TW202128996A - 用於活體內合成非天然多肽的組合物及方法 - Google Patents
用於活體內合成非天然多肽的組合物及方法 Download PDFInfo
- Publication number
- TW202128996A TW202128996A TW109134953A TW109134953A TW202128996A TW 202128996 A TW202128996 A TW 202128996A TW 109134953 A TW109134953 A TW 109134953A TW 109134953 A TW109134953 A TW 109134953A TW 202128996 A TW202128996 A TW 202128996A
- Authority
- TW
- Taiwan
- Prior art keywords
- natural
- amino
- acid
- cell
- unnatural
- Prior art date
Links
Images






































Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/43504—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from invertebrates
- C07K14/43595—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from invertebrates from coelenteratae, e.g. medusae
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/67—General methods for enhancing the expression
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/70—Vectors or expression systems specially adapted for E. coli
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/12—Transferases (2.) transferring phosphorus containing groups, e.g. kinases (2.7)
- C12N9/1241—Nucleotidyltransferases (2.7.7)
- C12N9/1247—DNA-directed RNA polymerase (2.7.7.6)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/93—Ligases (6)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P21/00—Preparation of peptides or proteins
- C12P21/02—Preparation of peptides or proteins having a known sequence of two or more amino acids, e.g. glutathione
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/20—Fusion polypeptide containing a tag with affinity for a non-protein ligand
- C07K2319/22—Fusion polypeptide containing a tag with affinity for a non-protein ligand containing a Strep-tag
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y207/00—Transferases transferring phosphorus-containing groups (2.7)
- C12Y207/07—Nucleotidyltransferases (2.7.7)
- C12Y207/07006—DNA-directed RNA polymerase (2.7.7.6)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y601/00—Ligases forming carbon-oxygen bonds (6.1)
- C12Y601/01—Ligases forming aminoacyl-tRNA and related compounds (6.1.1)
- C12Y601/01026—Pyrrolysine-tRNAPyl ligase (6.1.1.26)
Abstract
本發明揭示用於細胞將非天然胺基酸併入非天然多肽中之組合物、方法及套組。本文亦揭示用於增加該細胞合成之非天然多肽之活性及產率的組合物、方法及套組。
Description
天然遺傳密碼係由64個可能藉由遺傳字母表之四個字母製成之密碼子組成。使用三種密碼子作為終止密碼子,保留61個藉由轉送RNA (tRNA)識別之正義密碼子,其藉由具有20種蛋白質性胺基酸中之一者的同源胺基醯基tRNA合成酶(本文中亦簡稱為tRNA合成酶)而帶電。儘管典型胺基酸已使得活生物體具有顯著多樣性,但仍存在多種其並不提供之化學功能性及相關反應性。擴增遺傳密碼以包括非天然或非典型胺基酸(ncAA)之能力可能賦予蛋白質所需功能或活性且顯著促進諸如治療發展之蛋白質之許多已知及新興應用。合成含有非天然胺基酸之非天然蛋白質或非天然多肽之當前方法具有侷限性。值得注意的是,大部分方法僅能夠將單一非天然胺基酸或非天然胺基酸之一個物種之若干複本引入至非天然多肽中。此外,藉由當前可用之方法合成之非天然多肽常常具有降低的酶活性、溶解度或產率。
解決此等侷限性之一個替代解決方案為藉由無細胞或活體外表現系統合成非天然多肽。然而,此類表現系統不足以提供轉譯後修飾環境,其中完全實現非天然多肽之氧化還原特性及合成非天然多肽之其他轉譯後修飾。因此,仍需要用於活體內合成含有非天然胺基酸之非天然多肽的組合物及方法。
本文描述用於活體內合成非天然多肽或非天然蛋白質之組合物、方法、細胞(未經工程改造及經工程改造兩者)、半合成生物體(semi-synthetic organism;SSO)、試劑、遺傳物質、質體及套組,其中各非天然多肽或非天然蛋白質包含兩個或更多個藉由細胞解碼之非天然胺基酸。
本文所描述的為活體內合成非天然多肽之方法,其包含:提供至少一個包含至少四個非天然鹼基對之非天然去氧核糖核酸(DNA)分子;轉錄該至少一個非天然DNA分子以得到包含至少兩個非天然密碼子之信使核糖核酸(mRNA)分子;轉錄該至少一個非天然DNA分子以得到至少兩個各自包含至少一個非天然反密碼子之轉送RNA (tRNA)分子,其中於對應DNA中之該至少兩個非天然鹼基對處於序列背景中,使得mRNA分子之非天然密碼子於tRNA分子中之每一者之非天然反密碼子互補;以及藉由利用該至少兩個非天然tRNA分子轉譯非天然mRNA分子來合成非天然多肽,其中各非天然反密碼子引導非天然胺基酸位點特異性併入至非天然多肽中。在一些實施例中,至少兩個非天然鹼基對包含選自dCNMO-dTPT3、dNaM-dTPT3、dCNMO-dTAT1或dNaM-dTAT1之鹼基對。
在一些實施例中,提供合成非天然多肽之方法,其包含:提供至少一個包含至少四個非天然鹼基對之非天然去氧核糖核酸(DNA)分子,其中該至少一個非天然DNA分子編碼(i)包含至少第一及第二非天然密碼子之信使核糖核酸(mRNA)分子,及(ii)至少第一及第二轉送RNA (tRNA)分子,該第一tRNA分子包含第一非天然反密碼子且該第二tRNA分子包含第二非天然反密碼子,並且該至少一DNA分子中之至少四個非天然鹼基對處於序列背景中,使得該mRNA分子之第一及第二非天然密碼子分別與第一及第二非天然反密碼子互補;轉錄該至少一個非天然DNA分子以得到mRNA;轉錄該至少一個非天然DNA分子以得到至少第一及第二tRNA分子;以及藉由利用該至少第一及第二非天然tRNA分子轉譯該非天然mRNA分子來合成非天然多肽,其中至少第一及第二非天然反密碼子中之每一者引導非天然胺基酸位點特異性併入非天然多肽中。
在一些實施例中,該等方法包含至少兩個非天然密碼子,其各自包含位於密碼子之第一位置、第二位置或第三位置處之第一非天然核苷酸,視情況其中該第一非天然核苷酸位於密碼子之第二位置或第三位置處。在一些情況下,該等方法包含至少兩個非天然密碼子,其各自包含核酸序列NNX或NXN,且非天然反密碼子包含核酸序列XNN、YNN、NXN或NYN,以形成包含NNX-XNN、NNX-YNN或NXN-NYN之非天然密碼子-反密碼子對,其中N為任何天然核苷酸,X為第一非天然核苷酸且Y為不同於第一非天然核苷酸之第二非天然核苷酸,其中X-Y形成DNA中之非天然鹼基對(unnatural base pair;UBP)。
在一些實施例中,UBP係在mRNA之密碼子序列與tRNA之反密碼子序列之間形成以促進mRNA轉譯成非天然多肽。在一些情況下,密碼子-反密碼子UBP包含:密碼子序列,其包含mRNA之三個連續核酸讀段5'至3' (例如UUX);及反密碼子序列,其包含tRNA之三個連續核酸讀段5'至3' (例如YAA或XAA)。在一些實施例中,當mRNA密碼子為UUX時,tRNA反密碼子為YAA或XAA。在一些實施例中,當mRNA密碼子為UGX時,tRNA反密碼子為YCA或XCA。在一些實施例中,當mRNA密碼子為CGX時,tRNA反密碼子為YCG或XCG。在一些實施例中,當mRNA密碼子為AGX時,tRNA反密碼子為YCU或XCU。在一些實施例中,當mRNA密碼子為GAX時,tRNA反密碼子為YUC或XUC。在一些實施例中,當mRNA密碼子為CAX時,tRNA反密碼子為YUG或XUG。在一些實施例中,當mRNA密碼子為GXU時,tRNA反密碼子為AYC。在一些實施例中,當mRNA密碼子為CXU時,tRNA反密碼子為AYG。在一些實施例中,當mRNA密碼子為GXG時,tRNA反密碼子為CYC。在一些實施例中,當mRNA密碼子為AXG時,tRNA反密碼子為CYU。在一些實施例中,當mRNA密碼子為GXC時,tRNA反密碼子為GYC。在一些實施例中,當mRNA密碼子為AXC時,tRNA反密碼子為GYU。在一些實施例中,當mRNA密碼子為GXA時,tRNA反密碼子為UYC。在一些實施例中,當mRNA密碼子為CXC時,tRNA反密碼子為GYG。在一些實施例中,當mRNA密碼子為UXC時,tRNA反密碼子為GYA。在一些實施例中,當mRNA密碼子為AUX時,tRNA反密碼子為YAU或XAU。在一些實施例中,當mRNA密碼子為CUX時,tRNA反密碼子為XAG或YAG。在一些實施例中,當mRNA密碼子為UUX時,tRNA反密碼子為XAA或YAA。在一些實施例中,當mRNA密碼子為GUX時,tRNA反密碼子為XAC或YAC。在一些實施例中,當mRNA密碼子為UAX時,tRNA反密碼子為XUA或YUA。在一些實施例中,當mRNA密碼子為GGX時,tRNA反密碼子為XCC或YCC。
在一些實施例中,至少一個非天然DNA分子經轉錄為包含本文所描述之非天然鹼基(例如d5SICS、dNaM、dTPT3、dMTMO、dCNMO、dTAT1)之信使RNA (mRNA)。例示性mRNA密碼子係藉由包含三個連續去氧核糖核苷酸(NNN)之非天然DNA之例示性區域編碼,該等去氧核糖核苷酸包含TTX、TGX、CGX、AGX、GAX、CAX、GXT、CXT、GXG、AXG、GXC、AXC、GXA、CXC、TXC、ATX、CTX、TTX、GTX、TAX或GGX,其中X為附接至2'去氧核糖基部分之非天然鹼基。由轉錄例示性非天然DNA產生之例示性mRNA密碼子包含三個連續核糖核苷酸(NNN),其分別包含UUX、UGX、CGX、AGX、GAX、CAX、GXU、CXU、GXG、AXG、GXC、AXC、GXA、CXC、UXC、AUX、CUX、UUX、GUX、UAX或GGX,其中X為附接至核糖基部分之非天然鹼基。在一些實施例中,非天然鹼基在密碼子序列中之第一位置(X-N-N)中。在一些實施例中,非天然鹼基在密碼子序列中之第二(或中間)位置(N-X-N)中。在一些實施例中,非天然鹼基在密碼子序列中之第三(最末)位置(N-N-X)中。
在一些實施例中,方法包含含至少一個G之密碼子及含至少一個C之反密碼子。在一些情況下,方法包含X及Y,其中X及Y獨立地選自由以下組成之群:(i) 2-硫尿嘧啶、2'-去氧尿苷、4-硫-尿嘧啶、尿嘧啶-5-基、次黃嘌呤-9-基(I)、5-鹵基尿嘧啶;5-丙炔基-尿嘧啶、6-偶氮基-尿嘧啶、5-甲基胺基甲基尿嘧啶、5-甲氧基胺基甲基-2-硫尿嘧啶、假尿嘧啶、尿嘧啶-5-氧基乙酸甲酯、尿嘧啶-5-氧基乙酸、5-甲基-2-硫尿嘧啶、3-(3-胺基-3-N-2-羧丙基)尿嘧啶、5-甲基-2-硫尿嘧啶、4-硫尿嘧啶、5-甲基尿嘧啶、5'-甲氧基羧甲基尿嘧啶、5-甲氧基尿嘧啶、尿嘧啶-5-氧基乙酸、5-(羧基羥甲基)尿嘧啶、5-羧甲基胺基甲基-2-硫尿苷、5-羧甲基胺基甲基尿嘧啶或二氫尿嘧啶;(ii) 5-羥甲基胞嘧啶、5-三氟甲基胞嘧啶、5-鹵基胞嘧啶、5-丙炔基胞嘧啶、5-羥基胞嘧啶、環胞嘧啶、胞嘧啶阿拉伯糖苷(arabinoside)、5,6-二氫胞嘧啶、5-硝基胞嘧啶、6-偶氮基胞嘧啶、氮雜胞嘧啶、N4-乙基胞嘧啶、3-甲基胞嘧啶、5-甲基胞嘧啶、4-乙醯基胞嘧啶、2-硫胞嘧啶、啡㗁𠯤胞苷([5,4-b][l,4]苯并㗁𠯤-2(3H)-酮)、啡噻𠯤胞苷(1H-嘧啶并[5,4-b][l,4]苯并噻𠯤-2(3H)-酮)、啡㗁𠯤胞苷(9-(2-胺基乙氧基)-H-嘧啶并[5,4-b][l,4]苯并㗁𠯤-2(3H)-酮)、咔唑胞苷(2H-嘧啶并[4,5-b]吲哚-2-酮)或吡啶并吲哚胞苷(H-吡啶并[3',2':4,5]吡咯并[2,3-d]嘧啶-2-酮);(iii) 2-胺基腺嘌呤;2-丙基腺嘌呤;2-胺基-腺嘌呤;2-F-腺嘌呤;2-胺基-丙基-腺嘌呤;2-胺基-2'-去氧腺苷去氧腺苷;3-去氮腺嘌呤;7-甲基腺嘌呤;7-去氮-腺嘌呤;8-氮雜腺嘌呤;經8-鹵基、8-胺基、8-硫醇、8-硫烷基及8-羥基取代之腺嘌呤;N6-異戊烯基腺嘌呤;2-甲基腺嘌呤;2,6-二胺基嘌呤;2-甲基硫-N6-異戊烯基腺嘌呤或6-氮雜-腺嘌呤;(iv) 2-甲基鳥嘌呤;鳥嘌呤之2-丙基及烷基衍生物;3-去氮鳥嘌呤;6-硫-鳥嘌呤;7-甲基鳥嘌呤;7-去氮鳥嘌呤;7-去氮鳥苷;7-去氮-8-氮雜鳥嘌呤;8-氮雜鳥嘌呤;經8-鹵基、8-胺基、8-硫醇、8-硫烷基及8-羥基取代之鳥嘌呤;1-甲基鳥嘌呤;2,2-二甲基鳥嘌呤;7-甲基鳥嘌呤或6-氮雜-鳥嘌呤;以及(v)次黃嘌呤、黃嘌呤、1-甲基肌苷、Q核苷、β-D-半乳糖基Q核苷、肌苷、β-D-甘露糖基Q核苷、懷丁氧苷、羥基脲、(acp3)w、2-胺基吡啶或2-吡啶酮。在一些實施例中,X及Y獨立地選自由以下組成之群:
 。
在一些情況下,X為
。
在一些情況下,X為 。在一些實施例中。Y為
。在一些實施例中。Y為 。
。
在一些實施例中,本文所描述之方法包含非天然密碼子-反密碼子對NNX-XNN,其中NNX-XNN係選自由以下組成之群:UUX-XAA、UGX-XCA、CGX-XCG、AGX-XCU、GAX-XUC、CAX-XUG、AUX-XAU、CUX-XAG、GUX-XAC、UAX-XUA及GGX-XCC。在一些實施例中,本文所描述之方法包含非天然密碼子-反密碼子對NNX-YNN,其中NNX-YNN係選自由以下組成之群:UUX-YAA、UGX-YCA、CGX-YCG、AGX-YCU、GAX-YUC、CAX-YUG、AUX-YAU、CUX-YAG、GUX-YAC、UAX-YUA及GGX-YCC。在一些情況下,本文所描述之方法包含非天然密碼子-反密碼子對NXN-NYN,其中NXN-NYN係選自由以下組成之群:GXU-AYC、CXU-AYG、GXG-CYC、AXG-CYU、GXC-GYC、AXC-GYU、GXA-UYC、CXC-GYG及UXC-GYA。在一些實施例中,本文所描述之方法包含至少兩種各自包含不同的非天然反密碼子之非天然tRNA分子。在一些情況下,至少兩個非天然tRNA分子包含來自甲烷八聯球菌屬(Methanosarcina genus
)之吡咯離胺醯基tRNA及來自詹氏甲烷球菌(Methanocaldococcus jannaschii
)之酪胺醯基tRNA,或其衍生物。在一些實施例中,方法包含藉由胺基-醯基tRNA合成酶而裝載有至少兩個非天然tRNA分子。在一些情況下,tRNA合成酶係選自由嵌合PylRS (chPylRS)及詹氏甲烷球菌AzFRS (Mjp
AzFRS)組成之群。在一些實施例中,如本文所描述之方法包含藉由至少兩種不同tRNA合成酶而裝載有至少兩個非天然tRNA分子。在一些情況下,至少兩種不同tRNA合成酶包含嵌合PylRS (chPylRS)及詹氏甲烷球菌AzFRS (Mjp
AzFRS)。
在一些實施例中,本文描述活體內合成非天然多肽之方法。在一些實施例中,非天然多肽包含兩個、三個或更多個非天然胺基酸。在一些情況下,非天然多肽包含至少兩個相同的非天然胺基酸。在一些實施例中,非天然多肽包含至少兩個不同的非天然胺基酸。在一些情況下,非天然胺基酸包含:
離胺酸類似物;芳族側鏈;疊氮基;炔基;或醛基或酮基。在一些情況下,非天然胺基酸不包含芳族側鏈。在一些實施例中,非天然胺基酸係選自:N6-疊氮基乙氧基-羰基-L-離胺酸(AzK)、N6-炔丙基乙氧基-羰基-L-離胺酸(PraK)、N6-(炔丙氧基)-羰基-L-離胺酸(PrK)、對疊氮基-苯丙胺酸(pAzF)、BCN-L-離胺酸、降冰片烯離胺酸、TCO-離胺酸、甲基四𠯤離胺酸、烯丙氧基羰基離胺酸、2-胺基-8-側氧基壬酸、2-胺基-8-側氧基辛酸、對乙醯基-L-苯丙胺酸、對疊氮基甲基-L-苯丙胺酸(pAMF)、對碘-L-苯丙胺酸、間乙醯基苯丙胺酸、2-胺基-8-側氧基壬酸、對炔丙基氧基苯丙胺酸、對炔丙基-苯丙胺酸、3-甲基-苯丙胺酸、左旋多巴、氟化苯丙胺酸、異丙基-L-苯丙胺酸、對疊氮基-L-苯丙胺酸、對醯基-L-苯丙胺酸、對苯甲醯基-L-苯丙胺酸、對溴苯基丙胺酸、對胺基-L-苯丙胺酸、異丙基-L-苯丙胺酸、O-烯丙基酪胺酸、O-甲基-L-酪胺酸、O-4-烯丙基-L-酪胺酸、4-丙基-L-酪胺酸、膦醯基酪胺酸、三-O-乙醯基-GlcNAcp-絲胺酸、L-磷絲胺酸、膦醯基絲胺酸、L-3-(2-萘基)丙胺酸、2-胺基-3-((2-((3-(苯甲氧基)-3-側氧基丙基)胺基)乙基)硒烷基)丙酸、2-胺基-3-(苯基硒烷基)丙酸、硒半胱胺酸、N6-(((2-疊氮基苯甲基)氧基)羰基)-L-離胺酸、N6-(((3-疊氮基苯甲基)氧基)羰基)-L-離胺酸及N6-(((4-疊氮基苯甲基)氧基)羰基)-L-離胺酸。
在一些實施例中,如本文所描述之活體內合成非天然多肽之方法包含呈質體形式之至少一個非天然DNA分子。在一些情況下,將至少一個非天然DNA分子整合至細胞之基因體中。在一些實施例中,至少一個非天然DNA分子編碼非天然多肽。在一些實施例中,本文所描述之方法包含非天然DNA分子之活體內複製及轉錄以及經轉錄mRNA分子於細胞生物體中之活體內轉譯。在一些實施例中,細胞生物體為微生物。在一些實施例中,細胞生物體為原核生物。在一些實施例中,細胞生物體為細菌。在一些情況下,細胞生物體為革蘭氏陽性細菌。在一些實施例中,細胞生物體為革蘭氏陰性細菌。在一些情況下,細胞生物體為大腸桿菌。在一些實施例中,細胞生物體包含核苷三磷酸轉運子。在一些情況下,核苷三磷酸轉運子包含Pt
NTT2之胺基酸序列。在一些實施例中,核苷三磷酸轉運子包含Pt
NTT2之截短胺基酸序列。在一些替代方案中,Pt
NTT2之截短胺基酸序列與由SEQ ID NO.1編碼之Pt
NTT2至少80%一致。在一些實施例中,細胞生物體包含至少一個非天然DNA分子。在一些實施例中,至少一個非天然DNA分子包含至少一個質體。在一些實施例中,將至少一個非天然DNA分子整合至細胞之基因體中。在一些情況下,至少一個非天然DNA分子編碼非天然多肽。在一些情況下,本發明中所描述之方法可為活體外方法,其包含使用無細胞系統合成非天然多肽。
在一些實施例中,本文描述用於活體內合成非天然多肽之方法,其中非天然多肽包含非天然糖部分。在一些實施例中,非天然鹼基對包含至少一個包含非天然糖部分之非天然核苷酸。在一些實施例中,非天然糖部分係選自由以下組成之群:OH、經取代之低碳數烷基、烷芳基、芳烷基、O-烷芳基或O-芳烷基、SH、SCH3
、OCN、Cl、Br、CN、CF3
、OCF3
、SOCH3
、SO2
CH3
、ONO2
、NO2
、N3
、NH2
F;O-烷基、S-烷基、N-烷基;O-烯基、S-烯基、N-烯基;O-炔基、S-炔基、N-炔基;O-烷基-O-烷基、2'-F、2'-OCH3
、2'-O(CH2
)2
OCH3
,烷基、烯基及炔基可為經取代或未經取代之C1
-C10
烷基、C2
-C10
烯基、C2
-C10
炔基、-O[(CH2
)n
O]m
CH3
、-O(CH2
)n
OCH3
、-O(CH2
)n
NH2
、-O(CH2
)n
CH3
、-O(CH2
)n
-NH2
及-O(CH2
)n
ON[(CH2
)n
CH3
)]2
,其中n及m為1至約10;及/或於5'位置處之修飾:5'-乙烯基、5'-甲基(R或S);於4'位置處之修飾:4'-S、雜環烷基、雜環烷芳基、胺基烷胺基、聚烷基胺基、經取代之矽烷基、RNA裂解基團、報導基團、嵌入基團、用於改良寡核苷酸之藥物動力學特性之基團或用於改良寡核苷酸之藥效動力學特性之基團,及其任何組合。
在一些實施例中,本文描述用於活體內合成非天然多肽之細胞,該細胞包含:至少兩個不同的非天然密碼子-反密碼子對,其中各非天然密碼子-反密碼子對包含來自非天然信使RNA (mRNA)之非天然密碼子及來自非天然轉移核糖核酸(tRNA)之非天然反密碼子,該非天然密碼子包含第一非天然核苷酸且該非天然反密碼子包含第二非天然核苷酸;以及至少兩個不同的各自共價連接至對應非天然tRNA之非天然胺基酸。在一些情況下,細胞進一步包含至少一個包含至少四個非天然鹼基對(UBP)之非天然DNA分子。在一些實施例中,本文所描述用於活體內合成非天然多肽之細胞,該細胞包含至少一個包含至少四個非天然鹼基對之非天然DNA分子,其中至少一個非天然DNA分子編碼(i)編碼非天然多肽且包含至少第一及第二非天然密碼子之信使核糖核酸(mRNA)分子;以及(ii)至少第一及第二轉送RNA (tRNA)分子,第一tRNA分子包含第一非天然反密碼子且第二tRNA分子包含第二非天然反密碼子,並且至少一個DNA分子中之至少四個非天然鹼基對處於序列背景中,使得mRNA分子之第一及第二非天然密碼子分別與第一及第二非天然反密碼子互補。在一些實施例中,細胞進一步包含mRNA分子及至少第一及第二tRNA分子。在細胞之一些實施例中,至少第一及第二tRNA分子共價連接至非天然胺基酸。在一些實施例中,細胞進一步包含非天然多肽。
在一些實施例中,第一非天然核苷酸位於非天然密碼子之第二或第三位置處且與非天然反密碼子之第二非天然核苷酸互補地鹼基配對。在一些情況下,第一非天然核苷酸及第二非天然核苷酸包含獨立地選自由以下組成之群的第一及第二鹼基:
 ,視情況其中第二鹼基不同於第一鹼基。在一些實施例中,細胞進一步包含至少一個包含至少四個非天然鹼基對(UBP)之非天然DNA分子。在一些情況下,至少四個非天然鹼基對獨立地選自由以下組成之群:dCNMO/dTPT3、dNaM/dTPT3、dCNMO/dTAT1或dNaM/dTAT1。在一些情況下,至少一個非天然DNA分子包含至少一個質體。在一些實施例中,將至少一個非天然DNA分子整合至細胞之基因體中。在一些實施例中,至少一個非天然DNA分子編碼非天然多肽。在一些實施例中,如本文所描述之細胞表現核苷三磷酸轉運子。在一些替代方案中,核苷三磷酸轉運子包含Pt
NTT2之胺基酸序列。在一些情況下,核苷三磷酸轉運子包含Pt
NTT2之截短胺基酸序列,視情況其中Pt
NTT2之該截短胺基酸序列與由SEQ ID NO.1編碼之Pt
NTT2至少80%一致。在一些實施例中,細胞表現至少兩種tRNA合成酶。在一些實施例中,至少兩種tRNA合成酶包含嵌合PylRS (chPylRS)及詹氏甲烷球菌AzFRS (MjpAzFRS)。在一些實施例中,細胞包含非天然核苷酸,該等非天然核苷酸包含非天然糖部分。在一些情況下,非天然糖部分係選自由以下組成之群:於2'位置處之修飾:OH、經取代之低碳數烷基、烷芳基、芳烷基、O-烷芳基或O-芳烷基、SH、SCH3
、OCN、Cl、Br、CN、CF3
、OCF3
、SOCH3
、SO2
CH3
、ONO2
、NO2
、N3
、NH2
F;O-烷基、S-烷基、N-烷基;O-烯基、S-烯基、N-烯基;O-炔基、S-炔基、N-炔基;
O-烷基-O-烷基、2'-F、2'-OCH3
、2'-O(CH2
)2
OCH3
,其中烷基、烯基及炔基可為經取代或未經取代之C1
-C10
烷基、C2
-C10
烯基、C2
-C10
炔基、-O[(CH2
)n
O]m
CH3
、-O(CH2
)n
OCH3
、-O(CH2
)n
NH2
、-O(CH2
)n
CH3
、-O(CH2
)n
-NH2
及-O(CH2
)n
ON[(CH2
)n
CH3
)]2
,其中n及m為1至約10;及/或於5'位置處之修飾:5'-乙烯基、5'-甲基(R或S);於4'位置處之修飾:4'-S、雜環烷基、雜環烷芳基、胺基烷胺基、聚烷基胺基、經取代之矽烷基、RNA裂解基團、報導基團、嵌入基團、用於改良寡核苷酸之藥物動力學特性之基團或用於改良寡核苷酸之藥效動力學特性之基團,及其任何組合。在一些實施例中,細胞包含至少一個在轉錄期間由RNA聚合酶識別之非天然核苷酸鹼基。在一些實施例中,如本文所描述之細胞轉譯至少一個包含至少兩個非天然胺基酸之非天然多肽。在一些情況下,至少兩個非天然胺基酸獨立地選自由以下組成之群:N6-疊氮基乙氧基-羰基-L-離胺酸(AzK)、N6-炔丙基乙氧基-羰基-L-離胺酸(PraK)、N6-(炔丙氧基)-羰基-L-離胺酸(PrK)、對疊氮基-苯丙胺酸(pAzF)、BCN-L-離胺酸、降冰片烯離胺酸、TCO-離胺酸、甲基四𠯤離胺酸、烯丙氧基羰基離胺酸、2-胺基-8-側氧基壬酸、2-胺基-8-側氧基辛酸、對乙醯基-L-苯丙胺酸、對疊氮基甲基-L-苯丙胺酸(pAMF)、對碘-L-苯丙胺酸、間乙醯基苯丙胺酸、2-胺基-8-側氧基壬酸、對炔丙基氧基苯丙胺酸、對炔丙基-苯丙胺酸、3-甲基-苯丙胺酸、左旋多巴、氟化苯丙胺酸、異丙基-L-苯丙胺酸、對疊氮基-L-苯丙胺酸、對醯基-L-苯丙胺酸、對苯甲醯基-L-苯丙胺酸、對溴苯基丙胺酸、對胺基-L-苯丙胺酸、異丙基-L-苯丙胺酸、O-烯丙基酪胺酸、O-甲基-L-酪胺酸、O-4-烯丙基-L-酪胺酸、4-丙基-L-酪胺酸、膦醯基酪胺酸、三-O-乙醯基-GlcNAcp-絲胺酸、L-磷絲胺酸、膦醯基絲胺酸、L-3-(2-萘基)丙胺酸、2-胺基-3-((2-((3-(苯甲氧基)-3-側氧基丙基)胺基)乙基)硒烷基)丙酸、2-胺基-3-(苯基硒烷基)丙酸、硒半胱胺酸、N6-(((2-疊氮基苯甲基)氧基)羰基)-L-離胺酸、N6-(((3-疊氮基苯甲基)氧基)羰基)-L-離胺酸及N6-(((4-疊氮基苯甲基)氧基)羰基)-L-離胺酸。在一些情況下,如本文所描述之細胞為經分離細胞。在一些替代方案中,本文所描述之細胞為原核生物。在一些情況下,本文所描述之細胞包含細胞株。
,視情況其中第二鹼基不同於第一鹼基。在一些實施例中,細胞進一步包含至少一個包含至少四個非天然鹼基對(UBP)之非天然DNA分子。在一些情況下,至少四個非天然鹼基對獨立地選自由以下組成之群:dCNMO/dTPT3、dNaM/dTPT3、dCNMO/dTAT1或dNaM/dTAT1。在一些情況下,至少一個非天然DNA分子包含至少一個質體。在一些實施例中,將至少一個非天然DNA分子整合至細胞之基因體中。在一些實施例中,至少一個非天然DNA分子編碼非天然多肽。在一些實施例中,如本文所描述之細胞表現核苷三磷酸轉運子。在一些替代方案中,核苷三磷酸轉運子包含Pt
NTT2之胺基酸序列。在一些情況下,核苷三磷酸轉運子包含Pt
NTT2之截短胺基酸序列,視情況其中Pt
NTT2之該截短胺基酸序列與由SEQ ID NO.1編碼之Pt
NTT2至少80%一致。在一些實施例中,細胞表現至少兩種tRNA合成酶。在一些實施例中,至少兩種tRNA合成酶包含嵌合PylRS (chPylRS)及詹氏甲烷球菌AzFRS (MjpAzFRS)。在一些實施例中,細胞包含非天然核苷酸,該等非天然核苷酸包含非天然糖部分。在一些情況下,非天然糖部分係選自由以下組成之群:於2'位置處之修飾:OH、經取代之低碳數烷基、烷芳基、芳烷基、O-烷芳基或O-芳烷基、SH、SCH3
、OCN、Cl、Br、CN、CF3
、OCF3
、SOCH3
、SO2
CH3
、ONO2
、NO2
、N3
、NH2
F;O-烷基、S-烷基、N-烷基;O-烯基、S-烯基、N-烯基;O-炔基、S-炔基、N-炔基;
O-烷基-O-烷基、2'-F、2'-OCH3
、2'-O(CH2
)2
OCH3
,其中烷基、烯基及炔基可為經取代或未經取代之C1
-C10
烷基、C2
-C10
烯基、C2
-C10
炔基、-O[(CH2
)n
O]m
CH3
、-O(CH2
)n
OCH3
、-O(CH2
)n
NH2
、-O(CH2
)n
CH3
、-O(CH2
)n
-NH2
及-O(CH2
)n
ON[(CH2
)n
CH3
)]2
,其中n及m為1至約10;及/或於5'位置處之修飾:5'-乙烯基、5'-甲基(R或S);於4'位置處之修飾:4'-S、雜環烷基、雜環烷芳基、胺基烷胺基、聚烷基胺基、經取代之矽烷基、RNA裂解基團、報導基團、嵌入基團、用於改良寡核苷酸之藥物動力學特性之基團或用於改良寡核苷酸之藥效動力學特性之基團,及其任何組合。在一些實施例中,細胞包含至少一個在轉錄期間由RNA聚合酶識別之非天然核苷酸鹼基。在一些實施例中,如本文所描述之細胞轉譯至少一個包含至少兩個非天然胺基酸之非天然多肽。在一些情況下,至少兩個非天然胺基酸獨立地選自由以下組成之群:N6-疊氮基乙氧基-羰基-L-離胺酸(AzK)、N6-炔丙基乙氧基-羰基-L-離胺酸(PraK)、N6-(炔丙氧基)-羰基-L-離胺酸(PrK)、對疊氮基-苯丙胺酸(pAzF)、BCN-L-離胺酸、降冰片烯離胺酸、TCO-離胺酸、甲基四𠯤離胺酸、烯丙氧基羰基離胺酸、2-胺基-8-側氧基壬酸、2-胺基-8-側氧基辛酸、對乙醯基-L-苯丙胺酸、對疊氮基甲基-L-苯丙胺酸(pAMF)、對碘-L-苯丙胺酸、間乙醯基苯丙胺酸、2-胺基-8-側氧基壬酸、對炔丙基氧基苯丙胺酸、對炔丙基-苯丙胺酸、3-甲基-苯丙胺酸、左旋多巴、氟化苯丙胺酸、異丙基-L-苯丙胺酸、對疊氮基-L-苯丙胺酸、對醯基-L-苯丙胺酸、對苯甲醯基-L-苯丙胺酸、對溴苯基丙胺酸、對胺基-L-苯丙胺酸、異丙基-L-苯丙胺酸、O-烯丙基酪胺酸、O-甲基-L-酪胺酸、O-4-烯丙基-L-酪胺酸、4-丙基-L-酪胺酸、膦醯基酪胺酸、三-O-乙醯基-GlcNAcp-絲胺酸、L-磷絲胺酸、膦醯基絲胺酸、L-3-(2-萘基)丙胺酸、2-胺基-3-((2-((3-(苯甲氧基)-3-側氧基丙基)胺基)乙基)硒烷基)丙酸、2-胺基-3-(苯基硒烷基)丙酸、硒半胱胺酸、N6-(((2-疊氮基苯甲基)氧基)羰基)-L-離胺酸、N6-(((3-疊氮基苯甲基)氧基)羰基)-L-離胺酸及N6-(((4-疊氮基苯甲基)氧基)羰基)-L-離胺酸。在一些情況下,如本文所描述之細胞為經分離細胞。在一些替代方案中,本文所描述之細胞為原核生物。在一些情況下,本文所描述之細胞包含細胞株。
相關申請案之交叉參考
本申請案主張於2019年10月10日申請之美國臨時申請案第62/913,664號及於2020年3月12日申請之美國臨時申請案第62/988,882號之優先權,該等申請案中之每一者均以全文引用之方式併入本文中。序列表
本申請案含有序列表,該序列表已以ASCII格式以電子方式提交且其特此以全文引用之方式併入。在2010年10月6日產生之該ASCII複本命名為「36271-809_851_SL.txt」且大小為21千位元組。
關於聯邦贊助之研究之申明
本發明係在政府支持下,在由美國國家衛生研究院(National Institutes of Health)授予的批准號GM118178下進行。政府對本發明可具有某些權利。以引用方式之併入
本說明書中所提及之所有公開案、專利及專利申請案均以引用之方式併入本文中,其引用之程度如同各個別公開案、專利或專利申請案經特定且個別地指示以引用之方式併入一般。在以引用之方式併入之公開案及專利或專利申請案與本說明書中所含之揭示內容相抵觸之情況下,本說明書意欲替代及/或優先於任何此類相抵觸之材料。某些術語
除非另有定義,否則本文所使用之所有技術及科學術語具有與熟習所主張之主題的此項技術者通常所理解相同之含義。應理解,以上一般描述及以下詳細描述僅為例示性及解釋性的且不限制所主張之任何主題。在本申請案中,除非另外明確說明,否則單數之使用包括複數。必須指出,除非上下文另外清楚指定,否則如本說明書及隨附申請專利範圍中所使用之單數形式「一(a/an)」及「該」包括複數個指示物。在本申請案中,除非另外說明,否則「或」之使用意謂「及/或」。此外,術語「包括(including)」以及其他形式(諸如「包括(include/includes/included)」)之使用不具限制性。
如本文所使用,範圍及量可以「約」特定值或範圍表示。約亦包括精確量。因此,「約5 µL」意謂「約5 µL」且亦意謂「5 µL」。一般而言,術語「約」包括預期在實驗誤差內之量。
在合成方法之上下文中,如本文所使用之諸如「在適合於提供……之條件下」或「在足以產生……之條件下」或其類似者的片語係指在實驗者改變之一般技術範圍內之反應條件,諸如時間、溫度、溶劑、反應物濃度及其類似物,其提供適用量或產率之反應產物。所需反應產物不必為唯一反應產物或不必完全消耗起始材料,其限制條件為所需反應產物可經分離或另作他用。
「化學上可行的」意謂不違反一般理解的有機結構之法則的鍵結佈置或化合物;舉例而言,在某些情形中將含有自然界中不存在之五價碳原子的申請專利範圍定義中之結構應理解為不在申請專利範圍內。本文所揭示之結構在其所有實施例中意欲僅包括「化學上可行的」結構,且化學上不可行的任何所述結構(例如在展示有可變原子或基團之結構中)均不意欲在本文中揭示或主張。
如本文所使用之術語化學結構之「類似物」係指化學結構保留與母結構相當大的相似性,但其可能不易於以合成方式衍生自母結構。在一些實施例中,核苷酸類似物為非天然核苷酸。在一些實施例中,核苷類似物為非天然核苷。易於以合成方式衍生自母化學結構之相關化學結構稱為「衍生物」。
因此,如本文所使用之術語聚核苷酸係指DNA、RNA、DNA樣或RNA樣聚合物,諸如肽核酸(PNA)、鎖核酸(LNA)、硫代磷酸酯、非天然鹼基及其類似物,其為此項技術中熟知的。聚核苷酸可在自動合成器中合成,例如使用胺基偶磷酯化學方法或其他適於合成器使用之化學方法。
DNA包括(但不限於) cDNA及基因體DNA。DNA可藉由共價或非共價方式附接至另一生物分子,包括(但不限於)RNA及肽。RNA包括編碼RNA,例如,信使RNA (mRNA)。在一些實施例中,RNA為rRNA、RNAi、snoRNA、微小RNA、siRNA、snRNA、exRNA、piRNA、長ncRNA或其任何組合或雜合體。在一些情況下,RNA為核糖核酸酶之組分。DNA及RNA可呈任何形式,包括(但不限於):直鏈、環狀、超螺旋、單股及雙股形式。
肽核酸(PNA)為合成DNA/RNA類似物,其中肽樣主鏈替代DNA或RNA之糖磷酸主鏈。PNA寡聚物在與互補DNA結合中展示更高結合強度及更大特異性,其中PNA/DNA鹼基錯配比DNA/DNA雙螺旋體中之類似錯配更去穩定化。此結合強度及特異性亦適用於PNA/RNA雙螺旋體。PNA不易於由核酸酶或蛋白酶識別,此使得其對酶降解具有抗性。PNA亦在寬pH範圍內為穩定的。亦參見Nielsen PE, Egholm M, Berg RH, Buchardt O (1991年12月). 「Sequence-selective recognition of DNA by strand displacement with a thymine-substituted polyamide」,Science
254 (5037): 1497-500. 數位物件識別碼:10.1126/science.1962210. PMID 1962210;及Egholm M, Buchardt O, Christensen L, Behrens C, Freier SM, Driver DA, Berg RH, Kim SK, Nordén B及Nielsen PE (1993), 「PNA Hybridizes to Complementary Oligonucleotides Obeying the Watson-Crick Hydrogen Bonding Rules」.Nature
365 (6446): 566-8. 數位物件識別碼:10.1038/365566a0. PMID 7692304。
鎖核酸(LNA)為經修飾之RNA核苷酸,其中LNA核苷酸之核糖部分經連接2'氧及4'碳的額外橋修飾。橋「鎖定」3'-內(北)構形中之核糖,其常常見於A-形式雙螺旋體中。不論何時需要,LNA核苷酸可與寡核苷酸中之DNA或RNA殘基混合。此類寡聚物可經化學合成,且為可商購的。鎖核糖構形增強鹼基堆疊及主鏈預組織。參見例如Kaur, H;Arora, A;Wengel, J;Maiti, S (2006), 「Thermodynamic, Counterion, and Hydration Effects for the Incorporation of Locked Nucleic Acid Nucleotides into DNA Duplexes」,Biochemistry
45 (23): 7347-55. 數位物件識別碼:10.1021/bi060307w. PMID 16752924;Owczarzy R.;You Y., Groth C.L., Tataurov A.V. (2011), 「Stability and mismatch discrimination of locked nucleic acid-DNA duplexes.」,Biochem.
50 (43): 9352-9367. 數位物件識別碼:10.1021/bi200904e. PMC 3201676. PMID 21928795;Alexei A. Koshkin;Sanjay K. Singh, Poul Nielsen, Vivek K. Rajwanshi, Ravindra Kumar, Michael Meldgaard, Carl Erik Olsen, Jesper Wengel (1998), 「LNA (Locked Nucleic Acids): Synthesis of the adenine, cytosine, guanine, 5-methylcytosine, thymine and uracil bicyclonucleoside monomers, oligomerisation, and unprecedented nucleic acid recognition」,Tetrahedron
54 (14): 3607-30. 數位物件識別碼:10.1016/S0040-4020(98)00094-5;以及Satoshi Obika;Daishu Nanbu, Yoshiyuki Hari, Ken-ichiro Morio, Yasuko In, Toshimasa Ishida, Takeshi Imanishi (1997), 「Synthesis of 2'-O
,4'-C
-methyleneuridine and -cytidine. Novel bicyclic nucleosides having a fixed C3'-endo sugar puckering」,Tetrahedron Lett.
38 (50): 8735-8. 數位物件識別碼:10.1016/S0040-4039(97)10322-7。
分子信標或分子信標探針為可偵測特定核酸序列在均質溶液中之存在的寡核苷酸雜交探針。分子信標為具有內部淬滅螢光團之髮夾形分子,其螢光在其結合於目標核酸序列時恢復。參見例如Tyagi S, Kramer FR (1996), 「Molecular beacons: probes that fluoresce upon hybridization」,Nat Biotechnol.
14 (3): 303-8. PMID 9630890;Täpp I, Malmberg L, Rennel E, Wik M, Syvänen AC (2000年4月), 「Homogeneous scoring of single-nucleotide polymorphisms: comparison of the 5'-nuclease TaqMan assay and Molecular Beacon probes」,Biotechniques
28 (4): 732-8. PMID 10769752;以及Akimitsu Okamoto (2011), 「ECHO probes: a concept of fluorescence control for practical nucleic acid sensing」,Chem. Soc. Rev.
40: 5815-5828。
在一些實施例中,核鹼基一般為核苷之雜環鹼基部分。核鹼基可為天然存在的、可經修飾、可不帶有與天然鹼基之類似性,且可例如藉由有機合成來合成。在某些實施例中,核鹼基包含能夠在使用或不使用氫鍵之情況下與另一核酸之鹼基相互作用的任何原子或原子群。在某些實施例中,非天然核鹼基並非衍生自天然核鹼基。應注意,非天然核鹼基不一定具有鹼性特性,然而為簡單起見而將其稱為核鹼基。在一些實施例中,當提及核鹼基時,「(d)」指示核鹼基可附接至去氧核糖或核糖。
在一些實施例中,核苷為包含核鹼基部分及糖部分之化合物。核苷包括(但不限於):天然存在之核苷(如在DNA及RNA中發現)、無鹼基核苷、經修飾之核苷及具有模擬鹼基及/或糖基團之核苷。核苷包括包含任何種類取代基之核苷。核苷可為經由核酸鹼基與還原糖基團之間的醣苷連接形成的醣苷化合物。
在一些實施例中,如本發明所描述之非天然mRNA密碼子及非天然tRNA反密碼子可依據其DNA編碼序列書寫。舉例而言,非天然tRNA反密碼子可書寫為GYU或GYT。
本文所使用之章節標題僅出於組織目的且不應理解為限制所描述之主題。用於活體內合成非天然多肽之組合物及方法
本文揭示用於具有擴展遺傳字母表之非天然多肽之活體內合成的組合物及方法。在一些情況下,如本文所描述之組合物及方法包含編碼非天然多肽之非天然核酸分子,其中非天然多肽包含非天然胺基酸。在一些情況下,非天然多肽包含至少兩個非天然胺基酸。在一些情況下,非天然多肽包含至少三個非天然胺基酸。在一些情況下,非天然多肽包含兩個非天然胺基酸。在一些情況下,非天然多肽包含三個非天然胺基酸。在一些情況下,併入非天然多肽中之至少兩個非天然胺基酸可為相同或不同的非天然胺基酸。在一些情況下,將非天然胺基酸以位點特異性方式併入非天然多肽中。在一些情況下,非天然多肽為非天然蛋白質。
在一些情況下,如本文所描述之組合物及方法包含半合成生物體(SSO)。在一些情況下,方法包含將至少一個非天然鹼基對(UBP)併入至少一個非天然核酸分子中。在一些實施例中,方法包含將一個UBP併入至少一個非天然核酸分子中。在一些實施例中,方法包含將兩個UBP併入至少一個非天然核酸分子中。在一些實施例中,方法包含將三個UBP併入至少一個非天然核酸分子中。UBP鹼基對係藉由在兩個非天然核苷之非天然核鹼基之間配對而形成。在一些實施例中,非天然核酸分子為非天然DNA分子。
在一些實施例中,至少一個非天然核酸分子為一個分子或包含一個分子(例如質體或染色體)。在一些實施例中,至少一個非天然核酸分子為兩個分子或包含兩個分子(例如,兩個質體、兩個染色體或染色體及質體)。在一些實施例中,至少一個非天然核酸分子為三個分子或包含三個分子(例如,三個質體、兩個質體及染色體、質體及兩個染色體或三個染色體)。染色體之實例包括其中已整合UBP之基因體染色體及包含UBP之人工染色體(例如細菌人工染色體)。在一些實施例中,在使用包含至少四個非天然鹼基對之至少一個非天然DNA分子且該至少一個非天然DNA分子為兩個或更多個分子之情況下,至少四個非天然鹼基對可以任何可行的方式(例如一個在第一位置及三個在第二位置,兩個在第一位置及兩個在第二位置等)分佈於兩個或更多個分子中。
在一些情況下,至少一個視情況包括UBP之非天然核酸分子經轉錄以得到信使RNA分子,其包含至少一個含有至少一個非天然核苷酸之非天然密碼子。在一些實施例中,轉錄係指產生與DNA分子之一部分互補之一或多個RNA分子。在一些情況下,非天然核苷酸佔據非天然密碼子之第一、第二或第三密碼子位置,例如第二或第三密碼子位置。在一些情況下,兩個非天然核苷酸佔據非天然密碼子之第一及第二、第一及第三、第二及第三或第一及第三密碼子位置。在一些情況下,三個非天然核苷酸佔據非天然密碼子之所有三個密碼子位置。在一些情況下,攜帶非天然核苷酸之mRNA包含至少兩個非天然密碼子(在一些實施例中,表述「至少兩個非天然密碼子」與「至少第一及第二非天然密碼子」可互換)。在一些情況下,攜帶非天然核苷酸之mRNA包含兩個非天然密碼子。在一些情況下,攜帶非天然核苷酸之mRNA包含三個非天然密碼子。
在一些實施例中,視情況包括UBP之非天然核酸分子經轉錄以得到至少一個tRNA分子,其中tRNA分子包含攜帶至少一個非天然核苷酸之非天然反密碼子。在一些情況下,非天然核苷酸佔據非天然反密碼子之第一、第二或第三反密碼子位置。在一些情況下,兩個非天然核苷酸佔據非天然反密碼子之第一及第二、第一及第三、第二及第三或第一及第三反密碼子位置。在一些情況下,三個非天然核苷酸佔據非天然反密碼子之所有三個反密碼子位置。在一些情況下,視情況包括UBP之非天然核酸分子經轉錄以得到至少兩個包含至少兩個非天然反密碼子之tRNA。在情況下,至少兩個非天然反密碼子可相同或不同。在一些情況下,視情況包括UBP之非天然核酸分子經轉錄以得到兩個包含可相同或不同的非天然反密碼子之tRNA。在一些情況下,視情況包括UBP之非天然核酸分子經轉錄以得到三個包含三個可相同或不同的非天然反密碼子之tRNA。
在一些實施例中,由mRNA編碼之至少一個非天然密碼子可與tRNA之至少非天然反密碼子互補以形成非天然密碼子-反密碼子對。在一些情況下,本文所描述之組合物及方法包含用一個、兩個、三個或更多個非天然密碼子-反密碼子對來合成非天然多肽。在一些情況下,本文所描述之組合物及方法包含用兩個非天然密碼子-反密碼子對來合成非天然多肽。在一些情況下,本文所描述之組合物及方法包含用三個非天然密碼子-反密碼子對來合成非天然多肽。
在一些情況下,本文所描述之組合物及方法包含使用一個、兩個、三個或更多個非天然密碼子-反密碼子對合成具有一個、兩個、三個或更多個非天然胺基酸之非天然多肽。在一些情況下,本文所描述之組合物及方法包含使用兩個非天然密碼子-反密碼子對合成具有兩個非天然胺基酸之非天然多肽。在一些情況下,本文所描述之組合物及方法包含使用三個非天然密碼子-反密碼子對合成具有三個非天然胺基酸之非天然多肽。
在一些情況下,非天然密碼子包含核酸序列XNN、NXN、NNX、XXN、XNX、NXX或XXX,且非天然反密碼子包含核酸序列XNN、YNN、NXN、NYN、NNX、NNY、NXX、NYY、XNX、YNY、XXN、YYN或YYY,以形成非天然密碼子-反密碼子對。在一些情況下,非天然密碼子-反密碼子對由NNX-XNN、NNX-YNN或NXN-NYN構成,其中N為任何天然核苷酸,X為第一非天然核苷酸,且Y為第二非天然核苷酸。在一些實施例中,任何天然核苷酸包括具有標準鹼基(諸如腺嘌呤、胸腺嘧啶、尿嘧啶、鳥嘌呤或胞嘧啶)的核苷酸及具有天然存在之經修飾之鹼基(諸如假尿苷、5-甲基胞嘧啶等)的核苷酸。在一些實施例中,非天然密碼子-反密碼子對在密碼子中包含至少一個G且在反密碼子中包含至少一個C。在一些實施例中,非天然密碼子-反密碼子對在密碼子中包含至少一個G或C且在反密碼子中包含至少一個互補C或G。X及Y各自獨立地選自由以下組成之群:(i) 2-硫尿嘧啶、2'-去氧尿苷、4-硫-尿嘧啶、尿嘧啶-5-基、次黃嘌呤-9-基(I)、5-鹵基尿嘧啶;5-丙炔基-尿嘧啶、6-偶氮基-尿嘧啶、5-甲基胺基甲基尿嘧啶、5-甲氧基胺基甲基-2-硫尿嘧啶、假尿嘧啶、尿嘧啶-5-氧基乙酸甲酯、尿嘧啶-5-氧基乙酸、5-甲基-2-硫尿嘧啶、3-(3-胺基-3-N-2-羧丙基)尿嘧啶、5-甲基-2-硫尿嘧啶、4-硫尿嘧啶、5-甲基尿嘧啶、5'-甲氧基羧甲基尿嘧啶、5-甲氧基尿嘧啶、尿嘧啶-5-氧基乙酸、5-(羧基羥甲基)尿嘧啶、5-羧甲基胺基甲基-2-硫尿苷、5-羧甲基胺基甲基尿嘧啶、二氫尿嘧啶、5-羥甲基胞嘧啶、5-三氟甲基胞嘧啶、5-鹵基胞嘧啶、5-丙炔基胞嘧啶、5-羥基胞嘧啶、環胞嘧啶、胞嘧啶阿拉伯糖苷、5,6-二氫胞嘧啶、5-硝基胞嘧啶、6-偶氮基胞嘧啶、氮雜胞嘧啶、N4-乙基胞嘧啶、3-甲基胞嘧啶、5-甲基胞嘧啶、4-乙醯基胞嘧啶、2-硫胞嘧啶、啡㗁𠯤胞苷([5,4-b][l,4]苯并㗁𠯤-2(3H)-酮)、啡噻𠯤胞苷(1H-嘧啶并[5,4-b][l,4]苯并噻𠯤-2(3H)-酮)、啡㗁𠯤胞苷(9-(2-胺基乙氧基)-H-嘧啶并[5,4-b][l,4]苯并㗁𠯤-2(3H)-酮)、咔唑胞苷(2H-嘧啶并[4,5-b]吲哚-2-酮)、吡啶并吲哚胞苷(H-吡啶并[3',2':4,5]吡咯并[2,3-d]嘧啶-2-酮)、2-胺基腺嘌呤、2-丙基腺嘌呤、2-胺基-腺嘌呤、2-F-腺嘌呤、2-胺基-丙基-腺嘌呤、2-胺基-2'-去氧腺苷、3-去氮腺嘌呤、7-甲基腺嘌呤、7-去氮-腺嘌呤、8-氮雜腺嘌呤、經8-鹵基、8-胺基、8-硫醇、8-硫烷基及8-羥基取代之腺嘌呤、N6-異戊烯基腺嘌呤、2-甲基腺嘌呤、2,6-二胺基嘌呤、2-甲基硫-N6-異戊烯基腺嘌呤、6-氮雜-腺嘌呤、2-甲基鳥嘌呤;鳥嘌呤之2-丙基及烷基衍生物、3-去氮鳥嘌呤、6-硫-鳥嘌呤、7-甲基鳥嘌呤、7-去氮鳥嘌呤、7-去氮鳥苷、7-去氮-8-氮雜鳥嘌呤、8-氮雜鳥嘌呤、經8-鹵基、8-胺基、8-硫醇、8-硫烷基及8-羥基取代之鳥嘌呤、1-甲基鳥嘌呤、2,2-二甲基鳥嘌呤、7-甲基鳥嘌呤、6-氮雜-鳥嘌呤、次黃嘌呤、黃嘌呤、1-甲基肌苷、Q核苷、β-D-半乳糖基Q核苷、肌苷、β-D-甘露糖基Q核苷、懷丁氧苷、羥基脲、(acp3)w、2-胺基吡啶或2-吡啶酮。
在一些實施例中,X及Y獨立地選自由以下組成之群:
 。
。
在一些情況下,非天然密碼子-反密碼子對包含NNX-XNN,其中NNX-XNN係選自由以下組成之群:AAX-XUU、AUX-XAU、ACX-XGU、AGX-XCU、UAX-XUA、UUX-XAA、UCX-XGA、UGX-XCA、CAX-XUG、CUX-XAG、CCX-XGG、CGX-XCG、GAX-XUC、GUX-XAC、GCX-XGC及GGX-XCC。在一些情況下,非天然密碼子-反密碼子對包含NNX-YNN,其中NNX-YNN係選自由以下組成之群:AAX-YUU、AUX-YAU、ACX-YGU、AGX-YCU、UAX-YUA、UUX-YAA、UCX-YGA、UGX-YCA、CAX-YUG、CUX-YAG、CCX-YGG、CGX-YCG、GAX-YUC、GUX-YAC、GCX-YGC及GGX-YCC。在一些實施例中,非天然密碼子-反密碼子對包含NXN-NXN,其中NXN-NXN係選自由以下組成之群:AXA-UXU、AXU-AXU、AXC-GXU、AXG-CXU、UXA-UXA、UXU-AXA、UXC-GXA、UXG-CXA、CXA-UXG、CXU-AXG、CXC-GXG、CXG-CXG、GXA-UXC、GXU-AXC、GXC-GXC及GXG-CXC。在一些情況下,非天然密碼子-反密碼子對包含NXN-NYN,其中NXN-NYN係選自由以下組成之群:AXA-UYU、AXU-AYU、AXC-GYU、AXG-CYU、UXA-UYA、UXU-AYA、UXC-GYA、UXG-CYA、CXA-UYG、CXU-AYG、CXC-GYG、CXG-CYG、GXA-UYC、GXU-AYC、GXC-GYC及GXG-CYC。
在一些實施例中,非天然密碼子-反密碼子對包含XNN-NNX,其中XNN-NNX係選自由以下組成之群:XAA-UUX、XAU-AUX、XAC-AGX、XAG-CUX、XUA-UAX、XUU-AAX、XUC-GAX、XUG-CAX、XCA-UGX、XCU-AGX、XCC-GGX、XCG-CGX、XGA-UCX、XGU-ACX、XGC-GCX及XGG-CCX。在一些實施例中,非天然密碼子-反密碼子對包含XNN-NNY,其中XNN-NNY係選自由以下組成之群:XAA-UUY、XAU-AUY、XAC-AGY、XAG-CUY、XUA-UAY、XUU-AAY、XUC-GAY、XUG-CAY、XCA-UGY、XCU-AGY、XCC-GGY、XCG-CGY、XGA-UCY、XGU-ACY、XGC-GCY及XGG-CCY。
在一些實施例中,非天然密碼子-反密碼子對包含XXN-NXX,其中XXN-NXX係選自由以下組成之群:XXA-UXX、XXU-AXX、XXC-GXX及XXG-CXX。在一些實施例中,非天然密碼子-反密碼子對包含XXN-NYY,其中XXN-NYY係選自由以下組成之群:XXA-UYY、XXU-AYY、XXC-GYY及XXG-CYY。在一些替代方案中,非天然密碼子-反密碼子對包含XNX-XNX,其中XNX-XNX係選自由以下組成之群:XAX-XUX、XUX-XAX、XCX-XGX及XGX-XCX。在一些實施例中,非天然密碼子-反密碼子對包含XNX-YNY,其中XNX-YNY係選自由以下組成之群:XAX-YUY、XUX-YAY、XCX-YGY及XGX-YCY。在一些情況下,非天然密碼子-反密碼子對包含NXX-XXN,其中NXX-XXN係選自由以下組成之群:AXX-XXU、UXX-XXA、CXX-XXG及GXX-XXC。在一些情況下,非天然密碼子-反密碼子對包含NXX-YYN,其中NXX-YYN係選自由以下組成之群:AXX-YYU、UXX-YYA、CXX-YYG及GXX-YYC。在一些情況下,非天然密碼子-反密碼子對包含XXX-XXX或XXX-YYY。
在產生具有擴展遺傳字母表之非天然多肽(圖 2
)之方法的例示性工作流程100 (圖 1
)中,編碼各自包含互補非天然核鹼基(X,Y)之蛋白質102及tRNA 103之DNA 101經轉錄104,以產生tRNA 106及mRNA 107。X為第一非天然核苷酸且Y為第二非天然核苷酸。使tRNA裝載有非天然胺基酸105之後,mRNA 107經轉譯108,以產生包含一或多個非天然胺基酸109之蛋白質110。在一些情況下,本文所描述之方法及組合物允許非天然胺基酸以高保真度及產率位點特異性併入。本文亦描述包含擴展遺傳字母表之半合成生物體、用於使用半合成生物體以產生包括包含至少一個非天然胺基酸殘基之彼等產物的蛋白質產物的方法。
選擇非天然核鹼基使得本文所描述之方法中之一或多個步驟最佳化。舉例而言,針對高效率複製、轉錄及/或轉譯選擇核鹼基。在一些情況下,針對本文所描述之方法使用超過一個非天然核鹼基對。舉例而言,包含去氧核糖部分之第一組核鹼基用於DNA複製(諸如第一核鹼基及第二核鹼基,經組態以形成第一鹼基對),且第二組核鹼基(諸如第三核鹼基及第四核鹼基,其中第三及第四核鹼基附接至核糖,經組態以形成第二鹼基對)用於轉錄/轉譯。在一些情況下,第一組核鹼基與第二組核鹼基之間的互補配對允許轉錄基因以自包含來自第一組之核鹼基的DNA模板產生tRNA或蛋白質。在一些情況下,第二組核鹼基之間的互補配對(第二鹼基對)允許藉由匹配包含非天然核酸之tRNA及mRNA來進行轉譯。在一些情況下,將第一組中之核鹼基附接至去氧核糖部分。在一些情況下,將第一組中之核鹼基附接至核糖部分。在一些情況下,兩組核鹼基為獨特的。在一些情況下,至少一個核鹼基在兩組中為相同的。在一些情況下,第一核鹼基及第三核鹼基為相同的。在一些實施例中,第一鹼基對及第二鹼基對為不相同的。在一些情況下,第一鹼基對、第二鹼基對及第三鹼基對為不相同的。
在一些實施例中,藉由如本文所揭示之組合物及方法合成之非天然多肽或非天然蛋白質之產率與藉由其他方法合成之相同非天然多肽或非天然蛋白質之產率相比更高。在一些情況下,藉由如本文所揭示之組合物及方法合成之非天然多肽或非天然蛋白質之產率比藉由其他方法合成之相同非天然多肽或非天然蛋白質之產率高至少10%、至少20%、至少30%、至少40%或至少50%。其他方法之實例包括利用琥珀色密碼子抑制之方法。
在一些實例中,藉由如本文所揭示之組合物及方法合成之非天然多肽或非天然蛋白質之溶解度與藉由其他方法合成之相同非天然多肽或非天然蛋白質之溶解度相比更高。在一些情況下,藉由如本文所揭示之組合物及方法合成之非天然多肽或非天然蛋白質之溶解度比藉由其他方法合成之相同非天然多肽或非天然蛋白質高至少10%、至少20%、至少30%、至少40%或至少50%。在一些情況下,藉由如本文所揭示之組合物及方法合成之非天然蛋白質之生物活性與藉由其他方法合成之相同非天然蛋白質之生物活性相比更高。在一些情況下,藉由如本文所揭示之組合物及方法合成之非天然蛋白質之生物活性比藉由其他方法合成之相同非天然蛋白質之生物活性高至少10%、至少20%、至少30%、至少40%或至少50%。
在一些實施例中,如本文所描述之用於活體內合成非天然多肽之組合物及方法利用或包含半合成生物體(SSO)。在一些實施例中,SSO在非天然多肽之合成期間經歷純系擴增。在一些情況下,SSO在非天然多肽之合成期間並不純系擴增。在一些情況下,SSO可在非天然多肽之合成期間在細胞週期之任何階段停滯。在一些實施例中,如本文所描述之組合物及方法可活體外合成非天然多肽。在一些情況下,如本文所描述之組合物及方法可包含無細胞系統以合成非天然多肽。核酸分子
在一些實施例中,舉例而言,核酸(例如在本文中亦稱為所關注之核酸分子)係來自諸如DNA、cDNA、基因體DNA (gDNA)、RNA、短抑制性RNA (siRNA)、RNAi、tRNA、mRNA或核糖體RNA (rRNA)之任何來源或組合物,且呈任何形式(例如線性、圓形、超螺旋、單股、雙股及其類似形式)。在一些實施例中,核酸包含核苷酸、核苷或聚核苷酸。在一些情況下,核酸包含天然及非天然核酸。在一些情況下,核酸亦包含非天然核酸,諸如DNA或RNA類似物(例如含有鹼基類似物、糖類似物及/或非天然主鏈及其類似物)。應理解,術語「核酸」並不係指或表示特定長度之聚核苷酸鏈,因此聚核苷酸及寡核苷酸亦包括於定義中。例示性天然核苷酸包括(但不限於):ATP、UTP、CTP、GTP、ADP、UDP、CDP、GDP、AMP、UMP、CMP、GMP、dATP、dTTP、dCTP、dGTP、dADP、dTDP、dCDP、dGDP、dAMP、dTMP、dCMP及dGMP。例示性天然去氧核糖核苷酸包括dATP、dTTP、dCTP、dGTP、dADP、dTDP、dCDP、dGDP、dAMP、dTMP、dCMP及dGMP。例示性天然核糖核苷酸包括ATP、UTP、CTP、GTP、ADP、UDP、CDP、GDP、AMP、UMP、CMP及GMP。對於天然RNA,尿嘧啶鹼基為尿苷。核酸有時為載體、質體、噬質體、自主複製序列(autonomously replicating sequence;ARS)、著絲點、人工染色體、酵母人工染色體(例如YAC)或其他能夠在宿主細胞中複製或經複製的核酸。在一些情況下,非天然核酸為核酸類似物。在其他情況下,非天然核酸來自細胞外來源。在其他情況下,非天然核酸可用於本文所提供之生物體,例如經基因修飾之生物體之細胞內空間。在一些實施例中,非天然核苷酸不為天然核苷酸。在一些實施例中,不包含天然鹼基之核苷酸包含非天然核鹼基。非天然核酸
核苷酸類似物或非天然核苷酸包含核苷酸,該核苷酸含有對鹼基、糖或磷酸酯部分之某些類型的修飾。在一些實施例中,修飾包含化學修飾。在一些情況下,修飾在3'OH或5'OH基團處、在主鏈處、在糖組分處或在核苷酸鹼基處進行。在一些情況下,修飾視情況包括非天然存在之連接分子及/或股間或股內交聯。在一個態樣中,經修飾之核酸包含3'OH或5'OH基團、主鏈、糖組分或核苷酸鹼基中之一或多者之修飾,及/或非天然存在之連接分子之添加。在一個態樣中,經修飾之主鏈包含除磷酸二酯主鏈以外的主鏈。在一個態樣中,經修飾之糖包含除去氧核糖(在經修飾之DNA中)以外或除核糖(經修飾之RNA)以外的糖。在一個態樣中,經修飾之鹼基包含除腺嘌呤、鳥嘌呤、胞嘧啶或胸腺嘧啶(在經修飾之DNA中)以外的鹼基或除腺嘌呤、鳥嘌呤、胞嘧啶或尿嘧啶(在經修飾之RNA中)以外的鹼基。
在一些實施例中,核酸包含至少一個經修飾之鹼基。在一些情況下,核酸包含2、3、4、5、6、7、8、9、10、15、20或更多個經修飾之鹼基。在一些情況下,對鹼基部分之修飾包括對A、C、G及T/U以及不同嘌呤或嘧啶鹼基之天然及合成修飾。在一些實施例中,修飾為腺嘌呤、鳥嘌呤、胞嘧啶或胸腺嘧啶之經修飾形式(在經修飾之DNA中)或腺嘌呤、鳥嘌呤、胞嘧啶或尿嘧啶之經修飾形式(經修飾之RNA)。
非天然核酸之經修飾鹼基包括(但不限於):尿嘧啶-5-基、次黃嘌呤-9-基(I)、2-胺基腺嘌呤-9-基、5-甲基胞嘧啶(5-me-C)、5-羥甲基胞嘧啶、黃嘌呤、次黃嘌呤、2-胺基腺嘌呤、腺嘌呤及鳥嘌呤之6-甲基及其他烷基衍生物、腺嘌呤及鳥嘌呤之2-丙基及其他烷基衍生物、2-硫尿嘧啶、2-硫胸腺嘧啶及2-硫胞嘧啶、5-鹵基尿嘧啶及胞嘧啶、5-丙炔基尿嘧啶及胞嘧啶、6-偶氮基尿嘧啶、胞嘧啶及胸腺嘧啶、5-尿嘧啶(假尿嘧啶)、4-硫尿嘧啶、經8-鹵基、8-胺基、8-巰基、8-硫基烷基、8-羥基及其他8-取代之腺嘌呤及鳥嘌呤、經5-鹵基(尤其為5-溴、5-三氟甲基)及其他5-取代之尿嘧啶及胞嘧啶、7-甲基鳥嘌呤及7-甲基腺嘌呤、8-氮雜鳥嘌呤及8-氮雜腺嘌呤、7-去氮鳥嘌呤及7-去氮腺嘌呤以及3-去氮鳥嘌呤及3-去氮腺嘌呤。某些非天然核酸,諸如經5-取代之嘧啶、6-氮雜嘧啶及經N-2取代之嘌呤、經N-6取代之嘌呤、經O-6取代之嘌呤、2-胺基丙基腺嘌呤、5-丙炔基尿嘧啶、5-丙炔基胞嘧啶、5-甲基胞嘧啶、增加雙螺旋體形成之穩定性之彼等、通用核酸、疏水性核酸、混雜核酸、尺寸擴展核酸、氟化核酸、經5-取代之嘧啶、6-氮雜嘧啶以及經N-2、N-6及O-6取代之嘌呤,包括2-胺基丙基腺嘌呤、5-丙炔基尿嘧啶及5-丙炔基胞嘧啶。5-甲基胞嘧啶(5-me-C)、5-羥甲基胞嘧啶、黃嘌呤、次黃嘌呤、2-胺基腺嘌呤、6-甲基、腺嘌呤及鳥嘌呤之其他烷基衍生物、腺嘌呤及鳥嘌呤之2-丙基及其他烷基衍生物、2-硫尿嘧啶、2-硫胸腺嘧啶及2-硫胞嘧啶、5-鹵基尿嘧啶、5-鹵基胞嘧啶、5-丙炔基(-C≡C-CH3
)尿嘧啶、5-丙炔基胞嘧啶、嘧啶核酸之其他炔基衍生物、6-偶氮基尿嘧啶、6-偶氮基胞嘧啶、6-偶氮基胸腺嘧啶、5-尿嘧啶(假尿嘧啶)、4-硫尿嘧啶、經8-鹵基、8-胺基、8-巰基、8-硫基烷基、8-羥基及其他8-取代之腺嘌呤及鳥嘌呤、經5-鹵基(尤其5-溴、5-三氟甲基)、其他5-取代之尿嘧啶及胞嘧啶、7-甲基鳥嘌呤、7-甲基腺嘌呤、2-F-腺嘌呤、2-胺基-腺嘌呤、8-氮雜鳥嘌呤、8-氮雜腺嘌呤、7-去氮鳥嘌呤、7-去氮腺嘌呤、3-去氮鳥嘌呤、3-去氮腺嘌呤、三環嘧啶、啡㗁𠯤胞苷([5,4-b][l,4]苯并㗁𠯤-2(3H)-酮)、啡噻𠯤胞苷(1H-嘧啶并[5,4-b][l,4]苯并噻𠯤-2(3H)-酮)、G-夾鉗、啡㗁𠯤胞苷(例如9-(2-胺基乙氧基)-H-嘧啶并[5,4-b][l,4]苯并㗁𠯤-2(3H)-酮)、咔唑胞苷(2H-嘧啶并[4,5-b]吲哚-2-酮)、吡啶并吲哚胞苷(H-吡啶并[3',2':4,5]吡咯并[2,3-d]嘧啶-2-酮)、其中嘌呤或嘧啶鹼基經其他雜環置換之彼等、7-去氮-腺嘌呤、7-去氮鳥苷、2-胺基吡啶、2-吡啶酮、氮雜胞嘧啶、5-溴胞嘧啶、溴尿嘧啶、5-氯胞嘧啶、氯化胞嘧啶、環胞嘧啶、胞嘧啶阿拉伯糖苷、5-氟胞嘧啶、氟嘧啶、氟尿嘧啶、5,6-二氫胞嘧啶、5-碘胞嘧啶、羥基脲、碘尿嘧啶、5-硝基胞嘧啶、5-溴尿嘧啶、5-氯尿嘧啶、5-氟尿嘧啶及5-碘尿嘧啶、2-胺基-腺嘌呤、6-硫基-鳥嘌呤、2-硫基-胸腺嘧啶、4-硫基-胸腺嘧啶、5-丙炔基-尿嘧啶、4-硫基-尿嘧啶、N4-乙基胞嘧啶、7-去氮鳥嘌呤、7-去氮-8-氮雜鳥嘌呤、5-羥基胞嘧啶、2'-去氧尿苷、2-胺基-2'-去氧腺苷,以及美國專利第3,687,808號;第4,845,205號;第4,910,300號;第4,948,882號;第5,093,232號;第5,130,302號;第5,134,066號;第5,175,273號;第5,367,066號;第5,432,272號;第5,457,187號;第5,459,255號;第5,484,908號;第5,502,177號;第5,525,711號;第5,552,540號;第5,587,469號;第5,594,121號;第5,596,091號;第5,614,617號;第5,645,985號;第5,681,941號;第5,750,692號;第5,763,588號;第5,830,653號及第6,005,096號;WO 99/62923;Kandimalla等人, (2001) Bioorg. Med. Chem. 9:807-813;The Concise Encyclopedia of Polymer Science and Engineering, Kroschwitz, J.I.,編, John Wiley & Sons, 1990, 858- 859;Englisch等人, Angewandte Chemie, 國際版, 1991, 30, 613;以及Sanghvi, 第15章, Antisense Research and Applications, Crooke及Lebleu編, CRC出版社, 1993, 273-288中所描述之彼等。額外鹼基修飾可見於例如美國專利第3,687,808號;Englisch等人, Angewandte Chemie, 國際版, 1991, 30, 613中。在一些情況下,非天然核酸包含圖 3
之核鹼基。在一些情況下,非天然核酸包含圖 4A
之核鹼基。在一些情況下,非天然核酸包含圖 4B
之核鹼基。
包含各種雜環鹼基及各種糖部分(及糖類似物)之非天然核酸為此項技術中可用的,且在一些情況下,核酸包括除天然存在之核酸之主要五種鹼基組分以外的一種或若干種雜環鹼基。舉例而言,在一些情況下,雜環鹼基包括尿嘧啶-5-基、胞嘧啶-5-基、腺嘌呤-7-基、腺嘌呤-8-基、鳥嘌呤-7-基、鳥嘌呤-8-基、4-胺基吡咯并[2.3-d]嘧啶-5-基、2-胺基-4-側氧基吡咯并[2,3-d]嘧啶-5-基、2-胺基-4-側氧基吡咯并[2.3-d]嘧啶-3-基,其中嘌呤經由9-位置、嘧啶經由1-位置、吡咯并嘧啶經由7-位置且吡唑并嘧啶經由1-位置附接至核酸之糖部分。
在一些實施例中,以下描繪非天然核酸之經修飾之鹼基,其中波浪線或R鑑別附接至去氧核糖或核糖之點。


 。
。
在一些實施例中,核苷酸類似物亦在磷酸酯部分經修飾。經修飾之磷酸酯部分包括(但不限於)具有兩個核苷酸之間的鍵處之修飾且含有例如以下之彼等磷酸酯部分:硫代磷酸酯;對掌性硫代磷酸酯;二硫代磷酸酯;磷酸三酯;胺基烷基磷酸三酯;甲基及其他烷基膦酸酯,包括3'-伸烷基膦酸酯及對掌性膦酸酯;亞膦酸酯;胺基磷酸酯,包括3'-胺基胺基磷酸酯及胺基烷基胺基磷酸酯;硫羰基胺基磷酸酯;硫羰基烷基膦酸酯;硫羰基烷基磷酸三酯及硼烷磷酸酯。應理解,在兩個核苷酸之間的此等磷酸酯或經修飾之磷酸酯鍵可為經由3'-5'鍵或2'-5'鍵,且鍵含有反轉極性,諸如3'-5'至5'-3'或2'-5'至5'-2'。亦包括各種鹽、混合鹽及游離酸形式。大量美國專利教示如何製備及使用含有經修飾之磷酸酯之核苷酸,且包括(但不限於):3,687,808;4,469,863;4,476,301;5,023,243;5,177,196;5,188,897;5,264,423;5,276,019;5,278,302;5,286,717;5,321,131;5,399,676;5,405,939;5,453,496;5,455,233;5,466,677;5,476,925;5,519,126;5,536,821;5,541,306;5,550,111;5,563,253;5,571,799;5,587,361;及5,625,050。
在一些實施例中,非天然核酸包括2',3'-二去氧基-2',3'-二脫氫-核苷(PCT/US2002/006460)、經5'-取代之DNA及RNA衍生物(PCT/US2011/033961;Saha等人, J. Org Chem., 1995, 60, 788-789;Wang等人, Bioorganic & Medicinal Chemistry Letters, 1999, 9, 885-890;以及Mikhailov等人, Nucleosides & Nucleotides, 1991, 10(1-3), 339-343;Leonid等人, 1995, 14(3-5), 901-905;以及Eppacher等人, Helvetica Chimica Acta, 2004, 87, 3004-3020;PCT/JP2000/004720;PCT/JP2003/002342;PCT/JP2004/013216;PCT/JP2005/020435;PCT/JP2006/315479;PCT/JP2006/324484;PCT/JP2009/056718;PCT/JP2010/067560)或製成具有經修飾之鹼基的單磷酸酯之經5'-取代的單體(Wang等人, Nucleosides Nucleotides & Nucleic Acids, 2004, 23 (1及2), 317-337)。
在一些實施例中,非天然核酸包括在糖環之5'位置及2'位置處之修飾(PCT/US94/02993),諸如經5'-CH2
-取代之2'-O-保護核苷(Wu等人, Helvetica Chimica Acta, 2000, 83, 1127-1143及Wu等人, Bioconjugate Chem. 1999, 10, 921-924)。在一些情況下,非天然核酸包括醯胺連接之核苷二聚體(已經製備用於併入寡核苷酸中),其中在該二聚體(5'至3')中之3'連接核苷包含2'-OCH3
及5'-(S)-CH3
(Mesmaeker等人, Synlett, 1997, 1287-1290)。非天然核酸可包括經2'-取代之5'-CH2
(或O)修飾之核苷(PCT/US92/01020)。非天然核酸可包括5'-亞甲基膦酸酯DNA及RNA單體以及二聚體(Bohringer等人, Tet. Lett., 1993, 34, 2723-2726;Collingwood等人, Synlett, 1995, 7, 703-705;及Hutter等人, Helvetica Chimica Acta, 2002, 85, 2777-2806)。非天然核酸可包括具有2'-取代之5'-膦酸酯單體(US2006/0074035)及其他經修飾之5'-膦酸酯單體(WO1997/35869)。非天然核酸可包括經5'-修飾之亞甲基膦酸酯單體(EP614907及EP629633)。非天然核酸可包括在5'及/或6'-位置處包含羥基之5'或6'-膦酸酯核糖核苷之類似物(Chen等人, Phosphorus, Sulfur and Silicon, 2002, 777, 1783-1786;Jung等人, Bioorg. Med. Chem., 2000, 8, 2501-2509;Gallier等人, Eur. J. Org. Chem., 2007, 925-933;以及Hampton等人, J. Med. Chem., 1976, 19(8), 1029-1033)。非天然核酸可包括具有5'-磷酸酯基之5'-膦酸酯去氧核糖核苷單體及二聚體(Nawrot等人, Oligonucleotides, 2006, 16(1), 68-82)。非天然核酸可包括核苷,其具有6'-膦酸酯基,其中5'或/及6'-位置未經取代或經硫基-第三丁基(SC(CH3
)3
) (及其類似物);亞甲基胺基(CH2
NH2
) (及其類似物)或氰基(CN) (及其類似物)取代(Fairhurst等人, Synlett, 2001, 4, 467-472;Kappler等人, J. Med. Chem., 1986, 29, 1030-1038;Kappler等人, J. Med. Chem., 1982, 25, 1179-1184;Vrudhula等人, J. Med. Chem., 1987, 30, 888-894;Hampton等人, J. Med. Chem., 1976, 19, 1371-1377;Geze等人, J. Am. Chem. Soc, 1983, 105(26), 7638-7640;以及Hampton等人, J. Am. Chem. Soc, 1973, 95(13), 4404-4414)。
在一些實施例中,非天然核酸亦包括糖部分之修飾。在一些情況下,核酸含有一或多個核苷,其中糖基經修飾。此類糖修飾之核苷可賦予增強的核酸酶穩定性、增加的結合親和力或某些其他有益的生物特性。在某些實施例中,核酸包含經化學修飾之核呋喃糖環部分。經化學修飾之核呋喃糖環之實例包括(但不限於)添加取代基(包括5'及/或2'取代基;橋接兩個環原子以形成雙環核酸(BNA);用S、N(R)或C(R1
)(R2
) (R=H、C1
-C12
烷基或保護基)置換核糖基環氧原子;及其組合。經化學修飾之糖之實例可見於WO2008/101157、US2005/0130923及WO2007/134181中。
在一些情況下,經修飾之核酸包含經修飾之糖或糖類似物。因此,除核糖及去氧核糖以外,糖部分可為戊醣、去氧戊醣、己醣、去氧已醣、葡萄糖、阿拉伯糖、木糖、來蘇糖或糖「類似」環戊基。糖可呈哌喃糖基或呋喃糖基形式。糖部分可為核糖、去氧核糖、阿拉伯糖或2'-O-烷基核糖之呋喃糖苷,且糖可以[α]或[β]變旋異構組態附接至各別雜環鹼基。糖修飾包括(但不限於):2'-烷氧基-RNA類似物、2'-胺基-RNA類似物、2'-氟-DNA及2'-烷氧基-RNA/DNA嵌合體或胺基-RNA/DNA嵌合體。舉例而言,糖修飾可包括2'-O-甲基-尿苷或2'-O-甲基-胞嘧啶核苷。糖修飾包括2'-O-烷基-取代之去氧核糖核苷及2'-O-乙二醇樣核糖核苷。已知此等糖或糖類似物及各別「核苷」之製備,其中此類糖或類似物附接至雜環鹼基(核酸鹼基)。糖修飾亦可進行且與其他修飾組合。
對糖部分之修飾包括核糖及去氧核糖之天然修飾以及非天然修飾。糖修飾包括(但不限於):於2'位置處之修飾:OH;F;O-烷基、S-烷基或N-烷基;O-烯基、S-烯基或N-烯基;O-炔基、S-炔基或N-炔基;或O-烷基-O-烷基,其中烷基、烯基及炔基可為經取代或未經取代之C1
至C10
烷基或C2
至C10
烯基及炔基。2'糖修飾亦包括(但不限於):-O[(CH2
)n
O]m
CH3
、-O(CH2
)n
OCH3
、-O(CH2
)n
NH2
、-O(CH2
)n
CH3
、-O(CH2
)n
ONH2
及-O(CH2
)n
ON[(CH2
)n
CH3
)]2
,其中n及m為1至約10。
於2'位置處之其他修飾包括(但不限於):C1
至C10
低碳數烷基、經取代之低碳數烷基、烷芳基、芳烷基、O-烷芳基、O-芳烷基、SH、SCH3
、OCN、Cl、Br、CN、CF3
、OCF3
、SOCH3
、SO2
CH3
、ONO2
、NO2
、N3
、NH2
、雜環烷基、雜環烷芳基、胺基烷胺基、聚烷基胺基、經取代之矽烷基、RNA裂解基團、報導基團、嵌入基團、用於改良寡核苷酸之藥物動力學特性之基團或用於改良寡核苷酸之藥效動力學特性之基團及具有類似特性之其他取代基。類似修飾亦可在糖上之其他位置處進行,尤其3'末端核苷酸上之糖之3'位置或5'末端核苷酸之2'-5'連接之寡核苷酸及5'位置。經修飾之糖亦包括含有在橋環氧(諸如CH2
及S)處之修飾的彼等經修飾之糖。核苷酸糖類似物亦可具有糖模擬物(諸如環丁基部分)而非呋喃戊醣基糖。存在大量教示此類經修飾之糖結構之製備且詳述及描述一系列鹼基修飾的美國專利,諸如美國專利第4,981,957號;第5,118,800號;第5,319,080號;第5,359,044號;第5,393,878號;第5,446,137號;第5,466,786號;第5,514,785號;第5,519,134號;第5,567,811號;第5,576,427號;第5,591,722號;第5,597,909號;第5,610,300號;第5,627,053號;第5,639,873號;第5,646,265號;第5,658,873號;第5,670,633號;第4,845,205號;第5,130,302號;第5,134,066號;第5,175,273號;第5,367,066號;第5,432,272號;第5,457,187號;第5,459,255號;第5,484,908號;第5,502,177號;第5,525,711號;第5,552,540號;第5,587,469號;第5,594,121號;第5,596,091號;第5,614,617號;第5,681,941號及第5,700,920號,該等專利中之每一者均以全文引用之方式併入本文中。
具有經修飾之糖部分之核酸的實例包括(但不限於)包含5'-乙烯基、5'-甲基(R或S)、4'-S、2'-F、2'-OCH3
及2'-O(CH2
)2
OCH3
取代基之核酸。2'位置處之取代基亦可選自烯丙基、胺基、疊氮基、硫基、O-烯丙基、O-(C1
-C10
烷基)、OCF3
、O(CH2
)2
SCH3
、O(CH2
)2
-O-N(Rm
)(Rn
)及O-CH2
-C(=O)-N(Rm
)(Rn
),其中各Rm
及Rn
獨立地為H或經取代或未經取代之C1
-C10
烷基。
在某些實施例中,本文所描述之核酸包括一或多種雙環核酸。在某些此類實施例中,雙環核酸包含在4'核糖基環原子與2'核糖基環原子之間的橋接。在某些實施例中,本文所提供之核酸包括一或多種雙環核酸,其中橋接包含4'至2'雙環核酸。此類4'至2'雙環核酸之實例包括(但不限於)以下式中之一者:4'-(CH2
)-O-2' (LNA);4'-(CH2
)-S-2';4'-(CH2
)2
-O-2' (ENA);4'-CH(CH3
)-O-2'及4'-CH(CH2
OCH3
)-O-2',以及其類似物(參見美國專利第7,399,845號);4'-C(CH3
)(CH3
)-O-2'及其類似物(參見WO2009/006478;WO2008/150729;US2004/0171570;美國專利第7,427,672號;Chattopadhyaya等人, J. Org. Chem., 209, 74, 118-134及WO2008/154401)。亦參見例如:Singh等人, Chem. Commun., 1998, 4, 455-456;Koshkin等人, Tetrahedron, 1998, 54, 3607-3630;Wahlestedt等人, Proc. Natl. Acad. Sci. U. S. A., 2000, 97, 5633-5638;Kumar等人, Bioorg. Med. Chem. Lett., 1998, 8, 2219-2222;Singh等人, J. Org. Chem., 1998, 63, 10035-10039;Srivastava等人, J. Am. Chem. Soc., 2007, 129(26) 8362-8379;Elayadi等人, Curr. Opinion Invens. Drugs, 2001, 2, 558-561;Braasch等人, Chem. Biol, 2001, 8, 1-7;Oram等人, Curr. Opinion Mol. Ther., 2001, 3, 239-243;美國專利第4,849,513號;第5,015,733號;第5,118,800號;第5,118,802號;第7,053,207號;第6,268,490號;第6,770,748號;第6,794,499號;第7,034,133號;第6,525,191號;第6,670,461號;及第7,399,845號;國際公開案第WO2004/106356號、第WO1994/14226號、第WO2005/021570號、第WO2007/090071號及第WO2007/134181號;美國專利公開案第US2004/0171570號、第US2007/0287831號及第US2008/0039618號;美國臨時申請案第60/989,574號、第61/026,995號、第61/026,998號、第61/056,564號、第61/086,231號、第61/097,787號及第61/099,844號;及國際申請案第PCT/US2008/064591號、第PCT US2008/066154號、第PCT US2008/068922號及第PCT/DK98/00393號。
在某些實施例中,核酸包含連接之核酸。核酸可使用任何核酸間鍵連接在一起。兩個主要類別之核酸間連接基團係由存在或不存在磷原子來定義。代表性含磷核酸間鍵包括(但不限於):磷酸二酯、磷酸三酯、甲基磷酸酯、胺基磷酸酯及硫代磷酸酯(P=S)。代表性不含磷核酸間連接基團包括(但不限於):亞甲基甲基亞胺基(-CH2
-N(CH3
)-O-CH2
-)、硫代二酯(-O-C(O)-S-)、硫羰基胺基甲酸酯(-O-C(O)(NH)-S-);矽氧烷(-O-Si(H)2-
O-);及N,N*-二甲基肼(-CH2
-N(CH3
)-N(CH3
))。在某些實施例中,具有對掌性原子之核酸間鍵可製備為外消旋混合物、個別對映異構體(例如烷基膦酸酯及硫代磷酸酯)。非天然核酸可含有單一修飾。非天然核酸可含有在該等部分中之一者內或在不同部分之間的多個修飾。
對核酸之主鏈磷酸酯修飾包括(但不限於):甲基膦酸酯、硫代磷酸酯、胺基磷酸酯(橋接或非橋接)、磷酸三酯、二硫代磷酸酯(phosphorodithioate/phosphodithioate)及硼烷磷酸酯,且可以任何組合形式使用。亦可使用其他非磷酸酯鍵。
在一些實施例中,主鏈修飾(例如,膦酸甲酯、硫代磷酸酯、磷醯胺酸及二硫代磷酸酯核苷酸間鍵)可對經修飾之核酸賦予免疫調節活性及/或增強其活體內穩定性。
在一些情況下,磷衍生物(或經修飾之磷酸酯基)在以下中附接至糖或糖類似物部分且可為以下:單磷酸酯、二磷酸酯、三磷酸酯、膦酸烷基酯、硫代磷酸酯、二硫代磷酸酯、胺基磷酸酯或其類似物。含有經修飾之磷酸酯鍵或非磷酸酯鍵之例示性聚核苷酸可見於Peyrottes等人, 1996, Nucleic Acids Res. 24: 1841-1848;Chaturvedi等人, 1996, Nucleic Acids Res. 24:2318-2323;以及Schultz等人, (1996) Nucleic Acids Res. 24:2966-2973;Matteucci, 1997, Oligonucleotides as Therapeutic Agents中之「Oligonucleotide Analogs: an Overview」, (Chadwick及Cardew編) John Wiley and Sons, New York, NY;Zon, 1993, Protocols for Oligonucleotides and Analogs, Synthesis and Properties中之「Oligonucleoside Phosphorothioates」, Humana出版社, 第165-190頁;Miller等人, 1971, JACS 93:6657-6665;Jager等人, 1988, Biochem. 27:7247-7246;Nelson等人, 1997, JOC 62:7278-7287;美國專利第5,453,496號;以及Micklefield, 2001, Curr. Med. Chem. 8: 1157-1179中。
在一些情況下,主鏈修飾包含用替代部分(諸如陰離子、中性或陽離子基團)替代磷酸二酯鍵。此類修飾之實例包括:陰離子核苷間鍵;N3'至P5'胺基磷酸酯修飾;硼烷磷酸酯DNA;促寡核苷酸;中性核苷間鍵,諸如甲基膦酸酯;醯胺連接之DNA;亞甲基(甲基亞胺基)鍵;甲縮醛及硫代甲縮醛鍵;含有磺醯基之主鏈;(N-嗎啉基)寡聚物;肽核酸(PNA);以及帶正電去氧核糖核酸胍(DNG)寡聚物(Micklefield, 2001, Current Medicinal Chemistry 8: 1157-1179)。經修飾之核酸可包含嵌合或混合主鏈,其包含一或多個修飾,例如磷酸酯鍵之組合,諸如磷酸二酯鍵與硫代磷酸酯鍵之組合。
用於磷酸酯之取代物包括例如短鏈烷基或環烷基核苷間鍵、混合雜原子及烷基或環烷基核苷間鍵或一或多個短鏈雜原子或雜環核苷間鍵。此等物包括具有以下之彼等物:N-嗎啉基鍵(部分地由核苷之糖部分形成);矽氧烷主鏈;硫化物、亞碸及碸主鏈;甲醯基及硫代甲醯基主鏈;亞甲基甲醯基及硫代甲醯基主鏈;含有烯烴之主鏈;胺基磺酸酯主鏈;亞甲基亞胺基及亞甲基肼基主鏈;磺酸酯及磺醯胺主鏈;醯胺主鏈;以及具有混合N、O、S及CH2
組成部分之其他者。大量美國專利揭示如何製備及使用此等類型之磷酸酯替代物且包括(但不限於):美國專利第5,034,506號;第5,166,315號;第5,185,444號;第5,214,134號;第5,216,141號;第5,235,033號;第5,264,562號;第5,264,564號;第5,405,938號;第5,434,257號;第5,466,677號;第5,470,967號;第5,489,677號;第5,541,307號;第5,561,225號;第5,596,086號;第5,602,240號;第5,610,289號;第5,602,240號;第5,608,046號;第5,610,289號;第5,618,704號;第5,623,070號;第5,663,312號;第5,633,360號;第5,677,437號;以及第5,677,439號。亦應理解,在核苷酸取代物中,核苷酸之糖及磷酸酯部分兩者均可經例如醯胺類鍵(胺基乙基甘胺酸) (PNA)置換。美國專利第5,539,082號、第5,714,331號;以及第5,719,262號教示如何製備及使用PNA分子,該等專利中之每一者均以引用之方式併入本文中。亦參見Nielsen等人, Science, 1991, 254, 1497-1500。亦有可能將其他類型之分子(結合物)連接至核苷酸或核苷酸類似物以增強例如細胞吸收。結合物可化學連接至核苷酸或核苷酸類似物。此類結合物包括(但不限於):脂質部分,諸如膽固醇部分(Letsinger等人, Proc. Natl. Acad. Sci. USA, 1989, 86, 6553-6556);膽酸(Manoharan等人, Bioorg. Med. Chem. Let., 1994, 4, 1053-1060);硫醚,例如己基-S-三苯甲基硫醇(Manoharan等人, Ann. KY. Acad. Sci., 1992, 660, 306-309;Manoharan等人, Bioorg. Med. Chem. Let., 1993, 3, 2765-2770);硫代膽固醇(Oberhauser等人, Nucl. Acids Res., 1992, 20, 533-538);脂族鏈,例如十二烷二醇或十一基殘基(Saison-Behmoaras等人, EM5OJ, 1991, 10, 1111-1118;Kabanov等人, FEBS Lett., 1990, 259, 327-330;Svinarchuk等人, Biochimie, 1993, 75, 49-54);磷脂,例如二-十六基-外消旋-甘油或三乙銨l-二-O-十六基-外消旋-甘油基-S-H-膦酸酯(Manoharan等人, Tetrahedron Lett., 1995, 36, 3651-3654;Shea等人, Nucl. Acids Res., 1990, 18, 3777-3783);多元胺或聚乙二醇鏈(Manoharan等人, Nucleosides & Nucleotides, 1995, 14, 969-973);或金剛烷乙酸(Manoharan等人, Tetrahedron Lett., 1995, 36, 3651-3654);軟脂基部分(Mishra等人, Biochem. Biophys. Acta, 1995, 1264, 229-237);或十八基胺或己基胺基-羰基-羥膽固醇部分(Crooke等人, J. Pharmacol. Exp. Ther., 1996, 277, 923-937)。大量美國專利教示此類結合物之製備且包括(但不限於):美國專利第4,828,979號;第4,948,882號;第5,218,105號;第5,525,465號;第5,541,313號;第5,545,730號;第5,552,538號;第5,578,717號、第5,580,731號;第5,580,731號;第5,591,584號;第5,109,124號;第5,118,802號;第5,138,045號;第5,414,077號;第5,486,603號;第5,512,439號;第5,578,718號;第5,608,046號;第4,587,044號;第4,605,735號;第4,667,025號;第4,762,779號;第4,789,737號;第4,824,941號;第4,835,263號;第4,876,335號;第4,904,582號;第4,958,013號;第5,082,830號;第5,112,963號;第5,214,136號;第5,082,830號;第5,112,963號;第5,214,136號;第5,245,022號;第5,254,469號;第5,258,506號;第5,262,536號;第5,272,250號;第5,292,873號;第5,317,098號;第5,371,241號、第5,391,723號;第5,416,203號、第5,451,463號;第5,510,475號;第5,512,667號;第5,514,785號;第5,565,552號;第5,567,810號;第5,574,142號;第5,585,481號;第5,587,371號;第5,595,726號;第5,597,696號;第5,599,923號;第5,599,928號;以及第5,688,941號。
本文描述用於複製、轉錄、轉譯及將非天然胺基酸併入蛋白質中之組合物及方法中所使用之核鹼基。在一些實施例中,本文所描述之核鹼基包含以下結構: ,其中各X獨立地為碳或氮;R2
為視情況存在的且若存在,則獨立地為氫、烷基、烯基、炔基;甲氧基、甲硫醇、甲烷硒基、鹵素、氰基或疊氮基;其中各Y獨立地為硫、氧、硒或二級胺;其中各E獨立地為氧、硫或硒;且其中波浪線指示鍵結至核糖基、去氧核糖基或二去氧核糖基部分或其類似者之點,其中核糖基、去氧核糖基或二去氧核糖基部分或其類似者呈游離形式,連接至單-磷酸酯基、二磷酸酯基或三磷酸酯基,視情況包含α-硫代三磷酸酯基、β-硫代三磷酸酯基或γ-硫代三磷酸酯基,或包括於RNA或DNA中或包括於RNA類似物或DNA類似物中。在一些實施例中,R2
為低碳數烷基(例如C1
-C6
)、氫或鹵素。在本文所描述之核鹼基之一些實施例中,R2
為氟。在本文所描述之核鹼基之一些實施例中,X為碳。在本文所描述之核鹼基之一些實施例中,E為硫。在本文所描述之核鹼基之一些實施例中,Y為硫。在本文所描述之核鹼基之一些實施例中,核鹼基具有以下結構:
,其中各X獨立地為碳或氮;R2
為視情況存在的且若存在,則獨立地為氫、烷基、烯基、炔基;甲氧基、甲硫醇、甲烷硒基、鹵素、氰基或疊氮基;其中各Y獨立地為硫、氧、硒或二級胺;其中各E獨立地為氧、硫或硒;且其中波浪線指示鍵結至核糖基、去氧核糖基或二去氧核糖基部分或其類似者之點,其中核糖基、去氧核糖基或二去氧核糖基部分或其類似者呈游離形式,連接至單-磷酸酯基、二磷酸酯基或三磷酸酯基,視情況包含α-硫代三磷酸酯基、β-硫代三磷酸酯基或γ-硫代三磷酸酯基,或包括於RNA或DNA中或包括於RNA類似物或DNA類似物中。在一些實施例中,R2
為低碳數烷基(例如C1
-C6
)、氫或鹵素。在本文所描述之核鹼基之一些實施例中,R2
為氟。在本文所描述之核鹼基之一些實施例中,X為碳。在本文所描述之核鹼基之一些實施例中,E為硫。在本文所描述之核鹼基之一些實施例中,Y為硫。在本文所描述之核鹼基之一些實施例中,核鹼基具有以下結構: 。在本文所描述之核鹼基之一些實施例中,E為硫且Y為硫。在本文所描述之核鹼基之一些實施例中,波浪線指示鍵結至核糖基或去氧核糖基部分之點。在本文所描述之核鹼基之一些實施例中,波浪線指示鍵結至連接至三磷酸酯基之核糖基或去氧核糖基部分之點。在本文所描述之核鹼基之一些實施例中為核酸聚合物之組分。在本文所描述之核鹼基之一些實施例中,核鹼基為tRNA之組分。在本文所描述之核鹼基之一些實施例中,核鹼基為tRNA中之反密碼子之組分。在本文所描述之核鹼基之一些實施例中,核鹼基為mRNA之組分。在本文所描述之核鹼基之一些實施例中,核鹼基為mRNA之密碼子之組分。在本文所描述之核鹼基之一些實施例中,核鹼基為RNA或DNA之組分。在本文所描述之核鹼基之一些實施例中,核鹼基為DNA中之密碼子之組分。在本文所描述之核鹼基之一些實施例中,核鹼基與另一互補核鹼基形成核鹼基對。核酸鹼基配對性質
。在本文所描述之核鹼基之一些實施例中,E為硫且Y為硫。在本文所描述之核鹼基之一些實施例中,波浪線指示鍵結至核糖基或去氧核糖基部分之點。在本文所描述之核鹼基之一些實施例中,波浪線指示鍵結至連接至三磷酸酯基之核糖基或去氧核糖基部分之點。在本文所描述之核鹼基之一些實施例中為核酸聚合物之組分。在本文所描述之核鹼基之一些實施例中,核鹼基為tRNA之組分。在本文所描述之核鹼基之一些實施例中,核鹼基為tRNA中之反密碼子之組分。在本文所描述之核鹼基之一些實施例中,核鹼基為mRNA之組分。在本文所描述之核鹼基之一些實施例中,核鹼基為mRNA之密碼子之組分。在本文所描述之核鹼基之一些實施例中,核鹼基為RNA或DNA之組分。在本文所描述之核鹼基之一些實施例中,核鹼基為DNA中之密碼子之組分。在本文所描述之核鹼基之一些實施例中,核鹼基與另一互補核鹼基形成核鹼基對。核酸鹼基配對性質
在一些實施例中,非天然核苷酸在併入DNA或RNA中期間或之後與另一非天然核苷酸形成鹼基對(非天然鹼基對;UBP)。在一些實施例中,經穩定整合之非天然核酸為可與另一核酸(例如天然或非天然核酸)形成鹼基對之非天然核酸。在一些實施例中,經穩定整合之非天然核酸為可與另一非天然核酸形成鹼基對(非天然核酸鹼基對(UBP))之非天然核酸。舉例而言,第一非天然核酸可與第二非天然核酸形成鹼基對。舉例而言,可在併入核酸期間及之後鹼基配對之一對非天然核苷三磷酸包括(d)5SICS之三磷酸((d)5SICSTP)及(d)NaM之三磷酸((d)NaMTP)。其他實例包括(但不限於):(d)CNMO之三磷酸((d)CNMOTP)及(d)TPT3之三磷酸((d)TPT3TP)。此類非天然核苷酸可具有核糖或去氧核糖糖部分(由「(d)」指示)。舉例而言,可在併入核酸中時鹼基配對之一對非天然核苷三磷酸包括TAT1之三磷酸(TAT1TP)及NaM之三磷酸(NaMTP)。在一些實施例中,可在併入核酸中時鹼基配對之一對非天然核苷三磷酸包括dCNMO之三磷酸(dCNMOTP)及TAT1之三磷酸(TAT1TP)。在一些實施例中,可在併入核酸中時鹼基配對之一對非天然核苷三磷酸包括dTPT3之三磷酸(dTPT3TP)及NaM之三磷酸(NaMTP)。在一些實施例中,非天然核酸不實質上與天然核酸(A、T、G、C)形成鹼基對。在一些實施例中,經穩定整合之非天然核酸可與天然核酸形成鹼基對。
在一些實施例中,經穩定整合之非天然(去氧)核糖核苷酸為可形成UBP但不與任一天然(去氧)核糖核苷酸實質上形成鹼基對之非天然(去氧)核糖核苷酸。在一些實施例中,經穩定整合之非天然(去氧)核糖核苷酸為可形成UBP但不與一或多種天然核酸實質上形成鹼基對之非天然(去氧)核糖核苷酸。舉例而言,經穩定整合之非天然核酸可能不與A、T及C實質上形成鹼基對,但可與G形成鹼基對。舉例而言,經穩定整合之非天然核酸可能不與A、T及G實質上形成鹼基對,但可與C形成鹼基對。舉例而言,經穩定整合之非天然核酸可能不與C、G及A實質上形成鹼基對,但可與T形成鹼基對。舉例而言,經穩定整合之非天然核酸可能不與C、G及T實質上形成鹼基對,但可與A形成鹼基對。舉例而言,經穩定整合之非天然核酸可能不與A及T實質上形成鹼基對,但可與C及G形成鹼基對。舉例而言,經穩定整合之非天然核酸可能不與A及C實質上形成鹼基對,但可與T及G形成鹼基對。舉例而言,經穩定整合之非天然核酸可能不與A及G實質上形成鹼基對,但可與C及T形成鹼基對。舉例而言,經穩定整合之非天然核酸可能不與C及T實質上形成鹼基對,但可與A及G形成鹼基對。舉例而言,經穩定整合之非天然核酸可能不與C及G實質上形成鹼基對,但可與T及G形成鹼基對。舉例而言,經穩定整合之非天然核酸可能不與T及G實質上形成鹼基對,但可與A及G形成鹼基對。舉例而言,經穩定整合之非天然核酸可能不與G實質上形成鹼基對,但可與A、T及C形成鹼基對。舉例而言,經穩定整合之非天然核酸可能不與A實質上形成鹼基對,但可與G、T及C形成鹼基對。舉例而言,經穩定整合之非天然核酸可能不與T實質上形成鹼基對,但可與G、A及C形成鹼基對。舉例而言,經穩定整合之非天然核酸可能不與C實質上形成鹼基對,但可與G、T及A形成鹼基對。
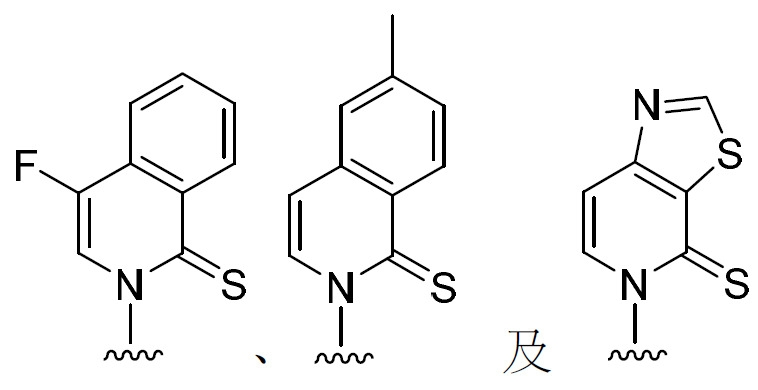
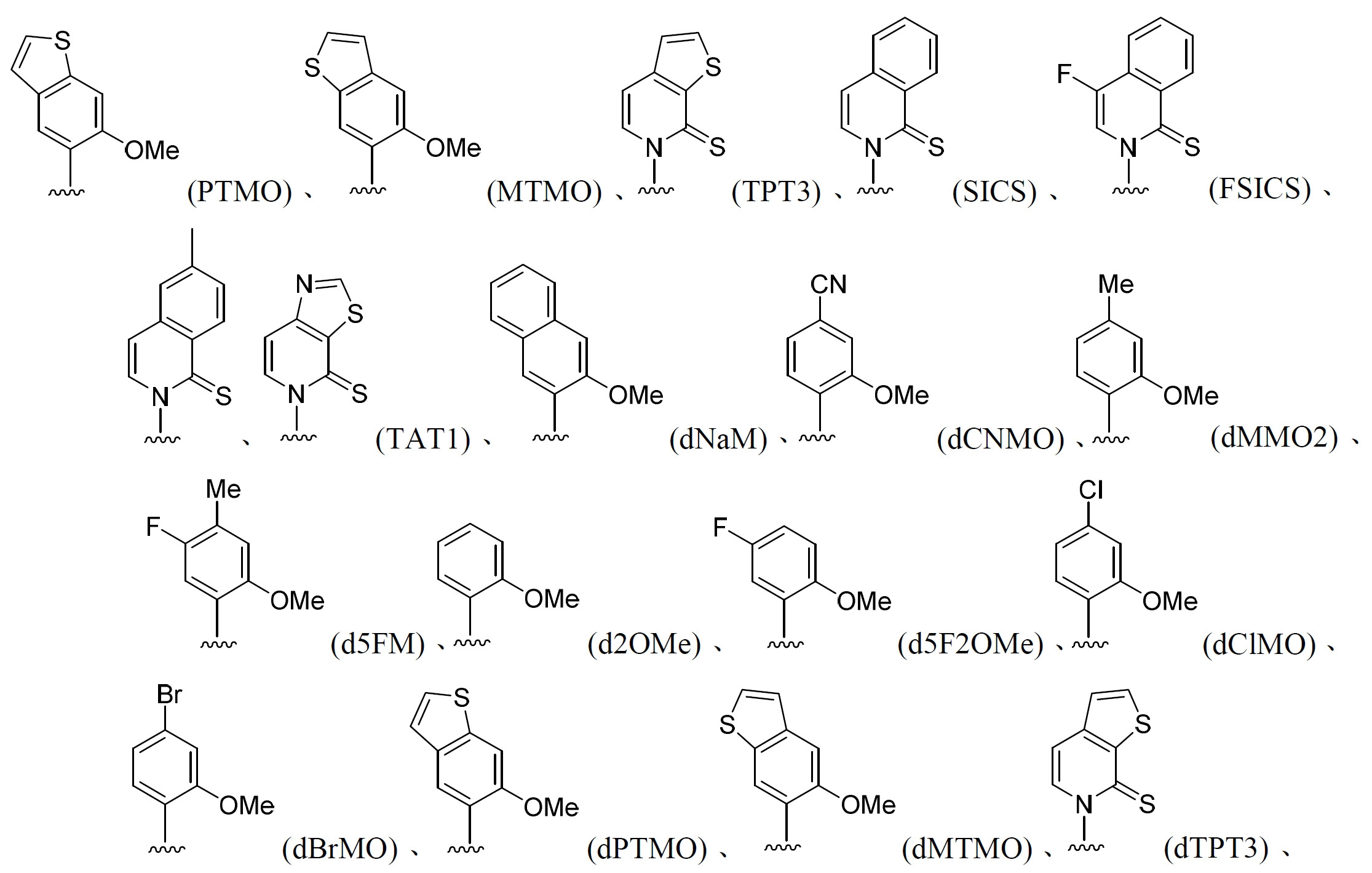
能夠在活體內條件下形成非天然DNA或RNA鹼基對(UBP)之例示性非天然核苷酸包括(但不限於):5SICS、d5SICS、NaM、dNaM、dTPT3、dMTMO、dCNMO、TAT1及其組合。在一些實施例中,非天然核苷酸鹼基對包括(但不限於): 。經工程改造之 生物體
。經工程改造之 生物體
在一些實施例中,本文所揭示之方法及質體進一步用於產生經工程改造之生物體,例如併入及複製非天然核苷酸或非天然核酸鹼基對(UBP)之生物體,且亦可使用含有非天然核苷酸之核酸來轉錄用於轉譯非天然多肽或含有至少一個非天然胺基酸殘基之非天然蛋白質之mRNA及tRNA。在一些情況下,非天然胺基酸殘基以位點特異性方式併入非天然多肽或非天然蛋白質中。在一些情況下,生物體為非人類半合成生物體(SSO)。在一些情況下,生物體為半合成生物體(SSO)。在一些情況下,SSO為細胞。在一些情況下,活體內方法包含半合成生物體(SSO)。在一些情況下,半合成生物體包含微生物。在一些情況下,生物體包含細菌。在一些情況下,生物體包含革蘭氏陰性細菌。在一些情況下,生物體包含革蘭氏陽性細菌。在一些情況下,生物體包含大腸桿菌。此類經修飾之生物體以各種方式包含額外組分,諸如DNA修復機制、經修飾之聚合酶、核苷酸轉運子或其他組分。在一些情況下,SSO包含大腸桿菌菌株YZ3。在一些情況下,SSO包含大腸桿菌菌株ML1或ML2,諸如Ledbetter等人.J. Am Chem. Soc.
2018, 140(2), 758之圖1(B-D)中所描述之彼等菌株。在一些情況下,SSO為細胞株。在一些情況下,細胞株為永生化細胞株。在一些情況下,細胞株包含初級細胞。在一些情況下,細胞株包含幹細胞。在一些情況下,SSO為類器官的。
在一些情況下,所採用之細胞用編碼異源蛋白質之表現卡匣(例如能夠將非天然核苷三磷酸轉運至細胞中之核苷三磷酸轉運子)及視情況選用之CRISPR/Cas9系統基因轉化以消除已喪失非天然核苷酸之DNA (例如大腸桿菌菌株YZ3、ML1或ML2)。在一些情況下,細胞進一步包含對於非天然核酸吸收之增強的活性。在一些情況下,細胞進一步包含對於非天然核酸導入之增強的活性。
在一些實施例中,將Cas9及適當的嚮導RNA (sgRNA)編碼於獨立質體上。在一些情況下,Cas9及sgRNA編碼於相同質體上。在一些情況下,編碼Cas9、sgRNA之核酸分子或包含非天然核苷酸之核酸分子位於一或多個質體上。在一些情況下,Cas9編碼於第一質體上且sgRNA及包含非天然核苷酸之核酸分子編碼於第二質體上。在一些情況下,Cas9、sgRNA及包含非天然核苷酸之核酸分子編碼於相同質體上。在一些情況下,核酸分子包含兩個或更多個非天然核苷酸。在一些情況下,將Cas9併入宿主生物體之基因體中且sgRNA編碼於質體上或生物體之基因體中。
在一些情況下,將編碼Cas9及sgRNA之第一質體及編碼包含非天然核苷酸之核酸分子之第二質體引入經工程改造之微生物中。在一些情況下,將編碼Cas9之第一質體及編碼sgRNA及包含非天然核苷酸之核酸分子之第二質體引入經工程改造之微生物中。在一些情況下,將編碼Cas9、sgRNA及包含非天然核苷酸之核酸分子之質體引入經工程改造之微生物中。在一些情況下,核酸分子包含兩個或更多個非天然核苷酸。
在一些實施例中,產生在其DNA (質體或基因體)內併有包含至少一個非天然鹼基對(UBP)之至少一個非天然核酸分子之活細胞。在一些情況下,至少一個非天然核酸分子包含一個、兩個、三個、四個或更多個UBP。在一些情況下,至少一個非天然核酸分子為質體。在一些情況下,將至少一個非天然核酸分子整合至細胞之基因體中。在一些實施例中,至少一個非天然核酸分子編碼非天然多肽或非天然蛋白質。在一些情況下,至少一個非天然核酸分子經轉錄以得到mRNA之非天然密碼子及tRNA之非天然反密碼子。在一些實施例中,至少一個非天然核酸分子為非天然DNA分子。
在一些情況下,非天然鹼基對包括一對非天然彼此鹼基配對之核苷酸,其能夠在非天然彼此鹼基配對之核苷酸(呈其各別三磷酸酯形式)藉由核苷酸三磷酸酯轉運子之作用吸收至細胞中時在活體內條件下形成非天然鹼基對。細胞可藉由編碼核苷酸三磷酸酯轉運子之表現卡匣經基因轉化,以使得核苷酸三磷酸酯轉運子經表現且可用於將非天然核苷酸轉運至細胞中。細胞可為原核或真核細胞,且一對非天然彼此鹼基配對之核苷酸(呈其各別三磷酸酯形式)可為dTPT3 (dTP3TP)之三磷酸酯及dNaM (dNaMTP)或dCNMO (dCNMOTP)之三磷酸酯。
在一些實施例中,細胞為具有核酸之經基因轉化之細胞,例如為編碼能夠將此類非天然核苷酸轉運至細胞中之核苷酸三磷酸酯轉運子之表現卡匣。細胞可包含異源核苷三磷酸轉運子,其中異源核苷三磷酸轉運子可將天然及非天然核苷三磷酸轉運至細胞中。
在一些情況下,本文所描述之方法亦包括在磷酸鉀及/或磷酸酶或核苷酸酶之抑制劑存在下使經基因轉化之細胞與各別三磷酸酯接觸。在此類接觸期間或之後,可將細胞置放於適用於細胞生長及複製之維持生命的培養基內。可將細胞維持在維持生命的培養基中,以使得非天然核苷酸之各別三磷酸酯形式併入細胞內之核酸中,且通過細胞之至少一個複製週期。一對非天然彼此鹼基配對之核苷酸(呈各別三磷酸酯形式)可包含dTPT3或(dTPT3TP)之三磷酸酯及dCNMO或dNaM (dCNOM或dNaMTP)之三磷酸酯,細胞可為大腸桿菌,且dTPT3TP及dNaMTP可藉由轉運子Pt
NTT2導入大腸桿菌中,其中大腸桿菌聚合酶(諸如Pol III或Pol II)可使用非天然三磷酸酯以複製含有UBP之DNA,從而將非天然核苷酸及/或非天然鹼基對併入細胞環境內之細胞核酸中。另外,在一些情況下,諸如NaMTP及TAT1TP、5FMTP及TPT3TP之核糖核苷酸藉由轉運子Pt
NTT2導入大腸桿菌中。在一些情況下,用於導入核糖核苷酸之Pt
NTT2為截短Pt
NTT2,其中截短Pt
NTT2具有與未截短Pt
NTT2之胺基酸序列至少60%、至少65%、至少70%、至少75%、至少80%、至少85%或至少90%一致之胺基酸序列。未截短Pt
NTT2之實例(NCBI寄存編號EEC49227.1,GI:217409295)具有胺基酸序列(SEQ ID NO: 1):

本文描述包含使用三個或更多個非天然鹼基配對之核苷酸的組合物及方法。在一些情況下,此類鹼基配對之核苷酸係經由使用核苷酸轉運子或經由此項技術中已知之標準核酸轉化方法(例如電穿孔、化學轉化或其他方法)進入細胞。在一些情況下,鹼基配對之非天然核苷酸作為聚核苷酸之一部分(諸如質體)進入細胞。作為聚核苷酸之一部分(RNA或DNA)進入細胞之一或多個鹼基配對之非天然核苷酸自身無需活體內複製。舉例而言,將雙股DNA質體或包含第一非天然去氧核糖核苷酸及具有經組態以形成第一非天然鹼基對之鹼基的第二非天然去氧核糖核苷酸之其他核酸電穿孔至細胞中。用第三非天然去氧核糖核苷酸、具有經組態以彼此形成第二非天然鹼基對之鹼基的第四非天然去氧核糖核苷酸處理細胞培養基,其中第一非天然去氧核糖核苷酸之鹼基與第三非天然去氧核糖核苷酸之鹼基形成第二非天然鹼基對,且其中第二非天然去氧核糖核苷酸之鹼基與第四非天然去氧核糖核苷酸鹼基形成第三非天然鹼基對。在一些情況下,最初轉化之雙股DNA質體的活體內複製產生包含第三非天然去氧核糖核苷酸及第四非天然去氧核糖核苷酸之後續複製之質體。替代或組合地,將第三非天然去氧核糖核苷酸及第四非天然去氧核糖核苷酸之核糖核苷酸變異體添加至細胞培養基中。在一些情況下,將此等核糖核苷酸併入RNA,諸如mRNA或tRNA中。在一些情況下,第一、第二、第三及第四去氧核苷酸包含不同的鹼基。在一些情況下,第一、第三及第四去氧核苷酸包含不同的鹼基。在一些情況下,第一及第三去氧核苷酸包含相同的鹼基。
藉由本發明之方法之實踐,一般技術者可獲得活的且繁殖的細胞群,該細胞群在維持於個別細胞中之至少一些內之至少一個核酸中具有至少一個非天然核苷酸及/或至少一個非天然鹼基對(UBP),其中至少一個核酸在細胞內經穩定繁殖,且其中細胞在與(例如,在以下存在下生長)適用於生物體生長及複製之維持生命的培養基中之一或多個非天然核苷酸接觸時,表現適用於提供一或多個非天然核苷酸之三磷酸酯形式之細胞吸收的核苷酸三磷酸酯轉運子。
在藉由核苷酸三磷酸酯轉運子轉運至細胞中之後,非天然鹼基配對之核苷酸藉由細胞機制併入細胞內之核酸中,該細胞機制例如細胞之自身DNA及/或RNA聚合酶、異源聚合酶或已使用定向進化而進化之聚合酶(Chen T, Romesberg FE, FEBS Lett. 2014年1月21日;588(2):219-29;Betz K等人, J Am Chem Soc. 2013年12月11日;135(49):18637-43)。非天然核苷酸可併入細胞核酸中,該等核酸諸如基因體DNA、基因體RNA、mRNA、tRNA、結構RNA、微小RNA及自主複製核酸(例如,質體、病毒或載體)。
在一些情況下,經基因工程改造之細胞係藉由將核酸(例如異源核酸)引入細胞中而產生。在一些情況下,引入細胞中之核酸呈質體形式。在一些情況下,將引入細胞中之核酸整合至細胞之基因體中。本文所描述之任何細胞均可為宿主細胞且可包含表現載體。在一個實施例中,宿主細胞為原核細胞。在另一實施例中,宿主細胞為大腸桿菌。在一些實施例中,細胞包含一或多個異源聚核苷酸。可使用各種技術將核酸試劑引入微生物中。用於將異源核酸引入各種生物體中之方法之非限制性實例包括:轉化、轉染、轉導、電穿孔、超音波介導之轉化、結合、粒子轟擊及其類似方法。在一些情況下,添加載體分子(例如,雙-苯并咪唑基化合物,例如參見美國專利第5,595,899號)可增加細胞中之DNA吸收,但其藉由習知方法通常難以轉化。習知轉化方法為技工容易地可用的,且可見於Maniatis, T., E. F. Fritsch及J. Sambrook (1982) Molecular Cloning: a Laboratory Manual;Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.中。
在一些情況下,遺傳轉化係使用在質體、病毒載體、病毒核酸、噬菌體核酸、噬菌體、黏質體及人工染色體(但不限於以上各者)中之表現卡匣直接轉移,或經由轉移細胞中之基因物質或載體(諸如陽離子脂質體)獲得。此類方法在此項技術中為可用的且容易地適用於本文所描述之方法中。轉移載體可為用於將基因遞送至細胞(例如質體)中之任何核苷酸構築體,或作為用以遞送基因之通用策略之一部分,例如作為重組逆轉錄病毒或腺病毒之一部分(Ram等人 Cancer Res. 53:83-88, (1993))。適當的轉染方式,包括病毒載體、化學轉染物或物理機械方法(諸如電穿孔及DNA之直接擴散)係由例如Wolff, J. A.等人, Science, 247, 1465-1468, (1990);及Wolff, J. A. Nature, 352, 815-818, (1991)描述。
舉例而言,可藉由任何方法將編碼核苷三磷酸轉運子或聚合酶表現卡匣及/或載體之DNA引入細胞中,該方法包括(但不限於):鈣介導之轉化、電穿孔、顯微注射、脂質體轉染、粒子轟擊及其類似方法。
在一些情況下,細胞包含併入細胞內之一或多個核酸中之非天然核苷三磷酸。舉例而言,細胞可為活細胞,其能夠將至少一個非天然核苷酸併入維持於細胞內之DNA或RNA內。細胞亦可在活體內條件下將至少一個包含一對非天然彼此鹼基配對之核苷酸之非天然鹼基對(UBP)併入細胞內之核酸中,其中非天然彼此鹼基配對之核苷酸(例如其各別三磷酸酯)係藉由核苷三磷酸轉運子之作用吸收至細胞中,其基因係藉由基因轉化呈現(例如引入)至細胞中。舉例而言,在併入維持於細胞內之核酸中後,dTPT3及dCNMO可形成穩定的非天然鹼基對,其可藉由生物體之DNA複製機制穩定繁殖,例如當在包含dTPT3TP及dCNMOTP之維持生命的培養基中生長時。
在一些情況下,細胞能夠複製含有非天然核苷酸之核酸。此類方法可包括用編碼核苷三磷酸轉運子之表現卡匣基因轉化之細胞,該核苷三磷酸轉運子能夠在活體內條件下將一或多個非天然核苷酸(其呈各別三磷酸酯形式)轉運至細胞中。替代地,可使用先前已用可表現經編碼之核苷三磷酸轉運子之表現卡匣經基因轉化的細胞。方法亦可包括在活體內條件下,將經基因轉化之細胞在適用於細胞生長及複製之維持生命的培養基中與磷酸鉀及至少一個非天然核苷酸(例如,能夠形成非天然鹼基對(UBP)之兩個彼此鹼基配對之核苷酸)之各別三磷酸酯形式接觸或暴露於磷酸鉀及至少一個非天然核苷酸之各別三磷酸酯形式,並且在至少一個非天然核苷酸(例如,能夠形成非天然鹼基對(UBP)之兩個彼此鹼基配對之核苷酸)之各別三磷酸酯形式存在下,將經轉化之細胞維持於維持生命的培養基中,通過至少一個細胞複製週期。
在一些實施例中,細胞包含穩定併入之非天然核酸。一些實施例包含細胞(例如呈大腸桿菌形式),其將除A、G、T及C以外的核苷酸穩定併入維持於細胞內之核酸內。舉例而言,除A、G、T及C以外的核苷酸可為d5SICS、dCNMO、dNaM及/或dTPT3,其在併入細胞之核酸中後,可在核酸內形成穩定的非天然鹼基對。在一個態樣中,當藉由用於三磷酸酯轉運子之基因轉化之生物體在包括磷酸鉀及d5SICS、dNaM、dCNMO及/或dTPT3之三磷酸酯之維持生命的培養基中生長時,非天然核苷酸及非天然鹼基對可藉由生物體之複製裝置穩定繁殖。
在一些情況下,細胞包含擴展遺傳字母表。細胞可包含經穩定併入之非天然核酸。在一些實施例中,具有擴展遺傳字母表之細胞包含含有可與另一非天然核苷酸配對之非天然核苷酸的非天然核酸。在一些實施例中,具有擴展遺傳字母表之細胞包含與另一核酸氫鍵結之非天然核酸。在一些實施例中,具有擴展遺傳字母表之細胞包含不與其所鹼基配對之另一核酸氫鍵結的非天然核酸。在一些實施例中,具有擴展遺傳字母表之細胞包含非天然核酸,其含有具有經由疏水性及/或填充相互作用與核鹼基或另一非天然核苷酸鹼基配對之核鹼基的非天然核苷酸。在一些實施例中,具有擴展遺傳字母表之細胞包含經由非氫鍵結相互作用與另一核酸鹼基配對之非天然核酸。具有擴展遺傳字母表之細胞可為複製同源核酸以形成包含非天然核酸之核酸的細胞。具有擴展遺傳字母表之細胞可為包含與另一非天然核酸配對(非天然核酸鹼基對(UBP))之非天然核酸鹼基之細胞。
在一些實施例中,細胞在活體內條件下由導入之非天然核苷酸形成非天然DNA鹼基對(UBP)。在一些實施例中,磷酸鉀及/或磷酸酶及/或核苷酸酶活性之抑制劑可促進非天然核苷酸之轉運。方法包括使用表現異源核苷三磷酸轉運子之細胞。當此類細胞與一或多個核苷酸三磷酸酯接觸時,核苷三磷酸經轉運至細胞中。細胞可處於磷酸鉀及/或磷酸酶及核苷酸酶之抑制劑之存在下。非天然核苷三磷酸可藉由細胞之天然機制(亦即聚合酶)而併入細胞內之核酸中並且例如彼此鹼基配對,以在細胞之核酸內形成非天然鹼基對。在一些實施例中,UBP在攜帶非天然鹼基之DNA與RNA核苷酸之間形成。
在一些實施例中,UBP可在暴露於非天然三磷酸酯時併入細胞或細胞群中。在一些實施例中,UBP可在實質上不斷地暴露於非天然三磷酸酯時併入細胞或細胞群中。
在一些實施例中,與在不具有異源基因之表現之誘導的細胞中的一或多個非天然三磷酸酯之生長及吸收相比,在細胞中之異源基因(例如,核苷三磷酸轉運子(NTT))之表現之誘導可導致較慢細胞生長及增加的非天然三磷酸酯吸收。吸收不同地包含將核苷酸轉運至細胞中,諸如經由擴散、滲透或經由轉運子之作用。在一些實施例中,與不具有異源基因之表現之誘導的細胞之生長及吸收相比,在細胞中之異源基因(例如NTT)之表現之誘導可導致增加的細胞生長及增加的非天然核酸吸收。
在一些實施例中,UBP係在對數生長期期間併入。在一些實施例中,UBP係在非對數生長期期間併入。在一些實施例中,UBP係在實質上線性生長期期間併入。在一些實施例中,UBP係在生長一段時間之後穩定地併入細胞或細胞群中。舉例而言,UBP可在生長至少約1、2、3、4、5、6,、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45或50或更多個複製之後穩定地併入細胞或細胞群中。舉例而言,UBP可在生長持續至少約1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23或24小時之生長之後穩定地併入細胞或細胞群中。舉例而言,UBP可在生長持續至少約1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30或31天之生長之後穩定地併入細胞或細胞群中。舉例而言,UBP可在生長持續至少約1、2、3、4、5、6、7、8、9、10、11或12個月之生長之後穩定地併入細胞或細胞群中。舉例而言,UBP可在生長持續至少約1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、50年之生長之後穩定地併入細胞或細胞群中。
在一些實施例中,細胞進一步利用RNA聚合酶以產生含有一或多個非天然核苷酸之mRNA。在一些情況下,細胞進一步利用聚合酶以產生含有包含一或多個非天然核苷酸之反密碼子的tRNA。在一些情況下,tRNA裝載有非天然胺基酸。在一些情況下,在轉譯以合成含有至少一個非天然胺基酸之非天然多肽或非天然蛋白質期間,tRNA之非天然反密碼子與mRNA之非天然密碼子配對。
天然及非天然胺基酸
如本文所使用,胺基酸殘基可指含有胺基及羧基兩者之分子。適合的胺基酸包括(但不限於):天然存在之胺基酸之D異構體及L異構體,以及藉由有機合成或任何其他方法製備之非天然存在之胺基酸。如本文所使用,術語胺基酸包括(但不限於):α-胺基酸、天然胺基酸、非天然胺基酸及胺基酸類似物。術語「α-胺基酸」可指含有與指定為α-碳之碳結合的胺基及羧基兩者的分子。例如: 。
。
術語「β-胺基酸」可指含有呈β組態之胺基及羧基兩者的分子。
「天然存在之胺基酸」可指通常見於自然界中合成之肽中且由一個字母縮寫A、R、N、C、D、Q、E、G、H、I、L、K、M、F、P、S、T、W、Y及V已知的二十種胺基酸中之任一者。
下表展示天然胺基酸之特性之概述:
| 胺基酸 | 3-字母代碼 | 1-字母代碼 | 側鏈極性 | 側鏈電荷 (pH 7.4) | 親水性指數 |
| 丙胺酸 | Ala | A | 非極性 | 中性 | 1.8 |
| 精胺酸 | Arg | R | 極性 | 正電荷 | −4.5 |
| 天冬醯胺酸 | Asn | N | 極性 | 中性 | −3.5 |
| 天冬胺酸 | Asp | D | 極性 | 負電荷 | −3.5 |
| 半胱胺酸 | Cys | C | 極性 | 中性 | 2.5 |
| 麩胺酸 | Glu | E | 極性 | 負電荷 | −3.5 |
| 麩醯胺酸 | Gln | Q | 極性 | 中性 | −3.5 |
| 甘胺酸 | Gly | G | 非極性 | 中性 | −0.4 |
| 組胺酸 | His | H | 極性 | 正電荷(10%) 中性(90%) | −3.2 |
| 異白胺酸 | Ile | I | 非極性 | 中性 | 4.5 |
| 白胺酸 | Leu | L | 非極性 | 中性 | 3.8 |
| 離胺酸 | Lys | K | 極性 | 正電荷 | −3.9 |
| 甲硫胺酸 | Met | M | 非極性 | 中性 | 1.9 |
| 苯丙胺酸 | Phe | F | 非極性 | 中性 | 2.8 |
| 脯胺酸 | Pro | P | 非極性 | 中性 | −1.6 |
| 絲胺酸 | Ser | S | 極性 | 中性 | −0.8 |
| 蘇胺酸 | Thr | T | 極性 | 中性 | −0.7 |
| 色胺酸 | Trp | W | 非極性 | 中性 | −0.9 |
| 酪胺酸 | Tyr | Y | 極性 | 中性 | −1.3 |
| 纈胺酸 | Val | V | 非極性 | 中性 | 4.2 |
「疏水性胺基酸」包括較小疏水性胺基酸及較大疏水性胺基酸。「較小疏水性胺基酸」可為甘胺酸、丙胺酸、脯胺酸及其類似物。「較大疏水性胺基酸」可為纈胺酸、白胺酸、異白胺酸、苯丙胺酸、甲硫胺酸、色胺酸及其類似物。「極性胺基酸」可為絲胺酸、蘇胺酸、天冬醯胺酸、麩醯胺酸、半胱胺酸、酪胺酸及其類似物。「帶電荷胺基酸」可為離胺酸、精胺酸、組胺酸、天冬胺酸、麩胺酸及其類似物。
「胺基酸類似物」可為在結構上與胺基酸類似且可在形成肽模擬物巨環中取代胺基酸的分子。胺基酸類似物包括(但不限於):β-胺基酸及其中胺基或羧基經類似反應基取代(例如,一級胺經二級胺或三級胺取代,或用酯取代羧基)之胺基酸。
非典型胺基酸(ncAA)或「非天然胺基酸」可為不係通常見於自然界中合成之肽中且由一個字母縮寫A、R、N、C、D、Q、E、G、H、I、L、K、M、F、P、S、T、W、Y及V已知的二十種胺基酸中之任一者的胺基酸。在一些情況下,非天然胺基酸為非典型胺基酸之子組。
胺基酸類似物可包括β-胺基酸類似物。β-胺基酸類似物之實例包括(但不限於)以下:環狀β-胺基酸類似物;β-丙胺酸;(R)-β-苯丙胺酸;(R)-1,2,3,4-四氫-異喹啉-3-乙酸;(R)-3-胺基-4-(1-萘基)-丁酸;(R)-3-胺基-4-(2,4-二氯苯基)丁酸;(R)-3-胺基-4-(2-氯苯基)-丁酸;(R)-3-胺基-4-(2-氰基苯基)-丁酸;(R)-3-胺基-4-(2-氟苯基)-丁酸;(R)-3-胺基-4-(2-呋喃基)-丁酸;(R)-3-胺基-4-(2-甲基苯基)-丁酸;(R)-3-胺基-4-(2-萘基)-丁酸;(R)-3-胺基-4-(2-噻吩基)-丁酸;(R)-3-胺基-4-(2-三氟甲基苯基)-丁酸;(R)-3-胺基-4-(3,4-二氯苯基)丁酸;(R)-3-胺基-4-(3,4-二氟苯基)丁酸;(R)-3-胺基-4-(3-苯并噻吩基)-丁酸;(R)-3-胺基-4-(3-氯苯基)-丁酸;(R)-3-胺基-4-(3-氰基苯基)-丁酸;(R)-3-胺基-4-(3-氟苯基)-丁酸;(R)-3-胺基-4-(3-甲基苯基)-丁酸;(R)-3-胺基-4-(3-吡啶基)-丁酸;(R)-3-胺基-4-(3-噻吩基)-丁酸;(R)-3-胺基-4-(3-三氟甲基苯基)-丁酸;(R)-3-胺基-4-(4-溴苯基)-丁酸;(R)-3-胺基-4-(4-氯苯基)-丁酸;(R)-3-胺基-4-(4-氰基苯基)-丁酸;(R)-3-胺基-4-(4-氟苯基)-丁酸;(R)-3-胺基-4-(4-碘苯基)-丁酸;(R)-3-胺基-4-(4-甲基苯基)-丁酸;(R)-3-胺基-4-(4-硝基苯基)-丁酸;(R)-3-胺基-4-(4-吡啶基)-丁酸;(R)-3-胺基-4-(4-三氟甲基苯基)-丁酸;(R)-3-胺基-4-五氟-苯基丁酸;(R)-3-胺基-5-己烯酸;(R)-3-胺基-5-己炔酸;(R)-3-胺基-5-苯基戊酸;(R)-3-胺基-6-苯基-5-己烯酸;(S)-1,2,3,4-四氫-異喹啉-3-乙酸;(S)-3-胺基-4-(1-萘基)-丁酸;(S)-3-胺基-4-(2,4-二氯苯基)丁酸;(S)-3-胺基-4-(2-氯苯基)-丁酸;(S)-3-胺基-4-(2-氰基苯基)-丁酸;(S)-3-胺基-4-(2-氟苯基)-丁酸;(S)-3-胺基-4-(2-呋喃基)-丁酸;(S)-3-胺基-4-(2-甲基苯基)-丁酸;(S)-3-胺基-4-(2-萘基)-丁酸;(S)-3-胺基-4-(2-噻吩基)-丁酸;(S)-3-胺基-4-(2-三氟甲基苯基)-丁酸;(S)-3-胺基-4-(3,4-二氯苯基)丁酸;(S)-3-胺基-4-(3,4-二氟苯基)丁酸;(S)-3-胺基-4-(3-苯并噻吩基)-丁酸;(S)-3-胺基-4-(3-氯苯基)-丁酸;(S)-3-胺基-4-(3-氰基苯基)-丁酸;(S)-3-胺基-4-(3-氟苯基)-丁酸;(S)-3-胺基-4-(3-甲基苯基)-丁酸;(S)-3-胺基-4-(3-吡啶基)-丁酸;(S)-3-胺基-4-(3-噻吩基)-丁酸;(S)-3-胺基-4-(3-三氟甲基苯基)-丁酸;(S)-3-胺基-4-(4-溴苯基)-丁酸;(S)-3-胺基-4-(4-氯苯基)丁酸;(S)-3-胺基-4-(4-氰基苯基)-丁酸;(S)-3-胺基-4-(4-氟苯基)丁酸;(S)-3-胺基-4-(4-碘苯基)-丁酸;(S)-3-胺基-4-(4-甲基苯基)-丁酸;(S)-3-胺基-4-(4-硝基苯基)-丁酸;(S)-3-胺基-4-(4-吡啶基)-丁酸;(S)-3-胺基-4-(4-三氟甲基苯基)-丁酸;(S)-3-胺基-4-五氟-苯基丁酸;(S)-3-胺基-5-己烯酸;(S)-3-胺基-5-己炔酸;(S)-3-胺基-5-苯基戊酸;(S)-3-胺基-6-苯基-5-己烯酸;1,2,5,6-四氫吡啶-3-甲酸;1,2,5,6-四氫吡啶-4-甲酸;3-胺基-3-(2-氯苯基)-丙酸;3-胺基-3-(2-噻吩基)-丙酸;3-胺基-3-(3-溴苯基)-丙酸;3-胺基-3-(4-氯苯基)-丙酸;3-胺基-3-(4-甲氧基苯基)-丙酸;3-胺基-4,4,4-三氟-丁酸;3-胺基己二酸;D-β-苯丙胺酸;β-白胺酸;L-β-高丙胺酸;L-β-高天冬胺酸γ-苯甲基酯;L-β-高麩胺酸δ-苯甲基酯;L-β-高異白胺酸;L-β-高白胺酸;L-β-高甲硫胺酸;L-β-高苯丙胺酸;L-β-高脯胺酸;L-β-高色胺酸;L-β-高纈胺酸;L-Nω-苯甲氧基羰基-β-高離胺酸;Nω-L-β-高精胺酸;O-苯甲基-L-β-高羥基脯胺酸;O-苯甲基-L-β-高絲胺酸;O-苯甲基-L-β-高蘇胺酸;O-苯甲基-L-β-高酪胺酸;γ-三苯甲基-L-β-高天冬醯胺酸;(R)-β-苯丙胺酸;L-β-高天冬胺酸γ-第三丁基酯;L-β-高麩胺酸δ-第三丁基酯;L-Nω-β-高離胺酸;Nδ-三苯甲基-L-β-高麩醯胺酸;Nω-2,2,4,6,7-五甲基-二氫苯并呋喃-5-磺醯基-L-β-高精胺酸;O-第三丁基-L-β-高羥基-脯胺酸;O-第三丁基-L-β-高絲胺酸;O-第三丁基-L-β-高蘇胺酸;O-第三丁基-L-β-高酪胺酸;2-胺基環戊烷甲酸;以及2-胺基環己烷甲酸。
胺基酸類似物可包括丙胺酸、纈胺酸、甘胺酸或白胺酸之類似物。丙胺酸、纈胺酸、甘胺酸及白胺酸之胺基酸類似物之實例包括(但不限於)以下:α-甲氧基甘胺酸;α-烯丙基-L-丙胺酸;α-胺基異丁酸;α-甲基-白胺酸;β-(1-萘基)-D-丙胺酸;β-(1-萘基)-L-丙胺酸;β-(2-萘基)-D-丙胺酸;β-(2-萘基)-L-丙胺酸;β-(2-吡啶基)-D-丙胺酸;β-(2-吡啶基)-L-丙胺酸;β-(2-噻吩基)-D-丙胺酸;β-(2-噻吩基)-L-丙胺酸;β-(3-苯并噻吩基)-D-丙胺酸;β-(3-苯并噻吩基)-L-丙胺酸;β-(3-吡啶基)-D-丙胺酸;β-(3-吡啶基)-L-丙胺酸;β-(4-吡啶基)-D-丙胺酸;β-(4-吡啶基)-L-丙胺酸;β-氯-L-丙胺酸;β-氰基-L-丙胺酸;β-環己基-D-丙胺酸;β-環己基-L-丙胺酸;β-環戊烯-1-基-丙胺酸;β-環戊基-丙胺酸;β-環丙基-L-Ala-OH.二環己基銨鹽;β-第三丁基-D-丙胺酸;β-第三丁基-L-丙胺酸;γ-胺基丁酸;L-α,β-二胺基丙酸;2,4-二硝基-苯基甘胺酸;2,5-二氫-D-苯基甘胺酸;2-胺基-4,4,4-三氟丁酸;2-氟-苯基甘胺酸;3-胺基-4,4,4-三氟-丁酸;3-氟-纈胺酸;4,4,4-三氟-纈胺酸;4,5-脫氫-L-leu-OH.二環己基銨鹽;4-氟-D-苯基甘胺酸;4-氟-L-苯基甘胺酸;4-羥基-D-苯基甘胺酸;5,5,5-三氟-白胺酸;6-胺基己酸;環戊基-D-Gly-OH.二環己基銨鹽;環戊基-Gly-OH.二環己基銨鹽;D-α,β-二胺基丙酸;D-α-胺基丁酸;D-α-第三丁基甘胺酸;D-(2-噻吩基)甘胺酸;D-(3-噻吩基)甘胺酸;D-2-胺基己酸;D-2-二氫茚基甘胺酸;D-烯丙基甘胺酸-二環己基銨鹽;D-環己基甘胺酸;D-正纈胺酸;D-苯基甘胺酸;β-胺基丁酸;β-胺基異丁酸;(2-溴苯基)甘胺酸;(2-甲氧基苯基)甘胺酸;(2-甲基苯基)甘胺酸;(2-噻唑基)甘胺酸;(2-噻吩基)甘胺酸;2-胺基-3-(二甲基胺基)-丙酸;L-α,β-二胺基丙酸;L-α-胺基丁酸;L-α-第三丁基甘胺酸;L-(3-噻吩基)甘胺酸;L-2-胺基-3-(二甲基胺基)-丙酸;L-2-胺基己酸二環己基-銨鹽;L-2-二氫茚基甘胺酸;L-烯丙基甘胺酸二環己基銨鹽;L-環己基甘胺酸;L-苯基甘胺酸;L-炔丙基甘胺酸;L-正纈胺酸;N-α-胺基甲基-L-丙胺酸;D-α,γ-二胺基丁酸;L-α,γ-二胺基丁酸;β-環丙基-L-丙胺酸;(N-β-(2,4-二硝基苯基))-L-α,β-二胺基丙酸;(N-β-1-(4,4-二甲基-2,6-二側氧基亞環己-1-基)乙基)-D-α,β-二胺基丙酸;(N-β-1-(4,4-二甲基-2,6-二側氧基亞環己-1-基)乙基)-L-α,β-二胺基丙酸;(N-β-4-甲基三苯甲基)-L-α,β-二胺基丙酸;(N-β-烯丙氧基羰基)-L-α,β-二胺基丙酸;(N-γ-1-(4,4-二甲基-2,6-二側氧基亞環己-1-基)乙基)-D-α,γ-二胺基丁酸;(N-γ-1-(4,4-二甲基-2,6-二側氧基亞環己-1-基)乙基)-L-α,γ-二胺基丁酸;(N-γ-4-甲基三苯甲基)-D-α,γ-二胺基丁酸;(N-γ-4-甲基三苯甲基)-L-α,γ-二胺基丁酸;(N-γ-烯丙氧基羰基)-L-α,γ-二胺基丁酸;D-α,γ-二胺基丁酸;4,5-脫氫-L-白胺酸;環戊基-D-Gly-OH;環戊基-Gly-OH;D-烯丙基甘胺酸;D-高環己基丙胺酸;L-1-芘基丙胺酸;L-2-胺基己酸;L-烯丙基甘胺酸;L-高環己基丙胺酸;以及N-(2-羥基-4-甲氧基-Bzl)-Gly-OH。
胺基酸類似物可包括精胺酸或離胺酸之類似物。精胺酸及離胺酸之胺基酸類似物之實例包括(但不限於)以下:瓜胺酸;L-2-胺基-3-胍基丙酸;L-2-胺基-3-脲基丙酸;L-瓜胺酸;Lys(Me)2
-OH;Lys(N3
)-OH;Nδ-苯甲氧基羰基-L-鳥胺酸;Nω-硝基-D-精胺酸;Nω-硝基-L-精胺酸;α-甲基-鳥胺酸;2,6-二胺基庚二酸;L-鳥胺酸;(Nδ-1-(4,4-二甲基-2,6-二側氧基-亞環己-1-基)乙基)-D-鳥胺酸;(Nδ-1-(4,4-二甲基-2,6-二側氧基-亞環己-1-基)乙基)-L-鳥胺酸;(Nδ-4-甲基三苯甲基)-D-鳥胺酸;(Nδ-4-甲基三苯甲基)-L-鳥胺酸;D-鳥胺酸;L-鳥胺酸;Arg(Me)(Pbf)-OH;Arg(Me)2
-OH (不對稱);Arg(Me)2
-OH (對稱);Lys(ivDde)-OH;Lys(Me)2
-OH.HCl;Lys(Me3
)-OH氯化物;Nω-硝基-D-精胺酸;以及Nω-硝基-L-精胺酸。
胺基酸類似物可包括天冬胺酸或麩胺酸之類似物。天冬胺酸及麩胺酸之胺基酸類似物之實例包括(但不限於)以下:α-甲基-D-天冬胺酸;α-甲基-麩胺酸;α-甲基-L-天冬胺酸;γ-亞甲基-麩胺酸;(N-γ-乙基)-L-麩醯胺酸;[N-α-(4-胺基苯甲醯基)]-L-麩胺酸;2,6-二胺基庚二酸;L-α-胺基辛二酸;D-2-胺基己二酸;D-α-胺基辛二酸;α-胺基庚二酸;亞胺基二乙酸;L-2-胺基己二酸;蘇-β-甲基-天冬胺酸;γ-羧基-D-麩胺酸γ,γ-二-第三丁基酯;γ-羧基-L-麩胺酸γ,γ-二-第三丁基酯;Glu(OAll)-OH;L-Asu(OtBu)-OH;以及焦麩胺酸。
胺基酸類似物可包括半胱胺酸及甲硫胺酸之類似物。半胱胺酸及甲硫胺酸之胺基酸類似物之實例包括(但不限於):Cys(法呢基)-OH、Cys(法呢基)-OMe、α-甲基-甲硫胺酸、Cys(2-羥乙基)-OH、Cys(3-胺基丙基)-OH、2-胺基-4-(乙硫基)丁酸、丁硫胺酸、丁硫胺酸磺醯亞胺、乙硫胺酸、氯化甲硫胺酸甲鋶、硒甲硫胺酸、氧化半胱胺酸、[2-(4-吡啶基)乙基]-DL-青黴胺、[2-(4-吡啶基)乙基]-L-半胱胺酸、4-甲氧基苯甲基-D-青黴胺、4-甲氧基苯甲基-L-青黴胺、4-甲基苯甲基-D-青黴胺、4-甲基苯甲基-L-青黴胺、苯甲基-D-半胱胺酸、苯甲基-L-半胱胺酸、苯甲基-DL-高半胱胺酸、胺甲醯基-L-半胱胺酸、羧乙基-L-半胱胺酸、羧甲基-L-半胱胺酸、二苯基甲基-L-半胱胺酸、乙基-L-半胱胺酸、甲基-L-半胱胺酸、第三丁基-D-半胱胺酸、三苯甲基-L-高半胱胺酸、三苯甲基-D-青黴胺、胱硫醚、高胱胺酸、L-高胱胺酸、(2-胺基乙基)-L-半胱胺酸、硒-L-胱胺酸、胱硫醚、Cys(StBu)-OH及乙醯胺基甲基-D-青黴胺。
胺基酸類似物可包括苯丙胺酸及酪胺酸之類似物。苯丙胺酸及酪胺酸之胺基酸類似物之實例包括β-甲基-苯丙胺酸、β-羥基苯基丙胺酸、α-甲基-3-甲氧基-DL-苯丙胺酸、α-甲基-D-苯丙胺酸、α-甲基-L-苯丙胺酸、1,2,3,4-四氫異喹啉-3-甲酸、2,4-二氯-苯丙胺酸、2-(三氟甲基)-D-苯丙胺酸、2-(三氟甲基)-L-苯丙胺酸、2-溴-D-苯丙胺酸、2-溴-L-苯丙胺酸、2-氯-D-苯丙胺酸、2-氯-L-苯丙胺酸、2-氰基-D-苯丙胺酸、2-氰基-L-苯丙胺酸、2-氟-D-苯丙胺酸、2-氟-L-苯丙胺酸、2-甲基-D-苯丙胺酸、2-甲基-L-苯丙胺酸、2-硝基-D-苯丙胺酸、2-硝基-L-苯丙胺酸、2,4,5-三羥基-苯丙胺酸、3,4,5-三氟-D-苯丙胺酸、3,4,5-三氟-L-苯丙胺酸、3,4-二氯-D-苯丙胺酸、3,4-二氯-L-苯丙胺酸、3,4-二氟-D-苯丙胺酸、3,4-二氟-L-苯丙胺酸、3,4-二羥基-L-苯丙胺酸、3,4-二甲氧基-L-苯丙胺酸、3,5,3'-三碘-L-甲狀腺胺酸、3,5-二碘-D-酪胺酸、3,5-二碘-L-酪胺酸、3,5-二碘-L-甲狀腺胺酸、3-(三氟甲基)-D-苯丙胺酸、3-(三氟甲基)-L-苯丙胺酸、3-胺基-L-酪胺酸、3-溴-D-苯丙胺酸、3-溴-L-苯丙胺酸、3-氯-D-苯丙胺酸、3-氯-L-苯丙胺酸、3-氯-L-酪胺酸、3-氰基-D-苯丙胺酸、3-氰基-L-苯丙胺酸、3-氟-D-苯丙胺酸、3-氟-L-苯丙胺酸、3-氟-酪胺酸、3-碘-D-苯丙胺酸、3-碘-L-苯丙胺酸、3-碘-L-酪胺酸、3-甲氧基-L-酪胺酸、3-甲基-D-苯丙胺酸、3-甲基-L-苯丙胺酸、3-硝基-D-苯丙胺酸、3-硝基-L-苯丙胺酸、3-硝基-L-酪胺酸、4-(三氟甲基)-D-苯丙胺酸、4-(三氟甲基)-L-苯丙胺酸、4-胺基-D-苯丙胺酸、4-胺基-L-苯丙胺酸、4-苯甲醯基-D-苯丙胺酸、4-苯甲醯基-L-苯丙胺酸、4-雙(2-氯乙基)胺基-L-苯丙胺酸、4-溴-D-苯丙胺酸、4-溴-L-苯丙胺酸、4-氯-D-苯丙胺酸、4-氯-L-苯丙胺酸、4-氰基-D-苯丙胺酸、4-氰基-L-苯丙胺酸、4-氟-D-苯丙胺酸、4-氟-L-苯丙胺酸、4-碘-D-苯丙胺酸、4-碘-L-苯丙胺酸、高苯丙胺酸、甲狀腺素、3,3-二苯丙胺酸、甲狀腺胺酸、乙基-酪胺酸及甲基-酪胺酸。
胺基酸類似物可包括脯胺酸之類似物。脯胺酸之胺基酸類似物之實例包括(但不限於):3,4-脫氫-脯胺酸、4-氟-脯胺酸、順-4-羥基-脯胺酸、噻唑啶-2-甲酸及反-4-氟-脯胺酸。
胺基酸類似物可包括絲胺酸及蘇胺酸之類似物。絲胺酸及蘇胺酸之胺基酸類似物之實例包括(但不限於):3-胺基-2-羥基-5-甲基己酸、2-胺基-3-羥基-4-甲基戊酸、2-胺基-3-乙氧基丁酸、2-胺基-3-甲氧基丁酸、4-胺基-3-羥基-6-甲基庚酸、2-胺基-3-苯甲氧基丙酸、2-胺基-3-苯甲氧基丙酸、2-胺基-3-乙氧基丙酸、4-胺基-3-羥基丁酸及α-甲基絲胺酸。
胺基酸類似物可包括色胺酸之類似物。色胺酸之胺基酸類似物之實例包括(但不限於)以下:α-甲基-色胺酸;β-(3-苯并噻吩基)-D-丙胺酸;β-(3-苯并噻吩基)-L-丙胺酸;1-甲基-色胺酸;4-甲基-色胺酸;5-苯甲氧基-色胺酸;5-溴-色胺酸;5-氯-色胺酸;5-氟-色胺酸;5-羥基-色胺酸;5-羥基-L-色胺酸;5-甲氧基-色胺酸;5-甲氧基-L-色胺酸;5-甲基-色胺酸;6-溴-色胺酸;6-氯-D-色胺酸;6-氯-色胺酸;6-氟-色胺酸;6-甲基-色胺酸;7-苯甲氧基-色胺酸;7-溴-色胺酸;7-甲基-色胺酸;D-1,2,3,4-四氫-正哈爾滿-3-甲酸;6-甲氧基-1,2,3,4-四氫正哈爾滿-1-甲酸;7-氮雜色胺酸;L-1,2,3,4-四氫-正哈爾滿-3-甲酸;5-甲氧基-2-甲基-色胺酸;以及6-氯-L-色胺酸。
胺基酸類似物可為外消旋的。在一些情況下,使用胺基酸類似物之D異構體。在一些情況下,使用胺基酸類似物之L異構體。在一些情況下,胺基酸類似物包含呈R或S組態之對掌性中心。有時,β-胺基酸類似物之一或多個胺基經保護基,例如第三丁氧基羰基(BOC基團)、9-茀基甲氧基羰基(FMOC)、甲苯磺醯基及其類似基團取代。有時,β-胺基酸類似物之羧酸官能基受保護,例如作為其酯衍生物。在一些情況下,使用胺基酸類似物之鹽。
在一些實施例中,非天然胺基酸為Liu C.C., Schultz, P.G.Annu. Rev. Biochem.
2010, 79, 413中所描述之非天然胺基酸。在一些實施例中,非天然胺基酸包含N6(2-疊氮基乙氧基) -羰基-L-離胺酸。
在一些實施例中,在與結合部分結合之前,本文所描述之胺基酸殘基(例如在蛋白質內)突變為非天然胺基酸。在一些情況下,針對非天然胺基酸之突變防止免疫系統之自身抗原反應或使免疫系統之自身抗原反應減至最小。如本文所使用,術語「非天然胺基酸」係指除20種天然存在於蛋白質中之胺基酸以外的胺基酸。非天然胺基酸之非限制性實例包括:對乙醯基-L-苯丙胺酸、對碘-L-苯丙胺酸、對甲氧基苯丙胺酸、O-甲基-L-酪胺酸、對炔丙氧基苯丙胺酸、對炔丙基-苯丙胺酸、L-3-(2-萘基)丙胺酸、3-甲基-苯丙胺酸、O-4-烯丙基-L-酪胺酸、4-丙基-L-酪胺酸、三-O-乙醯基-GlcNAcp-絲胺酸、左旋多巴、氟化苯丙胺酸、異丙基-L-苯丙胺酸、對疊氮基-L-苯丙胺酸、對疊氮基-苯丙胺酸、對苯甲醯基-L-苯丙胺酸、對二羥硼基苯丙胺酸、O-炔丙基酪胺酸、L-磷絲胺酸、膦醯基絲胺酸、膦醯基酪胺酸、對溴苯丙胺酸、硒半胱胺酸、對胺基-L-苯丙胺酸、異丙基-L-苯丙胺酸、N6-((炔丙氧基)-羰基)-L-離胺酸(PrK)、疊氮基-離胺酸(N6-疊氮基乙氧基-羰基-L-離胺酸;AzK)、N6-(((2-疊氮基苯甲基)氧基)羰基)-L-離胺酸、N6-(((3-疊氮基苯甲基)氧基)羰基)-L-離胺酸及N6-(((4-疊氮基苯甲基)氧基)羰基)-L-離胺酸、酪胺酸胺基酸之非天然類似物;麩醯胺酸胺基酸之非天然類似物;苯丙胺酸胺基酸之非天然類似物;絲胺酸胺基酸之非天然類似物;蘇胺酸胺基酸之非天然類似物;經烷基、芳基、醯基、疊氮基、氰基、鹵基、肼、醯肼、羥基、烯基、炔基、醚、硫醇、磺醯基、硒基、酯、硫代酸、硼酸酯、 酸鹽(boronate)、磷酸基、膦醯基、膦、雜環、烯酮、亞胺、醛、羥胺、酮基或胺基取代之胺基酸或其組合;具有光活化交聯劑之胺基酸;自旋標記之胺基酸;螢光胺基酸;金屬結合胺基酸;含金屬胺基酸;放射性胺基酸;光籠化及/或光異構化胺基酸;含生物素或生物素類似物之胺基酸;含酮基胺基酸;包含聚乙二醇或聚醚之胺基酸;經重原子取代之胺基酸;化學上可裂解或可光裂解之胺基酸;具有經延長側鏈之胺基酸;含有毒性基團之胺基酸;經糖取代之胺基酸;含有碳連接之糖的胺基酸;氧化還原活性胺基酸;含有α-羥基的酸;胺基硫基酸;經α,α二取代之胺基酸;β-胺基酸;除脯胺酸或組胺酸以外的環狀胺基酸及除苯丙胺酸、酪胺酸或色胺酸以外的芳族胺基酸。
酸鹽(boronate)、磷酸基、膦醯基、膦、雜環、烯酮、亞胺、醛、羥胺、酮基或胺基取代之胺基酸或其組合;具有光活化交聯劑之胺基酸;自旋標記之胺基酸;螢光胺基酸;金屬結合胺基酸;含金屬胺基酸;放射性胺基酸;光籠化及/或光異構化胺基酸;含生物素或生物素類似物之胺基酸;含酮基胺基酸;包含聚乙二醇或聚醚之胺基酸;經重原子取代之胺基酸;化學上可裂解或可光裂解之胺基酸;具有經延長側鏈之胺基酸;含有毒性基團之胺基酸;經糖取代之胺基酸;含有碳連接之糖的胺基酸;氧化還原活性胺基酸;含有α-羥基的酸;胺基硫基酸;經α,α二取代之胺基酸;β-胺基酸;除脯胺酸或組胺酸以外的環狀胺基酸及除苯丙胺酸、酪胺酸或色胺酸以外的芳族胺基酸。
在一些實施例中,非天然胺基酸包含選擇性反應基或用於位點選擇性標記目標蛋白或多肽之反應基。在一些情況下,化學反應係雙正交反應(例如生物相容及選擇性反應)。在一些情況下,化學反應係Cu(I)催化或「無銅」炔烴-疊氮化物三唑形成反應、施陶丁格連接(Staudinger ligation)、反向需電子狄爾斯-阿德(inverse-electron-demand Diels-Alder;IEDDA)反應、「光點擊(photo-click)」化學反應或金屬介導之方法,諸如烯烴複分解及鈴木-宮浦或薗頭交叉偶合(Suzuki-Miyaura or Sonogashira cross-coupling)。在一些實施例中,非天然胺基酸包含光反應性基團,其在用例如UV照射時交聯。在一些實施例中,非天然胺基酸包含光籠蔽胺基酸。在一些情況下,非天然胺基酸係經對位取代、經間位取代或經鄰位取代之胺基酸衍生物。
在一些情況下,非天然胺基酸包含對乙醯基-L-苯丙胺酸、對疊氮基甲基-L-苯丙胺酸(pAMF)、對碘-L-苯丙胺酸、O-甲基-L-酪胺酸、對甲氧基苯丙胺酸、對炔丙基氧基苯丙胺酸、對炔丙基-苯丙胺酸、L-3-(2-萘基)丙胺酸、3-甲基-苯丙胺酸、O-4-烯丙基-L-酪胺酸、4-丙基-L-酪胺酸、三-O-乙醯基-GlcNAcp-絲胺酸、左旋多巴、氟化苯丙胺酸、異丙基-L-苯丙胺酸、對疊氮基-L-苯丙胺酸、對醯基-L-苯丙胺酸、對苯甲醯基-L-苯丙胺酸、L-磷絲胺酸、膦醯基絲胺酸、膦醯基酪胺酸、對溴苯基丙胺酸、對胺基-L-苯丙胺酸或異丙基-L-苯丙胺酸。
在一些情況下,非天然胺基酸為3-胺基酪胺酸、3-硝基酪胺酸、3,4-二羥基-苯丙胺酸或3-碘酪胺酸。在一些情況下,非天然胺基酸為苯基硒半胱胺酸。在一些情況下,非天然胺基酸為含有二苯甲酮、酮、碘、甲氧基、乙醯基、苯甲醯基或疊氮基之苯丙胺酸衍生物。在一些情況下,非天然胺基酸為含有二苯甲酮、酮、碘、甲氧基、乙醯基、苯甲醯基或疊氮基之離胺酸衍生物。在一些情況下,非天然胺基酸包含芳族側鏈。在一些情況下,非天然胺基酸不包含芳族側鏈。在一些情況下,非天然胺基酸包含疊氮基。在一些情況下,非天然胺基酸包含邁克爾受體(Michael-acceptor)基團。在一些情況下,邁克爾受體基團包含能夠經由1,2-加成反應形成共價鍵之不飽和部分。在一些情況下,邁克爾受體基團包含缺電子烯烴或炔烴。在一些情況下,邁克爾受體基團包括(但不限於)α,β不飽和的:酮、醛、亞碸、碸、腈、亞胺或芳族物。在一些情況下,非天然胺基酸為脫氫丙胺酸。在一些情況下,非天然胺基酸包含醛基或酮基。在一些情況下,非天然胺基酸為包含醛或酮基團之離胺酸衍生物。在一些情況下,非天然胺基酸為在β、γ或δ位置處包含一或多個O、N、Se或S原子之離胺酸衍生物。在一些情況下,非天然胺基酸為在γ位置處包含O、N、Se或S原子之離胺酸衍生物。在一些情況下,非天然胺基酸為其中ε N原子經氧原子置換之離胺酸衍生物。在一些情況下,非天然胺基酸為並非天然存在之轉譯後修飾之離胺酸的離胺酸衍生物。
在一些情況下,非天然胺基酸為包含側鏈之胺基酸,其中來自α位置之第六原子包含羰基。在一些情況下,非天然胺基酸為包含側鏈之胺基酸,其中來自α位置之第六原子包含羰基,且來自α位置之第五原子為氮。在一些情況下,非天然胺基酸為包含側鏈之胺基酸,其中來自α位置之第七原子為氧原子。
在一些情況下,非天然胺基酸為包含硒之絲胺酸衍生物。在一些情況下,非天然胺基酸為硒絲胺酸(2-胺基-3-氫硒丙酸)。在一些情況下,非天然胺基酸為2-胺基-3-((2-((3-(苯甲氧基)-3-側氧基丙基)胺基)乙基)硒烷基)丙酸。在一些情況下,非天然胺基酸為2-胺基-3-(苯基硒烷基)丙酸。在一些情況下,非天然胺基酸包含硒,其中硒之氧化導致形成包含烯烴之非天然胺基酸。
在一些情況下,非天然胺基酸包含環辛炔基。在一些情況下,非天然胺基酸包含反環辛烯基。在一些情況下,非天然胺基酸包含降冰片烯基。在一些情況下,非天然胺基酸包含環丙烯基。在一些情況下,非天然胺基酸包含二氮環丙烯基。在一些情況下,非天然胺基酸包含四𠯤基。
在一些情況下,非天然胺基酸為離胺酸衍生物,其中側鏈氮經胺甲醯基化。在一些情況下,非天然胺基酸為離胺酸衍生物,其中側鏈氮經醯基化。在一些情況下,非天然胺基酸為2-胺基-6-{[(第三丁氧基)羰基]胺基}己酸。在一些情況下,非天然胺基酸為2-胺基-6-{[(第三丁氧基)羰基]胺基}己酸。在一些情況下,非天然胺基酸為N6-Boc-N6-甲基離胺酸。在一些情況下,非天然胺基酸為N6-乙醯基離胺酸。在一些情況下,非天然胺基酸為吡咯離胺酸。在一些情況下,非天然胺基酸為N6-三氟乙醯基離胺酸。在一些情況下,非天然胺基酸為2-胺基-6-{[(苯甲氧基)羰基]胺基}己酸。在一些情況下,非天然胺基酸為2-胺基-6-{[(對碘苯甲氧基)羰基]胺基}己酸。在一些情況下,非天然胺基酸為2-胺基-6-{[(對硝基苯甲氧基)羰基]胺基}己酸。在一些情況下,非天然胺基酸為N6-脯胺醯基離胺酸。在一些情況下,非天然胺基酸為2-胺基-6-{[(環戊氧基)羰基]胺基}己酸。在一些情況下,非天然胺基酸為N6-(環戊烷羰基)離胺酸。在一些情況下,非天然胺基酸為N6-(四氫呋喃-2-羰基)離胺酸。在一些情況下,非天然胺基酸為N6-(3-乙炔基四氫呋喃-2-羰基)離胺酸。在一些情況下,非天然胺基酸為N6-((丙-2-炔-1-基氧基)羰基)離胺酸。在一些情況下,非天然胺基酸為2-胺基-6-{[(2-疊氮基環戊氧基)羰基]胺基}己酸。在一些情況下,非天然胺基酸為N6-((2-疊氮基乙氧基)羰基)離胺酸。在一些情況下,非天然胺基酸為2-胺基-6-{[(2-硝基苯甲氧基)羰基]胺基}己酸。在一些情況下,非天然胺基酸為2-胺基-6-{[(2-環辛炔基氧基)羰基]胺基}己酸。在一些情況下,非天然胺基酸為N6-(2-胺基丁-3-炔醯基)離胺酸。在一些情況下,非天然胺基酸為2-胺基-6-((2-胺基丁-3-炔醯基)氧基)己酸。在一些情況下,非天然胺基酸為N6-(烯丙氧基羰基)離胺酸。在一些情況下,非天然胺基酸為N6-(丁烯基-4-氧基羰基)離胺酸。在一些情況下,非天然胺基酸為N6-(戊烯基-5-氧基羰基)離胺酸。在一些情況下,非天然胺基酸為N6-((丁-3-炔-1-基氧基)羰基)-離胺酸。在一些情況下,非天然胺基酸為N6-((戊-4-炔-1-基氧基)羰基)-離胺酸。在一些情況下,非天然胺基酸為N6-(噻唑啶-4-羰基)離胺酸。在一些情況下,非天然胺基酸為2-胺基-8-側氧基壬酸。在一些情況下,非天然胺基酸為2-胺基-8-側氧基辛酸。在一些情況下,非天然胺基酸為N6-(2-側氧乙醯基)離胺酸。在一些情況下,非天然胺基酸為N6-(((2-疊氮基苯甲基)氧基)羰基)-L-離胺酸。在一些情況下,非天然胺基酸為N6-(((3-疊氮基苯甲基)氧基)羰基)-L-離胺酸。在一些情況下,非天然胺基酸為N6-(((4-疊氮基苯甲基)氧基)羰基)-L-離胺酸。
在一些情況下,非天然胺基酸為N6-丙醯基離胺酸。在一些情況下,非天然胺基酸為N6-丁醯基離胺酸,在一些情況下,非天然胺基酸為N6-(丁-2-烯醯基)離胺酸,在一些情況下,非天然胺基酸為N6-((雙環[2.2.1]庚-5-烯-2-基氧基)羰基)離胺酸。在一些情況下,非天然胺基酸為N6-((螺[2.3]己-1-烯-5-基甲氧基)羰基)離胺酸。在一些情況下,非天然胺基酸為N6-(((4-(1-(三氟甲基)環丙-2-烯-1-基)苯甲基)氧基)羰基)離胺酸。在一些情況下,非天然胺基酸為N6-((雙環[2.2.1]庚-5-烯-2-基甲氧基)羰基)離胺酸。在一些情況下,非天然胺基酸為半胱胺醯基離胺酸。在一些情況下,非天然胺基酸為N6-((1-(6-硝基苯并[d][1,3]間二氧雜環戊烯-5-基)乙氧基)羰基)離胺酸。在一些情況下,非天然胺基酸為N6-((2-(3-甲基-3H-二氮雜環丙烯-3-基)乙氧基)羰基)離胺酸。在一些情況下,非天然胺基酸為N6-((3-(3-甲基-3H-二氮雜環丙烯-3-基)丙氧基)羰基)離胺酸。在一些情況下,非天然胺基酸為N6-((間硝基苯甲氧基)N6-甲基羰基)離胺酸。在一些情況下,非天然胺基酸為N6-((雙環[6.1.0]壬-4-炔-9-基甲氧基)羰基)-離胺酸。在一些情況下,非天然胺基酸為N6-((環庚-3-烯-1-基氧基)羰基)-L-離胺酸。
在一些情況下,非天然胺基酸為2-胺基-3-(((((苯甲氧基)羰基)胺基)甲基)硒烷基)丙酸。在一些實施例中,非天然胺基酸藉由重新設置之琥珀、蛋白石或赭石終止密碼子併入非天然多肽或非天然蛋白質中。在一些實施例中,非天然胺基酸藉由4-鹼基密碼子併入非天然多肽或非天然蛋白質中。在一些實施例中,非天然胺基酸藉由重新設置之罕見正義密碼子併入蛋白質中。
在一些實施例中,非天然胺基酸藉由包含非天然核苷酸之非天然密碼子併入非天然多肽或非天然蛋白質中。
在一些情況下,將非天然胺基酸併入蛋白質中係藉由正交的經修飾之合成酶/tRNA對介導。此類正交對包含天然或突變合成酶,其能夠使非天然tRNA裝載有特異性非天然胺基酸,常常同時使a)其他內源性胺基酸或替代非天然胺基酸裝至非天然tRNA上及b)裝載有任何其他(包括內源性) tRNA減至最少。此類正交對包含能夠藉由合成酶裝入,同時避免藉由內源性合成酶裝載有其他內源性胺基酸之tRNA。在一些實施例中,此類對係自諸如細菌、酵母、古細菌或人類來源之各種生物體鑑別出。在一些實施例中,正交合成酶/tRNA對包含來自單一生物體之組分。在一些實施例中,正交合成酶/tRNA對包含來自兩個不同生物體之組分。在一些實施例中,正交合成酶/tRNA對包含在修飾之前促進不同胺基酸之轉譯的組分。在一些實施例中,正交合成酶為經修飾之丙胺酸合成酶。在一些實施例中,正交合成酶為經修飾之精胺酸合成酶。在一些實施例中,正交合成酶為經修飾之天冬醯胺酸合成酶。在一些實施例中,正交合成酶為經修飾之天冬胺酸合成酶。在一些實施例中,正交合成酶為經修飾之半胱胺酸合成酶。在一些實施例中,正交合成酶為經修飾之麩醯胺酸合成酶。在一些實施例中,正交合成酶為經修飾之麩胺酸合成酶。在一些實施例中,正交合成酶為經修飾之丙胺酸甘胺酸。在一些實施例中,正交合成酶為經修飾之組胺酸合成酶。在一些實施例中,正交合成酶為經修飾之白胺酸合成酶。在一些實施例中,正交合成酶為經修飾之異白胺酸合成酶。在一些實施例中,正交合成酶為經修飾之離胺酸合成酶。在一些實施例中,正交合成酶為經修飾之甲硫胺酸合成酶。在一些實施例中,正交合成酶為經修飾之苯丙胺酸合成酶。在一些實施例中,正交合成酶為經修飾之脯胺酸合成酶。在一些實施例中,正交合成酶為經修飾之絲胺酸合成酶。在一些實施例中,正交合成酶為經修飾之蘇胺酸合成酶。在一些實施例中,正交合成酶為經修飾之色胺酸合成酶。在一些實施例中,正交合成酶為經修飾之酪胺酸合成酶。在一些實施例中,正交合成酶為經修飾之纈胺酸合成酶。在一些實施例中,正交合成酶為經修飾之磷絲胺酸合成酶。在一些實施例中,正交tRNA為經修飾之丙胺酸tRNA。在一些實施例中,正交tRNA為經修飾之精胺酸tRNA。在一些實施例中,正交tRNA為經修飾之天冬醯胺酸tRNA。在一些實施例中,正交tRNA為經修飾之天冬胺酸tRNA。在一些實施例中,正交tRNA為經修飾之半胱胺酸tRNA。在一些實施例中,正交tRNA為經修飾之麩醯胺酸tRNA。在一些實施例中,正交tRNA為經修飾之麩胺酸tRNA。在一些實施例中,正交tRNA為經修飾之丙胺酸甘胺酸。在一些實施例中,正交tRNA為經修飾之組胺酸tRNA。在一些實施例中,正交tRNA為經修飾之白胺酸tRNA。在一些實施例中,正交tRNA為經修飾之異白胺酸tRNA。在一些實施例中,正交tRNA為經修飾之離胺酸tRNA。在一些實施例中,正交tRNA為經修飾之甲硫胺酸tRNA。在一些實施例中,正交tRNA為經修飾之苯丙胺酸tRNA。在一些實施例中,正交tRNA為經修飾之脯胺酸tRNA。在一些實施例中,正交tRNA為經修飾之絲胺酸tRNA。在一些實施例中,正交tRNA為經修飾之蘇胺酸tRNA。在一些實施例中,正交tRNA為經修飾之色胺酸tRNA。在一些實施例中,正交tRNA為經修飾之酪胺酸tRNA。在一些實施例中,正交tRNA為經修飾之纈胺酸tRNA。在一些實施例中,正交tRNA為經修飾之磷絲胺酸tRNA。
在一些實施例中,非天然胺基酸可藉由胺基醯基(aaRS或RS)-tRNA合成酶-tRNA對併入非天然多肽或非天然蛋白質中。例示性aaRS-tRNA對包括(但不限於):詹氏甲烷球菌(Mj-Tyr
) aaRS/tRNA對、詹氏甲烷球菌(Methanococcus jannaschii/M. jannaschii
) TyrRS變異體p
AzFRS (Mjp
AzFRS)、大腸桿菌TyrRS (Ec-Tyr
)/嗜熱脂肪芽孢桿菌tRNACUA
對、大腸桿菌LeuRS (Ec-Leu
)/嗜熱脂肪芽孢桿菌tRNACUA
對及吡咯離胺醯基-tRNA對。在一些情況下,非天然胺基酸藉由Mj-Tyr
RS/tRNA對併入非天然多肽或非天然蛋白質中。可藉由Mj-Tyr
RS/tRNA對併入之例示性非天然胺基酸(UAA)包括(但不限於):經對位取代之苯丙胺酸衍生物,諸如對疊氮基-L-苯丙胺酸(p
AzF)、N6-(((2-疊氮基苯甲基)氧基)羰基)-L-離胺酸、N6-(((3-疊氮基苯甲基)氧基)羰基)-L-離胺酸、N6-(((4-疊氮基苯甲基)氧基)羰基)-L-離胺酸、對胺基苯丙胺酸及對甲基苯丙胺酸;經間位取代之酪胺酸衍生物,諸如3-胺基酪胺酸、3-硝基酪胺酸、3,4-二羥基苯丙胺酸及3-碘酪胺酸;苯基硒半胱胺酸;對溴苯丙胺酸;以及鄰硝基苯甲基酪胺酸。
在一些情況下,非天然胺基酸可藉由Ec-Tyr
/tRNACUA
或Ec-Leu
/tRNACUA
對併入非天然多肽或非天然蛋白質中。可藉由Ec-Tyr
/tRNACUA
或Ec-Leu
/tRNACUA
對併入之例示性UAA包括(但不限於):含有二苯甲酮、酮、碘或疊氮取代基之苯丙胺酸衍生物;O-炔丙基酪胺酸;α-胺基辛酸、O-甲基酪胺酸、O-硝基苯甲基半胱胺酸;以及3-(萘-2-基胺基)-2-胺基-丙酸。
在一些情況下,非天然胺基酸可藉由吡咯離胺醯基-tRNA對併入非天然多肽或非天然蛋白質中。在一些情況下,PylRS可自古細菌物種獲得,例如自產甲烷古細菌獲得。在一些情況下,PylRS可自巴氏甲烷八聯球菌(Methanosarcina barkeri
)、馬氏甲烷八聯球菌或嗜乙酸甲烷八聯球菌(Methanosarcina acetivorans
)。在一些情況下,PylRS可為嵌合PylRS。可藉由吡咯離胺醯基-tRNA對併入之例示性UAA包括(但不限於):經醯胺及胺基甲酸酯取代之離胺酸,諸如N6-(2-疊氮基乙氧基)-羰基-L-離胺酸(AzK)、N6-(((2-疊氮基苯甲基)氧基)羰基)-L-離胺酸、N6-(((3-疊氮基苯甲基)氧基)羰基)-L-離胺酸、N6-(((4-疊氮基苯甲基)氧基)羰基)-L-離胺酸、2-胺基-6-((R)-四氫呋喃-2-甲醯胺基)己酸、N-ε-D-脯胺醯基-L-離胺酸及N-ε-環戊基氧基羰基-L-離胺酸;N-ε-丙烯醯基-L-離胺酸;N-ε-[(1-(6-硝基苯并[d][1,3]間二氧雜環戊烯-5-基)乙氧基)羰基]-L-離胺酸;以及N-ε-(1-甲基環丙-2-烯甲醯胺基)離胺酸。
在一些情況下,如本文所描述之組合物及方法包含使用至少兩個tRNA合成酶將至少兩個非天然胺基酸併入非天然多肽或非天然蛋白質中。在一些情況下,至少兩個tRNA合成酶可相同或不同。在情況下,至少兩個非天然胺基酸可相同或不同。在一些情況下,併入非天然多肽中之至少兩個非天然胺基酸係不同的。在一些情況下,至少兩個不同非天然胺基酸可以位點特異性方式併入非天然多肽或非天然蛋白質中。
在一些情況下,非天然胺基酸可藉由US 9,988,619及US 9,938,516中所揭示之合成酶併入本文所描述之非天然多肽或非天然蛋白質中。可藉由此類合成酶併入之例示性UAA包括對甲基疊氮基-L-苯丙胺酸、芳烷基、雜環基、雜芳烷基非天然胺基酸及其他非天然胺基酸。在一些實施例中,此類UAA包含吡啶基、吡𠯤基、吡唑基、三唑基、㗁唑基、噻唑基、噻吩基或其他雜環。在一些實施例中,此類胺基酸包含疊氮基、四𠯤或諸如水溶性部分之能夠結合至偶合搭配物之其他化學基團。在一些實施例中,此類合成酶經表現且用於將UAA活體內併入蛋白質中。在一些實施例中,此類合成酶用以使用無細胞轉譯系統將UAA併入蛋白質中。
在一些情況下,非天然胺基酸可藉由天然存在之合成酶併入本文所描述之非天然多肽或非天然蛋白質中。在一些實施例中,非天然胺基酸藉由對於一或多個胺基酸具營養缺陷性之生物體併入非天然多肽或非天然蛋白質中。在一些實施例中,對應於營養缺陷型胺基酸之合成酶能夠使對應tRNA裝載有非天然胺基酸。在一些實施例中,非天然胺基酸為硒半胱胺酸或其衍生物。在一些實施例中,非天然胺基酸為硒甲硫胺酸或其衍生物。在一些實施例中,非天然胺基酸為芳族胺基酸,其中芳族胺基酸包含芳基鹵化物,諸如碘化物。在實施例中,非天然胺基酸在結構上與營養缺陷型胺基酸類似。在一些情況下,非天然胺基酸包含圖 5A
中所說明之非天然胺基酸。
在一些情況下,非天然胺基酸包含離胺酸或苯丙胺酸衍生物或類似物。在一些情況下,非天然胺基酸包含離胺酸衍生物或離胺酸類似物。在一些情況下,非天然胺基酸包含吡咯離胺酸(Pyl)。在一些情況下,非天然胺基酸包含苯丙胺酸衍生物或苯丙胺酸類似物。在一些情況下,非天然胺基酸為Wan等人, 「Pyrrolysyl-tRNA synthetase: an ordinary enzyme but an outstanding genetic code expansion tool」, Biocheim Biophys Aceta 1844(6): 1059-4070 (2014)中所描述之非天然胺基酸。在一些情況下,非天然胺基酸包含圖 5B
及圖 5C
中所說明之非天然胺基酸。
在一些實施例中,非天然胺基酸包含圖 5D- 圖 5G
中所說明之非天然胺基酸(改編自Dumas等人, Chemical Science 2015, 6, 50-69之表1)。
在一些實施例中,併入本文所描述之蛋白質中之非天然胺基酸揭示於US 9,840,493;US 9,682,934;US 2017/0260137;US 9,938,516;或US 2018/0086734中。可藉由此類合成酶併入之例示性UAA包括對甲基疊氮基-L-苯丙胺酸、芳烷基、雜環基及雜芳烷基,以及離胺酸衍生物非天然胺基酸。在一些實施例中,此類UAA包含吡啶基、吡𠯤基、吡唑基、三唑基、㗁唑基、噻唑基、噻吩基或其他雜環。在一些實施例中,此類胺基酸包含疊氮基、四𠯤或諸如水溶性部分之能夠結合至偶合搭配物之其他化學基團。在一些實施例中,UAA包含經由烷基連接子附接至芳族部分之疊氮基。在一些實施例中,烷基連接子為C1
-C10
連接子。在一些實施例中,UAA包含經由烷基連接子附接至芳族部分之四𠯤。在一些實施例中,UAA包含經由胺基附接至芳族部分之四𠯤。在一些實施例中,UAA包含經由烷胺基附接至芳族部分之四𠯤。在一些實施例中,UAA包含經由烷基鏈附接至胺基酸側鏈之末端氮(例如離胺酸衍生物之N6,或包含較短烷基側鏈之衍生物之N5、N4或N3)之疊氮基。在一些實施例中,UAA包含經由烷基鏈附接至胺基酸側鏈之末端氮之四𠯤。在一些實施例中,UAA包含經由烷基連接子附接至醯胺之疊氮基或四𠯤。在一些實施例中,UAA為3-胺基丙胺酸、絲胺酸、離胺酸或其衍生物之含有疊氮基或四𠯤之胺基甲酸酯或醯胺。在一些實施例中,此類UAA併入活體內蛋白質中。在一些實施例中,此類UAA併入無細胞系統中之蛋白質中。
細胞類型
在一些實施例中,許多類型之細胞/微生物用於例如轉化或基因工程改造。在一些實施例中,細胞為原核或真核細胞。在一些情況下,細胞為微生物,諸如細菌細胞、真菌細胞、酵母或單細胞原蟲。在其他情況下,細胞為真核細胞,諸如經培養之動物、植物或人類細胞。在其他情況下,細胞存在於生物體,諸如植物或動物中。
在一些實施例中,經工程改造之微生物為常常能夠分裂及增殖之單細胞生物體。微生物可包括以下特徵中之一或多者:好氧、厭氧、絲狀、非絲狀、單倍體、二倍體、營養缺陷型及/或非營養缺陷型。在某些實施例中,經工程改造之微生物為原核微生物(例如細菌),且在某些實施例中,經工程改造之微生物為非原核微生物。在一些實施例中,經工程改造之微生物為真核微生物(例如,酵母、真菌、阿米巴原蟲(amoeba))。在一些實施例中,經工程改造之微生物為真菌。在一些實施例中,經工程改造之生物體為酵母。
任何適合的酵母可經選為宿主微生物、經工程改造之微生物、經基因修飾之生物體或用於異源或經修飾之聚核苷酸之來源。酵母包括(但不限於):耶氏酵母(Yarrowia yeast) (例如解脂耶氏酵母(Y. lipolytica) (先前分類為解脂假絲酵母(Candida lipolytica))、假絲酵母(Candida yeast) (例如拉考夫假絲酵母(C. revkaufi)、維斯假絲酵母(C. viswanathii)、鐵紅假絲酵母(C. pulcherrima)、熱帶假絲酵母(C. tropicalis)、產朊假絲酵母(C. utilis))、紅酵母(Rhodotorula yeast) (例如黏紅酵母(R. glutinus)、牧草紅酵母(R. graminis))、紅冬孢酵母(Rhodosporidium yeast) (例如圓紅冬孢酵母(R. toruloides))、酵母屬酵母(Saccharomyces yeast) (例如,釀酒酵母(S. cerevisiae)、貝酵母(S. bayanus)、巴斯德酵母(S. pastorianus)、卡爾斯伯酵母(S. carlsbergensis))、隱球酵母(Cryptococcus yeast)、毛芽胞酵母(Trichosporon yeast) (例如茁牙毛芽胞酵母(T. pullans)、皮狀毛芽胞酵母(T. cutaneum))、畢赤酵母(Pichia yeast) (例如,巴斯德畢赤酵母(P. pastoris))及油脂酵母(Lipomyces yeast) (例如,斯達油脂酵母(L. starkeyii)、產油油脂酵母(L. lipoferus))。在一些實施例中,適合的酵母為蛛網黴屬(Arachniotus)、麴菌屬(Aspergillus)、短梗黴屬(Aureobasidium)、奧氏菌屬(Auxarthron)、芽生菌屬(Blastomyces)、假絲酵母、金孢子菌屬(Chrysosporuim)、德巴利酵母金孢子菌屬(Chrysosporuim Debaryomyces)、球孢子菌(Coccidiodes)、隱球酵母、裸囊菌屬(Gymnoascus)、漢森酵母(Hansenula)、組織漿菌屬(Histoplasma)、伊薩酵母屬(Issatchenkia)、克魯維酵母(Kluyveromyces)、油脂酵母、伊薩酵母屬(Lssatchenkia)、小芽孢菌屬(Microsporum)、地衣內生菌屬(Myxotrichum)、密氏菌屬(Myxozyma)、樹粉孢屬(Oidiodendron)、管囊酵母屬(Pachysolen)、青黴菌屬(Penicillium)、畢赤酵母、紅冬孢酵母屬、紅酵母、紅酵母、酵母、裂殖酵母屬(Schizosaccharomyces)、帚黴屬(Scopulariopsis)、瘤孢屬(Sepedonium)、毛芽胞酵母或耶氏酵母屬。在一些實施例中,適合的酵母為以下物種之酵母:弗氏蛛網黴菌(Arachniotus flavoluteus)、黃麴黴(Aspergillus flavus)、煙麯黴(Aspergillus fumigatus)、黑麴菌(Aspergillus niger)、出芽短梗黴菌(Aureobasidium pullulans)、薩氏奧氏菌(Auxarthron thaxteri)、皮炎芽生菌(Blastomyces dermatitidis)、白假絲酵母(Candida albicans)、都柏林假絲酵母(Candida dubliniensis)、無名假絲酵母(Candida famata)、光滑假絲酵母(Candida glabrata)、高里假絲酵母(Candida guilliermondii)、克菲假絲酵母(Candida kefyr)、克魯斯假絲酵母(Candida krusei)、郎比可假絲酵母(Candida lambica)、解脂假絲酵母、魯氏假絲酵母(Candida lustitaniae)、近平滑假絲酵母(Candida parapsilosis)、鐵紅假絲酵母、拉考夫假絲酵母、皺褶假絲酵母(Candida rugosa)、熱帶假絲酵母、高蛋白假絲酵母、維斯假絲酵母、西氏假絲酵母(Candida xestobii)、克拉氏金孢子菌(Chrysosporuim keratinophilum)、粗球黴菌(Coccidiodes immitis)、白色隱球菌液化變種(Cryptococcus albidus var. diffluens)、羅倫隱球酵母(Cryptococcus laurentii)、新型隱球酵母(Cryptococcus neofomans)、漢森德巴利酵母(Debaryomyces hansenii)、杜氏裸囊菌(Gymnoascus dugwayensis)、異常漢森酵母(Hansenula anomala)、莢膜組織胞漿菌(Histoplasma capsulatum)、西方伊薩酵母(Issatchenkia occidentalis)、東方伊薩酵母(Isstachenkia orientalis)、乳酸克魯維酵母(Kluyveromyces lactis)、馬克斯克魯維酵母(Kluyveromyces marxianus)、耐熱克魯維酵母(Kluyveromyces thermotolerans)、瓦爾提克魯維酵母(Kluyveromyces waltii)、產油油脂酵母、斯達油脂酵母、石膏狀小芽孢菌(Microsporum gypseum)、反折地衣內生菌(Myxotrichum deflexum)、刺樹粉孢菌(Oidiodendron echinulatum)、嗜鞣管囊酵母(Pachysolen tannophilis)、點青黴(Penicillium notatum)、異常畢赤酵母(Pichia anomala)、巴斯德畢赤酵母(Pichia pastoris)、樹幹畢赤酵母(Pichia stipitis)、圓紅冬孢酵母、黏紅酵母、牧草紅酵母、釀酒酵母、克魯維酵母(Saccharomyces kluyveri)、粟酒裂殖酵母(Schizosaccharomyces pombe)、枝頂孢帚黴菌(Scopulariopsis acremonium)、黃瘤孢菌(Sepedonium chrysospermum)、皮狀毛芽胞酵母、茁牙毛芽胞酵母、解脂耶氏酵母或解脂耶氏酵母(先前分類為解脂假絲酵母)。在一些實施例中,酵母為解脂耶氏酵母菌株,其包括(但不限於):ATCC20362、ATCC8862、ATCC18944、ATCC20228、ATCC76982及LGAM S(7)1菌株(Papanikolaou S.及Aggelis G., Bioresour. Technol. 82(1):43-9 (2002))。在某些實施例中,酵母為假絲酵母物種(亦即假絲酵母屬)酵母。任何適合的假絲酵母物種可用於及/或經基因修飾以用於產生脂肪二羧酸(例如,辛二酸、癸二酸、十二烷二酸、十四烷二酸、十六烷二酸、十八烷二酸、二十烷二酸)。在一些實施例中,適合的假絲酵母物種包括(但不限於):白假絲酵母、都柏林假絲酵母、無名假絲酵母、光滑假絲酵母、高里假絲酵母、克菲假絲酵母、克魯斯假絲酵母、郎比可假絲酵母、解脂假絲酵母、魯氏假絲酵母、近平滑假絲酵母、鐵紅假絲酵母、拉考夫假絲酵母、皺褶假絲酵母、熱帶假絲酵母、高蛋白假絲酵母、維斯假絲酵母、西氏假絲酵母及本文所描述之任何其他假絲酵母屬酵母。假絲酵母屬菌株之非限制性實例包括(但不限於):sAA001 (ATCC20336)、sAA002 (ATCC20913)、sAA003 (ATCC20962)、sAA496 (US2012/0077252)、sAA106 (US2012/0077252)、SU-2 (ura3-/ura3-)、H5343 (β氧化阻斷;美國專利第5648247號)菌株。來自假絲酵母屬酵母之任何適合的菌株均可用作用於基因修飾之親本菌株。
酵母屬、物種及菌株常常在基因內容上緊密相關以使得其可能難以區分、分類及/或命名。在一些情況下,解脂假絲酵母與解脂耶氏酵母之菌株可能難以區分、分類及/或命名,且在一些情況下,其可能視為相同的生物體。在一些情況下,熱帶假絲酵母及維斯假絲酵母之各種菌株可能難以區分、分類及/或命名(例如參見Arie等人, J. Gen. Appl.Microbiol., 46, 257-262 (2000)。某些自ATCC以及自其他市售或學術來源獲得之熱帶假絲酵母及維斯假絲酵母菌株可視為等效的且同樣適用於本文所描述之實施例。在一些實施例中,熱帶假絲酵母及維斯假絲酵母之某些親本菌株視為僅在名稱上不同。
任何適合的真菌可經選為宿主微生物、經工程改造之微生物或用於異源聚核苷酸之來源。真菌之非限制性實例包括(但不限於):麴菌屬真菌(例如寄生麴菌(A. parasiticus)、構巢麴菌(A. nidulans))、破囊壺菌屬(Thraustochytrium)真菌、裂殖壺菌(Schizochytrium)真菌及根黴菌屬(Rhizopus)真菌(例如鬚根黴菌(R. arrhizus)、米根黴菌(R. oryzae)、黑根黴菌(R. nigricans))。在一些實施例中,真菌為寄生麴菌菌株,其包括(但不限於)菌株ATCC24690,且在某些實施例中,真菌為構巢麴菌菌株,其包括(但不限於)菌株ATCC38163。
任何適合的原核生物可經選為宿主微生物、經工程改造之微生物或用於異源聚核苷酸之來源。可選擇革蘭氏陰性細菌或革蘭氏陽性細菌。細菌之實例包括(但不限於):芽孢桿菌屬(Bacillus)細菌(例如枯草芽孢桿菌(B. subtilis)、巨大芽孢桿菌(B. megaterium))、不動桿菌屬(Acinetobacter)細菌、奴卡菌屬(Norcardia)細菌、黃色桿菌屬(Xanthobacter)細菌、艾氏菌屬(Escherichia)細菌(例如,大腸桿菌(例如菌株DH10B、Stbl2、DH5-α、DB3、DB3.1)、DB4、DB5、JDP682及ccdA-over (例如,美國申請案第09/518,188號)))、鏈黴菌屬(Streptomyces)細菌、伊文氏桿菌屬(Erwinia)細菌、克雷伯氏菌屬(Klebsiella)細菌、沙雷菌屬(Serratia)細菌(例如黏質沙雷菌屬(S. marcessans))、假單胞菌屬(Pseudomonas)細菌(例如,銅綠假單胞菌(P. aeruginosa)、沙門氏菌屬(Salmonella)細菌(例如,鼠傷寒沙門氏菌(S. typhimurium)、傷寒沙門氏菌(S. typhi)、巨型球菌屬(Megasphaera)細菌(例如埃氏巨型球菌(Megasphaera elsdenii))。細菌亦包括(但不限於):光合細菌(例如,綠色非硫細菌(例如綠彎菌屬(Choroflexus)細菌(例如橙色綠彎菌(C. aurantiacus))、直立原絲體(Chloronema)細菌(例如吉氏直立原絲體(C. gigateum)))、綠色硫細菌(例如綠色硫黃細菌屬(Chlorobium)細菌(例如,泥生綠色硫黃細菌(C. limicola)、暗網菌屬(Pelodictyon)細菌(例如微黃暗網菌(P. luteolum))、紫色硫細菌(例如,紅色硫黃細菌屬(Chromatium)細菌(例如硫黃菌(C. okenii)))及紫色非硫細菌(例如,紅螺菌屬(Rhodospirillum)細菌(例如深紅紅螺菌(R. rubrum))、紅細菌屬(Rhodobacter)細菌(例如,類球紅細菌(R. sphaeroides)、莢膜紅細菌(R. capsulatus))及紅微菌屬(Rhodomicrobium)細菌(例如瓦氏紅微菌(R. vanellii))。
來自非微生物生物體之細胞可用作宿主微生物、經工程改造之微生物或用於異源聚核苷酸之來源。此類細胞之實例包括(但不限於):昆蟲細胞(例如果蠅屬(Drosophila) (例如黑腹果蠅(D. melanogaster))、夜蛾屬(Spodoptera) (例如草地夜蛾(S. frugiperda) Sf9或Sf21細胞)及粉夜蛾(Trichoplusa) (例如High-Five細胞)、線蟲細胞(例如秀麗隱桿線蟲(C. elegans)細胞);禽類細胞;兩棲動物細胞(例如,有爪蟾蜍(Xenopus laevis)細胞);爬行動物細胞;哺乳動物細胞(例如,NIH3T3、293、CHO、COS、VERO、C127、BHK、Per-C6、布茲(Bowes)黑素瘤及海拉(HeLa)細胞);及植物細胞(例如阿拉伯芥(Arabidopsis thaliana)、煙草(Nicotania tabacum)、腺泡葉萼距花(Cuphea acinifolia)、等大花瓣萼距花(Cuphea aequipetala)、狹葉萼距花(Cuphea angustifolia)、附蓋萼距花(Cuphea appendiculata)、阿維吉拉萼距花(Cuphea avigera)、阿維吉拉萼距花鐵紅變種(Cuphea avigera var. pulcherrima)、腋花萼距花(Cuphea axilliflora)、巴希萼距花(Cuphea bahiensis)、巴氏萼距花(Cuphea baillonis)、短柄萼距花(Cuphea brachypoda)、巴氏萼距花(Cuphea bustamanta)、囊距萼距花(Cuphea calcarata)、美葉萼距花(Cuphea calophylla)、美葉萼距花亞種美索萼距花(Cuphea calophylla subsp. mesostemon)、香膏萼距花(Cuphea carthagenensis)、露珠萼距花(Cuphea circaeoides)、密花萼距花(Cuphea confertiflora)、柯達他萼距花(Cuphea cordata)、厚瓣萼距花(Cuphea crassiflora)、藍萼距花(Cuphea cyanea)、十蕊萼距花(Cuphea decandra)、齒狀萼距花(Cuphea denticulata)、二籽萼距花(Cuphea disperma)、艾氏萼距花(Cuphea epilobiifolia)、長枝萼距花(Cuphea ericoides)、黃萼距花(Cuphea flava)、弗氏萼距花(Cuphea flavisetula)、紫斑葉萼距花(Cuphea fuchsiifolia)、高姆瑞萼距花(Cuphea gaumeri)、黏毛萼距花(Cuphea glutinosa)、異葉萼距花(Cuphea heterophylla)、虎克里亞納萼距花(Cuphea hookeriana)、細葉萼距花(Cuphea hyssopifolia) (墨西哥石南(Mexican-heather))、尖果萼距花(Cuphea hyssopoides)、火紅萼距花(Cuphea ignea)、臭味萼距花(Cuphea ingrata)、約魯萼距花(Cuphea jorullensis)、杉木萼距花(Cuphea lanceolata)、長葉萼距花(Cuphea linarioides)、拉維亞萼距花(Cuphea llavea)、羅氏萼距花(Cuphea lophostoma)、黃萼距花(Cuphea lutea)、金色萼距花(Cuphea lutescens)、美麗萼距花(Cuphea melanium)、梅爾維亞萼距花(Cuphea melvilla)、小花萼距花(Cuphea micrantha)、小瓣萼距花(Cuphea micropetala)、米姆羅德萼距花(Cuphea mimuloides)、光葉萼距花(Cuphea nitidula)、沼澤萼距花(Cuphea palustris)、同心結萼距花(Cuphea parsonsia)、巴斯庫洛姆萼距花(Cuphea pascuorum)、寡瓣萼距花(Cuphea paucipetala)、樸氏萼距花(Cuphea procumbens)、假蠅子草萼距花(Cuphea pseudosilene)、假越橘萼距花(Cuphea pseudovaccinium)、美花萼距花(Cuphea pulchra)、紅花萼距花(Cuphea racemosa)、匍匐萼距花(Cuphea repens)、柳葉萼距花(Cuphea salicifolia)、薩爾瓦多萼距花(Cuphea salvadorensis)、斯庫瑪妮萼距花(Cuphea schumannii)、無梗萼距花(Cuphea sessiliflora)、無柄萼距花(Cuphea sessilifolia)、刺芒萼距花(Cuphea setosa)、奇麗萼距花(Cuphea spectabilis)、闊葉萼距花(Cuphea spermacoce)、華麗萼距花(Cuphea splendida)、華麗萼距花變種綠黃萼距花(Cuphea splendida var. viridiflava)、斯氏萼距花(Cuphea strigulosa)、薩氏萼距花(Cuphea subuligera)、特氏萼距花(Cuphea teleandra)、賽姆德萼距花(Cuphea thymoides)、托卡尼卡萼距花(Cuphea tolucana)、尖葉萼距花(Cuphea urens)、膨脹萼距花(Cuphea utriculosa)、藍葉柄萼距花(Cuphea viscosissima)、沃森萼距花(Cuphea watsoniana)、來提萼距花(Cuphea wrightii)、杉木萼距花(Cuphea lanceolata)。
用作宿主生物體或異源聚核苷酸之來源的微生物或細胞為可商購的。本文所描述之微生物及細胞以及其他適合的微生物及細胞可獲自例如Invitrogen公司(Carlsbad, CA)、美國菌種保藏中心(American Type Culture Collection) (Manassas, Virginia)及農業研究菌種保藏中心(Agricultural Research Culture Collection) (NRRL;Peoria, Illinois)。宿主微生物及經工程改造之微生物可以任何適合的形式提供。舉例而言,此類微生物可以液體培養物或固體培養物(例如基於瓊脂之培養基)提供,其可為原始培養物或可傳代(例如,經稀釋及培養)一或多次。微生物亦可以冷凍形式或乾燥形式(例如凍乾)提供。微生物可以任何適合的濃度提供。
聚合酶
聚合酶之尤其適用的功能為使用現存核酸作為模板催化核酸股之聚合。其他適用的功能在本文中別處描述。適用的聚合酶之實例包括DNA聚合酶及RNA聚合酶。
在例如需要併入非天然核酸之多種情形中,包括擴增、定序、標記、偵測、選殖及諸多其他,將高度需要改良聚合酶非天然核酸之特異性、持續力或其他特徵之能力。
在一些情況下,本文所揭示之內容包括,例如在DNA擴增期間將非天然核酸併入生長模板複本中之聚合酶。在一些實施例中,聚合酶可經修飾以使得聚合酶之活性位點經修飾以減小非天然核酸至活性位點中之立體進入抑制。在一些實施例中,聚合酶可經修飾以提供與非天然核酸之一或多個非天然特徵之互補。此類聚合酶可在細胞中經表現或工程改造以用於將UBP穩定地併入細胞中。因此,本發明包括含異源或重組聚合酶之組合物及其使用方法。
可使用關於蛋白質工程改造之方法修飾聚合酶。舉例而言,可基於晶體結構進行分子模型化以鑑別可進行突變以修飾目標活性之聚合酶之位置。鑑別為替代目標之殘基可經使用能量最小化模型化、同源性模型化及/或保守胺基酸取代所選擇之殘基來置換,諸如Bordo等人, J Mol Biol 217: 721-729 (1991)及Hayes等人, Proc Natl Acad Sci, USA 99: 15926- 15931 (2002)中所描述。
各種聚合酶中之任一者可用於本文所闡述之方法或組合物中,包括例如自生物系統分離之蛋白質類酶及其功能變異體。除非另外指示,否則提及特定聚合酶(諸如下文所例示之彼等聚合酶)應理解為包括其功能變異體。在一些實施例中,聚合酶為野生型聚合酶。在一些實施例中,聚合酶為經修飾或突變之聚合酶。
亦可使用具有用於改良非天然核酸至活性位點區域中之進入及用於與非天然核苷酸在活性位點區域中配位之特徵的聚合酶。在一些實施例中,經修飾之聚合酶具有經修飾之核苷酸結合位點。
在一些實施例中,經修飾之聚合酶具有針對非天然核酸之特異性,其為野生型聚合酶針對非天然核酸之特異性的至少約10%、20%、30%、40%、50%、60%、70%、80%、90%、95%、97%、98%、99%、99.5%、99.99%。在一些實施例中,經修飾或野生型聚合酶具有針對包含經修飾之糖之非天然核酸的特異性,其為野生型聚合酶針對不具有經修飾之糖之天然核酸及/或非天然核酸之特異性的至少約10%、20%、30%、40%、50%、60%、70%、80%、90%、95%、97%、98%、99%、99.5%、99.99%。在一些實施例中,經修飾或野生型聚合酶具有針對包含經修飾之鹼基之非天然核酸的特異性,其為野生型聚合酶針對不具有經修飾之鹼基之天然核酸及/或非天然核酸之特異性的至少約10%、20%、30%、40%、50%、60%、70%、80%、90%、95%、97%、98%、99%、99.5%、99.99%。在一些實施例中,經修飾或野生型聚合酶具有針對包含三磷酸酯之非天然核酸的特異性,其為野生型聚合酶針對包含三磷酸酯之核酸及/或不具有三磷酸酯之非天然核酸的特異性的至少約10%、20%、30%、40%、50%、60%、70%、80%、90%、95%、97%、98%、99%、99.5%、99.99%。舉例而言,經修飾或野生型聚合酶可具有針對包含三磷酸酯之非天然核酸的特異性,其為野生型聚合酶針對具有二磷酸酯或單磷酸酯或無磷酸酯或其組合之非天然核酸之特異性的至少約10%、20%、30%、40%、50%、60%、70%、80%、90%、95%、97%、98%、99%、99.5%、99.99%。
在一些實施例中,經修飾或野生型聚合酶具有針對非天然核酸寬鬆的特異性。在一些實施例中,經修飾或野生型聚合酶具有針對非天然核酸之特異性及針對天然核酸之特異性,其為野生型聚合酶針對天然核酸之特異性的至少約10%、20%、30%、40%、50%、60%、70%、80%、90%、95%、97%、98%、99%、99.5%、99.99%。在一些實施例中,經修飾或野生型聚合酶具有針對包含經修飾之糖之非天然核酸的特異性及針對天然核酸之特異性,其為野生型聚合酶針對天然核酸之特異性的至少約10%、20%、30%、40%、50%、60%、70%、80%、90%、95%、97%、98%、99%、99.5%、99.99%。在一些實施例中,經修飾或野生型聚合酶具有針對包含經修飾鹼基之非天然核酸之特異性及針對天然核酸之特異性,其為野生型聚合酶針對天然核酸之特異性的至少約10%、20%、30%、40%、50%、60%、70%、80%、90%、95%、97%、98%、99%、99.5%、99.99%。
缺乏核酸外切酶活性可為野生型特徵或由變體聚合酶或經工程改造之聚合酶賦予之特徵。舉例而言,exo minus克列諾片段為不具有3'至5'校正核酸外切酶活性之克列諾片段之突變版本。
本發明之方法可用於擴展缺乏固有3至5'核酸外切酶校正活性或3至5'核酸外切酶校正活性已(例如經由突變)失能之任何DNA聚合酶的基質範圍。DNA聚合酶之實例包括polA、polB (參見,例如Parrel及Loeb, Nature Struc Biol 2001)、polC、polD、polY、polX及逆轉錄酶(RT),但較佳為前進型高保真聚合酶(PCT/GB2004/004643)。在一些實施例中,經修飾或野生型聚合酶實質上不具有3'至5'校正核酸外切酶活性。在一些實施例中,經修飾或野生型聚合酶實質上不具有針對非天然核酸之3'至5'校正核酸外切酶活性。在一些實施例中,經修飾或野生型聚合酶具有3'至5'校正核酸外切酶活性。在一些實施例中,經修飾或野生型聚合酶具有針對天然核酸之3'至5'校正核酸外切酶活性且實質上缺乏針對非天然核酸之3'至5'校正核酸外切酶活性。
在一些實施例中,經修飾之聚合酶具有3'至5'校正核酸外切酶活性,其為野生型聚合酶之校正核酸外切酶活性之至少約60%、70%、80%、90%、95%、97%、98%、99%、99.5%、99.99%。在一些實施例中,經修飾之聚合酶具有針對非天然核酸之3'至5'校正核酸外切酶活性,其為野生型聚合酶針對天然核酸之校正核酸外切酶活性的至少約60%、70%、80%、90%、95%、97%、98%、99%、99.5%、99.99%。在一些實施例中,經修飾之聚合酶具有針對非天然核酸之3'至5'校正核酸外切酶活性及針對天然核酸之3'至5'校正核酸外切酶活性,其為野生型聚合酶針對天然核酸之校正核酸外切酶活性的至少約60%、70%、80%、90%、95%、97%、98%、99%、99.5%、99.99%。在一些實施例中,經修飾之聚合酶具有針對天然核酸之3'至5'校正核酸外切酶活性,其為野生型聚合酶針對天然核酸之校正核酸外切酶活性的至少約60%、70%、80%、90%、95%、97%、98%、99%、99.5%、99.99%。
在一些實施例中,聚合酶係根據其自核酸解離之速率表徵。在一些實施例中,聚合酶具有對於一或多種天然及非天然核酸之相對較低的解離速率。在一些實施例中,聚合酶具有對於一或多種天然及非天然核酸之相對較高的解離速率。解離速率為可經調節以在本文所闡述之方法中調節反應速率之聚合酶活性。
在一些實施例中,聚合酶可根據其在與特定天然及/或非天然核酸或天然及/或非天然核酸之集合一起使用時之保真度來表徵。保真度通常係指在製備核酸模板之複本時聚合酶將正確核酸併入生長核酸鏈中之精確度。DNA聚合酶保真度可量測為在聚合酶-股-模板核酸二元複合物中,在相同位點處競爭股合成的正確與不正確天然及非天然核酸(當天然及非天然核酸存在時(例如在相同濃度下))併入之比率。DNA聚合酶保真度可計算為(kcat
/Km
)(針對天然及非天然核酸)與(kcat
/Km
) (針對不正確天然及非天然核酸)之比率;其中kcat
及Km
為在穩態酶動力學中之Michaelis-Menten參數(Fersht, A. R. (1985) Enzyme Structure and Mechanism, 第2版, 第350頁, W. H. Freeman & Co., New York.,以引用之方式併入本文中)。在一些實施例中,在具有或不具有校正活性之情況下,聚合酶具有至少約100、1000、10,000、100,000或1×106
之保真度值。
在一些實施例中,使用偵測具有特定結構之非天然核酸之併入的分析篩選來自其天然來源之聚合酶或其變異體。在一個實例中,聚合酶可針對併入非天然核酸或UBP之能力來篩選;例如d5SICSTP、dCNMOTP、d dTPT3TP、dNaMTP、dCNMOTP-dTPT3TP或d5SICSTP-dNaMTP UBP。可使用呈現與野生型聚合酶相比針對非天然核酸之經修飾的特性的聚合酶,例如異源聚合酶。舉例而言,經修飾之特性可為例如在非天然核酸(或天然存在之核苷酸)存在下之Km
、kcat
、Vmax
、聚合酶持續力;在非天然核酸存在下之藉由聚合酶之平均模板讀段-長度;聚合酶針對非天然核酸之特異性;非天然核酸之結合速率;產物(焦磷酸酯、三磷酸酯等)釋放之速率;分支化速率或其任何組合。在一個實施例中,經修飾之特性為針對非天然核酸之減小的Km
及/或針對非天然核酸之增加的kcat
/Km
或Vmax
/Km
。類似地,與野生型聚合酶相比,該聚合酶視情況具有增加的非天然核酸結合速率、增加的產物釋放速率及/或降低的分支化速率。
同時,聚合酶可將天然核酸(例如A、C、G及T)併入生長核酸複本中。舉例而言,聚合酶視情況呈現針對天然核酸之比活性,其為對應的野生型聚合酶之至少約5%高(例如5%、10%、25%、50%、75%、100%或更高);及在天然核酸存在下模板之持續合成能力,其為在天然核酸存在下野生型聚合酶之至少5%高(例如5%、10%、25%、50%、75%、100%或更高)。視情況,聚合酶呈現針對天然存在之核苷酸之kcat
/Km
或Vmax
/Km
,其為野生型聚合酶之至少約5%高(例如約5%、10%、25%、50%、75%或100%或更高)。
本文所使用之可具有併入具有特定結構之非天然核酸之能力的聚合酶亦可使用定向進化方法來產生。核酸合成分析可用於篩選具有針對多種非天然核酸中之任一者之特異性的聚合酶變異體。舉例而言,聚合酶變異體可針對在DNA模板中併入與非天然核苷酸相對之非天然核苷三磷酸之能力篩選;例如dTPT3TP相對dCNMO、dCNMOTP相對dTPT3、NaMTP相對dTPT3或TAT1TP相對dCNMO或dNaM。在一些實施例中,此類分析為例如使用重組聚合酶變異體之活體外分析。在一些實施例中,此類分析為例如在細胞中表現聚合酶變異體之活體內分析。此類定向進此類化技術可用於篩選任何適合的聚合酶之變異體對於本文所闡述的非天然核酸中之任一者之活性。在一些情況下,本文所使用之聚合酶具有將非天然核糖核苷酸併入諸如RNA之核酸中之能力。舉例而言,使用本文所描述之聚合酶將NaM或TAT1核糖核苷酸併入核酸中。
所描述之組合物之經修飾之聚合酶可視情況為經修飾及/或重組Φ29型DNA聚合酶。視情況,聚合酶可為經修飾及/或重組Φ29、B103、GA-1、PZA、Φ15、BS32、M2Y、Nf、G1、Cp-1、PRD1、PZE、SF5、Cp-5、Cp-7、PR4、PR5、PR722或L17聚合酶。
所描述之組合物之經修飾之聚合酶可視情況為經修飾及/或重組原核DNA聚合酶,例如DNA聚合酶II (Pol II)、DNA聚合酶III (Pol III)、DNA聚合酶IV (Pol IV)、DNA聚合酶V (Pol V)。在一些實施例中,經修飾之聚合酶包含介導跨越非指令破壞之核苷酸的DNA合成的聚合酶。在一些實施例中,編碼Pol I、Pol II (polB
)、Poll IV (dinB
)及/或Pol V (umuCD
)之基因在經工程改造之細胞或SSO中組成性表現或過度表現。在一些實施例中,Pol II之表現或過度表現增加促成經工程改造之細胞或SSO中之非天然鹼基對(UBP)之保留率增加。
通常適用於本發明之核酸聚合酶包括DNA聚合酶、RNA聚合酶、逆轉錄酶及其突變形式或變化形式。DNA聚合酶及其特性尤其詳細描述於DNA Replication第2版, Kornberg及Baker, W. H. Freeman, New York, N. Y. (1991)中。已知的適用於本發明之習知DNA聚合酶包括(但不限於):強烈火球菌(Pyrococcus furiosus;Pfu) DNA聚合酶(Lundberg等人, 1991, Gene, 108: 1, Stratagene);烏茲火球菌(Pyrococcus woesei;Pwo) DNA聚合酶(Hinnisdaels等人, 1996, Biotechniques, 20:186-8, Boehringer Mannheim);嗜熱棲熱菌(Thermus thermophilus;Tth) DNA聚合酶(Myers及Gelfand 1991, Biochemistry 30:7661);嗜熱脂肪芽孢桿菌(Bacillus stearothermophilus) DNA聚合酶(Stenesh及McGowan, 1977, Biochim Biophys Acta 475:32);海濱嗜熱球菌(Thermococcus litoralis;TIi) DNA聚合酶(亦稱為Vent™ DNA聚合酶, Cariello等人, 1991, Polynucleotides Res, 19: 4193, New England Biolabs);9°Nm™ DNA聚合酶(New England Biolabs)、Stoffel片段;Thermo Sequenase®
(Amersham Pharmacia Biotech UK);Therminator™ (New England Biolabs);海棲熱孢菌(Thermotoga maritima;Tma) DNA聚合酶(Diaz及Sabino, 1998 Braz J Med. Res, 31 :1239);水生棲熱菌(Thermus aquaticus;Taq) DNA聚合酶(Chien等人, 1976, J. Bacteoriol, 127: 1550);DNA聚合酶;鹿兒島火球菌(Pyrococcus kodakaraensis) KOD DNA聚合酶(Takagi等人, 1997, Appl. Environ. Microbiol. 63:4504);JDF-3 DNA聚合酶(來自熱球菌屬JDF-3,專利申請案WO 0132887);GB-D火球菌(PGB-D) DNA聚合酶(亦稱為Deep Vent™ DNA聚合酶,Juncosa-Ginesta等人, 1994, Biotechniques, 16:820, New England Biolabs);UlTma DNA聚合酶(來自嗜熱海棲熱孢菌(thermophile Thermotoga maritima);Diaz及Sabino, 1998 Braz J. Med. Res, 31 :1239; PE Applied Biosystems);Tgo DNA聚合酶(來自柳珊瑚熱球菌(thermococcus gorgonarius),Roche Molecular Biochemicals);大腸桿菌DNA聚合酶I (Lecomte及Doubleday, 1983, Polynucleotides Res. 11 :7505);T7 DNA聚合酶(Nordstrom等人, 1981, J Biol. Chem. 256:3112);及古細菌DP1I/DP2 DNA聚合酶II (Cann等人, 1998, Proc. Natl. Acad. Sci. USA 95:14250)。考慮嗜溫聚合酶及嗜熱聚合酶兩者。嗜熱DNA聚合酶包括(但不限於):ThermoSequenase®、9°Nm™、Therminator™、Taq、Tne、Tma、Pfu、TfI、Tth、TIi、Stoffel片段、Vent™及Deep Vent™ DNA聚合酶、KOD DNA聚合酶、Tgo、JDF-3及其突變體、變異體及衍生物。亦考慮作為3'核酸外切酶缺失型突變體之聚合酶。適用於本發明之逆轉錄酶包括(但不限於)來自以下之逆轉錄酶:HIV、HTLV-I、HTLV-II、FeLV、FIV、SIV、AMV、MMTV、MoMuLV及其他逆轉錄病毒(參見Levin, Cell 88:5-8 (1997);Verma, Biochim Biophys Acta. 473:1-38 (1977);Wu等人, CRC Crit Rev Biochem. 3:289- 347(1975))。聚合酶之其他實例包括(但不限於):9°N™ DNA聚合酶、Taq DNA聚合酶、Phusion® DNA聚合酶、Pfu DNA聚合酶、RB69 DNA聚合酶、KOD DNA聚合酶及VentR® DNA聚合酶(Gardner等人, (2004) 「Comparative Kinetics of Nucleotide Analog Incorporation by Vent DNA Polymerase」 J. Biol. Chem., 279(12), 11834-11842;Gardner及Jack 「Determinants of nucleotide sugar recognition in an archaeon DNA polymerase」 Nucleic Acids Research, 27(12) 2545-2553)。自非嗜熱性生物體分離之聚合酶可為可熱滅活的。實例為來自噬菌體之DNA聚合酶。應理解,來自各種來源中之任一者的聚合酶可經修飾以增加或降低其對高溫條件之耐受性。在一些實施例中,聚合酶可為嗜熱性的。在一些實施例中,嗜熱聚合酶可為可熱滅活的。嗜熱聚合酶典型地適用於高溫條件或熱循環條件,諸如用於聚合酶鏈反應(polymerase chain reaction;PCR)技術之熱循環條件之彼等嗜熱聚合酶。
在一些實施例中,聚合酶包含:Φ29、B103、GA-1、PZA、Φ15、BS32、M2Y、Nf、G1、Cp-1、PRD1、PZE、SF5、Cp-5、Cp-7、PR4、PR5、PR722、L17、ThermoSequenase®、9°Nm™、Therminator™ DNA聚合酶、Tne、Tma、TfI、Tth、TIi、Stoffel片段、Vent™及Deep Vent™ DNA聚合酶、KOD DNA聚合酶、Tgo、JDF-3、Pfu、Taq、T7 DNA聚合酶、T7 RNA聚合酶、PGB-D、UlTma DNA聚合酶、大腸桿菌DNA聚合酶I、大腸桿菌DNA聚合酶III、古細菌DP1I/DP2 DNA聚合酶II、9°N™ DNA聚合酶、Taq DNA聚合酶、Phusion® DNA聚合酶、Pfu DNA聚合酶、SP6 RNA聚合酶、RB69 DNA聚合酶、鳥成髓細胞瘤病毒(Avian Myeloblastosis Virus;AMV)逆轉錄酶、莫洛尼鼠類白血病病毒(Moloney Murine Leukemia Virus;MMLV)逆轉錄酶、SuperScript® II逆轉錄酶及SuperScript® III逆轉錄酶。
在一些實施例中,聚合酶為DNA聚合酶I (或克列諾片段)、Vent聚合酶、Phusion® DNA聚合酶、KOD DNA聚合酶、Taq聚合酶、T7 DNA聚合酶、T7 RNA聚合酶、Therminator™ DNA聚合酶、POLB聚合酶、SP6 RNA聚合酶、大腸桿菌DNA聚合酶I、大腸桿菌DNA聚合酶III、鳥成髓細胞瘤病毒(AMV)逆轉錄酶、莫洛尼鼠類白血病病毒(MMLV)逆轉錄酶、SuperScript® II逆轉錄酶或SuperScript® III逆轉錄酶。
核苷酸轉運子
核苷酸轉運體(NT)為促進核苷酸基質跨越細胞膜及囊泡轉移之一組膜轉運蛋白。在一些實施例中,存在兩種類型之NT,即集中型核苷轉運子及平衡型核苷轉運子。在一些情況下,NT亦涵蓋有機陰離子轉運子(OAT)及有機陽離子轉運子(OCT)。在一些情況下,核苷酸轉運子為核苷三磷酸轉運子(NTT)。
在一些實施例中,核苷三磷酸轉運子(NTT)係來自細菌、植物或藻類。在一些實施例中,核苷三磷酸轉運子為Tp
NTT1、Tp
NTT2、Tp
NTT3、Tp
NTT4、Tp
NTT5、Tp
NTT6、Tp
NTT7、Tp
NTT8 (假微型海鏈藻)、Pt
NTT1、Pt
NTT2、Pt
NTT3、Pt
NTT4、Pt
NTT5、Pt
NTT6 (三角褐指藻)、Gs
NTT (溫泉藻(Galdieria sulphuraria
))、At
NTT1、At
NTT2 (阿拉伯芥)、Ct
NTT1、Ct
NTT2 (沙眼披衣菌(Chlamydia trachomatis
))、Pam
NTT1、Pam
NTT2 (阿莫氏原披衣菌(Protochlamydia amoebophila
))、Cc
NTT (嗜核凱氏桿菌(Caedibacter caryophilus
))、Rp
NTT1 (普氏立克次體(Rickettsia prowazekii
))。在一些實施例中,NTT為CNT1、CNT2、CNT3、ENT1、ENT2、OAT1、OAT3或OCT1。在一些情況下,NTT為Pt
NTT1、Pt
NTT2、Pt
NTT3、Pt
NTT4、Pt
NTT5或Pt
NTT6。
在一些實施例中,NTT將非天然核酸導入生物體(例如細胞)中。在一些實施例中,NTT可經修飾以使得NTT之核苷酸結合位點經修飾以減小非天然核酸至核苷酸結合位點中之立體進入抑制。在一些實施例中,NTT可經修飾以提供與非天然核酸之一或多個天然或非天然特徵之增加的相互作用。此類NTT可在細胞中經表現或工程改造以用於將UBP穩定地導入細胞中。因此,本發明包括含異源或重組NTT之組合物及其使用方法。
NTT可使用關於蛋白質工程改造之方法修飾。舉例而言,可基於晶體結構進行分子模型化以鑑別NTT之位置,其中可進行突變以修飾目標活性或結合位點。鑑別為替代目標之殘基可經使用能量最小化模型化、同源性模型化及/或保守胺基酸取代所選擇之殘基來置換,諸如Bordo等人, J Mol Biol 217: 721-729 (1991)及Hayes等人, Proc Natl Acad Sci, USA 99: 15926- 15931 (2002)中所描述。
各種NTT中之任一者可用於本文所闡述之方法或組合物中,包括例如自生物系統分離之蛋白質類酶及其功能變異體。除非另外指示,否則提及特定NTT (諸如下文所例示之彼等NTT)應理解為包括其功能變異體。在一些實施例中,NTT為野生型NTT。在一些實施例中,NTT為經修飾或突變NTT。在一些實施例中,如本文所使用之經修飾或突變NTT為在N端、C端或N端與C端兩者處經截短的NTT。在一些實施例中,截短NTT與未截短NTT至少60%、至少65%、至少70%、至少75%、至少80%、至少85%或至少90%一致。在一些情況下,如本文所使用之NTT為Pt
NTT1、Pt
NTT2、Pt
NTT3、Pt
NTT4、Pt
NTT5或Pt
NTT6。在一些情況下,如本文所使用之Pt
NTT在N端、C端或N端與C端兩者處經截短。在一些實施例中,截短Pt
NTT與未截短Pt
NTT至少60%、至少65%、至少70%、至少75%、至少80%、至少85%或至少90%一致。在一些情況下,如本文所使用之NTT為截短Pt
NTT2,其中截短Pt
NTT2具有與未截短Pt
NTT2之胺基酸序列至少60%、至少65%、至少70%、至少75%、至少80%、至少85%或至少90%一致之胺基酸序列。未截短Pt
NTT2之實例(NCBI寄存編號EEC49227.1,GI:217409295)具有胺基酸序列SEQ ID NO: 1。
亦可使用具有用於改良非天然核酸至細胞中之進入及用於與非天然核苷酸在核苷酸結合區域中配位之特徵的NTT。在一些實施例中,經修飾之NTT具有經修飾之核苷酸結合位點。在一些實施例中,經修飾或野生型NTT具有針對非天然核酸之寬鬆的特異性。舉例而言,NTT視情況呈現針對非天然核苷酸之特異性導入活性,其為對應野生型NTT之至少約0.1%高(例如約0.1%、0.2%、0.5%、0.8%、1%、1.1%、1.2%、1.5%、1.8%、2%、3%、4%、5%、10%、25%、50%、75%、100%或更高)。視情況,NTT呈現針對非天然核苷酸之kcat
/Km
或Vmax
/Km
,其為野生型NTT之至少約0.1%高(例如約0.1%、0.2%、0.5%、0.8%、1%、1.1%、1.2%、1.5%、1.8%、2%、3%、4%、5%、10%、25%、50%、75%或100%或更高)。
NTT可根據其對於三磷酸酯之親和力(亦即Km)及/或導入速率(亦即Vmax)表徵。在一些實施例中,NTT具有對於一或多種天然及非天然三磷酸酯之相對Km或Vmax。在一些實施例中,NTT具有對於一或多種天然及非天然三磷酸酯之相對較高的Km或Vmax。
來自天然來源之NTT或其變異體可使用偵測三磷酸酯之量(若三磷酸酯經適當標記,則使用質譜或放射能)的分析篩選。在一個實例中,可針對導入非天然三磷酸酯之能力篩選NTT;例如dTPT3TP、dCNMOTP、d5SICSTP、dNaMTP、NaMTP及/或TPT1TP。可使用呈現與野生型NTT相比針對非天然核酸之經修飾的特性的NTT,例如異源NTT。舉例而言,經修飾之特性可為例如針對三磷酸酯導入之Km
、kcat
、Vmax
。在一個實施例中,經修飾之特性為針對非天然三磷酸酯之減小的Km
及/或針對非天然三磷酸酯之增加的kcat
/Km
或Vmax
/Km
。類似地,與野生型NTT相比,該NTT視情況具有增加的非天然三磷酸酯結合速率、增加的細胞內釋放速率及/或增加的細胞導入速率。
同時,NTT可將天然三磷酸酯(例如dATP、dCTP、dGTP、dTTP、ATP、CTP、GTP及/或TTP)導入細胞中。在一些情況下,NTT視情況呈現能夠支持複製及轉錄之針對天然核酸之特異性導入活性。在一些實施例中,NTT視情況呈現能夠支持複製及轉錄之針對天然核酸之kcat
/Km
或Vmax
/Km
。
本文所使用之可具有導入具有特定結構之非天然三磷酸酯之能力的NTT亦可使用定向進化方法來產生。核酸合成分析可用於篩選具有針對各種非天然三磷酸酯中之任一者之特異性的NTT變異體。舉例而言,可針對輸入非天然三磷酸酯之能力篩選NTT變異體;例如d5SICSTP、dNaMTP、dCNMOTP、dTPT3TP、NaMTP及/或TPT1TP。在一些實施例中,此類分析為例如使用重組NTT變異體之活體外分析。在一些實施例中,此類分析為例如在細胞中表現NTT變異體之活體內分析。此類技術可用於篩選任何適合的NTT之變異體針對本文所闡述之非天然三磷酸酯中之任一者的活性。核酸試劑及工具
與本文所描述之方法、細胞或經工程改造之微生物一起使用之核苷酸及/或核酸試劑(或聚核苷酸)包含一或多個具有或不具有非天然核苷酸之ORF。ORF可來自任何適合的來源,有時來自基因體DNA、mRNA、逆轉錄RNA或互補DNA (cDNA)或包含前述中之一或多者之核酸庫,且其來自任何含有所關注之核酸序列、所關注之蛋白質或所關注之活性的生物體物種。可自其獲得ORF之生物體之非限制性實例例如包括細菌、酵母、真菌、人類、昆蟲、線蟲、牛、馬、犬、貓、大鼠或小鼠。在一些實施例中,本文所描述之核苷酸及/或核酸試劑或其他試劑經分離或純化。可經由公開的活體外方法產生包括非天然核苷酸之ORF。在一些情況下,核苷酸或核酸試劑包含非天然核鹼基。
核酸試劑有時包含鄰近於ORF之核苷酸序列,其係與ORF結合轉譯且編碼胺基酸標籤。編碼標籤之核苷酸序列定位成核酸試劑中之ORF之3'及/或5',從而在藉由ORF所編碼之蛋白質或肽之C端或N端處編碼標籤。可利用任何不廢止活體外轉錄及/或轉譯之標籤且其可藉由技工適當選擇。標籤可促進所需ORF產物自培養物或醱酵培養基之分離及/或純化。在一些情況下,核酸試劑之庫與本文所描述之方法及組合物一起使用。舉例而言,至少100、1000、2000、5000、10,000或超過50,000個特有聚核苷酸之庫存在於庫中,其中各聚核苷酸包含至少一個非天然核鹼基。
具有或不具有非天然核苷酸之核酸或核酸試劑可包含某些元件,例如調節元件,常常根據核酸之預期用途來選擇。以下元件中之任一者可包括於核酸試劑中或自核酸試劑排除。核酸試劑例如可包括一或多個或所有以下核苷酸元件:一或多個啟動子元件、一或多個5'非轉譯區域(5'UTR)、一或多個可插入目標核苷酸序列之區域(「插入元件」)、一或多個目標核苷酸序列、一或多個3'非轉譯區域(3'UTR)及一或多個選擇元件。核酸試劑可具備此類元件中之一或多者,且其他元件可在核酸引入所需生物體中之前插入核酸中。在一些實施例中,所提供之核酸試劑包含啟動子、5'UTR、視情況存在之3'UTR及插入元件,目標核苷酸序列係藉由該插入元件插入(亦即殖於)核苷酸試劑中。在某些實施例中,所提供之核酸試劑包含啟動子、插入元件及視情況存在之3'UTR,且5' UTR/目標核苷酸序列係與視情況存在之3'UTR一起插入。該等元件可以任何適用於在所選表現系統中表現(例如在所選生物體中表現,或例如在無細胞系統中表現)的次序來配置,且在一些實施例中,核酸試劑在5'至3'方向上包含以下元件:(1)啟動子元件、5'UTR及插入元件;(2)啟動子元件、5'UTR及目標核苷酸序列;(3)啟動子元件、5'UTR、插入元件及3'UTR;以及(4)啟動子元件、5'UTR、目標核苷酸序列及3'UTR。在一些實施例中,UTR可經最佳化以改變或增加完全天然或含有非天然核苷酸之ORF之轉錄或轉譯。
核酸試劑(例如表現卡匣及/或表現載體)可包括多種調節元件,包括啟動子、強化子、轉譯起始序列、轉錄終止序列及其他元件。「啟動子」通常為在關於轉錄起始位點相對固定位置時起作用之一或多個DNA序列。舉例而言,啟動子可在核苷酸三磷酸酯轉運子核酸區段上游。「啟動子」含有RNA聚合酶與轉錄因子之基本相互作用所需的核心元件,且可含有上游元件及反應元件。「強化子」通常係指在距轉錄起始位點不固定距離處起作用之DNA序列,且可在轉錄單元之5'或3''。此外,強化子可在內含子內以及在編碼序列自身內。其長度通常在10與300之間,且其以順式起作用。強化子起作用以增加附近啟動子之轉錄。如同啟動子,強化子亦常常含有介導轉錄調節的反應元件。強化子常常決定表現之調節,且可用以改變或最佳化ORF表現,包括完全天然或含有非天然核苷酸之ORF。
如上文所指出,核酸試劑亦可包含一或多個5' UTR及一或多個3'UTR。舉例而言,用於真核宿主細胞(例如,酵母、真菌、昆蟲、植物、動物、人類或成核細胞)及原核宿主細胞(例如,病毒、細菌)之表現載體可含有傳導信號以用於終止轉錄的序列,其可影響mRNA表現。此等區域可經轉錄為編碼mRNA之組織因子蛋白之未轉譯部分中之聚腺苷酸化區段。3'非轉譯區域亦包括轉錄終止位點。在一些較佳實施例中,轉錄單元包含聚腺苷酸化區域。此區域之一個益處為其增加經轉錄單元將如同mRNA一般經處理及轉運之可能性。已明確聚腺苷酸化信號於表現構築體中之鑑別及使用。在一些較佳實施例中,同源聚腺苷酸化信號可用於轉殖基因構築體中。
5'UTR可包含其所起源之核苷酸序列內源之一或多個元件,且有時包括一或多個外源性元件。5'UTR可起源於例如來自任何適合的生物體(例如,病毒、細菌、酵母、真菌、植物、昆蟲或哺乳動物)之任何適合的核酸,諸如基因體DNA、質體DNA、RNA或mRNA。技工可基於所選表現系統(例如,在所選生物體中表現,或例如在無細胞系統中表現)來選擇用對於5' UTR適當的元件。5'UTR有時包含技工已知的以下元件中之一或多者:強化子序列(例如,經轉錄或轉譯的)、轉錄起始位點、轉錄因子結合位點、轉譯調節位點、轉譯起始位點、轉譯因子結合位點、附屬蛋白結合位點、反饋調節劑結合位點、Pribnow盒、TATA盒、-35元件、E-盒(螺旋-環-螺旋結合元件)、核糖體結合位點、複製子、內部核糖體進入位點(IRES)、靜止子元件及其類似者。在一些實施例中,啟動子元件可經分離以使得所有對於適當條件性調節所必需的5'UTR元件均含於啟動子元件片段中或含於啟動子元件片段之功能子序列內。
核酸試劑中之5' UTR可包含轉譯強化子核苷酸序列。轉譯強化子核苷酸序列常常定位於核酸試劑中之啟動子與目標核苷酸序列之間。轉譯強化子序列常常與核糖體結合,有時為18S rRNA結合核糖核苷酸序列(亦即,40S核糖體結合序列)且有時為內部核糖體進入序列(IRES)。IRES通常經由多個特異性分子間相互作用與精確置放之與40S核糖體子單元接觸的RNA三級結構形成RNA骨架。核糖體強化子序列之實例為已知的且可藉由技工鑑別(例如,Mignone等人, Nucleic Acids Research 33: D141-D146 (2005);Paulous等人, Nucleic Acids Research 31: 722-733 (2003);Akbergenov等人, Nucleic Acids Research 32: 239-247 (2004);Mignone等人, Genome Biology 3(3): reviews0004.1-0001.10 (2002);Gallie, Nucleic Acids Research 30: 3401-3411 (2002);Shaloiko等人, 數位物件識別碼: 10.1002/bit.20267;以及Gallie等人, Nucleic Acids Research 15: 3257-3273 (1987))。
轉譯強化子序列有時為真核序列,諸如克紮克(Kozak)共同序列或其他序列(例如,螅體(hydroid polyp)序列,GenBank寄存編號U07128)。轉譯強化子序列有時為原核序列,諸如夏因-達爾加諾(Shine-Dalgarno)共同序列。在某些實施例中,轉譯強化子序列為病毒核苷酸序列。轉譯強化子序列有時來自植物病毒之5' UTR,該植物病毒諸如煙草嵌紋病毒(Tobacco Mosaic Virus;TMV)、苜蓿嵌紋病毒(Alfalfa Mosaic Virus;AMV)、煙草蝕刻病毒(Tobacco Etch Virus;ETV);馬鈴薯病毒Y (Potato Virus Y;PVY)、蕪菁嵌紋(poty)病毒(Turnip Mosaic Virus)及豌豆種子源性嵌紋病毒(Pea Seed Borne Mosaic Virus)。在某些實施例中,來自TMV之長度為約67個鹼基的ω序列係以轉譯強化子序列(例如,不含鳥苷核苷酸且包括長度為25個核苷酸之聚(CAA)中心區域)之形式包括於核酸試劑中。
3'UTR可包含其所起源之核苷酸序列內源之一或多個元件,且有時包括一或多個外源性元件。3'UTR可起源於例如來自任何適合的生物體(例如,病毒、細菌、酵母、真菌、植物、昆蟲或哺乳動物)之任何適合的核酸,諸如基因體DNA、質體DNA、RNA或mRNA。技工可基於所選表現系統(例如,在所選生物體中表現)來選擇用對於3'UTR適當的元件。3'UTR有時包含技工已知的以下元件中之一或多者:轉錄調節位點、轉錄起始位點、轉錄終止位點、轉錄因子結合位點、轉譯調節位點、轉譯終止位點、轉譯起始位點、轉譯因子結合位點、核糖體結合位點、複製子、強化子元件、靜止子元件及聚腺苷尾。3'UTR常常包括聚腺苷尾且有時不包括聚腺苷尾,且若聚腺苷尾存在,則一或多個腺苷部分可經添加或自其缺失(例如,約5、約10、約15、約20、約25、約30、約35、約40、約45或約50個腺苷部分可經添加或減去)。
在一些實施例中,5' UTR及/或3' UTR之修飾用於改變(例如,增加、添加、降低或實質上消除)啟動子之活性。啟動子活性之變化可繼而改變肽、多肽或蛋白之活性(例如酶活性),其係藉由包含經修飾之5'或3' UTR之可操作地連接的啟動子元件改變所關注之一或多個核苷酸序列之轉錄。舉例而言,在某些實施例中,微生物可藉由基因修飾經工程改造以表現包含經修飾之5'或3'UTR之核酸試劑,該經修飾之5'或3'UTR可添加新穎活性(例如,通常不在宿主生物體中發現之活性)或藉由增加自可操作地連接於所關注之核苷酸序列(所關注之同源或異源核苷酸序列)之同源或異源啟動子之轉錄來增加現存活性之表現。在一些實施例中,微生物可藉由基因修飾經工程改造以表現包含經修飾之5'或3'UTR之核酸試劑,在某些實施例中,該經修飾之5'或3'UTR可藉由減小或實質上消除自可操作地連接於所關注之核苷酸序列之同源或異源啟動子之轉錄來降低活性的表現。
來自表現卡匣或表現載體之核苷酸三磷酸酯轉運子之表現可藉由任何能夠在原核細胞或真核細胞中表現之啟動子控制。啟動子元件典型地為DNA合成及/或RNA合成所需。啟動子元件常常包含可藉由提供用於對應於基因之RNA之合成的起始位點而促進特定基因之轉錄的DNA之區域。在一些實施例中,啟動子通常定位於靠近其所調節之基因處、定位於基因之上游(例如,基因之5'),且在與基因之有義股相同的DNA股上。在一些實施例中,啟動子元件可自基因或生物體分離,且經插入成與聚核苷酸序列功能性連接以允許經改變及/或調節之表現。用於核酸表現之非天然啟動子(例如,通常不與給定核酸序列相關之啟動子)常常稱為異源啟動子。在某些實施例中,異源啟動子及/或5'UTR可經插入成與聚核苷酸功能性連接,該聚核苷酸編碼具有如本文所描述之所需活性之多肽。如本文關於啟動子所使用之術語「可操作地連接」及「與……功能性連接」係指編碼序列與啟動子元件之間的關係。當來自編碼序列之經由轉錄之表現係藉由啟動子元件調節或控制時,啟動子與編碼序列可操作地連接或功能性連接。術語「可操作地連接」及「與……功能性連接」在本文中關於啟動子元件可互換地利用。
啟動子常常與RNA聚合酶相互作用。聚合酶為催化使用先前存在之核酸試劑的核酸合成之酶。當模板為DNA模板時,RNA分子在蛋白質合成之前經轉錄。具有適用於本發明方法之聚合酶活性之酶包括任何在具有所選模板以合成蛋白質之所選系統中具有活性之聚合酶。在一些實施例中,在本文中亦稱為啟動子元件之啟動子(例如異源啟動子)可經可操作地連接於核苷酸序列或開放閱讀框架(open reading frame;ORF)。自啟動子元件之轉錄可催化對應於可操作地連接於啟動子之核苷酸序列或ORF序列的RNA之合成,其繼而導致所要肽、多肽或蛋白質之合成。
啟動子元件有時展現對於調節控制之反應性。啟動子元件亦有時可藉由選擇性劑來調節。亦即,自啟動子元件之轉錄有時可回應於環境、營養或內部條件或信號(例如,熱誘導性啟動子、光調節啟動子、反饋調節啟動子、激素影響之啟動子、組織特異性啟動子、氧及pH影響之啟動子、回應於選擇性劑(例如康黴素(kanamycin))之啟動子及其類似者(例如))之變化而啟動、關閉、上調或下調。頻繁地受環境、營養或內部信號影響之啟動子受在啟動子處或靠近啟動子處結合且在某些條件下增加或降低目標序列之表現之信號(直接或間接)影響。如同本文所揭示之所有方法,包括天然或經修飾之啟動子可用於改變或最佳化完全天然ORF(例如NTT或aaRS)或含有非天然核苷酸(例如mRNA或tRNA)之ORF的表現。
影響自用於本文所描述之實施例中之啟動子之轉錄的選擇性或調節劑之非限制性實例包括(但不限於):(1)編碼提供針對以其他方式具有毒性的化合物之抗性之產物(例如,抗生素)的核酸片段;(2)編碼以其他方式不含受體細胞之產物(例如,必需產物、tRNA基因、營養缺陷型標記)之核酸片段;(3)編碼遏制基因產物之活性之產物的核酸片段;(4)編碼可容易地鑑別之產物(例如,表型標記,諸如抗生素(例如,β-內醯胺酶)、β-半乳糖、綠色螢光蛋白(GFP)、黃色螢光蛋白(YFP)、紅色螢光蛋白(RFP)、強化型藍色螢光蛋白(CFP)及細胞表面蛋白)之核酸片段;(5)結合以其他方式對細胞存活及/或功能有害的產物之核酸片段;(6)以其他方式抑制描述於上述第1-5號中之核酸片段中之任一者(例如,反義寡核苷酸)之活性的核酸片段;(7)結合改變基質之產物(例如,限制性核酸內切酶)之核酸片段;(8)可用於分離或鑑別所要分子(例如,特定蛋白質結合位點)之核酸片段;(9)編碼可以其他方式非功能性(例如,對於分子之亞群之PCR擴增)之特定核苷酸序列之核酸片段;(10)若不存在,則直接或間接地賦予對特定化合物之抗性或靈敏性之核酸片段;(11)編碼為毒性的或將相對無毒化合物轉化為受體細胞中之毒性化合物之產物(例如,單純疱疹胸苷酶胞嘧啶脫胺酶)的核酸片段;(12)抑制含有核酸片段之核酸分子之複製、分割或遺傳可能性的該等核酸片段;(13)編碼條件性複製功能之核酸片段,例如在某些宿主或宿主細胞菌株中或在某些環境條件(例如,溫度、營養條件及其類似條件)下之複製;及/或(14)編碼一或多個包含非天然核苷酸之mRNA或tRNA之核酸。在一些實施例中,可添加調節或選擇性劑以改變生物體所經受之現有生長條件(例如,在液體培養物中之生長、在醱酵槽中之生長、在固體養分培養盤上之生長及其類似者(例如))。
在一些實施例中,啟動子元件之調節可用於改變(例如,增加、添加、降低或實質上消除)肽、多肽或蛋白質之活性(例如酶活性)。舉例而言,在某些實施例中,微生物可藉由基因修飾經工程改造以表現核酸試劑,該核酸試劑可添加新穎活性(例如,通常不在宿主生物體中發現之活性)或藉由增加自可操作地連接於所關注之核苷酸序列(例如,所關注之同源或異源核苷酸序列)之同源或異源啟動子之轉錄來增加現存活性之表現。在一些實施例中,微生物可藉由基因修飾經工程改造以表現核酸試劑,在某些實施例中,該核酸試劑可藉由減小或實質上消除自可操作地連接於所關注之核苷酸序列之同源或異源啟動子之轉錄來降低活性之表現。
編碼核酸之異源蛋白質(例如核苷酸三磷酸酯轉運子)可插入任何適合的表現系統中或與其一起使用。在一些實施例中,核酸試劑有時經穩定地整合至宿主生物體之染色體中,或在某些實施例中,核酸試劑可缺失宿主染色體之一部分(例如,經基因修飾之生物體,其中宿主基因體之變化賦予選擇性地或較佳地維持攜帶基因修飾之所要生物體的能力)。可選擇此類核酸試劑(例如,其改變的基因體向生物體賦予可選性狀之核酸或基因修飾之生物體)以使用其導引所要蛋白質或核酸分子之產生的能力。當需要時,可改變核酸試劑以使得密碼子編碼(i)相同胺基酸,其使用不同於在天然序列中所規定的tRNA之tRNA,或(ii)不同於正常胺基酸之胺基酸,包括非習知或非天然胺基酸(包括經可偵測標記之胺基酸)。
使用可為載體(諸如質體)之一部分的表現卡匣有效地實現重組表現。載體可包括啟動子,其可操作地連接於編碼核苷酸三磷酸酯轉運子之核酸。載體亦可包括其他如本文所描述之轉錄及轉譯所需之元件。表現卡匣、表現載體及卡匣或載體中之序列可與非天然核苷酸所接觸之細胞異源。舉例而言,核苷酸三磷酸酯轉運子序列可與細胞異源。
可產生各種適用於攜帶、編碼及/或表現核苷酸三磷酸酯轉運子之原核及真核表現載體。此類表現載體包括例如pET、pET3d、pCR2.1、pBAD、pUC及酵母載體。該等載體可用於例如多種活體內及活體外情形中。可使用之原核啟動子之非限制性實例包括SP6、T7、T5、tac
、bla
、trp
、gal
、lac
或麥芽糖啟動子。可使用之真核啟動子之非限制性實例包括組成性啟動子,例如病毒啟動子,諸如CMV、SV40及RSV啟動子,以及可調節啟動子,例如誘導型或可抑制啟動子,諸如tet
啟動子、hsp70啟動子及藉由CRE調節之合成啟動子。用於細菌表現之載體包括pGEX-5X-3,且用於真核表現之載體包括pCIneo-CMV。可使用之病毒載體包括與慢病毒、腺病毒、腺相關病毒、疱疹病毒、痘瘡病毒、小兒麻痹病毒、AIDS病毒、神經元營養病毒、辛德比(Sindbis)及其他病毒相關之彼等病毒載體。亦適用的為與此等病毒共有使其適用作載體之特性的任何病毒家族。可用之逆轉錄病毒載體包括在Verma, American Society for Microbiology, 第229-232頁, Washington, (1985)中所描述之彼等逆轉錄病毒載體。舉例而言,此類逆轉錄病毒載體可包括鼠類馬洛尼白血病病毒(Murine Maloney Leukemia virus)、MMLV及其他表現所要特性之逆轉錄病毒。病毒載體典型地含有非結構早期基因、結構晚期基因、RNA聚合酶III轉錄物、複製及殼體化所必需之反轉末端重複序列及用以控制病毒基因體之轉錄及複製的啟動子。當經工程改造為載體時,病毒典型地具有經移除早期基因中之一或多者,且將基因或基因/啟動子卡匣插入病毒基因體中代替經移除之病毒核酸。選殖
此項技術中已知的任何適宜的選殖策略均可用以將元件(諸如ORF)併入核酸試劑中。已知方法可用以獨立於插入元件將元件插入模板中,諸如(1)在一或多個現存限制酶位點處裂解模板且接合所關注之元件,及(2)藉由使包括一或多個適合的限制酶位點之寡核苷酸引子雜交及藉由聚合酶鏈反應以擴增來將限制酶位點添加至模板(本文中更詳述地描述)。其他選殖策略利用一或多個存在的或經插入至核酸試劑中之插入位點,諸如用於PCR之寡核苷酸引子雜交位點(例如)及本文所描述之其他位點。在一些實施例中,選殖策略可與基因操作組合,該基因操作諸如重組(例如,將具有所關注之核酸序列之核酸試劑重組至待修飾之生物體之基因體中,如本文所進一步描述)。在一些實施例中,一或多個經選殖之ORF可藉由用一或多個所關注之ORF以工程改造微生物來產生(直接或間接)經修飾或野生型核苷酸三磷酸酯轉運子及/或聚合酶,微生物包含核苷酸三磷酸酯轉運子活性或聚合酶活性之變化的活性。
可藉由使核酸與一或多種特異性裂解劑接觸來使核酸特異性裂解。特異性裂解劑常常根據在特定位點處之特定核苷酸序列特異性地裂解。酶特異性裂解劑之實例包括(但不限於):核酸內切酶(例如,DNA酶(例如,DNA酶I、DNA酶II);RNA酶(例如,RNA酶E、RNA酶F、RNA酶H、RNA酶P);Cleavase™酶;Taq DNA聚合酶;大腸桿菌DNA聚合酶I及真核結構特異性核酸內切酶;鼠類FEN-1核酸內切酶;I型、II型或III型限制性核酸內切酶,諸如Acc I、Afl III、Alu I、Alw44 I、Apa I、Asn I、Ava I、Ava II、BamH I、Ban II、Bcl I、Bgl I、Bgl II、Bln I、BsaI、Bsm I、BsmBI、BssH II、BstE II、Cfo I、CIa I、Dde I、Dpn I、Dra I、EcIX I、EcoR I、EcoR I、EcoR II、EcoR V、Hae II、Hae II、Hind II、Hind III、Hpa I、Hpa II、Kpn I、Ksp I、Mlu I、MIuN I、Msp I、Nci I、Nco I、Nde I、Nde II、Nhe I、Not I、Nru I、Nsi I、Pst I、Pvu I、Pvu II、Rsa I、Sac I、Sal I、Sau3A I、Sca I、ScrF I、Sfi I、Sma I、Spe I、Sph I、Ssp I、Stu I、Sty I、Swa I、Taq I、Xba I、Xho I);醣苷酶(例如,尿嘧啶-DNA醣苷酶(UDG)、3-甲基腺嘌呤DNA醣苷酶、3-甲基腺嘌呤DNA醣苷酶II、嘧啶水合-DNA醣苷酶、FaPy-DNA醣苷酶、胸腺嘧啶錯配-DNA醣苷酶、次黃嘌呤-DNA醣苷酶、5-羥甲基尿嘧啶DNA醣苷酶(HmUDG)、5-羥甲基胞嘧啶DNA醣苷酶或1,N6-乙烯橋-腺嘌呤DNA醣苷酶);核酸外切酶(例如核酸外切酶III);核糖核酸酶及DNA酶。樣品核酸可用化學劑處理,或使用經修飾之核苷酸合成,且經修飾核酸可經裂解。在非限制性實例中,樣品核酸可用以下處理:(i)烷基化劑,諸如甲基亞硝脲,其產生若干烷基化鹼基,包括N3-甲基腺嘌呤及N3-甲基鳥嘌呤,其藉由烷基嘌呤DNA-醣苷酶識別及裂解;(ii)亞硫酸氫鈉,其導致DNA中之胞嘧啶殘基之去胺基化以形成可藉由尿嘧啶N-醣苷酶裂解之尿嘧啶殘基;以及(iii)將鳥嘌呤轉化為其氧化形式8-羥基鳥嘌呤之化學劑,該8-羥基鳥嘌呤可藉由甲醯胺基嘧啶DNA N-醣苷酶裂解。化學裂解方法之實例包括(但不限於):烷基化(例如,硫代磷酸酯-經修飾核酸之烷基化);含P3'-N5'-磷醯胺酸之核酸之酸不穩定性之裂解;及核酸之四氧化鋨及哌啶處理。
在一些實施例中,核酸試劑包括一或多個重組酶插入位點。重組酶插入位點為在核酸分子上之識別序列,其參與藉由重組蛋白之整合/重組反應。舉例而言,用於Cre重組酶之重組位點為loxP,其為34鹼基對序列,其由側接8鹼基對核心序列之兩個13鹼基對反轉重複序列(其充當重組酶結合位點)構成(例如,Sauer, Curr. Opin. Biotech. 5:521-527 (1994))。重組位點之其他實例包括attB、attP、attL及attR序列及其突變體、片段、變異體及衍生物,其係藉由重組蛋白λ Int且藉由輔助蛋白質整合宿主因子(IHF)、FIS及切除酶(Xis)識別(例如,美國專利第5,888,732號;第6,143,557號;第6,171,861號;第6,270,969號;第6,277,608號及第6,720,140號;美國專利申請案第09/517,466號及第09/732,914號;美國專利公開案第US2002/0007051號;以及Landy, Curr. Opin. Biotech. 3:699-707 (1993))。
選殖核酸之重組酶之實例在Gateway®系統(Invitrogen, California)中,其包括至少一個用於活體內或活體外選殖所要核酸分子之重組位點。在一些實施例中,系統利用含有至少兩個不同的位點特異性重組位點之載體,該等重組位點常常基於噬菌體λ系統(例如,att1及att2),且係自野生型(att0)位點突變而來。各突變位點具有對於相同類型之其同源搭配物att位點(亦即,其結合搭配物重組位點)之特有的特異性(例如,attB1與attP1,或attL1與attR1),且不會與另一突變類型之重組位點或與野生型att0位點交叉反應。不同的位點特異性使得所要分子之定向選殖或鍵聯,因此提供選殖分子之所要定向。藉由重組位點側接之核酸片段係使用Gateway®系統藉由替代可選標記(例如ccdB)而經選殖及次選殖,該可選標記係藉由受體質體分子(有時稱為目標載體(Destination Vector))上之att位點側接。所要選殖隨後藉由ccdB敏感宿主菌株之轉化及受體分子上之標記之正選擇而進行選擇。用於負選擇之類似策略(例如,使用毒性基因)可用於其他生物體,諸如哺乳動物及昆蟲中之胸苷激酶(TK)。
核酸試劑有時含有一或多個複製起點(ORI)元件。在一些實施例中,模板包含兩個或更多個ORI,其中一個在一種生物體(例如細菌)中有效地起作用,且另一個在另一生物體(例如,真核生物,如例如酵母)中有效地起作用。在一些實施例中,ORI可在一個物種(例如釀酒酵母)中有效地起作用,且另一ORI可在不同物種(例如,粟酒裂殖酵母)中有效地起作用。核酸試劑有時亦包括一或多個轉錄調節位點。
核酸試劑,例如表現卡匣或載體可包括編碼標記產物之核酸序列。標記產物係用於判定基因是否已遞送至細胞且在遞送後是否經表現。實例標記基因包括大腸桿菌lacZ基因,其編碼β-半乳糖苷酶及綠色螢光蛋白。在一些實施例中,標記可為可選標記。當此類可選標記成功地轉移至宿主細胞中時,經轉化宿主細胞可在置放於選擇性壓力下之情況下存活。存在兩種廣泛使用之不同類別之選擇性方案。第一類係基於細胞之代謝及使用缺乏獨立於補充培養基生長之能力的突變細胞株。第二類為顯性選擇,其係指用於任何細胞類型之選擇方案,且並不需要使用突變細胞株。此等方案典型地使用藥物以遏制宿主細胞之生長。具有新穎基因之彼等細胞將表現蛋白質輸送抗藥性且將在選擇中存活。此類顯性選擇之實例使用藥物新黴素(neomycin) (Southern等人, J. Molec. Appl. Genet. 1: 327 (1982))、黴酚酸(mycophenolic acid) (Mulligan等人, Science 209: 1422 (1980))或潮黴素(Sugden等人, Mol. Cell. Biol. 5: 410-413 (1985))。
核酸試劑可包括一或多個選擇元件(例如,用於核酸試劑之存在之選擇,且不用於可經選擇性調節之啟動子元件之活化的元件)。常常使用已知方法利用選擇元件以判定細胞中是否包括核酸試劑。在一些實施例中,核酸試劑包括兩個或更多個選擇元件,其中一個在一種生物體中有效地起作用,且另一個在另一生物體中有效地起作用。選擇元件之實例包括(但不限於):(1)編碼提供針對以其他方式具有毒性的化合物之抗性之產物(例如抗生素)的核酸片段;(2)編碼以其他方式不含受體細胞之產物(例如,必需產物、tRNA基因、營養缺陷型標記)之核酸片段;(3)編碼遏制基因產物之活性之產物的核酸片段;(4)編碼可容易地鑑別之產物(例如,表型標記,諸如抗生素(例如,β-內醯胺酶)、β-半乳糖、綠色螢光蛋白(GFP)、黃色螢光蛋白(YFP)、紅色螢光蛋白(RFP)、強化型藍色螢光蛋白(CFP)及細胞表面蛋白)之核酸片段;(5)結合以其他方式對細胞存活及/或功能有害的產物之核酸片段;(6)以其他方式抑制描述於上述第1-5號中之核酸片段中之任一者(例如,反義寡核苷酸)之活性的核酸片段;(7)結合改變基質之產物(例如,限制性核酸內切酶)之核酸片段;(8)可用於分離或鑑別所要分子(例如,特定蛋白質結合位點)之核酸片段;(9)編碼可以其他方式非功能性(例如,對於分子之亞群之PCR擴增)之特定核苷酸序列之核酸片段;(10)若不存在,則直接或間接地賦予對特定化合物之抗性或靈敏性之核酸片段;(11)編碼為毒性的或將相對無毒化合物轉化為受體細胞中之毒性化合物之產物(例如,單純疱疹胸苷酶胞嘧啶脫胺酶)的核酸片段;(12)抑制含有核酸片段之核酸分子之複製、分割或遺傳可能性的該等核酸片段;及/或(13)編碼條件性複製功能之核酸片段,例如在某些宿主或宿主細胞菌株中或在某些環境條件(例如,溫度、營養條件及其類似條件)下之複製。
核酸試劑可呈任何適用於活體內轉錄及/或轉譯之形式。核酸有時為質體(諸如超螺旋質體),有時為酵母人工染色體(例如YAC),有時為線性核酸(例如,由PCR或由限制性消化產生之線性核酸),有時為單股且有時為雙股。核酸試劑有時係藉由擴增方法製備,諸如聚合酶鏈反應(PCR)方法或轉錄介導之擴增方法(TMA)。在TMA中,兩種酶用於等溫反應以產生藉由光發射所偵測到之擴增產物(例如,Biochemistry 1996年6月25日;35(25):8429-38)。標準PCR方法為已知的(例如,美國專利第4,683,202號;第4,683,195號;第4,965,188號;以及第5,656,493號),且通常循環進行。各循環均包括熱變性,其中雜交核酸解離;冷卻,其中引子寡核苷酸雜交;及藉由聚合酶(亦即Taq聚合酶)之寡核苷酸延伸。PCR循環方法之一實例為在95℃下處理樣品5分鐘;重複四十五次以下循環:95℃持續1分鐘、59℃持續1分鐘10秒及72℃持續1分30秒;且隨後在72℃下處理樣品5分鐘。使用可商購的熱循環器頻繁地進行多次循環。PCR擴增產物在分析之前有時在較低溫度下(例如,在4℃下)儲存一定時間且有時經冷凍(例如,在-20℃下)。
類似於上文所描述之策略的彼等選殖策略可用以產生含有非天然核苷酸之DNA。舉例而言,在所要位置處含有非天然核苷酸之寡核苷酸係使用標準固相合成來合成且藉由HPLC純化。隨後使用利用選殖位點,諸如BsaI位點(但可使用上文所論述之其他位點)之選殖方法(諸如金門組裝(Golden Gate Assembly))將寡核苷酸插入含有所需序列情境(亦即,UTR及編碼序列)之質體中。套組及製品
在某些實施例中,本文揭示與本文所描述之一或多種方法一起使用的套組及製品。此類套組包括載體、封裝或容器,其經分隔以接收一或多個容器,諸如小瓶、試管及其類似容器,一或多個容器中之每一者包含待用於本文所描述之方法中的各別元件中之一者。適合的容器包括例如瓶子、小瓶、注射器及試管。在一個實施例中,容器係由多種材料(諸如玻璃或塑膠)形成。
在一些實施例中,套組包括適合的封裝材料以容納套組之內含物。在一些情況下,封裝材料係由熟知方法構築,較佳地提供無菌、無污染物的環境。本文所採用之封裝材料可包括例如在經出售以與核酸定序系統一起使用之商業套組中所常用的彼等封裝材料。例示性封裝材料包括(但不限於):玻璃、塑膠、紙、箔及其類似材料,其能夠將本文所闡述之組分保持在固定界限內。
封裝材料可包括標記,其指示組分之特定用途。在對於存在於套組中之組分之特定組合適當的情況下,藉由標記所指示之套組之用途可為本文所闡述的方法中之一或多者。舉例而言,標記可指示套組適用於合成聚核苷酸之方法或適用於測定核酸之序列的方法。
封裝試劑或組分之使用說明書亦可包括於套組中。說明書將通常包括描述反應參數之有形表述,諸如待摻合的套組組分及樣品之相對量、試劑/樣品摻合物之維持時間段、溫度、緩衝條件及其類似者。
應理解,並非所有對於特定反應所必需的組分均需存在於特定套組中。實際上,一或多種額外組分可由其他來源提供。與套組一起提供之說明書可鑑別待提供之一或多種額外組分及其可自何處獲得。
在一些實施例中,提供適用於例如使用本發明所提供之方法將非天然核酸穩定地併入細胞核酸中以製備經基因工程改造之細胞的套組。在一個實施例中,本文所描述之套組包括經基因工程改造之細胞及一或多種非天然核酸。
在其他實施例中,本文所描述之套組提供細胞及核酸分子,該核酸分子含有用於引入該細胞中之異源基因從而提供經基因工程改造之細胞,諸如包含此段落中上文所描述之實施例中之任一者之核酸的表現載體。
編號實施例 .
本發明包括以下非限制性編號實施例:
實施例1. 一種合成非天然多肽之方法,其包含:
a. 提供至少一個包含至少四個非天然鹼基對之非天然去氧核糖核酸(DNA)分子;
b. 轉錄該至少一個非天然DNA分子,以得到包含至少兩個非天然密碼子之信使核糖核酸(mRNA)分子;
c. 轉譯該至少一個非天然DNA分子以得到各自包含至少一個非天然反密碼子之至少兩個轉送RNA (tRNA)分子,其中對應DNA中之該至少兩個非天然鹼基對處於序列背景中,使得該mRNA分子之該等非天然密碼子與該等tRNA分子中之每一者之該非天然反密碼子互補;以及
d. 藉由利用該至少兩個非天然tRNA分子轉譯該非天然mRNA分子來合成該非天然多肽,其中各非天然反密碼子引導非天然胺基酸位點特異性併入該非天然多肽中。
實施例1.1.一種合成非天然多肽之方法,其包含:
a. 提供至少一個包含至少四個非天然鹼基對之非天然去氧核糖核酸(DNA)分子;
b. 轉錄該至少一個非天然DNA分子,以得到包含至少兩個非天然密碼子之信使核糖核酸(mRNA)分子;
c. 轉譯該至少一個非天然DNA分子以得到至少兩個各自包含至少一個非天然反密碼子之轉送RNA (tRNA)分子,其中對應DNA中之該至少兩個非天然鹼基對處於序列背景中,使得該mRNA分子之該等非天然密碼子中之一者與該等tRNA分子中之一者之該非天然反密碼子互補,且一或多個其他非天然密碼子中之至少一者與其他tRNA分子中之至少一者之該非天然反密碼子互補;以及
d.藉由利用該至少兩個非天然tRNA分子轉譯該非天然mRNA分子來合成該非天然多肽,其中各非天然反密碼子引導非天然胺基酸位點特異性併入該非天然多肽中。
實施例2. 一種合成非天然多肽之方法,其包含:
a. 提供至少一個包含至少四個非天然鹼基對之非天然去氧核糖核酸(DNA)分子,其中該至少一個非天然DNA分子編碼(i)包含至少第一及第二非天然密碼子之信使核糖核酸(mRNA)分子,及(ii)至少第一及第二轉送RNA (tRNA)分子,該第一tRNA分子包含第一非天然反密碼子且該第二tRNA分子包含第二非天然反密碼子,並且該至少一個DNA分子中之該至少四個非天然鹼基對處於序列背景中,使得該mRNA分子之該第一非天然密碼子及該第二非天然密碼子分別與該第一非天然反密碼子及該第二非天然反密碼子互補;
b. 轉錄該至少一個非天然DNA分子,以得到該mRNA;
c. 轉錄該至少一個非天然DNA分子,以得到該至少第一及第二tRNA分子;以及
d. 藉由利用該至少第一及第二非天然tRNA分子轉譯該非天然mRNA分子來合成該非天然多肽,其中該至少第一及第二非天然反密碼子中之每一者引導非天然胺基酸位點特異性併入該非天然多肽中。
實施例3. 如實施例1、1.1或2之方法,其中該至少兩個非天然密碼子各自包含位於該密碼子之第一位置、第二位置或第三位置處之第一非天然核苷酸,視情況其中該第一非天然核苷酸位於該密碼子之該第二位置或該第三位置處。
實施例4. 如前述實施例中任一項之方法,其中該至少兩個非天然密碼子各自包含核酸序列NNX或NXN,且該非天然反密碼子包含核酸序列XNN、YNN、NXN或NYN,以形成包含NNX-XNN、NNX-YNN或NXN-NYN之該非天然密碼子-反密碼子對,其中N為任何天然核苷酸,X為第一非天然核苷酸,且Y為不同於該第一非天然核苷酸之第二非天然核苷酸,其中X-Y或X-X形成DNA中之該非天然鹼基對。
實施例4.1. 如前述實施例中任一項之方法,其中該至少兩個非天然密碼子各自包含核酸序列XNN、NXN、NNX,且該非天然反密碼子包含核酸序列NNX、NNY、NXN、NYN、NNX或NNY,以形成包含XNN-NNX、XNN-NNY、NXN-NXN、NXN-NYN、NNX-XNN或NNX-YNN之該非天然密碼子-反密碼子對,其中N為任何天然核苷酸,X為第一非天然核苷酸,且Y為不同於該第一非天然核苷酸之第二非天然核苷酸,其中X-X或X-Y形成DNA中之該非天然鹼基對。
實施例5. 如實施例4之方法,其中該密碼子包含至少一個G或C且該反密碼子包含至少一個互補C或G。
實施例6. 如實施例4或5之方法,其中X及Y獨立地選自由以下組成之群:
(i) 2-硫尿嘧啶、2'-去氧尿苷、4-硫-尿嘧啶、尿嘧啶-5-基、次黃嘌呤-9-基(I)、5-鹵基尿嘧啶;5-丙炔基-尿嘧啶、6-偶氮基-尿嘧啶、5-甲基胺基甲基尿嘧啶、5-甲氧基胺基甲基-2-硫尿嘧啶、假尿嘧啶、尿嘧啶-5-氧基乙酸甲酯、尿嘧啶-5-氧基乙酸、5-甲基-2-硫尿嘧啶、3-(3-胺基-3-N-2-羧丙基)尿嘧啶、5-甲基-2-硫尿嘧啶、4-硫尿嘧啶、5-甲基尿嘧啶、5'-甲氧基羧甲基尿嘧啶、5-甲氧基尿嘧啶、尿嘧啶-5-氧基乙酸、5-(羧基羥甲基)尿嘧啶、5-羧甲基胺基甲基-2-硫尿苷、5-羧甲基胺基甲基尿嘧啶或二氫尿嘧啶;
(ii) 5-羥甲基胞嘧啶、5-三氟甲基胞嘧啶、5-鹵基胞嘧啶、5-丙炔基胞嘧啶、5-羥基胞嘧啶、環胞嘧啶、胞嘧啶阿拉伯糖苷、5,6-二氫胞嘧啶、5-硝基胞嘧啶、6-偶氮基胞嘧啶、氮雜胞嘧啶、N4-乙基胞嘧啶、3-甲基胞嘧啶、5-甲基胞嘧啶、4-乙醯基胞嘧啶、2-硫胞嘧啶、啡㗁𠯤胞苷([5,4-b][l,4]苯并㗁𠯤-2(3H)-酮)、啡噻𠯤胞苷(1H-嘧啶并[5,4-b][l,4]苯并噻𠯤-2(3H)-酮)、啡㗁𠯤胞苷(9-(2-胺基乙氧基)-H-嘧啶并[5,4-b][l,4]苯并㗁𠯤-2(3H)-酮)、咔唑胞苷(2H-嘧啶并[4,5-b]吲哚-2-酮)或吡啶并吲哚胞苷(H-吡啶并[3',2':4,5]吡咯并[2,3-d]嘧啶-2-酮);
(iii) 2-胺基腺嘌呤、2-丙基腺嘌呤;2-胺基-腺嘌呤;2-F-腺嘌呤;2-胺基-丙基-腺嘌呤;2-胺基-2'-去氧腺苷;3-去氮腺嘌呤;7-甲基腺嘌呤;7-去氮-腺嘌呤;8-氮雜腺嘌呤;經8-鹵基、8-胺基、8-硫醇、8-硫烷基及8-羥基取代之腺嘌呤;N6-異戊烯基腺嘌呤;2-甲基腺嘌呤;2,6-二胺基嘌呤;2-甲基硫-N6-異戊烯基腺嘌呤或6-氮雜-腺嘌呤;
(iv) 2-甲基鳥嘌呤;鳥嘌呤之2-丙基及烷基衍生物;3-去氮鳥嘌呤;6-硫-鳥嘌呤;7-甲基鳥嘌呤;7-去氮鳥嘌呤;7-去氮鳥苷;7-去氮-8-氮雜鳥嘌呤;8-氮雜鳥嘌呤;經8-鹵基、8-胺基、8-硫醇、8-硫烷基及8-羥基取代之鳥嘌呤;1-甲基鳥嘌呤;2,2-二甲基鳥嘌呤;7-甲基鳥嘌呤或6-氮雜-鳥嘌呤;以及
(v) 次黃嘌呤、黃嘌呤、1-甲基肌苷、Q核苷、β-D-半乳糖基Q核苷、肌苷、β-D-甘露糖基Q核苷、懷丁氧苷、羥基脲、(acp3)w、2-胺基吡啶或2-吡啶酮。
實施例7. 如實施例4或5之方法,其中包含X及Y中之每一者之該等鹼基獨立地選自由以下組成之群:
 。
實施例8. 如實施例7之方法,其中包含各X之該鹼基為
。
實施例8. 如實施例7之方法,其中包含各X之該鹼基為 。
實施例9. 如實施例7或8之方法,其中包含各Y之該鹼基為
。
實施例9. 如實施例7或8之方法,其中包含各Y之該鹼基為 。
實施例10. 如實施例4至9中任一項之方法,其中NNX-XNN係選自由以下組成之群:UUX-XAA、UGX-XCA、CGX-XCG、AGX-XCU、GAX-XUC、CAX-XUG、AUX-XAU、CUX-XAG、GUX-XAC、UAX-XUA及GGX-XCC。
實施例11. 如實施例4至9中任一項之方法,其中NNX-YNN係選自由以下組成之群:UUX-YAA、UGX-YCA、CGX-YCG、AGX-YCU、GAX-YUC、CAX-YUG、AUX-YAU、CUX-YAG、GUX-YAC、UAX-YUA及GGX-YCC。
實施例12. 如實施例4至9中任一項之方法,其中NXN-NYN係選自由以下組成之群:GXU-AYC、CXU-AYG、GXG-CYC、AXG-CYU、GXC-GYC、AXC-GYU、GXA-UYC、CXC-GYG及UXC-GYA。
實施例13. 如實施例12之方法,其中NXN-NYN係選自由以下組成之群:AXG-CYU、GXC-GYC、AXC-GYU、GXA-UYC、CXC-GYG及UXC-GYA。
實施例13.1. 如實施例4.1至9中任一項之方法,其中XNN-NNY係選自由以下組成之群:XUU-AAY、XUG-CAY、XCG-CGY、XAG-CUY、XGA-UCY、XCA-UGY、XAU-AUY、XCU-AGY、XGU-ACY、XUA-UAY、XUC-GAY、XCC-GGY、XAA-UUY、XAC-GUY、XGC-GCY、XGG-CCY及XGG-CCY。
實施例13.2. 如實施例4.1至9中任一者之方法,其中XNN-NNX係選自由以下組成之群:XUU-AAX、XUG-CAX、XCG-CGX、XAG-CUX、XGA-UCX、XCA-UGX、XAU-AUX、XCU-AGX、XGU-ACX、XUA-UAX、XUC-GAX、XCC-GGX、XAA-UUX、XAC-GUX、XGC-GCX、XGG-CCX及XGG-CCX。
實施例14. 如前述實施例中任一項之方法,其中該至少兩個非天然tRNA分子各自包含不同的非天然反密碼子。
實施例15. 如實施例14之方法,其中該至少兩個非天然tRNA分子包含來自甲烷八聯球菌屬之吡咯離胺醯基tRNA及來自詹氏甲烷球菌之酪胺醯基tRNA,或其衍生物。
實施例16. 如實施例13、14或15中任一項之方法,其包含藉由胺基-醯基tRNA合成酶而裝載有該至少兩個非天然tRNA分子。
實施例17. 如實施例16之方法,其中該胺基醯基tRNA合成酶係選自由嵌合PylRS (chPylRS)及詹氏甲烷球菌AzFRS (Mjp
AzFRS)組成之群。
實施例18. 如實施例14或15之方法,其包含藉由至少兩種不同tRNA合成酶而裝載有該至少兩個非天然tRNA分子。
實施例19. 如實施例18之方法,其中該至少兩種tRNA合成酶包含嵌合PylRS (chPylRS)及詹氏甲烷球菌AzFRS (Mjp
AzFRS)。
實施例20. 如實施例1至19中任一項之方法,其中該非天然多肽包含兩個、三個或更多個非天然胺基酸。
實施例21. 如實施例1至20中任一項之方法,其中該非天然多肽包含至少兩個相同的非天然胺基酸。
實施例22. 如實施例1至20中任一項之方法,其中該非天然多肽包含至少兩個不同的非天然胺基酸。
實施例23. 如實施例1至22中任一項之方法,其中該非天然胺基酸包含
離胺酸類似物;
芳族側鏈;
疊氮基;
炔基;或
醛基或酮基。
實施例24. 如實施例1至22中任一項之方法,其中該非天然胺基酸不包含芳族側鏈。
實施例25. 如實施例1至22中任一項之方法,其中該非天然胺基酸係選自:N6-疊氮基乙氧基-羰基-L-離胺酸(AzK)、N6-炔丙基乙氧基-羰基-L-離胺酸(PraK)、N6-(炔丙氧基)-羰基-L-離胺酸(PrK)、對疊氮基-苯丙胺酸(pAzF)、BCN-L-離胺酸、降冰片烯離胺酸、TCO-離胺酸、甲基四𠯤離胺酸、烯丙氧基羰基離胺酸、2-胺基-8-側氧基壬酸、2-胺基-8-側氧基辛酸、對乙醯基-L-苯丙胺酸、對疊氮基甲基-L-苯丙胺酸(pAMF)、對碘-L-苯丙胺酸、間乙醯基苯丙胺酸、2-胺基-8-側氧基壬酸、對炔丙基氧基苯丙胺酸、對炔丙基-苯丙胺酸、3-甲基-苯丙胺酸、左旋多巴、氟化苯丙胺酸、異丙基-L-苯丙胺酸、對疊氮基-L-苯丙胺酸、對醯基-L-苯丙胺酸、對苯甲醯基-L-苯丙胺酸、對溴苯基丙胺酸、對胺基-L-苯丙胺酸、異丙基-L-苯丙胺酸、O-烯丙基酪胺酸、O-甲基-L-酪胺酸、O-4-烯丙基-L-酪胺酸、4-丙基-L-酪胺酸、膦醯基酪胺酸、三-O-乙醯基-GlcNAcp-絲胺酸、L-磷絲胺酸、膦醯基絲胺酸、L-3-(2-萘基)丙胺酸、2-胺基-3-((2-((3-(苯甲氧基)-3-側氧基丙基)胺基)乙基)硒烷基)丙酸、2-胺基-3-(苯基硒烷基)丙酸、硒半胱胺酸、N6-(((2-疊氮基苯甲基)氧基)羰基)-L-離胺酸、N6-(((3-疊氮基苯甲基)氧基)羰基)-L-離胺酸及N6-(((4-疊氮基苯甲基)氧基)羰基)-L-離胺酸。
實施例26. 如前述實施例中任一項之方法,其中該至少一個非天然DNA分子呈質體形式。
實施例27. 如實施例1至26中任一項之方法,其中將該至少一個非天然DNA分子整合至細胞之基因體中。
實施例28. 如實施例26或27之方法,其中該至少一個非天然DNA分子編碼該非天然多肽。
實施例29. 如前述實施例中任一項之方法,其中該方法包含該非天然DNA分子之活體內複製及轉錄以及該經轉錄mRNA分子於細胞生物體中之活體內轉譯。
實施例30. 如實施例29之方法,其中該細胞生物體為微生物。
實施例31. 如實施例30之方法,其中該細胞生物體為原核生物。
實施例32. 如實施例31之方法,其中該細胞生物體為細菌。
實施例33. 如實施例32之方法,其中該細胞生物體為革蘭氏陽性細菌。
實施例34. 如實施例32之方法,其中該細胞生物體為革蘭氏陰性細菌。
實施例35. 如實施例34之方法,其中該細胞生物體為大腸桿菌。
實施例36. 如前述實施例中任一項之方法,其中該至少兩個非天然鹼基對包含選自dCNMO-dTPT3、dNaM-dTPT3、dCNMO-dTAT1或dNaM-dTAT1之鹼基對。
實施例37. 如實施例29至36中任一項之方法,其中該細胞生物體包含核苷三磷酸轉運子。
實施例38. 如實施例37之方法,其中該核苷三磷酸轉運子包含Pt
NTT2之胺基酸序列。
實施例39. 如實施例38之方法,其中該核苷三磷酸轉運子包含Pt
NTT2之截短胺基酸序列。
實施例40. 如實施例39之方法,其中Pt
NTT2之該截短胺基酸序列與藉由SEQ ID NO.1編碼之Pt
NTT2至少80%一致。
實施例41. 如實施例29至40中任一項之方法,其中該細胞生物體包含至少一個非天然DNA分子。
實施例42. 如實施例41之方法,其中該至少一個非天然DNA分子包含至少一個質體。
實施例43. 如實施例42之方法,其中將該至少一個非天然DNA分子整合至該細胞之該基因體中。
實施例44. 如實施例42或43之方法,其中該至少一個非天然DNA分子編碼該非天然多肽。
實施例45. 如實施例1至26中任一項之方法,其中該方法為活體外方法,其包含使用無細胞系統合成該非天然多肽。
實施例46. 如前述實施例中任一項之方法,其中該等非天然鹼基對包含至少一個包含非天然糖部分之非天然核苷酸。
實施例47. 如實施例46之方法,其中該非天然糖部分包含選自由以下組成之群的部分:OH、經取代之低碳烷基、烷芳基、芳烷基、O-烷芳基或O-芳烷基、SH、SCH3
、OCN、Cl、Br、CN、CF3
、OCF3
、SOCH3
、SO2
CH3
、ONO2
、NO2
、N3
、NH2
F;
O-烷基、S-烷基、N-烷基;
O-烯基、S-烯基、N-烯基;
O-炔基、S-炔基、N-炔基;
O-烷基-O-烷基、2'-F、2'-OCH3
、2'-O(CH2
)2
OCH3
,其中該烷基、烯基及炔基可為經取代或未經取代之C1
-C10
烷基、C2
-C10
烯基、C2
-C10
炔基、-O[(CH2
)n
O]m
CH3
、-O(CH2
)n
OCH3
、-O(CH2
)n
NH2
、-O(CH2
)n
CH3
、-O(CH2
)n
-NH2
及-O(CH2
)n
ON[(CH2
)n
CH3
)]2
,其中n及m為1至約10;
及/或於5'位置處之修飾:
5'-乙烯基、5'-甲基(R或S);
於4'位置處之修飾:
4'-S、雜環烷基、雜環烷芳基、胺基烷胺基、聚烷基胺基、經取代之矽烷基、RNA裂解基團、報導基團、嵌入基團、用於改良寡核苷酸之藥物動力學特性之基團或用於改良寡核苷酸之藥效動力學特性之基團及其任何組合。
實施例48. 一種細胞,其包含至少一個包含至少四個非天然鹼基對之非天然DNA分子,其中該至少一個非天然DNA分子編碼(i)編碼非天然多肽且包含至少第一及第二非天然密碼子之信使核糖核酸(mRNA)分子;以及(ii)至少第一及第二轉送RNA (tRNA)分子,該第一tRNA分子包含第一非天然反密碼子且該第二tRNA分子包含第二非天然反密碼子,並且該至少一個DNA分子中之該至少四個非天然鹼基對處於序列背景中,使得該mRNA分子之該第一非天然密碼子及該第二非天然密碼子分別與該第一非天然反密碼子及該第二非天然反密碼子互補。
實施例49. 如實施例48之細胞,其進一步包含該mRNA分子以及該至少第一及第二tRNA分子。
實施例50. 如實施例49之細胞,其中該至少第一及第二tRNA分子共價連接至非天然胺基酸。
實施例51. 如實施例50之細胞,其進一步包含該非天然多肽。
實施例52. 一種細胞,其包含:
a. 至少兩個不同的非天然密碼子-反密碼子對,其中各非天然密碼子-反密碼子對包含來自非天然信使RNA (mRNA)之非天然密碼子及來自非天然轉移核糖核酸(tRNA)之非天然反密碼子,該非天然密碼子包含第一非天然核苷酸且該非天然反密碼子包含第二非天然核苷酸;以及
b. 各自共價連接至對應非天然tRNA之至少兩個不同的非天然胺基酸。
實施例53. 如實施例52之細胞,其進一步包含至少一個包含至少四個非天然鹼基對(UBP)之非天然DNA分子。
實施例54. 如實施例48至53中任一項之細胞,其中該第一非天然核苷酸位於該非天然密碼子之第二或第三位置。
實施例54.1. 如實施例48至53中任一項之細胞,其中該第一非天然核苷酸位於該非天然密碼子之第一、第二或第三位置。
實施例55. 如實施例54或54.1之細胞,其中該第一非天然核苷酸與該非天然反密碼子之該第二非天然核苷酸互補地鹼基配對。
實施例56. 如實施例48至55中任一項之細胞,其中該第一非天然核苷酸及該第二非天然核苷酸分別包含第一及第二鹼基,其獨立地選自由以下組成之群:
。
實施例10. 如實施例4至9中任一項之方法,其中NNX-XNN係選自由以下組成之群:UUX-XAA、UGX-XCA、CGX-XCG、AGX-XCU、GAX-XUC、CAX-XUG、AUX-XAU、CUX-XAG、GUX-XAC、UAX-XUA及GGX-XCC。
實施例11. 如實施例4至9中任一項之方法,其中NNX-YNN係選自由以下組成之群:UUX-YAA、UGX-YCA、CGX-YCG、AGX-YCU、GAX-YUC、CAX-YUG、AUX-YAU、CUX-YAG、GUX-YAC、UAX-YUA及GGX-YCC。
實施例12. 如實施例4至9中任一項之方法,其中NXN-NYN係選自由以下組成之群:GXU-AYC、CXU-AYG、GXG-CYC、AXG-CYU、GXC-GYC、AXC-GYU、GXA-UYC、CXC-GYG及UXC-GYA。
實施例13. 如實施例12之方法,其中NXN-NYN係選自由以下組成之群:AXG-CYU、GXC-GYC、AXC-GYU、GXA-UYC、CXC-GYG及UXC-GYA。
實施例13.1. 如實施例4.1至9中任一項之方法,其中XNN-NNY係選自由以下組成之群:XUU-AAY、XUG-CAY、XCG-CGY、XAG-CUY、XGA-UCY、XCA-UGY、XAU-AUY、XCU-AGY、XGU-ACY、XUA-UAY、XUC-GAY、XCC-GGY、XAA-UUY、XAC-GUY、XGC-GCY、XGG-CCY及XGG-CCY。
實施例13.2. 如實施例4.1至9中任一者之方法,其中XNN-NNX係選自由以下組成之群:XUU-AAX、XUG-CAX、XCG-CGX、XAG-CUX、XGA-UCX、XCA-UGX、XAU-AUX、XCU-AGX、XGU-ACX、XUA-UAX、XUC-GAX、XCC-GGX、XAA-UUX、XAC-GUX、XGC-GCX、XGG-CCX及XGG-CCX。
實施例14. 如前述實施例中任一項之方法,其中該至少兩個非天然tRNA分子各自包含不同的非天然反密碼子。
實施例15. 如實施例14之方法,其中該至少兩個非天然tRNA分子包含來自甲烷八聯球菌屬之吡咯離胺醯基tRNA及來自詹氏甲烷球菌之酪胺醯基tRNA,或其衍生物。
實施例16. 如實施例13、14或15中任一項之方法,其包含藉由胺基-醯基tRNA合成酶而裝載有該至少兩個非天然tRNA分子。
實施例17. 如實施例16之方法,其中該胺基醯基tRNA合成酶係選自由嵌合PylRS (chPylRS)及詹氏甲烷球菌AzFRS (Mjp
AzFRS)組成之群。
實施例18. 如實施例14或15之方法,其包含藉由至少兩種不同tRNA合成酶而裝載有該至少兩個非天然tRNA分子。
實施例19. 如實施例18之方法,其中該至少兩種tRNA合成酶包含嵌合PylRS (chPylRS)及詹氏甲烷球菌AzFRS (Mjp
AzFRS)。
實施例20. 如實施例1至19中任一項之方法,其中該非天然多肽包含兩個、三個或更多個非天然胺基酸。
實施例21. 如實施例1至20中任一項之方法,其中該非天然多肽包含至少兩個相同的非天然胺基酸。
實施例22. 如實施例1至20中任一項之方法,其中該非天然多肽包含至少兩個不同的非天然胺基酸。
實施例23. 如實施例1至22中任一項之方法,其中該非天然胺基酸包含
離胺酸類似物;
芳族側鏈;
疊氮基;
炔基;或
醛基或酮基。
實施例24. 如實施例1至22中任一項之方法,其中該非天然胺基酸不包含芳族側鏈。
實施例25. 如實施例1至22中任一項之方法,其中該非天然胺基酸係選自:N6-疊氮基乙氧基-羰基-L-離胺酸(AzK)、N6-炔丙基乙氧基-羰基-L-離胺酸(PraK)、N6-(炔丙氧基)-羰基-L-離胺酸(PrK)、對疊氮基-苯丙胺酸(pAzF)、BCN-L-離胺酸、降冰片烯離胺酸、TCO-離胺酸、甲基四𠯤離胺酸、烯丙氧基羰基離胺酸、2-胺基-8-側氧基壬酸、2-胺基-8-側氧基辛酸、對乙醯基-L-苯丙胺酸、對疊氮基甲基-L-苯丙胺酸(pAMF)、對碘-L-苯丙胺酸、間乙醯基苯丙胺酸、2-胺基-8-側氧基壬酸、對炔丙基氧基苯丙胺酸、對炔丙基-苯丙胺酸、3-甲基-苯丙胺酸、左旋多巴、氟化苯丙胺酸、異丙基-L-苯丙胺酸、對疊氮基-L-苯丙胺酸、對醯基-L-苯丙胺酸、對苯甲醯基-L-苯丙胺酸、對溴苯基丙胺酸、對胺基-L-苯丙胺酸、異丙基-L-苯丙胺酸、O-烯丙基酪胺酸、O-甲基-L-酪胺酸、O-4-烯丙基-L-酪胺酸、4-丙基-L-酪胺酸、膦醯基酪胺酸、三-O-乙醯基-GlcNAcp-絲胺酸、L-磷絲胺酸、膦醯基絲胺酸、L-3-(2-萘基)丙胺酸、2-胺基-3-((2-((3-(苯甲氧基)-3-側氧基丙基)胺基)乙基)硒烷基)丙酸、2-胺基-3-(苯基硒烷基)丙酸、硒半胱胺酸、N6-(((2-疊氮基苯甲基)氧基)羰基)-L-離胺酸、N6-(((3-疊氮基苯甲基)氧基)羰基)-L-離胺酸及N6-(((4-疊氮基苯甲基)氧基)羰基)-L-離胺酸。
實施例26. 如前述實施例中任一項之方法,其中該至少一個非天然DNA分子呈質體形式。
實施例27. 如實施例1至26中任一項之方法,其中將該至少一個非天然DNA分子整合至細胞之基因體中。
實施例28. 如實施例26或27之方法,其中該至少一個非天然DNA分子編碼該非天然多肽。
實施例29. 如前述實施例中任一項之方法,其中該方法包含該非天然DNA分子之活體內複製及轉錄以及該經轉錄mRNA分子於細胞生物體中之活體內轉譯。
實施例30. 如實施例29之方法,其中該細胞生物體為微生物。
實施例31. 如實施例30之方法,其中該細胞生物體為原核生物。
實施例32. 如實施例31之方法,其中該細胞生物體為細菌。
實施例33. 如實施例32之方法,其中該細胞生物體為革蘭氏陽性細菌。
實施例34. 如實施例32之方法,其中該細胞生物體為革蘭氏陰性細菌。
實施例35. 如實施例34之方法,其中該細胞生物體為大腸桿菌。
實施例36. 如前述實施例中任一項之方法,其中該至少兩個非天然鹼基對包含選自dCNMO-dTPT3、dNaM-dTPT3、dCNMO-dTAT1或dNaM-dTAT1之鹼基對。
實施例37. 如實施例29至36中任一項之方法,其中該細胞生物體包含核苷三磷酸轉運子。
實施例38. 如實施例37之方法,其中該核苷三磷酸轉運子包含Pt
NTT2之胺基酸序列。
實施例39. 如實施例38之方法,其中該核苷三磷酸轉運子包含Pt
NTT2之截短胺基酸序列。
實施例40. 如實施例39之方法,其中Pt
NTT2之該截短胺基酸序列與藉由SEQ ID NO.1編碼之Pt
NTT2至少80%一致。
實施例41. 如實施例29至40中任一項之方法,其中該細胞生物體包含至少一個非天然DNA分子。
實施例42. 如實施例41之方法,其中該至少一個非天然DNA分子包含至少一個質體。
實施例43. 如實施例42之方法,其中將該至少一個非天然DNA分子整合至該細胞之該基因體中。
實施例44. 如實施例42或43之方法,其中該至少一個非天然DNA分子編碼該非天然多肽。
實施例45. 如實施例1至26中任一項之方法,其中該方法為活體外方法,其包含使用無細胞系統合成該非天然多肽。
實施例46. 如前述實施例中任一項之方法,其中該等非天然鹼基對包含至少一個包含非天然糖部分之非天然核苷酸。
實施例47. 如實施例46之方法,其中該非天然糖部分包含選自由以下組成之群的部分:OH、經取代之低碳烷基、烷芳基、芳烷基、O-烷芳基或O-芳烷基、SH、SCH3
、OCN、Cl、Br、CN、CF3
、OCF3
、SOCH3
、SO2
CH3
、ONO2
、NO2
、N3
、NH2
F;
O-烷基、S-烷基、N-烷基;
O-烯基、S-烯基、N-烯基;
O-炔基、S-炔基、N-炔基;
O-烷基-O-烷基、2'-F、2'-OCH3
、2'-O(CH2
)2
OCH3
,其中該烷基、烯基及炔基可為經取代或未經取代之C1
-C10
烷基、C2
-C10
烯基、C2
-C10
炔基、-O[(CH2
)n
O]m
CH3
、-O(CH2
)n
OCH3
、-O(CH2
)n
NH2
、-O(CH2
)n
CH3
、-O(CH2
)n
-NH2
及-O(CH2
)n
ON[(CH2
)n
CH3
)]2
,其中n及m為1至約10;
及/或於5'位置處之修飾:
5'-乙烯基、5'-甲基(R或S);
於4'位置處之修飾:
4'-S、雜環烷基、雜環烷芳基、胺基烷胺基、聚烷基胺基、經取代之矽烷基、RNA裂解基團、報導基團、嵌入基團、用於改良寡核苷酸之藥物動力學特性之基團或用於改良寡核苷酸之藥效動力學特性之基團及其任何組合。
實施例48. 一種細胞,其包含至少一個包含至少四個非天然鹼基對之非天然DNA分子,其中該至少一個非天然DNA分子編碼(i)編碼非天然多肽且包含至少第一及第二非天然密碼子之信使核糖核酸(mRNA)分子;以及(ii)至少第一及第二轉送RNA (tRNA)分子,該第一tRNA分子包含第一非天然反密碼子且該第二tRNA分子包含第二非天然反密碼子,並且該至少一個DNA分子中之該至少四個非天然鹼基對處於序列背景中,使得該mRNA分子之該第一非天然密碼子及該第二非天然密碼子分別與該第一非天然反密碼子及該第二非天然反密碼子互補。
實施例49. 如實施例48之細胞,其進一步包含該mRNA分子以及該至少第一及第二tRNA分子。
實施例50. 如實施例49之細胞,其中該至少第一及第二tRNA分子共價連接至非天然胺基酸。
實施例51. 如實施例50之細胞,其進一步包含該非天然多肽。
實施例52. 一種細胞,其包含:
a. 至少兩個不同的非天然密碼子-反密碼子對,其中各非天然密碼子-反密碼子對包含來自非天然信使RNA (mRNA)之非天然密碼子及來自非天然轉移核糖核酸(tRNA)之非天然反密碼子,該非天然密碼子包含第一非天然核苷酸且該非天然反密碼子包含第二非天然核苷酸;以及
b. 各自共價連接至對應非天然tRNA之至少兩個不同的非天然胺基酸。
實施例53. 如實施例52之細胞,其進一步包含至少一個包含至少四個非天然鹼基對(UBP)之非天然DNA分子。
實施例54. 如實施例48至53中任一項之細胞,其中該第一非天然核苷酸位於該非天然密碼子之第二或第三位置。
實施例54.1. 如實施例48至53中任一項之細胞,其中該第一非天然核苷酸位於該非天然密碼子之第一、第二或第三位置。
實施例55. 如實施例54或54.1之細胞,其中該第一非天然核苷酸與該非天然反密碼子之該第二非天然核苷酸互補地鹼基配對。
實施例56. 如實施例48至55中任一項之細胞,其中該第一非天然核苷酸及該第二非天然核苷酸分別包含第一及第二鹼基,其獨立地選自由以下組成之群:
 ,其中該第二鹼基不同於該第一鹼基。
實施例57. 如實施例48或50至56中任一項之細胞,其中該至少四個非天然鹼基對獨立地選自由以下組成之群:dCNMO-dTPT3、dNaM-dTPT3、dCNMO-dTAT1或dNaM-dTAT1。
實施例58. 如實施例48或50至57中任一項之細胞,其中該至少一個非天然DNA分子包含至少一個質體。
實施例59. 如實施例48或50至58中任一項之細胞,其中將該至少一個非天然DNA分子整合至該細胞之基因體中。
實施例60. 如實施例50至59中任一項之細胞,其中該至少一個非天然DNA分子編碼非天然多肽。
實施例61. 如實施例48至60中任一項之細胞,其中該細胞表現核苷三磷酸轉運子。
實施例62. 如實施例61之細胞,其中該核苷三磷酸轉運子包含Pt
NTT2之胺基酸序列。
實施例63. 如實施例62之細胞,其中該核苷三磷酸轉運子包含Pt
NTT2之截短胺基酸序列。
實施例64. 如實施例63之細胞,其中Pt
NTT2之該截短胺基酸序列與由SEQ ID NO.1編碼之Pt
NTT2至少80%一致。
實施例65. 如實施例48至64中任一項之細胞,其中該細胞表現至少兩種tRNA合成酶。
實施例66. 如實施例65之細胞,其中該至少兩種tRNA合成酶為嵌合PylRS (chPylRS)及詹氏甲烷球菌AzFRS (Mjp
AzFRS)。
實施例67. 如實施例48至66中任一項之細胞,其中該細胞包含非天然核苷酸,該等非天然核苷酸包含非天然糖部分。
實施例68. 如實施例67之細胞,其中該非天然糖部分係選自由以下組成之群:
於2'位置處之修飾:
OH、經取代之低碳數烷基、烷芳基、芳烷基、O-烷芳基或O-芳烷基、SH、SCH3
、OCN、Cl、Br、CN、CF3
、OCF3
、SOCH3
、SO2
CH3
、ONO2
、NO2
、N3
、NH2
F;
O-烷基、S-烷基、N-烷基;
O-烯基、S-烯基、N-烯基;
O-炔基、S-炔基、N-炔基;
O-烷基-O-烷基、2'-F、2'-OCH3
、2'-O(CH2
)2
OCH3
,其中該烷基、烯基及炔基可為經取代或未經取代之C1
-C10
烷基、C2
-C10
烯基、C2
-C10
炔基、-O[(CH2
)n
O]m
CH3
、-O(CH2
)n
OCH3
、-O(CH2
)n
NH2
、-O(CH2
)n
CH3
、-O(CH2
)n
-NH2
及-O(CH2
)n
ON[(CH2
)n
CH3
)]2
,其中n及m為1至約10;
及/或於5'位置處之修飾:
5'-乙烯基、5'-甲基(R或S);
於4'位置處之修飾:
4'-S、雜環烷基、雜環烷芳基、胺基烷胺基、聚烷基胺基、經取代之矽烷基、RNA裂解基團、報導基團、嵌入基團、用於改良寡核苷酸之藥物動力學特性之基團或用於改良寡核苷酸之藥效動力學特性之基團及其任何組合。
實施例69. 如實施例48至68中任一項之細胞,其中該至少一個非天然核苷酸鹼基在轉錄期間藉由RNA聚合酶識別。
實施例70. 如實施例48至69中任一項之細胞,其中該細胞轉譯包含該至少兩個非天然胺基酸之至少一個非天然多肽。
實施例71. 如實施例48至70中任一項之細胞,其中該至少兩個非天然胺基酸獨立地選自由以下組成之群:N6-疊氮基乙氧基-羰基-L-離胺酸(AzK)、N6-炔丙基乙氧基-羰基-L-離胺酸(PraK)、N6-(炔丙氧基)-羰基-L-離胺酸(PrK)、對疊氮基-苯丙胺酸(pAzF)、BCN-L-離胺酸、降冰片烯離胺酸、TCO-離胺酸、甲基四𠯤離胺酸、烯丙氧基羰基離胺酸、2-胺基-8-側氧基壬酸、2-胺基-8-側氧基辛酸、對乙醯基-L-苯丙胺酸、對疊氮基甲基-L-苯丙胺酸(pAMF)、對碘-L-苯丙胺酸、間乙醯基苯丙胺酸、2-胺基-8-側氧基壬酸、對炔丙基氧基苯丙胺酸、對炔丙基-苯丙胺酸、3-甲基-苯丙胺酸、左旋多巴、氟化苯丙胺酸、異丙基-L-苯丙胺酸、對疊氮基-L-苯丙胺酸、對醯基-L-苯丙胺酸、對苯甲醯基-L-苯丙胺酸、對溴苯基丙胺酸、對胺基-L-苯丙胺酸、異丙基-L-苯丙胺酸、O-烯丙基酪胺酸、O-甲基-L-酪胺酸、O-4-烯丙基-L-酪胺酸、4-丙基-L-酪胺酸、膦醯基酪胺酸、三-O-乙醯基-GlcNAcp-絲胺酸、L-磷絲胺酸、膦醯基絲胺酸、L-3-(2-萘基)丙胺酸、2-胺基-3-((2-((3-(苯甲氧基)-3-側氧基丙基)胺基)乙基)硒烷基)丙酸、2-胺基-3-(苯基硒烷基)丙酸、硒半胱胺酸、N6-(((2-疊氮基苯甲基)氧基)羰基)-L-離胺酸、N6-(((3-疊氮基苯甲基)氧基)羰基)-L-離胺酸及N6-(((4-疊氮基苯甲基)氧基)羰基)-L-離胺酸。
實施例72. 如實施例48至71中任一項之細胞,其中該細胞經分離。
實施例73. 如實施例48至72中任一項之細胞,其中該細胞為原核生物。
實施例74. 一種細胞株,其包含如實施例48至73中任一項之細胞。實例 實例 1. 初始密碼子篩選
,其中該第二鹼基不同於該第一鹼基。
實施例57. 如實施例48或50至56中任一項之細胞,其中該至少四個非天然鹼基對獨立地選自由以下組成之群:dCNMO-dTPT3、dNaM-dTPT3、dCNMO-dTAT1或dNaM-dTAT1。
實施例58. 如實施例48或50至57中任一項之細胞,其中該至少一個非天然DNA分子包含至少一個質體。
實施例59. 如實施例48或50至58中任一項之細胞,其中將該至少一個非天然DNA分子整合至該細胞之基因體中。
實施例60. 如實施例50至59中任一項之細胞,其中該至少一個非天然DNA分子編碼非天然多肽。
實施例61. 如實施例48至60中任一項之細胞,其中該細胞表現核苷三磷酸轉運子。
實施例62. 如實施例61之細胞,其中該核苷三磷酸轉運子包含Pt
NTT2之胺基酸序列。
實施例63. 如實施例62之細胞,其中該核苷三磷酸轉運子包含Pt
NTT2之截短胺基酸序列。
實施例64. 如實施例63之細胞,其中Pt
NTT2之該截短胺基酸序列與由SEQ ID NO.1編碼之Pt
NTT2至少80%一致。
實施例65. 如實施例48至64中任一項之細胞,其中該細胞表現至少兩種tRNA合成酶。
實施例66. 如實施例65之細胞,其中該至少兩種tRNA合成酶為嵌合PylRS (chPylRS)及詹氏甲烷球菌AzFRS (Mjp
AzFRS)。
實施例67. 如實施例48至66中任一項之細胞,其中該細胞包含非天然核苷酸,該等非天然核苷酸包含非天然糖部分。
實施例68. 如實施例67之細胞,其中該非天然糖部分係選自由以下組成之群:
於2'位置處之修飾:
OH、經取代之低碳數烷基、烷芳基、芳烷基、O-烷芳基或O-芳烷基、SH、SCH3
、OCN、Cl、Br、CN、CF3
、OCF3
、SOCH3
、SO2
CH3
、ONO2
、NO2
、N3
、NH2
F;
O-烷基、S-烷基、N-烷基;
O-烯基、S-烯基、N-烯基;
O-炔基、S-炔基、N-炔基;
O-烷基-O-烷基、2'-F、2'-OCH3
、2'-O(CH2
)2
OCH3
,其中該烷基、烯基及炔基可為經取代或未經取代之C1
-C10
烷基、C2
-C10
烯基、C2
-C10
炔基、-O[(CH2
)n
O]m
CH3
、-O(CH2
)n
OCH3
、-O(CH2
)n
NH2
、-O(CH2
)n
CH3
、-O(CH2
)n
-NH2
及-O(CH2
)n
ON[(CH2
)n
CH3
)]2
,其中n及m為1至約10;
及/或於5'位置處之修飾:
5'-乙烯基、5'-甲基(R或S);
於4'位置處之修飾:
4'-S、雜環烷基、雜環烷芳基、胺基烷胺基、聚烷基胺基、經取代之矽烷基、RNA裂解基團、報導基團、嵌入基團、用於改良寡核苷酸之藥物動力學特性之基團或用於改良寡核苷酸之藥效動力學特性之基團及其任何組合。
實施例69. 如實施例48至68中任一項之細胞,其中該至少一個非天然核苷酸鹼基在轉錄期間藉由RNA聚合酶識別。
實施例70. 如實施例48至69中任一項之細胞,其中該細胞轉譯包含該至少兩個非天然胺基酸之至少一個非天然多肽。
實施例71. 如實施例48至70中任一項之細胞,其中該至少兩個非天然胺基酸獨立地選自由以下組成之群:N6-疊氮基乙氧基-羰基-L-離胺酸(AzK)、N6-炔丙基乙氧基-羰基-L-離胺酸(PraK)、N6-(炔丙氧基)-羰基-L-離胺酸(PrK)、對疊氮基-苯丙胺酸(pAzF)、BCN-L-離胺酸、降冰片烯離胺酸、TCO-離胺酸、甲基四𠯤離胺酸、烯丙氧基羰基離胺酸、2-胺基-8-側氧基壬酸、2-胺基-8-側氧基辛酸、對乙醯基-L-苯丙胺酸、對疊氮基甲基-L-苯丙胺酸(pAMF)、對碘-L-苯丙胺酸、間乙醯基苯丙胺酸、2-胺基-8-側氧基壬酸、對炔丙基氧基苯丙胺酸、對炔丙基-苯丙胺酸、3-甲基-苯丙胺酸、左旋多巴、氟化苯丙胺酸、異丙基-L-苯丙胺酸、對疊氮基-L-苯丙胺酸、對醯基-L-苯丙胺酸、對苯甲醯基-L-苯丙胺酸、對溴苯基丙胺酸、對胺基-L-苯丙胺酸、異丙基-L-苯丙胺酸、O-烯丙基酪胺酸、O-甲基-L-酪胺酸、O-4-烯丙基-L-酪胺酸、4-丙基-L-酪胺酸、膦醯基酪胺酸、三-O-乙醯基-GlcNAcp-絲胺酸、L-磷絲胺酸、膦醯基絲胺酸、L-3-(2-萘基)丙胺酸、2-胺基-3-((2-((3-(苯甲氧基)-3-側氧基丙基)胺基)乙基)硒烷基)丙酸、2-胺基-3-(苯基硒烷基)丙酸、硒半胱胺酸、N6-(((2-疊氮基苯甲基)氧基)羰基)-L-離胺酸、N6-(((3-疊氮基苯甲基)氧基)羰基)-L-離胺酸及N6-(((4-疊氮基苯甲基)氧基)羰基)-L-離胺酸。
實施例72. 如實施例48至71中任一項之細胞,其中該細胞經分離。
實施例73. 如實施例48至72中任一項之細胞,其中該細胞為原核生物。
實施例74. 一種細胞株,其包含如實施例48至73中任一項之細胞。實例 實例 1. 初始密碼子篩選
諸如sfGFP之綠色螢光蛋白及變異體已用作用於ncAA併入(尤其在位置Y151處)之研究的模型系統,其已展示出包容各種天然及ncAA取代。構築質體以含有兩個dNaM-dTPT3 UBP,一個位於sfGFP之密碼子151內且另一個經定位以編碼馬氏甲烷八聯球菌tRNAPyl
之反密碼子(圖 6C
),其藉由PylRS選擇地裝載有ncAA N6-((2-疊氮基乙氧基)-羰基)-L-離胺酸(AzK) (圖 6B
)。構築質體以檢查六個密碼子之解碼,包括兩個第一位置非天然密碼子(XTC及XTG;X係指dNaM)、兩個第二位置非天然密碼子(AXC及GXA)及兩個非天然第三位置密碼子(AGX及CAX),以及相對股背景密碼子(YTC、YTG、AYC、GYA、AGY及CAY;Y係指dTPT3)。
儘管SSO之純系群體能夠產生較大量純非天然蛋白質,其可能係由於在活體外構築期間消除錯誤組裝之質體,以促進初始密碼子篩選蛋白質表現首先用非純系細胞群體進行探索,且在轉化之後立即分析蛋白質產生。使用質體轉化攜有編碼嵌合吡咯離胺醯基-tRNA合成酶(chPylRSIPYE
)之附屬質體的大腸桿菌ML2 (BL21(DE3) lacZYA:PtNTT2(66-575) ΔrecA polB++),且在生長至早期固定相之後,在補充有dNaMTP及dTPT3TP之選擇性培養基中,將細胞轉移至新制培養基中。在生長至中指數期之後,將培養物補充有NaMTP、TPT3TP及AzK,且添加異丙基-β-D-硫代半乳糖苷(IPTG)以誘導T7 RNA聚合酶(T7 RNAP)、chPylRSIPYE
及tRNAPyl
之表現。在額外生長1小時之後,添加無水四環素(aTc)以誘導sfGFP之表現,其藉由螢光監測。
第一位置密碼子在不存在或存在AzK之情況下展示無顯著螢光,不管解碼是否嘗試用雜配對或自配對反密碼子(例如,對於XTG分別為tRNAPyl
(CAY)或tRNAPyl
(CAX)) (圖 10
)。在第二位置處具有dNaM之密碼子在不存在AzK之情況下展示極少螢光,但在其存在下,當用雜配對之反密碼子tRNAPyl
(GYT)或tRNAPyl
(TYC)重新編碼之tRNAPyl
解碼時展示出顯著螢光,但自配對反密碼子tRNAPyl
(GXT)或tRNAPyl
(TXC)則沒有。在dTPT3處於第二位置之情況下,在添加或不添加AzK之情況下未觀測到螢光,不管是否用雜配對或自配對tRNA嘗試解碼。在不存在AzK之情況下,第三位置密碼子CAX及CAY展示較高螢光,且出人意料地展示添加較少螢光,不管是否用雜配對或自配對tRNAPyl
嘗試解碼。此結果表明對應第三位置非天然tRNA在核糖體處非產生性結合且藉由天然tRNA阻斷非天然密碼子通讀。在不存在AzK之情況下,AGX及AGY展示極少螢光,且在添加AzK之情況下,具有tRNAPyl
(XCT)之AGX展示螢光增加。
由於第一位置密碼子看起來無前景,因此進行第二位置密碼子之更全面篩選。因為初始分析指示僅密碼子中之NaM及反密碼子中之TPT3的潛在解碼,因此檢查NXN密碼子及同源tRNAPyl
(NYN)。由於對應序列背景未充分保留於SSO之DNA中,排除在16種可能密碼子中之CXA、CXG及TXG。與先前結果一致,在不存在AzK之情況下,密碼子AXC及GXC之使用導致幾乎沒有螢光,而在存在AzK之情況下,其導致顯著螢光(圖 6D
)。類似地,在GXT、CXC、TXC、GXG、GXA、CXT及AXG密碼子之情況下,相對於當AzK被保留時,添加AzK導致螢光顯著增加。其餘四密碼子AXA、AXT、TXA及TXT產生極少螢光,不管是否添加AzK,揭露對於至少一個G-C對之迫切需求。
為了篩選非天然蛋白質產生,sfGFP經由C端StrepII親和標籤純化且藉由四個PEG單元(DBCO-PEG4-TAMRA)與連接至若丹明染料(TAMRA)之二苯并環辛炔(DBCO)反應進行菌株促進之疊氮-炔烴環加成(SPAAC)反應。如先前所示,成功結合不僅用可偵測螢光團標記含有ncAA之蛋白質,且亦產生電泳遷移之可偵測移位,從而允許相對於所產生之總蛋白質定量含有AzK之蛋白質(亦即ncAA併入之保真度;圖 6D
)。與先前結果一致,密碼子GXC及AXC之使用導致產生大量具有AzK殘基之sfGFP。值得注意地,七個額外非天然密碼子,即GXT、CXC、TXC、GXG、GXA、CXT及AXG亦產生顯著含量之非天然蛋白質(圖 6D
,圖 11
)。
最後,進行第三位置密碼子之更全面篩選。因為在初始篩選中,僅AGX呈現經解碼,且隨後僅藉由自配對tRNAPyl
(XCT),在具有同源自配對tRNAPyl
(XNN)之密碼子之第三位置具有dNaM之密碼子(圖 6C
)進一步檢查。排除NCX密碼子,因為其產生NCXA之序列背景,如上文所指出,其並不充分保留於SSO之DNA中。與初始分析一致,在不存在AzK之情況下,此等密碼子通常產生比在第二位置密碼子下所觀測到的更多的螢光,但在存在AzK變數之情況下,觀測到螢光增加(圖 6D
)。無論如何,當如上文所描述分離且分析蛋白質時,使用CGX、ATX、CAX、AGX、GAX、TGX、CTX、TTX、GTX或TAX均導致顯著量之非天然蛋白質產生(圖 6D
,圖 11
)。密碼子GGX產生多個移位物種,表明tRNAPyl
(XCC)解碼一或多個天然密碼子。當使用密碼子AAX時未偵測到非天然蛋白質。實例 2. 純系 SSO 中之密碼子表徵
為選擇上文所描述之密碼子篩選中所鑑別之最有前景的密碼子/反密碼子對,比較在AzK存在下所觀測到之螢光於分離蛋白質(圖 6D
,插圖)中之誘導遷移移位。基於此分析,選擇七個非天然密碼子/反密碼子對,即GXC/GYC、GXT/AYC、AXC/GYT、AGX/XCT、CGX/XCG、TTX/XAA、TGX/XCA,以用於進一步表徵。在純系SSO中檢查此等密碼子/反密碼子對,其消除經錯誤組裝之質體或在活體外構築期間喪失UBP之質體轉化的細胞。純系SSO係藉由將轉化子劃線至含有dNaMTP及dTPT3TP之固體生長培養基上、選定個別群落及確定質體完整性及高UBP保留率獲得。使高保留純系再生長且誘導以產生如上文所描述之蛋白質。值得注意地,所觀測之螢光指示七個密碼子/反密碼子對中之每一者以與琥珀色抑制對照物有利地比較之水準產生蛋白質,且此外,凝膠移位分析表明幾乎所有sfGFP含有ncAA (圖 7A
,圖 12
)。使用密碼子/反密碼子AGX/XCT、CGX/XCG、TTX/XAA及TGX/XCA之解碼僅取決於表現培養基中之NaMTP且在添加及不添加TPT3TP之情況下,產生具有類似AzK含量之sfGFP (圖 13
)。
以上所分析之七個非天然密碼子/反密碼子對在核糖體處清楚地介導高效解碼;然而,當在純系SSO中分析時,來自初步非純系篩選之其他密碼子可能展示高效解碼。因此,探索在具有四個額外密碼子/反密碼子對TXC/GYA、GXG/CYC、CXC/GYG及AXT/AYT之純系SSO中之非天然蛋白質產生。儘管UBP保留率較高(表1),在具有或不具有AzK之情況下,AXT展示無螢光信號,進一步支持對具有第二位置密碼子之G-C對之要求。在添加AzK之情況下,對於TXC、CXC及GXG之螢光與七個最初表徵之密碼子之彼螢光類似,儘管在不存在AzK之情況下其略微較高(圖 7A
)。SPAAC凝膠移位分析揭露,儘管來自初步篩選之資料的相對較大誤差排除了定量比較,但CXC清楚地在純系SSO中產生的比在具有非純系SSO以及亦可能TXC及GXG之初步篩選中觀測到之蛋白質顯著更多的移位蛋白質(圖 7B
)。資料表明對於某些密碼子,篩選中之次佳效能至少部分由活體外質體構築中之序列依賴性差異產生。無論如何,結果鑑別兩個額外高保真度密碼子TXC及CXC,且表明仍可能鑑別更多可行的密碼子。
為了開始評估非天然密碼子/反密碼子對之正交性,選擇AXC/GYT、GXT/AYC及AGX/XCT且針對具有非天然密碼子與反密碼子之所有成對組合之純系SSO中之蛋白質產生進行檢查。在添加AzK之情況下,當各非天然密碼子與同源非天然反密碼子配對時觀測到顯著螢光,且當與非同源非天然反密碼子配對時,觀察到相比於背景幾乎無增加(圖 7B
)。因此,AXC/GYT、GXT/AYC及AGX/XCT為正交的且能夠在SSO中同時使用。實例 3. 兩個非天然密碼子之同時解碼 .
為探索多個密碼子之同時解碼,首先用位置190及200處之天然sf
GFP密碼子(分別經GXT及AXC,(sf
GFP190,200
(GXT,AXC)))構築質體。另外,質體編碼tRNAPyl
(AYC)及詹氏甲烷球菌RNApAzF
,其藉由詹氏甲烷球菌TyrRS (Mj
TyrRS)選擇性裝載有對疊氮基-L-苯丙胺酸(p
AzF;圖 6B
),且其反密碼子經重新編碼以識別AXC (tRNApAzF
(GYT);圖 8A
)。藉由含UBP之質體轉化攜帶編碼chPylRSIPYE
及Mjp
AzFRS兩者之附屬質體之大腸桿菌ML2,且獲得、生長及誘導SSO以產生如上文所描述之sfGFP。在提供AzK及p
AzF兩者之情況下,在與具有單一密碼子構築體之表現相同之時間標度內觀測到增加之細胞螢光(圖 8B
,圖 14
),儘管具有自sf
GFP190,200
(GXT,AXC)之表現之螢光水準略微小於使用sf
GFP190
(GXT)或sf
GFP200
(AXC)所觀測到之螢光水準,但明顯大於使用對應抑制因子tRNA解碼之來自琥珀色赭色對照物(sf
GFP190,200
(TAA,TAG))之螢光水準(圖 8C
,圖 14
)。在兩種情況下,當藉由SPAAC凝膠移位分析時,與針對併入單一ncAA所觀測到之遷移相比,未顯而易見未移位之帶且進一步延遲主要帶之遷移,表明實際上已併入兩種ncAA (圖 8D
)。為證實併入p
AzF及AzK兩者,使用定量完整蛋白質質譜(HRMS ESI-TOF)來分析純化蛋白質。與凝膠移位分析一致,此分析揭露91±1.1%之分離蛋白質含有p
AzF及AzK兩者,而1.7±0.4%含有單一p
AzF及7.5±0.78%含有單個AzK(圖 15
)。在兩種情況下,所鑑別之雜質之質量對應於與dX至dT突變一致之胺基酸取代,表明在複製期間ncAA併入保真度之大部分損失由dNaM或dTPT3損失引起,且不歸因於轉錄或轉譯期間之誤差。UBP之保留率基於抗生蛋白鏈菌素-生物素移位分析。保留率包含相對於ssDNA模板對照之相對移位而歸一化之相對移位(亦即,經移位帶之信號除以經移位及未經移位帶之總信號),不同之處在於不可進行tRNA p AzF
及tRNASer
之歸一化。展示平均值±標準差(表 1
)。表 1. 所報導之 SSO 中之鹼基對 (BP) 保留率
SSO產生16 ± 3.2 μg·ml-1
純化蛋白質,而琥珀色抑制對照物產生6.8 ± 1.1 μg·ml-1
。然而,應注意,SSO培養物與琥珀色赭色對照細胞相比生長至更低密度,且當針對OD600
歸一化時,SSO產生13 ± 1.6 μg·ml-1
純化蛋白質,而琥珀色赭色抑制產生2.8 ± 0.28 μg ml-1
,表明每OD600
,以超過4.5倍的蛋白質產生SSO。所有產量均藉由在親和純化期間使用過量Strep-Tactin XT珠粒之sfGFP捕獲來測定。在t=180分鐘表現下將產量歸一化至最終OD600
。展示平均值±標準差(表 2
)。因此,SSO有效地產生具有兩個ncAA之非天然蛋白質。表 2. sfGFP 表現之蛋白質產量
| 構築體 | ||||||
| 單一密碼子實驗 | 呈現於以下中 | n | 一或多個密碼子 | 一或多個UBP保留密碼子 | 一或多個反密碼子 | 一或多個UBP保留反密碼子 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 圖6A | 3 | AXC | 94±3 | GYT | 92±4 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 圖6A | 3 | GXC | 94±3 | GYC | 96±5 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 圖6A | 3 | GXT | 99±1 | AYC | 99±1 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 圖6A | 3 | AGX | 89±3 | XCT | 61±18 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 圖6A | 3 | CGX | 89±3 | XCG | 83±8 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 圖6A | 3 | TGX | 91±2 | XCA | 78±13 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 圖6A | 3 | TTX | 95±3 | XAA | 76±37 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 圖6A | 5 | CXC | 67±8 | GYG | 91±4 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 圖6A | 4 | GXG | 58±2 | CYC | 60±10 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 圖6A | 3 | TXC | 87±6 | GYA | 94±11 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 圖6A | 3 | AXT | 97±3 | AYT | 95±1 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 圖6B | 3 | AGX | 91±1 | AYC | 101±1 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 圖6B | 3 | AGX | 92±1 | GYT | 99±6 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 圖6B | 3 | AGX | 82±3 | XCT | 100±4 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 圖6B | 3 | AXC | 96±3 | AYC | 99±2 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 圖6B | 3 | AXC | 98±1 | GYT | 94±8 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 圖6B | 3 | AXC | 99±2 | XCT | 84±12 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 圖6B | 3 | GXT | 99±4 | AYC | 97±2 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 圖6B | 3 | GXT | 100±1 | GYT | 100±1 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 圖6B | 3 | GXT | 99±1 | XCT | 101±1 |
| 多密碼子密碼子實驗(包括對照) | ||||||
| sfGFP190 馬氏甲烷八聯球菌tRNAPyl | 圖7B | 3 | GXT | 103±4 | AYC | 101±4 |
| sfGFP200 詹氏甲烷球菌tRNA p AzF | 圖7B | 3 | AXC | 96±2 | GYT | >94±1 |
| sfGFP190,200 馬氏甲烷八聯球菌tRNAPyl 詹氏甲烷球菌tRNA p AzF | 圖7B | 3 | GXT、AXC | 98±3、86±2 | AYC、GYT | 96±1、>88±1 |
| sfGFP151,190,200 馬氏甲烷八聯球菌tRNAPyl 詹氏甲烷球菌tRNA p AzF 大腸桿菌tRNASer | 圖7B | 3 | AXC、GXT、AGX | 92±1、101±2、96±3 | XCT、GYT、AYC | 93±3、>87±3、>94±2 |
| 構築體 | n | 一或多個密碼子/一或多個反密碼子 | 蛋白質產量(μg/ml) | 歸一化蛋白質產量(μg/ml/OD600) |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 3 | TAC/- | 66±13 | 23±1.9 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 3 | TAG/CTA | 52±11 | 18±3.0 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 3 | AXC/GYT | 28±6.3 | 19±2.1 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 3 | GXC/GYC | 31±0.32 | 18±2.9 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 3 | GXT/AYC | 29±3.3 | 21±0.22 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 3 | AGX/XCT | 34±4.7 | 19±1.7 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 3 | CGX/XCG | 29±2.8 | 19±5.2 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 3 | TGX/XCA | 27±3.2 | 18±4.8 |
| sfGFP151 馬氏甲烷八聯球菌tRNAPyl | 3 | TTX/XAA | 27±4.1 | 19±4.6 |
| sfGFP190 、200 馬氏甲烷八聯球菌tRNAPyl 、詹氏甲烷球菌tRNA p AzF | 3 | TAA、TAG/TTA、CTA | 5.6±1.0 | 5.0±0.24 |
| sfGFP190 、200 馬氏甲烷八聯球菌tRNAPyl 、詹氏甲烷球菌tRNA p AzF | 3 | TAA、TAG/TTA、CTA | 6.8±1.1 | 2.8±0.28 |
| sfGFP190 、200 馬氏甲烷八聯球菌tRNAPyl 、詹氏甲烷球菌 tRNA p AzF | 3 | GXT、AXC/AYC、GYT | 16±3.2 | 13±1.6 |
| sfGFP151 、190 、200 馬氏甲烷八聯球菌tRNAPyl 、詹氏甲烷球菌tRNA p AzF 、大腸桿菌tRNASer | 3 | AXC、GXT、AGX/XCT、GYT、AYC | 12±1.9 | 7.8±1.1 |
為表徵具有不同官能基之ncAA之蛋白質表現,在如上所描述之SSO中表現sfGFP190,200
(GXT,AXC),但補充具有N6
-(炔丙氧基)-羰基-L-離胺酸之生長介質(PrK,圖 6B
),其亦藉由chPylRSIPYE
而非AzK識別。對於SSO或琥珀色赭色對照物,未藉由螢光觀測到對表現之實質性影響(圖 8E
)。在各情況下,經驗證,藉由SPAAC用TAMRA-PEG4
-DBCO校正PrK及p
AzF兩者之併入,繼之以使用TAMRA-PEG4
-疊氮基之銅催化之炔烴疊氮環加成(CuAAC),兩者均誘導電泳遷移之可觀測移位。藉由SSO產生之蛋白質以及琥珀色赭色對照物展示預期凝膠移位及TAMRA信號(圖 8F
)。實例 4. 三個非天然密碼子之同時解碼
為探索三個正交非天然密碼子之同時解碼,採用內源性絲胺酸tRNASer
、大腸桿菌SerT,其藉由不具有反密碼子識別之內源性SerRS裝入且預先重新編碼以解碼非天然密碼子。攜帶編碼chPylRSIPYE
及Mjp
AzFRS之附屬質體之大腸桿菌ML2用表現sfGFP151,190,200
(AXC,GXT,AGX)以及tRNAPyl
(XCT)、tRNA p AzF
(GYT)及tRNASer
(AYC)之質體轉化(圖 9A
),且純系SSO經製備、生長及誘導以產生如上文所描述之蛋白質。在AzK及p
AzF添加至培養基之情況下,觀測到顯著螢光,類似於上文針對兩個密碼子之同時解碼所獲得之結果(圖 9B
,圖 14
)。此等細胞產生12.1±1.9 μg ml-1
(7.8±1.1 μg ml-1
OD-1
)之分離蛋白質,其僅略微小於用兩個非天然密碼子之解碼分離之量(表2)。為證實p
AzF、AzK及Ser均已併入,經純化之蛋白經由定量完整蛋白質譜分析(HRMS ESI-TOF)來分析且發現96±0.63%之分離蛋白含有p
AzF、AzK及Ser,而主要雜質為僅含有AzK及Ser之sfGFP (3.5±0.63%)。不併入Ser之蛋白質幾乎不可偵測(0.20±0.087%),而對應於僅含有p
AzF及Ser之蛋白質之質量不可偵測(圖 9C
,圖 16
)。另外,未偵測到對應於Ser、AzK或p
AzF之多次插入之任何雜質。實例 5. 活體內表現非天然多肽材料之方法
所使用之寡核苷酸及質體之完整清單在表 3
中。天然ssDNA寡核苷酸及gBlock係購自IDT (San Diego, CA)。Genewiz (San Diego, CA)進行定序。所有DNA之純化均使用Zymo Research二氧化矽管柱套組來進行。所有選殖酶及聚合酶均購自New England Biolabs (Ipswich, MA)。所有生物結合試劑均購自Click Chemistry Tools (Scottsdale, AZ)。用於此研究之所有非天然核苷三磷酸及核苷胺基磷酸酯均獲自商業來源。除如文獻中所描述合成之sfGFP200
(AGX)以外,所有ssDNA dNaM模板亦獲自商業來源。表 3. 用於 PCR 及抗生蛋白鏈菌素 - 生物素移位分析中之單股 DNA 寡核苷酸
生長條件
| ID | 應用 | 序列(5'至3') | SEQ ID NO: |
| 用於UBP PCR之引子 | |||
| Efo309 | sfGFP Y151插入F | ATGGGTCTCACACAAACTCGAGTACAACTTTAACTCACAC | 2 |
| Efo310 | sfGFP Y151插入R | ATGGGTCTCGATTCCATTCTTTTGTTTGTCTGC | 3 |
| Efo296 | sfGFP Y200插入F | CATAATGGTCTCGCTGCTGCCCGATAACCAC | 4 |
| Efo297 | sfGFP Y200插入R | TGATATTGGTCTCGGTCTTTCGATAAAACACTCTGAGTAGAG | 5 |
| Efo311 | 馬氏甲烷八聯球菌tRNAPyl 插入F | ATGGGTCTCGAAACCTGATCATGTAGATCGAACGG | 6 |
| Efo312 | 馬氏甲烷八聯球菌tRNAPyl 插入R | ATGGGTCTCATCTAACCCGGCTGAACGG | 7 |
| Efo313 | 詹氏甲烷球菌tRNA p AzF 插入F | ATGGGTCTCCGGTAGTTCAGCAGGGCAGAACG | 8 |
| Efo314 | 詹氏甲烷球菌tRNA p AzF 插入R | ATGGGTCTCGGAGGGGATTTGAACCCCTGCCATG | 9 |
| Efo294 | sfGFP D190插入 | ATATTCGGTCTCGTCAGCAGAATACGCCGATTGG | 10 |
| Efo295 | sfGFP D190插入 | ACGCGTTGGTCTCGGTTATCGGGCAGCAGCACC | 11 |
| YZ401 | 大腸桿菌tRNASer 插入F | ATTGGTCTCGGCCGAGCGGTTGAAGGCAC | 12 |
| YZ403 | 大腸桿菌tRNASer 插入R | ATTGGTCTCTCTGGAACCCTTTCGGGTCG | 13 |
| 用於抗生蛋白鏈菌素-生物素移位分析之引子 | |||
| Efo251 | 位置Y151插入F | CTCGAGTACAACTTTAACTCACAC | 14 |
| Efo252 | 位置Y151插入R | GATTCCATTCTTTTGTTTGTCTGC | 15 |
| Efo294 | 位置D190插入F | ATATTCGGTCTCGTCAGCAGAATACGCCGATTGG | 10 |
| Efo295 | 位置D190插入R | ACGCGTTGGTCTCGGTTATCGGGCAGCAGCACC | 11 |
| Efo347 | 位置Y200插入F | GCTGCTGCCCGATAACCAC | 16 |
| Efo348 | 位置Y200插入R | GGTCTTTCGATAAAACACTCTGAGTAGAG | 17 |
| Efo343 | 馬氏甲烷八聯球菌tRNAPyl 插入F | GAAACCTGATCATGTAGATCGAACGG | 18 |
| Efo344 | 馬氏甲烷八聯球菌tRNAPyl 插入R | ATCTAACCCGGCTGAACGG | 19 |
| Efo313 | 詹氏甲烷球菌tRNA p AzF 插入F | ATGGGTCTCCGGTAGTTCAGCAGGGCAGAACG | 8 |
| Efo305 | 詹氏甲烷球菌tRNA p AzF 插入R | CCGCTGCCACTAGGAAGCTTATG | 20 |
| Efo119 | 大腸桿菌tRNASer 插入F | CCTCTAGAAAATCATTCCGGAAGTGTG | 21 |
| Efo162 | 大腸桿菌tRNASer 插入R | CTCTGGAACCCTTTCGGGTCGCCGGTTTGXTAGACCGGTGCCTTCAACCGCTCGGC | 22 |
| 用於UBP PCR之模板([NNN]指示任何指定密碼子/反密碼子三重組) | |||
| GFP151_[NNN] | sfGFP Y151插入 | CTCGAGTACAACTTTAACTCACACAATGTA[NNN]ATCACGGCAGACAAACAAAAGAATGGAATC | 23 |
| GFP190_GXT | sfGFP D190插入 | CAGCAGAATACGCCGATTGGCGXTGGCCCGGTGCTGCTGCCCGATAACC | 24 |
| GFP200_AXC | sfGFP Y200插入F | GCTGCTGCCCGATAACCACAXCCTCTCTACTCAGAGTGTTTTATCGAAAGACC | 25 |
| GFP200_opt_AGX | sfGFP Y200插入R | GCTGCCCGATAACCACAGXTTGTCTACTCAGAGTGTTTTATCG | 26 |
| tRNA_Pyl_[NNN] | 馬氏甲烷八聯球菌tRNAPyl 插入 | GAATCTAACCCGGCTGAACGGATT[NNN]AGTCCGTTCGATCTACATGATCAGG | 27 |
| tRNA_Mj_GYT | 馬氏甲烷八聯球菌tRNAPyl 插入 | GATTTGAACCCCTGCCATGCGGATTAXCAGTCCGCCGTTCTGCCCTGCTGAA | 28 |
| Trna_Eser_AYC | 大腸桿菌tRNASer 插入 | CTCTGGAACCCTTTCGGGTCGCCGGTTTGXTAGACCGGTGCCTTCAACCGCTCGGC | 22 |
所有細菌實驗均在補充有磷酸鉀(50 mM pH 7)之300 μl 2×YT (Fisher Scientific)培養基中進行。在37℃下,在200 r.p.m.震盪下(Infors HT Minitron),在平底48孔培養盤(CELLSTAR,Greiner Bio-One)進行生長。在以下濃度下使用抗生素(除非另外說明):氯黴素(5 μg/ml)、卡本西林(100 μg/ml)及吉歐黴素(50 μg/ml)。在以下濃度下使用非天然核苷三磷酸(除非另外說明):dNaMTP (150 μM)、dTPT3TP (10 μM)、NaMTP (250 μM)、TPT3TP (30 μM)。UBP培養基定義為含有dNaMTP及dTPT3TP之該2×YT培養基。質體構築
較大插入(>100 bp),即Mjp
AzFRS、tRNA或抗生素抗性卡匣之插入係藉由PCR擴增子或gBlock之吉布森組裝進行。在室溫下用DpnI處理擴增子隔夜,隨後在50℃下用組裝處理1.5小時。藉由將所需變化引入經設計以擴增整個質體之PCR引子懸垂物中來構築缺失或較小插入(<50 bp;例如密碼子或反密碼子突變誘發、移除限制位點或引入金閘目標位點)。在PCR之前使用T4 PNK使引子磷酸化,且在室溫下用DpnI處理所得PCR擴增子隔夜且使用T4 DNA連接酶重新循環。在初始組裝/接合之後,將質體轉移至電感受態XL-10金細胞中且在選擇性LB Lennox瓊脂(BP Difco)上生長。自個別群落分離質體且在使用之前藉由桑格定序(Sanger sequencing)來驗證。用於此研究中之所有質體可見於表 4
中。所有sfGFP
閱讀框係藉由PT7-tetO
控制且所有tRNA係藉由PT7-lacO
主鏈pSYN控制,該PT7-lacO
主鏈pSYN含有:ori(p15A)bleoR
。主鏈pGEX含有:ori(pBR322)ampR
。金閘目標位點(dest)係由識別序列BsaI-KpnI-BsaI構成。表 4. 用於實例中之質體
1
Zhang, Y等人. A semi-synthetic organism that stores and retrieves increased genetic information.Nature
551, 644-647 (2017)UBP 寡聚物之 PCR
| 主鏈 | 來源 | 應用 | 相關特性 |
| 超摺疊GFP表現質體 | |||
| pSYN | Zhang等人.1 | 天然表現質體 | sfGFP151 (TAG)、馬氏甲烷八聯球菌tRNAPyl (CTA) |
| pSYN | Zhang等人.1 | 天然表現質體 | sfGFP151 (TAC) |
| pSYN | 此研究 | 天然表現質體 | sfGFP190 (TAA)、馬氏甲烷八聯球菌tRNAPyl (TTA)、蛋白石終止密碼子 |
| pSYN | 此研究 | 天然表現質體 | sfGFP200 (TAG)、詹氏甲烷球菌tRNA p AzF (CTA) |
| pSYN | 此研究 | 天然表現質體 | sfGFP190,200 (TAA,TAG)、馬氏甲烷八聯球菌tRNAPyl (TTA)、詹氏甲烷球菌tRNA p AzF (CTA);蛋白石終止密碼子 |
| pSYN | Zhang等人.1 | UBP目標質體 | sfGFP151 (dest)、馬氏甲烷八聯球菌tRNAPyl (dest) |
| pSYN | 此研究 | UBP目標質體 | sfGFP190 (dest)、馬氏甲烷八聯球菌tRNAPyl (dest) |
| pSYN | 此研究 | UBP目標質體 | sfGFP200 (dest)、詹氏甲烷球菌tRNA p AzF (dest) |
| pSYN | 此研究 | UBP目標質體 | sfGFP190-200 (dest)、馬氏甲烷八聯球菌tRNAPyl (dest)、詹氏甲烷球菌tRNA p AzF (dest) |
| pSYN | 此研究 | UBP目標質體 | sfGFP151,190-200 (dest,dest)、馬氏甲烷八聯球菌tRNAPyl (dest)、詹氏甲烷球菌tRNA p AzF (dest)、大腸桿菌tRNASer (dest) |
| 輔助質體 | |||
| pGEX | 此研究 | 輔助質體 | PAmpR -tetR 、 PlacIq -lacI 、 Ptac-lacO -chPylRSIPYE |
| pGEX | 此研究 | 輔助質體 | PAmpR -tetR 、 PlacIq -lacI 、 PlacUV5 -lacO-MjpAzFRS 、 Ptac-lacO -chPylRSIPYE |
使用化學合成之含有ssDNA寡核苷酸(在清單B中)之dNaM作為模板,用引子(在清單A中)進行之PCR (OneTaq標準緩衝液1×、0.025單位/μl OneTaq、0.2 mM dNTP、0.1 mM dTPT3TP、0.1 mM dNaMTP、1.2 mM MgSO4
、1×SYBR綠、1.0 μM引子、約20 pM模板;循環:96℃ 0:30分鐘、96℃ 0:30分鐘、54℃ 0:30分鐘、68℃ 4:00分鐘、螢光讀數、轉至步驟2<24次)獲得含有UBP序列之雙股DNA插入物。使用與上文相同之條件,但其中兩種模板處於1 nM下,藉由重疊延伸組合用於位置sfGFP190
及sfGFP200
之插入。當SYBR綠追蹤達至平穩時,監測擴增且將反應物置於冰上。產物經由天然PAGE (6%丙烯醯胺:雙丙烯醯胺29:1;1×TBE中之SYBR金染色)分析以驗證單一擴增子,在旋轉管柱(Zymo Research)上純化且使用Qubit dsDNA HS (賽默飛世爾)定量。SSO 表現載體之金閘 組裝
含有UBP之插入物經由金閘組裝(Cutsmart緩衝液1×,1 mM ATP,6.67單位/μl T4 DNA連接酶,0.67單位/μl BsaI-HFv2、20 ng/μl進入載體DNA;循環:37℃ 10:00分鐘、37℃ 5:00分鐘、16℃ 5:00分鐘、22℃ 2:00分鐘,重複步驟2 39次,37℃ 20:00分鐘、55℃ 15:00分鐘、80℃ 30:00分鐘)在各插入物與進入載體之莫耳比為3:1下併入pSYN進入載體框架(表4)中。BsaI-HF用於圖 6
中之實驗。殘餘線性DNA及未消化之進入載體首先用KpnI-HF (在37℃下1小時,0.33 單位/μl)消化,接著用T5核酸外切酶(在37℃下30分鐘,0.17單位/μl)消化。在旋轉管柱上純化產物且使用Qubit dsDNA HS (ThermoFisher)定量。勝任起始細胞之製備
菌株ML2 (BL21(DE3)lac
ZYA::Pt
NTT2(66-57
5)ΔrecA polB++
)係用附屬pGEX質體轉化(表4)且接種於具有氯黴素及卡本西林之LB Lennox瓊脂上。如先前所描述藉由攝取放射性[α-32
P] dATP來挑選單一群落且驗證Pt
NTT2活性(Zhang等人2017)。用於UBP複製及轉譯之勝任細胞係藉由在具有檔板之培養物燒瓶中,在37℃ 250 r.p.m.下,在2×YT培養基中生長直至OD600
為0.25-0.30為止來製備。將培養物轉移至預先冷卻之50 mL法爾康管中且在冰水浴中溫和震盪2分鐘。細胞藉由離心(10分鐘,3200 r.p.m)粒化且在冷無菌水中洗滌,再次粒化及洗滌,隨後最後粒化且懸浮於50 μl 10%甘油/10 mL培養物中。細胞立即使用或在-80℃下冷凍以供隨後使用。非純系群體實驗
新鮮製備的勝任細胞用約0.4 ng金閘組裝產物電穿孔(2.5 kV)且立即懸浮於補充有磷酸鉀(50 mM pH 7)之950 μl 2×YT中,將其中10 μl稀釋至含有1.25X dNaMTP及dTPT3TP之40 μl UBP培養基中而無吉歐黴素。在37℃下回收細胞1小時之後,在具有吉歐黴素下將15 μl細胞懸浮於285 μl UBP培養基中且在37℃震盪下在48孔培養盤中生長。在OD600
約1下,將培養物轉移至冰中,隨後達至固定相,且儲存隔夜以用於蛋白質表現。純系 SSO 實驗
勝任細胞用金閘組裝產物(1-20 ng)電穿孔且如非純系群體實驗進行回收。藉由將10 μl回收培養物(及其稀釋液)擴散成含有氯黴素、卡本西林、勻黴素、dNaMTP及dTPT3TP之瓊脂小液滴(250 μl 2×YT 2%瓊脂50 mM磷酸鉀)來進行接種。在生長於培養盤(12-20小時;37℃)上之後,挑選直徑為大致0.5 mm之群落且將其懸浮於UBP培養基(300 μl)中。在OD約1下,將各培養物轉移至冰上之預冷卻管中,隨後達至固定相,且儲存隔夜以用於蛋白質表現。各培養物1)使用抗生蛋白鏈菌素生物素移位分析(如下文所描述)對於UBP保留率進行預先篩選,且2)藉由使培養物與已含有用於表現之組分(核苷三磷酸、ncAA、IPTG及無水四環素)的介質以1:4混合定性sfGFP
表現來進行預先篩選。若在37℃下培育2小時或在室溫下培育隔夜後添加適當ncAA時群落並不產生任何螢光信號,則丟棄該等群落。另外,丟棄在sfGFP
中具有<80% UBP保留率之群落。若超過三個群落滿足此等準則,則僅選擇具有最高UBP保留率之三個群落以限制材料費用。圖 7A
中之虛線右側之資料係經由略微修改之方法獲得。代替如上文所描述之預先篩選群落,對大量群落進行表現,但僅對在表現期間展示有前景螢光之培養物進行蛋白質分析。在表現期間,使用10 mM AzK。另外,在蛋白質純化期間使用緩衝液W2代替緩衝液W。預選殖 SSO 表現載體
在圖 7B
、圖 8
及圖 9
中之實驗中,分離來自預先篩選之群落的質體(Zymo Research Miniprep)以充當用於(預選殖)轉化之起始質體,以便易化群落預先篩選。針對來自具有一或多個適當ncAA之sfGFP表現之定性螢光,預篩選質體(如上文所描述)。用於圖 7B
中之資料之群落替代地在存在AzK之情況下在具有及不具有rNaMTP及rTPT3TP之情況下經預先篩選以定性地分別產生暗色及螢光信號。在sfGFP
中預先篩選所有預選殖質體之UBP保留率(>80%)。此外,此等質體使用標準OneTaq方案(New England Biolabs)進行PCR擴增,而無用於迫使dX至dN突變之非天然核苷三磷酸,且擴增子經桑格定序以驗證質體中之天然序列之完整性。在蛋白質編碼序列中允許沉默突變。UBP 蛋白質表現
當將核糖核苷酸三磷酸酯添加至250 μM NaMTP及30 μM TPT3TP中,同時添加5 mMp
AzF、20 mM AzK或10 mM PrK下之ncAA時,將培養物在UBP培養基中再新至OD600
0.10-0.15且37℃震盪直至OD 0.5-0.8。僅10 mM AzK用於雙/三密碼子實驗或其對照物中(圖 8
,圖 9
)。在進一步培育20分鐘之後,藉由添加IPTG (1 mM)引發預先誘導且培養物另外培育1小時。最終,藉由抑制tetO,藉由添加無水四環素(100 ng/μl)來誘導sfGFP表現。使用Perkin Elmer Envision 2103 Multilabel Reader (OD:590/20 nm過濾器;sfGFP:ex. 485/14 nm, em. 535/25 nm)監測(每30分鐘)OD600
及GFP螢光。在表現3小時之後,使培養物粒化且儲存在-80℃下用於隨後分析。用於 UBP 保留率之抗生蛋白鏈菌素 - 生物素移位分析
藉由PCR擴增使用非天然核苷三磷酸d5SICSTP以及生物素標記之dNaM類似物dMMO2Bio
TP測定質體DNA中之UBP保留率。經由標準小規模純化分離來自SSO之質體,產生SSO表現質體(pSYN)與附屬質體(pGEX)之混合物。總共2 ng質體混合物用作15 μl PCR反應中之模板(OneTaq標準緩衝液1×、0.018單位/μl OneTaq、0.007單位/μl DeepVent、0.4 mM dNTP、0.1 mM d5SICSTP、0.1 mM dMMO2Bio
TP、2.2 mM MgSO4
、1X SYBR綠、1.0 μM引子;循環:96℃ 2:00分鐘、96℃ 0:30分鐘、50℃ 0:10分鐘、68℃ 4:00分鐘、螢光讀數、68℃ 0:10分鐘,轉至步驟2<24次)。在各循環之最後步驟期間移除個別樣品,因為SYBR綠I追蹤展示擴增至平穩階段。所得生物素標記之擴增子補充有10 μg抗生蛋白鏈菌素(Promega)/1.5-2.0 μl粗PCR反應。抗生蛋白鏈菌素結合部分藉由6%天然-PAGE目測為移位,且藉由ImageStudioLite或Fiji定量移位及未移位兩者以產生相對原始的移位百分比。藉由將原始移位標準化為對照移位(藉由用化學合成寡核苷酸使PCR反應模板化來產生),評估總體UBP保留率。對於tRNApAzF
或tRNASer
標準化係不可能的,因為僅在引子黏合外部金閘插入物之情況下可靠的擴增係可能的,且因此並不黏合至對應對照寡核苷酸。蛋白質純化
使用BugBuster (100 μl;EMD Millipore;15分鐘;室溫;220 r.p.m.)溶解來自蛋白質表現實驗(200 μl)之細胞集結粒。隨後在緩衝液W (50 mM HEPES pH 8,150 mM NaCl,1 mM EDTA)中將細胞溶解物稀釋至等於500 μl減去所用親和性珠粒之體積的最終體積。以20 μl使用磁性Strep-Tactin XT珠粒(MagStrep「3型」XT珠粒、IBA Lifesciences之5% (v/v)懸浮液)用於常規純化且100 μl用於估算總表現量。將蛋白質與珠粒結合(30分鐘;4℃;溫和地旋轉),隨後將珠粒下拉且用緩衝液W (2×500 μl)洗滌。在針對HRMS分析之蛋白質純化中,替代地使用緩衝液W2 (50 mM HEPES pH 8,1 mM EDTA)。最後,在室溫下,在偶發性渦旋下,使用25 μl緩衝液BXT (50 mM HEPES pH 8,150 mM NaCl,1 mM EDTA,50 mM d-生物素)溶離蛋白質10分鐘。將蛋白質用緩衝液BXT2 (50 mM HEPES pH 8,1 mM EDTA,50 mM d-生物素)溶離以用於HRMS分析。使用Qubit蛋白質分析套組(ThermoFisher)進行定量。TAMRA 結合之 sfGFP 之西方墨點法
藉由在室溫下在黑暗中將33 ng/μl純蛋白質用0.1 mM TAMRA-PEG4
-DBCO (Click Chemistry Tools)培育隔夜來進行SPAAC。反應物與SDS-PAGE負載染料(250 mM Tris-HCl pH 6、30%甘油、5% βME、0.02%溴酚藍)以2:1混合且在95℃下變性5分鐘。凝膠為5%丙烯醯胺堆疊凝膠及當分析位置sfGFP151
時SDS-PAGE時為15%丙烯醯胺解析凝膠,以及當分析sfGFP190,200
時為17%丙烯醯胺解析凝膠(解析凝膠:15%或17%丙烯醯胺:雙丙烯醯胺29:1,0.1% (w/v) APS、0.04% TEMED、0.375 M Tris-HCl pH 8.8、0.1% (w/v) SDS;堆疊:5%丙烯醯胺:雙丙烯醯胺29:1,0.1% (w/v) APS、0.1% TEMED、0.125 M Tris-HCl pH 6.8、0.1% (w/v) SDS)。電泳在40 V下進行15分鐘,之隨後在120 V下對於15%凝膠操作約5小時且對於17%凝膠操作約6.5小時。每2小時更換操作緩衝液(25 mM Tris鹼、200 mM甘胺酸、0.1% (w/v) SDS)。在冷轉移緩衝液(20% (v/v) MeOH、50 mM Tris鹼、400 mM甘胺酸、0.0373% (w/v) SDS)中在90 V下使用濕式轉移將所得凝膠印跡至PVDF (EMD Millipore 0.45 μm PVDF-FL)上1小時。膜在4℃下在溫和攪動下使用含5%脫脂奶溶液之PBS-T (PBS pH 7.4,0.01% (v/v) Tween20)阻斷隔夜。將初級抗體(家兔α-Nterm-GFP Sigma Aldrich #G1544)施加於PBS-T (1:3,000)1小時(室溫;溫和攪動)。在PBS-T中洗滌墨點(5分鐘),隨後將二級抗體(山羊α-家兔-Alexa Fluor 647-結合抗體,ThermoFisher #A32733)施加於PBS-T (1:20,000)45分鐘(室溫;溫和攪動)。用PBS-T洗滌墨點,隨後(3×5分鐘)在50-100 μm解析度下,使用Typhoon 9410雷射掃描器(Typhoon Scanner Control v5 GE Healthcare Life Sciences)成像,首先掃描AlexaFluor 647 (Ex. 633 nm;Em. 670/30 nm;PMT 500 V),且隨後掃描TAMRA (Ex. 532 nm;Em. 580/30 nm;PMT 400 V)。PrK-p
AzF 標記之蛋白質之雙生物結合
使用BugBuster (100 μl;EMD Millipore;在室溫下15分鐘;220 r.p.m.)溶解來自1 mL培養物之細胞集結粒。將溶解物稀釋於緩衝液W (600 μl)中且添加MagStrep珠粒(200 μl)並且使其結合(30分鐘;4℃;溫和地旋轉)。使用磁鐵將珠粒拉下且用冷緩衝液W (2×1000 μl)洗滌,隨後懸浮於緩衝液W (200 μl)中。使用具有TAMRA-PEG4
-DBCO (0.5 mM)之此懸浮液之一半進行SPAAC 12-16小時(室溫;溫和地旋轉)。珠粒用無EDTA之緩衝液W (2×500 μl;HEPES 50 mM pH 7.4,150 mM NaCl)洗滌,隨後懸浮於無EDTA之緩衝液W (100 μl)中。使用具有疊氮基-PEG4-TAMRA (0.2 mM)以及硫酸銅(II) (0.5 mM)、參(苯甲基三唑基甲基)胺(2 mM;THPTA)及抗壞血酸鈉(15 mM)之此懸浮液中之一半進行CuAAC (1.5小時;室溫;溫和地旋轉)。用緩衝液W (2×500 μl)洗滌珠粒,隨後使用緩衝液BXT進行溶離(10分鐘;室溫;偶爾渦旋)。完整蛋白質高解析度質譜分析
藉由四次離心循環經由如前所描述之14,000 × g (3×10分鐘,且隨後1×18分鐘)下之10K Amicon超離心過濾器(EMD Millipore)將純化蛋白質(5 μg)去鹽至HPLC級水中(4×500 μl)。在回收蛋白質之後,將6 μl蛋白質注射至連接至Waters G2-XS TOF之Waters I類LC中。流動條件為0.4 mL/分鐘的50:50水:乙腈加上0.1%甲酸。藉由ESI+完成離子化且收集m/z 500-2000之資料。在質量峰之主部分上方進行光譜合併,且使用Waters MaxEnt1去捲積經合併光譜。藉由自動峰整合以及手動峰鑑別進行分析(圖 15
,圖 16
)。相對於鑑別為產物或雜質之所有物質之積分,將保真度計算為預期質量之積分,而不考慮技術雜質(例如,鹽加合物、精胺酸氧化)。
儘管已在本文中展示且描述本發明之較佳實施例,但對於熟習此項技術者應顯而易見,此類實施例僅以實例方式提供。在不脫離本發明之情況下,熟習此項技術者現將想到許多變化、改變及取代。應理解,本文所描述之本發明之實施例的各種替代方案可用於實踐本發明。預期以下申請專利範圍界定本發明之範疇,且由此涵蓋此等申請專利範圍及其同等者之範疇內的方法及結構。
100:例示性工作流程
101:DNA
102,110:蛋白質
103,106:tRNA
104:轉錄
105,109:非天然胺基酸
107:mRNA
108:轉譯
190:位置
200:位置
本發明之各種態樣詳細闡述於隨附申請專利範圍中。將參考闡述利用本發明原理之說明性實施例及其附圖的以下詳細描述來獲得對本發明之特徵及優點的較佳理解:
圖 1
說明使用非天然鹼基對(UBP)將非典型胺基酸(ncAA)位點特異性併入非天然多肽或非天然蛋白質中之工作流程,其使用非天然X-Y鹼基對。將三個ncAA併入非天然多肽或非天然蛋白質中僅展示為實例;可併入任何數目之ncAA。
圖 2
描繪例示性非天然核苷酸鹼基對(UBP)。
圖 3
描繪去氧核糖X類似物。為了清楚起見,已省略去氧核糖及磷酸酯。
圖 4A-B
說明核糖核苷酸類似物。為了清楚起見圖 4A
為已省略核糖及磷酸酯之核糖核苷酸X類似物之描述。為了清楚起見圖 4B
為已省略核糖及磷酸酯之核糖核苷酸Y類似物之描述。
圖 5A-G
說明例示性非天然胺基酸。圖 5A
係自Young等人, 「Beyond the canonical 20 amino acids: expanding the genetic lexicon」,J. of Biological Chemistry
285(15): 11039-11044 (2010)之圖2改編。圖 5B
為例示性非天然胺基酸離胺酸衍生物。圖 5C
為例示性非天然胺基酸苯丙胺酸衍生物。圖 5D-5G
說明例示性非天然胺基酸。此等非天然胺基酸(UAA)已基因編碼於蛋白質中(圖 5D
- UAA 1-42號;圖 5E
- UAA 43-89號;圖 5F
- UAA 90-128號;圖 5G
- UAA 129-167號)。圖 5D-5G
係自Dumas等人, Chemical Science
2015, 6, 50-69之表1改編。
圖 6A-D
說明使用非天然密碼子及反密碼子在非純系SSO中產生蛋白質。非天然密碼子及非天然反密碼子依據其DNA編碼序列書寫。圖 6A
為dNaM-dTPT3 UBP之化學結構。圖 6B
為ncAAs、AzK、PrK及p
AzF之化學結構。圖 6C
為用於表現sfGFP151
(NNN)及馬氏甲烷八聯球菌tRNAPyl
(NNN)之基因卡匣之示意圖,其中NNN係指任何指定密碼子或反密碼子。圖 6D
描繪在蛋白質表現端點(亦即,在添加aTc之後t=180分鐘)處,在培養基中在具有及不具有AzK之情況下,使用指定密碼子及反密碼子之來自非純系SSO培養物之歸一化螢光(a.u.,任意單位)。各複製培養物來源於用攜帶UBP之質體轉型之勝任SSO起始細胞之不同批料(n=3,生物複本)。展示平均值及個別數據點。經純化sfGFP之一個代表性裁剪西方墨點法展示以上各密碼子及反密碼子,其用來自SSO培養物之TAMRA-PEG4
-DBCO進行SPAAC (僅α-GFP通道)。圖 6D
插圖為在AzK(來自圖 6D
)存在下之平均端點螢光相對於藉由SPAAC誘導之定量相對蛋白質轉換之平均值之散點圖(n=3;生物複本)。選擇用於進一步分析之七個頂部密碼子經環繞。
圖 7A-B
說明蛋白產生及純系SSO中之密碼子正交性之分析。非天然密碼子及非天然反密碼子依據其DNA編碼序列書寫。圖 7A
描繪在七個頂部密碼子及反密碼子(左側
)以及四個其他所選密碼子(右側
)之蛋白質表現端點(亦即在添加aTc之後t=180分鐘)處,在具有及不具有AzK之情況下,來自純系SSO之歸一化螢光。各複製培養物係自個別SSO群落繁殖(左側
:n=3
,右側
:n
= [5 , 4 , 3 , 3
];生物複本)。展示平均值及個別數據點。展示經純化sfGFP之一個代表性裁剪西方墨點法,其用來自SSO培養物之TAMRA-PEG4
-DBCO進行SPAAC (僅α-GFP通道)。圖 7B
描繪在AXC、GXT及AGX密碼子及GYT、AYC及XCT反密碼子之表現端點處,來自純系SSO培養物之歸一化螢光。檢查在培養基中具有及不具有AzK以及在培養基中不具有核苷三磷酸NaMTP及TPT3TP兩者之情況下之所有成對組合。各培養物係自單一群落繁殖且指示平均值±標準差(黑色文字;n=3;生物複本)。
圖 8A-F
說明兩個非天然密碼子之同時解碼。非天然密碼子及非天然反密碼子依據其DNA編碼序列書寫。圖 8A
為含有sfGFP190,200
(GXT,AXC)、馬氏甲烷八聯球菌tRNAPyl
(AYC)及詹氏甲烷球菌tRNApAzF
(GYT)之基因卡匣之示意圖。圖 8B-C
,在指示ncAA存在下,在sfGFP
表現期間之歸一化螢光之時程曲線圖。在t=-60分鐘處添加IPTG且在t=0處添加aTc。在自個別SSO群落繁殖之培養物中進行各複製表現(n=3,生物複本)。展示平均值及個別數據點。圖 8B
說明在圖 8A
中之卡匣之純系SSO表現以及展示僅含有具有適當tRNA之單一密碼子之卡匣之表現的對照物。圖 8C
說明亦展示含有sfGFP190,200
(TAA,TAG)、馬氏甲烷八聯球菌tRNAPyl
(TTA)及詹氏甲烷球菌tRNApAzF
(CTA)之卡匣,以及含有具有適當抑制因子tRNA之單一終止密碼子之對照卡匣的純系表現。圖 8D
展示在具有及不具有藉由SPAAC結合至TAMRA-PEG4
-DBCO之情況下,來自圖 8B-C
之SSO之純化sfGFP之α-GFP及TAMRA螢光掃描之偽染色西方墨點法。影像係由相同墨點(UBP構築體及終止密碼子抑制因子)裁剪,但定位成對準未移位之帶以便容易比較電泳遷移。圖 8E
展示在添加PrK及p
AzF之情況下,在來自圖 8B-C
之雙密碼子/tRNA卡匣之純系表現期間的歸一化螢光之時程曲線圖。所展示平均值及個別數據點(n=3,生物複本)。圖 8F
展示在具有及不具有藉由SPAAC結合至TAMRA-PEG4
-DBCO及藉由CuAAC結合至TAMRA-PEG4
-疊氮之情況下,來自圖 8E
之SSO之純化sfGFP之α-GFP及TAMRA螢光掃描之偽染色西方墨點法。
圖 9A-C
說明三個非天然密碼子之同時解碼。非天然密碼子及非天然反密碼子依據其DNA編碼序列書寫。圖 9A
為含有sfGFP151,190,200
(AXC,GXT,AGX)、馬氏甲烷八聯球菌tRNAPyl
(XCT)、詹氏甲烷球菌tRNApAzF
(GYT)及大腸桿菌tRNASer
(AYC)之基因卡匣之示意圖。圖 9B
為在不存在或存在AzK及/或p
AzF之情況下,在sfGFP
表現期間之歸一化螢光之時程曲線圖。在t=-60分鐘處添加IPTG且在t=0處添加aTc。在自個別SSO群落繁殖之培養物中進行各複製表現(n=3,生物複本)。展示平均值及個別數據點。圖 9C
為來自圖 9B
中之SSO純化之完整sfGFP之HRMS分析的代表性去捲積質譜。峰值標記表示相對於其他相關物種之各峰值之分子量以及定量。使用標準單字母胺基酸代碼。針對此等物種中之每一者展示之平均值±標準差(n=3)。
圖 10
說明非純系SSO中之非天然密碼子之初始篩選。非天然密碼子及非天然反密碼子依據其DNA編碼序列書寫。用於選擇在密碼子之第一、第二或第三位置攜帶UBP之密碼子/反密碼子對的在蛋白質表現端點(亦即在補充aTc之後t=180分鐘)處,來自SSO細胞之歸一化螢光之配對帶狀圖。加/減指示將20 mM AzK添加至培養基。各複本來源於勝任SSO起始細胞之不同批料(n=3,生物複本)。
圖 11A-B
說明非純系SSO表現之西方墨點法及螢光掃描。非天然密碼子及非天然反密碼子依據其DNA編碼序列書寫。圖 11A
,在具有藉由SPAAC結合至TAMRA-PEG4
-DBCO之情況下,來自圖 6D
之培養物之純化sfGFP之α-GFP及TAMRA螢光掃描之偽染色西方墨點法。加/減號指示是否進行SPAAC。進行三個試驗(指示為1、2、3;生物複本)。將各組(NXN/NYN及NNX/XNN)之三個試驗平行處理。圖11B,針對指定密碼子/反密碼子對之西方墨點法(圖 11A
)中之相對位移之定量(亦即,經移位頻帶之信號除以經移位及未經移位頻帶兩者之總信號)。加/減號指示是否進行SPAAC。展示平均值±標準差以及個別數據點(n=3)。
圖 12A-B
說明純系SSO表現之西方墨點法及螢光掃描。非天然密碼子及非天然反密碼子依據其DNA編碼序列書寫。圖 12A
,在具有藉由SPAAC結合至TAMRA-PEG4
-DBCO之情況下,來自圖 7A
之培養物之純化sfGFP之α-GFP及TAMRA螢光掃描之偽染色西方墨點法。顯示之(裁剪)區域遷移於32 kDa與25 kDa標準蛋白質標記之間。圖 12B
,針對指定密碼子之西方墨點法(圖 12A
)中之相對位移之定量。展示平均值±標準差以及個別數據點(除CXC之n=5及GXG之n=4以外n=3)。
圖 13
說明在不存在TPT3TP之情況下,純系SSO表現。非天然密碼子及非天然反密碼子依據其DNA編碼序列書寫。在頂部四個自我配對密碼子/反密碼子之蛋白質表現端點(亦即在補充aTc之後t=180分鐘)處,來自純系SSO之歸一化螢光。在自個別群落繁殖之培養物中進行各複製表現,如圖 7A
中所進行(n=3,生物複本)。針對螢光及定量西方墨點蛋白質位移(亦即相對位移;凝膠未展示)以及螢光之個別數據點展示平均值±標準差。
圖 14
說明用於雙密碼子表現之對照。非天然密碼子及非天然反密碼子依據其DNA編碼序列書寫。在具有或不具有於培養基中之指示ncAA之情況下,在指定基因型之sfGFP表現期間之歸一化螢光之時程曲線圖。在t=-60分鐘處添加IPTG且在t=0處添加aTc。在自個別群落繁殖之培養物中進行各複製表現(n=3,生物複本)。展示平均值及個別數據點。
圖 15A-B
說明來自雙密碼子表現之蛋白質之HRMS分析。在培養基中具有AzK及p
AzF之情況下,由表現sfGFP151,190,200
(GXT,AXC)、tRNAPyl
(AYC)及tRNApAzF
(GYT)之SSO純化之完整sfGFP之HRMS分析,如圖 8B
中所展示(n=3,生物複本)。使用標準單字母胺基酸代碼。圖 15A
描繪具有相關峰值之標註及其彼此相對豐度之去捲積光譜。圖 15B
描繪峰值分配及解釋。
圖 16A-B
說明來自參密碼子表現之蛋白質之HRMS分析。在培養基中具有AzK及p
AzF之情況下,由表現sfGFP151,190,200
(AXC,GXT,AGX)、tRNAPyl
(XCT)、tRNApAzF
(GYT)及tRNASer
(AYC)之SSO純化之完整sfGFP之HRMS分析,如圖 9B
中所展示(n=3,生物複本)。使用標準單字母胺基酸代碼。圖 16A
描繪具有相關峰值之標註及其彼此相對豐度之去捲積光譜。圖 16B
描繪峰值分配及解釋。












100:例示性工作流程
101:DNA
102,110:蛋白質
103,106:tRNA
104:轉錄
105,109:非天然胺基酸
107:mRNA
108:轉譯
Claims (70)
- 一種合成非天然多肽之方法,其包含: a. 提供至少一種包含至少四個非天然鹼基對之非天然去氧核糖核酸(DNA)分子,其中該至少一種非天然DNA分子編碼(i)包含至少第一及第二非天然密碼子之信使核糖核酸(mRNA)分子,及(ii)至少第一及第二轉送RNA (tRNA)分子,該第一tRNA分子包含第一非天然反密碼子且該第二tRNA分子包含第二非天然反密碼子,並且該至少一種DNA分子中之至少四個非天然鹼基對於序列中使得該mRNA分子之第一非天然密碼子及第二非天然密碼子分別與該第一非天然反密碼子及該第二非天然反密碼子互補; b. 轉錄該至少一種非天然DNA分子以得到該mRNA; c. 轉錄該至少一種非天然DNA分子以得到該至少第一及第二tRNA分子;以及 d. 藉由利用該至少第一及第二非天然tRNA分子轉譯該非天然mRNA分子來合成該非天然多肽,其中該至少第一及第二非天然反密碼子各引導非天然胺基酸位點特異性併入該非天然多肽中。
- 如請求項1之方法,其中該至少兩個非天然密碼子各自包含位於該密碼子之第一位置、第二位置或第三位置之第一非天然核苷酸,視情況其中該第一非天然核苷酸位於該密碼子之第二位置或第三位置。
- 如前述請求項中任一項之方法,其中該至少兩個非天然密碼子各自包含核酸序列NNX或NXN,且該非天然反密碼子包含核酸序列XNN、YNN、NXN或NYN,形成包含NNX-XNN、NNX-YNN或NXN-NYN之非天然密碼子-反密碼子對,其中N為任何天然核苷酸,X為第一非天然核苷酸,且Y為不同於該第一非天然核苷酸之第二非天然核苷酸,其中X-Y形成DNA中之非天然鹼基對。
- 如請求項3之方法,其中該密碼子包含至少一個G或C且該反密碼子包含至少一個互補C或G。
- 如請求項3或4之方法,其中X及Y獨立地選自由以下組成之群: (i) 2-硫尿嘧啶、2'-去氧尿苷、4-硫-尿嘧啶、尿嘧啶-5-基、次黃嘌呤-9-基(I)、5-鹵基尿嘧啶;5-丙炔基-尿嘧啶、6-偶氮基-尿嘧啶、5-甲基胺基甲基尿嘧啶、5-甲氧基胺基甲基-2-硫尿嘧啶、假尿嘧啶、尿嘧啶-5-乙醛酸(oxacetic acid)甲酯、尿嘧啶-5-乙醛酸、5-甲基-2-硫尿嘧啶、3-(3-胺基-3-N-2-羧丙基)尿嘧啶、5-甲基-2-硫尿嘧啶、4-硫尿嘧啶、5-甲基尿嘧啶、5'-甲氧基羧甲基尿嘧啶、5-甲氧基尿嘧啶、尿嘧啶-5-氧基乙酸(oxyacetic acid)、5-(羧基羥甲基)尿嘧啶、5-羧甲基胺基甲基-2-硫尿苷、5-羧甲基胺基甲基尿嘧啶,或二氫尿嘧啶; (ii) 5-羥甲基胞嘧啶、5-三氟甲基胞嘧啶、5-鹵基胞嘧啶、5-丙炔基胞嘧啶、5-羥基胞嘧啶、環胞嘧啶、胞嘧啶阿拉伯糖苷、5,6-二氫胞嘧啶、5-硝基胞嘧啶、6-偶氮基胞嘧啶、氮雜胞嘧啶、N4-乙基胞嘧啶、3-甲基胞嘧啶、5-甲基胞嘧啶、4-乙醯基胞嘧啶、2-硫胞嘧啶、啡㗁𠯤胞苷([5,4-b][l,4]苯并㗁𠯤-2(3H)-酮)、啡噻𠯤胞苷(1H-嘧啶并[5,4-b][l,4]苯并噻𠯤-2(3H)-酮)、啡㗁𠯤胞苷(9-(2-胺基乙氧基)-H-嘧啶并[5,4-b][l,4]苯并㗁𠯤-2(3H)-酮)、咔唑胞苷(2H-嘧啶并[4,5-b]吲哚-2-酮),或吡啶并吲哚胞苷(H-吡啶并[3',2':4,5]吡咯并[2,3-d]嘧啶-2-酮); (iii) 2-胺基腺嘌呤、2-丙基腺嘌呤;2-胺基-腺嘌呤;2-F-腺嘌呤;2-胺基-丙基-腺嘌呤;2-胺基-2'-去氧腺苷;3-去氮腺嘌呤;7-甲基腺嘌呤;7-去氮-腺嘌呤;8-氮雜腺嘌呤;經8-鹵基、8-胺基、8-硫醇、8-硫烷基及8-羥基取代之腺嘌呤;N6-異戊烯基腺嘌呤;2-甲基腺嘌呤;2,6-二胺基嘌呤;2-甲硫基-N6-異戊烯基腺嘌呤,或6-氮雜-腺嘌呤; (iv) 2-甲基鳥嘌呤;鳥嘌呤之2-丙基及烷基衍生物;3-去氮鳥嘌呤;6-硫-鳥嘌呤;7-甲基鳥嘌呤;7-去氮鳥嘌呤;7-去氮鳥苷;7-去氮-8-氮雜鳥嘌呤;8-氮雜鳥嘌呤;經8-鹵基、8-胺基、8-硫醇、8-硫烷基及8-羥基取代之鳥嘌呤;1-甲基鳥嘌呤;2,2-二甲基鳥嘌呤;7-甲基鳥嘌呤,或6-氮雜-鳥嘌呤;以及 (v) 次黃嘌呤、黃嘌呤、1-甲基肌苷、Q核苷(queosine)、β-D-半乳糖基Q核苷、肌苷、β-D-甘露糖基Q核苷、懷丁氧苷(wybutoxosine)、羥基脲、(acp3)w、2-胺基吡啶,或2-吡啶酮。
- 如請求項4或5之方法,其中各包含X及Y之該等鹼基獨立地選自由以下組成之群:
 。
。
- 如請求項6之方法,其中包含X之鹼基為。
- 如請求項6或7之方法,其中包含Y之鹼基為。

- 如請求項3至8中任一項之方法,其中NNX-XNN係選自由以下組成之群:UUX-XAA、UGX-XCA、CGX-XCG、AGX-XCU、GAX-XUC、CAX-XUG、AUX-XAU、CUX-XAG、GUX-XAC、UAX-XUA及GGX-XCC。
- 如請求項3至8中任一項之方法,其中NNX-YNN係選自由以下組成之群:UUX-YAA、UGX-YCA、CGX-YCG、AGX-YCU、GAX-YUC、CAX-YUG、AUX-YAU、CUX-YAG、GUX-YAC、UAX-YUA及GGX-YCC。
- 如請求項3至8中任一項之方法,其中NXN-NYN係選自由以下組成之群:GXU-AYC、CXU-AYG、GXG-CYC、AXG-CYU、GXC-GYC、AXC-GYU、GXA-UYC、CXC-GYG及UXC-GYA。
- 如前述請求項中任一項之方法,其中該至少兩個非天然tRNA分子各自包含不同的非天然反密碼子。
- 如請求項12之方法,其中該至少兩個非天然tRNA分子包含來自甲烷八聯球菌屬(Methanosarcina genus)之吡咯離胺醯基tRNA及來自詹氏甲烷球菌(Methanocaldococcus jannaschii )之酪胺醯基tRNA,或其衍生物。
- 如請求項11至13中任一項之方法,其包含藉由胺基-醯基tRNA合成酶而裝載該至少兩個非天然tRNA分子。
- 如請求項14之方法,其中該tRNA合成酶係選自由嵌合PylRS (chPylRS)及詹氏甲烷球菌AzFRS (Mjp AzFRS)組成之群。
- 如請求項12或13之方法,其包含藉由至少兩種不同的tRNA合成酶而裝載該至少兩個非天然tRNA分子。
- 如請求項16之方法,其中該至少兩種不同tRNA合成酶包含嵌合PylRS (chPylRS)及詹氏甲烷球菌AzFRS (Mjp AzFRS)。
- 如請求項1至17中任一項之方法,其中該非天然多肽包含兩個、三個或更多個非天然胺基酸。
- 如請求項1至18中任一項之方法,其中該非天然多肽包含至少兩個相同的非天然胺基酸。
- 如請求項1至18中任一項之方法,其中該非天然多肽包含至少兩個不同的非天然胺基酸。
- 如請求項1至20中任一項之方法,其中該非天然胺基酸包含 離胺酸類似物; 芳族側鏈; 疊氮基; 炔基;或 醛基或酮基。
- 如請求項1至20中任一項之方法,其中該非天然胺基酸不包含芳族側鏈。
- 如請求項1至20中任一項之方法,其中該非天然胺基酸係選自N6-疊氮基乙氧基-羰基-L-離胺酸(AzK)、N6-炔丙基乙氧基-羰基-L-離胺酸(PraK)、N6-(炔丙氧基)-羰基-L-離胺酸(PrK)、對-疊氮基-苯丙胺酸(p AzF)、BCN-L-離胺酸、降冰片烯(norbornene)離胺酸、TCO-離胺酸、甲基四𠯤離胺酸、烯丙氧基羰基離胺酸、2-胺基-8-側氧基(oxo)壬酸、2-胺基-8-側氧基辛酸、對-乙醯基-L-苯丙胺酸、對-疊氮基甲基-L-苯丙胺酸(pAMF)、對碘-L-苯丙胺酸、間-乙醯基苯丙胺酸、2-胺基-8-側氧基壬酸、對-炔丙基氧基苯丙胺酸、對-炔丙基-苯丙胺酸、3-甲基-苯丙胺酸、左旋多巴、氟化苯丙胺酸、異丙基-L-苯丙胺酸、對-疊氮基-L-苯丙胺酸、對-醯基-L-苯丙胺酸、對-苯甲醯基-L-苯丙胺酸、對-溴苯基丙胺酸、對-胺基-L-苯丙胺酸、異丙基-L-苯丙胺酸、O-烯丙基酪胺酸、O-甲基-L-酪胺酸、O-4-烯丙基-L-酪胺酸、4-丙基-L-酪胺酸、膦醯基酪胺酸、三-O-乙醯基-GlcNAcp-絲胺酸、L-磷絲胺酸、膦醯基絲胺酸、L-3-(2-萘基)丙胺酸、2-胺基-3-((2-((3-(苯甲氧基)-3-側氧基丙基)胺基)乙基)硒烷基(selanyl))丙酸、2-胺基-3-(苯基硒烷基)丙酸、硒半胱胺酸、N6-(((2-疊氮基苯甲基)氧基)羰基)-L-離胺酸、N6-(((3-疊氮基苯甲基)氧基)羰基)-L-離胺酸,及N6-(((4-疊氮基苯甲基)氧基)羰基)-L-離胺酸。
- 如前述請求項中任一項之方法,其中該至少一種非天然DNA分子呈質體形式。
- 如請求項1至23中任一項之方法,其中該至少一種非天然DNA分子整合至細胞之基因體中。
- 如請求項24或25之方法,其中該至少一種非天然DNA分子編碼該非天然多肽。
- 如前述請求項中任一項之方法,其中該方法包含該非天然DNA分子之活體內複製及轉錄以及該經轉錄mRNA分子於細胞生物體中之活體內轉譯。
- 如請求項27之方法,其中該細胞生物體為微生物。
- 如請求項28之方法,其中該細胞生物體為原核生物。
- 如請求項29之方法,其中該細胞生物體為細菌。
- 如請求項30之方法,其中該細胞生物體為革蘭氏陽性細菌。
- 如請求項30之方法,其中該細胞生物體為革蘭氏陰性細菌。
- 如請求項32之方法,其中該細胞生物體為大腸桿菌(Escherichia coli )。
- 如前述請求項中任一項之方法,其中該至少兩個非天然鹼基對包含選自dCNMO-dTPT3、dNaM-dTPT3、dCNMO-dTAT1或dNaM-dTAT1之鹼基對。
- 如請求項27至34中任一項之方法,其中該細胞生物體包含核苷三磷酸轉運子。
- 如請求項35之方法,其中該核苷三磷酸轉運子包含Pt NTT2之胺基酸序列。
- 如請求項36之方法,其中該核苷三磷酸轉運子包含Pt NTT2之截短胺基酸序列,視情況其中Pt NTT2之該截短胺基酸序列與由SEQ ID NO.1編碼之Pt NTT2至少80%一致。
- 如請求項27至37中任一項之方法,其中該細胞生物體包含該至少一種非天然DNA分子。
- 如請求項38之方法,其中該至少一種非天然DNA分子包含至少一種質體。
- 如請求項38之方法,其中該至少一種非天然DNA分子整合至該細胞之基因體中。
- 如請求項39或40之方法,其中該至少一種非天然DNA分子編碼該非天然多肽。
- 如請求項1至24中任一項之方法,其中該方法為活體外方法,其包含使用無細胞系統合成該非天然多肽。
- 如前述請求項中任一項之方法,其中該等非天然鹼基對包含至少一種包含非天然糖部分之非天然核苷酸。
- 如請求項43之方法,其中該非天然糖部分包含選自由以下組成之群的部分: 於2'位置之修飾,其包含: OH、經取代之低碳數烷基、烷芳基、芳烷基、O-烷芳基或O-芳烷基、SH、SCH3 、OCN、Cl、Br、CN、CF3 、OCF3 、SOCH3 、SO2 CH3 、ONO2 、NO2 、N3 或NH2 F; O-烷基、S-烷基或N-烷基; O-烯基、S-烯基或N-烯基; O-炔基、S-炔基或N-炔基; O-烷基-O-烷基、2'-F、2'-OCH3 或2'-O(CH2 )2 OCH3 ,其中該烷基、烯基及炔基可為經取代或未經取代之C1 -C10 烷基、C2 -C10 烯基、C2 -C10 炔基、-O[(CH2 )n O]m CH3 、-O(CH2 )n OCH3 、-O(CH2 )n NH2 、-O(CH2 )n CH3 、-O(CH2 )n -NH2 或-O(CH2 )n ON[(CH2 )n CH3 )]2 ,其中n及m為1至約10; 於5'位置之修飾,其包含: 5'-乙烯基或5'-甲基(R或S);或 於4'位置之修飾,4'-S、雜環烷基、雜環烷芳基、胺基烷胺基、聚烷基胺基、經取代之矽基、RNA裂解基團、報導基團、嵌入基團(intercalator)、用於改良寡核苷酸之藥物動力性質之基團,或用於改良寡核苷酸之藥效動力性質之基團;或 其任何組合。
- 一種細胞,其包含至少一種包含至少四個非天然鹼基對之非天然DNA分子,其中該至少一種非天然DNA分子編碼(i)編碼非天然多肽且包含至少第一及第二非天然密碼子之信使核糖核酸(mRNA)分子;以及(ii)至少第一及第二轉送RNA (tRNA)分子,該第一tRNA分子包含第一非天然反密碼子且該第二tRNA分子包含第二非天然反密碼子,其中該至少一種DNA分子中之該至少四個非天然鹼基對於序列中使得該mRNA分子之該第一非天然密碼子及該第二非天然密碼子分別與該第一非天然反密碼子及該第二非天然反密碼子互補。
- 如請求項45之細胞,其進一步包含該mRNA分子以及該至少第一及第二tRNA分子。
- 如請求項46之細胞,其中該至少第一及第二tRNA分子共價連接至非天然胺基酸。
- 如請求項47之細胞,其進一步包含該非天然多肽。
- 一種細胞,其包含: a. 至少兩個不同的非天然密碼子-反密碼子對,其中各非天然密碼子-反密碼子對包含來自非天然信使RNA (mRNA)之非天然密碼子及來自非天然轉移核糖核酸(tRNA)之非天然反密碼子,該非天然密碼子包含第一非天然核苷酸且該非天然反密碼子包含第二非天然核苷酸;以及 b. 至少兩個不同的非天然胺基酸各自共價連接至對應非天然tRNA。
- 如請求項49之細胞,其進一步包含至少一種包含至少四個非天然鹼基對(UBP)之非天然DNA分子。
- 如請求項45至50中任一項之細胞,其中該第一非天然核苷酸位於該非天然密碼子之第二或第三位置。
- 如請求項51之細胞,其中該第一非天然核苷酸與該非天然反密碼子之第二非天然核苷酸互補地鹼基配對。
- 如請求項45至52中任一項之細胞,其中該第一非天然核苷酸及該第二非天然核苷酸分別包含第一及第二鹼基獨立地選自由以下組成之群:
 ,其中該第二鹼基不同於該第一鹼基。
,其中該第二鹼基不同於該第一鹼基。
- 如請求項45或47至53中任一項之細胞,其中該至少四個非天然鹼基對獨立地選自由以下組成之群:dCNMO/dTPT3、dNaM/dTPT3、dCNMO/dTAT1或dNaM/dTAT1。
- 如請求項45或47至54中任一項之細胞,其中該至少一種非天然DNA分子包含至少一種質體。
- 如請求項45或47至54中任一項之細胞,其中該至少一種非天然DNA分子整合至該細胞之基因體中。
- 如請求項47至56中任一項之細胞,其中該至少一種非天然DNA分子編碼非天然多肽。
- 如請求項45至57中任一項之細胞,其中該細胞表現核苷三磷酸轉運子。
- 如請求項58之細胞,其中該核苷三磷酸轉運子包含Pt NTT2之胺基酸序列。
- 如請求項59之細胞,其中該核苷三磷酸轉運子包含Pt NTT2之截短胺基酸序列,視情況其中Pt NTT2之該截短胺基酸序列與由SEQ ID NO.1編碼之Pt NTT2至少80%一致。
- 如請求項45至60中任一項之細胞,其中該細胞表現至少兩種tRNA合成酶。
- 如請求項61之細胞,其中該至少兩種tRNA合成酶為嵌合PylRS (chPylRS)及詹氏甲烷球菌AzFRS (Mjp AzFRS)。
- 如請求項45至62中任一項之細胞,其中該細胞包含非天然核苷酸,該等非天然核苷酸包含非天然糖部分。
- 如請求項63之細胞,其中該非天然糖部分係選自由以下組成之群: 於2'位置之修飾,其包含 OH、經取代之低碳數烷基、烷芳基、芳烷基、O-烷芳基或O-芳烷基、SH、SCH3 、OCN、Cl、Br、CN、CF3 、OCF3 、SOCH3 、SO2 CH3 、ONO2 、NO2 、N3 或NH2 F; O-烷基、S-烷基或N-烷基; O-烯基、S-烯基或N-烯基; O-炔基、S-炔基或N-炔基; O-烷基-O-烷基、2'-F、2'-OCH3 、2'-O(CH2 )2 OCH3 ,其中該烷基、烯基及炔基可為經取代或未經取代之C1 -C10 烷基、C2 -C10 烯基、C2 -C10 炔基、-O[(CH2 )n O]m CH3 、-O(CH2 )n OCH3 、-O(CH2 )n NH2 、-O(CH2 )n CH3 、-O(CH2 )n -NH2 或-O(CH2 )n ON[(CH2 )n CH3 )]2 ,其中n及m為1至約10; 於5'位置之修飾,其包含: 5'-乙烯基、5'-甲基(R或S);或 於4'位置之修飾,4'-S、雜環烷基、雜環烷芳基、胺基烷胺基、聚烷基胺基、經取代之矽基、RNA裂解基團、報導基團、嵌入基團、用於改良寡核苷酸之藥物動力性質之基團,或用於改良寡核苷酸之藥效動力性質之基團;或 其任何組合。
- 如請求項45至64中任一項之細胞,其中至少一個非天然核苷酸鹼基在轉錄期間由RNA聚合酶識別。
- 如請求項45至65中任一項之細胞,其中該細胞轉譯至少一種包含該至少兩個非天然胺基酸之非天然多肽。
- 如請求項45至66中任一項之細胞,其中該至少兩個非天然胺基酸獨立地選自由以下組成之群:N6-疊氮基乙氧基-羰基-L-離胺酸(AzK)、N6-炔丙基乙氧基-羰基-L-離胺酸(PraK)、N6-(炔丙氧基)-羰基-L-離胺酸(PrK)、對-疊氮基-苯丙胺酸(p AzF)、BCN-L-離胺酸、降冰片烯離胺酸、TCO-離胺酸、甲基四𠯤離胺酸、烯丙氧基羰基離胺酸、2-胺基-8-側氧基壬酸、2-胺基-8-側氧基辛酸、對-乙醯基-L-苯丙胺酸、對-疊氮基甲基-L-苯丙胺酸(pAMF)、對-碘-L-苯丙胺酸、間-乙醯基苯丙胺酸、2-胺基-8-側氧基壬酸、對-炔丙基氧基苯丙胺酸、對-炔丙基-苯丙胺酸、3-甲基-苯丙胺酸、左旋多巴、氟化苯丙胺酸、異丙基-L-苯丙胺酸、對-疊氮基-L-苯丙胺酸、對-醯基-L-苯丙胺酸、對-苯甲醯基-L-苯丙胺酸、對-溴苯基丙胺酸、對-胺基-L-苯丙胺酸、異丙基-L-苯丙胺酸、O-烯丙基酪胺酸、O-甲基-L-酪胺酸、O-4-烯丙基-L-酪胺酸、4-丙基-L-酪胺酸、膦醯基酪胺酸、三-O-乙醯基-GlcNAcp-絲胺酸、L-磷絲胺酸、膦醯基絲胺酸、L-3-(2-萘基)丙胺酸、2-胺基-3-((2-((3-(苯甲氧基)-3-側氧基丙基)胺基)乙基)硒烷基)丙酸、2-胺基-3-(苯基硒烷基)丙酸、硒半胱胺酸、N6-(((2-疊氮基苯甲基)氧基)羰基)-L-離胺酸、N6-(((3-疊氮基苯甲基)氧基)羰基)-L-離胺酸,及N6-(((4-疊氮基苯甲基)氧基)羰基)-L-離胺酸。
- 如請求項45至67中任一項之細胞,其中該細胞經分離。
- 如請求項45至68中任一項之細胞,其中該細胞為原核生物。
- 一種細胞株,其包含如請求項45至69中任一項之細胞。
Applications Claiming Priority (4)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201962913664P | 2019-10-10 | 2019-10-10 | |
| US62/913,664 | 2019-10-10 | ||
| US202062988882P | 2020-03-12 | 2020-03-12 | |
| US62/988,882 | 2020-03-12 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| TW202128996A true TW202128996A (zh) | 2021-08-01 |
Family
ID=75436820
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| TW109134953A TW202128996A (zh) | 2019-10-10 | 2020-10-08 | 用於活體內合成非天然多肽的組合物及方法 |
Country Status (12)
| Country | Link |
|---|---|
| US (1) | US20220243244A1 (zh) |
| EP (1) | EP4041249A4 (zh) |
| JP (1) | JP2022552271A (zh) |
| KR (1) | KR20220080136A (zh) |
| CN (1) | CN114761026A (zh) |
| AU (1) | AU2020363962A1 (zh) |
| BR (1) | BR112022006233A2 (zh) |
| CA (1) | CA3153855A1 (zh) |
| IL (1) | IL291663A (zh) |
| MX (1) | MX2022004316A (zh) |
| TW (1) | TW202128996A (zh) |
| WO (1) | WO2021072167A1 (zh) |
Families Citing this family (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP4079745A1 (en) | 2013-08-08 | 2022-10-26 | The Scripps Research Institute | A method for the site-specific enzymatic labelling of nucleic acids in vitro by incorporation of unnatural nucleotides |
| US11761007B2 (en) | 2015-12-18 | 2023-09-19 | The Scripps Research Institute | Production of unnatural nucleotides using a CRISPR/Cas9 system |
| US20200131555A1 (en) | 2017-07-11 | 2020-04-30 | Synthorx, Inc. | Incorporation of unnatural nucleotides and methods thereof |
| BR112021025130A2 (pt) * | 2019-06-14 | 2022-03-15 | Scripps Research Inst | Reagentes e métodos para replicação, transcrição e tradução em organismos semissintéticos |
| WO2024039516A1 (en) * | 2022-08-19 | 2024-02-22 | Illumina, Inc. | Third dna base pair site-specific dna detection |
Family Cites Families (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| MXPA03009563A (es) * | 2001-04-19 | 2004-12-06 | Scripps Research Inst | Incorporacion in vivo de aminoacidos no naturales. |
| US20080051317A1 (en) * | 2005-12-15 | 2008-02-28 | George Church | Polypeptides comprising unnatural amino acids, methods for their production and uses therefor |
| WO2019014262A1 (en) * | 2017-07-11 | 2019-01-17 | The Scripps Research Institute | INCORPORATION OF NON-NATURAL NUCLEOTIDES AND METHODS OF IN VIVO USE THEREOF |
| US20200131555A1 (en) * | 2017-07-11 | 2020-04-30 | Synthorx, Inc. | Incorporation of unnatural nucleotides and methods thereof |
| BR112021025130A2 (pt) * | 2019-06-14 | 2022-03-15 | Scripps Research Inst | Reagentes e métodos para replicação, transcrição e tradução em organismos semissintéticos |
-
2020
- 2020-10-08 TW TW109134953A patent/TW202128996A/zh unknown
- 2020-10-09 WO PCT/US2020/054947 patent/WO2021072167A1/en unknown
- 2020-10-09 JP JP2022521262A patent/JP2022552271A/ja active Pending
- 2020-10-09 KR KR1020227015123A patent/KR20220080136A/ko unknown
- 2020-10-09 AU AU2020363962A patent/AU2020363962A1/en active Pending
- 2020-10-09 MX MX2022004316A patent/MX2022004316A/es unknown
- 2020-10-09 CN CN202080083870.3A patent/CN114761026A/zh active Pending
- 2020-10-09 BR BR112022006233A patent/BR112022006233A2/pt unknown
- 2020-10-09 EP EP20874652.9A patent/EP4041249A4/en active Pending
- 2020-10-09 CA CA3153855A patent/CA3153855A1/en active Pending
-
2022
- 2022-03-24 IL IL291663A patent/IL291663A/en unknown
- 2022-04-08 US US17/716,848 patent/US20220243244A1/en active Pending
Also Published As
| Publication number | Publication date |
|---|---|
| EP4041249A1 (en) | 2022-08-17 |
| CA3153855A1 (en) | 2021-04-15 |
| AU2020363962A1 (en) | 2022-04-14 |
| BR112022006233A2 (pt) | 2022-06-21 |
| CN114761026A (zh) | 2022-07-15 |
| KR20220080136A (ko) | 2022-06-14 |
| WO2021072167A1 (en) | 2021-04-15 |
| US20220243244A1 (en) | 2022-08-04 |
| MX2022004316A (es) | 2022-05-11 |
| JP2022552271A (ja) | 2022-12-15 |
| IL291663A (en) | 2022-05-01 |
| EP4041249A4 (en) | 2024-03-27 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US20240117363A1 (en) | Production of unnatural nucleotides using a crispr/cas9 system | |
| US11466279B2 (en) | Import of unnatural or modified nucleoside triphosphates into cells via nucleic acid triphosphate transporters | |
| US11879145B2 (en) | Reagents and methods for replication, transcription, and translation in semi-synthetic organisms | |
| TW202128996A (zh) | 用於活體內合成非天然多肽的組合物及方法 | |
| JP7429642B2 (ja) | 非天然塩基対組成物および使用の方法 | |
| US20220228148A1 (en) | Eukaryotic semi-synthetic organisms | |
| RU2799441C2 (ru) | Композиции на основе неприродных пар оснований и способы их применения |