JP2004016131A - Dnaマイクロアレイおよびその解析方法 - Google Patents
Dnaマイクロアレイおよびその解析方法 Download PDFInfo
- Publication number
- JP2004016131A JP2004016131A JP2002177214A JP2002177214A JP2004016131A JP 2004016131 A JP2004016131 A JP 2004016131A JP 2002177214 A JP2002177214 A JP 2002177214A JP 2002177214 A JP2002177214 A JP 2002177214A JP 2004016131 A JP2004016131 A JP 2004016131A
- Authority
- JP
- Japan
- Prior art keywords
- dna
- probe
- carrier
- sequence
- binding
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
Images





Landscapes
- Apparatus Associated With Microorganisms And Enzymes (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
Abstract
【課題】DANマイクロアレイに代表されるプローブ担体により検体中の変異やプローブとのミスマッチを検出する際に、変異やミスマッチ部位の数が少ない場合でもこれを検出できる方法を提供すること。
【解決手段】プローブDNAと検体DNAとの間にある変異などに由来するミスマッチ部分の修飾に標識を有するヌクレオチドを使用して、ミスマッチ部分を検出可能とする。
【選択図】 図5
【解決手段】プローブDNAと検体DNAとの間にある変異などに由来するミスマッチ部分の修飾に標識を有するヌクレオチドを使用して、ミスマッチ部分を検出可能とする。
【選択図】 図5
Description
【0001】
【産業上の利用分野】
本発明はゲノムの多様性、遺伝子多型、SNPs等、変異またはミスマッチ塩基の検出方法に関するものである。
【0002】
【従来の技術】
従来、ゲノムの多様性を解析する方法の一つに、RFLPs法が上げられる。RFLPs法は、制限酵素で切断したDNA断片が、電気泳動で移動度の差として十分識別できるときは、複数のゲノム間または遺伝的背景を調べる上で、有効な方法であった。しかしながら、この方法では、例えば一塩基のミスマッチ等、移動度にまったく差が出ない場合などに十分な解析ができないことが多く、近年ではSNPs解析の重要性が広く認識されるなかで、その重要性は低いものとなっている。
【0003】
こうした中で、Affimetrix社のGeneChipなど、DNAマイクロアレイによる方法が近年注目されている。しかしながら、この方法は一つの変異塩基を検出するのに20個のマトリックスを使用してしまうため、その分のロスがある。また、ユーザ側にとっては、基板上のターゲットDNAが予め決められてしまうという制約もある。その他の方法としては、Nanogen社の印可電圧式ハイブリダイゼーション法や質量分析による方法もあるが、DNAマイクロアレイほど大規模な一括処理をすることができていない。
【0004】
【発明が解決しようとする課題】
本発明の目的は、DANマイクロアレイに代表されるプローブ担体により検体中の変異やプローブとのミスマッチを検出する際に、変異やミスマッチ部位の数が少ない場合でもこれを検出できる方法を提供することにある。
【0005】
【課題を解決するための手段】
本発明にかかる対照DNA配列に対する検体DNA配列における変化を検出するための方法は、
前記対照DNA配列から選択した配列に相補的な配列をプローブ配列とし、該プローブ配列を担体上の固定化領域に配置したプローブ担体を用意する工程と、
前記検体DNA配列を前記プローブ担体と反応させて前記プローブと該検体DNA配列とをハイブリダイズさせてこれらの配列から二本鎖体を得る工程と、
前記二本鎖体中に二本鎖を形成していない塩基がある場合に、該塩基の二本鎖体の一方の鎖にある側のヌクレオチドを該二本鎖体から除去して、鎖の切断部位を形成する工程と、
前記鎖の切断部位を検出可能な標識を有するヌクレオチドで修復する工程と、
前記修復された部位に挿入された標識を検出することで、前記検体DNA配列中における変化を検出する工程と
を有することを特徴とするものである。
【0006】
上記のプローブ配列としては、制限酵素の認識配列を介して担体に結合しているものが好適に利用でき、検体DNA配列が、プローブ配列の有する認識配列に対応する制限酵素によって処理された断片としてプローブ担体との反応に供給される方法を用いることができる。
【0007】
本発明にかかるプローブ結合用DNA担体の第1の態様は、担体上にDNAの固定化領域の複数を配置したDNA担体において、互いに異なる第1〜第nの制限酵素(n≧2)により形成される突出末端の配列と相補的な第1〜第nのプローブ結合用配列の各々を前記固定化領域ごとに配置したことを特徴とするプローブ結合用DNA担体である。
【0008】
本発明にかかるプローブ結合用DNA担体の第2の態様は、担体上にDNAの固定化領域の複数を配置したDNA担体において、任意の制限酵素により提供される突出末端の塩基数の範囲内でランダムに塩基を組み合わせた多数の塩基配列の各々を前記固定化領域ごとに配置したことを特徴とするプローブ結合用DNA担体である。
【0009】
また、本発明にかかる対照DNA配列に対する検体DNA配列における変化を検出するために用いるプローブ担体は、前記対照DNA配列に相補的配列を第1〜第nの制限酵素(n≧2)のそれぞれで処理して得られた第1〜第nのDNA断片の各々を対応する制限酵素の認識配列を介して担体に固定して各DNA断片ごとに固定化領域を形成したことを特徴とするプローブ担体である。
【0010】
本発明にかかるプローブ担体の製造方法の第1の態様は、上記のプローブ担体の製造方法であって、上記第1の態様にかかるプローブ結合用DNA担体の有するプローブ結合用配列に対応する第1〜第nの制限酵素のそれぞれを用いて前記対照DNA配列を有する二本鎖DNAを処理して得られるDNA断片をプローブとして用意する工程と、
これらのDNA断片を上記第1の態様にかかるプローブ結合用DNA担体と反応させ、各DNA断片の有する突出末端を、該プローブ結合用DNA担体上の対応するプローブ結合用配列とハイブリダイズさせて、該DNA断片を該担体上に固定する工程と、
該担体上に固定されたDNA断片を一本鎖化して、前記プローブ結合用配列を介して該担体に前記対照DNA配列由来の断片がプローブとして結合した構造を得る工程と
を有することを特徴とするプローブ担体の製造方法である。
【0011】
本発明にかかるプローブ担体の製造方法の第2の態様は、
上記第2の態様にかかるプローブ結合用DNA担体の有するプローブ結合用配列の塩基数に相当する突出末端を生じさせる制限酵素を用いて前記対照DNA配列を有する二本鎖DNAを処理して得られるDNA断片をプローブとして用意する工程と、
これらの上記第2の態様にかかるプローブ結合用DNA担体と反応させ、各DNA断片の有する突出末端を、該プローブ結合用DNA担体上の対応するプローブ結合用配列とハイブリダイズさせて、該DNA断片を該担体上に固定する工程と、
該担体上に固定されたDNA断片を一本鎖化して、前記プローブ結合用配列を介して該担体に前記対照DNA配列由来の断片がプローブとして結合した構造を得る工程と
を有することを特徴とするプローブ担体の製造方法である。
【0012】
本発明では、プローブ担体をDNAマイクロアレイとした場合の大規模一括処理という特長を活かすことができ、任意の制限酵素の認識切断部位と相補的な配列をターゲットDNAに用意し、ライゲーション反応によって、ある基準としたいDNA断片の鎖を継ぎ足すという工夫と、継ぎ足された新規のオリゴヌクレオチドと検体とのハイブリダイゼーション、および遺伝子の修復機構とを組み合わせ、応用することによって、たった1個のマトリックスで、変異またはミスマッチ塩基を検出することを可能とした。
【0013】
また従来のDNAマイクロアレイは大規模一括処理という特長はあっても、実際アレイを構成するターゲットDNA群をどのように選定するかについては、依然として難しい問題である。本発明による方法では、ランダムニックによる制限酵素を想定した場合、例えば突出末端部分を6塩基とすると、すべての場合の数を想定しても46=4096スポットにしかならず、現行の一般的なマイクロアレイサイズと比べても十分余裕で、尚且つその上にライゲーションによって実在する配列をすべて継ぎ足すことができるため、解析上、優位なマイクロアレイを提供することができる。
【0014】
なお、本発明においては、検体側のDNAをターゲット、マイクロアレイに固定化するDNAをプローブと表現した。
【0015】
【発明の実施の形態】
本発明にかかるプローブ担体は、検体に対する標準の配列を規定する対照DNA配列から選択された少なくとも1種(対照DNA配列全体あるいはこれから選択された部分配列)をプローブとして担体上に固定したもので、複数種のプローブをそれぞれ独立した領域、例えばドット状スポットとして担体表面に固定化したものである。このようなプローブの固定化領域が所定の間隔で配列されたものをプローブ・アレイという。このプローブ担体には、DNAマイクロアレイ、DNAチップ、プローブ・アレイと一般的に呼ばれている検査用等のプレートやチップが含まれる。
【0016】
一方、プローブは担体表面に結合可能な構造を有しており、担体上へのプローブの固定は固定用の官能基を用いて行われる好ましい。その際、プローブが有する担体表面に結合可能な構造は、アミノ基、スルフィドリル基、カルボキシル基、水酸基、酸ハライド化物(ハロホルミル基;−COX)、ハライド化物(−X)、アジリジン、マレイミド基、スクシイミド基、イソチオシアネート基、スルフォニルクロリド基(−SO2Cl)、アルデヒド基(ホルミル基;−CHO)、ヒドラジン、ヨウ化アセトアミド、チオール基などの有機官能基の少なくとも1種を導入する処理により形成されたものであることが好ましい。また、プローブ側の担体への結合に必要な構造に応じて、担体の表面に必要とされる処理を施してもよい。例えば、後述する実施例におけるような、基板側にマレイミド基を導入し、プローブ側にチオール基を導入してプローブの固定化を行う方法を好ましい方法として挙げることができる。
【0017】
本発明においては、固定化するプローブとしてのオリゴヌクレオチド(一本鎖)が、現在機能の知られている、入手可能な制限酵素の認識配列を介してプローブが担体に固定される。
【0018】
このプローブを担体に固定するためのプローブ結合用DNA担体は、多くの異なる制限酵素の認識配列、具体的には第1〜第nの制限酵素(n≧2)のそれぞれが生じる突出末端に相補的なプローブ結合用配列の複数を、好ましくはアレイ状に並べた構成とすることができる。そして、このプローブ結合用DNA担体に上記第1〜第nの制限酵素(n≧2)のそれぞれで対照DNA配列を含む二本鎖DNAを処理して得られた二本鎖DNA断片を反応させて、対応する第1〜第nの制限酵素(n≧2)のそれぞれが生じる突出末端に相補的なプローブ結合用配列とそれぞれ二本鎖を形成させ、必要に応じてリガーゼによる修復を行う。そして、この二本鎖を加熱および洗浄することによって一本鎖化することで、担体には制限酵素の認識配列を介して対照DNA配列に由来するプローブとしての配列が結合したプローブ担体を得ることができる。
【0019】
このプローブ担体を新しいDNAマイクロアレイ等として、これと比較したい検体を同様に制限酵素で処理したあと、これとハイブリダイゼーションを行う。もしもここで、2つの検体間にミスマッチがあった場合の検出方法は、以下のプロセスで構成される。
【0020】
ハイブリダイズ後、DNAグシコシラーゼを作用させてミスマッチ塩基を取り除く。次にAPエンドヌクレアーゼを作用させて鎖を切断する。予め蛍光標識されたdNTPsを混入させておき、DNAポリメラーゼβを作用させて、ミスマッチがあった所の塩基を蛍光標識された塩基に置き換えて修復することによって、蛍光検出できるようにした。
【0021】
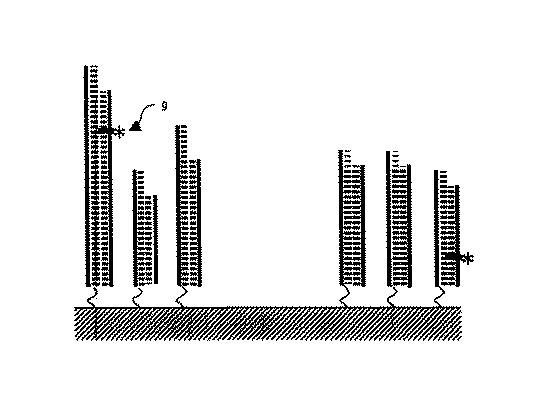
以下に、図を用いて本発明における各操作について具体的に説明する。図1〜5は本発明の実施形態を表したものである。まず、図1に示すとおり、オリゴヌクレオチド断片をいくつもスポット状に配置するための基板1(これはガラスや樹脂などが用いられ各スポットの形状はウェル状でも板状でもかまわない。例えばマイクロプレートや顕微鏡スライドガラス、シリコンウエハなどに相当する)の上に、あらかじめ用意された複数種のオリゴヌクレオチド断片を固相化する。オリゴヌクレオチド断片2とオリゴヌクレオチド断片3は、それぞれ異なるスポットを形成している。ここで各スポットのオリゴヌクレオチド断片は、図2(1)に図示のように、例えば、ある6カッターの制限酵素に相当する断片4−1、とある4カッターの制限酵素に相当する断片4−2・・・というように、現在機能の知られている制限酵素の認識部位に相補的なオリゴヌクレオチド断片で、複数のスポットが構成されることを特徴としたDNAアレイもしくはDNAマイクロアレイである。
【0022】
プローブ担体としてのアレイの構成にかかわる実施形態は、この他、DNase Iによるランダムニックに対応するように、まったく任意の配列、予め個々に決められた長さのオリゴヌクレオチド断片を複数のスポットに固相化したDNAアレイもしくはDNAマイクロアレイ(プローブ結合用担体の第1及び第2の実施態様を用いて形成されたもの)と、何の制約もない市販のDNAアレイもしくはDNAマイクロアレイと、アレイの構成に関して3つの実施形態が可能である。
【0023】
このようなDNAアレイに対して、mRNAを出発点としてds−cDNAを取得し、これを幾つかの制限酵素で切断し、これを該DNAアレイと図2の(1)に示されるような通常のライゲーション反応をT4 DNA リガーゼによって行う。もしもプロモーター領域や転写制御領域の解析が対象であるなら、プライマー伸長反応を併用することによってさらに上流のシークエンスを取得しても良いし、ゲノムを直接制限酵素で処理し、図2(1)のライゲーションに供給してもかまわない。ただしこのとき供給されるゲノムにはメチル化や変異によるウラシル化が予め刷り込まれている可能性があるので、解析が複雑である。図2(2)は、ライゲーション反応が終了した段階を示している。各スポットには、同じ切断部位認識配列をもつ、異なる長さのDNA断片が結合される。この場合の用途はスクリーニング的用途である。その他の実施形態としては、制限酵素で切断したDNAを電気泳動やHPLCにて分画したものを各スポットに反応させれば、同じ長さのDNA断片が結合される。図3の(1)は、十分なディネーチャーと洗浄操作により、一本鎖化したところを表した。この操作は、次の図3(2)(比較したいサンプルDNAの断片とのハイブリダイゼーション)と併せて行ってプロセスを短縮できるが、もともとのDNA断片と競合し、反応の確率が1/2になるので感度の点では不利である。サンプルDNAの断片は、組織、血液、培養細胞、菌体などから抽出し、さきほどのds−cDNAと同じ制限酵素処理によって得られたサンプルである。制限酵素の処理形態には、二つの実施形態がある。第一は、サンプルを予め小分けし、各制限酵素切断、完全に失活させたあと、もとの一つの容器に混合する方法で、この場合DNA断片の両側は同じ切断構造となるので、解析には有利な情報となる。第二は、幾つかの制限酵素はバッファーを共有できるため、一つの容器内で複数タイプの切断が可能である。さて、図3(2)では塩基配列のミスマッチや変異6を伴ったDNA断片がハイブリダイゼーションされたことを表している。未反応物を除去した後、グリコシラーゼ活性のある物質を使って、変異塩基、ミスマッチ塩基を除去する。ウラシル化による変異塩基では図4(1)に示されるようなabasicな状態、AP部位7を作るのにUracil−N−Glycosylaseが第一候補に上げられる。ミスマッチ塩基の場合には、第一候補としてExonucleaseIIIのグリコシラーゼ活性を利用できるが、その他にもAAG/MPG(alkil−adenine DNA gl
ycosylase)、TDG(Thymine(T:G)−missmatch DNA glycosylase)、hMYH(Adenin DNA glycosylase) 、hOGG1(8−oxo−guanine DNA glycosylase)、hNTH1(Thymineglycol DNA glycosylase)が利用できる。
【0024】
このようにしてできたAP部位に、APエンドヌクレアーゼ活性をもつHuman AP Endonuclease(APE)、もしくはExonucleaseIIIのAPエンドヌクレアーゼ活性を利用して、図4(2)に示されるような一塩基ギャップ8を作る。外れたフリーの塩基を分離後、次にHuman DNA Polymerase βと、蛍光標識されたdNTPs8を加えて、ギャップを埋めることによって、図5に示されるように、変異もしくはミスマッチを検出できるアッセイ系を構築することができる。蛍光標識されたdNTPsの蛍光色素は、たとえばCy Dyeシリーズ、FITC、ローダミン、テキサスレッドを使うことができる。また、DNA Polymerase β以外に、フリーの塩基を分離後、リガーゼを用いてリン酸ジエステル結合の鎖を修復することも可能である。この場合ペア塩基が何であれ、アミノアリルdUTPを使えば、上記の蛍光色素のほかにAlexaシリーズの蛍光色素も使える。ビオチン―アビジン結合を介したこれら蛍光標識物も同様に使用できる。またこれらの標識された塩基が蛍光色素ではなく、RI標識されたdNTPsであっても良い。
【0025】
【実施例】
これより実施例により本発明をより具体的に説明するが、DNAマイクロアレイにかかわる、cDNAの調製、PCR増幅、蛍光ラべリング、ハイブリダイゼーションといった基本的実験操作は、当業者によく知られた参考書、WEBサイト等に詳しく記載されている(参考:DNAマイクロアレイ実践マニュアル(羊土社、監修 林崎良英、編集 岡崎康司)、Molecular Biology Protocols(http://www.nwfsc.noaa.gov/protocols.htlm)など)ものであって、以下の説明により本発明は何等限定されるものではない。
【0026】
(A)切断部位認識配列の相補的な断片をもつDNAマイクロアレイの作成
表1に選択した制限酵素群に対応するアレイの構成を示した。5番から68番については、DNaseIを用い、塩基配列はAAAAから順番にすべての組み合わせに対応させた。表1の塩基配列をもつプローブDNAは、DNA自動合成機を用いて合成し、5’末端にはThiol−Modifier(GlenResearch社)を用いて−SH基を導入した。一方基板となるガラス板上には、シランカップリング剤とN−マレイミドカプロイロキシスクシンイミドによってマレイミド基を導入し、先ほどのプローブDNAを所定の濃度、Bufferに調製した後、インクジェットプリンタ(キャノン社)で基板上に打ち、化学的に結合、ブロッキング処理を行った。以上の工程の詳細は、特開平11−187900号公報の実施例を採用したものである。
【0027】
【表1】
 【0028】
【0028】
(B)制限酵素処理断片を継ぎ足したDNAマイクロアレイの作成
NHBE−Cell(気管支/気管上皮細胞、BioWhittaker社)からCell−to−DNAキット(Ambion社)を使ってDNAを抽出し、表1の5種類の制限酵素で処理できるよう小分けした。各試験管は、制限酵素に添付のBufferで溶解、切断した後、EDTA0.1mMを1滴滴下し、Mgイオンをマスキング、失活処理を70℃、30分行った。各制限酵素の処理時間および濃度は、カタログデータ完全分解に要する時間を参考にした。次にMicroSpin S−200 HR Columnsを用いて各溶液を脱塩、混合したあと、ライゲーション反応バッファー(Tris−HCl pH8.0、4mM MgCl2、10mM (NH4)2SO4、1.2mM EDTA、100uM NAD、0.005%BSA)の組成となるよう、20×濃縮バッファーを添加、E.coli DNA Ligaseを1Uになるように添加し、速やかに(A)のスライドグラス上で室温、45分間ライゲーションを行った。
【0029】
(C)制限酵素処理断片を継ぎ足した一本鎖DNAマイクロアレイの作成
ライゲーション終了後、スライドグラスを95℃の同バッファーに浸し、3〜4回液を交換しながらよく洗浄して一本鎖化したのち、4℃で保存した。
【0030】
(D)検体側の調整
Hep2−Cell(ヒト咽頭癌由来)からCell−to−DNAキット(Ambion社)を使ってDNAを抽出し、表1の5種類の制限酵素で処理できるよう小分けした。各試験管は、制限酵素に添付のBufferで溶解、切断した後、EDTA0.1mMを1滴滴下し、Mgイオンをマスキング、失活処理を70℃、30分行った。各制限酵素の処理時間および濃度は、カタログデータ完全分解に要する時間を参考にした。以上は(A)と同様に行った。
【0031】
(E)ハイブリダイゼーション
(D)の検体に、ホルムアミドが30%となるように添加し、95℃10分間一本鎖化を行った後、予め4℃に冷やしたスライドグラスの上に約数十〜数百ul滴下し、保湿反応チャンバー内で65℃2時間反応させた。反応終了後5×SSCバッファー(0.2%SDS)で洗浄し、そのあと1×SSCで3回洗浄した。
【0032】
(F)ミスマッチ塩基の除去
(E)のハイブリダイゼーション終了後のスライドグラスを予め10mM HEPES−KOH、pH7.4、10mM KCl、10mM EDTA の溶液に浸しておき、そこに1UnitとなるようにE.coli Endonuclease III(Trevigen Inc.)を加え、37℃1時間処理した。
【0033】
(G)1塩基ギャップ処理
(F)のミスマッチ塩基除去終了後のスライドグラスを予め50mM HEPES、pH7.5、50mM KCl、1ug/ml BSA、10mM MgCl2、0.05%TritonXの溶液に浸しておき、そこに1UnitとなるようにHuman AP Endonuclease(Trevigen Inc.)を加え、37℃1時間処理した。
【0034】
(H)蛍光標識修復
(G)の1塩基ギャップ処理終了後のスライドグラスを予め50mM Tris−HCl、pH8.8、10mM KCl、0.4mg/ml BSA、10mM MgCl2、1mM DTTの溶液に浸しておき、そこに1UnitとなるようにHuman DNA Polymerrase β(Trevigen Inc.)と、各50uMとなるようにCy5−dATP、dTTP、Cy3−dCTP、dGTPを加え、37℃1時間処理した。
【0035】
(I)洗浄および観察
(H)の蛍光標識修復終了後のスライドグラスを1×SSCで3回洗浄し、最後水に置換し、風乾したものを、GenePix4000Bで測定した。
【0036】
【発明の効果】
本発明のプローブ固定用の担体は、固定するプローブの種類の選択幅が広くユニバーサルなDNAチップを提供することを可能としている。すなわち、わざわざ専用のDNAチップを直接作るのではなく、もともとのわずかな配列にリガーゼで必要な配列を追加することで、任意のチップを作ることが出来る。また従来フルマッチと1塩基ミスマッチのハイブリダイゼーションは、殆ど差がなくその識別は大変困難なものであったが、DNAの修復機構を応用することによって、その識別を蛍光標識によって直接行うことが出来る。さらに応用として、ある塩基についてはDNAメチル化の修復が同じ方法で行われているので、適当な酵素を選択することによって、メチル化の有無、程度を知ることも本法で可能である。
【図面の簡単な説明】
【図1】プローブ担体調製用の基板の構成を示す図である。
【図2】(1)及び(2)は検体DNAの検出方法における各反応工程を示す図である。
【図3】(1)及び(2)は検体DNAの検出方法における各反応工程を示す図である。
【図4】(1)及び(2)は検体DNAの検出方法における各反応工程を示す図である。
【図5】検体DNAの検出方法における各反応工程を示す図である。
【符号の説明】
1 基板
2 スポット
3 スポット
4−1、4−2 制限酵素認識配列
5 対照DNA由来の制限酵素断片
6 ミスマッチ(変異)
7 AP部位
8 一塩基ギャップ
【産業上の利用分野】
本発明はゲノムの多様性、遺伝子多型、SNPs等、変異またはミスマッチ塩基の検出方法に関するものである。
【0002】
【従来の技術】
従来、ゲノムの多様性を解析する方法の一つに、RFLPs法が上げられる。RFLPs法は、制限酵素で切断したDNA断片が、電気泳動で移動度の差として十分識別できるときは、複数のゲノム間または遺伝的背景を調べる上で、有効な方法であった。しかしながら、この方法では、例えば一塩基のミスマッチ等、移動度にまったく差が出ない場合などに十分な解析ができないことが多く、近年ではSNPs解析の重要性が広く認識されるなかで、その重要性は低いものとなっている。
【0003】
こうした中で、Affimetrix社のGeneChipなど、DNAマイクロアレイによる方法が近年注目されている。しかしながら、この方法は一つの変異塩基を検出するのに20個のマトリックスを使用してしまうため、その分のロスがある。また、ユーザ側にとっては、基板上のターゲットDNAが予め決められてしまうという制約もある。その他の方法としては、Nanogen社の印可電圧式ハイブリダイゼーション法や質量分析による方法もあるが、DNAマイクロアレイほど大規模な一括処理をすることができていない。
【0004】
【発明が解決しようとする課題】
本発明の目的は、DANマイクロアレイに代表されるプローブ担体により検体中の変異やプローブとのミスマッチを検出する際に、変異やミスマッチ部位の数が少ない場合でもこれを検出できる方法を提供することにある。
【0005】
【課題を解決するための手段】
本発明にかかる対照DNA配列に対する検体DNA配列における変化を検出するための方法は、
前記対照DNA配列から選択した配列に相補的な配列をプローブ配列とし、該プローブ配列を担体上の固定化領域に配置したプローブ担体を用意する工程と、
前記検体DNA配列を前記プローブ担体と反応させて前記プローブと該検体DNA配列とをハイブリダイズさせてこれらの配列から二本鎖体を得る工程と、
前記二本鎖体中に二本鎖を形成していない塩基がある場合に、該塩基の二本鎖体の一方の鎖にある側のヌクレオチドを該二本鎖体から除去して、鎖の切断部位を形成する工程と、
前記鎖の切断部位を検出可能な標識を有するヌクレオチドで修復する工程と、
前記修復された部位に挿入された標識を検出することで、前記検体DNA配列中における変化を検出する工程と
を有することを特徴とするものである。
【0006】
上記のプローブ配列としては、制限酵素の認識配列を介して担体に結合しているものが好適に利用でき、検体DNA配列が、プローブ配列の有する認識配列に対応する制限酵素によって処理された断片としてプローブ担体との反応に供給される方法を用いることができる。
【0007】
本発明にかかるプローブ結合用DNA担体の第1の態様は、担体上にDNAの固定化領域の複数を配置したDNA担体において、互いに異なる第1〜第nの制限酵素(n≧2)により形成される突出末端の配列と相補的な第1〜第nのプローブ結合用配列の各々を前記固定化領域ごとに配置したことを特徴とするプローブ結合用DNA担体である。
【0008】
本発明にかかるプローブ結合用DNA担体の第2の態様は、担体上にDNAの固定化領域の複数を配置したDNA担体において、任意の制限酵素により提供される突出末端の塩基数の範囲内でランダムに塩基を組み合わせた多数の塩基配列の各々を前記固定化領域ごとに配置したことを特徴とするプローブ結合用DNA担体である。
【0009】
また、本発明にかかる対照DNA配列に対する検体DNA配列における変化を検出するために用いるプローブ担体は、前記対照DNA配列に相補的配列を第1〜第nの制限酵素(n≧2)のそれぞれで処理して得られた第1〜第nのDNA断片の各々を対応する制限酵素の認識配列を介して担体に固定して各DNA断片ごとに固定化領域を形成したことを特徴とするプローブ担体である。
【0010】
本発明にかかるプローブ担体の製造方法の第1の態様は、上記のプローブ担体の製造方法であって、上記第1の態様にかかるプローブ結合用DNA担体の有するプローブ結合用配列に対応する第1〜第nの制限酵素のそれぞれを用いて前記対照DNA配列を有する二本鎖DNAを処理して得られるDNA断片をプローブとして用意する工程と、
これらのDNA断片を上記第1の態様にかかるプローブ結合用DNA担体と反応させ、各DNA断片の有する突出末端を、該プローブ結合用DNA担体上の対応するプローブ結合用配列とハイブリダイズさせて、該DNA断片を該担体上に固定する工程と、
該担体上に固定されたDNA断片を一本鎖化して、前記プローブ結合用配列を介して該担体に前記対照DNA配列由来の断片がプローブとして結合した構造を得る工程と
を有することを特徴とするプローブ担体の製造方法である。
【0011】
本発明にかかるプローブ担体の製造方法の第2の態様は、
上記第2の態様にかかるプローブ結合用DNA担体の有するプローブ結合用配列の塩基数に相当する突出末端を生じさせる制限酵素を用いて前記対照DNA配列を有する二本鎖DNAを処理して得られるDNA断片をプローブとして用意する工程と、
これらの上記第2の態様にかかるプローブ結合用DNA担体と反応させ、各DNA断片の有する突出末端を、該プローブ結合用DNA担体上の対応するプローブ結合用配列とハイブリダイズさせて、該DNA断片を該担体上に固定する工程と、
該担体上に固定されたDNA断片を一本鎖化して、前記プローブ結合用配列を介して該担体に前記対照DNA配列由来の断片がプローブとして結合した構造を得る工程と
を有することを特徴とするプローブ担体の製造方法である。
【0012】
本発明では、プローブ担体をDNAマイクロアレイとした場合の大規模一括処理という特長を活かすことができ、任意の制限酵素の認識切断部位と相補的な配列をターゲットDNAに用意し、ライゲーション反応によって、ある基準としたいDNA断片の鎖を継ぎ足すという工夫と、継ぎ足された新規のオリゴヌクレオチドと検体とのハイブリダイゼーション、および遺伝子の修復機構とを組み合わせ、応用することによって、たった1個のマトリックスで、変異またはミスマッチ塩基を検出することを可能とした。
【0013】
また従来のDNAマイクロアレイは大規模一括処理という特長はあっても、実際アレイを構成するターゲットDNA群をどのように選定するかについては、依然として難しい問題である。本発明による方法では、ランダムニックによる制限酵素を想定した場合、例えば突出末端部分を6塩基とすると、すべての場合の数を想定しても46=4096スポットにしかならず、現行の一般的なマイクロアレイサイズと比べても十分余裕で、尚且つその上にライゲーションによって実在する配列をすべて継ぎ足すことができるため、解析上、優位なマイクロアレイを提供することができる。
【0014】
なお、本発明においては、検体側のDNAをターゲット、マイクロアレイに固定化するDNAをプローブと表現した。
【0015】
【発明の実施の形態】
本発明にかかるプローブ担体は、検体に対する標準の配列を規定する対照DNA配列から選択された少なくとも1種(対照DNA配列全体あるいはこれから選択された部分配列)をプローブとして担体上に固定したもので、複数種のプローブをそれぞれ独立した領域、例えばドット状スポットとして担体表面に固定化したものである。このようなプローブの固定化領域が所定の間隔で配列されたものをプローブ・アレイという。このプローブ担体には、DNAマイクロアレイ、DNAチップ、プローブ・アレイと一般的に呼ばれている検査用等のプレートやチップが含まれる。
【0016】
一方、プローブは担体表面に結合可能な構造を有しており、担体上へのプローブの固定は固定用の官能基を用いて行われる好ましい。その際、プローブが有する担体表面に結合可能な構造は、アミノ基、スルフィドリル基、カルボキシル基、水酸基、酸ハライド化物(ハロホルミル基;−COX)、ハライド化物(−X)、アジリジン、マレイミド基、スクシイミド基、イソチオシアネート基、スルフォニルクロリド基(−SO2Cl)、アルデヒド基(ホルミル基;−CHO)、ヒドラジン、ヨウ化アセトアミド、チオール基などの有機官能基の少なくとも1種を導入する処理により形成されたものであることが好ましい。また、プローブ側の担体への結合に必要な構造に応じて、担体の表面に必要とされる処理を施してもよい。例えば、後述する実施例におけるような、基板側にマレイミド基を導入し、プローブ側にチオール基を導入してプローブの固定化を行う方法を好ましい方法として挙げることができる。
【0017】
本発明においては、固定化するプローブとしてのオリゴヌクレオチド(一本鎖)が、現在機能の知られている、入手可能な制限酵素の認識配列を介してプローブが担体に固定される。
【0018】
このプローブを担体に固定するためのプローブ結合用DNA担体は、多くの異なる制限酵素の認識配列、具体的には第1〜第nの制限酵素(n≧2)のそれぞれが生じる突出末端に相補的なプローブ結合用配列の複数を、好ましくはアレイ状に並べた構成とすることができる。そして、このプローブ結合用DNA担体に上記第1〜第nの制限酵素(n≧2)のそれぞれで対照DNA配列を含む二本鎖DNAを処理して得られた二本鎖DNA断片を反応させて、対応する第1〜第nの制限酵素(n≧2)のそれぞれが生じる突出末端に相補的なプローブ結合用配列とそれぞれ二本鎖を形成させ、必要に応じてリガーゼによる修復を行う。そして、この二本鎖を加熱および洗浄することによって一本鎖化することで、担体には制限酵素の認識配列を介して対照DNA配列に由来するプローブとしての配列が結合したプローブ担体を得ることができる。
【0019】
このプローブ担体を新しいDNAマイクロアレイ等として、これと比較したい検体を同様に制限酵素で処理したあと、これとハイブリダイゼーションを行う。もしもここで、2つの検体間にミスマッチがあった場合の検出方法は、以下のプロセスで構成される。
【0020】
ハイブリダイズ後、DNAグシコシラーゼを作用させてミスマッチ塩基を取り除く。次にAPエンドヌクレアーゼを作用させて鎖を切断する。予め蛍光標識されたdNTPsを混入させておき、DNAポリメラーゼβを作用させて、ミスマッチがあった所の塩基を蛍光標識された塩基に置き換えて修復することによって、蛍光検出できるようにした。
【0021】
以下に、図を用いて本発明における各操作について具体的に説明する。図1〜5は本発明の実施形態を表したものである。まず、図1に示すとおり、オリゴヌクレオチド断片をいくつもスポット状に配置するための基板1(これはガラスや樹脂などが用いられ各スポットの形状はウェル状でも板状でもかまわない。例えばマイクロプレートや顕微鏡スライドガラス、シリコンウエハなどに相当する)の上に、あらかじめ用意された複数種のオリゴヌクレオチド断片を固相化する。オリゴヌクレオチド断片2とオリゴヌクレオチド断片3は、それぞれ異なるスポットを形成している。ここで各スポットのオリゴヌクレオチド断片は、図2(1)に図示のように、例えば、ある6カッターの制限酵素に相当する断片4−1、とある4カッターの制限酵素に相当する断片4−2・・・というように、現在機能の知られている制限酵素の認識部位に相補的なオリゴヌクレオチド断片で、複数のスポットが構成されることを特徴としたDNAアレイもしくはDNAマイクロアレイである。
【0022】
プローブ担体としてのアレイの構成にかかわる実施形態は、この他、DNase Iによるランダムニックに対応するように、まったく任意の配列、予め個々に決められた長さのオリゴヌクレオチド断片を複数のスポットに固相化したDNAアレイもしくはDNAマイクロアレイ(プローブ結合用担体の第1及び第2の実施態様を用いて形成されたもの)と、何の制約もない市販のDNAアレイもしくはDNAマイクロアレイと、アレイの構成に関して3つの実施形態が可能である。
【0023】
このようなDNAアレイに対して、mRNAを出発点としてds−cDNAを取得し、これを幾つかの制限酵素で切断し、これを該DNAアレイと図2の(1)に示されるような通常のライゲーション反応をT4 DNA リガーゼによって行う。もしもプロモーター領域や転写制御領域の解析が対象であるなら、プライマー伸長反応を併用することによってさらに上流のシークエンスを取得しても良いし、ゲノムを直接制限酵素で処理し、図2(1)のライゲーションに供給してもかまわない。ただしこのとき供給されるゲノムにはメチル化や変異によるウラシル化が予め刷り込まれている可能性があるので、解析が複雑である。図2(2)は、ライゲーション反応が終了した段階を示している。各スポットには、同じ切断部位認識配列をもつ、異なる長さのDNA断片が結合される。この場合の用途はスクリーニング的用途である。その他の実施形態としては、制限酵素で切断したDNAを電気泳動やHPLCにて分画したものを各スポットに反応させれば、同じ長さのDNA断片が結合される。図3の(1)は、十分なディネーチャーと洗浄操作により、一本鎖化したところを表した。この操作は、次の図3(2)(比較したいサンプルDNAの断片とのハイブリダイゼーション)と併せて行ってプロセスを短縮できるが、もともとのDNA断片と競合し、反応の確率が1/2になるので感度の点では不利である。サンプルDNAの断片は、組織、血液、培養細胞、菌体などから抽出し、さきほどのds−cDNAと同じ制限酵素処理によって得られたサンプルである。制限酵素の処理形態には、二つの実施形態がある。第一は、サンプルを予め小分けし、各制限酵素切断、完全に失活させたあと、もとの一つの容器に混合する方法で、この場合DNA断片の両側は同じ切断構造となるので、解析には有利な情報となる。第二は、幾つかの制限酵素はバッファーを共有できるため、一つの容器内で複数タイプの切断が可能である。さて、図3(2)では塩基配列のミスマッチや変異6を伴ったDNA断片がハイブリダイゼーションされたことを表している。未反応物を除去した後、グリコシラーゼ活性のある物質を使って、変異塩基、ミスマッチ塩基を除去する。ウラシル化による変異塩基では図4(1)に示されるようなabasicな状態、AP部位7を作るのにUracil−N−Glycosylaseが第一候補に上げられる。ミスマッチ塩基の場合には、第一候補としてExonucleaseIIIのグリコシラーゼ活性を利用できるが、その他にもAAG/MPG(alkil−adenine DNA gl
ycosylase)、TDG(Thymine(T:G)−missmatch DNA glycosylase)、hMYH(Adenin DNA glycosylase) 、hOGG1(8−oxo−guanine DNA glycosylase)、hNTH1(Thymineglycol DNA glycosylase)が利用できる。
【0024】
このようにしてできたAP部位に、APエンドヌクレアーゼ活性をもつHuman AP Endonuclease(APE)、もしくはExonucleaseIIIのAPエンドヌクレアーゼ活性を利用して、図4(2)に示されるような一塩基ギャップ8を作る。外れたフリーの塩基を分離後、次にHuman DNA Polymerase βと、蛍光標識されたdNTPs8を加えて、ギャップを埋めることによって、図5に示されるように、変異もしくはミスマッチを検出できるアッセイ系を構築することができる。蛍光標識されたdNTPsの蛍光色素は、たとえばCy Dyeシリーズ、FITC、ローダミン、テキサスレッドを使うことができる。また、DNA Polymerase β以外に、フリーの塩基を分離後、リガーゼを用いてリン酸ジエステル結合の鎖を修復することも可能である。この場合ペア塩基が何であれ、アミノアリルdUTPを使えば、上記の蛍光色素のほかにAlexaシリーズの蛍光色素も使える。ビオチン―アビジン結合を介したこれら蛍光標識物も同様に使用できる。またこれらの標識された塩基が蛍光色素ではなく、RI標識されたdNTPsであっても良い。
【0025】
【実施例】
これより実施例により本発明をより具体的に説明するが、DNAマイクロアレイにかかわる、cDNAの調製、PCR増幅、蛍光ラべリング、ハイブリダイゼーションといった基本的実験操作は、当業者によく知られた参考書、WEBサイト等に詳しく記載されている(参考:DNAマイクロアレイ実践マニュアル(羊土社、監修 林崎良英、編集 岡崎康司)、Molecular Biology Protocols(http://www.nwfsc.noaa.gov/protocols.htlm)など)ものであって、以下の説明により本発明は何等限定されるものではない。
【0026】
(A)切断部位認識配列の相補的な断片をもつDNAマイクロアレイの作成
表1に選択した制限酵素群に対応するアレイの構成を示した。5番から68番については、DNaseIを用い、塩基配列はAAAAから順番にすべての組み合わせに対応させた。表1の塩基配列をもつプローブDNAは、DNA自動合成機を用いて合成し、5’末端にはThiol−Modifier(GlenResearch社)を用いて−SH基を導入した。一方基板となるガラス板上には、シランカップリング剤とN−マレイミドカプロイロキシスクシンイミドによってマレイミド基を導入し、先ほどのプローブDNAを所定の濃度、Bufferに調製した後、インクジェットプリンタ(キャノン社)で基板上に打ち、化学的に結合、ブロッキング処理を行った。以上の工程の詳細は、特開平11−187900号公報の実施例を採用したものである。
【0027】
【表1】
(B)制限酵素処理断片を継ぎ足したDNAマイクロアレイの作成
NHBE−Cell(気管支/気管上皮細胞、BioWhittaker社)からCell−to−DNAキット(Ambion社)を使ってDNAを抽出し、表1の5種類の制限酵素で処理できるよう小分けした。各試験管は、制限酵素に添付のBufferで溶解、切断した後、EDTA0.1mMを1滴滴下し、Mgイオンをマスキング、失活処理を70℃、30分行った。各制限酵素の処理時間および濃度は、カタログデータ完全分解に要する時間を参考にした。次にMicroSpin S−200 HR Columnsを用いて各溶液を脱塩、混合したあと、ライゲーション反応バッファー(Tris−HCl pH8.0、4mM MgCl2、10mM (NH4)2SO4、1.2mM EDTA、100uM NAD、0.005%BSA)の組成となるよう、20×濃縮バッファーを添加、E.coli DNA Ligaseを1Uになるように添加し、速やかに(A)のスライドグラス上で室温、45分間ライゲーションを行った。
【0029】
(C)制限酵素処理断片を継ぎ足した一本鎖DNAマイクロアレイの作成
ライゲーション終了後、スライドグラスを95℃の同バッファーに浸し、3〜4回液を交換しながらよく洗浄して一本鎖化したのち、4℃で保存した。
【0030】
(D)検体側の調整
Hep2−Cell(ヒト咽頭癌由来)からCell−to−DNAキット(Ambion社)を使ってDNAを抽出し、表1の5種類の制限酵素で処理できるよう小分けした。各試験管は、制限酵素に添付のBufferで溶解、切断した後、EDTA0.1mMを1滴滴下し、Mgイオンをマスキング、失活処理を70℃、30分行った。各制限酵素の処理時間および濃度は、カタログデータ完全分解に要する時間を参考にした。以上は(A)と同様に行った。
【0031】
(E)ハイブリダイゼーション
(D)の検体に、ホルムアミドが30%となるように添加し、95℃10分間一本鎖化を行った後、予め4℃に冷やしたスライドグラスの上に約数十〜数百ul滴下し、保湿反応チャンバー内で65℃2時間反応させた。反応終了後5×SSCバッファー(0.2%SDS)で洗浄し、そのあと1×SSCで3回洗浄した。
【0032】
(F)ミスマッチ塩基の除去
(E)のハイブリダイゼーション終了後のスライドグラスを予め10mM HEPES−KOH、pH7.4、10mM KCl、10mM EDTA の溶液に浸しておき、そこに1UnitとなるようにE.coli Endonuclease III(Trevigen Inc.)を加え、37℃1時間処理した。
【0033】
(G)1塩基ギャップ処理
(F)のミスマッチ塩基除去終了後のスライドグラスを予め50mM HEPES、pH7.5、50mM KCl、1ug/ml BSA、10mM MgCl2、0.05%TritonXの溶液に浸しておき、そこに1UnitとなるようにHuman AP Endonuclease(Trevigen Inc.)を加え、37℃1時間処理した。
【0034】
(H)蛍光標識修復
(G)の1塩基ギャップ処理終了後のスライドグラスを予め50mM Tris−HCl、pH8.8、10mM KCl、0.4mg/ml BSA、10mM MgCl2、1mM DTTの溶液に浸しておき、そこに1UnitとなるようにHuman DNA Polymerrase β(Trevigen Inc.)と、各50uMとなるようにCy5−dATP、dTTP、Cy3−dCTP、dGTPを加え、37℃1時間処理した。
【0035】
(I)洗浄および観察
(H)の蛍光標識修復終了後のスライドグラスを1×SSCで3回洗浄し、最後水に置換し、風乾したものを、GenePix4000Bで測定した。
【0036】
【発明の効果】
本発明のプローブ固定用の担体は、固定するプローブの種類の選択幅が広くユニバーサルなDNAチップを提供することを可能としている。すなわち、わざわざ専用のDNAチップを直接作るのではなく、もともとのわずかな配列にリガーゼで必要な配列を追加することで、任意のチップを作ることが出来る。また従来フルマッチと1塩基ミスマッチのハイブリダイゼーションは、殆ど差がなくその識別は大変困難なものであったが、DNAの修復機構を応用することによって、その識別を蛍光標識によって直接行うことが出来る。さらに応用として、ある塩基についてはDNAメチル化の修復が同じ方法で行われているので、適当な酵素を選択することによって、メチル化の有無、程度を知ることも本法で可能である。
【図面の簡単な説明】
【図1】プローブ担体調製用の基板の構成を示す図である。
【図2】(1)及び(2)は検体DNAの検出方法における各反応工程を示す図である。
【図3】(1)及び(2)は検体DNAの検出方法における各反応工程を示す図である。
【図4】(1)及び(2)は検体DNAの検出方法における各反応工程を示す図である。
【図5】検体DNAの検出方法における各反応工程を示す図である。
【符号の説明】
1 基板
2 スポット
3 スポット
4−1、4−2 制限酵素認識配列
5 対照DNA由来の制限酵素断片
6 ミスマッチ(変異)
7 AP部位
8 一塩基ギャップ
Claims (8)
- 対照DNA配列に対する検体DNA配列における変化を検出するための方法であって、
前記対照DNA配列から選択した配列に相補的な配列をプローブ配列とし、該プローブ配列を担体上の固定化領域に配置したプローブ担体を用意する工程と、
前記検体DNA配列を前記プローブ担体と反応させて前記プローブと該検体DNA配列とをハイブリダイズさせてこれらの配列から二本鎖体を得る工程と、
前記二本鎖体中に二本鎖を形成していない塩基がある場合に、該塩基の二本鎖体の一方の鎖にある側のヌクレオチドを該二本鎖体から除去して、鎖の切断部位を形成する工程と、
前記鎖の切断部位を検出可能な標識を有するヌクレオチドで修復する工程と、
前記修復された部位に挿入された標識を検出することで、前記検体DNA配列中における変化を検出する工程と
を有することを特徴とする対照DNA配列に対する検体DNA配列における変化を検出するための方法。 - 前記プローブ配列が、制限酵素の認識配列を介して前記担体に結合しており、前記検体DNA配列が、該プローブ配列の有する認識配列に対応する制限酵素によって処理された断片として前記プローブ担体との反応に供給される請求項1に記載の方法。
- 担体上にDNAの固定化領域の複数を配置したDNA担体において、
互いに異なる第1〜第nの制限酵素(n≧2)により形成される突出末端の配列と相補的な第1〜第nのプローブ結合用配列の各々を前記固定化領域ごとに配置したことを特徴とするプローブ結合用DNA担体。 - 担体上にDNAの固定化領域の複数を配置したDNA担体において、
任意の制限酵素により提供される突出末端の塩基数の範囲内でランダムに塩基を組み合わせた多数の塩基配列の各々を前記固定化領域ごとに配置したことを特徴とするプローブ結合用DNA担体。 - 対照DNA配列に対する検体DNA配列における変化を検出するために用いるプローブ担体であって、
前記対照DNA配列に相補的配列を第1〜第nの制限酵素(n≧2)のそれぞれで処理して得られた第1〜第nのDNA断片の各々を対応する制限酵素の認識配列を介して担体に固定して各DNA断片ごとに固定化領域を形成したことを特徴とするプローブ担体。 - 請求項5に記載のプローブ担体の製造方法であって、
請求項2に記載されるプローブ結合用DNA担体の有するプローブ結合用配列に対応する第1〜第nの制限酵素のそれぞれを用いて前記対照DNA配列を有する二本鎖DNAを処理して得られるDNA断片をプローブとして用意する工程と、
これらのDNA断片を請求項2に記載されるプローブ結合用DNA担体と反応させ、各DNA断片の有する突出末端を、該プローブ結合用DNA担体上の対応するプローブ結合用配列とハイブリダイズさせて、該DNA断片を該担体上に固定する工程と、
該担体上に固定されたDNA断片を一本鎖化して、前記プローブ結合用配列を介して該担体に前記対照DNA配列由来の断片がプローブとして結合した構造を得る工程と
を有することを特徴とするプローブ担体の製造方法。 - 請求項5に記載のプローブ担体の製造方法であって、
請求項3に記載されるプローブ結合用DNA担体の有するプローブ結合用配列の塩基数に相当する突出末端を生じさせる制限酵素を用いて前記対照DNA配列を有する二本鎖DNAを処理して得られるDNA断片をプローブとして用意する工程と、
これらのDNA断片を請求項2に記載されるプローブ結合用DNA担体と反応させ、各DNA断片の有する突出末端を、該プローブ結合用DNA担体上の対応するプローブ結合用配列とハイブリダイズさせて、該DNA断片を該担体上に固定する工程と、
該担体上に固定されたDNA断片を一本鎖化して、前記プローブ結合用配列を介して該担体に前記対照DNA配列由来の断片がプローブとして結合した構造を得る工程と
を有することを特徴とするプローブ担体の製造方法。 - 請求項6または7に記載の製造方法により得られたことを特徴とするプローブ担体。
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2002177214A JP2004016131A (ja) | 2002-06-18 | 2002-06-18 | Dnaマイクロアレイおよびその解析方法 |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2002177214A JP2004016131A (ja) | 2002-06-18 | 2002-06-18 | Dnaマイクロアレイおよびその解析方法 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| JP2004016131A true JP2004016131A (ja) | 2004-01-22 |
Family
ID=31175299
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2002177214A Pending JP2004016131A (ja) | 2002-06-18 | 2002-06-18 | Dnaマイクロアレイおよびその解析方法 |
Country Status (1)
| Country | Link |
|---|---|
| JP (1) | JP2004016131A (ja) |
-
2002
- 2002-06-18 JP JP2002177214A patent/JP2004016131A/ja active Pending
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP6571895B1 (ja) | 核酸プローブ及びゲノム断片検出方法 | |
| US20190024141A1 (en) | Direct Capture, Amplification and Sequencing of Target DNA Using Immobilized Primers | |
| KR102307230B1 (ko) | 핵산의 다중 검출 | |
| US7892732B2 (en) | Method of performing PCR amplification on a microarray | |
| EP3129505B1 (en) | Methods for clonal replication and amplification of nucleic acid molecules for genomic and therapeutic applications | |
| US20070287151A1 (en) | Methods and Means for Nucleic Acid Sequencing | |
| US20130324419A1 (en) | Methods for nucleic acid capture and sequencing | |
| JP2007509629A (ja) | 二本鎖dnaの切断による複合核酸分析 | |
| US6692915B1 (en) | Sequencing a polynucleotide on a generic chip | |
| US20180073063A1 (en) | Reusable microarray compositions and methods | |
| CN116194591A (zh) | 序列转换反应 | |
| JP2004016131A (ja) | Dnaマイクロアレイおよびその解析方法 | |
| CN101280338B (zh) | 一种检测核酸多态性的核酸扩增方法 | |
| US11970736B2 (en) | Methods for accurate parallel detection and quantification of nucleic acids | |
| EP1207209A2 (en) | Methods using arrays for detection of single nucleotide polymorphisms | |
| JP2024035110A (ja) | 変異核酸の正確な並行定量するための高感度方法 | |
| WO2003070977A2 (en) | Method for detecting single nucleotide polymorphisms | |
| JP2002223761A (ja) | 標的核酸検出法およびそのための試薬 |