JP2006503088A - ターンライブラリーをファージ上にディスプレイするための構造化ペプチド骨格 - Google Patents
ターンライブラリーをファージ上にディスプレイするための構造化ペプチド骨格 Download PDFInfo
- Publication number
- JP2006503088A JP2006503088A JP2004544870A JP2004544870A JP2006503088A JP 2006503088 A JP2006503088 A JP 2006503088A JP 2004544870 A JP2004544870 A JP 2004544870A JP 2004544870 A JP2004544870 A JP 2004544870A JP 2006503088 A JP2006503088 A JP 2006503088A
- Authority
- JP
- Japan
- Prior art keywords
- peptide
- dna
- peptides
- hairpin
- protein
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Withdrawn
Links
Images




















Classifications
-
- C—CHEMISTRY; METALLURGY
- C40—COMBINATORIAL TECHNOLOGY
- C40B—COMBINATORIAL CHEMISTRY; LIBRARIES, e.g. CHEMICAL LIBRARIES
- C40B40/00—Libraries per se, e.g. arrays, mixtures
- C40B40/02—Libraries contained in or displayed by microorganisms, e.g. bacteria or animal cells; Libraries contained in or displayed by vectors, e.g. plasmids; Libraries containing only microorganisms or vectors
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K1/00—General methods for the preparation of peptides, i.e. processes for the organic chemical preparation of peptides or proteins of any length
- C07K1/04—General methods for the preparation of peptides, i.e. processes for the organic chemical preparation of peptides or proteins of any length on carriers
- C07K1/047—Simultaneous synthesis of different peptide species; Peptide libraries
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/705—Receptors; Cell surface antigens; Cell surface determinants
- C07K14/70503—Immunoglobulin superfamily
- C07K14/70514—CD4
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/705—Receptors; Cell surface antigens; Cell surface determinants
- C07K14/70503—Immunoglobulin superfamily
- C07K14/70535—Fc-receptors, e.g. CD16, CD32, CD64 (CD2314/705F)
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/705—Receptors; Cell surface antigens; Cell surface determinants
- C07K14/71—Receptors; Cell surface antigens; Cell surface determinants for growth factors; for growth regulators
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K7/00—Peptides having 5 to 20 amino acids in a fully defined sequence; Derivatives thereof
- C07K7/04—Linear peptides containing only normal peptide links
- C07K7/06—Linear peptides containing only normal peptide links having 5 to 11 amino acids
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K7/00—Peptides having 5 to 20 amino acids in a fully defined sequence; Derivatives thereof
- C07K7/04—Linear peptides containing only normal peptide links
- C07K7/08—Linear peptides containing only normal peptide links having 12 to 20 amino acids
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/10—Processes for the isolation, preparation or purification of DNA or RNA
- C12N15/1034—Isolating an individual clone by screening libraries
- C12N15/1037—Screening libraries presented on the surface of microorganisms, e.g. phage display, E. coli display
Landscapes
- Chemical & Material Sciences (AREA)
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Organic Chemistry (AREA)
- Genetics & Genomics (AREA)
- Biochemistry (AREA)
- Molecular Biology (AREA)
- Biophysics (AREA)
- Medicinal Chemistry (AREA)
- General Health & Medical Sciences (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Zoology (AREA)
- Immunology (AREA)
- Toxicology (AREA)
- Cell Biology (AREA)
- Engineering & Computer Science (AREA)
- Gastroenterology & Hepatology (AREA)
- Biomedical Technology (AREA)
- Biotechnology (AREA)
- Microbiology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Wood Science & Technology (AREA)
- General Engineering & Computer Science (AREA)
- Physics & Mathematics (AREA)
- Plant Pathology (AREA)
- Crystallography & Structural Chemistry (AREA)
- Virology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Analytical Chemistry (AREA)
- Peptides Or Proteins (AREA)
Abstract
本発明は、重要な生物学的プロセスに関与しているペプチド又はタンパク質分子の構造−活性分析のためのモデルシステムに関する。本発明によって提供されるものは、エネルギー安定化コンホメーションにとって好ましい配列を持つジスルフィド制約型環状ペプチドを含んでなるコンビナトリアルペプチドライブラリーである。本発明の一態様は、溶液中でβヘアピン構造を示す環状ペプチド骨格に関する。このようなペプチド骨格を選択し利用する方法がここに提供され、それらは、インビボでの分子相互作用の模倣及び治療的薬剤の設計に有用である。従って、本発明は生物学的研究や薬剤開発のための意味深い有用性を有する。
Description
本発明は、一般に、タンパク質構造活性相関の研究、特にコンホメーション制約型ペプチドのコンビナトリアルライブラリー及び生物学的及び製薬的利用のためのそのようなライブラリーの作製とスクリーニングの方法に関する。
構造活性相関(SAR)の研究は、タンパク質又はペプチドと他の生物学的に活性な分子との間の分子間相互作用の理解のために価値ある知見を提供する。その天然の環境下では、ペプチド又はタンパク質は、その結合パートナーを認識して結合し、それと分子複合体を形成するために、独特でコンホメーション的に制約を受けている構造をとり、それが特定の活性を誘導する。タンパク質−タンパク質結合パートナーの例には、酵素−基質、リガンド−レセプター、抗原−抗体が含まれる。従って、その天然形態のペプチドのコンホメーションを決定することは、そのインビボ活性を厳密に模倣し、薬剤として有用であろうその類似体を合理的に設計するために極めて重要になる。
殆どの小ペプチドは、高度に柔軟で、典型的には、独特の溶液コンホメーションをとらない;特に、それらは、同じ配列が天然タンパク質においてとる構造を維持しない。固定した構造の欠如は、ペプチドが標的(エントロピーの理由から)に対して有するであろう親和性を減じ、分子の活性コンホメーションの決定を極めて困難にする。この理由から、多くの戦略は、ペプチドへ制約を導入する(例えば、D−アミノ酸,ジスルフィド又は他の架橋)、又はペプチドの一部を更に強固な非ペプチド骨格と置き換えることであると記述されている。実際、そのようなペプチド模倣物は、特定のコンホメーションへ適応し、生物学的活性にとって重要なペプチド中の特定のアミノ酸又は官能基に関する情報を提供する体系的な構造−活性研究を行うために広く利用されている。
対象のタンパク質をコンホメーション制約型ドメインとして表すことが可能な幾つかの制約型タンパク質骨格が同定されていて、それらはミニボデイー構造(minibody structures)(Bianchiら (1994) J Mol Biol 236:649-659)、βシートターンのループ、コイルドコイル基部構造(Myszka及びChaiken (1994) Biochem 33:2363-2372)、ジンクフィンガードメイン、システイン結合(ジスルフィド)構造、トランスグルタミナーゼ結合構造、環状ペプチド、らせん状バレル又は束状構造、ロイシンジッパーモチーフ(Martinら (1994) EMBO J 13:5303-5309)等々である。同定されたタンパク質骨格の中では、βターンが、多くの生物学的活性ペプチドにおける分子認識にとって重要な部位であると考えられている。Smith及びPease(1980) CRC Crit Rev Biochem 8:315-300。従って、コンホメーション制約型βターンを含むペプチドが特に望ましい。同定されたβターン担持ペプチドの大部分は、天然基質の配列と類似したペプチドの環状化によって生成された環状ペプチドである。Milner-White (1989) Trends Pharmacol Sci. 10:70-74。しかしながら、これら環状ペプチドは、まだ顕著な柔軟性を保持しうる。このため、多くの研究は、βターンを模倣する強固な非ペプチド化合物を導入することを試みている。そのような非ペプチドβターン模倣物を有するペプチドは、薬剤発見のための有用な手掛かりを提供する。Ball及びAlewood(1990) J Mol Recog 3:55-64;国際公開94/03494(Kahn)。
薬剤発見における革命的進歩の一つは、コンビナトリアルライブラリーの開発である。コンビナトリアルライブラリーは、合成的に又は組換え法によって作製することができるペプチドのような、異なった分子の集合である。コンビナトリアルペプチドライブラリーは、全アミノ酸が無作為にペプチド配列の幾つかの位置又は全ての位置に導入されているペプチドを含む。このようなライブラリーは、標的分子へ効果的に結合するペプチド配列をスクリーニングし、そのような配列を同定するために、様々な方法で作製され、利用されている。
ペプチドライブラリーを作製するための多くの方法が開発され、記載されてきた。例えば、ペプチドライブラリーのメンバーは、Lamら (1991) Nature 354:82及びPCT公開WO92/00091によって記載されているように、ポリスチレン又はポリアクリルアミド樹脂のような固体支持体上で行われるスプリット合成 (split-synthesis)によって作製できる。Geysenら,米国特許第4833092号に開示されているその他の方法は、各ライブラリーメンバーのペプチドの配置が、そのペプチドの合成構造に関する情報を与えるように、秩序だっていて予め決まった様式でのペプチド合成を含む。
メンバーペプチドが、それらの天然対応物へより近いものを表すように、コンビナトリアルペプチドライブラリーへ構造的制約を導入することに、かなりの努力が捧げられている。Houstonらの米国特許第5824483号は、αらせんコンホメーションを特徴とし、よって互いにコイルドコイル二量体を形成することができるペプチドを含む合成ペプチドライブラリーについて記載している。McBrideら(1996) J Mol Biol 259:819-827は、同定されたプロテイナーゼ阻害剤の抗トリプシンループ領域を模倣する環状ペプチドの合成ライブラリーについて記載している。
ペプチドライブラリーに基づく先導的発見の補完的方法は、繊維状バクテリオファージでのライブラリーのディスプレイである。この方法は、合成によって調製されうるライブラリーよりも何桁も大きい1010−1012ほどの独特なペプチドメンバーのライブラリーの調製を可能にする。大きなライブラリーのサイズに加えて、ファージディスプレイの利点は、ライブラリー構築の容易さ(Kunkel突然変異誘発)、独特の識別子(そのDNA配列)へ結合体(ディスプレイされたペプチド)をカップリングすること、プール中の希な結合クローンの増幅のための選択プロトコール、及び生合成の高いフィデリティー(合成法との比較)である。更に、迅速で安価な選択プロトコールが、対象の標的へ結合するこれらのライブラリーメンバーを同定するために利用可能である。しかしながら、L−アミノ酸からなる天然ペプチドのみがファージにディスプレイされ、三次構造−活性相関を形成する問題は、非天然的に生じるペプチド又は非ペプチド化合物を含む制約型ペプチド模倣物の場合よりも更に困難である。この問題に対する一つの可能な解決法は、小さな可変ペプチドセグメントを提供するために、折り畳みタンパク質の構造上の制約を利用することである。実際に、幾つかの小さな安定タンパク質が、ペプチドディスプレイ骨格として提案されている。Nygren及びUhlen (1997) Curr. Opin. Struct. Biol. 7:463-469;Vitaら, (1998) Biopolymers 47:93-100;Vitaら, (1999) Proc. Natl. Acad. Sci. USA 96:13091-13096;Smithら, (1998) J. Mol. Biol. 277:317-332;Christmannら, (1999) Protein Engng. 12:797-806。残念ながら、このアプローチ法によって得られたタンパク質リガンドが小分子リード薬物へ転換できることは明らかではない。エピトープのタンパク質から小ペプチド又は非ペプチド小分子への移動は、極めて挑戦的な課題のままである。Cochran (2000) Chem. Biol. 7:R85-R94。
従って、天然ペプチドのコンホメーション優先度を支配している法則に関する広範な研究や幾つかのペプチドライブラリー系の存在にもかかわらず、天然ペプチドの構造的安定性に必要な特徴は理解が不十分のままである。特に、構造に対する残基置換及び非共有的相互作用の効果に関する体系的又は定量的評価は殆どなかった。
本発明は、確定したペプチド骨格の安定性への個々の残基の貢献度を評価し、コンビナトリアルペプチドライブラリーに提示された一連の置換を評価するための新規モデルシステムを提供する。本発明のペプチドは、ペプチド配列内の二つのシステイン間のジスルフィド結合を介して環状化される。様々な確定された残基部位でのアミノ酸置換は、環状ペプチドのコンホメーションとそれらの構造的安定性へ影響する。本発明は、また、ペプチドへ更なる構造的制約を提供する特異的二次構造であるβターンを持つ環状ペプチドをスクリーニングし分析する方法を提供する。βターンを有する環状ペプチドの集団を含んでなる主題ペプチドライブラリーは、分子結合アッセイによる生物学的に活性な候補分子のスクリーニングに利用できる。そのようなスクリーニングのための方法がまた本発明によって提供される。本発明の組成物と方法は、対象ペプチドの構造活性相関を分析し、よって、治療薬剤の合理的設計と同様に、特定の生物学的プロセスに関与している分子相互作用の研究のために有用な情報を提供するのに使用できる。
I.定義
「βターン」という用語は、ペプチド鎖に方向を逆にさせ、疑似10員環を形成する4' から1'の水素結合をしばしば含むテトラペプチド配列からなるタンパク質二次構造を意味する。βターンの様々なコンホメーションの最も広く受け入れられている分類法は、出典明示によってその開示がここに明示的に取り込まれるChou及びFasman (1977) J Mol Biol 115:135-175に記載されている。例えばI、I'、II、及びII'型を含む様々なβターン型が定義されている。本発明の目的に対して、「逆向ターン」という用語は、βターン、γターン、βヘアピン及びβバルジを含む、良く知られたタンパク質二次構造を包含する一般的な意味で使用される。
「細胞」、「細胞株」及び「細胞培養」はここでは相互に交換可能に用いられ、かかる標記は細胞又は細胞株の全ての子孫を含む。よって、例えば「形質転換体」や「形質転換細胞」のような用語には、初代対象細胞と何世代移行したかを問わずそれから由来した培養とを含む。全ての子孫が、意図的な突然変異あるいは意図せざる突然変異のため、正確に同一のDNA内容であるわけではないことも理解される。最初に形質転換された細胞に対してスクリーニングされたものと同じ機能あるいは生物的活性を有する突然変異子孫も含まれる。区別しての標記を意図している場合は、文脈から明らかなはずである。
「βターン」という用語は、ペプチド鎖に方向を逆にさせ、疑似10員環を形成する4' から1'の水素結合をしばしば含むテトラペプチド配列からなるタンパク質二次構造を意味する。βターンの様々なコンホメーションの最も広く受け入れられている分類法は、出典明示によってその開示がここに明示的に取り込まれるChou及びFasman (1977) J Mol Biol 115:135-175に記載されている。例えばI、I'、II、及びII'型を含む様々なβターン型が定義されている。本発明の目的に対して、「逆向ターン」という用語は、βターン、γターン、βヘアピン及びβバルジを含む、良く知られたタンパク質二次構造を包含する一般的な意味で使用される。
「細胞」、「細胞株」及び「細胞培養」はここでは相互に交換可能に用いられ、かかる標記は細胞又は細胞株の全ての子孫を含む。よって、例えば「形質転換体」や「形質転換細胞」のような用語には、初代対象細胞と何世代移行したかを問わずそれから由来した培養とを含む。全ての子孫が、意図的な突然変異あるいは意図せざる突然変異のため、正確に同一のDNA内容であるわけではないことも理解される。最初に形質転換された細胞に対してスクリーニングされたものと同じ機能あるいは生物的活性を有する突然変異子孫も含まれる。区別しての標記を意図している場合は、文脈から明らかなはずである。
「コンピテント細胞」や「エレクトロポレーションコンピテント細胞」という用語は、コンピテント(適格)な状態にあって、様々なソースからDNAを取り込むことが可能な細胞を意味する。この状態は一過性でも永久的でもよい。エレクトロポレーションコンピテント細胞は、エレクトロポレーションの間にDNAを取り込むことが可能である。
「制御配列」は、発現について言う場合、特定の宿主生物中での作用可能に結合しているコード配列の発現に必要なDNA配列を意味する。例えば原核生物に好適な制御配列には、プロモーター、場合によってはオペレータ配列、リボソーム結合部位、及びおそらくはまだよく理解されていない他の配列が含まれる。真核細胞は、プロモーター、ポリアデニル化シグナル及びエンハンサーを利用することが知られている。
「コートタンパク質」という用語は、少なくとも一部がウイルス粒子の表面に存在するタンパク質を意味する。機能的な観点からは、コートタンパク質は、宿主細胞におけるウイルス構築プロセスの間にウイルス粒子と会合する任意のタンパク質であり、他の宿主細胞に感染するまで構築ウイルスに付随したままである。コートタンパク質は、主要なコートタンパク質であってもよいし、又は小数のコートタンパク質であってもよい。「主要な」コートタンパク質とは、10コピーのタンパク質あるいはそれ以上のウイルスコートに存在しているコートタンパク質である。主要なコートタンパク質は、ビリオン(ウイルス粒子)当たり何十、何百又は何千のコピーさえも存在している場合もある。
「制御配列」は、発現について言う場合、特定の宿主生物中での作用可能に結合しているコード配列の発現に必要なDNA配列を意味する。例えば原核生物に好適な制御配列には、プロモーター、場合によってはオペレータ配列、リボソーム結合部位、及びおそらくはまだよく理解されていない他の配列が含まれる。真核細胞は、プロモーター、ポリアデニル化シグナル及びエンハンサーを利用することが知られている。
「コートタンパク質」という用語は、少なくとも一部がウイルス粒子の表面に存在するタンパク質を意味する。機能的な観点からは、コートタンパク質は、宿主細胞におけるウイルス構築プロセスの間にウイルス粒子と会合する任意のタンパク質であり、他の宿主細胞に感染するまで構築ウイルスに付随したままである。コートタンパク質は、主要なコートタンパク質であってもよいし、又は小数のコートタンパク質であってもよい。「主要な」コートタンパク質とは、10コピーのタンパク質あるいはそれ以上のウイルスコートに存在しているコートタンパク質である。主要なコートタンパク質は、ビリオン(ウイルス粒子)当たり何十、何百又は何千のコピーさえも存在している場合もある。
「エレクトロポレーション」及び「エレクトロポレーションをする」という用語は、細胞へ外来物質が取り込まれることを十分に可能にする条件下で、細胞へ電圧を印加することによって細胞へ外来物質(タンパク質、核酸など)を導入するプロセスを意味する。外来物質は、典型的にはDNAである。
「融合タンパク質」とは、二つの部分が共有結合的に結合したポリペプチドであり、その各部分は異なった性質を有するポリペプチドである。その性質とは、インビトロ又はインビボにおける活性のような生物学的性質である。また、その性質は、標的分子への結合や反応の触媒などのような、単純な化学的又は物理的性質でありうる。上記二つの部分は、単一のペプチド結合によって直接結合していてもよいし、又は一又は複数のアミノ酸残基を含むペプチドリンカーを介して結合していてもよい。一般には、二つの部分とリンカーは互いに読み枠にある。
「異種DNA」は、宿主細胞中に導入される任意のDNAである。DNAは、ゲノムDNA,cDNA、合成DNA、これらの融合体又は組合わせを含む様々なソースから誘導されうる。DNAには、宿主あるいは受容細胞と同じ細胞又は細胞型からのDNA、又は例えば哺乳類あるいは植物からの異なった細胞型からのDNAが含まれうる。場合によっては、DNAは、例えば抗生物質耐性遺伝子、温度耐性遺伝子などの選択遺伝子を含んでもよい。
「融合タンパク質」とは、二つの部分が共有結合的に結合したポリペプチドであり、その各部分は異なった性質を有するポリペプチドである。その性質とは、インビトロ又はインビボにおける活性のような生物学的性質である。また、その性質は、標的分子への結合や反応の触媒などのような、単純な化学的又は物理的性質でありうる。上記二つの部分は、単一のペプチド結合によって直接結合していてもよいし、又は一又は複数のアミノ酸残基を含むペプチドリンカーを介して結合していてもよい。一般には、二つの部分とリンカーは互いに読み枠にある。
「異種DNA」は、宿主細胞中に導入される任意のDNAである。DNAは、ゲノムDNA,cDNA、合成DNA、これらの融合体又は組合わせを含む様々なソースから誘導されうる。DNAには、宿主あるいは受容細胞と同じ細胞又は細胞型からのDNA、又は例えば哺乳類あるいは植物からの異なった細胞型からのDNAが含まれうる。場合によっては、DNAは、例えば抗生物質耐性遺伝子、温度耐性遺伝子などの選択遺伝子を含んでもよい。
「ライゲーション」とは、二つの核酸断片の間にリン酸ジエステル結合を形成するプロセスである。二つの断片のライゲーションのためには、断片の末端に互いに適合性がなければならない。ある場合には、末端はエンドヌクレアーゼ消化後に直接的に適合性がある。しかしながら、最初に、エンドヌクレアーゼ消化によって共通に生成されるねじれ末端を、ライゲーションために適合性があるように平滑末端へ変換することが必要である。末端の平滑化のためには、DNAを、4つのデオキシリボヌクレオチド三リン酸の存在下で、約10単位のDNAポリメラーゼI又はT4DNAポリメラーゼのクレノウフラグメントで、適切な緩衝液中、少なくとも15℃で15分間にわたって処理することができる。次に、DNAを、フェノール−クロロホルム抽出及びエタノール沈殿によって精製することができる。互いにライゲーションされることになるDNA断片は、約等モル量の溶液に入れられる。また、この溶液は、一般に、ATP、リガーゼ緩衝液、及び0.5μgのDNAあたり約10単位のT4DNAリガーゼのようなリガーゼを含む。DNAがベクターへライゲーションされるのであれば、ベクターは、最初に、適切な制限エンドヌクレアーゼによる消化によって直線化される。直線化された断片は、次に、ライゲーション工程中でのセルフライゲーションを防止するために、細菌アルカリホスファターゼ又はウシ腸ホスファターゼによって処理される。
「変異」とは、野生型配列のような基準ヌクレオチド配列に対するヌクレオチドの欠失、挿入、又は置換である。
核酸に言及する場合の「作用可能に結合」とは、核酸が、その他の核酸配列と機能的関連性で位置づけられていることを意味する。例えば、ポリペプチドの分泌に関与するプレタンパク質として発現される場合、プレ配列又は分泌性リーダーのDNAは、ポリペプチドのDNAへ作用可能に結合している:配列の転写へ影響を及ぼす場合、プロモーター又はエンハンサーは、コード化配列へ作用可能に結合している:又は、翻訳を促進するように位置する場合、リボゾーム結合部位は、コード化配列へ作用可能に結合している。一般に、「作用可能に結合」とは、結合しているDNA配列が隣接し、分泌性リーダーの場合には近接していて、読み取り段階にあることを意味する。しかしながら、エンハンサーは近接している必要はない。結合は、簡便な制限部位でのライゲーションによって達成される。そのような部位が存在しない場合は、合成オリゴヌクレオチドアダプター又はリンカーが、常套的な実務に従って使用される。
核酸に言及する場合の「作用可能に結合」とは、核酸が、その他の核酸配列と機能的関連性で位置づけられていることを意味する。例えば、ポリペプチドの分泌に関与するプレタンパク質として発現される場合、プレ配列又は分泌性リーダーのDNAは、ポリペプチドのDNAへ作用可能に結合している:配列の転写へ影響を及ぼす場合、プロモーター又はエンハンサーは、コード化配列へ作用可能に結合している:又は、翻訳を促進するように位置する場合、リボゾーム結合部位は、コード化配列へ作用可能に結合している。一般に、「作用可能に結合」とは、結合しているDNA配列が隣接し、分泌性リーダーの場合には近接していて、読み取り段階にあることを意味する。しかしながら、エンハンサーは近接している必要はない。結合は、簡便な制限部位でのライゲーションによって達成される。そのような部位が存在しない場合は、合成オリゴヌクレオチドアダプター又はリンカーが、常套的な実務に従って使用される。
「ファージディスプレイ」は、変異体ポリペプチドが、例えば、繊維状ファージ粒子のような、ファージの表面のコートタンパク質への融合タンパク質として表示される技術のことである。ファージディスプレイの有用性は、ランダム化されたタンパク質変異体の大きなライブラリーが、標的分子と高い親和性で結合する配列について、迅速かつ効果的に識別できることにある。ファージ上のペプチド及びタンパク質ライブラリーの表示は、特異的な結合特性を有するものを目的とした数百万のポリペプチドのスクリーニングに利用されている。多価ファージディスプレイ法は、繊維状ファージのgene III又はgene VIIIのどちらかへの融合を通して、小さなランダムペプチド及び小さなタンパク質を表示することに利用されている。Wells及びLowman(1992) Curr. Opin. Struct. Biol B:355-362及びそこに引用されている参考文献。一価のファージディスプレイでは、タンパク質又はペプチドライブラリーはgene III又はその一部分へ融合され、野生型gene IIIの存在下で低濃度で発現されるので、ファージ粒子は1コピーを表示するか融合タンパク質を表示しない。結合活性効果は、多価ファージに比例して低下するので、識別は内在性リガンド親和性に基づき、DNA操作を単純化するファージミドベクターが使用される。Lowman及びWells(1991) Methods: A companion to Methods in Enzymology 3:205-216。ファージディスプレイでは、表示されたポリペプチドを含むファージ粒子の表現型は、ファージ粒子内の遺伝子型、ファージコートタンパク質によって囲まれたDNAに対応する。
「ファージミド」とは、バクテリアの複製開始点、例えばColE1、及びバクテリオファージの遺伝子間領域のコピーを有するプラスミドベクターである。ファージミドは、繊維状バクテリオファージを含む、任意の既知のバクテリオファージに基づいてもよい。また、プラスミドは、一般に、抗生物質耐性の選択マーカーを含む。これらのベクターへクローニングされるDNAの断片はプラスミドとして増殖できる。これらのベクターを内に有する細胞へファージ粒子の生産に必要な全ての遺伝子が提供された場合、プラスミドの複製方式は、プラスミドDNAの一本鎖のコピーを生成しファージ粒子を包むローリングサークル複製へ変わる。ファージミドは、感染性又は非感染性のファージ粒子を形成しうる。この用語は、異種ポリペプチドがファージ粒子の表面に表示されるように、遺伝子融合体として異種ポリペプチド遺伝子へ結合したファージコートタンパク質遺伝子又はその断片を含むファージミドを含む。Sambrookら, 4.17。
「ファージベクター」とは、異種遺伝子を含み、複製が可能な二重鎖の複製型バクテリオファージを意味する。ファージベクターは、ファージ複製及びファージ粒子形成を可能にするファージ複製開始点を有する。ファージは、好ましくは、M13、f1、fd、pf3ファージ又はその誘導体のような繊維状バクテリオファージ、ラムダ、21、phi80、phi81、82、424、434などのようなラムドイドファージ、又はその誘導体、バキュロウイルス又はその誘導体、T4ファージ又はその誘導体、T7ファージウイルス又はその誘導体である。
「ファージベクター」とは、異種遺伝子を含み、複製が可能な二重鎖の複製型バクテリオファージを意味する。ファージベクターは、ファージ複製及びファージ粒子形成を可能にするファージ複製開始点を有する。ファージは、好ましくは、M13、f1、fd、pf3ファージ又はその誘導体のような繊維状バクテリオファージ、ラムダ、21、phi80、phi81、82、424、434などのようなラムドイドファージ、又はその誘導体、バキュロウイルス又はその誘導体、T4ファージ又はその誘導体、T7ファージウイルス又はその誘導体である。
細胞からのDNAの「調製」とは、宿主細胞の培養からのプラスミドDNAの単離を意味する。DNAの調製に一般的に利用されている方法は、Sambrookらのセクション1.25−1.33に記載されている大規模及び小規模プラスミド調製法である。DNAの調製の後、Sambrookらのセクション1.40に記載されているような当該分野で良く知られた方法によって精製することができる。
「オリゴヌクレオチド」は、既知の方法によって化学的に合成される短い単鎖又は二重鎖ポリデオキシリボヌクレオチドである(例えば、リン酸トリエステル、亜リン酸エステル、又はホスホラミダイト化学などの知られた方法によって、1988年5月4日に公開されたEP266032に記載されたもののような固相技術を用いて、あるいはFroehlerら,(1986) Nucl. Acids Res. 14: 5399-5407 に記載されたようなデオキシヌクレオシドH-ホスホネート中間体を介して化学的に合成される)。更なる方法は、下記に明記されるポリメラーゼ連鎖反応法、及び他のオートプライマー法、及び固体支持体上でのオリゴヌクレオチド合成を含む。これらの方法は全てEngelsら,(1989)Agnew. Chem. Int. Ed. Engl. 28:216-734に記載されている。これらの方法は、遺伝子の全核酸配列が知られている場合、又はコード鎖に対して相補的な核酸配列が入手可能な場合に使用が可能である。あるいは、標的であるアミノ酸配列が知られている場合、各アミノ酸残基の知られており好ましいコード化残基を使用して潜在的な核酸配列を推測することが可能である。次に、オリゴヌクレオチドは、ポリアクリルアミドゲル上で精製される。
「オリゴヌクレオチド」は、既知の方法によって化学的に合成される短い単鎖又は二重鎖ポリデオキシリボヌクレオチドである(例えば、リン酸トリエステル、亜リン酸エステル、又はホスホラミダイト化学などの知られた方法によって、1988年5月4日に公開されたEP266032に記載されたもののような固相技術を用いて、あるいはFroehlerら,(1986) Nucl. Acids Res. 14: 5399-5407 に記載されたようなデオキシヌクレオシドH-ホスホネート中間体を介して化学的に合成される)。更なる方法は、下記に明記されるポリメラーゼ連鎖反応法、及び他のオートプライマー法、及び固体支持体上でのオリゴヌクレオチド合成を含む。これらの方法は全てEngelsら,(1989)Agnew. Chem. Int. Ed. Engl. 28:216-734に記載されている。これらの方法は、遺伝子の全核酸配列が知られている場合、又はコード鎖に対して相補的な核酸配列が入手可能な場合に使用が可能である。あるいは、標的であるアミノ酸配列が知られている場合、各アミノ酸残基の知られており好ましいコード化残基を使用して潜在的な核酸配列を推測することが可能である。次に、オリゴヌクレオチドは、ポリアクリルアミドゲル上で精製される。
「ポリメラーゼ連鎖反応」又は「PCR」は、核酸、RNA及び/又はDNAの特定の切片の少量が1987年7月28日発行の米国特許第4683195号に記載されたようにして増幅される方法又は技術を意味する。一般に、対象とする領域の末端からの又はそれを越える配列情報は、オリゴヌクレオチドプライマーの設計を可能にするように利用可能でなければならない;これらのプライマーは増幅されるテンプレートの反対鎖の配列と同一又は類似している。2つのプライマーの5'末端ヌクレオチドは増幅された物質の末端と一致しうる。PCRを用いて、全細胞性RNA、バクテリオファージ、又はプラスミド配列等から転写されたcDNA、及び全ゲノムDNAから特異的RNA配列、特異的DNA配列を増幅することができる。一般的には、Mullisら, Cold Spring Harbor Symp. Quant Biol. 51: 263 (1987); Erlich, Ed., PCR Technology, (Stockton Press, NY, 1989)を参照のこと。ここで用いられる場合、PCRは、プライマーとして既知の核酸を、そして核酸の特定の切片を増幅又は生成するために核酸ポリメラーゼを使用することを含む核酸試験試料を増幅するための核酸ポリメラーゼ反応法の唯一ではないが一つの例であると考えられる。
DNAが非核酸不純物から分離された場合、DNAは「精製」される。不純物は、極性、非極性、イオン性などである。
DNAが非核酸不純物から分離された場合、DNAは「精製」される。不純物は、極性、非極性、イオン性などである。
制限酵素消化物からの与えられたDNA断片の「回収」又は「単離」とは、電気泳動によるポリアクリルアミドゲル又はアガロースゲル上での消化物の分離、分子量が知られているマーカーDNA断片との移動度の比較による対象断片の同定、所望する断片を含むゲル部分の除去、そしてDNAからのゲルの分離を意味する。この方法は一般的に知られている。例えば、Lawnら (1981) Nucleic Acids Res., 9:6103-6114、及びGoeddelら (1980) Nucleic Acids Res., 8:4057を参照のこと。
「転写制御エレメント」は、以下の構成物の一又は複数を含む:エンハンサーエレメント、プロモーター、オペレーター配列、レプレッサー遺伝子、及び転写終結配列。これらの構成物は、当該分野において良く知られている。米国特許第5667780号。
「形質転換体」とは、そのDNAに関連した表現型の発現(例えば、DNAによってコードされたタンパク質によって与えられる抗生物質耐性)によって証明されるように、DNAを取り入れて維持している細胞である。
「形質転換」又は「形質転換する」とは、細胞がDNAを取り込み、「形質転換体」になるプロセスを意味する。DNAの取り込みは、永久的あるいは一過性である。
「転写制御エレメント」は、以下の構成物の一又は複数を含む:エンハンサーエレメント、プロモーター、オペレーター配列、レプレッサー遺伝子、及び転写終結配列。これらの構成物は、当該分野において良く知られている。米国特許第5667780号。
「形質転換体」とは、そのDNAに関連した表現型の発現(例えば、DNAによってコードされたタンパク質によって与えられる抗生物質耐性)によって証明されるように、DNAを取り入れて維持している細胞である。
「形質転換」又は「形質転換する」とは、細胞がDNAを取り込み、「形質転換体」になるプロセスを意味する。DNAの取り込みは、永久的あるいは一過性である。
開始ポリペプチドの「変異体」又は「突然変異体」、例えば融合タンパク質又は異種ポリペプチド(ファージに対して異種)は、1)アミノ酸配列が、開始ポリペプチドのそれとは異なるものを有し、及び2)天然又は人工(人為的)突然変異誘発のどちらかによって開始ポリペプチドから誘導されたポリペプチドである。このような変異体は、例えば、対象ポリペプチドのアミノ酸配列内の残基の欠失、及び/又は挿入、及び/又は置換を含む。最終構成物(コンストラクト)が所望する機能的特徴を有するならば、欠失、挿入、及び置換のどの組み合わせでも、最終変異体又は突然変異体構成物に到達するように生成されうる。また、アミノ酸変化は、グリコシル化部位の数又は位置を変えるように、ポリペプチドの翻訳後プロセスを変えることができる。ポリペプチドのアミノ酸配列変異体を生成する方法は、出典明示によりここに明示的に取り込まれる米国特許第5534615号に記載されている。
「ペプチド類似体」という用語は、上に定義したように、アミノ酸からなり、その結合能及び/又は特異性に関して、特異的分子と類似する分子又はその一部を意味する。このようなペプチド類似体は、タンパク質工学技術によって発見あるいは構築され、そのような方法は当業者には良く知られている。あるいは、このようなペプチド類似体はスクリーニングプロセスによって見出すことができ、それは、例えば特定分子の天然結合パートナー(特定分子は、必ずしもタンパク質又はペプチドではない)、又はその一部が、ここに記載のように(すなわちキメラタンパク質において)、それへ結合する能力に関してペプチド化合物をスクリーニングするために使用される。二番目のスクリーニング段階では、新しく見出されたペプチド化合物(又はその一部)は、それ自体が、天然結合パートナーの類似体をスクリーニングするために、キメラタンパク質中で特定分子のペプチド類似体として利用される。ペプチド類似体を見出し又は作製する他の方法も当業者には明らかであろう。
「ペプチド類似体」という用語は、上に定義したように、アミノ酸からなり、その結合能及び/又は特異性に関して、特異的分子と類似する分子又はその一部を意味する。このようなペプチド類似体は、タンパク質工学技術によって発見あるいは構築され、そのような方法は当業者には良く知られている。あるいは、このようなペプチド類似体はスクリーニングプロセスによって見出すことができ、それは、例えば特定分子の天然結合パートナー(特定分子は、必ずしもタンパク質又はペプチドではない)、又はその一部が、ここに記載のように(すなわちキメラタンパク質において)、それへ結合する能力に関してペプチド化合物をスクリーニングするために使用される。二番目のスクリーニング段階では、新しく見出されたペプチド化合物(又はその一部)は、それ自体が、天然結合パートナーの類似体をスクリーニングするために、キメラタンパク質中で特定分子のペプチド類似体として利用される。ペプチド類似体を見出し又は作製する他の方法も当業者には明らかであろう。
「エピトープ」という用語は、抗原決定基として抗体と結合することができる抗体又はその一部を意味する。
「結合パートナー複合体」とは、特定の検出可能形式で互いに結合している二つ以上の分子の会合を意味している:従って、リガンドとレセプター、抗体と抗原、及びキメラタンパク質とそれが結合する化合物の結合である。
「直接又は間接的に標識化」という用語は、分子が、放射性同位体、染料、又は蛍光あるいは化学発光部分のような検出可能シグナルを発する標識成分、又はそのもの自体は検出されないが、ある付加的な反応を通して化合物の存在を示すことが可能な、ビオチン、酵素基質、エピトープ、又はヌクレオチド配列のような付着酵素、リガンドのような成分を含みうることを意味する。
「リガンド」とは、他の与えられた分子又は分子複合体と特異的に結合することができる分子又は多量体分子複合体を意味する。必ずしもそうではないが、しばしば、リガンドは可溶性である一方、細胞膜へ埋め込まれたアンカードメインによるように、その標的が固定化されている。
「結合パートナー複合体」とは、特定の検出可能形式で互いに結合している二つ以上の分子の会合を意味している:従って、リガンドとレセプター、抗体と抗原、及びキメラタンパク質とそれが結合する化合物の結合である。
「直接又は間接的に標識化」という用語は、分子が、放射性同位体、染料、又は蛍光あるいは化学発光部分のような検出可能シグナルを発する標識成分、又はそのもの自体は検出されないが、ある付加的な反応を通して化合物の存在を示すことが可能な、ビオチン、酵素基質、エピトープ、又はヌクレオチド配列のような付着酵素、リガンドのような成分を含みうることを意味する。
「リガンド」とは、他の与えられた分子又は分子複合体と特異的に結合することができる分子又は多量体分子複合体を意味する。必ずしもそうではないが、しばしば、リガンドは可溶性である一方、細胞膜へ埋め込まれたアンカードメインによるように、その標的が固定化されている。
「レセプター」という用語は、細胞膜に埋め込まれた他のアンカードメインを有し、与えられた分子又は分子複合体へ結合することができる少なくとも分子の一部又は多量体分子複合体を意味する。例えば二量体のように、レセプター又はリガンドのどちらか、あるいは両方がホモ又はヘテロ多量体型である場合、多くのレセプターは、特に高い親和性をリガンドに対して有している。
「固体支持体」という用語は、限定を受けずに、細胞又はバクテリオファージ粒子のような天然の生物学的なもの、又は、限定を受けずに、アクリルアミド誘導体、セルロース、ナイロン、シリカなどの合成的なもののどちらか、及び磁化粒子で、可溶性分子が結合あるいは連結しうる不溶性マトリックスを意味する。
「天然に生じる」とは、通常は自然に見出せることを意味する。一般には、化学物質は自然に発生しうるが、如何なる特定の場合でも天然源から生成又は誘導される必要はない。
「非天然生成」によってとは、殆ど又は決して自然に見出されることがなく、及び/又は有機合成法を利用して生成されることを意味する。
「修飾された」とは、非天然生成か又は天然に生じた化合物とは逸脱する形で改変されることを意味する。
「固体支持体」という用語は、限定を受けずに、細胞又はバクテリオファージ粒子のような天然の生物学的なもの、又は、限定を受けずに、アクリルアミド誘導体、セルロース、ナイロン、シリカなどの合成的なもののどちらか、及び磁化粒子で、可溶性分子が結合あるいは連結しうる不溶性マトリックスを意味する。
「天然に生じる」とは、通常は自然に見出せることを意味する。一般には、化学物質は自然に発生しうるが、如何なる特定の場合でも天然源から生成又は誘導される必要はない。
「非天然生成」によってとは、殆ど又は決して自然に見出されることがなく、及び/又は有機合成法を利用して生成されることを意味する。
「修飾された」とは、非天然生成か又は天然に生じた化合物とは逸脱する形で改変されることを意味する。
II.概略
本発明は、コンホメーション制約型ペプチド、及び生物活性分子の構造−活性分析及び薬剤リード化合物の発見のために有用なペプチドライブラリーに関する。本発明のペプチドは、互いにジスルフィド結合を形成することができる二つのシステイン残基を含む。従って、ペプチドは、溶液中でβヘアピン骨格の形成を容易にする環状形をとる。多くのペプチドの構造を制約するのには十分ではないものの、ジスルフィド環状化は有益である。ペプチドの残りの残基は更に選択されてヘアピン構造の形成へ有意にバイアスされる。更には、例えばβターンのような、生物学的プロセスにおける重要性が証明されている同定済みの二次構造を有する種々の生物活性ペプチドを模倣するための相対的な多様性を提供するために、本発明のペプチド内の残基のサブセットが変えられる。
一側面では、本発明は、構造制約型環状ペプチドの集合を含んでなるペプチドライブラリーを包含する。ライブラリーの各ペプチドメンバーは、アミノ酸配列C1-A1-A2-(A3)n-A4-A5-C2[配列番号:1]を含み、ここで、
A1、A2、A3、A4及びA5は、天然に生じるL−アミノ酸であり:
システインC1の末端は、場合によってはアミノ保護基によって保護され;
システインC2の末端は、場合によってはカルボキシ保護基によって保護され;
A1及びA5は、アミノ酸W、Y、F、H、I、V及びTからなる群から選択され;
A2及びA4は、アミノ酸W、Y、F、L、M、I、及びVからなる群から選択され;
A3は、任意の天然に生じるL-アミノ酸であり、nは3、4、5、6、7、8、9、10、11及び12からなる群から選択される整数であり;
C1及びC2はジスルフィド結合で互いに結合し、それによって環状ペプチドを形成している。
本発明は、コンホメーション制約型ペプチド、及び生物活性分子の構造−活性分析及び薬剤リード化合物の発見のために有用なペプチドライブラリーに関する。本発明のペプチドは、互いにジスルフィド結合を形成することができる二つのシステイン残基を含む。従って、ペプチドは、溶液中でβヘアピン骨格の形成を容易にする環状形をとる。多くのペプチドの構造を制約するのには十分ではないものの、ジスルフィド環状化は有益である。ペプチドの残りの残基は更に選択されてヘアピン構造の形成へ有意にバイアスされる。更には、例えばβターンのような、生物学的プロセスにおける重要性が証明されている同定済みの二次構造を有する種々の生物活性ペプチドを模倣するための相対的な多様性を提供するために、本発明のペプチド内の残基のサブセットが変えられる。
一側面では、本発明は、構造制約型環状ペプチドの集合を含んでなるペプチドライブラリーを包含する。ライブラリーの各ペプチドメンバーは、アミノ酸配列C1-A1-A2-(A3)n-A4-A5-C2[配列番号:1]を含み、ここで、
A1、A2、A3、A4及びA5は、天然に生じるL−アミノ酸であり:
システインC1の末端は、場合によってはアミノ保護基によって保護され;
システインC2の末端は、場合によってはカルボキシ保護基によって保護され;
A1及びA5は、アミノ酸W、Y、F、H、I、V及びTからなる群から選択され;
A2及びA4は、アミノ酸W、Y、F、L、M、I、及びVからなる群から選択され;
A3は、任意の天然に生じるL-アミノ酸であり、nは3、4、5、6、7、8、9、10、11及び12からなる群から選択される整数であり;
C1及びC2はジスルフィド結合で互いに結合し、それによって環状ペプチドを形成している。
好ましい一実施態様では、本発明のペプチドは、位置A1又はA5あるいは両方のアミノ酸残基のβ炭素に二つの非水素置換基を有するβ分岐残基を有する。発明の好ましいペプチドはA1又はA5又は両方に分岐脂肪族残基I又はV又はTを持つ。更に好ましくは、A1又はA5はスレオニン(T)である。更により好ましくは、A1及びA5の両方がスレオニン残基である。
その他の好ましい実施態様によると、ペプチドは、位置A1又はA5又はその両方に芳香族残基W、Y、F又はHを有する。更に好ましくは、A1又はA5はWである。好適な一ペプチドはA1にHを、A5にVを含む。
その他の好ましい実施態様では、本発明のペプチドは、位置A2又はA4あるいは両方に芳香族残基W、Y、又はFを有する。更に好ましくは、A2又はA4はWである:また更により好ましくは、A2及びA4はWである。その他の好ましい実施態様は、位置A2又はA4あるいは両方に非分枝脂肪族残基L又はMを含む:より好ましくは、A2又はA4はロイシンである。更なる他の好ましいペプチドは、位置A2又はA4あるいは両方に分枝脂肪族残基I又はVを有する。
その他の好ましい実施態様によると、ペプチドは、位置A1又はA5又はその両方に芳香族残基W、Y、F又はHを有する。更に好ましくは、A1又はA5はWである。好適な一ペプチドはA1にHを、A5にVを含む。
その他の好ましい実施態様では、本発明のペプチドは、位置A2又はA4あるいは両方に芳香族残基W、Y、又はFを有する。更に好ましくは、A2又はA4はWである:また更により好ましくは、A2及びA4はWである。その他の好ましい実施態様は、位置A2又はA4あるいは両方に非分枝脂肪族残基L又はMを含む:より好ましくは、A2又はA4はロイシンである。更なる他の好ましいペプチドは、位置A2又はA4あるいは両方に分枝脂肪族残基I又はVを有する。
本発明のペプチドでは、A3残基の数nは、3、4、5、6、7、8、9、10、11、又は12;好ましくは4、5、6、7、8、9又は10;より好ましくは4、5、又は6をとることが可能である。一実施態様では、nは4であり、生じるペプチドは十量体である。これらの十量体では、残基部位A1、A2、A4及びA5は、それぞれ上記に記したアミノ酸残基の選択群からのものであるが、中央の(A3)4 は、種々のアミノ酸のテトラペプチド配列である。本発明の一側面では、(A3)4 テトラペプチド配列は、限定されるものではないが、EGNK、ENGK、QGSF、VWQL及びGPLTを含むβターン構造を好ましくは形成するものから選択される。
一側面では、本発明のライブラリーは、少なくとも約102メンバーのペプチドを含み、それぞれは他に比べて少なくとも一のアミノ酸変異を有する。好ましくは、ライブラリーは、少なくとも約104ペプチド、より好ましくは約1010ペプチド、更により好ましくは少なくとも約1012ペプチドを含む。種々の実施態様によると、アミノ酸変異は、ペプチドの確定した位置で生じる。例えば、変異は、水素結合(HB)鎖部位(例えばA1/A5)又は非水素結合(NHB)鎖部位(例えばA2/A4)で生じうる;残基及びその交差ストランド対(例えばA1/A5又はA2/A4)は、同じ又は異なったアミノ酸を有することができる。変異は中央部位(A3)nでも生じ、A3は20の天然に生じるL-アミノ酸のどれでも可能である。
一側面では、本発明のライブラリーは、少なくとも約102メンバーのペプチドを含み、それぞれは他に比べて少なくとも一のアミノ酸変異を有する。好ましくは、ライブラリーは、少なくとも約104ペプチド、より好ましくは約1010ペプチド、更により好ましくは少なくとも約1012ペプチドを含む。種々の実施態様によると、アミノ酸変異は、ペプチドの確定した位置で生じる。例えば、変異は、水素結合(HB)鎖部位(例えばA1/A5)又は非水素結合(NHB)鎖部位(例えばA2/A4)で生じうる;残基及びその交差ストランド対(例えばA1/A5又はA2/A4)は、同じ又は異なったアミノ酸を有することができる。変異は中央部位(A3)nでも生じ、A3は20の天然に生じるL-アミノ酸のどれでも可能である。
環状ペプチドのカルボキシ末端及びアミノ末端は、(L)及び(D)型の両方において、従来のアミドペプチド結合を通して、任意の既知の保護基で保護されるか、又は他のアミノ酸残基(一般的には、天然に生じる残基)へ結合する。保護基及び付加的残基は、常套的なペプチド合成技術を利用して加えることが可能である。一般的に、1から約50、好ましくは1から20のアミノ酸残基が、各カルボキシ及びアミノ基末端位置に独立して存在しうる。これらの付加的残基は、対象となるβターンを含む既知のタンパク質の一部であってもよいし、又は任意の他の所望する残基配列であってもよい。これらの付加的残基は、ペプチド全体の構造に対するβターンの効果を決定するため、又はβターン環状ペプチドと対象タンパク質の結合に対する付加的残基の効果を決定するために加えてもよい。
あるいは、A1、A2、A4及び/又はA5の一又は複数が独立に固定され、残基A3が、ペプチドライブラリーを作製する既知の方法を用いて変化される本発明の環状ペプチドのライブラリーを調製することができる。ライブラリーを作製する好ましい方法は、ファージディスプレイである。その更なる詳細が下記に説明されているファージディスプレイのどの既知の方法でも本発明の方法に用いることができる。
あるいは、A1、A2、A4及び/又はA5の一又は複数が独立に固定され、残基A3が、ペプチドライブラリーを作製する既知の方法を用いて変化される本発明の環状ペプチドのライブラリーを調製することができる。ライブラリーを作製する好ましい方法は、ファージディスプレイである。その更なる詳細が下記に説明されているファージディスプレイのどの既知の方法でも本発明の方法に用いることができる。
他の側面では、本発明はβターンヘアピン構造を提示するための環状ペプチド骨格を包含する。環状ペプチド骨格は、アミノ酸配列C1-A1-A2-(A3)n-A4-A5-C2(配列番号:1)を含み、ここで、C1とC2はシステインであり;A1、A2、A3、A4及びA5は天然に生じるL−アミノ酸であり;システインC1の末端はアミノ保護基によって保護されていてもよく;システインC2の末端はカルボキシ保護基によって保護されていてもよく;A1とA5はアミノ酸W、Y、F、H、I、V及びTからなる群から選択され;A2とA4はアミノ酸W、Y、F、L、M、I、及びVからなる群から選択され;A3は任意の天然に生じるL-アミノ酸であり、nは3、4、5、6、7、8、9、10、11、又は12の整数であり;C1とC2はジスルフィド結合によって互いに結合して、環状ペプチドを形成する。
好適な一実施態様では、本発明のペプチド骨格は、位置A1又はA5又はその両方のアミノ酸残基のβ炭素に二つの非水素置換基を有するβ分岐残基を有する。本発明の好適なペプチド骨格はA1、A5又は両方に分岐脂肪族残基I、V又はTを有する。より好ましくは、A1又はA5はスレオニン(T)である。更により好ましくは、A1とA5の双方がスレオニン残基である。
好適な一実施態様では、本発明のペプチド骨格は、位置A1又はA5又はその両方のアミノ酸残基のβ炭素に二つの非水素置換基を有するβ分岐残基を有する。本発明の好適なペプチド骨格はA1、A5又は両方に分岐脂肪族残基I、V又はTを有する。より好ましくは、A1又はA5はスレオニン(T)である。更により好ましくは、A1とA5の双方がスレオニン残基である。
その他の実施態様によると、ペプチド骨格は、位置A1又はA5又はその両方に芳香族残基W、Y、F又はHを有する。更に好ましくは、A1又はA5はWである。好適な一ペプチドはA1にHを、A5にVを含む。
その他の好ましい実施態様では、本発明のペプチド骨格は、位置A2又はA4あるいは両方に芳香族残基W、Y、又はFを有する。更に好ましくは、A2又はA4はWである:また更により好ましくは、A2及びA4はWである。その他の好ましい実施態様は、位置A2又はA4あるいは両方に非分枝脂肪族残基L又はMを有するペプチド骨格を含む:より好ましくは、A2又はA4はロイシンである。更なる他の好ましいペプチドは、位置A2又はA4あるいは両方に分枝脂肪族残基I又はVを有する。
本発明のペプチドでは、A3残基の数nは、3、4、5、6、7、8、9、10、11、又は12;好ましくは4、5、6、7、8、9又は10;より好ましくは4、5、又は6をとることが可能である。一実施態様では、nは4であり、生じるペプチドは十量体である。これらの十量体では、残基部位A1、A2、A4及びA5は、それぞれ上記に記したアミノ酸残基の選択群からのものであるが、中央の(A3)4 は、種々のアミノ酸のテトラペプチド配列である。本発明の一側面では、(A3)4 テトラペプチド配列は、限定されるものではないが、EGNK、ENGK、QGSF、VWQL及びGPLTを含むβターン構造を好ましくは形成するものから選択される。
その他の好ましい実施態様では、本発明のペプチド骨格は、位置A2又はA4あるいは両方に芳香族残基W、Y、又はFを有する。更に好ましくは、A2又はA4はWである:また更により好ましくは、A2及びA4はWである。その他の好ましい実施態様は、位置A2又はA4あるいは両方に非分枝脂肪族残基L又はMを有するペプチド骨格を含む:より好ましくは、A2又はA4はロイシンである。更なる他の好ましいペプチドは、位置A2又はA4あるいは両方に分枝脂肪族残基I又はVを有する。
本発明のペプチドでは、A3残基の数nは、3、4、5、6、7、8、9、10、11、又は12;好ましくは4、5、6、7、8、9又は10;より好ましくは4、5、又は6をとることが可能である。一実施態様では、nは4であり、生じるペプチドは十量体である。これらの十量体では、残基部位A1、A2、A4及びA5は、それぞれ上記に記したアミノ酸残基の選択群からのものであるが、中央の(A3)4 は、種々のアミノ酸のテトラペプチド配列である。本発明の一側面では、(A3)4 テトラペプチド配列は、限定されるものではないが、EGNK、ENGK、QGSF、VWQL及びGPLTを含むβターン構造を好ましくは形成するものから選択される。
環状ペプチドのカルボキシ末端及びアミノ末端は、(L)及び(D)型の両方において、従来のアミドペプチド結合を通して、任意の既知の保護基で保護されるか、又は他のアミノ酸残基(一般的には、天然に生じる残基)へ結合する。保護基及び付加的残基は、常套的なペプチド合成技術を利用して加えることが可能である。一般的に、1から約50、好ましくは1から20のアミノ酸残基が、各カルボキシ及びアミノ基末端位置に独立して存在しうる。これらの付加的残基は、対象となるβターンを含む既知のタンパク質の一部であってもよいし、又は任意の他の所望する残基配列であってもよい。これらの付加的残基は、ペプチド全体の構造に対するβターンの効果を決定するため、又はβターン環状ペプチドと対象タンパク質の結合に対する付加的残基の効果を決定するために加えてもよい。
また、本発明は、a)上述のような本発明のコンビナトリアルライブラリーを提供し;b)特定位置A1、A2、A3、A4又はA5の一つのアミノ酸に相違のある少なくとも二つのペプチドをコンビナトリアルライブラリーから選択し;c)ペプチドのコンホメーションを確定し;d)ペプチドの相対的安定性を測定し、比較し;e)コンホメーション的に安定なβヘアピン骨格を有するペプチドを選択する工程を含んでなる、コンホメーション的に安定したβヘアピン骨格を有するペプチドをスクリーニングする方法を包含する。ペプチドのコンホメーション及び安定性は、NMR、分子モデリング、結晶学及び自由エネルギー計算などの当該分野で知られた多くの方法を用いて確定することができる。例えば、Cavanaghら (1995) Protein NMR Spectroscopy, Principles and Practice (Academic Press, San Diego)を参照のこと。ペプチドのコンホメーション及び安定性を確定する特定の方法は、実施例によって以下により詳しく記載する。本発明のペプチドを含むβターンは、その結合活性について天然生物活性タンパク質を模倣するのに有用でありうる。
また、本発明は、a)上述のような本発明のコンビナトリアルライブラリーを提供し;b)特定位置A1、A2、A3、A4又はA5の一つのアミノ酸に相違のある少なくとも二つのペプチドをコンビナトリアルライブラリーから選択し;c)ペプチドのコンホメーションを確定し;d)ペプチドの相対的安定性を測定し、比較し;e)コンホメーション的に安定なβヘアピン骨格を有するペプチドを選択する工程を含んでなる、コンホメーション的に安定したβヘアピン骨格を有するペプチドをスクリーニングする方法を包含する。ペプチドのコンホメーション及び安定性は、NMR、分子モデリング、結晶学及び自由エネルギー計算などの当該分野で知られた多くの方法を用いて確定することができる。例えば、Cavanaghら (1995) Protein NMR Spectroscopy, Principles and Practice (Academic Press, San Diego)を参照のこと。ペプチドのコンホメーション及び安定性を確定する特定の方法は、実施例によって以下により詳しく記載する。本発明のペプチドを含むβターンは、その結合活性について天然生物活性タンパク質を模倣するのに有用でありうる。
βターン残基A3の同一性は、既知のタンパク質構造を研究し、次に既知の構造配列を本発明の構造化されたβヘアピン化合物へ置換することによって確定できる。この実施態様では、残基A3は既知のタンパク質より得られるが、残基A1、A2、A4及びA5は本発明について記載されている通りである。このようにして、本発明の固定化残基は、対象のタンパク質から特定のターンを構造化するために利用することができ、これにより、既知のタンパク質結合パートナーと結合するために、又は関連タンパク質−タンパク質相互作用をアンタゴナイズするために、タンパク質のターンが十分かどうかを試験することが可能になる。
また、本発明は、a)上述のようなコンビナトリアルライブラリーを提供し;b)コンビナトリアルライブラリーを結合パートナーと接触させ;c)結合パートナーと非共有複合体を形成することが可能なペプチドをライブラリーから選択し;d)場合によっては前記ペプチドを単離する工程を含んでなる、特定の結合パートナーと結合することが可能なペプチドを同定する方法を含む。ペプチド結合活性を評価し対象ペプチドを単離するための方法及び技術は、当該分野で知られており、下記により詳しく記載される。
本発明のペプチドの結合パートナーは、ペプチドと結合し場合によっては生物活性を発揮することができる任意の既知又は未知のペプチド、タンパク質、他の高分子又は化学物質を含む任意の分子の少なくとも一部であることができる。レセプター、リガンド、抗原、抗体、酵素及び酵素基質とその断片又は一部のようなタンパク質分子は、「結合パートナー」に包含される。また、他の非タンパク質化学的化合物は、有機であれ無機であれ、ペプチドの結合パートナーとなりうる。
また、本発明は、a)上述のようなコンビナトリアルライブラリーを提供し;b)コンビナトリアルライブラリーを結合パートナーと接触させ;c)結合パートナーと非共有複合体を形成することが可能なペプチドをライブラリーから選択し;d)場合によっては前記ペプチドを単離する工程を含んでなる、特定の結合パートナーと結合することが可能なペプチドを同定する方法を含む。ペプチド結合活性を評価し対象ペプチドを単離するための方法及び技術は、当該分野で知られており、下記により詳しく記載される。
本発明のペプチドの結合パートナーは、ペプチドと結合し場合によっては生物活性を発揮することができる任意の既知又は未知のペプチド、タンパク質、他の高分子又は化学物質を含む任意の分子の少なくとも一部であることができる。レセプター、リガンド、抗原、抗体、酵素及び酵素基質とその断片又は一部のようなタンパク質分子は、「結合パートナー」に包含される。また、他の非タンパク質化学的化合物は、有機であれ無機であれ、ペプチドの結合パートナーとなりうる。
III.βヘアピンペプチド
本発明の一実施態様は、溶液中でβヘアピンコンホメーションをとる環状ペプチド骨格を含む。βヘアピン構造の構成成分は、対となった逆平行βストランド、好ましくはβターンを含む。βストランドにおける非水素結合部位のジスルフィド結合システイン対の好ましい置換が研究され、少なくともタンパク質においては、それは、統計的に好ましい交差ストランド残基の特定の対(水素結合又は非水素結合部位のどちらか)とされている。一つの研究は、近接する逆平行ストランドの水素結合部位へ種々の残基対が導入されている変異タンパク質の実験的安定性測定を記載している。Smith & Regan (1995) Science 270:980-982。βストランドの幾何学的性質に適したコンホメーションをとる為の個々のアミノ酸の内在的優先度を確定するための試みが、βストランドの残基内容の分析(Chou & Fassman(1978) Annu. Rev. Biochem. 47:251-276)、又は種々のアミノ酸をタンパク質のβストランドへ置換して取り入れ、変異体の相対的安定性を測定すること(Kim & Berg(1993) Nature 362:267-270;Minor & Kim(1994) Nature 367: 660-663;Minor & Kim(1993) Nature 371:264-267;Smithら, Biochemistry(1994) 33:5510-5517)のどちらかによって行われてきた。残基のコンホメーションを指定するための改良統計方法は、種々の実験的性向の規模との相関を改善した(Munoz & Serrano(1994) Proteins 20:301-311)。トリプトファンへ割り当てられる傾向は、全ての報告済みの規模の中で中程度である。
最近、幾つかの短い直鎖状ペプチド(4−16アミノ酸)では、ヘアピンコンホメーションが水溶液中で部分的に存在していることが示されている。設計されたペプチド及びタンパク質配列から得られたペプチドの両方がこの性質を示している。一般に、これらの研究は、統計的に強いターン配列(例えば、i+1、i+2においてasn-gly)を有するペプチドを含む。しかしながら、ヘアピンの総数は、水溶液中において40−50%を越えることは殆どない。
本発明の一実施態様は、溶液中でβヘアピンコンホメーションをとる環状ペプチド骨格を含む。βヘアピン構造の構成成分は、対となった逆平行βストランド、好ましくはβターンを含む。βストランドにおける非水素結合部位のジスルフィド結合システイン対の好ましい置換が研究され、少なくともタンパク質においては、それは、統計的に好ましい交差ストランド残基の特定の対(水素結合又は非水素結合部位のどちらか)とされている。一つの研究は、近接する逆平行ストランドの水素結合部位へ種々の残基対が導入されている変異タンパク質の実験的安定性測定を記載している。Smith & Regan (1995) Science 270:980-982。βストランドの幾何学的性質に適したコンホメーションをとる為の個々のアミノ酸の内在的優先度を確定するための試みが、βストランドの残基内容の分析(Chou & Fassman(1978) Annu. Rev. Biochem. 47:251-276)、又は種々のアミノ酸をタンパク質のβストランドへ置換して取り入れ、変異体の相対的安定性を測定すること(Kim & Berg(1993) Nature 362:267-270;Minor & Kim(1994) Nature 367: 660-663;Minor & Kim(1993) Nature 371:264-267;Smithら, Biochemistry(1994) 33:5510-5517)のどちらかによって行われてきた。残基のコンホメーションを指定するための改良統計方法は、種々の実験的性向の規模との相関を改善した(Munoz & Serrano(1994) Proteins 20:301-311)。トリプトファンへ割り当てられる傾向は、全ての報告済みの規模の中で中程度である。
最近、幾つかの短い直鎖状ペプチド(4−16アミノ酸)では、ヘアピンコンホメーションが水溶液中で部分的に存在していることが示されている。設計されたペプチド及びタンパク質配列から得られたペプチドの両方がこの性質を示している。一般に、これらの研究は、統計的に強いターン配列(例えば、i+1、i+2においてasn-gly)を有するペプチドを含む。しかしながら、ヘアピンの総数は、水溶液中において40−50%を越えることは殆どない。
統計的により共通のターン配列(MQIGVKNPDGTITLEV)を伴うことを除いて、ユビキチンタンパク質から誘導された16マー(16-mer)ペプチドは、水中(約80%)では、高頻度で存在するヘアピンを形成しなかったが、ヘアピンは天然タンパク質におけるのと同じストランドレジスターを持っていなかった(Searleら (1995)Nat.Struct.Biol. 2:999-1006)。別のグループは、ターン領域が幾つかの配列(MQIGVKSXXKTITLKV,ここでXX=pro-ala又はpro-gly:Haque & Gellman (1997) J. Am. Chem. Soc.119:2303-2304)で置き換えられた類似のペプチドを研究した。天然ストランドレジスターを伴うヘアピン構造の証拠は、L-アミノ酸配列ではなくD-アミノ酸を含むターンについて観察された。総数の推定は、この研究では与えられていない。
幾つかのグループは、本来はタンパク質tendamistatからの配列に基づくモデルペプチドを研究してきた。ペプチドYQNPDGSQAは、水中において小数のヘアピンの存在を示すNMRの証拠を示した(Blancoら (1993) J. Am. Chem. Soc. 115: 5887-5888;de Albaら (1995) Eur. J. Biochem. 233:282-292;Constantineら (1995) J. Am. Chem. Soc. 117:10841-10854;Friedrichsら J. Am. Chem. Soc. (1995)v 117, pp 10855-10864)。より高く期待されるβ性向(IYSNPDGTWT)の鎖残基をともなうこのペプチドの変異体は、異なるターン配列(IYSNSDGTWT)をともなう二番目のペプチドと比較された。両ペプチドは、NMRによって、水中において30%ヘアピンであると見積もられた(de Albe ら (1996)Fold. Des. 1: 133-144)。このペプチドのさらなる変異は、ほとんどがターン配列にあり、種々の構造及び混合集団のヘアピンを生じる。一般的に、50%を越すコンフォーマー集団は一つもない(de Albe ら (1997) J. Am. Chem. Soc. 119:175-183)。最後の研究では、ペプチドITSNSDGTWTの三つのN末端残基が種々の配列で置き換えられている。再び、混合コンフォーマーが頻繁に観察され、与えられたヘアピンコンフォーマーの集団は、一般的に50%より低かった:一つのペプチド(YITNSDGTWT)は、高頻度のレジスターシフトヘアピンを形成しなかった(80%:de Albe ら (1997) Protein Sci. 6:2584-2560)。これらの研究の著者らは、少なくともこれらの短いモデルペプチドにおいて、ターン残基のコンホメーションの好みが、ヘアピンの安定性の決定において交差ストランド相互作用を支配するものと結論付けた。
幾つかのグループは、本来はタンパク質tendamistatからの配列に基づくモデルペプチドを研究してきた。ペプチドYQNPDGSQAは、水中において小数のヘアピンの存在を示すNMRの証拠を示した(Blancoら (1993) J. Am. Chem. Soc. 115: 5887-5888;de Albaら (1995) Eur. J. Biochem. 233:282-292;Constantineら (1995) J. Am. Chem. Soc. 117:10841-10854;Friedrichsら J. Am. Chem. Soc. (1995)v 117, pp 10855-10864)。より高く期待されるβ性向(IYSNPDGTWT)の鎖残基をともなうこのペプチドの変異体は、異なるターン配列(IYSNSDGTWT)をともなう二番目のペプチドと比較された。両ペプチドは、NMRによって、水中において30%ヘアピンであると見積もられた(de Albe ら (1996)Fold. Des. 1: 133-144)。このペプチドのさらなる変異は、ほとんどがターン配列にあり、種々の構造及び混合集団のヘアピンを生じる。一般的に、50%を越すコンフォーマー集団は一つもない(de Albe ら (1997) J. Am. Chem. Soc. 119:175-183)。最後の研究では、ペプチドITSNSDGTWTの三つのN末端残基が種々の配列で置き換えられている。再び、混合コンフォーマーが頻繁に観察され、与えられたヘアピンコンフォーマーの集団は、一般的に50%より低かった:一つのペプチド(YITNSDGTWT)は、高頻度のレジスターシフトヘアピンを形成しなかった(80%:de Albe ら (1997) Protein Sci. 6:2584-2560)。これらの研究の著者らは、少なくともこれらの短いモデルペプチドにおいて、ターン残基のコンホメーションの好みが、ヘアピンの安定性の決定において交差ストランド相互作用を支配するものと結論付けた。
結晶構造におけるヘアピン配列の分析は、異なる一連のβヘアピンの設計を可能にする。標的構造は、三残基鎖が隣接するI'型ターンであった。arg-gly配列が、溶解度をを向上させるために末端に加えられた。ペプチド RGITVNGKTYGRは、部分的にヘアピンコンホメーション(約30%)へ折り畳まれることがNMRによって確定した(Ramirez-Alvaradoら (1996) Nat. Strruct. Biol. 3:604-612)。鎖残基の重要性は、ile及びval、lys及びtyr、又は4つ全ての残基をアラニンへ置き換えることによって示された。アラニン置換ペプチドは、どれもヘアピンを形成する傾向を示さなかった。同じ著者らは、位置i+1のターンが変えられた(asnをasp、ala、gly、又はserへ)第二シリーズの実験を報告している。asnをターンに有する最初の配列よりも構造化されたペプチドはなかった(Ramirez-Alvaradoら (1997) J.Mol. Biol. 273:898-912)。この研究を記載した総説は、gly-lys対をモデルペプチドの末端に加えることでヘアピンが安定化したことを示しているが、更なる詳細は提供しなかった(Ramirez-Alvaradoら (1999)Bioorg. Med. Chem. 7:93-103)。
もう一つのモデルペプチドシリーズ(RYVEVXGOrnKILQ)は、水中でのヘアピン形成の証拠を与えている。D-pro又はL-asnである残基Xは、特徴的なNOEs及びα-Hシフトを与えるが、L-proペプチドはアンフォールドである。集団推定値は与えられていないが、D-proはより安定なヘアピンを与えると思われる(Stanger & Gellman (1998) J. Am. Chem. Soc. 120:4236-4237)。後の研究では完全に折り畳まれたヘアピンのモデルとしてこのペプチドのジスルフィド環化及び骨格環化型が用いられ、未環化類似体におけるヘアピン集団の推定が可能になった(30−70%)。Syudら (1999) Am Chem Soc. 121:11577-11578。
もう一つのモデルペプチドシリーズ(RYVEVXGOrnKILQ)は、水中でのヘアピン形成の証拠を与えている。D-pro又はL-asnである残基Xは、特徴的なNOEs及びα-Hシフトを与えるが、L-proペプチドはアンフォールドである。集団推定値は与えられていないが、D-proはより安定なヘアピンを与えると思われる(Stanger & Gellman (1998) J. Am. Chem. Soc. 120:4236-4237)。後の研究では完全に折り畳まれたヘアピンのモデルとしてこのペプチドのジスルフィド環化及び骨格環化型が用いられ、未環化類似体におけるヘアピン集団の推定が可能になった(30−70%)。Syudら (1999) Am Chem Soc. 121:11577-11578。
metレプレッサーDNA結合領域に基づいて設計された16残基ペプチド(KKYTVSINGKKITVSI)は、303Kで50%の集団推定値で水中でヘアピン構造を形成した。程度はより低い(35%)が、一つの鎖の切断は、ターンが鎖の相互作用なしで集団化していたことを示している。ヘアピン形成の熱力学的パラメーターの分析は、298KにおいてΔG=0.08kcal/molで、フォールデイングはエンタルピー的には指向性は不利であり、エントロピー的には推進されることを示した(Maynard & Searle (1997) Chem. Commun. 1297-1298;Griffiths-Jonesら (1998) Chem. Commun. 789-790;Maynardradら (1998) J. Am. Chem. Soc. 120:1996-2007)。Griffiths-Jonesら (1999) J. Mol Biol. 292:1051-1069。
タンパク質G(GB1)のB1ドメインに由来する最終ヘアピンペプチド(GEWTYDDATKTFTVTE)は、本発明のペプチドに関連する幾つかの特徴を有する。上述のモデルヘアピンとは異なり、GB1は、一つのthr-thr交差ストランド対を含む、4つのスレオニン残基を鎖内の水素結合部位に有している。これは、好ましくない対形成であると一般には考えられている。更に、相互作用して小さな疎水性コアを形成するかもしれない近接する非水素結合部位に、trp-val及びtyr-phe対がある。報告されたデータは、GB1ペプチドが水中で良く集団化したヘアピン(約50%)を形成したことを示す。データは、天然の鎖対形成と一致する(Blancoら (1994) Nat. Struct. Biol. 1:584-590)。GB1ペプチドの変性研究は、273Kにおいて80%のヘアピンの推定値を与え、データの分析(ΔCp=0と仮定)は、ΔH=−11.6kcal/mol、ΔS=−39cal/molKを与えた:すなわちフォールディングは、エンタルピー的に推進され、エントロピー的に好ましくない(Munozら, Nature(1998)v390, pp.196-199)。エンタルピー及びエントロピーの相対的役割は、上述のmetレプレッサーペプチドに比較して逆である。GB1ペプチドの熱変性のNMR研究から同様の結果が報告されている。Hondaら (2000) J. Mol. Biol. 295:269-278。同じグループの突然変異誘発研究(Kobayashiら (2000) Biochemistry 39:6564-6571)ではヘアピン安定性に対して特に重要なものとして交差ストランドフェニルアラニン及びチロシン残基が同定された。最後に、このグループはGB1ペプチドのジスルフィド環化類似体を報告している(CEWTYDDATKTFTVTCK;Kobayashiら (1999) Biochemistry 38:3228-3243)。この類似体の構造は記述されていない;しかしながら、それは、未環化ペプチドよりもGB1タンパク質の残りに7倍強く結合した。
タンパク質G(GB1)のB1ドメインに由来する最終ヘアピンペプチド(GEWTYDDATKTFTVTE)は、本発明のペプチドに関連する幾つかの特徴を有する。上述のモデルヘアピンとは異なり、GB1は、一つのthr-thr交差ストランド対を含む、4つのスレオニン残基を鎖内の水素結合部位に有している。これは、好ましくない対形成であると一般には考えられている。更に、相互作用して小さな疎水性コアを形成するかもしれない近接する非水素結合部位に、trp-val及びtyr-phe対がある。報告されたデータは、GB1ペプチドが水中で良く集団化したヘアピン(約50%)を形成したことを示す。データは、天然の鎖対形成と一致する(Blancoら (1994) Nat. Struct. Biol. 1:584-590)。GB1ペプチドの変性研究は、273Kにおいて80%のヘアピンの推定値を与え、データの分析(ΔCp=0と仮定)は、ΔH=−11.6kcal/mol、ΔS=−39cal/molKを与えた:すなわちフォールディングは、エンタルピー的に推進され、エントロピー的に好ましくない(Munozら, Nature(1998)v390, pp.196-199)。エンタルピー及びエントロピーの相対的役割は、上述のmetレプレッサーペプチドに比較して逆である。GB1ペプチドの熱変性のNMR研究から同様の結果が報告されている。Hondaら (2000) J. Mol. Biol. 295:269-278。同じグループの突然変異誘発研究(Kobayashiら (2000) Biochemistry 39:6564-6571)ではヘアピン安定性に対して特に重要なものとして交差ストランドフェニルアラニン及びチロシン残基が同定された。最後に、このグループはGB1ペプチドのジスルフィド環化類似体を報告している(CEWTYDDATKTFTVTCK;Kobayashiら (1999) Biochemistry 38:3228-3243)。この類似体の構造は記述されていない;しかしながら、それは、未環化ペプチドよりもGB1タンパク質の残りに7倍強く結合した。
幾つかの設計された三重により合わさったシートが報告されている:それらの一つは、タンパク質に生じる通常の20のアミノ酸のみを含み、水中で折り畳まれる(Kortemmeら (1998) Science 281:253-256)。その設計の一側面は、非分岐アミノ酸の隣の鎖の二つの非水素結合残基をも変える一方で、trpを非水素結合位置へ加えることである(WWドメインとの類推)。著者は、trp側鎖が隣りの鎖へと密集することを、分岐残基が妨げると述べている。変性データの熱力学的分析は、278Kにおいて−0.6kcal/molのフォールデイング自由エネルギーを与える(フォールデイング集団の推定値=80−90%)。
タンパク質ヘアピンを模倣することを意図した、又はデノボ設計ヘアピンとしてのジスルフィド制約型ペプチドについての多くの例が報告されている。多くの場合、ジスルフィド結合幾何構造がヘアピンの交差ストランド幾何構造と適合しないと当初考えられていたので、設計は一又は両末端にD-システインを含む。しかしながら、L-cysを利用する幾つかの例がある。
ジスルフィド環状ペプチドの殆どの研究において、構造の証拠を欠いている。ここで列挙された例は、実験的に確定されてきたものか、生物学的アッセイにおいて正常でないアミノ酸を使用せずに、より大きなヘアピン含有タンパク質に近い効力を有しているものである。
タンパク質ヘアピンを模倣することを意図した、又はデノボ設計ヘアピンとしてのジスルフィド制約型ペプチドについての多くの例が報告されている。多くの場合、ジスルフィド結合幾何構造がヘアピンの交差ストランド幾何構造と適合しないと当初考えられていたので、設計は一又は両末端にD-システインを含む。しかしながら、L-cysを利用する幾つかの例がある。
ジスルフィド環状ペプチドの殆どの研究において、構造の証拠を欠いている。ここで列挙された例は、実験的に確定されてきたものか、生物学的アッセイにおいて正常でないアミノ酸を使用せずに、より大きなヘアピン含有タンパク質に近い効力を有しているものである。
ヘキサペプチド(Boc-CL-Aib-AVC-NMe)の構造が、結晶学的に決定され、II'型ターン及びβシート幾何学が明らかにされた(Karleら J. Am. Chem. Soc. (1998)v110, pp1958-1963)。同じシステイン間隔を有するオクタペプチド(ACSPGHCE)はNMRで研究がなされ、pro-glyの中心にあるターンと類似の構造を有している(Walseら (1996) J. Comput.-Aided Mol Des. 10:11-22)。Ac-CXPGXC-NHMe形のペプチドはジスルフィド交換平衡の測定によって評価され、1kcal/molと同程度のペプチド間のターン指向性を示した(Milburnら (1987) J. Am. Chem. Soc. 109:4486-4496)。
M13の遺伝子5タンパク質(gene 5 protein)に基づいた11残基環状ペプチド(CGVSRQGKPYC)は、水溶液中で安定的に構造化されることがNMR分析によって実証された。環状ペプチドは、対応するタンパク質ループへかなり類似の構造をとる。著者は、明確なβヘアピン構造は、これまではどの非保護ジスルフィド制約型環においても報告されていなかったと主張する(Rietmanら (1996) Eur. J. Biochem. 238:706-713)。このペプチドは、システインへ最も近接する非水素結合部位においてval-pro対を有する。
X線構造に基づくカブトガニ抗リポ多糖因子(LALF)のループに一致するペプチドの環状化は、強力なリピッドAバインダーを産する。これらペプチドの構造に関する証拠はない。幾つかのペプチドは、システインへ最も近接する非水素結合部位に芳香族-芳香族対を有する:しかしながら、最も強力なもの(GCKPTFRRLKWKYKCG)は、pro-tyr対を有する(Riedら (1996)J. Biol. Chem. 271:2820-28127)。
M13の遺伝子5タンパク質(gene 5 protein)に基づいた11残基環状ペプチド(CGVSRQGKPYC)は、水溶液中で安定的に構造化されることがNMR分析によって実証された。環状ペプチドは、対応するタンパク質ループへかなり類似の構造をとる。著者は、明確なβヘアピン構造は、これまではどの非保護ジスルフィド制約型環においても報告されていなかったと主張する(Rietmanら (1996) Eur. J. Biochem. 238:706-713)。このペプチドは、システインへ最も近接する非水素結合部位においてval-pro対を有する。
X線構造に基づくカブトガニ抗リポ多糖因子(LALF)のループに一致するペプチドの環状化は、強力なリピッドAバインダーを産する。これらペプチドの構造に関する証拠はない。幾つかのペプチドは、システインへ最も近接する非水素結合部位に芳香族-芳香族対を有する:しかしながら、最も強力なもの(GCKPTFRRLKWKYKCG)は、pro-tyr対を有する(Riedら (1996)J. Biol. Chem. 271:2820-28127)。
ウサギデフェンシンのヘアピン領域のジスルフィド環状化ペプチドは、その線状類似体を上回る抗菌活性(約5から10倍)を有する。円偏光二色性分光法は、リン酸緩衝液内の幾つかの非ランダム構造を示した。より強力なペプチド(CAGFMRIRGRIHPLCMRR)は、システインへ最も近接する非水素結合部位にgly-pro対を有する(Thennarasu & Nagaraj(1999) Biochem. Biophys. Res. Commun. 254:281-283)。
最後の研究は、ヒトCD4のドメイン1のループの幾つかのペプチドを記載する。ジスルフィド制約に加えて、著者らは、ペプチド末端へ環外芳香族アミノ酸を加えた。例えば、CD4残基39−44を覆うペプチドは、FCNQGSFLCYとして制約されている。構造に関する証拠は与えられていないが、一つの環状ペプチド(FCYICEVEDQCY)が、正常なCD4相互作用及びHIVによるCD-4媒介細胞侵入に関与しているCD4相互作用の両方と拮抗することが報告されている(Zhangら (1996) Nature Biotechnology 14:472-475;Zhangら (1997) Nature Biotechnology 15:150-154)。
最後の研究は、ヒトCD4のドメイン1のループの幾つかのペプチドを記載する。ジスルフィド制約に加えて、著者らは、ペプチド末端へ環外芳香族アミノ酸を加えた。例えば、CD4残基39−44を覆うペプチドは、FCNQGSFLCYとして制約されている。構造に関する証拠は与えられていないが、一つの環状ペプチド(FCYICEVEDQCY)が、正常なCD4相互作用及びHIVによるCD-4媒介細胞侵入に関与しているCD4相互作用の両方と拮抗することが報告されている(Zhangら (1996) Nature Biotechnology 14:472-475;Zhangら (1997) Nature Biotechnology 15:150-154)。
IV.ペプチドライブラリー
当該分野で知られているペプチドライブラリーを作製するための多くの方法は、本発明のライブラリーを作製するために利用することができる。一実施態様では、ペプチドライブラリーのメンバーは、Lamら,(1991) Nature 354:82及びPCT公開WO92/00091によって記載のようなポリスチレン又はポリアクリルアミド樹脂などの固体支持体上で行われるスプリット合成によって作製することができる。本発明の一側面では、環状ペプチドのライブラリーは、一つ以上の残基A1、A2、A4及び/又はA5が独立に固定され、残基A3が産するように調製できる。
本発明のライブラリーを作製する好ましい方法は、ファージディスプレイ法である。ファージディスプレイ法では、本発明の環状ペプチドは、融合タンパク質を形成するために、少なくともファージコートタンパク質の一部分に融合される。この融合タンパク質は、下記に記載のようなファージディスプレイの既知の技術を利用し、融合タンパク質をコードする融合遺伝子を発現することによって作製できる。融合タンパク質は、環状ペプチドの一つ以上のコピーが粒子の表面に表示されているファージ又はファージミド粒子の一部分を形成しうる。環状ペプチド又は融合ペプチドをコードする核酸を含む遺伝子は発明の範囲内である。
当該分野で知られているペプチドライブラリーを作製するための多くの方法は、本発明のライブラリーを作製するために利用することができる。一実施態様では、ペプチドライブラリーのメンバーは、Lamら,(1991) Nature 354:82及びPCT公開WO92/00091によって記載のようなポリスチレン又はポリアクリルアミド樹脂などの固体支持体上で行われるスプリット合成によって作製することができる。本発明の一側面では、環状ペプチドのライブラリーは、一つ以上の残基A1、A2、A4及び/又はA5が独立に固定され、残基A3が産するように調製できる。
本発明のライブラリーを作製する好ましい方法は、ファージディスプレイ法である。ファージディスプレイ法では、本発明の環状ペプチドは、融合タンパク質を形成するために、少なくともファージコートタンパク質の一部分に融合される。この融合タンパク質は、下記に記載のようなファージディスプレイの既知の技術を利用し、融合タンパク質をコードする融合遺伝子を発現することによって作製できる。融合タンパク質は、環状ペプチドの一つ以上のコピーが粒子の表面に表示されているファージ又はファージミド粒子の一部分を形成しうる。環状ペプチド又は融合ペプチドをコードする核酸を含む遺伝子は発明の範囲内である。
その他の実施態様では、本発明は、複数個の複製可能な発現ベクターを含むライブラリーを構築する工程を含んでなる方法で、各発現ベクターは、融合タンパク質をコードする遺伝子融合体へ作用可能に結合している転写制御エレメントを含み、その遺伝子融合体は、本発明の環状ペプチドをコードする最初の遺伝子とファージコートタンパク質の少なくとも一部をコードする二番目の遺伝子を含み、ライブラリーは変異体環状ペプチド融合タンパク質をコードする複数個の遺伝子を含む。変異体の一番目の遺伝子及び変異体環状ペプチドをコードするそのライブラリーは、下記により詳細に記載する既知の突然変異誘発技術を利用して調製される。
本発明は、更に、上述の融合遺伝子を含む発現ベクター、並びにこれらベクターのライブラリーを含む。ベクターのライブラリーは、DNAライブラリーの形態、融合遺伝子のライブラリーを含むウイルス(ファージ又はファージミド)粒子のライブラリー、又は発現ベクターあるいはウイルス粒子のライブラリーを含む宿主細胞のライブラリーの形態であってもよい。
本発明は、更に、上述の融合遺伝子を含む発現ベクター、並びにこれらベクターのライブラリーを含む。ベクターのライブラリーは、DNAライブラリーの形態、融合遺伝子のライブラリーを含むウイルス(ファージ又はファージミド)粒子のライブラリー、又は発現ベクターあるいはウイルス粒子のライブラリーを含む宿主細胞のライブラリーの形態であってもよい。
また、本発明に含まれるものは、(a)融合タンパク質をコードする遺伝子融合へ作用可能に結合している転写制御エレメントを含む変異体の複製可能な発現ベクターのライブラリーを構築する工程であって、遺伝子融合体は本発明の環状ペプチドをコードする一番目の遺伝子と、ファージコートタンパク質の少なくとも一部をコードする二番目の遺伝子を含み、変異体発現ベクターは変異体の一番目の遺伝子を含む工程:(b)ベクターで適切な宿主細胞を形質転換する工程:(c)粒子が一つ以上の融合タンパク質のコピーを粒子の表面に表示するように、少なくとも発現ベクターの一部分を含み、宿主細胞を形質転換することが可能な組み換え体ファージ又はファージミドウイルス粒子を形成するのに適切な条件下で形質転換細胞を培養する工程:(d)粒子を標的分子と接触させ、少なくとも粒子の一部が標的分子へ結合するようにする工程:及び(e)結合する粒子を結合しない粒子から分離する工程、を含んでなる新規結合ポリペプチドを選択する方法である。本発明の方法では、ファージコートタンパク質は、好ましくはM13のような繊維状ファージの遺伝子III(geneIII)又は遺伝子VIII(geneVIII)コートタンパク質である。さらに、好ましくは、形質転換宿主細胞の培養は、組換え体ファージ又はファージミド粒子の形成に適切な条件下で行われ、少量にすぎないファージ又はファージミドが粒子の表面に一つ以上の融合タンパク質を表示するように(一価の表示)条件が調整される。
また、本発明は、上述の工程(a)から(e)を利用し、ファージディスプレイライブラリーへ構造バイアスを導入する方法を含む。本発明は、更に、標的がベーターヘアピンペプチド構造へ結合することが知られていて、好ましくはタンパク質標的が結合することが知られている場合に、上述の(a)から(e)の工程を利用し、ファージディスプレイライブラリーからペプチド構造を形成するベーターヘアピンを選択する方法を含む。
バクテリオファージ(ファージ)ディスプレイは、変異体ポリペプチドが、バクテリオファージ粒子の表面のコートタンパク質へ融合タンパク質として表示される既知の技術である(Scott, J.K.及びSmith, G.P.(1990) Science 249:386)。ファージディスプレイの有用性は、選択的にランダム化されたタンパク質変異体(又はランダムにクローニングされたcDNA)の大きなライブラリーが、高い親和性で標的分子と結合するこれら配列のために迅速かつ効率良く分類できるという事実にある。ファージ上のペプチド(Cwrilaら (1990)Proc. Natl. Acad. Sci. USA 87:6378)又はファージ上のタンパク質(Lowmanら (1991)Biochemistry 30:10832;Clacksonら (1991) Nature 352:624;Marksら (1991) J.Mol. Biol.222:581;Kangら (1991) Proc.Natl. Acad. Sci. USA 88:8363)ライブラリーの表示は、特異的結合特性を持つものに対して、何百万ものポリペプチドのスクリーニングに利用されてきた(Smith.G.P. (1991) Current Opin. Biotechnology 2:668)。ランダム変異のファージライブラリーの分類は、非常に多くの変異体の構築及び増殖のための戦略、標的レセプターを利用したアフィニテイー精製の手順、及び結合濃縮の結果を評価する手段を必要とする。米国特許第5223409号;米国特許第5403484号;米国特許第5571689号;米国特許第5663143号。
バクテリオファージ(ファージ)ディスプレイは、変異体ポリペプチドが、バクテリオファージ粒子の表面のコートタンパク質へ融合タンパク質として表示される既知の技術である(Scott, J.K.及びSmith, G.P.(1990) Science 249:386)。ファージディスプレイの有用性は、選択的にランダム化されたタンパク質変異体(又はランダムにクローニングされたcDNA)の大きなライブラリーが、高い親和性で標的分子と結合するこれら配列のために迅速かつ効率良く分類できるという事実にある。ファージ上のペプチド(Cwrilaら (1990)Proc. Natl. Acad. Sci. USA 87:6378)又はファージ上のタンパク質(Lowmanら (1991)Biochemistry 30:10832;Clacksonら (1991) Nature 352:624;Marksら (1991) J.Mol. Biol.222:581;Kangら (1991) Proc.Natl. Acad. Sci. USA 88:8363)ライブラリーの表示は、特異的結合特性を持つものに対して、何百万ものポリペプチドのスクリーニングに利用されてきた(Smith.G.P. (1991) Current Opin. Biotechnology 2:668)。ランダム変異のファージライブラリーの分類は、非常に多くの変異体の構築及び増殖のための戦略、標的レセプターを利用したアフィニテイー精製の手順、及び結合濃縮の結果を評価する手段を必要とする。米国特許第5223409号;米国特許第5403484号;米国特許第5571689号;米国特許第5663143号。
典型的には、本発明の環状化合物のような変異体ペプチドは、遺伝子III(geneIII)タンパク質へ融合し、これがウイルス粒子の一末端に表示される。あるいは、変異体ペプチドは遺伝子VIII(geneVIII)タンパク質へ融合が可能で、これはウイルス粒子の主要なコートタンパク質である。このような多価ディスプレイライブラリーは、ファージ遺伝子III(geneIII)を遺伝子III(geneIII)タンパク質のアミノ末端へ融合した外来配列をコードするcDNAと置き換えることによって構築される。
一価ファージディスプレイは、タンパク質又はペプチド配列が遺伝子III(geneIII)タンパク質の一部分へ融合し、野生型遺伝子III(geneIII)タンパク質の存在下で低レベルで発現し、その結果、粒子が主に野生型遺伝子III(geneIII)タンパク質及び一つのコピーを表示し、あるいは融合タンパク質を全く表示しないプロセスである(Bassら (1990)Proteins 80:309;Lowman. H.B.及びWells. J.A.(1991) Methods: a Companion to Methods in Enzymology 3:205)。一価ディスプレイは、多価ファージディスプレイに対して利点があり、後代ファージミド粒子が全感染力を保持する。アビディティー効果は低下し、分類は内在性リガンド親和性に基礎を置き,DNA操作を単純化するファージミドベクターが利用される。米国特許第5750373号及び米国特許第5780279号を参照のこと。他者も、タンパク質、特に抗体を表示するためにファージミドを利用している。米国特許第5667988号;米国特許第5759817号;米国特許第5770356号;及び米国特許第5658727号。
一価ファージディスプレイは、タンパク質又はペプチド配列が遺伝子III(geneIII)タンパク質の一部分へ融合し、野生型遺伝子III(geneIII)タンパク質の存在下で低レベルで発現し、その結果、粒子が主に野生型遺伝子III(geneIII)タンパク質及び一つのコピーを表示し、あるいは融合タンパク質を全く表示しないプロセスである(Bassら (1990)Proteins 80:309;Lowman. H.B.及びWells. J.A.(1991) Methods: a Companion to Methods in Enzymology 3:205)。一価ディスプレイは、多価ファージディスプレイに対して利点があり、後代ファージミド粒子が全感染力を保持する。アビディティー効果は低下し、分類は内在性リガンド親和性に基礎を置き,DNA操作を単純化するファージミドベクターが利用される。米国特許第5750373号及び米国特許第5780279号を参照のこと。他者も、タンパク質、特に抗体を表示するためにファージミドを利用している。米国特許第5667988号;米国特許第5759817号;米国特許第5770356号;及び米国特許第5658727号。
ペプチドライブラリーを作製する方法、及びこれらのライブラリーのスクリーニングは、米国特許第5723286;米国特許第5432018;米国特許第5580717;米国特許第5427908;及び米国特許第5498530に開示されている。また、米国特許第5770434;米国特許第5734018;米国特許第5698426;米国特許第5763192;及び米国特許第5723323を参照のこと。
M13ファージ上に表示されるペプチドライブラリーから高親和性リガンドを選択するために、二工程法を利用してもよい。低親和性リード物質が、最初に主要コートタンパク質(タンパク質VIII)上に表示された未処置の多価ライブラリーから選択される。低親和性セレクタント(selectant)は、次に遺伝子IIIの小さなコートタンパク質へ移され、一価の型で高親和性へと成熟した。
殆どのファージディスプレイは繊維状ファージを利用しているが、ラムドイドファージディスプレイシステム(WO95/34683;米国特許第5627024号)、T4ファージディスプレイ(Renら (1998) Gene 215:439;Zhu (1997) CAN 33:534;Jiangら (1997)CAN 128:44380;Renら (1997) CAN 127:215644;Ren (1996) Protein Sci 5:1833;Efimovら (1995)Virus Genes 10:173)及びT7ファージディスプレイシステム(Smith & Scott (1993) Methods in Enzymology 217:228-257;米国特許第5766905号)は,また良く知られており、本発明の環状ペプチドのライブラリーの作製に利用が可能である。
本発明の環状ペプチドのディスプレイのための適切な遺伝子IIIは、fUSE5(Scott. J. K.,及びSmith G.P.(1990) Science 249:386-390);fAFF1(Cwirlaら (1990) Proc. Natl. Sci. U.S.A. 87:6378-6382);fd-CAT1(McCaffertyら (1990) Nature (London)348:552-554);m663(Fowlkesら (1992) Biotechniques 13:422-427);fdtetDOG,pHEN1(Hoogeboomら (1991) Nucleic Acids Res. 19:4133-4137);pComb3(Gramら (1992) Proc. Natl. Acad. Sci. U.S.A. 89:3576-3580);pCANTAB 5E(Pharmacia);及びLamdaSurfZap(Hogrefe(1993) Gene 137:85-91)を含む。
M13ファージ上に表示されるペプチドライブラリーから高親和性リガンドを選択するために、二工程法を利用してもよい。低親和性リード物質が、最初に主要コートタンパク質(タンパク質VIII)上に表示された未処置の多価ライブラリーから選択される。低親和性セレクタント(selectant)は、次に遺伝子IIIの小さなコートタンパク質へ移され、一価の型で高親和性へと成熟した。
殆どのファージディスプレイは繊維状ファージを利用しているが、ラムドイドファージディスプレイシステム(WO95/34683;米国特許第5627024号)、T4ファージディスプレイ(Renら (1998) Gene 215:439;Zhu (1997) CAN 33:534;Jiangら (1997)CAN 128:44380;Renら (1997) CAN 127:215644;Ren (1996) Protein Sci 5:1833;Efimovら (1995)Virus Genes 10:173)及びT7ファージディスプレイシステム(Smith & Scott (1993) Methods in Enzymology 217:228-257;米国特許第5766905号)は,また良く知られており、本発明の環状ペプチドのライブラリーの作製に利用が可能である。
本発明の環状ペプチドのディスプレイのための適切な遺伝子IIIは、fUSE5(Scott. J. K.,及びSmith G.P.(1990) Science 249:386-390);fAFF1(Cwirlaら (1990) Proc. Natl. Sci. U.S.A. 87:6378-6382);fd-CAT1(McCaffertyら (1990) Nature (London)348:552-554);m663(Fowlkesら (1992) Biotechniques 13:422-427);fdtetDOG,pHEN1(Hoogeboomら (1991) Nucleic Acids Res. 19:4133-4137);pComb3(Gramら (1992) Proc. Natl. Acad. Sci. U.S.A. 89:3576-3580);pCANTAB 5E(Pharmacia);及びLamdaSurfZap(Hogrefe(1993) Gene 137:85-91)を含む。
融合ポリペプチドをコードする遺伝子融合体へ作用可能に結合している転写制御エレメントを含む複製可能なベクター変異体のファミリーの構築し、適切な宿主細胞を形質転換し、ファージ粒子の表面に融合ポリペプチドを表示するファージ粒子を形成するために形質転換細胞を培養し、組換え体ファージ粒子を標的分子と接触させて、粒子の少なくとも一部分を標的と結合させ、結合する粒子を結合しない粒子から分離することを含む、タンパク質、ペプチド、及びその突然変異体のファージディスプレイ法は知られており、本発明の方法と共に利用することができる。米国特許第5750373号;WO97/09446;米国特許第5514548号;米国特許第5498538号;米国特許第5516637号;米国特許第5432018号;WO96/22393;米国特許第5658727号;米国特許第5627024号;WO97/29185;O'Boyleら (1997) Virology 236:338-347;Soumillionら (1994) Appl. Biochem. Biotech. 47:175-190;O'Neil 及び Hoess.(1995) Curr. Orin. Struct. Biol. 5:443-449;Makowski (1993) Gene 128:5-11;Dunn(1996) Curr. Opin. Struct. Biol. 7:547-553;Choo 及び Klug (1995) Curr. Opin. Struct. Biol. 6:431-436;Bradbury及びCattaneo (1995) TINS 18:242-249;Corteseら (1995) Curr. Opin. Struct. Biol. 6:73-80;Allenら (1995) TIBS 20:509-516;Lindquist及びNaderi(1995) FEMS Micro. Rev. 17:33-39;Clarkson及びWells (1994) Tibtech. 12:173-184;Barbas (1993) Curr. Opin. Biol. 4:526-530;McGregor (1996) Mol. Biotech. 6:155-162;Corteseら (1996) Curr. Opin. Biol. 7:616-621;McLaffertyら (1993) Gene 128:29-36を参照のこと。
ファージのコートタンパク質をコードする遺伝子と、本発明の融合タンパク質の所望する環状ポリペプチド部分(すなわち、ファージコートタンパク質の少なくとも一部へ融合している本発明の環状ペプチド)をコードする遺伝子は、当該分野において知られている方法によって得ることができる(一般的には、Sambrookらを参照のこと)。その遺伝子をコードしているDNAは、化学的に合成(Merrfield(1963) J.Am.Chem.Soc. 85:2149)した後に、下記に記載のような変異体のライブラリーを調製するために変異されてされてもよい。
遺伝子融合体を含む機能的ベクターを形成する目的でDNA断片をライゲーションするには、DNA断片の末端は互いに適合化させなければならない。幾つかの場合では、末端は、エンドヌクレアーゼ消化の後で直接に適合する。しかしながら、通常はエンドヌクレアーゼ消化によって生成される粘着末端を、それらをライゲーションにおいて適合せしめるために平滑末端へ最初に変換することが必要である。末端を平滑末端化するために、DNAは適切な緩衝液中にて少なくとも15分間、15℃で、4つのデオキシヌクレオチド三リン酸の存在下で10単位のDNAポリメラーゼIのクレノウ断片(クレノウ)によって処理される。DNAは、次にフェノール-クロロホルム抽出及びタノール沈殿又は他のDNA精製技術で精製される。
遺伝子融合体を含む機能的ベクターを形成する目的でDNA断片をライゲーションするには、DNA断片の末端は互いに適合化させなければならない。幾つかの場合では、末端は、エンドヌクレアーゼ消化の後で直接に適合する。しかしながら、通常はエンドヌクレアーゼ消化によって生成される粘着末端を、それらをライゲーションにおいて適合せしめるために平滑末端へ最初に変換することが必要である。末端を平滑末端化するために、DNAは適切な緩衝液中にて少なくとも15分間、15℃で、4つのデオキシヌクレオチド三リン酸の存在下で10単位のDNAポリメラーゼIのクレノウ断片(クレノウ)によって処理される。DNAは、次にフェノール-クロロホルム抽出及びタノール沈殿又は他のDNA精製技術で精製される。
切断DNA断片は、大きさによって分離し、DNAゲル電気泳動を利用して選択されうる。DNAは、アガロース又はポリアクリルアミドマトリックスのどちらかによって電気泳動してもよい。マトリックスの選択は、分離されるDNA断片の大きさに依存する。Sambrookらのセクション6.30−6.33に記載のように、電気泳動の後、DNAを電解溶出(electroelution)によってマトリックスから抽出するか、低融解アガロースがマトリックスとして利用されている場合には、アガロースを融解し、DNAをアガロースから抽出する。
互いにライゲーションされるべきDNA断片(その前に適した制限酵素で消化し、ライゲーションされるべき各断片の末端が適合性であるもの)は、約等モル量で水溶液に加えられる。また、この溶液は、ATP、ライゲーション緩衝液及びリガーゼ、例えば0.5μgのDNA当たり約10単位のT4DNAリガーゼを含む。DNA断片がベクターへライゲーションされるのであれば、ベクターは、最初に適した制限エンドヌクレアーゼによって切断されて直線化される。次に、直線化されたベクターは、アルカリホスファターゼ又はウシ腸ホスファターゼによって処理される。脱リン酸化は、ライゲーション工程でのベクターのセルフライゲーション(自己連結)を防ぐ。
互いにライゲーションされるべきDNA断片(その前に適した制限酵素で消化し、ライゲーションされるべき各断片の末端が適合性であるもの)は、約等モル量で水溶液に加えられる。また、この溶液は、ATP、ライゲーション緩衝液及びリガーゼ、例えば0.5μgのDNA当たり約10単位のT4DNAリガーゼを含む。DNA断片がベクターへライゲーションされるのであれば、ベクターは、最初に適した制限エンドヌクレアーゼによって切断されて直線化される。次に、直線化されたベクターは、アルカリホスファターゼ又はウシ腸ホスファターゼによって処理される。脱リン酸化は、ライゲーション工程でのベクターのセルフライゲーション(自己連結)を防ぐ。
ライゲーション後、外来遺伝子が挿入されたベクターは、精製され、適切な宿主細胞へ形質転換される。好ましい形質転換方法は、エレクトロポレーションである。エレクトロポレーションは、当該分野で知られ、例えば、米国特許第4910140;米国特許第5186800;米国特許第4849355;米国特許第5173158;米国特許第5098843;米国特許第5422272;米国特許第5232856;米国特許第5283194;米国特許第5128257;米国特許第5750373;米国特許第4956288に記載の方法、又は他の既知のバッチあるいは連続エレクトロポレーションプロセスを利用して実施することができる。一以上(複数)のエレクトロポレーションが、宿主細胞へ形質転換されるDNAの量を増加させるために行われてもよい。繰り返しエレクトロポレーションが、当該技術分野で記載されている通りに行われる。Vaughanら (1996) Nature Biotechnology 14:309-314を参照のこと。付加的エレクトロポレーションの数は、数個(2,3,4.....10)から十位(10,20,30,.....100)及びさらには百位(100,200,300,.....1000)まで、所望する通りに変化しうる。繰り返しエレクトロポレーションは、宿主細胞へ形質転換されるコンビナトリアルライブラリー、例えば抗体ライブラリーの大きさが増大させるために望ましい。
ライブラリーの構築のためには、好ましくは、DNAは25マイクログラム/mL又はそれ以上の濃度で存在する。より好ましくは、DNAは、約30マイクログラム/mL又はそれ以上の濃度で存在し、より好ましくは約70マイクログラム/mL又はそれ以上、更により好ましくは約100マイクログラム/mL又はそれ以上でさらに数百マイクログラム/mLまでの濃度でDNAは存在する。一般的には、エレクトロポレーションでは、約50から約500マイクログラム/mLの範囲のDNA濃度が使われる。3.0ミリ秒(ms)より大きなエレクトロポレーション中の時定数は、高い形質転換効率を生じる。
ライブラリーの構築のためには、好ましくは、DNAは25マイクログラム/mL又はそれ以上の濃度で存在する。より好ましくは、DNAは、約30マイクログラム/mL又はそれ以上の濃度で存在し、より好ましくは約70マイクログラム/mL又はそれ以上、更により好ましくは約100マイクログラム/mL又はそれ以上でさらに数百マイクログラム/mLまでの濃度でDNAは存在する。一般的には、エレクトロポレーションでは、約50から約500マイクログラム/mLの範囲のDNA濃度が使われる。3.0ミリ秒(ms)より大きなエレクトロポレーション中の時定数は、高い形質転換効率を生じる。
DNAは、好ましくは、夾雑物を除くために精製される。DNAは、どの既知の方法によっても精製されるが、好ましい精製方法はDNAアフィニテイー精製法の利用である。DNA結合樹脂及び親和性試薬を利用したDNAの精製、例えば組換え体プラスミドDNAは、よく知られており、その既知の方法の何れも本発明で利用できる(Vogelstein.B.及び Gillespie.D.(1979) Proc. Natl. Acad. Sci.USA 76:615;Callen.W.(1993) Strategies 6:52-53)。また、商業的に入手可能なDNA単離及び精製キットは、Stratagene(CLEARCUT Miniprep Kit)及びLife Technologies(GLASSMAX DNA Isolation Systems)を含む幾つかのソースから入手可能である。DNA精製の適切な非限定的方法は、カラムクロマトグラフィー(米国特許第5707812号)、ヒドロキシル化シリカポリマー(米国特許第5693785号)、再水和シリカゲル(米国特許第4923978号)、ホウ素化ケイ酸塩(米国特許第5674997号)、修飾ガラス繊維膜(米国特許第5650506号;米国特許第5438127号)、フッ化吸収剤(米国特許第5625054号;米国特許第5438129号)、珪藻土(米国特許第5075430号)、透析(米国特許第4921952号)、ゲルポリマー(米国特許第5106966号)及びDNA結合試薬をともなうカオトロピック剤の利用(米国特許第5234809号)を含む。精製後、DNAは、本発明での濃度でのエレクトロポレーションでの利用のために、溶出されるか、そうでなければ水、好ましくは蒸留水又は脱イオン化水に再懸濁される。低塩緩衝液溶液の利用も考慮される。
エレクトロポレーションによって形質転換できる任意の適切な細胞を、本発明の方法における宿主細胞として利用することができる。形質転換が可能な適切な宿主細胞は、大腸菌のようなグラム陰性細菌を含む。適切な大腸菌株は、JM101、大腸菌K12株294(ATCC番号31446)、大腸菌株W3110(ATCC番号27325)、大腸菌X1776(ATCC番号31537)、大腸菌XL-1Blue(Stratagene)、及び大腸菌Bを含むが、大腸菌の多くの他の株、例えばXL1-BlueMRF、SURE、ABLE C、ABLE K、WM1100、MC1061、HB101、CJ136、MV1190、JS4、JS5、NM522、NM538、及びNM539が同じく利用される。細胞は、既知の方法を利用してコンピテント(形質転換受容性)にされる。Sambrookら, 上掲. 1.76-1.81, 16.30。
約1010コロニー形成単位(cfu)/mL)及びそれ以上の濃度の生存可能細胞は、好まれてエレクトロポレーションに利用される。更に好ましくは、生存可能細胞は、約1x1011から4x1011cfu/mLへ濃縮される。この範囲へ濃縮してもよい好ましい細胞は、下記に記載のSS320細胞である。細胞は、場合によっては37℃で約6−48時間(又はOD600=0.6−0.8まで)の間、標準的培養液の培養で生育し、次に培養液は遠心分離されて上清を取り除かれる(例えば、容器を傾けて上清が移される)。最初の精製は、好ましくは細胞ペレットを緩衝液溶液(例えばHEPES pH7.4)へ再懸濁であり、再遠心分離及び上清の取り除きがその後に続く。生じた細胞ペレットは、希グリセロール(例えば5−20%v/v)に再懸濁し、細胞ペレットを形成するために再び遠心分離され、上清は取り除かれる。最終の細胞濃度は、細胞ペレットを水又は希グリセロールへ再懸濁して所望する濃度とすることによって得られる。
約1010コロニー形成単位(cfu)/mL)及びそれ以上の濃度の生存可能細胞は、好まれてエレクトロポレーションに利用される。更に好ましくは、生存可能細胞は、約1x1011から4x1011cfu/mLへ濃縮される。この範囲へ濃縮してもよい好ましい細胞は、下記に記載のSS320細胞である。細胞は、場合によっては37℃で約6−48時間(又はOD600=0.6−0.8まで)の間、標準的培養液の培養で生育し、次に培養液は遠心分離されて上清を取り除かれる(例えば、容器を傾けて上清が移される)。最初の精製は、好ましくは細胞ペレットを緩衝液溶液(例えばHEPES pH7.4)へ再懸濁であり、再遠心分離及び上清の取り除きがその後に続く。生じた細胞ペレットは、希グリセロール(例えば5−20%v/v)に再懸濁し、細胞ペレットを形成するために再び遠心分離され、上清は取り除かれる。最終の細胞濃度は、細胞ペレットを水又は希グリセロールへ再懸濁して所望する濃度とすることによって得られる。
エレクトロポレーションに特に好ましい受容細胞は、ファージF'エピソームを含むコンピテント大腸菌株である。菌株においてファージの複製を可能にするどんなF'エピソームでも本発明で利用することができる。適切なエピソームは、ATCCへ寄託された菌株より入手可能であるか、商業的に入手可能である(CJ236、CSH18、DH5alphaF'、JM101、JM103、JM105、JM107、JM109、JM110)、KS1000、XL1-BLUE、71−18及び他)。SS320株は、XL1-BLUEの稔性エピソーム(F'プラスミド)をMC1061細胞へ転移させるために十分な条件下で、MC1061細胞をXL1−BLUE細胞と交配させることにより調製された。一般的には、二つの細胞型の培養を混合し、37℃で約1時間で培養培地の混合物を生育させることは、交配を可能にし、エピソーム転移を起こすために十分である。新しく生じた大腸菌株は、ストレプトマイシン耐性染色体マーカーを有するMC1061の遺伝子型、及びテトラサイクリン耐性を与えるF'プラスミドの遺伝子型を有する。この交配の後代は、両抗生物質に対して耐性であり、ストレプトマイシン及びテトラサイクリンの存在下で選択的に生育することが可能である。SS320株は、1998年6月18日に、アメリカン・タイプ・カルチャー・コレクション(ATCC), 10801 University Boulevard, Manassas, Virginia, USAへ寄託され、寄託受託番号98795が割り当てられている。
この菌株SS320の寄託は、特許手続き上の微生物の寄託の国際的承認に関するブダペスト条約及びその規則(ブダペスト条約)の規定に従って行われた。これは、寄託の日付から30年間、寄託の生存可能な培養が維持されることを保証するものである。寄託物はブダペスト条約の条項に従い、またジェネンテック社とATCCとの間の合意に従い、ATCCから入手することができ、これは、どれが最初であろうとも、関連した米国特許の発行時又は任意の米国又は外国特許出願の公開時に、寄託培養物の後代を永久かつ非制限的に入手可能とすることを保証し、米国特許法第122条35及びそれに従う特許庁長官規則(特に参照番号886OG638の37CFR第1.14条を含む)に従って権利を有すると米国特許庁長官が決定した者に後代を入手可能とすることを保証するものである。
本出願の譲受人は、寄託した培養物が、適切な条件下で培養されていた場合に死亡もしくは損失又は破壊されたならば、材料は通知時に同一の他のもの即座に取り替えることに同意する。寄託物質の入手可能性は、特許法に従いあらゆる政府の権限下で認められた権利に違反して、本発明を実施するライセンスであるとみなされるものではない。
本出願の譲受人は、寄託した培養物が、適切な条件下で培養されていた場合に死亡もしくは損失又は破壊されたならば、材料は通知時に同一の他のもの即座に取り替えることに同意する。寄託物質の入手可能性は、特許法に従いあらゆる政府の権限下で認められた権利に違反して、本発明を実施するライセンスであるとみなされるものではない。
オリゴヌクレオチド媒介突然変異誘発は、本発明の置換、欠失、及び挿入変異体を調製するためにの好ましい方法である。この技術は、Zollerら (1987) Nucl. Acids Res., 10: 6487-6504に記載のように、当該分野においてよく知られている。要約すると、タンパク質融合又は異種ポリペプチドをコードする遺伝子は、所望する変異をコードするオリゴヌクレオチドをDNAテンプレートへハイブリダイゼーションすることで改変し、そのテンプレートは、遺伝子の未改変又は天然DNA配列を含むプラスミドの一本鎖型である。ハイブリダイゼーション後、DNAポリメラーゼは、オリゴヌクレオチドプライマーを取り入れ、遺伝子内の選択改変をコードする、テンプレートの完全な二番目の相補鎖を合成するために利用される。一般的に、少なくとも25ヌクレオチド長のオリゴヌクレオチドが利用される。最適なオリゴヌクレオチドは、変異をコードしているどちらかの側のヌクレオチド上のテンプレートに対して完全に相補的な12から15ヌクレオチドを有する。これは、オリゴヌクレオチドが、正確に一本鎖DNAテンプレート分子へハイブリダイゼーションすることを確かなものにする。オリゴヌクレオチドは、例えばCreaら (1978)Proc.Natl. Acad. Sci. USA 75:5765によって記載された当該分野で知られた技術を利用することで容易に合成される。
DNAテンプレートは、例えばバクテリオファージM13ベクター(商業的に入手可能なM13mp18及びM13mp19ベクターは適切)のようなファージディスプレイシステムで利用されているバクテリオファージより誘導されるベクター、又は一本鎖ファージの複製開始点を含むベクターによって生成される:例は、Vieraら (1987)Meth. Enzymol. 153:3によって記載されている。従って、変異されるDNAは、一本鎖テンプレートを生成するために、これらベクターのうちの一つへ挿入することが可能である。一本鎖テンプレートの生産は、Sambrookらのセクション4.21−4.41に記載されている。
天然DNA配列を改変するためには、適切なハイブリダイゼーション条件下で、オリゴヌクレオチドを一本鎖テンプレートへハイブリダイゼーションする。次に、通常はT7DNAポリメラーゼ又はDNAポリメラーゼIのクレノウ断片であるDNA重合化酵素が、合成のプライマーとしてオリゴヌクレオチドを用いてテンプレートの相補鎖を合成するために加えられる。従って、ヘテロ二重鎖は形成され、DNAの一つの鎖は変異型をコードし、他の鎖(元のテンプレート)は遺伝子の天然、未改変配列をコードする。このヘテロ二重鎖分子は、次に通常は大腸菌JM101のような原核生物である適切な宿主細胞へ形質転換される。細胞を生育させた後、細胞はアガロースプレートへ播かれ、変異DNAを含む細菌コロニーを同定するために、32-リン酸で放射標識されたオリゴヌクレオチドプライマーを用いてスクリーニングされる。
天然DNA配列を改変するためには、適切なハイブリダイゼーション条件下で、オリゴヌクレオチドを一本鎖テンプレートへハイブリダイゼーションする。次に、通常はT7DNAポリメラーゼ又はDNAポリメラーゼIのクレノウ断片であるDNA重合化酵素が、合成のプライマーとしてオリゴヌクレオチドを用いてテンプレートの相補鎖を合成するために加えられる。従って、ヘテロ二重鎖は形成され、DNAの一つの鎖は変異型をコードし、他の鎖(元のテンプレート)は遺伝子の天然、未改変配列をコードする。このヘテロ二重鎖分子は、次に通常は大腸菌JM101のような原核生物である適切な宿主細胞へ形質転換される。細胞を生育させた後、細胞はアガロースプレートへ播かれ、変異DNAを含む細菌コロニーを同定するために、32-リン酸で放射標識されたオリゴヌクレオチドプライマーを用いてスクリーニングされる。
直ぐ上に記載した方法は、プラスミドの両鎖が変異を含むホモ二重鎖分子が作製されるように改変されてもよい。この改変は以下の通りである:一本鎖オリゴヌクレオチドは、上述のように一本鎖テンプレートへアニールされる。デオキシリボアデノシン(dATP)、デオキシリボグアノシン(dGTP)、及びデオキシリボチミジン(dTTP)の三つのデオキシリボヌクレオチドの混合物は、dCTP-(aS)と呼ばれる修飾チオ-デオキシリボシステイン(Amershamより得ることができる)と混ぜられる。この混合物は、テンプレート−オリゴヌクレオチド複合体へ添加される。この混合物へのDNAポリメラーゼの添加によって、変異塩基を除くテンプレートと同一のDNAの鎖が生成される。さらには、このDNAの新しい鎖は、dCTPに代わってdCTP-(aS)を含み、これが新しい鎖を制限エンドヌクレアーゼ消化から保護する。二本鎖ヘテロ二重鎖のテンプレート鎖に、適した制限酵素で切れ目を入れた後、テンプレート鎖はExoIIIヌクレアーゼ、又は変異誘発される部位を含む領域を通過したその他の適切なヌクレアーゼによって消化されることが可能である。そして、反応は、単に部分的に一本鎖化された分子を遊離するように止められる。次に、完全な二本鎖DNAのホモ二重鎖は、全4つのデオキシリボヌクレオチド三リン酸、ATP、及びDNAリガーゼの存在下でDNAポリメラーゼを用いて形成される。そして、このホモ二重鎖分子を、上記されている大腸菌JM101のような適切な宿主細胞へ形質転換することが可能である。
一つ以上の置換されるアミノ酸をともなう変異は、幾つかの方法のうちの一つの方法で生成されうる。アミノ酸がポリペプチド鎖内に近接して位置している場合は、アミノ酸は、所望するアミノ酸置換のすべてをコードする一つのオリゴヌクレオチドを用いて同時に変異される。しかしながら、アミノ酸が各々からある距離で位置している場合は、所望する変異のすべてをコードする一つのオリゴヌクレオチドを生成することはより困難である。代わりに、二つの代替法のうちの一つを用いてもよい。
最初の方法では、置換される各々のアミノ酸に対する個々のオリゴヌクレオチドが生成される。次に、オリゴヌクレオチドは、一本鎖テンプレートDNAへ同時にアニールされ、テンプレートから合成されるDNAの二番目の鎖は、所望するアミノ酸置換のすべてをコードする。代替法は、所望する変異体生産するためのを二回以上の突然変異誘発を含む。一回目は、単一の突然変異体のために記載されている:野生型DNAはテンプレートのために使用される。最初の所望するアミノ酸置換をコードするオリゴヌクレオチドは、このテンプレートへアニールされ、ヘテロ二重鎖DNA分子が生成される。二回目の突然変異誘発は、一回目の突然変異誘発で生産された変異DNAをテンプレートとして利用する。従って、このテンプレートは、既に一つ以上の突然変異を含む。次に、さらなる所望のアミノ酸置換をコードするオリゴヌクレオチドは、このテンプレートへアニールされ、生じるDNA鎖は、今回は、一回目及び二回目の突然変異誘発の両方からの変異をコードする。この結果として生じたDNAは、三回目の突然変異誘発のテンプレートとして利用でき、以後同様である。
最初の方法では、置換される各々のアミノ酸に対する個々のオリゴヌクレオチドが生成される。次に、オリゴヌクレオチドは、一本鎖テンプレートDNAへ同時にアニールされ、テンプレートから合成されるDNAの二番目の鎖は、所望するアミノ酸置換のすべてをコードする。代替法は、所望する変異体生産するためのを二回以上の突然変異誘発を含む。一回目は、単一の突然変異体のために記載されている:野生型DNAはテンプレートのために使用される。最初の所望するアミノ酸置換をコードするオリゴヌクレオチドは、このテンプレートへアニールされ、ヘテロ二重鎖DNA分子が生成される。二回目の突然変異誘発は、一回目の突然変異誘発で生産された変異DNAをテンプレートとして利用する。従って、このテンプレートは、既に一つ以上の突然変異を含む。次に、さらなる所望のアミノ酸置換をコードするオリゴヌクレオチドは、このテンプレートへアニールされ、生じるDNA鎖は、今回は、一回目及び二回目の突然変異誘発の両方からの変異をコードする。この結果として生じたDNAは、三回目の突然変異誘発のテンプレートとして利用でき、以後同様である。
また、カセット突然変異誘発は、本発明の置換、欠失及び挿入変異体を調製するために好ましい方法である。この方法は、Wellsら (1985)Gene 34:315により記載された方法に基づく。出発材料は変異される遺伝子を含んでなるプラスミド(又は他のベクター)である。変異される遺伝子のコドンが同定される。同定される変異部位の各側に独特な制限エンドヌクレアーゼ部位がなければならない。そのような制限部位が存在しない場合は、遺伝子の適切な位置にそれを導入するために上述したオリゴヌクレオチド媒介突然変異誘発法を使用してこれを産生しうる。制限部位がプラスミドへ導入された後、プラスミドDNAはこれらの部位で切断されてそれを線形化する。制限部位間のDNA配列をコードするが所望の変異を含んでいる二本鎖オリゴヌクレオチドは常套的な手順を使用して合成される。オリゴヌクレオチドの二本のストランドが別個に合成され、ついで標準的な方法を使用して共にハイブリダイズされる。この二本鎖オリゴヌクレオチドはカセットと呼ばれる。このカセットは線形化されたプラスミドの末端と適合性である5'及び3'末端を有するように設計され、プラスミドに直接に結合できる。しかして、このプラスミドは遺伝子の変異DNA配列を含んでいる。変異された変異体を含むベクターは、上述の適切な宿主細胞へ形質転換されることが可能である。
形質転換細胞は、ベクターにtet及び/又はamp耐性遺伝子が存在していることによって耐性が付与されていることから、一般的には、抗生物質、通常はテトラサイクリン(tet)又はアンピシリン(amp)上で生育させて選択する。
本発明での使用に適切なファージ及びファージミドベクターは、ファージディスプレイのための全ての既知のベクターを含む。さらなる例は、pComb8(Gramら (1995) Proc. Natl. Acad. Sci. USA 89:3576-3580);pC89(Feliciら (1991)J.Mol.Biol 222:310-310);pIF4(Bianchiら (1995) J.Mol.Biol. 247:154-160);PM48、PM52、及びPM54(Iannolo.(1995) J.Mol.Biol. 248:835-844);fdH(Geenwoodら (1991)J. Mol. Biol. 220:821-827);pfd8SHU、pfd8SU、pfd8SY、及びfdISPLAY8(Malik & Perham(1996) Gene 171:49-51);「88」(Smith(1993) Gene 128:1-2);f88.4(Zhongら (1994)J. Biol. Chem.269:24183-24188);p8V5(Affymax);MB1、MB20、MB26、MB27、MB28、MB42、MB48、MB49、MB56:(Marklandら (1991) Gene 109:13-19)を含む。同様に、ファージディスプレイシステムでファージミドベクターが用いられた場合、任意の既知のヘルパーファージを利用してもよい。適切なヘルパーファージの例には、M13-KO7(Pharmacia)、M13-VCS(Stratagene)、及びR408(Statagene)が含まれる。
本発明での使用に適切なファージ及びファージミドベクターは、ファージディスプレイのための全ての既知のベクターを含む。さらなる例は、pComb8(Gramら (1995) Proc. Natl. Acad. Sci. USA 89:3576-3580);pC89(Feliciら (1991)J.Mol.Biol 222:310-310);pIF4(Bianchiら (1995) J.Mol.Biol. 247:154-160);PM48、PM52、及びPM54(Iannolo.(1995) J.Mol.Biol. 248:835-844);fdH(Geenwoodら (1991)J. Mol. Biol. 220:821-827);pfd8SHU、pfd8SU、pfd8SY、及びfdISPLAY8(Malik & Perham(1996) Gene 171:49-51);「88」(Smith(1993) Gene 128:1-2);f88.4(Zhongら (1994)J. Biol. Chem.269:24183-24188);p8V5(Affymax);MB1、MB20、MB26、MB27、MB28、MB42、MB48、MB49、MB56:(Marklandら (1991) Gene 109:13-19)を含む。同様に、ファージディスプレイシステムでファージミドベクターが用いられた場合、任意の既知のヘルパーファージを利用してもよい。適切なヘルパーファージの例には、M13-KO7(Pharmacia)、M13-VCS(Stratagene)、及びR408(Statagene)が含まれる。
形質転換細胞の選択後、これらの細胞は培養によって生育され、ついでベクターDNAが単離されうる。ファージ又はファージミドベクターDNAは、例えばSambrookら, Molecular Cloning: A Laboratory Manual, 2版(1989) Cold Spring Harbor Laboratory Press. Cold Spring Harbor, NYに記述されているように、当該分野において既知の方法を用いて単離することが可能である。
単離されたDNAは、上掲のSambrookらのセクション1.40に記述され、上述されたような当該分野において既知の方法によって精製することが可能である。ついで、精製されたDNAは、DNA塩基配列決定法によって分析される。DNA塩基配列決定法は、Messingら (1981)Nucleic Acids Res. 9:309の方法、Maxamら (1980)Meth. Enzymol. 65:499の方法、又はその他の既知の方法によって行ってもよい。
単離されたDNAは、上掲のSambrookらのセクション1.40に記述され、上述されたような当該分野において既知の方法によって精製することが可能である。ついで、精製されたDNAは、DNA塩基配列決定法によって分析される。DNA塩基配列決定法は、Messingら (1981)Nucleic Acids Res. 9:309の方法、Maxamら (1980)Meth. Enzymol. 65:499の方法、又はその他の既知の方法によって行ってもよい。
V.応用
本発明の種々の側面及び実施態様は、構造的特徴が明確化されているペプチドの合理的設計及び分析のための新規モデルシステムの利点を証明する。そのようなペプチド及びそれを利用する方法を含んでなるコンビナトリアルライブラリーは、殆どすべての生物学的分子相互作用に関与している基礎的構造−活性相関を探索するために、有用な情報及び手段を提供する。ここで開示された又は本発明の開示に従って生成されたペプチドは、限定されるものではないが、酵素阻害剤、リガンドアンタゴニスト、リガンドアゴニスト、毒素、及び免疫原を含む種々の生物学的又は治療的薬剤の候補となりうる。
次の実施例は、例示的な方法を提供するものであって、限定するものではない。ここで引用する文献の開示の全てが、出典明示によりその全体がここに取り込まれる。
本発明の種々の側面及び実施態様は、構造的特徴が明確化されているペプチドの合理的設計及び分析のための新規モデルシステムの利点を証明する。そのようなペプチド及びそれを利用する方法を含んでなるコンビナトリアルライブラリーは、殆どすべての生物学的分子相互作用に関与している基礎的構造−活性相関を探索するために、有用な情報及び手段を提供する。ここで開示された又は本発明の開示に従って生成されたペプチドは、限定されるものではないが、酵素阻害剤、リガンドアンタゴニスト、リガンドアゴニスト、毒素、及び免疫原を含む種々の生物学的又は治療的薬剤の候補となりうる。
次の実施例は、例示的な方法を提供するものであって、限定するものではない。ここで引用する文献の開示の全てが、出典明示によりその全体がここに取り込まれる。
実施例1:構造化されたジスルフィド制約型βヘアピンペプチド骨格の構築
この実施例では、我々はβターンディスプレイのための骨格としてCX8Cの形態のデカマー(decamer)のジスルフィド制約型βヘアピンを調べることを選んだ。我々の目的のためには、多くのターン配列と適合性のある構造を構築することが必須である。つまり、ターンの残基以外の残基は、ペプチド構造をヘアピンの方へかなり偏らせなければならない。ジスルフィド環化は、多くのペプチドを構造化させるのには充分ではないが、役に立つ。我々の初期の目的は、ジスルフィド結合が共有結合的制約としてのみならず、β鎖の更に広い範囲の相互作用の中核を成すようにするためにも使用されうるかどうかを決定することであった。
この実施例では、我々はβターンディスプレイのための骨格としてCX8Cの形態のデカマー(decamer)のジスルフィド制約型βヘアピンを調べることを選んだ。我々の目的のためには、多くのターン配列と適合性のある構造を構築することが必須である。つまり、ターンの残基以外の残基は、ペプチド構造をヘアピンの方へかなり偏らせなければならない。ジスルフィド環化は、多くのペプチドを構造化させるのには充分ではないが、役に立つ。我々の初期の目的は、ジスルフィド結合が共有結合的制約としてのみならず、β鎖の更に広い範囲の相互作用の中核を成すようにするためにも使用されうるかどうかを決定することであった。
材料と方法
ペプチドの合成。 ペプチドを、パイオニアシンセサイザー(PE Biosystems)で標準的Fmoc化学を用いて合成し、トリフルオロ酢酸(TFA)中の5%トリイソプロピルシランで樹脂から分離し、逆相HPLC(アセトニトリル/H2O/0.1%TFA)により精製した。ペプチド同一性は質量分析により確認した。ペプチドは、I2の飽和酢酸溶液を滴下することによって環状ジスルフィドに転換させ、HPLCによって再精製した。精製したペプチドはC18分析用カラム(0−40%アセトニトリルで40分)で単一の対称ピークとして溶出した。
ペプチドの合成。 ペプチドを、パイオニアシンセサイザー(PE Biosystems)で標準的Fmoc化学を用いて合成し、トリフルオロ酢酸(TFA)中の5%トリイソプロピルシランで樹脂から分離し、逆相HPLC(アセトニトリル/H2O/0.1%TFA)により精製した。ペプチド同一性は質量分析により確認した。ペプチドは、I2の飽和酢酸溶液を滴下することによって環状ジスルフィドに転換させ、HPLCによって再精製した。精製したペプチドはC18分析用カラム(0−40%アセトニトリルで40分)で単一の対称ピークとして溶出した。
システイン有効濃度の測定。 グルタチオン原液を、3定量の0.2Mの還元グルタチオン(GSH)を1定量の0.1Mの酸化グルタチオン(GSSG)と混合することにより調製した。分量したものを−80℃で保存したが、数ヶ月は安定であった:単一のバッチの使用により、全グルタチオン濃度の変動から生じうるΔΔG値のあらゆる誤差を排除した。チオールジスルフィド平衡は、50μLのペプチド貯蔵物(水中に、およそ3mM)を50μLのグルタチオン貯蔵物と混合し、ファイアストーンバルブから真空/アルゴン循環で酸性溶液を脱酸し、ついでシリンジで300μLの脱酸した緩衝液を添加した後(0.2Mトリス、pH8.0;1mMEDTA;グルタチオンを滴定するための67mMトリスベース)、更に混合物を脱酸することにより合成した。全反応混合物の最終pHは8.10±0.05であった。溶液を、アルゴン下で撹拌し、ウォーターバスに20℃で保った。1.5時間後、連続して分量物(100μL)を、気密性シリンジで取り出し、直ちに400μLの31mM HCl中に放出することでクエンチし、最小の遅れでHPLCにより分析した。Ceff値を、グルタチオンモノマーは全体で0.025Mとみなして(つまり、ペプチドと混合したジスルフィド中に存在する僅かな量(<1%)のグルタチオンを無視して)ペプチド及びグルタチオンモルの還元及び酸化型のモル比から計算した(ピーク面積比は、HPLCにより測定された吸光度の違いで補正した)、:
Ceff=([ペプチド酸化型]/[ペプチド還元型])×([GSH]2/[GSSG])
[GSH]+2[GSSG]=0.025M
[GSSG]=0.025M/{2+3.26(GSHピーク域 / GSSGピーク域)}
[ペプチド酸化型]/[ペプチド還元型]=平衡ピーク面積比 / 吸光度比
各反応混合物からの2又は3のサンプルを分析した;経時的な数の変化は無く、計算されたCeff値は、典型的に5%未満と変化した(ΔΔGで30cal/molにする確かさに相当する)。
Ceff=([ペプチド酸化型]/[ペプチド還元型])×([GSH]2/[GSSG])
[GSH]+2[GSSG]=0.025M
[GSSG]=0.025M/{2+3.26(GSHピーク域 / GSSGピーク域)}
[ペプチド酸化型]/[ペプチド還元型]=平衡ピーク面積比 / 吸光度比
各反応混合物からの2又は3のサンプルを分析した;経時的な数の変化は無く、計算されたCeff値は、典型的に5%未満と変化した(ΔΔGで30cal/molにする確かさに相当する)。
NMR分光法。 NMRサンプルは、92%H2O/8%D2O pH5.1中の5−10mMのペプチド及び化学シフト基準としての0.1mMの1,4-ジオキサンを含む。全てのスペクトルは、15℃でBrukerDRX-500又はVarian Unity-400スペクトル計で得られた。2QF-COSY、TOCSY及びROESYスペクトルは、(Cavanaghら (1995) Protein NMR Spectroscopy, Principles and Practices(Academic Press, サンディエゴ)に記載されるようにして、グラジェントコヒーレンス選択法(van Zijlら (1995)J. Magn. Reson. 113A:265-270)、又は水サプレッションに対して励起像スカルプティング法(Hwang及びShaka, (1995)J. Magn. Reson. 112A, 275-279)で取得した。プロトン共鳴は標準的な方法(Wuthrich (1986)NMR of Protains and Nucelic Acids (John Wiley and Sons, New York)により与えた。3JHN-Hαは、F2の高デジタル解像度で処理された2QF-COSYスペクトルのHN−Hαのピーク逆位相二重線にLorentzianラインにフィットさせることにより得た。3JHN-HαはペプチドのD2O溶液で得たCOSY-35スペクトルから導き出した。距離及び二面角拘束は、(Skeltonら (1994) Biochemistry 33:13581-13592)に記載されるようにして生成した。100の初期構造を、ハイブリッドディスタンスジオメトリー法/シュミレーテッドアニーリングプログラムDGII(Havelら (1991) Prog. Biophys. Mol. Biol. 56:43-78)を用いて計算した;これらのうち80は、既に記載されているように(Skeltonら (1994)Prog. Biophys. Mol. Biol. 56:43-78)DISCOVERで実施されるAMBER全原子フォースフィールドを用いて分子動力学により更に精製された。最も低い拘束バイオレーションエネルギーの20コンフォメーションを各ペプチドの溶液コンフォメーションを表すために選択した。
構造計算。 構造は、78のROE-誘導距離拘束(10の中間及び28の長距離拘束;5.4、4.3、3.4又は3.0Åの上限)及び12の二面角拘束で計算した。最終的な20構造は、それぞれ0.05±0.02Å及び0.7±0.2°の距離及び二面角の平均最大バイオレーションを有し;実験上の距離及び二面角からのRMS偏差は、それぞれ0.007±0.002Å及び0.29±0.08°であった。残基の75%がラマチャンドランプロットの最適な部分でΦΨ値を有する(否認又は容認された領域に何もない)一方、平均構造の平均RMSDは、残基Cys1-Cys10のN、Cα、及びC原子に対して0.28±0.04Åである(Laskowskiら (1993)J. Appl. Crystallogr. 26:283-291)。
NMR分析。CD4ペプチドのNMRサンプルは、化学シフト基準として50μMの3-(トリメチルシリル)-1-プロパン-1,1,2,2,3,3,-d6-スルホン酸(DSS)で92%のH2O/8%D2O、pH3.5中に〜2mMのペプチドを含有していた。スペクトルは前記のようにして得て、分析した。cd2の構造を、84の(13の中-及び23の長-距離を含む)ROE誘導距離拘束及び13の二面角拘束から計算した。距離及び二面角拘束の平均最大バイオレーションは、それぞれ0.05±0.01Å及び0.6±0.4°であり;実験上の距離及び二面角拘束からのRMSDは、それぞれ0.009±0.002Å及び0.2±0.1°である。共有幾何学は良好で、ΦΨ角の74%が最適の範囲で、ラマチャンドランプロットの否認又は容認された範囲には何もない(Laskowskiら (1993) J. Appl. Crystallogr. 26:283-291)。
側鎖回転異性体の分析。 6−9Hzの範囲の3J Hα-Hβ1及び3J Hα-Hβ2の両方の観察により、側鎖は単一の古典的な回転異性体(χ1=−60°、+60°又は180°)を占有しないことが示され、大体においてサンプルは3つ全てが回転異性体のくぼみ(well)を揺らした。これが、bhpWのTrp3及びLeu8及びcd2のGln4、Phe7及びLeu8の場合の状態である。これらの側鎖に見られるROEピークは、サンプリングしたχ1値の範囲に対する時間平均を表す。これらのコンフォメーションの全てが、すぐに観察できるROEピークを生じるわけではなく、従って、構造計算過程は、拘束が得られる回転異性体の方に偏りうる。例えば、cd2のPhe7からのROEは、逆ストランドのプロトンに見られ、従って+60°の回転異性体のくぼみにPhe7を強制的に位置させる。多くのバックボーン-バックボーンの距離及びΦ二面角拘束が与えられると、bhpW及びcd2に対して計算された構造は、いくつかの側鎖の方向性の過剰決定以外においては、これらのペプチドの溶液コンフォメーションを正確に示す。
結果
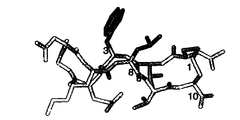
928の非余剰タンパク質構造の集合からのβシートの我々の調査では、隣接したストランドにおける残基の水素結合対と非水素結合対の間の平均Cβ-Cβ距離は、それぞれ4.82±0.58及び5.37±0.56Åであったが、ジスルフィド結合システインの平均Cβ-Cβの距離は、3.84Åであった。従って、逆平行鎖の逆残基のCβ原子は、通常ジスルフィド結合形成のためにはあまりにかけ離れている。それにもかかわらず、ジスルフィド架橋は、時々、βシートの非水素結合レジスターのシステインの間で見いだされている。我々は隣接した逆平行鎖を結合している23のジスルフィド結合システインを見つけた。23のうち14の場合において、ジスルフィドはシステインの1つ(又は両方)の前に2つの残基を疎水性側鎖に対してしっかりと詰め込む(図1a)。5つの更なる場合においては、この疎水性部位は極性又は荷電した残基によりβ及びγ-メチレンで(E、Q又はR)占有されていた。特に、ロイシン又は芳香族アミノ酸のどちらかの側鎖は、この特性を有するジスルフィドコンホメーションに良好な形の相補性を提供した。その結果、我々は、我々のモデルペプチドの残基8としてロイシンを選択し(図1b)、拡張した主鎖コンフォメーションを促進するために位置2及び9でトレオニンを含み、更に非常に強力なタイプII'βターンではなく、代わりにターン配列EGNKを選択した。ロイシンと対をなす最良の交差ストランド対合を決定するために、位置3が変えられた。
928の非余剰タンパク質構造の集合からのβシートの我々の調査では、隣接したストランドにおける残基の水素結合対と非水素結合対の間の平均Cβ-Cβ距離は、それぞれ4.82±0.58及び5.37±0.56Åであったが、ジスルフィド結合システインの平均Cβ-Cβの距離は、3.84Åであった。従って、逆平行鎖の逆残基のCβ原子は、通常ジスルフィド結合形成のためにはあまりにかけ離れている。それにもかかわらず、ジスルフィド架橋は、時々、βシートの非水素結合レジスターのシステインの間で見いだされている。我々は隣接した逆平行鎖を結合している23のジスルフィド結合システインを見つけた。23のうち14の場合において、ジスルフィドはシステインの1つ(又は両方)の前に2つの残基を疎水性側鎖に対してしっかりと詰め込む(図1a)。5つの更なる場合においては、この疎水性部位は極性又は荷電した残基によりβ及びγ-メチレンで(E、Q又はR)占有されていた。特に、ロイシン又は芳香族アミノ酸のどちらかの側鎖は、この特性を有するジスルフィドコンホメーションに良好な形の相補性を提供した。その結果、我々は、我々のモデルペプチドの残基8としてロイシンを選択し(図1b)、拡張した主鎖コンフォメーションを促進するために位置2及び9でトレオニンを含み、更に非常に強力なタイプII'βターンではなく、代わりにターン配列EGNKを選択した。ロイシンと対をなす最良の交差ストランド対合を決定するために、位置3が変えられた。
ジスルフィドの存在は、ヘアピン安定性のための簡便なプローブを提供する。チオール-ジスルフィド平衡を、基準チオールグルタチオンに関して測定し、システイン対の効果的濃度(Ceff)を得た。Ceffの大きい方の値は、増加した近似値を、システインチオールの平均値で示し、ヘアピン構造の形成と一致する。この方法はヘアピン安定性に対するβターン中の残基置換の効果を評価するために使用されている。Stroup及びGierasch (1990) Biochemistry 29:9765-9711。ペプチドCeff値は、位置3の異なる残基のためにかなり変化した(図2a)。著しく、位置3のトリプトファンは酸化型の方にペプチド平衡を強力にシフトした:この挙動はペプチド凝集によっては引き起こされない。アニリン類似物(-RT1n{Ceff,x/Ceff,ala})のCeff値のスケーリングは、折り畳みのためのペプチドの関連性質と解釈されうる>0.8kcal/molの範囲である自由エネルギー相違(図2b)を引き起こす。しかしながら、これらのデータはペプチドの折り畳まれた及び折り畳まれていない状態の効果の間で相違しない。例えば、与えられた置換は酸化ペプチドに詰め込まれている好ましい側鎖を促進しうるか、又は、還元したペプチドの拡張した主鎖コンフォメーションの方に単に偏る。
ペプチドが実際にβヘアピンを形成するかどうかを判断するために、それらのいくつかを、1H NMR分光法により評価した(表1)。トリプトファンペプチド(bhpW)は、強力につながったHα-HNNOE、多数の主鎖交差ストランドNOE及びストランド残基の大主鎖スカラーカップリング定数(3JHN-Hα>8.0Hz)の点で、高度に凝集したβヘアピンの特徴の全てを表す。Cys1及びCys10のHα化学シフトは、組織化されていないペプチドで得られる値に関して低磁場(downfield)であり、逆平行鎖はこれらの末端残基に含まれることが示された。他のペプチド研究では、少ない数のヘアピン構造を有することが判断された(表1参照)。興味深いことには、NMRデータはCeffと相関している(表1):従って、ジスルフィド交換アッセイは、酸化ペプチドのヘアピン構造の角度の有用な量を提供する。

実施例2:選択的テトラペプチドターン配列のヘアピン骨格への移動
実施例1によってbhpWに対して計算した構造は、タイプII'ターンと逆平行ヘアピンを形成するくぼみ(Gly5−Asn6)及びCys1、Trp3、Leu8及びCys10の側鎖の間の疎水性接触を明らかにした(図3)。bhpW安定性の熱力学分析は、高温又は化学変性剤の存在のどちらかで完全にほどくための酸化ペプチドの不足により、複雑であった。しかしながら、我々は、15℃でおそらくは>80%と、高い密度であるとヘアピンコンフォメーションを推定する。その構造安定性のため、我々はターンディスプレイ骨格としての研究のためにbhpWを選んだ。従って、我々は、異なるターン配列がbhpWストランド配列により構築できるできるかどうか試験した。
中和抗体及びヒトCD4に結合したHIVgp120の最近の結晶構造から接触表面の詳細が明らかになった(Kwongら, (1998) Nature 393:648-659.)。多数の突然変異研究から予想されるように、gp120結合に最も重要なCD4領域は、タンパク質表面から伸びる重要なPhe43側鎖を有する、C'-C"ヘアピンループ(残基37−46)である。実際、CD4残基40−48は境界面に埋まった表面積の63%に寄与し、全体の23%にPhe43が貢献している(Kwongら, (1998) Nature 393:648-659)。意外にも、Phe43結合部位の後にgp120には大きな穴があり、その内側には疎水性残基がある。C'-C"ターンに基づいて構築されたペプチドはgp120に結合しているかもしれず、もしそうであれば、結晶構造に見られる穴内に伸びるリガンドを設計するための開始点であるかもしれない。
実施例1によってbhpWに対して計算した構造は、タイプII'ターンと逆平行ヘアピンを形成するくぼみ(Gly5−Asn6)及びCys1、Trp3、Leu8及びCys10の側鎖の間の疎水性接触を明らかにした(図3)。bhpW安定性の熱力学分析は、高温又は化学変性剤の存在のどちらかで完全にほどくための酸化ペプチドの不足により、複雑であった。しかしながら、我々は、15℃でおそらくは>80%と、高い密度であるとヘアピンコンフォメーションを推定する。その構造安定性のため、我々はターンディスプレイ骨格としての研究のためにbhpWを選んだ。従って、我々は、異なるターン配列がbhpWストランド配列により構築できるできるかどうか試験した。
中和抗体及びヒトCD4に結合したHIVgp120の最近の結晶構造から接触表面の詳細が明らかになった(Kwongら, (1998) Nature 393:648-659.)。多数の突然変異研究から予想されるように、gp120結合に最も重要なCD4領域は、タンパク質表面から伸びる重要なPhe43側鎖を有する、C'-C"ヘアピンループ(残基37−46)である。実際、CD4残基40−48は境界面に埋まった表面積の63%に寄与し、全体の23%にPhe43が貢献している(Kwongら, (1998) Nature 393:648-659)。意外にも、Phe43結合部位の後にgp120には大きな穴があり、その内側には疎水性残基がある。C'-C"ターンに基づいて構築されたペプチドはgp120に結合しているかもしれず、もしそうであれば、結晶構造に見られる穴内に伸びるリガンドを設計するための開始点であるかもしれない。
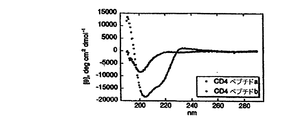
我々は、CD4ヘアピンの天然構造に基づいてジスルフィド拘束ペプチドを合成し(表2中のcd1残基38−45)、それが溶液中で本質的に構造化されていないことが分かった(図4a)。ついで我々は、bhpWの相当する残基(cd2、表2)に適合するように、置換G2T及びN3Wを作った;残基L8及びT9は既に天然CD4配列中に存在する。ペプチドcd2は、タイプII'ターンを有するヘアピン構造を取り入れて、きちんと整列している(図4)。図4Aに、cd2のピーク表示が見られた:cd1の対応するクロスピークの位置が示される。3JHN-Hα>8.3Hzを有するこれらのcd2残基に下線を引いた;全てのcd1残基が5.9と7.7Hzの間の骨格カップリング定数を有する。測定されたCeff値から、CD4ターン(QGSF)は、0.5kcal/molだけモデルヘアピン(EGNKターン)を不安定化し、我々はT2及びW3置換の両方が安定したヘアピン構造に必要であることを発見した(表2)。重要なことには、ペプチド構造をCD4のものと比較すると、骨格コンホメーションは、構造決定の不確実さの範囲内にあり、実質的に同じである(0.93ÅRMSD;図4B)。図4Bは、gp120結合CD4の結晶構造からCD4残基37−46(赤)上に重なって示されたcd2のNMR構造全体(20モデル;2つの垂直図)を示す。20モデルのRMSDは、残基1−10の骨格原子の平均座標に関して、0.50±0.09Åであり;結晶構造からのCD4の残基37−46を有する残基1−10の平均座標の比較は0.93ÅのRMSDを生じる。cd2のPhe7の3JHα-Hβカップリング定数は、側鎖が全体に見られる回転異性体を固定しないことを示し;Phe7側鎖は、共結晶構造で見られるものを確実にサンプリングして、溶液中の複数のコンフォメーションを取り入れることに留意のこと。これは、ペプチド骨格がCD4βターンを正しく示すことを実証する。
末端のセリン及びリジン残基は、そうしないと荷電していないCD4ペプチドのいくつかの変異体の溶解性を改善するために加えた。同様の修飾は、コントロールとしてのbhpWにも行った。bhpWとCD4の間で異なる非ターン残基には下線を引いた。還元ペプチドと酸化ペプチドの共溶離は、CD4ペプチドのT2、N3変異体のCeffの測定を妨げた。環状2色性スペクトルはAviv Instruments. Inc.のモデル202分光光度計で10℃にて得られ:ペプチド濃度は20mMリン酸カリウム、pH7.0中、20μMであった。
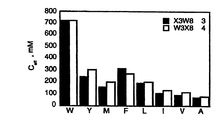
評価した2つの他のターン配列は、ヒトFc-ε-RIのドメイン2のF-GループからのVWQL、及びEPOアゴニストペプチドEMP1からのGPLTであった。3つのターンの全ては、trpペプチド骨格で、その配列が天然親ヘアピンループにより近接して適合している環化ペプチドで評価した。
SCGNQGSFLTCK-NH2 (配列番号:10) CD4ペプチド a
SCTWQGSFLTCK-NH2 (配列番号:11) b
Ac-CTKVWQLWTC-NH2 (配列番号:12) Fc-ε-RIペプチド c
SCTWVWQLLTCK-NH2 (配列番号:13) d
SCHFGPLTWVCK-NH2 (配列番号:14) EMP1ペプチド e
SCTWGPLTLTCK-NH2 (配列番号:15) f
SCGNQGSFLTCK-NH2 (配列番号:10) CD4ペプチド a
SCTWQGSFLTCK-NH2 (配列番号:11) b
Ac-CTKVWQLWTC-NH2 (配列番号:12) Fc-ε-RIペプチド c
SCTWVWQLLTCK-NH2 (配列番号:13) d
SCHFGPLTWVCK-NH2 (配列番号:14) EMP1ペプチド e
SCTWGPLTLTCK-NH2 (配列番号:15) f
環状2色性スペクトルは、それぞれの場合において、設計されたtrpヘアピン骨格が更に構築されたペプチドを生じることを示す(図5a-c)。NMRデータは、ペプチドで増加したヘアピン構造と一致し、骨格は構築状態の向けて様々な「異なる」ターンをバイアスし得ることを証明している。
ヘアピン骨格上に存在可能な他の通常のターンは、γターン(3アミノ酸)、バルジ(膨らんだ)ターン(5又は6アミノ酸)、及びより長いヘアピン(8アミノ酸)を含む。他のターンの長さは既知で、また骨格と適合する。
実施例1及び実施例2の結果は、小ジスルフィド拘束ヘアピンの1つのストランド位置の最適化が、非常に不完全に構築された分子を高度に構造化された(-ΔΔG>0.8kcal/mol)ものに置き換えるのに充分であることを実証している。構築されたヘアピンのステム位置、-CTW----LTC-は、最適化されたターン配列を必要としない;従って、βターンライブラリーのディスプレイにとって、またそうでなければ高度に集合していないかもしれない特定のターンの研究にとって適した骨格である。重要なことは、単なる天然アミノ酸が必要とされ、ターンライブラリーはファージに表示されうることである。
ヘアピン骨格上に存在可能な他の通常のターンは、γターン(3アミノ酸)、バルジ(膨らんだ)ターン(5又は6アミノ酸)、及びより長いヘアピン(8アミノ酸)を含む。他のターンの長さは既知で、また骨格と適合する。
実施例1及び実施例2の結果は、小ジスルフィド拘束ヘアピンの1つのストランド位置の最適化が、非常に不完全に構築された分子を高度に構造化された(-ΔΔG>0.8kcal/mol)ものに置き換えるのに充分であることを実証している。構築されたヘアピンのステム位置、-CTW----LTC-は、最適化されたターン配列を必要としない;従って、βターンライブラリーのディスプレイにとって、またそうでなければ高度に集合していないかもしれない特定のターンの研究にとって適した骨格である。重要なことは、単なる天然アミノ酸が必要とされ、ターンライブラリーはファージに表示されうることである。
我々がここで報告した置換エネルギーをβシートシステムの先の研究と比較することは興味深い。エネルギーの相違の大きさは同様であるが、我々が得た順序は、経験的なβ性向のスケールと、又は既知のβシートの得られた残基対頻度と相関していない(Hutchinsonら (1998) Protain Sci. 7:2287-2300, Wouters, M. A. & Curmi. P.M.G. (1995) Proteins 22:119-131)。特に、トリプトファンはそのようなスケールの例外ではない。これらの差は、典型的なタンパク質ドメインにおける平均的な傾向は、ほとんどの残基が高度に溶媒に暴露される小ペプチドには直接には適用できず、デノボ設計でのそのような情報の使用を複雑化していることを強調する。更に、ΔΔG順序は、側鎖の非極性表面積の増加に良好には相関しないが、好ましい残基は疎水性である。
最後に、ヘアピンステムは非常に小さいが、ジスルフィド及び交差ストランド3次接触の組合わせは、例えばCX4Cのようなジスルフィドのみのものを越える構造的偏りを与える。いくつかの特定の配列(例えばVVVV)(Milburnら (1998) Int. J. Peptide Protein Res. 31:311-321)が我々のヘアピンと適合するターンコンホメーションを取り入れることができないことが知られているが、またタンパク質に観察される非常に少ないターン配列が、単離されたペプチドに良好に定まったターンコンホメーションを取り入れることを示したのも真実である。我々はこの数を増加させる簡単な方法を示した。我々は、ランダム化されたターン配列(例えばXCTWX4LTCX)を有するヘアピンライブラリーは、結合決定基が小さい合成ターン擬態に直ぐに移ることができるか、または高度な処理能力の親和性最適化のための小分子リード体を同定するのに直接使用さえされる(Rohrerら (1998)science 282:737-740)構築リガンドを生じうると考える。
実施例3:ターン及び交差ストランド相互作用からの相対的貢献度の定量化
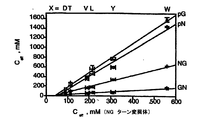
上記の実施例1において、置換はモデルペプチドbhp(ペプチド1)の位置3(X)に導入された。このゲスト部位は、タイプII'ターンに空間的にかなり近い(gly-asn、図1)。異なったターン配列及び幾何構造を有するヘアピンが異なる残基優先度をNHBゲスト部位に持つかどうかを更に調べるために、モデルペプチド1の中央gly-asn配列をタイプI'ターンasn-gly(ペプチド2)及びタイプII'ターンD-pro-asn及びD-pro-gly(ペプチド3及び4)と置き換える。位置3(X)での置換は、我々がgly-asn列で観察したヘアピン安定性の範囲をスパンするために選択した。Ceffは、実施例1に既に記載したようにして測定した。異なるターンから我々が得た値を図6で比較する。
CTXEGNKLTC 1 II' 配列番号:16
CTXENGKLTC 2 I' 配列番号:17
CTXEpNKLTC 3 II' 配列番号:18
CTXEpGKLTC 4 II' 配列番号:19
X=W,Y,L,V,T,D又はp=D-pro。
上記の実施例1において、置換はモデルペプチドbhp(ペプチド1)の位置3(X)に導入された。このゲスト部位は、タイプII'ターンに空間的にかなり近い(gly-asn、図1)。異なったターン配列及び幾何構造を有するヘアピンが異なる残基優先度をNHBゲスト部位に持つかどうかを更に調べるために、モデルペプチド1の中央gly-asn配列をタイプI'ターンasn-gly(ペプチド2)及びタイプII'ターンD-pro-asn及びD-pro-gly(ペプチド3及び4)と置き換える。位置3(X)での置換は、我々がgly-asn列で観察したヘアピン安定性の範囲をスパンするために選択した。Ceffは、実施例1に既に記載したようにして測定した。異なるターンから我々が得た値を図6で比較する。
CTXEGNKLTC 1 II' 配列番号:16
CTXENGKLTC 2 I' 配列番号:17
CTXEpNKLTC 3 II' 配列番号:18
CTXEpGKLTC 4 II' 配列番号:19
X=W,Y,L,V,T,D又はp=D-pro。
全ての場合に、位置3のトリプトファンは、与えられたターンの最も大きいCeff値を生じ、その安定化作用が一般的であることを証明している。異なる交差ストランド相互作用(横軸)とターン配列(縦軸)のCeffの大きな変化は、両方がこれらの環状ヘアピンペプチドにおける安定性に有意に貢献し得ることを示している。最後に、データセット間の著しい線形相関があり、ストランド位置3の置換とターン置換が環状ヘアピンの安定性に独立して貢献することを示している。これらのデータは、ヘアピンの折り畳みがかなりモジュラーであってもよく、ヘアピン設計を有意に単純化することを示唆している。
相対的なターンエネルギーは、ペプチドの適切な対のCeffを比較することにより計算することができる。しかしながら、図6の相関は、勾配からの相対ターンエネルギーの計算を可能にし、それは実験誤差に対して低い感度でなければならない。これらの値は表3に示す。asn-gly(タイプI')と比較して、gly-asn(タイプII')は安定性がより低いが、D-pro含有ターン(またタイプII')はヘアピン安定性を促進する。asn-gly対D-pro-glyで比較がなされてもよい一つの場合には、ここで得られたΔΔG値が、NMRにより得られたものと適度によく一致する。これは、Syudら (1999) J AM Chem Soc 121:11577によってあてがわれた基準状態及び2つの状態の折り畳みのその仮定がそのモデルシステムに適している;しかしながら、そのような基準状態を定めることが常に実現可能とは限らない。
相対的なターンエネルギーは、ペプチドの適切な対のCeffを比較することにより計算することができる。しかしながら、図6の相関は、勾配からの相対ターンエネルギーの計算を可能にし、それは実験誤差に対して低い感度でなければならない。これらの値は表3に示す。asn-gly(タイプI')と比較して、gly-asn(タイプII')は安定性がより低いが、D-pro含有ターン(またタイプII')はヘアピン安定性を促進する。asn-gly対D-pro-glyで比較がなされてもよい一つの場合には、ここで得られたΔΔG値が、NMRにより得られたものと適度によく一致する。これは、Syudら (1999) J AM Chem Soc 121:11577によってあてがわれた基準状態及び2つの状態の折り畳みのその仮定がそのモデルシステムに適している;しかしながら、そのような基準状態を定めることが常に実現可能とは限らない。
あるいは、ストランド位置に対する置換エネルギーは、残基X(図示せず)による代わりに、分類された同じデータをプロットすることによって得てもよい。相関性はまたすばらしく、勾配が自由エネルギー変化を生じている(表4)。エネルギーの範囲はペプチド1に関して実施例1で報告したものよりも大きい(1.42対0.85kcal・mol−1)。差異の多くは安定性スケールの底の置換までさかのぼる(特にasp)。gly-asnターンペプチドの安定性の低さは検知可能に構造化されておらず、Ceffアッセイはそれらの間の如何なる差異も記録しない。よって、より強いターン配列を持つペプチドにおいて得られるデータが、ストランド置換エネルギーのより完全な概要を提供する。
ターンのタイプが如何にヘアピン構造へ影響を及ぼすかを評価するために、2と3のトリプトファン類似体がNMR分析によって特徴付けされ、ペプチド1について実施例1に記載したようにして構造が算定された。図7の最小化平均構造の比較により、骨格及び側鎖コンホメーションが、存在するターンのタイプにかかわらず、非ターン残基(RMSD〜0.3Å)に対して非常に類似していることが明らかである。よって、Ceffの線形相関性と一致して(図6)、これらの三種のターンは隣接するストランドに対して如何なる構造的影響も及ぼさない。
ヘアピン構造に対するターン配列と良好な交差ストランド対合の重要性は多くのモデル研究において着目されてきた。しかし、残基置換の定量的なデータ又は体系的な評価は殆ど存在しない。我々のデータは、これらの簡単な環状ペプチドに対して、ストランド部位の置換とターンの置換は簡単な線形自由エネルギー関係と一致し、ヘアピン安定性に対して独立した相加効果を有していることを示している。これは、その構造の近接性が示され(図3及び7)、タンパク質固着の特徴に対しての算定ターンエネルギーの敏感性が報告されていることから(Mattosら(1994) J. Mol. Biol. 238:733)、意外なことである。しかしながら、これらのターンとストランドの間の対合は、それぞれが単独で及ぼす大きな影響に比較すると無視できるようである。これは、これらの成分の別個の解析によりβヘアピンの安定性が理解されうることを示唆している。
ヘアピン構造に対するターン配列と良好な交差ストランド対合の重要性は多くのモデル研究において着目されてきた。しかし、残基置換の定量的なデータ又は体系的な評価は殆ど存在しない。我々のデータは、これらの簡単な環状ペプチドに対して、ストランド部位の置換とターンの置換は簡単な線形自由エネルギー関係と一致し、ヘアピン安定性に対して独立した相加効果を有していることを示している。これは、その構造の近接性が示され(図3及び7)、タンパク質固着の特徴に対しての算定ターンエネルギーの敏感性が報告されていることから(Mattosら(1994) J. Mol. Biol. 238:733)、意外なことである。しかしながら、これらのターンとストランドの間の対合は、それぞれが単独で及ぼす大きな影響に比較すると無視できるようである。これは、これらの成分の別個の解析によりβヘアピンの安定性が理解されうることを示唆している。
実施例4:交差ストランド残基からのエネルギー寄与の定量化
上記の実施例の結果から、トリプトファンが、交差ストランドのロイシンと対合するとペプチド1の非水素結合(NHB)ストランド部位Xにおいてかなり安定化することが明らかになった。トリプトファンペプチド(bhpW)は水中で高度に構造化され、意図したヘアピン高次構造を採っている(図3)。ここでは、我々はNHB交差ストランド残基間の関係を調べる。顕著なことには、我々は、二種の構造的に等価でない部位に対する残基の優先傾向は同じであり、特異的な対の相互作用が単一部位の寄与からのほんの僅かな偏差をつくり出すことを見出す。従って、トリプトファン-トリプトファン交差ストランド対がヘアピン安定性に対して最適であるように思われる。
上記の実施例の結果から、トリプトファンが、交差ストランドのロイシンと対合するとペプチド1の非水素結合(NHB)ストランド部位Xにおいてかなり安定化することが明らかになった。トリプトファンペプチド(bhpW)は水中で高度に構造化され、意図したヘアピン高次構造を採っている(図3)。ここでは、我々はNHB交差ストランド残基間の関係を調べる。顕著なことには、我々は、二種の構造的に等価でない部位に対する残基の優先傾向は同じであり、特異的な対の相互作用が単一部位の寄与からのほんの僅かな偏差をつくり出すことを見出す。従って、トリプトファン-トリプトファン交差ストランド対がヘアピン安定性に対して最適であるように思われる。
トリプトファンからの安定化寄与の我々の知見は、効果が如何に一般的であるかを問うことを我々に促した。ペプチドbhpWのトリプトファン(図3)はII’型ターンの残基の側鎖と交差ストランドロイシンの双方に空間的に近接している。よって、トリプトファンの効果はこれらの他の残基との安定化接触に依存することがありうるように思われる。この問題に取り組むために、我々は疎水性対(ペプチド5)を逆にし、3位にロイシンを固定して8位(ジスルフィドに最も近い、図3)のアミノ酸を変化させた。システインチオールの有効濃度(Ceff)は我々の過去の研究にあるように決定した。
CTXEGNKLTC 1 配列番号:20
CTLEGNKXTC 5 配列番号:21
CTXEGNKWTC 6 配列番号:22
CTWEGNKXTC 7 配列番号:23
X=W,Y,F,L,M,I,V又はA
CTXEGNKLTC 1 配列番号:20
CTLEGNKXTC 5 配列番号:21
CTXEGNKWTC 6 配列番号:22
CTWEGNKXTC 7 配列番号:23
X=W,Y,F,L,M,I,V又はA
我々は、8位のトリプトファンが、試験したこれらの残基の中で最も安定化することを見出す(図8A)。重要なことに、Ceff値はtrp-leu対とleu-trp対とでかなり近いことで、この二つの配置がエネルギー的におよそ等価であることを示している。この結果は、ロイシンを持つ他の残基対にも成り立つと思われる:二つの群においてCeffの数値と順序は類似しているが、正確ではない(図8A)。
逆にした疎水性対の等価性がより一般的かどうかを試験するために、我々は、残基が代わりに交差ストランドトリプトファンと対合しているペプチド群6及び7を調べた(図8B)。ロイシン対の場合のように、密接な対応が、順序とCeffの数値の双方について二群のトリプトファンの間に見られる。我々は、ターンとジスルフィドに対する側鎖位置の差異にかかわらず、二つの交差ストランド部位が本質的に等価であると結論する。
逆にした疎水性対の等価性がより一般的かどうかを試験するために、我々は、残基が代わりに交差ストランドトリプトファンと対合しているペプチド群6及び7を調べた(図8B)。ロイシン対の場合のように、密接な対応が、順序とCeffの数値の双方について二群のトリプトファンの間に見られる。我々は、ターンとジスルフィドに対する側鎖位置の差異にかかわらず、二つの交差ストランド部位が本質的に等価であると結論する。
二つのロイシン群(1及び5)がトリプトファン群(6及び7)と比較されうる。二つのデータの傾向は顕著に類似しており(図8A及びB)、交差ストランド残基が独立して安定性に寄与していることを示唆している。この考えを探求するために、我々はペプチド群の各々においてその群の基準ペプチドに対しての、置換に対する自由エネルギー差を計算した(ΔΔG=−RTln(Ceff,X/Ceff,ref))。代表的な比較は図9にプロットする。
線形の自由エネルギー関係は4つのデータセットの間に存在する。これは、NHB部位3及び8の間で切り替えられる特定の交差ストランド対の比較と、同じ配向のleu対とのtrp対の比較にも見られる(図3)。(後者の相関性にはより多くの散乱性が存在する。)勾配(ρ)は各群の二つの異なる基準ペプチドに対して計られたΔΔG値を使用して決定した(X=ala及びtrp)。得られたρ値は大きくは異ならなかった(表5)。
線形の自由エネルギー関係は4つのデータセットの間に存在する。これは、NHB部位3及び8の間で切り替えられる特定の交差ストランド対の比較と、同じ配向のleu対とのtrp対の比較にも見られる(図3)。(後者の相関性にはより多くの散乱性が存在する。)勾配(ρ)は各群の二つの異なる基準ペプチドに対して計られたΔΔG値を使用して決定した(X=ala及びtrp)。得られたρ値は大きくは異ならなかった(表5)。
3位と8位が等価であるという考えと一致して、これらのデータを比較するプロットではρは1に近い。これに対して、leu対をtrp対と比較すると、ρは約0.4である。これは、与えられた残基対Xに対して、ヘアピン安定性の予想される差異が、交差ストランドパートナーとしてのtrpの〜2.5倍大きいことを意味している。これらの簡単な関係が与えられると、ΔΔGは、交差ストランドパートナーのρを置換エネルギーσxに乗じることにより基準対に対する任意の置換について計算されうる(以下を参照されたい)。これは、これらの残基が接触距離内にあるので驚くべきことであり、βヘアピンの設計に重要な意味を有している。
βシートタンパク質におけるHB及びNHB交差ストランド対の統計的解析では、多くの残基対が高い信頼性でもって正又は負に相関していることが見いだされる。主として統計的な優先性に従って、タンパク質の突然変異誘発の研究によりHB対間に1kcal・mol−1の大きさの相互作用エネルギーを特定した。タンパク質において好まれる交差ストランド対を含めることが、安定性を改善し、又は単離されたβヘアピンにストランドレジスターを固定するかもしれないことが提案される。
βシートタンパク質におけるHB及びNHB交差ストランド対の統計的解析では、多くの残基対が高い信頼性でもって正又は負に相関していることが見いだされる。主として統計的な優先性に従って、タンパク質の突然変異誘発の研究によりHB対間に1kcal・mol−1の大きさの相互作用エネルギーを特定した。タンパク質において好まれる交差ストランド対を含めることが、安定性を改善し、又は単離されたβヘアピンにストランドレジスターを固定するかもしれないことが提案される。
これらの効果を調べるために、我々は対相互作用エネルギーを計算した(図10)。Ceff比は単一の又は二重の置換に対してΔΔGをもたらす。典型的には、二重置換に対するΔΔGと単一の置換に対するΣΔΔGの差が相互作用エネルギーと見なされる。示された実施例では、これは、leu-leu基準状態に対するtrp-tyr対について−136cal・mol−1となろう。単一置換エネルギーが連続的に計算される場合は、第二工程でρの倍率をかけると、不一致は+41cal・mol−1のみである(これらの実験では有意ではない)。phe-trp対に対して(図2の上部と底部を比較されたい)、同様な分析によりρが含まれる場合、ΔΔΔG=−253cal・mol−1が得られる。この不一致は有意であり、(trpの一般的な優位性を越えて)trpとのpheの対合にある種の特定の構造的利点が存在しているかも知れないことを示唆している。単一の部位置換に対して見られる全エネルギー範囲と比較した場合、不一致が小さいことは興味深い(図9);我々は、このような対特異的効果(及び実験誤差)が我々の相関性における散在性を説明しうると考える。
我々の実験は、交差ストランド三次接触がヘアピン安定性を向上させることを明らかに示している。顕著なことは、trp-trp対の導入により、我々の元のペプチドbhpWと比較して大なる安定化が得られており(図3及び8)、我々は、タンパク質中で希であるにもかかわらず、trp-trpが単離されたヘアピンに対して最適なNHB対であると考える。大抵の場合、一般的な二重変異分析によって我々が得る対相互作用エネルギーはρの差によって十分に説明される。つまり、これらのエネルギーは単一の対に特異的ではないが、代わりに与えられた交差ストランドパートナーとは反対に全ての残基置換に対して程度の差はあれ感受性を示す。よって、我々は、組み合わせた単一部位優先性(σ及びρ)がヘアピン安定性を予想するのに最も重要であると結論する。顕著なことは、実験データの限られた基礎となる集合からこれらの予想を行うことが可能であろうことである。
実施例5:trpペプチド骨格に基づくファージディスプレイライブラリーの作製
糸状バクテリオファージM13の遺伝子8タンパク質に融合したランダムペプチドのライブラリーを、pS349の誘導体であるプラスミドpS1302b(出典明示によりここに取り込まれる米国特許出願第60/103514号及び同第60/134870号)のクンケル突然変異誘発により作製した。プラスミドpS1302bはpS349のtacプロモーターとmalEリーダー配列を含む。hGH配列とpS349のGly/Serリッチリンカー配列は次の配列によって置換した:
5'-TAA-TAA-TAA-ATG-GCT-GAT-CCG-AAC-CGT-TTC-CGC-GGT-AAA-GAT-CTG-GGT- GGC-GGT-ACT-CCA-AAC-GAC-CCG-CCA-ACC-ACT-CCA-CCA-ACT-GAT-AGC-CCA-GGC-GGT-3' (配列番号:24)
糸状バクテリオファージM13の遺伝子8タンパク質に融合したランダムペプチドのライブラリーを、pS349の誘導体であるプラスミドpS1302b(出典明示によりここに取り込まれる米国特許出願第60/103514号及び同第60/134870号)のクンケル突然変異誘発により作製した。プラスミドpS1302bはpS349のtacプロモーターとmalEリーダー配列を含む。hGH配列とpS349のGly/Serリッチリンカー配列は次の配列によって置換した:
5'-TAA-TAA-TAA-ATG-GCT-GAT-CCG-AAC-CGT-TTC-CGC-GGT-AAA-GAT-CTG-GGT- GGC-GGT-ACT-CCA-AAC-GAC-CCG-CCA-ACC-ACT-CCA-CCA-ACT-GAT-AGC-CCA-GGC-GGT-3' (配列番号:24)
挿入配列は3つの停止コドン、GDエピトープタグ、及びhGHの高レベルのディスプレイのために選択されたリンカーをコードする。プラスミドはまたpS349からのlacリプレッサー(lacIq)とアンピシリン耐性遺伝子を含む。ライブラリーを作製するために使用したオリゴヌクレオチドは次の通りであった:
5'-TCC-GCC-TCG-GCT-TAT-GCA-NNS-TGC-ACT-TGG-NNS-NNS-NNS-NNS-CTG-ACT- TGT-NNS-ATG-GCT-GAT-CCG-AAC-CGT-3' (配列番号:25)
ランダムペプチドの形態はよってXCTWX4LTCXであった。109から1010の個々の形質転換体のライブラリーを過去に記載されている方法(米国特許出願第60/103514号及び同第60/134870号)によって調製した。個々のクローンのおよそ三分の一が機能性ペプチド配列をコードしていた。残りは開始鋳型であり、停止コドンを含み、又は単一のヌクレオチド欠失を含んでいた。よって、ライブラリーのサイズはそれぞれの可能なランダム配列の幾つかのコピーを含むのに十分である。
5'-TCC-GCC-TCG-GCT-TAT-GCA-NNS-TGC-ACT-TGG-NNS-NNS-NNS-NNS-CTG-ACT- TGT-NNS-ATG-GCT-GAT-CCG-AAC-CGT-3' (配列番号:25)
ランダムペプチドの形態はよってXCTWX4LTCXであった。109から1010の個々の形質転換体のライブラリーを過去に記載されている方法(米国特許出願第60/103514号及び同第60/134870号)によって調製した。個々のクローンのおよそ三分の一が機能性ペプチド配列をコードしていた。残りは開始鋳型であり、停止コドンを含み、又は単一のヌクレオチド欠失を含んでいた。よって、ライブラリーのサイズはそれぞれの可能なランダム配列の幾つかのコピーを含むのに十分である。
実施例6:構造的にバイアスされたライブラリーからの結合ペプチドの選択
NuncのMaxiSorpプレートをPBS中2μg/mLのrhuFc-ε-RI-IgG融合体で終夜被覆した。ついで、プレートをPBS中0.5%のBSA(シグマのA-7638)で室温にて1時間の間ブロックした。0.5%のBSAだけで被覆することによって陰性のウェルを準備した。ファージ(1ウェル当たり1011ifu)を10の各正及び陰性のウェルに添加し、室温で20時間の間振とうしながらインキュベートした。非特異的に結合したファージを除去するために十分に洗浄した後、0.2Mのグリシン、pH2で5分間処理することによってバインダーを溶出させた。溶出されたファージをついでトリス塩基を添加して中和させ、大腸菌の培養物を感染させるために使用した(XL1-ブルー、ストラタジーン)。結合、溶出及び増幅の数サイクル(全体で3−5)を同様の条件下で実施した。192の個々のクローンを、ファージソーティングに対して記載されたようにして調製されたプレートでのファージ上清のインキュベーションにより、標的レセプターへの結合についてスクリーニングした。洗浄後、ウェルをα-M13-HRPコンジュゲート(ファーマシア・バイオテック27-9421-01)で処理し、結合した抗体をOPD基質(シグマP-9187)で検出した。プレート吸光度(A492−A405)を陽性プレートと陰性プレートの間で比較してレセプター結合に対して陽性のクローンを同定した。12のそのようなクローンを同定した。
NuncのMaxiSorpプレートをPBS中2μg/mLのrhuFc-ε-RI-IgG融合体で終夜被覆した。ついで、プレートをPBS中0.5%のBSA(シグマのA-7638)で室温にて1時間の間ブロックした。0.5%のBSAだけで被覆することによって陰性のウェルを準備した。ファージ(1ウェル当たり1011ifu)を10の各正及び陰性のウェルに添加し、室温で20時間の間振とうしながらインキュベートした。非特異的に結合したファージを除去するために十分に洗浄した後、0.2Mのグリシン、pH2で5分間処理することによってバインダーを溶出させた。溶出されたファージをついでトリス塩基を添加して中和させ、大腸菌の培養物を感染させるために使用した(XL1-ブルー、ストラタジーン)。結合、溶出及び増幅の数サイクル(全体で3−5)を同様の条件下で実施した。192の個々のクローンを、ファージソーティングに対して記載されたようにして調製されたプレートでのファージ上清のインキュベーションにより、標的レセプターへの結合についてスクリーニングした。洗浄後、ウェルをα-M13-HRPコンジュゲート(ファーマシア・バイオテック27-9421-01)で処理し、結合した抗体をOPD基質(シグマP-9187)で検出した。プレート吸光度(A492−A405)を陽性プレートと陰性プレートの間で比較してレセプター結合に対して陽性のクローンを同定した。12のそのようなクローンを同定した。
陽性のクローンの配列を、コード化DNAを配列決定することにより同定する。ディスプレイされた配列に対応するペプチド(すなわち、12マー)を標準的な固相法を使用して合成する。ついで、ペプチドを、適切な生物学的又は結合アッセイを使用してアッセイしてその効力を測定する。ペプチドは、上で概説した既知の方法:円偏光二色性、NMR、又はジスルフィド平衡の何れかを使用してヘアピン構造について評価することができる。ついで、ペプチドに置換を行って、選択されたターン残基の結合への相対寄与を決定することができる。理想的には、これらの置換は骨格構造を乱さない。結合モチーフの性質が理解されれば、ついで、ターン配列を更なる最適化のための好適な有機骨格上に移すことができる。
実施例7.特異的HB残基を持つ十量体bhp骨格の安定性
本実施例はbhp骨格の水素結合(HB)ストランド位置2(つまり、A1)及び9(つまり、A5)における置換を研究する。結果は、これらの部位における安定性決定要因は非水素結合(NHB)位置3(つまり、A2)及び8(つまり、A4)におけるものとは異なることを示している。位置3及び8の観察された等価性とは著しく異なり、位置2及び9は互いに非常に異なる残基優先度を有している。特に、環状ヘアピンに対する非常に大きな安定化は位置9のバリン残基から折り畳む;更に少ない安定化はVal2変異体で生じる。この効果は、自由エネルギー差及びVal9及びVal2bhp類似体のNMR構造分析から明らかである。更なる証拠は、bhpペプチドにおけるVal9の強い安定化効果が交差ストランドジスルフィドにそれが近接する結果であることを示唆している。興味深いことに、Val9はファージディスプレイペプチドライブラリーから選択されたジスルフィド環状化βヘアピンにしばしば存在しており、その安定化効果はこのフォールドに一般的であることを示唆している。
本実施例はbhp骨格の水素結合(HB)ストランド位置2(つまり、A1)及び9(つまり、A5)における置換を研究する。結果は、これらの部位における安定性決定要因は非水素結合(NHB)位置3(つまり、A2)及び8(つまり、A4)におけるものとは異なることを示している。位置3及び8の観察された等価性とは著しく異なり、位置2及び9は互いに非常に異なる残基優先度を有している。特に、環状ヘアピンに対する非常に大きな安定化は位置9のバリン残基から折り畳む;更に少ない安定化はVal2変異体で生じる。この効果は、自由エネルギー差及びVal9及びVal2bhp類似体のNMR構造分析から明らかである。更なる証拠は、bhpペプチドにおけるVal9の強い安定化効果が交差ストランドジスルフィドにそれが近接する結果であることを示唆している。興味深いことに、Val9はファージディスプレイペプチドライブラリーから選択されたジスルフィド環状化βヘアピンにしばしば存在しており、その安定化効果はこのフォールドに一般的であることを示唆している。
βヘアピンの水素結合ストランド位置における残基優先度を評価するために、以前に記載した環状ペプチドbhpWを宿主として使用した。このペプチドは明確に定まったヘアピンコンホメーションを採っており、そのNMR構造は図3に示されている。bhpWのジスルフィドはヘアピンコンホメーションを安定化させるのに役立ち、同時に構造のアッセイを提供する。ヘアピン安定性の変化はチオール-ジスルフィド平衡の位置の変化によって定量することができる。bhpWの残基2及び9は互いに交差するストランドであり、骨格水素結合を形成する。ついで、幾つかの置換をbhpWの位置2及び9に導入した:
bphW CTWEGNKLTC 配列番号:2
X2列 CXWEGNKLTC 配列番号:26
X9列 CTWEGNKLXC 配列番号:27
KGNEターン CTWKGNELTC 配列番号:28
V2H9対("VH") CVWEGNKLHC 配列番号:29
H2V9対("HV") CHWEGNKLVC 配列番号:30
HV,X3列 CHXEGNKLVC 配列番号:31
bphW CTWEGNKLTC 配列番号:2
X2列 CXWEGNKLTC 配列番号:26
X9列 CTWEGNKLXC 配列番号:27
KGNEターン CTWKGNELTC 配列番号:28
V2H9対("VH") CVWEGNKLHC 配列番号:29
H2V9対("HV") CHWEGNKLVC 配列番号:30
HV,X3列 CHXEGNKLVC 配列番号:31
bhpW変異体の相対的安定性をシステインチオールの有効濃度(Ceff)から決定し(実施例1の実験セクションを参照)、図11に示す。双方の位置での置換はヘアピン安定性に大きな効果を有している。安定性の範囲は位置2の置換に対して1kcal・mol−1であり(図11A)、位置9での同じ置換に対して0.7kcal・mol−1である(図11B)。これらの位置の各々における残基優先度の順番は、NHB位置3及び8に対してこれまでに観察されたもの(実施例4参照)とはかなり異なっている。有意なことに、位置2に対する安定性変化のパターンは位置9のものとは完全に異なっている。これに対して、残基優先度は位置3及び8に対して同じである。従って、残基2及び9の間の交差ストランド水素結合の影響に加えて、bhpヘアピンはこれらの部位における残基優先度分析の局在化した非対称性を示している。
位置2及び9の残基優先の非対称性は隣接する側鎖との密着性の差を反映する。バリン残基がスレオニンの代わりに交差ストランドヒスチジンと対をなしているペプチドを調製した。我々は側鎖が残基2及び9と同じヘアピン面にある残基4及び7をまた交換した。別のヘアピンモデル系では、ここのHB残基4及び9のようにストランド間のねじれによって近接して位置しているNHB残基の側鎖間の「対角」の相互作用がヘアピン安定性に影響を及ぼすために提案されている。Syudら (2001) J. Am. Chem. Soc. 123:8667-8677。何れの場合にも、位置9のバリンの強い安定化効果が残っている(図12)。従って、特定の側鎖-側鎖接触以外の何らかの構造的特徴がこの効果を生じせしめていると思われる。
NHBストランド位置のトリプトファン残基がβヘアピン構造の明確なねじれに関連している;これが局所的なコンホメーションの歪みをつくり出し、隣接位置の残基優先性に影響を及ぼしている。これを調べるために、我々は残基2及び9をより安定化させる「HV」の組合わせに固定した後、位置3を変えた。これらのペプチドの相対的安定性を図13の類似T2T9ペプチドのものと比較する。HVタンパク質は、そのより高いCeff値に示されるように、そのTT対応物よりも一律により安定である。注目すべきは、各列内の自由エネルギー差は線形に相関していることである。これは、バリン9の置換によりもたらされる大きな安定化は隣接するNHB部位における特定の残基の存在に依存しないことを証明している。興味深いことは、プロットの傾きは1ではないが、代わりに、HV宿主が位置3の置換に2倍、感応性が高いことを示している。これらを併せて考慮すると、図11−13のデータは、位置9のバリンによる強い非対称安定化がCX8Cジスルフィド制約ヘアピンの一般的性質であることを示唆している。
ヘアピンコンホメーションに対してHB部位の置換が有していた効果を評価するために、1H NMRデータに基づいてHV及びVHの構造を決定した。HVのNMRデータは、これが高度に構造化されたβヘアピンであることと完全に一致している:1H NMR二次化学シフト(観察されたシフトとランダムコイルシフト間の差異)はbhpWに対してよりも更に大きく、3JHα-Hβ結合定数値は3対のストランド残基(Cys1-Cys10、His2-Val9、Trp3-Leu8;表6)に対して明確な側鎖回転異性体を示している。特に、His2及びVal9は、疑いなく、それぞれ+60°及び180°のχ1回転異性体を採るように割り当てることができる。ペプチドVHに対しては、NMRデータから得られた拘束は明らかにコンホメーションを定めるが、Hα二次化学シフトはHVに対してよりも極端さは少ない。更に、3JHα-Hβ結合定数値は、Val2、His9、Trp3及びLeu8側鎖に対しての運動平均化を示している(表6)。これらの特徴の全てが、Ceff測定値と一致して、HVと比較したVHに対する安定性の低下を示している。HV及びVHの相対的構造安定性は、ヒスチジンの存在には依存していない;非常に類似したNMRパラメータが、ヒスチジンをスレオニンに置き換えたペプチドに対してもまた観察される(表6)。つまり、TVは明確に定まった側鎖コンホメーションを有しているが、VTは側鎖レベルでのコンホメーション平均化を示している。
HV及びVHに対して決定された構造は骨格レベルではかなり類似している(平均構造間で0.37ÅのRMS差):bhpWに対してこれまでに見られたように、双方のペプチドがII'型ターンを採り、かなり右巻きのストランドねじれを有している。ストランド間のねじれ角を計算すると、環化ヘアピンに対してねじれは一様ではなく、ジスルフィドに隣接したねじれは少なく(Cys1、Cys10対及び2、9対間でΘ<20°)、NHB対Trp3、Leu8の前後でねじれが大きい(Θ〜30°)ことが示される。これらの測定は両ストランドからの原子を含むので、各個々のストランドに存在するねじれ度はΘから明らかではない。あるいは、一残基からのψと続く残基からのφの合計(ψi+φi+1)が同じストランドにおける連続α炭素間のねじれの指標を付与し、非ねじれストランドはゼロ値で、右巻きねじれのストランドは正の値である。図14では、HV及びVHアンサンブルに対するねじれデータがbhpWと比較される。これらのデータは、大きなストランド間Θ値に寄与する大半のねじれがNHB疎水性残基のN末端側で生じる(Trp3及びGlu4間と、Leu8及びVal/His/Thr9間の<20°と比較して、His/Val/Thr2及びTrp3間と、Lys7及びLeu8間の45−65°)ことを明らかに示している。これらのアンサンブル内のψi+φi+1の範囲はオーバーラップしているが、図15のデータは、VHと比較して、HVがNHB部位の前に僅かにより顕著なねじれを有し、Cys1の廻りに顕著さが少ないねじれを有していることを示している(各アンサンブル内の2面角の正規分布を仮定すると、独立サンプルt-検定は、HVとVHに対する平均ψi+φi+1が、0.01未満のp値で、残基1−2、2−3、及び7−8に対して異なっていることを示している)。これは、His及びVal側鎖の位置の交換が骨格の幾何構造に微妙な影響を有していることを示唆する。
HV及びVHアンサンブル間の主な差異は側鎖次数の度合いである。Cys1、Trp3、Leu8及びCys10はHV及びVHにおいて同じχ1を採るけれども(Cys1、Trp3及びCys10に対して−60°;Leu8に対して180°)、3JHα-Hβの大きさは後者で極端さが少なく、VHの側鎖が交互のコンホメーションを一過性に採取していることを示唆している。より重要なことは、HVでは、His2及びVal9のχ1回転異性体が固定され(それぞれ+60°と180°)、これらの二つの側鎖を、交差ストランド水素結合上でファンデルワールス接触させる。ストランドのねじれにより、Val9のγ2メチル基がまたGlu4とLys7の双方の側鎖メチレン基とファンデルワールス接触させられる。しかし、VHでは、Val2が−60°及び180°の回転異性体ウェルの双方を集合させ、アンサンブルの構造の殆どがHis9に対して−60°のχ1を有し、側鎖をターンの向きに配向させる。更に、ストランドのねじれはVal2側鎖をペプチドの末端に向けている。従って、これらの側鎖間に接触は殆どないか全くなく、これらの残基間の骨格水素結合は溶媒によりさらされる。
HVペプチドに対して見られる非対称安定化がジスルフィド結合により課せられるストランド間ねじれの差に関係しているかどうかを評価するために、ペプチドHV及びVHを還元後にNMRによって調べた。驚いたことに、還元されたHVは観察可能な残基構造を有している:4の3JHN-Hα結合定数が8Hzを越え、Leu8メチル化学シフトはランダムコイルと比較して0.41ppm及び0.33ppm高磁場であり、3つの骨格交差ストランドNOEが観察される(Cys1 HαからCys10 Hα、His2 HNからVal9 HN及びGlu4 HNからLys7 HN)。これに対して、還元されたVHは好ましいコンホメーションを有しているという指標はない。おそらくは、Val9は酸化形態におけるように還元されたペプチドにおいて類似のコンホメーションを採る一方、位置3の前のストランドの強い右巻きねじれ(上記を参照)は何れの形態のVal2側鎖もC末端ストランドと相互作用させ得ないであろう。
併せると、データは、ジスルフィド環化βヘアピンを、C末端システインの直ぐ前のバリン残基によって顕著に安定化させることができ、イソロイシンがほぼ同様に安定化していると思われる。この効果はVal9とCys10の間の好ましいコンホメーション関係のためであろう。安定なβターンファージディスプレイに対しては、位置3又は8のTrp残基とここで報告されたまさに安定化しているHis-Val2,9対の組合わせが特に有用であろう。明確なヘアピン構造が検出可能な最も安定性が低い組合わせと比較すると、これらの置換は1.5kcal・mol−1だけ折り畳み構造を安定化させる。最適なW3-W8対の導入は安定性を更に増大させるべきで、構造化されるかもしれないβターンの範囲を拡大する。データは、固定されたCXnWVCモチーフを持つライブラリーが構造化されたヘアピンリガンドを生成させるのにまた有用であるかもしれないことを更に示唆している。
実施例8.bhpBR3ペプチドの構造と活性
この実施例はBR3レセプターのそのリガンドBAFFへの結合を模倣するターンペプチドを生成するためのbhp骨格の使用を記述する。TNFファミリーの最近明確になったメンバーであるBAFF(BLyS、TALL-1、zTNF4、THANK及びTNFS13Bとしても知られている)はマクロファージ、単球及び樹状細胞によって発現されるホモ三量体2型膜貫通タンパク質である。Mooreら (1999) Science 285:260-263;Schneiderら (1999) J. Exp. Med. 189:1747-1756。BAFFは末梢B細胞の発達と生存に対して重要であることが見出されている。Grossら (2001) Immunity 15:289-302;Schiemannら (2001) Science 293:2111-2114。興味深いことに、全身性エリテマトーデス(SLE)、関節リウマチ及び主に腎臓、関節及び唾液/涙腺にそれぞれ末端器官損傷があるシェーグレン症候群を含む自己免疫症候群に罹っているヒトでは、血清BAFFレベルが高い。更に、BAFFレベルは、疾患の重症度と相関しており、これらの障害を起こす病弊の原因における可能な役割と一致している。Cheemaら (2001) Arthritis Rheum. 44:1313-1319;Groomら (2002) J. Clin. Invest. 109:59-68;Zhangら (2001) J. Immunol. 166:6-10。
この実施例はBR3レセプターのそのリガンドBAFFへの結合を模倣するターンペプチドを生成するためのbhp骨格の使用を記述する。TNFファミリーの最近明確になったメンバーであるBAFF(BLyS、TALL-1、zTNF4、THANK及びTNFS13Bとしても知られている)はマクロファージ、単球及び樹状細胞によって発現されるホモ三量体2型膜貫通タンパク質である。Mooreら (1999) Science 285:260-263;Schneiderら (1999) J. Exp. Med. 189:1747-1756。BAFFは末梢B細胞の発達と生存に対して重要であることが見出されている。Grossら (2001) Immunity 15:289-302;Schiemannら (2001) Science 293:2111-2114。興味深いことに、全身性エリテマトーデス(SLE)、関節リウマチ及び主に腎臓、関節及び唾液/涙腺にそれぞれ末端器官損傷があるシェーグレン症候群を含む自己免疫症候群に罹っているヒトでは、血清BAFFレベルが高い。更に、BAFFレベルは、疾患の重症度と相関しており、これらの障害を起こす病弊の原因における可能な役割と一致している。Cheemaら (2001) Arthritis Rheum. 44:1313-1319;Groomら (2002) J. Clin. Invest. 109:59-68;Zhangら (2001) J. Immunol. 166:6-10。
BAFFに対する3つのレセプターのなかでBR3だけが特異的である;他の二つのTACIとBCMAはまた関連したリガンドAPRILにまた結合する。Grossら (2000) Nature 404:995-999;Thompsonら (2001) Science 293:2108-2111;Yanら (2001a) Curr. Biol. 11:1547-1552;Yanら (2000) Nat. Immunol. 1:37-41。TACIの細胞外ドメインは、TNFレセプターファミリーの特徴である二つのシステインリッチドメイン(CRD)を包含する特徴的なTNFR様構造を有している。これらのおよそ40残基の疑似反復は、6つの高度に保存されたシステインを含む3つの鎖内ジスルフィドを典型的には特徴とする独特に二次構造を有している。BCMAは、単一の限界CRDのみを含む点で普通ではない。しかしながら、BR3は、その細胞外ドメインが、これまでに特徴付けられている他のTNFRモジュールとは区別される間隙を持つ4つのシステイン残基を含む部分的なCRDのみから構成される点で更に一層相違する。Bodmerら (2002) Trends Biochem. Sci. 27:19-26;Naismith及びSprang (1998) Trends Biochem. Sci 23:74-79。TNFRファミリーの一般的なメンバーはリガンドの結合に対して二つのCRDを利用する;リガンド上の二つの別個の表面パッチと相互作用する各CRDからの類似のループから接点は主として由来する(Bodmerら, 2002において概説)。よって、BAFFへの高親和性結合がBR3のものと同様に単一の又は部分的なCRDのみによって如何にして達成されるかは明らかではない。BAFFの最近の結晶構造は幾つかの識別性のある特徴を有する三量体TNF様折り畳み構造を明らかにしている(Karpusasら (2002) J. Mol. Biol. 315:1145-1154;Liuら (2002) Cell 108:383-394;Orenら (2002) Nat. Struct. Biol. 9:288-292)が、何れのBAFFレセプターの構造も記載されていない。
方法と材料
ペプチド合成
ミニBR3及びbhpBR3を標準的なFmoc化学を使用してパイオニアペプチド合成機(PEバイオシステムズ)でC末端アミドとして合成した。その配列は次の通りである(図15Aにも示す):
ミニBR3: TPCVPAECFDLLVRHCVACGLLRTPR (配列番号32)
bhpBR3: CHWDLLVRHWVC (配列番号33)
ペプチド合成
ミニBR3及びbhpBR3を標準的なFmoc化学を使用してパイオニアペプチド合成機(PEバイオシステムズ)でC末端アミドとして合成した。その配列は次の通りである(図15Aにも示す):
ミニBR3: TPCVPAECFDLLVRHCVACGLLRTPR (配列番号32)
bhpBR3: CHWDLLVRHWVC (配列番号33)
ミニBR3に対しては、システイン19及び32の側鎖チオールを、トリフルオロ酢酸(TFA)安定アセトアミドメチル(Acm)誘導体として保護した。室温で1.5−4時間の間、TFA中5%トリイソプロピルシランで処理をして樹脂からペプチドを切断した。ロータリーエバポレーターによってTFAを除去した後、エチルエーテルを添加してペプチドを沈殿させた後、逆相HPLC(アセトニトリル/H2O/0.1%TFA)によって精製した。ペプチドの同定は熱スプレー質量分析法によって確認した。bhpBR3はHPLC画分にI2の飽和溶液(酢酸中)を滴下することによって環状ジスルフィドに転換させた。凍結乾燥後、酸化されたペプチドをHPLCによって精製した。還元されたミニBR3を含むHPLC画分をNH4OHを用いて〜9のpHまで調節した;ついで、システイン24及び35間にジスルフィドを、若干過剰のK3Fe(CN)6の添加によって形成し、酸化されたペプチドをHPLCによって精製した。若干過剰のI2を用いて〜4時間にわたってHPLC溶出液を処理することによって(第二のジスルフィドを同時形成すると共に)Acm基を除去した。酸化の進行を、分析用HPLCによってモニターし、最終生成物をHPLCによって再び精製した。ミニBR3は樹脂上にあるときにアミノ末端がビオチン化された後、未変性ペプチドについて上述したものと正確に同じようにして切断し、精製した。
NMR分光分析法
化学シフト基準として0.1mMのDSSと共に2.9mMのbhpBR3、pH4.5を含む試料に対して293KでBrukerのDRX-600を使用して、Starovasnikら, (1996) Biochemistry 35:15558-155569に記載されたようにして、二次元(2D)NMR実験装置を入手し分析した。距離の制約は2D NOESYスペクトルから誘導した(τm250ms);二面角制約は92%H2O/8%D2O中で獲得した2D DQF-COSYスペクトルと100%D2Oに溶解した試料で獲得された2D COSY-35スペクトルの分析から誘導した。共鳴割当と結合定数値は追加データに含める。
化学シフト基準として0.1mMのDSSと共に2.9mMのbhpBR3、pH4.5を含む試料に対して293KでBrukerのDRX-600を使用して、Starovasnikら, (1996) Biochemistry 35:15558-155569に記載されたようにして、二次元(2D)NMR実験装置を入手し分析した。距離の制約は2D NOESYスペクトルから誘導した(τm250ms);二面角制約は92%H2O/8%D2O中で獲得した2D DQF-COSYスペクトルと100%D2Oに溶解した試料で獲得された2D COSY-35スペクトルの分析から誘導した。共鳴割当と結合定数値は追加データに含める。
bhpBR3の三次元構造を、78のNOE誘導(26のロングレンジを含む)距離制約及び17の二面角制約に基づいて計算した。100の初期構造をDGIIを使用して計算した;これらの80を、記載されたようにして(上掲のStarovasnikら, (1996))、DISCOVERを使用して制約分子動力学によって更に精巧にした。最も低い制約バイオレーションエネルギーと良好な幾何構造を持つ20の構造がbhpBR3の溶液コンホメーションを示す。アンサンブルの平均座標に対して最低rms偏差(RMSD) のモデルを代表的な構造として選択した(PDBファイルのモデル1)。20のモデルの最終アンサンブルが入力データをよく満たし、0.1Å又は1°をそれぞれ越える距離又は二面角制約バイオレーションがない。構造はよく定まり、平均座標に対する平均骨格RMSDは0.42±0.07Åであり、PROCHECKによって判定して良好な共有的幾何構造を有している(φψ空間の最も好ましい領域で87%の残基、許容される領域で9%、広く許容される領域において4%)。Laskowskiら,(1993) J. Appl. Crystallogr. 26: 283-291。bhpBR3の構造はRCSBプロテインデータバンクから入手できるであろう(IDコードxxxx)。
競合置換ELISA
ヌンクMaxisorp96ウェルプレートをpH9.6の炭酸塩バッファー中BAFF2μg/mlの溶液を100μl用いて4℃で一晩被覆した。そのプレートをPBSで洗浄して、PBS中1%の脱脂乳でブロックした。BR3変異体の連続希釈物を、3ng/mlのビオチン化ミニBR3を含むPBS/0.05%Tween20中で調製した。PBS/Tweenでの洗浄後に、100μl/ウェルの各希釈物を移し、室温で1時間の間、インキュベートした。プレートを、PBS/Tweenで洗浄し、PBS/Tween中0.1U/mlのストレプトアビジン-POD(ベーリンガー・マンハイム)を100μl/ウェル用いて15分インキュベートした。PBS/Tweenの後、PBSによって洗浄した後、プレートを、0.8mg/mlのOPD(シグマ)及び0.01%のH2O2を含む100μlのPBS基質溶液を用いて5分インキュベートした。反応を100μl/ウェルの1M H3PO4を用いて停止させ、490nmでプレートを読み取った。IC50値は競合的ELISAシグナルの4パラメータフィットによって決定した。bhpBR3の初期原液溶液の濃度を記載されているようにして(Gill及びvon Hippel, (1989) Anal. Biochem. 182: 319-326)分光光度法で決定する一方、ミニBR3及びBR3細胞外ドメインの濃度を定量アミノ酸分析法によって決定した。
ヌンクMaxisorp96ウェルプレートをpH9.6の炭酸塩バッファー中BAFF2μg/mlの溶液を100μl用いて4℃で一晩被覆した。そのプレートをPBSで洗浄して、PBS中1%の脱脂乳でブロックした。BR3変異体の連続希釈物を、3ng/mlのビオチン化ミニBR3を含むPBS/0.05%Tween20中で調製した。PBS/Tweenでの洗浄後に、100μl/ウェルの各希釈物を移し、室温で1時間の間、インキュベートした。プレートを、PBS/Tweenで洗浄し、PBS/Tween中0.1U/mlのストレプトアビジン-POD(ベーリンガー・マンハイム)を100μl/ウェル用いて15分インキュベートした。PBS/Tweenの後、PBSによって洗浄した後、プレートを、0.8mg/mlのOPD(シグマ)及び0.01%のH2O2を含む100μlのPBS基質溶液を用いて5分インキュベートした。反応を100μl/ウェルの1M H3PO4を用いて停止させ、490nmでプレートを読み取った。IC50値は競合的ELISAシグナルの4パラメータフィットによって決定した。bhpBR3の初期原液溶液の濃度を記載されているようにして(Gill及びvon Hippel, (1989) Anal. Biochem. 182: 319-326)分光光度法で決定する一方、ミニBR3及びBR3細胞外ドメインの濃度を定量アミノ酸分析法によって決定した。
ターンペプチドbhpBR3はBAFFに競合的に結合する
BAFF-BR3相互作用を乱す薬剤の開発に向けての第一ステップとして、BR3の細胞外リガンド結合ドメインのNMR分析を実施した。興味深いことに、タンパク質の中央の三分の一だけが溶液中で安定な構造を採る;このコアはCysl9/Cys32とCys24/Cys35を連結する2つのジスルフィド結合によって安定化される。従って、1:3,2:4ジスルフィド結合パターンを導入して、26残基のミニBR3ペプチドを合成し、構造的に特徴付けした(図15A)。確かに、ミニBR3のNMRスペクトルは、このペプチドが完全長タンパク質の場合と本質的に同じ構造を採ることを示していた。重要なことに、ミニBR3はまたBAFFに完全長BR3と同じ親和性で結合する(〜70nM IC50;図16A)。
BAFF-BR3相互作用を乱す薬剤の開発に向けての第一ステップとして、BR3の細胞外リガンド結合ドメインのNMR分析を実施した。興味深いことに、タンパク質の中央の三分の一だけが溶液中で安定な構造を採る;このコアはCysl9/Cys32とCys24/Cys35を連結する2つのジスルフィド結合によって安定化される。従って、1:3,2:4ジスルフィド結合パターンを導入して、26残基のミニBR3ペプチドを合成し、構造的に特徴付けした(図15A)。確かに、ミニBR3のNMRスペクトルは、このペプチドが完全長タンパク質の場合と本質的に同じ構造を採ることを示していた。重要なことに、ミニBR3はまたBAFFに完全長BR3と同じ親和性で結合する(〜70nM IC50;図16A)。
高親和性BAFF結合が26残基コア内に含まれる場合、BR3のBAFF結合部を更に描写する試みを行った。他のTNF様リガンド/レセプター複合体の結晶構造において、BR3残基26DLLVRH31に類似のレセプターループがリガンドとの直接の接触点の形成に関与している。Bodmerら (2002) Trends Biochem. Sci 27:19-26。完全長及びミニBR3の双方において、NMR分析は、配列26DLLVRH31がAsp26とHis31の間に骨格水素結合を持つI型βターンを表していることを示唆していた。従って、BR3からの6の残基がジスルフィド結合βヘアピン(bhp)骨格内に埋め込まれた12残基のペプチドを合成した(図15A)。これまでに記載されているように、これらの骨格における強いストランド-ストランド相互作用は様々なβターンを構造化することができる。
ペプチドbhpBR3は高度の化学シフト分散、骨格及び側鎖結合定数の多くの極値、及びそのNMRスペクトル中に存在する多数のロングレンジNOEによって示されるように、溶液中で顕著に安定なコンホメーションを採る。bhpBR3の三次元構造は、予期されたように、BR3ターン配列がI型βターン構造を採るβヘアピンからなり、Arg30は正のφ角を採り、Asp26、Leu28、Val29、及びHis31の側鎖はβターンの面に突出している(図15B及び15C)。BR3が、TNFR及びDR5に対して観察されたもの(Hymowitzら, (1999) Mol. Cell 4 : 563-571)に相同の相互作用を使用してBAFFに結合するならば、この面(図15Cに示すターンの「底」面)がBAFFに接触することが予想される。
bhpBR3はBR3ターンを構造的に模倣するので、BAFFへの結合に対してミニBR3と競合するその能力を競合的置換ELISAで検査した(図16A)。顕著なことに、12残基ペプチドがより大きなコアドメインの結合を阻害し(IC50=15μM)、図15Cに示される6残基ターンに重要な結合決定因子が確かに存在していることを示している。更に、bhpBR3の活性を生物検定で確認した:bhpBR3は一次B細胞においてBAFF媒介NF-κB2/p52誘導をブロックしたが、コントロールヘアピンペプチドはブロックしなかった(図16B)。
BR3からのβターン構造がBAFFに対して有意な親和性を有しているという知見はその他のレセプターによるBAFFの認識に対して意味を持っている。TACI及びBCMAは、BR3のものに類似のターンコンホメーションを採ることが予想されるこのループ領域に相同的配列を共有している。従って、リガンドとのこのターンの相互作用はすべてのBAFF/レセプター複合体の保存された特徴であると思われる。そのような焦点の認識エピトープの驚くべき同定によりBAFF-BR3相互作用の小分子ペプチド模倣インヒビターを開発するための枠組みが提供されるであろう;これらのインヒビターは狼瘡のような自己免疫疾患の治療における治療薬となる可能性を有している。
本発明を必要的に好適な実施態様との関連で説明したが、当業者であれば、上述の明細書を読んだ後に、ここに記載された主題事項に、その精神と範囲から逸脱しないで、様々な変化、均等物との置換及び改変を施すことができるであろう。よって、本発明はここに特に記載したもの以外の方法で実施することができる。
Claims (18)
- βターンヘアピン構造を提示するための環状ペプチド骨格であって、アミノ酸配列C1-A1-A2-(A3)n-A4-A5-C2(配列番号:1)
(ここで、
C1とC2がシステインであり;
A1、A2、A3、A4及びA5が天然に生じるL−アミノ酸であり;
骨格のN'末端がアミノ保護基によって保護されていてもよく;
骨格のC'末端がカルボキシ保護基によって保護されていてもよく;
A1とA5がアミノ酸W、Y、F、H、I、V及びTからなる群から選択され;
A2とA4がアミノ酸W、Y、F、L、M、I、及びVからなる群から選択され;
A3が任意の天然に生じるL−アミノ酸であり、nが3、4、5、6、7、8、9、10、11、又は12の整数であり;
C1とC2がジスルフィド結合を形成して、環状ペプチドを形成する)
を含む環状ペプチド骨格。 - A1又はA5が、アミノ酸残基のβ炭素に二つの非水素置換基を有するβ分岐残基である、請求項1に記載の環状ペプチド骨格。
- A1又はA5がTである、請求項1に記載の環状ペプチド骨格。
- A1又はA5がアミノ酸W、F、H又はYである、請求項1に記載の環状ペプチド骨格。
- A1がHであり、A5がVである、請求項1に記載の環状ペプチド骨格。
- A2又はA4がアミノ酸W、F、又はYである、請求項1に記載の環状ペプチド骨格。
- A2とA4がWである、請求項6に記載の環状ペプチド骨格。
- A2とA4の双方がWである、請求項7に記載の環状ペプチド骨格。
- nが少なくとも4である、請求項1ないし8の何れか一項に記載の環状ペプチド骨格。
- nが10以下である、請求項9に記載の環状ペプチド骨格。
- nが4である、請求項9に記載の環状ペプチド骨格。
- (A3)4がEGNK、ENGK、QGSF又はVWQLである、請求項1ないし11の何れか一項に記載の環状ペプチド骨格。
- A1がT又はHであり、A2がW又はLであり、A4がW又はLであり、A5がT又はVである、請求項1に記載の環状ペプチド骨格。
- A1が、HとVからなる群から選択され、A5が、V、H及びIからなる群から選択される、請求項1に記載の環状ペプチド骨格。
- (A3)nがDLLVRHである、請求項1ないし14の何れか一項に記載の環状ペプチド骨格。
- アミノ酸配列がCHWDLLVRHWVC(配列番号33)である、請求項15に記載の環状ペプチド骨格。
- アミノ酸配列がCTWEGNKLTC(配列番号2)である、請求項13に記載の環状ペプチド骨格。
- アミノ酸配列がCHWEGNKLVC(配列番号30)である、請求項13に記載の環状ペプチド骨格。
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US10/271,343 US20030166003A1 (en) | 1999-06-14 | 2002-10-15 | Structured peptide scaffold for displaying turn libraries on phage |
| PCT/US2003/032450 WO2004035735A2 (en) | 2002-10-15 | 2003-10-14 | A structured peptide scaffold for displaying turn libraries on phage |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2006503088A true JP2006503088A (ja) | 2006-01-26 |
| JP2006503088A5 JP2006503088A5 (ja) | 2006-11-30 |
Family
ID=32106415
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2004544870A Withdrawn JP2006503088A (ja) | 2002-10-15 | 2003-10-14 | ターンライブラリーをファージ上にディスプレイするための構造化ペプチド骨格 |
Country Status (6)
| Country | Link |
|---|---|
| US (1) | US20030166003A1 (ja) |
| EP (1) | EP1558644A4 (ja) |
| JP (1) | JP2006503088A (ja) |
| AU (1) | AU2003301301A1 (ja) |
| CA (1) | CA2502243A1 (ja) |
| WO (1) | WO2004035735A2 (ja) |
Cited By (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2013513382A (ja) * | 2009-12-11 | 2013-04-22 | グワンジュ・インスティテュート・オブ・サイエンス・アンド・テクノロジー | 細胞内ターゲット結合用二座ペプチドバインダー |
| WO2014103203A1 (ja) | 2012-12-27 | 2014-07-03 | 独立行政法人産業技術総合研究所 | 微小タンパク質の骨格構造に基づく分子ライブラリ |
Families Citing this family (11)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20030095967A1 (en) | 1999-01-25 | 2003-05-22 | Mackay Fabienne | BAFF, inhibitors thereof and their use in the modulation of B-cell response and treatment of autoimmune disorders |
| UA83458C2 (uk) | 2000-09-18 | 2008-07-25 | Байоджен Айдек Ма Інк. | Виділений поліпептид baff-r (рецептор фактора активації в-клітин сімейства tnf) |
| AU2004233164B2 (en) | 2003-03-28 | 2009-10-08 | Biogen Ma Inc. | Truncated BAFF receptors |
| FR2898895B1 (fr) * | 2006-03-23 | 2012-04-06 | Univ Reims Champagne Ardenne | Cyclopeptide a activite anti-cancereuse derive du collagene de type iv |
| WO2008009085A1 (en) * | 2006-07-21 | 2008-01-24 | Cristália Produtos Químicos Farmacêuticos Ltda | Anti-inflammatory and antiallergic cyclic peptides |
| CA2657338C (en) * | 2006-07-21 | 2013-10-22 | Cristalia Produtos Quimicos Farmaceuticos Ltda. | Anti-inflammatory and antiallergic cyclic peptides |
| GB0913775D0 (en) * | 2009-08-06 | 2009-09-16 | Medical Res Council | Multispecific peptides |
| EP2764140B1 (en) | 2011-10-07 | 2017-08-30 | Bicycle Therapeutics Limited | Modulation of structured polypeptide specificity |
| GB201306623D0 (en) | 2013-04-11 | 2013-05-29 | Bicycle Therapeutics Ltd | Modulation of structured polypeptide specificity |
| EP3137482A1 (en) | 2014-05-02 | 2017-03-08 | MorphoSys AG | Peptide libraries |
| CA3159182A1 (en) * | 2019-11-21 | 2021-05-27 | Cameron PYE | Cell-permeable cyclic peptides and uses thereof |
Family Cites Families (16)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| NZ215865A (en) * | 1985-04-22 | 1988-10-28 | Commw Serum Lab Commission | Method of determining the active site of a receptor-binding analogue |
| US5223409A (en) * | 1988-09-02 | 1993-06-29 | Protein Engineering Corp. | Directed evolution of novel binding proteins |
| DK0564531T3 (da) * | 1990-12-03 | 1998-09-28 | Genentech Inc | Berigelsesfremgangsmåde for variantproteiner med ændrede bindingsegenskaber |
| US5885780A (en) * | 1991-07-19 | 1999-03-23 | University Of Utah | Method of obtaining small conformationally rigid conopeptides |
| US5830851A (en) * | 1993-11-19 | 1998-11-03 | Affymax Technologies N.V. | Methods of administering peptides that bind to the erythropoietin receptor |
| US5534615A (en) * | 1994-04-25 | 1996-07-09 | Genentech, Inc. | Cardiac hypertrophy factor and uses therefor |
| US5824483A (en) * | 1994-05-18 | 1998-10-20 | Pence Inc. | Conformationally-restricted combinatiorial library composition and method |
| US6100377A (en) * | 1994-06-10 | 2000-08-08 | The Trustees Of The University Of Pennsylvania | Constrained peptides |
| US5627024A (en) * | 1994-08-05 | 1997-05-06 | The Scripps Research Institute | Lambdoid bacteriophage vectors for expression and display of foreign proteins |
| US5556752A (en) * | 1994-10-24 | 1996-09-17 | Affymetrix, Inc. | Surface-bound, unimolecular, double-stranded DNA |
| US6475806B1 (en) * | 1995-06-07 | 2002-11-05 | Praecis Pharmaceuticals, Inc. | Anchor libraries and identification of peptide binding sequences |
| US6013458A (en) * | 1995-10-27 | 2000-01-11 | Molecumetics, Ltd. | Reverse-turn mimetics and methods relating thereto |
| US5866341A (en) * | 1996-04-03 | 1999-02-02 | Chugai Pharmaceutical Co., Ltd. | Compositions and methods for screening drug libraries |
| US5766905A (en) * | 1996-06-14 | 1998-06-16 | Associated Universities Inc. | Cytoplasmic bacteriophage display system |
| US6180343B1 (en) * | 1998-10-08 | 2001-01-30 | Rigel Pharmaceuticals, Inc. | Green fluorescent protein fusions with random peptides |
| CN1318084C (zh) * | 2000-05-26 | 2007-05-30 | 奥索-麦克尼尔药品公司 | 神经保护肽 |
-
2002
- 2002-10-15 US US10/271,343 patent/US20030166003A1/en not_active Abandoned
-
2003
- 2003-10-14 EP EP03808995A patent/EP1558644A4/en not_active Withdrawn
- 2003-10-14 AU AU2003301301A patent/AU2003301301A1/en not_active Abandoned
- 2003-10-14 JP JP2004544870A patent/JP2006503088A/ja not_active Withdrawn
- 2003-10-14 CA CA002502243A patent/CA2502243A1/en not_active Abandoned
- 2003-10-14 WO PCT/US2003/032450 patent/WO2004035735A2/en not_active Application Discontinuation
Cited By (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2013513382A (ja) * | 2009-12-11 | 2013-04-22 | グワンジュ・インスティテュート・オブ・サイエンス・アンド・テクノロジー | 細胞内ターゲット結合用二座ペプチドバインダー |
| WO2014103203A1 (ja) | 2012-12-27 | 2014-07-03 | 独立行政法人産業技術総合研究所 | 微小タンパク質の骨格構造に基づく分子ライブラリ |
| US9897611B2 (en) | 2012-12-27 | 2018-02-20 | National Institute Of Advanced Industrial Science And Technology | Molecule library constructed on the basis of backbone structure of microprotein |
| EP3489396A1 (en) | 2012-12-27 | 2019-05-29 | National Institute of Advanced Industrial Science and Technology | Molecule library constructed on the basis of backbone structure of microprotein |
Also Published As
| Publication number | Publication date |
|---|---|
| EP1558644A2 (en) | 2005-08-03 |
| WO2004035735A2 (en) | 2004-04-29 |
| CA2502243A1 (en) | 2004-04-29 |
| AU2003301301A1 (en) | 2004-05-04 |
| EP1558644A4 (en) | 2006-04-05 |
| WO2004035735A3 (en) | 2004-08-26 |
| AU2003301301A2 (en) | 2004-05-04 |
| US20030166003A1 (en) | 2003-09-04 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US20060110777A1 (en) | Structured peptide scaffold for displaying turn libraries on phage | |
| EP0610448B1 (en) | Peptide library and screening method | |
| JP4975324B2 (ja) | T細胞レセプターディスプレイ | |
| JP2008516210A (ja) | 治療、診断およびクロマトグラフィーに使用するためのタンパク質複合体 | |
| WO1991018980A1 (en) | Compositions and methods for identifying biologically active molecules | |
| US20080108789A1 (en) | DNA & protein binding miniature proteins | |
| JP2003159086A (ja) | 改良された提示ファージ | |
| JP2006503088A (ja) | ターンライブラリーをファージ上にディスプレイするための構造化ペプチド骨格 | |
| EP1689863B1 (en) | Template fixed beta-hairpin loop mimetics and their use in phage display | |
| US7229777B2 (en) | Hairpin peptides with a novel structural motif and methods relating thereto | |
| CA2576435A1 (en) | Determination of the 3d-structure of an extracellular domain of a b1 g-protein coupled receptor by nmr analysis | |
| Weiner et al. | Biological approaches to rational drug design | |
| US7943731B1 (en) | Dimerizing peptides | |
| Slusky et al. | Understanding Membrane Proteins. How to Design Inhibitors of Transmembrane Protein—Protein Interactions | |
| Egloff | Structure determination of neurotensin receptor 1 variants functionally expressed in e. coli | |
| EP1724347A2 (en) | DNA & protein binding miniature proteins | |
| Stewart | β-hairpin peptides and WW domains designed for selective recognition of oligonucleotides | |
| Chen | Construction and Characterization of Single-Chain DNA-Binding Proteins Derived from Phage 434 Repressor |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20061013 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20061013 |
|
| A761 | Written withdrawal of application |
Free format text: JAPANESE INTERMEDIATE CODE: A761 Effective date: 20070807 |