CN114729007A - 麻疹的联合抗病毒治疗 - Google Patents
麻疹的联合抗病毒治疗 Download PDFInfo
- Publication number
- CN114729007A CN114729007A CN202080077668.XA CN202080077668A CN114729007A CN 114729007 A CN114729007 A CN 114729007A CN 202080077668 A CN202080077668 A CN 202080077668A CN 114729007 A CN114729007 A CN 114729007A
- Authority
- CN
- China
- Prior art keywords
- peptide
- hrc
- fip
- antiviral
- fusion
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
- 230000000840 anti-viral effect Effects 0.000 title claims abstract description 203
- 201000005505 Measles Diseases 0.000 title claims abstract description 103
- 238000011282 treatment Methods 0.000 title description 14
- 108090000765 processed proteins & peptides Proteins 0.000 claims abstract description 358
- 239000000863 peptide conjugate Substances 0.000 claims abstract description 196
- 230000004927 fusion Effects 0.000 claims abstract description 189
- 208000015181 infectious disease Diseases 0.000 claims abstract description 125
- 108010068327 4-hydroxyphenylpyruvate dioxygenase Proteins 0.000 claims abstract description 122
- 238000000034 method Methods 0.000 claims abstract description 85
- 102000004196 processed proteins & peptides Human genes 0.000 claims abstract description 75
- 239000000203 mixture Substances 0.000 claims abstract description 66
- 210000004899 c-terminal region Anatomy 0.000 claims abstract description 56
- 230000002401 inhibitory effect Effects 0.000 claims abstract description 48
- 241000712079 Measles morbillivirus Species 0.000 claims description 263
- 229920001223 polyethylene glycol Polymers 0.000 claims description 168
- 239000002202 Polyethylene glycol Substances 0.000 claims description 161
- 210000004027 cell Anatomy 0.000 claims description 131
- 230000035772 mutation Effects 0.000 claims description 108
- 125000003275 alpha amino acid group Chemical group 0.000 claims description 69
- LYCAIKOWRPUZTN-UHFFFAOYSA-N Ethylene glycol Chemical group OCCO LYCAIKOWRPUZTN-UHFFFAOYSA-N 0.000 claims description 59
- HVYWMOMLDIMFJA-DPAQBDIFSA-N cholesterol Chemical group C1C=C2C[C@@H](O)CC[C@]2(C)[C@@H]2[C@@H]1[C@@H]1CC[C@H]([C@H](C)CCCC(C)C)[C@@]1(C)CC2 HVYWMOMLDIMFJA-DPAQBDIFSA-N 0.000 claims description 50
- 239000004471 Glycine Substances 0.000 claims description 45
- 239000012528 membrane Substances 0.000 claims description 43
- 230000004807 localization Effects 0.000 claims description 34
- 239000002105 nanoparticle Substances 0.000 claims description 31
- 235000012000 cholesterol Nutrition 0.000 claims description 25
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 claims description 21
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 claims description 21
- 102000003886 Glycoproteins Human genes 0.000 claims description 19
- 108090000288 Glycoproteins Proteins 0.000 claims description 19
- 230000002163 immunogen Effects 0.000 claims description 18
- 230000000069 prophylactic effect Effects 0.000 claims description 16
- 230000028993 immune response Effects 0.000 claims description 15
- 239000000539 dimer Substances 0.000 claims description 12
- GVJHHUAWPYXKBD-UHFFFAOYSA-N d-alpha-tocopherol Chemical group OC1=C(C)C(C)=C2OC(CCCC(C)CCCC(C)CCCC(C)C)(C)CCC2=C1C GVJHHUAWPYXKBD-UHFFFAOYSA-N 0.000 claims description 11
- 230000001939 inductive effect Effects 0.000 claims description 11
- 235000010384 tocopherol Nutrition 0.000 claims description 11
- 229960001295 tocopherol Drugs 0.000 claims description 11
- 229930003799 tocopherol Chemical group 0.000 claims description 11
- 239000011732 tocopherol Chemical group 0.000 claims description 11
- GVJHHUAWPYXKBD-IEOSBIPESA-N α-tocopherol Chemical group OC1=C(C)C(C)=C2O[C@@](CCC[C@H](C)CCC[C@H](C)CCCC(C)C)(C)CCC2=C1C GVJHHUAWPYXKBD-IEOSBIPESA-N 0.000 claims description 11
- 125000000913 palmityl group Chemical group [H]C([*])([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])[H] 0.000 claims description 9
- 230000002265 prevention Effects 0.000 claims description 9
- 230000034217 membrane fusion Effects 0.000 claims description 8
- COLNVLDHVKWLRT-QMMMGPOBSA-N phenylalanine group Chemical group N[C@@H](CC1=CC=CC=C1)C(=O)O COLNVLDHVKWLRT-QMMMGPOBSA-N 0.000 claims description 8
- 150000008574 D-amino acids Chemical class 0.000 claims description 7
- 125000001584 benzyloxycarbonyl group Chemical group C(=O)(OCC1=CC=CC=C1)* 0.000 claims description 7
- 239000006199 nebulizer Substances 0.000 claims description 4
- 238000007920 subcutaneous administration Methods 0.000 claims description 3
- 208000031886 HIV Infections Diseases 0.000 abstract description 6
- 208000037357 HIV infectious disease Diseases 0.000 abstract description 5
- 208000033519 human immunodeficiency virus infectious disease Diseases 0.000 abstract description 5
- 241000700605 Viruses Species 0.000 description 198
- 125000005647 linker group Chemical group 0.000 description 158
- 210000004556 brain Anatomy 0.000 description 92
- 210000002220 organoid Anatomy 0.000 description 78
- 108090000623 proteins and genes Proteins 0.000 description 70
- 230000003612 virological effect Effects 0.000 description 56
- 210000003169 central nervous system Anatomy 0.000 description 50
- 210000004379 membrane Anatomy 0.000 description 42
- 235000018102 proteins Nutrition 0.000 description 38
- 102000004169 proteins and genes Human genes 0.000 description 38
- 102000005962 receptors Human genes 0.000 description 37
- 108020003175 receptors Proteins 0.000 description 37
- 239000002552 dosage form Substances 0.000 description 29
- 239000003112 inhibitor Substances 0.000 description 29
- 239000008194 pharmaceutical composition Substances 0.000 description 28
- 238000002474 experimental method Methods 0.000 description 27
- 241000699670 Mus sp. Species 0.000 description 26
- 206010042297 Subacute sclerosing panencephalitis Diseases 0.000 description 26
- 230000014509 gene expression Effects 0.000 description 26
- 150000002632 lipids Chemical class 0.000 description 26
- 229940125777 fusion inhibitor Drugs 0.000 description 24
- 230000000694 effects Effects 0.000 description 23
- 235000001014 amino acid Nutrition 0.000 description 22
- 108010059722 Viral Fusion Proteins Proteins 0.000 description 21
- 150000001875 compounds Chemical class 0.000 description 21
- 101000633780 Homo sapiens Signaling lymphocytic activation molecule Proteins 0.000 description 20
- 241000699666 Mus <mouse, genus> Species 0.000 description 20
- 239000000562 conjugate Substances 0.000 description 20
- 238000001727 in vivo Methods 0.000 description 20
- 239000002609 medium Substances 0.000 description 20
- 102100029215 Signaling lymphocytic activation molecule Human genes 0.000 description 19
- 238000004458 analytical method Methods 0.000 description 19
- 150000001413 amino acids Chemical class 0.000 description 18
- 108020001507 fusion proteins Proteins 0.000 description 18
- 210000001519 tissue Anatomy 0.000 description 18
- 238000003556 assay Methods 0.000 description 17
- 102000037865 fusion proteins Human genes 0.000 description 17
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 16
- 230000008685 targeting Effects 0.000 description 16
- 241000725303 Human immunodeficiency virus Species 0.000 description 15
- 241000713772 Human immunodeficiency virus 1 Species 0.000 description 15
- 239000003795 chemical substances by application Substances 0.000 description 15
- 238000009472 formulation Methods 0.000 description 15
- 150000003573 thiols Chemical class 0.000 description 15
- 229960005486 vaccine Drugs 0.000 description 15
- 230000005764 inhibitory process Effects 0.000 description 14
- -1 linker compound Chemical class 0.000 description 14
- 239000000243 solution Substances 0.000 description 14
- 108700028369 Alleles Proteins 0.000 description 13
- 102100026189 Beta-galactosidase Human genes 0.000 description 13
- 108010074687 Signaling Lymphocytic Activation Molecule Family Member 1 Proteins 0.000 description 13
- 102000008115 Signaling Lymphocytic Activation Molecule Family Member 1 Human genes 0.000 description 13
- 108010005774 beta-Galactosidase Proteins 0.000 description 13
- 238000003559 RNA-seq method Methods 0.000 description 12
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 12
- 206010014599 encephalitis Diseases 0.000 description 12
- 230000002829 reductive effect Effects 0.000 description 12
- 239000007921 spray Substances 0.000 description 12
- 125000000101 thioether group Chemical group 0.000 description 12
- 230000014599 transmission of virus Effects 0.000 description 12
- 230000009385 viral infection Effects 0.000 description 12
- PEDCQBHIVMGVHV-UHFFFAOYSA-N Glycerine Chemical compound OCC(O)CO PEDCQBHIVMGVHV-UHFFFAOYSA-N 0.000 description 11
- 230000005540 biological transmission Effects 0.000 description 11
- 238000004113 cell culture Methods 0.000 description 11
- 230000008859 change Effects 0.000 description 11
- 239000003814 drug Substances 0.000 description 11
- 125000000524 functional group Chemical group 0.000 description 11
- 230000003993 interaction Effects 0.000 description 11
- 238000001890 transfection Methods 0.000 description 11
- 230000004913 activation Effects 0.000 description 10
- 239000004480 active ingredient Substances 0.000 description 10
- 125000000151 cysteine group Chemical group N[C@@H](CS)C(=O)* 0.000 description 10
- 239000003937 drug carrier Substances 0.000 description 10
- 125000005439 maleimidyl group Chemical group C1(C=CC(N1*)=O)=O 0.000 description 10
- 230000007246 mechanism Effects 0.000 description 10
- 239000013642 negative control Substances 0.000 description 10
- 210000001778 pluripotent stem cell Anatomy 0.000 description 10
- 239000013641 positive control Substances 0.000 description 10
- 239000003981 vehicle Substances 0.000 description 10
- 230000007502 viral entry Effects 0.000 description 10
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 10
- LFQSCWFLJHTTHZ-UHFFFAOYSA-N Ethanol Chemical compound CCO LFQSCWFLJHTTHZ-UHFFFAOYSA-N 0.000 description 9
- 239000000443 aerosol Substances 0.000 description 9
- 238000013459 approach Methods 0.000 description 9
- 230000000975 bioactive effect Effects 0.000 description 9
- 230000008878 coupling Effects 0.000 description 9
- 238000010168 coupling process Methods 0.000 description 9
- 238000005859 coupling reaction Methods 0.000 description 9
- 201000010099 disease Diseases 0.000 description 9
- 238000005516 engineering process Methods 0.000 description 9
- 108010048367 enhanced green fluorescent protein Proteins 0.000 description 9
- 230000012010 growth Effects 0.000 description 9
- 238000004519 manufacturing process Methods 0.000 description 9
- 239000000463 material Substances 0.000 description 9
- 150000007523 nucleic acids Chemical class 0.000 description 9
- 210000002345 respiratory system Anatomy 0.000 description 9
- 241001465754 Metazoa Species 0.000 description 8
- 241001529936 Murinae Species 0.000 description 8
- 210000003743 erythrocyte Anatomy 0.000 description 8
- 239000007788 liquid Substances 0.000 description 8
- 239000002773 nucleotide Substances 0.000 description 8
- 125000003729 nucleotide group Chemical group 0.000 description 8
- 238000012163 sequencing technique Methods 0.000 description 8
- 210000002966 serum Anatomy 0.000 description 8
- 108050001175 Connexin Proteins 0.000 description 7
- 102000010970 Connexin Human genes 0.000 description 7
- 108020004414 DNA Proteins 0.000 description 7
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Chemical compound OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 7
- 101000852870 Homo sapiens Interferon alpha/beta receptor 1 Proteins 0.000 description 7
- 102100036714 Interferon alpha/beta receptor 1 Human genes 0.000 description 7
- 241000282560 Macaca mulatta Species 0.000 description 7
- 238000011529 RT qPCR Methods 0.000 description 7
- 241000144282 Sigmodon Species 0.000 description 7
- 230000027455 binding Effects 0.000 description 7
- 210000005013 brain tissue Anatomy 0.000 description 7
- 229940079593 drug Drugs 0.000 description 7
- 239000010408 film Substances 0.000 description 7
- 230000001965 increasing effect Effects 0.000 description 7
- 108020004707 nucleic acids Proteins 0.000 description 7
- 102000039446 nucleic acids Human genes 0.000 description 7
- 239000013612 plasmid Substances 0.000 description 7
- 230000002064 post-exposure prophylaxis Effects 0.000 description 7
- 239000000047 product Substances 0.000 description 7
- 238000003762 quantitative reverse transcription PCR Methods 0.000 description 7
- 239000006228 supernatant Substances 0.000 description 7
- 239000004094 surface-active agent Substances 0.000 description 7
- 108090000626 DNA-directed RNA polymerases Proteins 0.000 description 6
- 102000004163 DNA-directed RNA polymerases Human genes 0.000 description 6
- 239000006144 Dulbecco’s modified Eagle's medium Substances 0.000 description 6
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 6
- 241000712003 Human respirovirus 3 Species 0.000 description 6
- 230000001154 acute effect Effects 0.000 description 6
- 239000002671 adjuvant Substances 0.000 description 6
- WQZGKKKJIJFFOK-VFUOTHLCSA-N beta-D-glucose Chemical compound OC[C@H]1O[C@@H](O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-VFUOTHLCSA-N 0.000 description 6
- 230000015572 biosynthetic process Effects 0.000 description 6
- FNXLCIKXHOPCKH-UHFFFAOYSA-N bromamine Chemical class BrN FNXLCIKXHOPCKH-UHFFFAOYSA-N 0.000 description 6
- 239000000969 carrier Substances 0.000 description 6
- 238000013461 design Methods 0.000 description 6
- 150000002148 esters Chemical group 0.000 description 6
- 230000002209 hydrophobic effect Effects 0.000 description 6
- 238000000338 in vitro Methods 0.000 description 6
- 238000002955 isolation Methods 0.000 description 6
- 231100000518 lethal Toxicity 0.000 description 6
- 230000001665 lethal effect Effects 0.000 description 6
- 239000002245 particle Substances 0.000 description 6
- 238000000746 purification Methods 0.000 description 6
- QFYXSLAAXZTRLG-UHFFFAOYSA-N pyrrolidine-2,3-dione Chemical group O=C1CCNC1=O QFYXSLAAXZTRLG-UHFFFAOYSA-N 0.000 description 6
- 239000003381 stabilizer Substances 0.000 description 6
- 230000000087 stabilizing effect Effects 0.000 description 6
- 230000004083 survival effect Effects 0.000 description 6
- 238000002560 therapeutic procedure Methods 0.000 description 6
- 102100034343 Integrase Human genes 0.000 description 5
- 238000000585 Mann–Whitney U test Methods 0.000 description 5
- 108010076504 Protein Sorting Signals Proteins 0.000 description 5
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 5
- 208000037065 Subacute sclerosing leukoencephalitis Diseases 0.000 description 5
- 230000006978 adaptation Effects 0.000 description 5
- 210000000170 cell membrane Anatomy 0.000 description 5
- 239000002299 complementary DNA Substances 0.000 description 5
- 239000004615 ingredient Substances 0.000 description 5
- 210000004072 lung Anatomy 0.000 description 5
- 230000001404 mediated effect Effects 0.000 description 5
- 210000002161 motor neuron Anatomy 0.000 description 5
- 230000001537 neural effect Effects 0.000 description 5
- 210000002569 neuron Anatomy 0.000 description 5
- 239000000546 pharmaceutical excipient Substances 0.000 description 5
- 229920000642 polymer Polymers 0.000 description 5
- 229920001184 polypeptide Polymers 0.000 description 5
- 238000002360 preparation method Methods 0.000 description 5
- 239000003755 preservative agent Substances 0.000 description 5
- 230000008569 process Effects 0.000 description 5
- 238000011321 prophylaxis Methods 0.000 description 5
- 108010054624 red fluorescent protein Proteins 0.000 description 5
- 230000002441 reversible effect Effects 0.000 description 5
- 230000011664 signaling Effects 0.000 description 5
- 239000007787 solid Substances 0.000 description 5
- 230000001225 therapeutic effect Effects 0.000 description 5
- 238000002255 vaccination Methods 0.000 description 5
- 239000013598 vector Substances 0.000 description 5
- 238000001262 western blot Methods 0.000 description 5
- GUBGYTABKSRVRQ-XLOQQCSPSA-N Alpha-Lactose Chemical compound O[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@H]1O[C@@H]1[C@@H](CO)O[C@H](O)[C@H](O)[C@H]1O GUBGYTABKSRVRQ-XLOQQCSPSA-N 0.000 description 4
- CURLTUGMZLYLDI-UHFFFAOYSA-N Carbon dioxide Chemical compound O=C=O CURLTUGMZLYLDI-UHFFFAOYSA-N 0.000 description 4
- 208000014912 Central Nervous System Infections Diseases 0.000 description 4
- 101150034814 F gene Proteins 0.000 description 4
- 241000282412 Homo Species 0.000 description 4
- GUBGYTABKSRVRQ-QKKXKWKRSA-N Lactose Natural products OC[C@H]1O[C@@H](O[C@H]2[C@H](O)[C@@H](O)C(O)O[C@@H]2CO)[C@H](O)[C@@H](O)[C@H]1O GUBGYTABKSRVRQ-QKKXKWKRSA-N 0.000 description 4
- 101710169105 Minor spike protein Proteins 0.000 description 4
- 101710081079 Minor spike protein H Proteins 0.000 description 4
- 241000714177 Murine leukemia virus Species 0.000 description 4
- 241001112090 Pseudovirus Species 0.000 description 4
- 229930006000 Sucrose Natural products 0.000 description 4
- CZMRCDWAGMRECN-UGDNZRGBSA-N Sucrose Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 CZMRCDWAGMRECN-UGDNZRGBSA-N 0.000 description 4
- 101800001690 Transmembrane protein gp41 Proteins 0.000 description 4
- 241000711975 Vesicular stomatitis virus Species 0.000 description 4
- 206010058874 Viraemia Diseases 0.000 description 4
- 208000036142 Viral infection Diseases 0.000 description 4
- 150000001408 amides Chemical group 0.000 description 4
- 239000000872 buffer Substances 0.000 description 4
- 230000001413 cellular effect Effects 0.000 description 4
- 230000002490 cerebral effect Effects 0.000 description 4
- 238000012512 characterization method Methods 0.000 description 4
- 238000006243 chemical reaction Methods 0.000 description 4
- 235000018417 cysteine Nutrition 0.000 description 4
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 4
- 230000001419 dependent effect Effects 0.000 description 4
- 239000012636 effector Substances 0.000 description 4
- 239000000839 emulsion Substances 0.000 description 4
- 238000010195 expression analysis Methods 0.000 description 4
- 239000013604 expression vector Substances 0.000 description 4
- 239000012091 fetal bovine serum Substances 0.000 description 4
- 230000002068 genetic effect Effects 0.000 description 4
- 239000008103 glucose Substances 0.000 description 4
- 235000011187 glycerol Nutrition 0.000 description 4
- 238000003306 harvesting Methods 0.000 description 4
- 238000004128 high performance liquid chromatography Methods 0.000 description 4
- 230000002458 infectious effect Effects 0.000 description 4
- 230000015788 innate immune response Effects 0.000 description 4
- NOESYZHRGYRDHS-UHFFFAOYSA-N insulin Chemical compound N1C(=O)C(NC(=O)C(CCC(N)=O)NC(=O)C(CCC(O)=O)NC(=O)C(C(C)C)NC(=O)C(NC(=O)CN)C(C)CC)CSSCC(C(NC(CO)C(=O)NC(CC(C)C)C(=O)NC(CC=2C=CC(O)=CC=2)C(=O)NC(CCC(N)=O)C(=O)NC(CC(C)C)C(=O)NC(CCC(O)=O)C(=O)NC(CC(N)=O)C(=O)NC(CC=2C=CC(O)=CC=2)C(=O)NC(CSSCC(NC(=O)C(C(C)C)NC(=O)C(CC(C)C)NC(=O)C(CC=2C=CC(O)=CC=2)NC(=O)C(CC(C)C)NC(=O)C(C)NC(=O)C(CCC(O)=O)NC(=O)C(C(C)C)NC(=O)C(CC(C)C)NC(=O)C(CC=2NC=NC=2)NC(=O)C(CO)NC(=O)CNC2=O)C(=O)NCC(=O)NC(CCC(O)=O)C(=O)NC(CCCNC(N)=N)C(=O)NCC(=O)NC(CC=3C=CC=CC=3)C(=O)NC(CC=3C=CC=CC=3)C(=O)NC(CC=3C=CC(O)=CC=3)C(=O)NC(C(C)O)C(=O)N3C(CCC3)C(=O)NC(CCCCN)C(=O)NC(C)C(O)=O)C(=O)NC(CC(N)=O)C(O)=O)=O)NC(=O)C(C(C)CC)NC(=O)C(CO)NC(=O)C(C(C)O)NC(=O)C1CSSCC2NC(=O)C(CC(C)C)NC(=O)C(NC(=O)C(CCC(N)=O)NC(=O)C(CC(N)=O)NC(=O)C(NC(=O)C(N)CC=1C=CC=CC=1)C(C)C)CC1=CN=CN1 NOESYZHRGYRDHS-UHFFFAOYSA-N 0.000 description 4
- 230000010468 interferon response Effects 0.000 description 4
- 210000003734 kidney Anatomy 0.000 description 4
- 239000008101 lactose Substances 0.000 description 4
- 239000002502 liposome Substances 0.000 description 4
- 238000004020 luminiscence type Methods 0.000 description 4
- 210000004962 mammalian cell Anatomy 0.000 description 4
- 239000003921 oil Substances 0.000 description 4
- 230000003389 potentiating effect Effects 0.000 description 4
- 239000000843 powder Substances 0.000 description 4
- 238000012545 processing Methods 0.000 description 4
- 238000011002 quantification Methods 0.000 description 4
- 238000003259 recombinant expression Methods 0.000 description 4
- 230000007480 spreading Effects 0.000 description 4
- 238000003892 spreading Methods 0.000 description 4
- 239000005720 sucrose Substances 0.000 description 4
- FBPFZTCFMRRESA-FSIIMWSLSA-N D-Glucitol Natural products OC[C@H](O)[C@H](O)[C@@H](O)[C@H](O)CO FBPFZTCFMRRESA-FSIIMWSLSA-N 0.000 description 3
- 108010010803 Gelatin Proteins 0.000 description 3
- 108010043121 Green Fluorescent Proteins Proteins 0.000 description 3
- 102000004144 Green Fluorescent Proteins Human genes 0.000 description 3
- 101710154606 Hemagglutinin Proteins 0.000 description 3
- 206010061598 Immunodeficiency Diseases 0.000 description 3
- 108060003951 Immunoglobulin Proteins 0.000 description 3
- 102000002227 Interferon Type I Human genes 0.000 description 3
- 108010014726 Interferon Type I Proteins 0.000 description 3
- 102000014150 Interferons Human genes 0.000 description 3
- 108010050904 Interferons Proteins 0.000 description 3
- 101710093908 Outer capsid protein VP4 Proteins 0.000 description 3
- 101710135467 Outer capsid protein sigma-1 Proteins 0.000 description 3
- 229920003171 Poly (ethylene oxide) Polymers 0.000 description 3
- DNIAPMSPPWPWGF-UHFFFAOYSA-N Propylene glycol Chemical compound CC(O)CO DNIAPMSPPWPWGF-UHFFFAOYSA-N 0.000 description 3
- 101710176177 Protein A56 Proteins 0.000 description 3
- 241000725643 Respiratory syncytial virus Species 0.000 description 3
- 241000282695 Saimiri Species 0.000 description 3
- 241000144290 Sigmodon hispidus Species 0.000 description 3
- VYPSYNLAJGMNEJ-UHFFFAOYSA-N Silicium dioxide Chemical compound O=[Si]=O VYPSYNLAJGMNEJ-UHFFFAOYSA-N 0.000 description 3
- 101710172711 Structural protein Proteins 0.000 description 3
- 108020000999 Viral RNA Proteins 0.000 description 3
- 230000002776 aggregation Effects 0.000 description 3
- 238000004220 aggregation Methods 0.000 description 3
- 125000003277 amino group Chemical group 0.000 description 3
- 239000003443 antiviral agent Substances 0.000 description 3
- 230000002238 attenuated effect Effects 0.000 description 3
- 230000000903 blocking effect Effects 0.000 description 3
- 150000001720 carbohydrates Chemical class 0.000 description 3
- 235000014633 carbohydrates Nutrition 0.000 description 3
- 150000001768 cations Chemical class 0.000 description 3
- 210000001638 cerebellum Anatomy 0.000 description 3
- 239000003153 chemical reaction reagent Substances 0.000 description 3
- 230000000295 complement effect Effects 0.000 description 3
- 230000001268 conjugating effect Effects 0.000 description 3
- 230000001086 cytosolic effect Effects 0.000 description 3
- 231100000135 cytotoxicity Toxicity 0.000 description 3
- 230000003013 cytotoxicity Effects 0.000 description 3
- 230000007423 decrease Effects 0.000 description 3
- 230000003247 decreasing effect Effects 0.000 description 3
- 230000002950 deficient Effects 0.000 description 3
- 238000001514 detection method Methods 0.000 description 3
- 238000010586 diagram Methods 0.000 description 3
- 230000009274 differential gene expression Effects 0.000 description 3
- 238000002050 diffraction method Methods 0.000 description 3
- 239000003623 enhancer Substances 0.000 description 3
- 230000026502 entry into host cell Effects 0.000 description 3
- 238000001914 filtration Methods 0.000 description 3
- 239000000499 gel Substances 0.000 description 3
- 239000008273 gelatin Substances 0.000 description 3
- 229920000159 gelatin Polymers 0.000 description 3
- 235000019322 gelatine Nutrition 0.000 description 3
- 235000011852 gelatine desserts Nutrition 0.000 description 3
- 239000005090 green fluorescent protein Substances 0.000 description 3
- 230000036541 health Effects 0.000 description 3
- 210000002865 immune cell Anatomy 0.000 description 3
- 230000003053 immunization Effects 0.000 description 3
- 238000002649 immunization Methods 0.000 description 3
- 238000003119 immunoblot Methods 0.000 description 3
- 102000018358 immunoglobulin Human genes 0.000 description 3
- 238000011534 incubation Methods 0.000 description 3
- 239000007924 injection Substances 0.000 description 3
- 238000002347 injection Methods 0.000 description 3
- 238000003780 insertion Methods 0.000 description 3
- 229940079322 interferon Drugs 0.000 description 3
- 230000000670 limiting effect Effects 0.000 description 3
- 238000010234 longitudinal analysis Methods 0.000 description 3
- 210000004698 lymphocyte Anatomy 0.000 description 3
- 230000004048 modification Effects 0.000 description 3
- 238000012986 modification Methods 0.000 description 3
- 210000000056 organ Anatomy 0.000 description 3
- 238000004806 packaging method and process Methods 0.000 description 3
- 230000008506 pathogenesis Effects 0.000 description 3
- 230000006320 pegylation Effects 0.000 description 3
- 230000010076 replication Effects 0.000 description 3
- 230000004044 response Effects 0.000 description 3
- 150000003839 salts Chemical class 0.000 description 3
- 238000012216 screening Methods 0.000 description 3
- 239000011780 sodium chloride Substances 0.000 description 3
- RYYKJJJTJZKILX-UHFFFAOYSA-M sodium octadecanoate Chemical compound [Na+].CCCCCCCCCCCCCCCCCC([O-])=O RYYKJJJTJZKILX-UHFFFAOYSA-M 0.000 description 3
- 239000002904 solvent Substances 0.000 description 3
- 239000000600 sorbitol Substances 0.000 description 3
- 235000010356 sorbitol Nutrition 0.000 description 3
- 238000006467 substitution reaction Methods 0.000 description 3
- 239000000725 suspension Substances 0.000 description 3
- 230000002195 synergetic effect Effects 0.000 description 3
- 238000003786 synthesis reaction Methods 0.000 description 3
- 238000012360 testing method Methods 0.000 description 3
- ARAIBEBZBOPLMB-UFGQHTETSA-N zanamivir Chemical compound CC(=O)N[C@@H]1[C@@H](N=C(N)N)C=C(C(O)=O)O[C@H]1[C@H](O)[C@H](O)CO ARAIBEBZBOPLMB-UFGQHTETSA-N 0.000 description 3
- 229960001028 zanamivir Drugs 0.000 description 3
- YBJHBAHKTGYVGT-ZKWXMUAHSA-N (+)-Biotin Chemical compound N1C(=O)N[C@@H]2[C@H](CCCCC(=O)O)SC[C@@H]21 YBJHBAHKTGYVGT-ZKWXMUAHSA-N 0.000 description 2
- VZQHRKZCAZCACO-PYJNHQTQSA-N (2s)-2-[[(2s)-2-[2-[[(2s)-2-[[(2s)-2-amino-5-(diaminomethylideneamino)pentanoyl]amino]propanoyl]amino]prop-2-enoylamino]-3-methylbutanoyl]amino]propanoic acid Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](C(C)C)NC(=O)C(=C)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCCNC(N)=N VZQHRKZCAZCACO-PYJNHQTQSA-N 0.000 description 2
- FPIPGXGPPPQFEQ-UHFFFAOYSA-N 13-cis retinol Natural products OCC=C(C)C=CC=C(C)C=CC1=C(C)CCCC1(C)C FPIPGXGPPPQFEQ-UHFFFAOYSA-N 0.000 description 2
- DQSIXGDDUJJEQH-QRPNPIFTSA-N 2-aminoacetic acid;(2s)-2-amino-3-phenylpropanoic acid Chemical compound NCC(O)=O.OC(=O)[C@@H](N)CC1=CC=CC=C1 DQSIXGDDUJJEQH-QRPNPIFTSA-N 0.000 description 2
- FWBHETKCLVMNFS-UHFFFAOYSA-N 4',6-Diamino-2-phenylindol Chemical compound C1=CC(C(=N)N)=CC=C1C1=CC2=CC=C(C(N)=N)C=C2N1 FWBHETKCLVMNFS-UHFFFAOYSA-N 0.000 description 2
- 108010085238 Actins Proteins 0.000 description 2
- 102000007469 Actins Human genes 0.000 description 2
- HJCMDXDYPOUFDY-WHFBIAKZSA-N Ala-Gln Chemical compound C[C@H](N)C(=O)N[C@H](C(O)=O)CCC(N)=O HJCMDXDYPOUFDY-WHFBIAKZSA-N 0.000 description 2
- 239000012103 Alexa Fluor 488 Substances 0.000 description 2
- 239000012110 Alexa Fluor 594 Substances 0.000 description 2
- 101000975750 Allium sativum Mannose-specific lectin Proteins 0.000 description 2
- CIWBSHSKHKDKBQ-JLAZNSOCSA-N Ascorbic acid Chemical compound OC[C@H](O)[C@H]1OC(=O)C(O)=C1O CIWBSHSKHKDKBQ-JLAZNSOCSA-N 0.000 description 2
- 239000012583 B-27 Supplement Substances 0.000 description 2
- 102000000905 Cadherin Human genes 0.000 description 2
- 108050007957 Cadherin Proteins 0.000 description 2
- 241000282465 Canis Species 0.000 description 2
- 102000000844 Cell Surface Receptors Human genes 0.000 description 2
- 108010001857 Cell Surface Receptors Proteins 0.000 description 2
- 108020004705 Codon Proteins 0.000 description 2
- 108091035707 Consensus sequence Proteins 0.000 description 2
- FBPFZTCFMRRESA-KVTDHHQDSA-N D-Mannitol Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-KVTDHHQDSA-N 0.000 description 2
- FBPFZTCFMRRESA-JGWLITMVSA-N D-glucitol Chemical compound OC[C@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-JGWLITMVSA-N 0.000 description 2
- 102000016911 Deoxyribonucleases Human genes 0.000 description 2
- 108010053770 Deoxyribonucleases Proteins 0.000 description 2
- RTZKZFJDLAIYFH-UHFFFAOYSA-N Diethyl ether Chemical compound CCOCC RTZKZFJDLAIYFH-UHFFFAOYSA-N 0.000 description 2
- 206010014612 Encephalitis viral Diseases 0.000 description 2
- 102100038132 Endogenous retrovirus group K member 6 Pro protein Human genes 0.000 description 2
- 102000004190 Enzymes Human genes 0.000 description 2
- 108090000790 Enzymes Proteins 0.000 description 2
- 101001066129 Homo sapiens Glyceraldehyde-3-phosphate dehydrogenase Proteins 0.000 description 2
- 101000978730 Homo sapiens Nephrin Proteins 0.000 description 2
- 101000687905 Homo sapiens Transcription factor SOX-2 Proteins 0.000 description 2
- 206010061218 Inflammation Diseases 0.000 description 2
- 102000004877 Insulin Human genes 0.000 description 2
- 108090001061 Insulin Proteins 0.000 description 2
- 102100034349 Integrase Human genes 0.000 description 2
- 101710203526 Integrase Proteins 0.000 description 2
- ONIBWKKTOPOVIA-BYPYZUCNSA-N L-Proline Chemical compound OC(=O)[C@@H]1CCCN1 ONIBWKKTOPOVIA-BYPYZUCNSA-N 0.000 description 2
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 2
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 2
- 102000003960 Ligases Human genes 0.000 description 2
- 108090000364 Ligases Proteins 0.000 description 2
- 108060001084 Luciferase Proteins 0.000 description 2
- 239000005089 Luciferase Substances 0.000 description 2
- 231100000002 MTT assay Toxicity 0.000 description 2
- 238000000134 MTT assay Methods 0.000 description 2
- 241000282567 Macaca fascicularis Species 0.000 description 2
- 241000124008 Mammalia Species 0.000 description 2
- 229930195725 Mannitol Natural products 0.000 description 2
- 102000018697 Membrane Proteins Human genes 0.000 description 2
- 108010052285 Membrane Proteins Proteins 0.000 description 2
- 208000005647 Mumps Diseases 0.000 description 2
- 239000012580 N-2 Supplement Substances 0.000 description 2
- 102100035486 Nectin-4 Human genes 0.000 description 2
- 240000002853 Nelumbo nucifera Species 0.000 description 2
- 235000006508 Nelumbo nucifera Nutrition 0.000 description 2
- 102100023195 Nephrin Human genes 0.000 description 2
- 108091028043 Nucleic acid sequence Proteins 0.000 description 2
- 108090001074 Nucleocapsid Proteins Proteins 0.000 description 2
- 229930040373 Paraformaldehyde Natural products 0.000 description 2
- 235000019483 Peanut oil Nutrition 0.000 description 2
- 108091005804 Peptidases Proteins 0.000 description 2
- 241000144300 Peromyscus gossypinus Species 0.000 description 2
- 102000007982 Phosphoproteins Human genes 0.000 description 2
- 108010089430 Phosphoproteins Proteins 0.000 description 2
- ONIBWKKTOPOVIA-UHFFFAOYSA-N Proline Natural products OC(=O)C1CCCN1 ONIBWKKTOPOVIA-UHFFFAOYSA-N 0.000 description 2
- 239000004365 Protease Substances 0.000 description 2
- 101710149136 Protein Vpr Proteins 0.000 description 2
- 206010037660 Pyrexia Diseases 0.000 description 2
- 108010092799 RNA-directed DNA polymerase Proteins 0.000 description 2
- 101710146873 Receptor-binding protein Proteins 0.000 description 2
- 108700008625 Reporter Genes Proteins 0.000 description 2
- 108010044012 STAT1 Transcription Factor Proteins 0.000 description 2
- 229920002684 Sepharose Polymers 0.000 description 2
- 238000012300 Sequence Analysis Methods 0.000 description 2
- 102100029904 Signal transducer and activator of transcription 1-alpha/beta Human genes 0.000 description 2
- PXIPVTKHYLBLMZ-UHFFFAOYSA-N Sodium azide Chemical compound [Na+].[N-]=[N+]=[N-] PXIPVTKHYLBLMZ-UHFFFAOYSA-N 0.000 description 2
- 229920002472 Starch Polymers 0.000 description 2
- 108010076818 TEV protease Proteins 0.000 description 2
- 102100024270 Transcription factor SOX-2 Human genes 0.000 description 2
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 2
- FPIPGXGPPPQFEQ-BOOMUCAASA-N Vitamin A Natural products OC/C=C(/C)\C=C\C=C(\C)/C=C/C1=C(C)CCCC1(C)C FPIPGXGPPPQFEQ-BOOMUCAASA-N 0.000 description 2
- ATBOMIWRCZXYSZ-XZBBILGWSA-N [1-[2,3-dihydroxypropoxy(hydroxy)phosphoryl]oxy-3-hexadecanoyloxypropan-2-yl] (9e,12e)-octadeca-9,12-dienoate Chemical compound CCCCCCCCCCCCCCCC(=O)OCC(COP(O)(=O)OCC(O)CO)OC(=O)CCCCCCC\C=C\C\C=C\CCCCC ATBOMIWRCZXYSZ-XZBBILGWSA-N 0.000 description 2
- 238000002679 ablation Methods 0.000 description 2
- 230000009471 action Effects 0.000 description 2
- 230000003044 adaptive effect Effects 0.000 description 2
- 210000001552 airway epithelial cell Anatomy 0.000 description 2
- FPIPGXGPPPQFEQ-OVSJKPMPSA-N all-trans-retinol Chemical compound OC\C=C(/C)\C=C\C=C(/C)\C=C\C1=C(C)CCCC1(C)C FPIPGXGPPPQFEQ-OVSJKPMPSA-N 0.000 description 2
- AWUCVROLDVIAJX-UHFFFAOYSA-N alpha-glycerophosphate Natural products OCC(O)COP(O)(O)=O AWUCVROLDVIAJX-UHFFFAOYSA-N 0.000 description 2
- 239000003242 anti bacterial agent Substances 0.000 description 2
- 229940088710 antibiotic agent Drugs 0.000 description 2
- 239000000427 antigen Substances 0.000 description 2
- 108091007433 antigens Proteins 0.000 description 2
- 102000036639 antigens Human genes 0.000 description 2
- 239000003963 antioxidant agent Substances 0.000 description 2
- 235000006708 antioxidants Nutrition 0.000 description 2
- 239000007864 aqueous solution Substances 0.000 description 2
- 125000004429 atom Chemical group 0.000 description 2
- 230000004888 barrier function Effects 0.000 description 2
- 210000000270 basal cell Anatomy 0.000 description 2
- 230000008901 benefit Effects 0.000 description 2
- 230000001588 bifunctional effect Effects 0.000 description 2
- 229960000074 biopharmaceutical Drugs 0.000 description 2
- 235000019282 butylated hydroxyanisole Nutrition 0.000 description 2
- 229910052799 carbon Inorganic materials 0.000 description 2
- 239000001569 carbon dioxide Substances 0.000 description 2
- 229910002092 carbon dioxide Inorganic materials 0.000 description 2
- 239000003729 cation exchange resin Substances 0.000 description 2
- 239000013553 cell monolayer Substances 0.000 description 2
- 238000004587 chromatography analysis Methods 0.000 description 2
- 238000002648 combination therapy Methods 0.000 description 2
- 238000004624 confocal microscopy Methods 0.000 description 2
- 230000021615 conjugation Effects 0.000 description 2
- 239000013078 crystal Substances 0.000 description 2
- 210000004292 cytoskeleton Anatomy 0.000 description 2
- 238000002784 cytotoxicity assay Methods 0.000 description 2
- 231100000263 cytotoxicity test Toxicity 0.000 description 2
- 210000004443 dendritic cell Anatomy 0.000 description 2
- 230000002074 deregulated effect Effects 0.000 description 2
- 230000001627 detrimental effect Effects 0.000 description 2
- 235000014113 dietary fatty acids Nutrition 0.000 description 2
- 230000004069 differentiation Effects 0.000 description 2
- 238000010790 dilution Methods 0.000 description 2
- 239000012895 dilution Substances 0.000 description 2
- 208000035475 disorder Diseases 0.000 description 2
- 239000006185 dispersion Substances 0.000 description 2
- 210000002889 endothelial cell Anatomy 0.000 description 2
- 238000001317 epifluorescence microscopy Methods 0.000 description 2
- 210000002919 epithelial cell Anatomy 0.000 description 2
- MMXKVMNBHPAILY-UHFFFAOYSA-N ethyl laurate Chemical compound CCCCCCCCCCCC(=O)OCC MMXKVMNBHPAILY-UHFFFAOYSA-N 0.000 description 2
- 239000000194 fatty acid Substances 0.000 description 2
- 229930195729 fatty acid Natural products 0.000 description 2
- 230000001605 fetal effect Effects 0.000 description 2
- 108091006047 fluorescent proteins Proteins 0.000 description 2
- 102000034287 fluorescent proteins Human genes 0.000 description 2
- 239000011888 foil Substances 0.000 description 2
- 230000000799 fusogenic effect Effects 0.000 description 2
- 102000034356 gene-regulatory proteins Human genes 0.000 description 2
- 108091006104 gene-regulatory proteins Proteins 0.000 description 2
- BRZYSWJRSDMWLG-CAXSIQPQSA-N geneticin Chemical compound O1C[C@@](O)(C)[C@H](NC)[C@@H](O)[C@H]1O[C@@H]1[C@@H](O)[C@H](O[C@@H]2[C@@H]([C@@H](O)[C@H](O)[C@@H](C(C)O)O2)N)[C@@H](N)C[C@H]1N BRZYSWJRSDMWLG-CAXSIQPQSA-N 0.000 description 2
- 238000012268 genome sequencing Methods 0.000 description 2
- 230000005182 global health Effects 0.000 description 2
- 230000001434 glomerular Effects 0.000 description 2
- 230000013595 glycosylation Effects 0.000 description 2
- 238000006206 glycosylation reaction Methods 0.000 description 2
- 239000008187 granular material Substances 0.000 description 2
- 239000000185 hemagglutinin Substances 0.000 description 2
- 229920001519 homopolymer Polymers 0.000 description 2
- 102000047486 human GAPDH Human genes 0.000 description 2
- 238000010166 immunofluorescence Methods 0.000 description 2
- 229940072221 immunoglobulins Drugs 0.000 description 2
- 239000002955 immunomodulating agent Substances 0.000 description 2
- 230000002584 immunomodulator Effects 0.000 description 2
- 229940121354 immunomodulator Drugs 0.000 description 2
- 238000002513 implantation Methods 0.000 description 2
- 230000001976 improved effect Effects 0.000 description 2
- 230000006872 improvement Effects 0.000 description 2
- 238000010348 incorporation Methods 0.000 description 2
- 230000006698 induction Effects 0.000 description 2
- 239000012678 infectious agent Substances 0.000 description 2
- 230000004054 inflammatory process Effects 0.000 description 2
- 229940125396 insulin Drugs 0.000 description 2
- 230000010354 integration Effects 0.000 description 2
- 210000003963 intermediate filament Anatomy 0.000 description 2
- 230000003834 intracellular effect Effects 0.000 description 2
- 230000009545 invasion Effects 0.000 description 2
- 239000008297 liquid dosage form Substances 0.000 description 2
- 239000006193 liquid solution Substances 0.000 description 2
- 210000001165 lymph node Anatomy 0.000 description 2
- 239000008176 lyophilized powder Substances 0.000 description 2
- 238000007726 management method Methods 0.000 description 2
- 239000000594 mannitol Substances 0.000 description 2
- 235000010355 mannitol Nutrition 0.000 description 2
- 239000003550 marker Substances 0.000 description 2
- 108010082117 matrigel Proteins 0.000 description 2
- 229910044991 metal oxide Inorganic materials 0.000 description 2
- 150000004706 metal oxides Chemical class 0.000 description 2
- 230000009456 molecular mechanism Effects 0.000 description 2
- 238000010172 mouse model Methods 0.000 description 2
- 208000010805 mumps infectious disease Diseases 0.000 description 2
- 210000000885 nephron Anatomy 0.000 description 2
- 230000002276 neurotropic effect Effects 0.000 description 2
- 231100000252 nontoxic Toxicity 0.000 description 2
- 230000003000 nontoxic effect Effects 0.000 description 2
- 210000004940 nucleus Anatomy 0.000 description 2
- 235000019198 oils Nutrition 0.000 description 2
- 239000002674 ointment Substances 0.000 description 2
- 229920002866 paraformaldehyde Polymers 0.000 description 2
- 244000052769 pathogen Species 0.000 description 2
- 230000001717 pathogenic effect Effects 0.000 description 2
- 230000037361 pathway Effects 0.000 description 2
- 238000003068 pathway analysis Methods 0.000 description 2
- 239000000312 peanut oil Substances 0.000 description 2
- 230000002688 persistence Effects 0.000 description 2
- 230000000144 pharmacologic effect Effects 0.000 description 2
- YBYRMVIVWMBXKQ-UHFFFAOYSA-N phenylmethanesulfonyl fluoride Chemical compound FS(=O)(=O)CC1=CC=CC=C1 YBYRMVIVWMBXKQ-UHFFFAOYSA-N 0.000 description 2
- 239000004033 plastic Substances 0.000 description 2
- 229920003023 plastic Polymers 0.000 description 2
- 210000000557 podocyte Anatomy 0.000 description 2
- 238000011533 pre-incubation Methods 0.000 description 2
- 238000001556 precipitation Methods 0.000 description 2
- 238000004321 preservation Methods 0.000 description 2
- 230000035755 proliferation Effects 0.000 description 2
- QELSKZZBTMNZEB-UHFFFAOYSA-N propylparaben Chemical compound CCCOC(=O)C1=CC=C(O)C=C1 QELSKZZBTMNZEB-UHFFFAOYSA-N 0.000 description 2
- 235000019419 proteases Nutrition 0.000 description 2
- 230000001681 protective effect Effects 0.000 description 2
- 210000000512 proximal kidney tubule Anatomy 0.000 description 2
- 238000011552 rat model Methods 0.000 description 2
- 230000008707 rearrangement Effects 0.000 description 2
- 230000009467 reduction Effects 0.000 description 2
- 230000001105 regulatory effect Effects 0.000 description 2
- 230000000241 respiratory effect Effects 0.000 description 2
- 238000012552 review Methods 0.000 description 2
- 238000001338 self-assembly Methods 0.000 description 2
- 239000008159 sesame oil Substances 0.000 description 2
- 235000011803 sesame oil Nutrition 0.000 description 2
- 108010061514 sialic acid receptor Proteins 0.000 description 2
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 2
- 239000003549 soybean oil Substances 0.000 description 2
- 235000012424 soybean oil Nutrition 0.000 description 2
- 235000019698 starch Nutrition 0.000 description 2
- 238000007619 statistical method Methods 0.000 description 2
- 239000000126 substance Substances 0.000 description 2
- 239000013589 supplement Substances 0.000 description 2
- 239000000829 suppository Substances 0.000 description 2
- 239000006188 syrup Substances 0.000 description 2
- 235000020357 syrup Nutrition 0.000 description 2
- 239000000454 talc Substances 0.000 description 2
- 229910052623 talc Inorganic materials 0.000 description 2
- 238000004448 titration Methods 0.000 description 2
- 231100000331 toxic Toxicity 0.000 description 2
- 230000002588 toxic effect Effects 0.000 description 2
- 238000003151 transfection method Methods 0.000 description 2
- 238000012546 transfer Methods 0.000 description 2
- 230000009466 transformation Effects 0.000 description 2
- 102000035160 transmembrane proteins Human genes 0.000 description 2
- 108091005703 transmembrane proteins Proteins 0.000 description 2
- 125000000430 tryptophan group Chemical group [H]N([H])C(C(=O)O*)C([H])([H])C1=C([H])N([H])C2=C([H])C([H])=C([H])C([H])=C12 0.000 description 2
- 210000005239 tubule Anatomy 0.000 description 2
- 210000003501 vero cell Anatomy 0.000 description 2
- 201000002498 viral encephalitis Diseases 0.000 description 2
- 230000029812 viral genome replication Effects 0.000 description 2
- 238000012800 visualization Methods 0.000 description 2
- 235000019155 vitamin A Nutrition 0.000 description 2
- 239000011719 vitamin A Substances 0.000 description 2
- 229940045997 vitamin a Drugs 0.000 description 2
- DGVVWUTYPXICAM-UHFFFAOYSA-N β‐Mercaptoethanol Chemical compound OCCS DGVVWUTYPXICAM-UHFFFAOYSA-N 0.000 description 2
- HDTRYLNUVZCQOY-UHFFFAOYSA-N α-D-glucopyranosyl-α-D-glucopyranoside Natural products OC1C(O)C(O)C(CO)OC1OC1C(O)C(O)C(O)C(CO)O1 HDTRYLNUVZCQOY-UHFFFAOYSA-N 0.000 description 1
- YYGNTYWPHWGJRM-UHFFFAOYSA-N (6E,10E,14E,18E)-2,6,10,15,19,23-hexamethyltetracosa-2,6,10,14,18,22-hexaene Chemical compound CC(C)=CCCC(C)=CCCC(C)=CCCC=C(C)CCC=C(C)CCC=C(C)C YYGNTYWPHWGJRM-UHFFFAOYSA-N 0.000 description 1
- WRIDQFICGBMAFQ-UHFFFAOYSA-N (E)-8-Octadecenoic acid Natural products CCCCCCCCCC=CCCCCCCC(O)=O WRIDQFICGBMAFQ-UHFFFAOYSA-N 0.000 description 1
- NWUYHJFMYQTDRP-UHFFFAOYSA-N 1,2-bis(ethenyl)benzene;1-ethenyl-2-ethylbenzene;styrene Chemical compound C=CC1=CC=CC=C1.CCC1=CC=CC=C1C=C.C=CC1=CC=CC=C1C=C NWUYHJFMYQTDRP-UHFFFAOYSA-N 0.000 description 1
- PZNPLUBHRSSFHT-RRHRGVEJSA-N 1-hexadecanoyl-2-octadecanoyl-sn-glycero-3-phosphocholine Chemical compound CCCCCCCCCCCCCCCCCC(=O)O[C@@H](COP([O-])(=O)OCC[N+](C)(C)C)COC(=O)CCCCCCCCCCCCCCC PZNPLUBHRSSFHT-RRHRGVEJSA-N 0.000 description 1
- JKMHFZQWWAIEOD-UHFFFAOYSA-N 2-[4-(2-hydroxyethyl)piperazin-1-yl]ethanesulfonic acid Chemical compound OCC[NH+]1CCN(CCS([O-])(=O)=O)CC1 JKMHFZQWWAIEOD-UHFFFAOYSA-N 0.000 description 1
- BFSVOASYOCHEOV-UHFFFAOYSA-N 2-diethylaminoethanol Chemical compound CCN(CC)CCO BFSVOASYOCHEOV-UHFFFAOYSA-N 0.000 description 1
- LQJBNNIYVWPHFW-UHFFFAOYSA-N 20:1omega9c fatty acid Natural products CCCCCCCCCCC=CCCCCCCCC(O)=O LQJBNNIYVWPHFW-UHFFFAOYSA-N 0.000 description 1
- XMTQQYYKAHVGBJ-UHFFFAOYSA-N 3-(3,4-DICHLOROPHENYL)-1,1-DIMETHYLUREA Chemical compound CN(C)C(=O)NC1=CC=C(Cl)C(Cl)=C1 XMTQQYYKAHVGBJ-UHFFFAOYSA-N 0.000 description 1
- YCAFMCMKNHYQFE-UHFFFAOYSA-N 4-nitro-2-[(2-phenylacetyl)amino]benzamide Chemical compound NC(=O)C1=CC=C([N+]([O-])=O)C=C1NC(=O)CC1=CC=CC=C1 YCAFMCMKNHYQFE-UHFFFAOYSA-N 0.000 description 1
- QSBYPNXLFMSGKH-UHFFFAOYSA-N 9-Heptadecensaeure Natural products CCCCCCCC=CCCCCCCCC(O)=O QSBYPNXLFMSGKH-UHFFFAOYSA-N 0.000 description 1
- 208000024893 Acute lymphoblastic leukemia Diseases 0.000 description 1
- 208000014697 Acute lymphocytic leukaemia Diseases 0.000 description 1
- 229920001817 Agar Polymers 0.000 description 1
- 229920000936 Agarose Polymers 0.000 description 1
- 102100027211 Albumin Human genes 0.000 description 1
- 108010088751 Albumins Proteins 0.000 description 1
- 102000002260 Alkaline Phosphatase Human genes 0.000 description 1
- 108020004774 Alkaline Phosphatase Proteins 0.000 description 1
- 241000416162 Astragalus gummifer Species 0.000 description 1
- 108091003079 Bovine Serum Albumin Proteins 0.000 description 1
- 239000004255 Butylated hydroxyanisole Substances 0.000 description 1
- 238000011740 C57BL/6 mouse Methods 0.000 description 1
- 241001678559 COVID-19 virus Species 0.000 description 1
- 108090000565 Capsid Proteins Proteins 0.000 description 1
- OKTJSMMVPCPJKN-UHFFFAOYSA-N Carbon Chemical compound [C] OKTJSMMVPCPJKN-UHFFFAOYSA-N 0.000 description 1
- 102000014914 Carrier Proteins Human genes 0.000 description 1
- 241000282693 Cercopithecidae Species 0.000 description 1
- 102100023321 Ceruloplasmin Human genes 0.000 description 1
- 241000282552 Chlorocebus aethiops Species 0.000 description 1
- 235000005979 Citrus limon Nutrition 0.000 description 1
- 244000131522 Citrus pyriformis Species 0.000 description 1
- 108091026890 Coding region Proteins 0.000 description 1
- 208000003322 Coinfection Diseases 0.000 description 1
- 208000035473 Communicable disease Diseases 0.000 description 1
- 229920002261 Corn starch Polymers 0.000 description 1
- 229920000742 Cotton Polymers 0.000 description 1
- 241000699800 Cricetinae Species 0.000 description 1
- 230000004543 DNA replication Effects 0.000 description 1
- 102000007260 Deoxyribonuclease I Human genes 0.000 description 1
- 108010008532 Deoxyribonuclease I Proteins 0.000 description 1
- LVGKNOAMLMIIKO-UHFFFAOYSA-N Elaidinsaeure-aethylester Natural products CCCCCCCCC=CCCCCCCCC(=O)OCC LVGKNOAMLMIIKO-UHFFFAOYSA-N 0.000 description 1
- 241000196324 Embryophyta Species 0.000 description 1
- 108010067770 Endopeptidase K Proteins 0.000 description 1
- 101710121417 Envelope glycoprotein Proteins 0.000 description 1
- 101710091045 Envelope protein Proteins 0.000 description 1
- 239000001856 Ethyl cellulose Substances 0.000 description 1
- ZZSNKZQZMQGXPY-UHFFFAOYSA-N Ethyl cellulose Chemical compound CCOCC1OC(OC)C(OCC)C(OCC)C1OC1C(O)C(O)C(OC)C(CO)O1 ZZSNKZQZMQGXPY-UHFFFAOYSA-N 0.000 description 1
- 244000187656 Eucalyptus cornuta Species 0.000 description 1
- 241000206602 Eukaryota Species 0.000 description 1
- 241000190598 Flexal mammarenavirus Species 0.000 description 1
- 101710160621 Fusion glycoprotein F0 Proteins 0.000 description 1
- 108700039691 Genetic Promoter Regions Proteins 0.000 description 1
- 108060003393 Granulin Proteins 0.000 description 1
- 239000007995 HEPES buffer Substances 0.000 description 1
- HTTJABKRGRZYRN-UHFFFAOYSA-N Heparin Chemical compound OC1C(NC(=O)C)C(O)OC(COS(O)(=O)=O)C1OC1C(OS(O)(=O)=O)C(O)C(OC2C(C(OS(O)(=O)=O)C(OC3C(C(O)C(O)C(O3)C(O)=O)OS(O)(=O)=O)C(CO)O2)NS(O)(=O)=O)C(C(O)=O)O1 HTTJABKRGRZYRN-UHFFFAOYSA-N 0.000 description 1
- 241000238631 Hexapoda Species 0.000 description 1
- 101000831616 Homo sapiens Protachykinin-1 Proteins 0.000 description 1
- 241000713340 Human immunodeficiency virus 2 Species 0.000 description 1
- 108010061833 Integrases Proteins 0.000 description 1
- 241000007938 Juquitiba virus Species 0.000 description 1
- XUJNEKJLAYXESH-REOHCLBHSA-N L-Cysteine Chemical compound SC[C@H](N)C(O)=O XUJNEKJLAYXESH-REOHCLBHSA-N 0.000 description 1
- 150000008575 L-amino acids Chemical class 0.000 description 1
- 241000713666 Lentivirus Species 0.000 description 1
- 240000007472 Leucaena leucocephala Species 0.000 description 1
- 235000010643 Leucaena leucocephala Nutrition 0.000 description 1
- 206010058467 Lung neoplasm malignant Diseases 0.000 description 1
- 235000007688 Lycopersicon esculentum Nutrition 0.000 description 1
- PEEHTFAAVSWFBL-UHFFFAOYSA-N Maleimide Chemical compound O=C1NC(=O)C=C1 PEEHTFAAVSWFBL-UHFFFAOYSA-N 0.000 description 1
- 241000712045 Morbillivirus Species 0.000 description 1
- GXCLVBGFBYZDAG-UHFFFAOYSA-N N-[2-(1H-indol-3-yl)ethyl]-N-methylprop-2-en-1-amine Chemical compound CN(CCC1=CNC2=C1C=CC=C2)CC=C GXCLVBGFBYZDAG-UHFFFAOYSA-N 0.000 description 1
- 101710043865 Nectin-4 Proteins 0.000 description 1
- 206010029350 Neurotoxicity Diseases 0.000 description 1
- 241000526636 Nipah henipavirus Species 0.000 description 1
- 108091092724 Noncoding DNA Proteins 0.000 description 1
- 102000011931 Nucleoproteins Human genes 0.000 description 1
- 108010061100 Nucleoproteins Proteins 0.000 description 1
- 239000005642 Oleic acid Substances 0.000 description 1
- ZQPPMHVWECSIRJ-UHFFFAOYSA-N Oleic acid Natural products CCCCCCCCC=CCCCCCCCC(O)=O ZQPPMHVWECSIRJ-UHFFFAOYSA-N 0.000 description 1
- 240000007594 Oryza sativa Species 0.000 description 1
- 235000007164 Oryza sativa Nutrition 0.000 description 1
- 101150084044 P gene Proteins 0.000 description 1
- 239000002033 PVDF binder Substances 0.000 description 1
- 108091059809 PVRL4 Proteins 0.000 description 1
- 241001111421 Pannus Species 0.000 description 1
- 241001579678 Panthea coenobita Species 0.000 description 1
- 208000037581 Persistent Infection Diseases 0.000 description 1
- 229940123066 Polymerase inhibitor Drugs 0.000 description 1
- 229920001213 Polysorbate 20 Polymers 0.000 description 1
- 208000006664 Precursor Cell Lymphoblastic Leukemia-Lymphoma Diseases 0.000 description 1
- 102100024304 Protachykinin-1 Human genes 0.000 description 1
- 108010001267 Protein Subunits Proteins 0.000 description 1
- 102000002067 Protein Subunits Human genes 0.000 description 1
- 101710188315 Protein X Proteins 0.000 description 1
- 229940122277 RNA polymerase inhibitor Drugs 0.000 description 1
- 238000013381 RNA quantification Methods 0.000 description 1
- 230000010799 Receptor Interactions Effects 0.000 description 1
- 206010057190 Respiratory tract infections Diseases 0.000 description 1
- 241000712907 Retroviridae Species 0.000 description 1
- 241000220010 Rhode Species 0.000 description 1
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 1
- 235000019485 Safflower oil Nutrition 0.000 description 1
- 229920005654 Sephadex Polymers 0.000 description 1
- 239000012507 Sephadex™ Substances 0.000 description 1
- 240000003768 Solanum lycopersicum Species 0.000 description 1
- 108010090804 Streptavidin Proteins 0.000 description 1
- UCKMPCXJQFINFW-UHFFFAOYSA-N Sulphide Chemical compound [S-2] UCKMPCXJQFINFW-UHFFFAOYSA-N 0.000 description 1
- 210000001744 T-lymphocyte Anatomy 0.000 description 1
- BHEOSNUKNHRBNM-UHFFFAOYSA-N Tetramethylsqualene Natural products CC(=C)C(C)CCC(=C)C(C)CCC(C)=CCCC=C(C)CCC(C)C(=C)CCC(C)C(C)=C BHEOSNUKNHRBNM-UHFFFAOYSA-N 0.000 description 1
- 241000723792 Tobacco etch virus Species 0.000 description 1
- 206010044221 Toxic encephalopathy Diseases 0.000 description 1
- 229920001615 Tragacanth Polymers 0.000 description 1
- 108091023040 Transcription factor Proteins 0.000 description 1
- 102000040945 Transcription factor Human genes 0.000 description 1
- 108700019146 Transgenes Proteins 0.000 description 1
- HDTRYLNUVZCQOY-WSWWMNSNSA-N Trehalose Natural products O[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@H](O)[C@@H](O)[C@@H](O)[C@@H](CO)O1 HDTRYLNUVZCQOY-WSWWMNSNSA-N 0.000 description 1
- LEHOTFFKMJEONL-UHFFFAOYSA-N Uric Acid Chemical compound N1C(=O)NC(=O)C2=C1NC(=O)N2 LEHOTFFKMJEONL-UHFFFAOYSA-N 0.000 description 1
- TVWHNULVHGKJHS-UHFFFAOYSA-N Uric acid Natural products N1C(=O)NC(=O)C2NC(=O)NC21 TVWHNULVHGKJHS-UHFFFAOYSA-N 0.000 description 1
- 241000221561 Ustilaginales Species 0.000 description 1
- 108700005077 Viral Genes Proteins 0.000 description 1
- 108010067390 Viral Proteins Proteins 0.000 description 1
- 108070000030 Viral receptors Proteins 0.000 description 1
- HCHKCACWOHOZIP-UHFFFAOYSA-N Zinc Chemical compound [Zn] HCHKCACWOHOZIP-UHFFFAOYSA-N 0.000 description 1
- WERKSKAQRVDLDW-ANOHMWSOSA-N [(2s,3r,4r,5r)-2,3,4,5,6-pentahydroxyhexyl] (z)-octadec-9-enoate Chemical compound CCCCCCCC\C=C/CCCCCCCC(=O)OC[C@H](O)[C@@H](O)[C@H](O)[C@H](O)CO WERKSKAQRVDLDW-ANOHMWSOSA-N 0.000 description 1
- DPXJVFZANSGRMM-UHFFFAOYSA-N acetic acid;2,3,4,5,6-pentahydroxyhexanal;sodium Chemical compound [Na].CC(O)=O.OCC(O)C(O)C(O)C(O)C=O DPXJVFZANSGRMM-UHFFFAOYSA-N 0.000 description 1
- 239000002253 acid Substances 0.000 description 1
- 230000002378 acidificating effect Effects 0.000 description 1
- 230000003213 activating effect Effects 0.000 description 1
- 230000033289 adaptive immune response Effects 0.000 description 1
- 230000004721 adaptive immunity Effects 0.000 description 1
- 239000000654 additive Substances 0.000 description 1
- 239000004964 aerogel Substances 0.000 description 1
- 238000001042 affinity chromatography Methods 0.000 description 1
- 239000008272 agar Substances 0.000 description 1
- 150000001298 alcohols Chemical class 0.000 description 1
- 235000010443 alginic acid Nutrition 0.000 description 1
- 239000000783 alginic acid Substances 0.000 description 1
- 229920000615 alginic acid Polymers 0.000 description 1
- 229960001126 alginic acid Drugs 0.000 description 1
- 150000004781 alginic acids Chemical class 0.000 description 1
- 229910045601 alloy Inorganic materials 0.000 description 1
- 239000000956 alloy Substances 0.000 description 1
- HDTRYLNUVZCQOY-LIZSDCNHSA-N alpha,alpha-trehalose Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 HDTRYLNUVZCQOY-LIZSDCNHSA-N 0.000 description 1
- 229940037003 alum Drugs 0.000 description 1
- WNROFYMDJYEPJX-UHFFFAOYSA-K aluminium hydroxide Chemical compound [OH-].[OH-].[OH-].[Al+3] WNROFYMDJYEPJX-UHFFFAOYSA-K 0.000 description 1
- 210000001132 alveolar macrophage Anatomy 0.000 description 1
- 150000001412 amines Chemical class 0.000 description 1
- 125000000539 amino acid group Chemical group 0.000 description 1
- 238000012870 ammonium sulfate precipitation Methods 0.000 description 1
- 230000003321 amplification Effects 0.000 description 1
- 239000003708 ampul Substances 0.000 description 1
- 210000004727 amygdala Anatomy 0.000 description 1
- 238000000540 analysis of variance Methods 0.000 description 1
- 239000003957 anion exchange resin Substances 0.000 description 1
- 230000008485 antagonism Effects 0.000 description 1
- 230000001093 anti-cancer Effects 0.000 description 1
- 239000007900 aqueous suspension Substances 0.000 description 1
- 235000010323 ascorbic acid Nutrition 0.000 description 1
- 229960005070 ascorbic acid Drugs 0.000 description 1
- 239000011668 ascorbic acid Substances 0.000 description 1
- 101150036080 at gene Proteins 0.000 description 1
- 239000012298 atmosphere Substances 0.000 description 1
- 230000001363 autoimmune Effects 0.000 description 1
- 210000003719 b-lymphocyte Anatomy 0.000 description 1
- 230000001580 bacterial effect Effects 0.000 description 1
- 108010058966 bacteriophage T7 induced DNA polymerase Proteins 0.000 description 1
- 210000000227 basophil cell of anterior lobe of hypophysis Anatomy 0.000 description 1
- 229960000686 benzalkonium chloride Drugs 0.000 description 1
- CADWTSSKOVRVJC-UHFFFAOYSA-N benzyl(dimethyl)azanium;chloride Chemical compound [Cl-].C[NH+](C)CC1=CC=CC=C1 CADWTSSKOVRVJC-UHFFFAOYSA-N 0.000 description 1
- 230000002457 bidirectional effect Effects 0.000 description 1
- 108091008324 binding proteins Proteins 0.000 description 1
- 229960002685 biotin Drugs 0.000 description 1
- 235000020958 biotin Nutrition 0.000 description 1
- 239000011616 biotin Substances 0.000 description 1
- 210000004369 blood Anatomy 0.000 description 1
- 239000008280 blood Substances 0.000 description 1
- 125000001246 bromo group Chemical group Br* 0.000 description 1
- 239000007853 buffer solution Substances 0.000 description 1
- 239000006172 buffering agent Substances 0.000 description 1
- CDQSJQSWAWPGKG-UHFFFAOYSA-N butane-1,1-diol Chemical compound CCCC(O)O CDQSJQSWAWPGKG-UHFFFAOYSA-N 0.000 description 1
- CZBZUDVBLSSABA-UHFFFAOYSA-N butylated hydroxyanisole Chemical compound COC1=CC=C(O)C(C(C)(C)C)=C1.COC1=CC=C(O)C=C1C(C)(C)C CZBZUDVBLSSABA-UHFFFAOYSA-N 0.000 description 1
- 229940043253 butylated hydroxyanisole Drugs 0.000 description 1
- 239000002775 capsule Substances 0.000 description 1
- 239000001768 carboxy methyl cellulose Substances 0.000 description 1
- 239000006143 cell culture medium Substances 0.000 description 1
- 230000030833 cell death Effects 0.000 description 1
- 230000007910 cell fusion Effects 0.000 description 1
- 239000013592 cell lysate Substances 0.000 description 1
- 239000006285 cell suspension Substances 0.000 description 1
- 229920002678 cellulose Polymers 0.000 description 1
- 239000001913 cellulose Substances 0.000 description 1
- 229920002301 cellulose acetate Polymers 0.000 description 1
- 239000003638 chemical reducing agent Substances 0.000 description 1
- 230000002759 chromosomal effect Effects 0.000 description 1
- 238000003776 cleavage reaction Methods 0.000 description 1
- 230000008045 co-localization Effects 0.000 description 1
- 229940110456 cocoa butter Drugs 0.000 description 1
- 235000019868 cocoa butter Nutrition 0.000 description 1
- 229940000425 combination drug Drugs 0.000 description 1
- 238000004891 communication Methods 0.000 description 1
- 230000000052 comparative effect Effects 0.000 description 1
- 230000001447 compensatory effect Effects 0.000 description 1
- 238000007906 compression Methods 0.000 description 1
- 230000006835 compression Effects 0.000 description 1
- 238000005094 computer simulation Methods 0.000 description 1
- 230000002153 concerted effect Effects 0.000 description 1
- 239000000356 contaminant Substances 0.000 description 1
- 230000001276 controlling effect Effects 0.000 description 1
- 235000005687 corn oil Nutrition 0.000 description 1
- 239000002285 corn oil Substances 0.000 description 1
- 239000008120 corn starch Substances 0.000 description 1
- 235000012343 cottonseed oil Nutrition 0.000 description 1
- 239000002385 cottonseed oil Substances 0.000 description 1
- 238000006880 cross-coupling reaction Methods 0.000 description 1
- 210000004748 cultured cell Anatomy 0.000 description 1
- 238000012258 culturing Methods 0.000 description 1
- DTPCFIHYWYONMD-UHFFFAOYSA-N decaethylene glycol Chemical compound OCCOCCOCCOCCOCCOCCOCCOCCOCCOCCO DTPCFIHYWYONMD-UHFFFAOYSA-N 0.000 description 1
- 238000012350 deep sequencing Methods 0.000 description 1
- 230000005860 defense response to virus Effects 0.000 description 1
- 230000003111 delayed effect Effects 0.000 description 1
- 239000008121 dextrose Substances 0.000 description 1
- 239000003085 diluting agent Substances 0.000 description 1
- 229940042399 direct acting antivirals protease inhibitors Drugs 0.000 description 1
- PRAKJMSDJKAYCZ-UHFFFAOYSA-N dodecahydrosqualene Natural products CC(C)CCCC(C)CCCC(C)CCCCC(C)CCCC(C)CCCC(C)C PRAKJMSDJKAYCZ-UHFFFAOYSA-N 0.000 description 1
- 238000003255 drug test Methods 0.000 description 1
- 239000005293 duran Substances 0.000 description 1
- 230000008030 elimination Effects 0.000 description 1
- 238000003379 elimination reaction Methods 0.000 description 1
- 239000003995 emulsifying agent Substances 0.000 description 1
- 210000000981 epithelium Anatomy 0.000 description 1
- 238000012869 ethanol precipitation Methods 0.000 description 1
- 235000019325 ethyl cellulose Nutrition 0.000 description 1
- 229920001249 ethyl cellulose Polymers 0.000 description 1
- LVGKNOAMLMIIKO-QXMHVHEDSA-N ethyl oleate Chemical compound CCCCCCCC\C=C/CCCCCCCC(=O)OCC LVGKNOAMLMIIKO-QXMHVHEDSA-N 0.000 description 1
- 229940093471 ethyl oleate Drugs 0.000 description 1
- 210000003527 eukaryotic cell Anatomy 0.000 description 1
- 238000011156 evaluation Methods 0.000 description 1
- 239000012526 feed medium Substances 0.000 description 1
- 239000000945 filler Substances 0.000 description 1
- 238000004108 freeze drying Methods 0.000 description 1
- 239000012737 fresh medium Substances 0.000 description 1
- 108700004026 gag Genes Proteins 0.000 description 1
- 101150098622 gag gene Proteins 0.000 description 1
- 238000005227 gel permeation chromatography Methods 0.000 description 1
- 238000002523 gelfiltration Methods 0.000 description 1
- 150000002334 glycols Chemical class 0.000 description 1
- 125000003630 glycyl group Chemical group [H]N([H])C([H])([H])C(*)=O 0.000 description 1
- 210000003714 granulocyte Anatomy 0.000 description 1
- 239000001963 growth medium Substances 0.000 description 1
- 229960002897 heparin Drugs 0.000 description 1
- 229920000669 heparin Polymers 0.000 description 1
- 108010037896 heparin-binding hemagglutinin Proteins 0.000 description 1
- 208000006454 hepatitis Diseases 0.000 description 1
- 231100000283 hepatitis Toxicity 0.000 description 1
- 210000001320 hippocampus Anatomy 0.000 description 1
- 239000005556 hormone Substances 0.000 description 1
- 229940088597 hormone Drugs 0.000 description 1
- 102000046537 human SLAMF1 Human genes 0.000 description 1
- 210000005260 human cell Anatomy 0.000 description 1
- 150000004677 hydrates Chemical class 0.000 description 1
- 239000000017 hydrogel Substances 0.000 description 1
- 229920001600 hydrophobic polymer Polymers 0.000 description 1
- WGCNASOHLSPBMP-UHFFFAOYSA-N hydroxyacetaldehyde Natural products OCC=O WGCNASOHLSPBMP-UHFFFAOYSA-N 0.000 description 1
- 238000010191 image analysis Methods 0.000 description 1
- 230000001900 immune effect Effects 0.000 description 1
- 210000000987 immune system Anatomy 0.000 description 1
- 230000006058 immune tolerance Effects 0.000 description 1
- 230000005847 immunogenicity Effects 0.000 description 1
- 238000001114 immunoprecipitation Methods 0.000 description 1
- 230000001771 impaired effect Effects 0.000 description 1
- 239000007943 implant Substances 0.000 description 1
- 238000000126 in silico method Methods 0.000 description 1
- 210000003000 inclusion body Anatomy 0.000 description 1
- 210000004263 induced pluripotent stem cell Anatomy 0.000 description 1
- 238000001802 infusion Methods 0.000 description 1
- 108091006086 inhibitor proteins Proteins 0.000 description 1
- 230000000977 initiatory effect Effects 0.000 description 1
- 230000000266 injurious effect Effects 0.000 description 1
- 239000002054 inoculum Substances 0.000 description 1
- 230000037431 insertion Effects 0.000 description 1
- 238000007918 intramuscular administration Methods 0.000 description 1
- 238000007912 intraperitoneal administration Methods 0.000 description 1
- 238000007915 intraurethral administration Methods 0.000 description 1
- 238000001990 intravenous administration Methods 0.000 description 1
- 238000005342 ion exchange Methods 0.000 description 1
- 238000004255 ion exchange chromatography Methods 0.000 description 1
- NPFOYSMITVOQOS-UHFFFAOYSA-K iron(III) citrate Chemical compound [Fe+3].[O-]C(=O)CC(O)(CC([O-])=O)C([O-])=O NPFOYSMITVOQOS-UHFFFAOYSA-K 0.000 description 1
- 230000002427 irreversible effect Effects 0.000 description 1
- QXJSBBXBKPUZAA-UHFFFAOYSA-N isooleic acid Natural products CCCCCCCC=CCCCCCCCCC(O)=O QXJSBBXBKPUZAA-UHFFFAOYSA-N 0.000 description 1
- 239000007951 isotonicity adjuster Substances 0.000 description 1
- 210000003292 kidney cell Anatomy 0.000 description 1
- 210000000265 leukocyte Anatomy 0.000 description 1
- 239000006194 liquid suspension Substances 0.000 description 1
- 238000011866 long-term treatment Methods 0.000 description 1
- 239000007937 lozenge Substances 0.000 description 1
- 201000005202 lung cancer Diseases 0.000 description 1
- 208000020816 lung neoplasm Diseases 0.000 description 1
- 210000003563 lymphoid tissue Anatomy 0.000 description 1
- 239000006166 lysate Substances 0.000 description 1
- 239000012139 lysis buffer Substances 0.000 description 1
- VTHJTEIRLNZDEV-UHFFFAOYSA-L magnesium dihydroxide Chemical compound [OH-].[OH-].[Mg+2] VTHJTEIRLNZDEV-UHFFFAOYSA-L 0.000 description 1
- 239000000347 magnesium hydroxide Substances 0.000 description 1
- 229910001862 magnesium hydroxide Inorganic materials 0.000 description 1
- 238000013507 mapping Methods 0.000 description 1
- 239000011159 matrix material Substances 0.000 description 1
- 229940041323 measles vaccine Drugs 0.000 description 1
- 238000005259 measurement Methods 0.000 description 1
- 230000010534 mechanism of action Effects 0.000 description 1
- 229910052751 metal Inorganic materials 0.000 description 1
- 239000002184 metal Substances 0.000 description 1
- 125000002496 methyl group Chemical group [H]C([H])([H])* 0.000 description 1
- 235000010270 methyl p-hydroxybenzoate Nutrition 0.000 description 1
- 239000004292 methyl p-hydroxybenzoate Substances 0.000 description 1
- 229960002216 methylparaben Drugs 0.000 description 1
- 239000011859 microparticle Substances 0.000 description 1
- 239000002480 mineral oil Substances 0.000 description 1
- 235000010446 mineral oil Nutrition 0.000 description 1
- 238000012544 monitoring process Methods 0.000 description 1
- 239000000178 monomer Substances 0.000 description 1
- 229940126619 mouse monoclonal antibody Drugs 0.000 description 1
- 239000002324 mouth wash Substances 0.000 description 1
- 229940051866 mouthwash Drugs 0.000 description 1
- 210000004400 mucous membrane Anatomy 0.000 description 1
- AFBIVMWJUFLUNW-UHFFFAOYSA-N n-(3-cyanophenyl)-2-phenylacetamide Chemical compound C=1C=CC(C#N)=CC=1NC(=O)CC1=CC=CC=C1 AFBIVMWJUFLUNW-UHFFFAOYSA-N 0.000 description 1
- 238000002663 nebulization Methods 0.000 description 1
- 210000005036 nerve Anatomy 0.000 description 1
- 210000001178 neural stem cell Anatomy 0.000 description 1
- 230000007135 neurotoxicity Effects 0.000 description 1
- 231100000228 neurotoxicity Toxicity 0.000 description 1
- 230000007935 neutral effect Effects 0.000 description 1
- 238000006386 neutralization reaction Methods 0.000 description 1
- 230000003472 neutralizing effect Effects 0.000 description 1
- 238000007481 next generation sequencing Methods 0.000 description 1
- 210000000633 nuclear envelope Anatomy 0.000 description 1
- 238000003199 nucleic acid amplification method Methods 0.000 description 1
- ZQPPMHVWECSIRJ-KTKRTIGZSA-N oleic acid Chemical compound CCCCCCCC\C=C/CCCCCCCC(O)=O ZQPPMHVWECSIRJ-KTKRTIGZSA-N 0.000 description 1
- 239000004006 olive oil Substances 0.000 description 1
- 235000008390 olive oil Nutrition 0.000 description 1
- 238000000424 optical density measurement Methods 0.000 description 1
- LXCFILQKKLGQFO-UHFFFAOYSA-N p-hydroxybenzoic acid methyl ester Natural products COC(=O)C1=CC=C(O)C=C1 LXCFILQKKLGQFO-UHFFFAOYSA-N 0.000 description 1
- 239000006174 pH buffer Substances 0.000 description 1
- 230000007170 pathology Effects 0.000 description 1
- 239000008188 pellet Substances 0.000 description 1
- 230000000149 penetrating effect Effects 0.000 description 1
- 230000035515 penetration Effects 0.000 description 1
- 239000000137 peptide hydrolase inhibitor Substances 0.000 description 1
- 239000003208 petroleum Substances 0.000 description 1
- 230000004526 pharmaceutical effect Effects 0.000 description 1
- UYWQUFXKFGHYNT-UHFFFAOYSA-N phenylmethyl ester of formic acid Natural products O=COCC1=CC=CC=C1 UYWQUFXKFGHYNT-UHFFFAOYSA-N 0.000 description 1
- 239000008363 phosphate buffer Substances 0.000 description 1
- 239000008055 phosphate buffer solution Substances 0.000 description 1
- WTJKGGKOPKCXLL-RRHRGVEJSA-N phosphatidylcholine Chemical compound CCCCCCCCCCCCCCCC(=O)OC[C@H](COP([O-])(=O)OCC[N+](C)(C)C)OC(=O)CCCCCCCC=CCCCCCCCC WTJKGGKOPKCXLL-RRHRGVEJSA-N 0.000 description 1
- 230000026731 phosphorylation Effects 0.000 description 1
- 238000006366 phosphorylation reaction Methods 0.000 description 1
- 239000000049 pigment Substances 0.000 description 1
- 108700004029 pol Genes Proteins 0.000 description 1
- 101150088264 pol gene Proteins 0.000 description 1
- 229920005862 polyol Polymers 0.000 description 1
- 150000003077 polyols Chemical class 0.000 description 1
- 239000000256 polyoxyethylene sorbitan monolaurate Substances 0.000 description 1
- 235000010486 polyoxyethylene sorbitan monolaurate Nutrition 0.000 description 1
- 239000000244 polyoxyethylene sorbitan monooleate Substances 0.000 description 1
- 235000010482 polyoxyethylene sorbitan monooleate Nutrition 0.000 description 1
- 229940068977 polysorbate 20 Drugs 0.000 description 1
- 229920000053 polysorbate 80 Polymers 0.000 description 1
- 229940068968 polysorbate 80 Drugs 0.000 description 1
- 229920001343 polytetrafluoroethylene Polymers 0.000 description 1
- 239000004810 polytetrafluoroethylene Substances 0.000 description 1
- 229920002981 polyvinylidene fluoride Polymers 0.000 description 1
- 239000011148 porous material Substances 0.000 description 1
- 230000004481 post-translational protein modification Effects 0.000 description 1
- 230000001323 posttranslational effect Effects 0.000 description 1
- 229920001592 potato starch Polymers 0.000 description 1
- 239000002243 precursor Substances 0.000 description 1
- 230000002335 preservative effect Effects 0.000 description 1
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 1
- 230000000750 progressive effect Effects 0.000 description 1
- 230000000644 propagated effect Effects 0.000 description 1
- 235000010232 propyl p-hydroxybenzoate Nutrition 0.000 description 1
- 239000004405 propyl p-hydroxybenzoate Substances 0.000 description 1
- 229960003415 propylparaben Drugs 0.000 description 1
- 230000004952 protein activity Effects 0.000 description 1
- 230000012846 protein folding Effects 0.000 description 1
- 230000004853 protein function Effects 0.000 description 1
- 230000009145 protein modification Effects 0.000 description 1
- 230000017854 proteolysis Effects 0.000 description 1
- 238000011084 recovery Methods 0.000 description 1
- 238000009877 rendering Methods 0.000 description 1
- 230000033458 reproduction Effects 0.000 description 1
- 238000011160 research Methods 0.000 description 1
- 239000011347 resin Substances 0.000 description 1
- 229920005989 resin Polymers 0.000 description 1
- 238000010839 reverse transcription Methods 0.000 description 1
- 238000004007 reversed phase HPLC Methods 0.000 description 1
- 235000009566 rice Nutrition 0.000 description 1
- 230000000630 rising effect Effects 0.000 description 1
- 102200070479 rs28933693 Human genes 0.000 description 1
- 235000005713 safflower oil Nutrition 0.000 description 1
- 239000003813 safflower oil Substances 0.000 description 1
- 230000007017 scission Effects 0.000 description 1
- 150000003335 secondary amines Chemical class 0.000 description 1
- 230000003248 secreting effect Effects 0.000 description 1
- 238000013207 serial dilution Methods 0.000 description 1
- 239000000741 silica gel Substances 0.000 description 1
- 229910002027 silica gel Inorganic materials 0.000 description 1
- 239000000377 silicon dioxide Substances 0.000 description 1
- 239000002356 single layer Substances 0.000 description 1
- 238000001542 size-exclusion chromatography Methods 0.000 description 1
- 235000020183 skimmed milk Nutrition 0.000 description 1
- 150000003384 small molecules Chemical class 0.000 description 1
- 235000019812 sodium carboxymethyl cellulose Nutrition 0.000 description 1
- 229920001027 sodium carboxymethylcellulose Polymers 0.000 description 1
- 239000012453 solvate Substances 0.000 description 1
- 239000008347 soybean phospholipid Substances 0.000 description 1
- 241000894007 species Species 0.000 description 1
- 238000001694 spray drying Methods 0.000 description 1
- 229940031439 squalene Drugs 0.000 description 1
- TUHBEKDERLKLEC-UHFFFAOYSA-N squalene Natural products CC(=CCCC(=CCCC(=CCCC=C(/C)CCC=C(/C)CC=C(C)C)C)C)C TUHBEKDERLKLEC-UHFFFAOYSA-N 0.000 description 1
- 230000010473 stable expression Effects 0.000 description 1
- 238000003153 stable transfection Methods 0.000 description 1
- 238000010561 standard procedure Methods 0.000 description 1
- 239000008107 starch Substances 0.000 description 1
- 210000000130 stem cell Anatomy 0.000 description 1
- 239000011550 stock solution Substances 0.000 description 1
- 238000003860 storage Methods 0.000 description 1
- 229960005322 streptomycin Drugs 0.000 description 1
- ADNPLDHMAVUMIW-CUZNLEPHSA-N substance P Chemical compound C([C@@H](C(=O)NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCSC)C(N)=O)NC(=O)[C@H](CC=1C=CC=CC=1)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@H]1N(CCC1)C(=O)[C@H](CCCCN)NC(=O)[C@H]1N(CCC1)C(=O)[C@@H](N)CCCN=C(N)N)C1=CC=CC=C1 ADNPLDHMAVUMIW-CUZNLEPHSA-N 0.000 description 1
- 239000000758 substrate Substances 0.000 description 1
- 235000000346 sugar Nutrition 0.000 description 1
- 150000008163 sugars Chemical class 0.000 description 1
- 238000013268 sustained release Methods 0.000 description 1
- 239000012730 sustained-release form Substances 0.000 description 1
- 208000024891 symptom Diseases 0.000 description 1
- 239000003826 tablet Substances 0.000 description 1
- 229940124597 therapeutic agent Drugs 0.000 description 1
- 238000011287 therapeutic dose Methods 0.000 description 1
- 125000003396 thiol group Chemical group [H]S* 0.000 description 1
- 230000009772 tissue formation Effects 0.000 description 1
- 231100000419 toxicity Toxicity 0.000 description 1
- 230000001988 toxicity Effects 0.000 description 1
- 235000010487 tragacanth Nutrition 0.000 description 1
- 239000000196 tragacanth Substances 0.000 description 1
- 229940116362 tragacanth Drugs 0.000 description 1
- 238000013518 transcription Methods 0.000 description 1
- 230000035897 transcription Effects 0.000 description 1
- 238000006276 transfer reaction Methods 0.000 description 1
- 230000009261 transgenic effect Effects 0.000 description 1
- 230000003363 transsynaptic effect Effects 0.000 description 1
- 239000013638 trimer Substances 0.000 description 1
- 239000000717 tumor promoter Substances 0.000 description 1
- 238000007492 two-way ANOVA Methods 0.000 description 1
- 241001430294 unidentified retrovirus Species 0.000 description 1
- 238000011144 upstream manufacturing Methods 0.000 description 1
- 229940116269 uric acid Drugs 0.000 description 1
- 229940125575 vaccine candidate Drugs 0.000 description 1
- 235000013311 vegetables Nutrition 0.000 description 1
- 230000007486 viral budding Effects 0.000 description 1
- 230000009447 viral pathogenesis Effects 0.000 description 1
- 210000002845 virion Anatomy 0.000 description 1
- 230000000007 visual effect Effects 0.000 description 1
- 239000001993 wax Substances 0.000 description 1
- 229910052725 zinc Inorganic materials 0.000 description 1
- 239000011701 zinc Substances 0.000 description 1
Images






























































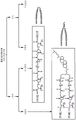


























































































Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K47/00—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient
- A61K47/50—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient the non-active ingredient being chemically bound to the active ingredient, e.g. polymer-drug conjugates
- A61K47/51—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient the non-active ingredient being chemically bound to the active ingredient, e.g. polymer-drug conjugates the non-active ingredient being a modifying agent
- A61K47/54—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient the non-active ingredient being chemically bound to the active ingredient, e.g. polymer-drug conjugates the non-active ingredient being a modifying agent the modifying agent being an organic compound
- A61K47/543—Lipids, e.g. triglycerides; Polyamines, e.g. spermine or spermidine
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/12—Viral antigens
- A61K39/21—Retroviridae, e.g. equine infectious anemia virus
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/12—Viral antigens
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/12—Viral antigens
- A61K39/155—Paramyxoviridae, e.g. parainfluenza virus
- A61K39/165—Mumps or measles virus
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K47/00—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient
- A61K47/50—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient the non-active ingredient being chemically bound to the active ingredient, e.g. polymer-drug conjugates
- A61K47/51—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient the non-active ingredient being chemically bound to the active ingredient, e.g. polymer-drug conjugates the non-active ingredient being a modifying agent
- A61K47/54—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient the non-active ingredient being chemically bound to the active ingredient, e.g. polymer-drug conjugates the non-active ingredient being a modifying agent the modifying agent being an organic compound
- A61K47/55—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient the non-active ingredient being chemically bound to the active ingredient, e.g. polymer-drug conjugates the non-active ingredient being a modifying agent the modifying agent being an organic compound the modifying agent being also a pharmacologically or therapeutically active agent, i.e. the entire conjugate being a codrug, i.e. a dimer, oligomer or polymer of pharmacologically or therapeutically active compounds
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/12—Antivirals
- A61P31/14—Antivirals for RNA viruses
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/12—Antivirals
- A61P31/14—Antivirals for RNA viruses
- A61P31/18—Antivirals for RNA viruses for HIV
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/005—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from viruses
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/60—Medicinal preparations containing antigens or antibodies characteristics by the carrier linked to the antigen
- A61K2039/6031—Proteins
- A61K2039/6075—Viral proteins
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/60—Medicinal preparations containing antigens or antibodies characteristics by the carrier linked to the antigen
- A61K2039/6093—Synthetic polymers, e.g. polyethyleneglycol [PEG], Polymers or copolymers of (D) glutamate and (D) lysine
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2760/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses negative-sense
- C12N2760/00011—Details
- C12N2760/18011—Paramyxoviridae
- C12N2760/18411—Morbillivirus, e.g. Measles virus, canine distemper
- C12N2760/18422—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2760/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses negative-sense
- C12N2760/00011—Details
- C12N2760/18011—Paramyxoviridae
- C12N2760/18411—Morbillivirus, e.g. Measles virus, canine distemper
- C12N2760/18433—Use of viral protein as therapeutic agent other than vaccine, e.g. apoptosis inducing or anti-inflammatory
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2760/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses negative-sense
- C12N2760/00011—Details
- C12N2760/18011—Paramyxoviridae
- C12N2760/18411—Morbillivirus, e.g. Measles virus, canine distemper
- C12N2760/18434—Use of virus or viral component as vaccine, e.g. live-attenuated or inactivated virus, VLP, viral protein
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Virology (AREA)
- General Health & Medical Sciences (AREA)
- Medicinal Chemistry (AREA)
- Pharmacology & Pharmacy (AREA)
- Veterinary Medicine (AREA)
- Public Health (AREA)
- Animal Behavior & Ethology (AREA)
- Epidemiology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Engineering & Computer Science (AREA)
- Organic Chemistry (AREA)
- Molecular Biology (AREA)
- Immunology (AREA)
- Communicable Diseases (AREA)
- Mycology (AREA)
- Microbiology (AREA)
- Biophysics (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Oncology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Genetics & Genomics (AREA)
- Gastroenterology & Hepatology (AREA)
- Biochemistry (AREA)
- Hematology (AREA)
- AIDS & HIV (AREA)
- Tropical Medicine & Parasitology (AREA)
- Pulmonology (AREA)
- Peptides Or Proteins (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Medicinal Preparation (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
本文描述了用抗病毒肽缀合物治疗麻疹或HIV感染的肽、组合物和方法,所述抗病毒肽缀合物包含缀合至C端七肽重复(HRC)肽的融合抑制肽(FIP)。本文还描述了稳定的可溶性麻疹F蛋白、组合物以及用稳定的F蛋白预防麻疹感染的方法,所述稳定的F蛋白可单独施用或与本文所述抗病毒肽缀合物结合施用。
Description
相关申请的交叉引用
本申请要求享有于2019年9月4日提交的美国申请号62/895,752;于2020年3月11日提交的美国申请号62/988,286;以及于2020年4月14日提交的美国申请号63/009,883的权益和优先权,三者均通过引用整体并入本文。
本文引用的所有专利、专利申请和出版物均通过引用整体并入本文。这些出版物的公开内容特此通过引用整体并入本申请。
本专利的公开内容包含受版权保护的材料。版权所有者不反对任何人对美国专利商标局专利档案或记录中呈现的专利文件或专利公开内容进行传真复制,但是除此以外保留任何和所有版权权利。
政府支持
本发明是在美国国立卫生研究院(National Institutes of Health)授予的资助NS091263、NS105699、AI121349和AI119762的政府支持下完成的。政府对本发明享有一定的权利。
背景技术
麻疹仍然是全球健康所面临的一项挑战。虽然1963年已推出麻疹疫苗,但对于已经感染的个体,尚无经FDA批准的抗病毒治疗。目前的方法防止麻疹病毒(“MV”或“MeV”)融合并进入宿主细胞的能力有限。目前的建议是在暴露后72小时内接种疫苗或免疫球蛋白(IG)。抗病毒化合物可以针对病毒活性的不同成分,例如,阻止病毒复制或阻止病毒进入宿主细胞。
发明内容
在某些方面,本文描述的本发明涉及抗病毒肽,该抗病毒肽包含C端七肽重复(HRC)肽和融合抑制肽(FIP)的组合。出乎意料的是,HRC和FIP的组合表现出协同作用,比单独使用任何一种方法都更有效。该组合整体防止MeV激活,并且也阻止已经激活病毒的重新折叠和融合。
在一个方面,本发明提供了一种抗病毒肽缀合物,该抗病毒肽缀合物包含融合抑制肽(FIP)和C端七肽重复(HRC)肽(FIP-HRC)。在一些实施方式中,抗病毒肽缀合物进一步包含膜定位部分区。在一些实施方式中,膜定位部分区包含选自胆固醇、生育酚和棕榈基的膜定位部分。在一些实施方式中,膜定位部分区缀合至HRC肽的C端。在一些实施方式中,抗病毒肽缀合物进一步包含连接体(linker)区。在一些实施方式中,连接体区包含聚乙二醇(polyethylene glycol,PEG)。在一些实施方式中,PEG的长度为4个乙二醇单元(PEG4)。在一些实施方式中,PEG的长度为11个乙二醇单元(PEG11)。
在一些实施方式中,连接体区与HRC肽的C端缀合。在一些实施方式中,抗病毒肽缀合物进一步包含膜定位部分区和连接体区。在一些实施方式中,连接体区与HRC肽的C端缀合,并且膜定位部分区与连接体区缀合。在一些实施方式中,连接体区包含聚乙二醇(PEG)。在一些实施方式中,PEG的长度为4个乙二醇单元(PEG4)。在一些实施方式中,PEG的长度为11个乙二醇单元(PEG11)。在一些实施方式中,PEG的长度为12个乙二醇单元(PEG12)。在一些实施方式中,PEG的长度为14个乙二醇单元(PEG14)。在一些实施方式中,抗病毒肽包含FIP区和HRC肽区的二聚体。在一些实施方式中,抗病毒肽包含缀合至连接体区的第一FIP-HRC肽和缀合至连接体区的第二FIP-HRC肽。在一些实施方式中,肽进一步包含丝氨酸-甘氨酸连接体。在一些实施方式中,丝氨酸-甘氨酸连接体位于FIP和HRC肽之间。在一些实施方式中,丝氨酸-甘氨酸连接体位于HRC肽的C端。在一些实施方式中,丝氨酸-甘氨酸连接体位于FIP区和HRC肽区之间,并且进一步包含位于HRC肽区的C端的第二丝氨酸-甘氨酸连接体。在一些实施方式中,丝氨酸-甘氨酸连接体包含氨基酸序列GSGSG。在一些实施方式中,抗病毒肽缀合物的FIP的第一个苯丙氨酸残基是D-氨基酸。在一些实施方式中,抗病毒肽缀合物的N端进一步包含苄氧羰基基团。在一些实施方式中,FIP肽包含氨基酸序列FFG。在一些实施方式中,HRC肽包含氨基酸序列PPISLERLDVGTNLGN AIAKLEDAKELLESSDQILR。在一些实施方式中,HRC肽缀合物包含氨基酸序列PPISLERLDVGTN。在一些实施方式中,抗病毒肽包含氨基酸序列FFGPPISLERLDVGTNLGNAIAKLEDAKELLESSDQILR。在一些实施方式中,抗病毒肽包含氨基酸序列FFGPPISLERLDVGTN。
在另一个方面,本发明提供了包含本文所述的任何抗病毒肽缀合物的纳米颗粒。在一些实施方式中,纳米颗粒具有约50nm至约150nm的直径。
在另一个方面,本发明提供了一种包含本文所述的任何抗病毒肽缀合物的组合物。
在另一个方面,本发明提供了一种预防性组合物,该预防性组合物包含抗病毒肽缀合物,该抗病毒肽缀合物包含融合抑制肽(FIP)和C端七肽重复(HRC)肽(FIP-HRC),其中,所述麻疹抗病毒肽缀合物防止麻疹病毒的膜融合。
在另一个方面,本发明提供了一种包含融合抑制(FIP)肽和C端七肽重复(HRC)肽(FIP-HRC)的纳米颗粒。
在另一个方面,本发明提供了一种感染后的麻疹预防(post-infection measlesprophylaxis)的方法,该方法包括向有需要的受试者施用包含融合抑制肽(FIP)和C端七肽(HRC)肽(FIP-HRC)的抗病毒肽缀合物。在一些实施方式中,抗病毒肽缀合物进一步包含连接体区。在一些实施方式中,抗病毒肽缀合物进一步包含膜定位部分区。在一些实施方式中,抗病毒肽缀合物进一步包含连接体区和膜定位部分区。在一些实施方式中,施用包括鼻内吸入或口腔吸入。在一些实施方式中,抗病毒肽缀合物通过选自喷雾器、气雾器和吸入器的装置施用。在一些实施方式中,施用包括皮下施用。在一些实施方式中,受试者已经暴露于麻疹病毒,该麻疹病毒包含野生型融合糖蛋白。在一些实施方式中,受试者已经暴露于麻疹病毒,该麻疹病毒包含选自N462K、L454W、T461I、E455G、E170G、G506E、M337L、D538G、G168R、S262G、A440P、R520C和L550P所组成的组中的融合糖蛋白的一种或多种突变。
在另一个方面,本发明提供了一种包含稳定的可溶性麻疹F蛋白的重组蛋白,该稳定的可溶性麻疹F蛋白包含E445G突变。
在另一个方面,本发明提供了一种包含稳定的可溶性麻疹F蛋白的重组蛋白,该稳定的可溶性麻疹F蛋白包含E170G和E455G双突变。
在另一个方面,本发明提供了一种包含氨基酸序列SEQ ID NO:3或SEQ ID NO:4的重组蛋白。
在另一个方面,本发明提供了一种包含氨基酸序列SEQ ID NO:5或SEQ ID NO:6的重组蛋白。
在另一个方面,本发明提供了一种免疫原性组合物,该免疫原性组合物包含本文所述的任何一种重组蛋白。在一些实施方式中,本发明提供了一种通过向受试者施用免疫原性组合物来在麻疹暴露之前预防麻疹感染的方法。在一些实施方式中,本发明提供了一种通过向受试者施用免疫原性组合物来诱导针对麻疹病毒的免疫反应的方法。
在另一个方面,本发明提供了一种免疫原性组合物,该免疫原性组合物包含氨基酸序列SEQ ID NO:1或SEQ ID NO:2,并且进一步包含本文所述的任何抗病毒肽缀合物。
在另一个方面,本发明提供了一种诱导针对麻疹病毒的免疫反应的方法,该方法通过向受试者施用本文所述的任何免疫原性组合物并进一步施用本文所述的任何抗病毒肽缀合物来进行。
在另一个方面,本发明提供了一种生产重组蛋白的方法,该重组蛋白包含氨基酸序列SEQ ID NO:3或SEQ ID NO:4。
在另一个方面,本发明提供了一种生产重组蛋白的方法,该重组蛋白包含氨基酸序列SEQ ID NO:5或SEQ ID NO:6。
在另一个方面,本发明提供了一种用于重组蛋白表达的细胞系的方法,该重组蛋白包含氨基酸序列SEQ ID NO:3或SEQ ID NO:4。
在另一个方面,本发明提供了一种用于重组蛋白表达的细胞系的方法,该重组蛋白包含氨基酸序列SEQ ID NO:5或SEQ ID NO:6。
在另一个方面,本发明提供了一种抗病毒肽缀合物,该抗病毒肽缀合物包含融合抑制肽(FIP)和C端七肽重复(HRC)肽(FIP-HRC),其中,HRC肽衍生自HIV-GP41(C34)。在一些实施方式中,HRC肽包含氨基酸序列WMEWDREINNYTSLIHSLIEESQNQQEKNEQELL。
在另一个方面,本发明提供了一种感染后的HIV预防的方法,该方法包括向有需要的受试者施用包含融合抑制肽(FIP)和C端七肽(HRC)肽(FIP-HRC)的抗病毒肽缀合物。在一些实施方式中,HRC肽衍生自HIV-GP41(C34)。
附图说明
本专利或申请文件包含至少一张彩色附图。为符合PCT专利申请的要求,本文中提供的许多附图是原始彩色图像的黑白表示。
图1示出了MV感染始于呼吸道。
图2示出了病毒血症和MV发病机制的出口(egress)。
图3示出了麻疹还可引起严重的并发症。
图4示出了患有MV脑炎的HIV感染患者的数据。
图5示出了麻疹病毒的示意图。
图6示出了两名患者的MV F基因含有相同的核苷酸突变。
图7示出了野生型麻疹病毒进入中枢神经系统(CNS)受到严格的控制。
图8示出了融合复合物的分析方法。
图9A-9C示出了融合水平。图9A-B示出了在存在已知受体的情况下,所有F蛋白都表现出相似的融合水平。图9C示出了F L454W在不存在已知受体的情况下诱导融合。
图10示出了F蛋白的两种热状态。
图11示出了由重组病毒诱导的细胞间融合(cell-to-cell fusion)。
图12示出了感染麻疹野生型F病毒和带有L454W F的病毒的90天大的脑类器官。
图13示出了来自小鼠的离体组织(无受体)。
图14示出了棉鼠的MV数据。
图15A-15B示出了小鼠的存活率数据。
图16示出了病毒进入作为治疗靶点。
图17示出了F糖蛋白衍生的肽。
图18示出了F糖蛋白衍生的肽抑制病毒进入。
图19示出了将HRC肽靶向脂质膜。
图20示出了提高HRC肽对F蛋白的亲和力(avidity)。
图21示出了将HRC肽靶向脂质膜的另一个实施方式。
图22A-22B示出了缀合的肽在体内是有效的。图22A中使用的模型是的棉鼠,图22B中使用的模型是SLAM:IFNARKO小鼠。
图23A-23C示出了MV HRC4肽阻断病毒的离体传播。
图24示出了MV HRC4的鼻内施用保护乳鼠免受带有L454W F的病毒的致命感染。
图25示出了肽粒径在纳摩尔范围内。
图26示出了两亲结构驱使自组装和纳米颗粒的形成。
图27示出了纳米颗粒在脂质膜界面处组装(解装配)并保留肽。
图28示出了纳米颗粒能够穿过HAE屏障并且在体内是生物可利用的。通过到达相关感染部位,这些纳米颗粒阻止麻疹病毒增殖。
图29示出了以6mg/kg经鼻内施用缀合的肽时,改善了棉鼠模型中的生物分布。
图30示出了组合策略,其中融合抑制肽(FIP)与融合蛋白结合并稳定麻疹F蛋白的融合前状态。
图31示出了等效线图曲线(isobologram curve)显示FIP和HRC肽之间的协同作用。
图32示出了添加至HRC区的FIP增强了抗病毒活性。
图33示出了当FIP和HRC结构相同时,效力超过这两种抑制剂加在一起的协同作用。
图34示出了当FIP和HRC结构相同时,效力超过这两种抑制剂加在一起的协同作用的额外数据。
图35示出了添加至HRC区的FIP增强了抗病毒活性。
图36示出了添加至HRC区的FIP增强了抗病毒活性的额外数据。
图37示出了存活比例。
图38示出了多种麻疹病毒(MV)。
图39示出了MV HRC4肽和RNA聚合酶抑制剂阻止人类运动神经元中的MV野生型感染。
图40示出了HRC4肽和RNA聚合酶抑制剂:野生型病毒和CNS适应病毒(CNS-adaptedviruses)。
图41示出了人类运动神经元中的MV HRC4肽和RNA聚合酶抑制剂对(vs.)CNS适应MV。
图42A-42B示出了进入的步骤。
图43A-43C也示出了进入的步骤。
图44A-44C示出了H-F相互作用在L454W F中发生了改变。
图45示出了MV HRC4的鼻内施用保护IFNAR KO小鼠免受致命的MV脑炎的影响。
图46示出了可以从多能干细胞生长的多种类器官以及所使用的发育信号的示意图。
图47A-47C示出了从多能干细胞产生的“迷你大脑(mini-brain)”。图47A示出了具有包含神经祖细胞(SOX2,红色)和神经元(TUJ1,绿色)的异质区的复杂形态是显而易见的。图47B示出了具有图案肾单元(patterned nephron)的从多能干细胞生长出的整个肾脏类器官的免疫荧光图像。形成肾小球的足细胞(NPHS1,黄色)、早期近端小管(莲藕凝集素(lotus tetragonolobus lectin),粉红色)和远端小管/收集管(E-钙粘蛋白,绿色)。图47C示出了人aSC来源的肺类器官中部的3D重建,对基底细胞(绿色)、肌动蛋白细胞骨架(红色)和细胞核(蓝色)的中间丝进行染色,并通过共聚焦显微镜成像。
图48示出了随着时间的推移,报告的麻疹病例。
图49示出了麻疹病毒的示意图。
图50示出了麻疹病毒进入细胞。
图51示出了麻疹病毒进入细胞的步骤。
图52示出了麻疹病毒进入作为潜在的治疗靶点。
图53示出了麻疹融合抑制肽(FIP)的结构。
图54示出了用于防止麻疹病毒进入细胞的组合策略。
图55示出了脂质-肽缀合物的设计。
图56示出了脂质-肽缀合物的优点。
图57-58示出了脂质-肽结构的示例。
图59-60示出了单体脂质-肽缀合物合成的反应方案。
图61示出了脂质-肽缀合物列表。
图62示出了脂质-肽缀合物的纯化和特征数据。
图63示出了基于β-半乳糖苷酶互补的融合试验的过程。
图64示出了麻疹脂质-肽缀合物在融合试验中的抑制活性的数据。
图65示出了基于来自融合试验数据的抑制活性的最佳脂质-肽缀合物候选物。
图66示出了MTT细胞毒性试验的过程。
图67示出了MTT试验数据。
图68示出了麻疹融合蛋白的热稳定性研究过程。
图69示出了来自麻疹融合蛋白的热稳定性研究的数据。
图70示出了麻疹脂质-肽缀合物的F稳定特性。
图71示出了基于麻疹脂质-肽缀合物的F稳定特性的最佳脂质-肽缀合物候选物。
图72示出了对融合激活的每个阶段进行量化。
图73-75示出了在多种浓度的不同脂质-肽缀合物存在的情况下,麻疹病毒与红细胞(RBC)的结合活性(图73:HRC;图74:FIP-HRC;图75:FIP)。
图76示出了麻疹脂质-肽缀合物的体内功效的示意图。
图77示出了麻疹脂质-肽缀合物的体内功效的数据。
图78示出了麻疹脂质-肽缀合物的粒径测量数据。
图79示出了脂质-肽缀合物之间相互作用的示意图。
图80示出了来自等效线图(isobologram)分析的数据。
图81示出了等效线图分析的量化。
图82图1示出了麻疹融合抑制缀合物的作用机制的示意图。
图83示出了用于不同脂质-肽缀合物的多种设计的示意图。
图84示出了体内功效的数据。
图85A-85C示出了FIP-HRC靶向MV F表达细胞。
图86示出了FIP-HRC将麻疹F稳定在其融合前状态。
图87示出了MeV肽的F-稳定特性。
图88-90示出了MeV肽对可溶性F的稳定特性。
图91示出了FIP-HRC防止F激活(与防止F重新折叠的HRC的机制完全不同)。
图92示出了FIP-HRC靶向MeV F表达细胞。
图93示出了来自三个不同实验的数据,表明FIP-HRC靶向MeV F表达细胞。
图94示出了MeV肽的稳定特性。
图95示出了MeV肽的细胞毒性。
图96示出了来自HRC+FIP缀合物等效线图分析的协同作用数据。
图97示出了衍生自麻疹HRC的带有12个氨基酸的FIP-HRC的效力。
图98示出了使用多种脂质-肽缀合物以及阳性和阴性对照对BG505(人体免疫缺陷病毒1型(HIV-1)毒株)的抑制数据。
图99示出了使用多种脂质-肽缀合物以及阳性和阴性对照对B41(HIV-1毒株)的抑制数据。
图100示出了使用多种脂质-肽缀合物以及阳性和阴性对照对16055(HIV-1毒株)的抑制数据。
图101示出了使用多种脂质-肽缀合物以及阳性和阴性对照对MN(HIV-1毒株)的抑制数据。
图102示出了使用多种脂质-肽缀合物以及阳性和阴性对照对水疱性口炎病毒(VSV)的抑制数据。
图103示出了使用多种脂质-肽缀合物对鼠白血病病毒(MLV)的抑制数据。
图104A-104B示出了来自CNS适应病毒的F蛋白内的替换位置。
图105A-105I示出了野生型(wt)病毒的离体感染与带有L454W F的病毒的离体感染:在器官型脑培养物(OBC)中,CNS适应病毒胜过wt病毒。
图106A-D示出了CNS适应的MeV变体在人多能干细胞(hiPSC)衍生的脑类器官中有效传播。
图107A-107H示出了带有指定突变的MeV融合(F)蛋白的融合活性和热稳定性,L454W F背景中的额外突变稳定了F蛋白的融合前状态。
图108示出了在高度易受影响的CD150xIFNARKO CNS中,抑制带有L454W F的病毒的传播。
图109A-109B示出了与野生型MeV相比,MeV F L454W在小鼠脑切片培养物中诱导干扰素刺激基因。
图110A-110B示出了在存在融合抑制剂的情况下感染人脑类器官。
图111A-111B示出了在感染融合蛋白突变体MeV的脑类器官中诱导宿主抗病毒基因。
图112示出了脑类器官和BrainSpan图集之间基因水平RPKM值的相关性热图。
图113示出了L454W感染后差异表达基因的KEGG通路分析。
图114示出了L454W和L454W/G506E融合蛋白突变病毒群的病毒等位基因纵向分析(LAVA)图。
图115示出了L454W和L454W/E455G融合蛋白突变病毒群的LAVA图。
图116示出了野生型融合蛋白病毒群的LAVA图。
图117A-117E示出了(A)可溶性MeV-FE170G-E455G已在使用或不使用FIP-HRC 1mM(不含胆固醇的二聚体)的情况下,在4℃或55℃下温育指定时间。温育后,使用小鼠融合前特异性抗体对F进行免疫沉淀。免疫沉淀的蛋白质在SDS-PAGE还原凝胶上运行,转移至PVDF膜并与α-MV-F-HRC生物素(1:1000)一起温育。链霉亲和素碱性磷酸酶缀合物已用作第二抗体(在磷酸盐缓冲液(Phosphate Buffer Saline,pbs)中的比例为1:1000)。(B)已使用ImageJ软件对(A)中描述的实验的蛋白质免疫印迹(Western Blot,WB)条带进行了量化。结果代表三个独立实验的平均值±SEM。(C)可溶性MeV-FE170G-E455G已在4℃、25℃或37℃下温育1周。如(A)中所述进行了免疫沉淀反应。(D)可溶性MeV-FWT或MeV-FE170G-E455G已在使用或不使用FIP-HRC、商购FIP、FIP二聚体或3g(1mM)的情况下,在4℃或55℃下温育60分钟或120分钟。(E)已使用ImageJ软件对(D)中描述的实验的蛋白质免疫印迹(WesternBlot,WB)条带进行了量化。结果代表三个独立实验的平均值±SEM。
具体实施方式
尽管50多年前推出了疫苗,麻疹仍然是全球健康所面临的一项挑战。对于已经感染的个体,没有经FDA批准的抗病毒治疗,并且目前的方法防止麻疹病毒(MeV)融合并进入宿主细胞的能力有限。在一个实施方式中,本文公开的主题涉及针对不同作用模式的两种现有MeV抗病毒方法的组合。第一种方法包括使用HRC衍生肽靶向MeV融合蛋白(F)的末端七肽重复(HRC)区,这干扰感染期间病毒融合所需的结构重排(例如,防止重新折叠)。第二种方法包括使用融合抑制肽(FIP)靶向MeV F的七肽重复B(HRB)区,这将MeV F稳定在其不能融合的融合前状态(例如,参见图52-32)。已经证明,在几种体外、离体和体内模型中,该组合方法比单独使用任何一种方法更有效。这种组合一起防止MeV激活,并且也阻止已经激活病毒的重新折叠和融合。
此外,在一个实施方式中,本文公开的主题涉及两种抗病毒方法的组合,以通过以下方式来防止麻疹病毒与细胞膜融合并进入细胞:使用HRC衍生肽靶向MeV F的C端七肽重复(HRC)区,这干扰感染期间病毒融合所需的结构重排;以及使用融合抑制肽(FIP)靶向MeVF的七肽重复B(HRB)区,这将MeV F稳定在激活前状态。该技术证明,在体外、离体以及体内,两种方法的组合比单独使用任何一种方法更有效。
本发明涉及用于通过呼吸道或皮下途径施用的麻疹抗病毒肽。本发明还涉及通过不同机制起作用以防止进入的两种不同的肽结构域的组合:(1)与麻疹F蛋白上的七肽重复序列互补的肽结构域(“HRC”),其对病毒进入的F-重新折叠步骤至关重要,一旦F蛋白被激活并插入靶细胞膜,即发生该F-重新折叠步骤。这种重新折叠是导致病毒-细胞膜融合和进入的原因,因此,通过HRC肽与F蛋白的结合来阻断它可以防止病毒进入和感染;(2)肽结构域(“FIP”),其与F蛋白的不同区(例如HRB区)结合并将F蛋白稳定在激活前的构象,防止F激活至能够融合的状态。如果F蛋白没有被激活到能够融合的状态,则不会发生任何后续的进入步骤。(1)和(2)的组合产生了肽,该肽首先将F蛋白稳定在其融合前的构象,使其不具备融合能力——然后,对于已被激活用于融合的F蛋白,该肽防止重新折叠和融合。该组合的功效已在体外和体内进行了研究,并且比单独使用任何一种方法都显著增强。
参考文献:
1.Plemper RK,Snyder JP.麻疹防治--麻疹病毒抑制剂是否能发挥作用?(Measles control–can measles virus inhibitors make a difference?)研究性药物的前沿观点(Curr Opin Investig Drugs).2009年8月;10(8):第811-20页.
2.Woo TM.疫苗可预防的疾病的暴露后管理(Postexposure management ofvaccine-preventable diseases).儿科保健杂志(J Pediatr Health Care).2016年3月;30(2):第173-82页.
3.Welsch JC,Talekar A,Mathieu C,Pessi A,Moscona A,Horvat B,Porotto M.脑穿透性融合抑制剂可预防致命的麻疹病毒感染(Fatal measles virus infectionprevented by brain-penetrant fusion inhibitors).病毒学杂志(JVirol).2013年12月;87(24):第13785-94页.
4.Hashiguchi T,Fukuda Y,Matsuoka R,Kuroda D,Kubota M,Shirogane Y,Watanabe S,Tsumoto K,Kohda D,Plemper RK,Yanagi Y.与抑制剂复合的麻疹病毒融合蛋白融合前形式的结构(Structures of the prefusion form of measles virus fusionprotein in complex with inhibitors).美国科学院院报(ProcNatl Acad Sci USA).2018年3月;115(10):第2496-501页.
抗病毒肽缀合物
在某些方面,本发明提供了包含融合抑制肽(FIP)和C端七肽重复(HRC)肽的抗病毒肽缀合物。
麻疹病毒属于麻疹病毒(Morbillivirus)属的副粘病毒。它是一种多形性病毒,直径范围为100nm至300nm。麻疹基因组由六个基因组成,每个基因编码一个结构蛋白:称为N(核衣壳蛋白)、P(磷蛋白)、M(基质蛋白)、F(融合蛋白)、H(血凝素蛋白)和L(大蛋白(LargeProtein))。这些基因之一的磷蛋白(P)基因还编码两种非结构蛋白(V和C)。基于核蛋白的核苷酸序列,存在不同的野生型麻疹病毒遗传谱系。
融合抑制肽(FIP)
在一些实施方式中,FIP是包含氨基酸序列苯丙氨酸-苯丙氨酸-甘氨酸(FFG)的融合抑制肽。在一些实施方式中,FIP是由氨基酸序列苯丙氨酸-苯丙氨酸-甘氨酸(FFG)组成的融合抑制肽。在一些实施方式中,麻疹FIP的第一个苯丙氨酸残基是D-氨基酸并且第二个苯丙氨酸残基和甘氨酸残基是L-氨基酸(即D-FFG)。D-氨基酸是其中氨基的立体碳α具有D-构型的氨基酸。在一些实施方式中,FIP的N末端进一步包含如图53所示的苄氧羰基基团(即,Z-D-FFG)。在一些实施方式中,FIP进一步包含一种或多种丝氨酸-甘氨酸连接体。在一些实施方式中,丝氨酸-甘氨酸连接体包含氨基酸序列GSGSG。在一些实施方式中,丝氨酸-甘氨酸连接体由氨基酸序列GSGSG组成。在一些实施方式中,FIP在FIP的C端包含丝氨酸-甘氨酸连接体(例如FFG-GSGSG、D-FFG-GSGSG、Z-D-GSGSG)。在一些实施方式中,FIP与MeV F的HRB区结合以防止病毒进入宿主细胞。
C端七肽重复(HRC)肽
在一些实施方式中,C端七肽重复(HRC)肽衍生自麻疹病毒F蛋白。HRC肽在麻疹毒株之间是保守的(conserved),并且可以衍生自任何麻疹毒株F蛋白。在一些实施方式中,麻疹病毒F蛋白衍生的C端七肽重复肽衍生自麻疹毒株B3。在一些实施方式中,麻疹病毒衍生的C端七肽重复肽衍生自麻疹毒株G954。麻疹病毒基因组的核苷酸和氨基酸序列以及其中被编码的蛋白质可在本领域技术人员已知的数据库中公开获得,例如但不限于GenBank和ViPR(www.viprbrc.org)。
在一些实施方式中,HRC是麻疹病毒F蛋白衍生的C端七肽重复肽。在一些实施方式中,HRC包含麻疹病毒衍生的F蛋白C端七肽重复肽的残基450至485。在一些实施方式中,HRC包含氨基酸序列PPISLERLDVGTNLGNAIAKLEDAKELLESSDQILR。在一些实施方式中,HRC由氨基酸序列PPISLERLDVGTNLGNAIAKLEDAKELLESSDQILR组成。在一些实施方式中,HRC包含氨基酸序列,该氨基酸序列包含与PPISLERLDVGTNLGNAIAKLEDAKELLESSDQILR约99%、98%、97%、96%、95%、94%、93%、92%、91%、90%、89%、88%、87%、86%、85%、84%、83%、82%、81%、80%、79%、78%、77%、76%或75%的序列同一性。在一些实施方式中,HRC包含麻疹病毒衍生的F蛋白C端七肽重复肽的残基450至462。在一些实施方式中,HRC包含氨基酸序列PPISLERLDVGTN。在一些实施方式中,HRC由氨基酸序列PPISLERLDVGTN组成。在一些实施方式中,HRC包含氨基酸序列,该氨基酸序列包含与PPISLERLDVGTN约99%、98%、97%、96%、95%、94%、93%、92%、91%、90%、89%、88%、87%、86%、85%、84%、83%、82%、81%、80%、79%、78%、77%、76%或75%的序列同一性。在一些实施方式中,HRC包含麻疹病毒衍生的C端七肽重复肽的残基450至463、464、465、466、467、468、469、470、471、472、473、474、475、476、477、478、479、480、481、482、483或484。在一些实施方式中,HRC包含长度在PPISLERLDVGTNLGNAIAKLEDAKELLESSDQILR和PPISLERLDVGTN之间的任何氨基酸序列。在一些实施方式中,HRC肽进一步包含一种或多种丝氨酸-甘氨酸连接体。在一些实施方式中,丝氨酸-甘氨酸连接体包含氨基酸序列GSGSG。在一些实施方式中,丝氨酸-甘氨酸连接体由氨基酸序列GSGSG组成。在一些实施方式中,HRC肽在HRC肽的C端处包含丝氨酸-甘氨酸连接体(例如PPISLERLDVGTNLGNAIAKLEDAKELLESSDQILR-GSGSG、PPISLERLDVGTN-GSGSG)。
人免疫缺陷病毒(HIV)是一种慢病毒,属于逆转录病毒科、正逆转录病毒亚科。根据遗传特征和病毒抗原的差异,将HIV分为1型和2型(HIV-1、HIV-2)。HIV基因组包括gag基因,该基因编码外核膜蛋白(MA,p17)、衣壳蛋白(CA,p24)、核衣壳(NC,p7)和更小的核酸稳定蛋白。gag阅读框的后面是pol阅读框,pol阅读框编码以下酶:蛋白酶(PR,p12)、逆转录酶(RT,p51)和核糖核酸酶H(p15)或RT核糖核酸酶H(共p66)和整合酶(IN,p32)。pol基因后面邻接的是env阅读框,由其衍生出两个包膜糖蛋白:gp120(表面蛋白,SU)和gp41(跨膜蛋白,TM)。除了结构蛋白外,HIV基因组还编码几种调节蛋白:Tat(反式激活蛋白)和Rev(RNA剪接调节剂)是HIV复制启动所必需的,而其他调节蛋白Nef(负调节因子)、Vif(病毒感染因子)、Vpr(病毒蛋白r)和Vpu(病毒蛋白独特)对病毒复制、病毒出芽和发病机制有影响。在一些实施方式中,C端七肽重复(HRC)肽衍生自HIV-1病毒gp41蛋白。gp41 HRC肽在HIV毒株之间是保守的,并且可以衍生自任何HIV-1毒株gp41。在一些实施方式中,gp41 HRC肽是如以下文献中所述的“C34”肽:Pessi等人,赋予天然融合蛋白衍生肽以有效的抗病毒活性的一般策略(A General Strategy to Endow Natural Fusion-protein-Derived Peptides withPotent Antiviral Activity),公共科学图书馆·综合(PLoS One),2012,7(5):e36833,该文献通过引用整体并入本文。HIV-1病毒基因组的核苷酸和氨基酸序列以及其中被编码的蛋白质可在本领域技术人员已知的数据库中公开获得,例如但不限于GenBank和洛斯阿拉莫斯国家实验室(Los Alamos National Laboratory)HIV序列数据库(www.hiv.lanl.gov)。
在一些实施方式中,HRC肽衍生自HIV-gp41(也称为“C34”肽)。
在一些实施方式中,HRC包含HIV-gp41的残基117至150。在一些实施方式中,HRC包含氨基酸序列WMEWDREINNYTSLIHSLIEESQNQQEKNEQELL。在一些实施方式中,HRC由氨基酸序列WMEWDREINNYTSLIHSLIE ESQNQQEKNEQELL组成。在一些实施方式中,HRC包含氨基酸序列,该氨基酸序列包含与WMEWDREINNYTSLIHSLIEESQNQQEKNEQELL约99%、98%、97%、96%、95%、94%、93%、92%、91%、90%、89%、88%、87%、86%、85%、84%、83%、82%、81%、80%、79%、78%、77%、76%或75%的序列同一性。在一些实施方式中,HRC肽进一步包含一种或多种丝氨酸-甘氨酸连接体。在一些实施方式中,丝氨酸-甘氨酸连接体包含氨基酸序列GSGSG。在一些实施方式中,丝氨酸-甘氨酸连接体由氨基酸序列GSGSG组成。在一些实施方式中,HRC肽在HRC肽的C端处包含丝氨酸-甘氨酸连接体(例如WMEWDREINNYTSLIHSLIEESQNQQEKNEQELL-GSGSG)。
在一些实施方式中,抗病毒肽缀合物包含融合抑制肽(FIP)和包含氨基酸序列FFG-PPISLERLDVGTNLGNAIAKLEDAKELLESSDQILR的C端七肽重复(HRC)肽。在一些实施方式中,抗病毒肽缀合物包含融合抑制肽(FIP)和由氨基酸序列FFG-PPISLERLDVGTNLGNAIAKLEDAKELLESSDQILR组成的C端七肽重复(HRC)肽。在一些实施方式中,抗病毒肽缀合物进一步包含一种或多种丝氨酸-甘氨酸连接体。在一些实施方式中,丝氨酸-甘氨酸连接体包含氨基酸序列GSGSG。在一些实施方式中,丝氨酸-甘氨酸连接体由氨基酸序列GSGSG组成。在一些实施方式中,抗病毒肽缀合物在FIP和HRC肽之间包含丝氨酸-甘氨酸连接体(例如FFG-GSGSG-PPISLERLDVGTNLGNAIAKLEDAKELLESSDQILR)。在一些实施方式中,抗病毒肽缀合物在HRC肽的C端处包含丝氨酸-甘氨酸连接体(例如FFG-PPISLERLDVGTNLGNAIAKLEDAKELLESSDQILR-GSGSG)。在一些实施方式中,抗病毒肽缀合物在FIP和HRC肽之间以及在HRC肽的C端处包含丝氨酸-甘氨酸连接体(例如FFG-GSGSG-PPISLERLDVGTNLGNAIAKLEDAKELLESSDQILR-GSGSG)。在一些实施方式中,抗病毒肽缀合物包含氨基酸序列,该氨基酸序列包含与FFG-PPISLERLDVGTNLGNAIAKLEDAKELLESSDQILR、FFG-GSGSG-PPISLERLDVGTNLGNAIAKLEDAKELLESSDQIL、FFG-PPISLERLDVGTNLGNAIAKLEDAKELLESSDQILR-GSGSG或FFG-GSGSG-PPISLERLDVGTNLGNAIAKLEDAKELLESSDQILR-GSGSG约99%、98%、97%、96%、95%、94%、93%、92%、91%、90%、89%、88%、87%、86%、85%、84%、83%、82%、81%、80%、79%、78%、77%、76%或75%的序列同一性。在一些实施方式中,FIP的第一个苯丙氨酸残基是D-氨基酸(即,D-FFG-……)。在一些实施方式中,抗病毒肽缀合物的N端进一步包含苄氧羰基基团(即,Z-D-FFG-……)。在一些实施方式中,抗病毒肽缀合物包含用于将FIP-HRC肽缀合至本文所述的连接体和膜定位部分的C端半胱氨酸残基(例如FFG-PPISLERLDVGTNLGNAIAKLEDAKELLESSDQILR-C、FFG-GSGSG-PPISLERLDVGTNLGNAIAKLEDAKELLESSDQIL-C、FFG-PPISLERLDVGTNLGNAIAKLEDAKELLESSDQILR-GSGSG-C或FFG-GSGSG-PPISLERLDVGTNLGNAIAKLEDAKELLESSDQILR-GSGSG-C)。
在一些实施方式中,抗病毒肽缀合物包含融合抑制肽(FIP)和包含氨基酸序列FFG-PPISLERDVGTN的C端七肽重复(HRC)肽。在一些实施方式中,抗病毒肽缀合物包含融合抑制肽(FIP)和由氨基酸序列FFG-PPISLERLDVGTN组成的C端七肽重复(HRC)肽。在一些实施方式中,抗病毒肽缀合物进一步包含一种或多种丝氨酸-甘氨酸连接体。在一些实施方式中,丝氨酸-甘氨酸连接体包含氨基酸序列GSGSG。在一些实施方式中,丝氨酸-甘氨酸连接体由氨基酸序列GSGSG组成。在一些实施方式中,抗病毒肽缀合物在FIP和HRC肽之间包含丝氨酸-甘氨酸连接体(例如FFG-GSGSG-PPISLERLDVGTN)。在一些实施方式中,抗病毒肽缀合物在HRC肽的C端包含丝氨酸-甘氨酸连接体(例如FFG-PPISLERLDVGTN-GSGSG)。在一些实施方式中,抗病毒肽缀合物在FIP和HRC肽之间以及在HRC肽的C端处包含丝氨酸-甘氨酸连接体(例如FFG-GSGSG-PPISLERLDVGTN-GSGSG)。在一些实施方式中,抗病毒肽缀合物包含的氨基酸包含与FFG-PPISLERLDVGTN、FFG-GSGSG-PPISLERLDVGTN、FFG-PPISLERLDVGTN-GSGSG或FFG-GSGSG-PPISLERLDVGTN-GSGSG约99%、98%、97%、96%、95%、94%、93%、92%、91%、90%、89%、88%、87%、86%、85%、84%、83%、82%、81%、80%、79%、78%、77%、76%或75%的序列同一性。在一些实施方式中,FIP的第一个苯丙氨酸残基是D-氨基酸(即,D-FFG-……)。在一些实施方式中,抗病毒肽缀合物的N端进一步包含苄氧羰基基团(即,Z-D-FFG-……)。在一些实施方式中,抗病毒肽缀合物包含用于将FIP-HRC肽缀合至本文所述的连接体和膜定位部分的C端半胱氨酸残基(例如FFG-PPISLERLDVGTN-C、FFG-GSGSG-PPISLERLDVGTN-C、FFG-PPISLERLDVGTN-GSGSG-C或FFG-GSGSG-PPISLERLDVGTN-GSGSG-C)。
在一些实施方式中,抗病毒肽缀合物包含融合抑制肽(FIP)和包含氨基酸序列FFG-WMEWDREINNYTSLIHSLIEESQNQQEKNEQELL的C端七肽重复(HRC)肽。在一些实施方式中,抗病毒肽缀合物包含融合抑制肽(FIP)和由氨基酸序列FFG-WMEWDREINNYTSLIHSLIEESQNQQEKNEQELL组成的C端七肽重复(HRC)肽。在一些实施方式中,抗病毒肽缀合物进一步包含一种或多种丝氨酸-甘氨酸连接体。在一些实施方式中,丝氨酸-甘氨酸连接体包含氨基酸序列GSGSG。在一些实施方式中,丝氨酸-甘氨酸连接体由氨基酸序列GSGSG组成。在一些实施方式中,抗病毒肽缀合物在FIP和HRC肽之间包含丝氨酸-甘氨酸连接体(例如FFG-GSGSG-WMEWDREINNYTSLIHSLIEESQNQQEKNEQELL)。在一些实施方式中,抗病毒肽缀合物在HRC肽的C端处包含丝氨酸-甘氨酸连接体(例如FFG-WMEWDREINNYTSLIHSLIEESQNQQEKNEQELL-GSGSG)。在一些实施方式中,抗病毒肽缀合物在FIP和HRC肽之间以及在HRC肽的C端处包含丝氨酸-甘氨酸连接体(例如FFG-GSGSG-WMEWDREINNYTSLIHSLIEESQNQQEKNEQELL-GSGSG)。在一些实施方式中,抗病毒肽缀合物包含的氨基酸包含与FFG-WMEWDREINNYTSLIHSLIEESQNQQEKNEQELL、FFG-GSGSG-WMEWDREINNYTSLIHSLIEESQNQQEKNEQELL、FFG-WMEWDREINNYTSLIHSLIEESQNQQEKNEQELL-GSGSG或FFG-GSGSG-WMEWDREINNYTSLIHSLIEESQNQQEKNEQELL-GSGSG约99%、98%、97%、96%、95%、94%、93%、92%、91%、90%、89%、88%、87%、86%、85%、84%、83%、82%、81%、80%、79%、78%、77%、76%或75%的序列同一性。在一些实施方式中,FIP的第一个苯丙氨酸残基是D-氨基酸(即,D-FFG-……)。在一些实施方式中,抗病毒肽缀合物的N端进一步包含苄氧羰基基团(即,Z-D-FFG-……)。在一些实施方式中,抗病毒肽缀合物包含用于将FIP-HRC肽缀合至本文所述的连接体和膜定位部分的C端半胱氨酸残基(例如FFG-WMEWDREINNYTSLIHSLIEESQNQQEKNEQELL-C、FFG-GSGSG-WMEWDREINNYTSLIHSLIEESQNQQEKNEQELL-C、FFG-WMEWDREINNYTSLIHSLIEESQNQQEKNEQELL-GSGSG-C或FFG-GSGSG-WMEWDREINNYTSLIHSLIEESQNQQEKNEQELL-GSGSG-C)。
无意受任何理论的束缚,病毒融合蛋白重新折叠并达到融合后状态(即,进入细胞)的能力依赖于位于蛋白质的N端和C端的两个互补的七肽重复(HR)区(分别为HRN和HRC)之间的相互作用。在一些实施方式中,包含FIP和HRC区的抗病毒肽缀合物稳定融合蛋白的融合前状态(例如,可逆结合,表明融合蛋白仍处于其融合前状态,参见图74)。在一些实施方式中,包含FIP和HRC区的抗病毒肽缀合物将麻疹融合蛋白稳定在融合前状态。在一些实施方式中,包含FIP和HRC区的抗病毒肽缀合物将HIV包膜蛋白稳定在融合前状态。
如本文所述,甘氨酸丝氨酸连接体包含氨基酸序列GSGSG。然而,可以使用更短或更长的甘氨酸丝氨酸连接体。在一个实施方式中,甘氨酸丝氨酸连接体具有式(GS)n,或G(SG)n,或S(GS)n,其中n为1、2、3、4、5、6、7、8、9和10。
连接体(Linker)
在一些实施方式中,抗病毒肽缀合物进一步包含连接体。在一些实施方式中,连接体是基于其记录的生物相容性、溶解性和低免疫原性或抗原性的组分。在一些实施方式中,连接体是聚乙二醇(PEG)连接体。PEG是指由重复的乙二醇单元组成的化合物。肽与PEG的共价缀合——称为聚乙二醇化——在本领域中是众所周知的。参见例如Turecek等人,生物制药的聚乙二醇化:已获批准药物的化学和非临床安全信息综述(PEGylation ofBiopharmaceuticals:A Review of Chemistry and Nonclinical Safety Informationof Approved Drugs),药物科学期刊(Journal of Pharmaceutical Sciences),105(2016,460-475,该文献通过引用整体并入本文)。可以通过与PEG部分和/或肽相连的官能团键合(linkage),将PEG连接体化合物与肽相连。可以使用生物活性分子的氨基作为连接位点来实现PEG(包括直链和支链PEG聚合物)与生物活性分子的共价连接。可以使用生物活性分子的硫醇基作为连接位点来实现PEG(包括直链和支链PEG聚合物)与生物活性分子的共价连接。可替代地,生物活性分子本身可以被修饰以包含官能团(例如胺、硫醇)以提供与直链和支链PEG聚合物的连接位点。
在一些实施方式中,PEG连接体是双官能的。双官能PEG连接体化合物具有两个官能团,因此可以通过每个官能团连接至两个生物活性分子(例如本文所述的抗病毒肽),以产生包含两种抗病毒肽的缀合物。在一些实施方式中,抗病毒肽缀合物的两种抗病毒肽是相同的(即,同型二价(homobivalent)缀合物)。在一些实施方式中,抗病毒肽缀合物的两种抗病毒肽是不同的(即,异型(heterobivalent)二价缀合物)。在一些实施方式中,PEG连接体可以是支链的和/或具有多个官能团,例如但不限于三个、四个、五个或更多个官能团。这样的多价PEG连接体可通过每个官能团连接至多个生物活性分子(例如本文所述的抗病毒肽),以产生包含多种抗病毒肽的缀合物,例如但不限于三种、四种、五种或更多种抗病毒肽。在一些实施方式中,多价抗病毒肽缀合物的抗病毒肽是相同的(即,同型多价缀合物)。在一些实施方式中,抗病毒肽缀合物的抗病毒肽是不同的(即,异型多价缀合物)。本发明的多价抗病毒肽缀合物可以包含FIP和HRC肽的多种组合(例如但不限于图83)。例如,在一个实施方式中,多价抗病毒肽缀合物包含FIP-HRC缀合物,进一步缀合至支链PEG连接体,可选地进一步包含如本文所述的膜定位部分。FIP-HRC缀合物可以是相同的或不同的。在另一个实施方式中,多价抗病毒肽缀合物包含缀合至支链PEG连接体的多个FIP,进一步缀合至HRC肽,可选地进一步包括膜定位部分。在一些实施方式中,多价抗病毒肽缀合物包含缀合至支链PEG连接体的多个FIP,可选地进一步包含膜定位部分。
在一些实施方式中,PEG连接体具有0至50个乙二醇单元(glycol unit)。在一些实施方式中,PEG连接体具有1个乙二醇单元(即,连接体是PEG1)。在一些实施方式中,PEG连接体具有2个乙二醇单元(即,连接体是PEG2)。在一些实施方式中,PEG连接体具有3个乙二醇单元(即,连接体是PEG3)。在一些实施方式中,PEG连接体具有4个乙二醇单元(即,连接体是PEG4)。在一些实施方式中,PEG连接体具有5个乙二醇单元(即,连接体是PEG5)。在一些实施方式中,PEG连接体具有6个乙二醇单元(即,连接体是PEG6)。在一些实施方式中,PEG连接体具有7个乙二醇单元(即,连接体是PEG7)。在一些实施方式中,PEG连接体具有8个乙二醇单元(即,连接体是PEG8)。在一些实施方式中,PEG连接体具有9个乙二醇单元(即,连接体是PEG9)。在一些实施方式中,PEG连接体具有10个乙二醇单元(即,连接体是PEG10)。在一些实施方式中,PEG连接体具有11个乙二醇单元(即,连接体是PEG11)。在一些实施方式中,PEG连接体具有12个乙二醇单元(即,连接体是PEG12)。在一些实施方式中,PEG连接体具有13个乙二醇单元(即,连接体是PEG13)。在一些实施方式中,PEG连接体具有14个乙二醇单元(即,连接体是PEG14)。在一些实施方式中,PEG连接体具有15个乙二醇单元(即,连接体是PEG15)。在一些实施方式中,PEG连接体具有16个乙二醇单元(即,连接体是PEG16)。在一些实施方式中,PEG连接体具有17个乙二醇单元(即,连接体是PEG17)。在一些实施方式中,PEG连接体具有18个乙二醇单元(即,连接体是PEG18)。在一些实施方式中,PEG连接体具有19个乙二醇单元(即,连接体是PEG19)。在一些实施方式中,PEG连接体具有20个乙二醇单元(即,连接体是PEG20)。在一些实施方式中,PEG连接体具有21个乙二醇单元(即,连接体是PEG21)。在一些实施方式中,PEG连接体具有22个乙二醇单元(即,连接体是PEG22)。在一些实施方式中,PEG连接体具有23个乙二醇单元(即,连接体是PEG23)。在一些实施方式中,PEG连接体具有24个乙二醇单元(即,连接体是PEG24)。在一些实施方式中,PEG连接体具有25个乙二醇单元(即,连接体是PEG25)。在一些实施方式中,PEG连接体具有26个乙二醇单元(即,连接体是PEG26)。在一些实施方式中,PEG连接体具有27个乙二醇单元(即,连接体是PEG27)。在一些实施方式中,PEG连接体具有28个乙二醇单元(即,连接体是PEG28)。在一些实施方式中,PEG连接体具有29个乙二醇单元(即,连接体是PEG29)。在一些实施方式中,PEG连接体具有30个乙二醇单元(即,连接体是PEG30)。在一些实施方式中,PEG连接体具有31个乙二醇单元(即,连接体是PEG31)。在一些实施方式中,PEG连接体具有32个乙二醇单元(即,连接体是PEG32)。在一些实施方式中,PEG连接体具有33个乙二醇单元(即,连接体是PEG33)。在一些实施方式中,PEG连接体具有34个乙二醇单元(即,连接体是PEG34)。在一些实施方式中,PEG连接体具有35个乙二醇单元(即,连接体是PEG35)。在一些实施方式中,PEG连接体具有36个乙二醇单元(即,连接体是PEG36)。在一些实施方式中,PEG连接体具有50个乙二醇单元(即,连接体是PEG50)。在一些实施方式中,PEG连接体具有4至12个乙二醇单元。在一些实施方式中,连接体具有4至24个乙二醇单元。在一些实施方式中,PEG连接体是PEG 5000,其是平均分子量约为5000Da的聚乙二醇聚合物。PEG 5000包含约114个乙二醇单元,因此,在一些实施方式中,PEG连接体具有约114个乙二醇单元(即,连接体是PEG114)。在一些实施方式中,PEG连接体是PEG 40,000,其是平均分子量约为40,000Da的聚乙二醇聚合物。PEG 40000包含约910个乙二醇单元,因此,在一些实施方式中,PEG连接体具有约910个乙二醇单元(即,连接体是PEG910)。
在一些实施方式中,抗病毒肽缀合物包含可选地缀合至PEG连接体的两个FIP-HRC肽,该缀合物具有下式:
[FIP]-Gx-[HRC]-Gx-L-Pn-L-Gx-[HRC]-Gx-[FIP]
其中,[FIP]包含如本文所述的融合抑制肽;
G包含如本文所述的GSGSG连接体,其中,任何一个G基团的x=0或1;
[HRC]包含如本文所述的C端七肽重复肽;
L是一个或多个官能团键合;以及
P是PEG部分,其中n是乙二醇单元的数量,并且n=0-50。在一些实施方式中,n是4。在一些实施方式中,n是11。在一些实施方式中,n是12。在一些实施方式中,n是24。在一些实施方式中,可以使用更短或更长的甘氨酸丝-氨酸连接体。在一些实施方式中,甘氨酸丝氨酸连接体具有式(GS)n,或G(SG)n,或S(GS)n,其中n为1、2、3、4、5、6、7、8、9和10。
在一些实施方式中,L包含一个或多个硫化物部分。在一些实施方式中,一个或多个硫化物部分源自一个或多个硫醇部分。在一些实施方式中,一个或多个硫化物部分源自半胱氨酸。在一些实施方式中,一个或多个硫化物部分缀合至HRC肽的C端。在一些实施方式中,一个或多个硫醇部分缀合至HRC肽的C端。在一些实施方式中,一个或多个半胱氨酸部分包含HRC肽的C端。在一些实施方式中,L包含一个或多个吡咯烷二酮部分。在一些实施方式中,一个或多个吡咯烷二酮部分源自一个或多个马来酰亚胺部分。在一些实施方式中,一个或多个吡咯烷二酮部分缀合至PEG。在一些实施方式中,一个或多个马来酰亚胺部分至缀合PEG。在一些实施方式中,通过将一个或多个硫醇部分与一个或多个马来酰亚胺部分偶联来形成L。在一些实施方式中,通过将缀合至HRC肽的C端的一个或多个硫醇部分与缀合至PEG的一个或多个马来酰亚胺部分偶联来形成L。在一些实施方式中,通过将HRC肽的C端处的一个或多个半胱氨酸部分与缀合至PEG的一个或多个马来酰亚胺部分偶联来形成L。
在一些实施方式中,未缀合的FIP-HRC肽的C端半胱氨酸残基终止于硫醇(通用结构“R-SH”,其中R是FIP-HRC肽)。硫醇和马来酰亚胺之间的交叉偶联反应将这两种组分连接起来以形成硫化物(通用结构“R1-S-R2”,其中R1是FIP-HRC肽,R2是PEG连接体)。

在一些实施方式中,L包含

在一些实施方式中,L包含

在一些实施方式中,抗病毒肽缀合物包含可选地缀合至PEG连接体的两个FIP-HRC肽,该缀合物具有下式:

其中,[FIP]包含如本文所述的融合抑制肽;
G包含如本文所述的GSGSG连接体,其中,任何一个G基团的x=0或1;[HRC]包含如本文所述的C端七肽重复肽;并且
其中n=0-50。在一些实施方式中,n是4。在一些实施方式中,n是11。在一些实施方式中,n是12。在一些实施方式中,n是24。在一些实施方式中,可以使用更短或更长的甘氨酸丝-氨酸连接体(linker)。在一些实施方式中,甘氨酸丝氨酸连接体具有式(GS)n,或G(SG)n,或S(GS)n,其中n为1、2、3、4、5、6、7、8、9和10。
膜定位部分(Membrane Localizing Moieties)/锚(Anchor)
在一些实施方式中,抗病毒肽缀合物包含如本文所述的融合抑制肽(FIP)和C端七肽重复(HRC)肽,并且进一步包含膜定位部分(在本文中也称为锚)。膜定位部分是增强肽定位在靶向治疗位置的能力的任何组分。在一些实施方式中,膜定位部分是疏水的(或以其他方式提高抗病毒肽缀合物的疏水性),这增加了肽定位在脂质膜处和/或插入到脂质膜中的趋势(例如,参见图56)。在一些实施方式中,膜定位部分是脂质。在一些实施方式中,膜定位部分是胆固醇、生育酚或棕榈基。在一些实施方式中,膜定位部分缀合至抗病毒肽的C端(参见图61和图64)。在一些实施方式中,膜定位部分缀合至连接体(例如PEG4或PEG11),并且该连接体缀合至肽的C端(参见图61和图64)。可以通过与膜定位部分和/或肽相连的官能团键合,将本发明的膜定位部分与肽相连。可以使用生物活性分子的氨基作为连接位点来实现膜定位部分与生物活性分子的共价连接。可替代地,生物活性分子本身可以被修饰以包含官能团,以提供与膜定位部分的连接位点。
在一些实施方式中,抗病毒肽缀合物包含可选地缀合至PEG连接体并缀合至膜定位部分的FIP-HRC肽,该缀合物具有下式:
[FIP]-Gx-[HRC]-Gx-L-Pn-L-MLM
其中,[FIP]包含如本文所述的融合抑制肽;
G包含如本文所述的GSGSG连接体,其中,任何一个G基团的x=0或1;[HRC]包含如本文所述的C端七肽重复肽;
L是一个或多个官能团键合;
P是PEG部分,其中,n是乙二醇单元的数量,n=0-50;并且MLM是膜定位部分。在一些实施方式中,n是4。在一些实施方式中,n是11。在一些实施方式中,n是12。在一些实施方式中,n是24。在一些实施方式中,MLM是胆固醇。在一些实施方式中,MLM是生育酚。在一些实施方式中,MLM是棕榈基。在一些实施方式中,可以使用更短或更长的甘氨酸丝-氨酸连接体(linker)。在一些实施方式中,甘氨酸丝氨酸连接体具有式(GS)n,或G(SG)n,或S(GS)n,其中n为1、2、3、4、5、6、7、8、9和10。
在一些实施方式中,L包含

在一些实施方式中,L包含

在一些实施方式中,L包含

在一些实施方式中,MLM包含

在一些实施方式中,MLM包含

在一些实施方式中,MLM包含

在一些实施方式中,抗病毒肽缀合物包含缀合至膜定位部分连接体的FIP-HRC肽,所述缀合物具有下式:
[FIP]-Gx-[HRC]-Gx-L-MLM
其中[FIP]包含如本文所述的融合抑制肽;
G包含如本文所述的GSGSG连接体,其中,任何一个G基团的x=0或1;[HRC]包含如本文所述的C端七肽重复肽;
L为一个或多个官能团键合;以及
MLM是膜定位部分。在一些实施方式中,MLM是胆固醇。在一些实施方式中,MLM是生育酚。在一些实施方式中,MLM是棕榈基。在一些实施方式中,可以使用更短或更长的甘氨酸丝-氨酸连接体(linker)。在一些实施方式中,甘氨酸丝氨酸连接体具有式(GS)n,或G(SG)n,或S(GS)n,其中n为1、2、3、4、5、6、7、8、9和10。
在一些实施方式中,L包含

在一些实施方式中,L包含

在一些实施方式中,L包含

在一些实施方式中,MLM包含

在一些实施方式中,MLM包含

在一些实施方式中,MLM包含

在一些实施方式中,L包含一个或多个硫化物部分。在一些实施方式中,一个或多个硫化物部分源自一个或多个硫醇部分。在一些实施方式中,一个或多个硫化物部分源自半胱氨酸。在一些实施方式中,一个或多个硫化物部分缀合至HRC肽的C端。在一些实施方式中,一个或多个硫醇部分缀合至HRC肽的C端。在一些实施方式中,一个或多个半胱氨酸部分包含HRC肽的C端。在一些实施方式中,L包含一个或多个酰胺部分。在一些实施方式中,一个或多个酰胺部分源自一个或多个溴酰胺。在一些实施方式中,L包含一个或多个酯部分。在一些实施方式中,一个或多个酯部分源自一个或多个溴酯。在一些实施方式中,一个或多个酰胺、溴酰胺、酯或溴酯部分缀合至MLM。在一些实施方式中,MLM缀合至PEG连接体,其继而包含一个或多个酰胺、溴酰胺、酯或溴酯部分。在一些实施方式中,通过将一个或多个硫醇部分与一个或多个溴酰胺或溴酯部分偶联来形成L。在一些实施方式中,通过将缀合至HRC肽的C端的一个或多个硫醇部分与缀合至MLM或PEG连接体的一个或多个溴酰胺或溴酯部分偶联来形成L。在一些实施方式中,通过将HRC肽的C端处的一个或多个半胱氨酸部分与缀合至MLM或PEG连接体的一个或多个溴酰胺或溴酯部分偶联来形成L。
在一些实施方式中,抗病毒肽缀合物包含可选地缀合至PEG连接体并缀合至膜定位部分的FIP-HRC肽,该缀合物具有下式:

其中,[FIP]包含如本文所述的融合抑制肽;
G包含如本文所述的GSGSG连接体,其中,任何一个G基团的x=0或1;[HRC]包含如本文所述的C端七肽重复肽;
其中n=0-50;并且MLM是膜定位部分。在一些实施方式中,n是4。在一些实施方式中,n是11。在一些实施方式中,n是12。在一些实施方式中,n是24。在一些实施方式中,MLM是胆固醇。在一些实施方式中,MLM是生育酚。在一些实施方式中,MLM是棕榈基。在一些实施方式中,可以使用更短或更长的甘氨酸丝-氨酸连接体(linker)。在一些实施方式中,甘氨酸丝氨酸连接体具有式(GS)n,或G(SG)n,或S(GS)n,其中n为1、2、3、4、5、6、7、8、9和10。
在一些实施方式中,MLM包含

在一些实施方式中,MLM包含

在一些实施方式中,MLM包含

在一些实施方式中,抗病毒肽缀合物包含缀合至膜定位部分连接体的FIP-HRC肽,所述缀合物具有下式:

其中,[FIP]包含如本文所述的融合抑制肽;
G包含如本文所述的GSGSG连接体,其中,任何一个G基团的x=0或1;[HRC]包含如本文所述的C端七肽重复肽;并且
MLM是膜定位部分。在一些实施方式中,MLM是胆固醇。在一些实施方式中,MLM是生育酚。在一些实施方式中,MLM是棕榈基。在一些实施方式中,可以使用更短或更长的甘氨酸丝-氨酸连接体(linker)。在一些实施方式中,甘氨酸丝氨酸连接体具有式(GS)n,或G(SG)n,或S(GS)n,其中n为1、2、3、4、5、6、7、8、9和10。
在一些实施方式中,MLM包含

在一些实施方式中,MLM包含

在一些实施方式中,MLM包含

在一些实施方式中,抗病毒肽缀合物包含两种FIP-HRC肽,每种FIP-HRC肽可选地缀合至PEG连接体并缀合至膜定位部分,该缀合物具有下式:

其中,[FIP]包含如本文所述的融合抑制肽;
G包含如本文所述的GSGSG连接体,其中,任何一个G基团的x=0或1;[HRC]包含如本文所述的C端七肽重复肽;
L为一个或多个官能团键合基团;
P是PEG部分,其中n是乙二醇单元的数量,n=0-50;以及
MLM是膜定位部分。在一些实施方式中,MLM是胆固醇。在一些实施方式中,MLM是生育酚。在一些实施方式中,MLM是棕榈基。在一些实施方式中,n是4。在一些实施方式中,n是11。在一些实施方式中,n是12。在一些实施方式中,n是24。在一些实施方式中,可以使用更短或更长的甘氨酸丝-氨酸连接体(linker)。在一些实施方式中,甘氨酸丝氨酸连接体具有式(GS)n,或G(SG)n,或S(GS)n,其中n为1、2、3、4、5、6、7、8、9和10。
在一些实施方式中,官能团键合包括

在一些实施方式中,官能团键合包括

在一些实施方式中,官能团键合包括

在一些实施方式中,官能团键合包括

在一些实施方式中,MLM包含

在一些实施方式中,MLM包含

在一些实施方式中,MLM包含

在一些实施方式中,L包含一个或多个硫化物部分。在一些实施方式中,一个或多个硫化物部分源自一个或多个硫醇部分。在一些实施方式中,一个或多个硫化物部分源自半胱氨酸。在一些实施方式中,一个或多个硫化物部分缀合至HRC肽的C端。在一些实施方式中,一个或多个硫醇部分缀合至HRC肽的C端。在一些实施方式中,一个或多个半胱氨酸部分包含HRC肽的C端。在一些实施方式中,L包含一个或多个吡咯烷二酮部分。在一些实施方式中,一个或多个吡咯烷二酮部分源自一个或多个马来酰亚胺部分。在一些实施方式中,一个或多个吡咯烷二酮部分缀合至一个或多个PEG,其继而可各自通过例如酯键或酰胺键缀合至支链连接体。在一些实施方式中,支链PEG连接体部分通过例如醚键缀合至MLM。在一些实施方式中,一个或多个马来酰亚胺部分缀合至一个或多个PEG或支链PEG连接体部分。在一些实施方式中,通过将一个或多个硫醇部分与一个或多个马来酰亚胺部分偶联来形成L。在一些实施方式中,通过将缀合至HRC肽的C端的一个或多个硫醇部分与缀合至一个或多个PEG或支链PEG连接体部分的一个或多个马来酰亚胺部分偶联来形成L。在一些实施方式中,通过将HRC肽的C端处的一个或多个半胱氨酸部分与缀合至一个或多个PEG或支链PEG连接体部分的一个或多个马来酰亚胺部分偶联来形成L。
在一些实施方式中,抗病毒肽缀合物包含两种FIP-HRC肽,每种FIP-HRC肽可选地缀合至PEG连接体并缀合至膜定位部分,该缀合物具有下式:

其中,[FIP]包含如本文所述的融合抑制肽;
G包含如本文所述的GSGSG连接体,其中,任何一个G基团的x=0或1;[HRC]包含如本文所述的C端七肽重复肽;
其中n=0-50;以及
MLM是膜定位部分。在一些实施方式中,MLM是胆固醇。在一些实施方式中,MLM是生育酚。在一些实施方式中,MLM是棕榈基。在一些实施方式中,n是4。在一些实施方式中,n是11。在一些实施方式中,n是12。在一些实施方式中,n是24。在一些实施方式中,可以使用更短或更长的甘氨酸丝-氨酸连接体(linker)。在一些实施方式中,甘氨酸丝氨酸连接体具有式(GS)n,或G(SG)n,或S(GS)n,其中n为1、2、3、4、5、6、7、8、9和10。
在一些实施方式中,MLM包含

在一些实施方式中,MLM包含

在一些实施方式中,MLM包含

在一些实施方式中,抗病毒肽缀合物包含如本文所述的单体形式的融合抑制肽(FIP)和C端七肽重复(HRC)肽(“FIP-HRC450-485”,参见图64)。
在一些实施方式中,抗病毒肽缀合物包含如本文所述的融合抑制肽(FIP)和C端七肽重复(HRC)肽,其呈单体形式并且进一步包含缀合至肽的C端的脂质(例如胆固醇)(“FIP-HRC450-485-chol”,参见图64)。
在一些实施方式中,抗病毒肽缀合物包含如本文所述的融合抑制肽(FIP)和C端七肽重复(HRC)肽,其呈单体形式并且进一步包含缀合至肽的C端的连接体(例如PEG4)和脂质(例如胆固醇)(“FIP-HRC450-485-peg4-chol”,参见图64)。
在一些实施方式中,抗病毒肽缀合物包含如本文所述的融合抑制肽(FIP)和C端七肽重复(HRC)肽,其呈二聚体形式并且进一步包含缀合至肽的C端的连接体(例如PEG4)和脂质(例如胆固醇)(“[FIP-HRC450-485-PEG4]2-chol”,参见图64)。
在一些实施方式中,抗病毒肽缀合物包含如本文所述的融合抑制肽(FIP)和C端七肽重复(HRC)肽,其呈二聚体形式并且进一步包含缀合至肽的C端的连接体(例如PEG11)(“[FIP-HRC450-485-peg11]2”,参见图64)。
抗病毒肽缀合物纳米颗粒
在某些方面,本发明提供了一种包含本文所述的抗病毒肽缀合物的纳米颗粒。在一些实施方式中,本发明的纳米颗粒具有约50nm至约150nm的直径。在一些实施方式中,本发明提供了包含多个纳米颗粒的组合物,该纳米颗粒包含多个本文所述的任何抗病毒肽缀合物。在一些实施方式中,纳米颗粒由抗病毒肽缀合物组成并且进一步包含其他融合脂质(fusogenic lipid)或天然脂质(例如1-棕榈酰-2-油酰-甘油-3-磷酸胆碱(POPC)和磷脂酰甘油(POPG))。在一些实施方式中,纳米颗粒的粒径适合在药物组合物中递送。在一些实施方式中,纳米颗粒被封装在水凝胶中,该水凝胶用于进行受控的、局部和缓慢的递送。在一些实施方式中,纳米颗粒的直径适合于吸入、鼻内施用或直接滴注至肺中(例如,使用通过吸入器、气雾器或喷雾器的递送)。
在一些实施方式中,当将抗病毒肽缀合物置于水溶液中时,它们自组装成纳米颗粒,使得肽的疏水区(例如膜定位部分)结合形成疏水核,而肽(例如FIP-HRC肽)的亲水区向外延伸(参见例如图26)。在一些实施方式中,当纳米颗粒接近双层脂质(例如宿主细胞膜)时,它们解装配,因为疏水区(例如膜定位部分)将与脂质膜相互作用,而亲水区(例如FIP-HRC肽)将面向水溶液(参见例如图27)。
稳定的可溶性麻疹F蛋白
之前,可溶性F总是转换至其融合后状态,除非经二硫键改造以维持融合前状态,就像呼吸道合胞病毒F疫苗候选物一样。本文描述的是麻疹F中的突变,其将可溶性F蛋白稳定在其融合前状态。在以可溶形式产生时,随着胞质内结构域和跨膜结构域的消融(ablation),人副流感病毒3型野生型(wt)F蛋白和麻疹wt F蛋白都在融合后状态下折叠,即使在C端添加了折叠体(foldomer)以保持三聚体低聚化。如果wt麻疹F蛋白用补充有本文所述的抗病毒肽缀合物(例如但不限于FIP-HRC-PEG11-二聚体)的培养基表达,则wt F蛋白保持在其融合前状态。在一些实施方式中,可溶性F蛋白包括SEQ ID NO:1或SEQ ID NO:2。在一些实施方式中,可溶性F蛋白由SEQ ID NO:1或SEQ ID NO:2构成。在一些实施方式中,可溶性F蛋白包含氨基酸序列,该氨基酸序列包含与SEQ ID NO:1或SEQ ID NO:2约99%、98%、97%、96%、95%、94%、93%、92%、91%、90%、89%、88%、87%、86%、85%、84%、83%、82%、81%、80%、79%、78%、77%、76%或75%的序列同一性。MGLKVNVSAIFMAVLLTLQTPTGQIHWGNLSKIGVVGIGSASYKVMTRSSHQSLVIKLMPNITLLNNCTRVEIAEYRRLLRTVLEPIRDALNAMTQNIRPVQSVASSRRHKRFAGVVLAGAALGVATAAQITAGIALHQSMLNSQAIDNLRASLETTNQAIEAIRQAGQEMILAVQGVQDYINNELIPSMNQLSCDLIGQKLGLKLLRYYTEILSLFGPSLRDPISAEISIQALSYALGGDINKVLEKLGYSGGDLLGILESRGIKARITHVDTESYFIVLSIAYPTLSEIKGVIVHRLEGVSYNIGSQEWYTTVPKYVATQGYLISNFDESSCTFMPEGTVCSQNALYPMSPLLQECLRGSTKSCARTLVSGSFGNRFILSQGNLIANCASILCKCYTTGTIINQDPDKILTYIAADHCPVVEVNGVTIQVGSRRYPDAVYLHRIDLGPPISLERLDVGTNLGNAIAKLEDAKELLESSDQILRSMKGLSSTS(SEQ ID NO:1)。
在一些实施方式中,可溶性F蛋白可选地包含烟草蚀纹病毒(Tobacco EtchVirus)蛋白酶位点(以斜体表示)、连接体(以粗体表示)、折叠体结构域(以双下划线表示)和6xHis标签(以粗斜体表示)。

在某些方面,本发明提供了一种包含可溶性麻疹F蛋白的重组多肽,该可溶性麻疹F蛋白包含一个或多个突变,该一个或多个突变产生稳定的蛋白而无需经改造的二硫键。在一些实施方式中,该突变是E455G。在一些实施方式中,一个或多个突变是E170G和E455G。在某些方面,该技术提供了编码这些重组多肽的核酸。
在一些实施方式中,包含突变E455G的稳定的可溶性F蛋白包含SEQ ID NO:3或SEQID NO:4。在一些实施方式中,包含突变E455G的稳定的可溶性F蛋白由SEQ ID NO:3或SEQID NO:4组成。在一些实施方式中,稳定的可溶性F蛋白包含氨基酸序列,该氨基酸序列包含与SEQ ID NO:3或SEQ ID NO:4约99%、98%、97%、96%、95%、94%、93%、92%、91%、90%、89%、88%、87%、86%、85%、84%、83%、82%、81%、80%、79%、78%、77%、76%或75%的序列同一性,其中位于455位的氨基酸是G。

在一些实施方式中,包含突变E455G的稳定的可溶性F蛋白可选地包含烟草蚀纹病毒蛋白酶位点(以斜体表示)、连接体(以粗体表示)、折叠体结构域(以双下划线表示)和6xHis标签(以粗斜体表示)。

在一些实施方式中,包含双突变体E170G和E455G的稳定的可溶性F蛋白包含SEQID NO:5或SEQ ID NO:6。在一些实施方式中,包含双突变体E170G和E455G的稳定的可溶性F蛋白由SEQ ID NO:5或SEQ ID NO:6组成。在一些实施方式中,包含双突变E170G和E455G的稳定的可溶性F蛋白包含氨基酸序列,该氨基酸序列包含与SEQ NO:5或SEQ ID NO:6约99%、98%、97%、96%、95%、94%、93%、92%、91%、90%、89%、88%、87%、86%、85%、84%、83%、82%、81%、80%、79%、78%、77%、76%或75%的序列同一性,其中,位于170位的氨基酸是G,位于455位的氨基酸是G。

在一些实施方式中,包含双突变E170G和E455G的稳定的可溶性F蛋白可选地包含烟草蚀纹病毒蛋白酶位点(以斜体表示)、连接体(以粗体表示)、折叠体结构域(以双下划线表示)和6xHis标签(以粗斜体表示)。

在某些方面,本发明提供了一种细胞,该细胞包含编码本发明的适用于重组表达的任何一种可溶性F蛋白的核酸(例如SEQ ID NO:1、2、3、4、5或6)。在某些方面,本发明提供了一种无性繁殖地衍生的细胞群,该细胞群包含编码本发明的适用于重组表达的任何一种可溶性F蛋白的核酸(例如SEQ ID NO:1、2、3、4、5或6)。在某些方面,本发明提供了稳定的细胞池(pool of cells),该细胞池包含编码本发明的适用于重组表达的任何一种可溶性F蛋白的核酸(例如SEQ ID NO:1、2、3、4、5或6)。在一些实施方式中,核酸序列经密码子优化以在宿主细胞(例如哺乳动物细胞)或任何其他合适的表达系统中实现最佳表达。
本领域技术人员理解,在本发明的可溶性F蛋白(例如SEQ ID No:1、2、3、4、5或6)的重组表达期间,存在于新合成的指定朝向分泌途径的大多数蛋白质的N端的信号肽(有时称为信号序列、靶向信号、定位信号、定位序列、转运肽、前导顺序或前导肽),可以被从最终的重组蛋白切割。在一些实施方式中,重组的可溶性F蛋白因此包含从第25个氨基酸开始或大约从第25个氨基酸开始的氨基酸(即,信号肽MGLKVNVSAIFMAVLLTLQTPTGQ从重组蛋白切割除去)。
在某些方面,本发明提供了一种用于生产处于融合前状态的麻疹F蛋白的方法。在某些方面,本发明提供了在培养基中培养表达本发明的任何一种可溶性F蛋白(例如SEQ IDNo:1、2、3、4、5或6)的细胞系,该培养基包含本文所述的任何抗病毒肽缀合物,从而产生处于融合前状态的稳定的可溶性F蛋白。
用于产生重组蛋白的多种表达系统在本领域中是已知的,并且包括原核(例如细菌)、植物、昆虫、酵母和哺乳动物表达系统。可以用含有本发明的可溶性F蛋白的编码序列的核酸转化、转导或转染合适的细胞系,以产生目标分子。可以通过本领域已知的方法引入含有这样的核酸序列的表达载体(vector),其可以以允许核苷酸序列在宿主细胞中的表达的方式连接至至少一个调节序列。本领域从业人员理解,设计表达载体可以取决于诸如待转染的宿主细胞的选择和/或待表达的所需蛋白质的类型和/或量等因素。增强子区是可见于非编码DNA区中的启动子区上游或下游的那些序列,其在优化表达中的重要性也是本领域中已知的。如果需要,可以使用来自病毒来源的复制起点,例如如果使用原核宿主来引入质粒DNA。然而,在真核生物中,染色体整合是DNA复制的常见机制。对于哺乳动物细胞的稳定转染,一小部分细胞可以将引入的DNA整合到它们的基因组中。所使用的表达载体和转染方法可能是促成成功整合事件的因素。对于所需蛋白质的稳定扩增和表达,含有编码目标蛋白质的DNA的载体稳定地整合到真核细胞(例如哺乳动物细胞)的基因组中,从而导致转染基因的稳定表达。可以将编码选择标记(例如对抗生素或药物的抗性)的基因与目标基因一起引入宿主细胞,以确定和选择稳定地表达编码目标蛋白质的基因的克隆。可以通过药物选择来确定含有目标基因的细胞,其中,掺入了选择标记基因的细胞将在药物存在的情况下存活。未掺入选择标记基因的细胞死亡。然后,可以筛选存活的细胞以产生所需的蛋白质分子。
也可选择调节插入序列的表达或以所需的特定方式修饰和加工核酸的宿主细胞株。蛋白质产物的此类修饰(例如糖基化和其他翻译后修饰)和加工(例如切割)对于蛋白质的功能可能是重要的。不同的宿主细胞株具有独特的和特定的机制用于蛋白质和基因产物的翻译后加工和修饰。因此,可以选择合适的宿主系统或细胞系以确保对所表达的外源蛋白进行正确的修饰和加工。因此,可以使用具有细胞机器的真核宿主细胞,该细胞机器用于对初级转录物的适当加工、对基因产物的糖基化和磷酸化。
对于正在培养的宿主细胞,可以使用多种培养参数。哺乳动物细胞的适当培养条件是本领域众所周知的(Cleveland WL等人,免疫学方法杂志(JImmunol Methods),1983,56(2):221-234),或可由技术人员确定(参见例如《动物细胞培养:实用方法(Animal CellCulture:A Practical Approach)》,第2版,编辑Rickwood,D.和Hames,B.D.(牛津大学出版社:纽约,1992年))。细胞培养条件可根据所选宿主细胞的类型而有所不同。可以使用市售的培养基。
本发明的可溶性F蛋白(例如SEQ ID NO:1、2、3、4、5或6)可以从表达多肽的任何人或非人细胞中纯化,包括已经用表达本发明的可溶性F蛋白的表达构建体转染的那些。对于蛋白质回收、分离和/或纯化,将细胞培养基或细胞裂解物(lysate)离心以去除颗粒细胞和细胞碎片。通过合适的纯化技术将所需的多肽分子从污染的可溶性蛋白质和多肽中分离或纯化。蛋白质的非限制性纯化方法包括:尺寸排阻色谱法;亲和色谱法;离子交换色谱法;乙醇沉淀;反相高效液相色谱法(HPLC);树脂(如二氧化硅或阳离子交换树脂(如DEAE))色谱;色谱聚焦;SDS-PAGE;硫酸铵沉淀;凝胶过滤,使用例如Sephadex G-75、琼脂糖(Sepharose);用于去除免疫球蛋白污染物的蛋白A琼脂糖色谱法等。其他添加剂、例如蛋白酶抑制剂(例如PMSF或蛋白酶K)可用于抑制纯化过程中的蛋白水解降解。也可以使用可以选择碳水化合物的纯化程序,例如离子交换软凝胶色谱法,或使用阳离子或阴离子交换树脂的HPLC,其中收集酸性更强的部分。
组合物
在一些实施方式中,抗病毒肽缀合物以包含抗病毒肽缀合物和药学上可接受的载体(carrier)的药物组合物的形式施用。在一些实施方式中,稳定的F蛋白以包含抗病毒肽缀合物和药学上可接受的载体的药物组合物的形式施用。在一些实施方式中,药物组合物是喷雾剂、气雾剂、凝胶、溶液、乳剂或混悬剂的形式。
该组合物优选地与药学上可接受的载体一起施用于哺乳动物。通常,在一些实施方式中,在制剂中使用适量的药学上可接受的盐,这在一些实施方式中可以使制剂等渗。
在某些实施方式中,抗病毒肽缀合物作为免疫原性组合物提供,该免疫原性组合物包含本文所述的任何一种抗病毒肽缀合物和药学上可接受的载体。在某些实施方式中,稳定的F蛋白作为免疫原性组合物提供,该免疫原性组合物包含本文所述的任何一种稳定的F蛋白和药学上可接受的载体。在某些实施方式中,免疫原性组合物进一步包含佐剂。
在一些实施方式中,药学上可接受的载体选自盐水、林格溶液、葡萄糖溶液以及这些的组合。考虑了本领域已知的其他合适的药学上可接受的载体。《雷明登药学大全(Remington's Pharmaceutical Sciences)》(2005年,马克出版公司(Mack PublishingCo.))中描述了合适的载体及其制剂。溶液的pH优选为约5至约8,更优选约7至约7.5。该制剂还可以包含冻干粉末。其他载体包括缓释制剂,例如固体疏水聚合物的半渗透基质,该基质呈成形制品的形式,例如薄膜、脂质体或微粒。对本领域技术人员而言显而易见的是,某些载体可能更优选,这取决于例如施用途径和所施用的抗病毒肽缀合物的浓度。
如本文所用的短语“药学上可接受的载体”是指本主题药剂从一个器官或身体部分携带或运输至另一个器官或身体部分所涉及的药学上可接受的材料、组合物或媒介(vehicle),例如液体或固体填充剂、稀释剂、赋形剂、溶剂或封装材料。与制剂的其他成分相容且对患者无害的每种载体都是可接受的。可用作药学上可接受的载体的材料的一些示例包括:糖类,例如乳糖、葡萄糖和蔗糖;淀粉,例如玉米淀粉和马铃薯淀粉;纤维素及其衍生物,如羧甲基纤维素钠、乙基纤维素和纤维素乙酸酯;粉状黄芪胶;麦芽;明胶;滑石;赋形剂,例如可可脂和栓剂蜡;油类,例如花生油、棉籽油、红花油、芝麻油、橄榄油、玉米油和大豆油;二醇类,例如丁二醇;多元醇,例如甘油、山梨糖醇、甘露糖醇和聚乙二醇;酯类,例如油酸乙酯和月桂酸乙酯;琼脂;缓冲剂,例如氢氧化镁和氢氧化铝;海藻酸;无热原水;等渗盐水;林格溶液;乙醇;磷酸盐缓冲溶液;以及其他用于药物制剂的无毒相容物质。术语“载体”表示天然的或合成的有机或无机成分,活性成分与其结合以促进应用。药物组合物的组分也能够以不存在会显著损害所需药效的相互作用的方式与本发明的化合物混合,并且能够相互混合。该组合物还可以包括额外的试剂,例如等渗剂、防腐剂、表面活性剂和二价阳离子(优选锌)。
该组合物还可以包括赋形剂或用于稳定抗病毒肽缀合物组合物的试剂,例如缓冲剂、还原剂、大体积蛋白(bulk protein)、氨基酸(例如甘氨酸或果仁糖)或碳水化合物。可用于配制抗病毒肽缀合物组合物的大体积蛋白包括白蛋白。用于配制抗病毒肽缀合物的典型碳水化合物包括但不限于蔗糖、甘露糖醇、乳糖、海藻糖或葡萄糖。
表面活性剂也可用于防止组合物中包含的肽或蛋白质的可溶性和不可溶性聚集和/或沉淀。合适的表面活性剂包括但不限于山梨糖醇三油酸酯、大豆卵磷脂和油酸。在某些情况下,优选使用乙醇等溶剂的溶液型气雾剂。因此,包括抗病毒肽缀合物或稳定的F蛋白的制剂还可以包括表面活性剂,该表面活性剂可以减少或防止在形成气雾剂中由溶液雾化引起的抗病毒肽缀合物或稳定的F蛋白的表面诱导聚集。可以使用多种常规的表面活性剂,例如聚氧乙烯脂肪酸酯和醇,以及聚氧乙烯山梨糖醇脂肪酸酯。用量通常在制剂的0.001重量%至4重量%之间。用于本发明目的的特别优选的表面活性剂是聚氧乙烯山梨糖醇单油酸酯、聚山梨醇酯80、聚山梨醇酯20。组合物中也可以包含本领域已知的其他试剂。
在一些实施方式中,药物组合物和剂型进一步包含一种或多种降低活性成分衰变或组合物性质改变速率的化合物。所谓的稳定剂或防腐剂可以包括但不限于氨基酸、抗氧化剂、pH缓冲剂或盐缓冲剂。抗氧化剂的非限制性示例包括丁基化羟基茴香醚(BHA)、抗坏血酸及其衍生物、生育酚及其衍生物、丁基化羟基茴香醚和半胱氨酸。防腐剂的非限制性示例包括对羟基苯甲酸酯,例如对羟基苯甲酸甲酯或丙酯和苯扎氯铵。氨基酸的其他非限制性示例包括甘氨酸或脯氨酸。
本发明还教导了使含有抗病毒肽缀合物的中性pH或低于中性pH的液体溶液稳定(防止或最大限度降低热或机械诱导的抑制剂蛋白的可溶性或不可溶性聚集和/或沉淀),其通过使用包括脯氨酸或甘氨酸的氨基酸(含或不含二价阳离子),产生在室温下稳定或优选地用于药物施用的澄清或几乎澄清的溶液。
在一个实施方式中,该组合物是单一单位或多单位剂型的药物组合物。本发明的单一单位或多单位剂型的药物组合物包含一种或多种组合物(例如本发明的化合物或其他预防剂或治疗剂)的预防或治疗有效量,所述一种或多种组合物通常为一种或多种媒介、载体或赋形剂、稳定剂和/或防腐剂。优选地,媒介、载体、赋形剂、稳定剂和防腐剂是药学上可接受的。
在一些实施方式中,药物组合物和剂型包括无水药物组合物和剂型。可以在低水分或低湿度条件下使用无水或低水分含量的成分来制备本发明的无水药物组合物和剂型。如果预期在制造、包装和/或储存期间与水分和/或湿气大量接触,则包含乳糖和至少一种包含伯胺或仲胺的活性成分的药物组合物和剂型优选是无水的。无水药物组合物的制备和储存应保持其无水性质。因此,优选地,使用已知防止暴露于水的材料对无水组合物进行包装,使得它们可包含在合适的处方试剂盒中。合适的包装的示例包括但不限于密封箔、塑料、单位剂量容器(例如小瓶)、泡罩包装和条形包装。
合适的媒介是药学领域技术人员所熟知的,并且合适的媒介的非限制性示例包括葡萄糖、蔗糖、淀粉、乳糖、明胶、大米、硅胶、甘油、滑石粉、氯化钠、脱脂奶粉、丙二醇、水、硬脂酸钠、乙醇和本领域熟知的类似物质。盐溶液以及葡萄糖水溶液和甘油溶液也可以用作液体载体。特定媒介是否适合掺入药物组合物或剂型取决于本领域熟知的多种因素,包括但不限于将剂型施用于患者的方式和剂型中的特定活性成分。药用媒介可以是无菌液体,例如水和油,包括石油、动物、植物或合成来源的那些油,例如花生油、大豆油、矿物油、芝麻油等。
本发明的药物组合物配制成与其预期的施用途径相容。下呼吸道内施用途径的示例包括但不限于经口或鼻腔吸入(例如,吸入足够小的、将明确地沉积在下呼吸道内的颗粒)。在多种实施方式中,药物组合物或单一单位剂型是无菌的并且为适合施用于受试者的形式,该受试者优选地为动物受试者,更优选地为哺乳动物受试者,并且最优选地为人类受试者。
本发明剂型的组合物、形状和类型通常会根据它们的用途而变化。剂型的非限制性示例包括粉剂;溶液剂;气雾剂(例如喷雾剂、定量或非定量雾化剂、口腔或鼻用吸入剂(包括定量吸入剂(MDI));适用于向患者黏膜施用的液体剂型,包括还可重构以提供适用于下呼吸道施用的液体剂型的混悬剂(例如水性或非水性液体混悬剂、水包油乳剂或油包水液体乳剂)、溶液剂和无菌固体剂(例如结晶或无定形固体)。可以使用例如混合器、流化床设备或喷雾干燥装备以常规方式使用上述成分来制备粉末或颗粒形式的制剂。
本发明还提供了,药物组合物可以包装在密封容器中,例如标明量的安瓿或小袋(sachette)。在一个实施方式中,药物组合物可以提供为适合施用至患者下呼吸道的递送装置中的干燥无菌的冻干粉末。如果需要,药物组合物可以呈现在可以包含一种或多种含有活性成分的单位剂型的包装或分配装置中。该包装可以例如包括金属或塑料箔,例如泡罩包装。包装或分配装置可以附有施用说明。
制备这些制剂或组合物的方法包括使本发明的化合物与载体和可选地一种或多种辅助成分结合的步骤。通常,通过以下步骤制备制剂:将本发明的化合物与液体载体或细散的固体载体或与这两者均匀且充分地混合,然后如果需要的话,使产物成形。
适用于施用的本发明的制剂可以是粉末、颗粒的形式,或作为在水性或非水性液体中的溶液剂或混悬剂,或作为水包油或油包水液体乳剂,或作为酏剂或糖浆,或作为锭剂(使用惰性基料,例如明胶和甘油,或蔗糖和阿拉伯胶)和/或作为漱口药等,每一种都含有预定量的本发明化合物(例如抗病毒肽缀合物)作为活性成分。
本文的液体组合物可以原样与递送装置一起使用,或者它们可以用于制备包含抗病毒肽缀合物的药学上可接受的制剂,该制剂例如通过喷雾干燥的方法制备。以下文献公开的用于药物施用的喷雾冷冻干燥蛋白的方法并入本文:Maa等人,当今药物生物技术杂志(Curr.Pharm.Biotechnol),2001,1,283-302。在另一个实施方式中,对本文的液体溶液进行冷冻喷雾干燥,并且将该喷雾干燥的产品收集为含有可分散抗病毒肽缀合物的粉末,该粉末施用到个体的下呼吸道中时具有疗效。
本发明的化合物和药物组合物可以用于联合治疗,即,该化合物和药物组合物可以与一个或多个其他所需的治疗或医疗程序同时、在其之前或之后施用。联合方案中将使用的特定的联合治疗(疗法或程序)将考虑到所需疗法和/或程序的相容性以及理想的治疗效果。还应当理解,所采用的疗法可以实现对相同紊乱的期望效果(例如本发明的化合物可以与另一种抗病毒剂同时施用)。
本发明还提供了一种药物包装或试剂盒,该药物包装或试剂盒包含一个或多个容器,该容器填充有本发明的药物组合物的一种或多种成分。可选地,与此类容器相关联的可以是监管药物或生物产品的生产、使用和销售的政府机构所规定的形式的通知,该通知反映所述机构批准用于人给药(human administration)的制造、使用或销售。
本发明提供了包含适用于治疗麻疹或HIV感染的抗病毒肽缀合物(例如FIP-HRC肽)的剂型。剂型可以配制为例如喷雾剂、气雾剂、纳米颗粒、脂质体或本领域技术人员已知的其他形式。参见例如《雷明登药学大全(Remington's Pharmaceutical Sciences)》;在前的《雷明登:药剂学科学与实践(Remington:The Science and Practice of Pharmacy)》;Howard C.、Ansel等人《药物剂型和施用系统(Pharmaceutical Dosage Forms and DrugDelivery Systems)》,利平科特·威廉斯·威尔金斯出版公司(Lippincott Williams&Wilkins);第7版(1999年10月1日)。
本发明还提供了包含适用于治疗或预防麻疹感染的稳定的F蛋白的剂型。剂型可以配制为例如喷雾剂、气雾剂、纳米颗粒、脂质体或本领域技术人员已知的其他形式。参见例如《雷明登药学大全(Remington's Pharmaceutical Sciences)》;在前的《雷明登:药剂学科学与实践(Remington:The Science and Practice of Pharmacy)》;Howard C.、Ansel等人《药物剂型和施用系统(Pharmaceutical Dosage Forms and Drug DeliverySystems)》,利平科特·威廉斯·威尔金斯出版公司(Lippincott Williams&Wilkins);第7版(1999年10月1日)。
通常,用于感染/紊乱的急救治疗的剂型可以比用于相同疾病的长期治疗的剂型包含更大量的一种或多种其所包含的活性成分。此外,预防和治疗有效的剂型可能因不同条件而异。例如,当旨在治疗已有的麻疹或HIV感染时,治疗有效的剂型可以包含具有适当的抗病毒作用的抗病毒肽缀合物。另一方面,当旨在将本发明的肽用作针对麻疹或HIV感染的预防剂(例如疫苗)时,不同的有效剂量可以包含具有适当的免疫原性作用的抗病毒肽缀合物。当旨在治疗现有的麻疹感染时,治疗有效的剂型可以包含具有适当的抗病毒作用的稳定的F蛋白。另一方面,当旨在将本发明的稳定的F蛋白用作麻疹感染的预防剂(例如疫苗)时,不同的有效剂量可以包含具有适当的免疫原性作用分的稳定的F蛋白。本发明所涵盖的具体剂型的这些和其他方式彼此不同,并且对于本领域技术人员而言是显而易见的。参见例如《雷明登药学大全((Remington's Pharmaceutical Sciences))》(2005年,马克出版公司(Mack Publishing Co.));Gennaro的《雷明登:药剂学科学与实践(Remington:TheScience and Practice of Pharmacy)》,利平科特·威廉斯·威尔金斯出版公司;第20版(2003年);Howard C.、Ansel等人《药物剂型和施用系统(Pharmaceutical Dosage Formsand Drug Delivery Systems)》,利平科特·威廉斯·威尔金斯出版公司;第7版(1999年10月1日);和《制剂技术百科全书(Encyclopedia of Pharmaceutical Technology)》,由Swarbrick,J.&JC Boylan编辑,马塞尔·德克尔公司(Marcel Dekker,Inc.),纽约,1988年,以上文献通过引用整体并入本文。
还可以调节药物组合物或剂型的pH以改善一种或多种活性成分的递送和/或稳定性。类似地,可以调节溶剂载体的极性、离子强度或张力以改善递送。还可以将诸如硬脂酸盐的化合物添加到药物组合物或剂型中以有利地改变一种或多种活性成分的亲水性或亲油性以改善递送。就此而言,硬脂酸盐也可以用作制剂的脂质媒介,作为乳化剂或表面活性剂,以及作为递送增强剂或渗透增强剂。活性成分的不同盐、水合物或溶剂化物可用于进一步调节所得组合物的性质。
可以使用技术将组合物与适当的载体和佐剂进行配制,以产生适用于免疫(immunization)的组合物。组合物可以包括佐剂,例如但不限于适用于免疫的明矾、聚肌胞苷酸(poly IC)、MF-59、基于角鲨烯的佐剂或基于脂质体的佐剂。
在一些实施方式中,组合物和方法包含可以调节宿主免疫耐受机制和诱导抗体释放的任何合适的试剂或免疫调节剂。在某些实施方式中,以足以瞬时调节受试者的免疫反应的时间和量施用免疫调节剂,以诱导包含针对麻疹F蛋白或HIV-1包膜的抗体的免疫反应。
治疗方法
在一个实施方式中,本文公开的主题涉及在暴露于MV之后开始的预防性医学治疗,以防止感染发生或恶化。在一个实施方式中,本文公开的主题涉及针对已经接触过MV或疑似接触过MV的受试者的预防。在一个实施方式中,可以对所述受试者施用包含本文所述的抗病毒肽缀合物或其药物组合物的暴露后预防。在一些实施方式中,抗病毒肽缀合物包含缀合至源自麻疹病毒的HRC肽的FIP,作为麻疹感染的预防剂。本发明考虑使用本文所述的任何抗病毒肽缀合物。在一些实施方式中,与FIP缀合的HRC肽是单体形式(例如“FIP-HRC450-485”,参见图64)。在一些实施方式中,与FIP缀合的HRC肽是单体形式并且进一步包含缀合至肽的C端的脂质(例如胆固醇)(例如“FIP-HRC450-485-chol”,参见图64)。在一些实施方式中,与FIP缀合的HRC肽是单体形式并且进一步包含缀合至肽的C端的连接体(例如PEG4)和脂质(例如胆固醇)(例如“FIP-HRC450-485-peg4-chol”,参见图64)。在一些实施方式中,与FIP缀合的HRC肽是二聚体形式并且进一步包含缀合至肽的C端的连接体(例如PEG4)和脂质(例如胆固醇)(例如“[FIP-HRC450-485-peg4]2-chol”,参见图64)。在一些实施方式中,与FIP缀合的HRC肽是二聚体形式并且进一步包含缀合至肽的C端的连接体(例如PEG4)(例如“[FIP-HRC450-485-peg11]2”,参见图64)。在一个实施方式中,本文所述的抗病毒肽缀合物可以以纳米颗粒的形式施用。在一个实施方式中,本文所述的抗病毒肽缀合物可以通过鼻内喷雾剂或本领域已知的任何其他合适的方法鼻内施用。在一个实施方式中,本文所述的抗病毒肽缀合物可以通过注射器或本领域已知的任何其他合适的方法皮下施用。在一个实施方式中,本文公开的主题可适用于并应用于除MV之外的副粘病毒(如腮腺炎)的暴露后预防。在一个实施方式中,本文公开的主题涉及通过抑制病毒融合来对任何病毒进行暴露后预防的方法。
在一个实施方式中,本文公开的主题涉及在暴露于HIV之后开始的预防性医学治疗,以防止感染发生或恶化。在一个实施方式中,本文公开的主题涉及针对已经接触过HIV或疑似接触过HIV的受试者的预防。在一个实施方式中,可以对所述受试者施用包含本文所述的抗病毒肽缀合物或其药物组合物的暴露后预防。在一些实施方式中,抗病毒肽缀合物包含缀合至衍生自HIV-gp41的HRC肽(“C34”)的FIP,作为HIV感染的预防剂。在一些实施方式中,抗病毒肽缀合物包含缀合至衍生自HIV-gp41的HRC肽(“C34”)的FIP,也可用作MV感染的预防剂。在一些实施方式中,与FIP缀合的HRC肽是单体形式并且进一步包含缀合至肽的C端的脂质(例如胆固醇)。在一些实施方式中,与FIP缀合的HRC肽是单体形式并且进一步包含缀合至肽的C端的连接体(例如PEG4)和脂质(例如胆固醇)。在一些实施方式中,与FIP缀合的HRC肽是二聚体形式并且进一步包含缀合至肽的C端的连接体(例如PEG4)和脂质(例如胆固醇)。在一些实施方式中,与FIP缀合的HRC肽是二聚体形式并且进一步包含缀合至肽的C端的连接体(例如PEG4)。在一个实施方式中,本文所述的抗病毒肽缀合物可以以纳米颗粒的形式施用。在一个实施方式中,所述抗病毒肽缀合物可以通过鼻内喷雾剂或本领域已知的任何其他合适的方法鼻内施用。在一个实施方式中,所述抗病毒肽缀合物可以通过注射器或本领域已知的任何其他合适的方法皮下施用。在一个实施方式中,本文公开的主题涉及通过抑制病毒融合来对任何病毒进行暴露后预防的方法。
在一些实施方式中,受试者已经暴露于包含野生型融合糖蛋白的麻疹病毒。在一些实施方式中,受试者已暴露于麻疹病毒,该麻疹病毒包含选自N462K、L454W、T461I、E455G、E170G、G506E、M337L、D538G、G168R、S262G、A440P、R520C和L550P的融合糖蛋白的一种或多种突变(例如,参见图104)。
本文公开的一种或多种化合物、或化合物的组合、或药物组合物可以通过任何合适的方式施用至细胞、哺乳动物或人。施用方法的非限制性示例包括(除其他外)(a)通过口服途径施用,其包括以胶囊、片剂、颗粒剂、喷雾剂、糖浆或其他此类形式施用;(b)通过眼内、鼻内、耳内、直肠、阴道、尿道内、经黏膜、口腔或经皮等非口服途径施用,包括作为水性悬浮液、油性制剂等或作为滴注剂、喷雾剂、栓剂、药膏、软膏等施用;(c)通过注射施用,包括皮下、腹膜内、静脉内、肌内、皮内、眶内、囊内、脊柱内、胸骨内等,包括输液泵递送;(d)局部施用(administration locally),例如通过直接在肾或心脏区注射,例如通过植入剂(depot)植入;(e)外敷施用(administration topically);如本领域技术人员认为合适的,用于使本文公开的化合物或化合物的组合与活组织接触;(f)通过吸入施用,包括通过气雾剂、雾化剂和粉状制剂;以及(g)通过植入施用。
将对本领域技术人员显而易见的是,待施用的有效体内剂量和特定的施用方式将根据所治疗对象的年龄、体重和物种以及本文公开的化合物或化合物的组合所用于的具体用途而变化。可以由本领域技术人员使用常规药理学方法来完成有效剂量水平的确定,即达到所需结果必需的剂量水平。通常,产品的人类临床应用以较低的剂量水平开始,然后增加剂量水平直到达到所需的效果。可替代地,可接受的体外研究可用于使用既定的药理学方法建立本方法所确定的组合物的有用剂量和施用途径。可以使用本领域已知的转换方法将来自体内研究的有效动物剂量(例如,在棉鼠中为6mg/kg;参见图22和图84)转换成合适的人类剂量(例如,参见Nair AB,Jacob S.动物和人类之间剂量转换的简明实践指南(Asimple practice guide for dose conversion between animals and human).基础与临床药学杂志(Journal of basic and clinical pharmacy).2016年3月;7(2):27)。
预防方法
不受理论的束缚,本文所述的抗病毒肽缀合物将可溶性麻疹F蛋白稳定在融合前状态。因此,在一些实施方式中,本发明的抗病毒肽缀合物可以与可溶性麻疹F蛋白(例如SEQ ID NO:1、2、3、4、5或6)一起使用作为疫苗,以促进针对融合前的麻疹F蛋白的免疫反应。在一些实施方式中,本发明的抗病毒肽缀合物可以与可溶性麻疹F蛋白(例如SEQ IDNO:1、2、3、4、5或6)一起施用,以引发针对麻疹的保护性免疫反应。在一些实施方式中,本发明提供了一种在受试者中诱导免疫反应的方法,所述方法包括施用免疫原性组合物,该免疫原性组合物包含本发明的任何一种抗病毒肽缀合物以及任何一种可溶性麻疹F蛋白(例如SEQ ID NO:1、2、3、4、5或6)。在一些实施方式中,本发明的抗病毒肽缀合物还可以与可溶性麻疹F蛋白(例如SEQ ID NO:1、2、3、4、5或6)一起使用作为对感染麻疹病毒的受试者的预防性治疗。
在一些实施方式中,稳定的可溶性F蛋白(例如SEQ ID NO:3、4、5或6)可用作疫苗以促进针对融合前的麻疹F蛋白的免疫反应(即,不添加本文所述的抗病毒肽缀合物)。在一些实施方式中,稳定的可溶性F蛋白(例如SEQ ID NO:3、4、5或6)可用于引发针对麻疹的保护性免疫反应。在一些实施方式中,本发明提供了一种在受试者中诱导免疫反应的方法,所述方法包括施用免疫原性组合物,该免疫原性组合物包含本文所述的任何一种稳定的可溶性F蛋白(例如SEQ ID NO:3、4、5或6)。
在一些实施方式中,稳定的F蛋白单独施用或与本文所述的任何抗病毒肽缀合物组合施用。在一些实施方式中,本发明的抗病毒肽缀合物可以与可溶性麻疹F蛋白(例如SEQID NO:1、2、3、4、5或6)一起使用作为对感染麻疹病毒的受试者的预防性治疗。
在一些实施方式中,本发明提供了用于诱导免疫反应、例如诱导针对麻疹病毒或针对HIV的抗体的组合物和方法。在一些实施方式中,抗体是广泛中和抗体。在一些实施方式中,该方法诱导针对麻疹F蛋白或HIV-1包膜的抗体。在一些实施方式中,该方法使用包含稳定的F蛋白和/或本文所述的任何抗病毒肽缀合物的组合物。在一些实施方式中,该方法进一步包括施用佐剂。在一些实施方式中,本发明提供了用于诱导针对包含野生型融合糖蛋白的麻疹病毒的免疫反应的组合物和方法。在一些实施方式中,本发明提供了用于诱导针对麻疹病毒的免疫反应的组合物和方法,该麻疹病毒包含选自N462K、L454W、T461I、E455G、E170G、G506E、M337L、D538G、G168R、S262G、A440P、R520C和L550P的融合糖蛋白的一种或多种突变(例如,参见图104)。
实施例
下文提供实施例,以促进更全面地理解本发明。以下实施例对制造和实践本发明的示例性模式进行了说明。然而,本发明的范围不限于这些实施例中公开的具体实施方式,这些实施例仅用于说明目的,因为可以使用替代方法来获得相似的结果。
实施例1
MeV感染可以始于呼吸道,如图1中所示。肺泡巨噬细胞和树突状细胞(DC)是表达MeV受体信号淋巴细胞激活分子(signaling lymphocyte activation molecule,SLAM,也称为CD150)的初始靶标。MeV受体结合蛋白血凝素(H)与CD150的结合导致这些细胞感染,然后,病毒传播到支气管相关淋巴组织和/或引流淋巴结。病毒在表达CD150的B和T淋巴细胞中增殖,随后出现病毒血症。粘附连接蛋白(PVRL4或连接素4(nectin 4))也可作为MeV受体,但存在于呼吸道上皮细胞的基底外侧表面;它与疾病后期的病毒传播有关。病毒血症和MeV发病机制的出口如图2中所示。如图3中所示,麻疹也可引起严重并发症。其中一些并发症影响中枢神经系统(CNS)。麻疹包涵体脑炎(MIBE)和亚急性硬化性全脑炎(SSPE)都是麻疹感染的致命并发症。MIBE可能在病毒感染后1-9个月发生。SSPE可能在感染后数年出现。在一个实施方式中,本文公开的主题聚焦来自MIBE和SSPE中枢神经系统并发症的临床分离株。在一个实施方式中,急性麻疹脑炎(AME)是MeV的另一并发症。过去,认为SSPE的发病率是1:100000例,但最近的数据显示,儿童出生第一年的感染可导致每600例感染者中有1例(Wendorf,K.A.等人,临床感染性疾病医学(Clin.Infect.Dis.),2017,65(2),226-232)。在严重免疫受损患者中,MIBE是麻疹的致命CNS表现。感染HIV的MeV脑炎患者的数据如图4中所示。患者8前面一年使用HAART。报告之时,患者1和患者8仍存活。所有患者最终均死亡(未公开的数据、个人通讯、戴安娜哈迪(Diana Hardie))。
MV病理学:中枢神经系统并发症
一个长期存在的问题是麻疹是如何在CNS中感染和传播的,因为CNS缺乏已知的MV受体。本文已经描述了副粘病毒的融合复合物。还描述了CNS临床分离株。因此,本文讨论的另一个问题是麻疹融合复合物在CNS中是如何适应的。包括F和H蛋白的麻疹病毒的示意图如图5中所示。本文公开的主题还涉及来自患者1和患者6的临床分离株序列的数据,如图4中所示。
在一个实施方式中,本文公开的主体涉及从在2009/2010年南非麻疹爆发期间罹患MIBE的患者的CNS回收的神经致病性的(neuropathogenic)MeV分离株的融合复合物的表征。MeV序列是从两名HIV感染患者的死后脑组织中分离出来的,这些患者在南非麻疹流行期间通过阳性MeV PCR被诊断为患有MIBE。病毒基因组测序显示,在这两个病例中,MeV F基因含有与图6中所示相同的核苷酸突变,其导致在第454位(L454W)亮氨酸替换为色氨酸。第一名患者是一名27岁的女性,她在患急性麻疹后3个月后患上MIBE。值得注意的是,从她的大脑中分离出的病毒中存在的L454W突变在她的早期血液样本(采集于她急性MV感染期间)的病毒中并不存在。第二名患者是一名34岁的女性,她在患急性MeV感染3周后出现典型的MIBE症状。野生型麻疹病毒进入CNS受到严格的控制,如图7中所示。野生型MeV的进入步骤需要连接素4或CD150受体。因此,出现的一个问题是,SSPE和MIBE的MeV毒株是如何进入CNS并在其中传播的?
在本文公开的主题的一个实施方式中,表征了从在2009/2010年南非麻疹爆发期间罹患MIBE的患者的CNS回收的神经致病性的MeV分离株的融合机器,并且提出的另一个问题是这种单一氨基酸突变是如何改变麻疹融合复合物的?病毒基因组测序显示,在这两个病例中,MeV F基因都含有相同的核苷酸突变,其导致在第454位(L454W)亮氨酸替换为色氨酸。融合复合物的分析方法如图8中所示。我们提出疑问,L454W核苷酸突变是否会改变MV融合机器的活性。
适应CNS的融合机器:改变了受体参与融合的要求
如图9A-B中所示,在存在已知受体的情况下,所有F蛋白都表现出相似的融合水平。如图9C中所示,F L454W在不存在已知受体的情况下诱导融合。
L454W突变会破坏F的稳定性吗?
本文中公开了野生型MeV F和适应CNS的MeV F的热稳定性。将表达所示MV F(x轴)的细胞在32℃下温育过夜,然后放置在指定的温度(T)下。然后将细胞与单克隆抗体(mAb)与融合前或融合后状态的MeV F在4℃下温育。y轴上的值表示相对发光单位(RLU)。F L454W的热稳定性低于wt F。这两种状态的示意图如图10中所示。带有CNS衍生的F的病毒可以在缺乏MeV受体的细胞中传播。重组病毒诱导细胞间融合,如图11中所示。MIBE衍生的F L454W能够在不存在已知MeV受体的情况下介导融合。MV F L454W的融合活性的增加超过F野生型,这与稳定性的降低相关。MV F L454W可以促进独立于H蛋白的融合。
带有L454W
F的病毒是否比野生型病毒更具致病性?
用于回答这个问题的模型包括但不限于人脑类器官,来自小鼠和棉鼠的离体组织。MeV感染棉鼠,并且可以对病毒滴度进行评估。还有一种CD150受体转基因乳鼠模型,它会导致野生型病毒的致命感染。
90天大的脑类器官被麻疹野生型F病毒和带有L454W F的病毒感染,如图12中所示。图13示出了来自小鼠的离体组织(不存在受体)。野生型病毒的离体感染与带有L454W F的病毒的离体感染:在器官型脑培养物中,CNS适应病毒胜过野生型病毒。来自IFNAR KO鼠脑的小脑切片使用带有EGFP基因的野生型病毒以5000PFU/片进行感染,持续4天。通过落射荧光显微镜观察到与24小时或96小时下的感染相关的红色荧光(比例尺=500μm)。带有L454W F的MeV在器官型脑培养物(OBC)中的传播。来自IFNAR KO鼠脑的小脑切片使用MeV-IC323-L454W F EGFP基因(绿色荧光)以及带有tdtomato的野生型病毒(红色荧光)以5000PFU/片进行感染,持续4天。通过落射荧光显微镜观察到与24小时(图13A)或72小时(图13B)下的感染相关的绿色荧光和红色荧光(比例尺=500μm)。
图14示出了棉鼠的MeV病毒数据。图15A-B示出了小鼠的存活率数据。图16示出了将病毒进入作为治疗靶标。如图17中所示的F糖蛋白衍生的肽抑制病毒进入,如图18中所示。图19示出了将HRC肽靶向脂质膜。图20示出了提高HRC肽对F蛋白的亲和力。图21示出了将HRC肽靶向脂质膜的另一个实施方式。图22A-B示出了缀合的肽在体内是有效的。所使用的模型是图22A中的棉鼠和图22B中的SLAM:IFNARKO小鼠。施用途径是鼻内施用,剂量为6mg/kg。图23A-C示出了MeV HRC4肽阻断病毒的离体传播。所使用的模型是来自感染带有L454W F的病毒的小鼠(无CD150)的离体组织。图24示出了MeV HRC4的鼻内施用保护乳鼠免受带有L454W F的病毒的致命感染。
表1-3提供了肽的非限制性示例。
表1提供了肽及其修饰的列表。a(氨基酸残基以单字母代码表示;Ac–乙酰化N端;GSGSG–含有五个氨基酸的连接体;D-FFG–D-氨基酸用于第一个苯丙氨酸残基;Z-D-FFG–Z是连接至D-FFG序列的N端的苄氧羰基;b(FIP–融合抑制肽;HRC450-485–始于第450位氨基酸并终于第485位氨基酸的麻疹HRC衍生肽序列);c(Peg–聚乙二醇;Chol–胆固醇);d(肽–HRC450-485/FIP-HRC450-485/FIP)。


表2示出了FIP、HRC和FIP-HRC肽在融合试验中的抑制活性(显著肽(salientpeptide))。进行了基于β-半乳糖苷酶互补的融合试验。简言之,在存在或不存在融合抑制肽的情况下,将用连接素-4和omega报告亚基瞬时转染的293T细胞(靶细胞)与共表达病毒糖蛋白(H和F)和α报告亚基的细胞(效应细胞)一起温育指定的时间。在不存在肽的情况下,靶细胞和效应细胞之间的融合导致β-半乳糖苷酶活性的重建,使用发光试剂盒Galacto-StarTMβ-半乳糖苷酶报告基因(赛默飞世尔(ThermoFisher),美国)对该活性进行量化。在肽的存在下,融合减少,并且β-半乳糖苷酶的活性也因此降低。数据来自至少三次独立实验。

表3示出了FIP、HRC和FIP-HRC肽在融合试验中的抑制活性(扩展版)。进行了基于β-半乳糖苷酶互补的融合试验,如前所述。简言之,在存在或不存在融合抑制肽的情况下,将用连接素-4或CD150和omega报告亚基瞬时转染的293T细胞(靶细胞)与共表达病毒糖蛋白(H和F)和α报告亚基的细胞(效应细胞)一起温育指定的时间。在不存在肽的情况下,靶细胞和效应细胞之间的融合导致β-半乳糖苷酶活性的重建,使用发光试剂盒Galacto-StarTMβ-半乳糖苷酶报告基因(赛默飞世尔(ThermoFisher),美国)对该活性进行量化。在肽的存在下,融合减少,并且β-半乳糖苷酶的活性也因此降低。数据来自三次独立实验(除了*=9次重复、§=6次重复和°=5次重复)。



肽缀合在溶液中诱导自缔合
图25示出了肽粒径在纳摩尔范围内。图26示出了两亲结构驱动自组装和纳米颗粒形成。图27示出了纳米颗粒在脂质膜界面处组装(解装配)并保留肽。图28示出了纳米颗粒能够穿过HAE屏障并且在体内是生物可利用的。通过到达相关感染部位,这些防止麻疹病毒增殖。图29示出了缀合的肽以6mg/kg经鼻内施用时,改善了棉鼠模型中的生物分布。图30示出了组合策略,其中融合抑制肽(FIP)与融合蛋白结合并稳定麻疹F蛋白的融合前状态。图31描绘了等效线图曲线(isobologram curve),该等效线图曲线示出了FIP和HRC肽之间的协同作用(组合药物测试:2种不同化合物的同时治疗)。图32示出了添加至HRC区的FIP增强了抗病毒活性。如图33和图34中所示,当FIP和HRC结构相同时,效力超过这两种抑制剂加在一起的协同作用。图35和图36示出了添加至HRC区的FIP增强了抗病毒活性。
针对MV IC323eGFP的肽抗病毒方法总结:
·病毒原液:IC323-EGFP,P3(ALa)滴度:2x 106pfu/ml
·肽原液:肽MeV HRC4浓度:50mg/ml
·治疗剂量为约1mg/kg或约0.1mg/kg,感染前24小时或感染前6小时
·小鼠:SLAM IFNAR KO(3.5至4周龄)
·鼻内感染(5uL/鼻孔=>10uL/小鼠)
·感染剂量=4x LD50
·20.000pfu/鼠
图37示出了存活比例。SLAM IFNAR KO小鼠的存活时间为3.5周。图38示出了多种MeV。图39示出了MV HRC4肽和RNA聚合酶抑制剂阻断人类运动神经元中的MeV野生型感染。感染天数:8天。绿色荧光表示MeV感染,它与病毒感染成正比。
MeV HRC4肽和RNA聚合酶抑制剂阻断病毒在运动神经元中的传播。有关聚合酶抑制剂的信息描述于《科学转化医学(Science Translational Medicine)》,2014年4月16日:第6卷,第232期,第232页。图40示出了HRC4肽和RNA聚合酶抑制剂:野生型病毒和CNS适应病毒。图41示出了人类运动神经元中的MV HRC4肽和RNA聚合酶抑制剂与CNS适应MV。随着时间的推移,聚合酶抑制剂的活性降低。
MeV HRC4肽仅在最高浓度下阻断病毒传播。图42A-B和图43A-C示出了进入的步骤。图44A-C示出了H-F相互作用在L454W F中发生改变。图45示出了MeV HRC4的鼻内施用保护IFNAR KO小鼠免受致命的MV脑炎。图46示出了可以从多能干细胞生长出的多种类器官的示意图以及所使用的发育信号。图47A-C示出了从多能干细胞产生的“迷你大脑”。图47A示出了具有包含神经祖细胞(SOX2,红色)和神经元(TUJ1,绿色)的异质区的复杂形态是显而易见的(Lancaster等人,2013)。图47B示出了具有图案肾单元(nephron)的从多能干细胞生长出的整个肾脏类器官的免疫荧光图像。形成肾小球的足细胞(NPHS1,黄色)、早期近端小管(莲藕凝集素(lotus tetragonolobus lectin),粉红色)和远端小管/收集管(E-钙粘蛋白,绿色)。图47C示出了人aSC来源的肺类器官的中间部分的3D重建,对基底细胞(绿色)、肌动蛋白细胞骨架(红色)和细胞核(蓝色)的中间丝进行染色,并通过共聚焦显微镜成像。
暴露后的预防
在一个实施方式中,本文公开的主题涉及在暴露于MeV之后开始的预防性医学治疗,以防止感染发生。在一个实施方式中,本文公开的主题涉及针对已经接触过MeV或疑似接触过MeV的受试者的预防。在一个实施方式中,可以对所述受试者施用由HRC肽纳米颗粒(例如HRC4肽纳米颗粒)组成的暴露后预防剂。在一个实施方式中,可以向受试者施用由FIP缀合的肽(例如FIP缀合的HRC4肽纳米颗粒)组成的暴露后预防剂。在一个实施方式中,该暴露后预防剂可以通过鼻内喷雾剂或本领域已知的任何其他合适的方法鼻内施用。在一个实施方式中,该暴露后预防剂可以通过注射器或本领域已知的任何其他合适的方法皮下施用。在一个实施方式中,本文公开的主题可适用于并应用于除MeV之外的副粘病毒(如腮腺炎)的暴露后预防。在一个实施方式中,本文公开的主题涉及通过抑制病毒融合来对任何病毒进行暴露后预防的方法。
实施例2
图48-83描述了本发明的进一步实施方式,包括某些脂质-肽缀合物,该脂质-肽缀合物由至少一种融合抑制肽、连接体(例如聚乙二醇(PEG))和膜定位部分组成。在一些实施方式中,融合抑制肽是麻疹融合抑制肽或麻疹HRC衍生肽。在一些实施方式中,PEG连接体重复或连接4-24次。在其他实施方式中,膜定位部分是胆固醇、生育酚或棕榈基。这些脂质-肽缀合物的其他实施方式可包含单体或二聚体、脂质或不含脂质。来自基于β-半乳糖苷酶互补的融合试验、MTT细胞毒性试验、热稳定性试验和F稳定性试验的体外和体内生物学测试数据表明,麻疹融合抑制缀合物抑制感染。
实施例3
图84-93对本发明作了进一步的描述。图84示出了体内功效数据。MeV衍生肽的鼻内施用保护棉鼠免受MV感染。用MeV“WTFb”毒株对棉鼠(n=4)进行鼻内感染。在感染前24小时和12小时以100μl(5m/kg剂量的肽)进行鼻内给药处理。感染4天后,对动物实施安乐死。肺匀浆的MeV滴定显示,[FIP-HRC450-485-peg4]2-胆固醇是最有效的抑制剂。与未经处理的动物相比,[HRC450-485-peg4]2-胆固醇降低了病毒滴度。
图85示出了FIP-HRC靶向MeV F表达细胞。流式细胞术——[FIP-HRC450-485-peg4]2-chol肽在HEK293T细胞中的定位。将表达MeV F的HEK293T细胞与肽(1μM)在37℃下温育60分钟。F蛋白和HRC-FIP肽分别用Alexa Fluor 488(绿色,x轴)和Alexa Fluor 594(红色,y轴)进行染色。代表三次独立实验。对于含有胆固醇的FIP-HRC,HRC信号与F信号成正比(A),表明FIP-HEC被引导至F表达细胞。单独的FIP(不含胆固醇)将HRC定位到F表达细胞,但HRC信号较低(B)。当脂质存在但FIP缺失时(C),HRC没有特异性地定位于F表达细胞。
图86示出了FIP-HRC将麻疹F稳定在其融合前状态。在FIP、HRC和FIP-HRC肽存在的情况下MeV F野生型的热稳定性。将表达MeV F(“WT”)的293T细胞在37℃下温育过夜。然后,在指定的肽的浓度增加的情况下将细胞置于55℃下10分钟。然后,将细胞在4℃下与融合前构象特异性小鼠mAb(77.4)一起温育。与Alexa 488缀合的抗小鼠第二抗体用于检测。使用细胞分析仪高内容图像系统识别染色细胞。y轴上的值指示阳性细胞与未处理细胞相比的百分比%,并表示构象抗体结合的百分比(反映融合前状态下F的百分比。)
这些值是来自三次实验的结果的平均值(±SE)。[FIP-HRC450-485-peg4]2-胆固醇肽在稳定融合前状态下的F方面最有效。
图87示出了MeV F(wt)在指定的肽存在下的热稳定性。在其融合前表位的浓度是未在55℃下温育10分钟的样本的50%(“稳定浓度50”或SC50)和90%(“稳定浓度90”或SC90)。这些值是来自至少三次实验的结果的平均值(±SE)。[FIP-HRC450-485-peg4]2-chol是最有效的F稳定剂。
图88-90示出了MeV肽的稳定特性(对于可溶性F)。参考图88,用两种可溶形式的MeV F转染细胞。使用了Wt F和突变体E455G(在融合前状态下内在稳定)。然后,在有或没有指定的肽的情况下,将细胞温育。转染后24小时,使用融合前构象特异性小鼠mAb(77.4)或抗组氨酸(HIS)抗体对等分的上清液进行免疫沉淀。使用抗HRC多克隆抗体进行检测。[FIP-HRC450-485-]2-peg11肽使wt可溶性F稳定(参见泳道(lane)1和泳道2,并与稳定的E455G F泳道5进行比较)。参见图89,用三种可溶形式的MeV F转染细胞。使用了Wt F、突变体E455G、双突变体E170G E455G(当在细胞上表达为跨膜蛋白时,两种突变体在融合前状态下均内在稳定)。然后,在有或没有指定的肽的情况下,将细胞在37℃下温育。转染后24小时,将每种组合的3份等分上清液转移至4℃、45℃或55℃下30分钟。然后,将样本转移到4℃并使用融合前构象特异性小鼠mAb(77.4)进行免疫沉淀。使用抗HRC多克隆抗体进行检测。[FIP-HRC450-485-]2-peg11肽在45℃和55℃下稳定所有可溶性F蛋白。即使在没有[FIP-HRC450-485-]2-peg11的情况下,带有E455G的突变体可溶性F以及带有E170G E455G突变的突变体可溶性F在融合前状态下在37℃下也是稳定的。
图90示出了蛋白质免疫印迹(Western blot)。在有(+)或没有(-)1uM指定的肽的情况下,将表达可溶形式的麻疹病毒(MeV)融合蛋白(F)wt、E455G或EG170-E455G的293T细胞培养24小时。收集培养细胞的上清液并在4℃、45℃或55℃下温育30分钟(30’)。然后将样本转移到4℃并使用识别融合前表位的融合前构象特异性小鼠mAb(77.4)进行免疫沉淀。(A)使用抗MeV FHRC对沉淀物进行蛋白质免疫印迹分析。(B)通过蛋白质免疫印迹分析检测到的免疫沉淀MV F蛋白的光密度测量。在存在[FIP-HRC450-485-]2-peg11的情况下,将蛋白质含量标准化为MeV F wt。(n=3,平均值±标准误差)。[FIP-HRC450-485-]2-peg11肽在45℃和55℃下稳定所有可溶性F蛋白。即使在没有[FIP-HRC450-485-]2-peg11的情况下,带有E455G的突变体可溶性F和带有E170G-E455G突变的突变体可溶性F在融合前状态下在37℃下也是稳定的。
图91示出了FIP-HRC防止F激活(与防止F重新折叠的HRC的机制不同)。允许共表达H-HN T193A(一种具有MeV茎部和HPIV3头部的嵌合结合蛋白,其结合唾液酸受体但触发MeVF)和MeV F(S262R,一种容易激活的F)的单层细胞在4℃下与唾液酸受体红细胞(RBC)结合。转移至37℃后,加入含有指定化合物或肽(1uM)的培养基60分钟。然后,加入10mM扎那米韦(zanamivir)以释放可逆结合(即,仅通过H-HN结合而不是通过F插入结合)的RBC。对通过HN受体相互作用可逆结合(橙色)、通过F插入不可逆结合(蓝色)或融合(白色)的RBC进行了量化。纵坐标值是来自一式三份进行的实验的结果的平均值(±SE)。[HRC450-485-peg4]2-chol在F插入靶细胞后阻断融合(不可逆结合,蓝色)。[FIP-HRC450-485-peg4]2-chol融合前状态进行阻断(可逆结合,橙色)。3G是将F稳定在其融合前状态的小分子(可逆结合的RBC,橙色)。在37℃温育开始时加入扎那米韦,释放所有RBC。
图85示出了FIP-HRC靶向MeV F表达细胞。图92示出了[FIP-HRC450-485-peg4]2-chol肽在HEK293T细胞中的定位。将HEK293T细胞培养物与肽(1μM)在37℃下温育60分钟。F蛋白和HRC-FIP肽分别用Alexa Fluor 488(绿色)和Alexa Fluor 594(红色)进行染色。合并后的图像示出了共定位。图93示出了FIP-HRC靶向MeV F表达细胞,来自三次独立实验。
图94示出了MeV F(WT)在指定的肽存在下的热稳定性。在其融合前表位的浓度是未在55℃下温育10分钟的样本的50%(“稳定浓度50”或SC50)和90%(“稳定浓度900”或SC90)。这些值是来自至少三次实验的结果的平均值(±SE)。[FIP-HRC450-485-peg4]2-chol是最有效的F稳定剂。
图95示出了MeV肽的细胞毒性。使用商业MTT试验在293T HEK细胞培养物中评估MeV肽的细胞毒性。肽没有毒性。
图96示出了协同作用。HRC+FIP的等效线图分析。对角线是加和线。位于该线下方、之上或上方的实验数据点(由点表示)分别表示协同作用、加和性或拮抗作用。红色虚线是由FIP和HRC在相同两个组分的不同比率下的贡献生成的曲线。蓝点表示HRC-4和FIP-PEG4-Chol-二聚体在IC50浓度下在FIP-HRC-PEG4-Chol-二聚体中的贡献。数据来自三次实验。该表示出了等效线图分析的结果。
图97示出了衍生自麻疹HRC的带有12个氨基酸的FIP-HRC的效力。替代衍生自麻疹F的HRC肽,使用了衍生自人类副流感病毒3(HPIV3)F的HRC肽。这种肽称为VIKI。对于HPIV3和Nipah病毒,VIKI肽非常有效,但是它是麻疹的弱抑制剂。图97示出了所述肽的IC50和IC90。FIP-VIKI HRC-PEG4-CHOL-二聚体的效力明显低于FIP-MV HRC-PEG4-CHOL-二聚体。这表明效力与氨基酸序列相关。修饰的FIP-MeV HRC(FIP-MeV HRC-Mod-PEG4-CHOL-二聚体)是带有12个氨基酸的FIP-HRC,衍生自麻疹HRC。带有来自麻疹HRC区的12aa的FIP-MeVHRC比FIP-VIKI更有效。
实施例4——FIP-HIV HRC
血清中和试验
在293T细胞中使用Effectine(Qiagen)试剂共转染假病毒env和NL-Luc-AM载体。16小时后更换培养基,分装上清液并且之后在-80°冷冻32小时。在TZM-BL细胞上进行病毒滴定(titration),以找到每秒产生100,000个荧光素酶计数的稀释度。血清在56°下解冻并热灭活1小时。未使用的样本储存在-20°。TZM-BL细胞在感染前16小时以1x104细胞/孔的浓度播种(seeded)在不透明的白细胞培养板中。将血清解冻并以最大速度旋转10分钟。血清首先以1:5稀释,并通过Spin-X(Costar)过滤管以最大速度旋转10分钟。(这不适用于纯化的IgG)。
血清再连续稀释五步,每步以1:4稀释。对照抑制剂以1:5连续稀释6步。将稀释的血清和对照抑制剂转移到预温育板中。将每个动物的血清转移到其各自的板中(每孔110ul)。每个板包含至少两个病毒对照孔和一个背景孔,此时它们仅是培养基。然后将假病毒解冻,稀释至先前确定可产生2,000,000CPS的浓度,并取(110ul)添加到除背景孔之外的所有接受培养基的孔中。板在37°下温育1小时。然后将预温育混合物与细胞混合。每个孔包含足够的体积进行两次重复(100ul的预温育混合物与100ul的细胞)。感染后16小时,可以从细胞中吸出培养基并在必要时更换。感染后第3天从细胞中吸出培养基,并将50ul Glo-Lysis缓冲液(Promega)添加到每个孔中。试验板在-80°下冷冻至少2小时。将试验板解冻,并用多通道移液器将每个孔混合。将等量(50ul)Bright-Glo底物(Promega)添加到每个孔中并检测荧光素酶计数。从样本数据中减去来自每个试验板内的背景孔的计数,并将计数绘制为抑制百分比,其中病毒对照孔设置为100%生长。
假病毒转染方案
该方案适用于在T175培养瓶中进行30ml转染。对于T75培养瓶,将所有的量减半。转染前一天:将293T细胞(ATCC,CRL-11268)播种到T175培养瓶中至约70%汇合度。注:以1:3拆分完全汇合的T175培养瓶可得到所需的细胞浓度。转染日:注:最好在当天晚些时候进行转染,以使得过夜温育时间更接近16小时。将1.5ml EC缓冲液(Effectene试剂盒–Qiagen,301427)添加到15ml组织培养管中。将12ug DNA添加至EC缓冲液中。对于假病毒,添加12ug env质粒和12ug骨架载体(backbone vector)(NL-Luc)。注:大多数测试的env以1:1的env:骨架主链转染最具感染性,然而,低滴度的env可根据具体情况通过经验确定它们的理想比例来增强(任一种质粒的比例不超过5倍)。注:与标准pNL4-3.Luc.R-E-(aidsreagent.org,#3418)相比,NL-Luc-AM骨架一直被证明能产生更具传染性的假病毒。添加100ul增强子(Effectene试剂盒)并通过旋转轻轻混合。在室温下温育5分钟。添加120ul的Effectene试剂(Effectene试剂盒)并通过旋转轻轻混合。在室温下温育10分钟。从293T培养瓶中轻轻吸出细胞系培养基。将10ml细胞系培养基(DMEM、10%FBS、Pen/Strep、Gln)快速添加到15ml管中的DNA复合物中。注:据称,没有必要将Opti-Mem与Effectene一起使用。从管中快速取出含有DNA复合物的细胞系培养基,然后轻轻加入293T培养瓶中。注:293T细胞未紧密附着在培养瓶上——不要将培养基直接吸取至细胞上面。将培养瓶在37度下温育过夜。转染后第1天:注:早晨第一件事是进行冲洗。从293T细胞培养瓶中轻轻吸出培养基。向培养瓶中轻轻加入30ml新的细胞系培养基。注:据称,Effectene转染方法对细胞没有毒性——作为预防措施,在转染假病毒时更换培养基。转染后第2天:注:在当天晚些时候进行收采,据称可以获得更高的滴度。收采也可推迟到第3天。从培养瓶中取出病毒上清液加至50ml管中。以1600RPM的速度将样本旋转10分钟。注:此时,可以标记冷冻管(VWR,#66021-996)。通过Steriflip(Fisher,SCGP00525)或等效装置无菌过滤病毒上清液。注:执行此步骤以去除任何剩余的细胞碎片,但据称这可以在过滤器上捕获一些病毒。对于低滴度病毒,此步骤可以省略。将病毒上清液快速等分至冷冻管中,每管1ml。立即将样本放至-80℃的冷冻冰箱内。注:病毒在没有细胞的情况下在室温下停留的时间越长,滴度就越低。不建议一次对超过一种或两种病毒进行这种收采。也可以在冰上进行等分。
表4:072920EF概要


图98-103示出了多种病毒的抑制数据。图98示出了使用多种脂质-肽缀合物以及阳性和阴性对照对BG505(HIV-1)毒株的抑制数据。图99示出了使用多种脂质-肽缀合物以及阳性和阴性对照对B41(HIV-1毒株)的抑制数据。图101示出了使用多种脂质-肽缀合物以及阳性和阴性对照对16055(HIV-1毒株)的抑制数据。图101示出了使用多种脂质-肽缀合物以及阳性和阴性对照对MN(HIV-1毒株)的抑制数据。图102示出了使用多种脂质-肽缀合物以及阳性和阴性对照对水疱性口炎病毒(VSV)的抑制数据。图103示出了使用多种脂质-肽缀合物对鼠白血病病毒(MLV)的抑制数据。
实施例5——有助于脑内感染和传播的麻疹病毒的病毒融合复合物的分子特征
摘要
在死于麻疹病毒(MeV)中枢神经系统(CNS)感染的患者中发现了一种MeV分离株,其融合蛋白(F)L454W中具有单一氨基酸变化。我们分析了这种突变是否比野生型病毒在CNS中具有适应性优势,从而导致这些患者患病。使用小鼠器官型脑培养物(OBC)和人脑类器官,我们示出了F中的特异性CNS适应性突变导致病毒的离体传播增加,这与增强的先天免疫反应有关。病毒在脑组织中的传播被靶向F的抑制肽阻断,这支持了F参与CNS内传播的观点。MeV F中的单个突变改变融合复合物,使病毒更具神经致病性,即使面对先天免疫反应,其也能在脑组织中传播。
引言
尽管有安全有效的麻疹病毒(MeV)疫苗可用,但MeV并未被根除,并且自2010年以来每年在全球造成100,000-140,000人死亡(Moss&Griffin,2012;Perry等人,2015;Simons等人,2012)。通过疫苗接种根除MeV的部分阻力因素是疫苗接种覆盖率低,这通常与父母对疫苗安全性的不合理担忧有关(Jansen等人,2003)。此外,该疫苗是减毒活病毒,不能用于免疫严重受损的人群。由于SARS-CoV-2大流行降低了儿童常规疫苗接种的覆盖率,并可能使更易感的个体暴露于感染,因此麻疹在全球范围内卷土重来,这种情况可能会恶化。
在初始感染时,MeV感染呼吸道中表达活化CD150(SLAM)的免疫细胞,从而得以进入免疫系统(Tatsuo、Ono、Tanaka和Yanagi,2000)。到达引流淋巴结后,病毒在表达CD150的淋巴细胞中增殖,并从那里继续引起病毒血症。在感染后期,MeV通过在这些细胞基底外侧膜上表达的连接素4感染呼吸道上皮细胞;MeV从这个位置离开宿主的呼吸道,并且可被传播(Mühlebach等人,2011;Noyce等人,2011)。
当MeV感染中枢神经系统(CNS)时,MeV会在感染急性期数天至数年后引起致命的并发症(Allen、McQuaid、McMahon、Kirk和McConnell,1996;Buchanan和Bonthius,2012;Hosoya,2006)。在一小部分病例中,亚急性硬化性全脑炎(SSPE)在初始感染数年后才发展。SSPE的特征是与超突变MeV基因组RNA和病毒转录体以及缺陷病毒颗粒组装相关的大脑持续感染(Cattaneo、Schmid、Billeter、Sheppard和Udem,1988;Rima和Duprex,2005;Schmid等人,1992)。麻疹包涵体脑炎(MIBE)发生在免疫功能不全的患者感染野生型(wt)病毒数周至数月后,并且在极少数情况下,在注射了先前类型的减毒MeV疫苗(不再使用)的情况下发生(Baldolli等人,2016;Buchanan和Bonthius,2012;Hughes、Jenney、Newton、Morris和Klapper,1993)。MIBE可能与介导融合而无需与细胞表面受体接合的病毒融合复合物相关,其称为超融合MeV融合复合物(Hardie、Albertyn、Heckmann和Smuts,2013;C.Mathieu等人,2015)。迄今为止,对控制在CNS中MeV感染和传播的机制仍然知之甚少,尽管CNS入侵似乎需要病毒融合(F)蛋白,从而可被融合抑制剂靶向(Hashiguchi等人,2018;Makhortova等人,2007;M.Watanabe等人,2016;Young和Rall,2009)。
MeV对细胞的感染始于与细胞表面受体的附着,然后由病毒表面上的MeV受体结合(H)和F蛋白的协同动作介导。因此,MeV的H/F复合物构成了促进进入宿主细胞的病毒融合机制(Chang和Dutch,2012;Harrison,2008)。受感染的细胞合成F作为前体(F0),该前体(F0)在细胞内被切割以产生融合前F复合物,该融合前F复合物由三个C端F1亚基组成,这些F1亚基通过二硫键与三个N端F2亚基联系。新的病毒颗粒显示了这种三聚体F结构在动力学上被困在病毒膜外表面的亚稳态构象中(Hashiguchi等人,2018)。通过靶细胞表面进入受体(即,野生型毒株的CD150或连接素4)接合H糖蛋白后,F准备进行融合激活(Mühlebach等人,2011;Noyce等人,2011;Tatsuo等人,2000)。在受体接合后,H触发融合前的F蛋白进行结构转变,从而延伸以将其疏水融合肽插入宿主细胞膜。然后F重新折叠成稳定的融合后6螺旋束结构,这将病毒和靶细胞膜结合在一起以启动融合孔的形成。F蛋白重新折叠并达到这种融合后状态的能力依赖于位于蛋白质的N端和C端的两个互补的七肽重复(HR)区(分别为HRN和HRC)之间的相互作用。该融合步骤可以被对应于这些HR区的肽抑制(Lambert等人,1996)。
已观察到几名患者的CNS感染MeV分离株,所述MeV分离株带有含有HRC结构域中的突变的F蛋白(Ayata等人,2010;Hardie等人,2013;Jurgens等人,2015;Watanabe等人,2013)。一些突变病毒在体内其他器官中的生长受损(S.Watanabe等人,2015)。从两名死于含有带有L454W突变的F的MIBE(Hardie等人,2013)的患者身上回收病毒序列,该突变赋予亚稳态F热不稳定性。我们之前已经证明,这种F突变影响进入靶细胞;表达绿色荧光蛋白并带有L454W F(MeV-IC323-EGFP-F L454W)的重组MeV(IC323毒株)在缺乏已知MeV受体的细胞中传播。在细胞间融合试验中,单独带有L454W突变的F独立于H蛋白介导融合(Jurgens等人,2015)。相比之下,其他超融合病毒依赖于H蛋白进行膜融合(Sato等人,2018)。
在MIBE患者中发现的F L454W突变可能是在CNS中从头合成的(Hardie等人,2013)或可能存在于wt病毒群中并在CNS中经历了阳性选择。该病毒的来源无法确定,并且带有该F的CNS分离株是否能感染其他组织尚不清楚。一份报告显示,带有L454W F的病毒可以在某些融合抑制剂的选择压力下出现(Ha等人,2017),这表明,带有这种神经致病性F蛋白的病毒可以在CNS之外找到。在最近的研究中,我们发现带有L454W F的病毒在棉鼠肺中比wt生长得好,在MeV CNS感染的哺乳期hCD150 tg小鼠模型中的致死率高10倍,并且比wt MeV更快地到达小鼠CNS(Mathieu等人,2019)。然而,就观察到的这种突变和大脑中改变的融合复合物的影响,尚未对其原因进行研究。
在这一系列实验中,我们提出疑问,在以下两个CNS感染的离体模型中,携带CNS适应融合复合物的MeV在生长和传播方面是否不同于wt MeV:小鼠器官型脑培养物和人脑类器官。在脑炎病例中观察到的超融合变体在这些模型中的传播比wt更有效。感染不需要任何已知的麻疹受体,并且感染程度与MeV F(wt或突变体)在其融合前状态下的稳定性成反比。病毒的传播被抑制F重新折叠的融合抑制剂阻断。诱发先天免疫反应,但这并不阻断病毒传播。上述的病毒的离体进化模型强调了MeV融合复合物进行CNS适应的功能要求。
结果
带有F糖蛋白L454W的麻疹病毒在细胞培养物中不稳定
我们以及其他人先前已描述过与从患者分离的或在实验环境中生成的神经致病性麻疹毒株相关联的MeV F糖蛋白中的突变(L454W、T461I和N462K)(Hardie等人,2013;Jurgens等人,2015;Mathieu等人,2019;Watanabe等人;2013)。这些突变降低了F蛋白融合前亚稳态的热稳定性。为了探索改变稳定性的分子决定因素,将这些突变映射到F蛋白的融合前构象(MeV F;PDB 5YXW;Hashiguchi等人,2018)和融合后构象(HPIV3 F;PDB 1ZTM;Yin等人,2005)的X射线结构上(图104)。三个突变(L454W、T461I和N462K)均位于HRC结构域内。基于融合前的结构,L454W突变可能会导致与同一原体内的T314和/或相邻原体内的L457的空间位阻。T461I和N462K突变发生在HRC结构域的良序α-螺旋区。在电脑模拟中(Insilico),这些残基的突变会导致与相邻原体的空间冲突。最值得注意的是,这三个突变发生在HRC结构域中融合前构象的头部和茎部区相交的部分。该交界处的相互作用可能对稳定融合前状态很重要,因此,正如我们之前的数据所表明的,该区的突变可能导致MeV-F融合前结构的稳定性降低(Jurgens等人,2015)。
在两个不同的临床病例中发现了MeV F L454W。该突变降低了融合蛋白的稳定性,产生了即使在没有已知受体的情况下,也允许MeV在Vero细胞中传播的超融合表型。为了评估这种突变对病毒适应性(fitness)的影响,我们生成了带有F L454W的MeV IC323-EGFP的重组感染性克隆体,并在37℃或32℃下在Vero-CD150细胞中培养重组病毒(更低的温度使F蛋白稳定在其融合前状态(Jurgens等人,2015))。在通过反向遗传学生成带有L454WF的重组病毒的过程中(认为该过程是常规过程),出现了一组意料之外的突变。与通常增长到5.106噬斑形成单位(pfu)/ml以上的wt病毒原液相比,两种病毒制剂的滴度保持较低,在32℃下仅达到2x105 pfu/ml以及在37℃下仅达到约5x105 pfu/ml。之前我们注意到,与wt病毒相比,突变体的病毒滴度在37℃下显著降低(Mathieu等人,2019),表明带有L454W突变的F蛋白对细胞培养物中的病毒生长不利。在3次传代后,从相同的初始质粒中取出但在不同的温度下生长的这两种病毒原液的下一代测序显示,两者的F蛋白中都有额外的突变:除了存在于100%序列中的L454W,在32℃下出现G506E突变,其等位基因频率为约36%,并且在37℃下出现E455G突变,等位基因频率为约22%。在图104中可以观察到E455G突变,但在可用的晶体结构中没有解析出G506。我们假设L454W F变体的不稳定性有利于细胞培养物中这两个新突变的出现。这些出人意料的突变提供了如下所述的显著调查工具。
在没有已知受体的情况下,脑组织中MeV-IC323-EGFP-F
L454W与wt病毒的生长
由于已经从CNS中分离出带有L454W F的MeV病毒,我们假设其在两种脑部感染模型(鼠和人)中可以很好地适应。我们假设这种突变会在大脑中受到阳性选择压力。我们之前曾使用来自IFNAR1敲除(KO)小鼠的小鼠小脑器官型脑培养物(OBC)来评估在CNS中存在的四种细胞类型中的病毒感染和传播(Welsch等人,2017),但缺少与1型IFN反应相关的抗病毒作用。来自表达人类CD150 F1转基因的小鼠的海马体和小脑OBC维持wt病毒感染和传播(J.C.Welsch等人,2013),但是当不存在已知的MeV受体时,wt病毒不传播(Ferren等人,2019;Welsch等人,2017)。在图105A-D所示的实验中,OBC源自不表达任何已知的麻疹受体的IFNAR1KO小鼠。wt病毒(表达增强型绿色荧光蛋白,EGFP)未能在96小时内传播(图106A和图106B)。在图105C-D中,来自IFNAR1KO小鼠的OBC与5000pfu的wt病毒共感染,该wt病毒表达不同的荧光蛋白(红色荧光蛋白tdTomato)和MeV-IC323-EGFP-F L454W(带有培养物中出现的额外G506E突变)。在24小时(图105C)和96小时(图105D)监测感染情况。虽然wt病毒没有(如预期)在组织中有效传播,但带有L454W(EGFP)的病毒感染并传播,并且G506E突变等位基因频率从约36%增加到约70%,表明对于F中的该突变的强大的阳性选择压力。
阻断融合抑制所有变体的传播
对MeV疫苗毒株感染的研究表明,干扰F蛋白功能可以在阻止CNS内的传播(Makhortova等人,2007)。我们之前示出了MeV F衍生的二聚体胆固醇缀合的融合抑制肽(称为HRC4,其阻断F介导的融合)阻断棉鼠和小鼠的体外、离体和体内wt MeV感染(Mathieu等人,2015、2019;Welsch等人,2013)。图105E-G中评估了HRC4肽阻断病毒在暴露于病毒后的OBC组织中传播的功效。本文中所使用的OBC获取自不表达任何已知MeV受体的IFNAR1KO(Welsch等人,2017)。
使用RT-qPCR对MeV N RNA拷贝的数量——反映实验结束时OBC中存在的病毒载量——进行了量化。图105F示出了,与未处理的样本中的病毒载量相比,用100nM HRC4肽处理的感染后培养物中的病毒载量显著降低2个对数去除率(log reduction)(**p=0.008,曼-惠特尼U检验(Mann-Whitney U-test)),并且用10nM处理的组中,MeV N RNA拷贝的数量显著降低4倍(*p=0.03,曼-惠特尼U检验)。在用较低浓度的融合抑制剂处理的组中,没有观察到病毒载量的显著变化。MeV IC323-EGFP-F L454W病毒侵入组织,感染四天后,在整个培养物中形成了广泛的融合区(图105G)。HRC4肽(100nM或10nM)在同一时间段内阻断MeV-IC323-EGFP-F L454W的扩散(图105G)。在所使用的最高浓度(100nM)下,仅观察到分离的单个感染细胞。更低浓度(10nM)仅是部分抑制性的,观察到几个焦点扩散区(图105G),并且更低浓度(低于10nM)和未经处理的组织之间没有发现显著差异(图105F)。由于融合抑制剂在感染后24小时添加,这些结果表明HRC4肽阻断了三维CNS组织中的细胞间扩散,这表明MeV在脑组织中的传播取决于融合蛋白的功能。即使使用来自表达人类CD150(SLAM)受体的IFNAR1KO小鼠的OBC——增强病毒在CNS中的传播,最高浓度的肽也阻断MeV IC323-EGFP-FL454W的传播,至少与WT病毒一样(图108)。
具有免疫能力的OBC中的带有L454W的病毒的进化
为了评估在完全合格的(fully competent)小鼠CNS存在的情况下,带有L454W加L454W/E455G或L454W/G506E双群的两种病毒的病毒进化,我们从wt小鼠中获得了OBC。来自C57BL/6乳鼠的OBC感染了MeV wt和L454W F病毒(使用额外的L454W/E455G或L454W/G506E)。我们之前观察到(Ferren等人,2019),wt病毒在这个模型中不传播,我们在本文中证实了这一发现。相比之下,带有L454W F的两种病毒均在C57BL/6OBC中有效传播,并且在7天后具有相似的感染水平(图105H)。7天后,裂解OBC进行测序,以评估转录组、病毒进化和病毒RNA量化。在C57BL/6OBC中,与感染wt病毒的OBC(传播最小)相比,带有L454W F的病毒诱导了与强干扰素信号相关的基因表达模式,其中该基因模式与未感染的OBC无法区分(参见图109差异基因表达分析)。尽管有这种强烈的先天免疫反应,然而,带有L454W F的病毒的传播虽然减少但并未停止(参见图105I)。然后,我们分析了这些OBC中的病毒序列,以确定L454W/E455G和L454W/G506E突变是否保守。序列分析示出于补充材料的LAVA图中(Lin等人,2019b);LAVA图#1示出了四份样本(CM005-8)中L454W/G506E的等位基因频率以及一份样本(CM017)中L454W/E455G的等位基因频率。输入L454W/G506E的病毒的双突变体的等位基因频率为约36%(在病毒制剂中观察到的),在7天后在离体组织中上升至97%、89%、96%和78%。对于L454W/E455G,输入病毒的双突变体等位基因频率为约22%,并且在离体序列中,双突变体降低至约4%。
人诱导性多能干细胞(hiPSC)衍生的脑类器官的感染:CNS适应变体与wt病毒
为了将CNS建模扩展到人类神经组织,我们将hiPSC(来自两名独立的供体,一名男性和一名女性,FA10和FA11)分化为脑类器官(Lancaster和Knoblich,2014)。90天大的类器官感染了wt病毒或带有L454W F的病毒。为了分析该模型中病毒传播的改变是否与F的特性有关而不是简单归因于个体突变,类器官也感染了带有含N462K突变(之前发现的在仓鼠大脑中有效生长的实验室适应突变)和T461I突变(之前在SSPE病例中发现)的F的MeV(Watanabe等人,2013)。图104中描述了这些突变。在10天期间内监测感染情况,并通过监测荧光来评估传播(图106A和图110示出了在第10天拍摄的照片)。为了确保用等量的病毒进行感染,在表达CD150的Vero细胞(Vero-CD150)中对用于感染人脑类器官的接种物进行了平行评估,如图106B和图110所示。所有病毒滴度均相似(pfu/ml),其中L454W突变体仅略低。在Vero-CD150中,所有病毒在3天内有效传播并破坏了细胞单层(数据未显示)。10天后,裂解感染和未感染的人脑类器官进行RNA测序,以评估转录组、监测类器官感染期间的病毒进化并量化病毒RNA(参见图107C-D、图112和补充材料)。
与带有wt F的病毒相比,带有L454W F的病毒在人脑类器官中的传播效率更高(图106A和106C),并且被感染后24小时添加的HRC4融合抑制剂有效阻断(图110)。带有T461I突变F的病毒(来自SSPE患者)也在脑类器官中传播。相比之下,与wt病毒相比,带有N462K F的病毒仅显示出适度的传播增加(图106A和106C)。
在图106D中,我们比较了人脑类器官中未感染或感染(wt病毒或带有L454W F蛋白的病毒)的细胞之间的差异基因表达。带有L454W F的MeV的每百万读数(reads permillion,RPM)值比MeV wt高出几倍(图106D)。与感染wt病毒的类器官和对照样本相比,带有L454W F的病毒诱导了与干扰素信号相关的基因表达模式(p=2.6x10-17,差异基因表达分析参见图110)。这种反应似乎可能是病毒生长差异的结果,因为干扰素反应似乎与独立于特定的F突变的病毒计数相关(图111示出了图106A中所示的所有MeV变体感染的差异表达)。对于两组人脑类器官,最高的基因水平表达系数与最年幼的胎儿发育阶段(受孕后8-13周)和源自杏仁核的脑组织相关(图112),如前所述(Luo等人,2016;Qian等人,2016)。对两种人脑类器官感染系列共有的差异表达基因的通路分析表明,人脑类器官中的麻疹感染引起了强烈的干扰素反应(图113)。
然后,我们提出疑问,这两个病毒群(L454W-L454W/E455G和L454W-L454W/G506E)是否在该模型中进化,就像它们在小鼠器官型脑切片中那样。序列分析于LAVA图中示出。在L454W-L454W/G506E群的情况下,值得注意的是从脑类器官中出现的病毒序列的F蛋白中的G506E变体,其等位基因频率从病毒原液中的约36%增加至FA10脑类器官中的约96%和FA11脑类器官中的约78%(参见图114中FA10的数据)。第二病毒原液(含有L454W和L454W/E455G,其中双突变体以22%的等位基因频率存在)用于感染第二组脑类器官(源自FA11iPSC)。L454W突变得以维持,并且类器官中E455G F的等位基因频率降低至约2%(参见图115)。野生型病毒未表现出任何显著的改变(参见图116)。
L454W
F背景中的额外突变稳定了F蛋白的融合前状态
在鼠和人体组织中,我们观察到F中G506E的阳性选择和E455G的阴性选择。为了理解这两种额外突变的不同选择的原因,我们使用采用指定F蛋白的cDNA转染的细胞评估带有额外突变(单独以及与L454W组合)的F的功能特性。在我们的功能试验中,带有L454W/G506E突变的F仍然明显不如wt F稳定,而带有L454W/E455G的F比wt F更稳定(图108A-B)。神经致病变体的标志是在没有已知病毒受体的情况下融合的能力,并且这种表型仍然存在:L454W/G506E F可以在没有任何已知受体(CD150/SLAM或连接素4)存在的情况下介导融合。然而,L454W/E455G F——带有在类器官生长期间频率显著降低的E455G突变——在没有已知受体的情况下不介导融合。
然后,基于图107A-B的结果,我们提出疑问,带有L454W/E455G F的同质病毒群在器官型脑切片和脑类器官模型中是否表现得像wt病毒,以及E455G是否会在这些神经组织中受到阴性选择压力。因此,我们试图生成两种带有L454W/E455G F或E455G F蛋白的新病毒,并分析它们在离体脑组织中的生长和传播。在Vero-CD150中,带有双突变体L454W/E455G F的重组病毒的生长与wt病毒相似。然而,带有单独突变的E455G F的重组病毒无法回收,这表明它在培养物中是不利的。为了确定双突变体在脑中是否具有优势,我们在OBC中共感染了wt(红色)和带有L454W/E455G(绿色)F的病毒,如图105D中所示。图107E示出了感染后96小时病毒传播的程度。与图105D所示结果(其中L454W病毒侵入整个组织,并且wt感染是有限的)形成鲜明对比,共感染导致wt和L454W/E455G病毒的传播相似(有限)。
为了确认人脑类器官中的L454W/E455G病毒在的表型,我们感染了90天大的类器官(图1107F-H)。在10天期间内监测感染情况,并且图107F示出了第10天的荧光病毒传播。为了确保用预期量的病毒(5000pfu/孔)进行感染,在Vero-CD150中对用于感染人脑类器官的接种物进行了平行评估,如图107G所示。带有L454W/E455G F的病毒在10天后在FA10和FA11hiPSC衍生的脑类器官中传播有限(图107F),但在Vero-CD150中传播并在2天内破坏细胞单层(图107G)。10天后,裂解受感染的人脑类器官进行RNA测序,以评估转录组、监测类器官感染期间的病毒进化、量化病毒RNA并评估病毒进化(参见图107H、图111和图113)。L454W/E455G感染的病毒基因组数量与我们观察到的wt病毒的情况相似(图106),并且10天后,双突变保持稳定(参见LAVA图#5)。为了证实这些发现,我们由hiPSC FA11分化出另一组脑类器官。用1000pfu/孔的wt、L454W F(L454W和L454W/E455G混合群)和带有L4545/E455GF的病毒感染共9个孔。感染后20天,裂解脑类器官并提取RNA用于病毒测序。数据示出于LAVA图#6中。在所有三份样本中,wt病毒仅在P基因(R77C)中发生变化,其等位基因频率为约30%。一份样本中带有L454W F的病毒(具有L454W和L454W/E455G混合群)完全消除了E455G突变。在第二份样本中,E455G保持在约24%,并且第三份样本丢失了E455G并获得了额外的突变(D538G),其频率为约23%。双突变体L454W/E455G保持稳定,没有显著变化,并且不能在脑类器官中很好地传播。
鉴于发现如G506E的F稳定型突变不一定干扰CNS适应,我们在体内寻找这种表型的证据。在最初发表关于F中的L454W突变(Hardie等人,2013)时,分离的患者病毒的F蛋白中带有一个额外的突变(M337L)。我们评估了M337L(单独和与L454W组合)对F的融合和稳定性的影响(图107A和107B)。表达的M337L F在没有受体的情况下不介导融合,其融合前状态比wt F更稳定。M337L/L454W F在没有受体的情况下促进融合,并且明显不如wt F耐热。体内进化(通过不同的突变残基)导致与G506E所引起的类似的功能改变。这种模式反映了CNS特异性适应模式。
讨论
在南非MeV爆发中,8名HIV感染者死于MeV CNS临床表现(Albertyn等人,2011)。我们表征了一种CNS适应的融合复合物(由H和F组成)的特性(Jurgens等人,2015)。来自MIBE患者的CNS的MeV融合复合物序列被改变,使得F在没有已知进入受体的情况下被激活。来自两名不同患者的分离株的F蛋白在第454位(L454W)处含有一个特定的氨基酸改变,这增加了F介导与任何异型粘附蛋白融合的能力,并显著降低了F的热稳定性(Jurgens等人,2015)。
在通过反向遗传学生成带有L454W F的重组病毒的技术方法中,意外地出现了一组出乎意料地提供有用信息的(unexpectedly informative)突变。在37℃下进行的标准方法产生带有L454W F或L454W/E455G F的混合病毒群。将病毒生产的温度降低至32℃以稳定F得到含有L454W和L454W/G506E的群。虽然E455位于HRC结构域中(参见图104),但残基G506位于跨膜结构域中并且在MeV F的可用晶体结构中没有解析出。将带电残基置于跨膜结构域中可能会对蛋白质折叠和活性产生巨大影响,但该突变在我们的离体模型中被阳性选择。我们的数据显示,HRC突变E455G导致F稳定性增加,并且L454W/E455G F(与L454W不同)要求H(MeV受体结合蛋白)与受体结合以介导融合。因此,该第二个HRC结构域突变对于H-受体相互作用具有要求。G506E跨膜结构域突变略微恢复了F的稳定性,但双突变体L454W/G506F仍然不如wt稳定,并在没有已知受体的情况下介导融合。尽管这些补偿性突变由于技术方法而产生,但它们提供了直接证据表明可以在没有已知受体的情况下融合的病毒在CNS中被阳性选择。
带有T461I和N462K F蛋白的病毒在细胞培养物中和离体情况下(数据未示出)均表现出遗传稳定性,然而,出人意料的是,携带N462K F的病毒没有在人脑类器官中像我们预期的那样令人满意地生长。双突变体L454W/G506E似乎已经达到了适于这两种培养条件的“平衡”。正如我们对于人类副流感病毒3型观察到的(Iketani等人,2018),从细胞培养中的临床标本中分离病毒为病毒进化提供了选择压力,这可掩盖临床毒株的真实特征。临床样本的直接测序避免产生这些细胞系伪影(Iketani等人,2018)。我们之前描述了来自SSPE病例的病毒,其中共有F(SSPE F)具有5个氨基酸变化(G168R、E170G、S262G、A440P、R520C、L550P)(Angius等人,2019)。SSPE F的热稳定性低于wt F,并且可以在没有已知受体的情况下介导融合。当单独评估突变的影响时,我们观察到单一的突变不能赋予融合特性。一些突变增加了融合前F的稳定性(例如E170G),而其他突变使其降低(例如S262G)。我们推测,临床病例中可能发生了导致完全适应CNS的SSPE F的体内进化。
对首次确定F中存在L454W突变的临床样本之一进行深度测序,揭示了F蛋白中存在其他几个突变。F蛋白中的一个额外突变(M337L)具有与L454W相似的等位基因频率,并且带有M337L/L454W的F蛋白与带有L454W/G506突变的F的融合表型相匹配,证明了病毒在体内向着与在脑类器官中观察到的相似的F功能进化。这些结果表明,在CNS中可能会阳性选择不同的突变,这些突变会得到类似的不稳定和不依赖受体的F。我们将M337L突变映射到MeV F的融合前结构上。该突变可能与L256和L257形成疏水性相互作用,以稳定MeV F的融合前构象。带有L454W/E455G F的病毒我们的实验中被成功回收,并且与wt病毒有相似的表现。脑类器官中的感染是有限的(对于wt病毒),并且在本文所呈现的两个独立实验的10-20天跨度中,其并未导致突变的阴性选择。更长时间的感染可能会导致E455G突变被消除,或导致引入额外的突变。尽管进行了多次尝试,但无法回收仅带有E455G F的病毒,并且我们认为E455G F稳定性的提高可能不利于适应。
附图注解
图104示出了来自CNS适应病毒的F蛋白内的替换位置。(A)带有融合肽(FP)、N端七肽重复(HRN)、C端七重复(HRC)、跨膜(TM)和细胞质(CT)结构域的MeV F的示意图。(B)融合前(左,MeV F;PDB5YXW)和融合后(右,HPIV3 F;PDB 1ZTM)构象的带状图。示出了F蛋白结构中的五个替换(M337L、L454W、E455G、T461I和N462K)。
图105示出了野生型(wt)病毒的离体感染与带有L454W F的病毒的离体感染:在器官型脑培养物(OBC)中,CNS适应病毒胜过wt病毒。(A-B)来自IFNARKO小鼠大脑的OBC使用5000噬斑形成单位(pfu)/切片的带有EGFP(绿色荧光)的wt病毒进行感染。(C,D)来自IFNARKO小鼠大脑的OBC使用5000噬斑形成单位(pfu)/切片的带有tdTomato(红色荧光)的wt病毒和MeV-IC323-L454W FEGFP(绿色荧光)(5000pfu/切片)进行共感染,并在96小时内进行监测。照片拍摄于24小时(C)和96小时(D)。比例尺=500μm。(E-G)MeV F衍生的融合抑制肽(HRC4)抑制带有L454WF的MeV在OBC中的传播。来自IFNARKO鼠脑的OBC使用5000pfu/切片的MeV-IC323-L454W FEGFP感染4天。通过在初始感染后24小时、48小时和72小时以指定的浓度添加HRC4融合抑制肽对OBC进行处理,或不作处理(NT对照)。(E)步骤示意图。(F)在感染后4天从器官型切片中收采总RNA,并通过RT-qPCR量化MeV N基因的表达水平。结果表示为来自5只不同小鼠的培养物的平均值±标准偏差(**,P<0.01;***,P<0.001[曼-惠特尼U检验])。(G)在以指定浓度处理的OBC中,通过落射荧光显微镜4dpi观察到与感染相关的绿色荧光(比例尺=500μm)。(H)在完全免疫合格的OBC中带有L454W F感染的离体病毒。来自C57/BL6鼠脑的OBC使用1000pfu/切片的带有L454W F的病毒(使用两种病毒制剂,一种在F中具有额外的E455G,另一种在F中具有G506E)感染7天。如图所示,照片拍摄于感染后4天。(I)带有L454W的病毒在wt和IFNARKO OBC中的生长。来自wt或IFNARKO鼠脑的OBC使用1000PFU/切片的MeV-IC323-L454WFEGFP感染7天。在感染后4天从OBC收采总RNA,并通过RT-qPCR量化MeV N基因的表达水平。结果表示为来自至少5只不同小鼠的培养物中的平均值±标准偏差(*,P<0.05;***,P<0.001[曼-惠特尼U检验])。
图106示出了CNS适应的MeV变体在人多能干细胞(hiPSC)衍生的脑类器官中有效传播。(A)两组独立的90天大的人脑类器官(源自两个hiPSC,FA10和FA11)使用带有指定的MeV融合(F)蛋白的重组MeV病毒(带有EGFP或tdTomato荧光蛋白)进行感染。对于每种病毒,对3个各自包含2-4个类器官的单独的孔进行感染(5,000pfu/孔)。随着时间的推移对脑类器官进行监测,并且本文所示的荧光反映了10天后的感染情况。比例尺=1000μm。(B)用于感染的接种物的病毒滴度在Vero CD150上评估,以pfu/ml(log)为单位。(C)在感染后10天从人脑类器官中收采总RNA,并通过RT-qPCR量化MeV N的基因表达水平。(D)脑类器官中wt病毒与带有L454WF的病毒的感染的RNA-Seq分析(来自三个独立实验的数据)。对未感染和MeV感染的脑类器官的七个重复进行了转录分析(n=6未感染,n=5WT,n=2带有L454W F的病毒)。每个热图下方描述了每份样本的MeV RPM值。在所有样本中对原始计数进行标准化,并进行差异表达分析。热图中描绘了L454W和未感染之间具有最低校正p值的50个基因,通过每份样本相对于每个基因的平均标准化计数的log2倍数变化着色。
图107示出了带有指定突变的MeV融合(F)蛋白的融合活性和热稳定性。(A)通过B-gal互补试验,对共表达指定MeV F蛋白和MeV wt血凝素(H)的HEK293T细胞与(没有任何已知的麻疹受体的)HEK293T细胞之间的细胞间融合进行评估。Y轴上的值表示为三个独立实验结果的相对发光单位(RLU)的平均值(标准误差,SE)。*p<0.05,**p<0.01,***p<0.001,****,p<0.0001(双向方差分析(2way ANOVA))。(B)在存在连接素4、CD150或没有受体的情况下,与wt F相比,带有指定突变的MeV F蛋白(IC323)的融合活性百分比。(C)用带有指定突变的MeV F蛋白转染HEK293T细胞,并在37℃下温育24小时,然后升至55℃指定的时间。Y轴上的值代表与指定的F蛋白结合的融合前构象特异性抗体的百分比(与零时的wtF蛋白相比)。这些值是三个独立实验的平均值。****,p<0.0001(图4D中总结了双向方差分析结果)。(D)与零时时的wt F(100%)相比,在55℃下将融合前表位的分数降低至50%(TS50)和降低至10%(TS10)的时间(分钟)。数据是三个实验的平均值+/-SE。(E)来自IFNARKO鼠脑的OBC使用5000pfu/切片的带有tdTomato(红色荧光)的wt病毒和MeV-IC323-L454W/E455G FEGFP(绿色荧光)共感染,并在96小时内进行监测。照片拍摄于96小时。比例尺=500μm。(F)两组独立的90天大的人脑类器官(源自两个hiPSC,FA10和FA11)使用带有L4545W/E455G F蛋白的重组MeV病毒进行感染。对包含2-4个类器官的三个单独的孔进行感染(5000pfu/孔)。随着时间的推移对脑类器官进行监测,并且本文所示的荧光反映了10天后的感染情况。比例尺=1000μm。(G)在Vero CD150(pfu/ml;log)上对用于感染的病毒接种物的病毒滴度进行评估。照片示出了2天后的感染程度(标明了pfu/孔)。(H)感染后10天,从人脑类器官中收采总RNA,并通过RT-qPCR量化MeV N的基因表达水平。
参考文献
1.Ader,N.,Brindley,M.,Avila,M., C.,Horvat,B.,Hiltensperger,G.,Schneider-Schaulies,J.,Vandevelde,M.,Zurbriggen,A.,Plemper,R.K.等人(2013).麻疹病毒属粘附蛋白触发主动膜融合的机制(Mechanism for active membrane fusiontriggering by morbillivirus attachment protein).病毒学杂志(J.Virol.),87,314–326.
C.,Horvat,B.,Hiltensperger,G.,Schneider-Schaulies,J.,Vandevelde,M.,Zurbriggen,A.,Plemper,R.K.等人(2013).麻疹病毒属粘附蛋白触发主动膜融合的机制(Mechanism for active membrane fusiontriggering by morbillivirus attachment protein).病毒学杂志(J.Virol.),87,314–326.
2.Albertyn,C.,van der Plas,H.,Hardie,D.,Candy,S.,Tomoka,T.,Leepan,E.B.和Heckmann,J.M.(2011).南非麻疹爆发的无记载的伤亡(Silent casualtiesfromthe measles outbreak in South Africa).南非医学杂志(S.Afr.Med.J.),101,313–314,316–317.
3.Allen,I.V,McQuaid,S.,McMahon,J.,Kirk,J.和McConnell,R.(1996).SSPE中麻疹病毒抗原和基因组在CNS中的分布对病毒传播和髓鞘脱失机制的意义(Thesignificance of measles virus antigen and genome distribution in the CNS inSSPE for mechanisms of viral spread and demyelination).神经病理学与实验神经病学杂志(Journal of Neuropathology and Experimental Neurology),55,471–480.
4.Angius,F.,Smuts,H.,Rybkina,K.,Stelitano,D.,Eley,B.,Wilmshurst,J.,Ferren,M.,Lalande,A.,Mathieu,C.,Moscona,A.等人(2019).对2009-2010年南非麻疹流行的亚急性硬化性全脑炎基因型B3病毒的分析表明,超融合F蛋白有助于脑中的麻疹病毒感染(Analysis of a Subacute Sclerosing Panencephalitis Genotype B3 Virus fromthe 2009-2010South African Measles Epidemic Shows That Hyperfusogenic FProteins Contribute to Measles Virus Infection in the Brain).病毒学杂志(J.Virol.),93.
5.Avila,M.,Alves,L.,Khosravi,M.,Ader-Ebert,N.,Origgi,F.,Schneider-Schaulies,J.,Zurbriggen,A.,Plemper,R.K.和Plattet,P.(2014).界定犬瘟热病毒融合前F复合物的触发范围的分子决定因素(Molecular determinants defining thetriggering range of prefusion F complexes of canine distemper virus).病毒学杂志(J.Virol.),88,2951–2966.
6.Ayata,M.,Takeuchi,K.,Takeda,M.,Ohgimoto,S.,Kato,S.,Sharma,L.B.,Tanaka,M.,Kuwamura,M.,Ishida,H.和Ogura,H.(2010).源自一例亚急性硬化性全脑炎的麻疹病毒的Osaka-2毒株的F基因是神经毒性的主要决定因素(The F gene of the Osaka-2strain of measles virus derived from a case of subacute sclerosingpanencephalitis is a major determinant of neurovirulence).病毒学杂志(J.Virol.),84,11189–11199.
7.Baldolli,A.,Dargère,S.,Cardineau,E.,Vabret,A.,Dina,J.,de LaBlanchardi ère,A.和Verdon,R.(2016).免疫功能不全患者的麻疹包涵体脑炎(MIBE)(Measles inclusion-body encephalitis(MIBE)in a immunocompromised patient).临床病毒学杂志(Journal of Clinical Virology),81,43–46.
8.Battles,M.B.,Langedijk,J.P.,Furmanova-Hollenstein,P.,Chaiwatpongsakorn,S.,Costello,H.M.,Kwanten,L.,Vranckx,L.,Vink,P.,Jaensch,S.,Jonckers,T.H.M.等人(2016).呼吸道合胞体病毒抑制剂的分子机制(Molecularmechanism of respiratory syncytial virus fusion inhibitors).自然化学生物(Nat.Chem.Biol.),12,87–93.
9.Buchanan,R.和Bonthius,D.J.(2012).麻疹病毒和相关的中枢神经系统后遗症(Measles Virus and Associated Central Nervous System Sequelae).儿科神经病学研讨会(Seminars in Pediatric Neurology),19,107–114.
10.Cattaneo,R.,Schmid,A.,Billeter,M.A.,Sheppard,R.D.和Udem,S.A.(1988).多重病毒突变而非宿主因素导致亚急性硬化性全脑炎细胞系中的麻疹病毒基因表达缺陷(Multiple viral mutations rather than host factors cause defectivemeasles virus gene expression in a subacute sclerosing panencephalitis cellline).病毒学杂志(J.Virol.),62,1388–1397.
11.Chang,A.和Dutch,R.E.(2012).副粘病毒的融合和进入:通向共同目标的多条路径(Paramyxovirus fusion and entry:multiple paths to a common end).病毒(Viruses),4,613–636.
12.Cosby,S.L.和Brankin,B.(1995).麻疹病毒感染脑内皮细胞以及对其粘附特性的影响(Measles virus infection of cerebral endothelial cells and effect ontheir adhesive properties).兽医微生物学(Vet.Microbiol),44,135–139.
13.Delhaye,S.,Paul,S.,Blakqori,G.,Minet,M.,Weber,F.,Staeheli,P.和Michiels,T.(2006).神经元在病毒性脑炎期间产生I型干扰素(Neurons produce type Iinterferon during viral encephalitis).美国科学院院报(Proc.Natl.Acad.Sci.U.S.A.),103,7835–7840.
14.Delpeut,S.,Sawatsky,B.,Wong,X.-X.,Frenzke,M.,Cattaneo,R.和vonMessling,V.(2017).连接素-4相互作用在一种新的发病机制模型中控制麻疹病毒毒性——松鼠猴(Saimiri sciureus)(连接素-4Interactions Govern Measles VirusVirulence in a New Model of Pathogenesis,the Squirrel Monkey(Saimirisciureus)).病毒学杂志(Journal of Virology),91,e02490-16.
15.Devaux,P.,Hodge,G.,McChesney,M.B.和Cattaneo,R.(2008).V-或C-缺陷麻疹病毒的减毒:通过恒河猴炎症和干扰素反应控制感染(Attenuation of V-or C-defective measles viruses:infection control by the inflammatory andinterferon responses of rhesus monkeys).病毒学杂志(Journal of Virology),82,5359–5367.
16.Devaux,P.,Hudacek,A.W.,Hodge,G.,Reyes-del Valle,J.,McChesney,M.B.和Cattaneo,R.(2011).一种无法拮抗STAT1功能的重组麻疹病毒无法控制恒河猴炎症并减弱(A Recombinant Measles Virus Unable To Antagonize STAT1 Function CannotControl Inflammation and Is Attenuated in Rhesus Monkeys).病毒学杂志(Journalof Virology),85,348–356.
17.Esolen,L.M.,Takahashi,K.,Johnson,R.T.,Vaisberg,A.,Moench,T.R.,Wesselingh,S.L.和Griffin,D.E.(1995).急性致死性麻疹患儿脑内皮细胞感染(Brainendothelial cell infection in children with acute fatal measles).临床研究杂志(J.Clin.Invest.),96,2478–2481.
18.Ferren,M.,Horvat,B.,and Mathieu,C.(2019).麻疹脑炎:迈向新疗法(Measles Encephalitis:Towards New Therapeutics).病毒(Viruses),11.
19.Frenzke,M.,Sawatsky,B.,Wong,X.X.,Delpeut,S.,Mateo,M.,Cattaneo,R.和von Messling,V.(2013).连接素-4依赖性麻疹病毒传播到食蟹猴气管上皮:感染的免疫细胞浸润固有层的作用(连接素-4-dependent measles virus spread to the cynomolgusmonkey tracheal epithelium:role of infected immune cells infiltrating thelamina propria).病毒学杂志(Journal of Virology),87,2526–2534.
20.Generous,A.R.,Harrison,O.J.,Troyanovsky,R.B.,Mateo,M.,Navaratnarajah,C.K.,Donohue,R.C.,Pfaller,C.K.,Alekhina,O.,Sergeeva,A.P.,Indra,I.等人(2019).由连接素引发的反式内吞作用在细胞之间转移细胞质货物,包括感染性物质(Trans-endocytosis elicited by连接素stransfers cytoplasmic cargo,including infectious material,between cells).细胞科学杂志(J.Cell.Sci.),132.
21.Griffin,D.E.,Lin,W.-H.和Pan,C.-H.(2012).麻疹病毒、免疫控制和持久性(Measles virus,immune control,and persistence).FEMS微生物学综述(FEMSMicrobiol.Rev.),36,649–662.
22.Ha,M.N.,Delpeut,S.,Noyce,R.S.,Sisson,G.,Black,K.M.,Lin,L.-T.,Bilimoria,D.,Plemper,R.K.,Privé,G.G.和Richardson,C.D.(2017).赋予对膜融合抑制剂苄氧羰基-d-Phe-l-Phe-Gly和4-硝基-2-苯乙酰氨基-苯甲酰胺的抗性的麻疹病毒融合蛋白的突变(Mutations in the Fusion Protein of Measles Virus That ConferResistance to the Membrane Fusion Inhibitors Carbobenzoxy-d-Phe-l-Phe-Gly和4-Nitro-2-Phenylacetyl Amino-Benzamide).病毒学杂志(J.Virol.),91.
23.Hardie,D.R.,Albertyn,C.,Heckmann,J.M.和Smuts,H.E.M.(2013).麻疹包涵体脑炎(MIBE)患者大脑中病毒的分子特征(Molecular characterisation of virus inthe brains of patients with measles inclusion body encephalitis (MIBE)).病毒学杂志(Virol.J),10,283.
24.Harrison,S.C.(2008).病毒膜融合(Viral membrane fusion).自然结构与分子生物学(Nat.Struct.Mol.Biol.),15,690–698.
25.Hashiguchi,T.,Fukuda,Y.,Matsuoka,R.,Kuroda,D.,Kubota,M.,Shirogane,Y.,Watanabe,S.,
26.Tsumoto,K.,Kohda,D.,Plemper,R.K.等人(2018).与抑制剂复合的麻疹病毒融合蛋白融合前形式的结构(Structures of the prefusion form of measles virusfusion protein in complex with inhibitors).美国科学院院报(Proc.Natl.Acad.Sci.U.S.A.),115,2496–2501.
27.Hosoya,M.(2006).麻疹脑炎:直接病毒入侵还是自身免疫介导的炎症?(Measles encephalitis:direct viral invasion or autoimmune-mediatedinflammation?),内科学(Intern.Med.),45,841–842.
28.Hughes,I.,Jenney,M.E.,Newton,R.W.,Morris,D.J.和Klapper,P.E.(1993).急性淋巴细胞白血病免疫抑制治疗期间的麻疹脑炎(Measles encephalitis duringimmunosuppressive treatment for acute lymphoblastic leukaemia).儿童疾病档案(Arch.Dis.Child),68,775–778.
29.Iketani,S.,Shean,R.C.,Ferren,M.,Makhsous,N.,Aquino,D.B.,desGeorges,A.,Rima,B.,Mathieu,C.,Porotto,M.,Moscona,A.等人(2018).人类健康所需的病毒进入特性因病毒分离过程中的快速基因组变化而丢失(Viral Entry PropertiesRequired for Fitness in Humans Are Lost through Rapid Genomic Change duringViral Isolation).MBio 9.
30.Jansen,V.a.A.,Stollenwerk,N.,Jensen,H.J.,Ramsay,M.E.,Edmunds,W.J.和Rhodes,C.J.(2003).麻疹在疫苗接种量下降的人群中暴发(Measles outbreaks in apopulation with declining vaccine uptake).科学(Science),301,804.
31.Jurgens,E.M.,Mathieu,C.,Palermo,L.M.,Hardie,D.,Horvat,B.,Moscona,A.和Porotto,M.(2015).麻疹融合机制在神经致病变体中失调(Measles fusionmachinery is dysregulated in neuropathogenic variants).MBio 6.
32.Lambert,D.M.,Barney,S.,Lambert,A.L.,Guthrie,K.,Medinas,R.,Davis,D.E.,Bucy,T.,Erickson,J.,Merutka,G.和Petteway,S.R.(1996).来自副粘病毒融合(F)蛋白保守区的肽是病毒融合的有效抑制剂(Peptides from conserved regions ofparamyxovirus fusion(F)proteins are potent inhibitors of viral fusion).美国科学院院报(Proc.Natl.Acad.Sci.U.S.A.),93,2186–2191.
33.Lancaster,M.A.,and Knoblich,J.A.(2014).从人类多能干细胞产生脑类器官(Generation of cerebral organoids from human pluripotent stem cells).自然实验手册(Nat Protoc),9,2329–2340.
34.Lemon,K.,de Vries,R.D.,Mesman,A.W.,McQuaid,S.,van Amerongen,G.,Yüksel,S.,Ludlow,M.,Rennick,L.J.,Kuiken,T.,Rima,B.K.等人(2011).非人灵长类动物气溶胶感染后麻疹病毒的早期靶细胞(Early target cells of measles virus afteraerosol infection of non-human primates).公共科学图书馆病原体(PLoSPathogens),7,e1001263.
35.Leonard,V.H.J.,Sinn,P.L.,Hodge,G.,Miest,T.,Devaux,P.,Oezguen,N.,Braun,W.,McCray,P.B.,McChesney,M.B.,Cattaneo,R.等人(2008).麻疹病毒无视其上皮细胞受体,在恒河猴中仍然具有毒性,但不能穿过气道上皮细胞并且不会脱落(Measlesvirus blind to its epithelial cell receptor remains virulent in rhesusmonkeys but cannot cross the airway epithelium and is not shed).临床研究杂志(The Journal of Clinical Investigation),118,2448–2458.
36.Leonard,V.H.J.,Hodge,G.,Reyes-Del Valle,J.,McChesney,M.B.和Cattaneo,R.(2010).麻疹病毒选择性地无视信号淋巴细胞激活分子(SLAM;CD150),在恒河猴中减弱并诱导强烈的适应性免疫反应(Measles virus selectively blind tosignaling lymphocytic activation molecule(SLAM;CD150)is attenuated andinduces strong adaptive immune responses in rhesus monkeys).病毒学杂志(Journal of Virology),84,3413–3420.
37.Lin,J.-Y.,Kuo,R.-L.和Huang,H.-I.(2019a).人神经干细胞中I型干扰素抗病毒反应的激活(Activation of type I interferon antiviral response in humanneural stem cells).干细胞研究与治疗(Stem Cell Res Ther),10,387.
38.Lin,M.J.,Shean,R.C.,Makhsous,N.和Greninger,A.L.(2019B).LAVA:用于病毒等位基因纵向分析的简化可视化工具(LAVA:a streamlined visualization tool forlongitudinal analysis of viral alleles).BioRxiv2019.12.17.879320.
39.Luo,C.,Lancaster,M.A.,Castanon,R.,Nery,J.R.,Knoblich,J.A.和Ecker,J.R.(2016).脑类器官概括了人类胎儿大脑的表观基因组特征(Cerebral OrganoidsRecapitulate Epigenomic Signatures of the Human Fetal Brain).细胞报告(CellRep),17,3369–3384.
40.Makhortova,N.R.,Askovich,P.,Patterson,C.E.,Gechman,L.A.,Gerard,N.P.和Rall,G.F.(2007).Neurokinin-1使麻疹病毒能够在神经元中跨突触传播(Neurokinin-1enables measles virus trans-synaptic spread in neurons).病毒学(Virology),362,235–244.
41.Mathieu,C.,Huey,D.,Jurgens,E.,Welsch,J.C.,DeVito,I.,Talekar,A.,Horvat,B.,Niewiesk,S.,Moscona,A.和Porotto,M.(2015).通过鼻内递送融合抑制肽预防麻疹病毒感染(Prevention of measles virus infection by intranasal delivery offusion inhibitor peptides).病毒学杂志(J.Virol.),89,1143–1155.
42.Mathieu,C.,Ferren,M.,Jurgens,E.,Dumont,C.,Rybkina,K.,Harder,O.,Stelitano,D.,Madeddu,S.,Sanna,G.,Schwartz,D.等人(2019).麻疹病毒携带麻疹包涵体脑炎-衍生融合蛋白经呼吸道感染后具有致病性(Measles Virus Bearing MeaslesInclusion Body Encephalitis-Derived Fusion Protein Is Pathogenic afterInfection via the Respiratory Route).病毒学杂志(J.Virol.),93:e01862-18.
43.Moss,W.J.和Griffin,D.E.(2012).麻疹(Measles).柳叶刀(Lancet),379,153–164.
44.Mühlebach,M.D.,Mateo,M.,Sinn,P.L.,Prüfer,S.,Uhlig,K.M.,Leonard,V.H.J.,Navaratnarajah,C.K.,Frenzke,M.,Wong,X.X.,Sawatsky,B.等人(2011).粘附连接蛋白连接素-4是麻疹病毒的上皮受体(Adherens junction protein连接素-4is theepithelial receptor for measles virus).自然(Nature),480,530–533.
45.Noyce,R.S.,Bondre,D.G.,Ha,M.N.,Lin,L.-T.,Sisson,G.,Tsao,M.-S.和Richardson,C.D.(2011).肿瘤细胞标志物PVRL4(连接素4)是麻疹病毒的上皮细胞受体(Tumor cell marker PVRL4(连接素4)is an epithelial cell receptor for measlesvirus).公共科学图书馆病原体(PLoS Pathog.),7,e1002240.
46.Perry,R.T.,Murray,J.S.,Gacic-Dobo,M.,Dabbagh,A.,Mulders,M.N.,Strebel,P.M.,Okwo-Bele,J.-M.,Rota,P.A.和Goodson,J.L.(2015).2000-2014年全球区域性消除麻疹的进展情况(Progress toward regional measleselimination-worldwide,2000-2014).发病率和死亡率周报(MMWR Morb.Mortal.Wkly.Rep.),64,1246–1251.
47.Plemper,R.K.,Doyle,J.,Sun,A.,Prussia,A.,Cheng,L.-T.,Rota,P.A.,Liotta,D.C.,Snyder,J.P.和Compans,R.W.(2005).一种对麻疹原发病毒株具有活性的小分子进入抑制剂的设计(Design of a small-molecule entry inhibitor with activityagainst primary measles virus strains).抗菌物和化学疗法(Antimicrob.AgentsChemother.),49,3755–3761.
48.Prussia,A.J.,Plemper,R.K.和Snyder,J.P.(2008).麻疹病毒进入抑制剂:作用机制和耐药性发展的结构性建议(Measles virus entry inhibitors:a structuralproposal for mechanism of action and the development of resistance).生物化学(Biochemistry),47,13573–13583.
49.Qian,X.,Nguyen,H.N.,Song,M.M.,Hadiono,C.,Ogden,S.C.,Hammack,C.,Yao,B.,Hamersky,G.R.,Jacob,F.,Zhong,C.等人(2016).使用微型生物反应器模拟ZIKV暴露的大脑区域特异性类器官(Brain-Region-Specific Organoids Using Mini-bioreactors for Modeling ZIKV Exposure).细胞(Cell),165,1238–1254.
50.Rima,B.K.和Duprex,W.P.(2005).麻疹病毒持续存在的分子机制(Molecularmechanisms of measles virus persistence).病毒研究(Virus Research),111,132–147.
51.Sato,Y.,Watanabe,S.,Fukuda,Y.,Hashiguchi,T.,Yanagi,Y.和Ohno,S.(2018).人类神经元之间的细胞间麻疹病毒传播依赖于血凝素和超融合蛋白(Cell-to-Cell Measles Virus Spread between Human Neurons Is Dependent on Hemagglutininand Hyperfusogenic Fusion Protein).病毒学杂志(J.Virol.),92:e02166-17.
52.Schmid,A.,Spielhofer,P.,Cattaneo,R.,Baczko,K.,ter Meulen,V.和Billeter,M.A.(1992).亚急性硬化性全脑炎的典型特征是持续存在的麻疹病毒的融合蛋白胞质结构域发生改变(Subacute sclerosing panencephalitis is typicallycharacterized by alterations in the fusion protein cytoplasmic domain of thepersisting measles virus).病毒学(Virology),188,910–915.
53.Simons,E.,Ferrari,M.,Fricks,J.,Wannemuehler,K.,Anand,A.,Burton,A.和Strebel,P.(2012).评估2010年全球麻疹死亡率降低目标:来自监测数据模型的结果(Assessment of the 2010global measles mortality reduction goal:results from amodel of surveillance data).柳叶刀(Lancet),379,2173–2178.
54.Sun,A.,Prussia,A.,Zhan,W.,Murray,E.E.,Doyle,J.,Cheng,L.-T.,Yoon,J.-J.,Radchenko,E.V.,Palyulin,V.A.,Compans,R.W.等人(2006).麻疹病毒进入的非肽抑制剂(Nonpeptide inhibitors of measles virus entry).医药化学杂志(J.Med.Chem.),49,5080–5092.
55.Tatsuo,H.,Ono,N.,Tanaka,K.和Yanagi,Y.(2000).SLAM(CDw150)是麻疹病毒的细胞受体(SLAM(CDw150)is a cellular receptor for measles virus).自然(Nature),406,893–897.
56.Watanabe,M.,Hashimoto,K.,Abe,Y.,Kodama,E.N.,Nabika,R.,Oishi,S.,Ohara,S.,Sato,M.,Kawasaki,Y.,Fujii,N.等人(2016).一种源自融合蛋白七肽重复序列的新型肽在体外和体内抑制亚急性硬化性全脑炎病毒的复制(A Novel Peptide Derivedfrom the Fusion Protein Heptad Repeat Inhibits Replication of SubacuteSclerosing Panencephalitis Virus In Vitro and In Vivo).公共科学图书馆·综合(PLoS One),11,e0162823.
57.Watanabe,S.,Shirogane,Y.,Suzuki,S.O.,Ikegame,S.,Koga,R.和Yanagi,Y.(2013).具有增强的融合活性的突变融合蛋白促进麻疹病毒在人类神经元细胞和哺乳仓鼠大脑中传播(Mutant fusion proteins with enhanced fusion activity promotemeasles virus spread in human neuronal cells and brains of sucklinghamsters).病毒学杂志(J.Virol.),87,2648–2659.
58.Watanabe,S.,Ohno,S.,Shirogane,Y.,Suzuki,S.O.,Koga,R.和Yanagi,Y.(2015).具有增强的融合活性的融合蛋白的麻疹病毒突变体在神经元细胞中有效传播,但在其他细胞中不然,不会引起强烈的细胞病理(Measles virus mutants possessing thefusion protein with enhanced fusion activity spread effectively in neuronalcells,but not in other cells,without causing strong cytopathology).病毒学杂志(J.Virol.),89,2710–2717.
59.Welsch,J.C.,Talekar,A.,Mathieu,C.,Pessi,A.,Moscona,A.,Horvat,B.和Porotto,M.(2013).脑穿透性融合抑制剂可预防致命的麻疹病毒感染(Fatal measlesvirus infection prevented by brain-penetrant fusion inhibitors).病毒学杂志(J.Virol.),87,13785–13794.
60.Welsch,J.C.,Lionnet,C.,Terzian,C.,Horvat,B.,Gerlier,D.和Mathieu,C.(2017).器官型脑培养物:研究CNS的嗜神经病毒感染和筛选抗病毒药物的框架(Organotypic Brain Cultures:A Framework for Studying CNS Infection byNeurotropic Viruses and Screening Antiviral Drugs).生物实验方案(Bio-Protocol)7,e2605.
61.Welsch,J.C.,Charvet,B.,Dussurgey,S.,Allatif,O.,Aurine,N.,Horvat,B.,Gerlier,D.和Mathieu,C.(2019).I型干扰素受体信号驱使星形胶质细胞和小神经胶质细胞在脑部感染期间对麻疹病毒的选择性许可(Type I Interferon Receptor SignalingDrives Selective Permissiveness of Astrocytes and Microglia to Measles Virusduring Brain Infection).病毒学杂志(J.Virol.),93:e00618-19.
62.Young,V.A.和Rall,G.F.(2009).进入突触:麻疹病毒在神经元内和之间的传播(Making it to the synapse:measles virus spread in and among_neurons).微生物学和免疫学前沿专题(Curr.Top.Microbiol.Immunol.),330,3–30.
方法
肽和化学品。之前已描述了MV F衍生的融合抑制肽(Mathieu等人,2015)。简言之,合成了源自MV F蛋白的C端七肽重复区的36-aa肽。在本研究中使用了肽的二聚胆固醇缀合(HRC4)形式。N-(3-氰基苯基)-2-苯乙酰胺(也称为3G)购自英国ZereneX分子有限公司(ZereneX Molecular Limited(UK))。通过高压液相色谱(HPLC)对3G的纯度进行测试,并显示纯度>95%。
质粒和试剂。MeV IC323H和F蛋白的基因经过密码子优化、合成和亚克隆到哺乳动物表达载体pCAGGS中。编码连接素4、CD150的质粒是商购的。
细胞。HEK293T(人肾上皮细胞)、293-3-46(Radecke和Billeter,1995;Radecke等人,1995)以及Vero和Vero-SLAM/CD150(非洲绿猴肾)细胞在补充有10%胎牛血清(FBS,生命技术公司(Life Technologies));赛默飞世尔(Thermo Fisher))和抗生素的杜氏改良Eagle培养基(Dulbecco’s modified Eagle’s medium(DMEM);生命技术公司;赛默飞世尔科技公司)中在37℃、在5%二氧化碳中生长。293-3-46和Vero-SLAM/CD150培养基中补充有1mg/ml遗传霉素(赛默飞世尔科技公司)。
重组病毒的生产和分析。MeV IC323-EGFP(Hashimoto等人,2002)是表达编码EGFP的基因的重组病毒。具有突变T461I、N462K和L454W的所有变体均使用反向遗传学在MeVIC323-EGFP背景中产生(使用编码MeV IC323-EGFP的质粒,由日本福冈九州大学的Yanagi友情提供)。MeV IC323-Td Tomato通过用编码tdTomato红色荧光蛋白的序列替换EGFP表达盒产生。如前所述,MeV IC323重组病毒在293-3-46细胞中取出(Radecke等人,1995)。带有L454W的病毒的病毒生产E37℃或32℃下进行。所有病毒都在Vero-SLAM/CD150细胞中繁殖和滴定。
结构建模。使用蛋白质同源服务器Phyre2为野生型(wt)麻疹病毒融合糖蛋白(MeVF)制作了20个模型(Kelley等人,2015)。通过程序XtalView(McRee,1999)手动修复MeV F融合前和融合后状态的每个模型的不良局部几何形状。随后,生成的模型通过CNS-1.3(Brünger等人,1998)分别针对5EVM和1ZTM的融合前和融合后状态的电子密度进行精炼。相同的方法用于制备所有MV-F突变蛋白的模型。所有结构性附图均使用PyMol(www.pymol.org/)制作。
基于β-半乳糖苷酶(Beta-Gal)互补的融合试验。进行了基于Beta-Gal互补的融合试验,如前文所述(Jurgens等人,2015)。简言之,在MV F HRC衍生的融合抑制肽存在或不存在的情况下,将用上述构建体和omega报告亚基瞬时转染的293T细胞与共表达病毒糖蛋白和α报告亚基的细胞一起孵育指定的时间段(Mathieu等人,2015)。
使用F-构象-特异性mAb对细胞表面进行染色。将用病毒糖蛋白构建体瞬时转染的293T细胞在37℃下在完全培养基(DMEM,10%FBS)中温育过夜。转染后20小时,将细胞转移至55℃,时间如图中所示。此后,将细胞与特异性检测融合前构象中的MeV F的小鼠单克隆抗体(mAb)(1:1,000)在冰上温育1小时。细胞用PBS洗涤,然后细胞用Alexa-488抗小鼠第二抗体(1:500;生命技术公司)在冰上温育1小时。细胞用PBS洗涤,然后细胞在冰上用4%多聚甲醛(PFA)用DAPI(4',6-二脒基-2-苯基吲哚(4′,6-diamidino-2-phenylindole);赛默飞世尔(Thermo Fisher))以1:1000稀释固定60分钟。对板进行洗涤,添加0.01%叠氮化钠,并通过使用活细胞图像分析系统(IN Cell Analyzer)对板进行成像。使用Cell Profiler确定阳性识别细胞的百分比。
器官型小脑培养物的制备和感染后的处理。小脑切片从IFNAR1KO(和SLAM/CD150tg x IFNAR1KO)或C57/BL6小鼠制备并保持在培养物中,如在其他地方所详述的(Welsch等人,2017)。简言之,从7日龄小鼠的脑中分离出小脑,并用McIlwain组织切片机(WPI公司-欧洲)切割以获得350μm厚的渐进切片。然后,将脑切片在冷的 -A/5g/L D-葡萄糖/1x犬尿酸缓冲液中解离,并放置在密理博(Millipore)细胞培养插入膜(Millicell细胞培养插入物,30mm,亲水性聚四氟乙烯,密理博(Millipore))上。随后,切片在补充有25%马血清、5g/L葡萄糖、1%HEPES(均来自赛默飞世尔科技公司)和0.1mg/L人重组胰岛素(R&D Systems公司)的GlutaMAX最低必需培养基(GlutaMAX minimal essentialmedium)中在37℃、5%二氧化碳的潮湿环境下培养。切片步骤之后,每天更换培养基。来自5只小鼠的切片在切片当天感染MeVIC323-EGFP-F L454W病毒(来自IFNAR1KO的5.103pfu/切片和来自SLAM/CD150tg x IFNAR1KO小鼠的100pfu/切片)。然后,从第1天到第4天每天用在Neurobasal培养基中的HRC4融合抑制剂的连续稀释液或用媒介(未处理的条件;“NT”)处理培养物。将2μl的10000nM、1000nM或100nM HRC4添加每个孔中5个切片的上面。几分钟后,含有肽的液滴被完全吸收并到达含有补料培养基的系统的下部隔层。培养基(1ml)中的最终浓度为100nM、10nM或1nM。在每个时间点,收集切片,提取RNA并如前所述进进行RT-qPCR(Welsch等人,2013)
-A/5g/L D-葡萄糖/1x犬尿酸缓冲液中解离,并放置在密理博(Millipore)细胞培养插入膜(Millicell细胞培养插入物,30mm,亲水性聚四氟乙烯,密理博(Millipore))上。随后,切片在补充有25%马血清、5g/L葡萄糖、1%HEPES(均来自赛默飞世尔科技公司)和0.1mg/L人重组胰岛素(R&D Systems公司)的GlutaMAX最低必需培养基(GlutaMAX minimal essentialmedium)中在37℃、5%二氧化碳的潮湿环境下培养。切片步骤之后,每天更换培养基。来自5只小鼠的切片在切片当天感染MeVIC323-EGFP-F L454W病毒(来自IFNAR1KO的5.103pfu/切片和来自SLAM/CD150tg x IFNAR1KO小鼠的100pfu/切片)。然后,从第1天到第4天每天用在Neurobasal培养基中的HRC4融合抑制剂的连续稀释液或用媒介(未处理的条件;“NT”)处理培养物。将2μl的10000nM、1000nM或100nM HRC4添加每个孔中5个切片的上面。几分钟后,含有肽的液滴被完全吸收并到达含有补料培养基的系统的下部隔层。培养基(1ml)中的最终浓度为100nM、10nM或1nM。在每个时间点,收集切片,提取RNA并如前所述进进行RT-qPCR(Welsch等人,2013)
人脑类器官分化。如前所述(Lancaster和Knoblich,2014)从两个iPSC系(FA0000010和FA0000011,简称为FA10和FA11)生成脑类器官。简言之,使用EB培养基(DMEM/F12(赛默飞世尔科技公司),cat.#11330-032),补充以20%血清替代物(Knockout SerumReplacement)(赛默飞世尔科技公司,cat.#10828-028)、3%ES优级批量测试的胎牛血清(赛默飞世尔科技公司,cat#10439024)、1%Glutamax(赛默飞世尔科技公司,cat.#35050-038)、1x非必需胺基酸、0.1mM 2-巯基乙醇、4ng/ml bFGF(R&D Systems公司,cat.#233FB01M)和50uM Y-27632,用Accutase细胞解离试剂(干细胞技术公司(Stem CellTechnologies),cat#7920)将细胞解离成单细胞悬浮液,并以4,500细胞/孔的浓度播种在96孔低粘附板(U形底)中。每隔一天更换一次新鲜培养基,直到第6天。在第6天,用神经诱导(NI)培养基(DMEM/F12、1x N2补充剂、1X Glutamax、1X NEAA和1μg/ml肝素(西格玛奥德里奇(Sigma-Aldrich),cat#H3149)替换EB培养基,并将类器官转移到60mm或100mm的低粘附板中。允许类器官形成神经上皮组织,直到第11-14天,每隔一天更换一次培养基。在第11-24天之间,类器官用Matrigel基质胶液滴包被,并通过在37℃下保持30分钟使其凝胶化。将Matrigel基质胶包被的类器官转移到分化培养基(1:1DMEM/F12:Neurobasal基础培养基、0.5%N2补充剂、2%不含维生素A的B27补充剂(生命技术公司,cat#12587010)、0.25%胰岛素溶液(生命技术公司,cat#12585014)、50uM 2-巯基乙醇、1%Glutamax、0.5%NEAA、1%青霉素-链霉素),持续4天。4天后,将类器官转移到含有维生素A的B27补充剂(赛默飞世尔科技公司,cat#17504-044)的分化培养基中。将脑类器官再培养60天,每7天更换一次培养基,然后在90天或270天用于进一步实验。
脑类器官RNA-Seq和分析。使用Direct-zoltm RNA MicroPrep(Zymo)从未感染和感染的脑类器官中提取RNA,并提交给JP Sulzberger哥伦比亚基因组中心(JP SulzbergerColumbia Genome Center)进行文库制备和测序。使用poly-A富集(poly-A enrichment)制备链特异性RNA-Seq文库,并在具有配对末端2x100读数的Illumina NovaSeq上进行测序。在质量和衔接头(adapter)修整后,以GRCh38为参考基因组、使用Kallisto版本0.44.0进行转录丰度量化(Bray等人,2016)。
为了了解脑类器官的发育阶段,我们使用了BrainSpan数据集(22031440)。因为BrainSpan提供基于Gencode v10注释的标准化RPKM表达值),我们使用bowtie2(22388286)将R1测序读数映射到人类基因组的GRCh37注释,并使用featureCounts对基因水平(gene-level)的RPKM水平进行量化(Liao等人,2014)。在所有539个RNA-Seq实验(524个BrainSpan,15个MeV感染)中,基于大于1000的任意基因水平(gene-level)RPKM总和对基因进行过滤。在R v3.6.2中生成了伪计数为1的log2对数转换过的(log2-transformed)RPKM值的所有相关矩阵。提取与未感染的FA10脑类器官复制1相关系数最高的前100个BrainSpan样本,并使用pheatmap(https://https://github.com/raivokolde/pheatmap)生成相关系数的热图。
使用Kallisto转录本丰度和R Bioconductor包(R Bioconductor package)DESeq2进行基因表达差异分析(Love等人,2014)。分析代码可在http://www.github.com/greninger-lab/MeV-brain-organoids获得。对每份样本平均少于一个计数的基因进行低计数过滤。将类器官之间的批次效应和生物学差异作为混杂因素纳入设计式,并使用DESeq2默认参数进行标准化和差异表达分析。在使用R包(R package)pheatmap生成的热图上绘制了在F454W和未感染的类器官之间具有最低本杰明-汉伯格(Benjamini-Hochberg)校正p值的50个基因的表达,并在R包ggplot中生成了条形图。使用R包ReactomePA(Yu和He,2016)评估在L454W和未感染的KEGG途径之间的差异表达基因(padj<0.0001,绝对值log2倍数变化>1)的富集。
为了计算MeV读数的每百万读数(RPM)值,使用Bowtie2使用默认参数(22388286),将每份样本与NC_001498MeV参考序列进行比对。使用生成的BAM文件中的映射读数的数量计算MeV RPM值。
小鼠脑切片的RNAseq。从脑切片中提取的RNA的制备和测序,如上述类器官一样。通过Kallisto v0.44(参考)将读数与小鼠参考转录组mmGRCm38进行伪比对。在DESeq2中进行差异表达分析,将批次效应纳入设计式。如上所述,为脑类器官生成了表达热图。按上述方法计算MeV RPM。
脑类器官基因组特异性RT-qPCR。根据制造商的说明,使用SuperScriptTM III第一链合成系统(赛默飞世尔(Thermofisher)),使用人GAPDH反向引物和FW 5’标记的MeV引物5'-gcagggcaatctcacaatcagg AAAACTGGTGTTCTACAACAA-3'(含有来自尼帕(Nipah)病毒的TAG序列的反基因组链的MeV序列),对500ng总RNA进行靶向MeV基因组链的特定逆转录。然后将获得的cDNA以1:10稀释。然后如前所述使用MeV Rev5’-TGAAGGCCACTGCATT-3’和TagFW 5’-gcagggcaatctcacaatcagg-3’引物进行QPCR(Iketani等人,2018)。所有结果都标准化为人GAPDH偏差。
mNGS和变体表达(calling)。如前所述进行mNGS(Iketani等人,2018)。简言之,使用Quick-RNA病毒试剂盒(Quick-RNA Viral Kit(Zymo))从50μL病毒培养物中提取RNA,并用TURBO DNase I(赛默飞世尔(Thermo Fisher))进行处理。使用Superscript IV逆转录酶(赛默飞世尔)和随机六聚体(IDT)从脱氧核糖核酸酶(DNase)处理的RNA生成cDNA,然后通过Sequenase 2.0DNA聚合酶进行第二链合成。然后,用DNA清洁和浓缩试剂盒(DNA Clean&Concentrator Kit)(Zymo)将得到的双链cDNA进行纯化。使用Nextera XT试剂盒(依诺米那(Illumina))从2μL的cDNA构建文库,并在1x192 bp Illumina MiSeq运行中进行测序。
使用Trimmomatic v0.38对测序读数进行衔接头和质量修整(Bolger等人,2014)。使用MeV参考基因组(NC_001498),用LAVA(github.com/greninger-lab/lava)识别出频率大于10%且覆盖率大于10x的变体。通过将测序读数映射到Geneious v11.1.4中相同的MeV参考毒株,手动确认所有变体(Kearse等人,2012)。存在于基质和融合蛋白之间的基因间区以及存在于均聚体(homopolymeric tract)中的那些变体排除在分析之外。可在美国国家生物技术信息中心(NCBI)BioProject编号PRJNA594952下获得测序读数。
统计分析:Mantel-Cox检验用于存活比较分析。所有其他统计比较均使用曼-惠特尼U检验进行。所有分析均在GraphPad Prism5软件中进行。RNAseq数据的统计分析在R中进行。
方法部分的参考文献
1.Bolger,AM,Lohse,M.和Usadel,B.(2014).Trimmomatic:用于Illumina序列数据的灵活修剪机(Trimmomatic:a flexible trimmer for Illumina sequence data)。生物信息学(英格兰牛津)(Bioinforma.Oxf.Engl.),30,2114–2120。
2.Bray,N.L.,Pimentel,H.,Melsted,P.和Pachter,L.(2016).近乎最优的概率性RNA-seq量化(Near-optimal probabilistic RNA-seq quantification).自然·生物技术(Nat.Biotechnol.),34,525–527.
3.Brünger,A.T.,Adams,P.D.,Clore,G.M.,DeLano,W.L.,Gros,P.,Grosse-Kunstleve,R.W.,Jiang,J.S.,Kuszewski,J.,Nilges,M.,Pannu,N.S.等人(1998).晶体学和核磁共振系统:用于大分子结构测定的新软件套件(Crystallography&NMR system:Anew software suite for macromolecular structure determination).晶体学学报D辑:生物晶体学(Acta Crystallogr.D Biol.Crystallogr.),54,905–921.
4.Hashimoto,K.,Ono,N.,Tatsuo,H.,Minagawa,H.,Takeda,M.,Takeuchi,K.和Yanagi,Y.(2002).表达绿色荧光蛋白的重组病毒揭示了独立于SLAM(CD150)的麻疹病毒进入(SLAM(CD150)-independent measles virus entry as revealed by recombinantvirus expressing green fluorescent protein).病毒学杂志(J.Virol.),76,6743–6749.
5.Iketani,S.,Shean,R.C.,Ferren,M.,Makhsous,N.,Aquino,D.B.,desGeorges,A.,Rima,B.,Mathieu,C.,Porotto,M.,Moscona,A.等人(2018).人类健康所需的病毒进入特性因病毒分离过程中的快速基因组变化而丢失(Viral Entry PropertiesRequired for Fitness in Humans Are Lost through Rapid Genomic Change duringViral Isolation).MBio 9.
6.Jurgens,E.M.,Mathieu,C.,Palermo,L.M.,Hardie,D.,Horvat,B.,Moscona,A.和Porotto,M.(2015).麻疹融合机制在神经致病变体中失调(Measles fusion machineryis dysregulated in neuropathogenic variants).MBio 6.
7.Kearse,M.,Moir,R.,Wilson,A.,Stones-Havas,S.,Cheung,M.,Sturrock,S.,Buxton,S.,Cooper,A.,Markowitz,S.,Duran,C.等人(2012).Geneious Basic:用于组织和分析序列数据的集成和可扩展桌面软件平台(Geneious Basic:an integrated andextendable desktop software platform for the organization and analysis ofsequence data).生物信息学(英格兰牛津)(Bioinforma.Oxf.Engl.),28,1647–1649.
8.Kelley,L.A.,Mezulis,S.,Yates,C.M.,Wass,M.N.和Sternberg,M.J.E.(2015).用于蛋白质建模、预测和分析的Phyre2门户网站(The Phyre2 web portal forprotein modeling,prediction and analysis).自然实验手册(Nat.Protoc.),10,845–858.
9.Lancaster,M.A.和Knoblich,J.A.(2014).从人类多能干细胞产生脑类器官(Generation of cerebral organoids from human pluripotent stem cells).自然实验手册(Nat.Protoc.),9,2329–2340.
10.Langmead,B.和Salzberg,S.L.(2012).使用Bowtie 2进行快速间隙读数对齐(Fast gapped-read alignment with Bowtie 2).自然方法学(Nat.Methods),9,357–359.
11.Liao,Y.,Smyth,G.K.和Shi,W.(2014).featureCounts:一个用于将序列读数分配给基因组特征的高效通用程序,(featureCounts:an efficient general purposeprogram for assigning sequence reads to genomic features).生物信息学(英格兰牛津)(Bioinforma.Oxf.Engl.),30,923–930.
12.Love,M.I.,Huber,W.和Anders,S.(2014).使用DESeq2对RNA-seq数据的倍数变化和分散进行适度估计(Moderated estimation of fold change and dispersion forRNA-seq data with DESeq2).基因组生物学(Genome Biol.),15,550.
13.Mathieu,C.,Huey,D.,Jurgens,E.,Welsch,J.C.,DeVito,I.,Talekar,A.,Horvat,B.,Niewiesk,S.,Moscona,A.和Porotto,M.(2015).通过鼻内递送融合抑制肽预防麻疹病毒感染(Prevention of measles virus infection by intranasal delivery offusion inhibitor peptides).病毒学杂志(J.Virol).89,1143–1155.
14.McRee,D.E.(1999).XtalView/Xfit--用于操作原子坐标和电子密度的多功能程序(XtalView/Xfit--A versatile program for manipulating atomic coordinatesand electron density).结构生物学杂志(J.Struct.Biol.),125,156–165.
15.Radecke,F.,and Billeter,M.A.(1995).附录:麻疹病毒反基因组和蛋白质共有序列(Appendix:measles virus antigenome and protein consensus sequences).微生物学和免疫学前沿专题(Curr.Top.Microbiol.Immunol.),191,181–192.
16.Radecke,F.,Spielhofer,P.,Schneider,H.,Kaelin,K.,Huber,M., C.,Christiansen,G.和Billeter,M.A.(1995).从克隆DNA中取出麻疹病毒(Rescue ofmeasles viruses from cloned DNA).欧洲分子生物学会杂志(EMBO J.),14,5773–5784.
C.,Christiansen,G.和Billeter,M.A.(1995).从克隆DNA中取出麻疹病毒(Rescue ofmeasles viruses from cloned DNA).欧洲分子生物学会杂志(EMBO J.),14,5773–5784.
17.Welsch,J.C.,Talekar,A.,Mathieu,C.,Pessi,A.,Moscona,A.,Horvat,B.和Porotto,M.(2013).脑穿透性融合抑制剂可预防致命的麻疹病毒感染(Fatal measlesvirus infection prevented by brain-penetrant fusion inhibitors).病毒学杂志(J.Virol).87,13785–13794.
18.Welsch,J.C.,Lionnet,C.,Terzian,C.,Horvat,B.,Gerlier,D.和Mathieu,C.(2017).器官型脑培养物:研究CNS的嗜神经病毒感染和筛选抗病毒药物的框架(Organotypic Brain Cultures:A Framework for Studying CNS Infection byNeurotropic Viruses and Screening Antiviral Drugs).Bio-Protoc.7,e2605.
实施例6——在融合前状态稳定的可溶性F蛋白
图117A-E示出了通过特异性突变E107G和E455G以及通过FIP-HRC(不含胆固醇的二聚体)稳定的麻疹融合蛋白的数据。处于融合前状态的可溶性F可用作疫苗。图117A-E示出了1)在FIP-HRC存在的情况下,稳定的F在55℃下保持其融合前状态温育长达两小时;2)稳定的F在37℃下保持其融合前状态长达一周;3)wt F被FIP-HRC肽部分稳定;和4)FIP-HRC的F稳定性优于市售的稳定剂(例如市售的FIP和3g)。
Claims (59)
1.一种抗病毒肽缀合物,所述抗病毒肽缀合物包含融合抑制肽(FIP)和C端七肽重复(HRC)肽(FIP-HRC)。
2.根据权利要求1所述的抗病毒肽缀合物,所述抗病毒肽缀合物进一步包含膜定位部分区。
3.根据权利要求2所述的抗病毒肽缀合物,其中,所述膜定位部分区包含选自胆固醇、生育酚和棕榈基所组成的组中的膜定位部分。
4.根据权利要求2所述的抗病毒肽缀合物,其中,所述膜定位部分区缀合至所述HRC肽的C端。
5.根据权利要求1所述的抗病毒肽缀合物,所述抗病毒肽缀合物进一步包含连接体区。
6.根据权利要求5所述的抗病毒肽缀合物,其中,所述连接体区包含聚乙二醇(PEG)。
7.根据权利要求6所述的抗病毒肽缀合物,其中,所述PEG的长度为4个乙二醇单元(PEG4)。
8.根据权利要求6所述的肽缀合物,其中,所述PEG的长度为11个乙二醇单元(PEG11)。
9.根据权利要求5所述的抗病毒肽缀合物,其中,所述连接体区缀合至所述HRC肽的C端。
10.根据权利要求1所述的抗病毒肽缀合物,所述抗病毒肽缀合物进一步包含膜定位部分区和连接体区。
11.根据权利要求10所述的抗病毒肽缀合物,其中,所述连接体区缀合至所述HRC肽的C端并且所述膜定位部分区缀合至所述连接体区。
12.根据权利要求11所述的抗病毒肽缀合物,其中,所述连接体区包含聚乙二醇(PEG)。
13.根据权利要求12所述的抗病毒肽缀合物,其中,所述PEG的长度为4个乙二醇单元(PEG4)。
14.根据权利要求12所述的抗病毒肽缀合物,其中,所述PEG的长度为11个乙二醇单元(PEG11)。
15.根据权利要求5-14中任一项所述的抗病毒肽缀合物,其中,所述抗病毒肽包含所述FIP区和所述HRC肽区的二聚体。
16.根据权利要求15所述的抗病毒肽缀合物,其中,所述抗病毒肽包含缀合至所述连接体区的第一FIP-HRC肽和缀合至所述连接体区的第二FIP-HRC肽。
17.根据权利要求1-16中任一项所述的抗病毒肽缀合物,其中,所述肽进一步包含丝氨酸-甘氨酸连接体。
18.根据权利要求17所述的抗病毒肽缀合物,其中,所述丝氨酸-甘氨酸连接体位于所述FIP和HRC肽之间。
19.根据权利要求18所述的抗病毒肽缀合物,其中,所述丝氨酸-甘氨酸连接体位于所述HRC肽的C端。
20.根据权利要求19所述的抗病毒肽缀合物,其中,所述丝氨酸-甘氨酸连接体位于所述FIP区和HRC肽区之间,并且所述抗病毒肽进一步包含位于所述HRC肽区的C端的第二丝氨酸-甘氨酸连接体。
21.根据权利要求17-20中任一项所述的抗病毒肽缀合物,其中,所述丝氨酸-甘氨酸连接体包含氨基酸序列GSGSG。
22.根据权利要求1-21中任一项所述的抗病毒肽缀合物,其中,所述FIP的第一个苯丙氨酸残基是D-氨基酸。
23.根据权利要求1-21中任一项所述的抗病毒肽缀合物,其中,所述肽的N端还包含苄氧羰基基团。
24.根据权利要求1-23中任一项所述的抗病毒肽缀合物,其中,所述FIP肽包含氨基酸序列FFG。
25.根据权利要求1-23中任一项所述的抗病毒肽缀合物,其中,所述HRC肽包含氨基酸序列PPISLERLDVGTNLGNAIAKLEDAKELLESSDQILR。
26.根据权利要求1-23中任一项所述的抗病毒肽缀合物,其中,所述HRC肽缀合物包含氨基酸序列PPISLERLDVGTN。
27.根据权利要求1-23中任一项所述的抗病毒肽缀合物,其中,所述抗病毒肽包含氨基酸序列FFGPPISLERLDVGTNLGNAIAKLEDAKELLESSDQILR。
28.根据权利要求1-23中任一项所述的抗病毒肽缀合物,其中,所述抗病毒肽包含氨基酸序列FFGPPISLERLDVGTN。
29.一种纳米颗粒,所述纳米颗粒包含根据权利要求1-28中任一项所述的抗病毒肽缀合物。
30.根据权利要求29所述的纳米颗粒,其中,所述纳米颗粒的直径在约50nm和约150nm之间。
31.一种组合物,所述组合物包含根据权利要求1-28中任一项所述的抗病毒肽缀合物。
32.一种预防性组合物,所述预防性组合物包含抗病毒肽缀合物,所述抗病毒肽缀合物包含融合抑制肽(FIP)和C端七肽重复(HRC)肽(FIP-HRC),其中,所述麻疹抗病毒肽缀合物防止麻疹病毒的膜融合。
33.一种纳米颗粒,所述纳米颗粒包含融合抑制肽(FIP)和C端七肽重复(HRC)肽(FIP-HRC)。
34.一种感染后的麻疹预防的方法,所述方法包括向有需要的受试者施用抗病毒肽缀合物,所述抗病毒肽缀合物包含融合抑制肽(FIP)和C端七肽(HRC)肽(FIP-HRC)。
35.根据权利要求34所述的方法,其中,所述抗病毒肽缀合物进一步包含连接体区。
36.根据权利要求34所述的方法,其中,所述抗病毒肽缀合物进一步包含膜定位部分区。
37.根据权利要求34所述的方法,其中,所述抗病毒肽缀合物进一步包含连接体区和膜定位部分区。
38.根据权利要求34所述的方法,其中,所述施用包括鼻内吸入或口腔吸入。
39.根据权利要求38所述的方法,其中,所述抗病毒肽缀合物通过选自喷雾器、气雾器和吸入器所组成的组中的装置施用。
40.根据权利要求34的方法,其中,所述施用包括皮下施用。
41.根据权利要求34所述的方法,其中,所述受试者已经暴露于麻疹病毒,所述麻疹病毒包含野生型融合糖蛋白。
42.根据权利要求34所述的方法,其中,所述受试者已经暴露于麻疹病毒,所述麻疹病毒包含选自N462K、L454W、T461I、E455G、E170G、G506E、M337L、D538G、G168R、S262G、A440P、R520C和L550P所组成的组中的融合糖蛋白的一种或多种突变。
43.一种重组蛋白,所述重组蛋白包含稳定的可溶性麻疹F蛋白,所述F蛋白包含E445G突变。
44.一种重组蛋白,所述重组蛋白包含稳定的可溶性麻疹F蛋白,所述F蛋白包含E170G和E455G双突变。
45.一种重组蛋白,所述重组蛋白包含氨基酸序列SEQ ID NO:3或SEQ ID NO:4。
46.一种重组蛋白,所述重组蛋白包含氨基酸序列SEQ ID NO:5或SEQ ID NO:6。
47.一种免疫原性组合物,所述免疫原性组合物包含根据权利要求43-46所述的任一重组蛋白。
48.一种在麻疹暴露之前预防麻疹感染的方法,所述方法通过向受试者施用根据权利要求47所述的免疫原性组合物。
49.一种诱导针对麻疹病毒的免疫反应的方法,所述方法通过向受试者施用根据权利要求47所述的免疫原性组合物。
50.一种免疫原性组合物,所述免疫原性组合物包含氨基酸序列SEQ ID NO:1或SEQ IDNO:2,并且进一步包含根据权利要求1-28中任一项所述的抗病毒肽缀合物。
51.一种诱导针对麻疹病毒的免疫反应的方法,所述方法通过向受试者施用根据权利要求47所述的免疫原性组合物,并进一步施用包含根据权利要求1-28中任一项所述的抗病毒肽缀合物的免疫原性组合物。
52.一种生产包含氨基酸序列SEQ ID NO:3或SEQ ID NO:4的重组蛋白的方法。
53.一种生产包含氨基酸序列SEQ ID NO:5或SEQ ID NO:6的重组蛋白的方法。
54.一种细胞系,所述细胞系表达包含氨基酸序列SEQ ID NO:3或SEQ ID NO:4的重组蛋白。
55.一种细胞系,所述细胞系表达包含氨基酸序列SEQ ID NO:5或SEQ ID NO:6的重组蛋白。
56.一种抗病毒肽缀合物,所述抗病毒肽缀合物包含融合抑制肽(FIP)和C端七肽重复(HRC)肽(FIP-HRC),其中,所述HRC肽衍生自HIV-gp41(C34)。
57.根据权利要求56所述的抗病毒肽缀合物,其中,所述HRC肽包含氨基酸序列WMEWDREINNYTSLIHSLIEESQNQQEKNEQELL。
58.一种感染后的HIV预防的方法,所述方法包括向有需要的受试者施用抗病毒肽缀合物,所述抗病毒肽缀合物包含融合抑制肽(FIP)和C端七肽重复(HRC)肽(FIP-HRC)。
59.根据权利要求58所述的方法,其中,所述HRC肽衍生自HIV-gp41(C34)。
Applications Claiming Priority (7)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201962895752P | 2019-09-04 | 2019-09-04 | |
| US62/895,752 | 2019-09-04 | ||
| US202062988286P | 2020-03-11 | 2020-03-11 | |
| US62/988,286 | 2020-03-11 | ||
| US202063009883P | 2020-04-14 | 2020-04-14 | |
| US63/009,883 | 2020-04-14 | ||
| PCT/US2020/049473 WO2021046398A2 (en) | 2019-09-04 | 2020-09-04 | Combination antiviral therapy for measles |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| CN114729007A true CN114729007A (zh) | 2022-07-08 |
Family
ID=74853434
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN202080077668.XA Pending CN114729007A (zh) | 2019-09-04 | 2020-09-04 | 麻疹的联合抗病毒治疗 |
Country Status (6)
| Country | Link |
|---|---|
| US (1) | US20230014151A1 (zh) |
| EP (1) | EP4025586A4 (zh) |
| JP (1) | JP2022551813A (zh) |
| CN (1) | CN114729007A (zh) |
| IL (1) | IL291091A (zh) |
| WO (1) | WO2021046398A2 (zh) |
Families Citing this family (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP4281096A1 (en) * | 2021-01-22 | 2023-11-29 | Decoy Therapeutics Inc. | Methods and compositions for treating infections |
| WO2023150375A2 (en) * | 2022-02-07 | 2023-08-10 | Decoy Therapeutics Inc. | Methods and compositions for treating covid infections |
Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20060280754A1 (en) * | 2003-11-04 | 2006-12-14 | Garry Robert F | Method of preventing virus: cell fusion by inhibiting the function of the fusion initiation region in rna viruses having class i membrane fusogenic envelope proteins |
| US20170216448A1 (en) * | 2014-05-07 | 2017-08-03 | Cornell University | Inhibitors of fusion between viral and cell membranes as well as compositions and methods of using them |
Family Cites Families (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP2603241A2 (en) * | 2010-08-11 | 2013-06-19 | Jv Bio Srl | Multimeric inhibitors of viral fusion and uses thereof |
-
2020
- 2020-09-04 WO PCT/US2020/049473 patent/WO2021046398A2/en unknown
- 2020-09-04 EP EP20861697.9A patent/EP4025586A4/en active Pending
- 2020-09-04 CN CN202080077668.XA patent/CN114729007A/zh active Pending
- 2020-09-04 JP JP2022515099A patent/JP2022551813A/ja active Pending
- 2020-09-04 US US17/640,327 patent/US20230014151A1/en active Pending
-
2022
- 2022-03-03 IL IL291091A patent/IL291091A/en unknown
Patent Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20060280754A1 (en) * | 2003-11-04 | 2006-12-14 | Garry Robert F | Method of preventing virus: cell fusion by inhibiting the function of the fusion initiation region in rna viruses having class i membrane fusogenic envelope proteins |
| US20170216448A1 (en) * | 2014-05-07 | 2017-08-03 | Cornell University | Inhibitors of fusion between viral and cell membranes as well as compositions and methods of using them |
Non-Patent Citations (2)
| Title |
|---|
| DENNIS HOFFMANN;WIBKE BAYER;OLIVER WILDNER;: "In situ tumor vaccination with adenovirus vectors encoding measles virus fusogenic membrane proteins and cytokines", WORLD JOURNAL OF GASTROENTEROLOGY, no. 22, 14 June 2007 (2007-06-14) * |
| TIAGO N. FIGUEIRA, ET AL: "Structure-Stability-Function Mechanistic Links in the Anti-Measles Virus Action of Tocopherol-Derivatized Peptide Nanoparticles", ACS NANO, vol. 12, no. 10, 19 September 2018 (2018-09-19), pages 1, XP093100901, DOI: 10.1021/acsnano.8b01422 * |
Also Published As
| Publication number | Publication date |
|---|---|
| EP4025586A2 (en) | 2022-07-13 |
| JP2022551813A (ja) | 2022-12-14 |
| US20230014151A1 (en) | 2023-01-19 |
| IL291091A (en) | 2022-05-01 |
| WO2021046398A3 (en) | 2021-04-08 |
| WO2021046398A2 (en) | 2021-03-11 |
| EP4025586A4 (en) | 2024-01-03 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US20220072697A1 (en) | Multivalent vaccines for rabies virus and coronaviruses | |
| AU2004230485A1 (en) | The severe acute respiratory syndrome coronavirus | |
| CA3202566A1 (en) | Neutralizing monoclonal antibodies against covid-19 | |
| WO1995004081A1 (en) | MONOCLONAL IgA ANTIBODY AGAINST RESPIRATORY SYNCYTIAL VIRUS | |
| EP4130047A1 (en) | Soluble ace2 and fusion protein, and applications thereof | |
| CN114729007A (zh) | 麻疹的联合抗病毒治疗 | |
| US20230235007A1 (en) | Humanized ace2-fc fusion protein for treatment and prevention of sars-cov-2 infection | |
| EA017957B1 (ru) | Композиции и способы, ингибирующие грипп | |
| US20240082386A1 (en) | Methods for immunizing pre-immune subjects against respiratory syncytial virus (rsv) | |
| WO2023020623A1 (zh) | 用于预防或治疗冠状病毒感染的融合蛋白、Spike蛋白纳米颗粒及其应用 | |
| WO2022051859A1 (en) | Pseudotyped retroviral particles for inducing immunity against coronavirus infections | |
| US20110305670A1 (en) | Nucleic acid encoding fusion polypeptides that prevent or inhibit hiv infection | |
| CN115772227A (zh) | 新型冠状病毒SARS-CoV-2的Delta变异株疫苗与应用 | |
| US20240101629A1 (en) | Use of interleukin-7 for the treatment of coronavirus | |
| TW201545755A (zh) | 重組呼吸道融合病毒及疫苗 | |
| JP2024502783A (ja) | コロナウイルスワクチン | |
| JP2023540705A (ja) | 修飾された抗ウイルス結合剤 | |
| US11186626B2 (en) | Broadly neutralizing antibody targeting the ebolavirus glycoprotein internal fusion loop | |
| US20060110740A1 (en) | Use of sendai virus as a human parainfluenza vaccine | |
| Ávila-Nieto et al. | Novel Spike-stabilized trimers with improved production protect K18-hACE2 mice and golden Syrian hamsters from the highly pathogenic SARS-CoV-2 Beta variant | |
| WO2024061188A1 (zh) | 一种冠状病毒多价疫苗及其应用 | |
| WO2023201304A2 (en) | Compositions for preventing or treating coronavirus infections | |
| WO2005063798A1 (en) | Identification of two linear epitopes on ebola or marburg virus glycoproteins critical for infection | |
| WO2022226060A1 (en) | Antibody cocktail for treatment of ebolavirus infections | |
| WO2022261343A1 (en) | Peptide vlp-based vaccines |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| PB01 | Publication | ||
| PB01 | Publication | ||
| SE01 | Entry into force of request for substantive examination | ||
| SE01 | Entry into force of request for substantive examination |