WO2008009895A2 - Matriptase-2 - Google Patents
Matriptase-2 Download PDFInfo
- Publication number
- WO2008009895A2 WO2008009895A2 PCT/GB2007/002598 GB2007002598W WO2008009895A2 WO 2008009895 A2 WO2008009895 A2 WO 2008009895A2 GB 2007002598 W GB2007002598 W GB 2007002598W WO 2008009895 A2 WO2008009895 A2 WO 2008009895A2
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- matriptase
- cells
- polypeptide
- nucleic acid
- acid molecule
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Ceased
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/113—Non-coding nucleic acids modulating the expression of genes, e.g. antisense oligonucleotides; Antisense DNA or RNA; Triplex- forming oligonucleotides; Catalytic nucleic acids, e.g. ribozymes; Nucleic acids used in co-suppression or gene silencing
- C12N15/1137—Non-coding nucleic acids modulating the expression of genes, e.g. antisense oligonucleotides; Antisense DNA or RNA; Triplex- forming oligonucleotides; Catalytic nucleic acids, e.g. ribozymes; Nucleic acids used in co-suppression or gene silencing against enzymes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/48—Hydrolases (3) acting on peptide bonds (3.4)
- C12N9/50—Proteinases, e.g. Endopeptidases (3.4.21-3.4.25)
- C12N9/64—Proteinases, e.g. Endopeptidases (3.4.21-3.4.25) derived from animal tissue
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y304/00—Hydrolases acting on peptide bonds, i.e. peptidases (3.4)
- C12Y304/21—Serine endopeptidases (3.4.21)
- C12Y304/21109—Matriptase (3.4.21.109)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/10—Type of nucleic acid
- C12N2310/12—Type of nucleic acid catalytic nucleic acids, e.g. ribozymes
Definitions
- the invention relates to a novel agent for treating prostate cancer; a composition including same; means for increasing the cellular levels of either exogenous or endogenous matriptase-2; anti-bodies to matriptase-2 and a method for treating prostate cancer which involves the use of matriptase-2.
- TTSP's type Il transmembrane serine proteases
- TTSP's are characterised by a short N-terminal cytoplasmic tail, a membrane-spanning region, potential ligand binding domains and a C-terminal serine protease domain. These proteolytic enzymes are therefore ideally positioned to interact with other cell surface proteins, as well as soluble proteins, extra-cellular matrix components and proteins on adjacent cells.
- Human members of the TTSP family include enteropeptidase/enterokinase, matriptase/membrane-type serine protease- 1/TADG-15, hepsin/TMPRSSI , TMPRSS3/TADG-12, and corin.
- Matriptase-2 (also known as TMPRSS6) is a new, highly conserved, membrane- bound enzyme that belongs to the TTSP family. Matriptase-2 shares structural similarities with matriptase-1 , however, matriptase-1 is located at 11q25, whereas the gene encoding matriptase-2 maps to chromosome 22q13.1. The majority of the identified TTSP's demonstrate restricted tissue distribution, but reports suggest TTSP expression is widely dys-regulated during tumour growth and progression. Matriptase-1 plays an important role in tumour invasion and metastasis as this protease possesses the ability to activate growth and angiogenic factors and degrade components of the extra-cellular matrix.
- matriptase-1 can activate hepatocyte growth factor (HGF), urokinase-type plasminogen activator (uPA) and protease-activated receptor 2 (PAR2) ' , importantly, these factors are known to enhance tumour cell invasion and metastasis.
- HGF hepatocyte growth factor
- uPA urokinase-type plasminogen activator
- PAR2 protease-activated receptor 2
- matriptase-1 The pro-tumorigenic nature of matriptase-1 is further demonstrated in several studies targeting this protease which show that the reduced expression or inhibition of matriptase-1 is associated with a decrease in in vitro cellular traits such as invasion, migration and scattering and with decreased tumour development in vivo.
- Matriptase-2 is related to matriptase-1 (35% homology) and appears to possess the same matrix-degrading properties. To date, the biological and biochemical roles of matriptase-2 are unknown, and it is not clear whether matriptase-2 has any function in the neoplastic process.
- Matriptase-1 Owing to its structural similarities to matriptase-1 , we anticipated a similar function between the two matripatases. Matriptase-1 has been indicated as an proteinase activator for pro-HGF, a cytokine that increase the invasiveness and aggressiveness of cancer cells and a known angiogenic factor. Pro-HGF, which is biological inactive, can be activated by matriptase-1 and become a pro-cancer active cytokine. All of these findings have led to a lot of interest being focused on Matriptase-1 , its involvement and role in various cancers, and its possible uses in future treatments.
- matriptase-2 we investigated the function of matriptase-2, by hypothesising that it may act in a similar fashion to matriptase-1 and thus act as a molecule that may promote the aggressiveness of cancer cells.
- matripase-2 had a very different pattern of expression and function in prostate cancer. Subsequently, we have obtained evidence to show that matriptase-2 is an inhibitor to the invasiveness of prostate cancer and the growth of prostate tumours. This makes matriptase-2 the first and only TMPRSS that has anti-cancer actions.
- Our invention therefore concerns the role of matriptase-2 in prostate cancer and more particularly the inhibitory role of matriptase-2 in prostate cancer.
- nucleic acid molecule encoding a polypeptide that has anti-cancer activity in or against prostate tumour cells and is characterised by the nucleic acid sequence shown in Figure 6, SEQ ID NO.1 ; or a homologue thereof; or a nucleic acid molecule that hybridises to the molecule shown in Figure 6 under stringent conditions; or a fragment thereof which fragment also has anti-cancer activity in prostate cells.
- Reference herein to stringent conditions includes reference to either increasing the temperature of incubation to above 25 0 C and more preferably above 5O 0 C and more preferably still up to 65 0 C and/or washing the annealed molecules using a salt solution having an ionic strength of 1.0N sodium chloride - 0.02N sodium chloride, and most preferably 0.5N sodium chloride - 0.02N sodium chloride and more preferably still 0.1 N sodium chloride to 0.02N sodium chloride.
- an anticancer agent for treating prostate cancer comprising a nucleic acid molecule encoding matriptase-2 as shown in Figure 6, SEQ ID NO:1.
- a polypeptide that has anti-cancer activity in or against treating prostate tumour cells comprising a polypeptide as shown in Figure 7, SEQ ID NO:2; or a homologue thereof; or a fragment thereof which fragment also has anti-cancer activity in or against prostate tumour tissue.
- an anti-cancer agent for treating prostate cancer comprising the polypeptide matriptase-2 as shown in Figure 7, SEQ ID NO:2.
- nucleic acid molecule encoding matriptase-2 and/or the corresponding aforementioned polypeptide for treating prostate cancer.
- a medicament comprising the aforementioned nucleic acid molecule encoding matriptase-2 and/or the corresponding aforementioned polypeptide.
- the medicament of the invention is most suitable for treating prostate cancer.
- the medicament of the invention includes the aforementioned nucleic acid molecule encoding matriptase-2 and/or the corresponding aforementioned polypeptide formulated with a suitable excipient, carrier or emollient.
- a method for treating prostate cancer comprising administering to an individual to be treated a medically effective amount of the aforementioned matriptase-2 nucleic acid molecule and/or its corresponding polypeptide.
- a method of treating prostate cancer comprising increasing the cellular levels of either exogenous or endogenous matriptase-2 polypeptide in prostate cells.
- Reference herein to the exogenous increase in cellular levels of matriptase-2 polypeptide includes reference to: introducing into prostate cells either externally manufactured matriptase-2 polypeptide; or introducing means for increasing the said cells production of matriptase-2 by, for example, increasing the number of copies of the matriptase-2 gene.
- Reference herein to the endogenous increase in cellular levels of matriptase-2 polypeptide includes introducing into prostate cells means that enables the prostate cells to increase native production of matriptase-2 polypeptide, for example, suitable means include supplementing the cells' manufacturing machinery to provide for the increased production of matriptase-2 such as by increasing the effectiveness or activity of the promoter controlling the expression of the matriptase-2 gene whereby the overall production of matriptase-2 is increased, or, additionally or alternatively, supplementing the cells' other cellular protein production pathways whereby the amount of matriptase-2 produced is increased.
- suitable means include supplementing the cells' manufacturing machinery to provide for the increased production of matriptase-2 such as by increasing the effectiveness or activity of the promoter controlling the expression of the matriptase-2 gene whereby the overall production of matriptase-2 is increased, or, additionally or alternatively, supplementing the cells' other cellular protein production pathways whereby the amount of matriptase-2 produced is
- a vector adapted to transfect or transform prostate cells wherein said vector includes:
- said vector is engineered to transfect or transform human prostate cells.
- an antibody selective for the aforementioned matriptase-2 polypeptide is provided.
- the antibody is selective for the human form of matriptase-2 and, ideally, is monoclonal.
- a process for the production of antibodies that selectively bind to matriptase-2 polypeptide comprising: (a) introducing matriptase-2 polypeptide into a mammalian animal;
- the matriptase-2 polypeptide comprises either the whole polypeptide or a fragment thereof and, most suitably, where a fragment is used the following fragment is preferred, GQGDGGDGEEAEPEGMFKAC. Ideally said animal is non-human.
- FIG. 1 shows staining of matriptase-1 (A and B) and matriptase-2 (C and D) in human prostate tissues. Prostate tumours over-expressed matriptase-1 protein
- matriptase-2 had a completely different staining pattern (C and D).
- Matriptase-2 is membranous protein in normal prostate epithelial cells (C).
- prostate cancer cells (D) loss the membranous pattern and reduced in its staining intensity.
- Figure 2 shows western blotting (A and B) and Q-RT-PCR (C and D) showing matriptase-2 expression levels in PC-3 (A and C) and DU-145 (B and D) cell lines;
- figure-2 we successfully over- expressed matriptase-2 in PC-3 (A and C) and DU-145 (B and D) cells. This was confirmed at protein level (A and B) and mRNA level (C and D) 1 using Western blotting and quantitative PCR analysis, respectively.
- Figure 3 shows matriptase-2's expression and cell growth in the PC-3 (A and B) and DU-145 cells (C and D);
- Figure 5 shows how matriptase-2 over-expressing prostate cancer cells had dramatically reduced in vivo growth.
- Figure 6 shows the nucleic acid molecule encoding matriptase-2 polypeptide.
- Figure 7 shows the amino acid sequence of the polypeptide matriptase-2.
- Figure 8 shows over-expression of matriptase-2 reduced cellular migration and motility, as assessed by a migration/wound healing assay, in the (A) PC-3 and (B) DU-145 cell lines.
- Both pc-3 Matriptase - 2 EXP and D U-145 Matriptase - 2 EXP cells were significantly less able to migrate into, and close a wound created in the cell monolayer than their respective pEF6 control cell lines.
- Figure 9 shows the localisation and expression of paxillin and FAK molecules in the presence and absence of HGF cytokine in PC-STM 1 , PC-3 PEF6 , and PC- 3 Matriptase"2 EXP cells.
- PC-3 and DU-145 cell lines were purchased from the American Type Culture Collection (ATCC 1 Rockville, Maryland, USA). These cells were maintained in Dubecco's Modified Eagle Medium (DMEM) (PAA Laboratories Ltd, Somerset, UK) supplemented with penicillin, streptomycin and 10% foetal calf serum (PAA Laboratories Ltd, Somerset, UK) and incubated at 37 0 C, 5% CO 2 and 95% humidity.
- DMEM Dubecco's Modified Eagle Medium
- PAA Laboratories Ltd, Somerset, UK supplemented with penicillin, streptomycin and 10% foetal calf serum
- Matriptase-2 Antibody is available commercially (Triple Point Biologies, Forest
- the matriptase peptide (GQGDGGDGEEAEPEGMFKAC. matrip2N20) was generated from the N-terminal domain of each protein, no overlapping between these two regions was ensured.
- the peptide was attached the KLH keyhole limpet Megathura crenulata and injected into rabbit with complete Freunds solution, following a standard procedure. Antisera were obtained after series injection and booster injections.
- Immunoglobulin was purified using a protein-A sepharose affinity column (Sigma, Poole, Dorset, England). The antibodies we obtained following a series of purification steps developed in our laboratories.
- matriptase-2 The specificity of matriptase-2 was verified using synthetic peptides and cell lysates.
- the cell lystates were from MCF-7, DU-145, HECV and MRC5 cell lines (see 'cell lines and tissues' for details).
- Matriptase peptides were blotted onto a nitrocellulose membrane, at different concentrations, to confirm that the purified matriptase antibodies specifically recognised the appropriate matriptase-1 or matriptase-2 peptide.
- a variety of human normal and cancer cell line lysates were also examined with Western Blot analysis to confirm the presence of the matriptase proteins.
- Human prostate, breast, endothelial cell and stromal fibroblast cell lines were purchased from ECACC and ATCC.
- Prostate cancer celll lines were: PC3,
- DU145, CA-HPV, breast cancer were MDA MB 231 , MCF-7, fibroblast was MRC5.
- a human endothelial cell line, HECV was from lnterlab Cell line Collection (ICLC), Naples, Italy.
- Frozen sections of prostate tumours were cut at a thickness of 6 ⁇ m using a cryostat.
- the sections were mounted on super frost plus microscope slides, air- dried and then fixed in a mixture of 50% Acetone and 50% methanol.
- the sections were then placed in "Optimax" wash buffer for 5 -10 minutes to rehydrate.
- Sections were incubated for 20 minutes in a 0.6% BSA blocking solution and probed with the primary antibodies.
- sections were incubated for 30 minutes in the secondary biotinylated antibody (Multilink Swine anti- goat/mouse/rabbit immunoglobulin, Dako Inc.).
- the Avidin Biotin Complex (Vector Laboratories) was then applied to the sections, followed by extensive washing steps.
- Diamino benzidine chromogen (Vector Labs) was then added to the sections, and incubated in the dark for 5 minutes. Sections were then counter stained in Gill's Haematoxylin and dehydrated in ascending grades of methanol before clearing in xylene and mounting under a cover slip
- Matriptase-2 The full sequence of Matriptase-2 was amplified from a cDNA bank generated in our laboratory from normal human tissues. PCR primers used were: forward 5'- ATGCCCGTGGCCGAGG -3' and reverse 5'- GGTCACCACTTGCTGGATCCA -
- This matriptase-2 sequence was then cloned into pEF6/V5-His-TOPO vector (Invitrogen, Paisley, UK).
- the technique used to generate this expression plasmid is outlined in greater depth in the recently published breast cancer study Parr et al 2007, CCR.
- the vector was then electroporated into the prostate cancer cell lines (PC-3 and DU-145) which were virtually negative in matriptase- 2 expression.
- Cells containing the matriptase-2 expression plasmid and displaying enhanced matriptase-2 expression were designated pc-3 Mat ⁇ ptase"2 EXP or DU-145 Matriptase"2 EXP , similarly cells containing a close pEF6 plasmid only (containing no expression sequence, to demonstrate the lack of effects exerted by the plasmid alone) were designated PC-3 pEF6 and DU-145 pEF6 whereas the unaltered wild type cells were simply labelled PC- ⁇ or 011-145 ⁇ .
- ribozyme insert was generated using the primers under a touch-down condition, the resulting products were T-A cloned into the pEF6/V5-His-TOPO vector.
- purified plasmid was then electroporated into the PLC-PRF-5 liver cancer cell line with the aim of suppressing the expression of matriptase-2 in the cell line were it is found most abundantly.
- the PLC-PRF-5 cells with matriptase-2 expression suppressed were called PLC-MAT-2-KO, and properties were assessed through a series of cell assays described below.
- PC-3 and DU-145 control cells The growth rates of PC-3 and DU-145 control cells was examined and compared to that of those containing the matriptase-2 expression plasmid using an in vitro growth assay.
- Cells were seeded in a 96 well plate at a density of 3,000 or 7,000 cells/well, and incubated at 37°C for 24, 48 or 72 hours. MTT was added in solution to the cells (200 ⁇ g/well) and incubated for 4 hours at 37°C, then the cells were lysed with Triton (10%) and the intensity of the colour released was determined by a plate reader (Titertek Multiskan, Eflab, Finland). The number of cells was shown as absorbance units.
- Tumour Cell Invasion Assay We quantified the invasive nature of the matriptase-2 modified cancer cells and control cells using the standard invasion assay procedure as described previously (Parr et al., 2001).
- Transwell chambers equipped with a 6.5mm diameter polycarbonate filter insert (pore size 8 ⁇ m)(Becton Dickinson, Labware, Oxford, UK), were pre-coated with 50 ⁇ g/insert of solubilised tissue basement membrane, Matrigel (Collaborative Research Products, Bedford, Massachusetts,
- the QPCR technique utilised the Amplifluor system (Intergen Inc, England) and Q-PCR master mix (ABgene, Surrey, England), in conjunction with a universal probe (UniPrimerTM). Real-time QPCR conditions were 95°C for 12 minutes, followed by 65 cycles at 95°C for15s, 55°C for 60s, 72°C for 20s. The results of the test molecules were normalised against levels of ⁇ -actin, using a ⁇ -actin quantitation kit from Perkin-Elmers (Perkin- Elmers, Surrey, England, UK).
- Cancer cells were grow to confluence, detached and lysed in HCMF buffer containing 0.5% SDS, 1% Triton X-100, 2 mM CaCI 2 , 100 ⁇ g/ml phenylmethylsulfonyl fluoride, 1 mg/ml leupeptin, 1 mg/ml aprotinin and, 10 mM sodium orthovanadate on a rotor wheel for 1 hour before being spun at 13,00Og to remove insolubles. Following lysis, protein levels were quantified using the
- Bio-Rad DC Protein Assay kit Bio-Rad laboratories, California, USA, standardised to 2mg/ml and diluted 1 :1 with Laemmli 2x concentrate sample buffer (Sigma, Dorset, UK). Samples were boiled for 5 minutes before loading into a 10% acrylamide gel and being subjected to electrophoretic separation. Once sufficient separation had occurred the proteins were blotted onto a Hybond-C Extra nitrocellulose membrane (Amersham Biosciences UK Ltd, Bucks, UK), blocked in 10% milk and subjected to specific antibody probing.
- Matriptase-2 expression was detected using both commercial antibodies available at the time of the study; RP-1 matriptase-2 recognising the cytoplasmic region and RP-2 matriptase-2 recognising the stem region of the matriptase-2 protein (both antibodies were obtained from Triple Point Biologies, Forest Grove, Oregon, USA), in addition to these commercially available antibodies a third antibody generated in house was also used. An anti ⁇ -actin antibody (Santa Cruz Biotechnology, Inc., California, USA) was also used to indicate uniform protein expression levels in the samples.
- Bound primary antibodies were then detected using peroxidise conjugated anti-rabbit (matriptase-2) or anti-goat ( ⁇ - actin) antibodies (Sigma, Dorset, UK) and visualised through the addition of Supersignal West Dura Extended Duration substrate chemi-luminescent system (Perbio Science UK Ltd, Cramlington, UK)
- RNA extraction and Reverse Transcription-Polymerase Chain Reaction RT-PCR
- RNA levels throughout samples were then standardised to 250ng for use in reverse transcription.
- Reverse transcription was carried out using a enhanced avain reverse transcriptase-PCR-100 kit with anchored oligo(dt) primers and in accordance with the supplied protocol (Sigma, Dorset, UK). The quality of the cDNA generated was tested using ⁇ -actin primers before probing for the expression of matriptase-2 using specific primers (full primer sequences are outlined in table, 1).
- PCR was performed in a T-Cy Thermocycler (Creacon Technologies Ltd, The Netherlands) using REDTaq® ReadyMixTM PCR Reaction Mix (Sigma, Dorset, UK). PCR conditions were as follows: denaturing at 94 0 C for 1 minute, annealing at 58 0 C for 2 minutes and extension at 72 0 C for 3 minutes. PCR was conducted over 38 cycles with an initial 5 minute denaturing step (94 0 C) and a final 10 minute extension step (72 0 C). Following PCR, products were loaded ⁇ on a 0.8% agarose gel, separated electrophoretically, stained in ethidium bromide and visualised under ultraviolet light.
- the adhesion of control and transfected cells to an artificial basement membrane was quantified using an in vitro Matrigel adhesion assay adapted form a previously described method [22]. Briefly, 45,000 cells were seeded into the wells of a 96 well plate that had been previously coated with 5 ⁇ g of Matrigel artificial basement membrane. Cells were incubated for 45 minutes before being subjected to vigorous washing in BSS 1 to remove non-adherent cells. Adherent cells were then fixed in 4% formaldehyde (v/v) and stained with 0.5% (w/v) crystal violet. The number of stained, adherent cells was counted in several random fields under x 40 objective magnification.
- Immuno-Fluorescent staining Cells for use in immuno-fluorescent staining were seeded at a density of 20,000 cells per well in a 16-well chamber slide (LAB-TEK Fisher Scientific UK, Longhborough, Leics, UK). Duplicate rows were seeded and the cells were cultured overnight to adhere and establish before one of the duplicate rows was treated with 40ng/ml HGF for 2 hours. Following treatment, the medium was aspirated and the cells were fixed in ice cold ethanol at -2O 0 C for 20 minutes.
- the cells were then rehydrated in BSS for 20 minutes at room temperature before being permeablized with a BSS solution containing 0.1% Triton X100 for 5 minutes.
- a blocking solution of MenaPath Autowash buffer (A. Menarini Diagnostics, Berkshire, UK), containing 2 drops of horse serum per 5ml, was then placed on the cells for 20 minutes to block non-specific binding.
- Cells were next washed with wash buffer twice before probing for specific expression of paxillin or FAK using anti-paxillin (Transduction Laboratories) and anti-FAK (Transduction Laboratories) primary antibodies made up in wash buffer at a 1 :100 concentration for 1 hour. The primary antibody was then completely removed by washing the cells three times in wash buffer.
- a FITC conjugated anti-mouse secondary antibody (Insight Biotechnology Ltd., Middlesex, UK) was then added and the slides were incubated on a shaker platform for 1 hour in the dark. Finally the slides were washed three times to remove un-bound secondary antibodies, mounted with Fluor-save (Calbiochem-Novabiochem Ltd., Nottingham, UK) and visualised under an Olympus BX51 microscope at X100 objective magnification.
- FIG. 1 shows antibody staining of normal prostate epithelial cells (A and C) and human prostate cancer tissues (B and D).
- Slides A and B shows staining of matriptase-1 and slides C and D show stainings of matriptase-2 in human prostate tissue.
- a comparison of slides A and B shows that tumours over- expressed matriptase-1 protein.
- slides C and D shows that matriptase-2 had a completely different staining pattern.
- matriptase-2 is membranous protein in normal prostate epithelial cells (C), but prostate cancer cells (D) lost the membranous pattern and reduced staining intensity.
- the transfection process appears to have been more efficient in the pc-3 Matriptase"2 EXP cell line than the DU-145 Matriptase -* EXP cell line, as forced expression was obvious using conventional RT-PCR in the pc-3 Mat ⁇ ptase'2 Exp cell line whereas Q-PCR was required to identify this trend in the DU- 145 Matriptase"2 EXP cell line.
- Matriptase-2's effects on the growth of PC-3 and DU-145 cells were examined using an in vitro tumour cell growth assay (see figure 3). Contrasting results for the effects of matriptase-2 expression on tumour cell growth were seen between the PC-3 and DU-145 cell lines. In the PC-3 cell line, forced-expression of matriptase-2 resulted in a significant decrease in the growth over a 5 day incubation period and a significant difference was seen between the PC-
- PC-3 pEF6 and DU-145 pEF6 This data is represented in Figure 4 where it can be seen that the expression of matriptase-2 in either of the cell lines investigated, i.e. PC-3 or DU-145, reduced cell invasiveness (A and B). However, the capability of both these cell lines to adhere to the extra cellular matrix was not affected (C and D).
- matriptase-2 was forcibly up-regulated in two prostate cancer cell lines, which normally have minimal expression levels of this molecule, using an expression plasmid. Transfection of these cell lines with the matriptase-2 expression plasmid affected a number of in vitro cellular traits directly linked to cancer aggression. In both of these prostate cancer cell lines, over-expression of matriptase-2 statistically reduced cellular migration and invasion, traits strongly linked to cancer invasion, dissemination and metastasis. These prostate studies suggest a role for matriptase-2 in the suppression of cellular invasion and migration, both key events in cancer metastasis, and imply a preventative effect of matriptase-2 in cancer progression.
Landscapes
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Engineering & Computer Science (AREA)
- Chemical & Material Sciences (AREA)
- Genetics & Genomics (AREA)
- Organic Chemistry (AREA)
- Zoology (AREA)
- Biomedical Technology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Wood Science & Technology (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Biotechnology (AREA)
- Biochemistry (AREA)
- Molecular Biology (AREA)
- Microbiology (AREA)
- Plant Pathology (AREA)
- Physics & Mathematics (AREA)
- Biophysics (AREA)
- Virology (AREA)
- Medicinal Chemistry (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Pharmaceuticals Containing Other Organic And Inorganic Compounds (AREA)
Abstract
The invention relates to the role of matriptase-2 in treating prostate cancer.
Description
Matriptase-2
The invention relates to a novel agent for treating prostate cancer; a composition including same; means for increasing the cellular levels of either exogenous or endogenous matriptase-2; anti-bodies to matriptase-2 and a method for treating prostate cancer which involves the use of matriptase-2.
Matriptases are a small family of serine proteases, implicated in tumour progression, that have been generating a high degree of interest in recent years. Members of this emerging group have been termed type Il transmembrane serine proteases (TTSP's). TTSP's are characterised by a short N-terminal cytoplasmic tail, a membrane-spanning region, potential ligand binding domains and a C-terminal serine protease domain. These proteolytic enzymes are therefore ideally positioned to interact with other cell surface proteins, as well as soluble proteins, extra-cellular matrix components and proteins on adjacent cells. Human members of the TTSP family include enteropeptidase/enterokinase, matriptase/membrane-type serine protease- 1/TADG-15, hepsin/TMPRSSI , TMPRSS3/TADG-12, and corin.
Matriptase-2 (also known as TMPRSS6) is a new, highly conserved, membrane- bound enzyme that belongs to the TTSP family. Matriptase-2 shares structural similarities with matriptase-1 , however, matriptase-1 is located at 11q25, whereas the gene encoding matriptase-2 maps to chromosome 22q13.1.
The majority of the identified TTSP's demonstrate restricted tissue distribution, but reports suggest TTSP expression is widely dys-regulated during tumour growth and progression. Matriptase-1 plays an important role in tumour invasion and metastasis as this protease possesses the ability to activate growth and angiogenic factors and degrade components of the extra-cellular matrix. Studies also report that matriptase-1 can activate hepatocyte growth factor (HGF), urokinase-type plasminogen activator (uPA) and protease-activated receptor 2 (PAR2)', importantly, these factors are known to enhance tumour cell invasion and metastasis. Matriptase-1 was originally identified in breast cancer cells, and has recently been gaining attention in human cancer-based studies as matriptase-1 is highly expressed in a variety of epithelial derived cancers including breast, colorectal, prostate, cervical and ovarian. Importantly, inhibition of matriptase-1 stopped both primary tumour growth and metastasis in murine models of prostate and ovarian cancer. Matriptase-1 expression also correlates with HGF and c-Met expression, while increased levels of matriptase-1 are associated with poor prognosis in breast cancer patients.
The pro-tumorigenic nature of matriptase-1 is further demonstrated in several studies targeting this protease which show that the reduced expression or inhibition of matriptase-1 is associated with a decrease in in vitro cellular traits such as invasion, migration and scattering and with decreased tumour development in vivo.
Matriptase-2 is related to matriptase-1 (35% homology) and appears to possess the same matrix-degrading properties. To date, the biological and biochemical roles of matriptase-2 are unknown, and it is not clear whether matriptase-2 has any function in the neoplastic process.
Owing to its structural similarities to matriptase-1 , we anticipated a similar function between the two matripatases. Matriptase-1 has been indicated as an proteinase activator for pro-HGF, a cytokine that increase the invasiveness and aggressiveness of cancer cells and a known angiogenic factor. Pro-HGF, which is biological inactive, can be activated by matriptase-1 and become a pro-cancer active cytokine. All of these findings have led to a lot of interest being focused on Matriptase-1 , its involvement and role in various cancers, and its possible uses in future treatments.
We investigated the function of matriptase-2, by hypothesising that it may act in a similar fashion to matriptase-1 and thus act as a molecule that may promote the aggressiveness of cancer cells.
However, to our surprise, matripase-2 had a very different pattern of expression and function in prostate cancer. Subsequently, we have obtained evidence to show that matriptase-2 is an inhibitor to the invasiveness of prostate cancer and the growth of prostate tumours. This makes matriptase-2 the first and only TMPRSS that has anti-cancer actions.
Statements of Invention
Our invention therefore concerns the role of matriptase-2 in prostate cancer and more particularly the inhibitory role of matriptase-2 in prostate cancer.
According to a first aspect of the invention there is therefore provided a nucleic acid molecule encoding a polypeptide that has anti-cancer activity in or against prostate tumour cells and is characterised by the nucleic acid sequence shown in Figure 6, SEQ ID NO.1 ; or a homologue thereof; or a nucleic acid molecule that hybridises to the molecule shown in Figure 6 under stringent conditions; or a fragment thereof which fragment also has anti-cancer activity in prostate cells.
Reference herein to stringent conditions includes reference to either increasing the temperature of incubation to above 250C and more preferably above 5O0C and more preferably still up to 650C and/or washing the annealed molecules using a salt solution having an ionic strength of 1.0N sodium chloride - 0.02N sodium chloride, and most preferably 0.5N sodium chloride - 0.02N sodium chloride and more preferably still 0.1 N sodium chloride to 0.02N sodium chloride.
According to a further aspect of the invention there is therefore provided an anticancer agent for treating prostate cancer comprising a nucleic acid molecule encoding matriptase-2 as shown in Figure 6, SEQ ID NO:1.
According to a further aspect of the invention there is provided a polypeptide that has anti-cancer activity in or against treating prostate tumour cells comprising a polypeptide as shown in Figure 7, SEQ ID NO:2; or a homologue thereof; or a fragment thereof which fragment also has anti-cancer activity in or against prostate tumour tissue.
According to a further aspect of the invention there is provided an anti-cancer agent for treating prostate cancer comprising the polypeptide matriptase-2 as shown in Figure 7, SEQ ID NO:2.
According to a further aspect of the invention there is provided the use of the aforementioned nucleic acid molecule encoding matriptase-2 and/or the corresponding aforementioned polypeptide for treating prostate cancer.
According to a further aspect of the invention there is provided a medicament comprising the aforementioned nucleic acid molecule encoding matriptase-2 and/or the corresponding aforementioned polypeptide.
The medicament of the invention is most suitable for treating prostate cancer.
Further, the medicament of the invention includes the aforementioned nucleic acid molecule encoding matriptase-2 and/or the corresponding aforementioned polypeptide formulated with a suitable excipient, carrier or emollient.
According to a further aspect of the invention there is provided a method for treating prostate cancer comprising administering to an individual to be treated a medically effective amount of the aforementioned matriptase-2 nucleic acid molecule and/or its corresponding polypeptide.
According to a further aspect of the invention there is provided a method of treating prostate cancer comprising increasing the cellular levels of either exogenous or endogenous matriptase-2 polypeptide in prostate cells.
Reference herein to the exogenous increase in cellular levels of matriptase-2 polypeptide includes reference to: introducing into prostate cells either externally manufactured matriptase-2 polypeptide; or introducing means for increasing the said cells production of matriptase-2 by, for example, increasing the number of copies of the matriptase-2 gene.
Reference herein to the endogenous increase in cellular levels of matriptase-2 polypeptide includes introducing into prostate cells means that enables the prostate cells to increase native production of matriptase-2 polypeptide, for example, suitable means include supplementing the cells' manufacturing machinery to provide for the increased production of matriptase-2 such as by increasing the effectiveness or activity of the promoter controlling the expression of the matriptase-2 gene whereby the overall production of matriptase-2 is increased, or, additionally or alternatively, supplementing the cells' other cellular protein production pathways whereby the amount of matriptase-2 produced is
increased. Those skilled in the art will be familiar with ways by which this can be achieved.
According to a yet further aspect of the invention there is provided a vector adapted to transfect or transform prostate cells wherein said vector includes:
(1) at least one copy of the matriptase-2 gene; and/or
(2) at least one over-expressing or constitutively active promoter which is either coupled to the matriptase-2 gene of part (1) and/or which is designed for insertion into a host genome upstream of the native matriptase-2 gene of said prostate cells whereby transfection or transformation of said cells with said vector results in the enhanced expression of matriptase-2.
Ideally, said vector is engineered to transfect or transform human prostate cells.
According to a further aspect of the invention there is provided an antibody selective for the aforementioned matriptase-2 polypeptide.
Most preferably, the antibody is selective for the human form of matriptase-2 and, ideally, is monoclonal.
According to a further aspect of the invention there is provided a process for the production of antibodies that selectively bind to matriptase-2 polypeptide comprising:
(a) introducing matriptase-2 polypeptide into a mammalian animal;
(b) obtaining anti-sera from said animal; and
(c) purifying the anti-sera in order to obtaining matriptase-2 antibody. In a preferred method of the invention the matriptase-2 polypeptide comprises either the whole polypeptide or a fragment thereof and, most suitably, where a fragment is used the following fragment is preferred, GQGDGGDGEEAEPEGMFKAC. Ideally said animal is non-human.
An embodiment of the invention will now be described by way of example only with reference to the following materials methods and results wherein:
Figure 1 shows staining of matriptase-1 (A and B) and matriptase-2 (C and D) in human prostate tissues. Prostate tumours over-expressed matriptase-1 protein
(B vs A);
However, matriptase-2 had a completely different staining pattern (C and D).
Matriptase-2 is membranous protein in normal prostate epithelial cells (C).
However, prostate cancer cells (D) loss the membranous pattern and reduced in its staining intensity.
Figure 2 shows western blotting (A and B) and Q-RT-PCR (C and D) showing matriptase-2 expression levels in PC-3 (A and C) and DU-145 (B and D) cell lines;
We successfully isolated full length human matriptase-2 from a cDNA of normal tissues and generated a mammalian expression construct. This was used to transfect prostate cancer cell lines PC-3 and DU-145, both had little expression of matriptase-2, figure 2 (A and B). As shown in figure-2, we successfully over- expressed matriptase-2 in PC-3 (A and C) and DU-145 (B and D) cells. This was confirmed at protein level (A and B) and mRNA level (C and D)1 using Western blotting and quantitative PCR analysis, respectively.
Figure 3 shows matriptase-2's expression and cell growth in the PC-3 (A and B) and DU-145 cells (C and D);
We tested the growth rate of prostate cancer cell lines that carried either matriptase-2 expression vector, a control vector or none (wild type). As shown in figure-3, over-expression of matriptase-2 resulted in significantly slower growth in PC-3 cells (figure 3 A and B), although not as significant in DU-145 cells, (C and D).
(A and C) Histogram representing the percentage increase at day 3 and day 5. Statistical comparison to the pEF6 control indicating a highly significant decrease in growth at day 5 (*** represents p < 0.001) following the forced expression of Matriptase-2 in the PC-3 cell line. Data show is the mean data of 3 individual experiments and error bars represent the standard error of the mean. (B and D) Line graph representing growth trends over the 5 days.
Figure 4 shows how over-expressing matriptase-2 resulted in significantly reduced in vitro invasiveness (A and B) but not matrix adhesiveness (C and D);
Prostate cancer cell lines that were forced to over-express matriptase-2, showed a significantly reduced in vitro invasiveness as shown in figure-4 (A-PC-3 cells,
B-DU-145 cells). However, the capability of both cells to extracellular matrix were not affected (Figure-4 C and D).
Significantly reduced invasiveness was seen in both the (A) PC-3 (*** represents p < 0.001) and (B) DU-145 (** represents p < 0.01) cell lines following forced expression of Matriptase-2. In contrast, no significant difference was seen with matrix adhesiveness in both cells.
Figure 5 shows how matriptase-2 over-expressing prostate cancer cells had dramatically reduced in vivo growth.
Using an in vivo model (athymic nude mice), it was shown that Matriptase-2 over-expression PC-3 cells had a dramatically reduced growth rate compared with control.
This reduced capacity was noticeable from the earliest experimental reading at 7 days and continued through the entire experiment to the experimental end point were pc-3Matπptase"2 EXP tumours were barely noticeable and were insufficient in size to be excised. In comparison to this, PC-3 cells containing an empty pEF6 plasmid (PC-3pEF6) grew rapidly and developed into noticeable tumours following
sub-cutaneously injection. The development of the PC-3pEF6 tumours continued steadily through the experiment and these tumours were obviously apparent upon reaching the experimental end point.
Data shows a significantly reduced capacity for PC-3 cells with forced expression of Matriptase-2 to growth and develop in the living environment (p<0.001 vs control).
Figure 6 shows the nucleic acid molecule encoding matriptase-2 polypeptide.
Figure 7 shows the amino acid sequence of the polypeptide matriptase-2.
Figure 8 shows over-expression of matriptase-2 reduced cellular migration and motility, as assessed by a migration/wound healing assay, in the (A) PC-3 and (B) DU-145 cell lines. Both pc-3Matriptase-2 EXP and DU-145Matriptase-2 EXP cells were significantly less able to migrate into, and close a wound created in the cell monolayer than their respective pEF6 control cell lines. *** p < 0.001 and ** p < 0.01 vs respective pEF6 control cells.
Figure 9 shows the localisation and expression of paxillin and FAK molecules in the presence and absence of HGF cytokine in PC-S™1, PC-3PEF6, and PC- 3Matriptase"2 EXP cells.
MATERIAL AND METHODS Human prostate tissues.
This was archival materials obtained from the operating theatre, immediately after surgery. PC-3 and DU-145 cell lines were purchased from the American Type Culture Collection (ATCC1 Rockville, Maryland, USA). These cells were maintained in Dubecco's Modified Eagle Medium (DMEM) (PAA Laboratories Ltd, Somerset, UK) supplemented with penicillin, streptomycin and 10% foetal calf serum (PAA Laboratories Ltd, Somerset, UK) and incubated at 370C, 5% CO2 and 95% humidity. Matriptase-2 Antibody is available commercially (Triple Point Biologies, Forest
Grove, Oregon, USA). However it can also be made using the following procedure.
Matriptase- 2 Antibody Production, Purification and Analysis of Specificity The matriptase peptide (GQGDGGDGEEAEPEGMFKAC. matrip2N20) was generated from the N-terminal domain of each protein, no overlapping between these two regions was ensured. The peptide was attached the KLH keyhole limpet Megathura crenulata and injected into rabbit with complete Freunds solution, following a standard procedure. Antisera were obtained after series injection and booster injections. Immunoglobulin was purified using a protein-A sepharose affinity column (Sigma, Poole, Dorset, England). The antibodies we obtained following a series of purification steps developed in our laboratories.
The specificity of matriptase-2 was verified using synthetic peptides and cell lysates. The cell lystates were from MCF-7, DU-145, HECV and MRC5 cell lines
(see 'cell lines and tissues' for details). Matriptase peptides were blotted onto a nitrocellulose membrane, at different concentrations, to confirm that the purified matriptase antibodies specifically recognised the appropriate matriptase-1 or matriptase-2 peptide. A variety of human normal and cancer cell line lysates were also examined with Western Blot analysis to confirm the presence of the matriptase proteins.
Cell lines and tissues
Human prostate, breast, endothelial cell and stromal fibroblast cell lines were purchased from ECACC and ATCC. Prostate cancer celll lines were: PC3,
DU145, CA-HPV, breast cancer were MDA MB 231 , MCF-7, fibroblast was MRC5. A human endothelial cell line, HECV, was from lnterlab Cell line Collection (ICLC), Naples, Italy.
Immuno-histochemical Staining of Breast Specimens
Frozen sections of prostate tumours were cut at a thickness of 6μm using a cryostat. The sections were mounted on super frost plus microscope slides, air- dried and then fixed in a mixture of 50% Acetone and 50% methanol. The sections were then placed in "Optimax" wash buffer for 5 -10 minutes to rehydrate. Sections were incubated for 20 minutes in a 0.6% BSA blocking solution and probed with the primary antibodies. Following extensive washings, sections were incubated for 30 minutes in the secondary biotinylated antibody (Multilink Swine anti- goat/mouse/rabbit immunoglobulin, Dako Inc.). Following washings, the Avidin Biotin Complex (Vector Laboratories) was then applied to
the sections, followed by extensive washing steps. Diamino benzidine chromogen (Vector Labs) was then added to the sections, and incubated in the dark for 5 minutes. Sections were then counter stained in Gill's Haematoxylin and dehydrated in ascending grades of methanol before clearing in xylene and mounting under a cover slip
Generation of a Matriptase-2-expressing prostate Cancer Cell Line
The full sequence of Matriptase-2 was amplified from a cDNA bank generated in our laboratory from normal human tissues. PCR primers used were: forward 5'- ATGCCCGTGGCCGAGG -3' and reverse 5'- GGTCACCACTTGCTGGATCCA -
3'
This matriptase-2 sequence was then cloned into pEF6/V5-His-TOPO vector (Invitrogen, Paisley, UK). The technique used to generate this expression plasmid is outlined in greater depth in the recently published breast cancer study Parr et al 2007, CCR. The vector was then electroporated into the prostate cancer cell lines (PC-3 and DU-145) which were virtually negative in matriptase- 2 expression. Cells containing the matriptase-2 expression plasmid and displaying enhanced matriptase-2 expression were designated pc-3Matπptase"2 EXP or DU-145Matriptase"2 EXP, similarly cells containing a close pEF6 plasmid only (containing no expression sequence, to demonstrate the lack of effects exerted by the plasmid alone) were designated PC-3pEF6 and DU-145pEF6 whereas the unaltered wild type cells were simply labelled PC-^ or 011-145^.
Knockout of Matriptase-2 Expression Using a Ribozvme Transgenes A Matriptase-2 ribozyme system was employed to knockout the expression of matriptase-2 in cancer cell lines, to examine the effect of its loss. The procedure was as we previously described (Jiang et al 2005, BGRC). The primers used for the generation of the ribozyme were forward 5'- CTGCAGCACTAC
AATTCCCGGCGGGTAACTGATGAGTCCGTGAGGA -3' and reverse 5'- ACTAGTTGTACTCAATCGCCACTTCTCCCAGGATTTCGTCCTCACGGA-S'. As described previously, ribozyme insert was generated using the primers under a touch-down condition, the resulting products were T-A cloned into the pEF6/V5-His-TOPO vector. Following selection and amplification in E. CoIi, purified plasmid was then electroporated into the PLC-PRF-5 liver cancer cell line with the aim of suppressing the expression of matriptase-2 in the cell line were it is found most abundantly. The PLC-PRF-5 cells with matriptase-2 expression suppressed were called PLC-MAT-2-KO, and properties were assessed through a series of cell assays described below.
Cell growth Assay
The growth rates of PC-3 and DU-145 control cells was examined and compared to that of those containing the matriptase-2 expression plasmid using an in vitro growth assay.
Cells were seeded in a 96 well plate at a density of 3,000 or 7,000 cells/well, and incubated at 37°C for 24, 48 or 72 hours. MTT was added in solution to the cells (200μg/well) and incubated for 4 hours at 37°C, then the cells were lysed with Triton (10%) and the intensity of the colour released was determined by a plate
reader (Titertek Multiskan, Eflab, Finland). The number of cells was shown as absorbance units.
Tumour Cell Invasion Assay We quantified the invasive nature of the matriptase-2 modified cancer cells and control cells using the standard invasion assay procedure as described previously (Parr et al., 2001). Transwell chambers, equipped with a 6.5mm diameter polycarbonate filter insert (pore size 8μm)(Becton Dickinson, Labware, Oxford, UK), were pre-coated with 50μg/insert of solubilised tissue basement membrane, Matrigel (Collaborative Research Products, Bedford, Massachusetts,
USA). 10,000 or 15,000 control and matriptase-2 modified cancer cells were seeded into each insert and allowed to invade for 3 days. Following incubation, cells that had invaded through the basement membrane were fixed (4% formaldehyde), and then stained with crystal violet 0.5%(w/v). For analysis, the cells were counted in 10 fields/insert (x40 magnification); to determine the mean number of invaded cancer cells.
Real-time Quantitative Polymerase Chain Reaction (QPCR)
Real-time Quantitative PCR was also used to assess the matriptase-2 mRNA levels present in the control and transfected cell lines.
The iCycler IQ system (BioRad, Camberley, UK), was employed to quantify the level (shown as copies/μl from internal standard) of matriptase-2 in each of the control and transfected cells. Results are given as number of transcripts/μl based on an internal standard and the results were further normalised using the expression of β actin in these samples. Matriptase-2 forward and reverse primers were δ'-CCGAGTACGAAGTGGAC-S' and 5'-
TAGCTGTAGCGGTAACAACC-3'. The QPCR technique utilised the Amplifluor system (Intergen Inc, England) and Q-PCR master mix (ABgene, Surrey, England), in conjunction with a universal probe (UniPrimer™). Real-time QPCR conditions were 95°C for 12 minutes, followed by 65 cycles at 95°C for15s, 55°C for 60s, 72°C for 20s. The results of the test molecules were normalised against levels of β-actin, using a β-actin quantitation kit from Perkin-Elmers (Perkin- Elmers, Surrey, England, UK).
SDS-PAGE and Western blotting
Cancer cells were grow to confluence, detached and lysed in HCMF buffer containing 0.5% SDS, 1% Triton X-100, 2 mM CaCI2, 100 μg/ml phenylmethylsulfonyl fluoride, 1 mg/ml leupeptin, 1 mg/ml aprotinin and, 10 mM sodium orthovanadate on a rotor wheel for 1 hour before being spun at 13,00Og to remove insolubles. Following lysis, protein levels were quantified using the
Bio-Rad DC Protein Assay kit (Bio-Rad laboratories, California, USA), standardised to 2mg/ml and diluted 1 :1 with Laemmli 2x concentrate sample buffer (Sigma, Dorset, UK). Samples were boiled for 5 minutes before loading into a 10% acrylamide gel and being subjected to electrophoretic separation.
Once sufficient separation had occurred the proteins were blotted onto a Hybond-C Extra nitrocellulose membrane (Amersham Biosciences UK Ltd, Bucks, UK), blocked in 10% milk and subjected to specific antibody probing. Matriptase-2 expression was detected using both commercial antibodies available at the time of the study; RP-1 matriptase-2 recognising the cytoplasmic region and RP-2 matriptase-2 recognising the stem region of the matriptase-2 protein (both antibodies were obtained from Triple Point Biologies, Forest Grove, Oregon, USA), in addition to these commercially available antibodies a third antibody generated in house was also used. An anti β-actin antibody (Santa Cruz Biotechnology, Inc., California, USA) was also used to indicate uniform protein expression levels in the samples. Bound primary antibodies were then detected using peroxidise conjugated anti-rabbit (matriptase-2) or anti-goat (β- actin) antibodies (Sigma, Dorset, UK) and visualised through the addition of Supersignal West Dura Extended Duration substrate chemi-luminescent system (Perbio Science UK Ltd, Cramlington, UK)
RNA extraction and Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
Cells were grown to confluence in a 25cm3 tissue culture flask; RNA was then extracted from these cells using the ABgene total RNA isolation reagent kit
(Advanced Biotechnologies Ltd, Surrey, UK) in accordance with the supplied protocol. Following extraction, cellular RNA was quantified through spectrophotometric measurement (WPA UV 1101 , Biotech Photometer, Cambridge, UK). RNA levels throughout samples were then standardised to
250ng for use in reverse transcription. Reverse transcription was carried out using a enhanced avain reverse transcriptase-PCR-100 kit with anchored oligo(dt) primers and in accordance with the supplied protocol (Sigma, Dorset, UK). The quality of the cDNA generated was tested using β-actin primers before probing for the expression of matriptase-2 using specific primers (full primer sequences are outlined in table, 1). PCR was performed in a T-Cy Thermocycler (Creacon Technologies Ltd, The Netherlands) using REDTaq® ReadyMix™ PCR Reaction Mix (Sigma, Dorset, UK). PCR conditions were as follows: denaturing at 940C for 1 minute, annealing at 580C for 2 minutes and extension at 720C for 3 minutes. PCR was conducted over 38 cycles with an initial 5 minute denaturing step (940C) and a final 10 minute extension step (720C). Following PCR, products were loaded < on a 0.8% agarose gel, separated electrophoretically, stained in ethidium bromide and visualised under ultraviolet light.
In vitro Matrigel adhesion assay
The adhesion of control and transfected cells to an artificial basement membrane was quantified using an in vitro Matrigel adhesion assay adapted form a previously described method [22]. Briefly, 45,000 cells were seeded into the wells of a 96 well plate that had been previously coated with 5μg of Matrigel artificial basement membrane. Cells were incubated for 45 minutes before being subjected to vigorous washing in BSS1 to remove non-adherent cells. Adherent cells were then fixed in 4% formaldehyde (v/v) and stained with 0.5% (w/v)
crystal violet. The number of stained, adherent cells was counted in several random fields under x 40 objective magnification.
In vitro migration/wound healing assay Migration rates of control and transfected cells were assessed using a wound healing/migration assay. This technique has been modified from a previously described method reported by our group [23], examining cell migration rates through their capacity to close an artificial wound. Briefly, cells were cultured in a 24 well plate, upon reaching confluence; the cell monolayer was wounded using a 21 G needle. The closure of this wound, through the migration of cells, was tracked and recorded over a 90 minute period using a time-lapsed video system (Panasonic, Japan) 90 minutes. Cellular migration was then calculated using the Optimas 6 motion analysis software.
In vivo development of prostate tumours.
Athymic nude mice (Nude CD-1) of 4-6 weeks old were purchased from Charles River, UK, and maintained in filter-toped units. 100μl of cell suspension (in 0.5mg/ml Matrigel) was injected subcutaneously at the left scapula area. Mice were weighed and tumour sizes measured twice weekly for 4 weeks. Mice with weight loss over 25% and tumour size larger than 1cm in any dimension were terminated according to the UK Home Office and UKCCCR guideline. The volume of the tumour was determined using the formula: tumour volume = 0.523*width2* length. At the conclusion of the experiment, animals were
terminally anaesthetised, primary tumours were dissected, weighed and frozen at -8O0C. Part of the primary tumours was fixed for histological examination.
Immuno-Fluorescent staining Cells for use in immuno-fluorescent staining were seeded at a density of 20,000 cells per well in a 16-well chamber slide (LAB-TEK Fisher Scientific UK, Longhborough, Leics, UK). Duplicate rows were seeded and the cells were cultured overnight to adhere and establish before one of the duplicate rows was treated with 40ng/ml HGF for 2 hours. Following treatment, the medium was aspirated and the cells were fixed in ice cold ethanol at -2O0C for 20 minutes.
The cells were then rehydrated in BSS for 20 minutes at room temperature before being permeablized with a BSS solution containing 0.1% Triton X100 for 5 minutes. A blocking solution of MenaPath Autowash buffer (A. Menarini Diagnostics, Berkshire, UK), containing 2 drops of horse serum per 5ml, was then placed on the cells for 20 minutes to block non-specific binding. Cells were next washed with wash buffer twice before probing for specific expression of paxillin or FAK using anti-paxillin (Transduction Laboratories) and anti-FAK (Transduction Laboratories) primary antibodies made up in wash buffer at a 1 :100 concentration for 1 hour. The primary antibody was then completely removed by washing the cells three times in wash buffer. A FITC conjugated anti-mouse secondary antibody (Insight Biotechnology Ltd., Middlesex, UK) was then added and the slides were incubated on a shaker platform for 1 hour in the dark. Finally the slides were washed three times to remove un-bound secondary antibodies, mounted with Fluor-save (Calbiochem-Novabiochem Ltd.,
Nottingham, UK) and visualised under an Olympus BX51 microscope at X100 objective magnification.
Results Figure 1 shows antibody staining of normal prostate epithelial cells (A and C) and human prostate cancer tissues (B and D). Slides A and B shows staining of matriptase-1 and slides C and D show stainings of matriptase-2 in human prostate tissue. A comparison of slides A and B shows that tumours over- expressed matriptase-1 protein. However a comparison of slides C and D shows that matriptase-2 had a completely different staining pattern. Indeed, matriptase-2 is membranous protein in normal prostate epithelial cells (C), but prostate cancer cells (D) lost the membranous pattern and reduced staining intensity.
The surprising results shown in Figure 1 were investigated further by determining the expression pattern of matriptase-2 in prostate cancer cell lines transfected with matriptase-2.
RT-PCR and Q-PCR demonstrated that both of the wild type cells (PC-β™1" and DU-Hδ^) and the pEF6 control cells (PC-3pEF6 and DU-145pEF6) showed minimal to no expression of the matriptase-2 gene. However, upon transfection with the expression plasmid an increase in matriptase-2 mRNA expression could be seen (figure 2A-D). The transfection process appears to have been more efficient in the pc-3Matriptase"2 EXP cell line than the DU-145Matriptase-* EXP cell line, as
forced expression was obvious using conventional RT-PCR in the pc-3Matπptase'2 Exp cell line whereas Q-PCR was required to identify this trend in the DU- 145Matriptase"2 EXP cell line.
Additionally, Western blotting was used to probe for matriptase-2 protein levels in both the control and transfected cell lines. Two commercially available antibodies and one antibody generated "in-house" were used to detect matriptase-2 protein expression (figure 2E). Similar to the trends seen at the mRNA level, protein expression of matriptase-2 was found to be minimal in all of the control cell lines (PC^, 011-145^, PC-3pEF6 and DU-145pEF6) and was enhanced in these cell lines following transfection with the matriptase-2 expression plasmid (PC-3Matriptase-2 EXP and DU-i45Matriptase-2 EXP).
However, when these cell lines were investigated for cell growth significantly slower growth was determined as illustrated in Figure 3.
Matriptase-2's effects on the growth of PC-3 and DU-145 cells were examined using an in vitro tumour cell growth assay (see figure 3). Contrasting results for the effects of matriptase-2 expression on tumour cell growth were seen between the PC-3 and DU-145 cell lines. In the PC-3 cell line, forced-expression of matriptase-2 resulted in a significant decrease in the growth over a 5 day incubation period and a significant difference was seen between the PC-
3Matriptase-2 EXP and pQ.βPEFβ ∞n ]]nQ afte|. 5 days (p < Q Q01 )
Even more surprising we discovered that this reduced growth also correlated with significantly reduced in vitro invasiveness without effecting matrix adhesiveness. Both cell lines containing the expression plasmid
 EXP and DU-145Matriptase"2 EXP) were found to have a significantly reduced capacity to invade through an artificial Matrigel basement membrane compared to their respective pEF6 control cell lines (PC-3pEF6 vs pc-3Matriptase'2 EXP p < 0.001 , DU- 145pEF6 vs DU-145Matriptase-2 EXP p = 0.003). This appears to highlight contrasting, roles between matriptase-1 and matriptase-2 in cellular invasion, indicating a suppressive role of matriptase-2 in prostate cancer cell invasion. The capacity of PC-3 and DU-145 prostate cancer cells to adhere to an artificial Matrigel basement membrane over a 45 minute period was examined using an in vitro Matrigel adhesion assay. Transfection of both prostate cancer cell lines with the matriptase-2 expression plasmid did not result in any noticeable change in the adhesive properties of the prostate cancer cell lines. No significant difference in adhesive capacity was seen between cells containing the expression plasmid
EXP and DU-145Matriptase"2 EXP) were found to have a significantly reduced capacity to invade through an artificial Matrigel basement membrane compared to their respective pEF6 control cell lines (PC-3pEF6 vs pc-3Matriptase'2 EXP p < 0.001 , DU- 145pEF6 vs DU-145Matriptase-2 EXP p = 0.003). This appears to highlight contrasting, roles between matriptase-1 and matriptase-2 in cellular invasion, indicating a suppressive role of matriptase-2 in prostate cancer cell invasion. The capacity of PC-3 and DU-145 prostate cancer cells to adhere to an artificial Matrigel basement membrane over a 45 minute period was examined using an in vitro Matrigel adhesion assay. Transfection of both prostate cancer cell lines with the matriptase-2 expression plasmid did not result in any noticeable change in the adhesive properties of the prostate cancer cell lines. No significant difference in adhesive capacity was seen between cells containing the expression plasmid
(pc_3Matriptase-2 EXP and DLM 45Matriptase-2 EXP) and thejf respectjve pEp6 cont|O|
cells (PC-3pEF6 and DU-145pEF6). This data is represented in Figure 4 where it can be seen that the expression of matriptase-2 in either of the cell lines investigated, i.e. PC-3 or DU-145, reduced cell invasiveness (A and B). However, the capability of both these cell lines to adhere to the extra cellular matrix was not affected (C and D).
The effect of matriptase-2 over-expression on cell migration was assessed using a migration/wound healing assay (see figure 8). Transfection with the matriptase-2 expression plasmid affected the closure of the artificial wound.
Both pc-3Matriptase-2 EXP and D|j-i45Matriptase-2 EXP cell lines were found to have
significantly reduced migratory rates compared to their respective pEF6 controls
(pc_3Matriptase-2 EXP vs pC_3pEF6 p = Q Q34 EXP vs DU.145PEF6 p _
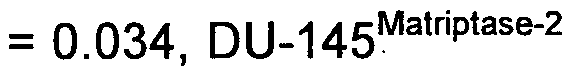
0.002).
In an in vivo environment, PC-3 cells grow rapidly and develop into large tumour masses. However, transfection of this aggressive prostate cancer cell line with the matriptase-2 expression plasmid was found to dramatically reduce in vivo growth and resulted in minimal to no tumour development (see figure 5). The effect of matriptase-2 over-expression was noticeable from the first experimental reading, where a significant difference in tumour development was seen between the pc-3Matriptase"2 EXP and PC-3pEF6 cell lines (p = 0.0002). This trend was continued over the course of the entire experiment, making the excision of the pc-3Matriptase-2 EXP tumour impossible due to its small size and lack of development. Similar in vivo effects demonstrated here using prostate cancer cells have also been found in the aggressive MDA-MB-231 cell line. Together this strongly indicates a role for matriptase-2 in the development of tumours in vivo.
The localisation and expression of the paxillin and FAK molecules in the presence and absence of the HGF cytokine were examined in PC-β™1", PC-3pEF6 and pc-3Matriptase-2 EXP cells (see figure 9). Minimal staining of both paxillin and FAK were apparent in the PC-S^ and PC-3pEF6 cell lines, both in the presence and absence of HGF. However pc-3Matriptase"2 EXP cells had a greater degree of
staining for both paxillin and FAK, and this staining was focused in the areas of the focal adhesion complexes at the cellular peripheries. Interestingly, treatment of these cells with HGF prior to fixation and staining seemed to reduce the degree of paxillin and FAK staining. This results suggests there is some interaction between these adhesion molecules and the matriptase-2 protease and that this interaction may be responsible for the changes in the migratory rates previously demonstrated in the pc-3Matriptase-2 EXP cells.
In this study, matriptase-2 was forcibly up-regulated in two prostate cancer cell lines, which normally have minimal expression levels of this molecule, using an expression plasmid. Transfection of these cell lines with the matriptase-2 expression plasmid affected a number of in vitro cellular traits directly linked to cancer aggression. In both of these prostate cancer cell lines, over-expression of matriptase-2 statistically reduced cellular migration and invasion, traits strongly linked to cancer invasion, dissemination and metastasis. These prostate studies suggest a role for matriptase-2 in the suppression of cellular invasion and migration, both key events in cancer metastasis, and imply a preventative effect of matriptase-2 in cancer progression.
In addition to the in vitro data, the results of the in vivo tumour growth and development assay again demonstrate the dramatic effects of matriptase-2 over- expression in tumour development in a complex living environment where numerous other factors and interactions can influence tumour growth. PC-3 cells containing the matriptase-2 expression plasmid developed very poorly in an
in vivo environment and were significantly smaller than PC-3 cells transfected with an empty pEF6 plasmid vector. This in vivo evidence further suggests that mariptase-2 may have an antagonistic effect on the development and progression of prostate cancer and highlights the potential of this protease for use in the development of new therapeutics for the treatment of cancer progression and metastasis.
References:
• Jiang WG, Davies G, Oystein Fodstad O.Com-1/p8 in oestrogen regulated growth of breast cancer cells, the ER-beta connection. Biochemical and Biophysical Research Communications, 2005, 330, 253-
262
• Parr C1 Davies G, Nakamura T, Matsumoto K, Mason MD, Jiang WG. A HGF/SF antagonist, NK4, regulates matrix-adhesion and paxillin phosphorylation in prostate cancer cells. Biochemical and Biophysical Research Communications, 2001 , 285, 1330-1337
• Parr, C, et al., Matriptase-2 inhibits breast growth and invasion and correlates with favorable prognosis for breast cancer patients. Clinical Cancer Research, 2007. 13: p. 3568 - 3576.
• Jiang, W.G., et al., Inhibition of invasion and motility of human colon cancer cells by gamma linolenic acid. British Journal of Cancer, 1995. 71 : p. 744 - 752.
• Jiang, W.G., et al., Antagonistic effect ofNK4, a novel hepatocyte growth factor variant, on in vitro angiogenesis of human vascular endothelial cells. Cancer Research, 1999. 5: p. 3695 - 3703.
Claims
1. A nucleic acid molecule encoding a polypeptide that has anticancer activity in prostate cells and is characterised by the nucleic acid sequence shown in Figure 6, SEQ ID NO:1 ; or a homologue thereof; or a nucleic acid molecule that hybridises to the molecule shown in Figure 6, SEQ ID NO:1 , under stringent conditions; or a fragment thereof which fragment also has anticancer activity in prostate cells.
2. A polypeptide that has anti-cancer activity in prostate cells comprising a polypeptide as shown in Figure 7, SEQ ID NO:2, or a homologue thereof; or a fragment thereof which fragment also has anti-cancer activity in prostate tissue.
3. A medicament comprising the nucleic acid molecule according to claim 1 and/or the polypeptide according to claim 2.
4. The use of a nucleic acid molecule according to claim 1 in the manufacture of a medicament for treating prostate cancer.
5. The use of the polypeptide according to claim 2 in the manufacture of a medicament for treating prostate cancer.
6. A medicament according to claims 3, 4 or 5 wherein the medicament is formulated with a suitable excipient, carrier or emollient.
7. A vector adapted to transfect or transform prostate cells wherein said vector includes:
(1) at least one copy of the matriptase-2 nucleic acid molecule according to claim 1 ; and/or
(2) . at least one over-expressing or constitutively active promoter which is either coupled to the nucleic acid molecule of part (1) and/or which is designed for insertion into a host genome upstream of the native matriptase-2 gene of said prostate cells whereby transfection or transformation of said cells with said vector results in the enhanced expression of matriptase-2.
8. An antibody selective for the polypeptide according to claim 2.
9. An antibody according to claim 8 wherein the antibody is selective for the human form of the said polypeptide.
10. A process for the production of antibodies that selectively bind to the polypeptide of claim 2 comprising: (a) introducing matriptase-2 polypeptide according to claim 2, or an immunogenic fragment thereof, into a non-human mammalian animal;
(b) obtaining anti-sera from said animal; and
(c) purifying the anti-sera in order to obtain matriptase-2 antibody.
11. A method according to claim 10 wherein the matriptase-2 polypeptide comprises at least the following fragment
GQGDGGDGEEAEPEGMFKAC.
12. An anti-cancer agent for treating prostate cancer comprising the nucleic acid molecule according to claim 1 and/or the polypeptide according to claim 2.
13. The use of a nucleic acid molecule according to claim 1 and/or the polypeptide according to claim 2 for treating prostate cancer.
14. A method for treating prostate cancer comprising administering to an individual to be treated a medically effective amount of the nucleic acid molecule according to claim 1 and/or the polypeptide molecule according to claim 2.
15. A method for treating prostate cancer comprising increasing cellular levels of either exogenous or endogenous matriptase-2 polypeptide according to claim 2 in prostate cells.
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| GB0614201.2 | 2006-07-18 | ||
| GB0614201A GB0614201D0 (en) | 2006-07-18 | 2006-07-18 | Matriptase-2 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| WO2008009895A2 true WO2008009895A2 (en) | 2008-01-24 |
| WO2008009895A3 WO2008009895A3 (en) | 2008-04-10 |
Family
ID=36955810
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/GB2007/002598 Ceased WO2008009895A2 (en) | 2006-07-18 | 2007-07-12 | Matriptase-2 |
Country Status (2)
| Country | Link |
|---|---|
| GB (1) | GB0614201D0 (en) |
| WO (1) | WO2008009895A2 (en) |
Cited By (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2023503513A (en) * | 2019-11-29 | 2023-01-30 | カイマブ・リミテッド | Treatment of physiological iron overload |
| JP2023520969A (en) * | 2020-04-07 | 2023-05-23 | マブウェル セラピューティクス インコーポレイテッド | Anti-TMPRSS6 antibody and use thereof |
Family Cites Families (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP1252300B1 (en) * | 2000-02-03 | 2011-01-19 | Dendreon Corporation | Nucleic acid molecules encoding transmembrane serine proteases, the encoded proteins and methods based thereon |
-
2006
- 2006-07-18 GB GB0614201A patent/GB0614201D0/en not_active Ceased
-
2007
- 2007-07-12 WO PCT/GB2007/002598 patent/WO2008009895A2/en not_active Ceased
Cited By (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2023503513A (en) * | 2019-11-29 | 2023-01-30 | カイマブ・リミテッド | Treatment of physiological iron overload |
| JP7709436B2 (en) | 2019-11-29 | 2025-07-16 | カイマブ・リミテッド | Treatment of Physiological Iron Overload |
| US12570740B2 (en) | 2019-11-29 | 2026-03-10 | Kymab Limited | Treatment for physiological iron overload with an anti-matriptase-2 antibody |
| JP2023520969A (en) * | 2020-04-07 | 2023-05-23 | マブウェル セラピューティクス インコーポレイテッド | Anti-TMPRSS6 antibody and use thereof |
| JP7813234B2 (en) | 2020-04-07 | 2026-02-12 | マブウェル セラピューティクス インコーポレイテッド | Anti-TMPRSS6 antibody and uses thereof |
Also Published As
| Publication number | Publication date |
|---|---|
| WO2008009895A3 (en) | 2008-04-10 |
| GB0614201D0 (en) | 2006-08-23 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| Pepper et al. | Upregulation of urokinase receptor expression on migrating endothelial cells | |
| Turck et al. | Matrix metalloproteinase 2 (gelatinase A) regulates glomerular mesangial cell proliferation and differentiation | |
| Devy et al. | The pro‐or antiangiogenic effect of plasminogen activator inhibitor 1 is dose dependent | |
| D'Anjou et al. | Molecular validation of PACE4 as a target in prostate cancer | |
| Cheng et al. | Hepatocyte growth factor activator inhibitor type 1 regulates epithelial to mesenchymal transition through membrane-bound serine proteinases | |
| Itahana et al. | Role of Id-2 in the maintenance of a differentiated and noninvasive phenotype in breast cancer cells | |
| Wang et al. | Pathway-related molecules of VEGFC/D-VEGFR3/NRP2 axis in tumor lymphangiogenesis and lymphatic metastasis | |
| Xiang et al. | Hepatocyte nuclear factor 4 alpha promotes the invasion, metastasis and angiogenesis of neuroblastoma cells via targeting matrix metalloproteinase 14 | |
| Singh et al. | Transcription factor C/EBP‐β mediates downregulation of dipeptidyl‐peptidase III expression by interleukin‐6 in human glioblastoma cells | |
| US9453050B2 (en) | Compositions for treating glioma | |
| JP2017511150A (en) | Kits and methods for reprogramming non-hepatocytes to hepatocytes | |
| Ahokas et al. | Matrix metalloproteinase-21 is expressed epithelially during development and in cancer and is up-regulated by transforming growth factor-β1 in keratinocytes | |
| Wang et al. | FOXF2 deficiency permits basal-like breast cancer cells to form lymphangiogenic mimicry by enhancing the response of VEGF-C/VEGFR3 signaling pathway | |
| Salvi et al. | In vitro c-met inhibition by antisense RNA and plasmid-based RNAi down-modulates migration and invasion of hepatocellular carcinoma cells | |
| Numa et al. | Syndecan-1 expression in cancer of the uterine cervix: association with lymph node metastasis | |
| Ghilardi et al. | Trypsinogen 4 boosts tumor endothelial cells migration through proteolysis of tissue factor pathway inhibitor-2 | |
| CN114457163A (en) | Marker of mesenchymal subtype glioblastoma and application of marker | |
| EP2588865A1 (en) | The role of fragile x mental retardation gene and protein in cancer metastasis | |
| Guttery et al. | Association of invasion-promoting tenascin-C additional domains with breast cancers in young women | |
| WO2008009895A2 (en) | Matriptase-2 | |
| KR101885982B1 (en) | Use of NAB2 as an oral cavity cancer prognostic marker | |
| Krizbai et al. | Growth factor-induced morphological, physiological and molecular characteristics in cerebral endothelial cells | |
| KR20160055923A (en) | Inhibiting cancer metastasis | |
| Jia et al. | CD147 depletion down-regulates matrix metalloproteinase-11, vascular endothelial growth factor-A expression and the lymphatic metastasis potential of murine hepatocarcinoma Hca-F cells | |
| Lin et al. | Alpha2 macroglobulin, a PSA binding protein, is expressed in human prostate stroma |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 07766186 Country of ref document: EP Kind code of ref document: A2 |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |
|
| NENP | Non-entry into the national phase |
Ref country code: RU |
|
| 122 | Ep: pct application non-entry in european phase |
Ref document number: 07766186 Country of ref document: EP Kind code of ref document: A2 |