therapeutic effect by modulating the function of the diseases or disorders described above, eg, diabetes. Any method involving gene therapy that can be used in the art may be used in the practice of the present invention. See, for example, Goldspiel et al. 1993, Clin Pharm 12: 488-505.
好ましい実施形態においては、治療は適当な宿主中、上記したPDXポリペプチドまたはそのフラグメント、誘導体または類縁体の1つ以上のいずれかを発現する発現ベクターの部分である核酸を含む。特定の実施形態においては、このような核酸はPDXポリペプチドのコーディング領域に作動可能に連結しているプロモーターを保有する。プロモーターは誘導型または構成型であってよく、場合により組織特異的である。プロモーターは例えばウィルスまたは哺乳類起源であってよい。別の特定の実施形態においては、コーディング配列(および何れかの他の所望の配列)がゲノム内の所望の部位において相同組み換えを誘発する領域にフランキングされており、これにより核酸の染色体内発現をもたらすような核酸分子を使用する。例えばKoller and Smithies,1989,Proc Natl Acad Sci USA 86:8932−8935を参照できる。更に別の実施形態においては、送達される核酸はエピソームにとどまり、そして内因性の、その他の面ではサイレントな遺伝子を誘導する。 In a preferred embodiment, the treatment comprises a nucleic acid that is part of an expression vector that expresses any one or more of the above-mentioned PDX polypeptides or fragments, derivatives or analogs thereof in a suitable host. In certain embodiments, such nucleic acids carry a promoter operably linked to the coding region of the PDX polypeptide. Promoters can be inducible or constitutive and are optionally tissue specific. The promoter may be of viral or mammalian origin, for example. In another specific embodiment, the coding sequence (and any other desired sequence) is flanked by regions that induce homologous recombination at the desired site in the genome, thereby chromosomal expression of the nucleic acid. Use nucleic acid molecules that result in See, for example, Koller and Smithies, 1989, Proc Natl Acad Sci USA 86: 8932-8935. In yet another embodiment, the delivered nucleic acid remains episomal and induces an endogenous, otherwise silent gene.
治療用の核酸の患者への送達は、直接(即ち患者を核酸または核酸含有ベクターに直接曝露する)または間接(即ち細胞をまず核酸にインビトロで接触させ、次に患者に移植する)のいずれかであってよい。これらの2つの方法はそれぞれインビボまたはエクスビボの遺伝子療法として知られている。本発明の特定の実施形態においては、核酸はインビボに直接投与し、そこで発現させてコードされた産物を生産する。これは当該分野で知られた多くの方法のいずれか、例えば該核酸を適切な核酸発現ベクターの部分として構築し、それを細胞内となるような態様で投与する(例えば欠損型または減衰型のレトロウィルスまたは他のウィルスベクターを用いた感染による;米国特許4,980,286参照);ネイキッドDNAを直接注射する;微粒子衝突を使用する(例えばGene Gun(登録商標);Biolistic,DuPont)を使用する;該核酸を脂質でコーティングする;関連する細胞表面受容体/トランスフェクション剤を使用する;リポソーム、微粒子またはマイクロカプセルに封入する;核に進入することがわかっているペプチドに連結させて投与する;または目的の受容体を特異的に発現する細胞型を「ターゲティング」するために使用できる受容体媒介エンドサイトーシスを起こしやすいリガンドに連結させて投与する(例えばWu and Wu,1987,J.Biol.Chem.262:4429−4432参照)等により達成される。 Delivery of therapeutic nucleic acid to the patient is either direct (ie, exposing the patient directly to the nucleic acid or nucleic acid-containing vector) or indirectly (ie, the cell is first contacted with the nucleic acid in vitro and then transplanted into the patient). It may be. These two methods are known as in vivo or ex vivo gene therapy, respectively. In certain embodiments of the invention, the nucleic acid is administered directly in vivo where it is expressed to produce the encoded product. This can be done in any of a number of ways known in the art, such as constructing the nucleic acid as part of a suitable nucleic acid expression vector and administering it in a manner that makes it intracellular (eg, a defective or attenuated form). By infection with retrovirus or other viral vectors; see US Pat. No. 4,980,286; direct injection of naked DNA; using microparticle bombardment (eg Gene Gun®; Biolistic, DuPont) Coating the nucleic acid with lipids; using relevant cell surface receptors / transfection agents; encapsulating in liposomes, microparticles or microcapsules; administering linked to peptides known to enter the nucleus Or “target” a cell type that specifically expresses the receptor of interest. Ligated to easily ligand cause receptor-mediated endocytosis can use to be administered (e.g., Wu and Wu, 1987, J.Biol.Chem.262: 4429-4432 reference) is achieved by like.
本発明の実施における遺伝子療法の別の方法ではエレクトロポレーション、リポフェクション、リン酸カルシウム媒介トランスフェクション、ウィルス感染等の方法によりインビトロの組織培養において細胞に遺伝子を転移させる。一般的に転移の方法は細胞への選択可能なマーカーの同時転移を包含する。次に転移された遺伝子を取り込んで発現している細胞の単離を容易にするために、細胞を淘汰圧(例えば抗生物質耐性)下に付す。次にこれらの細胞を患者に送達する。特定の実施形態においては、得られた組み換え細胞のインビボの投与の前に、核酸を当該分野で知られた方法、例えば目的の核酸配列を含有するウィルスまたはバクテリオファージベクターを用いたトランスフェクション、エレクトロポレーション、マイクロインジェクション、感染、細胞融合、染色体媒介遺伝子転移、マイクロセル媒介遺伝子転移、スフェロプラスト融合、および、レシピエント細胞の必要な発生生理機能が転移により損なわれない同様の方法により、細胞内に核酸を導入する。例えばLoeffler and Behr,1993,Meth Enzymol 217:599−618を参照できる。選択された手法は、細胞により核酸が発現され得るような、細胞への核酸の安定な転移をもたらすものでなければならない。好ましくは、該転移核酸は細胞の子孫に遺伝可能であり発現され得るものである。別の実施形態においては、転移した核酸はエピソームにとどまり、その他の面ではサイレントな内因性核酸の発現を誘導する。 In another method of gene therapy in the practice of the present invention, genes are transferred to cells in in vitro tissue culture by methods such as electroporation, lipofection, calcium phosphate mediated transfection, viral infection and the like. In general, the method of transfer involves the simultaneous transfer of selectable markers to cells. The cells are then subjected to selection pressure (eg, antibiotic resistance) to facilitate isolation of cells that have taken up and are expressing the transferred gene. These cells are then delivered to the patient. In certain embodiments, prior to in vivo administration of the resulting recombinant cells, the nucleic acid is transferred to a method known in the art, such as transfection with a virus or bacteriophage vector containing the nucleic acid sequence of interest, electro Cells by poration, microinjection, infection, cell fusion, chromosome-mediated gene transfer, microcell-mediated gene transfer, spheroplast fusion, and similar methods in which the necessary developmental physiology of the recipient cell is not impaired by the transfer Nucleic acid is introduced into the inside. See, for example, Loeffler and Behr, 1993, Meth Enzymol 217: 599-618. The technique selected should result in a stable transfer of the nucleic acid into the cell such that the nucleic acid can be expressed by the cell. Preferably, the transfer nucleic acid is inheritable and can be expressed in the progeny of the cell. In another embodiment, the transferred nucleic acid remains episomal and induces endogenous nucleic acid expression that is otherwise silent.
本発明の好ましい実施形態においては、得られた組み換え細胞を当該分野で知られた種々の方法、例えば上皮細胞の注射(例えば皮下)、患者への皮膚移植片としての組み換え皮膚細胞の適用、および組み換え血球(例えば造血幹細胞または前駆細胞)または肝細胞の静脈内注射により患者に送達する。使用予定となる細胞の総量は所望の作用、患者の状態等により変動し、そして当業者が決定してよい。 In a preferred embodiment of the present invention, the resulting recombinant cells are subjected to various methods known in the art, such as injection of epithelial cells (eg subcutaneously), application of recombinant skin cells as a skin graft to a patient, and Delivered to the patient by intravenous injection of recombinant blood cells (eg, hematopoietic stem or progenitor cells) or hepatocytes. The total amount of cells to be used will vary depending on the desired effect, patient condition, etc. and may be determined by one skilled in the art.
遺伝子療法の目的のために核酸を導入することができる細胞は何れかの所望の入手可能な細胞型を包含し、そして外因、異種、同種または自原のものであってよい。細胞型は例えば分化した細胞、例えば上皮細胞、内皮細胞、ケラチノサイト、線維芽細胞、筋細胞、肝細胞および血球、または、種々の幹細胞または前駆細胞、特に胚性心筋細胞、肝幹細胞(国際特許出願WO94/08598)、神経幹細胞(Stemple and Anderson,1992,Cell 71:973−985)、造血幹細胞または前駆細胞、例えば骨髄、臍帯血、末梢血、胎児肝臓から得られるもの等を包含する。好ましい実施形態においては、遺伝子療法のために使用される細胞は患者自家のものである。 Cells into which the nucleic acid can be introduced for gene therapy purposes include any desired available cell type and may be exogenous, xenogeneic, allogeneic or autologous. The cell types are eg differentiated cells such as epithelial cells, endothelial cells, keratinocytes, fibroblasts, myocytes, hepatocytes and blood cells, or various stem or progenitor cells, especially embryonic cardiomyocytes, hepatic stem cells (international patent applications) WO94 / 08598), neural stem cells (Steple and Anderson, 1992, Cell 71: 973-985), hematopoietic stem cells or progenitor cells such as those obtained from bone marrow, umbilical cord blood, peripheral blood, fetal liver and the like. In a preferred embodiment, the cells used for gene therapy are the patient's own.
遺伝子療法のためのDNAは患者に対し、非経口的、例えば静脈内、皮下、筋肉内および腹腔内に投与できる。DNAまたは誘導剤は製薬上許容しうる担体、即ち動物への投与に適する生体適合性のあるベヒクル、例えば生理食塩水中で投与する。治療有効量は治療される動物において医療上望ましい結果、例えばPDXまたは遺伝子産物の増大または低減をもたらすことができる量である。このような量は当業者により決定できる。医療分野で知られている通り、何れかの所定の患者に対する用量は多くの要因、例えば患者の体格、体表面積、年齢、投与する特定の化合物、性別、投与の時間および経路、全身状態および同時に投与する他剤に応じて変動する。用量は変動するが、DNAの静脈内投与のための好ましい用量はDNA分子約106〜1022コピーである。例えばDNAはkg当たり約2x1012ビリオンで投与する。 DNA for gene therapy can be administered to patients parenterally, for example intravenously, subcutaneously, intramuscularly and intraperitoneally. The DNA or inducer is administered in a pharmaceutically acceptable carrier, i.e. a biocompatible vehicle suitable for administration to animals, such as saline. A therapeutically effective amount is an amount that can result in a medically desirable result in the treated animal, such as an increase or decrease in PDX or gene product. Such amount can be determined by one skilled in the art. As is known in the medical field, the dose for any given patient will depend on many factors such as the patient's size, body surface area, age, the particular compound being administered, sex, time and route of administration, general condition and simultaneously. It varies depending on other drugs to be administered. Although the dose varies, the preferred dose for intravenous administration of DNA is about 10 6 to 10 22 copies of the DNA molecule. For example, DNA is administered at about 2 × 10 12 virions per kg.
(医薬組成物)
本発明の化合物、例えばPDXポリペプチド、PDXポリペプチドをコードする核酸、PDXポリペプチドをコードする核酸の発現を増大させる核酸(本明細書においては「活性化合物」とも称する)およびその誘導体、フラグメント、類縁体および相同体は投与に適する医薬組成物に配合できる。このような組成物は典型的には核酸分子、または蛋白および製薬上許容しうる担体を含む。本明細書においては、「製薬上許容しうる担体」とは医薬品投与に適合する溶媒、分散媒体、コーティング、抗細菌剤および抗カビ剤、等張性付与および吸収遅延剤等のいずれかおよび全てを包含する。適当な担体は参照により本明細書に組み込まれるこの分野の標準的な参考書であるRemington’s Pharmaceutical Sciencesの最新版に記載されている。このような担体または希釈剤の好ましい例は、水、生理食塩水、フィンガー溶液、デキストロース溶液、および5%ヒト血清アルブミンを包含する。リポソームおよび非水性のベヒクル、例えば固定油も使用してよい。薬学的に活性な物質のためのこのような媒体および試薬の使用は当該分野でよく知られている。何れかの従来の媒体または試薬が活性化合物と非適合でない限りにおいて、組成物中のその使用が意図される。補助的な活性化合物もまた組成物に配合できる。
(Pharmaceutical composition)
Compounds of the invention, eg, PDX polypeptides, nucleic acids encoding PDX polypeptides, nucleic acids that increase the expression of nucleic acids encoding PDX polypeptides (also referred to herein as “active compounds”) and derivatives, fragments thereof, Analogs and homologues can be formulated into pharmaceutical compositions suitable for administration. Such compositions typically comprise a nucleic acid molecule, or protein, and a pharmaceutically acceptable carrier. As used herein, “pharmaceutically acceptable carrier” refers to any and all of solvents, dispersion media, coatings, antibacterial and antifungal agents, isotonicity and absorption delaying agents, and the like that are compatible with pharmaceutical administration. Is included. Suitable carriers are described in the latest edition of Remington's Pharmaceutical Sciences, a standard reference book in this field, incorporated herein by reference. Preferred examples of such carriers or diluents include water, saline, finger solution, dextrose solution, and 5% human serum albumin. Liposomes and non-aqueous vehicles such as fixed oils may also be used. The use of such media and reagents for pharmaceutically active substances is well known in the art. As long as any conventional medium or reagent is not incompatible with the active compound, its use in the composition is contemplated. Supplementary active compounds can also be incorporated into the compositions.
本発明の医薬組成物はその投与の意図される経路に適合するように製剤される。投与経路の例は非経口、例えば静脈内、皮内、皮下、経口(例えば吸入)、経皮(局所)、経粘膜および直腸投与を包含する。非経口、経皮または皮下の適用のために使用される溶液または懸濁液は以下の成分、即ち滅菌希釈剤、例えば注射用水、生理食塩水、固定油、ポリエチレングリコール、グリセリン、プロピレングリコールまたは他の合成の溶媒、抗細菌剤、例えばベンジルアルコールまたはメチルパラベン、抗酸化剤、例えばアスコルビン酸または重亜硫酸ナトリウム、キレート剤、例えばエチレンジアミン4酢酸、緩衝剤、例えば酢酸塩、クエン酸塩またはリン酸塩、および、浸透圧を調節するための薬剤、例えば塩化ナトリウムまたはデキストロースを包含する。pHは酸または塩基、例えば塩酸または水酸化ナトリウムで調節できる。非経口製剤はガラスまたはプラスチック製のアンプル、使い捨てシリンジまたは多用量のバイアル中に充填できる。 A pharmaceutical composition of the invention is formulated to be compatible with its intended route of administration. Examples of routes of administration include parenteral, eg intravenous, intradermal, subcutaneous, oral (eg inhalation), transdermal (topical), transmucosal and rectal administration. Solutions or suspensions used for parenteral, transdermal or subcutaneous applications may contain the following ingredients: sterile diluents such as water for injection, saline, fixed oil, polyethylene glycol, glycerin, propylene glycol or others Synthetic solvents, antibacterial agents such as benzyl alcohol or methyl paraben, antioxidants such as ascorbic acid or sodium bisulfite, chelating agents such as ethylenediaminetetraacetic acid, buffers such as acetate, citrate or phosphate, And agents for regulating osmotic pressure, such as sodium chloride or dextrose. The pH can be adjusted with acids or bases, such as hydrochloric acid or sodium hydroxide. The parenteral preparation can be enclosed in ampoules, disposable syringes or multiple dose vials made of glass or plastic.
注射用に適した医薬組成物は滅菌水溶液(水溶性の場合)または分散液および滅菌注射溶液または分散液の即時調製用の滅菌粉末を含む。静脈内投与のためには、適当な担体は生理食塩水、殺菌水、CremophorEL(登録商標)(BASF,Parsippany,NJ)またはリン酸塩緩衝食塩水(PBS)を包含する。全ての場合において、組成物は滅菌されなければならず、そして注射針を容易に通過できる程度の流動性を有さなければならない。製造および保存条件下で安定であり、細菌やカビのような微生物による汚染に抵抗して保存されなければならない。担体は例えば水、エタノール、ポリオール(例えばグリセロール、プロピレングリコールおよび液体ポリエチレングリコール等)およびこれらの適当な混合物を含有する溶媒または分散媒体であることができる。適切な流動性は例えばレシチンのようなコーティングの使用により、分散液の場合は所望の粒径を維持することにより、そして、界面活性剤の使用により維持することができる。微生物の作用の防止は種々の抗細菌剤および抗カビ剤、例えばパラベン、クロロブタノール、フェノール、アスコルビン酸、チメロサール等により達成できる。多くの場合において、等張性付与剤、例えば糖類、多価アルコール、例えばマンニトール、ソルビトール、塩化ナトリウムを組成物中に含有させることが好ましい。注射用組成物の長時間持続する吸収は吸収を遅延させる物質、例えばモノステアリン酸アルミニウムおよびゼラチンを組成物に含有させることにより達成できる。 Pharmaceutical compositions suitable for injection include sterile aqueous solutions (where water soluble) or dispersions and sterile powders for the extemporaneous preparation of sterile injectable solutions or dispersion. For intravenous administration, suitable carriers include physiological saline, sterile water, Cremophor EL® (BASF, Parsippany, NJ) or phosphate buffered saline (PBS). In all cases, the composition must be sterile and must be fluid to the extent that easy syringability exists. It must be stable under the conditions of manufacture and storage and must be preserved against contamination by microorganisms such as bacteria and mold. The carrier can be a solvent or dispersion medium containing, for example, water, ethanol, polyol (for example, glycerol, propylene glycol, and liquid polyethylene glycol, and the like), and suitable mixtures thereof. The proper fluidity can be maintained, for example, by the use of a coating such as lecithin, by the maintenance of the desired particle size in the case of dispersion and by the use of surfactants. Prevention of the action of microorganisms can be achieved by various antibacterial and antifungal agents, for example, parabens, chlorobutanol, phenol, ascorbic acid, thimerosal, and the like. In many cases, it will be preferable to include isotonic agents, for example, sugars, polyalcohols such as mannitol, sorbitol, sodium chloride in the composition. Long-lasting absorption of injectable compositions can be achieved by including in the composition an agent that delays absorption, for example, aluminum monostearate and gelatin.
適当な注射用溶液は所望により上記した成分の1つまたは組み合わせと共に適切な溶媒中に必要量の活性化合物(例えばPDXポリペプチドまたはPDXコード核酸)を配合すること、ついで、濾過滅菌することにより調製できる。一般的に、分散液は基剤としての分散媒体および上記した物質のうち必要な他の成分を含有する滅菌ベヒクル内に活性化合物を配合することにより調製する。滅菌注射用溶液の調製のための滅菌粉末の場合は、調製方法は、あらかじめ滅菌濾過された溶液から得られる活性成分と別の所望の成分の粉末を与えるような真空乾燥または凍結乾燥である。 Appropriate injectable solutions are prepared by formulating the required amount of the active compound (eg PDX polypeptide or PDX encoding nucleic acid) in an appropriate solvent, optionally together with one or a combination of the above components, and then filter sterilizing. it can. Generally, dispersions are prepared by incorporating the active compound into a sterile vehicle that contains a dispersion medium as a base and the required other ingredients from those enumerated above. In the case of sterile powders for the preparation of sterile injectable solutions, the method of preparation is vacuum or lyophilized to give a powder of the active ingredient and another desired ingredient obtained from a pre-sterilized filtered solution.
経口用組成物は一般的に不活性希釈剤または食用担体を含有する。それらはゼラチンカプセルに封入するか、圧縮成型して錠剤とすることができる。経口投与治療の目的のためには、活性化合物は賦形剤に配合し、錠剤、トローチまたはカプセルの形態で使用する。経口用組成物はまた、液体担体中の化合物を口腔内に適用し、嗽をし、吐出または嚥下する洗口剤として使用するための液体担体を用いて調製できる。薬学的に適合する結合剤および/または補助的物質を組成物の部分として含有できる。錠剤、丸薬、カプセル、トローチ等は何れかの以下の成分または同様の性質の化合物、即ち、結合剤、例えば微結晶セルロース、トラガカントガムまたはゼラチン;賦形剤、例えば澱粉または乳糖、崩壊剤、例えばアルギン酸、プリモゲルまたはコーンスターチ;潤滑剤、例えばステアリン酸マグネシウムまたはSterote;滑剤、例えばコロイド状に酸化ケイ素;甘味剤、例えばスクロースまたはサッカリン;またはフレーバー剤、例えばペパーミント、サリチル酸メチルまたはオレンジフレーバーを含有できる。 Oral compositions generally contain an inert diluent or an edible carrier. They can be enclosed in gelatin capsules or compressed into tablets. For the purpose of oral therapeutic treatment, the active compound is incorporated with excipients and used in the form of tablets, troches, or capsules. Oral compositions can also be prepared using a liquid carrier for use as a mouthwash to apply the compound in a liquid carrier to the oral cavity, to wrinkle, expel or swallow. Pharmaceutically compatible binding agents, and / or adjuvant materials can be included as part of the composition. Tablets, pills, capsules, troches, etc. are any of the following ingredients or compounds of similar properties, ie binders such as microcrystalline cellulose, tragacanth gum or gelatin; excipients such as starch or lactose, disintegrants such as alginic acid , Primogel or corn starch; lubricants such as magnesium stearate or Sterote; lubricants such as colloidal silicon oxide; sweeteners such as sucrose or saccharin; or flavoring agents such as peppermint, methyl salicylate or orange flavor.
吸入による投与のためには、化合物は適当な高圧ガス、例えば二酸化炭素の入った加圧容器またはディスペンサー、または、ネブライザーからエアゾールスプレーの形態で送達される。 For administration by inhalation, the compounds are delivered in the form of an aerosol spray from pressured container or dispenser with a suitable high pressure gas, such as carbon dioxide, or a nebulizer.
全身投与は経粘膜または経皮の手段によっても達成される。経粘膜または経皮の投与のためには、浸透すべき障壁に対して適切である浸透剤を製剤中に使用する。このような浸透剤は当該分野で知られており、例えば経粘膜投与のためには、洗剤、胆汁酸塩およびフシジン酸の誘導体が挙げられる。経粘膜投与は鼻スプレーまたは座剤の使用により達成できる。経皮投与のためには、活性化合物は当該分野で一般的に知られるとおり軟膏、膏薬、ゲルまたはクリーム中に処方する。 Systemic administration can also be accomplished by transmucosal or transdermal means. For transmucosal or transdermal administration, penetrants appropriate to the barrier to be permeated are used in the formulation. Such penetrants are known in the art and include, for example, for transmucosal administration, detergents, bile salts and fusidic acid derivatives. Transmucosal administration can be accomplished through the use of nasal sprays or suppositories. For transdermal administration, the active compounds are formulated into ointments, salves, gels, or creams as generally known in the art.
化合物はまた、直腸投与のための座剤(例えば従来の座剤用基剤、例えばカカオ脂または他のグリセリドを使用)または保持浣腸剤の形態に調製できる。 The compounds can also be prepared in the form of suppositories (eg, using conventional suppository bases such as cocoa butter or other glycerides) or retention enemas for rectal administration.
1つの実施形態において、活性化合物は身体からの急速な排除に対して化合物を保護する担体と共に調製され、例えば、インプラントおよびマイクロカプセルの送達系などの制御放出製剤が挙げられる。生体分解性、生体適合性の重合体、例えばエチレンビニルアセテート、ポリ無水物、ポリグリコール酸、コラーゲン、ポリオルトエステルおよびポリ乳酸を使用できる。このような製剤の調製方法は当業者の知るとおりである。材料はまたAlza CorporationおよびNova Pharmaceuticals,Inc.より購入可能である。リポソーム懸濁液(ウィルス抗原に対するモノクローナル抗体を用いて感染細胞にターゲティングされたリポソームも含む)もまた製薬上許容しうる担体として使用できる。これらは当該分野で知られた方法に従って、例えば、参照によりその全体が本明細書に組み込まれる米国特許4,522,811に記載の通り調製できる。 In one embodiment, the active compounds are prepared with carriers that will protect the compound against rapid elimination from the body, including, for example, controlled release formulations such as implants and microcapsule delivery systems. Biodegradable, biocompatible polymers can be used, such as ethylene vinyl acetate, polyanhydrides, polyglycolic acid, collagen, polyorthoesters, and polylactic acid. Those skilled in the art know how to prepare such formulations. Materials are also available from Alza Corporation and Nova Pharmaceuticals, Inc. More can be purchased. Liposomal suspensions (including liposomes targeted to infected cells using monoclonal antibodies against viral antigens) can also be used as pharmaceutically acceptable carriers. These can be prepared according to methods known in the art, for example, as described in US Pat. No. 4,522,811, incorporated herein by reference in its entirety.
投与の容易さおよび用量の均一性のためには、経口または非経口の組成物を単位剤型で製剤することが特に好都合である。本明細書においては単位剤型とは治療すべき対象に対する統一された用量に適する物理的に個別の単位を指し、各単位は所望の製薬上許容しうる担体と共に所望の治療効果を得るために計算された活性化合物の所定量を含有する。本発明の単位剤型の詳細は活性化合物の独特の性質および達成すべき特定の治療効果により決定され、これに直接依存している。 It is especially advantageous to formulate oral or parenteral compositions in dosage unit form for ease of administration and uniformity of dosage. As used herein, a unit dosage form refers to a physically discrete unit suitable for a unified dosage for a subject to be treated, each unit together with a desired pharmaceutically acceptable carrier to obtain a desired therapeutic effect. Contains a predetermined amount of calculated active compound. The details of the unit dosage form of the present invention are determined by and depend directly on the unique nature of the active compound and the particular therapeutic effect to be achieved.
本発明の核酸分子はベクター内に挿入し、遺伝子療法ベクターとして使用できる。遺伝子療法ベクターは例えば米国特許5,703,055に記載の多くの経路の何れかにより対象に送達できる。即ち送達はまた、例えば静脈注射、局所投与(米国特許5,328,470参照)または定位注射(例えばChen et al.,(1994)PNAS 91:3054−3057参照)を包含できる。遺伝子療法ベクターの医薬品調製物は適当な希釈剤中に遺伝子療法ベクターを含有するか、または、遺伝子送達ベヒクルを包埋した緩徐放出マトリックスを含むことができる。或いは、完全な遺伝子送達ベクター、例えばレトロウィルスベクターを組み換え細胞から未損傷で製造できる場合は、医薬品製剤は遺伝子送達系を与える細胞1つ以上を含むことができる。 The nucleic acid molecules of the invention can be inserted into vectors and used as gene therapy vectors. Gene therapy vectors can be delivered to a subject by any of the many routes described, for example, in US Pat. Thus delivery can also include, for example, intravenous injection, topical administration (see US Pat. No. 5,328,470) or stereotaxic injection (see, eg, Chen et al., (1994) PNAS 91: 3054-3057). The gene therapy vector pharmaceutical preparation may contain the gene therapy vector in a suitable diluent or may include a slow release matrix embedded with the gene delivery vehicle. Alternatively, where a complete gene delivery vector, such as a retroviral vector, can be produced intact from recombinant cells, the pharmaceutical formulation can include one or more cells that provide a gene delivery system.
医薬組成物は、投与の説明書と共に、容器、パックまたはディスペンサー内に入れることができる。 The pharmaceutical composition can be placed in a container, pack or dispenser together with instructions for administration.
本発明は以下の実施例において更に説明するが、これは請求項に記載した本発明を制限するものではない。 The invention will be further described in the following examples, which do not limit the invention described in the claims.
(実施例1:一般的方法)
以下の一般的方法を用いて本明細書に記載した実験を実施した。
(Example 1: General method)
The experiments described herein were performed using the following general method.
(組み換えアデノウィルス)
AdCMVPDX−1はR.Seijffers et al.,Endocrinology 140:1133(1999)に記載の通り構築した。これはSTF−1 cDNA、pACCMVpLpAベクターのBamH1部位にライゲーションされたPDX−1のラット相同体を含んでいる。
(Recombinant adenovirus)
AdCMVPDX-1 is R.I. Seijffers et al. , Endocrinology 140: 1133 (1999). This includes the rat homologue of PDX-1 ligated to the BamH1 site of the STF-1 cDNA, pACCMVpLpA vector.
AdCMVβ−galはβ−ガラクトシダーゼの核局在化シグナルを含む。 AdCMV β-gal contains the nuclear localization signal of β-galactosidase.
AdCMV−hInsは非相同のサイトメガロウィルスプロモーターの制御下のヒトインスリンcDNAを含む。 AdCMV-hIns contains human insulin cDNA under the control of a heterologous cytomegalovirus promoter.
AdRIP−1−hInsはラットインスリンプロモーター−1(RIP−1)の制御下のヒトインスリンcDNAを含む。RIP−1はラットインスリン−1遺伝子の5’フランキングDNA領域の410塩基である。 AdRIP-1-hIns contains human insulin cDNA under the control of rat insulin promoter-1 (RIP-1). RIP-1 is 410 bases in the 5 'flanking DNA region of the rat insulin-1 gene.
全ての第1世代の組み換えアデノウィルスはBecker et al,39に従って構築した。目的の遺伝子はpACCMV.pLpAプラスミドにライゲーションし、その後アデノウィルスプラスミドpJM17と同時遺伝子導入し、その後HEK−293細胞から組み換えビリオンを回収した。Ad−CMV−PDX−1はPDX−1のラット相同体を有し、Ad−CMV−hInsはCMVプロモーターから遠位にヒトインスリンcDNAを有する40。Ad−RIP−GFP−CMV−PDX−1はGFP遺伝子発現を駆動するpACCMV.pLpAプラスミド内のウィルスCMVプロモーター(HindIII/PstI)の代わりにラットインスリン−1遺伝子(Dr.Larry Mossより提供)の5’−フランキング領域の410ヌクレオチド、ついで、pACCMV−PDX−1.pLpAプラスミド由来のCMV−mPDX−1(PDX−1のマウス相同体)を含むNotI/NotIインサートの挿入により調製される2官能性の組み換えアデノウィルスである40。組み換えウィルスの特定の機能は非β細胞系統と比較した場合のインスリノーマ細胞系統において分析した。ウィルス保存株の調製は以前に報告40されている通り実施した。 All first generation recombinant adenoviruses were constructed according to Becker et al, 39 . The gene of interest is pACCMV. Ligation into the pLpA plasmid followed by co-transfection with the adenovirus plasmid pJM17 followed by recovery of the recombinant virions from HEK-293 cells. Ad-CMV-PDX-1 has a rat homologue of PDX-1, Ad-CMV-hIns has human insulin cDNA distal from the CMV promoter 40 . Ad-RIP-GFP-CMV-PDX-1 is a pACCMV. 410 nucleotides of the 5′-flanking region of the rat insulin-1 gene (provided by Dr. Larry Moss) instead of the viral CMV promoter (HindIII / PstI) in the pLpA plasmid, followed by pACCMV-PDX-1. A bifunctional recombinant adenovirus prepared by insertion of a NotI / NotI insert containing CMV-mPDX-1 (a mouse homolog of PDX-1) from the pLpA plasmid 40 . The specific function of the recombinant virus was analyzed in an insulinoma cell line compared to a non-β cell line. Preparation of virus stocks was performed as previously reported 40 .
(細胞培養)
マウス膵誘導細胞系統β−TC−1、α−TC−1およびラット膵細胞系統RIN1046−38を以前に報告されている条件(21,22)に従って培養した。
(Cell culture)
Mouse pancreatic derived cell lines β-TC-1, α-TC-1 and rat pancreatic cell line RIN 1046-38 were cultured according to previously reported conditions (21, 22).
(動物および組み換えアデノウィルス)
マウスを空調された環境下、12時間の照明/消灯周期の下に飼育し、研究用動物の福祉に関する規制に従って取り扱った。Balb/cマウス(8〜9週齢、24〜27gr)に対し、尾静脈内への全身投与により所定の組み換えアデノウィルス1〜5x1010moiを投与した(生理食塩水200〜300μlの容量)。尾部より採血し、グルコース濃度を測定した(Accutrend(登録商標)GC,Boehringer Mannheim,Mannheim,Germany)。免疫組織化学的染色(4%ホルムアルデヒド固定、パラフィン包埋)用、遺伝子発現分析(全RNA)用、および、肝中の膵ホルモン含量測定用に肝を採取した。後者2分析については、肝標本を即座に液体窒素中で凍結し、−70℃で保存した。
(Animals and recombinant adenoviruses)
Mice were housed in an air-conditioned environment with a 12-hour light on / off cycle and handled according to regulations on the welfare of research animals. Balb / c mice (8-9 weeks old, 24-27 gr) were administered predetermined recombinant adenovirus 1-5 × 10 10 moi by systemic administration into the tail vein (volume of physiological saline 200-300 μl). Blood was collected from the tail and the glucose concentration was measured (Accutrend® GC, Boehringer Mannheim, Mannheim, Germany). Livers were collected for immunohistochemical staining (4% formaldehyde fixation, paraffin embedding), gene expression analysis (total RNA), and measurement of pancreatic hormone content in the liver. For the latter two analyses, liver specimens were immediately frozen in liquid nitrogen and stored at -70 ° C.
雄性NOD/LtJおよびNOD/Scidマウスを、Weizmann Institute of ScienceのAnimal Breeding Center内において病原体非存在条件下で飼育した。実験はAnimal Welfare Committeeの監督と指針の下に実施した。マウスは実験開始時には1ヶ月齢であった。 Male NOD / LtJ and NOD / Scid mice were bred under pathogen-free conditions in the Animal Breeding Center at Weizmann Institute of Science. The experiment was conducted under the direction and guidance of the Animal Welfare Committee. The mice were 1 month old at the start of the experiment.
(ヒト肝細胞)
成熟ヒト肝組織を4〜10歳の小児に由来する異なる肝臓移植手術8例および40歳超の患者2人より得た。
(Human hepatocytes)
Mature human liver tissue was obtained from 8 different liver transplant operations from 2-10 year old children and 2 patients over 40 years old.
胎児ヒト肝は妊娠20〜22週の異なる中絶4例より得た。成熟および胎児の肝組織は両方ともCommittee on Clinical Investigations(施設内治験審査委員会)の許可を得て使用した。 Fetal human liver was obtained from 4 different abortions at 20-22 weeks of gestation. Both mature and fetal liver tissue were used with the permission of the Committee on Clinical Investigations (Institutional Review Board).
(細胞の採取および培養の条件)
ヒト肝細胞の単離は以前に報告41されている通り実施した。要約すれば、肝試料を冷Hanks Balanced Salt溶液(HBSS)で灌流し、薄片(厚み1〜2mm)に切り出し、37℃で20分間0.03%コラゲナーゼI型(Worthington Biochemical Corp.)で消化した。解離した細胞を採取しHBSS+EGTA(5mM)で2回洗浄し、4℃で5分間500xgで遠心分離することにより収集した。細胞を10%FCS、100U/mlペニシリン、100μg/mlストレプトマイシンおよび250ng/mlアンホテリシンB(Biological Industries,Israel)を添加したDulbeccoの最小必須培地(1gr/Lグルコース)に再懸濁した。細胞の生存性と数を測定し、細胞をフィブロネクチン(3μg/cm2、Biological Industries,Israel)予備コーティングプレート上に、1〜2x105個/mlでプレーティングした。最初の3日間は毎日培地を交換することにより非付着細胞を除去した。コンフルエントとなった培養物は0.05%トリプシン−EDTA溶液を用いて1:3に分割した。細胞は5%CO2および95%空気の湿潤雰囲気下37℃に維持した。
(Cell collection and culture conditions)
Isolation of human hepatocytes was performed as previously reported 41 . In summary, liver samples were perfused with cold Hanks Balanced Salt solution (HBSS), cut into slices (thickness 1-2 mm) and digested with 0.03% collagenase type I (Worthington Biochemical Corp.) for 20 minutes at 37 ° C. . Dissociated cells were collected, washed twice with HBSS + EGTA (5 mM), and collected by centrifugation at 500 × g for 5 minutes at 4 ° C. The cells were resuspended in Dulbecco's minimal essential medium (1 gr / L glucose) supplemented with 10% FCS, 100 U / ml penicillin, 100 μg / ml streptomycin and 250 ng / ml amphotericin B (Biological Industries, Israel). Cell viability and number were measured and cells were plated at 1-2 × 10 5 cells / ml on fibronectin (3 μg / cm 2 , Biological Industries, Israel) pre-coated plates. Non-adherent cells were removed by changing the medium daily for the first 3 days. Confluent cultures were split 1: 3 using 0.05% trypsin-EDTA solution. Cells were maintained at 37 ° C. in a humidified atmosphere of 5% CO 2 and 95% air.
(ウィルス感染および成長因子の投与)
肝細胞を個別に示すとおり成長因子の存在下または非存在下(EGF20ng/ml,Cytolab Ltd.;ニコチンアミド10mM、Sigma)でDulbeccoの最小必須培地(1gr/Lグルコース)中培養し、5日間組み換えアデノウィルス(500moi)を感染させた。本試験で使用した組み換えアデノウィルスはAd−RIP−GFP−CMV−PDX−1、Ad−CMV−PDX−1、Ad−CMV−hIns40、Ad−CMV−GFP(Clontech,BD Biosciences)であった。
(Virus infection and administration of growth factors)
Hepatocytes were cultured individually in Dulbecco's minimal essential medium (1 gr / L glucose) in the presence or absence of growth factors (
(RNA単離およびRT−PCR分析)
全RNAをTri−Reagent(Molecular Research Center,Ohio)を用いて凍結組織から単離した。RNA試料を60分間RQ1 RNase非含有DNaseI(Promega)10単位で処理した。cDNAは4μgのDNA非含有全RNAおよび0.5μgオリゴ(dT)15を用いて逆転写(ネイティブAMV逆転写酵素、Chimerx)により調製した。PCRはT3サーモサイクラーを(Biometra,Gottingen,Germany)を用いて実施し、産物は1.8%アガロースゲル上で分離し、臭化エチジウムで可視化した。PCRのために使用したプライマーの配列および反応条件は表1および表3に示すとおりである。内因性マウスPDX−1の発現と異所性ラット相同体の間の判別のために、2セットの特異的オリゴヌクレオチドプライマーを設計した(表1および表3参照)ことに留意されたい。
(RNA isolation and RT-PCR analysis)
Total RNA was isolated from frozen tissue using Tri-Reagent (Molecular Research Center, Ohio). RNA samples were treated with 10 units of RQ1 RNase-free DNase I (Promega) for 60 minutes. cDNA was prepared by reverse transcription (native AMV reverse transcriptase, Chimerx) using 4 μg of DNA-free total RNA and 0.5 μg oligo (dT) 15 . PCR was performed using a T3 thermocycler (Biometra, Gottingen, Germany) and the products were separated on a 1.8% agarose gel and visualized with ethidium bromide. The primer sequences and reaction conditions used for PCR are as shown in Tables 1 and 3. Note that two sets of specific oligonucleotide primers were designed for discrimination between endogenous mouse PDX-1 expression and ectopic rat homologues (see Tables 1 and 3).
(リアルタイムPCR)
RT−PCRはSYBR−Green I染料を用いてLightCycler(Roche Applied Science, Mannheim,Germany)上で実施した。
(Real-time PCR)
RT-PCR was performed on a LightCycler (Roche Applied Science, Mannheim, Germany) using SYBR-Green I dye.
使用した増幅条件は初期の変性を95℃10分間、ついでマウスおよびラットPDX−1の両方につき55サイクルまたはβ−アクチンにつき30サイクルとした。両方のPDX−1相同体につき、各サイクルの内容は、変性95℃15秒、アニーリング59℃および伸長72℃15秒とした。β−アクチンのアニーリングは56℃で10分間行った。蛍光シグナルは各サイクル後に5秒間、β−アクチンでは90℃、マウスPDX−1では87℃およびラットPDX−1では88℃でモニタリングした。融解曲線プログラムは68℃で40秒行い、発生した産物の特異性を分析した。 The amplification conditions used were initial denaturation at 95 ° C. for 10 minutes, then 55 cycles for both mouse and rat PDX-1, or 30 cycles for β-actin. For both PDX-1 homologues, the content of each cycle was denaturation 95 ° C. for 15 seconds, annealing 59 ° C., and extension 72 ° C. for 15 seconds. Annealing of β-actin was performed at 56 ° C. for 10 minutes. The fluorescence signal was monitored for 5 seconds after each cycle at 90 ° C. for β-actin, 87 ° C. for mouse PDX-1 and 88 ° C. for rat PDX-1. The melting curve program was carried out at 68 ° C. for 40 seconds, and the specificity of the generated product was analyzed.
マウスPDX−1のRT−PCRは3回実施し、ラットPDX−1は2回実施した。 Mouse PDX-1 RT-PCR was performed 3 times and rat PDX-1 was performed twice.
ラットおよびマウスのPDX−1の濃度は共に同じ試料におけるそれぞれのβ−アクチンmRNA濃度に対して規格化した。 Both rat and mouse PDX-1 concentrations were normalized to their respective β-actin mRNA concentrations in the same sample.
或いは、ABI Prism 7000配列検出システム(Applied Biosystems)を用いて定量的リアルタイムRT−PCRを実施した。 Alternatively, quantitative real-time RT-PCR was performed using an ABI Prism 7000 sequence detection system (Applied Biosystems).
本試験において使用したTaqMan(登録商標)蛍光発生プローブ、Assay−On−Demand(Applied Biosystems)は、ヒトβ−アクチンHs99999903_ml、ヒトインスリンHs00355773_ml、ヒトグルカゴンHs00174967_ml、ヒトソマトスタチンHs00356144_ml、ヒトPDX−1Hs00173014_ml、ヒトGlut−2Hs00165775_ml、ヒトグルコキナーゼHs00175951_ml、ヒトPC2Hs00159922_ml、ヒトSCG2Hs00185761_ml、ヒトSGNE1Hs00161638_mlであった。 The TaqMan® fluorogenic probe, Assay-On-Demand (Applied Biosystems) used in this study was human β-actin Hs9999993_ml, human insulin Hs00355773_ml, human glucagon Hs00174967_ml, human somatostatin Hs00356144_ml0017, human lux14144 ml -2Hs00165775_ml, human glucokinase Hs00175951_ml, human PC2Hs00159922_ml, human SCG2Hs00185761_ml, human SGNE1Hs00161638_ml.
増幅条件は開始時50℃2分、変性95℃10分、ついで変性95℃15分およびアニーリング60℃1分の各サイクルを40サイクルとし、TaqMan PCR Master Mix(Applied Biosystems)を用いて行った。相対的な定量分析は2^(−ΔCt)等式を用いてABIプリズム7000SDSソフトウェアに従って行った。mRNA濃度はヒトβ−アクチンmRNAに対して補正した。 The amplification conditions were 50 ° C. for 2 minutes at the start, denaturation at 95 ° C. for 10 minutes, then denaturation at 95 ° C. for 15 minutes and annealing at 60 ° C. for 1 minute for 40 cycles, and TaqMan PCR Master Mix (Applied Biosystems) was used. Relative quantitative analysis was performed according to ABI Prism 7000 SDS software using the 2 ^ (-ΔCt) equation. The mRNA concentration was corrected for human β-actin mRNA.
(膵ホルモン免疫組織化学)
4μmのパラフィン包埋切片のスライドを脱パラフィン化し、3%H2O2中でインキュベートし、市販のHistomouse(登録商標)−SPキット(Zymed laboratories,South San Francisco,California)を用いてブロッキング溶液(インスリンおよびグルカゴンの検出の両方)中でインキュベートした。次に切片を共に1:200の希釈度のヒトインスリンおよびヒトグルカゴンに対するモノクローナル抗体(Sigma)と共に37℃で1時間インキュベートした。スライドを室温で30分間二次ビオチニル化IgGに曝露し、次にストレプトアビジン−パーオキシダーゼ、その後、色原体パーオキシド溶液中でインキュベートした。一次抗体は使用せず二次抗体のみを使用し、その後ストレプトアビジン−パーオキシダーゼおよび色原体パーオキシド溶液を使用した対照群を設けることにより系の考えられるバックグラウンドを排除した。
(Pancreatic hormone immunohistochemistry)
Slides of 4 μm paraffin-embedded sections were deparaffinized, incubated in 3% H 2 O 2 , and blocked using a commercially available Histomouse®-SP kit (Zymed laboratories, South San Francisco, Calif.). Incubation in both insulin and glucagon detection). The sections were then incubated for 1 hour at 37 ° C. with a 1: 200 dilution of human insulin and a monoclonal antibody to human glucagon (Sigma). Slides were exposed to secondary biotinylated IgG for 30 minutes at room temperature and then incubated in streptavidin-peroxidase followed by chromogenic peroxide solution. The possible background of the system was eliminated by providing a control group using only the secondary antibody and no streptavidin-peroxidase and chromogenic peroxide solution, without using the primary antibody.
(膵ホルモンに関するラジオイムノアッセイ(RIA))
膵臓および肝臓を単離し、即座に液体窒素中に凍結し、−70℃で保存した。凍結した組織は0.18NのHCl/35%エタノール中でホモゲナイズした。ホモジネートを攪拌しながら4℃で一夜抽出し、上澄みを凍結乾燥した。試料をプロテアーゼ阻害剤カクテル添加0.8mlRIAアッセイバッファー(Sigma)に溶解した。肝インスリンおよびグルカゴン濃度をラットラジオイムノアッセイ(RIA、カタログ番号SRI−13KおよびGL−32K、Linco,Missouri,USAおよびCoat−A−Count,DPC;CA,USA)を用いて測定した。ソマトスタチン濃度はRIA(Euro−diagnostica,Sweden)により測定した。膵ホルモンの肝含量は摘出した組織の重量に対して規格化した。
(Radioimmunoassay for pancreatic hormones (RIA))
Pancreas and liver were isolated and immediately frozen in liquid nitrogen and stored at -70 ° C. Frozen tissues were homogenized in 0.18N HCl / 35% ethanol. The homogenate was extracted overnight at 4 ° C. with stirring, and the supernatant was lyophilized. Samples were dissolved in 0.8 ml RIA assay buffer (Sigma) with protease inhibitor cocktail. Hepatic insulin and glucagon concentrations were measured using rat radioimmunoassay (RIA, catalog numbers SRI-13K and GL-32K, Linco, Missouri, USA and Coat-A-Count, DPC; CA, USA). Somatostatin concentration was measured by RIA (Euro-diagnostica, Sweden). The liver content of pancreatic hormone was normalized to the weight of the extracted tissue.
(肝機能の測定)
血清試料中のアルブミン、AST(アスパルテートアミノトランスフェラーゼ)、ALT(アラニンアミノトランスフェラーゼ)および総ビリルビンよりなる血清生化学数値をOlympus AU2700 Apparatus(Olympus,Germany)を用いて測定した。
(Measurement of liver function)
Serum biochemical values consisting of albumin, AST (aspartate aminotransferase), ALT (alanine aminotransferase) and total bilirubin in serum samples were measured using Olympus AU2700 Apparatus (Olympus, Germany).
(インスリンおよびC−ペプチドの検出)
インスリンおよびC−ペプチドの分泌および一次成熟肝細胞中の含量を初回ウィルスおよび成長因子投与の3日後に48時間の静的インキュベーションにより測定した。インスリンの培地への分泌はUltra Sensitive Human Insulin RIAキット(Linco Research)を用いてRIAにて測定し、そして、C−ペプチドの分泌はHuman C−Peptide RIAキット(Linco Research)により測定した。
(Insulin and C-peptide detection)
Insulin and C-peptide secretion and content in primary mature hepatocytes was measured by static incubation for 48
インスリン含量は細胞ペレットを0.18NのHCl、35%エタノール中でホモゲナイズした後に測定した。ホモジネートを攪拌しながら4℃で一夜抽出し、上澄みを凍結乾燥した。試料を0.2%BSAおよびプロテアーゼ阻害剤カクテル(Sigma)含有PBS0.5mlに溶解した。試料100μlをRIAに使用した。インスリン含量はBio−Rad蛋白試験キットにより測定された全細胞蛋白に対して規格化した。 Insulin content was determined after homogenizing the cell pellet in 0.18N HCl, 35% ethanol. The homogenate was extracted overnight at 4 ° C. with stirring, and the supernatant was lyophilized. Samples were dissolved in 0.5 ml PBS containing 0.2% BSA and protease inhibitor cocktail (Sigma). 100 μl of sample was used for RIA. Insulin content was normalized to total cellular protein measured with the Bio-Rad protein test kit.
(グルコース負荷試験)
成熟肝細胞にAd−CMV−PDX−1および成長因子を5日間投与した。細胞を105個/ウェルの密度で6穴プレートにプレーティングした。
(Glucose tolerance test)
Adult liver cells were administered Ad-CMV-PDX-1 and growth factors for 5 days. Cells were plated in 6-well plates at a density of 10 5 cells / well.
経時分析のために、細胞を0.1%BSA含有Krebs−Ringer緩衝液(KRB)中で2時間予備インキュベートし、次に2mMまたは25mMのグルコースを含有する培地中で所定時間インキュベートした。各時点において、培地試料中のインスリン(Ultra SensitiveヒトインスリンRIAキット−Linco Research)およびC−ペプチド(ヒトC−ペプチドRIAキット)を分析した。 For time-lapse analysis, cells were preincubated in Krebs-Ringer buffer (KRB) containing 0.1% BSA for 2 hours, and then incubated for a predetermined time in medium containing 2 mM or 25 mM glucose. At each time point, the medium samples were analyzed for insulin (Ultra Sensitive Human Insulin RIA Kit-Linco Research) and C-peptide (Human C-Peptide RIA Kit).
グルコース用量応答;細胞を0.1%BSA含有KRBで2時間予備インキュベートし、洗浄し、その後、漸増濃度のD−グルコースまたは2−デオキシグルコース(0〜25mM)を2時間負荷した。25mMグルコースにおけるインキュベート終了時に、細胞をKRBで洗浄し、2mMグルコース含有培地中更に2時間インキュベートした。 Glucose dose response; cells were preincubated with KRB containing 0.1% BSA for 2 hours, washed, and then loaded with increasing concentrations of D-glucose or 2-deoxyglucose (0-25 mM) for 2 hours. At the end of the incubation in 25 mM glucose, the cells were washed with KRB and incubated for a further 2 hours in medium containing 2 mM glucose.
(電子顕微鏡観察)
肝細胞を2.5%グルタルアルデヒドに固定し、オスミウム酸染色し、一連の等級のエタノールおよびプロピレンオキシドで脱水し、Araladite溶液(Polyscience Inc.)中に包埋した。ウルトラミクロトームで超薄片を切り出し、2%ウラニルアセテートおよびReynoldクエン酸鉛溶液で染色した。包埋後イムノゴールド反応のために、50〜90nmの肝切片をニッケルグリッド上においた。グリッドを一夜室温でインスリンに対する抗体(モルモットポリクローナル;7.8μg/ml、Dako)と共にインキュベートし、次に室温で1.5時間モルモットIgGに対するイムノゴールドコンジュゲート抗体(15nmゴールド;1:40希釈、Dako)と共にインキュベートした。切片を電子顕微鏡下に観察した(Jeol 1200EX2)。
(Electron microscope observation)
Hepatocytes were fixed in 2.5% glutaraldehyde, stained with osmate, dehydrated with a series of grades of ethanol and propylene oxide, and embedded in Araladite solution (Polyscience Inc.). Ultrathin sections were cut with an ultramicrotome and stained with 2% uranyl acetate and Reynold lead citrate solution. A 50-90 nm liver section was placed on a nickel grid for post-embedding immunogold reaction. The grid is incubated overnight at room temperature with an antibody against insulin (guinea pig polyclonal; 7.8 μg / ml, Dako), then immunogold-conjugated antibody against guinea pig IgG for 1.5 hours at room temperature (15 nm gold; 1:40 dilution, Dako) Incubated with. Sections were observed under an electron microscope (Jeol 1200EX2).
(細胞移植)
5週齢の非肥満糖尿病重度合併免疫不全(SCID−NOD)雄性マウス(Weizmann Institute,Israel)に180mg/kg体重のストレプトゾトシンを腹腔内注射することにより高血糖症とした。血中グルコース濃度が2回連続測定で約300mg/dlに達した時点で、マウスの腎皮膜下に30ゲージ針を用いてMatrigel(Sigma)50μl中、5日間に亘り、PDX−1および成長因子をあらかじめ投与した3x106ヒト肝細胞を移植した。血液を尾部から週2回採取し、グルコース濃度の測定に付した(Accutrend(登録商標)GC,Roche Applied Science)。製造元の取扱説明書に従って、マウスC−ペプチドに対して0%交差反応性のUltrasensitiveヒトC−ペプチドELISAキット(Mercodia)およびヒトインスリンに対して0%交差反応性のマウスインスリンELISAキット(Mercodia)を用いることにより、給餌マウスの血液中の血清をC−ペプチドおよびインスリン濃度の分析用に採取した。腎臓および膵臓は免疫組織化学的分析用に採取した。
(Cell transplantation)
Hyperglycemia was achieved by intraperitoneal injection of streptozotocin at 180 mg / kg body weight into 5-week-old non-obese diabetic severely combined immunodeficient (SCID-NOD) male mice (Weizmann Institute, Israel). When the blood glucose concentration reached approximately 300 mg / dl in two consecutive measurements, PDX-1 and growth factors were used for 5 days in 50 μl of Matrigel (Sigma) using a 30 gauge needle under the kidney capsule of mice. 3 × 10 6 human hepatocytes were administered in advance. Blood was collected from the tail twice a week and subjected to glucose concentration measurement (Accutrend® GC, Roche Applied Science). According to the manufacturer's instructions, an ultrasensitive human C-peptide ELISA kit (Mercodia) with 0% cross-reactivity to mouse C-peptide and a mouse insulin ELISA kit (Mercodia) with 0% cross-reactivity to human insulin By using, serum in the blood of fed mice was collected for analysis of C-peptide and insulin concentrations. Kidneys and pancreas were collected for immunohistochemical analysis.
(組織学的検討および染色)
腎臓および膵臓を4%ホルムアルデヒドに固定し、パラフィン包埋した。パラフィン包埋組織の厚み5m切片を脱パラフィン化し、3%H2O2中でインキュベートし、次にブロッキング溶液中でインキュベートする前に抗原回復のためにクエン酸塩緩衝液中でマイクロウエーブ処理する(PDX−1検出)か、または、Histomouse(登録商標)−SPキット(Zymed laboratories)を用いてブロッキング溶液に即座に曝露した(インスリン検出)。PDX−1の検出のためには、切片をカエルPDX−1のN末端部分に対して構築された抗血清(1:5000、C.V.E.Wrightより入手)と共に4℃で一夜インキュベートした。インスリンの検出のためには、切片をヒトインスリンに対するモノクローナル抗体(1:100、Sigma)と共に37℃で1時間インキュベートした。スライドを二次ビオチニル化IgGに30分間曝露し、次にストレプトアビジン−パーオキシダーゼ、ついで色原体パーオキシド溶液中でインキュベートした。
(Histological examination and staining)
Kidneys and pancreas were fixed in 4% formaldehyde and embedded in paraffin. Paraffin embedded tissue 5 m thick sections are deparaffinized and incubated in 3% H 2 O 2 and then microwaved in citrate buffer for antigen retrieval prior to incubation in blocking solution. (PDX-1 detection) or immediately exposed to blocking solution using Histomouse-SP kit (Zymed laboratories) (insulin detection). For detection of PDX-1, sections were incubated overnight at 4 ° C. with antisera constructed against the N-terminal portion of the frog PDX-1 (1: 5000, obtained from CVE Wright). . For detection of insulin, sections were incubated with a monoclonal antibody against human insulin (1: 100, Sigma) for 1 hour at 37 ° C. Slides were exposed to secondary biotinylated IgG for 30 minutes and then incubated in streptavidin-peroxidase followed by chromogenic peroxide solution.
(シクロホスファミド加速糖尿病(CAD))
糖尿病の発症を前述の通り(3)シクロホスファミド(Sigma,Rehovot,Israel)により加速した。雄性NODマウスに4週齢の時点でシクロホスファミド200mg/kgを腹腔内注射した。その後2週間以内にマウスが糖尿病にならない場合は、Cyの二回目の腹腔内注射(200mg/kg)を行い、1週間後に工程を再度反復した。糖尿病マウスをSPFから除き、12時間の照明/消灯周期の下に空調環境内において飼育し、そこで72時間馴化させた後に新たに血糖値を測定して糖尿病を確認した。
(Cyclophosphamide accelerated diabetes (CAD))
As described above, the onset of diabetes was accelerated by (3) cyclophosphamide (Sigma, Rehovot, Israel). Male NOD mice were injected intraperitoneally with 200 mg / kg of cyclophosphamide at the age of 4 weeks. If the mouse did not become diabetic within 2 weeks, a second intraperitoneal injection of Cy (200 mg / kg) was performed and the process was repeated again after 1 week. The diabetic mice were removed from the SPF, bred in an air-conditioned environment under a 12-hour lighting / extinction cycle, and after acclimatization for 72 hours, blood glucose levels were newly measured to confirm diabetes.
(高血糖症)
血中グルコースはAccutrend(登録商標)GC,Glucose Analyzed(Boehringer Mannheim,Mannheim,Germany)を用いて週二回測定した。血中グルコース濃度が2回の連続測定で300mg/dlより高値であればマウスを糖尿病と見なした。
(Hyperglycemia)
Blood glucose was measured twice a week using Accutrend® GC, Glucose Analyzed (Boehringer Mannheim, Mannheim, Germany). A mouse was considered diabetic if the blood glucose concentration was higher than 300 mg / dl in two consecutive measurements.
(ウィルス感染)
8〜10週齢の糖尿病NODマウス(体重20〜22gr)に対し、尾静脈内への全身投与により所定の組み換えアデノウィルス1.5x1010pfuを投与した。ウィルスはPBS中200〜300μlの容量で投与した。
(Virus infection)
Predetermined recombinant adenovirus 1.5 × 10 10 pfu was administered to 8-10 week old diabetic NOD mice (body weight 20-22 gr) by systemic administration into the tail vein. Virus was administered in a volume of 200-300 μl in PBS.
(膵臓組織の検討)
膵臓および肝臓を4%ホルムアルデヒド中に固定し、パラフィン包埋し、切り出して標準的なヘマトキシリンおよびエオシンにより染色した。免疫組織化学的に染色した4μmパラフィン包埋切片のスライドを脱パラフィン化し、ブロックして(21)に記載の通りHistomouse(登録商標)−SPキット(Zymed laboratories,South San Francisco,California)を用いて分析した。切片をヒトインスリンに対するモノクローナル抗体(Sigma,Rehovot,Israel)の1/100希釈物と共にインキュベートした。スライドをストレプトアビジン−パーオキシダーゼと組み合わせた抗マウスIgGビオチニル化抗体、ついで色原体パーオキシド溶液を用いて発色させた。対照スライドはインスリン特異的抗体を添加しない以外は同じプロトコルに従って並行して発色させた。
(Examination of pancreatic tissue)
The pancreas and liver were fixed in 4% formaldehyde, embedded in paraffin, excised and stained with standard hematoxylin and eosin. Immunohistochemically stained 4 μm paraffin-embedded slides were deparaffinized and blocked using Histomouse®-SP kit (Zymed laboratories, South San Francisco, Calif.) As described in (21). analyzed. Sections were incubated with a 1/100 dilution of a monoclonal antibody against human insulin (Sigma, Rehovot, Israel). The slides were developed with an anti-mouse IgG biotinylated antibody combined with streptavidin-peroxidase and then with a chromogenic peroxide solution. Control slides were developed in parallel according to the same protocol except that no insulin specific antibody was added.
(実施例2:異所性に同時送達されたインスリンプロモーターのPDX−1誘導内因性インスリン遺伝子の発現および活性化の測定)
肝における異所性PDX−1発現の作用を調べるために、雄性Balb/cおよびC57BL/6マウス(11〜14週齢)の尾部静脈にAd−CMV−PDX−1組み換えアデノウィルス2x109プラーク形成単位(0.2ml生理食塩水中)を注射した。対照として、AdCMV−β−galまたはAdCMV−hInsおよびAdRIP−I−hIns組み換えアデノウィルスをマウスに同様に注射した。動物は通常の非制限食餌において12時間照明/消灯周期下の空調環境下に飼育し、ウィルス投与後1週間に屠殺した。肝臓、脾臓、膵臓および腎臓を摘出し、迅速に液体窒素中で凍結し、−70℃で保存して全RNAの単離に付した。
Example 2: Measurement of PDX-1-induced endogenous insulin gene expression and activation of an ectopically co-delivered insulin promoter
To examine the effects of ectopic PDX-1 expression in the liver, Ad-CMV-PDX-1
PDX−1およびインスリンの遺伝子の発現はRT−PCRで測定した。全RNAをRNAzol(CINNA−BIOTEX)を用いて凍結組織から単離した。RNA試料は10ulのDNase I(Promega)で処理した。cDNAは1μgDNA非含有全RNAおよび0.5μgオリゴ(dT)15を用いて逆転写により調製した。RT反応液1.5μlを以下の表1に示すプライマーおよびPCR条件を用いて増幅した。PCRはGeneAmp PCRシステム2400(Perkin Elmer)で行い、産物は1.7%アガロースゲル上で分離した。別のPCR反応を逆転写酵素を用いることなく各RNA試料について実施することにより増幅された産物がDNA汚染によるものではないことを確認した。プライマーはマウス相同体ではなく異所性ラットPDX−1の発現のみを検出するように設計した。mI−2増幅用のプライマーは異なるエクソン上に位置する。試料変性の第1の工程は全ての増幅遺伝子について同一であり、94℃1分とした。 PDX-1 and insulin gene expression was measured by RT-PCR. Total RNA was isolated from frozen tissue using RNAzol (CINNA-BIOTEX). RNA samples were treated with 10 ul DNase I (Promega). cDNA was prepared by reverse transcription using 1 μg DNA-free total RNA and 0.5 μg oligo (dT) 15 . 1.5 μl of the RT reaction solution was amplified using the primers and PCR conditions shown in Table 1 below. PCR was performed on a GeneAmp PCR system 2400 (Perkin Elmer) and the products were separated on a 1.7% agarose gel. A separate PCR reaction was performed on each RNA sample without using reverse transcriptase to confirm that the amplified product was not due to DNA contamination. Primers were designed to detect only ectopic rat PDX-1 expression, not mouse homologues. Primers for mI-2 amplification are located on different exons. The first step of sample denaturation was the same for all amplified genes and was 94 ° C. for 1 minute.
全RNAの分析によればAdCMV−PDX−1の投与により主に肝においてPDX−1の発現が起こったことがわかる。同じマウスに由来する脾臓、膵臓および腎臓はPDX−1のラット相同体についてはRT−PCRによれば陰性であった。 The analysis of total RNA shows that PDCM-1 expression occurred mainly in the liver by administration of AdCMV-PDX-1. Spleen, pancreas and kidneys from the same mouse were negative for rat homologues of PDX-1 by RT-PCR.
異所性ラットPDX−1メッセージのついて陽性であったマウスの75%(35匹中25匹)はmI−2遺伝子を発現したのに対し、マウスの35%がmI−1遺伝子を発現した(図1)。mI−2とmI−1の間のこのような発現の相違がPDX−1発現肝細胞中に存在する転写因子の本質または量によりmI−1プロモーターが異なって影響されるためであるかどうかを明らかにするために、上記した通りマウスにAdRIP−I−hIns組み換えアデノウィルスをAdCMV−PDX−1と同時送達した。図1に示すとおり、PDX−1が内因性mI−2の発現のみを誘導した肝においては、これはラットインスリン−1プロモーター(RIP−1)も活性化した。このことはDNAメチル化の異なった水準または個別の染色質構造が肝におけるPDX−1発現による内因性mI−1発現の活性化の効率が低いことの原因である可能性を示唆している。更にまたこれらのデータはPDXと同時送達された場合の肝におけるβ細胞特異的インスリンプロモーターを活性化する能力を示している。 75% (25/35) of mice positive for ectopic rat PDX-1 message expressed mI-2 gene, whereas 35% of mice expressed mI-1 gene ( FIG. 1). Whether such expression differences between mI-2 and mI-1 are due to the mI-1 promoter being affected differently by the nature or amount of transcription factors present in PDX-1 expressing hepatocytes. For clarity, mice were co-delivered with AdRIP-I-hIns recombinant adenovirus with AdCMV-PDX-1 as described above. As shown in FIG. 1, in the liver where PDX-1 induced only endogenous mI-2 expression, it also activated the rat insulin-1 promoter (RIP-1). This suggests that different levels of DNA methylation or individual chromatin structures may be responsible for the low efficiency of activation of endogenous mI-1 expression by PDX-1 expression in the liver. Furthermore, these data indicate the ability to activate the β-cell specific insulin promoter in the liver when co-delivered with PDX.
内因性のマウスインスリンおよび異所性ヒトインスリンの遺伝子の発現はそれぞれ同じ濃度の対照組み換えアデノウィルスAdCMV−β−galまたはAdCMV−hInsおよびAdRIP−hInsの投与(n=20)では誘導されなかった。これらの結果はPDX−1は内因性インスリン遺伝子の発現を誘導するため、および、膵臓外の組織におけるRIP−1の活性化のために必須かつ十分であることを示している。 Expression of endogenous mouse insulin and ectopic human insulin genes was not induced by administration of the same concentration of control recombinant adenovirus AdCMV-β-gal or AdCMV-hIns and AdRIP-hIns, respectively (n = 20). These results indicate that PDX-1 is essential and sufficient for inducing the expression of the endogenous insulin gene and for activation of RIP-1 in tissues outside the pancreas.
(実施例3:PDX−1誘導のソマトスタチン遺伝子発現およびインビボの蛋白生産の測定)
実施例2に記載の通り動物に組み換えアデノウィルスを投与した。ソマトスタチンの遺伝子発現は表1に示す条件に従って実施例2に記載の通りRT−PCRで測定した。
Example 3: Measurement of PDX-1-induced somatostatin gene expression and in vivo protein production
Recombinant adenovirus was administered to animals as described in Example 2. The gene expression of somatostatin was measured by RT-PCR as described in Example 2 according to the conditions shown in Table 1.
図3に示すとおり、AdCMV−PDX−1を投与したマウスの肝はソマトスタチンの遺伝子発現を示した。AdCMV−PDX−1を投与したマウスは免疫化学的分析を行った肝組織においてはソマトスタチン蛋白について陽性に染色された。AdCMV−β−galを投与したマウスはソマトスタチンを発現しなかった。 As shown in FIG. 3, the liver of the mouse administered with AdCMV-PDX-1 showed gene expression of somatostatin. Mice administered with AdCMV-PDX-1 stained positive for somatostatin protein in the liver tissue subjected to immunochemical analysis. Mice administered AdCMV-β-gal did not express somatostatin.
(実施例4:PDX−1誘導グルカゴン遺伝子発現の測定)
実施例2に記載するとおりAdCMVPDX−1組み換えアデノウィルスを動物に投与した。グルカゴン遺伝子発現を表1に示す条件およびプライマーを用いて実施例2に記載の通りRT−PCRで測定した。
(Example 4: Measurement of PDX-1 induced glucagon gene expression)
The AdCMVPDX-1 recombinant adenovirus was administered to the animals as described in Example 2. Glucagon gene expression was measured by RT-PCR as described in Example 2 using the conditions and primers shown in Table 1.
図3に示すとおり、AdCMV−PDX−1を投与したマウスの肝はグルカゴン遺伝子を発現した。AdCMV−β−galを投与したマウスはグルカゴンを発現しなかった。 As shown in FIG. 3, the livers of mice administered with AdCMV-PDX-1 expressed the glucagon gene. Mice administered with AdCMV-β-gal did not express glucagon.
(実施例5:プロホルモン変換酵素1/3誘導遺伝子発現の測定)
実施例2に記載の通り組み換えアデノウィルスを動物に投与した。プロホルモン変換酵素1/3(PC1/3)遺伝子発現はオリゴ(dT)15の代わりに遺伝子特異的オリゴヌクレオチド(TCCAGGTGCCTACAGGATTCTCT)(配列番号1)を用いてcDNAを逆転写した以外は実施例2に記載の通りRT−PCRにより測定した。図3に示すとおり、PDX−1を投与した動物に由来する肝のみがKexinファミリープロテアーゼのメンバーであるPC1/3の発現を誘導し、PC1/3の発現は調節された分泌経路を有する内分泌および神経内分泌の細胞に限定されている。細胞内コンパートメント内に生産されたホルモンを貯留する能力と合わせると、ホルモンの生産、プロセシング、保存および分泌に関わる調節された経路の誘導を含む内分泌の表現型のPDX−1依存性の誘導が示唆される。
(Example 5: Measurement of
Recombinant adenovirus was administered to animals as described in Example 2.

実施例2に記載の通り組み換えアデノウィルスを動物に投与した。肝臓、脾臓、膵臓および腎臓を摘出した。組織の一部を4%ホルムアルデヒドに固定し、パラフィン包埋し、免疫組織化学的染色に付した。残余の肝および膵臓の組織を70%エタノール−0.18N塩酸中でホモゲナイズし、凍結乾燥し、PBS(リン酸塩緩衝食塩水)中に再懸濁してIRI含量のRIA測定に付した。
Recombinant adenovirus was administered to animals as described in Example 2. The liver, spleen, pancreas and kidney were removed. A portion of the tissue was fixed in 4% formaldehyde, embedded in paraffin, and subjected to immunohistochemical staining. The remaining liver and pancreatic tissues were homogenized in 70% ethanol-0.18N hydrochloric acid, lyophilized, resuspended in PBS (phosphate buffered saline) and subjected to RIA measurement of IRI content.
(免疫組織化学的検討)
パラフィン包埋組織の5μm切片を脱パラフィン化し、3%H2O2中でインキュベートし、次にブロッキング溶液中でインキュベートする前に抗原回復のためにクエン酸塩緩衝液中でマイクロウエーブ処理する(PDX−1検出)か、または、ブロッキング溶液に即座に曝露した(インスリン検出)(Histomouse(登録商標)−SPキット、Zymed laboratories、CA,USA)。
(Immunohistochemical study)
A 5 μm section of paraffin-embedded tissue is deparaffinized and incubated in 3% H 2 O 2 and then microwaved in citrate buffer for antigen retrieval prior to incubation in blocking solution ( PDX-1 detection) or immediately exposed to blocking solution (insulin detection) (Histomouse-SP kit, Zymed laboratories, CA, USA).
PDX−1の検出:切片をカエルPDX−1のN末端部分に対して構築された抗血清と共に4℃で一夜インキュベートした。 Detection of PDX-1: Sections were incubated overnight at 4 ° C. with antisera constructed against the N-terminal portion of the frog PDX-1.
インスリンの検出:切片をモノクローナル抗ヒトインスリン(Sigma,St−Louis MO)と共に37℃で1時間インキュベートした。 Insulin detection: Sections were incubated with monoclonal anti-human insulin (Sigma, St-Louis MO) for 1 hour at 37 ° C.
スライドを二次ビオチニル化IgGに30分間曝露し、ストレプトアビジン−パーオキシダーゼ、ついで色原体パーオキシド溶液中でインキュベートした。 Slides were exposed to secondary biotinylated IgG for 30 minutes and incubated in streptavidin-peroxidase followed by chromogenic peroxide solution.
PDX−1を投与したマウスの肝切片の免疫組織化学的分析によれば肝細胞核の30〜60%においてホメオプロテインの発現があり、肝細胞の0.1〜1%が(プロ)インスリンについて陽性に染色されたことがわかった。対照のAdCMVβ−gal投与肝は(プロ)インスリンについては陽性に染色されなかったが、β−ガラクトシダーゼ活性は核の50%で明らかとなった。AdCMV−hInsを投与したマウスの肝は肝切片においてはインスリンについて陽性に染色されなかったが、同じマウスの血清IRIはPDX−1投与マウスにおける血清中IRI濃度と同様、3倍高値であった。インスリンではなくPDX−1の異所性発現が(プロ)インスリンについて陽性に免疫染色されたという事実は、β細胞の表現型に移行していると考えられる、肝細胞の小さいサブ集団(肝細胞ではなく、内分泌細胞に特徴的な、調節経路に属する分泌ベシクル)におけるインスリンの貯留を支援する細胞モジュレーションの誘導を示唆している。 According to immunohistochemical analysis of liver sections of mice administered with PDX-1, there is homeoprotein expression in 30-60% of hepatocyte nuclei, and 0.1-1% of hepatocytes are positive for (pro) insulin. Was found to be stained. Control AdCMV β-gal treated livers did not stain positively for (pro) insulin, but β-galactosidase activity was evident in 50% of the nuclei. The liver of mice administered with AdCMV-hIns did not stain positively for insulin in liver sections, but the serum IRI of the same mice was three times higher, similar to the serum IRI concentration in PDX-1-treated mice. The fact that the ectopic expression of PDX-1 but not insulin was immunostained positively for (pro) insulin is the result of a small subpopulation of hepatocytes (hepatocytes) that appears to have shifted to the β-cell phenotype Rather, it suggests the induction of cell modulation that supports insulin storage in secretory vesicles belonging to regulatory pathways, characteristic of endocrine cells).
(ラジオイムノアッセイ)
肝インスリンmRNAが効率的に蛋白に翻訳されるかどうかを調べるために、ラジオイムノアッセイ(RIA)により肝組織に由来する抽出液中の免疫反応性インスリン(IRI)含量を試験した。RT−PCRによりインスリン遺伝子発現について陽性であったPDX−1投与マウスの肝(図1)は対照ウィルスを投与した動物の肝臓よりも約25倍高値のIRIを含有していた(表2)。PDX−1投与肝由来抽出液中の平均IRI濃度は20.7±3.97μU/mg蛋白であり、対照肝ではIRIは僅か0.78±0.25μU/mg蛋白であった。対照肝試料中で測定したインスリンのバックグラウンド濃度はおそらくはこの臓器中に大量に存在するその受容体に結合したインスリン(膵起源)を示している可能性がある。PDX−1投与肝抽出液中に検出されたIRIは膵抽出液中に検出された濃度の<0.1%であったが(表2)、PDX−1投与マウスにおける血清中IRI濃度は対照群と比較してほぼ3倍高値であり(それぞれ323±48.4対118.2±23.7μU/ml(表2))、インスリンが合成されておりその大部分が血流中に分泌されたことを示している。これらのデータは分子操作により誘導されたインスリン遺伝子発現がプロ/ホルモンの特異的肝生産に良好に翻訳されたことを示している。
(Radioimmunoassay)
In order to examine whether liver insulin mRNA is efficiently translated into protein, the immunoreactive insulin (IRI) content in the extract derived from liver tissue was examined by radioimmunoassay (RIA). The liver of PDX-1 treated mice that were positive for insulin gene expression by RT-PCR (FIG. 1) contained about 25 times higher IRI than the liver of animals that received control virus (Table 2). The average IRI concentration in the PDX-1-administered liver-derived extract was 20.7 ± 3.97 μU / mg protein, and in the control liver, the IRI was only 0.78 ± 0.25 μU / mg protein. The background concentration of insulin measured in the control liver sample may indicate insulin bound to its receptor (pancreatic origin), probably present in large quantities in this organ. The IRI detected in the PDX-1 treated liver extract was <0.1% of the concentration detected in the pancreatic extract (Table 2), but the serum IRI concentration in the PDX-1 administered mice was the control Almost three times higher than the group (323 ± 48.4 vs. 118.2 ± 23.7 μU / ml, respectively) (Table 2), insulin is synthesized and most of it is secreted into the bloodstream It shows that. These data indicate that insulin gene expression induced by molecular manipulation was successfully translated into pro / hormone specific liver production.
PDX−1投与肝において検出された免疫反応性のインスリンは膵抽出液中のIRI濃度の1%未満であった(表2)。肝抽出液中のラジオイムノアッセイ(RIA)により測定されたIRI値はこの臓器における実際のインスリン生産を過小評価していると考えられる。RIAのために本発明者等が使用した抗体はプロセシングされたホルモンに優先的に結合し、肝細胞に主に存在し、膵臓にははるかに低い濃度で存在すると推定されるプロインスリンとの交差反応性は僅か60%である。 Immunoreactive insulin detected in PDX-1 treated liver was less than 1% of the IRI concentration in the pancreatic extract (Table 2). The IRI value measured by radioimmunoassay (RIA) in the liver extract is considered to underestimate the actual insulin production in this organ. The antibodies we used for RIA bind preferentially to processed hormones, crossover with proinsulin presumed to be predominantly present in hepatocytes and in much lower concentrations in the pancreas The reactivity is only 60%.
(実施例7:血中グルコースおよび血清中インスリンの濃度)
実施例2に記載の通り組み換えアデノウィルスを動物に投与した。屠殺する前にグルコース濃度(Accutrend(登録商標)GC,Boehringer Mannheim,Mannheim,Germany)およびラジオイムノアッセイによるインスリン濃度(Coat−a−Count,DPC,Los Angeles,CA,USA,ラットインスリン標準物質(Linco)使用、使用した抗インスリン抗体はヒトプロインスリンとは交差反応性は僅か60%である)の測定用に、下大静脈より採血した。
(Example 7: Concentrations of blood glucose and serum insulin)
Recombinant adenovirus was administered to animals as described in Example 2. Before sacrifice, glucose concentration (Accutren® GC, Boehringer Mannheim, Mannheim, Germany) and insulin concentration by radioimmunoassay (Coat-a-Count, DPC, Los Angeles, CA, USA, rat insulin standard (Linco) Blood was collected from the inferior vena cava for the measurement of the anti-insulin antibody used and the cross-reactivity with human proinsulin being only 60%.
AdCMV−PDX−1組み換えアデノウィルスを投与したマウスは高血糖症ではなかったが、その血中グルコース濃度はAdCMV−β−galまたはAdCMV−Lucを投与したマウスよりも有意に低値であった[それぞれ197±11.2対228±15.74mg/dl、(表2)]。血漿中免疫反応性インスリン濃度は対照群と比較してPDX−1投与マウスで有意に高値であった[それぞれ323±48.4対118.2±23.7μU/ml、(表2)]。 Mice treated with AdCMV-PDX-1 recombinant adenovirus were not hyperglycemic, but their blood glucose levels were significantly lower than those administered AdCMV-β-gal or AdCMV-Luc [respectively 197 ± 11.2 vs. 228 ± 15.74 mg / dl, (Table 2)]. Plasma immunoreactive insulin levels were significantly higher in PDX-1 treated mice compared to the control group [323 ± 48.4 vs 118.2 ± 23.7 μU / ml, respectively (Table 2)].
PDX−1投与マウスにおける血清中IRI濃度の3倍上昇は、PDX−1投与肝抽出液中に示された肝IRI含量の25倍上昇(表2)をそれ自体では説明することができない。即ち、肝プロ/インスリン含量の増大は局所的生産に起因している。 A three-fold increase in serum IRI concentration in PDX-1-treated mice cannot itself explain the 25-fold increase in liver IRI content (Table 2) shown in the PDX-1-treated liver extract. That is, the increase in hepatic pro / insulin content is due to local production.
実施例2に記載するとおり組み換えアデノウィルスを動物に投与した。肝および膵臓を摘出し、70%エタノール−0.18N塩酸中でホモゲナイズし、凍結乾燥し、0.1M塩酸−0.1%BSA中に再懸濁し、HPLC分析に付した。
Recombinant adenovirus was administered to the animals as described in Example 2. The liver and pancreas were removed, homogenized in 70% ethanol-0.18N hydrochloric acid, lyophilized, resuspended in 0.1M hydrochloric acid-0.1% BSA, and subjected to HPLC analysis.
肝および膵の抽出液から得たインスリン関連ペプチドをLichrospher100R−18カラム(Merck、Darmstadt,Germany)およびGross et alの溶離条件を用いた逆相HPLCにより分解した。1ml画分を水中0.1%BSA0.1mlの入った試験管中に採取し、Speed−Vac装置で乾燥し、RIA緩衝液(PBS中0.1%BSA)1ml中に希釈再調製し、RIAによるペプチド測定に付した。モルモット抗ブタインスリン抗体(Linco,St Charles,MO)とラットまたはヒトインスリン標準物質のいずれかを用いてそれぞれマウスまたはヒトIRIを測定した。 Insulin-related peptides from liver and pancreatic extracts were resolved by reverse phase HPLC using a Lichrospher 100R-18 column (Merck, Darmstadt, Germany) and Gross et al elution conditions. 1 ml fraction was collected in a test tube containing 0.1 ml of 0.1% BSA in water, dried on a Speed-Vac device, reconstituted in 1 ml of RIA buffer (0.1% BSA in PBS) It was subjected to peptide measurement by RIA. Mouse or human IRI was measured using guinea pig anti-pig insulin antibody (Linco, St Charles, MO) and either rat or human insulin standards, respectively.
PDX−1投与マウスの肝IRI含量のHPLC分析によれば完全にプロセシングされたmI−1およびmI−2への変換は59±7%(n=3)であった。これと比較して、膵抽出液は85±5%(n=3)の成熟インスリンを含有していた(図2)。これに対し、ヒトインスリンの異所性発現(AdCMV−hIns)は、抽出されたIRIの大部分が未成熟インスリンであった肝一例を除き、肝細胞中のIRIの貯留をもたらさなかった。このことは、インスリン遺伝子産物の貯留が観察されずその大部分が構成性の分泌経路により分泌されたトランスフェクトFAO細胞における以前の観察結果と合致している。これらのデータは肝における異所性PDX−1発現は誘導されたプロホルモンのプロセシングが可能な内分泌組織に特徴的である細胞機序を誘導し、肝においてプロインスリンのみが異所性に発現される場合には誘導されないことを示している。即ち、異所性PDX−1発現による肝細胞における拡張されたβ細胞表現型の誘導を示す。 According to HPLC analysis of liver IRI content of mice treated with PDX-1, the conversion to fully processed mI-1 and mI-2 was 59 ± 7% (n = 3). In comparison, the pancreatic extract contained 85 ± 5% (n = 3) mature insulin (FIG. 2). In contrast, ectopic expression of human insulin (AdCMV-hIns) did not result in retention of IRI in hepatocytes, except in one case where the extracted IRI was mostly immature insulin. This is consistent with previous observations in transfected FAO cells in which no accumulation of insulin gene product was observed and most of it was secreted by a constitutive secretion pathway. These data indicate that ectopic PDX-1 expression in the liver induces a cellular mechanism characteristic of endocrine tissues capable of processing the induced prohormone, and only proinsulin is ectopically expressed in the liver The case shows that it is not induced. That is, it shows induction of an expanded β-cell phenotype in hepatocytes by ectopic PDX-1 expression.
(実施例9:肝プロ/インスリン生産の生物学的活性)
糖尿病マウスにおける血糖値を制御するPDX−1誘導肝インスリン生産の能力を試験した。C57BL/6マウスを200mg/kgの腹腔内STZ注射の24時間後にケトアシドーシスを伴った糖尿病(>600mg/dl)とした。STZ注射の24〜48時間後に、マウスには尾部静脈から生理食塩水中のAdCMV−PDX−1またはAdCMVβ−gal(対照)組み換えアデノウィルスを投与した。図4に示すとおり、AdCMV−PDX−1投与マウスは組み換えアデノウィルス投与の2日後から開始して、約600から200〜300mg/dlまでの血中グルコース値の緩やかな低下を示した。これとは対照的に、対照のAdCMVβ−gal投与マウスにおいては、高血糖症は持続し、死亡率の増大を伴い、試験した22匹中12匹が死亡し、アデノウィルス投与の1〜3日後には重度のケトアシドーシスを有していた。更にまた、両群とも高血糖症誘導後は体重減少を示し、屠殺時前までに回復しなかった。要すれば、データは、PDX−1の発現は損なわれたβ細胞機能を有するマウスにおける高血糖症を軽減する成熟した生物学的に活性なインスリンの生産を誘導するのに十分であることを示している。
Example 9: Biological activity of hepatic pro / insulin production
The ability of PDX-1 induced hepatic insulin production to control blood glucose levels in diabetic mice was tested. C57BL / 6 mice were diabetic (> 600 mg / dl) with
(実施例10:異所性PDX−1発現によるインスリンプロモーターのインビトロの活性化)
PDX−1はラットインスリン−1プロモーターによりヒトインスリンの発現を送達する組み換えアデノウィルスAdRip−1hInsと共に同時送達された場合にラットインスリン−1プロモーターを活性化する(実施例2および図1参照)。PDX−1はラット肝細胞においてラットインスリンプロモーター−1をインビトロで活性化するのに十分であることがわかっている。成熟および胎児の肝細胞の一次培養物を血清非含有の化学的に明らかにされている培地中、コラーゲン−1被覆組織培養皿上で培養した。プレーティングから2日後に、細胞にAdCMV−PDX−1&AdRIP−1hInsまたはAdCMVβ−gal&AdRIP−1hInsのいずれかを投与した。アデノウィルス投与の48時間後、全RNAを抽出し、プロインスリン遺伝子の発現を実施例2に記載したとおり調べた。
Example 10 In Vitro Activation of Insulin Promoter by Ectopic PDX-1 Expression
PDX-1 activates the rat insulin-1 promoter when co-delivered with recombinant adenovirus AdRip-1hIns that delivers human insulin expression via the rat insulin-1 promoter (see Example 2 and FIG. 1). PDX-1 has been found to be sufficient to activate rat insulin promoter-1 in rat hepatocytes in vitro. Primary cultures of mature and fetal hepatocytes were cultured on collagen-1 coated tissue culture dishes in chemically defined medium without serum. Two days after plating, cells were administered either AdCMV-PDX-1 & AdRIP-1 hIns or AdCMVβ-gal & AdRIP-1 hIns. 48 hours after adenovirus administration, total RNA was extracted and proinsulin gene expression was examined as described in Example 2.
PDX−1は異所性発現されたRIP−hIns(ラットインスリンプロモーター−1、このプロモーターの410bp、ヒトインスリン駆動、組み換えアデノウィルスにより導入)を活性化し、β−galはこのような能力を保有していなかった(図5)。 PDX-1 activates ectopically expressed RIP-hIns (rat insulin promoter-1, 410 bp of this promoter, human insulin driven, introduced by recombinant adenovirus), β-gal possesses such ability None (Figure 5).
(実施例11:肝細胞における内因性ソマトスタチン遺伝子発現のインビトロの誘導)
胎児(E14−Fisher−344ラット)から単離した肝細胞の一次培養物を培養し、実施例9に記載の通り組み換えアデノウィルスを投与した。ソマトスタチン遺伝子発現は表1に示すプライマーおよびRT−PCR条件を用いて、実施例2に記載の通り逆転写全RNA試料中で検出した。
Example 11: In vitro induction of endogenous somatostatin gene expression in hepatocytes
Primary cultures of hepatocytes isolated from fetuses (E14-Fisher-344 rats) were cultured and recombinant adenovirus was administered as described in Example 9. Somatostatin gene expression was detected in reverse transcribed total RNA samples as described in Example 2 using the primers and RT-PCR conditions shown in Table 1.
データはインビトロの肝細胞における異所性PDX−1発現は、インビトロで、肝細胞における内因性の、その他の面ではサイレントであるソマトスタチンの遺伝子の発現を誘導することを示している(図6)。 The data show that ectopic PDX-1 expression in hepatocytes in vitro induces expression of the somatostatin gene that is endogenous in hepatocytes and otherwise silent (FIG. 6). .
(実施例12:肝細胞における内因性インスリン遺伝子発現のインビトロの誘導)
実施例10に記載の通り胎児(E14−Fisher−344ラット)の一次培養物を培養し、組み換えアデノウィルスを投与した。ラットインスリン1遺伝子の発現は表1に示すプライマーおよびRT−PCR条件を用いて、実施例2に記載の通り逆転写全RNA試料中で検出した。
Example 12: In vitro induction of endogenous insulin gene expression in hepatocytes
Primary cultures of fetuses (E14-Fisher-344 rats) were cultured as described in Example 10, and recombinant adenovirus was administered. Rat
データはインビトロの胎児肝細胞の一次培養物における異所性PDX−1発現は、内因性の、その他の面ではサイレントであるインスリンの遺伝子の発現を誘導することを示している(図6)。 The data show that ectopic PDX-1 expression in primary cultures of fetal hepatocytes in vitro induces endogenous, otherwise silent gene expression of insulin (FIG. 6).
(実施例13:生産されたホルモンの貯留およびその調節された分泌を可能にする内分泌および神経内分泌の特徴である細胞内コンパートメントの肝細胞内異所性PDX−1発現による誘導)
実施例2に記載の通り、Ad−CMVhInsまたはAdCMVPDX−1のいずれかをマウスに投与した。投与により肝細胞によるヒトインスリンの生産を示す3倍の血清中IRI増大がもたらされた(図1)。免疫細胞化学試験によりインスリン蛋白が陽性であった細胞はAdCMVPDX投与のみで検出された。更にまた、肝抽出液のHPLC分析では肝抽出液中には僅か痕跡量のIRIのみしか検出されず、その全てはAd−CMVhIns投与マウスでは未プロセシングであり、これに対し、AdCMVPDX−1投与マウスでは25倍増大が観察された。更に、AdCMVPDX−1投与マウスにおいて生産されたインスリンの59%が、プロセシングされた。更に、AdCMVPDX−1を投与した肝のみが、インスリンのプロセシングの保存および調節された分泌のための調節された経路が可能である細胞にのみ特徴的であるプロホルモンプロセシング酵素PC1/3の誘導を示した。これらのデータは肝細胞におけるインスリンの調節された分泌をPDXが誘導することを示している。
Example 13: Induction of hepatocyte ectopic PDX-1 expression of intracellular compartments characteristic of endocrine and neuroendocrine enabling the retention of the produced hormone and its regulated secretion
Mice were administered either Ad-CMVhIns or AdCMVPDX-1 as described in Example 2. Administration resulted in a 3-fold increase in serum IRI indicating production of human insulin by hepatocytes (FIG. 1). Cells that were positive for insulin protein by immunocytochemistry were detected only by AdCMVPDX administration. Furthermore, in the HPLC analysis of the liver extract, only a trace amount of IRI was detected in the liver extract, all of which were unprocessed in the Ad-CMVhIns-administered mice, whereas the AdCMVPDX-1-administered mice A 25-fold increase was observed. Furthermore, 59% of the insulin produced in AdCMVPDX-1 treated mice was processed. Furthermore, only the liver administered with AdCMVPDX-1 shows induction of the prohormone processing enzyme PC1 / 3, which is characteristic only for cells that are capable of a regulated pathway for the preservation and regulated secretion of insulin processing. It was. These data indicate that PDX induces regulated secretion of insulin in hepatocytes.
(実施例14:PDXにより発現がモジュレートされる核酸の識別)
PDXによりモジュレートされる核酸は異所性PDX発現により識別される。膵組織と比較した場合に、対照投与の膵外島組織において発現されない核酸はPDXによりモジュレートされる核酸である。このようにして識別されたこれらの核酸は膵関連障害を治療するための治療用化合物として使用される。
(Example 14: Identification of nucleic acid whose expression is modulated by PDX)
Nucleic acids modulated by PDX are distinguished by ectopic PDX expression. Nucleic acids that are not expressed in control administered extrapancreatic islet tissue when compared to pancreatic tissue are nucleic acids that are modulated by PDX. These nucleic acids thus identified are used as therapeutic compounds for treating pancreatic related disorders.
標的遺伝子の識別はサブトラクティブのライブラリ、市販のマイクロアレイチップ(IncyteまたはAffimetrix)またはメンブレンハイブリダイゼーション(CLONTECH、Atlas(登録商標)発現アレイまたはMultiple Tissue Northern(MTN(登録商標))ブロット)の何れかにより実施する。投与組織からのRNAの単離、その精製およびcDNAプローブの合成は、製造元の取扱説明書に従って実施する。 Target gene identification can be done either by subtractive library, commercially available microarray chip (Incyte or Affimetrix) or membrane hybridization (CLONTECH, Atlas® expression array or Multiple Tissue Northern (MTN®) blot) carry out. Isolation of RNA from the administered tissue, its purification, and synthesis of the cDNA probe are performed according to the manufacturer's instructions.
PDX投与非膵島組織において発現され、そして膵島プローブされたメンブレンまたはチップ中にも存在するが対照投与非膵島組織には存在しない遺伝子は直接または非直接のPDX標的遺伝子であり、これは遺伝子発現の島細胞特徴的性質を示す。直接または間接の間の判別は、図7に示す電子移動性シフト試験(EMSA)による候補標的遺伝子プロモーター分析、および、プロモーターフットプリンティング(Sambrook et al.,Molecular Cloning:A Laboratory Manual,2nd ed.,Cold Spring Harbor Laboratory,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,N.Y.,1989に記載)により解明される。 A gene that is expressed in PDX-administered non-islet tissue and is also present in the islet-probeed membrane or chip but not in control-administered non-islet tissue is a direct or non-direct PDX target gene, which is a gene expression gene Shows islet cell characteristic properties. Discrimination between direct and indirect can be performed by analyzing the candidate target gene promoter by the electron mobility shift test (EMSA) shown in FIG. 7 and promoter footprinting (Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd ed.,). Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989).
(実施例15:宿主組織における所望の異所性発現遺伝子の調節発現の誘導)
本実施例はインスリン以外の何れかの受容体の調節された発現の誘導を説明する。PDXが非膵島組織においてインスリンプロモーターを活性化しそのグルコースおよび成長因子感知能力を媒介する場合、何らかの別のプロモーターがグルコースおよび成長因子により同様に調節される。即ち、本発明はインスリンプロモーターにより駆動され、これによりその転写および他の面では非内分泌性の組織からの調節された分泌を制御するインスリンプロモーターにより駆動される例えばグルカゴン、成長ホルモン、ステロイドホルモンを含む多くの分泌および/または非分泌の因子の発現を栄養的およびホルモン的に調節するために使用できる(図7)。
(Example 15: Induction of regulated expression of desired ectopic expression gene in host tissue)
This example illustrates the induction of regulated expression of any receptor other than insulin. When PDX activates the insulin promoter in non-islet tissue and mediates its glucose and growth factor sensing ability, some other promoter is similarly regulated by glucose and growth factor. That is, the invention includes eg glucagon, growth hormone, steroid hormone driven by an insulin promoter driven by an insulin promoter, thereby controlling its transcription and otherwise regulated secretion from non-endocrine tissue It can be used to nutritionally and hormonally regulate the expression of many secreted and / or non-secreted factors (Figure 7).
(実施例16:βまたは島細胞特異的転写因子の階層構造中のPDX位置の識別)
本実施例はβ細胞または島細胞に特異的な転写因子の階層構造におけるPDXの位置の識別を説明する。膵島において発現されるが肝における異所性のPDX−1の発現により誘導されない各転写因子は、肝のような非内分泌膵臓組織中のより包括的で完全または完全に近いβ細胞表現型の誘導のためにPDXと協調すると考えられる。肝における島細胞特異的転写因子の誘導された発現の検出は、表1に例示する適切なプライマーおよび条件を用いて実施例2に記載するとおり実施される。
(Example 16: Identification of PDX position in the hierarchical structure of β- or islet cell-specific transcription factor)
This example illustrates the identification of the position of PDX in the hierarchical structure of transcription factors specific to β cells or islet cells. Each transcription factor that is expressed in islets but not induced by ectopic PDX-1 expression in the liver induces a more comprehensive, complete or near-complete β-cell phenotype in non-endocrine pancreatic tissues such as liver For this reason, it is considered to cooperate with PDX. Detection of induced expression of islet cell-specific transcription factors in the liver is performed as described in Example 2 using the appropriate primers and conditions exemplified in Table 1.
転写因子の活性を分析するための別の方法はフットプリンティングにより実施され、そして以下の通り実施される。 Another method for analyzing transcription factor activity is performed by footprinting and is performed as follows.
電子移動性シフト試験(EMSA):核抽出液(蛋白3〜4μg)を10%グリセロール、15mMHepes(pH7.9)、150mMKCl、5mMDTTおよび0.3gのポリdIdC、ポリdAdT(SIGMA St Louis MO)を含有するDNA結合混合物中10分間氷上でインキュベートした。最初のインキュベーションの後、プローブ約0.2ngを添加し、更に25分間の氷上インキュベーションに付した。結合反応はネイティブの4%ポリアクリルアミドゲル上で分析した。 Electron mobility shift test (EMSA): Nuclear extract (3-4 μg protein) with 10% glycerol, 15 mM Hepes (pH 7.9), 150 mM KCl, 5 mM DTT and 0.3 g poly dIdC, poly dAdT (SIGMA St Louis MO) Incubated on ice for 10 minutes in the contained DNA binding mixture. After the initial incubation, approximately 0.2 ng of probe was added and subjected to an additional 25 minute incubation on ice. The binding reaction was analyzed on a native 4% polyacrylamide gel.
オリゴヌクレオチド(プローブ)。合成の2本鎖オリゴヌクレオチドをDNAポリメラーゼのクレノウフラグメントを用いて[α32P]ATPで末端標識する。インスリンプロモーター上のPDX−1結合部位(の1つ)に関する例であるオリゴヌクレオチドA3/A4の配列は5’GATCTGCCCCTTGTTAATAATCTAATG3’(配列番号24)である。A1(インスリンプロモーター上の別のPDX−1結合部位)の配列は5’GATCCGCCCTTAATGGGCCAAACGGCA−3’(配列番号25)である。図7に示すとおり、標識されたオリゴを電子移動性シフト試験のためのプローブとして使用する。PDX−1の識別は、プロモーター上のその同属体遺伝子座へのPDX−1の結合を防止するか、または、pdx−1+標識プローブのみを含有するものと比較してPAGE(抗体+pdx−1+プローブ)上で分離される複合体の分子量を増大させる特異的抗体を用いたスーパーシフトにより二重推定される(図7における最後2レーン)。 Oligonucleotide (probe). Synthetic double-stranded oligonucleotides are end-labeled with [α 32 P] ATP using Klenow fragment of DNA polymerase. The sequence of oligonucleotide A3 / A4, which is an example for (one of) the PDX-1 binding sites on the insulin promoter, is 5′GATCTGCCCCCTGTTAATAATCTAATG3 ′ (SEQ ID NO: 24). The sequence of A1 (another PDX-1 binding site on the insulin promoter) is 5′GATCCGCCCTTAATGGGCCAAACGGGCA-3 ′ (SEQ ID NO: 25). As shown in FIG. 7, labeled oligos are used as probes for electron mobility shift testing. Identification of PDX-1 prevents binding of PDX-1 to its homologous locus on the promoter, or PAGE (antibody + pdx-1 + probe compared to that containing only pdx-1 + labeled probe) ) Double presumed by supershift with a specific antibody that increases the molecular weight of the complex separated above (last 2 lanes in FIG. 7).
(実施例17:肝における膵内分泌および外分泌マーカーの異所性PDX−1発現による誘導)
インビボの成熟肝における異所性PDX−1発現は膵遺伝子の広範なレパートリーを活性化する。内分泌および外分泌の両方のマーカー、例えば外分泌膵転写因子p48は肝における異所性PDX−1発現に応答して独特の発現を示した(図8)。対照投与マウスは膵遺伝子発現に対して殆ど陰性であった。インスリン遺伝子発現は異所性PDX−1投与マウスのほぼ100%で誘導されたが、対照投与マウスの20%においても蛋白に翻訳されない極めて低い濃度で発現された。
(Example 17: Induction of pancreatic endocrine and exocrine markers in liver by ectopic PDX-1 expression)
Ectopic PDX-1 expression in mature liver in vivo activates a broad repertoire of pancreatic genes. Both endocrine and exocrine markers, such as exocrine pancreatic transcription factor p48, showed unique expression in response to ectopic PDX-1 expression in the liver (FIG. 8). Control-treated mice were almost negative for pancreatic gene expression. Insulin gene expression was induced in almost 100% of ectopic PDX-1 treated mice but was expressed at very low concentrations that were not translated into protein in 20% of control treated mice.
発生中の膵においてPDX−1は膵の関与と一時的に相関する早期分子マーカーとして作用する。このデータはPDX−1は肝のような成熟した完全に分化した組織において、膵臓器形成におけるその役割を反復していることを示している。 In the developing pancreas, PDX-1 acts as an early molecular marker that temporarily correlates with pancreatic involvement. This data indicates that PDX-1 repeats its role in pancreatic organogenesis in mature, fully differentiated tissues such as the liver.
(実施例18:肝から膵への発生のシフトの長時間持続過程に対するPDX−1による惹起)
マウスインビボ肝への初回単回アデノウィルス媒介PDX−1投与後6ヶ月のインスリン、グルカゴンおよびソマトスタチンの遺伝子の発現および蛋白の生産を調べた。
(Example 18: Induction by PDX-1 for a long-lasting process of developmental shift from liver to pancreas)
Insulin, glucagon and somatostatin gene expression and
8〜9週齢のマウスにAd−CMV−PDX−1、即ちCMVプロモーターの制御下にラットPDX−1遺伝子(STF−1)を担持する組み換えアデノウィルスを全身投与した。肝における膵遺伝子発現を齢マッチ対照マウス(Ad−CMV−β−ガラクトシダーゼ投与または未投与)との比較において分析した。 8- to 9-week-old mice were systemically administered with Ad-CMV-PDX-1, that is, a recombinant adenovirus carrying the rat PDX-1 gene (STF-1) under the control of the CMV promoter. Pancreatic gene expression in the liver was analyzed in comparison to age-matched control mice (Ad-CMV-β-galactosidase administered or not).
肝への遺伝子のアデノウィルス媒介送達により達成される予測された一過性のPDX−1発現にもかかわらず(組み換えPDX−1の発現はウィルス注射後30日〜56日の間に減衰する)、インスリンとソマトスタチンの発現はmRNA(図9)および蛋白のレベル(図11)の両方で6ヶ月間持続した。グルカゴンの遺伝子発現は最初の4ヶ月の間に顕著となった(図9および10)。重要なことであるが、インスリンIおよびインスリンIIの遺伝子の発現は初回PDX−1投与後6〜8ヶ月の時点でもPDX−1投与マウスの80〜100%において明らかであった。 Despite the predicted transient PDX-1 expression achieved by adenovirus-mediated delivery of genes to the liver (recombinant PDX-1 expression decays between 30 and 56 days after virus injection) Insulin and somatostatin expression persisted for 6 months at both mRNA (FIG. 9) and protein levels (FIG. 11). Glucagon gene expression became prominent during the first 4 months (FIGS. 9 and 10). Importantly, insulin I and insulin II gene expression was evident in 80-100% of PDX-1 treated mice even at 6-8 months after the initial PDX-1.
インスリンおよびグルカゴンの遺伝子発現の間の一時的相違は成熟肝における膵臓器形成を特徴付ける独特の現象を反映しており、βおよびδ細胞の表現型に向けてより安定な相互変換があることを示唆している。グルカゴン遺伝子は直接のPDX−1標的遺伝子ではなく、肝におけるその持続的発現はPDX−1がこの臓器において分化因子として機能していることを示唆している。 Transient differences between insulin and glucagon gene expression reflect a unique phenomenon that characterizes pancreatic organogenesis in the mature liver, suggesting that there is a more stable interconversion towards β and δ cell phenotypes is doing. The glucagon gene is not a direct PDX-1 target gene and its sustained expression in the liver suggests that PDX-1 functions as a differentiation factor in this organ.
(実施例19:インスリン、グルカゴンおよびソマトスタチンの定量分析
PDX−1投与肝のホルモン生産)
免疫組織化学的分析(図10)により、PDX−1異所性遺伝子送達後4〜6ヶ月においても主に中心静脈近接部においてインスリン生産細胞の位置が特定される(図10Aおよび10C)。グルカゴン陽性細胞もまた中心静脈近接部に局在するが(図10B)、逐次的スライド上で行ったこれら2種のホルモンの免疫組織化学的分析によれば、これらのホルモンは同じ細胞内には同時に局在しないことが示唆される。肝内の中心静脈に近接する領域に存在する肝細胞は成熟細胞に相当することがわかっている。
(Example 19: Quantitative analysis of insulin, glucagon and somatostatin hormone production in the liver treated with PDX-1)
Immunohistochemical analysis (FIG. 10) identifies the location of insulin-producing cells primarily in the central vein proximity even 4-6 months after PDX-1 ectopic gene delivery (FIGS. 10A and 10C). Although glucagon-positive cells are also localized in the central vein vicinity (FIG. 10B), according to immunohistochemical analysis of these two hormones performed on sequential slides, these hormones are not located in the same cell. It is suggested that they are not localized at the same time. It has been found that hepatocytes present in the region close to the central vein in the liver correspond to mature cells.
PDX−1投与マウスの肝内に保存された肝インスリンの定量的分析によれば、投与後4〜6ヶ月においても、肝インスリン含量は約30〜75ng/g組織であり、これに対し、齢マッチの対照肝では1〜9ng/g組織であった(図11A)。初回Ad−CMV−PDX−1投与後少なくとも4ヶ月まで肝プロ/グルカゴンおよびソマトスタチン含量の有意な2倍増量が観察された(図11BおよびC)。PDX−1投与後の他の2種の膵ホルモンと比較した場合のインスリンの肝含量の大きな相違は膵におけるこれらのホルモンの比に似ていると考えられる。肝インスリン生産にもかかわらず、正常な膵機能を有するPDX−1投与マウスにおける血清中インスリンおよびグルコースの濃度は実験期間中を通して正常であった(PDX−1投与と対照の比較において、それぞれ、インスリン:1.0±0.5対0.9±0.4ng/ml、および、グルカゴン:0.16±0.08対0.12±0.05ng/ml)。 According to the quantitative analysis of hepatic insulin stored in the liver of PDX-1 treated mice, the liver insulin content is about 30-75 ng / g tissue even after 4-6 months after administration, whereas In matched control livers, it was 1-9 ng / g tissue (FIG. 11A). Significant 2-fold increases in hepatic pro / glucagon and somatostatin content were observed for at least 4 months after the first Ad-CMV-PDX-1 administration (FIGS. 11B and C). The large difference in the liver content of insulin when compared to the other two pancreatic hormones after PDX-1 administration appears to be similar to the ratio of these hormones in the pancreas. Despite hepatic insulin production, serum insulin and glucose concentrations in PDX-1 treated mice with normal pancreatic function were normal throughout the experimental period (in the comparison of PDX-1 administration and control, respectively, : 1.0 ± 0.5 vs. 0.9 ± 0.4 ng / ml and glucagon: 0.16 ± 0.08 vs. 0.12 ± 0.05 ng / ml).
肝内の膵ホルモンの持続的生産は異所性PDX−1がPDX−1トランスジーンの持続的存在を必要としない事象のカスケードを惹起することを示唆している。 The sustained production of pancreatic hormones in the liver suggests that ectopic PDX-1 triggers a cascade of events that do not require the persistent presence of the PDX-1 transgene.
(実施例20:肝における内因性の他の面ではサイレントなPDX−1遺伝子の発現の異所性PDX−1による惹起)
一過性の異所性PDX−1発現により惹起される肝における持続性発生シフトを評価するために、トランスジーンが他の面ではサイレントな膵転写因子の発現を誘導するかどうか分析した。
Example 20: Ectopic PDX-1 Induction of Silent PDX-1 Gene Expression in Other Endogenous Endogenous Livers
To assess the persistent developmental shift in the liver caused by transient ectopic PDX-1 expression, we analyzed whether the transgene induces expression of otherwise silent pancreatic transcription factors.
異所性遺伝子による肝内における、内因性で他の面ではサイレントなPDX−1遺伝子の誘導を分析するために、ラットPDX−1相同体の発現を指向する組み換えアデノウィルスを全身送達によりマウスに投与し、そして、RT−PCRにより異所性PDX−1トランスジーン(ラット)mRNA(cDNA)と内因性マウスmRNAとを判別するために特定のオリゴヌクレオチドプライマーを使用した。 Recombinant adenovirus directed to the expression of rat PDX-1 homolog is administered to mice by systemic delivery to analyze endogenous and otherwise silent PDX-1 gene induction in the liver by ectopic genes And specific oligonucleotide primers were used to discriminate between ectopic PDX-1 transgene (rat) mRNA (cDNA) and endogenous mouse mRNA by RT-PCR.
Ad−CMV−PDX−1投与マウスの肝から単離したDNA試料のPCR分析によれば、アデノウィルス注射後30〜56日にウィルスコードトランスジーンは消失することが確認される(図12A)。 PCR analysis of DNA samples isolated from the livers of mice treated with Ad-CMV-PDX-1 confirms that the virus-coded transgene disappears 30-56 days after adenovirus injection (FIG. 12A).
図12Bは異所性ラットPDX−1発現は肝における送達されたウィルスDNAの観察された存在と並行し、そして1ヵ月後において消滅することも示している(図12A)。実験の全期間にわたり投与肝において発現された唯一のPDX−1の相同体は内因性でその他の面ではサイレントなマウス相同体である(図12B)。内因性PDX−1の発現はラットPDX−1トランスジーンを投与されたマウスに限定されており、異所性PDX−1投与マウスの75%(28匹中21匹)および分析した対照投与肝25例中0例で明らかとなった。リアルタイムPCRを使用して、肝におけるマウス対ラットのPDX−1遺伝子発現の識別および定量された相対量を、同一条件(ただし異なるプライマー)を用いて初回投与後の時間の関数として分析し、同じ試料内のβアクチンに対して規格化した。
FIG. 12B also shows that ectopic rat PDX-1 expression parallels the observed presence of delivered viral DNA in the liver and disappears after one month (FIG. 12A). The only PDX-1 homologue expressed in the treated liver for the entire duration of the experiment is an endogenous and otherwise silent mouse homologue (FIG. 12B). Endogenous PDX-1 expression is restricted to mice receiving rat PDX-1 transgene, 75% of ectopic PDX-1 treated mice (21 of 28) and analyzed
図12Cに示す通り、異所性ラットPDX−1をコードするmRNAは5日において最高値を示し、30日には85%低下し、その後は消失した。これとは対照的に、内因性マウスPDX−1は実験の全期間を通じてかなりの濃度で発現された。これらのデータを総括すると、内因性であるが他の面ではサイレントであるPDX−1の肝内における自己誘導が示唆されており、これは肝から膵への相互変換過程の長時間持続する様式を機序的に説明していることを示唆している。
As shown in FIG. 12C, mRNA encoding ectopic rat PDX-1 showed the highest value on
(実施例21:機能性でありSTZ誘導高血糖症を防止するPDX−1トランスジーン発現に応答した肝内で生産されるインスリン)
生物学的に活性なインスリンの長時間持続する生産をPDX−1遺伝子送達により誘導できるかどうかを調べるために、それがSTZ誘導糖尿病に対抗して保護作用をもたらすかどうかを分析した。初回のAd−CMV−PDX−1投与後8ヶ月に、マウスに220mg/kgのSTZを投与し、高血糖症の発生率を齢マッチ対照群と比較した。対照Balb/cマウスの60%(10匹中6匹)がSTZ注射後3〜5日以内に高血糖症を発症した。これとは対照的に、5匹のPDX−1投与マウスでは、それらがAd−CMV−PDX−1投与後8ヶ月に分析したという事実にもかかわらず、うち僅か1匹のみがSTZ投与に応答して高血糖症を発症していた(20%)。
Example 21: Insulin produced in the liver in response to PDX-1 transgene expression that is functional and prevents STZ-induced hyperglycemia
To investigate whether long-lasting production of biologically active insulin can be induced by PDX-1 gene delivery, it was analyzed whether it provided a protective effect against STZ-induced diabetes. Mice were administered 220 mg /
免疫組織化学的試験およびRIAによるインスリン含量の定量によれば、STZ投与に応答して膵β細胞は殆どが破壊され、そして対照糖尿病マウスと、重要なことであるが、PDX−1投与マウス(正常血糖のまま推移)の両方において、膵インスリン含量が95±1%低下したことが判った。これとは対照的に、PDX−1投与マウスにおける肝免疫反応性インスリン(IRI)含量はPDX−1を投与しなかった対照糖尿病マウスと比較して40倍高値であった(図13)。健常マウスにおいては、肝インスリンは膵内の生産量の僅か1%であったが、STZ投与に応答して肝インスリン生産は同じマウスのSTZ投与膵において生産された免疫反応性インスリンの量の25.6%となっている。 According to immunohistochemical tests and quantification of insulin content by RIA, most pancreatic β cells were destroyed in response to STZ administration, and, importantly, control diabetic mice and, more importantly, PDX-1 treated mice ( It was found that the pancreatic insulin content was reduced by 95 ± 1% in both cases. In contrast, hepatic immunoreactive insulin (IRI) content in PDX-1 treated mice was 40 times higher compared to control diabetic mice not administered PDX-1 (FIG. 13). In healthy mice, hepatic insulin was only 1% of production in the pancreas, but in response to STZ administration, hepatic insulin production was 25. of the amount of immunoreactive insulin produced in the same mouse STZ-administered pancreas. 6%.
これらの結果から、PDX−1誘導の発生シフトは長時間持続し、かつ機能的であり、肝により寄与された比較的中等度のIRI濃度は肝内で生産されたインスリンは、肝グルコース生産とグルコース廃棄の間のバランスの効率的調節によってもSTZ誘導高血糖症に対抗する保護作用を示すことが示唆される。重要な点は、これはまた肝内で発生シフトした細胞がβ細胞特異的毒素に抵抗性であることも示唆していることである。 From these results, the PDX-1-induced developmental shift is long-lasting and functional, and the relatively moderate IRI concentration contributed by the liver is that insulin produced in the liver It is suggested that efficient regulation of the balance between glucose disposal also shows a protective effect against STZ-induced hyperglycemia. Importantly, this also suggests that cells shifted in the liver are resistant to β-cell specific toxins.
重要なことであるが、初回ウィルス感染の6〜8ヵ月後であっても肝内で局所的インスリン生産が継続しているにもかかわらず、肝機能は損なわれていなかった(表4a)。アデノウィルス投与に応答して肝機能の一過性の変化が起こったが、肝機能は1〜2ヶ月以内に正常値を回復した。更にまた、血清中アミラーゼ濃度は内因性PDX−1および膵ホルモン発現にも関わらず全時点において増大しなかった(表4b)。PDX−1投与マウスの体重増加率は齢マッチ対照マウスと同様であった。 Importantly, liver function was not impaired despite local insulin production continuing in the liver even 6-8 months after the initial viral infection (Table 4a). Transient changes in liver function occurred in response to adenovirus administration, but liver function returned to normal within 1-2 months. Furthermore, serum amylase concentrations did not increase at all time points despite endogenous PDX-1 and pancreatic hormone expression (Table 4b). The rate of weight gain of PDX-1 treated mice was similar to age-matched control mice.
(細胞培養)
ケラチノサイトの培養は剥離厚みの皮膚の小型生検標本(2〜4cm2)から開始した。トリプシン−EDTA中一夜(ON)インキュベートした後、表皮を分離し、上皮をトリプシン−EDTA中で破砕して単細胞の懸濁液を形成した。細胞懸濁液をケラチノサイト培地(Nature 265:421−4,1977)中で培養し、細胞懸濁液をファルコン培養プレートに結合させ、2〜5継代で使用した。細胞が70%コンフルエントに達した時点で48〜96時間、以下に記載する所定の投与法により投与した。
(Cell culture)
Keratinocyte culture was initiated from small biopsy specimens (2-4 cm 2 ) of exfoliated skin. After overnight (ON) incubation in trypsin-EDTA, the epidermis was separated and the epithelium was disrupted in trypsin-EDTA to form a single cell suspension. The cell suspension was cultured in keratinocyte medium (Nature 265: 421-4, 1977) and the cell suspension was bound to a Falcon culture plate and used at passages 2-5. When the cells reached 70% confluence, they were administered for 48 to 96 hours according to the prescribed administration method described below.
(遺伝子発現)
遺伝子発現分析はTaqmanリアルタイムPCR(ABI)を用いて実施した。
(Gene expression)
Gene expression analysis was performed using Taqman real-time PCR (ABI).
(細胞の投与)
K1:EGF+KGF+NIC+PDX−1(100moi)
K2:EGF+KGF+NIC+PDX−1(10moi)
K3:EGF+KGF
K4:EGF+KGF+RGCI
全投与においてEGF、KGFは20ng/ml;NIC:10ng/ml。
(Administration of cells)
K1: EGF + KGF + NIC + PDX-1 (100moi)
K2: EGF + KGF + NIC + PDX-1 (10moi)
K3: EGF + KGF
K4: EGF + KGF + RGCI
EGF and KGF were 20 ng / ml in all administrations; NIC: 10 ng / ml.
(対照群)
対照群細胞は非関連性のAd−CMV−Hインスリン、即ちCMVプロモーターの制御下にヒトインスリン遺伝子の発現を行っている組み換えアデノウィルスを投与した。RGCIは2官能性の組み換えアデノウィルスコンストラクトAd−CMV−PDX−1−RIP−GFPであり、これはインスリン遺伝子発現に向けてPDX−1媒介転移分化を起こしている細胞を識別した。このウィルスにおけるPDX−1の発現はCMVプロモーターにより駆動されたのに対し、GFP発現はインスリンに関する組織特異的プロモーター(RIP)により駆動された。
(Control group)
Control group cells were administered unrelated Ad-CMV-H insulin, ie, recombinant adenovirus expressing human insulin gene under the control of CMV promoter. RGCI is a bifunctional recombinant adenovirus construct Ad-CMV-PDX-1-RIP-GFP, which identified cells undergoing PDX-1-mediated transdifferentiation towards insulin gene expression. PDX-1 expression in this virus was driven by a CMV promoter, whereas GFP expression was driven by a tissue specific promoter (RIP) for insulin.
(結果)
投与K1〜K4において、内因性の他の面ではサイレントな膵遺伝子がケラチノサイトにおいて発現された。興味深いことに、グルカゴンの遺伝子発現は低濃度のPDX−1(K4)により、そして、重要なことであるがEGF+KGF投与単独により、異所性PDX−1を必要とすることなく誘導された(図15)。
(result)
In administrations K1-K4, an otherwise endogenous pancreatic gene was expressed in keratinocytes. Interestingly, glucagon gene expression was induced by low concentrations of PDX-1 (K4) and, importantly, EGF + KGF administration alone, without the need for ectopic PDX-1 (FIG. 15).
(実施例23:ヒト肝細胞におけるインスリンプロモーターのPDX−1による活性化)
ヒト肝細胞を成人および胎児の組織から単離した。細胞は異種の表現型を示し、20継代までは培養物中効率的に増殖した。何れの事前の選別も行うことなく成人の完全に分化した肝臓から単離したヒト細胞が異所性PDX−1発現に応答して膵表現型に向けた発生再指向の過程を起こすかどうか分析した。膵特徴の第1の指標はその他の面では肝中で不活性であるインスリンプロモーターの活性化である。細胞に異種のCMVプロモーターの制御下にPDX−1を発現する2官能性組み換えアデノウィルスAd−RIP−GFP−CMV−PDX−1を投与し、インスリンプロモーターにGFP発現(図17a)を制御させ、これによりPDX−1「応答」細胞を緑色蛍光により識別した(図17b)。成人ヒト肝細胞が組み換えアデノウィルスに感染する総能力はAd−CMV−GFP感染により調べ、成人肝細胞の40±7%が継代1〜6においてAd−CMV−GFP感染に応答して緑色蛍光を示した。意外にも、これらの細胞の約半数(23±3.5%)がFACS分析(データ示さず)でも判明したとおり、膵プロモーターの活性化による異所性PDX−1発現に応答している(図17b)。部分的な応答が成人肝細胞の分化状態により影響され、制限されるかどうかを調べるために、分化程度が低くおそらくは成人ヒト肝細胞よりも多能細胞を多く含んでいる胎児ヒト肝細胞の発生再指向を誘導するPDX−1の能力を分析した。実際、培養物中の胎児ヒト肝細胞(妊娠22週で単離)の27±7.8%が膵プロモーターの活性化による異所性PDX−1発現に応答し、Ad−CMV−GFP形質導入へのその応答は成人肝から単離された細胞と同様であった(図17c)。応答細胞のこの中等度の増大は細胞の分化状態がPDX−1により誘導される発生シフト過程において限定された役割しか果たしていないことを示唆していると考えられる。
(Example 23: Activation of insulin promoter by PDX-1 in human hepatocytes)
Human hepatocytes were isolated from adult and fetal tissues. The cells showed a heterogeneous phenotype and grew efficiently in culture until
3つの重要な観察結果がヒト肝細胞の一次培養から得られる。第1に、成人および胎児の両方の細胞はインビトロで培養した場合、6ヶ月まで効率的に増殖し、そして、PDX−1投与に応答してインスリンプロモーターを活性化することができる。しかしながら、その感染および「転移分化」の能力は継代数が増加するに従って低下する(図17cおよびd)。第2に、胎児ヒト肝組織は成人肝より単離された細胞よりも多数の多能細胞よりなるが、それらは膵への転移分化過程を起こす同様の能力を有する(図17c)。第3に、異所性にPDX−1を発現するヒト肝細胞においてインスリンプロモーターを活性化する能力は、組み換えアデノウィルスにより感染可能な細胞の半数は培養の低継代ではPDX−1依存性の態様で異所性インスリンプロモーターをやはり活性化させるため、細胞の希少な集団においては生じない(図17d)。 Three important observations are obtained from the primary culture of human hepatocytes. First, both adult and fetal cells grow efficiently up to 6 months when cultured in vitro and can activate the insulin promoter in response to PDX-1 administration. However, its ability to infect and “metastasize” decreases as the number of passages increases (FIGS. 17c and d). Secondly, fetal human liver tissue consists of a greater number of pluripotent cells than cells isolated from adult liver, but they have a similar ability to undergo a metastatic differentiation process to the pancreas (FIG. 17c). Third, the ability to activate the insulin promoter in human hepatocytes ectopically expressing PDX-1 indicates that half of the cells that can be infected by recombinant adenovirus are PDX-1-dependent in low passage cultures. It also activates the ectopic insulin promoter and thus does not occur in a rare population of cells (FIG. 17d).
(実施例24:可溶性因子によるPDX−1誘導の肝から膵への転移分化の促進)
転移分化過程の範囲をより良好に識別することはPDX−1投与肝細胞における内因性の他面ではサイレントな膵遺伝子の発現の誘導を分析することである。
(Example 24: Promotion of PDX-1-induced liver-to-pancreas differentiation by a soluble factor)
A better discrimination of the extent of the metastatic differentiation process is to analyze the induction of endogenous pancreatic gene expression in PDX-1 treated hepatocytes.
3種の主要な膵ホルモン遺伝子発現の発現は、対照の未投与の肝細胞と比較して2桁高値のPDX−1により誘導した(図18)。 The expression of the three major pancreatic hormone gene expressions was induced by PDX-1 which was two orders of magnitude higher compared to control untreated hepatocytes (FIG. 18).
ニコチンアミドおよび表皮成長因子(EGF)は胚性膵臓器培養を含めて未分化の膵細胞の膵内分泌分化を促進することがわかっている。興味深いことに、PDX−1投与にニコチンアミドおよびEGF(総称してGFと称する)を添加した場合、膵ホルモン遺伝子発現は劇的に増大した。成人肝細胞の一次培養におけるインスリン遺伝子発現は対照未投与肝細胞と比較して7桁高値となった。ニコチンアミドおよびEGFは単独でも組み合わせた場合も肝細胞における膵遺伝子発現に対してPDX−1依存作用を示した。これらのデータはPDX−1は肝から膵への転移分化の過程に必要であるが、GFは非依存的に誘導するのには十分ではない過程に対して相乗作用を有することを示唆している。重要な点は、胎児および成人のヒト肝細胞は、両方の培養におけるインスリン遺伝子の発現のリアルタイムPCR定量により示されたとおり、異所性PDX−1発現およびGF投与に応答して膵遺伝子発現の同様の水準を示していることである(図18b)。 Nicotinamide and epidermal growth factor (EGF) have been shown to promote pancreatic endocrine differentiation of undifferentiated pancreatic cells, including embryonic pancreatic organ cultures. Interestingly, pancreatic hormone gene expression increased dramatically when nicotinamide and EGF (collectively referred to as GF) were added to PDX-1 administration. Insulin gene expression in primary cultures of adult hepatocytes was 7 orders of magnitude higher than in control untreated hepatocytes. Nicotinamide and EGF, both alone and in combination, showed a PDX-1 dependent effect on pancreatic gene expression in hepatocytes. These data suggest that PDX-1 is required for the process of metastatic differentiation from liver to pancreas, but GF has a synergistic effect on a process that is not sufficient to induce independently. Yes. Importantly, fetal and adult human hepatocytes showed pancreatic gene expression in response to ectopic PDX-1 expression and GF administration, as demonstrated by real-time PCR quantification of insulin gene expression in both cultures. It shows a similar level (FIG. 18b).
PDX−1投与の前数週間に亘りGFの存在下に肝細胞を培養したところ、PDX−1およびGFを同時に投与した細胞と比較してインスリン遺伝子発現の増大はもたらされなかった。これとは対照的に、予備投与培養からGFを除去した場合、インスリン遺伝子発現がPDX−1単独の場合と同様の濃度でもたらされた。総括すると、PDX−1誘導転移分化に対してGFが有している促進作用は、この過程の影響を受けやすい希少なサブ集団の増殖を誘発することではなく、むしろ、それらはいまだ未知の態様で寄与することによりPDX−1誘導過程をもたらす細胞内シグナル伝達を増強していることが示唆されている。Ad−CMV−hIns,即ちCMVプロモーター制御下にヒトプロインスリンcDNAの構成的異所性発現を起こす組み換えアデノウィルスの同様の感染多重度はPDX−1およびGF投与細胞の場合に匹敵する濃度のインスリン遺伝子発現をもたらした(PDX−1投与の場合のようにグルカゴンとソマトスタチンの遺伝子発現の誘導を伴わない)。Ad−CMV−hInsを投与した場合にインスリンを発現する細胞の数がPDX−1誘導内因性インスリン遺伝子を発現する細胞の数の二倍であること(細胞の僅か23%のみが転移分化過程を起こし、細胞の40%が異所性ヒトインスリン遺伝子を発現するため、図17cおよび18c参照)を考慮すると、PDX−1投与肝細胞における(内因性)インスリンプロモーターは非相同のCMVプロモーターと同様の活性および力価を有することが示唆される。 Culture of hepatocytes in the presence of GF for several weeks prior to PDX-1 administration did not result in increased insulin gene expression compared to cells administered PDX-1 and GF simultaneously. In contrast, removal of GF from the pre-dosing culture resulted in insulin gene expression at a concentration similar to PDX-1 alone. In summary, the facilitating effect of GF on PDX-1 induced metastatic differentiation does not induce the growth of a rare subpopulation that is susceptible to this process, but rather they are still in an unknown manner It is suggested that the intracellular signal transduction resulting in the PDX-1 induction process is enhanced by contributing in the above. A similar multiplicity of infection of Ad-CMV-hIns, a recombinant adenovirus that causes constitutive expression of human proinsulin cDNA under the control of the CMV promoter, is comparable to that of PDX-1 and GF treated cells. This resulted in expression (without induction of glucagon and somatostatin gene expression as in PDX-1 administration). The number of cells that express insulin when Ad-CMV-hIns is administered is twice the number of cells that express the PDX-1-induced endogenous insulin gene (only 23% of the cells undergo the metastatic differentiation process) Waking up and 40% of the cells express the ectopic human insulin gene (see FIGS. 17c and 18c), the (endogenous) insulin promoter in PDX-1 treated hepatocytes is similar to the heterologous CMV promoter It is suggested to have activity and potency.
(実施例25:PDX−1による成人ヒト肝細胞への内分泌特性の付与)
既に転移分化した成人ヒト肝細胞が内分泌細胞の特徴を獲得するかどうか分析するために、これらの細胞のPDX−1誘導インスリンを貯蔵してプロセシングする能力を分析した。図19はインスリンおよびC−ペプチドの分泌およびPDX−1投与細胞におけるインスリン含量を示す。PDX−1投与単独では免疫反応性インスリン(IRI)含量は34.5±4.5倍増大、IRI分泌は38.7±8.7倍増大、そして、C−ペプチド分泌は7.5±2.1倍増大となった。GFを培地に添加したところ過程に対するPDX−1の作用が大きく増強され、未投与肝細胞と比較して、IRI含量は91.3±20.3倍増大し、その分泌は74.5±33.3倍増大し、そしてC−ペプチドの分泌は33.9±14.6倍増大した。過程に対するPDX−1の作用をヒトプロインスリンの異所性発現の作用と比較した(Ad−CMV−hIns組み換えアデノウィルスを使用)。Ad−CMV−hInsを投与した細胞においては生産されたIRIの大部分が放出されたが、PDX−1投与細胞中のIRIの大半は細胞内に貯留された。Ad−CMV−hIns投与によるC−ペプチドの中等度の分泌は肝細胞中のフリンのような既存のエンドペプチダーゼに起因するものと考えられる。重要なことであるが、プロホルモン変換酵素2の誘導はPDX−1投与の場合のみ顕著であり、Ad−CMV−hIns投与肝細胞では観察されなかった(図21a)。
(Example 25: Giving endocrine properties to adult human hepatocytes by PDX-1)
In order to analyze whether adult human hepatocytes already metastasized differentiated to acquire endocrine cell characteristics, the ability of these cells to store and process PDX-1-induced insulin was analyzed. FIG. 19 shows insulin and C-peptide secretion and insulin content in PDX-1 treated cells. PDX-1 administration alone increased immunoreactive insulin (IRI) content by 34.5 ± 4.5 fold, IRI secretion increased by 38.7 ± 8.7 fold, and C-peptide secretion increased by 7.5 ± 2 Increased by a factor of 1. When GF was added to the medium, the effect of PDX-1 on the process was greatly enhanced, the IRI content increased by 91.3 ± 20.3 times compared to untreated hepatocytes, and its secretion was 74.5 ± 33. .3 fold increase and C-peptide secretion increased by 33.9 ± 14.6 fold. The effect of PDX-1 on the process was compared to the effect of ectopic expression of human proinsulin (using Ad-CMV-hIns recombinant adenovirus). Although most of the produced IRI was released in cells administered Ad-CMV-hIns, most of the IRI in PDX-1-treated cells was retained intracellularly. The moderate secretion of C-peptide by Ad-CMV-hIns administration is thought to be due to existing endopeptidases such as furin in hepatocytes. Importantly, induction of
インスリンに対する抗体を用いたイムノゴールド組織化学の電子顕微鏡分析によれば、インスリンは分泌顆粒に保存された(図20a)。これらの顆粒はインビボの未損傷の膵島の場合に用に特徴的な緻密コア部を含まないが、より低濃度のインスリン保存を有するβ細胞系統に存在するものと類似していた。内分泌表現型は神経内分泌ベシクル特異的遺伝子発現の特異的誘導に関連していた。SCG−2(セクレトグラニン−2)およびSGNE1(分泌顆粒ニューロエンドクリン−1、図21b)の特異的発現はPDX−1投与細胞のみで観察され、Ad−CMV−hIns投与では観察されなかった。 According to immunogold histochemical electron microscopic analysis using an antibody against insulin, insulin was stored in secretory granules (FIG. 20a). These granules do not contain the dense core that is characteristic for undamaged islets in vivo, but were similar to those present in beta cell lines with a lower concentration of insulin storage. The endocrine phenotype was associated with specific induction of neuroendocrine vesicle-specific gene expression. Specific expression of SCG-2 (secretogranin-2) and SGNE1 (secretory granule neuroendocrine-1, FIG. 21b) was observed only in cells treated with PDX-1, but not with Ad-CMV-hIns .
これらのデータを総括すれば、PDX−1および適当な因子を投与された成人ヒト肝細胞は、膵内分泌細胞に特徴的な多くの特性を模倣した膵ホルモン生産細胞への広範で効率的な転移分化を起こすことが示唆される。 To summarize these data, adult human hepatocytes administered PDX-1 and appropriate factors have extensive and efficient metastasis to pancreatic hormone-producing cells that mimic many of the characteristics characteristic of pancreatic endocrine cells. It is suggested that differentiation occurs.
(実施例26:転移分化した成人ヒト肝細胞のグルコース感知能力)
グルコース感知能力およびグルコース感知とインスリン分泌との間の関連は膵β細胞機能を保証するものである。
(Example 26: Glucose sensing ability of metastasized adult human hepatocytes)
Glucose sensing ability and the link between glucose sensing and insulin secretion guarantee pancreatic beta cell function.
PDX−1誘導転移分化した肝細胞はGLUT−2およびグルコキナーゼ(GK、図21a)遺伝子を発現し、グルコース調節の態様でインスリンを分泌することが示された。25mMグルコースへのPDX−1投与成人ヒト肝細胞の曝露はインスリン分泌の即座で強度の増大をもたらす(図21b)。グルコース刺激インスリン(図21b)およびC−ペプチド(図21c)の分泌の経時的分析によれば、膵β細胞の場合と同様の2相性の動的な特徴が明らかになり、これには即座でシャープな第1のピークとそれに続く延長した第2のピークの分泌が伴っていた。グルコース惹起が排除されると、インスリン分泌は即座に低下した(図21d)。初回グルコース惹起後60〜90分の細胞外インスリン濃度の低下(図21b)はC−ペプチドの場合(図21c)よりも強度であり、異種培養物中の肝細胞による分泌インスリンの広範な取り込みを表していると考えられる。グルコース用量応答によれば、正常な膵島における8〜16mMでの最大インスリン分泌と比較して最大C−ペプチド分泌が25mMグルコースで生じ(図21d)たことから、正常膵β細胞と比較して右側へのシフトが明らかになった。重要な点は、グルコース感知能力とインスリン分泌の間の関連が正常膵β細胞と同様の様式で転移分化肝細胞中で起こったことであり、グルコースはインスリン分泌に対するその作用を発揮するためには代謝されなければならない。非代謝性のグルコース類縁体である2−デオキシ−グルコース(2DOG)は転移分化肝細胞においてC−ペプチドの分泌を惹起しなかった(図21d)。予測された通り、構成性プロモーター(Ad−CMV−hIns)により駆動されるヒトインスリンの異所性発現はプロホルモンのグルコース調節分泌をもたらさなかった。これらのデータはグルコース感知能力およびインスリン分泌との関連性が転移分化過程の結果であることを示している。 PDX-1 induced metastatic hepatocytes expressed GLUT-2 and glucokinase (GK, FIG. 21a) genes and were shown to secrete insulin in a glucose-regulated manner. Exposure of adult human hepatocytes treated with PDX-1 to 25 mM glucose results in an immediate increase in insulin secretion (FIG. 21b). A time course analysis of glucose-stimulated insulin (FIG. 21b) and C-peptide (FIG. 21c) reveals a biphasic dynamic characteristic similar to that of pancreatic beta cells, which is immediate. It was accompanied by a sharp first peak followed by an extended second peak secretion. When glucose induction was eliminated, insulin secretion immediately declined (FIG. 21d). The decrease in extracellular insulin concentration 60-90 minutes after the initial glucose challenge (FIG. 21b) is more intense than in the case of C-peptide (FIG. 21c), indicating a broad uptake of secreted insulin by hepatocytes in heterogeneous cultures. It is thought that it represents. According to the glucose dose response, maximal C-peptide secretion occurred at 25 mM glucose compared to maximal insulin secretion at 8-16 mM in normal islets (FIG. 21d), so that the right side compared to normal pancreatic β cells. The shift to became clear. The important point is that the link between glucose sensing ability and insulin secretion occurred in metastatic differentiated hepatocytes in a manner similar to normal pancreatic β cells, and glucose must exert its effect on insulin secretion. Must be metabolized. 2-Deoxy-glucose (2DOG), a non-metabolic glucose analog, did not elicit secretion of C-peptide in metastatic differentiated hepatocytes (FIG. 21d). As expected, ectopic expression of human insulin driven by a constitutive promoter (Ad-CMV-hIns) did not result in glucose-regulated secretion of the prohormone. These data indicate that the relationship between glucose sensing ability and insulin secretion is a result of the metastatic differentiation process.
(実施例27:糖尿病マウスにおける高血糖症の転移分化成人ヒト肝細胞による軽減)
β細胞機能を代替する転移分化成人ヒト肝細胞の能力を調べるために、STZ投与により糖尿病とされた免疫不全SCID−NODマウスに細胞を移植した。図22は対照投与マウスは高血糖症のままであったのに対し、PDX−1投与成人ヒト肝細胞を移植されたマウスは血糖値の徐々であるが有意な低下を示したことを示している。免疫組織化学的分析によれば、これらのマウスの膵臓はインスリン枯渇であったが、腎被膜下に移植したヒト肝細胞はPDX−1およびインスリンについて陽性に染色された(図18b)。ヒトC−ペプチドはPDX−1投与ヒト肝細胞を移植されたSTZ投与マウスの血清中で検出された。ヒトC−ペプチド濃度は正常SCID−NODおよびSTZ投与対照マウスにおける0.04±0.02ng/ml(p<0.01)と比較して有意な6〜7倍上昇を示し、平均は0.26±0.03ng/mlであった(図22c)。ヒト細胞移植マウスにおける血清中マウスインスリン濃度は未変化のままであり、対照の正常血糖SCID−NOCマウス(0.45±0.03ng/ml)よりも有意に低値(0.16±0.03ng/ml)であった。これらを総括すると、これらの所見は移植マウスにおける高血糖症は転移分化ヒト肝細胞から分泌されたヒトインスリンにより正常化されたことを示している。これらの結果はPDX−1投与転移分化成人ヒト肝細胞がインビボのβ細胞を代替物として機能する能力を明らかにしている。
(Example 27: Reduction of hyperglycemia in diabetic mice by metastatic differentiated adult human hepatocytes)
To examine the ability of metastatic differentiated adult human hepatocytes to substitute for β-cell function, cells were transplanted into immunodeficient SCID-NOD mice rendered diabetic by STZ administration. FIG. 22 shows that control-treated mice remained hyperglycemic, whereas mice transplanted with adult human hepatocytes treated with PDX-1 showed a gradual but significant decrease in blood glucose levels. Yes. According to immunohistochemical analysis, the pancreas of these mice was insulin depleted, but human hepatocytes transplanted under the renal capsule stained positive for PDX-1 and insulin (FIG. 18b). Human C-peptide was detected in the serum of STZ-treated mice transplanted with PDX-1-treated human hepatocytes. Human C-peptide concentration showed a significant 6-7 fold increase compared to 0.04 ± 0.02 ng / ml (p <0.01) in normal SCID-NOD and STZ treated control mice, with an average of 0. 26 ± 0.03 ng / ml (FIG. 22c). Serum mouse insulin concentrations in human cell transplanted mice remain unchanged and are significantly lower (0.16 ± 0.00%) than control normoglycemic SCID-NOC mice (0.45 ± 0.03 ng / ml). 03 ng / ml). In summary, these findings indicate that hyperglycemia in transplanted mice was normalized by human insulin secreted from metastatic differentiated human hepatocytes. These results demonstrate the ability of PDX-1 administered metastatic differentiated adult human hepatocytes to function in vivo as a substitute for β cells.
(実施例28:PDX−1誘導の肝から膵への転移分化によるCAD−NODマウスにおける高血糖症の退行)
明らかな自己免疫糖尿病に対するPDX−1誘導の肝から膵への転移分化の作用を調べるために、糖尿病NODマウスにAd−CMV−PDX−1またはAd−Rip−βGalを前述のとおり投与した。1群のマウスは対照として未投与のままとした。グルコース代謝の調節を調べるために血中グルコース濃度および体重をモニタリングした。未投与マウスおよびAd−Rip−βGal投与マウスは高血糖症のまま存続し、体重を減少させ、投与後最初の2週間以内に死亡した(図24)。これとは対照的に、Ad−CMV−PDX−1を投与したマウスの65%(34匹中20匹)が投与後最初の5日間以内に正常血糖となった。しかしながら、この正常血糖は一部のマウスでは一過性であった。PDX1を投与したマウスの38%(13/34)は投与の1ヶ月後に正常血糖のままであり、Ad−CMV−PDX−1を投与した他の20%(7/34)は投与の10〜14日後に高血糖症(図24a)となったが、実験の全期間を通じて安定した体重を維持した(図24b)。
Example 28: Regression of hyperglycemia in CAD-NOD mice due to PDX-1-induced liver-to-pancreas metastatic differentiation
In order to examine the effect of PDX-1-induced liver-to-pancreas metastatic differentiation on apparent autoimmune diabetes, Adeno-CMV-PDX-1 or Ad-Rip-βGal was administered as described above to diabetic NOD mice. One group of mice remained untreated as a control. Blood glucose concentration and body weight were monitored to examine the regulation of glucose metabolism. Untreated mice and Ad-Rip-βGal treated mice remained hyperglycemic, lost weight and died within the first 2 weeks after administration (FIG. 24). In contrast, 65% (20/34) of mice receiving Ad-CMV-PDX-1 became normoglycemic within the first 5 days after administration. However, this normoglycemia was transient in some mice. 38% (13/34) of mice receiving PDX1 remained normoglycemic one month after administration, and the other 20% (7/34) receiving Ad-CMV-PDX-1 Hyperglycemia (FIG. 24a) occurred after 14 days, but maintained a stable weight throughout the experiment (FIG. 24b).
これらのデータはPDX−1により誘導された肝から膵への転移分化の過程は自己免疫糖尿病(例えば1型)を退行させることを示唆している。 These data suggest that the process of metastatic differentiation from liver to pancreas induced by PDX-1 regresses autoimmune diabetes (eg, type 1).
(実施例29:Ad−CMV−PDX−1投与糖尿病マウスにおけるインスリンの合成および調節)
膵および肝のインスリン発現の免疫組織化学的分析によれば、Ad−CMV−PDX−1投与マウスの肝におけるインスリン生産細胞の存在が明らかになったが、その膵には存在しなかった(図25)。肝インスリン生産細胞は前述の通り中心静脈に近接して位置していた。更にまた、肝インスリン含量および血清中インスリン濃度は対照群と比較してAd−CMV−PDX−1投与マウスでは有意に高値であった(図26)。即ちAd−CMV−PDX−1は肝におけるインスリンの発現およびその血中への分泌を誘導した。
(Example 29: Synthesis and regulation of insulin in diabetic mice treated with Ad-CMV-PDX-1)
Immunohistochemical analysis of pancreatic and hepatic insulin expression revealed the presence of insulin-producing cells in the liver of Ad-CMV-PDX-1 treated mice, but not in the pancreas (Fig. 25). Liver insulin-producing cells were located close to the central vein as described above. Furthermore, hepatic insulin content and serum insulin concentration were significantly higher in mice administered with Ad-CMV-PDX-1 than in the control group (FIG. 26). That is, Ad-CMV-PDX-1 induced the expression of insulin in the liver and its secretion into the blood.
血中グルコースによる肝インスリン放出の調節を分析するために、Ad−CMV−PDX−1投与後正常血糖となってから2〜3週間後の糖尿病マウス5匹におけるグルコース耐容性試験を実施した。対照として、Ad−CMV−βGal投与糖尿病マウスおよび健常BALB/cマウスを使用した。健常BALB/cおよびAd−CMV−PDX−1投与マウスとの間にはグルコースクリアランス速度に差は観察されず、肝転移分化細胞によるインスリン分泌は実際にグルコースにより調節されることを示している(図27)。これとは対照的に、糖尿病Ad−CMV−βGal投与マウスはグルコースクリアランスを示さず、試験中を通じて高血糖症のまま存続した。これらのデータを総括すると、PDX−1誘導の機能的な肝から膵への転移分化は自己免疫のマウスモデルにおいても起こっている。PDX−1誘導正常血糖は肝におけるインスリン生産の誘導およびグルコース調節態様におけるその放出と関連していた。
In order to analyze the regulation of hepatic insulin release by blood glucose, a glucose tolerance test was conducted in 5
(実施例30:骨髄細胞の発生シフトのPDX−1による誘導)
骨髄細胞から機能的膵細胞への発生シフトを誘導する異所性PDX−1の能力を以下の通り測定する。純度90%超の新鮮凍結ヒトBMから単離したAC133+細胞を(a)24穴の組織培養プラスチックプレートまたは(b)テフロン(登録商標)バッグのいずれかにおいて、IL−6、TPO、Flt−3リガンドおよびSCFの継続的存在下、TEPAの存在下または非存在下に3週間増殖させた。前駆細胞の組成および能力は投与終了時および長期インキュベーション培養後に検査する。
(Example 30: Induction of bone marrow cell development by PDX-1)
The ability of ectopic PDX-1 to induce a developmental shift from bone marrow cells to functional pancreatic cells is measured as follows. AC133 + cells isolated from fresh frozen human BM of greater than 90% purity were either IL-6, TPO, Flt-3 in either (a) a 24-well tissue culture plastic plate or (b) a Teflon bag. Growth was continued for 3 weeks in the presence or absence of TEPA in the continuous presence of ligand and SCF. The composition and capacity of progenitor cells is examined at the end of administration and after long incubation cultures.
感染および実験の条件を最適化するために、2官能性のアデノウィルス(AdRIP−GFP−CMV−PDX−1)を用いて培養物を感染させる。2〜103MOI(培養物中の細胞数でウィルス粒子数を割ったもの、MOI、感染多重度)での感染後の時間の関数としてのGFP−レポーター遺伝子の発現は以下の通りである。 In order to optimize infection and experimental conditions, the bifunctional adenovirus (AdRIP-GFP-CMV-PDX-1) is used to infect cultures. Expression of the GFP-reporter gene as a function of time after infection at 2-10 3 MOI (number of virus particles divided by number of cells in culture, MOI, multiplicity of infection) is as follows.
応答細胞(インスリンプロモーターを活性化し、これによりGFPを発現する)を分類し、非応答細胞から分離して培養し、その後の分析に付する。対照BM細胞(TEPA投与または未投与)の分離された培養物は転移分化を伴わないインスリン生産に関する対照としてAdCMV−インスリンを投与する。 Responder cells (which activate the insulin promoter and thereby express GFP) are sorted, cultured from non-responder cells and subjected to subsequent analysis. Isolated cultures of control BM cells (TEPA administered or not) receive AdCMV-insulin as a control for insulin production without metastatic differentiation.
遺伝子発現の後にRT−PCRを行う。遺伝子発現の濃度および発生シフト後の誘導後の時間を定量的試験においてリアルタイムPCR(ABI、USA)を用いてリアルタイムPCRにより分析する。膵ホルモン、インスリン、グルカゴン、ソマトスタチンおよびプロホルモン変換酵素および特定の膵転写因子、即ちBETA2、Isl−1、Nkx6.1、Pax4およびPax6および内因性PDX−1の発現を分析する。 RT-PCR is performed after gene expression. The concentration of gene expression and the time after induction after the developmental shift are analyzed by real-time PCR using real-time PCR (ABI, USA) in a quantitative test. The expression of pancreatic hormones, insulin, glucagon, somatostatin and prohormone converting enzyme and specific pancreatic transcription factors, namely BETA2, Isl-1, Nkx6.1, Pax4 and Pax6 and endogenous PDX-1, are analyzed.
ホルモン生産は特異的抗体、即ちモルモット抗ブタインスリン;ウサギ抗ヒトグルカゴン;ウサギ抗ヒトソマトスタチン(全てDAKO A/S、Glostrup,Denmark)を用いて免疫組織化学的に検出する。インスリンおよびグルカゴンの生産および分泌は市販のRIA特異的キットであるSensitiveヒトインスリンおよびC−ペプチドRIAキットおよびグルカゴンRIAキット(Linco resrarch ICN,Missouri、USA)により分析する。グルコースの用量応答およびプロ/インスリン合成の過程および成熟した生物学的に活性なインスリンへの変換は記載したとおり(2)逆相HPLCにより、そして、特異的ヒトc−ペプチドRIAキット(Linco)を用いた培地へのC−ペプチドの分泌を分析することにより分解する。異なるグルコース濃度における細胞内インスリン含量を分析する。 Hormone production is detected immunohistochemically using specific antibodies: guinea pig anti-pig insulin; rabbit anti-human glucagon; rabbit anti-human somatostatin (all DAKO A / S, Glostrup, Denmark). Insulin and glucagon production and secretion are analyzed by commercially available RIA-specific kits, Sensitive Human Insulin and C-peptide RIA kit and Glucagon RIA kit (Linco research ICN, Missouri, USA). Dose response of glucose and the process of pro / insulin synthesis and conversion to mature biologically active insulin was as described (2) by reverse phase HPLC and a specific human c-peptide RIA kit (Linco) Decompose by analyzing the secretion of C-peptide into the medium used. Analyze intracellular insulin content at different glucose concentrations.
PDX−1誘導BM細胞中のグルコースへのインスリン分泌応答の動的特徴をインビトロで分析し、異所性インスリン発現(AdCMV−hIns)の場合と比較する。細胞を6穴皿上種々の濃度のグルコース中でインキュベートし、培地へのIRI分泌の用量応答および経時変化をラジオイムノアッセイ(RIA)で測定する。フォルスコリン/IBMXによる増大したインスリン分泌はモジュレートされた肝細胞における蛋白の分泌に関わる調節の誘導を示している。2−DOCやL−グルコースに対してではなくグルコースへの感受性は、「モジュレート」された肝細胞における栄養代謝へのインスリン分泌経路の特異的な関連を示す。 The dynamic characteristics of the insulin secretory response to glucose in PDX-1 induced BM cells are analyzed in vitro and compared to the case of ectopic insulin expression (AdCMV-hIns). Cells are incubated in various concentrations of glucose on 6-well dishes, and the dose response and time course of IRI secretion into the medium is measured by radioimmunoassay (RIA). Increased insulin secretion by forskolin / IBMX indicates the induction of regulation related to protein secretion in modulated hepatocytes. Sensitivity to glucose but not to 2-DOC or L-glucose indicates a specific association of the insulin secretion pathway to nutrient metabolism in “modulated” hepatocytes.
増殖および転移分化した骨髄細胞が糖尿病SCIDマウスにおいて血中グルコース濃度を制御し、糖尿病を退行させる能力を分析することにより、それらが膵β細胞機能を模倣する能力を十分に調べることができる。 By analyzing the ability of proliferating and metastatic differentiated bone marrow cells to control blood glucose levels and diabetic diabetes in diabetic SCID mice, one can fully examine their ability to mimic pancreatic beta cell function.
(実施例31:PDX−1投与ヒト肝細胞のDNAマイクロアレイチップ分析)
図に示すとおり、PDX−1投与ヒト肝細胞のDNAマイクロアレイ分析によれば、対照細胞と比較して500超の遺伝子がアップレギュレートまたはダウンレギュレートされたことが判った。PDX−1投与に応答してモジュレートされた遺伝子は膵転写因子(表5参照)およびカタラーゼおよび肝ジスムターゼポリペプチド(表6参照)を包含する。
(Example 31: DNA microarray chip analysis of PDX-1-administered human hepatocytes)
As shown in the figure, DNA microarray analysis of PDX-1-administered human hepatocytes revealed that more than 500 genes were up-regulated or down-regulated compared to control cells. Genes modulated in response to PDX-1 administration include pancreatic transcription factors (see Table 5) and catalase and liver dismutase polypeptides (see Table 6).
(等価物)
本発明の特定の実施形態の上記した詳細な説明から、膵ホルモン生産を誘導する独特の方法が記載されていることは明らかである。本明細書においては特定の実施形態を詳述したが、これは説明を目的とするのみの例示であり、添付する請求項の範囲に関して制限することを意図するものではない。特に本発明者等は種々の置き換え、改変および修飾は請求項記載の本発明の精神および範囲から外れることなく本発明に対して行えることを意図している。
(Equivalent)
From the above detailed description of specific embodiments of the invention, it is evident that unique methods for inducing pancreatic hormone production have been described. Although specific embodiments have been described in detail herein, this is for illustrative purposes only and is not intended to be limiting with respect to the scope of the appended claims. In particular, it is intended by the inventors that various substitutions, alterations, and modifications may be made to the invention without departing from the spirit and scope of the invention as claimed.
Claims (43)
a.該非膵細胞において内因性PDX−1、胚マーカー、インスリン、グルカゴン、またはソマトスタチンの発現を誘導するための量、または、
b.該非膵細胞においてC/EBPβ、アルブミンまたはADH−1の発現を抑制するための量
のニューロDポリペプチドまたはベータセルリンポリペプチドをコードする核酸に該非膵細胞を接触させることを含む上記方法。 A method of converting non-pancreatic cells into pancreatic cells,
a. An amount to induce expression of endogenous PDX-1, an embryonic marker, insulin, glucagon, or somatostatin in the non-pancreatic cells, or
b. The above method comprising contacting the non-pancreatic cell with a nucleic acid encoding a neuro D polypeptide or betacellulin polypeptide in an amount to suppress the expression of C / EBPβ, albumin or ADH-1 in the non-pancreatic cell.
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US46971503P | 2003-05-12 | 2003-05-12 | |
| PCT/IB2004/001973 WO2004098646A1 (en) | 2003-05-12 | 2004-05-12 | Methods of inducing regulated pancreatic hormone production in non-pancreatic islet tissues |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2010261850A Division JP2011068660A (en) | 2003-05-12 | 2010-11-24 | Method of inducing regulated pancreatic hormone production in non-pancreatic islet tissue |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| JP2006525994A true JP2006525994A (en) | 2006-11-16 |
Family
ID=33435254
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2006506631A Pending JP2006525994A (en) | 2003-05-12 | 2004-05-12 | Methods for inducing regulated pancreatic hormone production in non-islet tissue |
| JP2010261850A Pending JP2011068660A (en) | 2003-05-12 | 2010-11-24 | Method of inducing regulated pancreatic hormone production in non-pancreatic islet tissue |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2010261850A Pending JP2011068660A (en) | 2003-05-12 | 2010-11-24 | Method of inducing regulated pancreatic hormone production in non-pancreatic islet tissue |
Country Status (4)
| Country | Link |
|---|---|
| EP (1) | EP1624898A1 (en) |
| JP (2) | JP2006525994A (en) |
| AU (1) | AU2004236573B2 (en) |
| WO (1) | WO2004098646A1 (en) |
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2019530439A (en) * | 2016-08-29 | 2019-10-24 | ハッケンサック ユニヴァーシティ メディカル センター | Compositions and methods for reprogramming adult cells with stemness of blood platelet-rich fractions containing platelet-like cells in humans |
Families Citing this family (13)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US8778899B2 (en) | 1999-06-01 | 2014-07-15 | Sarah Ferber | Methods of inducing regulated pancreatic hormone production in non-pancreatic islet tissues |
| US6774120B1 (en) | 1999-06-01 | 2004-08-10 | Sarah Ferber | Methods of inducing regulated pancreatic hormone production in non-pancreatic islet tissues |
| JP5250423B2 (en) | 2005-12-21 | 2013-07-31 | ユニヴァルシテ カソリック デ ルーバン | Isolated hepatic stem cells |
| US20080145937A1 (en) * | 2006-09-22 | 2008-06-19 | Baylor Research Institute | In Vivo Transformation of Pancreatic Acinar Cells into Insulin-Producing Cells |
| WO2014141165A1 (en) * | 2013-03-15 | 2014-09-18 | Universite De Geneve | Use of insulin signaling antagonists, optionally in combination of transfection of non-beta cells, for inducing insulin production |
| CA2915143C (en) | 2013-06-13 | 2021-08-03 | Orgenesis Ltd. | Cell populations, methods of transdifferentiation and methods of use thereof |
| US20160129047A1 (en) | 2013-07-05 | 2016-05-12 | Université Catholique de Louvain | Conditioned medium from human adult liver stem cells and its use in the treatment of liver disorders |
| MA41296A (en) * | 2014-12-30 | 2017-11-07 | Orgenesis Ltd | TRANSDIFFERENTIATION PROCESSES AND METHODS FOR USING THE SAME |
| US9957287B2 (en) | 2016-05-26 | 2018-05-01 | Cornell University | Methods and compositions to treat type-1 and type-2 diabetes |
| JP7177041B2 (en) | 2016-08-29 | 2022-11-22 | ハッケンサック ユニヴァーシティ メディカル センター | Compositions and methods for programming adult cells using a platelet-rich fraction of blood containing platelet-like cells |
| WO2018207179A1 (en) | 2017-05-08 | 2018-11-15 | Orgenesis Ltd. | Transdifferentiated cell populations and methods of use thereof |
| US11788062B2 (en) * | 2017-07-27 | 2023-10-17 | Board Of Supervisors Of Louisiana State University And Agricultural And Mechanical College | Functional feline pancreatic cells from adipose tissue |
| CN112899351B (en) * | 2021-02-03 | 2022-08-26 | 东华大学 | Double-fluorescence quantitative PCR detection kit for detecting cell infiltration state of PDX model mouse |
Citations (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2003500457A (en) * | 1999-06-01 | 2003-01-07 | サラ ファーバー, | Methods for inducing regulated pancreatic hormones in non-Langerhans islet tissues |
| WO2003033697A1 (en) * | 2001-10-18 | 2003-04-24 | Ixion Biotechnology, Inc. | Conversion of liver stem and progenitor cells to pancreatic functional cells |
| JP2003304878A (en) * | 2002-04-17 | 2003-10-28 | Susumu Kiyono | Induction of insulin-producing cell |
Family Cites Families (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| GB0206357D0 (en) * | 2002-03-18 | 2002-05-01 | Univ Bath | Cells |
-
2004
- 2004-05-12 JP JP2006506631A patent/JP2006525994A/en active Pending
- 2004-05-12 AU AU2004236573A patent/AU2004236573B2/en active Active
- 2004-05-12 EP EP04732369A patent/EP1624898A1/en not_active Ceased
- 2004-05-12 WO PCT/IB2004/001973 patent/WO2004098646A1/en active Application Filing
-
2010
- 2010-11-24 JP JP2010261850A patent/JP2011068660A/en active Pending
Patent Citations (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2003500457A (en) * | 1999-06-01 | 2003-01-07 | サラ ファーバー, | Methods for inducing regulated pancreatic hormones in non-Langerhans islet tissues |
| WO2003033697A1 (en) * | 2001-10-18 | 2003-04-24 | Ixion Biotechnology, Inc. | Conversion of liver stem and progenitor cells to pancreatic functional cells |
| JP2003304878A (en) * | 2002-04-17 | 2003-10-28 | Susumu Kiyono | Induction of insulin-producing cell |
Non-Patent Citations (1)
| Title |
|---|
| JPN6010028021, KOJIMA,H. et al, "Combined expression of pancreatic duodenal homeobox 1 and islet factor 1 induces immature enterocyte", Diabetes, 2002, Vol.51, No.5, p.1398−408 * |
Cited By (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2019530439A (en) * | 2016-08-29 | 2019-10-24 | ハッケンサック ユニヴァーシティ メディカル センター | Compositions and methods for reprogramming adult cells with stemness of blood platelet-rich fractions containing platelet-like cells in humans |
| JP7219211B2 (en) | 2016-08-29 | 2023-02-07 | ハッケンサック ユニヴァーシティ メディカル センター | Compositions and methods for reprogramming adult cells by stemness of platelet-rich fractions of blood containing platelet-like cells in humans |
Also Published As
| Publication number | Publication date |
|---|---|
| EP1624898A1 (en) | 2006-02-15 |
| AU2004236573B2 (en) | 2009-10-22 |
| WO2004098646A1 (en) | 2004-11-18 |
| WO2004098646B1 (en) | 2005-04-28 |
| AU2004236573A1 (en) | 2004-11-18 |
| JP2011068660A (en) | 2011-04-07 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP2011068660A (en) | Method of inducing regulated pancreatic hormone production in non-pancreatic islet tissue | |
| US9481894B2 (en) | Methods of inducing regulated pancreatic hormone production in non-pancreatic islet tissues | |
| US8119405B2 (en) | Methods of inducing regulated pancreatic hormone production in non-pancreatic islet tissues | |
| Miyatsuka et al. | Ectopically expressed PDX-1 in liver initiates endocrine and exocrine pancreas differentiation but causes dysmorphogenesis | |
| JP2003500457A5 (en) | ||
| KR102487883B1 (en) | Transdifferentiation method and method of use thereof | |
| Trinh et al. | Pax-6 activates endogenous proglucagon gene expression in the rodent gastrointestinal epithelium | |
| KR102181336B1 (en) | Cell populations, methods of transdifferentiation and methods of use thereof | |
| KR20080036015A (en) | Glucose inducible insulin expression and methods of treating diabetes | |
| US20030157071A1 (en) | Treatment or replacement therapy using transgenic stem cells delivered to the gut | |
| Zhang et al. | Genetically engineered K cells provide sufficient insulin to correct hyperglycemia in a nude murine model | |
| JP2003531581A (en) | Melanoma differentiation-related gene-5 and promoter and uses thereof | |
| US20160083733A1 (en) | Novel treatment of metabolic diseases | |
| JP2004520006A (en) | Nucleic acid comprising a region of rat PEG-3 promoter and use thereof | |
| AU2002344233A1 (en) | Treatment or replacement therapy using transgenic stem cells delivered to the gut | |
| AU2008200852A1 (en) | Treatment or replacement therapy using transgenic stem cells delivered to the gut |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20070511 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20100524 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20100823 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20100830 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20100916 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20100927 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20101022 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20101029 |
|
| A521 | Written amendment |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20101124 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20110221 |