KR20150028232A - 프로게스테론 수용체 서브타입의 유방암 예후, 예측 및 유전자 발현을 토대로 한 항프로게스틴 치료에 대한 반응성 예측 방법 - Google Patents
프로게스테론 수용체 서브타입의 유방암 예후, 예측 및 유전자 발현을 토대로 한 항프로게스틴 치료에 대한 반응성 예측 방법 Download PDFInfo
- Publication number
- KR20150028232A KR20150028232A KR20147033298A KR20147033298A KR20150028232A KR 20150028232 A KR20150028232 A KR 20150028232A KR 20147033298 A KR20147033298 A KR 20147033298A KR 20147033298 A KR20147033298 A KR 20147033298A KR 20150028232 A KR20150028232 A KR 20150028232A
- Authority
- KR
- South Korea
- Prior art keywords
- expression
- genes
- cancer
- gene
- progestin
- Prior art date
Links
Images






















































Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q1/00—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions
- C12Q1/68—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions involving nucleic acids
- C12Q1/6813—Hybridisation assays
- C12Q1/6834—Enzymatic or biochemical coupling of nucleic acids to a solid phase
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q1/00—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions
- C12Q1/68—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions involving nucleic acids
- C12Q1/6876—Nucleic acid products used in the analysis of nucleic acids, e.g. primers or probes
- C12Q1/6883—Nucleic acid products used in the analysis of nucleic acids, e.g. primers or probes for diseases caused by alterations of genetic material
- C12Q1/6886—Nucleic acid products used in the analysis of nucleic acids, e.g. primers or probes for diseases caused by alterations of genetic material for cancer
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K31/00—Medicinal preparations containing organic active ingredients
- A61K31/56—Compounds containing cyclopenta[a]hydrophenanthrene ring systems; Derivatives thereof, e.g. steroids
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P35/00—Antineoplastic agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P43/00—Drugs for specific purposes, not provided for in groups A61P1/00-A61P41/00
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P5/00—Drugs for disorders of the endocrine system
- A61P5/24—Drugs for disorders of the endocrine system of the sex hormones
- A61P5/32—Antioestrogens
-
- C—CHEMISTRY; METALLURGY
- C40—COMBINATORIAL TECHNOLOGY
- C40B—COMBINATORIAL CHEMISTRY; LIBRARIES, e.g. CHEMICAL LIBRARIES
- C40B40/00—Libraries per se, e.g. arrays, mixtures
- C40B40/04—Libraries containing only organic compounds
- C40B40/06—Libraries containing nucleotides or polynucleotides, or derivatives thereof
- C40B40/08—Libraries containing RNA or DNA which encodes proteins, e.g. gene libraries
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q2600/00—Oligonucleotides characterized by their use
- C12Q2600/106—Pharmacogenomics, i.e. genetic variability in individual responses to drugs and drug metabolism
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q2600/00—Oligonucleotides characterized by their use
- C12Q2600/158—Expression markers
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Organic Chemistry (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Wood Science & Technology (AREA)
- Zoology (AREA)
- Genetics & Genomics (AREA)
- Analytical Chemistry (AREA)
- Immunology (AREA)
- Pathology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Molecular Biology (AREA)
- Biochemistry (AREA)
- Medicinal Chemistry (AREA)
- Biophysics (AREA)
- Physics & Mathematics (AREA)
- General Engineering & Computer Science (AREA)
- Microbiology (AREA)
- Biotechnology (AREA)
- Public Health (AREA)
- Veterinary Medicine (AREA)
- Animal Behavior & Ethology (AREA)
- Pharmacology & Pharmacy (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Hospice & Palliative Care (AREA)
- Oncology (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Endocrinology (AREA)
- Diabetes (AREA)
- Epidemiology (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
- Pharmaceuticals Containing Other Organic And Inorganic Compounds (AREA)
- Apparatus Associated With Microorganisms And Enzymes (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
Abstract
본 발명은 PR 유발성 악성 종양을 앓고 있는 환자가 항-프로게스틴인 치료 화합물을 이용한 치료에 반응할 가능성이 있는지를 나타내는 유전자 발현 프로파일을 제공한다. 이러한 반응성을 동정함으로써, 치료 제공자는 이러한 치료가 유익한 환자를 사전에 확인할 수 있을 뿐만 아니라 무-반응자에 대한 대안 요법을 동정할 수 있다. 또한, 유전자 발현 프로파일의 이용 방법과 환자에서 유전자 발현 프로파일의 존재를 확인하기 위한 분석을 제공한다.
Description
본 출원은 2012년 4월 27일자 미국 가출원 번호 61/639,407에 대해 우선권을 주장하며, 이의 내용은 그 전체가 원용에 의해 본 명세서에 포함된다.
정부 권리에 대한 언급
본 발명은 국립 보건원으로부터 미국 지원 번호 CA1159712-01 하에 정부 지원을 받아 수행되었다. 정부는 본 발명에 대해 일정한 권리를 가진다.
현행 mRNA (예컨대, 유전자 발현에 기반을 둔) 예후적인 유방암 스크리닝 검사 (예, Oncotype DX)는, 유방암 진행에 관여하는 것으로 공지된, 한정된 수의 비-관련 유전자들의 발현을 분석하는 것이다. 유방암은 매우 혼성적인 질환 (heterogeneous disease)이기 때문에, 이러한 분석 방법으로는 빠르게 증가하는 기존 약물과 새로운 약물의 리스트를 포함하는 소정의 표적화된 치료에 가장 효과적일 것으로 보이는 환자를 선택하지 못한다. 그래서, 건강 관리 서비스 제공자는, 조합적인 치료가 임상 반응 또는 임상적인 혜택을 제공할 것이라는 기대로, 이용가능한 약물들을 무작위 조합하여 시도하게 된다. 또한, 이 전략으로는 임의의 유전자 발현 데이타 콜렉션의 발현과 이의 발현을 담당하는 임의의 규명된 기전(들)을 연결시킬 수가 없다 (즉, 타겟이 미지임).
Amit, I., et al. (2007). Nat Genet 39, 503-512.
Anderson, G.L., et al. (2012). Lancet Oncol.
Barski, A., et al. (2007). Cell 129, 823-837.
Beerli, R.R., and Hynes, N.E. (1996). J Biol Chem 271, 6071-6076.
Benjamini, Y., and Hochberg, Y. (1995). J Roy Stat Soc B Met 57, 289-300.
Blind and Garabedian (2008). J Steroid Biochem Mol Biol 109, 150-157.
Bonnefoi, H., et al. (2007). Lancet Oncol 8, 1071-1078.
Byron, S.A., et al. (2006). British journal of cancer 95, 1220-1228.
Carroll, J.S., et al. (2005). Cell 122, 33-43.
Cadepond, F., et al. (1997). Annu Rev Med, 48, 129-56.
Cartharius, K., et al. (2005). Bioinformatics 21, 2933-2942.
Chang, A.C., et al. (2003). Endocr Relat Cancer 10, 359-373.
Cheang, M.C., et al. (2009). JNCI 101, 736-750.
Chlebowski, R.T., et al. (2010). JAMA 304, 1684-1692.
Chlebowski, R.T., et al. (2003). Jama 289, 3243-3253.
Chlebowski, et al. (2009). The New England journal of medicine 360, 573-587.
Clemm, D.L., et al. (2000). Mol Endocrinol 14, 52-65.
Covington, K., and Parikh, A. (2011). The Red-R Journal 1, http://www.red-r.org/journal/published-articles/1-08082011-red-r-framework-integrated-discovery.
Crouch, S.P., et al. (1993). J Immunol Methods 160, 81-88.
Daniel, A.R., et al. (2007a). Mol Endocrinol 21, 2890-2906.
Daniel et al. (2011). Expert review of endocrinology & metabolism 6, 359-369.
Daniel, A.R., et al. (2009). Mol Cell Endocrinol 308, 47-52.
Daniel, A.R., and Lange, C.A. (2009). PNAS 106, 14287-14292.
Daniel, A.R., et al. (2007b). Steroids 72, 188-201.
De Vivo, I., et al. (2002). Proc Natl Acad Sci U S A 99, 12263-12268.
di Bari, M.G., et al. (2009). J Cell Physiol 219, 659-666.
Early Breast Cancer Trialists' Collaborative Group (2005). Lancet 365, 1687-1717.
ENCODE Project Consortium (2007). Nature 447, 799-816.
Gaujoux, R., and Seoighe, C. (2010). BMC bioinformatics 11, 367.
Geiss-Friedlander and Melchior, F. (2007). Nature reviews Molecular cell biology 8, 947-956.
Gentleman, R.C., et al. (2004). Genome Biol 5, R80.
Goss, P.E., et al. (2011). NEJM 364, 2381-2391.
Graham, J.D., et al. (2009). Endocrinology 150, 3318-3326.
Hagan, C.R., et al. (2011). Mol Cell Biol 31, 2439-2452.
Hager, G.L., et al. (2009). Molecular cell 35, 741-753.
Halford, S.E., and Marko, J.F. (2004). Nucleic Acids Res 32, 3040-3052.
He, H.H., et al. (2010). Nat Genet 42, 343-347.
Heintzman, N.D., et al. (2007). Nat Genet 39, 311-318.
Holmstrom, S.R., et al. (2008). Mol Endocrinol.
Horwitz, K.B., et al. (1982). Cell 28, 633-642.
Huang, E., et al. (2003). Nat Genet 34, 226-230.
Hurtado, A., et al. (2011). Nat Genet 43, 27-33.
Iniguez-Lluhi, J.A., and Pearce, D. (2000). Mol Cell Biol 20, 6040-6050.
Irizarry, R.A., et al. (2003). Nucleic Acids Res 31, e15.
Jacobsen, B.M., et al. (2005). Mol Endocrinol 19, 574-587.
Johnston, S., et al. (2009). J Clin Oncol 27, 5538-5546.
Kaufman, B., et al. (2009). J Clin Oncol 27, 5529-5537.
Labriola, L., et al. (2003). Mol Cell Biol 23, 1095-1111.
LaCroix, A.Z., et al. (2011). JAMA 305, 1305-1314.
Lambert, J.R., and Nordeen, S.K. (2003). Mol Endocrinol 17, 1085-1094.
Lange, C.A. (2008). Steroids 73, 914-921.
Lange, C.A., et al. (2000). Proc Natl Acad Sci U S A 97, 1032-1037.
Lanigan, F., et al. (2010). Breast Cancer Res 12, R59.
Lenferink, A.E., et al. (2001). Cancer Res 61, 6583-6591.
Li, X., et al. (2003). Mol Cell Biol 23, 3763-3773.
Liang, Y., et al. (2007). Cancer Res 67, 9929-9936.
Liu, S., et al. (2010). Breast Cancer Res Treat 119, 53-61.
Liu, W.M. (2004). Curr Med Chem 11, 2143-2151.
Locke, J.A., et al. (2008). Cancer Res 68, 6407-6415.
Loi, S., et al. (2007). J Clin Oncol 25, 1239-1246.
Lupien, M., et al. (2008). Cell 132, 958-970.
Man, J.-H., et al. (2006). Nucleic Acids Res 34, 5552-5566.
Meijsing, S.H., et al. (2009). Science 324, 407-410.
Melchior, F. (2000). Annu Rev Cell Dev Biol 16, 591-626.
Million Women Study Collaborators (2003). Lancet 362, 419-427.
Mo, R., et al. (2006). J Biol Chem 281, 15714-15720.
Moore, M.R., et al. (2000). Biochem Biophys Res Commun 277, 650-654.
Mootha, V.K., et al. (2003). Nat Genet 34, 267-273.
Musgrove, E.A., and Sutherland, R.L. (2009). Nat Rev Cancer 9, 631-643.
Nicholson, D.W., et al. (1995). Nature 376, 37-43.
Ogryzko, V.V., et al. (1996). Cell 87, 953-959.
Oliveros, J.C. (2007). http:// http://bioinfogp.cnb.csic.es/tools/venny/index.html.
Ong, C.T., and Corces, V.G. (2011). Nat Rev Genet 12, 283-293.
Paik, S., et al. (2004). NEJM 351, 2817-2826.
Parker, J.S., et al. (2009). J Clin Oncol 27, 1160-1167.
Perou, C.M., et al. (2000). Nature 406, 747-752.
Perrault, D., et al. (1996). J Clin Oncol, 14, 2709-12.
Pooley, K.A., et al. (2006). Cancer Epidemiol Biomarkers Prev 15, 675-682.
Prat, A., and Perou, C.M. (2011). Molecular oncology 5, 5-23.
Reginato, M.J., et al. (2005). Mol Cell Biol 25, 4591-4601.
Rhodes, D.R., et al. (2007). Neoplasia 9, 166-180.
Richer, J.K., et al. (2002). J Biol Chem 277, 5209-5218.
Salatino, M., et al. (2004). Oncogene 23, 5161-5174.
Sartorius, C.A., et al. (1994). Cancer Res 54, 3868-3877.
Satoh et al. (2004). Journal of mammary gland biology and neoplasia 9, 195-205.
Satoh, K., et al. (2007). Oncogene 26, 7526-7534.
Satokata, I., et al. (2000). Nat Genet 24, 391-395.
Siegel, R., et al. (2012). CA Cancer J Clin 62, 10-29.
Smalley, M.J., et al. (2007). Breast Cancer Res 9, R85.
Sorlie, T., et al. (2001). Proc Natl Acad Sci USA 98, 10869-10874.
Su, B., et al. (2011). J Steroid Biochem Mol Biol 123, 101-108.
Subramanian, A., et al. (2005). Proc Natl Acad Sci USA 102, 15545-15550.
Suzuki, T., et al. (2005). Endocr Relat Cancer 12, 701-720.
Takahashi, C., et al. (1997). Leukemia 11 Suppl 3, 340-343.
Takimoto, G.S., et al. (1996). J Biol Chem 271, 13308-13316.
Tang, Q., et al. (2011). Cancer Res.
Terry, K.L., et al. (2005). American journal of epidemiology 161, 442-451.
van 't Veer, L.J., et al. (2002). Nature 415, 530-536.
Verzi, M.P., et al. (2010). Dev Cell 19, 713-726.
Vicent, G.P., et al. (2011). Genes Dev 25, 845-862.
프로게스테론 수용체는 유방암 진행의 중요한 활성화 인자 (driver)인 것으로 확인되고 있다. 폐경기 여성에서 (에스트로겐과 조합하여 호르몬 대체 요법의 일환으로서) 프로게스틴을 이용한 치료는 이들의 유방암 위험성을 크게 높인다. 최근 연구에서는, 에스트로겐-단일 보충이 실제로 여성을 유방암으로부터 보호할 수 있는 것으로 제시되었다. 본원에는, 프로게스틴 작용에 대한 한가지 예시적인 원인 기전을 개시한다: 활성화된 탈SUMO일화된 (deSUMOylated) 포스포-프로게스테론 수용체 전사. 일 측면에서, 일차적으로 프로게스테론 수용체 (PR)에 의해 활성화되며, 따라서 항-에스테로겐 (예, 타목시펜), 항-프로게스틴 또는 아로마타제 저해제 요법에 감수성일 것으로 보이는 종양을 가진 유방암 환자를 동정하는데 사용할 수 있는, 독특한 유전자 시그니처를 규정한다. 다른 측면에서, PR 유전자 시그니처를 사용하여, 항-프로게스틴을 포함하는 치료에 적합한 후보자인 여성 개체군을 동정한다.
항-프로게스틴 치료와 관련된 주요 문제는 적절한 임상 치료 타겟인 활성화된 PR을 동정하는 방법에 관한 것이다. 일 측면에서, 본 발명의 예시적인 방법은 임상 조건에서 일상적으로 입수가능한 인간 종양 조직에서 기능적인 (활성화된) 상태로 존재하는 PR을 특정화하는 것을 목적으로 한다. 비-활성형 PR을 특이적인 항-프로게스틴으로 길항하는 것은 치료학적으로 무의미하기 때문에, 본 발명의 예시적인 방법은 항-프로게스틴을 이용한 환자 치료를 유도하기 위한 새롭고 중요한 정보를 제공해준다. 이러한 예후 진단 검사는, (1) 항-프로게스틴에 대한 치료 의사 결정을 뒷받침하는 일관된 방법을 제공하며, (2) 항-프로게스틴 치료에 반응할 것으로 보이는 개별 환자와 환자 개체군을 선별하는데 지침이 되며, (3) 항-프로게스틴 치료에 반응하거나 이러한 치료가 유익할 가능성이 거의 없는 개별 환자들을 제외한다.
본원에는, 예를 들어, 키나제 활성이 높은 암 세포에서, 프로게스테론 수용체에 의해 상향 조절되는 유전자 (예, 인간 PR 이소형 A 및/또는 B)를 동정하기 위한 예시적인 전략들과 방법들을 개시하며, 여기에서 PR-B는 (Ser294에서) 인산화되거나 또는 (Lys388에서) 탈SUMO일화되어, 전사적으로 과잉 활성형의 (핵 전사 인자) 수용체를 형성할 수 있다. PR의 전사 작용에 대한 지금까지의 지식은, 단백질 키나제 케스케이드와의 특이적인 상호작용의 결과로서 생기는 (PR-A에 대해 상대적인) PR-B의 고유한 전사 활성을 고려하지 못해, 어려움을 겪었다. 암 세포에서 탈SUMO일화된 (그리고, 인산화될 수 있는) PR-B에 의해 특이적으로 상향 또는 하향 조절되는 내인성 유전자들을 동정하기 위한 본원에 기술된 전략 및 방법이, 그런 유형으로는 최초이다.
현재, 스테로이드 호르몬 (SR) 양성 (내강형, luminal) 유방암을 가진 전체 여성의 절반 이상이 에스트로겐 생산 또는 에스트로겐 수용체 (ER) 작용을 차단하는 것을 목적으로 하는 내분비 요법에 적절하지 않다. SR 양성 또는 내강형 유방암 (전체 유방암 환자의 ~70%)에 걸린 환자에 대한 새로운 임상 스크리닝 (예측) 프로토콜의 일환으로서, 탈SUMO일화된 포스포-PR-유발성 유전자 시그니처의 발현을 이용함으로써, PR-유발성 (PR-driven) 증식 및 진행에서, 이용가능한 항-에스트로겐 및 아로마타제 저해제 치료에 반응하여 엔도크린-내성으로 진행될 가능성이 높은 종양을 가진 환자를 동정할 수 있다. 이런 환자는 항-프로게스틴을 포함하는 내분비 요법의 후보가 될 것이다. 비제한적인 예로, 미페프리스톤 (mifepristone) (RU486), 로나프리산 (Lonaprisan) (ZK-230211), 텔라프리스톤 (Telapristone) (Proellex 또는 CDB-4124), 오나프리스톤 (onapristone) (ZK-98299), 아소프리스닐 (asoprisnil), 울리프리스탈 아세테이트 (ulipristal acetate), 아글레프리스톤 (aglepristone), ZM172406, ZM172405 및 ZM150271과 같은, 항-프로게스틴/선택적인 PR 모듈레이터를 비롯하여, 몇몇은 신규한 것이지만, 선택적인 PR 모듈레이터가 존재한다.
본 발명은, 항-프로게스틴을 이용한 치료에 반응할 가능성이 있는 환자 (이러한 환자는 "반응자"로 지칭됨) 뿐만 아니라 이러한 치료가 유익할 가능성이 없는 환자 (이러한 환자는 "무-반응자"로 지칭됨)를 동정하기 위한, 유전자 발현 프로파일과 방법을 제공한다. 본 발명에 제시된 측면들은, 치료 제공자가, 제제를 투여하기 전에, 항-프로게스틴을 이용한 치료에 반응자인 환자와 이러한 치료에 무-반응자인 환자를 동정할 수 있게 한다.
또한, 본 발명은, 암을 앓고 있는 환자가 항-프로게스틴을 이용한 치료에 반응하는 성향을 나타내는, 유전자 발현 프로파일 ("유전자 시그니처"라고도 함)을 포함한다. 유전자 발현 프로파일은, 표 1a와 1b에 표시된 군으로부터 선택되는 유전자를 하나 이상, 바람직하게는 복수개를 포함한다. 이러한 유전자 군은, 본원에서, "항-프로게스틴 반응자 유전자"로 언급된다. 본 발명의 측면에서, 이들 유전자들은 일부 또는 전체가 항-프로게스틴 치료에 반응자인 환자에서 차별적으로 발현된다 (예, 상향-조절되거나 또는 하향-조절됨).
본 발명은, 또한, 암 환자가 항-프로게스틴 치료에 반응자 또는 무-반응자인지를 결정하는 방법을 포함한다. 일 측면에서, 본 방법은, 환자로부터 악성 조직 또는 세포의 샘플 (예, 종양 샘플, 순환성 종양 세포)을 수득하는 단계, 상기 샘플의 하나 이상의 유전자 발현 프로파일을 측정하는 단계, 및 예를 들어, 대조군 샘플의 하나 이상의 유전자 발현 프로파일과 비교함으로써, 항-프로게스틴 반응자 유전자들로부터 선택되는 하나 이상의 유전자가 상기 샘플에서 과다 또는 과소 발현되는 지를 하나 이상의 유전자 발현 프로파일에서 확인하는 단계를 포함한다. 이러한 정보를 통해, 치료 제공자는 항-프로게스틴 치료가 환자에게 유익할 것인 지를 확인할 수 있다.
다른 측면에서, 본 발명은 환자의 조직 샘플에서 유전자의 발현 프로파일을 측정하는 분석 방법과 이러한 분석 방법을 이용하기 위한 설명서를 추가로 포함한다.
일 구현예는, (a) 환자로부터 생물 샘플을 수득하는 단계; (b) 상기 생물 샘플에서 표 1a 및/또는 표 1b에 표시된 하나 이상의 유전자의 발현 수준을 측정하는 단계; 및 (c) 단계 (b)의 상기 발현 수준을 대조군에서 동일 유전자(들)의 발현 수준과 비교하는 단계를 포함하며, 표 1a 및/또는 표 1b의 하나 이상의 유전자의 수준이 상기 대조군과 비교하여 상기 생물 샘플에서 증가/상향 조절된 경우, 상기 환자는 항-프로게스틴 치료에 반응자인 것을 특징으로 하는, 암으로 진단받은 환자가 항-프로게스틴을 이용한 치료학적 치료에 반응할 가능성이 있는지를 확인하는 분석 방법을 제공한다. 다른 구현예에서, 하나 이상의 유전자의 발현 수준은 상기 생물 샘플에서 감소/하향-조절된다.
다른 구현예는, a. 환자의 생물 샘플에서 표 1a 및/또는 표 1b에 표시된 하나 이상의 유전자의 발현 수준을 측정하는 단계를 포함하며, b. 상기 생물 샘플에서의 하나 이상의 유전자의 발현 수준이 상기 개체가 항-프로게스틴 치료에 반응할 것임을 나타내는 지표인 것인, 유방암 환자가 항-프로게스틴 치료에 반응할 것인지를 확인하는 방법을 제공한다.
일 구현예에서, 유전자 발현 수준의 지표로서 mRNA 수준을 측정한다. 일 구현예에서, 복수의 mRNA들을 각각 측정한다. 다른 구현예에서, 복수의 mRNA들을 동시에 측정한다. 일 구현예에서, 하나 이상의 유전자의 발현 수준은 인 시츄 (in situ) 혼성화, 노던 블롯, 핵산 증폭, 마이크로어레이 분석 또는 이들의 조합으로 이루어진 군으로부터 선택되는 임의의 기법을 이용하여 측정할 수 있다. 일 구현예에서, 핵산 증폭 방법은 중합효소 연쇄 반응, 정량적인 중합효소 연쇄 반응, 역전사 중합효소 연쇄 반응, 리가제 연쇄 반응 또는 이들의 조합으로 이루어진 군으로부터 선택된다. 다른 구현예에서, 유전자 발현 수준은 마이크로어레이 분석을 통해 측정한다.
일 구현예에서, 표 1a 및/또는 표 1b에 표시된 유전자 2종 이상의 발현을 측정한다. 다른 구현예에서, 표 1a 및/또는 표 1b에 표시된 유전자 3종 이상의 발현을 측정한다. 일 구현예에서, 표 1a 및/또는 표 1b에 표시된 유전자 4종 이상의 발현을 측정한다. 다른 구현예에서, 표 1a 및/또는 표 1b에 표시된 유전자 6종 이상의 발현을 측정한다. 일 구현예에서, 표 1a 및/또는 표 1b에 표시된 유전자 9종 이상의 발현을 측정한다. 다른 구현예에서, 표 1a 및/또는 표 1b에 표시된 유전자 12종 이상의 발현을 측정한다. 다른 구현예에서, 표 1a에 표시된 유전자 15종 이상 및/또는 표 1b에 표시된 유전자 16종 이상의 발현을 측정한다.
일 구현예에서, 유전자(들)의 발현은 대조군에 비해 증가된다.
일 구현예에서, 생물 샘플은 조직 생검, 유관 세척물 (ductal lavage), 미세 바늘 흡입물 (fine needle aspiration), 외과적으로 적출된 조직의 단편, 순환하는 종양 세포, 순환성 DNA 또는 순환성 엑소좀이다. 다른 구현예에서, 대조군은 비-암성 조직의 샘플이다. 일 구현예에서, 비-암성 조직의 대조군은 환자로부터 유래된다. 다른 구현예에서, 대조군은 하나 이상의 유전자의 사전 결정된 대조군의 양 또는 농도이다. 일 구현예에서, 음성 대조군은 소정의 수치 값 또는 수치 값들의 대조군 범위이다.
일 구현예에서, 환자는 포유류이다. 다른 구현예에서, 포유류는 인간이다. 일 구현예에서, 의료 제공자는 정보를 제공받는다. 일 구현예에서, 환자는 유방암으로 치료받는다. 일 구현예에서, 환자는 하나 이상의 항-프로게스틴을 유효량으로 투여받는다. 다른 구현예에서, 치료는 하나 이상의 추가적인 치료학적 제제의 투여를 더 포함한다.
일 구현예는, 표 1a 및/또는 표 1b에 표시된 하나 이상의 유전자의 발현 수준이 상기 환자의 생물 샘플에서 대조군에 비해 증가/상향 조절된 환자에게, 항-프로게스틴을 단독으로 또는 다른 치료와 병행하여 투여하는 단계를 포함하는, 암 환자의 치료 방법을 제공한다.
일 구현예는, 표 1a에 표시된 하나 이상의 유전자의 발현 수준이 상기 환자의 생물 샘플에서 대조군에 비해 감소/하향 조절된 환자에게, 항-프로게스틴을 단독으로 또는 다른 치료와 병행하여 투여하는 단계를 포함하는, 암 환자의 치료 방법을 제공한다.
PR 유전자들 중 다수가 혈액과 같은 생물 샘플에서 검출될 수 있는 분비되는 인자이다. 따라서, 다른 구현예에서, 본원에 기술된 유전자 어레이 또는 이의 일부는 아직 진단받지 않은 개체에서 암을 조기 검출하기 위한 바이오마커 (예, mRNA 및 단백질 수준에서의 유전자 발현을 포함함)로서 사용할 수 있다.
일 구현예는 암으로 진단받은 환자가 활성형 프로게스테론 수용체 (KR)를 발현하는 세포를 포함하는 암을 앓고 있으며 항-프로게스틴을 이용한 치료학적 치료에 반응할 가능성이 있는지를 확인하기 위한 방법을 제공하며, 이 방법은 (a) 환자로부터 생물 샘플을 수득하는 단계; (b) KBTBD11, RBPMS2, PLA2G48, FL112684, SH2D4B, RASCD2, CLDN8 및 이들의 임의 조합으로 이루어진 군으로부터 선택되는 하나 이상의 유전자의 발현 수준을 상기 생물 샘플의 세포에서 측정하는 단계; 및 (c) 단계 (b)의 상기 발현 수준을 야생형 (WT) 대조군 샘플 및/또는 기준 샘플의 세포에서 상기 하나 이상의 유전자의 발현 수준과 비교하는 단계를 포함하며, 상기 생물 샘플에서의 상기 하나 이상의 유전자의 발현 수준이 상기 대조군/기준 샘플에 비해 감소된 경우, 상기 환자는 항-프로게스틴을 이용한 치료에 반응자이다. 다른 구현예는, (d) VCX, CHN2, AFAP1L2, PXMP4, THY1, ZNF26, CDH10, ZNF812 및 이들의 임의 조합으로 이루어진 군으로부터 선택되는 하나 이상의 유전자의 발현 수준을 상기 생물 샘플의 세포에서 측정하는 단계; 및 (e) 단계 (d)의 상기 발현 수준을 아생형 (WT) 대조군 샘플 및/또는 기준 샘플에서 상기 하나 이상의 유전자의 발현 수준과 비교하는 단계를 더 포함하며, 상기 생물 샘플에서의 상기 하나 이상의 유전자의 발현 수준이 상기 WT 대조군/기준 샘플에 비해 증가된 경우, 상기 환자는 항-프로게스틴을 이용한 치료에 반응자이다.
일 구현예에서, 암은 유방암, 난소암, 자궁내막암, 뇌암, 폐암, 전립선암, 수막종 또는 자궁암이다.
다른 구현예는 암 환자가 항-프로게스틴 치료에 반응할 것인지를 확인하는 방법을 제공하며, 이 방법은 a. 환자의 생물 샘플에서 KR78D11, RBPMS2, PLA2G48, FL112684, SH2D4B, RASCD2, CLDN8 및 이들의 임의 조합으로 이루어진 군으로부터 선택되는 하나 이상의 유전자의 발현 수준을 암 세포에서 측정하는 단계를 포함하며, b. 대조군 WT 샘플 및/또는 기준 샘플에서의 수준에 비해, 상기 생물 샘플에서 상기 하나 이상의 유전자의 발현 수준 감소는 상기 개체가 항-프로게스틴 치료에 반응할 것임을 나타내는 지표이다. 일 구현예는, (c) VCX, CHN2, AFAP1L2, PXMP4, THY1, ZNF26, CDH10, ZNF812 및 이들의 임의 조합으로 이루어진 군으로부터 선택되는 하나 이상의 유전자의 발현 수준을 상기 생물 샘플의 세포에서 측정하는 단계; 및 (d) 단계 (c)에서의 상기 발현 수준을 야생형 (WT) 대조군 샘플 및/또는 기준 샘플에서의 상기 하나 이상의 유전자의 발현 수준과 비교하는 단계를 더 포함하며, 생물 샘플에서 상기 하나 이상의 유전자의 발현 수준이 상기 WT 대조군 샘플 및/또는 기준 샘플에 비해 증가된 경우 (기준 대조군이 확립되어야 하며, 진단 장치를 기준 대조군(들)으로 보정함) 상기 환자는 항-프로게스틴을 이용한 치료에 반응자이다.
일 구현예에서, 하나 이상의 유전자의 mRNA 수준은 유전자 발현 수준의 지표로서 측정된다. 다른 구현예에서, 하나 이상의 유전자의 발현 수준은 1차 시기와 2차 시기에 측정된다. 일 구현예에서, 예를 들어, 유전자 KB7BD11의 발현은, 예컨대 서열번호 1의 프로브와의 혼성화에 의해 검출된다. 일 구현예에서, 복수의 mRNA들을 각각 측정한다. 다른 구현예에서, 복수의 mRNA들을 동시에 측정한다. 일 구현예에서, 프로브는 상기 유전자 2종 이상에 혼성하는 복수개의 고정된 프로브들 중 하나이다. 일 구현예에서, 상기 유전자들 중 하나 이상의 발현 수준의 측정은 인 시츄 혼성화, 노던 블롯, 핵산 증폭, 마이크로어레이 분석 또는 이들의 조합을 포함한다.
일 구현예에서, KR78D11, RBPMS2, PLA2G48, FL112684, SH2D4B, RASCD2, CLDN8, VCX, CHN2, AFAP1L2, PXMP4, THY1, ZNF26, CDH10, ZNF812 및 이들의 임의 조합으로 이루어진 군으로부터 선택되는 2종 이상의 유전자의 발현을 측정한다. 다른 구현예에서, KR78D11, RBPMS2, PLA2G48, FL112684, SH2D4B, RASCD2, CLDN8, VCX, CHN2, AFAP1L2, PXMP4, THY1, ZNF26, CDH10, ZNF812 및 이들의 임의 조합으로 이루어진 군으로부터 선택되는 3종 이상의 유전자의 발현을 측정한다. 다른 구현예에서, KR78D11, RBPMS2, PLA2G48, FL112684, SH2D4B, RASCD2, CLDN8, VCX, CHN2, AFAP1L2, PXMP4, THY1, ZNF26, CDH10, ZNF812 및 이들의 임의 조합으로 이루어진 군으로부터 선택되는 4종 이상의 유전자의 발현을 측정한다. 다른 구현예에서, KR78D11, RBPMS2, PLA2G48, FL112684, SH2D4B, RASCD2, CLDN8, VCX, CHN2, AFAP1L2, PXMP4, THY1, ZNF26, CDH10, ZNF812 및 이들의 임의 조합으로 이루어진 군으로부터 선택되는 6종 이상의 유전자의 발현을 측정한다. 다른 구현예에서, KR78D11, RBPMS2, PLA2G48, FL112684, SH2D4B, RASCD2, CLDN8, VCX, CHN2, AFAP1L2, PXMP4, THY1, ZNF26, CDH10, ZNF812 및 이들의 임의 조합으로 이루어진 군으로부터 선택되는 7, 8, 9, 10, 11, 12, 13, 14 또는 15종 이상의 유전자의 발현을 측정한다.
일 구현예에서, 생물 샘플은 조직 생검, 유관 세척물, 미세 바늘 흡입물, 외과적으로 적출된 종양의 단편, 순환하는 종양 세포, 순환하는 DNA 또는 순환하는 엑소좀이다. 다른 구현예에서, 대조군 샘플은 비-암성 조직의 샘플이다. 일 구현예에서, 대조군 샘플은 환자로부터 유래된다.
일 구현예는, 항-프로게스틴 요법을 개시 또는 중지하기 위한 조언을 건강 관리 서비스 제공자에게 제공한다. 다른 구현예는, 예를 들어, 하나 이상의 항-프로게스틴을 유효량으로 투여함으로써, 암 환자를 치료한다. 일 구현예에서, 치료는 하나 이상의 부가적인 치료학적 제제를 투여하는 단계를 더 포함한다.
일 구현예는, 암 환자를 치료하기 위해, 대조군과 비교하여, KR78D11, RBPMS2, PLA2G48, FL112684, SH2D4B, RASCD2, CLDN8 및 이들의 임의 조합으로 이루어진 군으로부터 선택되는 하나 이상의 유전자의 발현 수준이 감소되거나 및/또는 VCX, CHN2, AFAP1L2, PXMP4, THY1, ZNF26, CDH10, ZNF812 및 이들의 임의 조합으로 이루어진 군으로부터 선택되는 하나 이상의 유전자의 발현 수준이 증가된 환자에게, 항-프로게스틴을 단독으로 또는 다른 치료와 병행하여 투여하는 단계를 포함하는, 암 환자의 치료 방법을 제공한다. 다른 구현예는, (c) VCX, CHN2, AFAP1L2, PXMP4, THY1, ZNF26, CDH10, ZNF812 및 이들의 임의 조합으로 이루어진 군으로부터 선택되는 하나 이상의 유전자의 발현 수준을 상기 생물 샘플의 세포에서 측정하는 단계; 및 (d) 단계 (a)의 상기 발현 수준을 대조군에서의 상기 하나 이상의 유전자의 발현 수준과 비교하는 단계를 더 포함하며, 생물 샘플에서 상기 하나 이상의 유전자의 발현 수준이 상기 대조군에 비해 증가된 경우 상기 환자는 항-프로게스틴을 이용한 치료에 반응자이다.
일 구현예는 암으로 진단받은 환자가 활성형 프로게스테론 수용체 (KR)를 발현하는 세포를 포함하며, 항-프로게스틴을 이용한 치료학적 치료에 반응할 가능성이 있는지를 확인하는 방법을 제공하며, 이 방법은 (a) 환자로부터 생물 샘플을 수득하는 단계; (b) THY1, KLF9, SPINK5L.3, PHLDA1, MAP1A, SPRYD5, ATG12, PDK4, MSX2, TUBA3E, TSC22D1, TUBA3D, KHDRBS3, UTS2D, SLC35C1, KIAA0513 및 이들의 임의 조합으로 이루어진 군으로부터 선택되는 하나 이상의 유전자의 발현 수준을 상기 생물 샘플의 세포에서 측정하는 단계; 및 (c) 단계 (b)에서의 상기 발현 수준을 야생형 (WT) 대조군 샘플 및/또는 기준 샘플에서의 상기 하나 이상의 유전자의 발현 수준과 비교하는 단계를 포함하며, 생물 샘플에서 상기 하나 이상의 유전자의 발현 수준이 상기 WT 대조군 샘플 및/또는 기준 샘플에 비해 증가된 경우, 상기 환자는 항-프로게스틴을 이용한 치료에 반응자이다
일 구현예에서, 암은 유방암, 난소암, 자궁 내막암, 뇌암, 폐암, 전립선암, 수막종 또는 자궁암이다.
다른 구현예는 암 환자가 항-프로게스틴 치료에 반응할 것인 지를 확인하는 방법을 제공하며, 이 방법은 a. 환자의 생물 샘플에서 THY1, KLF9, SPINK5L.3, PHLDA1, MAP1A, SPRYD5, ATG12, PDK4, MSX2, TUBA3E, TSC22D1, TUBA3D, KHDRBS3, UTS2D, SLC35C1, KIAA0513 및 이들의 임의 조합으로 이루어진 군으로부터 선택되는 하나 이상의 유전자의 발현 수준을 측정하는 단계를 포함하며, b. 생물 샘플에서 하나 이상의 유전자의 발현 수준이 WT 대조군 샘플 및/또는 기준 샘플에서의 발현 수준에 비해 증가된 것은, 상기 개체가 항-프로게스틴 치료에 반응할 것임을 나타내는 지표이다.
일 구현예에서, mRNA 수준은 유전자 발현 수준의 지표로서 측정된다. 일 구현예에서, 유전자, 예를 들어, THY1의 발현은 예컨대 서열번호 16의 프로브와의 혼성화에 의해 검출한다.
일 구현예에서, 복수의 mRNA들을 각각 측정한다. 다른 구현예에서, 복수의 mRNA들을 동시에 측정한다. 일 구현예에서, 하나 이상의 유전자의 발현 수준의 측정은 인 시츄 혼성화, 노던 블롯, 핵산 증폭, 마이크로어레이 분석 또는 이들의 조합을 포함한다.
일 구현예에서, THY1, KLF9, SPINK5L.3, PHLDA1, MAP1A, SPRYD5, ATG12, PDK4, MSX2, TUBA3E, TSC22D1, TUBA3D, KHDRBS3, UTS2D, SLC35C1, KIAA0513 및 이들의 임의 조합으로 이루어진 군으로부터 선택되는 2종 이상의 유전자의 발현을 측정한다. 다른 구현예에서, THY1, KLF9, SPINK5L.3, PHLDA1, MAP1A, SPRYD5, ATG12, PDK4, MSX2, TUBA3E, TSC22D1, TUBA3D, KHDRBS3, UTS2D, SLC35C1, KIAA0513 및 이들의 임의 조합으로 이루어진 군으로부터 선택되는 3종 이상의 유전자의 발현을 측정한다. 다른 구현예에서, THY1, KLF9, SPINK5L.3, PHLDA1, MAP1A, SPRYD5, ATG12, PDK4, MSX2, TUBA3E, TSC22D1, TUBA3D, KHDRBS3, UTS2D, SLC35C1, KIAA0513 및 이들의 임의 조합으로 이루어진 군으로부터 선택되는 4종 이상의 유전자의 발현을 측정한다. 다른 구현예에서, THY1, KLF9, SPINK5L.3, PHLDA1, MAP1A, SPRYD5, ATG12, PDK4, MSX2, TUBA3E, TSC22D1, TUBA3D, KHDRBS3, UTS2D, SLC35C1, KIAA0513 및 이들의 임의 조합으로 이루어진 군으로부터 선택되는 6종 이상의 유전자의 발현을 측정한다. 다른 구현예에서, THY1, KLF9, SPINK5L.3, PHLDA1, MAP1A, SPRYD5, ATG12, PDK4, MSX2, TUBA3E, TSC22D1, TUBA3D, KHDRBS3, UTS2D, SLC35C1, KIAA0513 및 이들의 임의 조합으로 이루어진 군으로부터 선택되는 7종 이상의 유전자의 발현을 측정한다. 다른 구현예에서, THY1, KLF9, SPINK5L.3, PHLDA1, MAP1A, SPRYD5, ATG12, PDK4, MSX2, TUBA3E, TSC22D1, TUBA3D, KHDRBS3, UTS2D, SLC35C1, KIAA0513 및 이들의 임의 조합으로 이루어진 군으로부터 선택되는 8종 이상, 9종 이상, 10종 이상, 11종 이상, 12종 이상, 13종 이상, 14종 이상, 15종 이상 또는 16종 이상의 유전자의 발현을 측정한다.
일 구현예에서, 생물 샘플은 조직 생검, 유관 세척물, 미세 바늘 흡입물, 외과적으로 적출된 종양의 단편, 순환하는 종양 세포, 순환하는 DNA 또는 순환하는 엑소좀이다. 다른 구현예에서, 대조군 샘플은 비-암성 조직의 샘플, 예를 들어, 상기 환자로부터 유래된다.
일 구현예는 건강 관리 서비스 제공자가 항-프로게스틴 치료를 개시 또는 중지하기 조언을 제공한다. 다른 구현예는 환자에서 암을 치료하는 것을 포함한다. 일 구현예에서, 치료는 하나 이상의 항-프로게스틴을 유효량으로 투여하는 것을 포함한다. 다른 구현예에서, 치료는 하나 이상의 부가적인 치료학적 제제의 투여를 더 포함한다.
일 구현예는, 암 환자를 치료하기 위해, THY1, KLF9, SPINK5L.3, PHLDA1, MAP1A, SPRYD5, ATG12, PDK4, MSX2, TUBA3E, TSC22D1, TUBA3D, KHDRBS3, UTS2D, SLC35C1, KIAA0513 및 이들의 임의 조합으로 이루어진 군으로부터 선택되는 하나 이상의 유전자의 발현 수준이 대조군에 비해 증가된 환자에게, 항-프로게스틴을 단독으로 또는 다른 치료와 병행하여 투여하는 단계를 포함하는, 암 환자의 치료 방법을 제공한다.
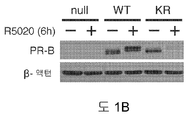
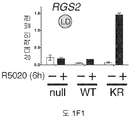
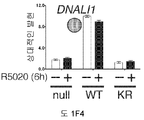
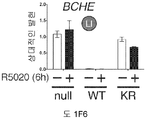
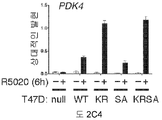
도 1. WT 또는 SUMO-결함성 PR을 안정적으로 발현하는 T47D 세포의, 6시간 동안 R5020 처리 또는 무처리시 유전자 발현 프로파일링. (A) 12종의 인간 유방 종양에서 전체 및 포스포-Ser294 PR 단백질 (로딩 대조군으로서 사용된 총 ERK1/2)을 보여주는 웨스턴 블롯. (B) 야생형 PR-B (WT), SUMO-결함성 돌연변이 K388R PR-B (KR) 또는 빈 벡터 (null) 대조군을 안정적으로 발현하는 T47D 세포에, PR-B에 대해 웨스턴 블롯팅하기 전, R5020을 처리하거나 무처리하였다. (C) 차별적으로 발현되는 전사체들에 대해 표준화된 발현 값들을 나타낸 히트 맵 (heat map) (1개 이상의 샘플에서 배수 변화 >8.0, BH 보정된 P <0.001). 각 치료군에서 생물 두플리케이트 (biological duplicate)를 나타내며, 주목할 만한 유전자 발현 카테고리 (우측에서 1-4로 번호를 매김)를 나타낸다 (결과 참조). (D) 프로게스틴 처리 후 상향 또는 하향 조절된 PR 타겟 유전자들을 나타낸 벤 다이아그램 (log2 배수 변화 >0.6, BH 보정된 P <0.01; 공통 배수 변화 >1.5). (E) PR-null 세포에 비해 상향 또는 하향 조절된 다수의 리간드-의존적인 PR 타겟 유전자들을 도시한 벤 다이아그램 (파트 D에서와 같음). (F) 벡터 대조군 (PR-null), WT 또는 KR PR을 안정적으로 발현하며, 6시간 동안 R5020으로 처리 또는 무처리된 T47D 세포에서 선택된 PR 타겟 유전자들의 (RT-qPCR에 의해 결정한) 상대적인 mRNA 발현; 리간드-의존성 (LD) or 리간드-비의존성 (LI) 벤 카테고리에서 선택한 유전자들을 나타냄 (매칭 유색 라벨 참조). 데이타는 n = 3 평균 +/- SD으로 나타낸다.
도 1.1. T47D 세포에서 유도성 PR 발현에 대한 동계 모델의 구축 및 검증. (A) 클론 유도성 세포주들은 재료 ? 방법에 기술된 바와 같이 개발하였으며, PR 단백질 발현은 2일간 유도체 분자 AP21967 및 1시간 동안 R5020으로 처리한 후 웨스턴 블롯팅에 의해 측정하였다. 프로게스틴-의존성 PR 인산화는 PR 포스포-Ser294 특이 항체를 이용하여 측정하였다. β-액틴 웨스턴 블롯팅을 로딩 대조군으로 수행하였다. 단기간 R5020 처리시 (총 PR에서 약간의 겔 업시프트에 의해 확인되는 바와 같이) 프로게스틴-의존성 PR 글로벌 인산화와 동일 수준의 리간드-의존성 Ser294 인산화가 확인되었다. (B) 2개의 독립적인 모델 시스템 및 플랫폼으로부터 유래된, 전체 게놈 발현 프로파일링 데이타 세트들의 유전자 세트 농화 분석 (GSEA) 비교: (i) Illumina HT-12v4 플랫폼을 이용한 WT 및 돌연변이 KR PR (-/+R5020)를 안정적으로 발현하는 T47D 세포 및 (ii) Affymetrix U133A 2.0 플랫폼을 이용한 유도성 WT 또는 돌연변이 KR PR (-/+AP21967, -/+R5020)을 발현하는 T47D 세포. WT +R5020 (또는 KR +R5020)에 의해 Illumina 데이타세트에서 가장 상향 조절된 유전자들은 가장 왼쪽에 나타내고, WT +R5020 (또는 KR +R5020)에 의해 가장 하향 조절된 유전자들은 가장 우측에 나타낸다. GSEA 어플리케이션을 이용하여, Affymetrix 유전자 (검정 수직 막대)들을 Illumina 데이타세트에 따라 위치시켰으며 (상향 조절된 유전자에서 하향 조절된 유전자), 통계 농화 스코어를 정하였다. Affymetrix와 Illumina 간의 치료군들 모두 통계적으로 유의하였다 (P <0.001). (C) iWT 및 iKR PR을 발현하는 T47D 세포주에서 2종의 PR 타겟 유전자 (MSX2 및 MAP1A)에 대해 유전자 발현 수준을 검증하였다. 세포에 AP21967을 처리하여 PR 발현을 유도하고, RT-qPCR 유전자 발현 분석을 수행하기 전에 RU486 및/또는 R5020을 공동-처리하였다. 데이타는 n=3의 평균 +/- SD으로 나타낸다.
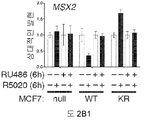
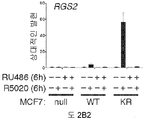
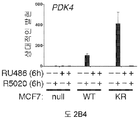
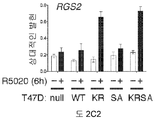
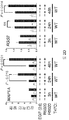
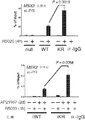
도 2. PR Ser294의 인산화는 MCF-7 및 T47D 세포들에서 SUMO-결함성 PR 유전자 발현 및 프로모터 선택성을 유발한다. (A) 환자 코호트의 조직 샘플에서 PR 타겟 유전자들의 상대적인 발현 수준 (카피수). (B) 빈 벡터 (PR-null), WT 또는 SUMO-결함성 K388R PR를 안정적으로 발현하는 MCF-7 세포에서 선택된 PR 타겟 유전자들의 상대적인 유전자 발현 수준. 세포에 합성 프로게스틴 R5020 및/또는 항-프로게스틴 RU486을 6시간 동안 공동-처리하고, mRNA 수준을 RT-qPCR (방법 참조)을 이용하여 측정하였다. (C) 동일한 PR 타겟 유전자들 (파트 A-B)의 상대적인 유전자 발현 수준을 PR을 안정적으로 발현하는 5종의 벡터-매칭된 T47D 세포주에서 RT-qPCR을 이용하여 측정하였다: 빈 벡터 (null), 야생형 (WT) PR, K388R 돌연변이 (KR) PR, S294A 돌연변이 (SA) PR, 및 K388R / S294A 이중 돌연변이 (KRSA) PR. 세포를 6시간 동안 R5020으로 처리하였다. (D) WT PR을 발현하는 T47D 세포에 2일간 상피 성장 인자 (EGF)를 처리하고, 3, 24 또는 48시간 동안 R5020을 처리하였다. 상대적인 MAP1A 및 RGS2 mRNA 수준을 RT-qPCR으로 측정하였다. (E) T47Dco 모 세포를, 20시간 동안 R5020로 처리하기 전 20분간 EGF를 전처리하였다. 상대적인 RGS2 mRNA 수준을 RT-qPCR로 측정하였다. 데이타는 n = 3의 평균 +/- SD로 나타내며, 스튜던트 t-검정을 이용하여 유의성을 계산하였다.
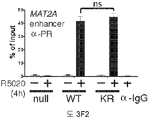
도 3. 프로모터 선택성은 인핸서 유전자좌에 SUMO-결함성 KR PR, CBP, MLL2 및 히스톤 테일 변형, H3K4me2의 동원을 높임으로써 달성된다. (A) 전사 개시부에서 15,094 bp 상류에 위치한 MSX2 유전자 PRE-함유 인핸서 영역을 도식적으로 도시한 것이다. (B) MSX2 인핸서 영역으로의 PR의 상대적인 동원을, 1 또는 4시간 동안 R5020으로 처리한 후, 구성적으로 PR null, WT 또는 KR을 발현하는 T47D 세포에서 ChIP-qPCR 분석을 통해 측정하였다. PR 동원 값들을 입력 염색질 DNA 값의 퍼센트로서 표준화하였다. 백그라운드 비-특이 항체 결합성을 제어하기 위해, 모든 샘플들로부터 유래된 혼합물에 함유된 염색질을 IgG 항체를 사용하여 면역침강시켰다. 유도성 PR을 발현하는 T47D 세포에서 (우측) 유사한 ChIP 결과들을 수득하였다. (C) MSX2 인핸서 영역으로의 CBP의 상대적인 동원을 파트 B에 기술된 바와 같이 측정하였다. (D) MSX2 인핸서에서 H3K4 다이메틸화 수준을 유도성 PR을 발현하는 세포주 (iWT 및 iKR)에서 확인하였다. H3K4me2의 존재는, 그 영역에 걸쳐있는 중첩되는 qPCR 산물들을 이용하여, PRE로부터 상류/하류 MSX2 인핸서에서 확인하였다. (E) MSX2 인핸서 영역으로의 MLL2 동원을 파트 B에 기술된 바와 같이 구성적인 PR과 유도성 PR 둘다를 발현하는 T47D 세포에서 확인하였다. (F) MAT2A 유전자 발현을 안정적인 WT 또는 SUMO-결함성 KR PR을 발현하는 T47D 세포에서 RT-qPCR에 의해 측정하였다. 부가적으로, 표준 ChIP-qPCR 분석에 의해 측정된 바와 같이, PR 및 MLL2 동원을 이들 세포에서 정량하였다. 데이타는 n = 3의 평균 +/- SD으로 나타내며, 유의성을 스튜던트의 t-검사를 이용하여 계산하였다. 또한, 도 3.1, 3.2를 참조한다.
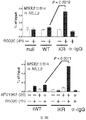
도. 3.1. 도 3과 관련하여, 선택된 PR 타겟 유전자 인핸서에 대한 WT 및 SUMO-결함성 PR 분자의 상대적인 동원을 보여주는 ChIP 분석. (A) (1시간 R5020 노출 후) RGS2, MAP1A 및 PDK4의 상류 프로모터/인핸서 영역들에서 컨센서스 PRE 서열로의 PR 분자의 동원을, WT (iWT) 및 KR (iKR) 수용체를 발현하는 T47D 세포의 유도성 모델에서 표준 ChIP 분석에 의해 측정하였다. HBB 유전자의 인트론 영역으로의 PR의 동원을 음성 대조군으로서 포함시켰다. (B) ChIP 분석을 파트 A와 같이 수행하여, WT 또는 SUMO-결함성 (KR) PR을 안정적으로 발현하는 T47D 세포에서 RGS2 인핸서로의 차별적인 PR 동원을 입증하였다. 데이타는 n = 3의 평균 +/- SD으로 나타낸다.
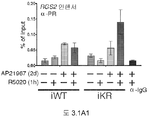
도 3.2. 포스포-Ser5 및 전체-RNA 중합효소 II의 동원에 대한 MSX2 근위 프로모터 영역에서의 ChIP 분석. (A) (1시간 R5020에 노출한 후) MSX2 근위 프로모터 영역으로의 전체 RNA 중합효소 II의 동원을, WT (iWT) 및 KR (iKR) 수용체를 발현하는 T47D 세포의 유도성 모델에서 표준 ChIP 분석에 의해 측정하였다. (B) ChIP 분석은, CTD Ser5 인산화 검출에 의해 측정한 바와 같이, 항체 타겟팅의 기능적으로 활성형인 RNA 중합효소 II를 이용하여, 파트 A에서와 같이 수행하였다. 데이타는 n = 3의 평균 +/- SD으로 나타낸다.
도 3.3. SUMO-결함성 PR은 창의적인 경로 분석에 의해 확인된 세포 증식에 관여하는 유전자들을 상향 조절한다. WT 또는 KR PR을 발현하는 세포에서, 프로게스틴에 의해 (log2 배수 변화 >1.0, BH 보정된 P <0.01; 공통 배수 변화 >2.0) 상향 조절되는 유전자들이 포함된 다중 세포 기능들 (x 축)의 유의한 발현 (y 축). 상향 조절된 유전자들이 상당한 수 포함된 생물학적 경로들을 수평선 위 막대로 나타내며, 이는 BH 보정된 P <0.05이다.
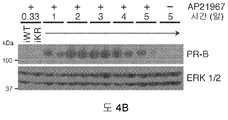
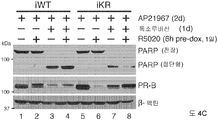
도 4. SUMO-결함성 프로게스테론 수용체는 세포 증식 증가 및 세포자살 감소를 촉진시킨다. (A) 유도성 PR을 발현하는 T47D 세포주의 증식 잠재력을 프로게스틴 (R5020) 및 유도인자, AP21967 (AP)의 존재 하에 MTT 분석으로 측정하였다. (B) 세포 배양 배지에 AP21967을 첨가한 후 5일 이상 동안 유도성 PR 발현이 지속됨을 보여주는 웨스턴 블롯, ERK1/2 웨스턴 블롯팅은 로딩 대조군으로서 수행하였다. (C) 유도성 PR을 발현하는 세포에서 발생하는 세포자살을, 폴리(ADP)-리보스 중합효소 1 (PARP) 절단에 대한 웨스턴 블롯팅에 의해 검출하였다. 세포에 프로게스틴 및/또는 독소루비신을 처리한 후, 단백질을 회수하였다. (D) 증식 및 세포자살을 세포 생존성 루시퍼라제 분석을 이용하여 PR을 구성적으로 발현하는 세포에서 측정하였으며, 이때 4일 발광성을 0일에 대해 표준화하였다. 수집한 데이타는 n = 6의 평균 +/- SD로 나타내며, 유의성을 스튜던트 t-검정을 이용하여 계산하였다.
도 4.1. 리간드-의존성 (LD) 및 리간드-비의존성 (LI) KR>WT 유전자 시그니처를 제시한다. LD (151종의 유전자) 및 LI (92종의 유전자) KR>WT 유전자 시그니처 리스트를 해당 프로브 ID와 더불어 전체를 제시한다.
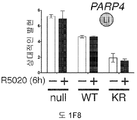
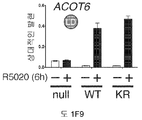
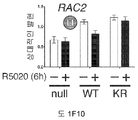
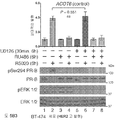
도 5. SUMO-결함성 PR 유전자 발현 시그니처는 HER2-양성의 인간 유방 종양과 관련있으며, 환자 생존성 감소를 나타낸다. (A) (본 발명의 LD KR>WT 유전자 시그니처에서 유전자들에 대해) 표준화된 유전자 발현 수준은 ERBB2 상태에 따라 정리된 환자 코호트에서 각 종양에 대해 제시된다 (Bonnefoi et al., 2007). (B) 유전자 발현 수준은 BT-474 인간 유방암 세포에서 CHN2 및 RGS2 (둘다 SUMO-결함성 PR에 의해 상향 조절되며 LD KR>WT 유전자 시그니처의 구성원임) 및 대조군 유전자 ACOT6 (WT 및 KR 수용체 둘다에서 동일하게 상향 조절됨)에 대한 RT-qPCR에 의해 측정하였다. 세포는 프로게스틴 또는 항-프로게스틴 공동-처리 전에 MEK 키나제 저해제 U0126을 전처리하였다. 총 PR, PR Ser294 인산화, 총 ERK1/2 및 ERK1/2 인산화에 대한 웨스턴 블롯팅에 의해 단백질 수준을 평가하였다. (C) 조합된 T47D 메타유전자 (WT 또는 KR, -/+R5020)를 발현하는 종양을 가진 환자에서, 이러한 메타유전자가 결핍된 환자 종양 대비, 멀리 전이하는 시간에 대한 카플란-메이어 생존 곡선. 환자 샘플에는 Loi et al. 데이타세트 (Loi et al., 2007)의 무처리 및 타목시펜-처리 ER-양성 종양이 포함된다. (D) 조합된 T47D 메타유전자 (KR -R5020, 또는 KR +R5020)를 발현하는 종양을 가진 환자에 대한, 이러한 메타유전자가 결핍된 환자 종양 대비, 파트 C에서와 같은 생존 곡선. 도 4.1을 참조한다.
도 5.1. 도에는 항체 정보와 RT- 및 ChIP-qPCR 분석에 사용된 프라이머 세트가 모두 포함되어 있다.
도 6. 프로게스테론 또는 항-프로게스틴으로 처리된 T47D 세포에서 조절된 상위 유전자. 임의의 가능한 샘플 비교 (예, 프로게스테론 대 에탄올)에서 상향 또는 하향 조절된 (>2배, BH 보정된 P <0.01) 임의의 전사체에 대한 표준화된 상대적인 발현 값들을 나타낸 히트 맵. 샘플을 6시간 동안 처리하였으며, 각 치료군에서 생물학적 트리플리케이트를 나타낸다. 유전자 (열)를 자율적인 계층 클러스터링을 토대로 그룹핑하였으며, 상향 조절된 발현 값은 적색으로, 하향 조절된 발현 값은 청색으로 나타낸다.
도 7. 프로게스테론-조절된 상위 유전자들은 또한 항-프로게스틴 RU486 및 아글레프리스톤으로 처리한 후 SUMO-결함의 PR을 발현하는 세포에서 상향 조절되지만, 오나프리스톤은 그렇지 않다. 임의의 세포주에서 프로게스테론 처리 (즉, 프로게스테론 대 에탄올) 후, 상향 조절된 (>2.5배, BH 보정된 P <0.01) 임의의 전사체에 대한 표준화된 상대적인 발현 값들을 나타낸 히트 맵. 세포주들을 각 개별 코호트에서 6시간 동안 처리하였으며, 각 치료군에서 생물학적 트리플리케이트를 나타낸다. 유전자 (열)를 자율적인 계층 클러스터링을 토대로 그룹핑하였으며, 상향 조절된 발현 값은 적색으로, 하향 조절된 발현 값은 청색으로 나타낸다.
도 8. 오나프리스톤으로 처리된 세포는 PR 발현성 세포에서 유전자 발현을 자극하지 않는다. 도 7의 처리군(세로열)의 자율적인 계층 클러스터링. 또한, 유전자 (열)를 자율적인 계층 클러스터링을 토대로 그룹핑하였으며, 상향 조절된 발현 값은 적색으로, 하향 조절된 발현 값은 청색으로 나타낸다.
도 9. WT 또는 KR PR을 발현하는 세포들을 구별할 수 있는 유전자 15종. 이들 유전자 15종은 3번의 독립적인 통계학적 방법 (방법 참조)을 수행하여 결정된 바와 같이, WT 및 KR 세포에서 독자적으로 조절된다. 각 전사체에 대한 표준화된 상대적인 발현 값들을 나타낸 히트 맵. 샘플을 6시간 동안 처리하고, 생물학적 트리플리케이트를 각 처리군에 대해 나타낸다 (에탄올 및 프로게스틴 군의 경우 n=5). 유전자 (열)를 자율적인 계층 클러스터링을 토대로 그룹핑하였으며, 상향 조절된 발현 값은 적색으로, 하향 조절된 발현 값은 청색으로 나타낸다.
도 10. 2가지의 독립적인 마이크로어레이 실험을 중복함으로써 동정된, KR 발현 세포에서 29종의 유전자들이 특이적으로 상향 조절된다. 프로게스테론 (P4) 처리 후, WT PR 발현 세포 대비, SUMO-결함성 PR (KR)을 발현하는 세포에서 특이적으로 상향 조절 (>1.5배, BH 보정된 P <0.01)된 모든 전사체들에 대한 표준화된 상대적인 발현 값들을 나타낸 히트 맵. 샘플들을 6시간 동안 처리하였으며, 각 치료군에서 생물학적 트리플리케이트를 나타낸다. 유전자 (열)를 자율적인 계층 클러스터링을 토대로 그룹핑하였으며, 상향 조절된 발현 값은 적색으로, 하향 조절된 발현 값은 청색으로 나타낸다.
도 11. 정제한 프로게스틴-의존성 KR>WT 유전자 시그니처. 오나프리스톤 처리 (단독 또는 P4와 조합)에 의해 특이적 (BH 보정된 P <0.01)으로 자극받은 도 10 유래 유전자들을 빼면, 유전자 16종이 남는다. 프로게스테론 처리 후, WT PR 발현 세포 대비, SUMO-결함성 PR (KR)을 발현하는 세포에서 특이적으로 상향 조절된 전사체 16종들에 대한 표준화된 상대적인 발현 값들을 나타낸 히트 맵. 샘플들을 6시간 동안 처리하였으며, 각 치료군에서 생물학적 트리플리케이트를 나타낸다. 유전자 (열)를 자율적인 계층 클러스터링을 토대로 그룹핑하였으며, 상향 조절된 발현 값은 적색으로, 하향 조절된 발현 값은 청색으로 나타낸다.
도 1.1. T47D 세포에서 유도성 PR 발현에 대한 동계 모델의 구축 및 검증. (A) 클론 유도성 세포주들은 재료 ? 방법에 기술된 바와 같이 개발하였으며, PR 단백질 발현은 2일간 유도체 분자 AP21967 및 1시간 동안 R5020으로 처리한 후 웨스턴 블롯팅에 의해 측정하였다. 프로게스틴-의존성 PR 인산화는 PR 포스포-Ser294 특이 항체를 이용하여 측정하였다. β-액틴 웨스턴 블롯팅을 로딩 대조군으로 수행하였다. 단기간 R5020 처리시 (총 PR에서 약간의 겔 업시프트에 의해 확인되는 바와 같이) 프로게스틴-의존성 PR 글로벌 인산화와 동일 수준의 리간드-의존성 Ser294 인산화가 확인되었다. (B) 2개의 독립적인 모델 시스템 및 플랫폼으로부터 유래된, 전체 게놈 발현 프로파일링 데이타 세트들의 유전자 세트 농화 분석 (GSEA) 비교: (i) Illumina HT-12v4 플랫폼을 이용한 WT 및 돌연변이 KR PR (-/+R5020)를 안정적으로 발현하는 T47D 세포 및 (ii) Affymetrix U133A 2.0 플랫폼을 이용한 유도성 WT 또는 돌연변이 KR PR (-/+AP21967, -/+R5020)을 발현하는 T47D 세포. WT +R5020 (또는 KR +R5020)에 의해 Illumina 데이타세트에서 가장 상향 조절된 유전자들은 가장 왼쪽에 나타내고, WT +R5020 (또는 KR +R5020)에 의해 가장 하향 조절된 유전자들은 가장 우측에 나타낸다. GSEA 어플리케이션을 이용하여, Affymetrix 유전자 (검정 수직 막대)들을 Illumina 데이타세트에 따라 위치시켰으며 (상향 조절된 유전자에서 하향 조절된 유전자), 통계 농화 스코어를 정하였다. Affymetrix와 Illumina 간의 치료군들 모두 통계적으로 유의하였다 (P <0.001). (C) iWT 및 iKR PR을 발현하는 T47D 세포주에서 2종의 PR 타겟 유전자 (MSX2 및 MAP1A)에 대해 유전자 발현 수준을 검증하였다. 세포에 AP21967을 처리하여 PR 발현을 유도하고, RT-qPCR 유전자 발현 분석을 수행하기 전에 RU486 및/또는 R5020을 공동-처리하였다. 데이타는 n=3의 평균 +/- SD으로 나타낸다.
도 2. PR Ser294의 인산화는 MCF-7 및 T47D 세포들에서 SUMO-결함성 PR 유전자 발현 및 프로모터 선택성을 유발한다. (A) 환자 코호트의 조직 샘플에서 PR 타겟 유전자들의 상대적인 발현 수준 (카피수). (B) 빈 벡터 (PR-null), WT 또는 SUMO-결함성 K388R PR를 안정적으로 발현하는 MCF-7 세포에서 선택된 PR 타겟 유전자들의 상대적인 유전자 발현 수준. 세포에 합성 프로게스틴 R5020 및/또는 항-프로게스틴 RU486을 6시간 동안 공동-처리하고, mRNA 수준을 RT-qPCR (방법 참조)을 이용하여 측정하였다. (C) 동일한 PR 타겟 유전자들 (파트 A-B)의 상대적인 유전자 발현 수준을 PR을 안정적으로 발현하는 5종의 벡터-매칭된 T47D 세포주에서 RT-qPCR을 이용하여 측정하였다: 빈 벡터 (null), 야생형 (WT) PR, K388R 돌연변이 (KR) PR, S294A 돌연변이 (SA) PR, 및 K388R / S294A 이중 돌연변이 (KRSA) PR. 세포를 6시간 동안 R5020으로 처리하였다. (D) WT PR을 발현하는 T47D 세포에 2일간 상피 성장 인자 (EGF)를 처리하고, 3, 24 또는 48시간 동안 R5020을 처리하였다. 상대적인 MAP1A 및 RGS2 mRNA 수준을 RT-qPCR으로 측정하였다. (E) T47Dco 모 세포를, 20시간 동안 R5020로 처리하기 전 20분간 EGF를 전처리하였다. 상대적인 RGS2 mRNA 수준을 RT-qPCR로 측정하였다. 데이타는 n = 3의 평균 +/- SD로 나타내며, 스튜던트 t-검정을 이용하여 유의성을 계산하였다.
도 3. 프로모터 선택성은 인핸서 유전자좌에 SUMO-결함성 KR PR, CBP, MLL2 및 히스톤 테일 변형, H3K4me2의 동원을 높임으로써 달성된다. (A) 전사 개시부에서 15,094 bp 상류에 위치한 MSX2 유전자 PRE-함유 인핸서 영역을 도식적으로 도시한 것이다. (B) MSX2 인핸서 영역으로의 PR의 상대적인 동원을, 1 또는 4시간 동안 R5020으로 처리한 후, 구성적으로 PR null, WT 또는 KR을 발현하는 T47D 세포에서 ChIP-qPCR 분석을 통해 측정하였다. PR 동원 값들을 입력 염색질 DNA 값의 퍼센트로서 표준화하였다. 백그라운드 비-특이 항체 결합성을 제어하기 위해, 모든 샘플들로부터 유래된 혼합물에 함유된 염색질을 IgG 항체를 사용하여 면역침강시켰다. 유도성 PR을 발현하는 T47D 세포에서 (우측) 유사한 ChIP 결과들을 수득하였다. (C) MSX2 인핸서 영역으로의 CBP의 상대적인 동원을 파트 B에 기술된 바와 같이 측정하였다. (D) MSX2 인핸서에서 H3K4 다이메틸화 수준을 유도성 PR을 발현하는 세포주 (iWT 및 iKR)에서 확인하였다. H3K4me2의 존재는, 그 영역에 걸쳐있는 중첩되는 qPCR 산물들을 이용하여, PRE로부터 상류/하류 MSX2 인핸서에서 확인하였다. (E) MSX2 인핸서 영역으로의 MLL2 동원을 파트 B에 기술된 바와 같이 구성적인 PR과 유도성 PR 둘다를 발현하는 T47D 세포에서 확인하였다. (F) MAT2A 유전자 발현을 안정적인 WT 또는 SUMO-결함성 KR PR을 발현하는 T47D 세포에서 RT-qPCR에 의해 측정하였다. 부가적으로, 표준 ChIP-qPCR 분석에 의해 측정된 바와 같이, PR 및 MLL2 동원을 이들 세포에서 정량하였다. 데이타는 n = 3의 평균 +/- SD으로 나타내며, 유의성을 스튜던트의 t-검사를 이용하여 계산하였다. 또한, 도 3.1, 3.2를 참조한다.
도. 3.1. 도 3과 관련하여, 선택된 PR 타겟 유전자 인핸서에 대한 WT 및 SUMO-결함성 PR 분자의 상대적인 동원을 보여주는 ChIP 분석. (A) (1시간 R5020 노출 후) RGS2, MAP1A 및 PDK4의 상류 프로모터/인핸서 영역들에서 컨센서스 PRE 서열로의 PR 분자의 동원을, WT (iWT) 및 KR (iKR) 수용체를 발현하는 T47D 세포의 유도성 모델에서 표준 ChIP 분석에 의해 측정하였다. HBB 유전자의 인트론 영역으로의 PR의 동원을 음성 대조군으로서 포함시켰다. (B) ChIP 분석을 파트 A와 같이 수행하여, WT 또는 SUMO-결함성 (KR) PR을 안정적으로 발현하는 T47D 세포에서 RGS2 인핸서로의 차별적인 PR 동원을 입증하였다. 데이타는 n = 3의 평균 +/- SD으로 나타낸다.
도 3.2. 포스포-Ser5 및 전체-RNA 중합효소 II의 동원에 대한 MSX2 근위 프로모터 영역에서의 ChIP 분석. (A) (1시간 R5020에 노출한 후) MSX2 근위 프로모터 영역으로의 전체 RNA 중합효소 II의 동원을, WT (iWT) 및 KR (iKR) 수용체를 발현하는 T47D 세포의 유도성 모델에서 표준 ChIP 분석에 의해 측정하였다. (B) ChIP 분석은, CTD Ser5 인산화 검출에 의해 측정한 바와 같이, 항체 타겟팅의 기능적으로 활성형인 RNA 중합효소 II를 이용하여, 파트 A에서와 같이 수행하였다. 데이타는 n = 3의 평균 +/- SD으로 나타낸다.
도 3.3. SUMO-결함성 PR은 창의적인 경로 분석에 의해 확인된 세포 증식에 관여하는 유전자들을 상향 조절한다. WT 또는 KR PR을 발현하는 세포에서, 프로게스틴에 의해 (log2 배수 변화 >1.0, BH 보정된 P <0.01; 공통 배수 변화 >2.0) 상향 조절되는 유전자들이 포함된 다중 세포 기능들 (x 축)의 유의한 발현 (y 축). 상향 조절된 유전자들이 상당한 수 포함된 생물학적 경로들을 수평선 위 막대로 나타내며, 이는 BH 보정된 P <0.05이다.
도 4. SUMO-결함성 프로게스테론 수용체는 세포 증식 증가 및 세포자살 감소를 촉진시킨다. (A) 유도성 PR을 발현하는 T47D 세포주의 증식 잠재력을 프로게스틴 (R5020) 및 유도인자, AP21967 (AP)의 존재 하에 MTT 분석으로 측정하였다. (B) 세포 배양 배지에 AP21967을 첨가한 후 5일 이상 동안 유도성 PR 발현이 지속됨을 보여주는 웨스턴 블롯, ERK1/2 웨스턴 블롯팅은 로딩 대조군으로서 수행하였다. (C) 유도성 PR을 발현하는 세포에서 발생하는 세포자살을, 폴리(ADP)-리보스 중합효소 1 (PARP) 절단에 대한 웨스턴 블롯팅에 의해 검출하였다. 세포에 프로게스틴 및/또는 독소루비신을 처리한 후, 단백질을 회수하였다. (D) 증식 및 세포자살을 세포 생존성 루시퍼라제 분석을 이용하여 PR을 구성적으로 발현하는 세포에서 측정하였으며, 이때 4일 발광성을 0일에 대해 표준화하였다. 수집한 데이타는 n = 6의 평균 +/- SD로 나타내며, 유의성을 스튜던트 t-검정을 이용하여 계산하였다.
도 4.1. 리간드-의존성 (LD) 및 리간드-비의존성 (LI) KR>WT 유전자 시그니처를 제시한다. LD (151종의 유전자) 및 LI (92종의 유전자) KR>WT 유전자 시그니처 리스트를 해당 프로브 ID와 더불어 전체를 제시한다.
도 5. SUMO-결함성 PR 유전자 발현 시그니처는 HER2-양성의 인간 유방 종양과 관련있으며, 환자 생존성 감소를 나타낸다. (A) (본 발명의 LD KR>WT 유전자 시그니처에서 유전자들에 대해) 표준화된 유전자 발현 수준은 ERBB2 상태에 따라 정리된 환자 코호트에서 각 종양에 대해 제시된다 (Bonnefoi et al., 2007). (B) 유전자 발현 수준은 BT-474 인간 유방암 세포에서 CHN2 및 RGS2 (둘다 SUMO-결함성 PR에 의해 상향 조절되며 LD KR>WT 유전자 시그니처의 구성원임) 및 대조군 유전자 ACOT6 (WT 및 KR 수용체 둘다에서 동일하게 상향 조절됨)에 대한 RT-qPCR에 의해 측정하였다. 세포는 프로게스틴 또는 항-프로게스틴 공동-처리 전에 MEK 키나제 저해제 U0126을 전처리하였다. 총 PR, PR Ser294 인산화, 총 ERK1/2 및 ERK1/2 인산화에 대한 웨스턴 블롯팅에 의해 단백질 수준을 평가하였다. (C) 조합된 T47D 메타유전자 (WT 또는 KR, -/+R5020)를 발현하는 종양을 가진 환자에서, 이러한 메타유전자가 결핍된 환자 종양 대비, 멀리 전이하는 시간에 대한 카플란-메이어 생존 곡선. 환자 샘플에는 Loi et al. 데이타세트 (Loi et al., 2007)의 무처리 및 타목시펜-처리 ER-양성 종양이 포함된다. (D) 조합된 T47D 메타유전자 (KR -R5020, 또는 KR +R5020)를 발현하는 종양을 가진 환자에 대한, 이러한 메타유전자가 결핍된 환자 종양 대비, 파트 C에서와 같은 생존 곡선. 도 4.1을 참조한다.
도 5.1. 도에는 항체 정보와 RT- 및 ChIP-qPCR 분석에 사용된 프라이머 세트가 모두 포함되어 있다.
도 6. 프로게스테론 또는 항-프로게스틴으로 처리된 T47D 세포에서 조절된 상위 유전자. 임의의 가능한 샘플 비교 (예, 프로게스테론 대 에탄올)에서 상향 또는 하향 조절된 (>2배, BH 보정된 P <0.01) 임의의 전사체에 대한 표준화된 상대적인 발현 값들을 나타낸 히트 맵. 샘플을 6시간 동안 처리하였으며, 각 치료군에서 생물학적 트리플리케이트를 나타낸다. 유전자 (열)를 자율적인 계층 클러스터링을 토대로 그룹핑하였으며, 상향 조절된 발현 값은 적색으로, 하향 조절된 발현 값은 청색으로 나타낸다.
도 7. 프로게스테론-조절된 상위 유전자들은 또한 항-프로게스틴 RU486 및 아글레프리스톤으로 처리한 후 SUMO-결함의 PR을 발현하는 세포에서 상향 조절되지만, 오나프리스톤은 그렇지 않다. 임의의 세포주에서 프로게스테론 처리 (즉, 프로게스테론 대 에탄올) 후, 상향 조절된 (>2.5배, BH 보정된 P <0.01) 임의의 전사체에 대한 표준화된 상대적인 발현 값들을 나타낸 히트 맵. 세포주들을 각 개별 코호트에서 6시간 동안 처리하였으며, 각 치료군에서 생물학적 트리플리케이트를 나타낸다. 유전자 (열)를 자율적인 계층 클러스터링을 토대로 그룹핑하였으며, 상향 조절된 발현 값은 적색으로, 하향 조절된 발현 값은 청색으로 나타낸다.
도 8. 오나프리스톤으로 처리된 세포는 PR 발현성 세포에서 유전자 발현을 자극하지 않는다. 도 7의 처리군(세로열)의 자율적인 계층 클러스터링. 또한, 유전자 (열)를 자율적인 계층 클러스터링을 토대로 그룹핑하였으며, 상향 조절된 발현 값은 적색으로, 하향 조절된 발현 값은 청색으로 나타낸다.
도 9. WT 또는 KR PR을 발현하는 세포들을 구별할 수 있는 유전자 15종. 이들 유전자 15종은 3번의 독립적인 통계학적 방법 (방법 참조)을 수행하여 결정된 바와 같이, WT 및 KR 세포에서 독자적으로 조절된다. 각 전사체에 대한 표준화된 상대적인 발현 값들을 나타낸 히트 맵. 샘플을 6시간 동안 처리하고, 생물학적 트리플리케이트를 각 처리군에 대해 나타낸다 (에탄올 및 프로게스틴 군의 경우 n=5). 유전자 (열)를 자율적인 계층 클러스터링을 토대로 그룹핑하였으며, 상향 조절된 발현 값은 적색으로, 하향 조절된 발현 값은 청색으로 나타낸다.
도 10. 2가지의 독립적인 마이크로어레이 실험을 중복함으로써 동정된, KR 발현 세포에서 29종의 유전자들이 특이적으로 상향 조절된다. 프로게스테론 (P4) 처리 후, WT PR 발현 세포 대비, SUMO-결함성 PR (KR)을 발현하는 세포에서 특이적으로 상향 조절 (>1.5배, BH 보정된 P <0.01)된 모든 전사체들에 대한 표준화된 상대적인 발현 값들을 나타낸 히트 맵. 샘플들을 6시간 동안 처리하였으며, 각 치료군에서 생물학적 트리플리케이트를 나타낸다. 유전자 (열)를 자율적인 계층 클러스터링을 토대로 그룹핑하였으며, 상향 조절된 발현 값은 적색으로, 하향 조절된 발현 값은 청색으로 나타낸다.
도 11. 정제한 프로게스틴-의존성 KR>WT 유전자 시그니처. 오나프리스톤 처리 (단독 또는 P4와 조합)에 의해 특이적 (BH 보정된 P <0.01)으로 자극받은 도 10 유래 유전자들을 빼면, 유전자 16종이 남는다. 프로게스테론 처리 후, WT PR 발현 세포 대비, SUMO-결함성 PR (KR)을 발현하는 세포에서 특이적으로 상향 조절된 전사체 16종들에 대한 표준화된 상대적인 발현 값들을 나타낸 히트 맵. 샘플들을 6시간 동안 처리하였으며, 각 치료군에서 생물학적 트리플리케이트를 나타낸다. 유전자 (열)를 자율적인 계층 클러스터링을 토대로 그룹핑하였으며, 상향 조절된 발현 값은 적색으로, 하향 조절된 발현 값은 청색으로 나타낸다.
프로게스테론 수용체 (PR)는 유방암 및 자궁내막암을 비롯한 특정 암의 증식과 생장에 중요한 역할을 한다. 유방암 세포에서 공통적인 인산화 현상은 PR의 전사 활성에 영향을 미친다. 포스포-Ser294 PR은 리간드-의존성 Lys388 SUMO일화 (즉, 억제성 변형)에 저항성을 나타낸다. 단백질 키나제에 의한 PR SUMO일화의 길항화는 PR 탈억제 (즉, 전사 활성화)를 위한 기전을 제공한다. 야생형 또는 K388R (SUMO일화-결함성) PR을 발현하는 유방암 세포에서 글로벌 유전자 발현 프로파일링을 통해, SUMO일화-결함성 PR이 본래 증식 및 생존에 도움이 되는 (pro-survival) 신호전달에 필요한 유전자들을 조절한다는 것이 드러났다. K388R PR은, 스테로이드 수용체 공동-활성인자, CBP 및 MLL2, 뉴클레오좀 리모델링의 매개 인자와 함께, "SUMO-민감성" 후보 유전자의 인핸서 영역으로 선호적으로 동원된다. SUMO-결함성 (포스포-Ser294) PR 유전자 시그니처는 ERBB2-과다발현성 유방 종양과 유의한 수준으로 연관되어 있으며, 조기 전이와 짧은 생존 예후를 나타낸다. 가역적인 PR SUMO일화/탈SUMO일화는 유방암 세포에서 타겟 유전자의 선택을 상당히 변화시킨다는 결론에 도달하였다. 포스포-PR에 의해 유발된 ER 양성 및/또는 PR 양성 종양을 가진 환자는 항-프로게스틴이 포함된 엔도크린 요법에서 효과가 있을 수 있다.
본원에 기술된 유전자 시그니처는, 암 진행에 기여하는 것으로 공지된 관련 유전자들의 콜렉션을 포함하며, 이들의 발현은 현재 활성화된 포스포-PR (탈SUMO일화된 PR-B)에 직접적으로 의존하는 것으로 공지되어 있다. 관련된 기전이 본원에 기술된 바와 같이 규명되었으므로, (PR 활성과, PR과 다른 악성 생장 및 증식 경로의 상호작용을 차단하기 위한 목적으로) 항-프로게스틴의 사용을 포함하는 치료가 유익할 것으로 보이는, PR-유동성 종양을 가진 유방암 환자가 검사를 통해 동정될 것 있다. 본원에 기술된 규명된 유전자 발현 패턴은 탈SUMO일화된 포스포-PR가 요인으로서, 약물 옵션 (항-프로게스틴 요법 단독 또는 다른 항암제와의 조합 사용)은 활성화된 PR 유전자 발현 패턴을 가진 사례에 대해 효과적인 치료 전략이 될 것이다. 이런 치료법은 항-프로게스틴 약물 + ER+ 유방 종양에 대한 현행 표준 엔도크린 케어 치료 (예, 항-프로게스틴과 조합된, 항-에스트로겐 또는 아로마스타제 저해제) 또는 항-프로게스틴 약물 + 그외 항암 화합물 (예, 에베롤리무스 (everolimus), 트라스투주맵 (trastuzumab), T-DM1, 항-HER2 약물, m-TOR 저해제, 항-VEGF 약물, 항-EGF 약물, 베박시주맵 (bevacizumab), 파클리탁셀, 독세탁셀 (docetaxel), 탁산, 독소루비신, 리포좀 독소루비신, 페길화된 리포좀 독소루비신, 안트라사이클린, 안트라센다이온, 카보플라틴 (carboplatin), 시스플라틴 (cisplatin), 5-FU, 겜시타빈, 사이클로포스파미드)을 포함한다. 즉, 항-프로게스틴을 이용한 치료시, 종양 억제 및 PR 유전자 시그니처가 반전될 것이다.
본 발명은 유전자 발현 프로파일과 암 치료에 대한 환자의 반응성을 예측하기 위한 이의 용도를 제공한다. 보다 구체적으로, 유전자 발현 프로파일은 유방암을 앓고 있는 환자가 항-프로게스틴을 포함하는 엔도크린 요법을 이용한 치료에 반응자인지 또는 무-반응자인지를 나타내는 지표이다.
유방암 치료 성과에 대해 상당한 개선이 이루어지고 있다. 하지만, 매번 정상 세포의 생장이 이런 치료에 영향을 받게 되어, 원치않거나 및/또는 좋지 못한 결과가 나타난다. 이런 기타 효과로는, 설사, 홍반, 여드름, 피부 건조, 구역질 (토할 것 같음) 및 구토, 식욕 감소, 체중 감소, 무력증, 소양증, 신경장애 및 복통을 포함할 수 있다. 본 발명의 측면들은, 항-프로게스틴 치료가 유익한 환자와 연관된 바이오마커를 제공한다. 따라서, 본 발명은, 항-프로게스틴 치료가 유익할 것으로 보이는 유방암 환자를 사전에 치료 제공자가 확인할 수 있으며, 무-반응자에 대해서는 다른 치료 옵션을 고려할 수 있게 한다.
본 발명의 측면들은 유방암 환자의 항-프로게스틴을 이용한 치료에 대한 반응 경향을 표시하는 유전자 발현 프로파일을 포함한다. 유전자 발현 프로파일은, 표 1a 및/또는 1b에 표시된 유전자 하나 이상, 바람직하게는 복수개를 포함한다. 이들 유전자 군은 본원에서 "항-프로게스틴 반응자 유전자"로 지칭된다. 본 발명의 측면들에서, 이들 유전자 일부 또는 전체가 항-프로게스틴을 이용한 치료에 반응자인 환자에서 차별적으로 발현 (예, 상향 조절 또는 하향 조절)된다. 이에, 환자가 환자의 조직으로부터 유전자 발현 프로파일을 수득하고, 항-프로게스틴 반응자 유전자들 중 하나 이상의 유전자가 상향 또는 하향 조절되었는지를 확인함으로써, 상기한 치료가 유효할 것인지를 사전에 확인할 수 있다.
일 구현예에서, 본 발명의 유전자 발현 프로파일은, 조절되는 항-프로게스틴 반응자 유전자를 약 4개 이상, 예컨대 약 4 - 약 9개, 약 9 - 15개 이상으로 포함한다. 일 구현예에서, 유전자 발현 프로파일은 조절되는 항-프로게스틴 반응자 유전자를 약 4개 이상, 예컨대 약 6 - 12개 포함한다.
본 발명의 유전자 발현 프로파일은, 항-프로게스틴을 이용한 치료에 대한 유방암 환자의 반응성을 예측하는데 사용할 수 있다. 일 측면에서, 본 방법은, (a) 유방암 환자의 생물 샘플 (조직 생검, 유관 세척물, 미세 바늘 흡입 샘플, 외과적으로 적출한 조직의 단편 또는 순환하는 종양 세포)로부터 유전자 발현 프로파일을 수득하는 단계; (b) 표 1a 및/또는 1b에 표시된 유전자들 중 하나 이상의 발현이 상향 또는 하향 조절 (과다 발현 또는 과소 발현)되는 지를 상기 유전자 발현 프로파일에서 확인하는 단계를 포함한다. 일 구현예에서, 이들 화합물에 대한 반응을 확인하는데 있어 유전자 프로파일의 예측 가치는 본 발명에서 상향 또는 하향 조절되는 것으로 확인된 관련 유전자들의 수에 따라 높아진다.
정의
달리 언급되지 않은 한, 본원에 사용된 모든 기술적 및 과학적 용어들은 본 발명이 속하는 당해 기술 분야의 당업자들이 통상적으로 이해하는 바와 동일한 의미를 가진다. 본원에 기술된 바와 유사하거나 또는 동일한 임의의 방법 및 물질들이 본 발명을 실시하거나 테스트하는데 사용될 수 있지만, 방법 및 재료에 대한 몇몇 구현예들을 본원에 개시한다. 본원에서, 아래 용어들은 각각 본 섹션에서 이와 관련된 의미를 가진다.
관사 ("a" 및 "an")는 본원에서 관사의 문법적인 대상 하나 또는 하나 이상 (즉, 하나 이상)을 지칭한다. 예를 들어, "(an) 인자"는 인자 하나 또는 2 이상의 인자를 의미한다.
"복수개"는 2 이상을 의미한다.
"개체" 또는 "환자"는 인간과 같은 포유류를 비롯한 척추동물이다. 포유류로는, 비제한적으로, 인간, 농장 동물, 스포츠 동물 및 애완 동물을 포함한다.
용어 "약"은 본원에서 대략, 정도, 대충 또는 쯤의 의미이다. 용어 "약"이 수치 범위와 함께 사용되었을 때, 이는 기술된 수치 값들의 상한 및 하한 경계를 확장함으로써 그 범위를 달라지게 한다. 일반적으로, 용어 "약"을 사용하여, 본원에서 언급된 수치에서 상하 10%의 가변성으로 수치 값을 확장시킨다. 일 측면에서, 용어 "약"은 사용되어진 숫자의 수치 값에서 ± 20%를 의미한다. 따라서, 약 50%는 45%-55%를 의미한다. 엔드포인트에 의해 본원에 기술된 수치 범위들은 그 범위에 속하는 모든 수와 분수를 포함한다 (예, 1 - 5는 1, 1.5, 2, 2.75, 3, 3.90, 4 및 5를 포함함). 또한, 모든 수와 분수는 용어 "약"에 의해 변경되는 것이 당연한 것으로 이해되어야 한다.
용어 "유전자"는 폴리펩타이드 또는 전구체를 만드는데 필요한 조절 서열과 코딩 서열을 포함하는 핵산 서열을 지칭한다. 폴리펩타이드는 전장 코딩 서열에 의해 또는 코딩 서열의 임의 일부에 의해 코딩될 수 있다. 유전자는 식물, 진균, 동물, 박테리아 게놈 또는 에피좀, 진핵생물, 핵 또는 플라스미드 DNA, cDNA, 바이러스 DNA 또는 화학적 합성 DNA 등의, 당해 기술 분야에 공지된 임의 소스로부터 전체 또는 일부가 유래될 수 있다. 유전자는 발현 산물의 생물학적 활성 또는 화학적 구조, 발현 속도 또는 발현 조적 방식에 영향을 미칠 수 있는, 비-번역 영역 또는 코딩 영역 어느 하나에 하나 이상의 변형을 포함할 수 있다. 이러한 변형으로는, 비제한적으로, 하나 이상의 뉴클레오티드의 돌연변이, 삽입, 결손 및 치환을 포함한다. 유전자는 연속적인 (uninterrupted) 코딩 서열로 구성되거나, 또는 적절한 스플라이스 정션에 의해 결합된 하나 이상의 인트론을 포함할 수 있다.
용어 "유전자 발현"은 뉴클레오티드 서열이 검출가능한 수준으로 발현되도록 핵산 서열에 대한 성공적인 전사 및/또는 번역이 수행되는 프로세스를 의미한다.
용어 "유전자 발현 프로파일" 또는 "유전자 시그니처"는 특정 세포 또는 조직 타입에 의해 발현되는 유전자들의 그룹을 지칭하며, 여기서 유전자들의 동시 존재 또는 이들 유전자들의 차별적인 발현이 특정 상황에 대한 지표/예측이다.
용어 "핵산"은, 본원에서, 한가지 이상의 뉴클레오티드, 즉, 리보뉴클레오티드, 데옥시리보뉴클레오티드, 또는 둘다로 구성된 분자를 지칭한다. 이 용어는 리보뉴클레오티드 및 데옥시리보뉴클레오티드의 모노머와 폴리머를 포함하며, 리보뉴클레오티드 및/또는 데옥시리보뉴클레오티드는 폴리머일 경우 5' -> 3' 결합을 통해 서로 결합되어 있다. 리보뉴클레오티드 및 데옥시리보뉴클레오티드 폴리머들은 단일 가닥이거나 이중 가닥일 수 있다. 그러나, 결합은, 예를 들어, 5' -> 3' 결합을 포함하는 핵산을 포함하여, 당해 기술 분야에 공지된 모든 결합을 포함할 수 있다. 아울러, 용어 "핵산 서열"은 상보적인 서열을 포함하며, 구체적으로 핵산 서열과 이의 상보체 둘다에 실질적으로 상보적인 임의의 핵산 서열을 포함한다.
용어 "어레이" 및 "마이크로어레이"는 어레이 상에 올리고뉴클레오티드로 유전자가 제시된 타입을 지칭하며, 여기서 어레이 상에 유전자가 제시된 타입은 어레이의 의도한 목적 (예, 인간 유전자의 발현을 모니터링하기 위해)에 따라 결정된다. 해당 어레이 상의 올리고뉴클레오티드는 동일한 타입, 카테고리 또는 그룹의 유전자에 해당될 수 있다. 유전자는, 이들이 기원 종 (예, 인간, 마우스, 랫); 질환 상태 (예, 암); 기능 (예, 단백질 키나제, 종양 억제인자); 또는 생물학적 프로세스 (예, 세포자살, 신호 전이, 세포 주기 조절, 증식, 분화) 등과 같이 일부 공통된 특징들을 공유한다면, 동일한 타입의 것으로 간주될 수 있다. 예를 들어, 하나의 어레이 타입은, 어레이의 각 올리고뉴클레오티드가 암과 관련된 유전자에 해당되는 "암 어레이"일 수 있다.
용어 "활성화"는 본원에서 예를 들어 기저 수준 이상으로의 증가, 저해된 상태로부터 기저 수준으로의 회복 및 기저 수준 이상으로의 경로의 자극을 비롯하여, 신호전달 경로나 생물학적 반응의 임의 변형을 지칭한다.
용어 "차별적인 발현"은 정상 인접 조직 대비 질환에 걸린 조직 또는 세포에서의 유전자의 일시적인 조직 발현 패턴의 정량적인 차이 뿐만 아니라 정성적인 차이 둘다를 지칭한다. 예를 들어, 차별적으로 발현된 유전자가 질환 상태에 비해 정상 상태에서 그 발현이 활성화되거나, 또는 일부 또는 완전히 불활성화되거나, 또는 정상 상태에 비해 질환 상태에서 상향 조절 (과다 발현)되거나 또는 하향 조절 (과소 발현)될 수 있다. 이러한 정성적으로 조절되는 유전자는, 해당 조직 또는 세포 타입에서, 대조군 또는 질환 상태에서는 검출가능하지만 2가지 상황 둘다에서는 검출할 수 없는, 발현 패턴을 나타낼 수 있다. 다르게 말하면, 환자의 질환에 걸린 조직이나 세포에서, 환자의 정상 (질환에 걸리지 않은) 조직 또는 세포 및/또는 대조군 조직 또는 세포에서의 발현 수준에 비해, 더 높거나 낮은 수준으로 유전자의 발현이 이루어질 때, 그 유전자는 차별적으로 발현된 것이다.
용어 "생물학적 샘플"은 유기체 (예, 인간 환자) 또는 유기체의 구성 성분 (예, 세포)로부터 수득되는 샘플을 지칭한다. 샘플은 임의의 생물학적 조직 또는 유체일 수 있다. 샘플은 환자로부터 유래된 샘플인 "임상 샘플"일 수 있다. 이러한 샘플로는, 비제한적으로, 객담, 혈액, 혈액 세포 (예, 백색 세포), 양수, 혈장, 정액, 골수, 순환하는 종양 세포, 순환하는 DNA 순환하는 엑소좀 및 조직 또는 미세 바늘 생검 샘플, 뇨, 복막액 및 흉수 또는 이로부터 유래된 세포를 포함한다. 또한, 생물 샘플은 조직학적인 목적으로 회수한 냉동 단편 또는 포르말린으로 고정된 파라핀 포매된 단편 등의 조직 단편을 포함할 수 있다. 또한, 생물 샘플은 "환자 샘플"로도 언급될 수 있다.
본원에서, "건강 관리 서비스 제공자"는 개체이거나, 또는 예방적, 치유적, 홍보성 또는 재활 건강 관리 서비스를 환자와 같은 개체에게 제공하는 기관 또는 개체를 포함한다. 일 구현예에서, 환자의 진단/치료에 사용할 수 있도록, 건강 관리 서비스 제공자에게 데이타가 제공된다.
용어 "표준"은, 본원에서, 대조군 또는 건강한 개체 등의 비교에 사용되는 것을 지칭한다.
용어 "포함한다", "포함하는" 등은 미국 특허법에서 간주된 의미를 가질 수 있으며, "함유한다", "비롯하는" 등을 의미할 수 있다. 본원에서, "비롯하는" 또는 "함유한다" 등은 비제한적인 예로 포함하는 것을 의미한다.
암
본원에 기술된 방법을 이용하여, PR-유발성 증식과 항-에스트로겐 또는 아로마타제 저해제 치료에 대한 엔도크린-내성으로의 진행을 이행할 것으로 보이는 암을 가진 환자를 동정할 수 있다. 이런 환자는 항-프로게스틴을 포함하는 엔도크린 요법의 후보가 될 수 있다. 본원에 기술된 유전자 시그니처는 폐암, 뇌암, 전립선암, 자궁내막암, 수막종, 전립선암, 난소암 및 자궁 육종/암과 같은 다수의 암들에서 이용될 수 있다. 본원에 기술된 유전자 시그니처는 림프관평활근종증 (lymphangioleiomyomatosis) 및 자궁근종 (자궁 근종)을 비롯한 다른 장애들에서도 이용될 수 있다.
유방암
유방암은 여성들에서 가장 흔히 진단되는 암으로서, 암 사망 원인의 2번째를 차지하고 있다. 유방암 (악성 유방 종양)은 유방 조직으로부터 기원하며, 가장 흔하게는 유관 또는 도관에 모유를 공급하는 소엽의 내부 라이닝으로부터 기원한 타입의 암이다. 유관으로부터 기원한 암은 유관암 (ductal carcinomas)이라고 하며; 소엽으로부터 기원한 암은 소엽 암종이라고 한다. 유방암은 인간 및 기타 포유류 질환으로서, 인간에서 가장 많은 사례는 여성이지만, 남성에서도 때때로 유방암이 발생할 수 있다.
종양의 크기, 단계, 증식 속도 및 그외 특징들은 치료의 종류를 결정하게 된다. 치료는 수술, 약물 (호르몬 치료 및 화학요법), 방사선 및/또는 면역요법을 포함할 수 있다. 종양을 외과적으로 제거하는 것이 단독으로도 가장 우수하 효과를 제공하며, 수술 만으로도 다수 사례들에서 치유를 달성할 수 있다. 장기간 질병없이 생존할 가능성을 다소 높이기 위해, 수술과 더불어 몇가지 화학요법의 용법들이 통상적으로 제공된다. 대부분의 화학요법 형태들은 체내 도처에서 빠르게 분열하는 세포를 사멸시키는 것으로, 그 결과 일시적으로 탈모, 골수 및 면역계의 손상, 및 소화성 교란이 유발된다. 방사선은 특히 유방 보존 수술 후에 처방되며, 실질적으로 국소 재발율과 또한 다수 사례들에서 전체 생존성을 개선시킨다. 일부 유방암은 에스트로겐 및/또는 프르게스테론과 같은 호르몬에 민감하므로, 이들 호르몬의 효과를 차단함으로써 이를 치료하는 것이 가능하다.
전세계적으로, 유방암은 여성의 전체 암 (비-흑색종 피부암을 제외하고)의 22.9%를 차지한다. 2008년, 세계적으로 유방암으로 인한 사망 건 수는 458,503건이었다 (여성에서의 암으로 인한 사망 건수의 13.7%를 차지함). 예후와 생존율은 암 타입, 단계 및 치료에 따라 크게 달라진다.
유방암에 대한 최초 인지 증상은 전형적으로 유방 조직의 다른 부분과 다르게 느껴지는 덩어리이다. 처음 유방암은 유방 촬영술 (mammogram)로 검출한다. 겨드랑이에 위치한 림프절에서 발견되는 덩어리도 유방암임을 나타낼 수 있다.
덩어리 이외에 유방암임을 나타내는 다른 표시는 유방 크기와 형태의 변화, 피부 딤플링 (dimpling), 유두 함몰 (nipple inversion) 또는 자연적인 단일-유두 분비를 포함할 수 있다. 통증 ("유방통")은 일반적으로 유방암 존재 또는 부재를 확인하는 불확실한 도구이지만, 유방 건강상의 다른 문제를 표현하는 것일 수 있다.
유방암은 일반적으로 수술로, 가능하게는 화학요법 또는 방사선요법, 또는 전술한 모든 요법들로 치료한다. 종합적인 접근이 바람직하다. 호르몬 양성 암을 장기간의 호르몬 차단 요법으로 치료한다. 예후에 따라 그리고 재발 위험성에 따라서 점차적으로 공격적인 치료가 제공된다. 1기 암 (및 DCIS)은 예후가 우수하며, 일반적으로 유방보존술로, 때로는 방사선으로 치료한다. HER2 양성 암은 트라스투주맵 (trastuzumab) (Herceptin®) 용법으로 치료할 수 있다. 1기 암 이외의 다른 타입들에서는 화학요법은 드물다. 예후가 점점 좋지 않으며 재발 위험성이 더 높은 2기 암과 3기 암은 일반적으로 수술 (유방보존술 또는 림프절 적출을 수반하거나 수반하지 않은 유방절제술), 화학요법 (HER2 양성 암의 경우, +트라스투주맵) 및 때로는 방사선 (특히 거대 암, 다중 양성 림프절 또는 유방보존술 이후에)으로 치료한다. 4기 전이암 (즉, 원거리 부위로 전파됨)은 예후가 좋지 않으며, 수술, 방사선, 화학요법 및 타겟화된 요법으로 유래된 모든 치료들을 다양하게 조합하여 관리한다. 10년 생존율은 무치료의 경우 5%이고, 최적 치료의 경우에는 10%이다.
수술 이후에, 그리고 수술과 함께 약물을 사용하는 것을 보강 요법이라 한다. 수술 전의 화학요법 또는 다른 타입의 요법들은 신보강 요법이라 한다.
현재 보강적인 유방암 치료에 사용되는 주된 치료 군에는 3가지가 있다: 호르몬 차단 요법, 화학요법 및 단일클론 항체.
호르몬 차단 요법: 일부 유방암은 계속 생장하기 위해 에스트로겐을 필요로 한다. 이는 그 표면에 에스트로겐 수용체 (ER 양성) 및 프로게스테론 수용체 (PR 양성)의 존재에 의해 구분할 수 있다 (때로는 호르몬 수용체로도 언급됨). 이들 ER 양성 암은 수용체를 차단하는 약물, 예컨대 타목시펜 (Nolvadex®), 랄록시펜 (raloxifene), 오르멜록시펜 (ormeloxifene) 또는 토레미펜 (toremifene), 또는 다른 예로 아로마타제 저해제를 이용하여 에스트로겐의 생산을 차단하는 약물, 예컨대 아나스트로졸 (anastrozole) (Arimidex®), 엑세메스탄 (exemestane) 또는 레트로졸 (Letrozole) (Femara®)을 사용하여 치료할 수 있다. 부가적으로, Iressa® /게피티닙 (gefitinib), 및 라파티닙 (lapatinib)과 같은 EGFR 저해제도 존재한다.
항-프로게스틴 제제도 요법에 사용될 수 있다. 항-프로게스틴 (호르몬 길항제)은 프로게스테론 (생리 주기와 임신에 작용하는 호르몬)을 생산하거나 이용하는 세포를 방지하는 물질이다. 항-프로게스틴은 일부 암 세포가 증식 못하록 정지시킬 수 있다. 항-프로게스틴으로는, 비제한적으로, 오나프리스톤 (onapristone), 로나프리산 (lonaprisan), PF-02413873, 릴로프리스톤 (lilopristone), ORG2058, 미페프리스톤 (mifepristone) (RU486), 아소프리스닐 (asoprisnil), 텔라프리스톤 (telapristone), 울리프리스탈 (ulipristal), 아글레프리스톤 (aglepristone), ZM172406, ZM172405 및 ZM150271을 포함한다.
아글레프리스톤 (8S,11R,13S,14S,17R)-11-(4-다이메틸아미노페닐)-17-하이드록시-13-메틸-17-[(Z)-프로프-1-에닐]-1,2,6,7,8,11,12,14,15,16-데카하이드로사이클로펜타[a]펜안트렌-3-온

오나프리스톤 (onapristone), (예, (8S,11R,13R,14S,17S)-11-[4-(다이메틸아미노)페닐]-17-하이드록시-17-(3-하이드록시프로필)-13-메틸-1,2,6,7,8,11,12,14,15,16-데카하이드로사이클로펜타[a]펜안트렌-3-온)은 아래 화학식을 가진다:

그외 항-프로게스틴으로는 다음과 같은 화합물을 포함한다: 프로게스테론의 3-(6,6-에틸렌-17B-하이드록시-3-옥소-17A-프레그나-4-엔-17A-일)프로피온산 G-락톤, 3-(6,6-에틸렌-17.β.-하이드록시-3-옥소-17.α.-프레그나-4-엔-17.알ㅍ.-y- l)프로피온산.감마.-락톤 및 하기의 것:
미페프리스톤
(10S,11S,14S,15S,17R)-17-[4-(다이메틸아미노)페닐]-14-하이드록시-15-메틸-14-(프로프-1-yn-1-일)테트라사이클로[8.7.0.0^{2,7}.0^{11,15}]헵타데카-1,6-다이엔-5-온

릴로프리스톤
(11-β,17-β,17(z))-로페닐);에스트라-4,9-다이엔-3-온,11-(4-(다이메틸아미노)페닐)-17-하이드록시-17-(3-하이드록시-1-p;11β-[4-(다이메틸아미노)페닐]-17β-하이드록시-17-[(Z)-3-하이드록시-1-프로페닐]에스트라-4,9-다이엔-3-온

ORG2058
(8R,9S,10R,13S,14S,16R,17S)-16-에틸-17-(2-하이드록시아세틸)-13-메틸-2,6,7,8,9,10,11,12,14,15,16,17-도데카하이드로-1H-사이클로펜타[a]펜안트렌-3-온

로나프리산
(8S,11R,13S,14S,17S)-11-(4-아세틸페닐)-17-하이드록시-13-메틸-17-(1,1,2,2,2-펜타플루오로에틸)-1,2,6,7,8,11,12,14,15,16-데카하이드로사이클로펜타[a]펜안트렌-3-온

아소프리스닐
(8S,11R,13S,14S,17S)-11-[4-[(E)-하이드록시이미노메틸]페닐]-17-메톡시-17-(메톡시메틸)-13-메틸-1,2,6,7,8,11,12,14,15,16-데카하이드로사이클로펜타[a]펜안트렌-3-온

울리프리스탈
(8S,11R,13S,14S,17R)-17-아세틸-11-[4-(다이메틸아미노)페닐]-17-하이드록시-13-메틸-1,2,6,7,8,11,12,14,15,16-데카하이드로사이클로펜타[a]펜안트렌-3-온

PF-2413873
4-[3-사이클로프로필-1-(메실메틸)-5-메틸-1H-피라졸-4-일]옥시,-2,6-다이메틸벤조니트릴

텔라프리스톤
[(8S,11R,13S,14S,17R)-11-[4-(다이메틸아미노)페닐]-17-(2-메톡시아세틸)-13-메틸-3-옥소-1,2,6,7,8,11,12,14,15,16-데카하이드로사이클로펜타[a]펜안트렌-17-일] 아세테이트

추가적인 항-프로게스틴으로는 하기의 화합물을 포함한다:
| ZM172406 (R)-N-(3-클로로-4-시아노페닐)-3,3,3-트리플루오로-2-하이드록시-2-메틸프로판아미드 |
 |
| ZM172405 (S)-N-)3-클로로-4-시아노페닐)-3,3,3-트리플루오로-2-하이드록시-2-메틸프로판아미드 |
 |
| ZM150271 N-(3-클로로-4-시아노페닐)-3,3,3-트리플루오로-2-하이드록시-2-메틸프로판아미드 |
 |
본원에 개술된 예시적인 시스템 및 방법을 이용하여, 항-프로게스틴 (예컨대, 오나프리스톤, 로나프리산, 미페프리스톤, PF-02413873, 텔라프리스톤, 릴로프리스톤, ORG2058, 아포프리스닐, 울리프리스탈, ZM172406, ZM150271, ZM172405 및 아글레프리스톤)에 의한 생장 저해에 감수성인 악성 종양을 가지는 것으로 의심되는 환자를 동정하고 치료할 수 있다. 일 측면에서, 항-프로게스틴을 이용한 생장 저해에 감수성인 악성 종양 (암)을 가진 것으로 의심되는 환자는 항-프로게스틴으로 치료할 수 있다. 다른 측면에서, 항-프로게스틴을 이용한 치료에 감수성인 종양으로는, 비제한적인 예로, 유방, 뇌암, 수막종, 전립선암, 난소암, 자궁내막암, 자궁 육종, 자궁 근종, 폐암 및 자궁 조직 암을 포함한다. 다른 측면에서, 항-프로게스틴은 환자에게 1일 당 약 10 mg - 약 200 mg으로 투여할 수 있다. 선택적으로, 항-종양 화합물 (예, 에베롤리무스, 트라스투주맵, T-DM1, 항-HER2 약물, m-TOR 저해제, 항-VEGF 약물, 항-EGF 약물, 베박시주맵 (bevacizumab), 파클리탁셀, 독세탁셀 (docetaxel), 탁산, 독소루비신, 리포좀 독소루비신, 페길화된 리포좀 독소루비신, 안트라사이클린, 안트라센디온, 카보플라틴 (carboplatin), 시스플라틴 (cisplatin), 5-FU, 겜시타빈, 사이클로포스파미드, 아로마타제 저해제 (아나스트로졸, 레트로졸 (Letrozole), 엑세메스탄 (exemestane), 보로졸 (vorozole), 포르메스탄 (formestane) 및 파드로졸 (fadrozole)), 항-에스트로겐 (풀베스트란트 (Fulvestrant)), 선택적인-에스트로겐 수용체 모듈레이터 (랄록시펜 (raloxifene), 타목시펜, 토레미펜, 라소폭시펜 (lasofoxifene), 아피목시펜 (afimoxifene), 아르족시펜 (arzoxifene) 및 바제독시펜 (bazedoxifene)), 안드로겐 수용체 차단제 (엔잘루타미드 (enzalutamide)) 또는 17 α-하이드록실라제/C17,20 리아제의 저해제 (아비라테론 (abiraterone))도 항-프로게스틴을 이용한 치료 전과 치료 후에 환자에게 병행 투여할 수 있다.
화학요법: 2-4기 질환에 주로 사용되며, 특히 에스트로겐 수용체-음성 (ER 음성) 질환에서 효과적이다. 이는 조합 요법으로 통상 3-6개월간 제공된다. 가장 일반적인 치료제 중 한가지는 AC로 알려진 사이클로포스파미드 + 독소루비신 (Adriamycin®)이다. 대부분의 화학요법 약물들은, 복제시 또는 다른 기전으로 DNA 손상을 야기함으로써 빠르게-증식하는 및/또는 빠르게-복제하는 암 세포를 파괴함으로써, 작용하며; 이들 약물은 또한 빠르게-증식하는 정상 세포를 손상시켜, 심각한 부작용을 유발한다. 심장 근육의 손상은 독소루비신의 가장 위험한 합병증이다. 때때로, 독세탁셀과 같은 탁산 약물이 추가되는데, 이 경우 이 용법을 CAT라고 하며; 탁산 (예, 독세탁셀 및 파클리탁셀)은 암 세포에서 미세소관을 공격한다. 동일한 결과를 나타내는 다른 일반적인 치료제로는 사이클로포스파미드, 메토트렉세이트 및 플루오로우라실 (CMF)이 있다. 화학요법은 일반적으로 임의의 약물을 지칭하는 것일 수 있다.
단일클론 항체: HER2에 대한 단일클론 항체인 트라스투주맵 (Herceptin®)은 1-3기의 HER2-양성 유방암을 5년 무질환 생존율 약 87%로 개선시킨다 (총 생존율 95%). 그러나, 트라스투주맵은 심장 독성과 관련되어 있으며, 환자의 약 2%가 유의한 심장 손상을 앓게 된다. 또한, 다른 단일클론 항체들도 임상 실험 중에 있다. 트라스투주맵은 종양에서 HER2가 증폭된 환자에게만 효과적이다.
방사선치료는 일반적으로 수술로 적출되지 못한 것일 수 있는 미세 종양 세포를 파괴하기 위해, 종양 전부위 (tumor bed) 영역 및 국지적인 림프절에 대한 수술 이후에 제공된다. 또한, 종양 미세환경에 효과적일 수 있다. 방사선 치료는 외부 빔 조사치료 (external beam radiotherapy) 또는 근접조사치료 (internal radiotherapy)으로서 전달될 수 있다. 전통적인 방사선요법은 유방암을 수술한 이후에 제공된다. 또한, 방사선은 유방암 수술 시에 - 수술 중에 - 제공될 수 있다. 방사선은 재발 위험성을 50-66%까지 낮출 수 있다 (위험성을 1/2 - 2/3 낮춤).
개시 및 진행을 초래하는 분자 인자들이 완전히 파악된 것은 아니다. 여성의 보건 발의 (WHI: Women's Health Initiative)에 의한 무작위 임상 실험을 통해, 에스트로겐 및 프로게스틴을 포함하는 (에스트로겐 단독이 아님) 호르몬 대체 요법 (HRT)이 폐경기 여성에서 침습적인 유방암의 발생 위험성을 현저하게 높인다는 것이 확인되었다 (Chlebowski et al., 2003; LaCroix et al., 2011). 유사한 결론은 여성 백만명을 대상으로 한 관찰 실험에서도 나타났다 (Million Women Study Collaborators, 2003). 이러한 결과는, HRT 처방의 급격한 감소로 이어졌으며, 그 결과 유방암 발생이 현저하게 감소하였다 (Chlebowski et al., 2009). WHI 데이타에 대한 추가적인 분석으로, 에스트로겐만을 포함하는 HRT를 처방받은 여성은 침습적인 유방암이 발생할 위험성이 낮은 것으로 입증되었다 (Anderson et al., 2012; LaCroix et al., 2011). PR 발현은 전통적으로 에스트로겐 수용체 (ER) 기능의 임상 지표로서 사용된다 (즉, PR은 ER 타겟 유전자임). 그러나, 논란은 많지만, 이러한 놀라운 병인적인 증거는 유방암 개시 및 조기 진행의 매개인자로서 프로게스테론 수용체 (PR)의 고유한 작용에 대한 추가적인 연구가 필요하다는 강력한 근거가 된다 (Lange, 2008의 리뷰).
전통적으로, PR은 효율적인 전사에 필요한 공동-조절성 분자들을 동원할 수 있는 다이머로서, 타겟 유전자 프로모터 또는 인핸서에 결합하는 리간드-활성화된 전사 인자로서 정의된다. 최근 들어, 단백질 키나제가 스테로이드 호르몬에 의해 (펩타이드 성장인자에 반응하여) 신속하여 활성화된다는 것이 널리 인지되게 되었다. 실제, 인산화 현상은 PR 작용에 대한 조절성 인풋 (input)을 제공한다 (Daniel et al., 2009에서 리뷰되었으며, 하기에 논의됨). PR에서의 몇가지 돌연변이가 암 위험성과 연계되어 있으며; 이는 PR 전사 활성에 영향을 주기 보다는 주로 PR의 발현 수준을 변형시키는 것으로 보인다 (De Vivo et al., 2002; Pooley et al., 2006; Terry et al., 2005). 2종의 PR 단백질 이소형, 즉 PR-A 및 PR-B는 유방암 조직에서 공동-발현된다. PR-B는 PR-A에 없는 N-말단 (B-상류 단편 또는 BUS로 칭해짐)에 아미노산 164개를 포함하는 전장 수용체이다. 이들 2종의 이소형 모두 번역 후 상당히 변형된다 (인산화, 유비퀴틴화, 아세틸화). PR의 N-말단은 주요 조절성 인산화 부위 (Ser294) 뿐만 아니라 본원에서 연구된 SUMO일화 부위 (Lys388)를 포함하고 있다. PR-A (예, 등재번호 NM_001202474.1 (GI:321117149)로서 NCBI 데이타베이스 참조)는 그렇지 않지만, PR-B (예, 등재번호 NM_000926.4 (GI:160358783)로서 NCBI 데이타베이스 참조)는 세포 배양시 및 생체내에서 Ser294에서 인산화된다 (Clemm et al., 2000). 리간드 결합시, PR 이소형 둘다 빠르게 (15분) Lys388에서 SUMO일화된다 (Daniel et al., 2007a). SUMO일화는, 작은 유비퀴틴-유사 변형제 (SUMO) 펩타이드 (~11.5 kD)가, 유비퀴틴화와 유사하게, ATP-의존성 효소 (3단계) 기전을 통해 주로 컨센서스 SUMO일화 모티프 (IKxE)에서 기질 분자의 라이신 잔기와 공유 결합함으로써 이루어진다 (Melchior, 2000). 기질의 SUMO일화는 종종 단백질-단백질 상호작용, 세포내 위치, 단백질 안정성 (즉, 유비퀴틴화에 반할 수 있음) 및/또는 효소 또는 전사 활성을 바꾼다 (Geiss-Friedlander and Melchior, 2007).
최근, Daniel 등은, 활성화된 미토겐 활성화된 단백질 키나제 (MAPK) 또는 세포 주기-의존성 단백질 키나제-2 (CDK2)에 반응하여, Ser294에서의 PR-B 인산화가, Lys388에서의 빠른 프로게스틴-유발성 SUMO일화를 방지한다는 것을 발견하였다 (Daniel et al., 2007a; Daniel and Lange, 2009). 아울러, Ser294 인산화-유발성 PR SUMO일화의 길항화가, 선택된 유방암-관련 유전자 프로모터, 즉 HBEGF (Daniel et al., 2007a), STC1 및 IRS1 (Daniel and Lange, 2009)를 탈억제(활성화)화함으로써; 유방암-관련 활성화 인자 STC1 및 IRS의 포스포-PR-의존적인 상향 조절이 프로게스틴 부재시에 발생하였다 (Daniel and Lange, 2009). 프로모터 구조 (즉, 호르몬 반응 인자들의 수)는 SUMO일화된 글루코코르티코이드 수용체 (GR)에 의해 리포터-유전자 프로모터를 인지하는 결정인자이지만 (Iniguez-Lluhi and Pearce, 2000), 스테로이드 수용체의 SUMO일화가 내인성 유전자의 조절을 (즉, 염색질에서) 변형시키는 방법에 대해서는 훨씬 덜 알려져 있다. 지금까지, 오직 수개의 내인성 유전자들만 PR SUMO일화에 민감한 것으로 알려져 있었다 (Daniel et al., 2007a; Daniel and Lange, 2009). 본원에서는, PR이 인간 유방암에서 흔히 상승되는 활성 미토겐성 단백질 키나제 (즉, MAPK 및 CDK2)에 대한 센서로서 작용하며; 상승된 Ser294 인산화의 영향으로 인해, 대신 PR SUMO일화에 민감한 (즉, 이에 의해 정상적으로 억제되는) 유전자가 유방암 세포의 증식과 생존 촉진 신호전달을 활성화하도록 협력할 수 있음을 개시한다. 포스포-PR (SUMO-결함성) 유전자 시그니처는 선택적인 항-프로게스틴을 포함하는 엔도크린 요법에 반응할 것으로 보이는 인간 유방암 환자 서브세트를 식별할 수 있다.
본원에서, 전사 이후의 다이나믹한 현상들과 관련된 PR 프로모터의 민감성 기전 (즉, PR Ser294 인산화가 Lys388의 탈SUMO일화와 커플링되어 있음)을 밝힌다. 전체 게놈 발현 분석을 이용하여, 야생형 (WT) 및 SUMO-결함성 (K388R) PR-B에 의해 차별적으로 조절되는 유전자를 동정하였으며, 변형된 PR 프로모터 선택성에 대한 기전을 조사하였다. 이러한 발견 사실은, SUMO-결함성 포스포-PR-B가 유방암 세포의 증식과 생존 촉진에 중요한 유전자의 선택적인 조절에 관련되어 있으며, 인산화 및 탈SUMO일화된 PR이 빠른 (내강) 유방암 종양 진행과 관련된 ERBB2-양성 표현형의 중요한 활성화 인자일 수 있음을 시사한다.
유전자 발현 프로파일 (마커) 및 유전자 발현 프로파일들의 측정
특정 유전자의 발현이 항-프로게스틴을 이용한 유방암 치료의 예측인자임을 본원에서 입증하였다. 이러한 유전자로는 하기 유전자 (또는 이의 상동체)를 포함한다:

"상동성"은, 본원에서, 2개의 폴리머 분자, 예컨대 2개의 핵산 분자, 예로, 2개의 DNA 분자 또는 2개의 RNA 분자, 또는 2개의 폴리펩타이드 분자 간의 서브유닛 서열의 유사성을 의미한다. 2개의 분자 양쪽에 서브유닛 위치가 동일한 모노머 서브유닛이 존재하는 경우, 예를 들어, 2개의 DNA 분자 각각에서의 위치에 아데닌이 있는 경우, 이들은 그 위치에서 상동적이다. 2개의 서열 간의 상동성은 일치되거나 상동적인 위치의 갯수에 대한 직접적인 함수이며, 예를 들어, 만일 2개의 컴파운드 화합물에서 위치들 중 절반 (예, 10개의 서브유닛으로 이루어진 폴리머에서 5곳)이 상동적이면, 이들 2 서열간의 상동성은 50%이며, 만일 위치들 중 90%, 예컨대 10곳 중 9곳이 일치하거나 상동적이면, 2개의 서열은 90%의 상동성을 공유한다. 예를 들어, DNA 서열 3'ATTGCC5'와 3'TATGGC는 50%의 상동성을 공유한다.
본원에서, "상동성"은 "동일성"과 동의어로 사용된다.
2개의 뉴클레오티드 서열 또는 아미노산 서열 간에 동일성 %를 결정하는 것은 수학적인 알고리즘을 이용하여 결정할 수 있다. 예를 들어, 2개의 서열을 비교하는데 사용가능한 수학적인 알고리즘으로는 Karlin and Altschul (1990)의 알고리즘이 있으며, Karlin and Altschul (1993)에서 수정되었다. 이 알고리즘은 Altschul 등의 NBLAST 및 XBLAST 프로그램에 통합되어 있으며, 예를 들어 국립 생물공합 정보 센터 (NCBI)의 월드 와이드 웹 사이트에서 접근할 수 있다. BLAST 뉴클레오티드 검색은 본원에 기술된 핵산과 상동적인 뉴클레오티드를 수득하기 위해 다음과 같은 파라미터를 이용하여 NBLAST 프로그램 (NCBI 웹사이트에서는 "blastn"으로 지칭됨)으로 수행할 수 있다: 갭 패널티 = 5; 갭 연장 패널티 = 2; 미스매치 패널티 = 3; 매치 보상 (match reward) = 1; 기대 값 (expectation value) 10.0; 및 글자 크기 = 11. BLAST 단백질 검색은, 본원에 기술된 단백질 분자에 상동적인 아미노산 서열을 수득하기 위해, 다음과 같은 파라미터를 이용하여 XBLAST 프로그램 (NCBI 웹사이트에서는 "blastn"로 지칭됨) 또는 NCBI "blastp" 프로그램으로 수행할 수 있다: 기대 값 10.0, BLOSUM62 스코링 매트릭스. 비교 목적으로 갭이 많은 정렬을 구하기 위해서는, Gapped BLAST를 Altschul 등에 기술된 바와 같이 사용할 수 있다. 다른 예로, PSI-Blast 또는 PHI-Blast를 사용하여, 공통 패턴을 공유한 분자들 간의 관련성과 분자들 간의 원위 관련성을 검출하는 반복 검색 (iterated search)을 수행할 수 있다. BLAST, Gapped BLAST, PSI-Blast 및 PHI-Blast 프로그램을 이용하는 경우, 각 프로그램 (예컨대, XBLAST 및 NBLAST)의 디폴트 파라미터를 사용할 수 있다.
2개의 서열 간의 동일성%는 전술한 바와 비슷한 기법으로 결정할 수 있으며, 갭이 허용되거나 허용되지 않을 수 있다. 동일성 % 계산시, 전형적으로 정확하게 일치되는 것을 계수한다.
본원에서, "실질적으로 상동적인 아미노산 서열" 또는 "실질적으로 동일한 아미노산 서열"은 기준 항체 체인의 아미노산 서열에 대해 약 92% 이상 또는 약 95% 이상의 상동성 또는 동일성, 예컨대, 약 96% 이상의 상동성 또는 동일성, 예컨대 약 97% 이상의 상동성 또는 동일성, 예컨대 약 98% 이상의 상동성 또는 동일성, 및 약 99% 이상의 상동성 또는 동일성을 가지는 아미노산 서열을 포함한다. 아미노산 서열의 유사성 또는 동일성은 BLAST (기본적인 국소 정렬 검색 툴) 2.0.14 알고리즘을 채택한 BLASTP 및 TBLASTN 프로그램을 이용함으로써 계산할 수 있다. 이들 프로그램에 사용된 디폴트 세팅이 본 발명의 목적에 따라 실질적으로 유사한 아미노산 서열을 동정하는데 적합하다.
본원에서, 용어 "보존적인 아미노산 치환"은 본원에서 하기 5개의 그룹들 중 하나의 그룹 내에서의 아미노산 치환으로 정의된다:
I. 소형 지방족, 무극성 또는 약간 극성의 잔기:
Ala, Ser, Thr, Pro, Gly;
II. 극성, 음으로 하전된 잔기 및 이들의 아미드:
Asp, Asn, Glu, Gln;
III. 극성, 양으로 하전된 잔기:
His, Arg, Lys;
IV. 크고, 지방족, 무극성 잔기:
Met Leu, Ile, Val, Cys
V. 크고, 방향족 잔기:
Phe, Tyr, Trp
"실질적으로 상동적인 핵산 서열" 또는 "실질적으로 동일한 핵산 서열"은, 대응 서열이 기준 핵산 서열에 의해 코딩되는 펩타이드와 실질적으로 동일한 구조와 기능을 가진 펩타이드를 코딩하도록, 기준 핵산 서열에 핵산 서열이 대응되는 것을 의미하며; 예를 들어, 아미노산에서 하나의 변화가 펩타이드 기능에 유의하게 작용하지 않는 경우를 의미한다. 일 구현예에서, 실질적으로 동일한 핵산 서열은 기준 핵산 서열에 의해 코딩되는 펩타이드를 코딩한다. 실질적으로 유사한 핵산 서열과 기준 핵산 서열 간의 동일성 %는 적어도 약 50%, 65%, 75%, 85%, 92%, 95%, 99% 또는 그 이상이다. 핵산 서열들의 실질적인 동일성은 2개의 서열들의 서열 동일성을, 예를 들어 물리적/화학적 방법 (즉, 혼성화) 또는 컴퓨터 알고리즘을 통한 서열 정렬을 통해 비교함으로써 결정할 수 있다.
뉴클레오티드 서열이 기준 뉴클레오티드 서열과 실질적으로 유사한 지를 확인하는데 적합한 핵산 혼성화 조건은 7% 소듐 도데실 설페이트 SDS, 0.5 M NaPO4, 1 mM EDTA 및 50℃이며, 세정은 2X 표준 식염수 사이트레이트 (SSC), 0.1% SDS 및 50℃이며; 바람직하게는 7% (SDS), 0.5 M NaPO4, 1 mM EDTA 및 50℃에서 혼성화하고, 1X SSC, 0.1% SDS 및 50℃에서 세정하는 것이며; 바람직하게는 7% SDS, 0.5 M NaPO4, 1 mM EDTA 및 50℃에서 혼성화하고, 0.5 x SSC, 0.1% SDS 및 50℃에서 세정하는 것이며; 더 바람직하게는 7% SDS, 0.5 M NaPO4, 1 mM EDTA 50℃에서 혼성화하고, 0.1X SSC, 0.1% SDS 및 65℃에서 세정하는 것이다. 2개의 핵산 간의 실질적인 유사성을 정하는데 적합한 컴퓨터 알고리즘으로는 GCS 프로그램 패키지를 포함한다. 이들 프로그램에 제공되는 디폴트 세팅은 본 발명의 목적에 따라 핵산 서열들의 실질적인 유사성을 결정하는데 적합하다.
발현 수준 측정
일 구현예에서, 대상 유전자의 mRNA 등의 핵산 발현을 측정한다. 사전 선택된 mRNA의 발현 수준은, 예를 들어, 인 시츄 혼성화, 노던 블롯, 핵산 증폭 기법 (예, PCR, 정량적인 PCR, 리가제 연쇄 반응 등), RNA Seq 및 마이크로어레이 분석 등의 다양한 임의의 기법에 의해 동정 및/또는 정량할 수 있다. mRNA 수준은 노던 블롯팅을 통해 정량적으로 측정할 수 있다. RNA 샘플을 아가로스 겔 상에서 분리시키고, 타겟 서열에 상보적인 방사선 표지된 RNA 프로브로 혼성화한다. 방사선 표지된 RNA를 그런 후 자동방사선촬영으로 검출한다.
mRNA 농도를 측정하는 다른 방법은 정량적인 역 전사중합효소 연쇄 반응이다. RT-PCR은 먼저 mRNA로부터 역전사에 의해 DNA 주형을 만들며, 이를 cDNA라고 한다. 이 cDNA 주형을 qPCR에 사용하며, 이때 DNA 증폭 공정이 진행됨에 따라 프로브의 형광에 변화가 발생한다. 표준 곡선을 이용하여 qPCR에서 전형적으로 균질화된 조직 나노리터 당 카피수 또는 세포당 카피수로서 mRNA의 카피수 등의 절대 측정값을 수득할 수 있다. qPCR은 민감도가 매우 우수하다 (mRNA 한 분자도 검출 가능함).
다른 방식으로, mRNA 한 분자를 형광 바코드 (나노스트링)로 각각 테그를 붙이는 것으로, 이로써 하나씩 검출할 수 있으며, 직접 디지털 정량화로 계수가능하다 (Krassen Dimitrov, NanoString Technologies). 또한, 여러가지 mRNA의 세포 농도에 대한 상대적인 척도를 제공할 수 있는 유전자 발현의 연속 분석 (SAGE)과 같은 "테그 기반의" 기법을 이용할 수 있다.
또한, DNA 마이크로어레이를 이용하여 다수 유전자의 전사체 수준을 한번에 확인할 수 있다 (발현 프로파일링). 최근 마이크로어레이 기술의 진보로 하나의 어레이에서 인간을 비롯한 여러 유기체의 게놈에서 모두 공지된 유전자에 의해 전사체 수준을 정량할 수 있다.
컴퓨터/프로세서
검출, 예후 예측 및/또는 진단 방법은 프로세스/컴퓨터 시스템의 이용을 채택할 수 있다. 예를 들어, 방법을 구현하기 위한 컴퓨터 프로그램 코드를 저장하는 프로그램 메모리, 작동 메모리 및 통상적인 컴퓨터 스크린, 키보드, 마우스 및 프린트와 같은 인터페이스, 뿐만 아니라 네트워크 인터페이스 및 데이타베이스 인터페이스 등의 소프트웨어 인터페이스와 커플링된 프로세서를 포함하는 일반적인 목적의 컴퓨터 시스템이 본원에 기술된 일 구현예에 사용된다.
컴퓨터 시스템은 키보드와 같은 데이타 입력 장치에서부터 사용자의 입력, 데이타 파일 입력 또는 네트워크 인터페이스, 또는 예를 들어 마이크로어레이 또는 PCR 데이타를 해석하는 시스템과 같은 다른 시스템을 허용하며, 프린터, 디스플레이, 네트워크 인터페이스 또는 데이타 저장 장치 등의 출력 장치로의 출력을 제공한다. 입력 장치, 예컨대 네트워크 인터페이스는 본원에 기술된 단백질/핵산의 검출 및/또는 이들 화합물들의 정량을 포함하는 입력값을 수신한다. 출력 장치는 디스플레이와 같은 출력을 제공하며, 이러한 것으로는 하나 이상의 숫자 및/또는 화합물의 검출 및/또는 정량화를 도시한 그래프를 포함한다.
컴퓨터 시스템은 본원에 기술된 방법에 의해 수득된 데이타를 저장하는 데이타 저장체와 커플링된다. 이 데이타는 각 측정 및/또는 각 개체를 저장하며; 선택적으로 이들 데이타 타입들 각각으로 구성된 복수의 세트들이 각 개체에 대해 저장된다. 하나 이상의 컴퓨터/프로세서가 예를 들어 분리된 장치로서, 예를 들어 네트워크 상의 컴퓨터 시스템에 연결된 분리된 장치로서 사용되거나, 또는 컴퓨트 시스템 상에서 구동되는 별개의 또는 통합된 프로그램을 포함할 수 있다. 어느 방법을 적용하던 간에, 이들 시스템은 데이타를 수신하며, 검출/진단에 대한 데이타를 다시 제시한다.
실시예
아래 실시예들은 본 발명의 특정 구현예들과 측면들을 기술하고 더욱 예시하기 위한 목적으로 제시되며, 본 발명의 범위를 한정하는 것으로 해석되지 않는다.
실시예 I
인산화된 SUMO-결함성 프로게스테론 수용체는 유방암이 진행되는 동안 증식성 유전자 시그니처를 유발한다
재료 및 방법
인간 유방 종양 샘플에서의 프로게스테론 수용체 발현
단백질 및 mRNA 분석을 위해 미국 미네소타 조직 입수 기관의 생물 물질 입수 네트워크 (BioNet)로부터 식별하지 않은 인간 유방 종양 샘플을 입수하였다. 냉동 조직 샘플들은 유관 암종, 침윤성 유관 암종, 소염 압종 또는 전이 암종으로 진단받은 환자로부터 유래된 것이다. 시료는 미네소타 대학의 임상 병리과에서 분석하였으며, 표준 임상 조직학적 방법을 이용하여 에스트로겐 수용체 (ER) 및 프로게스테론 수용체 (PR) 발현에 대해 스코어를 매겼다. 종양 샘플을 표준 방법들 (동결 조직의 분쇄, RIPA 완충제, 트리-시약)을 이용하여 단백질 또는 mRNA용으로 각각 회수하고, 총 PR, 포스포-Ser294 PR, 및 ERK1/2 단백질 발현 수준을 (후술하는) 웨스턴 블롯팅에 의해 측정하였다. 모든 시료들은 연구 목적으로 조직 샘플을 사용한다는 사전 고지를 제공받은 환자로부터 입수하였으며, 미네소타 대학의 기관감사위원회 (IRB)로부터 승인받았다.
세포 배양, 발현 벡터 및 웨스턴 블롯팅
T47Dco 모 세포주는 기존에 특징이 규명되었다 (Horwitz et al., 1982). PR을 안정적으로 발현하는 T47D 세포는, WT, K388R, S294A 또는 K388R/S294A PR을 코딩하는 cDNA를 pIRES-neo3 발현 벡터 (Clontech, catalog #631621)에 분자 클로닝한 다음, 이 벡터를 FuGENE HD (Roche, catalog #04709713001)를 이용하여 T47D-Y에 형질전환함으로써, 구축하였다 (Sartorius et al., 1994). 단세포 클론을 고 농도 G418 선별 (500 ug/ml) 하에 증식시키고, 저 농도 G418 선별 (200 ug/ml) 하에 유지시켰다 (EMD Chemicals, catalog #345810). 이들 세포는 5% 소 태아 혈청 (FBS), 1% 비-필수 아미노산 (NEAA), 1% 페니실린/스트렙토마이신, 6 ng/ml 인슐린 (CellGro, catalog #10-010-CV)이 첨가된 완전 최소 필수 배지 (cMEM)에서 유지시켰다. 유발성 PR을 발현하는 T47D 세포는 기존에 개시된 바 있다 (Hagan et al., 2011). 유발성 PR 발현은, 최소 처리 기간 2일 동안 세포 배양 배지에 AP21967 (10-9 M, Ariad Pharmaceuticals, Cambridge, MA)을 첨가함으로써 달성하였다. PR을 발현하는 MCF-7 세포주는, WT 또는 KR PR을 코딩하는 cDNA 삽입체를 포함하는 pIRES-neo3를 FuGENE HD를 이용하여 세포에 형질감염시킴으로써, 구축하였다. 단일 세포 클론을 고 농도 G418 선별 하에 증식시키고, 저 농도 G418 선별 하에 유지시켰다. MCF-7 세포는 5% FBS, 1% 페니실린/스트렙토마이신이 보충된 DMEM (Dulbecco's modification of Eagle's medium, CellGro, catalog #10-013-CV)에서 유지시켰다. BT-474 세포 (ATCC, Manassas, VA)는 10% FBS, 1% 페니실린/스트렙토마이신이 첨가된 RPMI 1640 배지 (Gibco, catalog #11875)에서 유지시켰다. SDS-PAGE를 8% 겔에서 수행하고, 기존에 기술된 바와 같이 웨스턴 블롯팅 분석을 수행하였다 (Daniel et al., 2007a). 항체 정보는 도 5.1을 참조한다.
유전자 발현 프로파일링
pIRES-neo3 빈 벡터, WT 또는 KR PR을 안정적으로 발현하는 T47D 세포를 하루 동안 변형된 IMEM (Gibco, catalog #A10488)에서 혈청 고갈시키고, 6시간 동안 R5020 (10-8 M) 또는 비히클 대조군을 처리한 다음, RNeasy 키트 (QIAgen, catalog #74104)를 이용하여 RNA를 추출하였다. 6시간 동안의 프로게스틴 처리로, 기존 연구와 비교하여 실질적인 PR-의존적인 유전자 발현이 가능하였다 (Jacobsen et al., 2005; Richer et al., 2002). 두플리케이트 실험에서 DNase I 처리한 (QIAgen, catalog #79254) RNA 샘플들은, 제조사의 프로토콜에 따라 Illumina HT-12v4 비드 칩 플랫폼을 이용하여 발현 분석을 위해 준비하였다. lumi로 지칭되는 Bioconductor (Gentleman et al., 2004) 패키지를 이용하여 R 소프트웨어로 데이타를 분석하였으며, 이때 세기에 대한 원 데이타를 log2로 변환하고, 분위 표준화하였다. 차별적으로 발현된 유전자들을 limma 패키지를 이용하여 분석하였으며, 이때 경험적 베이즈를 이용하여 유전자들의 분산을 예측하였다. 제시된 유전자 발현 데이타는 log2 표준화된 세기를 포함하며, 제시된 생물 비교 (예, R5020/비히클)는 Benjamini and Hochberg (BH) 조정된 P 값과 log2 배수 변화를 포함한다 (Benjamini and Hochberg, 1995). 도 1C에서 히트 맵을 작성하기 위해, 유전자의 자율적인 계층 클러스터링을 R 패키지 g 플롯에서 heatmap.2 기능을 이용하여 수행하였다. 클러스터링은 유클리드 거리 및 완전 연관 (Euclidean distance and complete linkage)을 이용하여 수행하였다. 열은, 평균 0 및 1과 같은 표준 편차를 가지는 것으로 변환하였다.
유발성 PR을 발현하는 T47D 세포에서의 유전자 발현 프로파일을 Affymetrix 마이크로어레이 플렛폼을 이용하여 측정하였다. PR 발현은 2일간 AP21967 (10-9 M)로 유발하였으며, 세포는 1일간 변형된 IMEM에서 혈청 고갈시킨 후, 6시간 동안 R5020 (10-8 M) 또는 비히클 대조군을 처리한 다음 RNeasy 키트를 이용하여 RNA를 추출하였다. DNase I을 처리한 샘플을 제조사의 프로토콜에 따라 Affymetrix U133A 2.0 마이크로어레이를 이용한 발현 분석을 위해 준비하였다. Raw Affymetrix CEL 파일들을 처리하고, Bioconductor (Gentleman et al., 2004) 패키지, affy 및 affyQCReport를 이용하여 R에서 표준화하였다. 데이타를 affy 패키지에 포함된 Robust Multi-array Average (Irizarry et al., 2003) 알고리즘을 이용하여 표준화하였다. MAS 5.0 알고리즘 (또한 affy 패키지에 포함됨)의 일부로서 윌콘슨이 지정한 순위 검사를 사용하여, 모든 프로브 세트들에 대한 존재/부재 판정을 실시하였다 (Liu, 2004). 선택된 조건 쌍들에 대한 표준화된 발현 수준을 log2 비율로서 계산하였다. 모든 유전자 발현 데이타는 NCBI 유전자 발현 옴니부스 (GEO) 데이타베이스 (등재 번호: GSE34149, http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE34149)에서 이용가능하다.
항-프로게스틴으로 처리된 T47D 세포주에서의 유전자 발현 프로파일을, 이들의 두드러진 차이를 제외하고는, 전술한 동일한 방법을 이용하여 입수하였다. 혈청 고갈 후, 빈 벡터, WT 또는 KR PR을 안정적으로 발현하는 세포를 6시간 동안 8가지의 잠재적인 조건들 중 한가지 조건 하에 처리하였다: (1) 에탄올 (비히클 대조군), (2) 프로게스테론 (10-8 M), (3) RU486 (10-7 M), (4) 아글레프리스톤 (10-7 M), (5) 오나프리스톤 (10-7 M), (6) RU486 (10-7 M) + 프로게스테론 (10-8 M), (7) 아글레프리스톤 (10-7 M) + 프로게스테론 (10-8 M), 또는 (8) 오나프리스톤 (10-7 M) + 프로게스테론 (10-8 M). 유전자 발현 수준을 측정하고, 표준화하고, 분석한 다음, 전술한 방법을 이용하여 히트 맵을 구축하였다.
활성화된 PR에 의해 유발된 종양에서 유전자 마커의 동정
2번의 독립적인 마이크로어레이 실험 (둘다 전술한 Illumina HT-12v4 플랫폼을 이용하여 수행됨)으로부터 수득한 마이크로어레이 원 데이타를 조합하고, lumi로 지칭되는 Bioconductor (Gentleman et al., 2004) 패키지를 이용하여 R 소프트웨어에서 표준화하였으며, 이때 원 데이타의 세기를 log2 변환하고, 분위 표준화 (quantile normalized)하였다.
샘플 크기 & 조성 : 1차 분석을 위해, 일차 실험에서 합성 프로게스틴인 R5020으로 자극한 2개의 KR 세포 리플리케이트를 사용하였으며, 2차 실험에서는 프로게스테론으로 자극한 3개의 KR 세포 리플리케이트를 사용하였다. 대응되는 WT 세포 리플리케이트에 대해서는 동일하게 수행하였다. 1차 분석과 관련하여, 2 그룹들 (KR & WT) 각각에 대해 n=5의 샘플 크기를 사용하였다. 2차 분석과 관련하여, 제2 실험에서 프로게스테론 + 오나프리스톤을 공동-처리한 KR 세포 리플리케이트 3개를 사용하였으며, 또한 제2 실험에서 프로게스테론 + 오나프리스톤을 공동-처리한 WT 세포 리플리케이트 3개를 사용하였다 (2개의 그룹 각각에 대해 n=3). 3차 분석과 관련하여, 제2 실험에서 프로게스테론 + RU486를 공동-처리한 KR 세포 리플리케이트 3개를 사용하였으며; 제2 실험에서도 역시 프로게스테론 + RU486로 공동-처리한 WT 세포 리플리케이트 3종을 사용하였다 (2개의 군 각각 n=3). 4차 분석과 관련하여, 프로게스테론 + 오나프리스톤을 공동-처리한 KR 세포 리플리케이트 3개를 사용하였으며; 비히클 대조군 (에탄올)을 처리한 WT 세포 리플리케이트 3종을 사용하였다 (2개의 군 각각 n=3). 5차 분석과 관련하여, 프로게스테론 + 오나프리스톤을 공동-처리한 WT 세포 리플리케이트 3개를 사용하였으며; 비히클 대조군 (에탄올)을 처리한 WT 세포 리플리케이트 3종을 사용하였다 (2개의 군 각각 n=3).
대조군 유전자: 가공한 (표준화된) 데이타를 정성 분석하기 위해, 하기 리스트의 대조군 유전자들을 사용하였다: TBP, GAPDH, ACTB, TRAP1, PPIB, FPGS, EEF1A1, UBC, TXN, B2M, HMBS 및 FARP1. 마이크로어레이 기술에서 널리 공지된 문제들을 감안하기 위해, 전술한 대조군 유전자를 타겟팅하는 이 칩 (Illumina HT-12v4)의 프로브들을 모두 동정하여, 이용하였다. 아래 괄호 안 숫자는, 특정 대조군 유전자를 타겟팅하는 이 칩 상의 상이한 프로브 갯수를 나타낸다: TBP (1), GAPDH (3), ACTB (3), TRAP1 (1), PPIB (1), FPGS (3), EEF1A1 (4), UBC (3), TXN (2), B2M (2), HMBS (3), 및 FARP1 (4).
통계학적 방법: 통계적 유의성을 분석하기 위해, 아래 3종의 다른 독립적인 방법들을 사용하였디.
1) P-값.
독립적인 t-검정은 모수적 유전자 변수 (parametric gene variable)들에 대해 사용하였으며 (분산 상태들의 정상성 (normality) 및 균등성 (equality) 둘다 충족되었음); 정상 상태는 총족하지만 분산 상태의 균등성은 충족시키지 않는 유전자 변수들에 대한 Aspin-Welch 비균등-분산 검정 (AW); 및 i) 정상 상태를 충족하지 않거나 또는 ii) 분산 상태의 정상성 및 균등성을 충족하지 않는, 비-모수적 유전자 변수 (non-parametric gene variable), 즉, 이들 변수에 대한 Mann-Whitney U 검정 (MW). Illumina HT-12v4 칩에 47,231개의 프로브 세트가 존재한다는 것을 감안하여, 본페로니 보정을 이용하면, 전체 실험에서 유의성 수준은 α = 1.06 x 10-6로 설정되었다. 따라서, 임의 변수에 대해 P-값 방법에 따라 유의성을 구하기 위해서는, 아래 상태를 충족하여야 한다: P < α. Mann-Whitney U 검정 (MW)과 관련하여, 비-모수적 변수가 독립적이라면 (하나의 그룹의 개체가 다른 그룹의 개체와 동일한 발현 값을 가진다면), 정확 확률 (exact probability)을 이용하며; 그렇지 않으면, 보정이 적용된 대략적인 확률 (approximated probability)을 이용하였다.
2) 배수 변화 (FC).
모든 유전자 변수들의 경우, 배수 변화 (FC)는 전자가 후자 보다 큰 경우 (과다 발현) WT 그룹의 평균 발현 값을 상회하는 KR 그룹의 평균 발현 값으로서 정의되며, 통계적 유의성은 FC ≥ 1.10에서 설정되었으며, 이는 log2 스케일에서 변화가 ≥ 10%인 것을 의미한다. KR 그룹의 평균 발현 값이 WT 그룹의 평균 발현 값 보다 낮은 경우 (과소 발현), FC는 KR 그룹의 평균 발현 값을 상회하는 WT 그룹의 평균 발현 값의 네거티브 비율로서 정의되며, 통계적 유의성은 FC ≤ -1.10에서 설정되었으며, 이는 log2 스케일애서 변화가 ≥ 10%인 것을 의미한다. 이러한 방법에 따라, 유전자 변수는 통계학적인 유의성을 구하기 위해 |FC| ≥ 1.10이어야 한다.
3) ROC 곡선 분석.
2가지 그룹에 대한 식별력을 분석하기 위해 ROC 곡선 분석을 모든 유전자 변수에 대해 수행하였다. 소형 샘플 크기의 효과를 가능한 많이 상쇄하기 위해, 통계학적 유의성을 ROC AUC = 1.00으로 설정하였다. ROC AUC = 1.00인 변수는 완벽한 식별력을 가지며, 즉, 변수에 대해 2개의 그룹이 완전히 분리되며, 2가지 그룹들 간에 중복이 없다 (AUC: 곡선 하 면적). 실험적 ROC 곡선을 본 분석에 사용하였다.
통계학적 유의성에 대한 전체 기준: 전술한 3가지의 독립적인 통계학적 유의성 평가 방법을 통합하여, 전체 유의성 기준을 다음과 같이 설정하였다: 임의의 변수가 가장 유의한 변수의 최종 리스트에 포함되도록 하기 위해서는, i) P < 1.06 x 10-6, ii) |FC| ≥ 1.10, 및 iii) ROC AUC = 1.00을 가지도록 해야 할 것이다. 아울러, 제1 방법의 경우에 위 음성인 경우를 최소화하기 위해, 제1 방법의 기준을 충족하지 못하지만 다른 2가지 방법들의 기준을 충족시키며, 변화가 log2 스케일에서 20% 보다 크다는 것을 의미하는 |FC| ≥ 1.20인 경우일 때만, 해당 변수가 통계적으로 유의한 것으로 하였다.
프로브 다중도: 통계적인 유의성의 3가지 기준을 모두 충족시키며 최종 리스트에 포함되는 모든 유전자 변수들을 대상으로, 복수의 프로브가 동일한 유전자 변수를 타겟팅할 가능성을 조사하였다. 소정의 유전자 변수를 타겟팅하는 (2 이상의) 복수의 프로브들이 존재하는 경우, 프로브들 모두 통계적 유의성에 대해 조사하였다. 이들 프로브 대부분이 유의성 기준 3가지 모두를 만족시킨다면, 주어진 유전자 변수를 가장 유의한 변수들로 구성된 최종 리스트에 유지시켰다. 프로브의 절반이 (3가지 방법들 모두에 의해) 유의한 것으로 결정되고, 나머지는 그렇지 않은 것으로 결정된 관계인 경우, 주어진 유전자 변수를 최종 리스트에서 제외하였다.
RT-qPCR
정량적인 역전사 중합효소 연쇄 반응 (RT-qPCR)을 분석하기 위해, 5 x 105 세포/웰로 6웰 디쉬에 접종하고, 처리 전 하루 동안 변형된 IMEM에서 혈청 고갈시켰다 (각 도면을 참조함). RNA를 TriPure 시약 (Roche, catalog #11667157001)을 이용하여 추출하고, 트랜스크립터 cDNA 제1 가닥 cDNA 합성 키트 (Roche, catalog #04897030001)를 이용하여 cDNA를 합성하였다. 상대적인 발현 수준을, SYBR 그린 마스터-믹스 (Roche, catalog #04887352001)를 이용하여 Roche LightCycler II에서 qPCR 분석을 수행하여, 측정하였다. 타겟 유전자의 정량 수준을 표준 하우스키퍼 유전자의 발현에 대해 표준화하였다: TBP, ACTB, 및/또는 GAPDH. 유도성 PR을 포함하는 세포의 경우, 리간드 처리를 제외하기 전에 세포를 2일간 AP21967 (10-9 M)로 유도하는 것을 제외하고는, 프로토콜은 상기와 동일하였다.
상피 성장 인자 (EGF) 처리를 수반하는 RT-qPCR 분석의 경우, 세포를 6웰 디쉬에 5 x 105 세포/웰로 접종하고, 변형된 IMEM에서 2일간 혈청 고갈시켰다. 세포에 R5020 (10-8 M)을 처리하기 전에 100 ng/ml EGF (Sigma, catalog #E9644)를 전처리하였다.
MEK 저해제를 이용한 실험들에서, BT-474 세포를 6웰 디쉬에 5 x 105 세포/웰로 접종하였다. 하루 후, 세포를 헹구고, IMEM에서 1일간 혈청 고갈시켰다. 이들 세포를 MEK 저해제 U0126 (5uM, EMD Chemicals, catalog #662005)으로 30분간 전처리하였다. 그런 후, 6시간 동안 R5020 (10-8 M) 및/또는 RU486 (10-7 M)을 세포 배양 웰에 넣은 후, RNA/단백질 분리하고, RT-qPCR/웨스턴 블롯팅을 전술한 바와 같이 수행하였다. 본 실험에 사용된 PCR 프라이머 세트를 도 5.1에 나타낸다.
정교한 경로 분석
정교한 경로 분석 (IPA)을 이용하여 2개의 개별 유전자 리스트를 비교하였다: SUMO-결함성 PR (+R5020/-R5020 log2 배수 변화 >1.0, BH 보정된 P <0.01)을 발현하는 세포에서 프로게스틴에 의해 상향 조절되는 유전자 대비, WT PR을 발현하는 T47D 세포에서 프로게스틴에 의해 상향 조절되는 유전자. 이들 유전자 리스트를, 각 유전자를 다양한 생물학적 기능 또는 네트워크 경로와의 관련성을 확인하기 위해, 코어 분석이 완료된 IPA 소프트웨어에 업로드하였다. IPA 비교 분석을 이용하여, 기능적으로 구분되는 경로들에서 상향 조절된 WT 또는 KR PR을 세포가 발현하는지 여부를 확인하였다. 분석은 유의성을 기준으로 점수를 매기고 (BH 보정된 P 값, 다중 가설 검정으로 보정함), 특정 생물 기능에 유의적으로 관여하는 유전자 리스트의 역치는 P <0.05 (또는 -log10 (BH 보정된 P value) >1.30)였다.
PR 발현 메타유전자 (metagene)의 동정
메타유전자 분석을 빈 벡터, WT PR 또는 K388R PR을 구성적으로 발현하는 세포주로부터 유전자 발현 마이크로어레이 데이타를 이용하여 수행하였으며, 비히클 또는 R5020 (도 5C-D)을 처리하였다. 각 샘플에서 메타유전자를 동정하는 전략을 비-음수 행렬 인수분해 (non-negative matrix factorization) (Gaujoux and Seoighe, 2010)를 이용하여 채택하였다. 이러한 전략은 메타유전자의 동정과 다른 데이타세트로의 적용을 용이하게 한다. 고 분산 하에 유전자에 대한 연구를 제한하고 메타유전자 피트 (metagene fit)의 계산에 사용되는 프로브의 수를 한정하기 위해, 상위 80 백분위에 속하는 프로브의 4분범위 (IQR)를 기반으로 한 메타유전자 분석으로 프로브들을 검토하였다. 데이타의 최적 랭킹을 8개로 계산하였으며, 따라서 8개의 메타유전자가 데이타에 존재한다. 이들 메타유전자들 중 3개는 모든 샘플들에서 고도로 발현되거나 또는 어떤 샘플에서도 발현되지 않는데, 이는 이들이 하우스키핑 또는 계속 발현되는 유전자들에 대한 메타유전자가 틀림없다는 것을 의미한다. 나머지 5개의 메타유전자는 빈 벡터 PR-null 샘플 (-R5020 및 +R5020 처리 사이에 구분이 없음), 및 R5020이 처리 또는 무처리된 WT 또는 KR PR의 쌍 조합에 해당된다. 따라서, 이들 분석을 통해 세포의 생물학적으로 관련된 아종들로부터 메타유전자를 동정하였다.
Loi 등의 인간 유방 종양 데이타세트 (Loi et al., 2007)는 수개의 데이타세트 전체에서 타목시펜 처리 및 무처리된 샘플 둘다에 대한 유전자 발현 데이타를 포함하고 있다. 이들 데이타를 함께 종합하였으며, 이는 유전자 발현 옴니부스 (GEO) (등재 번호 GSE6532)를 통해 이용가능하다. 데이타세트 (Loi et al., 2007)를 가공하기 위해 Red-R (Covington and Parikh, 2011)에 로딩하였다. 메타유전자 분석을 위한 기저 행렬을 재형상화하여, 유전자 심볼 전체를 모우고, 유전자의 각 프로브에 대한 메타유전자 값들을 평균내었다 (평균 값). 동일한 조작을 발현 데이타에 대해 수행하였다. 비-매칭 유전자 (메타유전자에는 존재하지만, 임상 발현 데이타에 존재하지 않거나, 또는 반대임)는 분석에서 제외하였다. 재형상화된 데이타를, (T47D 세포주 데이타에 대한 초기 메타유전자 피트를 구축하기 위해 행해진 바와 같이) 스코링하기 위해 비-음수 행렬 인수분해 (NMF) 패키지 함수 (fcnnls)에 적용하였다. Loi 등의 데이타는 z-스코어로서 제공되기 때문에, 데이타를 언-로깅(un-logged)하고, (정상 형태에서 음수를 포함하고 있기 때문에) fcnnls 알고리즘에 사용하였다. 샘플이 피팅된 계수 행렬 (스코어링 행렬)에서 비-0 값을 나타낸다면 샘플은 메타유전자를 발현하는 것으로 취하였다.
새로운 PR-타겟 유전자의 동정 및 유전자 발현 플랫폼의 비교 분석
기존에 공개된 2가지의 실험 (Jacobsen et al., 2005; Richer et al., 2002)에서 도출된 의존성 및 -비의존성 PR-타겟 유전자 리스트를 합하였다 (같은 것은 제거함). 본원에서 동정된 유전자들은, 양쪽 플랫폼 (Illumina 및 Affymetrix를 조합함)을 이용하여 측정된 바와 같이 상향 조절되었으며 (>1.5배, BH 보정된 P <0.01), 바이오인포메틱스 툴인 VENNY (Oliveros, 2007)를 이용하여 기존에 공지된 상향 조절된 유전자에 대한 벤 다이아그램 비교를 수행하기 전에 동일한 것은 제거하였다.
유전자 세트 농화 분석 (GSEA) 소프트웨어 (Mootha et al., 2003; Subramanian et al., 2005)를 사용하여, WT 또는 KR PR을 안정적으로 발현하는 세포에서 상향 또는 하향 조절된 유전자들을 유도성 iWT 또는 iKR PR을 발현하는 세포와 비교하였다. Affymetrix 발현 데이타를 이용하여, 4개의 유전자 세트들이 구축되었다: R5020 처리시 iWT에 의해 >2.0배 상향 또는 하향 조절된 유전자, 및 R5020 처리시 iKR에 의해 >2.0배 상향 또는 하향 조절된 유전자. 마찬가지로, 2개의 GSEA-형식의 데이타세트를 Illumina 발현 데이타로부터 구축하였다: 제1 데이타세트에서 2개의 표현형 (WT +R5020 vs WT -R5020)을 비교하고, 제2 데이타세트에서는 2개의 표현형 (KR +R5020 vs KR -R5020)을 비교하였다. 이들 Illumina 데이타세트를 이용하여 GSEA를 수행하였으며, Affymetrix 유전자 세트들의 농화에 대해 조회하였다. 순열 타입 (permutation type)을 순열 1000개인 유전자_세트로 설정한 것을 제외하고는, 디폴트 세팅을 이용하여 GSEA를 실행하였으며, 본 발명의 데이타세트에는 log 스케일의 데이타가 포함되어 있기 때문에 랭킹 유전자들의 행렬은 Diff_of_Classes로 설정하였다.
염색질 면역침강 (ChIP)
ChIP 분석을 ChIP-IT Express instruction manual (Active Motif, catalog # 53008)에 따라 수행하였다. 세포를 15 cm 배양 디쉬 당 세포 15 x 106개로 2일간 cMEM에 접종한 다음, 변형된 IMEM에서 2일간 혈청 고갈시켰다. 세포에 R5020 (10-8 M) 또는 비히클을 1시간 또는 4시간 처리하였다. 유도성 PR을 발현하는 T47D 세포의 경우, AP21967 (10-9M)를 고갈 단계에 첨가하였다. 염색질을 Bioruptor sonicator (Diagenode, model UCB-200)를 30분간 사용 (30초 온/오프)하여 절단시켰다. 면역침강은 절단된 염색질 60 ul, 항체 2 ug으로 준비하고, 밤새 면역침강시켰다. 정제된 ChIP 및 투입 DNA를 이용하여, 3세트의 qPCR에 의해 상대적인 동원을 측정하였다. 분석은 SYBR 그린 마스터-믹스를 이용하여 Roche LightCycler II에서 수행하였다. 타겟 유전자 좌 정량 값을 인풋 DNA 정량 값의 퍼센트로서 표준화하였다.
H3K4me2 수준을 분석하기 위해, 뉴클레오솜을 MNase (micrococcal nuclease)를 이용하여 분리하였다. 15 cm 디쉬에서, 세포 12 x 106를 cMEM 중에 접종하고, 변형된 IMEM에서 혈청 고갈시킨 후, 2일간 AP21967 (10-9 M) 처리로 유도하였다. 1일 후, 세포에 R5020 (10-8 M)을 4일간 처리하고, 전술한 바와 같이 면역침강시켰다 (Verzi et al.).
세포 증식 및 세포자살 분석
세포 증식을 MTT 분석 (3-[4,5-다이메틸티아졸-2-일]- 2,5-다이페닐테트라졸륨 브로마이드, Sigma catalog #M2128)으로 측정하였다. 24웰 플레이트에서, 1 x 104 세포/웰로 cMEM 중에 접종하고 (유도성 PR 발현은 2일간 AP21967 (10-9 M)을 이용하여 유도함), 세포를 세척한 후, 5% 덱스트란-코팅된 차콜 처리된 (DCC) FBS가 첨가된 변형된 IMEM에서 1일간 스테로이드를 고갈시킨 다음, R5020 (10-8 M)을 첨가하였다. 0, 2, 4, 6일째에, 각 0.5 ml 세포 배양 웰에 60 ul MTT (5 mg/ml)를 3시간 동안 첨가하여 세포 증식을 측정하였으며, 배지를 조심스럽게 제거한 다음, 가용화 용액 (90% v/v DMSO/PBS)을 첨가하여 세포를 용혈시켰다. 세포 용혈물의 흡광도 (650 및 570 nm)를 플레이트 판독기를 이용하여 측정하였다. 650 nm 측정값을 570 측정값에서 공제하고, 샘플 평균을 0일에 대해 표준화하였다.
폴리 (ADP)-리보스 중합효소 1 (PARP) 절단 분석을 사용하여, 독소루비신을 세포독성 농도로 처리한 후 세포 배양물에서의 세포자살 수준을 측정하였다. 유도성 PR을 발현하는 T47D 세포를 10 cm 디쉬 (2 x 106 cells/dish)내 cMEM에 접종하고, AP21967 (10-9 M)로 유도하였다. 세포를 헹구고, 유도한 다음 4일간 혈청 고갈시켰다. 그런 후, 세포에 R5020 (10-8 M)을 6시간 처리한 다음, 디쉬에 독소루비신 (8 uM)을 첨가하여 24시간 처리하였다. 단백질을 표준 RIPA 세포용혈 완충제를 사용하여 회수하고, SDS-PAGE한 다음 절단형-PARP 및 PR 항체를 이용하여 웨스턴 블롯팅하였다. β-액틴 웨스턴 블롯팅을 샘플 로딩 대조군에 대해 수행하였다.
세포독성을 나타내는 독소루비신으로 처리한 이후의 세포 생존성을, 살아있는 세포의 수와 직접적인 비례 관계인 아데노신 트리포스페이트 (ATP)의 농도를 Cell-Titer-Glo 생발광 분석 (Promega, catalog #G7571)을 이용하여 측정함으로써 결정하였다 (Crouch et al., 1993). WT 또는 KR PR을 발현하는 T47D 세포를 cMEM이 담긴 24웰 디쉬 (1 x 104 세포/웰)에 접종하였다. 세포를 헹구고, 5% DCC FBS가 첨가된 변형된 IMEM에서 1일간 스테로이드를 고갈시켰다. 세포에 R5020 (10-8 M)을 6시간 처리한 다음, 독소루비신 (6 uM)을 웰에 첨가하였다. 4일 후, 세포 생존성을 Cell-Titer-Glo 기질을 첨가함으로써 측정하고, 플레이트 판독기를 이용하여 발광성을 측정하였다. 샘플 평균을 0일에 대해 표준화하였다 (n=6, -/+ SD).
온코마인 (Oncomine) 데이타 분석
인간 유방 종양 샘플에서 개개 PR 타갯 유전자들의 상대적인 발현을, 온코마인 데이타베이스 (버전 4.4, 2011년 10월에 데이타가 공개됨)를 검색함으로써 확인하였다. 개별 PR 타겟 유전자 (예, RGS2)를 암 게놈 Atlas (TCGA) 유방 2 데이타세트에서 조회하였다. 온코마인 아웃풋 데이타를 분류하여, "암 대 정상" 조합으로 분리하고, 혈액, 정상 유방 및 유방 암종 샘플에 대한 카피수 단위 발현 값으로서 박스-및-위스커 플롯 (box-and-whiskers plot)을 이용하여 기록하였다 (도 2a) (점: max/min, 위스커: 90/10 백분위, 박스: 75/25 백분위, 라인: 모든 샘플들의 중간값). 각각의 분석을 위해, 각 유전자에 특이적인 특정 유방 암종은 다음과 같다: 침습성 소엽 유방 암종 (MSX2), 침습성 유관 및 소엽 암종 (RGS2), 유관내 사상형 (Intraductal Cribriform) 유방 선암종 (MAP1A), 및 점액 유방 암종 (PDK4).
온코마인 데이타베이스에 기술된 바와 같이, 다중 유방암 "컨셉 (concept)"을 리간드 의존적인 (LD) KR>WT 유전자 시그니처와 조합하였다. 온코마인에 따르면, 컨셉은 유전자 발현 마이크로어레이로부터 유래되거나, 또는 종앙 코호트 또는 암 세포주 실험들에서 유래된 유전자-카피수 데이타세트이다. 구체적으로, 컨셉은 몇가지 기준 (예, ERBB2-양성 유방 종양에서 발현된 유전자들 중 상위 5%)에 의해 규정된 다양한 공개된 데이타세트로부터 유래된 유전자들의 리스트이다. 리간드 의존적인 (LD) 유전자 시그니처는, R5020 처리군의 유전자 발현 값을 R5020 비-처리군에 대해 표준화한 다음, KR 및 WT PR 발현성 세포주들 간에 표준화된 배수 변화 값들을 비교함으로써, 구축하였다. 이러한 분석으로, WT PR 발현성 세포에 비해 SUMO-결함성 PR을 발현하는 세포에서 >1.5배 상향 조절된 LD 유전자 151개가 동정되었다. 리간드-비의존성 (LI) 유전자 시그니처는, WT 또는 KR 발현성 세포에서 R5020 비-처리군에서의 유전자 발현 값을 PR-null 발현성 세포에 대해 표준화한 다음, KR 및 WT 발현성 세포주들 간에 표준화된 배수 변화 값들을 비교함으로써, 구축하였다. 이러한 분석으로, WT PR 발현성 세포에 비해 SUMO-결함성 PR을 발현하는 세포에서 >1.5배 상향 조절된 LI 유전자 92개가 동정되었다. 이들 PR 유전자 시그니처를 온코마인 검색 프리미엄 편집 소프트웨어 (Compendia Bioscience, Ann Arbor, MI)에 업로드하고, 데이타베이스에서 조합된 컨셉을 검색하였다.
결과
PR SUMO일화는 T47D 유방암 세포에서 프로모터 선택성을 변형시킨다. 미공지된 이유로, 종양성 유방 조직에 비해, 정상적으로 PR-조절된 유전자들 간에는 거의 겹치지 않는다 (Graham et al., 2009). PR 기능의 명백한 다양성에 대한 한가지 기전은 변형된 신호 전달과 같은 유방암 발생시 초기 현상들과 관련있을 수 있다. 본 발명자들의 이전 연구들의 일부를 토대로 (Daniel et al., 2007a; Daniel and Lange, 2009; Daniel et al., 2007b), SUMO일화된 PR과 인산화된 (즉, 탈SUMO일화된) PR 간의 밸런스가 유방암에서 종종 변형되며, 이로써 PR 프로모터의 선택성에 변이와 유전자 발현 패턴에 변형이 생기는 것으로 추정되었다. PR 양성인 것으로 임상적으로 규정된 유방 종양 10개에 대한 스크리닝에서, 광범위한 총 PR mRNA (도시 안함) 및 단백질 발현 (도 1A)을 검출하였다. RT-qPCR 및 웨스턴 블롯팅에서 모두 PR 양성인 것으로 확인된 유방 종양들 중 (10개 중) 7개에서, 5개 이상의 샘플 (1, 3, 6, 8 및 9번 레인)이 포스포-Ser294 PR-B를 어느 정도의 수준으로 함유하고 있는 것으로 명확하게 나타났다 (도 1A). 특히, 종양 10개 중 2개 (레인 1번과 3번)는 포스포-Ser294 PR-B를 다량 함유하였다. 주목하게도, PR-B는, 주로 MAPK 및 CDK 패밀리들내에서, 프롤린-특이적인 단백질 키나제로 투입되는 프로게스틴 또는 펩타이드 성장인자에 반응하여 빨리 인산화되지만, PR-A는 그렇지 않았다 (Clemm et al., 2000). 이러한 사실에 부합되게, EGF는 프로게스틴-유발성 PR-B를 차단하지만, PR-A SUMO일화는 그렇지 않다 (Daniel et al., 2007a).
본원에서는 PR 타겟 유전자들이 PR SUMO일화 상태에 따라 차이가 있음을 개시한다. 임상 시료에서의 광범위한 PR 발현 (도 1A 및 (Liu et al., 2010))은, PR-의존적인 유전자 발현이 유방암 표현형에 대한 PR 기여를 나타내는 좀더 정확한 마커를 제공해줄 수 있음을 시사해주었다. 인산화된 및 SUMO-결함성 PR-B의 독특한 작용을 알기 위해, 야생형 (SUMO일화될 수 있음) 또는 SUMO-결함성 (K388R 돌연변이/포스포 모방) PR-B 분자 중 어느 하나를 안정적으로 발현하는 유방암 세포의 전사 프로파일을 전 게놈 발현 프로파일링을 이용하여 측정하였다. 복수의 클론들을 야생형 (WT) PR-B 또는 Lys388에서 SUMO로의 변형을 이행할 수 었는 돌연변이 K388R (KR) PR-B 중 어느 하나를 발현하는 벡터-매칭된 PR-null T47D 유방암 세포로 조작하였으며; 이러한 SUMO-결함성 수용체는 Ser294에서 영구적으로 인산화되는 PR-B의 기능성 모방체이다 (Daniel et al., 2007a; Lange et al., 2000). 포스포-Ser294 및 S294D 수용체는 WT PR에 비해 빠른 리간드-의존적인 (유비퀴틴-매개) 하향 조절을 이행하는 과잉 활성의 전사 인자이다 (Daniel et al., 2007b). WT 또는 KR PR-B 중 어느 하나를 발현하는 세포를 합성 프로게스틴, R5020 (10-8 M)으로 6시간 처리하였다 (도 1B). 리간드 결합시, PR는 복수의 부위에서, 약간 겔 상향 이동에 의해 확인되는 바와 같이 전체적으로 인산화된다 (Takimoto et al., 1996). 기존의 보고에 부합되게 (Daniel et al., 2007a; Daniel and Lange, 2009), 과잉 활성화된 KR PR은 WT PR에 비해 약간 더 빨리 리간드-유발에 의해 (유비퀴틴 프로테아좀-의존적임) 하향 조절되게 된다 (6시간째에 명확해짐) (Lange et al., 2000). 이러한 실험 조건을 이용하여, Illumina HT-12v4 전 게놈 유전자 발현 비드 어레이를 사용하여 글로벌 유전자 발편 프로파일을 동시에 측정하였다 (도 1C). 조절된 상위 유전자들을, 상향 또는 하향 조절된 유전자들을 나타낸 히트 맵으로 작성하였다 (하나 이상의 샘플에서, 배수 변화 >8.0, BH 보정된 P <0.001, 도 1C). 프로게스틴 처리시, 이들 세포는 다양한 발현 패턴을 나타내었으며; 복수의 PR-조절된 유전자 세트들이 신속하게 명확해졌다 (도 1C; 무처리 대조군 대비, 리간드-의존성 PR에 의해 상향 (1a) 또는 하향 (1b) 조절된 PR-조절된 유전자들의 그룹, PR-null 대조군 대비 리간드-비의존성 PR에 의해 상향 (2a) 또는 하향 (2b) 조절된 유전자, 및 WT(3) 대비 KR 또는 KR (4) 발현성 세포주 대비 WT에서 주로 상향 조절된 리간드-의존성 유전자 군들을 비교함).
리간드-의존적인 또는 리간드-비의존적인 방식으로 PR에 의해 >1.5배 상향 조절된 유전자가 동정되었으며, KR 또는 WT 수용체 중 어느 하나를 발현하는 세포들 간의 유전자 발현 중복이 확인된 유전자 뿐만 아니라 독특하게 조절된 유전자들의 아종이 동정되었다 (도 1D-E). 다음으로, RT-qPCR을 이용하여 이들 클래스들 유래 다수의 PR 타겟 유전자들에 대해 발현 프로파일들을 검증하였다 (도 1F). 놀랍게도, RGS2 발현 (주로 KR 수용체에 의해 상향 조절됨)이 기저/근상피 구획에서 과다 발현되고, 실질적으로 대부분의 유방 종양에서 상승되었다 (Smalley et al., 2007). 이와는 대조적으로, BCL2L11 (BIM)은 ERBB/MAPK-의존성 내강 세포 클리어링 (luminal cell clearing)에 참여하는 세포자살 촉진 매개인자 (pro-apoptotic mediator)로서 (Reginato et al., 2005), 이의 발현은 KR 수용체가 아닌 주로 WT 수용체에 의해 상향 조절된다. 이들 예에서 시사되는 바와 같이, 유전자 어레이로 PR 타겟 유전자들의 다양한 클래스들이 명확하게 동정되었으며, 여기에는 PR-매개 세포 및 생존 기전을 나타내는 유전자 발현 프로파일이 포함된다.
이들 결과는 기본적으로 유도성 벡터 시스템으로부터 유래된 WT 또는 KR PR을 발현하도록 조작된 T47D 세포에서도 반복되었다 (도 1.1). 이 모델에서, PS의 유도성 발현 (iWT 또는 iKR)은 오직 세포 배양 배지에 첨가된 소형 분자 이량체화제인 AP21967의 존재에 의존적이며; AP21967 처리시 동일 수준의 iWT 또는 iKR이 유도되었으며, 이들 수용체는 프로게스틴에 반응하여 Ser294를 동일하게 인산화하였다 (도 1.1A). 세포에 AP21967 (10-9 M) 및 R5020 (10-8 M)를 처리한 다음, ffymetrix U133A 2.0 마이크로어레이 플랫폼을 이용하여 유전자 발현의 변화를 분석하였다. PR을 안정적으로 발현하는 T47D 세포로부터 입수한 PR-의존적인 유전자 발현 프로파일들 (Illumina 플랫폼을 이용하여 분석함)은, PR을 유도에 의해 발현하는 동일 모 세포(T47D)로부터 입수한 유전자 어레이 데이타 (Affymetrix 플랫폼으로 분석함; 도 1.1B-C)와 상당히 비슷하였다. 요컨대, 어레이를 통해 기존 보고 (Jacobsen et al., 2005; Richer et al., 2002)에 비해 PR 조절된 유전자들이 더 많이 동정되었으며 (>1.5배, BH 보정된 P <0.01), 마이크로어레이 플랫폼은 현재 초기 기술에 비해 수천개 더 많은 수의 "리포터"를 포함한다. 기존에 공지된 PR 타겟 유전자의 70%가 동정되었지만, 새로운 PR 타겟 유전자들 수백개도 동정되었다 (데이타 도시 안함).
PR Ser294의 인산화는 SUMO-결함성 PR 유전자의 발현을 유도한다. "SUMO-민감한" PR-타겟 유전자의 조절 기전을 조사하기 위해, 마이크로어레이 분석을 통해 4개의 유전자를 향후 연구를 위해 선별하였다 (MSX2, RGS2, MAP1A 및 PDK4). 이들 특정한 유전자들은 KR 발현 세포에서는 상향 조절되지만, WT 수용체 발현 세포에서는 그렇지 않았다 (도 1D, 197 유전자 카테고리). 온코마인 데이타베이스의 쿼리는, 4개의 유전자 모두 정상 유방 조직과 혈액 보다는 유방 암종에서 증폭됨을 나타내었다 (도 2a). 추가적인 유방암 모델에서, PR 타겟 유전자 발현의 SUMO-의존적인 변화를 검증하기 위해, 벡터 대조군, WT 또는 KR 수용체를 (에스트로겐 부재시) 내인성 PR을 저농도로 발현하는 MCF-7 세포로 안정적으로 도입하였다. 이들 세포에 비히클 대조군 (에탄올) 또는 R5020 (10-8 M)을 PR 길항제, RU486 (10-7 M)의 존재 또는 부재 하에 6시간 동안 처리하였다 (도 2b). MCF-7 세포에서 프로게스틴-유발성 유전자 발현 프로파일들은, 본 T47D 세포 모델에서 수득한 것 (MSX2, RGS2, MAP1A 및 PDK4)과 거의 동일하였다. 아울러, 이들의 R5020-유도성 mRNA 발현은 RU486 첨가시 완전히 없어졌는데, 이는 이들 유전자의 조절이 거의 PR-의존적임을 의미한다.
기존에 SUMO-결함성 KR 수용체가 포스포-Ser294 (WT) PR 종들을 거의 유사하게 모방한다고 공지되었다 (Daniel et al., 2007a). 동일한 유전자 세트 (MSX2, RGS2, MAP1A, 및 PDK4)에서 PR 조절이 인산화-의존적임을 입증하기 위해, PR-null T47D 세포, 또는 WT, KR 또는 포스포-돌연변이 S294A (SA) PR-B를 안정적으로 발현하는 T47D 세포 (Lange et al., 2000)를 사용하였다. PR Ser294의 돌연변이로, 루시퍼라제 리포터 분석에서 측정된 바와 같이, 전사를 억제하는 상당히 SUMO일화된 수용체가 형성되었다 (Daniel et al., 2007a). 이러한 사실과 일관되게, 내인성 PR 타겟 유전자의 프로게스틴-유도성 상향 조절이, SUMO-결함성 KR PR을 발현하는 세포에 비해 S294A PR을 발현하는 세포에서 차단되었다 (도 2c). 프로게스틴-유도성 유전자 발현이, Ser294 및 Lys388 2곳 모두에 점 돌연변이를 가진 PR 이중 돌연변이 (KRSA)를 발현하는 세포에서 (즉, R5020-처리된 KR 세포에서 유도되는 수준과 유사하게) 복원되었는데, 이는 이들 인산화-의존적인 PR 타겟 유전자에서 PR 탈SUMO일화가 리간드-의존적인 상향 조절에 필요한 유력한 현상임을 시사해준다.
유방암 세포에 EGF를 처리하면, 명확한 PR Ser294 인산화와 탈SUMO일화가 유도된다 (Daniel et al., 2007a). 따라서, WT PR을 안정적으로 발현하는 T47에 EGF (100 ng/ml)를 전처리한 다음, 비히클 대조군 또는 R5020 (10-8 M)을 처리하였다. MAP1A 및 RGS2 둘다 2일의 시간 경과시 EGF 단독에 둔감하였다 (도 2D). 하지만, EGF의 전처리는 이들 유전자들의 프로게스틴 자극성 mRNA 발현을 유의하게 증가시켰다 (도 2D). 비슷한 결과는 6시간 처리한 모 (내인성 PR-A 및 PR-B 이소형 둘다를 발현하는) T47Dco 세포에서 RGS2에 대해서도 관찰되었지만, MAP1A 발현은 그렇지 않았다 (도 2E). 복수의 인자들 (즉, PR 인산화의 강도 및 지속 기간, 전사 활성 및 단백질 수준)들도 성장인자를 이용하게 광범위하게 자극받은 세포들에서 PR-조절성 MAP1A 발현 카이네틱스에 영향을 미칠 것으로 보인다. WT PR을 안정적으로 발현하는 T47D-B에서, MAP1A mRNA 발현은 프로게스틴 + 헤레굴린-eta1 3시간 처리한 직후 상승적으로 상향 조절되었으며; 프로게스틴-단독 처리시에는 이 수준에 도달하는데 24시간 걸렸다 (데이타 도시 안함). 요컨대, 이들 데이타는, PR이 인산화 및 SIMO일화 상태에 따라 복수의 내인성 유전자들을 역동적으로 조절하며; 성장인자는 탈억제된 전사 인자로서 작용하는 포스포-PR에 우호적이라는 것을 시사해준다.
PR SUMO 변형은 프로모터 선택 기전을 제공한다. 유전자 어레이 분석에 따르면, PR의 SUMO 변형은 선택된 프로모터에 대한 전사 반응의 정도를 변형시키며, 다른 PR 타겟 유전자의 조절은 PR의 SUMO일화에 완전히 둔감한 것으로 나타났다 (도 1). PR 프로모터 선택 기전을 조사하기 위해, 차별적으로 조절된 PR 타겟 유전자의 염색질로의 PR과 선택된 공동-조절인자들의 동원을 조사하였다. 먼저, MSX2에 실험 포커스를 두었다. 이 호메오박스 전사 인자는, PR-B와 마찬가지로, 유선 발생에 필수적이며, MSX2의 이식유전자 발현이 마우스에서 유관 과형성을 유발한다 (Satoh et al., 2007; Satokata et al., 2000). 기능 연구를 통해, MSX2가 사이클린 D1과 E1의 발현을 유도하는 것으로 확인되었으며 (Satoh et al., 2004), 이는 RAS-매개의 세포 형질전환에 관여하며 (Takahashi et al., 1997), 상피 마커의 하향 조절을 통해 상피에서 간엽으로의 전이를 유도한다 (di Bari et al., 2009). Lanigan et al. (Lanigan et al., 2010)은, 양호한 예후 (즉, ER 및 PR과 유사함)와 조합되어 있음에도 불구하고, MSX2 발현이 내강 B 및 HER2-농화된 분자 아종의 유방암 둘다에서 크게 상승한다는 것을 보여주었다. MSX2 전사 개시부의 상류 및 하류에 위치한 복수의 컨센서스 프로게스테론 반응 인자 (PRE) 서열들은 MatInspector 소프트웨어 (Cartharius et al., 2005)를 이용하여 동정되었다. 특히, 하나의 PRE는 (즉, 미국 메사추세츠, 보스톤에 위치한 하버드 대상의 밀레스 브라운에 의해 제공된 공개되지 않은 ChIP-chip 실험들로부터 유래된) PR 시스트롬을 토대로 공지된 PR 동원 영역과 정렬되었다. MSX2는 SUMO-결함성 PR을 안정적으로 또는 유도에 의해 (T47D) 발현하는 T47D 세포 또는 MCF-7 세포에서 프로게스틴 처리에 반응하여 전사적으로 상향 조절되지만, WT 수용체는 그렇지 않았다 (도 2b-C, 1.1C). MSX2의 PRE 인핸서 영역으로의 PR의 직접 동원을 조사하기 위해 (도 3A), WT 또는 KR PR을 구성적으로 (또는 유도에 의해) 발현하는 세포에 R5020 (10-8 M)을 처리하고, 염색질 면역침강 (ChIP) 분석을 수행하였다. 프로게스틴 처리 후, WT 및 KR PR 둘다 PRE 인핸서 영역에서 쉽게 검출되었지만 (도 3B, 좌측), WT PR을 발현하는 프로게스틴-처리 세포에서는 (RT-qPCR에 의해 측정한 mRNA 수준은) 전사 활성이 검출되지 않았다 (도 2b-C, 1.1C). 특히, WT PR에 비해 MSX2 인핸서 유전자 좌에 SUMO-결함성 KR PR가 상당히 많이 동원되었다. 이러한 사실은 유도성 PR를 발현하는 세포 (도 3B, 우측) 뿐만 아니라 SUMO-결함성 PR에 의해 상향 조절된 복수의 다른 유전자들의 PRE-함유 인핸서 (도 3.1)에서도 반복되었다. 다음으로, 공통 PR 전사 공동-활성인자, cAMP-반응 인자-결합 단백질 (CREB)-결합 단백질 (CBP)의 MSX2 인핸서 유전자 좌로 동원되는 지를 조사하였다. CBP는 복수의 핵 수용체와 상호작용하며, 전사 스캐폴드로서 기능하며, 히스톤 아세틸트랜스퍼라제 (HAT) 활성을 가진다 (Lambert and Nordeen, 2003; Li et al., 2003; Ogryzko et al., 1996). ChIP 분석을 이용한 결과, MSX2 유전자 좌로의 CBP의 동원이 WT PR에서는 그렇지 않았지만 SUMO-결함성 KR PR을 발현하는 세포에서 현저하게 상승하였다 (도 3C). KR PR과 조합된 이들 공동-활성 인자의 존재 증가와 일관되게, iWT PR 발현성 세포에 비해, iKR PR 발현성 세포의 프로게스틴 처리시, MSX2 근위 프로모터 영역으로의 기능적으로 활성형인 포스포-Ser5 RNA 중합효소 II의 전체적인 동원이 증가된 것으로 관찰되었다 (도 3.2). 이러한 데이타는, WT PR이 프로게스틴의 존재 시 이 영역으로 확실히 동원됨 (도 3B)에도 불구하고, 유의한 mRNA 발현이 이루어지지 않는 이유를 설명해 줄 수 있다 (도 2b-C, 1.1C). 내인성 유전자 좌에서 탈SUMO일화된 PR 및 SRC1의 구성적인 조합은 기존에 보고된 바 있다 (Daniel and Lange, 2009).
히스톤 테일 변형 (메틸화, 아세틸화, 인산화, 등)은 염색질 다이나믹스에 유의하게 영향을 미침으로써, 유전자 발현에 변화를 일으키는 것으로 공지된 후생적 변형이다 (Ong and Corces, 2011에서 리뷰됨). 일반적으로, 히스톤 H3 Lys4 다이메틸화 (H3K4me2)는 전사 활성화와 관련된 후생 마크이다 (Barski et al., 2007; He et al., 2010). H3K4me2는 전사 인자-촉매화된 쌍을 이룬 뉴클레오좀 포지셔닝 영역을 표시하며, 근처 유전자 활성화를 나타내는 표시이다 (He et al., 2010). MSX2 인핸서 유전자 좌에서 H3K4me2 수준을 측정하기 위해, 유도성 PR을 발현하는 T47D 세포 (iWT 및 iKR)에 R5020 (10-8 M)을 4시간 처리하고, MNase (micrococcal nuclease)로 분해하여 뉴클레오솜을 분리한 다음; 히스톤 메틸화를 ChIP 및 qPCR (도 3D, 좌측)으로 확인하였다. H3K4me2 수준은 iWT PR 발현 세포에 비해 iKR을 발현하는 프로게스틴 처리된 세포에서 증가하였다. MSX2 PRE 유전자 좌 주변 (중첩성 qPCR 산물을 이용한 상류 및 하류 약 500 pb)의 H3K4me2의 R5020-유도성 배수 변화를 측정하여, 국소 히스톤 다이메틸화 패턴을 가시화하였다 (도 3D, 우측). 프로게스틴-의존성 H3K4me2는 iWT 발현 세포에 비해 SUMO-결함성 iKR PR을 발현하는 세포에서 농화되었다. 실제, PRE 서열의 측면에서의 히스톤 메틸화 수준 증가는 기능적인 인핸서 영역에 전사 인자 복합체의 동원을 촉진시키는 뉴클레오솜 리모델링 및 전파의 결과인 것으로 보인다. (He et al., 2010).
이러한 결과들은, 하나 이상의 히스톤 메틸트랜스퍼라제가 iWT 또는 iKR PR을 발현하는 세포에서 MSX2 인핸서로 차별적으로 동원됨을 시사해준다. 최근, 서브유닛 혼성 계통 백혈병 2 (MLL2) 메틸트랜스퍼라제를 비롯한 염색질 리모델링 복합체가 프로게스틴-의존성 H3K4 트리메틸화에 관련 있는 것으로 시사되었다 (Vicent et al., 2011). 아울러, ER-α는 그것의 LXXLL 모티프를 통해 MLL2와 직접 상호작용하며, MLL2는 MCF-7 세포에서 에스트로겐-의존적인 전사 상향 조절을 매개한다 (Mo et al., 2006). 본원에서는, 안정적인 T47D 모델과 유도성 T47D 모델을 이용하여, MLL2가 SUMO-결함성 KR PR을 발현하는 프로게스틴이 처리된 세포에서 MSX2 인핸서로 현저하게 동원되지만, WT PR에서는 그렇지 않다는 것을 확인하였다 (도 3E).
마지막으로, PR SUMO일화 상태에 둔감한 대조군 PR-타겟 유전자인 MAT2A 근처 PRE-함유 인핸서 유전자 좌로의 PR의 상대적인 동원을 측정하였다 (도 1D, 중첩성 벤 카테고리). MAT2A mRNA 발현은 프로게스틴-처리된 WT 발현 세포 또는 KR PR 발현 세포에서 동일하게 상향 조절되었다 (도 3F, 좌측). 마찬가지로, MAT2A 인핸서에서 동일한 PRE-함유 영역으로의 PR 및 MLL2의 프로게스틴-의존적인 동원은 WT 또는 KR PR중 어느 하나를 발현하는 세포에서 매우 비슷하였다 (도 3F, 가운데 및 우측). 요컨대, 이 데이타는, (염색질에서) 인핸서/프로모터 구조가 PR SUMO일화와 조합되어, PR과 초기 염색질 리모델링의 매개인자 (MLL2)와의, 뿐만 아니라 CBP를 비롯한 주요 공동-조절인자 간의 중요한 상호작용을 차단하는 기능을 하며; 이들 인자들이 더 높은 농도로 SUMO-결함성 PR을 발현하는 세포에서 "민감한" PRE 영역과 특이적으로 조합되었다. 아마도, SUMO-민감성 인핸서 영역은 비교적 "폐쇄된" 영역 (즉, MSX2와 같은 유전자에 대해)에서 뉴클레오솜 포지셔닝화에 변화를 일으키기 위해, MLL2의 PR-의존적인 동원을 필요로 한다. 반면, 기존의 "열린" 영역은 (즉, MAT2A와 같은 유전자에 대해) PR SUMO 변형에 둔감할 수 있다. 아울러, SUMO-결함성 PR과 다른 인자 (즉, 파이오니어-타입의 전사 인자)와의 선호적인 조합이 PR 프로모터 선택에 기여할 수도 있으며; MSX2 인핸서 영역으로의 KR 동원은 프로게스틴 존재시 WT 수용체에 비해 현저하게 강화되었다 (도 3B). 이러한 의문들은 보다 상세한 글로벌 유전자 및 시스트롬 분석에서 다룬다 (고찰 참조).
SUMO-결함성 포스포-PR은 세포 증식을 증가시키고, 세포자살은 낮춘다. 정교한 경로 분석 (IPA, Ingenuity Systems) 소프트웨어에는 분자적으로 규명된 경로, 생물 기능 또는 질환 상태가 수동으로 지정되어 있으며, 현행 문헌을 기초로 유전자 데이타베이스가 포함되어 있다. 이 툴을 이용하여, WT 또는 KR 수용체를 안정적으로 발현하는 세포에서 리간드-의존적으로 상향 조절된 유전자들 (>2배, BH 보정된 P <0.01)을 비교하였다. 프로게스틴 처리시, SUMO-결함성 PR은 다중적인 증식성 및 생존 촉진성 생물 기능으로 지정된 유전자 세트들을 현저하게 상향 조절시켰지만, WT는 그렇지 않았다 (도 3.3). SUMO-결함성 PR을 안정적으로 발현하는 유방암 세포는 WT 또는 인산화-결함 S294A PR을 안정적으로 발현하는 세포에 비해, 소프트-아가에서 증식 증가를 나타내었다 (Daniel et al., 2007a; Daniel and Lange, 2009). 본원에서, MTT 증식 분석을 유도 모델을 이용하여 수행하였다 (도 4A). 이러한 이소제닉 시스템의 이점은 세포 생장/사망 비율과 안정적인 세포주 모델에서 발생할 수 있는 표현형 드리프트 (phenotypic drift)에서의 클론 편차를 없앤다는 것이다. 세포를 1일에 동일 밀도로 접종하고, PR 발현을 유도하기 위해 AP21967 화합물을 처리 또는 무처리한 다음, 비히클 (에탄올) 또는 R5020에 노출시켰다. R5020-처리한, iWT 또는 iKR PR 발현성 세포는 비-유도 또는 무-처리한 카운터파트들 보다 빨리 증식하였다. 그러나, AP21967 및 R5020 둘다에 지속적으로 노출시킨 6일까지, iWT 수용체를 발현하는 배양물 보다, iKR 발현성 배양물에 세포 수가 유의하게 더 많이 존재하였으며, 대조군들은 모두 비슷하게 유지되었다. 웨스턴 블롯팅을 통해, 세포 배양물에 AP21967을 첨가하는 경우 유도성 PR 발현이 지속되며, 상응하는 수준의 iWT 및 iKR PR 단백질이 발현되는 것으로 확인되었다 (도 4B).
MTT 분석은 시간 경과에 따른 생활성 (생존성) 세포를 측정하며, PR은 유방암 세포의 생존 촉진과 관련있는 것으로 보고된 바 있다 (Lange, 2008; Moore et al., 2000). 따라서, 세포자살의 간접 표시 인자로서 폴리(ADP-리보스) 중합효소 1 (PARP)의 절단을 측정하였다. PARP는 활성화된 카스파제-3에 의한 Asp214에서의 절단의 타겟으로서, 위임된 세포자살 신호전달에 대한 민감성 지표이다 (Nicholson et al., 1995). PR 발현을 AP21967 처리로 유도하고, 세포에 R5020을 6시간 전처리하여 각각의 iWT 또는 SUMO-결함성 iKR 유전자의 발현 프로그램을 활성화하였다. R5020 전처리 후, 세포 배양 배지에 독소루비신을 첨가하여, 1일 간 세포자살을 유도한 다음, 세포 용혈물을 회수한 후, 웨스턴 블롯팅에 의해 절단된 PARP의 상대적인 수준을 측정하였다 (도 4C). 특히, SUMO-결함성 iKR PR를 발현하는 독소루비신-처리된 세포는 iWT PR 발현 세포에 비해, 특히 R5020 전처리된 세포에서 PARP 절단 수준이 감소되었다 (레인 4와 8을 비교함). 독소루비신 처리는 WT 및 KR PR 단백질 발현을 둘다 감소시켰다 (도 4C, 1번 및 3번 레인을 비교하거나, 5번과 7번 레인을 비교함). 그러나, 단백질 발현 변화에 대해 표준화한 복수의 반복 실험들에서, iKR PR을 발현하는 세포는 iWT PR 발현성 세포에 비해 PARP 절단율이 일관적으로 낮게 나타났다. 이러한 사실은 PR을 안정적으로 발현하는 T47D 세포에서 검증되었다. PR-null 세포와, WT 또는 KR PR 중 어느 하나를 안정적으로 발현하는 세포를 완전 배지에 접종하고, 혈청 고갈 후, 독소루비신 처리 또는 무처리하면서 R5020을 처리하였다 (도 4D). 다시 말해, 프로게스틴-처리된 SUMO-결함성 KR PR을 발현하는 세포에서 현저한 세포 생존성 증가가 관찰되었다. 흥미롭게도, 이들 세포에 독소루비신을 세포독성 농도로 처리하였을 때, 생존성은 WT PR 발현성 세포에 비해 2배가 되었다 (도 4D). 이들 데이타는, SUMO-결함성 PR이 세포 증식 증가 및 생존 촉진 신호전달의 중요한 매개 인자라는 것을 시사해주며; 변형된 PR을 발현하는 세포는 이와 관련된 유전자 발현 프로파일과 일치되는 생물학적 프로세스를 이행한다 (도 1).
SUMO-결함성 PR 유전자 시그니처는 ERBB2 양성 유방암과 관련되어 있다. 인간 유방암은 종종 MAPK, AKT, 및/또는 CDK 단백질 및/또는 키나제 활성을 높은 수준으로 포함하고 있어, PR 억제에 우호적이다 (Daniel et al., 2007a; Daniel and Lange, 2009). 포스포-PR-유동성 (SUMO-결함) 병변을 시사하는 유전자 패턴의 증거로 공개된 인간 유방암 데이타베이스를 입증하기 위해, WT 수용체를 발현하는 세포에 상대적으로 KR 발현성 세포에서 발현 수준이 더 높은 유전자들로 구성된 고유한 PR 유전자 시그니처를 규명하였다 (발현 >1.5배, KR vs. WT, BH 보정된 P <0.01). 이들 유전자들은 KR 수용체를 발현하는 세포에서 주로 상향 조절되거나 및/또는 WT 수용체를 발현하는 세포에서만 하향 조절된다. 이러한 분석은, 리간드-의존성 및 리간드-비의존성 PR 타겟 유전자 둘다에 대해 수행하였다. 이러한 기준을 이용하여, 고유한 151- 및 92-유전자 시그니처를 구축하였으며, 각각 리간드-의존성 (LD) 및 리간드-비의존성 (LI) KR 수용체에 의해 (WT에 비해) 차별적으로 상향 조절된 PR-타겟 유전자로 정의하였다 (도 4.1).
이들 유전자 시그니처를 온코마인 검색 프리미움 에디션 (Compendia Bioscience, Ann Arbor, MI)에 업로딩하고, 관련 컨셉을 데이타베이스에서 조사하였다 (Rhodes et al., 2007에서 리뷰됨). 온코마인 컨셉은 특정 기준으로 정의된 유전자 리스트이다 (예, 특정 종양 코호트애서 과다 발현된 상위 유전자들). LD 151-유전자 시그니처는 복수의 유방암 컨셉과 연관되어 있었으며, 유의성 수준이 높았다 (P <0.0001, FDR <0.01) (데이타 도시 안함). 특히, 5개의 구분되는 ERBB2-양성 유방암 컨셉 (2개의 세포주 유래이고, 3개는 종양 코호트 유래임)은 독립적으로 LD PR-유전자 시그니처와 연관되어 있었다. 즉, SUMO-결함성 PR을 발현하는 세포에서 프로게스틴 존재시 특이적으로 상향 조절된 유전자들은, ERBB2-양성 유방암에서 고도로 과다 발현된 동일한 유전자 (상위 5-10%)들이었다 (도 5A, 데이타 도시 안함). 특히, LI 92-유전자 시그니처는 하나 이상의 ERBB2-양성 컨셉과 유의하게 연관되어 있었다 (Bonnefoi et al., 2007). 이들 데이타는, 리간드-의존성 및 -비의존성의 고유한 PR-조절된 유전자 세트들이 ERBB2-양성인 것으로 공지된 것을 비롯하여 단백질-키나제-유발성 종양에서 유의하게 상향 조절된다는 것을 보여준다 (도 5A).
이들 관련된 유전자 프로그램 (SUMO-결함성 PR 및 ERBB2 신호전달)의 발현은 세포가 증식 및 생존하도록 유방암 세포에 의해 이용되는 독립적인 수단을 의미하는 것일 수 있다. 실제, HER2-농화된 유방암은 대개 스테로이드 호르몬 수용체 (SR)에 음성이다 (Perou et al., 2000; Sorlie et al., 2001). 다른 예로, 이들 통계적으로 유의하게 연관된 컨셉들은 기능적으로 연결될 수도 있다. 내강 유방암은 주로 SR-양성이지만, 내강 A 종양의 대략 7%, 그리고 내강 B 종양의 20%가 HER2-농화된 형태이다 (Cheang et al., 2009; Prat and Perou, 2011). ERBB2 과다발현 (도 5A) 및 SUMO-민감성 (상기 기술됨)과 공동-연관된 선택 유전자들의 PR- 및 MAPK-의존성 조절을 HER-증폭된 유방암 세포에서 검사하였지만, 구성적으로 활성화된 MAPL를 포함하는 SR-양성의 BT-474 유방암 세포는 검사하지 않았다 (Lenferink et al., 2001). RU486 처리는 이종 모델에서 BT-474 종양의 증식을 현저하게 저해하였으며 (Liang et al., 2007), 6일간 시험관내에서 수행된 MTT 분석에서 BT-474 세포의 증식을 유의하게 차단하였으며; 비슷한 결과는 MEK 저해제, U0126을 이용한 경우에도 관찰되었다 (데이타 도시 안함). 먼저, WT PR이 아닌 주로 KR (및 ERBB2-연관된; 도 5A의 열들을 참조함)에 의해 조절되는 PR 타겟 유전자 (CHN2 및 RGS2)의 발현을, PR SUMO일화에 둔감한 대조군 유전자 (ACOT6; WT 및 SUMO-결합성 PR에 의해 동일하게 상향 조절됨, 도 1F)에 대해 상대적으로 측정하였다. 주목하게도, R5020 처리는 PR-B Ser294 인산화 증가 (2번 레인)와 BT-474 세포에서 CHN2 및 RGS2의 명확한 상향 조절을 유도하였으며; 각각 17배 및 26배였다 (도 5B). KR PR (~20배)에 비해 T47D WT PR 발현성 세포 (~2배)가 RGS2 발현에 대해 R5020 처리에 대해 덜 민감함을 상기하여야 한다 (도 1F). ACOT6 발현은 또한 R5020에 의해 유도되며; 3가지 유전자 모두 발현은 항-프로게스틴 RU486에 의해 완전히 차단되었다 (도 5B). CHN2 및 RGS2 mRNA 발현이 최대일 때 (+R5020; 1번과 2번 레인 비교), 포스포-Ser294 PR이 쉽게 검출되지만, 아마도 활성화된 PR 종들의 리간드-의존적인 (프로테아좀-매개) 하향 조절로 인해, 전체 PR 수준이 크게 감소하며, 검출불가능한 수준으로 보인다는 것에 유념하여야 한다 (Lange et al., 2000). 이들 세포에 MEK 키나제 저해제, U0126을 전처리하면 R5020-유도성 PR Ser294 인산화가 차단되며, 부분적으로, 그러나 유의한 수준으로 CHN2 및 RGS2 발현이 모두 감소된다 (6번 레인). 반면에, PR SUMO-상태에 영향을 받지 않는 대조군 유전자인 ACOT6는 MEK 키나제 저해제에 완전히 둔감하였다. 이러한 데이타는 본 발명의 가설을 뒷받침해주며, 인산화 현상이 비변형된 (즉, WT PR을 함유한) SR-양성 유방암 세포에서 SUMO-결함성 PR 유전자 시그니처의 발현과 PR-유발성 증식에 기여한다는 것을 입증해준다. CHN2 및 RGS2 (도 5B)와 마찬가지로, ERBB2 과다 발현성 내강 유방암에서 상향 조절된 상당수의 유전자들이 실제 PR-유발성인 것으로 추측된다.
전술한 사실들은, 본 발명의 세포주 모델에서 유래된 PR 유전자 시그니처가 공개된 인간 유방 종양 코호트에서 종양 단계, 림프절 양성 (node positivity) 및 환자 생존에 대한 예측 인자인지에 대한 검사를 촉구하였다. 예를 들어, Loi et al. Dataset (Loi et al., 2007)에서는 먼저 ER 양성/PR 양성인 유방 종양을 가진 환자에 대한 가장 큰 생존 데이타 콜렉션들 중 하나를 보여주었다. 메타유전자들 (Huang et al., 2003)을 각 샘플을 나타낸 (PR-null, WT PR, KR PR; R5020 처리 또는 무처리) 본 발명의 T47D 마이크로어레이 데이타세트로부터 분리하였다. 카플란-메이어의 생존 분석을 이용하여, PR-관련 메타유전자를 발현하는 환자 종양 (WT 또는 KR, -/+R5020)을 다른 전체 환자 종양과 비교하였다. 이 분석을 통해, 임의의 PR 유전자 시그니처 (즉, 전사 활성형의 PR임을 나타내는)를 발현하는 종양을 가진 이러한 종양 코호트의 환자들은 유의하게 감소된 무-전이성-생존 (P = 0.000785; 도 5C)한다는 것이 밝혀졌다. 특히, PR-관련 메타유전자를 발현하지 않는 환자 종양 (도 5C, 상위 곡선)은 장기 생존율이 ~80%였다. 추측컨대, 이 그룹의 종양은 PR을 다량 발현하지만, 이들 수용체는 상대적으로 불활성형이다. 이러한 사실과 일관되게, PR mRNA의 고수준은 양호한 성과와 연관되어 있다 (Loi et al., 2007). 이러한 사실은, (활성 보다는) PR 발현을 기초로한 종양의 분류는 오해의 소지가 있다는 것을 시사한다. 흥미롭게도, 종양의 유전자 시그니처가 KR을 발현하는 T47D 세포 +R5020와 비슷한 환자는, 성과가 좋지 않은 경향이 있다 (P <0.1). 리간드-비의존성 (KR) PR 타겟 유전자의 기여를 포함시키기 위해, 종양이 KR 메타유전자들 (KR -R5020, 또는 KR +R5020)을 모두 발현하는 환자들을 조합하였다. 이들 환자는 종양에서 2가지의 KR 메타유전자 중 어느 하나를 발현하지 않는 환자 보다 생존성이 유의하게 감소되었다 (P = 0.0261) (도 5D). 림프절의 상태와 원발성 종양 단계와 관련하여, 메타유전자의 발현과의 명확한 연관성은 나타나지 않았다. 이들 데이타는, PR-의존성 전사, 특히, 탈SUMO일화된 (포스포-Ser294) 수용체의 작용이 (내강) 유방암 환자 서브세트에서의 종양 진행과 좋지 않은 성과의 원인임을 시사해준다.
PR 길항제, RU486 및 아글레프리스톤은 SUMO-결함성 PR을 발현하는 세포에서만 유전자 발현을 자극한다.
리간드-의존성 PR 프로모터 선택성은 그 수용체의 인산화 및 SUMO일화 상태에 의존한다. 아울러, PR 리간드 구조 역시 PR 활성에 영향을 미쳐, 타겟 유전자 조절에 대해 작용성 또는 길항성 특성을 야기한다. PR 리간드 (프로게스테론 및 R5020)는 강력한 작용제인 반면, PR 길항제 (RU486, 아글레프리스톤 및 오나프리스톤)은 일반적으로 PR 전사 작용을 차단하지만; 이들 리간드는 고유한 작용 기전을 가지고 있으며, 여러가지 세포 배경 하에 다양한 수준의 작용효과/길항효과를 촉발할 수 있다 (Cadepond, 1997). 이에, 본 발명에서는 야생형 PR (WT), SUMO-결함성 PR (KR) 또는 빈 벡터 (PR-Null)를 발현하는 T47D 유방암 세포에서 이들 길항제들의 전사 효과를 조사하기 위해, 유전자 발현 프로파일링을 사용하였다.
각 세포주에 아래 잠재적인 조건 8가지 중 한가지를 6시간 처리한 후, Illumina HT-12v4 마이크로어레이 플랫폼을 이용하여 글로벌 유전자 발현 프로파일을 측정하였다: (1) 에탄올 (비히클 대조군), (2) 프로게스테론 (10-8 M), (3) RU486 (10-7 M), (4) 아글레프리스톤 (10-7 M), (5) 오나프리스톤 (10-7 M), (6) RU486 + 프로게스테론, (7) 아글레프리스톤 + 프로게스테론, 또는 (8) 오나프리스톤 + 프로게스테론. 히트 맵 분석은 상향 또는 하향 조절된 상위 유전자들을 나열한다 (도 6) (하나 이상의 샘플 비교에서 배수 변화 >2.0, BH 보정된 P <0.01). 빈 벡터 (PR-null)를 발현하는 세포에서, 리간드 노출에 의존적인 유전자 발현의 유의한 변화는 관찰되지 않았다. 즉, 빈 벡터 세포에서의 발현 프로파일링은, PR 및/또는 리간드 의존적인 발현 수준 차이를 명확하게 설명해줄 수 있는, 필연적인 베이스라인 대조군을 제공해주었다. 예상한 바와 같이, 다수의 유전자들이 프로게스테론 (P4) 처리 후 WT 및 KR 세포들에서 모두 유의하게 상향 조절되었다 (도 6, 10번 및 18번 레인). PR 길항제의 전사 영향력을 파악하기 위해, 프로게스테론 처리 후 WT 또는 KR 세포에서 상향 조절 (배수 변화 >2.5, BH P 값 <0.01)된 유전자들을 모두 분리하여, 모든 샘플들에 대한 발현 값들을 히트 맵을 이용하여 나타내었다 (도 7). 여기서, PR-null 세포는 임의의 리간드 노출시 조절되지 않았으며, 비히클 대조군 (에탄올)으로만 처리된 WT 및 KR 세포들 역시 조절되지 않았다. RU486 또는 아글레프리스톤의 처리는 다수의 PR 타겟 유전자들의 상향 조절을 유발하였으며, 특히 WT 세포가 아닌 SUMO-결함성 PR (KR) 세포에서 상향 조절을 유발하였다 (도 7, 11-14 및 19-22번 레인 비교). 반대로, KR 세포에 오나프리스톤 (또는 오나프리스톤 + P4)을 처리하면, 이들 PR 타겟 유전자의 상향 조절이 야기되지 않았다 (도 7, 19-22번 레인 및 23-24번 레인 비교). 도 7에서 모든 샘플들에 대한 자율적인 계층 클러스터링을 통해, 에탄올 또는 오나프리스톤 처리된 KR 샘플은 PR-null 샘플과 가장 관련성이 높은 위치로 설정되었는데, 이는 오나프리스톤으로 처리된 KR 세포가 PR 타겟 유전자들을 유의하게 상향 조절하지 않는다는 것을 의미한다 (도 8). 전체적으로, 이들 데이타는, 오나프리스톤이 과잉 활성의 PR (즉, 인산화되고 탈SUMO일화된 PR)을 발현하는 세포에서 PR 타겟 유전자의 발현을 성공적으로 저해할 수 있음을 시사해준다 (도 7, 18번 레인과 23-34번 레인 비교). 그러나, 항-프로게스틴 RU486 및 아글레프리스톤은 과잉 활성의 PR을 발현하는 세포에서 상당한 작용제 활성을 가진다 (도 7, 18번 레인과 19-22번 레인 비교). 따라서, 공격적인 종양을 가진 유방암 환자는 PR 길항제, 특히 오나프리스톤을 포함하는 치료로부터 상당히 효과를 얻을 수 있을 것으로 예상된다. 이들 데이타는 RU486을 유방암 치료제로 조사한 기존의 II상 임상 실험이, 아마도 과잉 활성의 PR을 발현하는 세포에서 RU486의 실질적인 작용성 특성으로 인해, 성공하지 못한 이유를 설명하는데 도움이 될 수 있기 때문에, 특별한 임상적인 의미를 가진다 (Perrault, 1996).
유전자 마커는 활성화된 PR에 의해 유발된 종양을 동정할 수 있다.
본원에서, 전사적으로 과잉 활성형의 PR (즉 인산화되고 탈SUMO화된 PR)이 유방암 세포 증식의 유발자이며 (도 4), 상승된 HER2 신호전달과 연관되어 있으며 (도 5), 감소된 무-전이 생존의 예측 인자 (도 5)라는 상당한 증거들이 제공되었다. 이에, WT 발현 세포와 활성화된 (KR) 발현 세포를, 높은 민감도와 특이성을 확보하기 위해 3가지의 독립적인 통계적 방법을 이용하여, 구별할 수 있는, PR-의존적인 유전자 마커 (유전자)들을 동정하고자 하였다.
전술한 바와 같이, 여러가지 실험적인 의문들을 해결하기 위해, 2가지의 독립적인 유전자 발현 마이크로어레이 실험을 수행하였다. 제1 실험에서는 프로게스틴-의존성 PR 타겟 유전자를 조사하였고, 제2 실험에서는 PR 발현성 세포에서 항-프로게스틴의 역할을 조사하였다. 이로써, 각 실험의 리플리케이트 샘플을 조합하고, WT 또는 KR 발현성 세포들 간의 유전적 차이를 프로게스틴 노출 조건 하에 조사하는 것이 가능하였다.
WT 또는 KR PR을 발현하는 세포들을 구별할 수 있는 유전자 (마커) 15개가 동정되었다 (표 1a). 이들 유전자를 파악하기 위해, WT 또는 KR 세포에서 발현된 유전자들의 리플리케이트 발현 값들을 비교하고, 프로게스틴 처리하고 (도 9, 10번과 18번 레인 비교), 미리 결정된 유의성 역치 P-value, 배수 변화 및 ROC (receiver operator characteristic) 곡선 분석을 통과한 유전자들을 분리하였다 (통계적 유의성에 필요한 기준에 대한 상세 내용은 방법 참조). 또한, 이들 그룹들 간에 임의의 유의성 검사를 통과하지 못한, 즉 이들 세포들 간의 유전자 발현 차이가 없음을 의미하는, 12개의 공지된 하우스키핑 유전자 (프로브 세트 30)들도 검증하였다 (즉, 어레이 샘플들 간에 기술적 에러는 없음) (데이타 도시 안함). 따라서, 이들 15종의 유전자들의 발현 수준이, 세포가 "활성화된 PR" (인산화된, 탈SUMO일화된 PR) 또는 WT PR에 의해 유발된 것인지를 정확하게 나타낸다는 점은 우수한 신뢰성을 가진다.
2차 분석에서, 유전자 차별적인 발현을, 모두 프로게스테론 + 항-프로게스틴인 오나프리스톤을 공동-처리한, KR 및 WT 세포들 간에 조사하였다. 표 2에서 알 수 있는 바와 같이, 오나프리스톤 처리는, 유전자 1종을 제외하고는, 전술한 15개의 바이오마커 유전자들의 유의한 차별적인 발현을 완전히 무효화하였다. 표 2는 KR (프로게스테론 + 오나프리스톤) 및 WT (프로게스테론 + 오나프리스톤) 세포들 간에 유의하게 차별적으로 발현되는 유전자 8종만을 나타낸다. 오리지날 유전자 15종들 중, 오직 하나, 즉 CDH10은 여전히 2가지 그룹들 간에 유의하게 차별적으로 발현되었다. 실험의 본 파트에서 하우스키핑 유전자 분석 (데이타 도시 안함)을 통해, 본 실험 및 분석 방법을 검증하였으며; 하우스키핑 유전자 12종을 타겟팅하는 프로브 30종 모두 본 발명의 전체 실험들에서 유의성 기준에 따라 유의한 것으로 확인되지 않았다 (방법 참조).

3차 분석에서, 프로게스테론 + 항-프로게스틴 RU486으로 공동 처리한 양쪽 KR 세포와 WT 세포 간의 차별적인 유전자 발현을 조사하였다. 표 3에서 알 수 있는 바와 같이, RU486 처리는, 유전자 1종을 제외하고는, 전술한 15개의 바이오마커 유전자들의 유의한 차별적인 발현을 완전히 무효화하였다. 표 3은 KR (프로게스테론 + RU486) 및 WT (프로게스테론 + RU486) 세포들 간에 유의하게 차별적으로 발현되는 유전자 7종만을 나타낸다. 오리지날 유전자 15종들 중, 오직 하나, 즉 CDH10은 여전히 2가지 그룹들 간에 유의하게 차별적으로 발현되었다. 실험의 본 파트에서 하우스키핑 유전자 분석 (데이타 도시 안함)을 통해, 본 실험 및 분석 방법을 검증하였으며; 하우스키핑 유전자 12종을 타겟팅하는 프로브 30종 모두 본 발명의 전체 실험들에서 유의성 기준에 따라 유의한 것으로 확인되지 않았다 (방법 참조).

다음으로, 항-프로게스틴인 오나프리스톤으로 KR 세포 처리시 전술한 전사 차이를 반전시킬 수 있는지를 테스트하였다 (표 1a). 동일한 엄격한 분석을 하기 2가지 그룹들을 비교하여 반복하였다: 오나프리스톤 + 프로게스테론 처리한 KR 발현성 세포와 비히클 대조군 (에탄올) 처리한 WT 발현성 세포. 2가지 그룹들이 유의하게 차별적으로 발현되는 유전자 5종 외에는 유전적으로 구분할 수 없다는 것을 확인하였다. 프로게스틴은 PR 타겟 유전자 상에서 수천개를 명확하게 활성화할 수 있지만, 이들 2가지 그룹들 간에 유의하게 차이가 있는 오직 유전자 5종만 동정되었다 (표 4). 이들 데이타는, 오나프리스톤 처리 (심지어 프로게스테론 노출시에도)가 활성화된 PR을 발현하는 세포에서 프로게스테론 노출 효과를 효과적으로 반전시킬 수 있다는 것을 시사해준다. 즉, 하우스키핑 대조군 유전자들은 이들 2가지 그룹들 간에 유의하게 차별적으로 발현되지 않았다 (데이타 도시 안함).

마지막으로, 항-프로게스틴인 오나프리스톤으로 KR 세포 처리시 전술한 (표 1a) 전사 차이를 반전시킬 수 있는지를 테스트하였다. (프로게스테론 + 오나프리스톤) 처리한 WT 세포와 에탄올 (비히클 대조군)만 처리한 WT 세포 간의 비교 분석을 수행하였다. 표 5에서 알 수 있는 바와 같이, 2가지 그룹들 간에 유의하게 차별적으로 발현된 상태로 유지되는 유전자는 오직 1종만 있었다. 이는, 항-프로게스틴인 오나프리스톤으로의 처리가 WT PR 수용체를 가진 세포에서 프로게스테론을 이용한 자극에 의해 유도된 전사 차이를 거의 완벽하게 반전시킬 수 있다는 것을 명확하게 입증해준다. 실험의 본 파트에서 하우스키핑 유전자 분석을 통해, 하우스키핑 유전자 12종을 타겟팅하는 프로브 30종 전체 실험들에서 유의성 기준에 따라 유의한 것으로 확인되지 않았다 (방법 참조).
| 유전자 | KR에서 상향 또는 하향 조절 |
WT (평균 ± SD)
n=5 |
KR (평균 ± SD)
n=5 |
P 값
α=1.06E-06 |
배수 변화
(KR/WT) 또는 -(WT/KR) |
ROC AUC | 프로브 ID | 등재 번호 | 프로브 서열 |
| GPR124 | 상향 | 6.651 ± 0.063 | 7.707 ± 0.068 | 5.27E-07* | 1.159* | 1.000* | ILMN_1773059 | NM_032777.6 | CTAGGGTTCCCTCCCAGTCTTCACATCACTCTGGCCTCATCACCAAGGTG 서열번호 52 |
| 표 5. 프로게스틴-자극 및 오나프리스톤-처리된 KR 세포와 비히클 대조군 (에탄올) 처리된 WT 세포 간에 유의하게 차벌적으로 발현되는 유전자 1종. (*) 해당 방법의 기준에 따라 통계적으로 유의함. | |||||||||
활성화된 PR에 의해 상향 조절된 유전자는 항-프로게스틴에 의해 억제된다.
전술한 통계적 분석은, 프로게스틴의 존재 하에 WT 또는 SUMO-결함성 PR을 발현하는 세포들을 서로 구분할 수 있는 유전자 15종을 신뢰할 수 있는 수준으로 동정하기 위한 강력한 민감도 및 특이성 기준을 제공해주었다. 그러나, 프로게스틴 처리에 반응하여 특이적으로 (즉, P4/에탄올 비)상향 조절되는 다른 PR 타겟 유전자들도 다수 있었다. 프로게스틴에 의해, 특히 활성화된 PR (KR)을 발현하는 세포에서, WT와 비교하여, 유도된 PR 타겟 유전자를 조사하였다. 이들 유전자 산물은 활성화된 PR 전사 프로그램의 활성화 인자일 것이다. 실제, 유전자 151종의 리간드-의존성 "KR>WT" 유전자 시그니처로서 이들 PR 유전자 타입들이 개시되었다 (도 2b, 4.1).
전술한 마이크로어레이 실험들은 각각 직접 비교할 수 있는 매우 유사한 처리 조건을 포함한다: WT 또는 KR PR을 발현하는, 비히클 또는 프로게스틴 처리 세포. 이들 실험들 간의 유일한 차이는 제1 실험에서는 프로게스틴 처리 R5020이었으며, 재2 실험에서는 천연 PR 리간드 프로게스테론 (P4)였다. 따라서, 거의 동일한 조건에서 제2 마이크로어레이 실험을 수행함으로써, 각 실험의 결과들을 비교하고, WT PR에 비해 SUMO-결함성 PR을 발현하는 세포에서 특이적으로 상향 조절되는 재현성이 높은 프로게스틴-의존적인 PR 타겟 유전자 세트로 수렴하는 기회가 생겼다. 그 결과, "활성화된 PR"을 발현하는 유방암 세포에서 사향 조절되는 확실한 유전자 리스트가 동정되었으며, 여기서 PR은 인산화 및 탈SUMO일화된 것이다.
제1 마이크로어레이 실험에서, WT PR에 비해 SUMO-결함성 PR을 발현하는 세포에서 특이적으로 상향 조절된 (배수 변화 >1.5, BH 보정된 P <0.01) 프로게스틴-의존성 PR 타겟 유전자 151종 (도 4.1)이 동정되었다. 본 분석을 제2 마이크로어레이 실험에서도 반복하였으며, 양쪽 실험으로부터 중복되는 유전자들을 분리하였다. 이로써, 유전자 151종 리스트는 유전자 29종으로 축소되었으며, 이들은 특히 WT PR에 비해 SUMO-결함성 PR을 발현하는 세포에서 프로게스틴 처리 (R5020 또는 프로게스테론)에 반응하여 상향 조절된 것이다 (도 10, 10번과 18번 레인 비교). 이들 유전자 29종은 프로게스틴에 반응하여 SUMO-결합성 PR 발현에 대한 재현성이 높은 마커인 것으로 생각된다 (즉, 2종의 PR 작용제, 즉 R5020과 프로게스테론을 처리한 2번의 독립적인 실험에서 유래된, 총 5개의 리플리케이트).
제2 마이크로어레이 실험으로부터 수득한 발현 데이타에 따르면, RU486과 아글레프리스톤은 SUMO-결함성 PR을 발현하는 세포에서 작용 특성을 가지는 반면, 오나프리스톤은 WT 또는 KR PR을 발현하는 세포에서 효과적인 길항제인 것으로 확인되었다. 따라서, "활성화된 PR" 유전자 리스트는, 오나프리스톤 처리 단독 (없음) 또는 오나프리스톤 + P4 처리 (유전자 13)에 의해 심지어 보통 수준으로 자극받은, 20종의 유전자들 중 임의의 것을 제거함으로써, 더 좁힐 수 있다. MSX2, MAP1A 및 PDK4가 포함된 최종 유전자 16종으로 이루어진 리스트가 수득되었다 (표 1b, 도 11). 이들 유전자 3종을 다중 유전자 발현 및 ChIP 실험에서 광범위하게 연구하였으며, SUMO-결함성 PR을 발현하는 세포에서의 특이적인 조절이 나타났다. 실제, 유전자 16종 중 7개는 정교한 경로 분석에 의해 확인된 바와 같이, 암 관련 기능에 참여한다. 이들 PR 타겟 유전자 16종은, PR이 인산화 및 탈SUMO일화되었을 때 프로게스틴에 의해 명확하게 상향 조절되며, 이들 유전자의 산물이 종양의 공격성을 높이도록 작용한다는 결론에 도달하였다 (도 2, 4, 5).
| 유전자 | KR에서 상향 또는 하향 조절 | 프로브 ID | 등재 번호 | 프로브 서열 |
| THY1 | 상향 | ILMN_1779875 | NM_006288.2 | CTGAGGCAAGCCATGGAGTGAGACCCAGGAGCCGGACACTTCTCAGGAAA 서열번호 16 |
| KLF9 | 상향 | ILMN_1778523 | NM_001206.2 | GCCCTTCACCATTGTGGAATGATGCCCTGGCTTTAAGGTTTAGCTCCACA 서열번호 17 |
| SPINK5L3 | 상향 | ILMN_1697543 | NM_001040129.2 | GCAGACTGCCCCAATGTGACAGCACCTGTTTGTGCCTCAAATGGCCACAC 서열번호 18 |
| PHLDA1 | 상향 | ILMN_1687978 | NM_007350.3 | AACAGTCTCTCCGCCCCGCACCAGATCAAGTAGTTTGGACATCACCCTAC 서열번호 19 |
| MAP1A | 상향 | ILMN_1701558 | NM_002373.4 | CCCAAGCAAGCCAGTGAGCAGCCCTGCCAGACTACTGCCAGACTGAGAAA 서열번호 20 |
| SPRYD5 | 상향 | ILMN_1753648 | NM_032681.1 | TCCCTGATATACACCATCCCCAATTGCTCCTTCTCACCTCCTCTCAGGCC 서열번호 21 |
| ATG12 | 상향 | ILMN_2188204 | NM_004707.2 | GAGTCGTGATTGTACCACTGCATTCCTGCTGAGCAACAGAGTGAGACCCC 서열번호 22 |
| PDK4 | 상향 | ILMN_1684982 | NM_002612.3 | CAGAAGTCCTAGACAGTGACATTTCTTAATGGTGGGAGTCCAGCTCATGC 서열번호 23 |
| MSX2 | 상향 | ILMN_1766951 | NM_002449.4 | AGGTACATTCATCCTCACAGATTGCAAAGGTGATTTGGGTGGGGGTTTAG 서열번호 24 |
| TUBA3E | 상향 | ILMN_1652464 | NM_207312.1 | GGTCCCCAAAGACGTCAATGCGGCCATCGCCACCATCAAGACCAAGCGCA 서열번호 25 |
| TSC22D1 | 상향 | ILMN_1692177 | NM_006022.2 | TCCCAATGGTGTAGACCAGTGGCGATGGATCTAGGAGTTTACCAACTGAG 서열번호 26 |
| TUBA3D | 상향 | ILMN_2215639 | NM_080386.1 | TCCCCTGCCACCCCCGGGATGGCTGCTTCCAAGTTGTTTGCAATTAAAGG 서열번호 27 |
| KHDRBS3 | 상향 | ILMN_1691747 | NM_006558.1 | AGGCACCTTCAGCGAGGACAGCAAAGGGCGTCTACAGAGACCAGCCATAT 서열번호 28 |
| UTS2D | 상향 | ILMN_2180232 | NM_198152.2 | GCTGGTATATCCAGTGCATTGTTGGCACCATGGGACCAGAAGGTGGTGAC 서열번호 29 |
| SLC35C1 | 상향 | ILMN_1680104 | NM_018389.3 | AGGGTGGCTTGCAGTCCCTGGCCCTTCTGGTGGGCATTTGGTATGTCCTT 서열번호 30 |
| KIAA0513 | 상향 | ILMN_1693233 | NM_014732.2 | CTTCTTGAACCTGGTGGCCCCCGTTGGAACTATCAGTGGCGTCTCCCATG 서열번호 31 |
| 표 1b. | ||||
고찰
본 실험에서, PR SUMO 변형이 전사 활성과 프로모터 선택에 영향을 미치는 방식을 더 잘 이해하기 위해, 유전자 발현 프로파일링을 수행하였다. 신규로 조작한 유방암 세포주 모델을 이용하여, 인간 종양에 존재하는 (탈SUMO일화된) PR-유발된 유전자 시그니처를 동정하였으며, 이는 환자 생존성 감소와 연관되어 있었다. 앞서, Ser294에서의 PR 인산화가 Lys388에서의 PR SUMO일화를 길항한다는 것이 입증되었다 (Daniel et al., 2007a). 본원에서, 새로운 데이타는, 유방암 세포가 세포 증식 및 생존 촉진 경로를 활성화하는 타겟 유전자 쪽으로 PR 전사 작용을 이동시키기 위해 이러한 기전을 활용할 수 있음을 암시해준다 (도 4, 5). 글로벌 유전자 발현 수준을 분석하기 위한 바이오인포메틱스를 이용하여 (도 1), 전사 반응에 있어 놀라운 차이가 WT PR과 탈SUMO일화된 PR 간에 확인되었으며, 이는 프로모터/인핸서 선택성에서의 변형으로서 ChIP 분석을 통해 추가로 특정화되었다 (도 3, 3.1). 아울러, 비변형된 유방암 세포 (또는 WT PR-B만 발현하는 세포)에의 EGF의 처리는, PR Ser294 인산화가 선택된 PR 타겟 유전자의 전사 탈억제에 관련되어 있음을 추가로 보여준다 (도 2). 특히, SUMO-결함성 PR (즉, 포스포-PR 유발된)에 의해 특이적으로 상향 조절되는 유전자들은 ERBB2-양성인 인간 유방 종양과 세포주들에서 다량 발현되는 유전자들과 유의한 연관성이 있으며; 연구들은 인산화된 (탈SUMO일화된) PR-B-특이적인 전사 작용과 ERBB2-관련 유전자들의 서브세트의 발현 간에 역학적인 연관성을 뒷받침해준다 (도 5). 요컨대, 데이타는, 엔도크린 내성으로의 유방암의 조기 및 신속한 진행에 포스포-PR 의존적인 전사 조절 기전과 이런 활성의 잠재적인 기여에 대한 추가적인 연구 필요성에 대한 강력한 근거를 제시한다.
유전자 발현 분석은 SUMO일화-민감성 PR 타겟 유전자들을 동정한다. PR SUMO일화가 HBEGF, IRS1, 및 STC1 등의 내인성 유전자 좌 몇몇을 전사적으로 억제한다는 것은 이미 보고되어 있으며 (Daniel et al., 2007a; Daniel and Lange, 2009); 이들 유전자 3종 모두 유방암 세포 증식에 기여하는 것으로 알려져 있다 (Beerli and Hynes, 1996; Byron et al., 2006; Chang et al., 2003). 본원에서, 포괄적인 실험 세트를 수행하여, WT PR 또는 SUMO-결함성 돌연변이 K388R PR (포스포-모방) 중 어느 하나를 발현하는 T47D 세포에서, 합성 프로게스틴인 R5020을 처리 또는 무처리하고, 전체 게놈 발현 프로파일링을 위한 현행 마이크로어레이 기법들을 이용함으로써, 내인성 PR 타겟 유전자의 조절을 측정하였다. 가역적인 PR SUMO일화의 역할을 조사하는 것과는 별개로, 이 마이크로어레이 데이타세트는 프로게스틴 처리에 따른 WT PR-B 전사 작용에 대한 최신의 잘-통제된 분석을 제공해준다 (새롭게 구축된 백터 일치시킨 세포주를 이용함). 구성적 (안정한) 또는 유발성 WT 또는 돌연변이 PR을 발현하는 추가적인 세포주 및 새로운 세포주 클론들을 이용하여 엄격한 독립적인 실험을 수행하였으며, 유전자 발현 수준은 별개의 마이크로어레이 플랫폼 (Illumina 및 Affymetrix)을 이용하여 측정하였다. 실제, 본 분석에서 기존에 동정된 PR 타겟 유전자들 중 70%가 검증되었지만 (Jacobsen et al., 2005; Richer et al., 2002), 또한 새로운 미공개 PR 타겟 유전자 수백개도 검증되었는데, 이들 다수가 리간드-의존적인 예에 해당되었다. 본 데이타세트는 리간드-의존성 및 리간드-비의존성의 PR-매개 전사 조절 기전을 조사하는 향후 연구에 중요한 자원을 제공한다.
특히, WT 대 KR PR에 의해 조절되는 유전자들의 비교를 통해, PR 조절된 유전자들 대부분이 상당히 중복되는 것으로 드러나, SUMO일화/탈SUMO일화에 의한 PR-B의 다이나믹 변형이 상대적으로 둔감한 것으로 보인다 (도 1D-E, 벤 카테고리의 중복). 그러나, 이들 카테고리에서, WT 또는 KR PR 중 어느 하나에 의해 조절되었을 때, 다수 유전자들은 중간 (바뀐) 발현 수준을 나타내었는데, 이는 PR 매개 전사에, 부분적으로는 PR SUMO일화 상태에 따라 다수 기전들이 작용한다는 것을 시사해준다. 역으로, 유전자들의 더 작은 서브세트들은 PR의 SUMO일화-상태에 매우 민감하다 (도 1D-E, 중복되는 영역을 제외한 모든 벤 카테고리). 놀랍게도, 이들 서브세트는, WT 대조군에 비해 KR PR에 의해 상향 조절 및 하향 조절된 유전자들이 포함되어 있어, PR-B의 SUMO일화가 프로모터 배경에 따라 억제 또는 활성화될 수 있는 것으로 시사된다. 예를 들어, 다수의 증식성 유전자들이 증가되는데, 공지된 종양 억제 유전자의 다수는 탈SUMO일화된 (KR) PR에 의해 억제되었다.
기존 연구를 토대로 (Daniel et al., 2007a; Daniel and Lange, 2009), 포스포-Ser294-PR (즉, 주로 탈SUMO일화된)이 암 세포 표현형에 상당한 영향을 미치는 유전자 조절에 변동을 야기하는 것으로 추측되었다. 따라서, 본원에서는 전체적으로 새로운 유방암 세포 모델을 이용하여, 이들 유전자들을 동정하고, (WT 및 KR PR에 의한) 차별적인 조절 기전을 파악하고자 하였다. Ser294에서 인산화될 수 없어 상당히 SUMO일화되는 수용체인 S294A PR을 안정적으로 발현하는 세포에서 (Daniel et al., 2007a; Lange et al., 2000), KR-상향 조절된 선택 유전자 (예, MSX2 등)은 발현이 전체적으로 차단되었으며; 전사의 상향 조절은 PR K388R/S294A 이중 돌연변이 (KRSA; 도 2c)를 발현하는 세포에서 회복되었다. 이들 데이타는, PR SUMO 변형이 인산화 현상에 반응하여 효과적으로 "탈억제"되는 PR 타겟 유전자를 전사적으로 우세적으로 억제함을 보여준다. 예를 들어, PR-의존성 MSX2 및 RGS2 mRNA 발현은 WT PR 발현성 세포에 EGF 처리시 상당히 증가하였다 (도 2D). PR 인산화와 탈SUMO일화가 유방암 세포에서 PR 전사 활성과 프로모터 선택성을 놀랍게도 변형시킴으로써 글로벌 유전자 발현 패턴에 영향을 미친다는 결론에 도달하였다.
PR 프로모터 선택성에 작용하는 기전들. 마이크로어레이 연구들을 통해, PR SUMO 변형이 광범위한 PR 타겟 유전자의 발현을 변형시키지만 다른 것에는 영향을 미치지 않는다는 것이 명확해졌다. 프로모터 선택성 기전에 대해서는 거의 알려져 있지 않다. 하지만, 이 의문은 다른 SR 패밀리 멤버들에서 해소되고 있다 (Tang et al., 2011). SR과 염색질의 상호작용은 상당히 다이나믹하며, 빠르고 지속적인 교환으로서 이루어진다 (Hager et al., 2009). 따라서, 전사 인자가 "결합하는" 밀집된 영역은 (ChIP로 측정함) 실제 그 영역에서 전사 인자 점유율을 높이는 방향으로의 평형 이동을 반영한다. 컨센서스 DNA 서열로의 SR 결합, 멀티-단백질 복합체로의 공동-자극 인자들의 참여 및/또는 특정 세포 위치로의 SR 격리 뿐만 아니라 염색질 접근성을 조절하는 히스톤 변형 등의 복수의 인자들이 이러한 평형에 영향을 미칠 수 있다. 아울러, 제한효소 연구를 통해, 분산을 통해 가능한 것 보다 최대 1000배 빨리 효소가 컨센서스 서열에 결합되도록 하는 기전이 밝혀졌는데, 이는 결합을 촉진시키는 보조 인자들의 존재를 시사한다 (Halford and Marko, 2004). 마찬가지로, 최근 연구에서는, "파이오니아 인자"로 칭해지는 특정 단백질이 발생 조직 또는 암 특이 조건에서 염색질의 리모델링과 SR 전사 인자의 근처 게놈 결합부 (인핸서)로의 국지화를 보조한다는 것이 확인되었다 (Carroll et al., 2005; Hurtado et al., 2011; Lupien et al., 2008).
SUMO 분자의 부가에 의한 단백질 기질의 변형은 단백질-단백질 상호작용에 영향을 미칠 수 있거나, 및/또는 단백질 안정성, 국지화 또는 전사 활성을 변경시킬 수 있다 (Geiss-Friedlander and Melchior, 2007의 리뷰). PR SUMO일화 (Lys388)는 대부분 PR의 전사 활성을 종종 억제하며 (그러나, 프로모터 의존적인 양상으로 이를 증가시킬 수 있음; 도 1F BCL2L11 및 DNALI1), 프로테아좀 매개 턴오버를 통해 리간드-의존적인 PR 하향 조절 속도를 늦추는 경향을 보이지만 (Daniel et al., 2007a), PR 위치를 바꾸는 지는 것은 명확하지 않다 (Man et al., 2006). 본 발병의 분석에서 다수의 유전자들이 MSX2처럼 행동하며; 발현이 SUMO-결함성 KR PR에 의해 실질적으로 상향 조절되었지만, WT PR에서는 그렇지 않았다 (도 2c). 아울러, KR PR은 WT 수용체 보다 2-3배 높은 수준으로 MSX2 인핸서를 점유하였다 (도 3B). KR PR 수준 증가가 이 유전자 좌로 동원되고 증가된 MSX2 mRNA 발현과 연관되어 있다는 사실은, PR SUMO일화 (SUMO-민감성 인핸서 영역과 염색질 상황에서)가 PR DNA 결합 수준에서 발생하는 조인자 상호작용을 변형시킨다는 것을 시사한다. 이러한 사실과 관련하여, PIAS3, 즉 PR SUMO일화 E3 리가제는 시험관내에서 PRE DNA 서열과의 PR 결합을 직접 저해한다 (Man et al., 2006). 따라서, (KR이 아닌) WT PR과의 PIAS3-매개 SUMO 접합은 선택된 PRE 서열과 수용체가 효과적으로 결합하는 것을 방지할 수 있으며, 따라서 후속적으로 그 유전자 좌에서 PR 점유와는 거리가 멀게 평형이 이동한다. 이런 기전이 서열 특이적인지 또는 프로모터 특이적인지는 미완으로 남아있다.
프로모터 구조는 PR 등의 SUMO일화된 전사 인자에 의한 프로모터 선택에 중요한 결정 인자일 가능성이 있다. Holmstrom 등 (Holmstrom et al., 2008)은, SUMO일화된 GR이 전사를 효과적으로 저해하기 위해서는 복수의 GR 결합부가 포함된 DNA와의 안정적인 상호작용을 필요로 한다는 것을 밝혀냈다. 흥미롭게도, 글루코코르티코이드 수용체 (GR)의 SUMO일화 역시 연관된 내인성 유전자들의 전사 유도에 선택적으로 작용한다 (Holmstrom et al., 2008). 이런 사실과 관련하여, 최근 염색질 변형 맵핑 연구들을 통해, 인핸서에서의 히스톤 H3 Lys4 모노- 및 다이메틸화(H3K4me1/2)가 전사적으로 활성형인 유전자들과 연관되어 있는 것으로 확인되었다 (He et al., 2010; Heintzman et al., 2007). 실제, DNA 반응 인자들에 전사 인자들이 접근가능한 영역들이 DNase 또는 MNase 과민감 부위로서 먼저 동정되었는데, 이들 영역은 비교적 점유된 뉴클레오솜으로부터 자유롭기 때문이었다 (ENCODE Project Consortium, 2007). H3K4me2는, 전사 인자가 결합하기 위한 영역의 뉴클레오솜 리모델링과 접근용이성을 촉진시키기 위해 추가적인 단백질 (파이오니아 인자)을 동원할 수 있는 기능성 인핸서로서, 후생 마커인 것으로 생각된다 (He et al., 2010). 본 발명자들은, PR 동원에서 이 파이오니아 인자들을 동정하지 않았지만, 본 실험에서, WT PR에 비해 SUMO-결함성 KR PR을 발현하는 세포에서 MSX2 인핸서에서의 H3K4 다이메틸화 증가를 관찰하게 되었다. 이 모델에서, 탈SUMO일화된 PR은 히스톤 메틸트랜스퍼라제, MLL2를 (즉, MSX2 인핸서로) 선호적으로 동원하여, SUMO일화된 수용체에 의해 정상적으로 억제되는 활성부에 생산적인 전사 복합체 형성을 가능하게 하는 지속적인 H3K4 다이메틸화를 형성할 수 있다.
마지막으로, SR의 DNA 결합 특이성 역시 서열 조성에 매우 의존적이다. GR 연구 실험에서, 컨센서스 GRE/PRE 서열내 한쌍의 염기 변화가 수용체 결합성과 조인자 상호작용에 상당한 영향을 미칠 수 있는 것으로 확인되었다 (Meijsing et al., 2009). 따라서, DNA 자체는 SR에 대한 서열 특이적인 알로스테릭 리간드인 것으로 보이며, 프로모터 선택성과 전사 결과에 직접 작용할 수 있다. SUMO일화된 GR는 거의-완전한 컨센서스 GR-결합부를 선호하는 것으로 보인다 (Holmstrom et al., 2008). 특히, PR이 그렇듯이, GR의 부위-특이적인 인산화 역시 이의 프로모터 선호성을 변형시킬 수 있다 (Blind and Garabedian, 2008). 현재, SUMO일화 대 탈SUMO일화된 PR들이 여러가지 PRE 서열들을 차별적으로 인지하는지에 대해서는 알려져 있지 않다 (즉, 모든 PR-결합부를 동정하기 위한 ChIP-seq 실험은 수행하지 않았음). 그러나, SUMO 변형이 기질 단백질의 형태를 상당히 변이시킬 수 있기 때문에, 이는 그럴듯하게 생각된다. 분명한 것은, 탈SUMO일화된 PR이 다량의 PR 공동-활성인자 (CBP, MLL2)를 인핸서 영역으로 동원할 수 있으며; 기능성 전사 복합체의 좀더 빠른 또는 안정적인 형성은 WT PR 대비 KR에 의한 선택된 프로모터의 사용 또는 "샘플링" 증가를 설명해 줄 수 있다 (도 3). 본 분석을 통해, SUMO일화된 또는 탈SUMO일화된 PR들 이상으로 선택된 인핸서 영역의 선호적인 억제 또는 활성화를 설명할 수 있는 명백한 글로벌 신호(들)은 드러나지 않았다.
탈SUMO일화된 PR 유전자 발현의 임상적인 의미. 내강 유방암에서 선택적인 ER 모듈레이터 (SERM [예, 타목시펜], 항-에스트로겐 [예, 풀베스트란트 (Fulvestrant)]) 및/또는 아로마타제 저해제 (예, 아나스트로졸, 레트로졸 (Letrozole), 또는 엑세메스탄 (exemestane))를 이용하여 ER 기능을 타겟팅하는 것은, 대부분의 여성에서 매우 효과적이다 (Early Breast Cancer Trialists' Collaborative Group, 2005; Goss et al., 2011). 실제, SR과 성장인자 신호전달 경로의 크로스-톡이 광범위하며, 종양이 상승된 성장인자 신호전달의 영향으로 엔도크린 내성으로 진행되는 경향이 있기 때문에, ER과 ERBB 수용체 둘다를 타겟팅하는 병용 요법이 무-진행성 생존율을 높인다 (Johnston et al., 2009; Kaufman et al., 2009). 본원에서는, 리간드-의존적인 (151개의 유전자) 및 리간드-비의존적인 조건 (92개의 유전자) 하에 탈SUMO일화된 (포스포-모방) PR 종들에 의해 상향 조절되거나 탈억제된 독특한 유전자 세트를 개시한다 (도 4.1). 이들 유전자들의 상승된 발현은, 과잉 활성의 포스포-PR(탈SUMO일화된) 종들에 의해, 특히 활성화된 성장인자 신호전달 케스케이드가 특징적인 암에서, 주로 유발되는 종양들에서 나타날 수 있다. 예를 들어, MAPK 및 CDK2 또는 CDK4/6는 일부 유방 종양들에서 영구적인 PR Ser294 인산화를 유도할 것으로 보이는 유방암 진행의 공지된 활성화 인자이다 (도 1A). 이 "포스포-PR" 유전자 시그니처를 발현하는 내강 타입 (ER 양성/PR 양성) 유방 종양을 가진 환자가 존재하는 것으로 추측되며 (도 1A 및 도 4.1 참조), 이 서브세트는, 조기에 동정된다면, 선택성이 높은 항-프로게스틴의 사용을 포함하는 엔도크린 요법을 통해, 아마도 현재 사용되는 항-에스트로겐 또는 아로마타제 저해제 및/또는 성장인자 경로 저해제와 병용하여, 효과를 볼 수 있을 것으로 예상된다.
실제, 많은 연구들이, PR이 기능성 ER 발현의 임상 마커일 뿐만 아니라 종양 진행의 중요한 독립 활성화 인자임을 밝혀냈다 (Daniel et al., 2011). 특히, SR 양성의 내강 A형 종양은 보다 공격적인 성장인자-고수준의 내강 B형 표현형으로 진행됨에 따라, SR 발현이 감소되기 시작하고, PR 감소가 시작된다. 이러한 예후가 좋지 않은 내강 B형 종양은 종종 임상적으로는 ER 양성/PR-저수준 또는 null로 특정화되며, 엔도크린 내성으로 될 가능성이 더 높다. 탈SUMO일화된 포스포-PR이 과잉 활성의 수용체로서 기능하지만, 또한 유비퀴틴-프로테아좀 경로를 통해 빠르게 턴오버한다는 것도, 기존에 확인되었다 (도 1B 및 (Lange et al., 2000)). 실제, 내인성 유전자 정보의 RT-qPCR (mRNA 수준, 도 5B에서와 같이) 또는 리포터 유전자를 이용하여 측정된 바와 같이, PR-의존적인 전사가 피크에 도달하였을 때, PR 단백질 수준은 실제 검출되지 않는다 (Daniel et al., 2007b). 이런 사실은, PR이 (즉, 임상 조건에서 총 단백질 검출 방법을 통해 통상적으로 측정한 바에 따라) PR-low 또는 null로 임상적으로 규정된 유방 종양의 서브세트에서 또한 과잉 활성형인지에 대한 중요한 의문을 갖게 한다. 흥미롭게도, 유방 종양은 새로운 (de novo) 프로게스테론 합성, 즉 성장인자-의존적인 신호전달에 의해 매개되는 공정을 수행할 수 있다 (Locke et al., 2008; Su et al., 2011; Suzuki et al., 2005). 종양 세포의 (국소) 프로게스테론 생산은 좀더 공격적인 ER 양성/PR 양성 종양들에서 (즉, 리간드-의존적인 유전자들에서) PR 작용을 연장시키는데 기여할 수 있다.
놀랍게도, 본원에서는, 탈SUMO일화된 포스포-PR을 발현하는 유방암 세포가, ERBB/MAPK 신호전달 경로의 파지티브 조절에 다수가 직접 관여하는 세포 증식 유전자들의 발현을 유발하며 (도 4.1), 따라서, 종양 진행과 분명하게 연관된 "피드-포워드"의 좋지 않은 사이클 타입을 확립한다는 것을 개시하게 되었다 (Amit et al., 2007; Prat and Perou, 2011). 본 데이타는, 본 발명에서 독자적으로 규정된 PR 시그니처와 유의한 유사성에 의해 확인되는 바와 같이, 포스포-PR이 이러한 이행 (transition) (즉, SR 감소에 앞서 발생할 수 있는 성장인자-유발성 경로의 획득 쪽으로의 종양 진행)의 활성화 인자로서 작용할 수 있음을 시사한다 (도 4.1). 이러한 사실은, HRT의 일부로서 프로게스틴을 복용하는 여성에서 기원한 유방 종양이 대조군에 비해 훨씬 빈번하고, 크며, 질병 단계가 높다는 것을 보여주는, 여성 보건 발의 및 백만 여성 실험으로부터 유래된 이용가능한 임상 데이타로부터 뒷받침된다 (Chlebowski et al., 2010; Million Women Study Collaborators, 2003). 특히, 최근 이들 데이타에 대한 분석을 통해, 에스트로겐-단일 HRT가 침습성 유방암으로부터 여성을 실제 보호할 수 있는 것으로 입증되었다 (Anderson et al., 2012; LaCroix et al., 2011). 다른 연구 (Labriola et al., 2003; Musgrove and Sutherland, 2009; Salatino et al., 2004)들과 종합하면, 이 데이타는, 유방암 환자에서의 PR 작용을 타겟팅하는 것이 특히 항-에스트로겐 또는 아로마타제 저해제에 내성이 되는 환자에게 매우 유익하다는 컨셉을 뒷받침해준다. 특히, 환자의 거의 40%는 에스트로겐 작용만을 타겟팅하는 엔도크린 요법에 실패하거나, 또는 궁극적으로는 내성을 발현하게 될 것이며, 이는 대규모의 취약 집단을 형성한다.
유방암의 분자 서브타입들을 둘러싼 집중적인 연구는, 이러한 혼성 암의 유전자 특징에 대한 훌륭한 통찰력을 제공해주게 되었지만 (Prat and Perou, 2011), 현행 타겟화된 요법은 여전히 적은 수의 임상-병리학적 마커들에만 집중되어 있다. 다양한 마커 (예, ER, PR 및 HER2)들의 상태에 대한 지식은 예후를 예측하는 유용성이 있으며 현행 요법들에 대한 정보를 제공할 수 있는 것이 사실이지만, 확장된 갯수의 관련 유전자들에 대한 mRNA 수준 측정 (즉, 유전자 시그니처)은 종양에서 활동하는 유전자 경로와 관련된 보다 민감하고 특이적인 정보를 제공해 줄 것이다. 이런 지식을 이용하여 특히 타겟화된 요법을 고려할 때 임상적인 결정에 정보를 제공할 수 있다. 따라서, 유방 종양을 분류하기 위한 예후 예측성 mRNA 발현에 기반한 분석법들이 빠르게 확장되고 있다 (Loi et al., 2007; Paik et al., 2004; Parker et al., 2009; van 't Veer et al., 2002). 하지만, 현재 이용가능한 예후를 나타내는 시그니처는 유전자 발현의 변화를 해당 종양에 존재하는 분자 활성화 인자와 연관짓지 못하고 있다. 본원에서는, 공경적인 종양의 특징을 나타낼 수 있는 가능성이 보다 높은 PR-의존적인 유전자 시그니처를 동정하게 되었다 (도 5D, 4.1). 실험들을 통해, 탈SUMO일화된 포스포-PR은 이러한 표현형의 주요 활성화 인자로서 관련되어 있는 것으로 확인된다. 동물 모델에서의 검증 실험들이 필요하지만 (진행 중임), 본 연구들은 최신 항-에스트로겐을 이용한 엔도크린 요법에 대한 유용한 부가로서 항-프로게스틴의 사용을 강력하게 뒷받침한다. PR-유발성 종양 (활성화된 PR 유전자 시그니처를 포함함)을 가진 환자의 동정은, 엔도크린 내성 발생을 방지하기 위한 개입을 가능케 하며, 환자에게 추가적인 임상적인 효과를 제공해준다.
요약
본원에서, PR은, 인산화 현상이 수용체를 탈SUMO일화된 상태로 유도하여, 세포 증식과 생존 촉진을 촉진하는, 급격하게 변형된 전사 프로그램을 발현시키는 미토겐 자극의 센서이므로, PR 전사 작용이 원래 생각했던 것 보다 더 복잡하다는 것을 확인하게 되었다. 탈SUMO일화된 포스포-PR 유전자 시그니처는, 유방암 세포 모델에서 과잉 활성의 PR 신호전달의 마커인 공지된 및 신규한 PR 타겟 유전자들 둘다를 동정하였으며; 이 시그니처는 실제 유방암 재발 환자의 서브세트에도 존재하고 있다 (도 1A 및 5D). 이러한 독특한 시그니처는 엔도크린-내성으로 (즉, 에스트로겐 타겟화된 요법에 대한 내성으로) 진행되거나 및/또는 내성이 될 가능성이 있는 종양을 가진 환자를 동정하기 위한 유용한 예측 수단을 제공할 수 있다.
간행물, 등재 번호로 표시된 뉴클레오티드 및 아미노산 서열, 특허 및 특허 출원들은 모두 원용에 의해 본 명세서에 포함된다. 전술한 내용에서 본 발명이 이의 특정한 바람직한 구현예를 들어 기술되고, 상세한 다수 내용들이 예시 목적으로 기술되어 있지만, 당해 기술 분야의 당업자라면, 본 발명이 첨부된 구현예예들을 허용하며 본원에 기술된 특정 상세 내용이 본 발명의 기본 개념으로부터 이탈되지 않는 한에서 상당히 변경될 수 있음은, 자명할 것이다.
본원에 기술된 구체적인 방법 및 조성물은 바람직한 구현예를 나타내고, 예시한 것일 뿐, 본 발명의 범위를 한정하는 것으로 의도되지 않는다. 당해 기술 분야의 당업자라면 본 명세서의 내용을 고찰함으로써 다른 목적, 측면 및 구현예들을 연상할 수 있을 것이며, 이것은 청구항 범위에 의해 규정되는 본 발명의 범위에 포함된다. 당해 기술 분야의 당업자라면, 다양한 치환 및 변형을 본원에 기술된 발명에 대해 본 발명의 범위 및 사상내에서 행할 수 있다는 것을 용이하게 인지할 것이다. 본원에 예시적으로 기술된 본 발명은 적절하게는 단계들의 순서를 달리하면서 수행할 수 있으며, 방법 및 공정이 본원 또는 청구항에 기술된 순서의 단계들로 반드시 한정되는 것은 아니다. 본원 및 첨부된 청구항에서, 단수형 ("a," "an" 및 "the")은 명백히 다르게 언급되지 않은 한 복수의 언급도 포함한다. 즉, 예를 들어, "핵산" 또는 "폴리팹타이드"라는 언급은 이러한 핵산 또는 폴리펩타이드 (예, 핵산 또는 폴리펩타이드의 용액, 또는 핵산 또는 폴리펩타이드 조제물 시리즈) 등의 복수도 포함한다. 본 문헌에서, 용어 "또는"은, 달리 언급되지 않은 한, 어느 하나만 지칭하는 것이 아니거나, 또는 "A 또는 B"는 "B는 아니고 A", "A는 아니고 B" 및 "A와 B"를 포함하는 것으로 사용된다.
어떤 상황에서도 본 특허가 본원에 구체적으로 기술된 특정 실시예, 구현예 또는 방법으로 한정되는 것으로 해석되지 않는다. 진술이 구체적이고 제한 또는 조건없이 출원인에 의해 작성된 의견서에서 명확하게 사용된 경우가 아닌 한, 어떤 경우에도 본 특허는 심사관 또는 특허상표청의 다른 사무관 또는 직원에 의해 행해진 어떠한 진술로 한정되는 것으로 해석되지 않는다.
사용된 용어들과 표현들은 설명하기 위해 사용된 것이며, 한정되지 않으며, 개시되고 설명된 특징 또는 이의 일부분에 대한 임의의 등가를 제외하기 위해 이들 용어 및 표현들을 사용한 것이 아니며, 청구된 본 발명의 범위 안에서 댜양한 수정이 가능한 것으로 인지된다. 따라서, 본 발명이 바람직한 구현예 및 선택적인 특징으로 구체적으로 기술되어 있더라도, 당해 기술 분야의 당업자라면 본원에 기술된 컨셉에 대한 수정 및 변형을 행할 수 있으며, 이러한 수정 및 변형은 본 발명의 첨부된 청구항 및 내용에 의해 정의되는 본 발명의 범위 안에 포함되는 것으로 이해될 것이다.
SEQUENCE LISTING
<110> Lange, Carol
Knutson, Todd
Nikas, Jason Basil
<120> BREAST CANCER PROGNOSIS
<130> 600.867WO1
<140> PCT/US2013/032677
<141> 2013-03-15
<150> US 61/639,407
<151> 2012-04-27
<160> 122
<170> FastSEQ for Windows Version 4.0
<210> 1
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 1
ggtaaactac acctgttgaa ggccaagttc agggcagctg ttgtgatctg 50
<210> 2
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 2
gaaccactga gtcaggagag ccaggtggag gaaccaccga gtcaggagag 50
<210> 3
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 3
ggccatttca gacttgggag atgaggcggc tgttgtcatt gctgatcctg 50
<210> 4
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 4
ccattggcac agggaggttt gacctcttcc ctgctattat ccctcctccc 50
<210> 5
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 5
gtgtaatcac ccaaaacccc ccggcctgtg cctgttttcc cttctgcgct 50
<210> 6
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 6
agcaggtctt accgagaatt cagctgccaa aaccctcctc tgagtgttcc 50
<210> 7
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 7
gggtcacgtg tctttggtga gtgagaagac ctaaactcct ggccatcatc 50
<210> 8
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 8
acgcattcct ggcggccttc ctcgggggta tcctggtgtt tggagaaaac 50
<210> 9
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 9
accagcagaa gccagcagag aggcatggga caggttcccc acaagcctta 50
<210> 10
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 10
ctgaggcaag ccatggagtg agacccagga gccggacact tctcaggaaa 50
<210> 11
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 11
tctcacccag gcacagcccc gccaccatgg atctccgtgt acactatcaa 50
<210> 12
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 12
tgtcaagggg ctttgcattc aaactgcttt tccagggcta tactcagaag 50
<210> 13
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 13
tggggtgctt cctgtggtag tgtctttcag gtatccgttc cactagctac 50
<210> 14
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 14
agcaacctca caaacaagcc gcttctgtta ggtacatgtc ctgcccttgc 50
<210> 15
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 15
ctcacccctt aatgttcacc tgcaaactca taccagagag aaagccctca 50
<210> 16
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 16
ctgaggcaag ccatggagtg agacccagga gccggacact tctcaggaaa 50
<210> 17
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 17
gcccttcacc attgtggaat gatgccctgg ctttaaggtt tagctccaca 50
<210> 18
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 18
gcagactgcc ccaatgtgac agcacctgtt tgtgcctcaa atggccacac 50
<210> 19
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 19
aacagtctct ccgccccgca ccagatcaag tagtttggac atcaccctac 50
<210> 20
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 20
cccaagcaag ccagtgagca gccctgccag actactgcca gactgagaaa 50
<210> 21
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 21
tccctgatat acaccatccc caattgctcc ttctcacctc ctctcaggcc 50
<210> 22
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 22
gagtcgtgat tgtaccactg cattcctgct gagcaacaga gtgagacccc 50
<210> 23
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 23
cagaagtcct agacagtgac atttcttaat ggtgggagtc cagctcatgc 50
<210> 24
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 24
aggtacattc atcctcacag attgcaaagg tgatttgggt gggggtttag 50
<210> 25
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 25
ggtccccaaa gacgtcaatg cggccatcgc caccatcaag accaagcgca 50
<210> 26
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 26
tcccaatggt gtagaccagt ggcgatggat ctaggagttt accaactgag 50
<210> 27
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 27
tcccctgcca cccccgggat ggctgcttcc aagttgtttg caattaaagg 50
<210> 28
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 28
aggcaccttc agcgaggaca gcaaagggcg tctacagaga ccagccatat 50
<210> 29
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 29
gctggtatat ccagtgcatt gttggcacca tgggaccaga aggtggtgac 50
<210> 30
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 30
agggtggctt gcagtccctg gcccttctgg tgggcatttg gtatgtcctt 50
<210> 31
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 31
cttcttgaac ctggtggccc ccgttggaac tatcagtggc gtctcccatg 50
<210> 32
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 32
agcaacctca caaacaagcc gcttctgtta ggtacatgtc ctgcccttgc 50
<210> 33
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 33
gccaggtgtc ctgactgtcc tacaatatca ttttcctggg agtgggagtc 50
<210> 34
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 34
ggtaaactac acctgttgaa ggccaagttc agggcagctg ttgtgatctg 50
<210> 35
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 35
ggagctcaag tgtcgggaac tgtctaactt caggttgtgt gagtgcgtta 50
<210> 36
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 36
atcactattc ctggttatct caccaacgaa ggctaggagg cggcgtcaga 50
<210> 37
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 37
ggtggaggaa ccactgagtc aggagagcga gatggaagaa ccactgagtc 50
<210> 38
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 38
ccccccaaaa ttatcagtgc tctgctttta gtcacgtgta ttttcattac 50
<210> 39
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 39
ccacatcgtc ttccctgtcc caatcgacca gtgtatcgac ggctgagtgc 50
<210> 40
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 40
cggcgcttcc cagcaccaac atgtaaccgg catgtttcca gcagaagaca 50
<210> 41
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 41
agcaacctca caaacaagcc gcttctgtta ggtacatgtc ctgcccttgc 50
<210> 42
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 42
cgatgttcag aggctgtttc ctgcagcatg tatttccatg gcccacacag 50
<210> 43
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 43
gacacagaac acagacgcct tactggcaac ctgctttcaa gacccctgtc 50
<210> 44
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 44
gtgtttatga tgagtcagag tgcttttcct cggtgggaca gttgctggcc 50
<210> 45
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 45
cagttctgca gtgtaatgga ggacgggcaa cgtgcatgtg caggctcacc 50
<210> 46
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 46
gagccaaagg ctcactcaaa ggcacaggta gatgcctggc agcaaattca 50
<210> 47
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 47
agcaacctca caaacaagcc gcttctgtta ggtacatgtc ctgcccttgc 50
<210> 48
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 48
tatagacctg tgtgaccagc ccccagttcc tcccccagtt cctcccagga 50
<210> 49
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 49
tctggtctac agtggaggga gagctggttt taaatgttgg ccgttgatgc 50
<210> 50
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 50
gatggaacca actttgtaca tcttggccat gtcactggtc attgtgtgaa 50
<210> 51
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 51
cgactggcag accgactact tgccctggtc atccaccctg aggaagatgt 50
<210> 52
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide probe
<400> 52
ctagggttcc ctcccagtct tcacatcact ctggcctcat caccaaggtg 50
<210> 53
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 53
ctgtctttca tccttcccag ag 22
<210> 54
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 54
tctttctgct tcttctccag tttag 25
<210> 55
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 55
ggcttatttc agatttgaag acctc 25
<210> 56
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 56
agaagcgcaa tactaggacc tttc 24
<210> 57
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 57
tgctcaacaa tgtcgattct g 21
<210> 58
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 58
ttccactccc attctgcttc 20
<210> 59
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 59
gtctgactct gactctcgga ctg 23
<210> 60
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 60
gaacttacat cagaaggttg ctttg 25
<210> 61
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 61
accagcctcc tatatggaaa tc 22
<210> 62
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 62
ctcccgagga caaatgattc 20
<210> 63
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 63
agaagaagca caatgaggag attc 24
<210> 64
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 64
catgtggaaa ttatcacttc tttgg 25
<210> 65
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 65
gccaaaaggg tcatcatctc tgcc 24
<210> 66
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 66
tggtggtgca ggaggcattg 20
<210> 67
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 67
tgctggatgc cctgctggag 20
<210> 68
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 68
accactcacg agtcacctcc g 21
<210> 69
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 69
aaatcccttg ttaaaggagg tctg 24
<210> 70
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 70
ctgagaggta ccataatgga aaatg 25
<210> 71
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 71
agcggcgtgg atgcaggaac 20
<210> 72
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 72
tgcgcggctt ccgattggtc 20
<210> 73
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 73
catactccac tgcaccaacg 20
<210> 74
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 74
agaaattggc aagccgtaac 20
<210> 75
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 75
tctcatcagc tacaccacca ac 22
<210> 76
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 76
acattggctg aatagttgtc aaac 24
<210> 77
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 77
agctgtcctc aaaagcaagg 20
<210> 78
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 78
tctgggcaat cagagttttg 20
<210> 79
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 79
ccgatgatca agatgtggag 20
<210> 80
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 80
caaaggcatt gatgaaggtg 20
<210> 81
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 81
ataacaagac ctctgccaga agaac 25
<210> 82
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 82
caggtgggtt tcctatgttc ag 22
<210> 83
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 83
agaagatgta gacttcctgc cctac 25
<210> 84
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 84
tttcgtttgg ataaacgtaa ttctg 25
<210> 85
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 85
tccaactcct aagccagtgc 20
<210> 86
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 86
tgctcctggg agtagattgg 20
<210> 87
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 87
gccagccaac tgctatcttc 20
<210> 88
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 88
ctgggagtgc tctggaagg 19
<210> 89
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 89
gttccctgag aacctcatta aacc 24
<210> 90
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 90
ggtctctgca gcctgttctg 20
<210> 91
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 91
cacatttctg tctgactctg aagc 24
<210> 92
<211> 26
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 92
ccacatttgc tagcttatta gttctg 26
<210> 93
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 93
gacgtccaga tcagaactaa taagc 25
<210> 94
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 94
ctgactttga caataggtcc tcaag 25
<210> 95
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 95
cttgaggacc tattgtcaaa gtcag 25
<210> 96
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 96
caagctcatg gacatcaaat agaag 25
<210> 97
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 97
attcttctat ttgatgtcca tgagc 25
<210> 98
<211> 26
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 98
tcagacccag tcactaacat ttctac 26
<210> 99
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 99
catgccaaga gtagaaatgt tagtg 25
<210> 100
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 100
aaatggtcca agagaatatg gtaag 25
<210> 101
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 101
ggatccattg tcaaggaaac ttac 24
<210> 102
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 102
agagattgcc atcagtacag gac 23
<210> 103
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 103
gtactgatgg caatctctgg ttc 23
<210> 104
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 104
atcattttgt tctgaaggat ttctc 25
<210> 105
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 105
aaatccttca gaacaaaatg atcc 24
<210> 106
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 106
taatcttctg gctctatcct tctcc 25
<210> 107
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 107
ctactggttg aggagaagga tagag 25
<210> 108
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 108
gagtttggaa ttcctattaa tgctc 25
<210> 109
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 109
aaactcagta tcagcagcct gtc 23
<210> 110
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 110
cactgatttg ctttctaacc gatac 25
<210> 111
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 111
tactgctgct gcttctaaca catc 24
<210> 112
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 112
cacaaggcaa ctggataatt aactg 25
<210> 113
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 113
ttaattatcc agttgccttg tgaag 25
<210> 114
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 114
agagagggca gacagatgta cc 22
<210> 115
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 115
tggaccattt aagggctgag 20
<210> 116
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 116
gactctgtgg gaagctgagg 20
<210> 117
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 117
gcatacattt ctttcacagg gatac 25
<210> 118
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 118
tggcttactt tacagttcca gaaag 25
<210> 119
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 119
cggagcccat agttctttct c 21
<210> 120
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 120
ttatttgtct ccccgcactc 20
<210> 121
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 121
aacaaataaa acttaatcaa ggaaactg 28
<210> 122
<211> 26
<212> DNA
<213> Artificial Sequence
<220>
<223> A synthetic oligonucleotide primer
<400> 122
cttgttgagt taggatttac cagaac 26
Claims (47)
- 암이 활성형 프로게스테론 수용체 (KR)를 발현하는 세포를 포함하는 지의 여부와 항-프로게스틴을 이용한 치료학적 치료에 대해 암이 반응할 가능성을 확인하기 위한 방법으로서,
(a) 암으로 진단받은 환자로부터 생물 샘플을 수득하는 단계;
(b) 상기 생물 샘플의 세포에서, KBTBD11, RBPMS2, PLA2G48, FL112684, SH2D4B, RASCD2, CLDN8 및 이들의 조합으로 이루어진 군으로부터 선택되는 1종 이상의 유전자의 하나 이상의 샘플 발현 수준을 측정하는 단계; 및
(c) 단계 (b)의 상기 하나 이상의 샘플 발현 수준을, 야생형 (WT) 대조군 샘플의 세포의 상기 하나 이상의 유전자의 하나 이상의 발현 수준과 비교하는 단계를 포함하며,
상기 샘플 발현 수준이 상기 대조군 샘플에 비해 감소된다면, 상기 암이 항-프로게스틴을 이용한 치료에 대한 반응성 암인 것을 특징으로 하는 방법. - 제1항에 있어서,
(d) VCX, CHN2, AFAP1L2, PXMP4, THY1, ZNF26, CDH10, ZNF812 및 이들의 조합으로 이루어진 군으로부터 선택되는 하나 이상의 유전자의 발현 수준을 상기 생물 샘플의 세포에서 측정하는 단계; 및 (e) 단계 (d)의 상기 발현 수준을 야생형 (WT) 대조군 샘플의 세포에서의 상기 하나 이상의 유전자의 하나 이상의 발현 수준과 비교하는 단계를 더 포함하며,
상기 생물 샘플에서 상기 하나 이상의 유전자의 발현 수준이 상기 WT 대조군 샘플 및/또는 기준 샘플에 비해 증가된다면, 상기 암이 항-프로게스틴을 이용한 치료에 대한 반응성 암인 것을 특징으로 하는 방법. - 제1항 또는 제2항에 있어서, 상기 암이 유방암, 난소암, 자궁내막암, 뇌암, 폐암, 전립선암, 수막종 또는 자궁암으로 이루어진 군으로부터 선택되는 것을 특징으로 하는 방법.
- 암 환자가 항-프로게스틴 치료에 반응할 것인지를 확인하는 방법으로서,
a. 상기 환자의 생물 샘플의 암 세포에서 KR78D11, RBPMS2, PLA2G48, FL112684, SH2D4B, RASCD2, CLDN8 및 이들의 조합으로 이루어진 군으로부터 선택되는 하나 이상의 유전자의 발현 수준을 측정하는 단계를 포함하며,
b. 대조군 WT 샘플 및/또는 기준 샘플에서의 수준과 비교하여, 상기 생물 샘플에서 상기 하나 이상의 유전자의 발현 수준 감소는, 상기 개체가 항-프로게스틴 치료에 반응할 것임을 나타내는 지표인 것을 특징으로 하는 방법. - 제4항에 있어서,
(c) VCX, CHN2, AFAP1L2, PXMP4, THY1, ZNF26, CDH10, ZNF812 및 이들의 조합으로 이루어진 군으로부터 선택되는 하나 이상의 유전자의 발현 수준을 상기 생물 샘플의 세포에서 측정하는 단계; 및 (d) 단계 (c)에서의 상기 발현 수준을 야생형 (WT) 대조군 샘플 및/또는 기준 샘플에서의 상기 하나 이상의 유전자의 발현 수준과 비교하는 단계를 더 포함하며,
상기 생물 샘플에서 상기 하나 이상의 유전자의 발현 수준이 상기 WT 대조군 샘플 및/또는 기준 샘플과 비교하여 증가되면, 상기 환자가 항-프로게스틴을 이용한 치료에 반응자임을 나타내는 것임을 특징으로 하는 방법. - 제1항 내지 제5항 중 어느 한항에 있어서, 상기 유전자의 mRNA 수준이 유전자 발현 수준의 지표로서 측정되는 것을 특징으로 하는 방법.
- 제1항 내지 제6항 중 어느 한항에 있어서, 상기 유전자 KB7BD11의 발현 수준은 서열번호 1의 프로브와의 혼성화에 의해 검출되는 것을 특징으로 하는 방법.
- 제7항에 있어서, 상기 프로브는 디바이스에 고정되는 것을 특징으로 하는 방법.
- 제8항에 있어서, 상기 디바이스가 마이크로어레이인 것을 특징으로 하는 방법.
- 제7항에 있어서, 상기 프로브가 상기 유전자들 중 2종 이상에 혼성하는 고정된 복수의 프로브들 중 하나인 것을 특징으로 하는 방법.
- 제1항 내지 제10항 중 어느 한항에 있어서, 상기 하나 이상의 유전자의 발현 수준 측정은 인 시츄 혼성화, 노던 블롯, 핵산 증폭, 마이크로어레이 분석 또는 이들의 조합을 포함하는 것을 특징으로 하는 방법.
- 제2항 내지 제6항 중 어느 한항에 있어서, KR78D11, RBPMS2, PLA2G48, FL112684, SH2D4B, RASCD2, CLDN8, VCX, CHN2, AFAP1L2, PXMP4, THY1, ZNF26, CDH10, ZNF812 및 이들의 조합으로 이루어진 군으로부터 선택되는 2종 이상의 유전자의 발현을 측정하는 것을 특징으로 하는 방법.
- 제12항에 있어서, KR78D11, RBPMS2, PLA2G48, FL112684, SH2D4B, RASCD2, CLDN8, VCX, CHN2, AFAP1L2, PXMP4, THY1, ZNF26, CDH10, ZNF812 및 이들의 조합으로 이루어진 군으로부터 선택되는 3종 이상의 유전자의 발현을 측정하는 것을 특징으로 하는 방법.
- 제13항에 있어서, KR78D11, RBPMS2, PLA2G48, FL112684, SH2D4B, RASCD2, CLDN8, VCX, CHN2, AFAP1L2, PXMP4, THY1, ZNF26, CDH10, ZNF812 및 이들의 조합으로 이루어진 군으로부터 선택되는 4종 이상의 유전자의 발현을 측정하는 것을 특징으로 하는 방법.
- 제14항에 있어서, KR78D11, RBPMS2, PLA2G48, FL112684, SH2D4B, RASCD2, CLDN8, VCX, CHN2, AFAP1L2, PXMP4, THY1, ZNF26, CDH10, ZNF812 및 이들의 조합으로 이루어진 군으로부터 선택되는 6종 이상의 유전자의 발현을 측정하는 것을 특징으로 하는 방법.
- 제15항에 있어서, KR78D11, RBPMS2, PLA2G48, FL112684, SH2D4B, RASCD2, CLDN8, VCX, CHN2, AFAP1L2, PXMP4, THY1, ZNF26, CDH10, ZNF812 및 이들의 조합으로 이루어진 군으로부터 선택되는 7, 8, 9, 10, 11, 12, 13, 14 또는 15종 이상의 유전자의 발현을 측정하는 것을 특징으로 하는 방법.
- 제1항 내지 제16항 중 어느 한항에 있어서, 상기 생물 샘플이 조직 생검, 유관 세척물 (ductal lavage) 또는 미세 바늘 흡입 샘플 (fine needle aspiration sample)인 것을 특징으로 하는 방법.
- 제1항 내지 제17항 중 어느 한항에 있어서, 상기 대조군 샘플이 비-암성 조직의 샘플인 것을 특징으로 하는 방법.
- 제18항에 있어서, 상기 대조군이 환자로부터 유래된 것을 특징으로 하는 방법.
- 제1항 내지 제19항 중 어느 한항에 있어서, 건강 관리 서비스 제공자에게 항-프로게스틴 요법의 개시 또는 중지에 대한 조언을 제공하는 단계를 더 포함하는 것을 특징으로 하는 방법.
- 제1항 내지 제20항 중 어느 한항에 있어서, 상기 환자에서 암을 치료하는 단계를 더 포함하는 것을 특징으로 하는 방법.
- 제21항에 있어서, 상기 치료는 하나 이상의 항-프로게스틴을 유효량으로 투여하는 단계를 포함하는 것을 특징으로 하는 방법.
- 제21항 또는 제22항에 있어서, 상기 치료는 하나 이상의 부가적인 치료학적 제제의 투여를 더 포함하는 것을 특징으로 하는 방법.
- 암 환자의 치료 방법으로서,
상기 암 환자를 치료하기 위해, 대조군과 비교하여, KR78D11, RBPMS2, PLA2G48, FL112684, SH2D4B, RASCD2, CLDN8 및 이들의 조합으로 이루어진 군으로부터 선택되는 1종 이상의 유전자의 수준이 감소되거나, 및/또는 VCX, CHN2, AFAP1L2, PXMP4, THY1, ZNF26, CDH10, ZNF812 및 이들의 조합으로 이루어진 군으로부터 선택되는 1종 이상의 유전자의 수준이 증가된 환자에게, 항-프로게스틴을 단독으로 또는 다른 치료와 병행하여 투여하는 단계를 포함하는 것을 특징으로 하는 방법. - 제24항에 있어서,
(c) VCX, CHN2, AFAP1L2, PXMP4, THY1, ZNF26, CDH10, ZNF812 및 이들의 조합으로 이루어진 군으로부터 선택되는 하나 이상의 유전자의 발현 수준을 상기 생물 샘플의 세포에서 측정하는 단계; 및 (d) 단계 (a)의 상기 발현 수준을 대조군에서의 상기 하나 이상의 유전자의 발현 수준과 비교하는 단계를 더 포함하며,
상기 생물 샘플에서 하나 이상의 유전자의 발현 수준이 상기 대조군에 비해 증가된다면, 상기 환자는 항-프로게스틴을 이용한 치료에 반응자인 것임을 특징으로 하는 방법. - 암으로 진단받은 환자가 활성형 프로게스테론 수용체 (KR)을 발현하는 세포를 포함하는지 여부와 항-프로게스틴을 이용한 치료학적 치료에 반응할 가능성이 있는지 여부를 확인하는 방법으로서,
(a) 환자로부터 생물 샘플을 수득하는 단계;
(b) THY1, KLF9, SPINK5L.3, PHLDA1, MAP1A, SPRYD5, ATG12, PDK4, MSX2, TUBA3E, TSC22D1, TUBA3D, KHDRBS3, UTS2D, SLC35C1, KIAA0513 및 이들의 조합으로 이루어진 군으로부터 선택되는 하나 이상의 유전자의 발현 수준을 상기 생물 샘플의 세포에서 측정하는 단계; 및
(c) 단계 (b)에서의 상기 발현 수준을 야생형 (WT) 대조군 샘플 및/또는 기준 샘플에서의 상기 하나 이상의 유전자의 발현 수준과 비교하는 단계를 포함하며,
상기 생물 샘플에서 상기 하나 이상의 유전자의 발현 수준이 상기 WT 대조군 샘플 및/또는 기준 샘플에 비해 증가된 경우, 상기 환자는 항-프로게스틴을 이용한 치료에 반응자인 것임을 특징으로 하는 방법. - 제26항에 있어서, 상기 암이 유방암, 난소암, 자궁내막암, 뇌암, 폐암, 전립선암, 수막종 또는 자궁암으로 이루어진 군으로부터 선택되는 것을 특징으로 하는 방법.
- 암 환자가 항-프로게스틴 치료에 반응할 것인지를 확인하는 방법으로서,
a. 환자의 생물 샘플에서 THY1, KLF9, SPINK5L.3, PHLDA1, MAP1A, SPRYD5, ATG12, PDK4, MSX2, TUBA3E, TSC22D1, TUBA3D, KHDRBS3, UTS2D, SLC35C1, KIAA0513 및 이들의 조합으로 이루어진 군으로부터 선택되는 하나 이상의 유전자의 발현 수준을 측정하는 단계를 포함하며,
b. 상기 생물 샘플에서 상기 하나 이상의 유전자의 발현이 WT 대조군 샘플에서의 상기 발현 수준에 비해 증가된 것은, 상기 개체가 항-프로게스틴 치료에 반응할 것임을 나타내는 지표인 것을 특징으로 하는 방법. - 제26항 내지 제28항 중 어느 한항에 있어서, mRNA 수준이 유전자 발현 수준의 지표로서 측정되는 것을 특징으로 하는 방법.
- 제26항에 있어서, 상기 유전자 THY1의 발현 수준은 서열번호 16의 프로브와의 혼성화에 의해 검출되는 것을 특징으로 하는 방법.
- 제30항에 있어서, 상기 프로브는 디바이스에 고정되는 것을 특징으로 하는 방법.
- 제31항에 있어서, 상기 디바이스가 마이크로어레이인 것을 특징으로 하는 방법.
- 제26항 내지 제32항 중 어느 한항에 있어서, 상기 하나 이상의 유전자의 발현 수준의 측정은 인 시츄 혼성화, 노던 블롯, 핵산 증폭, 마이크로어레이 분석 또는 이들의 조합을 포함하는 것을 특징으로 하는 방법.
- 제26항 내지 제29항 중 어느 한항에 있어서, THY1, KLF9, SPINK5L.3, PHLDA1, MAP1A, SPRYD5, ATG12, PDK4, MSX2, TUBA3E, TSC22D1, TUBA3D, KHDRBS3, UTS2D, SLC35C1, KIAA0513 및 이들의 조합으로 이루어진 군으로부터 선택되는 2종 이상의 유전자의 발현을 측정하는 것을 특징으로 하는 방법.
- 제34항에 있어서, THY1, KLF9, SPINK5L.3, PHLDA1, MAP1A, SPRYD5, ATG12, PDK4, MSX2, TUBA3E, TSC22D1, TUBA3D, KHDRBS3, UTS2D, SLC35C1, KIAA0513 및 이들의 조합으로 이루어진 군으로부터 선택되는 3종 이상의 유전자의 발현을 측정하는 것을 특징으로 하는 방법.
- 제35항에 있어서, THY1, KLF9, SPINK5L.3, PHLDA1, MAP1A, SPRYD5, ATG12, PDK4, MSX2, TUBA3E, TSC22D1, TUBA3D, KHDRBS3, UTS2D, SLC35C1, KIAA0513 및 이들의 임의 조합으로 이루어진 군으로부터 선택되는 4종 이상의 유전자의 발현을 측정하는 것을 특징으로 하는 방법.
- 제36항에 있어서, THY1, KLF9, SPINK5L.3, PHLDA1, MAP1A, SPRYD5, ATG12, PDK4, MSX2, TUBA3E, TSC22D1, TUBA3D, KHDRBS3, UTS2D, SLC35C1, KIAA0513 및 이들의 임의 조합으로 이루어진 군으로부터 선택되는 6종 이상의 유전자의 발현을 측정하는 것을 특징으로 하는 방법.
- 제37항에 있어서, THY1, KLF9, SPINK5L.3, PHLDA1, MAP1A, SPRYD5, ATG12, PDK4, MSX2, TUBA3E, TSC22D1, TUBA3D, KHDRBS3, UTS2D, SLC35C1, KIAA0513 및 이들의 임의 조합으로 이루어진 군으로부터 선택되는 7종 이상의 유전자의 발현을 측정하는 것을 특징으로 하는 방법.
- 제38항에 있어서, THY1, KLF9, SPINK5L.3, PHLDA1, MAP1A, SPRYD5, ATG12, PDK4, MSX2, TUBA3E, TSC22D1, TUBA3D, KHDRBS3, UTS2D, SLC35C1, KIAA0513 및 이들의 조합으로 이루어진 군으로부터 선택되는 8종 이상, 9종 이상, 10종 이상, 11종 이상, 12종 이상, 13종 이상, 14종 이상, 15종 이상 또는 16종 이상의 유전자의 발현을 측정하는 것을 특징으로 하는 방법.
- 제26항 내지 제39항 중 어느 한항에 있어서, 상기 생물 샘플이 조직 생검, 유관 세척물 또는 미세 바늘 흡입 샘플인 것을 특징으로 하는 방법.
- 제26항 내지 제40항 중 어느 한항에 있어서, 상기 대조군 샘플이 비-암성 조직의 샘플인 것을 특징으로 하는 방법.
- 제41항에 있어서, 상기 대조군이 환자로부터 유래된 것을 특징으로 하는 방법.
- 제26항 내지 제42항 중 어느 한항에 있어서, 건강 관리 서비스 제공자에게 항-프로게스틴 요법의 개시 또는 중지에 대한 조언을 제공하는 단계를 더 포함하는 것을 특징으로 하는 방법.
- 제26항 내지 제43항 중 어느 한항에 있어서, 상기 환자에서 암을 치료하는 단계를 더 포함하는 것을 특징으로 하는 방법.
- 제44항에 있어서, 상기 치료는 하나 이상의 항-프로게스틴을 유효량으로 투여하는 단계를 포함하는 것을 특징으로 하는 방법.
- 제45항에 있어서, 상기 치료는 하나 이상의 부가적인 치료학적 제제의 투여를 더 포함하는 것을 특징으로 하는 방법.
- 암 환자의 치료 방법으로서,
상기 암 환자를 치료하기 위해, 대조군과 비교하여, THY1, KLF9, SPINK5L.3, PHLDA1, MAP1A, SPRYD5, ATG12, PDK4, MSX2, TUBA3E, TSC22D1, TUBA3D, KHDRBS3, UTS2D, SLC35C1, KIAA0513 및 이들의 조합으로 이루어진 군으로부터 선택되는 1종 이상의 유전자의 수준이 증가된 환자에게, 항-프로게스틴을 단독으로 또는 다른 치료와 병행하여 투여하는 단계를 포함하는 것을 특징으로 하는 방법.
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201261639407P | 2012-04-27 | 2012-04-27 | |
| US61/639,407 | 2012-04-27 | ||
| PCT/US2013/032677 WO2013162776A1 (en) | 2012-04-27 | 2013-03-15 | Breast cancer prognosis, prediction of progesterone receptor subtype and|prediction of response to antiprogestin treatment based on gene expression |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| KR20150028232A true KR20150028232A (ko) | 2015-03-13 |
Family
ID=48045769
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR20147033298A KR20150028232A (ko) | 2012-04-27 | 2013-03-15 | 프로게스테론 수용체 서브타입의 유방암 예후, 예측 및 유전자 발현을 토대로 한 항프로게스틴 치료에 대한 반응성 예측 방법 |
Country Status (14)
| Country | Link |
|---|---|
| US (1) | US10501800B2 (ko) |
| EP (1) | EP2841594A1 (ko) |
| JP (1) | JP2015516155A (ko) |
| KR (1) | KR20150028232A (ko) |
| AU (1) | AU2013252913A1 (ko) |
| CA (1) | CA2871590A1 (ko) |
| HK (1) | HK1208057A1 (ko) |
| IL (1) | IL235351A0 (ko) |
| MX (1) | MX2014013044A (ko) |
| NZ (1) | NZ701652A (ko) |
| PH (1) | PH12014502410A1 (ko) |
| RU (1) | RU2014147625A (ko) |
| WO (1) | WO2013162776A1 (ko) |
| ZA (1) | ZA201407853B (ko) |
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US10501800B2 (en) | 2012-04-27 | 2019-12-10 | Regents Of The University Of Minnesota | Breast cancer prognosis |
Families Citing this family (9)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20140363425A1 (en) * | 2013-03-13 | 2014-12-11 | J. Dinny Graham | Systems and methods for identifying cancers having activated progesterone receptors |
| US20150276767A1 (en) * | 2014-03-12 | 2015-10-01 | The University Of Sydney | Systems and methods for identifying cancers having activated progesterone receptors |
| JP6655075B2 (ja) | 2014-11-17 | 2020-02-26 | コンテクスト バイオファーマ インコーポレイテッド | オナプリストン持続放出組成物および方法 |
| JP2018528944A (ja) | 2015-09-25 | 2018-10-04 | コンテクスト バイオファーマ インコーポレイテッド | オナプリストン中間体の製造方法 |
| AU2016370499B2 (en) | 2015-12-15 | 2022-06-30 | Context Biopharma Inc. | Amorphous onapristone compositions and methods of making the same |
| WO2018102369A1 (en) | 2016-11-30 | 2018-06-07 | Arno Therapeutics, Inc. | Methods for onapristone synthesis dehydration and deprotection |
| US10942183B2 (en) | 2017-04-05 | 2021-03-09 | Regents Of The University Of Minnesota | Methods of detecting progesterone receptor and of detecting an expression level |
| EP3822368A1 (en) | 2019-11-14 | 2021-05-19 | Koninklijke Philips N.V. | Assessment of pr cellular signaling pathway activity using mathematical modelling of target gene expression |
| JP2022062465A (ja) * | 2020-10-08 | 2022-04-20 | 藤倉化成株式会社 | 動脈病変を検出し、並びに、脳心臓血管障害、糖尿病、慢性腎臓病、又は固形癌を検出する、体液抗体バイオマーカー |
Family Cites Families (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US6750015B2 (en) * | 2000-06-28 | 2004-06-15 | Kathryn B. Horwitz | Progesterone receptor-regulated gene expression and methods related thereto |
| WO2002000618A2 (en) * | 2000-06-28 | 2002-01-03 | University Technology Corporation | Progesterone receptor-regulated gene expression and methods related thereto |
| US10501800B2 (en) | 2012-04-27 | 2019-12-10 | Regents Of The University Of Minnesota | Breast cancer prognosis |
-
2013
- 2013-03-15 US US13/843,482 patent/US10501800B2/en active Active
- 2013-03-15 EP EP13714144.6A patent/EP2841594A1/en not_active Withdrawn
- 2013-03-15 JP JP2015508975A patent/JP2015516155A/ja active Pending
- 2013-03-15 AU AU2013252913A patent/AU2013252913A1/en not_active Abandoned
- 2013-03-15 WO PCT/US2013/032677 patent/WO2013162776A1/en active Application Filing
- 2013-03-15 CA CA2871590A patent/CA2871590A1/en not_active Abandoned
- 2013-03-15 KR KR20147033298A patent/KR20150028232A/ko not_active Application Discontinuation
- 2013-03-15 RU RU2014147625A patent/RU2014147625A/ru not_active Application Discontinuation
- 2013-03-15 NZ NZ701652A patent/NZ701652A/en not_active IP Right Cessation
- 2013-03-15 MX MX2014013044A patent/MX2014013044A/es unknown
-
2014
- 2014-10-27 PH PH12014502410A patent/PH12014502410A1/en unknown
- 2014-10-27 IL IL235351A patent/IL235351A0/en unknown
- 2014-10-28 ZA ZA2014/07853A patent/ZA201407853B/en unknown
-
2015
- 2015-09-04 HK HK15108666.9A patent/HK1208057A1/xx unknown
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US10501800B2 (en) | 2012-04-27 | 2019-12-10 | Regents Of The University Of Minnesota | Breast cancer prognosis |
Also Published As
| Publication number | Publication date |
|---|---|
| HK1208057A1 (en) | 2016-02-19 |
| US20130316992A1 (en) | 2013-11-28 |
| US10501800B2 (en) | 2019-12-10 |
| CA2871590A1 (en) | 2013-10-31 |
| PH12014502410A1 (en) | 2015-01-12 |
| AU2013252913A1 (en) | 2014-11-27 |
| MX2014013044A (es) | 2015-06-23 |
| RU2014147625A (ru) | 2016-06-20 |
| NZ701652A (en) | 2016-09-30 |
| WO2013162776A1 (en) | 2013-10-31 |
| JP2015516155A (ja) | 2015-06-11 |
| EP2841594A1 (en) | 2015-03-04 |
| IL235351A0 (en) | 2014-12-31 |
| ZA201407853B (en) | 2015-12-23 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| Al-Othman et al. | Role of androgen and microRNA in triple-negative breast cancer | |
| KR20150028232A (ko) | 프로게스테론 수용체 서브타입의 유방암 예후, 예측 및 유전자 발현을 토대로 한 항프로게스틴 치료에 대한 반응성 예측 방법 | |
| JP6381519B2 (ja) | がん転移の予後診断および処置のための方法 | |
| Knutson et al. | Phosphorylated and sumoylation-deficient progesterone receptors drive proliferative gene signatures during breast cancer progression | |
| US8877445B2 (en) | Methods for identification of tumor phenotype and treatment | |
| JP6550045B2 (ja) | 乳がんに由来する骨の転移がんの予後診断および処置のための方法 | |
| Zhang et al. | Hypoxia-induced TPM2 methylation is associated with chemoresistance and poor prognosis in breast cancer | |
| Liedtke et al. | PIK3CA-activating mutations and chemotherapy sensitivity in stage II–III breast cancer | |
| Peiro et al. | Increased insulin-like growth factor-1 receptor mRNA expression predicts poor survival in immunophenotypes of early breast carcinoma | |
| Gu et al. | Targeting thyroid hormone receptor beta in triple-negative breast cancer | |
| Siegfried et al. | Expression of PAM50 genes in lung cancer: evidence that interactions between hormone receptors and HER2/HER3 contribute to poor outcome | |
| Stute et al. | Life stage differences in mammary gland gene expression profile in non-human primates | |
| Mohebi et al. | Expression analysis of vimentin and the related lncRNA network in breast cancer | |
| Suenaga et al. | Gene Polymorphisms in the CCL5/CCR5 Pathway as a Genetic Biomarker for Outcome and Hand–Foot Skin Reaction in Metastatic Colorectal Cancer Patients Treated With Regorafenib | |
| Eissa et al. | Prognostic and biological significance of microRNA-221 in breast cancer | |
| Clarke et al. | Overview of biomarkers in metastatic colorectal cancer: Tumour, blood and patient-related factors | |
| Guo et al. | Genetic variations in the PI3K/AKT pathway predict platinum-based neoadjuvant chemotherapeutic sensitivity in squamous cervical cancer | |
| Adams et al. | Identification of a novel diagnostic gene expression signature to discriminate uterine leiomyoma from leiomyosarcoma | |
| De Mattia et al. | Germline polymorphisms in the nuclear receptors PXR and VDR as novel prognostic markers in metastatic colorectal cancer patients treated with FOLFIRI | |
| Bredemeier et al. | Comparison of the PI3KCA pathway in circulating tumor cells and corresponding tumor tissue of patients with metastatic breast cancer | |
| Rizzolo et al. | Somatic alterations of targetable oncogenes are frequently observed in BRCA1/2 mutation negative male breast cancers | |
| Martinez-Becerra et al. | Up-regulation of FXR isoforms is not required for stimulation of the expression of genes involved in the lack of response of colon cancer to chemotherapy | |
| Waissengrin et al. | The effect of non-oncology drugs on clinical and genomic risk in early luminal breast cancer | |
| Hagiwara et al. | CHFR-promoter-methylation status is predictive of response to irinotecan-based systemic chemotherapy in advanced colorectal cancer | |
| Xiao et al. | Methylation-mediated silencing of ATF3 promotes thyroid cancer progression by regulating prognostic genes in the MAPK and PI3K/AKT pathways |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| WITN | Application deemed withdrawn, e.g. because no request for examination was filed or no examination fee was paid |