CN116033910A - 编辑b细胞中b2m基因座的方法和组合物 - Google Patents
编辑b细胞中b2m基因座的方法和组合物 Download PDFInfo
- Publication number
- CN116033910A CN116033910A CN202180047549.4A CN202180047549A CN116033910A CN 116033910 A CN116033910 A CN 116033910A CN 202180047549 A CN202180047549 A CN 202180047549A CN 116033910 A CN116033910 A CN 116033910A
- Authority
- CN
- China
- Prior art keywords
- cell
- cells
- modified
- gene
- nuclease
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
- 210000003719 b-lymphocyte Anatomy 0.000 title claims abstract description 260
- 238000000034 method Methods 0.000 title claims abstract description 119
- 239000000203 mixture Substances 0.000 title abstract description 29
- 210000004027 cell Anatomy 0.000 claims abstract description 289
- 108010081355 beta 2-Microglobulin Proteins 0.000 claims abstract description 198
- 102000015736 beta 2-Microglobulin Human genes 0.000 claims abstract description 195
- 101150076800 B2M gene Proteins 0.000 claims abstract description 128
- 210000002865 immune cell Anatomy 0.000 claims abstract description 72
- 108090000623 proteins and genes Proteins 0.000 claims abstract description 61
- 230000000735 allogeneic effect Effects 0.000 claims abstract description 50
- 230000001965 increasing effect Effects 0.000 claims abstract description 25
- 230000022534 cell killing Effects 0.000 claims abstract description 23
- 230000002147 killing effect Effects 0.000 claims abstract description 17
- 108090000765 processed proteins & peptides Proteins 0.000 claims description 129
- 101710163270 Nuclease Proteins 0.000 claims description 103
- 102000004196 processed proteins & peptides Human genes 0.000 claims description 96
- 229920001184 polypeptide Polymers 0.000 claims description 87
- 210000001744 T-lymphocyte Anatomy 0.000 claims description 86
- 108020005004 Guide RNA Proteins 0.000 claims description 70
- 210000000822 natural killer cell Anatomy 0.000 claims description 59
- 102000039446 nucleic acids Human genes 0.000 claims description 52
- 108020004707 nucleic acids Proteins 0.000 claims description 52
- 150000007523 nucleic acids Chemical class 0.000 claims description 52
- 230000008439 repair process Effects 0.000 claims description 50
- 239000002773 nucleotide Substances 0.000 claims description 41
- 125000003729 nucleotide group Chemical group 0.000 claims description 41
- 102000004169 proteins and genes Human genes 0.000 claims description 37
- 230000001225 therapeutic effect Effects 0.000 claims description 30
- 239000013603 viral vector Substances 0.000 claims description 30
- 238000001727 in vivo Methods 0.000 claims description 29
- 238000010459 TALEN Methods 0.000 claims description 26
- 102100028970 HLA class I histocompatibility antigen, alpha chain E Human genes 0.000 claims description 25
- 101000986085 Homo sapiens HLA class I histocompatibility antigen, alpha chain E Proteins 0.000 claims description 21
- 238000000338 in vitro Methods 0.000 claims description 18
- 108091033409 CRISPR Proteins 0.000 claims description 17
- 230000000295 complement effect Effects 0.000 claims description 17
- 230000001472 cytotoxic effect Effects 0.000 claims description 17
- 102100026277 Alpha-galactosidase A Human genes 0.000 claims description 16
- 108010017070 Zinc Finger Nucleases Proteins 0.000 claims description 16
- 231100000433 cytotoxic Toxicity 0.000 claims description 16
- 108010043645 Transcription Activator-Like Effector Nucleases Proteins 0.000 claims description 15
- 239000013598 vector Substances 0.000 claims description 15
- 210000001948 pro-b lymphocyte Anatomy 0.000 claims description 14
- 108010042407 Endonucleases Proteins 0.000 claims description 13
- 102000004127 Cytokines Human genes 0.000 claims description 10
- 108090000695 Cytokines Proteins 0.000 claims description 10
- 230000003612 virological effect Effects 0.000 claims description 10
- 102100022641 Coagulation factor IX Human genes 0.000 claims description 9
- 102000004190 Enzymes Human genes 0.000 claims description 9
- 108090000790 Enzymes Proteins 0.000 claims description 9
- 108010076282 Factor IX Proteins 0.000 claims description 9
- 102100028967 HLA class I histocompatibility antigen, alpha chain G Human genes 0.000 claims description 9
- 108010024164 HLA-G Antigens Proteins 0.000 claims description 9
- 102100033342 Lysosomal acid glucosylceramidase Human genes 0.000 claims description 9
- 229960004222 factor ix Drugs 0.000 claims description 9
- 210000003958 hematopoietic stem cell Anatomy 0.000 claims description 9
- 108020003175 receptors Proteins 0.000 claims description 9
- 102000005962 receptors Human genes 0.000 claims description 9
- 102100035765 Angiotensin-converting enzyme 2 Human genes 0.000 claims description 8
- 108090000975 Angiotensin-converting enzyme 2 Proteins 0.000 claims description 8
- 108010017544 Glucosylceramidase Proteins 0.000 claims description 8
- 108010030291 alpha-Galactosidase Proteins 0.000 claims description 8
- 239000000427 antigen Substances 0.000 claims description 8
- 108091007433 antigens Proteins 0.000 claims description 8
- 102000036639 antigens Human genes 0.000 claims description 8
- 210000000014 large pre-b cell Anatomy 0.000 claims description 8
- 210000000345 small pre-b cell Anatomy 0.000 claims description 8
- 230000008685 targeting Effects 0.000 claims description 8
- 210000001266 CD8-positive T-lymphocyte Anatomy 0.000 claims description 7
- 108010019670 Chimeric Antigen Receptors Proteins 0.000 claims description 7
- 230000027455 binding Effects 0.000 claims description 7
- 239000012634 fragment Substances 0.000 claims description 7
- 210000003297 immature b lymphocyte Anatomy 0.000 claims description 7
- 210000003519 mature b lymphocyte Anatomy 0.000 claims description 7
- 210000001806 memory b lymphocyte Anatomy 0.000 claims description 7
- 210000003720 plasmablast Anatomy 0.000 claims description 7
- 102100028966 HLA class I histocompatibility antigen, alpha chain F Human genes 0.000 claims description 6
- 101000986080 Homo sapiens HLA class I histocompatibility antigen, alpha chain F Proteins 0.000 claims description 6
- 238000001802 infusion Methods 0.000 claims description 6
- 210000002809 long lived plasma cell Anatomy 0.000 claims description 6
- 210000003826 marginal zone b cell Anatomy 0.000 claims description 6
- 241000124008 Mammalia Species 0.000 claims description 5
- 210000005260 human cell Anatomy 0.000 claims description 5
- 239000008194 pharmaceutical composition Substances 0.000 claims description 5
- 102000004533 Endonucleases Human genes 0.000 claims description 4
- 238000004519 manufacturing process Methods 0.000 claims description 3
- 239000000546 pharmaceutical excipient Substances 0.000 claims description 3
- 238000003780 insertion Methods 0.000 abstract description 16
- 230000037431 insertion Effects 0.000 abstract description 16
- 101000937544 Homo sapiens Beta-2-microglobulin Proteins 0.000 abstract description 7
- 230000002779 inactivation Effects 0.000 abstract description 7
- 102100027314 Beta-2-microglobulin Human genes 0.000 abstract 1
- 108020001507 fusion proteins Proteins 0.000 description 52
- 102000037865 fusion proteins Human genes 0.000 description 52
- 108091033319 polynucleotide Proteins 0.000 description 48
- 102000040430 polynucleotide Human genes 0.000 description 48
- 239000002157 polynucleotide Substances 0.000 description 48
- 238000010362 genome editing Methods 0.000 description 33
- 230000000415 inactivating effect Effects 0.000 description 31
- 239000002299 complementary DNA Substances 0.000 description 26
- 108020004414 DNA Proteins 0.000 description 24
- 210000004180 plasmocyte Anatomy 0.000 description 17
- 241000699666 Mus <mouse, genus> Species 0.000 description 15
- 239000013638 trimer Substances 0.000 description 13
- 238000012217 deletion Methods 0.000 description 11
- 230000037430 deletion Effects 0.000 description 11
- 238000002054 transplantation Methods 0.000 description 11
- 238000003556 assay Methods 0.000 description 10
- 238000011740 C57BL/6 mouse Methods 0.000 description 9
- 102100031780 Endonuclease Human genes 0.000 description 9
- 101000868279 Homo sapiens Leukocyte surface antigen CD47 Proteins 0.000 description 9
- 102100032913 Leukocyte surface antigen CD47 Human genes 0.000 description 9
- 238000003501 co-culture Methods 0.000 description 9
- 238000002744 homologous recombination Methods 0.000 description 9
- 230000006801 homologous recombination Effects 0.000 description 9
- 238000006467 substitution reaction Methods 0.000 description 9
- 108700018351 Major Histocompatibility Complex Proteins 0.000 description 8
- 238000002474 experimental method Methods 0.000 description 8
- 238000012986 modification Methods 0.000 description 8
- 230000004048 modification Effects 0.000 description 8
- 230000020382 suppression by virus of host antigen processing and presentation of peptide antigen via MHC class I Effects 0.000 description 8
- 238000012546 transfer Methods 0.000 description 8
- 102000004547 Glucosylceramidase Human genes 0.000 description 7
- 101000884271 Homo sapiens Signal transducer CD24 Proteins 0.000 description 7
- 102100024295 Maltase-glucoamylase Human genes 0.000 description 7
- 241000699670 Mus sp. Species 0.000 description 7
- 102100038081 Signal transducer CD24 Human genes 0.000 description 7
- 239000002253 acid Substances 0.000 description 7
- 108010028144 alpha-Glucosidases Proteins 0.000 description 7
- 230000001404 mediated effect Effects 0.000 description 7
- 230000004083 survival effect Effects 0.000 description 7
- 102100039341 Atrial natriuretic peptide receptor 2 Human genes 0.000 description 6
- 101710102159 Atrial natriuretic peptide receptor 2 Proteins 0.000 description 6
- 238000010453 CRISPR/Cas method Methods 0.000 description 6
- 102100030704 Interleukin-21 Human genes 0.000 description 6
- 102000047279 human B2M Human genes 0.000 description 6
- 108010074108 interleukin-21 Proteins 0.000 description 6
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 5
- 239000013607 AAV vector Substances 0.000 description 5
- 108700028369 Alleles Proteins 0.000 description 5
- 102000008949 Histocompatibility Antigens Class I Human genes 0.000 description 5
- 108010088652 Histocompatibility Antigens Class I Proteins 0.000 description 5
- 102000004389 Ribonucleoproteins Human genes 0.000 description 5
- 108010081734 Ribonucleoproteins Proteins 0.000 description 5
- -1 cas8 Proteins 0.000 description 5
- 230000034431 double-strand break repair via homologous recombination Effects 0.000 description 5
- 230000001717 pathogenic effect Effects 0.000 description 5
- 239000002243 precursor Substances 0.000 description 5
- 230000008569 process Effects 0.000 description 5
- 102000053602 DNA Human genes 0.000 description 4
- 102100028976 HLA class I histocompatibility antigen, B alpha chain Human genes 0.000 description 4
- 108010058607 HLA-B Antigens Proteins 0.000 description 4
- 101100395313 Homo sapiens HLA-E gene Proteins 0.000 description 4
- 108091028043 Nucleic acid sequence Proteins 0.000 description 4
- 206010052779 Transplant rejections Diseases 0.000 description 4
- 238000004458 analytical method Methods 0.000 description 4
- 230000005782 double-strand break Effects 0.000 description 4
- 238000000684 flow cytometry Methods 0.000 description 4
- 229920002521 macromolecule Polymers 0.000 description 4
- 230000006780 non-homologous end joining Effects 0.000 description 4
- 108091026890 Coding region Proteins 0.000 description 3
- 230000004568 DNA-binding Effects 0.000 description 3
- 241000702421 Dependoparvovirus Species 0.000 description 3
- 101100219622 Escherichia coli (strain K12) casC gene Proteins 0.000 description 3
- 101100382541 Escherichia coli (strain K12) casD gene Proteins 0.000 description 3
- 101100326871 Escherichia coli (strain K12) ygbF gene Proteins 0.000 description 3
- 101100005249 Escherichia coli (strain K12) ygcB gene Proteins 0.000 description 3
- 238000012413 Fluorescence activated cell sorting analysis Methods 0.000 description 3
- 241000282412 Homo Species 0.000 description 3
- 101000874179 Homo sapiens Syndecan-1 Proteins 0.000 description 3
- 108090001005 Interleukin-6 Proteins 0.000 description 3
- 101100219625 Mus musculus Casd1 gene Proteins 0.000 description 3
- 101100387128 Myxococcus xanthus (strain DK1622) devR gene Proteins 0.000 description 3
- 101100387131 Myxococcus xanthus (strain DK1622) devS gene Proteins 0.000 description 3
- 102100035721 Syndecan-1 Human genes 0.000 description 3
- 101100273269 Thermus thermophilus (strain ATCC 27634 / DSM 579 / HB8) cse3 gene Proteins 0.000 description 3
- 108010073062 Transcription Activator-Like Effectors Proteins 0.000 description 3
- 230000000890 antigenic effect Effects 0.000 description 3
- 101150117416 cas2 gene Proteins 0.000 description 3
- 101150055191 cas3 gene Proteins 0.000 description 3
- 101150111685 cas4 gene Proteins 0.000 description 3
- 101150049463 cas5 gene Proteins 0.000 description 3
- 101150106467 cas6 gene Proteins 0.000 description 3
- 101150044165 cas7 gene Proteins 0.000 description 3
- 101150055766 cat gene Proteins 0.000 description 3
- 230000015556 catabolic process Effects 0.000 description 3
- 230000002950 deficient Effects 0.000 description 3
- 238000006731 degradation reaction Methods 0.000 description 3
- 230000003828 downregulation Effects 0.000 description 3
- 230000001939 inductive effect Effects 0.000 description 3
- 230000010354 integration Effects 0.000 description 3
- 239000000463 material Substances 0.000 description 3
- 238000011002 quantification Methods 0.000 description 3
- 108091008146 restriction endonucleases Proteins 0.000 description 3
- 238000010361 transduction Methods 0.000 description 3
- 230000026683 transduction Effects 0.000 description 3
- 238000001890 transfection Methods 0.000 description 3
- NHBKXEKEPDILRR-UHFFFAOYSA-N 2,3-bis(butanoylsulfanyl)propyl butanoate Chemical compound CCCC(=O)OCC(SC(=O)CCC)CSC(=O)CCC NHBKXEKEPDILRR-UHFFFAOYSA-N 0.000 description 2
- 102000014914 Carrier Proteins Human genes 0.000 description 2
- 108020004705 Codon Proteins 0.000 description 2
- 230000007018 DNA scission Effects 0.000 description 2
- 108700024394 Exon Proteins 0.000 description 2
- 102100028972 HLA class I histocompatibility antigen, A alpha chain Human genes 0.000 description 2
- 108010075704 HLA-A Antigens Proteins 0.000 description 2
- 101100220044 Homo sapiens CD34 gene Proteins 0.000 description 2
- 108010002350 Interleukin-2 Proteins 0.000 description 2
- 102000018697 Membrane Proteins Human genes 0.000 description 2
- 108010052285 Membrane Proteins Proteins 0.000 description 2
- 206010027476 Metastases Diseases 0.000 description 2
- 241001465754 Metazoa Species 0.000 description 2
- 108020004682 Single-Stranded DNA Proteins 0.000 description 2
- HCHKCACWOHOZIP-UHFFFAOYSA-N Zinc Chemical compound [Zn] HCHKCACWOHOZIP-UHFFFAOYSA-N 0.000 description 2
- 101710185494 Zinc finger protein Proteins 0.000 description 2
- 102100023597 Zinc finger protein 816 Human genes 0.000 description 2
- 230000004913 activation Effects 0.000 description 2
- 239000011324 bead Substances 0.000 description 2
- 108091008324 binding proteins Proteins 0.000 description 2
- 238000004113 cell culture Methods 0.000 description 2
- 239000003153 chemical reaction reagent Substances 0.000 description 2
- 238000012258 culturing Methods 0.000 description 2
- 238000001514 detection method Methods 0.000 description 2
- 230000004069 differentiation Effects 0.000 description 2
- 239000003937 drug carrier Substances 0.000 description 2
- 230000008030 elimination Effects 0.000 description 2
- 238000003379 elimination reaction Methods 0.000 description 2
- 238000005206 flow analysis Methods 0.000 description 2
- 238000001943 fluorescence-activated cell sorting Methods 0.000 description 2
- 230000037433 frameshift Effects 0.000 description 2
- 230000006870 function Effects 0.000 description 2
- 230000004927 fusion Effects 0.000 description 2
- 102000054766 genetic haplotypes Human genes 0.000 description 2
- 230000028993 immune response Effects 0.000 description 2
- 108020004999 messenger RNA Proteins 0.000 description 2
- 230000009401 metastasis Effects 0.000 description 2
- 244000052769 pathogen Species 0.000 description 2
- 230000001681 protective effect Effects 0.000 description 2
- 230000002829 reductive effect Effects 0.000 description 2
- 210000000952 spleen Anatomy 0.000 description 2
- 239000011701 zinc Substances 0.000 description 2
- 229910052725 zinc Inorganic materials 0.000 description 2
- 102100021253 Antileukoproteinase Human genes 0.000 description 1
- 101100327165 Arabidopsis thaliana CCD8 gene Proteins 0.000 description 1
- 108091008875 B cell receptors Proteins 0.000 description 1
- 102100024222 B-lymphocyte antigen CD19 Human genes 0.000 description 1
- 102100035875 C-C chemokine receptor type 5 Human genes 0.000 description 1
- 101710149870 C-C chemokine receptor type 5 Proteins 0.000 description 1
- 108010029697 CD40 Ligand Proteins 0.000 description 1
- 102100032937 CD40 ligand Human genes 0.000 description 1
- 238000010356 CRISPR-Cas9 genome editing Methods 0.000 description 1
- 102100024965 Caspase recruitment domain-containing protein 11 Human genes 0.000 description 1
- 230000033616 DNA repair Effects 0.000 description 1
- 102100030442 Derlin-3 Human genes 0.000 description 1
- 102100021807 ER degradation-enhancing alpha-mannosidase-like protein 1 Human genes 0.000 description 1
- 108010054218 Factor VIII Proteins 0.000 description 1
- 102000001690 Factor VIII Human genes 0.000 description 1
- 102100028971 HLA class I histocompatibility antigen, C alpha chain Human genes 0.000 description 1
- 108010052199 HLA-C Antigens Proteins 0.000 description 1
- 102100031573 Hematopoietic progenitor cell antigen CD34 Human genes 0.000 description 1
- 208000031220 Hemophilia Diseases 0.000 description 1
- 208000009292 Hemophilia A Diseases 0.000 description 1
- 101000615334 Homo sapiens Antileukoproteinase Proteins 0.000 description 1
- 101000980825 Homo sapiens B-lymphocyte antigen CD19 Proteins 0.000 description 1
- 101001095043 Homo sapiens Bone marrow proteoglycan Proteins 0.000 description 1
- 101100382122 Homo sapiens CIITA gene Proteins 0.000 description 1
- 101000761179 Homo sapiens Caspase recruitment domain-containing protein 11 Proteins 0.000 description 1
- 101000842622 Homo sapiens Derlin-3 Proteins 0.000 description 1
- 101000895701 Homo sapiens ER degradation-enhancing alpha-mannosidase-like protein 1 Proteins 0.000 description 1
- 101000777663 Homo sapiens Hematopoietic progenitor cell antigen CD34 Proteins 0.000 description 1
- 101000840258 Homo sapiens Immunoglobulin J chain Proteins 0.000 description 1
- 101000840257 Homo sapiens Immunoglobulin kappa constant Proteins 0.000 description 1
- 101001130862 Homo sapiens Oligoribonuclease, mitochondrial Proteins 0.000 description 1
- 101000891028 Homo sapiens Peptidyl-prolyl cis-trans isomerase FKBP11 Proteins 0.000 description 1
- 101001131990 Homo sapiens Peroxidasin homolog Proteins 0.000 description 1
- 101000582986 Homo sapiens Phospholipid phosphatase-related protein type 3 Proteins 0.000 description 1
- 101000919297 Homo sapiens Protein disulfide isomerase CRELD2 Proteins 0.000 description 1
- 101001098824 Homo sapiens Protein disulfide-isomerase A4 Proteins 0.000 description 1
- 101000621057 Homo sapiens Serum paraoxonase/lactonase 3 Proteins 0.000 description 1
- 101000685001 Homo sapiens Stromal cell-derived factor 2-like protein 1 Proteins 0.000 description 1
- 101000914514 Homo sapiens T-cell-specific surface glycoprotein CD28 Proteins 0.000 description 1
- 102000026659 IL10 Human genes 0.000 description 1
- 102100029571 Immunoglobulin J chain Human genes 0.000 description 1
- 102100029572 Immunoglobulin kappa constant Human genes 0.000 description 1
- 108090000174 Interleukin-10 Proteins 0.000 description 1
- 108090000172 Interleukin-15 Proteins 0.000 description 1
- 102000003812 Interleukin-15 Human genes 0.000 description 1
- 108090000978 Interleukin-4 Proteins 0.000 description 1
- 102100026371 MHC class II transactivator Human genes 0.000 description 1
- 108700002010 MHC class II transactivator Proteins 0.000 description 1
- 101100437231 Mus musculus B2m gene Proteins 0.000 description 1
- 101000851433 Mus musculus Tumor necrosis factor ligand superfamily member 13B Proteins 0.000 description 1
- 206010028980 Neoplasm Diseases 0.000 description 1
- 102000011931 Nucleoproteins Human genes 0.000 description 1
- 108010061100 Nucleoproteins Proteins 0.000 description 1
- 108091034117 Oligonucleotide Proteins 0.000 description 1
- 102100032835 Oligoribonuclease, mitochondrial Human genes 0.000 description 1
- 102100024894 PR domain zinc finger protein 1 Human genes 0.000 description 1
- 102100040348 Peptidyl-prolyl cis-trans isomerase FKBP11 Human genes 0.000 description 1
- 102100034601 Peroxidasin homolog Human genes 0.000 description 1
- 108010009975 Positive Regulatory Domain I-Binding Factor 1 Proteins 0.000 description 1
- 102100029370 Protein disulfide isomerase CRELD2 Human genes 0.000 description 1
- 102100037089 Protein disulfide-isomerase A4 Human genes 0.000 description 1
- 235000011449 Rosa Nutrition 0.000 description 1
- 102100022833 Serum paraoxonase/lactonase 3 Human genes 0.000 description 1
- 108091081024 Start codon Proteins 0.000 description 1
- 101000898746 Streptomyces clavuligerus Clavaminate synthase 1 Proteins 0.000 description 1
- 101000879712 Streptomyces lividans Protease inhibitor Proteins 0.000 description 1
- 102100023183 Stromal cell-derived factor 2-like protein 1 Human genes 0.000 description 1
- 108091008874 T cell receptors Proteins 0.000 description 1
- 102000016266 T-Cell Antigen Receptors Human genes 0.000 description 1
- 102100027213 T-cell-specific surface glycoprotein CD28 Human genes 0.000 description 1
- 108700019146 Transgenes Proteins 0.000 description 1
- 241000700605 Viruses Species 0.000 description 1
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 1
- 238000011316 allogeneic transplantation Methods 0.000 description 1
- 230000005784 autoimmunity Effects 0.000 description 1
- 210000000649 b-lymphocyte subset Anatomy 0.000 description 1
- 230000008901 benefit Effects 0.000 description 1
- 230000008033 biological extinction Effects 0.000 description 1
- 230000007321 biological mechanism Effects 0.000 description 1
- 201000011510 cancer Diseases 0.000 description 1
- 238000002659 cell therapy Methods 0.000 description 1
- 230000036755 cellular response Effects 0.000 description 1
- 238000012937 correction Methods 0.000 description 1
- 210000004748 cultured cell Anatomy 0.000 description 1
- 230000001351 cycling effect Effects 0.000 description 1
- 230000003247 decreasing effect Effects 0.000 description 1
- 230000000593 degrading effect Effects 0.000 description 1
- 230000001419 dependent effect Effects 0.000 description 1
- 201000010099 disease Diseases 0.000 description 1
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 1
- 229940079593 drug Drugs 0.000 description 1
- 239000003814 drug Substances 0.000 description 1
- 210000004667 early pro-b cell Anatomy 0.000 description 1
- 230000000694 effects Effects 0.000 description 1
- 239000003623 enhancer Substances 0.000 description 1
- 229960000301 factor viii Drugs 0.000 description 1
- 238000010230 functional analysis Methods 0.000 description 1
- 238000001476 gene delivery Methods 0.000 description 1
- 238000010363 gene targeting Methods 0.000 description 1
- 238000010353 genetic engineering Methods 0.000 description 1
- 208000007345 glycogen storage disease Diseases 0.000 description 1
- 230000005745 host immune response Effects 0.000 description 1
- 238000011502 immune monitoring Methods 0.000 description 1
- 230000037451 immune surveillance Effects 0.000 description 1
- 210000000987 immune system Anatomy 0.000 description 1
- 230000036039 immunity Effects 0.000 description 1
- 230000016784 immunoglobulin production Effects 0.000 description 1
- 230000000091 immunopotentiator Effects 0.000 description 1
- 229960003444 immunosuppressant agent Drugs 0.000 description 1
- 239000003018 immunosuppressive agent Substances 0.000 description 1
- 238000011065 in-situ storage Methods 0.000 description 1
- 208000015181 infectious disease Diseases 0.000 description 1
- 208000027866 inflammatory disease Diseases 0.000 description 1
- 230000002401 inhibitory effect Effects 0.000 description 1
- 238000002372 labelling Methods 0.000 description 1
- 210000002202 late pro-b cell Anatomy 0.000 description 1
- 210000000265 leukocyte Anatomy 0.000 description 1
- 230000002197 limbic effect Effects 0.000 description 1
- 210000004698 lymphocyte Anatomy 0.000 description 1
- 239000003550 marker Substances 0.000 description 1
- 238000010172 mouse model Methods 0.000 description 1
- 239000013642 negative control Substances 0.000 description 1
- 210000003819 peripheral blood mononuclear cell Anatomy 0.000 description 1
- 230000008488 polyadenylation Effects 0.000 description 1
- 238000002360 preparation method Methods 0.000 description 1
- 230000006798 recombination Effects 0.000 description 1
- 238000005215 recombination Methods 0.000 description 1
- 238000011160 research Methods 0.000 description 1
- 230000002441 reversible effect Effects 0.000 description 1
- 238000000926 separation method Methods 0.000 description 1
- 210000001082 somatic cell Anatomy 0.000 description 1
- 230000000638 stimulation Effects 0.000 description 1
- 238000003786 synthesis reaction Methods 0.000 description 1
- 238000012360 testing method Methods 0.000 description 1
- 210000001519 tissue Anatomy 0.000 description 1
- 238000013518 transcription Methods 0.000 description 1
- 230000035897 transcription Effects 0.000 description 1
- 230000001131 transforming effect Effects 0.000 description 1
- 230000014621 translational initiation Effects 0.000 description 1
Images





















Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/705—Receptors; Cell surface antigens; Cell surface determinants
- C07K14/70503—Immunoglobulin superfamily
- C07K14/70539—MHC-molecules, e.g. HLA-molecules
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/46—Cellular immunotherapy
- A61K39/461—Cellular immunotherapy characterised by the cell type used
- A61K39/4612—B-cells
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/46—Cellular immunotherapy
- A61K39/462—Cellular immunotherapy characterized by the effect or the function of the cells
- A61K39/4621—Cellular immunotherapy characterized by the effect or the function of the cells immunosuppressive or immunotolerising
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/46—Cellular immunotherapy
- A61K39/462—Cellular immunotherapy characterized by the effect or the function of the cells
- A61K39/4622—Antigen presenting cells
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/46—Cellular immunotherapy
- A61K39/464—Cellular immunotherapy characterised by the antigen targeted or presented
- A61K39/4643—Vertebrate antigens
- A61K39/46434—Antigens related to induction of tolerance to non-self
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/113—Non-coding nucleic acids modulating the expression of genes, e.g. antisense oligonucleotides; Antisense DNA or RNA; Triplex- forming oligonucleotides; Catalytic nucleic acids, e.g. ribozymes; Nucleic acids used in co-suppression or gene silencing
- C12N15/1138—Non-coding nucleic acids modulating the expression of genes, e.g. antisense oligonucleotides; Antisense DNA or RNA; Triplex- forming oligonucleotides; Catalytic nucleic acids, e.g. ribozymes; Nucleic acids used in co-suppression or gene silencing against receptors or cell surface proteins
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/87—Introduction of foreign genetic material using processes not otherwise provided for, e.g. co-transformation
- C12N15/90—Stable introduction of foreign DNA into chromosome
- C12N15/902—Stable introduction of foreign DNA into chromosome using homologous recombination
- C12N15/907—Stable introduction of foreign DNA into chromosome using homologous recombination in mammalian cells
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K2239/00—Indexing codes associated with cellular immunotherapy of group A61K39/46
- A61K2239/26—Universal/off- the- shelf cellular immunotherapy; Allogenic cells or means to avoid rejection
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/60—Fusion polypeptide containing spectroscopic/fluorescent detection, e.g. green fluorescent protein [GFP]
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/10—Type of nucleic acid
- C12N2310/20—Type of nucleic acid involving clustered regularly interspaced short palindromic repeats [CRISPRs]
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2750/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssDNA viruses
- C12N2750/00011—Details
- C12N2750/14011—Parvoviridae
- C12N2750/14111—Dependovirus, e.g. adenoassociated viruses
- C12N2750/14141—Use of virus, viral particle or viral elements as a vector
- C12N2750/14143—Use of virus, viral particle or viral elements as a vector viral genome or elements thereof as genetic vector
Landscapes
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Genetics & Genomics (AREA)
- Cell Biology (AREA)
- Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Organic Chemistry (AREA)
- Microbiology (AREA)
- Immunology (AREA)
- Zoology (AREA)
- Biomedical Technology (AREA)
- Molecular Biology (AREA)
- Medicinal Chemistry (AREA)
- Mycology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biotechnology (AREA)
- Wood Science & Technology (AREA)
- General Engineering & Computer Science (AREA)
- Public Health (AREA)
- Epidemiology (AREA)
- Pharmacology & Pharmacy (AREA)
- Animal Behavior & Ethology (AREA)
- Veterinary Medicine (AREA)
- Biochemistry (AREA)
- Biophysics (AREA)
- Plant Pathology (AREA)
- Physics & Mathematics (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Gastroenterology & Hepatology (AREA)
- Toxicology (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
- Document Processing Apparatus (AREA)
- Virology (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Hematology (AREA)
- Developmental Biology & Embryology (AREA)
- Crystallography & Structural Chemistry (AREA)
Abstract
本文提供的方法和组合物的一些实施方案包括制备修饰的B细胞。在一些实施方案中,B细胞中的内源性β‑2微球蛋白(B2M)基因被修饰。一些实施方案涉及增加修饰的B细胞对同种异体免疫细胞杀伤的抗性。在一些实施方案中,所述内源性B2M基因失活的,增加了所述修饰的B细胞对同种异体免疫细胞杀伤的抗性。在一些实施方案中,将替代MHC‑I插入失活的内源性B2M基因中,增加了修饰的B细胞对同种异体免疫细胞杀伤的抗性。一些实施方案包括富集成功修饰的细胞。
Description
交叉引用
本申请请求保护于2020年11月16日提交的,申请号为63/114,131,标题为“编辑B细胞中B2M基因座的方法和组合物”的美国临时申请,以及于2020年7月3日提交的,申请号为63/0479,78,标题为“编辑B细胞中B2M基因座的方法和组合物”的美国临时申请的优先权,它们的全部内容通过引用并入本文。
序列表的引入
本申请与电子格式的序列表一起提交。序列表以名称为SCRI295WOSEQLIST的文件的提供,创建于2021年6月29日,大小约为4.5Kb。所述序列表的电子格式的信息通过引用全部并入本文。
技术领域
本文提供的方法和组合物的一些实施方案包括制备修饰的B细胞。在一些实施方案中,B细胞中的内源性β-2微球蛋白(B2M)基因被修饰。一些实施方案涉及增加修饰的B细胞对同种异体免疫细胞杀伤的抗性。在一些实施方案中,所述内源性B2M基因是失活的,增加了修饰的B细胞对同种异体免疫细胞杀伤的抗性。在一些实施方案中,将替代MHC-I插入失活的内源性B2M基因中,增加了修饰的B细胞对同种异体免疫细胞杀伤的抗性。一些实施方案包括对成功修饰的细胞进行富集。
背景技术
由于CRISPR和TALEN系统等最新工具的有效性和易用性,基因组编辑应用的次数有所增加。然而,临床相关的人类体细胞的基因组编辑仍然是一个挑战,例如,由于对该细胞的同种异体移植产生的不期望的宿主免疫应答。因此,亟需适合于同种异体移植的细胞,所述细胞可消除或减少对该细胞进行同种异体移植触发不期望的受体免疫应答的可能性。
发明内容
本文提供的方法和组合物的一些实施方案包括用于修饰的细胞中内源性β-2微球蛋白(B2M)基因的系统,所述系统包括:能够切割细胞基因组中内源性B2M基因中靶向基因座的核酸酶,或编码所述核酸酶的核酸;和任选地:包括与B2M基因互补的序列的向导RNA(gRNA),和/或修复模板,所述修复模板包括第一同源臂、第二同源臂和编码第一同源臂和第二同源臂之间有效载荷的核酸,其中所述第一同源臂和/或所述第二同源性臂与所述B2M基因中的序列具有同源性。
在一些实施方案中,所述gRNA包括:与SEQ ID NO:01-04中任一条的核苷酸序列具有至少85%、86%、87%、88%、89%、90%、95%、96%、97%、98%、99%或100%的序列一致性的核苷酸序列;SEQ ID NO:01-04中任一条的核苷酸序列;或其变体,所述变体与SEQ IDNO:01-04中的任一条相比具有不超过3个错配。
在一些实施方案中,所述gRNA适于使所述内源性B2M基因失活。
在一些实施方案中,所述靶向基因座位于B2M基因的第一编码外显子内。
在一些实施方案中,所述核酸酶包括Cas核酸酶。在一些实施方案中,所述Cas核酸酶包括Cas9核酸酶。
在一些实施方案中,所述核酸酶选自锌指核酸酶、转录激活因子样效应物核酸酶(TALEN)、归巢核酸内切酶(HE)和组合的TALEN-HE蛋白(megaTAL)。
在一些实施方案中,所述修复模板缺乏核酸酶靶向的序列,和/或缺乏能够与gRNA或其互补序列杂交的序列。在一些实施方案中,所述有效载荷缺乏核酸酶靶向的序列,和/或缺乏能够与gRNA或其互补序列杂交的序列。在一些实施方案中,所述第一同源臂和/或第二同源臂缺少核酸酶靶向的序列,和/或缺少能够与gRNA或其互补序列杂交的序列。
在一些实施方案中,所述有效载荷包括编码第一多肽的第一核酸。在一些实施方案中,所述有效载荷包括编码第二多肽的第二核酸。在一些实施方案中,所述编码核糖体跳跃序列的核酸位于所述第一核酸和所述第二核酸之间。
在一些实施方案中,所述第一多肽或第二多肽编码B2M cDNA。在一些实施方案中,所述第一多肽或第二多肽编码非聚合HLA多肽或治疗性多肽。在一些实施方案中,所述非聚合HLA多肽选自HLA-E、HLA-F或HLA-G。一些实施方案还包括编码由非聚合HLA多肽呈递的自体肽的第三多肽。在一些实施方案中,所述治疗性多肽包括酶、抗体或其抗原结合片段、受体、嵌合抗原受体或细胞因子。在一些实施方案中,所述治疗性多肽包括因子IX、血管紧张素转换酶2(Ace2)、β-葡萄糖脑苷脂酶(GBA)、α-半乳糖苷酶A(GLA)或酸性α-葡萄糖苷酶(GAA)。
在一些实施方案中,所述第一核酸可操作地与内源性B2M基因的启动子连接。
在一些实施方案中,所述第一多肽与所述内源性B2M基因编码的多肽符合读码框架。
在一些实施方案中,所述有效载荷包括异源启动子,并且所述第一核酸可操作地与所述异源启动子连接。在一些实施方案中,所述异源启动子是组成型启动子。在一些实施方案中,所述启动子选自MND启动子、EF1α启动子或PGK启动子。
在一些实施方案中,载体包括修复模板。在一些实施方案中,所述载体包括病毒载体。在一些实施方案中,所述病毒载体包括腺相关病毒(AAV)载体或慢病毒载体。
一些实施方案还包括细胞,其中所述细胞包括所述核酸酶;和任选地,所述gRNA和/或修复模板。在一些实施方案中,所述细胞包括所述核酸酶。在一些实施方案中,所述细胞包括所述gRNA和/或修复模板。在一些实施方案中,所述细胞选自B细胞、造血干细胞、早期祖B细胞(early pro-B cell)、晚期祖B细胞(late pro-B cell),大的前B细胞(largepre-B cell)、小的前B细胞(small pre-B cell)、不成熟B细胞、T1 B细胞、T2 B细胞、边缘区B细胞、成熟B细胞、初始B细胞、浆母(短寿命)细胞、GC B细胞、记忆B细胞和长寿命浆细胞。在一些实施方案中,所述细胞是B细胞。在一些实施方案中,所述细胞是人的细胞。
在一些实施方案中,所述细胞是受试者自体的细胞。
在一些实施方案中,所述细胞与受试者同种异体。
在一些实施方案中,所述细胞是体外的。
在一些实施方案中,所述细胞是体内的。
一些实施方案包括药物组合物,所述药物组合物包括本文提供的任何一种系统和药学上可接受的赋形剂。
一些实施方案包括制备修饰的细胞的方法,包括获得本文提供的任何一种系统;将所述核酸酶引入第一细胞;和任选地:将gRNA引入所述细胞,和/或将修复模板引入所述细胞,从而制备在修饰的细胞基因组中包括修饰的B2M基因座的修饰的细胞。
在一些实施方案中,所述修饰的B2M基因座包括失活的内源性B2M基因。在一些实施方案中,所述gRNA适于使内源性B2M基因失活。一些实施方案还包括选择修饰的细胞。在一些实施方案中,所述选择包括使所述修饰的细胞与免疫细胞接触。在一些实施方案中,所述免疫细胞与所述第一细胞同种异体。在一些实施方案中,所述免疫细胞包括与所述第一细胞的MHC-I不同的MHC-I。在一些实施方案中,所述免疫细胞选自T细胞,例如细胞毒性CD8+T细胞或自然杀伤细胞。
在一些实施方案中,所述免疫细胞是体内的。
在一些实施方案中,所述免疫细胞是体外的。
在一些实施方案中,所述修饰的B2M基因座表达活性B2M基因或B2McDNA。在一些实施方案中,所述修复模板缺乏被核酸酶靶向的序列,和/或缺乏能够与所述gRNA或其互补序列杂交的序列,使得修饰的B2M基因座缺乏被核酸酶靶向的序列,和/或缺少能够与所述gRNA或其互补序列杂交的序列。在一些实施方案中,所述修复模板包括B2M cDNA。一些实施方案还包括选择修饰的细胞。在一些实施方案中,所述选择包括使修饰的细胞与免疫细胞接触。在一些实施方案中,所述免疫细胞与第一细胞的自体的。在一些实施方案中,所述免疫细胞选自T细胞,例如细胞毒性CD8+T细胞或自然杀伤细胞。
在一些实施方案中,所述免疫细胞是体内的。
在一些实施方案中,所述免疫细胞是体外的。
在一些实施方案中,所述修饰的B2M基因座表达替换MHC-I。在一些实施方案中,所述修饰的B2M基因座包括失活的B2M内源性基因。在一些实施方案中,所述修复模板包括编码B2M cDNA、非聚合HLA多肽和/或由非聚合HLA肽呈递的自体肽的有效载荷。一些实施方案还包括选择修饰的细胞。在一些实施方案中,所述选择包括使修饰的细胞与免疫细胞接触。在一些实施方案中,所述免疫细胞与第一细胞同种异体。在一些实施方案中,所述免疫细胞选自T细胞,例如细胞毒性CD8+T细胞或自然杀伤细胞。
在一些实施方案中,所述免疫细胞是体内的。
在一些实施方案中,所述免疫细胞是体外的。
在一些实施方案中,所述第一细胞是B细胞。
一些实施方案包括通过本文提供的任一种方法制备的细胞。
一些实施方案包括用于富集对T细胞或自然杀伤细胞杀伤的抗性增加的修饰的B细胞的方法,包括:通过本文提供的任一种方法制备修饰的细胞,其中所述第一细胞是B细胞,并且其中所述修饰的B2M基因座是失活的;以及使所述修饰的细胞与T细胞或自然杀伤细胞接触,其中所述T细胞或自然杀伤细胞与所述第一细胞同种异体,并且其中所述修饰的B细胞与表达B2M基因的B细胞相比,对T细胞或自然杀伤细胞的杀伤具有增加的抗性。
一些实施方案包括富集修饰的B细胞的方法,包括通过本文提供的任一种方法制备修饰的细胞,其中修饰的B2M基因座表达活性B2M基因或B2McDNA;以及使所述修饰的细胞与T细胞或自然杀伤细胞接触,其中所述T细胞或自然杀伤细胞与所述第一细胞是自体的,并且其中不表达活性B2M基因或B2M cDNA的细胞被所述T细胞或自然杀伤细胞杀死。
一些实施方案包括用于富集修饰的B细胞的方法,包括通过本文提供的任一种方法制备修饰的细胞,其中修饰的B2M基因座表达替换的MHC-I,其中修复模板包括编码B2McDNA、非聚合HLA多肽和由非聚合HLA肽呈递的自体肽的有效载荷;以及使所述修饰的细胞与T细胞或自然杀伤细胞接触,其中所述T细胞或自然杀伤细胞与所述第一细胞同种异体。
一些实施方案包括制备用于同种异体输注的修饰的B细胞的方法,包括通过本文提供的任一种方法制备修饰的细胞,其中所述细胞是B细胞,并且所述修复模板包括编码B2M cDNA、非聚合HLA多肽和由非聚合HLA肽呈递的自体肽的有效载荷。一些实施方案还包括向受试者施用修饰的B细胞,其中所述B细胞与受试者同种异体。在一些实施方案中,所述非聚合HLA多肽选自HLA-E、HLA-F或HLA-G。在一些实施方案中,所述受试者是哺乳动物。在一些实施方案中,所述受试者是人。
本文提供的方法和组合物的一些实施方案包括制备修饰的B细胞的方法,包括:使B细胞中的内源性β-2微球蛋白(B2M)基因失活;以及将编码融合蛋白的多核苷酸引入所述失活的B2M基因中,从而获得修饰的B细胞,其中所述融合蛋白包括外源B2M多肽。
本文提供的方法和组合物的一些实施方案包括在自体受试者体内富集修饰的B细胞的方法,包括:使B细胞中的内源性β-2微球蛋白(B2M)基因失活;将编码融合蛋白的多核苷酸引入所述失活的B2M基因中,从而获得修饰的B细胞,其中所述融合蛋白包括外源B2M多肽;并向受试者施用所述修饰的B细胞,其中所述修饰的B细胞是所述受试者自体的细胞。
本文提供的方法和组合物的一些实施方案包括制备用于同种异体输注的修饰B细胞的方法,包括:使B细胞中的内源性β-2微球蛋白(B2M)基因失活;以及将编码融合蛋白的多核苷酸引入所述失活的B2M基因中,从而获得修饰的B细胞,其中所述融合蛋白包括:外源性B2M多肽、非聚合HLA多肽和由所述非聚合HLA肽呈递的自体肽。一些实施方案还包括向受试者施用所述修饰的B细胞,其中所述修饰的B细胞与受试者同种异体。在一些实施方案中,所述非聚合HLA多肽选自HLA-E或HLA-G。
在一些实施方案中,所述内源性B2M基因通过内源性B2M基因的至少一部分的插入、缺失或替换而失活。
在一些实施方案中,所述内源性B2M基因的失活包括将与Cas核酸酶偶联的成簇的规律间隔的短回文重复序列DNA(CRISPR)引入B细胞。在一些实施方案中,所述Cas核酸酶包括Cas9核酸酶。在一些实施方案中,所述使内源性B2M基因失活包括将向导RNA(gRNA)引入B细胞。在一些实施方案中,所述gRNA包括与SEQ ID NO:01-04中任一条的核苷酸序列具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%或99%的序列一致性的核苷酸序列。
在一些实施方案中,所述使内源性B2M基因失活包括将包括修复模板的病毒载体引入B细胞。在一些实施方案中,所述病毒载体包括腺相关病毒(AAV)载体。
在一些实施方案中,所述使内源性B2M基因失活包括将核酸酶引入B细胞,其中所述核酸酶选自锌指核酸酶、转录激活因子样效应物核酸酶(TALEN)、归巢核酸内切酶(HE)和组合的TALEN-HE蛋白(megaTAL)。
在一些实施方案中,将编码融合蛋白的所述多核苷酸引入失活的B2M基因中,使得所述融合蛋白与内源性B2M基因的外显子符合读码框架。
在一些实施方案中,所述多核苷酸包括同源臂、编码自切割肽的核酸和/或启动子。
在一些实施方案中,将编码融合蛋白的多核苷酸引入失活的B2M基因,所述失活的B2M基因将与Cas核酸酶偶联的成簇的规律间隔的短回文重复序列DNA(CRISPR)引入B细胞。在一些实施方案中,所述Cas核酸酶包括Cas9核酸酶。在一些实施方案中,将编码融合蛋白的多核苷酸引入所述失活的B2M基因包括将向导RNA(gRNA)引入B细胞。在一些实施方案中,所述gRNA包括与SEQ ID NO:01-04中任一条的核苷酸序列具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%或99%的序列一致性的核苷酸序列。
在一些实施方案中,所述将编码融合蛋白的多核苷酸引入所述失活的B2M基因包括将包括修复模板的病毒载体引入B细胞。在一些实施方案中,所述病毒载体包括腺相关病毒(AAV)载体。
在一些实施方案中,所述将编码融合蛋白的多核苷酸引入所述失活的B2M基因包括将核酸酶引入B细胞,其中所述核酸酶选自锌指核酸酶、转录激活因子样效应物核酸酶(TALEN)、归巢核酸内切酶(HE)和组合的TALEN-HE蛋白(megaTAL)。
在一些实施方案中,使B细胞中的内源性β-2微球蛋白(B2M)基因失活;将编码融合蛋白的多核苷酸引入所述失活的B2M基因中是顺序进行的。在一些实施方案中,使B细胞中的内源性β-2微球蛋白(B2M)基因失活;将编码融合蛋白的多核苷酸引入失活的B2M基因中是同时进行的。
在一些实施方案中,所述融合蛋白包括治疗性多肽或治疗性核酸。在一些实施方案中,所述治疗性多肽包括酶、抗体或其抗原结合片段、受体、嵌合抗原受体或细胞因子。在一些实施方案中,所述治疗性多肽包括因子IX、血管紧张素转换酶2(Ace2)、β-葡萄糖脑苷脂酶(GBA)、α-半乳糖苷酶A(GLA)或酸性α-葡萄糖苷酶(GAA)。
在一些实施方案中,所述B细胞选自造血干细胞、早期祖B细胞、晚期祖B细胞,大的前B细胞、小的前B细胞、不成熟B细胞、T1 B细胞、T2 B细胞、边缘区B细胞、成熟B细胞、初始B细胞、浆母(短寿命)细胞、GC B细胞、记忆B细胞和长寿命浆细胞。
在一些实施方案中,所述受试者是哺乳动物。在一些实施方案中,所述受试者是人。
本文提供的方法和组合物的一些实施方案包括通过上述任一种方法制备的修饰的B细胞。
本文提供的方法和组合物的一些实施方案包括药物组合物,所述药物组合物包括本文提供的任一种修饰的B细胞和药学上可接受的载体。
本文提供的方法和组合物的一些实施方案包括分离的核酸,所述分离的核酸包括与SEQ ID NO:01-04中任一条的核苷酸序列具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%或99%的序列一致性的核苷酸序列。
附图说明
图1描绘了在以下条件下培养的浆细胞(CD138+)中MHC-II表达的流式分析:(1)用MCD40L和CpG B或αCD180活化2天;(2)I期,用IL-21和IL-6或R848治疗3天;和(3)II期,用IL-21治疗3天。
图2描绘了小鼠B2M基因座的示意图,包括B2M基因,相应的表示外显子的B2MmRNA,表示为B2M cDNA的编码部分mRNA的相应蛋白质;还示出了向导RNA的位点:gRNA-1和gRNA_5。
图3示出了向导RNA:gRNA-1和gRNA_5及其反向序列的缺失频率(ICE评分)和B2M基因的预测失活(敲除(KO)评分)。
图4A描绘了用gRNA-1、gRNA_5或对照处理的小鼠B细胞的流式分析。
图4B描绘了用gRNA-1、gRNA_5或对照处理的细胞的表达H2Kb蛋白的细胞的百分比的条形图。
图4C描绘了用gRNA处理以靶向人B2M基因座(gRNA-1、gRNA-2)或对照的人B细胞的流式分析。
图5A描绘了表达TCR蛋白的细胞毒性CD8+T细胞,其识别错配的MHC多肽,杀伤具有错配的MHC-I蛋白的细胞(左图);缺乏MHC-I的细胞可能不会诱导细胞毒性CD8+T细胞的杀伤(右图)。
图5B描绘了混合细胞测定法,以确定缺乏MHC-I的B细胞是否会抵抗这种具有来自BALB/c小鼠的T细胞(HLA,D类)和来自C57BL/6小鼠(HLA、B类)的B细胞的细胞毒性CD8+T细胞的杀伤。
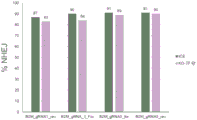
图6A描绘了用B2M gRNA_1RNP、Rosa26RNP或模拟物处理的B细胞和T细胞的混合培养物的流式分析。
图6B示出了混合培养物中CD45.1(B2M敲除B细胞)与CD45.2(对照B细胞)之比与T细胞与B细胞之比。
图7描绘了CD8+体外杀伤测定的时间线。
图8A描绘了针对CD45.2(靶向)和CD45.1(诱饵)C57/B6 B细胞的修饰的细胞在Balb/C CD8+T细胞共培养24小时和48小时的时间点的FACS分析。
图8B描绘了与来自图8A相关的实验的定量CD45.1:CD45.2比值相关的修饰的细胞的折线图,用不存在CD8+T细胞的共培养条件时的基础比例进行标准化。
图9A描绘了在与图8A相关的实验中,CD45.1 B细胞群体中H2Kb(C57/B6B细胞中相关MHC1单倍型)的修饰的细胞的FACS分析。
图9B描绘了修饰的细胞随着CD8+T细胞比例的增加在24小时和48小时的时间点在B2M编辑的CD45.1 B细胞中定量H2Kb敲除水平的折线图。
图10描绘了修饰B2M基因座的构建体(3307)的示意图,并包括编码5′和3′同源臂(HA)、可操作地与GFP多肽连接的MND启动子、P2A自切割肽和B2M cDNA的序列。
图11是通过B2M gRNA_1RNP递送和3307AAV转导的组合编辑的原代小鼠B细胞的流式分析。
图12A描绘了B2M基因座处的取代的策略,其中所述B2M基因座被修饰以使内源性B2M表达失活,并表达工程化的HLA-E。
图12B描绘了混合细胞测定法,以检测B细胞存活,所述B细胞被修饰以使内源性B2M表达失活并表达工程化的HLA-E。
图13描绘了编码5’和3’同源臂(HA)的HDR供体模板(3310);外源性MND启动子;融合蛋白,所述融合蛋白包括Qdm自体肽、B2M cDNA和通过柔性接头(linker)连接的非多态性HLA-Eα链;2A自切割肽;GFP标记;和SV40聚腺苷酸化序列。
图14是用图13所示的构建体编辑的B细胞和在H2Kb-中GFP表达、第7天分化的B细胞的流式分析。
图15A描绘了一项用于建立体内移植MHC-I敲除细胞的NK细胞杀伤参数的研究。
图15B描绘了用小鼠细胞进行同种异体B细胞的移植的小鼠模型的研究。
图16描绘了用人细胞进行同种异体B细胞的移植的异种移植模型的研究。
具体实施方式
本文提供的方法和组合物的一些实施方案包括制备修饰的B细胞。在一些实施方案中,B细胞中的内源性β-2微球蛋白(B2M)基因被修饰。一些实施方案涉及增加修饰的B细胞对同种异体免疫细胞杀伤的抗性。在一些实施方案中,所述内源性B2M基因是失活的,这增加了修饰的B细胞对同种异体免疫细胞杀伤的抗性。在一些实施方案中,将替代MHC-I插入失活的内源性B2M基因中,增加了修饰的B细胞对同种异体免疫细胞杀伤的抗性。一些实施方案包括对成功修饰的细胞进行富集。
本文提供的方法和组合物的一些实施方案包括制备修饰的B细胞。在一些实施方案中,B细胞中的内源性β-2微球蛋白(B2M)基因被修饰。在一些实施方案中,所述B细胞中的内源性B2M基因是失活的。在一些这样的实施方案中,将编码融合蛋白的多核苷酸引入失活的B2M基因中。一些实施方案包括在B2M基因座修饰以用于同种异体输注的B细胞的制备和使用、融合蛋白的表达和/或修饰的细胞的体内富集。
本文提供的一些实施方案包括在B2M基因座引入外源转基因和工程化的基因组DNA的方法,以改善B细胞治疗的递送。一些实施方案包括具有AAV递送的修复模板的CRISPR-Cas9重组核蛋白复合物(向导RNA和蛋白质)的使用。一些实施方案包括替代核酸酶(例如锌指核酸酶、转录激活因子样效应物核酸酶或组成的megaTAL)或修复模板(例如单链或双链DNA)平台在基因递送中的应用。在一些实施方案中,递送至B2M基因座的序列促进了工程化人B细胞的同种异体递送和/或富集。在一些实施方案中,递送至B2M基因座的序列通过体内富集工程化B细胞促进自体递送。
一些实施方案包括用于有效编辑B2M基因座,例如敲除/灭活MHC-I的人B2M的合成向导RNA。一些实施方案包括用于将外源序列引入B2M,同时也保持内源性B2M的表达的修复模板。这样的修复模板可以包括编码治疗蛋白,例如因子IX、血管紧张素转换酶2(Ace2)、β-葡萄糖脑苷脂酶(GBA)、α-半乳糖苷酶A(GLA)或酸性α-葡萄糖苷酶(GAA)的序列。一些实施方案包括用与HLA(例如,HLA-E或HLA-G)的非聚合型融合的B2M和抑制NK细胞杀伤的肽替代内源性B2M的修复模板。这些序列促进血浆细胞的同种异体移植,而不会被宿主免疫细胞(例如T和/或NK细胞)排斥。一些实施方案包括用与非多聚合HLA融合的B2M替换B2M并引入治疗蛋白的外源序列的修复模板。
一些实施方案涉及增加修饰的B细胞对同种异体免疫细胞杀伤的抗性。例如,在一些实施方案中,所述内源性B2M基因被失活,增加了修饰的B细胞对同种异体免疫细胞杀伤的抗性。一些这样的实施方案包括用于富集修饰的B细胞的组合物和方法,所述修饰的B细胞对同种异体T细胞或同种异体自然杀伤细胞的杀伤的抗性增加。一些这样的实施方案包括制备修饰的B细胞,其中所述细胞基因组的B2M基因座被失活,使所述修饰的细胞与免疫细胞,例如T细胞(例如CD8+T细胞)接触,其中所述免疫细胞与修饰的B细胞同种异体。在一些这样的实施方案中,所述修饰的B细胞和免疫细胞之间缺乏错配的MHC-I增加了修饰的B细胞对免疫细胞杀伤的抗性。
一些实施方案涉及增加修饰的B细胞对同种异体免疫细胞杀伤的抗性。例如,在一些实施方案中,将替代MHC-I插入失活的内源性B2M基因中,增加了修饰的B细胞对同种异体免疫细胞杀伤的抗性。一些这样的实施方案包括用于富集修饰的B细胞的组合物和方法,所述修饰的B细胞通过对细胞的MHC-I进行替代而对同种异体T细胞或同种异体自然杀伤细胞的杀伤有增加的抗性。一些这样的实施方案包括制备修饰的B细胞,其中所述修饰的B2M基因座表达替代MHC-I。在一些这样的实施方案中,所述内源性B2M基因座被失活,并被编码B2M cDNA、非聚合HLA多肽和由非聚合HLA肽呈递的自体肽的有效载荷取代。一些实施方案还包括将所述修饰的细胞与免疫细胞,例如T细胞(例如CD8+T细胞)接触,其中所述免疫细胞与所述免疫细胞同种异体。
一些这样的实施方案包括用于富集具有某些成功修饰的修饰的B细胞的组合物和方法。一些这样的实施方案包括制备修饰的B细胞,其中所述细胞的修饰的B2M基因座表达活性B2M基因或B2M cDNA,使所述修饰的细胞与免疫细胞,例如T细胞(例如CD8+T细胞)接触,其中所述免疫细胞是修饰的B细胞自体的。当修饰未导致活性B2M基因或B2M cDNA成功表达,修饰的细胞可被免疫细胞杀死、去除或清除。
可用于本文提供的方法和组合物的某些实施方案的某些方面被US.20180141992;Hung K.L.等人,(2018)Mol Ther 26:456-467;Voss J.E.等人,(2019)Elife 8:e42995;Hartweger H.,等人,(2019)J Exp Med 216:1301-1310;JohnsonM.J.等人,(2018)Scientific Reports 8:12144;和Moffett H.F.等人,(2019)Sci Immunol 4(35)所公开,其各自通过引用全部明确并入。
定义
如本文所用,“基因组编辑”在根据说明书理解时具有其通常的和普通的含义,可以包括但不限于,例如,包括在活体基因组中插入、缺失或替换DNA的基因工程方法的过程。编辑基因也称为基因编辑。在本文描述的一些替代方案中,提供了一种制备表达分子(例如大分子)的浆细胞或浆细胞前体的方法,其中B细胞或B细胞前体经过至少一轮基因组编辑。基因组编辑的方法可以包括但不限于在细胞基因组中插入、缺失或替换核酸。在一些替代方案中,使用核酸酶来实现这一过程。在一些替代方案中,所述核酸酶是工程化的。在一些替代方案中,所述方法包括诱导双链断裂,所述断裂可以通过非同源性末端接合(NHEJ)或同源重组(HR)修复。在一些替代方案中,通过引入单链核酸来进行基因组编辑步骤。在一些替代方案中,所述至少一轮基因组编辑还包括循环B细胞以将单链DNA寡核苷酸或重组腺相关病毒同源重组到候选的基因座中。在一些替代方案中,在没有病毒整合的情况下进行B细胞的基因组编辑以表达蛋白质。在一些替代方案中,进行第二轮基因组编辑以切除区域。在一些替代方案中,进行第三轮基因组编辑以使得药物激活的生长增强子表达。在本文的一些替代方案中,所述基因组编辑是通过定向同源重组非致病性AAV介导的编辑来进行的。
基因组编辑也可以使用基于RNA和蛋白质的转染。例如,CRISPR/Cas系统可以被修饰以编辑基因组。这项技术需要将与合成向导RNA(gRNA)复合的Cas核酸酶递送到细胞中,因此可以在特定位置切割所述细胞基因组,并允许移除现有基因和/或添加新基因。因此,CRISPR/Cas和相关的可编程核酸内切酶系统正迅速成为生物医学研究实验室的重要基因组编辑工具,其在基因破坏和/或基因靶向方面的应用在多种培养的细胞和模型生物系统中得到了证明。在本文描述的CRISPR/Cas系统的一些替代方案中,所述Cas核酸酶包括Cas1、Cas2、Cas3、Cas4、Cas5、Cas6、Cas7、Cas8或Cas9。
CRISPR/Cas系统的基本组成部分包括靶基因、原型间隔区相邻的基序(protospacer adjacent motif,PAM)、向导RNA、Cas核酸内切酶。应用CRISPR/Cas进行基因组编辑的一个重要方面是需要有效地将向导RNA递送到各种细胞类型的系统。例如,这可以涉及递送体外产生的向导RNA作为核酸(通过体外转录或化学合成产生的向导RNA)。在一些替代方案中,所述核酸可以通过引入修饰的碱基而具有核酸酶抗性。
所述CRISPR-Cas系统分为两类。1类系统具有用于降解外源核酸的多个Cas蛋白的复合物。2类系统具有单个大Cas蛋白,同样用于降解外源核酸。共有93个cas基因,分为35个家族。源于cas核心(cas core)的35个家族中的11个,包括蛋白家族CAS1至CAS9。如本文所述,Cas包括CAS1、Cas2、Cas3、Cas4、Cas5、Cas6、Cas7、Cas8或CAS9。
基因编辑也可以通过基于非核酸酶的新型基因编辑平台进行。以前从人类造血干细胞中分离出一个新的AAV家族。这些非致病性AAV天然存在于健康个体中,并可能具有独特的基因编辑和基因转移特性。该技术也被称为AAV介导的定向同源重组编辑(AmENDRTM)。这一过程是通过自然生物机制进行的同源重组,细胞利用这种机制来确保高度精确的DNA修复。
AAV通过定向同源重组介导的编辑是通过设计同源序列“臂”启动的,所述同源序列“臂”对基因组的某个区域具有特异性,并在施用于细胞时得到DNA的永久性校正。在本文的一些替代方案中,通过定向同源重组非致病性AAV介导的编辑来进行基因编辑。对新型AAV基因组的鉴定如Smith等人(Mol Ther.2014Sep;22(9):1625–1634;通过引用将其全部并入本文)中所述。Smith等人描述的新型AAV代表了一类新的操纵HSC基因组的基因载体。此外,这些载体可以极大地增强向靶组织和细胞(包括对基因转移不敏感的细胞)递送基因的能力,这规避了对AAV2普遍存在的预存免疫。在一些替代方案中,基因编辑由造血细胞中天然存在的非致病性AAV进行,其中所述编辑是通过AAV介导的编辑来进行的,所述通过AAV介导的编辑通过使用非致病性AAV的定向同源重组来进行,如Smith等人所述,。
如本文所用,“工程化的核酸酶”在根据说明书理解时具有其通常的和普通的含义,并且可以包括但不限于,例如,被工程化为杂交酶的酶,其可以用于特异性识别DNA序列并通过引入双链断裂有效地编辑基因组。非限制性的,四个优选的工程化核酸酶家族适用于本文所述的实施方案,包括归巢核酸内切酶、锌指蛋白核酸酶(ZFN)、转录激活因子样效应物核酸酶(TALEN)或CRISPR-Cas系统。
如本文所用,当根据说明书理解时,“归巢核酸内切酶”具有其通常的和普通的含义,可以包括但不限于,例如,以大识别位点(12至40个碱基对的双链DNA序列)为特征的脱氧核糖核酸内切酶。在制备表达分子如大分子的浆细胞或浆细胞前体的一些替代方法中,所述方法包括:(a)分离B细胞,(b)培养所述B细胞,(c)在没有病毒整合的情况下,对B细胞进行第一轮基因组编辑以进行蛋白表达,(d)扩增B细胞,(e)分化B细胞,任选地,在步骤(c)或(d)后,从而产生表达蛋白的浆细胞。在一些替代方案中,所述第一轮基因组编辑是通过基于RNA和蛋白质的转染进行的。在一些替代方案中,所述核酸酶是归巢核酸内切酶。
如本文所用,“锌指核酸酶(ZFN)”在根据说明书理解时具有其通常的和普通的含义,可以包括但不限于例如通过将锌指DNA结合结构域与DNA切割域融合而产生的工程化的限制性核酸内切酶。锌指结构域可以被工程化为靶向特定的目标DNA序列,这使得锌指核酸酶能够靶向复杂基因组中的特定序列。在制备表达分子(例如大分子)的浆细胞的一些替代方法中,所述方法包括:(a)分离B细胞,(b)培养所述B细胞,(c)在没有病毒整合的情况下,对B细胞进行第一轮基因组编辑以进行蛋白表达,(d)扩增B细胞,(e)分化B细胞,任选地,在步骤(c)或(d)后,从而产生表达蛋白的浆细胞。在一些替代方案中,所述第一轮基因组编辑是通过基于RNA和蛋白质的转染进行的。在一些替代方案中,所述核酸酶是锌指核酸酶。
如本文所用,“转录激活因子样效应物核酸酶”(TALEN)在根据说明书理解时具有其通常的和普通的含义,可以包括但不限于,例如,限制性核酸内切酶,可以被工程化以切割DNA中特定序列或位点。它们是通过将TAL效应物DNA结合结构域与DNA切割结构域(一种切割DNA链的核酸酶)融合而制成的。转录激活因子样效应物(TALE)可以被工程化为与目标DNA序列结合,因此当与核酸酶结合时,DNA可以在特定位置被切割。因此,所述限制性核酸内切酶可以被引入细胞,用于基因组编辑或原位基因组编辑(已知的利用工程化的核酸酶进行基因组编辑的技术)。TALEN的使用是本领域技术人员已知的。在本文所述的一些替代方案中,提供了一种制备表达分子(例如大分子,例如归巢核酸内切酶)的浆细胞或浆细胞前体的方法,其中B细胞或B细胞前体经过至少一轮基因组编辑。基因组编辑的方法可以包括但不限于在细胞基因组中插入、缺失或替换核酸。在一些替代方案中,使用核酸酶来实现这一过程。在一些替代方案中,所述核酸酶是工程化的。在一些替代方案中,所述方法包括诱导双链断裂,通过非同源末端接合(NHEJ)或同源重组(HR)修复双链断裂。在一些替代方案中,所述方法包括第一轮基因组编辑或基因组编辑。在一些替代方案中,所述第一轮基因组编辑包括递送核酸酶,其中所述核酸酶靶向B细胞中的至少一个基因座。在一些替代方案中,所述至少一个基因座包括JCHAIN、IGKC、IGMC、PON3、PRG2、FKBP11、SDC1、SLPI、DERL3、EDEM1、LY6C2、CRELD2、REXO2、PDIA4、PRDM1、CARD11、CCR5或SDF2L1。在一些替代方案中,所述核酸酶是锌指核酸酶、转录活化因子样效应物核酸酶(TALEN)、归巢核酸内切酶(HE)、组合的TALEN-HE蛋白(megaTAL)或靶向与CAS核酸酶偶联的CRISPR的合成的向导RNA。在一些替代方案中,所述Cas核酸酶包括Cas1、Cas2、Cas3、Cas4、Cas5、Cas6、Cas7、Cas8或Cas9。在一些替代方案中,所述第一轮基因组编辑包括用重组腺相关病毒载体转化B细胞,作为同源重组到候选基因座的供体模板。在一些替代方案中,所述重组腺相关病毒载体是单链、双链或自补的。
如本文所用,“敲除”包括以干扰靶向多核苷酸序列功能的方式删除全部或部分靶向多核苷酸序列。例如,通过在靶向多核苷酸序列的功能域(例如DNA结合域)中诱导靶向多核苷酸序列中的indel来改变靶多核苷酸,从而实现敲除。基于本文的具体实施方式,本领域技术人员容易理解如何使用本发明的CRISPR/Cas系统敲除靶向多核苷酸序列或其一部分。
制备修饰的B细胞的某些方法
本文提供的方法和组合物的一些实施方案包括制备修饰的B细胞的方法。一些这样的实施方案包括使B细胞中的内源性β-2微球蛋白(B2M)基因失活;以及将编码融合蛋白的多核苷酸引入所述失活的B2M基因中,从而获得修饰的B细胞,其中所述融合蛋白包括外源B2M多肽。
在一些实施方案中,所述内源性B2M基因通过插入、缺失或替换至少一部分内源性B2M蛋白而失活。在一些实施方案中,基因组(例如二倍体基因组)中B2M的每个等位基因均被失活。
在一些实施方案中,所述使内源性B2M基因失活包括将与Cas核酸酶偶联的CRISPR核酸引入B细胞。在一些实施方案中,所述Cas核酸酶包括Cas9核酸酶。在一些实施方案中,所述使内源性B2M基因失活包括将向导RNA(gRNA)引入B细胞。在一些实施方案中,所述gRNA包括与SEQ ID NO:01-04中任一条的核苷酸序列具有至少85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性的核苷酸序列,或与SEQ ID NO:01-04中任一条的核苷酸序列具有前述百分比中任意两个之间的任何百分比的序列一致性的核苷酸序列。
在一些实施方案中,所述使内源性B2M基因失活包括将病毒载体引入B细胞,所述病毒载体包括修复模板。在一些实施方案中,所述病毒载体包括腺相关病毒(AAV)载体。
在一些实施方案中,所述使内源性B2M基因失活包括将核酸酶引入B细胞,其中所述核酸酶选自锌指核酸酶、转录激活因子样效应物核酸酶(TALEN)、归巢核酸内切酶(HE)和组合的TALEN-HE蛋白(megaTAL)。
在一些实施方案中,将编码融合蛋白的多核苷酸引入失活的B2M基因中,使得融合蛋白与内源性B2M基因的外显子符合读码框架。
在一些实施方案中,所述多核苷酸包括同源臂、编码自切割肽的核酸和/或启动子。
在一些实施方案中,所述将编码融合蛋白的多核苷酸引入失活的B2M基因,包括将与Cas核酸酶偶联的CRISPR核酸引入B细胞。在一些实施方案中,所述Cas核酸酶包括Cas9核酸酶。在一些实施方案中,将编码融合蛋白的多核苷酸引入失活的B2M基因包括将gRNA引入B细胞。在一些实施方案中,所述gRNA包括与SEQ ID NO:01-04中任一条的核苷酸序列具有至少85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性的核苷酸序列,或与SEQ ID NO:01-04中任一条的核苷酸序列具有前述百分比中任意两个之间的任何百分比的序列一致性的核苷酸序列。
在一些实施方案中,所述将编码融合蛋白的多核苷酸引入失活的B2M基因包括将病毒载体引入B细胞,所述病毒载体包括修复模板。在一些实施方案中,所述病毒载体包括AAV载体。
在一些实施方案中,所述将编码融合蛋白的多核苷酸引入失活的B2M基因包括将核酸酶引入B细胞,其中所述核酸酶选自锌指核酸酶、TALEN、HE和megaTAL。
在一些实施方案中,所述使B细胞中的B2M基因失活;将编码融合蛋白的多核苷酸引入所述失活的B2M基因中是顺序进行的。在一些实施方案中,所述使B细胞中的内源性B2M基因失活;将编码融合蛋白的多核苷酸引入失活的B2M基因中是同时进行的。
在一些实施方案中,所述融合蛋白包括治疗性多肽或治疗性核酸。在一些实施方案中,所述治疗性多肽包括酶、抗体或其抗原结合片段、受体、嵌合抗原受体或细胞因子。在一些实施方案中,所述治疗性多肽包括因子IX、血管紧张素转换酶2(Ace2)、β-葡萄糖脑苷脂酶(GBA)、α-半乳糖苷酶A(GLA)或酸性α-葡萄糖苷酶(GAA)。
治疗性多肽或治疗性核酸的更多示例包括:(a)被设计用于抑制病原体感染的亲和试剂,包括治疗性抗体、免疫粘附素和双特异性T细胞结合蛋白(bi-specific T cellengaging protein);(b)用于治疗炎性疾病、移植物排斥和/或自身免疫的免疫抑制剂,包括细胞因子如IL10、抗体(如抗TNF)和肽;(c)被设计用于增加宿主细胞对癌症或病原体的应答的免疫增强剂,包括细胞因子、抗体(如抗PDL1)和双特异性T细胞结合蛋白;和(d)被设计用于治疗由缺失蛋白质(包括与糖原贮积症相关的蛋白质)或血友病(如因子IX或因子VIII)引起的疾病的酶。
在一些实施方案中,所述B细胞选自造血干细胞、早期祖B细胞、晚期祖B细胞,大的前B细胞、小的前B细胞、不成熟B细胞、T1 B细胞、T2 B细胞、边缘区B细胞、成熟B细胞、初始B细胞、浆母(短寿命)细胞、GC B细胞、记忆B细胞和长寿命浆细胞。
体内富集修饰的B细胞的某些方法
本文提供的方法和组合物的一些实施方案包括用于在受试者(例如自体受试者)体内富集修饰的B细胞的方法。一些这样的实施方案包括使B细胞中的内源性B2M基因失活;将编码融合蛋白的多核苷酸引入所述失活B2M基因中,从而获得修饰的B细胞,其中所述融合蛋白包括外源B2M多肽;以及向受试者施用所述修饰的B细胞,其中所述B细胞是所述受试者自体的。在一些这样的实施方案中,缺乏活性内源性B2M基因或外源性B2M多肽的修饰的细胞被受试者免疫系统清除的可能性增加,例如被自然杀伤细胞清除。
在一些实施方案中,所述内源性B2M基因通过插入、缺失或替换至少一部分所述内源性B2M蛋白而失活。在一些实施方案中,基因组(例如二倍体基因组)中B2M的每个等位基因均失活。
在一些实施方案中,所述使内源性B2M基因失活包括将与Cas核酸酶偶联的CRISPR核酸引入B细胞。在一些实施方案中,所述Cas核酸酶包括Cas9核酸酶。在一些实施方案中,所述使内源性B2M基因失活包括将gRNA引入B细胞。在一些实施方案中,所述gRNA包括与SEQID NO:01-04中任一条的核苷酸序列具有至少85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性的核苷酸序列,或与SEQ ID NO:01-04中任一条的核苷酸序列具有前述百分比中任意两个之间的任何百分比的序列一致性的核苷酸序列。
在一些实施方案中,所述使内源性B2M基因失活包括将病毒载体引入B细胞,所述病毒载体包括修复模板。在一些实施方案中,所述病毒载体包括AAV载体。
在一些实施方案中,所述使内源性B2M基因失活包括将核酸酶引入B细胞,其中所述核酸酶选自锌指核酸酶、TALEN、HE和megaTAL。
在一些实施方案中,所述将编码融合蛋白的多核苷酸引入所述失活的B2M基因中,使得融合蛋白与内源性B2M基因的外显子符合读码框架。
在一些实施方案中,所述多核苷酸包括同源臂、编码自切割肽的核酸和/或启动子。
在一些实施方案中,所述将编码融合蛋白的多核苷酸引入失活的B2M基因,包括将偶联至Cas核酸酶的CRISPR核酸引入B细胞。在一些实施方案中,所述Cas核酸酶包括Cas9核酸酶。在一些实施方案中,将编码融合蛋白的多核苷酸引入灭活的B2M基因包括将gRNA引入B细胞。在一些实施方案中,所述gRNA包括与SEQ ID NO:01-04中任一条的核苷酸序列具有至少85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性的核苷酸序列,或与SEQ ID NO:01-04中任一条的核苷酸序列具有前述百分比中任意两个之间的任何百分比的序列一致性的核苷酸序列。
在一些实施方案中,将编码融合蛋白的多核苷酸引入失活的B2M基因包括将病毒载体引入B细胞,所述病毒载体包括修复模板。在一些实施方案中,所述病毒载体包括AAV载体。
在一些实施方案中,将编码融合蛋白的多核苷酸引入失活的B2M基因包括将核酸酶引入B细胞,其中所述核酸酶选自锌指核酸酶、TALEN、HE和megaTAL。
在一些实施方案中,所述使B细胞中的B2M基因失活;将编码融合蛋白的多核苷酸引入失活的B2M基因中是顺序进行的。在一些实施方案中,所述使B细胞中的内源性B2M基因失活;将编码融合蛋白的多核苷酸引入失活的B2M基因中是同时进行的。
在一些实施方案中,所述融合蛋白包括治疗性多肽或治疗性核酸。在一些实施方案中,所述治疗性多肽包括酶、抗体或其抗原结合片段、受体、嵌合抗原受体或细胞因子。在一些实施方案中,所述治疗性多肽包括因子IX、血管紧张素转换酶2(Ace2)、β-葡萄糖脑苷脂酶(GBA)、α-半乳糖苷酶A(GLA)或酸性α-葡萄糖苷酶(GAA)。
在一些实施方案中,所述B细胞选自造血干细胞、早期祖B细胞、晚期祖B细胞,大的前B细胞、小的前B细胞、不成熟B细胞、T1 B细胞、T2 B细胞、边缘区B细胞、成熟B细胞、初始B细胞、浆母(短寿命)细胞、GC B细胞、记忆B细胞和长寿命浆细胞。
在一些实施方案中,所述受试者是哺乳动物。在一些实施方案中,所述受试者是人。
制备同种异体输注用修饰的B细胞的某些方法
本文提供的方法和组合物的一些实施方案包括制备用于同种异体输注的修饰B细胞的方法。一些这样的实施方案包括使B细胞中的内源性B2M基因失活;以及将编码融合蛋白的多核苷酸引入所述失活的B2M基因中,从而获得修饰的B细胞,其中所述融合蛋白包括:外源性B2M多肽、非聚合HLA多肽和由所述非聚合HLA肽呈递的自体肽。一些方法还包括向受试者施用所述修饰的B细胞,其中所述B细胞与受试者同种异体。在一些实施方案中,所述非聚合HLA多肽选自HLA-E或HLA-G。
在一些实施方案中,所述内源性B2M基因通过插入、缺失或替换至少一部分内源性B2M蛋白而失活。在一些实施方案中,基因组(例如二倍体基因组)中B2M的每个等位基因均失活。
在一些实施方案中,所述使内源性B2M基因失活包括将与Cas核酸酶偶联的CRISPR核酸引入B细胞。在一些实施方案中,所述Cas核酸酶包括Cas9核酸酶。在一些实施方案中,所述使内源性B2M基因失活包括将gRNA引入B细胞。在一些实施方案中,所述gRNA包括与SEQID NO:01-04中任一条的核苷酸序列具有至少85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性的核苷酸序列,或与SEQ ID NO:01-04中任一条的核苷酸序列具有前述百分比中任意两个之间的任何百分比的序列一致性的核苷酸序列。
在一些实施方案中,所述使内源性B2M基因失活包括将病毒载体引入B细胞,所述病毒载体包括修复模板。在一些实施方案中,所述病毒载体包括AAV载体。
在一些实施方案中,所述使内源性B2M基因失活包括将核酸酶引入B细胞,其中所述核酸酶选自锌指蛋白核酸酶、TALEN、HE和megaTAL。
在一些实施方案中,将编码融合蛋白的多核苷酸引入失活的B2M基因中,使得所述融合蛋白与内源性B2M基因的外显子符合读码框架。
在一些实施方案中,所述多核苷酸包括同源臂、编码自切割肽的核酸和/或启动子。
在一些实施方案中,所述将编码融合蛋白的多核苷酸引入失活的B2M基因,包括将与Cas核酸酶偶联的CRISPR核酸引入B细胞。在一些实施方案中,所述Cas核酸酶包括Cas9核酸酶。在一些实施方案中,所述将编码融合蛋白的多核苷酸引入失活的B2M基因包括将gRNA引入B细胞。在一些实施方案中,所述gRNA包括与SEQ ID NO:01-04中任一条的核苷酸序列具有至少85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性的核苷酸序列,或与SEQ ID NO:01-04中任一条的核苷酸序列具有前述百分比中任意两个之间的任何百分比的序列一致性的核苷酸序列。
在一些实施方案中,所述将编码融合蛋白的多核苷酸引入失活的B2M基因包括将病毒载体引入B细胞,所述病毒载体包括修复模板。在一些实施方案中,所述病毒载体包括AAV载体。
在一些实施方案中,所述将编码融合蛋白的多核苷酸引入失活的B2M基因包括将核酸酶引入B细胞,其中所述核酸酶选自锌指核酸酶、TALEN、HE和megaTAL。
在一些实施方案中,使B细胞中的B2M基因失活;将编码融合蛋白的多核苷酸引入失活的B2M基因中是顺序进行的。在一些实施方案中,使B细胞中的内源性B2M基因失活;将编码融合蛋白的多核苷酸引入失活的B2M基因中是同时进行的。
在一些实施方案中,所述融合蛋白包括治疗性多肽或治疗性核酸。在一些实施方案中,所述治疗性多肽包括酶、抗体或其抗原结合片段、受体、嵌合抗原受体或细胞因子。在一些实施方案中,所述治疗性多肽包括因子IX、血管紧张素转换酶2(Ace2)、β-葡萄糖脑苷脂酶(GBA)、α-半乳糖苷酶A(GLA)或酸性α-葡萄糖苷酶(GAA)。
在一些实施方案中,所述B细胞选自造血干细胞、早期祖B细胞、晚期祖B细胞,大的前B细胞、小的前B细胞、不成熟B细胞、T1 B细胞、T2 B细胞、边缘区B细胞、成熟B细胞、初始B细胞、浆母(短寿命)细胞、GC B细胞、记忆B细胞和长寿命浆细胞。
在一些实施方案中,所述受试者是哺乳动物。在一些实施方案中,所述受试者是人。
某些组合物和系统
本文提供的方法和组合物的一些实施方案包括通过本文提供的任何一种方法制备的修饰的B细胞。本文提供的方法和组合物的一些实施方案包括药物组合物,所述药物组合物包括本文提供的任何一种修饰的B细胞和药学上可接受的载体。本文提供的方法和组合物的一些实施方案包括分离的核酸,所述分离的核酸包括与SEQ ID NO:01-04中任一条的核苷酸序列具有至少85%、86%、87%、88%、89%、90%、95%、96%、97%、98%、99%、100%的序列一致性的核苷酸序列,或与SEQ ID NO:01-04中任一条的核苷酸序列具有前述百分比中任意两个之间的任何百分比的序列一致性的核苷酸序列。
方法和组合物的一些实施方案包括用于使细胞中内源性β-2微球蛋白(B2M)基因失活的系统。一些这样的实施方案包括能够切割细胞基因组中内源性B2M基因中的靶向基因座的核酸酶或编码所述核酸酶的核酸。一些实施方案还包括具有与B2M基因互补的序列的向导RNA(gRNA)、和/或修复模板,所述修复模板包括第一同源臂、第二同源臂和编码第一同源臂和第二同源臂之间有效载荷的核酸,其中所述第一同源臂和/或所述第二同源性臂与B2M基因中的序列具有同源性。在一些实施方案中,所述gRNA适于使内源性B2M基因失活。在一些实施方案中,所述gRNA包括:与SEQ ID NO:01-04中任一条的核苷酸序列具有至少85%、86%、87%、88%、89%、90%、95%、96%、97%、98%、99%或100%的序列一致性的核苷酸序列;或其变体,所述变体与SEQ ID NO:01-04中的任一条相比具有不超过3个错配。在一些实施方案中,所述靶向的基因座位于B2M基因的第一编码外显子内。
在一些实施方案中,所述核酸酶包括Cas核酸酶。在一些实施方案中,所述Cas核酸酶包括Cas9核酸酶。
在一些实施方案中,所述核酸酶选自锌指核酸酶、转录激活因子样效应物核酸酶(TALEN)、归巢核酸内切酶(HE)和组合的TALEN-HE蛋白(megaTAL)。
在一些实施方案中,所述修复模板缺乏核酸酶靶向的序列。在一些实施方案中,所述第一同源臂和/或所述第二同源臂缺少所述核酸酶靶向的序列,和/或缺少能够与所述gRNA或其互补序列杂交的序列。在一些实施方案中,所述有效载荷缺乏所述核酸酶靶向的序列,和/或缺乏能够与所述gRNA或其互补序列杂交的序列。在一些实施方案中,所述有效载荷包括编码第一多肽的第一核酸。在一些实施方案中,所述有效载荷包括编码第二多肽的第二核酸。在一些实施方案中,所述编码核糖体跳跃序列的核酸位于所述第一核酸和所述第二核酸之间。
在一些实施方案中,所述第一多肽或第二多肽编码B2M cDNA。
在一些实施方案中,所述第一多肽或第二多肽编码非聚合HLA多肽。在一些实施方案中,所述非聚合HLA多肽选自HLA-E、HLA-F或HLA-G。一些实施方案还包括编码由非聚合HLA多肽呈递的自体肽的第三多肽。
在一些实施方案中,所述第一多肽或所述第二多肽包括治疗性多肽。在一些实施方案中,所述治疗性多肽包括酶、抗体或其抗原结合片段、受体、嵌合抗原受体或细胞因子。在一些实施方案中,所述治疗性多肽包括因子IX、血管紧张素转换酶2(Ace2)、β-葡萄糖脑苷脂酶(GBA)、α-半乳糖苷酶A(GLA)或酸性α-葡萄糖苷酶(GAA)。
在一些实施方案中,所述第一核酸可操作地与所述内源性B2M基因的启动子连接。在一些实施方案中,所述第一多肽与所述内源性B2M基因编码的多肽符合读码框架。在一些实施方案中,所述有效载荷包括异源启动子,并且所述第一核酸可操作地与异源启动子连接。在一些实施方案中,所述异源启动子是组成型启动子。在一些实施方案中,所述启动子选自MND启动子、EF1α启动子或PGK启动子。
在一些实施方案中,载体包括修复模板。在一些实施方案中,所述载体包括病毒载体。在一些实施方案中,所述病毒载体包括腺相关病毒(AAV)载体。
一些实施方案还包括细胞,其中所述细胞包括核酸酶;和任选地,gRNA和/或修复模板。在一些实施方案中,所述细胞选自B细胞、造血干细胞、早期祖B细胞、晚期祖B细胞,大的前B细胞、小的前B细胞、不成熟B细胞、T1 B细胞、T2 B细胞、边缘区B细胞、成熟B细胞、初始B细胞、浆母(短寿命)细胞、GC B细胞、记忆B细胞和长寿命浆细胞。在一些实施方案中,所述细胞是B细胞。在一些实施方案中,所述细胞是人的细胞。在一些实施方案中,所述细胞是受试者自体的。在一些实施方案中,所述细胞与受试者同种异体。在一些实施方案中,所述细胞是体外的。在一些实施方案中,所述细胞细胞是体内的。
一些实施方案包括药物组合物,所述药物组合物包括本文提供的任何一种系统和药学上可接受的赋形剂。
一些实施方案包括制备修饰的细胞的方法,所述方法包括获得本文公开的任何一种系统;并将核酸酶引入第一细胞。一些实施方案还包括将gRNA引入所述细胞,和/或将修复模板引入细胞,从而制备在修饰的细胞基因组中包括修饰的B2M基因座的修饰的细胞。
在一些实施方案中,所述修饰的B2M基因座是失活的。在一些实施方案中,所述gRNA适于使内源性B2M基因失活。一些实施方案还包括选择修饰的细胞。在一些实施方案中,所述选择包括使所述修饰的细胞与免疫细胞接触。在一些实施方案中,所述免疫细胞与第一细胞同种异体。例如,在一些实施方案中,所述免疫细胞包括与第一细胞的MHC-I不同的MHC-Ⅰ。在一些实施方案中,所述免疫细胞选自T细胞,例如细胞毒性CD8+T细胞或自然杀伤细胞。在一些实施方案中,所述免疫细胞是体内的。在一些实施方案中,所述免疫细胞是体外的。
在一些实施方案中,所述修饰的B2M基因座表达活性B2M基因或B2McDNA。在一些实施方案中,所述修复模板缺乏核酸酶靶向的序列,和/或缺乏能够与gRNA或其互补序列杂交的序列,使得修饰的B2M基因座缺乏核酸酶的靶序列和/或缺少能够与gRNA或其互补序列杂交的序列。在一些实施方案中,所述修复模板包括B2M cDNA。一些实施方案还包括选择修饰的细胞。在一些实施方案中,所述选择包括使所述修饰的细胞与免疫细胞接触。在一些实施方案中,所述免疫细胞与第一细胞自体的。在一些实施方案中,所述免疫细胞选自T细胞,例如细胞毒性CD8+T细胞或自然杀伤细胞。在一些实施方案中,所述免疫细胞是体内的。在一些实施方案中,所述免疫细胞是体外的。
在一些实施方案中,所述修饰的B2M基因座表达替换MHC-I。在一些实施方案中,所述修复模板包括编码B2M cDNA、非聚合HLA多肽和/或由非聚合HLA肽呈递的自体肽的有效载荷。一些实施方案还包括选择修饰的细胞。在一些实施方案中,所述选择包括使修饰的细胞与免疫细胞接触。在一些实施方案中,所述免疫细胞与第一细胞同种异体。在一些实施方案中,所述免疫细胞选自T细胞,例如细胞毒性CD8+T细胞或自然杀伤细胞。在一些实施方案中,所述免疫细胞是体内的。在一些实施方案中,所述免疫细胞是体外的。在一些实施方案中,所述第一细胞是B细胞。
一些实施方案包括用于富集对T细胞或自然杀伤细胞杀伤的抗性增加的修饰B细胞的方法,包括通过获得本文公开的任何一种系统来制备的修饰的细胞;并将所述核酸酶引入第一细胞;和/或将修复模板引入所述细胞中,从而制备在修饰的细胞的基因组中包括修饰的B2M基因座的修饰的细胞。在一些这样的实施方案中,所述第一细胞是B细胞,并且其中所述修饰的B2M基因座是失活的。一些实施方案还包括使修饰的细胞与T细胞或自然杀伤细胞接触,其中所述T细胞或自然杀伤细胞与所述第一细胞同种异体,并且其中与表达B2M基因的B细胞相比,所述修饰的B细胞对T细胞或自然杀伤细胞的杀伤抗性增加。
一些实施方案包括用于富集修饰的B细胞的方法,包括通过获得本文公开的任何一种系统来制备的修饰的细胞;并将所述核酸酶引入第一细胞;和/或将修复模板引入所述细胞,从而制备在修饰的细胞的基因组中包括修饰的B2M基因座的修饰的细胞。在一些实施方案中,所述修饰的B2M基因座表达活性B2M基因或B2M cDNA。一些实施方案还包括使修饰的细胞与T细胞或自然杀伤细胞接触,其中所述T细胞或自然杀伤细胞与所述第一细胞是自体,并且其中不表达活性B2M基因或B2M cDNA的细胞被T细胞或自然杀伤细胞杀死。
一些实施方案包括用于富集修饰的B细胞的方法,包括通过获得本文公开的任何一种系统来制备的修饰的细胞;并将所述核酸酶引入第一细胞;和/或将修复模板引入所述细胞,从而制备在修饰的细胞的基因组中包括修饰的B2M基因座的修饰的细胞。在一些实施方案中,所述修饰的B2M基因座表达替换MHC-I,其中修复模板包括编码B2M cDNA、非聚合HLA多肽和由非聚合HLA肽呈递的自体肽的有效载荷。一些实施方案还包括使修饰的细胞与T细胞或自然杀伤细胞接触,其中所述T细胞或自然杀伤细胞与第一细胞同种异体。
一些实施方案包括通过本文提供的方法中的任一种制备的细胞。
实施例
实施例1-浆细胞中MHC II的下调
主要组织相容性复合体(MHC)-I和MHC-II在移植物排斥反应中各自发挥作用。每个MHC都呈递抗原肽,所述抗原肽可以触发免疫应答,最终导致呈递细胞的消亡。未分化B细胞和一些分化的B细胞亚群同时表达MHC-I和MHC-II。
为了研究分化的血浆细胞是否下调MHC-II,使用阴性选择分离小鼠脾脏B细胞,并与各种细胞因子和饲养细胞一起培养。培养条件包括:(1)用MCD40L和CpG B或αCD180活化2天;(2)I期,用IL-21和IL-6或R848处理3天;和(3)II期,用IL-21处理3天。培养8天后,如使用流式细胞术以检测与MHC-II结合的抗体(MHCII-B-BV650)所测量的,血浆细胞亚群下调MHC-II。如图1所示,包括MCD40L和CpG B;以及IL-21和IL-6或R848导致MHC-II水平降低的细胞的百分比增加(26.3和23.3)。使用某些培养条件将B细胞分化为分化的B细胞或分化的B细胞产物导致MHC-II的下调。体外分化的B细胞培养物中MHC-II的意外下调可以减轻移植物排斥反应,这可能是由于修饰的B细胞表达错配的MHC-II蛋白引起的。
实施例2-B细胞中B2M基因座的修饰
分化的和未分化的B细胞都表达MHC-I,并在MHC-I肽复合物中呈递抗原肽。MHC-I肽复合物可被T细胞定向识别,所述T细胞表达对MHC-I多肽复合物具有特异性的T细胞受体。供体MHC-I与受体MHC-I不同的情况下可导致移植物排斥。当T细胞识别移植物中MHC-I复合物中呈递的不同序列时,T细胞可以直接杀死移植物细胞。MHC-I分子包括B2M蛋白、人类白细胞抗原(HLA)蛋白和抗原肽。B2M是MHC-I复合物运输到细胞表面所必需的。
CRISPR靶向B2M被用于消除功能性B2M基因座。使用ccTop CRISPR/Cas9在线靶点预测(target online predictor)工具在小鼠B2M基因的编码区内开发了两个gRNA靶序列(B2M gRNA_1和B2M gRNA_5)(图2)。gRNA_1与小鼠B2M的第一外显子的起始密码子相邻(基因组组装GRCm39/mm39,第121978227-121978246位)。gRNA_5位于B2M的外显子2(基因组组装GRCm39/mm39,第121981407-121981388位)。两个向导均被预测为通过引入引入移码的核苷酸插入或缺失,或终止密码子,使转录的RNA易于无义介导的降解,导致内源性B2M基因的灭活/敲除。B2M gRNA_1接近翻译起始位点和内源性B2M启动子,这将有利于多核苷酸(例如编码蛋白质的多核苷酸)的符合读码框架的插入。B2M gRNA_1和B2M gRNA_5的序列如表1所示。
表1

将B2M gRNA_1和gRNA_5以及重组Cas9作为核糖核蛋白(RNP)复合物递送至原代小鼠B细胞。从所述细胞中制备基因组DNA,并进行序列反卷积分析。所述分析揭示了两种向导的高插入和缺失频率(ICE评分)和预测失活indel(KO评分)的高发生率(图3)。
在人类细胞中进行了一项基本类似的研究,以表1中列示的gRNA靶向人类B2M基因座。人向导_1位于人B2M的第一外显子(基因组组装GRCh38/hg38,第44711585-44711566位)。人向导_2位于人B2M的外显子2(基因组组装GRCh38/hg38,第44715531-44715512位)。两种向导均被预测为通过引入核苷酸插入或缺失来引入移码,或终止密码子,使RNA易于无义介导的降解,导致内源性B2M基因的失活/敲除。
实施例3-修饰的B细胞中的蛋白表达
HLA-B编码的蛋白质(H2kb)是C57BL/6小鼠表达的主要HLA分子。为了确定B2M敲除是否消除了MHC-I在细胞表面的表达,通过流式细胞术定量了B细胞中H2kb的表达。用B2MgRNA_1或gRNA_5-RNP复合物处理来自C57BL/6小鼠的原代B细胞培养物。处理4天后,通过流式细胞术分析细胞中H2Kb蛋白的表达。与对照相比,用gRNA_1或gRNA_5处理的表达H2kb的细胞的数量减少(图4A、图4B)。基于流式细胞术的分析表明,在75%以上的初始、活化和分化的B细胞中,BM2/MHC-I蛋白表达被敲除/失活:(a)总淋巴细胞,(b)CD19+、CD138-B细胞,以及(c)CD19mid、CD138+浆细胞。功能性B2M基因座的基因组消除消除了MHC-I复合物的表面表达。
在人B细胞中进行了基本类似的研究,以表1中列示的gRNA靶向人B2M基因座(图4C)。使用gRNA-1而不是gRNA-2,90%以上的编辑的B细胞中MHC-I复合物(HLA-A)的表达被消除。这些结果与在小鼠B细胞中B2M耗竭实验中观察到的结果基本相似。
实施例4-修饰的B细胞的功能分析
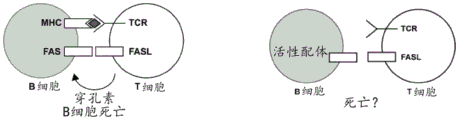
表达TCR蛋白的细胞毒性CD8+T细胞可以杀死具有错配MHC-I蛋白的细胞,所述TCR蛋白可以识别错配的MHC多肽。然而,细胞毒性CD8+T细胞可能不会杀死缺乏MHC-I的细胞,即使所述CD8+T和靶细胞来自错配供体(图5A)。用来自BALB/c小鼠(HLA,D类)的T细胞和来自C57BL/6小鼠(HLA,B类)的B细胞进行混合细胞试验,通过使B2M失活,以确定缺乏MHC-I的B细胞是否将对这种细胞毒性CD8+T细胞的杀伤有抗性,参见图5B。简而言之,在第0天,用抗CD3/CD28珠刺激Hkd+CD8+T细胞。第3天,从所述T细胞中移除刺激珠,并在IL-2中扩增T细胞。第3天从脾脏中分离Hkb+B细胞,并与MCD40L和CpG一起培养。在第4天,用RNP处理所述Hkb+B细胞以使B2M失活,或使作为阴性对照或模拟物的不相关基因座(Rosa26)失活。在第6天,与处理的B细胞和刺激的T细胞建立共培养组,并用MCD40-L、IL-4、IL-2、和无饲养支持物来处理。在第7天和第8天,通过FACS分析所有培养物。
使用B2M gRNA_1在原代C57BL/6(Hkb+)CD45.1 B细胞中诱导MHC-1敲除。将这些细胞与未编辑的C57BL/6CD45.2 B细胞混合,然后将其以不同比例与菌株错配的Balb/C(Hkd+)CD8+T细胞共培养。与用Rosa26RNP或模拟物处理的B细胞相比,用B2M gRNA_1RNP处理的B细胞群体具有更高的存活水平和更高的CD45.1表达水平(图6A)。在共培养24小时后,B2M编辑的CD45.1细胞优选地出现在与模拟物或Rosa26编辑的对照细胞相关的B细胞混合物中。B2M敲除细胞相对于Rosa26敲除细胞具有更高的生存优势。将CD45.1(B2M敲除B细胞)比CD45.2(对照B细胞)的比例与T细胞比B细胞的比例进行比较(图6B)。随着T细胞比B细胞比例的增加,CD8 T细胞杀伤试验中B2M敲除的保护作用更加明显。这些数据表明,功能性B2m的基因组消除保护了B细胞免于T细胞杀伤。
实施例5-B2M失活对CD8+T细胞杀伤的保护
进行CD8+体外杀伤试验。图7描绘了与实施例4中进行的测定基本类似的测定的时间线。
图8A描绘了针对CD45.2(靶向的)和CD45.1(诱饵)C57/B6 B细胞在与Balb/C CD8+T细胞共培养24小时和48小时的代表性FACS图。用靶向Rosa26安全港基因座的对照RNP或B2M RNP编辑的CD45.1诱饵细胞诱导MHC1敲除。图8A显示,在Rosa26对照中,CD45.2和CD45.1细胞的相对比例在两个时间点上都是稳定的,并且大致等于初始的靶向:诱饵为1:1的接种比例。相反,对于B2M编辑的CD42.1诱饵细胞,观察到向诱饵群体的显著倾斜,随着时间的推移,这种倾斜增加,这表明MHC1缺失具有保护作用。
图8B描述了来自图8A实验的CD45.1∶CD45.2比例的定量,以没有CD8+T细胞的共培养条件下的基础比例将其标准化。通过增加CD8+T细胞与B细胞的比例,绘制了共培养24小时(上图)和共培养48小时(下图)的时间点时的比例。图8B显示,对于Mock编辑的或Rosa26对照编辑的CD45.1诱饵B细胞,CD45.1∶CD45.2的比例在两个时间点都是稳定的,但是对于B2MCD45.1诱饵细胞,随着CD8+T细胞比例的增加而逐渐增加。每个数据点N=3。
图9A描述了在与图8A相关的实验中CD45.1 B细胞群体中H2Kb(C57/B6B细胞中的相关MHC1单倍型)的代表性FACS分析。示出了CD8+T细胞与B细胞的比例降低到1.5:1、1:1和1:2,共培养24小时(上排)和共培养48小时(下排)的时间点的结果。每个直方图覆盖了三种不同CD45.1诱饵种群:模拟物编辑的、Rosa26对照编辑的和B2M编辑的。每个直方图中显示的百分比值针对B2M编辑的组。图9A显示,B2M编辑的组中的MHC1敲除水平随着时间和CD8+T细胞比例的增加而增加,这表明在细胞毒性CD8+T细胞存在的情况下对该MHC1阴性群体的选择。
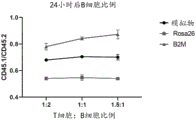
图9B描述了在24小时和48小时的时间点,随着CD8+T细胞比例的增加,B2M编辑的CD45.1 B细胞中H2Kb敲除水平的定量。图9B显示在两个时间点随着CD8+T细胞比例增加,MHC1敲除百分比逐渐增加。每个数据点N=3。
实施例6-B细胞中B2M表达的恢复
NK细胞进行免疫监测,以消除表面不表达MHC-I分子的潜在有害细胞。工程化B2M基因座的一个可能应用是使用CRISPR和同源性定向修复将cargo DNA引入B2M基因,并使用NK细胞的天然免疫监测功能来丰富工程化的细胞群体。在同源性定向修复实验中,CRISPR-gRNA被用于破坏基因组靶向基因座,AAV递送的修复模板被引入基因座,取代内源性序列。在一般的实验中,基因组破坏事件是高效的,同源性定向修复事件是低效的。由此产生的细胞群包括大多数只敲除等位基因的细胞和一小部分表达修复的等位基因的亚群。
在该实施例中,开发了包括HDR供体模板的构建体,以修饰B2M基因座,使其包括强MND启动子和GFP标记,从而得到的序列与内源性B2M编码序列符合读码框架(图10)。所述构建体包括B2M gRNA_1靶位点的突变版本。P2A自切割序列将GFP与B2M cDNA分离,允许在HDR编辑的细胞中表达可追踪标记,所述HDR编辑的细胞也表达人工重组的B2M编码序列。所述供体模板(命名为3307)被设计为作为腺相关病毒(AAV)被递送。
在B2M基因座先前被敲除/失活的B细胞中,用构建体成功修饰B2M基因基因座可以恢复B细胞MHC-I表达。相反,在B2M基因座先前被敲除/失活的B细胞中,用构建体对B2M基因的不成功修饰,例如通过在非B2M基因座的不正确插入或在不正确的部分插入,将不会恢复B细胞的MHC-I表达。简而言之,部分编辑的细胞是H2Kb-;通过NK细胞监测,这些细胞在体内死亡;H2Kb应在GFP+细胞中恢复;并且只有HDR和未编辑的细胞在体内存活。
B2M gRNA_1RNP递送和3307AAV转导的联合使用在原代小鼠B细胞中引发同源性定向修复。在接受RNP和病毒的细胞中,编辑后第3天观察到GFP+细胞群,GFP+细胞群是MHC-I阳性(图11)。在MHC-1阴性群体中未观察到GFP+细胞。接受不含RNP的3307病毒构建体的细胞全部为GFP阴性,这是意料之中的,因为AAV附加体在循环B细胞中不稳定,并且如果没有相关的B2M CRISPR试剂的共同递送,DNA序列将不会整合。这表明,细胞可以在体内成功修饰和选择B2M基因座,所述细胞内(1)未发生修饰;和(2)B2M基因座的一部分被修饰,使得所述基因座对gRNA抗性的失活,并表达B2M产物。
实施例7-B2M cDNA表达恢复的B细胞的存活
根据实施例6制备B细胞,其中内源性B2M基因座被敲除/失活,外源编码序列与B2M基因座处的B2M cDNA符合读码框架。
将从受试者制备的细胞群施用于同一受试者或具有匹配MHC-I序列的不同受试者。制备构建体适当修饰了B2M基因座的细胞,所述制备的细胞在B2M基因座表达B2M cDNA,并且对体内NK细胞杀伤具有抗性。相反,制备的B2M基因座未被构建体适当修饰且在B2M基因座上不表达B2M cDNA的细胞对体内NK细胞杀伤敏感,并且在体内被NK细胞清除。在一些实施方案中,可以使用类似的策略来选择体内工程化的B细胞,例如,受试者自体的工程化B细胞。
在另一个实施例中,从第一受试者获得B细胞。所述细胞被修饰。将所述修饰的细胞施用于所述第一受试者,或施用于具有与第一受试的MHC-I序列匹配的MHC-I序列的第二受试者。通过将B2M cDNA插入B2M基因组基因座,所述细胞在B2M基因组基因座处被修饰。表达B2M cDNA的修饰的细胞对体内NK细胞杀伤具有抗性。相反,不表达B2M cDNA的细胞在体内对NK细胞杀伤敏感,并在体内被NK细胞清除。
在另一个实施例中,根据实施例5制备B细胞,其中内源性B2M和/或CIITA被敲除/失活,并且插入外源编码序列以取代B2M。在一些实施方案中,B2M基因座被修饰以表达(a)三聚体,其包括工程化的HLA-E、可灭活NK细胞的常见自体肽和B2M;(b)CD47,一种能抑制NK细胞细胞毒性活性的表面蛋白;或(c)CD24,一种可以抑制NK细胞活性的表面蛋白。
实施例8-B细胞中B2M基因座的取代
HLA分子具有高度多态性,这意味着不同人之间HLA分子具有许多不同的序列。在MHC-I中,人表达一种类型的HLA。HLA-A、HLA-B和HLA-C被认为是高度多态性的,而HLA-E、HLA-F和HLA-G的是低多态性的。图12A中描绘了B2M基因座处的取代策略,其中B2M基因座被修饰以使内源性B2M表达失活,并表达由工程化HLA-E、一种通常在人中不突变的常见自体肽和B2M组成的三聚体。用这种非多态三聚体重组B2M基因座后,细胞在细胞表面表达所述三聚体,但不表达内源性的高度多态性HLA。表达这种来自不匹配个体的工程化三聚体的B细胞不会引发CD8+T细胞杀伤,但会抑制NK细胞免疫监视。图12B中描绘了用于测试修饰的B细胞的B细胞存活的混合细胞测定。
开发了包括HDR供体模板的构建体,以递送编码非多态性HLA-E融合多肽的多核苷酸,所述非多态性HLA-E融合多肽包括Qdm自体肽、B2M和HLA-Eα链,通过柔性接头序列连接并由外源MND启动子驱动(图13)。所述Qdm是由非多态性HLA-E呈递的自体肽。所述融合多肽与GFP荧光团配对用于追踪,并被设计用于诱导内源性B2M敲除。含有三聚体修复模板的构建体(命名为3310)被设计为作为腺相关病毒(AAV)递送。构建体3210的示例核苷酸序列如表2所示。
将B2M gRNA_1RNP递送和3310AAV转导的联合使用,编辑从C57BL/6小鼠分离的原代小鼠B细胞。在编辑后第7天,在经过同源性定向修复的细胞中,在CD19+、CD138-B细胞和CD19mid、CD138+血浆细胞中均可检测到GFP(图14)。重要的是,在两个群体中,GFP表达与HLA-B(Hk2b)表面表达的缺失完全同时发生,而HLA-B阳性细胞全部为GFP阴性,这表明HLA-E融合蛋白已经替换了这些细胞中内源性MHC-I的表达。
表2




实施例9-异体移植基因编辑的隐形(stealth)B细胞的体内存活
为了建立NK杀伤的参数,进行了与图15A和图15B所示研究基本相似的体内研究。从表达固定B细胞受体/抗体(SW-HEL)的C57BL/6小鼠分离原代小鼠B细胞。如实施例6和8所述对细胞进行修饰,使修饰的细胞表达HLA-E三聚体、CD47、CD24或含有HLA-E三聚物与CD47或CD24的组合。在表达小鼠BAFF、小鼠CD40L并补充可溶性R848或小鼠IL21的饲养细胞上扩增修饰的细胞后,约1000万修饰的细胞被转移到BalB/C小鼠中。细胞转移后,通过测量随时间的推移SW-HEL抗体的产生来跟踪移植。将移植与经编辑以在B2M基因座中表达GFP的C57BL/6细胞的移植以及在所有编辑策略中向C57BL/6小鼠的转移的所有细胞的移植进行比较。表达HLAE三聚体、CD47、CD24或其组合的同种异体B细胞在转移到不匹配的受体动物中后表达SW-HEL。转移的细胞的抗体表达的检测表明成功移植。相比之下,被工程化为表达GFP而不表达B2M的细胞,或被编辑为删除B2M的的细胞,由于NK细胞的清除,无法存活转移。由于T细胞的清除,未经编辑的细胞不能成功移植。用HLAE三聚体、CD47和/或CD24成功替代MHC 1类,表明修饰的B细胞群在转移到错配受体后存活。
实施例10-异体移植基因编辑的隐形B细胞的体内存活
进行与图16所示研究基本类似的研究。从外周血单核细胞分离人B细胞,并对其进行编辑,以使修饰的细胞表达HLA-E三聚体、CD47、CD24或含有HLA-E三聚物与CD47或CD24的组合。使用三阶段分化程序(如美国2018/0282692中披露的程序)在体外扩增和分化改性的细胞,所述程序通过引用全部并入。在扩增和分化后,1000万个工程化细胞被转移到免疫缺陷小鼠(NSG)中,所述小鼠被工程化以表达IL15,并预先移植了从不同的、错配的供体分离的人CD34细胞。通过转移人CD34细胞而人源化的NSG-IL15小鼠能够产生NK细胞,从而能够评估NK细胞的杀伤。细胞转移后,通过测量随时间的推移人IgG抗体的产生来跟踪移植。用CD34细胞人源化的NSG小鼠不产生IgG抗体。将同种异体编辑的细胞与编辑为在B2M基因座表达GFP的B细胞,及将所有编辑策略中向自体人源化NSG-IL15小鼠中转移的所有细胞的移植进行比较。
表达HLAE三聚体、CD47、CD24或其组合的同种异体B细胞在转移到错配的受体动物中后表达hIgG。转移的细胞中的抗体表达的检测表明成功移植。相比之下,被工程化为表达GFP而不表达B2M的细胞,或被编辑为删除B2M的细胞,由于NK细胞的清除,无法存活转移。由于T细胞的清除,未经编辑的细胞不能成功移植。用HLAE三聚体、CD47和/或CD24成功替代MHC 1类,表明修饰的B细胞群在转移到错配受体后存活。
本文所用术语“包含(comprsing)”与“包括(including)”、“含有(containing)”或“以……为特征”同义,并且是包含的或开放的,不排除额外的、未引用的元素或方法步骤。
上述描述公开了本发明的几种方法和材料。本发明易受方法和材料的改变以及制备方法和设备的改变的影响。考虑到本公开或本发明公开的实践,这些改变对于本领域技术人员将变得显而易见。因此,本发明并不局限于本文公开的具体实施方案,而是涵盖在本发明的真正范围和精神内的所有改变和替代方案。
本文引用的所有参考文献,包括但不限于已发表和未发表的申请、专利和文献参考文献,通过引用全部并入本文,并在此作为本说明书的一部分。如果通过引用并入的出版物和专利或专利申请与本说明书中包括的公开内容相矛盾,本说明书旨在取代和/或优先于任何此类矛盾的材料。
序列表
<110> 大卫·J罗林斯
理查德·G·詹姆斯
克莱尔·玛瑞亚·斯托弗斯
彼得·J库克
<120> 编辑B细胞中B2M基因座的方法和组合物
<130> SCRI.295WO
<150> 63/047978
<151> 2020-07-03
<160> 10
<170> FastSEQ for Windows Version 4.0
<210> 1
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 小鼠 B2M gRNA_1 (gRNA_1)
<400> 1
ctcggtgacc ctggtctttc 20
<210> 2
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 小鼠0B2M gRNA_5 (gRNA_5)
<400> 2
tcggcttccc attctccggt 20
<210> 3
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人类向导 1
<400> 3
gagtagcgcg agcacagcta 20
<210> 4
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人类向导 2
<400> 4
aagtcaactt caatgtcgga 20
<210> 5
<211> 301
<212> DNA
<213> 人工序列
<220>
<223> 5’同源臂
<400> 5
caccaaactc aagagcacac cctagatagt agggcaccaa gggtccagcc caggctgttt 60
gaaatatcac gggactttat aagaacatga aactgaaaat gggaaagtcc ctttgtaacc 120
tagttcagca tcaacagcta ggagactggt gacgacctcc ggatctgagt ccggattggc 180
tgtgagttca ggaactatat aagagcgcgc gccctggctg gctctcattt cagtggctgc 240
tactcggcgc ttcagtcgcg gtcgcttcag tcgtcagcat ggctcgctcg gtgaccctgg 300
t 301
<210> 6
<211> 347
<212> DNA
<213> 人工序列
<220>
<223> MND启动子
<400> 6
gaacagagaa acaggagaat atgggccaaa caggatatct gtggtaagca gttcctgccc 60
cggctcaggg ccaagaacag ttggaacagc agaatatggg ccaaacagga tatctgtggt 120
aagcagttcc tgccccggct cagggccaag aacagatggt ccccagatgc ggtcccgccc 180
tcagcagttt ctagagaacc atcagatgtt tccagggtgc cccaaggacc tgaaatgacc 240
ctgtgcctta tttgaactaa ccaatcagtt cgcttctcgc ttctgttcgc gcgcttctgc 300
tccccgagct ctatataagc agagctcgtt tagtgaaccg tcagatc 347
<210> 7
<211> 27
<212> DNA
<213> 人工序列
<220>
<223> Qdm肽
<400> 7
gctatggctc ccaggacact cctgctg 27
<210> 8
<211> 297
<212> DNA
<213> 人工序列
<220>
<223> beta-2微球蛋白
<400> 8
attcagaaga cacctcaaat ccaggtgtac tccagacacc cccccgagaa cggaaagcct 60
aatattctca attgttacgt gacccagttt cacccccccc atattgagat ccagatgctc 120
aaaaacggca agaaaattcc taaggtggag atgagcgaca tgagctttag caaggactgg 180
agcttctaca ttctcgccca cacagaattt acacccaccg agaccgacac ctatgcttgc 240
agggtgaaac atgccagcat ggctgagcct aagacagtgt actgggatag ggacatg 297
<210> 9
<211> 1011
<212> DNA
<213> 人工序列
<220>
<223> alpha链
<400> 9
agcccacact cgctgcggta tttcaccacc gccgtgtccc ggcccggcct cggggagccc 60
cggttcatca ttgtcggcta cgtggacgac acgcagttcg tgcgcttcga cagcgacgcg 120
gaaaatccga ggatggagcc tcgggcgcgg tggattgagc aggaggggcc ggagtattgg 180
gagcgggaga cttggaaagc cagggacatg gggaggaact tcagagtaaa cctgaggacc 240
ctgctcggct actacaatca gagtaacgac gaatctcaca cgctgcagtg gatgtacggc 300
tgcgacgtgg ggcccgatgg gcgcctgctc cgcgggtatt gtcaggaggc ctacgatggc 360
caggattaca tctccctgaa cgaggacctg cgttcctgga ccgcgaatga catagcctca 420
cagatctcta agcacaagtc agaggcagtc gatgaggccc accaacagag ggcatacctg 480
caaggtcctt gcgtggagtg gctccataga tacctacggc tgggaaatga gacactgcag 540
cgctcagacc ctccaaaggc acatgtgacc catcacccta gatctgaaga tgaagtcacc 600
ctgaggtgct gggccctggg cttctaccct gctgacatca ccctgacctg gcagttgaat 660
ggggaggagc tgacccagga catggagctt gtggagacca ggcctgcagg ggatggaacc 720
ttccagaagt gggcagctgt cgtggtgcct cttgggaagg agcagtatta cacatgccat 780
gtgtaccatg aggggctgcc tgagcccctc accctgagat gggagcctcc tccatccact 840
gtctccaaca tggtaatcat agctgttctg gttgtccttg gagctgtgat catccttgga 900
gctgtggtgg cttttgtgat gaagaggagg agacacatag gtgtaaaagg atgctatgct 960
catgttctag gcagcaagag cttccagacc tctgactggc ctcagaaggc a 1011
<210> 10
<211> 301
<212> DNA
<213> 人工序列
<220>
<223> 3’同源臂
<400> 10
ttctggtgct tgtctcactg accggcctgt atgctatcca gagtgagtgc ctctttcccc 60
tctcttggca ttaaattttt agttctcctt agttctcctt cccgacggga ttggccaggg 120
ctgcgtctct cgggaaagga gtgggcgtcc cgctgcttac agctttgagg ctacagggtg 180
gatgcgcctg tttgcgagtg tgctgacggt cagcgctgaa gatggggaag gcttctcttt 240
ttctcctctg ctggcgggag cccctagact ccctgagcgc aaaaggaggg tggggctgcc 300
t 301
Claims (77)
1.一种用于修饰细胞中内源性β-2微球蛋白(B2M)基因的系统,包括:
能够切割所述细胞基因组中内源性B2M基因中的靶向基因座的核酸酶或编码所述核酸酶的核酸;和可选地:
向导RNA(gRNA),所述向导RNA包括与B2M基因互补的序列,和/或
修复模板,所述修复模板包括第一同源臂、第二同源臂和编码所述第一同源臂和所述第二同源臂之间的有效载荷的核酸,其中所述第一同源臂和/或所述第二同源性臂与所述B2M基因中的序列具有同源性。
2.如权利要求1所述的系统,其中所述gRNA包括:与SEQIDNO:01-04中任一条的核苷酸序列具有至少85%、86%、87%、88%、89%、90%、95%、96%、97%、98%、99%或100%的序列一致性的核苷酸序列;SEQIDNO:01-04中任一条的核苷酸序列;或其变体,所述变体与SEQIDNO:01-04中的任一条相比具有不超过3个错配。
3.如权利要求1或2所述的系统,其中所述gRNA适于使所述内源性B2M基因失活。
4.如权利要求1-3中任一项所述的系统,其中所述靶向基因座位于所述B2M基因的第一编码外显子内。
5.如权利要求1-4中任一项所述的系统,其中所述核酸酶包括Cas核酸酶。
6.如权利要求5所述的系统,其中所述Cas核酸酶包括Cas9核酸酶。
7.如权利要求1所述的系统,其中所述核酸酶选自锌指核酸酶、转录激活因子样效应物核酸酶(TALEN)、归巢核酸内切酶(HE)和组合的TALEN-HE蛋白(megaTAL)。
8.如权利要求1-7中任一项所述的系统,其中所述修复模板缺少核酸酶靶向的序列,和/或缺少能够与所述gRNA或其互补序列杂交的序列。
9.如权利要求8所述的系统,其中所述有效载荷缺乏所述核酸酶靶向的序列,和/或缺乏能够与所述gRNA或其互补序列杂交的序列。
10.如权利要求8所述的系统,其中所述第一同源臂和/或所述第二同源臂缺少所述核酸酶靶向的序列,和/或缺少能够与所述gRNA或其互补序列杂交的序列。
11.如权利要求1-10中任一项所述的系统,其中所述有效载荷包括编码第一多肽的第一核酸。
12.如权利要求11所述的系统,其中所述有效载荷包括编码第二多肽的第二核酸。
13.如权利要求12所述的系统,其中编码核糖体跳跃序列的核酸位于所述第一核酸和所述第二核酸之间。
14.如权利要求11-13中任一项所述的系统,其中所述第一多肽或所述第二多肽编码B2McDNA。
15.如权利要求11-14中任一项所述的系统,其中所述第一多肽或所述第二多肽编码非聚合HLA多肽或治疗性多肽。
16.如权利要求15所述的系统,其中所述非聚合HLA多肽选自HLA-E、HLA-F或HLA-G。
17.如权利要求15或16所述的系统,还包括编码由所述非聚合HLA多肽呈递的自体肽的第三多肽。
18.如权利要求15所述的系统,其中所述治疗性多肽包括酶、抗体或其抗原结合片段、受体、嵌合抗原受体或细胞因子。
19.如权利要求15所述的系统,其中所述治疗性多肽包括因子IX、血管紧张素转换酶2(Ace2)、β-葡萄糖脑苷脂酶(GBA)、α-半乳糖苷酶A(GLA)或酸性-α-葡糖苷酶(GAA)。
20.如权利要求11-19中任一项所述的系统,其中所述第一核酸可操作地与所述内源性B2M基因的启动子连接。
21.如权利要求11-20中任一项所述的系统,其中所述第一多肽与所述内源性B2M基因编码的多肽符合读码框架。
22.如权利要求11-21中任一项所述的系统,其中所述有效载荷包括异源启动子,并且所述第一核酸可操作地与所述异源启动子连接。
23.如权利要求22所述的系统,其中所述异源启动子是组成型启动子。
24.如权利要求22或23所述的系统,其中所述启动子选自MND启动子、EF1α启动子或PGK启动子。
25.如权利要求1-24中任一项所述的系统,其中载体包括所述修复模板。
26.如权利要求25所述的系统,所述载体包括病毒载体。
27.如权利要求26所述的系统,其中所述病毒载体包括腺相关病毒(AAV)载体或慢病毒载体。
28.如权利要求1-27中任一项所述的系统,还包括细胞,其中所述细胞包括所述核酸酶;和任选地,所述gRNA和/或修复模板。
29.如权利要求28所述的系统,其中所述细胞包括所述核酸酶。
30.如权利要求28或29所述的系统;其中所述细胞包括所述gRNA和/或修复模板。
31.如权利要求28-30中任一项所述的系统,其中所述细胞选自B细胞、造血干细胞、早期祖B细胞、晚期祖B细胞,大的前B细胞、小的前B细胞、不成熟B细胞、T1B细胞、T2B细胞、边缘区B细胞、成熟B细胞、初始B细胞、浆母(短寿命)细胞、GCB细胞、记忆B细胞和长寿命浆细胞。
32.如权利要求28-31中任一项所述的系统,其中所述细胞是B细胞。
33.如权利要求28-32中任一项所述的系统,其中所述细胞是人的细胞。
34.如权利要求28-33中任一项所述的系统,其中所述细胞是受试者自体的。
35.如权利要求28-33中任一项所述的系统,其中所述细胞与受试者同种异体。
36.如权利要求28-35中任一项所述的系统,其中所述细胞是体外的。
37.如权利要求28-35中任一项所述的系统,其中所述细胞是体内的。
38.一种药物组合物,所述药物组合物包括权利要求1-37中任一项所述的系统和药学上可接受的赋形剂。
39.一种制备修饰的细胞的方法,包括:
获得权利要求1-37中任一项所述的系统;
将所述核酸酶引入第一细胞;并且可选地:
将所述gRNA引入所述细胞,和/或将修复模板引入所述细胞,从而制备在修饰的细胞基因组中包括修饰的B2M基因座的修饰的细胞。
40.如权利要求39所述的方法,其中所述修饰的B2M基因座包括失活的内源性B2M基因。
41.如权利要求39或40所述的方法,其中所述gRNA适于使所述内源性B2M基因失活。
42.如权利要求39-41中任一项所述的方法,还包括选择所述修饰的细胞。
43.如权利要求42所述的方法,其中所述选择包括使所述修饰的细胞与免疫细胞接触。
44.如权利要求43所述的方法,其中所述免疫细胞与所述第一细胞同种异体。
45.如权利要求43或44所述的方法,其中所述免疫细胞包括与所述第一细胞的MHC-I不同的MHC-Ⅰ。
46.如权利要求43-45中任一项所述的方法,其中所述免疫细胞选自T细胞,例如细胞毒性CD8+T细胞,或自然杀伤细胞。
47.如权利要求43-46中任一项所述的方法,其中所述免疫细胞是体内的。
48.如权利要求43-46中任一项所述的方法,其中所述免疫细胞是体外的。
49.如权利要求39所述的方法,其中所述修饰的B2M基因座表达活性B2M基因或B2McDNA。
50.如权利要求49所述的方法,其中所述修复模板缺少被所述核酸酶靶向的序列,和/或缺少能够与所述gRNA或其互补序列杂交的序列,使得所述修饰的B2M基因座缺少被所述核酸酶靶向的序列,和/或缺少能与所述gRNA序列或其互补杂交的序列。
51.如权利要求49或50所述的方法,其中所述修复模板包括B2McDNA。
52.如权利要求49-51中任一项所述的方法,还包括选择所述修饰的细胞。
53.如权利要求52所述的方法,其中所述选择包括使所述修饰的细胞与免疫细胞接触。
54.如权利要求53所述的方法,其中所述免疫细胞是所述第一细胞自体的。
55.如权利要求53或54所述的方法,其中所述免疫细胞选自T细胞,例如细胞毒性CD8+T细胞,或自然杀伤细胞。
56.如权利要求53-55中任一项所述的方法,其中所述免疫细胞是体内的。
57.如权利要求53-55中任一项所述的方法,其中所述免疫细胞是体外的。
58.如权利要求39所述的方法,其中所述修饰的B2M基因座表达替换的MHC-I。
59.如权利要求58所述的方法,其中所述修饰的B2M基因座包括失活的内源性B2M基因。
60.如权利要求58或59所述的方法,其中所述修复模板包括编码B2McDNA、非聚合HLA多肽和/或由所述非聚合HLA肽呈递的自体肽的有效载荷。
61.如权利要求58-60中任一项所述的方法,还包括选择所述修饰的细胞。
62.如权利要求61所述的方法,其中所述选择包括使所述修饰的细胞与免疫细胞接触。
63.如权利要求62所述的方法,其中所述免疫细胞与所述第一细胞同种异体。
64.如权利要求62或63所述的方法,其中所述免疫细胞选自T细胞,例如细胞毒性CD8+T细胞,或自然杀伤细胞。
65.如权利要求62-64中任一项所述的方法,其中所述免疫细胞是体内的。
66.如权利要求62-64中任一项所述的方法,其中所述免疫细胞是体外的。
67.如权利要求39-66中任一项所述的方法,其中所述第一细胞是B细胞。
68.如权利要求39-67中任一项所述的方法制备的细胞。
69.一种用于富集对T细胞或自然杀伤细胞杀伤的抗性增加的修饰的B细胞的方法,包括:
通过权利要求39所述的方法制备修饰的细胞,其中所述第一细胞是B细胞,并且其中所述修饰的B2M基因座被失活;和
使所述修饰的细胞与T细胞或自然杀伤细胞接触,其中所述T细胞或自然杀伤细胞与所述第一细胞同种异体,并且其中所述修饰的B细胞与表达B2M基因的B细胞相比,对T细胞或自然杀伤细胞杀伤的抗性增加。
70.一种用于富集修饰的B细胞的方法,包括:
通过权利要求39所述的方法制备修饰的细胞,其中所述修饰的B2M基因座表达活性B2M基因或B2McDNA;和
使所述修饰的细胞与T细胞或自然杀伤细胞接触,其中所述T细胞或自然杀伤细胞与所述第一细胞是自体的,并且其中不表达活性B2M基因或B2McDNA的细胞被所述T细胞或自然杀伤细胞杀死。
71.一种用于富集修饰的B细胞的方法,包括:
通过权利要求39所述的方法制备修饰的细胞,其中所述修饰的B2M基因座表达替换的MHC-I,其中所述修复模板包括编码B2McDNA、非聚合HLA多肽和由所述非聚合HLA肽呈递的自体肽的有效载荷;和
使所述修饰的细胞与T细胞或自然杀伤细胞接触,其中所述T细胞或自然杀伤细胞与所述第一细胞同种异体。
72.一种制备用于同种异体输注的修饰B细胞的方法,包括:
通过权利要求39所述的方法制备修饰的细胞,其中所述细胞是B细胞,并且所述修复模板包括编码B2McDNA、非聚合HLA多肽和由所述非聚合HLA肽呈递的自体肽的有效载荷。
73.如权利要求72所述的方法,还包括向受试者施用所述修饰的B细胞,其中所述B细胞与所述受试者同种异体。
74.如权利要求72或72所述的方法,其中所述非聚合HLA多肽选自HLA-E、HLA-F或HLA-G。
75.如权利要求72-74中任一项所述的方法,其中所述受试者是哺乳动物。
76.如权利要求72-75中任一项所述的方法,其中所述受试者是人。
77.一种分离的核酸,所述分离的核酸包括与SEQIDNO:01-04中任一条的核苷酸序列具有至少85%、86%、87%、88%、89%、90%、95%、96%、97%、98%、99%或100%的序列一致性的核苷酸序列;SEQIDNO:01-04中任一条的核苷酸序列;或其变体,所述变体与SEQIDNO:01-04中的任一条相比具有不超过3个错配。
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US202063047978P | 2020-07-03 | 2020-07-03 | |
| US63/047,978 | 2020-07-03 | ||
| US202063114131P | 2020-11-16 | 2020-11-16 | |
| US63/114,131 | 2020-11-16 | ||
| PCT/US2021/039953 WO2022006309A1 (en) | 2020-07-03 | 2021-06-30 | Methods and compositions for editing the b2m locus in b cells |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| CN116033910A true CN116033910A (zh) | 2023-04-28 |
Family
ID=79315579
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN202180047549.4A Pending CN116033910A (zh) | 2020-07-03 | 2021-06-30 | 编辑b细胞中b2m基因座的方法和组合物 |
Country Status (10)
| Country | Link |
|---|---|
| EP (1) | EP4175651A1 (zh) |
| JP (1) | JP2023532917A (zh) |
| KR (1) | KR20230035048A (zh) |
| CN (1) | CN116033910A (zh) |
| AU (1) | AU2021300358A1 (zh) |
| BR (1) | BR112022026534A2 (zh) |
| CA (1) | CA3186620A1 (zh) |
| IL (1) | IL299491A (zh) |
| MX (1) | MX2022015788A (zh) |
| WO (1) | WO2022006309A1 (zh) |
Families Citing this family (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2024030857A1 (en) * | 2022-08-01 | 2024-02-08 | Idexx Laboratories, Inc. | Human mouse fractional abundance assays |
Family Cites Families (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| IL297018A (en) * | 2015-12-18 | 2022-12-01 | Sangamo Therapeutics Inc | Directed cleavage of cell mhc receptor |
| EP3645036A1 (en) * | 2017-06-30 | 2020-05-06 | Cellectis | Cellular immunotherapy for repetitive administration |
-
2021
- 2021-06-30 WO PCT/US2021/039953 patent/WO2022006309A1/en active Application Filing
- 2021-06-30 AU AU2021300358A patent/AU2021300358A1/en active Pending
- 2021-06-30 EP EP21832140.4A patent/EP4175651A1/en active Pending
- 2021-06-30 CA CA3186620A patent/CA3186620A1/en active Pending
- 2021-06-30 JP JP2022581341A patent/JP2023532917A/ja active Pending
- 2021-06-30 MX MX2022015788A patent/MX2022015788A/es unknown
- 2021-06-30 CN CN202180047549.4A patent/CN116033910A/zh active Pending
- 2021-06-30 IL IL299491A patent/IL299491A/en unknown
- 2021-06-30 KR KR1020237001885A patent/KR20230035048A/ko unknown
- 2021-06-30 BR BR112022026534A patent/BR112022026534A2/pt unknown
Also Published As
| Publication number | Publication date |
|---|---|
| KR20230035048A (ko) | 2023-03-10 |
| EP4175651A1 (en) | 2023-05-10 |
| CA3186620A1 (en) | 2022-01-06 |
| MX2022015788A (es) | 2023-02-27 |
| BR112022026534A2 (pt) | 2023-04-18 |
| IL299491A (en) | 2023-02-01 |
| JP2023532917A (ja) | 2023-08-01 |
| WO2022006309A1 (en) | 2022-01-06 |
| AU2021300358A1 (en) | 2023-02-02 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US11180776B1 (en) | Universal donor cells | |
| JP7385281B2 (ja) | 低抗原性細胞の製造方法 | |
| US20230279351A1 (en) | Artificial antigen-specific immunoregulatory t (airt) cells | |
| US20220017715A1 (en) | Compositions and Methods for Efficacy Enhancement of T-Cell Based Immunotherapy | |
| CN116033910A (zh) | 编辑b细胞中b2m基因座的方法和组合物 | |
| AU2022283895A1 (en) | Gene editing in primary immune cells using cell penetrating crispr-cas system | |
| US20230272431A1 (en) | Methods and compositions for editing the b2m locus in b cells | |
| US20220279767A1 (en) | Transgenic swine, methods of making and uses thereof, and methods of making human immune system mice | |
| WO2023191063A1 (ja) | 遺伝子工学、細胞工学、および細胞医薬に適した細胞およびその製造方法 | |
| EP4347810A2 (en) | Ciita targeting zinc finger nucleases |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| PB01 | Publication | ||
| PB01 | Publication | ||
| SE01 | Entry into force of request for substantive examination | ||
| SE01 | Entry into force of request for substantive examination |