WO2000028072A1 - Biomarkers for oxidative stress - Google Patents
Biomarkers for oxidative stress Download PDFInfo
- Publication number
- WO2000028072A1 WO2000028072A1 PCT/US1999/026133 US9926133W WO0028072A1 WO 2000028072 A1 WO2000028072 A1 WO 2000028072A1 US 9926133 W US9926133 W US 9926133W WO 0028072 A1 WO0028072 A1 WO 0028072A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- antibody
- protein
- oxidative stress
- selenium
- proteineaceous
- Prior art date
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/76—Albumins
- C07K14/765—Serum albumin, e.g. HSA
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/46—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates
- C07K14/47—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates from mammals
- C07K14/4701—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates from mammals not used
- C07K14/4723—Cationic antimicrobial peptides, e.g. defensins
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/575—Hormones
- C07K14/62—Insulins
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/775—Apolipopeptides
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/68—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving proteins, peptides or amino acids
Definitions
- This invention relates generally to methods of detecting the presence of oxidatively damaged proteins and endogenous/autoantibodies to oxidatively damaged proteins.
- Chronic inflammation and oxidative stress are associated with a wide variety of diseases and disorders in human populations.
- diseases and disorders affect organs and systems including, but are not limited to, reproductive organs, immune system, lungs, cardiovascular system, nervous system, gastrointestinal system, as well as organs and systems controlling growth and development.
- diseases include, but are not limited to, coronary artery disease, renal disease, cancer, and psychiatric diseases.
- Inflammation and oxidative stress in animals result from interaction with the environment and involve exposure to a wide variety of physical, chemical and biological agents (Ames, B.N. et al. (1993), "Oxidants, antioxidants, and the degenerative diseases of aging,” Proc. Natl. Acad. Sci. USA 90:7915-7922; Sies, H. (ed.) (1991), Oxidative Stress, Oxidants and Antioxidants, New York: Academic Press). When these conditions become chronic, they can lead to changes in the normal cellular balance between antioxidants and oxidants that are associated with many different diseases in aging human populations (Grisham, M.G.
- Atherosclerosis is a chronic inflammatory disease that develops in response to metabolic, physical or environmental injuries such as hypercholesterolemia, hypertension or cigarette smoking (Munro, J.M and Cotran, R.S. (1988), "The pathogenesis of atherosclerosis: Atherogenesis and inflammation,” Lab. Invest. 58:249-261; Parthasarathy, S. et al.
- Sulfur moieties in amino acid residues in proteins are particularly susceptible to oxidation.
- cysteine residues which contain a thiol moiety
- cystine residues which contain a disulfide moiety
- cysteic acid cysteic acid
- cysteic acid cysteic acid
- cysteic acid cysteic acid
- cysteic acid cysteic acid
- selenium analogs of cysteine and cystine, selenocysteine and selenocystine which are more easily oxidized than their sulfur analogs, can be oxidized to selenocysteic acid (Huber, R. and Criddle, R. (1967),
- Reagents capable of oxidizing sulfur or selenium moieties in sulfur- or selenium- containing amino acids in proteins to sulfonic or selenocysteic acid moieties and sulfone or selenone moieties may be encountered directly in the environment (e.g., ozone), or may be generated endogenously, e.g., hypochlorous acid (HOC1) is generated by the myeloperoxidase (MPO)/hydrogen peroxide (H 2 O 2 )/chloride ion (Cl " ) system of activated phagocytic leukocytes during inflammation.
- MPO myeloperoxidase
- H 2 O 2 hydrogen peroxide
- Cl " chloride ion
- interaction with a wide variety of environmental oxidants may also contribute to oxidative stress in vivo and the formation of oxidized sulfur or selenium moieties in sulfur- or selenium-containing amino acids, e.g., cysteic acid in proteins.
- endogenous sources of strong oxidants include but are not limited to aerobic mitochondrial respiration, peroxisomes, and cytochrome P-450 enzymes (Ames, B.N. et al. (1993), "Oxidants, antioxidants, and the degenerative diseases of aging,” Proc. Natl. Acad. Sci.
- a major feature of this host defense function is a powerful oxygen-dependent, microbicidal, viricidal and tumoricidal system that utilizes two different peroxidases, myeloperoxidase (MPO) and eosinophilic peroxidase (EPO) (Henderson, W.R., Jr. (1991) "Eosinophil peroxidase: occurrence and biological function," in Peroxidases in Chemistry and Biology, Vol 1, Everse, J. et al. (eds.), CRC Press, Boca Raton, pp. 105-121; Klebanoff, S.J. (1992), "Oxygen metabolites from phagocytes," in Inflammation.
- MPO myeloperoxidase
- EPO eosinophilic peroxidase
- MPO Basic Principles and Clinical Correlates, J. I. Gallin et al. (eds.), Raven Press, NY, pp.391-444). MPO is found only in the granules of neutrophils and monocytes/macrophages, and is biochemically distinct from EPO (Bainton, D.F. (1992), "Developmental biology of neutrophils and eosinophils," in Inflammation: Basic Principles and Clinical Correlates, 2nd ed., Gallin, J.L. et al. (eds.), Raven Press, NY, pp. 303-324; Henderson, W.R., Jr.
- MPO utilizes H 2 O 2 and Cl “ (Carr, A.C. et al. (1996), “Peroxidase-mediated bromination of unsaturated fatty acids to form bromohydrins," Arch. Biochem. Biophys. 327:227-233; Thomas, E.L. and Learn, D.B.
- HOC1 In large interstitial inflammatory sites, the concentration of HOC1 has been estimated to be in the mM range (Weiss, S.J. (1989), “Tissue destruction by neutrophils,” N. Engl. J. Med. 320:365- 376). Reactivity of the MPO-H 2 O 2 -Cl " system is enhanced when phagocytes are activated on biological surfaces (Nathan, CF. (1987), "Neutrophil activation on biological surfaces. Massive secretion of hydrogen peroxide in response to products of macrophages and lymphocytes," J.
- EPO In mammalian cells, EPO also is able to catalyze the formation of HOC1, however, its preferred in vivo substrate is thought to be bromide and/or thiocyanate, both of which give rise to potent oxidants (HOBr and HOCN, respectively)
- HOBr and HOCN potent oxidants
- HOC1 can also react with superoxide radical anion in a metal-ion independent Haber- Weiss type reaction to form hydroxyl radical (OH) or with H 2 O 2 to form singlet oxygen (O 2 ) (Candeias, L.P et al. (1993), "Free hydroxyl radicals are formed on reaction between the neutrophil derived species superoxide anion and hypochlorous acid," FEBS Lett.333:151-153). Recent evidence suggests that Haber- Weiss chemistry also involves production of O 2 (Khan,
- HOC1 Under acidic conditions in the presence of chloride ion, HOC1 is in equilibrium with chlorine gas, and phagocytes have been shown to utilize this powerful oxidant at sites of inflammation and vascular disease (Hazen, S.L. et al. (1996), "Human neutrophils employ chlorine gas as an oxidant during phagocytosis,” J. Clin. Invest.98: 1283-1289). Both phagocytic and endothelial cells produce nitric oxide (NO) and reaction of NO with superoxide leads to formation of peroxynitrite (Halliwell, B. (1996), “Antioxidants in human health and disease," Annu. Rev. Nutr.
- ROS reactive oxygen species
- reactive non-oxygen species are strong enough to oxidize the sulfur or selenium moieties of sulfur- or selenium-containing amino acid residues in proteins to cysteic acid (cysteine sulfonic acid) or selenocysteic acid and to sulfone or selenone moieties such as methionine sulfone or selenone.
- albumin the main contributor to a plasma thiol concentration of 469 ⁇ M serves as a major antioxidant defense against HOC1 oxidation in vivo (Hu, M.-L. et al (1993), "Antioxidant protection against hypochlorous acid in human plasma,” J. Lab. Clin. Med. 121:257-262).
- HOC1 oxidation of low density lipoprotein was reported to result in loss of all thiols in the protein constituent, apolipoprotein B-100, (apoB) at a concentration of HOC1 that formed little, if any, chloramines with the many primary amino groups of lysine side chains or oxidation of lipid (Arnhold, J. etal. (1991), "Modification of low density lipoproteins by sodium hypochlorite," Biomed. Biochim. Acta 8:967-973; Hazell, LJ. and Stocker, R.
- LDL low density lipoprotein
- HOC1 oxidation of low density lipoproteins appears to produce sulfmic acids (Yang, E.Y. et al (1999), "Selective modification of apoB-100 in the oxidation of low density lipoproteins by myeloperoxidase in vitro," J. Lipid Res.40:686-98).
- HOC1 oxidation of immunoglobulins and several other proteins Naskalski, J.W. (1994), "Oxidative modification of protein structures under the action of myeloperoxidase and the hydrogen peroxide and chloride system," Ann. Biol. Clin.
- Oxidized sulfur-containing amino acid residues have been found to be associated with many diseases which affect mammals, particularly humans.
- cysteic acid has been reported to occur in proteins isolated from four sources known to be under oxidative stress: senile cataractous lens tissue (Garner, M.H. and Spector, A. (1980), "Selective oxidation of cysteine and methionine in normal and senile cataractous lenses,” Proc. Natl. Acad. Sci. USA
- a monoclonal antibody has been produced against human HOCl-oxidized LDL (oxLDL) that cross-reacts with other HOCl-oxidized proteins but was reported to not cross-react with LDL modified with reactive aldehyde products of lipid peroxidation or LDL that has been oxidized with copper (Cu 2+ ) (Malle, E. et al. (1995), "Immuno logic detection and measurement of hypochlorite- modified LDL with specific monoclonal antibodies," Arterioscler. Thromb. Vase. Biol. 15:982- 989).
- Protein carbonyl derivatives are also reported to be formed from oxidative injury.
- a method of derivatizing carbonyl groups using 2,4-dinitrophenylhydrazine (DNPH) and probing with a commercial biotinylated anti-DNP antibody followed by reacting with a streptavidin- linked horseradish peroxidase has been reported (Buss, H. et al (1997), "Protein carbonyl measurement by a sensitive ELISA method," Free Radic. Biol. Med. 23:361-366; Winterbourn. C. C. and Buss, I. H. (1999), "Protein Carbonyl Measurement by Enzyme-Linked Immunosorbent Assay," Methods in Enzymology 300:106-111).
- Cysteine (thiol) and cystine (disulfide) residues are generally less abundant in proteins than other amino acid side chains, but their importance in the structure and function of proteins is universally recognized. Oxidation or other chemical modification of protein thiols and disulfides usually leads to loss of biological activity (Albrich, J.M. et al. (1981), "Biological reactivity of hypochlorous acid: implications for microbicidal mechanisms of leukocyte myeloperoxidase," Proc. Natl. Acad. Sci. USA 78:210-214; Little, C. and O'Brien, P. (1967), "Products of Oxidation of a Protein Thiol Group after Reaction with Various Oxidizing Agents," Arch.
- Protein cysteic acid (cysteine sulfonic acid) is the stable end-product of oxidation ofthe functional sulfur moieties of cysteine and cystine and is not a normal constituent of naturally occurring mammalian proteins (Manneberg, M. et al. (1995), "Oxidation of cysteine and methionine residues during acid hydrolysis of proteins in the presence of sodium azide," Anal. Biochem. 224:122-127; Manneberg, M. et al. (1995), "Quantification of cysteine residues following oxidation to cysteic acid in the presence of sodium azide," Anal. Biochem. 231:349- 353).
- Selenocysteine is a functional residue in a number of enzymes that exhibit important antioxidant properties (Gallegos, A. et al. (1997), “Mechanisms ofthe Regulation of Thioredoxin Reductase Activity in Cancer Cells by the Chemopreventive Agent Selenium,” Cancer Res. 57:4965-4970): these include cellular and plasma glutathione peroxidases, phospholipid hydroperoxide glutathione peroxidase and thioredoxin reductase.
- Thioredoxin has been shown to regulate gene transcription by controlling the redox state of several transcription factors and their binding to DNA (Matthews, J.R. et al., 1992; Hirota, K. et al. (1997), "AP-1 transcriptional activity is regulated by a direct association between thioredoxin and Ref-1.” Proc. Natl. Acad. Sci. USA 94:3633-3638).
- the thioredoxin gene is overexpressed in human cancer and inhibits apoptosis (Baker, A. et al. (1997),
- the method appears to involve combining an internal standard which is a deuterium labeled oxidized sulfhydryl amino acid noted above, with a body fluid containing an oxidized sulfhydryl amino acid; then at least partially purifying the oxidized sulfhydryl amino acid and the internal standard from other components of the body fluid; quantifying the oxidized sulfhydryl amino acid concentrations by gas chromatography/mass spectrometry and correcting for losses in oxidized sulfhydryl amino acid by determining losses in the deuterium labeled oxidized sulfhydryl amino acid.
- an internal standard which is a deuterium labeled oxidized sulfhydryl amino acid noted above
- the method disclosed in the '038 patent appears to be quantifying unknown metabolites containing oxidized sulfhydryl amino acids.
- the oxidized sulfhydryl amino acid are not measured as concentrations in proteins, but rather as concentration in a body fluid and thus the proteins are not hydro lyzed to their constituent amino acid residues prior to analysis.
- the method uses gas chromatography/mass spectrometry to quantify the oxidized sulfhydryl amino acids.
- Biomarkers of oxidative stress are needed, among other reasons, to identify etiological relationships, to further define the pathophysiological mechanisms underlying diseases related to inflammation and oxidative stress, and to aid in assessing the efficacy of environmental, nutritional and therapeutic interventions.
- the present invention provides methods and compositions for detecting the presence of biomarkers of oxidative stress in proteins.
- the biomarker may be any amino acid that has undergone oxidation (or other modification, e.g. chloro-tyrosine, dityrosine).
- oxidized sulfur- or selenium-containing amino acids SSAA
- Oxidized SSAA are amino acids in which the sulfur or selenium moiety has been oxidized to some oxidation state.
- Oxidized SSAA include, but are not limited to, cysteine, cystine, methionine, selenomethionine, selenocystine and selenocysteine in their various possible oxidation states.
- the present invention provides methods to detect a biomarker of oxidative stress in a biological sample utilizing an antibody or antigen binding fragment thereof which binds said biomarker of oxidative stress, said method comprising the steps of:
- the biological samples used in the present invention include proteins, peptides or proteineaceous aggregates.
- the biological samples may be from any source, preferably an organism selected from the group consisting of: plants, bacteria, animals, viruses and fungi, and are most preferably mammalian or human.
- the antibody or antigen binding fragment may be bound to a solid phase support.
- the biomarkers may be oxidized sulfur- or selenium-containing amino acids or proteins, peptides or proteineaceous aggregates which include one or more oxidized sulfur or selenium-containing amino acids.
- the oxidized sulfur- or selenium-containing amino acid may be selected from the group consisting of the oxidation products of cysteine, cystine, methionine, selenocysteine, selenomethionine and selenocystine.
- Polyclonal and monoclonal antibodies both protein non-specific and protein specific for a protein, peptide or any proteineaceous aggregate that contains an oxidized sulfur- or selenium- containing amino acid are also provided.
- the polyclonal antibodies ofthe present invention may be obtained from a mouse, or other suitable source.
- Methods for detecting the presence of oxidative stress in an organism comprising detecting the presence of an antibody or antigen binding fragment thereof that binds an analyte comprising an oxidized sulfur- or selenium containing amino acid, whereby the presence of said antibody or antigen binding fragment thereof is indicative ofthe presence of oxidative stress in said organism, are also provided.
- the organisms are preferably selected from the group consisting of: plants, bacteria, animals, viruses and fungi, and are most preferably mammalian or human.
- Methods of measuring the amount of oxidative stress an organism has been exposed to comprising
- the organisms are preferably selected from the group consisting of: plants, bacteria, animals, viruses and fungi, and are most preferably mammalian or human.
- Methods of removing oxidatively damaged protein from a sample comprising contacting said sample with an antibody or antigen binding fragment thereof which is specific for oxidized sulfur- or selenium-containing amino acids, whereby at least a portion of oxidatively damaged protein is bound to said antibody or antigen binding fragment thereof, are also provided.
- the sample may be contacted with a solid support to which said antibody or antigen binding fragment thereof is attached.
- the samples used in these methods are preferably selected from the group consisting of plasma, biological fluids and cells, and mixtures thereof.
- the plasmas and biological fluids and cells used in these methods are preferably mammalian or human.
- Methods of detecting one or more selected oxidatively damaged proteins, peptides, or proteineaceous aggregate in a sample, or detecting the concentration of one or more selected oxidatively damaged proteins, peptides or proteineaceous aggregate in a sample comprising:
- the first antibody or antigen binding fragment thereof is specific for a selected oxidatively damaged protein, peptide or proteineaceous aggregate.
- the oxidatively damaged protein, peptide or proteineaceous aggregate preferably contains an oxidized sulfur- or selenium-containing amino acid.
- Methods for detecting or diagnosing the presence of a disease associated with oxidative stress in a mammalian subject comprising:
- step (b) comparing the level detected in step (a) to a level of biomarker of oxidative stress normally present in the mammalian subject; wherein an increase in the level of biomarker for oxidative stress as compared to normal levels indicates a disease associated with elevated levels of biomarker of oxidative stress, are also provided.
- Methods for monitoring the course of a disease associated with elevated levels of biomarker of oxidative stress in a mammalian subject comprising evaluating the level of biomarker of oxidative stress in a series of biological samples obtained at different time points from a mammalian subject according to the methods ofthe invention, wherein an increase in the level of biomarker of oxidative stress over time indicates progression ofthe disease, and wherein a decrease in the level of biomarker of oxidative stress over time indicates regression of the disease, are also provided.
- Methods for monitoring a therapeutic treatment of a disease associated with elevated levels of biomarker of oxidative stress comprising evaluating the level of biomarker of oxidative stress in a series of biological samples obtained at different time points from a mammalian subject undergoing a therapeutic treatment for a disease associated with elevated levels of biomarker of oxidative stress according to the methods ofthe invention, wherein a decrease in the level of biomarker of oxidative stress over time indicates an effective therapeutic outcome, are also provided.
- Monoclonal antibodies which bind a protein, peptide or any proteineaceous aggregate containing an oxidized sulfur- or selenium-containing amino acid are also provided. These monoclonal antibodies are produced by any means known in the art. Preferably, the monoclonal antibody is produced by hybridoma cell line K2.F1. The monoclonal antibodies may also be produced by hybridoma cell line K2.F1.6 deposited with the American Type Culture Collection (ATCC) on October 29, 1999, and assigned Patent Deposit Number PTA-897.
- ATCC American Type Culture Collection
- Polyclonal antibodies which bind proteins, peptides or proteineaceous aggregates containing oxidized sulfur- or selenium-containing amino acids are also provided.
- Polyclonal antibodies which bind biomarkers of oxidative stress, including protein, peptide or proteineaceous aggregate which contains an oxidized sulfur- or selenium-containing amino acid are also provided.
- Monoclonal and polyclonal antibody preparations wherein the monoclonal antibody is specific for oxidized sulfur- or selenium-containing amino acids in a protein, peptide or proteineaceous aggregate are also provided.
- the monoclonal antibody preparation is produced by hybridoma cell line K2.F 1.
- the polyclonal antibody preparation is produced by any animal capable of producing the polyclonal antibody preparation, preferably a mouse, rat, rabbit, chicken or goat, but other animals may be used.
- a hybridoma cell line producing a monoclonal antibody specific for oxidized sulfur- or selenium containing amino acids in a protein or proteineaceous aggregate is also provided.
- this hybridoma cell line is K2.F1.
- the hybridoma cell line is K2.F1.6.
- a test kit for measuring the presence of oxidative damage in an analyte comprising an antibody or an antigen binding fragment thereof, which antibody or antigen binding fragment is an antibody or antigen binding fragment thereof specific for a protein, peptide or any proteineaceous aggregate which contains oxidized sulfur- or selenium-containing amino acids, is also provided.
- a method for preparing a polyclonal antibody against a specific disease associated with an oxidized sulfur- or selenium-containing amino acid comprising:
- the selected disease may be coronary artery disease, renal disease, diabetes, or other disease associated with oxidative stress.
- the present invention also provides antibodies that bind to plasma proteins that contain oxidized sulfur- or selenium-containing amino acids.
- the nature ofthe oxidized proteins in a sample can be identified by the use of a secondary antibody specific for the non- modified portion of the proteins.
- the rate of production and removal of these damaged proteins can be determined in vivo.
- the immunoassay methods and antibodies provided in the present invention are useful, for example, in large population-based molecular epidemiologic studies aimed at the prevention and control of diseases or disorders associated with inflammation and oxidative stress.
- the methods and antibody compositions are useful in providing hypothesis-driven information on mechanisms of oxidative damage associated with a broad spectrum of human diseases.
- the present invention is also useful in assessing the efficacy of various environmental, nutritional and therapeutic antioxidant interventions, as well as in providing for the diagnosis, treatment and management of early stages of diseases associated with inflammation and/or oxidative stress.
- Specific applications ofthe present invention include, but are not limited to, study ofthe red cell membrane oxidation process; study of oxidative damage as a result of oxidative stress diseases, such as diabetes; and the study of mitochondria, which are proposed as a source for oxygen species that cause oxidation.
- the presence of endogenous antibody to oxidized protein can be correlated to the presence of a disease state.
- the partial oxidation of cysteine is believed to be involved in the redox control of gene expression (Claiborne, A. et al. (1993), "Protein-sulfenic acid stabilization and function in enzyme catalysis and gene regulation," FASEB J. 7:1483-1490), and the present invention can be used to further elucidate the mechanisms and species involved.
- Other specific applications of the present invention include, but are not limited to, the study of which plasma protein(s) is more susceptible to oxidative modification.
- the characteristics of the protein(s) that is (are) oxidatively modified can be determined. It is also possible, for a given sample, to determine the percent of a particular protein that may be oxidatively modified. The determination of the percent of this specific protein that may be oxidatively modified could reflect the severity ofthe disease as well as the efficacy of any invention designed to reduce . oxidative modification.
- Other specific applications of the present invention include, but are not limited to, the study ofthe in vivo rate of production and clearance ofthe oxidatively modified protein.
- the antibodies described in the present invention can be used to isolate the oxidatively modified protein in a sample.
- tracer methodology it is possible to assess the rate of removal of the tracer associated with the oxidatively modified protein.
- the determination of whether the presence of the damaged proteins is due to increased production or impaired removal of the oxidatively modified proteins may be important in selecting a proper management strategy for the patients.
- proteineaceous aggregate includes aggregates with protein, lipids, carbohydrates or nucleic acids.
- an “analyte” as used herein comprises a sample containing proteins, peptides or proteineaceous aggregates.
- biomarker of oxidative stress refers to SSAA which contain oxidized sulfur or selenium moieties and to epitopes containing such SSAA, i.e. short sequences of amino acids, preferably fewer than 50 amino acids, more preferably between 5 and 15 amino acids, which contain oxidized SSAA.
- the epitope could be a single oxidized SSAA in the protein.
- control value refers to a basal level of biomarker, i.e., SSAA which contain oxidized sulfur or selenium moieties and to epitopes containing such SSAA, that is normal, i.e., the amount present in a corresponding healthy cohort in the absence of any pathology (disease or disorder) which is associated with oxidative stress.
- the present invention provides methods and compositions for determining control values for oxidative stress. Such control values may need to account for age ofthe individual and therefore be directed to certain age ranges, as oxidative stress may accumulate over time. Such control values may additionally need to account for gender and race, and for environmental exposures, e.g., smoking, diet, etc.
- detection means determination that a substance, e.g., a biomarker, a particular oxidized SSAA, etc., is present.
- the methods and compositions of this invention can also be used to determine the amount of or concentration of a substance, e.g., biomarker, in a sample. Quantification and detection of biomarkers can be performed by any means known to those skilled in the art.
- Means of detection and quantification include but are not limited to precipitation ofthe protein containing the biomarker by an antibody which binds to the biomarker; Western immunoblotting in which the oxidized protein containing the biomarker (either as part of a mixture or contained in an immunoprecipitated complex) is separated by gel electrophoresis, transferred to a suitable support (e.g., nitrocellulose) and visualized by reaction with an antibody(ies); radioimmunoassay, in which the degree to which the protein competes with a radioactively labeled standard for binding to the antibody is used as a means of detecting and quantifying the protein; and enzyme-linked immuno-sorbant assay (ELISA).
- Western immunoblotting in which the oxidized protein containing the biomarker (either as part of a mixture or contained in an immunoprecipitated complex) is separated by gel electrophoresis, transferred to a suitable support (e.g., nitrocellulose) and visualized by reaction with an antibody(ies); radioimmun
- ELISA is a known technique for quantifying proteins in which, generally, an antibody against the protein of interest is immobilized on an inert solid, e.g., polystyrene.
- a sample to be assayed for the protein of interest is applied to the surface containing immobilized antibody.
- Protein binds the antibody, forming a complex.
- This complex is then contacted by a second antibody which binds the same protein and which is covalently bound to an easily assayed enzyme. After washing away any ofthe second antibody which is unbound, the enzyme in the immobilized complex is assayed, providing a measurement of the amount of protein in the sample.
- the ELISA procedure can be reversed, i.e., the antigen is immobilized on an inert support (e.g. 96-well microplate) and samples are probed for the presence of antibody to the immobilized antigen.
- the biomarker can also be detected and its localization determined in cells and tissues using immunohistochemical procedures.
- ELISA Western immunoblotting following electrophoretic separation of a protein mixture, and immunohistochemical procedures are the preferred methods of detecting and quantifying the biomarkers in proteins.
- Biomarkers can be detected and quantified in proteins taken from samples including, but not limited to, plasma, serum, cerebrospinal fluid, saliva, semen, pleural fluid, peritoneal fluid and amniotic fluid. These samples may be of human origin or they may be taken from animals other than humans, for example, avian species, but preferably mammals. As will be apparent to those skilled in the art, the subject methods and compositions can be used to detect and quantify oxidized SSAA in non-biological samples.
- Oxidatively damaged protein includes protein which has undergone oxidation and is intended to include all forms of modification that occur upon oxidation.
- the present invention includes an immunoassay utilizing an antibody for biomarkers of oxidative stress.
- immunoassay refers to a method of detecting or measuring antigens, in this case biomarkers of oxidative stress, by using antibodies as reagents.
- the antibodies can be polyclonal or, preferably, monoclonal.
- polyclonal antibodies and “monoclonal antibodies” have the standard meanings understood by those skilled in the art and refer to antibodies, either a mixture of different antibodies in the case of polyclonal antibodies, or a single antibody in the case of monoclonal antibodies, both of which are produced, in general, by immunization of an animal with an antigen.
- antibody-producing cells are selected from the animal and fused with myeloma cells. These cells are then cultured.
- the antibodies ofthe present invention detect oxidized SSAA in proteins to a desired level. Harlow, E. and Lane, D. (1988) Antibodies: A Laboratory Manual (Cold Spring Harbor Laboratory), which is inco ⁇ orated by reference in its entirety herein to the extent not inconsistent with the disclosure herewith teaches methods regarding the making and usage of antibodies.
- “Visualizing" the biomarker may be performed by any means known in the art, including, but not limited to, ELISA, radioimmunoassay, dot blots, Western immunoblotting combined with gel electrophoresis, immunohistochemistry at light and electron microscope levels, HPLC and mass spectrometry.
- oxidative stress refers to damage to biological molecules resulting from oxidation. Examples include but are not limited to oxidation of lipoproteins, membrane phospholipids; lipid peroxidation; protein damage, including cleavage of amino acid bonds and oxidation of functional groups; nucleic acid strand breaks; nucleic acid base modifications leading to point mutations; inhibition of RNA and protein synthesis; protein cross- linking; impaired maintenance of membrane ion gradients; and depletion of cellular levels of ATP, leading to cellular dysfunction and eventually to disease.
- the oxidant oxidizing reagent
- the methods and compositions of this invention provide means for detecting and measuring accumulated oxidative stress.
- accumulated oxidative stress refers to oxidative stress which is present in a biological molecule at the time of detection and measurement; such damage has not been repaired or otherwise removed.
- Oxidized sulfur- or selenium-containing amino acid includes an SSAA in any oxidation state or epitopes containing such SSAA, i.e., short sequences of amino acids, preferably fewer than 50 amino acids, more preferably between 5 and 15 amino acids, which contain oxidized SSAA.
- the epitope could be a single oxidized SSAA in the protein. This may be cysteic acid, or an intermediate oxidation state such as the thiosulfmate or thiosulfonate of cystine or the diselenide or mixtures thereof.
- the immunoassay methods disclosed herein include both protein nonspecific and protein specific detection.
- protein nonspecific refers to detection of biomarkers of oxidative stress in any protein(s) containing such biomarkers.
- protein specific refers to detection of biomarkers of oxidative stress in particular protein(s) containing such biomarkers.
- the present invention provides a monoclonal antibody directed against an oxidized moiety of protein cysteine/cystine residues that recognizes this biomarker in any protein.
- the present invention provides an ELISA assay for quantification of the biomarker.
- Monoclonal antibodies against other oxidized SSAA can be similarly prepared.
- An ELISA assay can be used in studies to assess variability in the basal levels of biomarkers of oxidative stress in plasma, serum, tissue or other bodily fluids from healthy, normal individuals of known age, gender, ethnicity and whatever parameters one wishes to consider (e.g., lifestyle or environmental exposures such as smoking, drinking, diet, etc.). These basal values for the biomarkers (e.g., mean values +/- variation and other relevant statistical parameters) can then be compared to individuals with pathophysiological conditions that are related to inflammation and oxidative stress, e.g., coronary artery disease, diabetes, etc.
- basal values for the biomarkers e.g., mean values +/- variation and other relevant statistical parameters
- Figure 1 is a chromatographic profile showing the HPLC separation, identification and quantification ofthe D and L isomers of cysteic acid (CA), L-cysteine sulfmic acid (L-CSA) and
- ASP D and L-aspartic acid
- APPA D-2 amino-3-phosphopropionic acid
- Figure 2 is a graph of cysteic acid (picomoles) versus LDL apoprotein (micrograms) showing a least squares linear regression plot of data from the hydrolysis of 3 concentrations of 2 different delipidated human LDL samples.
- Figure 3 is a graph of cysteic acid (picomoles)/protein (micrograms) versus sample number, samples 1-15 being from healthy humans, samples 16 and 17 from human patients with renal disease, samples 18-20 from human patients with unspecified diagnoses, samples 21-25 from human patients with coronary artery disease (CAD).
- CAD coronary artery disease
- Figure 4 is a graph of cysteic acid (picomoles)/protein (micrograms) versus FPLC fraction number, protein (milligram 100 microliters) versus FPLC fraction number, and cholesterol (milligram/milliliter) versus FPLC fraction number.
- Figure 5A is SDS-PAGE of oxidized and unoxidized proteins using murine antiserum raised against the oxidized A-chain of bovine insulin.
- Each lane contained 0.5 ⁇ g of the following proteins: (1) prestained M r markers; (2) unoxidized glyceraldehyde-3-phosphate dehydrogenase (GAPDH); (3) H 2 O 2 -oxidized GAPDH; (4) carbonic anhydrase; (5) oxidized A- chain of bovine insulin conjugated to ovalbumin, oxA-OVA; (6) OVA; (7) M r markers; (8) bovine insulin; (9) H 2 O 2 -oxidized BSA; (10) unoxidized BSA.
- GPDH unoxidized glyceraldehyde-3-phosphate dehydrogenase
- GAPDH H 2 O 2 -oxidized GAPDH
- carbonic anhydrase (5) oxidized A- chain of bovine insulin conjugated to ovalbumin, o
- Figure 5B is Western blotting of oxidized and unoxidized proteins using murine antiserum raised against the oxidized A-chain of bovine insulin. The lanes correspond to those in Figure 5A.
- Figure 6 is ELISA titration of mylein basic protein (MBP) with different dilutions of rabbit anti-MBP antiserum.
- MBP mylein basic protein
- Figure 7A is ELISA titration of oxidized A-chain of bovine insulin (oxA) conjugated to ovalbumin (oxA-OVA).
- Figure 7B is ELISA titration of oxidized A-chain of bovine insulin (oxA) conjugated to ovalbumin (oxA-OVA) and titration of performic acid oxidized ovalbumin (PAoxOVA) using mouse antiserum against performic acid oxidized bovine serum albumin (oxBSA).
- oxA oxidized A-chain of bovine insulin
- PAoxOVA performic acid oxidized ovalbumin
- oxBSA mouse antiserum against performic acid oxidized bovine serum albumin
- Figure 8 is ELISA analysis of H 2 O 2 -oxidized human serum albumin (oxHS A) containing different amounts of cysteic acid using murine antiserum raised against performic acid oxidized bovine serum albumin (oxBS A).
- Figure 9A is reducing SDS-PAGE (lanes 1-9) and non-denaturing PAGE (lane 10) of oxidized and unoxidized proteins using murine antiserum raised against oxidized bovine serum albumin (oxBSA).
- Figure 9B is immunoblotting of oxidized and unoxidized proteins using murine antiserum raised against oxidized bovine serum albumin (oxBSA). The lanes correspond to those in Figure 9A.
- oxBSA oxidized bovine serum albumin
- Figure 10 shows the reactivity ofthe monoclonal antibody against unoxidized BSA and PAoxBSA.
- Figure 11 shows the template of a 96-well plate of an experiment from which the data of
- Figures 12A-B show ELISA results on PAoxOVA of normal kids (aged 12-16).
- Figure 13 shows ELISA results for 62 human serum samples.
- Figure 14 shows ELISA results on patient sera on PAoxOVA in the absence and presence of oxOVA.
- Figure 15 shows PAoxOVA ELISA for plasma samples from 47 patients undergoing renal dialysis in the course of management of end stage renal disease.
- Figure 16 A illustrates an immunoassay that can determine the specificity ofthe oxidative modification process.
- Figure 16B illustrates the conventional immunoassay used to determine the total concentration of a protein in a sample.
- oxidized SSAA residues or partially oxidized residues in protein are biomarkers of irreversible protein oxidative damage, and the quantification thereof reflects the severity and/or duration of in vivo oxidative stress to which the protein has been subjected.
- Some of these oxidized SSAA residues include cysteic acid (cysteine sulfonic acid), methionine sulfone, methionine selenone or selenocysteic acid.
- cysteic acid cysteic acid
- methionine sulfone methionine selenone or selenocysteic acid.
- the presence of these biomarkers are diagnostic of oxidative stress associated with particular diseases. A decrease in these biomarkers with treatment or therapy, e.g., drug therapy, neutraceutical therapy, lifestyle changes, can be an indicator of effectiveness of treatment.
- Cysteine and cystine can be ultimately oxidized to cysteic acid, although intermediate oxidation states can occur.
- Methionine can be ultimately oxidized to methionine sulfone.
- Selenocysteine can be oxidized to selenocysteic acid.
- Selenomethionine can be oxidized to methionine selenone.
- a number of intermediate oxidation states may occur, depending on reaction conditions.
- cystine moieties ofthe protein can also be present as mixed disulfides, i.e., one of the contributing sulfur-containing moieties is not in peptide linkage with the protein, but is, e.g., a glutathione, cysteine, or cysteinylglycine moiety.
- the intermediate oxidation states of cysteine/cystine may undergo chemical reaction(s) to produce oligomeric proteins.
- SSAA biomarker because of its increased sensitivity for minimally oxidized protein. Little information is available on the in vivo turnover of HOCl-oxidized proteins (see Panzenboeck, U. et al., "Effects of reagent and enzymatically generated hypochlorite on physiochemical and metabolic properties of high density lipoproteins," J. Biol. Chem. 272:29711-29720), but such treatment can lead to fragmentation of polypeptide chains and cross-linking (Hazell, L.J. et al.
- oxidized protein can also be used to one's advantage.
- SSAA in proteins can be recognized by endogenous antibodies, e.g., antibodies made by the organism to recognize oxidatively damaged protein (ODP) containing oxidized SSAA.
- ODP oxidatively damaged protein
- one aspect of this invention provides for identifying such endogenous antibodies.
- One method for detecting endogenous antibodies to ODP containing oxidized SSAA is to use the ELISA assay developed to screen for a monoclonal antibody to ODP.
- the measurement of endogenous antibodies to oxidized SSAA in ODP can be used as an indicator of oxidative stress.
- monoclonal antibody to ODP may be used clinically to clear ODP from a patient's blood, for example, by passing the blood over a solid support to which the antibody has been attached.
- cysteine Under the conditions of strong acid (6N HCl) and high temperature (110°C) generally employed in art-known amino acid analysis of proteins, cysteine is unstable and is oxidized to the sulfmic and sulfonic acids (Inglis, A.S. and Liu, T.-Y. (1970), "The stability of cysteine and cystine during acid hydrolysis of proteins and peptides," J. Biol. Chem. 245:112-116).
- a chemical method for the detection of cysteic acid in protein which prevents production of cysteic acid by artifactual oxidation during hydrolysis is described herein. This method is further described below, in the Examples.
- disorders associated with inflammation/oxidative stress have been shown to have significantly elevated levels (as high as 10-20 fold higher) of LDL associated cysteic acid compared to healthy subjects.
- the protein cysteic acid was associated primarily with the plasma LDL fraction.
- the methods described in the invention are used to demonstrate the presence of oxidized SSAA in several naturally occurring proteins, including protein(s) associated with the low density lipoprotein (LDL) fraction of human plasma.
- HOC1 enzyme HOC1 or HOC1 generated by the MPO/H 2 O 2 /Cl " system
- the cysteic acid content of these proteins increased from 10 to 200-fold over background, depending on the protein and the concentration of HOC1.
- the methods ofthe present invention have shown as high as 20-fold differences between the levels of cysteic acid associated with the plasma lipoprotein fraction obtained from healthy individuals and patients with coronary artery and renal disease.
- the approach taken in this invention does not require derivatization ofthe ODP before detection.
- the ODP may be detected directly with the antibody approach of the present invention. This allows for the possibility of doing immunohistochemistry on tissue sections, e.g., atherosclerotic plaques and adjoining tissue (Johnston, J.A., et al. (1998) "Aggresomes: a cellular response to misfolded proteins," J. Cell. Biol. 143:1883-98), mitochondria and cells found in the blood and elsewhere, as well as cells in culture. See, for example, Lawrence, D.A., et al. (1996) "Surface thiols of human lymphocytes and their changes after in vitro and in vivo activation," J. Leukocyte Biol.
- Monoclonal and polyclonal antibodies have been prepared against phosphoserine (the phospho-analogue of cysteic acid) that recognize this residue in different proteins (i.e., it is protein nonspecific) as well as the free amino acid, but do not cross-react with phosphothreonine, phosphotyrosine, AMP or ATP (Abu-Lawi, K.I. and Sultzer, B.M.(1995), "Induction of serine and threonine protein phosphorylation by endotoxin-associated protein in murine resident peritoneal macrophages," Infect. Immun. 63:498-502; Hasegawa, M. et al.
- Protein nonspecific mAb(s) can be prepared which recognize the sulfonic acid moiety (-CH 2 -SO 3 -) of cysteic acid or other oxidation states highly specifically. Protein nonspecific antibodies are, in general, preferred for general screening pu ⁇ oses. Protein specific antibodies are, in general, preferred for detection of particular diseases or disorders.
- a mAb that is protein nonspecific in general increases the potential sensitivity of the biomarker as a measure of oxidative stress. Also, a protein nonspecific mAb is useful in identifying oxidative damage in any protein associated with any pathological condition, from any biological medium or tissue.
- Monoclonal antibodies are generally preferred to polyclonal antibodies for several reasons. Monoclonal antibodies have previously been successful at recognizing very small protein epitopes, e.g., phosphoserine (Abu-Lawi, K.I. and Sultzer, B.M.(1995), "Induction of serine and threonine protein phosphorylation by endotoxin-associated protein in murine resident peritoneal macrophages," Infect. Immun. 63:498-502; Hasegawa, M. et al. (1996), “Characterization of mAb AP422, a novel phosphorylation-dependent monoclonal antibody against tau protein," FEBS Lett. 384:25-30).
- phosphoserine Abu-Lawi, K.I. and Sultzer, B.M.(1995), "Induction of serine and threonine protein phosphorylation by endotoxin-associated protein in murine resident peritoneal macrophage
- Monoclonal antibodies provide a means of detecting oxidized sulfur- or selenium-containing amino acids with high specificity and low nonspecific signals, and thus low background signal.
- the hapten/antigen specificity can be characterized in detail.
- Monoclonal antibodies can be produced in unlimited quantity for an unlimited period of time.
- Measurements obtained by chemical methods for detecting oxidized SSAA can be compared to those obtained by the immunoassay-ELISA method to determine the sensitivity, valid concentration range and variability (both intra- and inter-assay) ofthe latter's measurement of protein SSAA.
- known methods of detecting oxidized or partially oxidized SSAA may be used to standardize the antibody.
- an immunochemical ELISA method for measuring protein carbonyl is calibrated/standardized using oxidized albumin in which the carbonyl content was determined using a colorimetric assay; the colorimetric results correlated well with the immunochemical method (Buss, H. et al. (1997), "Protein carbonyl measurement by a sensitive ELISA method," Free Rad. Biol. Med.
- EXAMPLE 1 Chemical Method for Detecting Cysteic Acid in Protein
- MSA Methanesulfonic acid
- Some commercial providers of methanesulfonic acid distribute methanesulfonic acid with 3-(2-aminoethyl)indole (about 0.2% v/v) added to it.
- 3-(2- aminoethyl)indole hinders oxidation of amino acid residues, particularly tryptophan; therefore, inclusion in the acid of 3-(2-aminoethyl)indole or another reagent which provides the same function is preferable.
- a strong reductant e.g., dithiothreitol
- the container holding the protein mixture preferably a glass hydrolysis tube, is then sealed under vacuum at room temperature. The container is evacuated to remove oxygen, to further prevent oxidation of amino acid residues. Following hydrolysis (at about 110°C, and about 18 h), samples are neutralized with
- the fluorescent amino acid derivatives can be quantified, e.g., using a fluorescence detection system coupled to a recorder with automated peak integration. Data can be preferably corrected for losses in handling using an internal standard, (D(-)-2-amino-3-phosphonopropionic acid (D-APPA).
- D-APPA D(-)-2-amino-3-phosphonopropionic acid
- Fig. 1 shows a representative chromatographic profile in which the D and L isomers of cysteic acid (CA), L-cysteine sulfmic acid (L-CSA) and D and L-aspartic acid (ASP) are well separated.
- CA cysteic acid
- L-CSA L-cysteine sulfmic acid
- ASP D and L-aspartic acid
- MSA may also be used that does not contain the antioxidant 3-(2-aminoethyl)indole.
- the sodium salt of 2-mercaptoethanesulfonic acid (MESNA) is added (as high as 48 mM final concentration) to all protein samples prior to hydrolysis. Addition of MESNA suppresses the conversion of intermediate oxidation products of cysteine/cystine to cysteic acid. MESNA has no effect on the recovery of cysteic acid as judged by the quantitative recovery of an added cysteic acid standard.
- MESNA is also used in place of N-acetyl-L-cysteine in the orthophthalaldehyde derivatization reaction. As a result, one does not obtain the D and L isomers ofthe amino acid derivatives. This increases the area under the curve ofthe cysteic acid (and the other amino acids), resulting in greater sensitivity.
- Example 1 A number of naturally occurring cysteine and/or cystine-containing proteins were analyzed using the chemical method described in Example 1 : bovine serum albumin (BSA); chicken ovalbumin (Oval); bovine spleen cathepsin B (Cat B); human myelin basic protein (MBP); chicken egg white lysozyme (LZ); and bovine pancreatic ribonuclease A (RNase).
- BSA bovine serum albumin
- Cat B chicken ovalbumin
- Cat B human myelin basic protein
- LZ chicken egg white lysozyme
- RNase bovine pancreatic ribonuclease A
- CatB was analyzed because a cysteic acid-containing peptide (active-site Cys- 29) has been isolated from this protein (Pohl, J. et al.
- the reaction was stopped by quenching with a 100- fold molar excess of methionine and allowed to remain at room temperature for 1-2 h to ensure that no protein-associated chloramines remained (Hazell, L.J. et al. (1994), "Oxidation of low- density lipoprotein by hypochlorite causes aggregation that is mediated by modification of lysine residues rather than lipid oxidation,” Biochem. J. 302:297-304).
- the samples were dialyzed to remove any small molecular weight oxidants, and reassayed for protein, since it is known that such treatment can lead to breakage of peptide bonds (Naskalski, J.W.
- EXAMPLE 5 Fractionation of Plasma Lipoproteins and Determination of Protein Cvsteic Acid The identity of the protein(s) responsible for the high levels of protein cysteic acid observed in the total plasma lipoprotein fraction from some individuals was analyzed. To do this, whole plasma samples were separated into different lipoprotein fractions using a gel exclusion (superose 6) fast protein liquid chromatographic procedure (FPLC) (Innis-Whitehouse, W. et al. , " An efficient chromatographic system for lipoprotein fractionation using whole plasma. Revised manuscript submitted for publication). Plasma was separated into 23 fractions containing lipoproteins of decreasing size, each of which was analyzed for cholesterol, triglycerides (Le, N.-A.
- FPLC fast protein liquid chromatographic procedure
- Fig. 4 shows the different lipoprotein fractions in a typical chromatographic profile in which cholesterol and total protein were measured.
- Fractions 21-23 contain variable amounts of human serum albumin (HSA) and immunoglobulins which overlap somewhat with the HDL fraction.
- HSA human serum albumin
- a number of plasma samples were fractionated and each of the 23 fractions was analyzed for cysteic acid at three different protein concentrations (see Fig. 2).
- the amount of cysteic acid per ⁇ g of protein was then plotted vs. fraction number and a representative result is shown in Fig. 4. It can be seen that the highest "specific level" of cysteic acid (— ⁇ — ⁇ — ⁇ — ) was associated with the LDL fraction.
- mice e.g. C57BL/10 or BALB/c
- mice are immunized s.c. or i.p. with 50 ⁇ g performic acid-oxidized A-chain of bovine insulin in complete Freund's adjuvant and boosted with antigen in Freund's incomplete adjuvant.
- This immunogen was chosen for two reasons: (a) beef insulin and the A chain fragment are highly immunogenic in these strains of mice (Keck, K. (1975), "Ir-gene control of immunogenicity of insulin and A-chain loop as a carrier determinant," Nature 254:78-79; Schroer, J.A. et al.
- mouse myeloma cells for example, mouse myeloma P3X3Ag8U.l (ATCC 1597)
- supematants are screened for mAbs that react with cysteic acid- containing polypeptides in ELISAs.
- mAbs that react with cysteic acid- containing polypeptides in ELISAs.
- two performic acid oxidized proteins that are different from the immunogen are used as screening antigens: human serum albumin (Hu, M.-L. et al (1993), "Antioxidant protection against hypochlorous acid in human plasma," J. Lab. Clin. Med.
- the fine-specificity ofthe candidate monoclonal antibodies are tested with the following related chemical moieties by competition in ELISAs: cysteic acid and homocysteic acid, taurine, ⁇ -alanine, phosphoserine, phosphothreonine and phosphotyrosein.
- cysteic acid and homocysteic acid cysteic acid and homocysteic acid, taurine, ⁇ -alanine, phosphoserine, phosphothreonine and phosphotyrosein.
- ascites are generated in mice.
- mice can be immunized with cysteic acid conjugated to Keyhole Limpet Hemocyanin (KLH), a procedure that was used in producing monoclonal antibodies to phosphoserine and phosphothreonine (Abu-Lawi, K.I. and Sultzer, B.M.(1995), "Induction of serine and threonine protein phosphorylation by endotoxin-associated protein in murine resident peritoneal macrophages," Infect. Immun. 63:498-502; Hasegawa, M. et al. (1996),
- the initial approach here was to immunize mice with the oxidized A-chain of bovine insulin (a 21 -mer obtained by performic acid oxidation), an immunogen of known structure that contains four cysteic acid residues.
- An HPLC procedure for screening sera against both oxidized proteins and unoxidized, control proteins that can detect as little as four picomoles cysteic acid per microgram oxidized protein was developed. These proteins were probed with mouse antisera or pre-non-immune sera, followed by a goat antimouse alkaline phosphatase conjugated antibody, and the signal is quantified and analyzed using an automated, computer-driven microplate reader (ELISA assay). Screening mouse sera for production of a significant antibody titer avoids beginning the time and labor-intensive procedures of cell fusions, selection and screening for positive hybridomas if the animals are not responding positively to the immunogen.
- ELISA assay automated, computer-driven microplate reader
- mice were screened against the oxidized A-chain of bovine insulin (ox A) that had been conjugated to chicken ovalbumin (OVA) using either the amine-reactive, homobifunctional cross-linking agent, Bis[sulfosuccinimidyl]suberate (BS 3 ) or the heterobifunctional (thiol-amine) cross-linking reagent, N[gamma-maleimidobutyryloxyl]sulfosuccinimido ester (sulfo-GMBS).
- OVA ovalbumin
- Vmax is the linear rate of change (mA 405 /min) in the ELISA assay. Values are the mean (range) of antisera from 5 or 10 different mice. Each well was coated with 1 ⁇ g ofthe sample protein.
- mice antiserum contained antibodies directed against the cysteic acid moieties of the oxidized A-chain, but not to other potentially oxidizable amino acids.
- this antiserum was tested against H 2 O 2 -oxidized bovine serum albumin (oxBSA), H 2 O 2 -oxidized human serum albumin (oxHSA) or performic acid oxidized OVA (PAoxOVA), all of which contained cysteic acid, no significant immunoreactivity was observed
- the oxidized A-chain of insulin was clearly antigenic, but may not provide a sufficient repertoire of cysteic acid epitopes to ensure reactivity against other heterologous cysteic-acid- containing proteins.
- performic acid oxidized BSA was used as the immunogen. Performic acid oxidation is reported to convert all thiols (1 cysteine) and disulfides (17 cystines) of BSA to sulfonic acid residues (35 cysteic acids) (94%). In practice, the yield of cysteic acid is never 100% since intermediate oxidation products are formed. This immunogen would be expected to present many different oxidation epitopes involving cysteine/cystine to the murine immune system.
- Hybridomas were prepared using immunocytes obtained from the draining lymph nodes of the immunized mice. Standard procedures were employed. Hybridoma supematants (SNs) were initially screened using a 96-well ELISA assay procedure. The wells were coated with performic acid oxidized chicken ovalbumin (PAoxOVA).
- PAoxOVA performic acid oxidized chicken ovalbumin
- Unoxidized OVA contains 4 free thiols/cysteines and one disulfide bond; based on sequence information, none ofthe oxidized peptides containing cysteine/cystine from BSA match those from OVA.
- Blocking was carried out using gelatin and detection and quantification of bound mouse antibody was accomplished using a goat anti-mouse IgG(whole)-alkaline phosphatase conjugate. Color was developed using p-nitrophenyl phosphate as a substrate and activity was measured using a 96-well microplate reader set up to measure the kinetics ofthe reaction at 405 nm. Activities in the accompanying figures/data are Vmax data given as milliabsorbance (OD) units/minute. Typically, values represent the means of 3-4 replicate wells.
- Figure 7 shows ELISA data testing the anti-oxBSA antiserum against oxA-OVA (Fig. 7A, Fraction 1) and performic acid oxidized OVA (PAoxOVA) (Fig. 7B).
- ELISA plate wells were coated with 2 ⁇ g of target protein and probed with increasing dilutions of murine anti- oxBSA antiserum. Reactivity was quantified using alkaline phosphatase-conjugated goat anti- mouse IgG (1 :30,000 dilution) and monitoring the absorbance at 405 nm.
- oxA-OVA fractions, 1,2 and 3 contained 37, 752 and 871 pmol cysteic acid per microgram protein.
- Figure 8 presents representative data from an ELISA analysis of antiserum obtained from these mice when tested against hydrogen peroxide oxidized HS A (oxHS A).
- ELISA plate wells were coated with 2 ⁇ g of either unoxidized HSA or hydrogen peroxide oxidized HSA containing
- polyclonal antibodies are also recognizing other oxidation epitopes of cysteine/cystine besides cysteic acid.
- FIG. 9 presents a set of representative composite data using the mouse antiserum raised against performic acid oxidized BSA.
- Fig. 9A shows the gels stained for protein
- Fig. 9 B shows these same lanes after immunoblotting with the mouse antiserum.
- Lanes 1-9 contained the following proteins: (1) prestained M r markers; (2) oxA- OVA, 4 ⁇ g; (3) OVA, 4 ⁇ g; (4) performic acid oxidized OVA, 0.5 ⁇ g (a low amount of protein that did not stain well was used because ofthe high immunoreactivity); (5) normal mouse serum, 2 ⁇ g; (6) unoxidized HSA, 1 ⁇ g; (7) H 2 O 2 -oxidized HSA, 1 ⁇ g; (8) human serum, 2 ⁇ g; (9) carbonic anhydrase, 0.5 ⁇ g. Lane 10 (Non-denaturing, native, PAGE), human serum, 2 ⁇ g..
- Lanes 6 and 7 show unoxidized and oxidized HSA, respectively. It can be seen that unoxidized HSA was unreactive, while oxidized HSA exhibited reactive bands with M r of about 90,000, 67,000 and 45,000-50,000. A lack of reactivity with unoxidized or oxidized HSA was only observed when the antiserum was first absorbed with BSA. Since the antiserum used for the blots in lanes 7 (oxHS A) and 8 (human serum) was not absorbed with BSA, the M r 67,000 albumin band showed reactivity. Of particular interest was the result with human serum (lane 8). Significant reactivity was seen with both the heavy and light chains of immunoglobulin (compare lanes 5 and 8).
- EXAMPLE 7 Synthesis of cvsteic acid-containing polypeptide of chemically defined sequence.
- mice immunized with the oxidized A-chain of bovine insulin produce antibodies that recognized the cysteic acid residues in the oxidized A-chain, but not those in other oxidized proteins.
- the use of performic acid oxidized BSA as an immunogen resulted in antisera that recognized oxidation-specific epitopes in heterologous oxidized proteins, including the oxidized A-chain of insulin that had been conjugated to chicken ovalbumin (oxA-OVA). Both the immunogen and some ofthe oxidized proteins used in the screen undoubtedly contained oxidized epitopes other than cysteic acid.
- a polypeptide that contains a single cysteic acid residue and no other amino acids that are capable of undergoing oxidation under standard conditions was synthesized.
- This chemically defined cysteic-acid peptide was chemically coupled to OVA and used to screen the antisera and hybridoma supematants.
- OVA oxidized B-chain of insulin
- the following 16-mer polypeptide that contains a single cysteine residue was synthesized: NH 2 -TAASCFQSQNPGVSTV-COOH (SEQ ID NO 1).
- This peptide constitutes residues 38-58 of CAP 37, a multifunctional human neutrophil protein important in host defense and inflammation.
- This peptide has been purified by HPLC and its purity and composition have been verified by MALDI mass spectrometric analysis and HPLC. Except for cysteine, none of the other amino acid residues undergo oxidation under standard conditions of oxidation, including performic acid oxidation. In principle, any peptide that contains at least one oxidizable sulfur- or selenium-containing amino acid may be used.
- the peptide is oxidized with performic acid according to a modification ofthe procedure of Moore which is routinely used in our laboratory for the oxidation of proteins and peptides.
- EXAMPLE 8 Screening with CAP-37 polypeptide Both unoxidized and oxidized CAP-37 polypeptide were coupled to OVA using standard procedures for use in ELISA screening of the hybridoma SNs.
- Figure 10 shows the reactivity ofthe monoclonal antibody against unoxodized BSA and PAoxBSA. The wells were coated with either 1 ⁇ g of PAoxBSA or l ⁇ g of unoxidized BSA.
- the monoclonal antibody reacts against PAoxOVA (see Table 5 below) and PAoxBSA, but does not react against unoxidized BSA (see Figure 10), unoxidized OVA or unoxidized OVA that has been reduced and carboxymethylated (data not shown). Further, the mAb does not react with performic acid oxidized human myelin basic protein (PAoxMBP). MBP does not contain cysteine/cystine but does contain all the other oxidizable amino acids: 10 histidine, 2 methionine, 19 serine, 8 threonine, 1 tryptophan and 4 tyrosine.
- PAoxMBP performic acid oxidized human myelin basic protein
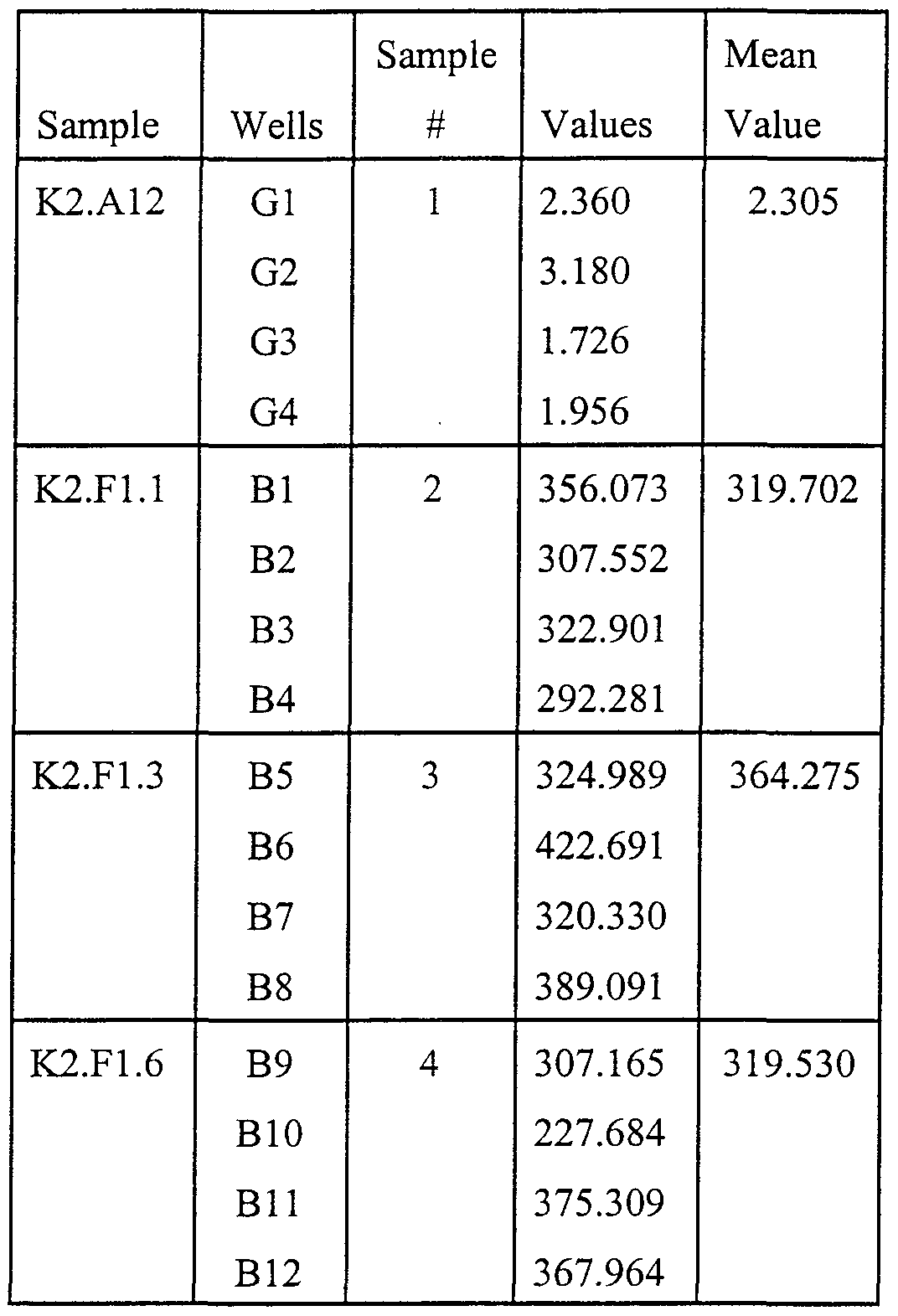
- Tables 3-5 show the activity expressed as Vmax values (milliAbsorbance(OD) units/minute at 405 nm) for 3 different clones ofthe mAb (K2.F1J, K2.F1.3 and K2.F1.6) and a non-producing clone used as a negative control (K2.A12).
- Table 3 shows the results for wells coated with 1 ⁇ g OVA-unoxCAP 37.
- Table 4 shows the results for wells coated with 1 ⁇ g OVA- oxCAP 37.
- Table 5 shows the results for wells coated with l ⁇ g PAox OVA.
- the cell line K2.F1.6 was deposited with the American Type Culture Collection (ATCC) on October 29, 1999, and assigned Patent Deposit Number PTA-897.
- the mAb did not show any significant reactivity against GSA conjugated to OVA, nor was the mAb activity against PAoxOVA significantly inhibited by preincubation with mM concentrations of GSA, cysteic acid, cysteine sulfinic acid, methionine sulfoxide or methionine sulfone.
- the mAb is not recognizing an oxidation product of cysteine but may be recognizing an intermediate oxidation product ofthe disulfide bond (oxides of cystine), e.g., cystine monoxide (thiosulfmate) and/or cystine dioxide (thiosulfonate).
- oxides of cystine e.g., cystine monoxide (thiosulfmate) and/or cystine dioxide (thiosulfonate).
- cystine monoxide thiosulfmate
- cystine dioxide thiosulfonate
- EXAMPLE 9 Human sera plasma testing for endogenous antibody to oxidized protein The ELISA assay developed to screen for mAb was used to look for the presence of endogenous antibody to oxidatively damaged protein (ODP) in the plasma of normal individuals and patients with coronary artery disease (CAD) and renal disease. 96-well microplates were coated with PAoxOVA and blocked with gelatin. Diluted plasma was added to the plates and the presence of endogenous antibody was detected and quantified by addition of a goat anti- human IgG(Fc)-alkaline phosphatase conjugate. As described above, the kinetic data are reported as Vmax values (milliAbsorbance(OD) units/minute at 405 nm). In most cases, these clinical data represent values from a single well.
- Vmax values milliAbsorbance(OD) units/minute at 405 nm
- Lpa a risk factor for development of CAD.
- the second set (samples 90-110) was from a group of American 12-13 year-olds who have been determined to be at risk for developing CAD (e.g. tendency toward obesity and/or have parents who are diabetic or obese or have other risk factors for CAD).
- the results of this study using plasma diluted 1 J00 are shown in Fig. 12A and Fig. 12B. Two points can be made. A number of these children have rather high levels of endogenous antibody to oxidatively damaged protein as measured by reactivity against PAoxOVA.
- Plasma samples were obtained from 47 patients undergoing renal dialysis in the course of management of end stage renal disease. These samples were diluted 1 :200 and examined in the PAoxOVA ELISA for the presence of endogenous antibody to ODP. The results are shown in Figure 15. Reactivity could be divided into three categories; high (Vmax > 50, 3 patients), intermediate (Vmax 10-50, 17 patients) and low (Vmax ⁇ 10, 27 patients), corresponding to 6%, 36% and 57% ofthe total number of samples. All three ofthe "high” patients had either CAD or diabetes, and nine ofthe 17 "intermediate” patients were diabetic. In marked contrast, 21 ofthe 27 "low” patients had no clinical history of CAD. These data also suggest that higher levels of endogenous antibody to ODP as defined by the PAoxOVA ELISA screen may be associated with CAD and/or diabetes, both diseases that have been strongly associated with inflammation and oxidative stress.
- LDL Low Density Lipoproteins
- Presence of oxidatively modified LDL has been demonstrated in human arteriosclerotic lesions (Avogaro, P. et al. (1988), "Presence of modified LDL in humans,” Arteriosclerosis 8:79-87).
- the major protein component of LDL is known as apolipoprotein B (apoB).
- Figure 16A illustrates an immunoassay that can determine the specificity of the oxidative modification process for apoB.
- the antibody described in this invention is used in the bottom ofthe well to trap all proteins exhibiting the modified epitope.
- the secondary antibody in this instance would be targeted against human apoB, but can be targeted against other proteins, as desired.
- Figure 16B illustrates the conventional immunoassay used to determine the total concentration of apoB in a sample (Le et al. (1999), "Lipid and apolipoprotein levels and distribution in patients with hypertriglyceridemia: Effect of triglyceride reductions with atorvastatin,” Metabolism, In press).
- the percent of apoB in a given sample that is oxidatively modified can be determined.
- This approach can be used, for example, in the clinical measurement of hemoglobin Ale, a glycosylated form of hemoglobin.
- the measurement of hemoglobin Ale is used in the management of diabetic patents (Bunn, H.F.
- hemoglobin Ale usually accounts for about 4-6% of the total hemoglobin, but in diabetics, hemoglobin Ale can increase to as much as 20% ofthe total hemoglobin. This method could be used for other proteins, as well.
- Immunoaffinity chromatography is a standard procedure for the isolation of specific protein based on the availability of a specific antibody (Le et al. (1986), "Direct determination of apoC-III specific activity using immunoaffinity chromatography,” Methods Enzymol. 129:457-469).
- the antibody is first coupled to a gel matrix and with the application of the sample containing the modified proteins, only the particles containing the oxidatively modified epitopes are retained on the column. By changing the condition ofthe column, the particles can be released from the antibody column.
- Monoclonal and polyclonal antibodies described herein were generated using performic acid oxidized bovine serum albumin. These antibodies were detected using an ELISA screening assay that employed performic acid oxidized chicken ovalbumin as the antigen.
- Oxidized proteins or proteineaceous aggregates comprising an oxidized sulfur- or selenium-containing amino acid from patients having specific diseases including, but not limited to, coronary artery disease (CAD; e.g., oxidized low density lipoprotein), renal disease, and diabetes may also be used for the development of disease-specific polyclonal and monoclonal antibodies.
- CAD coronary artery disease
- proteins or proteineaceous aggregates from normal, healthy individuals can be isolated and oxidized in vitro.
- oxidatively damaged proteins or proteineaceous aggregates are characteristic of the diseased tissue or the disease state.
- Such oxidized protein or proteineaceous aggregates may be used as the antigen in a screening ELISA assay to select antibodies that are specific to the disease.
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Organic Chemistry (AREA)
- Molecular Biology (AREA)
- Medicinal Chemistry (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Zoology (AREA)
- Genetics & Genomics (AREA)
- Biophysics (AREA)
- Gastroenterology & Hepatology (AREA)
- Toxicology (AREA)
- Engineering & Computer Science (AREA)
- Immunology (AREA)
- Hematology (AREA)
- Biomedical Technology (AREA)
- Urology & Nephrology (AREA)
- Microbiology (AREA)
- Cell Biology (AREA)
- Diabetes (AREA)
- Endocrinology (AREA)
- Biotechnology (AREA)
- Food Science & Technology (AREA)
- Physics & Mathematics (AREA)
- Analytical Chemistry (AREA)
- General Physics & Mathematics (AREA)
- Pathology (AREA)
- Peptides Or Proteins (AREA)
Abstract
Description







Claims
Priority Applications (4)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| EP99971864A EP1124985A4 (en) | 1998-11-06 | 1999-11-05 | Biomarkers for oxidative stress |
| AU12435/00A AU1243500A (en) | 1998-11-06 | 1999-11-05 | Biomarkers for oxidative stress |
| CA002348685A CA2348685A1 (en) | 1998-11-06 | 1999-11-05 | Biomarkers for oxidative stress |
| US09/831,123 US6953666B1 (en) | 1998-11-06 | 1999-11-05 | Biomarkers for oxidative stress |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US10740498P | 1998-11-06 | 1998-11-06 | |
| US60/107,404 | 1998-11-06 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| WO2000028072A1 true WO2000028072A1 (en) | 2000-05-18 |
| WO2000028072A9 WO2000028072A9 (en) | 2000-09-21 |
Family
ID=22316499
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/US1999/026133 WO2000028072A1 (en) | 1998-11-06 | 1999-11-05 | Biomarkers for oxidative stress |
Country Status (4)
| Country | Link |
|---|---|
| EP (1) | EP1124985A4 (en) |
| AU (1) | AU1243500A (en) |
| CA (1) | CA2348685A1 (en) |
| WO (1) | WO2000028072A1 (en) |
Cited By (12)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2001092879A1 (en) * | 2000-05-30 | 2001-12-06 | Danmarks Miljø Undersøgelser | An assay method and kit for testing biological material for exposure to stress using biomarkers |
| WO2002032445A2 (en) * | 2000-10-20 | 2002-04-25 | Beate Kehrel | Inhibition of the interaction between oxidized proteins and cd36 or the active mechanism thereof |
| JP2010183843A (en) * | 2009-02-10 | 2010-08-26 | National Institute Of Advanced Industrial Science & Technology | Antibody against oxidation type polypeptide containing oxidized sulfur-containing amino acid residue |
| US8034769B2 (en) | 2000-10-20 | 2011-10-11 | Hamburger Stiftung Zur Foerderung Von Wissenschaft Und Kultur | Oxidized proteins and oxidized protein inhibitor compositions and methods of use thereof |
| US8110347B2 (en) | 2005-04-15 | 2012-02-07 | Canimguide Therapeutics Ab | Diagnostic method for detecting cancer by measuring amount of cytokine like IL-6 |
| US8129191B2 (en) | 2003-12-18 | 2012-03-06 | Inverness Medical Switzerland Gmbh | Monitoring method and apparatus |
| US8133688B2 (en) | 2004-10-20 | 2012-03-13 | Canimguide Therapeutics Ab | Immunoregulation in cancer, chronic inflammatory and autoimmune diseases |
| US8182983B2 (en) | 2002-05-27 | 2012-05-22 | Canimguide Therapeutics Ab | Method for determining immune system affecting compounds |
| WO2014201451A1 (en) * | 2013-06-14 | 2014-12-18 | Wellmetris Llc | Stress and inflammation biomarker urine panel for dairy cows and beef cattle |
| US9120874B2 (en) | 2007-05-08 | 2015-09-01 | Canimguide Therapeutics Ab | Methods of immunoregulation by albumin neo-structures |
| US9638699B2 (en) | 2013-07-18 | 2017-05-02 | The University Of Sydney | Biomarkers of oxidative stress |
| US10161928B2 (en) | 2010-07-26 | 2018-12-25 | Wellmetris, Llc | Wellness panel |
Citations (6)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US4859613A (en) * | 1986-11-12 | 1989-08-22 | Lawrence David A | Immunoassays for glutathione and antibodies useful therein |
| US5585232A (en) * | 1992-07-06 | 1996-12-17 | President And Fellows Of Harvard College | Methods and diagnostic kits for determining toxicity utilizing E. coli stress promoters fused to reporter genes |
| US5605826A (en) * | 1994-06-10 | 1997-02-25 | Panorama Research, Inc. | 24 kilodalton cytoplasmic protease activating DNA fragmentation in apoptosis |
| US5667776A (en) * | 1986-11-21 | 1997-09-16 | Chiron Corporation | Treatment for biological damage using tumor necrosis factor and a free-radical scavenger |
| US5700654A (en) * | 1991-06-14 | 1997-12-23 | Vanderbilt Unversity | Method and compositions to assess oxidative stress in vivo |
| US5814300A (en) * | 1995-03-03 | 1998-09-29 | Cephalon, Inc. | Gene-targeted non-human mammals deficient in the SOD-1 gene |
Family Cites Families (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO1996004311A1 (en) * | 1994-08-03 | 1996-02-15 | University Of Alabama At Birmingham Research Foundation | Monoclonal antibody to nitrotyrosine, methods for diagnosis and methods for treatment of disease |
| AU7113696A (en) * | 1995-09-19 | 1997-04-09 | Cytochem, Inc. | Detection and quantitation of 8-oh-adenine using monoclonal antibodies |
| SK66198A3 (en) * | 1996-09-20 | 1999-09-10 | Atherogenics Inc | Diagnostics for and mediators of inflammatory disorders |
-
1999
- 1999-11-05 AU AU12435/00A patent/AU1243500A/en not_active Abandoned
- 1999-11-05 WO PCT/US1999/026133 patent/WO2000028072A1/en active Application Filing
- 1999-11-05 CA CA002348685A patent/CA2348685A1/en not_active Abandoned
- 1999-11-05 EP EP99971864A patent/EP1124985A4/en not_active Ceased
Patent Citations (7)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US4859613A (en) * | 1986-11-12 | 1989-08-22 | Lawrence David A | Immunoassays for glutathione and antibodies useful therein |
| US5667776A (en) * | 1986-11-21 | 1997-09-16 | Chiron Corporation | Treatment for biological damage using tumor necrosis factor and a free-radical scavenger |
| US5702697A (en) * | 1986-11-21 | 1997-12-30 | Chiron Corporation | Treatment for biological damage using a colony stimulating factor and a biological modifier |
| US5700654A (en) * | 1991-06-14 | 1997-12-23 | Vanderbilt Unversity | Method and compositions to assess oxidative stress in vivo |
| US5585232A (en) * | 1992-07-06 | 1996-12-17 | President And Fellows Of Harvard College | Methods and diagnostic kits for determining toxicity utilizing E. coli stress promoters fused to reporter genes |
| US5605826A (en) * | 1994-06-10 | 1997-02-25 | Panorama Research, Inc. | 24 kilodalton cytoplasmic protease activating DNA fragmentation in apoptosis |
| US5814300A (en) * | 1995-03-03 | 1998-09-29 | Cephalon, Inc. | Gene-targeted non-human mammals deficient in the SOD-1 gene |
Non-Patent Citations (5)
| Title |
|---|
| DATABASE BIOSIS, [online] LAINE M.L. ET AL.: "Novel polysaccharide capsular serotypes in Porphyromonas gingivalis", XP002948084, accession no. STN Database accession no. 1996:377715 * |
| DATABASE CAPLUS, [online] HANAZAWA ET AL.: "Monoclonal antibody against a serotype antigen of Porphyromonas (Bacteroides) endodontalis and characteristics of the antigen", XP002948085, accession no. STN Database accession no. 1990:530260 * |
| INFECTIOUS IMMUNOLOGY,, vol. 58, no. 8, 1990, pages 2542 - 2546 * |
| JOURNAL OF PERIODONTAL RESEARCH,, vol. 31, no. 4, 1996, pages 278 - 284 * |
| See also references of EP1124985A4 * |
Cited By (18)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2001092879A1 (en) * | 2000-05-30 | 2001-12-06 | Danmarks Miljø Undersøgelser | An assay method and kit for testing biological material for exposure to stress using biomarkers |
| WO2002032445A2 (en) * | 2000-10-20 | 2002-04-25 | Beate Kehrel | Inhibition of the interaction between oxidized proteins and cd36 or the active mechanism thereof |
| WO2002032445A3 (en) * | 2000-10-20 | 2003-04-17 | Beate Kehrel | Inhibition of the interaction between oxidized proteins and cd36 or the active mechanism thereof |
| EP1994941A3 (en) * | 2000-10-20 | 2012-07-25 | Hamburger Stiftung zur Förderung von Wissenschaft und Kultur | Medicine containing at least one oxidised protein |
| US8034769B2 (en) | 2000-10-20 | 2011-10-11 | Hamburger Stiftung Zur Foerderung Von Wissenschaft Und Kultur | Oxidized proteins and oxidized protein inhibitor compositions and methods of use thereof |
| US8182983B2 (en) | 2002-05-27 | 2012-05-22 | Canimguide Therapeutics Ab | Method for determining immune system affecting compounds |
| US8129191B2 (en) | 2003-12-18 | 2012-03-06 | Inverness Medical Switzerland Gmbh | Monitoring method and apparatus |
| US8361389B2 (en) | 2003-12-18 | 2013-01-29 | Inverness Medical Switzerland Gmbh | Monitoring methods and apparatus |
| US9689866B2 (en) | 2003-12-18 | 2017-06-27 | Inverness Medical Switzerland Gmbh | Monitoring method and apparatus |
| US8133688B2 (en) | 2004-10-20 | 2012-03-13 | Canimguide Therapeutics Ab | Immunoregulation in cancer, chronic inflammatory and autoimmune diseases |
| US9688746B2 (en) | 2004-10-20 | 2017-06-27 | Canimguide Therapeutics Ab | Immunoregulation in cancer, chronic inflammatory and autoimmune diseases |
| US8110347B2 (en) | 2005-04-15 | 2012-02-07 | Canimguide Therapeutics Ab | Diagnostic method for detecting cancer by measuring amount of cytokine like IL-6 |
| US9120874B2 (en) | 2007-05-08 | 2015-09-01 | Canimguide Therapeutics Ab | Methods of immunoregulation by albumin neo-structures |
| US9796777B2 (en) | 2007-05-08 | 2017-10-24 | Canimguide Therapeutics Ab | Methods of treating malignant tumors |
| JP2010183843A (en) * | 2009-02-10 | 2010-08-26 | National Institute Of Advanced Industrial Science & Technology | Antibody against oxidation type polypeptide containing oxidized sulfur-containing amino acid residue |
| US10161928B2 (en) | 2010-07-26 | 2018-12-25 | Wellmetris, Llc | Wellness panel |
| WO2014201451A1 (en) * | 2013-06-14 | 2014-12-18 | Wellmetris Llc | Stress and inflammation biomarker urine panel for dairy cows and beef cattle |
| US9638699B2 (en) | 2013-07-18 | 2017-05-02 | The University Of Sydney | Biomarkers of oxidative stress |
Also Published As
| Publication number | Publication date |
|---|---|
| AU1243500A (en) | 2000-05-29 |
| WO2000028072A9 (en) | 2000-09-21 |
| CA2348685A1 (en) | 2000-05-18 |
| EP1124985A4 (en) | 2003-01-15 |
| EP1124985A1 (en) | 2001-08-22 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US6953666B1 (en) | Biomarkers for oxidative stress | |
| Barelli et al. | Characterization of new polyclonal antibodies specific for 40 and 42 amino acid-long amyloid β peptides: Their use to examine the cell biology of presenilins and the immunohistochemistry of sporadic Alzheimer’s disease and cerebral amyloid angiopathy cases | |
| FI84107B (en) | HBA1C: S IMMUNOLOGICAL BESTAEMNINGSFOERFARANDE SOM INNEFATTAR DENATURERING. | |
| US4340581A (en) | Process for the immunological determination of basal membrane material and new basal membrane suitable therefor | |
| McDONALD et al. | Properties and distribution of the protein inhibitor (M r 17,000) of protein kinase C | |
| EP1124985A1 (en) | Biomarkers for oxidative stress | |
| Conrad et al. | Rat duodenal iron-binding protein mobilferrin is a homologue of calreticulin | |
| WO2009032722A1 (en) | Carbamylated proteins and risk of cardiovascular disease | |
| JP2006515561A (en) | Compositions and methods for assaying Apo-B48 and Apo-B100 | |
| WO2008055242A9 (en) | Early detection of diabetes | |
| EA002520B1 (en) | Isolated antibody and a fragment thereof, methods and a kit for the assessment of lipid peroxidation level in a biological sample, method for diagnosing diseases caused by a lipid peroxidation | |
| Nagnan-Le Meillour et al. | Binding specificity of native Odorant-binding protein isoforms is driven by phosphorylation and O-N-acetylglucosaminylation in the pig Sus scrofa | |
| Campos-Pinto et al. | Epitope mapping and characterization of 4-hydroxy-2-nonenal modified-human serum albumin using two different polyclonal antibodies | |
| CA2274776C (en) | Method for analyzing annexin v in urine and use thereof | |
| JP3259768B2 (en) | Testing methods for kidney disease | |
| US20030124616A1 (en) | Homocysteinylated transthyretin | |
| Li et al. | Differential expression of advanced glycosylation end-products in neurons of different species | |
| NO175639B (en) | ||
| MXPA06004234A (en) | Method of pretreating sample and immunological assay method using the same. | |
| JPH06239899A (en) | Antibody for human tau protein and determination of human tau protein in body fluid utilizing the same | |
| JP3978226B2 (en) | Immunoassay to identify alcoholics and monitor alcohol consumption | |
| KR101396300B1 (en) | Biomarkers for diagnosing the addiction of ethylbenzene | |
| Schifferli et al. | Two isotypes of human C4, C4A and C4B have different structure and function | |
| Kortesuo et al. | Characterization of two phospholipases A2 in serum of patients with sepsis and acute pancreatitis | |
| Schlaf et al. | A novel enzyme-linked immunosorbent assay for determination of synaptophysin as compared with other quantification procedures |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| ENP | Entry into the national phase |
Ref country code: AU Ref document number: 2000 12435 Kind code of ref document: A Format of ref document f/p: F |
|
| AK | Designated states |
Kind code of ref document: A1 Designated state(s): AE AL AM AT AU AZ BA BB BG BR BY CA CH CN CR CU CZ DE DK DM EE ES FI GB GD GE GH GM HR HU ID IL IN IS JP KE KG KP KR KZ LC LK LR LS LT LU LV MD MG MK MN MW MX NO NZ PL PT RO RU SD SE SG SI SK SL TJ TM TR TT UA UG US UZ VN YU ZA ZW |
|
| AL | Designated countries for regional patents |
Kind code of ref document: A1 Designated state(s): GH GM KE LS MW SD SL SZ TZ UG ZW AM AZ BY KG KZ MD RU TJ TM AT BE CH CY DE DK ES FI FR GB GR IE IT LU MC NL PT SE BF BJ CF CG CI CM GA GN GW ML MR NE SN TD TG |
|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application | ||
| AK | Designated states |
Kind code of ref document: C2 Designated state(s): AE AL AM AT AU AZ BA BB BG BR BY CA CH CN CR CU CZ DE DK DM EE ES FI GB GD GE GH GM HR HU ID IL IN IS JP KE KG KP KR KZ LC LK LR LS LT LU LV MD MG MK MN MW MX NO NZ PL PT RO RU SD SE SG SI SK SL TJ TM TR TT UA UG US UZ VN YU ZA ZW |
|
| AL | Designated countries for regional patents |
Kind code of ref document: C2 Designated state(s): GH GM KE LS MW SD SL SZ TZ UG ZW AM AZ BY KG KZ MD RU TJ TM AT BE CH CY DE DK ES FI FR GB GR IE IT LU MC NL PT SE BF BJ CF CG CI CM GA GN GW ML MR NE SN TD TG |
|
| COP | Corrected version of pamphlet |
Free format text: PAGES 5/17, 9/17, 10/17, 14/17 AND 16/17, DRAWINGS, REPLACED BY NEW PAGES 5/17, 9/17, 10/17, 14/17 AND 16/17 |
|
| DFPE | Request for preliminary examination filed prior to expiration of 19th month from priority date (pct application filed before 20040101) | ||
| ENP | Entry into the national phase |
Ref document number: 2348685 Country of ref document: CA Ref country code: CA Ref document number: 2348685 Kind code of ref document: A Format of ref document f/p: F |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 1999971864 Country of ref document: EP |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 09831123 Country of ref document: US |
|
| WWP | Wipo information: published in national office |
Ref document number: 1999971864 Country of ref document: EP |
|
| REG | Reference to national code |
Ref country code: DE Ref legal event code: 8642 |