CN110913881A - 工程化crispr cas9免疫隐身 - Google Patents
工程化crispr cas9免疫隐身 Download PDFInfo
- Publication number
- CN110913881A CN110913881A CN201880031835.XA CN201880031835A CN110913881A CN 110913881 A CN110913881 A CN 110913881A CN 201880031835 A CN201880031835 A CN 201880031835A CN 110913881 A CN110913881 A CN 110913881A
- Authority
- CN
- China
- Prior art keywords
- cas9
- protein
- aav
- proteins
- mhc
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
- 108091033409 CRISPR Proteins 0.000 title claims abstract description 233
- 238000000034 method Methods 0.000 claims abstract description 111
- 150000001413 amino acids Chemical class 0.000 claims abstract description 51
- 230000005847 immunogenicity Effects 0.000 claims abstract description 32
- 230000028993 immune response Effects 0.000 claims abstract description 25
- 108090000623 proteins and genes Proteins 0.000 claims description 226
- 102000004169 proteins and genes Human genes 0.000 claims description 201
- 108700018351 Major Histocompatibility Complex Proteins 0.000 claims description 108
- 230000020382 suppression by virus of host antigen processing and presentation of peptide antigen via MHC class I Effects 0.000 claims description 106
- 108091033319 polynucleotide Proteins 0.000 claims description 59
- 102000040430 polynucleotide Human genes 0.000 claims description 59
- 239000002157 polynucleotide Substances 0.000 claims description 59
- 239000013598 vector Substances 0.000 claims description 42
- 108020005004 Guide RNA Proteins 0.000 claims description 27
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 claims description 27
- 210000000234 capsid Anatomy 0.000 claims description 26
- 201000010099 disease Diseases 0.000 claims description 20
- 238000011282 treatment Methods 0.000 claims description 20
- 238000010362 genome editing Methods 0.000 claims description 19
- 239000013607 AAV vector Substances 0.000 claims description 18
- 241000193996 Streptococcus pyogenes Species 0.000 claims description 16
- 241000894007 species Species 0.000 claims description 15
- 241001634120 Adeno-associated virus - 5 Species 0.000 claims description 14
- 241000702421 Dependoparvovirus Species 0.000 claims description 14
- 230000004048 modification Effects 0.000 claims description 14
- 238000012986 modification Methods 0.000 claims description 14
- 108090000565 Capsid Proteins Proteins 0.000 claims description 13
- 102100023321 Ceruloplasmin Human genes 0.000 claims description 13
- 241001608472 Bifidobacterium longum Species 0.000 claims description 12
- 206010010356 Congenital anomaly Diseases 0.000 claims description 12
- 238000006467 substitution reaction Methods 0.000 claims description 10
- 241000191967 Staphylococcus aureus Species 0.000 claims description 9
- 238000010459 TALEN Methods 0.000 claims description 8
- 101710169336 5'-deoxyadenosine deaminase Proteins 0.000 claims description 7
- 102000055025 Adenosine deaminases Human genes 0.000 claims description 7
- 241000894006 Bacteria Species 0.000 claims description 7
- 102100026846 Cytidine deaminase Human genes 0.000 claims description 7
- 108010031325 Cytidine deaminase Proteins 0.000 claims description 7
- 229940009291 bifidobacterium longum Drugs 0.000 claims description 7
- 102000035118 modified proteins Human genes 0.000 claims description 7
- 108091005573 modified proteins Proteins 0.000 claims description 7
- 208000011580 syndromic disease Diseases 0.000 claims description 7
- 230000033228 biological regulation Effects 0.000 claims description 6
- 208000035475 disorder Diseases 0.000 claims description 6
- 108010017070 Zinc Finger Nucleases Proteins 0.000 claims description 5
- 241000801628 Odoribacter laneus Species 0.000 claims description 4
- 208000024827 Alzheimer disease Diseases 0.000 claims description 3
- 206010010099 Combined immunodeficiency Diseases 0.000 claims description 3
- 208000006682 alpha 1-Antitrypsin Deficiency Diseases 0.000 claims description 3
- 206010002026 amyotrophic lateral sclerosis Diseases 0.000 claims description 3
- 208000007442 rickets Diseases 0.000 claims description 3
- 108700019265 Aromatic amino acid decarboxylase deficiency Proteins 0.000 claims description 2
- 208000033810 Choroidal dystrophy Diseases 0.000 claims description 2
- 208000023105 Huntington disease Diseases 0.000 claims description 2
- 208000018737 Parkinson disease Diseases 0.000 claims description 2
- 201000009628 adenosine deaminase deficiency Diseases 0.000 claims description 2
- 208000036556 autosomal recessive T cell-negative B cell-negative NK cell-negative due to adenosine deaminase deficiency severe combined immunodeficiency Diseases 0.000 claims description 2
- 208000003571 choroideremia Diseases 0.000 claims description 2
- 201000006938 muscular dystrophy Diseases 0.000 claims description 2
- 201000004569 Blindness Diseases 0.000 claims 2
- 208000030162 Maple syrup disease Diseases 0.000 claims 2
- 208000000323 Tourette Syndrome Diseases 0.000 claims 2
- 208000016620 Tourette disease Diseases 0.000 claims 2
- 208000024393 maple syrup urine disease Diseases 0.000 claims 2
- 206010003694 Atrophy Diseases 0.000 claims 1
- 201000003883 Cystic fibrosis Diseases 0.000 claims 1
- 241000588722 Escherichia Species 0.000 claims 1
- 208000001914 Fragile X syndrome Diseases 0.000 claims 1
- 208000032007 Glycogen storage disease due to acid maltase deficiency Diseases 0.000 claims 1
- 206010053185 Glycogen storage disease type II Diseases 0.000 claims 1
- 208000031220 Hemophilia Diseases 0.000 claims 1
- 208000009292 Hemophilia A Diseases 0.000 claims 1
- 208000005176 Hepatitis C Diseases 0.000 claims 1
- 208000002972 Hepatolenticular Degeneration Diseases 0.000 claims 1
- 208000017604 Hodgkin disease Diseases 0.000 claims 1
- 208000010747 Hodgkins lymphoma Diseases 0.000 claims 1
- 101000986595 Homo sapiens Ornithine transcarbamylase, mitochondrial Proteins 0.000 claims 1
- 208000030673 Homozygous familial hypercholesterolemia Diseases 0.000 claims 1
- 206010061598 Immunodeficiency Diseases 0.000 claims 1
- 208000029462 Immunodeficiency disease Diseases 0.000 claims 1
- 102100033448 Lysosomal alpha-glucosidase Human genes 0.000 claims 1
- 208000002678 Mucopolysaccharidoses Diseases 0.000 claims 1
- 206010068871 Myotonic dystrophy Diseases 0.000 claims 1
- 208000003019 Neurofibromatosis 1 Diseases 0.000 claims 1
- 208000024834 Neurofibromatosis type 1 Diseases 0.000 claims 1
- 208000000599 Ornithine Carbamoyltransferase Deficiency Disease Diseases 0.000 claims 1
- 206010052450 Ornithine transcarbamoylase deficiency Diseases 0.000 claims 1
- 208000035903 Ornithine transcarbamylase deficiency Diseases 0.000 claims 1
- 102100028200 Ornithine transcarbamylase, mitochondrial Human genes 0.000 claims 1
- 201000011252 Phenylketonuria Diseases 0.000 claims 1
- 201000007737 Retinal degeneration Diseases 0.000 claims 1
- 206010045261 Type IIa hyperlipidaemia Diseases 0.000 claims 1
- 208000018839 Wilson disease Diseases 0.000 claims 1
- 206010001902 amaurosis Diseases 0.000 claims 1
- 230000037444 atrophy Effects 0.000 claims 1
- 201000004502 glycogen storage disease II Diseases 0.000 claims 1
- 208000002672 hepatitis B Diseases 0.000 claims 1
- 230000007813 immunodeficiency Effects 0.000 claims 1
- 208000002780 macular degeneration Diseases 0.000 claims 1
- 206010028093 mucopolysaccharidosis Diseases 0.000 claims 1
- 201000006417 multiple sclerosis Diseases 0.000 claims 1
- 201000011278 ornithine carbamoyltransferase deficiency Diseases 0.000 claims 1
- 208000030761 polycystic kidney disease Diseases 0.000 claims 1
- 230000004258 retinal degeneration Effects 0.000 claims 1
- 208000002320 spinal muscular atrophy Diseases 0.000 claims 1
- 108090000765 processed proteins & peptides Proteins 0.000 abstract description 102
- 102000004196 processed proteins & peptides Human genes 0.000 abstract description 62
- 230000002163 immunogen Effects 0.000 abstract description 28
- 238000002560 therapeutic procedure Methods 0.000 abstract description 20
- 238000010354 CRISPR gene editing Methods 0.000 abstract description 2
- 108010040467 CRISPR-Associated Proteins Proteins 0.000 description 41
- 230000027455 binding Effects 0.000 description 34
- 241000699670 Mus sp. Species 0.000 description 31
- 101150106478 GPS1 gene Proteins 0.000 description 30
- 208000002267 Anti-neutrophil cytoplasmic antibody-associated vasculitis Diseases 0.000 description 29
- 101000910035 Streptococcus pyogenes serotype M1 CRISPR-associated endonuclease Cas9/Csn1 Proteins 0.000 description 25
- 210000004027 cell Anatomy 0.000 description 24
- 125000003275 alpha amino acid group Chemical group 0.000 description 23
- 239000000427 antigen Substances 0.000 description 23
- 108091007433 antigens Proteins 0.000 description 23
- 102000036639 antigens Human genes 0.000 description 23
- 229920001184 polypeptide Polymers 0.000 description 20
- 210000001519 tissue Anatomy 0.000 description 19
- 101710172824 CRISPR-associated endonuclease Cas9 Proteins 0.000 description 17
- 230000000694 effects Effects 0.000 description 17
- 230000001225 therapeutic effect Effects 0.000 description 15
- 241001164825 Adeno-associated virus - 8 Species 0.000 description 14
- 238000003556 assay Methods 0.000 description 14
- 101150038500 cas9 gene Proteins 0.000 description 14
- 230000014509 gene expression Effects 0.000 description 14
- 239000000203 mixture Substances 0.000 description 14
- 108700028369 Alleles Proteins 0.000 description 13
- 241000700605 Viruses Species 0.000 description 13
- 238000004458 analytical method Methods 0.000 description 12
- 238000013459 approach Methods 0.000 description 12
- 230000001900 immune effect Effects 0.000 description 12
- 108020004414 DNA Proteins 0.000 description 11
- 238000005516 engineering process Methods 0.000 description 11
- 108091079001 CRISPR RNA Proteins 0.000 description 10
- 230000009260 cross reactivity Effects 0.000 description 10
- 230000002829 reductive effect Effects 0.000 description 10
- 101100275882 Emericella nidulans (strain FGSC A4 / ATCC 38163 / CBS 112.46 / NRRL 194 / M139) csnA gene Proteins 0.000 description 9
- 101100166134 Streptococcus thermophilus (strain ATCC BAA-491 / LMD-9) cas9-1 gene Proteins 0.000 description 9
- 101100166135 Streptococcus thermophilus (strain ATCC BAA-491 / LMD-9) cas9-2 gene Proteins 0.000 description 9
- 101150008672 csn-1 gene Proteins 0.000 description 9
- 238000002347 injection Methods 0.000 description 9
- 239000007924 injection Substances 0.000 description 9
- 239000000463 material Substances 0.000 description 9
- 244000005700 microbiome Species 0.000 description 9
- 238000004806 packaging method and process Methods 0.000 description 9
- 230000001177 retroviral effect Effects 0.000 description 9
- 101150101962 rplI gene Proteins 0.000 description 9
- 101710132601 Capsid protein Proteins 0.000 description 8
- 108010042407 Endonucleases Proteins 0.000 description 8
- 102000004533 Endonucleases Human genes 0.000 description 8
- 102000018713 Histocompatibility Antigens Class II Human genes 0.000 description 8
- 230000006870 function Effects 0.000 description 8
- 238000009163 protein therapy Methods 0.000 description 8
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 7
- 241000702423 Adeno-associated virus - 2 Species 0.000 description 7
- 101710197658 Capsid protein VP1 Proteins 0.000 description 7
- 102000043129 MHC class I family Human genes 0.000 description 7
- 108091054437 MHC class I family Proteins 0.000 description 7
- 108091054438 MHC class II family Proteins 0.000 description 7
- 241000699666 Mus <mouse, genus> Species 0.000 description 7
- 101710118046 RNA-directed RNA polymerase Proteins 0.000 description 7
- 101710108545 Viral protein 1 Proteins 0.000 description 7
- 210000001151 cytotoxic T lymphocyte Anatomy 0.000 description 7
- 210000000987 immune system Anatomy 0.000 description 7
- 238000001727 in vivo Methods 0.000 description 7
- 230000000670 limiting effect Effects 0.000 description 7
- 210000002966 serum Anatomy 0.000 description 7
- 230000003612 virological effect Effects 0.000 description 7
- 238000002965 ELISA Methods 0.000 description 6
- ZHNUHDYFZUAESO-UHFFFAOYSA-N Formamide Chemical compound NC=O ZHNUHDYFZUAESO-UHFFFAOYSA-N 0.000 description 6
- 239000000872 buffer Substances 0.000 description 6
- 239000003623 enhancer Substances 0.000 description 6
- 238000001415 gene therapy Methods 0.000 description 6
- 238000009396 hybridization Methods 0.000 description 6
- 238000011534 incubation Methods 0.000 description 6
- 230000035772 mutation Effects 0.000 description 6
- 210000000056 organ Anatomy 0.000 description 6
- 239000002245 particle Substances 0.000 description 6
- 239000000243 solution Substances 0.000 description 6
- 241000713813 Gibbon ape leukemia virus Species 0.000 description 5
- 108010027412 Histocompatibility Antigens Class II Proteins 0.000 description 5
- 241001465754 Metazoa Species 0.000 description 5
- 241000713869 Moloney murine leukemia virus Species 0.000 description 5
- 101710163270 Nuclease Proteins 0.000 description 5
- 108091028043 Nucleic acid sequence Proteins 0.000 description 5
- 241000194020 Streptococcus thermophilus Species 0.000 description 5
- 108091008874 T cell receptors Proteins 0.000 description 5
- 102000016266 T-Cell Antigen Receptors Human genes 0.000 description 5
- 210000001744 T-lymphocyte Anatomy 0.000 description 5
- 108700019146 Transgenes Proteins 0.000 description 5
- 238000004422 calculation algorithm Methods 0.000 description 5
- 108010078428 env Gene Products Proteins 0.000 description 5
- 238000002474 experimental method Methods 0.000 description 5
- 230000001404 mediated effect Effects 0.000 description 5
- 210000003071 memory t lymphocyte Anatomy 0.000 description 5
- 108020004999 messenger RNA Proteins 0.000 description 5
- 150000007523 nucleic acids Chemical group 0.000 description 5
- 230000001717 pathogenic effect Effects 0.000 description 5
- 239000013612 plasmid Substances 0.000 description 5
- 208000024891 symptom Diseases 0.000 description 5
- 230000008685 targeting Effects 0.000 description 5
- 241000702462 Akkermansia muciniphila Species 0.000 description 4
- 238000010356 CRISPR-Cas9 genome editing Methods 0.000 description 4
- 108010008532 Deoxyribonuclease I Proteins 0.000 description 4
- 102000007260 Deoxyribonuclease I Human genes 0.000 description 4
- 241000282412 Homo Species 0.000 description 4
- 241000186660 Lactobacillus Species 0.000 description 4
- 102000043131 MHC class II family Human genes 0.000 description 4
- 241000605939 Wolinella succinogenes Species 0.000 description 4
- 230000033289 adaptive immune response Effects 0.000 description 4
- 238000004364 calculation method Methods 0.000 description 4
- 230000001413 cellular effect Effects 0.000 description 4
- 230000008859 change Effects 0.000 description 4
- 239000003795 chemical substances by application Substances 0.000 description 4
- 230000000295 complement effect Effects 0.000 description 4
- 230000004927 fusion Effects 0.000 description 4
- 230000002068 genetic effect Effects 0.000 description 4
- 230000006698 induction Effects 0.000 description 4
- 230000017730 intein-mediated protein splicing Effects 0.000 description 4
- 238000001990 intravenous administration Methods 0.000 description 4
- 229940039696 lactobacillus Drugs 0.000 description 4
- 208000032839 leukemia Diseases 0.000 description 4
- 230000007246 mechanism Effects 0.000 description 4
- 102000039446 nucleic acids Human genes 0.000 description 4
- 108020004707 nucleic acids Proteins 0.000 description 4
- 244000052769 pathogen Species 0.000 description 4
- 230000010076 replication Effects 0.000 description 4
- 230000004044 response Effects 0.000 description 4
- 238000012360 testing method Methods 0.000 description 4
- 238000011746 C57BL/6J (JAX™ mouse strain) Methods 0.000 description 3
- 206010062346 Congenital neuropathy Diseases 0.000 description 3
- 102220605874 Cytosolic arginine sensor for mTORC1 subunit 2_D10A_mutation Human genes 0.000 description 3
- LFQSCWFLJHTTHZ-UHFFFAOYSA-N Ethanol Chemical compound CCO LFQSCWFLJHTTHZ-UHFFFAOYSA-N 0.000 description 3
- PEDCQBHIVMGVHV-UHFFFAOYSA-N Glycerine Chemical compound OCC(O)CO PEDCQBHIVMGVHV-UHFFFAOYSA-N 0.000 description 3
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 3
- 241000701024 Human betaherpesvirus 5 Species 0.000 description 3
- 241000725303 Human immunodeficiency virus Species 0.000 description 3
- 102100034353 Integrase Human genes 0.000 description 3
- 241000204031 Mycoplasma Species 0.000 description 3
- 241000588653 Neisseria Species 0.000 description 3
- 241000588650 Neisseria meningitidis Species 0.000 description 3
- 208000017442 Retinal disease Diseases 0.000 description 3
- 206010038923 Retinopathy Diseases 0.000 description 3
- 206010039710 Scleroderma Diseases 0.000 description 3
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 3
- 230000005867 T cell response Effects 0.000 description 3
- 102100034922 T-cell surface glycoprotein CD8 alpha chain Human genes 0.000 description 3
- 108010043645 Transcription Activator-Like Effector Nucleases Proteins 0.000 description 3
- 102100029613 Transient receptor potential cation channel subfamily V member 1 Human genes 0.000 description 3
- 108050004388 Transient receptor potential cation channel subfamily V member 1 Proteins 0.000 description 3
- 125000000539 amino acid group Chemical group 0.000 description 3
- 230000030741 antigen processing and presentation Effects 0.000 description 3
- 210000003719 b-lymphocyte Anatomy 0.000 description 3
- 239000000969 carrier Substances 0.000 description 3
- 230000036755 cellular response Effects 0.000 description 3
- 238000006243 chemical reaction Methods 0.000 description 3
- 239000002299 complementary DNA Substances 0.000 description 3
- 238000000205 computational method Methods 0.000 description 3
- 238000005094 computer simulation Methods 0.000 description 3
- 230000000875 corresponding effect Effects 0.000 description 3
- 238000011161 development Methods 0.000 description 3
- 230000018109 developmental process Effects 0.000 description 3
- 239000003085 diluting agent Substances 0.000 description 3
- 230000007613 environmental effect Effects 0.000 description 3
- 230000036039 immunity Effects 0.000 description 3
- 238000000126 in silico method Methods 0.000 description 3
- 230000015788 innate immune response Effects 0.000 description 3
- 230000003993 interaction Effects 0.000 description 3
- 230000007774 longterm Effects 0.000 description 3
- 210000004072 lung Anatomy 0.000 description 3
- 239000011159 matrix material Substances 0.000 description 3
- 210000001806 memory b lymphocyte Anatomy 0.000 description 3
- 238000010172 mouse model Methods 0.000 description 3
- 238000006386 neutralization reaction Methods 0.000 description 3
- 230000003472 neutralizing effect Effects 0.000 description 3
- 125000003729 nucleotide group Chemical group 0.000 description 3
- 230000037361 pathway Effects 0.000 description 3
- 230000008569 process Effects 0.000 description 3
- 238000003259 recombinant expression Methods 0.000 description 3
- 238000012546 transfer Methods 0.000 description 3
- 238000010200 validation analysis Methods 0.000 description 3
- 239000003981 vehicle Substances 0.000 description 3
- KDCGOANMDULRCW-UHFFFAOYSA-N 7H-purine Chemical compound N1=CNC2=NC=NC2=C1 KDCGOANMDULRCW-UHFFFAOYSA-N 0.000 description 2
- 108010079649 APOBEC-1 Deaminase Proteins 0.000 description 2
- 108010004483 APOBEC-3G Deaminase Proteins 0.000 description 2
- 241000186046 Actinomyces Species 0.000 description 2
- 102100036664 Adenosine deaminase Human genes 0.000 description 2
- 241000702460 Akkermansia Species 0.000 description 2
- 241000695953 Alcanivorax pacificus Species 0.000 description 2
- 241001147780 Alicyclobacillus Species 0.000 description 2
- 208000031277 Amaurotic familial idiocy Diseases 0.000 description 2
- 241001470561 Arracacha virus V Species 0.000 description 2
- 241000193399 Bacillus smithii Species 0.000 description 2
- 102100040397 C->U-editing enzyme APOBEC-1 Human genes 0.000 description 2
- 102100027221 CD81 antigen Human genes 0.000 description 2
- 101710094648 Coat protein Proteins 0.000 description 2
- 208000006992 Color Vision Defects Diseases 0.000 description 2
- 241001464948 Coprococcus Species 0.000 description 2
- 241000186216 Corynebacterium Species 0.000 description 2
- 102100040263 DNA dC->dU-editing enzyme APOBEC-3A Human genes 0.000 description 2
- 102100040262 DNA dC->dU-editing enzyme APOBEC-3B Human genes 0.000 description 2
- 102100040261 DNA dC->dU-editing enzyme APOBEC-3C Human genes 0.000 description 2
- 102100040264 DNA dC->dU-editing enzyme APOBEC-3D Human genes 0.000 description 2
- 102100040266 DNA dC->dU-editing enzyme APOBEC-3F Human genes 0.000 description 2
- 102100038076 DNA dC->dU-editing enzyme APOBEC-3G Human genes 0.000 description 2
- 230000004568 DNA-binding Effects 0.000 description 2
- 229920002307 Dextran Polymers 0.000 description 2
- 238000011510 Elispot assay Methods 0.000 description 2
- 241000194032 Enterococcus faecalis Species 0.000 description 2
- 241000186394 Eubacterium Species 0.000 description 2
- 241000605986 Fusobacterium nucleatum Species 0.000 description 2
- 241000968725 Gammaproteobacteria bacterium Species 0.000 description 2
- 241000193385 Geobacillus stearothermophilus Species 0.000 description 2
- 108050008753 HNH endonucleases Proteins 0.000 description 2
- 102000000310 HNH endonucleases Human genes 0.000 description 2
- 102000008949 Histocompatibility Antigens Class I Human genes 0.000 description 2
- 108010088652 Histocompatibility Antigens Class I Proteins 0.000 description 2
- 101000914479 Homo sapiens CD81 antigen Proteins 0.000 description 2
- 101000964378 Homo sapiens DNA dC->dU-editing enzyme APOBEC-3A Proteins 0.000 description 2
- 101000964385 Homo sapiens DNA dC->dU-editing enzyme APOBEC-3B Proteins 0.000 description 2
- 101000964383 Homo sapiens DNA dC->dU-editing enzyme APOBEC-3C Proteins 0.000 description 2
- 101000964382 Homo sapiens DNA dC->dU-editing enzyme APOBEC-3D Proteins 0.000 description 2
- 101000964377 Homo sapiens DNA dC->dU-editing enzyme APOBEC-3F Proteins 0.000 description 2
- 101000716102 Homo sapiens T-cell surface glycoprotein CD4 Proteins 0.000 description 2
- 102000007330 LDL Lipoproteins Human genes 0.000 description 2
- 108010007622 LDL Lipoproteins Proteins 0.000 description 2
- 241000282560 Macaca mulatta Species 0.000 description 2
- 101710081079 Minor spike protein H Proteins 0.000 description 2
- 241000713862 Moloney murine sarcoma virus Species 0.000 description 2
- 241001148552 Mycoplasma canis Species 0.000 description 2
- 241001148556 Mycoplasma ovipneumoniae Species 0.000 description 2
- 208000002537 Neuronal Ceroid-Lipofuscinoses Diseases 0.000 description 2
- 241000605156 Nitrobacter hamburgensis Species 0.000 description 2
- 241000385061 Oenococcus kitaharae Species 0.000 description 2
- 241000288906 Primates Species 0.000 description 2
- 108010026552 Proteome Proteins 0.000 description 2
- 241001135508 Ralstonia syzygii Species 0.000 description 2
- 241000190984 Rhodospirillum rubrum Species 0.000 description 2
- 241000714474 Rous sarcoma virus Species 0.000 description 2
- 101100166144 Staphylococcus aureus cas9 gene Proteins 0.000 description 2
- 241001134656 Staphylococcus lugdunensis Species 0.000 description 2
- 241000194023 Streptococcus sanguinis Species 0.000 description 2
- 229930006000 Sucrose Natural products 0.000 description 2
- CZMRCDWAGMRECN-UGDNZRGBSA-N Sucrose Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 CZMRCDWAGMRECN-UGDNZRGBSA-N 0.000 description 2
- 102100036011 T-cell surface glycoprotein CD4 Human genes 0.000 description 2
- 102000002689 Toll-like receptor Human genes 0.000 description 2
- 108020000411 Toll-like receptor Proteins 0.000 description 2
- 241001148134 Veillonella Species 0.000 description 2
- 241000193453 [Clostridium] cellulolyticum Species 0.000 description 2
- 201000000761 achromatopsia Diseases 0.000 description 2
- 210000005006 adaptive immune system Anatomy 0.000 description 2
- -1 antibody Proteins 0.000 description 2
- 230000004888 barrier function Effects 0.000 description 2
- 230000009286 beneficial effect Effects 0.000 description 2
- 230000008901 benefit Effects 0.000 description 2
- 230000004071 biological effect Effects 0.000 description 2
- 230000000903 blocking effect Effects 0.000 description 2
- 210000004204 blood vessel Anatomy 0.000 description 2
- 210000004556 brain Anatomy 0.000 description 2
- 238000003776 cleavage reaction Methods 0.000 description 2
- 201000007254 color blindness Diseases 0.000 description 2
- 239000002131 composite material Substances 0.000 description 2
- 230000007812 deficiency Effects 0.000 description 2
- 238000012217 deletion Methods 0.000 description 2
- 230000037430 deletion Effects 0.000 description 2
- 230000001419 dependent effect Effects 0.000 description 2
- LOKCTEFSRHRXRJ-UHFFFAOYSA-I dipotassium trisodium dihydrogen phosphate hydrogen phosphate dichloride Chemical compound P(=O)(O)(O)[O-].[K+].P(=O)(O)([O-])[O-].[Na+].[Na+].[Cl-].[K+].[Cl-].[Na+] LOKCTEFSRHRXRJ-UHFFFAOYSA-I 0.000 description 2
- 239000002552 dosage form Substances 0.000 description 2
- 229940032049 enterococcus faecalis Drugs 0.000 description 2
- 108700004025 env Genes Proteins 0.000 description 2
- 238000003114 enzyme-linked immunosorbent spot assay Methods 0.000 description 2
- 210000003527 eukaryotic cell Anatomy 0.000 description 2
- 238000001476 gene delivery Methods 0.000 description 2
- 210000002216 heart Anatomy 0.000 description 2
- 230000005745 host immune response Effects 0.000 description 2
- 230000028996 humoral immune response Effects 0.000 description 2
- 239000001257 hydrogen Substances 0.000 description 2
- 229910052739 hydrogen Inorganic materials 0.000 description 2
- 230000009851 immunogenic response Effects 0.000 description 2
- 238000000338 in vitro Methods 0.000 description 2
- 230000000977 initiatory effect Effects 0.000 description 2
- 210000000936 intestine Anatomy 0.000 description 2
- NBQNWMBBSKPBAY-UHFFFAOYSA-N iodixanol Chemical compound IC=1C(C(=O)NCC(O)CO)=C(I)C(C(=O)NCC(O)CO)=C(I)C=1N(C(=O)C)CC(O)CN(C(C)=O)C1=C(I)C(C(=O)NCC(O)CO)=C(I)C(C(=O)NCC(O)CO)=C1I NBQNWMBBSKPBAY-UHFFFAOYSA-N 0.000 description 2
- 229960004359 iodixanol Drugs 0.000 description 2
- FZWBNHMXJMCXLU-BLAUPYHCSA-N isomaltotriose Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1OC[C@@H]1[C@@H](O)[C@H](O)[C@@H](O)[C@@H](OC[C@@H](O)[C@@H](O)[C@H](O)[C@@H](O)C=O)O1 FZWBNHMXJMCXLU-BLAUPYHCSA-N 0.000 description 2
- 208000017476 juvenile neuronal ceroid lipofuscinosis Diseases 0.000 description 2
- 210000003734 kidney Anatomy 0.000 description 2
- 210000004185 liver Anatomy 0.000 description 2
- 230000004807 localization Effects 0.000 description 2
- 210000004698 lymphocyte Anatomy 0.000 description 2
- 238000004519 manufacturing process Methods 0.000 description 2
- 201000007607 neuronal ceroid lipofuscinosis 3 Diseases 0.000 description 2
- 230000006780 non-homologous end joining Effects 0.000 description 2
- 239000002773 nucleotide Substances 0.000 description 2
- 230000003287 optical effect Effects 0.000 description 2
- 210000000496 pancreas Anatomy 0.000 description 2
- 239000000816 peptidomimetic Substances 0.000 description 2
- 239000002953 phosphate buffered saline Substances 0.000 description 2
- 230000008488 polyadenylation Effects 0.000 description 2
- 230000023603 positive regulation of transcription initiation, DNA-dependent Effects 0.000 description 2
- 230000004853 protein function Effects 0.000 description 2
- 230000007420 reactivation Effects 0.000 description 2
- 230000007017 scission Effects 0.000 description 2
- 238000012216 screening Methods 0.000 description 2
- 210000003491 skin Anatomy 0.000 description 2
- 239000011780 sodium chloride Substances 0.000 description 2
- 238000007920 subcutaneous administration Methods 0.000 description 2
- 239000000758 substrate Substances 0.000 description 2
- 239000005720 sucrose Substances 0.000 description 2
- 210000001685 thyroid gland Anatomy 0.000 description 2
- 238000013518 transcription Methods 0.000 description 2
- 230000035897 transcription Effects 0.000 description 2
- 238000010361 transduction Methods 0.000 description 2
- 230000026683 transduction Effects 0.000 description 2
- 238000001890 transfection Methods 0.000 description 2
- 239000013603 viral vector Substances 0.000 description 2
- 238000005406 washing Methods 0.000 description 2
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Chemical compound O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 2
- 102000040650 (ribonucleotides)n+m Human genes 0.000 description 1
- NEWKHUASLBMWRE-UHFFFAOYSA-N 2-methyl-6-(phenylethynyl)pyridine Chemical compound CC1=CC=CC(C#CC=2C=CC=CC=2)=N1 NEWKHUASLBMWRE-UHFFFAOYSA-N 0.000 description 1
- UAIUNKRWKOVEES-UHFFFAOYSA-N 3,3',5,5'-tetramethylbenzidine Chemical compound CC1=C(N)C(C)=CC(C=2C=C(C)C(N)=C(C)C=2)=C1 UAIUNKRWKOVEES-UHFFFAOYSA-N 0.000 description 1
- 241001430193 Absiella dolichum Species 0.000 description 1
- 241000604451 Acidaminococcus Species 0.000 description 1
- 241001134630 Acidothermus cellulolyticus Species 0.000 description 1
- 241001437069 Acidovorax ebreus TPSY Species 0.000 description 1
- 241001655883 Adeno-associated virus - 1 Species 0.000 description 1
- 241000580270 Adeno-associated virus - 4 Species 0.000 description 1
- 229920001817 Agar Polymers 0.000 description 1
- 241000193412 Alicyclobacillus acidoterrestris Species 0.000 description 1
- GUBGYTABKSRVRQ-XLOQQCSPSA-N Alpha-Lactose Chemical compound O[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@H]1O[C@@H]1[C@@H](CO)O[C@H](O)[C@H](O)[C@H]1O GUBGYTABKSRVRQ-XLOQQCSPSA-N 0.000 description 1
- 241001621927 Aminomonas Species 0.000 description 1
- 241001621924 Aminomonas paucivorans Species 0.000 description 1
- 108091023037 Aptamer Proteins 0.000 description 1
- 241001478327 Azospirillum sp. Species 0.000 description 1
- 230000003844 B-cell-activation Effects 0.000 description 1
- 241000423333 Bacteroides fragilis NCTC 9343 Species 0.000 description 1
- 241001148536 Bacteroides sp. Species 0.000 description 1
- 241000927512 Barnesiella Species 0.000 description 1
- 241000260432 Barnesiella intestinihominis Species 0.000 description 1
- 241000611351 Bergeyella Species 0.000 description 1
- 241000186000 Bifidobacterium Species 0.000 description 1
- 241000589173 Bradyrhizobium Species 0.000 description 1
- 241000589171 Bradyrhizobium sp. Species 0.000 description 1
- 102100040399 C->U-editing enzyme APOBEC-2 Human genes 0.000 description 1
- 210000001266 CD8-positive T-lymphocyte Anatomy 0.000 description 1
- 241000589876 Campylobacter Species 0.000 description 1
- 241000589875 Campylobacter jejuni Species 0.000 description 1
- 241001453245 Campylobacter jejuni subsp. jejuni Species 0.000 description 1
- 241000589986 Campylobacter lari Species 0.000 description 1
- 241000222120 Candida <Saccharomycetales> Species 0.000 description 1
- 241000282465 Canis Species 0.000 description 1
- 241000283707 Capra Species 0.000 description 1
- 208000024172 Cardiovascular disease Diseases 0.000 description 1
- 102000053642 Catalytic RNA Human genes 0.000 description 1
- 108090000994 Catalytic RNA Proteins 0.000 description 1
- 241000761552 Catenibacterium mitsuokai DSM 15897 Species 0.000 description 1
- 241001478240 Coccus Species 0.000 description 1
- 108091026890 Coding region Proteins 0.000 description 1
- 235000021508 Coleus Nutrition 0.000 description 1
- 244000061182 Coleus blumei Species 0.000 description 1
- 241000064942 Corynebacterium diphtheriae C7 (beta) Species 0.000 description 1
- FBPFZTCFMRRESA-KVTDHHQDSA-N D-Mannitol Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-KVTDHHQDSA-N 0.000 description 1
- WQZGKKKJIJFFOK-QTVWNMPRSA-N D-mannopyranose Chemical compound OC[C@H]1OC(O)[C@@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-QTVWNMPRSA-N 0.000 description 1
- 102000053602 DNA Human genes 0.000 description 1
- 102100038050 DNA dC->dU-editing enzyme APOBEC-3H Human genes 0.000 description 1
- 101710082737 DNA dC->dU-editing enzyme APOBEC-3H Proteins 0.000 description 1
- 102000004163 DNA-directed RNA polymerases Human genes 0.000 description 1
- 108090000626 DNA-directed RNA polymerases Proteins 0.000 description 1
- 101710088194 Dehydrogenase Proteins 0.000 description 1
- 241000933091 Dinoroseobacter shibae DFL 12 = DSM 16493 Species 0.000 description 1
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 1
- 238000008157 ELISA kit Methods 0.000 description 1
- 102000004190 Enzymes Human genes 0.000 description 1
- 108090000790 Enzymes Proteins 0.000 description 1
- 241000186588 Erysipelatoclostridium ramosum Species 0.000 description 1
- 241000282326 Felis catus Species 0.000 description 1
- 241000192016 Finegoldia magna Species 0.000 description 1
- 241000589565 Flavobacterium Species 0.000 description 1
- 241000555689 Flavobacterium branchiophilum Species 0.000 description 1
- 241000589601 Francisella Species 0.000 description 1
- 241000589602 Francisella tularensis Species 0.000 description 1
- 241000589599 Francisella tularensis subsp. novicida Species 0.000 description 1
- 241000551711 Fructobacillus Species 0.000 description 1
- 108091006027 G proteins Proteins 0.000 description 1
- 102000030782 GTP binding Human genes 0.000 description 1
- 108091000058 GTP-Binding Proteins 0.000 description 1
- 101710177291 Gag polyprotein Proteins 0.000 description 1
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 1
- 239000004471 Glycine Substances 0.000 description 1
- 241000590002 Helicobacter pylori Species 0.000 description 1
- 101000964322 Homo sapiens C->U-editing enzyme APOBEC-2 Proteins 0.000 description 1
- 101000742736 Homo sapiens DNA dC->dU-editing enzyme APOBEC-3G Proteins 0.000 description 1
- 101000800426 Homo sapiens Putative C->U-editing enzyme APOBEC-4 Proteins 0.000 description 1
- 101000684820 Homo sapiens Sodium channel protein type 3 subunit alpha Proteins 0.000 description 1
- 101000654386 Homo sapiens Sodium channel protein type 9 subunit alpha Proteins 0.000 description 1
- 108090000144 Human Proteins Proteins 0.000 description 1
- 102000003839 Human Proteins Human genes 0.000 description 1
- 241000598436 Human T-cell lymphotropic virus Species 0.000 description 1
- 206010020772 Hypertension Diseases 0.000 description 1
- 241001398692 Ignavibacterium Species 0.000 description 1
- 241001203194 Ilyobacter polytropus DSM 2926 Species 0.000 description 1
- 101710172072 Kexin Proteins 0.000 description 1
- 241000917009 Lactobacillus rhamnosus GG Species 0.000 description 1
- GUBGYTABKSRVRQ-QKKXKWKRSA-N Lactose Natural products OC[C@H]1O[C@@H](O[C@H]2[C@H](O)[C@@H](O)C(O)O[C@@H]2CO)[C@H](O)[C@@H](O)[C@H]1O GUBGYTABKSRVRQ-QKKXKWKRSA-N 0.000 description 1
- 241001193656 Legionella pneumophila str. Paris Species 0.000 description 1
- 241000713666 Lentivirus Species 0.000 description 1
- 108010066345 MHC binding peptide Proteins 0.000 description 1
- 241000282553 Macaca Species 0.000 description 1
- 101710125418 Major capsid protein Proteins 0.000 description 1
- 241000124008 Mammalia Species 0.000 description 1
- 229930195725 Mannitol Natural products 0.000 description 1
- 102000018697 Membrane Proteins Human genes 0.000 description 1
- 108010052285 Membrane Proteins Proteins 0.000 description 1
- 201000009906 Meningitis Diseases 0.000 description 1
- 241000863391 Methylophilus Species 0.000 description 1
- 241000589354 Methylosinus Species 0.000 description 1
- 108090000143 Mouse Proteins Proteins 0.000 description 1
- 241000714177 Murine leukemia virus Species 0.000 description 1
- 101100113998 Mus musculus Cnbd2 gene Proteins 0.000 description 1
- 241001465821 Mycoplasma gallisepticum str. F Species 0.000 description 1
- 241000202942 Mycoplasma synoviae Species 0.000 description 1
- 241000588654 Neisseria cinerea Species 0.000 description 1
- 241000588649 Neisseria lactamica Species 0.000 description 1
- 101100166142 Neisseria meningitidis serogroup A / serotype 4A (strain DSM 15465 / Z2491) cas9 gene Proteins 0.000 description 1
- 206010028980 Neoplasm Diseases 0.000 description 1
- 241000772415 Neovison vison Species 0.000 description 1
- 241001178035 Odoribacter laneus YIT 12061 Species 0.000 description 1
- 240000007019 Oxalis corniculata Species 0.000 description 1
- 101150094724 PCSK9 gene Proteins 0.000 description 1
- 241000701945 Parvoviridae Species 0.000 description 1
- 101100166143 Pasteurella multocida (strain Pm70) cas9 gene Proteins 0.000 description 1
- 241000432806 Pasteurella multocida subsp. multocida str. Pm70 Species 0.000 description 1
- 241000374256 Peptoniphilus duerdenii Species 0.000 description 1
- 241000193804 Planococcus <bacterium> Species 0.000 description 1
- 241000605894 Porphyromonas Species 0.000 description 1
- 241001300940 Porphyromonas sp. Species 0.000 description 1
- 241000611831 Prevotella sp. Species 0.000 description 1
- 241000530934 Prevotella timonensis Species 0.000 description 1
- 102000004245 Proteasome Endopeptidase Complex Human genes 0.000 description 1
- 108090000708 Proteasome Endopeptidase Complex Proteins 0.000 description 1
- 241000192142 Proteobacteria Species 0.000 description 1
- 241000588769 Proteus <enterobacteria> Species 0.000 description 1
- 102100033091 Putative C->U-editing enzyme APOBEC-4 Human genes 0.000 description 1
- CZPWVGJYEJSRLH-UHFFFAOYSA-N Pyrimidine Chemical compound C1=CN=CN=C1 CZPWVGJYEJSRLH-UHFFFAOYSA-N 0.000 description 1
- 241000232299 Ralstonia Species 0.000 description 1
- 108020004511 Recombinant DNA Proteins 0.000 description 1
- 241000191025 Rhodobacter Species 0.000 description 1
- 241000187693 Rhodococcus rhodochrous Species 0.000 description 1
- 241001478306 Rhodovulum sp. Species 0.000 description 1
- 108091028664 Ribonucleotide Proteins 0.000 description 1
- 241000192031 Ruminococcus Species 0.000 description 1
- 241000192029 Ruminococcus albus Species 0.000 description 1
- 101150067145 SCARB1 gene Proteins 0.000 description 1
- 101150110009 SCN11A gene Proteins 0.000 description 1
- 101150010053 Scn10a gene Proteins 0.000 description 1
- 108020004682 Single-Stranded DNA Proteins 0.000 description 1
- 102100031367 Sodium channel protein type 9 subunit alpha Human genes 0.000 description 1
- 241001464874 Solobacterium moorei Species 0.000 description 1
- 241000639167 Sphaerochaeta globosa Species 0.000 description 1
- 241000713880 Spleen focus-forming virus Species 0.000 description 1
- 206010041660 Splenomegaly Diseases 0.000 description 1
- 241000191940 Staphylococcus Species 0.000 description 1
- 241000194017 Streptococcus Species 0.000 description 1
- 241000194019 Streptococcus mutans Species 0.000 description 1
- 241001267418 Streptococcus pasteuri Species 0.000 description 1
- 108090000787 Subtilisin Proteins 0.000 description 1
- 208000000389 T-cell leukemia Diseases 0.000 description 1
- 208000028530 T-cell lymphoblastic leukemia/lymphoma Diseases 0.000 description 1
- 108010062740 TRPV Cation Channels Proteins 0.000 description 1
- 241001313536 Thermothelomyces thermophila Species 0.000 description 1
- 108010073062 Transcription Activator-Like Effectors Proteins 0.000 description 1
- 108091023040 Transcription factor Proteins 0.000 description 1
- 102000040945 Transcription factor Human genes 0.000 description 1
- 241000589886 Treponema Species 0.000 description 1
- 241000589884 Treponema pallidum Species 0.000 description 1
- 101710159648 Uncharacterized protein Proteins 0.000 description 1
- HCHKCACWOHOZIP-UHFFFAOYSA-N Zinc Chemical compound [Zn] HCHKCACWOHOZIP-UHFFFAOYSA-N 0.000 description 1
- 241001531183 [Eubacterium] yurii Species 0.000 description 1
- 239000002253 acid Substances 0.000 description 1
- 230000004913 activation Effects 0.000 description 1
- 101150063416 add gene Proteins 0.000 description 1
- 239000002671 adjuvant Substances 0.000 description 1
- 239000008272 agar Substances 0.000 description 1
- 239000000956 alloy Substances 0.000 description 1
- 229910045601 alloy Inorganic materials 0.000 description 1
- WNROFYMDJYEPJX-UHFFFAOYSA-K aluminium hydroxide Chemical compound [OH-].[OH-].[OH-].[Al+3] WNROFYMDJYEPJX-UHFFFAOYSA-K 0.000 description 1
- 238000010171 animal model Methods 0.000 description 1
- 230000000692 anti-sense effect Effects 0.000 description 1
- 230000005875 antibody response Effects 0.000 description 1
- 238000009175 antibody therapy Methods 0.000 description 1
- 210000000612 antigen-presenting cell Anatomy 0.000 description 1
- 230000000890 antigenic effect Effects 0.000 description 1
- 239000003963 antioxidant agent Substances 0.000 description 1
- 230000003078 antioxidant effect Effects 0.000 description 1
- 210000000702 aorta abdominal Anatomy 0.000 description 1
- 238000013528 artificial neural network Methods 0.000 description 1
- 230000001580 bacterial effect Effects 0.000 description 1
- 230000003796 beauty Effects 0.000 description 1
- WQZGKKKJIJFFOK-VFUOTHLCSA-N beta-D-glucose Chemical compound OC[C@H]1O[C@@H](O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-VFUOTHLCSA-N 0.000 description 1
- 210000003445 biliary tract Anatomy 0.000 description 1
- 230000031018 biological processes and functions Effects 0.000 description 1
- 210000000746 body region Anatomy 0.000 description 1
- 239000007975 buffered saline Substances 0.000 description 1
- 239000006172 buffering agent Substances 0.000 description 1
- 230000003139 buffering effect Effects 0.000 description 1
- 201000011510 cancer Diseases 0.000 description 1
- 150000001720 carbohydrates Chemical class 0.000 description 1
- 235000014633 carbohydrates Nutrition 0.000 description 1
- 230000015556 catabolic process Effects 0.000 description 1
- 238000012512 characterization method Methods 0.000 description 1
- 239000002738 chelating agent Substances 0.000 description 1
- 239000003153 chemical reaction reagent Substances 0.000 description 1
- 210000000038 chest Anatomy 0.000 description 1
- 239000007979 citrate buffer Substances 0.000 description 1
- 239000011248 coating agent Substances 0.000 description 1
- 238000000576 coating method Methods 0.000 description 1
- 238000004737 colorimetric analysis Methods 0.000 description 1
- 230000000052 comparative effect Effects 0.000 description 1
- 150000001875 compounds Chemical class 0.000 description 1
- 108091036078 conserved sequence Proteins 0.000 description 1
- 238000007796 conventional method Methods 0.000 description 1
- 210000004087 cornea Anatomy 0.000 description 1
- 238000012937 correction Methods 0.000 description 1
- 230000002596 correlated effect Effects 0.000 description 1
- 230000001186 cumulative effect Effects 0.000 description 1
- 230000001461 cytolytic effect Effects 0.000 description 1
- 230000001086 cytosolic effect Effects 0.000 description 1
- 231100000433 cytotoxic Toxicity 0.000 description 1
- 230000001472 cytotoxic effect Effects 0.000 description 1
- 230000003013 cytotoxicity Effects 0.000 description 1
- 231100000135 cytotoxicity Toxicity 0.000 description 1
- 230000006378 damage Effects 0.000 description 1
- 230000000911 decarboxylating effect Effects 0.000 description 1
- 238000006731 degradation reaction Methods 0.000 description 1
- 239000008367 deionised water Substances 0.000 description 1
- 229910021641 deionized water Inorganic materials 0.000 description 1
- 229940124447 delivery agent Drugs 0.000 description 1
- 238000002716 delivery method Methods 0.000 description 1
- 210000004443 dendritic cell Anatomy 0.000 description 1
- 238000002298 density-gradient ultracentrifugation Methods 0.000 description 1
- 239000005547 deoxyribonucleotide Substances 0.000 description 1
- 125000002637 deoxyribonucleotide group Chemical group 0.000 description 1
- 238000013461 design Methods 0.000 description 1
- 206010012601 diabetes mellitus Diseases 0.000 description 1
- 238000010790 dilution Methods 0.000 description 1
- 239000012895 dilution Substances 0.000 description 1
- 238000009826 distribution Methods 0.000 description 1
- 230000005782 double-strand break Effects 0.000 description 1
- 239000003814 drug Substances 0.000 description 1
- 229940079593 drug Drugs 0.000 description 1
- 239000003937 drug carrier Substances 0.000 description 1
- 210000001198 duodenum Anatomy 0.000 description 1
- 239000012636 effector Substances 0.000 description 1
- 230000008030 elimination Effects 0.000 description 1
- 238000003379 elimination reaction Methods 0.000 description 1
- 230000002255 enzymatic effect Effects 0.000 description 1
- 229940088598 enzyme Drugs 0.000 description 1
- 150000002148 esters Chemical class 0.000 description 1
- HQPMKSGTIOYHJT-UHFFFAOYSA-N ethane-1,2-diol;propane-1,2-diol Chemical compound OCCO.CC(O)CO HQPMKSGTIOYHJT-UHFFFAOYSA-N 0.000 description 1
- 150000002170 ethers Chemical class 0.000 description 1
- 230000017188 evasion or tolerance of host immune response Effects 0.000 description 1
- 230000001747 exhibiting effect Effects 0.000 description 1
- 239000013604 expression vector Substances 0.000 description 1
- 239000012634 fragment Substances 0.000 description 1
- 229940118764 francisella tularensis Drugs 0.000 description 1
- 239000007789 gas Substances 0.000 description 1
- 230000030279 gene silencing Effects 0.000 description 1
- 238000010363 gene targeting Methods 0.000 description 1
- 108091006104 gene-regulatory proteins Proteins 0.000 description 1
- 102000034356 gene-regulatory proteins Human genes 0.000 description 1
- 239000008103 glucose Substances 0.000 description 1
- RWSXRVCMGQZWBV-WDSKDSINSA-N glutathione Chemical compound OC(=O)[C@@H](N)CCC(=O)N[C@@H](CS)C(=O)NCC(O)=O RWSXRVCMGQZWBV-WDSKDSINSA-N 0.000 description 1
- 229940037467 helicobacter pylori Drugs 0.000 description 1
- 238000002744 homologous recombination Methods 0.000 description 1
- 230000006801 homologous recombination Effects 0.000 description 1
- 102000054962 human APOBEC3G Human genes 0.000 description 1
- 230000004727 humoral immunity Effects 0.000 description 1
- 230000002209 hydrophobic effect Effects 0.000 description 1
- 210000003111 iliac vein Anatomy 0.000 description 1
- 230000003053 immunization Effects 0.000 description 1
- 238000003018 immunoassay Methods 0.000 description 1
- 230000000951 immunodiffusion Effects 0.000 description 1
- 230000006054 immunological memory Effects 0.000 description 1
- 238000012405 in silico analysis Methods 0.000 description 1
- 230000001939 inductive effect Effects 0.000 description 1
- 239000004615 ingredient Substances 0.000 description 1
- 230000002401 inhibitory effect Effects 0.000 description 1
- 230000005764 inhibitory process Effects 0.000 description 1
- 230000002452 interceptive effect Effects 0.000 description 1
- 238000001361 intraarterial administration Methods 0.000 description 1
- 238000000185 intracerebroventricular administration Methods 0.000 description 1
- 238000007918 intramuscular administration Methods 0.000 description 1
- 238000010255 intramuscular injection Methods 0.000 description 1
- 239000007927 intramuscular injection Substances 0.000 description 1
- 238000007912 intraperitoneal administration Methods 0.000 description 1
- 238000007913 intrathecal administration Methods 0.000 description 1
- 238000005304 joining Methods 0.000 description 1
- 229940059406 lactobacillus rhamnosus gg Drugs 0.000 description 1
- 239000008101 lactose Substances 0.000 description 1
- 230000021633 leukocyte mediated immunity Effects 0.000 description 1
- 238000012417 linear regression Methods 0.000 description 1
- 238000010801 machine learning Methods 0.000 description 1
- 239000000594 mannitol Substances 0.000 description 1
- 235000010355 mannitol Nutrition 0.000 description 1
- 230000015654 memory Effects 0.000 description 1
- 238000002493 microarray Methods 0.000 description 1
- 230000003278 mimic effect Effects 0.000 description 1
- 210000003205 muscle Anatomy 0.000 description 1
- 238000002703 mutagenesis Methods 0.000 description 1
- 231100000350 mutagenesis Toxicity 0.000 description 1
- SQMWSBKSHWARHU-SDBHATRESA-N n6-cyclopentyladenosine Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C2=NC=NC(NC3CCCC3)=C2N=C1 SQMWSBKSHWARHU-SDBHATRESA-N 0.000 description 1
- 210000005036 nerve Anatomy 0.000 description 1
- 230000007935 neutral effect Effects 0.000 description 1
- 239000003921 oil Substances 0.000 description 1
- 238000007911 parenteral administration Methods 0.000 description 1
- 230000036961 partial effect Effects 0.000 description 1
- 239000001814 pectin Substances 0.000 description 1
- 229920001277 pectin Polymers 0.000 description 1
- 235000010987 pectin Nutrition 0.000 description 1
- 230000006320 pegylation Effects 0.000 description 1
- 239000008177 pharmaceutical agent Substances 0.000 description 1
- 239000008194 pharmaceutical composition Substances 0.000 description 1
- 239000000546 pharmaceutical excipient Substances 0.000 description 1
- 210000002826 placenta Anatomy 0.000 description 1
- 108010089520 pol Gene Products Proteins 0.000 description 1
- 229920001993 poloxamer 188 Polymers 0.000 description 1
- 229920001223 polyethylene glycol Polymers 0.000 description 1
- 229920000642 polymer Polymers 0.000 description 1
- 239000003755 preservative agent Substances 0.000 description 1
- 230000002335 preservative effect Effects 0.000 description 1
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 1
- 238000012545 processing Methods 0.000 description 1
- 210000001236 prokaryotic cell Anatomy 0.000 description 1
- 230000016434 protein splicing Effects 0.000 description 1
- 238000010188 recombinant method Methods 0.000 description 1
- 230000006798 recombination Effects 0.000 description 1
- 238000005215 recombination Methods 0.000 description 1
- 238000007634 remodeling Methods 0.000 description 1
- 230000008439 repair process Effects 0.000 description 1
- 238000011160 research Methods 0.000 description 1
- 108091008146 restriction endonucleases Proteins 0.000 description 1
- 239000002336 ribonucleotide Substances 0.000 description 1
- 125000002652 ribonucleotide group Chemical group 0.000 description 1
- 108091092562 ribozyme Proteins 0.000 description 1
- 238000005070 sampling Methods 0.000 description 1
- 238000007423 screening assay Methods 0.000 description 1
- 230000019491 signal transduction Effects 0.000 description 1
- 230000005783 single-strand break Effects 0.000 description 1
- 238000004513 sizing Methods 0.000 description 1
- 229940126586 small molecule drug Drugs 0.000 description 1
- 150000003431 steroids Chemical class 0.000 description 1
- 238000011285 therapeutic regimen Methods 0.000 description 1
- 210000001541 thymus gland Anatomy 0.000 description 1
- 230000000699 topical effect Effects 0.000 description 1
- 239000011573 trace mineral Substances 0.000 description 1
- 235000013619 trace mineral Nutrition 0.000 description 1
- 238000003151 transfection method Methods 0.000 description 1
- 230000009261 transgenic effect Effects 0.000 description 1
- 230000001052 transient effect Effects 0.000 description 1
- 230000007704 transition Effects 0.000 description 1
- 238000011269 treatment regimen Methods 0.000 description 1
- 210000003954 umbilical cord Anatomy 0.000 description 1
- 241001430294 unidentified retrovirus Species 0.000 description 1
- 210000003932 urinary bladder Anatomy 0.000 description 1
- 235000013311 vegetables Nutrition 0.000 description 1
- 239000011701 zinc Substances 0.000 description 1
- 229910052725 zinc Inorganic materials 0.000 description 1
- NWONKYPBYAMBJT-UHFFFAOYSA-L zinc sulfate Chemical compound [Zn+2].[O-]S([O-])(=O)=O NWONKYPBYAMBJT-UHFFFAOYSA-L 0.000 description 1
Images

























Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/16—Hydrolases (3) acting on ester bonds (3.1)
- C12N9/22—Ribonucleases RNAses, DNAses
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/08—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from viruses
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K48/00—Medicinal preparations containing genetic material which is inserted into cells of the living body to treat genetic diseases; Gene therapy
- A61K48/005—Medicinal preparations containing genetic material which is inserted into cells of the living body to treat genetic diseases; Gene therapy characterised by an aspect of the 'active' part of the composition delivered, i.e. the nucleic acid delivered
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K49/00—Preparations for testing in vivo
- A61K49/0004—Screening or testing of compounds for diagnosis of disorders, assessment of conditions, e.g. renal clearance, gastric emptying, testing for diabetes, allergy, rheuma, pancreas functions
- A61K49/0008—Screening agents using (non-human) animal models or transgenic animal models or chimeric hosts, e.g. Alzheimer disease animal model, transgenic model for heart failure
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P35/00—Antineoplastic agents
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/12—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from bacteria
- C07K16/1267—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from bacteria from Gram-positive bacteria
- C07K16/1275—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from bacteria from Gram-positive bacteria from Streptococcus (G)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/111—General methods applicable to biologically active non-coding nucleic acids
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/85—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells
- C12N15/86—Viral vectors
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/10—Type of nucleic acid
- C12N2310/20—Type of nucleic acid involving clustered regularly interspaced short palindromic repeats [CRISPRs]
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2750/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssDNA viruses
- C12N2750/00011—Details
- C12N2750/14011—Parvoviridae
- C12N2750/14111—Dependovirus, e.g. adenoassociated viruses
- C12N2750/14141—Use of virus, viral particle or viral elements as a vector
- C12N2750/14143—Use of virus, viral particle or viral elements as a vector viral genome or elements thereof as genetic vector
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Engineering & Computer Science (AREA)
- Organic Chemistry (AREA)
- Genetics & Genomics (AREA)
- Molecular Biology (AREA)
- General Health & Medical Sciences (AREA)
- Biomedical Technology (AREA)
- Medicinal Chemistry (AREA)
- Biochemistry (AREA)
- Zoology (AREA)
- Biotechnology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Wood Science & Technology (AREA)
- Biophysics (AREA)
- Immunology (AREA)
- General Engineering & Computer Science (AREA)
- Microbiology (AREA)
- Animal Behavior & Ethology (AREA)
- Public Health (AREA)
- Veterinary Medicine (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Physics & Mathematics (AREA)
- Urology & Nephrology (AREA)
- Virology (AREA)
- Pharmacology & Pharmacy (AREA)
- Plant Pathology (AREA)
- Epidemiology (AREA)
- Hematology (AREA)
- Pathology (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Endocrinology (AREA)
- Diabetes (AREA)
- Rheumatology (AREA)
- Toxicology (AREA)
- Food Science & Technology (AREA)
- Cell Biology (AREA)
Abstract
本发明描述了一种在被施用需要Cas9的方案的受试者中避免免疫应答的方法,以优化和扩展包括施用免疫正交的Cas9的基于CRISPR的疗法的应用。本文还描述了通过将高免疫原性的肽或氨基酸与低免疫原性的等同物进行互换来修饰Cas9蛋白的方法。这些方法对于将Cas9库用于重复治疗特别有用。还提供了经修饰的的Cas9蛋白以降低免疫原性。
Description
相关申请的交叉引用
本申请根据35U.S.C第119(e)条要求于2017年3月14日提交的美国第62/471,267号和2018年1月8日提交的美国第62/614,875号的优先权,其全部内容在此通过引用并入本文。
背景技术
针对用于基因组工程目的的体内CRISPR/Cas9的免疫应答仍不明确。Cas9是一种外源蛋白,具有原核生物起源,并可能引起强烈的免疫应答,最终可能导致通过细胞毒性T细胞介导的免疫应答消除基因编辑的细胞或Cas9蛋白。
由于需要反复治疗,可能引起Cas9特异性细胞毒性细胞应答的原因有两个:1)当前体内CRISPRCas9介导的基因组编辑的整体功效较低,可能需要重复治疗;以及2)如果dCas9的基因组调控是一种参考的基因治疗方法,重复治疗对于持续的抑制/激活是必要的。另外,在诸如AAV介导的递送的某些递送系统下,Cas9可以具有长期表达,从而进一步增加Cas9特异性细胞毒性的细胞应答的可能性,进而妨碍了长期治疗效果。因此需要施用Cas9的新方法,以降低免疫原性,从而逃避免疫检测。本发明满足了这种需求并且还提供了相关的优势。
发明内容
避免对Cas9的免疫应答问题的新方法包括对每种处理使用直系同源的Cas9蛋白和/或工程化不会引起免疫应答的Cas9。本发明提供了在被施用需要Cas9的方案的受试者中避免免疫应答的方法,以优化和扩展基于包括施用免疫正交的Cas9的基于CRISPR的疗法的应用。本发明还提供了通过将高免疫原性的肽或氨基酸与低免疫原性的对应物进行交换来修饰Cas9蛋白的方法。这些方法对于将Cas9库(arsenal)用于重复治疗的应用特别有用。还提供了经修饰的以降低免疫原性的Cas9蛋白。
本发明的方面涉及一种产生蛋白质的方法,包括:鉴定对主要组织相容性复合体(MHC)具有亲和力的蛋白质的一个或多个区域,以及通过一个或多个氨基酸替代修饰对MHC具有亲和力的蛋白质的一个或多个区域,从而使得经修饰的区域对MHC没有亲和力,其中当所得的经修饰的蛋白质或编码该经修饰的蛋白质的多核苷酸施用于受试者时,该经修饰的蛋白质是免疫沉默的。在一些实施方案中,对MHC的亲和力是高亲和力。在一些实施方案中,至少一个经替代的氨基酸是不用作MHC蛋白质核心残基的氨基酸。在一些实施方案中,该蛋白质选自以下组:胞苷脱氨酶、腺苷脱氨酶、锌指核酸酶、转录激活因子样效应物核酸酶、Cas9、或AAV衣壳蛋白。在一些实施方案中,蛋白质是Cas9,任选地是SpCas9。
进一步的方面涉及根据以上公开的方法产生的经修饰的Cas9蛋白。更进一步的方面涉及一种经修饰的Cas9蛋白,包含一种或多种、两种或更多种、三种或更多种、四种或更多种、五种或更多种、六种或更多种、七种或更多种、八种或更多种、九种或更多种、十种或更多种、十五种或更多种、二十种或更多种表1中提供的氨基酸修饰。一些实施方案涉及编码经修饰的Cas9的分离的多核苷酸。进一步的实施方案涉及包含分离的多核苷酸的载体,任选地为AAV载体,以及还任选地为AAV5载体。另外的实施方案涉及包含所述载体的AAV衣壳。在一些实施方案中,一种或多种AAV衣壳蛋白已被修饰为免疫沉默的。
本发明的一些方面涉及一种鉴定免疫正交直系同源物的方法,该方法包括:确定蛋白质或其区域对多种主要组织相容性复合体(MHC)的亲和力集,比较所述蛋白质或其区域的亲和力集与所述蛋白质的直系同源物对所述多种MHC的亲和力集,以及基于非重叠的亲和力集确定免疫正交的直系同源物集。在一些实施方案中,对MHC的亲和力是高亲和力。在一些实施方案中,该蛋白质选自以下组:胞苷脱氨酶、腺苷脱氨酶、锌指核酸酶、转录激活因子样效应物核酸酶、Cas9或AAV衣壳蛋白。在一些实施方案中,蛋白质是Cas9,任选地是SpCas9或SaCas9。在一些实施方案中,直系同源的Cas9蛋白选自酿脓链球菌(S.pyogenes)Cas9(spCas9)、金黄色葡萄球菌(S.aureus)Cas9(saCas9)、长双歧杆菌(B.longum)Cas9、嗜黏蛋白艾克曼菌(A.muiciniphilia)Cas9、或O.laneus Cas9。
一些方面涉及一种在被施用需要蛋白的方案的受试者中避免免疫应答的方法,该方法包括:向受试者顺序施用免疫正交的两种或更多种蛋白质。在一些实施方案中,免疫正交的蛋白质不共享大于5个连续氨基酸的氨基酸序列。在一些实施方案中,免疫正交的蛋白质不共享对主要组织相容性复合体(MHC)的亲和力。在一些实施方案中,顺序施用三种或更多种、四种或更多种、五种或更多种、六种或更多种、七个或更多种、八种或更多种、九种或更多种、或十种或更多种免疫正交的蛋白质。
非限制性示例性方面涉及一种在被施用需要Cas9的方案的受试者中避免免疫应答的方法、和/或对受试者进行基因编辑或基因调控的方法、和/或治疗需要基因编辑或基因调控的受试者的方法,该方法包括:对受试者顺序施用两种或更多种免疫正交的Cas9蛋白。在一些实施方案中,免疫正交的Cas9蛋白不共享大于5个连续氨基酸的氨基酸序列。在一些实施方案中,免疫正交的Cas9蛋白不共享对主要组织相容性复合体(MHC)的亲和力。在一些实施方案中,顺序施用三种或更多种、四种或更多种、五种或更多种、六种或更多种、七种或更多种、八种或更多种、九种或更多种、或十种或更多种免疫正交的Cas9蛋白。在一些实施方案中,每种免疫正交的Cas9蛋白是衍生自不同种的细菌的Cas9。在一些实施方案中,免疫正交的Cas9蛋白选自酿脓链球菌(S.pyogenes)Cas9(spCas9)、金黄色葡萄球菌(S.aureus)Cas9(saCas9)、长双歧杆菌(B.longum)Cas9、嗜黏蛋白艾克曼菌(A.muiciniphilia)Cas9、或O.laneus Cas9。在一些实施方案中,免疫正交的Cas9蛋白包含spCas9和saCas9。在一些实施方案中,两种或更多种Cas9蛋白中的至少一种被修饰,以在施用于受试者时降低免疫原性。在一些实施方案中,根据以上公开的方法对两种或更多种Cas9蛋白中的至少一种进行修饰。在一些实施方案中,两种或更多种Cas9蛋白或编码所述Cas9蛋白的多核苷酸中的至少一种包含在AAV载体中。在一些实施方案中,AAV载体是AAV5载体。在一些实施方案中,AVV载体包含在AVV衣壳中。在一些实施方案中,两种或更多种Cas9蛋白或编码所述Cas9蛋白的多核苷酸包含在AAV载体中。在一些实施方案中,每种AAV载体包含在AAV衣壳中,任选地其中AAV衣壳彼此免疫正交。在一些实施方案中,该方法进一步包括对受试者施用一种或多种指导RNA。在一些实施方案中,选择指导RNA来治疗选自以下组的疾病、病症或病况:全色盲(achromatopsia)、腺苷脱氨酶缺乏症(adenosinedeaminase(ADA)deficiency)、α1-抗胰蛋白酶缺乏症(alpha-1-antitrypsindeficiency)、阿尔茨海默病(Alzheimer’s disease)、肌萎缩侧索硬化(amyotrophiclateral sclerosis)、芳香族氨基酸脱羧酶缺乏症(aromatic amino acid decarboxylasedeficiency)、巴顿病(Batten disease)、无脉络膜(choroideremia)、克里格勒-纳贾尔综合征(Crigler Najjar syndrome)、囊性纤维化(cystic fibrosis)、脆性X综合征(fragileX syndrome)、血友病(hemophilia)、乙型肝炎(hepatitis B)、丙型肝炎(hepatitis C)、纯合子家族性高胆固醇血症(homozygous familial hypercholesteremia)、亨廷顿病(Huntington’s Disease)、Leber先天性黑朦(Leber congenital amaurosis)、黄斑变性(macular degeneration)、枫糖尿病(maple syrup urine disease(MSUD))、粘多糖病(mucopolysarccharidosis)(I-IX)、多发性硬化(multiple sclerosis)、肌肉不良(muscular dystrophy)、强直性肌营养不良(myotonic dystrophy)、1型神经纤维瘤病(neurofibramotosis type 1)、鸟氨酸转氨甲酰酶缺乏症(ornithine transcarbamylasedeficiency)、先天性甲肥厚(pachyonychia congenita)、帕金森氏病(Parkinson’sdisease)、苯丙酮尿症(phenylketonuria)、多囊肾病(polycystickidney disease)、庞贝病(Pompe disease)、视网膜变性(retinal degeneration)、瑞特氏综合症(Rett’ssyndrome)、佝偻病(rickets)、脊髓性肌萎缩(spinal muscular atrophy)、重症联合免疫缺陷(severe combined immunodeficiency)、镰状细胞病(sickle cell disease)、史-莱-奥综合征(Smith-Lemli-Opitz syndrome)、Y连锁非梗阻性生精障碍(Y-linkednonobstructive spermatogenic failure)、地中海贫血(thalassemia)、泰-萨克斯病(Tay-Sachs disease)、威尔逊氏病(Wilson’s disease)、心血管疾病(cardiovasculardisease)、代谢综合征(metabolic syndrome)、疼痛管理(pain management)和X连锁视网膜劈裂症(X-linked retinoschisis)。
附图说明
图1是描述实施例1中描述的过程的流程图。
图2示出(A)免疫正交的蛋白质集,利用递归寻群(clique-finding)算法进行定位(粗体轮廓表示前4个正交的蛋白质集。颜色表示蛋白质对之间5-mer重叠的数量。该方法可以保证找到所有正交的蛋白质的最大集。酿脓链球菌属于5个相互正交的蛋白质集。)(B)包含每个蛋白质的最大群数,按大小细分(大小为4的群最常见)。
图3示出(A)由每个肽位置交换(swaps)引起的亲和力变化(数据示出的是酿脓链球菌中发现的98种高亲和力肽的平均值。)(B)交换后,按交换位置,每种亲和力类别中的肽的分布(交换出在高亲和力肽的开始处具有最大的效果的氨基酸。)(C)累积总数,示出具有至少一个无亲和力的交换选项(蓝色)、或至少一个无亲和力或低亲和力的交换选项(绿色)的肽的数量(在该蛋白质中有98种高亲和力肽(黑色虚线)。)。
图4示出由没有高亲和力肽重叠的Cas9的菌株构成的群,因此提供没有高亲和力肽重叠的五种Cas9蛋白的五个集。
图5A-图5H示出基于蛋白质的疗法引起适应性免疫应答:实验和经由计算机模拟(in silico)的分析:(图5A)蛋白质具有巨大的治疗潜力,但主要缺点是对治疗性蛋白质及其递送工具两者的免疫应答。(图5B)作为案例研究,我们探索了CRISPR-Cas9系统和基于AAV的相应递送工具。(图5C)经眼眶后向小鼠注射1012vg/小鼠的靶向PCSK9基因的AAV8-SaCas9或非靶向对照(空载体)。在接受AAV-SaCas9-PCSK9病毒的小鼠中可以看到PCSK9血清水平的降低,这是由于成功的基因打靶(每组中,n=6只小鼠)。(图5D)在用于SaCas9蛋白质的ELISA中检测到对有效载荷的免疫应答(n=12)。(图5E)在用于AAV8病毒衣壳的ELISA中检测到对递送工具的免疫应答(n=12只小鼠)。(图5F)用于找到免疫正交的蛋白同源物群的经由计算机模拟的工作流程。(图5G)通过检查一种蛋白质中的所有k-mer是否以零或一个错配存在于另一个蛋白质序列中来进行非免疫学获知(immunologicallyuninformed)的序列比较。x轴对应于k,而MHC I和MHC II示出仅仅预测为与MHC I类和MHCII类分子结合的肽的重叠。48%的Cas9对示出无6-mer重叠,而83%的Cas9对示出无重叠的MHC结合的肽。(图5H)与(g)相同,但适用于AAV VP1衣壳蛋白。所有AAV对都包含重叠的MHC结合的肽。
图6A-图6E示出Cas9和AAV免疫原性预测的实验验证。(图6A)以1012vg/小鼠经眼眶后注射使小鼠暴露于抗原。在注射前第0天以及在4-6周的过程中的多个点处收集血清。(图6B)注射SpCas9(n=6)和SaCas9(n=12)的小鼠中产生抗-SpCas9抗体,以及注射SpCas9(n=6)和SaCas9(n=12)的小鼠中产生抗-SaCas9抗体。(图6C)注射AAV8 SpCas9(n=12;左图)或AAVDJ SpCas9(n=12;右图)的小鼠产生抗-SpCas9和抗-SaCas9抗体。(图6D)针对注射AAV8或AAVDJ的小鼠产生的抗AAV8/DJ/2/5抗体(对于所有图,n=4)。(图6E)针对注射AAV2或AAV5的小鼠产生的抗AAV8/DJ/2/5抗体(对于所有图,n=5)。
图7描述了Cas9免疫正交群。描述了对应于6-mer重叠的群。突出显示了正交群的实例,包括酿脓链球菌(S.pyogenes)、金黄色葡萄球菌(S.aureus)、长双歧杆菌(B.longum)、嗜黏蛋白艾克曼菌(A.muiciniphilia)或O.laneus的Cas9。
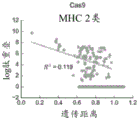
图8A-图8D示出Cas9和AAV直系同源物的免疫原性经由计算机模拟分析的结果以及比较。线性回归排除不重叠的对。(图8A)Cas9 MHC I类肽重叠与系统发育距离。(图8B)AAV MHC I类肽重叠与系统发育距离。(图8C)Cas9 MHC II类肽重叠与系统发育距离。(图8D)AAV MHC II类肽重叠与系统发育距离。
图9A-图9B显示主要的AAV血清型的组。(图9A)超过81个HLA等位基因的AAV免疫正交群。与其他血清型相比,AAV5的免疫扩散最大。不存在正交群。(图9B)AAV系统发育示出主要的血清型分组以及重构序列Anc80L65的位置。
图10示出通过ELISPOT测定法对MHC II肽的预测的实验验证。
图11示出极端微生物Cas9和来自共生的、致病的和环境的种的Cas9库的肽重叠的免疫正交群。
具体实施方式
在下文中将更全面地描述根据本发明的实施方案。然而,本发明的各个方面可以以不同的形式体现,并且不应被解释为仅限于本文所述的实施方案。相反,提供这些实施方案是为了使得本发明更加透彻和完整,并且将本发明的范围充分传达给本领域技术人员。在本文的描述中使用的术语仅出于描述特定实施方案的目的,而不旨在限制。
除非另有定义,否则本文中使用的所有术语(包括技术和科学术语)具有与本发明所属领域的普通技术人员通常所理解的相同含义。还将理解的是,诸如在常用词典中定义的那些的术语应被解释为具有与其在本申请的上下文和相关技术中的含义一致的含义,并且不应以理想化或过于正式的意义进行解释,除非本文明确定义。尽管下面没有明确定义,但这些术语应根据其通常的含义进行解释。
在本文的描述中使用的术语仅出于描述特定实施方案的目的,而不旨在限制本发明。本文提及的所有出版物、专利申请、专利和其他参考文献通过引用的方式整体并入本文。
除非另有说明,否则本技术的实践将采用本领域技术范围内的组织培养技术、免疫学、分子生物学、微生物学、细胞生物学和重组DNA的常规技术。
除非上下文另外指出,否则特别旨在可以以任何组合使用本文描述的本发明的各种特征。此外,本发明还预期在一些实施方案中,可以排除或省略本文所述的任何特征或特征的组合。为了说明,如果说明书指出复合物包含组分A、B和C,则特别旨在可以单独地或者组合地省略和具体放弃A、B或C中的任何一种或其组合。
除非另外明确指出,否则所有指定的实施方案、特征和术语旨在包括所列举的实施方案、特征或术语及其生物学等同物两种。
所有数字名称(例如pH、温度、时间、浓度和分子量,包括范围)均为近似值,可以根据需要以1.0或0.1的增量变化(+)或(-),或可选地以+/-15%、或可选地+/-10%,或可选地+/-5%,或可选地+/-2%的变量进行变化。应理解,尽管并不总是明确指出,但是所有数字名称之前均带有术语“约”。还应理解,尽管并不总是明确指出,本文所述的试剂仅是示例性的,并且其等同物是本领域已知的。
定义
如在本发明的说明书和所附权利要求书中所使用的,不定冠词“a”、“an”和定冠词“the”也旨在包括复数形式,除非上下文另外明确指出。
如本文所用,术语“约”当指可测量值如量或浓度等时,意在涵盖指定量的20%、10%、5%、1%、0.5%或甚至0.1%的变量。
当术语或“可接受的”、“有效的”或“足够的”用于描述本文公开的任何组分、范围、剂型等的选择时,意指所述组分、范围、剂型等适用于所公开的目的。
本文所用的术语“腺相关病毒”或“AAV”是指与此名称相关并属于细小病毒科,dependoparvovirus属的病毒类别的成员。已知该病毒的多种血清型适用于基因递送。所有已知的血清型均可以感染来自各种组织类型的细胞。现有技术公开了至少11或12(按顺序编号)种血清型。可用于本文公开的方法中的非限制性示例性血清型包括11或12种血清型(例如AAV2、AAV5和AAV8)或变体血清型(例如AAV-DJ)中的任一种。AAV结构颗粒由60个由VP1、VP2和VP3构成的蛋白质分子组成。每种颗粒包含以二十面体结构排列的约5个VP1蛋白、5个VP2蛋白和50个VP3蛋白。下面提供了可用于本文公开的方法中的非限制性示例性VP1序列。
AAT46339.1 AAV-11

pdb|4IOV|A AAV-rh32

ABI16639.1 AAV-12

NP_044927.1 AAV-4

YP_077178.1 AAV-7

YP_077180.1 AAV-8

AAT46337.1 AAV-10

AAS99264.1 AAV-9

NP_049542.1 AAV-1

AAB95450.1 AAV-6

NP_043941.1 AAV-3

ABZ10812.1 AAV-13

YP_680426.1 AAV-2

YP_068409.1 AAV-5

3J1Q_A AAV-DJ

AKU89595.1 Anc80

同样如本文中所使用的,“和/或”是指并且涵盖一个或多个相关联的所列项目的任何和所有可能的组合,以及当以可选方式(“或”)解释时缺乏组合。
如本文所用,术语“适配体”是指可以以高亲和力和特异性结合一个或多个选定目标的单链DNA或RNA分子。非限制性的示例性目标包括但不限于蛋白质或肽。
术语“Cas9”是指CRISPR相关的RNA指导的核酸内切酶(诸如,酿脓链球菌Cas9(spCas9))及其直系同源物及其生物等同物。Cas9的生物等同物包括但不限于来自嗜酸耐热菌(Alicyclobacillus acidoterrestris)的C2c1和来自包括氨基酸球菌属(Acidaminococcus spp.)和新凶手弗朗西丝菌(Francisella novicida)U112的各种细菌的Cpf1(执行与Cas9类似的切割功能)。Cas9可以指引起DNA中的双链断裂的核酸内切酶,导致DNA中的单链断裂的切口酶变体(诸如RuvC或HNH突变体),以及缺乏核酸内切酶活性的其他变体(诸如deadCas-9或dCas9)。Cas9也可以指“分裂Cas9”,其中CAs9分为两半:C-Cas9和N-Cas9,并与两个内含肽部分融合。参见,例如,美国专利第9,074,199 B1号;Zetsche等人,(2015)Nat Biotechnol.33(2):139-42;Wright等人,(2015)PNAS 112(10)2984-89。包含质粒的SpCas9的商用来源的非限制性实例可以在以下AddGene参考编号下找到:
42230:PX330;SpCas9和单指导RNA
48138:PX458;SpCas9-2A-EGFP和单指导RNA
62988:PX459;SpCas9-2A-Puro和单指导RNA
48873:PX460;SpCas9n(D10A切口酶)和单指导RNA
48140:PX461;SpCas9n-2A-EGFP(D10A切口酶)和单指导RNA
62987:PX462;SpCas9n-2A-Puro(D10A切口酶)和单指导RNA
48137:PX165;SpCas9
下表提供了Cas9的更多实例:













下面提供了本文公开的实施例中使用的那些Cas9序列。
YP_898402.1膜蛋白[土拉弗朗西丝菌(Francisella tularensis subsp.)novicida亚种U112]


ZP_05061364.1 CRISPR相关大蛋白质(暂时性的),推定[γ变形杆菌(gammaproteobacterium)HTCC5015]


ZP_08324662.1 CRISPR相关蛋白,CSX12家族[Parasutterellaexcrementihominis YIT 11859]

YP_122507.1假定蛋白lpp0160[嗜肺军团杆菌巴黎株(Legionella pneumophilastr.Paris)]

NP_907747.1假定蛋白WS1613[产琥珀酸沃林氏菌(Wolinella succinogenes)DSM1740]


ADX75954.1 CRISPR相关蛋白,Csn1家族[伪中间葡萄球菌(Staphylococcuspseudintermedius)ED99]


ZP_10206685.1 CRISPR相关蛋白,Csn1家族[南极动性球菌(Planococcusantarcticus)DSM 14505]


ZP_16930555.1 csn1家族的CRISPR-相关蛋白[血链球菌(Streptococcussanguinis)SK49]


AAK33936.1保守的假定蛋白[酿脓链球菌(Streptococcus pyogenes)M1 GAS]

YP_820832.1 CRISPR系统样蛋白[嗜热链球菌(Streptococcus thermophilus)LMD-9]

NP_721764.1假定蛋白SMU_1405c[变形链球菌(Streptococcus mutans)UA159]


YP_004373648.1 CRISPR相关蛋白,Csn1家族[球团科里氏杆菌(Coromerbacterium glomerans)PW2]


ZP_08576281.1可能的CRISPR相关蛋白[香肠乳杆菌(Lactobacillusfarciminis)KCTC 3681]


ZP_03683851.1假定蛋白CATMIT_02512,部分[Catenibacterium mitsuokai DSM15897]


YP_003171950.1 CRISPR相关蛋白Csn1[鼠李糖乳杆菌(Lactobacillusrhamnosus)GG]

YP_003937986.1 CRISPR相关蛋白[双歧杆菌(Bifidobacterium bifidum)S17]

EHN59352.1 CRISPR相关蛋白[北原酒球菌(Oenococcus kitaharae)DSM 17330]

ZP_08660870.1可能的CRISPR相关蛋白[果糖杆菌(Fructobacillus fructosus)KCTC 3544]

YP_001691366.1假定蛋白FMG_0058[大芬戈尔德菌(Finegoldia magna)ATCC29328]

ZP_07316256.1 CRISPR相关蛋白,Csn1家族[非典型韦荣氏球菌(Veillonellaatypica)ACS-134-V-Col7a]

ZP_08029929.1 CRISPR相关蛋白,Csn1家族[莫雷梭菌(Solobacterium moorei)F0204]

ZP_03989815.1 crispr相关蛋白[氨基酸球菌种(Acidaminococcussp.)D21]


ZP_07455288.1 csn1家族CRISPR相关蛋白[尤氏真杆菌玛格丽特亚种(Eubacterium yurii subsp.margaretiae)ATCC 43715]


CBK78998.1 CRISPR相关核酸内切酶,Csn1家族[灵巧粪球菌(Coprococcuscatus)GD/7]


ZP_00143587.1假定蛋白[具核梭杆菌文森特里亚种(Fusobacterium nucleatumsubsp.vincentii)ATCC 49256]


YP_005054169.1 CRISPR相关蛋白,Csn1家族[龈沟产线菌(Filifactor alocis)ATCC 35896]

ZP_07398877.1 csn1家族CRISPR相关蛋白[Peptoniphilus duerdenii ATCCBAA-1640]

NP_970941.1 CRISPR相关Cas5e[牙密螺旋体(Treponema denticola)ATCC35405]


ZP_07912707.1保守的假定蛋白[路邓葡萄球菌(Staphylococcus lugdunensis)M23590]


ZP_02077990.1假定蛋白EUBDOL_01797[细长真杆菌(Eubacterium dolichum)DSM3991]

YP_820161.1 CRISPR系统样蛋白[嗜热链球菌(Streptococcus thermophilus)LMD-9]

EFT93846.1 CRISPR相关蛋白,Csn1家族[粪肠球菌(Enterococcus faecalis)TX0012]


YP_002937591.1 CRISPR系统相关蛋白[直肠真杆菌(Eubacterium rectale)ATCC33656]


YP_015730.1假定蛋白MMOB0330[运动支原体(Mycoplasma mobile)163K]

ZP_09312133.1假定蛋白MoviS_00710[绵羊肺炎支原体(Mycoplasmaovipneumoniae)SC01]


ADC31648.1 Csn1家族CRISPR相关蛋白[鸡毒支原体F株(Mycoplasmagallisepticum str.F)]


YP_278700.1假定蛋白MS53_0582[滑液囊支原体(Mycoplasma synoviae)53]


EIE39736.1 Csn1家族CRISPR相关蛋白[犬支原体(Mycoplasma canis)PG 14]

NP_907605.1假定蛋白WS1445[产琥珀酸沃林氏菌(Wolinella succinogenes)DSM1740]


YP_002344900.1 CRISPR相关蛋白[空肠弯曲菌空肠亚种(Campylobacter jejunisubsp.Jejuni)NCTC 11168=ATCC 700819]

YP_003516037.1 CRISPR相关蛋白[雪貂螺杆菌(Helicobacter mustelae)12198]

ZP_06887976.1 CRISPR相关蛋白,Csn1家族[甲基弯菌(Methylosinustrichosporium)OB3b]


YP_003968716.1 CRISPR相关蛋白,Csn1家族(plasmid)[Ilyobacter polytropusDSM 2926]

ZP_09352959.1 CRISPR相关蛋白cas9/csn1,II/nmemi亚型[史氏芽胞杆菌(Bacillus smithii)7_3_47FAA]

YP_002507391.1 CRISPR相关蛋白,Csn1家族[解纤维梭菌(Clostridiumcellulolyticum)H10]


YP_002551549.1 crispr相关蛋白,csn1家族[Acidovorax ebreus TPSY]

YP_002342100.1假定蛋白NMA0631[脑膜炎奈瑟氏菌(Neisseria meningitidis)Z2491]

NP_246064.1假定蛋白PM1127[多杀性巴氏杆菌多杀性亚种Pm70株(Pasteurellamultocida subsp.multocida str.Pm70)]


ZP_07738815.1 CRISPR相关蛋白,Csn1家族[氨基单胞菌(Aminomonaspaucivorans)DSM 12260]

ZP_08574780.1 CRISPR相关蛋白,Csn1家族[棒状乳杆菌极曲亚种(Lactobacillus coryniformis subsp.torquens)KCTC 3535]

ZP_03755025.1假想蛋白ROSEINA2194_03455[食葡糖罗斯拜瑞氏菌(Roseburiainulinivorans)DSM 16841]


ZP_10953934.1 HNH核酸内切酶[Alicyclobacillus hesperidum URH17-3-68]

ADI19058.1细菌中保守的未表征蛋白[不可培养的β变形杆菌HF0070_07E19]

ZP_08157403.1 CRISPR相关蛋白,Csn1家族[白色瘤胃球菌(Ruminococcusalbus)8]


ZP_10010146.1 CRISPR相关蛋白Cas9/Csn1,II/NMEMI亚型[密螺旋体属(Treponema sp.)JC4]


ZP_11150502.1 CRISPR相关蛋白,Csn1家族[太平洋食烷菌(Alcanivoraxpacificus)W11-5]

ZP_18919511.1假定蛋白C882_0672[Caenispirillum salinarum AK4]


YP_425545.1 CRISPR相关核酸内切酶Csn1家族蛋白[深红红螺菌(Rhodospirillum rubrum)ATCC 11170]


CCA84553.1保守的假定蛋白[青枯雷尔氏菌(Ralstonia syzygii)R24]

ZP_10898214.1 CRISPR相关蛋白,Csn1家族[小红卵菌属(Rhodovulum sp.)PH10]

YP_004386148.1 CRISPR相关蛋白,Csn1家族[脱氮嗜脂环物菌(Alicycliphilusdenitrificans)K601]


YP_003552871.1 CRISPR相关蛋白,Csn1家族[Candidatus Puniceispirillummarinum IMCC1322]

YP_003448082.1 CRISPR相关蛋白,Csn1家族[偶氮螺菌(Azospirillum sp.)B510]

YP_571550.1假定蛋白Nham_4054(质粒)[汉民硝化细菌(Nitrobacterhamburgensis)X14]


YP_001239928.1假定蛋白BBta_3952[缓生根瘤菌属(Bradyrhizobiumsp.)BTAi1]


YP 001531750.1 CRISPR相关蛋白[Dinoroseobacter shibae DFL 12=DSM16493]

YP_001411379.1 CRISPR相关核酸内切酶Csn1家族蛋白[Parvibaculumlavamentivorans DS-1]


ZP_17295095.1 CRISPR相关蛋白cas9/csn1,II/nmemi亚型[动物溃疡伯格菌(Bergeyella zoohelcum)ATCC 43767]


ZP_07217791.1保守的假定蛋白[拟杆菌属(Bacteroides sp.)20_3]


YP_005848005.1假定蛋白IALB_3034[Ignavibacterium aibum JCM 16511]


YP_213533.1保守的假定蛋白[脆弱类杆菌(Bacteroides fragilis)NCTC 9343]

ZP_10895610.1 CRISPR相关蛋白Cas9/Csn1,II/NMEMI亚型[卟啉单胞菌属(Porphyromonas sp.)口服分类(oral taxon)279str.F0450]

ZP_11022414.1 CRISPR相关蛋白cas9/csn1,II/nmemi亚型[肠道巴恩斯氏菌(Barnesiella intestinihominis)YIT 11860]


ZP_09642280.1 CRISPR相关蛋白cas9/csn1,II/nmemi亚型[Odoribacter laneusYIT 12061]


YP_004843922.1推定的CRISPR相关(Cas)蛋白[嗜鳃黄杆菌(Flavobacteriumbranchiophilum)FL-15]


ZP_08837074.1假定蛋白HMPREF0666_03250[普雷沃菌属(Prevotella sp.)C561]


ZP_06288774.1 CRISPR相关蛋白,Csn1家族[Prevotella timonensis CRIS 5C-Bl]

YP_001875142.1 CRISPR相关核酸内切酶Csn1家族蛋白[Elusimicrobiumminutum Pei191]


YP_004248194.1 CRISPR相关蛋白,Csn1家族[Sphaerochaeta globosa Buddy株]


YP_873709.1 HNH核酸内切酶[嗜酸纤维素分解菌(Acidothermuscellulolyticus)11B]

ZP_07880770.1保守的假定蛋白[放线菌属(Actinomyces sp.)口服分类(oraltaxon)180 str.F0310]

ZP_03925169.1保守的假定蛋白[Actinomyces coleocanis DSM 15436]


YP_001955845.1限制性核酸内切酶[长双歧杆菌(Bifidobacterium longum)DJO10A]


YP_001878601.1假定蛋白Amuc_2010[嗜黏蛋白艾克曼菌(Akkermansiamuciniphila)ATCC BAA-835]

YP_004168469.1 CRISPR相关蛋白,csn1家族[Nitratifractor salsuginis DSM16511]


ZP_08015909.1假定蛋白HMPREF9464_01128[沃兹沃思萨特氏菌(Sutterellawadsworthensis)3_1_45B]


J7RUA5.1 CRISPR相关的核酸内切酶Cas9[金黄色葡萄球菌(Staphylococcusaureus)]

AEX66236.1 CRISPR相关的核酸内切酶[白喉杆菌(Corynebacteriumdiphtheriae)C7(β)]

WP_013852048.1 II型CRISPR RNA指导的核酸内切酶Cas9[巴氏链球菌(Streptococcus pasteurianus)]


EEZ71796.1 CRISPR相关蛋白,Csn1家族[灰色奈瑟球菌(Neisseria cinerea)ATCC 14685]


BAK69486.1推定的CRISPR相关蛋白[红嘴鸥弯曲杆菌(Campylobacter lari)]

OJI07263.1假定蛋白BK997_03320[Candidatus Micrarchaeum acidiphilumARMAN-1]


APG80630.1 CRISPR相关核酸内切酶Cas9[Candidatus Parvarchaeumacidiphilum ARMAN-4]

tr|I0AP30|I0AP30_IGNAJ CRISPR相关核酸内切酶Cas9 OS=Ignavibacteriumalbum(菌株DSM 19864/JCM 16511/NBRC 101810/Mat9-16)OX=945713 GN=cas9 PE=3SV=1

Ga0054994_10813嗜热脂肪芽孢杆菌(Geobacillus stearothermophilus)Cas9

WP_036475267.1 II型CRISPR RNA指导的核酸内切酶Cas9[乳糖奈瑟菌(Neisseria lactamica)]


本文所用的术语“细胞”可以指原核细胞或真核细胞,其任选地获自受试者或可商购的来源。
本文所用的术语“CRISPR”是指成簇的规律间隔的短回文重复(CRISPR)。CRISPR还可以指依赖于CRISPR途径的序列特异性基因操纵的技术或系统。可以使用CRISPR核酸内切酶和指导RNA对CRISPR重组表达系统进行编程以切割目标多核苷酸。CRISPR系统可以用于引起目标多核苷酸中的双链或单链断裂。CRISPR系统也可以用于募集蛋白质或标记目标多核苷酸。在一些方面,CRISPR介导的基因编辑利用非同源末端连接(NHEJ)或同源重组的途径来实施编辑。CRISPR技术的这些应用是本领域已知的并且广泛实践的。参见,例如,美国专利第8,697,359号和Hsu等(2014)Cell 156(6):1262-1278。
本文所用的术语“包括/包含”旨在表示所述组合物和方法包括所列举的要素,但不排除其他要素。如本文所用的,过渡短语“基本上由……组成”(和语法变体)应解释为涵盖所列举的实施方式的所列举的材料或步骤“以及实质上对基本特征和新颖特征没有影响的那些”。参见In re Herz,537 F.2d 549,551-52,190U.S.P.Q.461,463(CCPA 1976)(原文为重点);另请参见MPEP§2111.03。因此,本文所用的术语“基本上由……组成”不应解释为等同于“包括/包含”。“由……组成”应表示不包括更多的其他成分的痕量要素和用于施用本文所公开的组合物的实质性方法步骤。由这些过渡术语中的每一个所限定的方面都在本发明的范围内。
术语“编码”在应用于核酸序列时,是指在天然状态或通过本领域技术人员众所周知的方法操作时,多核苷酸如果可以被转录和/或转译来产生mRNA从而用于多肽和/或其片段,则该多核苷酸被认为“编码”多肽。反义链是这种核酸的互补物,并且编码的序列可以从其推导出。
当涉及特定分子、生物或细胞材料时,术语“等同物”或“生物等同物”可互换使用,并且意指具有最低同源性而仍保持所需结构或功能的那些。
本文所用的术语“表达”是指多核苷酸被转录成mRNA的过程和/或转录的mRNA随后被翻译成肽、多肽或蛋白质的过程。如果多核苷酸衍生自基因组DNA,则表达可以包括在真核细胞中mRNA的剪接。基因的表达水平可以通过测量细胞或组织样品中的mRNA或蛋白质的量来确定。此外,可以确定多个基因的表达水平以建立特定样本的表达谱。
本文所用的术语“功能性”可以用于修饰任何分子、生物或细胞材料,以使其实现特定的指定效果。
本文所用的术语“gRNA”或“指导RNA”是指用于靶向特定基因以利用CRISPR技术进行校正的指导RNA序列。设计用于靶向特异性的gRNA和供体治疗性多核苷酸的技术是本领域众所周知的。例如,Doench,J.,et al.Nature biotechnology 2014;32(12):1262-7,Mohr,S.et al.(2016)FEBS Journal 283:3232-38,和Graham,D.,et al.GenomeBiol.2015;16:260。gRNA包括融合多核苷酸或多核苷酸,或者可选地基本上由融合多核苷酸或多核苷酸组成,或者进一步由融合多核苷酸或多核苷酸组成,其中,所述融合多核苷酸包括CRISPRRNA(crRNA)和反式激活的CRIPSPRRNA(tracrRNA),所述多核苷酸包括CRISPRRNA(crRNA)和反式激活的CRIPSPR RNA(tracrRNA)的。在一些方面,gRNA是合成的(Kelley,M.et al.(2016)J of Biotechnology 233(2016)74-83)。
本文所用的术语“免疫正交”是指两种或多种抗原之间缺乏免疫交叉反应。在一些实施方案中,抗原是蛋白质(例如,Cas9)。在一些实施方案中,抗原是病毒(例如,AAV)。在一些实施方案中,免疫正交的抗原不共享大于5、大于6、大于7、大于8、大于9、大于10、大于11、大于12、大于13、大于14、大于15或大于16个连续氨基酸的氨基酸序列。在一些实施方案中,免疫正交的抗原不共享任何高免疫原性肽。在一些实施方案中,免疫正交的抗原不共享对主要组织相容性复合体(例如,MHCI类或II类)的亲和力。免疫正交的抗原适合连续施用以避开宿主的免疫系统。
术语“免疫沉默的”是指在施用后不引起宿主的免疫应答的抗原。在一些实施方案中,抗原不引起适应性免疫应答。在一些实施方案中,抗原不引起先天性免疫应答。在一些实施方案中,抗原不引起适应性免疫应答或先天性免疫应答。在一些实施方案中,免疫沉默的抗原具有降低的免疫原性。
术语“内含肽”是指能够通过蛋白质剪接将自身切除并连接蛋白质的其余部分的一类蛋白质。“分裂内含肽”来自两个基因。“分裂内含肽”的非限制性实例是最初衍生自点形念珠藻(N.punctiforme)的C-内含肽和N-内含肽序列。
本文所用的术语“分离的”是指基本上不含其他材料的分子、或生物制品、或细胞材料。
本文所用的术语“核酸序列”和“多核苷酸”可互换使用,指任何长度的核苷酸的聚合形式,即核糖核苷酸或脱氧核糖核苷酸。因此,该术语包括但不限于,单链、双链或多链DNA或RNA,基因组DNA,cDNA,DNA-RNA杂交体,或包含嘌呤和嘧啶碱基的聚合物,或其他天然、化学或生物化学修饰的非天然的或衍生的核苷酸碱基。
术语“主要组织相容性复合体”(MHC)是指负责将肽(包括自身和非自身(抗原))呈递给T细胞的蛋白质家族。T细胞识别抗原肽并触发一系列事件,从而导致病原体和感染细胞被破坏。MHC家庭分为三个子组:I类、II类和III类。MHC I类分子具有仅被CD8辅助受体识别的β2亚基。MHC II类分子具有仅被CD4辅助受体识别的β1和β2亚基。这样,MHC分子会伴随以高亲和力结合至给定的抗原的淋巴细胞类型,因为不同的淋巴细胞表达不同的T细胞受体(TCR)辅助受体。通常,MHC I类分子结合短肽,其N-端和C-端固定在位于肽结合槽的端部的袋中。尽管大多数的肽的长度为9个氨基酸残基,但更长的肽可以通过其中心部分的凸出部来容纳,从而形成长度为8-15的结合肽。与II类蛋白质结合的肽的大小不受限制,并且氨基酸长度可以从11变化到30个。MHC II类分子中的肽结合槽在两端是开放的,从而可以结合长度相对较长的肽。“核心”是指对肽的识别贡献最多的氨基酸残基。在一些实施方案中,核心的长度为九个氨基酸。除了核心之外,侧翼区域对于肽对MHC分子的特异性也很重要。
本文所用的术语“器官”是作为个体生物体的特定部分的结构,其中个体生物体的某个功能或多个功能是局部执行的并且在形态上是分开的。器官的非限制性实例包括皮肤、血管、角膜、胸腺、肾脏、心脏、肝脏、脐带、肠、神经、肺、胎盘、胰腺、甲状腺和脑。
术语“直系同源物”用于指另一种基因或蛋白质,并且意指从相同的祖先来源进化而来的所述基因或蛋白质的同源物。直系同源物可以或可以不保留与其直系同源的基因或蛋白质相同的功能。Cas9直系同源物的非限制性实例包括金黄色葡萄球菌(S.aureus)Cas9(“spCas9”)、嗜热链球菌(S.thermophiles)Cas9、嗜肺乳杆菌(L.pneumophilia)Cas9、乳酸奈瑟氏菌(N.lactamica)Cas9、脑膜炎奈瑟氏菌(N.meningitides)Cas9、长双歧杆菌(B.longum)Cas9、嗜黏蛋白艾克曼菌(A.muciniphila)Cas9和O.laneus Cas9。
本文所用的术语“启动子”是指调节诸如基因的编码序列的表达的任何序列。例如,启动子可以是组成型的、诱导型的、阻抑型的或组织特异性的。“启动子”是控制序列,其是多核苷酸序列中控制起始和转录速率的区域。它可以包含遗传元件,在该遗传元件处诸如RNA聚合酶和其他转录因子的调节蛋白和分子可以结合。非限制性的示例性启动子包括CMV启动子和U6启动子。
术语“蛋白质”、“肽”和“多肽”可互换使用,并且在其最广泛的意义上是指氨基酸、氨基酸类似物或拟肽物的两个或多个亚基的化合物。亚基可以通过肽键连接。在另一方面,亚基可以通过例如酯、醚等其他键连接。蛋白质或肽必须包含至少两个氨基酸,并且不限制于可以包括蛋白质序列或肽序列的氨基酸的最大数量。如本文所用的,术语“氨基酸”是指天然和/或非天然或合成的氨基酸,其包括甘氨酸以及D和L旋光异构体、氨基酸类似物和拟肽物。
本文所用的术语“重组表达系统”是指用于表达通过重组形成的某些遗传材料的遗传结构。
本文所用的术语“受试者”旨在表示任何动物。在一些实施方案中,受试者可以是哺乳动物;在进一步的实施方案中,所述受试者可以是牛、马、猫、鼠、猪、犬科、人或大鼠。
本文所用的术语“组织”用于指活的或死的生物体的组织,或源自或设计为模仿活的或死的生物体的任何组织。该组织可以是健康的、患病的和/或具有基因突变。生物组织可以包括任何单个组织(例如,可以相互连接的细胞的集合)或构成生物体的身体的器官或部分或区域的一组组织。该组织可以包括均质的细胞材料,或者可以是复合结构,诸如在包括胸腔的身体区域中出现的结构,例如可以包括肺组织、骨骼组织和/或肌肉组织。示例性组织包括,但不限于,源自肝、肺、甲状腺、皮肤、胰腺、血管、膀胱、肾脏、脑、胆道、十二指肠、腹主动脉、髂静脉、心脏和肠的组织,包括其任何组合。
如本文所用的,对受试者的疾病的“处理(treating)”或“治疗(treatment)”是指(1)防止症状或疾病出现在易患或尚未表现出疾病症状的受试者中;(2)抑制疾病或阻止其发展;(3)改善疾病或疾病症状或引起疾病或疾病症状的消退。如本领域中所理解的,“治疗”是一种用于获得有益的或期望的结果(包括临床结果)的方法。就本技术的目的而言,有益的或期望的结果可以包括一种或多种,但不限于,一种或多种症状的减轻或改善,病况(包括疾病)程度的减轻,病况(包括疾病)状态的稳定(即,不恶化),病况(包括疾病)的延缓或减慢,病况(包括疾病)、状态的进展、改善或缓和,以及缓解(无论是部分还是全部),无论是可检测的还是不可检测的。
如本文所用的,术语“载体”是指保留感染和转导未分裂和/或缓慢分裂细胞并将其整合到靶细胞基因组中的能力的重组载体。该载体可以衍生自或基于野生型病毒。本发明的方面涉及腺相关的病毒载体。
除非另有规定,否则在没有明确叙述的情况下可以推断出,当本发明涉及多肽、蛋白质、多核苷酸或抗体时,其等同物或生物等同物预期在本发明的范围内。如本文所用的,术语“其生物等同物”在提及参考蛋白质、抗体、多肽或核酸时旨在与“其等同物”同义,指具有最低同源性但同时仍保持所需结构或功能性的那些。除非本文具体叙述,否则预期本文提及的任何多核苷酸、多肽或蛋白质也包括其等同物。例如,等同物指至少约70%的同源性或同一性、或至少80%的同源性或同一性,以及可选地,或至少约85%,或可选地至少约90%,或可选地至少约95%、或可选地98%的同源性或同一性,并且表现出与参考蛋白质、多肽或核酸基本等同的生物学活性。可选地,当提及多核苷酸时,其等同物是在严格条件下与参考多核苷酸或其互补物杂交的多核苷酸。
申请人在本文中提供了用于下文所述的基因和蛋白质的转移和表达技术的多肽和/或多核苷酸序列。应当理解,尽管并非总是明确指出,本文提供的序列可用于提供表达产物以及基本相同的序列,其中,该基本相同的序列产生具有相同的生物学特性的蛋白质。这些“生物等同物”或“生物活性”多肽由本文所述的等效多核苷酸编码。当使用在默认条件下运行的序列同一性方法进行比较时,它们可以具有至少60%、或可选地至少65%、或可选地至少70%、或可选地至少75%、或可选地至少80%、或可选地至少85%、或可选地至少90%、或可选地至少95%、或可选地至少98%与参考多肽相同的一级氨基酸序列。提供了特定的多肽序列作为具体实施方案的实例。用具有类似电荷的替代氨基酸对氨基酸的序列进行修饰。另外,等效多核苷酸是在严格条件下与参考多核苷酸或其互补序列杂交的多核苷酸,关于多钛,该多肽由多核苷酸或其互补链编码,该多核苷酸在严格条件下与参考的编码多核苷酸杂交。可选地,等效多肽或蛋白质是由等效多核苷酸表达的多肽或蛋白质。
“杂交”是指其中一种或多种多核苷酸反应形成通过核苷酸残基的碱基之间的氢键而稳定的复合物的反应。氢键可以通过沃森-克里克(Watson-Crick)碱基配对、霍格斯坦(Hoogstein)结合或以任何其他特定序列的方式产生。该复合物可以包括形成双链结构的两条链、形成多链复合物的三条或多条链、自杂交的单链、或它们的任何组合。杂交反应可以构成更广泛过程中的步骤,例如启动PC反应或通过核酶对多核苷酸进行酶解。
严格杂交条件的实例包括:约25℃至约37℃的孵育温度;和约6x SSC至约10x SSC的杂交缓冲液浓度;约0%至约25%的甲酰胺浓度;以及约4x SSC至约8x SSC的洗涤溶液。中等杂交条件的实例包括:约40℃至约50℃的孵育温度;约9x SSC至2x SSC的缓冲液浓度;约30%至约50%的甲酰胺浓度;以及约5x SSC至约2x SSC的洗涤溶液。高严格条件的实例包括:约55℃至约68℃的孵育温度;约1x SSC至约0.1x SSC的缓冲液浓度;约55%至约75%的甲酰胺浓度;以及约1x SSC、0.1x SSC或去离子水的洗涤溶液。通常,杂交孵育时间为5分钟至24小时,具有1、2或更多个洗涤步骤,并且洗涤孵育时间为约1、2或15分钟。SSC是0.15MNaCl和15mM柠檬酸盐缓冲液。应当理解,可以采用使用其他缓冲系统的SSC的等同物。
“同源性”或“同一性”或“相似性”是指两个肽之间或两个核酸分子之间的序列相似性。同源性可以通过比较每个序列中可用于比较而被比对的位置来确定。当被比较的序列中的位置被相同的碱基或氨基酸占据时,则分子在该位置是同源的。序列之间的同源性的程度是根据序列共享的匹配位置或同源位置的数目。“无关的”或“非同源的”序列与本发明的序列之一共享小于40%的同一性,或可选地小于25%的同一性。
实施本发明的方法
产生免疫沉默的蛋白质和鉴定免疫正交的蛋白质的方法
本发明公开了鉴定或修饰蛋白质序列以降低免疫原性并且任选地至免疫沉默的方法。在一些方面,该方法包括鉴定主要组织相容性复合体(MHC)对蛋白质的一个或多个区域的亲和力,由所述鉴定组成,或基本上由所述鉴定组成。对MHC没有亲和力的那些蛋白质区域是免疫沉默的,而无需进一步修饰。相反,对MHC具有亲和力、任选地具有高亲和力的那些蛋白质区域可以通过一个或多个氨基酸替代来修饰,从而使得修饰的区域对MHC没有亲和力。在一些实施方案中,MHC是MHC I类。在一些实施方案中,MHC是MHC II类。
同时地或顺序地,可以任选地通过比对的方法或无比对的方法(例如k-mer分析)来鉴定蛋白质的直系同源物。因此,直系同源物的区域可以被靶向用于类似修饰或可以被认为是免疫沉默的,而无需基于上述结果进行进一步的修饰。可选地,可以基于直系同源物是免疫正交(例如,他们对与最初筛选的蛋白的MHC不同的MHC具有亲和力)的事实选择直系同源物,用于顺序施用。这种免疫正交的蛋白质的顺序施用,作为可选方案,图5F提供了鉴定和/或修饰这些蛋白质的工作流程的示例性示意图。
鉴定直系同源蛋白质的技术是本领域已知的,包括,但不限于传统的基于比对的方法和无比对的方法。此外,直系同源蛋白质的数据库是众所周知的,并且包括但不限于COG、eggNOG、InParanoid、OrthoDB、Ortholuge、CDD、Ensmbl Compara和KEGG。因此,应当理解,本领域普通技术人员可以容易地鉴别直系同源物。例如,k-mer分析是一种计算方法,其识别字符串(string)(例如序列)中包含的长度为k的所有可能的子字符串。k-mer的频率产生基础序列的“签名”,其继而可以用作比较序列并确定全面肽重叠的无比对手段。其他计算方法包括基于比对的计算方法,例如BLOSM(块替换矩阵)或PAM(点接受突变)矩阵。
确定MHC亲和力的方法在本领域中也是已知的,并且可以包括可以通过软件或公众可访问的数据库或“湿实验室”测定法获得的计算方法。预测MHC亲和力的计算方法的实例包括,但不限于,可通过IEDB分析资源获得的MHC结合预测模型(http://tools.immuneepitope.org/mhci/(MHC I)和http://tools.immuneepitope.org/mhcii/(MHC II))或NetMHC(http://www.cbs.dtu.dk/services/NetMHC/)。可选地或额外地,可以使用测定法来确定MHC亲和力或来验证其计算预测,所述测定法为诸如但不限于免疫测定法,诸如ELISA、微阵列、四聚体测定法和肽诱导的MHC稳定性测定法。使用这种测定法和计算方法可以进一步适用于对所治疗的特定受试者或患者的MHC概况进行解释。因此,可以优化蛋白质的修饰以使其对具体受试者或患者是免疫沉默的。类似地,比较可以是受宿主限制的,从而使得鉴定或修饰该蛋白质以使其对例如小鼠或人的具体宿主具有特异性。
申请人考虑将这种方法用于存在引发免疫应答风险的多种蛋白质。感兴趣的非限制性的示例性蛋白质包括胞苷脱氨酶,其可通过催化DNA碱基从C到T的变化来进行基因编辑(例如APOBEC——在许多物种中都是保守的,例如大鼠APOBEC3、大鼠APOBEC1、恒河猕猴(Resus Macaque)APOBEC3G、人APOBEC1(A1)、AID、APOBEC2(A2)、APOBEC3A(A3A)、APOBEC3B(A3B)、APOBEC3C(A3C)、APOBEC3DE(A3DE)、APOBEC3F(A3F)、APOBEC3G(A3G)、APOBEC3H(A3H)和APOBEC4(A4));腺苷脱氨酶,其可用于通过催化DNA碱基从A到G的变化来进行基因编辑(例如,ADA(DNA编辑器)——在几乎所有物种中是广泛保守的,和ADAR(RNA编辑器)——在大多数后生动物物种中是保守的);Zing Finger核酸酶(ZFN),其以与CRISPR/Cas9类似的方式用于基因组工程化,并且可以被工程化为由3-6个重复的锌指结构域组成的位点特异性核酸酶,该锌指结构域是一个广泛保守的DNA结合基序和核酸酶结构域;转录激活因子样效应物核酸酶(TALEN),其以与CRISPR/Cas9类似的方式用于基因组工程,并且类似于ZFN,因为它们被工程化为由TAL效应物DNA结合域(通常衍生自黄单胞变形杆菌(Xanthomonasproteobacteria)属)和核酸酶结构域组成的位点特异性核酸酶。上面提到的位点特异性酶(ZFN和TALEN)的结构域已得到充分表征,并且经过广泛地工程化以产生了期望的特异性。因此,存在这种蛋白质的许多变体。与MHC亲和力分析相关的其他蛋白质包括Cas9蛋白质和AAV衣壳,两者均用于基于CRISPR的基因编辑中。
本发明的方面涉及一种产生蛋白质的方法,该方法包括:鉴定对主要组织相容性复合体(MHC)具有亲和力的蛋白质的一个或多个区域,以及通过一个多个氨基酸替代修饰对该MHC具有亲和力的蛋白质的一个或多个区域,从而使得经修饰的区域对MHC没有亲和力,其中,所得到的经修饰的蛋白质在将该经修饰的蛋白质或编码该经修饰的蛋白质的多核苷酸施用于受试者时是免疫沉默的。在一些实施方案中,对MHC的亲和力是高亲和力。在一些实施方案中,至少一个经替代的氨基酸是不用作MHC蛋白核心残基的氨基酸。在一些实施方案中,该蛋白质选自胞苷脱氨酶、腺苷脱氨酶、锌指核酸酶、转录激活因子样效应物核酸酶、Cas9或AAV衣壳蛋白的组。在一些实施方案中,蛋白质是Cas9,任选地是SpCas9。
例如,为了优化和拓宽基于CRIPSR的疗法的应用,本发明人相应地开发了两种技术:1)通过将高免疫原性的结构域或肽与免疫原性相对较低的对应物进行交换来“人源化”Cas9蛋白。这对于将Cas9库(arsenal)用于重复治疗的应用尤为有用。在SpCas9中定位高度免疫原性肽之后,申请人计算了这些免疫原性肽中每个位置的单个氨基酸交换,这被预期会降低总体的免疫原性,而不会潜在地改变活性。本发明教导了要突变的区域和突变成的对象。另外,申请人鉴定了在免疫空间中正交的天然Cas9直系同源蛋白,即,不共享任何高免疫原性肽,并且因此适合顺序施用以逃避宿主的免疫系统并改善治疗方案。
因此,本发明的方面涉及用于免疫隐身(immune stealth)的经修饰的Cas9以及Cas9直系同源物在增强免疫逃避中的应用。对于需要体内递送的任何应用,经修饰的Cas9可以替代现有的野生型Cas9,其在重复使用后可能不会丧失功效。在免疫空间中直系同源的Cas9蛋白也可以用于体内应用,其中如果需要重复治疗,则可以顺序地利用在免疫空间中直系同源的Cas9蛋白。在下文描述与Cas9的这些非限制性方面。
本文公开的一些实施方案涉及一种产生经修饰的Cas9的方法,该方法包括:鉴定对主要组织相容性复合体(MHC)具有高亲和力的Cas9的一个或多个区域,以及通过一个或多个氨基酸替代修饰对该MHC具有高亲和力的Cas9的一个或多个区域,从而使得经修饰的区域对该MHC没有亲和力,其中,所得的经修饰的Cas9在施用于受试者时是免疫沉默的。在一些实施方案中,Cas9是SpCas9。进一步的实施方案涉及根据该方法产生的经修饰的Cas9。本文公开的一些实施方案涉及包含表1中提供的一种或多种氨基酸修饰的经修饰的SpCas9。本文公开的一些实施方案涉及在被施用需要Cas9的方案的受试者中避免免疫应答的方法,所述方法包括:顺序施用对主要组织相容性复合体(MHC)没有共享亲和力的一组直系同源的Cas9蛋白中的每一种。在一些实施方案中,Cas9蛋白的组选自图4中提供的Cas9蛋白的组。
在一些方面,本文提供了产生经修饰的Cas9的方法,所述方法包括以下步骤,由其组成,或基本上由其组成:鉴定对主要组织相容性复合体(MHC)具有亲和力的Cas9的一个或多个区域,以及通过一个或多个氨基酸替代修饰对该MHC具有亲和力的Cas9的一个或多个区域,从而使得经修饰的区域对该MHC没有亲和力,其中,所得的经修饰的Cas9在施用于受试者时具有降低的免疫原性。在一些实施方案中,对MHC的亲和力是高亲和力。在一些实施方案中,Cas9是SpCas9。在一些实施方案中,至少一个经替代的氨基酸是不用作MHC蛋白核心残基的氨基酸。在一些方面,本文提供了一种经修饰的Cas9,其通过鉴定对主要组织相容性复合体(MHC)具有亲和力的Cas9的一个或多个区域,并通过一个或多个氨基酸替代修饰对该MCH具有亲和力的Cas9的一个或多个区域,从而使得经修饰的区域对该MHC没有亲和力,其中所得的经修饰的Cas9在施用于受试者时具有降低的免疫原性。
在一些方面,本文提供了一种经修饰的Cas9,其包含一种或多种、两种或更多种、三种或更多种、四种或更多种、五种或更多种、六种或更多种、七种或更多种、八种或更多种、九种或更多种、十种或更多种、十五种或更多种、或二十种或更多种表1中提供的氨基酸修饰,由其组成,或基本上由其组成。
在一些方面,本文提供了编码经修饰的Cas9蛋白的分离的多核苷酸,其中,经修饰的Cas9通过以下方法产生:鉴定对主要组织相容性复合体(MHC)具有亲和力的Cas9的一个或多个区域,以及通过一个或多个氨基酸替代修饰对MHC具有亲和力的Cas9的一个或多个区域,从而使得经修饰的区域对MHC没有亲和力,其中所得的经修饰的Cas9在施用于受试者时具有降低的免疫原性。在一些方面,本文提供了编码经修饰的Cas9蛋白的分离的多核苷酸,其中,经修饰的Cas9包含一种或多种、两种或更多种、三种或更多种、四种或更多种、五种或更多种、六种或更多种、七种或更多种、八种或更多种、九种或更多种、十种或更多种、十五种或更多种、或20种或更多种表1中提供的氨基酸修饰物,由其组成,或基本上由其组成。在一些方面,本文提供了包含分离的多核苷酸的载体。在一些实施方案中,载体是AAV载体,任选地其中AAV载体是AAV5。
还应理解,根据相同方法可以将AAV衣壳修饰为免疫沉默的,即鉴定对主要组织相容性复合体(MHC)具有亲和力的一个或多个AAV衣壳蛋白的一个或多个区域,并通过一个或多个氨基酸替代修饰对该MHC具有亲和力的一个或多个AAV衣壳蛋白的一个或多个区域,从而使得经修饰的区域对MHC没有亲和力,其中,所得的包含一个或多个AAV衣壳蛋白的衣壳在施用于受试者时具有降低的免疫原性。根据该方法产生的经修饰的AAV可以用于本文公开的任何一个或多个实施方案中以逃避免疫系统。
此外,可以根据本文公开的方法鉴定免疫正交的AAV。因此,本文考虑了其中免疫正交的Cas9包含在免疫正交的AAV中的实施方案。
鉴定免疫正交的直系同源物的方法的其他方面包括:确定蛋白质或其区域对多种主要组织相容性复合体(MHC)的亲和力集,比较该蛋白质或其区域的亲和力集与该蛋白质的直系同源物对所述多种MHC的亲和力集,以及基于非重叠的亲和力集确定免疫正交的直系同源物集。在一些实施方案中,对MHC的亲和力是高亲和力。在一些实施方案中,该蛋白质选自胞苷脱氨酶、腺苷脱氨酶、锌指核酸酶、转录激活因子样效应物核酸酶、Cas9或AAV衣壳蛋白的组。在一些实施方案中,蛋白质是Cas9,任选地SpCas9或SaCas9。在一些实施方案中,Cas9蛋白直系同源物选自酿脓链球菌(S.pyogenes)Cas9(spCas9)、金黄色葡萄球菌(S.aureus)Cas9(saCas9)、长双歧杆菌(B.longum)Cas9、嗜黏蛋白艾克曼菌(A.muiciniphilia)Cas9或O.laneus Cas9。
不受理论的束缚,申请人认为,即使在MHC筛选之后,受试者仍可能具有先前存在的免疫反应的所有成分(repitoire),这会导致与蛋白质或其直系同源物的交叉反应。因此,在顺序施用免疫正交的蛋白质时存在一些混淆的风险。可能引起这种问题的非限制性的示例性蛋白质是源自受试者中致病的生物体的那些(例如,人中的金黄色葡萄球菌或酿脓链球菌)。因此,申请人建议鉴定极端微生物的这种蛋白质的免疫正交的直系同源物(并且因此在正常情况下不太可能与人类或其他受试者接触)和/或受试者免疫系统对其产生耐受性的高丰度的共生种。可以基于文献和/或基于对受试者群体或特定受试者的采样来确定正常微生物组或特定受试者的微生物组中的种丰度。在一些实施方案中,当建立耐受性时,共生种是存在于发育早期的种。
蛋白质和载体
其他方面涉及根据以上公开的方法产生的经修饰的Cas9蛋白。更进一步的方面涉及包含一种或更多种、两种或更多种、三种或更多种、四种或更多种、五种或更多种、六种或更多种、七种或更多种、八种或更多种、九种或更多种、十种或更多种、十五种或更多种、或二十种或更多种表1中提供的氨基酸修饰的经修饰的Cas9蛋白。一些实施方案涉及编码经修饰的Cas9的分离的多核苷酸。进一步的实施方案涉及包含该分离的多核苷酸的载体,任选地为AAV载体,以及还任选地为AAV5载体。另外的实施方案涉及包含所述载体的AAV衣壳。在一些实施方案中,一种或多种AAV衣壳蛋白已被修饰为免疫沉默的。
通常,将诸如RNA的遗传材料包装到一种或多种载体中的方法是本领域众所周知的。例如,可以使用包装载体和细胞系包装遗传材料,并通过传统的重组方法来引入。
在一些实施方案中,包装载体可以包括,但不限于,逆转录病毒载体、慢病毒载体、腺病毒载体和腺相关的病毒载体(任选地AAV8)。包装载体包含促进遗传材料向细胞内递送的元件和序列。例如,逆转录病毒构建体是包装质粒,该包装质粒包含至少一个逆转录病毒辅助DNA序列,其衍生自反式编码包装无复制能力的逆转录病毒载体所需的所有病毒体蛋白质的无复制能力的逆转录病毒基因组,并且用于产生高效价的能够包装无复制能力的逆转录病毒载体的病毒体蛋白,而不产生能复制的辅助病毒。逆转录病毒DNA序列缺少编码病毒的病毒5′LTR的天然增强子和/或启动子的区域,并且缺少负责包装辅助基因组和3′LTR的psi功能序列,但编码外源多腺苷酸化位点(例如,SV40多腺苷酸化位点)以及外源增强子和/或启动子,其在需要产生病毒的细胞类型中指导有效的转录。逆转录病毒是白血病病毒,诸如莫洛尼鼠白血病病毒(Moloney Murine Leukemia Virus,MMLV)、人类免疫缺陷病毒(Human Immunodeficiency Virus,HIV)或长臂猿白血病病毒(Gibbon Ape Leukemiavirus,GALV)。外源增强子和启动子可以是人巨细胞病毒(HCMV)即刻早期(IE)的增强子和启动子、莫洛尼鼠肉瘤病毒(MMSV)的增强子和启动子(U3区域)、劳斯肉瘤病毒(RousSarcoma Virus,RSV)的U3区域、脾聚焦形成病毒(Spleen Focus Forming Virus,SFFV)的U3区域、或加入天然的莫洛尼鼠白血病病毒(MMLV)启动子的HCMV IE增强子。
逆转录病毒包装质粒可以由基于质粒的表达载体编码的两个逆转录病毒辅助DNA序列组成,例如,其中第一辅助序列包含编码嗜亲性MMLV或GALV的gag蛋白和pol蛋白的cDNA,以及第二辅助序列包含编码env蛋白的cDNA。决定宿主范围的Env基因可以衍生自:编码以下蛋白的基因:嗜异性、双嗜性、嗜亲性、多变性(水貂病灶形成(mink focusforming))或10A1鼠白血病病毒env蛋白,或长臂猿白血病病毒(GALV)env蛋白,人类免疫缺陷病毒env(gp160)蛋白,泡状气孔病毒(VSV)G蛋白,人类T细胞白血病(HTLV)I型和II型env基因产物;衍生自一种或多种上述env基因的组合的嵌合包膜基因;或编码上述env基因产物的胞质和跨膜以及针对所需的靶细胞上的特定表面分子的单克隆抗体的嵌合的包膜基因。类似的基于载体的系统可以采用其他载体,诸如睡眠美容(sleeping beauty)载体或转座子元件。
然后可以通过适当的施用途径引入所得的包装表达系统,本文公开的方法的各个方面对其进行了详细讨论。
治疗方法
一些方面涉及在被施用需要蛋白质的方案的受试者中避免免疫应答的方法,该方法包括:向受试者顺序施用免疫正交的两种或更多种蛋白质。在一些实施方案中,免疫正交的蛋白质不共享大于5个连续氨基酸的氨基酸序列。在一些实施方案中,免疫正交的蛋白质不共享对主要组织相容性复合体(MHC)的亲和力。在一些实施方案中,顺序施用免疫正交的三种或更多种、四个或更多种、五种或更多种、六种或更多种、七种或更多种、八种或更多种、九种或更多种、或十种或更多种免疫正交的蛋白质。
非限制性的示例性方面涉及Cas9。在一些实施方案中,免疫正交的Cas9蛋白不共享大于5个连续氨基酸的氨基酸序列。在一些实施方案中,免疫正交的Cas9蛋白不共享对主要组织相容性复合体(MHC)的亲和力。在一些实施方案中,根据以上公开的方法修饰两种或更多种Cas9蛋白中的至少一种。在一些实施方案中,两种或更多种Cas9蛋白或编码所述Cas9蛋白的多核苷酸中的至少一种包含在AAV载体中。在一些实施方案中,AAV载体是AAV5载体。在一些实施方案中,AAV载体包含在AAV衣壳中。在一些实施方案中,两种或更多种Cas9蛋白或编码所述Cas9蛋白的多核苷酸包含在AAV载体中。在一些实施方案中,每个AAV载体包含在AAV衣壳中,任选地,其中该AAV衣壳彼此免疫正交。
本文公开了一种基因编辑的方法,其包括使细胞与两种或更多种免疫正交的Cas9或编码所述Cas9的多核苷酸(任选地包含在AAV衣壳中)顺序地接触。在一些实施方案中,包含Cas9或编码它们的多核苷酸中的每种的AAV衣壳可以是免疫正交的。在一些方面,接触是体外的。在其他方面,接触是体内的。在一些方面,接触是体内或体外的。在一些方面,多核苷酸中的至少一种包含编码指导RNA(gRNA)的多核苷酸,或基本上由其组成,或进一步由其组成。在一些方面,多核苷酸中的至少一种包含治疗性多肽,或可选地基本上由其组成,或进一步由其组成。
本文进一步公开了一种在有需要的受试者中进行基因编辑的方法,其包括向受试者顺序施用有效量的两种或更多种免疫正交的Cas9或编码所述Cas9的多核苷酸(任选地包含在AAV中)。在一些实施方案中,包含Cas9或编码它们的多核苷酸中的每种的AAV衣壳可以是免疫正交的。在一些方面,多核苷酸中的至少一种包含编码指导RNA(gRNA)的多核苷酸,或基本上由其组成,或进一步由其组成。在一些方面,多核苷酸中的至少一种包含治疗性多肽,或可选地基本上由其组成,或进一步由其组成。
在一些方面,编码gRNA的多核苷酸包含:包含CRISPR RNA(crRNA)和反式激活的CRIPSPR RNA(tracrRNA)的融合多肽;或包含CRISPRRNA(crRNA)和反式激活的CRIPSPRRNA(tracrRNA)的多肽,或可选地基本上由其组成,或进一步由其组成。一方面,编码gRNA的多核苷酸包含表2或表3中的一个或多个序列或其等同物,或由其组成。在一些方面,gRNA对受试者中需要进行基因编辑的DNA区域或需要其的细胞具有特异性。
在一些方面,本文提供了治疗需要基因编辑或基因调控的受试者的方法,该方法包括:对该受试者顺序施用两种或更多种免疫正交的Cas9蛋白。在一些实施方案中,免疫正交的Cas9蛋白不共享大于5个连续氨基酸的氨基酸序列。在一些实施方案中,免疫正交的Cas9蛋白不共享对主要组织相容性复合体(MHC)的亲和力。在一些实施方案中,顺序施用三种或更多种、四种或更多种、五种或更多种、六种或更多种、七种或更多种、八种或更多种、九种或更多种、或十种或更多种免疫正交的Cas9蛋白。在一些实施方案中,每种免疫正交的Cas9蛋白是衍生自不同种的细菌的Cas9。在一些实施方案中,免疫正交的Cas9蛋白选自酿脓链球菌Cas9(spCas9)、金黄色葡萄球菌Cas9(saCas9)、长双歧杆菌Cas9、嗜黏蛋白艾克曼菌Cas9或O.laneus Cas9。在具体实施方案中,免疫正交的Cas9蛋白包含spCas9和saCas9。在一些实施方案中,至少一种Cas9被修饰以在施用于受试者时降低免疫原性。在一些实施方案中,该方法进一步包括在AAV5载体中施用两种或更多种Cas9蛋白中的至少一种。在一些实施方案中,该方法进一步包括对受试者施用一种或多种指导RNA。
在一些实施方案中,选择指导RNA来治疗选自以下组的疾病、病症或病况:全色盲(achromatopsia)、腺苷脱氨酶缺乏症(adenosine deaminase(ADA)deficiency)、α1-抗胰蛋白酶缺乏症(alpha-1-antitrypsin deficiency)、阿尔茨海默病(Alzheimer’sdisease)、肌萎缩侧索硬化(amyotrophic lateral sclerosis)、芳香族氨基酸脱羧酶缺乏症(aromatic amino acid decarboxylase deficiency)、巴顿病(Batten disease)、无脉络膜(choroideremia)、克里格勒-纳贾尔综合征(CriglerNajjar syndrome)、囊性纤维化(cystic fibrosis)、脆性X综合征(fragile X syndrome)、血友病(hemophilia)、乙型肝炎(hepatitis B)、丙型肝炎(hepatitis C)、纯合子家族性高胆固醇血症(homozygousfamilial hypercholesteremia)、亨廷顿病(Huntington’s Disease)、Leber先天性黑朦(Leber congenital amaurosis)、黄斑变性(macular degeneration)、枫糖尿病(maplesyrup urine disease(MSUD))、粘多糖病(mucopolysarccharidosis(I-IX))、多发性硬化(multiple sclerosis)、肌肉不良(muscular dystrophy)、强直性肌营养不良(myotonicdystrophy)、1型神经纤维瘤病(neurofibramotosis type 1)、鸟氨酸转氨甲酰酶缺乏症(ornithine transcarbamylase deficiency)、先天性甲肥厚(pachyonychia congenita)、帕金森氏病(Parkinson’s disease)、苯丙酮尿症(phenylketonuria)、多囊肾病(polycystickidneydisease)、庞贝病(Pompe disease)、视网膜变性(retinaldegeneration)、瑞特氏综合症(Rett’s syndrome)、佝偻病(rickets)、脊髓性肌萎缩(spinal muscular atrophy)、重症联合免疫缺陷(severe combined immunodeficiency)、镰状细胞病(sickle cell disease)、史-莱-奥综合征(Smith-Lemli-Opitz syndrome)、Y连锁非梗阻性生精障碍(Y-linked nonobstructive spermatogenicfailure)、地中海贫血(thalassemia)和X连锁视网膜劈裂症(X-linked retinoschisis)。
在一些方面,设计和/或选择指导RNA以靶向或修复选自以下组的基因:Nav 1.7(SCN9A)、Nav 1.8(SCN10A基因)、1.9(SCN11A基因)和1.3(SCN3A基因);瞬时受体电位阳离子通道亚家族V成员1(TrpV1),也称为辣椒素受体和香草酸受体1;PRDM12;或HCN2。
本领域技术人员应当理解,可以产生用于靶向特异性的gRNA以靶向特定基因,任选地与疾病、病症或病况相关的基因。因此,与Cas9组合,指导RNA促进了CRISPR/Cas9系统的靶向特异性。如上所述,诸如启动子选择的其他方面可以提供实现靶向特异性的额外机制,例如,为编码指导RNA的多核苷酸选择启动子,以促进在具体器官或组织中的表达。因此,本文考虑针对具体疾病、病症或病况选择合适的gRNA。表2和表3分别提供了适用于人类和小鼠基因的gRNA的非限制性实例。
在整个治疗过程中以一剂量连续或间歇地施用经修饰的AAV或组合物就可以起到作用。可以通过任何合适的施用方式进行施用,包括但不限于:静脉内、动脉内、肌肉内、心脏内、鞘内、心室下、硬膜外、脑内、脑室内、视网膜下、玻璃体内、关节内、眼内、腹膜内、子宫内、皮内、皮下、经皮肤、经粘膜和吸入。
确定最有效的施用方式和剂量的方法是本领域技术人员已知的,并且将根据用于治疗的组合物、治疗的目的和所治疗的受试者而变化。可以通过治疗医师选择剂量水平和方式进行单次或多次施用。值得注意的是,剂量会受到施用途径的影响。合适的剂量和施用试剂的方法是本领域已知的。这种合适的剂量的非限制性实例可以低至每次施用1E+9载体基因组至多达1E+17载体基因组。
在另一方面,本发明的经修饰的病毒颗粒和组合物可以与其他治疗(例如,适用于具体疾病、病症或病况的那些经批准的治疗)联合施用。非限制性实例包括用经修饰的病毒颗粒和一种或多种类固醇的组合来治疗肌营养不良。
可以进行本发明的经修饰的病毒颗粒或组合物的这种施用,以产生所需疾病、病症或病况的动物模型,以用于实验和筛选测定。
可以通过任何合适的途径(例如静脉内、经皮、鼻内、口服、粘膜或其他递送方法)和/或通过单剂量或多剂量递送适用于本发明的剂量。应当理解,实际剂量可以根据所使用的重组表达系统(例如,AAV或慢病毒)、靶细胞、器官或组织、受试者以及所寻求的作用程度而变化。组织、器官和/或患者的大小和重量也会影响剂量。药剂可以进一步包括其他试剂,包括但不限于载体。本领域已知的合适载体的非限制性实例:例如水、盐水、乙醇、甘油、乳糖、蔗糖、葡聚糖、琼脂、果胶、植物来源的油、磷酸盐缓冲盐水和/或稀释剂。额外的材料(例如在WO 2017/070605的第[00533]段中公开的那些材料)可以适合于与本文公开的组合物一起使用。WO 2017/070605的第[00534]至[00537]段也提供了本文可以使用的CRISPR-Cas系统的给药惯例的非限制性实例。通常,给药考虑是本领域技术人员所熟知的。
组合物和试剂盒
本发明还提供了包含任何一种或多种免疫沉默的和/或免疫正交的蛋白质的组合物或试剂盒。一方面,载体是药学上可接受的载体。这些组合物可以如本文所述用于治疗,并且可以与其他已知疗法组合使用和/或根据本文所述的方法的方面使用。
简而言之,本发明的药物组合物可以包含免疫沉默的和/或免疫正交的Cas9或编码所述Cas9的多核苷酸(其任选地包含在AAV中,AAV任选地也是免疫正交的)与一种或多种药学或生理学上可接受的载体、稀释剂或赋形剂的组合。这种组合物可以包含缓冲剂,诸如中性缓冲盐水、磷酸盐缓冲盐水等;碳水化合物,诸如葡萄糖、甘露糖、蔗糖或葡聚糖、甘露醇;蛋白质;多肽或氨基酸,诸如甘氨酸;抗氧化剂;螯合剂,诸如EDTA或谷胱甘肽;佐剂(例如,氢氧化铝);和防腐剂。本发明的组合物可以配制为用于口服、静脉内、局部、肠内和/或肠胃外施用。在某些实施方案中,将本发明的组合物配制为用于静脉内施用。
实施例
以下实施例是非限制性且示例性的程序,其可以在使本发明生效的各个情况下使用。另外,本文公开的所有参考文献通过引用整体并入本文。
实施例1——Cas9蛋白的免疫原性
已经开发了几种经由计算机模拟的表位结合预测方法,其采用机器学习方法来预测肽-MHC I类结合亲和力。申请人已利用NetMHC 4.0 Server 4(一种基于神经网络和权矩阵的预测算法)来确定来自88菌株6的先前鉴定的Cas9蛋白序列中的肽在所有HLA等位基因超型上的免疫原性水平。
NetMHC以默认参数运行,预测每个等位基因在8到11个氨基酸的肽序列上的免疫原性评分。高免疫原性肽被定义为具有<50nM的亲和力得分,并且中间值为50nM500nM。
鉴定出最具免疫原性的肽之后,申请人利用两种经由计算机模拟的方法来确定降低SpCas9免疫原性所必需的修饰。
1)确定了每个高免疫原性肽中的单个氨基酸交换对降低免疫原性的影响
2)发现哪些Cas9直系同源物在其“免疫原性空间”中最接近,以确定哪些Cas9蛋白质可以顺序地用于重复治疗。
总体工作流程如图1所示。
实施例2——SpCas9中免疫原性肽中单个氨基酸交换的影响
在定位SpCas9中的高免疫原性肽之后,申请人在这些免疫原性肽中的每个位置处进行了单个氨基酸交换,以确定这些交换是否会降低该肽的总体的免疫原性。将这一新的肽列表首先提交给NetMHC服务器,以预测其免疫原性得分。目的是发现改变这种肽中的单个AA是否会显著更改亲和力。
计算免疫原性肽中每个单个氨基酸交换的亲和力得分。例如,位于原始蛋白质中的氨基酸位置327-334的肽“HHQDLTLL”具有32个无亲和力得分且具有单个氨基酸交换的肽(例如‘HHQDLTLK’、‘HHQDLTLN’、‘HHQDLTLD’)。得分最高的肽被定义为是在所有可能的肽交换中显示出最低亲和力值的肽。随后,“无”亲和力肽被提交给PROVEAN服务器,该服务器预测某些位置的单个氨基酸变化能够具有的对蛋白质功能性的影响。7导致“无”或“低”免疫原性且无害的单个氨基酸交换随后将用于SpCas9的实验诱变。这些突变列于表1中,其中,匹配的颜色对应于其免疫原性可以随着相同的AA交换而改变的肽。
然后可以将这种突变的SpCas9顺序地用于体内基因组治疗。不受理论的束缚,相信在重复治疗后,可以在不降低其功效的情况下实现这一点,而不引起免疫原性的应答。
实施例3——用于顺序给药以逃避宿主免疫系统的Cas9蛋白的正交性
目的是确定在‘免疫原性空间’中正交的Cas9直系同源物。这会使得申请人能够开具Cas9的顺序治疗方案,以用于治疗干预。分析表明,对于最保守的数据,总是至少存在相互正交且包括SpCas9的35种蛋白质的组。实施的方法如下:从一种蛋白质中选择高亲和力的肽,并确定那些确切的肽序列在整个其他序列中出现的次数。如果没有发现肽,则确定蛋白质是正交的。所选择的肽通常由8至11个氨基酸组成,被进一步分成长度为5至11的亚肽。这使得能够鉴定出蛋白质序列之间更细微的相似性。对每个可能的蛋白质对进行了这种分析。这里介绍的相互正交的蛋白质组没有平均长度为5的配对。Python软件包Networkx中提供了用于确定相互正交的算法‘寻群(find_cliques)’。
申请人创建了一个网络,其中两个蛋白质(节点)如果正交则通过一条边连接。申请人然后应用寻群算法来定位图中的所有最大群,其中最大群是完整的子图,从而使得在保持完整性的同时可以不添加其他节点。参见例如图4。
实施例4——小鼠实验
给两个月大的小鼠以6E+11GC/小鼠注射AAV病毒。申请人测试了两个不同的AAV衣壳(AAV8和AAVDJ),以及两个正交的Cas9蛋白(SpCas9和SaCas9),以测试使用不同衣壳或不同SpCas9蛋白进行的顺序AAV病毒注射是否由于免疫原性应答而对降低基因组编辑的功效有任何影响。
| 0周 | 3周 | 6周 |
| A1 | B2 | 测定(基线和AAV的作用) |
| A2 | A1 | 测定(基线和AAV的作用) |
| B1 | B2 | 测定(基线和AAV的作用) |
| B2 | B1 | 测定(基线和AAV的作用) |
| A1 | B2 | 测定(Cas9正交性) |
| B2 | A1 | 测定(Cas9正交性) |
| A2 | B1 | 测定(Cas9正交性) |
| B1 | A2 | 测定(Cas9正交性) |
说明:
A1:AAV8 SpCas9 CD81;A2:AAVDJ SpCas9 Scarb1;B1:AAV8 SaCas9 CD81;B2:AAVDJ SaCas9 Scarbl
实施例5——针对预测的肽确定记忆性T细胞群的存在
为针对任何预测的Cas9直系同源物的T细胞的存在,评估存在于人类群体中的记忆性T细胞群体。特别地,研究了金黄色葡萄球菌肽,因为大约30%的人类群体被这种病原体定殖。
实施例6——筛选“免疫正交的”直系同源物
基于蛋白质的治疗方法的主要障碍是与适应性免疫系统的相互作用,这可以导致被循环抗体中和以及被细胞毒性T淋巴细胞清除治疗细胞。避免这些问题的一种方法是使用人类或人源化的蛋白质,其通过自我识别来避免免疫应答。然而,这种方法将潜在的蛋白质疗法局限于源于人类的那些,排除了许多令人兴奋的效应物和递送载体,诸如CRISPR-Cas9和腺相关病毒(AAV)。为了解决这个问题,申请人在此提出了顺序地使用直系同源蛋白,直系同源蛋白的功能受到自然选择的限制,但是其结构因遗传漂移而具有多样性。原则上,这使得能够通过‘免疫正交的’直系同源物进行重复治疗,而不降低疗效,这是由于蛋白质之间缺乏免疫交叉反应。为了探索和验证这一构思,申请人选择了91种II型CRISPR-Cas9直系同源物和167种AAV衣壳蛋白质直系同源物,并开发了一条传递途径(pipeline)来比较总序列相似性以及与I类和II类主要组织相容性复合体(MHC)蛋白的预测结合。有趣的是,MHC结合的预测揭示了Cas9直系同源物集之间的广泛多样性,其中83%的配对被预测为具有非交叉反应性的免疫应答,而未观察到AAV血清型之间的总体免疫正交性。为了证实这些发现,申请人从酿脓链球菌和金黄色葡萄球菌中选择了两种Cas9直系同源物,它们在免疫空间中被预测为是正交的,并通过多种AAV血清型将它们递送到小鼠中。申请人在免疫小鼠的血清中观察到了针对AAV的交叉反应性的抗体,但未观察到针对Cas9直系同源物的交叉反应性的抗体,从而验证了这些蛋白质之间的计算预测的免疫正交性。展望未来,申请人期望该框架可以用于合理地设计蛋白质直系同源物之间的免疫正交性。
蛋白质疗法(包括基于蛋白质的基因疗法)对小分子药物具有多个优势。它们通常具有复杂的特定功能,并且对正常的生物过程具有最小的脱靶干扰。然而,任何基于蛋白质的疗法的基本挑战之一是与适应性免疫系统的相互作用。通过B细胞活化而被循环抗体中和以及通过CD8+细胞毒性T淋巴细胞(CTL)清除治疗细胞,为有效的蛋白质治疗创造了实质性障碍10。尽管对新型蛋白质的适应性免疫应答的延迟会使得有足够的时间以使初始剂量发挥作用,但由于存在记忆性T细胞和B细胞,后续药剂面临更快、更强的次级免疫应答。此外,基因转移研究表明,针对递送载体和/或治疗性转基因的宿主免疫应答可以消除治疗细胞,从而限制了治疗的功效11-16。
解决这些问题的常用方法是利用人类蛋白质,或利用被非人类成分替代的人源化蛋白质17,18。但是,这种方法局限于在人类或近亲物种中天然存在的一小部分治疗性蛋白质。此外,尽管蛋白质的人源化可以产生明显更少的免疫原性产物,但它们仍具有免疫风险18。规避对蛋白质疗法的免疫应答的另一种方法是去除免疫原性T细胞表位。19,20一旦鉴定了免疫原性T细胞表位,关键氨基酸被替代会降低蛋白质的免疫原性,因为关键的锚定残基处的氨基酸修饰可以消除与MHC分子的结合并防止抗原呈递。但是,由于HLA位点的巨大多样性,这可能很难做到。由于表位工程必须考虑到每个不同的HLA等位基因的底物特异性,因此疗法很可能必须对每个患者进行独特的改变。同样地,表位缺失已成功地应用于几种蛋白质,21但仅在限于不能代表完全多样性的少数HLA等位基因时才能保留蛋白质功能。还已知诸如PEG化的结构修饰通过干扰抗原处理机制来降低免疫原性。然而,有证据表明,在用PEG化医药用酶治疗的患者中诱发了PEG特异性抗体22-25。
此外,由于蛋白质的降解或治疗细胞的更新,或者在基因疗法的情况下转基因的表达降低,蛋白质疗法需要重复治疗26,27。这提供了更大的挑战,因为重复暴露于同一抗原会引起更强的次级免疫应答28,这会完全抑制后续剂量,甚至使免疫系统对首次暴露后残留的抗原敏感。为了促进有效的重复蛋白疗法,申请人提出使用直系同源蛋白质,其功能受到自然选择的限制,但其结构因遗传漂移而具有多样性。在给定足够的序列差异的情况下,直系同源物不会与暴露于其他直系同源物而产生的免疫应答发生交叉反应,使得重复药剂能够避免被现有抗体中和并避免治疗细胞被激活的CTL清除。
作为探索这种方法的案例研究,申请人专注于CRISPR-Cas9系统,这可能是用于基因编辑的最值得期待的疗法29-36。比较基因组学已证明,Cas9蛋白广泛分布于细菌种之间,并且在广泛的进化史上已经多样化37-39。申请人假设,这种多样性可以提供一种机制,在每种治疗中通过利用直系同源的Cas9蛋白来规避诱导免疫记忆。另外,还必须考虑由于Cas9和相关的指导RNA(gRNA)的递送载体或施用途径而引起的免疫原性。在这方面,腺相关病毒(AAV)已经成为基因递送的高度优选的载体,因为它们与低免疫原性和毒性相关14,15,这促进了长期的转基因表达40,41和治疗效果。尽管AAV载体的免疫原性相对较低,但仍然可以诱导针对衣壳和转基因的抗体42-46。此外,针对AAV的中和抗体(NAB)在人类群体中的流行率47和血清型之间的交叉反应性48仍然是有效的AAV疗法的障碍。尽管起初AAV由于其抗原呈递细胞(APC)的不良转导而被认为是非免疫原性的49,但现在人们知道它们可以转导树突状细胞(DC)50并通过Toll样受体(TLR)信号通路51触发先天性免疫应答。转导DC的能力取决于AAV血清型和基因组,并可以预测整体的免疫原性52。
为了评估AAV递送的CRISPR-Cas系统的免疫正交性,申请人分析了91种Cas9直系同源物和167种AAV VP1直系同源物。通过比较总序列相似性以及预测的与MHC I类和II类分子的结合强度,申请人构建了免疫交叉反应性图,并计算了在免疫原性谱图中正交的蛋白质的群。尽管MHC表位不能预测抗体表位,但更强大的记忆应答的诱导主要取决于记忆B细胞的再激活,其借助记忆T细胞通过在MHC II类分子上呈递抗原来实现。53,54最后,申请人通过测定治疗小鼠的蛋白质靶向抗体的诱导实验性地证实了这些免疫学预测。
对AAV和Cas9的体液免疫应答
顺序基因疗法治疗的主要障碍之一是存在由首次施用该疗法诱导的针对递送载体和转基因货物的中和抗体。为了确定对AAV-8衣壳和Cas9转基因的体液免疫应答动力学,申请人首先向C57BL/6J小鼠的眼眶后注射了1012vg的AAV-8-SaCas9,以靶向蛋白转化酶枯草杆菌蛋白酶/kexin9型(PCSK9),其是一种有前景的基因靶,在被破坏时可以降低低密度脂蛋白(LDL)的水平并预防心血管疾病。与先前的研究55一致,由于成功的SaCas9介导的基因编辑,小鼠在注射后的第一周就具有降低的PCSK9血清水平,这在整个实验期间(4周)得以维持(图5C)。值得注意的是,小鼠在注射后一周内对AAV8衣壳产生了体液免疫(图5D)。另外,申请人注意到,一部分小鼠产生了针对SaCas9蛋白的IgG1抗体(图5E)。为了评估多次给药AAV-Cas9的可行性,接下来申请人调查了是否存在AAV和Cas9直系同源物的免疫正交集。
鉴定免疫正交蛋白
自然选择产生了以直系同源基因形式的具有保守功能的多样的结构变体。申请人测定了这种多样性对91种II型Cas9直系同源物和167种AAV直系同源物的免疫交叉反应性的相关性:通过首先比较这91种II型Cas9直系同源物和167种AAV直系同源物的总体氨基酸序列相似性,并且其次使用了更特定的约束条件,即,其相应的氨基酸序列如何被预测与MHC I型和II型分子结合(图5F)。从这些分析中,申请人首先获得了对纯粹基于序列水平的Cas9和AAV直系同源物之间的综合免疫重叠的估计,其次基于预测的MHC结合获得对预测免疫重叠的更严格的估计。通过序列水平的聚类(clustering)和寻群方法,申请人定义了许多Cas9直系同源物集,其包含最多达9个成员且没有6-mer重叠(图7)。值得注意的是,基于MHC结合预测,申请人发现在Cas9直系同源物集中,有83%的配对被预测为具有非交叉反应的免疫应答,即,它们被预测为在免疫空间中是正交的(图5G)。相反,在AAV衣壳(VP1蛋白)直系同源物中,即使在用MHC-结合强度约束预测时,申请人也没有发现最高达16-mer水平的完全正交性(图5H),这可能反映了AAV的强序列保守性和较短的进化史56。该分析表明,与以前的观察结果一致57,58,暴露于一种AAV血清型可以诱导对所有AAV的广泛免疫,这对AAV递送平台提出了重大挑战,因为某些血清型在人类群体中很普遍。尽管差异最大的AAV血清型(AAV-5)显示出共享的免疫原性肽最少,但在VP1直系同源物内仍然存在完全保守的序列片段。如所期望的,尽管存在未被该回归(regression)捕获的显著变化,但是预测的免疫交叉反应与系统发育距离负相关(图8),这表明MHC结合预测可以改善仅系统发育距离之外的序列直系同源物的选择。
确认Cas9蛋白之间的体液免疫正交性
为了测试这些免疫学预测并建立该方法的实用性,申请人将范围缩小在包含普遍使用的酿脓链球菌Cas9以及充分表征的金黄色葡萄球菌Cas9的5个成员的群(图7)。为了确定这两种蛋白质是否具有交叉反应的抗体应答,申请人通过眼眶后注射向小鼠注射了1012vg的AAV8-SaCas9或AAV8-SpCas9,并在第0天(预注射)且在第4-6周定期收集血清(图6A)。在所有注射了SpCas9的小鼠(n=6)的血浆中检测到了SpCas9特异性抗体,值得注意的是注射SaCas9的小鼠(n=12)没有检测到该抗体(图6B)。尽管SaCas9似乎诱导了较弱的应答,但由于注射有SaCas9 AAV(n=12)的小鼠中只有一半对产生了针对SaCas9的可检测抗体,而注射有SpCas9 AAV(n=6)的小鼠均未产生针对SaCas9的抗体。这些结果在一项独立研究中得到了证实,其中在注射AAV-SpCas9的小鼠(n=12)血浆中检测出了SpCas9特异性抗体,但未检测到SaCas9特异性抗体。这些小鼠经眼眶后注射1012vg的AAV8-SpCas9或AAVDJ-SpCas9,并且在第4周还接受了另外的1011vg的肌内注射(图6C)。两者合计,该数据证实SpCas9和SaCas9具有体液免疫正交性。
AAV血清型之间广泛的交叉反应性
AAV由于能够避免诱导CD8+T细胞的强应答而成为一种首选的运载工具,然而,中和抗体的存在仍然是AAV治疗成功应用的重要障碍。与先前的结果一致,57申请人在所有各种人类AAV血清型中发现了共享的免疫原性肽(图9)。申请人证实两种血清型(AAV8和AAVDJ)缺乏正交性,其中申请人发现注射AAV8和AAVDJ的小鼠产生的抗体对AAV8和AAVDJ抗原两者都有反应(图6D)。这项分析表明,在所有HLA基因型中,还没有暴露于一种AVV可确保对另一种AVV没有免疫性的两种已知的AVV。但是,通过使用系统发育差异最大的血清型AAV5可以最小化免疫交叉反应58,59。这些预测仅确定了AAV5与小鼠模型中常用的AAV2和AAV8之间的单个共享的高度免疫原性肽(尽管存在其他几种具有轻度MHC亲和力的共享肽)。申请人通过ELISA证实了这一点,其中注射AAV2的小鼠未引发针对AAV5和AAV8的抗体,并且注射AAV5的小鼠未引发针对AAVDJ和AAV8的抗体(图6E)。
蛋白质疗法的使用需要规避宿主免疫应答的方法。例如,Cas9具有原核起源,并且可以引起T细胞应答,这可以导致转导细胞被清除。另外,循环的抗体可以中和AAV载体,并在重复给药后阻止有效的转导。抑制免疫力的药物可以缓解其中的这些方面中的一些,但并非没有明显的副作用,也不适合健康状况不佳的患者60-63。与癌症抗体疗法64相似,SpCas9蛋白也可以通过交换高免疫原性结构域来进行去免疫。这是一种有前景的方法,但是,由于申请人预期有数十种突变来实现隐身,因此它将是复杂且费力的,并且可能导致活性降低和整体疗效降低。
为了避免这个问题,申请人在这里开发了一个框架,通过检查蛋白质中所有k-mers是否存在于另一蛋白的滑动窗口,来比较蛋白质直系同源物及其与MHC I和MHC II的预测结合,专注于被预测为与至少一个MHC等位基因结合的肽。通过该分析,申请人鉴定了免疫正交的Cas9蛋白群。根据这些预测,来自一个直系同源物的特异性T细胞应答不会与同一个群的另一个直系同源物发生交叉反应,从而阻止了CD8+细胞毒性T细胞的再激活,并阻止了再激活记忆性B细胞所需要的CD4+T细胞的帮助。申请人通过ELISA证实了这些结果,并验证了两个充分表征的Cas9蛋白(即SpCas9和SaCas9)是免疫正交的。因此,申请人期望可以顺序使用属于同一群的蛋白质,而不会引起记忆性T细胞和B细胞应答。
由于AAV在基因疗法中作为递送剂的重要性,申请人还通过该MHC I和MHC II比较框架分析了AAV血清型,并证明没有两个AAV是相互免疫正交的。然而,利用已知的HLA基因型,可以使用当前定义的血清型来定义免疫正交的AAV的个性化方案。例如,AAV5的使用使得小鼠和灵长类动物的免疫交叉反应最小化,因为最近一项研究证实,其中嵌合AAV5免疫的小鼠和灵长类动物成功接受了第二剂AAV1的治疗59。然而,在人类环境中,申请人预测,AAV5与其他AAV之间将存在实质上更多的免疫重叠。该分析建议,创建一对用于人类应用的全面正交的AAV衣壳需要在两种蛋白质之一中进行10个突变[0053]。这种假设的正交AAV衣壳提出了重大的工程挑战,因为它需要对许多最保守的区域进行突变,以实现免疫正交性。
先前的工作已经证实,MHC亲和力高度依赖于结合袋(pocket)中的任一端的锚残基56。残基多样性在结合口袋的中心更容易被接受,尽管可能最影响抗原特异性的是这些残基,因为它们被认为是与T细胞受体(TCR)相互作用的核心。将9-mer空间中直系同源物对的数量与基于II类结合预测而预测的直系同源物配对的数量进行比较,表明即使在申请人在此探索的成千上万个HLA-2中,只有约65%的9-mer肽用作合适的MHC II类结合核心。MHC分子对肽空间的这种采样不足很可能反映了对疏水锚残基的需求,并通过将免疫原性肽突变为从未用作MHC结合核心的肽而留出了一些蛋白质去免疫的空间。然而,在保持蛋白质功能的同时实现这一点已被证明即使对于很少的HLA等位基因也是困难的,并且仍然是重大的蛋白质工程挑战。
申请人也注意到了这项工作的一些局限性。主要是,申请人使用了近亲交配的C57BL/6J作为小鼠模型,该模型具有非常有限的MHC多样性66,并且可能无法概括其他人类免疫学特征(诸如抗原处理和呈递的差异)。在这种情况下,申请人尝试用ELISPOT测定法测量所预测的一小部分MHC II肽的T细胞应答,并且确实证实了针对某些MHC II肽的免疫原性,尽管申请人也注意到,C57BL/6J小鼠对AAV-CRISPR总体上没有显示出强烈的应答(图10)。展望未来,可以使用其他小鼠模型(诸如表达人类HLA同种异型的小鼠)来潜在地重复这项工作,但是,这些模型面临着自身的技术挑战,诸如受限制的HLA等位基因(仅代表主要的MHC II亚组)以及受限制的TCR库66。另外,还可以预测B细胞表位并将其整合到免疫正交性分析中。但是,由于B细胞表位可以是线性和构象的,因此更难以预测。这些经由计算机模拟的模型的进展和进一步验证使得能够在将来进行更好的预测67-71。最后,最近的工作表明,MHC I类肽会对蛋白酶体处理产生的剪接的宿主和病原体衍生的肽具有重大贡献72。目前尚不清楚这会如何影响申请人预测是免疫正交的蛋白质的交叉识别。一方面,它提供了一种机制,使得剪接到相同宿主蛋白上的非常短的抗原序列可能导致对本质上不同的外来抗原的交叉识别,然而,申请人希望这是不可能的,因为抗原和整个宿主蛋白质组之间有大量可能的剪接肽。
总体而言,申请人认为,该框架为有效的基因疗法提供了潜在的解决方案,不仅可以用于Cas9介导的基因组工程,而且还可以用于可能需要重复治疗的其他蛋白质疗法。尽管使用这种方法仍然需要减轻初次免疫应答,特别是CTL清除,但申请人希望表位缺失和低免疫原性递送载体(诸如AAV)可以缓解此问题,并且重复给药的可能性将减少对非常高的第一给药效率的需求。
计算方法
对于Cas9,申请人选择了对Cas9蛋白的多样性进行分类的探索性研究中引用的91个直系同源物73,其中包括在实验上已经充分表征的几个。对于AAV,申请人分析了167个序列,重点研究了所有13种表征的人血清型,以及从恒河猴分离的一种(rh32)、一种经工程化的变体(DJ)和一种重构的祖先蛋白(Anc80L65)。然后,申请人比较了这些蛋白质之间的总序列相似性(非免疫学获知的),并预测了与MHC I类和MHC II类分子的结合(免疫学获知的)。通过检查蛋白质中所有连续k-mer是否以零或一个错配存在于另一蛋白质序列的滑动窗口来进行非免疫学获知的序列比较。免疫学获知的(immunologically informed)序列比较以类似的方式进行,但仅使用预测为对I类使用netMHC 4.074与81个HLA-1等位基因中的至少一个(等位基因可以在http://www.cbs.dtu.dk/services/NetMHC/MHC_allele_names.txt上找到)、以及对II类使用netMHCIIpan 3.175基于936个HLA-2等位基因与5620种可能的MHC II分子中的至少一个(等位基因可以在http://www.cbs.dtu.dk/services/NetMHCIIpan-3.1/alleles_name.list上找到)结合的那些k-mer。申请人比较netMHC与其他免疫表位预测平台(诸如免疫表位数据库(iedb.org)76的使用,发现整个软件之间的一致性非常强。最终,申请人选择了netMHC,因为它支持更多的HLA等位基因。如果预测的亲和力在软件指南中建议的400,000个随机肽的测试文库的排名前2%,则将序列定义为结合。使用Bron-Kerbosch算法进行免疫正交群的产生。简而言之,以每个直系同源物作为顶点来构建图,其中边缘由连接顶点之间共享的免疫原性肽的数量来定义。其中所述集中的每一对都是免疫正交的蛋白质集构成一个群。使用BLOSUM 62矩阵(不包括插入缺失)测量蛋白质序列之间的系统发育距离。所有软件、输入和输出文件都可以在GitHub上获得。
实验方法
AAV生产
使用HEK293T细胞通过三重转染方法生产AAV2/8、AAV2/2、AAV2/DJ病毒颗粒,并通过碘克沙醇梯度纯化(Grieger等,2006)。转染的融合率在80%到90%之间。转染前2小时,将培养基替换为预热的培养基。每种病毒均在5×15cm的平板中产生,其中每个平板均使用PEI(1ug/uL线性PEI,在1x DPBS pH 4.5,使用HCl),以4∶1的PEI∶DNA质量比,用7.5μg pXR衣壳(pXR-8、pXR-2、pXR-DJ)、7.5μg重组转移载体和22.5μg pAd5辅助载体进行转染。将混合物在室温下温育10分钟,并且然后滴加到培养基上。72小时后收获病毒,并使用碘克沙醇密度梯度超速离心法进行纯化。然后使用100kDa过滤器(Millipore)用补充有50mM NaCl和0.0001%的Pluronic F68(Thermo Fisher)的1x PBS(pH 7.2)对病毒进行透析,至最终体积约为~1ml,并使用针对ITR区域的特异性引物,对照标准(ATCC VR-1616)用QPCR进行定量。
AA V-ITR-F:5’-CGGCCTCAGTGAGCGA-3’和
AA V-ITR-R:5’-GGAACCCCTAGTGATGGAGTT-3’。
动物研究
所有的动物程序均按照加利福尼亚大学圣地亚哥分校的协会动物护理和使用委员会(IACUC)批准的协议进行的。所有小鼠都是从杰克逊实验室获得的。在成年C57BL/6J小鼠(10周)中,以1×1012vg/小鼠通过眼眶后注射进行AAV注射。
ELISA
PCSK9:根据制造商的指南,使用Mouse Proprotein Convertase 9/PCSK9Quantikine ELISA试剂盒(R&D Systems)测量血清PCSK9的水平。简而言之,将血清样品在Calibrator稀释剂中以1∶200稀释,并使其能够在预包被有捕获抗体的微孔板上结合2h。然后,用PCSK9偶联物、接着用PCSK9底物溶液顺序地温育样品,在每个步骤之间进行间歇性洗涤。使用标准酶标仪(BioRad iMark)比色法评估血清中PCSK9的含量。
Cas9和AAV:将重组SpCas9蛋白(PNA Bio,目录号CP01)或SaCas9蛋白(ABM good,目录号K144)稀释到1x包被缓冲液(Bethyl)中,并用0.5μg包被96孔Nunc MaxiSorp板(ab210903)中的每个孔在4℃下过夜。对于AAV实验,使用1x包被缓冲液中的109vg AAV-2、AAV-5,AAV-8或AAV-DJ包被96孔Nuc MaxiSorp板中的每个孔。将板用350μl的1x洗涤缓冲液(Bethyl)洗涤3次,每次5分钟,然后在室温下用300μl的1x BSA封闭溶液(Bethyl)封闭2h。重复洗涤程序。以1∶40的稀释度添加血清样品,并将板在振动下于4℃孵育5小时。将孔洗涤3次,每次5分钟,然后将100μl的HRP标记的山羊抗小鼠IgG1(Bethyl;在1%BSA封闭溶液中以1∶100,000稀释)加入到每个板中。在室温下孵育1小时后,将孔洗涤4次,每次5分钟,然后向每个孔中添加100μl的TMB底物(Behtyl)。使用酶标仪(BioRad iMark)测量450nm处的光密度(OD)。
实施例7——极端微生物Cas9
申请人探索了从极端微生物物种中选择另外的直系同源物(在该极端微生物正常情况下预期不会与人接触)和/或从常见物种中选择在正常的微生物组中是高丰度的(尤其是在发育的早期阶段),且免疫系统对其具有发展的耐受的直系同源物的策略。
申请人从符合极端微生物、共生菌群、病原菌群和非极端环境物种的这些物种中挖掘了Cas9序列。使用这些序列,申请人探索了这些类别中Cas9的正交性,以鉴定哪些是不与已有免疫发生交叉反应的良好候选者的直系同源物(图11)。尽管极端微生物Cas9之间存在广泛的正交性,但与较大的共生菌群、病原菌群和环境种进行比较时,观察到了一些重叠的肽。少数Cas9直系同源物没有显示出实质性的重叠,因此这些可能是用于表征、测试和将来使用的有用候选者。此外,在极端环境中探索Cas9直系同源物的多样性可能会为免疫正交性提供其他有希望的目标。
等同物
除非另有定义,否则本文所用的所有技术和科学术语具有与该技术所属领域的普通技术人员通常所理解的相同含义。
本文说明性地描述的本技术可以在不存在本文未具体公开的任何一个或多个要素、一个或多个限制的情况下适当地实践。因此,例如,术语“包含(comprising)”、“包括(including)”、“含有(containing)”等应被广泛地且不受限制地理解。另外,本文所采用的术语和表达已被用作术语的说明而非限制,并且不旨在使用这样的术语和表达来排除所示出和描述的特征或其部分的任何等同物,但是应当认识到,在所要求保护的本技术的范围内可以进行各种修改。
因此,应当理解,本文提供的材料、方法和实施例代表优选的方面,是示例性的,并且不旨在限制本技术的范围。
在此已经广泛且概括地描述了本技术。落入一般公开范围内的每个较窄的物种和亚属分组也形成本技术的一部分。这包括对本技术的一般性描述,其附带条件或否定限制从该属中去除了任何主题,无论所切除的材料是否在本文中进行了具体叙述。
另外,在根据马库什组来描述本技术的特征或方面的情况下,本领域技术人员会意识到,本技术由此也根据马库什组的任何单个成员或成员的子组来描述。
本文提及的所有出版物、专利申请、专利和其他参考文献均通过引用的方式明确地并入本文,其程度与单独引用并入的程度相同。在有冲突的情况下,以本说明书(包括定义)为准。
其他方面在所附权利要求书中描述。
参考文献
1.Chew W,et al.(2016)A multifunctional AAV-CRISPR-Cas9 and its hostresponse.Nature Methods,13(10):868-874.
2.Wang D,Mou H,Li S,Li Y,Hough S,Tran K,et al.AdenovirusMediatedSomatic Genome Editing of Pten by CRISPR/Cas9 in Mouse Liver in Spite ofCas9-Specific Immune Responses.Hum Gene Ther.2015;26
3.Riechmann L,et al.(1988)Reshaping human antibodies fortherrapy.Nature 332:323-327.
4.Lundegaard C,et al.(2010)“Major Histocompatibility Complec Class IBinding Predictions as a Tool in Epitope Discovery.”Immunology 130.3(2010):309-318.PMC.Web.7 Nov.2016.
5.Massimo A,et al.(2016)Gapped sequence alignment using artificialneural networks:application to the MHC class I system.Bioinformatics,32(4):5117.
6.Fonfara I,et al.(2014)Phylogeny of Cas9 Determines FunctionalExchangeability of Dual-RNA and Cas9 among Orthologous Type II CRISPR-CasSystems.Nucleic Acids Research 42.4:2577-2590.
7.Choi Y and Chan AP(2015)PROVEAN web server:a tool to predict thefunetional effect of amino acid substitutions and indels.Bioinformatics 31(16):27452747.
8.Massimo Andreatta and Morten Nielsen.Gapped sequence alignmentusing artificial neural netwvorks:application to the MHC class Isystem.Bioinformatics,Feb 15;32(4):51172016.
9.Tong,SYC et al.(2015)Staphylococcus aureus Infections:Epidemiology,Pathophysiology,Clinical Manifestations,and Management.Clinical MicrobiologyReviews.28:603661.
10.Mingozzi,F.&High,K.A.Immune responses to AAV vectors:overcomingbarriers to successful gene therapy.Blood 122,23-36(2013).
11.Mays,L.E.&Wilson,J.M.The Complex and Evolving Story of T cellActivation to AAV Vector-encoded Transgene Products.Mol.Ther.19,16-27(2011).
12.Basner-Tschakarjan,E.,Bijjiga,E.&Martino,A.T.Pre-clinicalassessment of immune responses to adeno-associated virus(AAV)vectors.Front.Immunol.5,(2014).
13.Ertl,H.C.J.&High,K.A.Impact of AAV Capsid-Specific T-CellResponses on Design and Outcome of Clinical Gene Transfer Trials withRecombinant Adeno-Associated Viral Vectors:An Evolving Controversy.Hum.GeneTher.28,328-337(2017).
14.Kotterman,M.A.,Chalberg,T.W.&Schaffer,D.V.Viral Vectors for GeneTherapy:Translational and Clinical Outlook.Annu.Rev.Biomed.Eng.17,63-89(2015).
15.Mingozzi,F.&High,K.A.Therapeutic in vivo gene transfer for geneticdisease using AAV:progress and challenges.Nat.Rev.Genet.12,341-355(2011).
16.Manno,C.S.et al.Successful transduction of liver in hemophilia byAAV-Factor IX and limitations imposed by the host immune response.Nat.Med.12,342-347(2006).
17.Sathish,J.G.et al.Challenges and approaches for the development ofsafer immunomodulatory biologics.Nat Rev Drug Discov 12,306-324(2013).
18.Harding,F.A.,Stickler,M.M.,Razo,J.&DuBridge,R.B.The immunogenicityof humanized and fully human antibodies:Residual immunogenicity resides inthe CDR regioas.MAbs 2,256-265(2010).
19.De Groot,a S.,Knopp,P.M.&Martin,W.De-immunization of therapeuticproteins by T-cell epitope modification.Dev.Biol.(Basel).122,171-194(2005).
20.Tangri,S.et al.Rationally Engineered Therapeutic Proteins withReduced Immunogenicity.J.Immunol.174,3187LP-3196(2005).
21.Salvat,R.S.,Choi,Y.,Bishop,A.,Bailey-Kellogg,C.&Griswold,K.E.Protein deimmunization via strucmre-based design enables efficientepitope deletion at high mutational loads.Biotechnol.Bioeng.112,1306-1318(2015).
22.Armstrong,J.K.et al.Antibody against poly(ethylene glycol)adversely affeets PEG-asparaginase therapy in acute lymphoblastic leukemiapatients.Cancer 110,103-111(2007).
23.Ganson,N.J.,Kelly,S.J.,Scarlett,E.,Sundy,J.S.&Hershfield,M.S.Control of hyperuricemia in subjects with refractory gout,and inductionof antibody against poly(ethylene glycol)(PEG),in a phase I trial ofsubcutaneous PEGylated urate oxidase.Arthritis Res.Ther.8,R12-R12(2006).
24.Veronese,F.M.&Mero,A.The impact of PEGylation on biologicaltherapies.BioDrugs 22,315-329(2008).
25. S.,Kunstelj,M.&Porekar,V.G.PEGylation of therapeuticproteins.Biotechnol.J.5,113-128(2010).
S.,Kunstelj,M.&Porekar,V.G.PEGylation of therapeuticproteins.Biotechnol.J.5,113-128(2010).
26.Jacobs,F.,Gordts,S.C.,Muthuramu,I.&De Geest,B.The liver as atarget organ for gene therapy:state of the art,challenges,and futureperspectives.Pharmaceuticals(Basel).5,1372-92(2012).
27.Kok,C.Y.et al.Adeno-associated Virus-mediated Rescue of NeonatalLethality in Argininosuccinate Synthetase-deficient Mice.Mol.Ther.21,1823-1831(2013).
28.Courtenay-Luck,N.S.,Epenetos,A.A.&Moore,R.Development of primaryand secondary immune responses to mouse monoclonal antibodies used in thediagnosis and therapy of malignant neoplasms.Cancer Res.46,6489-6493(1986).
29.Jinek,M.et al.A Programmable Dual-RNA-Guided DNA Endonuclease inAdaptice Bacterial Immunity.Science 337,816-822(2012).
30.Mali,P.et al.RNA-guided human genome engineering via Cas9.Science339,823-6(2013).
31.Gasiunas,G.,Barrangou,R.,Horvath,P.&Siksnys,V.Cas9-crRNAribonucleoprotein complex mediates specific DNA cleavage for adaptiveimmunity in bacteria.Proc.Natl.Acad.Sci.109,E2579-E2586(2012).
32.Cong,L.et al.Multiplex genome engineering using CRISPR/Cassystems.Science 339,819-23(2013).
33.Ran,F.A.et al.In vivo genome editing using Staphylococcus aureusCas9.Nature 520,186-190(2015).
34.Jinek,M.et al.RNA-programmed genome editing in human cells.Elife2013,(2013).
35.Mali,P.,Esvelt,K.M.&Church,G.M.Cas9 as a versatile tool forengineering biology.Nat.Methods 10,957-963(2013).
36.Hsu,P.D.,Lander,E.S.&Zhang,F.Development and applications ofCRISPR-Cas9 for genome engineering.Cell 157,1262-1278(2014).
37.Makarova,K.S.et al.An updated evolutionary classification ofCRISPR-Cas systems.Nat.Rev.Microbiol.13,722-736(2015).
38.Chylinski,K.,Makarova,K.S.,Charpentier,E.&Koonin,E.V.Classification and evolution of type II CRISPR-Cas systems.Nucleic AcidsResearch 42,6091-6105(2014).
39.Shmakov,S.et al.Diversity and evolution of class 2 CRISPR-Cassystems.Nat.Rev.Microbiol.15,169-182(2017).
40.Wagner,J.a gt al.Safety and biological efficacy ofan adeno-associated virus vector-cystic fibrosis transmembrane regulator(AAV-CFTR)inthe cystic fibrosis maxillary sinus.Laryngoscope 109,266-74(1999).
41.Song,S.et al.Sustained secretion of human alpha-1-antitrypsinfrommurine muscle transduced with adeno-associated virus vectors.Proc.Natl.Acad.Sci.U.S.A.95,14384-8(1998).
42.Chirmule,N.et al.Humoral Immunity to Adeno-Associated Virus Type 2Vectors following Administration to Murine and Nonhuman PrimateMusclc.J.Virol.74,2420-2425(2000).
43.Fields,P.a et al.Risk and prevention of anti-factor IX formationin AAV-mediated gene transfer in the context of a large deletion ofF9.Mol.Ther.4,201-210(2001).
44.Herzog,R.W.et al.Influence of vector dose on factor IX-specific Tand B cell responses in muscle-directed gene therapy.Hum.Gene Ther.13,1281-91(2002).
45.Lozier,J.N.,Tayebi,N.&Zhang,P.Mapping of genes that controltheantibody responso to human factor IX in mice.Blood 105,1029-1035(2005).
46.Zhang,H.G.et al.Genetic analysis of the antibody response to AAV2and factor IX.Mol.Ther.11,866-874(2005).
47.Benveniste,O.et al.Prevalence of Serum IgG and NeutralizingFactors Against Adeno-Associated Virus(AAV)Types 1,2,5,6,8,and 9 in theHealthy Population:Implications for Gene Therapy Using AAV Vectors.Hum.GeneTher.21,704-712(2010).
48.Gao,G.-P.et al.Novel adeno-associated viruses from rhesus monkeysas vectors for human gene therapy.Proc.Natl.Acsd.Sci.99,11854-11859(2002).
49.Jooss,K.,Yang,Y.,Fisher,K.J.&Wilson,J.M.Transduction of DendriticCells by DNA Viral Vectors Directs the Immune Response to Transgene Productsin Muscle Fibers.J.Virol.72,4212-4223(1998).
50.Gernoux,G.et al.Early Interaction of Adeno-Associated VirusSerotype 8 Vector with the Host Immune System Following IntramuscularDelivery Results in Weak but Detectable Lymphocyte and Dendritic CellTransduction.Hum.Gene Ther.26,1-13(2015).
51.Zhu,J.,Huang,X.&Yang,Y.The TLR9-MyD88 pathway is critical foradaptive immune responses to adeno-associated virus gene therapy vectors inmice.J.Clin.Invest.119,2388-2398(2009).
52.Gernoux,G.,Wilson,J.M.&Mueller,C.Regulatory and Exhausted T CellResponses to AAV Capsid.Hum.Gene Ther.28,338-349(2017).
53.Kurosaki,T.,Kometani,K.&Ise,W.Memory B cells.Nat.Rev.Immunol.15,149-159(2015).
54.Zabel,F.et al.Distinct T helper cell dependence of memory B-cellproliferation versus plasma cell differentiation.Immunology 150,329-342(2017).
55.Ding,Q.et al.Permanent Alteration of PCSK9 With In Vivo CRISPR-Cas9 Genome Editing.Circ.Res.115,488-492(2014).
56.Zinn,E.et al.In Silico Reconstruction of the Viral EvolutionaryLineage Yields a Potent Gene Therapy Vector.Cell Rep.12,1056-1068(2017).
57.Calcedo,R.&Wilson,J.M.AAV Natural Infection Induces Broad Cross-Neutralizing Antibody Responses to Multiple AAV Serotypes inChimpanzees.Hum.Gene Ther.Clin.Dev.27,79-82(2016).
58.Harbison,C.E.et al.Examining the cross-reactivity andneutralization mechanisms of a panel of mabs against adeno-associated virusserotypes l and 5.J.Gen.Virol.93,(2012).
59.Majowicz,A.et al.Successful Repeated Hepatic Gene Delivery in Miceand Non-human Primates Achieved by Sequential Administration of AAV5ch andAAV1.Mol.Ther.25,1831-1842(2017).
60.McIntosh,J.H.et al.Successful attenuation of humoral immunity toviral capsid and transgenic protein following AAV-mediated gene transfer witha non-depleting CD4 antibody and cyclosporine.Gene Ther 19,78-85(2012).
61.Mingozzi,F.et al.Prevalence and pharmacological modulation ofhumoral immunity to AAV vectors in gene transfer to synovial tissue.Gene Ther20,417-424(2013).
62.Mingozzi,F.et al.Pharmacological Modulation of Humoral Immunity ina Nonhuman Primate Model of AAV Gene Transfer for Hemophilia B.Mol.Ther.20,1410-1416(2017).
63.Unzu,C.et al.Transient and intensive pharmacologicalimmunosuppression fails to improve AAV-based livet gene transfer in non-humanprimates.J.Transl.Med.10,122(2012).
64.Riechmann,L.,Clark,M.,Waldmann,H.&Winter,G.Reshaping humanantibodies for therapy.Nature 332,323-7(1988).
65.Rupperr,J.et al.Prominent role of secondary anchor residues inpeptide binding to HLA-A2.1 molecules.Cell 74,929-937(2017).
66.Baker,M.P.,Reynolds,H.M.,Lumicisi,B.&Bryson,C.J.Immunogenicity ofprotein therapeutic5:The key causes,consequences and challenges.Self Nonself1,314-322(2010).
67.EL-Manzalawy,Y.,Dobbs,D.&Honavar,V.Predicting linear B-cellepitopes using string kernels.J.Mol.Recognit.21,243-255(2008).
68.Larsen,J.E.P.,Lund,O.&Nielsen,M.Improved method for predictinglinear B-cell epitopes.Immunome Res.2,2(2006).
69.Sollner,J.et al.Analysis and prediction of protective continuousB-cell epitopes on pathogen proteins.Immunome Res.4,1(2008).
70.Dalkas,G.A.&Rooman,M.SEPIa,a knowledge-driven algorithm forpredicting conformational B-cell epitopes from the amino acid sequence.BMCBioinformatics 18,95(2017).
71.Sun,P.et al.Bioinformatics resources and tools for conformationalB-cell epitope prediction.Computational and Mathematical Methods in Medicine2013,(2013).
72.Liepe,J.et al.A large fraction of HLA class I ligands areproteasome-generated spliced peptides.Science(80-.).354,(2016).
73.Fonfara,I.et al.Phylogeny of Cas9 determines functionalexchangeability of dual-RNA and Cas9 among orthologous type II CRISPR-Cassystems.Nucleic Acids Res.42,2577-2590(2014).
74.Andreatta,M.&Nielsen,M.Gapped sequence alignment using artificialneural networks:application to the MHC class I system.Bioinformatics 32,511-517(2015).
75.Andreatta,M.et al.Accurate pan-specific prediction of peptide-MHCclass II binding affinity with improved binding coreidentification.Immunogenetics 67,641-650(2015).
76.Vita,R.et al.The immune epitope database(IEDB)3.0.Nucleic AcidsRes.43,D405-12(2015).
77.Gdell,M.,Yang,L.&Church,G.M.Genome editing assessment using CRISPRGenome Analyzer(CRISPR-GA).Bioinformatics 30,2968-2970(2014).

表2





表3




Claims (31)
1.一种产生蛋白质的方法,包括:
鉴定对主要组织相容性复合体(MHC)具有亲和力的蛋白质的一个或多个区域,以及
通过一个或多个氨基酸替代修饰所述对MHC具有亲和力的蛋白质的一个或多个区域,从而使得经修饰的区域对所述MHC没有亲和力,
其中,当将所得的经修饰的蛋白质或编码所述经修饰的蛋白质的多核苷酸施用于受试者时,所述经修饰的蛋白质是免疫沉默的。
2.根据权利要求1所述的方法,其中,对所述MHC的亲和力是高亲和力。
3.根据权利要求1或2所述的方法,其中,至少一个经替代的氨基酸是不用作MHC蛋白核心残基的氨基酸。
4.根据权利要求1-3中任一项所述的方法,其中,所述蛋白质选自以下组:胞苷脱氨酶、腺苷脱氨酶、锌指核酸酶、转录激活因子样效应物核酸酶、Cas9或AAV衣壳蛋白。
5.根据权利要求4所述的方法,其中,所述蛋白质是Cas9。
6.根据权利要求5所述的方法,其中,所述Cas9是SpCas9。
7.一种根据权利要求1-6中任一项所述的方法产生的经修饰的Cas9蛋白。
8.一种经修饰的Cas9蛋白,包含一种或更多种、两种或更多种、三种或更多种、四种或更多种、五种或更多种、六种或更多种、七种或更多种、八种或更多种、九种或更多种、十种或更多种、十五种或更多种、二十种或更多种表1中提供的氨基酸修饰。
9.一种分离的多核苷酸,编码权利要求7或8所述的经修饰的Cas9蛋白。
10.一种载体,包含权利要求9所述的分离的多核苷酸。
11.根据权利要求10所述的载体,其中,所述载体是AAV载体,任选地其中所述AAV载体是AAV5。
12.一种AVV衣壳,包含权利要求11所述的载体。
13.根据权利要求12所述的AAV衣壳,其中,所述AAV衣壳蛋白中的一种或多种已经根据权利要求1-4中任一项所述的方法进行了修饰。
14.一种在被施用需要Cas9的方案的受试者中避免免疫应答的方法,所述方法包括:对所述受试者顺序施用两种或更多种免疫正交的Cas9蛋白。
15.一种对受试者进行基因编辑或基因调控的方法,所述方法包括:对所述受试者顺序施用两种或更多种免疫正交的Cas9蛋白。
16.一种治疗需要基因编辑或基因调控的受试者的方法,所述方法包括:对所述受试者顺序施用两种或更多种免疫正交的Cas9蛋白或编码所述Cas9蛋白的多核苷酸。
17.根据权利要求14-16中任一项所述的方法,其中,所述免疫正交的Cas9蛋白不共享大于5个连续氨基酸的氨基酸序列。
18.根据权利要求14-17中任一项所述的方法,其中,所述免疫正交的Cas9蛋白不共享对主要组织相容性复合体(MHC)的亲和力。
19.根据权利要求14-18中任一项所述的方法,其中,顺序施用三种或更多种、四种或更多种、五种或更多种、六种或更多种、七种或更多种、八种或更多种、九种或更多种、或十种或更多种免疫正交的Cas9蛋白。
20.根据权利要求14-19中任一项所述的方法,其中,每种免疫正交的Cas9蛋白是衍生自不同种的细菌的Cas9。
21.根据权利要求20所述的方法,其中,所述免疫正交的Cas9蛋白选自酿脓链球菌(S.pyogenes)Cas9(spCas9)、金黄色葡萄球菌(S.aureus)Cas9(saCas9)、长双歧杆菌(B.longum)Cas9、嗜黏蛋白艾克曼菌(A.muiciniphilia)Cas9或O.laneus Cas9。
22.根据权利要求21所述的方法,其中,所述免疫正交的Cas9蛋白包括spCas9和saCas9。
23.根据权利要求14-22中任一项所述的方法,其中,所述两种或更多种Cas9蛋白中的至少一种被修饰,以在施用于所述受试者时降低免疫原性。
24.根据权利要求23所述的方法,其中,所述两种或更多种Cas9蛋白中的至少一种是根据权利要求1-6中任一项所述的方法修饰的。
25.根据权利要求14-24中任一项所述的方法,其中,所述两种或更多种Cas9蛋白或编码所述Cas9蛋白的多核苷酸中的至少一种包含在AAV载体中。
26.根据权利要求25所述的方法,其中,所述AAV载体是AAV5载体。
27.根据权利要求25或26所述的方法,其中,所述AAV载体包含在AAV衣壳中。
28.根据权利要求25-27中任一项所述的方法,其中,两种或更多种Cas9蛋白或编码所述Cas9蛋白的多核苷酸包含在AAV载体中。
29.根据权利要求28所述的方法,其中,每种AAV载体包含在AAV衣壳中,任选地其中所述AAV衣壳彼此免疫正交。
30.根据权利要求14-29中任一项所述的方法,还包括向所述受试者施用一种或多种指导RNA。
31.根据权利要求30所述的方法,其中,所述指导RNA被选择用于治疗选自以下的疾病、病症或病况:全色盲、腺苷脱氨酶缺乏症、α1-抗胰蛋白酶缺乏症、阿尔茨海默病、肌萎缩侧索硬化、芳香族氨基酸脱羧酶缺乏症、巴顿病、无脉络膜、克里格勒-纳贾尔综合征、囊性纤维化、脆性X综合征、血友病、乙型肝炎、丙型肝炎、纯合子家族性高胆固醇血症、亨廷顿病、Leber先天性黑朦、黄斑变性、枫糖尿病(MSUD)、粘多糖病(I-IX)、多发性硬化、肌肉不良、强直性肌营养不良、1型神经纤维瘤病、鸟氨酸转氨甲酰酶缺乏症、先天性甲肥厚、帕金森氏病、苯丙酮尿症、多囊肾病、庞贝病、视网膜变性、瑞特氏综合症、佝偻病、脊髓性肌萎缩、重症联合免疫缺陷、镰状细胞病、史-莱-奥综合征、Y连锁非梗阻性生精障碍、地中海贫血、泰-萨克斯病、威尔逊氏病、心血管疾病、代谢综合征、疼痛管理和X连锁视网膜劈裂症。
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201762471267P | 2017-03-14 | 2017-03-14 | |
| US62/471,267 | 2017-03-14 | ||
| US201862614875P | 2018-01-08 | 2018-01-08 | |
| US62/614,875 | 2018-01-08 | ||
| PCT/US2018/022258 WO2018170015A1 (en) | 2017-03-14 | 2018-03-13 | Engineering crispr cas9 immune stealth |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| CN110913881A true CN110913881A (zh) | 2020-03-24 |
Family
ID=63522593
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN201880031835.XA Pending CN110913881A (zh) | 2017-03-14 | 2018-03-13 | 工程化crispr cas9免疫隐身 |
Country Status (7)
| Country | Link |
|---|---|
| US (1) | US11332727B2 (zh) |
| EP (1) | EP3595694A4 (zh) |
| JP (1) | JP2020513814A (zh) |
| CN (1) | CN110913881A (zh) |
| GB (1) | GB2574561A (zh) |
| IL (1) | IL268829A (zh) |
| WO (1) | WO2018170015A1 (zh) |
Cited By (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN113717960A (zh) * | 2021-08-27 | 2021-11-30 | 电子科技大学 | 一种新Cas9蛋白、CRISPR-Cas9基因组定向编辑载体及基因组编辑方法 |
| CN116751764A (zh) * | 2023-05-08 | 2023-09-15 | 珠海舒桐医疗科技有限公司 | 一种Cas9蛋白、II型CRISPR/Cas9基因编辑系统及应用 |
Families Citing this family (36)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CA2853829C (en) | 2011-07-22 | 2023-09-26 | President And Fellows Of Harvard College | Evaluation and improvement of nuclease cleavage specificity |
| US20150044192A1 (en) | 2013-08-09 | 2015-02-12 | President And Fellows Of Harvard College | Methods for identifying a target site of a cas9 nuclease |
| US9359599B2 (en) | 2013-08-22 | 2016-06-07 | President And Fellows Of Harvard College | Engineered transcription activator-like effector (TALE) domains and uses thereof |
| US9388430B2 (en) | 2013-09-06 | 2016-07-12 | President And Fellows Of Harvard College | Cas9-recombinase fusion proteins and uses thereof |
| US9737604B2 (en) | 2013-09-06 | 2017-08-22 | President And Fellows Of Harvard College | Use of cationic lipids to deliver CAS9 |
| US9228207B2 (en) | 2013-09-06 | 2016-01-05 | President And Fellows Of Harvard College | Switchable gRNAs comprising aptamers |
| US11053481B2 (en) | 2013-12-12 | 2021-07-06 | President And Fellows Of Harvard College | Fusions of Cas9 domains and nucleic acid-editing domains |
| US10077453B2 (en) | 2014-07-30 | 2018-09-18 | President And Fellows Of Harvard College | CAS9 proteins including ligand-dependent inteins |
| CA2963820A1 (en) | 2014-11-07 | 2016-05-12 | Editas Medicine, Inc. | Methods for improving crispr/cas-mediated genome-editing |
| EP3294896A1 (en) | 2015-05-11 | 2018-03-21 | Editas Medicine, Inc. | Optimized crispr/cas9 systems and methods for gene editing in stem cells |
| JP7396783B2 (ja) | 2015-06-09 | 2023-12-12 | エディタス・メディシン、インコーポレイテッド | 移植を改善するためのcrispr/cas関連方法および組成物 |
| US11667911B2 (en) | 2015-09-24 | 2023-06-06 | Editas Medicine, Inc. | Use of exonucleases to improve CRISPR/CAS-mediated genome editing |
| IL294014B2 (en) | 2015-10-23 | 2024-07-01 | Harvard College | Nucleobase editors and their uses |
| US11597924B2 (en) | 2016-03-25 | 2023-03-07 | Editas Medicine, Inc. | Genome editing systems comprising repair-modulating enzyme molecules and methods of their use |
| EP4047092A1 (en) | 2016-04-13 | 2022-08-24 | Editas Medicine, Inc. | Cas9 fusion molecules, gene editing systems, and methods of use thereof |
| IL308426A (en) | 2016-08-03 | 2024-01-01 | Harvard College | Adenosine nuclear base editors and their uses |
| US11661590B2 (en) | 2016-08-09 | 2023-05-30 | President And Fellows Of Harvard College | Programmable CAS9-recombinase fusion proteins and uses thereof |
| US11542509B2 (en) | 2016-08-24 | 2023-01-03 | President And Fellows Of Harvard College | Incorporation of unnatural amino acids into proteins using base editing |
| SG11201903089RA (en) | 2016-10-14 | 2019-05-30 | Harvard College | Aav delivery of nucleobase editors |
| WO2018119359A1 (en) | 2016-12-23 | 2018-06-28 | President And Fellows Of Harvard College | Editing of ccr5 receptor gene to protect against hiv infection |
| EP4095263A1 (en) | 2017-01-06 | 2022-11-30 | Editas Medicine, Inc. | Methods of assessing nuclease cleavage |
| US11898179B2 (en) | 2017-03-09 | 2024-02-13 | President And Fellows Of Harvard College | Suppression of pain by gene editing |
| EP3592777A1 (en) | 2017-03-10 | 2020-01-15 | President and Fellows of Harvard College | Cytosine to guanine base editor |
| JP7191388B2 (ja) | 2017-03-23 | 2022-12-19 | プレジデント アンド フェローズ オブ ハーバード カレッジ | 核酸によってプログラム可能なdna結合蛋白質を含む核酸塩基編集因子 |
| WO2018201086A1 (en) | 2017-04-28 | 2018-11-01 | Editas Medicine, Inc. | Methods and systems for analyzing guide rna molecules |
| US11560566B2 (en) | 2017-05-12 | 2023-01-24 | President And Fellows Of Harvard College | Aptazyme-embedded guide RNAs for use with CRISPR-Cas9 in genome editing and transcriptional activation |
| WO2018227114A1 (en) | 2017-06-09 | 2018-12-13 | Editas Medicine, Inc. | Engineered cas9 nucleases |
| US11866726B2 (en) | 2017-07-14 | 2024-01-09 | Editas Medicine, Inc. | Systems and methods for targeted integration and genome editing and detection thereof using integrated priming sites |
| CN111801345A (zh) | 2017-07-28 | 2020-10-20 | 哈佛大学的校长及成员们 | 使用噬菌体辅助连续进化(pace)的进化碱基编辑器的方法和组合物 |
| US11319532B2 (en) | 2017-08-30 | 2022-05-03 | President And Fellows Of Harvard College | High efficiency base editors comprising Gam |
| CN111757937A (zh) | 2017-10-16 | 2020-10-09 | 布罗德研究所股份有限公司 | 腺苷碱基编辑器的用途 |
| BR112020020266A2 (pt) * | 2018-04-03 | 2021-01-19 | Stridebio, Inc. | Vetores de vírus com evasão de anticorpos |
| WO2020191243A1 (en) | 2019-03-19 | 2020-09-24 | The Broad Institute, Inc. | Methods and compositions for editing nucleotide sequences |
| CN110499334A (zh) * | 2019-08-08 | 2019-11-26 | 复旦大学 | CRISPR/SlugCas9基因编辑系统及其应用 |
| DE112021002672T5 (de) | 2020-05-08 | 2023-04-13 | President And Fellows Of Harvard College | Vefahren und zusammensetzungen zum gleichzeitigen editieren beider stränge einer doppelsträngigen nukleotid-zielsequenz |
| WO2024112479A1 (en) * | 2022-11-22 | 2024-05-30 | The Regents Of The University Of California | Crispr-cas effector polypeptides, lipid nanoparticles, and methods of use thereof |
Citations (6)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN1523996A (zh) * | 2001-07-13 | 2004-08-25 | Ĭ��ר������˾ | 降低多肽免疫原性的方法 |
| CN1524090A (zh) * | 2001-03-20 | 2004-08-25 | Ĭ��ר������˾ | 免疫原性减弱的经修饰胰岛素 |
| US20140271550A1 (en) * | 2013-03-14 | 2014-09-18 | The Trustees Of The University Of Pennsylvania | Constructs and Methods for Delivering Molecules via Viral Vectors with Blunted Innate Immune Responses |
| WO2015153789A1 (en) * | 2014-04-01 | 2015-10-08 | Editas Medicine, Inc. | Crispr/cas-related methods and compositions for treating herpes simplex virus type 1 (hsv-1) |
| US20160017295A1 (en) * | 2013-05-31 | 2016-01-21 | The Regents Of The University Of California | Adeno-associated virus variants and methods of use thereof |
| WO2016033246A1 (en) * | 2014-08-27 | 2016-03-03 | Caribou Biosciences, Inc. | Methods for increasing cas9-mediated engineering efficiency |
Family Cites Families (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US6730507B1 (en) | 1993-06-24 | 2004-05-04 | Merck & Co., Inc. | Use of helper-dependent adenoviral vectors of alternative serotypes permits repeat vector administration |
| WO2002066514A2 (en) | 2001-02-19 | 2002-08-29 | Merck Patent Gmbh | Artificial fusion proteins with reduced immunogenicity |
| DK2800811T3 (en) * | 2012-05-25 | 2017-07-17 | Univ Vienna | METHODS AND COMPOSITIONS FOR RNA DIRECTIVE TARGET DNA MODIFICATION AND FOR RNA DIRECTIVE MODULATION OF TRANSCRIPTION |
| KR20160056869A (ko) * | 2013-06-17 | 2016-05-20 | 더 브로드 인스티튜트, 인코퍼레이티드 | 바이러스 구성성분을 사용하여 장애 및 질환을 표적화하기 위한 crispr-cas 시스템 및 조성물의 전달, 용도 및 치료 적용 |
| CN105517579B (zh) * | 2013-07-10 | 2019-11-15 | 哈佛大学校长及研究员协会 | 用于RNA向导的基因调节和编辑的正交Cas9蛋白 |
-
2018
- 2018-03-13 WO PCT/US2018/022258 patent/WO2018170015A1/en unknown
- 2018-03-13 EP EP18768009.5A patent/EP3595694A4/en not_active Withdrawn
- 2018-03-13 JP JP2019550861A patent/JP2020513814A/ja active Pending
- 2018-03-13 GB GB1913821.3A patent/GB2574561A/en not_active Withdrawn
- 2018-03-13 CN CN201880031835.XA patent/CN110913881A/zh active Pending
- 2018-03-13 US US16/494,269 patent/US11332727B2/en active Active
-
2019
- 2019-08-21 IL IL26882919A patent/IL268829A/en unknown
Patent Citations (6)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN1524090A (zh) * | 2001-03-20 | 2004-08-25 | Ĭ��ר������˾ | 免疫原性减弱的经修饰胰岛素 |
| CN1523996A (zh) * | 2001-07-13 | 2004-08-25 | Ĭ��ר������˾ | 降低多肽免疫原性的方法 |
| US20140271550A1 (en) * | 2013-03-14 | 2014-09-18 | The Trustees Of The University Of Pennsylvania | Constructs and Methods for Delivering Molecules via Viral Vectors with Blunted Innate Immune Responses |
| US20160017295A1 (en) * | 2013-05-31 | 2016-01-21 | The Regents Of The University Of California | Adeno-associated virus variants and methods of use thereof |
| WO2015153789A1 (en) * | 2014-04-01 | 2015-10-08 | Editas Medicine, Inc. | Crispr/cas-related methods and compositions for treating herpes simplex virus type 1 (hsv-1) |
| WO2016033246A1 (en) * | 2014-08-27 | 2016-03-03 | Caribou Biosciences, Inc. | Methods for increasing cas9-mediated engineering efficiency |
Cited By (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN113717960A (zh) * | 2021-08-27 | 2021-11-30 | 电子科技大学 | 一种新Cas9蛋白、CRISPR-Cas9基因组定向编辑载体及基因组编辑方法 |
| CN116751764A (zh) * | 2023-05-08 | 2023-09-15 | 珠海舒桐医疗科技有限公司 | 一种Cas9蛋白、II型CRISPR/Cas9基因编辑系统及应用 |
| CN116751764B (zh) * | 2023-05-08 | 2024-01-30 | 珠海舒桐医疗科技有限公司 | 一种Cas9蛋白、II型CRISPR/Cas9基因编辑系统及应用 |
Also Published As
| Publication number | Publication date |
|---|---|
| IL268829A (en) | 2019-10-31 |
| GB201913821D0 (en) | 2019-11-06 |
| EP3595694A4 (en) | 2021-06-09 |
| JP2020513814A (ja) | 2020-05-21 |
| EP3595694A1 (en) | 2020-01-22 |
| US20200299657A1 (en) | 2020-09-24 |
| WO2018170015A1 (en) | 2018-09-20 |
| GB2574561A (en) | 2019-12-11 |
| US11332727B2 (en) | 2022-05-17 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| CN110913881A (zh) | 工程化crispr cas9免疫隐身 | |
| Moreno et al. | Immune-orthogonal orthologues of AAV capsids and of Cas9 circumvent the immune response to the administration of gene therapy | |
| JP7498665B2 (ja) | 新規アデノ随伴ウイルス(aav)ベクター、低減されたカプシド脱アミド化を有するaavベクター、およびその使用 | |
| US11466258B2 (en) | Methods of predicting ancestral virus sequences and uses thereof | |
| US20200407750A1 (en) | Novel adeno-associated virus (aav) vectors, aav vectors having reduced capsid deamidation and uses therefor | |
| Herrmann et al. | A robust and all-inclusive pipeline for shuffling of adeno-associated viruses | |
| US20220298500A1 (en) | Compositions for regulating and self-inactivating enzyme expression and methods for modulating off-target activity of enzymes | |
| US20230002788A1 (en) | Aav3b variants with improved production yield and liver tropism | |
| EP4256046A1 (en) | Engineering immune orthoganol aav and immune stealth crispr-cas | |
| Moreno | Engineering efficient and safe in situ genome regulation via CRISPR-Cas9 for enabling gene therapies | |
| Wang et al. | Delivery of CRISPR/Cas9 system by AAV as vectors for gene therapy | |
| WO2024015966A2 (en) | Recombinant aav having aav clade d and clade e capsids and compositions containing same | |
| RU2774874C2 (ru) | Печень-специфичные конструкции, экспрессионные кассеты фактора viii и способы их применения | |
| Raghavan | Engineering minimally immunogenic cargos and delivery modalities for gene therapy | |
| CN116875597A (zh) | 修正lmna心肌病致病突变的载体及其应用 | |
| Weinstein | New methods in engineering adeno-associated virus (AAV) for improved gene delivery |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| PB01 | Publication | ||
| PB01 | Publication | ||
| SE01 | Entry into force of request for substantive examination | ||
| SE01 | Entry into force of request for substantive examination | ||
| WD01 | Invention patent application deemed withdrawn after publication |
Application publication date: 20200324 |
|
| WD01 | Invention patent application deemed withdrawn after publication |