KR101593224B1 - 신경세포 축약 모델링 방법 - Google Patents
신경세포 축약 모델링 방법 Download PDFInfo
- Publication number
- KR101593224B1 KR101593224B1 KR1020140070324A KR20140070324A KR101593224B1 KR 101593224 B1 KR101593224 B1 KR 101593224B1 KR 1020140070324 A KR1020140070324 A KR 1020140070324A KR 20140070324 A KR20140070324 A KR 20140070324A KR 101593224 B1 KR101593224 B1 KR 101593224B1
- Authority
- KR
- South Korea
- Prior art keywords
- cell body
- cell
- dendrite
- dendritic
- dendrites
- Prior art date
Links
Images






Classifications
-
- G—PHYSICS
- G06—COMPUTING; CALCULATING OR COUNTING
- G06F—ELECTRIC DIGITAL DATA PROCESSING
- G06F17/00—Digital computing or data processing equipment or methods, specially adapted for specific functions
- G06F17/10—Complex mathematical operations
-
- G—PHYSICS
- G16—INFORMATION AND COMMUNICATION TECHNOLOGY [ICT] SPECIALLY ADAPTED FOR SPECIFIC APPLICATION FIELDS
- G16H—HEALTHCARE INFORMATICS, i.e. INFORMATION AND COMMUNICATION TECHNOLOGY [ICT] SPECIALLY ADAPTED FOR THE HANDLING OR PROCESSING OF MEDICAL OR HEALTHCARE DATA
- G16H50/00—ICT specially adapted for medical diagnosis, medical simulation or medical data mining; ICT specially adapted for detecting, monitoring or modelling epidemics or pandemics
- G16H50/50—ICT specially adapted for medical diagnosis, medical simulation or medical data mining; ICT specially adapted for detecting, monitoring or modelling epidemics or pandemics for simulation or modelling of medical disorders
-
- G—PHYSICS
- G06—COMPUTING; CALCULATING OR COUNTING
- G06F—ELECTRIC DIGITAL DATA PROCESSING
- G06F2111/00—Details relating to CAD techniques
- G06F2111/10—Numerical modelling
-
- G—PHYSICS
- G06—COMPUTING; CALCULATING OR COUNTING
- G06F—ELECTRIC DIGITAL DATA PROCESSING
- G06F30/00—Computer-aided design [CAD]
- G06F30/20—Design optimisation, verification or simulation
Landscapes
- Engineering & Computer Science (AREA)
- Data Mining & Analysis (AREA)
- Public Health (AREA)
- Health & Medical Sciences (AREA)
- Medical Informatics (AREA)
- Physics & Mathematics (AREA)
- Databases & Information Systems (AREA)
- Mathematical Physics (AREA)
- General Physics & Mathematics (AREA)
- Theoretical Computer Science (AREA)
- Biomedical Technology (AREA)
- Epidemiology (AREA)
- Pathology (AREA)
- General Health & Medical Sciences (AREA)
- Primary Health Care (AREA)
- Mathematical Analysis (AREA)
- General Engineering & Computer Science (AREA)
- Software Systems (AREA)
- Pure & Applied Mathematics (AREA)
- Mathematical Optimization (AREA)
- Computational Mathematics (AREA)
- Algebra (AREA)
- Measurement And Recording Of Electrical Phenomena And Electrical Characteristics Of The Living Body (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
- Apparatus Associated With Microorganisms And Enzymes (AREA)
- Investigating Or Analysing Biological Materials (AREA)
Abstract
세포체(soma)와 수상돌기(dendrite) 사이의 비대칭적인 신호 전달과 수상돌기 흥분성 사이의 관계를 기반으로 복잡한 구조를 갖는 실제 신경세포를 생물리학적으로 타당하게 축약하여 사실적인 시험분석을 가능하게 하는 모델링 방법이 개시된다. 상기 신경세포 축약 모델링 방법은, 세포체 및 수상돌기를 포함하는 신경세포의 축약 모델을 구현하는 방법으로서, 상기 신경세포의 해부학적 모델로부터 상기 세포체와 상기 수상돌기의 신호 전달 특성을 나타내는 전압 감쇠 인자를 결정하는 단계-상기 전압 감쇠 인자는 상기 세포체로부터 경로 길이(Dpath)의 함수로 결정됨-; 및 상기 전압 감쇠 인자를 포함하는 상기 해부학적 모델의 시스템 파라미터를 이용하여 특정 경로 길이에서 결정된 수동적 파라미터를 갖는 세포체 구획 및 수상돌기 구획으로 구성된 상기 축약 모델을 결정하는 단계를 포함한다.
Description
본 발명은 신경세포 축약 모델링 방법에 관한 것으로, 더욱 상세하게는 세포체(soma)와 수상돌기(dendrite) 사이의 비대칭적인 신호 전달 특성과 수상돌기 흥분성 사이의 관계를 이용하여, 복잡한 구조를 갖는 실제 신경세포를 생물리학적으로 타당하게 축약하여 사실적인 시험분석을 가능하게 하는 모델링 방법에 관한 것이다.
중추 신경계에서 많은 신경세포들이 그들의 수상돌기에 전압 게이트 이온 채널(voltage gated ion channel: VGIC)을 갖는다. 전압 게이트 이온 채널의 활성화는 위치에 민감하며, 수상돌기 신호 및 발화 패턴(firing pattern)에 대한 기능적 효과로 이어진다(London M, Hausser M (2005) Dendritic computation. Annu Rev Neurosci 28: 503-532, Oviedo H, Reyes AD (2005) Variation of input-output properties along the somatodendritic axis of pyramidal neurons. J Neurosci 25: 4985-4995, Gidon A, Segev I (2012) Principles governing the operation of synaptic inhibition in dendrites. Neuron 75: 330-341, Lee RH, Kuo JJ, Jiang MC, Heckman CJ (2003) Influence of active dendritic currents on input-output processing in spinal motoneurons in vivo. J Neurophysiol 89: 27-39). 따라서, 수상돌기의 전압 게이트 이온 채널의 활성화에 대한 수상돌기 구조의 영향을 정확하게 이해하는 것이 필요하다. 다시 말하면, 수상돌기 구조는 수상돌기의 수동적 흥분성뿐만 아니라 그의 능동적 흥분성에 영향을 미친다(여기서, 수상돌기 흥분성이라는 용어는 수상돌기 전압 게이트 이온 채널을 포함하는 능동적 요소를 주로 지칭한다).
수상돌기 전압 게이트 이온 채널의 활성화는 세포체에서 전압 또는 전류 클램프(clamp)를 이용하여 연구될 수 있다. 세포체에서 인가되는 명령 신호(전압 또는 전류)는 수상돌기 트리(dendritic tree)의 경로를 따라 전파되므로, 이들의 케이블 특성(cable property)(Rall W (1962) Theory of physiological properties of dendrites. Ann N Y Acad Sci 96: 1071-1092)으로 인해 더 먼 전압 게이트 이온 채널을 활성화하는데 더 높은 명령 신호가 요구된다. 이러한 효과는 수상돌기 흥분성의 해석을 위해 잠재적 혼란 인자(confounding factor)로서 공간적 클램프(space-clamp)를 발생시킨다(Bar-Yehuda D, Korngreen A (2008) Space-clamp problems when voltage clamping neurons expressing voltage-gated conductances. J Neurophysiol 99: 1127-1136; Muller W, Lux HD (1993) Analysis of voltage-dependent membrane currents in spatially extended neurons from point-clamp data. J Neurophysiol 69: 241-247). 신경세포 모델을 이용하는 많은 연구들이 전압 게이트 이온 채널 활성화의 위치 의존도에 대한 핵심 결정 요인으로써 세포체(cell body)로부터 수상돌기로의 신호 전달 크기, 즉 결합 컨덕턴스(coupling conductance)를 특정함으로써 이 혼란 인자를 설명하였다(Carlin KP, Bui TV, Dai Y, Brownstone RM (2009) Staircase currents in motoneurons: insight into the spatial arrangement of calcium channels in the dendritic tree. J Neurosci 29: 5343-5353; Mainen ZF, Sejnowski TJ (1996) Influence of dendritic structure on firing pattern in model neocortical neurons. Nature 382: 363-366; Zomorrodi R, Kroger H, Timofeev I (2008) Modeling thalamocortical cell: impact of ca channel distribution and cell geometry on firing pattern. Front Comput Neurosci 2: 5).
그러나, 수상돌기 해부에 초점을 맞춘 연구들은 세포체와 수상돌기 사이에 더 복잡한 신호 전달 과정이 존재함을 입증하였다. 실험적 증거(Hausser M, Spruston N, Stuart GJ (2000) Diversity and dynamics 509 of dendritic signaling. Science 290: 739-744)와 계산적 분석(Segev I, London M (2000) Untangling dendrites with quantitative models. Science 290: 744-750; Rall W, Rinzel J (1973) Branch input resistance and steady attenuation for input to one branch of a dendritic neuron model. Biophys J 13: 648-687)는 모두 수상돌기 내 전기적 신호의 전달이 전달 방향(즉, 세포체에서 수상돌기로 및 그 반대로)에 대해 비대칭적이며 주파수(직류(DC) 또는 교류(AC) 전류)에 의해 조정된다는 점을 강조하였다. 이러한 방향과 주파수 의존적인 수상돌기 신호는 2-포트 회로 이론(Carnevale NT, Johnston D (1982) Electrophysiological characterization of remote chemical synapses. J Neurophysiol 47: 606-621) 및 수상돌기 해부학적 구조의 형태 전기긴장성 변환(morphoelectrotonic transformation)(Zador AM, Agmon-Snir H, Segev I (1995) The morphoelectrotonic transform: a graphical approach to dendritic function. J Neurosci 15: 1669-1682)을 이용하여 분석되었다. 이러한 연구들은, 수상돌기 흥분성의 이론적인 검토 및 이해가 세포체에서 수상돌기로의 직류 신호 전달의 기존 정성적 또는 현상학적인 재현보다 앞서야 한다고 제안하였다. 더욱이, 세포체와 수상돌기의 단일 지점 사이의 수상돌기 신호에 대한 이전의 분석은, 전압 게이트 이온 채널 또는 시냅스 입력(synaptic input)이 많은 수상돌기에 위치하는 더욱 자연스런 조건을 반영하도록 확장되어야 한다.
신경세포의 세포체(soma)와 수상돌기(dendrite) 사이의 전기생리학적(electrophysiological) 신호의 전달은 비대칭적이라는 점이 알려져 있으나, 이러한 비대칭성이 전압-게이트 수상돌기 채널(voltage-gated dendritic channel) 및 결과적인 신경세포 행동의 활성화에 얼마나 영향을 미치는 지는 불명확한 것으로 남아있다.
더욱이, 이와 같은 기존의 신경 세포 모델링 방법들은 추상적으로 개발되어 다양한 생물리학적 성질을 갖는 개별 신경세포들을 사실적으로 모델링하는데 한계가 있었다.
상기와 같은 종래 기술의 문제점을 해결하기 위해, 본 발명은 실험적으로 모델 변수들을 결정하는 수학적 기법을 개발하여 특정 신경 세포의 주요 생물리학적 성질들을 사실적으로 반영하는 신경세포의 축약 모델링 방법을 제공하고자 한다.
위와 같은 과제를 해결하기 위해, 본 발명의 한 특징에 따른 신경세포 축약 모델링 방법은 모델링 대상 신경 세포에 대하여, 상기 신경세포의 세포체와 수상돌기 사이의 비대칭적 신호 전달 특성을 확인하는 단계; 상기 신경 세포의 수상돌기 흥분성을 확인하는 단계; 상기 비대칭적 신호 전달 특성과 상기 수상돌기 흥분성 사이의 관계를 확인하는 단계; 및 상기 상기 비대칭적 신호 전달 특성과 상기 수상돌기 흥분성 사이의 관계를 이용하여 모델링 대상 신경 세포에 대하여 신경 세포 축약 모델링 방법을 결정하는 단계;를 포함하여 구성된다.
여기에서, 상기 비대칭적 신호 전달 특성과 상기 수상돌기 흥분성 사이의 관계는 상기 세포체로부터 상기 수상돌기 방향으로 신호감쇠의 증가가 상기 수상 돌기의 흥분성 역치를 증가시키고, 상기 수상돌기로부터 상기 세포체 방향으로 신호감쇠의 증가가 상기 수상 돌기의 흥분성 역치를 감소시키는 것을 포함한다.
여기에서, 상기 비대칭적 신호 전달 특성은 3개의 전압 감쇠 파라미터인 세포체에 주입된 일정 전류에 반응하여 수상돌기로 전파되는 신경세포 세포막 전압의 공간적 감쇠 프로파일(VASD DC), 기시분절 및 세포체에서 생성되어 수상돌기로 전파되는 활동전위(action potential)의 공간적 감쇠 프로파일(VASD AC), 및 수상돌기의 VGIC 및 일정한 시냅스 입력에 의해 생성된 고원전위의 공간적 감쇠 프로파일(VADS DC)를 포함한다.
여기에서, 상기 세포체에 주입된 일정 전류에 반응하여 수상돌기로 전파되는 세포막 전압의 공간적 감쇠 프로파일(VASD DC), 상기 기시분절 및 세포체에서 생성되어 수상돌기로 전파되는 활동전위의 공간적 감쇠 프로파일(VASD AC), 및 상기 수상돌기의 VGIC 및 일정한 시냅스 입력에 의해 생성된 고원전위의 공간적 감쇠 프로파일(VADS DC)은 각각 식 1 내지 3에 의해 상기 세포체로부터 경로 길이 인자로 결정된다.
[식 1]
[식 2]
[식 3]

여기에서, 상기 비대칭적 신호 전달 특성은 상기 세포체와 상기 수상돌기 단일 지점 사이에서 DC 전압 감쇠의 비율(VASD DC/VADS DC)에 의해 아래와 같이 표현될 수 있다. 아래 식에서 RN,D은 수상돌기의 동일 위치(site)에서 입력 저항을 나타낸다.
본 발명의 또 다른 특징에 따른 신경세포 축약 모델링 방법은 신호 전달 특성을 나타내며, 상기 세포체로부터 경로 길이(Dpath)의 함수인 전압 감쇠 인자를 결정하는 단계; 및 상기 전압 감쇠 인자를 포함하는 상기 해부학적 모델의 시스템 파라미터를 이용하여 특정 경로 길이에서 결정된 복수개의 수동적 파라미터 관계식을 결정하는 단계; 를 포함한다.
여기에서, 상기 전압 감쇠 인자는 상기 세포체에서 상기 수상돌기로 향하는 DC 성분의 VASD DC 및 AC 성분의 VASD AC 및 상기 수상돌기에서 상기 세포체로 향하는 DC 성분의 VADS DC를 포함할 수 있다.
여기에서, 상기 VASD DC는 세포체에 위치해있는 안정된 시냅스 입력 및 지속 내부 전류(persistent inward current), 그리고 DC 자극전류에 대한 수상돌기의 전파특성으로 측정될 수 있다.
여기에서, 상기 VADS DC는 세포체로부터 동일 거리에 있는 안정된 시냅스 입력 및 수상돌기의 모든 가지에 분포된 VGIC에 의해 생성된 지속 내부 전류(persistent inward current)에 대한 수상돌기의 전파특성으로 측정될 수 있다.
여기에서, 상기 VASD AC는 세포체에서 생성되는 활동전위에 대한 수상돌기의 전파특성으로 측정될 수 있다.
여기에서, 상기 VASD DC 및 VASD AC는 상기 세포체로부터 경로 길이(Dpath)의 함수로서 전압 감쇠 상수(각각 λSD DC 및 λSD AC)를 갖는 단일 지수 함수로 피팅될 수 있다.
여기에서, 상기 VASD DC 및 VASD AC는 하기 식 1 및 식 2에 의해 결정될 수 있다.
[식 1]
[식 2]
여기에서, 상기 VADS DC는 상기 세포체로부터 경로 길이(Dpath)의 함수로서 두 개의 파라미터(α1 및 α2, α1은 VADS DC=0.5에서 거리를 개략적으로 나타내며, α2는 α1에서 VADS DC의 역-시그모이드(inverse-sigmoid) 곡선의 기울기 변화를 나타냄)를 갖는 수정된 볼츠만 방정식으로 피팅될 수 있다.
여기에서, 상기 VADS DC는 하기 식 3에 의해 결정될 수 있다.
[식 3]

여기에서, 상기 해부학적 모델의 시스템 파라미터는 상기 전압 감쇠 인자 및 상기 신경세포의 입력저항인 RN 과 상기 신경세포의 막 시상수인 τm를 포함하며, 상기 축약 모델을 결정하는 단계는, 상기 해부학적 모델의 시스템 파라미터에 대해 역 방정식을 풂으로써 세포체로부터 상기 특정 경로 길이에서 상기 수동적 파라미터를 결정할 수 있다.
여기에서, 상기 축약 모델을 결정하는 단계는, 하기 식 4 내지 식 8에 의해 상기 수동적 파라미터를 결정하는 단계를 포함할 수 있다.
[식 4]
[식 5]
[식 6]
[식 7]

[식 8]
(VASD DC: 상기 세포체에서 상기 수상돌기로 향하는 DC 성분의 전압 감쇠 인자, VASD AC: 상기 세포체에서 상기 수상돌기로 향하는 AC 성분의 전압 감쇠 인자, VADS DC: 상기 수상돌기에서 상기 세포체로 향하는 DC 성분의 전압 감쇠 인자, rN: 상기 체세포 구획(somatic compartment)의 표면적으로 정규화된 입력 저항(RN), : 활동 전위(action potential)에서 최대 주파수 성분이며, p: 상기 축약 모델의 전체 면적에 대한 체세포의 비율)
: 활동 전위(action potential)에서 최대 주파수 성분이며, p: 상기 축약 모델의 전체 면적에 대한 체세포의 비율)
본 발명의 신경세포 축약 모델링 방법은 세포체로부터 모든 거리에서 신호 전달 비대칭성 및 수상돌기 흥분성 사이의 밀접한 관계를 규명하여 이를 통해 수립되어, 모델링 대상 신경세포에 대하여 신호 전달 비대칭성과 수상 돌기 흥분성의 관계를 이용하여 신경 세포를 모델링 할 수 있다.
본 발명에서 신경세포에 대한 신호 전달 비대칭성은 전압 감쇠 인자를 통해 설명되는 바, 전압 감쇠 인자와 수상 돌기 흥분성을 이용하여 신경 세포를 모델링 할 수 있다.
또한, 본 발명에 따르면, 기존 사용된 전기적 영역(길이 상수 λ)이 아닌 물리적 영역(Dpath) 상에서 축소 신경세포 모델을 통해 수상돌기 흥분성을 생리학적으로 나타내기 위한 이론적인 기반을 제공할 수 있다.
이로써 수상돌기 흥분성의 공간적 이질성(heterogeneity)을 지배하는 전압 감쇠 인자의 식별이 해부학적으로 재구성된 모델과 축소 신경세포 모델 사이의 차이를 연결할 수 있다.
이러한 본 발명의 신경 세포 축약 모델링 방법을 이용하면, 생체 신경망 모사 계산 알고리즘과 같은 차세대 정보 처리 분야, 생체 신경계 손상이나 질환으로 인해 생긴 운동 기능 장애를 복구하기 위한 신경-기계 인터페이스 기술 및 첨단 보철 장치과 같은 재활 분야, 및 생체 신경계로부터 신경 신호를 측정하여 생체 신경망의 기능 상태를 진단하는 의료진단장치 개발 분야 및 신약 개발을 위한 시험 평가 분야와 같은 의료 분야에 큰 기술 진전을 이룩할 수 있다.
도 1은 수상돌기의 전압 감쇠 특성을 도시한 도면이다.
도 2는 해부학적 운동 신경세포 모델(청색)과 축소 운동 신경세포 모델(적색)을 비교한 도면이다.
도 3은 수상돌기 내 다른 위치의 PIC 채널에 대한 해부학적 모델 및 축약 모델의 세포 흥분성을 도시한다.
도 4는 VA 인자에 대한 세포 흥분성의 민감도를 도시한다.
도 5는 VADS DC 및 VASD AC에 대한 셀 흥분성의 공간 의존도를 도시한다.
도 6은 수상돌기에서의 입력 저항의 DC VA 인자에 대한 의존성을 도시한다.
도 2는 해부학적 운동 신경세포 모델(청색)과 축소 운동 신경세포 모델(적색)을 비교한 도면이다.
도 3은 수상돌기 내 다른 위치의 PIC 채널에 대한 해부학적 모델 및 축약 모델의 세포 흥분성을 도시한다.
도 4는 VA 인자에 대한 세포 흥분성의 민감도를 도시한다.
도 5는 VADS DC 및 VASD AC에 대한 셀 흥분성의 공간 의존도를 도시한다.
도 6은 수상돌기에서의 입력 저항의 DC VA 인자에 대한 의존성을 도시한다.
본 발명의 연구자들은 중추 신경계를 구성하는 주요 신경세포(primary neuron)에서 신호 전달 방향 및 주파수에 대한 수상돌기 신호전파의 의존성을 연구하였다. 이를 위해, 본 발명의 연구자들은 신경세포 종류 중 하나(척추 운동 신경세포(spinal motoneuron)를 선택하였다.
구체적으로, 척추 운동 신경세포 종류 전체에서 이러한 일관된 비대칭성의 특징에 기반하여, 비대칭적 신호 전달 및 수상돌기 흥분성 사이의 관계를 확인하도록 방향 및 주파수 의존성 수상돌기 신호의 영향을 조직적으로 시험 가능하게 하는 간략화된 모델링 기법을 개발하고자 하였다.
특히, 본 발명은 수상돌기의 복잡한 신호 특성이 수상돌기 흥분성을 생리학적으로 표현하는데 필수적인 것인지를 설명하며, 이러한 신호 특성의 변화가 수상돌기 트리에서 전압 반응성 채널(voltage sensitive channel) 활성화의 일반적 패턴에 얼마나 영향을 미치는지 조사하였다.
이러한 연구 개발을 통해, 본 발명자들은 신경세포에서 신호 전달의 비대칭성의 분석에 기반하여, 운동 신경세포를 비대칭 신호 전달을 보존한 두 구획(compartment)의 표현으로 축약 모델링하여 재구성하는 것이 가능함을 발견하여 본 발명에 이르렀다.
본 발명의 축약 모델은 수상돌기 흥분성(dendritic excitability) 및 신호 전달 비대칭성을 이용한다. 본 발명에서 신호 전달 비대칭성은 수상 돌기들로 분산되는 지속적 내부 전류로 표현되며, 그에 따라 본 발명의 축약 모델은 수상 돌기 흥분성과 수상돌기들로 분산되는 지속적 내부 전류(persistent inward current: PIC)를 포함하는 해부학상 모델의 다이내믹스(dynamics)을 정확하게 모사했다.
본 발명의 축약 모델은 비대칭 신호 전달과 수상돌기 흥분성 사이의 관계를 이해하는데 사용되었다. 세포체에서 수상돌기로 신호감쇠의 증가는 PIC의 활성화 역치(threshold)를 증가시키는(저흥분성(hypo-excitability)) 반면 수상돌기에서 세포체로 신호감쇠의 증가는 PIC의 활성화 역치를 감소시킨다는(과흥분성(hyper-excitability)) 것에 기초한다.
이러한 효과는 매우 강력해서, 세포체와 수상돌기 사이의 정상적인 비대칭 신호전달 특성을 반전시키는 것이 PIC 역치와 세포체로부터 이 전류원의 거리 사이의 상관관계를 반전시킨다. 따라서, 세포의 정상적인 생리학적 기능뿐만 아니라 수상돌기 흥분성에 대한 생물물리학적으로 현실적인 시뮬레이션을 위한 수상돌기 신호전달의 정상적인 비대칭성을 유지하는 것이 매우 중요하게 된다.
이하, 첨부된 도면 및 실시예를 참조하여 본 발명을 보다 상세하게 설명한다. 그러나, 본 발명의 실시예는 여러 가지 다른 형태로 변형될 수 있으며, 본 발명의 범위가 이하 설명되는 실시예로 한정되는 것은 아니다. 본 발명의 실시예는 본 발명이 속하는 기술분야에서 통상의 지식을 가진 자에게 본 발명을 보다 완전하게 설명하기 위해서 제공되는 것이다. 또한, 본 발명을 설명함에 있어서, 정의되는 용어들은 본 발명에서의 기능을 고려하여 정의 내려진 것으로, 이는 당 분야에 종사하는 기술자의 의도 또는 관례 등에 따라 달라질 수 있으므로, 본 발명의 기술적 구성요소를 한정하는 의미로 이해되어서는 아니 될 것이다.
실시예
대상 신경세포의 준비
여섯 개의 고양이 척추 α-운동 신경세포(Burke 아카이브(archive)의 Vemoto 1-6)의 형태학적(morphological) 데이터가 ‘http://NeuroMorpho.Org’ (Ascoli GA (2006) Mobilizing the base of neuroscience data: the case of neuronal morphologies. Nat Rev Neurosci 7: 318-324)로부터 다운로드 되었으며, Import3D 툴을 이용하여 NEURON 시뮬레이션 환경 v6.1.1(Hines ML, Carnevale NT (1997) The NEURON simulation environment. Neural Comput 9: 1179-1209)으로 변환되었다. 개별적으로 불러들인 운동 신경세포에 대해, 기시분절(initial segment)/축삭소구(axon hillock)가 세포체의 사이즈를 이전 보고된 치수(Kim H, Major LA, Jones KE (2009) Derivation of cable parameters for a reduced model that retains asymmetric voltage attenuation of reconstructed spinal motor neuron dendrites. J Comput Neurosci 27: 321-336, Cullheim S, Fleshman JW, Glenn LL, Burke RE (1987) Membrane area and dendritic structure in type-identified triceps surae alpha motoneurons. J Comp Neurol 255: 68-81)로 수정한 후 세포체에 부가될 수 있다.
수동적 막 특성(passive membrane property)의 결정(assignment)
균일한 세포질 저항(Ra=70 Ω·㎝) 및 고유 막 커패시턴스(Cm=1 ㎌/㎠)를 가정하여, 운동 신경세포의 고유 막 저항(specific membrane resistivity: Rm) 값과 결과적인 전기 긴장성 특성이 모두 본 발명의 발명자에 의한 이전 연구(Kim H, Major LA, Jones KE (2009) Derivation of cable parameters for a reduced model that retains asymmetric voltage attenuation of reconstructed spinal motor neuron dendrites. J Comput Neurosci 27: 321-336)에서 다루어졌다. 운동 신경세포(즉, Vemoto6)의 Rm은 세포체에서 25 Ω·㎠이고 수상돌기에서 11 ㏀·㎠)로 설정되었다.
전압 감쇠 곡선 및 전압 감쇠 상수의 생성
수상돌기의 신호 전달을 나타내는 전압 감쇠(VA) 인자는 전류 주입 위치에서의 전압에 대한 측정 위치에서의 전압 비로 정의 된다(Rall W, Rinzel J (1973) Branch input resistance and steady attenuation for input to one branch of a dendritic neuron model. Biophys J 13: 648-687, Carnevale NT, Johnston D (1982) Electrophysiological characterization of remote chemical synapses. J Neurophysiol 47: 606-621). 세 개의 VA 인자(세포체에서 주사된(somatically injected) DC(VASD DC) 및 250 Hz-AC(VASD AC)를 포함하는 세포체 밖으로 향하는(somatofugal) 방향에 대한 두 개의 VA 인자 및 세포체에서 동일 거리에 있는 수상돌기의 모든 지점에 대해 동시적 DC 입력을 갖는 세포체 중심으로 향하는(somatocentric) 방향에 대한 하나(VADS DC))는 NEURON 소프트웨어(Hines ML, Carnevale NT (1997) The NEURON simulation environment. Neural Comput 9: 1179-1209)에서 임피던스 클래스 툴을 사용하여 해부학적으로 재구성된 신경세포의 수동적 수상돌기(passive dendrite)에서 연산되었다.
특히, VADS DC는 안정된 시냅스 입력 및 세포체로부터 동일 거리에서 수상돌기의 모든 가지에 분포된 VGIC에 의해 생성된 지속 내부 전류(persistent inward current)의 전달에 대해 측정된다. VADS DC에 대한 전류 주입량은 각 지점이 포함되는 수상돌기 세그먼트의 표면적(surface area)에 비례하여 수상돌기의 개별 지점으로 차등적으로 공급된다. 250 Hz의 AC 주파수는 운동 신경세포에 대해 2ms(주기는 4ms)의 평균 스파이크 폭으로부터 산출되었다. 낮은 주파수(<250Hz)를 갖는 AC 신호에 대한 VASD AC는 본 발명의 축약 모델링 프레임워크에서 잘 보존되는 것으로 나타났다. 개별 VA 데이터는 세포체로부터 경로 길이(즉, Dpath)의 함수로서 표시되었다.
거리에 따른 감쇠율을 계량화하기 위해, VASD DC 및 VASD AC는 전압 감쇠 상수(λSD DC 및 λSD AC)를 갖는 단일 지수 함수로 산출되었으며, VADS DC는 두 개의 파라미터(α1 및 α2)를 갖는 수정된 볼츠만 방정식으로 산출되었다. 여기서, α1은 VADS DC=0.5에서 거리를 개략적으로 나타내며, α2는 α1에서 VADS DC의 역-시그모이드(inverse-sigmoid) 곡선의 기울기 변화를 나타낸다(상세한 형상은 도 1을 참조). 개별 피팅 함수의 파라미터 값은 MATLAB에서 비선형 회귀 함수를 이용하여 결정된다.
[식 1]
[식 2]
[식 3]

이와 같이, 운동 신경세포(Vemoto6)에 대해 특정된 네 개의 전압 감쇠 상수(상기 식 1-3에서 λSD DC, λSD AC, α1 및 α2)는 본 발명의 축약 모델에 대한 세 가지 VA 인자의 값을 결정하는데 사용되었다.
본 발명의 축약 모델에 대한 VA 인자의 초기값은 600 ㎛의 Dpath에서 특정되었다. 이 값은, 해부학적 모델(Elbasiouny SM, Bennett DJ, Mushahwar VK (2005) Simulation of dendritic CaV1.3 channels in cat lumbar motoneurons: spatial distribution. J Neurophysiol 94: 3961-3974, Bui TV, Ter-Mikaelian M, Bedrossian D, Rose PK (2006) Computational estimation of the distribution of L-type Ca(2+) channels in motoneurons based on variable threshold of activation of persistent inward currents. J Neurophysiol 95: 225-241)에서 실험적으로 관찰된 비선형 발화 패턴을 생성하기 위한 PIC 채널의 분포에 대해 이전에 보고된 거리 간격(300-850 ㎛)의 평균값을 이용하였다.
신경세포 축약 모델
상기 해부학적 모델로부터 정의된 다섯 가지의 시스템 파라미터(RN, τm,  ,
,  , 및
, 및  )가 본 발명의 두 구획 모델링 프레임워크에 분석적으로 포함되었다. 축약 모델의 다섯 가지 수동적 파라미터(세포체의 세포막 전도도(Gm,S), 수상돌기의 세포막 전도도(Gm,D), 세포체의 세포막 전정용량(Cm,S), 수상돌기의 세포막 전정용량(Cm,D) 및 세포체와 수상돌기 사이의 전도도(GC)) 모두는 다섯 가지 시스템 파라미터 모두에 대해 세포체로부터 특정 거리(Dpath)의 함수로 역 방정식을 풂으로써 결정하였다.
)가 본 발명의 두 구획 모델링 프레임워크에 분석적으로 포함되었다. 축약 모델의 다섯 가지 수동적 파라미터(세포체의 세포막 전도도(Gm,S), 수상돌기의 세포막 전도도(Gm,D), 세포체의 세포막 전정용량(Cm,S), 수상돌기의 세포막 전정용량(Cm,D) 및 세포체와 수상돌기 사이의 전도도(GC)) 모두는 다섯 가지 시스템 파라미터 모두에 대해 세포체로부터 특정 거리(Dpath)의 함수로 역 방정식을 풂으로써 결정하였다.
[식 4]
[식 5]
[식 6]
[식 7]

[식 8]
여기서, rN은 세포체 구획(somatic compartment)의 표면적으로 정규화된 입력 저항(RN)이고,  는 활동 전위(action potential)에서 최대 주파수 성분이며, p는 축약 모델의 전체 면적에 대한 세포체의 비율이다.
는 활동 전위(action potential)에서 최대 주파수 성분이며, p는 축약 모델의 전체 면적에 대한 세포체의 비율이다.
본 발명에서, 축약 모델의 세포체 표면적(315759.2 ㎛2)은 해부학적으로 재구성된 운동 신경세포(즉, Vomoto6)에서 Dpath=600㎛까지 세포체로부터 누적된 전체 표면적을 나타낸다. rN 및 p는 Vemoto6의 RN(1.29 ㏁) 및 전체 표면적(641786.9 ㎛2)으로부터 산출될 수 있었다. rN(0.407 Ω·㎠),  (7.2 ms), p(0.492) 및
(7.2 ms), p(0.492) 및  (2ms의 스파이크 폭을 가정할 때 4ms의 주기로부터 산출된 2π×250 Hz)의 초기값은 모든 거리(즉, Dpath)에서 상수로서 설정되었다.
(2ms의 스파이크 폭을 가정할 때 4ms의 주기로부터 산출된 2π×250 Hz)의 초기값은 모든 거리(즉, Dpath)에서 상수로서 설정되었다.
능동적 막 특성(active membrane property)의 결정(assignment)
여섯 가지 해부학적 모델 중 급속 경련(fast twitch)-피로 내성(fatigue resistant) 타입(FR-타입) 운동 신경세포(Vemoto6)이 축약 모델과 비교를 위해 선택되었다.
동일한 타입 및 운동역학의 전압 게이트 이온 채널 및 흥분성(excitatory) 시냅스가 해부학적 모델 및 축소 운동 모델에 모두 사용되었다. 간략히 설명하면, 활동 전위는 다양한 활동 전류에 의해 생성되었다. 이 다양한 활동 전류는 세포체에서 INa,f, INa,p, IK,Dr, ICa,N, IK(Ca)와, 해부학적 모델의 기시분절(initial segment)/축삭소구(axon hillock)에서 INa,f, INa,p, IK,Dr, 및 축약 모델의 세포체 구획에서 INa,f, INa,p, IK,Dr, ICa,N, IK(Ca)를 포함한다. 고원 전위(plateau potential)가 양 모델의 수상돌기 내 L-타입 Ca2+ 채널의 영향을 받는 전류(ICa,L)에 의해 생성되었다. ICa,L이 존재하지 않는 경우, 스파이크 생성을 담당하는 운동 채널 밀도(GNa,f [mS/cm2], GNa,p [mS/cm2], GK,Dr [mS/cm2], GCa,N [mS/cm2] 및 GK(Ca) [mS/cm2])는 축약 모델 및 해부학적 모델 모두에서 동일한 전위(92.3 mv)의 활동 전위 및 동일한 기전류(rheobase current)(10.5 nA)에서의 후과분극(afterhyperpolarization: AHP) 특성(98.5 ms의 지속시간 및 3.1 mV의 깊이)을 갖도록 조정되었다. 축약 모델의 수상돌기에서 ICa,L에 대한 GCa,L [mS/cm2]은 세포체에서 모든 활동 전류를 차단한 상태에서 삼각 전압 클램프 시뮬레이션이 진행되는 동안 해부학적 모델에서 측정된 피크 전류(-22 nA)에 맞게 결정되었다(Lee RH, Heckman CJ (1999) Paradoxical effect of QX-314 on persistent inward currents and bistable behavior in spinal motoneurons in vivo. J Neurophysiol 82: 2518-2527).
시뮬레이션 프로토콜
세포체 입력-세포체 출력 관계가 느리게 증가 및 감소하는 전압(20초 동안 -70 mV에서 시작하여 30mV의 피크를 갖도록) 및 전류(10초 동안 0에서 시작하여 20 nA의 피크를 갖도록) 클램프를 세포체에 인가함으로써 평가될 수 있다. 수상돌기 입력-세포체 출력 관계가, 삼각파형으로 수상돌기 내 흥분성 시냅스 수용체(excitatory synaptic receptor)의 최대 컨덕턴스(Gsyn)를 천천히 증가시키고 감소시키는 동안(20초 동안 0에서 시작하여 해부학적 케이스에 대해서는 1.2 mS/cm2의 피크를 갖고 축소 케이스에 대해서는 1.2 mS/cm2의 피크를 갖도록) 조사될 수 있다. 축소모델에서 Gsyn의 피크는 고원 전위의 활동 타이밍(onset timing)(5 ms) 및 피크값(-13.6 mV)을 해부학적 케이스와 같이 매칭하도록 조정되었다.
결과
신경세포에서 수상돌기 신호를 특정하는 세 가지 전압 감쇠 파라미터
수상돌기의 신호 전달은 세가지 전압 감쇠(VA) 인자에 의해 완전하게 재구성된 신경세포 모델에서 특정된다. 세가지 전압 감쇠는, 세포체에 대해 DC(VASD DC) 및 AC(VASD AC) 입력을 포함하는 세포체에서 수상돌기 방향의 두 가지 및 세포체로부터 동일 거리의 수상돌기 내 모든 지점에 대해 동시 DC 입력을 갖는 수상돌기에서 세포체 방향에 대한 한 가지(VADS DC)를 포함한다.
이 세가지 VA 인자는, 세포체에 주입된 일정 전류에 반응하는 신경세포 전압 프로파일(VASD DC), 기시분절 및 세포체로부터 수상돌기로의 활동 전위 전달(VASD AC), 및 수상돌기에서 VGIC에 의해 생성된 일정한 시냅스 입력 및 고원 전위(VADS DC)를 정의한다.
도 1은 수상돌기의 전압 감쇠 특성을 도시한다. 도 1의 (a) 내지 (e)에서 왼쪽 열은 운동 신경세포의 형태를 각각 도시하며, 오른쪽 열은 세포체로부터 모든 거리(Dpath)에서 측정된 세가지 전압 감쇠 인자(VASD DC는 흑색, VASD AC는 회색, 및 VADS DC는 보라색)를 도시한다. 단일 지수 함수(exp(-Dpath/λ)가 VASD DC(도 1의 (a) 내지 (e)에서 λ=2678.87, 3085.6, 2763.7, 1945.5 및 2156.4) 및 VASD AC(도 1의 (a) 내지 (e)에서 λ=420.1, 437.1, 402.3, 373.1, 464.7)를 산출하는데 사용되었으며, 수정된 볼츠만 방정식( )이 VADS DC(도 1의 (a) 내지 (e)에서 α1&α2=1020.8&307.7, 635.2&439.5, 327.9&469.6, 374.2&504.8, 861.9&268.3)를 산출하는데 사용되었다.
)이 VADS DC(도 1의 (a) 내지 (e)에서 α1&α2=1020.8&307.7, 635.2&439.5, 327.9&469.6, 374.2&504.8, 861.9&268.3)를 산출하는데 사용되었다.
보라색 실선은 동일 거리에 있는 세포체와 수상돌기의 모든 점 사이의 VADS DC를 나타내는 반면(즉, 점 대 모든 지점), 보라색 점선(도 1의 (a) 내지 (e)에서 λ=225, 143.6, 118.5, 144, 193.7일 때 exp(-Dpath/λ))은 세포체와 세포체에서 모든 개별 수상돌기 트리로의 경로를 따른 단일 지점 사이의 VADS DC를 나타낸다(즉, 점 대 점).
도 1의 (f)는 0.4 에서 4.0 ㏁으로 입력저항(RN)이 변화할 때 여섯 가지 종류의 확인된 해부학적으로 재구성된 운동 신경세포에서 측정된 VADS DC를 도시한다. α1(VADS DC가 50% 감소한 거리) 파라미터는 α2 파라미터(α1 일 때의 곡선의 기울기 측정)와 비교하여 RN에 더 민감하다. 삽입된 도면은 0.4 및 4.0 ㏁의 극단값에서 VADS DC의 예를 도시한다. 동일한 RN에서 포인트(α1, α2)의 분포는 타입에 특정되지 않는다는 점을 주목해야 한다(FR- 타입 운동 신경세포에 대해 회색 및 흑색으로 채워진 원으로 표시). S, FR 및 FF는 각각 느린 경련, 급속 경련-피로 내성, 및 급속 경련-급속 피로 타입의 운동 신경세포 형식을 나타낸다.
도 1의 (a) 내지 (e)는 해부학적으로 재구성된 다섯 가지의 척추 운동 신경세포(즉, Vemoto1-4, 및 6) 에서 세포체로부터 거리(Dpath)의 함수로 전압 감쇠를 도시한다. 이 도면의 각 포인트는 표시된 경로 거리에서 수상돌기 가지(dendritic branch) 내 단일 포인트에서의 전압 감쇠를 나타낸다. 다섯 가지의 모든 해부학적 운동 신경세포 모델에서, 개별 VA 인자의 공간 프로파일은 수상돌기 신호의 방향 및 주파수 의존성을 나타내도록 구별 가능하게 다르다. 거리에 대한 개별 VA 인자의 다른 의존성은 격심한 분지 구조(branching structure)에 기인한 세포체와 수상돌기 사이의 전기 부하의 불균형 및 케이블 특성(cable property)에 기인한 저역 통과 필터링 효과로부터 유래한 것이다. VADS DC에 대한 데이터의 수직 정렬은 50 ㎛의 해상도에서 공간적인 샘플링에 의해 도출된다. 이 VADS DC 데이터는, 이 파라미터가 세포체 및 수상돌기 내 단일 지점 사이에서 산출될 때(Kim H, Major LA, Jones KE (2009) Derivation of cable parameters for a reduced model that retains asymmetric voltage attenuation of reconstructed spinal motor neuron dendrites. J Comput Neurosci 27: 321-336) VADS DC에 대해 종래에 피팅되었던 단일 지수함수(보라색 점선) 보다는 역 시그모이드 곡선(보라색 실선)으로 피팅된다. 새로운 기법을 이용하여 산출될 때 수상돌기에서 세포체로 향하는 DC 전압의 감쇠차이는 다수의 수상돌기 가지에 입력된 DC 전류가 세포체에서 합해져서 세포체의 세포막 전압을 결정하기 때문이다.
다른 두 가지 전압 감쇠 파라미터의 데이터는 단일 지수함수에 의해 양호하게 피팅된다. 개별 VA 데이터를 피팅하는 방정식이 축소 운동 신경세포 모델에 대한 세 가지 VA 인자의 값을 산출하는데 사용되었다. 도 1(a)에서 개별 VA 인자의 공간적 프로파일은 다른 타입(Vemoto1-5)의 운동 신경세포 모델에 걸쳐 일관되게 나타났다.
본 발명의 발명자는 모든 해부학적 모델이 세포체에서 동일한 입력 저항(RN)을 갖도록 고유 막 저항(specific membrane resistivity)을 변경하였으며, 계수 α1 및 α2을 찾기 위해 VADS DC 데이터를 역 시그모이드 커브로 재산출하고 피팅하였다. α2 파라미터와 비교하여 α1 파라미터는 RN의 변화에 더 민감하였다.
여섯 가지의 모든 운동 신경세포 형태는 RN과 α1 및 α2의 변화 사이에 유사한 관계를 가졌으며, 이는 여섯가지 모델의 형태학적 차이점이 VADS DC 작용에 큰 역할을 하지 못하였음을 나타낸다.
이러한 모든 결과는 전압 감쇠 특성이 신경세포의 종류에 특정되지 않으며, 수상돌기 트리의 가지 구조의 포괄적 특성에 특정됨을 나타낸다.
수상돌기 흥분성에 대한 VA 인자를 포함하는 축소 신경세포 모델
척추 운동 신경세포는 크고 많은 갈래로 나뉜 수상돌기 및 강한 지속적 내부 전류(persistent inward current: PIC)를 생성하는 VGIC를 가지므로(Schwindt PC, Crill WE (1980) Properties of a persistent inward current in normal and TEA-injected motoneurons. J Neurophysiol 43: 1700-1724), 신호 감쇠 특성과 수상돌기 흥분성 사이의 관계를 평가하기 위해 척추 운동 신경세포가 선택되었다. 운동 신경세포 발화 패턴에 대한 PIC의 영향은 매우 크고 거북이(Hounsgaard J, Kiehn O (1989) Serotonin-induced bistability of turtle motoneurones caused by a nifedipine-sensitive calcium plateau potential. J Physiol 414: 265-282), 쥐(rat)( Li Y, Bennett DJ (2003) Persistent sodium and calcium currents cause plateau potentials in motoneurons of chronic spinal rats. J Neurophysiol 90: 857-869), 생쥐(mouse)(Meehan CF, Sukiasyan N, Zhang M, Nielsen JB, Hultborn H (2010) Intrinsic properties of mouse lumbar motoneurons revealed by intracellular recording in vivo. J Neurophysiol 103: 2599-2610) 및 고양이(Lee RH, Heckman CJ (1998) Bistability in spinal motoneurons in vivo: systematic variations in persistent inward currents. J Neurophysiol 80: 583-593)를 포함하는 다양한 종에서 충분하게 조사되어 왔다.
도 2는 해부학적 운동 신경세포 모델(청색)과 축소 운동 신경세포 모델(적색)을 비교한 도면이다. 도 2의 (a)는 해부학적 수상돌기를 축소한 것이다. 도 2(a)에서 상부에 도시된 도면은 세포체로부터 경로 길이(Dpath)에 대한 운동 신경세포(vemoto6)의 덴드로그램이며, 하부에 도시된 도면은 축약 모델의 두 구획을 도시한다. 좌측 화살표로 지시된 부분은 세포체/기시분절(initial segment)/축삭소구(axon hillock)를 나타내며, 우측 화살표로 지시된 부분은 원 세포의 수상돌기의 협대역(평균 600 ㎛이고 표준편차 4 ㎛) 내 모든 지점을 나타낸다. 해부학적 모델에서 수상돌기 내 개별 지점에서 막 전위는 축약 모델과의 비교를 위해 평균 연산됨을 주지하여야 한다. 도 2(b)내지 2(e)는 수동적 다이내믹스를 도시한다. 축약 모델의 다섯 가지 수동적 파라미터들 모두(Gm,S,=0.143 mS/cm2, Gm,D=0.211 mS/cm2, Cm,S=1.058 mS/cm2, Cm,D=0.915 mS/cm2, 및 GC=0.211 mS/cm2)는 해부학적 모델으로부터 측정된 다섯 가지 시스템 특성을 유지하도록 해부학적으로 결정되었다. 다섯 가지 시스템 특성은, 세포체에서 필링(peeling) 분석에 의해 획득되는 입력 저항(RN= 1.29 ㏁) 및 막 시상수( =7.2 ms)와, Dpath=600 ㎛에서의 세 가지 전압 감쇠 인자(VASD DC=0.76, VADS DC=0.75, VASD AC=0.27)를 포함한다. 도 2(b)는 RN을 연산하기 위해 세포체에서 주사되는 스텝 전류(Isoma)에 반응하는 막 전위(Vm)의 시간 경과를 도시한다. 도 2(c)는 τm 및 τ1(등화 시상수)를 측정하기 위해 축소 모델 보다 해부학적 모델의 세포체에 더 작은 진폭을 갖는 짧은(brief) 전류 펄스를 입력하여 세포막 전압의 과도 응답을 세미로그(semilog) 플롯(실선)으로 도시한다. τm 및 τ1(등화 시상수)는 이 곡선(실선)의 꼬리부분을 피팅한 선형 회귀 라인(점선)의 기울기의 역수에 상응한다. 도 2(d)는 DC 입력에 응답하는 Vm의 시간 경과를 도시한다. 하부 그래프에서 실선과 점선은 세포체와 수상돌기에 입력된 DC 전류를 각각 나타내며, 상부 그래프는 세포체(실선)로부터 수상돌기(점선) 방향의 VASD DC에 대한 Vm 및 그 반대 방향의 VADS DC에 대한 Vm이다. 도 2(e)는 세포체에 주사되는 250 Hz의 AC 전류(Isoma)에 대한 세포막 전압의 반응(VASD AC)에 대해 세포체(점선)와 수상돌기(실선)에서 최대 진폭으로 정규화된 Vm의 시간 경과를 도시한다. 도 2(f) 및 (g)는 두 모델의 수상돌기에 대한 부가된 ICa,L에 따른 PIC 활성화를 도시한다. 도 2(f)의 상부 그래프는 중간 그래프에 도시된 세포체에 적용된 삼각 전압 클램프(Vm)에 대한 전체 전류(ITotal)로부터 누설 전류(ILeak)를 감산함으로써 획득된 유효 PIC(IPIC)의 시간 경과를 도시하며, 하부 그래프는 ITotal-Vm 관계를 도시한다. 도 2(g)에서 상부 그래프는 중간 그래프에 도시된 세포체에 적용된 삼각 전류 클램프(Isoma)에 반응하는 세포체(실선) 및 수상돌기(점선)에서의 Vm의 시간 경과를 도시하며, 하부 그래프는 Vm-Isoma 관계를 도시한다. 도 2(f) 및 (g)에서 화살표는 삼각 전압 및 삼각 전류의 상승 및 하강 위상(phase)을 나타낸다. ICa,L에 대한 최대 컨덕턴스(GCa,L)는 해부학적 케이스의 경우 1.37 mS/cm2이고 축소 케이스의 경우 0.124 mS/cm2였다. 도 2(h) 및 (i)는 두 모델에서 세포체에 INa,f, INa,p, IK,Dr, ICa,N, 및 IK(Ca)가 부가되는 경우 세포 흥분성을 도시한다. 도 2(h)에서 상부의 네 그래프는 세포체에서 삼각 전류 자극(Isoma)에 반응하는 즉석 발화 빈도(F)와 Vm의 시간 경과를 도시하며(청색 및 적색은 세포체에 대한 것이고 회색은 수상돌기에 대한 것임), 최하부 그래프는 F-Isoma 곡선이다. 해부학적 케이스에서 활성 전류에 대한 최대 컨덕턴스는 세포체에서 GNa,f=0.71 [S/cm2], GK,Dr=0.23 [S/cm2], GCa,N=0.01 [S/cm2], GK(Ca)=0.0258 [S/cm2] 였고, 기시분절(initial segment) 및 축삭소구(axon hillock)에서 GNa,f=2.7 [S/cm2], GNa,p=0.033*10-3 [S/cm2], GK,Dr=0.17 [S/cm2] 였으며, 수상돌기에서 GCa,L=1.37 [mS/cm2] 였다. 축약 모델에서는, 세포체에서 GNa,f=26.75[mS/cm2], GNa,p=0.00086 [mS/cm2], GK,Dr=6.2 [mS/cm2], GCa,N=0.008 [mS/cm2], GK(Ca)= 0.54 [mS/cm2] 였고, 수상돌기에서 GCa,L=0.124 [mS/cm2]였다. 도 2(i)에서, 상부의 네 그래프는 Dpath=600 ㎛의 모든 지점에 위치한 시냅스 수용체에 대한 최대 컨덕턴스(Gsyn) 변동에 반응하는 F와 Vm의 시간 경과를 도시하며(청색 및 적색은 세포체에 대한 것이고 회색은 수상돌기에 대한 것임), 최하부 그래프는 F-Gsyn 곡선이다. 도 2(i)에서 화살표는 Gsyn의 상승 위상 및 하강 위상을 나타낸다.
=7.2 ms)와, Dpath=600 ㎛에서의 세 가지 전압 감쇠 인자(VASD DC=0.76, VADS DC=0.75, VASD AC=0.27)를 포함한다. 도 2(b)는 RN을 연산하기 위해 세포체에서 주사되는 스텝 전류(Isoma)에 반응하는 막 전위(Vm)의 시간 경과를 도시한다. 도 2(c)는 τm 및 τ1(등화 시상수)를 측정하기 위해 축소 모델 보다 해부학적 모델의 세포체에 더 작은 진폭을 갖는 짧은(brief) 전류 펄스를 입력하여 세포막 전압의 과도 응답을 세미로그(semilog) 플롯(실선)으로 도시한다. τm 및 τ1(등화 시상수)는 이 곡선(실선)의 꼬리부분을 피팅한 선형 회귀 라인(점선)의 기울기의 역수에 상응한다. 도 2(d)는 DC 입력에 응답하는 Vm의 시간 경과를 도시한다. 하부 그래프에서 실선과 점선은 세포체와 수상돌기에 입력된 DC 전류를 각각 나타내며, 상부 그래프는 세포체(실선)로부터 수상돌기(점선) 방향의 VASD DC에 대한 Vm 및 그 반대 방향의 VADS DC에 대한 Vm이다. 도 2(e)는 세포체에 주사되는 250 Hz의 AC 전류(Isoma)에 대한 세포막 전압의 반응(VASD AC)에 대해 세포체(점선)와 수상돌기(실선)에서 최대 진폭으로 정규화된 Vm의 시간 경과를 도시한다. 도 2(f) 및 (g)는 두 모델의 수상돌기에 대한 부가된 ICa,L에 따른 PIC 활성화를 도시한다. 도 2(f)의 상부 그래프는 중간 그래프에 도시된 세포체에 적용된 삼각 전압 클램프(Vm)에 대한 전체 전류(ITotal)로부터 누설 전류(ILeak)를 감산함으로써 획득된 유효 PIC(IPIC)의 시간 경과를 도시하며, 하부 그래프는 ITotal-Vm 관계를 도시한다. 도 2(g)에서 상부 그래프는 중간 그래프에 도시된 세포체에 적용된 삼각 전류 클램프(Isoma)에 반응하는 세포체(실선) 및 수상돌기(점선)에서의 Vm의 시간 경과를 도시하며, 하부 그래프는 Vm-Isoma 관계를 도시한다. 도 2(f) 및 (g)에서 화살표는 삼각 전압 및 삼각 전류의 상승 및 하강 위상(phase)을 나타낸다. ICa,L에 대한 최대 컨덕턴스(GCa,L)는 해부학적 케이스의 경우 1.37 mS/cm2이고 축소 케이스의 경우 0.124 mS/cm2였다. 도 2(h) 및 (i)는 두 모델에서 세포체에 INa,f, INa,p, IK,Dr, ICa,N, 및 IK(Ca)가 부가되는 경우 세포 흥분성을 도시한다. 도 2(h)에서 상부의 네 그래프는 세포체에서 삼각 전류 자극(Isoma)에 반응하는 즉석 발화 빈도(F)와 Vm의 시간 경과를 도시하며(청색 및 적색은 세포체에 대한 것이고 회색은 수상돌기에 대한 것임), 최하부 그래프는 F-Isoma 곡선이다. 해부학적 케이스에서 활성 전류에 대한 최대 컨덕턴스는 세포체에서 GNa,f=0.71 [S/cm2], GK,Dr=0.23 [S/cm2], GCa,N=0.01 [S/cm2], GK(Ca)=0.0258 [S/cm2] 였고, 기시분절(initial segment) 및 축삭소구(axon hillock)에서 GNa,f=2.7 [S/cm2], GNa,p=0.033*10-3 [S/cm2], GK,Dr=0.17 [S/cm2] 였으며, 수상돌기에서 GCa,L=1.37 [mS/cm2] 였다. 축약 모델에서는, 세포체에서 GNa,f=26.75[mS/cm2], GNa,p=0.00086 [mS/cm2], GK,Dr=6.2 [mS/cm2], GCa,N=0.008 [mS/cm2], GK(Ca)= 0.54 [mS/cm2] 였고, 수상돌기에서 GCa,L=0.124 [mS/cm2]였다. 도 2(i)에서, 상부의 네 그래프는 Dpath=600 ㎛의 모든 지점에 위치한 시냅스 수용체에 대한 최대 컨덕턴스(Gsyn) 변동에 반응하는 F와 Vm의 시간 경과를 도시하며(청색 및 적색은 세포체에 대한 것이고 회색은 수상돌기에 대한 것임), 최하부 그래프는 F-Gsyn 곡선이다. 도 2(i)에서 화살표는 Gsyn의 상승 위상 및 하강 위상을 나타낸다.
수상돌기 흥분성의 위치 의존도에 대한 세 가지 VA 인자의 영향을 명확하게 조사하기 위해, 해부학적으로 재구성된 운동 신경세포는 원 세포의 VA 인자뿐만 아니라 입력 저항(RN) 및 시스템 시상수(τm)와 같은 세포체에서 측정된 세포 특성을 해부학적으로 유지하는 본 발명의 두 구획(two-compartment) 모델을 이용하여 축소되었다(도 2(a)). 해부학적 모델과 축약 모델 사이의 체계적인 비교는 두 단계로 이루어진다. 첫째로 세포막의 수동적인 반응특성에 대한 비교이고 둘째로 PIC 채널을 갖는 수상돌기의 흥분성 및 세포체의 발화 출력과 관련된 세포 흥분성에 대한 비교이며, Dpath=600 ㎛에서 모든 수준의 입력/출력 관계가 비교된다. 이어, 본 발명의 발명자들은 다수의 2-구획 모델을 구성하였으며 해부학적 모델의 수상돌기를 따라 Dpath값에 의존하는 수상돌기의 흥분성을 매칭시켰다.
축약 모델과 전체 모델의 수동적 다이내믹스(passive dynamics) 비교.
축약 모델의 수동적 반응특성을 설명하는데 사용된 다섯 가지 운동 파라미터(Dpath=600 ㎛인 경우, RN, τm, VASD DC, VADS DC 및 VASD AC)는 해부학적 모델의 수동적 막 특성을 타당하게 모사하기에 충분하였다(도 2(b), 2(c)). 세포체에서 길게 지속되는 스텝 전류에 반응하는 탈분극(depolarization) 및 재분극(repolarization)의 시간 경과는 양 케이스에서 매우 유사한 반면(도 2(a)), 제2 시상수(또는 등화(equalizing) 시상수, 도 2(c)에서 τ1)은 미소한 차이를 나타내었다. 비교를 위해, 도 2(c)에서 해부학적 케이스에 대한 전압 반응의 하향 전이를 일으키도록 축소 케이스(적색) 보다 더 작은 진폭을 갖는 짧은 전류 펄스가 해부학적 케이스(청색)에 대해 세포체로 인가되었다. VASD DC의 산출을 위해 측정된 세포체 및 수상돌기 막 전위는 동일한 반면, VADS DC의 산출을 위해 측정된 막전위는 정상 상태에서 해부학적 모델과 비교하여 축약 모델에서 약 0.09% 가량 약간 크게 나타났다(도 2(d)). 두 모델에서 수상돌기 탈분극의 유사성은, 동일한 수상돌기 주입 전류에 의해, Dpath=600 ㎛인 경우 축약 모델의 수상돌기에서 입력 저항이 해부학적 모델의 복잡한 수상돌기 네트워크의 입력 저항을 획득한다는 점을 나타낸다. 세포체에 대한 250 Hz의 AC 입력에 응답하는 세포체 막 전위 및 수상돌기 막 전위 진폭 비율(VASD AC)은 양 모델에서 동일하였다(도 2(e)).
수상돌기 PIC 활성화의 비교.
세포체에서 생성된 모든 활성 전류(active current)를 차단한 상태에서 느린 경사의 전압 또는 전류 클램프가 세포체로 인가되는 동안 수상돌기 PIC 활성화에 대한 비교가 이루어졌다. 이 방법에서, 본 발명의 발명자들은 AC 신호 감쇠(즉, VASD AC)의 그것으로부터 PIC 활성화에 대한 DC 신호 감쇠(즉, VASD DC 및 VADS DC)의 영향을 차단할 수 있었다. 축약 모델의 수상돌기에서 Ca 채널의 최대 컨덕턴스는 삼각 전압 클램프 자극(triangular voltage clamp stimulation) 동안 해부학적 모델에서 특정된 피크 전류에 피팅되었다(도 2(f)에서 IPIC,Peak). 삼각 전압 클램프의 상승 위상 동안 두 모델은 유사한 N자 형상의 동력의 총 전류를 생성하였다(도 2(f)에서 최상부 그래프의 ITotal). PIC 활성화의 결과는 세포체에 대한 삼각 전류 자극(triangular current stimulation) 동안 세포체 및 수상돌기 모두에서 길게 지속되는 탈분극(도 2(g)에서 최상부 그래프의 고원 전위)이라는 것이다. 전압 클램프의 상승 위상과 비교하여 전압 클램프의 하강 위상 동안 PIC의 지연된 오프셋은 전류/전압 관계의 히스테리시스(hysteresis)를 발생시킨다(도 2(f) 및 (g)의 최하부 그래프). 두 모델 간의 차이는, Dpath=600 ㎛에서 개별 수상돌기 가지의 입력 저항의 변동에 기인한 것으로 예상되는, 해부학적 수상돌기에서 PIC의 더 빠른 온셋과 더 느린 오프셋이다. 그러나, 이러한 차이는 고원 전위의 활성화 동안 전류 클램프 조건에서 무시될 수 있다(도 2(g)).
발화 반응(firing response)의 비교.
수상돌기에서 PIC 채널의 활성화는 수상돌기로 전달되는 활동 전위에 의해 영향을 받으므로(Kim H, Jones KE (2012) The retrograde frequency response of passive dendritic trees constrains the nonlinear firing behaviour of a reduced neuron model. PLoS One 7: e43654), 본 발명의 발명자들은 해부학적 모델과 축약 모델 사이에서 세포체에 대한 삼각 전류 자극에 반응하는 발화 패턴(firing pattern)을 비교하였다. Ca PIC 채널의 존재하는 경우, 두 모델은 유사한 비선형 발화 양상을 나타냈다(도 2(h)에서 최하부 그래프). 즉, 스파이크 시작(spike initiation)에 대한 전류 역치 아래에서 지속적인 발화를 갖는 반시계방향 히스테리시스 주파수-전류(F-Isoma)를 나타냈다. 전류 주입의 하강 위상의 끝에서 발화 빈도(firing rate)는 해부학적 모델에서 더 크게 나타났다. 이는 PIC의 더 느린 오프셋이 더 내부로의 전류를 발생시키기 때문이다. 전체적으로, 두 모델의 발화 빈도 양상은 잘 일치되었다. 도 2(i)는 토닉 시냅스 입력(tonic synaptic input)이 수상돌기 내 PIC 채널과 상호작용하는 경우에도 축약 모델이 전체 모델에 잘 일치됨을 나타낸다(Bennett DJ, Hultborn H, Fedirchuk B, Gorassini M (1998) Synaptic activation 550 of plateaus in hindlimb otoneurons of decerebrate cats. J Neurophysiol 80: 2023-2037). 세포체 및 수상돌기에서 막 전위의 진폭은 Gsyn의 삼각 변동 동안 두 모델 사이에 유사하였다. 세포체 주입 전류와의 F-Isoma 관계에 일치하게(도 2(i)의 최하부 그래프), 해부학적 모델은 축약 모델보다 더 빠른 PIC 채널의 온셋 및 스파이킹에 대한 역치 아래에서 유지되는 발화의 더 긴 지속시간을 갖는다.
PIC 위치에 대한 거리 변화의 효과 비교.
상술한 결과들은 전체 모델 및 축약 모델에서 세포체로부터 600 마이크로미터의 거리에 PIC 채널이 위치한 케이스에 적용되었다. 다음으로, 본 발명의 발명자들은 PIC를 생성하는 이온 채널의 위치를 변경하였다. 각 위치에서, 새로운 2-구획 모델이 생성되었으며, 이 2-구획 모델은 해부학적 모델의 수동적 특성(RN, τm, 및 세 가지 VAS)에 매칭되었다. 이는 축약 모델의 VA 파라미터에서 거리 의존성 변화를 필요로 하였다. 두 모델의 PIC 채널 밀도는 Dpath=600 ㎛에서 결정된 IPIC의 일정한 피크를 유지하도록 조정되었다(Lee RH, Heckman CJ (1999) Paradoxical effect of QX-314 on persistent inward currents and bistable behavior in spinal motoneurons in vivo. J Neurophysiol 82: 2518-2527).
도 3은 수상돌기 내 다른 위치의 PIC 채널에 대한 해부학적 모델 및 축약 모델의 세포 흥분성을 도시한다. 도 3에서 신경세포 모델과 자극 프로토콜은 수상돌기 내 PIC 채널 위치의 변동을 제외하면 도 2(h) 및 (i)에 사용된 것과 동일하다. 도 3(a) 내지 (d)는 세포체에 주사한 삼각 전류 자극(Isoma)과 이에 반응하는 발화 빈도(F)의 관계를 도시한다. 도 3(e) 내지 (h)는 수상돌기에서 흥분성 시냅스 수용체의 최대 컨덕턴스(Gsyn)를 삼각형태로 증가 및 감소하였을 때 반응하는 발화 빈도(F)를 도시한다. 도 3의 도면들에서 화살표는 삼각 전류 자극 및 Gsyn의 삼각 변화에서 상승 위상 및 하강 위상을 나타낸다. 거리와 입력 역치 간에는 부의 관계(negative relation)가 형성됨을 주지하여야 한다. 수상돌기 내 다른 거리에서, VA 인자는 VASD DC={0.91, 0.77, 0.69, 0.63}, VADS DC={0.96, 0.79, 0.57, 0.38} 및 VASD AC ={0.65, 0.31, 0.18, 0.12}이고, Ca PIC 채널의 밀도는 해부학적 모델에서 GCa,L={1.14, 1.27, 1.95, 4.1 mS/cm2}이고 축약 모델에서 GCa,L={0.11, 0.122, 0.132, 0.189 mS/cm2}이며, 축약 모델의 수동적 파라미터는 Gm,S={0.078, 0.132, 0.174, 0.2 mS/cm2}, Gm,D={0.179, 0.143, 0.1, 0.07 mS/cm2}, GC={0.918, 0.244, 0.114, 0.06 mS/cm2}, Cm,S={0.609, 1.077, 1.211, 1.302 μF/cm2}, Cm,D={1.239, 0.903, 0.764, 0.62 μF/cm2} 였다. 삽입된 파라미터 값은 200에서 1000 ㎛까지 증가된 거리를 나타낸다.
일반적으로, 축약 모델은 세포체 전류 자극 또는 시냅스 입력 동안 세포체로부터 모든 거리에서 해부학적 모델의 전체 발화 패턴을 예측할 수 있다(도 3). PIC 채널이 수상돌기 터미널으로 이동할 때, 역치 전류 및 고원 전위의 활성화에 대한 시냅스 컨덕턴스는 감소한다(도 3의 회색 화살표). 이 결과는, 수상돌기에서 PIC 채널이 더 멀수록 셀은 더 높은 흥분성을 갖는다는 것을 나타낸다.
VA 인자에 대한 수상돌기 흥분성의 의존도
상술한 결과들은, 2-구획 모델에 대한 VA 인자의 조직적 변화가 복잡한 운동 신경세포 수상돌기의 경로를 따라 고원 전위를 활성화하기 위한 역치에서 공간적 변화를 나타내는 것을 허용함을 보여주었다. 따라서, 다음으로 본 발명의 발명자는 VA 인자가 어떻게 수상돌기 흥분성에 영향을 미치는지 평가하는데 2-구획 축약 모델을 이용하였다. 이를 위해, 수상돌기 PIC 활성화에 대한 VA 인자의 영향의 민감도 분석이 세포체로부터 다른 거리에 위치한 PIC 채널에 대해 이루어졌다.
도 4는 VA 인자에 대한 세포 흥분성의 민감도를 도시한다. 도 2(h) 및 (i)에 사용된 것과 동일한 축약 모델 및 램프 자극 프로토콜을 이용하여, F-Isoma 및 F-Gsyn 관계가 개별 VA 인자가 개별적으로 변화되는 동안 시뮬레이션 되었다. 도 4(a) 및 (b)는 원래값(0.76)에서 4.7% 감소 및 20.4% 증가된 VASD DC에서 F-Isoma 곡선을 도시한다. 수직의 회색선에 연결된 원 내의 문자, IS1(8.1 nA) 및 IS2(19.8 nA)는, VASD DC 변화의 결과로서 초기 발화 가속에 대해 감소되고 증가된 전류 역치를 나타낸다. 흑색 화살표는 세포체로의 삼각 전류 자극의 상승 위상 및 하강 위상을 나타낸다. 도 4(c) 및 (d)는 도 4(a) 및 (b)에서 인가된 것과 동일한 변화율(%)로 변경된 VASD DC에서의 F-Gsyn 곡선을 도시한 것이다. 수직의 회색 화살표는 VASD DC를 변경시킨 결과로서 발화 가속도에 대한 증가(약 0.055 mS/cm2, 왼쪽) 및 감소된(약 0.029 mS/cm2, 오른쪽) Gsyn 역치를 나타낸다. 흑색 화살표는 수상돌기에서 Gsyn의 삼각 변화의 상승 위상 및 하강 위상을 나타낸다. 도 4(e)는 IS1 또는 IS2에서 발화 가속을 생성하기 위한 개별 VA 인자(VASD DC, VADS DC, 및 VASD AC)의 퍼센트 변화를 도시한다. 0%는 VA 인자의 원래 값을 나타내며, 좌표에서 마이너스 부호는 VA 인자 값의 감소를 나타낸다. 도 4(f)는 발화 가속 및 DC VA 인자의 퍼센트 변화에 대한 역치 전류(IS)의 관계를 도시한다. 데이터 포인트들은 선형 회귀선으로 피팅되었다.
VA 인자에 대한 세포 흥분성의 민감도.
주파수-전류(F-Isoma) 곡선을 생성하기 위해 삼각 전류가 세포체에 주입되었으며, PIC 활성화에 대한 역치가 발화의 가속으로부터 측정되었다. VASD DC가 원래 값(original value)에서 4.7% 감소한 경우, 높은 역시(high threshold)는 19.8 nA에서 검출되었으며, IS2로 표시되었다(도 4(a)). 이와 유사하게, VASD DC가 20.4% 증가한 경우, 이 가속에 대한 낮은 전류 역치는 8.1 nA로 검출되었고 IS1로 표시되었다(도 4(b)). IS2에서 IS1로의 시프트는 흥분성의 증가를 지시하는, 역치의 과분극 시프트를 나타낸다. 세 가지 전압 감쇠 인자 각각의 민감도는 IS1과 IS2 간 점화의 가속을 변경하는데 요구되는 변화량을 측정함으로써 추정되었다.
본 발명의 발명자들은 목표 역치를 달성하기 위해 작은 변화가 필요한 경우 VA 인자에 대한 높은 민감도를 추정하였다. 전화 가속(firing acceleration)에 대한 역치는 AC VA 인자 보다 DC에서의 변화에 대해 더 민감하였다(도 4(e)). DC VA 인자의 변화는 반대 방향으로 역치 목표를 변경하였다. VADS DC의 감소에 의해 수상돌기에서 세포체로의 감쇠를 증가시키는 것은 흥분성의 증가를 가져온다. 반대로, 반대 방향으로 감쇠, 즉 VASD DC를 증가시키는 것은 흥분성의 감소를 가져온다. 그러므로, 본 발명의 발명자들은 VASD DC가 흥분성과 양의 상관 관계를 가지며 반대로 VADS DC가 흥분성과 음의 상관 관계를 갖는다는 결론을 도출하였다(도 4(f)). 회귀선의 기울기는 세포체에서 수상돌기로의 전압 감쇠에 대해 전류 역치가 가장 민감함을 나타낸다. 또한, VASD AC에 대한 상대적으로 크지 않은 변동의 효과는 흥분성을 증가시킨다.
개별 전압 감쇠 인자를 변경하는 유사한 효과가 수상돌기로 인가된 시냅스 컨덕턴스에 의해 PIC가 활성화되는 경우에 발견되었다(도 4(c) 및 (d)). 세 가지 전압 감쇠 인자는 세포체로의 전류 입력으로서 시냅스 입력에 대한 흥분성에 관한 동일한 상관 관계를 나타내었다. 즉, VASD DC의 증가는 흥분성을 증가시켰고, 반면 VADS DC 또는 VASD AC의 증가는 흥분성을 감소시켰다.
수상돌기 흥분성의 공간 프로파일에서 VA 인자의 역할.
다음으로, 본 발명의 발명자들은 전압 감쇠 인자들이 수상돌기 상의 위치 및 그 활성화 역치 사이의 관계에 어떻게 영향을 미치는지 연구하였다.
도 5는 VADS DC 및 VASD AC에 대한 셀 흥분성의 공간 의존도를 도시한다. 도 5의 모든 시뮬레이션 조건은 VADS DC 및 VASD AC의 값을 조직적으로 변경하였다는 점을 제외하고 도 4에 사용된 것과 동일하였다. 생리학적 케이스(도 3)과 비교하여 거리가 증가함에 따라 발화 패턴은 역순으로 나타남을 주지하여야 한다. 도 5(a)-(d)는 F-Isoma의 관계를 도시하며, 도 5(e)-(h)는 F-Gsyn 관계를 도시한다. 흑색 화살표는 삼각 전류 자극 및 Gsyn 변화의 상승 위상 및 하강 위상을 나타낸다. 회색 화살표는 세포체로부터 다른 거리에서 활성화 고원 전압에 대한 전류 및 Gsyn 역치를 나타낸다. 다른 위치에서 VADS DC={0.47, 0.6, 0.78, 0.876} 및 VASD AC ={0.65, 0.31, 0.18, 0.5}였다. 축약 모델의 수동적 파라미터 값은 Gm,S={0.228, 0.183, 0.117, 0.067 mS/cm2}, Gm,D={0.019, 0.079, 0.181, 0.276 mS/cm2}, GC={0.099, 0.135, 0.204, 0.234 mS/cm2}, Cm,S={1.644, 1.387, 0.766, 1.617 μF/cm2}, Cm,D={0.134, 0.499, 1.373, 0.352 μF/cm2} 였다. 삽입된 파라미터 값은 200에서 1000 ㎛까지 증가된 거리를 나타낸다.
세 가지 VA 인자 모두가 해부학적 운동 신경세포 모델(도 1의 (a))에 의한 시뮬레이션으로부터 획득된 생물물리학적 값들에 의해 제한될 때, PIC 위치를 세포체에 가깝게 이동하는 것은 PIC 활성화 역치의 증가를 가져온다: 더 가까운 PIC 일수록 그 활성화에 더 큰 전류(주입되거나 시냅스)가 요구된다(도 3). 본 발명에 따른 분석에서, 본 발명의 발명자들은 각 PIC 거리에 대한 그 생리학적 값으로 VASD DC를 유지하였고, VADS DC 및 VASD AC를 변경하였다. 이러한 변경의 결과는, 도 5에 도시된 것과 같이 생리학적 케이스(도 3)에서 관찰된 PIC 활성화의 공간 차수(spatial order)가 전도되는(inverted) 결과를 나타내었다. 즉, PIC 역치 활성화는 세포체에 더 가까이 이동함에 따라 더 낮은 값으로 시프트하였다.
결과적으로, 세 가지 VA 인자들이 모두 비생리학적 위치에서 PIC와 함께 그 생리학적 범위(도 3)로 제한되는 경우, VADS DC 및 VASD AC의 변동은 발화 패턴에 매칭되는 것이 발견될 수 있다. 예를 들어, 생리학적 케이스에서 Dpath=1000 ㎛의 F-Isoma 곡선(도 3 (d) 및 (h))은, VADS DC를 0.96에서 0.47로 단지 감소시킴으로써 Dpath=200 ㎛에서 재생산될 수 있다. 반면, VADS DC (0.63 ->0.876)및 VASD AC(0.12->0.5) 모두의 증가는, Dpath=1000 ㎛일 때 생리학적 케이스(도 3(a))의 Dpath=200 ㎛에서의 F-Isoma 곡선과 동일한 곡선을 도출할 수 있다(도 5(d)). 수상돌기 모든 지점에서 PIC의 강한 의존성 및 VADS DC 및 VASD AC에 대한 관련 발화 패턴의 강한 의존성은 수상돌기 흥분성의 생화학적인 실제의 시뮬레이션에 요구된다.
수상돌기 시스템의 높은 차원(dimensionality)은 수상돌기 내 전압 게이트 이온 채널의 활성화에 대해 복잡한(방향 및 주파수 의존의) 수상돌기 신호가 기여하는지 여부 또는 어떻게 기여하는지를 확인하는 것을 어렵게 한다. 본 발명의 발명자들은 현실적인 축소 운동 신경세포 모델을 이용하여 세포체와 수상돌기 간 전압 전달의 비대칭성이 전압 반응 수상돌기 채널의 공간 활성화 패턴과 관련된 세포체 발화 출력을 결정하는데 중요한 역할을 한다는 것을 입증하였다. 본 발명은 다양한 종류의 척추 운동 신경세포들에서 전압 전달 특성이 현저하게 유사한 비대칭적 프로파일을 갖는다는 것을 보여주었으며, 일반적인 기능에 대한 이러한 비대칭 신호전달의 중요한 역할이 신경세포에 광범위하게 적용될 수 있음을 제안하였다. 이러한 모든 결과들은 수상돌기 신호 전달에서의 비대칭성에 생물물리학적으로 기반한 결론이 수상돌기 흥분성을 생리학적으로 나타내기 위해 신경세포의 축약 모델에서 유지되어야 한다.
수상돌기 흥분성과 전압 감쇠 인자의 상호 작용
수상돌기 신호의 비대칭성은 세포체와 수상돌기 단일 지점 사이에서 DC 전압 감쇠의 비율(VASD DC/VADS DC)에 의해 계량화 되었다. 이러한 비대칭성 인덱스는 수상돌기의 동일 위치(site)에서 입력 저항(RN,D)에 비례하는 것으로 이론적으로 나타났으며, 다음의 식으로 나타난다:  .
.
본 발명에서, VADS DC는 동일 거리에서 세포체와 수상돌기의 모든 지점 사이에서 특정되었다. 나아가 본 발명은 방정식을 이용하여 예견된 입력 저항이 해부학적 모델에서 직접 측정된 것과 잘 일치된다는 것을 확인하였다(도 6).
도 6은 수상돌기에서의 입력 저항의 DC VA 인자에 대한 의존성을 도시한다. 도 6에서 입력 저항(RN,D, 회색 점들)은 수상돌기에서 주사되는 전체 전류에 대하여 동일 거리의 운동 신경세포 수상돌기의 개별 지점에 측정되었다(VADS DC에 대해 사용된 자극 프로토콜을 참조). 측정된 RN,D,는 방정식( , 여기서 RN은 세포체(Dpath=0)에서의 입력 저항)을 사용하여 DC VA 데이터에 대한 피팅 곡선으로부터 예측된 흑색 실선에 의해 중첩될 수 있다. 수상돌기에서 예측된 입력 저항과 측정된 입력 저항이 일치함을 주목하여야 한다. 먼 거리에서 잘 일치하지 않는 것은 VADS DC 데이터와 그 피팅 곡선 사이의 오차(도 1(a))에 기인한 것이다.
, 여기서 RN은 세포체(Dpath=0)에서의 입력 저항)을 사용하여 DC VA 데이터에 대한 피팅 곡선으로부터 예측된 흑색 실선에 의해 중첩될 수 있다. 수상돌기에서 예측된 입력 저항과 측정된 입력 저항이 일치함을 주목하여야 한다. 먼 거리에서 잘 일치하지 않는 것은 VADS DC 데이터와 그 피팅 곡선 사이의 오차(도 1(a))에 기인한 것이다.
이 결과는 ‘지점 대 모든 지점’의 경우 수상돌기 에서 복잡한 RN,D 분포가 ‘지점 대 지점’의 케이스에 대한 것과 동일한 방성식에 의해 획득될 수 있음을 나타낸다. 이 방정식에 기반하여, 생리학적 범위에서 VASD DC를 유지하는 동안 VADS DC의 증가로 인해 고원 전위에 대한 입력 역치와 거리사이의 역 관계(도 5)가 RN,D를 감소시킴에 의해 설명될 수 있다. 이러한 모든 결과들은 수상돌기 입력 저항을 결정함을 통한 수상돌기에서 VGIC의 활성화에 대한 DC VA 특성의 직접적인 영향을 나타낸다.
본 발명에서, AC 신호의 감쇠는 수상돌기에 대한 토닉 시냅스 입력을 가정하는 경우 세포체에서 수상돌기로의 전달에 대해서만 고려되었다. 그 이유는 본 축약 모델링 기법의 한계뿐만 아니라 수상돌기 흥분성에 대한 후방전파 활동 전위(back-propagating action potential)의 효과 때문이다(도 2(g) 및 2(h)의 수상돌기에서 고원 전위에 대한 전류 역치를 비교). 수상돌기로 수동적으로 전달되는 활동 전위는 스파이크 및 후과분극(afterhyperpolarization: AHP) 단계에 의한 PIC 채널의 활성화에 대한 흥분성 및 억제성 효과를 모두 가질 수 있다. 활동 전위의 후방전파에 대한 수동적 수상돌기의 기여도는 신호 주파수에 대해 네거티브 하게 관련된 VASD AC를 이용하여 특정될 수 있다. AHP(100ms)의 시간 간격은 운동 신경세포에서의 스파이크(2ms)보다 훨씬 길기 때문에 후방전파 활동 전위의 억제성 효과는 흥분성 효과에 비해 더 우세적일 것으로 예상된다(도 2(g) 및 2(h)의 PIC 활성화에 대한 전류 역치 비교). 이러한 억제성 효과는 올-오어-넌(all-or-none) 방식으로 활성화 되는 것을 방지하는 고원 전위의 단계적인(graded) 활성화를 형성하는데 중요한 것으로 나타났다(Elbasiouny SM, Bennett DJ, Mushahwar VK (2006) Simulation of Ca2+ persistent inward currents in spinal motoneurones: mode of activation and integration of synaptic inputs. J Physiol 570: 355-374). 거리 증가에 따른 VASD AC의 지수적 감소로 인해, 후방전파 활동 전위는 세포체 주변 PIC 채널의 활성화를 더욱 효과적으로 제한할 것이다. 이는 더 가까운(proximal) 수상돌기 위치에서 VGIC가 먼 위치(distal site)에 비해 활성화되기 더 어렵다는 것을 나타낸다. 먼 수상돌기보다 세포체 주변에서 더 낮은, 수상돌기 흥분성의 비균일성은, 세포체로부터 거리 함수인 세가지 VA 인자의 유사한 프로파일을 나타내므로 다른 주된 종류의 신경세포에서도 발생할 것으로 예측된다(도 1).
운동 신경세포의 비선형 발화 패턴에 대한 많은 컴퓨터를 이용한 연구가, 비선형 발화 패턴을 생성하기 위해 고원 전위를 생성하는 L-타입 Ca+ 채널이 세포체가 아닌 수상돌기에서 집단화(cluster) 되어야 한다는 점을 제안하였다(F-Isoma의 이력 관계, 도 2(h)의 최하부 그래프)(Carlin KP, Jones KE, Jiang Z, Jordan LM, Brownstone RM (2000) Dendritic L-type calcium currents in mouse spinal motoneurons: implications for bistability. Eur J Neurosci 12: 1635-1646, Booth V, Rinzel J (1995) A minimal, compartmental model for a dendritic origin of bistability of motoneuron firing patterns. J Comput Neurosci 2: 299-312). 그러나, 최근의 면역조직 화학적(immunohistochemical) 연구는 세포체로부터 가까운 위치(<100 ㎛) 및 먼 위치(450~650 ㎛) 모두에서 집단화된 CaV 1.3의 높은 밀도를 나타내었다(Ballou EW, Smith WB, Anelli R, Heckman CJ (2006) Measuring dendritic distribution of membrane proteins. J Neurosci Methods 156: 257-266). 더하여, 수상돌기 없이 고립된 세포체로부터 유핵 패치 고정 기록(nucleated patch clamp recordings)이 체포체에서 Ca PIC 채널의 존재를 나타내었다(Moritz AT, Newkirk G, Powers RK, Binder MD (2007) Facilitation of somatic calcium channels can evoke prolonged tail currents in rat hypoglossal motoneurons. J Neurophysiol 98: 1042-1047). 그러나, 가까운 위치에 비해 먼 수상돌기 영역에서의 더 높은 흥분성 때문에, 세포체에 대한 전류 자극이 발생하는 동안 발화 패턴을 결정하는데 있어, 먼 거리의 PIC 채널이 우세하다(도 3 및 도 6).
수상돌기에서 위치 의존적인 흥분성은 모델 운동 신경세포의 입출력 관계와 관련이 있었다(도 3). PIC 채널의 위치가 변동되는 동안, 운동 신경세포는 운동 신경세포에서 실험적으로 관찰된 모든 비선형 점화 패턴을 재 생산할 수 있었다. 네 가지 발화 타입(타입 Ⅰ-Ⅳ)은 세포체에 대한 삼각 전류 자극이 발생하는 동안 F-I 관계에 기반하여 분류되었다(Bennett DJ, Li Y, Siu M (2001) Plateau potentials in sacrocaudal motoneurons of chronic spinal rats, recorded in vitro. J Neurophysiol 86: 1955-1971). 타입 Ⅰ 발화(자기 지속 발화(self-sustained firing) 없이 선형적으로 중첩하는 F-I 관계) 및 타입 Ⅱ 발화(자기 지속 발화 없이 시계방향의 F-I 관계)가 가까운 세포체에서(<300 ㎛) 관찰된 반면, 타입 Ⅲ 발화(자기 지속 발화를 갖는 선형적으로 중첩하는 F-I 관계) 및 타입 Ⅳ 발화(자기 지속 발화를 갖는 반시계 방향 F-I 관계)는 세포체로부터 더 멀리 이격된 수상돌기 영역에서(>600 ㎛) 발생하는 경향이 있었다. 운동 신경세포들 사이의 발화 패턴의 변동은 활성 태널 특성의 변동 대신 수상돌기에서 PIC 채널의 다른 분포 때문일 수 있다.
전압 감쇠 특성 및 수상돌기 흥분성의 변동성
수상돌기의 거리 의존적 VA 특성은 신경세포 네트워크의 일반적인 동태 동안 잠재적으로 조정될 수 있다. 고양이의 내측 비복근(medial gastrocnemius muscle)의 운동 신경세포 풀(pool)에서, RN, τm 및 AHP 특성을 포함하는 전기적 특성들은 연속적인 방법으로 운동 신경세포의 형식(S-, FR- 및 FF- 타입)과 조직적으로 관련되어 있음이 보고되었다(Zengel JE, Reid SA, Sypert GW, Munson JB (1985) Membrane electrical properties and prediction of motor-unit type of medial gastrocnemius motoneurons in the cat. J Neurophysiol 53: 1323-1344). RN은 개별 VA 인자들(VASD DC(Kim H, Major LA, Jones KE (2009) Derivation of cable parameters for a reduced model that retains asymmetric voltage attenuation of reconstructed spinal motor neuron dendrites. J Comput Neurosci 27: 321-336), VASD AC(Kim H, Jones KE (2012) The retrograde frequency response of passive dendritic trees constrains the nonlinear firing behaviour of a reduced neuron model. PLoS One 7: e43654), VADS DC(도 1(e)))의 공간 프로파일을 변화 시킴으로써 전압 감쇠의 감소에 대한 포지티브한 효과를 나타내었다. τm 및 AHP 특성은 활동 전위(action potential)의 형상을 결정하는 중요 파라미터이므로, τm 및 AHP 특성은 주파수 의존적인 VASD AC에 더 영향을 미칠 것이다. 더하여, 개별 신경세포의 막 전기적 특성(membrane electrical property)은 백그라운드 활동성의 다른 레벨 또는 수상돌기 트리를 충격하는(bombard) 시냅시스에 의한 션팅 효과(shunting effect)에 의해 모듈화 될 수 있다(Destexhe A, Pare D (1999) Impact of network activity on the integrative properties of neocortical pyramidal neurons in vivo. J Neurophysiol 81: 1531-1547, Williams SR (2004) Spatial compartmentalization and functional 570 impact of conductance in pyramidal neurons. Nat Neurosci 7: 961-967). 나아가, 케이블 특성을 결정하는 누설 전류 채널의 특성은 다양한 확산성 신경전달물질(diffusive neurotransmitter)에 의해 공간 수상돌기 흥분성을 조정하기 위한 대상이 될 수 있다(Goldstein SA, Bockenhauer D, O'Kelly I, Zilberberg N (2001) Potassium leak channels and the KCNK family of two-P-domain subunits. Nat Rev Neurosci 2: 175-184, Nicoll RA, Malenka RC, Kauer JA (1990) Functional comparison of neurotransmitter receptor subtypes in mammalian central nervous system. Physiol Rev 70: 513-565). 더하여, 수상돌기 형태는 생리학적 조건에 따라 고정적이지 않고 동적으로 변동된다. 또한, 형태의 미묘한 변화가 병리학적 조건(Yamada M, Wada Y, Tsukagoshi H, Otomo E, Hayakawa M (1988) A quantitative Golgi study of basal dendrites of hippocampal CA1 pyramidal cells in senile dementia of Alzheimer type. J Neurol Neurosurg Psychiatry 51: 1088-1090, Teskey GC, Monfils MH, Silasi G, Kolb B (2006) Neocortical kindling is associated with opposing alterations in dendritic morphology in neocortical layer V and striatum from neocortical layer III. Synapse 59: 1-9, Sousa N, Lukoyanov NV, Madeira MD, Almeida OF, Paula-Barbosa MM (2000) Reorganization of the morphology of hippocampal neurites and synapses after stress-induced damage correlates with behavioral improvement. Neuroscience 97: 253-266) 또는 물리적 손상(Krenz NR, Weaver LC (1998) Changes in the morphology of sympathetic preganglionic neurons parallel the development of autonomic dysreflexia after spinal cord injury in rats. Neurosci Lett 243: 61-64)에 의해 발생할 수 있다. 수상돌기의 VA 특성에 대한 이러한 모든 전기적 또는 형태학적 영향은 수상돌기 흥분성의 공간적 프로파일을 현저하게 변동시킬 수 있으며, 수상돌기에서 VGIC의 물리적 위치 또는 활성화 특성을 변화시키지 않고서 신경망 내 신경세포의 입력/출력 기능의 레퍼토리를 풍부하게 할 수 있다.
수상돌기 신경세포의 축약 모델링의 응용
복잡한 수상돌기 트리는 많은 축소 신경세포 모델에서 다중 등가 케이블에서 하나의 구획으로 넓은 범위의 레벨로 붕괴되었다. 축약 모델에서 수상돌기 흥분성은 표면적의 보존(Clements JD, Redman SJ (1989) Cable properties of cat spinal motoneurones measured by combining voltage clamp, current clamp and intracellular staining. J Physiol 409: 63-87) 또는 축방향 저항(axial resistance)(Bush PC, Sejnowski TJ (1993) Reduced compartmental models of 592 neocortical pyramidal cells. J Neurosci Methods 46: 159-166)과 같은 축소 기법에 의존한다. 그러나, 축소된 수상돌기에서 위치에 따른 흥분성이 해부학적 모델과 비교하여 어느 정도인지 보여주지 못하였으며, 해부학적 모델 및 축약 모델 사이의 추적 가능성의 희생을 가져왔다(Hendrickson EB, Edgerton JR, Jaeger D (2011) The capabilities and limitations of conductance-based compartmental neuron models with reduced branched or unbranched morphologies and active dendrites. J Comput Neurosci 30: 301-321). 본 발명에서, 발명자들은 세포체로부터 모든 거리에서 전압 감쇠 인자 및 수상돌기 흥분성 사이의 밀접한 관계를 보여주었다(도 3 및 도 6). 이러한 발견은 지금까지 사용된 전기적 영역(길이 상수 λ)이 아닌 물리적 영역(Dpath) 상에서 축소 신경세포 모델로 수상돌기 흥분성을 생리학적으로 나타내기 위한 이론적인 기반을 제공할 수 있다. 이로써 수상돌기 흥분성의 공간적 이질성(heterogeneity)을 지배하는 전압 감쇠 인자의 식별이 해부학적으로 재구성된 모델과 축소 신경세포 모델 사이의 차이를 연결할 수 있다.
Claims (16)
- 신경세포의 세포체와 수상돌기 사이의 신호 전달을 모델링하는 방법에 있어서,
신경세포의 세포체와 수상돌기 사이의 신호 전달에 대하여,
1) 상기 신경세포의 세포체에서 상기 수상돌기로 향하는 DC 성분의 전압 감쇠 인자(VASD DC),
2) 상기 세포체에서 상기 수상돌기로 향하는 AC 성분의 전압 감쇠 인자(VASD AC), 및
3) 상기 수상돌기에서 상기 세포체로 향하는 DC 성분의 전압 감쇠 인자(VADS DC)의 3가지의 전압 감쇠 인자를 확인하는 단계, 및
상기 전압 감쇠 인자를 이용하여 세포체의 세포막 전도도(Gm,S), 수상돌기의 세포막 전도도(Gm,D), 세포체의 세포막 전정용량(Cm,S), 수상돌기의 세포막 전정용량(Cm,D), 및 세포체와 수상돌기 사이의 전도도(Gc)의 신경세포 모델의 5가지 수동적 파라미터 중 하나 이상을 결정하는 단계,
를 포함하는 것을 특징으로 하는 신경세포 축약 모델링 방법. - 제 1항에 있어서,
상기 신경세포의 세포체와 수상돌기 사이의 신호 전달은 상기 세포체로부터 상기 수상돌기의 방향으로 신호감쇠의 증가가 상기 수상돌기의 흥분성 역치를 증가시키고,
상기 수상돌기로부터 상기 세포체의 방향으로 신호감쇠의 증가가 상기 수상돌기의 흥분성 역치를감소시키는 비대칭적 신호 전달 특성을 가지는 것을 특징으로 하는 신경세포 축약 모델링 방법. - 삭제
- 제1항 또는 제2항에 있어서,
상기 VASD DC, 상기 VASD AC, 및 상기 VADS DC가 각각 식 1 내지 3에 의해 상기 세포체로부터 경로 길이 인자로 결정되는 것을 특징으로 하는 신경세포 축약 모델링 방법.
[식 1]

[식 2]

[식 3]

(여기서, λSD DC 및 λSD AC -는 전압 감쇠 상수이며, α1은 VASD DC=0.5 에서의 거리, α2는 α1에서 VADS DC의 역-시그모이드 곡선의 기울기 변화이고, Dpath는 세포체로부터의 경로 길이이다.) - 제2항에 있어서,
상기 비대칭적 신호 전달 특성은 상기 세포체와 상기 세포체로부터 동일거리에 있는 상기 수상돌기 위의 모든 지점들 사이에서 식 4에 의해 표현되는 DC 전압 감쇠의 비율로 표현되는 것을 특징으로 하는 신경 세포 축약 모델링 방법.
[식 4]
.
(여기서, RN,D는 세포체로부터 동일거리(Dpath)에 있는 수상돌기 위의 모든 지점에 대한 입력 저항이고, RN은 세포체(Dpath=0)에서의 입력 저항이다.) - 삭제
- 삭제
- 제1항에 있어서,
상기 VASD DC는 상기 세포체에 있는 안정된 시냅스 입력 및 지속 내부 전류(persistent inward current), 그리고 DC 자극전류에 대한 상기 수상돌기의 전파특성으로 측정하는 것을 특징으로 하는 신경세포 축약 모델링 방법. - 제1항에 있어서,
상기 VADS DC는 상기 세포체로부터 동일 거리에 있는 안정된 시냅스 입력 및 상기 수상돌기의 모든 가지에 분포된 VGIC에 의해 생성된 지속 내부 전류(persistent inward current)에 대한 상기 수상돌기의 전파특성으로 측정하는 것을 특징으로 하는 신경세포 축약 모델링 방법. - 제1항에 있어서,
상기 VASD AC는 상기 세포체에서 생성되는 활동전위(action potential)에 대한 상기 수상돌기의 전파특성으로 측정하는 것을 특징으로 하는 신경세포 축약 모델링 방법. - 삭제
- 삭제
- 삭제
- 삭제
- 삭제
- 제1항에 있어서,
상기 세포체의 세포막 전도도(Gm,S), 수상돌기의 세포막 전도도(Gm,D), 세포체의 세포막 전정용량(Cm,S), 수상돌기의 세포막 전정용량(Cm,D), 및 세포체와 수상돌기 사이의 전도도(Gc)의 신경세포 모델의 5가지 수동적 파라미터는 하기 식5 내지 식 9에 의해 각각 결정되는 것을 특징으로 하는 신경세포 축약 모델링 방법.
[식 5]
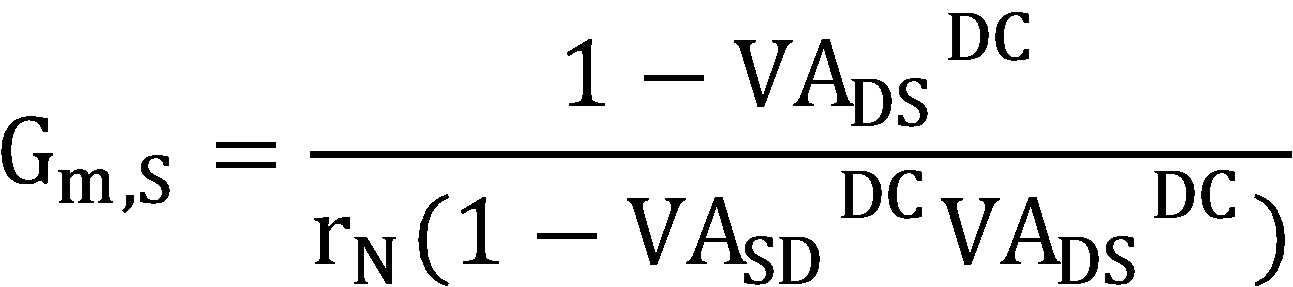
[식 6]

[식 7]

[식 8]

[식 9]

(여기서, rN은 세포체 구획(somatic compartment)의 표면적으로 정규화된 입력 저항(RN)이고, ω는 활동 전위(action potential)에서 최대 주파수 성분이며, p는 축약 모델의 전체 면적에 대한 세포체의 비율이다.)
Priority Applications (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| KR1020140070324A KR101593224B1 (ko) | 2014-06-10 | 2014-06-10 | 신경세포 축약 모델링 방법 |
| PCT/KR2015/000195 WO2015190664A1 (ko) | 2014-06-10 | 2015-01-08 | 신경세포 축약 모델링 방법 |
| US14/679,383 US11068561B2 (en) | 2014-06-10 | 2015-04-06 | Reduced modeling method for neurons |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| KR1020140070324A KR101593224B1 (ko) | 2014-06-10 | 2014-06-10 | 신경세포 축약 모델링 방법 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| KR20150142145A KR20150142145A (ko) | 2015-12-22 |
| KR101593224B1 true KR101593224B1 (ko) | 2016-02-12 |
Family
ID=54769778
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020140070324A KR101593224B1 (ko) | 2014-06-10 | 2014-06-10 | 신경세포 축약 모델링 방법 |
Country Status (3)
| Country | Link |
|---|---|
| US (1) | US11068561B2 (ko) |
| KR (1) | KR101593224B1 (ko) |
| WO (1) | WO2015190664A1 (ko) |
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| KR20190097482A (ko) * | 2018-02-12 | 2019-08-21 | 재단법인대구경북과학기술원 | 이질적 신경세포 집단 모델링 방법 |
Families Citing this family (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN110472733B (zh) * | 2019-07-22 | 2023-05-02 | 天津大学 | 一种基于神经形态学的在体神经元建模方法 |
Family Cites Families (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US6723698B2 (en) * | 1991-03-11 | 2004-04-20 | Curis, Inc. | Methods and compositions for the treatment of motor neuron injury and neuropathy |
| US8906360B2 (en) * | 2005-07-22 | 2014-12-09 | The Board Of Trustees Of The Leland Stanford Junior University | Light-activated cation channel and uses thereof |
| BRPI1012842A2 (pt) * | 2009-05-27 | 2019-09-24 | Easyglide Ltd | sistema de endoscópio com limpeza de fluido. |
| US9427581B2 (en) * | 2013-04-28 | 2016-08-30 | ElectroCore, LLC | Devices and methods for treating medical disorders with evoked potentials and vagus nerve stimulation |
| JP2014239871A (ja) * | 2013-05-07 | 2014-12-25 | 安東 秀夫 | 生体活動検出方法、生体活動測定装置、生体活動検出信号の転送方法および生体活動情報を利用したサービスの提供方法 |
-
2014
- 2014-06-10 KR KR1020140070324A patent/KR101593224B1/ko active IP Right Grant
-
2015
- 2015-01-08 WO PCT/KR2015/000195 patent/WO2015190664A1/ko active Application Filing
- 2015-04-06 US US14/679,383 patent/US11068561B2/en active Active
Non-Patent Citations (2)
| Title |
|---|
| BMC NEUROSCIENCE |
| J COMPUT NEUROSCI (2009) |
Cited By (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| KR20190097482A (ko) * | 2018-02-12 | 2019-08-21 | 재단법인대구경북과학기술원 | 이질적 신경세포 집단 모델링 방법 |
| KR102028946B1 (ko) | 2018-02-12 | 2019-10-07 | 재단법인대구경북과학기술원 | 이질적 신경세포 집단 모델링 방법 |
Also Published As
| Publication number | Publication date |
|---|---|
| WO2015190664A1 (ko) | 2015-12-17 |
| US20150356267A1 (en) | 2015-12-10 |
| US11068561B2 (en) | 2021-07-20 |
| KR20150142145A (ko) | 2015-12-22 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| ElBasiouny et al. | Simulation of dendritic CaV1. 3 channels in cat lumbar motoneurons: spatial distribution | |
| Arlotti et al. | Axon terminal polarization induced by weak uniform DC electric fields: a modeling study | |
| Miyasho et al. | Low-threshold potassium channels and a low-threshold calcium channel regulate Ca2+ spike firing in the dendrites of cerebellar Purkinje neurons: a modeling study | |
| Hounsgaard | Motor neurons | |
| Herrera-Valdez et al. | Relating ion channel expression, bifurcation structure, and diverse firing patterns in a model of an identified motor neuron | |
| Rathour et al. | Influence fields: a quantitative framework for representation and analysis of active dendrites | |
| Santamaria et al. | Modulatory effects of parallel fiber and molecular layer interneuron synaptic activity on Purkinje cell responses to ascending segment input: a modeling study | |
| KR101593224B1 (ko) | 신경세포 축약 모델링 방법 | |
| Kim et al. | Asymmetric electrotonic coupling between the soma and dendrites alters the bistable firing behaviour of reduced models | |
| Kim et al. | Asymmetry in signal propagation between the soma and dendrites plays a key role in determining dendritic excitability in motoneurons | |
| Elbasiouny | Development of modified cable models to simulate accurate neuronal active behaviors | |
| Dai et al. | Motoneuron output regulated by ionic channels: a modeling study of motoneuron frequency-current relationships during fictive locomotion | |
| Balbi et al. | Axon-somatic back-propagation in detailed models of spinal alpha motoneurons | |
| Grande et al. | Estimates of the location of L-type Ca2+ channels in motoneurons of different sizes: a computational study | |
| Kurian et al. | Motoneuron model of self-sustained firing after spinal cord injury | |
| Poznanski et al. | A dendritic cable model for the amplification of synaptic potentials by an ensemble average of persistent sodium channels | |
| Maex et al. | Temporal integration and 1/f power scaling in a circuit model of cerebellar interneurons | |
| Mousa et al. | Dendritic distributions of L-type Ca2+ and SKL channels in spinal motoneurons: a simulation study | |
| KR102028946B1 (ko) | 이질적 신경세포 집단 모델링 방법 | |
| Mahapatra et al. | Simulation of In Vitro-Like Electrical Activities in Urinary Bladder Smooth Muscle Cells | |
| Pfeiffer et al. | A dynamic clamp protocol to artificially modify cell capacitance | |
| Kim | Impact of the localization of dendritic calcium persistent inward current on the input-output properties of spinal motoneuron pool: a computational study | |
| Mahapatra et al. | A Mathematical Model of Stochastic Synaptic Noise Dynamics Based Spontaneous Action Potential in Non-neural Cell | |
| Fan et al. | Electric field effects on neuronal input–output relationship by regulating NMDA spikes | |
| Kim et al. | Foundational dendritic processing that is independent of the cell type-specific structure in model primary neurons |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A201 | Request for examination | ||
| E902 | Notification of reason for refusal | ||
| E701 | Decision to grant or registration of patent right | ||
| GRNT | Written decision to grant | ||
| FPAY | Annual fee payment |
Payment date: 20181212 Year of fee payment: 4 |
|
| FPAY | Annual fee payment |
Payment date: 20191203 Year of fee payment: 5 |