JP2022550141A - 肺線維症モデルおよびそれを使用する方法 - Google Patents
肺線維症モデルおよびそれを使用する方法 Download PDFInfo
- Publication number
- JP2022550141A JP2022550141A JP2022519551A JP2022519551A JP2022550141A JP 2022550141 A JP2022550141 A JP 2022550141A JP 2022519551 A JP2022519551 A JP 2022519551A JP 2022519551 A JP2022519551 A JP 2022519551A JP 2022550141 A JP2022550141 A JP 2022550141A
- Authority
- JP
- Japan
- Prior art keywords
- cells
- pats
- alveolar
- lung
- aec1
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
- 238000000034 method Methods 0.000 title claims abstract description 73
- 208000005069 pulmonary fibrosis Diseases 0.000 title claims abstract description 27
- 210000004027 cell Anatomy 0.000 claims abstract description 414
- 210000002950 fibroblast Anatomy 0.000 claims abstract description 40
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 claims abstract description 34
- 201000010099 disease Diseases 0.000 claims abstract description 30
- 230000002685 pulmonary effect Effects 0.000 claims abstract description 15
- 206010014561 Emphysema Diseases 0.000 claims abstract description 11
- 210000004072 lung Anatomy 0.000 claims description 168
- 210000001519 tissue Anatomy 0.000 claims description 91
- 230000014509 gene expression Effects 0.000 claims description 71
- 208000027418 Wounds and injury Diseases 0.000 claims description 61
- 208000014674 injury Diseases 0.000 claims description 61
- 239000003795 chemical substances by application Substances 0.000 claims description 59
- 108090000623 proteins and genes Proteins 0.000 claims description 56
- 230000006378 damage Effects 0.000 claims description 55
- 108010006654 Bleomycin Proteins 0.000 claims description 51
- 229960001561 bleomycin Drugs 0.000 claims description 51
- OYVAGSVQBOHSSS-UAPAGMARSA-O bleomycin A2 Chemical group N([C@H](C(=O)N[C@H](C)[C@@H](O)[C@H](C)C(=O)N[C@@H]([C@H](O)C)C(=O)NCCC=1SC=C(N=1)C=1SC=C(N=1)C(=O)NCCC[S+](C)C)[C@@H](O[C@H]1[C@H]([C@@H](O)[C@H](O)[C@H](CO)O1)O[C@@H]1[C@H]([C@@H](OC(N)=O)[C@H](O)[C@@H](CO)O1)O)C=1N=CNC=1)C(=O)C1=NC([C@H](CC(N)=O)NC[C@H](N)C(N)=O)=NC(N)=C1C OYVAGSVQBOHSSS-UAPAGMARSA-O 0.000 claims description 51
- NKANXQFJJICGDU-QPLCGJKRSA-N Tamoxifen Chemical compound C=1C=CC=CC=1C(/CC)=C(C=1C=CC(OCCN(C)C)=CC=1)/C1=CC=CC=C1 NKANXQFJJICGDU-QPLCGJKRSA-N 0.000 claims description 48
- 210000002220 organoid Anatomy 0.000 claims description 48
- 201000009794 Idiopathic Pulmonary Fibrosis Diseases 0.000 claims description 47
- 208000036971 interstitial lung disease 2 Diseases 0.000 claims description 47
- 210000000130 stem cell Anatomy 0.000 claims description 47
- 102000000802 Galectin 3 Human genes 0.000 claims description 46
- 108010001517 Galectin 3 Proteins 0.000 claims description 46
- 230000011664 signaling Effects 0.000 claims description 44
- 238000002679 ablation Methods 0.000 claims description 42
- 102100038447 Claudin-4 Human genes 0.000 claims description 40
- 101000882890 Homo sapiens Claudin-4 Proteins 0.000 claims description 40
- 108091008606 PDGF receptors Proteins 0.000 claims description 40
- 102000011653 Platelet-Derived Growth Factor Receptors Human genes 0.000 claims description 40
- 238000001727 in vivo Methods 0.000 claims description 38
- 108010043471 Core Binding Factor Alpha 2 Subunit Proteins 0.000 claims description 37
- 206010016654 Fibrosis Diseases 0.000 claims description 31
- 230000004761 fibrosis Effects 0.000 claims description 31
- 239000003550 marker Substances 0.000 claims description 30
- 108010053187 Diphtheria Toxin Proteins 0.000 claims description 28
- 102000016607 Diphtheria Toxin Human genes 0.000 claims description 28
- 239000003112 inhibitor Substances 0.000 claims description 27
- 101000723509 Homo sapiens 14-3-3 protein sigma Proteins 0.000 claims description 26
- 101001130862 Homo sapiens Oligoribonuclease, mitochondrial Proteins 0.000 claims description 26
- 108010038512 Platelet-Derived Growth Factor Proteins 0.000 claims description 26
- 102000010780 Platelet-Derived Growth Factor Human genes 0.000 claims description 25
- 102100025373 Runt-related transcription factor 1 Human genes 0.000 claims description 24
- 229960001603 tamoxifen Drugs 0.000 claims description 24
- 101001126417 Homo sapiens Platelet-derived growth factor receptor alpha Proteins 0.000 claims description 19
- 102100030485 Platelet-derived growth factor receptor alpha Human genes 0.000 claims description 19
- 102100027881 Tumor protein 63 Human genes 0.000 claims description 19
- 101710140697 Tumor protein 63 Proteins 0.000 claims description 19
- 239000003446 ligand Substances 0.000 claims description 19
- 210000000651 myofibroblast Anatomy 0.000 claims description 19
- 102000015775 Core Binding Factor Alpha 1 Subunit Human genes 0.000 claims description 18
- 108010024682 Core Binding Factor Alpha 1 Subunit Proteins 0.000 claims description 18
- 108010079362 Core Binding Factor Alpha 3 Subunit Proteins 0.000 claims description 18
- 230000002068 genetic effect Effects 0.000 claims description 18
- 102100037596 Platelet-derived growth factor subunit A Human genes 0.000 claims description 17
- 239000001963 growth medium Substances 0.000 claims description 17
- 208000020816 lung neoplasm Diseases 0.000 claims description 16
- 101000998027 Homo sapiens Keratin, type I cytoskeletal 17 Proteins 0.000 claims description 15
- 102100033511 Keratin, type I cytoskeletal 17 Human genes 0.000 claims description 15
- 206010058467 Lung neoplasm malignant Diseases 0.000 claims description 14
- 241000700605 Viruses Species 0.000 claims description 14
- 239000000556 agonist Substances 0.000 claims description 14
- 210000002919 epithelial cell Anatomy 0.000 claims description 14
- 201000005202 lung cancer Diseases 0.000 claims description 14
- 102100027833 14-3-3 protein sigma Human genes 0.000 claims description 13
- 230000004071 biological effect Effects 0.000 claims description 13
- 241000233866 Fungi Species 0.000 claims description 12
- 241000894006 Bacteria Species 0.000 claims description 11
- 208000004852 Lung Injury Diseases 0.000 claims description 11
- 206010069363 Traumatic lung injury Diseases 0.000 claims description 11
- 230000000694 effects Effects 0.000 claims description 11
- 231100000515 lung injury Toxicity 0.000 claims description 11
- 102000006990 Core Binding Factors Human genes 0.000 claims description 9
- 108010072732 Core Binding Factors Proteins 0.000 claims description 9
- 238000003501 co-culture Methods 0.000 claims description 9
- 102000004169 proteins and genes Human genes 0.000 claims description 9
- 101000975496 Homo sapiens Keratin, type II cytoskeletal 8 Proteins 0.000 claims description 8
- 102100023972 Keratin, type II cytoskeletal 8 Human genes 0.000 claims description 8
- XWNMORIHKRROGW-UHFFFAOYSA-N Ro 5-3335 Chemical group C12=CC(Cl)=CC=C2NC(=O)CN=C1C1=CC=CN1 XWNMORIHKRROGW-UHFFFAOYSA-N 0.000 claims description 8
- 102100033601 Collagen alpha-1(I) chain Human genes 0.000 claims description 7
- 101000642514 Homo sapiens Transcription factor SOX-4 Proteins 0.000 claims description 7
- 208000029523 Interstitial Lung disease Diseases 0.000 claims description 7
- 102100036693 Transcription factor SOX-4 Human genes 0.000 claims description 7
- 108010029483 alpha 1 Chain Collagen Type I Proteins 0.000 claims description 7
- 230000003472 neutralizing effect Effects 0.000 claims description 7
- 208000035143 Bacterial infection Diseases 0.000 claims description 6
- 208000006545 Chronic Obstructive Pulmonary Disease Diseases 0.000 claims description 6
- 108010037462 Cyclooxygenase 2 Proteins 0.000 claims description 6
- 101000998011 Homo sapiens Keratin, type I cytoskeletal 19 Proteins 0.000 claims description 6
- 101000990912 Homo sapiens Matrilysin Proteins 0.000 claims description 6
- 101000685726 Homo sapiens Protein S100-A2 Proteins 0.000 claims description 6
- 101000848014 Homo sapiens Trypsin-2 Proteins 0.000 claims description 6
- 102100033420 Keratin, type I cytoskeletal 19 Human genes 0.000 claims description 6
- 102100030417 Matrilysin Human genes 0.000 claims description 6
- 102100038280 Prostaglandin G/H synthase 2 Human genes 0.000 claims description 6
- 102100023089 Protein S100-A2 Human genes 0.000 claims description 6
- 102100034392 Trypsin-2 Human genes 0.000 claims description 6
- 208000036142 Viral infection Diseases 0.000 claims description 6
- 208000022362 bacterial infectious disease Diseases 0.000 claims description 6
- 210000000270 basal cell Anatomy 0.000 claims description 6
- 206010006475 bronchopulmonary dysplasia Diseases 0.000 claims description 6
- 239000006143 cell culture medium Substances 0.000 claims description 6
- 201000000306 sarcoidosis Diseases 0.000 claims description 6
- 210000003491 skin Anatomy 0.000 claims description 6
- WJBSSBFGPKTMQQ-UHFFFAOYSA-N 6-(trifluoromethoxy)-2-[5-[2-[2-[6-[6-(trifluoromethoxy)-1h-benzimidazol-2-yl]pyridin-3-yl]oxyethoxy]ethoxy]pyridin-2-yl]-1h-benzimidazole Chemical compound C1=C(OC(F)(F)F)C=C2NC(C3=CC=C(C=N3)OCCOCCOC3=CC=C(N=C3)C3=NC4=CC=C(C=C4N3)OC(F)(F)F)=NC2=C1 WJBSSBFGPKTMQQ-UHFFFAOYSA-N 0.000 claims description 5
- 206010001052 Acute respiratory distress syndrome Diseases 0.000 claims description 5
- 101100351501 Mus musculus Cbfb gene Proteins 0.000 claims description 5
- 102100040990 Platelet-derived growth factor subunit B Human genes 0.000 claims description 5
- 208000013616 Respiratory Distress Syndrome Diseases 0.000 claims description 5
- 206010040047 Sepsis Diseases 0.000 claims description 5
- 201000000028 adult respiratory distress syndrome Diseases 0.000 claims description 5
- 210000005058 airway cell Anatomy 0.000 claims description 5
- 210000001185 bone marrow Anatomy 0.000 claims description 5
- 238000012258 culturing Methods 0.000 claims description 5
- 238000000338 in vitro Methods 0.000 claims description 5
- 210000000936 intestine Anatomy 0.000 claims description 5
- 210000003734 kidney Anatomy 0.000 claims description 5
- 210000004185 liver Anatomy 0.000 claims description 5
- QHRCVGXBBIRPTE-UHFFFAOYSA-N 5-ethyl-4-(4-methoxyphenyl)-1,3-thiazol-2-amine Chemical compound S1C(N)=NC(C=2C=CC(OC)=CC=2)=C1CC QHRCVGXBBIRPTE-UHFFFAOYSA-N 0.000 claims description 4
- 101710103506 Platelet-derived growth factor subunit A Proteins 0.000 claims description 4
- 101710103494 Platelet-derived growth factor subunit B Proteins 0.000 claims description 4
- 101150106167 SOX9 gene Proteins 0.000 claims description 4
- 210000000233 bronchiolar non-ciliated Anatomy 0.000 claims description 4
- 210000002216 heart Anatomy 0.000 claims description 4
- 210000004263 induced pluripotent stem cell Anatomy 0.000 claims description 4
- 210000004412 neuroendocrine cell Anatomy 0.000 claims description 4
- 238000012216 screening Methods 0.000 claims description 4
- 210000002955 secretory cell Anatomy 0.000 claims description 4
- 101100058960 Arabidopsis thaliana CALS1 gene Proteins 0.000 claims description 3
- 101000693933 Arabidopsis thaliana Fructose-bisphosphate aldolase 8, cytosolic Proteins 0.000 claims description 3
- 102100024436 Caldesmon Human genes 0.000 claims description 3
- 102100032215 Cathepsin E Human genes 0.000 claims description 3
- 102100040896 Growth/differentiation factor 15 Human genes 0.000 claims description 3
- 101000910297 Homo sapiens Caldesmon Proteins 0.000 claims description 3
- 101000869031 Homo sapiens Cathepsin E Proteins 0.000 claims description 3
- 101000893549 Homo sapiens Growth/differentiation factor 15 Proteins 0.000 claims description 3
- 101000840577 Homo sapiens Insulin-like growth factor-binding protein 7 Proteins 0.000 claims description 3
- 101000990990 Homo sapiens Midkine Proteins 0.000 claims description 3
- 101000652736 Homo sapiens Transgelin Proteins 0.000 claims description 3
- 101000658574 Homo sapiens Transmembrane 4 L6 family member 1 Proteins 0.000 claims description 3
- 102100029228 Insulin-like growth factor-binding protein 7 Human genes 0.000 claims description 3
- 102100030335 Midkine Human genes 0.000 claims description 3
- 101100005010 Mus musculus Ca8 gene Proteins 0.000 claims description 3
- 102100031013 Transgelin Human genes 0.000 claims description 3
- 102100034902 Transmembrane 4 L6 family member 1 Human genes 0.000 claims description 3
- 239000010425 asbestos Substances 0.000 claims description 3
- 229910052895 riebeckite Inorganic materials 0.000 claims description 3
- 230000009385 viral infection Effects 0.000 claims description 3
- 230000003612 virological effect Effects 0.000 claims description 3
- 206010017533 Fungal infection Diseases 0.000 claims description 2
- 208000031888 Mycoses Diseases 0.000 claims description 2
- 231100000196 chemotoxic Toxicity 0.000 claims description 2
- 230000002604 chemotoxic effect Effects 0.000 claims description 2
- 239000000126 substance Substances 0.000 claims description 2
- 208000011580 syndromic disease Diseases 0.000 claims description 2
- 102000012666 Core Binding Factor Alpha 3 Subunit Human genes 0.000 claims 5
- 102100036732 Actin, aortic smooth muscle Human genes 0.000 claims 2
- 101000929319 Homo sapiens Actin, aortic smooth muscle Proteins 0.000 claims 2
- 206010038687 Respiratory distress Diseases 0.000 claims 1
- 230000001154 acute effect Effects 0.000 claims 1
- 230000000266 injurious effect Effects 0.000 claims 1
- 230000008929 regeneration Effects 0.000 abstract description 16
- 238000011069 regeneration method Methods 0.000 abstract description 16
- 208000019693 Lung disease Diseases 0.000 abstract description 7
- 238000007877 drug screening Methods 0.000 abstract description 6
- 238000007876 drug discovery Methods 0.000 abstract description 3
- 210000002821 alveolar epithelial cell Anatomy 0.000 description 208
- 241000699670 Mus sp. Species 0.000 description 62
- 102100025064 Cellular tumor antigen p53 Human genes 0.000 description 56
- 108010078814 Tumor Suppressor Protein p53 Proteins 0.000 description 55
- 239000005090 green fluorescent protein Substances 0.000 description 33
- 238000012744 immunostaining Methods 0.000 description 29
- 238000004458 analytical method Methods 0.000 description 28
- 102100029599 Advanced glycosylation end product-specific receptor Human genes 0.000 description 26
- 108010045108 Receptor for Advanced Glycation End Products Proteins 0.000 description 26
- 230000004069 differentiation Effects 0.000 description 26
- 230000003176 fibrotic effect Effects 0.000 description 26
- 238000012174 single-cell RNA sequencing Methods 0.000 description 26
- -1 ACTA2 Proteins 0.000 description 24
- IAZDPXIOMUYVGZ-UHFFFAOYSA-N Dimethylsulphoxide Chemical compound CS(C)=O IAZDPXIOMUYVGZ-UHFFFAOYSA-N 0.000 description 20
- 230000005778 DNA damage Effects 0.000 description 18
- 231100000277 DNA damage Toxicity 0.000 description 18
- 241000699666 Mus <mouse, genus> Species 0.000 description 18
- 238000012360 testing method Methods 0.000 description 17
- BDUHCSBCVGXTJM-WUFINQPMSA-N 4-[[(4S,5R)-4,5-bis(4-chlorophenyl)-2-(4-methoxy-2-propan-2-yloxyphenyl)-4,5-dihydroimidazol-1-yl]-oxomethyl]-2-piperazinone Chemical compound CC(C)OC1=CC(OC)=CC=C1C1=N[C@@H](C=2C=CC(Cl)=CC=2)[C@@H](C=2C=CC(Cl)=CC=2)N1C(=O)N1CC(=O)NCC1 BDUHCSBCVGXTJM-WUFINQPMSA-N 0.000 description 14
- 230000009758 senescence Effects 0.000 description 14
- 102000002664 Core Binding Factor Alpha 2 Subunit Human genes 0.000 description 13
- 102100025369 Runt-related transcription factor 3 Human genes 0.000 description 13
- 230000037361 pathway Effects 0.000 description 13
- 108010017843 platelet-derived growth factor A Proteins 0.000 description 13
- 230000019491 signal transduction Effects 0.000 description 13
- 108010016788 Cyclin-Dependent Kinase Inhibitor p21 Proteins 0.000 description 12
- 102100033270 Cyclin-dependent kinase inhibitor 1 Human genes 0.000 description 12
- 239000002158 endotoxin Substances 0.000 description 12
- 229920006008 lipopolysaccharide Polymers 0.000 description 12
- 101000612671 Homo sapiens Pulmonary surfactant-associated protein C Proteins 0.000 description 11
- 241001465754 Metazoa Species 0.000 description 11
- 102100040971 Pulmonary surfactant-associated protein C Human genes 0.000 description 11
- 239000005557 antagonist Substances 0.000 description 11
- 108010051779 histone H3 trimethyl Lys4 Proteins 0.000 description 10
- 230000001965 increasing effect Effects 0.000 description 10
- 230000002688 persistence Effects 0.000 description 10
- 230000001052 transient effect Effects 0.000 description 10
- FWBHETKCLVMNFS-UHFFFAOYSA-N 4',6-Diamino-2-phenylindol Chemical compound C1=CC(C(=N)N)=CC=C1C1=CC2=CC=C(C(N)=N)C=C2N1 FWBHETKCLVMNFS-UHFFFAOYSA-N 0.000 description 9
- 238000006243 chemical reaction Methods 0.000 description 9
- 210000000056 organ Anatomy 0.000 description 9
- 238000010186 staining Methods 0.000 description 9
- 101150091877 Ccn2 gene Proteins 0.000 description 8
- 101150023976 Krt19 gene Proteins 0.000 description 8
- 102100030742 Transforming growth factor beta-1 proprotein Human genes 0.000 description 8
- 230000002159 abnormal effect Effects 0.000 description 8
- 230000004913 activation Effects 0.000 description 8
- 230000010094 cellular senescence Effects 0.000 description 8
- 230000002950 deficient Effects 0.000 description 8
- 238000012217 deletion Methods 0.000 description 8
- 230000037430 deletion Effects 0.000 description 8
- 102000005962 receptors Human genes 0.000 description 8
- 108020003175 receptors Proteins 0.000 description 8
- OPIFSICVWOWJMJ-AEOCFKNESA-N 5-bromo-4-chloro-3-indolyl beta-D-galactoside Chemical compound O[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@H]1OC1=CNC2=CC=C(Br)C(Cl)=C12 OPIFSICVWOWJMJ-AEOCFKNESA-N 0.000 description 7
- 101150106169 LGALS3 gene Proteins 0.000 description 7
- 238000000692 Student's t-test Methods 0.000 description 7
- 230000032683 aging Effects 0.000 description 7
- 238000002487 chromatin immunoprecipitation Methods 0.000 description 7
- 108010033040 Histones Proteins 0.000 description 6
- 102000001393 Platelet-Derived Growth Factor alpha Receptor Human genes 0.000 description 6
- 108010068588 Platelet-Derived Growth Factor alpha Receptor Proteins 0.000 description 6
- 238000002474 experimental method Methods 0.000 description 6
- 238000010172 mouse model Methods 0.000 description 6
- 230000001575 pathological effect Effects 0.000 description 6
- 230000002062 proliferating effect Effects 0.000 description 6
- 238000011002 quantification Methods 0.000 description 6
- 230000004044 response Effects 0.000 description 6
- 239000000523 sample Substances 0.000 description 6
- 102100026189 Beta-galactosidase Human genes 0.000 description 5
- 238000001353 Chip-sequencing Methods 0.000 description 5
- 101100043050 Mus musculus Sox4 gene Proteins 0.000 description 5
- 101150038994 PDGFRA gene Proteins 0.000 description 5
- 108090001012 Transforming Growth Factor beta Proteins 0.000 description 5
- 108010005774 beta-Galactosidase Proteins 0.000 description 5
- 230000001413 cellular effect Effects 0.000 description 5
- 230000003436 cytoskeletal effect Effects 0.000 description 5
- 238000009826 distribution Methods 0.000 description 5
- 239000003623 enhancer Substances 0.000 description 5
- 230000006870 function Effects 0.000 description 5
- 230000003284 homeostatic effect Effects 0.000 description 5
- 230000000977 initiatory effect Effects 0.000 description 5
- 239000007928 intraperitoneal injection Substances 0.000 description 5
- 239000002609 medium Substances 0.000 description 5
- 108010054624 red fluorescent protein Proteins 0.000 description 5
- 230000022532 regulation of transcription, DNA-dependent Effects 0.000 description 5
- 230000008439 repair process Effects 0.000 description 5
- 230000028617 response to DNA damage stimulus Effects 0.000 description 5
- 230000017423 tissue regeneration Effects 0.000 description 5
- 230000007704 transition Effects 0.000 description 5
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 4
- 101150014060 Ager gene Proteins 0.000 description 4
- 101150025841 CCND1 gene Proteins 0.000 description 4
- 108050002772 E3 ubiquitin-protein ligase Mdm2 Proteins 0.000 description 4
- 102000012199 E3 ubiquitin-protein ligase Mdm2 Human genes 0.000 description 4
- WZUVPPKBWHMQCE-UHFFFAOYSA-N Haematoxylin Chemical compound C12=CC(O)=C(O)C=C2CC2(O)C1C1=CC=C(O)C(O)=C1OC2 WZUVPPKBWHMQCE-UHFFFAOYSA-N 0.000 description 4
- 102100033636 Histone H3.2 Human genes 0.000 description 4
- 101150073145 SFTPC gene Proteins 0.000 description 4
- 238000009825 accumulation Methods 0.000 description 4
- 230000015572 biosynthetic process Effects 0.000 description 4
- 230000000903 blocking effect Effects 0.000 description 4
- 230000024245 cell differentiation Effects 0.000 description 4
- 208000035475 disorder Diseases 0.000 description 4
- 239000003814 drug Substances 0.000 description 4
- 238000001943 fluorescence-activated cell sorting Methods 0.000 description 4
- 238000003306 harvesting Methods 0.000 description 4
- 230000013632 homeostatic process Effects 0.000 description 4
- 238000010166 immunofluorescence Methods 0.000 description 4
- 108700016226 indium-bleomycin Proteins 0.000 description 4
- 239000000463 material Substances 0.000 description 4
- 230000007246 mechanism Effects 0.000 description 4
- 230000004660 morphological change Effects 0.000 description 4
- 210000001087 myotubule Anatomy 0.000 description 4
- 230000001717 pathogenic effect Effects 0.000 description 4
- 230000000144 pharmacologic effect Effects 0.000 description 4
- 108090000765 processed proteins & peptides Proteins 0.000 description 4
- 230000035755 proliferation Effects 0.000 description 4
- 230000001172 regenerating effect Effects 0.000 description 4
- 230000001105 regulatory effect Effects 0.000 description 4
- 230000007480 spreading Effects 0.000 description 4
- 238000003892 spreading Methods 0.000 description 4
- 230000002103 transcriptional effect Effects 0.000 description 4
- 241000008904 Betacoronavirus Species 0.000 description 3
- 108091003079 Bovine Serum Albumin Proteins 0.000 description 3
- 108010025464 Cyclin-Dependent Kinase 4 Proteins 0.000 description 3
- 102000013701 Cyclin-Dependent Kinase 4 Human genes 0.000 description 3
- 108020004414 DNA Proteins 0.000 description 3
- 230000005971 DNA damage repair Effects 0.000 description 3
- 239000006144 Dulbecco’s modified Eagle's medium Substances 0.000 description 3
- LFQSCWFLJHTTHZ-UHFFFAOYSA-N Ethanol Chemical class CCO LFQSCWFLJHTTHZ-UHFFFAOYSA-N 0.000 description 3
- 101000635938 Homo sapiens Transforming growth factor beta-1 proprotein Proteins 0.000 description 3
- 101000635958 Homo sapiens Transforming growth factor beta-2 proprotein Proteins 0.000 description 3
- 101000759453 Homo sapiens YY1-associated protein 1 Proteins 0.000 description 3
- 101150090494 KRT8 gene Proteins 0.000 description 3
- 239000003798 L01XE11 - Pazopanib Substances 0.000 description 3
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 3
- 239000004472 Lysine Substances 0.000 description 3
- 206010028980 Neoplasm Diseases 0.000 description 3
- 101150057744 PDGFA gene Proteins 0.000 description 3
- 229930040373 Paraformaldehyde Natural products 0.000 description 3
- 102000040945 Transcription factor Human genes 0.000 description 3
- 108091023040 Transcription factor Proteins 0.000 description 3
- 102100030737 Transforming growth factor beta-2 proprotein Human genes 0.000 description 3
- 102100023267 YY1-associated protein 1 Human genes 0.000 description 3
- 239000012190 activator Substances 0.000 description 3
- 210000004504 adult stem cell Anatomy 0.000 description 3
- 210000002383 alveolar type I cell Anatomy 0.000 description 3
- 210000002588 alveolar type II cell Anatomy 0.000 description 3
- 230000008512 biological response Effects 0.000 description 3
- 201000011510 cancer Diseases 0.000 description 3
- 230000030833 cell death Effects 0.000 description 3
- 150000001875 compounds Chemical class 0.000 description 3
- 230000007423 decrease Effects 0.000 description 3
- 230000003247 decreasing effect Effects 0.000 description 3
- 238000010586 diagram Methods 0.000 description 3
- 229940079593 drug Drugs 0.000 description 3
- 210000000981 epithelium Anatomy 0.000 description 3
- 239000012091 fetal bovine serum Substances 0.000 description 3
- 238000007490 hematoxylin and eosin (H&E) staining Methods 0.000 description 3
- 230000003993 interaction Effects 0.000 description 3
- 230000003902 lesion Effects 0.000 description 3
- 210000005265 lung cell Anatomy 0.000 description 3
- 230000001404 mediated effect Effects 0.000 description 3
- 210000004940 nucleus Anatomy 0.000 description 3
- 229920002866 paraformaldehyde Polymers 0.000 description 3
- 230000036961 partial effect Effects 0.000 description 3
- 229960000639 pazopanib Drugs 0.000 description 3
- CUIHSIWYWATEQL-UHFFFAOYSA-N pazopanib Chemical compound C1=CC2=C(C)N(C)N=C2C=C1N(C)C(N=1)=CC=NC=1NC1=CC=C(C)C(S(N)(=O)=O)=C1 CUIHSIWYWATEQL-UHFFFAOYSA-N 0.000 description 3
- 102000004196 processed proteins & peptides Human genes 0.000 description 3
- 238000000159 protein binding assay Methods 0.000 description 3
- 230000008458 response to injury Effects 0.000 description 3
- 239000000243 solution Substances 0.000 description 3
- CCEKAJIANROZEO-UHFFFAOYSA-N sulfluramid Chemical group CCNS(=O)(=O)C(F)(F)C(F)(F)C(F)(F)C(F)(F)C(F)(F)C(F)(F)C(F)(F)C(F)(F)F CCEKAJIANROZEO-UHFFFAOYSA-N 0.000 description 3
- 210000001578 tight junction Anatomy 0.000 description 3
- 230000005945 translocation Effects 0.000 description 3
- RFHPWPWVGFSLOP-ZROIWOOFSA-N (3z)-5-bromo-3-(4,5,6,7-tetrahydro-1h-indol-2-ylmethylidene)-1h-indol-2-one Chemical compound C1CCCC(N2)=C1C=C2\C=C1/C(=O)NC2=CC=C(Br)C=C21 RFHPWPWVGFSLOP-ZROIWOOFSA-N 0.000 description 2
- DEEOXSOLTLIWMG-UHFFFAOYSA-N 1-[2-[5-(2-methoxyethoxy)-1-benzimidazolyl]-8-quinolinyl]-4-piperidinamine Chemical compound C1=NC2=CC(OCCOC)=CC=C2N1C(N=C12)=CC=C1C=CC=C2N1CCC(N)CC1 DEEOXSOLTLIWMG-UHFFFAOYSA-N 0.000 description 2
- WPHKIQPVPYJNAX-UHFFFAOYSA-N 1-[4-[4-amino-7-[1-(2-hydroxyethyl)pyrazol-4-yl]thieno[3,2-c]pyridin-3-yl]phenyl]-3-(3-fluorophenyl)urea Chemical compound C1=2SC=C(C=3C=CC(NC(=O)NC=4C=C(F)C=CC=4)=CC=3)C=2C(N)=NC=C1C=1C=NN(CCO)C=1 WPHKIQPVPYJNAX-UHFFFAOYSA-N 0.000 description 2
- SRSGVKWWVXWSJT-ATVHPVEESA-N 5-[(z)-(5-fluoro-2-oxo-1h-indol-3-ylidene)methyl]-2,4-dimethyl-n-(2-pyrrolidin-1-ylethyl)-1h-pyrrole-3-carboxamide Chemical compound CC=1NC(\C=C/2C3=CC(F)=CC=C3NC\2=O)=C(C)C=1C(=O)NCCN1CCCC1 SRSGVKWWVXWSJT-ATVHPVEESA-N 0.000 description 2
- 101150033916 ACTN1 gene Proteins 0.000 description 2
- 101150063781 AKAP5 gene Proteins 0.000 description 2
- 102000007469 Actins Human genes 0.000 description 2
- 108010085238 Actins Proteins 0.000 description 2
- 108700028369 Alleles Proteins 0.000 description 2
- 241000004176 Alphacoronavirus Species 0.000 description 2
- 102100038778 Amphiregulin Human genes 0.000 description 2
- 238000011740 C57BL/6 mouse Methods 0.000 description 2
- 101150052583 CALM1 gene Proteins 0.000 description 2
- 101150065209 CCNG1 gene Proteins 0.000 description 2
- 208000025721 COVID-19 Diseases 0.000 description 2
- 241001678559 COVID-19 virus Species 0.000 description 2
- 101100356682 Caenorhabditis elegans rho-1 gene Proteins 0.000 description 2
- 101150079049 Ccnd2 gene Proteins 0.000 description 2
- 101150069156 Cdkn2b gene Proteins 0.000 description 2
- 102100037313 Core-binding factor subunit beta Human genes 0.000 description 2
- 101710145030 Core-binding factor subunit beta Proteins 0.000 description 2
- 108010058546 Cyclin D1 Proteins 0.000 description 2
- 230000012746 DNA damage checkpoint Effects 0.000 description 2
- 101100239628 Danio rerio myca gene Proteins 0.000 description 2
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 2
- 101150084967 EPCAM gene Proteins 0.000 description 2
- 102000004190 Enzymes Human genes 0.000 description 2
- 108090000790 Enzymes Proteins 0.000 description 2
- 102000018651 Epithelial Cell Adhesion Molecule Human genes 0.000 description 2
- 108010066687 Epithelial Cell Adhesion Molecule Proteins 0.000 description 2
- 102100024165 G1/S-specific cyclin-D1 Human genes 0.000 description 2
- 101150029115 HOPX gene Proteins 0.000 description 2
- 102100032742 Histone-lysine N-methyltransferase SETD2 Human genes 0.000 description 2
- 108010077223 Homer Scaffolding Proteins Proteins 0.000 description 2
- 102000010029 Homer Scaffolding Proteins Human genes 0.000 description 2
- 241000282412 Homo Species 0.000 description 2
- 101000809450 Homo sapiens Amphiregulin Proteins 0.000 description 2
- 101001046870 Homo sapiens Hypoxia-inducible factor 1-alpha Proteins 0.000 description 2
- 101000669513 Homo sapiens Metalloproteinase inhibitor 1 Proteins 0.000 description 2
- 101000606506 Homo sapiens Receptor-type tyrosine-protein phosphatase eta Proteins 0.000 description 2
- 208000002979 Influenza in Birds Diseases 0.000 description 2
- MIJPAVRNWPDMOR-ZAFYKAAXSA-N L-ascorbic acid 2-phosphate Chemical compound OC[C@H](O)[C@H]1OC(=O)C(OP(O)(O)=O)=C1O MIJPAVRNWPDMOR-ZAFYKAAXSA-N 0.000 description 2
- 239000002138 L01XE21 - Regorafenib Substances 0.000 description 2
- 101150018665 MAPK3 gene Proteins 0.000 description 2
- 101150039798 MYC gene Proteins 0.000 description 2
- 238000000585 Mann–Whitney U test Methods 0.000 description 2
- 102100039364 Metalloproteinase inhibitor 1 Human genes 0.000 description 2
- 208000025370 Middle East respiratory syndrome Diseases 0.000 description 2
- 108700027649 Mitogen-Activated Protein Kinase 3 Proteins 0.000 description 2
- 102100024192 Mitogen-activated protein kinase 3 Human genes 0.000 description 2
- 108010057466 NF-kappa B Proteins 0.000 description 2
- 102000003945 NF-kappa B Human genes 0.000 description 2
- 101150071967 Nupr1 gene Proteins 0.000 description 2
- 241000283973 Oryctolagus cuniculus Species 0.000 description 2
- 101150111584 RHOA gene Proteins 0.000 description 2
- 241000700159 Rattus Species 0.000 description 2
- 102100039808 Receptor-type tyrosine-protein phosphatase eta Human genes 0.000 description 2
- 241000725643 Respiratory syncytial virus Species 0.000 description 2
- 235000011449 Rosa Nutrition 0.000 description 2
- 101150001462 SFTPB gene Proteins 0.000 description 2
- 201000003176 Severe Acute Respiratory Syndrome Diseases 0.000 description 2
- 241000282887 Suidae Species 0.000 description 2
- 239000003819 Toceranib Substances 0.000 description 2
- 101150018417 VIM gene Proteins 0.000 description 2
- 102000013814 Wnt Human genes 0.000 description 2
- 108050003627 Wnt Proteins 0.000 description 2
- 101100459258 Xenopus laevis myc-a gene Proteins 0.000 description 2
- RJURFGZVJUQBHK-UHFFFAOYSA-N actinomycin D Natural products CC1OC(=O)C(C(C)C)N(C)C(=O)CN(C)C(=O)C2CCCN2C(=O)C(C(C)C)NC(=O)C1NC(=O)C1=C(N)C(=O)C(C)=C2OC(C(C)=CC=C3C(=O)NC4C(=O)NC(C(N5CCCC5C(=O)N(C)CC(=O)N(C)C(C(C)C)C(=O)OC4C)=O)C(C)C)=C3N=C21 RJURFGZVJUQBHK-UHFFFAOYSA-N 0.000 description 2
- 230000003321 amplification Effects 0.000 description 2
- 238000010171 animal model Methods 0.000 description 2
- 239000002246 antineoplastic agent Substances 0.000 description 2
- 206010064097 avian influenza Diseases 0.000 description 2
- 229960000074 biopharmaceutical Drugs 0.000 description 2
- 210000002459 blastocyst Anatomy 0.000 description 2
- 210000004369 blood Anatomy 0.000 description 2
- 239000008280 blood Substances 0.000 description 2
- 230000037396 body weight Effects 0.000 description 2
- 239000000872 buffer Substances 0.000 description 2
- 230000025084 cell cycle arrest Effects 0.000 description 2
- 230000032823 cell division Effects 0.000 description 2
- 230000005754 cellular signaling Effects 0.000 description 2
- 230000008859 change Effects 0.000 description 2
- 238000010205 computational analysis Methods 0.000 description 2
- 229940127089 cytotoxic agent Drugs 0.000 description 2
- 238000003745 diagnosis Methods 0.000 description 2
- 238000002224 dissection Methods 0.000 description 2
- 238000005516 engineering process Methods 0.000 description 2
- 229940088598 enzyme Drugs 0.000 description 2
- 230000012010 growth Effects 0.000 description 2
- 239000003102 growth factor Substances 0.000 description 2
- 230000036541 health Effects 0.000 description 2
- 229950000568 ilorasertib Drugs 0.000 description 2
- 238000010185 immunofluorescence analysis Methods 0.000 description 2
- 238000007901 in situ hybridization Methods 0.000 description 2
- 238000011534 incubation Methods 0.000 description 2
- 208000015181 infectious disease Diseases 0.000 description 2
- 239000007924 injection Substances 0.000 description 2
- 238000002347 injection Methods 0.000 description 2
- 238000011813 knockout mouse model Methods 0.000 description 2
- 208000037841 lung tumor Diseases 0.000 description 2
- 210000002540 macrophage Anatomy 0.000 description 2
- 238000012423 maintenance Methods 0.000 description 2
- 108010082117 matrigel Proteins 0.000 description 2
- 108020004999 messenger RNA Proteins 0.000 description 2
- DZFZXPPHBWCXPQ-UHFFFAOYSA-N n-[5-[2-(cyclopropanecarbonylamino)imidazo[1,2-b]pyridazin-6-yl]oxy-2-methylphenyl]-2,5-dimethylpyrazole-3-carboxamide Chemical compound CN1N=C(C)C=C1C(=O)NC1=CC(OC2=NN3C=C(NC(=O)C4CC4)N=C3C=C2)=CC=C1C DZFZXPPHBWCXPQ-UHFFFAOYSA-N 0.000 description 2
- 239000002858 neurotransmitter agent Substances 0.000 description 2
- 238000003199 nucleic acid amplification method Methods 0.000 description 2
- 239000012188 paraffin wax Substances 0.000 description 2
- 239000004031 partial agonist Substances 0.000 description 2
- 230000008506 pathogenesis Effects 0.000 description 2
- 230000007170 pathology Effects 0.000 description 2
- 238000003068 pathway analysis Methods 0.000 description 2
- 239000008188 pellet Substances 0.000 description 2
- 230000009038 pharmacological inhibition Effects 0.000 description 2
- 238000007747 plating Methods 0.000 description 2
- 230000008569 process Effects 0.000 description 2
- 238000012545 processing Methods 0.000 description 2
- 230000000750 progressive effect Effects 0.000 description 2
- 230000000306 recurrent effect Effects 0.000 description 2
- 229960004836 regorafenib Drugs 0.000 description 2
- FNHKPVJBJVTLMP-UHFFFAOYSA-N regorafenib Chemical compound C1=NC(C(=O)NC)=CC(OC=2C=C(F)C(NC(=O)NC=3C=C(C(Cl)=CC=3)C(F)(F)F)=CC=2)=C1 FNHKPVJBJVTLMP-UHFFFAOYSA-N 0.000 description 2
- 238000011160 research Methods 0.000 description 2
- 208000000649 small cell carcinoma Diseases 0.000 description 2
- 150000003384 small molecules Chemical class 0.000 description 2
- 238000007619 statistical method Methods 0.000 description 2
- 239000000758 substrate Substances 0.000 description 2
- 229960005048 toceranib Drugs 0.000 description 2
- 210000004881 tumor cell Anatomy 0.000 description 2
- DWYRIWUZIJHQKQ-SANMLTNESA-N (1S)-1-(4-fluorophenyl)-1-[2-[4-[6-(1-methylpyrazol-4-yl)pyrrolo[2,1-f][1,2,4]triazin-4-yl]piperazin-1-yl]pyrimidin-5-yl]ethanamine Chemical compound Cn1cc(cn1)-c1cc2c(ncnn2c1)N1CCN(CC1)c1ncc(cn1)[C@@](C)(N)c1ccc(F)cc1 DWYRIWUZIJHQKQ-SANMLTNESA-N 0.000 description 1
- KCOYQXZDFIIGCY-CZIZESTLSA-N (3e)-4-amino-5-fluoro-3-[5-(4-methylpiperazin-1-yl)-1,3-dihydrobenzimidazol-2-ylidene]quinolin-2-one Chemical compound C1CN(C)CCN1C1=CC=C(N\C(N2)=C/3C(=C4C(F)=CC=CC4=NC\3=O)N)C2=C1 KCOYQXZDFIIGCY-CZIZESTLSA-N 0.000 description 1
- UAKWLVYMKBWHMX-PTNGSMBKSA-N (3z)-3-[[4-(dimethylamino)phenyl]methylidene]-1h-indol-2-one Chemical compound C1=CC(N(C)C)=CC=C1\C=C/1C2=CC=CC=C2NC\1=O UAKWLVYMKBWHMX-PTNGSMBKSA-N 0.000 description 1
- PQPBIMIFGPFTNF-FYOMJGPWSA-N (z)-but-2-enedioic acid;5-[(z)-(5-fluoro-2-oxo-1h-indol-3-ylidene)methyl]-n-[(2s)-2-hydroxy-3-morpholin-4-ylpropyl]-2,4-dimethyl-1h-pyrrole-3-carboxamide Chemical compound OC(=O)\C=C/C(O)=O.C([C@@H](O)CNC(=O)C=1C(C)=C(\C=C/2C3=CC(F)=CC=C3NC\2=O)NC=1C)N1CCOCC1 PQPBIMIFGPFTNF-FYOMJGPWSA-N 0.000 description 1
- DSCFFEYYQKSRSV-UHFFFAOYSA-N 1L-O1-methyl-muco-inositol Natural products COC1C(O)C(O)C(O)C(O)C1O DSCFFEYYQKSRSV-UHFFFAOYSA-N 0.000 description 1
- YSDNWNOGHQYWPK-UHFFFAOYSA-N 2-[(3,4-dichlorophenyl)carbamoyl]bicyclo[2.2.1]hept-5-ene-3-carboxylic acid Chemical compound OC(=O)C1C(C=C2)CC2C1C(=O)NC1=CC=C(Cl)C(Cl)=C1 YSDNWNOGHQYWPK-UHFFFAOYSA-N 0.000 description 1
- TWYCZJMOEMMCGC-UHFFFAOYSA-N 2-[4-(6,7-dimethoxyquinazolin-4-yl)oxyphenyl]-n-(1-propan-2-ylpyrazol-4-yl)acetamide Chemical compound C=12C=C(OC)C(OC)=CC2=NC=NC=1OC(C=C1)=CC=C1CC(=O)NC=1C=NN(C(C)C)C=1 TWYCZJMOEMMCGC-UHFFFAOYSA-N 0.000 description 1
- SEMWODVUFFPLLJ-BYYHNAKLSA-N 2-[4-[(e)-1,2-diphenylbut-1-enyl]phenoxy]-n,n-diethylethanamine Chemical compound C1=CC(OCCN(CC)CC)=CC=C1C(\C=1C=CC=CC=1)=C(/CC)C1=CC=CC=C1 SEMWODVUFFPLLJ-BYYHNAKLSA-N 0.000 description 1
- CMYHZFCJPORPHY-UHFFFAOYSA-N 2-methylsulfanyl-6-[4-[3-[3-(trifluoromethyl)anilino]-1h-1,2,4-triazol-5-yl]phenoxy]pyrimidin-4-amine Chemical compound CSC1=NC(N)=CC(OC=2C=CC(=CC=2)C=2NC(NC=3C=C(C=CC=3)C(F)(F)F)=NN=2)=N1 CMYHZFCJPORPHY-UHFFFAOYSA-N 0.000 description 1
- BUOYTFVLNZIELF-UHFFFAOYSA-N 2-phenyl-1h-indole-4,6-dicarboximidamide Chemical compound N1C2=CC(C(=N)N)=CC(C(N)=N)=C2C=C1C1=CC=CC=C1 BUOYTFVLNZIELF-UHFFFAOYSA-N 0.000 description 1
- YNFBMDWHEHETJW-UHFFFAOYSA-N 2-pyridin-2-yl-1h-benzimidazole Chemical compound N1=CC=CC=C1C1=NC2=CC=CC=C2N1 YNFBMDWHEHETJW-UHFFFAOYSA-N 0.000 description 1
- NXERGIJHYVUXHM-UHFFFAOYSA-N 3-(3-fluorophenyl)-5,5-dimethyl-4-(4-methylsulfonylphenyl)furan-2-one Chemical compound CC1(C)OC(=O)C(C=2C=C(F)C=CC=2)=C1C1=CC=C(S(C)(=O)=O)C=C1 NXERGIJHYVUXHM-UHFFFAOYSA-N 0.000 description 1
- JNDVEAXZWJIOKB-JYRVWZFOSA-N 3-[(3-(2-carboxyethyl)-4-methylpyrrol-2-yl)methylene]-2-indolinone Chemical compound CC1=CNC(\C=C/2C3=CC=CC=C3NC\2=O)=C1CCC(O)=O JNDVEAXZWJIOKB-JYRVWZFOSA-N 0.000 description 1
- NHFDRBXTEDBWCZ-ZROIWOOFSA-N 3-[2,4-dimethyl-5-[(z)-(2-oxo-1h-indol-3-ylidene)methyl]-1h-pyrrol-3-yl]propanoic acid Chemical compound OC(=O)CCC1=C(C)NC(\C=C/2C3=CC=CC=C3NC\2=O)=C1C NHFDRBXTEDBWCZ-ZROIWOOFSA-N 0.000 description 1
- APYYTEJNOZQZNA-MOSHPQCFSA-N 3-[2,4-dimethyl-5-[(z)-(2-oxo-6-phenyl-1h-indol-3-ylidene)methyl]-1h-pyrrol-3-yl]propanoic acid Chemical compound OC(=O)CCC1=C(C)NC(\C=C/2C3=CC=C(C=C3NC\2=O)C=2C=CC=CC=2)=C1C APYYTEJNOZQZNA-MOSHPQCFSA-N 0.000 description 1
- XXJWYDDUDKYVKI-UHFFFAOYSA-N 4-[(4-fluoro-2-methyl-1H-indol-5-yl)oxy]-6-methoxy-7-[3-(1-pyrrolidinyl)propoxy]quinazoline Chemical compound COC1=CC2=C(OC=3C(=C4C=C(C)NC4=CC=3)F)N=CN=C2C=C1OCCCN1CCCC1 XXJWYDDUDKYVKI-UHFFFAOYSA-N 0.000 description 1
- AQSSMEORRLJZLU-UHFFFAOYSA-N 4-chloro-3-[5-methyl-3-[4-(2-pyrrolidin-1-ylethoxy)anilino]-1,2,4-benzotriazin-7-yl]phenol Chemical compound N1=C2C(C)=CC(C=3C(=CC=C(O)C=3)Cl)=CC2=NN=C1NC(C=C1)=CC=C1OCCN1CCCC1 AQSSMEORRLJZLU-UHFFFAOYSA-N 0.000 description 1
- 102100032500 40S ribosomal protein S27-like Human genes 0.000 description 1
- YBBAOKYVJCNJIV-UHFFFAOYSA-N 5,7-dimethoxy-3-pyridin-4-ylquinoline;dihydrochloride Chemical compound Cl.Cl.C1=NC2=CC(OC)=CC(OC)=C2C=C1C1=CC=NC=C1 YBBAOKYVJCNJIV-UHFFFAOYSA-N 0.000 description 1
- CTNPALGJUAXMMC-PMFHANACSA-N 5-[(z)-(5-fluoro-2-oxo-1h-indol-3-ylidene)methyl]-n-[(2s)-2-hydroxy-3-morpholin-4-ylpropyl]-2,4-dimethyl-1h-pyrrole-3-carboxamide Chemical compound C([C@@H](O)CNC(=O)C=1C(C)=C(\C=C/2C3=CC(F)=CC=C3NC\2=O)NC=1C)N1CCOCC1 CTNPALGJUAXMMC-PMFHANACSA-N 0.000 description 1
- 101150096982 ACTG1 gene Proteins 0.000 description 1
- 108010042708 Acetylmuramyl-Alanyl-Isoglutamine Proteins 0.000 description 1
- 101150020966 Acta2 gene Proteins 0.000 description 1
- 102100027308 Apoptosis regulator BAX Human genes 0.000 description 1
- 108050006685 Apoptosis regulator BAX Proteins 0.000 description 1
- 201000002909 Aspergillosis Diseases 0.000 description 1
- 208000036641 Aspergillus infections Diseases 0.000 description 1
- 102100034691 Astrocytic phosphoprotein PEA-15 Human genes 0.000 description 1
- MLDQJTXFUGDVEO-UHFFFAOYSA-N BAY-43-9006 Chemical compound C1=NC(C(=O)NC)=CC(OC=2C=CC(NC(=O)NC=3C=C(C(Cl)=CC=3)C(F)(F)F)=CC=2)=C1 MLDQJTXFUGDVEO-UHFFFAOYSA-N 0.000 description 1
- 101150063177 Basp1 gene Proteins 0.000 description 1
- 102100021573 Bcl-2-binding component 3, isoforms 3/4 Human genes 0.000 description 1
- 102100026596 Bcl-2-like protein 1 Human genes 0.000 description 1
- 101150008012 Bcl2l1 gene Proteins 0.000 description 1
- 241000588832 Bordetella pertussis Species 0.000 description 1
- 101150114882 CALM2 gene Proteins 0.000 description 1
- 101150014024 CAPN2 gene Proteins 0.000 description 1
- 102100027217 CD82 antigen Human genes 0.000 description 1
- 101150046141 CLDN3 gene Proteins 0.000 description 1
- 101150068199 CSRP1 gene Proteins 0.000 description 1
- 101150093222 CTTN gene Proteins 0.000 description 1
- 241000282472 Canis lupus familiaris Species 0.000 description 1
- 241000283707 Capra Species 0.000 description 1
- 101150056960 Casp8 gene Proteins 0.000 description 1
- 102100026548 Caspase-8 Human genes 0.000 description 1
- 108050001278 Cdc42 Proteins 0.000 description 1
- 102000011068 Cdc42 Human genes 0.000 description 1
- 108091007854 Cdh1/Fizzy-related Proteins 0.000 description 1
- 102000038594 Cdh1/Fizzy-related Human genes 0.000 description 1
- 241000282693 Cercopithecidae Species 0.000 description 1
- 108010077544 Chromatin Proteins 0.000 description 1
- 101150057694 Cldn7 gene Proteins 0.000 description 1
- 108010035532 Collagen Proteins 0.000 description 1
- 102000008186 Collagen Human genes 0.000 description 1
- 108060005980 Collagenase Proteins 0.000 description 1
- 102000029816 Collagenase Human genes 0.000 description 1
- 208000035473 Communicable disease Diseases 0.000 description 1
- 241000711573 Coronaviridae Species 0.000 description 1
- 208000001528 Coronaviridae Infections Diseases 0.000 description 1
- 241000699802 Cricetulus griseus Species 0.000 description 1
- 102000009512 Cyclin-Dependent Kinase Inhibitor p15 Human genes 0.000 description 1
- 108010009356 Cyclin-Dependent Kinase Inhibitor p15 Proteins 0.000 description 1
- 108010009392 Cyclin-Dependent Kinase Inhibitor p16 Proteins 0.000 description 1
- 102100024458 Cyclin-dependent kinase inhibitor 2A Human genes 0.000 description 1
- 101150100287 DDX39B gene Proteins 0.000 description 1
- 108010061982 DNA Ligases Proteins 0.000 description 1
- 102000012410 DNA Ligases Human genes 0.000 description 1
- 102100033195 DNA ligase 4 Human genes 0.000 description 1
- 230000033616 DNA repair Effects 0.000 description 1
- 102100022204 DNA-dependent protein kinase catalytic subunit Human genes 0.000 description 1
- 108010092160 Dactinomycin Proteins 0.000 description 1
- 102000007260 Deoxyribonuclease I Human genes 0.000 description 1
- 108010008532 Deoxyribonuclease I Proteins 0.000 description 1
- 101100028719 Dictyostelium discoideum pats1 gene Proteins 0.000 description 1
- 101100203525 Drosophila melanogaster snf gene Proteins 0.000 description 1
- 102100023877 E3 ubiquitin-protein ligase RBX1 Human genes 0.000 description 1
- 101710095156 E3 ubiquitin-protein ligase RBX1 Proteins 0.000 description 1
- 101150014084 EI24 gene Proteins 0.000 description 1
- 101150022145 Eif4ebp1 gene Proteins 0.000 description 1
- 101150056578 Emp2 gene Proteins 0.000 description 1
- 108010067770 Endopeptidase K Proteins 0.000 description 1
- 241000709661 Enterovirus Species 0.000 description 1
- 101150031329 Ets1 gene Proteins 0.000 description 1
- 102100022466 Eukaryotic translation initiation factor 4E-binding protein 1 Human genes 0.000 description 1
- 108010037362 Extracellular Matrix Proteins Proteins 0.000 description 1
- 102000010834 Extracellular Matrix Proteins Human genes 0.000 description 1
- 101150114877 Ezr gene Proteins 0.000 description 1
- 101150065562 F11R gene Proteins 0.000 description 1
- 102100037362 Fibronectin Human genes 0.000 description 1
- 108010067306 Fibronectins Proteins 0.000 description 1
- 101150033270 Gadd45a gene Proteins 0.000 description 1
- 208000007569 Giant Cell Tumors Diseases 0.000 description 1
- 102100020972 Glutamine amidotransferase-like class 1 domain-containing protein 3, mitochondrial Human genes 0.000 description 1
- 241001064231 Goat enterovirus Species 0.000 description 1
- 102100031150 Growth arrest and DNA damage-inducible protein GADD45 alpha Human genes 0.000 description 1
- 108700039143 HMGA2 Proteins 0.000 description 1
- 241000606768 Haemophilus influenzae Species 0.000 description 1
- HTTJABKRGRZYRN-UHFFFAOYSA-N Heparin Chemical compound OC1C(NC(=O)C)C(O)OC(COS(O)(=O)=O)C1OC1C(OS(O)(=O)=O)C(O)C(OC2C(C(OS(O)(=O)=O)C(OC3C(C(O)C(O)C(O3)C(O)=O)OS(O)(=O)=O)C(CO)O2)NS(O)(=O)=O)C(C(O)=O)O1 HTTJABKRGRZYRN-UHFFFAOYSA-N 0.000 description 1
- 101800001649 Heparin-binding EGF-like growth factor Proteins 0.000 description 1
- 102100028999 High mobility group protein HMGI-C Human genes 0.000 description 1
- 101150073387 Hmga2 gene Proteins 0.000 description 1
- 102100032827 Homeodomain-interacting protein kinase 2 Human genes 0.000 description 1
- 101000731896 Homo sapiens 40S ribosomal protein S27-like Proteins 0.000 description 1
- 101000734668 Homo sapiens Astrocytic phosphoprotein PEA-15 Proteins 0.000 description 1
- 101000971203 Homo sapiens Bcl-2-binding component 3, isoforms 1/2 Proteins 0.000 description 1
- 101000971209 Homo sapiens Bcl-2-binding component 3, isoforms 3/4 Proteins 0.000 description 1
- 101000868273 Homo sapiens CD44 antigen Proteins 0.000 description 1
- 101000914469 Homo sapiens CD82 antigen Proteins 0.000 description 1
- 101000927810 Homo sapiens DNA ligase 4 Proteins 0.000 description 1
- 101000619536 Homo sapiens DNA-dependent protein kinase catalytic subunit Proteins 0.000 description 1
- 101000678280 Homo sapiens Eukaryotic translation initiation factor 4E-binding protein 1 Proteins 0.000 description 1
- 101001002170 Homo sapiens Glutamine amidotransferase-like class 1 domain-containing protein 3, mitochondrial Proteins 0.000 description 1
- 101001066158 Homo sapiens Growth arrest and DNA damage-inducible protein GADD45 alpha Proteins 0.000 description 1
- 101001066401 Homo sapiens Homeodomain-interacting protein kinase 2 Proteins 0.000 description 1
- 101001044927 Homo sapiens Insulin-like growth factor-binding protein 3 Proteins 0.000 description 1
- 101001055222 Homo sapiens Interleukin-8 Proteins 0.000 description 1
- 101000880398 Homo sapiens Metalloreductase STEAP3 Proteins 0.000 description 1
- 101000733743 Homo sapiens Phorbol-12-myristate-13-acetate-induced protein 1 Proteins 0.000 description 1
- 101000595923 Homo sapiens Placenta growth factor Proteins 0.000 description 1
- 101000611888 Homo sapiens Platelet-derived growth factor C Proteins 0.000 description 1
- 101000876829 Homo sapiens Protein C-ets-1 Proteins 0.000 description 1
- 101001116548 Homo sapiens Protein CBFA2T1 Proteins 0.000 description 1
- 101000639763 Homo sapiens Regulator of telomere elongation helicase 1 Proteins 0.000 description 1
- 101001095783 Homo sapiens Ribonucleoside-diphosphate reductase subunit M2 B Proteins 0.000 description 1
- 101000695043 Homo sapiens Serine/threonine-protein kinase BRSK1 Proteins 0.000 description 1
- 101000729945 Homo sapiens Serine/threonine-protein kinase PLK2 Proteins 0.000 description 1
- 101000691614 Homo sapiens Serine/threonine-protein kinase PLK3 Proteins 0.000 description 1
- 101000739911 Homo sapiens Sestrin-3 Proteins 0.000 description 1
- 101000795222 Homo sapiens TP53-regulated inhibitor of apoptosis 1 Proteins 0.000 description 1
- 101000659879 Homo sapiens Thrombospondin-1 Proteins 0.000 description 1
- 101001087394 Homo sapiens Tyrosine-protein phosphatase non-receptor type 1 Proteins 0.000 description 1
- 101000894590 Homo sapiens Uncharacterized protein C20orf85 Proteins 0.000 description 1
- 101000742579 Homo sapiens Vascular endothelial growth factor B Proteins 0.000 description 1
- 101000742596 Homo sapiens Vascular endothelial growth factor C Proteins 0.000 description 1
- 101000723893 Homo sapiens Zinc finger matrin-type protein 3 Proteins 0.000 description 1
- 101000802094 Homo sapiens mRNA decay activator protein ZFP36L1 Proteins 0.000 description 1
- 101001127470 Homo sapiens p53 apoptosis effector related to PMP-22 Proteins 0.000 description 1
- 101150054249 Hspa4 gene Proteins 0.000 description 1
- 101150050268 ITGB1 gene Proteins 0.000 description 1
- 241000712431 Influenza A virus Species 0.000 description 1
- 241000713196 Influenza B virus Species 0.000 description 1
- 102100022708 Insulin-like growth factor-binding protein 3 Human genes 0.000 description 1
- 102100026236 Interleukin-8 Human genes 0.000 description 1
- 206010072877 Intestinal fibrosis Diseases 0.000 description 1
- 101150078927 KRT18 gene Proteins 0.000 description 1
- 101150105104 Kras gene Proteins 0.000 description 1
- 101150042912 Krt7 gene Proteins 0.000 description 1
- DSCFFEYYQKSRSV-MBXCVVGISA-N L-Quebrachitol Chemical compound COC1[C@H](O)[C@@H](O)C(O)[C@H](O)[C@H]1O DSCFFEYYQKSRSV-MBXCVVGISA-N 0.000 description 1
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 1
- 239000005517 L01XE01 - Imatinib Substances 0.000 description 1
- 239000002147 L01XE04 - Sunitinib Substances 0.000 description 1
- 239000005511 L01XE05 - Sorafenib Substances 0.000 description 1
- 239000005536 L01XE08 - Nilotinib Substances 0.000 description 1
- 239000002137 L01XE24 - Ponatinib Substances 0.000 description 1
- 108010001831 LDL receptors Proteins 0.000 description 1
- 101000844802 Lacticaseibacillus rhamnosus Teichoic acid D-alanyltransferase Proteins 0.000 description 1
- 101150057799 Lamp3 gene Proteins 0.000 description 1
- 102100024640 Low-density lipoprotein receptor Human genes 0.000 description 1
- 101150040099 MAP2K2 gene Proteins 0.000 description 1
- 101150079856 MSN gene Proteins 0.000 description 1
- 101150037751 MYH9 gene Proteins 0.000 description 1
- 101150064183 MYL6 gene Proteins 0.000 description 1
- 101150056121 MYL9 gene Proteins 0.000 description 1
- FYYHWMGAXLPEAU-UHFFFAOYSA-N Magnesium Chemical compound [Mg] FYYHWMGAXLPEAU-UHFFFAOYSA-N 0.000 description 1
- 101150060694 Mapk13 gene Proteins 0.000 description 1
- 102100037653 Metalloreductase STEAP3 Human genes 0.000 description 1
- 206010027476 Metastases Diseases 0.000 description 1
- 229930192392 Mitomycin Natural products 0.000 description 1
- 241000588655 Moraxella catarrhalis Species 0.000 description 1
- 241001529936 Murinae Species 0.000 description 1
- 101100490443 Mus musculus Acvr1 gene Proteins 0.000 description 1
- 101100338311 Mus musculus H2-T23 gene Proteins 0.000 description 1
- 101100365659 Mus musculus Scgb3a1 gene Proteins 0.000 description 1
- NWIBSHFKIJFRCO-WUDYKRTCSA-N Mytomycin Chemical compound C1N2C(C(C(C)=C(N)C3=O)=O)=C3[C@@H](COC(N)=O)[C@@]2(OC)[C@@H]2[C@H]1N2 NWIBSHFKIJFRCO-WUDYKRTCSA-N 0.000 description 1
- NWVOHJOHQOHBMB-UHFFFAOYSA-N N-[2-(dimethylamino)ethyl]-N-[[4-[[4-methyl-3-[(4-pyridin-3-ylpyrimidin-2-yl)amino]phenyl]carbamoyl]phenyl]methyl]pyridine-3-carboxamide Chemical compound CN(C)CCN(Cc1ccc(cc1)C(=O)Nc1ccc(C)c(Nc2nccc(n2)-c2cccnc2)c1)C(=O)c1cccnc1 NWVOHJOHQOHBMB-UHFFFAOYSA-N 0.000 description 1
- 101150079463 NBL1 gene Proteins 0.000 description 1
- 108010018525 NFATC Transcription Factors Proteins 0.000 description 1
- 102000002673 NFATC Transcription Factors Human genes 0.000 description 1
- 101150097297 Nedd4 gene Proteins 0.000 description 1
- 241000588650 Neisseria meningitidis Species 0.000 description 1
- 102000007530 Neurofibromin 1 Human genes 0.000 description 1
- 108010085793 Neurofibromin 1 Proteins 0.000 description 1
- 108010015832 Non-Receptor Type 2 Protein Tyrosine Phosphatase Proteins 0.000 description 1
- 102000005650 Notch Receptors Human genes 0.000 description 1
- 108010070047 Notch Receptors Proteins 0.000 description 1
- 101150030755 Ocln gene Proteins 0.000 description 1
- 108091034117 Oligonucleotide Proteins 0.000 description 1
- 240000007594 Oryza sativa Species 0.000 description 1
- 235000007164 Oryza sativa Nutrition 0.000 description 1
- 101150109086 PDGFD gene Proteins 0.000 description 1
- 229910019142 PO4 Inorganic materials 0.000 description 1
- 101150022681 PPP1CA gene Proteins 0.000 description 1
- 101150083342 PPP3CA gene Proteins 0.000 description 1
- 201000010630 Pancoast tumor Diseases 0.000 description 1
- 208000015330 Pancoast tumour Diseases 0.000 description 1
- 241001494479 Pecora Species 0.000 description 1
- 102100033716 Phorbol-12-myristate-13-acetate-induced protein 1 Human genes 0.000 description 1
- 102000004160 Phosphoric Monoester Hydrolases Human genes 0.000 description 1
- 108090000608 Phosphoric Monoester Hydrolases Proteins 0.000 description 1
- 108010022233 Plasminogen Activator Inhibitor 1 Proteins 0.000 description 1
- 102100039418 Plasminogen activator inhibitor 1 Human genes 0.000 description 1
- 102000018967 Platelet-Derived Growth Factor beta Receptor Human genes 0.000 description 1
- 108010051742 Platelet-Derived Growth Factor beta Receptor Proteins 0.000 description 1
- 102100040681 Platelet-derived growth factor C Human genes 0.000 description 1
- 101710179684 Poly [ADP-ribose] polymerase Proteins 0.000 description 1
- 229920000776 Poly(Adenosine diphosphate-ribose) polymerase Polymers 0.000 description 1
- 102100033762 Proheparin-binding EGF-like growth factor Human genes 0.000 description 1
- 101800004937 Protein C Proteins 0.000 description 1
- 102100035251 Protein C-ets-1 Human genes 0.000 description 1
- 102100024952 Protein CBFA2T1 Human genes 0.000 description 1
- 108010029485 Protein Isoforms Proteins 0.000 description 1
- 102000001708 Protein Isoforms Human genes 0.000 description 1
- 108010019674 Proto-Oncogene Proteins c-sis Proteins 0.000 description 1
- 101150117834 Prpf19 gene Proteins 0.000 description 1
- 241000589517 Pseudomonas aeruginosa Species 0.000 description 1
- 101150020518 RHEB gene Proteins 0.000 description 1
- 101710178916 RING-box protein 1 Proteins 0.000 description 1
- 101150046080 RPL26 gene Proteins 0.000 description 1
- 239000005464 Radotinib Substances 0.000 description 1
- 102000003901 Ras GTPase-activating proteins Human genes 0.000 description 1
- 108090000231 Ras GTPase-activating proteins Proteins 0.000 description 1
- 102000046951 Ras Homolog Enriched in Brain Human genes 0.000 description 1
- 108700019578 Ras Homolog Enriched in Brain Proteins 0.000 description 1
- 102100034469 Regulator of telomere elongation helicase 1 Human genes 0.000 description 1
- 102100038013 Ribonucleoside-diphosphate reductase subunit M2 B Human genes 0.000 description 1
- 101150090785 Rps27l gene Proteins 0.000 description 1
- 101150093156 Rras gene Proteins 0.000 description 1
- 241000315672 SARS coronavirus Species 0.000 description 1
- 108010005173 SERPIN-B5 Proteins 0.000 description 1
- 101150089938 SKP1 gene Proteins 0.000 description 1
- 101150093801 SLC9A3R1 gene Proteins 0.000 description 1
- 102100036546 Salivary acidic proline-rich phosphoprotein 1/2 Human genes 0.000 description 1
- 101800001700 Saposin-D Proteins 0.000 description 1
- 101150049354 Scgb1a1 gene Proteins 0.000 description 1
- 102100028623 Serine/threonine-protein kinase BRSK1 Human genes 0.000 description 1
- 102100031462 Serine/threonine-protein kinase PLK2 Human genes 0.000 description 1
- 102100026209 Serine/threonine-protein kinase PLK3 Human genes 0.000 description 1
- 102100030333 Serpin B5 Human genes 0.000 description 1
- 102100037575 Sestrin-3 Human genes 0.000 description 1
- 101150007091 Shisa5 gene Proteins 0.000 description 1
- 206010050207 Skin fibrosis Diseases 0.000 description 1
- 101150092978 Slc25a4 gene Proteins 0.000 description 1
- 108020004459 Small interfering RNA Proteins 0.000 description 1
- 101150101311 Sqstm1 gene Proteins 0.000 description 1
- 241000191967 Staphylococcus aureus Species 0.000 description 1
- 201000005010 Streptococcus pneumonia Diseases 0.000 description 1
- 241000193998 Streptococcus pneumoniae Species 0.000 description 1
- 241000193996 Streptococcus pyogenes Species 0.000 description 1
- 101710172711 Structural protein Proteins 0.000 description 1
- CZMRCDWAGMRECN-UGDNZRGBSA-N Sucrose Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 CZMRCDWAGMRECN-UGDNZRGBSA-N 0.000 description 1
- 229930006000 Sucrose Natural products 0.000 description 1
- 101710137500 T7 RNA polymerase Proteins 0.000 description 1
- SAQSTQBVENFSKT-UHFFFAOYSA-M TCA-sodium Chemical compound [Na+].[O-]C(=O)C(Cl)(Cl)Cl SAQSTQBVENFSKT-UHFFFAOYSA-M 0.000 description 1
- 101150000629 TGFB1 gene Proteins 0.000 description 1
- 101150072275 TGFB2 gene Proteins 0.000 description 1
- 102100029669 TP53-regulated inhibitor of apoptosis 1 Human genes 0.000 description 1
- 101150027463 TUBA1A gene Proteins 0.000 description 1
- 101150052918 TUBA1B gene Proteins 0.000 description 1
- 101150078072 TUBA1C gene Proteins 0.000 description 1
- 229940123237 Taxane Drugs 0.000 description 1
- 102100036034 Thrombospondin-1 Human genes 0.000 description 1
- 102000000591 Tight Junction Proteins Human genes 0.000 description 1
- 108010002321 Tight Junction Proteins Proteins 0.000 description 1
- 101150107801 Top2a gene Proteins 0.000 description 1
- 108700009124 Transcription Initiation Site Proteins 0.000 description 1
- 239000007983 Tris buffer Substances 0.000 description 1
- 108090000631 Trypsin Proteins 0.000 description 1
- 102000004142 Trypsin Human genes 0.000 description 1
- 108010091356 Tumor Protein p73 Proteins 0.000 description 1
- 102000015098 Tumor Suppressor Protein p53 Human genes 0.000 description 1
- 102100030018 Tumor protein p73 Human genes 0.000 description 1
- 102100033001 Tyrosine-protein phosphatase non-receptor type 1 Human genes 0.000 description 1
- 102100033019 Tyrosine-protein phosphatase non-receptor type 11 Human genes 0.000 description 1
- 101710116241 Tyrosine-protein phosphatase non-receptor type 11 Proteins 0.000 description 1
- 102100033141 Tyrosine-protein phosphatase non-receptor type 2 Human genes 0.000 description 1
- 102100021442 Uncharacterized protein C20orf85 Human genes 0.000 description 1
- 101150044482 VDAC2 gene Proteins 0.000 description 1
- 108010073919 Vascular Endothelial Growth Factor D Proteins 0.000 description 1
- 108010019530 Vascular Endothelial Growth Factors Proteins 0.000 description 1
- 102000005789 Vascular Endothelial Growth Factors Human genes 0.000 description 1
- 102100038217 Vascular endothelial growth factor B Human genes 0.000 description 1
- 102100038232 Vascular endothelial growth factor C Human genes 0.000 description 1
- 102100038234 Vascular endothelial growth factor D Human genes 0.000 description 1
- 101150099479 Vdac3 gene Proteins 0.000 description 1
- 101150096461 ZFP36L1 gene Proteins 0.000 description 1
- 102100028482 Zinc finger matrin-type protein 3 Human genes 0.000 description 1
- 101150098162 abcA3 gene Proteins 0.000 description 1
- 108010076089 accutase Proteins 0.000 description 1
- 230000000397 acetylating effect Effects 0.000 description 1
- RJURFGZVJUQBHK-IIXSONLDSA-N actinomycin D Chemical compound C[C@H]1OC(=O)[C@H](C(C)C)N(C)C(=O)CN(C)C(=O)[C@@H]2CCCN2C(=O)[C@@H](C(C)C)NC(=O)[C@H]1NC(=O)C1=C(N)C(=O)C(C)=C2OC(C(C)=CC=C3C(=O)N[C@@H]4C(=O)N[C@@H](C(N5CCC[C@H]5C(=O)N(C)CC(=O)N(C)[C@@H](C(C)C)C(=O)O[C@@H]4C)=O)C(C)C)=C3N=C21 RJURFGZVJUQBHK-IIXSONLDSA-N 0.000 description 1
- 230000003213 activating effect Effects 0.000 description 1
- 239000004480 active ingredient Substances 0.000 description 1
- 208000009956 adenocarcinoma Diseases 0.000 description 1
- 230000001270 agonistic effect Effects 0.000 description 1
- 230000004075 alteration Effects 0.000 description 1
- 210000001132 alveolar macrophage Anatomy 0.000 description 1
- 125000000539 amino acid group Chemical group 0.000 description 1
- 239000000427 antigen Substances 0.000 description 1
- 108091007433 antigens Proteins 0.000 description 1
- 102000036639 antigens Human genes 0.000 description 1
- 229950009576 avapritinib Drugs 0.000 description 1
- 229960003005 axitinib Drugs 0.000 description 1
- RITAVMQDGBJQJZ-FMIVXFBMSA-N axitinib Chemical compound CNC(=O)C1=CC=CC=C1SC1=CC=C(C(\C=C\C=2N=CC=CC=2)=NN2)C2=C1 RITAVMQDGBJQJZ-FMIVXFBMSA-N 0.000 description 1
- 108700000707 bcl-2-Associated X Proteins 0.000 description 1
- 108700000711 bcl-X Proteins 0.000 description 1
- 239000011324 bead Substances 0.000 description 1
- 230000009286 beneficial effect Effects 0.000 description 1
- 230000008901 benefit Effects 0.000 description 1
- 230000031018 biological processes and functions Effects 0.000 description 1
- 230000033228 biological regulation Effects 0.000 description 1
- 239000000090 biomarker Substances 0.000 description 1
- 210000003969 blast cell Anatomy 0.000 description 1
- 238000004364 calculation method Methods 0.000 description 1
- 230000009787 cardiac fibrosis Effects 0.000 description 1
- 229960002412 cediranib Drugs 0.000 description 1
- 229960000590 celecoxib Drugs 0.000 description 1
- RZEKVGVHFLEQIL-UHFFFAOYSA-N celecoxib Chemical compound C1=CC(C)=CC=C1C1=CC(C(F)(F)F)=NN1C1=CC=C(S(N)(=O)=O)C=C1 RZEKVGVHFLEQIL-UHFFFAOYSA-N 0.000 description 1
- 230000021164 cell adhesion Effects 0.000 description 1
- 230000008568 cell cell communication Effects 0.000 description 1
- 230000010261 cell growth Effects 0.000 description 1
- 230000006037 cell lysis Effects 0.000 description 1
- 239000008004 cell lysis buffer Substances 0.000 description 1
- 230000009028 cell transition Effects 0.000 description 1
- 230000007248 cellular mechanism Effects 0.000 description 1
- 210000003483 chromatin Anatomy 0.000 description 1
- 230000001684 chronic effect Effects 0.000 description 1
- 208000019425 cirrhosis of liver Diseases 0.000 description 1
- 229920001436 collagen Polymers 0.000 description 1
- 229960002424 collagenase Drugs 0.000 description 1
- 208000011829 combined pulmonary fibrosis-emphysema syndrome Diseases 0.000 description 1
- 238000004891 communication Methods 0.000 description 1
- 229950009240 crenolanib Drugs 0.000 description 1
- DYNHJHQFHQTFTP-UHFFFAOYSA-N crenolanib Chemical compound C=1C=C2N(C=3N=C4C(N5CCC(N)CC5)=CC=CC4=CC=3)C=NC2=CC=1OCC1(C)COC1 DYNHJHQFHQTFTP-UHFFFAOYSA-N 0.000 description 1
- 101150114348 cycs gene Proteins 0.000 description 1
- 210000004292 cytoskeleton Anatomy 0.000 description 1
- 229960000640 dactinomycin Drugs 0.000 description 1
- 238000013079 data visualisation Methods 0.000 description 1
- 230000003412 degenerative effect Effects 0.000 description 1
- 229960003964 deoxycholic acid Drugs 0.000 description 1
- KXGVEGMKQFWNSR-LLQZFEROSA-N deoxycholic acid Chemical compound C([C@H]1CC2)[C@H](O)CC[C@]1(C)[C@@H]1[C@@H]2[C@@H]2CC[C@H]([C@@H](CCC(O)=O)C)[C@@]2(C)[C@@H](O)C1 KXGVEGMKQFWNSR-LLQZFEROSA-N 0.000 description 1
- 238000001514 detection method Methods 0.000 description 1
- 230000009274 differential gene expression Effects 0.000 description 1
- 108010007093 dispase Proteins 0.000 description 1
- 229950005778 dovitinib Drugs 0.000 description 1
- 230000009977 dual effect Effects 0.000 description 1
- 230000004064 dysfunction Effects 0.000 description 1
- 210000001671 embryonic stem cell Anatomy 0.000 description 1
- 210000002889 endothelial cell Anatomy 0.000 description 1
- 230000002708 enhancing effect Effects 0.000 description 1
- 238000010201 enrichment analysis Methods 0.000 description 1
- YQGOJNYOYNNSMM-UHFFFAOYSA-N eosin Chemical compound [Na+].OC(=O)C1=CC=CC=C1C1=C2C=C(Br)C(=O)C(Br)=C2OC2=C(Br)C(O)=C(Br)C=C21 YQGOJNYOYNNSMM-UHFFFAOYSA-N 0.000 description 1
- SEACYXSIPDVVMV-UHFFFAOYSA-L eosin Y Chemical compound [Na+].[Na+].[O-]C(=O)C1=CC=CC=C1C1=C2C=C(Br)C(=O)C(Br)=C2OC2=C(Br)C([O-])=C(Br)C=C21 SEACYXSIPDVVMV-UHFFFAOYSA-L 0.000 description 1
- 230000007275 epithelial homeostasis Effects 0.000 description 1
- 230000007705 epithelial mesenchymal transition Effects 0.000 description 1
- 210000003743 erythrocyte Anatomy 0.000 description 1
- 210000003238 esophagus Anatomy 0.000 description 1
- 229960004945 etoricoxib Drugs 0.000 description 1
- MNJVRJDLRVPLFE-UHFFFAOYSA-N etoricoxib Chemical compound C1=NC(C)=CC=C1C1=NC=C(Cl)C=C1C1=CC=C(S(C)(=O)=O)C=C1 MNJVRJDLRVPLFE-UHFFFAOYSA-N 0.000 description 1
- 231100000776 exotoxin Toxicity 0.000 description 1
- 239000002095 exotoxin Substances 0.000 description 1
- 238000013401 experimental design Methods 0.000 description 1
- 238000010195 expression analysis Methods 0.000 description 1
- 210000002744 extracellular matrix Anatomy 0.000 description 1
- 230000001605 fetal effect Effects 0.000 description 1
- 238000001914 filtration Methods 0.000 description 1
- 210000001650 focal adhesion Anatomy 0.000 description 1
- 108020001507 fusion proteins Proteins 0.000 description 1
- 102000037865 fusion proteins Human genes 0.000 description 1
- SDUQYLNIPVEERB-QPPQHZFASA-N gemcitabine Chemical compound O=C1N=C(N)C=CN1[C@H]1C(F)(F)[C@H](O)[C@@H](CO)O1 SDUQYLNIPVEERB-QPPQHZFASA-N 0.000 description 1
- 229960005277 gemcitabine Drugs 0.000 description 1
- 238000009499 grossing Methods 0.000 description 1
- 229960002897 heparin Drugs 0.000 description 1
- 229920000669 heparin Polymers 0.000 description 1
- 239000005556 hormone Substances 0.000 description 1
- 229940088597 hormone Drugs 0.000 description 1
- 238000009396 hybridization Methods 0.000 description 1
- 229960002411 imatinib Drugs 0.000 description 1
- KTUFNOKKBVMGRW-UHFFFAOYSA-N imatinib Chemical compound C1CN(C)CCN1CC1=CC=C(C(=O)NC=2C=C(NC=3N=C(C=CN=3)C=3C=NC=CC=3)C(C)=CC=2)C=C1 KTUFNOKKBVMGRW-UHFFFAOYSA-N 0.000 description 1
- 229960003685 imatinib mesylate Drugs 0.000 description 1
- YLMAHDNUQAMNNX-UHFFFAOYSA-N imatinib methanesulfonate Chemical compound CS(O)(=O)=O.C1CN(C)CCN1CC1=CC=C(C(=O)NC=2C=C(NC=3N=C(C=CN=3)C=3C=NC=CC=3)C(C)=CC=2)C=C1 YLMAHDNUQAMNNX-UHFFFAOYSA-N 0.000 description 1
- 238000003125 immunofluorescent labeling Methods 0.000 description 1
- 230000002055 immunohistochemical effect Effects 0.000 description 1
- 230000002401 inhibitory effect Effects 0.000 description 1
- 108010044426 integrins Proteins 0.000 description 1
- 102000006495 integrins Human genes 0.000 description 1
- 230000035992 intercellular communication Effects 0.000 description 1
- 229940125425 inverse agonist Drugs 0.000 description 1
- 238000011835 investigation Methods 0.000 description 1
- 230000002427 irreversible effect Effects 0.000 description 1
- 238000002372 labelling Methods 0.000 description 1
- 208000003849 large cell carcinoma Diseases 0.000 description 1
- 229960003784 lenvatinib Drugs 0.000 description 1
- WOSKHXYHFSIKNG-UHFFFAOYSA-N lenvatinib Chemical compound C=12C=C(C(N)=O)C(OC)=CC2=NC=CC=1OC(C=C1Cl)=CC=C1NC(=O)NC1CC1 WOSKHXYHFSIKNG-UHFFFAOYSA-N 0.000 description 1
- 238000007169 ligase reaction Methods 0.000 description 1
- 210000000982 limb bud Anatomy 0.000 description 1
- 229950002216 linifanib Drugs 0.000 description 1
- MPVGZUGXCQEXTM-UHFFFAOYSA-N linifanib Chemical compound CC1=CC=C(F)C(NC(=O)NC=2C=CC(=CC=2)C=2C=3C(N)=NNC=3C=CC=2)=C1 MPVGZUGXCQEXTM-UHFFFAOYSA-N 0.000 description 1
- 230000007774 longterm Effects 0.000 description 1
- 208000026807 lung carcinoid tumor Diseases 0.000 description 1
- 102100034702 mRNA decay activator protein ZFP36L1 Human genes 0.000 description 1
- 239000011777 magnesium Substances 0.000 description 1
- 229910052749 magnesium Inorganic materials 0.000 description 1
- 229940049920 malate Drugs 0.000 description 1
- BJEPYKJPYRNKOW-UHFFFAOYSA-N malic acid Chemical compound OC(=O)C(O)CC(O)=O BJEPYKJPYRNKOW-UHFFFAOYSA-N 0.000 description 1
- 208000015179 malignant superior sulcus neoplasm Diseases 0.000 description 1
- 210000005075 mammary gland Anatomy 0.000 description 1
- 238000004519 manufacturing process Methods 0.000 description 1
- 238000013507 mapping Methods 0.000 description 1
- 239000011159 matrix material Substances 0.000 description 1
- 230000035800 maturation Effects 0.000 description 1
- 230000002503 metabolic effect Effects 0.000 description 1
- 239000002207 metabolite Substances 0.000 description 1
- 230000009401 metastasis Effects 0.000 description 1
- 229960004857 mitomycin Drugs 0.000 description 1
- 238000012986 modification Methods 0.000 description 1
- 230000004048 modification Effects 0.000 description 1
- 230000009456 molecular mechanism Effects 0.000 description 1
- 230000000877 morphologic effect Effects 0.000 description 1
- BSOQXXWZTUDTEL-ZUYCGGNHSA-N muramyl dipeptide Chemical compound OC(=O)CC[C@H](C(N)=O)NC(=O)[C@H](C)NC(=O)[C@@H](C)O[C@H]1[C@H](O)[C@@H](CO)O[C@@H](O)[C@@H]1NC(C)=O BSOQXXWZTUDTEL-ZUYCGGNHSA-N 0.000 description 1
- 230000035772 mutation Effects 0.000 description 1
- 206010028537 myelofibrosis Diseases 0.000 description 1
- LBWFXVZLPYTWQI-IPOVEDGCSA-N n-[2-(diethylamino)ethyl]-5-[(z)-(5-fluoro-2-oxo-1h-indol-3-ylidene)methyl]-2,4-dimethyl-1h-pyrrole-3-carboxamide;(2s)-2-hydroxybutanedioic acid Chemical compound OC(=O)[C@@H](O)CC(O)=O.CCN(CC)CCNC(=O)C1=C(C)NC(\C=C/2C3=CC(F)=CC=C3NC\2=O)=C1C LBWFXVZLPYTWQI-IPOVEDGCSA-N 0.000 description 1
- AUMMSMBRVDYUME-UHFFFAOYSA-N n-[4-[(5-tert-butyl-1,2-oxazol-3-yl)carbamoylamino]phenyl]-5-(1-ethyl-2,2,6,6-tetramethylpiperidin-1-ium-4-yl)oxypyridine-2-carboxamide;methanesulfonate Chemical compound CS([O-])(=O)=O.C1C(C)(C)[NH+](CC)C(C)(C)CC1OC1=CC=C(C(=O)NC=2C=CC(NC(=O)NC3=NOC(=C3)C(C)(C)C)=CC=2)N=C1 AUMMSMBRVDYUME-UHFFFAOYSA-N 0.000 description 1
- JVCWPUFNLFSKFS-UHFFFAOYSA-N n-[4-[(5-tert-butyl-1,2-oxazol-3-yl)carbamoylamino]phenyl]-5-(1-ethyl-2,2,6,6-tetramethylpiperidin-4-yl)oxypyridine-2-carboxamide Chemical compound C1C(C)(C)N(CC)C(C)(C)CC1OC1=CC=C(C(=O)NC=2C=CC(NC(=O)NC3=NOC(=C3)C(C)(C)C)=CC=2)N=C1 JVCWPUFNLFSKFS-UHFFFAOYSA-N 0.000 description 1
- 229940052778 neisseria meningitidis Drugs 0.000 description 1
- 201000011519 neuroendocrine tumor Diseases 0.000 description 1
- 229960001346 nilotinib Drugs 0.000 description 1
- HHZIURLSWUIHRB-UHFFFAOYSA-N nilotinib Chemical compound C1=NC(C)=CN1C1=CC(NC(=O)C=2C=C(NC=3N=C(C=CN=3)C=3C=NC=CC=3)C(C)=CC=2)=CC(C(F)(F)F)=C1 HHZIURLSWUIHRB-UHFFFAOYSA-N 0.000 description 1
- 229960004378 nintedanib Drugs 0.000 description 1
- XZXHXSATPCNXJR-ZIADKAODSA-N nintedanib Chemical compound O=C1NC2=CC(C(=O)OC)=CC=C2\C1=C(C=1C=CC=CC=1)\NC(C=C1)=CC=C1N(C)C(=O)CN1CCN(C)CC1 XZXHXSATPCNXJR-ZIADKAODSA-N 0.000 description 1
- 208000002154 non-small cell lung carcinoma Diseases 0.000 description 1
- 238000010606 normalization Methods 0.000 description 1
- 102000039446 nucleic acids Human genes 0.000 description 1
- 108020004707 nucleic acids Proteins 0.000 description 1
- 150000007523 nucleic acids Chemical class 0.000 description 1
- 238000003305 oral gavage Methods 0.000 description 1
- 229950006354 orantinib Drugs 0.000 description 1
- 210000001672 ovary Anatomy 0.000 description 1
- 230000036542 oxidative stress Effects 0.000 description 1
- 102100030898 p53 apoptosis effector related to PMP-22 Human genes 0.000 description 1
- 244000052769 pathogen Species 0.000 description 1
- MQHIQUBXFFAOMK-UHFFFAOYSA-N pazopanib hydrochloride Chemical compound Cl.C1=CC2=C(C)N(C)N=C2C=C1N(C)C(N=1)=CC=NC=1NC1=CC=C(C)C(S(N)(=O)=O)=C1 MQHIQUBXFFAOMK-UHFFFAOYSA-N 0.000 description 1
- 230000006611 pharmacological activation Effects 0.000 description 1
- NBIIXXVUZAFLBC-UHFFFAOYSA-K phosphate Chemical compound [O-]P([O-])([O-])=O NBIIXXVUZAFLBC-UHFFFAOYSA-K 0.000 description 1
- 239000010452 phosphate Substances 0.000 description 1
- 229920001184 polypeptide Polymers 0.000 description 1
- 229960001131 ponatinib Drugs 0.000 description 1
- PHXJVRSECIGDHY-UHFFFAOYSA-N ponatinib Chemical compound C1CN(C)CCN1CC(C(=C1)C(F)(F)F)=CC=C1NC(=O)C1=CC=C(C)C(C#CC=2N3N=CC=CC3=NC=2)=C1 PHXJVRSECIGDHY-UHFFFAOYSA-N 0.000 description 1
- 230000003334 potential effect Effects 0.000 description 1
- 101150108030 ppiD gene Proteins 0.000 description 1
- 239000002243 precursor Substances 0.000 description 1
- 238000002360 preparation method Methods 0.000 description 1
- 230000002265 prevention Effects 0.000 description 1
- 230000002206 pro-fibrotic effect Effects 0.000 description 1
- 230000001737 promoting effect Effects 0.000 description 1
- 210000002307 prostate Anatomy 0.000 description 1
- 229960000856 protein c Drugs 0.000 description 1
- 230000004850 protein–protein interaction Effects 0.000 description 1
- 230000005855 radiation Effects 0.000 description 1
- 229950004043 radotinib Drugs 0.000 description 1
- DUPWHXBITIZIKZ-UHFFFAOYSA-N radotinib Chemical compound C1=NC(C)=CN1C1=CC(NC(=O)C=2C=C(NC=3N=C(C=CN=3)C=3N=CC=NC=3)C(C)=CC=2)=CC(C(F)(F)F)=C1 DUPWHXBITIZIKZ-UHFFFAOYSA-N 0.000 description 1
- 230000002829 reductive effect Effects 0.000 description 1
- 201000002793 renal fibrosis Diseases 0.000 description 1
- 238000010839 reverse transcription Methods 0.000 description 1
- 230000002441 reversible effect Effects 0.000 description 1
- 238000012552 review Methods 0.000 description 1
- 235000009566 rice Nutrition 0.000 description 1
- 229940121487 ripretinib Drugs 0.000 description 1
- CEFJVGZHQAGLHS-UHFFFAOYSA-N ripretinib Chemical compound O=C1N(CC)C2=CC(NC)=NC=C2C=C1C(C(=CC=1F)Br)=CC=1NC(=O)NC1=CC=CC=C1 CEFJVGZHQAGLHS-UHFFFAOYSA-N 0.000 description 1
- 238000005096 rolling process Methods 0.000 description 1
- 210000003079 salivary gland Anatomy 0.000 description 1
- 238000012163 sequencing technique Methods 0.000 description 1
- 108091006024 signal transducing proteins Proteins 0.000 description 1
- 210000002460 smooth muscle Anatomy 0.000 description 1
- 238000002791 soaking Methods 0.000 description 1
- 239000001509 sodium citrate Substances 0.000 description 1
- NLJMYIDDQXHKNR-UHFFFAOYSA-K sodium citrate Chemical compound O.O.[Na+].[Na+].[Na+].[O-]C(=O)CC(O)(CC([O-])=O)C([O-])=O NLJMYIDDQXHKNR-UHFFFAOYSA-K 0.000 description 1
- 229960003787 sorafenib Drugs 0.000 description 1
- 206010041823 squamous cell carcinoma Diseases 0.000 description 1
- 230000006641 stabilisation Effects 0.000 description 1
- 238000011105 stabilization Methods 0.000 description 1
- 229940076156 streptococcus pyogenes Drugs 0.000 description 1
- 230000035882 stress Effects 0.000 description 1
- 239000005720 sucrose Substances 0.000 description 1
- 229960001796 sunitinib Drugs 0.000 description 1
- WINHZLLDWRZWRT-ATVHPVEESA-N sunitinib Chemical compound CCN(CC)CCNC(=O)C1=C(C)NC(\C=C/2C3=CC(F)=CC=C3NC\2=O)=C1C WINHZLLDWRZWRT-ATVHPVEESA-N 0.000 description 1
- 229960002812 sunitinib malate Drugs 0.000 description 1
- 239000004094 surface-active agent Substances 0.000 description 1
- 238000001356 surgical procedure Methods 0.000 description 1
- 230000008961 swelling Effects 0.000 description 1
- 101150005655 syf-2 gene Proteins 0.000 description 1
- 208000024891 symptom Diseases 0.000 description 1
- 230000008685 targeting Effects 0.000 description 1
- 108010057210 telomerase RNA Proteins 0.000 description 1
- 229940124597 therapeutic agent Drugs 0.000 description 1
- 230000030968 tissue homeostasis Effects 0.000 description 1
- 208000037816 tissue injury Diseases 0.000 description 1
- 230000001256 tonic effect Effects 0.000 description 1
- 231100000167 toxic agent Toxicity 0.000 description 1
- 239000003440 toxic substance Substances 0.000 description 1
- 201000007363 trachea carcinoma Diseases 0.000 description 1
- 238000013518 transcription Methods 0.000 description 1
- 230000035897 transcription Effects 0.000 description 1
- 238000011222 transcriptome analysis Methods 0.000 description 1
- 230000026683 transduction Effects 0.000 description 1
- 238000010361 transduction Methods 0.000 description 1
- 230000009466 transformation Effects 0.000 description 1
- 230000001131 transforming effect Effects 0.000 description 1
- 238000011830 transgenic mouse model Methods 0.000 description 1
- 238000002054 transplantation Methods 0.000 description 1
- LENZDBCJOHFCAS-UHFFFAOYSA-N tris Chemical compound OCC(N)(CO)CO LENZDBCJOHFCAS-UHFFFAOYSA-N 0.000 description 1
- 239000012588 trypsin Substances 0.000 description 1
- 208000029729 tumor suppressor gene on chromosome 11 Diseases 0.000 description 1
- 230000007306 turnover Effects 0.000 description 1
- OUYCCCASQSFEME-UHFFFAOYSA-N tyrosine Natural products OC(=O)C(N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-UHFFFAOYSA-N 0.000 description 1
- 210000003932 urinary bladder Anatomy 0.000 description 1
- 101150101430 vdac-1 gene Proteins 0.000 description 1
- 238000012800 visualization Methods 0.000 description 1
Images






















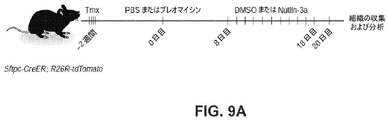




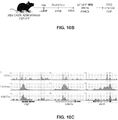



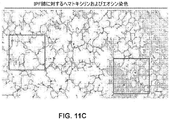

















Classifications
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/5005—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells
- G01N33/5008—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells for testing or evaluating the effect of chemical or biological compounds, e.g. drugs, cosmetics
- G01N33/502—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells for testing or evaluating the effect of chemical or biological compounds, e.g. drugs, cosmetics for testing non-proliferative effects
- G01N33/5023—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells for testing or evaluating the effect of chemical or biological compounds, e.g. drugs, cosmetics for testing non-proliferative effects on expression patterns
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/06—Animal cells or tissues; Human cells or tissues
- C12N5/0602—Vertebrate cells
- C12N5/0688—Cells from the lungs or the respiratory tract
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K35/00—Medicinal preparations containing materials or reaction products thereof with undetermined constitution
- A61K35/12—Materials from mammals; Compositions comprising non-specified tissues or cells; Compositions comprising non-embryonic stem cells; Genetically modified cells
- A61K35/42—Respiratory system, e.g. lungs, bronchi or lung cells
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P11/00—Drugs for disorders of the respiratory system
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/5005—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells
- G01N33/5008—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells for testing or evaluating the effect of chemical or biological compounds, e.g. drugs, cosmetics
- G01N33/5082—Supracellular entities, e.g. tissue, organisms
- G01N33/5088—Supracellular entities, e.g. tissue, organisms of vertebrates
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/10—Growth factors
- C12N2501/135—Platelet-derived growth factor [PDGF]
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2502/00—Coculture with; Conditioned medium produced by
- C12N2502/27—Lung cells, respiratory tract cells
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2503/00—Use of cells in diagnostics
- C12N2503/04—Screening or testing on artificial tissues
Landscapes
- Health & Medical Sciences (AREA)
- Engineering & Computer Science (AREA)
- Life Sciences & Earth Sciences (AREA)
- Biomedical Technology (AREA)
- Chemical & Material Sciences (AREA)
- Immunology (AREA)
- Cell Biology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biotechnology (AREA)
- General Health & Medical Sciences (AREA)
- Organic Chemistry (AREA)
- Zoology (AREA)
- Hematology (AREA)
- Urology & Nephrology (AREA)
- Molecular Biology (AREA)
- Biochemistry (AREA)
- Microbiology (AREA)
- Wood Science & Technology (AREA)
- Medicinal Chemistry (AREA)
- Genetics & Genomics (AREA)
- Pulmonology (AREA)
- Pathology (AREA)
- General Physics & Mathematics (AREA)
- Analytical Chemistry (AREA)
- Physics & Mathematics (AREA)
- Food Science & Technology (AREA)
- Tropical Medicine & Parasitology (AREA)
- Toxicology (AREA)
- General Engineering & Computer Science (AREA)
- Public Health (AREA)
- Animal Behavior & Ethology (AREA)
- Veterinary Medicine (AREA)
- Pharmacology & Pharmacy (AREA)
- Virology (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Physiology (AREA)
- Developmental Biology & Embryology (AREA)
- General Chemical & Material Sciences (AREA)
- Epidemiology (AREA)
- Chemical Kinetics & Catalysis (AREA)
Abstract
Description
本出願は、2020年2月12日出願の米国仮特許出願第62/975,294号および2019年9月26日出願の米国仮特許出願第62/906,241号に対する優先権を主張し、これらの各々の内容は、それらの全体がこれにより参照により本明細書に組み込まれる。
本発明は、National Institutes of Health,National Heart,Lung,and Blood Institute Grant Nos. R00HL127181;R01HL153375の下で、政府の支援によりなされた。連邦政府は、本発明に対して一定の権利を有する。
分野
成体幹細胞は、組織損傷に応答して動的な変化を受ける。これらの変化は、静止状態または平衡を保った状態からの復活、増殖の開始、新たな遺伝子発現の活性化、およびホメオスタシスへの復帰を含む。多くの場合、修復には、例えば、損傷または裸出のエリアをカバーする一過的な伸展および拡張を介した、上皮細胞形状における変化もまた関与する。幹細胞再生の研究は、通常、ニッチからのシグナルに応答して新たな分化プログラムを細胞がどのように選択するかを理解することに焦点を当てている。しかし、細胞形状および広がりなどのパラメーターにおける変化の意義、ならびにそれらにDNA修復および老化を含む病理学的状態に通常関連する遺伝子の一過的な緊密に制御された発現が関与するかどうかについては、知られていることははるかに少ない。
本開示は、肺疾患の理解、薬物発見および/または薬物スクリーニングのための使用のためのモデルに役立つ移行細胞状態が肺胞再生において存在するという、本発明者らによる発見に一部基づく。
本開示の原理の理解を促進することを目的として、ここで、好ましい実施形態に対して参照がなされ、特定の言語がそれを記載するために使用される。それにもかかわらず、本開示の範囲の限定はそれによって意図されないことが理解され、本明細書で示される本開示のかかる変更およびさらなる改変は、本開示が関連する分野の当業者に通常想起されることが企図される。
材料および方法
単一細胞トランスクリプトミクスは、ex vivoオルガノイドにおける以前には未知であった肺胞上皮細胞状態を明らかにする
最近の研究は、AEC2が、肺傷害に応答して増殖し、AEC1を生じることを示している。さらに、AEC2は、肺胞オルガノイドにおいてAEC1を自発的に生成する。しかし、AEC2のAEC1への分化の根底にある分子機構および移行細胞状態は、あまり理解されていない。これらの問題に対処するために、本発明者らは、AEC2およびPDGFRα+線維芽細胞を精製して、肺胞オルガノイドを樹立した。単一細胞トランスクリプトーム分析を、10日目のオルガノイド培養物から単離した細胞に対して実施した(図1A)。均一マニホールド近似投影(uniform manifold approximation and projection)(UMAP)は、Epcam+上皮細胞およびVim+Pdgfra+線維芽細胞からなる2つの主要なクラスターを同定した(図1B)。次に、本発明者らは、上皮細胞集団をさらにデコンボリューションおよび可視化し、それらを、リポポリサッカリド(LPS)で傷害された肺胞上皮細胞の公に入手可能な単一細胞RNA配列決定(scRNA-seq)データセットと比較した14。オルガノイド由来上皮細胞は、それらのin vivo対応物と重複し、これは、転写的類似性および細胞状態類似性を明らかにしている(図1C)。本発明者らは、オルガノイド由来上皮細胞において、以下の複数のサブクラスターを観察した:Sftpc(AEC2のマーカー)を発現する細胞、Ager(AEC1のマーカー)を発現する細胞およびSftpc+Mki67+増殖性AEC2(図1D)。
PATS細胞は、肺胞傷害後にin vivoで出現する。
次いで、PATS細胞が、ホメオスタシスおよび再生中の肺胞組織においてin vivoで観察され得るかどうかを調査した。これを試験するために、本発明者らは、LPSで処置したマウス肺および対照マウス肺からのscRNA-seqデータをUMAPプロットでレンダリングし、LPS傷害に独自の集団を見出した。UMAPプロットは、肺胞オルガノイドscRNA-seqデータセット(4,573細胞)およびLPSで処置した肺または対照肺(13,204細胞)における、AEC2(LampおよびSftpb)、AEC2増殖性(Mki67およびTop2a)、AEC1(AgerおよびAkap5)および新規肺胞上皮細胞状態における遺伝子の発現を示した。重要なことに、この集団は、Cldn4、Sox4、Lgals3、S100a14およびFn1を含む、PATSにおいて発現される遺伝子について富化されている。
系列追跡は、PATS細胞がAEC2に起源することを明らかにする。
オルガノイドおよび傷害モデルから先に提示されたデータは、PATS細胞がAEC2に起源することを示唆した。この仮説を実験的に試験するために、本発明者らは、タモキシフェンの投与が、AEC2およびその子孫において特異的に、tdTomato発現を恒久的に誘導する、Sftpc-CreER;R26R-tdTomato(本明細書で以後Sftpc-tdT+と呼ぶ)マウスモデルを使用した(図5A)。ブレオマイシン投与後の10日目に、損傷した肺胞領域において、LGALS3、SFNおよびCLDN4を共発現する系列標識された(tdTomato+、tdt+)細胞が観察されたが、対照肺では観察されなかった。AGERを共発現するtdt+細胞は、後の時点では、平たく薄く細長い形態を示し、AEC1へのAEC2移行が確認された。興味深いことに、SFN、CLDN4およびLGALS3を含むPATSマーカーを共発現する系列標識された細胞は、修復プロセスの間に、別個の形態学的特色(円形の、「伸展した」、および細長い)を有した(図5B)。これを定量化するために、本発明者らは、CLDN4+および/またはLGALS3+発現ならびにそれらの形状に従って、系列陽性細胞をスコア付けした(図5C)。3つの別個のパターンのマーカー発現、CLDN4+、CLDN4+LGALS3+およびLGALS3+が観察され、これらがそれぞれ、円形、伸展したおよび細長い形態と相関したことが見出された(図5Cおよび図5D)。さらに、本発明者らは、ブレオマイシンで誘導された傷害の後、8日目のCLDN4+細胞の、10日目のCLDN4+LGALS3+細胞の、および12日目のLGALS3+細胞の、実質的な富化を観察した(図2l)。scRNA-seqデータセットにおけるCLDN4+およびCLDN4+LGALS3+細胞状態が注目されたが、別個のLGALS3+集団は注目されなかった。類似のPATSマーカー発現が、肺切除術で誘導された「生理学的」再生の間に、AEC2系列標識されたマウスにおいて検出された(図5E)。scRNA-seqにおけるLGALS3の発現 対 免疫染色におけるLGALS3の発現との間の食い違いは、最終AEC1におけるLGALS3タンパク質の永続に起因し得る。合わせると、この分析により、AEC2のAEC1への分化が、2つの別個の状態を通過することが明らかになった(図5F)。
系列追跡分析は、PATS細胞がAEC1を生成することを明らかにする
先に示された実験からの複数ラインのデータは、PATS細胞がAEC1に向かう途中であることを示唆している。これを試験するために、スプライシングされたメッセンジャーRNAとスプライシングされていないメッセンジャーRNAとの間の比率に基づいた細胞分化トラジェクトリーの予測を可能にする、Velocytoと呼ばれるアルゴリズムツール18を使用した。PATS1 Ctgf+集団に起源して、PATS2 Lgals3+集団を介してAEC1への、強いRNA速度(トラジェクトリー)が観察された(図6Aおよび図6B)。次いで、R26R-tdTomatoと組み合わせた、PATSにおいて富化される遺伝子であるKrt19のCreER対立遺伝子(Krt19-tdt)を使用して、ブレオマイシン傷害後の系列追跡を実施して、同じトラジェクトリーが、再生中の肺胞においてin vivoで生じるかどうかを試験した。最初に、マウスを、ブレオマイシンで傷害し、タモキシフェンを7日後に投与して、Krt19発現細胞およびそれらの子孫を標識した(図6C)。次いで、AEC2、AEC1およびPATSマーカーについての免疫蛍光を実施した。
PATS中の保存された転写プログラムおよび経路。
AEC2のAEC1への分化は、立方体状から平たく薄いものへの、細胞の形状および構造における劇的な変化に関連する。かかる移行には、典型的には、シグナル伝達タンパク質および構造タンパク質の発現における多くの変化が伴う。したがって、これらの変化がPATS細胞において生じると考えられた。これらの変化がPATS細胞において生じるかどうかを決定するために、オルガノイド培養物(4,573細胞)およびLPSで誘導された傷害モデル(13,204細胞)からのscRNA-seqデータを分析した。以下を含む多数の遺伝子が、PATS細胞間で保存されていた:TGFβシグナル伝達経路中の遺伝子(Cdkn2b、Mapk3、Myc、Nbl1、Ppp2r1a、Rbx1、Rhoa、Skp1a、Tgfb1、Tgif1およびThbs1)、p53シグナル伝達経路中の遺伝子(Bax、Casp8、Ccnd1、Ccnd2、Ccng1、Cdk4、Cdkn1a、Cycs、Ei24、Gadd45a、Gadd45b、Sesn3、Sfn、Shisa5、Thbs1およびTrp53)、細胞老化に関与する遺伝子(Calm1、Calm2、Capn2、Ccnd1、Ccnd2、Cdk4、Cdkn1a、Cdkn2b、Eif4ebp1、Ets1、Gadd45b、H2-D1、H2-K1、h2-Q4、H2-Q6、H2-Q7、H2-T22、H2-T23、Kras、Map2k2、Mapk13、Mapk3、Myc、Ppid、Ppp1ca、Ppp3ca、Rbbp4、Rheb、Rras、Sqstm1、Slc25a4、Tgfb2、Trp53、Vdac1、Vdac2、Vdac3およびZfp36l1)、タイトジャンクションシグナル伝達遺伝子(Actb、Actg1、Actn1、Actn4、Actr3、Ccnd1、Cdc42、Cdk4、Cldn3、Cldn4、Cldn7、Cldrn18、cttn、Ezr、F11r、Hspa4、Itgb1、Jun、Msn、Myh9、Myl6、Myl9、Myl12a、Myl12b、Myn12a、Nedd4、Ocln、Ppp2r1a、Rac1、Rap1a、Rhoa、Slc9a3r1、Tuba1a、Tuba1b、Tuba1cおよびVasp)、およびDNA損傷チェックポイント遺伝子(Ccnd1、Ccng1、Cdkn1a、Ddx39b、ler3、Prpf19、Rpl26、Rps27l、Sox4、Syf2およびTrp53)。
PATS細胞は、in vivoでの肺胞再生の間に、DNA損傷および老化を天然に示す。
これらのデータは、DNA損傷関連遺伝子および老化関連遺伝子が、PATSにおいて高度に富化されることを示した。これらの知見を検証するために、ブレオマイシンで処置したマウスおよびPBSで処置したマウスの肺を、老化細胞のバイオマーカーとして機能するβ-ガラクトシダーゼ活性およびDNA損傷のマーカーであるγH2AXについて、10日後にアッセイした。興味深いことに、ブレオマイシンで傷害された肺胞において、5-ブロモ-4-クロロ-3-インドリル-β-d-ガラクトピラノシド(X-gal)由来の青色の沈着に基づいて、多数のβ-ガラクトシダーゼ活性細胞が検出されたが、対照肺胞では検出されなかった(図8A)。これは、ブレオマイシンで処置した肺において特異的な、老化マーカーCDKN1A(p21)の発現によってさらに支持された(図8B)。CDKN1A発現は、伸展したtdt+細胞において観察されたが、AGER+細胞では観察されず、これは、老化がAEC1において観察されないことを示している(図8B)。同様に、γH2AX斑の蓄積もまた、ブレオマイシンで処置した肺において、伸展したSftpc-tdt+細胞の核において検出されたが、AGER+細胞では検出されなかった(図8C)。さらなる分析により、ブレオマイシンで傷害された肺において、LGALS3を共発現するγH2AX+Sftpc-tdt+細胞の顕著な数が明らかになったが、対照では明らかにならず、これは、PATS細胞が肺胞再生の間にDNA損傷を受けることを示している(図8D)。
PATS細胞は、機械的伸展で誘導されたDNA損傷に対して脆弱である
以前の研究は、電離放射線、酸化ストレス、ならびに細胞の遊走および伸展の間に生じる機械的な力が、全てDNA損傷を引き起こし得ることを示している。経路分析により、酸化ストレスではなく細胞骨格変化に関与する遺伝子についての富化が明らかになった。AEC1へのAEC2分化が広範な細胞の広がりおよび細胞骨格動態を要求することを考慮して、PATS細胞が、DNA損傷をもたらし得る機械的伸展を経験すると仮説を立てた。これを試験するために、AEC2を、SFTPC-GFPマウスから精製し、AEC2が広がりAEC1へと分化する条件であるプラスチック表面上でそれらを培養した(二次元(2D)培養)(図8Hb)。プレーティング後5日以内に、AEC2のほとんどは伸展し、GFP発現は喪失した(GFP-)または下方調節された(GFPlo)。これらの細胞は、伸展の間中、その表面積を増加させ、AEC1マーカーを発現し始めた。興味深いことに、AGER+細胞は、DNA損傷マーカーγH2AXについても陽性であった(図8Hおよび図8I)。プレーティング後の2日目には、細胞においてDNA損傷マーカーが観察されなかったことに留意されたい(図8I)。合わせると、これらのデータは、大きく薄いAEC1へと分化する立方体状AEC2が、DNA損傷修復を天然に経験し、一過的老化を受けることを示している。
TP53シグナル伝達の遺伝的機能喪失および薬理学的活性化は、肺胞傷害-修復の間のPATSを介したAEC2~AEC1の分化を異常調節する
これらのデータは、TP53シグナル伝達の活性化が、傷害された肺におけるPATSに関連することを示唆している。他の組織における以前の研究は、TP53シグナル伝達が分化を促進し、幹細胞自己更新を抑制することを示唆している。TP53シグナル伝達の増強された活性化が、傷害後のPATSが関与するAEC1へのAEC2分化を増加させるかどうかを決定するために、Sftpc-CreER;R26-tdTomatoマウスにおいてブレオマイシンで誘導された傷害を実施し、その後、傷害後の8日目に開始するNutlin-3a(TP53活性化因子)またはジメチルスルホキシド(DMSO;対照)投与を実施し、20日目に組織収集した(図9A)。AGERについての免疫染色により、Nultin-3a処置が、DMSO処置と比較して、Sftpc-tdt+細胞のAEC1への有意に高い分化をもたらしたことが明らかになった(図9B)。Nutlin-3aを受けた未傷害の肺では、AGER+Sftpc-tdt+細胞の存在は観察されず、これは、Nutlin-3aで誘導されたTP53活性化単独では、AEC2のAEC1への分化を誘導するのに不十分であることを示唆している(図9B)。さらに、肺胞オルガノイド中のAEC2に対するNutlin-3aの影響を試験した。オルガノイドを、7日目に開始して、15日目の回収まで、Nutlin-3aまたはDMSOで処置した(図9C)。Nutlin-3aで処置したオルガノイドは、対照と比較して、AGER+細胞の数および強度における増加、ならびにKi67+増殖性細胞の数における減少を示した(図9D)。これらのデータは、TP53シグナル伝達の異所的活性化が、AEC2分化をin vivoおよびex vivoで増強することをさらに支持している。
TP53は、PATS関連遺伝子座に結合し、それらの発現を制御する
PATS細胞において潜在的に活性な転写因子モジュールを同定するために、クロマチン免疫沈降(ChIP)分析を、ヒストンH3K4me3、H3K36me3およびH3K27acについて実施して、それぞれ、活性なプロモーター、転写している遺伝子およびエンハンサーを同定した。ブレオマイシンで処置したCtgf-GFPおよび対照マウス由来のPATS細胞およびAEC2をそれぞれ単離し、ヒストンマーク分析に使用した。H3K4me3富化は、細胞型特異的プロモーターに対応する遺伝子(AEC2ではSftpc、PATSではFn1)において見出された。さらなる分析により、対照由来のAEC2と比較して、ブレオマイシン損傷で誘導された肺由来のPATS細胞に特異的な転写物の転写開始部位およびプロモーターと重複する多数のH3K4me3ピークの富化が明らかになった(図10A)。H3K4me3ピークのモチーフ分析は、TP53、ETS1、NF1、ATF3およびSOX4 - これらは全て、PATSで富化された経路に関与している - を含む転写因子についての結合部位の富化を予測した。AEC2と比較したPATSにおける、公知のTP53標的遺伝子座(FasおよびMdm2)におけるH3K4me3、H3K27acおよびH3K36me3についての富化。TP53について、予測された結合部位が、Mdm2エンハンサーおよびFasプロモーター中に見出され、これらは、TP53の2つの周知の直接的標的である。合わせると、これらのデータは、PATS特異的遺伝子の転写制御におけるTP53の直接的な役割を暗示している。
PATS関連遺伝子発現シグネチャーおよびシグナル伝達経路は、ヒト線維化肺において富化される
最近の研究は、特発性肺線維症(IPF)において線維化病巣を裏打ちする肺胞上皮細胞が、老化、成長停止および分化遮断の特色を示すことを示唆している。肺胞前駆体が、非許容的な病理学的微小環境に応答してその分化プロセスをPATS様状態で留め置くかどうかを試験するために、IPFを有するヒト患者の肺からの、最近報告されたscRNA-seqデータセット29を分析した。最初に、全ての非上皮および気道細胞を、さらなる分析から排除した。健康な肺およびIPF肺の両方において、UMAPプロットにおける見かけの重複を伴って、多数のAEC2およびAEC1が観察された(図11A)。対照的に、IPF試料において高度に富化され、AEC1またはAEC2のいずれとも重複しなかった、別個の細胞クラスターが見出された(図11B)。これらの細胞は、それらのマーカー遺伝子発現によって、KRT5-KRT17+と以前にアノテーションされた。特に、差次的遺伝子発現の分析により、マウスオルガノイドおよび傷害モデル由来のIPFで富化された細胞クラスターとPATS細胞との間で、Sfn、Sox4およびFn1、ならびにまたS100A2、PTGS2、KRT17、KRT8、CALS1、CLDN4、MMP7、PRSS2、IGFBP7、COL1A1、MDK、TAGLN、GDF15、TM4SF1およびCTSEを含む転写物の、著しい類似点が明らかになった。したがって、ヒトIPF特異的クラスターを「PATS様」細胞と命名した(図11Aおよび図11B)。PATS様集団において高度に富化された他の転写物は、CALD1、PRSS2、MMP7およびS100A2である。scRNA-seqデータを検証するために、免疫蛍光分析を、健康なヒト、ならびにIPFを有する肺の線維化領域および非線維化領域に対して実施した(図11C)。PATS様細胞(SFN、CLDN4およびKRT17)、AEC2(SFTPCおよびHTII-280)、AEC1(AGER)および筋線維芽細胞(ACTA2)の共免疫蛍光分析により、IPF肺の線維化領域中において特異的な、PATS様マーカーの発現が明らかになったが、健康な対照中および健康な外観を有するIPF肺の領域中では明らかにならなかった(図11D、図11E、図11F、図11G、図11H、図11I)。興味深いことに、KRT17は、正常な肺の基底細胞およびIPF肺中の基底様細胞において発現されることが最近示された(Habermann et al., 2019, doi.org/10.1101/753806;Adams et al., 2019, doi.org/10.1101/759902)。IPF肺の線維化領域において特異的な、基底細胞の別のマーカーであるTP63の類似の発現パターンもまた見出されたが、非線維化領域では見出されなかった(図11H、図11I)。PATS様マーカーは、高レベルのCOL1A1、および筋線維芽細胞の蓄積(図7D)と一致して、重症線維症の領域において高度に富化されることが見出された(図11D~図11I)。PATSマーカー(SFN、CLDN4、KRT17およびTP63)の定量化が、これらの知見をさらに支持した(図11E、図11Gおよび図11I)。さらに、PATS細胞は、健康な肺における立方体状AEC2とは対照的に、IPF肺において細長い形態を獲得したことが観察された(図11D、図11Fおよび図11G)。AGER発現が、多くのPATS様細胞を有するIPF肺中の領域には存在しなかったことに留意されたい(図11F)。遺伝子オントロジーおよび経路分析により、p53シグナル伝達(CDKN1A、CDKN2AおよびMDM2、ならびにCCND1、CCND1、SNF、GADD45A、BID、BAX、PMAIP1、BBC3、BCL2L1、PERP、ZMAT3、IGFBP3、CD82、THBS1、SERPINB5、RRM2B、SESN3、STEAP3、TP73)、DNA損傷チェックポイント(RPS27LおよびPLK3、ならびにBRSK1、HMGA2、PEA15、PLK2、PRKDC、SOX4、TRIAP1)および細胞老化(TGFB2およびHIPK2、ならびにCDKN2B、CXCL8、EIF4EBP1、ES1、NFATC1、SERPINE1、TGFB1、ZFP36L1)の構成要素についての富化が明らかになった(図11J)。さらに、PATS様クラスターは、接着斑、タイトジャンクション、およびアクチン細胞骨格の調節の構成要素について富化され、これは、IPFにおけるPATS様細胞とオルガノイドおよび再生中の肺胞におけるPATS様細胞との間の著しい類似点を示している。線維症の公知の調節因子であるAREG、TGFB1、TGFB2およびTIMP1についての有意な富化が、PATS様細胞において見出された(図11K)。次に、細胞老化(β-ガラクトシダーゼ活性およびCDKN1A)およびDNA損傷応答(γH2AX)のマーカーを、IPF肺および健康な肺において分析した。このデータにより、IPF中のPATS様集団におけるこれらのマーカーの特異的発現が明らかになったが、健康な肺では明らかにならなかった(図11Lおよび図11M)。合わせると、この分析により、再生中の組織由来のPATS細胞と病理学的線維化肺に特異的なPATS様細胞との間での類似性が明らかになった。
新規の調節可能な肺傷害モデルは、一過的な線維症をin vivoで再現する
肺線維症は、劇的な細胞外マトリックス変化および組織リモデリングを全てが次に生じる、上皮、間葉および免疫集団における変更が関与する、複雑な疾患である。これらの複雑な相互作用の結果として、線維症の開始はあまり理解されておらず、これらの個々の因子の全ての組合せの結果であると仮定されている。今のところ、単一の細胞型への損傷が線維化開始を生じ得るかどうかについては、いまだ未知である。
PATSの持続は、リポ線維芽細胞をin vivoで筋線維芽細胞へと駆動する
単一ラウンドのAEC1アブレーションが、一過的で可逆的な線維症、線維筋細胞移行のみを生じ、筋線維芽細胞を生じなかったことを考慮すると、反復AEC1アブレーション(反復肺胞傷害の代用物)が、線維症をさらに駆動し得ると考えられた(図14A)。反復AEC1アブレーション後のマウス肺の予備的組織学的分析により、実際に、肺胞における増加したACTA2発現、ならびに星型から「帯状」形態への、ACTA2+細胞における形態学的変化が明らかになった(図14B、図14Cおよび図14D)。さらに、反復アブレーションは、線維化病巣の同定を可能にし、マッソントリクローム分析は、これらの肺におけるECM産生における増加を実証した。
拡張可能な共培養モデルは、肺線維症をex vivoで再現する
集合的に、in vivoデータは、PATSが、筋線維芽細胞変換において直接的な役割を果たすことを示唆した。この細胞間コミュニケーションを決定的に実証するために、上皮およびリポ線維芽細胞の両方の成長を支持し、それらの正常な表現型を維持する培養条件を確立した。次いで、リポ線維芽細胞を、AEC2細胞(SFTPC-GFPマウスに由来する)またはPATS(ブレオマイシン傷害後にCTGF-GFPマウス(GFP+/EpCAM+)細胞から単離した)と共に7日間共培養し、免疫染色によって分析した。これらの条件下で、リポ線維芽細胞は、PATSの存在下でのみACTA2を自発的に発現したが、AEC2とでは発現しなかった。これらのデータは、PATSが、線維化促進性因子を産生し、リポ線維芽細胞の筋線維芽細胞への変換を誘導することをさらに示唆している(図15)。
リポ線維芽細胞の正体は、強直性のAEC1由来PDGFAシグナル伝達によって維持される
しかし、これら3つの傷害モデルからのデータは、PATSの存在がさらなる線維形成を駆動することを示唆しているが、単回AEC1アブレーションでは、ネイティブ間葉におけるaSMA発現がPATSの出現に先行することが観察された。これは、AEC1の孤立した喪失が、リポ線維芽細胞の筋線維芽細胞への移行を誘導するのに十分であることを示唆しており、したがって、AEC1が、細胞間コミュニケーションを介してリポ線維芽細胞の正体を維持すると仮説を立てた。
PDGFAシグナル伝達の崩壊は、筋線維形成の開始に十分である
PDGFAシグナル伝達軸の喪失が筋線維芽細胞変換に十分であるかどうかを試験するために、PDGFRA、PDGFRA-CreER/PDGFRAfloxの細胞型特異的条件付きノックアウトモデルを生成した。このマウス系は、PDGFRAの単一の機能的対立遺伝子のみを有し、タモキシフェン投与の際に、PDGFRA+細胞において排他的に、PDGFRA発現の完全な喪失を生じる。特に、PDGFRA欠失のたった2日後に、これらのマウスは、肺間葉においてaSMAを発現し始め、これは、PDGFRAシグナル伝達軸の喪失の際の迅速な筋線維芽細胞変換を示す(図17)。これらのデータに基づいて、PDGFAがリポ線維芽細胞の維持のために必要かつ十分であると仮説を立てた。
Runx1は、筋線維形成(myofibrogenic)運命のマスター転写調節因子である
リポ線維芽細胞から筋線維芽細胞への移行をもたらすマスター調節因子を決定するために、傷害後条件において優先的に上方調節される転写因子を調べた。複数の臓器における線維化進行に関連付けられることが公知の転写因子であるRunx1は、AEC1アブレーション後およびブレオマイシン傷害後に強く上方調節されることが見出された(図18)。重要なことに、Runx1発現は、AEC1アブレーション、反復AEC1アブレーションおよびPDGFRA欠失を含む、本明細書に記載される線維化傷害モデルの全てのモデルにわたって増加する(図19)。
RUNX1の遺伝学的および薬理学的阻害は、TGFbにより誘導される線維形成を遮断する
この培養システムを使用して、筋線維形成におけるRunx1の直接的な役割を決定した。培養物中のTGF-Bで処置した野生型リポ線維芽細胞は、aSMAを発現し、PDGFRAを下方調節する。Runx1flox/floxマウス由来のリポ線維芽細胞を培養し、Adeno-Cre-GFPウイルスを投与したところ、培養物における効率的なRunx1欠失が生じた(図20A)。Runx1欠失の後、TGF-Bを使用して筋線維形成を誘導した。しかし、野生型線維芽細胞とは異なり、Runx1が欠失した線維芽細胞は、aSMAを発現せず、PDGFRA発現を維持し、これは、Runx1が筋線維形成に必要であることを示している(図20B)。遺伝的欠失に加えて、これらの実験を、薬物Ro 5-3335によるRunx1の薬理学的阻害を用いて反復した。リポ線維芽細胞を、Ro 5-3335およびTGF-Bの存在下で48時間培養した。Runx1欠失と同様に、Ro 5-3335で処置したリポ線維芽細胞は、PDGFRA発現およびそれらの星型形状を維持し、aSMA発現および筋線維化(myofibrotic)変換は事実上存在しなかった。
Claims (37)
- 肺傷害オルガノイドモデルを生成する方法であって、
i)細胞の共培養物を、前記細胞の培養のために適合させた培養培地中に提供するステップであって、前記共培養物が、プレ1型肺胞移行細胞状態(PATS)細胞および肺胞線維芽細胞を含む、ステップ;
ii)前記培養培地を、1種または複数の作用物質と接触させるステップ;ならびに
iii)前記作用物質と接触されていない対照細胞培養培地と比較した、前記PATS細胞および/または前記肺胞線維芽細胞の少なくとも1つの発現マーカーに対する前記作用物質の生物学的影響を分析するステップ
を含む、方法。 - 前記PATS細胞が、罹患組織から単離される、請求項1に記載の方法。
- 前記罹患組織が、慢性閉塞性肺疾患(COPD)肺組織、肺線維症肺組織、特発性肺線維症肺組織、肺気腫肺組織、肺がん組織、サルコイドーシス肺組織、間質性肺炎肺組織、敗血症肺組織、ウイルス感染および細菌感染を有する肺組織、急性呼吸促迫症候群肺組織、ならびに気管支肺異形成症肺組織である、請求項2に記載の方法。
- 前記PATS細胞が、肺上皮細胞をin vivoまたはin vitroで傷害を引き起こす作用物質に曝露させることによって生成される、請求項1に記載の方法。
- 前記傷害を引き起こす作用物質が、ブレオマイシン、ジフテリア毒素(DT)、タモキシフェン、照射、ウイルス、細菌または真菌である、請求項4に記載の方法。
- 前記肺胞線維芽細胞が、リポ線維芽細胞である、請求項1に記載の方法。
- 前記肺胞線維芽細胞が、疾患状態特徴を有する、請求項1に記載の方法。
- 前記疾患状態特徴が、ACTA2を発現するリポ線維芽細胞、筋線維芽細胞の存在、および/または星型形状から帯状形状への細胞変化を含む、請求項7に記載の方法。
- 前記培養培地が、血小板由来増殖因子受容体(PDGFR)リガンドを含む、請求項1に記載の方法。
- 前記PDGFRリガンドが、血小板由来増殖因子サブユニットA(PDGFA)、血小板由来増殖因子サブユニットB(PDGFB)、血小板由来増殖因子サブユニットC(PDGFC)および/または血小板由来増殖因子サブユニットD(PDGFD)である、請求項9に記載の方法。
- 前記培養培地が、PDGFRA発現を維持することが可能な作用物質を含む、請求項1に記載の方法。
- 前記作用物質が、Ro 5-3335、AI-10-49、またはRUNXもしくはコア結合因子(CBF)タンパク質を調節する他の分子である、請求項11に記載の方法。
- 肺線維症のためのex vivoモデルを生成する方法であって、
i)in vivoでアブレーションを受けたAEC1細胞の培養物を、前記細胞の培養のために適合させた培養培地中に提供するステップ;
ii)前記培養培地を、1種または複数の作用物質と接触させるステップ;ならびに
iii)前記作用物質と接触されていない対照細胞培養培地と比較した、前記AEC1細胞の少なくとも1つの発現マーカーに対する前記作用物質の生物学的影響を分析するステップ
を含む、方法。 - 前記AEC1細胞が、少なくとも1ラウンドのアブレーションを受けている、請求項13に記載の方法。
- 前記AEC1細胞が、単独のまたは他の傷害モデルと組み合わせたアブレーションを受けている、請求項13に記載の方法。
- 前記AEC1細胞が、反復アブレーションを受けている、請求項13に記載の方法。
- 前記反復アブレーションが、約3日間~約2週間の過程にわたって行われる、請求項16に記載の方法。
- 前記AEC1細胞が、ブレオマイシン、ジフテリア毒素(DT)、アスベスト、化学的毒性作用物質、タモキシフェン、ウイルス、細菌または真菌に曝露されることによるアブレーションを受けている、請求項13に記載の方法。
- 前記AEC1細胞が、aSMAおよび/またはACTA2を発現する、請求項13に記載の方法。
- 前記AEC1細胞が、線維筋細胞の特徴を示す、請求項13に記載の方法。
- プレ1型肺胞移行細胞状態(PATS)細胞またはPATS様細胞を同定する方法であって、
i)肺胞細胞を得るステップ;および
ii)前記細胞を、1つまたは複数のマーカーについてスクリーニングするステップ
を含み、前記1つまたは複数のマーカーの存在が、PATS細胞またはPATS様細胞の存在を示す、方法。 - 前記肺胞細胞が、気管基底細胞、細気管支分泌細胞(クラブ細胞またはクララ細胞としても公知)、クラブバリアント細胞、肺胞上皮前駆(AEP)細胞、クララバリアント細胞、遠位肺前駆体、p63+Krt5-気道細胞、系列陰性上皮前駆体、気管支肺胞上皮幹細胞(BASC)、Sox9+p63+細胞、神経内分泌前駆細胞、遠位気道幹細胞、粘膜下腺管細胞、誘導多能性幹細胞由来肺幹細胞および2型肺胞上皮(AEC2)細胞からなる群より選択される、請求項21に記載の方法。
- 前記肺胞細胞が、慢性閉塞性肺疾患(COPD)、肺線維症、特発性肺線維症、肺気腫、肺がん、サルコイドーシス、間質性肺炎、敗血症、ウイルス感染、細菌感染、真菌感染、急性呼吸促迫症候群および/または気管支肺異形成症を患う患者の肺から得られる、請求項21に記載の方法。
- 前記肺胞細胞を作用物質と接触させるステップをさらに含む、請求項21に記載の方法。
- 前記作用物質が、ブレオマイシン、ジフテリア毒素(DT)、タモキシフェン、ウイルス、細菌または真菌である、請求項23に記載の方法。
- 前記1つまたは複数のマーカーが、CLDN4、KRT19、SFN、LGALS3、SOX4、S100A2、PTGS2、KRT17、KRT8、CALS1、MMP7、PRSS2、IGFBP7、COL1A1、MDK、TAGLN、GDF15、TM4SF1 TP63および/またはCTSEを含む、請求項21に記載の方法。
- 前記1つまたは複数のマーカーが、CLDN4、LGALS3およびLGALS3である、請求項21に記載の方法。
- 前記1つまたは複数のマーカーが、CLDN4、KRT19およびSFNである、請求項21に記載の方法。
- 前記1つまたは複数のマーカーが、CALD1、PRSS2、MMP7およびS100A2である、請求項21に記載の方法。
- in vivo組織線維症モデルを生成する方法であって、
i)被験体においてPDGFリガンドおよび/もしくはPDGF受容体(PDGFR)シグナル伝達の遺伝的喪失を導入するステップ;ならびに/あるいは
ii)PDGF-PDGFRシグナル伝達をモジュレートすることができる作用物質を被験体に投与するステップ;ならびに
iii)前記被験体由来の細胞を分析して、対照被験体と比較した、PDGFリガンドおよび/もしくはPDGF受容体(PDGFR)シグナル伝達の遺伝的喪失の生物学的影響ならびに/またはPDGF-PDGFRシグナル伝達をモジュレートすることができる前記作用物質の生物学的影響を決定するステップ
を含む、方法。 - PDGF-PDGFRシグナル伝達をモジュレートすることができる前記作用物質が、PDGFR阻害剤、PDGFRアゴニスト、PDGF阻害剤またはPDGFアゴニストを含む、請求項30に記載の方法。
- PDGF-PDGFRシグナル伝達をモジュレートすることができる前記作用物質が、PDGFまたはPDGFRに対する中和抗体を含む、請求項30に記載の方法。
- in vivo組織線維症モデルを生成する方法であって、
i)被験体においてRUNX1、RUNX2、RUNX3もしくはCBFbの遺伝的喪失を導入するステップ;ならびに/あるいは
ii)RUNX1阻害剤、RUNX2阻害剤、RUNX3阻害剤もしくはCBFb阻害剤を前記被験体に投与するステップ;ならびに
iii)前記被験体由来の細胞を分析して、対照被験体と比較した、RUNX1、RUNX2、RUNX3もしくはCBFβの前記遺伝的喪失の生物学的影響および/または前記RUNX1阻害剤、前記RUNX2阻害剤、前記RUNX3阻害剤もしくは前記CBFβ阻害剤の生物学的影響を決定するステップ
を含む、方法。 - 前記阻害剤が、Ro 5-3335、AI-10-49、またはRUNX1、RUNX2、RUNX3および/もしくはCBFβに対する中和抗体である、請求項33に記載の方法。
- 前記被験体がマウスである、請求項30または33に記載の方法。
- 前記細胞が、前記被験体の肺、心臓、腎臓、肝臓、腸、皮膚または骨髄に由来する、請求項30または33に記載の方法。
- 前記モデルが、肺、肝臓、腎臓、皮膚、腸または骨髄における線維症のための疾患モデルである、請求項30または33に記載の方法。
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201962906241P | 2019-09-26 | 2019-09-26 | |
| US62/906,241 | 2019-09-26 | ||
| US202062975294P | 2020-02-12 | 2020-02-12 | |
| US62/975,294 | 2020-02-12 | ||
| PCT/US2020/053164 WO2021062412A1 (en) | 2019-09-26 | 2020-09-28 | Lung fibrosis model and methods of using the same |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2022550141A true JP2022550141A (ja) | 2022-11-30 |
| JPWO2021062412A5 JPWO2021062412A5 (ja) | 2023-10-05 |
Family
ID=75166322
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2022519551A Pending JP2022550141A (ja) | 2019-09-26 | 2020-09-28 | 肺線維症モデルおよびそれを使用する方法 |
Country Status (5)
| Country | Link |
|---|---|
| US (1) | US20220341915A1 (ja) |
| EP (1) | EP4034142A4 (ja) |
| JP (1) | JP2022550141A (ja) |
| CA (1) | CA3156163A1 (ja) |
| WO (1) | WO2021062412A1 (ja) |
Families Citing this family (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2019099560A1 (en) | 2017-11-14 | 2019-05-23 | The Schepens Eye Research Institute, Inc. | Runx1 inhibition for treatment of proliferative vitreoretinopathy and conditions associated with epithelial to mesenchymal transition |
| CA3180984A1 (en) * | 2020-04-23 | 2021-10-28 | The Schepens Eye Research Institute, Inc. | Methods and materials for treatment of fibrosis |
| CN118234852A (zh) * | 2021-10-29 | 2024-06-21 | 特伦托大学 | 用于追踪和/或消融静止细胞的遗传构建体 |
| CN115772492B (zh) * | 2023-02-13 | 2023-05-02 | 淇嘉科技(天津)有限公司 | 利用博来霉素体外构建肺纤维化疾病模型的方法 |
-
2020
- 2020-09-28 US US17/763,991 patent/US20220341915A1/en active Pending
- 2020-09-28 JP JP2022519551A patent/JP2022550141A/ja active Pending
- 2020-09-28 EP EP20868029.8A patent/EP4034142A4/en active Pending
- 2020-09-28 CA CA3156163A patent/CA3156163A1/en active Pending
- 2020-09-28 WO PCT/US2020/053164 patent/WO2021062412A1/en unknown
Also Published As
| Publication number | Publication date |
|---|---|
| CA3156163A1 (en) | 2021-04-01 |
| US20220341915A1 (en) | 2022-10-27 |
| EP4034142A4 (en) | 2023-11-15 |
| WO2021062412A1 (en) | 2021-04-01 |
| EP4034142A1 (en) | 2022-08-03 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| Kobayashi et al. | Persistence of a regeneration-associated, transitional alveolar epithelial cell state in pulmonary fibrosis | |
| Lynch et al. | Submucosal gland myoepithelial cells are reserve stem cells that can regenerate mouse tracheal epithelium | |
| Paris et al. | STAT3–BDNF–TrkB signalling promotes alveolar epithelial regeneration after lung injury | |
| Schutgens et al. | Tubuloids derived from human adult kidney and urine for personalized disease modeling | |
| JP2022550141A (ja) | 肺線維症モデルおよびそれを使用する方法 | |
| Wang et al. | Single-cell analysis of murine fibroblasts identifies neonatal to adult switching that regulates cardiomyocyte maturation | |
| Choi et al. | Release of Notch activity coordinated by IL-1β signalling confers differentiation plasticity of airway progenitors via Fosl2 during alveolar regeneration | |
| Pu et al. | Bipotent transitional liver progenitor cells contribute to liver regeneration | |
| Ruan et al. | Capillarized liver sinusoidal endothelial cells undergo partial endothelial-mesenchymal transition to actively deposit sinusoidal ECM in liver fibrosis | |
| Moghieb et al. | Time-resolved proteome profiling of normal lung development | |
| Biermann et al. | Epigenetic priming of human pluripotent stem cell-derived cardiac progenitor cells accelerates cardiomyocyte maturation | |
| Li et al. | Acquisition of cholangiocarcinoma traits during advanced hepatocellular carcinoma development in mice | |
| Inagaki et al. | Use of mouse liver mesothelial cells to prevent postoperative adhesion and promote liver regeneration after hepatectomy | |
| Ó hAinmhire et al. | A conditionally immortalized Gli1-positive kidney mesenchymal cell line models myofibroblast transition | |
| Summers et al. | Resident mesenchymal vascular progenitors modulate adaptive angiogenesis and pulmonary remodeling via regulation of canonical Wnt signaling | |
| Kobayashi et al. | Collagen type I-mediated mechanotransduction controls epithelial cell fate conversion during intestinal inflammation | |
| Wang et al. | An organoid library of salivary gland tumors reveals subtype-specific characteristics and biomarkers | |
| Yeo et al. | Chromatin accessibility dynamics of neurogenic niche cells reveal defects in neural stem cell adhesion and migration during aging | |
| Zhou et al. | MKI67 as a potential diagnostic biomarker in pulmonary hypertension | |
| Reinecke et al. | Aberrant activation of wound healing programs within the metastatic niche facilitates lung colonization by osteosarcoma cells | |
| He et al. | Stiffness regulates intestinal stem cell fate | |
| van Bree et al. | Development of an orthotopic medulloblastoma zebrafish model for rapid drug testing | |
| US20210322454A1 (en) | Hippo regulation of cardiac vascularity, fibrosis, and inflammation | |
| Myszczyszyn et al. | Mice with renal-specific alterations of stem cell-associated signaling develop symptoms of chronic kidney disease but surprisingly no tumors | |
| JP2022546253A (ja) | 細胞外マトリクスの調節 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20220809 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20230927 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20230927 |
|
| A977 | Report on retrieval |
Free format text: JAPANESE INTERMEDIATE CODE: A971007 Effective date: 20240830 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20241003 |