特許法第30条第2項適用 (刊行物等1)公開者:Ganesh D、Torigoe K、Kumano-Kuramochi M、Machida S、Kobori T.、「Microplate-Based Assay for Screening of Advanced Glycation End Products Binding to Its Receptor.」、公開日:平成31年1月11日、Analytical Sciences、2019 Jan 11において早期公開された論文のコピー (刊行物等2)公開者:小堀俊郎、「食品研究部門 単年度試験研究概要書」公開日:平成30年2月22日、食品研究部門 単年度試験研究概要書(平成29年度(2017)食品試験研究成績・計画概要集(国立研究法人編)、国立研究開発法人農業・食品産業技術総合研究機構食品研究部門発行の表紙及び第91-92頁のコピー (刊行物等3)公開者:町田幸子、「食品研究部門 単年度試験研究概要書」公開日:平成30年2月22日、食品研究部門 単年度試験研究概要書(平成29年度(2017)食品試験研究成績・計画概要集(国立研究法人編)、国立研究開発法人農業・食品産業技術総合研究機構食品研究部門発行の表紙及び第85-86頁のコピー
以下、本発明を説明する。本明細書の全体にわたり、単数形の表現は、特に言及しない限り、その複数形の概念をも含むことが理解されるべきである。従って、単数形の冠詞(例えば、英語の場合は「a」、「an」、「the」など)は、特に言及しない限り、その複数形の概念をも含むことが理解されるべきである。また、本明細書において使用される用語は、特に言及しない限り、当該分野で通常用いられる意味で用いられることが理解されるべきである。したがって、他に定義されない限り、本明細書中で使用されるすべての専門用語および科学技術用語は、本発明の属する分野の当業者によって一般的に理解されるのと同じ意味を有する。矛盾する場合、本明細書(定義を含めて)が優先する。
(用語の定義)
以下に本明細書において特に使用される用語の定義を列挙する。
本明細書において、「約」とは、示される値の±10%を意味する。
本明細書において使用される場合、用語「終末糖化産物」または「後期糖化反応生成物」(いずれも英文では、Advanced Glycation End Products)とは、AGEsともいわれ、タンパク質の糖化産物であり多様な構造体の総称である。食品の加工過程で生じ、食味向上に重要である一方、生体内でも生成し、一部は生体に機能不全を誘導し加齢性疾患の引き金となる。糖尿病患者の生活の質を損ねる元凶である血管合併症として知られる糖尿病性血管障害の発症・進展への関与も知られている。血管合併症による眼、神経、腎臓の障害は、それぞれ糖尿病網膜症、神経症、腎症(あわせて三大合併症)とよばれており、糖尿病患者に特徴的な病態である。グルコースに代表される還元糖は、タンパク質、アミノ酸のアミノ基と非酵素的に反応して、シッフ塩基またはアマドリ転位化合物などの糖化生成物を形成する。ここまでの反応は可逆的であり、前期反応とよばれている。その後、さらに縮合、開裂、架橋形成などの複雑かつ不可逆的な反応を経て、終末糖化産物を形成する。このような一連の反応は、糖化反応と称される。AGEsはまた、このような過程を経て生成された構造物の総称である。生体中に存在するAGEs構造としては、カルボキシメチルリジン(CML)、カルボキエチルリジン(CEL)、ペントシジン、ピラリン、イミダゾリン、メチルグリオキサール、クロスリンなどが挙げられるが、これに限定されない。血漿中に存在するアルブミン、イムノグロブリン、オボアルブミンなどが上記の糖化を受けた産物もAGEであり、AGEとして実験系に汎用されている。さらに、インビトロ実験系では、BSA(ウシ血清アルブミン)に糖化処理を施したもの、例えば、R-AGE(リボースにより糖化処理をしたBSA)、F-AGE(フルクトースにより処理をしたBSA);G-AGE(グルコースにより糖化処理をしたBSA)なども汎用されている。血糖コントロールの指標として用いられているヘモグロビンA1cはアマドリ転移化合物であるが、AGEsに包含される。また、任意のタンパク質も、AGEsに変換可能である。例えば、AGEsに包含されるCMLアルブミンおよびCELアルブミンは、いずれもアルブミンが糖化を受けたAGEsである。このようなAGEs生成反応は、生体内において循環血液中、細胞外マトリクス、細胞内のいずれでも起こり得る。例えば、糖尿病患者の血管に存在するAGEsとしては、:蛍光性で架橋構造を有するもの(ペントシジン、クロスリンなど)および蛍光も架橋もないもの(カルボキシメチルリジン、ピラリン、メチルグリオキサール(MG)-イミダゾロンなど)の2つに大別できる。AGEsが異常値を示す場合、細小血管症(腎症、網膜症、神経症など)、大血管障害(虚血性心疾患、脳血管疾患、閉塞性動脈硬化症のような疾患が予想される。一般に使用される検査方法では、基準物質としては、ピラリン(正常範囲:血漿中23pmol/mL未満)、ペントシジン(正常範囲:血漿中0.00915~0.0431μg/mL(ELISAで測定した場合))などが使用される(「今日の臨床検査 2007-2008」発行所 株式会社 南江堂、参照)。
本明細書において、「刺激性の終末糖化産物」または「刺激性のAGEs(刺激性AGEs)」とは、疾患との関連性が高いAGEsを指し、sRAGEと強く結合する性質を有している。以前は、血中のグルコースによる糖化が主であると考えられていたが、グルコースによる糖化は長時間を要する上、グルコース糖化AGEsは生体への刺激性も弱いことが示唆され始めた。過剰なグルコースは、ポリオール代謝系で代謝されグリセルアルデヒド(Glycer)を生じるほか、酸化反応によりグリオキサール(GO)、グリコールアルデヒド(Glycol)などを生じるが、これらは、反応性が高く、短時間でAGEsを生成する上、その糖化産物は生体刺激性が高いことが報告されている(図1)。
本明細書において「捕捉用(または競合用)AGEs」とは、本発明の競合アッセイ系において、測定対象となる刺激性AGEsのsRAGEへの結合と競合するAGEsであって、測定対象のAGEsよりもsRAGEへの結合能が低いものを指す。例えば、グルコース(G)-AGEsは、sRAGEへの結合能がどのAGEsよりも低いため、グルコース(G)-AGEsよりもsRAGEへの結合能力が高いあらゆる刺激性AGEsを検出することができる。この場合、試料中の刺激性AGEsを網羅的に検出したい場合に有効である。また、フルクトース(F)-AGEsは、G-AGEsよりもsRAGEに対する結合能が高いため、G-AGEsを除く、F-AGEsよりもsRAGEに対する結合能が高い刺激性AGEsを検出することが可能である。このように、捕捉用AGEsは、測定対象となるAGEsの種類に応じて、捕捉用AGEsを適宜選択することができる。捕捉用AGEsの代表例としては、例えば、グルコース(G)-AGEsまたはフルクトース(F)-AGEs、ガラクトース-AGEs等を挙げることができ、さらに具体的な例としては、ウシ血清アルブミン、ヒト血清アルブミン、オボアルブミン、コラーゲン、クリスタリン、チューブリン、リゾチーム、RNase、カルモジュリン、ヘモグロビン、筋繊維タンパク質、アンチトロンビンIII、フェリチン、フィブリン、フィブリノーゲン、高密度リポタンパク質(HDL)、免疫グロブリン、低密度リポタンパク質(LDL)、インシュリン、ケラチン、オステオカルシン、アルコールデヒドロゲナーゼ、アルドースレダクターゼ、カテプシンB、スパーオキシドジスムダーゼ、ミオシンATPase、赤血球スペクトリン、またはミエリンのAGEsを挙げることができるがこれらに限定されない。
本明細書において「固定化」とは、AGEs等の物質が、対象となる別の実体(例えば、基板(基材、固相))に対して、動かなくさせることをいう。固定化は、種々の手段により達成され、例えば、炭酸-重炭酸緩衝液の利用、タグ(例えばビオチン)と結合する物質(例えばアビジン)でコーティングしたプレートの使用等を挙げることができるがこれらに限定されない。
本明細書において使用される場合、「複合体」とは、2以上の部分を含む任意の構成体を意味する。例えば、一方の部分がポリペプチドである場合は、他方の部分は、ポリペプチドであってもよく、それ以外の物質(例えば、糖、脂質、核酸、他の炭化水素等)であってもよい。本明細書において複合体を構成する2以上の部分は、共有結合で結合されていてもよくそれ以外の結合(例えば、水素結合、イオン結合、疎水性相互作用、ファンデルワールス力等)で結合されていてもよい。2以上の部分がポリペプチドの場合は、キメラポリペプチドとも称しうる。したがって、本明細書において「複合体」は、ポリペプチド、ポリヌクレオチド、脂質、糖、低分子などの分子が複数種連結してできた分子を含む。
本明細書において使用される場合、用語「リガンド」とは、特異的な受容体または受容体のファミリーに対する結合パートナーである。リガンドは、受容体に対する内因性のリガンドであるか、またはその代わりに、薬剤、薬剤候補、もしくは薬理学的手段のような受容体に対する合成リガンドであり得る。
本明細書において「AGEs分子」とは、上記AGEに含まれる任意の分子をいう。例えば、AGEsとしては、Lys-AGE(グルタルアルデヒド修飾リジン修飾AGE)、グルコース修飾AGE(G-AGE)、リボース修飾AGE(R-AGE)、フルクトース修飾AGE(F-AGE)またはそれらの改変体あるいはそれらの複合体を挙げることができるがそれらに限定されない。
本明細書において「AGEs様活性を示す分子」とは、少なくとも上述のAGEsの活性(本明細書において「AGEs様活性」という。)の一つを有する分子をいう。そのようなAGEs様活性としては、RAGEに対する結合活性(リガンド活性)を挙げることができるが、それに限定されない。
本明細書において使用される場合、用語「AGE受容体(Receptor for AGE)」とは、RAGEともいわれ、(1)配列番号8に示されるアミノ酸配列からなるポリペプチド;(2)上記配列番号8に示されるアミノ酸配列において1または数個のアミノ酸の置換、付加および/または欠失を含むアミノ酸配列を含み、かつ天然型RAGEの活性を示すポリペプチド;(3)上記配列番号8に示されるアミノ酸配列と少なくとも90%の配列同一性を有するアミノ酸配列を含み、かつ天然型RAGEの活性を示すポリペプチド;(4)上記配列番号8に示されるアミノ酸配列と少なくとも80%の配列相同性を有するアミノ酸配列を含み、かつ天然型RAGEの活性を示すポリペプチド;(5)配列番号7に示される核酸分子によってコードされるアミノ酸配列を含むポリペプチド;(6)上記配列番号7に示される核酸配列に対して相補的な核酸配列とストリンジェントな条件下でハイブリダイズする核酸分子によってコードされるアミノ酸配列を含み、かつ天然型RAGEの活性を示すポリペプチド;(7)上記配列番号7に示される核酸配列において1または数個のヌクレオチドの置換、付加および/または欠失を有する核酸分子によってコードされるアミノ酸配列を含み、かつ天然型RAGEの活性を示すポリペプチド;(8)上記配列番号7に示される核酸配列と少なくとも90%の配列同一性を有する核酸分子によってコードされるアミノ酸配列を含み、かつ天然型RAGEの活性を示すポリペプチド;および(9)上記配列番号7に示される核酸配列と少なくとも80%の配列相同性を有する核酸分子によってコードされるアミノ酸配列を含み、かつ天然型RAGEの活性を示すポリペプチド、のうちの1つである。上記の同一性または相同性は、配列分析用ツールであるBLAST(NCBIのBLAST 2.2.9(2004.5.12 発行))を用いてデフォルトパラメータを用いて算出される。ストリンジェントな条件は配列に依存して変化し、このような条件の決定は、当業者の技術範囲内である。RAGEはまた、1992年に、ウシ肺から同定され、AGEと結合するイムノグロブリンスーパーファミリーに属する、分子量約35kDaのI型膜タンパク質(糖鎖修飾を受けた完全なRAGEは、分子量55kDa)である。RAGEsの細胞外ドメインは、1つのV型イムノグロブリンドメイン、続いて、2つのC型イムノグロブリンドメイン(C1領域およびC2領域)の、3つのイムノグロブリンフォールド構造を取るドメインが結合した構造を取っている。RAGEはまた、細胞膜1回貫通型のドメインおよび43アミノ酸の細胞質ドメインを含む。RAGEは、多様なクラスのリガンド(AGEs、S100/カルグラニュリン、アンフォテリンおよびアミロイド-βペプチド)と相互作用する。Vドメインは、リガンド結合に必須の部位であり、細胞質ドメインは、RAGE媒介性細胞内シグナル伝達に必須である。RAGEはまた、各ドメイン内でジスルフィド結合を有するので、本発明の変異RAGE-界面活性剤複合体は、配列番号3のアミノ酸配列における38位、99位、144位、208位、259位および301位に対応するシステイン残基を保持していることが好ましい。RAGEは、正常組織および血管系においては低レベルでしか発現されない。しかし、この受容体は、そのリガンドが蓄積した場所においてアップレギュレートされる。RAGEの発現は、糖尿病血管系において内皮細胞、平滑筋細胞、周皮細胞、腎メサンギウム細胞および浸潤性単核食細胞で増加している。また、AGEsが蓄積している動脈硬化巣のような病的部位においても、RAGEの発現が増加している。AGEs-RAGE相互作用は、血管系ホメオスタシスにおいて重要な細胞の特性を変化させる。例えば、RAGEがAGEsと結合した後、血管内皮細胞は、VCAM-1、組織因子、およびIL-6の発現、ならびに高分子へのそれらの透過性を増加させる。単核食細胞において、RAGEは、サイトカインおよび増殖因子の発現を活性化し、可溶性AGEsに応じて細胞移動を誘導するのに対して、走触性は、固定リガンドで起こる。
本明細書では、「RAGEリガンド認識領域」または「sRAGE(soluble Receptorfor Advanced Glycation End products)」とは、交換可能に使用され、RAGEリガンドが認識する領域をいう。詳細には、sRAGEすなわちRAGEリガンド認識領域は、RAGEの細胞外領域の全部または一部をいう。sRAGEは、代表的には、配列番号3または配列番号8の22~332位で構成されるがこれに限定されない。
本明細書において使用される場合、用語「RAGE様ポリペプチド」とは、「RAGE8」、「mRAGE8」、「RAGE1」、「mRAGE1」、「RAGE2」、「mRAGE2」、「RAGE3」、「mRAGE3」、「RAGE4」、「mRAGE4」、「RAGE7」、「mRAGE7」、「RAGE143」、「mRAGE143」、「RAGE223」、「mRAGE223」、「RAGE226」および「mRAGE226」と称されるポリペプチドまたはこれらの変異体を包含する。これらの説明は、特開2013-209330等に開示されており、適宜本明細書において参考としてその内容を援用する。
本明細書において使用される場合、用語「RAGE分子」とは、RAGE様ポリペプチドの他、それらの任意の複合体を含むことが理解される。したがって、RAGE分子には、RAGE様ポリペプチド等、例えば、RAGE(全長)、RAGE細胞外領域(配列番号8の22-332位)、RAGE143、RAGE223、RAGE226等が包含されることが理解される。また、RAGE分子は、RAGEを構成する3つのドメインのうちあるドメイン全体、またはあるドメインの一部を欠いたRAGE(ミニRAGE)も包含される。また、ミニRAGEは、RAGE様ポリペプチドのミニRAGEも包含する。
本明細書では、「RAGEリガンド認識領域」を含む分子は、「RAGE分子」のうち全長RAGE以外のもの(「RAGE様ポリペプチド」を含む)、例えば、「RAGE8」、「mRAGE8」、「RAGE1」、「mRAGE1」、「RAGE2」、「mRAGE2」、「RAGE3」、「mRAGE3」、「RAGE4」、「mRAGE4」、「RAGE7」、「mRAGE7」、「RAGE143」、「mRAGE143」、「RAGE223」、「mRAGE223」、「RAGE226」および「mRAGE226」、RAGE細胞外領域(配列番号8の22-332位)、などを挙げることができる。
なお、上記RAGE様ポリペプチドは、天然型RAGEの活性が保持されている限り、非天然アミノ酸を含んでいてもよいし、アミノ酸アナログ、アミノ酸誘導体などを含んでいてもよい。
上記のRAGE様ポリペプチドにおいても、分子内ジスルフィド結合を形成することは重要であるので、配列番号8のアミノ酸配列の38位、99位、144位、208位、259位および301位に対応するシステインは保持されていることが好ましい。
本明細書において、「リガンド」とは、特異的な受容体または受容体のファミリーに対する結合パートナーである。リガンドは、受容体に対する内因性のリガンドであるか、またはその代わりに、薬剤、薬剤候補、もしくは薬理学的手段のような受容体に対する合成リガンドであり得る。
本明細書において「抗体」は、広義には、ポリクローナル抗体、モノクローナル抗体、多重特異性抗体、キメラ抗体、および抗イディオタイプ抗体、ならびにそれらの機能性フラグメント(例えば、F(ab’)2、およびFabフラグメント)、ならびにその他の組換えにより生産された結合体または機能的等価物(例えば、キメラ抗体、ヒト化抗体、多機能抗体、二重特異性またはオリゴ特異性(oligospecific)抗体、単鎖抗体、一本鎖抗体(scFV)、ダイアボディー(diabody)、sc(Fv)2(single chain (Fv)2)、scFv-Fc)を含む。さらにこのような抗体を、酵素、例えばアルカリホスファターゼ、西洋ワサビペルオキシダーゼ、αガラクトシダーゼなど、に共有結合させまたは組換えにより融合させてよい。さらにこのような抗体を、酵素、例えばアルカリホスファターゼ、西洋ワサビペルオキシダーゼ、αガラクトシダーゼなど、に共有結合させまたは組換えにより融合させてよい。狭義に使用する場合は、抗体は、全長抗体(例えば、ポリクローナル抗体、モノクローナル抗体等)を指し、その他は改変体または抗原結合フラグメントと称することがある。本発明で用いられる抗体は、その標的に結合すればよく、その由来、種類、形状などは問われない。具体的には、非ヒト動物の抗体(例えば、マウス抗体、ラット抗体、ラクダ抗体)、ヒト抗体、キメラ抗体、ヒト化抗体などの公知の抗体をもとに製造することができる。本発明においては、一本鎖抗体が使用される。抗体の標的への結合は識別的なあるいは特異的な結合であることが好ましい。抗体の改変体は、抗体と、例えばポリエチレングリコール等の各種分子が結合していてもよい。抗体の改変体は、抗体に公知の手法を用いて化学的な修飾を施すことによって得ることができる。
本明細書において「一本鎖抗体」とは、「scFv(single chain Fv)」ともいい、抗体の重鎖および軽鎖の可変部領域(VHおよびVL)を適当なリンカーペプチドで連結させたものに相当する。このようなコンストラクトを遺伝子レベルで構築し、タンパク質発現用ベクターを用いて大腸菌に導入することで一本鎖抗体タンパク質を発現させることができる。
本明細書において、「フラグメント」とは、全長のポリペプチドまたはポリヌクレオチド(長さがn)に対して、1~n-1までの配列長を有するポリペプチドまたはポリヌクレオチドをいう。フラグメントの長さは、その目的に応じて、適宜変更することができ、例えば、その長さの下限としては、ポリペプチドの場合、3、4、5、6、7、8、9、10、15、20、25、30、40、50、およびそれ以上のアミノ酸が挙げられ、ここの具体的に列挙していない整数で表される長さ(例えば、11など)もまた、下限として適切であり得る。また、ポリヌクレオチドの場合、5、6、7、8、9、10、15、20、25、30、40、50、75、100およびそれ以上のヌクレオチドが挙げられ、ここの具体的に列挙していない整数で表される長さ(例えば、11など)もまた、下限として適切であり得る。本明細書において、ポリペプチドおよびポリヌクレオチドの長さは、上述のようにそれぞれアミノ酸または核酸の個数で表すことができるが、上述の個数は絶対的なものではなく、同じ機能を有する限り、上限または下限としての上述の個数は、その個数の上下数個(または例えば上下10%)のものも含むことが意図される。本明細書において有用なフラグメントの長さは、そのフラグメントの基準となる全長タンパク質の機能のうち少なくとも1つの機能が保持されているかどうかによって決定され得る。
本明細書において遺伝子の「相同性」とは、2以上の遺伝子配列の、互いに対する同一性の程度をいう。従って、ある2つの遺伝子の相同性が高いほど、それらの配列の同一性または類似性は高い。2種類の遺伝子が相同性を有するか否かは、配列の直接の比較、または核酸の場合ストリンジェントな条件下でのハイブリダイゼーション法によって調べられ得る。2つの遺伝子配列を直接比較する場合、その遺伝子配列間でDNA配列が、代表的には少なくとも50%同一である場合、好ましくは少なくとも70%同一である場合、より好ましくは少なくとも80%、90%、95%、96%、97%、98%または99%同一である場合、それらの遺伝子は相同性を有する。
アミノ酸は、その一般に公知の3文字記号か、またはIUPAC-IUB Biochemical Nomenclature Commissionにより推奨される1文字記号のいずれかにより、本明細書中で言及され得る。ヌクレオチドも同様に、一般に認知された1文字コードにより言及され得る。
本明細書では、アミノ酸配列および塩基配列の類似性、同一性および相同性の比較は、配列分析用ツールであるBLASTを用いてデフォルトパラメータを用いて算出される。同一性の検索は例えば、NCBIのBLAST 2.2.9(2004.5.12 発行)を用いて行うことができる。本明細書における同一性の値は通常は上記BLASTを用い、デフォルトの条件でアラインした際の値をいう。ただし、パラメータの変更により、より高い値が出る場合は、最も高い値を同一性の値とする。複数の領域で同一性が評価される場合はそのうちの最も高い値を同一性の値とする。
本明細書において、「改変体」とは、もとのポリペプチドまたはポリヌクレオチドなどの物質に対して、一部が変更されているものをいう。そのような改変体としては、置換改変体、付加改変体、欠失改変体、短縮(truncated)改変体、対立遺伝子変異体などが挙げられる。対立遺伝子(allele)とは、 同一遺伝子座に属し、互いに区別される遺伝的改変体のことをいう。従って、「対立遺伝子変異体」とは、ある遺伝子に対して、対立遺伝子の関係にある改変体をいう。「種相同体またはホモログ(homolog)」とは、ある種の中で、ある遺伝子とアミノ酸レベルまたはヌクレオチドレベルで、相同性(好ましくは、60%以上の相同性、より好ましくは、80%以上、85%以上、90%以上、95%以上の相同性)を有するものをいう。そのような種相同体を取得する方法は、本明細書の記載から明らかである。「オルソログ(ortholog)」とは、オルソロガス遺伝子(orthologous gene)ともいい、二つの遺伝子がある共通祖先からの種分化に由来する遺伝子をいう。例えば、多重遺伝子構造をもつヘモグロビン遺伝子ファミリーを例にとると、ヒトとマウスのαヘモグロビン遺伝子はオルソログであるが,ヒトのαヘモグロビン遺伝子とβヘモグロビン遺伝子はパラログ(遺伝子重複で生じた遺伝子)である。オルソログは、分子系統樹の推定に有用であることから、オルソログもまた、本発明において有用であり得る。
本明細書において「保存的(に改変された)改変体」は、アミノ酸配列および核酸配列の両方に適用される。特定の核酸配列に関して、保存的に改変された改変体とは、同一のまたは本質的に同一のアミノ酸配列をコードする核酸をいい、核酸がアミノ酸配列をコードしない場合には、本質的に同一な配列をいう。このような塩基配列の改変法としては、制限酵素などによる切断、DNAポリメラーゼ、Klenowフラグメント、DNAリガーゼなどによる処理等による連結等の処理、合成オリゴヌクレオチドなどを用いた部位特異的塩基置換法(特定部位指向突然変異法;MarkZollerand Michael Smith,Methods in Enzymology,100,468-500(1983))が挙げられるが、この他にも通常分子生物学の分野で用いられる方法によって改変を行うこともできる。遺伝コードの縮重のため、多数の機能的に同一な核酸が任意の所定のタンパク質をコードする。例えば、コドンGCA、GCC、GCG、およびGCUはすべて、アミノ酸アラニンをコードする。したがって、アラニンがコドンにより特定される全ての位置で、そのコドンは、コードされたポリペプチドを変更することなく、記載された対応するコドンの任意のものに変更され得る。このような核酸の変動は、保存的に改変された変異の1つの種である「サイレント改変(変異)」である。ポリペプチドをコードする本明細書中のすべての核酸配列はまた、その核酸の可能なすべてのサイレント変異を記載する。当該分野において、核酸中の各コドン(通常メチオニンのための唯一のコドンであるAUG、および通常トリプトファンのための唯一のコドンであるTGGを除く)が、機能的に同一な分子を産生するために改変され得ることが理解される。したがって、ポリペプチドをコードする核酸の各サイレント変異は、記載された各配列において暗黙に含まれる。好ましくは、そのような改変は、ポリペプチドの高次構造に多大な影響を与えるアミノ酸であるシステインの置換を回避するようになされ得る。
あるアミノ酸は、相互作用結合能力の明らかな低下または消失なしに、例えば、リガンド分子の結合部位のようなタンパク質構造において他のアミノ酸に置換され得る。あるタンパク質の生物学的機能を規定するのは、タンパク質の相互作用能力および性質である。従って、特定のアミノ酸の置換がアミノ酸配列において、またはそのDNAコード配列のレベルにおいて行われ得、置換後もなお、もとの性質を維持するタンパク質が生じ得る。従って、生物学的有用性の明らかな損失なしに、種々の改変が、本明細書において開示されたペプチドまたはこのペプチドをコードする対応するDNAにおいて行われ得る。
このような核酸は、周知のPCR法により得ることができ、化学的に合成することもできる。これらの方法に、例えば、部位特異的変位誘発法、ハイブリダイゼーション法などを組み合わせてもよい。
上記のような改変を設計する際に、アミノ酸の疎水性指数が考慮され得る。タンパク質における相互作用的な生物学的機能を与える際の疎水性アミノ酸指数の重要性は、一般に当該分野で認められている(Kyte.JおよびDoolittle,R.F.J.Mol.Biol.157(1):105-132,1982)。アミノ酸の疎水的性質は、生成したタンパク質の二次構造に寄与し、次いでそのタンパク質と他の分子(例えば、酵素、基質、受容体、DNA、抗体、抗原など)との相互作用を規定する。各アミノ酸は、それらの疎水性および電荷の性質に基づく疎水性指数を割り当てられる。それらは:イソロイシン(+4.5);バリン(+4.2);ロイシン(+3.8);フェニルアラニン(+2.8);システイン/シスチン(+2.5);メチオニン(+1.9);アラニン(+1.8);グリシン(-0.4);スレオニン(-0.7);セリン(-0.8);トリプトファン(-0.9);チロシン(-1.3);プロリン(-1.6);ヒスチジン(-3.2);グルタミン酸(-3.5);グルタミン(-3.5);アスパラギン酸(-3.5);アスパラギン(-3.5);リジン(-3.9);およびアルギニン(-4.5))である。
あるアミノ酸を、同様の疎水性指数を有する他のアミノ酸により置換して、そして依然として同様の生物学的機能を有するタンパク質(例えば、リガンド結合能において等価なタンパク質)を生じさせ得ることが当該分野で周知である。このようなアミノ酸置換において、疎水性指数が±2以内であることが好ましく、±1以内であることがより好ましく、および±0.5以内であることがさらにより好ましい。疎水性に基づくこのようなアミノ酸の置換は効率的であることが当該分野において理解される。米国特許第4,554,101号に記載されるように、以下の親水性指数がアミノ酸残基に割り当てられている:アルギニン(+3.0);リジン(+3.0);アスパラギン酸(+3.0±1);グルタミン酸(+3.0±1);セリン(+0.3);アスパラギン(+0.2);グルタミン(+0.2);グリシン(0);スレオニン(-0.4);プロリン(-0.5±1);アラニン(-0.5);ヒスチジン(-0.5);システイン(-1.0);メチオニン(-1.3);バリン(-1.5);ロイシン(-1.8);イソロイシン(-1.8);チロシン(-2.3);フェニルアラニン(-2.5);およびトリプトファン(-3.4)。アミノ酸が同様の親水性指数を有しかつ依然として生物学的等価体を与え得る別のものに置換され得ることが理解される。このようなアミノ酸置換において、親水性指数が±2以内であることが好ましく、±1以内であることがより好ましく、および±0.5以内であることがさらにより好ましい。
本発明において、「保存的置換」とは、アミノ酸置換において、元のアミノ酸と置換されるアミノ酸との親水性指数または/および疎水性指数が上記のように類似している置換をいう。保存的置換の例は、当業者に周知であり、例えば、次の各グループ内での置換:アルギニンおよびリジン;グルタミン酸およびアスパラギン酸;セリンおよびスレオニン;グルタミンおよびアスパラギン;ならびにバリン、ロイシン、およびイソロイシン、などが挙げられるがこれらに限定されない。
本明細書中において、機能的に等価なポリペプチドを作製するために、アミノ酸の置換のほかに、アミノ酸の付加、欠失、または修飾もまた行うことができる。アミノ酸の置換とは、もとのペプチドを1つ以上、例えば、1~10個、好ましくは1~5個、より好ましくは1~3個のアミノ酸で置換することをいう。アミノ酸の付加とは、もとのペプチド鎖に1つ以上、例えば、1~10個、好ましくは1~5個、より好ましくは1~3個のアミノ酸を付加することをいう。アミノ酸の欠失とは、もとのペプチドから1つ以上、例えば、1~10個、好ましくは1~5個、より好ましくは1~3個のアミノ酸を欠失させることをいう。アミノ酸修飾は、アミド化、カルボキシル化、硫酸化、ハロゲン化、アルキル化、リン酸化、水酸化、アシル化(例えば、アセチル化)などを含むが、これらに限定されない。置換、または付加されるアミノ酸は、天然のアミノ酸であってもよく、非天然のアミノ酸、またはアミノ酸アナログでもよい。天然のアミノ酸が好ましい。
本明細書において、ポリペプチドまたはポリヌクレオチドの「置換、付加および/または欠失」とは、もとのポリペプチドまたはポリヌクレオチドに対して、それぞれアミノ酸もしくはその代替物、またはヌクレオチドもしくはその代替物が、置き換わること、付け加わること、または取り除かれることをいう。このような置換、付加および/または欠失の技術は、当該分野において周知であり、そのような技術の例としては、部位特異的変異誘発技術などが挙げられる。基準となる核酸分子またはポリペプチドにおけるこれらの変化は、目的とする機能(例えば、RAGEの認識能など)が保持される限り、この核酸分子の5’末端もしくは3’末端で生じ得るか、またはこのポリペプチドを示すアミノ酸配列のアミノ末端部位もしくはカルボキシ末端部位で生じ得るか、またはそれらの末端部位の間のどこにでも生じ得、基準配列中の残基間で個々に散在する。置換、付加または欠失は、1つ以上であれば任意の数でよく、そのような数は、その置換、付加または欠失を有する改変体において目的とする機能が保持される限り、多くすることができる。例えば、そのような数は、1または数個であり得、そして好ましくは、全体の長さの20%以内、15%以内、10%以内、5%以内、または150個以下、100個以下、50個以下、25個以下などであり得る。
本明細書において使用される場合、用語「タグ配列」とは、受容体-リガンドのような特異的認識機構により分子を選別するための物質、より具体的には、特定の物質を結合するための結合パートナーの役割を果たす物質(例えば、ビオチン-アビジン、ビオチン-ストレプトアビジンのような関係を有する)をいう。よって、例えば、タグ配列が結合した特定の物質は、タグ配列の結合パートナーを結合させた基材を接触させることで、この特定の物質を選別することができる。このようなタグ配列は、当該分野で周知である。代表的なタグ配列としては、mycタグ、Hisタグ、HA、Aviタグなどが挙げられるが、これらに限定されない。
本明細書において、「検出剤」とは、広義には、目的の物質または状態(例えば、AGEs)を検出できるあらゆる因子(剤)をいう。
本明細書において「タンパク質」、「ポリペプチド」、「オリゴペプチド」および「ペプチド」は、本明細書において同じ意味で使用され、任意の長さのアミノ酸のポリマーをいう。このポリマーは、直鎖であっても分岐していてもよく、環状であってもよい。アミノ酸は、天然のものであっても非天然のものであってもよく、改変されたアミノ酸であってもよい。この用語はまた、複数のポリペプチド鎖の複合体へとアセンブルされたものを包含し得る。この用語はまた、天然または人工的に改変されたアミノ酸ポリマーも包含する。そのような改変としては、例えば、ジスルフィド結合形成、グリコシル化、脂質化、アセチル化、リン酸化または任意の他の操作もしくは改変(例えば、標識成分との結合体化)。この定義にはまた、例えば、アミノ酸の1または2以上のアナログを含むポリペプチド(例えば、非天然アミノ酸などを含む)、ペプチド様化合物(例えば、ペプトイド)および当該分野において公知の他の改変が包含される。
本明細書において、「アミノ酸」は、本発明の目的を満たす限り、天然のものでも非天然のものでもよい。本明細書において「アミノ酸誘導体」または「アミノ酸アナログ」とは、天然に存在するアミノ酸とは異なるがもとのアミノ酸と同様の機能を有するものをいう。そのようなアミノ酸誘導体およびアミノ酸アナログは、当該分野において周知である。本明細書では、アミノ酸誘導体およびアミノ酸アナログは、アミノ酸と同じ生物学的機能を提供し得る限り代替として使用され得ることが理解される。本明細書において「天然のアミノ酸」とは、天然のアミノ酸のL-異性体を意味する。天然のアミノ酸は、グリシン、アラニン、バリン、ロイシン、イソロイシン、セリン、メチオニン、トレオニン、フェニルアラニン、チロシン、トリプトファン、システイン、プロリン、ヒスチジン、アスパラギン酸、アスパラギン、グルタミン酸、グルタミン、γ-カルボキシグルタミン酸、アルギニン、オルニチン、およびリジンである。特に示されない限り、本明細書でいう全てのアミノ酸はL体であるが、D体のアミノ酸を用いた形態もまた本発明の範囲内にある。本明細書において「非天然アミノ酸」とは、タンパク質中で通常は天然に見出されないアミノ酸を意味する。非天然アミノ酸の例として、ノルロイシン、パラ-ニトロフェニルアラニン、ホモフェニルアラニン、パラ-フルオロフェニルアラニン、3-アミノ-2-ベンジルプロピオン酸、ホモアルギニンのD体またはL体およびD-フェニルアラニンが挙げられる。本明細書において「アミノ酸アナログ」とは、アミノ酸ではないが、アミノ酸の物性および/または機能に類似する分子をいう。アミノ酸アナログとしては、例えば、エチオニン、カナバニン、2-メチルグルタミンなどが挙げられる。アミノ酸模倣物とは、アミノ酸の一般的な化学構造とは異なる構造を有するが、天然に存在するアミノ酸と同様な様式で機能する化合物をいう。
アミノ酸は、その一般に公知の3文字記号か、またはIUPAC-IUB Biochemical Nomenclature Commissionにより推奨される1文字記号のいずれかにより、本明細書中で言及され得る。ヌクレオチドも同様に、一般に認知された1文字コードにより言及され得る。
本明細書において「ポリヌクレオチド」、「オリゴヌクレオチド」および「核酸」は、本明細書において同じ意味で使用され、任意の長さのヌクレオチドのポリマーをいう。この用語はまた、「オリゴヌクレオチド誘導体」または「ポリヌクレオチド誘導体」を含む。「オリゴヌクレオチド誘導体」または「ポリヌクレオチド誘導体」とは、ヌクレオチドの誘導体を含むか、またはヌクレオチド間の結合が通常とは異なるオリゴヌクレオチドまたはポリヌクレオチドをいい、互換的に使用される。そのようなオリゴヌクレオチドとして具体的には、例えば、2’-O-メチル-リボヌクレオチド、オリゴヌクレオチド中のリン酸ジエステル結合がホスホロチオエート結合に変換されたオリゴヌクレオチド誘導体、オリゴヌクレオチド中のリン酸ジエステル結合がN3’-P5’ホスホロアミデート結合に変換されたオリゴヌクレオチド誘導体、オリゴヌクレオチド中のリボースとリン酸ジエステル結合とがペプチド核酸結合に変換されたオリゴヌクレオチド誘導体、オリゴヌクレオチド中のウラシルがC-5プロピニルウラシルで置換されたオリゴヌクレオチド誘導体、オリゴヌクレオチド中のウラシルがC-5チアゾールウラシルで置換されたオリゴヌクレオチド誘導体、オリゴヌクレオチド中のシトシンがC-5プロピニルシトシンで置換されたオリゴヌクレオチド誘導体、オリゴヌクレオチド中のシトシンがフェノキサジン修飾シトシン(phenoxazine-modified cytosine)で置換されたオリゴヌクレオチド誘導体、DNA中のリボースが2’-O-プロピルリボースで置換されたオリゴヌクレオチド誘導体およびオリゴヌクレオチド中のリボースが2’-メトキシエトキシリボースで置換されたオリゴヌクレオチド誘導体などが例示される。他にそうではないと示されなければ、特定の核酸配列はまた、明示的に示された配列と同様に、その保存的に改変された改変体(例えば、縮重コドン置換体)および相補配列を包含することが企図される。具体的には、縮重コドン置換体は、1またはそれ以上の選択された(または、すべての)コドンの3番目の位置が混合塩基および/またはデオキシイノシン残基で置換された配列を作成することにより達成され得る(Batzerら、Nucleic Acid Res.19:5081(1991);Ohtsukaら、J.Biol.Chem.260:2605-2608(1985);Rossoliniら、Mol.Cell.Probes 8:91-98(1994))。
本明細書において「ヌクレオチド」は、天然のものでも非天然のものでもよい。「ヌクレオチド誘導体」または「ヌクレオチドアナログ」とは、天然に存在するヌクレオチドとは異なるがもとのヌクレオチドと同様の機能を有するものをいう。そのようなヌクレオチド誘導体およびヌクレオチドアナログは、当該分野において周知である。そのようなヌクレオチド誘導体およびヌクレオチドアナログの例としては、ホスホロチオエート、ホスホルアミデート、メチルホスホネート、キラルメチルホスホネート、2’-O-メチルリボヌクレオチド、ペプチド型核酸(PNA)が含まれるが、これらに限定されない。
本明細書において「複合分子」とは、ポリペプチド、ポリヌクレオチド、脂質、糖、低分子などの分子が複数種連結してできた分子をいう。そのような複合分子としては、例えば、糖脂質、糖ペプチドなどが挙げられるがそれらに限定されない。本明細書では、配列番号3等で例示されるアミノ酸を有するポリペプチドまたはその改変体もしくはフラグメントであって、診断に関与する生物学的な活性性を有する限り、それぞれの改変体もしくはフラグメントなどをコードする核酸分子も使用することができる。また、そのような核酸分子を含む複合分子も使用することができる。
本明細書において「核酸」はまた、遺伝子、cDNA、mRNA、オリゴヌクレオチド、およびポリヌクレオチドと互換可能に使用される。特定の核酸配列はまた、「スプライス改変体」を包含する。同様に、核酸によりコードされた特定のタンパク質は、その核酸のスプライス改変体によりコードされる任意のタンパク質を暗黙に包含する。その名が示唆するように「スプライス改変体」は、遺伝子のオルタナティブスプライシングの産物である。転写後、最初の核酸転写物は、異なる(別の)核酸スプライス産物が異なるポリペプチドをコードするようにスプライスされ得る。スプライス改変体の産生機構は変化するが、エキソンのオルタナティブスプライシングを含む。読み過し転写により同じ核酸に由来する別のポリペプチドもまた、この定義に包含される。スプライシング反応の任意の産物(組換え形態のスプライス産物を含む)がこの定義に含まれる。
本明細書において「遺伝子」とは、遺伝形質を規定する因子をいう。通常染色体上に一定の順序に配列している。タンパク質の一次構造を規定する遺伝子を構造遺伝子といい、その発現を左右する遺伝子を調節遺伝子という。本明細書では、「遺伝子」は、「ポリヌクレオチド」、「オリゴヌクレオチド」および「核酸」ならびに/あるいは「タンパク質」「ポリペプチド」、「オリゴペプチド」および「ペプチド」をさすことがある。
本明細書において「ストリンジェントな条件でハイブリダイズするポリヌクレオチド」とは、当該分野で慣用される周知の条件をいう。本発明のポリヌクレオチド中から選択されたポリヌクレオチドをプローブとして、コロニー・ハイブリダイゼーション法、プラーク・ハイブリダイゼーション法あるいはサザンブロットハイブリダイゼーション法などを用いることにより、そのようなポリヌクレオチドを得ることができる。具体的には、コロニーあるいはプラーク由来のDNAを固定化したフィルターを用いて、0.7~1.0MのNaCl存在下、65℃でハイブリダイゼーションを行った後、0.1~2倍濃度のSSC(saline-sodium citrate)溶液(1倍濃度のSSC溶液の組成は、150mM 塩化ナトリウム、15mM クエン酸ナトリウムである)を用い、65℃条件下でフィルターを洗浄することにより同定できるポリヌクレオチドを意味する。ハイブリダイゼーションは、Molecular Cloning 2nd ed.,Current Protocols in Molecular Biology,Supplement 1~38、DNA Cloning 1:Core Techniques,A PRac1tical Approach,Second Edition,Oxford University Press(1995)などの実験書に記載されている方法に準じて行うことができる。ここで、ストリンジェントな条件下でハイブリダイズする配列からは、好ましくは、A配列のみまたはT配列のみを含む配列が除外される。従って、本発明において使用されるポリペプチド(例えば、RAGEなど)には、本発明で特に記載されたポリペプチドをコードする核酸分子に対して、ストリンジェントな条件下でハイブリダイズする核酸分子によってコードされるポリペプチドも包含される。
本明細書において「ハイブリダイズ可能なポリヌクレオチド」とは、上記ハイブリダイズ条件下で別のポリヌクレオチドにハイブリダイズすることができるポリヌクレオチドをいう。ハイブリダイズ可能なポリヌクレオチドとして具体的には、配列番号3などで表されるアミノ酸配列を有するポリペプチドをコードするDNAの塩基配列と少なくとも60%以上の相同性を有するポリヌクレオチド、好ましくは80%以上の相同性を有するポリヌクレオチド、さらに好ましくは95%以上の相同性を有するポリヌクレオチドを挙げることができる。
DNA二重鎖の安定性に影響を与える因子としては、塩基の組成、長さおよび塩基対不一致の程度が挙げられる。ハイブリダイゼーション条件は、当業者によって調整され得、これらの変数を適用させ、そして異なる配列関連性のDNAがハイブリッドを形成するのを可能にする。完全に一致したDNA二重鎖の融解温度は、以下の式によって概算され得る。
Tm(℃)=81.5+16.6(log[Na+])+0.41(%G+C)-600/N-0.72(%ホルムアミド)
ここで、Nは、形成される二重鎖の長さであり、[Na+]は、ハイブリダイゼーション溶液または洗浄溶液中のナトリウムイオンのモル濃度であり、%G+Cは、ハイブリッド中の(グアニン+シトシン)塩基のパーセンテージである。不完全に一致したハイブリッドに関して、融解温度は、各1%不一致(ミスマッチ)に対して約1℃ずつ減少する。
本明細書において、「リフォールディング」とは、異常な折り畳みを有するためにその本来有する機能を失っているポリペプチドの異常な構造を解きほぐし、界面活性剤により再凝集を防ぎつつ、シクロアミロースの包接能を活用したり、あるいは、酸化還元電位制御下での希釈透析により、そのポリペプチドの本来の正しい構造に再折りたたみすることをいう。
本明細書において、「精製用カラム」とは、タンパク質などの分子(例えば、抗体)と親和性を有するか、または結合することができる固体または半固体の支持体をいい、例えば、金属キレートアガロースカラム等を挙げることができる。分子ごとの親和性または結合力の違いにより、各分子を分離することができる。精製する分子の特性に応じて、当業者であれば適切な支持体が選択される。たとえば、TALONカラムでは、Hisタグ等を付加したタンパク質を効率よく精製することができる。
本明細書において、「TALON」とは、Coを使用した金属キレートアフィニティレジンをいう。
本明細書において、「リン酸緩衝化生理食塩水(PBS)とは、NaCl、KCl、Na2HPO4、およびKH2PO4を含むpH7~8の水溶液である。各成分の濃度およびpHは、用途に応じて好適に調節することができる。本明細書において、「PBS(+)」は、カルシウムイオンおよびマグネシウムイオンを含むことを意味しており、「PBS(-)」は、カルシウムイオンおよびマグネシウムイオンを含まないことを意味しているが、本明細書では、明示的に記載しない限り、「PBS」は「PBS(-)」を意味するものとする。本明細書では、代表的には、Dulbecco’s PBS(-)を使用することができる。Dulbecco’s PBS(-)の組成は、NaCl 8g,KCl 0.2g,Na2HPO4 1.15g,KH2PO4 0.2g/L、(pH 7.4)である。
本明細書において、「セルロースエステル膜チューブ」とは、酢酸セルロースの高分子混合物を押し出し成型することにより製造された透析チューブをいう。
本明細書において使用される場合、用語「受容体」とは、1個以上のリガンドと可逆的、かつ特異的に複合体化する1個以上の結合ドメインを備える生物学的な構造であって、ここで、この複合体化は生物学的な構造を有する。受容体は、完全に細胞の外部(細胞外の受容体)、細胞膜の中(しかし、受容体の部分を細胞外部の環境および細胞質ゾルに向けている)、または完全に細胞の中(細胞内の受容体)に存在し得る。これらはまた、細胞と独立的に機能し得る。細胞膜中の受容体は、細胞を、その境界の外部の空間と連絡(例えば、シグナル伝達)させ、そして細胞の内側および外側への分子およびイオンの輸送において機能させることを可能とする。本明細書において使用する場合、受容体は、受容体全長であっても、受容体のフラグメントであってもよい。
本明細書において、「抗原抗体反応」とは、当該分野で使用される最も広い意味で用いられ、特に、抗原と抗体との間の特異的結合に基づく反応をいう。検出系としてイムノブロット(ウェスタンブロット)形式を用いることによって、試料中の抗原を検出し、そして定量化するための試薬および方法も提供する。
本明細書中で使用される用語「結合」は、2つのタンパク質もしくは化合物または関連するタンパク質もしくは化合物の間、あるいはそれらの組み合わせの間での、物理的相互作用または化学的相互作用を意味する。結合には、イオン結合、非イオン結合、水素結合、ファンデルワールス結合、疎水性相互作用などが含まれる。物理的相互作用(結合)は、直接的または間接的であり得、間接的なものは、別のタンパク質または化合物の効果を介するかまたは起因する。直接的な結合とは、別のタンパク質または化合物の効果を介してもまたはそれらに起因しても起こらず、他の実質的な化学中間体を伴わない、相互作用をいう。
本明細書において第一の物質または因子が第二の物質または因子に「特異的に相互作用する」とは、第一の物質または因子が、第二の物質または因子に対して、第二の物質または因子以外の物質または因子(特に、第二の物質または因子を含むサンプル中に存在する他の物質または因子)に対するよりも高い親和性で相互作用することをいう。物質または因子について特異的な相互作用としては、例えば、核酸におけるハイブリダイゼーション、タンパク質における抗原抗体反応、リガンド-受容体反応、酵素-基質反応など、核酸およびタンパク質の両方が関係する場合、転写因子とその転写因子の結合部位との反応など、タンパク質-脂質相互作用、核酸-脂質相互作用などが挙げられるがそれらに限定されない。従って、物質または因子がともに核酸である場合、第一の物質または因子が第二の物質または因子に「特異的に相互作用する」ことには、第一の物質または因子が、第二の物質または因子に対して少なくとも一部に相補性を有することが包含される。また例えば、物質または因子がともにタンパク質である場合、第一の物質または因子が第二の物質または因子に「特異的に相互作用する」こととしては、例えば、抗原抗体反応による相互作用、受容体-リガンド反応による相互作用、酵素-基質相互作用などが挙げられるがそれらに限定されない。2種類の物質または因子がタンパク質および核酸を含む場合、第一の物質または因子が第二の物質または因子に「特異的に相互作用する」ことには、転写因子と、その転写因子が対象とする核酸分子の結合領域との間の相互作用が包含される。したがって、本明細書においてポリヌクレオチドまたはポリペプチドなどの生物学的因子に対して「特異的に相互作用する因子」とは、そのポリヌクレオチドまたはそのポリペプチドなどの生物学的因子に対する親和性が、他の無関連の(特に、同一性が30%未満の)ポリヌクレオチドまたはポリペプチドに対する親和性よりも、代表的には同等またはより高いか、好ましくは有意に(例えば、統計学的に有意に)高いものを包含する。そのような親和性は、例えば、ハイブリダイゼーションアッセイ、結合アッセイなどによって測定することができる。
本明細書中で使用される「接触(させる)」とは、化合物を、直接的または間接的のいずれかで、本発明のポリペプチドまたはポリヌクレオチドに対して物理的に近接させることを意味する。ポリペプチドまたはポリヌクレオチドは、多くの緩衝液、塩、溶液などに存在し得る。接触とは、核酸分子またはそのフラグメントをコードするポリペプチドを含む、例えば、ビーカー、マイクロタイタープレート、細胞培養フラスコまたはマイクロアレイ(例えば、遺伝子チップ)などに化合物を置くことが挙げられる。
本発明において酵素結合イムノソルベント検定法(ELISA)のような機構を使用する場合、一般に、固相(基材または基板)としては、マイクロタイタープレートが使用される。本明細書において使用される場合、用語「固相」とは、本明細書中において「基板」および「基材」と互換的に使用され、本発明のデバイスが構築される材料をいう。抗体のような分子が固定され得る平面状の支持体をいう。基板の材料としては、共有結合かまたは非共有結合のいずれかで、本発明において使用される生体分子に結合する特性を有するかまたはそのような特性を有するように誘導体化され得る、任意の固体材料が挙げられる。適切な基材としては、ビーズ、金粒子、プレート(例えば、マイクロタイタープレート)、試験管、チップ、磁性粒子、膜、繊維、スライドガラス、金属薄膜、フィルター、チューブ、ボール、ダイアモンド様炭素被膜ステンレスなどが挙げられるが、これらに限定されない。
固相および基板として使用するためのそのような材料としては、固体表面を形成し得る任意の材料が使用され得るが、例えば、ガラス、シリカ、シリコーン、セラミック、二酸化珪素、プラスチック、金属(合金も含まれる)、天然および合成のポリマー(例えば、ポリスチレン、セルロース、アミロース、キトサン、デキストラン、およびナイロン)以下が挙げられるがそれらに限定されない。基板は、複数の異なる材料の層から形成されていてもよい。例えば、ガラス、石英ガラス、アルミナ、サファイア、フォルステライト、炭化珪素、酸化珪素、窒化珪素などの無機絶縁材料を使用できる。また、ポリエチレン、エチレン、ポリプロピレン、ポリイソブチレン、ポリエチレンテレフタレート、不飽和ポリエステル、含フッ素樹脂、ポリ塩化ビニル、ポリ塩化ビニリデン、ポリ酢酸ビニル、ポリビニルアルコール、ポリビニルアセタール、アクリル樹脂、ポリアクリロニトリル、ポリスチレン、アセタール樹脂、ポリカーボネート、ポリアミド、フェノール樹脂、ユリア樹脂、エポキシ樹脂、メラミン樹脂、スチレン・アクリロニトリル共重合体、アクリロニトリルブタジエンスチレン共重合体、シリコーン樹脂、ポリフェニレンオキサイド、ポリスルホン等の有機材料を用いることができる。本発明においてはまた、ナイロン膜、ニトロセルロース膜、PVDF膜など、ブロッティングに使用される膜を用いることもできる。高密度のものを解析する場合は、ガラスなど硬度のあるものを材料として使用することが好ましい。基板として好ましい材質は、測定機器などの種々のパラメータによって変動し、当業者は、上述のような種々の材料から適切なものを適宜選択することができる。
「サンドイッチELISA」とは、ELISAの特定の形式を言い、抗体または抗原に特異的に結合できる分子(例えばRAGE)が固体材料に結合され、抗原を含有する試料に供され、その後、非結合抗原を取り除くべく、その固体材料表面が洗われその後、標識抗体(例えば、酵素が連結された標識抗体)が、結合している状態の抗原(存在する場合)に結合され、抗体-抗原-抗体のサンドイッチを形成するような形式をいう。抗体に連結され得る酵素の例は、アルカリホスファターゼ、セイヨウワサビペルオキシダーゼ、ルシフェラーゼ、ウレアーゼおよびβ-ガラクトシダーゼである。酵素が連結された抗体は、基質と反応し、測定され得る呈色反応生成物を発生させる。標識抗体の代わりに、標識されていない抗体もまた使用することができる。この場合、標識されていない抗体に、標識された二次抗体を結合させることによって、検出することができる。
本明細書において、「カイコ」とは、通常の意味のカイコ(蚕)を意味し、チョウ目(鱗翅目)・カイコガ科に属する昆虫の一種であるものとする。正式和名はカイコガ(学名:Bombix mori)で、カイコはこの幼虫の名称だが、一般的にはこの種全般をも指す。クワ(桑)を食餌とし、絹を産生して蛹の繭を作る。カイコは家蚕(かさん)とも呼ばれ、野生に生息する昆虫ではない。カイコの祖先は東アジアに生息するクワコ(Bombyxmandarina)であると考えられている。カイコとクワコは学問的には別種とされるが、これらの雑種は生殖能力をもつ。本明細書では、カイコには、クワコが含まれるものとする。本明細書において「カイコと同様の糖鎖を付与する生物」とは、カイコと同様の糖鎖を付加する能力を有する生物をいい、カイコと同様の糖鎖を付加する酵素をコードする遺伝子等でトランスジェニック生物等を含みうる。
本明細書において、「絹糸腺」とは、熟蚕の体内に存在する左右1対の器官であり、クワの葉から摂取した多量のタンパク質(アミノ酸)を2種類の絹タンパク質(フィブロイン、セリシン)に変える器官を意味する。絹糸腺は左右一対となっており、マユ糸の原料となる液状絹を分泌する。絹糸腺は、後部絹糸腺、中部絹糸腺、前部絹糸腺の3つの部分に分けられる。本発明では、いずれの絹糸腺を用いても合成可能であるが、合成後の取り扱いを考慮し、後部絹糸腺、中部絹糸腺が通常使用され、好ましくは中部絹糸腺が使用されるが、それに限定されるものではない。また、全身に発現させて全身から回収することも可能であるし、マユを形成させた後にマユから回収することも可能である
後部絹糸腺とは、最後部にある細長い部分で、伸ばすと約20cmになる。ここでは後にマユ糸の中心となるフィブロインタンパク質を合成する。
中部絹糸腺とは、中央部分にあるS字に曲がった太い部分で、伸ばすと約6cmになる。後部絹糸腺から送られてきたフィブロインタンパク質を濃縮して蓄え、繊維にしやすい形に整える。またもうひとつの絹タンパク質であるセリシンも分泌する。マユ糸を吐きだすとき、フィブロインタンパク質をまとめる接着剤の役割をする。
前部絹糸腺とは、長さは約4cmの吐糸口につながる細い管で、先に行くほど細くなる。液状のフィブロインタンパク質の分子が引き伸ばされて一定方向に揃えられ、互いに集合することでさらに水分が除かれる。管の先端でもう一対の管と一本に合流し、吐糸口から吐きだされて一本のマユ糸になる。
カイコは5齢の終わり頃にクワを食べるのをやめる(熟蚕)。熟蚕の体の中は、マユ糸の原料となる水飴のような液(液状絹)をため込んだ一対の器官(絹糸腺)でいっぱいになっている。絹糸腺は、細い吐糸管をつうじてカイコの口元にある吐糸口につながっている。液状絹は細い吐糸管を通ることで引き伸ばされて固まり、マユ糸となる。さらには、幼虫が吐糸管から吐きだした糸を近くのものに貼りつけ、頭と胸を8の字状に動かし、引っぱるという一連の運動で、マユ糸は次々と絹糸腺から引きだされるのである。
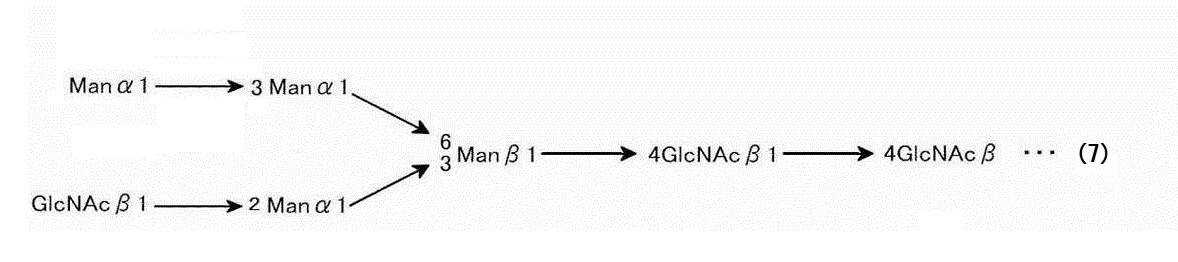
本明細書において「カイコ型糖鎖」とは、カイコで生産される糖タンパク質において特有の糖鎖構造をいい、代表的にはトリマンノシルコア(自体)、オリゴマンノース型糖鎖および複合型糖鎖、あるいはそのハイブリッド型がある。本発明では、中部絹糸腺を用いてカイコ型糖タンパク質を生産していることから、特に断らない限り、「カイコ型糖鎖」とは、この中部絹糸腺で生産される特有の糖鎖型をいう。このようなカイコ型糖鎖としては、例えば、アスパラギン(Asn)に結合したNアセチルグルコサミン(GlcNAc)が2個結合した後、マンノース(Man)が3分子結合したもの(トリマンノシルコアといい、下記(1)式で示される)をコアとして、そこから分岐した構造をとり、さらに種々の糖鎖が結合している。
本明細書において「精製された」生物学的因子(例えば、核酸またはタンパク質など)とは、その生物学的因子に天然に随伴する因子の少なくとも一部が除去されたものをいう。したがって、通常、精製された生物学的因子におけるその生物学的因子の純度は、その生物学的因子が通常存在する状態よりも高い(すなわち濃縮されている)。
本明細書中で使用される用語「精製された」は、好ましくは少なくとも75重量%、より好ましくは少なくとも85重量%、よりさらに好ましくは少なくとも95重量%、そして最も好ましくは少なくとも98重量%の、同型の生物学的因子が存在することを意味する。
本明細書において「対応する」アミノ酸または核酸とは、あるポリペプチド分子またはポリヌクレオチド分子において、比較の基準となるポリペプチドまたはポリヌクレオチドにおける所定のアミノ酸またはヌクレオチドと同様の作用を有するか、または有することが予測されるアミノ酸またはヌクレオチドをいい、特に酵素分子にあっては、活性部位中の同様の位置に存在し触媒活性に同様の寄与をするアミノ酸をいう。例えば、アンチセンス分子であれば、そのアンチセンス分子の特定の部分に対応するオルソログにおける同様の部分であり得る。対応するアミノ酸は、例えば、システイン化、グルタチオン化、S-S結合形成、酸化(例えば、メチオニン側鎖の酸化)、ホルミル化、アセチル化、リン酸化、糖鎖付加、ミリスチル化などがされる特定のアミノ酸であり得る。あるいは、対応するアミノ酸は、二量体化を担うアミノ酸であり得る。このような「対応する」アミノ酸または核酸は、一定範囲にわたる領域またはドメインであってもよい。従って、そのような場合、本明細書において「対応する」領域またはドメインと称される。
本明細書において「対応する」遺伝子(例えば、ポリペプチド分子またはポリヌクレオチド分子)とは、ある種において、比較の基準となる種における所定の遺伝子と同様の作用を有するか、または有することが予測される遺伝子(例えば、ポリペプチド分子またはポリヌクレオチド分子)をいい、そのような作用を有する遺伝子が複数存在する場合、進化学的に同じ起源を有するものをいう。従って、ある遺伝子に対応する遺伝子は、その遺伝子のオルソログであり得る。従って、マウス、ラットのRAGE(または可溶性形態のsRAGE)は、それぞれ、ヒトにおいて、対応するRAGE(sRAGEまたは可溶性形態のsRAGE)を見出すことができる。そのような対応する遺伝子は、当該分野において周知の技術を用いて同定することができる。したがって、例えば、ある動物(例えば、マウス)における対応する遺伝子は、対応する遺伝子の基準となる遺伝子(例えば、RAGEまたは可溶性形態のsRAGE)は、ある動物の配列をクエリ配列として用いてその動物(例えばヒト、ラット)の配列データベースを検索することによって見出すことができる。
本明細書中で使用される「異種」とは、異なる配列または対応しない配列、あるいは異なる種由来の配列である、ヌクレオチド配列またはアミノ酸配列をいう。例えば、ヒトのヌクレオチド配列またはアミノ酸配列は、マウスのヌクレオチド配列またはアミノ酸配列に対して異種であり、そしてヒトRAGEの核酸配列またはアミノ酸配列は、ヒトアルブミンのヌクレオチド配列またはアミノ酸配列に対して異種である。
本明細書において「生物学的機能」とは、ある遺伝子またはそれに関する核酸分子もしくはポリペプチドについて言及するとき、その遺伝子、核酸分子またはポリペプチドが生体内において有し得る特定の機能をいい、これには、例えば、特異的な抗体の生成、酵素活性、抵抗性の付与等を挙げることができるがそれらに限定されない。本発明においては、例えば、RAGEがヘモペキシン等のマーカーを認識する機能などを挙げることができるがそれらに限定されない。本明細書において、生物学的機能は、「生物学的活性」によって発揮され得る。本明細書において「生物学的活性」とは、ある因子(例えば、ポリヌクレオチド、タンパク質など)が、生体内において有し得る活性のことをいい、種々の機能(例えば、転写促進活性)を発揮する活性が包含され、例えば、ある分子との相互作用によって別の分子が活性化または不活化される活性も包含される。2つの因子が相互作用する場合、その生物学的活性は、その二分子との間の結合およびそれによって生じる生物学的変化、例えば、一つの分子を抗体を用いて沈降させたときに他の分子も共沈するとき、2分子は結合していると考えられる。したがって、そのような共沈を見ることが一つの判断手法として挙げられる。例えば、ある因子が酵素である場合、その生物学的活性は、その酵素活性を包含する。別の例では、ある因子がリガンドである場合、そのリガンドが対応する受容体への結合を包含する。そのような生物学的活性は、当該分野において周知の技術によって測定することができる。
したがって、「活性」は、結合(直接的または間接的のいずれか)を示すかまたは明らかにするか;応答に影響する(すなわち、いくらかの曝露または刺激に応答する測定可能な影響を有する)、種々の測定可能な指標をいい、例えば、本発明のポリペプチドまたはポリヌクレオチドに直接結合する化合物の親和性、または例えば、いくつかの刺激後または事象後の上流または下流のタンパク質の量あるいは他の類似の機能の尺度が、挙げられる。
本明細書において「マーカー(物質)」とは、ある状態(AGEsに関連する糖尿病、糖尿病性腎症、糖尿病性網膜症、糖尿病性神経症等の糖尿病合併症などの疾患)に罹患しているかまたはその危険性があるかどうかを追跡する示標となる物質をいう。このようなマーカーとしては、遺伝子、遺伝子産物、代謝物質、酵素などを挙げることができる。本発明において、糖尿血管障害、非アルコール性脂肪肝炎、アルツハイマー型認知症、などのAGEsに関連する疾患、糖尿病、高血糖、高血圧症、高血圧性腎硬化症、動脈硬化、虚血再灌流傷害、血管バルーン傷害後のような病態についての診断、予備的検出、予測または事前診断は、その状態に関連するマーカーに特異的な薬剤、剤、因子または手段、あるいはそれらを含む組成物、キットまたはシステム等を用いて実現することができる。さらに、本発明において、ある状態(例えば、糖尿病、糖尿病性腎症、糖尿病性網膜症、糖尿病性神経症等の糖尿病合併症などの疾患)についての診断、予備的検出、予測または事前診断、(人工透析などの)治療効果の評価は、その状態に関連するマーカーに特異的な薬剤、剤、因子または手段、あるいはそれらを含む組成物、キットまたはシステム等を用いて実現することもできる。
本明細書において、「遺伝子産物」とは、遺伝子によってコードされるタンパク質またはmRNAをいう。本明細書では、糖代謝に直接関連しない遺伝子産物(すなわち、インスリンなどの糖代謝に関連しないタンパク質など)が糖尿病の指標として使用可能であることが見出された。
本明細書において「被験体」とは、本発明の診断または検出等の対象となる生物(例えば、ヒト)をいう。
本明細書において「サンプル」とは、被験体等から得られた任意の物質をいい、例えば、体液(血液、唾液、尿、涙液、脳脊髄液等)が含まれる。好ましくは血液、尿、涙液が使用され、マーカーによって、特異性に相違があるが、当業者は本明細書の記載をもとに適宜好ましいサンプルを選択することができる。
本明細書において「薬剤」、「剤」または「因子」(いずれも英語ではagentに相当する)は、広義には、交換可能に使用され、意図する目的を達成することができる限りどのような物質または他の要素(例えば、光、放射能、熱、電気などのエネルギー)でもあってもよい。そのような物質としては、例えば、タンパク質、ポリペプチド、オリゴペプチド、ペプチド、ポリヌクレオチド、オリゴヌクレオチド、ヌクレオチド、核酸(例えば、cDNA、ゲノムDNAのようなDNA、mRNAのようなRNAを含む)、ポリサッカリド、オリゴサッカリド、脂質、有機低分子(例えば、ホルモン、リガンド、情報伝達物質、有機低分子、コンビナトリアルケミストリで合成された分子、医薬品として利用され得る低分子(例えば、低分子リガンドなど)など)、これらの複合分子が挙げられるがそれらに限定されない。ポリヌクレオチドに対して特異的な因子としては、代表的には、そのポリヌクレオチドの配列に対して一定の配列相同性を(例えば、70%以上の配列同一性)もって相補性を有するポリヌクレオチド、プロモーター領域に結合する転写因子のようなポリペプチドなどが挙げられるがそれらに限定されない。ポリペプチドに対して特異的な因子としては、代表的には、そのポリペプチドに対して特異的に指向された抗体またはその誘導体あるいはその類似物(例えば、単鎖抗体)、そのポリペプチドが受容体またはリガンドである場合の特異的なリガンドまたは受容体、そのポリペプチドが酵素である場合、その基質などが挙げられるがそれらに限定されない。
本明細書において「相互作用」とは、2つの物質についていうとき、一方の物質と他方の物質との間で力(例えば、分子間力(ファンデルワールス力)、水素結合、疎水性相互作用など)を及ぼしあうこという。通常、相互作用をした2つの物質は、会合または結合している状態にある。
したがって、本明細書においてポリヌクレオチドまたはポリペプチドなどの生物学的因子に対して「特異的に相互作用する因子」とは、そのポリヌクレオチドまたはそのポリペプチドなどの生物学的因子に対する親和性が、他の無関連の(特に、同一性が30%未満の)ポリヌクレオチドまたはポリペプチドに対する親和性よりも、代表的には同等またはより高いか、好ましくは有意に(例えば、統計学的に有意に)高いものを包含する。そのような親和性は、例えば、ハイブリダイゼーションアッセイ、結合アッセイなどによって測定することができる。
本明細書において第一の物質または因子が第二の物質または因子に「特異的に相互作用する」とは、第一の物質または因子が、第二の物質または因子に対して、第二の物質または因子以外の物質または因子(特に、第二の物質または因子を含むサンプル中に存在する他の物質または因子)に対するよりも高い親和性で相互作用することをいう。物質または因子について特異的な相互作用としては、例えば、リガンド-受容体反応、核酸におけるハイブリダイゼーション、タンパク質における抗原抗体反応、酵素-基質反応など、核酸およびタンパク質の両方が関係する場合、転写因子とその転写因子の結合部位との反応など、タンパク質-脂質相互作用、核酸-脂質相互作用などが挙げられるがそれらに限定されない。従って、物質または因子がともに核酸である場合、第一の物質または因子が第二の物質または因子に「特異的に相互作用する」ことには、第一の物質または因子が、第二の物質または因子に対して少なくとも一部に相補性を有することが包含される。また例えば、物質または因子がともにタンパク質である場合、第一の物質または因子が第二の物質または因子に「特異的に相互作用する」こととしては、例えば、抗原抗体反応による相互作用、受容体-リガンド反応による相互作用、酵素-基質相互作用などが挙げられるがそれらに限定されない。2種類の物質または因子がタンパク質および核酸を含む場合、第一の物質または因子が第二の物質または因子に「特異的に相互作用する」ことには、転写因子と、その転写因子が対象とする核酸分子の結合領域との間の相互作用が包含される。
本明細書において「手段」とは、ある目的を達成する任意の道具となり得るものをいい、特に、本明細書では、「選択的に認識する手段」とは、ある対象を他のものとは異なって認識することができる手段をいう。
本明細書中で使用される用語「結合」は、2つのタンパク質もしくは化合物または関連するタンパク質もしくは化合物の間、あるいはそれらの組み合わせの間での、物理的相互作用または化学的相互作用を意味する。結合には、イオン結合、非イオン結合、水素結合、ファンデルワールス結合、疎水性相互作用などが含まれる。物理的相互作用(結合)は、直接的または間接的であり得、間接的なものは、別のタンパク質または化合物の効果を介するかまたは起因する。直接的な結合とは、別のタンパク質または化合物の効果を介してもまたはそれらに起因しても起こらず、他の実質的な化学中間体を伴わない、相互作用をいう。
本明細書中で使用される「接触(させる)」とは、化合物を、直接的または間接的のいずれかで、本発明のマーカー、リガント等として機能しうるポリペプチドまたはポリヌクレオチドに対して物理的に近接させることを意味する。ポリペプチドまたはポリヌクレオチドは、多くの緩衝液、塩、溶液などに存在させることができる。接触とは、核酸分子またはそのフラグメントをコードするポリペプチドを含む、例えば、ビーカー、マイクロタイタープレート、細胞培養フラスコまたはマイクロアレイ(例えば、遺伝子チップ)などに化合物を置くことが挙げられる。
本明細書において「標識」とは、目的となる分子または物質を他から識別するための存在(たとえば、物質、エネルギー、電磁波など)をいう。そのような標識方法としては、RI(ラジオアイソトープ)法、蛍光法、ビオチン法、化学発光法等を挙げることができる。本発明のマーカーまたはそれを捕捉する因子または手段を複数、蛍光法によって標識する場合には、蛍光発光極大波長が互いに異なる蛍光物質によって標識を行う。蛍光発光極大波長の差は、10nm以上であることが好ましい。リガンドを標識する場合、機能に影響を与えないものならば何れも用いることができるが、AGEsを標識する場合は、蛍光物質としては、Alexa Fluorが望ましい。Alexa Fluorは、クマリン、ローダミン、フルオレセイン、シアニンなどを修飾して得られた水溶性の蛍光色素であり、広範囲の蛍光波長に対応したシリーズであり、他の該当波長の蛍光色素に比べ、非常に安定で、明るく、またpH感受性が低い。蛍光極大波長が10nm以上ある蛍光色素の組み合わせとしては、Alexa555とAlexa633の組み合わせ、Alexa488とAlexa555の組み合わせ等を挙げることができる。核酸を標識する場合は、その塩基部分と結合できるものであれば何れも用いることができるが、シアニン色素(例えば、CyDyeTMシリーズのCy3、Cy5等)、ローダミン6G試薬、N-アセトキシ-N2-アセチルアミノフルオレン(AAF)、AAIF(AAFのヨウ素誘導体)等を使用することが好ましい。蛍光発光極大波長の差が10nm以上である蛍光物質としては、例えば、Cy5とローダミン6G試薬との組み合わせ、Cy3とフルオレセインとの組み合わせ、ローダミン6G試薬とフルオレセインとの組み合わせ等を挙げることができる。本発明では、このような標識を利用して、使用される検出手段に検出され得るように目的とする対象を改変することができる。そのような改変は、当該分野において公知であり、当業者は標識におよび目的とする対象に応じて適宜そのような方法を実施することができる。
本明細書において「診断」とは、被験体における疾患、障害、状態などに関連する種々のパラメータを同定し、そのような疾患、障害、状態の現状または未来を判定することをいう。本発明の方法、装置、システムを用いることによって、体内の状態を調べることができ、そのような情報を用いて、被験体における疾患、障害、状態、投与すべき処置または予防のための処方物または方法などの種々のパラメータを選定することができる。本明細書において、狭義には、「診断」は、現状を診断することをいうが、広義には「予測診断」「事前診断」等を含む。早期の診断は「早期診断」ということもある。
本明細書において特に、「予測診断」または「事前診断」とは、交換可能に使用され、sRAGE、AGEsを認識することができる分子等を用いて、糖尿病、糖尿病性腎症、糖尿病性網膜症、糖尿病性神経症等の糖尿病合併症等について言及する場合、糖尿病、糖尿病性腎症、糖尿病性網膜症、糖尿病性神経症等の糖尿病合併症等の発症前の段階を検出することをいい、将来の発症リスクを判定すること、糖尿病、糖尿病性腎症、糖尿病性網膜症、糖尿病性神経症等の糖尿病合併症等の予防を目的として糖尿病に罹患するおそれの有無を判定することを含む。本発明の方法、キット、組成物、検出剤、診断剤、システム等を用いることによって、体内の状態を事前に調べることができ、そのような情報を用いて、被験体における疾患、障害、状態、投与すべき処置または予防のための処方物または方法などの種々のパラメータを選定することができる。本明細書では、「予測診断」または「事前診断」は、他の従来の方法では診断できない段階での診断をも包含することから、「早期診断」の概念と一部重なって用いられる。
本発明の診断方法は、原則として、身体から出たものを利用することができることから、医師などの医療従事者の手を離れて実施することができることから、産業上有用である。本明細書において、医師などの医療従事者の手を離れて実施することができることを明確にするために、特に「予測診断、事前診断もしくは診断」を「支援」すると称することがある。
本明細書において使用される場合、用語「検出剤」とは、広義には、目的の物質(例えば、変性LDL、AGEsなど)を検出できるあらゆる因子をいう。
本明細書において使用される場合、用語「診断剤」とは、広義には、目的の状態(例えば、疾患など)を診断できるあらゆる因子をいう。
本明細書において「治療」とは、ある疾患または障害について、そのような状態になった場合に、そのような疾患または障害の悪化を防止、好ましくは、現状維持、より好ましくは、軽減、さらに好ましくは消退させることをいう。
本明細書において「予防」とは、ある疾患または障害について、そのような状態になる前に、そのような状態にならないようにすることをいう。本発明の予測診断または事前診断を行うことによって、糖尿血管障害、非アルコール性脂肪肝炎、アルツハイマー型認知症、などのAGEsに関連する疾患、の予防をするか、あるいは予防のための対策を講じることができる。本発明の予測診断または事前診断を行うことによって、糖尿病性腎症、糖尿病性網膜症、糖尿病性神経症等の糖尿病合併症等の予防をするか、あるいは予防のための対策を講じることもできる。
(好ましい実施形態の説明)
以下に好ましい実施形態の説明を記載するが、この実施形態は本発明の例示であり、本発明の範囲はそのような好ましい実施形態に限定されないことが理解されるべきである。当業者はまた、以下のような好ましい実施例を参考にして、本発明の範囲内にある改変、変更などを容易に行うことができることが理解されるべきである。これらの実施形態について、当業者は適宜、任意の実施形態を組み合わせ得る。
<アッセイ系1:捕捉用AGEsが基板に固定化されている系>
本発明者らは、AGEsのsRAGEとの反応性が、AGEsの種類によって異なることを見出した。この知見に基づき、発明者らは、捕捉用物質として反応性が低いものAGEs(捕捉用AGEs(競合用AGEsともいう))と、試料中のより反応性が高いAGEs(刺激性AGEs)とを競合的にsRAGEに結合させることによって、試料中の刺激性のAGEsを検出することができるアッセイ系を構築した。より反応性が高い刺激性AGEsは、sRAGEに強く結合し、捕捉用AGEsの結合量が低下するため、捕捉用AGEsからのまたは捕捉用AGEsと結合したsRAGEからのシグナル(例えば、発色)が低下し、この低下の度合いから、試料中の刺激性AGEsの量を算出することが可能である。シグナルとしては、比色、蛍光、化学発光、ラジオアイソトープなどが挙げられる。AGEsの量の算出は、あらかじめ検量線を作成しておき、吸光度(OD値)を濃度に変換することで算出することが可能である。
したがって、一態様において、本発明は、試料中の刺激性の終末糖化産物(AGEs)を検出するためのキットであって、該キットは、捕捉用AGEsが固定化された基板と、配列番号3と少なくとも約90%の同一性を有するアミノ酸配列を含む終末糖化産物受容体(sRAGE)とを含む、キットを提供する。図14に示されるように、捕捉用AGEsを基板に固定化し、標識(例えば、ビオチン化)されたsRAGEを検出することで、捕捉用AGEsとsRAGEとの結合を検出することができる。この系では、図4のようなsRAGEを基板に固定化した系で使用される競合用AGEsに対する一次抗体および一次抗体を認識する二次抗体を使用する必要がなく有利であるが、いずれの場合でも、検出能力に大きな差はなく、同様に使用することができる。sRAGEを基板に固定化した系については、<アッセイ系:sRAGEが基板に固定化されている系>を参照のこと。
また、基板に捕捉用AGEsを固定化した場合、sRAGEによって認識される部位が表面に現れず、結合が阻害される可能性が懸念されるが、予想外にも、基板に固定化されたAGEsとsRAGEとの結合は阻害されることはなかった。
捕捉用AGEsは、任意のAGEsを使用することができる。例えば、フルクトース(F)-AGEsは、G-AGEsよりもsRAGEに対する結合能が高いため、F-AGEsを捕捉用AGEsとして使用した場合、G-AGEsを除く、F-AGEsよりもsRAGEに対する結合能が高い刺激性AGEsを検出することが可能である。このように測定対象に応じて、捕捉用AGEsを適宜選択することができる。刺激性AGEsを網羅的に検出する系では、好ましくは、捕捉用AGEsは、グルコース(G)-AGEsまたはフルクトース(F)-AGEsであり、より好ましくは、グルコース(G)-AGEsである。
捕捉用AGEsの基板への固定化は、当該分野で周知の技術に基づいて行うことが可能であるが、炭酸-重炭酸緩衝液(炭酸ナトリウム-重炭酸ナトリウム緩衝液)を使用して基板に固定化されるのが好ましい。固定化に使用され得る炭酸-重炭酸緩衝液は、約9.2~約10.6の範囲内のpHを有し、例えば、約9.2~約9.6、約9.2~約9.8、約9.2~約10.0、約9.2~約10.2、約9.2~約10.4、約9.4~約9.6、約9.4~約9.8、約9.4~約10.0、約9.4~約10.2、約9.4~約10.4、約9.6~約9.8、約9.6~約10.0、約9.6~約10.2、または約9.6~約10.4の範囲内のpHを有し、好ましくは、約9.4~約9.8の範囲内のpHを有し、より好ましくは、約9.6のpHを有し得る。
前記刺激性AGEsとしては、グリオキサール(GO)-AGEs、グリコールアルデヒド(Glycol)-AGEs、グリセルアルデヒド(Glycer)-AGEs、3-デオキシグルコソン(3-DG)-AGEs、メチルグリオキサール(MGO)-AGEs、アセトアルデヒド(AA)-AGEs、リボース-AGEs、キシロース-AGEs、アラビノース-AGEs、フルクトース(F)-AGEs、ソルビトール-AGEs、ガラクトーズーAGEsまたはこれらの任意の組合せが挙げられるが、これらに限定されない。特に、本発明のアッセイ系では、グリセルアルデヒド(Glycer)-AGEs、およびグリコールアルデヒド(Glycol)-AGEsを高感度で検出することが可能である。
いくつかの実施形態では、前記捕捉用AGEsは、ウシ血清アルブミン、ヒト血清アルブミン、オボアルブミン、コラーゲン、クリスタリン、チューブリン、リゾチーム、RNase、カルモジュリン、ヘモグロビン、筋繊維タンパク質、アンチトロンビンIII、フェリチン、フィブリン、フィブリノーゲン、高密度リポタンパク質(HDL)、免疫グロブリン、低密度リポタンパク質(LDL)、インシュリン、ケラチン、オステオカルシン、アルコールデヒドロゲナーゼ、アルドースレダクターゼ、カテプシンB、スパーオキシドジスムダーゼ、ミオシンATPase、赤血球スペクトリン、またはミエリンのAGEsであり得る。好ましい実施形態では、捕捉用AGEsは、アルブミンのAGEsである。
上記捕捉用AGEsが基板に固定化されている態様において、本発明のキットは、捕捉用AGEsに結合したsRAGEを検出するための検出剤をさらに含んでもよい。いくつかの実施形態において、sRAGEは、ビオチン化されていてもよい。検出剤としては、ストレプトアビジン、抗sRAGE抗体などが挙げられる。その他の実施形態において、sRAGEは、mycタグ、flagタグ、HAタグなどが付加されていてもよく、抗myc抗体、抗flag抗体、抗HA抗体などの検出剤を使用して検出することもできる。
本発明のアッセイ系は、非常に幅広い濃度域の刺激性AGEsを検出することができ、約100ng/ml以上、例えば約100ng/ml~約100、000ng/mlの濃度域、特に約10ng/ml以上、例えば約10ng/ml~約10,000ng/mlの濃度域の試料中の刺激性AGEsを検出することが可能である。高濃度の試料は、希釈してから測定すればよく、測定濃度域の上限は上記に限定されない。
本発明のキットにおいて使用され得るsRAGEは、配列番号3に示されるアミノ酸配列を有し得るが、AGEsを認識する能力を有するものである限り、配列番号3と少なくとも約80%、少なくとも約85%、少なくとも約90%、少なくとも約95%、少なくとも約96%、少なくとも約97%、少なくとも約98%、少なくとも約99%同一であるアミノ酸配列を有していてもよい。いくつかの実施形態では、sRAGEは、当該分野で周知の発現系、例えば、大腸菌、酵母、昆虫細胞、動物細胞等により発現され得る。好ましい実施形態では、本発明のキットは、カイコ発現系を使用したカイコ型sRAGEを含み得る。カイコ発現系を用いることで、カイコ型の糖鎖修飾がされ、安定性の高いsRAGEを生成することができる。カイコ型sRAGEの製造方法については、<sRAGE(カイコ型)およびその製造法>で詳述する。カイコ型の糖鎖修飾による安定化の他にsRAGEを安定化する方法としては、糖、グリセロール等の安定化剤を添加するなど当該分野で周知の手法を使用することができる。
<アッセイ系2:sRAGEが基板に固定化されている系>
別の態様において、本発明は、sRAGEが基板に固定化された系も提供する。より具体的には、本発明は、試料中の刺激性の終末糖化産物(AGEs)を検出するためのキットであって、該キットは、競合用AGEsと、配列番号3と少なくとも約90%の同一性を有するアミノ酸配列を含む終末糖化産物受容体(sRAGE)が固定化されている基板とを含む、キットを提供する。この態様においては、sRAGEと結合した競合用AGEsに特異的な検出剤を用いて検出が行われる。いくつかの実施形態では、本発明のキットは、競合用AGEsに特異的に結合する検出剤をさらに含み得る。検出剤は、使用したAGEsの種類に応じて、適宜選択することができる。例えば、競合用AGEsとして、ウシ血清アルブミン(BSA)を使用した場合、抗アルブミン抗体を検出剤として使用することができる。その他の種々の実施形態は、上記<アッセイ系1:捕捉用AGEsが基板に固定化されている系>、その他に記載の実施形態を適宜使用し得る。
<刺激性AGEs検出用基板>
さらなる態様において、本発明は、試料中の刺激性の終末糖化産物(AGEs)を検出するため基板であって、捕捉用AGEsが固定化されている、基板を提供する。固定化に関する種々の実施形態は、上記<アッセイ系1:捕捉用AGEsが基板に固定化されている系>、その他に記載の実施形態を適宜使用し得る。
<刺激性AGEsを検出する方法>
さらなる態様において、本発明は、試料中の刺激性の終末糖化産物(AGEs)を検出する方法を提供する。上記アッセイ系1を使用する態様においては、本方法は、捕捉用AGEsが固定化された基板に、刺激性AGEsを含むことが予想される試料と、本明細書に記載のsRAGEとを接触させる工程と、sRAGEの検出剤を添加する工程とを含み得る。本方法は、吸光度を測定する工程と、吸光度を試料中の刺激性AGEsの濃度に変換する工程とをさらに含んでもよい。刺激性AGEsの濃度への変換は、検量線を用いて行うことができる。その他の種々の実施形態は、上記<アッセイ系1:捕捉用AGEsが基板に固定化されている系>、その他に記載の実施形態を適宜使用し得る。
1つの実施形態において、本発明の検出方法において用いられる固相としては、固定される分子、例えば、AGEsまたはsRAGEが固定されうる限り任意のものを利用することができる。1つの実施形態では、酵素結合イムノソルベント検定法(ELISA)のような機構を使用する場合、一般に、固相(基材)としては、マイクロタイタープレートが使用される。基板の材料としては、共有結合かまたは非共有結合のいずれかで、本発明において使用される生体分子に結合する特性を有するかまたはそのような特性を有するように誘導体化され得る、任意の固体材料が挙げられる。その材料としては、例えば、ポリエチレン、エチレン、ポリプロピレン、ポリイソブチレン、ポリエチレンテレフタレート、不飽和ポリエステル、含フッ素樹脂、ポリ塩化ビニル、ポリ塩化ビニリデン、ポリ酢酸ビニル、ポリビニルアルコール、ポリビニルアセタール、アクリル樹脂、ポリアクリロニトリル、ポリスチレン、アセタール樹脂、ポリカーボネート、ポリアミド、フェノール樹脂、ユリア樹脂、エポキシ樹脂、メラミン樹脂、スチレン・アクリロニトリル共重合体、アクリロニトリルブタジエンスチレン共重合体、シリコーン樹脂、ポリフェニレンオキサイド、ポリスルホン等の有機材料を用いることができる。
本明細書において使用される場合、用語「固相」とは、本明細書中において「基板」および「基材」と互換的に使用され、本発明のデバイスが構築される材料をいう。AGEsまたはsRAGEのような分子が固定され得る平面状の支持体をいう。本発明において表面プラズモン共鳴の原理を用いて検出する場合、固相は、金、銀またはアルミニウムを含む金属薄膜を片面に持つガラス基板の基材であることが好ましい。本発明において酵素結合イムノソルベント検定法(ELISA)のような機構を使用する場合、一般に、固相(基材)としては、マイクロタイタープレートが使用される。基板の材料としては、共有結合かまたは非共有結合のいずれかで、本発明において使用される生体分子に結合する特性を有するかまたはそのような特性を有するように誘導体化され得る、任意の固体材料が挙げられる。適切な基材としては、ビーズ、金粒子、半導体ナノ粒子(例えば、CdTeナノ粒子、CdSeナノ粒子、GaNナノ粒子、ZnSナノ粒子、InPナノ粒子など)、シリカナノ粒子、ポリスチレンナノ粒子、アクリル系ナノ粒子、ラテックスナノ粒子、カーボンナノ粒子、プレート(例えば、マイクロタイタープレート)、試験管、チップ、磁性粒子、膜、繊維、スライドガラス、金属薄膜、フィルター、チューブ、ボール、ダイアモンド様炭素被膜ステンレスなどが挙げられるが、これらに限定されない。
固相および基板として使用するためのそのような材料としては、固体表面を形成し得る任意の材料が使用され得るが、例えば、ガラス、シリカ、シリコーン、セラミック、二酸化珪素、プラスチック、金属(合金も含まれる)、天然および合成のポリマー(例えば、ポリスチレン、セルロース、アミロース、キトサン、デキストラン、およびナイロン)以下が挙げられるがそれらに限定されない。基板は、複数の異なる材料の層から形成されていてもよい。例えば、ガラス、石英ガラス、アルミナ、サファイア、フォルステライト、炭化珪素、酸化珪素、窒化珪素などの無機絶縁材料を使用できる。また、ポリエチレン、エチレン、ポリプロピレン、ポリイソブチレン、ポリエチレンテレフタレート、不飽和ポリエステル、含フッ素樹脂、ポリ塩化ビニル、ポリ塩化ビニリデン、ポリ酢酸ビニル、ポリビニルアルコール、ポリビニルアセタール、アクリル樹脂、ポリアクリロニトリル、ポリスチレン、アセタール樹脂、ポリカーボネート、ポリアミド、フェノール樹脂、ユリア樹脂、エポキシ樹脂、メラミン樹脂、スチレン・アクリロニトリル共重合体、アクリロニトリルブタジエンスチレン共重合体、シリコーン樹脂、ポリフェニレンオキサイド、ポリスルホン等の有機材料を用いることができる。本発明においてはまた、ナイロン膜、ニトロセルロース膜、PVDF膜など、ブロッティングに使用される膜を用いることもできる。高密度のものを解析する場合は、ガラスなど硬度のあるものを材料として使用することが好ましい。基板として好ましい材質は、測定機器などの種々のパラメータによって変動し、当業者は、上述のような種々の材料から適切なものを適宜選択することができる。
本明細書においてポリヌクレオチドまたはポリペプチド発現の「検出」または「定量」は、例えば、mRNAの測定および免疫学的測定方法を含む適切な方法を用いて達成され得る。分子生物学的測定方法としては、例えば、ノーザンブロット法、ドットブロット法またはPCR法などが例示される。免疫学的測定方法としては、例えば、方法としては、マイクロタイタープレートを用いるELISA法、RIA法、蛍光抗体法、ウェスタンブロット法、免疫組織染色法などが例示される。また、定量方法としては、ELISA法またはRIA法などが例示される。アレイ(例えば、DNAアレイ、プロテインアレイ)を用いた遺伝子解析方法によっても行われ得る。DNAアレイについては、(秀潤社編、細胞工学別冊「DNAマイクロアレイと最新PCR法」)に広く概説されている。プロテインアレイについては、NatGenet.2002 Dec;32 Suppl:526-32に詳述されている。遺伝子発現の分析法としては、上述に加えて、RT-PCR、RACE法、SSCP法、免疫沈降法、two-hybridシステム、インビトロ翻訳などが挙げられるがそれらに限定されない。そのようなさらなる分析方法は、例えば、ゲノム解析実験法・中村祐輔ラボ・マニュアル、編集・中村祐輔羊土社(2002)などに記載されており、本明細書においてそれらの記載はすべて参考として援用される。
本明細書において「検索」とは、電子的にまたは生物学的あるいは他の方法により、ある核酸塩基配列を利用して、特定の機能および/または性質を有する他の核酸塩基配列を見出すことをいう。電子的な検索としては、BLAST(Altschul et al.,J.Mol.Biol.215:403-410(1990))、FASTA(Pearson & Lipman,Proc.Natl.Acad.Sci.,USA 85:2444-2448(1988))、Smith and Waterman法(Smith and Waterman,J.Mol.Biol.147:195-197(1981))、およびNeedleman and Wunsch法(Needleman and Wunsch,J.Mol.Biol.48:443-453(1970))などが挙げられるがそれらに限定されない。生物学的な検索としては、ストリンジェントハイブリダイゼーション、ゲノムDNAをナイロンメンブレン等に貼り付けたマクロアレイまたはガラス板に貼り付けたマイクロアレイ(マイクロアレイアッセイ)、PCRおよびin situハイブリダイゼーションなどが挙げられるがそれらに限定されない。本明細書において、本発明において使用される遺伝子には、このような電子的検索、生物学的検索によって同定された対応遺伝子も含まれるべきであることが意図される。
本明細書において「キット」とは、通常2つ以上の区画に分けて、提供されるべき部分(例えば、抗体、標識など)が提供されるユニットをいう。混合されて提供されるべきでなく、使用直前に混合して使用することが好ましいような組成物の提供を目的とするときに、このキットの形態は好ましい。そのようなキットは、好ましくは、提供される部分(例えば、試薬をどのように処理すべきかを記載する指示書または説明書を備えていることが有利である。本明細書においてキットが試薬キットとして使用される場合、キットには、通常、抗体の使い方などを記載した指示書などが含まれる。
本明細書において「指示書」は、本発明を使用する方法を医師または他の使用者に対する説明を記載したものである。この指示書は、本発明の検出方法、診断薬の使い方、または医薬などを投与することを指示する文言が記載されている。また、指示書には、投与部位として、骨格筋に投与(例えば、注射などによる)することを指示する文言が記載されていてもよい。この指示書は、本発明が実施される国の監督官庁(例えば、日本であれば厚生労働省、米国であれば食品医薬品局(FDA)など)が規定した様式に従って作成され、その監督官庁により承認を受けた旨が明記される。指示書は、いわゆる添付文書(package insert)であり、通常は紙媒体で提供されるが、それに限定されず、例えば、電子媒体(例えば、インターネットで提供されるホームページ、電子メール)のような形態でも提供され得る。
本明細書において「生体内」または「インビボ」(in vivo)とは、生体の内部をいう。特定の文脈において、「生体内」は、目的とする物質が配置されるべき位置をいう。
本明細書において「インビトロ」とは、種々の研究目的のために生体の一部分が「生体外に」(例えば、試験管内に)摘出または遊離されている状態をいう。インビボと対照をなす用語である。
本明細書において「エキソビボ」とは、ある処置について、体外で行われるがその後体内に戻されることが意図される場合、一連の動作をエキソビボという。
本発明の体外診断薬等の医薬等としての処方手順は、当該分野において公知であり、例えば、日本薬局方、米国薬局方、他の国の薬局方などに記載されている。従って、当業者は、本明細書の記載があれば、過度な実験を行うことなく、使用すべき量を決定することができる。
<sRAGE(カイコ型)およびその製造法>
1つの局面において、本発明は、カイコ型糖鎖を有する、配列番号3に示すアミノ酸配列またはその改変体を含む再構築終末糖化産物受容体(sRAGE)を提供する。
好ましい実施形態では、本発明において前記カイコ型糖鎖は、トリマンノシルコア、複合型糖鎖、オリゴマンノース型糖鎖またはハイブリッド型糖鎖の糖鎖を含む。
1つの実施形態では、前記カイコ型糖鎖は、トリマンノシルコア(アスパラギン残基からGlcNAc-β1,4-GlcNAc-β1,4-Man(-α1,6-Man)-α1,3-Man)の構造に加え、1分子あたりGlcNAc0~2分子およびMan0~4分子のうちの0~4分子が結合した糖鎖を含む。別の実施形態では、本発明のRAGEの前記カイコ型糖鎖は、トリマンノシルコア(アスパラギン残基からGlcNAc-β1,4-GlcNAc-β1,4-Man(-α1,6-Man)-α1,3-Man)の構造に加え、2分子あたりGlcNAc0~4分子およびMan0~8分子のうちの0~8分子が結合した糖鎖を含む。別の実施形態では、sRAGEのカイコ型糖鎖としては、上記(1)~(8)([化1]~[化8])の構造を有する糖鎖が挙げられるが、これらに限定されない。
1つの実施形態では、カイコ型sRAGEの糖鎖の組成比が、オリゴマンノース型が90%以上を占め、複合型およびハイブリッド型が5%未満である。カイコ型sRAGEの糖組成は、(Man)5(GlcNAc)2は、46%~56%であり、(Man)7(GlcNAc)2は、23%~33%であり、(Man)6(GlcNAc)2は、7%~15%であり、(Man)3(GlcNAc)2は、1%~5%であり、(Man)3(GlcNAc)3は、2%~8%であり、(Man)4(GlcNAc)3は、1%~5%である。好ましい実施形態において、糖組成は、(Man)5(GlcNAc)2は、48%~54%であり、(Man)7(GlcNAc)2は、25%~31%であり、(Man)6(GlcNAc)2は、9%~13%であり、(Man)3(GlcNAc)2は、2%~4%であり、(Man)3(GlcNAc)3は、3%~6%であり、(Man)4(GlcNAc)3は、2%~4%である。より好ましい実施形態において、糖組成は、(Man)5(GlcNAc)2は、51.4%であり、(Man)7(GlcNAc)2は、27.5%であり、(Man)6(GlcNAc)2は、10.8%であり、(Man)3(GlcNAc)2は、2.8%であり、(Man)3(GlcNAc)3は、4.6%であり、(Man)4(GlcNAc)3は、2.5%である。これらの詳細な情報は、本明細書において参考として援用される、WO2016/051808の図54等を参照することができる。
上記糖鎖または上記糖鎖にさらなる糖が付加した糖鎖の付加を受けたsRAGEは、安定性に優れ、かつ多様な構造のAGEsに対する認識能を有する。
1つの実施形態では、前記カイコ型糖鎖は、配列番号3の3位のアスパラギンおよび/または59位のアスパラギンに結合する。
別の実施形態では、本発明のsRAGEは、ビオチン化されたものである。ビオチン化を行うことによって、バイオアッセイにおいて、高感度で測定結果を得ることができたり、ストレプトアビジンを介してビーズや膜上に一定の方向で高密度の集積が可能になる。
1つの実施形態において、本発明のsRAGEは、終末期糖化生成物(AGEs)を検出するための組成物として用いることができる。したがって、本発明は、本発明のカイコ型糖鎖を含むsRAGEを含む、AGEsを検出するための組成物を提供する。
理論に束縛されることを望まないが、本発明のsRAGEを用いることによって、分析の対象となるAGEsを従来の1割程度からほぼすべてに該当するレベルにまで引き上げることができ、より正確な診断をできるようになったという点で顕著な効果を奏するものである。また、従来の再構築RAGE(sRAGE)による検出方法でも、広範なAGEsを検出可能であるが、sRAGEは一ヶ月半程度で断片化し、認識能を失うなどの実用化上の問題があったが、本発明のsRAGEを用いることによって、これをも克服し得たという点でも顕著である。本発明の改良型sRAGEは、糖鎖付加をうけており、非常に安定な分子であり、4℃での保存で長期にわたり、約1年以上認識活性を維持していた。また、本発明の改良型sRAGEにより微量なAGEsの濃縮、多様な構造のAGEsなどの検出が可能であった。
したがって、別の局面において、本発明はまた、sRAGEの製造方法であって、A)カイコまたはカイコと同様の糖鎖を付与する生物に、sRAGEをコードする核酸分子を発現可能に組み込む工程;B)該遺伝子が発現する条件下に該カイコまたはカイコと同様の糖鎖を付与する生物を配置し、該sRAGEを発現させる工程;およびC)該sRAGEを得る工程を包含する、方法を提供する。
本発明は、カイコで製造されるものであるが、カイコ型糖鎖を付与することができる限り、どのような生物を用いてもよく、カイコと同様の糖鎖を付与する任意の生物が使用されることができることが理解される。
1つの実施形態では、前記発現は、前記カイコまたはカイコと同様の糖鎖を付与する生物の中部絹糸腺において行われる。
本発明の好ましい実施形態では、前記A)工程は、前記sRAGEをコードする核酸分子を含む発現ベクターをマイクロインジェクションすることで達成される。もちろん、マイクロインジェクション以外でも、本明細書において説明し、あるいは他の公知の任意の手法で発現ベクターを導入することができることが理解される。
1つの実施形態では、前記核酸分子は、配列番号2に示す核酸配列またはその改変体を含む。
1つの実施形態では、本発明のタンパク質の製造方法では、中部絹糸腺特異的に発現するタンパク質をコードする核酸配列のプロモーター、および該プロモーターによって直接的または間接的に発現制御される任意のタンパク質をコードする核酸配列を有するトランスジェニックカイコまたはカイコと同様の糖鎖を付与する生物であって、該任意のタンパク質を絹糸腺で発現するあるいは繭糸に分泌するトランスジェニックカイコまたはカイコと同様の糖鎖を付与する生物を製造する工程;および(b)製造されたトランスジェニックカイコカイコと同様の糖鎖を付与する生物から、該任意のタンパク質を回収する工程によって達成されてもよい。
1つの例示的な実施形態では、本発明のカイコまたはカイコと同様の糖鎖を付与する生物を生産する工程では、まず、中部絹糸腺特異的に発現するタンパク質をコードする核酸配列のプロモーター、および該プロモーターによって直接的または間接的に発現制御される任意のタンパク質をコードする核酸配列を有するカイコ卵を生産する。次いで、製造されたカイコ卵から生じたカイコまたはカイコと同様の糖鎖を付与する生物の中から、任意のタンパク質を発現するトランスジェニックカイコまたはカイコと同様の糖鎖を付与する生物を選択することによって実現することができる。
1つの例示的な実施形態では、トランスジェニックカイコまたはカイコと同様の糖鎖を付与する生物を選択するには、例えば、選択マーカーを用いて行うことができる。本発明における選択マーカーとしては、当業者において一般的に使用されるマーカー、例えば、CFP、GFP、YFP、DsRed等の蛍光タンパク質を使用することができる。これらのマーカーを用いることにより、実体蛍光顕微鏡で観察するだけでトランスジェニックカイコを検出することができる。また、蛍光色が異なるので、複数のマーカーを同時に使用することもできる。
さらに別の実施形態では、製造されたトランスジェニックカイコまたはカイコと同様の糖鎖を付与する生物から、任意のタンパク質を回収する方法としては、例えば、トランスジェニックカイコまたはカイコと同様の糖鎖を付与する生物が 吐糸した繭から、任意のタンパク質を回収する方法が挙げられる。回収方法としては、当業者に周知の方法、例えば、繭を60%LiSCNで溶かし、20mM Tris、5M ureaで透析することによって回収する方法(Inoue,S.,Tsuda,H.,Tanaka,H.,Magoshi,Y and Mizuno(2001)Sericologia 4,157-163.)を使用することができる。また、その他のタンパク質回収法としては、例えば界面活性剤を用いる方法や水溶液で溶かす方法等が可能である。
また、中部絹糸腺特異的に発現するタンパク質をコードする核酸配列のプロモーター、および該プロモーターによって間接的に発現制御される任意のタンパク質をコードする核酸配列を有するカイコ卵またはカイコと同様の糖鎖を付与する生物の卵としては、例えば、(i)中部絹糸腺特異的に発現するタンパク質をコードする核酸配列のプロモーターの下流に、転写制御因子をコードする核酸配列が機能的に結合した核酸配列、および(ii)該転写制御因子の標的プロモーターの下流に、任意のタンパク質をコードする核酸配列が作動可能に連結される核酸配列を有するカイコ卵またはカイコと同様の糖鎖を付与する生物の卵が挙げられる。
中部絹糸腺特異的に発現するタンパク質をコードする核酸配列のプロモーター、および該プロモーターによって直接的に発現制御される任意のタンパク質をコードする核酸配列を有するカイコ卵またはカイコと同様の糖鎖を付与する生物の卵としては、例えば、中部絹糸腺特異的に発現するタンパク質をコードする核酸配列のプロモーターの下流に、任意のタンパク質をコードする核酸配列が作動可能に連結される核酸配列を有するカイコ卵またはカイコと同様の糖鎖を付与する生物の卵が挙げられる。このようなカイコ卵は、中部絹糸腺特異的に発現するタンパク質をコードする核酸配列のプロモーターの下流に、任意のタンパク質をコードする核酸配列が作動可能に連結される核酸配列を卵に導入することで製造できる。
本明細書において「作動可能に連結される」とは、目的のタンパク質の発現が実現されるように連結されていることをいい、代表的に、プロモーターに転写制御因子が結合することにより、プロモーターの下流に存在する核酸配列の発現が誘導されるように、該プロモーターと該核酸配列とが結合していることをいう。転写制御因子と標的配列の組み合わせとしては、GAL4とUAS、TetRとTREなどが挙げられる。GAL4とUAS、またはTetRとTREを用いることにより、目的とする遺伝子の発現部位や時期、量を正確に制御でき、多くの組織で容易に発現させることができる。また、発現させる遺伝子が致死性の遺伝子でも系統の作出が可能である。本発明で使用される核酸配列は、ハイブリダイゼーション技術やポリメラーゼ連鎖反応(PCR)技術、site-directedmutagenesis法、DNA合成等の方法により調製することが可能である。調製された核酸配列がプロモーター活性を有するか否かは、当業者においてはレポーター遺伝子を用いた周知のレポーターアッセイ等により検討することが可能である。使用され得るレポーター遺伝子は本明細書の別の箇所において説明されており、任意の公知のものを使用することができる。
本発明の糖タンパク質としては、絹糸中で不可逆的な変性が生じないタンパク質であることが好ましい。また、このようなタンパク質としては、絹糸腺細胞から絹糸腺内腔への分泌シグナルを有さないタンパク質を例示することができる。
カイコ卵またはカイコと同様の糖鎖を付与する生物の卵へのDNAの導入は、例えば、発生初期卵へ、トランスポゾンをベクターとして注射する方法(Tamura, T., Thibert, C., Royer ,C., Kanda, T., Abraham, E., Kamba, M., Komoto, N., Thomas, J.-L., Mauchamp, B., Chavancy, G., Shirk, P., Fraser, M., Prudhomme, J.-C. and Couble, P., 2000, Nature Biotechnology 18, 81-84)に従って行うことができる。
また、例えば、トランスポゾンの逆位末端反復配列(Handler AM, McCombs SD, Fraser MJ, Saul SH.(1998) Proc. Natl. Acad. Sci. U.S.A. 95(13):7520-5)の間に上記DNAを挿入したベクターとともに、トランスポゾン転移酵素をコードする核酸配列を有するベクター(ヘルパーベクター)をカイコ卵またはカイコと同様の糖鎖を付与する生物の卵に導入することもできる。ヘルパーベクターとしては、pHA3PIG (Tamura, T., Thibert, C., Royer ,C., Kanda, T., Abraham, E., Kamba, M., Komoto, N., Thomas, J.-L., Mauchamp, B., Chavancy, G., Shirk, P., Fraser, M., Prudhomme, J.-C. and Couble, P., 2000, Nature Biotechnology 18, 81-84)が挙げられるが、これに限定されるものではない。
本発明におけるトランスポゾンとしては、piggyBacが好ましいが、これに限定されるものではなく、マリーナ(mariner)、ミノス(minos)等を用いることもできる(Shimizu, K., Kamba, M., Sonobe, H., Kanda, T., Klinakis, A. G., Savakis, C. and Tamura, T. (2000) Insect Mol. Biol., 9, 277-281;Wang W, Swevers L, Iatrou K.(2000) Insect Mol Biol 9(2):145-55)。
また、本発明では、バキュロウイルスベクターを使用することによりトランスジェニックカイコまたはカイコと同様の糖鎖を付与する生物を作出することも可能である(Yamao, M., N. Katayama, H. Nakazawa, M. Yamakawa, Y. Hayashi et al., 1999, Genes Dev 13: 511-516)。
また、本発明におけるカイコまたはカイコと同様の糖鎖を付与する生物としては、特に制限はないが、また、目的の糖タンパク質の大量生産のためには、フィブロインタンパク質などの絹糸を構成するタンパク質をコードする遺伝子領域(コード領域、プロモーター領域、非翻訳領域を含む)の変異によって、絹糸を構成するタンパク質の生産が抑制されているカイコまたはカイコと同様の糖鎖を付与する生物を用いることが好ましい。このようなカイコまたはカイコと同様の糖鎖を付与する生物としては、絹糸を構成するタンパク質をコードする遺伝子領域の変異によって絹糸を構成するタンパク質の生産が抑制されている変異系統のカイコまたはカイコと同様の糖鎖を付与する生物、好ましくは該変異によって絹糸を構成するタンパク質の生産が抑制されている裸蛹系統のカイコまたはカイコと同様の糖鎖を付与する生物、より好ましくはNd-sDが挙げられるが、絹糸を構成するタンパク質の生産抑制の原因が、人為的か否か、また、自然界において生じた変異に依存するか否かに関わらず、絹糸を構成するタンパク質の生産が抑制されているカイコまたはカイコと同様の糖鎖を付与する生物であればよい。このようなカイコまたはカイコと同様の糖鎖を付与する生物は、セリシンカイコとして当業者には周知のカイコである。セリシンカイコを利用することによって、染色体に導入された任意の遺伝子から合成されるタンパク質の精製がさらに容易になる。
また、本発明におけるカイコまたはカイコと同様の糖鎖を付与する生物としては、非休眠卵を産下する性質を有するカイコだけでなく、休眠卵を産下する性質を有するカイコ(例えば実用品種であるぐんま、200、春嶺、鐘月、錦秋、鐘和等)も使用することができる。ここで、休眠卵とは産卵後胚発生が一時的に停止する卵を言い、非休眠卵とは産卵後胚発生が停止せず、幼虫が孵化する卵を言う。
休眠卵を産下する性質を有するカイコを用いる場合は、非休眠卵を産下させ、該非休眠卵にDNAを導入する。非休眠卵を産下させる方法としては、例えばぐんまにおいては、休眠卵を15℃~21℃で培養することで該休眠卵から生じた成虫に非休眠卵を産下させる方法、好ましくは休眠卵を16℃~20℃で培養することで該休眠卵から生じた成虫に非休眠卵を産下させる方法、より好ましくは休眠卵を18℃で培養することで該休眠卵から生じた成虫に非休眠卵を産下させる方法、最も好ましくは休眠卵を18℃で培養することで該休眠卵から生じた幼虫を全明で飼育し、生育した成虫に非休眠卵を産下させる方法を挙げることができる。また、200においては、休眠卵を15℃~21℃で培養することで該休眠卵から生じた成虫に非休眠卵を産下させる方法、好ましくは休眠卵を16℃~20℃で培養することで該休眠卵から生じた成虫に非休眠卵を産下させる方法、より好ましくは休眠卵を18℃で培養することで該休眠卵から生じた成虫に非休眠卵を産下させる方法、または休眠卵から生じた幼虫を全明で飼育し、生育した成虫に非休眠卵を産下させる方法、最も好ましくは休眠卵を25℃で培養することで該休眠卵から生じた幼虫を全明で飼育し、生育した成虫に非休眠卵を産下させる方法が挙げられる。卵の培養は、例えば、18℃~25℃のインキュベーター、または定温の部屋に入れることによって行うことができ、幼虫の飼育は20℃~29℃の飼育室で人工飼料を用いて行うことができる。
本発明において、日長条件とは卵の培養、または幼虫の飼育中における一日毎の明暗サイクルを意味する。このような条件としては明条件と暗条件があり、特に全明条件とは暗の無い24時間明の条件を意味する。日長条件は、品種に応じて変化させることができる。本発明の上記休眠卵の培養は、当業者においては、一般的なカイコ卵の培養法に従って行うことができる。例えば、「文部省(1978)蚕種製造.pp193、実教出版社、東京」に記載の方法に従って培養を行う。また、本発明におけるカイコ幼虫の飼育は、当業者においては、周知の方法によって行うことができる。例えば、「文部省(1978)蚕種製造.pp193、実教出版社、東京」に記載の方法に従って飼育を行う。
本発明において、産卵された卵が非休眠卵であるか否かは、卵の色で判定することができる。一般に、休眠卵は濃い茶褐色に着色し、非休眠卵は黄白色であることが知られている。よって、本発明においては、濃い茶褐色ではないこと、より好ましくは黄白色であることをもって産卵された卵が非休眠卵であると判定する。
別の局面では、本発明は、sRAGEをコードする核酸分子を発現可能に組み込んだカイコまたはカイコと同様の糖鎖を付与する生物を提供する。
この局面の実施形態では、本発明で用いられる前記核酸分子は、配列番号2に示す核酸配列またはその改変体を含む。
1つの実施形態では、本発明のカイコまたはカイコと同様の糖鎖を付与する生物は、中部絹糸腺特異的に発現するsRAGEをコードする核酸配列のプロモーター、および該プロモーターによって直接的または間接的に発現制御される任意のタンパク質をコードする核酸配列を有するトランスジェニックカイコまたはカイコと同様の糖鎖を付与する生物であって、該sRAGEを絹糸腺で発現するあるいは繭糸に分泌するトランスジェニックカイコまたはカイコと同様の糖鎖を付与する生物を提供する。本発明のトランスジェニックカイコまたはカイコと同様の糖鎖を付与する生物の状態に特に制限はなく、例えば卵の状態であってもよい。本発明のトランスジェニックカイコまたはカイコと同様の糖鎖を付与する生物を使用することで、sRAGEを大量に生産することができる。
本発明のトランスジェニックカイコまたはカイコと同様の糖鎖を付与する生物としては、好ましくは、(i)中部絹糸腺特異的に発現するsRAGEをコードする核酸配列のプロモーターの下流に、作動可能に連結される転写制御因子をコードする核酸配列、および(ii)該転写制御因子の標的プロモーターの下流に、作動可能に連結されるsRAGEをコードする核酸配列を有するトランスジェニックカイコまたはカイコと同様の糖鎖を付与する生物、または中部絹糸腺特異的に発現するsRAGEをコードする核酸配列のプロモーターの下流に、sRAGEをコードする核酸配列が作動可能に連結される核酸配列を有するトランスジェニックカイコまたはカイコと同様の糖鎖を付与する生物である。
他の実施形態において、sRAGEは、後部絹糸腺(セリシンの代わりにフィブロインを産生する)またはマユから抽出され得る。 さらに、1つの別の実施形態では、本発明は、本発明のトランスジェニックカイコまたはカイコと同様の糖鎖を付与する生物(絹糸腺または繭を生産するもの)の絹糸腺または繭を提供する。このような絹糸腺または繭は、sRAGEを大量に含有する絹糸腺または繭として有用である。
また、本発明は、本発明の方法に用いるための核酸配列または核酸分子(例えば、DNA)を提供する。このような核酸配列または核酸分子としては、(a)セリシンをコードする核酸配列のプロモーターの下流に、作動可能に連結される転写制御因子をコードする核酸配列、(b)該転写制御因子の標的プロモーターの下流に、作動可能に連結される任意のタンパク質をコードする核酸配列、(c)セリシンをコードする核酸配列のプロモーターの下流に、sRAGEをコードする核酸配列が作動可能に連結される核酸配列、などが挙げられ、これらの組み合わせからなるキットとして提供してもよい。また、本発明は、トランスポゾンの逆位末端反復配列の間に、(a)~(c)の核酸配列を挿入したベクターを提供する。さらに、該ベクターとトランスポゾン転移酵素をコードする核酸配列を有するベクター(ヘルパーベクター)を含むキットを提供する。
<糖鎖の構造>
糖鎖は、糖鎖の結合様式により、アスパラギンと結合する糖鎖(N-グリコシド結合糖鎖という)ならびに、セリン、スレオニンなどと結合する糖鎖(0-グリコシド結合糖鎖という)の2種類に大別される。上述のN-グリコシド結合糖鎖は、様々な構造を有しているが[生物化学実験法23-糖タンパク質糖鎖研究法(学会出版センター)高橋禮子編(1989年)]、いずれの場合も下記に示す基本となる共通のコア構造を有することが好ましい。もっとも、この点は、上述の糖タンパク質が抗体ではない場合にも、同様である。
カイコ型のものでは以下のとおりである(Iizuka et al. FEBS Journal 276(2009)5806-5820)。すなわち、上述の生産方法において、上述のカイコ型糖タンパク質には、下記化学式(1)で示される糖鎖構造(トリマンノシルコアとも呼ばれる。)
を含むN-グリコシド結合糖鎖(トリマンノシルコア)を有する糖タンパク質が含まれることが好ましい。理論に束縛されることを望まないが、上記糖鎖構造はカイコ型糖鎖に共通の特徴であるところ、実施例等で示されるように、安定性に優れていること、および変性LDLまたはAGEsを広く感度よく検出することができることが判明していることから、この糖鎖が一定の役割を果たしているものと推測されるからである。本明細書において、糖鎖(1)は、N4-1または糖鎖000.1とも表示される。
そして、上記の構造において、アスパラギンと結合する糖鎖の末端を還元末端、反対側を非還元末端という。還元末端のN-アセチルグルコサミンへのフコースの結合としては、α1,3結合、α1,6結合などがあげられる。
N-グリコシド結合糖鎖には、コア構造の非還元末端にマンノースのみが結合するハイマンノース型(オリゴマンノース型)、コア構造の非還元末端側にガラクトース-N-アセチルグルコサミン(以下、Gal-GlcNAcと称す。)の枝を並行して1ないしは複数本有し、更にGal-GlcNAcの非還元末端側にシアル酸、バイセクティングのN-アセチルグルコサミンなどの構造を有する複合型(コンプレックス型ともいい、両者は同義である。)、コア構造の非還元末端側にハイマンノース型(オリゴマンノース型)と複合型(コンプレックス型)の両方の枝を持つハイブリッド型などがあげられる。
本実施形態では、上述の糖タンパク質には、下記化学式(2)、(3)または(4)で示される糖鎖構造
この型は、アスパラギン残基からみて「GlcNAc-β1,4-GlcNAc-β1,4-Man(-α1,6-Man-β1,2-GlcNAc)-α1,3-Man-β1,2-GlcNAc」とも記載される。本明細書において、糖鎖(2)は、糖鎖200.1とも表示される。
(この型は、アスパラギン残基からみて「GlcNAc-β1,4-GlcNAc-β1,4-Man(-α1,6-Man-β1,2-GlcNAc)-α1,3-Man」とも記載され、複合型に分類されることもある。)本明細書において、糖鎖(3)は、糖鎖100.1とも表示される。
(この型は、アスパラギン残基からみて「GlcNAc-β1,4-GlcNAc-β1,4-Man(-α1,6-Man)-α1,3-Man-β1,2-GlcNAc」とも記載され、複合型に分類されることもある。)
を含むN-グリコシド結合糖鎖を有する糖タンパク質が含まれることが好ましい。カイコ型の糖鎖では、上述のような複合型糖鎖が含まれていることが知られているからである。本明細書において、糖鎖(4)は、N4-2または糖鎖100.2とも表示される。
そして、本発明では、(2)が代表的に認められるものである。上述の糖タンパク質のうち、化学式(3)または(4)で示される糖鎖構造を含むN-グリコシド結合糖鎖を有する糖タンパク質を含んでいてもよい。
化学式(2)、(3)または(4)で示される糖鎖構造を含むN-グリコシド結合糖鎖を有する糖タンパク質の割合が10モル%以上、20モル%以上、または30モル%以上であれば、その糖タンパク質のN-グリコシド結合糖鎖は、カイコ型を生産しない一般の細胞によって生産される糖タンパク質のN-グリコシド結合糖鎖に比べて、カイコ型の糖鎖により付与される特徴が強化されるからである。
(オリゴマンノース型)
この構造は、アスパラギン残基からみて「GlcNAc-β1,4-GlcNAc-β1,4-Man(-α1,3-Man)-α1,6-Man(-α1,3-Man)-α1,6-Man」とも記載される。本明細書において、糖鎖(5)は、N3または糖鎖M5.1とも表示される。本発明では、(2)のほかこの(5)も代表的に認められるものである。
この構造は、アスパラギン残基からみて「GlcNAc-β1,4-GlcNAc-β1,4-Man(-α1,3-Man-α1,2-Man)-α1,6-Man(-α1,3-Man)-α1,6-Man」とも記載される。本明細書において、糖鎖(6)は、N2または糖鎖M6.1とも表示される。本発明では、(2)のほかこの(6)も代表的に認められるものである。
もちろん、これらのオリゴマンノース型に加え、さらなる糖が付加したものも本発明の糖鎖として含まれ得ることが理解される。
(sRAGEの糖鎖)
sRAGEの糖鎖型として、上記(1)~(6)([化1]~[化6])に加えて以下のものも挙げられる。
(この型は、アスパラギン残基からみて「GlcNAc-β1,4-GlcNAc-β1,4-Man(-α1,6-Man-α1,3-Man)-α1,3-Man-β1,2-GlcNAc」とも記載され、ハイブリッド型に分類されることもある。)本明細書において、糖鎖(7)は、N4-3または糖鎖H4.11とも表示される。
(この構造は、アスパラギン残基からみて「GlcNAc-β1,4-GlcNAc-β1,4-Man(-α1,3-Man-α1,2-Man)-α1,6-Man(-α1,3-Man)-α1,6-Man-α1,2-Man」とも記載され、オリゴマンノース型に分類されることもある。)本明細書において、糖鎖(8)は、N1または糖鎖M7.2とも表示される。
sRAGEの糖鎖構造解析により、約90%以上がオリゴマンノース型糖鎖であり、数%の複合型およびハイブリッド型糖鎖を含むことを明らかにした。さらに、糖鎖付加により安定性が向上するという優れた効果を有する。
本発明に含まれ得るsRAGEの糖鎖型としては、参照により本明細書に組み込まれるWO2016/051808の図20および図54を参照することができる。
1つの実施形態では、本発明のRAGEの前記カイコ型糖鎖は、トリマンノシルコア(アスパラギン残基からGlcNAc-β1,4-GlcNAc-β1,4-Man(-α1,6-Man)-α1,3-Man)の構造に加え、1分子あたりGlcNAc0~2分子およびMan0~4分子のうちの0~4分子が結合した糖鎖を含む。別の実施形態では、本発明のRAGEの前記カイコ型糖鎖は、トリマンノシルコア(アスパラギン残基からGlcNAc-β1,4-GlcNAc-β1,4-Man(-α1,6-Man)-α1,3-Man)の構造に加え、2分子あたりGlcNAc0~4分子およびMan0~8分子のうちの0~8分子が結合した糖鎖を含む。
本発明のsRAGEは、糖鎖付加を受け得る部位が2か所存在し、糖鎖は、2か所のうち一方にのみ付加されていてもよいし、両方に付加されていてもよい。好ましい実施形態において、2か所の糖鎖付加を受け得る部位のうち両方に糖鎖が付加されている。2か所両方に糖鎖が付加されている場合、付加される糖鎖は同一の糖鎖であってもよいし、異なっていてもよい。付加される糖鎖は、上記(1)~(8)([化1]~[化8])の構造を有する糖鎖が挙げられる。
上述の通り、カイコ型の糖鎖は、オリゴマンノース型、複合型、ハイブリッド型に分類される。sRAGEの糖鎖構造は、87%~97%がオリゴマンノース型であり、2%~8%が複合型であり、1%~5%がハイブリッド型である。好ましくは、90%~94%がオリゴマンノース型であり、3%~6%が複合型であり、2%~4%がハイブリッド型である。最も好ましくは、92.5%がオリゴマンノース型であり、4.6%が複合型であり、2.5%がハイブリッド型である。
1つの実施形態において、カイコ型sRAGEの糖組成は、(Man)5(GlcNAc)2(参照により本明細書に組み込まれるWO2016/051808図54のN3)は、46%~56%であり、(Man)7(GlcNAc)2は、23%~33%であり、(Man)6(GlcNAc)2は、7%~15%であり、(Man)3(GlcNAc)2は、1%~5%であり、(Man)3(GlcNAc)3は、2%~8%であり、(Man)4(GlcNAc)3は、1%~5%である。好ましくは、糖組成は、(Man)5(GlcNAc)2は、48%~54%であり、(Man)7(GlcNAc)2は、25%~31%であり、(Man)6(GlcNAc)2は、9%~13%であり、(Man)3(GlcNAc)2は、2%~4%であり、(Man)3(GlcNAc)3は、3%~6%であり、(Man)4(GlcNAc)3は、2%~4%である。最も好ましくは、糖組成は、(Man)5(GlcNAc)2は、51.4%であり、(Man)7(GlcNAc)2は、27.5%であり、(Man)6(GlcNAc)2は、10.8%であり、(Man)3(GlcNAc)2は、2.8%であり、(Man)3(GlcNAc)3は、4.6%であり、(Man)4(GlcNAc)3は、2.5%である。これらの詳細な説明は、参照により本明細書に組み込まれるWO2016/051808の図54を参照することができる。
糖タンパク質に結合している糖鎖を成す糖の種類はそれほど多くなく、よく見られるものは、グルコース、ガラクトース、マンノース、フコース、N-アセチルグルコサミン、N-アセチルガラクトサミン、N-アセチルノイラミン酸、キシロースなど7~8種程度である。構造様式もある程度限られており、その中のわずかな構造の違いが識別され、精密に認識されて様々な生命現象が制御されている。
本明細書では、「フコース」とは、通常のフコース(fucose)を意味し、デオキシ糖の一種である6-デオキシ-ガラクトースで、化学式はC6H12O5、分子量164.16、融点163℃、比旋光度-76゜で六炭糖、単糖に分類される。天然にはL型がL-フコシドの形で、動植物に幅広く存在する。哺乳動物および植物では細胞表面のN結合糖鎖上で見つかる。
本明細書では、「N-アセチルグルコサミン」とは、通常のN-アセチルグルコサミン(N-アセチル-D-グルコサミン、GlcNAc、NAG)を意味し、グルコースから誘導された単糖で、いくつかの生化学的機構にとって重要な物質である。化学的にはこの物質はグルコサミンと酢酸の間のアミドである。N-アセチルグルコサミンは、哺乳動物においては、糖タンパク質、ヒアルロン酸などグリコサミノグリカン(ムコ多糖)の成分となっている。N-アセチルグルコサミンは、アスパラギンにマンノースを中心とするオリゴ糖鎖が結合するN結合型糖タンパク質の骨格をなすほか(キトビオース構造)、更に複雑構造を持つ糖鎖の主要構成糖である。
本明細書では、「ガラクトース」とは、通常のガラクトース(Galactose)を意味し、分子式、分子量はグルコースと同じC6H12O6、180である。立体配置は2位(Fischer投影式で上から2番目)、5位の-OHが同じ方向3位、4位が反対方向であり、D-ガラクトースは5位D-グリセルアルデヒドと同じ配向をもっている。グルコースの4-エピマーである。天然ではD-ガラクトースがほとんどである。
本明細書では、「マンノース」とは、通常のマンノース(Mannose)を意味し、アルドヘキソースに分類される単糖の一種であり、分子式、分子量はグルコースと同じC6H12O6、180である。立体配置は2位(Fischer投影式で上から2番目)、3位の-OHが同じ方向4位、5位が反対方向であり、D-マンノースは5位D-グリセルアルデヒドと同じ配向をもっている。グルコースの2-エピマーである。天然ではD-マンノースがほとんどである。
<糖鎖の分析方法>
中性糖・アミノ糖組成分析
糖鎖は、代表的に、ガラクトース、マンノース、フコースなどの中性糖、N-アセチルグルコサミンなどのアミノ糖、シアル酸などの酸性糖から構成されている。糖鎖の組成分析は、トリフルオロ酢酸等で、糖鎖の酸加水分解を行うことにより、中性糖またはアミノ糖を遊離し、その組成比を分析することができる。具体的な方法として、Dionex社製糖組成分析装置(BioLC)を用いる方法が挙げられる。BioLCはHPAEC-PAD(high performance anion-exchange chromatography-pulsed amperometric detection)法[J.Liq.Chromatogr.,6,1577(1983)]によって糖組成を分析する装置である。
また、2-アミノピリジンによる蛍光標識化法でも組成比を分析することができる。具体的には、公知の方法[Agric.Biol.Chem.,55(1),283-284(1991)]に従って酸加水分解した試料を2-アミノピリジル化で蛍光標識し、HPLC分析して組成比を算出することができる。
糖鎖構造解析
糖鎖の構造解析は、2次元糖鎖マップ法[Anal.Biochem.,171,73(1988)、生物化学実験法23-糖タンパク質糖鎖研究法(学会出版センター)高橋禮子編(1989年)]により行うことができる。2次元糖鎖マップ法は、例えば、X軸には逆相クロマトグラフィー糖鎖の保持時間または溶出位置を、Y軸には順相クロマトグラフィーによる糖鎖の保持時間または溶出位置を、それぞれプロットし、既知糖鎖のそれらの結果と比較することにより、糖鎖構造を推定する方法である。
具体的には、糖タンパク質をヒドラジン分解して、糖タンパク質から糖鎖を遊離し、2-アミノピリジン(以下、PAと略記する)による糖鎖の蛍光標識[J.Biochem.,95,197(1984)]を行った後、ゲルろ過により糖鎖を過剰のPA化試薬などと分離し、逆相クロマトグラフィーを行う。次いで、分取した糖鎖の各ピークについて順相クロマトグラフィーを行う。これらの結果をもとに、2次元糖鎖マップ上にプロットし、糖鎖スタンダード(TaKaRa社製)、文献[Anal.Biochem.,171,73(1988)]とのスポットの比較より糖鎖構造を推定することができる。さらに各糖鎖のMALDI-TOF-MSなどの質量分析を行い、2次元糖鎖マップ法により推定される構造を確認することができる。
<糖タンパク質の生産方法>
本実施形態では、カイコ等の鱗翅目に分類される昆虫の絹糸腺細胞において、糖タンパク質を発現させる。
その際、上述の昆虫は、カイコであることが好ましい。なぜなら、カイコの場合には、養蚕業において飼育方法が確立しており、さらに絹糸腺から繭をはき出す能力が優れているため、上述の糖タンパク質を大量に効率よく得ることができるからである。
カイコは、代表的に一頭あたり0.3~0.5gの繭を作るが、このほとんどは、後部絹糸腺で合成されるフィブロインや、中部絹糸腺で合成されるセリシンなどの絹タンパク質である。このように、カイコは優れたタンパク質合成能力を有した生物であり、この能力を利用すれば、分析・評価用の組換えタンパク質等を大量に、かつ、安価に生産することができるからである。
さらに、上述の絹糸腺細胞は、中部絹糸腺細胞であることが好ましい。なぜなら、中部絹糸腺ではセリシンが合成されるため、セリシン中に上述の糖タンパク質を発現させれば、フィブロインの周りに存在し、比較的水に溶けやすいセリシン層に、組換えタンパク質を局在させるために、中部絹糸腺で効率良く組換えタンパク質を発現させることができ、繭に含まれる組換えタンパク質を、その立体構造を変性させることなく容易に抽出することが可能となるからである。
すなわち、本実施形態における糖タンパク質の生産方法は、糖タンパク質を発現させる工程が、本発明の糖タンパク質を発現するように遺伝的に改変されてなる上述の昆虫(例えば、カイコ)個体の絹糸腺において本発明の糖タンパク質を含む繭を生成させる工程を含むことが好ましい。ここで、通常は、昆虫個体の絹糸腺において上述の糖タンパク質を含む繭を生成させるということは、代表的に、昆虫の絹糸腺細胞において上述の糖タンパク質を発現させることになる。
そして、目的の糖タンパク質を抽出する工程は、上述の絹糸腺から前記糖タンパク質を含む繭を吐出させる工程と、上述の吐出された繭から上述の糖タンパク質を抽出する工程と、を含むことが好ましい。なぜなら、絹糸腺ではフィブロインまたはセリシンが合成されるため、フィブロインまたはセリシン中に上述の糖タンパク質を発現させれば、フィブロイン層またはセリシン層に、組換えタンパク質を局在させるために、絹糸腺で効率良く組換えタンパク質を発現させることができ、繭に含まれる組換えタンパク質を抽出することが可能となるからである。
1つの実施形態では、上述の糖タンパク質を発現させる工程は、上述の昆虫個体において、上述の糖タンパク質をコードする遺伝子が上述の昆虫のゲノムにおいてセリシンプロモーターの下流に発現可能に設けられている昆虫個体を用いる工程と、上述の繭として、セリシン部に上述の糖タンパク質を含む繭を生成させる工程と、を含むことが好ましい。そして、この場合には、上述の糖タンパク質を抽出する工程が、上述の繭を抽出液に浸して、上述の繭の上述のセリシン部から上述の抽出液中に上述の糖タンパク質を抽出する工程を含むことが好ましい。なぜなら、繭を構成する絹糸は、中心に存在するフィブロイン層の周りにセリシン層が存在する構造をしている。絹タンパク質を合成している絹糸腺は、機能的および形態的に、後部絹糸腺、中部絹糸腺、および前部絹糸腺に区別されるが、セリシン層を構成するセリシンは中部絹糸腺で合成され、フィブロイン層を構成するフィブロインは後部絹糸腺で合成される。糖タンパク質を発現する組織が中部絹糸腺であった場合、糖タンパク質は絹糸のセリシン層へ分泌される。一方、糖タンパク質を発現する組織が後部絹糸腺であった場合、糖タンパク質は絹糸のフィブロイン層へ分泌される。そして、セリシン層に糖タンパク質を分泌するトランスジェニックカイコを作製するためには、ベクターにおいて、中部絹糸腺で糖タンパク質を発現させるために、糖タンパク質の構造遺伝子に加えて、例えばそれぞれの構造遺伝子の上流に、中部絹糸腺細胞で遺伝子発現を引き起こすプロモーターを組めばよい。中部絹糸腺細胞で遺伝子発現を引き起こすプロモーターとしては、例えば、セリシン1遺伝子またはセリシン2遺伝子のプロモーターをあげることができる。中部絹糸腺または後部絹糸腺は通常使用される合成経路であり、本発明でも同様に利用することができる。あるいは、全身に発現させてもよい。なお、絹糸腺で発現させた場合、中部(および後部)絹糸腺を摘出して抽出する方法と、繭から抽出する方法がある。また、全身で発現させた場合、全身を磨砕したり、体液を絞ったりする方法がある。
このような発現ベクターを用いると、一回の遺伝子導入操作で、セリシン層に糖タンパク質を分泌するトランスジェニックカイコを作製することができる。一方、セリシン層に糖タンパク質を分泌するトランスジェニックカイコは複数回の遺伝子導入操作によって作製することもできる。
もっとも、フィブロイン層に糖タンパク質を分泌するトランスジェニックカイコを作製するという方法を排除する趣旨ではない。後部絹糸腺で糖タンパク質を発現させるために、糖タンパク質の構造遺伝子に加えて、それぞれの構造遺伝子の上流に、例えば後部絹糸腺細胞で遺伝子発現を引き起こすプロモーターが組み込まれているベクターを用いてフィブロイン層にタンパク質を分泌するトランスジェニックカイコを作製してもかまわない。後部絹糸腺細胞で遺伝子発現を引き起こすプロモーターとしては、例えば、フィブロインL鎖、フィブロインH鎖、またはフィブロヘキサマリンなどの遺伝子のプロモーターをあげることができる。
セリシン層に糖タンパク質を分泌するトランスジェニックカイコを作製する場合と同様に、フィブロイン層に糖タンパク質を分泌するトランスジェニックカイコを作製する場合においても、上述の発現ベクターセットを用いて、複数回の遺伝子導入操作によりトランスジェニックカイコを作製しても良い。
これらの発現ベクターは、カイコ染色体へ遺伝子を導入するための機能を有している。例えば、昆虫由来のDNA型トランスポゾンの部分配列が組み込まれていれば、遺伝子をカイコ染色体に導入することが可能である。具体的には、DNA型トランスポゾンの末端に存在する一対の逆向き反復配列を有したプラスミドベクターであり、一対の逆向き反復配列に挟まれた領域に、染色体に挿入される遺伝子配列、すなわち、糖タンパク質の遺伝子とプロモーターが組み込まれている発現ベクターである。昆虫由来DNA型トランスポゾンとしては、piggyBac、mariner(Insect Mol.Biol.9,145-155,2000)、およびMinos(Insect Mol.Biol.9,277-281,2000)などが知られているが、この中でもpiggyBac由来の配列は、最も頻繁に利用されている。トランスジェニックカイコを作製するためには、このプラスミドを、piggyBacのトランスポゼース発現ベクター(ヘルパープラスミド)と共にカイコ卵へ微量注入する。このヘルパープラスミドは、piggyBacの逆向き反復配列の片方または両方を欠いた、実質的にはpiggyBacのトランスポゼース遺伝子領域のみが組み込まれている組換えプラスミドベクターである。ヘルパープラスミドにおいて、トランスポゼースを発現させるためのプロモーターは、内在性のトランスポゼースプロモーターをそのまま利用しても良いし、あるいは、カイコ・アクチンプロモーターやショウジョウバエHSP70プロモーター等を利用してもよい。次世代カイコのスクリーニングを容易にするために、挿入するポリヌクレオチドを組み込んだベクター内に同時にマーカー遺伝子を組み込んでおくこともできる。
本明細書において、プロモーターに転写制御因子が結合することにより、プロモーターの下流に存在する構造遺伝子の核酸配列の発現が誘導されるように、該プロモーターと該核酸配列とが結合させることもできる。上記転写制御因子と標的配列の組み合わせとしては、GAL4とUAS、TetRとTREなどが挙げられる。GAL4とUAS、又はTetRとTREを用いることにより、目的とする遺伝子の発現部位や時期、量を正確に制御でき、多くの組織で容易に発現させることができる。また、発現させる遺伝子が致死性の遺伝子でも系統の作出が可能である。
1つの実施形態において、本発明の方法によって製造される糖タンパク質は、本発明の方法によって製造される限りなんら限定されず、シグナル配列(例えば、フィブロインHシグナルペプチド)、エンテロキナーセ認識部位(例えば、DDDDK(配列番号1))、ビオチン化部位(例えばBioEaseタグ)、またはタグ配列(例えば、BioEaseタグ、Hisタグ等)を有していても、有していなくてもよい。即ち、本発明の糖タンパク質の製造方法によって製造される糖タンパク質には、シグナル配列を有している糖タンパク質、シグナル配列を有していない糖タンパク質の両方が含まれる。
本発明における絹糸腺特異的に発現するタンパク質をコードする核酸配列のプロモーター、および該プロモーターによって直接的に発現制御される糖タンパク質をコードする核酸配列を有するカイコ卵としては、例えば、絹糸腺特異的に発現するタンパク質をコードする核酸配列のプロモーターの下流に、目的とする糖タンパク質をコードする核酸配列が作動可能に連結される核酸配列を有するカイコ卵が挙げられる。このようなカイコ卵は、絹糸腺特異的に発現するタンパク質をコードする核酸配列のプロモーターの下流に、目的とする糖タンパク質をコードする核酸配列が作動可能に連結される核酸配列をカイコ卵に導入することで製造できる。
また、本発明における絹糸腺特異的に発現するタンパク質をコードする核酸配列のプロモーター、および該プロモーターによって間接的に発現制御される組換え糖タンパク質をコードする核酸配列を有するカイコ卵としては、例えば、(i)絹糸腺特異的に発現するタンパク質をコードする核酸配列のプロモーターの下流に、転写制御因子をコードする核酸配列が機能的に結合した核酸配列、および(ii)該転写制御因子の標的プロモーターの下流に、組換え抗体をコードする核酸配列が作動可能に連結される核酸配列、を有するカイコ卵が挙げられる。なお本発明においては、絹糸腺特異的に発現するタンパク質をコードする核酸配列のプロモーターによって直接的又は間接的に発現制御される組換え抗体をコードする核酸配列は、抗体の分泌を促進し回収量を多くするためシグナル配列を有するものが好ましい。シグナル配列の具体的な態様は、本明細書において他の場所において説明しており、任意の公知のものを使用することができる。
1つの例示的な製法として、ベクターを微量注入したカイコ卵から孵化した幼虫(F0世代)を飼育する。得られた全F0世代のカイコを野生型カイコと、あるいはF0カイコ同士で交配し、次世代(F1世代)のカイコからトランスジェニックカイコを選抜する。トランスジェニックカイコの選抜は、例えばPCR法やサザンブロット法を用いて行う。また、マーカー遺伝子を組み込んだ場合には、その表現形質を利用して選抜することも可能である。例えばマーカー遺伝子としてGFP等の蛍光タンパク質遺伝子を利用した場合には、F1世代のカイコ卵や幼虫に励起光を照射し、蛍光タンパク質の発する蛍光を検出することにより行うことができる。以上のような方法によりトランスジェニックカイコを作出することができる。
トランスジェニックカイコの繭から、目的とする糖タンパク質を回収する際には、糖タンパク質がセリシン層に局在する場合はセリシン層から、フィブロイン層に含まれている場合はフィブロイン層から回収する。セリシン層からの糖タンパク質の回収は、特に容易に実施することができる。セリシン層を構成するセリシンは親水性であるため、この層に局在する組換えタンパク質は、タンパク質を変性させてしまう溶液を用いることなく抽出することが可能である。セリシン層から目的の糖タンパク質を抽出するための抽出液は、その糖タンパク質の抽出が可能であるものならば特段の制限はない。例えば、中性の塩類溶液であっても良いし、界面活性剤や、その他、抽出を効率的に行うための試薬などが含まれる溶液であっても良い。これらの抽出液を使って繭から糖タンパク質を抽出するには、例えば、断片化した繭を抽出液に浸し攪拌するなどの方法を用いることができる。また、抽出の前に、繭を微粉末化する処理を行っても良いし、抽出の際に超音波処理を行うなどの機械的処理を併用しても良い。また、上述のカイコの絹糸腺細胞は、さらなる糖鎖付加がされるように適宜転移酵素(例えば、βガラクトース転移酵素、N-アセチルグルコサミン転移酵素等)を発現させるように形質転換されていてもよい。
本発明の実施において、発現を確認するためにレポーター遺伝子を組み込んでもよい。このようなレポーター遺伝子としては、その発現が検出可能なものであれば特に制限されず、例えば、当業者において一般的に使用されるCAT遺伝子、lacZ遺伝 子、ルシフェラーゼ遺伝子、β-グルクロニダーゼ遺伝子(GUS)およびGFP遺伝子等を挙げることができる。レポーター遺伝子の発現レベルは、該レポーター遺伝子の種類に応じて、当業者に公知の方法により測定することができる。例えば、レポーター遺伝子がCAT遺伝子である場合には、該遺伝子産物によるクロラムフェニコールのアセチル化を検出することによって、レポーター遺伝子の発現レベルを測定することができる。レポーター遺伝子がlacZ遺伝子である場合には、該遺伝子発現産物の触媒作用による色素化合物の発色を検出することにより、また、ルシフェラーゼ遺伝子である場合には、該遺伝子発現産物の触 媒作用による蛍光化合物の蛍光を検出することにより、また、β-クロニダーゼ遺伝子(GUS)である場合には、該遺伝子発現産物の触媒作用によるGlucuron(ICN社)の発光や5-ブロモ-4-クロロ-3-インドリル-β-グルクロニド(X-Gluc)の発色を検出することにより、さらに、GFP遺伝子である場合には、GFPタンパク質による蛍光を検出することにより、レポーター遺伝子の発現レベルを測定することができる。
本発明の組換え糖タンパク質の製造方法は、カイコ体内で合成された目的の糖タンパク質を回収する工程を含む。合成された目的の糖タンパク質は、不溶化せずに、活性のある状態で、中部絹糸腺又は後部絹糸腺に分泌される。従って、目的の糖タンパク質は、中部絹糸腺又は後部絹糸腺から回収することが可能である。目的の糖タンパク質を中部絹糸腺又は後部絹糸腺から回収する方法としては、例えば、吐糸期になったカイコを解剖し、中部絹糸腺又は後部絹糸腺を20mM Tris-HCl pH7.4中に摘出し、絹糸腺をピンセットやメスで傷をいれることにより絹糸腺中の目的の糖タンパク質を回収できる。
また、本発明の目的の糖タンパク質は、例えば、トランスジェニックカイコが吐糸した繭から回収することも可能である。回収方法としては、当業者に周知の方法、例えば、繭を60%LiSCNで溶かし、20mMTris、5M尿素で透析することによって回収する方法(Inoue,S.,Tsuda,H.,Tanaka,H.,Magoshi,Y and Mizuno(2001)Sericologia 4,157-163.)を使用することができる。また、その他のタンパク質回収法としては、例えば界面活性剤を用いる方法や水溶液で溶かす方法等が可能である。
また、本発明におけるカイコとしては、特に制限はないが、目的の糖タンパク質の大量生産のためには、フィブロインタンパク質などの絹糸を構成するタンパク質をコードする核酸配列領域(コード領域、プロモーター領域、非翻訳領域を含む)の変異によって、絹糸を構成するタンパク質の生産が抑制されているカイコを用いることが好ましい。このようなカイコとしては、絹糸を構成するタンパク質をコードする核酸配列領域の変異によって絹糸を構成するタンパク質の生産が抑制されている変異系統のカイコ、好ましくは該変異によって絹糸を構成するタンパク質の生産が抑制されている裸蛹系統のカイコ、より好ましくはNd-sDが挙げられるが、絹糸を構成するタンパク質の生産抑制の原因が、人為的か否か、また、自然界において生じた変異に依存するか否かに関わらず、絹糸を構成するタンパク質の生産が抑制されているカイコであればよい。
このようなカイコの1つの態様は、セリシンカイコとして当業者には周知のカイコである。セリシンカイコを利用することによって、中部絹糸腺における目的の糖タンパク質の大量生産が可能となり、染色体に導入された組み換え抗体をコードする核酸配列から合成されるタンパク質の精製も容易になる。また、後部絹糸腺において組換え抗体を産生させる際にも、生産量の点でセリシンカイコを用いることが好ましい。
また、本発明におけるカイコとしては、非休眠卵を産下する性質を有するカイコ、休眠卵を産下する性質を有するカイコ(例えば実用品種であるぐんま、200、春嶺、鐘月、錦秋、鐘和等)を使用することができる。ここで、休眠卵とは産卵後胚発生が一時的に停止する卵を言い、非休眠卵とは産卵後胚発生が停止せず、幼虫が孵化する卵を言う。休眠卵を産下する性質を有するカイコを用いる場合は、非休眠卵を産下させ、該非休眠卵にDNAを導入する。非休眠卵を産下させる方法としては、例えばぐんまにおいては、休眠卵を15℃~21℃で培養することで該休眠卵から生じた成虫に非休眠卵を産下させる方法、好ましくは休眠卵を16℃~20℃で培養することで該休眠卵から生じた成虫に非休眠卵を産下させる方法、より好ましくは休眠卵を18℃で培養することで該休眠卵から生じた成虫に非休眠卵を産下させる方法、最も好ましくは休眠卵を18℃で培養することで該休眠卵から生じた幼虫を全明で飼育し、生育した成虫に非休眠卵を産下させる方法を挙げることができる。また、200においては、休眠卵を15℃~21℃で培養することで該休眠卵から生じた成虫に非休眠卵を産下させる方法、好ましくは休眠卵を16℃~20℃で培養することで該休眠卵から生じた成虫に非休眠卵を産下させる方法、より好ましくは休眠卵を18℃で培養することで該休眠卵から生じた成虫に非休眠卵を産下させる方法、又は休眠卵から生じた幼虫を全明で飼育し、生育した成虫に非休眠卵を産下させる方法、最も好ましくは休眠卵を25℃で培養することで該休眠卵から生じた幼虫を全明で飼育し、生育した成虫に非休眠卵を産下させる方法が挙げられる。
卵の培養は、例えば、18℃~25℃のインキュベーター、又は定温の部屋に入れることによって行うことができ、幼虫の飼育は20℃~29℃の飼育室で人工飼料を用いて行うことができる。
本発明の上記休眠卵の培養は、当業者においては、一般的なカイコ卵の培養法に従って行うことができる。例えば、「文部省(1978)蚕種製造.pp193、実教出版社、東京」に記載の方法に従って培養を行う。また、本発明におけるカイコ幼虫の飼育は、当業者においては、周知の方法によって行うことができる。例えば、「文部省(1978)蚕種製造.pp193、実教出版社、東京」に記載の方法に従って飼育を行う。
本発明において、産卵された卵が非休眠卵であるか否かは、卵の色で判定することができる。一般に、休眠卵は濃い茶褐色に着色し、非休眠卵は黄白色であることが知られている。よって、本発明においては、濃い茶褐色ではないこと、より好ましくは黄白色であることをもって産卵された卵が非休眠卵であると判定する。
以下、カイコ卵への核酸分子の導入方法の具体例を記すが、本発明におけるカイコ卵への核酸分子の導入方法は、この方法に限定されるものでない。例えば、カイコ卵へ核酸分子注入用の管を使用して直接卵内へ核酸分子を導入することが可能であるが、好ましい態様としては、前もって物理的又は化学的に卵殻に穴を空け、該穴から核酸分子を導入する。この際、核酸分子注入用の管を挿入角度が該卵の腹側の側面に対してほぼ垂直となるように該穴から卵内に挿入することができる。
本発明において、物理的に卵殻に穴を空ける方法としては、例えば針、微小レーザー等を用いて穴を空ける方法が挙げられる。好適には針を用いた方法によって卵殻に穴を空けることができる。該針は、カイコの卵殻に穴を空けることができるものであれば、その針の材質、強度等は、特に制限されない。なお、本発明における針とは、通常、先端が尖った棒状の針を指すが、この形状に限定されず、卵殻に穴を空けることができるものであれば、全体の形状は特に制限されない。例えば、先端の尖ったピラミッド型の物質、又は先端の尖った三角錐の形状の物質もまた、本発明の「針」に含まれる。本発明においては、タングステン針を好適に使用することができる。本発明の針の太さ(直径)は、後述のキャピラリーが通過可能な穴を空けることができる程度の太さであればよく、通常2~20μm、好ましくは5~10μmである。一方、化学的に卵殻に穴を空ける方法としては、例えば薬品(次亜塩素酸等)等を用いて穴を空ける方法が挙げられる。
本発明において、穴を空ける位置としては、該穴から核酸分子注入用の管を挿入した場合に卵の腹側の側面に対する挿入角度を、ほぼ垂直にできる位置ならば特に制限はないが、好ましくは腹側の側面又はその反対側であり、より好ましくは腹側の側面であり、よりさらに好ましくは卵の腹側側面のやや後端よりの中央部である。
本発明において、「ほぼ垂直」とは、70°~120°を意味し、好ましくは80°~100°を意味する。本発明において、「将来的に生殖細胞になる位置」としては、通常、卵の腹側の卵表に近い位置(通常、卵表から0.01mm~0.05mmの位置)であり、好ましくは、卵の腹側中央の卵表に近い位置でやや後極よりの位置である。
本発明の核酸分子注入用の管は、その管の材質、強度、内径等は特に制限されないが、核酸分子注入用の管を挿入する前に、物理的又は化学的に卵殻に穴を空ける場合は、空けられた穴を通過できる太さ(外径)であることが好ましい。本発明の核酸分子注入用の管としては、例えば、ガラスキャピラリー等を挙げることができる。
本発明の核酸分子の導入方法において、好ましい態様としては、上記のカイコ卵に物理的又は化学的に穴を空け、核酸分子入用の管を挿入角度が該卵の腹側の側面に対してほぼ垂直となるように該穴から卵内に挿入し、核酸分子を注入する工程を、針と核酸分子注入用の管が一体型となったマニュピュレーターを使用して行う。好ましい実施形態では、本発明は、通常、該マニュピュレーターを構成要素の1つとする装置を使用して実施される。
このような装置としては、解剖顕微鏡、照明装置、可動式のステージ、顕微鏡に金具で固定した粗動マニュピュレーター、このマニュピュレーターに付けたマイクロマニュピュレーター、核酸分子を注射するための空気圧を調整するインジェクターから構成されている。インジェクターに用いる圧力は窒素ボンベから供給され、圧力のスイッチはフットスイッチによっていれることができる。注射はガラススライド等の基板上に固定した卵に対して行い、卵の位置は移動式のステージによって決める。また、マイクロマニュピュレーターのガラスキャピラリーは4本のチューブで繋がれた操作部によって操作する。実際の手順は、卵に対するタングステン針の位置を粗動マニュピュレーターで決め、ステージのレバーで水平方向に卵を動かし穴を空ける。続いて、マイクロマニュピュレーターの操作部のレバーを操作して、穴の位置にガラスキャピラリーの先端を誘導し、再びステージのレバーによりキャピラリーを卵に挿入する。この場合、卵の腹側の側面に対し垂直にガラスキャピラリーが挿入される必要がある。フットスイッチを入れ核酸分子を注射し、レバーを操作して卵からキャピラリーを抜く。空けた穴を瞬間接着剤等でふさぎ、一定の温度および、一定の湿度のインキュベーターで保護する。本発明に使用される装置としては、好適には、特許第1654050号に記載の装置又は該装置を改良した装置が挙げられる。
また、本発明の1つの実施形態においては、核酸分子の導入に用いるカイコ卵が基板に固定されていてもよい。本発明の基板として、例えば、スライドグラス、プラスチック板等を用いることができるが、これらに特に制限されない。本発明の上記態様においては、カイコ卵内の将来的に生殖細胞になる位置に正確に核酸分子を注射するために、卵の方向を揃えて固定することが望ましい。また、上記態様においては、基板へ固定するカイコ卵の数には、特に制限はない。また、複数個のカイコ卵を用いる場合、カイコ卵を基板へ固定する方向性としては、好ましくは背腹の向きが一定となるような方向である。本発明の上記カイコ卵の基板への固定は、例えば、水性の糊をあらかじめ塗布した市販の台紙(バラ種台紙)の上に産卵させ、台紙に水を加えて卵をはがし、次いで濡れた状態の卵を基板に整列させ、風乾することによって行う。卵はスライドグラス上に卵の方向を揃えて固定することが好ましい。また、卵の基盤への固定は両面テープや接着剤等を用いることによっても可能である。
カイコ卵に核酸分子が導入されたか否かは、例えば、注射した核酸分子を卵から再度抽出して測定する方法(Nagaraju, J., Kanda, T., Yukuhiro, K., Chavancy, G., Tamura, T. and Couble, P.(1996)Attempt of transgenesis of the silkworm(Bombyx mori L) by egg-injection of foreign DNA. Appl. Entomol. Zool., 31, 589-598)や、注射した核酸分子の卵内での発現を見る方法(Tamura, T., Kanda, T., Takiya, S., Okano, K. and Maekawa, H. (1990). Transient expression of chimeric CAT genes injected into early embryos of the domesticated silkworm, Bombyx mori. Jpn. J. Genet., 65, 401-410)等によって確認することができる。
1つの局面では、本発明は、本発明の方法を実現するために、システムを構築することができる。
本明細書において、「システム」とは、検出、予測診断、事前診断、診断等を行うための任意の系をいい、一般に、1または複数の構成要素からなり、複数の構成要素がある場合それらの要素は互いに作用・関連し合っており、全体として調和のとれた挙動・機能を示すという3条件を満足する系をいう。システムは、装置、組成物、診断薬など任意の形態であり得る。従って、システムは、例えば、測定装置を備える大掛かりなシステムから、クロマトグラフィーを備えるシステム、免疫反応を利用したキット、抗体を含む組成物(すなわち、マーカーのモノクローナル抗体を含む、体外医薬品である診断薬)などを包含することが理解される。
1つの実施形態では、本発明は、被験体由来のサンプル中のAGEsを検出するためのsRAGEを含む、被験体がAGEsに関連する疾患(例えば、糖尿病、糖尿病性腎症、糖尿病性網膜症、糖尿病性神経症等の糖尿病合併症等)であるかどうか事前診断または診断するためのシステムまたは組成物を提供する。これらの組成物またはシステムは、上記AGEsを同定することができる限り、任意の因子または手段を用いることができることが理解され得る。従って、本明細書において具体的に記載された因子または手段のみならず、当該分野において公知の任意の等価の因子または手段を用いることができることが理解される。
1つの実施形態では、本発明において使用される因子は、核酸分子、ポリペプチド、脂質、糖鎖、有機低分子およびそれらの複合分子からなる群より選択され、好ましくは、因子は、タンパク質または複合分子(例えば、糖タンパク質、脂質タンパク質など)である。好ましくは、因子は、抗体(例えば、ポリクローナル抗体またはモノクローナル抗体)である。このような因子は、標識されるか、または標識可能であることが好ましい。なぜなら、診断することが容易となるからである。
本発明の好ましい実施形態において、使用される手段は、質量分析装置、核磁気共鳴測定装置、X線解析装置、SPR、クロマトグラフィー(例えば、HPLC、薄層クロマトグラフィー、ガスクロマトグラフィー)、免疫学的手段(例えば、ウェスタンブロッティング、ELISA、RIA)、生化学的手段(例えば、pI電気泳動、サザンブロッティング、二次元電気泳動)、電気泳動機器、化学的分析機器、蛍光二次元ディファレンシャル電気泳動法(2DE-DIGE)、同位体標識法(ICAT)、タンデムアフィニティ精製法(TAP法)、物理学的手段、レーザーマイクロダイセクションおよびこれらの組み合わせからなる群より選択される。
本発明の好ましい実施形態では、本発明のシステムは、さらに、AGEsの標準を含む。このような標準は、AGEsの検出手段が正常に機能しているかどうかを確認するために用いることが好ましい。
好ましい実施形態では、本発明では、対象となるサンプルを精製する手段をさらに備え得る。このような精製手段としては、例えば、クロマトグラフィーなどを挙げることができる。精製することによって、診断の精度を上げることができることから、好ましい実施形態において使用され得るが、これは必須ではない。
本発明において、被験体は、哺乳動物を含み、1つの実施形態では、被験体は、齧歯類を含む。このような齧歯類(例えば、ラット、マウスなど)は、モデル動物、特に、糖尿病または糖尿病性腎症のモデル動物(例えば、ストレプトゾシン(Stz)マウス等)が作製されていることから好ましい。別の好ましい実施形態では、被験体は、ヒトを含む。
1つの実施形態では、本発明において使用される因子または手段は、本発明のAGEの定量をする能力を有する。このような定量は、標準曲線を描いたときに、検量線がきちんと描ける手段または因子であるものがよい。好ましくは、例えば、抗体、質量分析、クロマトグラフィー分析などを挙げることができる。従って、ある実施形態では、本発明のシステムは、AGEsの定量を行うための定量手段をさらに備える。
1つの実施形態では、定量手段は、標準曲線と測定結果とを比較して前記AGEsが正常値の範囲内かどうかを判定する判定手段を含む。このような判定手段は、コンピュータを用いて実現することができる。本発明の検出または診断において、AGEsの濃度を測定する方法は、そのAGEsの濃度を特異的に測定できる方法であれば、タンパク質の定量に一般に用いられている方法をそのまま用いることができる。例えば、各種のイムノアッセイ、質量分析(MS)、クロマトグラフィー、電気泳動等を用いることができる。
本発明の検出または診断における好ましい実施形態の一つは、AGEsを担体上に捕捉し、その捕捉されたAGEsの濃度を測定することである。すなわち、AGEsに対する親和性を有する物質を担体に固定化し、その親和性を有する物質を介してAGEsを担体上に捕捉する。本実施形態によれば、サンプル中に含まれる夾雑物質の影響を低減させることができ、より高感度かつ高精度でAGEsの濃度を測定することができる。検出の方法としては、公知技術、例えばEIA(エンザイムイムノアッセイ)、RIA(ラジオイムノアッセイ)、ELISA(酵素連結イムノソルベントアッセイ)等が挙げられる。
本実施形態においてAGEsの測定方法にイムノアッセイを用いる場合は、抗体を固定化した担体を用いることが好ましい。このようにすれば、担体に固定化された抗体を1次抗体としたイムノアッセイの系を簡単に構築することができる。例えば、AGEsまたはsRAGEに特異的でエピトープの異なる2種類の抗体を用意し、一方を1次抗体として担体に固定化し、他方を2次抗体として酵素標識し、サンドイッチEIAの系を構築することができる。その他、結合阻止法や競合法によるイムノアッセイの系も構築可能である。さらに、担体として基板を用いる場合は、抗体チップによるイムノアッセイが可能である。抗体チップによれば、複数のマーカーの濃度を同時に測定でき、迅速な測定が可能である。したがって、本発明は、sRAGEが基板に固定化されている試薬を提供することができる。このような試薬を用いて、AGEsを検出するための組成物も提供することができる。
一方、本実施形態においてAGEsの測定方法に質量分析を用いる場合は、抗体の他、イオン結合や疎水性相互作用によってAGEを担体に捕捉することもできる。イオン結合や疎水性相互作用は抗原と抗体等のバイオアフィニティほどの特異性がなく、AGEs以外の物質も捕捉されるが、質量分析によれば分子量を反映した質量分析計スペクトルによって定量するので、問題はない。特に、担体として基板を使用したプロテインチップを用い、表面エンハンス型レーザー脱離イオン化(surface-enhanced laser desorption/ionization)-飛行時間質量分析(time-of-flight mass spectrometry)(以下、「SELDI-TOF-MS」と称する)を行えば、AGEsの濃度をより正確に測定することができる。使用できる基板の種類としては、陽イオン交換基板、陰イオン交換基板、順相基板、逆相基板、金属イオン基板、抗体基板等を用いることができるが、陽イオン交換基板、特に弱陽イオン交換基板と、金属イオン基板が好ましく用いられる。
<ビオチン化タンパク質の製造>
(ビオチンリガーゼを共発現するカイコ)
本発明の1つの局面において、標的タンパク質をコードする核酸分子およびビオチンリガーゼを共発現可能に組み込んだカイコまたはカイコと同様の糖鎖を付与する生物を提供する。
標的タンパク質、例えばsRAGEをコードする核酸配列には、ビオチン化タグ配列が含まれているため、ビオチンリガーゼを共発現させることによって、標的タンパク質がビオチン化を受ける。1つの実施形態において、ビオチンリガーゼは、BirA(配列番号10)である。ビオチン化を受けるタグ配列としては、BioEase.tag、Avi.tag、ビオチン化を受け得る任意の配列が挙げられるが、これらに限定されない。これらのカイコはビオチンの経口投与によりビオチン化sRAGEを効率的に製造することができる。また、得られたsRAGEの糖鎖構造解析により、(Man)5(GlcNAc)2を主とするオリゴマンノース型糖鎖が90%以上を占め、数%のコンプレックス型、および、ハイブリッド型の糖鎖を含むことを明らかにした。さらに糖鎖付加により安定性が向上し(1年以上安定)、ビオチンを介した方向性を維持した固定化により、多様な構造のAGEsなどの検出が可能である。
(ビオチンリガーゼを共発現するカイコよるビオチン化タンパク質の製造)
本発明の別の局面において、ビオチン化タンパク質の製造方法であって、A)カイコまたはカイコと同様の糖鎖を付与する生物に、ビオチンリガーゼおよびタンパク質をコードする核酸分子を共発現可能に組み込む工程;B)該核酸分子が発現する条件下に該カイコまたはカイコと同様の糖鎖を付与する生物を配置し、該ビオチンリガーゼおよび該タンパク質を発現させる工程;およびC)該生物にビオチンを投与し、ビオチン化された該タンパク質を得る工程を包含する、方法を提供する。本発明のカイコ型ビオチン化タンパク質の製造方法は、目的のタンパク質(例えば、sRAGEなど)を可溶性タンパク質として、高いビオチン化効率でビオチン化された状態で、数日以内に大量に発現させることが可能であるという優れた効果を有する。さらに、カイコまたはカイコと同様の糖鎖を付与する生物を用いて発現された糖鎖付加タンパク質は、糖鎖付加が行われない大腸菌発現系を用いたタンパク質よりも、pH安定性に優れ、脱塩処理によって凝集する率が低く、種々の緩衝液を適用することができるため、活用範囲が広い。また、得られたsRAGEの糖鎖構造解析により、(Man)5(GlcNAc)2を主とするオリゴマンノース型糖鎖が90%以上を占め、数%のコンプレックス型、および、ハイブリッド型の糖鎖を含むことを明らかにした。さらに糖鎖付加により安定性が向上し(1年以上安定)、ビオチンを介した方向性を維持した固定化により、多様な構造のAGEsなどの検出が可能である。
カイコへのビオチンの投与は、ビオチンを添加した餌の経口投与が好ましいが、注射などの方法により投与することも可能である。
本発明の好ましい実施形態では、前記A)工程は、ビオチンリガーゼおよびタンパク質をコードする核酸分子を含む発現ベクターをマイクロインジェクションすることで達成される。もちろん、マイクロインジェクション以外でも、本明細書において説明し、あるいは他の公知の任意の手法で発現ベクターを導入することができることが理解される。
(カイコ型ビオチン化sRAGE)
別の局面において、本発明は、高い効率でビオチン化されたsRAGEを提供する。高効率のビオチン化に起因して、ビオチンを介した方向性を維持した基板等への固定が可能であり、多様な構造のAGEsの検出が可能である。1つの実施形態において、本発明のsRAGEのビオチン化効率は、約10%以上、約20%以上、約30%以上、約40%以上、約50%以上、または約60%以上である。好ましい実施形態では、本発明のsRAGEのビオチン化効率は、約60%以上である。得られたsRAGEの糖鎖構造解析により、(Man)5(GlcNAc)2を主とするオリゴマンノース型糖鎖が90%以上を占め、数%のコンプレックス型、および、ハイブリッド型の糖鎖を含むことを明らかにした。さらに糖鎖付加により安定性が向上し(1年以上安定)、ビオチンを介した方向性を維持した固定化により、多様な構造のAGEsなどの検出が可能である。
1つの実施形態において、本発明のsRAGEは、カイコ型糖鎖付加を受けている。本発明の特定の実施形態では、本発明のsRAGEは、糖鎖付加を受け得る部位が2か所存在する配列番号3のアミノ酸配列を有する。これらの糖鎖付加部位に付加される糖鎖は同一であってもよいし、異なってもよい。糖鎖が異なる場合は、付加される糖鎖は、上記(1)~(8)([化1]~[化8])の糖鎖の任意の組み合わせであり得る。
<カイコによる一本鎖抗体の生産>
1つの実施形態において、本発明によって製造されたタンパク質は、カイコ型糖鎖の付加を受け得る。付加される糖鎖としては、上記トリマンノシルコア、複合型、オリゴマンノース型およびハイブリッド型が挙げられ、好ましくは、上記(1)~(8)([化1]~[化8])の構造のいずれかを有する糖鎖である。
(一般技術)
本明細書において用いられる分子生物学的手法、生化学的手法、微生物学的手法は、当該分野において周知であり慣用されるものであり、例えば、Sambrook J.et al.(1989).Molecular Cloning: A Laboratory Manual,Cold Spring Harborおよびその3rd Ed.(2001);Ausubel,F.M.(1987).Current Protocols in Molecular Biology,Greene Pub.Associates and Wiley-Interscience;Ausubel,F.M.(1989).Short Protocols in Molecular Biology: A Compendium of Methods from Current Protocols in Molecular Biology,Greene Pub.Associates and Wiley-Interscience;Innis,M.A.(1990).PCR Protocols: A Guide to Methods and Applications,Academic Press;Ausubel,F.M.(1992).Short Protocols in Molecular Biology: A Compendium of Methods from Current Protocols in Molecular Biology,Greene Pub.Associates;Ausubel,F.M.(1995).Short Protocols in Molecular Biology: A Compendium of Methods from Current Protocols in Molecular Biology,Greene Pub.Associates;Innis,M.A.et al.(1995).PCR Strategies,Academic Press;Ausubel,F.M.(1999).Short Protocols in Molecular Biology: A Compendium of Methods from Current Protocols in Molecular Biology,Wiley,and annual updates;Sninsky,J.J.et al.(1999).PCR Applications: Protocols for Functional Genomics,Academic Press、別冊実験医学「遺伝子導入&発現解析実験法」羊土社、1997などに記載されており、これらは本明細書において関連する部分(全部であり得る)が参考として援用される。
人工的に合成した遺伝子を作製するためのDNA合成技術および核酸化学については、例えば、Gait,M.J.(1985).Oligonucleotide Synthesis:A Practical Approach,IRLPress;Gait,M.J.(1990).Oligonucleotide Synthesis:A Practical Approach,IRL Press;Eckstein,F.(1991).Oligonucleotides and Analogues:A Practical Approac,IRL Press;Adams,R.L.et
al.(1992).The Biochemistry of the Nucleic Acids,Chapman&Hall;Shabarova,Z.et al.(1994).Advanced Organic Chemistry of Nucleic Acids,Weinheim;Blackburn,G.M.et al.(1996).Nucleic Acids in Chemistry and Biology,Oxford University Press;Hermanson,G.T.(I996).Bioconjugate Techniques,Academic Pressなどに記載されており、これらは本明細書において関連する部分が参考として援用される。
本明細書において引用された、科学文献、特許、特許出願などの参考文献は、その全体が、各々具体的に記載されたのと同じ程度に本明細書において参考として援用される。
以上、本発明を、理解の容易のために好ましい実施形態を示して説明してきた。以下に、実施例に基づいて本発明を説明するが、上述の説明および以下の実施例は、例示の目的のみに提供され、本発明を限定する目的で提供したものではない。したがって、本発明の範囲は、本明細書に具体的に記載された実施形態にも実施例にも限定されず、特許請求の範囲によってのみ限定される。
(製造例1:遺伝子組換えカイコによる中部絹糸腺におけるビオチン化sRAGEの生産)
手短に述べると、sRAGEベクターをカイコ卵にマイクロインジェクションを行い、RAGE発現カイコを得た。さらに、中部絹糸腺で発現可能な系統と交配し、中部絹糸腺(抽出が容易で純度も高い)にsRAGEを発現する遺伝子組換えカイコを作出した。さらにビオチンリガーゼを共発現させた遺伝子組換えカイコにビオチンを含む餌を与えることによりビオチン化sRAGEを生産する。遺伝子組換えカイコによるsRAGEの生産方法は、参照により本明細書に組み込まれる国際公開第2016/051808号にも詳細に記載されている。
(材料および方法)
(系統作製、発現等)
pBac[Ser-UAS/3xP3EGFP](Ken-ichiro Tatematsu, Isao Kobayashi, Keiro Uchino, Hideki Sezutsu, Tetsuya Iizuka, Naoyuki Yonemura, Toshiki Tamura, Transgenic Research, 19, 473 (2010))のUAS(Upstream Activation Sequence)配列の下流に、PCRで増幅したフィブロインH鎖のシグナルペプチドをコードする断片およびsRAGEをコードする断片を挿入することによりUASベクターを構築した。この発現ベクターを用いて遺伝子組換えカイコを作出した。農業生物資源研究所で維持されている白眼・白卵・非休眠系統のw1-pnd系統を宿主系統として用いた。得られた遺伝子組換えカイコを中部絹糸腺でGAL4を発現する系統(Ken-ichiro Tatematsu, Isao Kobayashi, Keiro Uchino, Hideki Sezutsu, Tetsuya Iizuka, Naoyuki Yonemura, Toshiki Tamura, Transgenic Research, 19, 473 (2010))のUAS(Upstream Activation Sequence)と交配した。得られた次世代カイコのうち、GAL4コンストラクトとUASコンストラクトを共に持つ個体を選抜マーカーにより選抜した。5齢6日目の幼虫を解剖し、中部絹糸腺を摘出した。1本当たり1mLのPBS+1% Triton X-100の抽出液で、4℃、2時間振とうすることにより、タンパク質を抽出した。
(精製)
凍結融解処理により中部絹糸腺抽出物からセリシンを除去した溶解液をPBSで平衡化したTALON レジン(Coを使用した金属キレートアフィニティクロマトグラフィレジン)に結合させ、PBS中のイミダゾール濃度を段階的に上げることによりsRAGEを溶出した。50mM~500mMイミダゾール溶出画分を収集しPBS(-)で透析した。さらにPBSで平衡化したMutein Matrixに結合させ、1.5mM~3mMのビオチンを含むPBSで溶出させた画分をPBS透析し、ビオチン化sRAGEを精製した。
(配列)
本実施例で製造したsRAGEの配列は、配列番号3に示される。カイコの中部絹糸腺発現タンパク質のN結合型糖鎖の構造は、以下の通りであり、sRAGEはカイコ中部絹糸腺に特異的に発現させている。中部絹糸腺に発現させた糖タンパク質の糖鎖は、中部絹糸腺以外では認められるバウチマンノース型(フコースあり)糖鎖や高マンノース型糖鎖は殆ど無く、複合型糖鎖、ハイブリッド型糖鎖もしくは、オリゴマンノース型糖鎖である。
(製造例2:遺伝子組換えカイコによる後部絹糸腺におけるsRAGEの生産)
本製造例では、後部絹糸腺においてsRAGEを発現させた。後部絹糸腺、または、後部/中部絹糸腺両者からのsRAGEの製造は、製造例1に準じて行ったが、絹糸腺1本当たりの抽出使用するバッファー量を5-10倍にすることにより、抽出量の増大が確認された。
(結果)
結果を図2および図3に示す。図2に示されるように、後部絹糸腺においてsRAGEが十分に発現されたことが示された。また、図3に示されるように、sRAGEがビオチン化されていることが示された。
(実施例1:刺激性AGEsの検出)
本実施例では、代表的な刺激性AGEs(GO、MGO、Glycer、Glycol処理OVA)の検出を行った。実施例1におけるアッセイ系の概要を図4に示す。
(材料および方法)
本実施例1で使用するアッセイ系では、sRAGEがウェル上に固定化されている。試料中の競合用AGEsと刺激性AGEsが競合的に固定化されたsRAGEに結合し、刺激性AGEsが多いほど、競合用AGEsの結合量が減少する。競合用AGEsに特異的な検出試薬を用いて発色させ、吸光度が少ないほど(競合用AGEsが少ないほど)、試料中の刺激性AGEsが多きことを示す。あらかじめ、検量線を作製しておき、吸光度を濃度に換算することで、試料中の刺激性AGEsの濃度を決定することができる。
・固定化(50μl/ウェル)
上記製造例において製造されたsRAGE(B. mori sRAGE)(5μl/ウェル/TBS)を4℃で一晩固定化した。アルカリ側のpHに長時間放置するとsRAGEが失活するため、固定化におけるバッファーとしてPBS(-)を使用した。その後、TBST(200μl/ウェル)で3回洗浄した。
・ブロッキング(250μl/ウェル)
Protein free blocking buffer(Thermo fisher scientific)を使用して、室温で2時間ブロッキングを行った。その後、TBST(200μl/ウェル)で3回洗浄した。
・競合的結合
試料および競合用AGEsをウェルに添加し(100μl/ウェル)、室温で2時間インキュベートした。試料は、各種還元糖処理OVA(最終濃度1~1500ng/ml PBS(-))を含む。競合用AGEsは、フルクトースまたはグルコース糖化BSA(最終濃度100ng/ml PBS(-))を使用した。その後、TBST(200μl/ウェル)で5回洗浄した。
競合用AGEsに対する一次抗体(100μl/ウェル)を添加し、室温で1時間インキュベートした。一次抗体は、抗BSA(マウスモノクローナル)(Pierceブロッキングバッファーで1:6000希釈)を用いた。次いで、TBST(200μl/ウェル)で5回洗浄した。さらにHRPで標識された二次抗体(100μl/ウェル)を添加し室温で1時間インキュベートした。次いで、TBST(200μl/ウェル)で5回洗浄した。
最後に、HRPの基質であるTMB(50μl/ウェル)を添加して25℃で10分インキュベートして発色させ、1N HCl(50μl/ウェル)により反応を停止した。450nmのO.D.値を測定した。
(結果)
図5に結果を示す。競合法のため刺激性AGEs量が多いほど、競合用AGEsがsRAGEに結合する量が少なくなりOD値は低くなる。G-BSAとの競合により、いずれのAGEsも100ng/ml以上の濃度で十分に検出可能なことが確認された(GlycerおよびGlycol処理AGEsは、10ng/ml程度でも検出可能であった。過去の報告から、ヒト検体中のAGEsの検出には、500~3000ng/mlの測定範囲が必要と思われ(村本弘昭、武藤寿生、竹内正義著、透析会誌46(5), 467-473;およびTakeuch et al., PLoS One. 2015; 10(3): e0118652.)、本発明のアッセイ系では、非常に低い濃度で感度良く検出することができることが明らかになった。
(実施例2:試料中の成分がアッセイ系に与える影響の確認)
本実施例では、血漿または血清中のAGEs以外の成分が、アッセイ系の能力に影響を与えないかを確認した。本実施例では、実施例1のアッセイ系を使用した。また、試料として希釈血漿または血清を使用した。
(結果)
図6に示されるように、希釈率1/20~1/1000において、OD値がほぼ一定であり、血漿を含まない試料と検出能力にほとんど差がないことが明らかになった。血清を使用した場合でも、同様の結果が得られた。このように、本発明のアッセイ系は、生体試料中においても感度良くAGEsを検出することができる。
(実施例3:血清存在下での代表的な刺激性AGEsの検出)
本実施例では、20倍希釈したベトナム人血清存在下で、代表的な刺激性AGEsの測定範囲/検出感度を確認した。検出は、実施例1と同様に行った。試料は、各還元糖処理OVA(GO、Glycer、MGO、Glycol処理OVA)を、PBS(-)で20倍に希釈したヒト血清中に溶解することによって調製された(100μl/ウェル)。
(結果)
結果を図7に示す。図7の横軸の各AGEsの濃度は、アッセイ系における最終濃度を示す。本発明のアッセイ系は、ヒト血清中のAGEsの検出にも使用可能であることが示された。
(実施例4:捕捉用AGEsを固定化したアッセイ系の構築)
実施例1では、sRAGEをウェルに固定化した系で検出を行ったが、本実施例では、捕捉用AGEsをウェルに固定化した場合のアッセイ系で検出を行うことができるか検討した。
(材料および方法)
・固定化(100μl/ウェル)
PGS(-)で希釈された捕捉用AGEs(G-BSAまたはF-BSA、0.1μg/ml)を4℃で一晩固定化し、TBS-0.05%Tween20(TBST)(200μl/ウェル)で3回洗浄した。
・ブロッキング(250μl/ウェル)
Protein free blocking buffer(Thermo fisher scientific)を使用して、37℃で2時間ブロッキングを行った。その後、TBST(200μl/ウェル)で3回洗浄した。
・競合的結合
試料(100μl/ウェル)およびビオチン化B.mori sRAGE(0.05μg/ml/20倍希釈血清)をウェルに添加し、4℃で一晩インキュベートした。試料は、各種AGEs(20倍希釈血清)を含む。次いで、TBST(200μl/ウェル)で5回洗浄した。
Pierceブロッキングバッファーで希釈されたHRPコンジュゲートストレプトアビジン(50μl/ウェル)を添加し、室温で30分間インキュベートし、TBST(200μl/ウェル)で5回洗浄した。HRPの基質であるTMB(50μl/ウェル)を添加し室温で10分間インキュベートして発色させ、1N HCl(50μl/ウェル)により反応を停止した。450nmのO.D.値を測定した。
(結果)
捕捉用AGEsをウェルに固定化した系においても、実施例1と同様、刺激性AGEsを検出することが可能であることが示された(図8A~図8D)。
(実施例5:マウス血清/血漿のアッセイ系に与える影響の確認)
本実施例では、実験動物(マウス)血清/血漿のアッセイ系に与える影響の確認を行った。
(材料および方法)
実験動物として、7週齢のマウス(雄、雌)を使用した。ヘパリン処理した注射筒を用い、心臓採血後に血液を2分した。半量は血漿分離財を用いて血漿の調製を行い、残りは、30分間静置し血液を凝固させた後に遠心して血清を調製した。調製した血漿および血清は、使用時まで-80℃で保存した(試料は凍結・融解を繰り返さない)。
・固定化(100μl/ウェル)
PGS(-)で希釈された捕捉用AGEs(G-BSAまたはF-BSA、0.1μg/ml)を4℃で一晩固定化し、TBS-0.05%Tween20(TBST)(200μl/ウェル)で3回洗浄した。
・ブロッキング(250μl/ウェル)
Protein free blocking buffer(Thermo fisher scientific)を使用して、37℃で2時間ブロッキングを行った。その後、TBST(200μl/ウェル)で3回洗浄した。
・競合的結合
試料(100μl/ウェル)およびビオチン化B.mori sRAGE(0.05μg/ml/20倍希釈血清)をウェルに添加し、4℃で一晩インキュベートした。試料は、PBS(-)で希釈したマウス血清または血漿である。次いで、TBST(200μl/ウェル)で5回洗浄した。
Pierceブロッキングバッファーで希釈されたHRPコンジュゲートストレプトアビジン(50μl/ウェル)を添加し、室温で30分間インキュベートし、TBST(200μl/ウェル)で5回洗浄した。HRPの基質であるTMB(50μl/ウェル)を添加し室温で10分間インキュベートして発色させ、1N HCl(50μl/ウェル)により反応を停止した。450nmのO.D.値を測定した。
(結果)
血清を使用した場合、200倍希釈でわずかな検出の阻害効果が見られたが、この阻害効果を考慮して、血清中の刺激性AGEsの検出が可能であることが示された(図9A)。一方、血漿を使用した場合、200倍希釈でも顕著な阻害が観察されたため、200倍以上の希釈についても試験した(図9B)。1000倍以上の希釈率の場合、阻害効果を考慮して血漿中の刺激性AGEsの検出が可能であることが示された。
(実施例6:牛乳および乳酸醗酵飲料を対象にした検出)
本実施例では、AGEsの生成が予想される牛乳およびその加工品である乳酸発酵飲料AおよびBを対象に検出を行った。使用したアッセイ系は、実施例4に準ずる。試料は、ビオチン化B.mori sRAGE(0.05μg/ml/20倍希釈血清)およびPBS(-)で希釈した牛乳または乳酸発酵飲料を含む。
(結果)
動物由来の食品中のAGEsの検出が可能であることが示唆された(図10)。食品の加熱調理・加工過程でAGEsが生じることはメイラード反応として以前から知られていた。食品中のAGEsは風味などの付加価値として捉えられていたが、近年、食品中のAGEsの一部が加齢性疾患の原因となることが指摘されていることから、風味は変わらず、加齢性疾患の原因となるAGEsのみを減らすような加工法の開発、AGEsを減らす効果のある食材の活用などが求められている。本発明のアッセイ系は、その際の食品中、加工過程の刺激性AGEsの検出、抗糖化能が期待される農林水産物などの抗糖化能の評価への応用が可能であることが示された。
(実施例7:アッセイ系の条件の最適化)
本実施例では、よりよい再現性のために、アッセイ系の条件の最適化を行った。
固相化するタンパク質の量、固相化後の安定性は、アッセイの感度、ダイナミックレンジを決める重要な要素とされている。アルカリ側のpHで処理した方が、汎用されている全ての基板の材質(ポリスチレンなど)に依らず、タンパク質の固相化量が向上し、かつ、固相化後の剥離が生じ難いとされており、抗体などの比較的安定なタンパク質を固相化する場合には、炭酸バッファー(pH9.2~10.6)が汎用されている。最も重要なのは、固相化後のタンパク質が、検出作業の過程で剥離する等の問題がなく、固相化されたタンパク質が安定していることである。一方、固相化量は多ければ良い訳ではなく、固相化反応時のタンパク質濃度を検討する等により、最適な固相化量(方法)を決定する必要がある。
AGEsは、抗体と異なり、アルカリ処理に対して構造が安定か、アルカリ処理後に活性を失わず機能するか不明であったため、実施例4では、AGEsの構造/機能に影響を与えないことを確認されたPBS(-)で希釈して固相化反応を行った。刺激性AGEsの検出には十分に耐えうるものであったが、基板に固相化されたAGEsが一定ではない(固相化量が異なる、検出過程でAGEsが剥離する)ことが予測された。そこで、固相化条件を検討したところ、アルカリ条件(例えば、pH9.6)にて固相化が強固に行われ、検出作業過程での剥離が生じ難くなり、1晩程度アルカリ条件に曝されても、固相化後にPBS(-)にて洗浄することにより固相化されたAGEsは構造(機能)を維持していることが確認された。本実施例では、炭酸ナトリウムバッファー(pH9.6)中で固相化反応を行った。炭酸ナトリウムバッファー(pH9.6)を用いた場合、G-BSAが強固に固定化されよりよい再現性が示された。また、図11に示すように、pH9.6での固定化を行った場合、pH7.4で固定化を行った場合と比べて、検量線の近似曲線の相関係数「Rの二乗」が1に近づき、また、フィッティングが改善していることが確認された。このように、pH9.6での固定化を行った場合、測定値を得るのにより適した検量線が得られた。
また、ビオチン化sRAGEの濃度は、0.1μg/ml(反応液中の最終濃度)とした場合に、固相化したG-BSAとの最適な競合反応を行うことができ、よりよい再現性が示された(図12)。また、捕捉用AGEsは、0.9μg/mlとした場合に、測定値を得るのにより適した検量線が得られた(図13)。当業者であれば、アッセイ系におけるsRAGEおよび捕捉用AGEsの濃度は、適宜調節できるものであり、上記濃度に限定されるものではない。
(材料および方法)
本実施例におけるアッセイ系の概要を図14に示す。
・コーティング(100μl/ウェル)
炭酸バッファー(pH9.6)で希釈された0.9μg/mlの捕捉用AGEs(G-BSAまたはF-BSA)を4℃で一晩固定化した。200μl/ウェルのTBS-0.05%Tween20(TBST)で3回洗浄した。
・ブロッキング(250μl/ウェル)
Protein free blocking buffer(Thermo fisher scientific)を使用して、37℃で2時間ブロッキングを行った。その後、TBST(200μl/ウェル)で3回洗浄した。
・競合的結合
試料(100μl/ウェル)をウェルに添加し、4℃で一晩インキュベートした。試料は、ビオチン化B.mori sRAGE(0.1μg/ml/PBS)とマウス血清またはglycer-BSAを含む。次いで、TBST(200μl/ウェル)で5回洗浄した。
Pierceブロッキングバッファーで希釈されたHRPコンジュゲートストレプトアビジン(50μl/ウェル)を添加し、室温で30分間インキュベートし、TBST(200μl/ウェル)で5回洗浄した。HRPの基質であるTMB(50μl/ウェル)を添加し室温で10分間インキュベートして発色させ、1N HCl(50μl/ウェル)により反応を停止した。450nmのO.D.値を測定した。
本アッセイ系を使用して、実験動物(マウス)の血清中のAGEs濃度を追跡した。本実施例では、特に加齢性疾患との関係が高いと考えられるグリセルアルデヒド-BSA濃度当量(μg/ml)で表示した(図15)。グリセルアルデヒド処理BSAを用いて検量線を作成し、試料中の刺激性AGEsの濃度は、グリセルアルデヒド処理BSA濃度に換算して示すことにより、各試料中の刺激性AGEs濃度(μg/ml)とした。ヒト血清に関しては、15~50倍希釈で検量線の直線性のある部分に測定値が得られることが確認された(健常者、疾患の進行により異なるため、2段階希釈でデータを取得した)。
(結果)
図16Aおよび図16Bに示されるように、若年期、壮年期、老年期群において、マウス血清、血漿中のAGEsの濃度の測定が可能なことが明らかになった。さらに、経時的な変動の追跡が可能であることが示唆された。
(実施例8:動物、植物由来の食品、調理品中のAGEs測定)
実施例7のアッセイ系を用いて動物、植物由来の食品、調理品中のAGEsの測定が可能かどうかを検証した。
(材料および方法)
(1)豚肉の焼成及びサンプリング
ブタ肩ロース(各6枚)を各々非焼成群(3枚)と焼成群(3枚)の2群に分けて刺激性AGEsの定量試験を実施した。焼成試料の調製に先立ち、フライパン(和平フレイズ製、COM-9784)をIHヒーター(Panasonic製、KZ-PH33)に載せ、放射温度計(堀場製作所製、IT-550)をフライパン上部約50cmの高さの位置に固定した。これにより、フライパン表面温度が一定温度(約195℃)で安定したことを確認してから3枚の豚肉を順次焼成した。その際、試料をフライパン中央に静置して表面を3分間焼成してから、裏面を3分間焼成し、その後改めて表面及び裏面を各1分間焼成した。なお、焼成は素焼きとし、乾燥を防止するためにフライパンを蓋で覆って焼成した。加えて、合計6枚の焼成群試料を断続的に焼成するにあたり、試料交換時にフライパンに残存した油分をキッチンペーパーで除去した。
次に、焼成群及び非焼成群の試料1枚あたり9か所から革ポンチ(φ=12mm)を用いて肉片を採取した。採取した肉片は混合してからナイフで細断することによって均一化した。
(2)豚肉からのペプチドの抽出
均一化した試料から以下の手順でペプチドを抽出した。まず、均一化試料から1g程度を正確に秤量して、1gあたり9mLの8M尿素溶液(生理食塩水に溶解)を添加した。室温で1時間静置した後、試料懸濁液はホモジナイザー(IKA Ultra-Turrax, DT-20)を9000rpmで2分間駆動させることを2回繰り返すことにより破砕した。破砕試料から100μLを分け取ってから900μLの生理食塩水を添加して尿素濃度を低下させたのち、20mg/mLトリプシン(和光純薬)を10μL添加して、ローテーターで撹拌しながら37℃で終夜反応させた。反応終了後、試料を95℃で5分間静置することによってトリプシンを失活させ、遠心分離(15000rpm、15分、室温)を行うことにより、残渣を沈殿画分として除去した。一方、ペプチドが含まれる上清画分のうち850μLを回収し、フィルター(ADVANTEC、DISMIC-13、pore size 0.2μm)に通して清浄化した。こうして得られた試料に生理食塩水を加えて総量を1.2mLとし、そのうち1mLをHPLC(島津製作所)に供した。超純水/0.2%ギ酸で平衡化した逆相HPLCカラム(InertSustain C18カラム、GLサイエンス)に供して吸着させた後、移動相を100%アセトニトリル/0.2%ギ酸に交換することで溶出した画分を分取した。溶出画分は遠心濃縮後に200μLの超純水に溶解し、BCAタンパク質定量キット(タカラバイオ、T9300A)を用いて濃度検定を行ったところ、各試料から数mg/mLのペプチドが得られた。
得られたペプチドを実施例7に準ずるAGEs検出系に添加し焼成により生成するAGEsを測定した。
(結果)
図17に示されるように、Glycer-BSA当量に換算することにより、タレなどの添加の無い単なる焼成では、肉中のAGEs量に大きな差が無いことが確認できる。同様に、青汁、牛乳などの液体試料においても、刺激性AGEsの測定が可能であった。素焼きの場合、調理により加齢性疾患の原因となる刺激性AGEsが生成することは確認されなかった。
(実施例9:各種還元糖で糖化したBSAの測定)
本実施例では、各種還元糖(グルコース、フルクトース、ガラクトース、マルトース、ラクトース、キシロース、リボース、グリコールアルデヒド、グリオキサール、メチルグリオキサール)で糖化したBSAを測定した。
4μg/mLのBSA-AGEsと0.1μg/mlのsRAGEを競合させ、ウェルに固定化されたグルコース-BSAに結合したsRAGEを定量した。測定は、実施例7のアッセイ系に準じて行った。また、各糖化BSAをトリプシン処理し糖化ペプチドにした際の刺激性AGEsの定量も行った。トリプシン処理は、タンパク質の全体の構造がRAGEの認識に必須なのではなく、還元糖により修飾されたペプチドでもRAGE結合能(生体への刺激性)があるか否かを確認するために行った。
(結果)
結果を図18Aおよび図18Bに示す。糖化処理に用いる還元糖の種類により、sRAGEとの反応性の異なるAGEsが生じること、また、実施例7のアッセイ系により、各種還元糖で糖化したAGEsをGlycer-BSA当量で示すことが可能なことが確認された。また、トリプシン処理された糖化ペプチドも測定可能であることが確認できた。結果から、sRAGEの各種還元糖で糖化されたAGEsとの結合力は、おおよそ以下のとおりである:グルコース≒マルトース≒ラクトース<フルクトース<ガラクトース<リボース≒グリオキサール<キシロース≒グリコールアルデヒド≒メチルグリオキサール。
(注記)
以上のように、本発明の好ましい実施形態を用いて本発明を例示してきたが、本発明は、この実施形態に限定して解釈されるべきものではない。本発明は、特許請求の範囲によってのみその範囲が解釈されるべきであることが理解される。当業者は、本発明の具体的な好ましい実施形態の記載から、本発明の記載および技術常識に基づいて等価な範囲を実施することができることが理解される。本明細書において引用した特許、特許出願および文献は、その内容自体が具体的に本明細書に記載されているのと同様にその内容が本明細書に対する参考として援用されるべきであることが理解される。