FR2851773A1 - New polynucleotides encoding proteins involved in spiramycin biosynthesis, useful for improving synthesis of macrolide antibiotics or for generating new hybrid macrolides - Google Patents
New polynucleotides encoding proteins involved in spiramycin biosynthesis, useful for improving synthesis of macrolide antibiotics or for generating new hybrid macrolides Download PDFInfo
- Publication number
- FR2851773A1 FR2851773A1 FR0302439A FR0302439A FR2851773A1 FR 2851773 A1 FR2851773 A1 FR 2851773A1 FR 0302439 A FR0302439 A FR 0302439A FR 0302439 A FR0302439 A FR 0302439A FR 2851773 A1 FR2851773 A1 FR 2851773A1
- Authority
- FR
- France
- Prior art keywords
- sep
- gene
- seq
- microorganism
- sequence
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/195—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria
- C07K14/36—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria from Actinomyces; from Streptomyces (G)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N1/00—Microorganisms, e.g. protozoa; Compositions thereof; Processes of propagating, maintaining or preserving microorganisms or compositions thereof; Processes of preparing or isolating a composition containing a microorganism; Culture media therefor
- C12N1/20—Bacteria; Culture media therefor
- C12N1/205—Bacterial isolates
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P19/00—Preparation of compounds containing saccharide radicals
- C12P19/44—Preparation of O-glycosides, e.g. glucosides
- C12P19/60—Preparation of O-glycosides, e.g. glucosides having an oxygen of the saccharide radical directly bound to a non-saccharide heterocyclic ring or a condensed ring system containing a non-saccharide heterocyclic ring, e.g. coumermycin, novobiocin
- C12P19/62—Preparation of O-glycosides, e.g. glucosides having an oxygen of the saccharide radical directly bound to a non-saccharide heterocyclic ring or a condensed ring system containing a non-saccharide heterocyclic ring, e.g. coumermycin, novobiocin the hetero ring having eight or more ring members and only oxygen as ring hetero atoms, e.g. erythromycin, spiramycin, nystatin
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12R—INDEXING SCHEME ASSOCIATED WITH SUBCLASSES C12C - C12Q, RELATING TO MICROORGANISMS
- C12R2001/00—Microorganisms ; Processes using microorganisms
- C12R2001/01—Bacteria or Actinomycetales ; using bacteria or Actinomycetales
- C12R2001/465—Streptomyces
Abstract
Description
<Desc/Clms Page number 1> <Desc / Clms Page number 1>
POLYPEPTIDES IMPLIQUES DANS LA BIOSYNTHESE DES SPIRAMYCINES. SEQUENCES NUCLEOTIDIOUES CODANT CES POLYPEPTIDES ET LEURS APPLICATIONS
La présente invention concerne l'isolement et l'identification de nouveaux gènes de la voie de biosynthèse des spiramycines et de nouveaux polypeptides impliqués dans cette biosynthèse. Elle est également relative à l'utilisation de ces gènes dans le but d'augmenter les taux de production et la pureté de la spiramycine produite. POLYPEPTIDES INVOLVED IN THE BIOSYNTHESIS OF SPIRAMYCINS. NUCLEOTIDIC SEQUENCES ENCODING THESE POLYPEPTIDES AND THEIR APPLICATIONS
The present invention relates to the isolation and identification of novel genes of the biosynthetic pathway of spiramycins and novel polypeptides involved in this biosynthesis. It also relates to the use of these genes in order to increase production rates and the purity of spiramycin produced.
L'invention a également trait à l'utilisation de ces gènes pour la construction de mutants pouvant conduire à la synthèse de nouveaux antibiotiques ou à des formes dérivées de spiramycines. L'invention concerne encore les molécules produites grâce à l'expression de ces gènes et enfin des compositions pharmacologiquement actives d'une molécule produite grâce à l'expression de tels gènes. The invention also relates to the use of these genes for the construction of mutants which can lead to the synthesis of new antibiotics or forms derived from spiramycins. The invention also relates to the molecules produced by the expression of these genes and finally pharmacologically active compositions of a molecule produced by the expression of such genes.
Les Streptomyces sont des bactéries filamenteuses Gram positif du sol. Elles jouent un rôle important dans la décomposition et la minéralisation des matières organiques grâce à la grande diversité des enzymes dégradatives qu'elles sécrètent. Elles présentent des phénomènes de différenciations morphologiques uniques chez les procaryotes s'accompagnant d'une différenciation métabolique caractérisée par la production de métabolites secondaires présentant une extraordinaire diversité de structures chimiques et d'activités biologiques. Parmi ces métabolites, on retrouve les spiramycines produites à l'état naturel par la bactérie Streptomyces ambofaciens. Streptomyces are Gram positive filamentous soil bacteria. They play an important role in the decomposition and mineralization of organic matter thanks to the great diversity of the degradative enzymes that they secrete. They present phenomena of morphological differentiation unique in prokaryotes accompanied by a metabolic differentiation characterized by the production of secondary metabolites with an extraordinary diversity of chemical structures and biological activities. These metabolites include spiramycins naturally produced by the bacterium Streptomyces ambofaciens.
La spiramycine est un antibiotique de la famille des macrolides utile aussi bien en médecine vétérinaire qu'en médecine humaine. Les macrolides sont caractérisés par la présence d'un cycle lactone sur lequel sont greffés un ou plusieurs sucres. Streptomyces ambofaciens produit à l'état naturel la spiramycine I, II et III (cf. Figure 1), cependant l'activité antibiotique de la spiramycine 1 est nettement supérieure à celle des Spiramycin is a macrolide antibiotic useful in both veterinary medicine and human medicine. Macrolides are characterized by the presence of a lactone ring on which are grafted one or more sugars. Streptomyces ambofaciens naturally produces spiramycin I, II and III (see Figure 1), however the antibiotic activity of spiramycin 1 is significantly higher than that of
<Desc/Clms Page number 2><Desc / Clms Page number 2>
spiramycines II et III (Liu et al., 1999). La molécule de spiramycine 1 est constituée d'un macro-cycle lactonique, appelle platénolide et de trois sucres : la forosamine, le mycaminose et le mycarose (cf. Figure 1). L'activité antibiotique des spiramycines est due à une inhibition de la synthèse protéique chez les procaryotes par un mécanisme impliquant la liaison de l'antibiotique au ribosome bactérien. Spiramycins II and III (Liu et al., 1999). The spiramycin 1 molecule consists of a lactone macro-cycle called platenolide and three sugars: forosamine, mycaminosis and mycarose (see Figure 1). The antibiotic activity of spiramycins is due to an inhibition of protein synthesis in prokaryotes by a mechanism involving the binding of the antibiotic to the bacterial ribosome.
Certains composés membres de la famille des macrolides et possédant également un cycle lactone ont donné lieu à des utilisations hors du domaine des antibiotiques. Certain compounds belonging to the macrolide family and also having a lactone ring have given rise to uses outside the field of antibiotics.
Ainsi le produit FK506 a des effets immunosuppresseurs et offre des perspectives d'application thérapeutique dans le domaine de la transplantation d'organe, de l'arthrite rhumatoïde et plus généralement dans des pathologies liées à des réactions autoimmunes. D'autres macrolides comme l'avermectine ont des activités insecticides et anti-helminthiques. Thus the product FK506 has immunosuppressive effects and offers prospects for therapeutic application in the field of organ transplantation, rheumatoid arthritis and more generally in pathologies related to autoimmune reactions. Other macrolides such as avermectin have insecticidal and helminthic activities.
De nombreuses voies de biosynthèse, concernant des antibiotiques appartenant à des classes variées ainsi que d'autres métabolites secondaires (pour une revue, Chater K. Many biosynthetic pathways, concerning antibiotics belonging to various classes as well as other secondary metabolites (for a review, Chater K.
1990), ont à ce jour déjà été étudiées chez les Streptomycètes. Cependant, la connaissance des voies de biosynthèse des spiramycines n'est que très partielle à cejour. 1990) have so far been studied in Streptomycetes. However, knowledge of the biosynthetic pathways of spiramycins is only very partial today.
La biosynthèse des spiramycines est un processus complexe comportant de nombreuses étapes et impliquant de nombreuses enzymes (Omura et al., 1979a, Omura et al., 1979b). Les spiramycines appartiennent à la large classe des polyketides qui regroupe des molécules complexes particulièrement abondantes dans les microorganismes du sol. Ces molécules sont regroupées non pas par analogie de structure mais par une certaine similarité au niveau d'étapes de leur voie de biosynthèse. The biosynthesis of spiramycins is a complex process involving many steps and involving many enzymes (Omura et al., 1979a, Omura et al., 1979b). Spiramycins belong to the broad class of polyketides which groups complex molecules particularly abundant in microorganisms of the soil. These molecules are grouped together not by analogy of structure but by a certain similarity in the steps of their biosynthetic pathway.
En effet, les polyketides sont produits par une série complexe de réactions mais possèdent en commun d'avoir dans leur voie de biosynthèse une série de réactions catalysées par une ou des enzymes appelées polyketides synthases (PKS). Chez Streptomyces ambofaciens, la biosynthèse du macro-cycle lactonique des spiramycines (platénolide) est réalisée par une série de huit modules codés par cinq gènes PKS différents (Kuhstoss S., 1996, brevet US 5,945,320). Les spiramycines sont obtenues à partir de ce cycle lactone. Cependant, les diverses étapes et enzymes impliquées dans la Indeed, the polyketides are produced by a complex series of reactions but have in common to have in their path of biosynthesis a series of reactions catalyzed by one or more enzymes called polyketide synthases (PKS). In Streptomyces ambofaciens, the biosynthesis of the lactam cyclic cycle of spiramycins (platenolide) is carried out by a series of eight modules coded by five different PKS genes (Kuhstoss S., 1996, US Patent 5,945,320). Spiramycins are obtained from this lactone ring. However, the various steps and enzymes involved in the
<Desc/Clms Page number 3><Desc / Clms Page number 3>
synthèse des sucres, ainsi que leur liaison au cycle lactone et la modification de ce cycle pour l'obtention des spiramycines restent encore inconnues à cejour. synthesis of sugars, as well as their linkage to the lactone cycle and the modification of this cycle to obtain spiramycins are still unknown today.
Le brevet US 5,514,544 décrit le clonage d'une séquence appelée srmR chez Streptomyces ambofaciens. Il est fait l'hypothèse dans ce brevet que le gène srmR code une protéine régulatrice de la transcription des gènes impliqués dans la biosynthèse des macrolides. US Patent 5,514,544 describes the cloning of a sequence called srmR in Streptomyces ambofaciens. It is hypothesized in this patent that the srmR gene encodes a protein regulating the transcription of genes involved in macrolide biosynthesis.
En 1987, Richardson et collaborateurs (Richardson et al., 1987) ont montré que la résistance à la spiramycine de S. ambofaciens est conférée par au moins trois gènes, ces derniers ont été appelés srmA, srmB et srmC. Le brevet US 4,886,757 décrit plus particulièrement la caractérisation d'un fragment d'ADN de S. ambofaciens contenant le gène srmC. Cependant la séquence de ce gène n'a pas été divulguée. En 1990, Richardson et collaborateurs (Richardson et al., 1990) ont posé l'hypothèse de l'existence de trois gènes de biosynthèse de la spiramycine proche de srmB. Le brevet US 5,098,837 rapporte le clonage de cinq gènes potentiellement impliqués dans la biosynthèse de la spiramycine. Ces gènes ont été baptisés srmD, srmE, srmF, srmG et srmH. In 1987, Richardson et al. (Richardson et al., 1987) showed that spiramycin resistance in S. ambofaciens is conferred by at least three genes, the latter called srmA, srmB and srmC. US Patent 4,886,757 describes more particularly the characterization of a DNA fragment of S. ambofaciens containing the srmC gene. However, the sequence of this gene has not been disclosed. In 1990, Richardson et al. (Richardson et al., 1990) hypothesized the existence of three spiramycin biosynthetic genes near srmB. US Patent 5,098,837 reports the cloning of five genes potentially involved in the biosynthesis of spiramycin. These genes have been named srmD, srmE, srmF, srmG and srmH.
Une des difficultés majeures dans la production de composés tels que les spiramycines réside dans le fait que de très grands volumes de fermentation sont nécessaires à la production d'une quantité relativement faible de produit. Il est donc souhaitable de pouvoir augmenter l'efficacité de production de telles molécules afin d'en diminuer les coûts de production. One of the major difficulties in producing compounds such as spiramycins is that very large volumes of fermentation are required to produce a relatively small amount of product. It is therefore desirable to be able to increase the production efficiency of such molecules in order to reduce their production costs.
La voie de biosynthèse des spiramycines est un processus complexe et il serait souhaitable d'identifier et de supprimer les réactions parasites qui pourraient exister au cours de ce processus. Le but d'une telle manipulation est d'obtenir un antibiotique plus pur et/ou une amélioration de la productivité. A ce sujet, Streptomyces ambofaciens produit à l'état naturel la spiramycine I, II et III (cf. Figure 1), cependant l'activité antibiotique de la spiramycine 1 est nettement supérieure à celle des spiramycines II et The biosynthetic pathway of spiramycins is a complex process and it would be desirable to identify and suppress the spurious reactions that may occur during this process. The purpose of such manipulation is to obtain a purer antibiotic and / or an improvement in productivity. In this regard, Streptomyces ambofaciens naturally produces spiramycin I, II and III (see Figure 1), however the antibiotic activity of spiramycin 1 is significantly higher than that of spiramycins II and
<Desc/Clms Page number 4><Desc / Clms Page number 4>
III (Liu et al., 1999). Il serait donc souhaitable de pouvoir disposer de souches ne produisant que de la spiramycine I. III (Liu et al., 1999). It would therefore be desirable to have strains producing only spiramycin I.
En raison de l'intérêt commercial des antibiotiques macrolides, l'obtention de nouveaux dérivés, notamment d'analogues de spiramycines ayant des propriétés avantageuses est intensivement recherchée. Il serait souhaitable de pouvoir générer en quantité suffisante les intermédiaires de biosynthèse de la voie de biosynthèse des spiramycines ou des dérivés des spiramycines notamment dans le but de fabriquer des antibiotiques hybrides dérivés des spiramycines. Because of the commercial interest of macrolide antibiotics, obtaining new derivatives, especially spiramycin analogues having advantageous properties is intensively sought. It would be desirable to be able to generate in sufficient quantity the biosynthetic intermediates of the biosynthetic pathway of spiramycins or derivatives of spiramycins in particular for the purpose of producing hybrid antibiotics derived from spiramycins.
Description générale de l'invention
La présente invention résulte du clonage de gènes dont le produit intervient dans la biosynthèse des spiramycines. L'invention concerne tout d'abord de nouveaux gènes de la voie de biosynthèse des spiramycines et de nouveaux polypeptides impliqués dans cette biosynthèse. General description of the invention
The present invention results from the cloning of genes whose product is involved in the biosynthesis of spiramycins. The invention firstly relates to novel genes of the spiramycin biosynthetic pathway and novel polypeptides involved in this biosynthesis.
Les gènes de la voie de biosynthèse et les séquences codantes associées ont été clonés et la séquence d'ADN de chacun a été déterminée. Les séquences codantes
![]()
clonées seront désignées par la suite o/7*c, orf2*c, orf3 *c, orf4*c, or/3* or/6*, orf7*c, or/8*, or/9*, orflO*, or/7, or/2, or/3, orf4, orf5, or/6, orf7, or/8, orj9c, or/10, orf11c, or/12, orfl3c, orfl4, orf15c, or/16, or/17, or/18, or/19, or/20, or/2 le, orf22c, orj23c,
![]()
or/24c, orj25c, or/26, or/27, orj28c, or/29, orf30c, or/31 et orj32c. La fonction des protéines codées par ces séquences dans la voie de biosynthèse des spiramycines est développée dans la discussion ci-après qui est illustrée par les figures 4, 5, 6 et 8. The genes of the biosynthetic pathway and the associated coding sequences were cloned and the DNA sequence of each was determined. Coding sequences
![]()
cloned will be referred to as o / 7 * c, orf2 * c, orf3 * c, orf4 * c, or / 3 * or / 6 *, orf7 * c, or / 8 *, or / 9 *, orflO *, gold / 7, gold / 2, gold / 3, orf4, orf5, gold / 6, gold7, gold / 8, gold9c, gold / 10, gold11c, gold / 12, goldfl3c, goldfl4, gold15c, gold / 16, gold / 17, gold / 18, gold / 19, gold / 20, gold / 2 le, orf22c, orj23c,
![]()
or / 24c, orj25c, or / 26, or / 27, orj28c, or / 29, orf30c, or / 31 and orj32c. The function of the proteins encoded by these sequences in the spiramycin biosynthetic pathway is developed in the following discussion which is illustrated by Figures 4, 5, 6 and 8.
1) Un premier objet de l'invention concerne un polynucléotide codant un polypeptide impliqué dans la biosynthèse des spiramycines caractérisé en ce que la séquence dudit polynucléotide est : 1) A first subject of the invention relates to a polynucleotide encoding a polypeptide involved in spiramycin biosynthesis, characterized in that the sequence of said polynucleotide is:
<Desc/Clms Page number 5><Desc / Clms Page number 5>
(a) l'une des séquences SEQ ID N 3,5, 7,9, 11, 13,15, 17, 19, 21, 23, 25, 28, 30, 34, 36, 40, 43, 45, 47, 49, 53, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 107, 109, 111, 113,115, 118, et 120, (b) l'une des séquences constituées par les variants des séquences (a), (c) l'une des séquences dérivées des séquences (a) et (b) en raison de la dégénérescence du code génétique. (a) one of SEQ ID NO: 3.5, 7.9, 11, 13, 15, 17, 19, 21, 23, 25, 28, 30, 34, 36, 40, 43, 45, 47 , 49, 53, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 107, 109, 111, 113, 115, 118, and 120, (b) one of sequences constituted by the variants of the sequences (a), (c) one of the sequences derived from the sequences (a) and (b) because of the degeneracy of the genetic code.
2) La présente invention a également pour objet un polynucléotide s'hybridant dans des conditions d'hybridation de forte stringence à au moins l'un des polynucléotides selon le paragraphe 1) ci-dessus. 2) The subject of the present invention is also a polynucleotide hybridizing under hybridization conditions of high stringency to at least one of the polynucleotides according to paragraph 1) above.
3) L'invention concerne également un polynucléotide présentant au moins 70%, 80%, 85%, 90%, 95% ou 98% d'identité en nucléotides avec un polynucléotide comprenant au moins 10,12, 15, 18, 20 à 25, 30, 40, 50,60, 70,80, 90,100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000, 1050, 1100, 1150, 1200, 1250, 1300,1350, 1400,1450, 1500,1550, 1600,1650, 1700, 1750, 1800,1850 ou 1900 nucléotides consécutifs d'un polynucléotide selon le paragraphe 1) ci dessus. 3) The invention also relates to a polynucleotide having at least 70%, 80%, 85%, 90%, 95% or 98% nucleotide identity with a polynucleotide comprising at least 10, 12, 15, 18, 20 to 25, 30, 40, 50,60, 70,80, 90,100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000, 1050, 1100, 1150, 1200, 1250, 1300, 1350, 1400, 1450, 1500, 1550, 1600, 1650, 1700, 1750, 1800, 1850 or 1900 consecutive nucleotides of a polynucleotide according to paragraph 1) above .
4) L'invention concerne également un polynucléotide selon le pragraphe 2) ou 3) ci-dessus caractérisé en ce qu'il est isolé à partir d'une bactérie du genre Streptomyces. 4) The invention also relates to a polynucleotide according to paragraph 2) or 3) above characterized in that it is isolated from a bacterium of the genus Streptomyces.
5) L'invention concerne également un polynucléotide selon le pragraphe 2), 3) ou 4) ci-dessus caractérisé en ce qu'il code une protéine impliquée dans la biosynthèse d'un macrolide. 5) The invention also relates to a polynucleotide according to paragraph 2), 3) or 4) above characterized in that it encodes a protein involved in the biosynthesis of a macrolide.
6) L'invention concerne également un polynucléotide selon le pragraphe 2), 3) ou 4) ci-dessus caractérisé en ce qu'il code une protéine ayant une activité similaire à la protéine codée par le polynucléotide avec lequel il s'hybride ou il présente l'identité. 6) The invention also relates to a polynucleotide according to paragraph 2), 3) or 4) above, characterized in that it encodes a protein having a similar activity to the protein encoded by the polynucleotide with which it hybridizes or he presents the identity.
7) L'invention concerne également un polypeptide résultant de l'expression d'un polynucléotide selon le pragraphe 1), 2), 3), 4), 5) ou 6) ci-dessus. 7) The invention also relates to a polypeptide resulting from the expression of a polynucleotide according to paragraph 1), 2), 3), 4), 5) or 6) above.
<Desc/Clms Page number 6> <Desc / Clms Page number 6>
8) Un autre aspect de l'invention concerne un polypeptide impliqué dans la biosynthèse des spiramycines caractérisée en ce que la séquence dudit polypeptide est : (a) l'une des séquences SEQ ID N 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 27, 29,31,32,33,35,37,38,39,41,42,44,46,48,50,51,52,54,55,56,57,58,59,61, 63, 65, 67, 69, 71, 73, 75, 77,-79, 81, 83, 85, 108, 110, 112, 114, 116, 117, 119 et 121, (b) l'une des séquences telles que définies en (a) excepté que tout au long de ladite séquence un ou plusieurs acides aminés ont été substitués, insérés ou délétés sans en affecter les propriétés fonctionnelles, (c) l'une des séquences constituées par les variants des séquences (a) et (b). 8) Another aspect of the invention relates to a polypeptide involved in spiramycin biosynthesis characterized in that the sequence of said polypeptide is: (a) one of SEQ ID N 4, 6, 8, 10, 12, 14 , 16, 18, 20, 22, 24, 26, 27, 29,31,32,33,35,37,38,39,41,42,44,46,48,50,51,52,54,55 , 56, 57, 58, 59, 61, 63, 65, 67, 69, 71, 73, 75, 77, -79, 81, 83, 85, 108, 110, 112, 114, 116, 117, 119 and 121, (b) one of the sequences as defined in (a) except that throughout said sequence one or more amino acids have been substituted, inserted or deleted without affecting its functional properties, (c) one of sequences consisting of the variants of sequences (a) and (b).
9) Un autre objet de l'invention concerne un polypeptide présentant au moins 70%, 80%, 85%, 90%, 95% ou 98% d'identité en acides aminés avec un polypeptide comprenant au moins 10,15, 20,30 à 40,50, 60,70, 80,90,100, 120,140, 160, 180, 200, 220, 240, 260, 280, 300, 320, 340, 360, 380, 400, 420, 440, 460, 480,500, 520, 540,560, 580,600, 620 ou 640 acides aminés consécutifs d'un polypeptide selon le paragraphe 8) ci-dessus. 9) Another subject of the invention relates to a polypeptide having at least 70%, 80%, 85%, 90%, 95% or 98% amino acid identity with a polypeptide comprising at least 10, 15, 20, 30 to 40.50, 60.70, 80.90, 100, 120, 140, 160, 180, 200, 220, 240, 260, 280, 300, 320, 340, 360, 380, 400, 420, 440, 460, 480, 500, 520, 540,560, 580,600, 620 or 640 consecutive amino acids of a polypeptide according to paragraph 8) above.
10) Un autre aspect de l'invention concerne un polypeptide selon le paragraphe 9) ci-dessus caractérisé en ce qu'il est isolé à partir d'une bactérie du genre Streptomyces. Another aspect of the invention relates to a polypeptide according to paragraph 9) above characterized in that it is isolated from a bacterium of the genus Streptomyces.
11) Un autre aspect de l'invention concerne un polypeptide selon le paragraphe 9) ou 10) ci-dessus caractérisé en ce qu'il code une protéine impliquée dans la biosynthèse d'un macrolide. 11) Another aspect of the invention relates to a polypeptide according to paragraph 9) or 10) above characterized in that it encodes a protein involved in the biosynthesis of a macrolide.
12) Un autre aspect de l'invention concerne un polypeptide selon le pragraphe 9), 10) ou 11) ci-dessus caractérisé en ce qu'il a une activité similaire à celle du polypeptide avec lequel il partage l'identité. 12) Another aspect of the invention relates to a polypeptide according to paragraph 9), 10) or 11) above characterized in that it has an activity similar to that of the polypeptide with which it shares the identity.
13) Un autre aspect de l'invention concerne un ADN recombinant caractérisé en ce qu'il comprend au moins un polynucléotide selon l'un des paragraphes 1), 2), 3), 4), 5) ou 6) ci-dessus. 13) Another aspect of the invention relates to a recombinant DNA characterized in that it comprises at least one polynucleotide according to one of paragraphs 1), 2), 3), 4), 5) or 6) above .
<Desc/Clms Page number 7> <Desc / Clms Page number 7>
14) Un autre aspect de l'invention concerne un ADN recombinant selon le pragraphe 13) ci-dessus caractérisé en ce que le dit ADN recombinant est compris dans un vecteur. 14) Another aspect of the invention relates to a recombinant DNA according to paragraph 13) above characterized in that said recombinant DNA is included in a vector.
15) Un autre aspect de l'invention concerne un ADN recombinant selon le paragraphe 14) ci-dessus caractérisé en ce que ledit vecteur est choisi parmi les bactériophages, les plasmides, les phagemides, les vecteurs intégratifs, les fosmides, les cosmides, les vecteurs navettes, les BAC ou les PAC. Another aspect of the invention relates to a recombinant DNA according to paragraph 14) above characterized in that said vector is selected from bacteriophages, plasmids, phagemids, integrative vectors, fosmids, cosmids, shuttle vectors, BACs or PACs.
16) Un autre aspect de l'invention concerne un ADN recombinant selon le pragraphe 15) ci-dessus caractérisé en ce qu'il est choisi parmi pOS49.1, pOS49.11, pOSC49. 12, pOS49. 14, pOS49. 16, pOS49. 28, pOS44. 1, pOS44. 2, pOS44. 4, pSPM5, pSPM7, pOS49. 67, pOS49. 88, pOS49. 106, pOS49. 120, pOS49. 107, pOS49.32, pOS49. 43, pOS49. 44, pOS49. 50, pOS49. 99, pSPM17, pSPM21, pSPM502, pSPM504, pSPM507, pSPM508, pSPM509, pSPMl, pBXLllll, pBXL1112, pBXL1113, pSPM520, pSPM521, pSPM522, pSPM523, pSPM524, pSPM525, pSPM527, pSPM528, pSPM34, pSPM35, pSPM36, pSPM37, pSPM38, pSPM39, pSPM40, pSPM41, pSPM42, pSPM43, pSPM44, pSPM45, pSPM47, pSPM48, pSPM50, pSPM51, pSPM52, pSPM53, pSPM55, pSPM56, pSPM58, pSPM72, pSPM73, pSPM515, pSPM519, pSPM74 et pSPM75. 16) Another aspect of the invention relates to a recombinant DNA according to paragraph 15) above, characterized in that it is selected from pOS49.1, pOS49.11, pOSC49. 12, pOS49. 14, pOS49. 16, pOS49. 28, pOS44. 1, pOS44. 2, pOS44. 4, pSPM5, pSPM7, pOS49. 67, pOS49. 88, pOS49. 106, pOS49. 120, pOS49. 107, pOS49.32, pOS49. 43, pOS49. 44, pOS49. 50, pOS49. 99, pSPM17, pSPM21, pSPM502, pSPM504, pSPM507, pSPM508, pSPM509, pSPM1, pBXLllll, pBXL1112, pBXL1113, pSPM520, pSPM521, pSPM522, pSPM523, pSPM524, pSPM525, pSPM527, pSPM528, pSPM34, pSPM35, pSPM36, pSPM37, pSPM38, pSPM39, pSPM40, pSPM41, pSPM42, pSPM43, pSPM44, pSPM45, pSPM47, pSPM48, pSPM50, pSPM51, pSPM52, pSPM53, pSPM55, pSPM56, pSPM58, pSPM72, pSPM73, pSPM515, pSPM519, pSPM74 and pSPM75.
17) Un autre aspect de l'invention concerne un vecteur d'expression caractérisé en ce qu'il comprend au moins une séquence d'acide nucléique codant un polypeptide selon le pragraphe 7), 8), 9), 10), 11) ou 12) ci-dessus. 17) Another aspect of the invention relates to an expression vector characterized in that it comprises at least one nucleic acid sequence encoding a polypeptide according to paragraph 7), 8), 9), 10), 11) or 12) above.
18) L'invention est également relative à un système d'expression comprenant un vecteur d'expression approprié et une cellule hôte permettant l'expression d'un ou plusieurs polypeptides selon le paragraphe 7), 8), 9), 10), 11) ou 12) ci-dessus. 18) The invention also relates to an expression system comprising an appropriate expression vector and a host cell for the expression of one or more polypeptides according to paragraph 7), 8), 9), 10), 11) or 12) above.
19) L'invention est également relative à un système d'expression selon le paragraphe 18 ci-dessus caractérisé en ce qu'il est choisi parmi les systèmes d'expression procaryotes ou les systèmes d'expression eucaryotes. 19) The invention also relates to an expression system according to paragraph 18 above, characterized in that it is chosen from prokaryotic expression systems or eukaryotic expression systems.
<Desc/Clms Page number 8> <Desc / Clms Page number 8>
20) L'invention est également relative à un système d'expression selon le paragraphe 19) ci-dessus caractérisé en ce qu'il est choisi parmi les systèmes d'expression chez la bactérie E. coli, les sytèmes d'expression baculovirus permettant une expression dans des cellules d'insectes, les sytèmes d'expression permettant une expression dans des cellules de levures, les sytèmes d'expression permettant une expression dans des cellules de mammifères. 20) The invention also relates to an expression system according to paragraph 19) above characterized in that it is selected from the expression systems in the bacterium E. coli, the baculovirus expression systems allowing expression in insect cells, expression systems allowing expression in yeast cells, expression systems allowing expression in mammalian cells.
21) L'invention est également relative à une cellule hôte dans laquelle a été introduit au moins un polynucléotide et/ou au moins un ADN recombinant et/ou au moins un vecteur d'expression selon l'un des pragraphes 1), 2), 3), 4), 5), 6), 13), 14), 15), 16) ou 17) ci-dessus. 21) The invention also relates to a host cell into which at least one polynucleotide and / or at least one recombinant DNA and / or at least one expression vector has been introduced according to one of the following steps: 1), 2) , 3), 4), 5), 6), 13), 14), 15), 16) or 17) above.
22) L'invention est également relative à un procédé de production d'un polypeptide selon le paragraphe 7), 8), 9), 10), 11) ou 12) ci-dessus caractérisé en ce que ledit procédé comprend les étapes suivantes: a) insérer au moins un acide nucléique codant ledit polypeptide dans un vecteur approprié ; b) cultiver, dans un milieu de culture approprié, une cellule hôte préalablement transformée ou transfectée avec le vecteur de l'étape a) ; c) récupérer le milieu de culture conditionné ou un extrait cellulaire; d) séparer et purifier à partir dudit milieu de culture ou encore à partir de l'extrait cellulaire obtenu à l'étape c), ledit polypeptide ; e) le cas échéant, caractériser le polypeptide recombinant produit. 22) The invention also relates to a method for producing a polypeptide according to paragraph 7), 8), 9), 10), 11) or 12) above characterized in that said method comprises the following steps a) inserting at least one nucleic acid encoding said polypeptide into a suitable vector; b) culturing, in a suitable culture medium, a host cell previously transformed or transfected with the vector of step a); c) recovering the conditioned culture medium or a cell extract; d) separating and purifying from said culture medium or from the cell extract obtained in step c), said polypeptide; e) where appropriate, characterizing the recombinant polypeptide produced.
23) Un autre aspect de l'invention concerne un microorganisme bloqué dans une étape de la voie de biosynthèse d'au moins un macrolide. 23) Another aspect of the invention relates to a microorganism blocked in a step of the biosynthetic pathway of at least one macrolide.
24) Un autre aspect de l'invention concerne un microorganisme selon le paragraphe 23) ci-dessus caractérisé en ce qu'il est obtenu en inactivant la fonction d'au moins une protéine impliquée dans la biosynthèse de ce ou de ces macrolides dans un microorganisme producteur de ce ou de ces macrolide. 24) Another aspect of the invention relates to a microorganism according to paragraph 23) above characterized in that it is obtained by inactivating the function of at least one protein involved in the biosynthesis of this or these macrolides in a microorganism producing this or these macrolide.
<Desc/Clms Page number 9> <Desc / Clms Page number 9>
25) Un autre aspect de l'invention concerne un microorganisme selon le paragraphe 24) ci-dessus caractérisé en ce que l'inactivation de cette ou de ces protéines est effectuée par mutagénèse dans le ou les gènes codant ladite ou lesdites protéines ou par expression d'un ou de plusieurs ARN anti-sens complémentaires de l'ARN ou des ARN messagers codant ladite ou lesdites protéines. 25) Another aspect of the invention relates to a microorganism according to paragraph 24) above characterized in that the inactivation of this or these proteins is carried out by mutagenesis in the gene or genes encoding said protein or by expression one or more antisense RNAs complementary to the RNA or messenger RNAs encoding said one or more proteins.
26) Un autre aspect de l'invention concerne un microorganisme selon le paragraphe 25) ci-dessus caractérisé en ce que l'inactivation de cette ou de ces protéines est effectuée par mutagénèse par irradiation, par action d'agent chimique mutagène, par mutagénèse dirigée ou par remplacement de gène. 26) Another aspect of the invention relates to a microorganism according to paragraph 25) above characterized in that the inactivation of this or these proteins is carried out by mutagenesis by irradiation, by action of mutagenic chemical agent, by mutagenesis directed or by gene replacement.
27) Un autre aspect de l'invention concerne un microorganisme selon le paragraphe 25) ou 26) ci-dessus caractérisé en ce que la ou les mutagénèses sont effectuées in vitro ou in situ, par suppression, substitution, délétion et/ou addition d'une ou plusieurs bases dans le ou les gènes considérés ou par inactivation génique. 27) Another aspect of the invention relates to a microorganism according to paragraph 25) or 26) above characterized in that the mutagenesis or mutagenesis are performed in vitro or in situ, by deletion, substitution, deletion and / or addition of one or more bases in the gene (s) considered or by gene inactivation.
28) Un autre aspect de l'invention concerne un microorganisme selon le paragraphe 23), 24), 25), 26) ou 27) ci-dessus caractérisé en ce que ledit microorganisme est une bactérie du genre Streptomyces. 28) Another aspect of the invention relates to a microorganism according to paragraph 23), 24), 25), 26) or 27) above characterized in that said microorganism is a bacterium of the genus Streptomyces.
29) Un autre aspect de l'invention concerne un microorganisme selon le paragraphe 23), 24), 25), 26), 27) ou 28) ci-dessus caractérisé en ce que le macrolide est la spiramycine. 29) Another aspect of the invention relates to a microorganism according to paragraph 23), 24), 25), 26), 27) or 28) above characterized in that the macrolide is spiramycin.
30) Un autre aspect de l'invention concerne un microorganisme selon le paragraphe 23), 24), 25), 26), 27), 28) ou 29) ci-dessus caractérisé en ce que ledit microorganisme est une souche de S. ambofaciens. Another aspect of the invention relates to a microorganism according to paragraph 23), 24), 25), 26), 27), 28) or 29) above characterized in that said microorganism is a strain of S. ambofaciens.
31) Un autre aspect de l'invention concerne un microorganisme selon le paragraphe 23), 24), 25), 26), 27), 28), 29) ou 30) ci-dessus caractérisé en ce que la mutagénèse est effectuée dans au moins un gène comprenant une séquence selon l'un des paragraphes 1), 2), 3), 4), 5) ou 6) ci-dessus. 31) Another aspect of the invention relates to a microorganism according to paragraph 23), 24), 25), 26), 27), 28), 29) or 30) above characterized in that the mutagenesis is carried out in at least one gene comprising a sequence according to one of paragraphs 1), 2), 3), 4), 5) or 6) above.
<Desc/Clms Page number 10> <Desc / Clms Page number 10>
32) Un autre aspect de l'invention concerne un microorganisme selon le paragraphe 25), 26), 27), 28), 29), 30) ou 31) ci-dessus caractérisé en ce que la mutagénèse est effectuée dans un ou plusieurs gènes comprenant une des séquences correspondant à une ou plusieurs des séquences SEQ ID N 3, 5, 7,9, 11, 13,15, 17,19, 21, 23, 25, 28, 30, 34, 36, 40, 43, 45, 47, 49, 53, 60, 62, 64, 66, 68,70, 72,74, 76,78, 80, 82, 84, 107,109, 111, 113, 115,118, et 120. 32) Another aspect of the invention relates to a microorganism according to paragraph 25), 26), 27), 28), 29), 30) or 31) above characterized in that the mutagenesis is carried out in one or more genes comprising one of the sequences corresponding to one or more of the sequences SEQ ID N 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 28, 30, 34, 36, 40, 43 , 45, 47, 49, 53, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 107, 109, 111, 113, 115, 118, and 120.
33) Un autre aspect de l'invention concerne un microorganisme selon le paragraphe 25), 26), 27), 28), 29), 30), 31) ou 32) ci-dessus caractérisé en ce que la mutagénèse consiste en l'inactivation génique d'un gène comprenant une séquence correspondant à la séquence SEQ ID N 13. 33) Another aspect of the invention relates to a microorganism according to paragraph 25), 26), 27), 28), 29), 30), 31) or 32) above characterized in that mutagenesis consists of gene inactivation of a gene comprising a sequence corresponding to the sequence SEQ ID N 13.
34) Un autre aspect de l'invention concerne une souche de Streptomyces ambofaciens caractérisée en ce qu'il s'agit d'une souche choisie parmi : - l'une des souches déposées auprès de la Collection Nationale de Cultures de Microorganismes (CNCM) le 10 juillet 2002 sous le numéro d'enregistrement 1-2909, 1- 2911,1-2912,1-2913,1-2914,1-2915,1-2916 ou 1-2917 - la souche SPM510. 34) Another aspect of the invention relates to a strain of Streptomyces ambofaciens characterized in that it is a strain selected from: - one of the strains deposited with the National Collection of Cultures of Microorganisms (CNCM) on July 10, 2002 under registration number 1-2909, 1- 2911,1-2912,1-2913,1-2914,1-2915,1-2916 or 1-2917 - strain SPM510.
35) Un autre aspect de l'invention concerne un procédé de préparation d'un intermédiaire de biosynthèse de macrolide caractérisé en ce qu'il comprend les étapes suivantes : a) cultiver, dans un milieu de culture approprié, un microorganisme selon l'un des paragraphes 23), 24), 25), 26), 27), 28), 29), 30), 31), 32), 33) ou 34) ci dessus, b) récupérer le milieu de culture conditionné ou un extrait cellulaire, c) séparer et purifier à partir dudit milieu de culture ou encore à partir de l'extrait cellulaire obtenu à l'étape b), ledit intermédiaire de biosynthèse. 35) Another aspect of the invention relates to a process for preparing a macrolide biosynthesis intermediate, characterized in that it comprises the following steps: a) cultivating, in a suitable culture medium, a microorganism according to one of of paragraphs 23), 24), 25), 26), 27), 28), 29), 30), 31), 32), 33) or 34) above, b) recovering the conditioned culture medium or a cell extract, c) separating and purifying from said culture medium or from the cell extract obtained in step b), said biosynthesis intermediate.
36) Un autre aspect de l'invention concerne un procédé de préparation d'une molécule dérivée d'un macrolide caractérisé en ce que l'on prépare un intermédiaire de biosynthèse selon le procédé du paragraphe 35) ci-dessus et que l'on modifie l'intermédiaire ainsi produit. 36) Another aspect of the invention relates to a method for preparing a molecule derived from a macrolide characterized in that a biosynthetic intermediate is prepared according to the method of paragraph 35) above and that one modifies the intermediate thus produced.
<Desc/Clms Page number 11> <Desc / Clms Page number 11>
37) Un autre aspect de l'invention concerne un procédé de préparation selon le paragraphe 36) ci-dessus caractérisé en ce que l'on modifie ledit intermédiaire par voie chimique, biochimique, enzymatique et/ou microbiologique. 37) Another aspect of the invention relates to a method of preparation according to paragraph 36) above characterized in that one modifies said intermediate chemically, biochemically, enzymatically and / or microbiologically.
38) Un autre aspect de l'invention concerne un procédé de préparation selon le paragraphe 36) ou 37) ci-dessus caractérisé en ce que l'on introduit dans ledit microorganisme un ou plusieurs gènes codant des protéines susceptibles de modifier l'intermédiaire en s'en servant comme substrat. 38) Another aspect of the invention relates to a method of preparation according to paragraph 36) or 37) above characterized in that one introduces into said microorganism one or more genes encoding proteins capable of modifying the intermediate by using it as a substrate.
39) Un autre aspect de l'invention concerne un procédé de préparation selon le paragraphe 36), 37) ou 38) ci-dessus caractérisé en ce que le macrolide est la spiramycine. 39) Another aspect of the invention relates to a method of preparation according to paragraph 36), 37) or 38) above characterized in that the macrolide is spiramycin.
40) Un autre aspect de l'invention concerne un procédé de préparation selon le paragraphe 36), 37), 38) ou 39) ci-dessus caractérisé en ce que le microorganisme utilisé est une souche de S. ambofaciens. 40) Another aspect of the invention relates to a method of preparation according to paragraph 36), 37), 38) or 39) above characterized in that the microorganism used is a strain of S. ambofaciens.
41) Un autre aspect de l'invention concerne un microorganisme produisant de la spiramycine I mais ne produisant pas de spiramycine II et III
42) Un autre aspect de l'invention concerne un microorganisme selon le paragraphe 41) ci-dessus caractérisé en ce qu'il comprend l'ensemble des gènes nécessaires à la biosynthèse de la spiramycine 1 mais que le gène comprenant la séquence SEQ ID N 13 ou l'un de ses variants ou l'une des séquences dérivées de celles-ci en raison de la dégénérescence du code génétique et codant un polypeptide de séquence SEQ ID N 14 ou l'un de ses variants n'est pas exprimé ou a été rendu inactif. 41) Another aspect of the invention relates to a microorganism producing spiramycin I but not producing spiramycin II and III
42) Another aspect of the invention relates to a microorganism according to paragraph 41) above, characterized in that it comprises all the genes necessary for the biosynthesis of spiramycin 1 but that the gene comprising the sequence SEQ ID N 13 or one of its variants or one of the sequences derived from them because of the degeneracy of the genetic code and encoding a polypeptide of sequence SEQ ID No. 14 or one of its variants is not expressed or has been rendered inactive.
43) Un autre aspect de l'invention concerne un microorganisme selon le paragraphe 42) ci-dessus caractérisé en ce que la dite inactivation est réalisée par mutagénèse dans le gène codant ladite protéine ou par l'expression d'un ARN anti-sens complémentaire de l'ARN messager codant ladite protéine. 43) Another aspect of the invention relates to a microorganism according to paragraph 42) above characterized in that said inactivation is carried out by mutagenesis in the gene encoding said protein or by the expression of a complementary antisense RNA messenger RNA encoding said protein.
<Desc/Clms Page number 12> <Desc / Clms Page number 12>
44) Un autre aspect de l'invention concerne un microorganisme selon le paragraphe 43) ci-dessus caractérisé en ce que la dite mutagénèse est effectuée dans le promoteur de ce gène, dans la séquence codante ou dans une séquence non codante de manière à rendre inactive la protéine codée ou à en empêcher son expression ou sa traduction. 44) Another aspect of the invention relates to a microorganism according to paragraph 43) above characterized in that said mutagenesis is carried out in the promoter of this gene, in the coding sequence or in a non-coding sequence so as to render inactivates the encoded protein or prevents its expression or translation.
45) Un autre aspect de l'invention concerne un microorganisme selon la revendication 43) ou 44) ci-dessus caractérisé en ce que la mutagénèse est effectuée par irradiation, par action d'agent chimique mutagène, par mutagénèse dirigée ou par remplacement de gène. 45) Another aspect of the invention relates to a microorganism according to claim 43) or 44) above characterized in that the mutagenesis is carried out by irradiation, by action of mutagenic chemical agent, by site-directed mutagenesis or by gene replacement .
46) Un autre aspect de l'invention concerne un microorganisme selon le paragraphe 43), 44) ou 45) ci-dessus caractérisé en ce que la mutagénèse est effectuée in vitro ou in situ, par suppression, substitution, délétion et/ou addition d'une ou plusieurs bases dans le gène considéré ou par inactivation génique. 46) Another aspect of the invention relates to a microorganism according to paragraph 43), 44) or 45) above characterized in that the mutagenesis is carried out in vitro or in situ, by deletion, substitution, deletion and / or addition of one or more bases in the gene under consideration or by gene inactivation.
47) Un autre aspect de l'invention concerne un microorganisme selon le paragraphe 41) ou 42) ci-dessus caractérisé en ce que ledit microorganisme est obtenu en exprimant les gènes de la voie de biosynthèse de la spiramycine sans que ceux-ci ne comprennent le gène comprenant la séquence correspondant à SEQ ID N 13 ou l'un de ses variants ou l'une des séquences dérivées de celles-ci en raison de la dégénérescence du code génétique et codant un polypeptide de séquence SEQ ID N 14 ou l'un de ses variants. 47) Another aspect of the invention relates to a microorganism according to paragraph 41) or 42) above, characterized in that said microorganism is obtained by expressing the genes of the spiramycin biosynthetic pathway without these genes being understood. the gene comprising the sequence corresponding to SEQ ID No. 13 or one of its variants or one of the sequences derived from them because of the degeneracy of the genetic code and encoding a polypeptide of sequence SEQ ID No. 14 or the one of its variants.
48) Un autre aspect de l'invention concerne un microorganisme selon le paragraphe 41), 42), 43), 44), 45), 46) ou 47) ci-dessus caractérisé en ce que ledit microorganisme est une bactérie du genre Streptomyces. 48) Another aspect of the invention relates to a microorganism according to paragraph 41), 42), 43), 44), 45), 46) or 47) above characterized in that said microorganism is a bacterium of the genus Streptomyces .
49) Un autre aspect de l'invention concerne un microorganisme selon le paragraphe 41), 42), 43), 44), 45), 46), 47) ou 48) ci-dessus caractérisé en ce que ledit microorganisme est obtenu à partir d'une souche de départ produisant les spiramycines I, II et III. 49) Another aspect of the invention relates to a microorganism according to paragraph 41), 42), 43), 44), 45), 46), 47) or 48) above characterized in that said microorganism is obtained at from a starting strain producing spiramycins I, II and III.
<Desc/Clms Page number 13> <Desc / Clms Page number 13>
50) Un autre aspect de l'invention concerne un microorganisme selon le paragraphe 41), 42), 43), 44), 45), 46), 47), 48) ou 49) ci-dessus caractérisé en ce qu'il est obtenu par mutagénèse dans un gène comprenant la séquence correspondant à SEQ ID N 13 ou l'un de ses variants ou l'une des séquences dérivées de celles-ci en raison de la dégénérescence du code génétique et codant un polypeptide de séquence SEQ ID N 14 ou l'un de ses variants ayant la même fonction. Another aspect of the invention relates to a microorganism according to paragraph 41), 42), 43), 44), 45), 46), 47), 48) or 49) above, characterized in that is obtained by mutagenesis in a gene comprising the sequence corresponding to SEQ ID No. 13 or one of its variants or one of the sequences derived from them because of the degeneracy of the genetic code and encoding a polypeptide of sequence SEQ ID N 14 or one of its variants having the same function.
51) Un autre aspect de l'invention concerne un microorganisme selon le paragraphe 41), 42), 43), 44), 45), 46), 47), 48), 49) ou 50) ci-dessus caractérisé en ce que ledit microorganisme est obtenu à partir d'une souche de S. ambofaciens produisant les spiramycines I, II et III, dans laquelle est effectuée une inactivation génique du gène comprenant la séquences correspondant à SEQ ID N 13 ou l'une des séquences dérivées de celle-ci en raison de la dégénérescence du code génétique. 51) Another aspect of the invention relates to a microorganism according to paragraph 41), 42), 43), 44), 45), 46), 47), 48), 49) or 50) above characterized in that that said microorganism is obtained from a strain of S. ambofaciens producing spiramycins I, II and III, in which gene inactivation of the gene comprising the sequence corresponding to SEQ ID No. 13 or one of the sequences derived from this one because of the degeneracy of the genetic code.
52) Un autre aspect de l'invention concerne une souche de S. ambofaciens caractérisée en ce qu'il s'agit de la souche déposée auprès de la Collection Nationale de Cultures de Microorganismes (CNCM) le 10 juillet 2002 sous le numéro d'enregistrement 1-2910. 52) Another aspect of the invention relates to a strain of S. ambofaciens characterized in that it is the strain deposited with the National Collection of Cultures of Microorganisms (CNCM) July 10, 2002 under the number of registration 1-2910.
53) Un autre aspect de l'invention concerne un procédé de production de spiramycine I, caractérisé en ce qu'il comprend les étapes suivantes : (a) cultiver, dans un milieu de culture approprié, un microorganisme selon l'un des paragraphes 41), 42), 43), 44), 45), 46), 47), 48), 49), 50), 51) ou 52) ci-dessus, (b) récupérer le milieu de culture conditionné ou un extrait cellulaire, (c) séparer et purifier à partir dudit milieu de culture ou encore à partir de l'extrait cellulaire obtenu à l'étape b), la spiramycine I. 53) Another aspect of the invention relates to a method for producing spiramycin I, characterized in that it comprises the following steps: (a) cultivating, in a suitable culture medium, a microorganism according to one of the paragraphs 41 ), 42), 43), 44), 45), 46), 47), 48), 49), 50), 51) or 52) above (b) recovering the conditioned culture medium or an extract (c) separating and purifying from said culture medium or from the cell extract obtained in step b), spiramycin I.
54) Un autre aspect de l'invention concerne l'utilisation d'un polynucléotide selon l'un des paragraphes 1), 2), 3), 4), 5) ou 6) ci-dessus pour améliorer la production en macrolides d'un microorganisme. 54) Another aspect of the invention relates to the use of a polynucleotide according to one of paragraphs 1), 2), 3), 4), 5) or 6) above to improve the production of macrolide d a microorganism.
<Desc/Clms Page number 14> <Desc / Clms Page number 14>
55) Un autre aspect de l'invention concerne un microorganisme mutant producteur de macrolide caractérisé en ce qu'il possède une modification génétique dans au moins un gène comprenant une séquence selon l'un des paragraphes 1), 2), 3), 4), 5) ou 6) ci-dessus et/ou qu'il surexprime au moins un gène comprenant une séquence selon l'un des paragraphes 1), 2), 3), 4), 5) ou 6) ci-dessus. 55) Another aspect of the invention relates to a mutant macrolide-producing microorganism characterized in that it possesses a genetic modification in at least one gene comprising a sequence according to one of the paragraphs 1), 2), 3), 4 ), 5) or 6) above and / or overexpressing at least one gene comprising a sequence according to one of paragraphs 1), 2), 3), 4), 5) or 6) above .
56) Un autre aspect de l'invention concerne un microorganisme mutant selon le paragraphe 55) ci-dessus caractérisé en ce que la modification génétique consiste en une suppression, une substitution, une délétion et/ou une addition d'une ou plusieurs bases dans le ou les gènes considérés dans le but d'exprimer une ou des protéines ayant une activité supérieure ou d'exprimer un niveau plus élevé de cette ou de ces protéines. 56) Another aspect of the invention relates to a mutant microorganism according to paragraph 55) above characterized in that the genetic modification consists of a deletion, a substitution, a deletion and / or an addition of one or more bases in the gene or genes considered for the purpose of expressing one or more proteins having a higher activity or of expressing a higher level of this or these proteins.
57) Un autre aspect de l'invention concerne un microorganisme mutant selon le paragraphe 55) ci-dessus caractérisé en ce que la surexpression du gène considéré est obtenue en augmentant le nombre de copie de ce gène et/ou en plaçant un promoteur plus actif que le promoteur sauvage. 57) Another aspect of the invention relates to a mutant microorganism according to paragraph 55) above characterized in that the overexpression of the gene of interest is obtained by increasing the number of copies of this gene and / or by placing a more active promoter as the wild promoter.
58) Un autre aspect de l'invention concerne un microorganisme mutant selon le paragraphe 55) ou 57) ci-dessus caractérisé en ce que la surexpression du gène considéré est obtenue en transformant un microorganisme producteur de macrolide par une construction d'ADN recombinant selon le paragraphe 13,14 ou 17 ci-dessus, permettant la surexpression de ce gène. 58) Another aspect of the invention relates to a mutant microorganism according to paragraph 55) or 57) above characterized in that overexpression of the gene of interest is obtained by transforming a macrolide-producing microorganism with a recombinant DNA construct according to 13.14 or 17 above, allowing overexpression of this gene.
59) Un autre aspect de l'invention concerne un microorganisme mutant selon le paragraphe 55), 56), 57) ou 58) ci-dessus caractérisé en ce que la modification génétique est effectuée dans un ou plusieurs gènes comprenant une des séquences correspondant à une ou plusieurs des séquences SEQ ID N 3,5, 7,9, 11,13, 15,17, 19, 21, 23, 25, 28, 30, 34, 36, 40, 43, 45, 47, 49, 53, 60, 62, 64, 66, 68, 70,72, 74,76, 78, 80, 82, 84, 107, 109, 111, 113, 115, 118, et 120, ou l'un de ses variants ou l'une des séquences dérivées de celles-ci en raison de la dégénérescence du code génétique. 59) Another aspect of the invention relates to a mutant microorganism according to paragraph 55), 56), 57) or 58) above characterized in that the genetic modification is carried out in one or more genes comprising one of the sequences corresponding to one or more of the sequences SEQ ID N 3.5, 7.9, 11, 13, 15, 17, 19, 21, 23, 25, 28, 30, 34, 36, 40, 43, 45, 47, 49, 53, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 107, 109, 111, 113, 115, 118, and 120, or one of its variants or one of the sequences derived from them because of the degeneracy of the genetic code.
<Desc/Clms Page number 15> <Desc / Clms Page number 15>
60) Un autre aspect de l'invention concerne un microorganisme mutant selon le paragraphe 55), 56), 57), 58) ou 59) ci-dessus caractérisé le microorganisme surexprime un ou plusieurs gènes comprenant une des séquences correspondant à une ou plusieurs des séquences SEQ ID N 3,5, 7,9, 11, 13,15, 17, 19, 21, 23, 25, 28,30, 34,36, 40, 43, 45, 47, 49, 53, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 107, 109, 111, 113, 115, 118, et 120, ou l'un de ses variants ou l'une des séquences dérivées de celles-ci en raison de la dégénérescence du code génétique. 60) Another aspect of the invention relates to a mutant microorganism according to paragraph 55), 56), 57), 58) or 59) above characterized the microorganism overexpresses one or more genes comprising one of the sequences corresponding to one or more sequences SEQ ID N 3.5, 7.9, 11, 13, 15, 17, 19, 21, 23, 25, 28, 30, 34, 36, 40, 43, 45, 47, 49, 53, 60 , 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 107, 109, 111, 113, 115, 118, and 120, or one of its variants or the one of the sequences derived from these because of the degeneracy of the genetic code.
61) Un autre aspect de l'invention concerne un microorganisme mutant selon le paragraphe 55), 56), 57), 58), 59) ou 60) ci-dessus caractérisé en ce que ledit microorganisme est une bactéries du genre Streptomyces. 61) Another aspect of the invention relates to a mutant microorganism according to paragraph 55), 56), 57), 58), 59) or 60) above characterized in that said microorganism is a bacterium of the genus Streptomyces.
62) Un autre aspect de l'invention concerne un microorganisme mutant selon le paragraphe 55), 56), 57), 58), 59), 60) ou 61) ci-dessus caractérisé en ce que le macrolide est la spiramycine. 62) Another aspect of the invention relates to a mutant microorganism according to paragraph 55), 56), 57), 58), 59), 60) or 61) above characterized in that the macrolide is spiramycin.
63) Un autre aspect de l'invention concerne un microorganisme mutant selon le paragraphe 55), 56), 57), 58), 59), 60), 61) ou 62) ci-dessus caractérisé en ce que ledit microorganisme est une souches de S. ambofaciens. 63) Another aspect of the invention relates to a mutant microorganism according to paragraph 55), 56), 57), 58), 59), 60), 61) or 62) above characterized in that said microorganism is a strains of S. ambofaciens.
64) Un autre aspect de l'invention concerne un procédé de production de macrolides, caractérisé en ce qu'il comprend les étapes suivantes : (a) cultiver, dans un milieu de culture approprié, un microorganisme selon l'un des paragraphes 55), 56), 57), 58), 59), 60), 61), 62), 63) ou 64) ci-dessus, (b) récupérer le milieu de culture conditionné ou un extrait cellulaire, (c) séparer et purifier à partir dudit milieu de culture ou encore à partir de l'extrait cellulaire obtenu à l'étape b), le ou lesdits macrolides produits. 64) Another aspect of the invention relates to a method for producing macrolides, characterized in that it comprises the following steps: (a) cultivating, in a suitable culture medium, a microorganism according to one of paragraphs 55) , 56), 57), 58), 59), 60), 61), 62), 63) or 64) above, (b) recovering the conditioned culture medium or cell extract, (c) separating and purifying from said culture medium or from the cell extract obtained in step b), the macrolide or macrolides produced.
65) Un autre aspect de l'invention concerne l'utilisation d'une séquence et/ou d'un ADN recombinant et/ou d'un vecteur selon l'un des paragraphes 1), 2), 3), 4), 5), 6), 7), 8), 9), 10), 11), 12), 13), 14), 15), 16) ou 17) ci-dessus pour la préparation d'antibiotiques hybrides. 65) Another aspect of the invention relates to the use of a sequence and / or a recombinant DNA and / or a vector according to one of paragraphs 1), 2), 3), 4), 5), 6), 7), 8), 9), 10), 11), 12), 13), 14), 15), 16) or 17) above for the preparation of hybrid antibiotics.
<Desc/Clms Page number 16> <Desc / Clms Page number 16>
66) Un autre aspect de l'invention concerne l'utilisation d'au moins un polynucléotide et/ou au moins un ADN recombinant et/ou au moins un vecteur d'expression et/ou au moins un polypeptide et/ou au moins une cellule hôte selon l'un des paragraphes 1), 2), 3), 4), 5), 6), 7), 8), 9), 10), 11), 12), 13), 14), 15), 16), 17) ou 21) ci-dessus pour réaliser une ou plusieurs bioconversions. 66) Another aspect of the invention relates to the use of at least one polynucleotide and / or at least one recombinant DNA and / or at least one expression vector and / or at least one polypeptide and / or at least one host cell according to one of paragraphs 1), 2), 3), 4), 5), 6), 7), 8), 9), 10), 11), 12), 13), 14), 15), 16), 17) or 21) above to perform one or more bioconversions.
67) Un autre aspect de l'invention concerne un polynucléotide caractérisé en ce qu'il s'agit d'un polynucléotide complémentaire de l'un des polynucléotides selon le paragraphe 1), 2), 3), 4), 5), ou 6) ci-dessus. 67) Another aspect of the invention relates to a polynucleotide characterized in that it is a polynucleotide complementary to one of the polynucleotides according to paragraph 1), 2), 3), 4), 5), or 6) above.
68) Un autre aspect de l'invention concerne un microorganisme producteur d'au moins une spiramycine caractérisé en ce qu'il surexprime : - un gène susceptible d'être obtenu par amplification en chaîne par polymérase (PCR) en utilisant le couple d'amorces de séquence suivante : 5' AAGCTTGTGTGCCCGGTGTACCTGGGGAGC 3' (SEQ ID N 138) et 5' GGATCCCGCGACGGACACGACCGCCGCGCA 3' (SEQ ID N 139) et comme matrice le cosmide pSPM36 ou l'ADN total de Streptomyces ambofaciens, - ou un gène dérivé de celui-ci en raison de la dégénérescence du code génétique. 68) Another aspect of the invention relates to a microorganism producing at least one spiramycin characterized in that it overexpresses: a gene that can be obtained by polymerase chain reaction (PCR) using the pair of primers of the following sequence: 5 'AAGCTTGTGTGCCCGGTGTACCTGGGGAGC 3' (SEQ ID N 138) and 5 'GGATCCCGCGACGGACACGACCGCCGCGCA 3' (SEQ ID N 139) and as a template the cosmid pSPM36 or the total DNA of Streptomyces ambofaciens, - or a gene derived from that because of the degeneracy of the genetic code.
69) Un autre aspect de l'invention concerne un microorganisme selon le paragraphe 68 caractérisé en ce qu'il s'agit d'une bactérie du genre Streptomyces. 69) Another aspect of the invention relates to a microorganism according to paragraph 68 characterized in that it is a bacterium of the genus Streptomyces.
70) Un autre aspect de l'invention concerne un microorganisme selon le paragraphe 68 ou 69 caractérisé en ce qu'il s'agit d'une bactérie de l'espèce Streptomyces ambofaciens. 70) Another aspect of the invention relates to a microorganism according to paragraph 68 or 69 characterized in that it is a bacterium of the species Streptomyces ambofaciens.
71) Un autre aspect de l'invention concerne un microorganisme selon le paragraphe 68,69 ou 70 caractérisé en ce que la surexpression dudit gène est obtenue par transformation dudit microorganisme par un vecteur d'expression. 71) Another aspect of the invention relates to a microorganism according to paragraph 68, 69 or 70 characterized in that the overexpression of said gene is obtained by transformation of said microorganism by an expression vector.
<Desc/Clms Page number 17> <Desc / Clms Page number 17>
72) Un autre aspect de l'invention concerne une souche de Streptomyces ambofaciens caractérisée en ce qu'il s'agit de la souche OSC2/pSPM75(l) ou de la souche OSC2/pSPM75(2). 72) Another aspect of the invention relates to a strain of Streptomyces ambofaciens characterized in that it is the strain OSC2 / pSPM75 (1) or strain OSC2 / pSPM75 (2).
73) Un autre aspect de l'invention concerne un ADN recombinant caractérisé en ce qu'il comprend : - un polynucléotide susceptible d'être obtenu par amplification en chaîne par polymérase en utilisant le couple d'amorces de séquence suivante : 5' AAGCTTGTGTGCCCGGTGTACCTGGGGAGC 3' (SEQ ID N 138) et 5' GGATCCCGCGACGGACACGACCGCCGCGCA 3' (SEQ ID N 139) et comme matrice le cosmide pSPM36 ou l'ADN total de Streptomyces ambofaciens, - ou un fragment d'au moins 10,12, 15,18, 20 à 25, 30, 40, 50,60, 70,80, 90,100, 150,200, 250,300, 350,400, 450,500, 550,600, 650,700, 750,800, 850, 900, 950, 1000, 1050, 1100, 1150, 1200, 1250, 1300, 1350, 1400, 1450, 1460, 1470, 1480, 1490 ou 1500 nucléotides consécutifs de ce polynucléotide. 73) Another aspect of the invention relates to a recombinant DNA characterized in that it comprises: a polynucleotide capable of being obtained by polymerase chain reaction using the following pair of primers: 5 'AAGCTTGTGTGCCCGGTGTACCTGGGGAGC 3 (SEQ ID NO: 138) and 5 'GGATCCCGCGACGGACACGACCGCCGCGCA 3' (SEQ ID N 139) and as a template the cosmid pSPM36 or the total DNA of Streptomyces ambofaciens, or a fragment of at least 10, 12, 15, 18, 20 to 25, 30, 40, 50.60, 70.80, 90.100, 150.200, 250.300, 350.400, 450.500, 550.600, 650.700, 750.800, 850, 900, 950, 1000, 1050, 1100, 1150, 1200, 1250, 1300, 1350, 1400, 1450, 1460, 1470, 1480, 1490 or 1500 consecutive nucleotides of this polynucleotide.
74) Un autre aspect de l'invention concerne un ADN recombinant selon le paragraphe 73 caractérisé en ce qu'il s'agit d'un vecteur. 74) Another aspect of the invention relates to a recombinant DNA according to paragraph 73 characterized in that it is a vector.
75) Un autre aspect de l'invention concerne un ADN recombinant selon le paragraphe 73 ou 74 caractérisé en ce qu'il s'agit d'un vecteur d'expression. Another aspect of the invention relates to a recombinant DNA according to paragraph 73 or 74 characterized in that it is an expression vector.
76) Un autre aspect de l'invention concerne une cellule hôte dans laquelle a été introduit au moins un ADN recombinant selon l'un des paragraphes 73,74 ou 75. 76) Another aspect of the invention relates to a host cell into which at least one recombinant DNA has been introduced according to one of paragraphs 73, 74 or 75.
77) Un autre aspect de l'invention concerne un procédé de production d'un polypeptide caractérisé en ce que ledit procédé comprend les étapes suivantes: a) transformer une cellule hôte par au moins un vecteur d'expression selon le paragraphe 75; b) cultiver, dans un milieu de culture approprié, ladite cellule hôte; c) récupérer le milieu de culture conditionné ou un extrait cellulaire; 77) Another aspect of the invention relates to a method of producing a polypeptide characterized in that said method comprises the following steps: a) transforming a host cell with at least one expression vector according to paragraph 75; b) culturing, in a suitable culture medium, said host cell; c) recovering the conditioned culture medium or a cell extract;
<Desc/Clms Page number 18><Desc / Clms Page number 18>
d) séparer et purifier à partir dudit milieu de culture ou encore à partir de l'extrait cellulaire obtenu à l'étape c), ledit polypeptide ; e) le cas échéant, caractériser le polypeptide recombinant produit. d) separating and purifying from said culture medium or from the cell extract obtained in step c), said polypeptide; e) where appropriate, characterizing the recombinant polypeptide produced.
78) Un autre aspect de l'invention concerne un microorganisme selon le paragraphe 51 caractérisé en ce que l'inactivation génique est effectuée par délétion en phase du gène ou d'une partie du gène comprenant la séquence correspondant à SEQ ID N 13 ou l'une des séquences dérivées de celle-ci en raison de la dégénérescence du code génétique. 78) Another aspect of the invention relates to a microorganism according to paragraph 51 characterized in that the gene inactivation is carried out by deletion in phase of the gene or a part of the gene comprising the sequence corresponding to SEQ ID No. 13 or one of the sequences derived from it because of the degeneracy of the genetic code.
79) Un autre aspect de l'invention concerne un microorganisme selon l'un des paragraphes 41,42, 43,44, 45,46, 47,48, 49,50, 51 ou 78 caractérisé en ce en ce qu'il surexprime en outre : - un gène susceptible d'être obtenu par amplification en chaîne par polymérase en utilisant le couple d'amorces de séquence suivante : 5' AAGCTTGTGTGCCCGGTGTACCTGGGGAGC 3' (SEQ ID N 138) et 5' GGATCCCGCGACGGACACGACCGCCGCGCA 3' (SEQ ID N 139) et comme matrice le cosmide pSPM36 ou l'ADN total de Streptomyces ambofaciens. 79) Another aspect of the invention relates to a microorganism according to one of paragraphs 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51 or 78, characterized in that it overexpresses in addition: a gene which can be obtained by polymerase chain reaction using the following pair of primers: 5 'AAGCTTGTGTGCCCGGTGTACCTGGGGAGC 3' (SEQ ID N 138) and 5 'GGATCCCGCGACGGACACGACCGCCGCGCA 3' (SEQ ID N 139 ) and as a template the cosmid pSPM36 or the total DNA of Streptomyces ambofaciens.
- ou un gène dérivé de celui-ci en raison de la dégénérescence du code génétique. or a gene derived from it because of the degeneracy of the genetic code.
80) Un autre aspect de l'invention concerne un vecteur d'expression caractérisé en ce que le polynucléotide de séquence SEQ ID N 47 ou un polynucléotide dérivé de celui-ci en raison de la dégénérescence du code génétique est placé sous le contrôle d'un promoteur permettant l'expression de la protéine codée par le dit polynucléotide chez Streptomyces ambofaciens. 80) Another aspect of the invention relates to an expression vector characterized in that the polynucleotide of sequence SEQ ID No. 47 or a polynucleotide derived therefrom due to the degeneracy of the genetic code is placed under the control of a promoter allowing the expression of the protein encoded by said polynucleotide in Streptomyces ambofaciens.
81) Un autre aspect de l'invention concerne un vecteur d'expression selon le paragraphe 80 caractérisé en ce qu'il s'agit du plasmide pSPM524 ou pSPM525. 81) Another aspect of the invention relates to an expression vector according to paragraph 80 characterized in that it is the plasmid pSPM524 or pSPM525.
82) Un autre aspect de l'invention concerne une souche de Streptomyces ambofaciens transformée par un vecteur selon le paragraphe 80 ou 81. 82) Another aspect of the invention relates to a strain of Streptomyces ambofaciens transformed by a vector according to paragraph 80 or 81.
<Desc/Clms Page number 19> <Desc / Clms Page number 19>
83) Un autre aspect de l'invention concerne un microorganisme selon l'un des paragraphes 41,42, 43,44, 45, 46, 47, 48, 49, 50, 51,78 ou 79 caractérisé en ce qu'il surexprime en outre le gène de séquence codante SEQ ID N 47 ou d'une séquence codante dérivée de celle-ci en raison de la dégénérescence du code génétique. 83) Another aspect of the invention relates to a microorganism according to one of paragraphs 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 78 or 79, characterized in that it overexpresses in addition, the coding sequence gene SEQ ID No. 47 or a coding sequence derived therefrom due to the degeneracy of the genetic code.
84) Un autre aspect de l'invention concerne un microorganisme selon le paragraphe 83 caractérisé en ce qu'il s'agit de la souche SPM502 pSPM525 déposée auprès de la Collection Nationale de Cultures de Microorganismes (CNCM) Institut Pasteur, 25, rue du Docteur Roux 75724 Paris Cedex 15, France, le 26 février 2003 sous le numéro d'enregistrement 1-2977. 84) Another aspect of the invention relates to a microorganism according to paragraph 83 characterized in that it is the SPM502 pSPM525 strain deposited with the National Collection of Cultures of Microorganisms (CNCM) Institut Pasteur, 25, rue du Dr. Roux 75724 Paris Cedex 15, France, February 26, 2003 under the registration number 1-2977.
85) Un autre aspect de l'invention concerne un procédé de production de spiramycine(s), caractérisé en ce qu'il comprend les étapes suivantes : (a) cultiver, dans un milieu de culture approprié, un microorganisme selon l'un des paragraphes 68, 69, 70,71, 72,78, 79,82, 83 ou 84, (b) récupérer le milieu de culture conditionné ou un extrait cellulaire, (c) séparer et purifier à partir dudit milieu de culture ou encore à partir de l'extrait cellulaire obtenu à l'étape b), les spiramycines. 85) Another aspect of the invention relates to a method for producing spiramycin (s), characterized in that it comprises the following steps: (a) cultivating, in a suitable culture medium, a microorganism according to one of the 68, 69, 70, 71, 72, 78, 79, 82, 83 or 84, (b) recovering the conditioned culture medium or cell extract, (c) separating and purifying from said culture medium, or from the cell extract obtained in step b), spiramycins.
86) Un autre aspect de l'invention concerne un polypeptide caractérisé en ce que sa séquence comprend la séquence SEQ ID N 112. 86) Another aspect of the invention relates to a polypeptide characterized in that its sequence comprises the sequence SEQ ID N 112.
87) Un autre aspect de l'invention concerne un polypeptide caractérisé en ce que sa séquence correspond à la séquence traduite de la séquence codante : - d'un gène susceptible d'être obtenu par amplification en chaîne par polymérase (PCR) en utilisant le couple d'amorces de séquence suivante : 5' AAGCTTGTGTGCCCGGTGTACCTGGGGAGC 3' (SEQ ID N 138) et 5' GGATCCCGCGACGGACACGACCGCCGCGCA 3' (SEQ ID N 139) et comme matrice le cosmide pSPM36 ou l'ADN total de Streptomyces ambofaciens, -ou d'un gène dérivé de celui-ci en raison de la dégénérescence du code génétique. 87) Another aspect of the invention relates to a polypeptide characterized in that its sequence corresponds to the translated sequence of the coding sequence of: - a gene capable of being obtained by polymerase chain reaction (PCR) using the pair of primers of the following sequence: 5 'AAGCTTGTGTGCCCGGTGTACCTGGGGAGC 3' (SEQ ID N 138) and 5 'GGATCCCGCGACGGACACGACCGCCGCGCA 3' (SEQ ID N 139) and as a template the cosmid pSPM36 or the total DNA of Streptomyces ambofaciens, or of a gene derived from it because of the degeneracy of the genetic code.
<Desc/Clms Page number 20> <Desc / Clms Page number 20>
88) Un autre aspect de l'invention concerne un vecteur d'expression permettant l'expression d'un polypeptide selon le paragraphe 86 ou 87 chez Streptomyces ambofaciens. 88) Another aspect of the invention relates to an expression vector for the expression of a polypeptide according to paragraph 86 or 87 in Streptomyces ambofaciens.
89) Un autre aspect de l'invention concerne un vecteur d'expression selon le paragraphe 88 caractérisé en ce qu'il s'agit du plasmide pSPM75. 89) Another aspect of the invention relates to an expression vector according to paragraph 88 characterized in that it is the plasmid pSPM75.
Définitions générales
Le terme " isolé " au sens de la présente invention désigne un matériel biologique (acide nucléique ou protéine) qui a été soustrait à son environnement originel (l'environnement dans lequel il est localisé naturellement). General Definitions
The term "isolated" in the sense of the present invention means a biological material (nucleic acid or protein) that has been removed from its original environment (the environment in which it is naturally located).
Par exemple un polynucléotide présent à l'état naturel dans une plante ou un animal n'est pas isolé. Le même polynucléotide séparé des acides nucléiques adjacents au sein desquels il est naturellement inséré dans le génome de la plante ou l'animal est considéré comme " isolé ". For example, a polynucleotide naturally occurring in a plant or animal is not isolated. The same polynucleotide separated from the adjacent nucleic acids in which it is naturally inserted into the genome of the plant or animal is considered "isolated".
Un tel polynucléotide peut être inclus dans un vecteur et/ou un tel polynucléotide peut être inclus dans une composition et demeurer néanmoins à l'état isolé du fait que le vecteur ou la composition ne constitue pas son environnement naturel. Such a polynucleotide may be included in a vector and / or such a polynucleotide may be included in a composition and still remain in the isolated state because the vector or composition does not constitute its natural environment.
Le terme " purifié " ne nécessite pas que le matériel soit présent sous une forme de pureté absolue, exclusive de la présence d'autres composés. Il s'agit plutôt d'une définition relative. The term "purified" does not require that the material be present in a form of absolute purity, exclusive of the presence of other compounds. It is rather a relative definition.
Un polynucléotide est à l'état " purifié " après purification du matériel de départ ou du matériel naturel d'au moins un ordre de grandeur, de préférence 2 ou 3 et préférentiellement 4 ou 5 ordres de grandeur. A polynucleotide is in the "purified" state after purification of the starting material or natural material of at least one order of magnitude, preferably 2 or 3 and preferably 4 or 5 orders of magnitude.
<Desc/Clms Page number 21> <Desc / Clms Page number 21>
Aux fins de la présente invention le terme ORF ( Open Reading Frame , c'est à dire cadre ouvert de lecture) a été employé pour désigner en particulier la séquence codante d'un gène. For the purposes of the present invention the term ORF (Open Reading Frame) has been used to refer in particular to the coding sequence of a gene.
Aux fins de la présente description, l'expression " séquence nucléotidique " peut être employée pour désigner indifféremment un polynucléotide ou un acide nucléique. For purposes of this disclosure, the term "nucleotide sequence" may be used to denote either a polynucleotide or a nucleic acid.
L'expression " séquence nucléotidique " englobe le matériel génétique lui-même et n'est donc pas restreinte à l'information concernant sa séquence. The term "nucleotide sequence" encompasses the genetic material itself and is therefore not restricted to information about its sequence.
Les termes " acide nucléique ", " polynucléotide ", " oligonucléotide " ou encore séquence nucléotidique " englobent des séquences d'ARN, d'ADN, d'ADNc ou encore des séquences hybrides ARN/ADN de plus d'un nucléotide, indifféremment sous la forme simple chaîne ou sous la forme de duplex. The terms "nucleic acid", "polynucleotide", "oligonucleotide" or nucleotide sequence "encompass RNA, DNA or cDNA sequences or hybrid RNA / DNA sequences of more than one nucleotide, regardless of whether the simple form chain or in the form of duplex.
Le terme " nucléotide " désigne à la fois les nucléotides naturels (A, T, G, C) ainsi que des nucléotides modifiés qui comprennent au moins une modification telle que (1) un analogue d'une purine, (2) un analogue d'une pyrimidine, ou (3) un sucre analogue, des exemples de tels nucléotides modifiés étant décrits par exemple dans la demande PCT N WO 95/04 064. The term "nucleotide" refers to both naturally occurring nucleotides (A, T, G, C) as well as modified nucleotides that include at least one modification such as (1) a purine analogue, (2) an analogue of a purine, a pyrimidine, or (3) a similar sugar, examples of such modified nucleotides being described for example in the PCT application N WO 95/04 064.
Aux fins de la présente invention, un premier polynucléotide est considéré comme étant " complémentaire " d'un second polynucléotide lorsque chaque base du premier polynucléotide est appariée à la base complémentaire du second polynucléotide dont l'orientation est inversée. Les bases complémentaires sont A et T (ou A et U), ou C et G. For purposes of the present invention, a first polynucleotide is considered to be "complementary" to a second polynucleotide when each base of the first polynucleotide is matched to the complementary base of the second polynucleotide whose orientation is reversed. The complementary bases are A and T (or A and U), or C and G.
Le terme gènes de la voie de biosynthèse des spiramycines comprend également les gènes régulateurs et les gènes conférant la résistance aux microorganismes producteurs. The term genes of the spiramycin biosynthetic pathway also includes regulatory genes and genes conferring resistance to producing microorganisms.
On entendra par " fragment " d'un acide nucléique de référence selon l'invention, une séquence nucléotidique de longueur réduite par rapport à l'acide nucléique de référence et comprenant, sur la partie commune, une séquence en nucléotides identique à l'acide nucléique de référence. The term "fragment" of a reference nucleic acid according to the invention, a nucleotide sequence of reduced length relative to the reference nucleic acid and comprising, on the common part, a nucleotide sequence identical to the acid reference nuclei.
<Desc/Clms Page number 22> <Desc / Clms Page number 22>
Un tel " fragment " d'acide nucléique selon l'invention peut être le cas échéant, compris dans un polynucléotide plus grand duquel il est constitutif. Such a "fragment" of nucleic acid according to the invention may optionally be included in a larger polynucleotide of which it is constitutive.
De tels fragments comprennent ou alternativement consistent en, des polynucléotides de longueur allant de 8,10, 12,15, 18, 20 à 25, 30, 40, 50,60, 70,80, 90, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950,1000, 1050,1100, 1150,1200, 1250, 1300, 1350,1400, 1450,1500, 1550,1600, 1650,1700, 1750,1800, 1850 ou 1900 nucléotides consécutifs d'un acide nucléique selon l'invention. Such fragments comprise or alternatively consist of polynucleotides of length ranging from 8, 10, 12, 15, 18, 20 to 25, 30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250. , 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000, 1050, 1100, 1150, 1200, 1250, 1300, 1350, 1400, 1450, 1500 , 1550, 1600, 1650, 1700, 1750, 1800, 1850 or 1900 consecutive nucleotides of a nucleic acid according to the invention.
Par " fragment " d'un polypeptide selon l'invention, on entendra un polypeptide dont la séquence d'acides aminés est plus courte que celle du polypeptide de référence et qui comprend sur toute la partie commune avec ces polypeptides de référence, une séquence en acides aminés identique. The term "fragment" of a polypeptide according to the invention will be understood to mean a polypeptide whose amino acid sequence is shorter than that of the reference polypeptide and which comprises over all the part common to these reference polypeptides, a sequence of amino acids identical.
De tels fragments peuvent, le cas échéant, être compris au sein d'un polypeptide plus grand duquel ils font partie. Such fragments may, where appropriate, be included within a larger polypeptide of which they are part.
De tels fragments d'un polypeptide selon l'invention peuvent avoir une longueur de 10, 15,20, 30 à 40,50, 60,70, 80,90,100, 120,140, 160,180, 200,220, 240,260, 280, 300, 320, 340, 360, 380, 400, 420, 440, 460, 480, 500, 520, 540, 560, 580, 600, 620 ou 640 acides aminés. Such fragments of a polypeptide according to the invention may have a length of 10, 15,20, 30 to 40,50, 60,70, 80,90,100, 120,140, 160,180, 200,220, 240,260, 280, 300, 320, 340, 360, 380, 400, 420, 440, 460, 480, 500, 520, 540, 560, 580, 600, 620 or 640 amino acids.
Par " conditions d'hybridation de forte stringence " au sens de la présente invention, on entendra des conditions d'hybridation défavorisant l'hybridation de brins d'acides nucléiques non homologues. Des conditions d'hybridations de forte stringence peuvent être par exemple décrites comme des conditions d'hybridation dans le tampon décrit par Church & Gilbert (Church & Gilbert, 1984) à une température comprise entre 55 C et 65 C, de manière préférée la température d'hyridation est de 55 C, de manière encore plus préférée la températue d'hybridation est de 60 C et de manière tout à fait ' préférée la température d'hybridation est de 65 C, suivi d'un ou plusieurs lavages effectué en tampon 2X SSC (le tampon IX SSC correspond à une solution aqueuse Highly stringent hybridization conditions within the meaning of the present invention will be understood to mean hybridization conditions that discriminate against the hybridization of non-homologous nucleic acid strands. Highly stringent hybridization conditions can be described, for example, as hybridization conditions in the buffer described by Church & Gilbert (Church & Gilbert, 1984) at a temperature of between 55 ° C. and 65 ° C., preferably the temperature The hybridization temperature is 60 ° C. and more preferably the hybridization temperature is 65 ° C., followed by one or more washings carried out in a buffer. 2X SSC (the IX SSC buffer corresponds to an aqueous solution
<Desc/Clms Page number 23><Desc / Clms Page number 23>
0,15M NaCl, 15 mM de citrate de sodium) à une température comprise entre 55 C et 65 C, de manière préférée cette température est de 55 C, de manière encore plus préférée cette températue est de 60 C et de manière tout à fait préférée cette température est de 65 C, suivi d'un ou plusieurs lavages en tampon 0.5X SSC à une température comprise entre 55 C et 65 C, de manière préférée cette température est de 55 C, de manière encore plus préférée cette températue est de 60 C et de manière tout à fait préférée cette température est de 65 C. 0.15 M NaCl, 15 mM sodium citrate) at a temperature of between 55 ° C. and 65 ° C., preferably this temperature is 55 ° C., more preferably still this temperature is 60 ° C. and quite exactly preferred this temperature is 65 C, followed by one or more washes in 0.5X SSC buffer at a temperature between 55 C and 65 C, preferably this temperature is 55 C, even more preferably this temperature is of 60 C and quite preferably this temperature is 65 C.
Il va sans dire que les conditions d'hybridation ci-dessus décrites peuvent être adaptées en fonction de la longueur de l'acide nucléique dont l'hybridation est recherchée ou du type de marquage choisi, selon des techniques connues de l'homme du métier. Les conditions convenables d'hybridation peuvent par exemple être adaptées selon l'ouvrage de F. Ausubel et al (2002). It goes without saying that the hybridization conditions described above can be adapted as a function of the length of the nucleic acid whose hybridization is sought or of the type of marking chosen, according to techniques known to those skilled in the art. . Suitable hybridization conditions may for example be adapted according to the work of F. Ausubel et al (2002).
Par " variant " d'un acide nucléique selon l'invention, on entendra un acide nucléique qui diffère d'une ou plusieurs bases par rapport au polynucléotide de référence. Un acide nucléique variant peut être d'origine naturelle, tel qu'un variant allélique retrouvé naturellement, ou peut être aussi un variant non naturel obtenu par exemple par des techniques de mutagénèse. By "variant" of a nucleic acid according to the invention is meant a nucleic acid which differs from one or more bases relative to the reference polynucleotide. A variant nucleic acid may be of natural origin, such as a naturally occurring allelic variant, or may also be a non-natural variant obtained for example by mutagenesis techniques.
En général, les différences entre l'acide nucléique de référence et l'acide nucléique variant sont réduites de telle sorte que les séquences nucléotidiques de l'acide nucléique de référence et de l'acide nucléique variant sont très proches et, dans de nombreuses régions, identiques. Les modifications de nucléotides présentes dans un acide nucléique variant peuvent être silencieuses, ce qui signifie qu'elles n'altèrent pas les séquences d'aminoacides codées par ledit acide nucléique variant. In general, the differences between the reference nucleic acid and the variant nucleic acid are reduced so that the nucleotide sequences of the reference nucleic acid and the variant nucleic acid are very close and, in many regions , identical. The nucleotide modifications present in a variant nucleic acid can be silent, which means that they do not alter the amino acid sequences encoded by said variant nucleic acid.
Cependant, les changements de nucléotides dans un acide nucléique variant peuvent aussi résulter de substitutions, additions, délétions dans le polypeptide codé par l'acide nucléique variant par rapport aux peptides codés par l'acide nucléique de référence. En outre, des modifications de nucléotides dans les régions codantes peuvent However, nucleotide changes in a variant nucleic acid may also result from substitutions, additions, deletions in the polypeptide encoded by the variant nucleic acid relative to the peptides encoded by the reference nucleic acid. In addition, nucleotide modifications in the coding regions may
<Desc/Clms Page number 24><Desc / Clms Page number 24>
produire des substitutions, conservatives ou non conservatives dans la séquence d'aminoacides. produce conservative or non-conservative substitutions in the amino acid sequence.
De préférence, les acides nucléiques variants selon l'invention codent des polypeptides qui conservent sensiblement la même fonction ou activité biologique que le polypeptide de l'acide nucléique de référence ou encore la capacité à être reconnus par des anticorps dirigés contre les polypeptides codés par l'acide nucléique initial. Preferably, the variant nucleic acids according to the invention encode polypeptides which retain substantially the same function or biological activity as the polypeptide of the reference nucleic acid or the capacity to be recognized by antibodies directed against the polypeptides encoded by the initial nucleic acid.
Certains acides nucléiques variants coderont ainsi des formes mutées des polypeptides dont l'étude systématique permettra de déduire des relations structure activité des protéines en question. Some variant nucleic acids will thus encode mutated forms of the polypeptides, the systematic study of which will make it possible to deduce the structure-activity relationships of the proteins in question.
Par " variant " d'un polypeptide selon l'invention, on entendra principalement un polypeptide dont la séquence d'acides aminés contient une ou plusieurs substitutions, additions ou délétions d'au moins un résidu d'acide aminé, par rapport à la séquence d'acides aminés du polypeptide de référence, étant entendu que les substitutions d'aminoacides peuvent être indifféremment conservatives ou non conservatives. By "variant" of a polypeptide according to the invention is meant mainly a polypeptide whose amino acid sequence contains one or more substitutions, additions or deletions of at least one amino acid residue, with respect to the sequence of amino acids of the reference polypeptide, it being understood that the amino acid substitutions may be indifferently conservative or non-conservative.
De préférence, les polypeptides variants selon l'invention conservent sensiblement la même fonction ou activité biologique que le polypeptide de référence ou encore la capacité à être reconnus par des anticorps dirigés contre les polypeptides initiaux. Preferably, the variant polypeptides according to the invention retain substantially the same function or biological activity as the reference polypeptide or the ability to be recognized by antibodies directed against the initial polypeptides.
Par polypeptide ayant " une activité similaire " à un polypeptide de référence au sens de l'invention, on entend un polypeptide ayant une activité biologique proche, mais pas nécessairement identique, de celle du polypeptide de référence tel que mesuré dans' un essai biologique convenable à la mesure de l'activité biologique du polypeptide de référence. By polypeptide having "similar activity" to a reference polypeptide in the sense of the invention is meant a polypeptide having a biological activity similar to, but not necessarily identical to, that of the reference polypeptide as measured in a suitable biological assay. to the extent of the biological activity of the reference polypeptide.
Par " antibiotique hybride " au sens de l'invention on entend un composé, généré par la construction d'une voie de biosynthèse artificielle utilisant la technologie de l'ADN recombinant. By "hybrid antibiotic" within the meaning of the invention is meant a compound generated by the construction of an artificial biosynthetic pathway using recombinant DNA technology.
<Desc/Clms Page number 25> <Desc / Clms Page number 25>
Description détaillée de l'invention
La présente invention a plus particulièrement pour objet, de nouveaux gènes de la voie de biosynthèse des spiramycines et de nouveaux polypeptides impliqués dans cette biosynthèse tels que présentés dans la description détaillée ci-dessous. Detailed description of the invention
The present invention more particularly relates to new genes of the biosynthetic pathway of spiramycins and new polypeptides involved in this biosynthesis as presented in the detailed description below.
Les gènes de la voie de biosynthèse ont été clonés et la séquence d'ADN de ces gènes a été déterminée. Les séquences obtenues ont été analysées grâce au programme FramePlot (Ishikawa J & Hotta K. 1999). On a identifié, parmi les phases ouvertes de lecture, les phases ouvertes de lecture présentant un usage des codons typiques de Streptomyces. Cette analyse a montré que cette région comporte 42 ORFs localisées de part et d'autre de cinq gènes codant l'enzyme polyketide synthase (PKS). De part et d'autre de ces cinq gènes codant des PKS, respectivement 10 et 32 ORFs ont été identifiées en aval et en amont (l'aval et l'amont étant définis par l'orientation des 5 gènes de PKS tous orientés dans le même sens) (cf. figure 3 et 37). Ainsi, les 32 phases ouvertes de lecture de ce type, occupant une région d'environ 40 kb (cf. SEQ ID N 1 présentant une première région de 31 kb contenant 25 ORFs et SEQ ID N 106 présentant une région d'environ 10,3 kb dont 1402 pb chevauchent la séquence précédente (SEQ ID N 1) et environ 9 kb correspondent à la suite de la séquence (avec cependant un région d'environ 450 nucléotides dont la séquence n'a pas été déterminée (région symbolisée par des N dans ladite séquence)), cette dernière partie de 9 kb contenant 7 ORFs supplémentaires (dont un ORF de séquence partielle, dûe à la région indéterminée), cf. également figure 3 et 37 et ci-dessous), ont été identifiées en amont des 5 gènes codant la PKS et 10 occupant une région d'environ Il,1 kb (SEQ ID N 2 et figure 3), ont été identifiées en aval des gènes de la PKS. Les 10 gènes situés en aval des 5 gènes PKS ont ainsi été nommés orfl *c, orf2*c, orf3*c, orf4*c, orf5*, orf6*, orf7*c, or/8* or/9* or/10* (SEQ ID N 3, 5, 7,9, 11, 13, 15,17, 19 et 21). Le c ajouté dans le nom du gène signifiant pour l'ORF en question que la séquence codante est dans l'orientation inverse (le brin codant est donc le brin complémentaire de la séquence donnée en SEQ ID N 2 pour ces gènes). En utilisant la même nomenclature, les 32 ORFs en amont des gènes de la PKS ont été nommés or/7, or/5, or/3, orf4, orf5, orf6, The genes of the biosynthetic pathway were cloned and the DNA sequence of these genes was determined. The sequences obtained were analyzed using the FramePlot program (Ishikawa J & Hotta K. 1999). Among the open reading phases, open reading frames with a typical codon usage of Streptomyces have been identified. This analysis showed that this region has 42 ORFs located on either side of five genes encoding the enzyme polyketide synthase (PKS). On either side of these five genes coding for PKS, respectively 10 and 32 ORFs were identified downstream and upstream (downstream and upstream being defined by the orientation of the PKS genes all oriented in the same meaning) (see Figure 3 and 37). Thus, the 32 open reading frames of this type, occupying a region of approximately 40 kb (see SEQ ID No. 1 having a first region of 31 kb containing 25 ORFs and SEQ ID N 106 having a region of about 10, 3 kb of which 1402 bp overlap the previous sequence (SEQ ID No. 1) and about 9 kb correspond to the remainder of the sequence (with however a region of about 450 nucleotides whose sequence has not been determined (region symbolized by N in said sequence)), this last part of 9 kb containing 7 additional ORFs (including a partial sequence ORF, due to the indeterminate region), see also FIGS. 3 and 37 and below), have been identified upstream 5 genes encoding PKS and occupying a region of about 11.1 kb (SEQ ID N 2 and Figure 3) were identified downstream of the PKS genes. The 10 genes located downstream of the 5 PKS genes have been named orfl * c, orf2 * c, orf3 * c, orf4 * c, orf5 *, orf6 *, orf7 * c, or / 8 * or / 9 * or / 10 * (SEQ ID NO: 3, 5, 7.9, 11, 13, 15, 17, 19 and 21). The c added in the name of the gene means for the ORF in question that the coding sequence is in the reverse orientation (the coding strand is therefore the complementary strand of the sequence given in SEQ ID No. 2 for these genes). Using the same nomenclature, the 32 ORFs upstream of the PKS genes were named gold / 7, or / 5, or / 3, orf4, orf5, orf6,
<Desc/Clms Page number 26><Desc / Clms Page number 26>
orf7, orf8, orj9c, or/70, orfllc, orfl2, orfi3c, orfl4, orfl5c, orfl6, orfl7, orfl8, orfl9,
![]()
or/20, o7c, orf22c, orf23c, orf24c, orf25c, orf26, orj27, orj28c, orf29, orf30c, orf31 et orj32c (SEQ ID N 23,25, 28,30, 34,36, 40,43, 45,47, 49,53, 60,62, 64,66, 68, 70,72, 74,76, 78,80, 82, 84, 107, 109, 111, 113, 115, 118, et 120) (cf. figure 3 et 37). orf7, orf8, orj9c, or / 70, orfllc, orfl2, orfi3c, orfl4, orfl5c, orfl6, orfl7, orfl8, orfl9,
![]()
or / 20, o7c, orf22c, orf23c, orf24c, orf25c, orf26, orj27, orj28c, orf29, orf30c, orf31 and orj32c (SEQ ID N 23,25, 28,30, 34,36, 40,43, 45,47 , 49.53, 60.62, 64.66, 68, 70.72, 74.76, 78.80, 82, 84, 107, 109, 111, 113, 115, 118, and 120) (see FIG. 3 and 37).
Les séquences protéiques déduites de ces phases ouvertes de lecture ont été comparées avec celles présentes dans différentes bases de données grâce à différents programmes : BLAST (Altschul et al., 1990) (Altschul et al., 1997), CD-search, COGs (Cluster of Orthologous Groups) (ces trois programmes sont accessibles notamment auprès du National Center for Biotechnology Information (NCBI) (Bethesda, Maryland, USA)), FASTA ((Pearson W. R & D. J. Lipman, 1988) et (Pearson W. R., 1990), BEAUTY (Worley K. C.. et al., 1995)), (ces deux programmes sont accessibles notamment auprès du centre de ressources INFOBIOGEN, Evry, France). Ces comparaisons ont permis de formuler des hypothèses sur la fonction des produits de ces gènes et d'identifier ceux susceptibles d'être impliqués dans la biosynthèse des spiramycines. The protein sequences deduced from these open reading phases were compared with those present in different databases through different programs: BLAST (Altschul et al., 1990) (Altschul et al., 1997), CD-search, COGs ( Cluster of Orthologous Groups) (these three programs are available from the National Center for Biotechnology Information (NCBI) (Bethesda, Maryland, USA)), FASTA (Pearson W. R & DJ Lipman, 1988) and (Pearson WR, 1990). BEAUTY (Worley KC et al., 1995)), (these two programs are accessible in particular from the INFOBIOGEN resource center, Evry, France). These comparisons made it possible to formulate hypotheses on the function of the products of these genes and to identify those likely to be involved in the biosynthesis of spiramycins.
Gènes situés en aval des gènes codant les PKS
Une représentation schématique de l'organisation de la région est présentée en figure 3. Ainsi qu'il sera démontré ci-dessous, sur les 10 gènes identifiés en aval des gènes codant les PKS 9 semblent impliqués dans la biosynthèse ou la résistance aux
![]()
spiramycines. Il s'agit des 9 gènes suivants : orfl *c, orf2*c, orf3*c, orf4*c, orf5*, or/6*, orf7*c, or/8* et or/9*. Genes located downstream of genes coding for PKS
A schematic representation of the organization of the region is presented in FIG. 3. As will be demonstrated below, on the 10 genes identified downstream of the genes encoding the PKS 9 appear to be involved in the biosynthesis or resistance to
![]()
spiramycin. These are the following 9 genes: orfl * c, orf2 * c, orf3 * c, orf4 * c, orf5 *, or / 6 *, orf7 * c, or / 8 * and or / 9 *.
Dans le tableau 1 suivant sont présentées les références à la séquence en ADN et en acides aminés des 10 gènes identifiés en aval des 5 gènes PKS. In the following Table 1 are presented references to the DNA and amino acid sequence of the genes identified downstream of the PKS genes.
Tableau 1

<tb>
<tb> Gène <SEP> Position <SEP> dans <SEP> la <SEP> Séquence <SEP> ADN <SEP> Séquences <SEP> polypeptidiques
<tb> séquence <SEP> SEQ <SEP> ID <SEP> N <SEP> 2
<tb> orf1*c <SEP> 10882 <SEP> à <SEP> 10172 <SEP> SEQ <SEP> ID <SEP> N <SEP> 3 <SEP> SEQ <SEP> ID <SEP> N <SEP> 4
<tb> <Tb>
<tb> Gene <SEP> Position <SEP> in <SEP><SEP> Sequence <SEP> DNA <SEP> Sequences <SEP> polypeptides
<tb> sequence <SEP> SEQ <SEP> ID <SEP> N <SEP> 2
<tb> orf1 * c <SEP> 10882 <SEP> to <SEP> 10172 <SEP> SEQ <SEP> ID <SEP> N <SEP> 3 <SEP> SEQ <SEP> ID <SEP> N <SEP> 4
<Tb>
<Desc/Clms Page number 27> <Desc / Clms Page number 27>

<tb>
<tb> orf2*c <SEP> 10052 <SEP> à <SEP> 8781 <SEP> SEQ <SEP> ID <SEP> N <SEP> 5 <SEP> SEQ <SEP> ID <SEP> N <SEP> 6 <SEP>
<tb> orf3*c <SEP> 8741 <SEP> à <SEP> 7476 <SEP> SEQ <SEP> ID <SEP> N <SEP> 7 <SEP> SEQ <SEP> ID <SEP> N <SEP> 8
<tb> orf4*c <SEP> 7459 <SEP> à <SEP> 6100 <SEP> SEQ <SEP> ID <SEP> N <SEP> 9 <SEP> SEQ <SEP> ID <SEP> N <SEP> 10 <SEP>
<tb> orf5* <SEP> 5302 <SEP> à <SEP> 5976 <SEP> SEQ <SEP> ID <SEP> N <SEP> 11 <SEP> SEQ <SEP> ID <SEP> N <SEP> 12
<tb> orf6* <SEP> 4061 <SEP> à <SEP> 5305 <SEP> SEQ <SEP> ID <SEP> N <SEP> 13 <SEP> SEQ <SEP> ID <SEP> N <SEP> 14
<tb> orf7*c <SEP> 3665 <SEP> à <SEP> 2817 <SEP> SEQ <SEP> ID <SEP> N <SEP> 15 <SEP> SEQ <SEP> ID <SEP> N <SEP> 16
<tb> orf8* <SEP> 1925 <SEP> à <SEP> 2755 <SEP> SEQ <SEP> ID <SEP> N <SEP> 17 <SEP> SEQ <SEP> ID <SEP> N <SEP> 18
<tb> orf9* <SEP> 1007 <SEP> à <SEP> 1888 <SEP> SEQ <SEP> ID <SEP> N <SEP> 19 <SEP> SEQ <SEP> ID <SEP> N <SEP> 20
<tb> orf10* <SEP> 710 <SEP> à <SEP> 937 <SEP> SEQ <SEP> ID <SEP> N <SEP> 21 <SEP> SEQ <SEP> ID <SEP> N <SEP> 22
<tb> <Tb>
<tb> orf2 * c <SEP> 10052 <SEP> to <SEP> 8781 <SEP> SEQ <SEP> ID <SEP> N <SEP> 5 <SEP> SEQ <SEP> ID <SEP> N <SEP> 6 <September>
<tb> orf3 * c <SEP> 8741 <SEP> to <SEP> 7476 <SEP> SEQ <SEP> ID <SEP> N <SEP> 7 <SEP> SEQ <SEP> ID <SEP> N <SEP> 8
<tb> orf4 * c <SEP> 7459 <SEP> to <SEP> 6100 <SEP> SEQ <SEP> ID <SEP> N <SEP> 9 <SEP> SEQ <SEP> ID <SEP> N <SEP> 10 <September>
<tb> orf5 * <SEP> 5302 <SEP> to <SEP> 5976 <SEP> SEQ <SEP> ID <SEP> N <SEP> 11 <SEP> SEQ <SEP> ID <SEP> N <SEP> 12
<tb> orf6 * <SEP> 4061 <SEP> to <SEP> 5305 <SEP> SEQ <SEP> ID <SEP> N <SEP> 13 <SEP> SEQ <SEP> ID <SEP> N <SEP> 14
<tb> orf7 * c <SEP> 3665 <SEP> to <SEP> 2817 <SEP> SEQ <SEP> ID <SEP> N <SEP> 15 <SEP> SEQ <SEP> ID <SEP> N <SEP> 16
<tb> orf8 * <SEP> 1925 <SEP> to <SEP> 2755 <SEP> SEQ <SEP> ID <SEP> N <SEP> 17 <SEP> SEQ <SEP> ID <SEP> N <SEP> 18
<tb> orf9 * <SEP> 1007 <SEP> to <SEP> 1888 <SEP> SEQ <SEP> ID <SEP> N <SEP> 19 <SEP> SEQ <SEP> ID <SEP> N <SEP> 20
<tb> orf10 * <SEP> 710 <SEP> to <SEP> 937 <SEP> SEQ <SEP> ID <SEP> N <SEP> 21 <SEP> SEQ <SEP> ID <SEP> N <SEP> 22
<Tb>
Le c ajouté dans le nom du gène indique que la séquence codante est dans l'orientation inverse (le brin codant est donc le brin complémentaire de la séquence donnée en SEQ ID N 2 pour ces gènes). The c added in the name of the gene indicates that the coding sequence is in the reverse orientation (the coding strand is therefore the complementary strand of the sequence given in SEQ ID No. 2 for these genes).
Dans le but de déterminer la fonction des polypeptides identifiés, trois types d'expériences ont été menés : la comparaison des séquences identifiées avec des séquences de fonctions connues, des expériences d'inactivation de gènes, conduisant à la construction de souches mutantes et des analyses de la production en spiramycines et en intermédiaires de biosynthèse des spiramycines par ces souches mutantes. In order to determine the function of the identified polypeptides, three types of experiments were conducted: the comparison of identified sequences with known function sequences, gene inactivation experiments, leading to the construction of mutant strains and assays production of spiramycins and spiramycin biosynthesis intermediates by these mutant strains.
Les séquences protéiques déduites de ces phases ouvertes de lecture ont tout d'abord été comparées avec celles présentes dans différentes bases de données grâce à différents programmes : BLAST (Altschul et al., 1990) (Altschul et al., 1997), CDsearch, COGs (Cluster of Orthologous Groups), FASTA ((Pearson W. R & D. J. The protein sequences deduced from these open reading phases were first compared with those present in different databases through different programs: BLAST (Altschul et al., 1990) (Altschul et al., 1997), CDsearch, COGs (Cluster of Orthologous Groups), FASTA (Pearson W. R & DJ
Lipman, 1988) et (Pearson W. R., 1990), BEAUTY (Worley K. C.. et al., 1995)). Ces comparaisons ont permis de formuler des hypothèses sur la fonction des produits de ces gènes et d'identifier ceux susceptibles d'être impliqués dans la biosynthèse de spiramycine. Le tableau 2 montre les protéines présentant une forte similitude avec les produits 10 gènes situés en aval des 5 gènes PKS. Lipman, 1988) and (Pearson W.R., 1990), BEAUTY (Worley K.C. et al., 1995)). These comparisons made it possible to formulate hypotheses on the function of the products of these genes and to identify those likely to be involved in spiramycin biosynthesis. Table 2 shows the proteins showing strong similarity to the gene products located downstream of the PKS genes.
<Desc/Clms Page number 28> <Desc / Clms Page number 28>
Tableau 2
![]()
![]()
<tb>
<tb> Produit <SEP> Protéine <SEP> présentant <SEP> une <SEP> Numéro <SEP> d'accès <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> de <SEP> gène <SEP> similitude <SEP> significative <SEP> GenBank <SEP> BLAST*
<tb> <Tb>
<tb> Product <SEP> Protein <SEP> presenting <SEP> a <SEP> Access <SEP> Number <SEP> Score <SEP> Reported <SEP> Function
<tb> of <SEP> gene <SEP> similarity <SEP> significant <SEP> GenBank <SEP> BLAST *
<Tb>
![]()
orfl *c TylMI(orf3 *) (S. fradiae) CAA57473 287 N-

![]()
orfl * c TylMI (orf3 *) (S. fradiae) CAA57473 287 N-
<tb>
<tb> méthyltransférase
<tb> orf2*c <SEP> dnrQ <SEP> gene <SEP> product <SEP> AAD <SEP> 15266 <SEP> 153 <SEP> Inconnue
<tb> (Streptomyces <SEP> peucetius)
<tb> orf3 <SEP> *c <SEP> TylM#(orf2*) <SEP> (S. <SEP> fradiae) <SEP> CAA57472 <SEP> 448 <SEP> Glycosyltransféras
<tb> e
<tb> orf4*c <SEP> Crotonyl-CoA <SEP> réductase <SEP> NP~630556 <SEP> 772 <SEP> Crotonyl-CoA
<tb> (S. <SEP> coelicolor) <SEP> réductase
<tb> or/5* <SEP> MdmC <SEP> (S. <SEP> mycarofaciens) <SEP> B42719 <SEP> 355 <SEP> Ométhyltransférase
<tb> orf6* <SEP> 3-O-acyltransférase <SEP> (S. <SEP> Q00718 <SEP> 494 <SEP> Acyltransférase
<tb> mycarofaciens)
<tb> orf7*c <SEP> MdmA <SEP> (S. <SEP> A60725 <SEP> 380 <SEP> Protéine <SEP> impliquée
<tb> mycarofaciens) <SEP> dans <SEP> la <SEP> résistance
<tb> à <SEP> la <SEP> midécamycine
<tb> orj8* <SEP> ABC-transporteur <SEP> (S. <SEP> CAC22119 <SEP> 191 <SEP> ABC-transporteur
<tb> griseus)
<tb> orf9* <SEP> ABC-transporteur <SEP> (S. <SEP> CAC22118 <SEP> 269 <SEP> ABC-transporteur
<tb> griseus)
<tb> orf10* <SEP> putative <SEP> small <SEP> conserved <SEP> NP-627432 <SEP> 109 <SEP> Inconnue
<tb> hypothetical <SEP> protein <SEP> (S.
<tb> coelicolor)
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> methyltransferase
<tb> orf2 * c <SEP> dnrQ <SEP> gene <SEP> product <SEP> ADF <SEP> 15266 <SEP> 153 <SEP> Unknown
<tb> (Streptomyces <SEP> peucetius)
<tb> orf3 <SEP> * c <SEP> TylM # (orf2 *) <SEP> (S. <SEP> fradiae) <SEQ> CAA57472 <SEQ> 448 <SEP> Glycosyltransferase
<tb> e
<tb> orf4 * c <SEP> Crotonyl-CoA <SEP> reductase <SEP> NP ~ 630556 <SEQ> 772 <SEP> Crotonyl-CoA
<tb> (S. <SEP> coelicolor) <SEP> reductase
<tb> or / 5 * <SEP> MdmC <SEP> (S. <SEP> Mycarofaciens) <SEP> B42719 <SEP> 355 <SEP> Omethyltransferase
<tb> orf6 * <SEP> 3-O-acyltransferase <SEP> (S. <SEP> Q00718 <SEP> 494 <SEP> Acyltransferase
<tb> mycarofaciens)
<tb> orf7 * c <SEP> MdmA <SEP> (S. <SEP> A60725 <SEP> 380 <SEP> Protein <SEP> involved
<tb> mycarofaciens) <SEP> in <SEP> the <SEP> resistance
<tb> to <SEP> the <SEP> midecamycin
<tb> orj8 * <SEP> ABC-transporter <SEP> (S. <SEP> CAC22119 <SEP> 191 <SEP> ABC-transporter
<tb> griseus)
<tb> orf9 * <SEP> ABC-transporter <SEP> (S. <SEP> CAC22118 <SEP> 269 <SEP> ABC-transporter
<tb> griseus)
<tb> orf10 * <SEP> putative <SEP> small <SEP> conserved <SEP> NP-627432 <SEQ> 109 <SEP> Not known
<tb> hypothetical <SEP> protein <SEP> (S.
<tb> coelicolor)
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Des expériences d'inactivation de gènes ont été réalisées pour confirmer ces résultats. Gene inactivation experiments were performed to confirm these results.
Les méthodes utilisées consistent à effectuer un remplacement de gène. Le gène cible à interrompre est remplacé par une copie de ce gène interrompue par une cassette conférant la résistance à un antibiotique (par exemple l'apramycine, la généticine ou The methods used consist in performing a gene replacement. The target gene to be interrupted is replaced by a copy of this gene interrupted by a cassette conferring resistance to an antibiotic (for example apramycin, geneticin or
<Desc/Clms Page number 29><Desc / Clms Page number 29>
l'hygromycine). Les cassettes utilisées sont bordées de part et d'autre par des codons de terminaison de la traduction dans toutes les phases de lecture et par des terminateurs de transcription actifs chez Streptomyces. L'insertion de la cassette dans le gène cible peut s'accompagner ou non d'une délétion dans ce gène cible. La taille des régions flanquant la cassette peut aller de quelques centaines à plusieurs milliers de paires de bases. Un deuxième type de cassettes peut être utilisé pour l'inactivation de gènes : des cassettes dites cassettes excisables . Ces cassettes présentent l'avantage de pouvoir être excisées chez Streptomyces par un événement de recombinaison de site spécifique après avoir été introduites dans le génome de S. ambofaciens. Le but est d'inactiver certains gènes dans des souches de Streptomyces sans laisser dans la souche finale de marqueurs de sélection ou de grandes séquences d'ADN n'appartenant pas à la souche. Après excision il subsiste uniquement une courte séquence d'une trentaine de paires de bases (appelé site cicatriciel ) dans le génome de la souche (cf. figure 10). La mise en oeuvre de ce système consiste, dans un premier temps, au remplacement de la copie sauvage du gène cible (grâce à deux événements de recombinaison homologue, cf. figure 9) par une construction dans laquelle une cassette excisable a été insérée dans ce gène cible. L'insertion de cette cassette est accompagnée d'une délétion dans le gène cible (cf. figure 9). Dans un deuxième temps, l'excision de la cassette excisable du génome de la souche est provoquée. La cassette excisable fonctionne grâce à un système de recombinaison site spécifique et a pour avantage de permettre l'obtention de mutants de Streptomyces ne portant finalement pas de gène de résistance. On s' affranchit également d'éventuels effets polaires sur l'expression des gènes situés en aval du ou des gènes inactivés (cf. figure 10). Les souches ainsi construites ont été testées pour leur production en spiramycines.
![]()
![]()
L'inactivation des gènes orfl *c, orf2*c, orf3*c et orf4*c n'a pas été effectuée car les expériences de comparaison de séquence ont permis de déterminer que ces gènes avaient une similitude relativement importante avec des gènes impliqués dans la biosynthèse d'un antibiotique relativement proche. Ainsi, le gène orfl *c code une protéine présentant une identité de 66% (déterminée grâce au programme BLAST) avec la protéine codée par gène tylMI qui code une N-méthyltransférase impliquée dans la Inactivation of the orfl * c, orf2 * c, orf3 * c and orf4 * c genes was not performed because the sequence comparison experiments determined that these genes had a relatively high similarity to genes involved in the biosynthesis of a relatively close antibiotic. Thus, the orfl * c gene encodes a protein having an identity of 66% (determined by the BLAST program) with the tylMI gene encoded protein which encodes an N-methyltransferase involved in the
<Desc/Clms Page number 30><Desc / Clms Page number 30>
biosynthèse de tylosine et catalysant la 3 N-méthylation durant la production du mycaminose chez Streptomyces fradiae (Gandecha,A.R. et al., 1997 ; Numéro d'accès GenBank: CAA57473 ; Score BLAST: 287). Cette similitude avec une protéine impliquée dans la voie de biosynthèse d'un autre antibiotique relativement proche et plus particulièrement dans la biosynthèse du mycaminose suggère que le gène orfl*c code une N-methyltransférase responsable d'une N-méthylation lors de la biosynthèse de la forosamine ou du mycaminose (cf. figure 5 et 6). Cette hypothèse est étayée par le fait que la protéine codée par le gène orf1*c présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 3). tylosin biosynthesis and catalyzing 3-N-methylation during mycaminosis production in Streptomyces fradiae (Gandecha, A.R. et al., 1997; GenBank Accession Number: CAA57473; BLAST Score: 287). This similarity with a protein involved in the biosynthetic pathway of another relatively close antibiotic and more particularly in the biosynthesis of mycaminosis suggests that the orfl * c gene encodes an N-methyltransferase responsible for N-methylation during the biosynthesis of forosamine or mycaminosis (see Figure 5 and 6). This hypothesis is supported by the fact that the protein encoded by the orf1 * c gene has a strong similarity with other proteins of similar function in other organisms (see Table 3).
Tableau 3

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> méthyltransferase <SEP> (S. <SEP> antibioticus) <SEP> CAA05643 <SEP> 277 <SEP> méthyltransférase
<tb> N,N-diméthyltransferase <SEP> (S. <SEP> venezuelae) <SEP> AAC68678 <SEP> 268 <SEP> N,Ndiméthyltransférase
<tb> probable <SEP> N-méthylase <SEP> snogX <SEP> (S. <SEP> T46679 <SEP> 243 <SEP> N-méthyltransférase
<tb> nogalater)
<tb>
@ * une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> methyltransferase <SEP> (S. <SEP> antibioticus) <SEP> CAA05643 <SEP> 277 <SEP> Methyltransferase
<tb> N, N-dimethyltransferase <SEP> (S. <SEP> venezuelae) <SEP> AAC68678 <SEP> 268 <SEP> N, N-dimethyltransferase
<tb> probable <SEP> N-methylase <SEP> snogX <SEP> (S. <SEP> T46679 <SEP> 243 <SEP> N-methyltransferase
<tb> nogalater)
<Tb>
@ * greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Le gène orf2*c code une protéine présentant une relative forte similitude (35% d'identité) avec une protéine codée par le gène tylMIII codant une NDP hexose 3,4 isomérase impliquée dans la biosynthèse de la tylosine chez Streptomyces fradiae (Gandecha,A.R., et al., 1997 ; Numéro d'accès GenBank : CAA57471 ; Score BMAST :
130). Cette similitude avec une protéine impliquée dans la voie de biosynthèse d'un autre antibiotique proche et plus particulièrement dans la biosynthèse du mycaminose The orf2 * c gene encodes a protein having a relatively high similarity (35% identity) with a protein encoded by the tylMIII gene encoding a NDP hexose 3,4 isomerase involved in the biosynthesis of tylosin in Streptomyces fradiae (Gandecha, AR , et al., 1997; GenBank Access Number: CAA57471; BMAST Score:
130). This similarity with a protein involved in the biosynthetic pathway of another close antibiotic and more particularly in the biosynthesis of mycaminosis
<Desc/Clms Page number 31><Desc / Clms Page number 31>
suggère fortement que le gène orf*2c code une NDP hexose 3,4 isomérase responsable d'isomérisation lors de la biosynthèse du mycaminose (cf. figure 5). strongly suggests that the orf * 2c gene encodes a NDP hexose 3,4 isomerase responsible for isomerization during the biosynthesis of mycaminosis (see Figure 5).
Le gène orj3*c code une protéine présentant une relative forte similitude (59% d'identité) avec une protéine codée par le gène tylMII codant une glycosyltransférase impliquée dans la biosynthèse de la tylosine chez Streptomycesfradiae (Gandecha,A.R. et al., 1997; Numéro d"accès GenBank: CAA57472; Score BLAST : 448). Cette similitude avec une protéine impliquée dans la voie de biosynthèse d'un autre antibiotique proche suggère fortement que le gène orf*3c code une glycosyltransférase. The orj3 * c gene encodes a protein with relatively high similarity (59% identity) with a protein encoded by the tylMII gene encoding a glycosyltransferase involved in the biosynthesis of tylosin in Streptomycesfradiae (Gandecha, AR et al., 1997; GenBank Accession Number: CAA57472; BLAST Score: 448) This similarity to a protein involved in the biosynthetic pathway of another nearby antibiotic strongly suggests that the orf * 3c gene encodes a glycosyltransferase.
Cette hypothèse est étayée par le fait que la protéine codée par le gène or/3 *c présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 4). This hypothesis is supported by the fact that the protein encoded by the or / 3 * c gene has a strong similarity with other proteins of similar function in other organisms (see Table 4).
Tableau 4

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> Glycosyl <SEP> transférase <SEP> (S. <SEP> venezuelae) <SEP> AAC68677 <SEP> 426 <SEP> Glycosyltransférase
<tb> Glycosyltransférase <SEP> (S. <SEP> antibioticus) <SEP> CAA05642 <SEP> 425 <SEP> Glycosyltransférase
<tb> Glycosyltransférase <SEP> CAA74710 <SEP> 395 <SEP> Glycosyltransférase
<tb> (Saccharopolyspora <SEP> erythraea)
<tb> Glycosyltransférase <SEP> (S. <SEP> antibioticus) <SEP> CAA05641 <SEP> 394 <SEP> Glycosyltransférase
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> Glycosyl <SEP> transferase <SEP> (S. <SEP> venezuelae) <SEP> AAC68677 <SEP> 426 <SEP> Glycosyltransferase
<tb> Glycosyltransferase <SEP> (S. <SEP> antibioticus) <SEP> CAA05642 <SEP> 425 <SEP> Glycosyltransferase
<tb> Glycosyltransferase <SEP> CAA74710 <SEP> 395 <SEP> Glycosyltransferase
<tb> (Saccharopolyspora <SEP> erythraea)
<tb> Glycosyltransferase <SEP> (S. <SEP> antibioticus) <SEP> CAA05641 <SEP> 394 <SEP> Glycosyltransferase
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Le gène orf4 *c code une protéine présentant une relative forte similitude avec plusieurs crotonyl-CoA réductases. Notamment, la protéine codée par orf4*c possède une similitude importante avec une crotonyl CoA réductase de chez Streptomyces coelicolor (Redenbach,M et al., 1996 ; Numéro d'accès GenBank : NP~630556 ; Score The orf4 * c gene encodes a protein with relatively strong similarity to several crotonyl-CoA reductases. Notably, the protein encoded by orf4 * c has a significant similarity to a crotonyl CoA reductase from Streptomyces coelicolor (Redenbach, M et al., 1996; GenBank Accession Number: NP ~ 630556;
<Desc/Clms Page number 32><Desc / Clms Page number 32>
BLAST : 772). Cette similitude avec une protéine impliquée dans la voie de biosynthèse d'un autre antibiotique proche suggère fortement que le gène orf4*c code également une crotonyl-CoA réductase. Cette hypothèse est étayée par le fait que la protéine codée par le gène orf4*c présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 5). BLAST: 772). This similarity to a protein involved in the biosynthetic pathway of another nearby antibiotic strongly suggests that the orf4 * c gene also encodes a crotonyl-CoA reductase. This hypothesis is supported by the fact that the protein encoded by the orf4 * c gene has a strong similarity with other proteins of similar function in other organisms (see Table 5).
Tableau 5

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> trans-2-énoyl-CoA <SEP> réductase <SEP> S72400 <SEP> 764 <SEP> trans-2-énoyl-CoA
<tb> (EC1.3.1.38) <SEP> (S. <SEP> collinus) <SEP> réductase
<tb> Crotonyl <SEP> CoA <SEP> réductase <SEP> (S. <SEP> fradiae) <SEP> CAA57474 <SEP> 757 <SEP> Crotonyl <SEP> CoA <SEP> réductase
<tb> Crotonyl-CoA <SEP> réductase <SEP> (S. <SEP> AAD53915 <SEP> 747 <SEP> Crotonyl-CoA <SEP> réductase
<tb> cinnamonensis)
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> trans-2-enoyl-CoA <SEP> reductase <SEP> S72400 <SEP> 764 <SEP> trans-2-enoyl-CoA
<tb> (EC1.3.1.38) <SEP> (S. <SEP> collinus) <SEP> reductase
<tb> Crotonyl <SEP> CoA <SEP> reductase <SEP> (S. <SEP> fradiae) <SEP> CAA57474 <SEP> 757 <SEP> Crotonyl <SEP> CoA <SEP> reductase
<tb> Crotonyl-CoA <SEP> reductase <SEP> (S. <SEP> AAD53915 <SEP> 747 <SEP> Crotonyl-CoA <SEP> reductase
<tb> cinnamonensis)
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Le gène orf6* présente une certaine similitude avec le gène mdmB présent chez Streptomyces mycarofaciens (Hara et Hutchisnson, 1992 ; numéro d'accès GenBank : A42719 ; Score BLAST : producteur d'antibiotique macrolide. Chez ce producteur, le gène est impliqué dans l'acylation du cycle lactone. Le gène or/6* coderait donc une acyltransférase. Cette hypothèse est étayée par le fait que la protéine codée par le gène orf6* présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 6). The orf6 * gene has some similarity to the mdmB gene present in Streptomyces mycarofaciens (Hara and Hutchisnson, 1992, GenBank accession number: A42719, BLAST score: producer of macrolide antibiotic, in this producer the gene is involved in acylation of the lactone cycle The or / 6 * gene would therefore encode an acyltransferase This assumption is supported by the fact that the protein encoded by the orf6 * gene has a strong similarity with other proteins of similar function in other organisms (see Table 6).
<Desc/Clms Page number 33> <Desc / Clms Page number 33>
Tableau 6

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> similitude <SEP> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> AcyA <SEP> (Streptomyces <SEP> J4001 <SEP> 450 <SEP> macrolide <SEP> 3-O-acyltransférase
<tb> thermotolerans)
<tb> Midécamycine <SEP> 4"-O- <SEP> BAA09815 <SEP> 234 <SEP> Midécamycine <SEP> 4"-O-propiony
<tb> propionyl <SEP> transférase <SEP> (S. <SEP> transférase
<tb> mycarofaciens)
<tb> <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> similarity <SEP> significant <SEP> of access <SEP> BLAST *
<tb> GenBank
<tb> AcyA <SEP> (Streptomyces <SEP> J4001 <SEP> 450 <SEP> macrolide <SEP> 3-O-acyltransferase
<tb> thermotolerans)
<tb> Midecamycin <SEP> 4 "-O- <SEP> BAA09815 <SEP> 234 <SEP> Midecamycin <SEP>4" -O-propiony
<tb> propionyl <SEP> transferase <SEP> (S. <SEP> transferase
<tb> mycarofaciens)
<Tb>
![]()
MycaroseO-acyltransférase AAG13909 189 Mycarose O-acyltransférase
![]()
![]()
MycaroseO-acyltransferase AAG13909 189 Mycarose O-acyltransferase
![]()
<tb>
<tb> (Micromonospora
<tb> megalomicea <SEP> subsp. <SEP> Nigra)
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> (Micromonospora
<tb> megalomicea <SEP> subsp. <SEP> Nigra)
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
L'inactivation du gène or/6* a été réalisée par une délétion/insertion en phase et il a été montré que la souche résultante ne produit plus de spiramycine II et III mais uniquement de la spiramycine I (cf figure 1). Ceci confirme que le gène orf6* est bien impliqué dans la synthèse de spiramycine II et III. L' enzyme codée par ce gène est responsable de la formation de spiramycine II et III par fixation d'un groupement acétyl ou butyryl sur le carbone 3. Les souches n'exprimant plus la protéine codée par le gène orf6* sont particulièrement intéressantes puisqu'elles ne produisent plus de spiramycine II et III mais seulement de la spiramycine I. Comme il a été précisé ci-dessus, l'activité antibiotique de la spiramycine I est nettement supérieure à celle des spiramycines II et III (Liu et al., 1999). The inactivation of the or / 6 * gene was carried out by a deletion / insertion in phase and it was shown that the resulting strain no longer produces spiramycin II and III but only spiramycin I (see FIG. 1). This confirms that the orf6 * gene is well implicated in the synthesis of spiramycin II and III. The enzyme encoded by this gene is responsible for the formation of spiramycin II and III by attachment of an acetyl or butyryl group on carbon 3. The strains that no longer express the protein encoded by the orf6 * gene are of particular interest since they no longer produce spiramycin II and III but only spiramycin I. As noted above, the antibiotic activity of spiramycin I is significantly greater than spiramycin II and III (Liu et al., 1999 ).
Le gène or/?*code une protéine présentant une relative forte similitude avec plusieurs O-méthyltransférase. Notamment, la protéine codée par orf5* possède une similitude importante avec une O-méthyltransférase (EC 2. 1.1.-) MdmC de The or /? * Gene encodes a protein with relatively strong similarity to several O-methyltransferases. In particular, the protein encoded by orf5 * has a significant similarity with an O-methyltransferase (EC 2. 1.1.-) MdmC of
<Desc/Clms Page number 34><Desc / Clms Page number 34>
Streptomyces mycarofaciens (Hara & Hutchinson, 1992 ; Numéro d'accès GenBank :B42719; Score BLAST : 355). Cette similitude avec une protéine impliquée dans la voie de biosynthèse d'un autre antibiotique suggère fortement que le gène or/5* code également une O-méthyltransférase. Le gène orf5* serait impliqué dans la formation de précurseurs incorporés dans le cycle lactone. En effet, d'après les comparaisons de séquences, le produit du gène orf5 est également relativement proche de FkbG qui est responsable de la méthylation de l'hydroxymalonyl-ACP d'après (Wu et al., 2000 ; Hoffmeister et al., 2000 ; Numéro d'accès GenBank : AAF86386 ; Score BLAST : 247) (cf. figure 8). Cette hypothèse est étayée par le fait que la protéine codée par le gène or/5* présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 7). Streptomyces mycarofaciens (Hara & Hutchinson, 1992, GenBank accession number: B42719, BLAST score: 355). This similarity to a protein involved in the biosynthetic pathway of another antibiotic strongly suggests that the gold / 5 * gene also encodes an O-methyltransferase. The orf5 * gene would be involved in the formation of precursors incorporated into the lactone ring. In fact, according to the sequence comparisons, the product of the orf5 gene is also relatively close to FkbG which is responsible for the methylation of hydroxymalonyl-ACP according to (Wu et al., 2000, Hoffmeister et al. 2000, GenBank Access Number: AAF86386, BLAST Score: 247) (see Figure 8). This hypothesis is supported by the fact that the protein encoded by the or / 5 * gene shows strong similarity with other proteins of similar function in other organisms (see Table 7).
Tableau 7

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> Probable <SEP> O-méthyltransferase <SEP> (EC <SEP> T18553 <SEP> 223 <SEP> O-méthyltransférase
<tb> 2.1.1.-) <SEP> safC <SEP> (Myxococcus <SEP> xanthus)
<tb> 4-O-méthyltransferase <SEP> (EC <SEP> 2.1.1.-) <SEP> - <SEP> JC4004 <SEP> 222 <SEP> O-méthyltransférase
<tb> (Streptomyces <SEP> sp.)
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> Likely <SEP> O-methyltransferase <SEP> (EC <SEP> T18553 <SEP> 223 <SEP> O-methyltransferase
<tb> 2.1.1.-) <SEP> safC <SEP> (Myxococcus <SEP> xanthus)
<tb> 4-O-methyltransferase <SEP> (EC <SEP> 2.1.1.-) <SEP> - <SEP> JC4004 <SEP> 222 <SEP> O-methyltransferase
<tb> (Streptomyces <SEP> sp.)
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Grâce à l'effet polaire de l'insertion d'une cassette non excisée dans le gène or/6*, il a pu être déterminé que le gène orf 5* est essentiel à la voie de biosynthèse des spiramycines. En effet, l'insertion de la cassette excisable dans la partie codante du gène orf 6* entraîne un arrêt total de la production en spiramycines. Cependant, une fois que la cassette insérée a été excisée (et donc lorsque seul le gène orf6* est inactivé, cf exemples 14 et 15), on retrouve une production en spiramycine I. Ceci montre que le Thanks to the polar effect of inserting an uncut cassette into the or / 6 * gene, it has been determined that the orf 5 * gene is essential for the spiramycin biosynthetic pathway. Indeed, the insertion of the excisable cassette in the coding part of the orf 6 * gene entails a total cessation of spiramycin production. However, once the inserted cassette has been excised (and thus when only the orf6 * gene is inactivated, see Examples 14 and 15), spiramycin I production is found. This shows that the
<Desc/Clms Page number 35><Desc / Clms Page number 35>
gène orf 5* est essentiel à la voie de biosynthèse des spiramycines puisque son inactivation entraîne un arrêt total de la production en spiramycines. orf 5 * gene is essential to the spiramycin biosynthetic pathway since its inactivation leads to a total cessation of spiramycin production.
Le gène orf7*c code une protéine présentant une relative forte similitude avec une protéine codée par le gène mdmA de Streptomyces mycarofaciens, ce dernier codant une protéine impliquée dans la résistance à la midécamycine chez cet organisme producteur (Hara et al., 1990 ; Numéro d'accès GenBank : A60725 ; Score BLAST : 380). Cette similitude avec une protéine impliquée dans la voie de biosynthèse d'un autre antibiotique suggère fortement que le gène orf7*c code également une protéine impliquée dans la résistance à la spiramycine. Plus particulièrement, l'enzyme codée par le gène orf7*c possède une activité méthyltransférase et est impliquée dans la résistance à la spiramycine chez Streptomyces ambofaciens. Il a été démontré que ce gène confère une résistance de type MLS I, résistance dont on sait qu'elle est dûe à la monométhylation en position A2058 de l'ARN ribosomal 23S (Pernodet et al., 1996). Cette hypothèse est étayée par le fait que la protéine codée par le gène orf7*c présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 8). The orf7 * c gene encodes a protein with relatively strong similarity to a protein encoded by the mdmA gene of Streptomyces mycarofaciens, the latter encoding a protein involved in midecamycin resistance in this producing organism (Hara et al., 1990; GenBank: A60725; BLAST Score: 380). This similarity to a protein involved in the biosynthetic pathway of another antibiotic strongly suggests that the orf7 * c gene also encodes a protein involved in spiramycin resistance. More particularly, the enzyme encoded by the orf7 * c gene has methyltransferase activity and is involved in spiramycin resistance in Streptomyces ambofaciens. This gene has been shown to confer MLS I resistance, a resistance known to be due to monomethylation in the A2058 position of 23S ribosomal RNA (Pernodet et al., 1996). This hypothesis is supported by the fact that the protein encoded by the orf7 * c gene has a strong similarity with other proteins of similar function in other organisms (see Table 8).
Tableau 8
![]()
![]()
<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<Tb>
![]()
maerolide-lincosamide-streptograminB JC5319 238 23S rRNA

![]()
monerolide-lincosamide-streptograminB JC5319 238 23S rRNA
<tb>
<tb> résistance <SEP> déterminant <SEP> (S. <SEP> fradiae) <SEP> méthyltransférase
<tb> 23S <SEP> ribosomal <SEP> RNA <SEP> methyltransferase <SEP> AAL68827 <SEP> 119 <SEP> 23S <SEP> rRNA
<tb> ErmML <SEP> (Micrococcus <SEP> luteus) <SEP> méthyltransférase
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> resistance <SEP> determining <SEP> (S. <SEP> fradiae) <SEP> methyltransferase
<tb> 23S <SEP> ribosomal <SEP> RNA <SEP> methyltransferase <SEP> AAL68827 <SEP> 119 <SEP> 23S <SEP> rRNA
<tb> ErmML <SEP> (Micrococcus <SEP> luteus) <SEP> methyltransferase
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
<Desc/Clms Page number 36> <Desc / Clms Page number 36>
Le gène orf8* code une protéine présentant une relative forte similitude avec une protéine de type transporteur ABC chez Streptomyces griseus (Campelo, 2002, Numéro d'accés GenBank : CAC22119 ; Score BLAST : 191). Cette similitude avec une protéine de type transporteur ABC suggère fortement que le gène orj8* code également une protéine de type transporteur ABC pouvant être impliquée dans la résistance à la spiramycine. Cette hypothèse est étayée par le fait que la protéine codée par le gène orf8* présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 9). The orf8 * gene encodes a protein with relatively strong similarity to an ABC transporter protein in Streptomyces griseus (Campelo, 2002, GenBank Accession Number: CAC22119, BLAST Score: 191). This similarity with an ABC transporter protein strongly suggests that the orj8 * gene also encodes an ABC transporter-like protein that may be involved in spiramycin resistance. This hypothesis is supported by the fact that the protein encoded by the orf8 * gene has a strong similarity with other proteins of similar function in other organisms (see Table 9).
Tableau 9

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> d'accès <SEP> Score <SEP> Fonction
<tb> significative <SEP> GenBank <SEP> BLAST* <SEP> rapportée
<tb> AcrW <SEP> (Streptomyces <SEP> galilaeus) <SEP> BAB72060 <SEP> 94 <SEP> Transporteur
<tb> ABC
<tb> Daunorubicin <SEP> résistance <SEP> transmembrane <SEP> P32011 <SEP> 89 <SEP> Résistance <SEP> à <SEP> la
<tb> protéine <SEP> (Streptomyces <SEP> peucetius) <SEP> daunorubicine
<tb> Probable <SEP> ABC-transporter, <SEP> NP~626506 <SEP> 89 <SEP> Transporteur
<tb> transmembrane <SEP> component. <SEP> ABC
<tb> (Streptomyces <SEP> coelicolor)
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> of access <SEP> Score <SEP> Function
<tb> Significantly <SEP> GenBank <SEP> BLAST * <SEP> reported
<tb> AcrW <SEP> (Streptomyces <SEP> Galilaeus) <SEP> BAB72060 <SEP> 94 <SEP> Carrier
<tb> ABC
<tb> Daunorubicin <SEP> resistance <SEP> transmembrane <SEP> P32011 <SEP> 89 <SEP> Resistance <SEP> to <SEP> la
<tb> protein <SEP> (Streptomyces <SEP> peucetius) <SEP> daunorubicin
<tb> Likely <SEP> ABC-transporter, <SEP> NP ~ 626506 <SEP> 89 <SEP> Carrier
<tb> transmembrane <SEP> component. <SEP> ABC
<tb> (Streptomyces <SEP> coelicolor)
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Le gène or/9* code une protéine présentant une relative forte similitude avec une protéine de type transporteur ABC chez Streptomyces griseus (Campelo, 2002, Numéro d'accès GenBank : CAC22118 ; Score BLAST : 269). Cette similitude avec une protéine de type transporteur ABC suggère fortement que le gène or/9* code également une protéine de type transporteur ABC pouvant être impliquée dans la résistance à la spiramycine. Cette hypothèse est étayée par le fait que la protéine codée par le gène The or / 9 * gene encodes a protein with relatively strong similarity to an ABC transporter protein in Streptomyces griseus (Campelo, 2002, GenBank accession number: CAC22118, BLAST score: 269). This similarity to an ABC transporter protein strongly suggests that the or / 9 * gene also encodes an ABC transporter-like protein that may be involved in spiramycin resistance. This hypothesis is supported by the fact that the protein encoded by the gene
<Desc/Clms Page number 37><Desc / Clms Page number 37>
orf8* présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 10). orf8 * has a strong similarity with other proteins of similar function in other organisms (see Table 10).
Tableau 10

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> significative <SEP> Numéro <SEP> Score <SEP> Fonction
<tb> d'accès <SEP> BLAST* <SEP> rapportée
<tb> GenBank
<tb> Probable <SEP> ABC-type <SEP> transport <SEP> protein, <SEP> ATP- <SEP> NP~626505 <SEP> 231 <SEP> Transporteur
<tb> binding <SEP> component <SEP> (S. <SEP> coelicolor) <SEP> ABC
<tb> Putative <SEP> ABC <SEP> transporter <SEP> ATP-binding <SEP> NP~627624 <SEP> 228 <SEP> Transporteur
<tb> component <SEP> (Streptomyces <SEP> Coelicolor) <SEP> ABC
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> significant <SEP> Number <SEP> Score <SEP> Function
<tb> access <SEP> BLAST * <SEP> reported
<tb> GenBank
<tb> Likely <SEP> ABC-type <SEP> transport <SEP> protein, <SEP> ATP- <SEP> NP ~ 626505 <SE> 231 <SEP> Carrier
<tb> binding <SEP> component <SEP> (SE <sep> coelicolor) <SEP> ABC
<tb> Putative <SEP> ABC <SEP> Carry <SEP> ATP-binding <SEP> NP ~ 627624 <SEP> 228 <SEP> Carrier
<tb> component <SEP> (Streptomyces <SEP> Coelicolor) <SEP> ABC
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Le gène orflO* code une protéine présentant une relative forte similitude avec une protéine de fonction inconnue. Cependant des gènes similaires à orf10* sont retrouvés au milieu de plusieurs groupes de gènes impliqués dans la biosynthèse d'antibiotiques. The orflO * gene encodes a protein with relatively high similarity to a protein of unknown function. However, genes similar to orf10 * are found in the middle of several groups of genes involved in the biosynthesis of antibiotics.
Ainsi un gène proche de or/10* est retrouvé chez S. coelicolor (Redenbach et al., 1996, numéro d'accès GenBank : NP~627432, score BLAST : 109). Un gène proche (CouY) est également retrouvé chez S. rishiriensis (Wang et al., 2000, numéro d'accès GenBank : AAG29779, score BLAST 97). Thus a gene close to gold / 10 * is found in S. coelicolor (Redenbach et al., 1996, GenBank accession number: NP ~ 627432, BLAST score: 109). A close gene (CouY) is also found in S. rishiriensis (Wang et al., 2000, GenBank accession number: AAG29779, BLAST score 97).
Gènes situés en amont des gènes codant les PKS
Dans la séquence d'ADN située en amont des gènes codant les PKS (l'aval et l'amont étant défini par l'orientation des 5 gènes de PKS tous orientés dans le même sens) (cf. figure 3), 32 ORFs ont été identifiées (cf. ci-dessus). Ainsi, les 32 phases ouvertes de lecture de ce type, occupent une région d'environ 40 kb (cf. SEQ ID N 1 présentant une première région de 31 kb contenant 25 ORFs et SEQ ID N 106 Genes located upstream of genes coding for PKS
In the DNA sequence located upstream of the genes coding for PKS (downstream and upstream being defined by the orientation of the PKS genes all oriented in the same direction) (see FIG. 3), 32 ORFs were identified (see above). Thus, the 32 open reading frames of this type occupy a region of about 40 kb (see SEQ ID No. 1 having a first region of 31 kb containing 25 ORFs and SEQ ID N 106
<Desc/Clms Page number 38><Desc / Clms Page number 38>
présentant une région d'environ 10,3 kb dont 1402 pb chevauchent la séquence précédente (SEQ ID N 1) et environ 9 kb correspondent à la suite de la séquence (avec cependant un région d'environ 450 nucléotides dont la séquence n'a pas été déterminée (région symbolisée par des N dans ladite séquence)), cette dernière partie de 9 kb contient 7 ORFs supplémentaires (dont une ORF de séquence partielle, dûe à la séquence indéterminée (orf28c)), cf. également figure 3 et 37 et ci-dessous). Une représentation schématique de l'organisation de la région est présentée en figure 5 et 37. having a region of about 10.3 kb, of which 1402 bp overlap the preceding sequence (SEQ ID No. 1) and about 9 kb correspond to the rest of the sequence (with however a region of about 450 nucleotides whose sequence has not has not been determined (region symbolized by N in said sequence)), this last part of 9 kb contains 7 additional ORFs (including a partial sequence ORF, due to the indeterminate sequence (orf28c)), cf. also Figure 3 and 37 and below). A schematic representation of the organization of the region is presented in Figure 5 and 37.
Les 32 gènes identifiés ont été nommés : or/7, or/2, or/3, orf4, orf5, orf6, or/7, orf8,
![]()
orj9c, orfl0, orj11c, orfl2, orj73c, orfl4, orfi5c, orj76, orj77, orfl8, orfl9, orf20, orfllc, orJ22c, orf23c, orf24c, orfl5c, orf26, orfl7, orf28c, orfl9, orj30c, orj3l et orf32c. The 32 identified genes have been named: gold / 7, gold / 2, gold / 3, orf4, orf5, orf6, or / 7, orf8,
![]()
orj9c, orfl0, orj11c, orfl2, orj73c, orfl4, orfi5c, orj76, orj77, orfl8, orfl9, orf20, orfllc, orJ22c, orf23c, orf24c, orfl5c, orf26, orfl7, orf28c, orfl9, orj30c, orj3l and orf32c.
Dans le tableau 11suivant sont présentées les références à la séquence en ADN et en acides aminés des 32 gènes identifiés en amont des 5 gènes PKS. In the following table are presented the references to the DNA and amino acid sequence of the 32 genes identified upstream of the PKS genes.
Tableau 11

<tb>
<tb> Gène' <SEP> Position <SEP> dans <SEP> la <SEP> Séquence <SEP> ADN <SEP> Séquence(s) <SEP> polypeptidique(s)2
<tb> séquence <SEP> SEQ <SEP> ID <SEP> N <SEP> 1
<tb> orf1 <SEP> 658 <SEP> à <SEP> 1869 <SEP> SEQ <SEP> ID <SEP> N <SEP> 23 <SEP> SEQ <SEP> ID <SEP> N <SEP> 24
<tb> orf2 <SEP> 1866 <SEP> à <SEP> 2405 <SEP> SEQ <SEP> ID <SEP> N <SEP> 25 <SEP> SEQ <SEP> ID <SEP> N <SEP> 26 <SEP> et <SEP> 27
<tb> or/3 <SEP> 2402 <SEP> à <SEP> 3568 <SEP> SEQ <SEP> ID <SEP> N <SEP> 28 <SEP> SEQ <SEP> ID <SEP> N <SEP> 29
<tb> orf4 <SEP> 3565 <SEP> à <SEP> 4473 <SEP> SEQ <SEP> ID <SEP> N <SEP> 30 <SEP> SEQ <SEP> ID <SEP> N <SEP> 31, <SEP> 32 <SEP> et <SEP> 33 <SEP>
<tb> orf5 <SEP> 4457 <SEP> à <SEP> 5494 <SEP> SEQ <SEP> ID <SEP> N <SEP> 34 <SEP> SEQ <SEP> ID <SEP> N <SEP> 35
<tb> orf6 <SEP> 5491 <SEP> à <SEP> 6294 <SEP> SEQ <SEP> ID <SEP> N <SEP> 36 <SEP> SEQ <SEP> ID <SEP> N <SEP> 37, <SEP> 38 <SEP> et <SEP> 39
<tb> orf7 <SEP> 6296 <SEP> à <SEP> 7705 <SEP> SEQ <SEP> ID <SEP> N <SEP> 40 <SEP> SEQ <SEP> ID <SEP> N <SEP> 41 <SEP> et <SEP> 42
<tb> orf8 <SEP> 8011 <SEP> 1 <SEP> à <SEP> 9258 <SEP> SEQ <SEP> ID <SEP> N <SEP> 43 <SEP> SEQ <SEP> ID <SEP> N <SEP> 44
<tb> orj9c <SEP> 10081 <SEP> à <SEP> 9362 <SEP> SEQ <SEP> ID <SEP> N <SEP> 45 <SEP> SEQ <SEP> ID <SEP> N <SEP> 46
<tb> <Tb>
<tb> Gene '<SEP> Position <SEP> in <SEP><SEP> Sequence <SEP> DNA <SEP> Sequence (s) <SEP> polypeptide (s) 2
<tb> sequence <SEP> SEQ <SEP> ID <SEP> N <SEP> 1
<tb> orf1 <SEP> 658 <SEP> to <SEP> 1869 <SEP> SEQ <SEP> ID <SEP> N <SEP> 23 <SEP> SEQ <SEP> ID <SEP> N <SEP> 24
<tb> orf2 <SEP> 1866 <SEP> to <SEP> 2405 <SEP> SEQ <SEP> ID <SEP> N <SEP> 25 <SEP> SEQ <SEP> ID <SEP> N <SEP> 26 <SEP > and <SEP> 27
<tb> or / 3 <SEP> 2402 <SEP> to <SEP> 3568 <SEP> SEQ <SEP> ID <SEP> N <SEP> 28 <SEP> SEQ <SEP> ID <SEP> N <SEP> 29
<tb> orf4 <SEP> 3565 <SEP> to <SEP> 4473 <SEP> SEQ <SEP> ID <SEP> N <SEP> 30 <SEP> SEQ <SEP> ID <SEP> N <SEP> 31, <SEP> 32 <SEP> and <SEP> 33 <SEP>
<tb> orf5 <SEP> 4457 <SEP> to <SEP> 5494 <SEP> SEQ <SEP> ID <SEP> N <SEP> 34 <SEP> SEQ <SEP> ID <SEP> N <SEP> 35
<tb> orf6 <SEP> 5491 <SEP> to <SEP> 6294 <SEP> SEQ <SEP> ID <SEP> N <SEP> 36 <SEP> SEQ <SEP> ID <SEP> N <SEP> 37, <SEP> 38 <SEP> and <SEP> 39
<tb> orf7 <SEP> 6296 <SEP> to <SEP> 7705 <SEP> SEQ <SEP> ID <SEP> N <SEP> 40 <SEP> SEQ <SEP> ID <SEP> N <SEP> 41 <SEP > and <SEP> 42
<tb> orf8 <SEP> 8011 <SEP> 1 <SEP> to <SEP> 9258 <SEP> SEQ <SEP> ID <SEP> N <SEP> 43 <SEP> SEQ <SEP> ID <SEP> N <SEP > 44
<tb> orj9c <SEP> 10081 <SEP> to <SEP> 9362 <SEP> SEQ <SEP> ID <SEP> N <SEP> 45 <SEP> SEQ <SEP> ID <SEP> N <SEP> 46
<Tb>
<Desc/Clms Page number 39> <Desc / Clms Page number 39>

<tb>
<tb> orflO <SEP> 10656 <SEP> à <SEP> 12623 <SEP> SEQ <SEP> ID <SEP> N <SEP> 47 <SEP> SEQ <SEP> ID <SEP> N <SEP> 48 <SEP>
<tb> orfllc <SEP> 14482 <SEP> à <SEP> 12734 <SEP> SEQ <SEP> ID <SEP> N <SEP> 49 <SEP> SEQ <SEP> ID <SEP> N <SEP> 50, <SEP> 51 <SEP> et <SEP> 52
<tb> orfl2 <SEP> 14601 <SEP> à <SEP> 16031 <SEP> SEQ <SEP> ID <SEP> N <SEP> 53 <SEP> SEQ <SEP> ID <SEP> N <SEP> 54,55, <SEP> 56,57, <SEP> 58 <SEP> et <SEP> 59
<tb> orfl3c <SEP> 17489 <SEP> à <SEP> 16092 <SEP> SEQ <SEP> ID <SEP> N <SEP> 60 <SEP> SEQ <SEP> ID <SEP> N <SEP> 61
<tb> or/14 <SEP> 17809 <SEP> à <SEP> 18852 <SEP> SEQ <SEP> ID <SEP> N <SEP> 62 <SEP> SEQ <SEP> ID <SEP> N <SEP> 63
<tb> orfl5c <SEP> 20001 <SEP> à <SEP> 18961 <SEP> SEQ <SEP> ID <SEP> N <SEP> 64 <SEP> SEQ <SEP> ID <SEP> N <SEP> 65
<tb> or)76 <SEP> 20314 <SEP> à <SEP> 21552 <SEP> SEQ <SEP> ID <SEP> N <SEP> 66 <SEP> SEQ <SEP> ID <SEP> N <SEP> 67 <SEP> ~~~~~~~~~~
<tb> or/77 <SEP> 21609 <SEP> à <SEP> 22879 <SEP> SEQ <SEP> ID <SEP> N <SEP> 68 <SEP> SEQ <SEP> ID <SEP> N <SEP> 69 <SEP> ~~~~~~~
<tb> orfl8 <SEP> 22997 <SEP> à <SEP> 24175 <SEP> SEQ <SEP> ID <SEP> N <SEP> 70 <SEP> SEQ <SEP> ID <SEP> N <SEP> 71
<tb> orfl9 <SEP> 24177 <SEP> à <SEP> 25169 <SEP> SEQ <SEP> ID <SEP> N <SEP> 72 <SEP> SEQ <SEP> ID <SEP> N <SEP> 73 <SEP>
<tb> or/20 <SEP> 25166 <SEP> à <SEP> 26173 <SEP> SEQ <SEP> ID <SEP> N <SEP> 74 <SEP> SEQ <SEP> ID <SEP> N <SEP> 75
<tb> <Tb>
<tb> orflO <SEP> 10656 <SEP> to <SEP> 12623 <SEP> SEQ <SEP> ID <SEP> N <SEP> 47 <SEP> SEQ <SEP> ID <SEP> N <SEP> 48 <SEP >
<tb> orfllc <SEP> 14482 <SEP> to <SEP> 12734 <SEP> SEQ <SEP> ID <SEP> N <SEP> 49 <SEP> SEQ <SEP> ID <SEP> N <SEP> 50, <SEP> 51 <SEP> and <SEP> 52
<tb> orfl2 <SEP> 14601 <SEP> to <SEP> 16031 <SEQ> SEQ <SEP> ID <SEQ> N <SEP> 53 <SEQ> SEQ <SEQ> ID <SEQ> N <SEP> 54.55 , <SEP> 56.57, <SEP> 58 <SEP> and <SEP> 59
<tb> orfl3c <SEQ> 17489 <SEP> to <SEP> 16092 <SEQ> SEQ <SEP> ID <SEP> N <SEP> 60 <SEQ> SEQ <SEP> ID <SEP> N <SEP> 61
<tb> or / 14 <SEP> 17809 <SEP> to <SEP> 18852 <SEQ> SEQ <SEP> ID <SEP> N <SEP> 62 <SEQ> SEQ <SEP> ID <SEP> N <SEP> 63
<tb> orfl5c <SEP> 20001 <SEP> to <SEP> 18961 <SEP> SEQ <SEP> ID <SEP> N <SEP> 64 <SEP> SEQ <SEP> ID <SEP> N <SEP> 65
<tb> or) 76 <SEP> 20314 <SEP> to <SEP> 21552 <SEP> SEQ <SEP> ID <SEP> N <SEP> 66 <SEP> SEQ <SEP> ID <SEP> N <SEP> 67 <SEP> ~~~~~~~~~~
<tb> or / 77 <SEP> 21609 <SEP> to <SEP> 22879 <SEP> SEQ <SEP> ID <SEP> N <SEP> 68 <SEP> SEQ <SEP> ID <SEP> N <SEP> 69 <SEP> ~~~~~~~
<tb> orfl8 <SEP> 22997 <SEP> to <SEP> 24175 <SEP> SEQ <SEP> ID <SEP> N <SEP> 70 <SEP> SEQ <SEP> ID <SEP> N <SEP> 71
<tb> orfl9 <SEP> 24177 <SEP> to <SEP> 25169 <SEP> SEQ <SEP> ID <SEP> N <SEP> 72 <SEP> SEQ <SEP> ID <SEP> N <SEP> 73 <SEP >
<tb> or / 20 <SEP> 25166 <SEP> to <SEP> 26173 <SEP> SEQ <SEP> ID <SEP> N <SEP> 74 <SEP> SEQ <SEP> ID <SEP> N <SEP> 75
<Tb>
![]()
orj21c 27448 à 26216 SEQ ID N 76 SEQ ID N 77 ~~~~~~~~~~~

![]()
orj21c 27448 to 26216 SEQ ID N 76 SEQ ID N 77 ~~~~~~~~~~~
<tb>
<tb> orj22c <SEP> 28560 <SEP> à <SEP> 27445 <SEP> SEQ <SEP> ID <SEP> N <SEP> 78 <SEP> SEQ <SEP> ID <SEP> N <SEP> 79 <SEP> ~~~~~~~~~~~
<tb> orj23c <SEP> 29770 <SEP> à <SEP> 28649 <SEP> SEQ <SEP> ID <SEP> N <SEP> 80 <SEP> SEQ <SEP> ID <SEP> N <SEP> 81
<tb> orj24c <SEP> 30074 <SEP> à <SEP> 29763 <SEP> SEQ <SEP> ID <SEP> N <SEP> 82 <SEP> SEQ <SEP> ID <SEP> N <SEP> 83
<tb> orj25c <SEP> 30937 <SEP> à <SEP> 30071 <SEP> SEQ <SEP> ID <SEP> N <SEP> 84 <SEP> SEQ <SEP> ID <SEP> N <SEP> 85
<tb> Gène1 <SEP> Position <SEP> dans <SEP> la <SEP> Séquence <SEP> ADN <SEP> Séquence(s) <SEP> polypeptidique(s)2
<tb> séquence <SEP> SEQ <SEP> ID <SEP> NO <SEP> : <SEP>
<tb> 106
<tb> or/26 <SEP> 1647 <SEP> à <SEP> 2864 <SEP> SEQ <SEP> ID <SEP> N <SEP> 107 <SEP> SEQ <SEP> ID <SEP> N <SEP> 108
<tb> or/27 <SEP> 2914 <SEP> à <SEP> 3534 <SEP> SEQ <SEP> ID <SEP> N <SEP> 109 <SEP> SEQ <SEP> ID <SEP> N <SEP> 110
<tb> orf28c <SEP> débute <SEP> à <SEP> la <SEP> position <SEP> 4973 <SEP> SEQ <SEP> ID <SEP> N <SEP> 111 <SEP> SEQ <SEP> ID <SEP> N <SEP> 112
<tb> or/29 <SEP> 5662 <SEP> à <SEP> 6669 <SEP> SEQ <SEP> ID <SEP> N <SEP> 113 <SEP> SEQ <SEP> ID <SEP> N <SEP> 114
<tb> orf30c <SEP> 7729 <SEP> à <SEP> 6692 <SEP> SEQ <SEP> ID <SEP> N <SEP> 115 <SEP> SEQ <SEP> ID <SEP> N <SEP> 116 <SEP> et <SEP> 117
<tb> or/31 <SEP> 7760 <SEP> à <SEP> 8734 <SEP> SEQ <SEP> ID <SEP> N <SEP> 118 <SEP> SEQ <SEP> ID <SEP> N <SEP> 119
<tb> <Tb>
<tb> orj22c <SEP> 28560 <SEP> to <SEP> 27445 <SEQ> SEQ <SEP> ID <SEP> N <SEP> 78 <SEQ> SEQ <SEP> ID <SEP> N <SEP> 79 <SEP > ~~~~~~~~~~~
<tb> orj23c <SEP> 29770 <SEP> to <SEP> 28649 <SEQ> SEQ <SEP> ID <SEP> N <SEP> 80 <SEQ> SEQ <SEP> ID <SEP> N <SEP> 81
<tb> orj24c <SEP> 30074 <SEP> to <SEP> 29763 <SEP> SEQ <SEP> ID <SEP> N <SEP> 82 <SEP> SEQ <SEP> ID <SEP> N <SEP> 83
<tb> orj25c <SEP> 30937 <SEP> to <SEP> 30071 <SEQ> SEQ <SEP> ID <SEP> N <SEP> 84 <SEQ> SEQ <SEP> ID <SEP> N <SEP> 85
<tb> Gene1 <SEP> Position <SEP> in <SEP><SEP> Sequence <SEP> DNA <SEP> Sequence (s) <SEP> polypeptide (s) 2
<tb> sequence <SEP> SEQ <SEP> ID <SEP> NO <SEP>: <SEP>
<tb> 106
<tb> or / 26 <SEP> 1647 <SEP> to <SEP> 2864 <SEP> SEQ <SEP> ID <SEP> N <SEP> 107 <SEP> SEQ <SEP> ID <SEP> N <SEP> 108
<tb> or / 27 <SEP> 2914 <SEP> to <SEP> 3534 <SEP> SEQ <SEP> ID <SEP> N <SEP> 109 <SEP> SEQ <SEP> ID <SEP> N <SEP> 110
<tb> orf28c <SEP> starts <SEP> at <SEP><SEP> position <SEP> 4973 <SEP> SEQ <SEP> ID <SEP> N <SEP> 111 <SEP> SEQ <SEP> ID <SEP > N <SEP> 112
<tb> or / 29 <SEP> 5662 <SEP> to <SEP> 6669 <SEP> SEQ <SEP> ID <SEP> N <SEP> 113 <SEP> SEQ <SEP> ID <SEP> N <SEP> 114
<tb> orf30c <SEP> 7729 <SEP> to <SEP> 6692 <SEP> SEQ <SEP> ID <SEP> N <SEP> 115 <SEP> SEQ <SEP> ID <SEP> N <SEP> 116 <SEP > and <SEP> 117
<tb> or / 31 <SEP> 7760 <SEP> to <SEP> 8734 <SEP> SEQ <SEP> ID <SEP> N <SEP> 118 <SEP> SEQ <SEP> ID <SEP> N <SEP> 119
<Tb>
<Desc/Clms Page number 40> <Desc / Clms Page number 40>

<tb>
<tb> orf32c <SEP> 10209 <SEP> à <SEP> 8983 <SEP> SEQIDN <SEP> 120 <SEP> SEQIDN <SEP> 121
<tb> <Tb>
<tb> orf32c <SEP> 10209 <SEP> to <SEP> 8983 <SEP> SEQIDN <SEP> 120 <SEP> SEQIDN <SEP> 121
<Tb>
1 Le c ajouté dans le nom du gène indique que la séquence codante est dans l'orientation inverse (le brin codant est donc le brin complémentaire de la séquence donnée en SEQ ID N 1 ou SEQ ID N 106 pour ces gènes). The c added in the gene name indicates that the coding sequence is in the reverse orientation (the coding strand is therefore the complementary strand of the sequence given in SEQ ID No. 1 or SEQ ID No. 106 for these genes).
2 Lorsque plusieurs séquences protéiques sont indiquées pour une seule orf, les protéines correspondantes sont issues de plusieurs sites possibles de démarrage de la traduction. When several protein sequences are indicated for a single orf, the corresponding proteins come from several possible sites for starting the translation.
Dans le but de déterminer la fonction des polypeptides identifiés dans le tableau 11 ci-dessus, trois types d'expériences ont été menées : la comparaison de l'identité des séquences identifiées avec des séquences de fonctions connues, des expériences d'inactivation de gènes et des analyses de la production en spiramycines de ces souches mutantes. In order to determine the function of the polypeptides identified in Table 11 above, three types of experiments were conducted: the comparison of the identity of the identified sequences with known sequences of functions, gene inactivation experiments and analyzes of the spiramycin production of these mutant strains.
Les séquences protéiques déduites de ces phases ouvertes de lecture ont tout d'abord été comparées avec celles présentes dans différentes bases de données grâce à différents programmes : BLAST (Altschul et al., 1990) (Altschul et al., 1997), CDsearch, COGs (Cluster of Orthologous Groups), FASTA ((Pearson W. R & D. J. The protein sequences deduced from these open reading phases were first compared with those present in different databases through different programs: BLAST (Altschul et al., 1990) (Altschul et al., 1997), CDsearch, COGs (Cluster of Orthologous Groups), FASTA (Pearson W. R & DJ
Lipman, 1988) et (Pearson W. R., 1990), BEAUTY (Worley K. C.. et al., 1995)), (cf. cidessus). Ces comparaisons ont permis de formuler des hypothèses sur la fonction des produits de ces gènes et d'identifier ceux susceptibles d'être impliqués dans la biosynthèse de spiramycine. Le tableau 12 montre les protéines présentant une forte similitude avec les 32 gènes situés en amont des 5 gènes PKS. Lipman, 1988) and (Pearson W.R., 1990), BEAUTY (Worley K.C. et al., 1995)), (see above). These comparisons made it possible to formulate hypotheses on the function of the products of these genes and to identify those likely to be involved in spiramycin biosynthesis. Table 12 shows the proteins showing a strong similarity with the 32 genes located upstream of the 5 PKS genes.
Tableau 12
![]()
![]()
<tb>
<tb> Gène <SEP> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction
<tb> significative <SEP> d'accès <SEP> BLAST* <SEP> rapportée
<tb> GenBank
<tb> <Tb>
<tb> Gene <SEP> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function
<tb> significant <SEP> of access <SEP> BLAST * <SEP> reported
<tb> GenBank
<Tb>
![]()
orfl Cytochrome P450 tylI (Streptomyces S49051 530 Cytochrome
![]()
![]()
orfl Cytochrome P450 tylI (Streptomyces S49051 530 Cytochrome
![]()
<tb>
<tb> fradiae) <SEP> P450
<tb> <Tb>
<tb> fradiae) <SEP> P450
<Tb>
<Desc/Clms Page number 41> <Desc / Clms Page number 41>
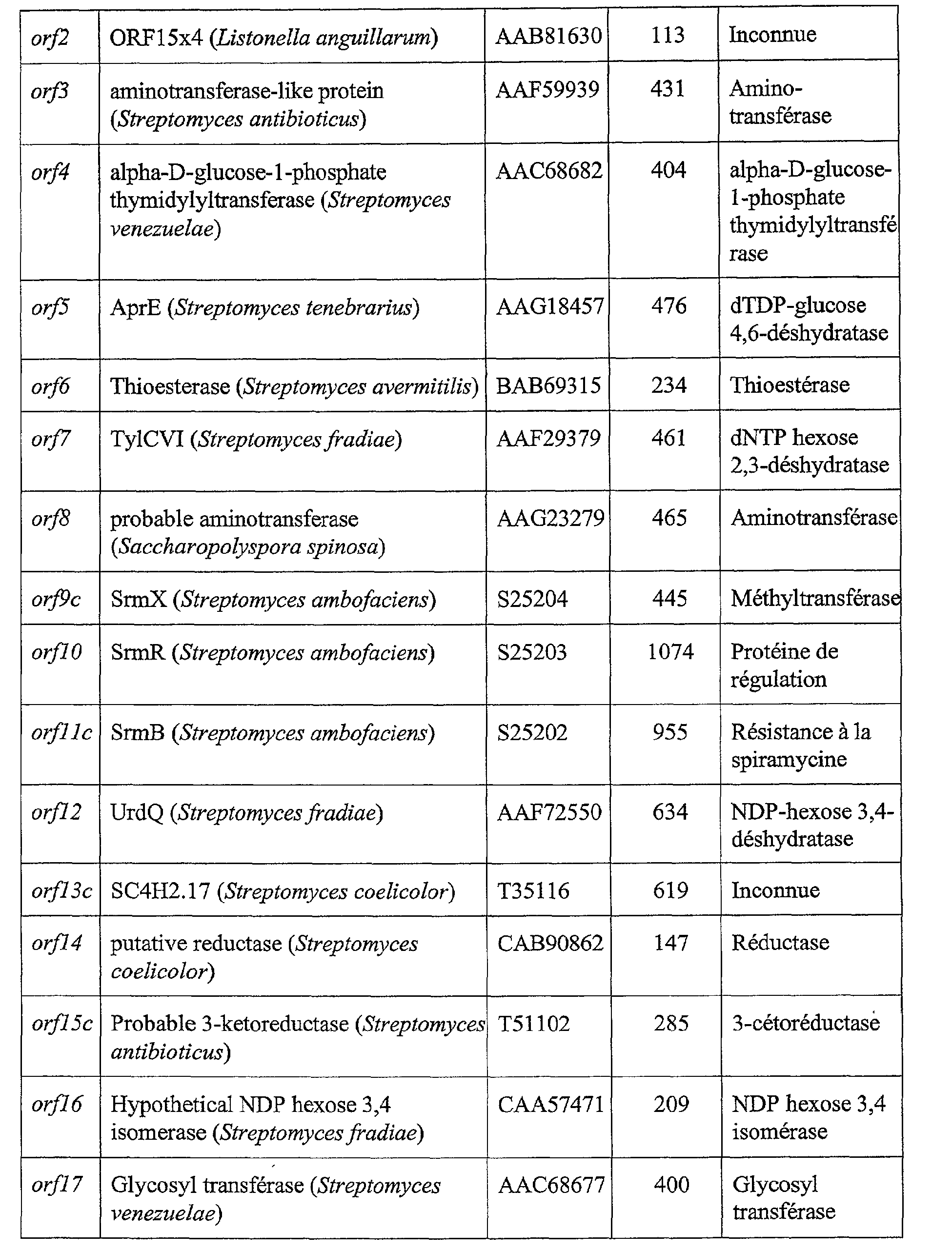
<tb>
<tb> orf2 <SEP> ORF15x4 <SEP> (Listonella <SEP> anguillarum) <SEP> AAB81630 <SEP> 113 <SEP> Inconnue
<tb> or/3 <SEP> aminotransferase-like <SEP> protein <SEP> AAF59939 <SEP> 431 <SEP> Amino-
<tb> (Streptomyces <SEP> antibioticus) <SEP> transférase
<tb> orf4 <SEP> alpha-D-glucose-1-phosphate <SEP> AAC68682 <SEP> 404 <SEP> alpha-D-glucosethymidylyltransferase <SEP> (Streptomyces <SEP> 1-phosphate
<tb> venezuelae) <SEP> thymidylyltransfé
<tb> rase
<tb> orf5 <SEP> AprE <SEP> (Streptomyces <SEP> tenebrarius) <SEP> AAG18457 <SEP> 476 <SEP> dTDP-glucose
<tb> 4,6-déshydratase
<tb> orf6 <SEP> Thioesterase <SEP> (Streptomyces <SEP> avermitilis) <SEP> BAB69315 <SEP> 234 <SEP> Thioestérase
<tb> orf7 <SEP> TylCVI <SEP> (Streptomyces <SEP> fradiae) <SEP> AAF29379 <SEP> 461 <SEP> dNTP <SEP> hexose <SEP>
<tb> 2,3-déshydratase
<tb> orf8 <SEP> probable <SEP> aminotransferase <SEP> AAG23279 <SEP> 465 <SEP> Aminotransférase
<tb> (Saccharopolyspora <SEP> spinosa)
<tb> orj9c <SEP> SrmX <SEP> (Streptomyces <SEP> ambofaciens) <SEP> S25204 <SEP> 445 <SEP> Méthyltransférase
<tb> orf10 <SEP> SrmR <SEP> (Streptomyces <SEP> ambofaciens) <SEP> S25203 <SEP> 1074 <SEP> Protéine <SEP> de
<tb> régulation
<tb> orf11c <SEP> SrmB <SEP> (Streptomyces <SEP> ambofaciens) <SEP> S25202 <SEP> 955 <SEP> Résistance <SEP> à <SEP> la
<tb> spiramycine
<tb> or/72 <SEP> UrdQ <SEP> (Streptomyces <SEP> fradiae) <SEP> AAF72550 <SEP> 634 <SEP> NDP-hexose <SEP> 3,4déshydratase
<tb> orfl3c <SEP> SC4H2.17 <SEP> (Streptomyces <SEP> coelicolor) <SEP> T35116 <SEP> 619 <SEP> Inconnue
<tb> orfl4 <SEP> putative <SEP> reductase <SEP> (Streptomyces <SEP> CAB90862 <SEP> 147 <SEP> Réductase
<tb> coelicolor)
<tb> orf15c <SEP> Probable <SEP> 3-ketoreductase <SEP> (Streptomyces <SEP> T51102 <SEP> 285 <SEP> 3-cétoréductase
<tb> antibioticus)
<tb> or/26 <SEP> Hypothetical <SEP> NDP <SEP> hexose <SEP> 3,4 <SEP> CAA57471 <SEP> 209 <SEP> NDP <SEP> hexose <SEP> 3,4 <SEP>
<tb> isomerase <SEP> (Streptomyces <SEP> fradiae) <SEP> isomérase
<tb> orf17 <SEP> Glycosyl <SEP> transférase <SEP> (Streptomyces <SEP> AAC68677 <SEP> 400 <SEP> Glycosyl
<tb> venezuelae) <SEP> transférase
<tb> <Tb>
<tb> orf2 <SEP> ORF15x4 <SEP> (Listonella <SEP> anguillarum) <SEQ> AAB81630 <SEP> 113 <SEP> Not known
<tb> or / 3 <SEP> aminotransferase-like <SEP> protein <SEP> AAF59939 <SEQ> 431 <SEP> Amino
<tb> (Streptomyces <SEP> antibioticus) <SEP> transferase
<tb> orf4 <SEP> alpha-D-glucose-1-phosphate <SEP> AAC68682 <SEP> 404 <SEP> alpha-D-glucosethymidylyltransferase <SEP> (Streptomyces <SEP> 1-phosphate
<tb> venezuelae) <SEP> thymidylyltransfé
<tb> shaves
<tb> orf5 <SEP> After <SEP> (Streptomyces <SEP> tenebrarius) <SEP> AAG18457 <SEQ> 476 <SEQ> dTDP-glucose
<tb> 4,6-dehydratase
<tb> orf6 <SEP> Thioesterase <SEP> (Streptomyces <SEP> avermitilis) <SEP> BAB69315 <SEP> 234 <SEP> Thioesterase
<tb> orf7 <SEP> TylCVI <SEP> (Streptomyces <SEP> fradiae) <SEP> AAF29379 <SEQ> 461 <SEP> dNTP <SEP> hexose <SEP>
<tb> 2,3-dehydratase
<tb> orf8 <SEP> probable <SEP> aminotransferase <SEP> AAG23279 <SEQ> 465 <SEP> Aminotransferase
<tb> (Saccharopolyspora <SEP> spinosa)
<tb> orj9c <SEP> SrmX <SEP> (Streptomyces <SEP> ambofaciens) <SEP> S25204 <SEQ> 445 <SEP> Methyltransferase
<tb> orf10 <SEP> SrmR <SEP> (Streptomyces <SEP> ambofaciens) <SEP> S25203 <SEQ> 1074 <SEP> Protein <SEP> from
<tb> regulation
<tb> orf11c <SEP> SrmB <SEP> (Streptomyces <SEP> ambofaciens) <SEP> S25202 <SEP> 955 <SEP> Resistance <SEP> to <SEP> la
<tb> spiramycin
<tb> or / 72 <SEP> UrdQ <SEP> (Streptomyces <SEP> fradiae) <SEP> AAF72550 <SEP> 634 <SEP> NDP-hexose <SEP> 3,4 dehydratase
<tb> orfl3c <SEP> SC4H2.17 <SEP> (Streptomyces SEP> coelicolor) <SEP> T35116 <SEP> 619 <SEP> Unknown
<tb> orfl4 <SEP> putative <SEP> reductase <SEP> (Streptomyces <SEP> CAB90862 <SEP> 147 <SEP> Reductase
<tb> coelicolor)
<tb> orf15c <SEP> Likely <SEP> 3-ketoreductase <SEP> (Streptomyces <SEP> T51102 <SEP> 285 <SEP> 3-ketoreductase
<tb> antibioticus)
<tb> or / 26 <SEP> Hypothetical <SEP> NDP <SEP> hexose <SEP> 3,4 <SEP> CAA57471 <SEP> 209 <SEP> NDP <SEP> hexose <SEP> 3,4 <SEP>
<tb> isomerase <SEP> (Streptomyces <SEP> fradiae) <SEP> isomerase
<tb> orf17 <SEP> Glycosyl <SEP> transferase <SEP> (Streptomyces <SEP> AAC68677 <SEP> 400 <SEP> Glycosyl
<tb> venezuelae) <SEP> transferase
<Tb>
<Desc/Clms Page number 42> <Desc / Clms Page number 42>

<tb>
<tb> orfl8 <SEP> Glycosyl <SEP> transférase <SEP> (Streptomyces <SEP> AAG29785 <SEP> 185 <SEP> Glycosyl
<tb> rishiriensis) <SEP> transférase
<tb> orfl9 <SEP> NDP-hexose <SEP> 4-cétoreductase <SEP> TylCIV <SEP> AAD41822 <SEP> 266 <SEP> NDP-hexose <SEP> 4-
<tb> (Streptomyces <SEP> fradiae) <SEP> cétoréductase
<tb> or/20 <SEP> EryBII <SEP> (Saccharopolyspora <SEP> erythraea) <SEP> AAB84068 <SEP> 491 <SEP> aldocéto
<tb> réductase
<tb> or/2 <SEP> le <SEP> TylCIII <SEP> (Streptomyces <SEP> fradiae) <SEP> AAD41823 <SEP> 669 <SEP> NDP-hexose <SEP> 3-Cméthyltransférase
<tb> orj22c <SEP> FkbH <SEP> (Streptomyces <SEP> hygroscopicus) <SEP> AAF86387 <SEP> 463 <SEP> Impliqué <SEP> dans <SEP> la
<tb> biosynthèse <SEP> du
<tb> méthoxymalonyl
<tb> orf23c <SEP> FkbI <SEP> (Streptomyces <SEP> hygroscopicus) <SEP> AAF86388 <SEP> 387 <SEP> Acyl <SEP> CoA
<tb> déshydrogénase
<tb> orj24c <SEP> FkbJ <SEP> (Streptomyces <SEP> hygroscopicus) <SEP> AAF86389 <SEP> 87 <SEP> Impliqué <SEP> dans <SEP> la
<tb> biosynthèse <SEP> du
<tb> méthoxymalonyl
<tb> orf25c <SEP> FkbK <SEP> (Streptomyces <SEP> hygroscopicus) <SEP> AAF86390 <SEP> 268 <SEP> Acyl <SEP> CoA
<tb> déshydrogénase
<tb> orj26 <SEP> TylCV <SEP> (Streptomyces <SEP> fradiae) <SEP> AAD41824 <SEP> 471 <SEP> Mycarosyl
<tb> transferase
<tb> or/27 <SEP> TylCVII <SEP> (Streptomyces <SEP> fradiae) <SEP> AAD41825 <SEP> 243 <SEP> NDP-hexose <SEP> 3,5-
<tb> (ou <SEP> 5-) <SEP> épimerase
<tb> orf28c <SEP> AcyB2 <SEP> (Streptomyces <SEP> thermotolerans) <SEP> JC2032 <SEP> 329 <SEP> protéine
<tb> régulatrice
<tb> or/29 <SEP> Béta-manannase <SEP> (Sorangium <SEP> AAK19890 <SEP> 139 <SEP> Glycosyl
<tb> cellulosum) <SEP> hydrolase
<tb> orf30c <SEP> Epimérase <SEP> de <SEP> sucre-nucléoside- <SEP> NP~600590 <SEP> 89 <SEP> Epimérase <SEP> de
<tb> diphosphate <SEP> (Corynebacterium <SEP> sucre-nucléosideglutamicum) <SEP> diphosphate
<tb> <Tb>
<tb> orf18 <SEP> Glycosyl <SEP> transferase <SEP> (Streptomyces <SEP> AAG29785 <SEP> 185 <SEP> Glycosyl
<tb> rishiriensis) <SEP> transferase
<tb> orf19 <SEP> NDP-hexose <SEP> 4-ketoreductase <SEP> TylCIV <SEP> AAD41822 <SEP> 266 <SEP> NDP-hexose <SEP> 4-
<tb> (Streptomyces <SEP> fradiae) <SEP> Cetoreductase
<tb> or / 20 <SEP> EryBII <SEP> (Saccharopolyspora <SEP> erythraea) <SEP> AAB84068 <SEQ> 491 <SEP> aldoketo
<tb> reductase
<tb> or / 2 <SEP> the <SEP> TylCIII <SEP> (Streptomyces <SEP> fradiae) <SEP> AAD41823 <SEP> 669 <SEP> NDP-hexose <SEP> 3-C-methyltransferase
<tb> orj22c <SEP> FkbH <SEP> (Streptomyces <SEP> hygroscopicus) <SEP> AAF86387 <SEP> 463 <SEP> Involved <SEP> in <SEP> la
<tb> biosynthesis <SEP> of
<tb> methoxymalonyl
<tb> orf23c <SEP> FkbI <SEP> (Streptomyces <SEP> hygroscopicus) <SEP> AAF86388 <SEP> 387 <SEP> Acyl <SEP> CoA
<tb> dehydrogenase
<tb> orj24c <SEP> FkbJ <SEP> (Streptomyces <SEP> hygroscopicus) <SEP> AAF86389 <SEP> 87 <SEP> Involved <SEP> in <SEP>
<tb> biosynthesis <SEP> of
<tb> methoxymalonyl
<tb> orf25c <SEP> FkbK <SEP> (Streptomyces <SEP> hygroscopicus) <SEP> AAF86390 <SEQ> 268 <SEP> Acyl <SEP> CoA
<tb> dehydrogenase
<tb> orj26 <SEP> TylCV <SEP> (Streptomyces <SEP> fradiae) <SEP> AAD41824 <SEP> 471 <SEP> Mycarosyl
<tb> transferase
<tb> or / 27 <SEP> TylCVII <SEP> (Streptomyces <SEP> fradiae) <SEP> AAD41825 <SEP> 243 <SEP> NDP-hexose <SEP> 3.5-
<tb> (or <SEP> 5-) <SEP> epimerase
<tb> orf28c <SEP> AcyB2 <SEP> (Streptomyces <SEP> thermotolerans) <SEP> JC2032 <SEP> 329 <SEP> Protein
<tb> regulator
<tb> or / 29 <SEP> Beta-Mananase <SEP> (Sorangium <SEP> AAK19890 <SEQ> 139 <SEP> Glycosyl
<tb> cellulosum) <SEP> hydrolase
<tb> orf30c <SEP> Epimerase <SEP> of <SEP> sugar-nucleoside- <SEP> NP ~ 600590 <SEP> 89 <SEP> Epimerase <SEP> of
<tb> diphosphate <SEP> (Corynebacterium <SEP> sugar-nucleosideglutamicum) <SEP> diphosphate
<Tb>
<Desc/Clms Page number 43> <Desc / Clms Page number 43>

<tb>
<tb> or/37 <SEP> Oxidoréductase <SEP> (Streptomyces <SEP> NP~631148 <SEP> 261 <SEP> Oxidoréductase
<tb> coelicolor)
<tb> orf32c <SEP> Protéine <SEP> régulatrice <SEP> de <SEP> la <SEP> famille <SEP> GntR <SEP> NP~625576 <SEP> 229 <SEP> Protéine
<tb> (Streptomyces <SEP> coelicolor) <SEP> régulatrice
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> or / 37 <SEP> Oxidoreductase <SEP> (Streptomyces <SEP> NP ~ 631148 <SEP> 261 <SEQ> Oxidoreductase
<tb> coelicolor)
<tb> orf32c <SEP> Regulatory <SEP> Protein <SEP> of <SEP><SEP> Family <SEP> GntR <SEP> NP ~ 625576 <SEQ> 229 <SEP> Protein
<tb> (Streptomyces <SEP> coelicolor) <SEP> regulator
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Des expériences d'inactivation de gènes ont été réalisées pour confirmer ces résultats. Les méthodes utilisées consistent à effectuer un remplacement de gène. Le gène cible à interrompre est remplacé par une copie de ce gène interrompue par une cassette conférant la résistance à un antibiotique (par exemple l'apramycine ou l'hygromycine). Les cassettes utilisées sont bordées de part et d'autre par des codons de terminaison de la traduction dans toutes les phases de lecture et par des terminateurs de transcription actifs chez Streptomyces. L'insertion de la cassette dans le gène cible peut s'accompagner ou non d'une délétion dans ce gène cible. La taille des régions flanquant la cassette peut aller de quelques centaines à plusieurs milliers de paires de bases. Un deuxième type de cassettes peut être utilisé pour l'inactivation de gènes : des cassettes dites cassettes excisables (cf. ci dessus). Les souches ainsi construites ont été testées pour leur production en spiramycines. Gene inactivation experiments were performed to confirm these results. The methods used consist in performing a gene replacement. The target gene to be interrupted is replaced by a copy of this gene interrupted by a cassette conferring resistance to an antibiotic (for example apramycin or hygromycin). The cassettes used are bordered on both sides by translation termination codons in all the reading phases and by active transcription terminators in Streptomyces. The insertion of the cassette into the target gene may or may not be accompanied by a deletion in this target gene. The size of the regions flanking the cassette can range from a few hundred to several thousand base pairs. A second type of cassette can be used for the inactivation of genes: cassettes called excisable cassettes (see above). The strains thus constructed were tested for their production in spiramycins.
Le gène orf1 code une protéine présentant une relative forte similitude avec plusieurs cytochrome P450. Notamment, la protéine codée par orfl possède une similitude importante avec la protéine codée par le gène tylI impliquée dans la biosynthèse de tylosine chez Streptomyces fradiae (Merson-Davies,L.A. et al., 1994 ; Numéro d'accès GenBank : S49051 ; Score BLAST : 530). Cette similitude avec une protéine impliquée dans la voie de biosynthèse d'un autre antibiotique proche suggère fortement que le gène orfl code également une cytochrome P450. Cette hypothèse est étayée par le fait que la protéine codée par le gène orf1 présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 13). The orf1 gene encodes a protein with relatively strong similarity to several cytochrome P450s. Notably, the orf-encoded protein has a significant similarity to the protein encoded by the tylI gene involved in tylosin biosynthesis in Streptomyces fradiae (Merson-Davies, LA et al., 1994; GenBank Accession Number: S49051; BLAST Score). : 530). This similarity to a protein involved in the biosynthetic pathway of another nearby antibiotic strongly suggests that the orf1 gene also encodes a cytochrome P450. This hypothesis is supported by the fact that the protein encoded by the orf1 gene shows strong similarity with other proteins of similar function in other organisms (see Table 13).
<Desc/Clms Page number 44> <Desc / Clms Page number 44>
Tableau 13

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> Putative <SEP> cytochrome <SEP> P450 <SEP> YJIB <SEP> 034374 <SEP> 248 <SEP> Cytochrome <SEP> P450
<tb> (Bacillus <SEP> subtilis)
<tb> Cytochrome <SEP> P450 <SEP> 113AI <SEP> P48635 <SEP> 237 <SEP> Cytochrome <SEP> P450
<tb> (Saccharopolyspora <SEP> erythraea)
<tb> Cytochrome <SEP> P-450 <SEP> hydroxylase <SEP> AAC46023 <SEP> 208 <SEP> Cytochrome <SEP> P-450
<tb> homolog <SEP> (Streptomyces <SEP> caelestis) <SEP> hydroxylase
<tb> Cytochrome <SEP> P450 <SEP> monooxygénase <SEP> AAC64105 <SEP> 206 <SEP> Cytochrome <SEP> P450
<tb> (Streptomyces <SEP> venezuelae) <SEP> monooxygénase
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> Putative <SEP> Cytochrome <SEP> P450 <SEQ> YJIB <SEP> 034374 <SEQ> 248 <SEP> Cytochrome <SEP> P450
<tb> (Bacillus <SEP> subtilis)
<tb> Cytochrome <SEP> P450 <SEQ> 113AI <SEQ> P48635 <SEQ> 237 <SEP> Cytochrome <SEP> P450
<tb> (Saccharopolyspora <SEP> erythraea)
<tb> Cytochrome <SEP> P-450 <SEP> hydroxylase <SEP> AAC46023 <SEP> 208 <SEP> Cytochrome <SEP> P-450
<tb> homolog <SEP> (Streptomyces <SEP> caelestis) <SEP> hydroxylase
<tb> Cytochrome <SEP> P450 <SEP> Monoxygenase <SEP> AAC64105 <SEP> 206 <SEP> Cytochrome <SEP> P450
<tb> (Streptomyces <SEP> venezuelae) <SEP> monooxygenase
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Le gène or/2 code une protéine présentant une similitude avec plusieurs protéines de fonctions inconnues (cf. Tableau 14). Notamment, la protéine codée par or/2 possède une similitude relativement importante avec la protéine codée par le gène ORF15x4 de Listonella anguillarum (numéro d'accès GenBank : AAB81630 ; Score BLAST : 113) de fonction inconnue. The or / 2 gene encodes a protein with similarity to several proteins of unknown function (see Table 14). In particular, the protein encoded by or / 2 has a relatively high similarity with the protein encoded by the ORF15x4 gene of Listonella anguillarum (GenBank accession number: AAB81630; BLAST score: 113) of unknown function.
Tableau 14
<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> unknown(Campylobacterjejuni) <SEP> AAK12959 <SEP> 106 <SEP> Inconnue
<tb> <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> unknown (Campylobacterjejuni) <SEP> AAK12959 <SEP> 106 <SEP> Unknown
<Tb>
<Desc/Clms Page number 45> <Desc / Clms Page number 45>

<tb>
<tb> WcgF <SEP> (Bacteroides <SEP> fragilis) <SEP> AA <SEP> D56738 <SEP> 105 <SEP> Inconnue
<tb> unknown <SEP> (Leptospira <SEP> borgpetersenii) <SEP> AAD12964 <SEP> 96 <SEP> Inconnue
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> WcgF <SEP> (Bacteroides <SEP> fragilis) <SEQ> AA <SEP> D56738 <SEQ> 105 <SEP> Unknown
<tb> unknown <SEP> (Leptospira <SEP> borgpetersenii) <SEP> AAD12964 <SEP> 96 <SEP> Unknown
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Le gène or/2 n'ayant pas d'homologue de fonction connue, son inactivation a été réalisée. Il a pu être montré que la souche résultante ne produit plus de spiramycines. Since the or / 2 gene does not have a homologue of known function, its inactivation has been performed. It could be shown that the resulting strain no longer produces spiramycins.
Ceci confirme que le gène or/2 est bien impliqué dans la biosynthèse des spiramycines. La protéine codée par ce gène régule ou est responsable d'une étape de bioconversion essentielle à la biosynthèse des spiramycines. This confirms that the or / 2 gene is well involved in the biosynthesis of spiramycins. The protein encoded by this gene regulates or is responsible for a bioconversion step essential for the biosynthesis of spiramycins.
Le gène or/3 code une protéine présentant une relative forte similitude avec plusieurs aminotransférases. Notamment, la protéine codée par or/3 possède une similitude importante avec une aminotransférase de Streptomyces antibioticus impliquée dans la biosynthèse de l'oléandomycine (Draeger, G., et al.,1999 ; Numéro d'accès GenBank: AAF59939 ; Score BLAST: 431). Cette similitude avec une protéine impliquée dans la voie de biosynthèse d'un autre antibiotique proche suggère fortement que le gène orf3 code une 3 amino-transférase responsable de la réaction de transamination nécessaire à la biosynthèse d'un des sucres aminés des spiramycines (cf. figure 5). Cette hypothèse est étayée par le fait que la protéine codée par le gène or/3 présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 15). The or / 3 gene encodes a protein with relatively strong similarity to several aminotransferases. Notably, the gold / 3 encoded protein has a significant similarity to an aminotransferase of Streptomyces antibioticus involved in the biosynthesis of oleandomycin (Draeger, G., et al., 1999; GenBank Accession Number: AAF59939; BLAST Score: 431). This similarity with a protein involved in the biosynthetic pathway of another nearby antibiotic strongly suggests that the orf3 gene encodes a 3-aminotransferase responsible for the transamination reaction necessary for the biosynthesis of one of the amino sugars of spiramycins (cf. Figure 5). This hypothesis is supported by the fact that the protein encoded by the or / 3 gene has a strong similarity with other proteins of similar function in other organisms (see Table 15).
Tableau 15

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> Aminotransférase <SEP> (Streptomyces <SEP> T51111 <SEP> 429 <SEP> Aminotransférase
<tb> antibioticus)
<tb> <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> Aminotransferase <SEP> (Streptomyces <SEP> T51111 <SEP> 429 <SEP> Aminotransferase
<tb> antibioticus)
<Tb>
<Desc/Clms Page number 46> <Desc / Clms Page number 46>

<tb>
<tb> Transaminase <SEP> (Streptomyces <SEP> AAC68680 <SEP> 419 <SEP> Transaminase
<tb> venezuelae)
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> Transaminase <SEP> (Streptomyces <SEP> AAC68680 <SEQ> 419 <SEP> Transaminase
<tb> venezuelae)
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
L'inactivation du gène or/3 a été réalisée. Il a ainsi pu être montré que la souche résultante ne produit plus de spiramycines. Ceci confirme que le gène or/3 est bien impliqué dans la biosynthèse des spiramycines. L'enzyme codée par ce gène est donc bien responsable d'une étape de bioconversion essentielle à la biosynthèse des spiramycines. La production de spiramycines peut être complémentée par l'expression de la protéine TylB de S. fradiae (cf exemple 23). Ceci démontre que le gène or/3 code une 3 amino-transférase responsable de la réaction de transamination nécessaire à la biosynthèse du mycaminose (cf. figure 5). Comme le mycaminose est le premier sucre à être fixé sur le platénolide, il est attendu que la souche interrompue dans orf3 (OS49. 67) accumule du platénolide. Inactivation of the or / 3 gene was performed. It has thus been shown that the resulting strain no longer produces spiramycins. This confirms that the or / 3 gene is well involved in the biosynthesis of spiramycins. The enzyme encoded by this gene is therefore responsible for a bioconversion step essential for the biosynthesis of spiramycins. The production of spiramycins may be supplemented by the expression of the TylB protein of S. fradiae (cf Example 23). This demonstrates that the or / 3 gene encodes a 3-aminotransferase responsible for the transamination reaction necessary for the biosynthesis of mycaminosis (see Figure 5). Since mycaminosis is the first sugar to be fixed on platenolide, the interrupted strain in orf3 (OS49,67) is expected to accumulate platenolide.
Les intermédaires de biosynthèse de la souche interrompue dans le gène or/3 ont été étudiés (cf. exemple 20). Ces expériences ont permis de démontrer que cette souche produit deux formes de platénolide : le platénolide A et le platénolide B, la structure déduite de ces deux molécules est présentée en figure 36. Intermediates of biosynthesis of the interrupted strain in the or / 3 gene were studied (see Example 20). These experiments made it possible to demonstrate that this strain produces two forms of platenolide: platenolide A and platenolide B, the structure deduced from these two molecules is presented in FIG.
Le gène orf4 code une protéine présentant une relative forte similitude avec
![]()
plusieurs NDP-glucose synthétases. Notamment, la protéine codée par orf4 possède une similitude importante avec une alpha-D-glucose-1-phosphate thymidylyltransférase de Streptomyces venezuelae (Xue Y et al.,1998 ; Numéro d'accès GenBank : AAC68682 ; Score BLAST 404). Cette similitude avec une protéine impliquée dans la voie de biosynthèse d'un autre antibiotique proche suggère fortement que le gène orf4 code une NDP-glucose syntéthase responsable de la synthèse du NDP-glucose nécessaire à la biosynthèse des trois sucres atypiques incorporés dans la molécule de spiramycine (cf. figure 4,5 et 6). Cette hypothèse est étayée par le fait que la protéine codée par le gène The orf4 gene encodes a protein with a relatively strong similarity with
![]()
several NDP-glucose synthetases. Notably, the orf4-encoded protein has a significant similarity to Streptomyces venezuelae alpha-D-glucose-1-phosphate thymidylyltransferase (Xue Y et al., 1998; GenBank Accession Number: AAC68682; BLAST 404 Score). This similarity with a protein involved in the biosynthetic pathway of another closely related antibiotic strongly suggests that the orf4 gene encodes an NDP-glucose synthase responsible for the synthesis of NDP-glucose required for the biosynthesis of the three atypical sugars incorporated into the molecule. spiramycin (see Figures 4,5 and 6). This hypothesis is supported by the fact that the protein encoded by the gene
<Desc/Clms Page number 47><Desc / Clms Page number 47>
orf4 présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 16). orf4 has a strong similarity with other proteins of similar function in other organisms (see Table 16).
Tableau 16

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> Glucose-1-phosphate <SEP> BAA84594 <SEP> 402 <SEP> Glucose-1-phosphate
<tb> thymidyltransférase <SEP> (Streptomyces <SEP> thymidyltransférase
<tb> avermitilis)
<tb> AclY <SEP> (Streptomyces <SEP> galilaeus) <SEP> BAB72036 <SEP> 400 <SEP> dTDP-1-glucose
<tb> synthétase
<tb> Putative <SEP> glucose-1-phosphate <SEP> AAK83289 <SEP> 399 <SEP> glucose-1-phosphate
<tb> thymidyltransférase <SEP> thymidyltransférase
<tb> (Saccharopolyspora <SEP> spinosa)
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> Glucose-1-Phosphate <SEP> BAA84594 <SEP> 402 <SEP> Glucose-1-Phosphate
<tb> Thymidyltransferase <SEP> (Streptomyces <SEP> thymidyltransferase
<tb> avermitilis)
<tb> AclY <SEP> (Streptomyces <SEP> galilaeus) <SEP> BAB72036 <SEQ> 400 <SEQ> dTDP-1-glucose
<tb> synthetase
<tb> Putative <SEP> glucose-1-phosphate <SEP> AAK83289 <SEP> 399 <SEP> glucose-1-phosphate
<tb> Thymidyltransferase <SEP> thymidyltransferase
<tb> (Saccharopolyspora <SEP> spinosa)
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Le gène orf5 code une protéine présentant une relative forte similitude avec plusieurs glucose déshydratases. Notamment, la protéine codée par orf5 possède une similitude importante avec une dTDP-glucose 4,6-déshydratase de Streptomyces tenebrarius (Li,T.B. et a/.,2001; Numéro d'accès GenBank : AAG18457, Score BLAST : 476). Cette similitude avec une protéine impliquée dans la voie de biosynthèse d'un autre antibiotique proche suggère fortement que le gène orf5 code une NDP glucose déshydratase nécessaire à la biosynthèse des trois sucres atypiques incorporés dans la molécule de spiramycine (cf. figure 4,5 et 6). Cette hypothèse est étayée par le fait que la protéine codée par le gène orf5 présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 17). The orf5 gene encodes a protein with relatively strong similarity to several glucose dehydratases. Notably, the orf5 encoded protein has a significant similarity to Streptomyces tenebrarius dTDP-glucose 4,6-dehydratase (Li, T.B., et al., 2001, GenBank Accession Number: AAG18457, BLAST Score: 476). This similarity with a protein involved in the biosynthetic pathway of another close antibiotic strongly suggests that the orf5 gene encodes an NDP glucose dehydratase necessary for the biosynthesis of the three atypical sugars incorporated in the spiramycin molecule (see Figure 4,5 and 6). This hypothesis is supported by the fact that the protein encoded by the orf5 gene has a strong similarity with other proteins of similar function in other organisms (see Table 17).
<Desc/Clms Page number 48><Desc / Clms Page number 48>
Tableau 17

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> DTDP-glucose <SEP> 4,6-déshydratase <SEP> AAK83290 <SEP> 464 <SEP> DTDP-glucose <SEP> 4,6-
<tb> (Saccharopolyspora <SEP> spinosa) <SEP> déshydratase
<tb> thymidine <SEP> diphosphoglucose <SEP> 4,6- <SEP> AAA68211 <SEP> 445 <SEP> thymidine
<tb> déshydratase <SEP> (Saccharopolyspora <SEP> diphosphoglucose
<tb> erythraea) <SEP> 4,6-déshydratase
<tb> dTDPglucose <SEP> 4,6-déshydratase <SEP> (EC <SEP> S49054 <SEP> 443 <SEP> dTDPglucose <SEP> 4,6-
<tb> 4.2.1.46) <SEP> - <SEP> (Streptomyces <SEP> fradiae) <SEP> déshydratase
<tb> TDP-glucose-4,6-déshydratase <SEP> AAC68681 <SEP> 421 <SEP> TDP-glucose-4,6-
<tb> (Streptomyces <SEP> venezuelae) <SEP> déshydratase
<tb> SgcA <SEP> (Streptomyces <SEP> globisporus) <SEP> AAF13998 <SEP> 418 <SEP> dNDP-glucose-4,6déshydratase
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> DTDP-glucose <SEP> 4,6-dehydratase <SEP> AAK83290 <SEP> 464 <SEP> DTDP-glucose <SEP> 4,6-
<tb> (Saccharopolyspora <SEP> spinosa) <SEP> dehydratase
<tb> thymidine <SEP> diphosphoglucose <SEP> 4,6- <SEP> AAA68211 <SEP> 445 <SEP> thymidine
<tb> dehydratase <SEP> (Saccharopolyspora <SEP> diphosphoglucose
<tb> erythraea) <SEP> 4,6-dehydratase
<tb> dTDP glucose <SEP> 4,6-dehydratase <SEP> (EC <SEP> S49054 <SEP> 443 <SEP> dTDPglucose <SEP> 4,6-
<tb> 4.2.1.46) <SEP> - <SEP> (Streptomyces <SEP> fradiae) <SEP> dehydratase
<tb> TDP-glucose-4,6-dehydratase <SEP> AAC68681 <SEP> 421 <SEP> TDP-glucose-4,6-
<tb> (Streptomyces <SEP> venezuelae) <SEP> dehydratase
<tb> SgcA <SEP> (Streptomyces <SEP> globisporus) <SEP> AAF13998 <SEP> 418 <SEP> dNDP-glucose-4,6 dehydratase
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Le gène orf6 code une protéine présentant une relative forte similitude avec plusieurs thioestérases. Notamment, la protéine codée par orf6 possède une similitude importante avec une thioestérase de Streptomyces avermitilis (Omura, S. et al., 2001 ;
Numéro d'accès GenBank : BAB69315 ; Score BLAST : 234). Cette similitude avec une protéine impliquée dans la voie de biosynthèse d'un autre antibiotique proche suggère fortement que le gène orf6 code également une thioestérase. Cette hypothèse est étayée par le fait que la protéine codée par le gène orf6 présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 18). The orf6 gene encodes a protein with relatively strong similarity to several thioesterases. In particular, the protein encoded by orf6 has a significant similarity with Streptomyces avermitilis thioesterase (Omura, S. et al., 2001;
GenBank Access Number: BAB69315; BLAST score: 234). This similarity to a protein involved in the biosynthetic pathway of another nearby antibiotic strongly suggests that the orf6 gene also encodes a thioesterase. This hypothesis is supported by the fact that the protein encoded by the orf6 gene has a strong similarity with other proteins of similar function in other organisms (see Table 18).
<Desc/Clms Page number 49> <Desc / Clms Page number 49>
Tableau 18

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> RifR <SEP> (Amycolatopsis <SEP> mediterranei) <SEP> AAG52991 <SEP> 216 <SEP> Thioestérase
<tb> Thioestérase <SEP> - <SEP> (Streptomyces <SEP> fradiae) <SEP> S49055 <SEP> 215 <SEP> Thioestérase
<tb> Thioestérase <SEP> (Streptomyces <SEP> BAB69188 <SEP> 213 <SEP> Thioestérase
<tb> avermitilis)
<tb> Thioestérase <SEP> II <SEP> (EC <SEP> 3.1.2.-) <SEP> - <SEP> T17413 <SEP> 203 <SEP> Thioestérase
<tb> (Streptomyces <SEP> venezuelae)
<tb> Piml <SEP> protein <SEP> (Streptomyces <SEP> natalensis) <SEP> CAC20922 <SEP> 201 <SEP> Thioestérase
<tb> Thioestérase <SEP> (Streptomyces <SEP> griseus) <SEP> CAC22116 <SEP> 200 <SEP> Thioestérase
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> RifR <SEP> (Amycolatopsis <SEP> mediterranei) <SEP> AAG52991 <SEP> 216 <SEP> Thioesterase
<tb> Thioesterase <SEP> - <SEP> (Streptomyces <SEP> fradiae) <SEP> S49055 <SEP> 215 <SEP> Thioesterase
<tb> Thioesterase <SEP> (Streptomyces <SEP> BAB69188 <SEP> 213 <SEP> Thioesterase
<tb> avermitilis)
<tb> Thioesterase <SEP> II <SEP> (EC <SEP> 3.1.2.-) <SEP> - <SEP> T17413 <SEP> 203 <SEP> Thioesterase
<tb> (Streptomyces <SEP> venezuelae)
<tb> Piml <SEP> protein <SEP> (Streptomyces <SEP> natalensis) <SEP> CAC20922 <SEP> 201 <SEP> Thioesterase
<tb> Thioesterase <SEP> (Streptomyces <SEP> griseus) <SEP> CAC22116 <SEP> 200 <SEP> Thioesterase
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Le gène orf7 code une protéine présentant une relative forte similitude avec plusieurs hexose déshydratases. Notamment, la protéine codée par orf6 possède une similitude importante avec une dNTP hexose 2,3-déshydratase (codée par lé gène TylCVI) de Streptomyces fradiae impliquée dans la biosynthèse de la tylosine (MersonDavies,L.A. et al., 1994 ; Numéro d'accès GenBank : AAF29379 ; Score BLAST : 461). The orf7 gene encodes a protein with relatively strong similarity to several hexose dehydratases. Notably, the orf6 encoded protein has a significant similarity to a dNTP hexose 2,3-dehydratase (encoded by the TylCVI gene) of Streptomyces fradiae involved in the biosynthesis of tylosin (MersonDavies, LA et al., 1994; GenBank access: AAF29379; BLAST score: 461).
Cette similitude avec une protéine impliquée dans la voie de biosynthèse d'un autre antibiotique proche suggère fortement que le gène orf7 code également une hexose 2-3 déshydratase nécessaire à la biosynthèse de deux sucres atypiques incorporés dans la molécule de spiramycine (cf. figure 4 et 6). Cette hypothèse est étayée par le fait que la protéine codée par le gène orf7 présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 19). This similarity with a protein involved in the biosynthetic pathway of another close antibiotic strongly suggests that the orf7 gene also encodes a hexose 2-3 dehydratase necessary for the biosynthesis of two atypical sugars incorporated in the spiramycin molecule (see Figure 4). and 6). This hypothesis is supported by the fact that the protein encoded by the orf7 gene has a strong similarity with other proteins of similar function in other organisms (see Table 19).
<Desc/Clms Page number 50> <Desc / Clms Page number 50>
Tableau 19

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> Rifl8 <SEP> (Amycolatopsis <SEP> mediterranei) <SEP> AAG52988 <SEP> 459 <SEP> Hexose <SEP> déshydratase
<tb> SimB3 <SEP> (Streptomyces <SEP> antibioticus) <SEP> AAK06810 <SEP> 444 <SEP> dNDP-4-keto-6-déoxyglucose-2,3-déshydratase
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> Rifl8 <SEP> (Amycolatopsis <SEP> mediterranei) <SEP> AAG52988 <SEP> 459 <SEP> Hexose <SEP> dehydratase
<tb> SimB3 <SEP> (Streptomyces <SEP> antibioticus) <SEP> AAK06810 <SEQ> 444 <SEP> dNDP-4-keto-6-deoxyglucose-2,3-dehydratase
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Le gène orf8 code une protéine présentant une relative forte similitude avec plusieurs aminotransférases. Notamment, la protéine codée par or/8 possède une similitude importante avec une aminotransférase probablement impliquée dans la biosynthèse de la forosamine chez Saccharopolyspora spinosa (Waldron,C. et al., 2001 ; Numéro d'accès GenBank : AAG23279 ; score BLAST : Cette similitude avec une protéine impliquée dans la voie de biosynthèse d'un autre antibiotique proche suggère fortement que le gène orf8 code une 4 amino-transférase responsable de la réaction de transamination nécessaire à la biosynthèse de la forosamine (cf. figure 6). Cette hypothèse est étayée par le fait que la protéine codée par le gène or/8 présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 20). The orf8 gene encodes a protein with relatively strong similarity to several aminotransferases. In particular, the protein encoded by or / 8 has a significant similarity with an aminotransferase probably involved in the biosynthesis of forosamine in Saccharopolyspora spinosa (Waldron, C. et al., 2001; GenBank Accession Number: AAG23279; BLAST score: This similarity with a protein involved in the biosynthetic pathway of another close antibiotic strongly suggests that the orf8 gene encodes a 4-amino transferase responsible for the transamination reaction necessary for the biosynthesis of forosamine (see Figure 6). is supported by the fact that the protein encoded by the or / 8 gene has a strong similarity with other proteins of similar function in other organisms (see Table 20).
<Desc/Clms Page number 51><Desc / Clms Page number 51>
Tableau 20

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> Putative <SEP> amino-sugar <SEP> biosynthesis <SEP> CAA07666 <SEP> 213 <SEP> Proteine <SEP> impliquée <SEP> dans <SEP> la
<tb> protein <SEP> (Bordetella <SEP> bronchiseptica) <SEP> biosynthèse <SEP> d'amino-sucre
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> Putative <SEP> amino-sugar <SEP> biosynthesis <SEP> CAA07666 <SEP> 213 <SEP> Protein <SEP> involved <SEP> in <SEP> la
<tb> protein <SEP> (Bordetella <SEP> bronchiseptica) <SEP> biosynthesis <SEP> of amino-sugar
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
L'inactivation du gène or/8 a été réalisée. Il a pu ainsi être montré que la souche résultante ne produit plus de spiramycines. Ceci confirme que le gène or/8 est bien impliqué dans la biosynthèse des spiramycines. L'enzyme codée par ce gène est donc bien responsable d'une étape de bioconversion essentielle à la biosynthèse des spiramycines. Inactivation of the or / 8 gene was performed. It could thus be shown that the resulting strain no longer produces spiramycins. This confirms that the or / 8 gene is well involved in the biosynthesis of spiramycins. The enzyme encoded by this gene is therefore responsible for a bioconversion step essential for the biosynthesis of spiramycins.
Le gène orf9c a déjà été identifié chez Streptomyces ambofaciens et a été désigné srmX par Geistlich et al. (Geistlich,M., et al., 1992). La similitude de la protéine codée par ce gène avec plusieurs méthyltransférases impliquées dans la voie de biosynthèse d'autres antibiotiques proches suggère fortement que le gène or9c code une méthyltransférase responsable de la réaction de méthylation nécessaire à la biosynthèse du mycaminose ou de la forosamine (cf. figure 5 et 6). Cette hypothèse est étayée par le fait que la protéine codée par le gène orf9c présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 21). The orf9c gene has already been identified in Streptomyces ambofaciens and has been designated srmX by Geistlich et al. (Geistlich, M., et al., 1992). The similarity of the protein encoded by this gene to several methyltransferases involved in the biosynthetic pathway of other closely related antibiotics strongly suggests that the or9c gene encodes a methyltransferase responsible for the methylation reaction required for the biosynthesis of mycaminosis or forosamine (cf. Figure 5 and 6). This hypothesis is supported by the fact that the protein encoded by the orf9c gene has a strong similarity with other proteins of similar function in other organisms (see Table 21).
Tableau 21

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> N,N-diméthyltransférase <SEP> AAC68678 <SEP> 240 <SEP> N,N-
<tb> (Streptomyces <SEP> venezuelae) <SEP> diméthyltransférase
<tb> <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> N, N-dimethyltransferase <SEP> AAC68678 <SEP> 240 <SEP> N, N-
<tb> (Streptomyces <SEP> venezuelae) <SEP> dimethyltransferase
<Tb>
<Desc/Clms Page number 52> <Desc / Clms Page number 52>

<tb>
<tb> Méthyltransférase <SEP> (Streptomyces <SEP> CAA05643 <SEP> 232 <SEP> Méthyltransférase
<tb> antibioticus)
<tb> Putative <SEP> amino <SEP> methylase <SEP> AAF01819 <SEP> 219 <SEP> Aminométhylase
<tb> (Streptomyces <SEP> nogalater)
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> Methyltransferase <SEP> (Streptomyces <SEP> CAA05643 <SEP> 232 <SEP> Methyltransferase
<tb> antibioticus)
<tb> Putative <SEP> Amino <SEP> Methylase <SEP> AAF01819 <SEP> 219 <SEP> Aminomethylase
<tb> (Streptomyces <SEP> nogalater)
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Le gène orflO a déjà été identifié chez Streptomyces ambofaciens et a été désigné srmR par Geistlich et al. (Geistlich, M., et al., 1992). La protéine codée par ce gène est impliquée dans la régulation de la voie de biosynthèse des spiramycines chez Streptomyces ambofaciens. L'inactivation du gène orf10 a été réalisée. Il a pu ainsi être montré que la souche résultante ne produit plus de spiramycines. Ceci confirme que le gène or/10 est bien impliqué dans la biosynthèse des spiramycines. La protéine codée par ce gène est donc bien essentielle à la biosynthèse des spiramycines. The orflO gene has already been identified in Streptomyces ambofaciens and has been designated srmR by Geistlich et al. (Geistlich, M., et al., 1992). The protein encoded by this gene is involved in the regulation of the spiramycin biosynthetic pathway in Streptomyces ambofaciens. Inactivation of the orf10 gene was performed. It could thus be shown that the resulting strain no longer produces spiramycins. This confirms that the gold / 10 gene is well involved in the biosynthesis of spiramycins. The protein encoded by this gene is therefore essential for the biosynthesis of spiramycins.
D'autre part ; le point de démarrage de la traduction de orf 10 a été déterminé et il a pu être montré que la surexpression de ce gène entraine une amélioration de la production en spiramycines. Le site de démarrage de la traduction correspond à un ATG situé en amont de l'ATG proposé par Geistlich et al. (Geistlich, M., et al., 1992). Il a en outre été démontré que cette extrémité 5' est esentielle à la fonction de OrflO puisqu'un messager tronqué en 5' est inactif (cf. exemple 17). Pour obtenir l'effet recherché sur la production en spiramycines, il est donc essentiel que la surexpression de orflO soit réalisée en prenant garde de ne pas exprimer un messager tronqué en 5' de or/70. On the other hand ; the starting point of the translation of orf 10 has been determined and it has been shown that the overexpression of this gene leads to an improvement in spiramycin production. The translation start site corresponds to an ATG located upstream of the ATG proposed by Geistlich et al. (Geistlich, M., et al., 1992). It has furthermore been demonstrated that this 5 'end is essential to the OrflO function since a truncated 5' messenger is inactive (see Example 17). To obtain the desired effect on spiramycin production, it is therefore essential that overexpression of orflO be performed with care not to express a truncated 5 'gold / 70 messenger.
Le gène orfllc a déjà été identifié chez Streptomyces ambofaciens et a été désigné srmB par Geistlich et al. (Geistlich,M. et al., 1992) et Schoner et al. (Schoner B et al., 1992). La protéine codée par ce gène est impliquée dans la résistance à la spiramycine chez Streptomyces ambofaciens et est un transporteur de type ABC. The orf11c gene has already been identified in Streptomyces ambofaciens and has been designated srmB by Geistlich et al. (Geistlich, M. et al., 1992) and Schoner et al. (Schoner B et al., 1992). The protein encoded by this gene is involved in spiramycin resistance in Streptomyces ambofaciens and is an ABC transporter.
<Desc/Clms Page number 53> <Desc / Clms Page number 53>
Le gène or/72 code une protéine présentant une relative forte similitude avec plusieurs hexose déshydratases. Notamment, la protéine codée par orfl2 possède une similitude importante avec une NDP-hexose 3,4-déshydratase codée par le gène UrdQ de Streptomyces fradiae et impliquée dans la biosynthèse de la tylosine (Hoffmeister,D. et al., 2000 ; Numéro d'accès GenBank : AAF72550 ; Score BLAST : 634). Cette similitude avec une protéine impliquée dans la voie de biosynthèse d'un autre antibiotique proche suggère fortement que le gène orfl2 code une 3,4 déshydratase responsable de la réaction de déshydratation nécessaire à la biosynthèse de la forosamine (cf. figure 6). Cette hypothèse est étayée par le fait que la protéine codée par le gène orfl2 présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 22). The or / 72 gene encodes a protein with relatively strong similarity to several hexose dehydratases. In particular, the protein encoded by orf21 has a significant similarity with a NDP-hexose 3,4-dehydratase encoded by the UrdQ gene of Streptomyces fradiae and involved in the biosynthesis of tylosin (Hoffmeister, D. et al., 2000; GenBank access: AAF72550; BLAST score: 634). This similarity to a protein involved in the biosynthetic pathway of another nearby antibiotic strongly suggests that the orfl2 gene encodes a 3,4 dehydratase responsible for the dehydration reaction necessary for the biosynthesis of forosamine (see Figure 6). This hypothesis is supported by the fact that the protein encoded by the orfl2 gene has a strong similarity with other proteins of similar function in other organisms (see Table 22).
Tableau 22

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> AknP <SEP> (Streptomyces <SEP> galilaeus) <SEP> AAF73452 <SEP> 625 <SEP> 3-déshydratase
<tb> NDP-hexose <SEP> 3,4-dehydratase <SEP> homolog <SEP> AAD13547 <SEP> 624 <SEP> NDP-hexose <SEP> 3,4
<tb> (Streptomyces <SEP> cyanogenus) <SEP> déshydratase
<tb> RdmI <SEP> (Streptomyces <SEP> purpurascens) <SEP> AAL24451 <SEP> 608 <SEP> hexose-C-3déshydratase
<tb> Probable <SEP> CDP-4-keto-6-deoxyglucose- <SEP> T46528 <SEP> 602 <SEP> CDP-4-céto-6-
<tb> 3-dehydratase <SEP> (El) <SEP> (Streptomyces <SEP> déoxyglucose-3violaceoruber) <SEP> déshydratase
<tb> Probable <SEP> NDP-hexose-3,4-dehydratase <SEP> AAG23278 <SEP> 582 <SEP> NDP-hexose-3,4-
<tb> (Saccharopolyspora <SEP> spinosa) <SEP> déshydratase
<tb> <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> AknP <SEP> (Streptomyces <SEP> galilaeus) <SEP> AAF73452 <SEP> 625 <SEP> 3-dehydratase
<tb> NDP-hexose <SEP> 3,4-dehydratase <SEP> homolog <SEP> AAD13547 <SEP> 624 <SEP> NDP-hexose <SEP> 3,4
<tb> (Streptomyces <SEP> cyanogenus) <SEP> dehydratase
<tb> RdmI <SEP> (Streptomyces <SEP> purpurascens) <SEP> AAL24451 <SEP> 608 <SEP> hexose-C-3deshydratase
<tb> Likely <SEP> CDP-4-keto-6-deoxyglucose- <SEP> T46528 <SEP> 602 <SEP> CDP-4-keto-6-
<tb> 3-dehydratase <SEP> (EI) <SEP> (Streptomyces <SEP> Deoxyglucose-3violaceoruber) <SEP> Dehydratase
<tb> Likely <SEP> NDP-hexose-3,4-dehydratase <SEP> AAG23278 <SEP> 582 <SEP> NDP-3,4-hexose
<tb> (Saccharopolyspora <SEP> spinosa) <SEP> dehydratase
<Tb>
<Desc/Clms Page number 54> <Desc / Clms Page number 54>

<tb>
<tb> dNTP-hexose <SEP> déshydratase <SEP> AAC01730 <SEP> 576 <SEP> dNTP-hexose
<tb> (Amycolatopsis <SEP> mediterranei) <SEP> déshydratase
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> dNTP-hexose <SEP> dehydratase <SEP> AAC01730 <SEP> 576 <SEP> dNTP-hexose
<tb> (Amycolatopsis <SEP> mediterranei) <SEP> dehydratase
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
L'inactivation du gène or/72 a été réalisée. Il a pu être montré que la souche résultante ne produit plus de spiramycines. Ceci confirme que le gène or/72 est bien impliqué dans la biosynthèse de la spiramycine. L'enzyme codée par ce gène est donc bien responsable d'une étape de bioconversion essentielle à la biosynthèse de la spiramycine. Inactivation of the or / 72 gene was performed. It could be shown that the resulting strain no longer produces spiramycins. This confirms that the or / 72 gene is well involved in the biosynthesis of spiramycin. The enzyme encoded by this gene is therefore responsible for a bioconversion step essential for the biosynthesis of spiramycin.
Le gène orf13c code une protéine présentant une relative forte similitude avec une protéine de fonction inconnue chez Streptomyces coelicolor. Cette protéine a été nommée SC4H2. 17 (numéro d'accès GeneBank : T35116; Score BLAST: 619). La protéine codée par le gène orf13c présente également une forte similitude avec d'autres protéines d'autres organismes (cf. tableau 23). The orf13c gene encodes a protein with relatively strong similarity to a protein of unknown function in Streptomyces coelicolor. This protein has been named SC4H2. 17 (GeneBank access number: T35116; BLAST score: 619). The protein encoded by the orf13c gene is also highly similar to other proteins of other organisms (see Table 23).
Tableau 23

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> hflX <SEP> protein <SEP> (Mycobacterium <SEP> leprae) <SEP> S72938 <SEP> 473 <SEP> Inconnue
<tb> Possible <SEP> ATP/GTP-binding <SEP> protein <SEP> NP~301739 <SEP> 470 <SEP> Protéine <SEP> fixant
<tb> (Mycobacterium <SEP> leprae) <SEP> l'ATP/GTP
<tb> GTP-binding <SEP> protein <SEP> (Mycobacterium <SEP> AAK47114 <SEP> 468 <SEP> Protéine <SEP> fixant <SEP> le
<tb> tuberculosis <SEP> CDC1551) <SEP> GTP
<tb> <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> hflX <SEP> protein <SEP> (Mycobacterium <SEP> leprae) <SEP> S72938 <SEQ> 473 <SEP> Unknown
<tb> Possible <SEP> ATP / GTP-binding <SEP> protein <SEP> NP ~ 301739 <SEQ> 470 <SEP> Protein <SEP> fixing
<tb> (Mycobacterium <SEP> leprae) <SEP> ATP / GTP
<tb> GTP-binding <SEP> protein <SEP> (Mycobacterium <SEP> AAK47114 <SEQ> 468 <SEP> Protein <SEP> fixing <SEP>
<tb> tuberculosis <SEP> CDC1551) <SEP> GTP
<Tb>
<Desc/Clms Page number 55> <Desc / Clms Page number 55>
![]()
![]()
<tb>
<tb> ATP/GTP-bindingprotein <SEP> T44592 <SEP> 388 <SEP> Protéine <SEP> fixant
<tb> <Tb>
<tb> ATP / GTP-bindingprotein <SEP> T44592 <SEP> 388 <SEP> Protein <SEP> Fixing
<Tb>
![]()
(Streptomyces fradiae) l'ATP/GTP * une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). ![]()
(Streptomyces fradiae) ATP / GTP * greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
* Aucune fonction précise n'a été assignée aux protéines proches de celle codée par orf13c. L'inactivation du gène orfl3c a été réalisée dans le but d'étudier la fonction de ce gène dans la voie de biosynthèse des spiramycines chez Streptomyces ambofaciens. Il a pu être montré que la souche résultante produit des spiramycines. Ceci indique que le gène orfl3c n'est pas essentiel à la biosynthèse des spiramycines et qu'il n'est pas essentiel à la survie de la bactérie. L'enzyme codée par ce gène n'est donc pas responsable d'une étape de bioconversion essentielle à la biosynthèse des spiramycines. * No specific function has been assigned to proteins close to that encoded by orf13c. Inactivation of the orfl3c gene was performed in order to study the function of this gene in the spiramycin biosynthetic pathway in Streptomyces ambofaciens. It has been shown that the resulting strain produces spiramycins. This indicates that the orfl3c gene is not essential for the biosynthesis of spiramycins and is not essential for the survival of the bacterium. The enzyme encoded by this gene is therefore not responsible for a bioconversion step essential for spiramycin biosynthesis.
Le gène or/74 code une protéine présentant une relative forte similitude avec une réductase putative (Redenbach,M., et al.,1996 ; Bentley et al., 2002 ; numéro d'accès GenBank : CAB90862 ; Score BLAST : 147). The or / 74 gene encodes a protein with relatively strong similarity to a putative reductase (Redenbach, M, et al., 1996, Bentley et al., 2002, GenBank accession number: CAB90862, BLAST score: 147).
L'inactivation du gène orfl4 a été réalisée. Il a pu ainsi être montré que la souche résultante ne produit plus de spiramycines. Ceci confirme que le gène orfl4 est bien impliqué dans la biosynthèse des spiramycines. L'enzyme codée par ce gène est donc bien responsable d'une étape de bioconversion essentielle à la biosynthèse des spiramycines. Les intermédaires de biosynthèse de la souche interrompue dans le gène or/24 ont été étudiés (cf. exemple 20). Ces expériences ont permis de démontrer que cette souche produit du platénolide A mais ne produit pas de platénolide B (cf. figure 36). Inactivation of the orflu4 gene was performed. It could thus be shown that the resulting strain no longer produces spiramycins. This confirms that the orfl4 gene is well implicated in the biosynthesis of spiramycins. The enzyme encoded by this gene is therefore responsible for a bioconversion step essential for the biosynthesis of spiramycins. Intermediates of biosynthesis of the interrupted strain in the gold / 24 gene were studied (see Example 20). These experiments made it possible to demonstrate that this strain produces platenolide A but does not produce platenolide B (see FIG.
Le gène orf15c code une protéine présentant une relative forte similitude avec plusieurs cétoréductases. Notamment, la protéine codée par orfl5c possède une similitude importante avec une 3-cétoréductase chez Streptomyces antibioticus (Numéro d'accès GenBak : T51102, Score BLAST : Cette similitude suggère fortement que The orf15c gene encodes a protein with relatively strong similarity to several ketoreductases. Notably, the orf15c-encoded protein has a significant similarity to a 3-ketoreductase in Streptomyces antibioticus (GenBak Accession Number: T51102, BLAST Score: This similarity strongly suggests that
<Desc/Clms Page number 56><Desc / Clms Page number 56>
le gène orf15c code une 3 céto-réductase responsable de la réaction de réduction nécessaire à la biosynthèse de la forosamine (cf. figure 6). Cette hypothèse est étayée par le fait que la protéine codée par le gène orf15c présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 24). the orf15c gene encodes a 3-keto reductase responsible for the reduction reaction necessary for the biosynthesis of forosamine (see FIG. This hypothesis is supported by the fact that the protein encoded by the orf15c gene shows strong similarity with other proteins of similar function in other organisms (see Table 24).
Tableau 24

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> oxidoreductase <SEP> homolog <SEP> (Streptomyces <SEP> AAD13550 <SEP> 272 <SEP> Oxidoréductase
<tb> cyanogenus)
<tb> D-oliose <SEP> 4-cétoreductase <SEP> CAB96550 <SEP> 265 <SEP> D-oliose <SEP> 4.
<tb> <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> oxidoreductase <SEP> homolog <SEP> (Streptomyces <SEP> AAD13550 <SEP> 272 <SEP> Oxidoreductase
<tb> cyanogenus)
<tb> D-oliose <SEP> 4-ketoreductase <SEP> CAB96550 <SEP> 265 <SEP> D-oliose <SEP> 4.
<Tb>
(Streptomyces <SEP> argillaceus) <SEP> cétoréductase
<tb> AknQ <SEP> (Streptomyces <SEP> galilaeus) <SEP> AAF73453 <SEP> 263 <SEP> putative <SEP> 3.
<tb> cétoréductase
<tb> Probable <SEP> NDP-hexose-3-ketoreductase <SEP> AAG23275 <SEP> 253 <SEP> NDP-hexose-3-
<tb> (Saccharopolyspora <SEP> spinosa) <SEP> cétoréductase
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). (Streptomyces <SEP> argillaceus) <SEP> Cetoreductase
<tb> AknQ <SEP> (Streptomyces <SEP> galilaeus) <SEP> AAF73453 <SEP> 263 <SEP> putative <SEP> 3.
<tb> cetoreductase
<tb> Likely <SEP> NDP-hexose-3-ketoreductase <SEP> AAG23275 <SEP> 253 <SEP> NDP-hexose-3
<tb> (Saccharopolyspora <SEP> spinosa) <SEP> Cetoreductase
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Le gène orfl6 code une protéine présentant une relative forte similitude avec plusieurs isomérases. Notamment, la protéine codée par orfl6 possède une similitude importante avec une NDP hexose 3,4 isomérase chez Streptomyces fradiae (Gandecha et al., 1997 ; Numéro d'accès GenBak : CAA57471, Score BLAST : Cette similitude suggère fortement que le gène orfl6 code une NDP hexose 3,4 isomérase responsable de la réaction d'isomérisation nécessaire à la biosynthèse du mycarose (cf. figure 5). Cette hypothèse est étayée par le fait que la protéine codée par le gène orf16 présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 25). The orflu6 gene encodes a protein with relatively strong similarity to several isomerases. In particular, the protein encoded by orflu6 has a significant similarity with a NDP hexose 3,4 isomerase in Streptomyces fradiae (Gandecha et al., 1997; GenBak Accession Number: CAA57471, BLAST Score: This similarity strongly suggests that the orfl6 gene code a NDP hexose 3,4 isomerase responsible for the isomerization reaction necessary for the mycarose biosynthesis (see Figure 5) This hypothesis is supported by the fact that the protein encoded by the orf16 gene has a strong similarity with others proteins of similar function in other organisms (see Table 25).
<Desc/Clms Page number 57> <Desc / Clms Page number 57>
Tableau 25

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> Putative <SEP> tautomerase <SEP> (Streptomyces <SEP> AAC68676 <SEP> 145 <SEP> Tautomérase
<tb> venezuelae)
<tb> TDP-4-céto-6-déoxyhexose <SEP> 3,4- <SEP> AAG13907 <SEP> 112 <SEP> TDP-4-céto-6isomérase <SEP> (Micromonospora <SEP> déoxyhexose <SEP> 3,4megalomicea <SEP> subsp. <SEP> nigra) <SEP> isomérase
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> Putative <SEP> tautomerase <SEP> (Streptomyces <SEP> AAC68676 <SEQ> 145 <SEP> Tautomerase
<tb> venezuelae)
<tb> TDP-4-keto-6-deoxyhexose <SEP> 3,4- <SEP> AAG13907 <SEP> 112 <SEP> TDP-4-keto-6 isomerase <SEP> (Micromonospora <SEP> deoxyhexose <SEP> 3 , 4megalomicea <SEP> subsp. <SEP> nigra) <SEP> isomerase
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Le gène orfl7 code une protéine présentant une relative forte similitude avec plusieurs glycosyl transférases. Notamment, la protéine codée par or/17 possède une similitude importante avec une glycosyl transférase de Streptomyces venezuelae (Xue,Y. et al.,1998 ; Numéro d'accès GenBank : AAC68677; score BLAST: 400). La similitude de la protéine codée par le gène orfl7 avec plusieurs glycosyl transférases impliquées dans la voie de biosynthèse d'autres antibiotiques proches suggère fortement que ce gène code également une glycosyl transférase. Cette hypothèse est étayée par le fait que la protéine codée par le gène or/77 présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 26). The orf17 gene encodes a protein with relatively strong similarity to several glycosyl transferases. Notably, the gold / 17 encoded protein has a significant similarity to Streptomyces venezuelae glycosyl transferase (Xue, Y. et al., 1998; GenBank Accession Number: AAC68677; BLAST score: 400). The similarity of the protein encoded by the orfl7 gene with several glycosyl transferases involved in the biosynthetic pathway of other closely related antibiotics strongly suggests that this gene also encodes a glycosyl transferase. This hypothesis is supported by the fact that the protein encoded by the or / 77 gene has a strong similarity with other proteins of similar function in other organisms (see Table 26).
Tableau 26

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> Glycosyltransférase <SEP> (Streptomyces <SEP> CAA57472 <SEP> 399 <SEP> Glycosyltransférase
<tb> fradiae)
<tb> <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> Glycosyltransferase <SEP> (Streptomyces <SEP> CAA57472 <SEP> 399 <SEP> Glycosyltransferase
<tb> fradiae)
<Tb>
<Desc/Clms Page number 58> <Desc / Clms Page number 58>

<tb>
<tb> Glycosyltransférase <SEP> (Streptomyces <SEP> CAA05642 <SEP> 378 <SEP> Glycosyltransférase
<tb> antibioticus)
<tb> Glycosyltransférase <SEP> (Streptomyces <SEP> CAA05641 <SEP> 360 <SEP> Glycosyltransférase
<tb> antibioticus)
<tb> Glycosyltransférase <SEP> CAA74710 <SEP> 344 <SEP> Glycosyltransférase
<tb> (Saccharopolyspora <SEP> erythraea)
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> Glycosyltransferase <SEP> (Streptomyces <SEP> CAA05642 <SEP> 378 <SEP> Glycosyltransferase
<tb> antibioticus)
<tb> Glycosyltransferase <SEP> (Streptomyces <SEP> CAA05641 <SEP> 360 <SEP> Glycosyltransferase
<tb> antibioticus)
<tb> Glycosyltransferase <SEP> CAA74710 <SEP> 344 <SEP> Glycosyltransferase
<tb> (Saccharopolyspora <SEP> erythraea)
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Le gène orfl8 code une protéine présentant une relative forte similitude avec plusieurs glycosyl transférases. Notamment, la protéine codée par or/75 possède une similitude importante avec une glycosyl transférase de Streptomyces rishiriensis (Wang et a/.,2000; Numéro d'accès GenBank : AAG29785; score BLAST: 185). La similitude de la protéine codée par le gène orfl8 avec plusieurs glycosyl transférases impliquées dans la voie de biosynthèse d'autres antibiotiques proches suggère fortement que ce gène code également une glycosyl transférase. Cette hypothèse est étayée par le fait que la protéine codée par le gène orfl8 présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 27). The orf18 gene encodes a protein with relatively strong similarity to several glycosyl transferases. Notably, the gold / 75 encoded protein has a significant similarity to Streptomyces rishiriensis glycosyltransferase (Wang et al., 2000, GenBank Accession Number: AAG29785, BLAST score: 185). The similarity of the protein encoded by the orfl8 gene with several glycosyl transferases involved in the biosynthetic pathway of other nearby antibiotics strongly suggests that this gene also encodes a glycosyl transferase. This hypothesis is supported by the fact that the protein encoded by the orfl8 gene has a strong similarity with other proteins of similar function in other organisms (see Table 27).
Tableau 27

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> NovM <SEP> (Streptomyces <SEP> spheroides) <SEP> AAF67506 <SEP> 184 <SEP> Glycosyltransférase
<tb> probable <SEP> glycosyl <SEP> transferase <SEP> T46519 <SEP> 169 <SEP> Glycosyltransférase
<tb> (Streptomyces <SEP> violaceoruber)
<tb> <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> NovM <SEP> (Streptomyces <SEP> Spheroids) <SEP> AAF67506 <SEQ> 184 <SEP> Glycosyltransferase
<tb> probable <SEP> glycosyl <SEP> transferase <SEP> T46519 <SEQ> 169 <SEP> Glycosyltransferase
<tb> (Streptomyces <SEP> violaceoruber)
<Tb>
<Desc/Clms Page number 59> <Desc / Clms Page number 59>

<tb>
<tb> Glycosyl <SEP> transferasehomolog <SEP> AAD13553 <SEP> 167 <SEP> Glycosyltransférase
<tb> (Streptomyces <SEP> cyanogenus)
<tb> Glycosyl <SEP> transferase <SEP> homolog <SEP> AAD13555 <SEP> 163 <SEP> Glycosyltransférase
<tb> (Streptomyces <SEP> cyanogenus)
<tb> dNTP-hexose <SEP> glycosyl <SEP> transférase <SEP> AAC01731 <SEP> 160 <SEP> Glycosyltransférase
<tb> (Amycolatopsis <SEP> mediterranei)
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> Glycosyl <SEP> transferasehomolog <SEP> AAD13553 <SEP> 167 <SEP> Glycosyltransferase
<tb> (Streptomyces <SEP> cyanogenus)
<tb> Glycosyl <SEP> transferase <SEP> homolog <SEP> AAD13555 <SEP> 163 <SEP> Glycosyltransferase
<tb> (Streptomyces <SEP> cyanogenus)
<tb> dNTP-hexose <SEP> glycosyl <SEP> transferase <SEP> AAC01731 <SEP> 160 <SEP> Glycosyltransferase
<tb> (Amycolatopsis <SEP> mediterranei)
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Le gène or/79 code une protéine présentant une relative forte similitude avec plusieurs cétoréductases. Notamment, la protéine codée par orfl9 possède une similitude importante avec une NDP-hexose 4-cétoréductase (TylCIV) de Streptomyces fradiae (Bate et al.,2000 ; Numéro d'accès GenBank : AAD41822 ; score BLAST : 266). La similitude de la protéine codée par le gène orfl9 avec cette cétoréductase impliquée dans la voie de biosynthèse d'un antibiotique proche suggère fortement que ce gène code également une 4-cétoréductase responsable de la réaction de réduction nécessaire à la biosynthèse du mycarose (cf. figure 4). Cette hypothèse est étayée par le fait que la protéine codée par le gène orfl9 présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 28). The or / 79 gene encodes a protein with relatively strong similarity to several ketoreductases. In particular, the protein encoded by orf19 has a significant similarity with a Streptomyces fradiae NDP-hexose 4-ketoreductase (TylCIV) (Bate et al., 2000 GenBank accession number: AAD41822, BLAST score: 266). The similarity of the protein encoded by the orfl9 gene with this ketoreductase involved in the biosynthetic pathway of a nearby antibiotic strongly suggests that this gene also encodes a 4-ketoreductase responsible for the reduction reaction necessary for mycarose biosynthesis (cf. Figure 4). This hypothesis is supported by the fact that the protein encoded by the orfl9 gene has a strong similarity with other proteins of similar function in other organisms (see Table 28).
Tableau 28

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> NDP-4-céto-6-déoxyhexose <SEP> 4- <SEP> AAL14256 <SEP> 251 <SEP> NDP-4-céto-6cétoreductase <SEP> (Streptomyces <SEP> venezuelae) <SEP> déoxyhexose <SEP> 4cétoréductase
<tb> <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> NDP-4-keto-6-deoxyhexose <SEP> 4- <SEP> AAL14256 <SEP> 251 <SEP> NDP-4-keto-6-ketoreductase <SEP> (Streptomyces <SEP> venezuelae) <SEP> deoxyhexose <MS> 4cororeductase
<Tb>
<Desc/Clms Page number 60> <Desc / Clms Page number 60>

<tb>
<tb> EryBIV <SEP> (Saccharopolyspora <SEP> erythraea) <SEP> AAB84071 <SEP> 249 <SEP> oxidoréductase
<tb> TDP-4-céto-6-déoxyhexose <SEP> 4- <SEP> AAG13916 <SEP> 218 <SEP> TDP-4-céto-6cétoreductase <SEP> (Micromonospora <SEP> déoxyhexose <SEP> 4megalomicea <SEP> subsp. <SEP> Nigra) <SEP> cétoréductase
<tb> dTDP-4-céto-6-déoxy-L-hexose <SEP> 4- <SEP> BAA84595 <SEP> 212 <SEP> dTDP-4-céto-6réductase <SEP> (Streptomyces <SEP> avermitilis) <SEP> déoxy-L-hexose <SEP> 4réductase
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> EryBIV <SEP> (Saccharopolyspora <SEP> erythraea) <SEP> AAB84071 <SEP> 249 <SEP> oxidoreductase
<tb> TDP-4-keto-6-deoxyhexose <SEP> 4- <SEP> AAG13916 <SEP> 218 <SEP> TDP-4-keto-6-ketoreductase <SEP> (Micromonospora <SEP> deoxyhexose <SEP> 4megalomicea <SEP > subsp. <SEP> Nigra) <SEP> Cetoreductase
<tb> dTDP-4-keto-6-deoxy-L-hexose <SEP> 4- <SEP> BAA84595 <SEP> 212 <SEP> dTDP-4-keto-6reductase <SEP> (Streptomyces <SEP> avermitilis) <MS> deoxy-L-hexose <SEP> 4reductase
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Le gène or/20 code une protéine présentant une relative forte similitude avec plusieurs hexose réductases. Notamment, la protéine codée par or/20 possède une similitude importante avec le gène EryBII de Saccharopolyspora erythraea qui code une dTDP-4-céto-L-6-déoxy-hexose 2,3-réductase (Summers,R.G., et al.,1997), Numéro d'accès GenBank : AAB84068 ; Score BLAST : 491). La similitude de la protéine codée par le gène or/20 avec plusieurs hexose réductases impliquées dans la voie de biosynthèse d'autres antibiotiques proches suggère fortement que ce gène code une 2-3 réductase responsable de la réaction de réduction nécessaire à la biosynthèse du mycarose (cf. figure 4). Cette hypothèse est étayée par le fait que la protéine codée par le gène or/20 présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 29). The gold / 20 gene encodes a protein with relatively strong similarity to several hexose reductases. Notably, the gold / 20 encoded protein has a significant similarity to the EryBII gene of Saccharopolyspora erythraea which encodes a dTDP-4-keto-L-6-deoxy-hexose 2,3-reductase (Summers, RG, et al. 1997), GenBank Access Number: AAB84068; BLAST score: 491). The similarity of the protein encoded by the or / 20 gene with several hexose reductases involved in the biosynthetic pathway of other closely related antibiotics strongly suggests that this gene encodes a 2-3 reductase responsible for the reduction reaction required for mycarose biosynthesis. (see Figure 4). This hypothesis is supported by the fact that the protein encoded by the gold / 20 gene has a strong similarity with other proteins of similar function in other organisms (see Table 29).
Tableau 29

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> TylCII <SEP> (Streptomyces <SEP> fradiae) <SEP> AAD41821 <SEP> 464 <SEP> NDP-hexose <SEP> 2,3-
<tb> énoyl <SEP> réductase
<tb> <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> TylCII <SEP> (Streptomyces <SEP> fradiae) <SEP> AAD41821 <SEP> 464 <SEP> NDP-hexose <SEP> 2,3-
<tb> enoyl <SEP> reductase
<Tb>
<Desc/Clms Page number 61> <Desc / Clms Page number 61>

<tb>
<tb> TDP-4-céto-6-deoxyhexose <SEP> 2,3- <SEP> AAG13914 <SEP> 446 <SEP> TDP-4-céto-6réductase <SEP> (Micromonospora <SEP> déoxyhexose <SEP> 2,3megalomicea <SEP> subsp. <SEP> Nigra) <SEP> réductase
<tb> dTDP-4-céto-6-déoxy-L-hexose <SEP> 2,3- <SEP> BAA84599 <SEP> 377 <SEP> dTDP-4-céto-6réductase <SEP> (Streptomyces <SEP> avermitilis) <SEP> déoxy-L-hexose
<tb> 2,3-réductase
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> TDP-4-keto-6-deoxyhexose <SEP> 2,3- <SEP> AAG13914 <SEP> 446 <SEP> TDP-4-keto-6reductase <SEP> (Micromonospora <SEP> deoxyhexose <SEP> 2 , 3megalomicea <SEP> subsp. <SEP> Nigra) <SEP> reductase
<tb> dTDP-4-keto-6-deoxy-L-hexose <SEP> 2,3- <SEP> BAA84599 <SEP> 377 <SEP> dTDP-4-keto-6reductase <SEP> (Streptomyces <SEP> avermitilis ) <SEP> deoxy-L-hexose
<tb> 2,3-reductase
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Le gène orf21c code une protéine présentant une relative forte similitude avec plusieurs hexose méthyltransférases. Notamment, la protéine codée par orf21c possède une similitude importante avec le gène TylCIII de Streptomyces fradiae qui code une NDP-hexose 3-C-méthyltransférase (Bate,N et al., 2000 ; Numéro d'accès GenBank : AAD41823 ; Score BLAST : 669). La similitude de la protéine codée par le gène orf21c avec plusieurs hexose méthyltransférases impliquées dans la voie de biosynthèse d'autres antibiotiques proches suggère fortement que ce gène code une hexose méthyltransférase responsable de la réaction de méthylation nécessaire à la biosynthèse du mycarose (cf. figure 4). Cette hypothèse est étayée par le fait que la protéine codée par le gène orf21c présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 30). The orf21c gene encodes a protein with relatively strong similarity to several hexose methyltransferases. Notably, the orf21c-encoded protein has a significant similarity to the Streptomyces fradiae TylCIII gene that encodes an NDP-hexose 3-C-methyltransferase (Bate, N et al., 2000; GenBank Accession Number: AAD41823; BLAST Score: 669). The similarity of the protein encoded by the orf21c gene with several hexose methyltransferases involved in the biosynthetic pathway of other nearby antibiotics strongly suggests that this gene encodes a hexose methyltransferase responsible for the methylation reaction required for the mycarose biosynthesis (see figure 4). This hypothesis is supported by the fact that the protein encoded by the orf21c gene has a strong similarity with other proteins of similar function in other organisms (see Table 30).
Tableau 30
<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> eryH <SEP> (Saccharopolyspora <SEP> erythraea) <SEP> 228448 <SEP> 592 <SEP> Gène <SEP> de
<tb> biosynthèse <SEP> de
<tb> l' <SEP> érythromycine <SEP>
<tb> <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> eryH <SEP> (Saccharopolyspora <SEP> erythraea) <SEP> 228448 <SE> 592 <SEP> Gene <SEP> of
<tb> biosynthesis <SEP> of
<tb><SEP> erythromycin <SEP>
<Tb>
<Desc/Clms Page number 62> <Desc / Clms Page number 62>

<tb>
<tb> S-adenosyl-dependent <SEP> methyl <SEP> AAK71270 <SEP> 358 <SEP> Méthyltransferase
<tb> transferase <SEP> (Coxiella <SEP> burnetii)
<tb> NovU <SEP> (Streptomyces <SEP> spheroides) <SEP> AAF67514 <SEP> 184 <SEP> C-méthyltransférase
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> S-adenosyl-dependent <SEP> methyl <SEP> AAK71270 <SEP> 358 <SEP> Methyltransferase
<tb> transferase <SEP> (Coxiella <SEP> burnetii)
<tb> NovU <SEP> (Streptomyces <SEP> spheroids) <SEP> AAF67514 <SEP> 184 <SEP> C-methyltransferase
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Le gène orf22c code une protéine présentant une relative forte similitude avec la protéine codée par le gène fkbH de Streptomyces hygroscopicus var. ascomyceticus qui code une enzyme impliquée dans la biosynthèse du méthoxymalonyl (Wu,K. et al.,2000; Numéro d'accès GenBank : AAF86387 ; Score BLAST : 463). La similitude de la protéine codée par le gène orj22c avec cette protéine impliquée dans la voie de biosynthèse d'un autre macrolide proche suggère fortement que ce gène code également une enzyme impliquée dans la biosynthèse du méthoxymalonyl chez Streptomyces ambofaciens (cf. figure 8). The orf22c gene encodes a protein with relatively high similarity to the protein encoded by the fkbH gene of Streptomyces hygroscopicus var. ascomyceticus that encodes an enzyme involved in the biosynthesis of methoxymalonyl (Wu, K et al., 2000; GenBank Accession Number: AAF86387; BLAST Score: 463). The similarity of the protein encoded by the orj22c gene with this protein involved in the biosynthetic pathway of another nearby macrolide strongly suggests that this gene also encodes an enzyme involved in the biosynthesis of methoxymalonyl in Streptomyces ambofaciens (see Figure 8).
Le gène orj23c code une protéine présentant une relative forte similitude avec la protéine codée par le gène fkbI de Streptomyces hygroscopicus var. ascomyceticus qui code une acyl CoA déshydrogénase impliquée dans la biosynthèse du méthoxymalonyl (Wu,K. et al., 2000 ; Numéro d'accès GenBank : AAF86388 ; Score BLAST 387). La similitude de la protéine codée par le gène orj23c avec plusieurs acyl CoA déshydrogénases impliquées dans la voie de biosynthèse d'autres antibiotiques proches suggère fortement que ce gène code une acyl CoA déshydrogénase impliquée dans la biosynthèse du méthoxymalonyl (cf. figure 8). Cette hypothèse est étayée par le fait que la protéine codée par le gène orf23c présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 31). The orj23c gene encodes a protein with relatively strong similarity to the protein encoded by the fkbI gene of Streptomyces hygroscopicus var. ascomyceticus which encodes an acyl CoA dehydrogenase involved in the biosynthesis of methoxymalonyl (Wu, K et al., 2000; GenBank Accession Number: AAF86388; BLAST Score 387). The similarity of the protein encoded by the orj23c gene with several acyl CoA dehydrogenases involved in the biosynthetic pathway of other nearby antibiotics strongly suggests that this gene encodes an acyl CoA dehydrogenase involved in the biosynthesis of methoxymalonyl (see Figure 8). This hypothesis is supported by the fact that the protein encoded by the orf23c gene has a strong similarity with other proteins of similar function in other organisms (see Table 31).
<Desc/Clms Page number 63><Desc / Clms Page number 63>
Tableau 31

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> Acyl-CoA <SEP> déshydrogénase <SEP> AAK19892 <SEP> 171 <SEP> Acyl-CoA
<tb> (Polyangium <SEP> cellulosum) <SEP> déshydrogénase
<tb> Probable <SEP> acyl-CoA <SEP> dehydrogenase <SEP> - <SEP> T36802 <SEP> 160 <SEP> acyl-CoA
<tb> (Streptomyces <SEP> coelicolor) <SEP> déshydrogénase
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> Acyl-CoA <SEP> dehydrogenase <SEP> AAK19892 <SEP> 171 <SEP> Acyl-CoA
<tb> (Polyangium <SEP> cellulosum) <SEP> dehydrogenase
<tb> Likely <SEP> acyl-CoA <SEP> dehydrogenase <SEP> - <SEP> T36802 <SEP> 160 <SEP> acyl-CoA
<tb> (Streptomyces <SEP> coelicolor) <SEP> dehydrogenase
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Le gène orf24c code une protéine présentant une relative forte similitude avec la protéine codée par le gène fkbJ de Streptomyces hygroscopicus var. ascomyceticus qui coderait la protéine de liaison du groupement acyl (Acyl Carrier Protein (ACP)) impliquée dans la biosynthèse du méthoxymalonyl (Wu,K., et al., 2000 ; Numéro d'accès GenBank : AAF86389 ; Score BLAST : 87). La similitude de la protéine codée par le gène orf24c avec cette protéine impliquées dans la voie de biosynthèse d'un autre macrolide proche suggère fortement que ce gène code une protéine impliquée dans la biosynthèse du méthoxymalonyl chez Streptomyces ambofaciens (cf. figure 8). The orf24c gene encodes a protein with relatively strong similarity to the protein encoded by the fkbJ gene of Streptomyces hygroscopicus var. ascomyceticus that would encode the acyl-binding protein (Acyl Carrier Protein (ACP)) involved in the biosynthesis of methoxymalonyl (Wu, K., et al., 2000; GenBank Accession Number: AAF86389; BLAST Score: 87). The similarity of the protein encoded by the orf24c gene with this protein involved in the biosynthetic pathway of another close macrolide strongly suggests that this gene encodes a protein involved in the biosynthesis of methoxymalonyl in Streptomyces ambofaciens (see Figure 8).
Le gène orj25c code une protéine présentant une relative forte similitude avec la protéine codée par le gène fkbK de Streptomyces hygroscopicus var. ascomyceticus qui code une acyl CoA déshydrogénase impliquée dans la biosynthèse du méthoxymalonyl (Wu,K., et al., 2000 ; Numéro d'accès GenBank : AAF86390 ; score BLAST : 268). La similitude de la protéine codée par le gène orj25c avec plusieurs acyl CoA déshydrogénases impliquées dans la voie de biosynthèse d'autres antibiotiques proches suggère fortement que ce gène code une acyl CoA déshydrogénase impliquée dans la biosynthèse du méthoxymalonyl (cf. figure 8). Cette hypothèse est étayée par le fait que la protéine codée par le gène orf25c présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 32). The orj25c gene encodes a protein with relatively strong similarity to the protein encoded by the fkbK gene of Streptomyces hygroscopicus var. ascomyceticus which encodes an acyl CoA dehydrogenase involved in the biosynthesis of methoxymalonyl (Wu, K., et al., 2000; GenBank Accession Number: AAF86390; BLAST score: 268). The similarity of the protein encoded by the orj25c gene with several acyl CoA dehydrogenases involved in the biosynthetic pathway of other nearby antibiotics strongly suggests that this gene encodes an acyl CoA dehydrogenase involved in the biosynthesis of methoxymalonyl (see Figure 8). This hypothesis is supported by the fact that the protein encoded by the orf25c gene shows strong similarity with other proteins of similar function in other organisms (see Table 32).
<Desc/Clms Page number 64><Desc / Clms Page number 64>
Tableau 32

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> Probable <SEP> 3-Hydroxybutyryl-CoA <SEP> P45856 <SEP> 177 <SEP> 3-HydroxybutyrylDehydrogenase <SEP> (Bacillus <SEP> subtilis) <SEP> CoA <SEP> déshydrogénase
<tb> 3-hydroxybutyryl-CoA <SEP> dehydrogenase <SEP> AAL32270 <SEP> 174 <SEP> 3-hydroxybutyrylprotein <SEP> (Bacillus <SEP> thuringiensis <SEP> serovar <SEP> CoA <SEP> déshydrogénase
<tb> kurstaki)
<tb> 3-hydroxybutyryl-CoA <SEP> dehydrogenase <SEP> NP~294792 <SEP> 167 <SEP> 3-hydroxybutyryl-
<tb> (Deinococcus <SEP> radiodurans) <SEP> CoA <SEP> déshydrogénase
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> Likely <SEP> 3-Hydroxybutyryl-CoA <SEP> P45856 <SEP> 177 <SEP> 3-HydroxybutyrylDehydrogenase <SEP> (Bacillus <SEP> subtilis) <SEP> CoA <SEP> dehydrogenase
<tb> 3-hydroxybutyryl-CoA <SEP> dehydrogenase <SEP> AAL32270 <SEQ> 174 <SEP> 3-hydroxybutyrylprotein <SEP> (Bacillus <SEP> thuringiensis <SEP> serovar <SEP> CoA <SEP> dehydrogenase
<tb> kurstaki)
<tb> 3-hydroxybutyryl-CoA <SEP> dehydrogenase <SEP> NP ~ 294792 <SEP> 167 <SEP> 3-hydroxybutyryl-
<tb> (Deinococcus <SEP> radiodurans) <SEP> CoA <SEP> dehydrogenase
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Le gène or/26 code une protéine présentant une identité de 65% (déterminée grâce au programme BLAST) avec la protéine codée par le gène tylCV qui code une mycarosyl transferase impliquée dans la biosynthèse de tylosine chez Streptomyces fradiae (Bate,N. et al., 2000 ; Numéro d'accès GenBank : AAD41824, Score BLAST : 471). Plus particulièrement, TylCV est une glycosyl-transférase liant la molécule de mycarose lors de la synthèse de la tylosine. Cette similitude avec une protéine impliquée dans la voie de biosynthèse d'un autre antibiotique relativement proche et plus particulièrement dans le transfert du mycarose, suggère que le gène or/26 code une glycosyl transferase. Cette hypothèse est étayée par le fait que la protéine codée par le gène or/26 présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 33). The or / 26 gene encodes a protein having a 65% identity (determined by the BLAST program) with the protein encoded by the tylCV gene which encodes a mycarosyl transferase involved in tylosin biosynthesis in Streptomyces fradiae (Bate, N. et al. ., 2000; GenBank Access Number: AAD41824, BLAST Score: 471). More particularly, TylCV is a glycosyl transferase binding the mycarose molecule during the synthesis of tylosin. This similarity with a protein involved in the biosynthetic pathway of another relatively close antibiotic, and more particularly in the transfer of mycarose, suggests that the gold / 26 gene encodes a glycosyl transferase. This hypothesis is supported by the fact that the protein encoded by the gold / 26 gene has a strong similarity with other proteins of similar function in other organisms (see Table 33).
<Desc/Clms Page number 65><Desc / Clms Page number 65>
Tableau 33

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> Glycosyl <SEP> transferase <SEP> (Streptomyces <SEP> BAA84592 <SEP> 218 <SEP> Glycosyl <SEP> transferase
<tb> avermitilis)
<tb> CalG4 <SEP> (Micromonospora <SEP> echinospora) <SEP> AAM70365 <SEP> 217 <SEP> Glycosyl <SEP> transferase
<tb> CalG2 <SEP> (Micromonospora <SEP> echinospora) <SEP> AAM70348 <SEP> 197 <SEP> Glycosyl <SEP> transferase
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> Glycosyl <SEP> transferase <SEP> (Streptomyces <SEP> BAA84592 <SEP> 218 <SEP> Glycosyl <SEP> transferase
<tb> avermitilis)
<tb> CalG4 <SEP> (Micromonospora <SEP> echinospora) <SEP> AAM70365 <SEP> 217 <SEP> Glycosyl <SEP> transferase
<tb> CalG2 <SEP> (Micromonospora <SEP> echinospora) <SEP> AAM70348 <SEP> 197 <SEP> Glycosyl <SEP> transferase
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Le gène orf27 code une protéine présentant une identité de 70% (déterminée grâce au programme BLAST) avec la protéine codée par le gène tylCVII qui code une NDPhexose 3,5- (ou 5-) épimerase impliquée dans la biosynthèse de tylosine chez Streptomyces fradiae (Bate,N. et al., 2000 ; Numéro d'accès GenBank : AAD41825, Score BLAST : 243). Plus particulièrement, TylCVII est une hexose 3,5- (ou 5-) épimerase impliquée dans la biosynthèse du mycarose. Cette similitude avec une protéine impliquée dans la voie de biosynthèse d'un autre antibiotique relativement proche et plus particulièrement dans la biosynthèse du mycarose, suggère que le gène or/27 code une épimérase. Cette hypothèse est étayée par le fait que la protéine codée par le gène or/27 présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 34). Une analyse des séquences proches obtenues grâce au programme BLAST suggère fortement que le gène orf27 code une 5 épimérase responsable de la réaction d'épimérisation nécessaire à la biosynthèse du mycarose (cf. figure 4). The orf27 gene encodes a protein having an identity of 70% (determined by means of the BLAST program) with the protein encoded by the tylCVII gene which encodes a NDPhexose 3,5- (or 5-) epimerase involved in the biosynthesis of tylosin in Streptomyces fradiae (Bate, N .; et al., 2000; GenBank Access Number: AAD41825, BLAST Score: 243). More particularly, TylCVII is a hexose 3,5- (or 5-) epimerase involved in the biosynthesis of mycarosis. This similarity with a protein involved in the biosynthetic pathway of another relatively close antibiotic and more particularly in the biosynthesis of mycarose suggests that the or / 27 gene encodes an epimerase. This hypothesis is supported by the fact that the protein encoded by the gold / 27 gene has a strong similarity with other proteins of similar function in other organisms (see Table 34). An analysis of the close sequences obtained by the BLAST program strongly suggests that the orf27 gene encodes an epimerase responsible for the epimerization reaction necessary for mycarose biosynthesis (see FIG. 4).
<Desc/Clms Page number 66><Desc / Clms Page number 66>
Tableau 34

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> LanZl <SEP> (Streptomyces <SEP> cyanogenus) <SEP> AAD13558 <SEP> 172 <SEP> NDP-hexose <SEP> 3,5epimerase
<tb> Epi <SEP> (Saccharopolyspora <SEP> spinosa) <SEP> AAK83288 <SEP> 169 <SEP> TDP-4-keto-6deoxyglucose <SEP> 3,5
<tb> epimerase
<tb> dNTP-hexose <SEP> 3,5 <SEP> epimerase <SEP> AAC01732 <SEP> 166 <SEP> dNTP-hexose <SEP> 3,5
<tb> (Amycolatopsis <SEP> mediterranei) <SEP> epimerase
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> LanZl <SEP> (Streptomyces <SEP> cyanogenus) <SEP> AAD13558 <SEP> 172 <SEP> NDP-hexose <SEP> 3,5epimerase
<tb> Epi <SEP> (Saccharopolyspora <SEP> Spinosa) <SEP> AAK83288 <SEP> 169 <SEP> TDP-4-keto-6-deoxyglucose <SEP> 3,5
<tb> epimerase
<tb> dNTP-hexose <SEP> 3.5 <SEP> epimerase <SEP> AAC01732 <SEP> 166 <SEP> dNTP-hexose <SEP> 3.5
<tb> (Amycolatopsis <SEP> mediterranei) <SEP> epimerase
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
La séquence d'orf28c est partielle, puisque la séquence d'une région d'environ 450 paires de bases n'a pas été déterminé (région symbolisée par des N dans la séquence SEQ ID N 106). La séquence partielle de cette ORF a néanmoins été utilisée pour l'analyse avec les différents programmes informatiques comme expliqué ci-dessus. Il a ainsi pû être déterminé que le gène orf28c code une protéine présentant une identité de 64% sur la séquence déterminée (SEQ ID N 112, qui est la séquence partielle de la protéine Orf28c) (déterminée grâce au programme BLAST) avec la protéine codée par le gène acyB2 qui code une protéine régulatrice impliquée dans la biosynthèse de carbomycine chez Streptomyces thermotolerans (Arisawa,A., et al., 1993 ; Numéro d'accès GenBank : JC2032, Score BLAST : 329). Cette similitude avec une protéine impliquée dans la voie de biosynthèse d'un antibiotique relativement proche suggère que le gène orf28c code une protéine régulatrice impliquée dans la biosynthèse des spiramycines. Cette hypothèse est étayée par le fait que la protéine codée par le gène orf28c présente également une forte similitude avec la protéine TylR qui est une The sequence of orf28c is partial, since the sequence of a region of approximately 450 base pairs has not been determined (region symbolized by N in the sequence SEQ ID N 106). The partial sequence of this ORF was nevertheless used for the analysis with the different computer programs as explained above. It has thus been possible to determine that the orf28c gene encodes a protein having an identity of 64% on the determined sequence (SEQ ID No. 112, which is the partial sequence of the Orf28c protein) (determined by means of the BLAST program) with the encoded protein. by the acyB2 gene which encodes a regulatory protein involved in the biosynthesis of carbomycin in Streptomyces thermotolerans (Arisawa, A., et al., 1993; GenBank Accession Number: JC2032, BLAST Score: 329). This similarity with a protein involved in the biosynthetic pathway of a relatively close antibiotic suggests that the orf28c gene encodes a regulatory protein involved in the biosynthesis of spiramycins. This hypothesis is supported by the fact that the protein encoded by the orf28c gene also has a strong similarity with the TylR protein which is a
<Desc/Clms Page number 67><Desc / Clms Page number 67>
protéine régulatrice impliquée dans la biosynthèse de tylosine chez Streptomycesfradiae (Bate,N. et al., 1999 ; Numéro d'accès GenBank : AAF29380, Score BLAST : 167). regulatory protein involved in tylosin biosynthesis in Streptomycesfradiae (Bate, N .; et al., 1999; GenBank Accession Number: AAF29380, BLAST Score: 167).
Le gène orf28c a pu être amplifié grace à des oligonucléotides situés de part et d'autre de la séquence non déterminée et sous cloné dans un vecteur d'expression. Il a pu ainsi être démontré que la surexpression du gène orf28c augmente significativement la production en spiramycines de la souche OSC2 (cf. exemple 24). Ceci démontre que la surexpression de orf28c conduit à une augmentation de la production en spiramycines et confirme son rôle en tant que régulateur de la voie de biosynthèse des spiramycines. The orf28c gene could be amplified by means of oligonucleotides located on either side of the undetermined sequence and subcloned into an expression vector. It has thus been demonstrated that the overexpression of the orf28c gene significantly increases the spiramycin production of the strain OSC2 (see Example 24). This demonstrates that the overexpression of orf28c leads to an increase in spiramycin production and confirms its role as a regulator of the spiramycin biosynthetic pathway.
Le gène orj29 code une protéine présentant une identité de 31 % (déterminée grâce au programme BLAST) avec une probable glycosyl hydrolase localisée dans le groupe de gènes impliqué dans la biosynthèse de soraphen A (un antifongique de la classe des polyketides) chez Sorangium cellulosum (Ligon,J., et al., 2002 ; Numéro d'accès GenBank: AAK19890, Score BLAST: 139). Cette similitude avec une protéine impliquée dans la voie de biosynthèse d'une molécule relativement proche suggère que le gène or/29 code une protéine ayant une activité glycosyl hydrolase. Cette hypothèse est étayée par le fait que la protéine codée par le gène or/29 présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 35). The orj29 gene encodes a protein having an identity of 31% (determined by the BLAST program) with a probable glycosyl hydrolase localized in the group of genes involved in the biosynthesis of soraphen A (a polyketide class antifungal) in Sorangium cellulosum ( Ligon, J., et al., 2002; GenBank Access Number: AAK19890, BLAST Score: 139). This similarity to a protein involved in the biosynthetic pathway of a relatively close molecule suggests that the or / 29 gene encodes a protein with glycosyl hydrolase activity. This hypothesis is supported by the fact that the protein encoded by the or / 29 gene has a strong similarity with other proteins of similar function in other organisms (see Table 35).
Une analyse de la séquence de la protéine codée par or/29 grâce au programme CDsearch (cf. ci-dessus) suggère également que le gène or/29 code une glycosyl hydrolase. An analysis of the sequence of the gold / 29 encoded protein using the CDResearch program (see above) also suggests that the or / 29 gene encodes a glycosyl hydrolase.
Tableau 35

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> ManA <SEP> (Caldicellulosiruptor <SEP> AAC44232 <SEP> 136 <SEP> beta-1,4-mannanase
<tb> saccharolyticus)
<tb> <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> ManA <SEP> (Caldicellulosiruptor <SEP> AAC44232 <SEP> 136 <SEP> beta-1,4-mannanase
<tb> saccharolyticus)
<Tb>
<Desc/Clms Page number 68> <Desc / Clms Page number 68>
![]()
![]()
<tb>
<tb> ManA <SEP> (Dictyoglomus <SEP> thermophilum) <SEP> AAB82454 <SEP> 129 <SEP> beta-mannanase
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> ManA <SEP> (Dictyoglomus <SEP> thermophilum) <SEP> AAB82454 <SEP> 129 <SEP> beta-mannanase
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
Le gène orf30c code une protéine présentant une identité de 31% (déterminée grâce au programme BLAST) avec une épimérase de sucre-nucléoside-diphosphate chez
Corynebacterium glutamicum (Numéro d'accès GenBank : NP~600590, Score BLAST :
89). Cette similitude suggère que le gène orf30c code une épimérase. Cette hypothèse est étayée par le fait qu'une analyse de la séquences grâce au programme CD-search (cf. ci-dessus) suggère également que le gène orf30c code une épimérase. The orf30c gene encodes a protein having an identity of 31% (determined by the BLAST program) with a sugar-nucleoside-diphosphate epimerase at
Corynebacterium glutamicum (GenBank Accession Number: NP ~ 600590, BLAST Score:
89). This similarity suggests that the orf30c gene encodes an epimerase. This hypothesis is supported by the fact that a sequence analysis using the CD-search program (see above) also suggests that the orf30c gene encodes an epimerase.
Le gène or/37 code une protéine présentant une identité de 52% (déterminée grâce au programme BLAST) avec une oxidoreductase chez Streptomyces coelicolor (Numéro d'accès GenBank : NP~631148, Score BLAST : 261). Cette similitude suggère que le gène or/37 code une réductase. Cette hypothèse est étayée par le fait qu'une analyse de la séquences grâce au programme CD-search (cf. ci-dessus) suggère également que le gène or/31 code une réductase. Cette hypothèse est également étayée par le fait que la protéine codée par le gène or/37 présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 36). The or / 37 gene encodes a protein having an identity of 52% (determined by the BLAST program) with an oxidoreductase in Streptomyces coelicolor (GenBank Accession Number: NP ~ 631148, BLAST Score: 261). This similarity suggests that the gold / 37 gene encodes a reductase. This hypothesis is supported by the fact that a sequence analysis using the CD-search program (see above) also suggests that the or / 31 gene encodes a reductase. This hypothesis is also supported by the fact that the protein encoded by the or / 37 gene has a strong similarity with other proteins of similar function in other organisms (see Table 36).
Tableau 36

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> Putative <SEP> oxidoreductase <SEP> (Streptomyces <SEP> BAB79295 <SEP> 173 <SEP> Oxidoreductase
<tb> griseus)
<tb> MocA <SEP> (Xanthomonas <SEP> axonopodis) <SEP> NP~640644 <SEP> 171 <SEP> Oxidoreductase
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> Putative <SEP> oxidoreductase <SEP> (Streptomyces <SEP> BAB79295 <SEP> 173 <SEP> Oxidoreductase
<tb> griseus)
<tb> MocA <SEP> (Xanthomonas <SEP> axonopodis) <SEP> NP ~ 640644 <SEQ> 171 <SEP> Oxidoreductase
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
<Desc/Clms Page number 69> <Desc / Clms Page number 69>
Le gène orf32c code une protéine présentant une identité de 47% (déterminée grâce au programme BLAST) avec une protéine régulatrice de la famille GntR chez
Streptomyces coelicolor (Numéro d'accès GenBank : NP~625576, Score BLAST : 229).
Cette similitude suggère que le gène orf32c code un régulateur transcriptionel de la famille GntR. Cette hypothèse est étayée par le fait que la protéine codée par le gène orf32c présente une forte similitude avec d'autres protéines de fonction similaire chez d'autres organismes (cf. tableau 37). The orf32c gene encodes a protein having an identity of 47% (determined by the BLAST program) with a regulatory protein of the GntR family in
Streptomyces coelicolor (GenBank accession number: NP ~ 625576, BLAST score: 229).
This similarity suggests that the orf32c gene encodes a transcriptional regulator of the GntR family. This hypothesis is supported by the fact that the protein encoded by the orf32c gene shows strong similarity with other proteins of similar function in other organisms (see Table 37).
Tableau 37

<tb>
<tb> Protéine <SEP> présentant <SEP> une <SEP> similitude <SEP> Numéro <SEP> Score <SEP> Fonction <SEP> rapportée
<tb> significative <SEP> d'accès <SEP> BLAST*
<tb> GenBank
<tb> SC5G8.04 <SEP> (Streptomyces <SEP> coelicolor) <SEP> NP~628991 <SEP> 209 <SEP> Protéine <SEP> régulatrice
<tb> de <SEP> la <SEP> famille <SEP> GntR
<tb> transcriptional <SEP> regulator <SEP> (Streptomyces <SEP> AAFOI064 <SEP> 176 <SEP> Régulateur
<tb> venezuelae) <SEP> transcriptionel
<tb> BH0578 <SEP> (Bacillus <SEP> halodurans) <SEP> NP~241444 <SEP> 172 <SEP> Régualteur
<tb> transcriptionl <SEP> de <SEP> la
<tb> famille <SEP> GntR
<tb>
* une plus grande similitude de séquence est associée à un score BLAST plus élevé (Altschul et al., 1990). <Tb>
<tb> Protein <SEP> presenting <SEP> a <SEP> similarity <SEP> Number <SEP> Score <SEP> Function <SEP> reported
<tb> Significant <SEP> of Access <SEP> BLAST *
<tb> GenBank
<tb> SC5G8.04 <SEP> (Streptomyces <SEP> coelicolor) <SEP> NP ~ 628991 <SEP> 209 <SEP> Regulating Protein <SEP>
<tb> of <SEP> the <SEP> family <SEP> GntR
<tb> transcriptional <SEP> regulator <SEP> (Streptomyces <SEP> AAFOI064 <SEP> 176 <SEP> Regulator
<tb> venezuelae) <SEP> transcriptional
<tb> BH0578 <SEP> (Bacillus <SEP> halodurans) <SEP> NP ~ 241444 <SEQ> 172 <SEP> Regulator
<tb> transcriptionl <SEP> of <SEP> la
<tb> family <SEP> GntR
<Tb>
* greater sequence similarity is associated with a higher BLAST score (Altschul et al., 1990).
La présente invention a également pour objet des polynucléotides s'hybridant dans des conditions d'hybridation de forte stringence à au moins l'un des polynucléotides de séquence SEQ ID N 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 28, 30, 34, 36, 40, 43, 45, 47, 49, 53, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 107, 109, 111,113, 115,118, et 120, ou l'un de ses variants ou l'une des séquences dérivées de celles-ci en raison de la dégénérescence du code génétique. Préférentiellement, cesdit polynucléotides sont isolés à partir d'une bactérie du genre Streptomyces, plus The present invention also relates to polynucleotides hybridizing under high stringency hybridization conditions to at least one of the polynucleotides of sequence SEQ ID N 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 28, 30, 34, 36, 40, 43, 45, 47, 49, 53, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 107, 109, 111, 113, 115, 118, and 120, or one of its variants or one of the sequences derived therefrom due to the degeneracy of the genetic code. Preferably, these polynucleotides are isolated from a bacterium of the genus Streptomyces, plus
<Desc/Clms Page number 70><Desc / Clms Page number 70>
préférentiellement, ces polynucléotides codent des protéines impliquées dans la biosynthèse d'un macrolide et encore plus préférentiellement, ces polynucléotides codent une protéine ayant une activité similaire à la protéine codée par les polynucléotides avec lesquels ils s'hybrident. Les conditions d'hybridation de forte stringence peuvent être définies comme des conditions d'hybridation défavorisant l'hybridation de brins d'acides nucléiques non homologues. Des conditions d'hybridations de forte stringence peuvent être par exemple décrites comme des conditions d'hybridation dans le tampon décrit par Church & Gilbert (Church & Gilbert, 1984) à une température comprise entre 55 C et 65 C, de manière préférée la température d'hyridation est de 55 C, de manière encore plus préférée la températue d'hybridation est de 60 C et de manière tout à fait préférée la température d'hybridation est de 65 C, suivi d'un ou plusieurs lavages effectué en tampon 2X SSC à une température comprise entre 55 C et 65 C, de manière préférée cette température est de 55 C, de manière encore plus préférée cette températue est de 60 C et de manière tout à fait préférée cette température est de 65 C, suivi d'un ou plusieurs lavages en tampon 0. 5X SSC à une température comprise entre 55 C et 65 C, de manière préférée cette température est de 55 C, de manière encore plus préférée cette températue est de 60 C et de manière tout à fait préférée cette température est de 65 C. Les conditions d'hybridation ci-dessus décrites peuvent être adaptées en fonction de la longueur de l'acide nucléique dont l'hybridation est recherchée ou du type de marquage choisi, selon des techniques connues de l'homme du métier. Les conditions convenables d'hybridation peuvent par exemple être adaptées selon l'ouvrage de F.Ausubel et al., 2002. preferentially, these polynucleotides encode proteins involved in the biosynthesis of a macrolide and even more preferably, these polynucleotides encode a protein having a similar activity to the protein encoded by the polynucleotides with which they hybridize. High stringency hybridization conditions can be defined as hybridization conditions that discriminate against the hybridization of non-homologous nucleic acid strands. Highly stringent hybridization conditions can be described, for example, as hybridization conditions in the buffer described by Church & Gilbert (Church & Gilbert, 1984) at a temperature of between 55 ° C. and 65 ° C., preferably the temperature The hybridization temperature is 60 ° C. and more preferably the hybridization temperature is 65 ° C., followed by one or more washes carried out in 2 × buffer. SSC at a temperature between 55 C and 65 C, preferably this temperature is 55 C, even more preferably this temperature is 60 C and very preferably this temperature is 65 C, followed by one or more washings in a buffer of 0.5 × SSC at a temperature of between 55 ° C. and 65 ° C., preferably this temperature is 55 ° C., even more preferably, this temperature is 60 ° C. and more preferably It is quite preferred that this temperature is 65 ° C. The hybridization conditions described above can be adapted as a function of the length of the nucleic acid whose hybridization is sought or of the type of marking chosen, according to the techniques described. known to those skilled in the art. Suitable hybridization conditions may for example be adapted according to the work of F.Ausubel et al., 2002.
L'invention concerne également un polynucléotide ayant au moins 70%, de façon plus préférée 80%, de façon plus préférée 85%, de façon encore plus préférée 90%, de façon encore plus préférée 95% et de manière tout à fait préférée 98% d'identité en nucléotides avec un polynucléotide comprenant au moins 10,12, 15,18, 20 à 25,30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750,800, 850,900, 950,1000, 1050,1100, 1150,1200, 1250,1300, 1350,1400, 1450, 1500,1550, 1600,1650, 1700,1750, 1800,1850 ou 1900 nucléotides consécutifs d'un The invention also relates to a polynucleotide having at least 70%, more preferably 80%, more preferably 85%, more preferably 90%, more preferably 95% and most preferably 98%. % nucleotide identity with a polynucleotide comprising at least 10,12, 15,18, 20 to 25,30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000, 1050, 1100, 1150, 1200, 1250, 1300, 1350, 1400, 1450, 1500, 1550, 1600, 1650, 1700, 1750, 1800, 1850 or 1900 consecutive nucleotides of a
<Desc/Clms Page number 71><Desc / Clms Page number 71>
polynucléotide choisi dans le groupe constitué des séquences nucléotidiques SEQ ID N 3,5, 7,9, 11, 13,15, 17, 19, 21, 23, 25, 28, 30, 34, 36, 40, 43, 45, 47, 49, 53,60, 62,64, 66,68, 70,72, 74,76, 78,80, 82,84, 107,109, 111,113, 115,118, et 120, ou l'un de ses variants ou l'une des séquences dérivées de celles-ci en raison de la dégénérescence du code génétique, ou un polynucléotide de séquence complémentaire. Préférentiellement, ces dits polynucléotides sont isolés à partir d'une bactérie du genre Streptomyces, plus préférentiellement, ces polynucléotides codent des protéines impliquées dans la biosynthèse d'un macrolide et encore plus préférentiellement, ces polynucléotides codent des protéines ayant des activités similaires aux protéines codées par les polynucléotides avec lesquels ils présentent l'identité. De façon tout à fait préférée, un polynucléotide selon l'invention est choisi dans le groupe constitué des séquences nucléotidiques SEQ ID N 3,5, 7,9, 11, 13,15, 17,19, 21,23, 25,28, 30, 34, 36, 40, 43, 45, 47, 49, 53, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 107, 109, 111,113, 115, 118, et 120, ou un polynucléotide de séquence complémentaire. polynucleotide selected from the group consisting of nucleotide sequences SEQ ID N 3.5, 7.9, 11, 13, 15, 17, 19, 21, 23, 25, 28, 30, 34, 36, 40, 43, 45, 47, 49, 53.60, 62.64, 66.68, 70.72, 74.76, 78.80, 82.84, 107.109, 111, 113, 115, 118, and 120, or one of its variants or one of the sequences derived therefrom due to the degeneracy of the genetic code, or a complementary sequence polynucleotide. Preferably, said polynucleotides are isolated from a bacterium of the genus Streptomyces, more preferably these polynucleotides encode proteins involved in the biosynthesis of a macrolide, and even more preferentially, these polynucleotides encode proteins having activities similar to the encoded proteins. by the polynucleotides with which they present the identity. Most preferably, a polynucleotide according to the invention is chosen from the group consisting of the nucleotide sequences SEQ ID N 3.5, 7.9, 11, 13, 15, 17, 19, 21, 23, 25, 28. , 30, 34, 36, 40, 43, 45, 47, 49, 53, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 107, 109, 111, 113 , 115, 118, and 120, or a polynucleotide of complementary sequence.
L'alignement optimal des séquences pour la comparaison peut être réalisé de manière informatique à l'aide d'algorithmes connus, par exemple ceux du package FASTA (Pearson W. R & D. J. Lipman, 1988) et (Pearson W. R., 1990), accessible notamment auprès du centre de ressources INFOBIOGEN, Evry, France. À titre d'illustration, le pourcentage d'identité de séquence pourra être déterminé à l'aide du logiciel LFASTA (Chao K.-M. et al., 1992) ou LALIGN (Huang X. et Miller W., 1991). Les programmes LFASTA et LALIGN font partie du package FASTA. LALIGN retourne les alignements locaux optimaux : ce programme est plus rigoureux, mais aussi plus lent que LFASTA. The optimal alignment of the sequences for comparison can be performed in a computer manner using known algorithms, for example those of the FASTA package (Pearson W. R & DJ Lipman, 1988) and (Pearson WR, 1990), accessible especially at the INFOBIOGEN resource center, Evry, France. By way of illustration, the percentage of sequence identity can be determined using LFASTA software (Chao K.-M. et al., 1992) or LALIGN (Huang X. and Miller W., 1991). The LFASTA and LALIGN programs are part of the FASTA package. LALIGN returns the optimal local alignments: this program is more rigorous, but also slower than LFASTA.
Un autre aspect de l'invention concerne un polypeptide résultant de l'expression d'une séquence d'acide nucléique telle que définie ci-dessus. Préférentiellement, les polypeptides selon l'invention présentent au moins 70%, de façon plus préférée 80%, de façon plus préférée 85%, de façon encore plus préférée 90%, de façon encore plus préférée 95% et de manière tout à fait préférée 98% d'identité en acides aminés avec un polypeptide comprenant au moins 10, 15, 20, 30 à 40, 50, 60, 70, 80, 90,100, 120, 140, Another aspect of the invention relates to a polypeptide resulting from the expression of a nucleic acid sequence as defined above. Preferably, the polypeptides according to the invention have at least 70%, more preferably 80%, more preferably 85%, even more preferably 90%, even more preferably 95% and most preferably 98% amino acid identity with a polypeptide comprising at least 10, 15, 20, 30 to 40, 50, 60, 70, 80, 90, 100, 120, 140,
<Desc/Clms Page number 72><Desc / Clms Page number 72>
160,180, 200,220, 240,260, 280,300, 320,340, 360,380, 400,420, 440,460, 480, 500,520, 540,560, 580,600, 620 ou 640 acides aminés consécutifs d'un polypeptide choisi parmi SEQ ID N 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24,26, 27,29, 31,32, 33,35, 37,38, 39,41, 42,44, 46, 48, 50,51, 52,54, 55,56, 57,58, 59,61, 63,65, 67,69, 71, 73,75, 77,79, 81,83, 85,108, 110,112, 114,116, 117,119 et 121, ou l'une de ces séquences excepté que tout au long de ladite séquence un ou plusieurs acides aminés ont été substitués, insérés ou délétés sans en affecter les propriétés fonctionnelles ou l'un des variant de ces séquences. Préferentiellement les polypeptides selon l'invention sont exprimés à l'état naturel par une bactérie du genre Streptomyces, plus préférentiellement, ces polypeptides sont impliquées dans la biosynthèse d'un macrolide et encore plus préférentiellement, ces polypeptides ont une activité similaire à celle du polypeptide avec lequel il partage l'identité. De façon préférée, un polypeptide selon l'invention est choisi dans le groupe constitué des séquences polypeptidiques SEQ ID N 4,6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 27, 29, 31, 32, 33, 35,37, 38,39, 41,42, 44,46, 48, 50, 51, 52,54, 55,56, 57,58, 59,61, 63,65, 67,69, 71, 73, 75, 77, 79, 81, 83, 85, 108, 110, 112, 114, 116, 117,119 et 121, ou l'une de ces séquences excepté que tout au long de ladite séquence un ou plusieurs acides aminés ont été substitués, insérés ou délétés sans en affecter les propriétés fonctionnelles ou l'un des variant de ces séquences. De façon tout à fait préférée, un polypeptide selon l'invention est choisi dans le groupe constitué des séquences polypeptidiques SEQ ID N 4,6, 8,10, 12, 14,16, 18, 20, 22, 24, 26, 27, 29, 31, 32, 33, 35, 37, 38, 39, 41, 42, 44, 46, 48, 50,51, 52,54, 55,56, 57,58, 59, 61, 63, 65,67, 69,71, 73,75, 77,79, 81, 83, 85, 108, 110, 112, 114, 116, 117, 119 et 121. 160,180, 200,220, 240,260, 280,300, 320,340, 360,380, 400,420, 440,460, 480, 500,520, 540,560, 580,600, 620 or 640 consecutive amino acids of a polypeptide selected from SEQ ID N 4, 6, 8, 10, 12, 14 , 16, 18, 20, 22, 24, 26, 27, 29, 31, 32, 33, 35, 37, 38, 39, 41, 42, 44, 46, 48, 50, 51, 52, 54, 55 , 56, 57, 58, 59, 61, 63, 65, 67, 69, 71, 73, 75, 77, 79, 81, 83, 85, 108, 110, 112, 114, 116, 117, 119 and 121, or one of these sequences except that throughout said sequence one or more amino acids have been substituted, inserted or deleted without affecting the functional properties or any variant thereof. Preferably, the polypeptides according to the invention are expressed in the natural state by a bacterium of the genus Streptomyces, more preferentially, these polypeptides are involved in the biosynthesis of a macrolide and even more preferentially, these polypeptides have an activity similar to that of the polypeptide. with whom he shares the identity. Preferably, a polypeptide according to the invention is chosen from the group consisting of the polypeptide sequences SEQ ID N 4,6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 27, 29, 31, 32, 33, 35.37, 38.39, 41.42, 44.46, 48, 50, 51, 52.54, 55.56, 57.58, 59.61, 63.65, 67, 69, 71, 73, 75, 77, 79, 81, 83, 85, 108, 110, 112, 114, 116, 117, 119 and 121, or one of these sequences except that throughout said one or more sequence Amino acids have been substituted, inserted or deleted without affecting the functional properties or any of the variants of these sequences. Most preferably, a polypeptide according to the invention is chosen from the group consisting of the polypeptide sequences SEQ ID N 4,6, 8,10, 12, 14, 16, 18, 20, 22, 24, 26, 27 , 29, 31, 32, 33, 35, 37, 38, 39, 41, 42, 44, 46, 48, 50.51, 52.54, 55.56, 57.58, 59, 61, 63, 65 , 67, 69, 71, 73, 75, 77, 79, 81, 83, 85, 108, 110, 112, 114, 116, 117, 119 and 121.
L'alignement optimal des séquences pour la comparaison peut être réalisé de manière informatique à l'aide d'algorithmes connus, par exemple ceux du package FASTA (Pearson W. R & D. J. Lipman, 1988) et (Pearson W. R., 1990), accessible notamment auprès du centre de ressources INFOBIOGEN, Evry, France. À titre d'illustration, le pourcentage d'identité de séquence pourra être déterminé à l'aide du logiciel LFASTA (Chao K.-M. et al., 1992) ou LALIGN (Huang X. et Miller W., The optimal alignment of the sequences for comparison can be performed in a computer manner using known algorithms, for example those of the FASTA package (Pearson W. R & DJ Lipman, 1988) and (Pearson WR, 1990), accessible especially at the INFOBIOGEN resource center, Evry, France. By way of illustration, the percentage of sequence identity can be determined using software LFASTA (Chao K.-M. et al., 1992) or LALIGN (Huang X. and Miller W.,
<Desc/Clms Page number 73><Desc / Clms Page number 73>
1991), en utilisant les paramètres par défaut tel que défini par le centre de ressources INFOBIOGEN, Evry, France. Les programmes LFASTA et LALIGN font partie du package FASTA. LALIGN retourne les alignements locaux optimaux : ce programme est plus rigoureux, mais aussi plus lent que LFASTA. 1991), using the default settings as defined by the INFOBIOGEN Resource Center, Evry, France. The LFASTA and LALIGN programs are part of the FASTA package. LALIGN returns the optimal local alignments: this program is more rigorous, but also slower than LFASTA.
Un autre aspect de l'invention concerne un ADN recombinant comprenant au moins un polynucléotide tel que décrit ci-dessus. Préférentiellement ; cet ADN recombinant est un vecteur. Encore plus préférentiellement, le vecteur est choisi parmi les bactériophages, les plasmides, les phagemides, les vecteurs intégratifs, les fosmides, les cosmides, les vecteurs navettes, les BAC (Bacterial Artificial Chromosome) ou les PAC (Pl-derived Artificial Chromosome). A titre illustratif on peut citer comme bactériophages le phage lambda et le phage M13. Comme plasmides on peut citer les plasmides se répliquant chez E. coli par exemple pBR322 et ses dérivés, pUC18 et ses dérivés, pUC19 et ses dérivés, pGB2 et ses dérivés (Churchward G. et al., 1984), pACYC177 (numéro d'accès GenBank : X06402) et ses dérivés, pACYC184 (numéro d'accès GenBank : X06403) et ses dérivés. On peut également citer les plasmides se répliquant chez Streptomyces comme par exemple pIJ101 et ses dérivés, pSG5 et ses dérivés, SLP1et ses dérivés, SCP2* et ses dérivés (Kieser et al. 2000). Comme phagemides on peut citer à titre illustratif pBluescript II et ses dérivés (commercialisés notamment par la société Stratagene (LaJolla, Californie, USA)), pGEM-T et ses dérivés (commercialisé par la société Promega (Madison, Wisconcin, USA)), #ZAPII et ses dérivés (commercialisés notamment par la société Stratagene (LaJolla, Californie, USA)). Comme vecteurs intégratifs on peut citer à titre illustratif, les vecteurs intégratifs chez Streptomyces comme par exemple ceux dérivés de SLP1(Kieser et al, 2000), ceux dérivés de pSAM2 (Kieser et al, 2000), les vecteurs utilisant les systèmes d'intégration des phages PhiC31 (Kieser et al, 2000) (par exemple pSET152 (Bierman et al., 1992)) ou de VWB (Van Mellaert L, et al. 1998), ainsi que les vecteurs utilisant le système d'intégration de IS 117 (Kieser et al. 2000). Comme fosmides on peut citer à titre illustratif le fosmide pFOSl (commercialisé par la société New England Bioloabs Inc., Beverly, Massachussetts, USA) et ses dérivés. Comme cosmides on peut citer à titre illustratif le cosmide SuperCos et ses dérivés (commercialisés notamment par la Another aspect of the invention relates to a recombinant DNA comprising at least one polynucleotide as described above. Preferentially; this recombinant DNA is a vector. Even more preferentially, the vector is chosen from bacteriophages, plasmids, phagemids, integrative vectors, fosmids, cosmids, shuttle vectors, BACs (Bacterial Artificial Chromosome) or PACs (Pl-derived Artificial Chromosome). By way of illustration, lambda phage and phage M13 may be mentioned as bacteriophages. Plasmids that may be mentioned include plasmids that replicate in E. coli, for example pBR322 and its derivatives, pUC18 and its derivatives, pUC19 and its derivatives, pGB2 and its derivatives (Churchward G. et al., 1984), pACYC177 (accession number GenBank Access: X06402) and its derivatives, pACYC184 (GenBank Access Number: X06403) and its derivatives. Streptomyces replicating plasmids such as, for example, pIJ101 and its derivatives, pSG5 and its derivatives, SLP1 and its derivatives, SCP2 * and its derivatives (Kieser et al., 2000) may also be mentioned. As illustrative pBluescript II and its derivatives (sold in particular by Stratagene (LaJolla, California, USA)), pGEM-T and its derivatives (marketed by Promega (Madison, Wisconcin, USA)), #ZAPII and its derivatives (marketed in particular by Stratagene (LaJolla, California, USA)). Integrative vectors that may be mentioned by way of illustration include integrative vectors in Streptomyces, for example those derived from SLP1 (Kieser et al., 2000), those derived from pSAM2 (Kieser et al., 2000), vectors using integration systems. PhiC31 phages (Kieser et al., 2000) (e.g., pSET152 (Bierman et al., 1992)) or VWB (Van Mellaert L, et al., 1998), as well as vectors using the IS integration system. (Kieser et al., 2000). As examples, fosmide pFOS1 (marketed by New England Bioloabs Inc., Beverly, Mass., USA) and its derivatives are exemplary. As cosmids, the cosmid SuperCos and its derivatives (marketed in particular by
<Desc/Clms Page number 74><Desc / Clms Page number 74>
société Stratagene (LaJolla, Californie, USA)), le cosmide pWED15 (Wahl et al., 1987) et ses dérivés. Comme vecteurs navettes on peut citer à titre illustratif les plasmides navettes E. coli / Streptomyces comme par exemple pIJ903 et ses dérivés, la série des plasmides pUWL, pCA0106, pWHM3, pOJ446 et leurs dérivés (Kieser et al. 2000), les BAC navettes E. coli / Streptomyces comme par exemple ceux décrits dans la demande de brevet WO 01/40497. Comme BAC (Bacterial Artificial Chromosome) on peut citer à titre illustratif le BAC pBeloBACll (numéro d'accès GenBank :U51113). Comme PAC (Pl-derived Artificial Chromosome) on peut citer à titre illustratif le vecteur pCYPAC6 (numéro d'accès GenBank : AF133437). De façon tout à fait préférée, un vecteur selon l'invention est choisi parmi pOS49.1, pOS49.11, pOSC49. 12, pOS49.14, pOS49. 16, pOS49. 28, pOS44.1, pOS44. 2, pOS44. 4, pSPM5, pSPM7, pOS49.67, pOS49. 88, pOS49.106, pOS49. 120, pOS49. 107, pOS49. 32, pOS49. 43, pOS49. 44, pOS49. 50, pOS49. 99, pSPM17, pSPM21, pSPM502, pSPM504, pSPM507, pSPM508, pSPM509, pSPMl, pBXLllll, pBXL1112, pBXL1113, pSPM520, pSPM521, pSPM522, pSPM523, pSPM524, pSPM525, pSPM527, pSPM528, pSPM34, pSPM35, pSPM36, pSPM37, pSPM38, pSPM39, pSPM40, pSPM41, pSPM42, pSPM43, pSPM44, pSPM45, pSPM47, pSPM48, pSPM50, pSPM51, pSPM52, pSPM53, pSPM55, pSPM56, pSPM58, pSPM72, pSPM73, pSPM515, pSPM519, pSPM74 et pSPM75. Stratagene company (LaJolla, California, USA)), cosmid pWED15 (Wahl et al., 1987) and its derivatives. As shuttle vectors, the E. coli / Streptomyces shuttle plasmids, for example pIJ903 and its derivatives, the plasmid series pUWL, pCA0106, pWHM3, pOJ446 and their derivatives (Kieser et al. E. coli / Streptomyces as for example those described in the patent application WO 01/40497. As BAC (Bacterial Artificial Chromosome) can be cited by way of illustration the BAC pBeloBACll (GenBank access number: U51113). As PAC (Pl-derived Artificial Chromosome) may be mentioned by way of illustration the vector pCYPAC6 (GenBank access number: AF133437). Most preferably, a vector according to the invention is selected from pOS49.1, pOS49.11, pOSC49. 12, pOS49.14, pOS49. 16, pOS49. 28, pOS44.1, pOS44. 2, pOS44. 4, pSPM5, pSPM7, pOS49.67, pOS49. 88, pOS49.106, pOS49. 120, pOS49. 107, pOS49. 32, pOS49. 43, pOS49. 44, pOS49. 50, pOS49. 99, pSPM17, pSPM21, pSPM502, pSPM504, pSPM507, pSPM508, pSPM509, pSPM1, pBXLllll, pBXL1112, pBXL1113, pSPM520, pSPM521, pSPM522, pSPM523, pSPM524, pSPM525, pSPM527, pSPM528, pSPM34, pSPM35, pSPM36, pSPM37, pSPM38, pSPM39, pSPM40, pSPM41, pSPM42, pSPM43, pSPM44, pSPM45, pSPM47, pSPM48, pSPM50, pSPM51, pSPM52, pSPM53, pSPM55, pSPM56, pSPM58, pSPM72, pSPM73, pSPM515, pSPM519, pSPM74 and pSPM75.
Un autre aspect de l'invention concerne un système d'expression comprenant un vecteur d'expression approprié et une cellule hôte permettant l'expression d'un ou plusieurs polypeptides tels que décrits ci-dessus dans un système biologique. Les vecteurs d'expression selon l'invention comprennent une séquence d'acide nucléique codant un ou plusieurs polypeptides tels que décrits ci-dessus et peuvent être destinés à l'expression des différents polypeptides selon l'invention dans diverses cellules hôtes bien connues de l'homme du métier. A titre d'exemple, on peut citer les systèmes d'expression procaryote comme les systèmes d'expression chez la bactérie E. coli, les sytèmes d'expression eucaryotes, comme le système d'expression baculovirus permettant une expression dans des cellules d'insectes, les sytèmes d'expression permettant une expression dans des cellules de levures, les sytèmes d'expression Another aspect of the invention relates to an expression system comprising an appropriate expression vector and a host cell for the expression of one or more polypeptides as described above in a biological system. The expression vectors according to the invention comprise a nucleic acid sequence encoding one or more polypeptides as described above and may be intended for the expression of the various polypeptides according to the invention in various well-known host cells of the invention. skilled person. By way of example, mention may be made of prokaryotic expression systems such as expression systems in E. coli bacteria, eukaryotic expression systems, such as the baculovirus expression system allowing expression in cells of insects, expression systems that allow expression in yeast cells, expression systems
<Desc/Clms Page number 75><Desc / Clms Page number 75>
permettant une expression dans des cellules de mammifères, notamment des cellules humaines. Les vecteurs d'expression utilisables dans de tels systèmes sont bien connus de l'homme du métier, en ce qui concerne les cellules procaryotes, on peut citer à titre illustratif les vecteurs d'expression chez E. coli par exemple, de la famille pET commercialisés par la société Stratagene (LaJolla, Californie, USA), les vecteurs de la famille GATEWAY commercialisés par la société Invitrogen (Carlsbad, Californie, USA), les vecteurs de la famille pBAD commercialisés par la société Invitrogen (Carlsbad, Californie, USA), les vecteurs de la famille pMAL commercialisés par la société New England Bioloabs Inc. (Beverly, Massachussetts, USA), les vecteurs d'expression inductibles par le rhamnose cités dans la publication Wilms B et al., 2001 et leurs dérivés, on peut également citer les vecteurs d'expression chez Streptomyces comme par exemple les vecteurs pIJ4123, pIJ6021, pPM927, pANT849, pANT 850, pANT 851, pANT1200, pANT1201, pANT1202 et leurs dérivés (Kieser et al., 2000). allowing expression in mammalian cells, especially human cells. The expression vectors that can be used in such systems are well known to those skilled in the art, with regard to prokaryotic cells, for example illustrative expression vectors in E. coli, for example, of the pET family. sold by Stratagene (LaJolla, California, USA), the GATEWAY family of vectors marketed by Invitrogen (Carlsbad, California, USA), the pBAD family vectors marketed by Invitrogen (Carlsbad, California, USA) , the vectors of the pMAL family sold by New England Bioloabs Inc. (Beverly, Massachussetts, USA), the rhamnose-inducible expression vectors cited in the publication Wilms B et al., 2001 and their derivatives, can be used. Streptomyces expression vectors such as pIJ4123, pIJ6021, pPM927, pANT849, pANT850, pANT851, pANT1200, pANT1201, pANT1202 and their derivatives (Kieser et al. 2000).
En ce qui concerne les cellules de levures, on peut citer à titre illustratif le vecteur pESC commercialisé par la société Stratagene (LaJolla, Californie, USA). En ce qui concerne le système d'expression baculovirus permettant une expression dans des cellules d'insectes on peut citer à titre illustratif le vecteur BacPAK6 commercialisé par la société BD Biosciences Clontech, (Palo Alto, Californie, USA). En ce qui concerne les cellules de mammifères, on peut citer à titre illustratif des vecteurs comprenant le promoteur des gènes précoces du virus CMV (Cytomegalovirus) (par exemple le vecteur pCMV et ses dérivés commercialisés par la société Stratagene (LaJolla, Californie, USA), ou le promoteur des gènes immédiats (SV40 early promoter) du virus simien vacuolisant SV40 (par exemple le vecteur pSG5 commercialisé par la société Stratagene (LaJolla, Californie, USA). As regards the yeast cells, the vector pESC marketed by the company Stratagene (LaJolla, California, USA) can be cited by way of illustration. As regards the baculovirus expression system allowing expression in insect cells, mention may be made, for example, of the BacPAK6 vector marketed by BD Biosciences Clontech, (Palo Alto, California, USA). As regards mammalian cells, mention may be made, for example, of vectors comprising the promoter of the early genes of the CMV (Cytomegalovirus) virus (for example the pCMV vector and its derivatives marketed by Stratagene (LaJolla, California, USA). or the immediate promoter (SV40 early promoter) of simian SV40 vacuolizing virus (for example the vector pSG5 marketed by Stratagene (LaJolla, California, USA).
L'invention est également relative à un procédé de production d'un polypeptide tel que décrit ci-dessus ledit procédé comprenant les étapes suivantes: a) insérer un acide nucléique codant ledit polypeptide dans un vecteur approprié The invention also relates to a process for producing a polypeptide as described above, said method comprising the steps of: a) inserting a nucleic acid encoding said polypeptide into a suitable vector
<Desc/Clms Page number 76><Desc / Clms Page number 76>
b) cultiver, dans un milieu de culture approprié, une cellule hôte préalablement transformée ou transfectée avec le vecteur de l'étape a) ; c) récupérer le milieu de culture conditionné ou un extrait cellulaire, par exemple par sonication ou par choc osmotique ; d) séparer et purifier à partir dudit milieu de culture ou encore à partir de l'extrait cellulaire obtenu à l'étape c), ledit polypeptide ; e) le cas échéant, caractériser le polypeptide recombinant produit. b) culturing, in a suitable culture medium, a host cell previously transformed or transfected with the vector of step a); c) recovering the conditioned culture medium or a cell extract, for example by sonication or osmotic shock; d) separating and purifying from said culture medium or from the cell extract obtained in step c), said polypeptide; e) where appropriate, characterizing the recombinant polypeptide produced.
Un polypeptide recombinant selon l'invention peut être purifié par passage sur une série appropriée de colonnes de chromatographie, selon les méthodes connues de l'homme de l'art et décrites par exemple dans F. Ausubel et al (2002). On peut citer à titre illustratif la technique du Tag-Histidine , qui consiste à ajouter une courte séquence poly-histidine au polypeptide à produire, ce dernier pouvant ensuite être purifié sur colonne de nickel. Un polypeptide selon l'invention peut être également préparé par les techniques de synthèse in vitro. A titre illustratif de telles techniques, un polypeptide selon l'invention pourra être préparé grâce au système rapid translation system (RTS) , commercialisé notamment par la société Roche Diagnostics France S.A, Meylan, France. A recombinant polypeptide according to the invention may be purified by passage over a suitable series of chromatography columns, according to the methods known to those skilled in the art and described for example in F. Ausubel et al (2002). By way of illustration, mention may be made of the Tag-Histidine technique, which consists in adding a short poly-histidine sequence to the polypeptide to be produced, which can then be purified on a nickel column. A polypeptide according to the invention may also be prepared by in vitro synthesis techniques. As an illustration of such techniques, a polypeptide according to the invention may be prepared using the rapid translation system (RTS) system, marketed in particular by Roche Diagnostics France SA, Meylan, France.
Un autre aspect de l'invention concerne des cellules hôtes dans laquelle a été introduit au moins un polynucléotide et/ou au moins un ADN recombinant et/ou au moins un vecteur d'expression selon l'invention. Another aspect of the invention relates to host cells into which at least one polynucleotide and / or at least one recombinant DNA and / or at least one expression vector according to the invention has been introduced.
Un autre aspect de l'invention concerne des microorganismes bloqués dans une étape de la voie de biosynthèse d'au moins un macrolide. L'intérêt réside d'une part dans l'étude de la fonction des protéines mutées et d'autre part dans la réalisation de microorganismes produisant des intermédiaires de biosynthèse. Ces intermédiaires peuvent être modifiés, éventuellement après séparation, soit par ajout de composants particuliers dans les milieux de production, soit par introduction dans les microorganismes ainsi mutés d'autres gènes codant des protéines susceptibles de Another aspect of the invention relates to microorganisms blocked in a step of the biosynthetic pathway of at least one macrolide. The interest lies in the study of the function of mutated proteins and in the production of microorganisms producing biosynthesis intermediates. These intermediates can be modified, optionally after separation, either by adding particular components to the production media, or by introducing into the microorganisms thus mutated other genes encoding proteins capable of
<Desc/Clms Page number 77><Desc / Clms Page number 77>
modifier l'intermédiaire en s'en servant comme substrat. Ces intermédiaires peuvent ainsi être modifiés par voie chimique, biochimique, enzymatique et/ou microbiologique. modify the intermediary by using it as a substrate. These intermediates can thus be modified chemically, biochemically, enzymatically and / or microbiologically.
Les microorganismes bloqués dans une étape de la voie de biosynthèse de macrolides peuvent être obtenus en inactivant la fonction d'une ou plusieurs protéines impliquées dans la biosynthèse de ce ou de ces macrolides dans des microorganismes producteurs de ce ou de ces macrolide. Selon la ou les protéines inactivées, on pourra ainsi obtenir des microorganismes bloqués dans les différentes étapes de la voie de biosynthèse de ce ou de ces macrolides. L'inactivation de cette ou de ces protéines peut être effectuée par toute méthode connue de l'homme du métier, par exemple par mutagénèse dans le ou les gènes codant ladite ou lesdites protéines ou par l'expression d'un ou de plusieurs ARN anti-sens complémentaires de l'ARN ou des ARN messagers codant ladite ou lesdites protéines. La mutagénèse peut, par exemple, être effectuée par irradiation, par action d'agent chimique mutagène, par mutagénèse dirigée, par remplacement de gène ou toute autre méthode connue de l'homme du métier. Les conditions convenables d'une telle mutagénèse peuvent par exemple être adaptées selon l'enseignement contenu dans les ouvrages de Kieser, T et al (2000) et Ausubel et al., (2002). La mutagénèse peut être effectuée in vitro ou in situ, par suppression, substitution, délétion et/ou addition d'une ou plusieurs bases dans le gène considéré ou par inactivation génique. Microorganisms blocked in a step of the macrolide biosynthetic pathway can be obtained by inactivating the function of one or more proteins involved in the biosynthesis of this or these macrolides in microorganisms producing this macrolide. Depending on the inactivated protein (s), it will thus be possible to obtain microorganisms blocked in the different steps of the biosynthetic pathway of this or these macrolides. The inactivation of this or these proteins can be carried out by any method known to those skilled in the art, for example by mutagenesis in the gene (s) encoding said protein (s) or by the expression of one or more anti-RNA. -following complementary RNA or messenger RNAs encoding said one or more proteins. The mutagenesis may, for example, be carried out by irradiation, by mutagenic chemical action, by site-directed mutagenesis, by gene replacement or any other method known to those skilled in the art. The suitable conditions for such mutagenesis can, for example, be adapted according to the teaching contained in the works of Kieser, T et al (2000) and Ausubel et al., (2002). Mutagenesis can be carried out in vitro or in situ by deletion, substitution, deletion and / or addition of one or more bases in the gene of interest or by gene inactivation.
Cette mutagénèse peut être effectuée dans un gène comprenant une séquence telle que décrite ci-dessus. Préférentiellement, les microorganismes bloqués dans une étape de la voie de biosynthèse de macrolides sont des bactéries du genre Streptomyces. Plus préférentiellement, l'inactivation de la fonction d'une ou plusieurs protéines impliquées dans la biosynthèse du ou des macrolides en question est effectuée par mutagénèse. This mutagenesis can be carried out in a gene comprising a sequence as described above. Preferentially, the microorganisms blocked in a step of the macrolide biosynthetic pathway are bacteria of the genus Streptomyces. More preferably, the inactivation of the function of one or more proteins involved in the biosynthesis of the macrolide (s) in question is carried out by mutagenesis.
Encore plus préférentiellement, le macrolide en question est la spiramycine et les microorganismes dans lesquels sont effectuées la ou les mutagénèses sont des souches de S. ambofaciens. Plus préférentiellement, la mutagénèse est effectuée dans un ou plusieurs gènes comprenant une des séquences correspondant à une ou plusieurs des séquences SEQ ID N 3,5, 7,9, 11, 13,15, 17,19, 21,23, 25,28, 30,34, 36,40, 43, 45, 47, 49, 53, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 107, 109, 111, 113, 115, 118, et 120. De façon préférée, la ou les mutagénèses sont effectuées par inactivation Even more preferentially, the macrolide in question is spiramycin and the microorganisms in which the mutagenesis (s) are carried out are strains of S. ambofaciens. More preferably, the mutagenesis is carried out in one or more genes comprising one of the sequences corresponding to one or more of the sequences SEQ ID N 3.5, 7.9, 11, 13, 15, 17, 19, 21, 23, 25, 28, 30, 34, 36, 40, 43, 45, 47, 49, 53, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 107, 109, 111, 113, 115, 118, and 120. Preferably, the mutagenesis (s) are carried out by inactivation.
<Desc/Clms Page number 78><Desc / Clms Page number 78>
génique. De façon tout à fait préférée, la mutagénèse consiste en l'inactivation génique d'un gène comprenant une séquence correspondant à la séquence SEQ ID N 13. gene. Most preferably, the mutagenesis consists in the gene inactivation of a gene comprising a sequence corresponding to the sequence SEQ ID No. 13.
A titre illustratif, les microorganismes suivants peuvent être cités à titre d'exemple de tels microorganismes : OS49. 16 (or/3:: #hyg, cf. exemple 2), OS49.67 (orf3délétion en phase, cf exemple 6), OS49. 107 (or/8:: #hyg, cf. exemple 7), OS49.50

(or/10:: f2hyg, cf. exemple 8), SPM21 (oYfZ:: att3S2aac-, cf. exemple 10), SPM22 (orfZ: : att3 délétion en phase, cf. exemple 10), SPM501 (of6*: : attl S2hyg+, cf. exemple 14), SPM502 (orf6*:: attl délétion en phase, cf. exemple 14), SPM507 (orfl2::att3slaac-, cf. exemple 11), SPM508 (orfl3c: : att3S2aac-, cf. exemple 12), SPM509 (ofl4:: att3S2aac-, cf. exemple 13). As an illustration, the following microorganisms can be cited as examples of such microorganisms: OS49. 16 (or / 3 :: #hyg, see example 2), OS49.67 (orf3deletion in phase, cf example 6), OS49. 107 (or / 8 :: #hyg, see example 7), OS49.50
(or / 10 :: f2hyg, see example 8), SPM21 (oYfZ :: att3S2aac-, see example 10), SPM22 (orfZ:: att3 in-phase deletion, see example 10), SPM501 (of6 *:: attn S2hyg +, see example 14), SPM502 (orf6 * :: attl deletion in phase, see example 14), SPM507 (orf1c :: att3S2aac-, cf. Example 12), SPM509 (ofl4 :: att3S2aac-, see Example 13).
Un autre aspect de l'invention concerne un procédé de préparation d'un intermédiaire de biosynthèse de macrolide utilisant les microorganismes bloqués dans une étape de la voie de biosynthèse de macrolides, tels que décrits ci-dessus. Le procédé consiste à cultiver, dans un milieu de culture approprié, un microorganisme bloqué dans une étape de la voie de biosynthèse de macrolides, tels que décrits ci-dessus, récupérer le milieu de culture conditionné ou un extrait cellulaire, par exemple par sonication ou par choc osmotique, séparer et purifier à partir dudit milieu de culture ou encore à partir de l'extrait cellulaire obtenu à l'étape précédente, ledit intermédiaire de biosynthèse. Another aspect of the invention relates to a process for preparing a macrolide biosynthesis intermediate using the microorganisms blocked in a step of the macrolide biosynthetic pathway as described above. The method comprises culturing, in a suitable culture medium, a microorganism blocked in a step of the macrolide biosynthetic pathway, as described above, recovering the conditioned culture medium or a cell extract, for example by sonication or by osmotic shock, separate and purify from said culture medium or from the cell extract obtained in the preceding step, said biosynthesis intermediate.
Les conditions de culture de tels microorganismes pourront être déterminées selon des techniques bien connues de l'homme du métier. Le milieu de culture pourra par exemple être le milieu MP5 ou le milieu SL11pour les Streptomyces et notamment pour Streptomyces ambofaciens (Pernodet et al., 1993). L'homme du métier pourra notamment se référer à l'ouvrage de Kieser et al. (2000) en ce qui concerne la culture des Streptomyces. L'intermédiaire produit peut être récupéré par toutes techniques connues de l'homme du métier. L'homme du métier pourra se référer, par exemple, aux techniques enseignées dans le brevet américain US 3,000,785 et plus particulièrement aux méthodes d'extraction des spiramycines décrites dans ce brevet. The culture conditions of such microorganisms may be determined according to techniques well known to those skilled in the art. The culture medium may for example be MP5 medium or SL11 medium for Streptomyces and in particular for Streptomyces ambofaciens (Pernodet et al., 1993). Those skilled in the art may in particular refer to the work of Kieser et al. (2000) with regard to the cultivation of Streptomyces. The product intermediate can be recovered by any techniques known to those skilled in the art. Those skilled in the art may refer, for example, to the techniques taught in US Pat. No. 3,000,785 and more particularly to the spiramycin extraction methods described in this patent.
<Desc/Clms Page number 79> <Desc / Clms Page number 79>
Un autre objet de l'invention concerne un procédé de préparation d'une molécule dérivée d'un macrolide utilisant les microorganismes bloqués dans une étape de la voie de biosynthèse de ce macrolide, tels que décrits ci-dessus. Le procédé consiste à obtenir un intermédiaire de biosynthèse selon le procédé ci-dessus et à modifier l'intermédiaire ainsi produit, éventuellement après séparation du milieu de culture. Les conditions de culture de tels microorganismes pourront être déterminées selon des techniques bien connues de l'homme du métier. Le milieu de culture pourra par exemple être le milieu MP5 ou le milieu SL11pour les Streptomyces et notamment pour Streptomyces ambofaciens (Pernodet et al., 1993). L'homme du métier pourra notamment se référer à l'ouvrage de Kieser et al. 2000 en ce qui concerne la culture des Streptomyces. Les intermédiaires produits peuvent être modifiés, éventuellement après séparation, soit par ajout de composants appropriés dans les milieux de production, soit par introduction dans les microorganismes d'autres gènes codant des protéines susceptibles de modifier l'intermédiaire en s'en servant comme substrat. Ces intermédiaires peuvent ainsi être modifiés par voie chimique, biochimique, enzymatique et/ou microbiologique. Plus préférentiellement, le macrolide en question est la spiramycine et les microorganismes dans lesquels sont effectuées la ou les mutagénèses sont des souches de S. ambofaciens. Another object of the invention relates to a process for preparing a molecule derived from a macrolide using the microorganisms blocked in a step of the biosynthetic pathway of this macrolide, as described above. The process consists of obtaining a biosynthesis intermediate according to the above process and modifying the intermediate thus produced, possibly after separation from the culture medium. The culture conditions of such microorganisms may be determined according to techniques well known to those skilled in the art. The culture medium may for example be MP5 medium or SL11 medium for Streptomyces and in particular for Streptomyces ambofaciens (Pernodet et al., 1993). Those skilled in the art may in particular refer to the work of Kieser et al. 2000 with regard to the cultivation of Streptomyces. The intermediates produced can be modified, optionally after separation, either by adding appropriate components to the production media, or by introducing into the microorganisms other genes encoding proteins capable of modifying the intermediate by using it as a substrate. These intermediates can thus be modified chemically, biochemically, enzymatically and / or microbiologically. More preferably, the macrolide in question is spiramycin and the microorganisms in which the mutagenesis (s) are carried out are strains of S. ambofaciens.
L'invention est également relative à un microorganisme produisant de la spiramycine I mais ne produisant pas de spiramycine II et III. Ce microorganisme comprend l'ensemble des gènes nécessaires à la biosynthèse de la spiramycine I mais ne produit pas de spiramycine II et III car le gène comprenant la séquence correspondant à SEQ ID N 13 ou l'un de ses variants ou l'une des séquences dérivées de celles-ci en raison de la dégénérescence du code génétique et codant un polypeptide de séquence SEQ ID N 14 ou l'un de ses variants n'est pas exprimé ou a été rendu inactif. The invention also relates to a microorganism producing spiramycin I but not producing spiramycin II and III. This microorganism comprises all the genes necessary for the biosynthesis of spiramycin I but does not produce spiramycin II and III because the gene comprising the sequence corresponding to SEQ ID No. 13 or one of its variants or one of the sequences derived from them because of the degeneracy of the genetic code and encoding a polypeptide of sequence SEQ ID N 14 or one of its variants is not expressed or has been rendered inactive.
L'inactivation de cette protéine peut être effectuée par toute méthode connue de l'homme du métier, par exemple par mutagénèse dans le gène codant ladite protéine ou par l'expression d'un ARN anti-sens complémentaire de l'ARN messager codant ladite protéine, étant entendu que si l'expression de orf5* est affectée par cette manipulation, il sera nécessaire de procéder à une autre modification pour que le gène orf5* soit The inactivation of this protein can be carried out by any method known to those skilled in the art, for example by mutagenesis in the gene encoding said protein or by the expression of an antisense RNA complementary to the messenger RNA encoding said protein. protein, provided that if the expression of orf5 * is affected by this manipulation, it will be necessary to make another modification so that the orf5 * gene is
<Desc/Clms Page number 80><Desc / Clms Page number 80>
correctement exprimé. La mutagénèse peut être effectuée dans la séquence codante ou dans une séquence non codante de manière à rendre inactive la protéine codée ou à en empêcher son expression ou sa traduction. La mutagénèse peut être effectuée par mutagénèse dirigée, par remplacement de gène ou toute autre méthode connue de l'homme du métier. Les conditions convenables d'une telle mutagénèse peuvent par exemple être adaptées selon l'enseignement contenu dans les ouvrages de Kieser, T et al (2000) et Ausubel et al., 2002. La mutagénèse peut être effectuée in vitro ou in situ, par suppression, substitution, délétion et/ou addition d'une ou plusieurs bases dans le gène considéré ou par inactivation génique. Le microorganisme peut être également obtenu en exprimant les gènes de la voie de biosynthèse de la spiramycine sans que ceux-ci ne comprennent le gène comprenant la séquence SEQ ID N 13 ou l'un de ses variants ou l'une des séquences dérivées de celles-ci en raison de la dégénérescence du code génétique et codant un polypeptide de séquence SEQ ID N 14 ou l'un de ses variants. Préférentiellement, le microorganisme est une bactérie du genre Streptomyces. correctly expressed. Mutagenesis may be performed in the coding sequence or in a non-coding sequence so as to render the encoded protein inactive or to prevent its expression or translation. Mutagenesis can be performed by site-directed mutagenesis, gene replacement or any other method known to those skilled in the art. The suitable conditions for such mutagenesis can, for example, be adapted according to the teaching contained in the works of Kieser, T et al (2000) and Ausubel et al., 2002. Mutagenesis can be performed in vitro or in situ, by deletion, substitution, deletion and / or addition of one or more bases in the gene of interest or by gene inactivation. The microorganism can also be obtained by expressing the genes of the spiramycin biosynthetic pathway without these comprising the gene comprising the sequence SEQ ID No. 13 or one of its variants or one of the sequences derived from those because of the degeneracy of the genetic code and encoding a polypeptide of sequence SEQ ID No. 14 or one of its variants. Preferably, the microorganism is a bacterium of the genus Streptomyces.
Plus préférentiellement, le microorganisme produisant de la spiramycine 1 mais ne produisant pas de spiramycine II et III est obtenu à partir d'un microorganisme de départ produisant les spiramycines I, II et III. Encore plus préférentiellement, le microorganisme est obtenu par mutagénèse dans un gène comprenant la séquence correspondant à SEQ ID N 13 ou l'un de ses variants ou l'une des séquences dérivées de celles-ci en raison de la dégénérescence du code génétique et codant un polypeptide de séquence SEQ ID N 14 ou l'un de ses variants ayant la même fonction. Encore plus préférentiellement, cette mutagénèse est effectuée par inactivation génique. De manière préférée le microorganisme est obtenu à partir d'une souche de S. ambofaciens produisant les spiramycines I, II et III dans laquelle est effectuée une inactivation génique du gène comprenant la séquence correspondant à SEQ ID N 13 ou l'une des séquences dérivées de celle-ci en raison de la dégénérescence du code génétique. De manière tout à fait préférée, l'inactivation génique est effectuée par délétion en phase du gène ou d'une partie du gène comprenant la séquence correspondant à SEQ ID N 13 ou l'une des séquences dérivées de celle-ci en raison de la dégénérescence du code génétique. A titre illustratif, la souche SPM502 (orf6*::attl, cf. exemple 14) peut être More preferably, the microorganism producing spiramycin 1 but not producing spiramycin II and III is obtained from a starting microorganism producing spiramycin I, II and III. Even more preferentially, the microorganism is obtained by mutagenesis in a gene comprising the sequence corresponding to SEQ ID No. 13 or one of its variants or one of the sequences derived from these because of the degeneracy of the genetic code and coding a polypeptide of sequence SEQ ID No. 14 or one of its variants having the same function. Even more preferentially, this mutagenesis is carried out by gene inactivation. Preferably, the microorganism is obtained from a strain of S. ambofaciens producing spiramycins I, II and III in which gene inactivation of the gene comprising the sequence corresponding to SEQ ID No. 13 or one of the derived sequences is carried out. of it because of the degeneracy of the genetic code. Most preferably, the gene inactivation is carried out by phase deletion of the gene or a part of the gene comprising the sequence corresponding to SEQ ID No. 13 or one of the sequences derived therefrom due to the degeneration of the genetic code. By way of illustration, the strain SPM502 (orf6 * :: attl, see Example 14) can be
<Desc/Clms Page number 81><Desc / Clms Page number 81>
citée comme un microorganisme produisant de la spiramycine I mais ne produisant pas de spiramycine II et III. cited as a microorganism producing spiramycin I but not producing spiramycin II and III.
L'invention est également relative à un microorganisme produisant de la spiramycine I mais ne produisant pas de spiramycine II et III tel que décrit ci-dessus caractérisé en ce en ce qu'il surexprime en outre : - un gène susceptible d'être obtenu par amplification en chaîne par polymérase en utilisant le couple d'amorces de séquence suivante : 5' AAGCTTGTGTGCCCGGTGTACCTGGGGAGC 3' (SEQ ID N 138) et 5' GGATCCCGCGACGGACACGACCGCCGCGCA 3' (SEQ ID N 139) et comme matrice le cosmide pSPM36 ou l'ADN total de Streptomyces ambofaciens - ou un gène dérivé de celui-ci en raison de la dégénérescence du code génétique. The invention also relates to a microorganism producing spiramycin I but not producing spiramycin II and III as described above, characterized in that it overexpresses in addition: a gene that can be obtained by polymerase chain reaction using the pair of primers of the following sequence: 5 'AAGCTTGTGTGCCCGGTGTACCTGGGGAGC 3' (SEQ ID N 138) and 5 'GGATCCCGCGACGGACACGACCGCCGCGCA 3' (SEQ ID N 139) and as template the cosmid pSPM36 or the total DNA of Streptomyces ambofaciens - or a gene derived from it because of the degeneracy of the genetic code.
Un exemple de séquence d'un tel gène est donnée en SEQ ID N 111(ADN), cependant cette séquence est partielle puisqu'elle ne comprend pas la partie 3' de la séquence codante correspondante. La traduction en protéine de cette partie de séquence codante est donnée en SEQ ID N 112. L'homme du métier pourra aisément la compléter notamment en utilisant l'enseignement présenté dans l'exemple 24. Il est ainsi présenté dans cet exemple une méthode pour cloner le gène orf28c et pour fabriquer un vecteur d'expression permettant l'expression de orf28c. Il est également montré dans cet exemple que la surexpression du gène orf28c dans la souche OSC2 conduit à l'amélioration de la production en spiramycines de cette souche. La surexpression du gène orf28c peut être obtenue en augmentant le nombre de copies de ce gène et/ou en plaçant un promoteur plus actif que le promoteur sauvage. De façon préférée, la surexpression dudit gène est obtenue en apportant dans le microorganisme une construction d'ADN recombinant, permettant la surexpression de ce gène. De façon préférée, cette construction d'ADN recombinant augmente le nombre de copie dudit gène et permet d'obtenir une surexpression dudit gène. Dans cette construction d'ADN recombinant, la séquence codante du gène peut être placée sous le contrôle d'un promoteur plus actif que le promoteur sauvage. On peut citer à titre d'illustration le promoteur ptrc actif chez Streptomyces ambofaciens (Amann, E. et al., 1988) ainsi que An example of a sequence of such a gene is given in SEQ ID No. 111 (DNA), however this sequence is partial since it does not include the 3 'part of the corresponding coding sequence. The protein translation of this portion of coding sequence is given in SEQ ID No. 112. Those skilled in the art can easily complete it, in particular by using the teaching presented in Example 24. It is thus presented in this example a method for clone the orf28c gene and to make an expression vector allowing the expression of orf28c. It is also shown in this example that the overexpression of the orf28c gene in the OSC2 strain leads to the improvement of the spiramycin production of this strain. Overexpression of the orf28c gene can be achieved by increasing the copy number of this gene and / or placing a more active promoter than the wild-type promoter. Preferably, the overexpression of said gene is obtained by providing the microorganism a recombinant DNA construct, allowing the overexpression of this gene. Preferably, this recombinant DNA construct increases the number of copies of said gene and makes it possible to obtain an overexpression of said gene. In this recombinant DNA construct, the coding sequence of the gene may be placed under the control of a more active promoter than the wild-type promoter. For example, the ptrc promoter active in Streptomyces ambofaciens (Amann, E. et al., 1988) and
<Desc/Clms Page number 82><Desc / Clms Page number 82>
le promoteur ermE*. Ainsi, de façon préférée, la ou les copies du gène orf28c apportée sont placées sous le contrôle du promoteur ermE* comme c'est le cas dans la construction pSPM75 (cf. exemple 24). the promoter ermE *. Thus, preferably, the copy or copies of the orf28c gene provided are placed under the control of the ermE * promoter as is the case in the pSPM75 construct (see Example 24).
L'invention est également relative à un microorganisme produisant de la spiramycine 1 mais ne produisant pas de spiramycine II et III tel que décrit ci-dessus caractérisé en ce en ce qu'il surexprime en outre le gène de séquence codante SEQ ID N 47 ou d'une séquence codante dérivée de celle-ci en raison de la dégénérescence du code génétique. Préférentiellement, ce microorganisme est caractérisé en ce qu'il s'agit de la souche SPM502 pSPM525 déposée auprès de la Collection Nationale de Cultures de Microorganismes (CNCM) Institut Pasteur, 25, rue du Docteur Roux 75724 Paris Cedex 15, France, le 26 février 2003 sous le numéro d'enregistrement I- 2977. The invention also relates to a microorganism producing spiramycin 1 but not producing spiramycin II and III as described above, characterized in that it also overexpresses the coding sequence gene SEQ ID N 47 or of a coding sequence derived therefrom due to the degeneracy of the genetic code. Preferably, this microorganism is characterized in that it is the SPM502 strain pSPM525 deposited with the National Collection of Cultures of Microorganisms (CNCM) Institut Pasteur, 25, rue du Docteur Roux 75724 Paris Cedex 15, France, on the 26th February 2003 under the registration number I-2977.
L'invention est également relative à un procédé de production de spiramycine I, le procédé consiste à cultiver dans un milieu de culture approprié un microorganisme produisant de la spiramycine 1 mais ne produisant pas de spiramycine II et III tel que décrit ci-dessus, récupérer le milieu de culture conditionné ou un extrait cellulaire, séparer et purifier à partir dudit milieu de culture ou encore à partir de l'extrait cellulaire obtenu à l'étape précédente la spiramycine I. Les conditions de culture de tel microorganisme pourront être déterminées selon des techniques bien connues de l'homme du métier. Le milieu de culture pourra par exemple être le milieu MP5 ou le milieu SL11 pour les Streptomyces et notamment pour Streptomyces ambofaciens (Pernodet et al., 1993). L'homme du métier pourra notamment se référer à l'ouvrage de Kieser et al. 2000 en ce qui concerne la culture des Streptomyces. La spiramycine 1 produite peut être récupérée par toutes techniques connues de l'homme du métier. The invention also relates to a process for the production of spiramycin I, the process consists in cultivating in a suitable culture medium a microorganism producing spiramycin 1 but not producing spiramycin II and III as described above, recovering the conditioned culture medium or a cell extract, separating and purifying from said culture medium or from the cell extract obtained in the preceding step spiramycin I. The culture conditions for such a microorganism may be determined according to techniques well known to those skilled in the art. The culture medium may for example be MP5 medium or SL11 medium for Streptomyces and in particular for Streptomyces ambofaciens (Pernodet et al., 1993). Those skilled in the art may in particular refer to the work of Kieser et al. 2000 with regard to the cultivation of Streptomyces. The spiramycin 1 produced can be recovered by any techniques known to those skilled in the art.
L'homme du métier pourra se référer, par exemple, aux techniques enseignées dans le brevet américain US 3,000,785 et plus particulièrement aux méthodes d'extraction des spiramycines décrites dans ce brevet. Those skilled in the art may refer, for example, to the techniques taught in US Pat. No. 3,000,785 and more particularly to the spiramycin extraction methods described in this patent.
<Desc/Clms Page number 83> <Desc / Clms Page number 83>
Un autre aspect de l'invention concerne l'utilisation d'une séquence nucléotidique selon l'invention pour améliorer la production en macrolides d'un microorganisme. Another aspect of the invention relates to the use of a nucleotide sequence according to the invention for improving the production of macrolides of a microorganism.
Ainsi, l'invention concerne un microorganisme mutant producteur de macrolide caractérisé en ce qu'il possède une modification génétique dans au moins un gène comprenant une séquence telle que définie ci-dessus et/ou qu'elle surexprime au moins un gène comprenant une séquence telle que définie ci-dessus. La modification génétique peut consister en une suppression, une substitution, une délétion et/ou une addition d'une ou plusieurs bases dans le ou les gènes considérés dans le but d'exprimer une ou des protéines ayant une activité supérieure ou d'exprimer un niveau plus élevé de cette ou de ces protéines. La surexpression du gène considéré peut être obtenue en augmentant le nombre de copies de ce gène et/ou en plaçant un promoteur plus actif que le promoteur sauvage. On peut citer à titre d'illustration le promoteur ptrc actif chez Streptomyces ambofaciens (Amann, E. et al., 1988) ainsi que le promoteur ermE* (Bibb et al., 1985), (Bibb et al., 1994). Ainsi la surexpression du gène considéré peut être obtenue en apportant dans un microorganisme producteur du macrolide considéré une construction d'ADN recombinant selon l'invention, permettant la surexpression de ce gène. En effet, certaines étapes de la biosynthèse des macrolides sont limitantes et si l'on exprime une ou plusieurs protéines plus actives ou un niveau d'expression plus important de la ou des protéines sauvages impliquées dans ces étapes limitantes, on peut améliorer la production du ou des macrolides concernés. Une augmentation du taux de production de la tylosine a par exemple été obtenue chez Streptomyces fradiae par duplication du gène codant une methyl transférase limitante convertissant la macrocine en tylosine (Baltz R, 1997). L'obtention de l'expression d'une protéine plus active peut être obtenue notamment par mutagénèse, l'homme du métier pourra par exemple se référer à ce sujet à l'ouvrage de F.Ausubel et al (2002). Préférentiellement, ces microorganismes mutants améliorés pour leur production en macrolides sont des bactéries du genre Streptomyces. Thus, the invention relates to a macrolide-producing mutant microorganism characterized in that it has a genetic modification in at least one gene comprising a sequence as defined above and / or that it overexpresses at least one gene comprising a sequence as defined above. The genetic modification may consist of the deletion, substitution, deletion and / or addition of one or more bases in the gene (s) considered for the purpose of expressing one or more proteins having a higher activity or expressing a higher level of this or these proteins. Overexpression of the gene of interest can be achieved by increasing the copy number of this gene and / or placing a more active promoter than the wild-type promoter. The active ptrc promoter in Streptomyces ambofaciens (Amann, E. et al., 1988) and the ermE * promoter (Bibb et al., 1985) (Bibb et al., 1994) can be cited as an example. Thus, the overexpression of the gene in question can be obtained by supplying a microorganism producing the macrolide considered a recombinant DNA construct according to the invention, allowing the overexpression of this gene. In fact, certain stages of macrolide biosynthesis are limiting and if one or more more active proteins or a higher level of expression of the wild protein or proteins involved in these limiting steps are expressed, the production of the macrolide can be improved. or macrolides involved. For example, an increase in the production rate of tylosin was obtained in Streptomyces fradiae by duplication of the gene encoding a limiting methyl transferase converting macrocin to tylosin (Baltz R, 1997). Obtaining the expression of a more active protein can be obtained in particular by mutagenesis, the person skilled in the art can for example refer to this subject in the work of F.Ausubel et al (2002). Preferentially, these mutant microorganisms improved for their production in macrolides are bacteria of the genus Streptomyces.
Plus préférentiellement, le macrolide en question est la spiramycine et les microorganismes dans lesquels sont effectuées la ou les mutagénèses sont des souches de S. ambofaciens. Plus préférentiellement, la modification génétique est effectuée dans un ou plusieurs gènes comprenant une des séquences correspondant à une ou plusieurs More preferably, the macrolide in question is spiramycin and the microorganisms in which the mutagenesis (s) are carried out are strains of S. ambofaciens. More preferably, the genetic modification is carried out in one or more genes comprising one of the sequences corresponding to one or more
<Desc/Clms Page number 84><Desc / Clms Page number 84>
des séquences SEQ ID N 3,5, 7, 9, Il, 13, 15, 17, 19,21,23,25,28,30,34,36,40,43, 45, 47, 49, 53, 60, 62, 64, 66, 68,70, 72,74, 76,78, 80,82, 84,107, 109,111, 113,115, 118, et 120, ou l'un de ses variants ou l'une des séquences dérivées de celles-ci en raison de la dégénérescence du code génétique. De façon préférée, le microorganisme surexprime un ou plusieurs gènes comprenant une des séquences correspondant à une ou plusieurs des séquences SEQ ID N 3,5, 7,9, 11,13, 15,17, 19, 21, 23, 25, 28, 30,34, 36,40, 43,45, 47,49, 53,60, 62,64, 66,68, 70,72, 74,76, 78,80, 82,84, 107, 109, 111,113, 115,118, et 120, ou l'un de ses variants ou l'une des séquences dérivées de celles-ci en raison de la dégénérescence du code génétique. De façon préférée, le microorganisme surexprime le gène comprenant une séquence correspondant à SEQ ID N 111, ou l'un de ses variants ou l'une des séquences dérivées de celles-ci en raison de la dégénérescence du code génétique. La séquence donnée en SEQ ID N 111 est partielle, cependant l'homme du métier pourra aisément la compléter notamment en utilisant l'enseignement présenté dans l'exemple 24. Il est ainsi présenté dans cet exemple une méthode pour cloner le gène orf28c et pour fabriquer un vecteur d'expression permettant l'expression de orf28c. Il est également montré dans cet exemple que la surexpression du gène orf28c dans la souche OSC2 conduit à l'amélioration de la production en spiramycines de cette souche. La surexpression du gène orf28c peut être obtenue en augmentant le nombre de copies de ce gène et/ou en plaçant un promoteur plus actif que le promoteur sauvage. De façon préférée, la surexpression du gène orf28c est obtenue en apportant dans le microorganisme une construction d'ADN recombinant, permettant la surexpression de ce gène. De façon préférée, cette construction d'ADN recombinant augmente le nombre de copie du gène orj28c et permet d'obtenir une surexpression du gène orf28c. Dans cette construction d'ADN recombinant, la séquence codante de orf28c peut être placée sous le contrôle , d'un promoteur plus actif que le promoteur sauvage. On peut citer à titre d'illustration le promoteur ptrc actif chez Streptomyces ambofaciens (Amann, E. et al., 1988) ainsi que le promoteur ermE*. Ainsi, de façon préférée, la ou les copies du gène orf28c apportée sont placées sous le contrôle du promoteur ermE* comme c'est le cas dans la construction pSPM75 (cf. exemple 24). sequences SEQ ID N 3.5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 28, 30, 34, 36, 40, 43, 45, 47, 49, 53, 60 , 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 107, 109, 111, 113, 115, 118, and 120, or one of its variants or one of the sequences derived from those because of the degeneracy of the genetic code. Preferably, the microorganism overexpresses one or more genes comprising one of the sequences corresponding to one or more of the sequences SEQ ID N 3.5, 7.9, 11, 13, 15, 17, 19, 21, 23, 25, 28. ,,,, , 115, 118, and 120, or one of its variants or one of the sequences derived therefrom due to the degeneracy of the genetic code. Preferably, the microorganism overexpresses the gene comprising a sequence corresponding to SEQ ID No. 111, or one of its variants or one of the sequences derived from these because of the degeneracy of the genetic code. The sequence given in SEQ ID No. 111 is partial, however the person skilled in the art will easily be able to complete it, in particular by using the teaching presented in Example 24. It is thus presented in this example a method for cloning the orf28c gene and for to make an expression vector allowing the expression of orf28c. It is also shown in this example that the overexpression of the orf28c gene in the OSC2 strain leads to the improvement of the spiramycin production of this strain. Overexpression of the orf28c gene can be achieved by increasing the copy number of this gene and / or placing a more active promoter than the wild-type promoter. Preferably, the overexpression of the orf28c gene is obtained by providing a recombinant DNA construct in the microorganism, allowing the overexpression of this gene. Preferably, this recombinant DNA construct increases the number of copies of the orj28c gene and makes it possible to obtain overexpression of the orf28c gene. In this recombinant DNA construct, the orf28c coding sequence may be under the control of a more active promoter than the wild-type promoter. For example, the ptrc promoter active in Streptomyces ambofaciens (Amann, E. et al., 1988) and the ermE * promoter can be cited. Thus, preferably, the copy or copies of the orf28c gene provided are placed under the control of the ermE * promoter as is the case in the pSPM75 construct (see Example 24).
<Desc/Clms Page number 85> <Desc / Clms Page number 85>
Un autre aspect de l'invention concerne un procédé de production de macrolides utilisant les microorganismes décrits au paragraphe précédent. Ce procédé consiste à cultiver dans un milieu de culture approprié un microorganisme défini dans le paragraphe précédent, récupérer le milieu de culture conditionné ou un extrait cellulaire, séparer et purifier à partir dudit milieu de culture ou encore à partir de l'extrait cellulaire obtenu à l'étape précédente le ou les macrolides produits. Les conditions de culture de tels microorganismes pourront être déterminées selon des techniques bien connues de l'homme du métier. Le milieu de culture pourra par exemple être le milieu MP5 ou le milieu SL11 pour les Streptomyces et notamment pour Streptomyces ambofaciens (Pernodet et al., 1993). L'homme du métier pourra notamment se référer à l'ouvrage de Kieser et al. 2000 en ce qui concerne la culture des Streptomyces. Le ou les macrolides produits peuvent être récupérés par toutes techniques connues de l'homme du métier. L'homme du métier pourra se référer, par exemple, aux techniques enseignées dans le brevet américain US 3,000,785 et plus particulièrement aux méthodes d'extraction des spiramycines décrites dans ce brevet. Préférentiellement, les microorganismes utilisés dans un tel procédé sont des bactéries du genre Streptomyces. Another aspect of the invention relates to a process for producing macrolides using the microorganisms described in the preceding paragraph. This method consists of cultivating in a suitable culture medium a microorganism defined in the preceding paragraph, recovering the conditioned culture medium or a cell extract, separating and purifying from said culture medium or from the cell extract obtained at the previous step the macrolide (s) produced. The culture conditions of such microorganisms may be determined according to techniques well known to those skilled in the art. The culture medium may for example be MP5 medium or SL11 medium for Streptomyces and in particular for Streptomyces ambofaciens (Pernodet et al., 1993). Those skilled in the art may in particular refer to the work of Kieser et al. 2000 with regard to the cultivation of Streptomyces. The macrolide (s) produced can be recovered by any technique known to those skilled in the art. Those skilled in the art may refer, for example, to the techniques taught in US Pat. No. 3,000,785 and more particularly to the spiramycin extraction methods described in this patent. Preferably, the microorganisms used in such a process are bacteria of the genus Streptomyces.
Plus préférentiellement, le macrolide en question est la spiramycine et les microorganismes mutants améliorés pour leur production en spiramycines sont des souches de S. ambofaciens. More preferably, the macrolide in question is spiramycin and the mutant microorganisms improved for their production in spiramycins are strains of S. ambofaciens.
Un autre aspect de l'invention concerne l'utilisation d'une séquence et/ou d'un vecteur selon l'invention pour la préparation d'antibiotiques hybrides. En effet, les polynucléotides selon l'invention peuvent servir à l'obtention de microorganisme exprimant une ou plusieurs protéines mutantes donnant lieu à une modification dans la spécificité du substrat ou encore être exprimés dans de multiples microorganismes producteurs d'antibiotiques dans le but de générer des antibiotiques hybrides. Ainsi les polynucléotides selon l'invention peuvent permettre, par transfert des gènes entre microorganismes producteurs, de fabriquer des antibiotiques hybrides ayant des propriétés pharmacologiquement intéressantes (Hopwood et al.,1985a, Hopwood et al.,1985b, Hutchinson et al., 1989). Le principe par lequel l'ingénierie génétique peut conduire à la production d'antibiotiques hybrides a été tout d'abord proposé par Another aspect of the invention relates to the use of a sequence and / or a vector according to the invention for the preparation of hybrid antibiotics. Indeed, the polynucleotides according to the invention can be used to obtain microorganisms expressing one or more mutant proteins giving rise to a modification in the specificity of the substrate or else to be expressed in multiple antibiotic-producing microorganisms in order to generate hybrid antibiotics. Thus the polynucleotides according to the invention can make it possible, by gene transfer between producer microorganisms, to produce hybrid antibiotics having pharmacologically interesting properties (Hopwood et al., 1985a, Hopwood et al., 1985b, Hutchinson et al., 1989). . The principle by which genetic engineering can lead to the production of hybrid antibiotics was first proposed by
<Desc/Clms Page number 86><Desc / Clms Page number 86>
Hopwood (Hopwood 1981). Il a ainsi été proposé que les enzymes participant à la biosynthèse d'antibiotiques acceptent souvent des substrats reliés structurellement, mais différents de leur substrat naturel. Il est généralement admis (Hopwood 1981, Hutchinson 1988, Robinson 1988) que les enzymes codées par les gènes de la voie de biosynthèse d'antibiotiques ont une spécificité de substrat moins stricte que les enzymes du métabolisme primaire. Il a ainsi pu être montré qu'un grand nombre de substrats nonnaturels sont transformés par des microorganismes producteurs d' antibiotique, leur mutants ou des enzymes purifiées de la voie de biosynthèse de ces antibiotiques (Hutchinson 1988). En utilisant cet enseignement, l'homme du métier pourra construire des microorganismes exprimant une ou plusieurs protéines mutantes donnant lieu à une modification dans la spécificité du substrat dans le but de générer des antibiotiques hybrides. Hopwood (Hopwood 1981). It has been proposed that enzymes involved in antibiotic biosynthesis often accept structurally related substrates, but different from their natural substrate. It is generally accepted (Hopwood 1981, Hutchinson 1988, Robinson 1988) that the enzymes encoded by genes in the antibiotic biosynthetic pathway have a less stringent substrate specificity than primary metabolic enzymes. It has thus been shown that a large number of natural substrates are transformed by antibiotic-producing microorganisms, mutants or purified enzymes of the biosynthetic pathway of these antibiotics (Hutchinson 1988). Using this teaching, those skilled in the art can construct microorganisms expressing one or more mutant proteins giving rise to a modification in the specificity of the substrate in order to generate hybrid antibiotics.
L'invention concerne également l'utilisation d'au moins un polynucléotide et/ou au moins un ADN recombinant et/ou au moins un vecteur d'expression et/ou au moins un polypeptide et/ou au moins une cellule hôte selon l'invention pour réaliser une ou plusieurs bioconversions. Ainsi l'invention permet de construire des souches bactériennes ou fongiques dans lesquelles on exprime, sous le contrôle de signaux d'expression appropriés, une ou plusieurs protéines selon l'invention. De telles souches peuvent alors être utilisées pour réaliser une ou des bioconversions. Ces bioconversions peuvent se faire soit à l'aide de cellules entières, soit à l'aide d'extraits acellulaires des dites cellules. Ces bioconversions peuvent permettre de transformer une molécule en une forme dérivée, avec une enzyme d'une voie de biosynthèse. Par exemple, Carreras et al. décrit l'utilisation de souche de Saccharopolyspora erythraea et de Streptomyces coelicolor pour la production de nouveau dérivés d'érithromycines (Carreras et al., 2002). Walczar et al. décrit l'utilisation d'une P450 mono-oxygénase de Streptomyces pour la bioconversion de désacétyladriamycine (un analogue de l'anthracycline) en de nouvelles anthracyclines (Walczar et al., 2001). Olonao et al. décrit l'utilisation d'une souche de Streptomyces lividans modifiée pour la bioconversion de rhodomycinone- The invention also relates to the use of at least one polynucleotide and / or at least one recombinant DNA and / or at least one expression vector and / or at least one polypeptide and / or at least one host cell according to the invention. invention for performing one or more bioconversions. Thus, the invention makes it possible to construct bacterial or fungal strains in which one or more proteins according to the invention are expressed under the control of appropriate expression signals. Such strains can then be used to carry out one or more bioconversions. These bioconversions can be done either with whole cells or with acellular extracts of said cells. These bioconversions can make it possible to transform a molecule into a derived form, with an enzyme of a biosynthetic pathway. For example, Carreras et al. describes the use of Saccharopolyspora erythraea strain and Streptomyces coelicolor strain for the production of new erithromycin derivatives (Carreras et al., 2002). Walczar et al. describes the use of Streptomyces P450 monooxygenase for the bioconversion of deacetyladriamycin (an anthracycline analogue) to novel anthracyclines (Walczar et al., 2001). Olonao et al. describes the use of a strain of Streptomyces lividans modified for rhodomycinone bioconversion.
<Desc/Clms Page number 87><Desc / Clms Page number 87>
epsilon en rhodomycine D (Olonao et al., 1999). L'homme du métier pourra appliquer ce principe à tout intermédiaire de biosynthèse. epsilon to rhodomycin D (Olonao et al., 1999). Those skilled in the art can apply this principle to any biosynthetic intermediate.
L'invention concerne également un ADN recombinant caractérisé en ce qu'il comprend : - un polynucléotide susceptible d'être obtenu par amplification en chaîne par polymérase en utilisant le couple d'amorces de séquence suivante : 5' AAGCTTGTGTGCCCGGTGTACCTGGGGAGC 3' (SEQ ID N 138) et 5' GGATCCCGCGACGGACACGACCGCCGCGCA 3' (SEQ ID N 139) et comme matrice le cosmide pSPM36 ou l'ADN total de Streptomyces ambofaciens - ou un fragment d'au moins 10,12, 15, 18, 20 à 25, 30, 40, 50,60, 70,80, 90,100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000,1050, 1100,1150, 1200,1250, 1300,1350, 1400,1450, 1460,1470, 1480, 1490 ou 1500 nucléotides consécutifs de ce polynucléotide. The invention also relates to a recombinant DNA characterized in that it comprises: a polynucleotide capable of being obtained by polymerase chain reaction using the following pair of primers: 5 'AAGCTTGTGTGCCCGGTGTACCTGGGGAGC 3' (SEQ ID N 138) and 5 'GGATCCCGCGACGGACACGACCGCCGCGCA 3' (SEQ ID N 139) and as a template the cosmid pSPM36 or the total DNA of Streptomyces ambofaciens - or a fragment of at least 10,12, 15, 18, 20 to 25, 30, 40, 50,60, 70,80, 90,100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000, 1050, 1100, 1150, 1200, 1250, 1300, 1350, 1400, 1450, 1460, 1470, 1480, 1490 or 1500 consecutive nucleotides of this polynucleotide.
Préférentiellement ; cet ADN recombinant est un vecteur. Encore plus préférentiellement, le vecteur est choisi parmi les bactériophages, les plasmides, les phagemides, les vecteurs intégratifs, les fosmides, les cosmides, les vecteurs navettes, les BAC (Bacterial Artificial Chromosome) ou les PAC (Pl-derived Artificial Chromosome). A titre illustratif on peut citer comme bactériophages le phage lambda et le phage M13. Comme plasmides on peut citer les plasmides se répliquant chez E. coli par exemple pBR322 et ses dérivés, pUC18 et ses dérivés, pUC19 et ses dérivés, pGB2 et ses dérivés (Churchward G. et al., 1984), pACYC177 (numéro d'accès GenBank : X06402) et ses dérivés, pACYC184 (numéro d'accès GenBank : X06403) et ses dérivés. On peut également citer les plasmides se répliquant chez Streptomyces comme par exemple pIJ101 et ses dérivés, pSG5 et ses dérivés, SLP1 et ses dérivés, SCP2* et ses dérivés (Kieser et al. 2000). Comme phagemides on peut citer à titre illustratif pBluescript II et ses dérivés (commercialisés notamment par la société Stratagene (LaJolla, Californie, USA)), pGEM-T et ses dérivés (commercialisé par la société Promega (Madison, Wisconcin, USA)), #ZAPII et ses dérivés (commercialisés notamment par la société Stratagene (LaJolla, Californie, USA)). Preferentially; this recombinant DNA is a vector. Even more preferentially, the vector is chosen from bacteriophages, plasmids, phagemids, integrative vectors, fosmids, cosmids, shuttle vectors, BACs (Bacterial Artificial Chromosome) or PACs (Pl-derived Artificial Chromosome). By way of illustration, lambda phage and phage M13 may be mentioned as bacteriophages. Plasmids that may be mentioned include plasmids that replicate in E. coli, for example pBR322 and its derivatives, pUC18 and its derivatives, pUC19 and its derivatives, pGB2 and its derivatives (Churchward G. et al., 1984), pACYC177 (accession number GenBank Access: X06402) and its derivatives, pACYC184 (GenBank Access Number: X06403) and its derivatives. Streptomyces replicating plasmids such as, for example, pIJ101 and its derivatives, pSG5 and its derivatives, SLP1 and its derivatives, SCP2 * and its derivatives can also be mentioned (Kieser et al., 2000). As illustrative pBluescript II and its derivatives (sold in particular by Stratagene (LaJolla, California, USA)), pGEM-T and its derivatives (marketed by Promega (Madison, Wisconcin, USA)), #ZAPII and its derivatives (marketed in particular by Stratagene (LaJolla, California, USA)).
<Desc/Clms Page number 88> <Desc / Clms Page number 88>
Comme vecteurs intégratifs on peut citer à titre illustratif, les vecteurs intégratifs chez Streptomyces comme par exemple ceux dérivés de SLP1 1 (Kieser et al, 2000), ceux dérivés de pSAM2 (Kieser et al, 2000), les vecteurs utilisant les systèmes d'intégration des phages PhiC31 (Kieser et al, 2000) (par exemple pSET152 (Bierman et al., 1992)) ou de VWB (Van Mellaert L, et al. 1998), ainsi que les vecteurs utilisant le système d'intégration de IS117 (Kieser et al. 2000). Comme fosmides on peut citer à titre illustratif le fosmide pFOSl (commercialisé par la société New England Bioloabs Inc., Beverly, Massachussetts, USA) et ses dérivés. Integrative vectors that may be mentioned by way of illustration include integrative vectors in Streptomyces, for example those derived from SLP1 (Kieser et al., 2000), those derived from pSAM2 (Kieser et al., 2000), vectors using integration of PhiC31 phages (Kieser et al., 2000) (eg pSET152 (Bierman et al., 1992)) or VWB (Van Mellaert L, et al., 1998), as well as vectors using the IS117 integration system (Kieser et al., 2000). As examples, fosmide pFOS1 (marketed by New England Bioloabs Inc., Beverly, Mass., USA) and its derivatives are exemplary.
Comme cosmides on peut citer à titre illustratif le cosmide SuperCos et ses dérivés (commercialisés notamment par la société Stratagene (LaJolla, Californie, USA)), le cosmide pWED15 (Wahl et al., 1987) et ses dérivés. Comme vecteurs navettes on peut citer à titre illustratif les plasmides navettes E. coli / Streptomyces comme par exemple pIJ903 et ses dérivés, la série des plasmides pUWL, pCA0106, pWHM3, pOJ446 et leurs dérivés (Kieser et al. 2000), les BAC navettes E. coli / Streptomyces comme par exemple ceux décrits dans la demande de brevet WO 01/40497. Comme BAC (Bacterial Artificial Chromosome) on peut citer à titre illustratif le BAC pBeloBACll (numéro d'accès GenBank :U51113). Comme PAC (Pl-derived Artificial Chromosome) on peut citer à titre illustratif le vecteur pCYPAC6 (numéro d'accès GenBank : AF133437). Plus préférentiellement, cet ADN recombinant est un vecteur d'expression. Les vecteurs d'expression utilisables dans différents systèmes d'expression sont bien connus de l'homme du métier, en ce qui concerne les cellules procaryotes, on peut citer à titre illustratif les vecteurs d'expression chez E. coli par exemple, de la famille pET commercialisés par la société Stratagene (LaJolla, Californie, USA), les vecteurs de la famille GATEWAY commercialisés par la société Invitrogen (Carlsbad, Californie, USA), les vecteurs de la famille pBAD commercialisés par la société Invitrogen (Carlsbad, Californie, USA), les vecteurs de la famille pMAL commercialisés par la société New England Bioloabs Inc. (Beverly, Massachussetts, USA), les vecteurs d'expression inductibles par le rhamnose cités dans la publication Wilms B et al., 2001 et leurs dérivés, on peut également citer les vecteurs d'expression chez Streptomyces comme par exemple les vecteurs pIJ4123, As cosmids, the cosmid SuperCos and its derivatives (sold in particular by the company Stratagene (LaJolla, California, USA)), the cosmid pWED15 (Wahl et al., 1987) and its derivatives are exemplary. As shuttle vectors, the E. coli / Streptomyces shuttle plasmids, for example pIJ903 and its derivatives, the plasmid series pUWL, pCA0106, pWHM3, pOJ446 and their derivatives (Kieser et al. E. coli / Streptomyces as for example those described in the patent application WO 01/40497. As BAC (Bacterial Artificial Chromosome) can be cited by way of illustration the BAC pBeloBACll (GenBank access number: U51113). As PAC (Pl-derived Artificial Chromosome) may be mentioned by way of illustration the vector pCYPAC6 (GenBank access number: AF133437). More preferably, this recombinant DNA is an expression vector. The expression vectors that can be used in different expression systems are well known to those skilled in the art, with regard to prokaryotic cells, for example, the expression vectors in E. coli, for example pET family marketed by Stratagene (LaJolla, California, USA), GATEWAY family vectors marketed by Invitrogen (Carlsbad, California, USA), the pBAD family vectors marketed by Invitrogen (Carlsbad, California, USA), the pMAL family vectors marketed by New England Bioloabs Inc. (Beverly, Mass., USA), the rhamnose-inducible expression vectors cited in Wilms B et al., 2001 and their derivatives, Streptomyces expression vectors can also be mentioned, for example pIJ4123 vectors,
<Desc/Clms Page number 89><Desc / Clms Page number 89>
pIJ6021, pPM927, pANT849, pANT 850, pANT 851, pANT1200, pANT1201, pANT1202 et leurs dérivés (Kieser et al., 2000). En ce qui concerne les cellules de levures, on peut citer à titre illustratif le vecteur pESC commercialisé par la société Stratagene (LaJolla, Californie, USA). En ce qui concerne le système d'expression baculovirus permettant une expression dans des cellules d'insectes on peut citer à titre illustratif le vecteur BacPAK6 commercialisé par la société BD Biosciences Clontech, (Palo Alto, Californie, USA). En ce qui concerne les cellules de mammifères, on peut citer à titre illustratif des vecteurs comprenant le promoteur des gènes précoces du virus CMV (Cytomegalovirus) (par exemple le vecteur pCMV et ses dérivés commercialisés par la société Stratagene (LaJolla, Californie, USA), ou le promoteur des gènes immédiats (SV40 early promoter) du virus simien vacuolisant SV40 (par exemple le vecteur pSG5 commercialisé par la société Stratagene (LaJolla, Californie, USA). Un autre aspect de l'invention concerne des cellules hôtes dans laquelle a été introduit au moins un ADN recombinant décrit dans ce paragraphe. pIJ6021, pPM927, pANT849, pANT 850, pANT 851, pANT1200, pANT1201, pANT1202 and their derivatives (Kieser et al., 2000). As regards the yeast cells, the vector pESC marketed by the company Stratagene (LaJolla, California, USA) can be cited by way of illustration. As regards the baculovirus expression system allowing expression in insect cells, mention may be made, for example, of the BacPAK6 vector marketed by BD Biosciences Clontech, (Palo Alto, California, USA). As regards mammalian cells, mention may be made, for example, of vectors comprising the promoter of the early genes of the CMV (Cytomegalovirus) virus (for example the pCMV vector and its derivatives marketed by Stratagene (LaJolla, California, USA). or the SV40 early promoter of simian SV40 vacuolizing virus (eg, the vector pSG5 marketed by Stratagene (LaJolla, Calif., USA). Another aspect of the invention relates to host cells in which at least one recombinant DNA described in this paragraph has been introduced.
Un autre aspect de l'invention concerne un procédé de production d'un polypeptide caractérisé en ce que ledit procédé comprend les étapes suivantes: a) transformer une cellule hôte par au moins un vecteur d'expression tel que décrit dans le paragraphe ci-dessus; b) cultiver, dans un milieu de culture approprié, ladite cellule hôte; c) récupérer le milieu de culture conditionné ou un extrait cellulaire; d) séparer et purifier à partir dudit milieu de culture ou encore à partir de l'extrait cellulaire obtenu à l'étape c), ledit polypeptide ; e) le cas échéant, caractériser le polypeptide recombinant produit. Another aspect of the invention relates to a method for producing a polypeptide characterized in that said method comprises the following steps: a) transforming a host cell with at least one expression vector as described in the paragraph above ; b) culturing, in a suitable culture medium, said host cell; c) recovering the conditioned culture medium or a cell extract; d) separating and purifying from said culture medium or from the cell extract obtained in step c), said polypeptide; e) where appropriate, characterizing the recombinant polypeptide produced.
Le polypeptide recombinant ainsi produit peut être purifié par passage sur une série appropriée de colonnes de chromatographie, selon les méthodes connues de l'homme de l'art et décrites par exemple dans F. Ausubel et al (2002). On peut citer à titre illustratif la technique du Tag-Histidine , qui consiste à ajouter une courte séquence poly-histidine au polypeptide à produire, ce dernier pouvant ensuite être purifié sur colonne de nickel. Ce polypeptide peut également être préparé par les The recombinant polypeptide thus produced can be purified by passage over a suitable series of chromatography columns, according to methods known to those skilled in the art and described for example in F. Ausubel et al (2002). By way of illustration, mention may be made of the Tag-Histidine technique, which consists in adding a short poly-histidine sequence to the polypeptide to be produced, which can then be purified on a nickel column. This polypeptide can also be prepared by
<Desc/Clms Page number 90><Desc / Clms Page number 90>
techniques de synthèse in vitro. A titre illustratif de telles techniques, le polypeptide pourra être préparé grâce au système rapid translation system (RTS) , commercialisé notamment par la société Roche Diagnostics France S.A, Meylan, France. in vitro synthesis techniques. As an illustration of such techniques, the polypeptide may be prepared by means of the rapid translation system (RTS), marketed in particular by Roche Diagnostics France SA, Meylan, France.
Un autre aspect de l'invention concerne un microorganisme producteur d'au moins une spiramycine caractérisé en ce qu'il surexprime : -un gène susceptible d'être obtenu par amplification en chaîne par polymérase (PCR) en utilisant le couple d'amorces de séquence suivante : 5' AAGCTTGTGTGCCCGGTGTACCTGGGGAGC 3' (SEQ ID N 138) et 5' GGATCCCGCGACGGACACGACCGCCGCGCA 3' (SEQ ID N 139) et comme matrice le cosmide pSPM36 ou l'ADN total de Streptomyces ambofaciens, - ou un gène dérivé de celui-ci en raison de la dégénérescence du code génétique. Another aspect of the invention relates to a microorganism producing at least one spiramycin characterized in that it overexpresses: a gene capable of being obtained by polymerase chain reaction (PCR) using the pair of primers of following sequence: 5 'AAGCTTGTGTGCCCGGTGTACCTGGGGAGC 3' (SEQ ID N 138) and 5 'GGATCCCGCGACGGACACGACCGCCGCGCA 3' (SEQ ID N 139) and as a template the cosmid pSPM36 or the total DNA of Streptomyces ambofaciens, - or a gene derived therefrom because of the degeneracy of the genetic code.
Un exemple de séquence d'un tel gène est donné en SEQ ID N 111 (ADN), cependant cette séquence est partielle puisqu'elle ne comprend pas la partie 3' de la séquence codante correspondante. La traduction en protéine de cette partie de séquence codante est donnée en SEQ ID N 112. L'homme du métier pourra aisément la compléter notamment en utilisant l'enseignement présenté dans l'exemple 24. Il est ainsi présenté dans cet exemple une méthode pour cloner le gène orj28c et pour fabriquer un vecteur d' expression permettant l' expression de orj28c. Il est également montré dans cet exemple que la surexpression du gène orj28c dans la souche OSC2 conduit à l'amélioration de la production en spiramycines de cette souche. De façon préférée, le microorganisme surexprimant - un gène susceptible d'être obtenu par amplification en chaîne par polymérase (PCR) en utilisant le couple d'amorces de séquence suivante : 5' AAGCTTGTGTGCCCGGTGTACCTGGGGAGC 3' (SEQ ID N 138) et 5' GGATCCCGCGACGGACACGACCGCCGCGCA 3' (SEQ ID N 139) et comme matrice le cosmide pSPM36 ou l'ADN total de Streptomyces ambofaciens, - ou un gène dérivé de celui-ci en raison de la dégénérescence du code génétique est une bactérie du genre Streptomyces, de manière encore plus préférée, il s'agit d'une bactérie de l'espèce Streptomyces ambofaciens. De façon préférée, la surexpression An example of a sequence of such a gene is given in SEQ ID No. 111 (DNA), however this sequence is partial since it does not include the 3 'portion of the corresponding coding sequence. The protein translation of this portion of coding sequence is given in SEQ ID No. 112. Those skilled in the art can easily complete it, in particular by using the teaching presented in Example 24. It is thus presented in this example a method for cloning the orj28c gene and to make an expression vector allowing the expression of orj28c. It is also shown in this example that the overexpression of the orj28c gene in the OSC2 strain leads to the improvement of the spiramycin production of this strain. Preferably, the microorganism overexpressing - a gene capable of being obtained by polymerase chain reaction (PCR) using the pair of primers with the following sequence: ## STR2 ## 3 '(SEQ ID N 139) and as a template the cosmid pSPM36 or the total DNA of Streptomyces ambofaciens, - or a gene derived from it because of the degeneracy of the genetic code is a bacterium of the genus Streptomyces, still more preferably, it is a bacterium of the species Streptomyces ambofaciens. Preferably, overexpression
<Desc/Clms Page number 91><Desc / Clms Page number 91>
dudit gène est obtenue par transformation dudit microorganisme par un vecteur d'expression, de manière tout à fait préféré, la souche de microorganisme est la souche OSC2/pSPM75(l) ou de la souche OSC2/pSPM75(2). said gene is obtained by transformation of said microorganism with an expression vector, most preferably the microorganism strain is the strain OSC2 / pSPM75 (1) or strain OSC2 / pSPM75 (2).
Un autre aspect de l'invention concerne un procédé de production de spiramycine(s) utilisant les microorganismes décrits au paragraphe précédent. Ce procédé consiste à cultiver dans un milieu de culture approprié un microorganisme défini dans le paragraphe précédent, récupérer le milieu de culture conditionné ou un extrait cellulaire, séparer et purifier à partir dudit milieu de culture ou encore à partir de l'extrait cellulaire obtenu à l'étape précédente le ou les spiramycines produites. Les conditions de culture de tels microorganismes pourront être déterminées selon des techniques bien connues de l'homme du métier. Le milieu de culture pourra par exemple être le milieu MP5 ou le milieu SL11pour les Streptomyces et notamment pour Streptomyces ambofaciens (Pernodet et al., 1993). L'homme du métier pourra notamment se référer à l'ouvrage de Kieser et al. 2000 en ce qui concerne la culture des Streptomyces. Le ou les spiramycines produites peuvent être récupérés par toutes techniques connues de l'homme du métier. L'homme du métier pourra se référer, par exemple, aux techniques enseignées dans le brevet américain US 3,000,785 et plus particulièrement aux méthodes d'extraction des spiramycines décrites dans ce brevet. Préférentiellement, les microorganismes utilisés dans un tel procédé sont des bactéries du genre Streptomyces. Plus préférentiellement, les microorganismes sont des souches de S. ambofaciens. Another aspect of the invention relates to a process for producing spiramycin (s) using the microorganisms described in the preceding paragraph. This method consists of cultivating in a suitable culture medium a microorganism defined in the preceding paragraph, recovering the conditioned culture medium or a cell extract, separating and purifying from said culture medium or from the cell extract obtained at the preceding step or the spiramycins produced. The culture conditions of such microorganisms may be determined according to techniques well known to those skilled in the art. The culture medium may for example be MP5 medium or SL11 medium for Streptomyces and in particular for Streptomyces ambofaciens (Pernodet et al., 1993). Those skilled in the art may in particular refer to the work of Kieser et al. 2000 with regard to the cultivation of Streptomyces. The spiramycin (s) produced can be recovered by any technique known to those skilled in the art. Those skilled in the art may refer, for example, to the techniques taught in US Pat. No. 3,000,785 and more particularly to the spiramycin extraction methods described in this patent. Preferably, the microorganisms used in such a process are bacteria of the genus Streptomyces. More preferably, the microorganisms are strains of S. ambofaciens.
Un autre aspect de l'invention concerne un vecteur d'expression caractérisé en ce que le polynucléotide de séquence SEQ ID N 47 ou un polynucléotide dérivé de celui-ci en raison de la dégénérescence du code génétique est placé sous le contrôle d'un promoteur permettant l'expression de la protéine codée par le dit polynucléotide chez Streptomyces ambofaciens. Des exemples de vecteurs d'expression utilisables chez Streptomyces ont été donnés ci-dessus. Another aspect of the invention relates to an expression vector characterized in that the polynucleotide of sequence SEQ ID No. 47 or a polynucleotide derived therefrom due to the degeneracy of the genetic code is placed under the control of a promoter. allowing expression of the protein encoded by said polynucleotide in Streptomyces ambofaciens. Examples of expression vectors that can be used in Streptomyces have been given above.
Préférentiellement un tel vecteur d'expression est caractérisé en ce qu'il s'agit du plasmide pSPM524 ou pSPM525. Preferably, such an expression vector is characterized in that it is the plasmid pSPM524 or pSPM525.
<Desc/Clms Page number 92> <Desc / Clms Page number 92>
Un autre aspect de l'invention concerne une souche de Streptomyces ambofaciens transformée par un vecteur défini dans le paragraphe précédent. Another aspect of the invention relates to a strain of Streptomyces ambofaciens transformed by a vector defined in the preceding paragraph.
Un autre aspect de l'invention concerne un polypeptide caractérisé en ce que sa séquence comprend la séquence SEQ ID N 112. L'invention est également relative à un polypeptide caractérisé en ce que sa séquence correspond à la séquence traduite de la séquence codante : - d'un gène susceptible d'être obtenu par amplification en chaîne par polymérase (PCR) en utilisant le couple d'amorces de séquence suivante : 5' AAGCTTGTGTGCCCGGTGTACCTGGGGAGC 3' (SEQ ID N 138) et 5' GGATCCCGCGACGGACACGACCGCCGCGCA 3' (SEQ ID N 139) et comme matrice le cosmide pSPM36 ou l'ADN total de Streptomyces ambofaciens, -ou d'un gène dérivé de celui-ci en raison de la dégénérescence du code génétique. Another aspect of the invention relates to a polypeptide characterized in that its sequence comprises the sequence SEQ ID No. 112. The invention also relates to a polypeptide characterized in that its sequence corresponds to the translated sequence of the coding sequence: a gene which can be obtained by polymerase chain reaction (PCR) using the pair of primers of the following sequence: 5 'AAGCTTGTGTGCCCGGTGTACCTGGGGAGC 3' (SEQ ID N 138) and 5 'GGATCCCGCGACGGACACGACCGCCGCGCA 3' (SEQ ID N 139) and as a template the cosmid pSPM36 or the total DNA of Streptomyces ambofaciens, or a gene derived from it because of the degeneracy of the genetic code.
Préferentiellement ces polypeptides sont exprimés à l'état naturel par une bactérie du genre Streptomyces, plus préférentiellement, ces polypeptides sont impliqués dans la biosynthèse des spiramycines. Preferably these polypeptides are expressed in the natural state by a bacterium of the genus Streptomyces, more preferably, these polypeptides are involved in the biosynthesis of spiramycins.
Un autre aspect de l'invention concerne un vecteur d'expression permettant l'expression d'un polypeptide tel que défini dans le paragraphe précédent chez Streptomyces ambofaciens. Des exemples de vecteurs d'expression utilisables chez Streptomyces ont été donnés ci-dessus. Préférentiellemnt, le vecteur d'expression en question est le plasmide pSPM75. Another aspect of the invention relates to an expression vector for the expression of a polypeptide as defined in the preceding paragraph in Streptomyces ambofaciens. Examples of expression vectors that can be used in Streptomyces have been given above. Preferentially, the expression vector in question is the plasmid pSPM75.
LISTE DES FIGURES Figure 1 : Structure chimique des spiramycines I, II et III. LIST OF FIGURES Figure 1: Chemical structure of spiramycins I, II and III.
Figure 2 : Cosmides utilisés pour le séquençage de la région. Figure 2: Cosmids used for sequencing the region.
Figure 3 : Organisation d'un groupe de gènes impliqués dans la voie de biosynthèse des spiramycines. Figure 3: Organization of a group of genes involved in the spiramycin biosynthetic pathway.
Figure 4 : Voie de biosynthèse proposée du mycarose. Figure 4: Proposed biosynthetic pathway of mycarosis.
Figure 5 : Voie de biosynthèse proposée du mycaminose. Figure 5: Proposed biosynthetic pathway of mycaminosis.
<Desc/Clms Page number 93> <Desc / Clms Page number 93>
Figure 6 : Voie de biosynthèse proposée de la forosamine. Figure 6: Proposed biosynthetic pathway of forosamine.
Figure 7 : Ordre préférentiel d'ajout des sucres dans la molécule de spiramycine et intermédiaires. Figure 7: Preferred order of addition of sugars in the spiramycin molecule and intermediates.
Figure 8 : Voie de biosynthèse proposée du méthoxymalonyl chez S. ambofaciens. Cette voie est proposée par analogie avec la biosynthèse du méthoxymalonyl chez Streptomyces hygroscopicus var. ascomyceticus (Wu,K. et al.,2000). Figure 8: Proposed biosynthetic pathway of methoxymalonyl in S. ambofaciens. This pathway is proposed by analogy with the biosynthesis of methoxymalonyl in Streptomyces hygroscopicus var. ascomyceticus (Wu, K et al., 2000).
Figure 9 : Etapes conduisant à l'inactivation d'un gène:
A) Clonage du gène cible dans un vecteur se répliquant chez E. coli et pas chez
Streptomyces;
B) Insertion de la cassette dans le gène cible (par clonage ou recombinaison entre courtes séquences identiques);
C) Introduction du plasmide chez Streptomyces ambofaciens (par transformation ou conjugaison) et sélection des clones ayant intégré la cassette puis criblage de clones ayant perdu la partie vecteur pour avoir un remplacement de gène;
D) Souche mutante dans laquelle le gène cible est inactivé. Figure 9: Steps leading to the inactivation of a gene:
A) Cloning of the target gene in a replicating vector in E. coli and not in
Streptomyces;
B) Insertion of the cassette into the target gene (by cloning or recombination between short identical sequences);
C) Introduction of the plasmid into Streptomyces ambofaciens (by transformation or conjugation) and selection of the clones having integrated the cassette and then screening of clones having lost the vector part to have a gene replacement;
D) Mutant strain in which the target gene is inactivated.
Figure 10 : optionnelles suivant l'inactivation d'un gène selon la méthode décrite en figure 9, pouvant être conduites si une cassette excisable a été employée pour interrompre le gène cible :
E) Introduction dans la souche mutante du plasmide pOSV508 portant les gènes xis et int de pSAM2 dont les produits vont permettre l'excision efficace par recombinaison spécifique de site de la cassette;
F) Obtention de clones ayant perdu la cassette excisable et sensibles à l'antibiotique à laquelle la cassette donnait la résistance;
G) Après croissance et sporulation sur milieu solide sans antibiotique le plasmide pOSV508 est perdu à haute fréquence. On peut obtenir des clones sensibles au thiostrepton dans lesquels le gène cible contient une délétion en phase. La séquence du gène cible délété peut être contrôlée par PCR et séquençage du produit PCR. Figure 10: optional following the inactivation of a gene according to the method described in Figure 9, which can be conducted if an excisable cassette has been used to interrupt the target gene:
E) Introduction into the mutant strain of plasmid pOSV508 carrying the xis and int genes of pSAM2 whose products will allow efficient excision by site-specific recombination of the cassette;
F) Obtaining clones having lost the excisable cassette and sensitive to the antibiotic to which the cassette gave the resistance;
G) After growth and sporulation on solid medium without antibiotic plasmid pOSV508 is lost at high frequency. Thiostrepton-sensitive clones in which the target gene contains an in-phase deletion can be obtained. The sequence of the deleted target gene can be monitored by PCR and sequencing of the PCR product.
<Desc/Clms Page number 94> <Desc / Clms Page number 94>
Figure 11 : Amplification de la cassette excisable dans le but de l'utiliser pour une expérience de recombinaison homologue. La technique de recombinaison homologue grâce à de courtes séquences homologues a été décrite par (Chaveroche et al., 2000). Les 39 ou 40 déoxy-nucléotides situés à l'extrémité 5' de ces oligonucléotides comportent une séquence correspondant à une séquence du gène à inactiver et les 20 déoxynucléotides situés le plus en 3' correspondent à la séquence d'une des extrémités de la cassette excisable. Figure 11: Amplification of the excisable cassette for the purpose of using it for a homologous recombination experiment. The homologous recombination technique using short homologous sequences has been described by (Chaveroche et al., 2000). The 39 or 40 deoxy nucleotides located at the 5 'end of these oligonucleotides comprise a sequence corresponding to a sequence of the gene to be inactivated and the 3' most deoxynucleotides correspond to the sequence of one end of the cassette. excisable.
Figure 12 : Obtention d'une construction pour l'inactivation d'un gène cible grâce à la technique décrite par (Chaveroche et al., 2000). Figure 12: Obtaining a construct for the inactivation of a target gene using the technique described by (Chaveroche et al., 2000).
Figure 13 : Carte du plasmide pWHM3. Strepto ori : origine de réplication Streptomyces. Figure 13: Map of plasmid pWHM3. Strepto ori: origin of replication Streptomyces.
Figure 14 : Carte du plasmide pOSV508. Figure 14: Map of plasmid pOSV508.
Figure 15 : Exemple de structure d'une cassette excisable. Celle-ci est constituée de la cassette #hyg (Blondelet-Rouault et al., 1997) bordée des sites attR et attL (Raynal et al., 1998), entre lesquels un événement de recombinaison permettra l'excision de la cassette, grâce à l'expression des gènes xis et int. Figure 15: Example of structure of an excisable cassette. This consists of the #hyg cassette (Blondelet-Rouault et al., 1997) bordered by the attR and attL sites (Raynal et al., 1998), between which a recombination event will allow the excision of the cassette, thanks to to the expression of the genes xis and int.
Figure 16 : Carte du plasmide pBXLl 111. Figure 16: Map of plasmid pBXL 111.
Figure 17 : Carte du plasmide pBXL1112. Figure 17: Map of plasmid pBXL1112.
Figure 18 : Test microbiologique de production en spiramycine, basé sur la sensibilité d'une souche de Micrococcus luteus à la spiramycine. La souche de Micrococcus luteus utilisée est une souche naturellement sensible à la spiramycine mais résistante à la congocidine. Les différentes souches de Streptomyces à tester ont été cultivées en erlenmeyers de 500 ml contenant 70 ml de milieu MP5, inoculés à une concentration initiale de 2.5 106 spores/ml et mises à pousser à 27 C sous agitation orbitale de 250 rpm. Des prélèvements de moûts de fermentation ont été réalisés après 48,72 et 96 heures de culture et centrifugés. Une dilution au dixième de ces surnageants dans du milieu de culture stérile est utilisée pour le test. La souche indicatrice de Micrococcus luteus résistante à la congocidine mais sensible à la spiramycine (Gourmelen et al., Figure 18: Microbiological test of spiramycin production, based on the sensitivity of a strain of Micrococcus luteus to spiramycin. The strain of Micrococcus luteus used is a strain naturally sensitive to spiramycin but resistant to congocidin. The different strains of Streptomyces to be tested were cultured in 500 ml Erlenmeyer flasks containing 70 ml of MP5 medium, inoculated at an initial concentration of 2.5 106 spores / ml and grown at 27 ° C. with orbital shaking of 250 rpm. Samples of fermentation musts were made after 48.72 and 96 hours of culture and centrifuged. A tenth dilution of these supernatants in sterile culture medium is used for the test. The indicator strain of Micrococcus luteus resistant to congocidine but sensitive to spiramycin (Gourmelen et al.,
<Desc/Clms Page number 95><Desc / Clms Page number 95>
1998) a été cultivée dans une boîte carrée de 12x12 cm. Des disques en papier Whatman AA ont été imbibés de 70 l de la dilution au dixième de chaque surnageant et déposés sur la surface de la boîte. Des disques imbibés d'une solution de spiramycine de différentes concentrations (2-4-8 ug/ml dans du milieu de culture MP5) sont utilisés comme gamme étalon. Les boîtes sont incubées à 37 C pendant 24 à 48h. Si le disque contient de la spiramycine, celle-ci diffuse dans l'agar et inhibe la croissance de la souche indicatrice de Micrococcus luteus. Cette inhibition crée un halo autour du disque, ce halo reflétant la zone où la souche de Micrococcus luteus n'a pas poussé. La présence de ce halo est donc une indication de la présence de spiramycine et permet de déterminer si la souche de S. ambofaciens correspondant au disque en question est productrice ou non productrice de spiramycine. Une comparaison avec les diamètres d'inhibition obtenus pour la gamme étalon permet d'obtenir une indication de la quantité de spiramycine produite par cette souche. 1998) was grown in a 12x12 cm square box. Whatman AA paper disks were soaked with 70 l of the tenth dilution of each supernatant and deposited on the surface of the box. Disks imbibed with a solution of spiramycin of different concentrations (2-4-8 μg / ml in MP5 culture medium) are used as a standard range. The dishes are incubated at 37 ° C. for 24 to 48 hours. If the disc contains spiramycin, it diffuses into the agar and inhibits the growth of the indicator strain of Micrococcus luteus. This inhibition creates a halo around the disc, this halo reflecting the area where the strain of Micrococcus luteus did not grow. The presence of this halo is therefore an indication of the presence of spiramycin and makes it possible to determine whether the S. ambofaciens strain corresponding to the disc in question is producing or not producing spiramycin. A comparison with the diameters of inhibition obtained for the standard range makes it possible to obtain an indication of the amount of spiramycin produced by this strain.
Figure 19: Chromatogramme CLHP du surnageant filtré du milieu de culture de la souche OSC2. Figure 19: HPLC chromatogram of the filtered supernatant of the culture medium of the strain OSC2.
Figure 20 : Chromatogramme CLHP du surnageant filtré du milieu de culture de la souche SPM501. Figure 20: HPLC chromatogram of the filtered supernatant of the culture medium of strain SPM501.
Figure 21 : Chromatogramme CLHP du surnageant filtré du milieu de culture de la souche SPM502. Figure 21: HPLC chromatogram of the filtered supernatant of the culture medium of strain SPM502.
Figure 22 : Chromatogramme CLHP du surnageant filtré du milieu de culture de la souche SPM507. Figure 22: HPLC chromatogram of the filtered supernatant of the culture medium of strain SPM507.
Figure 23 : Chromatogramme CLHP du surnageant filtré du milieu dé culture de la souche SPM508. FIG. 23: HPLC chromatogram of the filtered supernatant of the culture medium of strain SPM508.
Figure 24 : Chromatogramme CLHP du surnageant filtré du milieu de culture de la souche SPM509. FIG. 24: HPLC chromatogram of the filtered supernatant of the culture medium of strain SPM509.
Figure 25 : Alignement de la protéine Orf3 (SEQ ID N 29) avec la protéine TylB (SEQ ID N 87) de S.fradiae réalisé grâce au programme FASTA. Figure 25: Alignment of the protein Orf3 (SEQ ID No. 29) with the protein TylB (SEQ ID No. 87) S.fradiae achieved through the FASTA program.
Figure 26 : Alignement de la protéine MdmA de S. mycarofaciens (SEQ ID N 88) avec la protéine SrmD (SEQ ID N 16) réalisé grâce au programme FASTA. Figure 26: Alignment of the MdmA protein of S. mycarofaciens (SEQ ID No. 88) with the SrmD protein (SEQ ID No. 16) produced using the FASTA program.
<Desc/Clms Page number 96><Desc / Clms Page number 96>
Figure 27 : Exemple de séquences des sites résiduels après excision de la cassette excisable. En gras est indiqué le site att26 minimum tel que défini dans Raynal et al., 1998. La séquence de la phase 1 (attl) de 33 nucléotides est présentée en SEQ ID N 104, la séquence de la phase 2 (att2) de 34 nucléotides est présentée en SEQ ID N 105, la séquence de la phase 3 (att3) de 35 nucléotides est présentée en SEQ ID N 95.Figure 28 : Représentation schématique du gène orflO, localisation des amorces PCR utlisées et construction obtenue avec chaque couple d'amorces. Figure 27: Example of sequences of the residual sites after excision of the excisable cassette. In bold is indicated the minimum att26 site as defined in Raynal et al., 1998. The sequence of the 33 nucleotide phase (attl) is presented in SEQ ID N 104, the phase 2 (att2) sequence of 34. nucleotides is presented in SEQ ID N 105, the sequence of phase 3 (att3) of 35 nucleotides is presented in SEQ ID N 95.Figure 28: Schematic representation of the orflO gene, location of the PCR primers utlisés and construction obtained with each pair of d primers.
Figure 29 : Construction de la cassette pac-oritT. Figure 29: Construction of the pac-oritT cassette.
Figure 30 : Carte du cosmide pWED2. Figure 30: Cosmid map pWED2.
Figure 31 : Représentation schématique du groupe de gènes impliqués dans la voie de biosynthèse des spiramycines et de la localisation des trois sondes utilisées pour isoler les cosmides de la banque d'ADN génomique de la souche OSC2 de Streptomyces ambofaciens chez E. coli (cf. exemple 19).Figure 32 : Localisation des inserts des cosmides isolés de la banque d'ADN génomique de la souche OSC2 de Streptomyces ambofaciens chez E. coli (cf. exemple 19).Figure 33 : Sous clonage du fragment PstIPstl du cosmide pSPM36 (insert du plasmide (pSPM58), sous clonage du fragment Stul-Stul du cosmide pSPM36 (insert du plasmide pSPM72) et sous clonage d'un fragment EcoRI-StuI (insert du plasmide pSPM73). Figure 31: Schematic representation of the group of genes involved in the spiramycin biosynthesis pathway and the location of the three probes used to isolate the cosmids of the genomic DNA library of Streptomyces ambofaciens strain OSC2 in E. coli (cf. Example 19) Figure 32: Location of inserts of cosmids isolated from the genomic DNA library of Streptomyces ambofaciens strain OSC2 in E. coli (see Example 19) .Figure 33: Subcloning of the PstIPst1 fragment of cosmid pSPM36 (FIG. plasmid insert (pSPM58), subcloning the Stul-Stul fragment of cosmid pSPM36 (insert of plasmid pSPM72) and subcloning an EcoRI-StuI fragment (insert of plasmid pSPM73).
Figure 34 : Localisation des phases ouvertes de lecture indentifiées dans l'insert PstIPstI du plasmide pSPM58 et dans l'insert EcoRI-Stul du plasmide pSPM73. Figure 34: Location of the open reading frames identified in the PstIPstI insert of plasmid pSPM58 and in the EcoRI-Stul insert of plasmid pSPM73.
Figure 35 : Superposition des chromatogrammes CLHP du surnageant filtré du milieu de culture de la souche OS49. 67 réalisés à 238 et 280nm (haut) et spectres UV des molécules élués à 33,4 minutes et 44,8 minutes (bas). Figure 35: Superposition of HPLC chromatograms of the filtered supernatant of the culture medium of strain OS49. 67 at 238 and 280 nm (high) and UV spectra of the eluted molecules at 33.4 minutes and 44.8 minutes (low).
Figure 36 : Structure moléculaire des molécules platénolide A et platénolide B. Figure 36: Molecular Structure of Platenolide A and Platenolide B Molecules
Figure 37 : Organisation du groupe de gènes impliqués dans la voie de biosynthèse des spiramycines. Figure 37: Organization of the gene cluster involved in the spiramycin biosynthetic pathway.
<Desc/Clms Page number 97> <Desc / Clms Page number 97>
La présente invention est illustrée à l'aide des exemples qui suivent, qui doivent être considérés comme illustratifs et non limitatifs. The present invention is illustrated with the aid of the following examples, which should be considered as illustrative and not limiting.
En résumé, les gènes de la voie de biosynthèse des spiramycines ont été isolés à partir d'une banque d'ADN génomique de Streptomyces ambofaciens. Cette banque a été obtenue par digestion partielle de l'ADN génomique de Streptomyces ambofaciens, par l'enzyme de restriction BamHI. De larges fragments d'ADN, de 35 à 45 kb en moyenne, ont été clonés dans le cosmide pWEDl (Gourmelen et al., 1998) dérivé du cosmide pWED15 (Wahl et al., 1987). Ces cosmides ont été introduits chez E. coli grâce à des particules phagiques. La banque ainsi obtenue a été hybridée avec une sonde (de séquence SEQ ID N 86) correspondant à une partie du gène tylB de S. fradiae (MersonDavies & Cundliffe, 1994, numéro d'accès GenBank : U08223). Après hybridation, un cosmide sur les 4 s'hybridant avec la sonde a été plus particulièrement sélectionné. Ce cosmide nommé pOS49. 1 a ensuite été digéré par SacI et un fragment de 3,3 kb contenant la région s'hybridant avec la sonde a été sous cloné dans le vecteur pUC19 et séquence. Quatre phases ouvertes de lectures ont été identifiées et l'une d'entre elles code une protéine (SEQ ID N 29) présentant de fortes similitudes de séquence avec la protéine TylB de S. fradiae (SEQ ID N 87) (cf. figure 25). Ce gène a été nommé or/3 (SEQ ID N 28) et a été inactivé chez S. ambofaciens. Il a pu être démontré que les clones inactivés dans le gène or/3 ne produisent plus de spiramycines. Ceci montre l'implication du gène orf3 ou de gènes situés en aval dans la biosynthèse des spiramycines. In summary, the genes of the spiramycin biosynthetic pathway were isolated from a genomic library of Streptomyces ambofaciens. This library was obtained by partial digestion of the genomic DNA of Streptomyces ambofaciens by the BamHI restriction enzyme. Large DNA fragments, averaging 35-45 kb, were cloned into cosmid pWED1 (Gourmelen et al., 1998) derived from cosmid pWED15 (Wahl et al., 1987). These cosmids were introduced into E. coli by means of phage particles. The library thus obtained was hybridized with a probe (of sequence SEQ ID No. 86) corresponding to part of the tylB gene of S. fradiae (MersonDavies & Cundliffe, 1994, GenBank accession number: U08223). After hybridization, a cosmid on the 4 hybridizing with the probe was more particularly selected. This cosmid named pOS49. 1 was then digested with SacI and a 3.3 kb fragment containing the hybridizing region with the probe was subcloned into the vector pUC19 and sequenced. Four open reading runs were identified and one of them encodes a protein (SEQ ID No. 29) with strong sequence similarities to the S. fradiae TylB protein (SEQ ID N 87) (see FIG. ). This gene was named gold / 3 (SEQ ID N 28) and was inactivated in S. ambofaciens. It has been shown that inactivated clones in the or / 3 gene no longer produce spiramycins. This shows the implication of the orf3 gene or genes located downstream in the biosynthesis of spiramycins.
Une fois cette confirmation obtenue, une plus grande région du cosmide pOS49.1 a été séquencée, de part et d'autre du fragment SacI précédemment étudié. Ainsi, à partir du cosmide pOS49. 1, la séquence d'une région comportant sept phases ouvertes de lecture entières et deux autres incomplètes, situées de part et d'autre de ces sept phases ouvertes complètes, a pu être obtenue. Grâce à une recherche dans les bases de données, il a pu être montré que l'une des phases ouvertes de lecture incomplète correspondait au locus srmG (région codant une enzyme appelée polyketide synthase (PKS)). Les gènes correspondants ont été clonés par Burgett S. et al. en 1996 (Brevet américain US Once this confirmation was obtained, a larger region of the cosmid pOS49.1 was sequenced on either side of the SacI fragment previously studied. Thus, from the cosmid pOS49. 1, the sequence of a region comprising seven whole open reading and two incomplete incomplete phases, located on either side of these seven complete open phases, could be obtained. Through a database search, it was shown that one of the open incomplete reading stages corresponded to the srmG locus (a region encoding an enzyme called polyketide synthase (PKS)). The corresponding genes were cloned by Burgett S. et al. in 1996 (US Patent US
<Desc/Clms Page number 98><Desc / Clms Page number 98>
5,945,320). Par ailleurs, les autres phases ouvertes de lecture, les sept ORFs entières nommées : or/7, orf2, or/3, orf4, or/?, orf6, orf7 (SEQ ID N 23,25, 28,30, 34,36, 40) et le début de la huitième ORF nommé or/8 (séquence SEQ ID N 43) n'ont pas été retrouvées dans les bases de données. 5945320). On the other hand, the other open reading phases, the seven entire ORFs named: or / 7, orf2, or / 3, orf4, or / ?, orf6, orf7 (SEQ ID N 23,25, 28,30, 34,36 , 40) and the beginning of the eighth ORF named or / 8 (sequence SEQ ID N 43) were not found in the databases.
Dans un deuxième temps et dans le but de cloner d'autres gènes impliqués dans la biosynthèse des spiramycines, d'autres cosmides comportant des fragments du génome de S. ambofaciens dans cette même région ont été isolés. Pour cela une nouvelle série d'hybridations sur colonies a été effectuée en utilisant trois sondes. La première sonde correspond à un fragment d'ADN de 3,7kb contenant orfl, orf2 et le début de or/3, sous cloné à partir de pOS49.1. La deuxième sonde correspond à un fragment de 2 kb d'ADN contenant une partie de orf7 et une partie de orf8 et sous cloné à partir de pOS49. 1. Une troisième sonde a également été utilisée. Cette dernière correspond à un fragment d'ADN de 1.8kb contenant le gène srmD. Le gène srmD est un gène isolé de S. ambofaciens capable de conférer la résistance à la spiramycine. En effet, des travaux antérieurs avaient permis le clonage de plusieurs déterminants de résistance de S. ambofaciens, conférant la résistance à la spiramycine à une souche de S. griseofuscus (souche sensible à la spiramycine) (Pernodet et al., 1993) (Pernodet et al., 1999). Pour l'isolement de gènes de résistances, une banque cosmidique de l'ADN génomique de la souche S. ambofaciens avait été réalisée dans le cosmide pKC505 (Richardson MA et al., 1987). Ce pool de cosmides avait été introduit chez S. griseofuscus, naturellement sensible à la spiramycine. Cinq cosmides capables de conférer la résistance à l'apramycine et la spiramycine chez S. griseofuscus avaient ainsi été obtenus. Parmi ces 5 cosmides, un cosmide nommé pOS44. 1 possède dans son insert le gène srmD qui code une protéine présentant une certaine similitude avec la protéine codée par le gène mdmA de Streptomyces mycarofaciens et impliquée dans la résistance à la midécamycine chez cet organisme producteur (Hara et al., 1990, numéro d'accès Genbank : A60725) (figure 26). La troisième sonde utilisée pour localiser les gènes de biosynthèse de spiramycine a été un insert d' environ 1.8kb comprenant le gène srmD. In a second time and in order to clone other genes involved in the biosynthesis of spiramycins, other cosmids containing fragments of the genome of S. ambofaciens in this same region were isolated. For this, a new series of hybridizations on colonies was carried out using three probes. The first probe corresponds to a 3.7kb DNA fragment containing orf1, orf2 and the beginning of gold / 3, subcloned from pOS49.1. The second probe corresponds to a 2 kb fragment of DNA containing part of orf7 and part of orf8 and subcloned from pOS49. 1. A third probe has also been used. The latter corresponds to a 1.8 kb DNA fragment containing the srmD gene. The srmD gene is an isolated gene of S. ambofaciens capable of conferring resistance to spiramycin. Indeed, previous work had allowed the cloning of several resistance determinants of S. ambofaciens, conferring resistance to spiramycin to a strain of S. griseofuscus (spiramycin-sensitive strain) (Pernodet et al., 1993) (Pernodet et al., 1999). For the isolation of resistance genes, a cosmid library of genomic DNA of the S. ambofaciens strain was made in the cosmid pKC505 (Richardson MA et al., 1987). This cosmid pool had been introduced into S. griseofuscus, naturally sensitive to spiramycin. Five cosmids capable of conferring resistance to apramycin and spiramycin in S. griseofuscus had thus been obtained. Among these 5 cosmids, a cosmid named pOS44. 1 has in its insert the srmD gene which encodes a protein having a certain similarity with the protein encoded by the mdmA gene of Streptomyces mycarofaciens and involved in resistance to midecamycin in this producing organism (Hara et al., 1990, issue number Genbank access: A60725) (Figure 26). The third probe used to locate the spiramycin biosynthetic genes was an insert of approximately 1.8 kb including the srmD gene.
<Desc/Clms Page number 99> <Desc / Clms Page number 99>
Ces trois sondes ont été utilisées pour hybrider la banque d'ADN génomique décrite ci-dessus et ont permis de choisir deux cosmides (pSPM7 et pSPM5) susceptibles de contenir les inserts les plus longs et n'ayant pas de bandes communes. These three probes were used to hybridize the genomic DNA library described above and made it possible to choose two cosmids (pSPM7 and pSPM5) capable of containing the longest inserts and having no common bands.
Le cosmide pSMP5 hybridait avec la première et la deuxième sonde, mais n'hybridait pas avec la troisième sonde, alors que le cosmide pSPM7 hybridait avec la troisième sonde seulement. Ces deux cosmides ont été séquences en totalité par la technique du séquençage en aveugle ( shotgun sequencing ). Les séquences des inserts de ces deux cosmides : pSPM7 et pSPM5 ont pu être assemblées car bien que ne se chevauchant pas, chacun des inserts comprenait une séquence connue à l'une de ses extrémités. En effet, chacun de ces inserts comportait un fragment de la séquence d'un des gènes codant une enzyme appelée polyketide synthase (PKS). Ces 5 gènes ont été clonés par Burgett S. et al. en 1996 (Brevet américain US 5,945,320) (cf. figure 2). Ainsi, il a pu être déterminé une séquence unique d'ADN génomique de S. ambofaciens. Une séquence de 30943 nucléotides partant, en 5', d'un site EcoRI situé dans le premier gène PKS et allant jusqu'à un site BamHI en 3' est présentée en SEQ ID N 1. Cette séquence correspond à la région amont des gènes de la PKS (cf. figure 2 et 3). Une deuxième région de 11171 nucléotides, partant d'un site PstI en 5' et allant jusqu'à un site BstEII en 3' situé dans le cinquième gène de la PKS est présentée en SEQ ID N 2. Cette région est la région aval des gènes de la PKS (l'aval et l'amont étant définis par l'orientation des 5 gènes de PKS tous orientés dans le même sens) (cf. figures 2 et 3). The cosmid pSMP5 hybridized with the first and second probes, but did not hybridize with the third probe, whereas the cosmid pSPM7 hybridized with the third probe only. These two cosmids were completely sequenced by the shotgun sequencing technique. The sequences of the inserts of these two cosmids: pSPM7 and pSPM5 could be assembled because although not overlapping, each of the inserts included a known sequence at one of its ends. Indeed, each of these inserts included a fragment of the sequence of one of the genes encoding an enzyme called polyketide synthase (PKS). These 5 genes were cloned by Burgett S. et al. in 1996 (US Patent 5,945,320) (see Figure 2). Thus, it has been possible to determine a unique genomic DNA sequence of S. ambofaciens. A sequence of 30943 nucleotides starting at 5 'from an EcoRI site located in the first PKS gene and going up to a 3' BamHI site is presented in SEQ ID No. 1. This sequence corresponds to the upstream region of the genes. of the PKS (see Figure 2 and 3). A second region of 11171 nucleotides, starting from a 5 'PstI site and going up to a 3' BstEII site located in the fifth PKS gene, is presented in SEQ ID No. 2. This region is the downstream region of the PKS genes (downstream and upstream are defined by the orientation of PKS genes all oriented in the same direction) (see Figures 2 and 3).
Dans un troisième temps et dans le but de cloner d'autres gènes impliqués dans la biosynthèse des spiramycines, d'autres cosmides comportant des fragments du génome de S. ambofaciens dans cette même région ont été isolés (cf. exemple 18 et 19). In a third step, and in order to clone other genes involved in the biosynthesis of spiramycins, other cosmids comprising fragments of the S. ambofaciens genome in the same region were isolated (see Examples 18 and 19).
<Desc/Clms Page number 100> <Desc / Clms Page number 100>
EXEMPLE 1 : Construction d'une banque d'ADN génomique de la souche ATCC23877 de Streptomyces ambofaciens chez E. coli 1. 1. Extraction de l'ADN génomique de la souche ATCC23877 de Streptomyces ambofaciens. EXAMPLE 1 Construction of a genomic DNA library of Streptomyces ambofaciens strain ATCC23877 in E. coli 1. Extraction of genomic DNA of Streptomyces ambofaciens strain ATCC23877
La souche ATCC23877 de Streptomyces ambofaciens (accessible notamment auprès de l'American Type Culture Collection (ATCC) (Manassas, Virginie, USA), sous le numéro 23877) a été cultivée en milieu YEME (Yeast Extract-Malt Extract) ((Kieser, T, et al.,2000) et l'ADN génomique de cette souche a été extrait et purifié selon les techniques standards de lyse et précipitation (Kieser T. et al., 2000). Strain ATCC23877 from Streptomyces ambofaciens (available especially from the American Type Culture Collection (ATCC) (Manassas, Virginia, USA), under the number 23877) was grown in YEME (Yeast Extract-Malt Extract) medium (Kieser, T, et al., 2000) and the genomic DNA of this strain was extracted and purified according to standard lysis and precipitation techniques (Kieser T. et al., 2000).
1.2. Construction de la banque d'ADNgénomique
L'ADN génomique de la souche ATCC23877 de Streptomyces ambofaciens tel qu'isolé ci-dessus a été digéré partiellement par l'enzyme de restriction BamHI de manière à obtenir des fragments d'ADN de taille comprise entre environ 35 et 45 kb. 1.2. Construction of the genomic DNA library
The genomic DNA of Streptomyces ambofaciens strain ATCC23877 as isolated above was partially digested with BamHI restriction enzyme so as to obtain DNA fragments between about 35 and 45 kb in size.
Ces fragments ont été clonés dans le cosmide pWEDl (Gourmelen et al., 1998) digéré au préalable par BamHI. Le cosmide pWEDl est dérivé du cosmide pWE15 (Wahl, et al., 1987) par délétion du fragment HpaI-HpaI de 4. 1 kb contenant le module d'expression actif chez les mammifères (Gourmelen et al., 1998). Le mélange de ligation a ensuite été encapsidé in vitro dans des particules de phages lambda grâce au système Packagene Lambda DNA packaging system commercialisé par la société Prômëgà en suivant les recommandations du fabriquant. Les particules phagiques obtenues ont été utilisées pour infecter la souche SURE@ d'E. coli commercialisée par la société Stratagene (LaJolla, Californie, USA). La séléction des clones a été effectuée sur milieu LB + ampicilline (50 g/ml) puisque le cosmide pWEDl confère la résistance à l'ampicilline. These fragments were cloned into cosmid pWED1 (Gourmelen et al., 1998) pre-digested with BamHI. The cosmid pWED1 is derived from the cosmid pWE15 (Wahl, et al., 1987) by deletion of the 4.1 kb HpaI-HpaI fragment containing the active expression module in mammals (Gourmelen et al., 1998). The ligation mixture was then packaged in vitro in lambda phage particles by means of the Packagene Lambda DNA packaging system marketed by Promegae according to the manufacturer's recommendations. The phage particles obtained were used to infect the strain SURE @ E. coli marketed by Stratagene (LaJolla, California, USA). The selection of the clones was carried out on LB medium + ampicillin (50 g / ml) since the cosmid pWED1 confers resistance to ampicillin.
<Desc/Clms Page number 101> <Desc / Clms Page number 101>
EXEMPLE 2 : Isolement et caractérisation de gènes impliqués dans la biosynthèse de spiramycines chez Streptomyces ambofaciens 2. 1 Hybridation sur colonie des clones d'E. coli de la banque génomique de Streptomyces ambofaciens ATCC23877
Environ 2000 clones d'E. coli de la banque obtenue ci-dessus, ont été transférés sur filtre pour hybridation sur colonies. La sonde utilisée pour l'hybridation est un fragment NaeI-NaeI d'ADN (SEQ ID N 86) comportant une partie du gène tylB de Streptomycesfradiae. Ce fragment correspond aux nucléotides 2663 à 3702 du fragment d'ADN décrit par Merson-Davies, L. A. & Cundliffe E. (Merson-Davies, L. A. & Cundliffe E., 1994, numéro d'accès GenBank : U08223), dans lequel la séquence codante du gène tylB correspond aux nucléotides 2677 à 3843. EXAMPLE 2 Isolation and Characterization of Genes Involved in the Biosynthesis of Spiramycins in Streptomyces Ambofaciens 2.1 Colony Hybridization of E. coli genome library of Streptomyces ambofaciens ATCC23877
About 2000 clones of E. coli from the library obtained above, were transferred to filter for colony hybridization. The probe used for the hybridization is an NaeI-NaeI fragment of DNA (SEQ ID No. 86) comprising part of the tylB gene of Streptomycesfradiae. This fragment corresponds to nucleotides 2663 to 3702 of the DNA fragment described by Merson-Davies, LA & Cundliffe E. (Merson-Davies, LA & Cundliffe E., 1994, GenBank accession number: U08223), in which the sequence coding of the tylB gene corresponds to nucleotides 2677 to 3843.
Le fragment NaeI-NaeI d'ADN portant une partie du gène tylB de Streptomyces fradiae (SEQ ID N 86) a été marquée au 32P par la technique du random priming (Kit commercialisé par la société Roche) et utilisé comme sonde pour l'hybridation des 2000 clones de la banque, transférés sur filtre. Les membranes utilisées sont des membranes nylon Hybond N commercialisées par la société Amersham (Amersham Biosciences, Orsay, France) et l'hybridation a été effectuée à 55 C dans le tampon décrit par Church & Gilbert (Church & Gilbert, 1984). Un lavage a été effectué en 2X SSC à 55 C pendant 15 minutes et deux lavages successifs en 0. 5X SSC ont ensuite été effectués à 55 C, d'une durée de 15 minutes chacun. Dans ces conditions d'hybridation et de lavage, 4 clones parmi les 2000 hybridés présentaient un fort signal d'hybridation. The NaeI-NaeI fragment of DNA bearing part of the tylB gene of Streptomyces fradiae (SEQ ID No. 86) was labeled with 32P by the random priming technique (Kit marketed by Roche) and used as a probe for hybridization. 2000 clones of the bank, transferred to filter. The membranes used are Hybond N nylon membranes sold by Amersham (Amersham Biosciences, Orsay, France) and the hybridization was carried out at 55 ° C. in the buffer described by Church & Gilbert (Church & Gilbert, 1984). Washing was carried out in 2X SSC at 55 ° C. for 15 minutes and two successive 0 × 5 × SSC washes were then carried out at 55 ° C. for a duration of 15 minutes each. Under these hybridization and washing conditions, 4 of the 2000 hybridized clones had a strong hybridization signal.
Ces 4 clones ont été cultivés en milieu LB+ ampicilline (50 ng/ml) et les 4 cosmides correspondants ont été extraits par lyse alcaline standard (Sambrook et al., 1989). Il a ensuite été vérifié que l'hybridation était bien due à un fragment d'ADN présent dans l'insert de ces quatre cosmides. Pour cela, les cosmides ont été digérés indépendamment par plusieurs enzymes (BamHI, PstI et SacI). Les produits de digestions ont été séparés sur gel d'agarose, transférés sur membrane Nylon et hybridés par le fragment NaeI-NaeI d'ADN comportant une partie du gène tylB de Streptomycesfradiae (cf. ci-dessus) dans These 4 clones were cultured in LB + ampicillin medium (50 ng / ml) and the 4 corresponding cosmids were extracted by standard alkaline lysis (Sambrook et al., 1989). It was then verified that the hybridization was due to a DNA fragment present in the insert of these four cosmids. For this, the cosmids were independently digested with several enzymes (BamHI, PstI and SacI). The digests were separated on agarose gel, transferred to nylon membrane and hybridized with the NaeI-NaeI fragment of DNA containing part of the tylB gene of Streptomycesfradiae (see above) in
<Desc/Clms Page number 102><Desc / Clms Page number 102>
les mêmes conditions que ci-dessus. Les quatres cosmides ont pu être validés et un de ces cosmides a été plus particulièrement retenu et a été nommé pOS49.1. the same conditions as above. The four cosmids could be validated and one of these cosmids was more particularly retained and was named pOS49.1.
2. 2 Vérification de l'implication de la région identifiée et séquençage de l'insert du cosmide pOS49.1
Plusieurs fragments de l'insert du cosmide pOS49.1 ont été sous-clonés et leurs séquences ont été déterminées. Le cosmide pOS49. 1 a été digéré par l'enzyme SacI et il a été montré par Southern blot, dans les conditions décrites précédemment, qu'un fragment de 3,3 kb contient la région s'hybridant avec la sonde tylB. Ce fragment de 3,3 kb a été isolé par électroélution à partir d'un gel d'agarose 0. 8% puis cloné dans le vecteur pUC19 (numéro d'accès GenBank M77789) et séquence. Le plasmide ainsi obtenu a été nommé pOS49.11. Quatre phases ouvertes de lecture présentant un usage des codons typique de Streptomyces ont pu être identifiées dans ce fragment (deux complètes et deux tronquées), grâce au programme FramePlot (Ishikawa J & Hotta K. 2. 2 Verification of the involvement of the identified region and sequencing of the cosmid insert pOS49.1
Several fragments of the cosmid insert pOS49.1 were subcloned and their sequences determined. The cosmid pOS49. 1 was digested with SacI enzyme and it was shown by Southern blot, under the conditions previously described, that a 3.3 kb fragment contains the hybridizing region with the tylB probe. This 3.3 kb fragment was isolated by electroelution from an 0.8% agarose gel and then cloned into the pUC19 vector (GenBank accession number M77789) and sequenced. The plasmid thus obtained was named pOS49.11. Four open reading runs with typical Streptomyces codon usage could be identified in this fragment (two complete and two truncated), using the FramePlot program (Ishikawa J & Hotta K.
1999). Des comparaisons de séquences grâce au programme FASTA (cf.. (Pearson W. R & D. J. Lipman, 1988) et (Pearson W. R., 1990), accessible notamment auprès du centre de ressources INFOBIOGEN, Evry, France) ont permis de montrer que la protéine déduite de l'une de ces 4 phases ouvertes de lecture présente de fortes similitudes de séquence avec la protéine TylB (SEQ ID N 87 ; Numéro d'accès GenBank : U08223) de S. fradiae (cf. figure 25). Cette protéine a été nommée Orf3 (SEQ ID N 29). 1999). Sequence comparisons using the FASTA program (see Pearson W. R & DJ Lipman, 1988) and (Pearson WR, 1990), available in particular from the INFOBIOGEN resource center, Evry, France, have shown that The protein deduced from one of these 4 open reading phases has strong sequence similarities with the S. fradiae TylB protein (SEQ ID N 87, GenBank accession number: U08223) (see Figure 25). This protein has been named Orf3 (SEQ ID N 29).
Dans le but de tester si le gène correspondant (le gène or/3 (SEQ ID N 28)) était impliqué dans la biosynthèse des spiramycines chez S. ambofaciens, ce gène a été interrompu par une cassette #hyg (Blondelet-Rouault M-H. et al.,1997, numéro d'accès GenBank : X99315). Pour cela, le plasmide pOS49.11 a été digéré par l'enzyme XhoI et le fragment contenant les quatre phases ouvertes de lecture (deux complètes et deux tronquées, dont or/3 en entier) a été sous-cloné au niveau du site XhoI du vecteur pBC SK+ commercialisé par la société Stratagene (LaJolla, Califonnie, USA). Le plasmide ainsi obtenu a été nommé pOS49. 12. Dans le but d'inactiver or/3, un fragment PmlI- BstEII interne à or/3 a été remplacé par la cassette #hyg par clonage bout franc dans ce In order to test whether the corresponding gene (the or / 3 gene (SEQ ID No. 28)) was involved in spiramycin biosynthesis in S. ambofaciens, this gene was interrupted by a #hyg cassette (Blondelet-Rouault MH. et al., 1997, GenBank Access Number: X99315). For this, the plasmid pOS49.11 was digested with the enzyme XhoI and the fragment containing the four open reading phases (two complete and two truncated, including or / 3 in full) was subcloned at the XhoI site pBC SK + vector marketed by the company Stratagene (LaJolla, Califonnie, USA). The plasmid thus obtained was named pOS49. 12. In order to inactivate or / 3, an internal PmlI-BstEII fragment at or / 3 has been replaced by the #hyg cassette by free-end cloning in this
<Desc/Clms Page number 103><Desc / Clms Page number 103>
dernier plasmide. Pour cela, le plasmide pOS49. 12 a été digéré par les enzymes PmlI et BstEII dont le site unique se trouve dans la séquence codante du gène or/3. Les extrémités du fragment correspondant au vecteur ont été rendues bouts francs par un traitement-par l'enzyme de Klenow (grand fragment de la DNA polymerase I). La cassette Qhyg a été obtenue par digestion par l'enzyme BamHI du plasmide pHP45 #hyg (Blondelet-Rouault et al., 1997, numéro d'accès GenBank : X99315). Le fragment correspondant à la cassette #hyg a été récupéré sur gel d'agarose et ses extrémités ont été rendues bouts francs par un traitement par l'enzyme de Klenow. Les deux fragments bouts francs ainsi obtenus (la cassette Qhyg et le plasmide pOS49.12) ont été ligués et le produit de ligation a été utilisé pour la transformation de bactéries E. coli. Le plasmide ainsi obtenu a été nommé pOS49. 14 et contient le gène or/3 interrompu par la cassette Qhyg. last plasmid. For this, the plasmid pOS49. 12 was digested with PmlI and BstEII enzymes whose unique site is in the coding sequence of the or / 3 gene. The ends of the fragment corresponding to the vector were blunted by treatment with the Klenow enzyme (large fragment of DNA polymerase I). The Qhyg cassette was obtained by digestion with the BamHI enzyme of the pHP45 #hyg plasmid (Blondelet-Rouault et al., 1997, GenBank accession number: X99315). The fragment corresponding to the #hyg cassette was recovered on agarose gel and its ends were blunted by treatment with the Klenow enzyme. The two free ends fragments thus obtained (the Qhyg cassette and the plasmid pOS49.12) were ligated and the ligation product was used for the transformation of E. coli bacteria. The plasmid thus obtained was named pOS49. 14 and contains the or / 3 gene interrupted by the Qhyg cassette.
L'insert du plasmide pOS49. 14, sous la forme d'un fragment Xhol-Xhol dont les extrémités ont été rendues non cohésives par un traitement par l'enzyme de Klenow, a été cloné au niveau du site EcoRV du plasmide pOJ260 (le plasmide pOJ260 est un plasmide conjugatif capable de se répliquer chez E. coli mais incapable de se répliquer chez S. ambofaciens (Bierman M et al., 1992). Ce plasmide confère la résistance à l'apramycine chez E. coli et Streptomyces). Le plasmide obtenu (insert du plasmide pOS49. 14 cloné dans le plasmide pOJ260) a été nommé pOS49. 16. Ce dernier a été transféré dans la souche ATCC23877 de S. ambofaciens par conjugaison à l'aide de la souche conjugative E. coli S 17-1, comme décrit par Mazodier et al. (Mazodier et al., 1989). La souche E. coli S17-1 est dérivée de la souche E. coli 294 (Simon et al., 1983) (Simon, et al., 1986). Des clones transconjugants possédant le marqueur de résistance à l'hygromycine porté par la cassette Qhyg et ayant perdu le marqueur de résistance à l'apramycine porté par le vecteur pOJ260 ont pu être obtenus. Pour cela, après conjugaison, les clones sont sélectionnés pour leur résistance à l'hygromycine. Les clones résistants à l'hygromycine sont ensuite repiqués respectivement sur milieu avec hygromycine (antibiotique B) et sur milieu avec apramycine (antibiotique A) (cf. figure 9). Les clones résistants à l'hygromycine (HygR) et sensibles à l'apramycine (ApraS) The insert of plasmid pOS49. 14, in the form of an XhoI-XhoI fragment whose ends were made non-cohesive by treatment with Klenow enzyme, was cloned at the EcoRV site of plasmid pOJ260 (plasmid pOJ260 is a conjugative plasmid capable of to replicate in E. coli but unable to replicate in S. ambofaciens (Bierman M et al., 1992) This plasmid confers resistance to apramycin in E. coli and Streptomyces). The resulting plasmid (plasmid insert pOS49, cloned into plasmid pOJ260) was named pOS49. 16. The latter was transferred to S. ambofaciens strain ATCC23877 by conjugation with the conjugative strain E. coli S 17-1, as described by Mazodier et al. (Mazodier et al., 1989). E. coli strain S17-1 is derived from E. coli strain 294 (Simon et al., 1983) (Simon, et al., 1986). Transconjugant clones possessing the hygromycin resistance marker carried by the Qhyg cassette and having lost the apramycin resistance marker carried by the vector pOJ260 could be obtained. For this, after conjugation, the clones are selected for their resistance to hygromycin. The hygromycin-resistant clones are then transplanted respectively on medium with hygromycin (antibiotic B) and on medium with apramycin (antibiotic A) (see FIG. 9). Hygromycin-resistant clones (HygR) and apramycin-sensitive clones (ApraS)
<Desc/Clms Page number 104><Desc / Clms Page number 104>
sont en principe ceux où un double événement de recombinaison s'est produit et qui possèdent donc le gène orf3 interrompu par la cassette Qhyg. Le remplacement de la copie sauvage de orf3 par la copie interrompue a été vérifié par deux hybridations successives. Ainsi, l'ADN total des clones obtenus, a été digéré par différentes enzymes, séparé sur gel d'agarose, transféré sur membrane et hybridé avec une sonde correspondant à la cassette #hyg (cf. ci-dessus)) pour vérifier la présence de la cassette dans l'ADN génomique des clones obtenus. Une deuxième hybridation a été effectuée en utilisant comme sonde l'insert XhoI-XhoI du plasmide pOS49.11 contenant les quatre phases ouvertes de lecture (deux complètes et deux tronquées, dont orf3 en entier). La vérification du génotype peut également être réalisée par toute méthode connue de l'homme du métier et notamment par PCR en utilisant les oligonucléotides appropriés et séquençage du produit de PCR. Un des clones or/3:: #hyg ainsi obtenu et dont le génotype a été vérifié, a été choisi et a été appelé OS49.16. are in principle those where a double recombination event has occurred and therefore have the orf3 gene interrupted by the Qhyg cassette. Replacing the wild copy of orf3 with the interrupted copy was verified by two successive hybridizations. Thus, the total DNA of the clones obtained, was digested with various enzymes, separated on agarose gel, transferred to the membrane and hybridized with a probe corresponding to the #hyg cassette (see above) to verify the presence of the cassette in the genomic DNA of the clones obtained. A second hybridization was carried out using as a probe the XhoI-XhoI insert of plasmid pOS49.11 containing the four open reading phases (two complete and two truncated, including orf3 in full). Verification of the genotype can also be carried out by any method known to those skilled in the art and in particular by PCR using the appropriate oligonucleotides and sequencing of the PCR product. One of the clones gold / 3 :: #hyg thus obtained and whose genotype was verified, was chosen and was called OS49.16.
La production en spiramycine du clone OS49. 16 ainsi obtenu a été testée grâce au test de production décrit plus bas (cf. exemple 15). Il a ainsi pu être démontré que cette souche ne produit plus de spiramycine, confirmant l'implication de or/3 et/ou des gènes situés en aval comme par exemple orf4 dans la biosynthèse des spiramycines. The spiramycin production of clone OS49. 16 thus obtained was tested by the production test described below (see Example 15). It has thus been demonstrated that this strain no longer produces spiramycin, confirming the involvement of or / 3 and / or downstream genes such as for example orf4 in the biosynthesis of spiramycins.
Une fois cette confirmation obtenue, une plus grande région du cosmide pOS49.1 a été séquencée, de part et d'autre du fragment SacI précédemment étudié. Ainsi, à partir du cosmide pOS49.1, la séquence d'une région comportant sept phases ouvertes de lecture entières et deux autres incomplètes, situées de part et d'autre de ces sept phases ouvertes complètes, a pu être obtenue. Grâce à une recherche dans les bases de données, il a pu être montré que l'une des phases ouvertes de lecture incomplète correspondait au locus srmG (région codant une enzyme appelée polyketide synthase (PKS)). Les gènes correspondants ont été clonés par Burgett S. et al. en 1996 (Brevet américain US 5,945,320). Par ailleurs, les autres phases ouvertes de lecture : sept ORFs entières nommées : orfl, orf2, orf3, orf4, or/5, orf6, or/7 (SEQ ID N 23,25, 28,30, 34,36 et 40) et le début de la huitième ORF nommée or/8 (la séquence entière de cette orf est donnée en SEQ ID N 43) n'ont pas été retrouvées dans les bases de données. Once this confirmation was obtained, a larger region of the cosmid pOS49.1 was sequenced on either side of the SacI fragment previously studied. Thus, from the cosmid pOS49.1, the sequence of a region comprising seven whole open reading and two incomplete incomplete phases located on either side of these seven complete open phases could be obtained. Through a database search, it was shown that one of the open incomplete reading stages corresponded to the srmG locus (a region encoding an enzyme called polyketide synthase (PKS)). The corresponding genes were cloned by Burgett S. et al. in 1996 (US Patent 5,945,320). On the other hand, the other open reading phases: seven full ORFs named: orf1, orf2, orf3, orf4, or5, orf6, or / 7 (SEQ ID N 23,25, 28,30, 34,36 and 40) and the beginning of the eighth ORF named or / 8 (the entire sequence of this orf is given in SEQ ID N 43) have not been found in the databases.
<Desc/Clms Page number 105> <Desc / Clms Page number 105>
EXEMPLE 3 : Isolement et caractérisation d'autres gènes impliqués dans la biosynthèse de spiramycines chez Streptomyces ambofaciens
Dans un deuxième temps et dans le but de cloner d'autres gènes impliqués dans la biosynthèse des spiramycines, d'autres cosmides comportant des fragments du génome de S. ambofaciens dans cette même région ont été isolés. Pour cela il a été procédé à une nouvelle série d'hybridations sur colonies en utilisant trois sondes : - La première sonde utilisée correspond à un fragment d'ADN BamHI-PstI de 3,7kb contenant un fragment du gène de la PKS (Les gènes correspondant à la PKS ont été clonés par Burgett S. et al. en 1996 (Brevet américain US 5,945,320)), orfl, orf2 et le début de or/3, sous cloné à partir de pOS49.1 et allant d'un site BamHI situé 1300 paires de bases en amont du site EcoRI définissant la position 1 de SEQ ID N 1, jusqu'au site Pstl situé en position 2472 (SEQ ID N 1). Ce fragment BamHI-Pstl a été sous cloné à partir de pOS49. 1 dans le plasmide pBC SK+, ce qui a permis d'obtenir le plasmide pOS49. 28. EXAMPLE 3 Isolation and Characterization of Other Genes Involved in the Biosynthesis of Spiramycins in Streptomyces ambofaciens
In a second time and in order to clone other genes involved in the biosynthesis of spiramycins, other cosmids containing fragments of the genome of S. ambofaciens in this same region were isolated. For this purpose, a new series of hybridizations on colonies was carried out using three probes: the first probe used corresponds to a 3.7 kb BamHI-PstI DNA fragment containing a fragment of the PKS gene (genes corresponding to PKS were cloned by Burgett S. et al., in 1996 (US Patent 5,945,320), orf1, orf2, and start of gold / 3, subcloned from pOS49.1 and from a BamHI site. located 1300 base pairs upstream of the EcoRI site defining the position 1 of SEQ ID No. 1, to the PstI site located at position 2472 (SEQ ID No. 1). This BamHI-PstI fragment was subcloned from pOS49. 1 in the plasmid pBC SK +, which made it possible to obtain the plasmid pOS49. 28.
- La deuxième sonde utilisée correspond à un fragment d'ADN PstI-BamHI d'environ 2kb contenant un fragment d'or/7 et d'orf8, sous cloné à partir de pOS49. 1 et allant d'un site PstI situé en position 6693 de SEQ ID N 1 jusqu'au site BamHI situé en position 8714 de SEQ ID N 1. Ce fragment PstI-BamHI a été sous cloné à partir de pOS49.1 dans le plamside pBC SK+, ce qui a permis d'obtenir le plasmide pOS49.76. The second probe used corresponds to a PstI-BamHI DNA fragment of approximately 2 kb containing a fragment of gold / 7 and orf8, subcloned from pOS49. 1 and ranging from a PstI site located at position 6693 of SEQ ID No. 1 to the BamHI site located at position 8714 of SEQ ID No. 1. This PstI-BamHI fragment was subcloned from pOS49.1 into plamside pBC SK +, which made it possible to obtain the plasmid pOS49.76.
- Une troisième sonde a également été utilisée. Cette dernière correspond à un fragment d'ADN de 1.8kb EcoRI-HindIII contenant le gène srmD. Le gène srmD est un gène isolé de S. ambofaciens capable de conférer la résistance à la spiramycine. En effet, des travaux antérieurs avaient permis le clonage de plusieurs déterminants de résistance de S. ambofaciens, conférant la résistance à la spiramycine à une souche de S. griseofuscus (souche sensible à la spiramycine) (Pernodet et al., 1993) (Pernodet et al., 1999). Pour isoler des gènes de résistance, une banque cosmidique de l'ADN génomique de la souche S. ambofaciens ATCC23877 avait été réalisée dans le cosmide pKC505 - A third probe was also used. The latter corresponds to a 1.8kb EcoRI-HindIII DNA fragment containing the srmD gene. The srmD gene is an isolated gene of S. ambofaciens capable of conferring resistance to spiramycin. Indeed, previous work had allowed the cloning of several resistance determinants of S. ambofaciens, conferring resistance to spiramycin to a strain of S. griseofuscus (spiramycin-sensitive strain) (Pernodet et al., 1993) (Pernodet et al., 1999). To isolate resistance genes, a cosmid library of the genomic DNA of the S. ambofaciens strain ATCC23877 had been made in the cosmid pKC505
<Desc/Clms Page number 106><Desc / Clms Page number 106>
(Richardson MA et al., 1987). Pour cela l'ADN génomique de la souche S. ambofaciens ATCC23877 avait été digéré partiellement par Sau3AI de manière à obtenir des fragments de taille comprise entre environ 30 et 40 kb. L'ADN génomique ainsi digéré (3 g) avait été ligué avec 1 g du pKC505 préalablement digéré par l'enzyme BamHI (Pernodet et al., 1999). Le mélange de ligation avait ensuite été encapsidé in vitro dans des particules phagiques. Les particules phagiques obtenues avaient été utilisées pour infecter la souche d'E. coli HB101 (accessible notamment auprès de l'American Type Culture Collection (ATCC) (Manassas, Virginie, USA), sous le numéro 33694). Environ 20 000 clones d'E. coli résistant à l'apramycine avaient été poolés et les cosmides de ces clones avaient été extraits. Ce pool de cosmides avait été introduit par transformation de protoplastes dans la souche DSM 10191 de S. griseofuscus (Cox KL & Baltz RH. 1984), naturellement sensible à la spiramycine (Rao RN et al., 1987, cette souche est disponible notamment auprès de la German Collection of Microorganisms and Cell Cultures (Deutsche Sammlung von Mikro-organismen und Zellkulturen GmbH, DSMZ), (Braunschweig, Allamagne), sous le numéro DSM 10191). Les transformants avaient été sélectionnés sur un milieu contenant de l'apramycine. 1300 des clones poussant sur milieu contenant de l'apramycine avaient été transférés sur milieu contenant 5 g/ml de spiramycine. Plusieurs clones résistants à l'apramycine avaient également poussé sur milieu contenant de la spiramycine et les cosmides de ces colonies avaient été extraits et utilisés pour transformer E. coli et S. griseofuscus (Pemodet et al., 1999). Cinq cosmides capables de conférer la résistance à l'apramycine à E. coli et la co-résistance à l'apramycine et la spiramycine chez S. griseofuscus avaient ainsi été obtenus. Parmi ces 5 cosmides, il a été déterminé qu'un cosmide nommé pOS44.1 possède dans son insert un gène (SEQ ID N 15) qui code une protéine (SEQ ID N 16) présentant une certaine similitude avec une protéine codée par le gène mdmA de Streptomyces mycarofaciens (SEQ ID N 88), ce gène a été nommé srmD (cf. alignement présenté en figure 26 réalisé grâce au programme FASTA (cf.. (Pearson W. R & D. J. Lipman, 1988) et (Pearson W. R., 1990), accessibles notamment auprès du centre de ressources INFOBIOGEN, Evry, France). (Richardson MA et al., 1987). For this, the genomic DNA of the S. ambofaciens ATCC23877 strain had been partially digested with Sau3AI so as to obtain fragments of size between about 30 and 40 kb. The genomic DNA thus digested (3 g) was ligated with 1 g of pKC505 previously digested with the BamHI enzyme (Pernodet et al., 1999). The ligation mixture was then packaged in vitro in phage particles. The phage particles obtained were used to infect the E. coli strain. coli HB101 (available in particular from the American Type Culture Collection (ATCC) (Manassas, Virginia, USA) under number 33694). About 20,000 clones of E. apramycin-resistant E. coli had been pooled and the cosmids of these clones had been extracted. This cosmid pool was introduced by transformation of protoplasts into S. griseofuscus strain DSM 10191 (Cox KL & Baltz RH, 1984), naturally sensitive to spiramycin (Rao RN et al., 1987, this strain is available especially from from the German Collection of Microorganisms and Cell Cultures, (Braunschweig, Germany), under the number DSM 10191). The transformants were selected on a medium containing apramycin. 1300 clams growing on medium containing apramycin had been transferred to medium containing 5 g / ml of spiramycin. Several apramycin-resistant clones had also grown on spiramycin-containing medium and the cosmids of these colonies had been extracted and used to transform E. coli and S. griseofuscus (Pemodet et al., 1999). Five cosmids capable of conferring resistance to apramycin to E. coli and co-resistance to apramycin and spiramycin in S. griseofuscus were thus obtained. Among these 5 cosmids, it has been determined that a cosmid named pOS44.1 has in its insert a gene (SEQ ID No. 15) which encodes a protein (SEQ ID No. 16) having a certain similarity with a protein coded by the gene. mdmA of Streptomyces mycarofaciens (SEQ ID N 88), this gene was named srmD (see alignment presented in FIG. 26 realized thanks to the FASTA program (cf. (Pearson W. R & DJ Lipman, 1988) and (Pearson WR, 1990), accessible in particular from the INFOBIOGEN resource center, Evry, France).
<Desc/Clms Page number 107> <Desc / Clms Page number 107>
Pour isoler le déterminant de résistance contenu dans le plasmide pOS44. 1, celuici a été digéré partiellement par l'enzyme de restriction Sau3AI de manière à obtenir des fragments de taille d'environ 1,5 à 3 kb, ces fragments ont été ligués dans le vecteur pIJ486 linéarisé par l'enzyme BamHI (Ward et al., 1986). Un plasmide a été sélectionné pour sa capacité à conférer la résistance à la spiramycine chez la souche DSM 10191 de S. griseofuscus (Rao RN et al., 1987), naturellement sensible à la spiramycine (cf. cidessus). Pour cela, le pool de plasmides correspondant aux fragment Sau3AI de pOS44.1 ligués dans le vecteur pIJ486 (cf. ci-dessus), a été introduit par transformation de protoplastes dans la souche DSM 10191 et les transformants ont été sélectionnés pour leur résistance au thiostrepton (due au gène tsr porté par pIJ486). Les clones poussant sur milieu contenant du thiostrepton ont été transférés sur milieu contenant de la spiramycine. Plusieurs clones résistants au thiostrepton ont également poussé sur milieu contenant de la spiramycine et les plasmides de ces colonies ont été extraits. Un plasmide conférant la résistance et contenant un insert d'environ 1.8kb a été sélectionné et nommé pOS44. 2. Cet insert de 1. 8kb peut être sorti facilement grâce à un site HindIII et un site EcoRI présents dans le vecteur de part et d'autre de l'insert. Cet insert de 1.8kb HindIII-EcoRI a été séquence et le gène de résistance qu'il renferme a été nommé srmD. Ce fragment contenant le gène srmD a ainsi pu être facilement sous-cloné dans le vecteur pUC19 (numéro d'accès GenBank M77789) ouvert par EcoRI-HindIII, le plasmide obtenu a été nommé pOS44. 4. L'insert de 1.8kb HindIII-EcoRI, contenant le gène srmD, de ce plasmide a été utilisé comme sonde pour localiser les gènes de biosynthèse de spiramycine (cf. ci-dessous). To isolate the resistance determinant contained in plasmid pOS44. 1, which was partially digested with the Sau3AI restriction enzyme so as to obtain fragments of size of about 1.5 to 3 kb, these fragments were ligated into the vector pIJ486 linearized by the enzyme BamHI (Ward et al. al., 1986). A plasmid was selected for its ability to confer spiramycin resistance in S. griseofuscus strain DSM 10191 (Rao RN et al., 1987), naturally sensitive to spiramycin (see above). For this, the plasmid pool corresponding to the Sau3AI fragments of pOS44.1 ligated into the pIJ486 vector (see above), was introduced by transformation of protoplasts into strain DSM 10191 and the transformants were selected for their resistance to thiostrepton (due to the tsr gene carried by pIJ486). Clones growing on medium containing thiostrepton were transferred to medium containing spiramycin. Several thiostrepton resistant clones also grew on spiramycin-containing medium and plasmids from these colonies were extracted. A resistance conferring plasmid containing an insert of about 1.8 kb was selected and named pOS44. 2. This insert of 1. 8kb can be easily extracted thanks to a HindIII site and an EcoRI site present in the vector on either side of the insert. This 1.8kb HindIII-EcoRI insert was sequenced and the resistance gene it contains was named srmD. This fragment containing the srmD gene was thus easily subcloned into the vector pUC19 (access number GenBank M77789) opened by EcoRI-HindIII, the plasmid obtained was named pOS44. 4. The 1.8kb HindIII-EcoRI insert, containing the srmD gene, of this plasmid was used as a probe to locate the spiramycin biosynthetic genes (see below).
Un échantillon d'une souche Escherichia Coli DH5a contenant le plasmide pOS44. 4 a été déposé auprès de la Collection Nationale de Cultures de Microorganismes (CNCM) Institut Pasteur, 25, rue du Docteur Roux 75724 Paris Cedex 15, France, le 10 juillet 2002 sous le numéro d'enregistrement 1-2918. A sample of an Escherichia Coli strain DH5a containing the plasmid pOS44. 4 has been deposited with the National Collection of Cultures of Microorganisms (CNCM) Pasteur Institute, 25, rue du Docteur Roux 75724 Paris Cedex 15, France, July 10, 2002 under the registration number 1-2918.
Environ 2000 clones de la banque, obtenus ci-dessus (cf. exemple 1), ont été transférés sur filtre pour hybridation sur colonies selon les techniques classiques (Sambrook et al., 1989). About 2000 clones of the library, obtained above (see Example 1), were transferred to a filter for hybridization on colonies according to standard techniques (Sambrook et al., 1989).
<Desc/Clms Page number 108> <Desc / Clms Page number 108>
Les trois sondes décrites ci-dessus ont été marquées au 32P par la technique du random priming (Kit commercialisé par Roche) et utilisées pour l'hybridation des 2000 clones de la banque, transférés sur filtre. L'hybridation a été effectuée à 65 C dans le tampon décrit par Church & Gilbert (Church & Gilbert, 1984). Un lavage a été effectué en 2X SSC à 65 C pendant 15 minutes et deux lavages successifs ont ensuite été effectués en 0.5X SSC à 65 C, d'une durée de 15 minutes chacun. Dans ces conditions d'hybridation et de lavage, 16 clones parmi les 2000 hybridés présentaient un fort signal d'hybridation avec au moins une des sondes. Cependant, aucun cosmide n'hybridait avec les trois sondes. Les 16 cosmides ont été extraits et digérés par l'enzyme de restriction BamHI. La comparaison des profils de restriction de ces différents cosmides entre eux, a conduit à choisir deux cosmides susceptibles de contenir les inserts les plus longs de la région et n'ayant pas de bandes communes. Ainsi deux cosmides l'un nommé pSPM5 et l'autre pSPM7 ont été choisis. Le cosmide pSPM5 hybridait avec les sondes orfl à orf4 et la sonde or/8, mais n'hybridait pas avec la sonde srmD. pSPM7 hybridait seulement avec la sonde srmD et pas avec les deux autres sondes. The three probes described above were labeled with 32P by the technique of random priming (Kit commercialized by Roche) and used for the hybridization of the 2000 clones of the bank, transferred to filter. Hybridization was performed at 65 ° C in the buffer described by Church & Gilbert (Church & Gilbert, 1984). A washing was carried out in 2X SSC at 65 ° C. for 15 minutes and two successive washes were then carried out in 0.5 × SSC at 65 ° C., with a duration of 15 minutes each. Under these hybridization and washing conditions, 16 of the hybridized 2000 clones had a strong hybridization signal with at least one of the probes. However, no cosmid hybridized with the three probes. The 16 cosmids were extracted and digested with BamHI restriction enzyme. The comparison of the restriction profiles of these different cosmids between them led to the choice of two cosmids likely to contain the longest inserts of the region and having no common bands. Thus two cosmids one named pSPM5 and the other pSPM7 were chosen. The cosmid pSPM5 hybridized with the orf1 probes at orf4 and the or8 probe, but did not hybridize with the srmD probe. pSPM7 hybridized only with the srmD probe and not with the other two probes.
Ces deux cosmides ont été séquences en totalité par la technique du séquençage en aveugle ( shotgun sequencing ). Les séquences des inserts de ces deux cosmides : pSPM7 et pSPM5 ont pu être assemblées car bien que ne se chevauchant pas, chacun des inserts comprenait une séquence connue à l'une de ses extrémités. En effet, chacun de ces inserts comportait un fragment de la séquence d'un des gènes codant une enzyme appelée polyketide synthase (PKS). Ces 5 gènes ont été clonés par Burgett S. et al. en 1996 (Brevet américain US 5,945,320) (cf. figure 2). Ainsi, il a pu être déterminé une séquence unique d'ADN génomique de S. ambofaciens. Une séquence de 30943 nucléotides partant en 5' d'un site EcoRI situé dans le premier gène PKS et allant jusqu'à un site BamHI en 3' est présentée en SEQ ID N 1. Cette séquence correspond à la région amont des gènes de la PKS (cf. figure 2 et 3). Une deuxième région de 11171 nucléotides, partant d'un site PstI en 5' et allant jusqu'à un site NcoI en 3' situé dans le cinquième gène de la PKS est présentée en SEQ ID N 2. Cette deuxième région est la These two cosmids were completely sequenced by the shotgun sequencing technique. The sequences of the inserts of these two cosmids: pSPM7 and pSPM5 could be assembled because although not overlapping, each of the inserts included a known sequence at one of its ends. Indeed, each of these inserts included a fragment of the sequence of one of the genes encoding an enzyme called polyketide synthase (PKS). These 5 genes were cloned by Burgett S. et al. in 1996 (US Patent 5,945,320) (see Figure 2). Thus, it has been possible to determine a unique genomic DNA sequence of S. ambofaciens. A sequence of 30943 nucleotides starting 5 'from an EcoRI site located in the first PKS gene and going to a 3' BamHI site is presented in SEQ ID No. 1. This sequence corresponds to the upstream region of the genes of the PKS (see Figure 2 and 3). A second region of 11171 nucleotides, starting from a 5 'PstI site and going up to a 3' NcoI site located in the fifth gene of the PKS, is presented in SEQ ID No. 2. This second region is the
<Desc/Clms Page number 109><Desc / Clms Page number 109>
région aval des gènes de la PKS (l'aval et l'amont étant défini par l'orientation des 5 gènes de PKS tous orientés dans le même sens) (cf. figure 2 et 3). downstream region of the PKS genes (downstream and upstream being defined by the orientation of the PKS genes all oriented in the same direction) (see Figure 2 and 3).
EXEMPLE 4 : Analyse des séquences nucléotidiques, détermination des phases ouvertes de lecture et caractérisation des gènes impliqués dans la biosynthèse des spiramycines. EXAMPLE 4 Analysis of the Nucleotide Sequences, Determination of the Open Reading Phases and Characterization of the Genes Involved in the Biosynthesis of Spiramycins
Les séquences obtenues ont été analysées grâce au programme FramePlot (Ishikawa J & Hotta K. 1999). Ceci a permis d'identifier, parmi les phases ouvertes de lecture, les phases ouvertes de lecture présentant un usage des codons typique de Streptomyces. Cette analyse a permis de déterminer que cette région comporte 35 ORFs localisées de part et d'autre de cinq gènes codant l'enzyme polyketide synthase (PKS). Respectivement 10 et 25 ORFs ont été identifiées en aval et en amont de ces gènes (l'aval et l'amont étant défini par l'orientation des 5 gènes de PKS tous orientés dans le même sens) (cf. figure 3). Ainsi, les 25 phases ouvertes de lecture de ce type, occupant une région d'environ 31 kb (SEQ ID N 1 et figure 3) ont été identifiées en amont des 5 gènes codant la PKS et 10 occupant une région d'environ 11,1kb (SEQ ID N 2 et figure 3), ont été identifiées en aval des gènes de la PKS. Les gènes de la région amont ont été nommés or/7, or/2, or/3, orf4, orf5, orf6, orf7, orf8, orf9c, or/70, orfllc,
![]()
orfl2, orfl3c, orfl4, orf]5c, orj76, orfl7, orfl8, orfl9, orflo, or/2 le, orf22c, orf23c, orf24c et orf25c (SEQ ID N 23,25, 28,30, 34,36, 40,43, 45,47, 49,53, 60,62, 64, 66,68, 70,72, 74,76, 78,80, 82 et 84). Les gènes de la région aval ont été nommés
![]()
orfl *c, orf2*c, orf3 *c, orf4 *c, or/?*, orf6*, orf7*c, orf8*, or/9* or/70* (SEQ ID N 3, 5,7, 9, 11, 13,15, 17,19 et 21). Le c ajouté dans le nom du gène signifiant pour l'ORF en question que la séquence codante est dans l'orientation inverse (le brin codant est donc le brin complémentaire de la séquence donnée en SEQ ID N 1 ou SEQ ID N 2 pour ces gènes) (cf. figure 3). The sequences obtained were analyzed using the FramePlot program (Ishikawa J & Hotta K. 1999). This made it possible to identify, among the open reading phases, the open reading stages with typical codon usage of Streptomyces. This analysis made it possible to determine that this region comprises 35 ORFs located on either side of five genes encoding the enzyme polyketide synthase (PKS). Respectively 10 and 25 ORFs were identified downstream and upstream of these genes (downstream and upstream being defined by the orientation of the PKS genes all oriented in the same direction) (see Figure 3). Thus, the 25 open reading frames of this type, occupying a region of about 31 kb (SEQ ID N 1 and FIG. 3) were identified upstream of the 5 genes encoding PKS and occupying a region of about 11, 1kb (SEQ ID No. 2 and FIG. 3) were identified downstream of the PKS genes. The genes in the upstream region have been named gold / 7, gold / 2, gold / 3, orf4, orf5, orf6, orf7, orf8, orf9c, or / 70, orfllc,
![]()
orf2, orf13c, orf1f, orf1fc, orf76, orf17, orf18, orf19, orf1, orf2c, orf22c, orf23c, orf24c and orf25c (SEQ ID N 23,25, 28,30, 34,36, 40, 43, 45.47, 49.53, 60.62, 64, 66.68, 70.72, 74.76, 78.80, 82 and 84). Genes in the downstream region have been named
![]()
## STR1 ## wherein , 11, 13, 15, 17, 19 and 21). The c added in the name of the gene means for the ORF in question that the coding sequence is in the reverse orientation (the coding strand is therefore the complementary strand of the sequence given in SEQ ID N 1 or SEQ ID N 2 for these genes) (see Figure 3).
Les séquences protéiques déduites de ces phases ouvertes de lecture ont été comparées avec celles présentes dans différentes bases de données grâce à différents The protein sequences deduced from these open reading phases were compared with those present in different databases thanks to different
<Desc/Clms Page number 110><Desc / Clms Page number 110>
programmes : BLAST (Altschul et al., 1990) (Altschul et al., 1997), CD-search, COGs (Cluster of Orthologous Groups) (ces trois programmes sont accessibles notamment auprès du National Center for Biotechnology Information (NCBI) (Bethesda, Maryland, USA)), FASTA ((Pearson W. R & D. J. Lipman, 1988) et (Pearson W. R., 1990), BEAUTY (Worley K. C.. et al., 1995)), (ces deux programmes sont accessibles notamment auprès du centre de ressources INFOBIOGEN, Evry, France). Ces comparaisons ont permis de formuler des hypothèses sur la fonction des produits de ces gènes et d'identifier ceux susceptibles d'être impliqués dans la biosynthèse de spiramycine. programs: BLAST (Altschul et al., 1990) (Altschul et al., 1997), CD-search, Cluster of Orthologous Groups (COGs) (these three programs are available in particular from the National Center for Biotechnology Information (NCBI) (Bethesda , Maryland, USA)), FASTA (Pearson W. R & DJ Lipman, 1988) and (Pearson WR, 1990), BEAUTY (Worley KC et al., 1995), (these two programs are accessible in particular from INFOBIOGEN resource center, Evry, France). These comparisons made it possible to formulate hypotheses on the function of the products of these genes and to identify those likely to be involved in spiramycin biosynthesis.
EXEMPLE 5 : Inactivation de gène : principe de la construction d'une souche de Streptomyces ambofaciens interrompue
Les méthodes utilisées consistent à effectuer un remplacement de gène. Le gène cible à interrompre est remplacé par une copie de ce gène interrompue par une cassette conférant la résistance à un antibiotique (par exemple l'apramycine ou l'hygromycine), comme l'illustre la figure 9. Les cassettes utilisées sont bordées de part et d'autre par des codons de terminaison de la traduction dans toutes les phases de lecture et par des terminateurs de transcription actifs chez Streptomyces. EXAMPLE 5 Gene Inactivation: Principle of the Construction of a Strain of Streptomyces Ambofaciens Interrupted
The methods used consist in performing a gene replacement. The target gene to be interrupted is replaced by a copy of this gene interrupted by a cassette conferring resistance to an antibiotic (for example apramycin or hygromycin), as illustrated in FIG. 9. The cassettes used are lined on the and other by translation termination codons in all reading phases and by active transcription terminators in Streptomyces.
L'insertion de la cassette dans le gène cible peut s'accompagner ou non d'une délétion dans ce gène cible. La taille des régions flanquant la cassette peut aller de quelques centaines à plusieurs milliers de paires de bases. The insertion of the cassette into the target gene may or may not be accompanied by a deletion in this target gene. The size of the regions flanking the cassette can range from a few hundred to several thousand base pairs.
Les constructions nécessaires à l'inactivation du gène par la cassette ont été obtenues chez E. coli, organisme de référence pour l'obtention de constructions d'ADN recombinant. Le gène interrompu a été obtenu dans un plasmide se répliquant chez E. coli mais ne pouvant pas se répliquer chez Streptomyces. The constructs necessary for inactivation of the gene by the cassette were obtained from E. coli, a reference organism for obtaining recombinant DNA constructs. The interrupted gene was obtained in a plasmid replicating in E. coli but unable to replicate in Streptomyces.
Les constructions ont ensuite été sous-clonées dans des vecteurs pour permettre la transformation et l'inactivation du gène voulu chez S. ambofaciens. Pour cela, deux plasmides ont été utilisés : The constructs were then subcloned into vectors to allow transformation and inactivation of the desired gene in S. ambofaciens. For this, two plasmids were used:
<Desc/Clms Page number 111><Desc / Clms Page number 111>
- pOJ260 (Bierman M. et al., 1992) (cf. exemple 2) qui confère la résistance à l'apramycine chez E. coli et Streptomyces et qui a été employé quand le gène cible a été interrompu par une cassette conférant la résistance à l'hygromycine. pOJ260 (Bierman M. et al., 1992) (see Example 2) which confers resistance to apramycin in E. coli and Streptomyces and which was used when the target gene was interrupted by a cassette conferring resistance. with hygromycin.
- pOSK1205 (4726pb). Ce plasmide dérive du plasmide pBK-CMV (commercialisé par la société Stratagene (LaJolla, Californie, USA)) dans lequel un fragment
Avril contenant la séquence codant la résistance à la Néomycine/Kanamycine a été remplacé par une séquence codant la résistance à l'hygromycine, tout en conservant le promoteur P SV40. Pour cela, le plasmide pHP45-#hyg (Blondelet-
Rouault et al., 1997) a été digéré par les enzymes NotI et PflmI et le fragment conférant la résistance à l'hygromycine a été sous-cloné au niveau du site Avril du vecteur pBK-CMV après que toutes les extrémités aient été rendues bout franc par traitement à l'enzyme de Klenow. Dans pOSK1205, la cassette qui confère la résistance à l'hygromycine est précédée du promoteur pSV40. Ce plasmide confère la résistance à l'hygromycine chez E. coli et Streptomyces et a été employé quand le gène cible a été interrompu par une cassette conférant la résistance à l'apramycine. - pOSK1205 (4726bp). This plasmid derives from the plasmid pBK-CMV (sold by the company Stratagene (LaJolla, California, USA)) in which a fragment
April containing the sequence encoding resistance to Neomycin / Kanamycin was replaced by a sequence encoding hygromycin resistance, while retaining the promoter P SV40. For this, the plasmid pHP45- # hyg (Blondelet-
Rouault et al., 1997) was digested with the NotI and PflmI enzymes and the hygromycin resistance conferring fragment was subcloned at the April site of the pBK-CMV vector after all the ends were made end-to-end. Frank by treatment with Klenow's enzyme. In pOSK1205, the cassette that confers hygromycin resistance is preceded by the pSV40 promoter. This plasmid confers hygromycin resistance in E. coli and Streptomyces and was used when the target gene was interrupted by a cassette conferring resistance to apramycin.
Les cassettes ont été introduites dans le gène cible soit par clonage en utilisant des sites de restriction présents dans le gène cible, soit par recombinaison entre de courtes séquences identiques comme décrit par exemple par Chaveroche et al (Chaveroche MK et al., 2000). The cassettes were introduced into the target gene either by cloning using restriction sites present in the target gene, or by recombination between short identical sequences as described for example by Chaveroche et al (Chaveroche MK et al., 2000).
Le plasmide portant le gène interrompu par la cassette peut ensuite être introduit chez Streptomyces ambofaciens, par exemple par conjugaison entre E. coli et Streptomyces (Mazodier P, et al., 1989). Cette technique a été utilisée dans le cas où le vecteur de base est le vecteur pOJ260. Une deuxième technique peut être utilisée : la technique de la transformation de protoplastes après dénaturation par traitement alcalin de l'ADN (Kieser, T et al., 2000), pour augmenter la fréquence de recombinaison comme décrit par exemple par Oh & Chater (Oh & Chater, 1997). Cette technique a été utilisée dans le cas où le vecteur de base est pOJ260 ou pOSK1205 (cf. ci-dessous). Les transformants sont ensuite sélectionnés avec l'antibiotique correspondant à la cassette présente dans le The plasmid carrying the gene interrupted by the cassette can then be introduced into Streptomyces ambofaciens, for example by conjugation between E. coli and Streptomyces (Mazodier P, et al., 1989). This technique was used in the case where the base vector is the vector pOJ260. A second technique can be used: the technique of protoplast transformation after denaturation by alkaline treatment of DNA (Kieser, T et al., 2000), to increase the recombination frequency as described for example by Oh & Chater (Oh & Chater, 1997). This technique was used in the case where the base vector is pOJ260 or pOSK1205 (see below). The transformants are then selected with the antibiotic corresponding to the cassette present in the
<Desc/Clms Page number 112><Desc / Clms Page number 112>
gène cible (cf. figure 9, antibiotique B). On sélectionne ainsi un mélange de clones parmi lesquels il y a eu intégration par un seul ou par deux événements de recombinaison. Dans un deuxième temps on recherche les clones sensibles à l'antibiotique pour lequel le gène de résistance est présent dans le vecteur (hors de la cassette de recombinaison) (cf. figure 9, antibiotique A). On peut ainsi sélectionner les clones pour lesquels il y a eu en principe deux événements de recombinaison aboutissant au remplacement du gène sauvage par la copie interrompue par la cassette. Ces étapes sont schématisées figure 9. target gene (see Figure 9, antibiotic B). A mixture of clones from which there has been integration by one or two recombination events is thus selected. In a second step, the antibiotic-sensitive clones for which the resistance gene is present in the vector (outside the recombination cassette) are sought (see FIG. 9, antibiotic A). Clones for which there have been in principle two recombination events leading to the replacement of the wild-type gene by the copy interrupted by the cassette can thus be selected. These steps are shown schematically in Figure 9.
Plusieurs cassettes peuvent être utilisées pour l'interruption des gènes cibles. On peut par exemple utiliser la cassette Qhyg qui confère la résistance à l'hygromycine (Blondelet-Rouault et al., 1997, numéro d'accès GenBank : X99315). Several cassettes can be used for the interruption of target genes. For example, the Qhyg cassette which confers resistance to hygromycin may be used (Blondelet-Rouault et al., 1997, GenBank accession number: X99315).
EXEMPLE 6: Construction d'une souche de Streptomyces ambofaciens interrompue en phase dans le gène or/3
Le gène orf 3 avait été interrompu par la cassette Qhyg (cf. exemple 2. 2) et il a pu être démontré qu'une souche or/3:: #hyg ne produit plus de spiramycine, confirmant l'implication d'un ou plusieurs gènes de la région clonée dans la biosynthèse des spiramycines (cf. exemple 2. 2). Au vu de leur orientation, une cotranscription des ORFs 1 à 7 est probable (cf. figure 3) et le phénotype observé (non producteur de spiramycines) peut être dû à l'inactivation d'un ou plusieurs des gènes co-transcrits avec or/3. Pour confirmer l'implication de orf3 dans la biosynthèse de spiramycines, une nouvelle inactivation du gène or/3 a été effectuée, cette dernière étant réalisée en phase. EXAMPLE 6 Construction of a Strain of Streptomyces Ambofaciens Interrupted in Phase in the Gold / 3 Gene
The orf3 gene had been interrupted by the Qhyg cassette (see Example 2.2) and it could be shown that a strain of gold / 3 :: #hyg no longer produces spiramycin, confirming the involvement of one or several genes of the cloned region in spiramycin biosynthesis (see example 2. 2). In view of their orientation, a cotranscription of ORFs 1 to 7 is likely (see Figure 3) and the observed phenotype (non-producer of spiramycins) may be due to the inactivation of one or more genes co-transcribed with gold. / 3. To confirm the involvement of orf3 in the biosynthesis of spiramycins, a new inactivation of the or / 3 gene was performed, the latter being performed in phase.
Pour cela, un fragment DraIII interne à or/3 de 504 paires de bases a été délété. Un fragment d'ADN obtenu à partir de pOS49. 1, allant du site EcoRI situé en position 1 (SEQ ID N 1) jusqu'au site SacI situé en position 5274 (SEQ ID N 1) et qui comporte une délétion entre les deux sites DraIII aux positions 2563 et 3067 (504 nucléotides retirés) a été cloné dans le plasmide pOJ260 (Bierman M. et al., 1992). Le plasmide ainsi obtenu a été nommé pOS49.67. For this, an internal DraIII fragment at or / 3 of 504 base pairs has been deleted. A DNA fragment obtained from pOS49. 1, from the EcoRI site located at position 1 (SEQ ID No. 1) to the SacI site located at position 5274 (SEQ ID No. 1) and which has a deletion between the two DraIII sites at positions 2563 and 3067 (504 nucleotides removed ) was cloned into plasmid pOJ260 (Bierman M. et al., 1992). The plasmid thus obtained was named pOS49.67.
<Desc/Clms Page number 113> <Desc / Clms Page number 113>
L'insert de pOS49. 67 est donc constitué par un fragment d'ADN de S. ambofaciens contenant les gènes orfl, or/5, or/3 avec la délétion en phase, orf4 et une partie de orf5. The insert of pOS49. 67 thus consists of an S. ambofaciens DNA fragment containing the orf1, or5, or / 3 genes with in-phase deletion, orf4 and part of orf5.
Le vecteur dans lequel cet insert a été sous-cloné est pOJ260, le plasmide pOS49.67 confère donc la résistance à l'apramycine et a été introduit par transformation de protoplastes dans la souche OS49.16 (cf. exemple 2). La souche OS49. 16 étant résistante à l'hygromycine, des transformants hygR et apraR ont été obtenus. Après deux passages de tels clones sur milieu non sélectif, les clones sensibles à l'apramycine et à l'hygromycine (apraS et hygS) ont été recherchés. Dans certains de ces clones, on s'attend en effet à ce qu'un événement de recombinaison entre séquences homologues conduise au remplacement de la copie de orf3 interrompue par la cassette #hyg (contenu dans le génome de la souche OS49. 16) par la copie de or/3 avec la délétion en phase présente sur le vecteur. Les clones résultant de cette recombinaison sont attendus comme apraS et hygS après l'élimination des séquences du vecteur. Le génotype des souches ainsi obtenues peut être vérifié par hybridation ou par PCR et séquençage du produit de PCR (pour vérifier que seule une copie de or/3 délétée en phase est présente dans le génome des clones obtenus). Des clones n'ayant que la copie de or/3 avec une délétion en phase ont ainsi été obtenus et leur génotype a été vérifié. Un clone présentant les caractéristiques recherchées a été plus particulièrement sélectionné et a été nommé OS49. 67. The vector in which this insert was subcloned is pOJ260, the plasmid pOS49.67 therefore confers resistance to apramycin and was introduced by transformation of protoplasts in the strain OS49.16 (see Example 2). The strain OS49. Since it was resistant to hygromycin, hygR and apraR transformants were obtained. After two passages of such clones on nonselective medium, the clones sensitive to apramycin and hygromycin (apraS and hygS) were searched. In some of these clones, it is indeed expected that a recombination event between homologous sequences lead to the replacement of the copy of orf3 interrupted by the #hyg cassette (contained in the genome of the strain OS49.16) by the copy of gold / 3 with the phase deletion present on the vector. The clones resulting from this recombination are expected as apraS and hygS after the elimination of the vector sequences. The genotype of the strains thus obtained can be verified by hybridization or by PCR and sequencing of the PCR product (to verify that only one copy of gold / 3 deleted in phase is present in the genome of the clones obtained). Clones having only the copy of gold / 3 with a deletion in phase were thus obtained and their genotype was verified. A clone with the desired characteristics was particularly selected and was named OS49. 67.
Un échantillon de la souche OS49. 67 a été déposé auprès de la Collection' Nationale de Cultures de Microorganismes (CNCM) Institut Pasteur, 25, rue du Docteur Roux 75724 Paris Cedex 15, France, le 10 juillet 2002 sous le numéro d'enregistrement 1-2916. A sample of the strain OS49. 67 has been deposited with the National Collection of Cultures of Microorganisms (CNCM) Pasteur Institute, 25, rue du Docteur Roux 75724 Paris Cedex 15, France, July 10, 2002 under the registration number 1-2916.
EXEMPLE 7: Construction d'une souche de Streptomyces ambofaciens interrompue dans le gène orf8
Pour réaliser l'inactivation du gène or/8, une construction dans laquelle la cassette #hyg a été introduite dans la séquence codante de or/8 a été obtenue. Pour cela, le EXAMPLE 7 Construction of a Strain of Streptomyces Ambofaciens Interrupted in the Orf8 Gene
To achieve the inactivation of the or / 8 gene, a construct in which the #hyg cassette was introduced into the or / 8 coding sequence was obtained. For that, the
<Desc/Clms Page number 114><Desc / Clms Page number 114>
plasmide pOS49. 88 a tout d'abord été construit. Le plasmide pOS49. 88 est dérivé du plasmide pUC19 (numéro d'accès GenBank M77789) par insertion d'un fragment de 3.7 kb (fragment PstI-EcoRI obtenu à partir du cosmide pSPM5), contenant la fin de or/7, or/8 et le début de or/9, cloné dans les sites PstI-EcoRI de pUC19. La cassette #hyg (sous forme d'un fragment BamHI, rendu bout franc par traitement à l'enzyme de Klenow) a été clonée au niveau du site unique Sali de pOS49. 88 situé dans or/8 après que toutes les extrémités aient été rendues bout franc par traitement à l'enzyme de Klenow. plasmid pOS49. 88 was first built. The plasmid pOS49. 88 is derived from plasmid pUC19 (GenBank accession number M77789) by insertion of a 3.7 kb fragment (PstI-EcoRI fragment obtained from cosmid pSPM5), containing the end of gold / 7, or / 8 and the beginning of gold / 9, cloned into the PstI-EcoRI sites of pUC19. The #hyg cassette (as a BamHI fragment, blunted by Klenow enzyme treatment) was cloned at the unique SalI site of pOS49. 88 located in or / 8 after all extremities were made blunt by treatment with Klenow enzyme.
* Le clonage étant bout franc, deux types de plasmides ont été obtenus selon le sens d'insertion de la cassette : pOS49. 106 dans lequel les gènes hyg et orf8 sont dans la même orientation et pOS49. 120 dans lequel les gènes hyg et orf8 sont dans des orientations opposées. L'insert du plasmide pOS49. 106 a ensuite été sous-cloné dans le plasmide pOJ260 pour donner pOS49. 107. Pour cela, le plasmide pOS49. 106 a été digéré par l'enzyme Asp718I et les extrémités ont été rendues bout franc par traitement à l'enzyme de Klenow, ce produit de digestion a été redigéré par l'enzyme PstI et le fragment contenant le gène or/8 dans lequel la cassette #hyg a été insérée, a été cloné dans le vecteur pOJ260 (cf. ci dessus). Pour cela, le vecteur pOJ260 a été digéré par les enzymes EcoRV et PstI et utilisé pour la ligation. Cette manipulation permet donc d'obtenir une ligation orientée puisque chacun des deux fragments est bout franc d'un côté et PstI de l'autre. Le plasmide obtenu a été nommé pOS49.107. * Cloning being a blunt end, two types of plasmids were obtained according to the direction of insertion of the cassette: pOS49. 106 in which the genes hyg and orf8 are in the same orientation and pOS49. 120 in which the genes hyg and orf8 are in opposite orientations. The insert of plasmid pOS49. 106 was then subcloned into plasmid pOJ260 to give pOS49. 107. For this, the plasmid pOS49. 106 was digested with the enzyme Asp718I and the ends were made blunt by treatment with Klenow enzyme, this digest was redigested by the enzyme PstI and the fragment containing the gene or / 8 in which the cassette #hyg was inserted, was cloned in the vector pOJ260 (see above). For this, the vector pOJ260 was digested with the enzymes EcoRV and PstI and used for ligation. This manipulation thus makes it possible to obtain an oriented ligation since each of the two fragments is frank on one side and PstI on the other. The resulting plasmid was named pOS49.107.
Le plasmide pOS49. 107 a été introduit dans la souche ATCC23877 de S. ambofaciens par transformation de protoplastes (Kieser, T et al., 2000). Après transformation des protoplastes, les clones sont sélectionnés pour leur résistance à l'hygromycine. Les clones résistants à l'hygromycine sont ensuite repiqués respectivement sur milieu avec hygromycine (antibiotique B) et sur milieu avec apramycine (antibiotique A) (cf. figure 9). Les clones résistants à l'hygromycine (HygR) et sensibles à l'apramycine (ApraS) sont en principe ceux où un double événement de crossing over s'est produit et qui possèdent le gène orf8 interrompu par la cassette Qhyg. The plasmid pOS49. 107 was introduced into S. ambofaciens strain ATCC23877 by protoplast transformation (Kieser, T et al., 2000). After transformation of the protoplasts, the clones are selected for their resistance to hygromycin. The hygromycin-resistant clones are then transplanted respectively on medium with hygromycin (antibiotic B) and on medium with apramycin (antibiotic A) (see FIG. 9). The hygromycin-resistant (HygR) and apramycin-sensitive (ApraS) -resistant clones are in principle those where a double crossing-over event has occurred and which have the orf8 gene interrupted by the Qhyg cassette.
<Desc/Clms Page number 115> <Desc / Clms Page number 115>
Le remplacement de la copie sauvage de orf8 par la copie interrompue par la cassette #hyg a été vérifié par Southern Blot. Ainsi, l'ADN total des clones obtenus, a été digéré par plusieurs enzymes, séparé sur gel d'agarose, transféré sur membrane et hybridé avec une sonde correspondant à la cassette Qhyg pour vérifier la présence de la cassette dans l'ADN génomique des clones obtenus. Une deuxième hybridation a été effectuée en utilisant comme sonde l'insert PstI-EcoRI contenant la fin de or/7, or/8 et le début de or/9 d'une taille d'environ 3,7 kb du plasmide pOS49. 88. La vérification du génotype peut être réalisée par toute méthode connue de l'homme du métier et notamment par PCR en utilisant les oligonucléotides appropriés et séquençage du produit de PCR. Replacing the wild copy of orf8 with the copy interrupted by the #hyg cassette has been checked by Southern Blot. Thus, the total DNA of the clones obtained was digested with several enzymes, separated on agarose gel, transferred to the membrane and hybridized with a probe corresponding to the Qhyg cassette to check the presence of the cassette in the genomic DNA of the cells. clones obtained. A second hybridization was carried out using as probe the PstI-EcoRI insert containing the end of gold / 7, or / 8 and the beginning of gold / 9 with a size of about 3.7 kb of plasmid pOS49. 88. Verification of the genotype can be carried out by any method known to those skilled in the art and in particular by PCR using the appropriate oligonucleotides and sequencing of the PCR product.
Un clone orj8::Qhyg a été choisi et nommé OS49.107. Un échantillon de la souche OS49. 107 a été déposé auprès de la Collection Nationale de Cultures de Microorganismes (CNCM) Institut Pasteur, 25, rue du Docteur Roux 75724 Paris Cedex 15, France, le 10 juillet 2002 sous le numéro d'enregistrement 1-2917. An orj8 :: Qhyg clone was chosen and named OS49.107. A sample of the strain OS49. 107 has been deposited with the National Collection of Cultures of Microorganisms (CNCM) Pasteur Institute, 25, rue du Docteur Roux 75724 Paris Cedex 15, France, July 10, 2002 under the registration number 1-2917.
EXEMPLE 8: Construction d'une souche de Streptomyces ambofaciens interrompue dans le gène orf10
Un fragment d'ADN de 1,5 kb interne au gène or/70 a été obtenu par PCR en utilisant comme matrice, l'ADN génomique de S. ambofaciens et les amorces suivantes : SRMRl : 5' CTGCCAGTCCTCTCCCAGCAGTACG 3' (SEQ ID N 89) SRMR2 : 5' TGAAGCTGGACGTCTCCTACGTCGG 3' (SEQ ID N 90) Ce fragment d'ADN issu de la PCR a été cloné dans le vecteur pCR2.1 commercialisé par la société Invitrogen (Carlsbad, Californie, USA). Le plasmide ainsi obtenu a été nommé pOS49. 32. La cassette Qhyg (sous forme d'un fragment BamHI, cf. ci-dessus) a été clonée au niveau du site unique BstEII interne au fragment du gène orf10, après que toutes les extrémités aient été rendues bout franc par traitement à l' enzyme de Klenow. EXAMPLE 8 Construction of a Strain of Streptomyces Ambofaciens Interrupted in the Orf10 Gene
A 1.5 kb DNA fragment internal to the or / 70 gene was obtained by PCR using as template the genomic DNA of S. ambofaciens and the following primers: SRMR1: 5 'CTGCCAGTCCTCTCCCAGCAGTACG 3' (SEQ ID N 89) SRMR2: 5 'TGAAGCTGGACGTCTCCTACGTCGG 3' (SEQ ID N 90) This DNA fragment resulting from the PCR was cloned into the vector pCR2.1 sold by the company Invitrogen (Carlsbad, California, USA). The plasmid thus obtained was named pOS49. 32. The Qhyg cassette (in the form of a BamHI fragment, see above) was cloned at the unique internal BstEII site at the orf10 gene fragment, after all the ends were made blunt by Klenow enzyme.
<Desc/Clms Page number 116> <Desc / Clms Page number 116>
Le clonage étant bout franc, deux types de plasmides ont été obtenus selon le sens d'insertion de la cassette : pOS49. 43 dans lequel les gènes hyg et or/70 sont dans la même orientation et pOS49. 44 dans lequel les gènes hyg et or/10 sont dans des orientations opposées. L'insert du plasmide pOS49. 43 a été transféré (sous forme d'un fragment Asp718I-XbaI dont les extrémités ont été rendues bout franc par traitement à l'enzyme de Klenow) dans le site EcoRV du plasmide pOJ260 ce qui a permis d'obtenir le plasmide pOS49. 50. Le plasmide pOS49. 50 contenant le fragment du gène orf10 interrompu par la cassette Qhyg a été introduit dans la souche ATCC23877 de Streptomyces ambofaciens. Après transformation, les clones sont sélectionnés pour leur résistance à l'hygromycine. Les clones résistants à l'hygromycine sont ensuite repiqués respectivement sur milieu avec hygromycine (antibiotique B) et sur milieu avec apramycine (antibiotique A) (cf. figure 9). Les clones résistants à l'hygromycine (HygR) et sensibles à l'apramycine (ApraS) sont en principe ceux où un double événement de crossing over s'est produit et qui possèdent le gène orflO interrompu par la cassette Qhyg. Des clones qui possédaient le marqueur de résistance à l'hygromycine porté par la cassette et qui avaient perdu le marqueur de résistance à l'apramycine porté par le vecteur pOJ260 ont ainsi été obtenus. L'événement de remplacement de la copie sauvage de or/70 par la copie interrompue orflO::Qhyg a été vérifié par Southern Blot. Ainsi, l'ADN génomique total des clones obtenus, a été digéré par plusieurs enzymes, séparé sur gel d'agarose, transféré sur membrane et hybridé avec une sonde correspondant à la cassette #hyg pour vérifier la présence de la cassette dans l'ADN génomique des clones obtenus. Une deuxième hybridation a été effectuée en utilisant comme sonde le produit PCR de 1,5 kb interne au gène orf10 (cf. ci-dessus). Cloning being a blunt end, two types of plasmids were obtained according to the direction of insertion of the cassette: pOS49. 43 in which the genes hyg and or / 70 are in the same orientation and pOS49. 44 in which the genes hyg and or / 10 are in opposite orientations. The insert of plasmid pOS49. 43 was transferred (as an Asp718I-XbaI fragment whose ends were made blunt by treatment with Klenow enzyme) in the EcoRV site of plasmid pOJ260 which made it possible to obtain the plasmid pOS49. 50. The plasmid pOS49. 50 containing the fragment of the orf10 gene interrupted by the Qhyg cassette was introduced into Streptomyces ambofaciens strain ATCC23877. After transformation, the clones are selected for their resistance to hygromycin. The hygromycin-resistant clones are then transplanted respectively on medium with hygromycin (antibiotic B) and on medium with apramycin (antibiotic A) (see FIG. 9). The hygromycin-resistant (HygR) and apramycin-sensitive (ApraS) -resistant clones are in principle those where a double crossing-over event has occurred and which have the orflO gene interrupted by the Qhyg cassette. Clones which possessed the hygromycin resistance marker carried by the cassette and which had lost the apramycin resistance marker carried by the vector pOJ260 were thus obtained. The replacement event of the wild copy of gold / 70 by the interrupted copy orflO :: Qhyg has been verified by Southern Blot. Thus, the total genomic DNA of the clones obtained was digested with several enzymes, separated on agarose gel, transferred to the membrane and hybridized with a probe corresponding to the #hyg cassette to check the presence of the cassette in the DNA. genomic clones obtained. A second hybridization was carried out using as probe the 1.5 kb PCR product internal to the orf10 gene (see above).
Un clone présentant les caractéristiques attendues (orf10::#hyg) a été plus particulièrement sélectionné et nommé OS49.50. Il a pu en effet être vérifié grâce aux deux hybridations que la cassette Qhyg était bien présente dans le génome de ce clone et que l'on obtient bien le profil de digestion attendu dans le cas d'un remplacement, à la suite d'un double événement de recombinaison, du gène sauvage par la copie interrompue par la cassette Qhyg dans le génome de ce clone. La vérification du A clone with the expected characteristics (orf10 :: # hyg) was specifically selected and named OS49.50. It has indeed been possible to verify, thanks to the two hybridizations, that the Qhyg cassette was indeed present in the genome of this clone and that the expected digestion profile is obtained in the case of a replacement, following a double recombination event of the wild-type gene by the copy interrupted by the Qhyg cassette in the genome of this clone. Verification of
<Desc/Clms Page number 117><Desc / Clms Page number 117>
génotype peut également être réalisée par toute méthode connue de l'homme du métier et notamment par PCR en utilisant les oligonucléotides appropriés et séquençage du produit de PCR. genotype can also be performed by any method known to those skilled in the art and in particular by PCR using the appropriate oligonucleotides and sequencing of the PCR product.
EXEMPLE 9 : Inactivation de gènes : principe de la construction d'une souche de Streptomyces ambofaciens interrompue selon la technique des cassettes excisables (cf. Figure 9 et 10)
Un deuxième type de cassettes peut être utilisé pour l'inactivation de gènes : les cassettes dites cassettes excisables . Ces cassettes présentent l'avantage de pouvoir être excisées chez Streptomyces par un événement de recombinaison spécifique de site après avoir été introduites dans le génome de S. ambofaciens. Le but est d'inactiver certains gènes dans des souches de Streptomyces sans laisser dans la souche finale de marqueurs de sélection ou de grandes séquences d'ADN n'appartenant pas à la souche. EXAMPLE 9 Inactivation of Genes: Principle of the Construction of an Streptomyces Ambofaciens Strain Interrupted by the Technique of Excisable Cassettes (see FIGS. 9 and 10)
A second type of cassette can be used for the inactivation of genes: cassettes called excisable cassettes. These cassettes have the advantage of being excised in Streptomyces by a site-specific recombination event after being introduced into the S. ambofaciens genome. The goal is to inactivate certain genes in Streptomyces strains without leaving in the final strain of selection markers or large DNA sequences not belonging to the strain.
Après excision il subsiste uniquement une courte séquence d'environ une trentaine de paires de bases (appelé site cicatriciel ) dans le génome de la souche (cf. figure 10). After excision there remains only a short sequence of about thirty base pairs (called scarred site) in the genome of the strain (see Figure 10).
La mise en oeuvre de ce système consiste, dans un premier temps, au remplacement de la copie sauvage du gène cible (grâce à deux événements de recombinaison homologue, cf. figure 9) par une construction dans laquelle une cassette excisable a été insérée dans ce gène cible. L'insertion de cette cassette est accompagnée d'une délétion dans le gène cible (cf. figure 9). Dans un deuxième temps, l'excision de la cassette excisable du génome de la souche est provoquée. La cassette excisable fonctionne grâce à un système de recombinaison spécifique de site et a pour avantage de permettre l'obtention de mutants de Streptomyces ne portant finalement pas de gène de résistance. On s'affranchit également d'éventuels effets polaires sur l'expression des gènes situés en aval du ou des gènes inactivés (cf. figure 10). The implementation of this system consists, first of all, in the replacement of the wild-type copy of the target gene (by virtue of two homologous recombination events, see FIG. 9) by a construction in which an excisable cassette has been inserted in this system. target gene. The insertion of this cassette is accompanied by a deletion in the target gene (see Figure 9). In a second step, the excision of the excisable cassette of the genome of the strain is caused. The excisable cassette operates by means of a site-specific recombination system and has the advantage of allowing the production of mutants of Streptomyces not ultimately carrying a resistance gene. It also avoids possible polar effects on the expression of genes located downstream of the inactivated gene (s) (see Figure 10).
<Desc/Clms Page number 118> <Desc / Clms Page number 118>
L'emploi de cassettes excisables a été décrit et utilisé chez de nombreux organismes dont les cellules de mammifère, de levures et chez E. coli (Bayley et al., 1992 ; Brunelli et Pall, 1993 ; Camilli et al., 1994 ; Dale et Ow, 1991 ; Russell et al., 1992 ; Lakso et al., 1992). Ces cassettes excisables utilisent toutes la recombinase spécifique de site Cre agissant sur les sites lox. Ce système de recombinaison provient du bactériophage PI . The use of excisable cassettes has been described and used in many organisms including mammalian, yeast and E. coli cells (Bayley et al., 1992, Brunelli and Pall, 1993, Camilli et al., 1994; and Ow, 1991, Russell et al., 1992, Lakso et al., 1992). These excisable cassettes all use the Cre site-specific recombinase acting on the lox sites. This recombination system comes from the bacteriophage PI.
Pour la construction d'un système de type cassette excisable chez Streptomyces, il a été tiré partie du système de recombinaison spécifique de site décrit pour l'élément génétique mobile pSAM2 de Streptomyces ambofaciens (Boccard et al., 1989a et b). Le système mis en place consiste dans un premier temps à construire un vecteur recombinant comportant le gène à interrompre dans lequel est insérée une cassette excisable. L'insertion dans le gène cible de la cassette excisable s'accompagne d'une délétion dans le gène cible. Elle peut être réalisée par clonage en utilisant des sites de restriction présents dans le gène cible ou par recombinaison entre de courtes séquences identiques comme décrit par exemple par Chaveroche et al (Chaveroche MK et al., 2000). La cassette excisable peut être construite en utilisant par exemple la cassette Qhyg (Blondelet Rouault et al., 1997). Cette cassette a été bordée par des séquences attR et attL qui flanquent normalement la copie intégrée de pSAM2 (cf. figure 15). Les séquences attL et attR contiennent tous les sites nécessaires à la recombinaison sitespécifique permettant l'excision de pSAM2 ou de tout fragment d'ADN situé entre ces deux régions (Sezonov et al., 1997, Raynal et al.,1998). La construction d'une telle cassette n'est évidemment pas limitée à l'utilisation d'une cassette Qhyg, mais d'autres cassettes de résistances peuvent servir de base à la construction de cette cassettte (par exemple la cassette #aac ou Qvph (Blondelet Rouault et al., 1997)). For the construction of a Streptomyces excisable cassette-type system, the site-specific recombination system described for the Streptomyces ambofaciens mobile genetic element pSAM2 was used (Boccard et al., 1989a and b). The implemented system consists first of all in constructing a recombinant vector comprising the gene to be interrupted into which an excisable cassette is inserted. Insertion into the target gene of the excisable cassette is accompanied by a deletion in the target gene. It can be performed by cloning using restriction sites present in the target gene or by recombination between short identical sequences as described for example by Chaveroche et al (Chaveroche MK et al., 2000). The excisable cassette can be constructed using, for example, the Qhyg cassette (Blondelet Rouault et al., 1997). This cassette was bordered by attR and attL sequences that normally flank the integrated copy of pSAM2 (see Figure 15). The attL and attR sequences contain all the sites necessary for site specific recombination allowing the excision of pSAM2 or any DNA fragment located between these two regions (Sezonov et al., 1997, Raynal et al., 1998). The construction of such a cassette is obviously not limited to the use of a cassette Qhyg, but other cassettes of resistors can serve as a basis for the construction of this cassette (for example the cassette #aac or Qvph ( Blondelet Rouault et al., 1997)).
Après l'obtention de cette construction, la souche de Streptomyces est transformée avec le plasmide recombinant. Les transformants sont ensuite sélectionnés avec l'antibiotique correspondant à la cassette présente dans le gène cible (cf. figure 9, antibiotique B, il s'agit par exemple d'une sélection par l'hygromycine si la cassette excisable dérive de la cassette Qhyg). On sélectionne ainsi un mélange de clones parmi lesquels il y a eu intégration par un seul ou par deux événements de recombinaison. After obtaining this construct, the Streptomyces strain is transformed with the recombinant plasmid. The transformants are then selected with the antibiotic corresponding to the cassette present in the target gene (see FIG. 9, antibiotic B, it is for example a selection by hygromycin if the excisable cassette derives from the Qhyg cassette. ). A mixture of clones from which there has been integration by one or two recombination events is thus selected.
<Desc/Clms Page number 119> <Desc / Clms Page number 119>
Dans un deuxième temps on recherche les clones sensibles à l'antibiotique pour lesquels le gène de résistance est présent dans le vecteur (hors de la cassette de recombinaison) (cf. figure 9, antibiotique A). On peut ainsi sélectionner les clones pour lesquel il y a eu en principe deux événements de recombinaison aboutissant au remplacement du gène sauvage par la copie interrompue par la cassette. Ces étapes sont schématisées figure 9, le génotype des clones ainsi obtenus est vérifié par Southern blot et une souche présentant les caractéristiques voulues (le remplacement du gène sauvage par la copie interrompue par la cassette excisable) est sélectionnée. In a second step, the antibiotic-sensitive clones for which the resistance gene is present in the vector (outside the recombination cassette) are sought (see FIG. 9, antibiotic A). It is thus possible to select the clones for which, in principle, there have been two recombination events leading to the replacement of the wild-type gene by the copy interrupted by the cassette. These steps are shown schematically in Figure 9, the genotype of the clones thus obtained is verified by Southern blot and a strain having the desired characteristics (the replacement of the wild-type gene by the copy interrupted by the excisable cassette) is selected.
Dans un deuxième temps, la souche sélectionnée ci-dessus, est transformée par un plasmide permettant l'expression des gènes xis et int qui sont tous deux nécessaires à la recombinaison spécifique de site entre les sites attR et attL. Cette recombinaison entraîne le départ de la cassette excisable du génome de la souche, grâce à un événement de recombinaison (cf. figure 10) (Raynal et al., 1998). Il est intéressant de choisir le vecteur portant les gènes xis et int parmi les vecteurs relativement instables chez Streptomyces (par exemple dérivé du vecteur Streptomyces pWHM3, (Vara et al., 1989)), ceci permet d'obtenir une souche ayant perdu ce dernier vecteur après quelques cycles de sporulation en absence de pression de sélection. In a second step, the strain selected above is transformed by a plasmid allowing the expression of the xis and int genes which are both necessary for site-specific recombination between the attR and attL sites. This recombination leads to the departure of the excisable cassette from the genome of the strain, thanks to a recombination event (see FIG. 10) (Raynal et al., 1998). It is interesting to choose the vector carrying the genes xis and int among the relatively unstable vectors in Streptomyces (for example derived from the vector Streptomyces pWHM3, (Vara et al., 1989)), this makes it possible to obtain a strain having lost the latter. vector after a few cycles of sporulation in the absence of selection pressure.
Pour exciser la cassette, on peut par exemple utiliser le plasmide pOSV508 (cf. figure 14) qui est introduit par transformation de protoplastes dans la souche de S. ambofaciens contenant un gène interrompu par la cassette excisable. Le plasmide pOSV508 est dérivé du plasmide pWHM3 (Vara J et al.,1989) (cf. figure 13) dans lequel ont été ajoutés les gènes xis et int de pSAM2 (Boccard F. et al., 1989b) placés sous le contrôle du promoteur ptrc (Amann, E. et al., 1988). Les gènes xis et int placés sous le contrôle du promoteur ptrc ont été sous clonés dans le plasmide pWHM3 à partir du plasmide pOSint3 (Raynal et al., 1998) (cf. figure 14). L'introduction dans la souche mutante du plasmide pOSV508 portant les gènes xis et int de pSAM2 va permettre l'excision efficace par recombinaison spécifique de site de la cassette excisable entre les sites attL et attR flanquant cette cassette (Raynal A. et al., 1998) (figure 10). Parmi les transformants, sélectionnés pour leur résistance au thiostrepton due au gène tsr porté par To excise the cassette, it is possible, for example, to use the plasmid pOSV508 (FIG. 14), which is introduced by transformation of protoplasts into the S. ambofaciens strain containing a gene interrupted by the excisable cassette. The plasmid pOSV508 is derived from the plasmid pWHM3 (Vara J et al., 1989) (see FIG. 13) in which the xis and int genes of pSAM2 (Boccard F. et al., 1989b) placed under the control of ptrc promoter (Amann, E. et al., 1988). The xis and int genes placed under the control of the ptrc promoter were subcloned in the plasmid pWHM3 from the plasmid pOSint3 (Raynal et al., 1998) (see FIG. 14). The introduction into the mutant strain of plasmid pOSV508 carrying the xis and int genes of pSAM2 will allow efficient excision by site-specific recombination of the excisable cassette between the attL and attR sites flanking this cassette (Raynal A. et al. 1998) (Figure 10). Of the transformants selected for their thiostrepton resistance due to the tsr gene carried by
<Desc/Clms Page number 120><Desc / Clms Page number 120>
pOSV508, on choisit ceux devenus sensibles à l'antibiotique auquel la présence de la cassette confère la résistance (cf. figure 10). L'excision est efficace et il a été observé que plus de 90% des transformants sont de ce type. Après un ou plusieurs cycles de croissance et sporulation sur un milieu solide dépourvu de thiostrepton, on obtient des clones ayant perdu le plasmide pOSV508. Ces clones sont repérables par leur sensibilité au thiostrepton. La séquence du gène cible délété peut être vérifiée par PCR et séquençage du produit PCR. pOSV508, one chooses those become sensitive to the antibiotic to which the presence of the cassette confers the resistance (cf figure 10). Excision is effective and it has been observed that more than 90% of the transformants are of this type. After one or more cycles of growth and sporulation on a solid medium devoid of thiostrepton, clones having lost the plasmid pOSV508 are obtained. These clones are identifiable by their sensitivity to thiostrepton. The sequence of the deleted target gene can be verified by PCR and sequencing of the PCR product.
Finalement, la souche résultante porte au niveau du gène inactivé (délétion interne par exemple) un site att cicatriciel correspondant au site attB minimal (Raynal et al., 1998), issu de la recombinaison entre les sites attR et attL. Ce site attB minimal qui subsiste est semblable à celui présent naturellement dans les souches de Streptomyces ambofaciens, Streptomyces pristinaespiralis et Streptomyces lividans (Sezonov et al., 1997). Finally, the resulting strain carries at the level of the inactivated gene (internal deletion, for example) a scar scar site corresponding to the minimal attB site (Raynal et al., 1998), resulting from the recombination between the attR and attL sites. This remaining minimal attB site is similar to that found naturally in strains of Streptomyces ambofaciens, Streptomyces pristinaespiralis and Streptomyces lividans (Sezonov et al., 1997).
Le gène que l'on veut inactiver peut être cotranscrit avec d'autres gènes situés en aval. Pour éviter que l'inactivation d'un des gènes ait un effet polaire sur l'expression des gènes en aval dans l'opéron, il est important d'obtenir une délétion en phase après excision de la cassette. Le système des cassettes excisables tel que décrit ci-dessus permet de répondre à une telle exigence. L'homme du métier pourra, en effet, aisément construire trois cassettes excisables distinctes, celles-ci laissant après excision une séquence de 33, 34 ou 35 nucléotides respectivement, sans codon stop quelle que soit la phase de lecture. Connaissant la séquence du gène cible et la taille de la délétion associée à l'insertion de la cassette, il est possible de choisir entre ces trois cassettes excisables de façon à ce que l'excision conduise à une délétion en phase. Sur les 33,34 ou 35 nucléotides rajoutés, 26 correspondent à la séquence attB minimale (cf. figure 27). The gene that one wants to inactivate can be cotranscribed with other genes located downstream. To prevent the inactivation of one gene from having a polar effect on downstream gene expression in the operon, it is important to obtain a phase deletion after excision of the cassette. The system of excisable cassettes as described above makes it possible to meet such a requirement. Those skilled in the art may, in fact, easily construct three distinct excisable cassettes, these leaving after excision a sequence of 33, 34 or 35 nucleotides respectively, without a stop codon whatever the reading phase. Knowing the sequence of the target gene and the size of the deletion associated with the insertion of the cassette, it is possible to choose between these three excisable cassettes so that the excision leads to an in-phase deletion. Of the 33,34 or 35 nucleotides added, 26 correspond to the minimal attB sequence (see Figure 27).
Dans le cas de la présente demande, deux cassettes excisables ont été utilisées. Ces deux cassettes sont les suivantes : att1#hyg+ (SEQ ID N 91) et att3Qaac- (SEQ ID N 92), ces cassette laissent respectivement 33 et 35 nucléotides après excision. Elles comportent respectivement la cassette Qhyg ou la cassette Qaac, les signes + et correspondant à l'orientation de la cassette de résistance. Ces deux cassettes ont été In the case of the present application, two excisable cassettes were used. These two cassettes are the following: att1 # hyg + (SEQ ID N 91) and att3Qaac- (SEQ ID N 92), these cassettes leave respectively 33 and 35 nucleotides after excision. They include the Qhyg cassette or the Qaac cassette respectively, the + signs and corresponding to the orientation of the resistance cassette. These two cassettes were
<Desc/Clms Page number 121><Desc / Clms Page number 121>
construites et clonées au niveau du site EcoRV du vecteur pBC SK+ dont le site HindIII a été suprimé au préalable. Les plasmides obtenus ont été nommés patt1#hyg+ et patt3Qaac- respectivement. Les cassettes excisables peuvent être facilement sorties par une digestion du plasmide par EcoRV. constructed and cloned at the EcoRV site of the pBC SK + vector whose HindIII site was previously deleted. The resulting plasmids were named patt1 # hyg + and patt3Qaac- respectively. The excisable cassettes can easily be removed by digestion of the plasmid with EcoRV.
EXEMPLE 10: Construction d'une souche de Streptomyces ambofaciens interrompue dans le gène orf2
L'inactivation du gène orf2 a été réalisée grâce à la technique des cassettes excisables (cf. ci-dessus). La souche de départ utilisée est la souche Streptomyces ambofaciens OSC2 qui dérive de la souche ATCC23877. Cependant, la souche OSC2 diffère de la souche ATCC23877 en ce qu'elle a perdu l'élément génétique mobile pSAM2 (Boccard et al., 1989a et b). Cet élément mobile peut être perdu de manière spontanée au cours de la protoplastisation (action du lysozyme pour digérer la paroi bactérienne et fragmenter le mycélium (Kieser et al., 2000)) et de la régénération des protoplastes de la souche ATCC23877. Pour sélectionner les clones ayant perdu l'élément pSAM2, il a été mis en place un crible, basé sur la répression du gènepra par le répresseur de transcription KorSA (Sezonov et al., 1995) (Sezonov G. et al., 2000). EXAMPLE 10 Construction of a Strain of Streptomyces Ambofaciens Interrupted in the Orf2 Gene
Inactivation of the orf2 gene was performed using the excisable cassettes technique (see above). The starting strain used is the strain Streptomyces ambofaciens OSC2 which derives from strain ATCC23877. However, strain OSC2 differs from strain ATCC23877 in that it lost the mobile genetic element pSAM2 (Boccard et al., 1989a and b). This mobile element can be lost spontaneously during protoplastization (action of lysozyme to digest the bacterial wall and fragment mycelium (Kieser et al., 2000)) and regeneration of protoplasts of strain ATCC23877. To select the clones having lost the pSAM2 element, a screen was put in place, based on the repression of the genepra by the KorSA transcription repressor (Sezonov et al., 1995) (Sezonov G. et al., 2000) .
Pour cela, un fragment d'ADN contenant le promoteur du gène pra placé en amont du gène aph (conférant la résistance à la kanamycine et dépourvu de son propre promoteur) a été cloné dans le vecteur instable pWHM3Hyg, ce dernier dérivant du plasmide pWHM3 (Vara et al., 1989) dans lequel le gène tsr a été remplacé par le gène hyg (conférant la résistance à l'hygromycine). Le plasmide ainsi obtenu a été nommé pOSV510. La souche ATCC23877 a été transformée après protoplastisation par le plasmide pOSV510. Le promoteur Pra est un promoteur réprimé par le répresseur KorSA, le gène codant ce dernier étant situé au sein de l'élément mobile pSAM2 (Sezonov G. et al., 2000). Après transformation par le plasmide pOSV510, les bactéries transformées sont sélectionnées pour leur résistance à la kanamycine (due au gène aph porté par pOSV510). Les clones ayant perdu l'élément intégratif pSAM2 ont perdu le répresseur KorSA et le promoteur Pra n'est donc plus réprimé et permet l'expression du For this, a DNA fragment containing the promoter of the pra gene placed upstream of the aph gene (conferring resistance to kanamycin and devoid of its own promoter) was cloned into the unstable vector pWHM3Hyg, the latter derived from the plasmid pWHM3 ( Vara et al., 1989) in which the tsr gene has been replaced by the hyg gene (conferring resistance to hygromycin). The plasmid thus obtained was named pOSV510. The strain ATCC23877 was transformed after protoplastisation with the plasmid pOSV510. The Pra promoter is a promoter repressed by the KorSA repressor, the gene encoding the latter being located within the mobile element pSAM2 (Sezonov G. et al., 2000). After transformation with the plasmid pOSV510, the transformed bacteria are selected for their resistance to kanamycin (due to the aph gene carried by pOSV510). Clones having lost the integrative element pSAM2 have lost the KorSA repressor and the Pra promoter is no longer repressed and allows the expression of the
<Desc/Clms Page number 122><Desc / Clms Page number 122>
gène de résistance à la kanamycine aph. Une sélection par la kanamycine après transformation par le plasmide pOSV510 permet donc de sélectionner les clones ayant perdu l'élément intégratif pSAM2 (et donc KorSA) et possédant le plasmide pOSV510. kanamycin resistance gene aph. Selection by kanamycin after transformation with the plasmid pOSV510 thus makes it possible to select the clones having lost the integrative element pSAM2 (and thus KorSA) and possessing the plasmid pOSV510.
Le plasmide pOSV510 étant instable, après quelques cycles de sporulation sans antibiotique, des clones isolés sont repiqués sur milieu avec kanamycine, sur milieu avec hygromycine et sur milieu sans antibiotique. Les clones sensibles à la kanamycine et à l'hygromycine ont perdu pOSV510. La perte de l'élément pSAM2 a été vérifiée par hybridation et PCR. Un clone présentant les caractéristiques souhaitées a été sélectionné et a été nommé OSC2. Plasmid pOSV510 being unstable, after a few antibiotic-free sporulation cycles, isolated clones are subcultured on medium with kanamycin, on medium with hygromycin and on medium without antibiotic. Clones sensitive to kanamycin and hygromycin lost pOSV510. The loss of the pSAM2 element was verified by hybridization and PCR. A clone with the desired characteristics was selected and was named OSC2.
Un échantillon de la souche OSC2 a été déposé auprès de la Collection Nationale de Cultures de Microorganismes (CNCM) Institut Pasteur, 25, rue du Docteur Roux 75724 Paris Cedex 15, France, le 10 juillet 2002 sous le numéro d'enregistrement I- 2908. A sample of the OSC2 strain has been deposited with the National Collection of Microorganism Cultures (CNCM) Pasteur Institute, 25, rue du Docteur Roux 75724 Paris Cedex 15, France, July 10, 2002 under the registration number I-2908 .
L'inactivation du gène or/2 a été réalisée grâce à la technique des cassettes excisables (cf. ci-dessus et figure 10). Pour cela, un insert de 4,5 kb dont la séquence part du site EcoRI situé en position 1 jusqu'au site BamHI situé en position 4521 (SEQ ID N 1) a été sous-cloné au niveau des sites EcoRI et BamHI du plasmide pUC19 (numéro d'accès GenBank M77789) à partir du cosmide pSPM5. Le plasmide ainsi obtenu a été nommé pOS49.99. The inactivation of the or / 2 gene was carried out using the technique of excisable cassettes (see above and FIG. 10). For this, a 4.5 kb insert whose sequence starts from the EcoRI site located in position 1 to the BamHI site located at position 4521 (SEQ ID No. 1) was subcloned at the EcoRI and BamHI sites of the plasmid. pUC19 (GenBank access number M77789) from the cosmid pSPM5. The plasmid thus obtained was named pOS49.99.
Ce plasmide a été introduit dans la souche E. coli KS272 qui contenait déjà le plasmide pKOBEG (Chaveroche et al., 2000) (cf. figure 12). This plasmid was introduced into the E. coli strain KS272 which already contained the plasmid pKOBEG (Chaveroche et al., 2000) (see FIG. 12).
En parallèle, la cassette excisable att3Qaac- (SEQ ID N 92, cf. ci dessus) a été amplifiée par PCR en utilisant comme matrice le plasmide pOSK1102 (Le plasmide pOSK1102 est un plasmide dérivé du vecteur pGP704Not (Chaveroche et al., 2000) (Miller VL & Mekalanos JJ, 1988) dans lequel la cassette att3Qaac- a été clonée comme un fragment EcoRV dans le site unique EcoRV de pGP704Not) et à l'aide des amorces suivantes : In parallel, the excisable cassette att3Qaac- (SEQ ID No. 92, see above) was amplified by PCR using the plasmid pOSK1102 as template (The plasmid pOSK1102 is a plasmid derived from the vector pGP704Not (Chaveroche et al., 2000) (Miller VL & Mekalanos JJ, 1988) in which the att3Qaac- cassette was cloned as an EcoRV fragment into the unique EcoRV site of pGP704Not) and using the following primers:
<Desc/Clms Page number 123><Desc / Clms Page number 123>
ORF2A 5' CCCGCGCGGCAGCCTCTCCGTGATCGAGTCCGGCGTGACCATCGCGCGCGCTTCGTTCGG-3' (SEQ ID N 93) ORF2B 5' GCTCCGTGCGTCATGCAGGAAGGTGTCGTAGTCGCGGTAGATCTGCCTCTTCGTCCCGAA-3' (SEQ ID N 94)
Les 40 déoxy-nucléotides situés à l'extrémité 5' de ces oligonucléotides comportent une séquence correspondant à une séquence dans le gène cible (orj2 dans le cas présent) et les 20 déoxy-nucléotides situés le plus en 3' (figuré en gras ci-dessus) correspondent à la séquence d'une des extrémités de la cassette excisable att3Qaac- (cf. figure 11). ORF2A 5 'CCCGCGCGGCAGCCTCTCCGTGATCGAGTCCGGCGTGACCATCGCGCGCGCTTCGTTCGG-3' (SEQ ID N 93) ORF2B 5 'GCTCCGTGCGTCATGCAGGAAGGTGTCGTAGTCGCGGTAGATCTGCCTCTTCGTCCCGA-3' (SEQ ID N 94)
The 40 deoxy nucleotides located at the 5 'end of these oligonucleotides comprise a sequence corresponding to a sequence in the target gene (orj2 in the present case) and the 3' most deoxy-nucleotides located (shown in bold). above) correspond to the sequence of one end of the excisable cassette att3Qaac- (see Figure 11).
Le produit de PCR ainsi obtenu a été utilisé pour transformer la souche E. coli contenant les plasmides pKOBEG et pOS49. 99 comme décrit (Chaveroche et al., 2000) (cf. figure 12). Ainsi, les bactéries ont été transformées par électroporation et sélectionnées pour leur résistance à l'apramycine. Les plasmides des clones obtenus ont été extraits et digérés par plusieurs enzymes de restriction, dans le but de vérifier que le profil de digestion obtenu correspond au profil attendu s'il y a eu insertion de la cassette (att3Qaac-) dans le gène cible (or/2), c'est à dire si il y a bien eu recombinaison homologue entre les extrémités du produit PCR et le gène cible (Chaveroche et al., 2000). La vérification de la construction peut également être réalisée par toute méthode connue de l'homme du métier et notamment par PCR en utilisant les oligonucléotides appropriés et séquençage du produit de PCR. Un clone dont le plasmide possède le profil attendu a été sélectionné et le plasmide correspondant a été nommé pSPM17. Ce plasmide dérive de pOS49. 99 dans lequel orf2 est interrompue par la cassette apramycine (cf. figure 12). The PCR product thus obtained was used to transform the E. coli strain containing plasmids pKOBEG and pOS49. 99 as described (Chaveroche et al., 2000) (see Figure 12). Thus, the bacteria were transformed by electroporation and selected for their resistance to apramycin. The plasmids of the clones obtained were extracted and digested with several restriction enzymes, with the aim of verifying that the obtained digestion profile corresponds to the expected profile if there was insertion of the cassette (att3Qaac-) into the target gene ( or / 2), ie if there was indeed homologous recombination between the ends of the PCR product and the target gene (Chaveroche et al., 2000). Verification of the construction can also be carried out by any method known to those skilled in the art and in particular by PCR using the appropriate oligonucleotides and sequencing of the PCR product. A clone whose plasmid has the expected profile was selected and the corresponding plasmid was named pSPM17. This plasmid derives from pOS49. 99 in which orf2 is interrupted by the apramycin cassette (see Figure 12).
L'insertion de la cassette s'accompagne d'une délétion dans or/2, entre les nucléotides 211 et 492 de la partie codante de or/2. The insertion of the cassette is accompanied by a deletion in or2 between nucleotides 211 and 492 of the coding part of or2.
Le plasmide pSPM17 a été digéré par l'enzyme EcoRI puis les extrémités ont été rendues bout franc par traitement à l'enzyme de Klenow, ce produit de digestion a été ensuite digéré par l'enzyme XbaI et l'insert contenant le gène orf2 délété a été cloné dans le vecteur pOSK1205 (cf. ci dessus). Pour cela, le vecteur pOSK1205 a été digéré par l'enzyme BamHI puis les extrémités ont été rendues bout franc par traitement à The plasmid pSPM17 was digested with the enzyme EcoRI and then the ends were made blunt by treatment with Klenow enzyme, this digestion product was then digested with the enzyme XbaI and the insert containing the gene orf2 deleted. was cloned into the vector pOSK1205 (see above). For this purpose, the vector pOSK1205 was digested with the BamHI enzyme and then the ends were made blunt by treatment with
<Desc/Clms Page number 124><Desc / Clms Page number 124>
l'enzyme de Klenow, ce produit a été ensuite digéré par l'enzyme XbaI, et utilisé pour la ligation avec l'insert obtenu à partir de pSPM17 comme ci-dessus. Cette manipulation permet donc d'obtenir une ligation orientée puisque chacun des deux fragments est bout franc d'un côté et Xbal de l'autre. Le plasmide ainsi obtenu a été nommé pSPM21, il porte un gène de résistance à l'hygromycine (partie vecteur) et un insert dans lequel le gène orf2 délété est remplacé par la cassette att3Qaac-. the Klenow enzyme, this product was then digested with the enzyme XbaI, and used for ligation with the insert obtained from pSPM17 as above. This manipulation therefore makes it possible to obtain an oriented ligation since each of the two fragments is frank on one side and XbaI on the other. The plasmid thus obtained was named pSPM21, it carries a hygromycin resistance gene (vector part) and an insert in which the deleted orf2 gene is replaced by the att3Qaac- cassette.
Le vecteur pSPM21 a été introduit dans la souche Streptomyces ambofaciens OSC2 (cf. ci dessus) par transformation de protoplastes (Kieser, T et al., 2000). Après transformation, les clones sont sélectionnés pour leur résistance à l'apramycine Les clones résistants à l'apramycine sont ensuite repiqués respectivement sur milieu avec apramycine (antibiotique B) et sur milieu avec hygromycine (antibiotique A) (cf. figure 9). Les clones résistants à l'apramycine (ApraR) et sensibles à l'hygromycine (HygS) sont en principe ceux où un double événement de crossing over s'est produit et qui possèdent le gène or/2 interrompu par la cassette att3#aac-. Ces clones ont été sélectionnés et le remplacement de la copie sauvage de orf2 par la copie interrompue par la cassette a été vérifié. La présence de la cassette att3Qaac- a été vérifiée par PCR sur colonie. Une hybridation a également été effectuée. Pour cela, l'ADN total des clones obtenus, a été digéré par plusieurs enzymes, séparé sur gel d'agarose, transféré sur membrane et hybridé avec une sonde de 3 kb correspondant à un fragment EcoRl- BamHI de l'insert du plasmide pOS49.99 (cf. ci-dessus). La vérification du génotype peut également être réalisée par toute méthode connue de l'homme du métier et notamment par PCR en utilisant les oligonucléotides appropriés et séquençage du produit de PCR. The vector pSPM21 was introduced into the strain Streptomyces ambofaciens OSC2 (see above) by protoplast transformation (Kieser, T et al., 2000). After transformation, the clones are selected for their resistance to apramycin The apramycin-resistant clones are then subcultured respectively on medium with apramycin (antibiotic B) and on medium with hygromycin (antibiotic A) (see FIG. 9). The apramycin-resistant (ApraR) and hygromycin-sensitive (HygS) -resistant clones are in principle those where a double crossing-over event has occurred and which have the or / 2 gene interrupted by the att3 # aac- . These clones were selected and the replacement of the wild copy of orf2 by the copy interrupted by the cassette was verified. The presence of the att3Qaac- cassette was verified by colony PCR. Hybridization was also performed. For this, the total DNA of the clones obtained, was digested with several enzymes, separated on agarose gel, transferred to the membrane and hybridized with a 3 kb probe corresponding to an EcoRI-BamHI fragment of the insert of plasmid pOS49. .99 (see above). Verification of the genotype can also be carried out by any method known to those skilled in the art and in particular by PCR using the appropriate oligonucleotides and sequencing of the PCR product.
Un clone présentant les caractéristiques attendues a été sélectionné et nommé SPM21. Il a pu être vérifié grâce à la PCR et à l'hybridation que la cassette att3#aac- était bien présente dans le génome de ce clone et que l'on obtient bien le profil de digestion attendu dans le cas d'un remplacement, à la suite d'un double événement de recombinaison, du gène sauvage par la copie interrompue par la cassette att3Qaac- dans le génome de ce clone. Ce clone possède donc le génotype : orf2::att3#aac-. A clone with the expected characteristics was selected and named SPM21. It has been verified by PCR and hybridization that the att3 # aac- cassette was indeed present in the genome of this clone and that the expected digestion profile is obtained in the case of a replacement, as a result of a double recombination event, the wild-type gene by the copy interrupted by the att3Qaac- cassette in the genome of this clone. This clone therefore has the genotype: orf2 :: att3 # aac-.
<Desc/Clms Page number 125> <Desc / Clms Page number 125>
Un échantillon de la souche SPM21 a été déposé auprès de la Collection Nationale de Cultures de Microorganismes (CNCM) Institut Pasteur, 25, rue du Docteur Roux 75724 Paris Cedex 15, France, le 10 juillet 2002 sous le numéro d'enregistrement I- 2914. A sample of strain SPM21 has been deposited with the National Collection of Microorganism Cultures (CNCM) Pasteur Institute, 25, rue du Docteur Roux 75724 Paris Cedex 15, France, July 10, 2002 under the registration number I-2914 .
La souche SPM21 a été transformée par le vecteur pOSV508 par transformation de protoplastes pour provoquer l'excision de la cassette (cf. figure 14). Le plasmide pOSV508 est dérivé du plasmide pWHM3 (Vara J et al.,1989) (cf. figure 13) dans lequel il a été ajouté les gènes xis et int de pSAM2 (Boccard F. et al., 1989b) placé sous le contrôle du promoteur ptrc (Amann, E. et al., 1988) (cf. figure 14). L'introduction dans la souche SPM21 du plasmide pOSV508 portant les gènes xis et int de pSAM2 permet l'excision efficace par recombinaison spécifique de site de la cassette excisable entre les sites attL et attR flanquant cette cassette (Raynal A. et al., 1998) (figure 10). The strain SPM21 was transformed by the vector pOSV508 by transformation of protoplasts in order to excise the cassette (see FIG. 14). The plasmid pOSV508 is derived from the plasmid pWHM3 (Vara J et al., 1989) (see FIG. 13) in which the genes xis and int of pSAM2 (Boccard F. et al., 1989b) placed under control were added. ptrc promoter (Amann, E. et al., 1988) (see Figure 14). The introduction into the SPM21 strain of plasmid pOSV508 carrying the xis and int genes of pSAM2 allows the efficient excision by site-specific recombination of the excisable cassette between the attL and attR sites flanking this cassette (Raynal A. et al., 1998). ) (Figure 10).
Parmi les transformants sélectionnés pour leur résistance au thiostrepton due au gène tsr porté par pOSV508, on choisit ceux devenus sensibles à l'apramycine dont le gène de résistance est porté par la cassette att3#aac-, l'excision entraîne en effet la perte de ce gène de résistance (cf. figure 10). Le plasmide pOSV508 est instable et après deux passages successifs sur milieu sans antibiotique des clones isolés sont repiqués sur milieu avec thiostrepton et sur milieu sans thiostrepton. Les clones sensibles au thiostrepton ont perdu pOSV508. Il a été vérifié que l'excision de la cassette aboutit bien à une délétion en phase dans le gène orf2 par PCR et séquençage du produit PCR, l'excision de la cassette laisse en effet une séquence cicatricielle att3 caractéristique (qui est similaire au site attB d'origine après recombinaison entre les sites attL et attR) : 5' ATCGCGCGCGCTTCGTTCGGGACGAAGAGGTAGAT 3' (SEQ ID N 95). Among the transformants selected for their thiostrepton resistance due to the tsr gene carried by pOSV508, those which have become sensitive to apramycin whose resistance gene is carried by the att3 # aac- cassette are chosen, the excision in fact causes the loss of this resistance gene (see Figure 10). The plasmid pOSV508 is unstable and after two successive passages on medium without antibiotic isolated clones are subcultured on medium with thiostrepton and medium without thiostrepton. Thiostrepton-sensitive clones lost pOSV508. It has been verified that the excision of the cassette results in an in-phase deletion in the orf2 gene by PCR and sequencing of the PCR product, the excision of the cassette leaves indeed a characteristic scarring sequence att3 (which is similar to the site). original attB after recombination between attL and attR sites): 5 'ATCGCGCGCGCTTCGTTCGGGACGAAGAGGTAGAT 3' (SEQ ID NO: 95).
La souche ainsi obtenue et possédant le génotype voulu (orf2::att3) a été nommée SPM22. The strain thus obtained and having the desired genotype (orf2 :: att3) was named SPM22.
Un échantillon de la souche SPM22 a été déposé auprès de la Collection Nationale de Cultures de Microorganismes (CNCM) Institut Pasteur, 25, rue du Docteur Roux A sample of the strain SPM22 has been deposited with the National Collection of Cultures of Microorganisms (CNCM) Institut Pasteur, 25, rue du Docteur Roux
<Desc/Clms Page number 126><Desc / Clms Page number 126>
75724 Paris Cedex 15, France, le 10 juillet 2002 sous le numéro d'enregistrement I- 2915. 75724 Paris Cedex 15, France, July 10, 2002 under the registration number I-2915.
EXEMPLE 11: Construction d'une souche de Streptomyces ambofaciens interrompue dans le gène orfl2
Pour l'inactivation de or/12, orf13c et orfl4 un même plasmide de départ (pSPM504) a été employé pour introduire à différentes positions une cassette de type cassette excisable . Ce plasmide possède un insert de 15,1 kb qui correspond à la région de or/7 à orfl7. Pour construire ce plasmide un fragment BglII de 15,1 kb provenant de la digestion du cosmide pSPM7 (cf. ci-dessus) a été cloné dans le plasmide pMBL18 (Nakano et al., 1995) digéré par BamHI. Les extrémités BamHI et BglII étant compatibles, on obtient après ligation, le plasmide pSPM502. La totalité de l'insert de pSPM502 a ensuite été sous-cloné (sous forme d'un fragment HindIII/NheI) dans le plasmide pOSK1205 (digéré par HindIII/NheI) ce qui a permis d'obtenir le plasmide pSPM504. EXAMPLE 11 Construction of a Strain of Streptomyces Ambofaciens Interrupted in the Orfl2 Gene
For the inactivation of gold / 12, orf13c and orf1, the same starting plasmid (pSPM504) was used to introduce at different positions an excisable cassette type cassette. This plasmid has a 15.1 kb insert that corresponds to the region from or / 7 to orf17. To construct this plasmid a 15.1 kb BglII fragment from the digestion of cosmid pSPM7 (see above) was cloned into the plasmid pMBL18 (Nakano et al., 1995) digested with BamHI. The BamHI and BglII ends being compatible, the plasmid pSPM502 is obtained after ligation. The entire pSPM502 insert was then subcloned (as a HindIII / NheI fragment) into the plasmid pOSK1205 (digested with HindIII / NheI), which made it possible to obtain the plasmid pSPM504.
Ce plasmide a été introduit dans la souche E. coli KS272 qui contenait déjà le plasmide pKOBEG (Chaveroche et al., 2000) (cf. figure 12). This plasmid was introduced into the E. coli strain KS272 which already contained the plasmid pKOBEG (Chaveroche et al., 2000) (see FIG. 12).
En parallèle, la cassette excisable att3Qaac- a été amplifiée par PCR en utilisant comme matrice le plasmide pOSK1102 (le plasmide pOSKl 102 est un plasmide dérivé du vecteur pGP704Not (Chaveroche et al., 2000) (Miller VL & Mekalanos JJ, 1988) dans lequel la cassette att3#aac- a été clonée comme un fragment EcoRV dans le site unique EcoRV de pGP704Not), les amorces utilisées sont les suivantes : EDR8:5' CGGGATGATCGCTTGTCCGGCGGCCGGATGCCTAGCCTCATCGCGCGCGCTTCGTTCGG 3' (SEQ ID N 96) In parallel, the excisable cassette att3Qaac- was amplified by PCR using the plasmid pOSK1102 as a template (the plasmid pOSK1 102 is a plasmid derived from the pGP704Not vector (Chaveroche et al., 2000) (Miller VL & Mekalanos JJ, 1988) in wherein the att3 # aac- cassette was cloned as an EcoRV fragment into the unique EcoRV site of pGP704Not), the primers used are as follows: EDR8: 5 'CGGGATGATCGCTTGTCCGGCGGCCGGATGCCTAGCCTCATCGCGCGCGCTTCGTTCGG 3' (SEQ ID N 96)
<Desc/Clms Page number 127><Desc / Clms Page number 127>
EDR9:5' CCCGATCCAGAACGTCTGGTCGGTGATCAGGTCGCTGTTCATCTGCCTCTTCGTCCCGAA 3' (SEQ ID N 97)
Les 40 (seulement 39 pour EDR8) déoxy-nucléotides situés à l'extrémité 5' de ces oligonucléotides comportent une séquence correspondant à une séquence dans le gène cible (or/72 dans le cas présent) et les 20 déoxy-nucléotides situés le plus en 3' (figuré en gras ci-dessus) correspondent à la séquence d'une des extrémités de la cassette excisable att3Qaac- (cf. figure 11). EDR9: 5 'CCCGATCCAGAACGTCTGGTCGGTGATCAGGTCGCTGTTCATCTGCCTCTTCGTCCCGAA 3' (SEQ ID N 97)
The 40 (only 39 for EDR8) deoxy nucleotides located at the 5 'end of these oligonucleotides have a sequence corresponding to a sequence in the target gene (or / 72 in this case) and the deoxynucleotides located the most in 3 '(shown in bold above) correspond to the sequence of one end of the excisable cassette att3Qaac- (see Figure 11).
Le produit de PCR ainsi obtenu a été utilisé pour transformer la souche E. coli KS272 contenant les plasmides pKOBEG et pSPM504 (cf ci-dessus), comme décrit par Chaveroche et al. (Chaveroche et al., 2000) (cf. figure 12 pour le principe, le plasmide pOS49. 99 doit être remplacé par le plasmide pSPM504 et le plasmide obtenu n'est plus pSPM17 mais pSPM507). Ainsi, les bactéries ont été transformées par électroporation par ce produit de PCR et les clones ont été sélectionnés pour leur résistance à l'apramycine. Les plasmides des clones obtenus ont été extraits et digérés par plusieurs enzymes de restriction, dans le but de vérifier que le profil de digestion obtenu correspond au profil attendu s'il y a eu insertion de la cassette (att3Qaac-) dans le gène cible (orf12), c'est à dire si il y a bien eu recombinaison homologue entre les extrémités du produit PCR et le gène cible (Chaveroche et al., 2000). La vérification de la construction peut également être réalisée par toute méthode connue de l'homme du métier et notamment par PCR en utilisant les oligonucléotides appropriés et séquençage du produit de PCR. Un clone dont le plasmide possède le profil attendu a été sélectionné et le plasmide correspondant a été nommé pSPM507. Ce plasmide dérive de pSPM504 dans lequel or/12 est interrompue par la cassette att3Qaac- (cf. figure 12). L'insertion de la cassette s'accompagne d'une délétion dans le gène orfl2, l'interruption commence au niveau du trentième codon de orfl2. Il reste après la cassette les 46 derniers codons de or/12. The PCR product thus obtained was used to transform the E. coli KS272 strain containing the plasmids pKOBEG and pSPM504 (cf above), as described by Chaveroche et al. (Chaveroche et al., 2000) (see Figure 12 for the principle, the plasmid pOS49.99 must be replaced by the plasmid pSPM504 and the plasmid obtained is no longer pSPM17 but pSPM507). Thus, the bacteria were transformed by electroporation with this PCR product and the clones were selected for their resistance to apramycin. The plasmids of the clones obtained were extracted and digested with several restriction enzymes, with the aim of verifying that the obtained digestion profile corresponds to the expected profile if there was insertion of the cassette (att3Qaac-) into the target gene ( orf12), ie if there was indeed homologous recombination between the ends of the PCR product and the target gene (Chaveroche et al., 2000). Verification of the construction can also be carried out by any method known to those skilled in the art and in particular by PCR using the appropriate oligonucleotides and sequencing of the PCR product. A clone whose plasmid has the expected profile was selected and the corresponding plasmid was named pSPM507. This plasmid derives from pSPM504 in which /12 is interrupted by the att3Qaac- cassette (see FIG. 12). The insertion of the cassette is accompanied by a deletion in the orfl2 gene, the interruption begins at the thirtieth codon of orfl2. There remains after the cassette the last 46 codons of gold / 12.
Le vecteur pSPM507 a été introduit dans la souche Streptomyces ambofaciens OSC2 (cf. ci-dessus) par transformation de protoplastes (Kieser, T et al., 2000). Après The vector pSPM507 was introduced into the strain Streptomyces ambofaciens OSC2 (see above) by protoplast transformation (Kieser, T et al., 2000). After
<Desc/Clms Page number 128><Desc / Clms Page number 128>
transformation, les clones sont sélectionnés pour leur résistance à l'apramycine. Les clones résistants à l'apramycine sont ensuite repiqués respectivement sur milieu avec apramycine (antibiotique B) et sur milieu avec hygromycine (antibiotique A) (cf. figure 9). Les clones résistants à l'apramycine (ApraR) et sensibles à l'hygromycine (HygS) sont en principe ceux où un double événement de crossing over s'est produit et qui possèdent le gène orfl2 interrompu par la cassette att3Qaac-. Ces clones ont été plus particulièrement sélectionnés et le remplacement de la copie sauvage de orfl2 par la copie interrompue par la cassette a été vérifié par hybridation. Ainsi, l'ADN total des clones obtenus, a été digéré par plusieurs enzymes, séparé sur gel d'agarose, transféré sur membrane et hybridé avec une sonde correspondant à la cassette att3Qaac- pour vérifier la présence de la cassette dans l'ADN génomique des clones obtenus. Une deuxième hybridation a été effectuée en utilisant comme sonde un fragment d'ADN obtenu par PCR et correspondant à une très large partie de la séquence codante du gène orf12. transformation, the clones are selected for their resistance to apramycin. The apramycin-resistant clones are then subcultured respectively on medium with apramycin (antibiotic B) and on medium with hygromycin (antibiotic A) (see FIG. 9). The apramycin-resistant (ApraR) and hygromycin-sensitive (HygS) -resistant clones are in principle those where a double crossing-over event has occurred and which have the orfl2 gene interrupted by the att3Qaac- cassette. These clones were more particularly selected and the replacement of the wild copy of orfl2 by the copy interrupted by the cassette was verified by hybridization. Thus, the total DNA of the clones obtained, was digested with several enzymes, separated on agarose gel, transferred to the membrane and hybridized with a probe corresponding to the cassette att3Qaac- to check the presence of the cassette in the genomic DNA. clones obtained. A second hybridization was carried out using as probe a DNA fragment obtained by PCR and corresponding to a very large part of the coding sequence of the orf12 gene.
La vérification du génotype peut également être réalisée par toute méthode connue de l'homme du métier et notamment par PCR en utilisant les oligonucléotides appropriés et séquençage du produit de PCR.
![]()
![]()
Un clone présentant les caractéristiques attendues (orfl2::att3SZaac-) a été plus particulièrement sélectionné et nommé SPM507. Il a pu en effet être vérifié grâce aux deux hybridations que la cassette att3#aac- était bien présente dans le génome de ce clone et que l'on obtient bien le profil de digestion attendu dans le cas d'un remplacement, à la suite d'un double événement de recombinaison, du gène sauvage par la copie interrompue par la cassette att3Qaac- dans le génome de ce clone. Ce clone
![]()
possède donc le génotype : orfl2::att3s2aac- et a été nommé SPM507. Il est inutile de procéder à l'excision de la cassette pour l'étude de l'effet de l'inactivation de orfl2, au vu de l'orientation des gènes (cf. figure 3). En effet, le fait que orf13c soit orienté en sens opposé à orfl2 montre que ces gènes ne sont pas co-transcrits. L'utilisation d'une cassette excisable permet en revanche d'avoir la possibilité de se débarrasser du marqueur de sélection à tout moment, notamment par transformation par le plasmide pOSV508. Un échantillon de la souche SPM507 a été déposé auprès de la Collection A clone with the expected characteristics (orfl2 :: att3SZaac-) was more particularly selected and named SPM507. It could indeed be verified by means of the two hybridizations that the att3 # aac- cassette was indeed present in the genome of this clone and that the expected digestion profile is obtained in the case of a replacement, following a double recombination event, the wild-type gene by the copy interrupted by the att3Qaac- cassette in the genome of this clone. This clone
![]()
therefore has the genotype: orfl2 :: att3s2aac- and has been named SPM507. There is no need to excise the cassette for the study of the effect of the inactivation of orfl2, given the orientation of the genes (see Figure 3). Indeed, the fact that orf13c is oriented in the opposite direction to orfl2 shows that these genes are not co-transcribed. The use of an excisable cassette makes it possible, on the other hand, to be able to get rid of the selection marker at any time, in particular by transformation with the plasmid pOSV508. A sample of strain SPM507 has been deposited with the Collection
<Desc/Clms Page number 129><Desc / Clms Page number 129>
Nationale de Cultures de Microorganismes (CNCM) Institut Pasteur, 25, rue du Docteur Roux 75724 Paris Cedex 15, France, le 10 juillet 2002 sous le numéro d'enregistrement 1-2911. National Microorganism Cultures (CNCM) Pasteur Institute, 25, rue du Docteur Roux 75724 Paris Cedex 15, France, July 10, 2002 under the registration number 1-2911.
EXEMPLE 12: Construction d'une souche de Streptomyces ambofaciens interrompue dans le gène orfl3c
La cassette excisable att3Qaac- a été amplifiée par PCR en utilisant comme matrice le plasmide pOSK1102 (cf. ci-dessus) à l'aide des amorces suivantes : EDR3:5' ACCGGGGCGGTCCTCCCCTCCGGGGCGTCACGGCCGCGGAATCTGCCTCTTCGTCCCGAA 3' (SEQ ID N 98) EDR4:5' CACGCAGCGAGCCGACGCACTGATGGACGACACGATGGCCATCGCGCGCGCTTCGTTCGG 3' (SEQ ID N 99)
Les 40 déoxy-nucléotides situés à l'extrémité 5' de ces oligonucléotides comportent une séquence correspondant à une séquence dans le gène cible (orf13c dans le cas présent) et les 20 déoxy-nucléotides situés le plus en 3' (figuré en gras ci-dessus) correspondent à la séquence d'une des extrémités de la cassette excisable att3Qaac- (cf figure 11). EXAMPLE 12 Construction of a Strain of Streptomyces Ambofaciens Interrupted in the Orfl3c Gene
The excisable cassette att3Qaac- was amplified by PCR using as template the plasmid pOSK1102 (see above) using the following primers: EDR3: 5 'ACCGGGGCGGTCCTCCCCTCCGGGGCGTCACGGCCGCGGAATCTGCCTCTTCGTCCCGAA 3' (SEQ ID N 98) EDR4: 5 'CACGCAGCGAGCCGACGCACTGATGGACGACACGATGGCCATCGCGCGCGCTTCGTTCGG 3 '(SEQ ID N 99)
The 40 deoxy nucleotides located at the 5 'end of these oligonucleotides comprise a sequence corresponding to a sequence in the target gene (orf13c in the present case) and the 3' most deoxy-nucleotides (shown in bold). above) correspond to the sequence of one end of the att3Qaac- excisable cassette (see FIG. 11).
Le produit de PCR ainsi obtenu a été utilisé pour transformer la souche E. coli KS272 contenant les plasmides pKOBEG et pSPM504 (cf. ci-dessus), comme décrit par Chaveroche et al. (Chaveroche et al., 2000) (cf. figure 12 pour le principe, le plasmide pOS49. 99 doit être remplacé par le plasmide pSPM504 et le plasmide obtenu n'est plus pSPM17 mais pSPM508). Ainsi, les bactéries ont été transformées par électroporation par le produit de PCR et les clones ont été sélectionnés pour leur résistance à l'apramycine. Les plasmides des clones obtenus ont été extraits et digérés par plusieurs enzymes de restriction, dans le but de vérifier que le profil de digestion obtenu correspond au profil attendu s'il y a eu insertion de la cassette (att3Qaac-) dans le gène The PCR product thus obtained was used to transform E. coli strain KS272 containing plasmids pKOBEG and pSPM504 (see above), as described by Chaveroche et al. (Chaveroche et al., 2000) (see Figure 12 for the principle, the plasmid pOS49.99 must be replaced by the plasmid pSPM504 and the plasmid obtained is no longer pSPM17 but pSPM508). Thus, the bacteria were transformed by electroporation with the PCR product and the clones were selected for their resistance to apramycin. The plasmids of the clones obtained were extracted and digested with several restriction enzymes, in order to verify that the digestion profile obtained corresponds to the expected profile if there was insertion of the cassette (att3Qaac-) into the gene.
<Desc/Clms Page number 130><Desc / Clms Page number 130>
cible (orfl3c), c'est à dire si il y a bien eu recombinaison homologue entre les extrémités du produit PCR et le gène cible (Chaveroche et al., 2000). La vérification de la construction peut également être réalisée par toute méthode connue de l'homme du métier et notamment par PCR en utilisant les oligonucléotides appropriés et séquençage du produit de PCR. Un clone dont le plasmide possède le profil attendu a été sélectionné et le plasmide correspondant a été nommé pSPM508. Ce plasmide dérive de pSPM504 dans lequel orfl3c est interrompue par la cassette apramycine (cf. figure 12). L'insertion de la cassette s'accompagne d'une délétion dans le gène orfl3c, l'interruption commence au niveau du sixième codon de orfl3c. Il reste après la cassette les 3 derniers codons de orfl3c. target (orf13c), that is to say whether there has been homologous recombination between the ends of the PCR product and the target gene (Chaveroche et al., 2000). Verification of the construction can also be carried out by any method known to those skilled in the art and in particular by PCR using the appropriate oligonucleotides and sequencing of the PCR product. A clone whose plasmid has the expected profile was selected and the corresponding plasmid was named pSPM508. This plasmid derives from pSPM504 in which orf13c is interrupted by the apramycin cassette (see FIG. The insertion of the cassette is accompanied by a deletion in the orfl3c gene, the interruption begins at the sixth codon of orfl3c. There remains after the cassette the last 3 codons of orfl3c.
Le vecteur pSPM508 a été introduit dans la souche Streptomyces ambofaciens OSC2 (cf. ci-dessus) par transformation de protoplastes (Kieser, T et al., 2000). Après transformation, les clones sont sélectionnés pour leur résistance à l'apramycine. Les clones résistants à l'apramycine sont ensuite repiqués respectivement sur milieu avec apramycine (antibiotique B) et sur milieu avec hygromycine (antibiotique A) (cf. figure 9). Les clones résistants à l'apramycine (ApraR) et sensibles à l'hygromycine (HygS) sont en principe ceux où un double événement de crossing over s'est produit et qui possèdent le gène orfl3c interrompu par la cassette att3Qaac-. Ces clones ont été plus particulièrement sélectionnés et le remplacement de la copie sauvage de orfl3c par la copie interrompue par la cassette a été vérifié par hybridation. Ainsi, l'ADN total des clones obtenus, a été digéré par plusieurs enzymes, séparé sur gel d'agarose, transféré sur membrane et hybridé avec une sonde correspondant à la cassette att3Qaac- pour vérifier la présence de la cassette dans l'ADN génomique des clones obtenus. Une deuxième hybridation a été effectuée en utilisant comme sonde un produit de PCR correspondant à une séquence débordant d'une centaine de paires de base en amont et en aval de la séquence codante de orfl3c. La vérification du génotype peut également être réalisée par toute méthode connue de l'homme du métier et notamment par PCR en utilisant les oligonucléotides appropriés et séquençage du produit de PCR. The vector pSPM508 was introduced into the strain Streptomyces ambofaciens OSC2 (see above) by protoplast transformation (Kieser, T et al., 2000). After transformation, the clones are selected for their resistance to apramycin. The apramycin-resistant clones are then subcultured respectively on medium with apramycin (antibiotic B) and on medium with hygromycin (antibiotic A) (see FIG. 9). The apramycin-resistant (ApraR) and hygromycin-sensitive (HygS) -resistant clones are in principle those where a double crossing-over event has occurred and which have the orfl3c gene interrupted by the att3Qaac- cassette. These clones were more particularly selected and the replacement of the wild copy of orfl3c by the copy interrupted by the cassette was verified by hybridization. Thus, the total DNA of the clones obtained, was digested with several enzymes, separated on agarose gel, transferred to the membrane and hybridized with a probe corresponding to the cassette att3Qaac- to check the presence of the cassette in the genomic DNA. clones obtained. A second hybridization was carried out using as probe a PCR product corresponding to a sequence overflowing a hundred base pairs upstream and downstream of the coding sequence of orf13c. Verification of the genotype can also be carried out by any method known to those skilled in the art and in particular by PCR using the appropriate oligonucleotides and sequencing of the PCR product.
<Desc/Clms Page number 131> <Desc / Clms Page number 131>
![]()
Un clone présentant les caractéristiques attendues (orfl3c::att3Qaac-) a été plus particulièrement sélectionné et nommé SPM508. Il a pu en effet être vérifié grâce aux deux hybridations que la cassette att3Qaac- était bien présente dans le génome de ce clone et que l'on obtient bien le profil de digestion attendu dans le cas d'un remplacement, à la suite d'un double événement de recombinaison, du gène sauvage par la copie interrompue par la cassette att3Qaac- dans le génome de ce clone. Ce clone
![]()
possède donc le génotype : orfl3c::att3P,.aac- et a été nommé SPM508. Il est inutile de procéder à l'excision de la cassette pour l'étude de l'effet de l'inactivation de orfl3c, au vu de l'orientation des gènes (cf. figure 3), le fait que orfl4 soit orienté en sens opposé à orfl3c montre que ces gènes ne sont pas co-transcrits. L'utilisation d'une cassette excisable permet en revanche d'avoir la possibilité de se débarrasser du marqueur de sélection à tout moment. Un échantillon de la souche SPM508 a été déposé auprès de la Collection Nationale de Cultures de Microorganismes (CNCM) Institut Pasteur, 25, rue du Docteur Roux 75724 Paris Cedex 15, France, le 10 juillet 2002 sous le numéro d'enregistrement 1-2912. ![]()
A clone with the expected characteristics (orfl3c :: att3Qaac-) was more particularly selected and named SPM508. It has indeed been possible to verify, thanks to the two hybridizations, that the att3Qaac- cassette was indeed present in the genome of this clone and that the expected digestion profile is obtained in the case of a replacement, as a result of a double recombination event of the wild-type gene by the copy interrupted by the att3Qaac- cassette in the genome of this clone. This clone
![]()
therefore has the genotype: orfl3c :: att3P, .aac- and has been named SPM508. There is no need to excise the cassette to study the effect of the inactivation of orfl3c, in view of the orientation of the genes (see Figure 3), the fact that orfl4 is oriented in the direction of opposite to orfl3c shows that these genes are not co-transcribed. Using an excisable cassette, however, allows you to get rid of the selection marker at any time. A sample of the SPM508 strain has been deposited with the National Collection of Microorganism Cultures (CNCM) Pasteur Institute, 25, rue du Docteur Roux 75724 Paris Cedex 15, France, July 10, 2002 under the registration number 1-2912 .
EXEMPLE 13 Construction d'une souche de Streptomyces ambofaciens interrompue dans le gène orfl4
La cassette excisable att3Qaac- a été amplifiée par PCR en utilisant comme matrice le plasmide pOSK1102 (cf. ci-dessus) à l'aide des amorces suivantes : EDR5: 5'GGGCGTGAAGCGGGCGAGTGTGGATGTCATGCGAGTACTCATCGCGCGCGCTTCGTTCGG 3' (SEQ ID N 100) EDR6: 5'CGGGAAACGGCGTCGCACTCCTCGGGGGCCGCGTCAGCCCATCTGCCTCTTCGTCCCGAA 3' (SEQ ID N 101) EXAMPLE 13 Construction of a Strain of Streptomyces Ambofaciens Interrupted in the Orfl4 Gene
The excisable cassette att3Qaac- was amplified by PCR using as template the plasmid pOSK1102 (see above) using the following primers: EDR5: 5'GGGCGTGAAGCGGGCGAGTGTGGATGTCATGCGAGTACTCATCGCGCGCGCTTCGTTCGG 3 '(SEQ ID NO 100) EDR6: 5'CGGGAAACGGCGTCGCACTCCTCGGGGGCCGCGTCAGCCCATCTGCCTCTTCGTCCCGAA 3 '(SEQ ID N 101)
<Desc/Clms Page number 132> <Desc / Clms Page number 132>
Les 40 déoxy-nucléotides situés à l'extrémité 5' de ces oligonucléotides comportent une séquence correspondant à une séquence dans le gène cible (or/74 dans le cas présent) et les 20 déoxy-nucléotides situés le plus en 3' (figuré en gras ci-dessus) correspondent à la séquence d'une des extrémités de la cassette excisable att3Qaac- (cf. figure 11). The 40 deoxy nucleotides at the 5 'end of these oligonucleotides have a sequence corresponding to a sequence in the target gene (gold / 74 in this case) and the 3' most deoxy nucleotides (shown in FIG. bold above) correspond to the sequence of one end of the att3Qaac- excisable cassette (see Figure 11).
Le produit de PCR ainsi obtenu a été utilisé pour transformer la souche E. coli KS272 contenant les plasmides pKOBEG et pSPM504 (cf. ci-dessus), comme décrit par Chaveroche et al. (Chaveroche et al., 2000) (cf. figure 12 pour le principe, le plasmide pOS49. 99 doit être remplacé par le plasmide pSPM504 et le plasmide obtenu n'est plus pSPM17 mais pSPM509). Ainsi, les bactéries ont été transformées par électroporation par le produit de PCR et les clones ont été sélectionnés pour leur résistance à l'apramycine. Les plasmides des clones obtenus ont été extraits et digérés par plusieurs enzymes de restriction, dans le but de vérifier que le profil de digestion obtenu correspond au profil attendu s'il y a eu insertion de la cassette (att3Qaac-) dans le gène cible (orfl4), c'est à dire si il y a bien eu recombinaison homologue entre les extrémités du produit PCR et le gène cible (Chaveroche et al., 2000). La vérification de la construction peut également être réalisée par toute méthode connue de l'homme du métier et notamment par PCR en utilisant les oligonucléotides appropriés et séquençage du produit de PCR. Un clone dont le plasmide possède le profil attendu a été sélectionné et le plasmide correspondant a été nommé pSPM509. Ce plasmide dérive de pSPM504 dans lequel orfl4 est interrompu par la cassette apramycine (cf. figure 12). L'insertion de la cassette s'accompagne d'une délétion dans le gène orfl4, l'interruption commence au niveau du quatrième codon de orfl4. Il reste après la cassette le dernier codon de orfl4. The PCR product thus obtained was used to transform E. coli strain KS272 containing plasmids pKOBEG and pSPM504 (see above), as described by Chaveroche et al. (Chaveroche et al., 2000) (see Figure 12 for the principle, the plasmid pOS49.99 must be replaced by the plasmid pSPM504 and the plasmid obtained is no longer pSPM17 but pSPM509). Thus, the bacteria were transformed by electroporation with the PCR product and the clones were selected for their resistance to apramycin. The plasmids of the clones obtained were extracted and digested with several restriction enzymes, with the aim of verifying that the obtained digestion profile corresponds to the expected profile if there was insertion of the cassette (att3Qaac-) into the target gene ( orfl4), ie if there was indeed homologous recombination between the ends of the PCR product and the target gene (Chaveroche et al., 2000). Verification of the construction can also be carried out by any method known to those skilled in the art and in particular by PCR using the appropriate oligonucleotides and sequencing of the PCR product. A clone whose plasmid has the expected profile was selected and the corresponding plasmid was named pSPM509. This plasmid derives from pSPM504 in which orf14 is interrupted by the apramycin cassette (see FIG. The insertion of the cassette is accompanied by a deletion in the orfl4 gene, the interruption begins at the fourth codon of orfl4. There remains after the cassette the last codon of orfl4.
Le vecteur pSPM509 a été introduit dans la souche Streptomyces ambofaciens OSC2 (cf. ci-dessus) par transformation de protoplastes (Kieser, T et al., 2000). Après transformation, les clones sont sélectionnés pour leur résistance à l'apramycine. Les clones résistants à l'apramycine sont ensuite repiqués respectivement sur milieu avec apramycine (antibiotique B) et sur milieu avec hygromycine (antibiotique A) (cf. figure 9). Les clones résistants à l'apramycine (ApraR) et sensibles à l'hygromycine (HygS) The vector pSPM509 was introduced into the strain Streptomyces ambofaciens OSC2 (see above) by protoplast transformation (Kieser, T et al., 2000). After transformation, the clones are selected for their resistance to apramycin. The apramycin-resistant clones are then subcultured respectively on medium with apramycin (antibiotic B) and on medium with hygromycin (antibiotic A) (see FIG. 9). Apramycin-resistant clones (ApraR) and hygromycin-sensitive clones (HygS)
<Desc/Clms Page number 133><Desc / Clms Page number 133>
sont en principe ceux où un double événement de crossing over s'est produit et qui possèdent le gène orfl4 interrompu par la cassette att3Qaac-. Ces clones ont été plus particulièrement sélectionnés et le remplacement de la copie sauvage de orfl4 par la copie interrompue par la cassette a été vérifié par hybridation. Ainsi, l'ADN total des clones obtenus, a été digéré par plusieurs enzymes, séparé sur gel d'agarose, transféré sur membrane et hybridé avec une sonde correspondant à la cassette att3#aac- pour vérifier la présence de la cassette dans l'ADN génomique des clones obtenus. Une deuxième hybridation a été effectuée en utilisant comme sonde un produit de PCR correspondant à une séquence débordant d'une centaine de paires de base en amont et en aval de la séquence codante de orfl4. La vérification du génotype peut également être réalisée par toute méthode connue de l'homme du métier et notamment par PCR en utilisant les oligonucléotides appropriés et séquençage du produit de PCR.
![]()
![]()
Un clone présentant les caractéristiques attendues (orfl4.: att3d2aac-) a été plus particulièrement sélectionné et nommé SPM509. Il a pu en effet être vérifié grâce aux deux hybridations que la cassette att3Qaac- était bien présente dans le génome de ce clone et que l'on obtient bien le profil de digestion attendu dans le cas d'un remplacement, à la suite d'un double événement de recombinaison, du gène sauvage par la copie interrompue par la cassette att3Qaac- dans le génome de ce clone. Ce clone possède donc le génotype : orf14::att3#aac- et a été nommé SPM509. Il est inutile de procéder à l'excision de la cassette pour l'étude de l'effet de l'inactivation de orfl4, au vu de l'orientation des gènes (cf. figure 3), le fait que orf15c soit orienté en sens opposé à orfl4 montre que ces gènes ne sont pas co-transcrits. L'utilisation d'une cassette excisable permet en revanche d'avoir la possibilité de se débarrasser du marqueur de sélection à tout moment. Un échantillon de la souche SPM509 a été déposé auprès de la Collection Nationale de Cultures de Microorganismes (CNCM) Institut Pasteur, 25, rue du Docteur Roux 75724 Paris Cedex 15, France, le 10 juillet 2002 sous le numéro d'enregistrement 1-2913. A clone with the expected characteristics (orfl4 .: att3d2aac-) was more particularly selected and named SPM509. It has indeed been possible to verify, thanks to the two hybridizations, that the att3Qaac- cassette was indeed present in the genome of this clone and that the expected digestion profile is obtained in the case of a replacement, as a result of a double recombination event of the wild-type gene by the copy interrupted by the att3Qaac- cassette in the genome of this clone. This clone therefore has the genotype: orf14 :: att3 # aac- and has been named SPM509. There is no need to excise the cassette to study the effect of the inactivation of orfl4, given the orientation of the genes (see Figure 3), the fact that orf15c is oriented in the direction of opposite to orfl4 shows that these genes are not co-transcribed. Using an excisable cassette, however, allows you to get rid of the selection marker at any time. A sample of the SPM509 strain has been deposited with the National Collection of Microorganism Cultures (CNCM) Pasteur Institute, 25, rue du Docteur Roux 75724 Paris Cedex 15, France, July 10, 2002 under the registration number 1-2913 .
<Desc/Clms Page number 134><Desc / Clms Page number 134>
EXEMPLE 14: Construction d'une souche de Streptomyces ambofaciens interrompue dans le gène orf6*
L'inactivation du gène orf6* a été réalisée grâce à la technique des cassettes excisables (cf. ci-dessus et figure 10). Pour cela, le cosmide pSPM7 a été utilisé comme matrice pour amplifier un fragment du gène orf6* grâce aux oligonucléotides suivants : C9583 : 5' CTGCAGGTGCTCCAGCGCGTCGATCT 3' (oligo sens) (SEQ ID N 102) C9584 : 5' CTGCAGACGGAGGCGGACCTGCGGCT 3' (oligo antisens) (SEQ ID N 103)
Les 20 déoxy-nucléotides situés à l'extrémité 3' de ces oligonucléotides correspondent à une séquence située dans la partie codante du gène orf6* (SEQ ID N 13) et les 6 déoxy-nucléotides situés les plus en 5' correspondent à la séquence d'un site PstI permettant de faciliter le clonage par la suite. Le fragment d'ADN amplifié fait une taille d'environ 1,11 kb. Ce produit de PCR est cloné dans le vecteur pGEM-T Easy (commercialisé par la société Promega (Madison, Wisconcin, USA)) ce qui a permis d'obtenir le plasmide nommé pBXLl 111 (cf. figure 16). EXAMPLE 14 Construction of a Strain of Streptomyces Ambofaciens Interrupted in the Orf6 Gene
The inactivation of the orf6 * gene was performed using the excisable cassettes technique (see above and FIG. 10). For this, the cosmid pSPM7 was used as a template to amplify a fragment of the orf6 * gene by virtue of the following oligonucleotides: C9583: 5 'CTGCAGGTGCTCCAGCGCGTCGATCT 3' (trace oligo) (SEQ ID No. 102) C9584: 5 'CTGCAGACGGAGGCGGACCTGCGGCT 3' (oligo antisense) (SEQ ID NO: 103)
The deoxy nucleotides located at the 3 'end of these oligonucleotides correspond to a sequence located in the coding part of the orf6 * gene (SEQ ID No. 13) and the 6 most deoxy-nucleotides located in 5' correspond to the sequence a PstI site to facilitate cloning later. The amplified DNA fragment is about 1.11 kb in size. This PCR product is cloned into the vector pGEM-T Easy (marketed by the company Promega (Madison, Wisconcin, USA)), which made it possible to obtain the plasmid named pBXL 111 (see FIG. 16).
La cassette excisable att1#hyg+ a ensuite été introduite dans la séquence codante du gène orf6*. Pour cela, le plasmide pBXL1111 a été digéré par les enzymes de restriction SmaI et Asp718I et le produit de digestion a été traité par l'enzyme de Klenow. Cette manipulation permet de réaliser une délétion interne de 120 pb dans la séquence codante du gène orf6* (cf. figure 15). De plus, de part et d'autre des sites de restrictions, il reste respectivement 511 pb et 485 pb de la séquence de orf6* qui permettront la recombinaison homologue pour l'inactivation du gène orf6*. La cassette
![]()
excisable attl2hyg+ a été préparée à partir du plasmide pattlS2hyg+ (cf. ci-dessus) par digestion de ce plasmide par EcoRV. Cette dernière a ensuite été sous-clonée dans le vecteur pBXLl 111 préalablement préparé comme décrit ci-dessus (digestion SmaI et Asp718I puis traitement à l' enzyme de Klenow). Le plasmide obtenu a été nommé The excisable cassette att1 # hyg + was then introduced into the coding sequence of the orf6 * gene. For this, the plasmid pBXL1111 was digested with the restriction enzymes SmaI and Asp718I and the digestion product was treated with Klenow enzyme. This manipulation makes it possible to carry out an internal deletion of 120 bp in the coding sequence of the orf6 * gene (see FIG. In addition, on either side of the restriction sites, 511 bp and 485 bp of the orf6 * sequence remain, which will allow homologous recombination for the inactivation of the orf6 * gene. The tape
![]()
excisable attl2hyg + was prepared from the plasmid pattlS2hyg + (see above) by digestion of this plasmid with EcoRV. The latter was then subcloned into the vector pBXL111 previously prepared as described above (SmaI and Asp718I digestion followed by treatment with the Klenow enzyme). The plasmid obtained was named
<Desc/Clms Page number 135><Desc / Clms Page number 135>
pBXL1112 (cf. figure 17). Dans cette construction, le gène or/6* comporte une délétion de 120pb et est interrompu par la cassette att1#hyg+. pBXL1112 (see Figure 17). In this construct, the or / 6 * gene has a deletion of 120pb and is interrupted by the att1 # hyg + cassette.
Le plasmide pBXL1112 a été ensuite digéré par l'enzyme PstI (site bordant la cassette puisque présent dans les oligonucléotides PCR) et l'insert PstI de 3. 7 kb comprenant une partie de orf6* interrompu par la cassette att1#hyg+ a ensuite été cloné au niveau du site PstI du plasmide pOJ260 (cf. ci-dessus). Le plasmide ainsi obtenu a été nommé pBXLl 113. The plasmid pBXL1112 was then digested with the enzyme PstI (site bordering the cassette since present in the PCR oligonucleotides) and the PstI insert of 3.7 kb comprising a portion of orf6 * interrupted by the cassette att1 # hyg + was then cloned at the PstI site of plasmid pOJ260 (see above). The plasmid thus obtained was named pBXL113.
Le vecteur pBXL1113 a été introduit dans la souche Streptomyces ambofaciens OSC2 (cf. ci-dessus) par transformation de protoplastes (Kieser, T et al., 2000). Après transformation, les clones sont sélectionnés pour leur résistance à l'hygromycine. Les clones résistants à l'hygromycine sont ensuite repiqués respectivement sur milieu avec hygromycine (antibiotique B) et sur milieu avec apramycine (antibiotique A) (cf. figure 9). Les clones résistants à l'hygromycine (HygR) et sensibles à l'apramycine (ApraS) sont en principe ceux où un double événement de crossing over s'est produit et qui possèdent le gène or/0* interrompu par la cassette att1#hyg+. Ces clones ont été plus particulièrement sélectionnés et le remplacement de la copie sauvage de orf6* par la copie interrompue par la cassette a été vérifié par la technique du Southern blot. Ainsi, l'ADN total des clones obtenus, a été digéré par plusieurs enzymes, séparé sur gel d'agarose, transféré sur membrane et hybridé avec une sonde correspondant au gène hyg (obtenu par PCR) pour vérifier la présence de la cassette dans l'ADN génomique des clones obtenus. Une deuxième hybridation a été effectuée en utilisant comme sonde l'insert PstI-PstI contenant le gène orf6*, d'une taille d'environ 1,1 kb et obtenu à partir du plasmide pBXLllll (cf. ci-dessus et figure 16). La vérification du génotype peut également être réalisée par toute méthode connue de l'homme du métier et notamment par PCR en utilisant les oligonucléotides appropriés et séquençage du produit de PCR. The vector pBXL1113 was introduced into the strain Streptomyces ambofaciens OSC2 (see above) by protoplast transformation (Kieser, T et al., 2000). After transformation, the clones are selected for their resistance to hygromycin. The hygromycin-resistant clones are then transplanted respectively on medium with hygromycin (antibiotic B) and on medium with apramycin (antibiotic A) (see FIG. 9). The hygromycin-resistant (HygR) and apramycin-sensitive (ApraS) -resistant clones are in principle those where a double crossing-over event has occurred and which have the or / 0 * gene interrupted by the cassette att1 # hyg + . These clones were more particularly selected and the replacement of the wild copy of orf6 * by the copy interrupted by the cassette was verified by the Southern blot technique. Thus, the total DNA of the clones obtained, was digested with several enzymes, separated on agarose gel, transferred to membrane and hybridized with a probe corresponding to the gene hyg (obtained by PCR) to check the presence of the cassette in the Genomic DNA of the clones obtained. A second hybridization was carried out using as probe the PstI-PstI insert containing the orf6 * gene, of a size of about 1.1 kb and obtained from the plasmid pBXLllll (see above and FIG. 16). . Verification of the genotype can also be carried out by any method known to those skilled in the art and in particular by PCR using the appropriate oligonucleotides and sequencing of the PCR product.
Un clone présentant les caractéristiques attendues (orf6*::attl2hyg+) a été plus particulièrement sélectionné et nommé SPM501. Il a pu en effet être vérifié grâce aux deux hybridations que la cassette att1#hyg+ était bien présente dans le génome de ce A clone with the expected characteristics (orf6 * :: attl2hyg +) was more particularly selected and named SPM501. It could indeed be verified thanks to the two hybridizations that the cassette att1 # hyg + was present in the genome of this
<Desc/Clms Page number 136><Desc / Clms Page number 136>
clone et que l'on obtient bien le profil de digestion attendu dans le cas d'un remplacement, à la suite d'un double événement de recombinaison, du gène sauvage par la copie interrompue par la cassette att1#hyg+ dans le génome de ce clone. Ce clone possède donc le génotype : orf6*::att1#hyg+ et a été nommé SPM501. Un échantillon de la souche SPM501 a été déposé auprès de la Collection Nationale de Cultures de Microorganismes (CNCM) Institut Pasteur, 25, rue du Docteur Roux 75724 Paris Cedex 15, France, le 10 juillet 2002 sous le numéro d'enregistrement 1-2909. clone and that we obtain the expected digestion profile in the case of a replacement, as a result of a double recombination event, of the wild-type gene by the copy interrupted by the cassette att1 # hyg + in the genome of this gene. clone. This clone therefore has the genotype: orf6 * :: att1 # hyg + and has been named SPM501. A sample of the strain SPM501 has been deposited with the National Collection of Cultures of Microorganisms (CNCM) Institut Pasteur, 25, rue du Docteur Roux 75724 Paris Cedex 15, France, July 10, 2002 under the registration number 1-2909 .
La souche SPM501 a été transformée par le vecteur pOSV508 par transformation de protoplastes pour provoquer l'excision de la cassette (cf. figure 14). Le plasmide pOSV508 est dérivé du plasmide pWHM3 (Vara J et al.,1989) (cf. figure 13) dans lequel les gènes xis et int de pSAM2 (Boccard F. et al., 1989b) placés sous le contrôle du promoteur ptrc (Amann, E. et al., 1988) ont été ajoutés (cf. figure 14). L'introduction dans la souche SPM501 du plasmide pOSV508 portant les gènes xis et int de pSAM2 permet l'excision efficace par recombinaison site spécifique de la cassette excisable entre les sites attL et attR flanquant cette cassette (Raynal A. et al., 1998) (figure 10). Parmi les transformants, sélectionnés pour leur résistance au thiostrepton due au gène tsr porté par pOSV508, on choisit ceux devenus sensibles à l'hygromycine dont le gène de résistance est porté par la cassette att1#hyg+, l'excision entraîne en effet la perte de ce gène de résistance (cf. figure 10). Le plasmide pOSV508 est instable et après deux passages successifs sur milieu sans antibiotique des clones isolés sont repiqués sur milieu avec thiostrepton et sur milieu sans thiostrepton. Les clones sensibles au thiostrepton ont perdu pOSV508. Il a été vérifié que l'excision de la cassette a bien eu lieu en phase dans le gène orf6* par PCR et séquençage du produit PCR. L'interruption commence au 158tème codon, 40 codons sont délétés (120pb) et l'excision de la cassette laisse une séquence cicatricielle attl caractéristique de 33pb : 5' ATCGCGCGCTTCGTTCGGGACGAAGAGGTAGAT 3' (SEQ ID N 104). The SPM501 strain was transformed by the vector pOSV508 by protoplast transformation to excise the cassette (see FIG. 14). Plasmid pOSV508 is derived from the plasmid pWHM3 (Vara J et al., 1989) (see FIG. 13) in which the xis and int genes of pSAM2 (Boccard F. et al., 1989b) placed under the control of the ptrc promoter ( Amann, E. et al., 1988) have been added (see Figure 14). The introduction into the SPM501 strain of plasmid pOSV508 carrying the xis and int genes of pSAM2 allows efficient excision by specific site-specific recombination of the excisable cassette between the attL and attR sites flanking this cassette (Raynal A. et al., 1998). (Figure 10). Among the transformants selected for their thiostrepton resistance due to the tsr gene carried by pOSV508, those which have become sensitive to hygromycin whose resistance gene is carried by the cassette att1 # hyg + are chosen, the excision in fact causes the loss of this resistance gene (see Figure 10). The plasmid pOSV508 is unstable and after two successive passages on medium without antibiotic isolated clones are subcultured on medium with thiostrepton and medium without thiostrepton. Thiostrepton-sensitive clones lost pOSV508. It was verified that the excision of the cassette did occur in the orf6 * gene by PCR and sequencing of the PCR product. The disruption begins at the 158th codon, 40 codons are deleted (120pb), and excision of the cassette leaves a characteristic 33bb att binding sequence: 5 'ATCGCGCGCTTCGTTCGGGACGAAGAGGTAGAT 3' (SEQ ID N 104).
La souche ainsi obtenue et possédant le génotype voulu (orf6*::attl) a été nommée SPM502. The strain thus obtained and having the desired genotype (orf6 * :: attl) was named SPM502.
<Desc/Clms Page number 137><Desc / Clms Page number 137>
Un échantillon de la souche SPM502 a été déposé auprès de la Collection Nationale de Cultures de Microorganismes (CNCM) Institut Pasteur, 25, rue du Docteur Roux 75724 Paris Cedex 15, France, le 10 juillet 2002 sous le numéro d'enregistrement 1-2910. A sample of the SPM502 strain has been deposited with the National Collection of Microorganism Cultures (CNCM) Pasteur Institute, 25, rue du Docteur Roux 75724 Paris Cedex 15, France, July 10, 2002 under the registration number 1-2910 .
EXEMPLE 15 : Analyse des souches de Streptomyces ambofaciens interrompue dans les gènes or/2, orf3, or/8, orflO, orfl2, orfl3c, orfl4, orf6*
Pour tester la production en spiramycine des diverses souches obtenues, un test microbiologique basé sur la sensibilité d'une souche de Micrococcus luteus a été mis au point (cf. (Gourmelen A. et al., 1998)). La souche de Micrococcus luteus utilisée est une souche dérivée de la souche DSM1790 naturellement sensible à la spiramycine (cette souche est disponible notamment auprès de la German Collection of Microorganisms and Cell Cultures (Deutsche Sammlung von Mikro-organismen und Zellkulturen GmbH, DSMZ), (Braunschweig, Allemagne), sous le numéro DSM1790), la souche utilisée dans le présent test diffère de la souche DSM1790 en ce qu'elle est résistante à la congocidine. Cette souche est un mutant spontané obtenu par sélection sur milieu contenant des doses croissantes de congocidine. Une telle souche a été sélectionnée du fait que Streptomyces ambofaciens produit à la fois de la spiramycine et de la congocidine. Le but étant de doser la production en spiramycine des diverses souches obtenues grâce à un test microbiologique basé sur la sensibilité d'une souche de Micrococcus luteus, il est nécessaire de disposer d'une souche résistante à la congocidine. EXAMPLE 15 Analysis of strains of Streptomyces ambofaciens interrupted in the genes gold / 2, orf3, or / 8, orfl0, orf12, orf13c, orf1, orf6 *
To test the spiramycin production of the various strains obtained, a microbiological test based on the sensitivity of a strain of Micrococcus luteus was developed (see Gourmelen A. et al., 1998). The strain of Micrococcus luteus used is a strain derived from the strain DSM1790 naturally sensitive to spiramycin (this strain is available in particular from the German Collection of Microorganisms and Cell Cultures (Deutsche Sammlung von Mikro-organismen und Zellkulturen GmbH, DSMZ), ( Braunschweig, Germany), under the number DSM1790), the strain used in this test differs from the strain DSM1790 in that it is resistant to congocidin. This strain is a spontaneous mutant obtained by selection on medium containing increasing doses of congocidine. Such a strain has been selected because Streptomyces ambofaciens produces both spiramycin and congocidin. The aim is to measure the spiramycin production of the various strains obtained by means of a microbiological test based on the sensitivity of a strain of Micrococcus luteus, it is necessary to have a strain resistant to congocidine.
Les différentes souches de Streptomyces à tester ont été cultivées dans des erlenmeyers à baffles (erlenmeyers chicanés) de 500 ml contenant 70 ml de milieu MP5 (Pernodet et al., 1993). Les erlenmeyers chicanés ont été inoculés à une concentration initiale de 2.5 106 spores/ml des différentes souches de S. ambofaciens et mises à pousser à 27 C sous agitation orbitale de 250 rpm. Des prélèvements de 2 ml de suspensions ont été réalisés après 48,72 et 96 heures de culture et centrifugés. Les The different Streptomyces strains to be tested were cultured in 500 ml baffled Erlenmeyer flasks (500 ml Erlenmeyer flasks) containing 70 ml of MP5 medium (Pernodet et al., 1993). The quenched Erlenmeyer flasks were inoculated at an initial concentration of 2.5 106 spores / ml of the different strains of S. ambofaciens and grown at 27 ° C. with orbital shaking of 250 rpm. Samples of 2 ml of suspensions were made after 48.72 and 96 hours of culture and centrifuged. The
<Desc/Clms Page number 138><Desc / Clms Page number 138>
différents surnageants ont ensuite été congelés à -20 C. Une dilution au dixième de ces surnageants dans du milieu de culture stérile est utilisée pour le test (cf. figure 18). various supernatants were then frozen at -20 C. A tenth dilution of these supernatants in sterile culture medium is used for the test (see Figure 18).
La souche indicatrice de Micrococcus luteus résistante à la congocidine mais sensible à la spiramycine a été cultivée dans du milieu 2TY (Sambrook et al., 1989) contenant de la congocidine à 5 g/ml pendant 48h à 37 C. La densité optique (DO) de la culture est mesurée et cette culture est diluée de façon à ajuster la densité optique à 4. The spirocynin-sensitive, but conocidin-resistant Micrococcus luteus indicator strain was cultured in 2TY medium (Sambrook et al., 1989) containing 5 g / ml congocidin for 48 h at 37 C. Optical density (OD) ) of the culture is measured and this culture is diluted so as to adjust the optical density to 4.
0,4 ml de cette pré-culture est dilué dans 40 ml de milieu DAM5 (Difco Antibiotic Médium 5, commercialisé par la société Difco), préalablement porté à une température d'environ 45 C. Ce milieu est ensuite coulé dans une boîte carrée de 12x12 cm et laissé à reposer à température ambiante. 0.4 ml of this preculture is diluted in 40 ml of DAM5 medium (Difco Antibiotic Medium 5, marketed by the company Difco), previously heated to a temperature of about 45 C. This medium is then poured into a square box 12x12 cm and allowed to rest at room temperature.
Une fois le milieu refroidi et solidifié, des disques en papier Whatman AA (cf. Once the medium has cooled and solidified, Whatman AA paper discs (cf.
Gourmelen A. et al., 1998) de 12 mm de diamètre ont été imbibés de 70 III de la dilution au dixième de chaque surnageant et déposés sur la surface de la boîte. Des disques imbibés d'une solution de spiramycine de différentes concentrations (2-4-8 g/ml dans du milieu de culture MP5) sont utilisés comme gamme étalon. Les boîtes sont laissées à 4 C pendant 2 h de façon à permettre la diffusion des antibiotiques dans l'agar puis sont incubées à 37 C pendant 24 à 48h. Gourmelen A. et al., 1998) 12 mm in diameter were impregnated with 70 III of the tenth dilution of each supernatant and deposited on the surface of the can. Disks imbibed with a solution of spiramycin of different concentrations (2-4-8 g / ml in MP5 culture medium) are used as the standard range. The dishes are left at 4 ° C. for 2 hours so as to allow the antibiotics to diffuse into the agar and are then incubated at 37 ° C. for 24 to 48 hours.
Si le disque contient de la spiramycine, celle-ci diffuse dans l'agar et inhibe la croissance de la souche indicatrice de Micrococcus luteus. Cette inhibition crée un halo autour du disque, ce halo reflétant la zone où la souche de Micrococcus luteus n'a pas poussé. La présence de ce halo est donc une indication de la présence de spiramycine et permet de déterminer si la souche de S. ambofaciens correspondant au disque en question est productrice ou non productrice de spiramycine. Une comparaison avec les diamètres d'inhibition obtenus pour la gamme étalon permet d'obtenir une indication de la quantité de spiramycine produite par cette souche. If the disc contains spiramycin, it diffuses into the agar and inhibits the growth of the indicator strain of Micrococcus luteus. This inhibition creates a halo around the disc, this halo reflecting the area where the strain of Micrococcus luteus did not grow. The presence of this halo is therefore an indication of the presence of spiramycin and makes it possible to determine whether the S. ambofaciens strain corresponding to the disc in question is producing or not producing spiramycin. A comparison with the diameters of inhibition obtained for the standard range makes it possible to obtain an indication of the amount of spiramycin produced by this strain.
Les différentes souches décrites dans les exemples précédents ont été utilisées dans ce test pour détecter leur production en spiramycine. Les résultats obtenus ont été regroupés dans le Tableau 38. The different strains described in the preceding examples were used in this test to detect their spiramycin production. The results obtained are summarized in Table 38.
<Desc/Clms Page number 139> <Desc / Clms Page number 139>
Tableau 3

<tb>
<tb> Souche <SEP> Gène <SEP> inactivé <SEP> Exemple <SEP> dans <SEP> Phénotype <SEP> : <SEP>
<tb> lequel <SEP> la <SEP> Producteur <SEP> (+) <SEP> ou
<tb> souche <SEP> est <SEP> non-producteur <SEP> (-) <SEP> de
<tb> décrite <SEP> spiramycine
<tb> ATCC23877 <SEP> Aucun <SEP> 1 <SEP> (+)
<tb> OS49. <SEP> 16 <SEP> orf3::#hyg <SEP> 2 <SEP> (-)
<tb> OS49. <SEP> 67 <SEP> orf3délétion <SEP> en <SEP> phase <SEP> 6 <SEP> (-)
<tb> OS49. <SEP> 107 <SEP> orf8::#hyg <SEP> 7 <SEP> (-)
<tb> OS49. <SEP> 50 <SEP> orf10:: <SEP> #hyg <SEP> 8 <SEP> (-)
<tb> OSC2 <SEP> Aucun <SEP> 10 <SEP> (+)
<tb> SPM21 <SEP> orf2::att3#aac- <SEP> 10 <SEP> (-)
<tb> SPM22 <SEP> orf2::att3 <SEP> délétion <SEP> en <SEP> phase <SEP> 10 <SEP> (-)
<tb> SPM501 <SEP> orf6*::att1#hyg+ <SEP> 14 <SEP> (-)
<tb> SPM502 <SEP> orf6*::attl <SEP> délétion <SEP> en <SEP> phase <SEP> 14 <SEP> (+)
<tb> <Tb>
<tb> Strain <SEP> Gene <SEP> inactivated <SEP> Example <SEP> in <SEP> Phenotype <SEP>: <SEP>
<tb> which <SEP> the <SEP> Producer <SEP> (+) <SEP> or
<tb> strain <SEP> is <SEP> non-producer <SEP> (-) <SEP> of
<tb> described <SEP> spiramycin
<tb> ATCC23877 <SEP> None <SEP> 1 <SEP> (+)
<tb> OS49. <SEP> 16 <SEP> orf3 :: # hyg <SEP> 2 <SEP> (-)
<tb> OS49. <SEP> 67 <SEP> orf3deletion <SEP> in <SEP> phase <SEP> 6 <SEP> (-)
<tb> OS49. <SEP> 107 <SEP> orf8 :: # hyg <SEP> 7 <SEP> (-)
<tb> OS49. <SEP> 50 <SEP> orf10 :: <SEP>#hyg<SEP> 8 <SEP> (-)
<tb> OSC2 <SEP> None <SEP> 10 <SEP> (+)
<tb> SPM21 <SEP> orf2 :: att3 # aac- <SEP> 10 <SEP> (-)
<tb> SPM22 <SEP> orf2 :: att3 <SEP> deletion <SEP> in <SEP> phase <SEP> 10 <SEP> (-)
<tb> SPM501 <SEP> orf6 * :: att1 # hyg + <SEP> 14 <SEP> (-)
<tb> SPM502 <SEP> orf6 * :: attl <SEP> deletion <SEP> in <SEP> phase <SEP> 14 <SEP> (+)
<Tb>

SPM507 orfl2: : att3.l.aac- 11 (-) SPM508 orfl3c: : att3.aac- 12 (+)
![]()
SPM507 orfl2:: att3.l.aac- 11 (-) SPM508 orfl3c:: att3.aac- 12 (+)
![]()
<tb>
<tb> SPM509 <SEP> orf4::att3#aac- <SEP> 13 <SEP> (-)
<tb> <Tb>
<tb> SPM509 <SEP> orf4 :: att3 # aac- <SEP> 13 <SEP> (-)
<Tb>
Ces résultats permettent de tirer un certain nombre de conclusions en ce qui concerne la fonction des différents gènes impliqués dans la biosynthèse de la These results make it possible to draw a number of conclusions regarding the function of the different genes involved in the biosynthesis of the
<Desc/Clms Page number 140><Desc / Clms Page number 140>
spiramycine. Ainsi le gène or/3 est essentiel à la biosynthèse de la spiramycine. En effet une inactivation en phase dans ce gène conduit à une souche (OS49.67, (cf. exemple 6)) ne produisant plus de spiramycine. L'inactivation en phase permet d'écarter l'hypothèse d'une éventuelle influence de la cassette introduite sur l'expression des gènes situés en aval de or/3. spiramycin. Thus the or / 3 gene is essential for the biosynthesis of spiramycin. In fact a phase inactivation in this gene leads to a strain (OS49.67, (see Example 6)) no longer producing spiramycin. Inactivation in phase makes it possible to rule out the hypothesis of a possible influence of the introduced cassette on the expression of genes located downstream of / 3.
De même, les gènes orf8 et orf10 codent des protéines essentielles à la biosynthèse de la spiramycine puisque les souches OS49.107 et OS49.50 ont un phénotype non producteur. De plus, dans ces deux dernières souches, c'est bien l'inactivation du gène correspondant qui est responsable de ce phénotype non producteur, puisqu'au vu de l'orientation des différentes orfs (cf. figure 3), la construction introduite ne peut avoir d'effet polaire. Similarly, the orf8 and orf10 genes encode proteins essential for the biosynthesis of spiramycin since strains OS49.107 and OS49.50 have a nonproductive phenotype. Moreover, in these last two strains, it is the inactivation of the corresponding gene which is responsible for this non-producing phenotype, since in view of the orientation of the various orfs (see FIG. can have polar effect.
L'étude des souches possédant une cassette excisable permet également de tirer un certain nombre de conclusion en ce qui concerne la fonction des gènes interrompus. La
![]()
souche SPM507 possède le génotype : orfl2: : att3SZaac-. Il est inutile de procéder à l'excision de la cassette pour l'étude de l'effet de l'inactivation de orfl2, au vu de l'orientation des gènes (cf. figure 3). Le fait que orfl3c soit orienté en sens opposé à orfl2 montre que ces gènes ne sont pas co-transcrits. L'utilisation d'une cassette excisable permet en revanche d'avoir la possibilité de se débarrasser du marqueur de sélection à tout moment. Le phénotype de la souche SPM507 est non producteur, on peut donc en conclure que le gène orfl2 est un gène essentiel à la biosynthèse de la spiramycine chez S. ambofaciens.
![]()
![]()
strain SPM507 has the genotype: orfl2:: att3SZaac-. There is no need to excise the cassette for the study of the effect of the inactivation of orfl2, given the orientation of the genes (see Figure 3). The fact that orfl3c is oriented in the opposite direction to orfl2 shows that these genes are not co-transcribed. Using an excisable cassette, however, allows you to get rid of the selection marker at any time. The phenotype of the SPM507 strain is nonproductive, so it can be concluded that the orfl2 gene is a gene essential for the biosynthesis of spiramycin in S. ambofaciens.
![]()
La souche SPM508 possède le génotype : orfl3c: : att3S2aac-. Il est inutile de procéder à l'excision de la cassette pour l'étude de l'effet de l'inactivation de orfl3c, au vu de l'orientation des gènes (cf. figure 3). Le fait que orfl4 soit orienté en sens opposé à orfl3c montre que ces gènes ne sont pas co-transcrits. L'utilisation d'une cassette excisable permet en revanche d 'avoir la possibilité de se débarrasser du marqueur de sélection à tout moment. Le phénotype de la souche SPM508 est producteur, on peut The SPM508 strain has the genotype: orfl3c :: att3S2aac-. There is no need to excise the cassette for the study of the effect of the inactivation of orfl3c, given the orientation of the genes (see Figure 3). The fact that orfl4 is oriented in the opposite direction to orfl3c shows that these genes are not co-transcribed. Using an excisable cassette, however, allows you to get rid of the selection marker at any time. The phenotype of strain SPM508 is a producer, one can
<Desc/Clms Page number 141><Desc / Clms Page number 141>
donc en conclure que le gène orf13c n'est pas un gène essentiel à la biosynthèse de la spiramycine chez S. ambofaciens.
![]()
La souche SPM509 possède le génotype : orfl4: : att3Saac-. Il est inutile de procéder à l'excision de la cassette pour l'étude de l'effet de l'inactivation de or/14, au vu de l'orientation des gènes (cf. figure 3), le fait que orfl5c soit orienté en sens opposé à or/14 montre que ces gènes ne sont pas co-transcrits. L'utilisation d'une cassette excisable permet en revanche d'avoir la possibilité de se débarrasser du marqueur de selection à tout moment. Le phénotype de la souche SPM509 est non producteur, on peut donc en conclure que le gène or/14 est un gène essentiel à la biosynthèse de la spiramycine chez S. ambofaciens. therefore conclude that the orf13c gene is not a gene essential for the biosynthesis of spiramycin in S. ambofaciens.
![]()
The strain SPM509 has the genotype: orfl4 :: att3Saac-. There is no need to excise the cassette for the study of the effect of gold / 14 inactivation, given the orientation of the genes (see Figure 3), the fact that orfl5c is oriented opposite to gold / 14 shows that these genes are not co-transcribed. Using an excisable cassette, however, allows you to get rid of the selection marker at any time. The phenotype of the SPM509 strain is nonproductive, so it can be concluded that the or / 14 gene is a gene essential for the biosynthesis of spiramycin in S. ambofaciens.
La souche SPM21 possède le génotype : orf2::att3#aac-. Le phénotype de cette souche est non producteur de spiramycines. Cependant, l'orientation des gènes orf1 à orf8 (cf. figure 3) laisse penser que ces gènes sont cotranscrits. Aussi, le phénotype observé peut être dû à un effet polaire de la cassette introduite dans or/2 sur l'expression de gènes situés en aval dans l'opéron. La souche SPM22 possède le génotype orf2::att3 et a été obtenue après excision en phase de la cassette introduite. L'excision de la cassette laisse seulement une séquence cicatricielle caractéristique (cf. exemple 10). The strain SPM21 has the genotype: orf2 :: att3 # aac-. The phenotype of this strain is non-producer of spiramycins. However, the orientation of the orf1 to orf8 genes (see Figure 3) suggests that these genes are cotranscribed. Also, the phenotype observed may be due to a polar effect of the cassette introduced in or / 2 on the expression of genes located downstream in the operon. The strain SPM22 has the genotype orf2 :: att3 and was obtained after excision in the introduced cassette. Excision of the cassette leaves only a characteristic cicatricial sequence (see Example 10).
La souche SPM22 étant également de phénotype non producteur, on peut en conclure que le gène or/2 est un gène essentiel à la biosynthèse de la spiramycine chez S. ambofaciens. On observe ici uniquement l'effet dû à l'inactivation de or/2.
![]()
![]()
La souche SPM501 possède le génotype : orf6*:: attl.s2hyg+. Le phénotype de cette souche est non producteur de spiramycines. Cependant, les gènes orf5* et orf6* (cf. figure 3) ayant la même orientation, le phénotype observé peut être dû à un effet polaire de la cassette introduite dans orf6* sur l'expression de orf5*. La disposition de ces gènes laisse penser qu'ils peuvent être cotranscrits. Pour répondre à cette question, la souche SPM502 a été obtenue après excision en phase de la cassette introduite. Dans cette souche, on observe uniquement l'effet de l'inactivation de orf6*. Cette souche The SPM501 strain has the genotype: orf6 * :: attl.s2hyg +. The phenotype of this strain is non-producer of spiramycins. However, since the orf5 * and orf6 * genes (see FIG. 3) have the same orientation, the observed phenotype may be due to a polar effect of the cassette introduced into orf6 * on the expression of orf5 *. The arrangement of these genes suggests that they can be cotranscribed. To answer this question, strain SPM502 was obtained after phase excision of the introduced cassette. In this strain, only the effect of the inactivation of orf6 * is observed. This strain
<Desc/Clms Page number 142><Desc / Clms Page number 142>
possède le génotype orf6*::attl (cf. exemple 14). L'excision de la cassette laisse seulement une séquence cicatricielle en phase (cf. exemple 14). La souche SPM502 possède un phénotype producteur (cette souche ne produit cependant que de la spiramycine I (cf*. exemple 16)). On peut donc conclure que le gène orf5*est un gène essentiel à la biosynthèse de la spiramycine chez S. ambofaciens, puisque son inactivation indirecte dans la souche SPM501 conduit à un phénotype non producteur. Par contre ; le gène orf6* n'est pas un gène essentiel à la biosynthèse de la spiramycine I chez S. ambofaciens (par contre il est essentiel à la production de spiramycine II et III (cf. exemple 16)). has the genotype orf6 * :: attl (see example 14). Excision of the cassette leaves only a cicatricial sequence in phase (see Example 14). The strain SPM502 has a producing phenotype (this strain, however, only produces spiramycin I (cf Example 16)). It can therefore be concluded that the orf5 * gene is a gene essential for the biosynthesis of spiramycin in S. ambofaciens, since its indirect inactivation in the SPM501 strain leads to a non-producing phenotype. On the other hand ; the orf6 * gene is not a gene essential for the biosynthesis of spiramycin I in S. ambofaciens (on the other hand it is essential for the production of spiramycin II and III (see Example 16)).
EXEMPLE 16. Dosage de la production des spiramycines I, II et III dans les souches mutantes obtenues
Les différentes souches à tester ont été cultivées chacune dans 7 erlensmeyers chicanés de 500 mL contenant 70 ml de milieu MP5 (Pernodet et al., 1993). Les erlens ont été inoculés par 2.5 106 spores/ml des différentes souches de S. ambofaciens et mises à pousser à 27 C sous agitation orbitale de 250 rpm pendant 72 heures. Les cultures correspondant au même clone sont rassemblées, éventuellement filtrées sur filtre plissé, et centrifugées 15 min à 7000 tr/min. Les différents surnageants ont ensuite été stockés à-30 C. EXAMPLE 16 Determination of the Production of Spiramycins I, II and III in Mutant Strains Obtained
The different strains to be tested were each cultured in 7 500 ml baffled erlenmeyers containing 70 ml of MP5 medium (Pernodet et al., 1993). The erlens were inoculated with 2.5 106 spores / ml of the different strains of S. ambofaciens and grown at 27 ° C. with orbital shaking at 250 rpm for 72 hours. The cultures corresponding to the same clone are pooled, possibly filtered on a pleated filter, and centrifuged for 15 min at 7000 rpm. The different supernatants were then stored at -30 C.
Les dosages ont été effectués par Chromatographie Liquide à Haute Performance (CLHP) par appariement d'ions. L'analyse CLHP du milieu de culture permet de déterminer précisément la concentration des trois formes de spiramycine. La colonne utilisée (Macherey-Nagel) est remplie avec une phase Nucleosil de silice greffée octyle. Assays were performed by high performance liquid chromatography (HPLC) by ion pairing. The HPLC analysis of the culture medium makes it possible to precisely determine the concentration of the three forms of spiramycin. The column used (Macherey-Nagel) is filled with a Nucleosil phase of octyl grafted silica.
La granulométrie est de 5 m et la taille des pores 100 . Le diamètre interne de la colonne est de 4,6mm et sa longueur 25cm. La phase mobile est un mélange de tampon H3PO4 (pH2,2) et d'acétonitrile 70/30 (v/v) contenant 6,25g/L de perchlorate de NaC104. L'analyse est réalisée en régime isocratique avec un débit fixé à 1 ml/min. La The particle size is 5 m and the pore size 100. The internal diameter of the column is 4.6mm and its length 25cm. The mobile phase is a mixture of buffer H3PO4 (pH2.2) and acetonitrile 70/30 (v / v) containing 6.25 g / l of NaClO4 perchlorate. The analysis is carried out under isocratic conditions with a flow rate set at 1 ml / min. The
<Desc/Clms Page number 143><Desc / Clms Page number 143>
colonne est thermorégulée à 23 C. La détection est assurée par spectrophotométrie UV à 238nm. L'échantillon est réfrigéré à +10 C et la quantification est déterminée à partir de l'aire des pics (par étalonnage externe). Dans ces conditions, les temps de rétention de la spiramycine I, II et III sont respectivement d'environ 17 ; et 30 minutes, comme il a pu être vérifié grâce à un échantillon commercial comprenant les trois formes de spiramycine. column is thermoregulated at 23 C. The detection is performed by UV spectrophotometry at 238 nm. The sample is refrigerated at +10 C and quantification is determined from the peak area (by external calibration). Under these conditions, the retention times of spiramycin I, II and III are respectively about 17; and 30 minutes, as could be verified by a commercial sample comprising all three forms of spiramycin.
La souche OSC2 possède un phénotype producteur de spiramycine. C'est la souche parentale utilisée pour l'obtention des souches possédant une cassette excisable (cf. exemple 15). Cette souche a donc été utilisée comme témoin positif de production des trois formes de spiramycine. Cette souche produit bien les trois formes de spiramycines comme il a été vérifié par CLHP (cf. figure 19). The OSC2 strain has a spiramycin-producing phenotype. This is the parental strain used to obtain strains possessing an excisable cassette (see Example 15). This strain was therefore used as a positive control of production of the three forms of spiramycin. This strain produces all three spiramycin forms as verified by HPLC (see Figure 19).
L'étude des souches possédant une cassette excisable permet de tirer un certain nombre de conclusions en ce qui concerne la fonction des gènes interrompus. La souche
![]()
SPM507 possède le génotype : of f12:: att3,aac-. Le phénotype de la souche SPM507 est non producteur (cf. exemple 15), on peut donc en conclure que le gène orfl2 est un gène essentiel à la biosynthèse de la spiramycine chez S. ambofaciens. Cette souche ne produit plus de spiramycines comme il a été vérifié par CLHP (cf. figure 22). Ce résultat confirme le caractère essentiel du gène or/72 dans la biosynthèse de la spiramycine.
![]()
![]()
SPM507 has the genotype: of f12 :: att3, aac-. The phenotype of the strain SPM507 is non-producer (see Example 15), it can therefore be concluded that the orfl2 gene is a gene essential for the biosynthesis of spiramycin in S. ambofaciens. This strain no longer produces spiramycins as verified by HPLC (see Figure 22). This result confirms the essential character of the or / 72 gene in the biosynthesis of spiramycin.
![]()
La souche SPM508 possède le génotype : orfl3c: : att3S2aac-. Le phénotype de la souche SPM508 est producteur de spiramycine (cf. exemple 15), on peut donc en conclure que le gène orfl3c n'est pas un gène essentiel à la biosynthèse de la spiramycine chez S. ambofaciens. Cette souche produit de la spiramycine I, II et III comme il a été vérifié par CLHP (cf. figure 23). Ce résultat confirme que le gène orfl3c n'est pas un gène essentiel à la biosynthèse des spiramycines I, II et III ches S. ambofaciens. The SPM508 strain has the genotype: orfl3c :: att3S2aac-. The phenotype of the SPM508 strain produces spiramycin (see Example 15), it can therefore be concluded that the orfl3c gene is not a gene essential for the biosynthesis of spiramycin in S. ambofaciens. This strain produces spiramycin I, II and III as verified by HPLC (see Figure 23). This result confirms that the orfl3c gene is not a gene essential for the biosynthesis of spiramycins I, II and III of S. ambofaciens.
La souche SPM509 possède le génotype : orf14::att3#aac-. Le phénotype de la souche SPM509 est non producteur, on peut donc en conclure que le gène orfl4 est un The SPM509 strain has the genotype: orf14 :: att3 # aac-. The phenotype of strain SPM509 is nonproductive, so it can be concluded that the orfl4 gene is a
<Desc/Clms Page number 144><Desc / Clms Page number 144>
gène essentiel à la biosynthèse de la spiramycine chez S. ambofaciens. Cette souche ne produit plus de spiramycines comme il a été vérifié par CLHP (cf. figure 24). Ce résultat confirme l'essentialité du gène or/14 dans la biosynthèse de la spiramycine.
![]()
![]()
La souche SPM501 possède le génotype : orf6*: : attl S2hyg+. Le phénotype de cette souche est non producteur de spiramycines. Cette souche ne produit plus de spiramycines comme il a été vérifié par CLHP (cf. figure 20). Cependant, les gènes orf5* et orf6* (cf. figure 3) ayant la même orientation, le phénotype observé peut être dû à un effet polaire de la cassette introduite dans orf6*sur l'expression de orf5* dans l'opéron. Ceci laisse penser que ces gènes sont cotranscrits. Pour répondre à cette question, la souche SPM502 a été obtenue par excision de la cassette introduite, produisant une délétion en phase dans le gène or6* et restaurant l'expression de orf5*. The SPM501 strain has the genotype: orf6 *:: attl S2hyg +. The phenotype of this strain is non-producer of spiramycins. This strain no longer produces spiramycins as verified by HPLC (see Figure 20). However, since the orf5 * and orf6 * genes (see FIG. 3) have the same orientation, the observed phenotype may be due to a polar effect of the cassette introduced into orf6 * on the expression of orf5 * in the operon. This suggests that these genes are cotranscribed. To answer this question, strain SPM502 was obtained by excision of the introduced cassette, producing a phase deletion in the or6 * gene and restoring the expression of orf5 *.
Cette souche possède le génotype orf6*::attl (cf. exemple 14 et 15). L'excision de la cassette laisse seulement une séquence att cicatricielle en phase (cf. exemple 14). La souche SPM502 possède un phénotype producteur de spiramycine. Cependant, comme il a été prouvé par CLHP, cette souche ne produit que de la spiramycine I et ne produit pas de spiramycine II et III (cf. figure 21). On peut donc conclure de ces résultats que le gène or/5* est un gène essentiel à la biosynthèse de la spiramycine chez S. ambofaciens, puisque son inactivation indirecte dans la souche SPM501 conduit à un phénotype non producteur de spiramycine (cf. figure 20). Par contre ; le gène orf6* n'est pas un gène, essentiel à la biosynthèse de la spiramycine I chez S. ambofaciens, puisque l'inactivation de ce gène conduit à un phénotype producteur de spiramycine I (cf. figure 21). This strain has the genotype orf6 * :: attl (see example 14 and 15). Excision of the cassette leaves only a scarring phase in phase (see Example 14). The strain SPM502 has a phenotype producing spiramycin. However, as proved by HPLC, this strain produces only spiramycin I and does not produce spiramycin II and III (see Figure 21). It can therefore be concluded from these results that the or / 5 * gene is an essential gene for the biosynthesis of spiramycin in S. ambofaciens, since its indirect inactivation in the SPM501 strain leads to a non-spiramycin-producing phenotype (see Figure 20). ). On the other hand ; the orf6 * gene is not a gene essential for the biosynthesis of spiramycin I in S. ambofaciens, since the inactivation of this gene leads to a spiramycin I-producing phenotype (see Figure 21).
Cependant, orf6* est essentiel à la production de spiramycine II et III (cf. exemple 16)). However, orf6 * is essential for the production of spiramycin II and III (see Example 16)).
Le gène orf6* code donc bien une acyl-transférase responsable de la modification du platénolide à la position 3 (cf. figure 1). The orf6 * gene therefore encodes an acyl transferase responsible for the modification of platenolide at position 3 (see FIG.
<Desc/Clms Page number 145><Desc / Clms Page number 145>
EXEMPLE 17 : Détermination du point de démarrage de la traduction de orf 10 et amélioration de la production en spiramycines
17. 1 Construction des plasmides pSPM523. pSPM524 et pSPM525 :
Le gène orf10 a été identifié chez Streptomyces ambofaciens et a été désigné srmR par Geistlich et al. (Geistlich, M., et al., 1992). L'inactivation du gène orf10 a été réalisée (cf. exemple 8). Il a pu ainsi être montré que la souche résultante ne produit plus de spiramycines (cf. exemple 15). Ceci confirme que le gène or/70 est bien impliqué dans la biosynthèse des spiramycines. La protéine codée par ce gène est donc bien essentielle à la biosynthèse des spiramycines. Cependant, l'analyse de la séquence montre que deux codons ATG situés dans la même phase de lecture peuvent être utilisés pour la traduction de orf10 (cf. Figure 28). Un des deux codons possible (le codons le plus en amont) démarre à la position 10656 de la séquence donnée en SEQ ID N 1, alors que l'autre codon possible situé plus en aval démarre à la position 10809 (cf. SEQ ID N 1). Avant de tester un éventuel effet de la surexpression de srmR sur la production de spiramycine, il est important de déterminer dans un premier temps le point de démarrage de la traduction. EXAMPLE 17 Determination of the starting point of translation of orf 10 and improvement of spiramycin production
17. 1 Construction of Plasmids pSPM523. pSPM524 and pSPM525:
The orf10 gene has been identified in Streptomyces ambofaciens and has been designated srmR by Geistlich et al. (Geistlich, M., et al., 1992). Inactivation of the orf10 gene was performed (see Example 8). It was thus possible to show that the resulting strain no longer produces spiramycins (see Example 15). This confirms that the or / 70 gene is well implicated in the biosynthesis of spiramycins. The protein encoded by this gene is therefore essential for the biosynthesis of spiramycins. However, the analysis of the sequence shows that two ATG codons located in the same reading phase can be used for the translation of orf10 (see Figure 28). One of the two possible codons (the most upstream codon) starts at the position 10656 of the sequence given in SEQ ID N 1, while the other possible codon situated further downstream starts at the position 10809 (see SEQ ID N 1). Before testing a possible effect of srmR overexpression on spiramycin production, it is important to first determine the start point of the translation.
Dans le but de déterminer le site de démarrage de la traduction, trois constructions comportant trois formes d'orf10 ont été réalisées. Ces dernières ont été obtenues par PCR en utilisant des oligonucléotides comportant soit un site de restriction HindIII soit un site de restriction BamHI. In order to determine the start site of the translation, three constructions comprising three forms of orf10 have been realized. The latter were obtained by PCR using oligonucleotides comprising either a HindIII restriction site or a BamHI restriction site.
Le premier couple utilisé pour l'amplification correspond aux oligonucléotides suivants : EDR39 : 5'CCCAAGCTTGAGAAGGGAGCGGACATTCATGGCCCGCGCCGAACGC3' (SEQ ID N 122) (le site HindIII figure en gras) EDR42 : 5'CGGGATCCGGCTGACCATGGGAGACGGGCGCATCGCCGAGTTCAGC3' (SEQ ID N 123) (le site BamHI figure en gras) The first pair used for the amplification corresponds to the following oligonucleotides: EDR39: 5'CCCAAGCTTGAGAAGGGAGCGGACATTCATGGCCCGCGCCGAACGC3 '(SEQ ID N 122) (the HindIII site is in bold) EDR42: 5'CGGGATCCGGCTGACCATGGGAGACGGGCGCATCGCCGAGTTCAGC3' (SEQ ID NO: 123) (the BamHI site FIG. in bold)
<Desc/Clms Page number 146><Desc / Clms Page number 146>
Le couple d'amorces EDR39-EDR42 permet l'amplification d'un fragment d'or/70 comportant l'ATG situé le plus en 3' (position 10809 dans la séquence donnée en SEQ ID N 1) (cf. Figure 28). Le fragment obtenu fait une taille d'environ 2 kb et sera appelé par la suite "orf10 court", il ne contient pas le promoteur de or/70. Ce fragment de 2kb a été cloné dans le vecteur pGEM-T easy pour donner naissance au plasmide pSPM520. The primer pair EDR39-EDR42 allows the amplification of a gold / 70 fragment comprising the most 3 'ATG (position 10809 in the sequence given in SEQ ID No. 1) (see FIG. 28). . The fragment obtained has a size of about 2 kb and will be called thereafter "orf10 short", it does not contain the promoter of gold / 70. This 2kb fragment was cloned into the pGEM-T easy vector to give rise to plasmid pSPM520.
Le deuxième couple utilisé pour l'amplification correspond aux oligonucléotides suivants : EDR40 : 5'CCCAAGCTTGAGAAGGGAGCGGACATTCAATGCTTTGGTAAAGCAC3' (SEQ ID N 124) (le site HindIII figure en gras) EDR42 : 5'CGGGATCCGGCTGACCATGGGAGACGGGCGCATCGCCGAGTTCAGC3' (SEQ ID N 123) (le site BamHI figure en gras) Le couple d'amorces EDR40-EDR42 permet l'amplification d'un fragment d'orf10 comportant l'ATG situé le plus en 5' (position 10656 dans la séquence donnée en SEQ ID N 1) (cf. Figure 28). Ce fragment de 2,2 kb appelé par la suite "orf10 long" a été cloné dans le vecteur pGEM-T easy pour donner naissance au plasmide pSPM521, ce plasmide ne contient pas le promoteur de or/70. The second pair used for the amplification corresponds to the following oligonucleotides: EDR40: 5'CCCAAGCTTGAGAAGGGAGCGGACATTCAATGCTTTGGTAAAGCAC3 '(SEQ ID N 124) (the HindIII site is in bold) EDR42: 5'CGGGATCCGGCTGACCATGGGAGACGGGCGCATCGCCGAGTTCAGC3' (SEQ ID N 123) (the BamHI site appears in bold) The primer pair EDR40-EDR42 allows the amplification of a fragment of orf10 comprising the most 5 'ATG (position 10656 in the sequence given in SEQ ID No. 1) (see FIG. ). This 2.2 kb fragment, hereinafter referred to as "orf10 long", was cloned into the pGEM-T easy vector to give rise to the plasmid pSPM521, this plasmid does not contain the gold / 70 promoter.
Le troisième couple utilisé pour l'amplification correspond aux oligonucléotides suivants : EDR41 : 5'-CCCAAGCTTTCAAGGAACGACGGGGTGGTCAGTCAAGT-3' (SEQ ID N 125) (le site HindIII figure en gras) EDR42 : 5'CGGGATCCGGCTGACCATGGGAGACGGGCGCATCGCCGAGTTCAGC3' (SEQ ID N 123) (le site BamHI figure en gras) Le couple d'amorces EDR41-EDR42 permet l'amplification du gène orf10 avec les deux ATG, ainsi que son propre promoteur (cf. Figure 28). Ce fragment de 2,8 kb The third pair used for the amplification corresponds to the following oligonucleotides: EDR41: 5'-CCCAAGCTTTCAAGGAACGACGGGGTGGTCAGTCAAGT-3 '(SEQ ID N 125) (the HindIII site is in bold) EDR42: 5'CGGGATCCGGCTGACCATGGGAGACGGGCGCATCGCCGAGTTCAGC3' (SEQ ID N 123) (the BamHI site is in bold) The primer pair EDR41-EDR42 allows amplification of the orf10 gene with both ATGs, as well as its own promoter (see Figure 28). This fragment of 2.8 kb
<Desc/Clms Page number 147><Desc / Clms Page number 147>
appelé par la suite "orf10 pro" a été cloné dans le vecteur pGEM-T easy pour donner naissance au plasmide pSPM522. hereinafter referred to as "orf10 pro" was cloned into the pGEM-T easy vector to give rise to plasmid pSPM522.
Le fragment "orf10 pro" a été obtenu en utilisant comme matrice l'ADN chromosomique de la souche OSC2. Les fragments "orf10 court" et "orf10 long" ont quant à eux été obtenus en utilisant comme matrice l'ADN du fragment "orf10 pro" préalablement purifié. The "orf10 pro" fragment was obtained using as template the chromosomal DNA of the OSC2 strain. The fragments "short orf10" and "orf10 long" were obtained by using as template the DNA of the fragment "orf10 pro" previously purified.
Les inserts HindIII-BamHI des plasmides pSPM520, pSPM521 et pSPM522 ont ensuite été sous-clonés dans le vecteur pUWL201 (plamside dérivé du plasmide pUWL199 (Wehmeier UF, 1995) dans lequel le fragment KpnI-BamHI de la région du promoteur de ermE (cf. Bibb et al., 1985, notamment figure 2) portant une mutation augmentant la force du promoteur (promoteur ermE*) (Bibb et al., 1994) a été introduit (cf. Doumith et al., 2000)) préalablement digéré par les enzymes HindIII-BamHI. Ainsi, il a été obtenu trois plasmides : pSPM523 (dérivé du pUWL201 avec la forme "orf10 court" comme insert), pSPM524 (dérivé du pUWL201 avec la forme "orf10 long" comme insert) et pSPM525 (dérivé du pUWL201 avec la forme "orf10 pro") (Figure 28). The HindIII-BamHI inserts of the plasmids pSPM520, pSPM521 and pSPM522 were then subcloned in the vector pUWL201 (plasmid derived from the plasmid pUWL199 (Wehmeier UF, 1995) in which the KpnI-BamHI fragment of the ermE promoter region (cf. Bibb et al., 1985, especially Figure 2) carrying a mutation increasing the strength of the promoter (ermE * promoter) (Bibb et al., 1994) was introduced (see Doumith et al., 2000)) previously digested with HindIII-BamHI enzymes. Thus, three plasmids were obtained: pSPM523 (derived from pUWL201 with the form "short orf10" as insert), pSPM524 (derived from pUWL201 with the form "orf10 long" as insert) and pSPM525 (derived from pUWL201 with the form " orf10 pro ") (Figure 28).
17. 2 Transformation de la souche OS49. 50 par les constructions pSPM523, pSPM524 et pSPM525 :
La souche OS49. 50 (souche interrompue dans le gène orf10, cf. exemple 8) a été transformée indépendamment par transformation de protoplastes (Kieser, T et al., 2000) par chacun des plasmides pSPM523, pSPM524 et pSPM525. Un témoin négatif a également été réalisé en transformant la souche OS49.50 par le plasmide pUWL201 sans insert. Après transformation des protoplastes, les clones sont sélectionnés pour leur résistance au au thiostrepton. La transformation des clones par chacun des plamides est vérifiée par extraction de ces plasmides. Ainsi quatre nouvelles souches ont été obtenues : la souche OSC49.50 pUWL201, issue de la transformation par le plasmide pUWL201 sans insert, la souche OSC49. 50 pSPM523, issue de la transformation par le plasmide pSPM523, la souche OSC49. 50 pSPM524, issue de la transformation par le 17. 2 Transformation of the OS49 strain. 50 by the constructions pSPM523, pSPM524 and pSPM525:
The strain OS49. 50 (strain interrupted in the orf10 gene, see Example 8) was transformed independently by protoplast transformation (Kieser, T et al., 2000) with each of the plasmids pSPM523, pSPM524 and pSPM525. A negative control was also carried out by transforming the strain OS49.50 by the plasmid pUWL201 without insert. After transformation of the protoplasts, the clones are selected for thiostrepton resistance. The transformation of the clones by each of the plasmids is verified by extraction of these plasmids. Thus four new strains were obtained: the strain OSC49.50 pUWL201, resulting from the transformation with the plasmid pUWL201 without insert, the strain OSC49. PSPM523, resulting from the transformation with plasmid pSPM523, strain OSC49. 50 pSPM524, resulting from the transformation by the
<Desc/Clms Page number 148><Desc / Clms Page number 148>
plasmide pSPM524 et enfin la souche OS49. 50 pSPM525, issue de la transformation par le plasmide pSPM525. plasmid pSPM524 and finally the strain OS49. PSPM525, resulting from transformation with plasmid pSPM525.
La production de spiramycines de chacune de ces quatre souches a été testée par CLHP. Pour cela, les différentes souches de Streptomyces à tester ont été cultivées dans des erlenmeyers à baffles (erlenmeyers chicanés) de 500 ml contenant 70 ml de milieu MP5 (Pernodet et al., 1993). Lorsque la souche contient le plasmide pUWL201 ou l'un de ses dérivés, il est ajouté 5 g/ml de thiostrepton. Les erlenmeyers chicanés ont été inoculés à une concentration initiale de 2.5 106 spores/ml des différentes souches de S. ambofaciens et les cultures ont été incubées à 27 C sous agitation orbitale de 250 rpm pendant 96 heures. Les cellules ont ensuite été séparées du milieu par centrifugation et le surnageant a été analysé par CLHP (cf. exemple 16) pour déterminer la quantité de spiramycine produite. Grâce à un échantillon étalon et à la mesure de l'air des pics, il a été possible de déterminer la quantité de chacune des spiramycines produites par ces souches. Les résultats de cette analyse sont présentés dans le tableau 40, les données sont exprimées en mg par litre de surnageant. Les résultats correspondent à la production totale en spiramycines (obtenue en additionnant la production en spiramycine I, II et III). The spiramycin production of each of these four strains was tested by HPLC. For this, the different Streptomyces strains to be tested were cultured in 500 ml baffled Erlenmeyer flasks (500 ml Erlenmeyer flasks) containing 70 ml of MP5 medium (Pernodet et al., 1993). When the strain contains the plasmid pUWL201 or one of its derivatives, 5 g / ml of thiostrepton is added. The quenched Erlenmeyer flasks were inoculated at an initial concentration of 2.5 106 spores / ml of the different strains of S. ambofaciens and the cultures were incubated at 27 ° C. with orbital shaking at 250 rpm for 96 hours. The cells were then removed from the medium by centrifugation and the supernatant was analyzed by HPLC (see Example 16) to determine the amount of spiramycin produced. Using a standard sample and measuring peak air, it was possible to determine the amount of each of the spiramycins produced by these strains. The results of this analysis are shown in Table 40, the data are expressed in mg per liter of supernatant. The results correspond to the total production of spiramycins (obtained by adding spiramycin I, II and III production).
Tableau 40. Production en spiramycines des souches dérivées de OS49.50, (résultats exprimés en mg/1).

<tb>
<tb> <Tb>
<Tb>
Souche <SEP> Production <SEP> en <SEP> Spiramycines
<tb> OS49.50 <SEP> 0
<tb> pUWL201
<tb> OS49.50 <SEP> 0
<tb> pSPM523
<tb> OS49. <SEP> 50 <SEP> 93
<tb> pSPM524
<tb> Strain <SEP> Production <SEP> in <SEP> Spiramycine
<tb> OS49.50 <SEP> 0
<tb> pUWL201
<tb> OS49.50 <SEP> 0
<tb> pSPM523
<tb> OS49. <SEP> 50 <SEP> 93
<tb> pSPM524
<Tb>
<Desc/Clms Page number 149> <Desc / Clms Page number 149>

<tb>
<tb> OS49. <SEP> 50 <SEP> 149
<tb> pSPM525
<tb> <Tb>
<tb> OS49. <SEP> 50 <SEP> 149
<tb> pSPM525
<Tb>
Comme le montre les résultats donnés dans le tableau 40, le témoin négatif (souche OS49. 50 transformée par le plasmide pUWL201) ne produit pas de spiramycine. Lorsque le plasmide pSPM523 (qui contient la forme "orflO court" ) est introduit dans la souche OS49. 50, aucune production de spiramycine n'est observée. As shown in the results given in Table 40, the negative control (strain OS49.50 transformed with plasmid pUWL201) does not produce spiramycin. When the plasmid pSPM523 (which contains the form "short orflO") is introduced into the strain OS49. 50, no production of spiramycin is observed.
En revanche, la présence du plasmide pSPM524 (qui contient la forme "orf10 long") et du plasmide pSPM525 (qui contient la forme "orf10 pro") restaure la production de spiramycine dans la souche hôte OS49. 50. Ainsi, seuls les fragments de orf10 contenant l'ATG le plus en amont permettent de restaurer la synthèse de spiramycine. In contrast, the presence of plasmid pSPM524 (which contains the "long orf10" form) and plasmid pSPM525 (which contains the "orf10 pro" form) restores the production of spiramycin in the OS49 host strain. 50. Thus, only the fragments of orf10 containing the most upstream ATG make it possible to restore spiramycin synthesis.
Dans le but de confirmer ces résultats, le plasmide pSPM521 (plasmide pGEM-T easy contenant la forme "orflO long") a été digéré par l'enzyme de restriction XhoI (cette enzyme possède un site unique dans ce plasmide, localisé entre les deux ATG (cf. Figure 28)). Les extrémités XhoI ont ensuite été rendues bout franc par traitement à l'enzyme de Klenow. Le plasmide a alors été refermé sur lui-même par action de la T4 DNA ligase pour donner naissance au plasmide pSPM527. Si l'ATG le plus en amont (position 10656 dans la séquence donnée en SEQ ID N 1) est bien utilisé comme site d'initiation de la traduction, ce traitement entrainera un décalage du cadre de lecture au niveau du site XhoI et aura pour effet la production d'une protéine ne présentant pas d'activité activatrice. Par contre si l'initiation de la traduction a lieu au niveau de l'ATG plus en aval (position 10809 dans la séquence donnée en SEQ ID N 1), ce traitement devrait avoir peu ou pas d'effet sur l'expression de OrflO (compte tenu de la localisation du point de démarrage de la transcription) et aucun effet sur la protéine produite. In order to confirm these results, the plasmid pSPM521 (plasmid pGEM-T easy containing the form "long orflO") was digested with the restriction enzyme XhoI (this enzyme has a unique site in this plasmid, located between the two ATG (see Figure 28)). The XhoI ends were then blunted by Klenow enzyme treatment. The plasmid was then closed on itself by the action of T4 DNA ligase to give rise to the plasmid pSPM527. If the most upstream ATG (position 10656 in the sequence given in SEQ ID No. 1) is well used as a translation initiation site, this processing will cause a shift of the reading frame at the XhoI site and will have effect the production of a protein not exhibiting activating activity. On the other hand, if the initiation of the translation takes place at the level of the ATG further downstream (position 10809 in the sequence given in SEQ ID No. 1), this treatment should have little or no effect on the expression of OrflO. (considering the location of the start point of transcription) and no effect on the protein produced.
L'insert BamHI-HindIII de pSPM527 a ensuite été sous-cloné dans le vecteur pUWL201 pour donner naissance au plasmide pSPM528. Ce plasmide a été introduit dans la souche OS49. 50 et un clone possédant le plasmide voulu a plus particulièrement été sélectionné. La production de spiramycine de la souche The BamHI-HindIII insert of pSPM527 was then subcloned into the vector pUWL201 to give rise to the plasmid pSPM528. This plasmid was introduced into the strain OS49. 50 and a clone possessing the desired plasmid was more particularly selected. Spiramycin production of the strain
<Desc/Clms Page number 150><Desc / Clms Page number 150>
résultante a ensuite été testée par CLHP (Cf. exemple 16 et ci-dessus). Contrairement à ce qui avait été observé avec le plasmide pSPM524 (cf. tableau 40), la présence du plasmide pSPM528 dans la souche OS49.50 ne rétablit pas la production de spiramycine. Ceci confirme que le démarrage de la traduction du gène orf10 est l'ATG situé le plus en aval (ATG 1 cf. Figure 28). The resultant was then tested by HPLC (see Example 16 and above). Contrary to what was observed with plasmid pSPM524 (see Table 40), the presence of plasmid pSPM528 in strain OS49.50 does not restore the production of spiramycin. This confirms that the start of the translation of the orf10 gene is the most downstream ATG (ATG 1 see Figure 28).
17. 3 Amélioration de la production en spiramycines de la souche OSC2 de S. ambofaciens :
Afin de tester l'effet de la surexpression du gène orf10 sur la production de spiramycines, les plasmides pSPM523, pSPM524, pSPM525 et pSPM528 ont été introduits dans la souche OSC2. Pour cela, des protoplastes de la souche OSC2 ont été transformés (Kieser, T et al., 2000) indépendamment par chacun des plasmides pSPM523, pSPM524, pSPM525 et pSPM528. Un témoin négatif a également été réalisé en transformant la souche OSC2 par le plasmide pUWL201 sans insert. Après transformation des protoplastes, les clones sont sélectionnés pour leur résistance au au thiostrepton. Ainsi cinq nouvelles souches ont été obtenues : la souche OSC2 pUWL201, issue de la transformation par le plasmide pUWL201 sans insert, la souche OSC2 pSPM523, issue de la transformation par le plasmide pSPM523, la souche OSC2 pSPM524, issue de la transformation par le plasmide pSPM524, la souche OSC2 pSPM525, issue de la transformation par le plasmide pSPM525 et enfin la souche OSC2 pSPM528, issue de la transformation par le plasmide pSPM528. La production en spiramycines de ces souches a alors été analysée par CLHP (de la même manière que dans l'exemple 17. 2). L'analyse de la production en spiramycine de la souche OSC2 a également été effectée en parallèle pour comparaison. Les résultats de cette analyse sont présentés dans le tableau 41, les données sont exprimées en mg par litre de surnageant. Les résultats correspondent à la production totale en spiramycines (obtenue en additionnant la production en spiramycine I, II et III). 17. 3 Improvement of spiramycin production of S. ambofaciens strain OSC2:
In order to test the effect of the overexpression of the orf10 gene on the production of spiramycins, the plasmids pSPM523, pSPM524, pSPM525 and pSPM528 were introduced into the strain OSC2. For this, protoplasts of strain OSC2 were transformed (Kieser, T et al., 2000) independently by each of the plasmids pSPM523, pSPM524, pSPM525 and pSPM528. A negative control was also carried out by transforming the strain OSC2 by the plasmid pUWL201 without insert. After transformation of the protoplasts, the clones are selected for thiostrepton resistance. Thus, five new strains were obtained: the OSC2 pUWL201 strain, resulting from the transformation by the plasmid pUWL201 without insert, the OSC2 strain pSPM523, resulting from the transformation with the plasmid pSPM523, the OSC2 strain pSPM524, resulting from the transformation with the plasmid pSPM524, the strain OSC2 pSPM525, resulting from the transformation with the plasmid pSPM525 and finally the OSC2 strain pSPM528, resulting from the transformation with the plasmid pSPM528. The spiramycin production of these strains was then analyzed by HPLC (in the same manner as in Example 17.2). Analysis of spiramycin production of strain OSC2 was also performed in parallel for comparison. The results of this analysis are shown in Table 41, the data are expressed in mg per liter of supernatant. The results correspond to the total production of spiramycins (obtained by adding spiramycin I, II and III production).
<Desc/Clms Page number 151><Desc / Clms Page number 151>
Tableau 41. Production en spiramycines des souches dérivées de OSC2, (résultats exprimés en mg/1).

<tb>
<tb> <Tb>
<Tb>
Souche <SEP> Production <SEP> en <SEP> spiramycines
<tb> OSC2 <SEP> 69
<tb> OSC2 <SEP> 103
<tb> pUWL201
<tb> OSC2 <SEP> 19
<tb> pSPM523
<tb> OSC2 <SEP> 135
<tb> pSPM524
<tb> OSC2 <SEP> 278
<tb> pSPM525
<tb> OSC2 <SEP> 68
<tb> pSPM528
<tb> Strain <SEP> Production <SEP> in <SEP> spiramycins
<tb> OSC2 <SEP> 69
<tb> OSC2 <SEP> 103
<tb> pUWL201
<tb> OSC2 <SEP> 19
<tb> pSPM523
<tb> OSC2 <SEP> 135
<tb> pSPM524
<tb> OSC2 <SEP> 278
<tb> pSPM525
<tb> OSC2 <SEP> 68
<tb> pSPM528
<Tb>
Ainsi, il est observé que la présence du plasmide pSPM524 ou du plasmide pSPM525 augmente significativement la production en spiramycines de la souche OSC2. Ceci démontre bien que la surexpression de OrflO a un effet positif sur la production en spiramycines. Le plasmide pSPM528 n'a par contre pas d'effet sur la production en spiramycines. Thus, it is observed that the presence of the plasmid pSPM524 or the plasmid pSPM525 significantly increases the spiramycin production of the strain OSC2. This demonstrates that overexpression of OrflO has a positive effect on spiramycin production. The plasmid pSPM528 has no effect on the production of spiramycins.
Les plasmides pSPM525 et pUWL201 ont de la même manière été introduit dans la souche SPM502 (cf. exemple 14). Ainsi deux nouvelles souches ont été obtenues : la souche SPM502 pUWL201, issue de la transformation par le plasmide Plasmids pSPM525 and pUWL201 were likewise introduced into strain SPM502 (see Example 14). Thus two new strains were obtained: the strain SPM502 pUWL201, resulting from the transformation with the plasmid
<Desc/Clms Page number 152><Desc / Clms Page number 152>
pUWL201 sans insert, et la souche SPM502 pSPM525, issue de la transformation par le plasmide pSPM525. pUWL201 without insert, and the strain SPM502 pSPM525, resulting from the transformation with the plasmid pSPM525.
Un échantillon de la souche SPM502 pSPM525 (cette souche contient le plasmide pSPM525, cf. ci-dessus) a été déposé auprès de la Collection Nationale de Cultures de Microorganismes (CNCM) Institut Pasteur, 25, rue du Docteur Roux 75724 Paris Cedex 15, France, le 26 février 2003 sous le numéro d'enregistrement 1-2977. A sample of the SPM502 pSPM525 strain (this strain contains the plasmid pSPM525, see above) has been deposited with the National Collection of Microorganism Cultures (CNCM) Institut Pasteur, 25, rue du Docteur Roux 75724 Paris Cedex 15, France, February 26, 2003 under the registration number 1-2977.
La production en spiramycines des souches SPM502 pUWL201 et SPM502 pSPM525 a été analysée par CLHP (de la même manière que dans l'exemple 17.2). The spiramycin production of strains SPM502 pUWL201 and SPM502 pSPM525 was analyzed by HPLC (in the same manner as in Example 17.2).
L' analyse de la production en spiramycines de la souche SPM502 a également été effectée en parallèle pour comparaison. Les résultats de cette analyse sont présentés dans le tableau 42, les données sont exprimées en mg par litre de surnageant. Les résultats correspondent à la production en spiramycine I. En effet, aucune de ces souches ne produit de spiramycine II et III. Analysis of the spiramycin production of the SPM502 strain was also performed in parallel for comparison. The results of this analysis are shown in Table 42, the data are expressed in mg per liter of supernatant. The results correspond to spiramycin I production. Indeed, none of these strains produce spiramycin II and III.
Tableau 42. Production en spiramycine 1 des souches dérivées de SPM502, (résultats exprimés en mg/1).

<tb>
<tb> <Tb>
<Tb>
Souche <SEP> Spiramycine <SEP> 1
<tb> SPM502 <SEP> 47
<tb> SPM502 <SEP> pUWL201 <SEP> 72
<tb> SPM502 <SEP> pSPM525 <SEP> 130
<tb> Strain <SEP> Spiramycin <SEP> 1
<tb> SPM502 <SEP> 47
<tb> SPM502 <SEP> pUWL201 <SEP> 72
<tb> SPM502 <SEP> pSPM525 <SEP> 130
<Tb>
Ainsi, il a pu être observé que la surexpression du gène orf10 dans la souche SPM502 augmente de facon importante la production de spiramycine I. Thus, it has been observed that the overexpression of the orf10 gene in the SPM502 strain significantly increases the production of spiramycin I.
<Desc/Clms Page number 153><Desc / Clms Page number 153>
EXEMPLE 18 : Construction d'une banque d'ADN génomique de la souche OSC2 de Streptomyces ambofaciens chez E. coli dans la cosmide pWED2. EXAMPLE 18 Construction of a genomic DNA library of Streptomyces ambofaciens strain OSC2 in E. coli in the cosmid pWED2.
18. 1 Construction du cosmide pWED2 :
Dans le but de faciliter l'inactivation de gènes chez Streptomyces, un cosmide portant la séquence oriT du plasmide RK2 (ce qui permet son introduction par conjugaison dans Streptomyces à partir d'une souche appropriée de E. coli) et portant également un gène de résistance à un antibiotique conférant un phénotype repérable chez Streptomyces, a été construit. Un tel cosmide contenant de grands inserts d'ADN génomique de Streptomyces ambofaciens peut être utilisé dans des expériences d'inactivation de gènes. 18. 1 Construction of the cosmid pWED2:
In order to facilitate the inactivation of genes in Streptomyces, a cosmid bearing the oriT sequence of plasmid RK2 (which allows its introduction by conjugation into Streptomyces from a suitable strain of E. coli) and also carrying a gene of resistance to an antibiotic conferring a detectable phenotype in Streptomyces, was constructed. Such a cosmid containing large genomic DNA inserts of Streptomyces ambofaciens can be used in gene inactivation experiments.
Pour construire ce vecteur, une cassette pac-oriT (fragment EcoRV) a été introduite dans le cosmide pWEDl (Gourmelen et al., 1998) , dérivé du cosmide pWED15 (Wahl et al., 1987), au niveau du site unique HpaI. La cassette pac-oriT a été obtenue par PCR. Pour cela, le gène pac a été amplifié par PCR à partir du plasmide pVF 10.4 (Vara et al., 1985 ; Lacalle et al., 1989) en utilisant comme première amorce, l'amorce A (de séquence 5'CCAGTAGATATCCCGCCAACCCGGAGCTGCAC-3' (SEQ ID N 126), le site de restriction EcoRV a été souligné et la séquence en gras de 20 nucléotides correspond à une région en amont du promoteur du gène pac) et comme deuxième amorce, l'amorce B (de séquence 5'GAAAAGATCCGTCATGGGGTCGTGCGCTCCTT-3' (SEQ ID N 127), qui comporte à son extrémité 5' une séquence de 12 nucléotides correspondant au début de la séquence oriT (souligné double) et une séquence de 20 nucléotides (en gras) correspondant à la fin du gènepac (cf. Figure 29, 1 ère PCR). To construct this vector, a pac-oriT cassette (EcoRV fragment) was introduced into cosmid pWED1 (Gourmelen et al., 1998), derived from the cosmid pWED15 (Wahl et al., 1987), at the unique HpaI site. The pac-oriT cassette was obtained by PCR. For this, the pac gene was amplified by PCR from the plasmid pVF 10.4 (Vara et al., 1985, Lacalle et al., 1989) using as primer the primer A (sequence 5'CCAGTAGATATCCCGCCAACCCGGAGCTGCAC-3 (SEQ ID NO: 126), the EcoRV restriction site was underlined and the 20 nucleotide fat sequence corresponds to a region upstream of the pac gene promoter) and as the second primer, the B (5 'sequence) primer GAAAAGATCCGTCATGGGGTCGTGCGCTCCTT-3 '(SEQ ID N 127), which comprises at its 5' end a sequence of 12 nucleotides corresponding to the beginning of the oriT sequence (double underlined) and a sequence of 20 nucleotides (in bold) corresponding to the end of the genepac (see Figure 29, 1st PCR).
Le gène oriT a quant à lui été amplifié par PCR à partir du plasmide pPM803 (Mazodier,P. et al., 1989) en utilisant comme première amorce, l'amorce C (de séquence 5'-CACGACCCCATGACGGATCTTTTCCGCTGCAT-3' (SEQ ID N 128)), qui comporte à son extrémité 5' une séquence de 12 nucléotides correspondant à une séquence en aval de la séquence codante du gène pac (en gras) et une séquence de The oriT gene was in turn amplified by PCR from plasmid pPM803 (Mazodier, P. et al., 1989) using, as first primer, primer C (sequence 5'-CACGACCCCATGACGGATCTTTTCCGCTGCAT-3 '(SEQ ID N 128)), which comprises at its 5 'end a sequence of 12 nucleotides corresponding to a sequence downstream of the coding sequence of the pac gene (in bold) and a sequence of
<Desc/Clms Page number 154><Desc / Clms Page number 154>
20 nucléotides correspondant au début de la séquence on?) et comme deuxième amorce, l'amorce D (de séquence 5'GAGCCGGATATCATCGGTCTTGCCTTGCTCGT-3' (SEQ ID N 129)) qui comporte le site de restriction EcoRV (souligné simple) et une séquence de 20 nucléotides correspondant à la fin de la séquence oriT (souligné double) (cf. Figure 29, 2ième PCR). 20 nucleotides corresponding to the beginning of the sequence on?) And as a second primer, the primer D (of sequence 5'GAGCCGGATATCATCGGTCTTGCCTTGCTCGT-3 '(SEQ ID N 129)) which comprises the restriction site EcoRV (underlined simple) and a sequence 20 nucleotides corresponding to the end of the oriT sequence (double underlined) (see Figure 29, 2nd PCR).
Le produit d'amplification obtenu avec les amorces A et B et celui obtenu avec les amorces C et D ont à une de leur extrémité, une séquence commune de 24 nucléotides. Une troisième PCR a été réalisée en mélangeant les deux produits d'amplification obtenus précédemment et en utilisant comme amorces, les amorces A et D (cf. figure 29, 3ième PCR). Ceci a permis d'obtenir un produit d'amplification correspondant à l'ensemble pac+oriT. Ce fragment pac-oriT a alors été clone dans le vecteur pGEM-T Easy (commercialisé par la société Promega (Madison, Wisconcin, USA)), ce qui a permis d'obtenir le plasmide pGEM-T-pac-oriT. L'insert de ce plasmide a ensuite été sous cloné dans le cosmide pWEDl. Pour cela le plasmide pGEM-pac-oriT a été digéré par l'enzyme EcoRV et l'insert EcoRV contenant l'ensemble pac+oriT a été inséré dans le cosmide pWEDl ouvert au préalable par l'enzyme HpaI. Le cosmide ainsi obtenu a été baptisé pWED2 (cf. Figure 30). The amplification product obtained with primers A and B and that obtained with primers C and D have at one end a common sequence of 24 nucleotides. A third PCR was performed by mixing the two amplification products obtained previously and using primers A and D as primers (see FIG. 29, 3rd PCR). This made it possible to obtain an amplification product corresponding to the pac + oriT set. This pac-oriT fragment was then cloned into the vector pGEM-T Easy (marketed by Promega (Madison, Wisconcin, USA)), which made it possible to obtain the plasmid pGEM-T-pac-oriT. The insert of this plasmid was then subcloned into cosmid pWED1. For this, the plasmid pGEM-pac-oriT was digested with the enzyme EcoRV and the EcoRV insert containing the pac + oriT set was inserted into the cosmid pWED1 previously opened with the enzyme HpaI. The cosmid thus obtained was named pWED2 (see Figure 30).
Ce cosmide permet de faciliter l'inactivation de gènes chez Streptomyces. This cosmid makes it easier to inactivate genes in Streptomyces.
En effet, il porte la séquence oriT (ce qui permet son introduction par conjugaison dans Streptomyces à partir d'une souche appropriée de E. coli) mais également un gène de résistance à un antibiotique conférant un phénotype repérable chez Streptomyces. Un tel cosmide contenant de grands inserts d'ADN génomique de Streptomyces ambofaciens peut être utilisé dans des expériences d'inactivation de gènes. Indeed, it carries the oriT sequence (which allows its introduction by conjugation into Streptomyces from a suitable strain of E. coli) but also an antibiotic resistance gene conferring a detectable phenotype in Streptomyces. Such a cosmid containing large genomic DNA inserts of Streptomyces ambofaciens can be used in gene inactivation experiments.
Ainsi un cosmide dérivé de pWED2 contenant le gène cible pourra par exmeple être introduit dans la souche de E. coli KS272 contenant le plasmide pKOBEG (cf. Chaveroche et al. 2000) et une cassette sera introduite dans le gène cible selon la technique décrite par Chaveroche et al. 2000. Le cosmide obtenu par cette technqiue (cosmide dans lequel le gène cible est inactivé), pourra ensuite être Thus a cosmid derived from pWED2 containing the target gene may, for example, be introduced into the E. coli strain KS272 containing the plasmid pKOBEG (see Chaveroche et al., 2000) and a cassette will be introduced into the target gene according to the technique described by Chaveroche et al. 2000. The cosmid obtained by this technic (cosmid in which the target gene is inactivated), can then be
<Desc/Clms Page number 155><Desc / Clms Page number 155>
introduit dans une souche de E. coli telle que la souche S 17.1 ou toute autre souche permettant de transférer par conjugaison des plasmides contenant la séquence oriT vers Streptomyces. introduced into a strain of E. coli such as strain S 17.1 or any other strain which makes it possible to transfer, by conjugation, plasmids containing the oriT sequence to Streptomyces.
Après conjugaison entre E. coli et Streptomyces, des clones de Streptomyces dans lesquelles la copie sauvage du gène cible aura été remplacée par la copie interrompue pourront être obtenues comme décrit dans l'exemple 2. After conjugation between E. coli and Streptomyces, Streptomyces clones in which the wild-type copy of the target gene has been replaced by the interrupted copy can be obtained as described in Example 2.
Le gène de résistance exprimé chez Streptomyces présent sur ce nouveau cosmide est le gène pac de Streptomyces alboniger (Vara, J et al., 1985 ; Lacalle et al., 1989) qui code la puromycine N-acétyl transférase et qui confère la résistance à la puromycine. Lors d'expériences d'inactivation de gène, on cherchera les clones dans lesquels un double événement de recombinaison aura eu lieu. Ces clones auront éliminer le cosmide pWED2 et seront donc redevenus sensibles à la puromycine. The resistance gene expressed in Streptomyces present on this new cosmid is the pac gene of Streptomyces alboniger (Vara, J et al., 1985, Lacalle et al., 1989) which encodes puromycin N-acetyltransferase and which confers resistance to puromycin. In gene inactivation experiments, we will look for clones in which a double recombination event has occurred. These clones will have eliminated cosmid pWED2 and will therefore become sensitive to puromycin again.
18. 2 Construction d'une banque d'ADN génomique de la souche OSC2 de Streptomyces ambofaciens chez E. coli dans le cosmide pWED2
L'ADN génomique de la souche OSC2 de Streptomyces ambofaciens a été digéré partiellement par l'enzyme de restriction BamHI de manière à obtenir des fragments d'ADN de taille comprise entre environ 35 et 45 kb. Ces fragments ont ensuite été clonés dans le cosmide pWED2, ce dernier ayant été au préalable digéré par BamHI, puis traité par la phosphatase alcaline. Le mélange de ligation a ensuite été encapsidé in vitro dans des particules de phages lambda grâce au système Packagene Lambda DNA packaging système commercialisé par la société Promega (Madison, Wisconcin, USA) en suivant les recommandations du fabriquant. Les particules phagiques obtenues ont été utilisées pour infecter la souche SURE@ d'E. coli commercialisée par la société Stratagene (LaJolla, Californie, USA). La séléction des clones a été effectuée sur milieu LB + ampicilline (50 g/ml), le cosmide pWED2 conférant la résistance à l'ampicilline. 18. 2 Construction of a genomic DNA library of the OSC2 strain of Streptomyces ambofaciens in E. coli in the cosmid pWED2
The genomic DNA of Streptomyces ambofaciens strain OSC2 was partially digested with BamHI restriction enzyme to obtain DNA fragments of size between about 35 and 45 kb. These fragments were then cloned into cosmid pWED2, the latter having been digested beforehand with BamHI and then treated with alkaline phosphatase. The ligation mixture was then packaged in vitro in lambda phage particles using the Packagene Lambda DNA packaging system sold by Promega (Madison, Wisconcin, USA) according to the manufacturer's recommendations. The phage particles obtained were used to infect the strain SURE @ E. coli marketed by Stratagene (LaJolla, California, USA). The selection of the clones was carried out on LB + ampicillin medium (50 g / ml), the cosmid pWED2 conferring resistance to ampicillin.
<Desc/Clms Page number 156> <Desc / Clms Page number 156>
EXEMPLE 19 : Isolement de cosmides de la nouvelle banque couvrant la région de la voie de biosynthèse des spiramycines
19. 1 Hybridation sur colonies de la banque génomique de Streptomyces ambofaciens OSC2 :
Des cosmides de la nouvelle banque de Streptomyces ambofaciens OSC2 (cf. exemple 18) couvrant les orfl * à orf10* ou une partie ou la totalité des orfl à orf25c, ou une région plus en amont de l'orf25c ont été isolés. Pour cela, des hybridations successives sur les colonies d'E. coli obtenues selon l'exemple 18 ont été réalisées en utilisant les 3 sondes suivantes (cf. figure 31): - La première sonde utilisée correspond à un fragment d'ADN d'environ 0,8kb amplifié par PCR en utilisant comme matrice le cosmide pSPM5 et les amorces suivantes : ORF23c: 5'-ACGTGCGCGGTGAGTTCGCCGTTGC-3' (SEQ ID N 130) et ORF25c : 5'-CTGAACGACGCCATCGCGGTGGTGC-3' (SEQ ID N 131). EXAMPLE 19 Isolation of cosmids from the new library covering the region of the spiramycin biosynthetic pathway
19. 1 Colony hybridization of the genomic library of Streptomyces ambofaciens OSC2:
Cosmids from the new Streptomyces ambofaciens OSC2 library (see Example 18) covering orf10 * or a part or all orf25c or a region further upstream of orf25c were isolated. For this, successive hybridizations on E. coli colonies. coli obtained according to Example 18 were carried out using the following 3 probes (see FIG. 31): the first probe used corresponds to a DNA fragment of approximately 0.8 kb amplified by PCR using the cosmid as matrix pSPM5 and the following primers: ORF23c: 5'-ACGTGCGCGGTGAGTTCGCCGTTGC-3 '(SEQ ID NO: 130) and ORF25c: 5'-CTGAACGACGCCATCGCGGTGGTGC-3' (SEQ ID NO: 131).
Le produit de PCR ainsi obtenu contient un fragment du début de l'orf23c, l'orf24c en entier et la fin de l'orf25c (cf. figure 31, sonde I). The PCR product thus obtained contains a fragment of the beginning of orf23c, orf24c in its entirety and the end of orf25c (see Figure 31, probe I).
- La deuxième sonde utilisée correspond à un fragment d'ADN d'environ 0,7kb amplifié par PCR en utilisant comme matrice l'ADN total de la souche S. ambofaciens ATCC23877 et les amorces suivantes : ORFl*c: 5'-GACCACCTCGAACCGTCCGGCGTCA-3' (SEQ ID N 132) et ORF2*c : 5'-GGCCCGGTCCAGCGTGCCGAAGC-3' (SEQ ID N 133). The second probe used corresponds to a DNA fragment of approximately 0.7 kb amplified by PCR using as template the total DNA of the S. ambofaciens strain ATCC23877 and the following primers: ORF1 * c: 5'-GACCACCTCGAACCGTCCGGCGTCA- 3 '(SEQ ID NO: 132) and ORF2 * c: 5'-GGCCCGGTCCAGCGTGCCGAAGC-3' (SEQ ID NO: 133).
Le produit de PCR ainsi obtenu contient un fragment de la fin de l'orfl *c et du début de l'orf2*c (cf. figure 31, sonde II). The PCR product thus obtained contains a fragment of the end of the orfl * c and the beginning of orf2 * c (see Figure 31, probe II).
- La troisième sonde utilisée correspond à un fragment EcoRI-BamHI d'environ 3kb contenant les orfl, or/2 et orf3 et obtenu par digestion du plasmide pOS49.99 (cf. figure 31, sonde III). The third probe used corresponds to an EcoRI-BamHI fragment of approximately 3 kb containing the orfl, or / 2 and orf3 and obtained by digestion of the plasmid pOS49.99 (see FIG. 31, probe III).
<Desc/Clms Page number 157> <Desc / Clms Page number 157>
Environ 2000 clones de la banque obtenue dans l'exemple 18. 2 ont été transférés sur filtre pour hybridation sur colonies selon les techniques classiques (Sambrook et al., 1989). About 2000 clones of the library obtained in Example 18. 2 were transferred to a colony hybridization filter according to standard techniques (Sambrook et al., 1989).
La première sonde (cf. figure 31, sonde I) a été marquée au 32P par la technique du random priming (Kit commercialisé par la société Roche) et utilisées pour l'hybridation sur 2000 clones de la banque, après transfert sur filtre. L'hybridation a été effectuée à 65 C dans le tampon décrit par Church & Gilbert (Church & Gilbert, 1984). Deux lavages ont été effectués en 2x SSC, 0,1% SDS à 65 C, le premier pendant 10 minutes et le deuxième pendant 20 minutes et un troisième lavage d'une durée de 30 minutes a ensuite été effectué en 0. 2X SSC, 0,1% SDS à 65 C. Dans ces conditions d'hybridation et de lavage, 20 clones parmi les 2000 hybridés présentaient un fort signal d'hybridation avec la première sonde. Ces 20 clones ont été cultivés en milieu LB+ ampicilline (50 g/ml) et les 20 cosmides correspondants ont été extraits par lyse alcaline (Sambrook et al., 1989) puis ils ont été digérés par l'enzyme de restriction BamHI. Les produits de digestions ont ensuite été séparés sur gel d'agarose, transférés sur membrane de Nylon et hybridés par la première sonde (cf. ci-dessus : le produit de PCR ORF23c-ORF25c, sonde I) dans les mêmes conditions que ci-dessus. Douze cosmides possédaient un fragment BamHI hybridant fortement avec la sonde utilisée. The first probe (see FIG. 31, probe I) was labeled with 32P by the random priming technique (Kit marketed by Roche) and used for hybridization on 2000 clones of the library, after transfer to a filter. Hybridization was performed at 65 ° C in the buffer described by Church & Gilbert (Church & Gilbert, 1984). Two washes were performed at 2x SSC, 0.1% SDS at 65 ° C, the first for 10 minutes and the second for 20 minutes and a third 30-minute wash was then performed at 0. 2 × SSC, 0.1% SDS at 65 ° C. Under these hybridization and washing conditions, 20 of the 2000 hybridized clones showed a strong hybridization signal with the first probe. These clones were cultured in LB + ampicillin medium (50 g / ml) and the corresponding cosmids were extracted by alkaline lysis (Sambrook et al., 1989) and then digested with BamHI restriction enzyme. The digests were then separated on agarose gel, transferred onto a nylon membrane and hybridized with the first probe (see above: the ORF23c-ORF25c PCR product, probe I) under the same conditions as above. above. Twelve cosmids had a BamHl fragment hybridizing strongly with the probe used.
Ces 12 cosmides ont été nommés pSPM34, pSPM35, pSPM36, pSPM37, pSPM38, pSPM39, pSPM40, pSPM41, pSPM42, pSPM43, pSPM44 et pSPM45. Les profils, après digestion par BamHI, de ces 12 cosmides ont été comparés entre eux et avec celui du cosmide pSPM5. De plus des expériences d'amplification par PCR en utilisant différentes amorces correspondant à différents gènes déjà identifiés dans la région orflorf25c ont permis de positionner l'insert de certains de ces cosmides les uns par rapport aux autres et de déterminer également la localisation de ces inserts dans la région orflorf25c déjà connue (cf. Figure 32). Le cosmide pSPM36 a plus particulièrement été choisi car il était susceptible de contenir une grande région en amont de l'orf25c (cf. These 12 cosmids were named pSPM34, pSPM35, pSPM36, pSPM37, pSPM38, pSPM39, pSPM40, pSPM41, pSPM42, pSPM43, pSPM44 and pSPM45. The profiles, after digestion with BamHI, of these 12 cosmids were compared with each other and with that of cosmid pSPM5. Moreover PCR amplification experiments using different primers corresponding to different genes already identified in the orflorf25c region have made it possible to position the insert of some of these cosmids with respect to each other and also to determine the location of these inserts. in the orflorf25c region already known (see Figure 32). The cosmid pSPM36 was particularly chosen because it was likely to contain a large region upstream of the orf25c (cf.
Figure31 et 32). Figure31 and 32).
<Desc/Clms Page number 158> <Desc / Clms Page number 158>
Dans un deuxième temps, et en utilisant les mêmes conditions que celles décrites ci-dessus, les 2000 clones de la banque de Streptomyces ambofaciens OSC2 ont été hybridés avec la deuxième sonde correspondant au produit de PCR : ORF1*c- ORF2*c (cf. figure 31, sonde II). Cette hybridation a permis d'isoler des cosmides dont l'insert est situé dans la région de orfl*c à orf10*c. Dans les conditions d'hybridation utilisés, 16 clones parmi les 2000 hybridés présentaient un fort signal d'hybridation avec cette deuxième sonde. Ces 16 clones ont été cultivés en milieu LB+ ampicilline (50 p,g/ml) et les 16 cosmides correspondants ont été extraits par lyse alcaline (Sambrook et al., 1989) puis ils ont été digérés par l'enzyme de restriction BamHI. Les profils de digestion (après digestion par BamHI) de ces 16 cosmides ont été comparés entre eux et avec celui du cosmide pSPM7. Des expériences d'amplication par PCR avec les amorces ORFl*c et ORF2*c ont permis de choisir deux cosmides qui renfermaient bien les gènes orf1*c et orf2*c et dont les profils possédaient des bandes communes mais aussi des bandes différentes. De plus d'autres expériences d'amplification par PCR en utilisant différentes amorces correspondant à différents gènes déjà identifiés dans la région orf1*c à orfl0*c ont permis de positionner l'insert de ces cosmides les uns par rapport aux autres et de déterminer également la localisation de ces inserts dans la région orfl *c à orfl0*c déjà connue (cf. Figure 32). Les deux cosmides plus particulièrement sélectionnés ont été nommés pSPM47 et pSPM48 (cf. Figure 32). In a second step, and using the same conditions as those described above, the 2000 clones of the Streptomyces ambofaciens OSC2 library were hybridized with the second probe corresponding to the PCR product: ORF1 * c-ORF2 * c (cf. Figure 31, probe II). This hybridization made it possible to isolate cosmids whose insert is located in the orfl * c region at orf10 * c. Under the hybridization conditions used, 16 of the 2000 hybridized clones showed a strong hybridization signal with this second probe. These 16 clones were cultured in LB + ampicillin medium (50 μg / ml) and the corresponding 16 cosmids were extracted by alkaline lysis (Sambrook et al., 1989) and then digested with BamHI restriction enzyme. The digestion profiles (after digestion with BamHI) of these 16 cosmids were compared with each other and with that of cosmid pSPM7. PCR amplification experiments with the ORF1 * c and ORF2 * c primers made it possible to choose two cosmids which contained the orf1 * c and orf2 * c genes and whose profiles had common bands but also different bands. In addition to other PCR amplification experiments using different primers corresponding to different genes already identified in the region orf1 * c to orfl0 * c have made it possible to position the insert of these cosmids with respect to each other and to determine also the location of these inserts in the region orfl * c to orfl0 * c already known (see Figure 32). The two most particular cosmids selected were named pSPM47 and pSPM48 (see Figure 32).
En utilisant les mêmes conditions que celles décrites ci-dessus, les 2000 clones de la banque de Streptomyces ambofaciens OSC2 ont également été hybridés avec la troisième sonde correspondant au fragment d'ADN EcoRI-BamHI du plasmide pOS49. 99 (cf. figure 31 sonde ni). Cette hybridation a permis d'isoler les cosmides contenant la région de l'orfl jusqu'à l'orf3 et susceptibles de contenir soit une grande partie des gènes des PKS, soit une grande partie des gènes orfl à orf25c de la voie de biosynthèse des spiramycines. Dans ces conditions d'hybridation, 35 clones parmi les 2000 hybridés présentaient un fort signal d'hybridation avec la troisième sonde. Ces 35 clones ont été cultivés en milieu LB+ ampicilline (50 g/ml) et les 35 cosmides correspondants ont été extraits par lyse alcaline (Sambrook et al., 1989) puis digérés par Using the same conditions as those described above, the 2000 clones of the Streptomyces ambofaciens OSC2 library were also hybridized with the third probe corresponding to the EcoRI-BamHI DNA fragment of the plasmid pOS49. 99 (see figure 31 probe ni). This hybridization made it possible to isolate the cosmids containing the orf3 orf3 region and likely to contain either a large part of the PKS genes, or a large part of the orf25c genes of the biosynthesis pathway. spiramycin. Under these hybridization conditions, 35 of the hybridized 2000 clones showed a strong hybridization signal with the third probe. These 35 clones were cultured in LB + ampicillin medium (50 g / ml) and the corresponding cosmids were extracted by alkaline lysis (Sambrook et al., 1989) and then digested with
<Desc/Clms Page number 159><Desc / Clms Page number 159>
l'enzyme de restriction BamHI. Les profils, après digestion par BamHI, de ces 35 cosmides ont été comparés entre eux et avec celui du cosmide pSPM5. De plus des expériences d'amplification par PCR en utilisant différentes amorces correspondant à différents gènes déjà identifiés dans la région orfl à orf25c ont permis de vérifier que ces cosmides renfermaient bien des inserts provenant de la région orfl à orf25c et de positionner les inserts de ces cosmides les uns par rapport aux autres et par rapport à la la région orf1 à orf25c déjà connue (cf. Figure 32). Cinq cosmides ont été plus particulièrement sélectionnés, ils ont été nommés pSPM50, pSPM51, pSPM53, pSPM55 et pSPM56 (cf. Figure 32). the BamHI restriction enzyme. The profiles, after digestion with BamHI, of these 35 cosmids were compared with each other and with that of the cosmid pSPM5. In addition, PCR amplification experiments using different primers corresponding to different genes already identified in the orf1 to c25c region, made it possible to verify that these cosmids contained well inserts from the orf1c to c25c region and to position the inserts of these inserts. cosmid with respect to each other and with respect to the region orf1 to orf25c already known (see Figure 32). Five cosmids were more particularly selected, they were named pSPM50, pSPM51, pSPM53, pSPM55 and pSPM56 (see Figure 32).
19. 2 Sous clonage et séquençage d'une partie de l'insert du cosmide pSPM36. 19. 2 Subcloning and sequencing of a portion of the cosmid insert pSPM36.
La sonde d'environ 0,8 kb obtenue par PCR avec les amorces ORF23c et ORF25c (cf. ci-dessus et figure 31, sonde I) a également été utilisée dans des expériences de Southern Blot sur l'ADN total de S. ambofaciens OSC2 digéré par l'enzyme PstI. Dans les conditions d'hybridation décrite ci-dessus, cette sonde révèle un fragment unique PstI d'environ 6 kb lorsque hybridé sur l'ADN total de S. ambofaciens OSC2 digéré par l'enzyme PstI. Il existe un site Pstl au niveau de l'orf23c (cf. SEQ ID N 80) mais aucun autre site Pstl jusqu'à l'extrémité (site BamHI) de la séquence connue (cf. SEQ ID N 1). Ce fragment PstI-BamHI a une taille d'environ 1. 4 kb. Le fragment PstI de 6 kb hybridé sur l'ADN total de S. ambofaciens digéré par l'enzyme PstI contient donc une région d'environ 4,6 kb située en amont de orj25c. Cette région est susceptible de contenir d'autres gènes dont les produits sont impliqués dans la voie de biosynthèse de la spiramycine. Il a été vérifié par digestion que le cosmide pSPM36 contenait bien ce fragment Pstl de 6 kb. Ce fragment a été isolé à partir de pSPM36, dans le but de déterminer la séquence plus en amont de orf25c. Pour cela, le cosmide pSPM36 a été digéré par l'enzyme de restriction PstI Le fragment Pstl-Pstl d'une taille d'environ 6kb a été isolé par électroélution à partir d'un gel d'agarose 0. 8% puis cloné dans le vecteur pBK-CMV (4512bp) (commercialisé par la société Stratagene (La Jolla, Californie, USA)). Le plasmide ainsi obtenu a été nommé pSPM58 (cf. Figure 33) et la séquence de son insert a été déterminée. La séquence de cet insert est présentée en SEQ ID N 134. The probe of approximately 0.8 kb obtained by PCR with primers ORF23c and ORF25c (see above and FIG. 31, probe I) was also used in Southern blot experiments on the total DNA of S. ambofaciens. OSC2 digested with PstI enzyme. Under the hybridization conditions described above, this probe reveals a unique PstI fragment of about 6 kb when hybridized to the total DNA of S. ambofaciens OSC2 digested with the enzyme PstI. There is a PstI site at the level of orf23c (see SEQ ID No. 80) but no other PstI site up to the end (BamHI site) of the known sequence (see SEQ ID No. 1). This PstI-BamHI fragment has a size of about 1.4 kb. The 6 kb PstI fragment hybridized to the total DNA of S. ambofaciens digested with the PstI enzyme therefore contains a region of approximately 4.6 kb located upstream of orj25c. This region is likely to contain other genes whose products are involved in the spiramycin biosynthetic pathway. It was verified by digestion that cosmid pSPM36 did indeed contain this 6 kb PstI fragment. This fragment was isolated from pSPM36, in order to determine the sequence further upstream of orf25c. For this, the cosmid pSPM36 was digested with the PstI restriction enzyme. The PstI-PstI fragment of a size of approximately 6 kb was isolated by electroelution from a 0.8% agarose gel and then cloned into pBK-CMV vector (4512bp) (marketed by Stratagene (La Jolla, California, USA)). The plasmid thus obtained was named pSPM58 (see FIG. 33) and the sequence of its insert was determined. The sequence of this insert is presented in SEQ ID N 134.
<Desc/Clms Page number 160> <Desc / Clms Page number 160>
Toutefois, toute la séquence n'a pas été déterminée et il subsite un trou d'environ 450 nucléotides, la partie de la séquence non déterminée a été notée par une succession de N dans la séquence correspondante. However, the entire sequence was not determined and it subsite a hole of about 450 nucleotides, the part of the undetermined sequence was noted by a succession of N in the corresponding sequence.
19. 3 Analyse des nouvelles séquences nucléotidiques, détermination des phases ouvertes de lecture et caractérisation des gènes impliqués dans la biosynthèse des spiramycines. 19. 3 Analysis of new nucleotide sequences, determination of open reading phases and characterization of genes involved in the biosynthesis of spiramycins.
La séquence de l'insert du cosmide pSPM58 obtenue a été analysée grâce au programme FramePlot (Ishikawa J & Hotta K. 1999). Ceci a permis d'identifier, parmi les phases ouvertes de lecture, les phases ouvertes de lecture présentant un usage des codons typique de Streptomyces. Cette analyse a permis de déterminer que cet insert comporte 4 nouvelles ORFs en amont de l'orf25c (cf. figure 34). Ces gènes ont été nommés or/26 (SEQ ID N 107), or/27 (SEQ ID N 109), orj28c (SEQ ID N 111, la séquence de cette orf n'a pas été déterminée complètement puisqu'un trou d'environ 450 nucléotide subsite lors du séquençage de l'insert de pSPM58, ces 450 nucléotides figurent sous la forme d'une sére de N dans la séquence SEQ ID N 111) et orf29 (la séquence de cette dernière orf était incomplète dans cette insert). Le c ajouté dans le nom du gène signifiant pour l'ORF en question que la séquence codante est dans l'orientation inverse (cf. figure 34). The sequence of the cosmid insert pSPM58 obtained was analyzed using the FramePlot program (Ishikawa J & Hotta K. 1999). This made it possible to identify, among the open reading phases, the open reading stages with typical codon usage of Streptomyces. This analysis determined that this insert has 4 new ORFs upstream of the orf25c (see Figure 34). These genes were named gold / 26 (SEQ ID N 107), or / 27 (SEQ ID N 109), orj28c (SEQ ID N 111, the sequence of this orf was not completely determined since a hole of about 450 nucleotide subsite during the sequencing of the insert of pSPM58, these 450 nucleotides are in the form of a segre of N in the sequence SEQ ID N 111) and orf29 (the sequence of this latter orf was incomplete in this insert) . The c added in the name of the gene means for the ORF in question that the coding sequence is in the reverse orientation (see Figure 34).
Les séquences protéiques déduites de ces phases ouvertes de lecture ont été comparées avec celles présentes dans différentes bases de données grâce à différents programmes : BLAST (Altschul et al., 1990) (Altschul et al., 1997), CD-search, (ces deux programmes sont accessibles notamment auprès du National Center for Biotechnology Information (NCBI) (Bethesda, Maryland, USA)), FASTA ((Pearson W. The protein sequences deduced from these open reading phases were compared with those present in different databases through different programs: BLAST (Altschul et al., 1990) (Altschul et al., 1997), CD-search, (these two programs are available from the National Center for Biotechnology Information (NCBI) (Bethesda, Maryland, USA), FASTA (Pearson W.
R & D. J. Lipman, 1988) et (Pearson W. R., 1990) (ce programme est accessible notamment auprès du centre de ressources INFOBIOGEN, Evry, France). Ces comparaisons ont permis de formuler des hypothèses sur la fonction des produits de ces gènes et d'identifier ceux susceptibles d'être impliqués dans la biosynthèse de spiramycines. R & D. J. Lipman, 1988) and (Pearson W. R., 1990) (this program is accessible in particular from the INFOBIOGEN resource center, Evry, France). These comparisons made it possible to formulate hypotheses on the function of the products of these genes and to identify those likely to be involved in the biosynthesis of spiramycins.
<Desc/Clms Page number 161> <Desc / Clms Page number 161>
19. 4 Sous clonage et séquençage d'une autre partie de l'insert du cosmide pSPM36. 19. 4 Subcloning and sequencing another part of cosmid insert pSPM36.
La sonde d'environ 0,8 kb obtenue par PCR avec les amorces ORF23c et ORF25c (cf. ci-dessus et figure 31, sonde 1) a également été utilisée dans des expériences de Southern Blot sur l'ADN total de la souche OSC2 digéré par l'enzyme StuI. Dans les conditions d'hybridation décrite ci-dessus pour cette sonde, cette sonde révèle un fragment unique StuI d'environ 10 kb lorsque hybridé sur l'ADN total de la souche OSC2 digéré par l'enzyme StuI. Compte tenu de la présence d'un site StuI dans l'orf23c (cf. SEQ ID N 80) et de la localisation de ce site par rapport au site PstI, ce fragment de 10 kb inclut la totalité du fragment PstI précédemment étudié (insert de pSPM58) et permet d'avoir accès à une région non encore étudiée d'environ 4 kb. (cf figure 33). Il a été vérifié par digestion que le cosmide pSPM36 contenait bien ce fragment StuI de 10 kb. Ce fragment a été isolé à partir du cosmide pSPM36, dans le but de déterminer la séquence de la fin d'or/29 et d'autres gènes plus en amont de orfl9. Pour cela, le cosmide pSPM36 a été digéré par l'enzyme de restriction StuI. Le fragment StuI-StuI d'une taille d'environ 10 kb a été isolé par électroélution à partir d'un gel d'agarose 0. 8% puis cloné dans le vecteur pBK-CMV (4512bp) (commercialisé par la société Stratagene (La Jolla, Californie, USA)). Le plasmide ainsi obtenu a été nommé pSPM72 (cf. Figure 33). Ce dernier a ensuite été digéré par EcoRI (site EcoRI dans l'insert de pSPM58) et HindIII (site HindIII dans le site de clonage multiple du vecteur, immédiatement après le site StuI de l'extrémité de l'insert) (cf. Figure 33). Le fragment d'ADN EcoRI-HindIII ainsi obtenu, correspond à un fragment de l'insert du plasmide pSPM72 (cf. figure 33) et a été sous-cloné dans le vecteur pBC-SK+ (commercialisé par la société Stratagene (La Jolla, Californie, USA)) digéré au préalable par EcoRI et HindIII. Le plasmide ainsi obtenu a été nommé pSPM73 et la séquence de son insert a été déterminée. La séquence de cet insert est présentée en SEQ ID N 135. The probe of about 0.8 kb obtained by PCR with primers ORF23c and ORF25c (see above and FIG. 31, probe 1) was also used in Southern blot experiments on the total DNA of the OSC2 strain. digested with StuI enzyme. Under the hybridization conditions described above for this probe, this probe reveals a unique StuI fragment of about 10 kb when hybridized to the total DNA of the StuI enzyme-digested OSC2 strain. Given the presence of a StuI site in orf23c (see SEQ ID No. 80) and the location of this site with respect to the PstI site, this 10 kb fragment includes all of the previously studied PstI fragment (insert of pSPM58) and allows access to an untested region of about 4 kb. (see Figure 33). It was verified by digestion that the cosmid pSPM36 contained this StuI fragment of 10 kb. This fragment was isolated from the cosmid pSPM36, in order to determine the sequence of the late / 29 and other genes further upstream of orfl9. For this, the cosmid pSPM36 was digested with the StuI restriction enzyme. The StuI-StuI fragment of a size of approximately 10 kb was isolated by electroelution from an 0.8% agarose gel and then cloned into the vector pBK-CMV (4512bp) (sold by the company Stratagene ( La Jolla, California, USA)). The plasmid thus obtained was named pSPM72 (see Figure 33). The latter was then digested with EcoRI (EcoRI site in the insert of pSPM58) and HindIII (HindIII site in the multiple cloning site of the vector, immediately after the StuI site of the end of the insert) (see FIG. 33). The EcoRI-HindIII DNA fragment thus obtained corresponds to a fragment of the insert of the plasmid pSPM72 (see FIG. 33) and was subcloned into the vector pBC-SK + (sold by the company Stratagene (La Jolla, California, USA)) previously digested with EcoRI and HindIII. The plasmid thus obtained was named pSPM73 and the sequence of its insert was determined. The sequence of this insert is presented in SEQ ID N 135.
<Desc/Clms Page number 162> <Desc / Clms Page number 162>
19. 5 Analyse des nouvelles séquences nucléotidiques, détermination des phases ouvertes de lecture et caractérisation des gènes impliqués dans la biosynthèse des spiramycines. 19. 5 Analysis of new nucleotide sequences, determination of open reading phases and characterization of genes involved in spiramycin biosynthesis.
La séquence partielle de l'insert du cosmide pSPM73 obtenue a été analysée grâce au programme FramePlot (Ishikawa J & Hotta K. 1999). Ceci a permis d'identifier, parmi les phases ouvertes de lecture, les phases ouvertes de lecture présentant un usage des codons typique de Streptomyces. Cette analyse a permis de déterminer que cet insert comporte 4 ORFs, une incomplètes et trois complètes (cf. figure 34). L'ORF incomplète correspond à la partie 3' de la séquence codante de or/29 ce qui à permis de compléter la séquence de ce gène grâce à la séquence partielle de ce même orf obtenue lors du séquençage de l'insert du plasmide pSPM58 (cf. exemple 19. 2 et 19. 3), l'ensemble des deux séquences à ainsi permis d'obtenir la séquence complète de orj29. Les 4 gènes ont ainsi été nommés or/29 (SEQ ID N 113), orf30c (SEQ ID N 115), or/31 (SEQ ID N 118) et orf32c (SEQ ID N 120). Le c ajouté dans le nom du gène signifiant pour l'ORF en question que la séquence codante est dans l'orientation inverse (cf. figure 34). The partial sequence of the cosmid insert pSPM73 obtained was analyzed using the FramePlot program (Ishikawa J & Hotta K. 1999). This made it possible to identify, among the open reading phases, the open reading stages with typical codon usage of Streptomyces. This analysis determined that this insert has 4 ORFs, one incomplete and three complete (see Figure 34). The incomplete ORF corresponds to the 3 'portion of the or / 29 coding sequence which made it possible to complete the sequence of this gene by means of the partial sequence of this same orf obtained during the sequencing of the insert of plasmid pSPM58 ( see example 19. 2 and 19. 3), the combination of the two sequences thus made it possible to obtain the complete sequence of orj29. The 4 genes have been named gold / 29 (SEQ ID No. 113), orf30c (SEQ ID No. 115), or / 31 (SEQ ID No. 118) and orf32c (SEQ ID No. 120). The c added in the name of the gene means for the ORF in question that the coding sequence is in the reverse orientation (see Figure 34).
Les séquences protéiques déduites de ces phases ouvertes de lecture ont été comparées avec celles présentes dans différentes bases de données grâce à différents programmes : BLAST (Altschul et al., 1990) (Altschul et al., 1997), CD-search, (ces deux programmes sont accessibles notamment auprès du National Center for Biotechnology Information (NCBI) (Bethesda, Maryland, USA)), FASTA ((Pearson W. R & D. J. Lipman, 1988) et (Pearson W. R., 1990) (ce programme est accessible notamment auprès du centre de ressources INFOBIOGEN, Evry, France). Ces comparaisons ont permis de formuler des hypothèses sur la fonction des produits de ces gènes et d'identifier ceux susceptibles d'être impliqués dans la biosynthèse de spiramycines. The protein sequences deduced from these open reading phases were compared with those present in different databases through different programs: BLAST (Altschul et al., 1990) (Altschul et al., 1997), CD-search, (these two programs are available from the National Center for Biotechnology Information (NCBI) (Bethesda, Maryland, USA), FASTA (Pearson W. R & DJ Lipman, 1988) and (Pearson WR, 1990). These comparisons made it possible to formulate hypotheses on the function of the products of these genes and to identify those likely to be involved in spiramycin biosynthesis.
<Desc/Clms Page number 163> <Desc / Clms Page number 163>
EXEMPLE 20 : Analyse de la production d'intermédiaires de biosynthèse de spiramycines :
20. 1 Préparation des échantillons :
Les différentes souches à tester ont été cultivées chacune dans 7 erlenmeyers chicanés de 500 ml contenant 70 ml de milieu MP5 (Pemodet et al., 1993). Les erlens ont été inoculés par 2.5 106spores/ml des différentes souches de S. ambofaciens et mises à pousser à 27 C sous agitation orbitale de 250 rpm pendant 96 heures. Les cultures correspondant au même clone sont rassemblées, éventuellement filtrées sur filtre plissé, et centrifugées 15 min à 7000 tr/min. EXAMPLE 20 Analysis of the Production of Spiramycin Biosynthetic Intermediates:
20. 1 Sample preparation:
The different strains to be tested were each cultured in 7 500 ml flasked Erlenmeyer flasks containing 70 ml of MP5 medium (Pemodet et al., 1993). The erlens were inoculated with 2.5 106 s / ml of the different strains of S. ambofaciens and grown at 27 C with orbital shaking at 250 rpm for 96 hours. The cultures corresponding to the same clone are pooled, possibly filtered on a pleated filter, and centrifuged for 15 min at 7000 rpm.
Le pH du moût est ensuite ajusté à 9 avec de la soude et le surnageant est extrait par de la méthyl iso-butyryl cétone (MIBK). La phase organique (MIBK) est alors récupérée et évaporée. L'extrait sec est ensuite repris dans 1ml d'acétonitrile, puis dilué au 1/10 (100 l qsp lml avec de l'eau) avant d'être utilisé pour les analyses en chromatographie liquide couplée à la spectrométrie de masse (CL/SM). The pH of the must is then adjusted to 9 with sodium hydroxide and the supernatant is extracted with methyl iso-butyryl ketone (MIBK). The organic phase (MIBK) is then recovered and evaporated. The dry extract is then taken up in 1 ml of acetonitrile, then diluted to 1/10 (100 l qsp lml with water) before being used for analyzes in liquid chromatography coupled to mass spectrometry (CL / MS).
20. 2 Analyse des échantillons en CL/SM :
Les échantillons ont été analyés en CL/SM dans le but de déterminer la masse des différents produits synthétisés par les souches à tester. 20. 2 Analysis of CL / MS samples:
The samples were analyzed in LC / MS in order to determine the mass of the different products synthesized by the strains to be tested.
La colonne de chromatographie liquide haute performance utilisée est une colonne Kromasil C8 150*4,6mmm, 5mmm, 100 . The high performance liquid chromatography column used is a Kromasil C8 150 * 4.6mmm, 5mmm, 100 column.
La phase mobile est un gradient constitué d'un mélange d'acétonitrile et d'une solution aqueuse d'acide trifluoroacétique à 0. 05%, le débit est fixé à 1 ml/min. La température du four colonne est maintenue à 30 C. The mobile phase is a gradient consisting of a mixture of acetonitrile and an aqueous solution of trifluoroacetic acid at 0. 05%, the flow rate is set at 1 ml / min. The temperature of the column oven is maintained at 30 C.
La détection UV en sortie de la colonne a été effectuée à deux longueurs d'onde différentes : 238 nm et 280 nm. The UV detection at the outlet of the column was carried out at two different wavelengths: 238 nm and 280 nm.
Le spectrométre de masse couplé à la colonne de chromatographie est un appareil Simple Quadripole commercialisé par la société Agilent, avec des tensions de cone à 30 et à 70V. The mass spectrometer coupled to the chromatography column is a Simple Quadripole device marketed by Agilent, with cone voltages at 30 and 70V.
<Desc/Clms Page number 164> <Desc / Clms Page number 164>
20. 3 Analyse des intermédiaires de biosynthèse produits par la souche OS49.67 :
La souche OS49. 67 dans laquelle le gène or/3 est inactivé par une délétion en phase ne produit pas de spiramycines (cf. exemple 6 et 15). 20. 3 Analysis of biosynthetic intermediates produced by strain OS49.67:
The strain OS49. 67 in which the or / 3 gene is inactivated by an in-phase deletion does not produce spiramycins (see Examples 6 and 15).
Un échantillon a été préparé selon la méthode décrite ci-dessus (cf. paragraphe 20.1) et a été analysé par CL/SM comme décrit ci-dessus (cf. le paragraphe 20.2). A sample was prepared according to the method described above (see section 20.1) and analyzed by LC / MS as described above (see section 20.2).
Plus particulièrement, l'analyse par chromatographie a été réalisée en gradient de solvant avec comme phase mobile : 20% d'acétonitrile du temps T=0 à 5min puis montée linéaire à 30% à T=35 minutes, suivie d'un palierjusqu'à T=50 minutes. More particularly, the analysis by chromatography was carried out in a solvent gradient with 20% of acetonitrile at time T = 0 to 5 min and then linear rise at 30% at T = 35 minutes, followed by a plateau of about 20 minutes. at T = 50 minutes.
Dans ces conditions, deux produits ont plus particulièrement été observés : un absorbant à 238 nm (temps de rétention de 33.4 min) et un absorbant à 280 nm (temps de rétention de 44.8 min) (cf. figure 35). La figure 35 montre la superposition des chromatogrammes CLHP réalisés à 238 et 280nm (haut) ainsi que les spectres UV des molécules élués à 33,4 minutes et 44,8 minutes (bas). Under these conditions, two products were observed more particularly: an absorbent at 238 nm (retention time of 33.4 min) and an absorbent at 280 nm (retention time of 44.8 min) (see Figure 35). Figure 35 shows the superposition of the HPLC chromatograms made at 238 and 280 nm (high) as well as the UV spectra of the eluted molecules at 33.4 minutes and 44.8 minutes (low).
Les conditions d'analyses en spéctrométrie de masse couplée étaient les suivantes : le balayage se fait en mode scan, couvrant une gamme de masse comprise entre 100 et 1000 Da. Le gain de l'électro-multiplicateur était de IV. Concernant la source électrospray, la pression du gaz de nébulisation était de 35 psig, le débit du gaz séchant de 12.0 1.min-1, la température du gaz séchant de 350 C, la tension du capilaire était portée à +/- 3000 V. Ces expériences ont permis de déterminer la masse des deux produits séparés. Ces masses sont respectivement de 370 g/mole pour le produit élué en premier ([M-H2O]+=353 produit majoritaire) et 368 g/mole pour le second produit ([MH2O]+=351 produit majoritaire). The analysis conditions in coupled mass spectrometry were as follows: the scanning is done in scan mode, covering a mass range between 100 and 1000 Da. The gain of the electro-multiplier was IV. For the electrospray source, the nebulization gas pressure was 35 psig, the drying gas flow rate was 12.0 1.min-1, the drying gas temperature was 350 C, the capillary tension was raised to +/- 3000 V These experiments made it possible to determine the mass of the two separated products. These masses are respectively 370 g / mol for the product eluted first ([M-H2O] + = 353 majority product) and 368 g / mol for the second product ([MH2O] + = 351 majority product).
Afin d'obtenir la structure, les produits mentionnés ci-dessus ont été isolés et purifiés dans les conditions suivantes : la phase mobile est un mélange d'une solution aqueuse d'acide trifluoroacétique à 0. 05% et d'acétonitrile 70/30 (v/v). La chromatographie est réalisée en régime isocratique avec un débit fixé de 1 ml/min. Dans ces conditions, les temps de rétention des 2 produits sont respectivement de 8 et 13.3 minutes. De plus dans ce cas, l'échantillon préparé (cf. paragraphe 20.1) n'est pas dilué dans l'eau avant injecton de 10 l. In order to obtain the structure, the above-mentioned products were isolated and purified under the following conditions: the mobile phase is a mixture of an aqueous solution of 0.05% trifluoroacetic acid and 70/30 acetonitrile (v / v). The chromatography is carried out in isocratic regime with a fixed flow rate of 1 ml / min. Under these conditions, the retention times of the two products are respectively 8 and 13.3 minutes. Moreover, in this case, the sample prepared (see paragraph 20.1) is not diluted in water before the 10 l injection.
<Desc/Clms Page number 165> <Desc / Clms Page number 165>
Les 2 produits sont récupérés à la sortie de la colonne chromatographique et isolés dans les conditions suivantes : une cartouche Oasis HLB 1 cc 30mg (Waters) est conditionnée séquentiellement par 1 ml d'acétonitrile, puis 1 ml d'eau/acétonitrile (20v/80v) et 1ml d'eau/acétonitrile 80/20. L'échantillon est ensuite introduit et la cartouche lavée successivement par 1 ml d'eau/acétonitrile (95/5), lml d'eau/acétonitrile deutéré (95/5), puis éluée avec 600ul d'eau/acétonitrile deutéré 40/60. The 2 products are recovered at the outlet of the chromatographic column and isolated under the following conditions: an Oasis HLB cartridge 1 cc 30mg (Waters) is conditioned sequentially with 1 ml of acetonitrile, then 1 ml of water / acetonitrile (20 v / 80v) and 1ml of water / acetonitrile 80/20. The sample is then introduced and the cartridge washed successively with 1 ml of water / acetonitrile (95/5), 1 ml of water / deuterated acetonitrile (95/5) and then eluted with 600 μl of water / deuterated acetonitrile 40 / 60.
La solution récupérée est ensuite directement analysée en RMN. The recovered solution is then directly analyzed by NMR.
Les spectres RMN obtenus pour ces deux composés sont les suivants : - Premier produit élue: Platenolide A : (Spectre 9646V) Spectre 1H dans CD3CN (déplacements chimiques en ppm): 0,90 (3H, t, J=6Hz), 0,93 (3H, d, J=5Hz), 1,27 (3H, d, J=5Hz), entre 1,27 et 1,40 (3H, m), 1,51 (1H, m), 1,95 (1H, m), 2,12 (1H, m), 2,30 (1H, d, J=12Hz), 2,50 (1H, d, J=llHz), 2,58 (1H, dd, J=9 et 12 Hz), 2,96 (1H, d, J=7Hz), 3,43 (3H, s), 3,70 (1H, d, J=9Hz), 3,86 (1H, d, J=7Hz), 4,10 (1H, m), 5,08 (1H, m), 5,58 (1H, dt, J=3 et 12Hz), 5,70 (1H, dd, J=8 et 12 Hz), 6,05 (1H, dd, J=9 et 12 Hz), 6,24 (1H, dd, J=9 et 12Hz). The NMR spectra obtained for these two compounds are as follows: First eluted product: Platenolide A: (Spectrum 9646V) Spectrum 1H in CD3CN (chemical shifts in ppm): 0.90 (3H, t, J = 6Hz), 0, 93 (3H, d, J = 5Hz), 1.27 (3H, d, J = 5Hz), between 1.27 and 1.40 (3H, m), 1.51 (1H, m), 1.95 (1H, m), 2.12 (1H, m), 2.30 (1H, d, J = 12Hz), 2.50 (1H, d, J = 11Hz), 2.58 (1H, dd, J). = 9 and 12 Hz), 2.96 (1H, d, J = 7 Hz), 3.43 (3H, s), 3.70 (1H, d, J = 9 Hz), 3.86 (1H, d, J = 7Hz), 4.10 (1H, m), 5.08 (1H, m), 5.58 (1H, dt, J = 3 and 12Hz), 5.70 (1H, dd, J = 8 and 12 Hz), 6.05 (1H, dd, J = 9 and 12 Hz), 6.24 (1H, dd, J = 9 and 12Hz).
-Deuxième produit élue: Platenolide B : (Spectre 9647V) Spectre 1H dans CD3CN (déplacements chimiques en ppm): 0,81 (3H, t, J=6Hz), 0,89 (1H, m), 1,17 (3H, d, J=5Hz), 1,30 (4H, m), 1,47 (2H, m), 1,61 (1H, t, J=10Hz), 2,20 (1H, m), 2,38 (1H, d, J=13Hz), 2,52 (1H, m), 2,58 (1H, m), 2,68 (1H, dd, J=8 et 13Hz), 3,10 (1H, d, J=7Hz), 3,50 (3H, s), 3,61 (1H, d, J=8Hz), 3,82 (1H, d, J=7Hz), 5,09 (1H, m), 6,20 (1H, m), 6,25 (1H, dd, J=9 et 12 Hz), 6,58 (1H, d, J=12 Hz), 7,19 (1H, dd, J=9 et 12Hz). -Second product eluted: Platenolide B: (Spectrum 9647V) Spectrum 1H in CD3CN (chemical shifts in ppm): 0.81 (3H, t, J = 6Hz), 0.89 (1H, m), 1.17 (3H , d, J = 5Hz), 1.30 (4H, m), 1.47 (2H, m), 1.61 (1H, t, J = 10Hz), 2.20 (1H, m), 2, 38 (1H, d, J = 13Hz), 2.52 (1H, m), 2.58 (1H, m), 2.68 (1H, dd, J = 8 and 13Hz), 3.10 (1H, m.p. d, J = 7Hz), 3.50 (3H, s), 3.61 (1H, d, J = 8Hz), 3.82 (1H, d, J = 7Hz), 5.09 (1H, m). , 6.20 (1H, m), 6.25 (1H, dd, J = 9 and 12Hz), 6.58 (1H, d, J = 12Hz), 7.19 (1H, dd, J = 9 and 12Hz).
Ces expériences ont ainsi permis de déterminer la structure de ces deux composés. These experiments made it possible to determine the structure of these two compounds.
Le premier produit élué est le platénolide A et le deuxième est le platénolide B, la structure déduite de ces deux méolécules est présentée en figure 36. The first eluted product is platenolide A and the second is platenolide B, the structure deduced from these two meolecules is shown in FIG.
<Desc/Clms Page number 166> <Desc / Clms Page number 166>
20.4 Analyse des intermédiaires de biosynthèse produits par la souche SPM509 :
![]()
La souche SPM509 dans laquelle le gène orfl4 est inactivé (orfl4:: att3,slaac-) ne produit pas de spiramycines (cf. exemple 13,15 et 16). Un échantillon a été préparé selon la méthode décrite ci-dessus (cf. paragraphe 20. 1) et a été analysé par CL/SM comme décrit ci-dessus (cf. le paragraphe 20. 2 et 20.3). L'analyse des intermédiaires de biosynthèse présents dans le surnageant de culture de la souche SPM509 cultivée en milieu MP5 a montré que cette souche ne produisait que la forme B du platénolide ( platénolide B , cf. figure 36) mais pas la forme A ( platénolide A , cf. figure 36). 20.4 Analysis of Biosynthesis Intermediates Produced by SPM509:
![]()
The strain SPM509 in which the orflu4 gene is inactivated (orf14 :: att3, slaac-) does not produce spiramycins (see Example 13, 15 and 16). A sample was prepared according to the method described above (see paragraph 20. 1) and analyzed by LC / MS as described above (see paragraph 20.2 and 20.3). Analysis of the biosynthetic intermediates present in the culture supernatant of the strain SPM509 grown in MP5 medium showed that this strain only produced the B form of platenolide (platenolide B, see FIG. 36) but not the A form (platenolide). A, see Figure 36).
EXEMPLE 21 : Interruption du gène orfl4 dans une souche interrompue dans le gène orf3 (OS49.67)
Le produit du gène orfl4 est indispensable à la biosynthèse des spiramycines (cf. exemple 13,15 et 16 : la souche SPM509 dans laquelle ce gène est interrompu ne produit plus de spiramycines). L'analyse des intermédiaires de biosynthèse présents dans le surnageant de culture de la souche SPM509 cultivée en milieu MP5 a montré que cette souche ne produisait que la forme B du platénolide mais pas la forme A (cf. exemple 20). Une des hypothèses pouvant expliquer cette observation est que le produit de orfl4 soit impliqué dans la conversion de la forme B du platénolide en forme A par une étape enzymatique de réduction. Pour tester cette hypothèse, le gène orfl4 a été inactivé dans un mutant ne produisant plus de spiramycine, mais produisant les formes A et B du platénolide. C'est le cas de la souche OS49. 67 (cf. exemple 6 et 20) dans laquelle le gène or/3 est inactivé par une délétion en phase (#orf3). Pour inactiver le gène orfl4 dans cette souche le plasmide pSPM509 a été introduit par transformation de protoplastes de la souche OS49. 67 (Kieser, T et al., 2000). L'inactivation du gène or/74 a déjà été décrite dans le cas de la souche OSC2 (cf. exemple 13) et il a été procédé de la même manière pour l'inactivation du gène orfl4 dans la souche OS49. 67. Après transformation par le plasmide pSPM509, les clones sont sélectionnés pour leur résistance à l'apramycine. Les clones résistants à EXAMPLE 21: Interruption of the Orfl4 Gene in a Strain Interrupted in the Orf3 Gene (OS49.67)
The orfl4 gene product is essential for spiramycin biosynthesis (see Example 13, 15 and 16: the SPM509 strain in which this gene is interrupted no longer produces spiramycins). Analysis of the biosynthesis intermediates present in the culture supernatant of the strain SPM509 grown in MP5 medium showed that this strain produced only the B form of platenolide but not the A form (see Example 20). One of the hypotheses that may explain this observation is that the orfl4 product is involved in the conversion of form B of platenolide to form A by an enzymatic reduction step. To test this hypothesis, the orfl4 gene was inactivated in a mutant no longer producing spiramycin, but producing platenolide forms A and B. This is the case of the strain OS49. 67 (see Example 6 and 20) in which the or / 3 gene is inactivated by a phase deletion (# orf3). To inactivate the orfl4 gene in this strain, the plasmid pSPM509 was introduced by transformation of protoplasts of the strain OS49. 67 (Kieser, T et al., 2000). The inactivation of the or / 74 gene has already been described in the case of the OSC2 strain (see Example 13) and the same procedure was used for the inactivation of the orf14 gene in the OS49 strain. 67. After transformation with the plasmid pSPM509, the clones are selected for their resistance to apramycin. Clones resistant to
<Desc/Clms Page number 167><Desc / Clms Page number 167>
l'apramycine sont ensuite repiqués respectivement sur milieu avec apramycine et sur milieu avec hygromycine. Les clones résistants à l'apramycine (ApraR) et sensibles à l'hygromycine (HygS) sont en principe ceux où un double événement de crossing over s'est produit et dans lesquels le gène orfl4 a été remplacé par une copie de orfl4 interrompue par la cassette att3#aac-. Ces clones ont été plus particulièrement sélectionnés et le remplacement de la copie sauvage de orfl4 par la copie interrompue par la cassette a été vérifié par hybridation. Ainsi, l'ADN total des clones obtenus, a été digéré par plusieurs enzymes, séparé sur gel d'agarose, transféré sur membrane et hybridé avec une sonde correspondant à la cassette att3#aac- pour vérifier que le remplacement de gène avait bien eu lieu. La vérification du génotype peut également être réalisée par toute méthode connue de l'homme du métier et notamment par PCR en utilisant les oligonucléotides appropriés et séquençage du produit PCR.
![]()
![]()
Un clone présentant les caractéristiques attendues (Aorf3, orfl4::att3S2aac-) a été plus particulièrement sélectionné et nommé SPM510. Il a pu en effet être vérifié grâce aux deux hybridations que la cassette att3#aac- était bien présente dans le génome de ce clone et que l'on obtient bien le profil de digestion attendu dans le cas d'un remplacement, à la suite d'un double événement de recombinaison, du gène sauvage orfl4 par la copie interrompue par la cassette att3#aac- dans le génome de ce clone. A clone with the expected characteristics (Aorf3, orfl4 :: att3S2aac-) was more particularly selected and named SPM510. It could indeed be verified by means of the two hybridizations that the att3 # aac- cassette was indeed present in the genome of this clone and that the expected digestion profile is obtained in the case of a replacement, following a double recombination event, wild-type gene orfl4 by the copy interrupted by the cassette att3 # aac- in the genome of this clone.
EXEMPLE 22 : Complémentation fonctionnelle de l'interruption du gène orfl4
22. 1 Construction du plasmide pSPM519 :
Le gène orfl4 a été amplifié par PCR en utilisant le couple d'oligonucléotides suivant : EDR31 : 5' CCCAAGCTTCTGCGCCCGCGGGCGTGAA 3' (SEQ ID N 136) et EDR37 : 5' GCTCTAGAACCGTGTAGCCGCGCCCCGG 3' (SEQ ID N 137) et comme matrice l'ADN chromosomique de la souche OSC2. Les EXAMPLE 22 Functional Complementation of the Orfl4 Gene Interruption
22. 1 Construction of the plasmid pSPM519:
The orflu4 gene was amplified by PCR using the following pair of oligonucleotides: EDR31: 5 'CCCAAGCTTCTGCGCCCGCGGGCGTGAA 3' (SEQ ID N 136) and EDR37: 5 'GCTCTAGAACCGTGTAGCCGCGCCCCGG 3' (SEQ ID N 137) and as DNA template chromosome of the OSC2 strain. The
<Desc/Clms Page number 168><Desc / Clms Page number 168>
oligonucléotides EDR31 et EDR37 portent respectivement le site de restriction HindIII et Xbal (séquence en gras). Le fragment de 1,2 kb ainsi obtenu a été cloné dans le vecteur pGEM-T easy (commercialisé par la société Promega (Madison, Wisconcin, USA)) pour donner naissance au plasmide pSPM515. Ce plasmide a ensuite été digéré par les enzymes de restriction HindIII et XbaI. L'insert HindIII/XbaI de 1,2 kb obtenu a été cloné dans le vecteur pUWL201 (cf. exemple 17. 1) préalablement digéré par les mêmes enzymes. Le plasmide ainsi obtenu a été nommé pSPM519. oligonucleotides EDR31 and EDR37 respectively carry the restriction site HindIII and XbaI (sequence in bold). The 1.2 kb fragment thus obtained was cloned into the vector pGEM-T easy (marketed by Promega (Madison, Wisconcin, USA)) to give rise to plasmid pSPM515. This plasmid was then digested with the restriction enzymes HindIII and XbaI. The 1.2 kb HindIII / XbaI insert obtained was cloned into the vector pUWL201 (see Example 17.1) previously digested with the same enzymes. The plasmid thus obtained was named pSPM519.
22. 1 Transformation des souches SPM509 et SPM510 par le plasmide pSPM519 :
Le plasmide pSPM519 a été introduit dans les souches SPM509 (cf. exemple 13) et SPM510 (cf. exemple 17) par transformation de protoplastes (Kieser, T et al., 2000). Après transformation, les clones sont sélectionnés pour leur résistance au thiostrepton. Les clones sont ensuite repiqués sur un milieu contenant du thiostrepton. 22. 1 Transformation of strains SPM509 and SPM510 by the plasmid pSPM519:
Plasmid pSPM519 was introduced into strains SPM509 (see Example 13) and SPM510 (see Example 17) by protoplast transformation (Kieser, T et al., 2000). After transformation, the clones are selected for thiostrepton resistance. The clones are then subcultured on a medium containing thiostrepton.
La souche SPM509 est une souche non productrice de spiramycines (cf. exemple 13,15 et 16 et figure 24). La production en spiramycines de la souche SPM509 transformée par le vecteur pSPM519 (souche nommée SPM509 pSPM519) a été analysée en cultivant cette souche dans du milieu MP5 en présence de thiostrepton. Les surnageants de culture ont ensuite été analysés par CLHP (cf. exemple 16 et 17). Les résultats de cette analyse sont présentés dans le tableau 43, les données sont exprimées en mg par litre de surnageant. Les résultats correspondent à la production totale en spiramycines (obtenue en additionnant la production en spiramycine I, II et III). Il a été observé que la présence du vecteur pSPM519 dans la souche SPM509 restaure la production de spiramycines (cf. tableau 43). The strain SPM509 is a strain which does not produce spiramycins (see Example 13, 15 and 16 and FIG. 24). The spiramycin production of the SPM509 strain transformed with the vector pSPM519 (strain named SPM509 pSPM519) was analyzed by cultivating this strain in MP5 medium in the presence of thiostrepton. The culture supernatants were then analyzed by HPLC (see Examples 16 and 17). The results of this analysis are shown in Table 43, the data are expressed in mg per liter of supernatant. The results correspond to the total production of spiramycins (obtained by adding spiramycin I, II and III production). It has been observed that the presence of the vector pSPM519 in strain SPM509 restores the production of spiramycins (see Table 43).
<Desc/Clms Page number 169><Desc / Clms Page number 169>
Tableau 43. Production en spiramycines de la souche SPM509 transformée par le vecteur pSPM519, (résultats exprimés en mg/1 de surnageant).

<tb>
<tb> <Tb>
<Tb>
Souche <SEP> Production <SEP> en <SEP> spyramicines
<tb> SPM509 <SEP> pSPM519 <SEP> 58
<tb> Strain <SEP> Production <SEP> in <SEP> spyramicines
<tb> SPM509 <SEP> pSPM519 <SEP> 58
<Tb>
La souche SPM510 transformée par le plasmide pSPM519 a été nommée SPM510 pSPM509. The SPM510 strain transformed with the plasmid pSPM519 was named SPM510 pSPM509.
EXEMPLE 23 : fonctionnelle de l'interruption du gène orf3 par le gène tylB de S. fradiae
23. 1 Construction du plasmide pOS49.52 :
Le plasmide pOS49. 52 correspond à un plasmide permettant l'expression de la protéine TylB chez S. ambofaciens. Pour le construire, la séquence codante du gène tylB de S. fradiae (Merson-Davies & Cundliffe, 1994, numéro d'accès GenBank: U08223 (séquence de la région), SFU08223 (séquence ADN) et AAA21342 (séquence protéique)) a été introduite dans le plasmide pKC1218 (Bierman et al., 1992, Kieser et al., 2000, une souche de E. coli contenant ce plasmide est accessible notamment auprès de l'ARS (NRRL) Agricultural Research Service Culture Collection) (Peoria, minois, USA), sous le numéro B-14790). De plus cette séquence codante à été placée sous le contrôle du promoteur ermE* (cf. cidessus, notamment exemple 17.1, Bibb et al., 1985, Bibb et al., 1994). EXAMPLE 23: functional of the interruption of the orf3 gene by the tylB gene of S. fradiae
23. 1 Construction of Plasmid pOS49.52:
The plasmid pOS49. 52 corresponds to a plasmid allowing the expression of the TylB protein in S. ambofaciens. To construct it, the coding sequence of the S. fradiae tylB gene (Merson-Davies & Cundliffe, 1994, GenBank accession number: U08223 (region sequence), SFU08223 (DNA sequence) and AAA21342 (protein sequence)) was introduced into the plasmid pKC1218 (Bierman et al., 1992, Kieser et al., 2000, an E. coli strain containing this plasmid is accessible in particular from the ARS (NRRL) Agricultural Research Service Culture Collection) (Peoria, Minois, USA), under number B-14790). In addition, this coding sequence has been placed under the control of the ermE * promoter (see above, in particular Example 17.1, Bibb et al., 1985, Bibb et al., 1994).
23. 1 Transformation de la souches OS49. 67 par le plasmide pOS49.52 :
La souche OS49. 67 dans laquelle le gène or/3 est inactivé par une délétion en phase ne produit pas de spiramycines (cf. exemple 6 et 15). Le plasmide pOS49. 52 a été introduit dans la souche OS49. 67 par transformation de protoplastes (Kieser, T et al., 2000). Après transformation, les clones sont sélectionnés pour leur résistance à 23. 1 Transformation of the OS49 strain. 67 by the plasmid pOS49.52:
The strain OS49. 67 in which the or / 3 gene is inactivated by an in-phase deletion does not produce spiramycins (see Examples 6 and 15). The plasmid pOS49. 52 was introduced in strain OS49. 67 by protoplast transformation (Kieser, T et al., 2000). After transformation, the clones are selected for their resistance to
<Desc/Clms Page number 170><Desc / Clms Page number 170>
l'apramycine. Les clones sont ensuite repiqués sur un milieu contenant de l'apramycine. Un clone a été plus particulièrement sélectionné et a été nommé OS49.67pOS49.52. apramycin. The clones are then subcultured on a medium containing apramycin. A clone was particularly selected and was named OS49.67pOS49.52.
Comme il a été démontré ci-dessus, la souche OS49.67 ne produit pas de spiramycines (cf. exemple 6 et 15). La production en spiramycines de la souche OS49. 67 transformée par le vecteur pOS49.52 a été analysée par la technique décrite dans l'exemple 15. Il a ainsi pu être démontré que cette souche possède un phénotype producteur de spiramycines. Ainsi la protéine TylB permet la complémentation fonctionnelle de l'interruption du gène orf3. As demonstrated above, strain OS49.67 does not produce spiramycins (see Examples 6 and 15). The spiramycin production of strain OS49. Transformed by the vector pOS49.52 was analyzed by the technique described in Example 15. It was thus possible to demonstrate that this strain has a phenotype producing spiramycins. Thus the TylB protein allows the functional complementation of the interruption of the orf3 gene.
EXEMPLE 24 : Amélioration de la production en spiramycines par surexpression du gène orfl8c
24. 1 Construction du plasmide pSPM75 :
Le gène orf28c a été amplifié par PCR en utilisant un couple d'oligonucléotides comportant un site de restriction HindIII ou un site de restriction BamHI. Ces amorces ont la séquence suivante : KF30 : 5' AAGCTTGTGTGCCCGGTGTACCTGGGGAGC 3' (SEQ ID N 138) avec un site de restriction HindIII (qui figure en gras) KF31 : 5' GGATCCCGCGACGGACACGACCGCCGCGCA 3' (SEQ ID N 139) avec le site de restriction BamHI (qui figure en gras) Les amorces KF30 et KF31 portent respectivement le site de restriction HindIII et BamHI (séquence en gras). Le couple d'amorces KF30 et KF31 permet d'amplifier un fragment d'ADN d'une taille d'environ 1,5 kb contenant le gène orj28c en utilisant comme matrice le cosmide pSPM36 (cf. ci-dessus). Le fragment de 1,5 kb ainsi obtenu a été cloné dans le vecteur pGEM-T easy (commercialisé par la société Promega (Madison, Wisconcin, USA)) pour donner naissance au plasmide pSPM74. Le plasmide pSPM74 a ensuite été digéré par les enzymes de restriction HindIII et BamHI et l'insert HindIII/BamHI d'environ 1,5 kb obtenu a été sous cloné dans le vecteur pUWL201 (cf. EXAMPLE 24: Improvement of spiramycin production by overexpression of the orfl8c gene
24. 1 Construction of the plasmid pSPM75:
The orf28c gene was amplified by PCR using an oligonucleotide pair having a HindIII restriction site or a BamHI restriction site. These primers have the following sequence: KF30: 5 'AAGCTTGTGTGCCCGGTGTACCTGGGGAGC 3' (SEQ ID NO: 138) with a HindIII restriction site (shown in bold) KF31: 5 'GGATCCCGCGACGGACACGACCGCCGCGCA 3' (SEQ ID NO: 139) with the restriction site BamHI (which appears in bold) The KF30 and KF31 primers carry the HindIII and BamHI restriction site respectively (sequence in bold). The pair of primers KF30 and KF31 makes it possible to amplify a DNA fragment of a size of approximately 1.5 kb containing the orj28c gene by using the cosmid pSPM36 as a template (see above). The 1.5 kb fragment thus obtained was cloned into the vector pGEM-T easy (marketed by Promega (Madison, Wisconcin, USA)) to give rise to plasmid pSPM74. The plasmid pSPM74 was then digested with the restriction enzymes HindIII and BamHI and the HindIII / BamHI insert of approximately 1.5 kb obtained was subcloned in the vector pUWL201 (cf.
<Desc/Clms Page number 171><Desc / Clms Page number 171>
exemple 17.1) préalablement digéré par les mêmes enzymes. Le plasmide ainsi obtenu a été nommé pSPM75, il contient l'ensemble de la séquence codante de orf28c placée sous le contrôle du promoteur ermE*. Example 17.1) previously digested with the same enzymes. The plasmid thus obtained was named pSPM75, it contains the entire coding sequence of orf28c placed under the control of the ermE * promoter.
* 24. 2 Transformation de la souche OSC2 par le plasmide pSPM75 :
Le plasmide pSPM75 a été introduit dans les souches OSC2 par transformation de protoplastes (Kieser, T et al., 2000). Après transformation des protoplastes, les clones sont sélectionnés pour leur résistance au thiostrepton. Les clones sont ensuite repiqués sur un milieu contenant du thiostrepton et la transformation par le plasmide est vérifiée par extraction de plasmides. Deux clones ont plus particulièrement été sélectionnés et nommés OSC2/pSPM75(l) et OSC2/pSPM75 (2). * 24. 2 Transformation of the strain OSC2 by the plasmid pSPM75:
Plasmid pSPM75 was introduced into OSC2 strains by protoplast transformation (Kieser, T et al., 2000). After transformation of the protoplasts, the clones are selected for thiostrepton resistance. The clones are then subcultured on a thiostrepton-containing medium and the transformation with the plasmid is verified by plasmid extraction. Two clones were more particularly selected and named OSC2 / pSPM75 (1) and OSC2 / pSPM75 (2).
Afin de tester l'effet de la surexpression du gène orj28c sur la production de spiramycines, la production en spiramycines des clones OSC2/pSPM75(l) et OSC2/pSPM75 (2) a été testée par la technique décrite dans l'exemple 15. L'analyse de la production en spiramycines de la souche OSC2 a également été effectée en parallèle pour comparaison. Il a ainsi pu être observé que la présence du plasmide pSPM75 augmente significativement la production en spiramycines de la souche OSC2. Ceci démontre que la surexpression de orf28c conduit à une augmentation de la production en spiramycines et confirme son rôle en tant que régulateur. In order to test the effect of overexpression of the orj28c gene on the production of spiramycins, the spiramycin production of clones OSC2 / pSPM75 (1) and OSC2 / pSPM75 (2) was tested by the technique described in Example 15. Analysis of spiramycin production of strain OSC2 was also performed in parallel for comparison. It has thus been observed that the presence of the plasmid pSPM75 significantly increases the spiramycin production of the strain OSC2. This demonstrates that the overexpression of orf28c leads to an increase in spiramycin production and confirms its role as a regulator.
La production en spiramycines des clones OSC2/pSPM75(l) et OSC2/pSPM75 (2) a également été analysée par CLHP (de la même manière que dans l'exemple 17. 2). L'analyse de la production en spiramycines de la souche OSC2 a également été effectée en parallèle pour comparaison. Les résultats de cette analyse sont présentés dans le tableau 44, les données sont exprimées en mg par litre de surnageant. Les résultats correspondent à la production totale en spiramycines (obtenue en additionnant la production en spiramycine I, II et III). Spiramycin production of clones OSC2 / pSPM75 (1) and OSC2 / pSPM75 (2) was also analyzed by HPLC (in the same manner as in Example 17.2). Analysis of spiramycin production of strain OSC2 was also performed in parallel for comparison. The results of this analysis are shown in Table 44, the data are expressed in mg per liter of supernatant. The results correspond to the total production of spiramycins (obtained by adding spiramycin I, II and III production).
<Desc/Clms Page number 172><Desc / Clms Page number 172>
Tableau 44. Production en spiramycines des souches dérivées de OSC2 transformé par pSPM75, (résultats exprimés en mg/1).

<tb>
<tb> <Tb>
<Tb>
Souche <SEP> Spiramycines
<tb> OSC2 <SEP> 50
<tb> OSC2/pSPM75(l) <SEP> 120
<tb> OSC2/pSPM75 <SEP> (2) <SEP> 155
<tb> Strain <SEP> Spiramycine
<tb> OSC2 <SEP> 50
<tb> OSC2 / pSPM75 (l) <SEP> 120
<tb> OSC2 / pSPM75 <SEP> (2) <SEP> 155
<Tb>
Ainsi, il est observé que la présence du plasmide pSPM75 augmente significativement la production en spiramycines de la souche OSC2. Ceci démontre bien que la surexpression de orf28c a un effet positif sur la production en spiramycines. Thus, it is observed that the presence of the plasmid pSPM75 significantly increases the spiramycin production of the strain OSC2. This demonstrates that overexpression of orf28c has a positive effect on spiramycin production.
<Desc/Clms Page number 173> <Desc / Clms Page number 173>
Liste des constructions décrites dans la présente demande Liste des abréviations : Am : Ampicilline ; Hyg : Hygromycine ; Sp : Spiramycine ; Ts : Thiostrepton ; Cm : Chloramphénicol. Kn : Kanamycine, Apra : apramycine.
![]()
![]()
<tb>
<tb> Nom <SEP> de <SEP> la <SEP> Marqueur <SEP> Principales <SEP> caractéristiques <SEP> Référence
<tb> construction <SEP> de <SEP> sélection
<tb> <Tb>
<tb> Name <SEP> of <SEP><SEP> Marker <SEP> Main <SEP> characteristics <SEP> Reference
<tb> construction <SEP> of <SEP> selection
<Tb>
![]()
pWE15 Am (Wahl, et al., I987)

![]()
pWE15 Am (Wahl, et al., I987)
<tb>
<tb> pWED1 <SEP> Am <SEP> pWE15 <SEP> dans <SEP> lequel <SEP> un <SEP> (Gourmelen <SEP> et <SEP> al.,
<tb> fragment <SEP> HpaI-HpaI <SEP> de <SEP> 4.1 <SEP> kb <SEP> 1998)
<tb> a <SEP> été <SEP> délété
<tb> pOJ260 <SEP> Apra <SEP> Conjugatif, <SEP> non <SEP> réplicatif <SEP> chez <SEP> (Bierman <SEP> et <SEP> al.,
<tb> Streptomyces <SEP> 1992)
<tb> pHP45 <SEP> Qhyg <SEP> Hyg <SEP> Cassette <SEP> Qhyg <SEP> dans <SEP> pHP45. <SEP> (Blondelet-Rouault
<tb> et <SEP> al., <SEP> 1997)
<tb> pKC505 <SEP> Apra <SEP> Cosmide <SEP> (Richardson <SEP> MA <SEP> et
<tb> al., <SEP> 1987)
<tb> pIJ486 <SEP> Ts <SEP> Plasmide <SEP> réplicatif <SEP> (Ward <SEP> et <SEP> al., <SEP> 1986)
<tb> multicopies <SEP> Streptomyces
<tb> pOSint3 <SEP> Am <SEP> ptrc-xis-int <SEP> dans <SEP> pTrc99A <SEP> (Raynal <SEP> et <SEP> al., <SEP> 1998)
<tb> pWHM3 <SEP> Am, <SEP> Ts <SEP> Vecteur <SEP> navette <SEP> réplicatif <SEP> E. <SEP> (Vara <SEP> et <SEP> al., <SEP> 1989)
<tb> coli/Streptomyces.
<tb> pKOBEG <SEP> Cm <SEP> (Chaveroche <SEP> et <SEP> al.,
<tb> 2000)
<tb> pGP704Not <SEP> Am <SEP> (Chaveroche <SEP> et <SEP> al.,
<tb> 2000)
<tb> pMBL18 <SEP> Am <SEP> (Nakano <SEP> et <SEP> al.,
<tb> 1995)
<tb> pGEM-T <SEP> Easy <SEP> Am <SEP> vecteur <SEP> E. <SEP> coli <SEP> pour <SEP> clonage <SEP> Mezei <SEP> et <SEP> al., <SEP> 1994
<tb> de <SEP> produits <SEP> PCR
<tb> <Tb>
<tb> pWED1 <SEP> Am <SEP> pWE15 <SEP> in <SEP> which <SEP> a <SEP> (Gourmelen <SEP> and <SEP> al.,
<tb> fragment <SEP> HpaI-HpaI <SEP> from <SEP> 4.1 <SEP> kb <SEP> 1998)
<tb> a <SEP> summer <SEP> deleted
<tb> pOJ260 <SEP> Apra <SEP> Conjugative, <SEP> no <SEP> replicative <SEP> in <SEP> (Bierman <SEP> and <SEP> al.,
<tb> Streptomyces <SEP> 1992)
<tb> pHP45 <SEP> Qhyg <SEP> Hyg <SEP> Cassette <SEP> Qhyg <SEP> in <SEP> pHP45. <SEP> (Blondelet-Rouault
<tb> and <SEP> al., <SEP> 1997)
<tb> pKC505 <SEP> Apra <SEP> Cosmid <SEP> (Richardson <SEP> MA <SEP>)
<tb> al., <SEP> 1987)
<tb> pIJ486 <SEP> Ts <SEP> Replicative <SEP> Plasmid <SEP> (Ward <SEP> and <SEP> Al., <SEP> 1986)
<tb> multicopies <SEP> Streptomyces
<tb> pOSint3 <SEP> Am <SEP> ptrc-xis-int <SEP> in <SEP> pTrc99A <SEP> (Raynal <SEP> and <SEP> al., <SEP> 1998)
<tb> pWHM3 <SEP> Am, <SEP> Ts <SEP> Vector <SEP> replication shuttle <SEP><SEP> E. <SEP> (Vara <SEP> and <SEP> al., <SEP> 1989)
<tb> coli / Streptomyces.
<tb> pKOBEG <SEP> Cm <SEP> (Chaveroche <SEP> and <SEP> al.,
<tb> 2000)
<tb> pGP704Not <SEP> Am <SEP> (Chaveroche <SEP> and <SEP> al.,
<tb> 2000)
<tb> pMBL18 <SEP> Am <SEP> (Nakano <SEP> and <SEP> al.,
<tb> 1995)
<tb> pGEM-T <SEP> Easy <SEP> Am <SEP> vector <SEP> E. <SEP> coli <SEP> for <SEP> cloning <SEP> Mezei <SEP> and <SEP> al., <SEP> 1994
<tb> of <SEP> products <SEP> PCR
<Tb>
<Desc/Clms Page number 174> <Desc / Clms Page number 174>

<tb>
<tb> pOS49. <SEP> 1 <SEP> Am <SEP> pWED <SEP> avec <SEP> insert <SEP> au <SEP> niveau <SEP> Exemple <SEP> 2
<tb> du <SEP> site <SEP> BamHI
<tb> pOS49. <SEP> 11 <SEP> Am <SEP> Fragment <SEP> SacI <SEP> de <SEP> pOS49. <SEP> 1 <SEP> Exemple <SEP> 2
<tb> dans <SEP> pUC <SEP> 19. <SEP>
<tb> pOSC49. <SEP> 12 <SEP> Ch <SEP> Fragment <SEP> XhoI <SEP> de <SEP> pOS49. <SEP> 11 <SEP> Exemple <SEP> 2
<tb> dans <SEP> pBC <SEP> SK+
<tb> pOS49. <SEP> 14 <SEP> Cm,Hyg <SEP> pOSC49.12 <SEP> avec <SEP> le <SEP> gène <SEP> or/3 <SEP> Exemple <SEP> 2
<tb> interrompu <SEP> par <SEP> la <SEP> cassette
<tb> #hyg
<tb> pOS49. <SEP> 16 <SEP> Apra, <SEP> Hyg <SEP> Insert <SEP> de <SEP> pOS49. <SEP> 14 <SEP> dans <SEP> Exemple <SEP> 2
<tb> pOJ260
<tb> Cm <SEP> Fragment <SEP> BamHI-PstI <SEP> de <SEP> Exemple <SEP> 3
<tb> pOS49. <SEP> 28 <SEP> 3,7kb <SEP> de <SEP> pOS49. <SEP> 1 <SEP> dans <SEP> pBC
<tb> SK+
<tb> pOS44. <SEP> 1 <SEP> Apra, <SEP> Sp <SEP> pKC505 <SEP> contenant <SEP> un <SEP> insert <SEP> (Pernodet <SEP> et <SEP> al.,
<tb> conférant <SEP> la <SEP> résistance <SEP> la <SEP> 1999)
<tb> spiramycine <SEP> chez <SEP> S.
<tb> griseofuscus
<tb> pOS44. <SEP> 2 <SEP> Ts, <SEP> Sp <SEP> Fragment <SEP> Sau3AI <SEP> de <SEP> 1,8 <SEP> kb <SEP> Exemple <SEP> 3
<tb> pOS44. <SEP> 1 <SEP> dans <SEP> pIJ486 <SEP>
<tb> pOS44. <SEP> 4. <SEP> Am <SEP> Insert <SEP> de <SEP> pOS44. <SEP> 2 <SEP> dans <SEP> Exemple <SEP> 3
<tb> pUC19
<tb> pSPM5 <SEP> Am <SEP> pWED <SEP> avec <SEP> insert <SEP> d'ADN <SEP> de <SEP> Exemple <SEP> 3
<tb> S. <SEP> ambofaciens <SEP> au <SEP> niveau <SEP> du
<tb> site <SEP> BamHI
<tb> pSPM7 <SEP> Am <SEP> pWEDl <SEP> avec <SEP> insert <SEP> d'ADN <SEP> de <SEP> Exemple <SEP> 3
<tb> S. <SEP> ambofaciens <SEP> au <SEP> niveau <SEP> du
<tb> site <SEP> BamHI
<tb> <Tb>
<tb> pOS49. <SEP> 1 <SEP> Am <SEP> pWED <SEP> with <SEP> insert <SEP> at <SEP> level <SEP> Example <SEP> 2
<tb> of the <SEP><SEP> BamHI site
<tb> pOS49. <SEP> 11 <SEP> Am <SEP> Fragment <SEP> SacI <SEP> of <SEP> pOS49. <SEP> 1 <SEP> Example <SEP> 2
<tb> in <SEP> pUC <SEP> 19. <SEP>
<tb> pOSC49. <SEP> 12 <SEP> Ch <SEP> Fragment <SEP> XhoI <SEP> of <SEP> pOS49. <SEP> 11 <SEP> Example <SEP> 2
<tb> in <SEP> pBC <SEP> SK +
<tb> pOS49. <SEP> 14 <SEP> Cm, Hyg <SEP> pOSC49.12 <SEP> with <SEP> the <SEP> gene <SEP> or / 3 <SEP> Example <SEP> 2
<tb> interrupted <SEP> with <SEP> the <SEP> cassette
<tb>#hyg
<tb> pOS49. <SEP> 16 <SEP> Apra, <SEP> Hyg <SEP> Insert <SEP> of <SEP> pOS49. <SEP> 14 <SEP> in <SEP> Example <SEP> 2
<tb> pOJ260
<tb> Cm <SEP> Fragment <SEP> BamHI-PstI <SEP> of <SEP> Example <SEP> 3
<tb> pOS49. <SEP> 28 <SEP> 3.7kb <SEP> from <SEP> pOS49. <SEP> 1 <SEP> in <SEP> pBC
<tb> SK +
<tb> pOS44. <SEP> 1 <SEP> Apra, <SEP> Sp <SEP> pKC505 <SEP> containing <SEP> a <SEP> insert <SEP> (Pernodet <SEP> and <SEP> al.,
<tb> conferring <SEP><SEP> resistance <SEP><SEP> 1999)
<tb> spiramycin <SEP> in <SEP> S.
<tb> griseofuscus
<tb> pOS44. <SEP> 2 <SEP> Ts, <SEP> Sp <SEP> Fragment <SEP> Sau3AI <SEP> of <SEP> 1.8 <SEP> kb <SEP> Example <SEP> 3
<tb> pOS44. <SEP> 1 <SEP> in <SEP> pIJ486 <SEP>
<tb> pOS44. <SEP> 4. <SEP> Am <SEP> Insert <SEP> of <SEP> pOS44. <SEP> 2 <SEP> in <SEP> Example <SEP> 3
<tb> pUC19
<tb> pSPM5 <SEP> Am <SEP> pWED <SEP> with <SEP> insert <SEP> of DNA <SEP> of <SEP> Example <SEP> 3
<tb> S. <SEP> ambofaciens <SEP> at <SEP> level <SEP> of
<tb> site <SEP> BamHI
<tb> pSPM7 <SEP> Am <SEP> pWEDl <SEP> with <SEP> insert <SEP> of DNA <SEP> of <SEP> Example <SEP> 3
<tb> S. <SEP> ambofaciens <SEP> at <SEP> level <SEP> of
<tb> site <SEP> BamHI
<Tb>
<Desc/Clms Page number 175> <Desc / Clms Page number 175>

<tb>
<tb> pOSK1205 <SEP> Hyg <SEP> pBK-CMV <SEP> dans <SEP> lequel <SEP> hyg <SEP> Exemple <SEP> 5 <SEP>
<tb> remplace <SEP> neo
<tb> pOS49. <SEP> 67 <SEP> Apra <SEP> Fragment <SEP> EcoRI-SacI <SEP> de <SEP> Exemple <SEP> 6
<tb> pOS49. <SEP> 1, <SEP> comportant <SEP> une
<tb> délétion <SEP> interne <SEP> de <SEP> 504
<tb> nucléotides, <SEP> dans <SEP> pOJ260
<tb> pOS49. <SEP> 88 <SEP> Am <SEP> Fragment <SEP> de <SEP> 3. <SEP> 7 <SEP> kb <SEP> PstI- <SEP> Exemple <SEP> 7 <SEP>
<tb> EcoRI <SEP> de <SEP> pOS49. <SEP> 1 <SEP> dans
<tb> pUC19
<tb> pOS49. <SEP> 106 <SEP> Am <SEP> p049.88 <SEP> avec <SEP> hyg <SEP> dans <SEP> or/8 <SEP> Exemple <SEP> 7
<tb> (hyg <SEP> et <SEP> or/8 <SEP> dans <SEP> la <SEP> même
<tb> orientation)
<tb> pOS49. <SEP> 120 <SEP> Am <SEP> pOS49.88 <SEP> avec <SEP> hyg <SEP> dans <SEP> or/8 <SEP> Exemple <SEP> 7
<tb> (hyg <SEP> et <SEP> or/8 <SEP> dans <SEP> des
<tb> orientations <SEP> opposées)
<tb> pOS49. <SEP> 107 <SEP> Apra, <SEP> Hyg <SEP> Insert <SEP> de <SEP> pOS49.106 <SEP> dans <SEP> Exemple <SEP> 7
<tb> pOJ260
<tb> pOS49. <SEP> 32 <SEP> Am, <SEP> Kn <SEP> Fragment <SEP> de <SEP> 1,5 <SEP> kb <SEP> interne <SEP> à <SEP> Exemple <SEP> 8 <SEP>
<tb> orf10 <SEP> dans <SEP> pCR2.1-TOPO
<tb> pOS49. <SEP> 43 <SEP> Am, <SEP> Kn <SEP> pOS49. <SEP> 32 <SEP> avec <SEP> hyg <SEP> dans <SEP> or/70 <SEP> Exemple <SEP> 8
<tb> (hyg <SEP> et <SEP> orflO <SEP> dans <SEP> la <SEP> même
<tb> orientation)
<tb> pOS49. <SEP> 44 <SEP> Am, <SEP> Kn <SEP> pOS49. <SEP> 32 <SEP> avec <SEP> hyg <SEP> dans <SEP> or/76' <SEP> Exemple <SEP> 8 <SEP>
<tb> (hyg <SEP> et <SEP> or/70 <SEP> dans <SEP> des
<tb> orientations <SEP> opposées)
<tb> pOS49. <SEP> 50 <SEP> Apra, <SEP> Hyg <SEP> Insert <SEP> de <SEP> pOS49. <SEP> 43 <SEP> dans <SEP> Exemple <SEP> 8 <SEP>
<tb> pOJ260
<tb> pWHM3Hyg <SEP> Am, <SEP> Hyg <SEP> pWHM3 <SEP> dans <SEP> lequel <SEP> tsr <SEP> est <SEP> Exemple <SEP> 10
<tb> remplacé <SEP> par <SEP> hyg
<tb> <Tb>
<tb> pOSK1205 <SEP> Hyg <SEP> pBK-CMV <SEP> in <SEP> which <SEP> hyg <SEP> Example <SEP> 5 <SEP>
<tb> replaces <SEP> neo
<tb> pOS49. <SEP> 67 <SEP> Apra <SEP> Fragment <SEP> EcoRI-SacI <SEP> of <SEP> Example <SEP> 6
<tb> pOS49. <SEP> 1, <SEP> with <SEP> one
<tb> internal <SEP> deletion <SEP> of <SEP> 504
<tb> nucleotides, <SEP> in <SEP> pOJ260
<tb> pOS49. <SEP> 88 <SEP> Am <SEP> Fragment <SEP> of <SEP> 3. <SEP> 7 <SEP> kb <SEP> PstI- <SEP> Example <SEP> 7 <SEP>
<tb> EcoRI <SEP> of <SEP> pOS49. <SEP> 1 <SEP> in
<tb> pUC19
<tb> pOS49. <SEP> 106 <SEP> Am <SEP> p049.88 <SEP> with <SEP> hyg <SEP> in <SEP> or / 8 <SEP> Example <SEP> 7
<tb> (hyg <SEP> and <SEP> or / 8 <SEP> in <SEP> the <SEP> same
<tb> orientation)
<tb> pOS49. <SEP> 120 <SEP> Am <SEP> pOS49.88 <SEP> with <SEP> hyg <SEP> in <SEP> or / 8 <SEP> Example <SEP> 7
<tb> (hyg <SEP> and <SEP> or / 8 <SEP> in <SEP> of
<tb> opposite <SEP> orientations)
<tb> pOS49. <SEP> 107 <SEP> Apra, <SEP> Hyg <SEP> Insert <SEP> of <SEP> pOS49.106 <SEP> in <SEP> Example <SEP> 7
<tb> pOJ260
<tb> pOS49. <SEP> 32 <SEP> Am, <SEP> Kn <SEP> Fragment <SEP> of <SEP> 1.5 <SEP> kb <SEP> internal <SEP> to <SEP> Example <SEP> 8 <SEP>
<tb> orf10 <SEP> in <SEP> pCR2.1-TOPO
<tb> pOS49. <SEP> 43 <SEP> Am, <SEP> Kn <SEP> pOS49. <SEP> 32 <SEP> with <SEP> hyg <SEP> in <SEP> or / 70 <SEP> Example <SEP> 8
<tb> (hyg <SEP> and <SEP> orflO <SEP> in <SEP> the same <SEP>
<tb> orientation)
<tb> pOS49. <SEP> 44 <SEP> Am, <SEP> Kn <SEP> pOS49. <SEP> 32 <SEP> with <SEP> hyg <SEP> in <SEP> or / 76 '<SEP> Example <SEP> 8 <SEP>
<tb> (hyg <SEP> and <SEP> or / 70 <SEP> in <SEP> of
<tb> opposite <SEP> orientations)
<tb> pOS49. <SEP> 50 <SEP> Apra, <SEP> Hyg <SEP> Insert <SEP> of <SEP> pOS49. <SEP> 43 <SEP> in <SEP> Example <SEP> 8 <SEP>
<tb> pOJ260
<tb> pWHM3Hyg <SEP> Am, <SEP> Hyg <SEP> pWHM3 <SEP> in <SEP> where <SEP> tsr <SEP> is <SEP> Example <SEP> 10
<tb> replaced <SEP> with <SEP> Hyg
<Tb>
<Desc/Clms Page number 176> <Desc / Clms Page number 176>
![]()
![]()
<tb>
<tb> pOSV508 <SEP> Am, <SEP> Ts <SEP> ptrc-xis-int <SEP> dans <SEP> pWHM3 <SEP> Exemple <SEP> 9
<tb> <Tb>
<tb> pOSV508 <SEP> Am, <SEP> Ts <SEP> ptrc-xis-int <SEP> in <SEP> pWHM3 <SEP> Example <SEP> 9
<Tb>
![]()
pattlS2hyg+ Cm, Hyg Cassette att1Ohyg+ dans pBC Exemple 9
![]()
![]()
pattlS2hyg + Cm, Hyg Cassette att1Ohyg + in pBC Example 9
![]()
<tb>
<tb> SK+ <SEP> dont <SEP> le <SEP> site <SEP> HindIII <SEP> a <SEP> été
<tb> supprimé
<tb> <Tb>
<tb> SK + <SEP> of which <SEP> the <SEP> site <SEP> HindIII <SEP> has <SEP> been
<tb> deleted
<Tb>
![]()
patt3naac- Cm, Gn Cassette att32aac- dans pBC Exemple 9

![]()
patt3naac- Cm, Gn Cassette att32aac- in pBC Example 9
<tb>
<tb> SK+ <SEP> dont <SEP> le <SEP> site <SEP> HindIII <SEP> a <SEP> été
<tb> supprimé
<tb> pOSV510 <SEP> Am, <SEP> Hyg <SEP> pro <SEP> pra-Amh <SEP> dans <SEP> Exemple <SEP> 10
<tb> pWHM3Hyg
<tb> pOS49. <SEP> 99 <SEP> Am <SEP> Fragment <SEP> EcoRI-BamHI <SEP> de <SEP> Exemple <SEP> 10
<tb> 4,5 <SEP> kb <SEP> de <SEP> pSPM5 <SEP> dans <SEP> pUC19
<tb> pOSK1102 <SEP> Am, <SEP> Apra <SEP> pGP704Not <SEP> contenant <SEP> la <SEP> Exemple <SEP> 10
<tb> cassette <SEP> att3QaacpSPM17 <SEP> Am, <SEP> Apra <SEP> pOS49. <SEP> 99 <SEP> dans <SEP> lequel <SEP> or/2 <SEP> est <SEP> Exemple <SEP> 10
<tb> interrompue <SEP> par <SEP> la <SEP> cassette
<tb> att3QaacpSPM21 <SEP> Hyg, <SEP> Apra <SEP> pOSK1205 <SEP> contenant <SEP> l'insert <SEP> Exemple <SEP> 10
<tb> EcoRI-XbaI <SEP> de <SEP> pSPM17 <SEP> (dans
<tb> lequel <SEP> orf2 <SEP> est <SEP> interrompue
<tb> par <SEP> la <SEP> cassette <SEP> att3#aac-)
<tb> pSPM502 <SEP> Am <SEP> Fragment <SEP> BglII <SEP> de <SEP> 15,1 <SEP> kb <SEP> de <SEP> Exemple <SEP> 11
<tb> pSPM7danspMBL18
<tb> pSPM504 <SEP> Hyg <SEP> Insert <SEP> de <SEP> pSPM502 <SEP> dans <SEP> Exemple <SEP> 11
<tb> pOSK1205
<tb> pSPM507 <SEP> Hyg, <SEP> Apra <SEP> pSPM504 <SEP> dans <SEP> lequel <SEP> or/72 <SEP> Exemple <SEP> 11
<tb> est <SEP> interrompue <SEP> par <SEP> la <SEP> cassette
<tb> att3#aacpSPM508 <SEP> Hyg, <SEP> Apra <SEP> pSPM504 <SEP> dans <SEP> lequel <SEP> orfl3c <SEP> Exemple <SEP> 12
<tb> est <SEP> interrompue <SEP> par <SEP> la <SEP> cassette
<tb> att3Qaac-
<tb> <Tb>
<tb> SK + <SEP> of which <SEP> the <SEP> site <SEP> HindIII <SEP> has <SEP> been
<tb> deleted
<tb> pOSV510 <SEP> Am, <SEP> Hyg <SEP> pro <SEP> pra-Amh <SEP> in <SEP> Example <SEP> 10
<tb> pWHM3Hyg
<tb> pOS49. <SEP> 99 <SEP> Am <SEP> Fragment <SEP> EcoRI-BamHI <SEP> of <SEP> Example <SEP> 10
<tb> 4.5 <SEP> kb <SEP> of <SEP> pSPM5 <SEP> in <SEP> pUC19
<tb> pOSK1102 <SEP> Am, <SEP> Apra <SEP> pGP704Not <SEP> containing <SEP><SEP> Example <SEP> 10
<tb> tape <SEP> att3QaacpSPM17 <SEP> Am, <SEP> Apra <SEP> pOS49. <SEP> 99 <SEP> in <SEP> where <SEP> or / 2 <SEP> is <SEP> Example <SEP> 10
<tb> interrupted <SEP> with <SEP> the <SEP> cassette
<tb> att3QaacpSPM21 <SEP> Hyg, <SEP> Apra <SEP> pOSK1205 <SEP> containing <SEP> insert <SEP> Example <SEP> 10
<tb> EcoRI-XbaI <SEP> of <SEP> pSPM17 <SEP> (in
<tb> which <SEP> orf2 <SEP> is <SEP> interrupted
<tb> by <SEP> the <SEP> cassette <SEP> att3 # aac-)
<tb> pSPM502 <SEP> Am <SEP> Fragment <SEP> BglII <SEP> of <SEP> 15.1 <SEP> kb <SEP> of <SEP> Example <SEP> 11
<tb> pSPM7danspMBL18
<tb> pSPM504 <SEP> Hyg <SEP> Insert <SEP> of <SEP> pSPM502 <SEP> in <SEP> Example <SEP> 11
<tb> pOSK1205
<tb> pSPM507 <SEP> Hyg, <SEP> Apra <SEP> pSPM504 <SEP> in <SEP> which <SEP> or / 72 <SEP> Example <SEP> 11
<tb> is <SEP> interrupted <SEP> with <SEP> the <SEP> cassette
<tb> att3 # aacpSPM508 <SEP> Hyg, <SEP> Apra <SEP> pSPM504 <SEP> in <SEP> which <SEP> orfl3c <SEP> Example <SEP> 12
<tb> is <SEP> interrupted <SEP> with <SEP> the <SEP> cassette
<tb> att3Qaac-
<Tb>
<Desc/Clms Page number 177> <Desc / Clms Page number 177>

<tb>
<tb> pSPM509 <SEP> Hyg, <SEP> Apra <SEP> pSPM504 <SEP> dans <SEP> lequel <SEP> orfl4 <SEP> Exemple <SEP> 13
<tb> est <SEP> interrompue <SEP> par <SEP> la <SEP> cassette
<tb> att3QaacpBXLllll <SEP> Am <SEP> Fragment <SEP> de <SEP> 1,11 <SEP> kb <SEP> Exemple <SEP> 14
<tb> contenant <SEP> or/6* <SEP> amplifié <SEP> par
<tb> PCR <SEP> à <SEP> partir <SEP> de <SEP> pSPM7, <SEP> dans
<tb> le <SEP> vecteur <SEP> pGEM-T <SEP> Easy
<tb> pBXL1112 <SEP> Am, <SEP> Hyg <SEP> pBXLllll <SEP> dans <SEP> lequel <SEP> la <SEP> Exemple <SEP> 14
<tb> cassette <SEP> attlQhyg+ <SEP> a <SEP> été
<tb> introduite <SEP> après <SEP> délétion <SEP> de
<tb> 120 <SEP> pb <SEP> dans <SEP> la <SEP> séquence
<tb> codante <SEP> du <SEP> gène <SEP> or/6*
<tb> pBXLI <SEP> 113 <SEP> Apra, <SEP> Hyg <SEP> Insert <SEP> PstI <SEP> de <SEP> 3. <SEP> 7 <SEP> kb <SEP> de <SEP> Exemple <SEP> 14
<tb> pBXLl <SEP> 112 <SEP> dans <SEP> pOJ260
<tb> pSPM520 <SEP> Am <SEP> Fragment <SEP> de <SEP> PCR <SEP> amplifié <SEP> par <SEP> Exemple <SEP> 17
<tb> les <SEP> oligonucléotides <SEP> EDR39EDR42 <SEP> dans <SEP> pGEM-T <SEP> Easy
<tb> pSPM521 <SEP> Am <SEP> Fragment <SEP> de <SEP> PCR <SEP> amplifié <SEP> par <SEP> Exemple <SEP> 17
<tb> les <SEP> oligonucléotides <SEP> EDR40EDR42 <SEP> dans <SEP> pGEM-T <SEP> Easy
<tb> pSPM522 <SEP> Am <SEP> Fragment <SEP> de <SEP> PCR <SEP> amplifié <SEP> par <SEP> Exemple <SEP> 17
<tb> les <SEP> oligonucléotides <SEP> EDR41EDR42 <SEP> dans <SEP> pGEM-T <SEP> Easy
<tb> pUWL201 <SEP> Am,Ts <SEP> (Doumith <SEP> et <SEP> al.,
<tb> 2000)
<tb> pSPM523 <SEP> Am, <SEP> Ts <SEP> Fragment <SEP> HindIII-BamHI <SEP> de <SEP> Exemple <SEP> 17
<tb> l'insert <SEP> du <SEP> plasmide <SEP> pSPM520
<tb> dans <SEP> le <SEP> vecteur <SEP> pUWL201
<tb> pSPM524 <SEP> Am, <SEP> Ts <SEP> Fragment <SEP> HindIII-BamHI <SEP> de <SEP> Exemple <SEP> 17
<tb> l'insert <SEP> du <SEP> plasmide <SEP> pSPM521
<tb> <Tb>
<tb> pSPM509 <SEP> Hyg, <SEP> Apra <SEP> pSPM504 <SEP> in <SEP> which <SEP> orfl4 <SEP> Example <SEP> 13
<tb> is <SEP> interrupted <SEP> with <SEP> the <SEP> cassette
<tb> att3QaacpBXLllll <SEP> Am <SEP> Fragment <SEP> of <SEP> 1.11 <SEP> kb <SEP> Example <SEP> 14
<tb> containing <SEP> or / 6 * <SEP> amplified <SEP> by
<tb> PCR <SEP> to <SEP> from <SEP> of <SEP> pSPM7, <SEP> in
<tb> the <SEP> vector <SEP> pGEM-T <SEP> Easy
<tb> pBXL1112 <SEP> Am, <SEP> Hyg <SEP> pBXLllll <SEP> in <SEP> which <SEP> the <SEP> Example <SEP> 14
<tb> cassette <SEP> attlQhyg + <SEP> a <SEP> summer
<tb> introduced <SEP> after <SEP> deletion <SEP> of
<tb> 120 <SEP> pb <SEP> in <SEP> the <SEP> sequence
<tb> coding <SEP> of <SEP> gene <SEP> or / 6 *
<tb> pBXLI <SEP> 113 <SEP> Apra, <SEP> Hyg <SEP> Insert <SEP> PstI <SEP> of <SEP> 3. <SEP> 7 <SEP> kb <SEP> of <SEP> Example <SEP> 14
<tb> pBXLl <SEP> 112 <SEP> in <SEP> pOJ260
<tb> pSPM520 <SEP> Am <SEP> Fragment <SEP> of <SEP> PCR <SEP> amplified <SEP> by <SEP> Example <SEP> 17
<tb> the <SEP> oligonucleotides <SEP> EDR39EDR42 <SEP> in <SEP> pGEM-T <SEP> Easy
<tb> pSPM521 <SEP> Am <SEP> Fragment <SEP> of <SEP> PCR <SEP> amplified <SEP> by <SEP> Example <SEP> 17
<tb> the <SEP> oligonucleotides <SEP> EDR40EDR42 <SEP> in <SEP> pGEM-T <SEP> Easy
<tb> pSPM522 <SEP> Am <SEP> Fragment <SEP> of <SEP> PCR <SEP> amplified <SEP> by <SEP> Example <SEP> 17
<tb> the <SEP> oligonucleotides <SEP> EDR41EDR42 <SEP> in <SEP> pGEM-T <SEP> Easy
<tb> pUWL201 <SEP> Am, Ts <SEP> (Doumith <SEP> and <SEP> al.,
<tb> 2000)
<tb> pSPM523 <SEP> Am, <SEP> Ts <SEP> Fragment <SEP> HindIII-BamHI <SEP> of <SEP> Example <SEP> 17
<tb> the <SEP> insert of the <SEP> plasmid <SEP> pSPM520
<tb> in <SEP> the <SEP> vector <SEP> pUWL201
<tb> pSPM524 <SEP> Am, <SEP> Ts <SEP> Fragment <SEP> HindIII-BamHI <SEP> of <SEP> Example <SEP> 17
<tb> the <SEP> insert of the <SEP> plasmid <SEP> pSPM521
<Tb>
<Desc/Clms Page number 178> <Desc / Clms Page number 178>

<tb>
<tb> dans <SEP> le <SEP> vecteur <SEP> pUWL201
<tb> pSPM525 <SEP> Am, <SEP> Ts <SEP> Fragment <SEP> HindIII-BamHI <SEP> de <SEP> Exemple <SEP> 17
<tb> l'insert <SEP> du <SEP> plasmide <SEP> pSPM522
<tb> dans <SEP> le <SEP> vecteur <SEP> pUWL201
<tb> pSPM527 <SEP> Am <SEP> pSPM521 <SEP> avec <SEP> décalage <SEP> du <SEP> Exemple <SEP> 17
<tb> cadre <SEP> de <SEP> lecture <SEP> au <SEP> niveau <SEP> du
<tb> site <SEP> XhoI <SEP>
<tb> pSPM528 <SEP> Am <SEP> , <SEP> Ts <SEP> Fragment <SEP> HindIII-BamHI <SEP> de <SEP> Exemple <SEP> 17
<tb> l'insert <SEP> du <SEP> plasmide <SEP> pSPM527
<tb> dans <SEP> le <SEP> vecteur <SEP> pUWL201 <SEP>
<tb> pVF <SEP> 10.4 <SEP> (Vara <SEP> et <SEP> al., <SEP> 1985 <SEP> ;
<tb> Lacalle <SEP> et <SEP> al., <SEP> 1989)
<tb> pPM803 <SEP> Ts <SEP> (Mazodier,P. <SEP> et <SEP> al.,
<tb> 1989)
<tb> pGEM-T-pac- <SEP> Am <SEP> Cassette <SEP> pac-oriT <SEP> (amplifiée <SEP> Exemple <SEP> 18
<tb> oriT <SEP> par <SEP> PCR <SEP> à <SEP> partir <SEP> de <SEP> pVF <SEP> 10.4
<tb> et <SEP> pPM803) <SEP> dans <SEP> dans <SEP> pGEMT <SEP> Easy <SEP>
<tb> pWED2 <SEP> Am <SEP> Cassette <SEP> pac-oriT <SEP> obtenue <SEP> à <SEP> Exemple <SEP> 18
<tb> partir <SEP> de <SEP> pGEM-T-pac-oriT
<tb> insérée <SEP> dans <SEP> pWEDl
<tb> pSPM34 <SEP> Am <SEP> pWED2 <SEP> avec <SEP> insert <SEP> au <SEP> niveau <SEP> Exemple <SEP> 19
<tb> du <SEP> site <SEP> BamHI
<tb> pSPM35 <SEP> Am <SEP> pWED2 <SEP> avec <SEP> insert <SEP> au <SEP> niveau <SEP> Exemple <SEP> 19
<tb> du <SEP> site <SEP> BamHI
<tb> pSPM36 <SEP> Am <SEP> pWED2 <SEP> avec <SEP> insert <SEP> au <SEP> niveau <SEP> Exemple <SEP> 19
<tb> du <SEP> site <SEP> BamHI
<tb> pSPM37 <SEP> Am <SEP> pWED2 <SEP> avec <SEP> insert <SEP> au <SEP> niveau <SEP> Exemple <SEP> 19
<tb> du <SEP> site <SEP> BamHI
<tb> <Tb>
<tb> in <SEP> the <SEP> vector <SEP> pUWL201
<tb> pSPM525 <SEP> Am, <SEP> Ts <SEP> Fragment <SEP> HindIII-BamHI <SEP> of <SEP> Example <SEP> 17
<tb> the <SEP> insert of the <SEP> plasmid <SEP> pSPM522
<tb> in <SEP> the <SEP> vector <SEP> pUWL201
<tb> pSPM527 <SEP> Am <SEP> pSPM521 <SEP> with <SEP><SEP> offset <SEP> Example <SEP> 17
<tb> frame <SEP> from <SEP> read <SEP> at <SEP> level <SEP> from
<tb> site <SEP> XhoI <SEP>
<tb> pSPM528 <SEP> Am <SEP>, <SE> Ts <SEP> Fragment <SEP> HindIII-BamHI <SEP> of <SEP> Example <SEP> 17
<tb> the <SEP> insert of the <SEP> plasmid <SEP> pSPM527
<tb> in <SEP> the <SEP> vector <SEP> pUWL201 <SEP>
<tb> pVF <SEP> 10.4 <SEP> (Vara <SEP> and <SEP> al., <SEP> 1985 <SEP>;
<tb> Lacalle <SEP> and <SEP> al., <SEP> 1989)
<tb> pPM803 <SEP> Ts <SEP> (Mazodier, P. <SEP> and <SEP> al.,
<tb> 1989)
<tb> pGEM-T-pac- <SEP> Am <SEP> Cassette <SEP> pac-oriT <SEP> (amplified <SEP> Example <SEP> 18
<tb> oriT <SEP> by <SEP> PCR <SEP> to <SEP> from <SEP> of <SEP> pVF <SEP> 10.4
<tb> and <SEP> pPM803) <SEP> in <SEP> in <SEP> pGEMT <SEP> Easy <SEP>
<tb> pWED2 <SEP> Am <SEP> Cassette <SEP> pac-oriT <SEP> obtained <SEP> to <SEP> Example <SEP> 18
<tb> from <SEP> of <SEP> pGEM-T-pac-oriT
<tb> inserted <SEP> in <SEP> pWEDl
<tb> pSPM34 <SEP> Am <SEP> pWED2 <SEP> with <SEP> insert <SEP> at <SEP> level <SEP> Example <SEP> 19
<tb> of the <SEP><SEP> BamHI site
<tb> pSPM35 <SEP> Am <SEP> pWED2 <SEP> with <SEP> insert <SEP> at <SEP> level <SEP> Example <SEP> 19
<tb> of the <SEP><SEP> BamHI site
<tb> pSPM36 <SEP> Am <SEP> pWED2 <SEP> with <SEP> insert <SEP> at <SEP> level <SEP> Example <SEP> 19
<tb> of the <SEP><SEP> BamHI site
<tb> pSPM37 <SEP> Am <SEP> pWED2 <SEP> with <SEP> insert <SEP> at <SEP> level <SEP> Example <SEP> 19
<tb> of the <SEP><SEP> BamHI site
<Tb>
<Desc/Clms Page number 179> <Desc / Clms Page number 179>

<tb>
<tb> pSPM38 <SEP> Am <SEP> pWED2 <SEP> avec <SEP> insert <SEP> au <SEP> niveau <SEP> Exemple <SEP> 19
<tb> du <SEP> site <SEP> BamHI
<tb> pSPM39 <SEP> Am <SEP> pWED2 <SEP> avec <SEP> insert <SEP> au <SEP> niveau <SEP> Exemple <SEP> 19
<tb> du <SEP> site <SEP> BamHI
<tb> pSPM40 <SEP> Am <SEP> pWED2 <SEP> avec <SEP> insert <SEP> au <SEP> niveau <SEP> Exemple <SEP> 19
<tb> du <SEP> site <SEP> BamHI
<tb> pSPM41 <SEP> Am <SEP> pWED2 <SEP> avec <SEP> insert <SEP> au <SEP> niveau <SEP> Exemple <SEP> 19
<tb> du <SEP> site <SEP> BamHI
<tb> pSPM42 <SEP> Am <SEP> pWED2 <SEP> avec <SEP> insert <SEP> au <SEP> niveau <SEP> Exemple <SEP> 19
<tb> du <SEP> site <SEP> BamHI
<tb> pSPM43 <SEP> Am <SEP> pWED2 <SEP> avec <SEP> insert <SEP> au <SEP> niveau <SEP> Exemple <SEP> 19
<tb> du <SEP> site <SEP> BamHI
<tb> pSPM44 <SEP> Am <SEP> pWED2 <SEP> avec <SEP> insert <SEP> au <SEP> niveau <SEP> Exemple <SEP> 19
<tb> du <SEP> site <SEP> BamHI
<tb> pSPM45 <SEP> Am <SEP> pWED2 <SEP> avec <SEP> insert <SEP> au <SEP> niveau <SEP> Exemple <SEP> 19
<tb> du <SEP> site <SEP> BamHI
<tb> pSPM47 <SEP> Am <SEP> pWED2 <SEP> avec <SEP> insert <SEP> au <SEP> niveau <SEP> Exemple <SEP> 19
<tb> du <SEP> site <SEP> BamHI
<tb> pSPM48 <SEP> Am <SEP> pWED2 <SEP> avec <SEP> insert <SEP> au <SEP> niveau <SEP> Exemple <SEP> 19
<tb> du <SEP> site <SEP> BamHI
<tb> pSPM50 <SEP> Am <SEP> pWED2 <SEP> avec <SEP> insert <SEP> au <SEP> niveau <SEP> Exemple <SEP> 19
<tb> du <SEP> site <SEP> BamHI
<tb> pSPM51 <SEP> Am <SEP> pWED2 <SEP> avec <SEP> insert <SEP> au <SEP> niveau <SEP> Exemple <SEP> 19
<tb> du <SEP> site <SEP> BamHI
<tb> pSPM52 <SEP> Am <SEP> pWED2 <SEP> avec <SEP> insert <SEP> au <SEP> niveau <SEP> Exemple <SEP> 19
<tb> du <SEP> site <SEP> BamHI
<tb> pSPM53 <SEP> Am <SEP> pWED2 <SEP> avec <SEP> insert <SEP> au <SEP> niveau <SEP> Exemple <SEP> 19
<tb> du <SEP> site <SEP> BamHI
<tb> pSPM55 <SEP> Am <SEP> pWED2 <SEP> avec <SEP> insert <SEP> au <SEP> niveau <SEP> Exemple <SEP> 19
<tb> <Tb>
<tb> pSPM38 <SEP> Am <SEP> pWED2 <SEP> with <SEP> insert <SEP> at <SEP> level <SEP> Example <SEP> 19
<tb> of the <SEP><SEP> BamHI site
<tb> pSPM39 <SEP> Am <SEP> pWED2 <SEP> with <SEP> insert <SEP> at <SEP> level <SEP> Example <SEP> 19
<tb> of the <SEP><SEP> BamHI site
<tb> pSPM40 <SEP> Am <SEP> pWED2 <SEP> with <SEP> insert <SEP> at <SEP> level <SEP> Example <SEP> 19
<tb> of the <SEP><SEP> BamHI site
<tb> pSPM41 <SEP> Am <SEP> pWED2 <SEP> with <SEP> insert <SEP> at <SEP> level <SEP> Example <SEP> 19
<tb> of the <SEP><SEP> BamHI site
<tb> pSPM42 <SEP> Am <SEP> pWED2 <SEP> with <SEP> insert <SEP> at <SEP> level <SEP> Example <SEP> 19
<tb> of the <SEP><SEP> BamHI site
<tb> pSPM43 <SEP> Am <SEP> pWED2 <SEP> with <SEP> insert <SEP> at <SEP> level <SEP> Example <SEP> 19
<tb> of the <SEP><SEP> BamHI site
<tb> pSPM44 <SEP> Am <SEP> pWED2 <SEP> with <SEP> insert <SEP> at <SEP> level <SEP> Example <SEP> 19
<tb> of the <SEP><SEP> BamHI site
<tb> pSPM45 <SEP> Am <SEP> pWED2 <SEP> with <SEP> insert <SEP> at <SEP> level <SEP> Example <SEP> 19
<tb> of the <SEP><SEP> BamHI site
<tb> pSPM47 <SEP> Am <SEP> pWED2 <SEP> with <SEP> insert <SEP> at <SEP> level <SEP> Example <SEP> 19
<tb> of the <SEP><SEP> BamHI site
<tb> pSPM48 <SEP> Am <SEP> pWED2 <SEP> with <SEP> insert <SEP> at <SEP> level <SEP> Example <SEP> 19
<tb> of the <SEP><SEP> BamHI site
<tb> pSPM50 <SEP> Am <SEP> pWED2 <SEP> with <SEP> insert <SEP> at <SEP> level <SEP> Example <SEP> 19
<tb> of the <SEP><SEP> BamHI site
<tb> pSPM51 <SEP> Am <SEP> pWED2 <SEP> with <SEP> insert <SEP> at <SEP> level <SEP> Example <SEP> 19
<tb> of the <SEP><SEP> BamHI site
<tb> pSPM52 <SEP> Am <SEP> pWED2 <SEP> with <SEP> insert <SEP> at <SEP> level <SEP> Example <SEP> 19
<tb> of the <SEP><SEP> BamHI site
<tb> pSPM53 <SEP> Am <SEP> pWED2 <SEP> with <SEP> insert <SEP> at <SEP> level <SEP> Example <SEP> 19
<tb> of the <SEP><SEP> BamHI site
<tb> pSPM55 <SEP> Am <SEP> pWED2 <SEP> with <SEP> insert <SEP> at <SEP> level <SEP> Example <SEP> 19
<Tb>
<Desc/Clms Page number 180> <Desc / Clms Page number 180>

<tb>
<tb> du <SEP> site <SEP> BamHI
<tb> pSPM56 <SEP> Am <SEP> pWED2 <SEP> avec <SEP> insert <SEP> au <SEP> niveau <SEP> Exemple <SEP> 19
<tb> du <SEP> site <SEP> BamHI
<tb> pSPM58 <SEP> Kn <SEP> Fragment <SEP> PstI-PstI <SEP> d'environ <SEP> Exemple <SEP> 19
<tb> 6kb <SEP> de <SEP> l'insert <SEP> de <SEP> pSPM36
<tb> dans <SEP> pBK-CMV
<tb> pSPM72 <SEP> Kn <SEP> Fragment <SEP> StuI-StuI <SEP> d'environ <SEP> Exemple <SEP> 19
<tb> 10 <SEP> kb <SEP> de <SEP> l'insert <SEP> de <SEP> pSPM36
<tb> cloné <SEP> dans <SEP> pBK-CMV
<tb> pSPM73 <SEP> Cm <SEP> Fragment <SEP> EcoRI-HindIII <SEP> de <SEP> Exemple <SEP> 19
<tb> l'insert <SEP> de <SEP> pSPM72 <SEP> dans <SEP> pBCSK+
<tb> pSPM515 <SEP> Am <SEP> Fragment <SEP> de <SEP> PCR <SEP> amplifié <SEP> par <SEP> Exemple <SEP> 22
<tb> EDR31-EDR37 <SEP> dans <SEP> pGEM-T
<tb> easy
<tb> pSPM519 <SEP> Am, <SEP> Ts <SEP> Insert <SEP> HindIII/XbaI <SEP> de <SEP> Exemple <SEP> 22
<tb> pSPM515 <SEP> dans <SEP> pUWL201
<tb> pOS49. <SEP> 52 <SEP> Apra <SEP> Séquence <SEP> codante <SEP> de <SEP> tylB <SEP> sous <SEP> Exemple <SEP> 23
<tb> contrôle <SEP> du <SEP> promoteur <SEP> ermE*
<tb> dans <SEP> le <SEP> plasmide <SEP> pKC1218
<tb> pSPM74 <SEP> Am <SEP> Fragment <SEP> de <SEP> PCR <SEP> amplifié <SEP> par <SEP> Exemple <SEP> 24
<tb> KF30-KF31 <SEP> dans <SEP> pGEM-T
<tb> easy
<tb> pSPM75 <SEP> Am, <SEP> Ts <SEP> Insert <SEP> HindIII/BamHI <SEP> de <SEP> Exemple <SEP> 24
<tb> pSPM74 <SEP> dans <SEP> pUWL201
<tb> <Tb>
<tb> of the <SEP><SEP> BamHI site
<tb> pSPM56 <SEP> Am <SEP> pWED2 <SEP> with <SEP> insert <SEP> at <SEP> level <SEP> Example <SEP> 19
<tb> of the <SEP><SEP> BamHI site
<tb> pSPM58 <SEP> Kn <SEP> Fragment <SEP> PstI-PstI <SEP> of about <SEP> Example <SEP> 19
<tb> 6kb <SEP> of <SEP> the <SEP> insert of <SEP> pSPM36
<tb> in <SEP> pBK-CMV
<tb> pSPM72 <SEP> Kn <SEP> Fragment <SEP> StuI-StuI <SEP> of about <SEP> Example <SEP> 19
<tb> 10 <SEP> kb <SEP> of <SEP> the <SEP> insert of <SEP> pSPM36
<tb> cloned <SEP> in <SEP> pBK-CMV
<tb> pSPM73 <SEP> Cm <SEP> Fragment <SEP> EcoRI-HindIII <SEP> of <SEP> Example <SEP> 19
<tb> the <SEP> insert of <SEP> pSPM72 <SEP> in <SEP> pBCSK +
<tb> pSPM515 <SEP> Am <SEP> Fragment <SEP> of <SEP> PCR <SEP> amplified <SEP> by <SEP> Example <SEP> 22
<tb> EDR31-EDR37 <SEP> in <SEP> pGEM-T
<tb> easy
<tb> pSPM519 <SEP> Am, <SEP> Ts <SEP> Insert <SEP> HindIII / XbaI <SEP> of <SEP> Example <SEP> 22
<tb> pSPM515 <SEP> in <SEP> pUWL201
<tb> pOS49. <SEP> 52 <SEP> Apra <SEP><SEP><SEP> coding sequence of <SEP> tylB <SEP> under <SEP> Example <SEP> 23
<tb><SEP> control of <SEP> promoter <SEP> ermE *
<tb> in <SEP> the <SEP> plasmid <SEP> pKC1218
<tb> pSPM74 <SEP> Am <SEP> Fragment <SEP> of <SEP> PCR <SEP> amplified <SEP> by <SEP> Example <SEP> 24
<tb> KF30-KF31 <SEP> in <SEP> pGEM-T
<tb> easy
<tb> pSPM75 <SEP> Am, <SEP> Ts <SEP> Insert <SEP> HindIII / BamHI <SEP> of <SEP> Example <SEP> 24
<tb> pSPM74 <SEP> in <SEP> pUWL201
<Tb>
<Desc/Clms Page number 181><Desc / Clms Page number 181>
Dépôt de matériel biologique
Les organismes suivants ont été déposés le 10 juillet 2002 auprès de la Collection Nationale de Cultures de Microorganismes (CNCM), 25 rue du Docteur Roux, 75724 Paris Cedex 15, France, selon les dispositions du Traité de Budapest. Deposit of biological material
The following organizations were registered on July 10, 2002 with the National Collection of Cultures of Microorganisms (CNCM), 25 rue du Docteur Roux, 75724 Paris Cedex 15, France, according to the provisions of the Budapest Treaty.
- Souche OSC2 sous le numéro d'enregistrement 1-2908. - Strain OSC2 under the registration number 1-2908.
- Souche SPM501 sous le numéro d'enregistrement 1-2909. Strain SPM501 under the registration number 1-2909.
- Souche SPM502 sous le numéro d'enregistrement 1-2910. - strain SPM502 under the registration number 1-2910.
- Souche SPM507 sous le numéro d'enregistrement 1-2911. Strain SPM507 under the registration number 1-2911.
- Souche SPM508 sous le numéro d'enregistrement 1-2912. Strain SPM508 under the registration number 1-2912.
- Souche SPM509 sous le numéro d'enregistrement 1-2913. - strain SPM509 under the registration number 1-2913.
- Souche SPM21 sous le numéro d'enregistrement 1-2914. Strain SPM21 under the registration number 1-2914.
- Souche SPM22 sous le numéro d'enregistrement 1-2915. - strain SPM22 under the registration number 1-2915.
- Souche OS49. 67 sous le numéro d'enregistrement 1-2916. - OS49 strain. 67 under the registration number 1-2916.
- Souche OS49. 107 sous le numéro d'enregistrement 1-2917. - OS49 strain. 107 under the registration number 1-2917.
- Souche Escherichia Coli DH5a contenant le plasmide pOS44. 4 sous le numéro d'enregistrement 1-2918. Escherichia coli strain DH5a containing the plasmid pOS44. 4 under the registration number 1-2918.
Les organismes suivants ont été déposés le 26 février 2003 auprès de la
Collection Nationale de Cultures de Microorganismes (CNCM), 25 rue du Docteur
Roux, 75724 Paris Cedex 15, France, selon les dispositions du Traité de Budapest. The following organizations were registered on February 26, 2003 with the
National Collection of Cultures of Microorganisms (CNCM), 25 rue du Docteur
Roux, 75724 Paris Cedex 15, France, according to the provisions of the Budapest Treaty.
- Souche SPM502 pSPM525 sous le numéro d'enregistrement 1-2977. Strain SPM502 pSPM525 under the registration number 1-2977.
<Desc/Clms Page number 182> <Desc / Clms Page number 182>
Bibliographie : - Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. (1990). 215 (3) :403-410. Bibliography: - Altschul SF, Gish W, W Miller, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. (1990). 215 (3): 403-410.
- Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ. - Altschul SF, Madden TL, AA Schaffer, Zhang J, Z Zhang, Miller W, Lipman DJ.
Gapped BLAST and PSI-BLAST : a new generation of protein database search programs. Nucleic Acids Res. (1997). 25 (17):3389-402. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. (1997). 25 (17): 3389-402.
- Amann,E., Ochs, B. and Abel,K.J. Tightly regulated tac promoter vectors useful for the expression of unfused and fused proteins in Escherichia coli Gene (1988)
69 (2), 301-315. - Amann, E., Ochs, B. and Abel, KJ Tightly regulated tac promoter vectors for the expression of unfused and fused proteins in Escherichia coli Gene (1988)
69 (2), 301-315.
- Arisawa,A., Kawamura,N., Tsunekawa,H., Okamura,K., Tone,H. and
Okamoto, R. Cloning and nucleotide séquences of two genes involved in the 4"-
O-acylation of macrolide antibiotics from Streptomyces thermotolerans. Biosci. - Arisawa, A., Kawamura, N., Tsunekawa, H., Okamura, K., Tone, H. and
Okamoto, R. Cloning and nucleotide sequences of two genes involved in the 4 "-
O-acylation of macrolide antibiotics from Streptomyces thermotolerans. Biosci.
Biotechnol. Biochem. (1993). 57 (12) : 2020-2025. Biotechnol. Biochem. (1993). 57 (12): 2020-2025.
- Arisawa A, Kawamura N, Takeda K, Tsunekawa H, Okamura K, Okamoto R. - Arisawa A, Kawamura N, Takeda K, Tsunekawa H, Okamura K, Okamoto R.
Cloning of the macrolide antibiotic biosynthesis gene acyA, which encodes 3-0- acyltransferase, from Streptomyces thermotolerans and its use for direct fermentative production of a hybrid macrolide antibiotic. Appl Environ
Microbiol. (1994). 60 (7) :2657-2660. Cloning of the macrolide antibiotic biosynthesis gene acyA, which encodes 3-0-acyltransferase, from Streptomyces thermotolerans and its use for direct fermentative production of a hybrid macrolide antibiotic. Appl Environ
Microbiol. (1994). 60 (7): 2657-2660.
- August,P.R., Tang,L., Yoon,Y.J., Ning,Streptomyces, Mueller, R., Yu,T.W.,
Taylor, M., Hoffmann,D., Kim, C.G., Zhang,X., Hutchinson,C.R. et Floss,H.G. - August, PR, Tang, L., Yoon, Y., Ning, Streptomyces, Mueller, R., Yu, TW,
Taylor, M., Hoffmann, D., Kim, CG, Zhang, X., Hutchinson, CR and Floss, HG
Biosynthesis of the ansamycin antibiotic rifamycin: deductions from the molecular analysis of the rif biosynthetic gene cluster of Amycolatopsis mediterranei S699. Chem. Biol. (1998) 5 (2), 69-79. Biosynthesis of the ansamycin antibiotic rifamycin: deductions from the molecular analysis of the biosynthetic rif gene cluster of Amycolatopsis mediterranei S699. Chem. Biol. (1998) 5 (2), 69-79.
- Ausubel Fred M., Brent Roger, Kingston Robert E., Moore David D., Seidman
J. G., Smith John A., Struhl Kevin (Editeurs). Current Protocols in Molecular
Biology, publié par John Wiley & Sons Inc. Current Protocols Customer Service,
605 Third Avenue, 9th Floor New York, NY 10158 USA (édition mise à jour de
Mars 2002). - Ausubel Fred M., Brent Roger, Robert E. Kingston, Moore David D., Seidman
JG, Smith John A., Struhl Kevin (Editors). Current Protocols in Molecular
Biology, published by John Wiley & Sons Inc. Current Protocols Customer Service,
605 Third Avenue, New York 9th Floor, NY 10158 USA (updated edition of
March 2002).
- Baltz RH, McHenney MA, Cantwell CA, Queener SW, Solenberg PJ. - Baltz RH, McHenney MA, Cantwell CA, Queener SW, Solenberg PJ.
Applications oftransposition mutagenesis in antibiotic producing streptomycetes. Applications oftransposition mutagenesis in antibiotic producing streptomycetes.
Antonie Van Leeuwenhoek. (1997). 71 (1-2) : 179-187. Antonie Van Leeuwenhoek. (1997). 71 (1-2): 179-187.
- Bate N, Butler AR, Gandecha AR, Cundliffe E. Multiple regulatory genes in the tylosin biosynthetic cluster of Streptomyces fradiae. Chem Biol. (1999). 6 (9):
617-624. Bate N, Butler AR, Gandecha AR, Cundliffe E. Multiple regulatory genes in the tylosin biosynthetic cluster of Streptomyces fradiae. Chem Biol. (1999). 6 (9):
617-624.
<Desc/Clms Page number 183> <Desc / Clms Page number 183>
- Bate,N., Butler, A.R., Smith,LP. et Cundliffe,E. The mycarose-biosynthetic genes of Streptomyces fradiae, producer of tylosin. Microbiology (2000). 146 (Pt 1),
139-146. - Bate, N., Butler, AR, Smith, LP. and Cundliffe, E. The mycarose-biosynthetic genes of Streptomyces fradiae, producer of tylosin. Microbiology (2000). 146 (Pt 1),
139-146.
- Bayley C., Morgan, Dale E. C., Ow D. W. Exchange of gene activity in transgenic plants catalysed by the Cre-lox site spécifie system. Plant Mol. Biol (1992). 18 :353-361. - Bayley C., Morgan, Dale E.C., D. Ow D. Exchange of gene activity in transgenic plants catalysed by the Cre-lox site specifies system. Plant Mol. Biol (1992). 18: 353-361.
- Bentley,S.D., Chater,K.F., Cerdeno-Tarraga,A.M., Challis,G.L., Thomson,N.R.,
James,K.D., Harris,D.E., Quail,M.A., Kieser,H., Harper,D., Bateman,A.,
Brown,S., Chandra,G., Chen,C.W., Collins,M., Cronin,A., Fraser,A., Goble, A.,
Hidalgo, J., Homsby,T., Howarth,S., Huang,C.H., Kieser, T., Larke, L., Murphy,L.,
Oliver,K., O'Neil,S., Rabbinowitsch,E., Rajandream,M.A., Rutherford,K.,
Rutter, S., Seeger, K., Saunders,D., Sharp, S., Squares,R., Squares, S., Taylor, K.,
Warren,T., Wietzorrek, A., Woodward, J., Barrell,B.G., Parkhill,J. et
Hopwood, D.A. Complete genome séquence of the model actinomycete
Streptomyces coelicolor A3(2). Nature (2002). 417 (6885), 141-147. Bentley, SD, Chater, KF, Cerdeno-Tarraga, AM, Challis, GL, Thomson, NR,
James, KD, Harris, DE, Quail, MA, Kieser, H., Harper, D., Bateman, A.,
Brown, S., Chandra, G., Chen, CW, Collins, M., Cronin, A., Fraser, A., Goble, A.,
Hidalgo, J., Homsby, T., Howarth, S., Huang, CH, Kieser, T., Larke, L., Murphy, L.,
Oliver, K., O'Neil, S., Rabbinowitsch, E., Rajandream, MA, Rutherford, K.,
Rutter, S., Seeger, K., Saunders, D., Sharp, S., Squares, R., Squares, S., Taylor, K.,
Warren, T., Wietzorrek, A., Woodward, J., Barrell, BG, Parkhill, J. and
Hopwood, Complete DNA sequence of the model actinomycete
Streptomyces coelicolor A3 (2). Nature (2002). 417 (6885), 141-147.
- Bibb MJ, Findlay PR, Johnson MW. The relationship between base composition and codon usage in bacterial genes and its use for the simple and reliable identification of protein-coding séquences. Gène. (1984) 30 (1-3):157-166. - Bibb MJ, Findlay PR, Johnson MW. The relationship between base composition and codon use in bacterial genes and its use for the simple and reliable identification of protein-coding sequences. Gene. (1984) 30 (1-3): 157-166.
- Bibb MJ, Janssen GR, Ward JM. Cloning and analysis of the promoter region of the erythromycin résistance gene (ermE) of Streptomyces erythraeus. Gene. - Bibb MJ, Janssen GR, Ward JM. Cloning and analysis of the promoter region of the erythromycin resistance gene (ermE) of Streptomyces erythraeus. Gene.
(1985). 38 (1-3) : 215-26. (1985). 38 (1-3): 215-26.
- Bibb MJ, White J, Ward JM, Janssen GR. The mRNA for the 23S rRNA methylase encoded by the ermE gene of Saccharopolyspora erythraea is translated in the absence of a conventional ribosome-binding site. Mol Microbiol. - Bibb MJ, White J, Ward JM, Janssen GR. The mRNA for the 23S rRNA methylase encoded by the ermE gene of Saccharopolyspora erythraea is translated into the absence of a standard ribosome-binding site. Mol Microbiol.
(1994). 14 (3) : 533-45. (1994). 14 (3): 533-45.
- Bierman M, Logan R, O'Brien K, Seno ET, Rao RN, Schoner BE. Plasmid cloning vectors for the conjugal transfer of DNA from Escherichia coli to
Streptomyces spp. Gene. 1992;116(1):43-9. - Bierman M, Logan R, O'Brien K, Seno ET, Rao RN, Schoner BE. Plasmid cloning vectors for the transfer of DNA from Escherichia coli to
Streptomyces spp. Gene. 1992; 116 (1): 43-9.
- Blondelet-Rouault M-H., Weiser J., Lebrihi A., Branny P. and Pemodet J-L. - Blondelet-Rouault M-H., Weiser J., Lebrihi A., Branny P. and Pemodet J-L.
Antibiotic résistance gene cassettes derived from the W interposon for use in E. coli and Streptomyces. Gène (1997) 190 : 315-317. Antibiotic resistance gene cassettes derived from the interposon for use in E. coli and Streptomyces. Gene (1997) 190: 315-317.
- Boccard F., Smokvina T., J.-L., Pemodet Fridmann A., and Guérineau M.,
Structural analysis of loci involved in pSAM2 site-specific integration in
Streptomyces. Plasmid, (1989a). 21 : 59-70. Boccard F., Smokvina T., J.-L., Pemodet Fridmann A., and Guerineau M.,
Structural analysis of loci involved in pSAM2 site-specific integration in
Streptomyces. Plasmid, (1989a). 21: 59-70.
- Boccard F., Smokvina T., J.-L., Pemodet Fridmann A., and Guérineau M.. The integrated conjugative plasmpid pSAM2 of Streptomyces ambofaciens is related to temperate bacteriophages. EMBO J. (1989b) 8 : 973-980. Boccard F., Smokvina T., J.-L., Pemodet A. Fridmann, and Guérineau M. The integrated conjugative plasmpid pSAM2 of Streptomyces ambofaciens is related to bacteriophage. EMBO J. (1989b) 8: 973-980.
<Desc/Clms Page number 184> <Desc / Clms Page number 184>
- Brunelli J. P., Pall M.L., A series of Yeast/ E. coli lambda expression vectors designed for directional cloning of cDNA and cre/lox mediated plasmid excision. - Brunelli J.P., Pall M.L., A series of Yeast / E. coli lambda expression vectors designed for the directional cloning of cDNA and cre / lox mediated plasmid excision.
Yeast. (1993) 9: 1309-1318. Yeast. (1993) 9: 1309-1318.
- Camilli A., Beattie D. T. et Mekalanos J. J. Use of genetic recombination as a reporter of gene expression. Proc. Natl. Acad. Sci. USA (1994) 91 (7), -Campelo, A.B. and Gil,J.A. The candicidin gene cluster from Streptomyces griseus IMRU 3570. Microbiology (2002) 148 (Pt 1), 51-59. A. Camilli, Beattie D. T. and J. Mekalanos J. Use of genetic recombination as a reporter of gene expression. Proc. Natl. Acad. Sci. USA (1994) 91 (7), -Campelo, A.B. and Gil, J.A. The Candicidal Gene Cluster from Streptomyces griseus IMRU 3570. Microbiology (2002) 148 (Pt 1), 51-59.
- Carreras C, Frykman S, Ou S, Cadapan L, Zavala S, Woo E, Leaf T, Carney J,
Burlingame M, Patel S, Ashley G, Licari P. Saccharopolyspora erythraea- catalyzed bioconversion of 6-deoxyerythronolide B analogs for production of novel erythromycins. JBiotechnol. (2002). 92 (3): - Chao K.-M., Pearson W. R., Miller W. Aligning two séquences within a specified diagonal band. Comput. Appl. Biosci. (1992) 8 :481-487. - Carreras C, Frykman S, Or S, Cadapan L, Zavala S, Woo E, Leaf T, Carney J,
Burlingame M, Patel S, Ashley G, Licari P. Saccharopolyspora erythraea-catalyzed bioconversion of 6-deoxyerythronolide B analogs for production of novel erythromycins. JBiotechnol. (2002). 92 (3): - Chao K.-M., Pearson WR, Miller W. Aligning two sequences within a specified diagonal band. Comput. Appl. Biosci. (1992) 8: 481-487.
- Chater K. F. The improving prospects for yield increase by genetic engineering in antibiotic-producing Streptomycetes. Biotechnology. (1990). 8 (2) : 115-121. - Chater K. F. The improving prospects for yield increase by genetic engineering in antibiotic-producing Streptomycetes. Biotechnology. (1990). 8 (2): 115-121.
- Chaveroche MK, Ghigo JM, d'Enfert C. A rapid method for efficient gene replacement in the filamentous fungus Aspergillus nidulans. Nucleic Acids Res. - Chaveroche MK, Ghigo JM, from Enfert C. A rapid method for efficient gene replacement in the filamentous fungus Aspergillus nidulans. Nucleic Acids Res.
2000 ; 28 (22) : E97. 2000; 28 (22): E97.
- Church GM, Gilbert W. Genomic sequencing. Proc Natl Acad Sci U S A. (1984)
81 (7):1991-5. - GM Church, Gilbert W. Genomic sequencing. Proc Natl Acad Sci US A. (1984)
81 (7): 1991-5.
- Churchward G, Belin D, Nagamine Y. A pSC101-derived plasmid which shows no séquence homology to other commonly used cloning vectors. Gene. (1984) 31 (1-3): 165-71. - Churchward G, Belin D, Nagamine Y. pSC101-derived plasmid which shows no sequence homology to other commonly used cloning vectors. Gene. (1984) 31 (1-3): 165-71.
- Comstock,L.E., Coyne,M.J., Tzianabos,A.O. and Kasper, D.L. Interstrain variation of the polysaccharide B biosynthesis locus of Bacteroides fragilis: characterization of the region from strain 638R J. Bacteriol. (1999). 181 (19) :
6192-6196. - Comstock, LE, Coyne, MJ, Tzianabos, AO and Kasper, DL Interstrain Variation of the Polysaccharide B biosynthesis locus of Bacteroides fragilis: Characterization of the region from strain 638R J. Bacteriol. (1999). 181 (19):
6192-6196.
- Cox KL, Baltz RH. Restriction of bacteriophage plaque formation in
Streptomyces spp. JBacteriol. (1984) 159 (2): - Dale E. C., Ow D.W., Gene transfer with subsequent removal of the selection gene from the host genome. Proc. Natl. Acad. Sci. USA (1991). 88 :
10562. - Cox KL, Baltz RH. Restriction of bacteriophage plaque formation in
Streptomyces spp. JBacteriol. (1984) 159 (2): Dale EC, Ow DW, Gene transfer with subsequent removal of the gene selection from the host genome. Proc. Natl. Acad. Sci. USA (1991). 88:
10562.
- Doumith M, Weingarten P, Wehmeier UF, Salah-Bey K, Benhamou B,
Capdevila C, Michel JM, Piepersberg W, Raynal MC. Analysis of genes - Doumith M, Weingarten P, Wehmeier UF, Salah-Bey K, Benhamou B,
Capdevila C, Michel JM, Piepersberg W, Raynal MC. Analysis of genes
<Desc/Clms Page number 185><Desc / Clms Page number 185>
involved in 6-deoxyhexose biosynthesis and transfer in Saccharopolyspora erythraea. Mol Gen Genet. (2000) 264 (4): -Draeger,G., Park,Streptomyces-H.H. et Floss,H.G. Mechanism of the 2- deoxygenation step in the biosynthesis of the deoxyhexose moieties of the antibiotics granaticin and oleandomycin J. Am. Chem. Soc. (1999) 121 : 2611-
2612. in 6-deoxyhexose biosynthesis and transfer in Saccharopolyspora erythraea. Mol Gen Genet. (2000) 264 (4): -Draeger, G., Park, Streptomyces-HH and Floss, HG Mechanism of the Deoxygenation-2 deoxygenation in the biosynthesis of the deoxyhexose moieties of the antibiotics granaticin and oleandomycin J. Am. Chem. Soc. (1999) 121: 2611-
2612.
Gandecha,A.R., Large, S.L. et Cundliffe,E. Analysis of four tylosin biosynthetic genes from the tylLM region of the Streptomyces fradiae genome. Gene. (1997). Gandecha, A. R., Large, S. L. and Cundliffe, E. Analysis of four types of biosynthetic genes from the region of the Streptomyces fradiae genome. Gene. (1997).
184 (2) :197-203. 184 (2): 197-203.
- Geistlich, M., Losick,R., Turner,J.R. et Rao, R.N. Characterization of a novel regulatory gene goveming the expression of a polyketide synthase gene in
Streptomyces ambofaciens. Mol. Microbiol. (1992). 6 (14) : 2019-2029. - Geistlich, M., Losick, R., Turner, JR and Rao, RN Characterization of a novel regulatory gene goveming the expression of a polyketide synthase gene in
Streptomyces ambofaciens. Mol. Microbiol. (1992). 6 (14): 2019-2029.
- Gourmelen A, Blondelet-Rouault MH, Pemodet JL. Characterization of a glycosyl transferase inactivating macrolides, encoded by gimA from
Streptomyces ambofaciens. Antimicrob Agents Chemother. (1998). 42(10): 2612-
9. - Gourmelen A, Blondelet-Rouault MH, Pemodet JL. Characterization of a glycosyl transferase inactivating macrolide, encoded by gimA from
Streptomyces ambofaciens. Antimicrob Agents Chemother. (1998). 42 (10): 2612-
9.
- Huang X. et Miller W. A time-efficient, linear-space local similarity algorithm. - Huang X. and Miller W. A time-efficient, linear-space local similarity algorithm.
Adv. Appl. Math. (1991). 12 : 337-357. Adv. Appl. Math. (1991). 12: 337-357.
- Hara 0 et Hutchinson CR. Cloning ofmidecamycin(MLS)-resistance genes from
Streptomyces mycarofaciens, Streptomyces lividans and Streptomyces coelicolor
A3 (2). JAntibiot (Tokyo). (1990). 43 (8):977-991. - Hara 0 and Hutchinson CR. Cloning ofmidecamycin (MLS) -resistance genes from
Streptomyces mycarofaciens, Streptomyces lividans and Streptomyces coelicolor
A3 (2). JAntibiot (Tokyo). (1990). 43 (8): 977-991.
- Hara 0 et Hutchinson CR. A macrolide 3-O-acyltransferase gene from the midecamycin-producing species Streptomyces mycarofaciens. J Bacteriol. - Hara 0 and Hutchinson CR. Macrolide 3-O-acyltransferase gene from the midecamycin-producing species Streptomyces mycarofaciens. J Bacteriol.
(1992). 174(15) : 5141-5144. (1992). 174 (15): 5141-5144.
- Hoffmeister D, Ichinose K, Domann S, Faust B, Trefzer A, Drager G, Kirschning
A, Fischer C, Kunzel E, Bearden D, Rohr J et Bechthold A. The NDP-sugar co- substrate concentration and the enzyme expression level influence the substrate specificity of glycosyltransferases: cloning and characterization of deoxysugar biosynthetic genes of the urdamycin biosynthetic gene cluster. Chem Biol. - Hoffmeister D, Ichinose K, Domann S, Faust B, Trefzer A, Drager G, Kirschning
A, Fischer C, Kunzel E, Bearden D, Rohr J and Bechthold A. The NDP-sugar co-substrate concentration and the enzyme expression level influence the substrate specificity of glycosyltransferases: cloning and characterization of deoxysugar biosynthetic genes of the urdamycin biosynthetic gene cluster . Chem Biol.
(2000).7(11):821-831. (2000) .7 (11): 821-831.
- Hopwood, D.A. Future possibilities for the discovery of new antibiotics by genetic engineering. In Beta-lactam antibiotics. Edited by M.R.J. Salton and G.D. - Hopwood, D.A. Future possibilities for the discovery of new antibiotics by genetic engineering. In Beta-lactam antibiotics. Edited by M.R.J. Salton and G.D.
Shockman. New York: Academic Press. (1981). 585-598. Shockman. New York: Academic Press. (nineteen eighty one). 585-598.
- Hopwood D. A., Malpartida F., Kieser H. M., Ikeda H., Duncan J., Fujii I., Rudd
A. M., Floss H. G. et Omura S. Production of 'hybrid' antibiotics by genetic engineering. Nature. (1985a). 314 (6012) : 642-644. - Hopwood DA, Malpartida F., Kieser HM, Ikeda H., Duncan J., Fujii I., Rudd
AM, Floss HG and Omura S. Production of 'hybrid' antibiotics by genetic engineering. Nature. (1985a). 314 (6012): 642-644.
<Desc/Clms Page number 186> <Desc / Clms Page number 186>
- Hopwood D. A., Malpartida F., Kieser H. M., Ikeda H. et Omura S. (1985b) In
Microbiology (ed S. Silver). American Society for Microbiology, Washington D. - Hopwood DA, Malpartida F., Kieser HM, Ikeda H. and Omura S. (1985b) In
Microbiology (ed S. Silver). American Society for Microbiology, Washington D.
C., 409-413. C., 409-413.
- Houben Weyl, 1974, in Meuthode der Organischen Chemie, E. Wunsch Ed.,
Volume 15-1 et 15-II, - Hutchinson, C. R. Prospects for the discovery of new (hybrid) antibiotics by genetic engineering of antibiotic-producing bacteria. Med Res Rev. (1988). 8 (4) :
557-567. Houben Weyl, 1974, in Meuthode der Organischen Chemie, E. Wunsch Ed.,
Volume 15-1 and 15-II, Hutchinson, CR Prospects for the discovery of new (hybrid) antibiotics by genetic engineering of antibiotic-producing bacteria. Med Res Rev. (1988). 8 (4):
557-567.
- Hutchinson C. R., Borell C. W., Otten S. L., Stutzman-Engwall K. J. et Wang Y. - Hutchinson C. R., Borell C. W., Otten S. L., Stutzman-Engwall K. J. and Wang Y.
Drug discovery and development through the genetic engineering of antibiotic- producing microorganisms J. Med. Chem. (1989) 32 (5) : 929-937. Drug discovery and development through the genetic engineering of antibiotic-producing microorganisms J. Med. Chem. (1989) 32 (5): 929-937.
Ishikawa J, Hotta K. FramePlot: a new implementation of the frame analysis for predicting protein-coding regions in bacterial DNA with a high G + C content. Ishikawa J, Hotta K. FramePlot: a new implementation of the frame for predicting protein-coding regions in bacterial DNA with a high G + C content.
FEMSMicrobiol Lett. (1999) 174(2):251-3. FEMSMicrobiol Lett. (1999) 174 (2): 251-3.
- Kieser, T, Bibb, MJ, Buttner MJ, Chater KF, Hopwood DA. Practical
Streptomyces Genetics. 2000. The John Innés Foundation, Norwich UK. - Kieser, T, Bibb, MJ, Buttner MJ, KF Chater, Hopwood DA. Practical
Streptomyces Genetics. 2000. The John Innés Foundation, Norwich UK.
Kuhstoss et al. Production of a novel polyketide through the construction of a hybrid polyketide synthase. Gene. (1996). 183(1-2): 231-236. Kuhstoss et al. Production of a novel polyketide through the construction of a hybrid polyketide synthase. Gene. (1996). 183 (1-2): 231-236.
- Lacalle RA, Pulido D, Vara J, Zalacain M, Jimenez A. Molecular analysis of the pac gene encoding a puromycin N-acetyl transferase from Streptomyces alboniger. Gene. (1989). 79(2):375-380. - Lacalle RA, Pulido D, Vara J, Zalacain M, Jimenez A. Molecular analysis of the pac gene encoding puromycin N-acetyl transferase from Streptomyces alboniger. Gene. (1989). 79 (2): 375-380.
- Lakso M, Sauer B, Mosinger B Jr, Lee EJ, Manning RW, Yu SH, Mulder KL,
Westphal H. Targeted oncogene activation by site-specific recombination in transgenic mice. Proc Natl Acad Sci USA. (1992) 89 (14) :6232-6. - Lakso M, Sauer B, Mosinger B Jr, Lee EJ, RW Manning, Yu SH, Mulder KL,
Westphal H. Targeted oncogene activation by site-specific recombination in transgenic mice. Proc Natl Acad Sci USA. (1992) 89 (14): 6232-6.
- Li,T.B., Shang,G.D., Xia, H.Z. and Wang,Y.G. Cloning of the sugar related biosynthesis gene cluster from Streptomyces tenebrarius H6. Sheng Wu Gong
ChengXue Bao (2001). 17 (3) : 329-331. Li, TB, Shang, GD, Xia, HZ and Wang, Cloning of the sugar related biosynthesis cluster gene from Streptomyces tenebrarius H6. Sheng Wu Gong
ChengXue Bao (2001). 17 (3): 329-331.
- Ligon,J., Hill, S., Beck, J., Zirkle, R., Molnar,I., Zawodny,J., Money,S. and
Schupp, T. Characterization of the biosynthetic gene cluster for the antifungal polyketide soraphen A from Sorangium cellulosum So ce26. Gene (2002). 285 (1-2), 257-267. Ligon, J., Hill, S., Beck, J., Zirkle, R., Molnar, I., Zawodny, J., Money, S. and
Schupp, T. Characterization of the biosynthetic cluster gene for the antifungal polyketide soraphen A from Sorangium cellulosum So ce26. Gene (2002). 285 (1-2), 257-267.
- Liu L, Saevels J, Louis P, Nelis H, Rico S, Dierick K, Guyomard S, Roets E,
Hoogmartens J. Interlaboratory study comparing the microbiological potency of spiramycins I, II and III. JPharm Biomed Anal. (1999). 20 (1-2):217-24. Liu L, Saevels J, Louis P, Nelis H, Rico S, Dierick K, Guyomard S, Roets E,
Hoogmartens J. Interlaboratory study comparing the microbiological potency of spiramycins I, II and III. JPharm Biomed Anal. (1999). 20 (1-2): 217-24.
<Desc/Clms Page number 187> <Desc / Clms Page number 187>
- Mazodier P, Petter R, Thompson C. Intergeneric conjugation between
Escherichia coli and Streptomyces species.JBacteriol. (1989). 171 (6):3583-5. - Mazodier P, Petter R, Thompson C. Intergeneric conjugation between
Escherichia coli and Streptomyces species.JBacteriol. (1989). 171 (6): 3583-5.
- Merrifield RB, 1965a, Nature, 207 (996): 522-523. Merrifield RB, 1965a, Nature, 207 (996): 522-523.
- Merrifield RB., 1965b, Science, 150 (693): 178-185. - Merrifield RB., 1965b, Science, 150 (693): 178-185.
- Merson-Davies,L.A. et Cundliffe,E. Analysis of five tylosin biosynthetic genes from the tylIBA region of the Streptomyces fradiae genome, Molecular microbiology. (1994). 13 (2) : 349-355. - Merson-Davies, L.A. and Cundliffe, E. Analysis of five tylosin biosynthetic genes from the tylIBA region of the Streptomyces fradiae genome, Molecular microbiology. (1994). 13 (2): 349-355.
- Miller VL, Mekalanos JJ. A novel suicide vector and its use in construction of insertion mutations: osmoregulation of outer membrane proteins and virulence déterminants in Vibrio cholerae requires toxR. J Bacteriol. (1988) 170(6):2575-
83. - Miller VL, Mekalanos JJ. A novel suicide vector and its use in construction of insertion mutations: osmoregulation of outer membrane proteins and virulence determinants in Vibrio cholerae requires toxR. J Bacteriol. (1988) 170 (6): 2575-
83.
- Nakano, Y., Yoshida, Y., Yamashita, Y. & Koga, T. Construction of a series of pACYC-derived plasmid vectors. Gène (1995). 162 : Oh SH, Chater KF. Denaturation of circular or linear DNA facilitates targeted integrative transformation of Streptomyces coelicolor A3 (2): possible relevance to other organisms. JBacteriol. 1997; 179 (1):122-7. - Nakano, Y., Yoshida, Y., Yamashita, Y. & Koga, T. Construction of a series of pACYC-derived plasmid vectors. Gene (1995). 162: Oh SH, Chater KF. Denaturation of circular or linear DNA facilitates targeted integrative transformation of Streptomyces coelicolor A3 (2): possible relevance to other organisms. JBacteriol. 1997; 179 (1): 122-7.
- Olano C, Lomovskaya N, Fonstein L, Roll JT, Hutchinson CR. A two-plasmid system for the glycosylation of polyketide antibiotics: bioconversion of epsilon- rhodomycinone to rhodomycin D. Chem Biol. (1999). 6 (12) :845-55. - Olano C, Lomovskaya N, Fonstein L, Roll JT, Hutchinson CR. A two-plasmid system for the glycosylation of polyketide antibiotics: bioconversion of epsilon-rhodomycinone to rhodomycin D. Chem Biol. (1999). 6 (12): 845-55.
Omura S, Kitao C, Hamada H, Ikeda H. Bioconversion and biosynthesis of 16- membered macrolide antibiotics. X. Final steps in the biosynthesis of spiramycin, using enzyme inhibitor: cerulenin. Chem Pharm Bull (Tokyo). (1979a). 27(1):
176-82. Omura S, Kitao C, Hamada H, Ikeda H. Bioconversion and biosynthesis of 16-membered macrolide antibiotics. X. Final steps in the biosynthesis of spiramycin, using enzyme inhibitor: cerulenin. Chem Pharm Bull (Tokyo). (1979a). 27 (1):
176-82.
Omura S, Ikeda H, Kitao C. Isolation and properties of spiramycin I 3-hydroxyl acylase from Streptomyces and ambofaciens. JBiochem (Tokyo). (1979b). 86 (6):
1753-8. Omura S, Ikeda H, Kitao C. Isolation and properties of spiramycin I 3-hydroxyl acylase from Streptomyces and ambofaciens. JBiochem (Tokyo). (1979b). 86 (6):
1753-8.
- Omura,S., Ikeda,H., Ishikawa,J., Hanamoto,A., Takahashi,C., Shinose,M.,
Takahashi, Y., Horikawa, H., Nakazawa,H., Osonoe,T., Kikuchi, H., Shiba, T.,
Sakaki,Y. and Hattori, M. Genome séquence of an industrial microorganism
Streptomyces avermitilis: Deducing the ability of producing secondary metabolites. Proc. Natl. Acad. Sci. U.S.A. (2001). 98 (21), 12215-12220. - Omura, S., Ikeda, H., Ishikawa, J., Hanamoto, A., Takahashi, C., Shinose, M.,
Takahashi, Y., Horikawa, H., Nakazawa, H., Osonoe, T., Kikuchi, H., Shiba, T.,
Sakaki Y. and Hattori, M. Genome sequence of an industrial microorganism
Streptomyces avermitilis: Deducing the ability of producing secondary metabolites. Proc. Natl. Acad. Sci. USA (2001). 98 (21), 12215-12220.
- Pearson W. R. and D. J. Lipman. Improved Tools for Biological Sequence
Analysis. Proc. Natl. Acad. Sci. USA 1988, 85: 2444-2448. - Pearson WR and DJ Lipman. Improved Tools for Biological Sequence
Analysis. Proc. Natl. Acad. Sci. USA 1988, 85: 2444-2448.
- Pearson W. R. Rapid and Sensitive Sequence Comparison with FASTP and
FASTA. Methods in Enzymology. 1990, 183: 63-98. Pearson WR Rapid and Sensitive Sequence Comparison with FASTP
FASTA. Methods in Enzymology. 1990, 183: 63-98.
<Desc/Clms Page number 188> <Desc / Clms Page number 188>
- Pemodet JL, Simonet JM, Guerineau M. Plasmids in différent strains of
Streptomyces ambofaciens: free and integrated form of plasmid pSAM2. Mol
Gen Genet. (1984) ;198 (1) :35-41. - Pemodet JL, Simonet JM, Guerineau M. Plasmids in different strains of
Streptomyces ambofaciens: free and integrated form of plasmid pSAM2. mol
Gen Genet. (1984) 198 (1): 35-41.
- Pernodet JL, Alegre MT, Blondelet-Rouault MH, Guerineau M. Resistance to spiramycin in Streptomyces ambofaciens, the producer organism, involves at least two différent mechanisms. J Gen Microbiol. (1993) ; 139 ( Pt 5): 1003-11. Pernodet JL, Alegre MT, Blondelet-Rouault MH, Raspberry M. Resistance to spiramycin in Streptomyces ambofaciens, the producer organism, involves at least two different mechanisms. J Gen Microbiol. (1993); 139 (Pt 5): 1003-11.
- Pemodet JL, Fish S, Blondelet-Rouault MH, Cundliffe E. The macrolide- lincosamide-streptogramin B résistance phenotypes characterized by using a specifically deleted, antibiotic-sensitive strain of Streptomyces lividans. Pemodet JL, Fish S, Blondelet-Rouault MH, Cundliffe E. The macrolide-lincosamide-streptogramin B resistance phenotypes characterized by using a specific deleted antibiotic-sensitive strain of Streptomyces lividans.
Antimicrob Agents Chemother. (1996), 40 (3) : 581-5. Antimicrob Agents Chemother. (1996), 40 (3): 581-5.
- Pernodet JL, Gourmelen A, Blondelet-Rouault MH, Cundliffe E. Dispensable ribosomal résistance to spiramycin conferred by srmA in the spiramycin producer
Streptomyces ambofaciens. Microbiology. (1999), 145 (Pt 9):2355-64. - Pernodet JL, Gourmelen A, Blondelet-Rouault MH, Cundliffe E. Dispensable ribosomal resistance to spiramycin conferred by srmA in the spiramycin producer
Streptomyces ambofaciens. Microbiology. (1999), 145 (Pt 9): 2355-64.
- Rao RN, Richardson MA, Kuhstoss S. Cosmid shuttle vectors for cloning and analysis of Streptomyces DNA. Methods Enzymol. (1987) ; 153 : 166-98. - Rao RN, Richardson MA, Kuhstoss, S. Cosmid shuttle vectors for cloning and analysis of Streptomyces DNA. Methods Enzymol. (1987); 153: 166-98.
- Raynal A, Tuphile K, Gerbaud C, Luther T, Guerineau M, and Pemodet JL.. - Raynal A, Tuphile K, Gerbaud C, Luther T, Guerineau M, and Pemodet JL ..
Structure of the chromosomal insertion site for pSAM2 : functional analysis in
Escherichia coli. Molecular Microbiology (1998) 28 (2) 333-342. Structure of the chromosomal insertion site for pSAM2: functional analysis in
Escherichia coli. Molecular Microbiology (1998) 28 (2) 333-342.
- Redenbach,M., Kieser,H.M., Denapaite,D., Eichner,A., Cullum,J., Kinashi,H. et
Hopwood,D.A. A set of ordered cosmids and a detailed genetic and physical map for the 8 Mb Streptomyces coelicolor A3 (2) chromosomeMol. Microbiol. - Redenbach, M., Kieser, HM, Denapaite, D., Eichner, A., Cullum, J., Kinashi, H. and
Hopwood, DA A set of ordered cosmids and a detailed genetic and physical map for the 8 Mb Streptomyces coelicolor A3 (2) chromosomeMol. Microbiol.
(1996). 21 (1) : 77-96. (1996). 21 (1): 77-96.
- Richardson MA, Kuhstoss S, Solenberg P, Schaus NA, Rao RN. A new shuttle cosmid vector, pKC505, for streptomycetes : use in the cloning of three différent spiramycin-resistance genes from a Streptomyces ambofaciens library. - Richardson MA, Kuhstoss S, Solenberg P, NA Schaus, Rao RN. A new shuttle cosmid vector, pKC505, for streptomycetes: use in the cloning of three different spiramycin-resistance genes from a Streptomyces ambofaciens library.
Gene. (1987) ; 61 (3):231-41. Gene. (1987); 61 (3): 231-41.
- Robinson, J.A. Enzymes of Secondary Metabolism in Microorganisms. Chem
Soc Rev. (1988). 17 :383-452. Robinson, JA Enzymes of Secondary Metabolism in Microorganisms. Chem
Soc. Rev. (1988). 17: 383-452.
- Russell SH, Hoopes JL, Odell JT. Directed excision of a transgene from the plant genome. Mol Gen Genet. (1992). 234 (1): 49-59. - Russell SH, Hoopes JL, Odell JT. Directed excision of a transgene from the plant genome. Mol Gen Genet. (1992). 234 (1): 49-59.
- Sambrook J, Frisch EF, Maniatis T . Molecular Cloning : a laboratory manual 2nd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York USA (1989). - Sambrook J, Frisch EF, Maniatis T. Molecular Cloning: a laboratory manual 2nd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York USA (1989).
- Schoner B, Geistlich M, Rosteck P Jr, Rao RN, Seno E, Reynolds P, Cox K,
Burgett S et Hershberger C. Sequence similarity between macrolide-resistance - Schoner B, Geistlich M, Rosteck P Jr, Rao RN, Seno E, Reynolds P, Cox K,
Burgett S and Hershberger C. Sequence similarity between macrolide-resistance
<Desc/Clms Page number 189><Desc / Clms Page number 189>
determinants and ATP-binding transport proteins. Gene. (1992). 115 (1-2), 93-
96. determinants and ATP-binding transport proteins. Gene. (1992). 115 (1-2), 93-
96.
- Sezonov G, Hagege J, Pernodet JL, Friedmann A and Guerineau M. - Sezonov G, Hagege J, Pernodet JL, Friedmann A and Guerineau M.
Characterization of pra, a gene for replication control in pSAM2, the integrating élément of Streptomyces ambofaciens. Mol Microbiol. (1995). 17(3): 533-44. Characterization of pra, a gene for replication control in pSAM2, the integrating element of Streptomyces ambofaciens. Mol Microbiol. (1995). 17 (3): 533-44.
- Sezonov G., Blanc V., Bamas-Jacques N., Friedmann A., Pernodet J.L. and M. Sezonov G., White V., Bamas-Jacques N., Friedmann A., Pernodet J.L. and M.
Guerineau, Complete conversion of antibiotic precursor to pristinamycin IIA by over expression of Streptomyces pristinaespiralis biosynthetic genes, Nature
Biotechnology. (1997) 15 : 349-353.Sezonov G, Possoz C, Friedmann A, Pemodet JL, Guerineau M. KorSA from the
Streptomyces integrative element pSAM2 is a central transcriptional repressor : target genes and binding sites. JBacteriol. (2000). 182(5):1243-50. Guerineau, Complete conversion of precursor antibiotic to pristinamycin IIA by the expression of Streptomyces pristinaespiralis biosynthetic genes, Nature
Biotechnology. (1997) 15: 349-353.Sezonov G, Possoz C, Friedmann A, Pemodet JL, Guerineau M. KorSA from the
Streptomyces integrative element pSAM2 is a central transcriptional repressor: target genes and binding sites. JBacteriol. (2000). 182 (5): 1243-1250.
Simon R., Priefer U and Pühler. A broad host range mobilisation system for in vivo genetic engeneering : transposon mutagenesis in gram negative bacteria. Simon R., Priefer U and Pühler. A broad host range mobilization system for in vivo genetic engeneering: transposon mutagenesis in gram negative bacteria.
BiolTechnology (1983). 1 : 784-791. BiolTechnology (1983). 1: 784-791.
Simon, et al., Plasmid vectors for the genetic analysis and manipulation of rhizobia and other gram-negative bacteria, p. 640-659. In A. Weissbach, and H. Simon, et al., Plasmid vectors for the genetic analysis and manipulation of rhizobia and other gram-negative bacteria, p. 640-659. In A. Weissbach, and H.
Weissbach (eds. ), Methods in enzymology, vol 118, Academic Press, Inc.,
Orlando, 1986 - Summers,R.G., Donadio,Streptomyces, Staver, M.J., Wendt-Pienkowski,E.,
Hutchinson,C.R. et Katz,L. Sequencing and mutagenesis of genes from the erythromycin biosynthetic gene cluster of Saccharopolyspora erythraea that are involved in L-mycarose and D-desosamine production. Microbiology (1997). 143 (Pt 10) : 3251-3262. Weissbach (eds.), Methods in Enzymology, Vol 118, Academic Press, Inc.,
Orlando, 1986 - Summers, RG, Donadio, Streptomyces, Staver, MJ, Wendt-Pienkowski, E.,
Hutchinson, CR and Katz, L. Sequencing and mutagenesis of genes from the erythromycin biosynthetic gene cluster of Saccharopolyspora erythraea that are involved in L-mycarose and D-desosamine production. Microbiology (1997). 143 (Pt 10): 3251-3262.
- Van Mellaert L, Mei L, Lammertyn E, Schacht S, Anne J. Site-specific integration of bacteriophage VWB genome into Streptomyces venezuelae and construction of a VWB-based integrative vector. Microbiology. (1998). 144 ( Pt
12): 3351-8. - Van Mellaert L, Mei L, Lammertyn E, Schacht S, Anne J. Site-specific integration of bacteriophage VWB genome into Streptomyces venezuelae and construction of a VWB-based integrative vector. Microbiology. (1998). 144 (Pt
12): 3351-8.
- Vara J, Lewandowska-Skarbek M, Wang YG, Donadio S, Hutchinson
CR.Cloning of genes goveming the deoxysugar portion of the erythromycin biosynthesis pathway in Saccharopolyspora erythraea (Streptomyces erythreus). J
Bacteriol. (1989). 171(11): 5872-81. - Vara J, Lewandowska-Skarbek M, Wang YG, Donadio S, Hutchinson
CR.Cloning of genes goveming the deoxysugar portion of the erythromycin biosynthesis pathway in Saccharopolyspora erythraea (Streptomyces erythreus). J
Bacteriol. (1989). 171 (11): 5872-81.
- Vara J, Malpartida F, Hopwood DA, Jimenez A. Cloning and expression of a puromycin N-acetyl transferase gene from Streptomyces alboniger in
Streptomyces lividans and Escherichia coli. Gene. (1985). 33 (2): - Vara J, Malpartida F, Hopwood DA, Jimenez A. Cloning and expression of a puromycin N-acetyl transferase gene from Streptomyces alboniger
Streptomyces lividans and Escherichia coli. Gene. (1985). 33 (2):
<Desc/Clms Page number 190><Desc / Clms Page number 190>
- Wahl, G. M., K. A. Lewis, J. C. Ruiz, B. Rothenberg, J. Zhao, and G. A. Evans. Wahl, G.M., K.A. Lewis, J.C. Ruiz, B. Rothenberg, J.Zhao, and G.A. Evans.
Cosmid vectors for rapid genomic walking, restriction mapping, and gene transfer. Proc. Natl. Acad. Sci. USA (1987). 84 : -Waldron,C., Matsushima,P., Rosteck,P.R., Broughton,M.C., Turner,J.,
Madduri, K., Crawford,K.P., Merlo,D.J. et Baltz,R.H. Cloning and analysis of the spinosad biosynthetic gene cluster of Saccharopolyspora spinosa(l) Chem. Biol. Cosmid vectors for rapid genomic walking, restriction mapping, and gene transfer. Proc. Natl. Acad. Sci. USA (1987). 84: -Waldron, C., Matsushima, P., Rosteck, PR, Broughton, MC, Turner, J.,
Madduri, K., Crawford, KP, Merlo, DJ and Baltz, RH Cloning and Spinosad Analysis of the Biosynthetic Gene Cluster of Saccharopolyspora Spinosa (l) Chem. Biol.
(2001) 8 (5) : 487-499. (2001) 8 (5): 487-499.
- Walczak RJ, Hines JV, Strohl WR, Priestley ND.Bioconversion of the anthracycline analogue desacetyladriamycin by recombinant DoxA, a P450- monooxygenase from Streptomyces sp. strain C5. Org Lett. (2001). 3 (15):2277-
9. - Walczak RJ, Hines JV, Strohl WR, Priestley ND.Bioconversion of the anthracycline analog desacetyladriamycin by recombinant DoxA, a P450-monooxygenase from Streptomyces sp. strain C5. Org Lett. (2001). 3 (15): 2277-
9.
- Wang,Z.X., Li, S.M. et Heide,L. Identification of the coumermycin A(l) biosynthetic gene cluster of Streptomyces rishiriensis DSM 40489. Antimicrob. Wang, Z.X., Li, S.M. and Heide, L. Identification of the coumermycin A (l) biosynthetic cluster gene of Streptomyces rishiriensis DSM 40489. Antimicrob.
Agents Chemother. (2000) 44 (11), 3040-3048. Agents Chemother. (2000) 44 (11), 3040-3048.
- Ward, J.M., Janssen, G. R., Kieser, T, Bibb, M.J., Buttner, M.J. & Bibb, M.J. - Ward, J.M., Janssen, G.R., Kieser, T, Bibb, M.J., Buttner, M.J. & Bibb, M.J.
Construction and characterization of a series of multi-copy promoter-probe plasmid vectors for Streptomyces using the aminoglycoside phosphotransferase from Tn5 as indicator. Mol Gen Genet (1986). 203 : 468-478. Construction and characterization of a multi-copy promoter-probe plasmid vectors for Streptomyces using the aminoglycoside phosphotransferase from Tn5 as indicator. Mol Gen Genet (1986). 203: 468-478.
- Wehmeier UF. New multifunctional Escherichia coli-Streptomyces shuttle vectors allowing blue-white screening on XGal plates. Gene. (1995). 165(1):
149-150. - Wehmeier UF. New multifunctional Escherichia coli-Streptomyces shuttle vectors permitting blue-white screening on XGal plates. Gene. (1995). 165 (1):
149-150.
- Wilms B, Hauck A, Reuss M, Syldatk C, Mattes R, Siemann M, Altenbuchner J. - Wilms B, Hauck A, Reuss M, Syldatk C, Mattes R, Siemann M, Altenbuchner J.
High-cell-density fermentation for production of L-N-carbamoylase using an expression system based on the Escherichia coli rhaBAD promoter. Biotechnol
Bioeng. (2001). 73 (2) - Worley K. C., Wiese B. A., and Smith R. F. BEAUTY: An enhanced BLAST- based search tool that integrates multiple biological information resources into séquence similarity search results. Genome Research (1995). 5: 173-184. High-cell-density fermentation for production of LN-carbamoylase using an expression system based on Escherichia coli rhaBAD promoter. Biotechnol
Bioeng. (2001). 73 (2) - Worley KC, Wiese BA, and Smith RF BEAUTY: An enhanced BLAST-based search tool that integrates multiple biological information resources into sequence similarity search results. Genome Research (1995). 5: 173-184.
- Wu K, Chung L, Revill WP, Katz L et Reeves CD. The FK520 gene cluster of
Streptomyces hygroscopicus var. ascomyceticus (ATCC 14891) contains genes for biosynthesis of unusual polyketide extender units. Gene. (2000). 251 (1): 81-
90. - Wu K, Chung L, Revill WP, Katz L and Reeves CD. The FK520 gene cluster of
Streptomyces hygroscopicus var. ascomyceticus (ATCC 14891) contains genes for biosynthesis of unusual polyketide extender units. Gene. (2000). 251 (1): 81-
90.
Xue Y, Zhao L, Liu HW et Sherman DH. A gene cluster for macrolide antibiotic biosynthesis in Streptomyces venezuelae: architecture of metabolic diversity. Xue Y, Zhao L, Liu HW and Sherman DH. Gene cluster for macrolide antibiotic biosynthesis in Streptomyces venezuelae: architecture of metabolic diversity.
Proc Natl Acad Sci USA. (1998). 95 (21) : 12111-12116. Proc Natl Acad Sci USA. (1998). 95 (21): 12111-12116.
Claims (89)
Priority Applications (19)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| FR0302439A FR2851773A1 (en) | 2003-02-27 | 2003-02-27 | New polynucleotides encoding proteins involved in spiramycin biosynthesis, useful for improving synthesis of macrolide antibiotics or for generating new hybrid macrolides |
| US10/680,860 US7579167B2 (en) | 2002-10-08 | 2003-10-07 | Polypeptides involved in the biosynthesis of spiramycins, nucleotide sequences encoding these polypeptides and applications thereof |
| CA2501445A CA2501445C (en) | 2002-10-08 | 2003-10-08 | Polypeptides involved in spiramycin biosynthesis, nucleotide sequences encoding said polypeptides and uses thereof |
| HUE07019306A HUE026468T2 (en) | 2002-10-08 | 2003-10-08 | Polypeptides involved in the biosynthesis of spiramycins, nucleotide sequences coding these polypeptides and their uses |
| AU2003300479A AU2003300479B2 (en) | 2002-10-08 | 2003-10-08 | Polypeptides involved in spiramycin biosynthesis, nucleotide sequences encoding said polypeptides and uses thereof |
| BRPI0314543A BRPI0314543B8 (en) | 2002-10-08 | 2003-10-08 | host cells, recombinant DNA, their use, processes for the production of a polypeptide involved in the biosynthesis of spiramycins, the production of macrolides and the production of spiramycin(s), and macrolide-producing mutant transgenic microorganism |
| MXPA05002852A MXPA05002852A (en) | 2002-10-08 | 2003-10-08 | Polypeptides involved in spiramycin biosynthesis, nucleotide sequences encoding said polypeptides and uses thereof. |
| JP2005500990A JP5042497B2 (en) | 2002-10-08 | 2003-10-08 | Polypeptides involved in the biosynthesis of spiramycin, nucleotide sequences encoding these polypeptides, and uses thereof |
| EP03807883A EP1551975A2 (en) | 2002-10-08 | 2003-10-08 | Polypeptides involved in spiramycin biosynthesis, nucleotide sequences encoding said polypeptides and uses thereof |
| PCT/FR2003/002962 WO2004033689A2 (en) | 2002-10-08 | 2003-10-08 | Polypeptides involved in spiramycin biosynthesis, nucleotide sequences encoding said polypeptides and uses thereof |
| KR1020057006111A KR101110175B1 (en) | 2002-10-08 | 2003-10-08 | Polypeptides involved in spiramycin biosynthesis, nucleotide sequences encoding said polypeptides and uses thereof |
| CN2008100823963A CN101302528B (en) | 2002-10-08 | 2003-10-08 | Polypeptides involved in the biosynthesis of spiramycins, nucleotide sequences coding these polypeptides and their uses |
| EP07019306.5A EP1905833B1 (en) | 2002-10-08 | 2003-10-08 | Polypeptides involved in the biosynthesis of spiramycins, nucleotide sequences coding these polypeptides and their uses |
| ES07019306.5T ES2552912T3 (en) | 2002-10-08 | 2003-10-08 | Polypeptides involved in the biosynthesis of spiramycins, nucelotide sequences that encode these polypeptides and their applications |
| DK07019306.5T DK1905833T3 (en) | 2002-10-08 | 2003-10-08 | Polypeptides involved in the biosynthesis of spiramyciner, nucleotide sequences encoding these polypeptides, and uses thereof |
| RU2005113874/13A RU2377304C2 (en) | 2002-10-08 | 2003-10-08 | Polynucleotide coding acyltransferase that responds for modification of platenolid in position 3 (versions), polypeptide that represents acyltransferase, which is responsible for modification of platenolid in position 3 (versions), bacterial expression vector (versions), bacterial expression system, bacterial host cell, method for production of polypeptide, strain streptomyces ambofaciens, application of polynucleotide (versions), cell of bacterium streptomyces ambofaciens, host cell streptomyces ambofaciens and method of polypeptide production |
| IL167854A IL167854A (en) | 2002-10-08 | 2005-04-04 | Nucleotide sequences encoding polypeptides involved in spiramycin biosynthesis, expression vector comprising same and uses thereof |
| NO20052160A NO335995B1 (en) | 2002-10-08 | 2005-05-02 | Polypeptides involved in spiramycin biosynthesis, nucleotide sequences encoding said polypeptides and uses thereof. |
| HK08113536.6A HK1122330A1 (en) | 2002-10-08 | 2008-12-12 | Polypeptides involved in spiramycin biosynthesis, nucleotide sequences encoding said polypeptides and uses thereof |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| FR0302439A FR2851773A1 (en) | 2003-02-27 | 2003-02-27 | New polynucleotides encoding proteins involved in spiramycin biosynthesis, useful for improving synthesis of macrolide antibiotics or for generating new hybrid macrolides |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| FR2851773A1 true FR2851773A1 (en) | 2004-09-03 |
Family
ID=32843051
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| FR0302439A Pending FR2851773A1 (en) | 2002-10-08 | 2003-02-27 | New polynucleotides encoding proteins involved in spiramycin biosynthesis, useful for improving synthesis of macrolide antibiotics or for generating new hybrid macrolides |
Country Status (1)
| Country | Link |
|---|---|
| FR (1) | FR2851773A1 (en) |
Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP0791656A2 (en) * | 1996-02-22 | 1997-08-27 | Eli Lilly And Company | Platenolide synthase gene |
| EP0791655A2 (en) * | 1996-02-22 | 1997-08-27 | Eli Lilly And Company | Polyketide synthase genes |
| WO1999005283A2 (en) * | 1997-07-25 | 1999-02-04 | Hoechst Marion Roussel | Biosynthesis genes and transfer of 6-desoxy-hexoses in saccharopolyspora erythraea and in streptomyces antibioticus and their use |
| WO2003010193A2 (en) * | 2001-07-26 | 2003-02-06 | Ecopia Biosciences Inc. | Genes and proteins for the biosynthesis of rosaramicin |
-
2003
- 2003-02-27 FR FR0302439A patent/FR2851773A1/en active Pending
Patent Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP0791656A2 (en) * | 1996-02-22 | 1997-08-27 | Eli Lilly And Company | Platenolide synthase gene |
| EP0791655A2 (en) * | 1996-02-22 | 1997-08-27 | Eli Lilly And Company | Polyketide synthase genes |
| WO1999005283A2 (en) * | 1997-07-25 | 1999-02-04 | Hoechst Marion Roussel | Biosynthesis genes and transfer of 6-desoxy-hexoses in saccharopolyspora erythraea and in streptomyces antibioticus and their use |
| WO2003010193A2 (en) * | 2001-07-26 | 2003-02-06 | Ecopia Biosciences Inc. | Genes and proteins for the biosynthesis of rosaramicin |
Non-Patent Citations (5)
| Title |
|---|
| DATABASE EMBL [online] 8 January 1997 (1997-01-08), GANDECHA A R: "Streptomyces fradiae tylMIII(orf1*), tylMII(orf2*), tylMI(orf3*) and ccr(orf4*) genes.", XP002250640, Database accession no. X81885; P95748 * |
| GANDECHA A R ET AL: "Analysis of four tylosin biosynthetic genes from the tylLM region of the Streptomyces fradiae genome", GENE: AN INTERNATIONAL JOURNAL ON GENES AND GENOMES, ELSEVIER SCIENCE PUBLISHERS, BARKING, GB, vol. 184, no. 2, 1997, pages 197 - 203, XP004017240, ISSN: 0378-1119 * |
| GEISTLICH M ET AL: "CHARACTERIZATION OF A NOVEL REGULATORY GENE GOVERNING THE EXPRESSION OF A POLYKETIDE SYNTHASE GENE IN STREPTOMYCES-AMBOFACIENS", MOLECULAR MICROBIOLOGY, vol. 6, no. 14, 1992, pages 2019 - 2029, XP008020560, ISSN: 0950-382X * |
| PERNODET JEAN-LUC ET AL: "Dispensable ribosomal resistance to spiramycin conferred by srmA in the spiramycin producer Streptomyces ambofaciens.", MICROBIOLOGY (READING), vol. 145, no. 9, 1999, pages 2355 - 2364, XP002250639, ISSN: 1350-0872 * |
| RICHARDSON M A ET AL: "CLONING OF SPIRAMYCIN BIOSYNTHETIC GENES AND THEIR USE IN CONSTRUCTING STREPTOMYCES-AMBOFACIENS MUTANTS DEFECTIVE IN SPIRAMYCIN BIOSYNTHESIS", JOURNAL OF BACTERIOLOGY, vol. 172, no. 7, 1990, pages 3790 - 3798, XP008020563, ISSN: 0021-9193 * |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| Oliynyk et al. | Analysis of the biosynthetic gene cluster for the polyether antibiotic monensin in Streptomyces cinnamonensis and evidence for the role of monB and monC genes in oxidative cyclization | |
| Waldron et al. | Cloning and analysis of the spinosad biosynthetic gene cluster of Saccharopolyspora spinosa1 | |
| Trefzer et al. | Biosynthetic gene cluster of simocyclinone, a natural multihybrid antibiotic | |
| Fouces et al. | The tylosin biosynthetic cluster from Streptomyces fradiae: genetic organization of the left region | |
| Madduri et al. | Rhamnose biosynthesis pathway supplies precursors for primary and secondary metabolism in Saccharopolyspora spinosa | |
| Liu et al. | Genes for production of the enediyne antitumor antibiotic C-1027 in Streptomyces globisporus are clustered with the cagA gene that encodes the C-1027 apoprotein | |
| KR100588436B1 (en) | Biosynthetic Genes For Spinosyn Insecticide Production | |
| Bihlmaier et al. | Biosynthetic gene cluster for the polyenoyltetramic acid α-lipomycin | |
| Metsä-Ketelä et al. | Partial activation of a silent angucycline-type gene cluster from a rubromycin β producing Streptomyces sp. PGA64 | |
| JP2002537833A (en) | Mitomycin biosynthesis gene cluster | |
| EP1905833B1 (en) | Polypeptides involved in the biosynthesis of spiramycins, nucleotide sequences coding these polypeptides and their uses | |
| CA2501445C (en) | Polypeptides involved in spiramycin biosynthesis, nucleotide sequences encoding said polypeptides and uses thereof | |
| RU2377304C2 (en) | Polynucleotide coding acyltransferase that responds for modification of platenolid in position 3 (versions), polypeptide that represents acyltransferase, which is responsible for modification of platenolid in position 3 (versions), bacterial expression vector (versions), bacterial expression system, bacterial host cell, method for production of polypeptide, strain streptomyces ambofaciens, application of polynucleotide (versions), cell of bacterium streptomyces ambofaciens, host cell streptomyces ambofaciens and method of polypeptide production | |
| JP2006516885A (en) | Genes and proteins for biosynthesis of glycopeptide antibiotic A40926 | |
| FR2851773A1 (en) | New polynucleotides encoding proteins involved in spiramycin biosynthesis, useful for improving synthesis of macrolide antibiotics or for generating new hybrid macrolides | |
| JP2020500561A (en) | Compositions and methods for enhancing enduracidin production in genetically modified strains of Streptomyces fungicidus | |
| FR2845394A1 (en) | New polynucleotides encoding proteins involved in spiramycin biosynthesis, useful for improving synthesis of macrolide antibiotics or for generating new hybrid macrolides | |
| WO2002097082A2 (en) | Engineered biosynthesis of novel polyenes | |
| US7070980B2 (en) | Midecamycin biosynthetic genes | |
| US7109019B2 (en) | Gene cluster for production of the enediyne antitumor antibiotic C-1027 | |
| US7105491B2 (en) | Biosynthesis of enediyne compounds by manipulation of C-1027 gene pathway | |
| Caffrey et al. | New Glycosylated Polyene Macrolides: Refining the Ore from Genome Mining. Antibiotics 2022, 11, 334 | |
| WO2000040596A1 (en) | Gene cluster for production of the enediyne antitumor antibiotic c-1027 | |
| Fiedler et al. | Biosynthetic Gene Cluster of | |
| Clustered | Genes for Production of the Enediyne |