JP4132810B2 - Na + / H + antiporter gene and improvement of salt tolerance by expression of the gene - Google Patents
Na + / H + antiporter gene and improvement of salt tolerance by expression of the gene Download PDFInfo
- Publication number
- JP4132810B2 JP4132810B2 JP2001390491A JP2001390491A JP4132810B2 JP 4132810 B2 JP4132810 B2 JP 4132810B2 JP 2001390491 A JP2001390491 A JP 2001390491A JP 2001390491 A JP2001390491 A JP 2001390491A JP 4132810 B2 JP4132810 B2 JP 4132810B2
- Authority
- JP
- Japan
- Prior art keywords
- antiporter
- apnhap
- cells
- dna
- activity
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Expired - Fee Related
Links
Images












Landscapes
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Peptides Or Proteins (AREA)
- Breeding Of Plants And Reproduction By Means Of Culturing (AREA)
Description
【0001】
【発明の属する技術分野】
本発明は、Na+/H+アンチポーター遺伝子、特に、耐塩性藍藻(らん色細菌:Aphanothece halopytica) 由来Na+/H+アンチポーター遺伝子、及び該遺伝子の発現による耐塩性の向上に関する。
【0002】
【従来の技術】
今日、世界中の耕作地の20%、灌漑地のほぼ半分が高塩度の被害を受けているといわれている。高塩度は植物のイオン不均衡および高浸透圧によるストレスを誘発する。高塩度環境で生き延びる生物は内部浸透圧を調節する特異な機序を有している。そのような機序のひとつが砂糖、ある種のアミノ酸、第四アンモニウム化合物などの低分子量有機化合物の溶質を蓄える能力である。高塩度に適応するもうひとつの機序はNa+/H+アンチポーターおよびNa+-ATPアーゼの一機能として提唱されている細胞からのNa+排出である。塩ストレスは、このような毒性作用のほかに、酸化ストレスも誘発する。従って、近年、遺伝子工学を利用して植物の耐塩性の改善が試みられている。
【0003】
Na+/H+アンチポーターは細胞膜を介してNa+をH+に交換する反応を触媒し(Na+/H+アンチポーター活性)、細胞質内のpH、細胞容積、細胞質内ナトリウム濃度の制御など種々の機能を果たしている(1-3)。Escherichia coliでは、3種のアンチポーター(NhaA, NhaB, ChaA)が知られており、その機能的特徴も十分に解明されている(1)。動物では6種の相同Na+/H+アンチポーター(交換)(NHE1-6)が判明している(2)。植物および酵母では、空胞型(NHX1およびAtNHX1)(4-6)と形質膜型(SOD2およびSOS1)(7,8)のNa+/H+アンチポーターが報告されている。E. coliアンチポーターと真核細胞アンチポーターの相同性はきわめて低く、Na+/H+アンチポーター遺伝子は別々に進化が独立したことが示唆される。
【0004】
このようなNa+/H+ アンチポーター遺伝子を用いると、植物の耐塩性が向上するという報告が、最近(1999年、Science、2001年Nature Biotechnology)にカナダBlumbald他によりなされた。Blumbald他はアラビドプシスの液胞膜のNa+/H+ アンチポーター遺伝子を、アラビドプシスの液胞膜(Science 1999)、トマトの液胞膜(2001 Nature Biotechnology)で発現させることにより、ともに0.2M NaCl中でも生育することを報告している。これは、細胞の中にある液胞膜の内側にNaClを蓄積させることにより耐塩性を向上させるものである。
【0005】
このように、遺伝子工学の技術によって植物の耐塩性を改善する少なからぬ努力が行われているが、実用化にこぎつけるほど十分な結果は得られていない。
さて、植物に比べると単細胞のらん食細胞は単純であり、光合成生物における耐塩性を担う因子を解明するためには適切なモデルである。藍藻は太陽エネルギーおよび無機体炭素CO2を利用して増殖できる酸素を発生する光合成生物である。このような生物はきわめて低濃度から海水よりも高濃度まで塩を含有する水中などほぼすべての生息地で確認される。しかし、どの遺伝子が異なる塩度への適応に関与するのか、不明である。そこで、高塩度に対応するマスター調節遺伝子を同定することは、藍藻の耐塩性の機序を理解するばかりでなく、作物の収穫量を向上させる点で植物への応用にも重要である。
【0006】
本発明者等はこれまでの試験で(9)、藍藻Synechocystis sp. PCC6803にはNa+/H+アンチポーターSynNhaP1が存在し、真核細胞および原核細胞(Pseudomonas aeruginosa由来NhaP)のアンチポーターと相同であるが、NhaA, NhaBおよびChaAとは相同でないことを明らかにしてきた。さらに、SynNhaPには膜貫通(TM)領域に保存されたAsp138と比較的長いC末端親水性テールがあり、いずれも担体活性に重要であることも示してきた。長いC末端テールは動物の輸送活性を制御する働きがあると考えられている(2,10)。このような知見から、藍藻のアンチポーターは真核細胞のNa+/H+アンチポーターの構造的特性と機能的特性を試験するためのモデル系になることがわかる。
【0007】
現在までに、Na+/H+アンチポーター蛋白質ではごくわずかな機能性アミノ酸残基、特に陽イオン輸送に関わるアミノ酸残基が特定されている。SynNhaPのAsp138(9)、NhaAのAsp133, Asp163, Asp164(11,12)の重要性が報告されている。Na+/H+アンチポーターはH+とNa+またはLi+との間を特異的に交換するが、H+とLi+との交換活性が弱いアンチポーターもいくつかある(13)。E. Coli ChaAはアルカリ性pHでCa2+とH+、Na+とH+の交換活性を有すると報告されているが、詳細に調べられたことがない。どの因子によってイオン特異性が決まるのか不明である。Na+/H+アンチポーターの陽イオン輸送のための保存配列を特定するためには、新たなイオン特異性を有するアンチポーターのクローニングが欠かせない。
【0008】
Aphanothece halophyticaは0.25〜3.0M NaClと広範な塩度条件下で増殖可能な耐塩性藍藻である(15,16)。A. halophyticaは高塩度で浸透圧保護物質グリシンベタインを蓄積し(15)、A. halophyticaのDnaK蛋白は、他のDnaK/Hsp70ファミリー(17)より長いC末端セグメントを有しており、高塩度できわめて強い蛋白質折りたたみ活性を示すことが報告されている(18)。A. halophytica由来DnaK蛋白によるタバコの形質転換によって、耐塩性が増強されることも判明している(19)。
【0009】
【発明が解決しようとする課題】
従って、本発明の目的は、遺伝子工学の技術によって植物の耐塩性を向上させることのできる有用な遺伝子を見出し、該遺伝子を利用して、耐塩性作物を開発、創出して食料生産の向上に寄与する方法を提供することである。
【0010】
【課題を解決するための手段】
そこで、本発明者は、A. halophyticaに独特のNa+/H+アンチポーター(ApNhaP)があるかどうか研究した結果、真核細胞に相同であるが、新たなイオン特異性を有するNa+/H+アンチポーター遺伝子を単離することに成功し、更にそのイオン特異性がC末端領域によって変化することも明らかにした。
【0011】
更に、淡水藍藻Synechococcus sp. PCC7942において8種のプラスミドを過剰発現させ、耐塩性を試験した。その結果、ベタイン合成、カタラーゼおよびシャペロンに関与する遺伝子を比較して、耐塩性藍藻由来のNa+/H+アンチポーター遺伝子を過剰発現することによって、このSynechococcus細胞を0.5M NaClを含有するBG11培地でも海水中でも増殖することが判明し、淡水藍藻の耐塩性を大きく改善することに成功した。
【0012】
即ち、本発明は、以下の各態様に係る。
(1)耐塩性藍藻 (Aphanothece halopytica) 由来Na+/H+アンチポーターをコードするDNA。
(2)全pH領域においてLi+/H+アンチポーター活性を実質的に示さず、中性及びアルカリ性pH領域においてCa2+/H+アンチポーター活性を示すことを特徴とする、藍藻由来Na+/H+アンチポーターをコードするDNA。
(3)藍藻が耐塩性藍藻 (Aphanothece halopytica)である、2記載のDNA。
(4)以下の(a)又は(b)のタンパク質をコードする塩基配列又はその部分配列を含有するDNA:
(a)配列番号2に示されるアミノ酸配列から成るタンパク質、
(b)アミノ酸配列(a)において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、Na+/H+アンチポーター活性を有するタンパク質。
(5)以下の(a)又は(b)の塩基配列又はその部分配列を含むDNA:
(a)配列番号1における第796〜2361番目の塩基対からなる塩基配列、
(b)塩基配列(a)からなるDNAとストリンジェントな条件下でハイブリダイズし、且つ、Na+/H+アンチポーター活性を有するタンパク質をコードする塩基配列。
(6)1乃至5のいずれか一項に記載のDNAを含有する組換え発現ビークル。
(7)大腸菌由来プラスミドベクターである6に記載の組換え発現ビークル。
(8)大腸菌/淡水藍藻シャトルベクターである6に記載の組換え発現ビークル。
(9)6、7又は8に記載の発現ビークルによって形質転換された形質転換体。
(10)細菌である9に記載の形質転換体。
(11)大腸菌である10に記載の形質転換体。
(12)大腸菌がアンチポーター欠損株である10に記載の形質転換体。
(13)Na+感受性表現型及びCa2+感受性表現型が相補された12に記載の形質転換体。
(14)藍藻である9に記載の形質転換体。
(15)藍藻が淡水性藍藻である14に記載の形質転換体。
(16)海水中で増殖可能な淡水性藍藻である15に記載の形質転換体。
(17)植物である9に記載の形質転換体。
(18)植物がイネ及びタバコから成る群から選択される、17に記載の形質転換体。
(19)Na+/H+アンチポーターを過剰発現することを特徴とする、14乃至18のいずれか一項に記載の形質転換体。
(20)Na+/H+アンチポーター活性が増加した、14乃至18のいずれか一項に記載の形質転換体。
(21)1乃至5のいずれか一項に記載のDNAを宿主細胞に導入し、該DNAを発現させることから成る、該宿主の耐塩性を向上させる方法。
(22)1乃至5のいずれか一項に記載のDNAを含有する組換え発現ビークルによって宿主を形質転換し、該DNAを宿主細胞内で発現させることから成る、該宿主の耐塩性を向上させる方法。
(23)細胞外にNaClを排出させることにより耐塩性を向上させる、21又は22に記載の方法。
(24)宿主が植物である、21、22又は23に記載の方法。
(25)植物がイネ及びタバコから成る群から選択される、24に記載の方法。
(26)1乃至5のいずれか一項に記載のDNAを宿主細胞に導入し、該DNAを発現させることから成る、耐塩性生物の創出方法。
(27)1乃至5のいずれか一項に記載のDNAを含有する組換え発現ビークルによって宿主を形質転換し、該DNAを宿主細胞内で発現させることから成る、耐塩性生物の創出方法。
(28)宿主が植物である、26又は27に記載の方法。
(29)植物がイネ及びタバコから成る群から選択される、28に記載の形質転換体。
(30)6乃至20に記載の形質転換体を培養し、得られる培養物からNa+/H+アンチポーターを単離及び精製することから成る、Na+/H+アンチポーターの調製方法。
(31)1乃至5のいずれか一項に記載のDNAにコードされるタンパク質。
(32)30に記載の調製方法で得られる組換えNa+/H+アンチポータータンパク質。
【0013】
【発明の実施の形態】
本発明の耐塩性藍藻 (Aphanothece halopytica) 由来Na+/H+アンチポーターをコードするDNAは、以下の実施例に示すように、公知である複数のNa+/H+アンチポーター間に高度に保存された2つのポリペプチド領域をもとにプライマーを作成し、一方、当業者に公知の方法でA. halophytica培養し、それから単離したゲノムを鋳型として、当業者に周知のPCRを用いて調製することが出来る。
【0014】
本発明DNAにコードされるNa+/H+アンチポーターは、全pH領域においてLi+/H+アンチポーター活性を実質的に示さず、中性及びアルカリ性pH領域においてCa2+/H+アンチポーター活性を示すことを特徴とする。
【0015】
より具体的には、本発明DNAは、以下の(a)又は(b)のタンパク質をコードする塩基配列又はその部分配列を含有するDNA:
(a)配列番号2に示されるアミノ酸配列から成るタンパク質、
(b)アミノ酸配列(a)において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、Na+/H+アンチポーター活性を有するタンパク質、である。
更に、本発明DNAは、以下の(a)又は(b)の塩基配列又はその部分配列を含むDNA:
(a)配列番号1における第796〜2361番目の塩基対からなる塩基配列、
(b)塩基配列(a)からなるDNAとストリンジェントな条件下でハイブリダイズし、且つ、Na+/H+アンチポーター活性を有するタンパク質をコードする塩基配列、である。
ここで、部分配列とは、例えば、上記Na+/H+アンチポーターのC末端領域(配列番号2に示されるアミノ酸配列における402番目のアミノ酸残基Gluから521番目のアミノ酸残基Gluまでの領域)に該当する配列を意味し、この部分はイオン特異性に関与する部分であることが本発明によって明らかにされた領域である。
【0016】
当業者であれば、本明細書で開示された配列情報に基づいて、当該技術分野における周知手段を用いた化学合成等によっても調製することが可能である。更に、当業者であれば、Na+/H+アンチポーター活性が実質的に保持されるように、配列番号2における特定のアミノ酸配列における1若しくは数個のアミノ酸を欠失、置換若しくは付加することも、当該技術分野における周知手段を用いて容易に実施することができる。Na+/H+アンチポーター活性が実質的に保持されるためには、アミノ酸配列における相同性が、65%以上、好ましくは80%以上、より好ましくは90%以上、更に好ましくは98%以上、特に好ましくは99.5%より高いものを挙げることができる
【0017】
本発明において、特定の遺伝子又はDNAは、当業者に周知の緩衝液中で、適当な温度及び塩濃度等の諸条件下のストリンジェントな条件でハイブリダイズさせることができる。このようなストリンジェントな条件下で本発明の遺伝子又はDNAとハイブリダイズし、且つ、Na+/H+アンチポーター活性と実質的に同等の活性を有する蛋白質をコードするDNAの例として、例えば、それぞれの対応する遺伝子と相同性が、90%以上、好ましくは98%以上、より好ましくは99.5%より高いようなDNAを挙げることができる。
【0018】
更に、本発明は別な態様として、上記の本発明遺伝子の少なくとも一つの遺伝子を含有する組換え発現ビークルに係る。組換え発現ビークルには各種ベクター等の当業者に公知の任意のものが含まれるが、特に、大腸菌由来の各種プラスミドベクター、大腸菌/淡水らん食細菌シャトルベクター、及びアグロバクテリウムのTiプラスミドベクターが好適である。
該組換え発現ビークルには、遺伝子組換え操作において当業者に公知である様々な配列、例えば、原核細胞である大腸菌等が有する転写因子である各種σサブユニットに対する結合ドメインである各種プロモーター、及びエンハンサーなどの各種転写調節要素、制限酵素部位、並びにカナマイシン耐性マーカー等の選択マーカー(マーカー酵素等)遺伝子を任意に含むことが出来、当業者に公知の方法によって容易に調製することが出来る。
【0019】
本発明の発現ビークルによって形質転換される宿主としては、特に、大腸菌及び藍藻等の各種細菌、並びにイネ、タバコ等の植物を挙げることが出来る。これらの形質転換体には、本発明のDNAが発現された結果、Na+感受性表現型及びCa2+感受性表現型が相補されたものや、Na+/H+アンチポーターが過剰発現された結果、Na+/H+アンチポーター活性が増加したもの等がある。
【0020】
本発明のDNAを宿主細胞に導入し、該DNAを発現させることによって、該宿主の耐塩性を向上させることが出来る。耐塩性の向上は、主に、Na+/H+アンチポーターによって細胞外にNaClが排出されることによるものである。導入方法としては、例えば、上記組換え発現ビークルによって宿主を形質転換することによって、本発明DNAを導入することが出来る。他にも、燐酸カルシウム共沈法、エレクトロポレーション法、及びパーティクルガン法等、当業者に公知の任意の手段及び条件を用いることが出来る。
【0021】
このようにして、本発明のDNAを宿主細胞に導入し、該DNAを発現させることによって、耐塩性生物、例えば、海水中でも増殖することが出来る淡水藍藻を遺伝子工学的手法によって創出することが出来る。
【0022】
更に、上記の形質転換体を培養し、得られる培養物からNa+/H+アンチポーターを単離及び精製することによって、Na+/H+アンチポーターを調製することが出来る。培養、単離及び精製等に関する諸条件は、使用するベクター、宿主及びその他の各因子の種類等に応じて当業者が適宜選択することが出来る。
【0023】
本発明は、上記の本発明DNAにコードされるタンパク質にかかる。このようなタンパク質は、当業者に公知の方法で、A. halophytica培養物から精製及び単離することができる。或いは、上記調製方法によって、得られる組換えタンパク質として、Na+/H+アンチポーターを得ることも可能である。
【0024】
【実施例】
以下、実施例により本発明をより具体的に説明するが、本発明はこれら実施例により何ら限定されるものではない。
【0025】
【実施例1】
A. halophytica 培養
A. halophytica細胞はBG11培地に18 mM NaNO3のほか、Turk Island塩溶液を加えて、光独立栄養的に増殖させた。Turk Islandの塩溶液は、培地のNaCl濃度を適宜0.25〜2.5Mの範囲に調節したことを除き、これまでに発表されている方法で調製した(15-17)。培地200 mlを入れて綿栓をした500 mlの三角フラスコを用いて、濃縮CO2を補充することなく往復振盪機で振盪した。連続して白色蛍光を照射しながら培養フラスコを28℃でインキュベートした(30 microeinsteins m−2s−1)。
【0026】
A. halophytica のNa + / H + アンチポーター単離
A.halophyticaゲノムDNAは発表されている方法で単離した(17)。複数のNa+/H+アンチポーター間の高度に保存された2つのポリペプチド領域をもとに、部分的に変性したオリゴヌクレオチドを特定した(9)。正方向のプライマーであるNP-Fはポリペプチドの伸張部Phe-Leu-Pro-Pro-Leu-Leu-Phe-Glu-Alaの後に特定された(SynNhaPのアミノ酸残基73〜81)。逆方向のプライマーであるNP-Rには、Glu-Gly-Glu-Ser-Leu-Phe-Asn-Asp-Glyに相当する相補配列を含む(SynNhaPのアミノ酸残基160〜168)。全プライマーの配列を以下の表1に示す。予測した大きさ(〜0.3 kb)の増幅DNA断片を入手し、配列決定した。決定したヌクレオチド配列を用いて、逆転写ポリメラーゼチェーン反応(PCR)法によって未知の隣接DNA領域を増幅した(20)。そのために、Sau3AIによる部分消化やEcoRIまたはAseIによる完全消化で得たDNA断片はアガロースゲル電気泳動で大きさごとに分画した。1〜5 kbの断片はライゲーションキット(宝酒造、滋賀県)を用いて自己連結し、特異なDNA断片を増幅して配列決定した。A. halophyticaのNa+/H+アンチポーター遺伝子(apnhaP)の配列全体をカバーする約2.4 kbのヌクレオチド配列を決定した。DNA配列はABI310遺伝子分析機(Applied Biosystems, Foster City, CA)を用いて決定し、DNASISプログラム(日立ソフトウェアエンジニアリング、神奈川県)によって解析した。
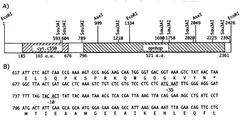
【0027】
その結果、配列番号1に示される、全体が2431 bpのヌクレオチドが配列決定された(図2)。この配列分析からオープンリーディングフレームと思われる796〜2361に位置するapnhaPが明らかになった。この予測遺伝子産物は配列番号2に示される521個のアミノ酸からなり、分子量は56,881 Daである(ApNhaP)。最初のMetの上流配列には-10(ATGAAT)および-35(TACACT)のコンセンサス配列の存在が明らかになった(図2)。さらに上流領域である185〜676には、光化学系II(チトクロムc550)に関与する低電位チトクロムcをコード化する別のオープンリーディングフレームがみつかった。ヌクレオチド配列から推定されたチトクロムc550のアミノ酸配列は、ラン色細菌、藻類および植物から得られた配列と相同性が高かった。
【0028】
図3に示されるように、ホモロジー検索によってApNhaPが真核細胞(SOS1, NHEs, AtNHX1およびNHX1)と原核細胞(NhaP, SynNhaPおよびSynNhaP2)に由来するNa+/H+アンチポーターにきわめて相同していることが明らかになった。SynNhaP2はSynechocystis sp. PCC 6803で確認された2つめのNhaP型アンチポーターである(9)。ApNhaPはSynNhaPと最も相同性が高かった(アミノ酸が61%まで同一)。親水性プロット解析(23)およびTM予測プログラムTopPredII(24)によって、ApNhaPの推定上のTMセグメントを11個特定した。ApNhaPのAsp139は膜貫通領域に保存されたイオン化アミノ酸であり、交換活性に対するその重要性はSynNhaPの突然変異誘発による分析で示されている(9)。尚、ApNhaPには長い親水性C末端テールがあるが、これは真核細胞のNa+/H+アンチポーターの場合であり、P. aeruginosa由来NhaPには認められない。このようなデータは、耐塩性ラン色細菌A. halophyticaに真核細胞に相同なNa+/H+アンチポーターがあることを示唆している。
【0029】
【表1】

【実施例2】
発現プラスミドの構築
apnhaPのコーディング領域をPCR反応により単離した。正方向のプライマーであるANP-NcoIは開始コドンATGとNcoI切断部位を含む。逆方向のプライマーであるANP-HindIIIにはHindIII制限部位が含まれる。増幅断片はpTrcHis2CプラスミドのNcoI/HindIII切断部位に連結した。得られたプラスミドpANhaPはインフレームでヒスチジンヘキサマー融合ApNhaPをコード化し、まずE. coli DH5α細胞に導入し、次にnhaA, nhaB, chaA遺伝子が欠失するTO114細胞に導入した(21)。
【0031】
このpANhaPのほかpSNhaPをNhaA, NhaB, ChaAが欠失したE. coli宿主細胞(TO114)に発現させた(9,21)。膜分画をウェスタンブロッティング法で分析し、pANhaPおよびpSNhaPで形質転換したE. coli細胞に〜53 kDaに相当する交叉反応を示すバンドがひとつあることが明らかになったが、ベクター単独(pTrcHis2C)で形質転換したE. coli細胞はまったくバンドがなかった。このような結果はApNhaPが発現し、E. coliの膜に集っている可能性を示唆するものである。 尚、SDS-PAGEおよび免疫ブロッティングは公知の方法(17)によって実施し、Lowryらの方法によって蛋白質を定量した(25)。ヒスチジンヒキサマー(His6タグ)に対して産生される抗体はR&D Systems Inc.(Minneapolis, MN)から得た。
【0032】
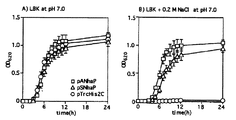
pANhaPで形質転換したE. coli TO114細胞がpH 7.0 37℃のLBK培地のなかでpTrcHis2CおよびpSNhaPで形質転換したTO114細胞とほぼ同じ速度で増殖したことを図4(A)に示す。しかし、TO114細胞にはNa+/H+アンチポーター遺伝子(nhaA, nhaB, chaA)が欠損するため、pTrcHis2Cで形質転換したE. coli細胞は0.2 M NaClの存在下で増殖することができない(図4(B))。これに対して、pANhaPまたはpSNhaPで形質転換したE.coli細胞は増殖可能であった(図4(B))。興味深いことに、pANhaPで形質転換したTO114細胞はpSNhaPで形質転換した細胞に比べて若干速く増殖した(図4(B))。
【0033】
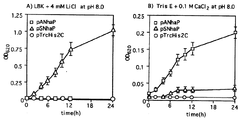
更に、ApNhaPとSynNhaPとでは相補する力が異なることが図4(B)からわかる。2種のアンチポーターにおける相補性の差をさらに明確にするために他の条件でも試験した。図5(A)では増殖温度を30℃にして、pANhaPで形質転換したTO114細胞がLBK培地のアルカリ性pHのもとpSNhaPで形質転換した細胞よりも若干速く増殖できたことを示す。このような条件下、増殖培地に0.2 M NaClを含む場合、ApNhaPはTO114細胞の塩感受性表現型を相補するが、SynNhaPは相補できない(図5(B)。0.5 M NaClの存在下でも、pANhaPで形質転換したTO114細胞は増殖可能であった(図5(C))。相補性の差はpHが小さいと観察されなかった。上のような結果はApNhaPがアルカリ性pHでSynNhaPに比して効果的に相補できることを示唆する。
【0034】
【実施例3】
Na + / H + アンチポーター活性
これまでに発表された方法(9,22)でLBK培地(NaClの代わりにKClを用いるLuriaブロス)で増殖させた細胞から調製した反転膜小胞によって、Na+/H+アンチポーター活性を調べた。簡潔に説明すると、3000×gで4℃、10分間遠心分離してE. coli細胞を収集し、浮遊用緩衝液TCDS(10 mMトリスHCl(pH7.5)、0.14 M塩化コリン、0.5 mMジチオスレイトール、0.25 Mスクロース)で洗浄した。このペレットをTCDS 緩衝液10 mlに浮遊させ、フレンチプレスにかけた(4000 p.s.i.)。次に、この溶液を12,000×gで4℃、10分間遠心分離した。この上清を110,000×gで4℃、60分間遠心分離し、ペレットをTCDS緩衝液600μlに浮遊させた。 アンチポーター活性は、10mMトリス緩衝液(規定pHになるまでHClを加えた)、5 mM MgCl2、0.14 M塩化コリン、1μMアクリジンオレンジ、膜小胞(蛋白質50μg)を含む2 mlの反応混合物に塩を加えて、ΔpH(膜内外のpH勾配)の変化によって評価した。ΔpHは励起492 nmおよび発光525 nmで得られるアクリジンオレンジの蛍光によってモニタリングした。塩を加える前に、呼吸による蛍光クエンチング(Q)を惹起するためにトリス-DL-乳酸(2 mM)を加えた。乳酸は小胞を賦活化し、小胞内にH+を蓄積させるが、これは色素の蓄積と乳酸誘導性蛍光クエンチング(Q)を誘発する。塩(5 mM)の追加によって、アンチポーターによるH+の排出のために蛍光性が増強し、クエンチングしていない蛍光、即ち、塩誘導性蛍光デクエンチング(ΔQ)をモニタリングした。次にNH4Cl(25 mM)を加えてΔpHを放散させた。
【0035】
その結果、図6上段に示すように、ApNhaP発現細胞にNaClを加えるとデクエンチング(ΔQ×100/Q)が観察されたが、対照(pTrcHis2C)細胞では観察されなかった。これはApNhaPがNa+/H+アンチポーター活性を有することを示している。図6(A)は、pH5〜9の広範囲にわたりApNhaPによる蛍光デクエンチングが観察されたことを示しており、これはSynNhaPによるデクエンチング(9)とほぼ同じであるが、E. coli NhaAによるデクエンチングとはまったく異なる。E. coli NhaAではアンチポーター活性がpH7.5以下で観察されることはなく、pHが上昇するにつれて活性が増強した(1)。
【0036】
更に、ApNhaPのイオン特異性を調べるために、異なる陽イオンを加えてデクエンチングを測定した。Li+とH+との交換を触媒できるほとんどのNa+/H+アンチポーターに比べて、ApNhaPは種々のpH値でLi+/H+アンチポーターの活性をほとんど示さなかった(図6(B))。又、ApNhaPが中性及びアルカリ性のpHでCa2+アンチポーター活性を示すことがわかったが(図6(C))、K+/H+ではごくわずかなアンチポーター活性を示し、Mg2+/H+ではまったく示さなかった。このような結果はApNhaPが新たなイオン特異性を有するアンチポーターであることを示唆している。
【0037】
【実施例4】
上記結果から、ApNhaPがCa2+とH+の交換活性を有するが、Li+とH+の交換活性はないことが示唆されるために、このようなイオン特異性がE. coli変異体細胞の塩感受性表現型を相補するかどうか、さらに試験した。その結果、図7(A)に示されるように、pH 8で4 mM LiClを含むLBK培地でApNhaPを発現するTO114細胞は増殖できないが、SynNhaPを発現するTO114は増殖できた。これに対して図7(B)に示すように、pH 8.0で0.1 M CaCl2を含むトリスE培地ではApNhaPを発現するTO114細胞が良好に増殖するが(26)、SynNhaPを発現するTO114は緩やかに増殖した。いずれも、対照細胞(ベクターのみ)では増殖不能であった。Ca2+含有培地でのSynNhaP発現細胞の増殖速度はSynNhaPのCa2+/H+交換活性から予測される速度より遅いが(図7(B)及び図6(C))、このような結果は、実施例3で得られたApNhaPとSynNhaPのLi+/H+およびCa2+/H+アンチポーター活性の結果に一致する(図6(B)及び図6(C))。
【0038】
E. coli ChaAはNa+またはCa2+によるプロトン/カチオン交換活性を有するが、Li+またはK+による活性はないと報告されているが(14,21)、これは基本的にApNhaPと同じイオン特異性である。ただし、ChaAはApNhaPと何ら相同性が認めらなかった。ChaAにはカルセケストリン、カルレティキュリンなど複数のCa2+結合蛋白やNa+/Ca2+イオン交換体に保存される酸性モチーフGlu200-His-Glu-Asp-Asp-Ser-Asp-Asp-Asp-Asp209を有するが(14)、ApNhaPにはこの酸性モチーフが欠けている。ChaAの親水性プロットによると、ApNhaPでは長い親水性C末端尾部がないことがわかる。ApNhaPは酵母(27)および植物(28)由来の空胞Ca2+/H+交換体とまったく相同していなかった。このようなすべてのデータは、ApNhaPがChaAとは異なる構造を有するNa+/H+アンチポーターであることを示している。
【0039】
【実施例5】
ApNhaP と SynNhaP のキメラ遺伝子構築
これまでに、C末端テールが一部欠失するとSynNhaPのNa+/H+対向輸送活性が減弱することがわかっている(9)。ApNhaPはSynNhaPに相同性が最も高いが、この2種のアンチポーターのC末端テールにかかる電荷は有意に異なり、ApNhaPは22個の塩基性アミノ酸と14個の酸性アミノ酸、SynNhaPは15個の塩基性アミノ酸と24個の酸性アミノ酸である。そのためわれわれは、図8に示されるようなASNhaPとSANhaPのキメラアンチポーターを構築し、そのなかでApNhaPの長いC末端テールをSynNhaPのテールに、SynNhaPの長いC末端テールをApNhaPのテールにそれぞれ置換した。反転膜小胞を用いてキメラの交換活性を評価した。
【0040】
即ち、ApNhaPのTM領域(アミノ酸残基Met1−Ile401)とSynNhaPのC末端領域細胞質領域(アミノ酸残基Gln401−Ser527)をコード化するキメラ遺伝子を次のように構築した(図1)。ApNhaPのTM領域とSynNhaPのC末端領域に相当するヌクレオチドをそれぞれ正方向/逆方向プライマーセットであるANP-NcoI/ANP-N-RおよびSNP-C-F/SNP-C-Rによって増幅した。前者ヌクレオチドの鋳型はpANhaPであり、後者ヌクレオチドの鋳型はsynnhaP遺伝子がpTrcHis2CのNcoI/EcoRI部位に結合したpSNhaPであった(9)。正方向のプライマーであるSNP-C-Fにはポリペプチドの伸長部であるApNhaPのGln398-Thr-Val-Ile401とSynNhaPのGln401-Phe-Val-Leu404に相当する配列が含まれており、逆方向のプライマーであるANP-N-RはSNP-C-Fの相補配列である。逆方向のプライマーSNP-C-RにはpSNhaPの終止コドンの直前にEcoRI切断部位がある。両方のPCR産物を合わせて、正方向のANP-NcoIプライマーと逆方向のSNP-C-Rプライマーを用いてキメラ遺伝子apsynnhaPを増幅し、プラスミドpASNhaPを作製したpTrcHis2CのNcoI/EcoRI消化部位に結合させた。
【0041】
次に、SynNhaPのTM領域(アミノ酸残基Met1−Thr400)とApNhaPのC末端領域(アミノ酸残基Glu402−Glu521)をコード化するキメラ遺伝子構築のために、SNP-N-F/SNP-N-RおよびANP-C-F/ANP-HindIIIの正方向/逆方向プライマーセットによって各ヌクレオチドを増幅した。本実施例で用いる逆方向プライマーであるSNP-N-RはANP-C-Fの相補的配列である。両方のPCR産物を合わせて、正方向SNP-N-Fおよび逆方向ANP-HindIIIプライマーを用いてキメラ遺伝子synapnhaPを増幅させ、プラスミドpSANhaPを作製したpTrcHis2CのNcoI/HindIII消化部位に結合させた。
【0042】
その結果、図9に示すように、ASNhaPキメラは図6に示す親ApNhaPのNa+/H+、Li+/H+、Ca2+/H+交換活性に匹敵する活性を示した。興味深いことに、ApNhaPではまず検出不能なLi+/H+交換活性が(図6(B))、ASNhaPキメラでは明らかに観察された(図9(B))。その一方でSANhaPキメラは親SynNhaPに比して、Na+/H+およびLi+/H+の両方の交換活性が減弱した(図9と図6)。更に、SANhaPキメラのCa2+/H+交換活性はpH依存的で、pHが中性またはアルカリ性の場合に活性の増強が観察されるようである(図9(C))。これはpHには関係なく相対的に活性が変化しない親SynNhaPでの観察とは対照的である(図6(C))。活性化のデータには矛盾もみられるが、このような結果はC末端領域がApNhaPおよびSynNhaPのイオン特異性にある種の働きをすることを示唆している。たとえば、SANhaPキメラはApNhaPおよびSynNhaPに比べてNa+/H+交換活性が低く(図9(A)と図6(A))、ASNhaPキメラのCa2+/H+交換活性はpH依存的であるが、これは親ApNhaPと同じであるがSynNhapとは異なる(図9(C))。このような結果は、ApNhaPおよびSynNhaPのイオン特異性もTM領域の構造によって変化し、ほとんどのNa+/H+アンチポーターにも同じことが考えられることを示唆している(1)。
【0043】
キメラの交換活性データは相補性解析によってさらに実証された。図10に示されるように、ASNhaPキメラはE. coli変異体のNa+、Li+およびCa2+感受性表現型を相補するが、SANhaPキメラがE. coli変異体のNa+およびLi+感受性表現型を相補することはまずない。SANhaPを発現するE. coli変異体細胞の増殖速度はSANhaPキメラのCa2+/H+交換活性から予測される速度よりも遅かった(図10(C)及び図9(C))。
【0044】
【実施例6】
次に、淡水らん色細菌Synechococcus sp. PCC7942において8種のプラスミドを過剰発現させ、耐塩性を試験した。
即ち、耐塩性らん色細菌Aphanothece halophytica(A. halophytica)由来のNa+/H+対向輸送体ApNhaPの発現プラスミドpUC303-ApNhaP(33)、A. halophytica由来の分子シャペロンDnaKの発現プラスミドpUC303-DnaK(34)、E. coli由来のカタラーゼの発現プラスミドpUC303-KatE(35,36)、E. coli由来のベタイン合成遺伝子の発現プラスミドpUC303-Bet(37,38)と、それを組み合わせたものである。これらはそれぞれイオンホメオスタシス、蛋白質の折りたたみ、活性酸素のクエンチ、浸透圧保護物質として機能すると考えられている。E. coliでは4つの遺伝子がベタイン合成に関与しており(37)、betA, betB、betTがそれぞれコリンデヒドロゲナーゼ、ベタインアルデヒドデヒドロゲナーゼ、コリン輸送体をコード化し、betIは調節遺伝子をコード化する。発現プラスミドpUC303-DnaK(34)ではgrpEおよびdnaJが部分遺伝子であり、dnaKは機能性蛋白質をコード化する。このすべての遺伝子が自らのプロモーター配列を使用する。Synechococcus sp. PCC7942細胞はpUC303-Bm(対照)、pUC303-ApNhaP, pUC303-KatE, pUC303-Bet, pUC303-DnaK, pUC303-KatE/Bet, pUC303-ApNhaP/KatE, pUC303-ApNhap/KatE/Betプラスミドで形質導入された(図11)。
【0045】
発現ベクターの構築
発現プラスミドpUC303-DnaKの構築のために、A. halophytica由来dnaK遺伝子(34)を含む3kbpの断片をSspIおよびEcoRVで消化して平滑断端にし、pUC303-BmをEcoRI-およびSalI-二重消化して平滑断端にした部位へ結合させた。耐塩性らん色細菌A. halophytica由来のNa+/H+対向輸送体の発現プラスミドで、自身のプロモーターを含有するプラスミドを次のように調製した。テンプレートとしてpANhaPを用いるPCR法で、apnhaP遺伝子の539位にあるEcoRI切断部位からpTrHis2CのHisタグ配列の直後にある終止コドンまでのNa+/H+対向輸送体断片を増幅した(33)。正方向のプライマーであるApNhEcF1 5’-TGGGAATTCCCCTCGGAA-3’にはEcoRI部位が含まれ、逆方向のプライマーであるApNhBmR1 5’-GTGGATCCTCAATGATGATGATGATG-3’には終止コドンの後にBamHI切断部位が含まれる。増幅断片をpBluescript IISK+のEcoRV部位に結合する。-194からEcoRI切断部位がある539までの別のNa+/H+対向輸送体断片はA. halophyticaをテンプレートとしてゲノム遺伝子を用いたPCR法で増幅した。正方向のApNhEcF 2,5’-CTGAATTCGATCGCGGCTCATATT-3’にはEcoRI切断部位が含まれ、逆方向のプライマーApNhEcR2 5’-GGGAATTCCCACTAATAGA-3’にも539にEcoRI切断部位がある。amnhaP遺伝子のC末端部がすでに結合しているpBluescript II SK+のEcoRI消化部位に増幅断片を結合させた。配向を確認した後、正しく結合したプラスミドpBApNhaPをBamHIによって消化し、E.coli/SynechococcusシャトルベクターpUC303-BmのBamHI消化部位に結合させた(38)。これで得られたプラスミドをpUC303-ApNhaPと呼ぶ。apnhaPおよびkatE遺伝子の共発現のために、E. coli由来のkatE遺伝子をSaII/BamHI消化部位に結合させて、プラスミドpUC303-KatEを用いた。KatE遺伝子はカナダのManitoba UniversityのP.C. Loewen教授から分譲していただいたものである(35)。プラスミドpUC303-KatEをXbaIで消化し、平滑断端にした。apnhaP遺伝子はBamHIでpUC303-ApNhaPを消化し、平滑断端して調製した。得られたapnhaP遺伝子はXbaI消化pUC303-KatEに結合させ、プラスミドpUC303-ApNhaP/KatEを調製した。apnhaP, katEおよびbet遺伝子の共発現のために、プラスミドpUC303-KatE/BeTを用いた。pUC303-KatE/BeTはXbaIで消化して平滑断端してから、そこにBamHIで切断して平滑断端にしたapnhaP遺伝子を結合させた。これで得られたプラスミドをpUC303-ApNhaP/KatE/BeTと呼ぶ。Synechococcus細胞を形質変換するためにこのプラスミドを用いた(38)。
【0046】
ストレス下でのらん色細菌の増殖
連続白色蛍光を照射(30μEm−2s−1)の下、10μgml−1ストレプトマイシンを補充したBG11液体培地に気泡を送りながら、Synechococcus細胞を30℃で継代培養した(359)。bet遺伝子が発現すると、100μMコリンを添加した。塩ストレスをかけるために、種々の濃度のNaCl(0〜0.6M)を含有する新鮮培地に後期対数増殖期の細胞を移した。この培養物を数日間インキュベートした。この期間中に、Shimadzu UV-160Aを用いて730nmの吸収を測定しながら、細胞増殖をモニタリングした。SDS-PAGEおよび免疫ブロッティング法をこれまでに発表された方法で実施した(34)。6-ヒスチジン(6×Hisタグ)に対して産生される抗体をR&D Systems, Inc.(USA)から得た。DnaK蛋白を免疫ブロッティング法で検出した(34)。TOF-MS(モデルKOMPACT MALDI IV tDE Shimdzu/Kratos)を用いてグリシンベタインを測定し、活性染色によってカタラーゼ活性を評価した(35)。
【0047】
その結果、すべての形質転換体がほぼ同じ速度で増殖でき、この形質転換体が低塩度条件下で増殖を損なわないことが示された(図12A)。高塩度では、淡水らん色細菌Synechococcus PCC7942(対照細胞)の増殖速度が低下し、0.375M NaClより高濃度では明らかに増殖が認められなくなった。0.4M NaClでは、対照細胞およびdnaK発現細胞は増殖せず、betまたはkatE発現細胞は増殖できたが、その速度はapnhaP発現細胞よりも遅かった(図12B)。katEおよびbetを共発現するSynechococcus細胞はkat発現細胞またはBet単独よりも若干増殖が速かった。このような結果はこれまでの報告と一致しており(35,38,39)、betまたはkatEの過剰発現によって塩ストレスに対する耐性を獲得したことを示している。apnhaP, katEおよびbetを共発現した細胞はapnhaP発現細胞に比べて増殖速度が若干低かった。katE(35), bet(38)およびdnaK(34)の発現は上記要領で確認した。NaClの濃度を0.5Mに上げると、種々の形質転換体でさらに増殖速度の低下が観察された。対照およびdnaK発現細胞のほかに、katEおよび/またはbetを発現する細胞も増殖できなかった(図12C)。apnhaP発現細胞のみが、apnhaPおよびkatE共発現細胞やapnhaP, katEおよびbet共発現細胞に比べて増殖速度が速かった。このようなデータから、ApNhaPの過剰発現によってSynechococcus細胞の耐塩性が向上し、他の関連蛋白によるApNhaPの共発現ではApNhaP発現細胞の耐塩性が向上しなかった。
【0048】
Na + / H + 対向輸送体活性の測定
ApNhaP蛋白はapnhaP発現細胞で検出されており、0.3M NaClで増殖した対照細胞では検出されなかった(図13A)。1日でApNhaPの最高濃度に達した。従って、apnhaP発現細胞の高塩度に対する耐性がApNhaP蛋白の活性によるものと考えられる。
そこで、次に、実際に、Na+/H+活性を基本的にNakamuraら(40)の方法にしたがって測定した。簡潔に説明すると、50mMジエタノールアミン-HClを含有する0.15MNaCl(pH9.3)にSynechococcus sp.PCC 7942細胞を浮遊させた。この処置によって、細胞内K+はK+/H+対向輸送体の働きによってジエタノールアミンに代わった。20mM HEPES-NaOH緩衝液を含む0.15M NaCl(pH8.5)を用いて、アミン負荷細胞を洗浄した。この段階で細胞内部はジエタノールアミンの放出によって酸性になり、Na+はNa+/H+対向輸送体によってH+に交換しようとする負荷がかかった。Na+/H+対向輸送体活性を測定する前に、0.2%グルコースを添加して細胞を賦活化し、次に50mMのジエタノールアミン-HCl(pH8.5)を加えた。適宜、細胞を濾過によって回収し、Shimdzu Personal Ion Analyzer PIA-1000によってフィルター上のNa+イオンを測定した(35)。
【0049】
その結果得られた、ジエタノールアミンの添加によるNa+負荷細胞のNa+排出活性を図13B〜Dに示す840)。中性ジエタノールアミンは細胞を貫通し、プロトン化した。必要なプロトンはNa+/H+対向輸送体を介して、Na+の交換によって、細胞外から取り込まなければならない。このようにNa+の排出活性はNa+/H+対向輸送体活性を反映している。対照とApNhaP発現細胞が異なる塩度条件下で異なるNa+排出活性を示すかどうか調べた。図13B〜Dに、ジエタノールアミンを添加して細胞内Na+が最初の数分できわめて急速に減少し、その後緩徐に減少していった。細胞内Na+が最も大きく減少したのは、0.3M NaClを含有するBG11培地で増殖する細胞(図13C)であり、これに続いて何も添加しない培地(図13B)、次に0.5M NaClを含有する培地(図13D)で増殖する細胞であった。対照とNaClが不在で増殖したApNhaP発現細胞との間にNa+排出活性の差はなかった(図13B)。0.3M NaClで増殖させた場合、対照細胞に比して、ApNhaP発現細胞ではやや強いNa+排出活性が観察され(図13C)、0.5M NaClで増殖させた場合さらに大きい差が観察された(図13D)。このようなすべての結果から、ApNhaP発現細胞が対照細胞に比して高塩度で対向輸送体活性が高いことが示された。
【0050】
次に、ApNhaP発現細胞の増殖速度に対する塩の刺激作用を調べた。塩刺激(0.65M NaCl)によって、ApNhaP発現細胞は10時間後まで増殖可能であったが、対照細胞はそれより遅い速度で3時間までしか増殖しなかった(図14A)。細胞死はApNhaP発現細胞に比べて対照細胞で高速かつ早期に検出された。1.2時間の塩ショック後に、ジエタノールアミンを添加すると対照細胞に比してApNhaP発現細胞で強いNa+排出活性が観察された(30%と75%、15分)(図14B)。
【0051】
光合成活性の評価
塩ストレスは数種の光合成生物で光合成活性を損なうことがわかっている(39)。ApNhaPの発現は塩刺激後の光合成活性を増強させるかどうかを調べるために、現在作用している光化学系II(PS II)の割合を以下にしめすように、R値で表して測定した。
即ち、光化学系II(PSII)における見かけ上の量子収量をPAM 蛍光光度計(Walz, Effeltrich,ドイツ)を用いて測定した(35)。650nmのピーク発光で光発光ダイオードからパルス変調励起を得た。変調した蛍光をλ>710nmで測定した(Schott RG9ロングパスフィルタ)。PAM-102およびPAM-103の付属モジュールはそれぞれ照射光(actinic light)と飽和パルスの管理に用いた。測定前に、10分間細胞を暗室におき、連続光(200μEm−2s−1)のもとPSIIの量子収量を測定した。PSIIの量子収量をF/Fm’=(Fm’-F)/Fm’の式で計算した。式中、FおよびFm’はそれぞれ飽和フラッシュ前後の照射下の蛍光レベルを表す。フラッシュの強度と持続時間は、それぞれ7000μEm−2s−1と800msであった。
その結果、図14Cに示すように、塩刺激前は対照のR値もApNhaP発現細胞のR値もほぼ同じ0.90と0.85であった。しかし、塩刺激から1.2時間後、この値はそれぞれ0.32と0.64に減少した。これを図14Bの結果と総合すると、光合成活性を強く維持する細胞の力は有効なNa+排出に左右されることが判った。
【0052】
ApNhaP 発現細胞の海水中での増殖
更に、図15Aに示すように、ApNhaP発現細胞が海水中で増殖できるということを実際に確認した。海水は愛知県三河地方で採取したものであり、その塩度は30.4psuであった。Betおよび/またはKatEのいずれかを発現する形質転換細胞は海水中で増殖できなかった。ApNhaPおよびKatEの共発現はApNhaP発現細胞の増殖速度に比べて速かったが、0.5M NaClを含有するBG11を増殖培地として用いた図12Cの結果とは対照的である。ApNhaP発現細胞に比べて増殖速度は遅かったものの、ApNhaP, KatEおよびBetを共発現するSynechococcus細胞は海水中でも増殖を維持することができた。このような結果から、ApNhaPの過剰発現によって淡水らん色細菌Synechococcus細胞が海水中で増殖できるほどの耐塩性を獲得したことは明確である。海水中でSynechococcus細胞が増殖する間、対照とApNhaP発現細胞との吸収スペクトルの差が認められた。図15Bに示すように、ApNhaP発現細胞は630 〜680nmの領域で典型的な吸収速度を示した。これに対して、対照細胞は680nm領域に比べて630nm領域での吸収性は有意に低いことがわかる。この吸収ピークはそれぞれフィコビリソームとクロロフィルを表しているので、上記データは対照細胞のフィコビリソームの変性がクロロフィルよりも早く生じたことを示しており、フィコビリソームが高塩度による損傷を受けやすいターゲットであることがわかる。
【0053】
Synechococcus細胞は低塩度でのNa+とH+の交換能が十分であることが上記データから判明した。高塩度では、Na+排出活性が弱いためにSynechococcus細胞がNa+流入にうまく対応できず、増殖が中止した。Synechococcus細胞でのApNhaPの過剰発現によって、高塩度培地および海水中において増殖を再開できた。これは淡水らん色細菌Synechococcus sp.PCC7942が遺伝子工学によって海水中で生存できたことを示す最初の報告である。
【0054】
【発明の効果】
本発明によって、新たなイオン特異性を有するNa+/H+アンチポーター遺伝子を単離することに成功し、該遺伝子で形質転換したアンチポーター欠損株である大腸菌においてNa+感受性表現型及びCa2+感受性表現型が相補されることが判明した。更に、該遺伝子で形質転換された淡水らん色細菌Synechococcus細胞内におけるApNhaPの過剰発現によって、淡水らん色細菌が海水中で増殖できるほどの耐塩性を獲得出来ることが判明した。
従って、本発明は、該遺伝子を利用して、耐塩性作物を開発、創出して食料生産の向上に寄与することの出来る、有力な方法を提供するものである。
【0055】
引用文献
1. Padan, E., and Schuldiner, S.(1996) in Handbook of Biological Physics (Konings, W. N., Kaback, H. R., and Lolkema, J. S., eds) vol.2, pp.501-531, Elsevier Science, Amsterdam
2. Orlowski, J., and Grinstein, S.(1997) J. Biol. Chem. 272, 22373-22376
3. Blumwald, E.,Aharon, G. S., and Apse, M. P.(2000) Biochim. Biophys. Acta 1465, 140-151
4. Nass, R., Cunningham, K. W., Rao, R. (1997) J. Biol. Chem. 272, 26145-26152
5. Nass, R., and Rao, R. (1998) J. Biol. Chem. 274, 21054-21060
6. Gaxiola, R. A., Rao, R., Scherman, A., Grisafi, P., Alper, S. L., and fink, G. R. (1999) Proc. Natl. Acad. Sci. U.S.A. 96,1480-1485
7. Jia, Z. P., McCullough, N., Martel, R., Hemmingsen, S., and Young, P. G.(1992) EMBO J. 11,1631-1640
8. Shi, H., Ishitani, M., Kim, C., and Zhu, J. K. (2000) Proc. Natl. Acad. Sci. U.S.A. 97, 6896-6901
9. Hamada, A., Hibino, T., Nakamura, T., and Takabe. T. (2001) Plant Physiol. 125, 437-446
10. Wakabayashi, S., Fafournoux, P., Sardet, C., and Pouyssegur, J. (1992) Proc. Natl. Acad. Sci. U.S.A. 89, 2424-2428
11. Inoue, H., Noumi, T., Tsuchiya, T., and Kanazawa, H. (1995) FEBS Lwtt.363, 264-268
12. Nakamura, T., Komano, Y., and Unemoto, R. (1995) Biochim. Biophys. Acta 1230, 170-176
13. Utsugi, J., Inaba, K., Kuroda, T., Tsuda, M., and Tsuchiya, T. (1998) Biochim. Biophys. Acta 1398, 330-334
14. Ivey, D. M., Guffanti, A. A., Zemsky. J., Pinner, E., Karpel, R., Padan, E., Schuldiner, S., and Krulwich, A. (1994) J. Biol. Chem. 268.11296-11303
15. Ishitani, M., Takabe, T., Kojima, K., and Takabe, T. (1993) Aust. J. Plant Physiol. 20, 693-703
16. Takabe, T., Incharoensakdi, A., Arakawa, K., and Takabe, T. (1997) Plant Physiol. 88, 1120-1124
17. Lee, B. H., Hibino, T., Jo, J. Viale, A. M., and Takabe, T. (1997) Plant Mol. Biol. 35, 763-775
18. Hibino, T., Kaku, N., Yoshikawa, H., Takabe, T., and Takabe, T. (1999) Plant Mol. Biol. 40, 409-418
19. Sugino, M., Hibino, T., Tanaka, Y., Nii, N., Takabe, T. and Takabe, T. (1999) Plant Sci, (Shannon) 137, 81-88
20. Triglia, T., Peterson, M. G., and Kemp, D. J. (1998) Nucleic Acids Res. 25, 8186
21. Ohyama, T., Igarashi, K., and Kobayashi, H. (1995) J. Bacteriol. 176, 4311-4315
22. Rosen, B. P. (1986) Methods Enzymol.125, 328-336
23. Kyte, J., and Doolittle, R. F. (1982) J. Mol. Biol 157, 105-132
24. Hofmann, K., and Stoffel, W. (1992) Comput. Appl. Biosci. 8, 331-337
25. Lowry, O.H., Rosebrough, N.J., Farr, A. L., and Randall, R.J. (1951) J. Bio;. Chem. 193, 265-275
26. Brockman, R. W., and Heppel, L. A. (1968) Biochemistry 7, 2554-2562
27. Cunningham, K. W., and Fink, G. R. (1996) Mol. Cell. Biol. 16, 2226-2237
28. Ueoka-Nakanishi, H., Nakanishi, Y., Tanaka, Y., and Maeshima, M. (1999) Eur. J. Biochem. 262, 417-425
29. Biemesderfer, D., DeGray, B., and Aronson, P. S. (1998) J. Biol. Chem. 273, 12391-12396
30. Wells, K. M., and Rao, R. (2001) J. Biol. Chem. 276, 3401-3407
31. Ono, K., Hibino, T., Kohinata, T., Suzuki, S., Tanaka, Y., Nakamura, T., Takabe, T., and Takabe, T. (2001) Plant Sci. (Shannon) 160, 455-461
32. Apse, M. P., Aharon, G. S., Snedden, W. A., and Blumwald, E. (1999) Science 285, 1256-1258
33. Waditee,R.et al. Halotolerant cyanobacterium Aphanothece halophytica,cotains an Na+/H+ antiporter, homologous to eukaryoticones,with novel ion specificity affected by C-terminal tail.(2001) J.Biol.Chem.in press
34. Lee, B. H., Hibino, T., Jo, J., Viale, A. M. and takabe, T. (1997) Plant. Mol. Biol. 35, 763-775
35. Kaku, N. et al (2000) Plant Science 159, 281-288
36. Ossowski, I., Mulvey, M. R., Leco, P.A., Borys, A. and Loewen, P. C. (1991) J. Bacteriol. 173 514-520
37. Lamark, T., Kaasen, I., Eshoo, M. W., McDougall, J. and Strom, A. P. (1991) Mol. Microbiol. 5, 1049-1064
38. Nomura, M., Ishitani, M., Takabe, T., Rai, A. K. and Takabe, T. (1995) Plant Physiol. 107, 703-708
39. Deshnium, P., Los, D. A., Hayashi, H., Mustardy, L. and Murata, N. (1995) Plant Mol Biol. 29, 897-907
40. Nakamura, T., Komano, Y. and Unemoto, T. (1995) Biochim. Biophys. Acta. 1230, 170-176
41. Kaku, N. et al. (1999) Plant Cell Physiol. 40, 557-564
42. Sugino, M., Hibino, T., Nii, N., Takabe, T., and Takabe, T. (1999) Plant Science 146, 81-88
43. Ono, K. et al. (2001) Plant Science 160, 455-461
44. Kishitani, S. et al. (2000) Plant Cell Environ. 23, 107-114
45. Takabe, T. et al. (1998) Glycinebetaine and the genetic engineering ofsalinity tolerance in plants. In Stress Responses of Photosynthetic Organisms. Edited by Satoh, K. and Murata, N. pp.115-131. Elsevier Science, Amsterdam.
46. Tanaka, T. et al. (1999) Plant Science 148, 131-138
【0056】
【配列表】
【図面の簡単な説明】
【図1】 pASNhaPとpSANhaPのキメラプラスミド構築に関する概略図(B)を示す。
【図2】 A. halophyticaのapnhaP座のゲノム構造 A:遺伝子と制限部位 B:apnhaPとチトクロムc550の接合部のヌクレオチド配列。apnhaPとチトクロムc550のヌクレオチド配列から演繹したapnhaPの推定上の-10(ATGAAT)および-35(TACACT)コンセンサス配列とアミノ酸配列も示す。
【図3】 Na+/H+アンチポーターの推定アミノ酸配列の比較 A:8種の生物に由来するNa+/H+アンチポーターの推定アミノ酸配列のアラインメント。この配列はプログラムClustalWによって整列化した。このアラインメントはSOS1、ヒトNHE1、酵母NHX1のN末端の550個のアミノ酸残基をもとにしている。すべての配列に保存されたアミノ酸残基を黒で強調表示し、保存的置換を点で示した。保存Asp(ApNhaPのAsp139)をアスタリスクで示す。予測される膜貫通領域をアラインメントの上に示す。 B:8個のNa+/H+アンチポーターの系統学的解析。複数の配列アラインメントと系統樹の作製はそれぞれClustalWおよびTreeViewソフトを用いた。
【図4】 3種のE. coli細胞の増殖速度に対するNaClの作用 LBK培地における対数増殖期の対照細胞と形質転換細胞を指示されたNaCl濃度を含む新鮮LBK培地(pH 7.0)に移した。 A:LBK培地における増殖の推移 B:0.2 M NaClを含有するLBKにおける増殖の推移 丸:対照細胞 四角:ApNhaP発現細胞
三角:SynNhaP発現細胞。各値は3つの測定値の平均である。
【図5】 3種のE. coli細胞の増殖速度に対するpHとNaClの作用 LBK培地における対数増殖期の対照細胞と形質転換細胞を指示されたNaCl濃度を含み、指示されたpHの新鮮LBK培地に移した。増殖温度は30℃であった。0 M NaCl(A)、0.2 M NaCl(B)0.5 M NaCl(C)を含むLBK培地での増殖の推移。白四角:ApNhaP発現細胞、pH 8.0。網掛けした四角:pH 8.5。黒四角:pH 9.0。白三角:SynNhaP発現細胞、pH 8.0。網掛けした三角:pH 8.5。黒三角:pH9.0。白丸:対照細胞、pH 8.0。網掛けした丸:ph 8.5。黒丸:pH 9.0。各値は3つの測定値の平均である。
【図6】 アクリジンオレンジ蛍光クエンチング法で測定したNa+/H+アンチポーター活性 対照E. coli細胞とApNhaPおよびSynNhaPを発現するE. coli細胞を塩を追加することなくLBK培地で増殖させた。この細胞から反転膜小胞を調製した。アンチポーター活性は実施例に記載した要領で測定した。上段にはApNhaP発現細胞と対照細胞(pTrcHis2C)の典型的なクエンチングパターンを示す。A, B, Cにはそれぞれ、Na+/H+, Li+/H+,Ca2+/H+アンチポーター活性のpH依存性を示した。塩(NaCl, LiCl, CaCl2)の最終濃度は5 mMであった。各値は3つの個別測定値の平均である。
【図7】 3種のE. coli細胞の増殖速度に対するLiClおよびCaCl2の作用 規定のLiClまたはCaCl2濃度のLBKまたはトリスE培地に、LBK培地で対数増殖期にある対照細胞および形質転換細胞をそれぞれ移した。他の実験条件は図4の説明に記載したとおりである。各値は3つの個別測定値の平均である。
【図8】 キメラNa+/H+アンチポーターの副次的構造モデル キメラNa+/H+アンチポーターを実施例に記載の要領で構築した。C末端親水性尾部にかかる陽性電荷と陰性電荷の数を示す。
【図9】 キメラアンチポーターの活性 ASNhaPキメラとSANhaPキメラを発現するE. coli細胞は塩を追加することなくLBK培地で増殖させた。この細胞から反転膜小胞を調製した。アンチポーター活性は実施例に記載の要領で構築した。各値は3つの個別測定値の平均である。
【図10】 ASNhaPキメラとSANhaPキメラを発現するE. coli細胞の増殖速度に対するNaCl, LiCl, CaCl2の作用 LBK培地で対数増殖期にあるASNhaPキメラとSANhaPキメラを発現するE. coli細胞を0.2 M NaClを含むLBK培地、0.4 mM LiClを含むLBK培地、0.1 M CaCl2を含むトリスE培地に移した。他の実験条件は図4の説明に記載した要領と同じである。各値は3つの個別測定値の平均である。
【図11】 発現ベクターの概略図を示す。
【図12】 対照と形質転換Synechococcus細胞の増殖速度に対する塩ストレスの作用。規定のNaCl濃度を含む新鮮培地にBG11培地で対数増殖期にある対照と形質転換細胞を含浸して、塩ストレスをかけた。
【図13】 ApNhaPの蓄積量(A)とNa+負荷細胞からのNa+排出量(B,C,D)。Aでは、対照とApNhaP発現細胞をBG11培地で増殖し、0.3M NaCl含有培地に移した。規定時間インキュベートした後、細胞を回収し、免疫学的分析を行った。B,C,Dでは、対照(白丸)とApNhaP発現細胞(黒丸)を規定濃度のNaClを含むBG11培地のなかで1日増殖させた。次に、Na+排出を誘発するジエタノールアミン(EA)を測定した。
【図14】 対照とApNhaP発現Synechococcus細胞における増殖速度(A)、Na+排出活性(B)、光合成電子輸送活性(C)に対する塩刺激の作用。BG11培地で対数増殖期の対照およびApNhaP発現細胞を回収し、0.65M NaClを含有する新たな培地に移した。
【図15】 海水中の対照と形質変換Synechococcus細胞の増殖曲線(A)と吸収スペクトル(B)。BG11培地で対数増殖期の対照および形質転換細胞を海水に移した。海水に移して3日後に、対照およびApNhaP発現細胞の吸収スペクトル(B)を測定した。[0001]
BACKGROUND OF THE INVENTION
The present invention relates to Na+/ H+Antiporter genes, especially Na from salt-tolerant cyanobacteria (Aphanothece halopytica)+/ H+The present invention relates to an antiporter gene and improvement of salt tolerance by expression of the gene.
[0002]
[Prior art]
Today, 20% of cultivated land around the world and almost half of irrigated land are said to be damaged by high salinity. High salinity induces stress due to plant ionic imbalance and high osmotic pressure. Organisms that survive in high salinity environments have a unique mechanism that regulates internal osmotic pressure. One such mechanism is the ability to store solutes of low molecular weight organic compounds such as sugar, certain amino acids, and quaternary ammonium compounds. Another mechanism to adapt to high salinity is Na+/ H+Antiporter and Na+-Na from cells proposed as a function of ATPase+It is discharge. In addition to such toxic effects, salt stress also induces oxidative stress. Therefore, in recent years, attempts have been made to improve the salt tolerance of plants using genetic engineering.
[0003]
Na+/ H+The antiporter is activated by Na through the cell membrane.+H+Catalyzed the exchange reaction (Na+/ H+It has various functions such as antiporter activity), pH in cytoplasm, cell volume, and sodium concentration in cytoplasm (1-3). In Escherichia coli, three types of antiporters (NhaA, NhaB, ChaA) are known, and their functional characteristics are well elucidated (1). In animals, 6 homologous Na+/ H+Antiporter (replacement) (NHE1-6) is known (2). In plants and yeast, vacuolar (NHX1 and AtNHX1) (4-6) and plasma membrane (SOD2 and SOS1) Na (7,8) Na+/ H+Antiporters have been reported. E. coli antiporter and eukaryotic antiporter have very low homology, Na+/ H+It is suggested that the evolution of the antiporter gene is independent.
[0004]
Such Na+/ H+ A recent report (1999, Science, 2001 Nature Biotechnology) by Canadian Blumbald et al. Reported that the use of antiporter genes improves plant salt tolerance. Blumbald et al. Describe Nab in the vacuolar membrane of Arabidopsis.+/ H+ It has been reported that both anti-porter genes grow in 0.2M NaCl by expressing them in Arabidopsis vacuolar membrane (Science 1999) and tomato vacuolar membrane (2001 Nature Biotechnology). This enhances salt tolerance by accumulating NaCl inside the vacuole membrane in the cell.
[0005]
As described above, considerable efforts have been made to improve the salt tolerance of plants by genetic engineering techniques. However, sufficient results have not been obtained for practical use.
Compared with plants, single-cell phagocytic cells are simple and are an appropriate model for elucidating factors responsible for salt tolerance in photosynthetic organisms. Cyanobacteria are solar energy and inorganic carbon CO2It is a photosynthetic organism that generates oxygen that can grow using Such organisms are found in almost all habitats, including salt-containing waters from very low to higher concentrations than seawater. However, it is unclear which genes are involved in adaptation to different salinities. Therefore, identification of a master regulatory gene corresponding to high salinity is important not only for understanding the mechanism of cyanobacterial salt tolerance, but also for plant applications in terms of improving crop yield.
[0006]
In the previous test (9), the present inventors have found that the cyanobacterium Synechocystis sp. PCC6803 contains Na.+/ H+Antiporter SynNhaP1Has been found to be homologous to the antiporters of eukaryotic and prokaryotic cells (NhaP from Pseudomonas aeruginosa), but not to NhaA, NhaB and ChaA. Furthermore, SynNhaP contains Asp conserved in the transmembrane (TM) region.138And a relatively long C-terminal hydrophilic tail, both of which have been shown to be important for carrier activity. The long C-terminal tail is thought to serve to control animal transport activity (2,10). From these findings, cyanobacterial antiporter is eukaryotic Na+/ H+It turns out to be a model system for testing the structural and functional properties of antiporters.
[0007]
To date, Na+/ H+Anti-porter proteins have identified very few functional amino acid residues, especially amino acid residues involved in cation transport. SynNhaP Asp138(9) NhaA Asp133, Asp163, Asp164The importance of (11,12) has been reported. Na+/ H+Antiporter is H+And Na+Or Li+Specifically exchanges with H+And Li+There are also some antiporters that have weak exchange activity (13). E. Coli ChaA is alkaline pH and Ca2+And H+, Na+And H+Have been reported to have exchange activity, but have not been investigated in detail. It is unclear which factor determines the ion specificity. Na+/ H+In order to specify a conserved sequence for cation transport of the antiporter, cloning of an antiporter having a new ion specificity is indispensable.
[0008]
Aphanothece halophytica is a salt-tolerant cyanobacteria that can grow under a range of 0.25-3.0M NaCl and a wide range of salinity conditions (15, 16). A. halophytica accumulates the osmoprotective substance glycine betaine with high salinity (15), and the DnaK protein of A. halophytica is another DnaK / Hsp70It has a C-terminal segment longer than that of the family (17), and has been reported to show a very strong protein folding activity at high salinity (18). It has also been found that transformation of tobacco with DnaK protein from A. halophytica enhances salt tolerance (19).
[0009]
[Problems to be solved by the invention]
Accordingly, an object of the present invention is to find a useful gene capable of improving plant salt tolerance by genetic engineering technology, and to develop and create a salt tolerant crop using the gene to improve food production. It is to provide a way to contribute.
[0010]
[Means for Solving the Problems]
Therefore, the present inventor has developed a unique Na in A. halophytica.+/ H+As a result of studying whether or not there is an antiporter (ApNhaP), Na is homologous to eukaryotic cells but has new ion specificity+/ H+We succeeded in isolating the antiporter gene, and further revealed that its ion specificity varies depending on the C-terminal region.
[0011]
Furthermore, 8 plasmids were overexpressed in the freshwater cyanobacteria Synechococcus sp. PCC7942, and the salt tolerance was tested. As a result, by comparing the genes involved in betaine synthesis, catalase and chaperone, Na derived from salt-tolerant cyanobacteria+/ H+By overexpressing the antiporter gene, this Synechococcus cell was found to grow in BG11 medium containing 0.5 M NaCl in seawater, and succeeded in greatly improving the salt tolerance of freshwater cyanobacteria.
[0012]
That is, the present invention relates to the following aspects.
(1) Na from salt-tolerant cyanobacteria (Aphanothece halopytica)+/ H+DNA encoding an antiporter.
(2) Li in the entire pH range+/ H+It exhibits virtually no antiporter activity and Ca in neutral and alkaline pH regions2+/ H+Cyanobacteria-derived Na, characterized by exhibiting antiporter activity+/ H+DNA encoding an antiporter.
(3) The DNA according to 2, wherein the cyanobacterium is a salt-tolerant cyanobacterium (Aphanothece halopytica).
(4) DNA containing a base sequence encoding the following protein (a) or (b) or a partial sequence thereof:
(A) a protein comprising the amino acid sequence shown in SEQ ID NO: 2,
(B) consisting of an amino acid sequence in which one or several amino acids are deleted, substituted or added in the amino acid sequence (a), and Na+/ H+Protein with antiporter activity.
(5) DNA comprising the following base sequence (a) or (b) or a partial sequence thereof:
(A) a base sequence consisting of the 796 to 2361th base pairs in SEQ ID NO: 1,
(B) hybridizes with DNA comprising the base sequence (a) under stringent conditions, and Na+/ H+A base sequence encoding a protein having antiporter activity.
(6) A recombinant expression vehicle containing the DNA according to any one of 1 to 5.
(7) The recombinant expression vehicle according to 6, which is an Escherichia coli-derived plasmid vector.
(8) The recombinant expression vehicle according to 6, which is an E. coli / freshwater cyanobacteria shuttle vector.
(9) A transformant transformed with the expression vehicle according to 6, 7, or 8.
(10) The transformant according to 9, which is a bacterium.
(11) The transformant according to 10, which is Escherichia coli.
(12) The transformant according to 10, wherein Escherichia coli is an antiporter-deficient strain.
(13) Na+Sensitive phenotype and Ca2+The transformant according to 12, wherein the sensitivity phenotype is complemented.
(14) The transformant according to 9, which is a cyanobacteria.
(15) The transformant according to 14, wherein the cyanobacterium is a freshwater cyanobacterium.
(16) The transformant according to 15, which is a freshwater cyanobacteria capable of growing in seawater.
(17) The transformant according to 9, which is a plant.
(18) The transformant according to 17, wherein the plant is selected from the group consisting of rice and tobacco.
(19) Na+/ H+The transformant according to any one of 14 to 18, wherein the anti-porter is overexpressed.
(20) Na+/ H+The transformant according to any one of 14 to 18, wherein the antiporter activity is increased.
(21) A method for improving the salt tolerance of the host, which comprises introducing the DNA according to any one of 1 to 5 into a host cell and expressing the DNA.
(22) The host is transformed with the recombinant expression vehicle containing the DNA according to any one of 1 to 5, and the DNA is expressed in the host cell, and the salt tolerance of the host is improved. Method.
(23) The method according to 21 or 22, wherein the salt tolerance is improved by discharging NaCl out of the cell.
(24) The method according to 21, 22, or 23, wherein the host is a plant.
(25) The method according to 24, wherein the plant is selected from the group consisting of rice and tobacco.
(26) A method for creating a salt-tolerant organism, which comprises introducing the DNA according to any one of 1 to 5 into a host cell and expressing the DNA.
(27) A method for creating a salt tolerant organism, comprising transforming a host with a recombinant expression vehicle containing the DNA according to any one of 1 to 5 and expressing the DNA in a host cell.
(28) The method according to 26 or 27, wherein the host is a plant.
(29) The transformant according to 28, wherein the plant is selected from the group consisting of rice and tobacco.
(30) The transformant according to 6 to 20 is cultured, and Na is obtained from the resulting culture.+/ H+Na consisting of isolating and purifying the antiporter+/ H+Preparation method of antiporter.
(31) A protein encoded by the DNA according to any one of 1 to 5.
(32) Recombinant Na obtained by the preparation method according to 30+/ H+Anti-porter protein.
[0013]
DETAILED DESCRIPTION OF THE INVENTION
Na from salt-tolerant cyanobacteria (Aphanothece halopytica) of the present invention+/ H+As shown in the following examples, the DNA encoding the antiporter contains a plurality of known Na+/ H+Primers are made based on two polypeptide regions highly conserved between antiporters, while A. halophytica is cultured by a method known to those skilled in the art, and then the genome isolated therefrom is used as a template for those skilled in the art. It can be prepared using well-known PCR.
[0014]
Na encoded by the DNA of the present invention+/ H+The anti-porter is Li in the entire pH range.+/ H+It exhibits virtually no antiporter activity and Ca in neutral and alkaline pH regions2+/ H+It exhibits antiporter activity.
[0015]
More specifically, the DNA of the present invention contains a base sequence encoding a protein of the following (a) or (b) or a partial sequence thereof:
(A) a protein comprising the amino acid sequence shown in SEQ ID NO: 2,
(B) consisting of an amino acid sequence in which one or several amino acids are deleted, substituted or added in the amino acid sequence (a), and Na+/ H+A protein having antiporter activity.
Furthermore, the DNA of the present invention comprises the following base sequence (a) or (b) or a partial sequence thereof:
(A) a base sequence consisting of the 796 to 2361th base pairs in SEQ ID NO: 1,
(B) hybridizes with DNA comprising the base sequence (a) under stringent conditions, and Na+/ H+A base sequence encoding a protein having antiporter activity.
Here, the partial sequence is, for example, the Na+/ H+This means a sequence corresponding to the C-terminal region of the antiporter (region from the 402th amino acid residue Glu to the 521st amino acid residue Glu in the amino acid sequence shown in SEQ ID NO: 2). This is a region that has been revealed by the present invention to be an involved part.
[0016]
Those skilled in the art can also prepare by chemical synthesis or the like using well-known means in the art based on the sequence information disclosed in the present specification. Furthermore, those skilled in the art will recognize Na+/ H+It is also easy to delete, substitute, or add one or several amino acids in the specific amino acid sequence in SEQ ID NO: 2 so as to substantially retain the antiporter activity using well-known means in the art. Can be implemented. Na+/ H+In order to substantially retain the antiporter activity, the homology in the amino acid sequence is 65% or more, preferably 80% or more, more preferably 90% or more, still more preferably 98% or more, particularly preferably 99. Can be higher than 5%
[0017]
In the present invention, a specific gene or DNA can be hybridized in a buffer solution well known to those skilled in the art under stringent conditions under various conditions such as an appropriate temperature and salt concentration. Hybridizes with the gene or DNA of the present invention under such stringent conditions, and Na+/ H+Examples of DNA encoding a protein having substantially the same activity as the antiporter activity include, for example, homology with each corresponding gene.90%more than,Preferably98% or more,ThanPreferably, the DNA is higher than 99.5%.
[0018]
Furthermore, this invention relates to the recombinant expression vehicle containing at least 1 gene of said gene of this invention as another aspect. The recombinant expression vehicle includes any vector known to those skilled in the art, such as various vectors, and in particular, various plasmid vectors derived from E. coli, E. coli / freshwater phagocytic shuttle vector, and Agrobacterium Ti plasmid vector. Is preferred.
The recombinant expression vehicle includes various promoters that are binding domains for various sequences known to those skilled in the art of genetic recombination, for example, various σ subunits that are transcription factors possessed by prokaryotic Escherichia coli, etc., and Various transcriptional regulatory elements such as enhancers, restriction enzyme sites, and selection marker (marker enzyme etc.) genes such as kanamycin resistance markers can be optionally included and can be easily prepared by methods known to those skilled in the art.
[0019]
Examples of the host transformed with the expression vehicle of the present invention include various bacteria such as Escherichia coli and cyanobacteria, and plants such as rice and tobacco. In these transformants, as a result of the expression of the DNA of the present invention, Na+Sensitive phenotype and Ca2+Those with complementary phenotypes, Na+/ H+As a result of overexpression of antiporter, Na+/ H+Some have increased antiporter activity.
[0020]
By introducing the DNA of the present invention into a host cell and expressing the DNA, the salt tolerance of the host can be improved. The improvement in salt tolerance is mainly due to Na+/ H+This is because NaCl is discharged out of the cell by the antiporter. As an introduction method, for example, the DNA of the present invention can be introduced by transforming a host with the above recombinant expression vehicle. In addition, any means and conditions known to those skilled in the art such as calcium phosphate coprecipitation method, electroporation method, and particle gun method can be used.
[0021]
Thus, by introducing the DNA of the present invention into a host cell and expressing the DNA, a salt-tolerant organism, for example, a freshwater cyanobacteria that can grow in seawater can be created by genetic engineering techniques. .
[0022]
Further, the above transformant is cultured, and the resulting culture is subjected to Na.+/ H+By isolating and purifying the antiporter, Na+/ H+An antiporter can be prepared. Various conditions relating to culture, isolation, purification, and the like can be appropriately selected by those skilled in the art according to the type of vector, host, and other factors used.
[0023]
The present invention relates to a protein encoded by the above-described DNA of the present invention. Such proteins can be purified and isolated from A. halophytica cultures by methods known to those skilled in the art. Alternatively, as a recombinant protein obtained by the above preparation method, Na+/ H+It is also possible to obtain an antiporter.
[0024]
【Example】
EXAMPLES Hereinafter, although an Example demonstrates this invention more concretely, this invention is not limited at all by these Examples.
[0025]
[Example 1]
A. halophytica culture
A. halophytica cells in 18 mM NaNO in BG11 medium3In addition, Turk Island salt solution was added to grow photoautotrophically. Turk Island's salt solution was prepared by methods published so far (15-17), except that the NaCl concentration of the medium was adjusted to the appropriate range of 0.25-2.5M. Using a 500 ml Erlenmeyer flask with 200 ml of medium and a cotton stopper, concentrate CO2It was shaken with a reciprocating shaker without replenishing. The culture flask was incubated at 28 ° C with continuous white fluorescence (30 microeinsteins m-2s-1).
[0026]
A. halophytica Na + / H + Antiporter isolation
A. halophytica genomic DNA was isolated by published methods (17). Multiple Na+/ H+Based on two highly conserved polypeptide regions between antiporters, partially denatured oligonucleotides were identified (9). The forward primer NP-F was identified after the polypeptide extension Phe-Leu-Pro-Pro-Leu-Leu-Phe-Glu-Ala (amino acids 73-81 of SynNhaP). The reverse primer NP-R contains a complementary sequence corresponding to Glu-Gly-Glu-Ser-Leu-Phe-Asn-Asp-Gly (amino residues 160 to 168 of SynNhaP). The sequences of all primers are shown in Table 1 below. An amplified DNA fragment of the expected size (˜0.3 kb) was obtained and sequenced. Using the determined nucleotide sequence, an unknown flanking DNA region was amplified by reverse transcription polymerase chain reaction (PCR) (20). Therefore, DNA fragments obtained by partial digestion with Sau3AI or complete digestion with EcoRI or AseI were fractionated according to size by agarose gel electrophoresis. The 1-5 kb fragment was self-ligated using a ligation kit (Takara Shuzo, Shiga Prefecture), and the specific DNA fragment was amplified and sequenced. A. halophytica Na+/ H+An approximately 2.4 kb nucleotide sequence covering the entire sequence of the antiporter gene (apnhaP) was determined. The DNA sequence was determined using an ABI310 gene analyzer (Applied Biosystems, Foster City, Calif.) And analyzed by the DNASIS program (Hitachi Software Engineering, Kanagawa Prefecture).
[0027]
As a result, the entire nucleotide of 2431 bp shown in SEQ ID NO: 1 was sequenced (FIG. 2). This sequence analysis revealed apnhaP located at 796-2361, which appears to be an open reading frame. This predicted gene product consists of 521 amino acids shown in SEQ ID NO: 2 and has a molecular weight of 56,881 Da (ApNhaP). The presence of -10 (ATGAAT) and -35 (TACACT) consensus sequences in the upstream sequence of the first Met was revealed (FIG. 2). Furthermore, in the
[0028]
As shown in FIG. 3, ApNhaP was derived from eukaryotic cells (SOS1, NHEs, AtNHX1 and NHX1) and prokaryotic cells (NhaP, SynNhaP and SynNhaP2) by homology search.+/ H+It became clear that it was very similar to the antiporter. SynNhaP2 is the second NhaP antiporter identified in Synechocystis sp. PCC 6803 (9). ApNhaP was most homologous to SynNhaP (amino acids are identical up to 61%). Eleven TM segments on ApNhaP were identified by hydrophilic plot analysis (23) and TM prediction program TopPredII (24). ApNhaP Asp139Is an ionized amino acid conserved in the transmembrane domain, and its importance for exchange activity has been shown by analysis of SynNhaP by mutagenesis (9). ApNhaP has a long hydrophilic C-terminal tail, which is eukaryotic Na.+/ H+This is the case for antiporters and is not observed in NhaP from P. aeruginosa. Such data indicate that the salt-tolerant orchid bacterium A. halophytica is Na homologous to eukaryotic cells.+/ H+It suggests that there is an antiporter.
[0029]
[Table 1]
[Example 2]
Construction of expression plasmid
The coding region of apnhaP was isolated by PCR reaction. ANP-NcoI, a forward primer, contains an initiation codon ATG and an NcoI cleavage site. The reverse primer, ANP-HindIII, contains a HindIII restriction site. The amplified fragment was ligated to the NcoI / HindIII cleavage site of the pTrcHis2C plasmid. The resulting plasmid pANhaP encoded histidine hexamer-fused ApNhaP in-frame and was first introduced into E. coli DH5α cells and then into TO114 cells lacking the nhaA, nhaB, and chaA genes (21).
[0031]
In addition to this pANhaP, pSNhaP was expressed in E. coli host cells (TO114) lacking NhaA, NhaB, and ChaA (9, 21). Membrane fractions were analyzed by Western blotting, and it was revealed that E. coli cells transformed with pANhaP and pSNhaP had a single cross-reactive band corresponding to ~ 53 kDa, but the vector alone (pTrcHis2C) E. coli cells transformed with had no bands at all. These results suggest the possibility that ApNhaP is expressed and collected in the membrane of E. coli. SDS-PAGE and immunoblotting were performed by a known method (17), and the protein was quantified by the method of Lowry et al. (25). Histidine Hixamer (His6The antibody produced against (tag) was obtained from R & D Systems Inc. (Minneapolis, Minn.).
[0032]
FIG. 4 (A) shows that E. coli TO114 cells transformed with pANhaP grew at about the same rate as TO114 cells transformed with pTrcHis2C and pSNhaP in LBK medium at pH 7.0 37 ° C. However, TO114 cells have Na+/ H+Since the antiporter genes (nhaA, nhaB, chaA) are deficient, E. coli cells transformed with pTrcHis2C cannot grow in the presence of 0.2 M NaCl (FIG. 4B). In contrast, E. coli cells transformed with pANhaP or pSNhaP were able to grow (FIG. 4B). Interestingly, TO114 cells transformed with pANhaP grew slightly faster than cells transformed with pSNhaP (FIG. 4B).
[0033]
Furthermore, it can be seen from FIG. 4B that ApNhaP and SynNhaP have different complementary forces. Other conditions were also tested to further clarify the complementarity differences between the two antiporters. FIG. 5 (A) shows that TO114 cells transformed with pANhaP at a growth temperature of 30 ° C. were able to grow slightly faster than cells transformed with pSNhaP under the alkaline pH of LBK medium. Under these conditions, when the growth medium contains 0.2 M NaCl, ApNhaP complements the salt-sensitive phenotype of TO114 cells, but SynNhaP cannot complement (Figure 5 (B). Even in the presence of 0.5 M NaCl, pANhaP. TO114 cells transformed with A. were able to grow (Figure 5 (C)). The difference in complementarity was not observed at low pH.The above results indicate that ApNhaP is alkaline pH compared to SynNhaP. This suggests that they can be complemented effectively.
[0034]
[Example 3]
Na + / H + Antiporter activity
Inverted membrane vesicles prepared from cells grown in LBK medium (Luria broth using KCl instead of NaCl) by previously published methods (9,22)+/ H+Antiporter activity was examined. Briefly, E. coli cells were collected by centrifugation at 3000 xg for 10 minutes at 4 ° C, and the suspension buffer TCDS (10 mM Tris HCl (pH 7.5), 0.14 M choline chloride, 0.5 mM dithio). Washed with threitol, 0.25 M sucrose). The pellet was suspended in 10 ml of TCDS buffer and applied to a French press (4000 p.s.i.). The solution was then centrifuged at 12,000 xg for 10 minutes at 4 ° C. The supernatant was centrifuged at 110,000 × g for 60 minutes at 4 ° C., and the pellet was suspended in 600 μl of TCDS buffer. Anti-porter activity is 10 mM Tris buffer (HCl added to the specified pH), 5 mM MgCl2Salt was added to a 2 ml reaction mixture containing 0.14 M choline chloride, 1 μM acridine orange, membrane vesicles (50 μg protein) and evaluated by changes in ΔpH (pH gradient inside and outside the membrane). ΔpH was monitored by acridine orange fluorescence obtained at excitation 492 nm and emission 525 nm. Prior to the addition of salt, Tris-DL-lactic acid (2 mM) was added to initiate fluorescence quenching (Q) by respiration. Lactic acid activates vesicles, and H in the vesicles+Which induces dye accumulation and lactate-induced fluorescence quenching (Q). Add salt (5 mM) to add anti-porter H+Fluorescence was enhanced due to the excretion of the unmonitored fluorescence, ie salt-induced fluorescence dequenching (ΔQ). Then NH4Cl (25 mM) was added to dissipate ΔpH.
[0035]
As a result, as shown in the upper part of FIG. 6, when NaCl was added to ApNhaP-expressing cells, dequenching (ΔQ × 100 / Q) was observed, but not in control (pTrcHis2C) cells. ApNhaP is Na+/ H+It shows having antiporter activity. FIG. 6 (A) shows that fluorescence dequenching by ApNhaP was observed over a wide range of pH 5-9, which is almost the same as that by SynNhaP (9), but what is dequenching by E. coli NhaA? Completely different. In E. coli NhaA, the antiporter activity was not observed below pH 7.5, and the activity increased as the pH increased (1).
[0036]
Furthermore, in order to investigate the ion specificity of ApNhaP, the dequenching was measured by adding different cations. Li+And H+Most Na that can catalyze exchange with+/ H+Compared to anti-porters, ApNhaP is Li at different pH values.+/ H+Almost no antiporter activity was shown (FIG. 6B). In addition, ApNhaP is a neutral and alkaline pH Ca2+It was found to show antiporter activity (FIG. 6C), but K+/ H+Shows very little antiporter activity, Mg2+/ H+Did not show at all. These results suggest that ApNhaP is an antiporter with a new ion specificity.
[0037]
[Example 4]
From the above results, ApNhaP is Ca2+And H+Exchange activity, but Li+And H+It was further tested whether such ion specificity would complement the salt-sensitive phenotype of E. coli mutant cells. As a result, as shown in FIG. 7 (A), TO114 cells expressing ApNhaP could not grow in an LBK medium containing 4 mM LiCl at
[0038]
E. coli ChaA is Na+Or Ca2+Proton / cation exchange activity by Li but with Li+Or K+It has been reported that there is no activity by (14, 21), which is basically the same ion specificity as ApNhaP. However, ChaA did not show any homology with ApNhaP. ChaA has multiple Ca such as calsequestrin and calreticulin.2+Binding protein and Na+/ Ca2+Acid motif conserved in ion exchangers Glu200-His-Glu-Asp-Asp-Ser-Asp-Asp-Asp-Asp209(14), but ApNhaP lacks this acidic motif. The hydrophilic plot of ChaA shows that ApNhaP does not have a long hydrophilic C-terminal tail. ApNhaP is a vacuolar Ca from yeast (27) and plants (28)2+/ H+There was no homology with the exchanger. All such data show that ApNhaP has a different structure than ChaA.+/ H+Indicates that it is an anti-porter.
[0039]
[Example 5]
ApNhaP When SynNhaP Chimera gene construction
So far, when the C-terminal tail is partially deleted, SynNhaP Na+/ H+It has been found that the oncoming transport activity is attenuated (9). ApNhaP has the highest homology to SynNhaP, but the charge on the C-terminal tails of the two antiporters is significantly different, ApNhaP has 22 basic amino acids and 14 acidic amino acids, and SynNhaP has 15 bases Sex amino acids and 24 acidic amino acids. Therefore, we constructed a chimeric antiporter of ASNhaP and SANhaP as shown in Figure 8, in which the long C-terminal tail of ApNhaP was replaced with the tail of SynNhaP, and the long C-terminal tail of SynNhaP was replaced with the tail of ApNhaP. did. Chimera exchange activity was assessed using inverted membrane vesicles.
[0040]
That is, the TM region of ApNhaP (amino acid residue Met1−Ile401) And SynNhaP C-terminal region cytoplasmic region (amino acid residue Gln)401−Ser527A chimeric gene encoding) was constructed as follows (FIG. 1). Nucleotides corresponding to the ApNhaP TM region and the SynNhaP C-terminal region were amplified by the forward / reverse primer sets ANP-NcoI / ANP-N-R and SNP-C-F / SNP-C-R, respectively. The former nucleotide template was pANhaP, and the latter nucleotide template was pSNhaP with the synnhaP gene bound to the NcoI / EcoRI site of pTrcHis2C (9). The forward primer, SNP-C-F, has a Gln of ApNhaP, which is a polypeptide extension.398-Thr-Val-Ile401And SynNhaP Gln401-Phe-Val-Leu404ANP-N-R, which is a primer in the reverse direction, is a complementary sequence of SNP-C-F. The reverse primer SNP-C-R has an EcoRI cleavage site immediately before the stop codon of pSNhaP. Both PCR products were combined to amplify the chimeric gene apsynnhaP using the forward ANP-NcoI primer and the reverse SNP-C-R primer, and ligated to the NcoI / EcoRI digestion site of pTrcHis2C, which produced the plasmid pASNhaP.
[0041]
Next, SynNhaP TM region (amino acid residue Met1−Thr400) And ApNhaP C-terminal region (amino acid residue Glu)402−Glu521Each nucleotide was amplified with forward / reverse primer sets of SNP-N-F / SNP-N-R and ANP-C-F / ANP-HindIII. SNP-N-R, which is a reverse primer used in this example, is a complementary sequence of ANP-C-F. Both PCR products were combined to amplify the chimeric gene synapnhaP using forward SNP-N-F and reverse ANP-HindIII primers and ligated to the NcoI / HindIII digestion site of pTrcHis2C, which generated plasmid pSANhaP.
[0042]
As a result, as shown in FIG. 9, the ASNhaP chimera has Na of the parent ApNhaP shown in FIG.+/ H+, Li+/ H+, Ca2+/ H+The activity was comparable to the exchange activity. Interestingly, ApNhaP does not detect Li+/ H+Exchange activity (FIG. 6B) was clearly observed with the ASNhaP chimera (FIG. 9B). On the other hand, the SANhaP chimera is Na compared to the parent SynNhaP.+/ H+And Li+/ H+Both exchange activities were attenuated (FIGS. 9 and 6). In addition, SANhaP chimera Ca2+/ H+The exchange activity is pH dependent, and it appears that enhanced activity is observed when the pH is neutral or alkaline (FIG. 9 (C)). This is in contrast to the observation with the parent SynNhaP, whose activity does not change relatively regardless of pH (FIG. 6 (C)). Although there are discrepancies in activation data, these results suggest that the C-terminal region plays a role in the ion specificity of ApNhaP and SynNhaP. For example, SANhaP chimeras are Na compared to ApNhaP and SynNhaP.+/ H+Exchange activity is low (FIG. 9 (A) and FIG. 6 (A)), and ASNhaP chimera Ca2+/ H+The exchange activity is pH dependent, which is the same as the parent ApNhaP but different from SynNhap (FIG. 9 (C)). These results indicate that the ion specificity of ApNhaP and SynNhaP also varies depending on the structure of the TM region, and most Na+/ H+This suggests that the same can be said for antiporters (1).
[0043]
The chimera exchange activity data was further verified by complementation analysis. As shown in FIG. 10, the ASNhaP chimera is a Na of the E. coli mutant.+, Li+And Ca2+The SANhaP chimera complements the sensitive phenotype, but the E. coli mutant Na+And Li+It is unlikely to complement the sensitive phenotype. The growth rate of E. coli mutant cells expressing SANhaP is the Ca of SANhaP chimera.2+/ H+The rate was slower than expected from the exchange activity (FIGS. 10C and 9C).
[0044]
[Example 6]
Next, eight kinds of plasmids were overexpressed in a freshwater orange-colored bacterium Synechococcus sp. PCC7942, and the salt tolerance was tested.
That is, Na derived from the salt-tolerant orange bacterium Aphanothece halophytica (A. halophytica)+/ H+Opposite transporter ApNhaP expression plasmid pUC303-ApNhaP (33), A. halophytica derived molecular chaperone DnaK expression plasmid pUC303-DnaK (34), E. coli derived catalase expression plasmid pUC303-KatE (35, 36), This is a combination of the expression plasmid pUC303-Bet (37,38) of betaine synthesis gene derived from E. coli and it. These are thought to function as ion homeostasis, protein folding, active oxygen quenching, and osmoprotective substances, respectively. In E. coli, four genes are involved in betaine synthesis (37), betA, betB and betT encode choline dehydrogenase, betaine aldehyde dehydrogenase and choline transporter, respectively, and betI encodes a regulatory gene. In the expression plasmid pUC303-DnaK (34), grpE and dnaJ are partial genes, and dnaK encodes a functional protein. All these genes use their own promoter sequences. Synechococcus sp. PCC7942 cells are pUC303-Bm (control), pUC303-ApNhaP, pUC303-KatE, pUC303-Bet, pUC303-DnaK, pUC303-KatE / Bet, pUC303-ApNhaP / KatE, pUC303-ApNhap / KatE / Bet Transduced (Figure 11).
[0045]
Construction of expression vector
To construct the expression plasmid pUC303-DnaK, a 3 kbp fragment containing the dnaK gene (34) from A. halophytica was digested with SspI and EcoRV to blunt ends, and pUC303-Bm was EcoRI- and SalI-double digested. Then, it was bonded to the site where the blunt end was made. Na from the salt-tolerant orange-colored bacterium A. halophytica+/ H+A plasmid containing its own promoter was prepared as follows with the expression plasmid of the opposite transporter. In PCR using pANhaP as a template, Na from the EcoRI cleavage site at position 539 of the apnhaP gene to the stop codon immediately after the His tag sequence of pTrHis2C+/ H+The antiporter fragment was amplified (33). The forward primer ApNhEcF1 5'-TGGGAATTCCCCTCGGAA-3 'contains an EcoRI site, and the reverse primer ApNhBmR1 5'-GTGGATCCTCAATGATGATGATGATG-3' contains a BamHI cleavage site after the stop codon. The amplified fragment is ligated to the EcoRV site of pBluescript IISK +. -194 to another 539 with EcoRI cleavage site+/ H+The opposite transporter fragment was amplified by PCR using genomic genes using A. halophytica as a template. The
[0046]
Growth of lanthanum bacteria under stress.
Irradiation with continuous white fluorescence (30μEm-2s-1) Under 10μgml-1Synechococcus cells were subcultured at 30 ° C. while sending bubbles to BG11 liquid medium supplemented with streptomycin (359). When the bet gene was expressed, 100 μM choline was added. To apply salt stress, cells in late logarithmic growth phase were transferred to fresh medium containing various concentrations of NaCl (0-0.6M). This culture was incubated for several days. During this period, cell proliferation was monitored while measuring absorbance at 730 nm using Shimadzu UV-160A. SDS-PAGE and immunoblotting were performed as previously published (34). Antibodies raised against 6-histidine (6 × His tag) were obtained from R & D Systems, Inc. (USA). DnaK protein was detected by immunoblotting (34). Glycine betaine was measured using TOF-MS (model KOMPACT MALDI IV tDE Shimdzu / Kratos), and catalase activity was evaluated by activity staining (35).
[0047]
As a result, it was shown that all the transformants can grow at almost the same rate, and this transformant does not impair the growth under low salinity conditions (FIG. 12A). At high salinity, the growth rate of the freshwater orange-colored bacterium Synechococcus PCC7942 (control cell) decreased, and at higher concentrations than 0.375 M NaCl, growth was clearly not observed. At 0.4 M NaCl, control cells and dnaK expressing cells did not grow, and bet or katE expressing cells could grow, but at a slower rate than apnhaP expressing cells (FIG. 12B). Synechococcus cells co-expressing katE and bet grew slightly faster than kat expressing cells or Bet alone. These results are consistent with previous reports (35,38,39), indicating that tolerance to salt stress was acquired by overexpression of bet or katE. Cells that co-expressed apnhaP, katE, and bet had a slightly slower growth rate than apnhaP-expressing cells. The expression of katE (35), bet (38) and dnaK (34) was confirmed as described above. When the NaCl concentration was increased to 0.5M, further reduction in growth rate was observed with various transformants. In addition to control and dnaK expressing cells, cells expressing katE and / or bet could not grow (FIG. 12C). Only apnhaP-expressing cells had a faster growth rate than apnhaP and katE co-expressing cells and apnhaP, katE and bet co-expressing cells. From these data, the salt tolerance of Synechococcus cells was improved by overexpression of ApNhaP, and the salt tolerance of ApNhaP-expressing cells was not improved by co-expression of ApNhaP with other related proteins.
[0048]
Na + / H + Measurement of antiporter activity
ApNhaP protein was detected in apnhaP-expressing cells, but not in control cells grown in 0.3M NaCl (FIG. 13A). The highest concentration of ApNhaP was reached in one day. Therefore, it is considered that the resistance of apnhaP-expressing cells to high salinity is due to the activity of ApNhaP protein.
So, next, actually, Na+/ H+The activity was measured basically according to the method of Nakamura et al. (40). Briefly, Synechococcus sp. PCC 7942 cells were suspended in 0.15 M NaCl (pH 9.3) containing 50 mM diethanolamine-HCl. By this treatment, intracellular K+Is K+/ H+Diethanolamine was replaced by the action of the oncoming transporter. Amine-loaded cells were washed with 0.15 M NaCl (pH 8.5) containing 20 mM HEPES-NaOH buffer. At this stage, the cell interior becomes acidic by the release of diethanolamine, and Na+Is Na+/ H+H by oncoming transporter+There was a load to try to replace. Na+/ H+Prior to measuring countertransporter activity, 0.2% glucose was added to activate the cells, followed by 50 mM diethanolamine-HCl (pH 8.5). If appropriate, cells are harvested by filtration and Nad on the filter by Shimdzu Personal Ion Analyzer PIA-1000.+Ions were measured (35).
[0049]
The resulting Na by addition of diethanolamine.+Na of loaded cells+Excretion activity is shown in FIGS. 13B-D 840). Neutral diethanolamine penetrated the cell and became protonated. Necessary proton is Na+/ H+Na through the opposite transporter+Must be taken up from outside the cell. In this way Na+Excretion activity of Na+/ H+Reflects oncoming transporter activity. Control and ApNhaP expressing cells differ under different salinity conditions+It was investigated whether or not it showed excretion activity. 13B to 13D, diethanolamine was added to add intracellular Na.+Decreased very rapidly in the first few minutes and then slowly decreased. Intracellular Na+Was the largest decrease in cells growing in BG11 medium containing 0.3M NaCl (FIG. 13C), followed by medium without any addition (FIG. 13B), followed by medium containing 0.5M NaCl. (FIG. 13D). Na between the control and ApNhaP expressing cells grown in the absence of NaCl+There was no difference in excretion activity (Figure 13B). When grown in 0.3 M NaCl, ApNhaP-expressing cells have a slightly stronger Na compared to control cells.+Excretion activity was observed (FIG. 13C), and a greater difference was observed when grown on 0.5M NaCl (FIG. 13D). All these results indicated that ApNhaP-expressing cells have higher salinity and higher countertransporter activity than control cells.
[0050]
Next, the stimulating effect of salt on the growth rate of ApNhaP-expressing cells was examined. Salt stimulation (0.65 M NaCl) allowed ApNhaP-expressing cells to grow up to 10 hours later, while control cells only grew up to 3 hours at a slower rate (FIG. 14A). Cell death was detected faster and earlier in control cells than in ApNhaP expressing cells. After 1.2 hours of salt shock, the addition of diethanolamine increases the Na concentration in ApNhaP-expressing cells compared to control cells.+Excretion activity was observed (30% and 75%, 15 minutes) (Figure 14B).
[0051]
Evaluation of photosynthetic activity
Salt stress has been shown to impair photosynthetic activity in several photosynthetic organisms (39). In order to investigate whether ApNhaP expression enhances the photosynthetic activity after salt stimulation, the ratio of currently acting photosystem II (PS II) was expressed as an R value as shown below.
That is, the apparent quantum yield in Photosystem II (PSII) was measured using a PAM fluorometer (Walz, Effeltrich, Germany) (35). Pulse modulated excitation was obtained from a light emitting diode with a peak emission at 650 nm. Modulated fluorescence was measured at λ> 710 nm (Schott RG9 long pass filter). The attached modules of PAM-102 and PAM-103 were used to manage actinic light and saturation pulse, respectively. Prior to measurement, place the cells in a dark room for 10 minutes and continuously light (200 μEm-2s-1) And the quantum yield of PSII was measured. The quantum yield of PSII was calculated by the formula F / Fm ′ = (Fm′−F) / Fm ′. In the formula, F and Fm ′ represent fluorescence levels under irradiation before and after the saturation flash, respectively. The intensity and duration of the flash is 7000μEm each-2s-1And 800ms.
As a result, as shown in FIG. 14C, before the salt stimulation, the R value of the control and the R value of ApNhaP expressing cells were approximately the same 0.90 and 0.85. However, 1.2 hours after salt stimulation, this value decreased to 0.32 and 0.64, respectively. When this is combined with the result of FIG. 14B, the power of the cell to maintain the photosynthetic activity strongly is effective Na.+It was found that it depends on the discharge.
[0052]
ApNhaP Proliferation of expressed cells in seawater
Furthermore, as shown in FIG. 15A, it was actually confirmed that ApNhaP-expressing cells can grow in seawater. Seawater was collected in the Mikawa region of Aichi Prefecture, and its salinity was 30.4psu. Transformed cells expressing either Bet and / or KatE failed to grow in seawater. ApNhaP and KatE co-expression was faster than the growth rate of ApNhaP-expressing cells, in contrast to the results in FIG. 12C using BG11 containing 0.5M NaCl as the growth medium. Although the growth rate was slower than ApNhaP expressing cells, Synechococcus cells co-expressing ApNhaP, KatE and Bet were able to maintain growth in seawater. From these results, it is clear that overexpression of ApNhaP has acquired salt tolerance sufficient for freshwater amber bacteria Synechococcus cells to grow in seawater. During the growth of Synechococcus cells in seawater, a difference in absorption spectrum between the control and ApNhaP expressing cells was observed. As shown in FIG. 15B, ApNhaP-expressing cells showed a typical absorption rate in the region of 630 to 680 nm. In contrast, it can be seen that the control cells have significantly lower absorbency in the 630 nm region than in the 680 nm region. These absorption peaks represent phycobilisomes and chlorophyll, respectively, so the above data indicate that the phycobilisome degeneration of the control cells occurred earlier than chlorophyll, and that phycobilisomes are a vulnerable target due to high salinity I understand.
[0053]
Synechococcus cells are Na at low salinity+And H+It was found from the above data that the exchange capacity of was sufficient. At high salinity, Na+Synechococcus cells are Na+Inability to respond to the inflow, proliferation stopped. Overexpression of ApNhaP in Synechococcus cells could resume growth in high salinity medium and seawater. This is the first report that the freshwater orange bacterium Synechococcus sp.PCC7942 was able to survive in seawater by genetic engineering.
[0054]
【The invention's effect】
According to the present invention, Na having a new ion specificity+/ H+In an Escherichia coli which is an antiporter-deficient strain transformed with the gene and succeeded in isolating the antiporter gene,+Sensitive phenotype and Ca2+It was found that the sensitivity phenotype was complemented. Furthermore, it has been found that overexpression of ApNhaP in the freshwater orange bacterium Synechococcus cells transformed with the gene can acquire salt tolerance sufficient for the freshwater orange bacterium to grow in seawater.
Accordingly, the present invention provides a powerful method that can contribute to the improvement of food production by developing and creating salt-tolerant crops using the gene.
[0055]
Cited references
1. Padan, E., and Schuldiner, S. (1996) in Handbook of Biological Physics (Konings, W. N., Kaback, H. R., and Lolkema, J. S., eds) vol.2, pp.501-531, Elsevier Science, Amsterdam
2. Orlowski, J., and Grinstein, S. (1997) J. Biol. Chem. 272, 22373-22376
3. Blumwald, E., Aharon, G. S., and Apse, M. P. (2000) Biochim. Biophys. Acta 1465, 140-151
4. Nass, R., Cunningham, K. W., Rao, R. (1997) J. Biol. Chem. 272, 26145-26152
5. Nass, R., and Rao, R. (1998) J. Biol. Chem. 274, 21054-21060
6. Gaxiola, R. A., Rao, R., Scherman, A., Grisafi, P., Alper, S. L., and fink, G. R. (1999) Proc. Natl. Acad. Sci. U.S.A. 96, 1480-1485
7. Jia, Z. P., McCullough, N., Martel, R., Hemmingsen, S., and Young, P. G. (1992) EMBO J. 11,1631-1640
8. Shi, H., Ishitani, M., Kim, C., and Zhu, J. K. (2000) Proc. Natl. Acad. Sci. U.S.A. 97, 6896-6901
9. Hamada, A., Hibino, T., Nakamura, T., and Takabe. T. (2001) Plant Physiol. 125, 437-446
10. Wakabayashi, S., Fafournoux, P., Sardet, C., and Pouyssegur, J. (1992) Proc. Natl. Acad. Sci. U.S.A. 89, 2424-2428
11. Inoue, H., Noumi, T., Tsuchiya, T., and Kanazawa, H. (1995) FEBS Lwtt. 363, 264-268
12. Nakamura, T., Komano, Y., and Unemoto, R. (1995) Biochim. Biophys. Acta 1230, 170-176
13. Utsugi, J., Inaba, K., Kuroda, T., Tsuda, M., and Tsuchiya, T. (1998) Biochim. Biophys. Acta 1398, 330-334
14. Ivey, DM, Guffanti, AA, Zemsky. J., Pinner, E., Karpel, R., Padan, E., Schuldiner, S., and Krulwich, A. (1994) J. Biol. Chem. 268.11296 -11303
15. Ishitani, M., Takabe, T., Kojima, K., and Takabe, T. (1993) Aust. J. Plant Physiol. 20, 693-703
16. Takabe, T., Incharoensakdi, A., Arakawa, K., and Takabe, T. (1997) Plant Physiol. 88, 1120-1124
17. Lee, B. H., Hibino, T., Jo, J. Viale, A. M., and Takabe, T. (1997) Plant Mol. Biol. 35, 763-775
18. Hibino, T., Kaku, N., Yoshikawa, H., Takabe, T., and Takabe, T. (1999) Plant Mol. Biol. 40, 409-418
19. Sugino, M., Hibino, T., Tanaka, Y., Nii, N., Takabe, T. and Takabe, T. (1999) Plant Sci, (Shannon) 137, 81-88
20. Triglia, T., Peterson, M. G., and Kemp, D. J. (1998) Nucleic Acids Res. 25, 8186
21. Ohyama, T., Igarashi, K., and Kobayashi, H. (1995) J. Bacteriol. 176, 4311-4315
22. Rosen, B. P. (1986) Methods Enzymol. 125, 328-336
23. Kyte, J., and Doolittle, R. F. (1982) J. Mol. Biol 157, 105-132
24. Hofmann, K., and Stoffel, W. (1992) Comput. Appl. Biosci. 8, 331-337
25. Lowry, O.H., Rosebrough, N.J., Farr, A. L., and Randall, R.J. (1951) J. Bio ;. Chem. 193, 265-275
26. Brockman, R. W., and Heppel, L. A. (1968)
27. Cunningham, K. W., and Fink, G. R. (1996) Mol. Cell. Biol. 16, 2226-2237
28. Ueoka-Nakanishi, H., Nakanishi, Y., Tanaka, Y., and Maeshima, M. (1999) Eur. J. Biochem. 262, 417-425
29. Biemesderfer, D., DeGray, B., and Aronson, P. S. (1998) J. Biol. Chem. 273, 12391-12396
30. Wells, K. M., and Rao, R. (2001) J. Biol. Chem. 276, 3401-3407
31. Ono, K., Hibino, T., Kohinata, T., Suzuki, S., Tanaka, Y., Nakamura, T., Takabe, T., and Takabe, T. (2001) Plant Sci. (Shannon ) 160, 455-461
32. Apse, M. P., Aharon, G. S., Snedden, W. A., and Blumwald, E. (1999) Science 285, 1256-1258
33. Waditee, R.D. et al. Halotolerant cyanobacterium Aphanothece halophytica, contains an Na + / H + antiporter, homologous to eukaryoticones, with novel ion specificity affected by C-terminal tail. Biol. Chem. in press
34. Lee, B. H., Hibino, T., Jo, J., Viale, A. M. and takabe, T. (1997) Plant. Mol. Biol. 35, 763-775
35. Kaku, N. et al (2000) Plant Science 159, 281-288
36. Ossowski, I., Mulvey, M. R., Leco, P.A., Borys, A. and Loewen, P. C. (1991) J. Bacteriol. 173 514-520
37. Lamark, T., Kaasen, I., Eshoo, M. W., McDougall, J. and Strom, A. P. (1991) Mol. Microbiol. 5, 1049-1064
38.Nomura, M., Ishitani, M., Takabe, T., Rai, A. K. and Takabe, T. (1995) Plant Physiol. 107, 703-708
39. Deshnium, P., Los, D. A., Hayashi, H., Mustardy, L. and Murata, N. (1995) Plant Mol Biol. 29, 897-907
40. Nakamura, T., Komano, Y. and Unemoto, T. (1995) Biochim. Biophys. Acta. 1230, 170-176
41. Kaku, N. et al. (1999) Plant Cell Physiol. 40, 557-564
42. Sugino, M., Hibino, T., Nii, N., Takabe, T., and Takabe, T. (1999) Plant Science 146, 81-88
43. Ono, K. et al. (2001) Plant Science 160, 455-461
44. Kishitani, S. et al. (2000) Plant Cell Environ. 23, 107-114
45.Takabe, T. et al. (1998) Glycinebetaine and the genetic engineering ofsalinity tolerance in plants.In Stress Responses of Photosynthetic Organisms.Edited by Satoh, K. and Murata, N. pp.115-131.Elsevier Science, Amsterdam .
46. Tanaka, T. et al. (1999) Plant Science 148, 131-138
[0056]
[Sequence Listing]
[Brief description of the drawings]
FIG. 1 shows a schematic diagram (B) regarding the construction of chimeric plasmids of pASNhaP and pSANhaP.
[Figure 2] Genomic structure of the apnhaP locus of A. halophytica A: Gene and restriction site B: apnhaP and cytochrome c550Nucleotide sequence of the junction. apnhaP and cytochrome c550Also shown are the putative -10 (ATGAAT) and -35 (TACACT) consensus and amino acid sequences of apnhaP deduced from the nucleotide sequence of.
[Figure 3] Na+/ H+Comparison of deduced amino acid sequences of antiporters A: Na derived from 8 organisms+/ H+Antiporter deduced amino acid sequence alignment. This sequence was aligned by the program ClustalW. This alignment is based on the N-terminal 550 amino acid residues of SOS1, human NHE1, and yeast NHX1. Amino acid residues conserved in all sequences are highlighted in black and conservative substitutions are indicated by dots. Save Asp (ApNhaP Asp139) Is indicated by an asterisk. The predicted transmembrane region is shown above the alignment. B: 8 Na+/ H+Phylogenetic analysis of antiporters. ClustalW and TreeView software were used to create multiple sequence alignments and phylogenetic trees, respectively.
FIG. 4: Effect of NaCl on the growth rate of three E. coli cells. Control cells and transformed cells in logarithmic growth phase in LBK medium were transferred to fresh LBK medium (pH 7.0) containing the indicated NaCl concentration. A: Transition of growth in LBK medium B: Transition of growth in LBK containing 0.2 M NaCl Circle: Control cells Square: ApNhaP expressing cells
Triangle: SynNhaP expressing cells. Each value is the average of three measurements.
FIG. 5: Effect of pH and NaCl on the growth rate of three E. coli cells Fresh LBK medium at indicated pH, containing indicated concentrations of control and transformed cells in logarithmic growth phase in LBK medium. Moved to. The growth temperature was 30 ° C. Transition of growth in LBK medium containing 0 M NaCl (A), 0.2 M NaCl (B) and 0.5 M NaCl (C). Open squares: ApNhaP expressing cells, pH 8.0. Shaded square: pH 8.5. Black square: pH 9.0. Open triangle: SynNhaP expressing cells, pH 8.0. Shaded triangle: pH 8.5. Black triangle: pH 9.0. Open circle: control cells, pH 8.0. Shaded circle: ph 8.5. Black circle: pH 9.0. Each value is the average of three measurements.
FIG. 6 Na measured by acridine orange fluorescence quenching method+/ H+Antiporter activity Control E. coli cells and E. coli cells expressing ApNhaP and SynNhaP were grown in LBK medium without the addition of salt. Inverted membrane vesicles were prepared from these cells. Antiporter activity was measured as described in the examples. The upper row shows typical quenching patterns of ApNhaP-expressing cells and control cells (pTrcHis2C). A, B, and C are Na+/ H+, Li+/ H+, Ca2+/ H+The pH dependence of the antiporter activity was shown. Salt (NaCl, LiCl, CaCl2) Final concentration was 5 mM. Each value is the average of three individual measurements.
FIG. 7 LiCl and CaCl versus growth rate of three E. coli cells.2Specified LiCl or CaCl2Control cells and transformed cells in logarithmic growth phase were transferred to LBK or Tris E medium at the respective concentrations. Other experimental conditions are as described in the explanation of FIG. Each value is the average of three individual measurements.
FIG. 8 Chimeric Na+/ H+Secondary structure model of antiporter Chimera Na+/ H+An antiporter was constructed as described in the examples. Shows the number of positive and negative charges on the C-terminal hydrophilic tail.
FIG. 9 Chimeric antiporter activity E. coli cells expressing ASNhaP chimera and SANhaP chimera were grown in LBK medium without the addition of salt. Inverted membrane vesicles were prepared from these cells. Antiporter activity was constructed as described in the Examples. Each value is the average of three individual measurements.
FIG. 10 shows NaCl, LiCl, CaCl with respect to the growth rate of E. coli cells expressing ASNhaP chimera and SANhaP chimera.2E. coli cells expressing ASNhaP chimera and SANhaP chimera in logarithmic growth phase in LBK medium are treated with LBK medium containing 0.2 M NaCl, LBK medium containing 0.4 mM LiCl, 0.1 M CaCl.2To Tris E medium containing Other experimental conditions are the same as those described in the explanation of FIG. Each value is the average of three individual measurements.
FIG. 11 shows a schematic diagram of an expression vector.
FIG. 12. Effect of salt stress on the growth rate of control and transformed Synechococcus cells. A fresh medium containing a prescribed NaCl concentration was impregnated with control and transformed cells in the logarithmic growth phase with BG11 medium and subjected to salt stress.
FIG. 13: ApNhaP accumulation (A) and Na+Na from loaded cells+Emissions (B, C, D). In A, control and ApNhaP-expressing cells were grown in BG11 medium and transferred to medium containing 0.3M NaCl. After incubation for a specified time, cells were collected and subjected to immunological analysis. In B, C, and D, control (open circles) and ApNhaP-expressing cells (filled circles) were grown for one day in a BG11 medium containing a prescribed concentration of NaCl. Next, Na+Diethanolamine (EA) was measured to induce excretion.
FIG. 14 Growth rate in control and ApNhaP expressing Synechococcus cells (A), Na+Effects of salt stimulation on excretion activity (B) and photosynthetic electron transport activity (C). Logarithmic growth phase control and ApNhaP-expressing cells were collected in BG11 medium and transferred to fresh medium containing 0.65 M NaCl.
FIG. 15 Growth curve (A) and absorption spectrum (B) of control and transformed Synechococcus cells in sea water. Logarithmic growth phase controls and transformed cells were transferred to seawater in BG11 medium. Three days after transferring to seawater, absorption spectra (B) of control and ApNhaP-expressing cells were measured.
Claims (15)
(a)配列番号2に示されるアミノ酸配列から成るタンパク質、
(b)アミノ酸配列(a)において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、Na+/H+アンチポーター活性を有するタンパク質。DNA containing a base sequence encoding the following protein (a) or (b):
(A) a protein comprising the amino acid sequence shown in SEQ ID NO: 2,
(B) A protein having an Na + / H + antiporter activity, comprising an amino acid sequence in which one or several amino acids are deleted, substituted or added in the amino acid sequence (a).
(a)配列番号1における第796〜2361番目の塩基対からなる塩基配列、
(b)塩基配列(a)からなるDNAとストリンジェントな条件下でハイブリダイズし、且つ、Na+/H+アンチポーター活性を有するタンパク質をコードする塩基配列。DNA comprising the following base sequence (a) or (b):
(A) a base sequence consisting of the 796 to 2361th base pairs in SEQ ID NO: 1,
(B) A nucleotide sequence that hybridizes with a DNA comprising the nucleotide sequence (a) under a stringent condition and encodes a protein having Na + / H + antiporter activity.
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2001390491A JP4132810B2 (en) | 2001-12-21 | 2001-12-21 | Na + / H + antiporter gene and improvement of salt tolerance by expression of the gene |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2001390491A JP4132810B2 (en) | 2001-12-21 | 2001-12-21 | Na + / H + antiporter gene and improvement of salt tolerance by expression of the gene |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2003180373A JP2003180373A (en) | 2003-07-02 |
| JP4132810B2 true JP4132810B2 (en) | 2008-08-13 |
Family
ID=27598393
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2001390491A Expired - Fee Related JP4132810B2 (en) | 2001-12-21 | 2001-12-21 | Na + / H + antiporter gene and improvement of salt tolerance by expression of the gene |
Country Status (1)
| Country | Link |
|---|---|
| JP (1) | JP4132810B2 (en) |
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPH0574923U (en) * | 1992-03-13 | 1993-10-12 | 関東自動車工業株式会社 | Clips for car mall mounting |
-
2001
- 2001-12-21 JP JP2001390491A patent/JP4132810B2/en not_active Expired - Fee Related
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPH0574923U (en) * | 1992-03-13 | 1993-10-12 | 関東自動車工業株式会社 | Clips for car mall mounting |
Also Published As
| Publication number | Publication date |
|---|---|
| JP2003180373A (en) | 2003-07-02 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US8168864B2 (en) | Transgenic pollen expressing exogenous plant vacuolar pyrophosphatase and methods for increasing seed production in plants | |
| Lopez et al. | Accumulation of a 22‐kDa protein and its mRNA in the leaves of Raphanus sativus in response to salt stress or water deficit | |
| MX2007007567A (en) | The transcription factor gene osnacx from rice and use for improving tolerance of plants to drought and salt thereof. | |
| AU2010283592B2 (en) | Stress tolerant plants | |
| US8586830B2 (en) | Ioslated TT1 polynucleotide, encoded proteins and vectors for increasing tolerance of plants and microbes to abiotic stresses and the use thereof | |
| BRPI0618328A2 (en) | method for improving plant growth characteristics over corresponding wild type plants, construction, host cell, method for producing a transgenic plant, plant part or plant cell having improved plant growth characteristics over wild type plants corresponding, and, uses of a construct and a nucleic acid | |
| DE112008003224T5 (en) | Transgenic plants with increased stress tolerance and increased yield | |
| WO2001006006A1 (en) | Environmental stress resistance gene | |
| CN101969759A (en) | Plants having enhanced yield-related traits and a method for making the same | |
| NL2005620C2 (en) | Plants with increased tolerance to metal ions. | |
| CA2390719A1 (en) | Method of transforming plants with vacuolar pyrophosphatase and stress-resistant oversized plants produced by the method | |
| CN104610439B (en) | Protein, its encoding gene and the purposes related to abiotic stress reaction | |
| KR100440097B1 (en) | Sodium/proton countertransporters, DNAs encoding the same, methods for preparing thereof and transformant plants having the DNAs | |
| CN111979253B (en) | TrFQR1 gene, cloning thereof, expression vector construction method and application | |
| CN107012167A (en) | The expression of the transcription regulaton factor of heat tolerance can be provided | |
| Roychoudhury et al. | Structural aspects and functional regulation of late embryogenesis abundant (LEA) genes and proteins conferring abiotic stress tolerance in plants | |
| CA2559093C (en) | Proteins imparting boron-tolerance and genes thereof | |
| Hossain et al. | Root specific expression of Na+/H+ antiporter gene from Synechocystis sp. PCC 6803 confers salt tolerance of tobacco plant | |
| JP4132810B2 (en) | Na + / H + antiporter gene and improvement of salt tolerance by expression of the gene | |
| CN113880926B (en) | Plant salt tolerance related protein and related biological material and application thereof | |
| CN107266542A (en) | Thick boisiana IpLEA genes and its encoding proteins and application | |
| US7816582B2 (en) | Arabinogalactan protein having activity of improving tolerance to heat or moisture stress | |
| KR20140063310A (en) | Transgenic plant with enhanced tolerance to cold stress by introducing grxc gene and preparation method thereof | |
| CN113046375A (en) | SpCPK33 gene and application of encoding protein thereof in regulation and control of tomato drought tolerance | |
| CN115322248B (en) | Potassium ion channel protein AlAKT1, coding gene and application thereof |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A711 | Notification of change in applicant |
Free format text: JAPANESE INTERMEDIATE CODE: A712 Effective date: 20031031 |
|
| RD03 | Notification of appointment of power of attorney |
Free format text: JAPANESE INTERMEDIATE CODE: A7423 Effective date: 20040129 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20040908 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20070213 |
|
| A521 | Written amendment |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20070416 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20071106 |
|
| A521 | Written amendment |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20080107 |
|
| A711 | Notification of change in applicant |
Free format text: JAPANESE INTERMEDIATE CODE: A711 Effective date: 20080229 |
|
| A521 | Written amendment |
Free format text: JAPANESE INTERMEDIATE CODE: A821 Effective date: 20080303 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20080520 |
|
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20080602 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20110606 Year of fee payment: 3 |
|
| R150 | Certificate of patent or registration of utility model |
Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20110606 Year of fee payment: 3 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20140606 Year of fee payment: 6 |
|
| LAPS | Cancellation because of no payment of annual fees |