BR112015000836B1 - Método para identificar um polipeptídeo tendo uma funcionalidade ou especificidade de ligação desejadas, biblioteca de moléculas de polinucleotídeo, método para gerar uma biblioteca de polinucleotídeos e método para gerar uma população de células hospedeiras - Google Patents
Método para identificar um polipeptídeo tendo uma funcionalidade ou especificidade de ligação desejadas, biblioteca de moléculas de polinucleotídeo, método para gerar uma biblioteca de polinucleotídeos e método para gerar uma população de células hospedeiras Download PDFInfo
- Publication number
- BR112015000836B1 BR112015000836B1 BR112015000836-4A BR112015000836A BR112015000836B1 BR 112015000836 B1 BR112015000836 B1 BR 112015000836B1 BR 112015000836 A BR112015000836 A BR 112015000836A BR 112015000836 B1 BR112015000836 B1 BR 112015000836B1
- Authority
- BR
- Brazil
- Prior art keywords
- cells
- binding
- sequence
- antibody
- expression
- Prior art date
Links
- 230000027455 binding Effects 0.000 title claims abstract description 132
- 238000009739 binding Methods 0.000 title claims abstract description 129
- 229920000023 polynucleotide Polymers 0.000 title claims abstract description 70
- 239000002157 polynucleotide Substances 0.000 title claims abstract description 70
- 229920001184 polypeptide Polymers 0.000 title claims abstract description 44
- 230000014509 gene expression Effects 0.000 claims abstract description 266
- 102000004965 antibodies Human genes 0.000 claims abstract description 210
- 108090001123 antibodies Proteins 0.000 claims abstract description 210
- 229920003013 deoxyribonucleic acid Polymers 0.000 claims abstract description 119
- 241000251539 Vertebrata <Metazoa> Species 0.000 claims abstract description 71
- 102000004190 Enzymes Human genes 0.000 claims abstract description 43
- 108090000790 Enzymes Proteins 0.000 claims abstract description 43
- 239000000427 antigen Substances 0.000 claims abstract description 41
- 102000038129 antigens Human genes 0.000 claims abstract description 41
- 108091007172 antigens Proteins 0.000 claims abstract description 41
- 230000017105 transposition Effects 0.000 claims abstract description 38
- 210000004027 cells Anatomy 0.000 claims description 393
- 240000007019 Oxalis corniculata Species 0.000 claims description 76
- 229920000970 Repeated sequence (DNA) Polymers 0.000 claims description 36
- 239000012528 membrane Substances 0.000 claims description 31
- 241000714175 Abelson murine leukemia virus Species 0.000 claims description 16
- 230000003321 amplification Effects 0.000 claims description 16
- 102000018358 Immunoglobulins Human genes 0.000 claims description 15
- 108060003951 Immunoglobulins Proteins 0.000 claims description 15
- 238000003199 nucleic acid amplification method Methods 0.000 claims description 14
- 210000004962 mammalian cells Anatomy 0.000 claims description 12
- 210000003719 B-Lymphocytes Anatomy 0.000 claims description 11
- 101700021280 PREB Proteins 0.000 claims description 10
- 238000002703 mutagenesis Methods 0.000 claims description 9
- 231100000350 mutagenesis Toxicity 0.000 claims description 9
- 210000002940 B-Lymphoid Precursor Cells Anatomy 0.000 claims description 8
- 238000003757 reverse transcription PCR Methods 0.000 claims description 8
- 210000000170 Cell Membrane Anatomy 0.000 claims description 7
- 102000006496 Immunoglobulin Heavy Chains Human genes 0.000 claims description 5
- 108010019476 Immunoglobulin Heavy Chains Proteins 0.000 claims description 5
- 210000004698 Lymphocytes Anatomy 0.000 claims description 5
- 241000283984 Rodentia Species 0.000 claims description 5
- 239000003446 ligand Substances 0.000 claims description 5
- 230000035693 Fab Effects 0.000 claims description 4
- 102000013463 Immunoglobulin Light Chains Human genes 0.000 claims description 4
- 108010065825 Immunoglobulin Light Chains Proteins 0.000 claims description 4
- 108010070144 Single-Chain Antibodies Proteins 0.000 claims description 3
- 102000005632 Single-Chain Antibodies Human genes 0.000 claims description 3
- 230000001413 cellular Effects 0.000 claims description 3
- 239000007787 solid Substances 0.000 claims description 3
- 239000000232 Lipid Bilayer Substances 0.000 claims description 2
- 241001024304 Mino Species 0.000 claims description 2
- 238000001943 fluorescence-activated cell sorting Methods 0.000 claims 1
- 238000009114 investigational therapy Methods 0.000 claims 1
- 102000004169 proteins and genes Human genes 0.000 abstract description 73
- 108090000623 proteins and genes Proteins 0.000 abstract description 73
- 229920001405 Coding region Polymers 0.000 abstract description 70
- 238000010367 cloning Methods 0.000 abstract description 47
- 238000001890 transfection Methods 0.000 abstract description 35
- 238000001712 DNA sequencing Methods 0.000 abstract description 15
- 238000000034 method Methods 0.000 abstract description 14
- 238000005457 optimization Methods 0.000 abstract description 8
- 230000001052 transient Effects 0.000 abstract description 8
- 108091007521 restriction endonucleases Proteins 0.000 description 75
- 235000018102 proteins Nutrition 0.000 description 64
- 102000004851 Immunoglobulin G Human genes 0.000 description 60
- 238000003752 polymerase chain reaction Methods 0.000 description 56
- 102000008579 Transposases Human genes 0.000 description 46
- 108010020764 Transposases Proteins 0.000 description 46
- 108091005938 enhanced green fluorescent protein Proteins 0.000 description 26
- 229920001850 Nucleic acid sequence Polymers 0.000 description 23
- 238000011144 upstream manufacturing Methods 0.000 description 22
- 229920002287 Amplicon Polymers 0.000 description 19
- 150000001413 amino acids Chemical group 0.000 description 17
- 108060000885 BCL2 Proteins 0.000 description 16
- 102100013894 BCL2 Human genes 0.000 description 16
- KTUFNOKKBVMGRW-UHFFFAOYSA-N Imatinib Chemical compound C1CN(C)CCN1CC1=CC=C(C(=O)NC=2C=C(NC=3N=C(C=CN=3)C=3C=NC=CC=3)C(C)=CC=2)C=C1 KTUFNOKKBVMGRW-UHFFFAOYSA-N 0.000 description 16
- 101710040533 TNFRSF8 Proteins 0.000 description 16
- 102100009538 TNFRSF8 Human genes 0.000 description 16
- 102000005133 glutamate 5-kinase Human genes 0.000 description 16
- 108020001979 glutamate 5-kinase Proteins 0.000 description 16
- 229960002411 imatinib Drugs 0.000 description 16
- 239000000203 mixture Substances 0.000 description 15
- 239000002773 nucleotide Substances 0.000 description 14
- 125000003729 nucleotide group Chemical group 0.000 description 14
- 230000004069 differentiation Effects 0.000 description 13
- 238000001976 enzyme digestion Methods 0.000 description 13
- 230000001177 retroviral Effects 0.000 description 13
- 239000005517 L01XE01 - Imatinib Substances 0.000 description 12
- 239000003550 marker Substances 0.000 description 12
- 235000001014 amino acid Nutrition 0.000 description 11
- 230000015572 biosynthetic process Effects 0.000 description 11
- 238000003786 synthesis reaction Methods 0.000 description 11
- 230000002194 synthesizing Effects 0.000 description 11
- 230000029087 digestion Effects 0.000 description 10
- 238000004520 electroporation Methods 0.000 description 10
- 230000001580 bacterial Effects 0.000 description 9
- 230000001225 therapeutic Effects 0.000 description 9
- 230000003612 virological Effects 0.000 description 9
- AVKUERGKIZMTKX-NJBDSQKTSA-N Ampicillin Chemical compound C1([C@@H](N)C(=O)N[C@H]2[C@H]3SC([C@@H](N3C2=O)C(O)=O)(C)C)=CC=CC=C1 AVKUERGKIZMTKX-NJBDSQKTSA-N 0.000 description 8
- 229940080856 Gleevec Drugs 0.000 description 8
- 229920002459 Intron Polymers 0.000 description 8
- 229960000723 ampicillin Drugs 0.000 description 8
- 102100019449 CD79B Human genes 0.000 description 7
- 101700045471 CD79B Proteins 0.000 description 7
- 229920000665 Exon Polymers 0.000 description 7
- 102000024070 binding proteins Human genes 0.000 description 7
- 108091007650 binding proteins Proteins 0.000 description 7
- 238000007374 clinical diagnostic method Methods 0.000 description 7
- 238000010790 dilution Methods 0.000 description 7
- 238000002826 magnetic-activated cell sorting Methods 0.000 description 7
- 230000001404 mediated Effects 0.000 description 7
- 238000010186 staining Methods 0.000 description 7
- 102100005826 CD19 Human genes 0.000 description 6
- 101700087100 CD19 Proteins 0.000 description 6
- 108020004705 Codon Proteins 0.000 description 6
- 229920002676 Complementary DNA Polymers 0.000 description 6
- 108010044023 Ki-1 Antigen Proteins 0.000 description 6
- 238000004458 analytical method Methods 0.000 description 6
- 239000003153 chemical reaction reagent Substances 0.000 description 6
- 239000002299 complementary DNA Substances 0.000 description 6
- 230000001965 increased Effects 0.000 description 6
- 238000002955 isolation Methods 0.000 description 6
- 102000005614 monoclonal antibodies Human genes 0.000 description 6
- 108010045030 monoclonal antibodies Proteins 0.000 description 6
- 241000894007 species Species 0.000 description 6
- QXRNAOYBCYVZCD-BQBZGAKWSA-N (2S)-6-amino-2-[[(2S)-2-aminopropanoyl]amino]hexanoic acid Chemical compound C[C@H](N)C(=O)N[C@H](C(O)=O)CCCCN QXRNAOYBCYVZCD-BQBZGAKWSA-N 0.000 description 5
- 102100019443 CD79A Human genes 0.000 description 5
- 101700037975 CD79A Proteins 0.000 description 5
- 210000004011 Plasma Cells Anatomy 0.000 description 5
- 238000010276 construction Methods 0.000 description 5
- 230000001809 detectable Effects 0.000 description 5
- 230000000694 effects Effects 0.000 description 5
- 238000005516 engineering process Methods 0.000 description 5
- 210000004602 germ cell Anatomy 0.000 description 5
- 108091008154 B cell receptors Proteins 0.000 description 4
- 108020004999 Messenger RNA Proteins 0.000 description 4
- 206010035226 Plasma cell myeloma Diseases 0.000 description 4
- 102100012830 RAG2 Human genes 0.000 description 4
- 210000003705 Ribosomes Anatomy 0.000 description 4
- 108010032099 V(D)J recombination activating protein 2 Proteins 0.000 description 4
- 101700082685 ampR Proteins 0.000 description 4
- 230000003115 biocidal Effects 0.000 description 4
- 230000024245 cell differentiation Effects 0.000 description 4
- 230000000295 complement Effects 0.000 description 4
- 230000001808 coupling Effects 0.000 description 4
- 238000010168 coupling process Methods 0.000 description 4
- 238000005859 coupling reaction Methods 0.000 description 4
- 238000001514 detection method Methods 0.000 description 4
- 238000011161 development Methods 0.000 description 4
- 230000018109 developmental process Effects 0.000 description 4
- 238000003745 diagnosis Methods 0.000 description 4
- 239000003814 drug Substances 0.000 description 4
- 239000001963 growth media Substances 0.000 description 4
- 230000001939 inductive effect Effects 0.000 description 4
- 229920002106 messenger RNA Polymers 0.000 description 4
- 230000035772 mutation Effects 0.000 description 4
- 201000000050 myeloid neoplasm Diseases 0.000 description 4
- 239000002245 particle Substances 0.000 description 4
- 238000005215 recombination Methods 0.000 description 4
- 238000006467 substitution reaction Methods 0.000 description 4
- UZOVYGYOLBIAJR-UHFFFAOYSA-N 4-isocyanato-4'-methyldiphenylmethane Chemical compound C1=CC(C)=CC=C1CC1=CC=C(N=C=O)C=C1 UZOVYGYOLBIAJR-UHFFFAOYSA-N 0.000 description 3
- 241000588724 Escherichia coli Species 0.000 description 3
- 241000282412 Homo Species 0.000 description 3
- 241000124008 Mammalia Species 0.000 description 3
- 102000018120 Recombinases Human genes 0.000 description 3
- 108010091086 Recombinases Proteins 0.000 description 3
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 3
- 102000000852 Tumor Necrosis Factor-alpha Human genes 0.000 description 3
- 108010001801 Tumor Necrosis Factor-alpha Proteins 0.000 description 3
- 241000700605 Viruses Species 0.000 description 3
- 238000005119 centrifugation Methods 0.000 description 3
- 108091006028 chimera Proteins 0.000 description 3
- 230000002950 deficient Effects 0.000 description 3
- 238000009509 drug development Methods 0.000 description 3
- 230000002068 genetic Effects 0.000 description 3
- 238000010353 genetic engineering Methods 0.000 description 3
- 230000005017 genetic modification Effects 0.000 description 3
- 235000013617 genetically modified food Nutrition 0.000 description 3
- 230000001976 improved Effects 0.000 description 3
- 238000003780 insertion Methods 0.000 description 3
- 230000000670 limiting Effects 0.000 description 3
- 238000004519 manufacturing process Methods 0.000 description 3
- 210000003519 mature B lymphocyte Anatomy 0.000 description 3
- 239000002609 media Substances 0.000 description 3
- 230000004481 post-translational protein modification Effects 0.000 description 3
- 229920000160 (ribonucleotides)n+m Polymers 0.000 description 2
- DGVVWUTYPXICAM-UHFFFAOYSA-N 2-mercaptoethanol Chemical compound OCCS DGVVWUTYPXICAM-UHFFFAOYSA-N 0.000 description 2
- 241000251468 Actinopterygii Species 0.000 description 2
- 206010059512 Apoptosis Diseases 0.000 description 2
- 102000019260 B-Cell Antigen Receptors Human genes 0.000 description 2
- 108010012919 B-Cell Antigen Receptors Proteins 0.000 description 2
- 101710040446 CD40 Proteins 0.000 description 2
- 102100013137 CD40 Human genes 0.000 description 2
- 108010047041 Complementarity Determining Regions Proteins 0.000 description 2
- 108010043121 Green Fluorescent Proteins Proteins 0.000 description 2
- 108010002162 IGK Proteins 0.000 description 2
- 102100007034 Immunoglobulin kappa light chain Human genes 0.000 description 2
- JTEDVYBZBROSJT-UHFFFAOYSA-N Indole-3-butyric acid Natural products C1=CC=C2C(CCCC(=O)O)=CNC2=C1 JTEDVYBZBROSJT-UHFFFAOYSA-N 0.000 description 2
- 108020004391 Introns Proteins 0.000 description 2
- 239000007759 RPMI Media 1640 Substances 0.000 description 2
- 108060007275 SCAI Proteins 0.000 description 2
- 241000710960 Sindbis virus Species 0.000 description 2
- 241000700618 Vaccinia virus Species 0.000 description 2
- 239000012190 activator Substances 0.000 description 2
- 230000001270 agonistic Effects 0.000 description 2
- 230000004075 alteration Effects 0.000 description 2
- 125000000539 amino acid group Chemical group 0.000 description 2
- 230000006907 apoptotic process Effects 0.000 description 2
- 238000006243 chemical reaction Methods 0.000 description 2
- 230000000875 corresponding Effects 0.000 description 2
- 229940079593 drugs Drugs 0.000 description 2
- 238000001962 electrophoresis Methods 0.000 description 2
- 239000002158 endotoxin Substances 0.000 description 2
- 238000002474 experimental method Methods 0.000 description 2
- -1 factors Proteins 0.000 description 2
- 230000002349 favourable Effects 0.000 description 2
- 238000000684 flow cytometry Methods 0.000 description 2
- 125000002485 formyl group Chemical group [H]C(*)=O 0.000 description 2
- 230000036039 immunity Effects 0.000 description 2
- 238000000338 in vitro Methods 0.000 description 2
- 230000002452 interceptive Effects 0.000 description 2
- 238000004806 packaging method and process Methods 0.000 description 2
- 230000002093 peripheral Effects 0.000 description 2
- 238000002823 phage display Methods 0.000 description 2
- OZAIFHULBGXAKX-UHFFFAOYSA-N precursor Substances N#CC(C)(C)N=NC(C)(C)C#N OZAIFHULBGXAKX-UHFFFAOYSA-N 0.000 description 2
- 238000000575 proteomic Methods 0.000 description 2
- 108020003175 receptors Proteins 0.000 description 2
- 102000005962 receptors Human genes 0.000 description 2
- 230000000638 stimulation Effects 0.000 description 2
- 230000013819 transposition, DNA-mediated Effects 0.000 description 2
- 238000005406 washing Methods 0.000 description 2
- 241000269350 Anura Species 0.000 description 1
- 239000004475 Arginine Substances 0.000 description 1
- 241000271566 Aves Species 0.000 description 1
- 102100015655 BCL2L1 Human genes 0.000 description 1
- 101710032374 BCL2L1 Proteins 0.000 description 1
- 229960001561 Bleomycin Drugs 0.000 description 1
- 108010006654 Bleomycin Proteins 0.000 description 1
- 210000001185 Bone Marrow Anatomy 0.000 description 1
- 210000000349 Chromosomes Anatomy 0.000 description 1
- 229920002839 Cis-regulatory element Polymers 0.000 description 1
- GRRNUXAQVGOGFE-UHFFFAOYSA-N Destomysin Chemical compound OC1C(NC)CC(N)C(O)C1OC1C2OC3(C(C(O)C(O)C(C(N)CO)O3)O)OC2C(O)C(CO)O1 GRRNUXAQVGOGFE-UHFFFAOYSA-N 0.000 description 1
- 229920002307 Dextran Polymers 0.000 description 1
- 241000255581 Drosophila <fruit fly, genus> Species 0.000 description 1
- 241000206602 Eukaryota Species 0.000 description 1
- 210000004700 Fetal Blood Anatomy 0.000 description 1
- 102000003688 G-protein coupled receptors Human genes 0.000 description 1
- 108090000045 G-protein coupled receptors Proteins 0.000 description 1
- 241000287828 Gallus gallus Species 0.000 description 1
- 229960002743 Glutamine Drugs 0.000 description 1
- 102000004144 Green Fluorescent Proteins Human genes 0.000 description 1
- 210000004408 Hybridomas Anatomy 0.000 description 1
- 229940097277 Hygromycin B Drugs 0.000 description 1
- 101700031293 IGG1 Proteins 0.000 description 1
- 108010054477 Immunoglobulin Fab Fragments Proteins 0.000 description 1
- 102000001706 Immunoglobulin Fab Fragments Human genes 0.000 description 1
- 102000012745 Immunoglobulin Subunits Human genes 0.000 description 1
- 108010079585 Immunoglobulin Subunits Proteins 0.000 description 1
- 102100009987 Immunoglobulin gamma-1 heavy chain Human genes 0.000 description 1
- 102000004310 Ion Channels Human genes 0.000 description 1
- 108090000862 Ion Channels Proteins 0.000 description 1
- XUJNEKJLAYXESH-REOHCLBHSA-N L-cysteine Chemical compound SC[C@H](N)C(O)=O XUJNEKJLAYXESH-REOHCLBHSA-N 0.000 description 1
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 1
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 1
- 101700029975 LIG4 Proteins 0.000 description 1
- 102100000794 LIG4 Human genes 0.000 description 1
- 210000001165 Lymph Nodes Anatomy 0.000 description 1
- 239000004472 Lysine Substances 0.000 description 1
- 101710034453 MT-CO3 Proteins 0.000 description 1
- 241001465754 Metazoa Species 0.000 description 1
- 102000005431 Molecular Chaperones Human genes 0.000 description 1
- 108010006519 Molecular Chaperones Proteins 0.000 description 1
- 241001182492 Nes Species 0.000 description 1
- 229920002332 Noncoding DNA Polymers 0.000 description 1
- 229920000272 Oligonucleotide Polymers 0.000 description 1
- 101700061400 PRKDC Proteins 0.000 description 1
- 102100003225 PRKDC Human genes 0.000 description 1
- RXWNCPJZOCPEPQ-NVWDDTSBSA-N PUROMYCIN Chemical compound C1=CC(OC)=CC=C1C[C@H](N)C(=O)N[C@H]1[C@@H](O)[C@H](N2C3=NC=NC(=C3N=C2)N(C)C)O[C@@H]1CO RXWNCPJZOCPEPQ-NVWDDTSBSA-N 0.000 description 1
- 229950010131 PUROMYCIN Drugs 0.000 description 1
- 210000002741 Palatine Tonsil Anatomy 0.000 description 1
- 210000002381 Plasma Anatomy 0.000 description 1
- 208000007452 Plasmacytoma Diseases 0.000 description 1
- 102000001253 Protein Kinases Human genes 0.000 description 1
- 108060006633 Protein Kinases Proteins 0.000 description 1
- 102000004022 Protein-Tyrosine Kinases Human genes 0.000 description 1
- 108090000412 Protein-Tyrosine Kinases Proteins 0.000 description 1
- 101800001295 Putative ATP-dependent helicase Proteins 0.000 description 1
- 101800001006 Putative helicase Proteins 0.000 description 1
- 102000007312 Recombinant Proteins Human genes 0.000 description 1
- 108010033725 Recombinant Proteins Proteins 0.000 description 1
- 241000282849 Ruminantia Species 0.000 description 1
- 210000002966 Serum Anatomy 0.000 description 1
- 210000000952 Spleen Anatomy 0.000 description 1
- 108010090804 Streptavidin Proteins 0.000 description 1
- 229940014598 TAC Drugs 0.000 description 1
- 102000009332 Tetraspanins Human genes 0.000 description 1
- 108050000196 Tetraspanins Proteins 0.000 description 1
- QORWJWZARLRLPR-UHFFFAOYSA-H Tricalcium phosphate Chemical compound [Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O QORWJWZARLRLPR-UHFFFAOYSA-H 0.000 description 1
- 102100009955 XRCC4 Human genes 0.000 description 1
- 108060009531 XRCC4 Proteins 0.000 description 1
- 101700029350 XRCC5 Proteins 0.000 description 1
- 102100009956 XRCC5 Human genes 0.000 description 1
- 101700032709 XRCC6 Proteins 0.000 description 1
- 102100009954 XRCC6 Human genes 0.000 description 1
- 240000008042 Zea mays Species 0.000 description 1
- 235000016383 Zea mays subsp huehuetenangensis Nutrition 0.000 description 1
- 235000002017 Zea mays subsp mays Nutrition 0.000 description 1
- 239000002253 acid Substances 0.000 description 1
- 230000003213 activating Effects 0.000 description 1
- 238000004873 anchoring Methods 0.000 description 1
- 230000003042 antagnostic Effects 0.000 description 1
- 230000002424 anti-apoptotic Effects 0.000 description 1
- 230000000890 antigenic Effects 0.000 description 1
- 229960000070 antineoplastic Monoclonal antibodies Drugs 0.000 description 1
- 238000002819 bacterial display Methods 0.000 description 1
- 239000011324 bead Substances 0.000 description 1
- 238000010009 beating Methods 0.000 description 1
- 238000003766 bioinformatics method Methods 0.000 description 1
- 230000031018 biological processes and functions Effects 0.000 description 1
- 230000033228 biological regulation Effects 0.000 description 1
- YBJHBAHKTGYVGT-ZKWXMUAHSA-N biotin Chemical compound N1C(=O)N[C@@H]2[C@H](CCCCC(=O)O)SC[C@@H]21 YBJHBAHKTGYVGT-ZKWXMUAHSA-N 0.000 description 1
- 229960002685 biotin Drugs 0.000 description 1
- 235000020958 biotin Nutrition 0.000 description 1
- 239000011616 biotin Substances 0.000 description 1
- OYVAGSVQBOHSSS-UAPAGMARSA-O bleomycin A2 Chemical compound N([C@H](C(=O)N[C@H](C)[C@@H](O)[C@H](C)C(=O)N[C@@H]([C@H](O)C)C(=O)NCCC=1SC=C(N=1)C=1SC=C(N=1)C(=O)NCCC[S+](C)C)[C@@H](O[C@H]1[C@H]([C@@H](O)[C@H](O)[C@H](CO)O1)O[C@@H]1[C@H]([C@@H](OC(N)=O)[C@H](O)[C@@H](CO)O1)O)C=1N=CNC=1)C(=O)C1=NC([C@H](CC(N)=O)NC[C@H](N)C(N)=O)=NC(N)=C1C OYVAGSVQBOHSSS-UAPAGMARSA-O 0.000 description 1
- 108091005941 blue fluorescent protein Proteins 0.000 description 1
- 108010006025 bovine growth hormone Proteins 0.000 description 1
- 229910000389 calcium phosphate Inorganic materials 0.000 description 1
- 239000001506 calcium phosphate Substances 0.000 description 1
- 235000011010 calcium phosphates Nutrition 0.000 description 1
- 230000003197 catalytic Effects 0.000 description 1
- 239000002458 cell surface marker Substances 0.000 description 1
- 239000006285 cell suspension Substances 0.000 description 1
- 230000004186 co-expression Effects 0.000 description 1
- 230000001010 compromised Effects 0.000 description 1
- 230000002596 correlated Effects 0.000 description 1
- 235000018417 cysteine Nutrition 0.000 description 1
- 125000000151 cysteine group Chemical group N[C@@H](CS)C(=O)* 0.000 description 1
- 230000001419 dependent Effects 0.000 description 1
- 201000009910 diseases by infectious agent Diseases 0.000 description 1
- 230000002708 enhancing Effects 0.000 description 1
- 210000003527 eukaryotic cell Anatomy 0.000 description 1
- 108091006031 fluorescent proteins Proteins 0.000 description 1
- 102000034387 fluorescent proteins Human genes 0.000 description 1
- 230000004927 fusion Effects 0.000 description 1
- 108020001507 fusion proteins Proteins 0.000 description 1
- 102000037240 fusion proteins Human genes 0.000 description 1
- 239000005090 green fluorescent protein Substances 0.000 description 1
- 230000012010 growth Effects 0.000 description 1
- 238000002744 homologous recombination Methods 0.000 description 1
- 230000002779 inactivation Effects 0.000 description 1
- 230000006882 induction of apoptosis Effects 0.000 description 1
- 239000003112 inhibitor Substances 0.000 description 1
- 230000002401 inhibitory effect Effects 0.000 description 1
- 230000003993 interaction Effects 0.000 description 1
- 238000002372 labelling Methods 0.000 description 1
- 238000003475 lamination Methods 0.000 description 1
- 239000002502 liposome Substances 0.000 description 1
- 239000006249 magnetic particle Substances 0.000 description 1
- 235000009973 maize Nutrition 0.000 description 1
- 238000005259 measurement Methods 0.000 description 1
- 239000003226 mitogen Substances 0.000 description 1
- 229960000060 monoclonal antibodies Drugs 0.000 description 1
- 231100000243 mutagenic effect Toxicity 0.000 description 1
- 229960004927 neomycin Drugs 0.000 description 1
- 239000002547 new drug Substances 0.000 description 1
- 230000003287 optical Effects 0.000 description 1
- 210000000056 organs Anatomy 0.000 description 1
- 230000002018 overexpression Effects 0.000 description 1
- 239000000825 pharmaceutical preparation Substances 0.000 description 1
- 239000003757 phosphotransferase inhibitor Substances 0.000 description 1
- 230000001402 polyadenylating Effects 0.000 description 1
- 230000001323 posttranslational Effects 0.000 description 1
- 230000003389 potentiating Effects 0.000 description 1
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 1
- 230000035755 proliferation Effects 0.000 description 1
- 230000000644 propagated Effects 0.000 description 1
- 238000002818 protein evolution Methods 0.000 description 1
- 210000001938 protoplasts Anatomy 0.000 description 1
- 238000003908 quality control method Methods 0.000 description 1
- 238000011084 recovery Methods 0.000 description 1
- 108010054624 red fluorescent protein Proteins 0.000 description 1
- 238000010839 reverse transcription Methods 0.000 description 1
- 238000010187 selection method Methods 0.000 description 1
- 238000000926 separation method Methods 0.000 description 1
- 231100000486 side effect Toxicity 0.000 description 1
- 230000001743 silencing Effects 0.000 description 1
- 239000012128 staining reagent Substances 0.000 description 1
- 230000003068 static Effects 0.000 description 1
- 239000000126 substance Substances 0.000 description 1
- 230000004083 survival Effects 0.000 description 1
- 239000000725 suspension Substances 0.000 description 1
- 238000004114 suspension culture Methods 0.000 description 1
- 238000002560 therapeutic procedure Methods 0.000 description 1
- 230000035897 transcription Effects 0.000 description 1
- 230000002103 transcriptional Effects 0.000 description 1
- 230000001131 transforming Effects 0.000 description 1
- 238000003146 transient transfection Methods 0.000 description 1
- 230000014616 translation Effects 0.000 description 1
- 230000001960 triggered Effects 0.000 description 1
- 230000010415 tropism Effects 0.000 description 1
- 241001430294 unidentified retrovirus Species 0.000 description 1
- 230000035899 viability Effects 0.000 description 1
- 108091005946 yellow fluorescent protein Proteins 0.000 description 1
Images
















Abstract
MÉTODO PARA IDENTIFICAR UM POLIPEPTÍDEO TENDO UMA FUNCIONALIDADE OU ESPECIFICIDADE DE LIGAÇÃO DESEJADAS, BIBLIOTECA DE MOLÉCULAS DE POLINUCLEOTÍDEO, MÉTODO PARA GERAR UMA BIBLIOTECA DE POLINUCLEOTÍDEOS, VETOR, CÉLULA HOSPEDEIRA, ANTICORPO E MÉTODO PARA GERAR UMA POPULAÇÃO DE CÉLULAS HOSPEDEIRAS. A presente invenção refere-se a um método que oferece eficiência, flexibilidade, utilidade e velocidade incomparáveis para a descoberta e otimização de polipeptídeos com funcionalidade e/ou especificidade de ligação desejadas, incluindo moléculas de ligação a antígeno, tais como anticorpos e fragmentos dos mesmos, para fenótipos funcionais e/ou de ligação desejados. O novo método baseia-se em construções de transposição e bibliotecas de ADN diversificadas clonadas em vetores de transposição e em sua transfeção em células hospedeiras por expressão transitória concomitante de uma enzima transponase funcional. Isto assegura uma introdução eficiente e estável de vetores de expressão baseados na transposição de células hospedeiras de vertebrados em uma etapa, que podem depois ser classificados em relação a um fenótipo funcional ou de ligação desejado das proteínas expressas, após o que as sequências codificantes relevantes para as proteínas expressas, incluindo anticorpos e fragmentos dos mesmos, podem ser identificadas por técnicas de clonagem e de sequenciação de ADN convencionais.
Description
[0001] A descoberta de proteínas específicas de alvo, incluindo anticorpos e fragmentos dos mesmos, é de significante interesse comercial, pelo fato de que a seleção de proteínas funcionais altamente seletivas ou proteínas de ligação, incluindo anticorpos e fragmentos dos mesmos, têm um alto potencial para o desenvolvimento de novas entidades biológicas (NBEs) com novas propriedades terapêuticas que muito especificamente integram, ou interferem com os processos biológicos, e portanto são previstos exibir perfis de efeitos colaterais mais baixos do que as novas entidades químicas convencionais (NCEs). Em relação a isso, particularmente o desenvolvimento de anticorpos terapêuticos altamente específicos um alvo, e terapêuticos com base em anticorpo, abriram o caminho para terapias completamente novas com aprimorada eficácia. Como uma consequência, anticorpos monoclonais terapêuticos representam o segmento de mais rápido desenvolvimento no desenvolvimento de novos fármacos das últimas décadas, e atualmente geram cerca de 50 bilhões de dólares de receita, o que é responsável por uma parte significante do mercado total global de produtos de fármacos.
[0002] Portanto, tecnologias eficientes e inovadoras, que permitem a descoberta de proteínas terapêuticas altamente potentes, mas também bem toleradas, em particular terapêuticos com base em anticorpo, estão em alta demanda.
[0003] De modo a identificar uma proteína com uma funcionalidade desejada ou uma propriedade de ligação específica, como é o caso para os anticorpos, é necessário se gerar, se expresssr de modo funcional e se fazer a leitura de coleções diversas e grandes, ou bibliotecas de proteínas, incluindo anticorpos e fragmentos dos mesmos, para as desejadas propriedades funcionais ou especificidade de ligação um alvo. Uma série de tecnologias tem sido desenvolvida nos últimos vinte anos, as quais permitem a expressão de diversas bibliotecas de proteínas seja em células hospedeiras, ou em partículas virais e de fago e métodos para a leitura de alta produtividade e/ou bateamento em direção da desejada propriedade funcional, ou fenótipo de ligação.
[0004] Tecnologias padrão do estado da técnica para alcançar a identificação de ligantes ou proteínas específicas de alvo com as desejadas propriedades funcionais incluem, por exemplo, as tecnologias de fago-display, display retroviral, display bacteriano, display de levedura e várias tecnologias de display de célula de mamífero, em combinação com ligação a superfície sólida (bateamento) e/ou outras tecnologias de enriquecimento. Todas as referidas tecnologias são cobertas por várias patentes e pedidos de patentes pendentes.
[0005] Embora os sistemas de fago display e de display procariótico tenham sido estabelecidos e são amplamente adotados na indústria de biotecnologia e em academia para a identificação de ligantes específicos um alvo, incluindo fragmentos de anticorpo (Hoogenboom, Nature Biotechnology 23, 1105-1116 (2005)), os mesmos sofrem de uma variedade de limitações, incluindo a incapacidade de expressar as versões de comprimento total das proteínas maiores, incluindo anticorpos de comprimento total, a falta de adequada modificação pós-translacional, a falta de adequado dobramento por vertebrados companheiros, e, no caso de anticorpos, uma combinação de cadeia leve e pesada artificialmente forçada. Portanto, no caso de descoberta de anticorpo pelos referidos métodos, “reformatar” em anticorpos de comprimento total e expressão em célula de mamífero é necessário. Em virtude das limitações acima mencionadas isso com frequência resulta em anticorpos com propriedades biofísicas desfavoráveis (por exemplo, baixa estabilidade, tendência a se agregar, vermelhouzida afinidade), limitando o potencial terapêutico e de diagnóstico das referidas proteínas. Isso, por um lado, leva a significantes coeficientes de atrito no desenvolvimento de moléculas de liderança geradas pelos referidos métodos, e, por outro lado, requer esforço significante para corrigir as viabilidades biofísicas e moleculares nas referidas proteínas para o desenvolvimento de fármaco adicional à jusante fármaco.
[0006] Portanto, uma tecnologia de descoberta de proteína e anticorpo tem sido desenvolvida usando eucarióticos inferiores (por exemplo, levedura) e, mais recentemente, também a expressão em sistemas de células de mamífero para a identificação de proteínas com as desejadas propriedades, na medida em que as referidas tecnologias permitem (i) a expressão de proteínas de comprimento total maiores, incluindo anticorpos de comprimento total, (ii) melhor ou normal modificação pós-translacional, e, (iii) em caso de anticorpos, o adequado pareamento de cadeias pesadas-leves (Beerli & Rader, mAbs 2, 365-378 (2010)). Isso, em agregado, seleciona as proteínas com propriedades biofísicas favoráveis que têm um maior potencial em desenvolvimento de fármaco e uso terapêutico.
[0007] Embora a expressão e o classificação de proteínas em células de vertebrado seja mais desejável, pelo fato das células de vertebrado (por exemplo, as células CHO de Hamster, HEK-293 humana, ou DT40 de galinha) sejam preferidas os sistemas de expressão para a produção de proteínas terapêuticas maiores, tal como anticorpos, as referidas tecnologias são atualmente também associadas com uma série de limitações, o que levou a uma lenta adoção das referidas tecnologias em academias e na indústria.
[0008] Primeiro, as células de vertebrado não são modificadas geneticamente e de modo estável com tanta eficiência, como, por exemplo, as células procarióticas ou eucarióticas inferiores tais como as leveduras. Portanto, permanence ainda um desafio de gerar diversas células de vertebrados (complexo) com base em bibliotecas de proteínas, a partir da qual candidatos com as desejadas propriedades ou as mais altas afinidades de ligação podem ser identificados. Segundo, de modo a isolar de modo eficiente as proteínas com as desejadas propriedades, em geral rodadas interativas de enriquecimento celular são necessárias. A expressão de vertebrados seja por transfecção transitória de plasmídeos (Higuchi et al J. Immunol. Methods 202, 193204(1997)), ou por sistemas de expressão viral transitória, tal como o sindbis ou vírus de vaccínia (Beerli et al. PNAS 105, 14336-14341 (2008), e pedido de patente internacional publicado sob o No. WO 02102885) não permitem múltiplas rodadas de seleção celular necessária para enriquecer de modo eficiente as proteínas altamente específicas, e os referidos métodos são portanto ou restritos ao classificação de pequenas bibliotecas de proteínas pré-enriquecidas, ou os mesmos requerem ciclos tediosos de isolamento de vírus/reinfecção de célula.
[0009] De modo a alcançar a expressão estável de proteínas de ligação e anticorpos em células de vertebrado, que permitem múltiplas rodadas de seleções com base no acoplamento estável de genótipo-fenótipo, tecnologias têm sido desenvolvidas, utilizando recombinases específicas (sistema de recombinase de flp/frt, Zhou et al. mAbs 5, 508-518 (2010)), ou vetores retrovirais (pedido de patente internacional publicado sob o No. WO 2009109368). Entretanto, a recombinação de flp/frt é um sistema de baixa eficiência para a integração estável de genes nos genomas de células hospedeiras de vertebrado e portanto, mais uma vez, apenas aplicável a pequenas bibliotecas pré-selecionadas, ou à otimização de proteína selecionada ou candidatos de anticorpo.
[0010] Em comparação com o sistema de recombinase flp/frt, os vetores retrovirais permitem uma modificação genética estável mais eficiente de células hospedeiras de vertebrado e a geração de bibliotecas celulares mais complexas. Entretanto, (i) as mesmas são restritas a apenas linhagens celulares selecionadas permissíveis, (ii) as mesmas representam um risco de biossegurança, quando células humanas são utilizadas, (iii) vetores de expressão retroviral são submetidos a mutagênese indesejada das sequências de biblioteca em virtude de transcrição reversa de baixa fidelidade, (iv) vetores retrovirais não permitem a integração de cassetes de expressão genômica com estrutura intacta de intron/exon, em virtude de splicing do genoma retroviral antes de empacotamento do vetor dentro das partículas retrovirais, (v) os retrovírus são submetidos a recombinação homóloga descontrolável e desfavorável de sequências de biblioteca durante o empacotamento dos genomas virais, (vi) são submetidos a silenciamento retroviral, e (vii) requer um procedimento tedioso de duas etapas de transfecção e empacotamento-célula/infecção de célula hospedeira. Todas as referidas limitações representam um desafio significante e limitações, e introduzir complexidades significantes para a utilidade do vetor retroviral com base em abordagens em gerar bibliotecas de células de vertebrado de alta qualidade/alta complexidade para eficiente descoberta de proteína específica de alvo, ou anticorpo.
[0011] Portanto, existe claramente uma necessidade para uma tecnologia mais eficiente, mais controlável e direta que permita a geração de células de vertebrados de alta qualidade e altamente complexas com base em bibliotecas que expressam diversas bibliotecas de proteínas incluindo anticorpos e fragmentos dos mesmos a partir dos quais as proteínas com função altamente específica e/ou propriedades de ligação e altas afinidades podem ser isoladas.(b) Transposases/Transposição:
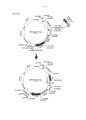
[0012] Transposons, ou elementos transponíveis (TEs), são elementos genéticos com a capacidade de se integrar de modo estável nos genomas das células hospedeiras, um processo que é chamado de transposição (Ivics et al. Mobile DNA 1, 25 (2010)) (incorporado aqui a título de referência em sua totalidade). TEs já foram postulados nos anos 1950s por Barbara McClintock em estudos genéticos com milho, mas os primeiros modelos funcionais para a transposição foram descritos para TEs bacterianos no final dos anos 1970s (Shapiro, PNAS 76, 1933-1937 (1979)) (incorporado aqui a título de referência em sua totalidade).
[0013] Nesse meio tempo está claro que os TEs estão presentes no genoma de cada organismo, e o sequenciamento genômico revelou que aproximadamente 45% do genoma humano é derivado de transposon (International Human Genome Sequencing Consortium Nature 409: 860-921 (2001)) (incorporado aqui a título de referência em sua totalidade). Entretanto, como diferente dos invertebrados, onde o TEs funcional (ou autônomo) foram identificados (figura 1a), os seres humanos e a maior parte dos vertebrados superiores não contêm TEs funcional. Foi levantada a hipótese de que a pressão seletiva evolucionaria contra o potencial mutagênico de TEs levou a inativação funcional dos mesmos há milhões de anos atrás durante a evolução.
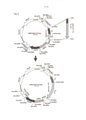
[0014] TEs autônomos compreendem DNA que codifica a enzima transposase localizada entre duas sequências de repetição terminal invertida (ITRs), que são reconhecidas pela enzima transposase codificada entre as ITRs e que pode catalisar a transposição do TE em qualquer sequência de DNA de dupla fita (figura 1a). Há duas diferentes classes de transposons: classe I, ou retrotransposons, que mobilizam via um Intermediário de RNA e um mecanismo de “copiar e colar” (figura 2b), e classe II, ou DNA transposons, que mobilizam via excisão-integração, ou um mecanismo de “cortar e colar” (figura 2a) (Ivics et al. Nat. Methods 6, 415-422(2009)) (incorporado aqui a título de referência em sua totalidade).
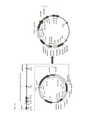
[0015] Eucarióticos inferiores bacterianos (por exemplo, levedura) e transposons de invertebrados parecem ser grandemente específicos de espécie, e não podem ser usados para a eficiente transposição de DNA em células de vertebrado. Apenas, após um primeiro transposon ativo ter sido artificialmente reconstruído por embaralhamento de sequência dos TEs inativos de peixe, que foi portanto chamado de “Sleeping Beauty” (Ivics et al. Cell 91, 501-510 (1997)) (incorporado aqui a título de referência em sua totalidade), se tornou possível se alcançar com sucesso a integração de DNA por transposição em células de vertebrado, incluindo células humanas. Sleeping Beauty é um transposon de DNA de classe II que pertence à family de transposons Tc1/mariner (Ni et al. Briefings Funct. Genomics Proteomics 7, 444-453 (2008)) (incorporado aqui a título de referência em sua totalidade). Nesse meio tempo, transposons funcionais adicionais foram identificados ou reconstruídos a partir de diferentes espécies, incluindo Drosophila, sapo e mesmo genomas humanos, que todos foram mostrados para permitir a Transposição de DNA em vertebrados e também genomas humanos das células hospedeiras (figura 3). Cada um dos referidos transposons, tem vantagens e desvantagens que estão relacionadas à eficiência da transposição, estabilidade de expressão, capacidade de carga genética, etc.
[0016] Até o presente, as tecnologias mediadas a transposon para a expressão de diversas bibliotecas de proteínas, incluindo anticorpos e fragmentos dos mesmos, em células hospedeiras de vertebrado para o isolamento específico de alvo, proteínas funcionais de ligação, incluindo anticorpos e fragmentos dos mesmos, não foram descritos na técnica anterior.
[0017] O método descrito aqui descreve uma nova tecnologia que oferece eficiência sem paralelo, flexibilidade, utilidade e velocidade para a descoberta e a otimização de polipeptídeos tendo a desejada especificidade e/ou funcionalidade de ligação, incluindo moléculas de ligação a antígeno tal como anticorpos e fragmentos dos mesmos, para os desejados fenótipos funcionais e/ou de ligação. O novo método é com base em construtores transponíveis e diversas bibliotecas de DNA clonados em vetores transponíveis e a transfecção dos mesmos em células hospedeiras por expressão transitória concomitante de uma enzima transponase funcional. Isso garante uma introdução eficiente, e estável dos vetores de expressão com base em transposon nas células hospedeiras de vertebrado em uma etapa, que pode então ser classificada para o desejado fenótipo de ligação ou funcional das proteínas expressas, após o que as sequências de codificação relevantes para as proteínas expressas, incluindo anticorpos e fragmentos dos mesmos, podem ser identificados por técnicas de clonagem padrão e de sequenciamento de DNA.
[0018] Em uma modalidade, a presente invenção é amplamente direcionada a um método para identificar um polipeptídeo tendo a desejada especificidade ou funcionalidade de ligação, que compreende:
[0019] (i) gerar uma coleção diversa de polinucleotídeos que codificam polipeptídeos tendo diferentes especificidades ou funcionalidades de ligação, em que os referidos polinucleotídeos compreendem uma sequência que codifica um polipeptídeo disposto entre as primeira e segunda sequências de repetição terminal invertida que são reconhecidas por e funcionais com pelo menos uma enzima transponase;
[0020] (ii) introduzir a coleção diversa de polinucleotídeos de (i) nas células hospedeiras;
[0021] (iii) expressar pelo menos uma enzima transposase funcional com as referidas sequências de repetição terminal invertidas nas referidas células hospedeiras de modo que a referida coleção diversa de polinucleotídeos é integrada no genoma das células hospedeiras para proporcionar a população das células hospedeiras que expressa a referida coleção diversa de polinucleotídeos que codificam polipeptídeos tendo diferentes especificidades ou funcionalidades de ligação;
[0022] (iv) rastrear as referidas células hospedeiras para identificar as células hospedeiras que expressam um polipeptídeo tendo a desejada especificidade ou funcionalidade de ligação; e
[0023] (v) isolar a sequência de polinucleotídeo que codifica o referido polipeptídeo a partir das referidas células hospedeiras.
[0024] Em uma modalidade preferida, os polinucleotídeos são moléculas de DNA. Em uma modalidade, a coleção diversa de polinucleotídeos compreende uma sequência de ligação a ligante de um receptor, ou uma sequência de ligação um alvo de uma molécula de ligação. Em uma modalidade preferida, os polinucleotídeos compreendem a sequência que codifica uma molécula de ligação a antígeno, tal como um domínio VH ou VL de anticorpo, ou um fragmento de ligação a antígeno dos mesmos, ou cadeias pesadas ou leves de anticorpo que são de comprimento total (isto é, que inclui a região constante). Em determinadas modalidades, os polinucleotídeos podem compreender uma sequência que codifica ambas as regiões VH e VL, ou ambas as cadeias pesada e leve de anticorpo. Em outra modalidade, os polinucleotídeos compreendem a sequência que codifica um domino de cadeia única Fv ou Fab.
[0025] Em uma modalidade, a coleção diversa de polinucleotídeos é gerada por submeter as sequências de gene da região V a PCR sob condições de mutagênese, por exemplo, por amplificação de PCR dos repertórios da região V a partir de células B de vertebrados. Em outra modalidade, a coleção diversa de polinucleotídeos é gerada por síntese de gene (por exemplo, por randomização das sequências que codificam um polipeptídeo tendo especificidade e/ou funcionalidade de ligação conhecida). Em uma modalidade útil, a coleção diversa de polinucleotídeos compreende vetores de plasmídeo. Em outra modalidade útil, a coleção diversa de polinucleotídeos compreende amplicons de PCR de DNA de dupla fita. Os vetores de plasmídeo podem compreender a sequência que codifica um marcador de gene, tal como um marcador fluorescente, um marcador de superfície celular, ou um marcador selecionável. A sequência de marcador de gene pode estar à montante ou à jusante da sequência que codifica o polipeptídeo tendo a especificidade ou funcionalidade de ligação, mas entre as sequências de repetição terminal invertidas. Alternativamente, a sequência de marcador de gene pode estar à jusante da referida sequência que codifica um polipeptídeo tendo especificidade ou funcionalidade de ligação e separado por um campo de entrada de ribossmo interno.
[0026] Em algumas modalidades, a coleção diversa de polinucleotídeos codifica uma pluralidade de moléculas de ligação a antígeno de um vertebrado, tal como um mamífero, por exemplo, um ser humano.
[0027] Em uma modalidade, a etapa (ii) do método compreende introduzir nas células hospedeiras polinucleotídeos que compreendem sequências que codificam as regiões VH ou VL de imunoglobulina, ou fragmento de ligação a antígenos das mesmas, e em que as referidas sequências de regiões VH e VL são codificadas em vetores separados. Em outra modalidade, a etapa (ii) do método da presente invenção compreende introduzir nas células hospedeiras polinucleotídeos que compreendem as sequências que codificam de cadeias pesadas ou leves de imunoglobulina de comprimento total, ou fragmento de ligação a antígenos dos mesmos, em que said sequências de cadeias pesadas e leves de comprimento total são em vetores separados. Os vetores podem ser introduzidos nas células hospedeiras simultaneamente ou em sequência. Em outra modalidade, as sequências que codificam as regiões VH e VL ou cadeias pesadas e leves de comprimento total são introduzidas nas células hospedeiras no mesmo vetor. No caso em que as sequências VH e VL ou as sequências de cadeias pesadas e leves de anticorpo de comprimento total são introduzidos nas células hospedeiras em diferentes vetores, é útil para as sequências de repetição terminal invertidas em cada vetor ser reconhecida por e funcional com diferentes enzimas transponases.
[0028] As células hospedeiras são preferivelmente células de vertebrado, e preferivelmente células de mamífero, tal como células de roedor ou células humanas. Células linfóides, por exemplo, células B, são particularmente úteis. As células B podem ser células B progenitoras ou células B precursoras tal como, por exemplo, células B progenitoras ou células B precursoras transformadas pelo vírus da Leucemia Abelson-Murine e precoe, células proB e preB humanas transformadas EBV imunoglobulina-null. Outras células hospedeiras úteis incluem linhagens de célula B tais como as células Sp2/0, células NS0, células X63, e células Ag8653, ou linhagens de célula de mamífero comum tal como as células CHO, células Per.C6 células, células BHK, e células 293.
[0029] Em uma modalidade do método da presente invenção, a etapa de expressar (iii) compreende introduzir nas referidas células hospedeiras um vetor de expressão que codifica a enzima transposase que reconhece e é funcional com pelo menos uma sequência de repetição terminal invertida nos polinucleotídeos. O vetor que codifica a enzima transposase pode ser introduzido nas células hospedeiras concomitantemente com ou antes ou subsequente a coleção diversa de polinucleotídeos. Em uma modalidade, a enzima transposase é transitoriamente expressa nas referidas células hospedeiras. Alternativamente, a etapa de expressar (iii) podem compreender induzir um sistema de expressão induzível que é integrado de modo estável no genoma das células hospedeiras, tal como, por exemplo, um sistema induzível por tetraciclina ou induzível por tamoxifeno. Em uma modalidade preferida, a etapa (iii) compreende expressar nas células hospedeiras(s) um vetor que compreende uma transponase funcional Sleeping Beauty ou uma transponase funcional PiggyBac. Em uma modalidade útil, a etapa (iii) compreende expressar nas referidas células hospedeiras um vetor que compreende SEQ ID NO:11. Em outra modalidade útil, o vetor codifica SEQ ID NO:12, ou a sequência com pelo menos 95% de homologia de sequência de amino ácido e tendo a mesma especificidade ou especificidade similar a da sequência de repetição terminal invertida.
[0030] Em outra modalidade útil, a etapa (iii) compreende expressar nas referidas células hospedeiras um vetor que compreende SEQ ID NO:17. Em outra modalidade útil, o vetor codifica SEQ ID NO:18, ou a sequência com pelo menos 95% de homologia de sequência de amino ácido e tendo a mesma especificidade ou especificidade similar a da sequência de repetição terminal invertida.
[0031] Em uma modalidade do método da presente invenção, a etapa de classificação (iv) compreende classificar células ativadas magnéticas (MACS), classificar células ativadas por fluorescência (FACS), bateamento contra moléculas imobilizadas em um bateamento de superfície sólida, seleção para ligação a moléculas associadas a membrana celular incorporadas em uma membrana celular de dupla camada de lipídeos, natural ou artificialmetne reconstituída, ou a classificação de alta produtividade de clones de células individuais em formato de múltiplas cavidades para o desejado fenótipo de ligação ou funcional. Em uma modalidade, a etapa de classificação (iv) compreende classificar para identificar polipeptídeos tendo a desejada especificidade ou funcionalidade de ligação um alvo. Em uma modalidade preferida, a etapa de classificação (iv) compreende classificação para identificar moléculas de ligação a antígeno tendo a desejada especificidade a antígeno. Em uma modalidade útil, a etapa de classificação adicionalmente compreende classificação para identificar moléculas de ligação a antígeno tendo uma ou mais desejadas propriedades funcionais. A etapa de classificação (iv) pode compreender múltiplos ciclos de enriquecimento de célula com a expansão das células hospedeiras entre os ciclos de enriquecimento de célula individuais.
[0032] Em uma modalidade do método da presente invenção, a etapa (v) de isolar a sequência de polinucleotídeo que codifica o polipeptídeo tendo a desejada especificidade ou funcionalidade de ligação compreende a genômica ou a amplificação de RT-PCR ou o sequenciamento profundo de próxima geração. Em uma modalidade útil, a sequência de polinucleotídeo isolada na etapa (v) é submetida a otimizacao de afinidade. Isso pode ser realizado ao se submeter a sequência de polinucleotídeo isolada a PCR ou RT-PCR sob condições de mutagênese. Em outra modalidade útil, a sequência que sofreu mutagênese é então adicionalmente submetida a etapas (i)-(v) do método da presente invenção. Em uma modalidade preferida, a sequência de polinucleotídeo obtida em (v) compreende a sequência que codifica a região VH ou VL de um anticorpo, ou um fragmento de ligação a antígeno dos mesmos, e em que a referida otimização de anticorpo compreende introduzir uma ou mais mutações na região de determinação de complementaridade ou região de framework da referida VH ou VL.
[0033] Em uma modalidade útil, as sequências de repetição terminal invertida são a partir do sistema transposon PiggyBac e são reconhecidas por e funcionais com a transponase PiggyBac. Em uma modalidade, a sequência que codifica a sequência de repetição terminal invertida PiggyBac à montante compreende SEQ ID NO:1. Em outra modalidade, a sequência que codifica a sequência de repetição terminal invertida PiggyBac à jusante compreende SEQ ID NO:2.
[0034] Em outra modalidade útil, as sequências de repetição terminal invertida são a partir do sistema detransposon Sleeping Beauty e são reconhecidas por e funcionais com a transponase transposase de Sleeping Beauty. Em uma modalidade, a sequência que codifica a sequência de repetição terminal invertida Sleeping Beauty à montante compreende SEQ ID NO:14. Em outra modalidade, a sequência que codifica a sequência de repetição terminal invertida Sleeping Beauty à jusante compreende SEQ ID NO:15.
[0035] Em uma modalidade da presente invenção, os polinucleotídeos compreendem sequências de região VH ou VL que codificam a sequência derivada a partir de um anticorpo anti-TNF alfa humano. Em uma modalidade, o anticorpo anti-TNF alfa humano é D2E7.
[0036] Em uma modalidade útil, a etapa (iii) compreende introduzir nas referidas células hospedeiras um vetor que compreende a sequência que codifica a transponase funcional PiggyBac. Em uma modalidade o vetor compreende SEQ ID NO:11. Em outra modalidade, o vetor codifica SEQ ID NO:12, ou uma sequência com pelo menos 95% de homologia de sequência de amino ácido e tendo a mesma especificidade ou especificidade similar a da sequência de repetição terminal invertida.
[0037] Em outra modalidade útil, a etapa (iii) compreende expressar nas referidas células hospedeiras um vetor que compreende SEQ ID NO:17. Em outra modalidade útil, o vetor codifica SEQ ID NO:18, ou uma sequência com pelo menos 95% de homologia de sequência de amino ácido e tendo a mesma especificidade ou especificidade similar a da sequência de repetição terminal invertida.
[0038] Em modalidades preferidas, as sequências de repetição terminal invertida são reconhecidas por e funcionais com pelo menos uma transposase selecionada a partir do grupo que consiste de: PiggyBac, Sleeping Beauty, Frog Prince, Himar1, Passport, Minos, hAT, Tol1, Tol2, Ac/Ds, PIF, Harbinger, Harbinger3-DR, e Hsmar1.
[0039] A presente invenção é adicionalmente direcionada a uma biblioteca de moléculas de polinucleotídeo que codificam polipeptídeos tendo diferentes especificidades ou funcionalidades de ligação, que compreende uma pluralidade de moléculas de polinucleotídeo, em que as referidas moléculas de polinucleotídeo compreendem uma sequência que codifica um polipeptídeo tendo a especificidade ou funcionalidade de ligação disposta entre as sequências de repetição terminal invertida que são reconhecidas por e funcionais com pelo menos uma enzima transponase. Preferivelmente os polinucleotídeos são moléculas de DNA e compreendem uma sequência de ligação a ligante de um receptor ou uma sequência de ligação um alvo da molécula de ligação. Em uma modalidade particularmente preferida, a biblioteca compreende polinucleotídeos, em que cada polinucleotídeo compreende a sequência que codifica uma sequência de ligação a antígeno de um anticorpo. Em uma modalidade, a biblioteca compreende polinucleotídeos que codificam a reigão VH ou VL de um anticorpo ou um fragmento de ligação a antígeno das mesmas. Alternativamente, os polinucleotídeos podem codificar a região VH e a região VL. Em uma modalidade preferida, os polinucleotídeos da biblioteca compreendem uma sequência que codifica a cadeia pesada ou leve de anticorpo de comprimento total (isto é, incluindo a região constante) ou um fragmento de ligação a antígeno da mesma. Alternativamente, os polinucleotídeos podem codificar ambas as cadeias pesada e leve de imunoglobulina de comprimento total. Em outras modalidades, os polinucleotídeos da biblioteca compreendem a sequência que codifica um um domínio Fv ou FAb de cadeia única. Em modalidades preferidas, os polinucleotídeos da biblioteca são na forma de plasmídeos ou amplicons de PCR de DNA de dupla fita. Em determinadas modalidades, os plasmídeos da biblioteca compreendem um marcador de gene. Em outra modalidade, os plasmídeos compreendem uma sequência que codifica a enzima transposase que reconhece e é funcional com as sequências de repetição terminal invertida. Em uma modalidade, a biblioteca da presente invenção compreende polinucleotídeos que codificam a a cadeia pesada da imunoglobulina de comprimento total incluindo a estrutura natural de intron/exon de uma cadeia pesada de anticorpo. A cadeia pesada da imunoglobulina de comprimento total pode compreender o domínio de ancoragem da membrana endógena.
[0040] A presente invenção é também direcionada a um método para gerar uma biblioteca de polinucleotídeos transponíveis que codificam polipeptídeos tendo diferentes especificidades ou funcionalidades de ligação, que compreende (i) gerar uma coleção diversa de polinucleotídeos que compreendem as sequências que codificam polipeptídeos tendo diferentes especificidades ou funcionalidades de ligação, em que os referidos polinucleotídeos compreendem a sequência que codifica o polipeptídeo tendo a especificidade ou funcionalidade de ligação disposta entre as sequências de repetição terminal invertida que são reconhecidas por e funcionais com pelo menos uma enzima transponase.
[0041] A presente invenção é também direcionada a um vetor que compreende a sequência que codifica a região VH ou VL de um anticorpo, ou porção de ligação a antígeno do mesmo, disposta entre as sequências de repetição terminal invertida que são reconhecidas por e funcionais com pelo menos uma enzima transponase. Em determinadas modalidades, o vetor codifica a cadeia pesada ou leve de comprimento total de uma imunoglobulina. Preferivelmente, a sequência que codifica a VH ou a VL ou a cadeia pesada ou leve é uma sequência randomizada gerada, por exemplo, por amplificação de PCR sob condições de mutagênese ou síntese de gene. Em uma modalidade, o vetor compreende as sequências de repetição terminal invertida que são reconhecidas por e funcionais com a transponase PiggyBac. Em outra modalidade, as sequências de repetição terminal invertida são reconhecidas por e funcionais com a transponase transposase de Sleeping Beauty. Em uma modalidade, o vetor compreende a sequência de região VH ou VL derivada a partir de um anticorpo anti-TNF alfa tal como, por exemplo, D2E7. Em determinadas modalidades, o vetor compreende pelo menos uma sequência selecionada a partir do grupo que consiste de: SEQ ID NO:5, SEQ ID NO:8, SEQ ID NO:9, SEQ ID NO:10, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:16, SEQ ID NO:17, e SEQ ID NO:19, SEQ ID NO: 20, SEQ ID NO:21, SEQ ID NO:22, SEQ ID NO:23, SEQ ID NO:24, SEQ ID NO:32, SEQ ID NO:33, SEQ ID NO:34, SEQ ID NO:35, SEQ ID NO:36, SEQ ID NO:37, SEQ ID NO:38, SEQ ID NO:39, SEQ ID NO:40, e SEQ ID NO:41.
[0042] A presente invenção é também direcionada a a células hospedeiras que compreende um vetor da presente invenção como descrito acima. Em uma modalidade preferida, as células hospedeiras adicionalmente compreendem um vetor de expressão que compreende a sequência que codifica a transposase que reconhece e é funcional com pelo menos uma sequência de repetição terminal invertida no vetor que codificam a referida sequência da região VH ou VL.
[0043] A presente invenção é ainda adicionalmente direcionada a moléculas de ligação a antígeno, por exemplo, anticorpos, produzidos por um método que compreende a reivindicação 1.
[0044] A presente invenção é também direcionada a um método para gerar a população de células hospedeiras capazes de expressar polipeptídeos tendo diferentes especificidades ou funcionalidades de ligação, que compreende:
[0045] (i) gerar uma coleção diversa de polinucleotídeos que compreendem as sequências que codificam polipeptídeos tendo diferentes especificidades ou funcionalidades de ligação, em que os referidos polinucleotídeos compreendem a sequência que codifica um polipeptídeo tendo a especificidade ou funcionalidade de ligação disposta entre as sequências de repetição terminal invertida que são reconhecidas por e funcionais com pelo menos uma enzima transponase; e
[0046] (ii) introduzir a referida coleção diversa de polinucleotídeos nas células hospedeiras.
[0047] Figura 1: a.) Esse desenho ilustra a configuração de um element transponível autônomo (TE), que pode transpor ou “pular” dentro de qualquer sequência de DNA alvo. Os componentes chave de um TE são uma enzima transposase ativa que reconhece as repetições terminais invertidas (ITRs) que margeiam a enzima transposase em si à montante e à jusante de sua sequência. TEs catalisa ou a cópia ou a excisão do TE, e a integração em sequências de DNA alvo não relacionadas. b.) Esse desenho ilustra a configuração de um sistema de vetor de transposon, no qual a expressão de uma enzima transposase ativa é efetuada por um vetor de expressão que não é acoplado ao TE em si. Em vez disso, o TE pode conter qualquer sequência(s), ou gene(s) de interesse que é/são clonado entre as ITRs à montante e à jusante. A integração do TE contendo qualquer sequência(s), ou gene(s) de interesse (por exemplo, uma biblioteca de DNA que codifica uma biblioteca de proteínas) pode se integrar nas sequências de DNA alvo não relacionadas, se a expressção de enzima transposase for proporcionada em trans, por exemplo, por um construtor de expressão de transponase separado, como ilustrado aqui.
[0048] Figura 2: a) Esse desenho ilustra os dois diferentes modos de como os TEs podem “pular” ou transpor em um DNA alvo não relacionado. Para o grupo II de transposons, a enzima transposase em uma primeira etapa reconhece as ITRs do elemento transponível e catalisa a excisão do TE a partir do DNA. Em uma segunda etapa, o TE excisado é inserido na sequência de DNA alvo não relacionada, que é também catalisada pela enzima transponase. Isso resulta em um mecanismo de transposição de “cortar e colar”. Para o grupo I de transposons (mostrado em b.) a informação de codificação do TE é primeiro replicada (por exemplo, transcrita e transcrita reversa, no caso dos retrotransposons) e o TE replicado então se integra na sequência de DNA alvo não relacionada, que é catalisada pela enzima transponase. Isso resulta em um mecanismo de transposição de “copiar e colar”.
[0049] Figura 3: A referida Figura proporciona uma visão geral de uma enzima transponase ativa que foi identificada e/ou reconstruída a partir de TEs inativos, dormentes, e que foram mostrados ser capazes de conferir transposição em vários vertebrados e também células humanas, como proporcionado na tabela. A tabela foi adaptada a partir da Tabela I da publicação de Ni et al. Briefings Funcional Genomics Proteomics 7, 444-453 (2008) (incorporado aqui a título de referência em sua totalidade).
[0050] Figura 4: Essa Figura esboça o princípio do método descrito aqui, para o isolamento de informação de codificação para proteínas, incluindo os anticorpos e os fragmentos das mesmas, com a desejada função, por exemplo, a ligação a um alvo de interesse, como ilustrado aqui. O(s) gene(s) de interesse, por exemplo, as proteínas que codificam a biblioteca de DNA transponível diversa, incluindo as cadeias de polipeptídeo de anticorpo ou fragmentos do mesmo, que é clonado entre as repetições terminais invertidas (ITRs) de um construtor transponível é introduzido em células hospedeiras de vertebrados jutno com um vetor de expressão para uma enzima transposase ativa (veja a parte superior do desenho). A expressão da enzima de transposon nas referidas células hospedeiras em trans e a presença do(s) gene(s) de interesse clonado(s) entre as ITRs que podem ser reconhecidas pela enzima transposase permite a integração estável do(s) gene(s) flanqueados pela ITR de interesse dentro do geonma das células hospedeiras, que pode então expressar de modo estável a(s) proteína(s) de interesse codificada(s) pelos genes de interesse. A biblioteca celular que expressa a(s) proteína(s) de interesse pode então ser classificada para a desejada funcionalidade das proteínas expressas, por exemplo, mas não limitada à ligação a uma proteína alvo de interesse, como ilustrado aqui. Por meio das técnicas de separação de célula conhecidas na técnica, por exemplo, MACS ou FACS, as células que expressam a(s) proteína(s) de interesse com o desejado fenótipo e que portanto contêm o genótipo correspondente, podem ser isoladas e a informação de codificação para o(s) gene(s) de interesse pode ser recuperada a partir das células isoladas por técnicas de clonagem conhecidas na técnica, por exemplo, mas não limitada a clonagem de PCR genômico, como ilustrado aqui.
[0051] Figuras 5a) e 5b): Esse desenho esboça a estratégia de clonagem para a geração de um vetor de expressão de cadeia leve kappa (LC) de imunoglobulina humana transponível(Ig), como descrito no Exemplo 1. A Figura 5 a.) ilustra a estratégia de clonagem para a inserção das ITRs 5’- e 3’ a partir do transposon PiggyBac no vetor de expressão de mamífero pIRES-EGFP (Invitrogen, Carlsbad, CA, USA), que já contém o forte elemento promotor de célula de mamífero pCMV(IE) (promotor precoce imediato de CMV), e intron/poliA sinaliza para a forte expressão de células hospedeiras de mamífero. Adicionalmente, à jusante de ClaI, EcoRV, NotI, EcoRI contendo múltiplos campos de clonagem, dentro dos quais o(s) gene(s) de interesse podem ser clonados, pIRES-EGFP contém um campo de entrada de ribossmo interno (IRES) com uma ORF à jusante de proteína fluorescente verde aumentada (EGFP), que efetua o acoplamento de expressão de gene(s) de interesse clonado à montante dos IRES. Elementos funcionais bacterianos (gene de resistência a ampicilina, ampR) e uma origem bacteriana de replicação (Col E1) para amplificação e seleção do plasmídeo em E. coli são ilustrados também. As ITRs resultants contendo plasmídeo PiggyBac é designada pIRES-EGFP-T1T2. FIG 5b) então ilustra a inserção de um gene LC kappa de Ig humano sintetizado em um único campo de enzima de restrição EcoRV de pIRES- EGFP-T1-T2, que posiciona o LC kappa de Ig humano à montante do cassette IRES-EGFP, e desse modo acopla a expressão de LC kappa de Ig humano a expressão de marcador de gene EGFP. A inserção de LC kappa de Ig humano resulta em um vetor de expressão de LC kappa de Ig humano transponível pIRES-EGFP-T1T2-IgL. Os desenhos mostram os campos de enzima de restrição únicos selecionados nos plasmídeos, assim como os campos duplicados selecionados que resultam a partir das etapas de clonagem.
[0052] Figura 6: Esse desenho esboça a clonagem de um vetor de expressão de cadeia pesada (HC) de imunoglobulina humana transponível (Ig) gama 1, que pode ser gerado por troca de LC kappa da Estrutura de leitura aberta de Ig humana (ORF) contra a ORF para a Ig humana gama 1 HC ORF. O desenho da Ig gama 1 final HC ORF é similar, também com relação a engenharia de um único Campo de enzima de restrição Eco47III que separa a variável (V) a partir das regiões de codificação constantes (C), que permite a troca de uma única região que codifica o anticorpo V contra as diversas regiões que codificam a biblioteca de anticorpo V, como descrito no Exemplo 3.
[0053] A Figura 7: Esse desenho ilustra a clonagem de um vetor de expressão da enzima transponase PiggyBac de mamífero, como descrito no Exemplo 4, usando pCDNA3.1(+) higro como a estrutura do vetor de expressão de mamífero, dentro dos quais o gene sintetizado ORF a partir de Transposase PiggyBac é clonado no único campo de enzima de restrição EcoRV de pCDNA3.1(+) higro, que resulta no vetor de expressão de transposon PiggyBac pCDNA3.1(+) higro-PB.expressão vetor pCDNA3.1(+) higro-PB. Também nesse desenho a posição relativa de outros elementos funcionais de mamífero (promotor CMV-IE, sinal BGH-poliA, segmento SV40- poliA, higromicinB ORF) e elementos funcionais bacterianos (gene de resistência a ampicilina, ampR, origem de replicação, ColE1), assim como os camposd e restrição de enzima selecionados relevantes são mostrados.
[0054] Figura 8: Esse desenho ilustra a clonagem de um vetor de expressão de imunoglobulina humana transponívelde cadeia leve (Ig-kappa LC) de Sleeping Beauty, como descrito no Exemplo 5. A clonagem pode ser realizada por substituir em sequência as ITRs 5’ e 3’ de PiggyBac com as ITRs 5’ e 3’ de Sleeping Beauty no construtor pIRES-EGFP-T1T2-IgL. Também nesse desenho a posição relativa de outros elementos funcionais de mamífero (Promotor CMV-IE, sinal BGH-poliA, segmento BGH-poliA, higromicinB ORF) e elementos funcionais bacterianos (gene de resistência a ampicilina, ampR, origem de replicação, ColE1), assim como campos de reconhecimento de enzima de restrição relevantes selecioandas são mostrados.
[0055] A Figura 9: Esse desenho ilustra a clonagem de um vetor de expressão de enzima transposase de Sleeping Beauty de mamífero, como descrito no Exemplo 6, usando pCDNA3.1(+) higro como a estrutura do vetor de expressão de mamífero, dentro do qual o gene sintetizado ORF a partir de transposase de Sleeping Beauty é clonado no único campo de enzima de restrição EcoRV de pCDNA3.1(+) higro, que resulta no vetor de expressão de transposon de Sleeping Beauty pCDNA3.1(+) higro-SB. Também nesse desenho a posição relativa de outros elementos funcionais de mamífero (Promotor CMV-IE, sinal BGH-poliA, segmento BGH-poliA, higromicinB ORF) e elementos funcionais bacterianos (gene de resistência a ampicilina, ampR, origem de replicação, ColE1), assim como os campos de reconhecimento de enzima de restrição relevantes selecioandas são mostrados.
[0056] Figura 10: Esse desenho mostra o arranjo dos elementos funcionais e posição dos campos de enzima de restrição única selecionada dentro de the fragmentos de DNA de gene sintetizado 1.) e 2.) que foram utilizados no Exemplo 4, de modo a clonar ambos os vetores de expressão de cadeia IgH “vazios” que permitem a transposição utilizando seja PiggyBac ou transposase de Sleeping Beauty. A origem dos elementos funcionais é descrita em detalhes na descrição do Exemplo.
[0057] A Figura 11: Esse desenho mostra o desenho final e o mapa de plasmídeo dos vetores de expressão transponíveis para ser humano, cadeias pesadas de Ig-gama1 ligado a membrana (esquerda) e cadeias leves de kappa de Ig humana (direita). Para o vetor de expressão IgH, a região de codificação de VH pode ser substituída pelas regiões de codificação de VH de qualquer outro anticorpo monoclonal, ou pela biblioteca de gene VH, usando os únicos campos de enzima de restrição NotI e NheI, que margeiam a região de codificação VH no referido vetor. Paara o vetor de expressão IgL, a região de codificação VL pode ser substituída por Regiões de codificação VL de qualquer outro anticorpo monoclonal, ou pela Biblioteca de gene de VL, usando únicos campos de enzima de restrição NotI e BsiWI, que margeiam a região de codificação VL no referido vetor. As construções de vetor 8 para os vetores IgH e IgL transponíveis para PiggyBac e Sleeping Beauty, descritos em detalhes no Exemplo 4 todos compartilham esse desenho geral. Os dois mapas de vetor exibidos aqui correspondem aos mapas de vetor de pPB-EGFP-HC-Ac10 (esquerda) e pPB-EGFP-LC-Ac10 (direita), e os vetores adicionais para a cadeia pesada de hBU12 (HC) ou cadeia leve (LC), contendo seja a ITRs PiggyBac ou Sleeping Beauty, são proporcionados nas tabelas abaixo. As sequências de todos os vetores nessa Figura são proporcionadas no Exemplo 4.
[0058] Figura 12: Essa Figura mostra dois gráficos de ponto de FACS bidimensional, nos quais a expressão de superfície de IgG humano a partir de vetores de expressão de IgHC e IgLC transfectados e transpostos é detectada na superfície de células proB de murino transformads por 63-12 A-MuLV derivada a partir de camundongos deficientes RAG-2. d2 pós TF quer dizer que as análises de FACS foram realizadas 2 dias após a transfecção de construtores de vetor nas células 63-12. Os gráficos de FACS na coluna da esquerda representam os controles negativos e positivos para a transfecção. NC=mock eletroporação de células sem plasmídeo DNA. pEGFP-N3=controle de transfecção com vetor de controle pEGFP-N3, que controla a eficiência da transfecção por tornar verde as células transfectadas. A segunda coluna a partir da esquerda mostra gráficos de FACS a partir de células 63-12 co-transfectadas seja com o vetor de transponase de PiggyBac, vetores pPB-EGFP-HC-Ac10, pPB-EGFP-LC-Ac10 (fileira de topo), ou vetor de transponase de PiggyBac, vetores pPB-EGFP-HC-hBU12, pPB-EGFP-LC-hBU12 (fileira do meio), ou com vetor de transponase de Sleeping Beauty, vetores pSB-EGFP-HC-Ac10, pSB-EGFP-LC- Ac10 (fileira de fundo). A segunda coluna da esquerda marcada “d2 pós TF” mostra a análise para células que expressam IgG na superfície da célula (Eixo Y) e a expressão de EGFP (Eixo X) dois dias pós a co-transfecção dos vetores como mencionado acima. IgG de superfície e células positivas duplas EGFP foram classificadas por FACS como indicado pelo portão retangular. A segunda coluna da direita marcada “d9 1x classificada” mostra a análise de IgG de superfície e expressão de EGFP na população celular que foi classificada no dia 2 após a transfecção, analisada do mesmo modo. Portões de classificação para a segunda classificação de FACS são também proporcionados como portões retangulares. A coluna mais a direita marcada “d16 2x classificada” mostra a análise de IgG de superfície e a expressão de EGFP das populações celulares que foram reclassificadas no dia 9 após a transfecção, e analisada do mesmo modo para IgG de superfície e para a expressão de EGFP como nos experimentos anteriores.
[0059] Figura 13: Essa Figura ilustra a demonstração de que as células proB que expressam IgG específico de CD30 na superfície de células 63-12 podem especificamente ser coradas e detectadas por antígeno CD30, e que as células específicas de CD30 são detectadas e re-isolada a partir da grande população de células que expressam IgG de superfície da especificidade não relacionada (aqui CD19), na qual as células específicas de CD30 foram enriquecidas com menor frequência. O graficod e pontos de FACS em cima mostra a detecção de IgG (via coloração anti-kappaLC) e ligação a CD30 (via coloração de antígeno CD30) na superfície das células de controle positivo, que são as células 63-12 estavelmente transpostas e 2x classificadas para expressar IgG anti-CD30 humano, clone Ac10 na superfície da célula. Como esperado, uma população relativamente pura (97,3%) de IgG-positivo/CD30-reativo foi detectável no quadrante superior direito do gráfico de pontos de FACs. Os números em cima de cada gráfico de FACs indica o número de células vivas com base em portao FSC/SSC que foram adquiridos em cada experimento. A fileira do meio mostra as análises de FACS para as células IgG-positivo/CD30 reativo detectáveis em um fundo de células específicas de IgG positivo/CD19. O número acima do número de eventos indica o fator de diluição de células positivas deIgG específica de anti-CD30 que foram usadas para a geração da população “enriquecida” de células positivas de IgG anti-CD30 mAb em um fundo de células positivas de IgG anti-CD19 mAb. Os portões de classificação são indicados que foram usados para especificamente isolar as células reativas de IgG- positivo/antígeno CD30 a partir das populações enriquecidas. Números maiores de eventos precisam ser adquiridos de modo a permitir a detecção e o isolamento das células IgG- positivo/CD30 em diluições mais elevadas. A fileria mais baixa dos gráficos de FACS então mostra a reanálise de células classificadas após as células terem sido expandidas por 12 dias pelos mesmos parâmetros (IgG-expressão & CD30 especificidade a antígeno).
[0060] Figura 14: Essa Figura mostra a clonagem de um vetor transponível para uma cadeia pesada de Ig-gama1 humano (HC) na configuração genômica. O fragmento linear em cima representa os exon e introns gama1 humano para Ig-gama1-HC ligado a membrana, com que margeiam os campos de restrição NheI e BstBI adicionados para permitir a ligação em Ig-gama1 HC cDNA vetor pPB-EGFP-HC-Ac10. H-designa a região de dobra de exon, M1 e M2 representam os exons que codificam a região de transmembrana da cadeia pesada de IgG expressa na superfície. Com uma simples ligação uma etapa da região cDNA C-gama1 do vetor de cadeia pesada humana transponível é substituída pela sua contraparte genômica como indicado na Figura. Usando essa estratégia, a região de codificação VH será ligada na estrutura ao exon que codifica a CH1 de C-gama1 humano.
[0061] Figura 15: Essa Figura mostra a sequência e a configuração geral da biblioteca de cadeia leve kappa. A região de codificação de CDR3 está sublinhada. Os campos de restrição úteis são indicados.
[0062] Figura 16: Essa Figura mostra a sequência e a configuração geral da biblioteca de cadeia pesada gama, mostrando como um exemplo a biblioteca de fragmento randomizadas usando a estratégia de randomização de NNK4. A biblioteca de fragmentos randomizados de cadeia pesada gama usando as estratégias de randomização NNK6, NNK8 e NNK10 diferem apenas no número de resíduos de amnio ácidos randomizados na região HCDR3. A região HA de codificação de CDR3 é sublinhada. O códon ARG codifica Lisina e Arginina. Os campos de restrição úteis são indicados.
[0063] Figura 17: Essa Figura mostra a digestão de modelos de PCR antes da amplificação com os primers. (A) A digestão de pUC57_Jkappa2-Ckappa com a endonuclease de restrição ScaI produz um fragmentod e DNA de terminação cega ideal para priming com o primer LCDR3-NNK6-F. (B) A digestão de pUC57_JH4 com a endonuclease de restrição DrdI produz um fragmento de DNA ideal para priming com os primers HCDR3-NNK4-F, HCDR3-NNK6-F, HCDR3- NNK8-F, e HCDR3-NNK10-F
[0064] Figura 18: Essa Figura mostra the eletroferogramas abrangendo as regiões LCDR3 e HCDR3 randomizadas dos amplicons de PCR gerados para diversificar a região LCDR3 pela abordagem de NNK-6 para Vkappa (A), e a região de HCDR3 pela abordagem de NNK4-para VH, como descrito nos Exemplos 12 e 13, respectivamente.
[0065] Como usado aqui, “coleção diversa” quer dizer uma pluralidade de variantes ou mutantes de proteínas de ligação ou funcionais particulares que exibem diferenças nas sequências de codificação de nucleotídeo ou nas sequências de amino ácido primário, que definem diferentes funcionalidades ou propriedades de ligação.
[0066] Como usado aqui, “biblioteca” quer dizer uma pluralidade de polinucleotídeos que codificam polipeptídeos tendo diferentes especificidades e/ou funcionalidades de ligação. Em determinadas modalidades, a biblioteca pode compreender polinucleotídeos que codificam pelo menos 102, pelo menos 103, pelo menos 104, pelo menos 105, pelo menos 106, pelo menos 107, pelo menos 108, ou pelo menos 109 unique polipeptídeos, tal como, por exemplo, cadeias pesadas ou leves de comprimento total de domínios de anticorpo ou VH ou VL.
[0067] Como usado aqui, “sequência de repetição terminal invertida” ou “ITR” quer dizer uma sequência identificada na terminação 5’ ou 3’ dos elementos transponíveis que são reconhecidos por transposases e que mediam a transposição das ITRs incluindo informação de codificação interveniente a partir de uma DNA construtor ou locus para outro DNA construtor ou locus.
[0068] Como usado aqui, “transposase” quer dizer uma enzima que tem a capacidade de reconhecer e de se ligar a ITRs e para mediar a mobilização de um elemento transponível a partir de uma sequência de DNA alvo para outra sequência de DNA alvo.
[0069] Como usado aqui, “molécula de ligação a antígeno” re em seu sentido mais amplo a uma molécula que se liga especificamente a um determinante antigênico. Um exemplo não limitante de uma molécula de ligação a antígeno é um anticorpo ou fragmento do mesmo que retém ligação específica a antígeno. Por “especificamente se liga” se quer dizer que a ligação é seletiva para o antígeno e pode ser discriminada a partir de interações indesejadas ou não específicas.
[0070] Como usado aqui, o termo “anticorpo” pretende incluir moléculas de anticoprpo total, incluindo anticporpos monoclonal, policlonal e multiespecíficas (por exemplo, biespecíficas), assim como os fragmentos de anticorpo tendo uma região Fc e retendo especificidade de ligação, e proteínas de fusão que incluem uma região equivalente à região Fc de uma imunoglobulina e que retém a especificidade de ligação. Também englobados estão os fragmentos de anticorpo que retêm a especificidade de ligação incluindo, mas não limitados a, fragmentos VH, fragmentos VL, fragmentos Fab, fragmentos F(ab’)2, fragmentos scFv, fragmentos Fv, minicorpos, diacorpos, triacorpos, e tetracorpos (vide, por exemplo, Hudson e Souriau, Nature Med. 9: 129-134 (2003)) (incorporado a título de referência em sua totalidade).
[0071] Uma modalidade da presente invenção descrita aqui é um método para a identificação de polipeptídeos de ligação específica ou funcional, incluindo, mas não limitados a cadeias de anticorpo ou fragmentos dos mesmos (figura 4), que compreende:
[0072] clonar as diversas bibliotecas transponíveis de DNA que codificam proteínas, incluindo cadeias de polipeptídeo de anticorpo ou fragmentos dos mesmos, entre as repetições terminais invertidas (ITRs) derivadas a partir de elementos transponíveis e reconhecíveis por e funcionais com pelo menos uma enzima transponase,
[0073] introduzir uma ou mais bibliotecas diversas transponíveis de DNA da etapa (i) nas células hospedeiras de vertebrado por métodos padrão conhecidos na técnica,
[0074] proporcionar a expressão temporária de pelo menos uma enzima transposase funcional nas referidas células hospedeiras de vertebrado em trans, de modo que as referidas uma ou mais bibliotecas diversas transponíveis de DNA são integradas de modo estável nos genomas das células hospedeiras de vertebrado, desse modo proporcionar a população das células hospedeiras de vertebrados que então expressam de modo estável as bibliotecas diversas de proteínas, incluindo cadeias de anticorpo ou fragmentos dos mesmos,
[0075] classificar as referidas diversas bibliotecas de células, que expressam de modo estável as proteínas, incluindo anticorpos ou fragmentos dos mesmos, para um desejado fenótipo de ligação ou funcional por métodos conhecidos na técnica,
[0076] opcionalmente, incluir ciclos de enriquecimento interativo com as células hospedeiras de vertebrado geneticametne modificadas de modoe stável para o desejado fenótipo de ligação ou funcional, e
[0077] isolar os genes correspondentes a partir das células hospedeiras enriquecidas que codificam o desejado fenótipo de ligação ou funcional por métodos de clonagem padrão, conhecidos na técnica, por exemplo, mas não limitados a, PCR (reação de cadeia de polimerase), usando primers específicos para as sequências contidas nos um ou mais construtores de bibliotecas de DNA transpostos.
[0078] A modalidade preferida da etapa (i) é de gerar bibliotecas diversas transponíveis de DNA seja por síntese de gene, ou por reação de cadeia de polimerase (PCR) usando primers adequados para a amplificação de diversas regiões que codificam proteína, e modelos de DNA que compreendem uma diversidade de proteínas de ligação, incluindo anticorpos, ou fragmentos dos mesmos, por métodos conhecidos na técnica.
[0079] Para a geração de diversas bibliotecas de anticorpos, uma coleção diversa de sequências de cadeia pesada e leve de anticorpos pode ser gerada por síntese padrão de gene na qual as sequências que codificam a região V podem ser randomizadas em determinadas posições, por exemplo, mas não limitadas a, qualquer uma ou toda a região de determinação de complementaridades (CDRs) das regiões V de cadeia pesada ou leve de anticorpo. A diversidade pode ser restrita a CDRs individuais das regiões V, ou a uma posição particular ou a diversas posições da estrutura, e/ou a posições particulares em uma ou mais das regiões de CDR. As regiões V com variações projetadas, como descrito acima, podem ser sintetizadas como um fragmento que codifica todas as cadeias pesadas ou leves de anticorpo que são margeadas por repetições terminais invertidas funcionais por pelo menos uma enzima transponase desejada. Preferivelmente, a biblioteca de DNA contendo diversos domínios variáveis que codificam as regiões V para as cadeias pesadas ou leves de anticorpo é gerada, e margeada por campos de clonagem apropriados, incluindo mas não limitados aos campos de reconhecimento de enzima de restrição, que são compatíveis com os campos de clonagem em vetores de expressão de cadeia pesada ou leve de anticorpo. Sistemas de expressão de transposon úteis para o uso nos métodos da presente invenção incluem, por exemplo, o sistema de transposon PiggyBac como descrito, por exemplo, em US Pat. Nos. 6,218,185; 6,551,825; 6,962,810; 7,105,343; e 7,932,088 (o conteúdo total dos quais está aqui incorporado a título de referência em sua totalidade) e o sistema de transposon de Sleeping Beauty como descrito nas patentes norte-americanas Nos. Nos. US 6,489,458; US 7,148,203; US 7,160,682; US 2011 117072; US 2004 077572; e US 2006 252140 (o conteúdo total dos quais está aqui incorporado a título de referência em sua totalidade.)
[0080] Diversas bibliotecas de cadeia leve e pesada de anticorpo podem também ser obtidas a partir da População de célula B isolada a partir das espécies desejadas de vertebrados, preferivelmente seres humanos, e preferivelmente a partir de compartimentos celulares contendo células B, por exemplo, mas não limitado a sangue periférico ou do cordão, e órgãos linfóides tais como medula óssea, baço, amidalas e tecidos de nodos linfáticos. Nesse caso, diversas sequências de região V de anticorpo para as cadeias pesada e leve de anticorpo podem ser isoladas por RT-PCR ou por PCR genômico usando pares de primers de PCR degenerados específicos de cadeuia leve e pesada de anticorpo, que pode amplificar a maioria das famílias da região V por proporcionar primers à montante que se ligam às sequências homólogas à montante de, ou dentro das sequências lideres, à montante de ou dentro das estruturas da região V, e por proporcionar primers à jusante que se ligam em regiões de homologia dentro de ou à jusante do segmento de gene de junção J de regiões de codificação de domínio variável, ou dentro de ou à jusante das regiões de codificação da região constantes de cadeias pesadas ou leves de anticorpo.
[0081] Os conjuntos de primers de PCR utilizados para a amplificação de diversas regiões de codificação variáveis pode ser margeada por campos de clonagem apropriados, por exemplo, mas não limitados aos campos de reconhecimento de enzima de restrição, que são compatíveis com os campos de clonagem em vetores de expressão de cadeia pesada ou leve de anticorpo.
[0082] As bibliotecas transponíveis de DNA da etapa (i) que codificam diversas proteínas, incluindo os anticorpos e os fragmentos de anticorpo dos mesmos, podem ser proporcionadas na forma de bibliotecas de plasmídeos, nas quais as bibliotecas de gene sintetizado ou de PCR amplificado transponíveis de DNA são clonados usando os campos de clonagem apropriados, como mencionado acima. Alternativamente, as bibliotecas transponíveis de DNA que codificam bibliotecas diversas de proteínas de ligação, tal como anticorpos e fragmentos dos mesmos, podem ser proporcionadas em forma de linear, construtores de DNA de dupla fita, diretamente como um resultado da síntese de DNA, ou como um resultado da amplificação de PCR. A última abordagem de proporcionar as bibliotecas transponíveis de DNA como amplicons lineares de PCR de DNA de dupla fita, que não foram clonados em vetores de expressão ou plasmídeos (em comparação a todos os outros sistemas de expressão de célula de vertebrados) tem a vantagem de que a máxima complexidade molecular das bibliotecas transponíveis de DNA é mantida e não comprometida pela limitada eficiência de clonagem ou de ligação em um vetor de expressão. De modo diferente, clonagem por ligação, ou de outro modo, no plasmídeo expressão ou vetores de transporte é um intermediário necessário para todos os outros sistemas de expressão em células de vertebrados com base em plasmídeo ou com base em vetor viral.
[0083] Entretanto, o uso de vetores de expressão de transposom com base em plasmídeo contendo as bibliotecas diversas transponíveis de DNA que codificam diversas proteínas de ligação, incluindo anticorpos e fragmentos de anticorpo dos mesmos, têm a vantagem de que os referidos vetores de expressão pode ser trabalhado por engenharia para conter elementos funcionais adicionais, que permitem a classificação, ou, alternativamente, a seleção de células hospedeiras de vertebrado transpostas de modo estável para a integração estável do vetor de expressão de transposon em células hospedeiras de vertebrado transpostas.
[0084] Isso é alcançado ao se proporcionar uma ligação operável às bibliotecas diversas transponíveis de DNA, isto é clonado nos vetores de expressão de transposon em cis, os cassetes de expressão para o marcador de genes incluindo, mas não limitados a, proteínas de marcação fluorescentes (por exemplo, verde, amarelo, vermelho, ou proteínas fluorescentes azuis, e as versões ampliadas das mesmas, como conhecido na técnica), ou cassetes de expressão para marcadores de superfície celular incluindo, mas não limitados a, marcadores CD, contra os quais anticorpos específicos de diagnóstico ou outras ferramentas de diagnóstico estão disponíveis.
[0085] Alternativamente, os cassetes de expressão para marcadores selecionáveis, que permitem a seleção de células hospedeiras de vertebrado transpostas para resistência a antibiótico, incluindo, mas não limitados a, puromicina, higromicinaB, bleomicina, resistência a neomicina, pode ser proporcionada em ligação operável às bibliotecas diversas transponíveis de DNA, isto é clonado nos vetores de expressão de transposon em cis.
[0086] A ligação operável pode ser alcançada por clonagem dos referidos cassetes de expressão para marcador de genes ou marcadores de resistência a antibiótico, seja à montante ou à jusante das regiões de codificação que compreendem as referidas bibliotecas diversas transponíveis de DNA, mas dentro das repetições terminais invertidas do vetor de transposon.
[0087] Alternativamente, a ligação operável pode ser alcançada por clonagem das regiões de codificação para o referido marcador ou genes de resistência a antibiótico à jusante das regiões de codificação que compreendem as referidas bibliotecas diversas transponíveis de DNA, mas separados por sequências de campo de entrada de ribossmo interno (IRES), que garantem o acoplamento de transcrição da expressão das referidas bibliotecas diversas transponíveis de DNA com o referido marcador ou genes de resistência a antibiótico, e desse modo permitindo a classificação para ou a seleção de células hospedeiras de vertebrado transpostas de modo estável.
[0088] Na etapa (ii) do método descrito aqui, as referidas bibliotecas diversas transponíveis de DNA que codificam as bibliotecas diversas de proteínas, incluindo anticorpos e fragmentos dos mesmos, são introduzidas nas células hospedeiras de vertebrado desejadas por métodos conhecidos na técnica para transferir de modo eficiente DNA através das membranas de células de vertebrados, incluindo, mas não limitados a, transfecção de DNA usando lipossomos, fosfatod e cálcio, DEAE-dextrano, partículas magnéticas de polietilenoimida (PEI), ou por fusão de protoplasto, transfecção mecânica, incluindo física, ou métodos balísticos(pistola de gene), ou por nucleofecção. Qualquer um dos métodos acima mencionados e outros métodos adequados para transferir DNA nas células hospedeiras de vertebrado pode ser usado individualmente, ou em combinação para a etapa (ii) do método descrito aqui.
[0089] No caso de proteínas diméricas, incluindo, mas não limitados a, anticorpos e fragmentos dos mesmos, é uma modalidade útil do método descrito aqui se introduzir bibliotecas diversas transponíveis de DNA e/ou vetores de transposon para cadeias pesadas ou leves de anticorpo contidas em vetores separados transponíveis, que podem independentemente ser introduzidos nas células hospedeiras de vertebrado. Isso ou permite a introdução em sequencia das bibliotecas diversas transponíveis de DNA para cadeias pesadas ou leves de anticorpo nas referidas células, ou a sua introdução simultânea das bibliotecas diversas transponíveis de DNA para as cadeias pesadas ou leves de anticorpo, o que, em qualquer caso, permite o transporte aleatório de qualquer anticorpo de cadeia pesada com qualquer anticorpo de cadeia leve codificado pelas pelo menos duas bibliotecas separadas diversas transponíveis de DNA.
[0090] Outra modalidade útil da modalidade anterior é de utilizar vetores de transposon separados e/ou diversas bibliotecas de DNA transponíveis para cadeias pesada e leve de anticorpo, onde os referidos construtores ou bibliotecas são contidos nos vetores transponíveis reconhecidos por diferentes enzimas transponases (figura 3). Isso permite a independente transposição de construtores de cadeia pesada de anticorpo e leve de anticorpo sem interferência entre as duas diferentes enzimas transponases, na medida em que um vetor transponível é apenas reconhecido e transposto por sua enzima transponase específica. No caso de transposição em sequência de vetores transponíveis ou de bibliotecas de DNA que codificam cadeias pesadas ou leves de anticorpo, a vantagem de utilizar diferentes enzimas transponases com diferentes sequências de ITR é, que com o segundo evento de transposição, o primeiro construtor já transposto de modoe stavel não é mais uma vez mobilizado para transposição adicional.
[0091] Essa modalidade também permite a descoberta de anticorpos pelo método de seleção guiada (Guo-Qiang et al. Methods Mol. Biol. 562, 133-142 (2009)) (incorporado aqui a título de referência em sua totalidade). Seleção guiada pode, por exemplo, ser usada para a conversão de qualquer anticorpo não humumo específico para a desejada especificidade de alvo/epítopo e com a desejada funcionalidade em um anticorpo totalmente humano, onde a mesma especificidade e funcionalidade alvo/epítopo é preservada. O princípio de seleção guiada acarreta na expressão de uma única cadeia de anticorpo (cadeia leve ou pesada) do anticorpo de referência (o “guia”), em combinação com a diversa biblioteca de cadeias de anticorpo complementares (isto é cadeia leve, ou cadeia pesada, respectivamente), e a classificação das referidas combinações de cadeia leve - pesada para o desejado fenótipo de ligação ou funcional. Desse modo, a primeira cadeia de anticorpo, “guia” a seleção de uma ou mais cadeias de anticorpo complementares a partir das diversas bibliotecas para o desejado fenótipo de ligação ou funcional. Uma vez que as uma ou mais novas cadeias de anticorpo complementares são isoladas, as mesmas podem ser clonadas em vetores de expressão e mais uma vez ser usadas para “guiar” a seleção da segunda cadeia de anticorpo complementar a partir de uma diversa de cadeias de anticorpos. O resultado final desse processo de duas eetapas é que ambas as cadeias pesada e leve originais de anticorpo de um anticorpo de referência são substituídas por sequências de cadeia de anticorpo novas e não relacionadas a partir das bibliotecas diversas, mas onde a nova combinação de cadeia leve e cadeia pesada de anticorpo exhibe as mesmas, ou similares propriedades de ligação ou funcionais do anticorpo de referência original. Portanto, o referido método requer a habilidade de independentemente expressar construtores de anticorpo de cadeia leve e cadeia pesada ou bibliotecas nas células hospedeiras de vertebrado, que podem ser alcançadas pela modalidade preferida para proporcionar cassetes de expressão de cadeia leve e cadeia pesada de anticorpo em diferentes sistemas de vetor transponível, reconhecidos por diferentes enzimas de transposons.
[0092] Entretanto, bibliotecas diversas transponíveis de DNA podem também ser construídas em um modo, que as regiões de codificação para proteínas multiméricas, incluindo anticorpos e fragmentos dos mesmos, são contidos no mesmo vetor de transposon, isto é onde a expressão das pelo menos duas diferentes subunidades de uma proteína multimérica, por exemplo, as regiões VH e VL ou cadeias pesadas e leves de comprimento total, são ligadas de modo operacional por clonagem de os respectivos cassetes de expressão ou regiões de codificação no mesmo vetor transponível.
[0093] Células hospedeiras de vertebrado úteis para a introdução de construtores transponíveis e/ou bibliotecas transponíveis de DNA da etapa (ii) são células a partir de espécies de vertebrados que podem ser ou que são imortalizadas e que podem ser cultivadas em meio de cultura celular apropriado e sob condições conhecidas na técnica. As mesmas incluem, mas não são limitados a, células a partir, por exemplo, de sapos, peixe, aves, mas preferivelmente a partir de espécies de mamíferos, incluindo, mas não limitados a, células a partir de roedores, ruminantes, espécies de primatas não humanos e seres humanos, com células a partir de roedor ou de origem humana sendo as preferidas.
[0094] Tipos de células úteis a partir das espécies acima mencioinadas incluem, mas não são limitadas a células da linhagem linfóide, que podem ser cultivadas em suspensão e em altas densidades, com as células derivadas da linhagem B sendo as preferidas, na medida em que as mesmas expressam endogenamente todas as necessárias proteínas, fatores, chaperones, e enzimas pós-translacionais para a ótima expressão de muitas proteínas, em particular de anticorpos, ou proteínas com base em anticorpos. Das células de vertebrado derivadas da linhagem B, são preferidas as que representam os estágios de diferenciação precoce, e são conhecidas como ceclulas B progenitoras (pro) ou precursoras (pre), pelo fatod e que as referidas células pro- ou preB na maioria dos casos não expressam as cadeias de anticorpo endógenas que podem interferir com a expressão de cadeia de anticorpo exógena ou heteróloga que são parte do método descrito aqui.
[0095] Células da linhagem pro- e pre- B úteis a partir de origem de roedor são as do vírus Abelson-Murine Leukemia (A-MuLV) células proB e preB transformadas (Alt et al. Cell 27, 381-390(1981) (incorporado aqui a título de referência em sua totalidade)) que expressam todos os necessários componentes para a expressão do anticorpo e também par a sua adequada deposição em superfície, incluindo os componentes receptores de célula B Ig-alfa (CD79a, ou mb-1), e Ig-beta (CD79b, ou B-29) (Hombach et al. Nature 343, 760-762 (1990)) (incorporado aqui a título de referência em sua totalidade), mas como mencionado acima, a sua maioria falta da expressão de anticorpo endógeno ou de cadeias de imunoglobulina. Aqui, células pro- e preB transformadas A- MuLV são preferidas que são derivadas a partir de mutantes de rato, incluindo, mas não limitadas a, mutantes de rato com defeito no gene-1 de ativação de recombinação (RAG-1), ou gene-2 de ativação de recombinação (RAG-2), ou animais que portam outras mutações em genes necessários para a recombinação V(D)J, por exemplo, XRCC4, DNA-ligase IV, Ku70, ou Ku80, Artemis, proteína quinase dependenten de DNA, subunidade catalítica (DNA-PKcs), e assim são desprovidos da habilidade de normalmente expressar os polipeptídeos endógenos de anticorpo.
[0096] Tipos úteis adicionais de células da linhagem B progenitoras (pro) e precursoras (pre) são as células precoces proB e preB humanas transformadas EBV imunoglobulina-nula (Ig-null) (Kubagawa et al. PNAS 85, 875-879(1988)) (incorporado aqui a título de referência em sua totalidade) que também expressam todos os fatores necessários para a expressão, modificação pós-translacional e expressão de superfície de anticorpos exógenos (incluindo CD79a e CD79b).
[0097] Outras células hospedeiras da linhagem B podem ser usadas, que representem os estágios de diferenciação da célula de plasma da linhagem de célula B, preferivelmente, mas não limitadas a myeloma linhagens célula de mieloma Ig-null, tais como Sp2/0, NSO, X63, Ag8653, e outras células de mieloma e de plasmacitoma, conhecidas na técnica. Opcionalmente, as referidas linhagens de células podem ser transfectadas de modo estável ou geneticamente modificadas de modo estável por outro meio diferente de transfecção, de modo a superexpressar os componentes receptores de célula B Ig-alfa (CD79a, ou mb-1), e Ig-beta (CD79b, ou B-29), em caso de uma ótima deposição de superfície dos anticorpos expressos exogenamente ser desejada.
[0098] Outras, células não linfóides de linhagens de mamífero, incluindo mas não limitados a, células hospedeiras de expressão de anticorpo de padrão industrial, incluindo, mas não limitados a, células CHO, células Per.C6, células BHK e células 293 podem ser usadas como as células hospedeiras para o método descrito aqui, e cada uma das referidas células pode opcionalmente também ser transfectada de modo estável ou geneticamente modificadas de modo estável para superexpressar os Componentes receptores de célula B Ig-alfa (CD79a, ou mb-1), e Ig-beta (CD79b, ou B-29), em caso de uma ótima deposição de superfície dos anticorpos expressos exogenamente ser desejada.
[0099] Essencialmente, quaisquer células hospedeiras de vertebrados, que seja transfectável, pode ser usada para o método descrito aqui, o que representa uma maior vantagem em comparação a quaisquer sistemas de expressão viral, tal como, mas não limitado aos sistemas de expressão do vírus de vaccínia, retroviral, adenoviral, ou vírus sindbis, pelo fatod e que o método descrito aqui não exibe restrição de células hospedeiras em virtude de tropismo de vírus para determinadas espécies ou tipos de célula, e adicionalmentemore pode ser usado com todas as células de vertebrado, incluindo células humanas, no niveld e biossegurança mais baixo, acrescentando à sua utilidade geral.
[00100] A Etapa (iii) do método descrito aqui resulta na modificação genética estável de células hospedeiras de vertebrado desejadas com os construtores transponíveis transfectados da etapa (ii) pela expressão temporária ou transitória da enzima transponase funcional, de modo que uma população estável de células hospedeiras de vertebrado é gerada que expressa bibliotecas diversas de proteínas codificadas pelos referidos construtores.
[00101] Uma modalidade útil da etapa (iii) é de se introduzir de modo transitório nas células hospedeiras, preferivelmente por co-transfecção, como descrito acima, um vetor de expressão de vertebrado que codifica a enzima transposase funcional junto com said a referida pelo menos uma biblioteca de DNA transponível diversa. Deve ser entendido que a co-transfecção transitória ou a co-integração de um vetor de expressão de transponase pode ser realizada ou simultaneamente, ou logo antes ou após a transferência dos construtores transponíveis e/ou das bibliotecas diversas transponíveis de DNA nas células hospedeiras de vertebrado, de modo que a transponase expressa transitoriamente pode usar de modo ótimo os vetores transponíveis transitoriamente introduzidos da etapa (ii) para a integração da biblioteca de DNA transponível no genoma das células hospedeiras de vertebrados.
[00102] Outra modalidade útil da etapa (iii) é de efetuar a integração estável dos vetores transponíveis e/ou bibliotecas transponíveis de DNA introduzidos da etapa (ii) por transitoriamente expressar a enzima transposase funcional por meio de um sistema de expressão induzível conhecido na técnica, que já é integrado de modo estável no genoma das células hospedeiras de vertebrados. A referida expressão induzível e transitória de uma transposase funcional pode ser alcançada, por exemplo, por sistemas promotores induzivela a tetraciclina (tet- on/tet-off) ou induzíveis a tamoxifeno conhecidas na técnica. Nesse caso, apenas os um ou mais de vetor transponível ou de biblioteca de DNA precisa ser introduzido no genoma das células hospedeiras, e a transposição estável dos construtores e a expressão estável das proteínas codificadas pelo um ou mais de vetor transponível ou de biblioteca de DNA são efetuadas pela alteração transitória na expressão da enzima transposase funcional nas células hospedeiras.
[00103] A Etapa (iv) do método descrito aqui efetua o isolamento de células hospedeiras de vertebrado transpostas que expressam proteínas com a desejada funcionalidade ou fenótipo de ligação.
[00104] A modalidade preferida da etapa (iv) é de classificar e de isolar as células hospedeiras transpostas da etapa (iii) que expressam as desejadas proteínas, incluindo anticorpos e fragmentos dos mesmos, com testes de ligação a alvo e por meio de tecncias padrão de separação de célula, tal como classificar células ativadas magnéticas (MACS) ou classificar em alta velocidade as células ativadas por fluorescência (FACS) conhecidas na técnica. Especialmente, em uma primeira etapa de enriquecimento de uma específica população de células hospedeiras de vertebrado transpostas, onde um grande número de células precisa ser processado, é preferido se isolar células específicas de alvo a partir de um grande número de células não específicas por técnicas com base em MACS.
[00105] Particularmente, para ciclos adicionais e iterativos de enriquecimento de célula, enriquecimento de FACS é preferido, na medida em que potencialmente menos números de células precisam ser processados, e pelo fato de que a citometria de fluxo de múltiplos canais permite o simultâneo enriquecimento das funcionalidades, incluindo, mas não limitados a, ligação específica alvo de mais do que uma espécie, ou a específica classificação para epítopos particulares usando anticorpos de competição específicos para epítopo na leitura de FACS.
[00106] Se proteínas, incluindo anticorpos e fragmentos dos mesmos, têm que ser descobertas as quais interagem com as parceiras de ligação solúveis, os referidos parceiros de ligação são preferivelmente marcados com marcações ou tags específicas, tais como mas não limitadas a biotina, myc, ou HA-tags conhecidas na técnica, que podem ser detectadas por reagentes secundários, por exemplo, mas não limitados a, estreptavidina ou anticorpos, que por si só são marcados magneticamente (para enriquecimento de célula com base em MACS) ou com fluorocromos (para enriquecimento de célula com base em FACS), de modo que as técnicas de separação de célula podem ser aplicadas.
[00107] Se proteínas, incluindo anticorpos e fragmentos dos mesmos têm que ser descobertas contra proteínas de ligação a membrana, que não podem ser expressas com facilidade como proteínas solúveis, tal como, por exemplo, mas não limitados a, tetraspaninas, spannsers de 7-transmembrana (tais como os receptores acoplados a proteína G acoplada), ou canais de íon, os referidos podem ser expressos em partículas virais, ou superexpressos em específica linhagens de células, que são então usadas para a marcação ou os métodos de bateamento conhecidos na técnica, que podem enriquecer as células hospedeiras de vertebrado que expressam as proteínas a partir dos construtores transpostos, incluindo anticorpos e fragmentos dos mesmos.
[00108] Em virtude do acoplamento estável de genótipo-fenótipo na população das células hospedeiras de vertebrados transpostas de modo estável, uma modalidade útil da etapa (v) é de repetir os ciclos de enriquecimento de célula para o desejado fenótipo de ligação ou funcional, até que uma população distinta de células é obtida a qual é associada com o desejado fenótipo de ligação ou funcional. Opcionalmente, os clones de células individuais podem ser isolados, por exemplo, mas não limitados a, por classificação de célula única usando tecnologia de citometria de fluxo, ou por limitar a diluição, de modo a recuperar a informação de DNA transposta a partir de clones de células individuais que são acoplados a um fenótipo de ligação ou funcional particular desejado.
[00109] Para a identificação de anticorpos funcionais específicos de alvo é com frequência favorável não só rastrear e selecionar um fenótipo de ligação particular, mas também adicionalmente rastrear por propriedades funcionais adicionais de anticorpos específicos de alvo, em particular os efeitos antagonísticos ou agonísticos em testes biológicos.
[00110] Portanto, é desejável se ser capaz de “alterar” de modo eficiente a expressão do anticorpo ligada a membrana celular para a expressão do anticorpo secretada nas células hospedeiras de vertebrado com rendimentos suficientes, de modo a produzir suficiente quantidade de um clone de anticorpo particular para os testes funcionais.
[00111] Em células de linhagem B naturais a alteração a partir da expressaod e anticorpo ligado a membrana para expressão do anticorpo secretado na membrana ocorre por meio de um mecanismo de splicing alternativo, no qual em células preB e células B um doador de splice alternativo próximo da terminação 3’ do último exon de região constante de cadeia pesada é preferivelmente spliced em um aceptor de splice do exon de ancoragem a membrana à jusante dos exons constantes da região de cadeia pesada. Desse modo, uma cadeia pesada de anticorpo é produzida nas células B com um C-terminal estendido, o domínio abrangendo a membrana, que ancora a cadeia pesada e desse modo todo o anticorpo contendo cadeia leve-pesada na membrana da célula. O domínio que abrange a membrana em C-terminal, também interage não covalentemente com a membrana abrangendo os componentes Ig-alfa (CD79a ou mb-1) e Ig-beta (CD79b ou B29), que resultam em melhor ancoragem na membrana e expressão mais elevada de imunoglobulina de superfície em linhagens de células B.
[00112] Uma vez que a célula B se diferencia adicionalmente para o estágio de célula plasmática, o splicing alternativo não ocorre mais e o doador de splice alternativo próximo da terminação 3’ da última região constante de cadeia pesada não é mais reconhecida ou utilizada, e o modelo de mRNA é terminado à jusante do códon de parada da região constante de cadeia pesada, e a cadeia pesada de um anticorpo secretado é traduzida.
[00113] De modo a explorar esse mecanismo natural de splicing alternativo e “alteração” a partir da expressão de ligado a membrana para a expressão secretada dos anticorpos expressos, é uma modalidade útil do método descrito aqui se construir os vetores transponíveis e bibliotecas diversas de DNA que codificam proteínas, incluindo anticorpos ou fragmentos dos mesmos, de tal modo que a estrutura natural de intron/exon de um anticorpo constante de cadeia pesada, incluindo os exons que codificam os domínios abrangendo a membrana é mantida. A referida modalidade representa uma clara vantagem em relação aos sistemas de expressão retrovirais, na medida em que o genoma do vetor retroviral já é spliced antes do mesmo ser empacotado em uma partícula retroviral e transduzido de modo estável dentro do geonma das células hospedeiras.
[00114] Outros sistemas de vetor viral podem ser restritos no comprimento do insersor de DNA que pode ser incorporado nos vetores, desse modo evitando a clonagem de regiões genômicas maiores dentro dos referidos vetores de expressão e desse modo evitando a exploração da “alteração” natural a partir da expressão ligada a membranapara a expressão secretada do anticorpo por splicing alternativo. Determinados transposons (por exemplo, Tol2, vide a Figura 3), foram caracterizados para serem capazes de transpor de modo eficiente mais do que fragmentos de DNA de 10 kb nas células hospedeiras de vertebrado sem qualquer perda na eficiência da transposição (Kawakami Genoma Biol. 8, Suppl I, S7 (2007)) (incorporado aqui a título de referência em sua totalidade). Portanto, é uma modalidade útil do método descrito aqui se construir vetores de expressão transponíveis que compreendam estruturas genômicas de exon/intron para expressão melhor e adequada e para a regulação natural da alteração a partir de expressão ligada a membrana para a expressão secretada do anticorpo. Os métodos da presente invenção são úteis para transpor os fragmentos de DNA pelo menos 5kb, pelo menos 6kb, pelo menos 7kb, pelo menos 8kb, pelo menos 9 kb, pelo menos 10 kb de tamanho em genomas das células hospedeiras.
[00115] A diferenciação de estágio de diferenciação de linhagem de célula B precoce que favorece a expressão ligada a membrana do anticorpo, para um estágio posterior, o estágio de célula plasmática, que favorece a expressão secretada do anticorpo pode ser induzida por fatores de diferenciação de célula B, tais como, mas não limitados a, engatilhamento de CD40/IL4, ou estimulação por mitógenos, tais como, mas não limitados a, lipopolissacarídeo (LPS), ou outros ativadores policlonais, ativadores de cepa de Staph. aureus Cowan (SAC), e nucleotídeos CpG, ou qualquer combinação dos mesmos.
[00116] Preferivelmente, a referida diferenciação é efetuada em células transformadas, nas quais a proliferação pode ser inibida de modo artificial, de modo que a adequada diferenciação de célula B pode mais uma vez ocorrer, como foi descrito para células preB de murino transformadas por A-MuLV, nas quais a tirosina quinase Abelson é especificamente inibida pelo inibidor de tirosina Gleevec (Muljo et al. Nat. Immunol 4, 31-37 (2003)) (incorporado aqui a título de referência em sua totalidade). Portanto, é uma modalidade preferida se utilizar células preB de murino transformadas a A-MuLV Ig-null para o método, que por tratamento com Gleevec, pode mais uma vez diferenciar em estágios de célula B mais maduras, incluindo células plasmáticas, que então secretam suficientes quantidades de anticorpo secretado para teste funcional adicional com base de splicing alternativo dos construtores da expressão de cadeia pesada genômica. É uma modalidade preferida do método descrito aqui, se aprimorar adicionalmente a referida diferenciação de linhagem de célula B pela superexpressao estável de fatores anti-apoptóticos, conhecidas na técnica, incluindo, mas não limitados a, bcl-2 ou bcl-xL.
[00117] Após a etapa (iv), o enriquecimento de células hospedeiras de vertebrado transpostas como descrito acima foi realizado, opcionalmente, o enriquecimento adicional de células de acordo com os métodos acima mencionados pode ser realizado (etapa (v)), até que população de células, ou células individuais sejam isoladas expressando proteínas, incluindo anticorpos e fragmentos dos mesmos, com as desejadas propriedades de ligação e/ou funcionais.
[00118] A Etapa (vi) do método descrito aqui é então realizada de modo a isolar a informação relevante de codificação contida nas células hospedeiras de vertebrado transpostas, isoladas para a desejada propriedade de ligação e/ou funcional.
[00119] Uma modalidade útil da etapa (vi) para o isolamento clonagem e sequenciamento da informação relevante de codificação para a desejada proteína de ligação ou funcional, incluindo um anticorpo ou fragmento de anticorpo dos mesmos, contido nas células isoladas, é de utilizar amplificação de RT-PCR ou genômica com pares de primers específicos para a informação relevante de codificação compreendida nos DNA construtores de DNA transpostos, e para sequenciar os amplicons genomicos ou RT-PCR seja diretamente, ou após sub-clonar nos vetores de sequenciamento, conhecidas na técnica, por exemplo, mas não limitados nos vetores de clonagem TA- ou Gateway.
[00120] A clonagem e o sequenciamento da informação relevante de codificação para a desejada proteína de ligação ou funcional, incluindo um anticorpo ou fragmento de anticorpo dos mesmos, como descrito no paragrafo anterior por amplificação de RT-PCR ou genômica pode também ser realizada com IgH transponível e vetor de expressão IgL, de modo que a região de codificação de proteína de ligação não pode apenas ser identificada, mas ao mesmo tempo também ser expressa com a transposição estável em células hospedeiras de mamífero como descrito aqui. Para a expressão de anticorpos secretados isso necessitaria apenas do uso de vetores de expressão de cadeia pesada Ig transponível desprovidos de região de codificação abrangendo a transmembrana IgH.
[00121] Outra modalidade útil da etapa (vi) é de submeter as populações celulares enriquecidas das etapas (iv) ou (v), que exibem o desejado fenótipo de ligação ou funcional na próxima geração de sequenciamento (“profundo”) (Reddy et al. Nat. Biotech. 28, 965-969 (2010)) (incorporado aqui a título de referência em sua totalidade), de modo a recuperar diretamente e em uma etapa representativa de diversos milhares de sequências para a informação de codificação contida nos construtores de DNA dos DNAs transpostos. Com base em uma análise de bioinformática da frequência relativa de sequências identificadas a partir das the populações celulares enriquecidas, permite a previsão sobre a qual as sequências codificadas daa proteína de ligação ou funcional, incluindo um anticorpo ou fragmento dos mesmos (Reddy et al. Nat. Biotech. 28, 965-969 (2010)) (incorporado aqui a título de referência em sua totalidade). Estatisticamente as sequências super representadas são então resintetizadas e clonados no vetor de expressão para a expressão como proteínas recombinantes, anticorpos ou fragmentos dos mesmos, de modo a caracterizar as mesmas de modo funcional e por suas propriedades de ligação. O referido método pode acelerar significativamente a identificação de sequências relevantes dentro da população celular funcionalmente e fenotipicamente enriquecida, que expressa as proteínas com propriedades funcionais ou específica de alvo.
[00122] Ainda outra modalidade útil do método descrito aqui é de utilizar a expressão de proteínas de células de vertebrados mediadas a transposição, incluindo anticorpos ou fragmentos dos mesmos, para a mutagênese e otimização das proteínas desejadas, incluindo a otimizacao da afinidade dos anticorpos e fragmentos dos mesmos.
[00123] Isso pode ser alcançado por se isolar os genes que codificam as proteínas, incluindo cadeias de anticorpo ou fragmentos dos mesmos, a partir de populações de células de vertebrados transpostas enriquecidas para o desejado fenótipo de ligação ou funcional de acordo com os métodos descritos na etapa (iv), tal como mas não limitados a, por PCR genômico ou amplificação de RT-PCR sob condições de mutagênese, conhecidos na técnica. A sequência que sofreu mutagêneses pode então ser re-clonada em vetores de transposição e então mais uma vez ser transposta nas células hospedeiras de vertebrado, de modo a submeter as mesmas a classificação de acordo com os métodos descrito aqui, para aprimoradas propriedades de ligação ou funcionais.
[00124] Em uma modalidade útil da referida abordagem, primers específicos são usados os quais permitem a amplificação de PCR sob condições de mutagênese dos construtores transpostos completos, incluindo as ITRs que margeiam.
[00125] Pelo referido método um amplicon de PCR que sofreu mtagênese contendo uma frequência média definida de mutações aleatórias é gerado a partir de células fucionalmente ou fenotipicamente selecionadas transpostas. O referido amplicon de PCR com mutações controladas (variações) dos modelos originais pode agora ser diretamente re-transposto em novas células hospedeiras de vertebrado, de acordo com as modalidades preferidas como descrito nos métodos applicable na etapa (ii).
[00126] A vantagem principal do referido método em relação as outras abordagens de células de vertebrado geneticamente modificadas é, que com essa tecnologia nenhuma reclonagem demorada do amplicon de PCR que sofreu mtagênese e controle de qualidade demorado da sequência que sofreu mutagêneses em vetores de expressão é necessária, o que é uma necessidade mandatória em todos os outros sistemas de expressão com base em plasmídeo ou virais, se a sequência que sofreu mutagênese tiver que ser submetida a outra rodada de classificação.
[00127] Pelo fato da transposição de DNA necessitar apenas da presença de ITRs que margeiam a codificação da região dos genes de interesse, amplicon amplificado por PCR do PCR que sofreu mtagêneses pode ser diretamente reintroduzido e retransposto em novas células hospedeiras de vertebrado para a expressão e a classificação para propriedades aprimoradas e/ou mutantes maduros de afinidade.
[00128] Juntos, os métodos descritos aqui, de utilizar TEs para a modificação genética estável de células hospedeiras de vertebrado com construtores transponíveis e/ou bibliotecas diversas transponíveis de DNA que codificam proteínas, incluindo anticorpos e fragmentos dos mesmos, oferecem uma eficiência sem paralelo, flexibilidade, utilidade e velocidade para a descoberta e a otimizacao das referidas proteínas para os ótimos e desejados fenótipos de ligação ou funcional.
[00129] Exemplo 1: Instrução para a clonagem de vetor de expressão de cadeia leve transponível básica de PiggyBac paara cadeias leves kappa de anticorpo humano compatíveis com a enzima transponase PiggyBac
[00130] Um vetor de expressão transponível básico para cadeias leves kappa humanas pode ser gerado por clonagem das ITRs a partir do transposon PiggyBac à montante e à jusante de um cassete de expressão de cadeia leve kappa de imunoglobulina humana.
[00131] Para isso, como uma primeira etapa, as sequências mínimas para as ITRs à montante e à jusante do transposon PiggyBac podem ser derivadas a partir de pXLBacII (publicada em US 7,105,343) (incorporado aqui a título de referência em sua totalidade) e podem ser gene sintetizados com campos de enzima de restrição de borda para clonar em vetor de expressão de mamífero pIRES-EGFP (PT3157-5, ordem #6064-1, Invitrogen-Life Technologies, Carlsbad, CA, USA)
[00132] A sequência ITR à montante PiggyBac com a repetição no terminal 5’ tem que ser gene sintetizado com sequência de enzima de restrição MunI de borda, compatível com um único Campo de enzima de restrição MunI em pIRES-EGFP, e quatro nucleotídeos aleatórios adicionais (em letras minúsculas) permitem a adequada digestão da enzima de restrição. A referida sequência é proporcionada em Seq-ID1.
[00133] A sequência de ITR à jusante PiggyBac com a repetição em terminal 3’ tem que ser gene sintetizada com sequência de enzima de restrição XhoI de borda compatível com o único campo de enzima de restrição XhoI em pIRES-EGFP, e quatro nucleotídeos aleatórios adicionais permitem a digestão adequada da enzima de restrição. A referida sequência é proporcionada em Seq-ID2. Com a digestão da enzima de restrição MunI do gene sintetizado Seq- ID1, o fragmento de DNA pode ser ligado em MunI linearized pIRES-EGFP, gerando pIRES-EGFP- TR1 de acordo com os métodos padrão, conhecidos na técnica. A orientação adequada do elemento de inserção pode ser verificada pela diagnóstico de digestão da enzima de restrição, e/ou por sequenciamento de DNA do construtor clonado (figura 5a).
[00134] Na próxima etapa o gene sintetizado e o fragmento de DNA digerido por XhoI Seq-ID2, pode ser ligado em XhoI linearizado pIRES-EGFP-T1 (figura 5a) por métodos padrão conhecidos na técnica, de modo a gerar pIRES-EGFP-T1T2, contendo não só ITRs de PiggyBac à montante mas também à jusante do cassete de expressão IRES-EGFP (figura 5b). A orientação adequada do elemento de inserção pode ser verificada por diagnóstico de digestão da enzima de restrição, e/ou por sequenciamento de DNA do construtor clonado (figura 5b).
[00135] Para a clonagem de uma cadeia leve kappa de imunoglobulina humana no vetor pIRES-EGFP-T1T2, a cadeia leve kappa de Ig humana a partir de anticorpo específico anti- TNF-alfa humano D2E7 pode ser sintetizado, que pode ser recuperado a partir do pedido de patente europeia EP 1 285 930 A2 (incorporado aqui a título de referência em sua totalidade).
[00136] A região de codificação para a cadeia leve kappa de Ig humana de anticorpo específico anti-TNF-alfa humano D2E7, na qual a região V de D2E7 é fundida na estrutura à sequência líder Vk1-27 (Entrada no banco de genes: X63398.1, que é o membro da família V- kappa da linha germinal mais próxima V-kappa de D2E7), e para a região constante kappa humana (Entrada no banco de genes: J00241) tem a sequência de nucleotídeo a seguir, que é proporcionada em Seq-ID3.
[00137] A sequência de nucleotídeo de Seq-ID3 se traduz em sequência de amino ácido Seq-ID4. O fragmento de DNA Seq-ID3 que codifica a cadeia leve kappa Ig D2E7 pode ser gene sintetizado e diretamente ligado por ligação de extremdiade cega no único campo de enzima de restrição EcoRV (que é também um cortador cego), por métodos conhecidos na técnica, que resultam no construtor pIRES-EGFP-T1T2-IgL (figura 5b)
[00138] Seq-ID3 foi trabalhado por engenharia para conter o único campo de enzima de restrição Eco47III entre as regiões de codificação V-kappa e C-kappa (realçadas em negrito e sublinhada), que permite a substituição das regiões de V-kappa no referido construtor contra outras regiões de V-kappa ou bibliotecas de V-kappa, usando a única enzima de restrição à montante da região de V-kappa de codificação no construtor, junto com Eco47III. A orientação adequada do elemento de inserção de cadeia leve kappa pode ser verificada por diagnóstico de digestão da enzima de restrição, e/ou por sequenciamento de DNA do construtor clonado (figura 5b).
[00139] Toda a sequência para o vetor de cadeia leve kappa de anticorpo humano transponível pIRES-EGFP-T1T2-IgL é proporcionada em sequência Seq-ID5.
[00140] Seq-ID1 (sequência de 5'-ITR de PiggyBac de 327 bp de comprimento. O campo de enzima de restrição MunIs em cada terminação é sublinhado e escrito em negrito, as adições de nucleotídeo aleatório aos términos são impressas em letra minúscula)
[00141] Seq- ID2 (sequência de 3'-ITR de PiggyBac de 264 bp de comprimento. O campo de enzima de restrição XhoIs em cada terminação é sublinhado e escrito em negrito, as adições de nucleotídeo aleatório nos términos são impressas em letra minúscula)
[00142] Seq-ID3 (região de codificação Ig-kappaLC de 711 bp de comprimento de mAb D2E7 específico para anti-TNF-alfa)
[00143] Seq-ID4 (sequência de mAb D2E7 específico para anti-TNF-alfa de 236 amino ácidos de comprimento)
[00144] Seq-ID5 (sequência de DNA 6436 bp de comprimento do vetor de expressão Ig-kappa-LC transponível para PiggyBac pIRES-EGFP-T1T2-IgL)
[00145] Exemplo 2: Instrução para a clonagem de um vetor de expressão básico de cadeia pesada transponível para PiggyBac para as cadeias pesadas gama1 de anticorpo humumo abrangendo a membrana
[00146] De modo a clonar um vetor de expressão de cadeia pesada Ig transponível, a cadeia leve kappa ORF a partir de pIRES-EGFP- T1T2-IgL (Seq-ID5) precisa ser trocada com um ORF que codifica uma região de codificação de cadeia pesada de IgG1 completamente humana.
[00147] Para a substituição da cadeia leve kappa humana no vetor pIRES-EGFP- T1T2-IgL pela cadeia pesada gama-1 de imunoglobulina humana, a região VH do anticorpo D2E7, que é específica para TNF-alfa humana (vide: EP 1 285 930 A2) (incorporado aqui a título de referência em sua totalidade) pode ser sintetizada. Para isso, uma sequência líder de um membro da família da região VH3 próximo à linha germinal é fundida na estrutura à região VH de anticorpo D2E7, que então é fundida na estrutura à região de codificação de uma região constante gama1 humana (Genbank: J00228) incluindo a membrana abrangendo exons (Genbank: X52847). De modo a ser capaz de substituir a cadeia leve kappa de Ig humana a partir de pIRES-EGFP- T1T2- IgL, campos únicos ClaI e NotI de enzima de restrição precisam estar presentes na terminação 5’ e na terminação 3’ da sequência (sublinhada), respectivamente. Adicionalmente, quatro nucleotídeos que margeiam os campos de enzima de restrição (realçados em letra minúscula nas terminações da sequência) em a adequada digestão da enzima de restrição do fragmento de DNA do gene sintetizado e a ligação na estrutura ClaI-NotI linearizada pIRES-EGFP- T1T2-IgL, de acordo com métodos padrão. A sequência que precisa ser gene sintetizada é proporcionada em Seq-ID6.
[00148] A partir do início do códon na posição 11 de Seq-ID6 (realçada em negrito), a referida sequência de nucleotídeo se traduz na cadeia pesada de IgG1 humano anti-TNF-alfa específica clone D2E7 (vide: EP 1 285 930 A2) (incorporado aqui a título de referência em sua totalidade), mas incluindo os exons de transmembrana gama1 humanos M1 e M2. A tradução da proteína de Seq-ID6 é proporcionada em Seq-ID7. O fragmento de DNA Seq-ID6 que codifica a cadeia pesada D2E7 Ig gama-1 pode então ser duplamente digerida pelas enzimas de restrição ClaI e NotI e direccionalmente ligadas em ClaI e NotI linearized pIRES-EGFP- T1T2-IgL, que resulta no construtor pIRES-EGFP- T1T2-IgH (figura 6)
[00149] Seq-ID6 foi também trabalhada por engenharia genética para conter o único campo de enzima de restrição Eco47III entre as regiões de codificação constantes V-pesada variável e a C-gama1 (realçadas em negrito e sublinhada em Seq-ID7), que permite a substituição de regiões V-pesada no referido construtor contra outras regiões V-pesadas ou bibliotecas V- pesadas, usando a única enzima de restrição à montante da região de codificação V-pesada no construtor, junto com Eco47III. A ligação correta do elemento de inserção pode ser verificada por diagnóstico de digestão da enzima de restrição, e/ou por sequenciamento de DNA do construtor clonado (figura 6).
[00150] Toda a sequência para o vetor de cadeia pesada gama1 de anticorpo humano transponível pIRES-EGFP- T1T2-IgH é proporcionada em sequência Seq-ID8
[00151] Os Exemplos 1 e 2 proporcionam instruçãos 'para a clonagem de vetores de expressão transponíveis básicos de PiggyBac para cadeia leve kappa de anticorpo humano e cadeia pesadas gama-1 de anticoprpo humano (forma ligada a membrana) e portanto para o IgG1 humano ligado a membrana de comprimento total, que pode ser utilizado para a redução tica da presente invenção.
[00152] Seq-ID6 (fragmento DNA 1642 bp de comprimento contendo a região de codificação para Ig-gama1-HC ligado a membrana de mAb D2E7 específico para anti-TNF-alfa)
[00153] Seq-ID7 (sequência de 539 amino ácidos de comprimento de Ig-gama1- HC ligado a membrana de anti-TNFalfa anticorpo
[00154] Seq-ID8 (sequência DNA de 7341 bp de comprimento de vetor de expressão Ig-gama1-membrana-HC humano transponível de PiggyBac pIRES-EGFP-T1T2-IgH)
[00155] Exemplo 3: Instruçãos para a clonagem de vetor de expressão cadeia leve transponível básico para cadeias leves kappa de anticorpo humano compatíveis com a enzima transponase Sleeping Beauty
[00156] De modo a transpor vetores de expressão de cadeia leve e pesada de imunoglobulina humana contida em um vetor transponível independentemente nas células hospedeiras, um construtor de cadeia leve de imunoglobulina transponível com diferentes sequências de repetição terminais invertidas (ITR) pode ser construído que são reconhecidos pela transposase de Sleeping Beauty.
[00157] Para isso, o vetor de expressão de cadeia leve Ig-kappa humano pIRES- EGFP-T1T2-IgL (Seq-ID-5) do exemplo 1 pode ser usado para substituir as ITRs 5’ e 3’ do sistema de transposon PiggyBac, contido no referido vetor, com as ITRs 5’ e 3’ do sistema de transposon de Sleeping Beauty. As sequências das 5’ITR e 3’ITR de Sleeping Beauty, reconhecidas e funcionais com a transposase de Sleeping Beauty, podem ser recuperadas a partir do documento de patente US 7160682B1/US 2003154500A1.
[00158] A sequência ITR à montante de Sleeping beauty com a repetição no terminal 5’ tem que ser gene sintetizado com a sequência de enzima de restrição MunI de bordas, permitindo a substituição de the MunI margeada PiggyBac 5’ ITR no construtor pIRES-EGFP-T1T2- IgL (Seq-ID-5) do exemplo 1 pela sequência 5’ ITR de Sleeping Beauty. A referida sequência é proporcionada como Seq-ID14 abaixo, no final desse Exemplo.
[00159] A sequência ITR à jusante de Sleeping beauty com a repetição na terminação 3’ (também publicada em US 7160682B1/US 2003154500A1) tem que ser gene sintetizado com as sequências de enzima de restrição XhoI de borda, permitindo a substituição de XhoI margeada PiggyBac 3’ ITR no construtor pIRES-EGFP-T1T2-IgL (Seq-ID-5) do exemplo 1 pela sequência 3’ ITR de Sleeping beauty. A referida sequência é como proporcionado em Seq-ID15 abaixo, no final desse Exemplo (campos de enzima de restrição XhoI são realçados em negrito e 4 nucleotídeos adicionais aleatórios de borda, permitindo a adequada digestão do fragmento de enzima de restrição do gene sintetizado, são indicados em letra minúscula):
[00160] Em uma primeira etapa, a 5’ ITR de MunI-margeada de PiggyBac de construtor pIRES-EGFP-T1T2-IgL (Seq-ID-5) tem que ser substituída pela 5’ ITR de Sleeping Beauty por digerir pIRES-EGFP-T1T2-IgL (Seq-ID-5) com a enzima de restrição MunI e por ligar o fragmento gene sintetizado digerido por MunI a partir de Seq-ID-14 na estrutura de vetor linearizada de MunI de pIRES-EGFP-T1T2-IgL (Seq-ID-5). A correta orientação de 5’ ITR de Sleeping Beauty pode ser checada por diagnóstico de digestão da enzima de restriçãos e/ou sequenciamento de DNA. O plasmídeo resultante é chamado de pIRES-EGFP-sbT1-pbT2-IgL (figura 8).
[00161] Em uma segunda etapa, a 3’ ITR de PiggyBac de borda XhoI de constructor ainda contida em pIRES-EGFP-sbT1-pbT2-IgL tem que ser substituída por the 3’ ITR de Sleeping Beauty por digerir pIRES-EGFP-sbT1-pbT2-IgL com a enzima de restrição XhoI e por ligar o fragmento de gene sintetizado digerido XhoI a partir de Seq-ID-15 na estrutura do vetor linearizado XhoI de pIRES-EGFP-sbT1-pbT2-IgL. A orientação correta de 3’ ITR de Sleeping Beauty pode ser checada por diagnóstico de digestão da enzima de restriçãos e/ou sequenciamento de DNA. O plasmídeo resultante é chamado de pIRES-EGFP-sbT1T2-IgL (figura 8).
[00162] Toda a sequência do vetor de expressão Ig-kappa LC pIRES-EGFP- sbT1T2-IgL transponível pela transposase de Sleeping Beauty é proporcionado em Seq-ID-16 abaixo, no final desse Exemplo.
[00163] Seq-ID14 (fragmento de DNA de 246 bp de comprimento contendo a 5' ITR do sistema de transponase de Sleeping Beauty. As bordas do campo de enzima de restrição MunIs são impressas em negrito e sublinhadas)
[00164] Seq-ID15 (fragmento de DNA de 248 bp de comprimento contendo a 3' ITR do sistema de transponase de Sleeping Beauty)
[00165] Seq-ID16 (sequência de DNA de 6339 bp de comprimento do vetor de expressão Ig-kappa-LC transponível de Sleeping Beauty pIRES-EGFP-sbT1T2-IgL)
[00166] Exemplo 4: Clonagem de vetores transponíveis de PiggyBac e Sleeping Beauty para IgG1 humano ligado a membrana
[00167] Além das instruções de clonagem para os vetores de expressão IgH e IgL transponíveis básicos de PiggyBac proporcionadas nos Exemplos 1 e 2, e a construção de um vetor de expressão transponível IgL básico de Sleeping Beauty proporcionada no Exemplo 3, a clonagem adicional dos aprimorados vetores de expressão transponíveis de PiggyBac e Sleeping Beauty IgH e IgL para um anti-humanCD30 mAb quimérico e para um anti-humanCD19 mAb humanizado foi realizada, de modo a reduzir a presente invenção na prática.
[00168] Para isso, em uma primeira etapa, os dois fragmentos de gene a seguir foram sintetizados (comissionado a Genscript, Piscataway, NJ, USA):
[00169] 1.) Um fragmento de DNA de 4975 bp contendo um cassete de expressão, no qual a expressão de uma cadeia pesada de IgG1 ligado a membrana humana é acionado pelo promotor EF1-alfa (pares de base 1-1335 de vetor de expressão Clontech pEF1-alfa-IRES, Cat-No. #631970), e no qual a expressão de cadeias de Ig é ligada a expressão de EGFP via um campo de entrada de ribossmo interno (IRES). As sequências de DN para as regiões de IRES e EGFP foram derivadas a partir de pIRES-EGFP (pares de base 1299-1884 e 1905-2621, respectivamente, do vetor de expressão Clontech pIRES-EGFP (Cat.-No. #6064-1, Life Technologies). Adicionalmente, o fragmento de DNA sintetizado continha um intron quimérico posicionado entre a região de codificação constente de Ig e a sequência IRES, cuja sequência foi derivada a partir do vetor de expressão de mamífero pCI (pares de base 857-989, de Promega, Cat.-No. #E1731). Na terminação 3’ de um cassete de expressão o fragmento sintetizado continha um sinal de poliadenilação de hormônio de crescimento bovino (BGH-poliA), cuja sequência foi derivada a partir de pCDNA3.1-higro(+) vetor de expressão (pares de base 1021-1235 de Invitrogen-Life Technologies, Cat.-No. #V870-20). Um cassete de expressão foi margeado à montante - e à jusante por ITRs de transposon PiggyBac já descrito em Seq-ID1 e Seq-ID2 adicionalmente acima.
[00170] Um mapa dos elementos e os seus arranjos no fragmento de gene de DNA sintetizado é proporcionado na Figura 10, incluindo adicionalmente adicionados únicos campos de enzima de restrição que podem ser usados para cortar ou para substituir qualquer um dos elementos funcionais de um cassete de expressão.
[00171] Uma sequência de fragmento de gene sintetizado de 4975 bp de comprimento é proporcionada como Seq-ID20 abaixo, no final desse Exemplo.
[00172] Deve ser observado aqui que o cassete de expressão do gene sintetizado para as cadeias de IgH humano proporcionadas em Seq-ID20, propositalmente, ainda não continham a região de codificação para um domínio VH, de modo que o construtor pode ser usado para a inserção de qualquer desejada região de codificação VH e/ou biblioteca de gene de codificação de VH usando os únicos campos de enzima de restrição NotI e NheI. O referido construtor portanto é designado cassete de expressão Ig-gama1-HC”vazio”.
[00173] 2.) De modo a proporcionar uma estrutura de plasmídeo para o cassete de expressão transponível de Seq-ID20, um fragmento de DNA de 2774 bp de comprimento foi gene sintetizado (realizado por Genscript, Piscataway, NJ, USA) que continha a ColE1 ori bacteriana e um gene de resistência a ampicilina. A informação de sequência para os referidos componentes de estrutura de plasmídeo foi derivada a partir da estrutura de plasmídeo de vetor de expressão pCI (Promega, Cat.-No. #E1731). O fragmento de gene sintético adicionalmente continha as ITRs 5' e 3' do transposon de Sleeping Beauty, já descrito em Seq-ID14 e Seq-ID15, respectivamente. O referido fragmento precisou ser circularizado e pode ser propagado em E.coli como um plasmídeo autônomo, em virtude da presença de ColE1 ori e do gene de resistência a ampicilina.
[00174] Um mapa dos elementos e do seu arranjo no fragmento de gene de DNA sintetizado é proporcionado na Figura 10, incluindo posição de únicos campos de enzima de restrição adicionados que podem ser usados para cortar ou para substituir qualquer um dos elementos funcionais de um vetor de expressão.
[00175] Uma sequência de fragmento de gene sintetizado de 2774 bp de comprimento é proporcionada como Seq-ID21 abaixo no final desse Exemplo:
[00176] Dois dos referidos fragmentos de gene permitiram a construção de ambos os vetores transponíveis de PiggyBac e Sleeping Beauty por ligaar os fragmentos a partir dos referidos vetores, com a digestão com diferentes enzimas de restrição, seguido pela ligação, como a seguir:
[00177] O vetor transponível PiggyBac foi clonado por ligar EcoRI-ClaI fragmentos a partir de Seq-ID20 e Seq-ID21, de modo que o construtor resultante contém todo o cassete de expressão ITR-margeada de PiggyBac de Seq-ID20, e a estrutura contendo ColE1-amp sem as ITRs de Sleeping Beauty de Sq-ID21. De modo oposto, a ligação de fragmentos XbaI-MluI a partir de Seq-ID20 e Seq-ID21 resulou na ligação de um cassete de expressão sem as ITRs de PiggyBac into em uma estrutura de plasmídeo linearizada de Seq-ID21 still contendo as ITRs de Sleeping Beauty. Plasmídeos miniprep que resultam a partir das duas ligaçãos foram analisados por diagnóstico de digestão de enzima de restrição usando uma mistura de XhoI-NheI-BamHI e adicionalmente com enzimas de restrição PvuI, de modo a identificar os plasmídeos corretamente ligados. Um clone de DNA selecionado de cada ligação foi retransformado em E.coli para gerar um maxiprep DNA, que foi verificado por sequenciamento de DNA usando primers de sequenciamento permitindo o sequenciamento de toda a sequência de plasmídeo.
[00178] Todas os vetores transponíveis das sequências de PiggyBac e Sleeping beauty (contendo o cassete de expressão gama1-HC humano “vasio”) gerados como descrito acima e verificados por sequenciamento de DNA são proporcionados como Seq-ID22 (vetor transponível de PiggyBac) e Seq-ID23 (vetor transponível de Sleeping Beauty) abaixo, no final desse Exemplo.
[00179] As regiões de codificação VL e VH do anticorpo CD30 anti-humano quimerico brentuximab (clone Ac10) podem ser recuperadas a partir das sequências 1 e 9 do pedido de patente US 2008213289A1, e são proporcionadas abaixo como Seq-ID24 e Seq-ID25, respectivamente.
[00180] As regiões de codificação VL e VH do anticorpo anti-humano CD19 humanizado hBU12 foram recuperadas a partir do documento de patente US 8,242,252 B2 como as sequências variantes HF e LG, respectivamente, e são proporcionadas em Seq-ID26 e Seq-ID27 adicionalmente abaixo, no final do Exemplo.
[00181] De modo a permitir a construção de final vetores de expressão anti-CD30 e anti-CD19 IgHC transponível de PiggyBac e Sleeping Beauty, os fragmentos de DNA para os domínios VH foram designados ter campos de enzima de restrição NheI e NotI de borda. A sequência de nucleotídeo que codifica a VH do anticorpo anti-CD30 brentuximab (clone Ac10) foi adicionalmente modificado para também conter a sequência líder para a expressão em célula de mamífero. As sequências de DNA dos fragmentos NotI-NheI que codificam a VH de anti-CD30 e anti-CD19 mAbs são proporcionados em Seq-ID28 e Seq-ID29 no final desse Exemplo. Os fragmentos de DNA foram gene sintetizados por GeneArt, Regensburg, Germany (os campos NotI e NheI são sublinhados).
[00182] De modo a gerar vetores de expressão de cadeia anti-CD30 e anti-CD19 IgH que são transponíveis ou com transposase de Sleeping Beauty ou de PiggyBac, os fragmentos digeridos NotI-NheI digeridos de Seq-ID28 Seq-ID29 foram ligados nos vetores linearizados NotI- NheI descritos em Seq-ID22 ou SeqID23, respectivamente. Isso resultrou na geração de quatro vetores contendo uma cadeia pesada (HC) completamente funcional de anti-CD30 mAb brentuximab (clone Ac10) e de anti-CD19 mAb hBU12 e os construtores foram designados: pPB- EGFP-HC-Ac10, pPB-EGFP-HC-hBU12, pSB-EGFP-HC-Ac10, e pSB-EGFP-HC-hBU12 e os seus mapas de vetor são proporcionados na Figura 11. Os referidos vetores foram especificamente projetados para permitir a expressão de superfície das cadeias pesadas, e, com a coexpressão de cadeias leves, expressão de IgG de superfície. Entretanto, a simples omissão da região de codificação da membrana abrangendo a região de teh Ig cadeia pesadas resulta em vetores de expressão transponíveis para IgG secreted IgG.
[00183] De modo a gerar vetores de expressão de cadeia IgL anti-CD30 e anti- CD19 que são transponíveis seja com a transponase de PiggyBac ou transposase de Sleeping Beauty, os genes de região constante de IgH a partir dos vetores descritos em Seq-ID22 e Seq- ID23 precisam ser substituídos com regiões de codificação de cadeia IgL de anticorpos anti-CD30 e anti-CD19. Isso foi alcançado por fragmentos de gene de síntese de gene contendo as regiões de codificação VL como descrito em Seq-ID25 e Seq-ID27 fundidas na estrutura para uma região de codificação de cadeia leve kappa constante humana, com a sequência líder na terminação 5’ e margeada por campos de clonagem NotI-BstBI que permite a ligação do fragmento digerido NotI- BstBI nos vetores linearizados NotI-BstBI descritos em Seq-ID22 e Seq-ID23, desse modo substituindo a região de codificação IgH constante de Seq-ID22 e Seq-ID23 com as regiões de codificação IgL de anti-CD30 mAb Ac10 e anti-CD19 mAb hBU12.
[00184] Os fragmentos de gene contendo as regiões de codificação IgL de anti- CD30 mAb Ac10 e anti-CD19 mAb hBU12, com a sequência líder e margeada pelos campos de clonagem NotI-BstBI são descritos em Seq-ID30 e Seq-ID31 abaixo, no final do Exemplo. A síntese de gene dos referidos fragmentos de DNA foi realizada por Genscript (Piscataway, NJ, USA).
[00185] De modo um gerar vetores de expressão de cadeia de IgL de anti-CD30 e anti-CD19 que são transponíveis seja com a transponase de PiggyBac ou com a transposase de Sleeping Beauty, fragmentos digeridos de NotI-BstBI de Seq-ID30 e Seq-ID31 foram ligados nos vetores linearizados NotI-BstBI descritos em Seq-ID22 ou SeqID23. Os quatro vetores resultantes foram chamados de: pPB-EGFP-LC-Ac10, pPB-EGFP-LC-hBU12, pSB-EGFP-LC-Ac10, e pSB- EGFP-LC-hBU12 e os seus mapas de vetor são proporcionados na Figura 11.
[00186] As sequências compleltas dos construtores IgH e IgL de PiggyBac e de Sleeping beauty anti-CD30 e anti-CD19 (oito combinações) são proporcionadas em Seq-ID-32 (pPB-EGFP-HC-Ac10), Seq-ID-33 (pPB-EGFP-HC-hBU12), Seq-ID-34 (pSB-EGFP-HC-Ac10), Seq-ID-35 (pSB-EGFP-HC-hBU12), e em Seq-ID-36 (pPB-EGFP-LC-Ac10), Seq-ID-37 (pPB- EGFP-LC-hBU12), Seq-ID-38 pSB-EGFP-LC-Ac10), e Seq-ID-39 (pSB-EGFP-LC-hBU12) abaixo, no final desse Exemplo
[00187] Seq-ID20 (sequência de DNA de 4975 bp de comprimento contendo um cassete de expressão para PiggyBac ITR-margeada para membrana abrangendo as cadeias pesadas de Ig-gama1 humano)
[00188] Seq-ID21 (sequência de DNA de 2774 bp de comprimento contendo componentes da estrutura de vetor ColE1 e resistência a ampicilina margeada por ITRs 5' e 3' de Sleeping Beauty)
[00189] Seq-ID22 (sequência de PiggyBac transponível de 7242 bp de comprimento de vetor “vazio” de gama1-HC humano)
[00190] Seq-ID23 (sequência de Sleeping Beauty de 7146 bp de comprimento transponível “vazia” de vetor gama1-HC humano)
[00191] SeqID-24 (VH região de codificação de 351 bp de comprimento de anticorpo anti-humano CD30 brentuximab)
[00192] Seq-ID25 (região de codificação VL de 333 bp de comprimento de anticorpo anti-humano CD30 brentuximab)
[00193] Seq-ID26 (VH região de codificação de 417 bp de comprimento de mAb huB12 anti-humano CD19, incluindo o líder)
[00194] Seq-ID27 (região de codificação VL de 375 bp de comprimento de mAb huB12 anti-humano CD19, incluindo o líder)
[00195] Seq-ID28 (Fagmento de DNA de 423 bp de comprimento, contendo a região de codificação de VH NotI-NheI-margeada do domínio VH de anti-humano CD30 mAb brentuximab)
[00196] Seq-ID29 (Fagmento de DNA de432 bp de comprimento, contendo a região de codificação de VH NotI-NheI-margeada do domínio VH de mAb hBU12anti-humano CD19)
[00197] Seq-ID30 (fragmento de DNA de 733 bp de comprimento contendo região de codificação IgL de anti-CD30 mAb Ac10 e margeada por campos de enzima de restrição NotI e BstBI)
[00198] Seq-ID31 (fragmento de DNA de 718 bp de comprimento contendo a região de codificação IgL de anti-CD19 mAb hBU12 e margeada por campos de enzima de restrição NotI e BstBI)
[00199] Seq-ID32 (7645 bp de sequência de pPB-EGFP-HC-Ac10)
[00200] Seq-ID33 (7654 bp de sequência de pPB-EGFP-HC-hBU12)
[00201] Seq-ID34 (7549 bp de sequência de pSB-EGFP-HC-Ac10)
[00202] Seq-ID35 (7558 bp de sequência de pSB-EGFP-HC-hBU12)
[00203] Seq-ID36 (6742 bp de sequência de pPB-EGFP-LC-Ac10)
[00204] Seq-ID37 (6727 bp de sequência de pPB-EGFP-LC-hBU12)
[00205] Seq-ID38 (6646 bp de comprimento sequência de pSB-EGFP-LC-Ac10)
[00206] Seq-ID39 (6631 bp de sequência de pSB-EGFP-LC-hBU12)
[00207] Exemplo 5: Instruçãos para a clonagem de um vetor de expressão de Transposase de PiggyBac
[00208] A ORF de enzima transposase PiggyBac funcional pode ser recuperada a partir da Patente US No. 7,105,343 B1(incorporada aqui por referência em sua totalidade) e é proporcionada em Seq-ID11 abaixo, no final desse Exemplo. A sequência de DNA de Seq-ID11 se traduz no amino ácido Seq-ID12 também proporcionada no final desse Exemplo.
[00209] De modo um gerar um vetor de expressão para células vertebrados para a enzima transponase PiggyBac, o referido ORF pode ser gene sintetizado e clonado como um DNA de terminação cega no único, campo de enzima de restrição de corte cega EcoRV no vetor de expressão padrão de células de vertebrados pCDNA3.1-higro(+) (catálogo # V870-20, Invitrogen, Carlsbad, CA, USA), por métodos conhecidos na técnica. A correta ligação de PiggyBac ORF, com relação ao promotor pCDNA3 pode ser verificada por diagnóstico de digestão da enzima de restrição, e/ou por sequenciamento de DNA do construtor de expressão PiggyBac clonado pCDNA3.1-higro(+)-PB (figura 7). A construção de um vetor de expressão PiggyBac foi realizada como descrito aqui e o vetor design foi verificado por diagnóstico de digestão da enzima de restrição, e sequenciamento de DNA.
[00210] A sequência do construtor de expressão de PiggyBac pCDNA3.1-higro(+)- PB é proporcionada como Seq-ID 13, abaixo no final desse Exemplo.
[00211] Seq-ID11 (ORF de Transposase de PiggyBac)
[00212] Seq-ID12 (sequência de amino ácido de Transposase PiggyBac)
[00213] Seq-ID13 (vetor de expressão pCDNA3.1-higro(+)-PiggyBac)
[00214] Exemplo 6: Instruçãos para a clonagem de um vetor de expressão de transposase de Sleeping Beauty
[00215] A estrutura de leitura aberta (ORF) da enzima transponase de Sleeping Beauty pode ser encontrada na referência de patente US 7160682B1/US 2003154500A1. A sequência é proporcionada em Seq-ID17, abaixo no final desse Exemplo. A referida sequência de DNA de Seq-ID17 se traduz em uma sequência de amino ácido de Seq-ID18, também proporcionada no final desse Exemplo, adicionalmente abaixo.
[00216] De modo a gerar um vetor de expressão de célula de vertebrado par a enzima transponase de Sleeping Beauty, a referida ORF pode ser gene sintetizado e clonado como um DNA de terminação cega em um único campo de enzima de restrição de corte cega EcoRV no vetor de expressão de célula de vertebrado padrão pCDNA3.1-higro(+) (catálogo # V870-20, Invitrogen, Carlsbad, CA, USA), por métodos conhecidos na técnica. A correta ligação da ORF de Sleeping Beauty, com relação ao promotor pCDNA3 pode ser verificada por diagnóstico de digestão da enzima de restrição, e/ou por sequenciamento de DNA do construtor de expressão clonado de Sleeping Beauty pCDNA3.1-higro(+)-SB (figura 9).
[00217] A sequência do construtor de expressão de Sleeping Beauty pCDNA3.1- higro(+)-SB é proporcionada em Seq-ID19, abaixo, no final desse Exemplo.
[00218] A construção de um vetor de expressão de Sleeping Beauty foi realizada como descrito aqui e o desenho do vetor foi verificado por diagnóstico de digestão da enzima de restrição, e sequenciamento de DNA. As regiões de codificação para enzimas transponases de PiggyBac e Sleeping Beauty foram gene sintetizadas por Genscript, Piscataway, NJ. com os oito diferentes vetores de expressão transponíveis IgH e IgL para a transponase de PiggyBac e transposase de Sleeping Beauty, e os vetores de expressão pCDNA3.1-higro(+)paara as enzimas transponases de PiggyBac e Sleeping Beauty, todos os vetores foram gerados que permitiram a expressão de anticorpos anti-CD30 e anti-CD19 na superfície da célula de células de mamífero.
[00219] Seq-ID17 (ORF de enzima transponase Sleeping Beauty)
[00220] Seq-ID18 (sequência de amino ácido de transposase de Sleeping Beauty)
[00221] Seq-ID19 (sequência de DNA de vetor de expressão de Sleeping Beauty pCDNA3.1-higro(+)-SB)
[00222] Exemplo 7: Geração de células preB de murino que expressam de modo estávela o IgG humano ligado a membrana a partir de vetores de expressão estavelmente transpostos
[00223] De modo a demonstrar a expressão estável dos anticorpos de IgG humano em células de mamífero, construtores expressão de IgH e IgL humano transponíveis foram transfectados em linhagem de célula proB 63-12 transformadas por vírus de leucemia de murinho Abelson (A-MuLV), originalmetne derivadas a partir de camundongos com deficiência de RAG-2 (Shinkai et al. (1992) Cell 68, 855-867) e portanto incapaz de iniciar a recombinação V(D)J. a referida linhagem de células hospedeiras representa o tipo de célula de linfócito de linhagem de célula B que expressa todos os componentes celulares para a ótima expressão ligada a membrana do anticorpo, incluindo os cofatores deceptores de célula B Ig-alfa (CD79a ou mb-1) e Ig-beta (CD79b ou B29) que interagem com a amino ácidos abrangendo a transmembrana de imunoglobulina ligada a membrana. Portanto, as referidas células ancoram de modo ótimo as moléculas de IgG com a região abrangendo a transmembrana em uma superfície da membrana da célula. As células 63-12 foram desenvolvidas em um meio de cultura estática em suspensão usando meio IMDM suplementado com 2 % de FCS, 0,03% de PrimatoneTM RL-UF (Sheffield Bioscience), 2 mM de L-glutamina, 50 μM de 2-mercaptoetanol, a 37°C em uma incubadora umdificada e uma atmosfera com 10% de CO2. Para a cotransfecção dos vetores de expressão IgH e IgL transponíveis (Exemplo 4) com um vetor de expressão de transposon (Exemplos 5 ou 6), as células foram passadas 24 horas antes de transfecção e semeadas em uma densidade de 5x105 células/ml, de modo a allow as células to enter into log-phase growth until the time-point de transfecção.
[00224] No dia da transfecção, células 63-12 foram harvested por centrifugação e resuspended em RPMI 1640 medium sem qualquer supplements ou serum at a densidade de 5x106 células/mL. 400 μL da referida suspensão de célula (que corresponde a 2x106 células) foram transferidos em 0,4 cm de cubetas de eletroporação (BioRad ordem #165-2081) e misturadas com 400 μL de meio RPMI 1640 contendo o DNA de plasmídeo desejado (ou uma mistura de plasmídeos). Células foram então transfectadas usando a BioRad Gene Pulser II em ajustes de 950μF/300V e incubadas por 5 min a temepratura ambiente após um único pulso de eletroporação. Após isso, as células foram transferidas ára 5 mL de meio de desenvolvimento com base em IMDM e as células foram centrifugadas uma vez, de modo a remover os resíduos de célula e DNA a partir da eletroporação, antes das células serem transferidas em um meio de desenvolvimento com base em IMDM para a recuperação e a expressão de proteínas a partir de plasmídeos transfectados.
[00225] Os ajustes de eletroporação foram determinados como as condições ótimas de transfecção para a linha celular 63-12 de célula proB transformadas por A-MuLV que rotineiramente resultou em eficiência de transfecção transitória que varia entre 30-40%. O resultado da referida transfecção por eletroporação é documentado nas análises de FACS ilustradas na Figura 12, onde os controles de transfecção, dois dias após a transfecção, são ilustrados nos painéis da coluna da esquerda. O controle negativo (marcado NC), que foi mock-eletroporado sem DNA, como esperado, não mostra quaisquer cellas fluorescentes verdes, enquanto que o controle de transfecção que foi transfectado com 15 μg de pEGFP-N3 plasmídeo (Clontech, ordem #6080- 1), mostrou que 38,8% das células foram transitoriamente transfectadas, como detectada por células que expressam a proteína fluorescente verde aumentada (vide célula no quadrante inferior direito). Como esperado, os controles de transfecção não mostram qualquer sinal de Ig-kappa, pelo fato de que nenhum dos controles de transfecção foi transfectado com um construtor de expressão de Ig.
[00226] Para a transposição dos vetores de expressão IgH e IgL, células 63-12 foram também transfectadas por eletroporação com uma mistura de 5 μg cada de um vetor de expressão IgH transponível, 5 μg de um vetor de expressão transponível IgL, e 5 μg de um vetor de expressão permitindo a expressão de uma transposase de mediação da transposição de the IgH e vetor de expressão IgL. O resultado dessa transfecção é também mostrado na Figura 12.
[00227] A expressão de IgG humano na superfície das células foi detectada pela biotinylated anti-cadeia leve kappa humana anticorpo específico de cadeia leve kappa anti-humana biotinilado (Affymetrix, ebioscience, ordem #13-9970-82) detectada com estreptavidina- aloficocianina (strep-APC) (Affymetrix, ebioscience, ordem #17-4317-82), e é mostrado no Eixo Y dos gráficos de pontos de FACS. Como pode ser visto na Figura 10, as medições ilustradas na segunda coluna a partir da esquerda mostram a análise de células transfectadas com vetores de expressão de transposase IgH+IgL+ dois dias após a eletroporação (marcada “d2 pós TF”). Como as análises de FACS após dois dias de transfecção mostraram que entre 1,8% e 2,8% das células expressam IgH+IgL humana na superfície da célula, pelo fato de que a expressão de IgL na superfície da célula pode apenas ser detectada, se as cadeias de IgH são co-expressas nas células, de modo que um IgG completo pode ser expresso na superfície das células.
[00228] A partir desses dados pode ser inferido que se ca. 38% das células são transitoriamente transfectadas, ca. 5-7,5% das referidas células foram co-transfectadas com ambos os vetores de expressão IgH e IgL. A partir desse experimento é concluído que os construtores de expressão IgH e IgL transponíveis permitem um alto nível de expressão de IgG humano na superfície de células transformadas proB de murino A-MuLV, que é comparável aos sinais de IgG de superfície obtidos por coloração de linfócitos B humanos periféricos com os mesmos reagentes de coloração de anticorpo (dados não mostrados).
[00229] Como esperado as células mostrando a expressão de IgG também exibiu a expressão de EGFP, pelo fato de que a expressão de EGFP foi transcripcionalmente acoplada à expressão de IgH ou IgL por meio das sequências IRES. Entretanto, a expressão de EGFP foi significantemente mais baixa, em comparação à expressão de EGFP a partir do plasmídeo de controle pEGFP-N3, que é esperado, como a expressão de EGFP em pEGFP-N3 é diretamente acionado pelo forte promotor constitutivo, enquanto que a expressão de EGFP nos vetores de expressão IgH e IgL transponíveisé efetuada pelo acoplamento de transcrição à região de codificação de IgH e IgL usando um campo de entrada de ribossmo interno (IRES). No entanto, como esperado, células que exibem maior expressão de IgG também exibem maior sinal de EGFP (o que leva a uma população de Ig-kappa+/EGFP+ relativamente diagonal), o que demonstra claramente, que ambos os níveis de expressão são acoplados.
[00230] Quando as células foram analisadas sem rastreamento celular após uma semana de transfecção, o sinal de EGFP nas transfecções de controle pEGFP-N3 não foi mais detectável (dados não mostrados), mostrando que as células não integraram de modoe stavel os construtores de expressão em qualquer frequência significante. De modo diferente, uma baixa ca. 1-2% de população de células de IgG-EGFP duplo positivo foi mantida em células que foram co- transfectadas com vetores IgH&IgL transponíveis junto com um vetor de expressão de transponase, já indicando que a ca. 3-6% de células transfectadas transitoriamente por célula integram de modo estável simultaneamente os vetores de expressão IgH e IgL transponíveis em seu genoma (dados não mostrados).
[00231] De modo a enriquecer as referidas células transpostas de modo estável, cadeia leve Ig-kappa e células positivas duplas EGFP foram classificadas por FACS no dia 2 após a transfecção, como indicado pelos portões de classificação (retângulos pretos na segunda coluna da esquerda gráfico de pontos de FACS). Cada 5’000 células que falham nesse portao foram classificadas a partir das transposições PiggyBac com IgH&IgL de anti-CD30 mAb Ac10 (fileira de topo), e de anti-CD19 mAb hBU12 (fileira do meio), e 3'000 células foram classificadas a partir da transposição de Sleeping Beauty com IgH&IgL de anti-CD30 mAb Ac10 (fileira de fundo), como indicado na Figura 12.
[00232] As células classificadas por FACS foram expandidas por uma semana (representando o dia 9 após a transfecção), e foram reanalisadas de novo por expressão de IgG de superfície por detecção de IgG com um anticorpo cadeia leve anti-kappa, como descrito acima. Como pode ser visto na Figura 10 (a segunda coluna a partir da direita), acima de 30%, 50% e 5% das células classificadas uma vez expressam de modo estáveled IgG na superfície da célula, enquanto as referidas células, como esperado, foram também EGFP-positivo. Isso demonstra que um percentual significante de co-células IgH & IgL transitoriamente transfectadas de modo estável mantêm a expressão de IgG. As transposições mediadas por PiggyBac nesse cenario experimental parece ter ocorrido cerca de 6-10-vezes mais eficiência do que as transposições mediadas por Sleeping Beauty.
[00233] Um par de conclusões adicionais pode ser traçado a partir do referido experimento de transposição: primeiro, a partir das células IgG/EGFP classificadas como duplo positivo no dia dois após a transfecção, cerca de 75% das células permaneceram estáveis EGFP+ nas transposições de PiggyBac, as ca. 40% e 30% da transposição PB-Ac10 e PB-hBU12 gera células EGFP+ desprovidas de expressão IgG de superfície. As referidas células mais provavelmente transpuseram de modo estável apenas um dos dois vetores de expressão IgH e IgL transponíveis, que não permite a expressão de IgG de superfície, mas suficiente para tornar as células EGFP-positivo. Isso também quer dizer que a partir de ca. 38% das células transitoriamente transfectadas originalmente, pelo menos 5% são transpostas de modo estável com pelo menos um vetor de expressão Ig transponível.
[00234] Os números de células transpostas de modo estável para a transposição de Sleeping Beauty foram mais baixos, do que os da transposição de PiggyBac, e após uma primeira rodada de FACS classificando as co-células transfectadas IgH+IgL+transposase, apenas cerca de 5% de células que expressam IgG de modo estável foi obtido. Entretanto, se também as células EGFP positivo estáveis são consideradas, cerca de 9% de células transpostas de modo estável foram obtidas após o primeiro ciclo de classificação por FACS usando transposase de Sleeping Beauty.
[00235] Quando as referidas células das células transpostas de modo estável de IgG-positivo e IgH/IgL foram classificadas por FACS de novo, cerca de 99% de células que expressam IgG de modo estável foram obtidas (figura 12, coluna mais a direita), e a expressão estável do fenótipo foi mantido por cerca de quatro semanas, sem qualquer mudança no percentual de células IgG+ (dados não mostrados). Portanto, é concluído que os vetores de expressão transponíveis para as cadeias de IgH e IgL, como descrito na presente invenção, são funcionais e podem ser integradas de modoe stavel em um genoma de mamífero das células hospedeiras com alta eficiência.
[00236] Exemplo 8: Enriquecimento de células que expressam IgG e IgG transpostos de modo estável por meio de ligação a antígeno específica
[00237] De modo a demonstrar que as células proB que expressam IgG humano, geradas por transposição dos vetores de expressão IgH e IgL, como descrito no Exemplo 7 acima, pode ser usado para o isolamento de células específicas de antígeno, reduzindo os números de linhagem de célula proB 63-12, que expressam anti-CD30 mAb (vide d16, 2x classificada, 63- 12+PB-Ac10, da Figura 12) foram misturadas com linhagem de célula proB 63-12 que expressa anti-CD19 mAb (vide d16, 2x classificada, 63-12+PB-hBU12, de A Figura 12) at ratios 10-2, 10-3, 10-4, 10-5, 10-6 (vide A Figura 13). Um total de 107 células foram coradas em 1 mL de PBS, suplementado com 2% de FCS, por 30 min em gelo, com os reagentes a seguir:
[00238] 0,1 μg de 6xHis-tagged, CD30 humana recombinante (Sino Biological Inc., Bejing, China, ordem #10777-H08H), e
[00239] 10 μL de anticorpo Ig-kappaLC-APC anti-humano de rato marcado (Life Tecnologias, Invitrogen, ordem #MH10515).
[00240] Após os referidos reagentes principais foram removidos a partir das células, por centrifugação e lavagem em PBS, 2% de FCS, uma coloração secundária foi realizada em 1 mL de PBS, suplementado com 2% de FCS, por 30 min em gelo, com:
[00241] 0,1 μg de anticorpo anti-His-tag biotinilado (IBA Life Sciences, Gottingen, Germany, ordem #2-1590-001)
[00242] Após o referido reagente secundário foi removido a partir das células, por centrifugação e lavagem em PBS, 2% de FCS, a combinação CD30-6xHis/anti-His-tag-bio foi detectada por coloração em 1 mL de PBS, 2% de FCS, por 30 min em gelo, com:
[00243] estreptavidina-ficoeritrina diluída de 1/500 (strep-PE) reagente (Affymetrix ebioscience, ordem #12-4317-87).
[00244] Após a coloração final de FACS, as células foram mais uma vez lavadas duas vezes em PBS gelado, 2% de FCS, e então resuspensas em 1,0 mL de PBS, 2% de FCS, após o que as células foram submetidas a análise de FACS e o rastreio de célula de Ig- kappaLC/células CD30 positivas (vide a Figura 13).
[00245] Como pode ser visto a partir dos resultados descritos na Figura 13, a população específica de células IgG positivo e anti-CD30 reativo é detectável no quadrante superior direito dos gráficos de FACs do controle positivo, e como esperado a intensidade do sinal de FACS para IgG de superfície (detectada via anti-Ig-kappa-APC) correlaciona com o sinal de FACS para anti-CD30 que resulta em um padrão de coloração diagonal para essa população.
[00246] Nas misturas de células que expressam anti-CD30 mAb com células que expressam anti-CD19 mAb, o nível de diluição das celulals que expressam anti-CD30 mAb específica é muito bem refletido pela frequência de células específicas de antígeno CD30 no quadrante superior direito e o portão de triagem de FACS estritamente definido (quadrado petro no quadrante superior direito). Os eventos bastante raros que correspondem às células CD30- reativo/IgG positivo com o aumento da diluição das células específicas (1:10'000, 1:100'000, 1:1'000'000) são dificilmente visíveis nas impressões do gráfico de pontos de FACss, mesmo, se maiores números de eventos forem adquiridos, como indicado acima nos gráficos de pontos individuais. Entretanto, a frequência de células CD30 detectáveis correlacionou bem com a frequência das mesmas como esperado a partir do fator de diluição. A partir desse resultado é concluído que a exibição e a detecção de células específicas de antígeno que expressam um anticorpo específico de antígeno por meio de IgG humano mediado a transposição na superfície de mamíferos e células proB, como mostrado aqui, podem ser realizadas de modo confiável.
[00247] A fileira de fundo do gráfico de pontos FACS na Figura 13 mostra a reanálise das células classificadas por FACS a partir dos diferentes experimentos de diluição de pico. Como pode ser visto a partir dos resultados, a re-análise das células classificadas por FACS a partir de diluição de 1:100, 1:1'000 e 1:10'000 resultou em quase a nesna população celular sendo enriquecida, que mostrou ca. 90-95% de células reativas a antígeno. As células classificadas por FACS a partir da diluição 1:100'000 continham uma pequena população adicional que não se encontrou dentro do portão de celulals IgG-positivo/CD30 reativo, mas também nesse experimento ca. 85% das células classificadas por FACS foram células que expressaram IgG específico de antígeno. Surpreendentemente, a mais alta pureza de células, com relação a reatividade de CD30 e expressão de IgG, resultou a partir da triagem de FACS, em que apenas 1 em 1'000'000 foi específica para antígeno CD30, e onde apenas ca. 14 células foram classificadas. Isso pode apenas ser explicado que os seus efeitos quase de clone precisam ser considerados de modo que a classificação não seja uma mistura a partir de células IgG-positivo-celulas CD30 reativo, mas em vez disso uns poucos clones que todos representaram clones de células IgG-positivo-CD30 reativo.
[00248] No entanto, os resultados da coloração específica mediada a antígeno e a identificação de anticorpo especifico de antígeno que expressa células e seu enriquecimento bem sucedido por triagem de célula mediada a FACS preparativa claramente demonstra a possibilidade do método descrito aqui para o isolamento de células que expressam anticorpos com o desejado fenótipo de ligação.
[00249] Exemplo 9: Instrução para a geração e o uso de vetores de expressão IgH transponíveis que podem ser usados para alternar a partir de expressão ligada a membrana para a expressão secretada do anticorpo
[00250] Os vetores de expressão Ig transponíveis descritos nos Exemplos 1 a 4 apenas permitem a expressão de IgG humano na superfície de células de mamífero, de modo que o fenótipo de ligação de anticorpos pode ser prontamente identificado e enriquecido por ligação de antígeno às células, por meio de FACS, como exemplificado no Exemplo 8, ou por bateamento de célula ou métodos de enriquecimento de lote (por exemplo, classificação de célula ativada por conta magnética, MACS). Entretanto, é com frequência desejado se analisar rapidamente as as propriedades de ligação do antígeno de um determinado anticorpo exibido pela célula também como um anticorpo secretado em solução. Embora seja possível se amplificar por PCR as regiões de codificação VL e VH relevantes de um clone de célula específica de antígeno em vetores de expressão para a expressão de IgG secretado, essa abordagem é demorada e trabalhosa.
[00251] Na descrição detalhada da presente invenção já é descrito que os construtores expressão de IgH transponíveis podem ser empregados que exploram a “alternância” natural a partir de expressão ligada a membrana para a expressão secretada do anticorpo, com base em splicing alternativo dos construtores de cadeia de IgH genômico.
[00252] Essa alternância a partir de expressão ligada a membrana para a expressão secretada do anticorpo pode ser alcançada como a seguir:
[00253] Em vez de um cassete de expressão com base em cDNA para a cadeia pesada de Ig-gama1 humanos, a organização genômica original do locus do gene Ig-gama1 humano precisa ser clonado nos vetores de expressão IgH como descrito antes no Exemplo 4. A sequência de todo o locus do gene de imunoglobulina na configuração da linha germinal pode ser recuperada a partir de contig NT_010168 do projeto genoma humano, que cobre o locus de cadeia pesada de Ig humana localizada no cromossomo 14. O locus de gene Ig-gama1 humano de cadeia pesada iniciando a partir do primeiro amino ácido do domínio CH1 na terminação 5’ a 500 bp à jusante do último códon de parada do segundo exon abrangendo a membrana gama1-M2 na terminação 3’ tem um comprimento de 5807 pares de base e não exibe campos internos NheI ou BstBI. Portanto, o referido locus de gene pode ser sintetizado com campos NheI e BstBI de borda, que podem ser usados para clonagem direcional. O referido fragmento de gene sintetizado pode então ser diretamente usado para substituir a região de codificação de cDNA da região de codificação gama1-constante ligada a membrana em pPB-EGFP-HC-Ac10 (Seq-ID32)
[00254] A sequência de DNA de um fragmento genômico Ig-gama1 humano a ser sintetizado é proporcionada em Seq-ID40 abaixo, no final desse Exemplo (os campos 5'-NheI e 3'- BstBI são ressaltados em negrito).
[00255] A organização dos exon e introns do locus da linha germinal de Ig-gama1- cadeia pesada humana, incluindo os seus exons que abrangem a membrana M1 e M2 é ilustrada na Figura 14. As regiões de codificação e as regiões de não codificação nesse fragmento de gene genômico esquerda em sua configuração genômica original são supostas conter todos os elementos necessários cis-regulatórios elementos permitir o splicing alternativo de um mRNA Ig- gama1 dependendo do estágio de diferenciação das células de linhagem B, nas quais o mRNA é processado (Peterson et al. (2002) Mol. Cell. Biol. 22, 5606-5615). A clonagem de fragmento Seq- ID40 em um vetor de expressão Ig-gama1 HC transponível pode ser realizada por substituir a região de codificação C-gama1 em pPB-EGFP-HC-Ac10 por digerir pPB-EGFP-HC-Ac10 com enzimas de restrição NheI e BstBI e ligar o fragmento genômico de Seq-ID40 como o fragmento NheI-BstBI digerido no fragmento de vetor linearizado NheI-BstBI de pPB-EGFP-HC-Ac10.
[00256] O resultado dessa ligação é mostrado de modo esquemático na Figura 14, e a sequência do construtor é proporcionada em Seq-ID41 abaixo no final desse Exemplo.
[00257] Células transformadas proB A-MuLV, como as células 63-12, representam uma linhagem celular adequada para explorar o mecanismo de splicing natural alternativo de um construtor genômico Ig-gama1 HC, como é possível para efetuar a diferenciação fenotípica das referidas células para as linhagens de células B mais maduras, se a a atividade de transformação da Abl-quinase for inibida. Isso pode especificamente ser alcançado com o inibidor de Abl-quinase Gleevec (também conhecido como Imatinib, ou STI-571) (Muljo e Schlissel (2003) Nature Immunol. 4, 31-37). Entretanto, se células proB transformadas A-MuLV são tratadas com Gleevec, as mesmas não só iniciam a diferenciação fenotípica a estágios de linhagem B mais madura, mas esse processo está também associado com uma indução de apoptose (observação não publicada). Isso pode ser evitado por primeiro se estabelecer uma linhagem de células transformadas 63-12 A-MuLV que é transfectada de modo estável com um vetor de expressão bcl-2.
[00258] A sequência de mRNA de rato bcl-2 pode ser encontrada em NCBI- Genbank NM_009741, e tem a sequência a seguir Seq-ID42, mostrado abaixo no final desse Exemplo. Esse quadro de leitura aberta se traduz na sequência de amino ácido a seguir Seq-ID43, também proporcionada abaixo, no final do Exemplo.
[00259] De modo a gerar um vetor de expressão bcl-2 de mamífero, a região de codificação bcl-2 de murinho pode ser gene sintetizado com KpnI de borda e campos de enzima de restrição XhoI que não estão presentes na região de codificação de bcl-2 e um fragmento de DNA sintetizado de duplo gene KpnI-XhoI digerido pode ser ligado em pCDNA3.1-higro(+) descrito adicionalmente acima de modo a gerar um vetor de expressão de mamífero para bcl-2 que pode ser transfectado de mod estável em células 63-12 de modo a selecionar transfectantes estáveis bcl-2.
[00260] Toda a sequência do vetor de expressão pCDNA3.1-higro(+) contendo o gene bcl-2 de murino inserido nos campos de restrição KpnI e XhoI de the múltiplos campos de clonagem é proporcionada como Seq-ID44 abaixo, no final desse Exemplo.
[00261] De modo a facilitar a geração de transfectantes estáveis, o referido vetor pode, por exemplo, ser linearizado fora de cassetes de expressão para bcl-2 ou higromicinB usando a enzima FspI, que lineariza o vetor no gene bacteriano de resistência a ampicilina. 20 μg do referido vetor linearizado pode ser transfectadas em into 2x106 células 63-12 por eletroporação a 950μF/300V exatamente como descrito adicionalmente acima para a transfecção de vetores transponíveis. Em seguida da eletroporação, as células podem então ser diluidas em 100 mL de meio de crescimento e laminads em cinco placas de 96 cavidades com cada 200 μL/cavidade, o que irá resultar na laminação de ca. 4x103 células/cavidade.
[00262] Os transfectantes estáveis podem então ser selecionados por adicionar 800 μg/mL de higromicinB 48 horas após a transfecção. Os clones de células individuais transfectadas de modo estável, das quais 20-100 podem ser esperadas a partir do referido experimento, pode então ser obtida 2-3 semanas mais tarde. Clones de células transfectadas bcl- 2 estáveis são melhor fucionalmente testadas para a sua capacidade de proteger as células a partir de apoptose, por medir a sobrevivência de clones individuais com a exposição de 0,1 a 1 mM Gleevec (Imatinib, ou STI-571). Uma vez que um transfectante bcl-2 estavel 63-12 é identificado que tem alta resistência a Gleevec (Imatinib, ou STI-571), esse clone pode ser utilizado como as células hospedeiras para a expressão de IgG humano a partir de Ig-gama1 HC genômico transponível e vetores de expressão Ig-kappa LC, por exemplo, utilizando os vetores Seq-ID41 e Seq-ID36.
[00263] Os referidos vetores podem ser co-transfectados com vetor de expressão PiggyBac (Seq-ID13) em células 63-12 transfectadas Bcl-2 de modo estável, e transpostas de modo estável e células que expressam IgG podem ser estabelecidas como descrito adicionalmente acima (equivalente ao Exemplo 7). Pelo fato das células proB representarem ume stágio de diferenciação, onde a imunoglobulina endógena é expressa como uma imunoglobulina ligada a membrana, pode ser esperado que também a Ig-HC expressa a partir de um vetor de expressão Ig-gama1 HC transponível em configuração genômica será expressa como a versão ligada a membrana.
[00264] Entretanto, se as células são tratadas com 0,1 a 1 mM Gleevec (Imatinib, ou STI-571), a Abl-quinase codificada por A-MuLV é especificamente inibida, as células não são mais transformadas e continuam o seu programa de diferenciação intrínseca para estágios de diferenciação de célula B mais madura. Em vitro, a referida diferenciação é independente de proteínas Ig expressas fucionalmente (Grawunder et al. (1995) Int. Immunol. 7, 1915-1925). Foi mostrado que a diferenciação in vitro de células não transformadas proB as torna responsivas a anti-IL4 derivado de célula T e estimulação de CD40, com o que as células mesmo se diferenciam em células do estágio plasmático que sofrem recombinação de troca de classe e onde as mesmas podem ser fundidas com células de mieloma para gerar hibridomas (Rolink et al. (1996) Immunity 5, 319-330).
[00265] Isso quer dizer que também as células 63-12 transformadas por A-MuLV, que são tornadas resistentes a apoptose por expressão estável de bcl-2 podem ser diferenciadas nas células doe stágio de célula plasmática com o tratamento com Gleevec e incubação simultânea com 10 μg/mL de anticorpo anti-CD40 agonístico, e 100 U/mL de IL4 recombinante, exatamente como descrito em Rolink et al. (1996) Immunity 5, 319-330.
[00266] O referido tratamento irá induzir uma mudança no programa de diferenciação celular, que mudará o programa de splicing alternativo celular a partir da expressão IgG ligado a membrana para a expressão de IgG secretada a partir de um construtor de expressão Ig HC em organização genômica. Isso irá permitir a produção de anticorpo secretado a partir de clones de células laminadas replica identificados e isolados por exibição de superfície e ligação a antígeno, sem a necessidade de re-clonar as Regiões de codificação VL e VH a partir de clones de células selecionadas e sem a necessidade de se ligar as mesmas em vetores de expressão para os anticorpos IgG secretados. Isso é uma característica funcional dos vetores que não podem ser facilmente incorporados na maioria dos sistemas de expressão em célula de mamífero, particularmente não em muitos sistemas de expressão com base em vírus, em quie os referidos extended genomic vetores de expressão genômicos estendidos não podem ser facilmente inseridos.
[00267] Seq-ID40 (gene Ig-gama1 de cadeia pesada genômico humano de 5812 bp de comprimento)
[00268] Seq-ID41 (vetor de expressão Ig-gama1-HC transponível na configuração genômica)
[00269] Seq-ID42 (região de codificacao bcl-2 de murinho)
[00270] Seq-ID43 (sequência de amino ácido de proteína Bcl-2 de murinho)
[00271] Seq-ID44 (etor de expressão de mamíferopCDNA3.1-higro(+)-bcl2 v)
[00272] Exemplo 10: Instrução para a geração de vetores que codificam bibliotecas humanas básicas de cadeia leve e pesada de anticorpo como vetores PiggyBac transponíveis.
[00273] De modo a gerar simples bibliotecas transponíveis de DNA que codificam bibliotecas humanas de cadeia leve e pesada de anticorpo, apenas as regiões VL e VH a partir de vetores transponíveis pIRES-EGFP-T1T2-IgL do Exemplo 2 e pIRES-EGFP-T1T2-IgH de Exemplo 3, respectivamente, precisam ser substituídas. Isso pode ser realizado pore sibntese de gene humano das Regiões de codificação VL e VH margeadas por ClaI e campos de enzima de restrição Eco47III, e por permitir variações de nucleotídeo em determinadas posições de HCDR e LCDR, como proporcionado em Seq-ID9, que codifica bibliotecas para os domínios variáveis de cadeia pesada, e Seq-ID-10, que codifica bibliotecas para domínios variáveis de cadeia leve, e que são proporcionadas no final desse Exemplo. Ambas as referidas sequências contêm um alongamento de N-sequências em HCDR3 (negrito), e LCDR3 (negrito), respectivamente. Ambas as sequencias Seq-ID9 e Seq-ID10 são margeadas por enzimas de restrição ClaI e Eco47III (sublinhadas), respectivamente, incluindo quatro nucleotídeos que margeiam os campos de enzima de restrição (realçados em letra minúscula nas terminações da sequência), permitindo a digestão adequada da enzima de restrição dos fragmentos de DNA de gene sintetizado e a ligação direcionada nas estruturas ClaI-Eco47III linearized pIRES-EGFP-T1T2-IgH e pIRES-EGFP-T1T2-IgL, respectivamente.
[00274] Desse modo, bibliotecas diversas transponíveis de DNA, que codificam cadeias pesada e leve de anticorpo on vetores separados, nas quais a expressão de cadeias de anticorpo são transcripcionalmente e portanto operacionalmente ligada a uma proteína de mercacao fluorescente verde pode ser gerada.
[00275] Seq-ID9 (região de codificação de domínio VL com variação de N- sequência variável em posições que codificam LCDR3)
[00276] Seq-ID10 (região de codificação de domínio VH com variação de N- sequência variável em posições que codificam HCDR3)
[00277] Exemplo 11: Instruçãos para a geração de uma biblioteca de expressão básica de cadeia leve Ig-kappa transponível humana de Sleeping Beauty
[00278] De modo a gerar uma biblioteca diversa de DNA transponível de Sleeping Beauty que codifica bibliotecas de cadeia leve de anticorpo humano, a região VL do vetor transponível de Sleeping Beauty pIRES-EGFP-sbT1T2-IgL do Exemplo 5 precisa ser substituída com um repertorio de gene VL diverso. Isso pode ser realizado por síntese de gene de regiões de codificação VL humana margeadas por ClaI e campo de enzima de restrição Eco47IIIs, e por permitir variações de nucleotídeo em determinadas posições HCDR e LCDR, como já proporcionado em Seq-ID-10 acima. A sequência Seq-ID10 é margeada por enzimas de restrição ClaI e Eco47III permitindo a ligação orientada em ClaI-Eco47III linearizadp pIRES-EGFP-sbT1T2- IgL. Desse modo a biblioteca de DNA transponível de Sleeping Beauty que codifica diversos anticorpos humanos de cadeia leve pode ser gerada.
[00279] Desse modo, bibliotecas diversas transponíveis de DNA, que codificam cadeias pesada e leve de anticorpo em vetores separados, nos quais a expressão de cadeias de anticorpo é transcripcionalmente e portanto operacionalmente ligada a uma proteína flurorescente de marcador verde pode ser gerada.
[00280] Exemplo 12: Clonagem de uma biblioteca de expressão de cadeia de IgL transponível
[00281] Uma biblioteca de cadeia leve V-kappa com região LCDR3 randomizada foi construída como descrito abaixo. Seis resíduos de amino ácidos foram randomizados, isto é, codificados pelo códon NNK (N = qualquer nucleotídeo; K = T ou G), que acomoda cada um dos 20 amino ácidos. A biblioteca foi com base em segmentos de gene Vkappa1-5 e Jkappa2 humanos de linha germinal e foi randomizada entre a cisteina conservada na terminação da região 3 da estrutura e a região de estrutura 4 com base em Jkappa2 como a seguir: Gln-Gln-(NNK)6-Thr. A sequência e a configuração geral da biblioteca de cadeia leve kappa são mostrados na Figura 15.
[00282] A molécula de DNA linear que codifica a biblioteca de cadeia leve kappa foi gerada por PCR. Para isso, dois modelos foram gerados por síntese de gene total (realizada por GenScript, Piscataway, NJ, USA). Por um lado, um construtor sintético foi gerado que compreende o segmento de gene Vkappa1-5 clonado no campo EcoRV de pUC57 (Genscript ordem #SD1176), pUC57_Vkappa1-5 (SEQ-ID45); por outro lado, um construtor sintético foi gerado que compreende o segmento de gene Jkappa2 fundido à região de codificação Ckappa clonada no campo EcoRV de pUC57, pUC57_Jkappa2-Ckappa (SEQ-ID46).
[00283] Um primeiro DNA linear que compreende o segmentod e gene Vkappa1-5 foi amplificado por PCR a partir de pUC57_Vkappa1-5 usando os primers pUC57-1 (5’-CGT TGT AAA ACG ACG GCC AG-3’) e LCDR3-B (5’-CTG TTG GCA GTA ATA AGT TGC-3’). Um segundo DNA linear que compreende a região de CDR3 randomizada (Gln-Gln-(NNK)6-Thr), o segmetnod e gene Jkappa2 e a região constante Ckappa foi amplificada a partir de pUC57_Jkappa2-Ckappa usando os primers LCDR3-NNK6-F (5’-GCA ACT TAT TAC TGC CAA CAG NNK NNK NNK NNK NNK NNK ACT TTT GGC CAG GGG ACC AAG-3’) e pUC57-2 (5’-TCA CAC AGG AAA CAG CTA TG-3’). Para evitar a introdução da orientação da sequência em virtude de priming da região randomizada do primer LCDR3-NNK6-F on pUC57_Jkappa2-Ckappa, o plasmídeo foi primeiro linearizado pela digestão com a enzima de restrição ScaI (Figura 17A).
[00284] As moléculas de DNA resultantes (SEQ-ID47 e SEQ-ID48) exibidas na sobreposição de 21bp e foram montadas por extensão de sobreposição de PCR usando os primers pUC57-1 e pUC57-2, gerando uma molécula de DNA que compreende a biblioteca de cadeia leve kappa margeada pelos campos de restrição NotI e AsuII (=BstBI) como mostrado na Figura 15. O amplicon PCR da biblioteca de cadeia leve V-kappa foi submetida a sequenciamento de fragmento de PCR, e o resultado mostrado na Figura 18, demonstra que de fato a diversidade esperada foi introduzida como projetado nas posições de LCDR3, como evidenciado por sinais de eletroferograma de sobreposição nas posições randomizadas. Esse fragmento de PCR foi digerido com endonucleases de restrição NotI e BstBI (um isosquizomero de AsuII) e clonados no PiggyBac- vetor transponível pPB-EGFP_HC-g1 (SEQ-ID049, que resulta em uma biblioteca que consiste de 5.2x 107 clones independentes. O tamanho da referida biblioteca pode ser facilmente aumentado em um fator 10 por aumentar a reação de ligação.
[00285] Bibliotecas de cadeia leve incorporando desenhos de randomização distinta, ou que compreendem segmentos de gene Vkappa e Jkappa diferentes dos usados no prsente exemplo, podem ser produzidas do mesmo modo. Da mesma forma, a estratégia descrita aqui pode ser empregada para a produção de bibliotecas Vlambda de cadeia leve.
[00286] SEQ-ID45 (pUC57_Vkappa1-5)
[00287] SEQ-ID46 (pUC57_Jkappa2-C-kappa)
[00288] SEQ-ID47 (produto de PCR Vkappa1-5)
[00289] SEQ-ID48 (produto de PCR NNK6-Jkappa2-C-kappa)
[00290] Exemplo 13: Clonagem de bibliotecas de expressão de adeia de IgH transponível com HCDR3 de comprimento variável
[00291] Uma biblioteca de cadeia pesada gama1 humana com região de HCDR3 randomizada foi construída como descrito abaixo. Diversos resíduos de amino ácido foram randomizados, isto é codificados pelo códon NNK (N = qualquer nucleotídeo; K = T ou G), que acomoda cada um dos 20 amino ácidos. A biblioteca foi com base nos segmentos de gene VH3-30 e JH4 e foi randomizada entre o resíduo de cisteína conservada na terminação da região 3 da estrutura e na região 4 da estrutura com base em JH4 como a seguir: Ala-Lys/Arg-(NNK)n-Asp- NNK. Vários comprimentos de HCDR3 foram explorados, com n = 4, 6, 8, ou 10 (randomização de NNK4, NNK6, NNK8, e NNK10). A sequência e a configuração geral da biblioteca de cadeia pesada gama é mostrada na Figura 16.
[00292] A molécula DNA linear que codifica a biblioteca de região variável de cadeia pesada (VH) foi gerada por PCR. Para isso, dois modelos foram gerados por síntese total de gene (realizada por GenScript, Piscataway, NJ, USA). Por um lado, um construtor sintético foi gerado que compreende o segmento de gene VH3-30 clonado no campo EcoRV de pUC57, pUC57_VH3-30 (SEQ-ID49); por outro lado, um construtor sintético foi gerado que compreende o segmento de gene JH4 clonado no campo EcoRV de pUC57, pUC57_JH4 (SEQ-ID50).
[00293] Um primeiro DNA linear que compreende o segmentod e gene VH3-30 foi amplificado por PCR a partir de pUC57_VH3-30 usando os primers VH3-30-F (5’-GAT ATC CAA TGC GGC CGC ATG -3’) e HCDR3-B (5’-CGC ACA GTA ATA CAC AGC CGT G -3’). Moléculas de DNA lineares adicionais que compreendem as regiões HCDR3 randomizadas Ala-Lys/Arg-(NNK)4- Asp-NNK, Ala-Lys/Arg-(NNK)6-Asp-NNK, Ala-Lys/Arg-(NNK)8-Asp-NNK, ou Ala-Lys/Arg-(NNK)10- Asp-NNK fundidas ao segmentod e gene JH4 foram amplificadas a partir de pUC57_JH4 usando, respectivamente, os primers HCDR3-NNK4-F (5’-CAC GGC TGT GTA TTA CTG TGC GAR GNN KNN KNN KNN KGA CNN KTG GGG CCA AGG AAC CCT GGT C-3’), HCDR3-NNK6-F (5’-CAC GGC TGT GTA TTA CTG TGC GAR GNN KNN KNN KNN KNN KNN KGA CNN KTG GGG CCA AGG AAC CCT GGT C-3’), HCDR3-NNK8-F (5’-CAC GGC TGT GTA TTA CTG TGC GAR GNN KNN KNN KNN KNN KNN KNN KNN KGA CNN KTG GGG CCA AGG AAC CCT GGT C-3’), ou HDR3-NNK10-F (5’-CAC GGC TGT GTA TTA CTG TGC GAR GNN KNN KNN KNN KNN KNN KNN KNN KNN KNN KGA CNN KTG GGG CCA AGG AAC CCT GGT C-3’) em combinação com o primer pUC57-3 (5’-CAG GTT TCC CGA CTG GAA AG-3’). Para evitar a introdução de uma orientação de sequência em virtude de priming da região randomizada dos primers HCDR3-NNK4-F, HCDR3- NNK6-F, HCDR3-NNK8-F e HCDR3-NNK10-F em pUC57_JH4, o plasmídeo foi primeiro linearizado pela digestão com a enzima de restrição DrdI (Figura 17B).
[00294] O produto de PCR resultante VH3-30 (SEQ-ID51) exibiu uma sobreposição de 22bp com os produtos de PCR NNK4-JH4, NNK6-JH4, NNK8-JH4 e NNK10-JH4 (SEQ-ID52 a 55), e foi montado com cada por extensão de sobreposição de PCR em 4 reaçoes separadas, usando os primers VH3-30-F e pUC57-3. As moléculas de DNA resultantes compreendidas da biblioteca VH margeada por campos de restrição NotI e NheI como mostrado na Figura 16. Todos os amplicons de PCr obtidos a partir dos PCRs que emprgaram os oligo degenerados NNK4-JH4, NNK6-JH4, NNK8-JH4 e NNK10-JH4 foram submetidos a sequenciamento direto de DNA, e foi confirmado que a randomization projetada das opsições de HCDR3 foi obtida, como esperado. Isso é mostrado por meio de exemplo na Figura 18 (B), onde o eletroferograma da região abrangendo a HCDR3 é proporcionado. As posições randomizadas mostram as esperadas sobreposições dos picos de sequência demonstrando a diversidade do nucleotídeo nas referidas posições (figura 18). Os 4 diferentes DNAs da biblioteca de VH foram misturados em relações equimolares, digeridos com endonucleases de restrição NotI e NheI e clonados no vetor transponível PiggyBac- pPB- EGFP_HC-gama1 (SEQ-ID22), à montante da região constante de cadeia pesada gama1, que resulta em uma biblioteca consistindo de 3.7x 107 clones independentes. O tamanho da referida biblioteca pode ser facilmente aumentado em um fator 10 ao aumentar a reação de ligação.
[00295] Bibliotecas de cadeia pesada que incorporam desenhhos de randomização distintos, ou que compreendem segmentos de genes de VH e JH diferentes dos usados nesse exemplo, podem ser produzidos da mesma maneira.
[00296] SEQ-ID49 (pUC57_VH3-30)
[00297] SEQ-ID50 (pUC57_JH4)
[00298] SEQ-ID51 (produto de PCR de VH3-30)
[00299] SEQ-ID52 (produto de PCR de NNK4-JH4)
[00300] SEQ-ID53 (produto de PCR de NNK6-JH4)
[00301] SEQ-ID54 (produto de PCR de NNK8-JH4)
[00302] SEQ-ID55 (produto de PCR de NNK10-JH4)
[00303] Exemplo 14: Identificação de regiões de codificação de cadeia leve e cadeia pesada variáveis a partir de células hospedeiras transpostas de modo estável enriquecidas com antígeno reativo
[00304] Em virtude da integração estável dos vetores de expressão transponíveis que codificam cadeias pesada e leve de anticorpo no hospedeiro, as regiões de codificação de cadeia pesada e cadeia leve variáveis podem ser re-isoladas em um modo direto por amplificação de PCR padrão seguido de sequenciamento direto dos amplicons de PCR ou, com a reclonagem, a partir de vetores de plasmídeo re-clonado. Para isso, células isoladas ou clones de célula, expressando anticorpos específicos de antígeno são centrifugados por 5 minutos a 1200x g. O RNA total é isolado a partir das referidas células usando reagente TRIzol (Sigma-Aldrich). Primeira fita de cDNA pode ser sintetizada com PowerScript (Clontech-Life Technologies) usando um primer oligio-dT. As regiões de codificação de cadeia leve podem então ser amplificadas por PCR usando os primers SP-F (5’-GAG GAG GAG GCG GCC GCC ATG AAT TTT GGA C-3’) e CK-rev (5’-GAG GAG GAG TTC GAA AGC GCT AAC ACT CTC-3’), que resultará em um amplicon de PCR de ca. 740 bp, dependendo do comprimento da região V-kappa contida no amplicon de PCR. Se desejado, o referido amplicon de PCR pode ser digerido com endonucleases de restrição NotI e BstBI, e clonados no vetor pPB-EGFP_HC-g1 (Seq-ID22), de modo a subclonar os clones individuais do amplicon de PCR para a identificação adicional da sequência. Clones de região V-kappa individuais podem então ser submetidos a sequenciamento usando o primer pPB-seq13 (5’-GGC CAG CTT GGC ACT TGA TG-3’), ligando no promotor EF1-alfa, à montante da região de V-codificação clonada.
[00305] As regiões variáveis de cadeia pesada podem ser amplificadas por PCR no cDNA, gerado como acima, usando os primers SP-F (5’-GAG GAG GAG GCG GCC GCC ATG AAT TTT GGA C-3’) e CG-revseq-1 (5’- GTT CGG GGA AGT AGT CCT TG-3’) que resultarão em um amplicon de PCR de ca. 530 bp de tamanho esperado, dependendo do comprimento da região VH contida no amplicon PCR. Se desejado, o referido amplicon de PCR pode ser digerido com endonucleases de restrição NotI e NheI, e clonados no vetor pPB-EGFP_HC-g1 (Seq-ID22). Clones individuais são então submetidos a sequenciamento da região VH- usando o primer pPB-seq13 (5’- GGC CAG CTT GGC ACT TGA TG-3’), que se liga no promotor EF1-alfa, à montante da região de V clonada.
Claims (14)
1. Método para identificar um polipeptídeo tendo uma funcionalidade ou especificidade de ligação desejadas, caracterizado pelo fato de que compreende: (i) gerar uma coleção diversificada de polinucleotídeos, preferivelmente vetores plasmídeos ou amplificações de PCR de DNA de cadeia dupla, que codificam polipeptídeos tendo diferentes funcionalidades ou especificidades de ligação, em que referidos polinucleotídeos compreendem uma sequência que codifica um polipeptídeo disposto entre uma primeira e uma segunda sequências de repetição terminal invertidas que são reconhecidas por e funcionais com pelo menos uma enzima transponase; (ii) introduzir a coleção diversificada de polinucleotídeos a partir de (i) em células hospedeiras; (iii) expressar pelo menos uma enzima transponase funcional com as referidas sequências de repetição terminal invertidas nas referidas células hospedeiras de modo que a referida coleção diversificada de polinucleotídeos é integrada no genoma da célula hospedeira para providenciar uma população de células hospedeiras que expressam a referida coleção diversificada de polinucleotídeos que codificam polipeptídeos tendo diferentes funcionalidades ou especificidades de ligação; (iv) investigar referidas células hospedeiras para identificar a célula hospedeira que expressa um polipeptídeo tendo uma funcionalidade ou especificidade de ligação desejada; e (v) isolar a sequência de polinucleotídeos que codifica referido polipeptídeo a partir da referida célula hospedeira.
2. Método de acordo com a reivindicação 1, caracterizado pelo fato que referidos polinucleotídeos compreendem a) uma sequência de ligação ao ligante de um receptor ou uma sequência de ligação a alvo de uma molécula de ligação; b) uma sequência de ligação a antígeno de um anticorpo; c) uma sequência que codifica uma região VH ou VL de um anticorpo ou um fragmento de ligação a antígeno da mesma; d) uma sequência que codifica uma região VH do anticorpo e uma região VL do anticorpo; e) uma sequência que codifica uma cadeia pesada ou uma cadeia leve de imunoglobulina de comprimento total, ou um fragmento de ligação a antígeno da mesma; e/ou f) uma sequência que codifica um domínio de Fab ou um Fv de cadeia simples.
3. Método de acordo com qualquer uma das reivindicações anteriores, caracterizado pelo fato que gerar a referida coleção diversificada de polinucleotídeos compreende submeter as sequências de gene da região V a PCR sob condições de mutagênese.
4. Método de acordo com qualquer uma das reivindicações anteriores, caracterizado pelo fato que a etapa (ii) compreende introduzir nas referidas células hospedeiras polinucleotídeos compreendendo sequências que codificam a) regiões VH ou VL de imunoglobulina, ou fragmentos de ligação a antígeno das mesmas, em que as referidas sequências das regiões VH e VL são codificadas em vetores separados; e/ou b) cadeias pesadas ou leves de imunoglobulina de comprimento total, ou fragmentos de ligação a antígeno das mesmas, em que as referidas sequências das cadeias pesadas e leves de comprimento inteiro estão em vetores separados; e c) um vetor compreendendo sequências que codificam cadeias VH e VL do anticorpo. d) um vetor compreendendo sequências que codificam uma cadeia pesada de imunoglobulina de comprimento inteiro e uma cadeia leve de imunoglobulina de comprimento inteiro.
5. Método de acordo com qualquer uma das reivindicações anteriores, caracterizado pelo fato que (a) referida etapa de expressar (iii) compreende introduzir nas referidas células hospedeiras um vetor de expressão que codifica a enzima transponase que reconhece e é funcional com pelo menos uma sequência de repetição terminal invertida, em que a referida enzima transponase é preferivelmente expressa de forma transitória na referida célula hospedeira, (b) referida etapa de investigar (iv) compreende uma seleção das células ativadas por meios magnéticos (MACS), uma seleção das células ativadas por fluorescência (FACS), uma seleção contra moléculas imobilizadas em uma superfície sólida, uma seleção para a ligação a moléculas associadas a membrana celular incorporadas em uma membrana de dupla camada de lipídeo celular, reconstituída natural ou artificialmente, ou uma investigação de alta produtividade de clones de células individuais em formato de múltiplas camadas para um fenótipo de ligação ou funcional desejado, (c) referida etapa (v) de isolar a sequência de polinucleotídeo que codifica o polipeptídeo tendo uma funcionalidade ou especificidade de ligação desejada compreende a amplificação de RT-PCR ou genômica ou sequenciamento profundo de próxima geração.
6. Método de acordo com qualquer uma das reivindicações anteriores, caracterizado pelo fato que a) as referidas sequências de repetição terminal invertidas são a partir do sistema de transposon PiggyBac ou a partir do sistema de transposon Sleeping Beauty; e/ou b) a etapa (iii) compreende introduzir em referida célula hospedeira um vetor compreendnendo uma sequência que codifica uma transponase PiggyBac funcional ou transponase Sleeping Beauty.
7. Método de acordo com qualquer uma das reivindicações anteriores, caracterizado pelo fato que referidas sequências de repetição terminal invertidas são reconhecidas por e funcionais com pelo menos uma transponase selecionada a partir do grupo que consiste de: PiggyBac, Sleeping Beauty, Frog Prince, Himar1, Passport, Minos, hAT, Tol1, Tol2, Ac/Ds, PIF, Harbinger, Harbinger3-DR, e Hsmar
8. Biblioteca de moléculas de polinucleotídeo que codificam polipeptídeos tendo diferentes funcionalidades ou especificidades de ligação, caracterizada pelo fato que compreende uma pluralidade de moléculas de polinucleotídeo, em que as referidas moléculas de polinucleotídeo compreendem uma sequência que codifica um polipeptídeo tendo uma funcionalidade ou especificidade de ligação disposta entre sequências de repetição terminal invertidas que são reconhecidas por e funcionais com pelo menos uma enzima transponase.
9. Biblioteca de acordo com a reivindicação 8, caracterizada pelo fato que referidos polinucleotídeos compreendem: a) pelo menos uma sequência que codifica uma sequência de ligação a antígeno de um anticorpo; b) uma sequência que codifica uma região VH ou VL de um anticorpo ou um fragmento de ligação a antígeno da mesma; c) uma sequência que codifica uma região VH do anticorpo e um região VL do anticorpo; d) uma sequência que codifica uma cadeia pesada ou uma cadeia leve de imunoglobulina de comprimento total, ou um fragmento de ligação a antígeno da mesma; e) uma sequência que codifica um domínio de Fab ou um FV de cadeia única.
10. Biblioteca de acordo com as reivindicações 8 ou 9, , caracterizada pelo fato que referidos plasmídeos ou amplificações por PCR de ADN de cadeia dupla adicionalmente compreendem uma sequência que codifica uma enzima transponase que reconhece e é funcional com as sequências de repetição terminal invertidas.
11. Método para gerar uma biblioteca de polinucleotídeos de transposição que codificam polipeptídeos tendo diferentes funcionalidades ou especificidades de ligação, caracterizado pelo fato de que compreende gerar uma coleção diversificada de polinucleotídeos compreendendo sequências que codificam polipeptídeos com diferentes funcionalidades ou especificidades de ligação, em que referidos polinucleotídeos compreendem uma sequência que codifica um polipeptídeo tendo uma funcionalidade ou especificidade de ligação disposta entre sequências de repetição terminal invertidas que são reconhecidas por e funcionais com pelo menos uma enzima transponase.
12. Método para gerar uma população de células hospedeiras capazes de expressar polipeptídeos com diferentes funcionalidades ou especificidades de ligação, caracterizado pelo fato que compreende: (i) gerar uma coleção diversificada de polinucleotídeos compreendendo sequências que codificam polipeptídeos tendo diferentes funcionalidades ou especificidades de ligação, em que os referidos polinucleotídeos compreendem uma sequência que codifica um polipeptídeo tendo uma funcionalidade ou especificidade de ligação disposta entre sequências de repetição terminal invertidas que são reconhecidas por e funcionais com pelo menos uma enzima transponase; e (ii) introduzir a referida coleção diversificada de polinucleotídeos em células hospedeiras.
13. Método ou vetor de acordo com as reivindicações anteriores, caracterizados pelo fato que o vetor compreendendo a sequência VH compreende sequências de repetição terminal invertidas que são reconhecidas por uma enzima transponase diferente daquelas das sequências de repetição terminal invertidas no vetor compreendendo a sequência VL.
14. Método ou célula hospedeira de acordo com as reivindicações anteriores, caracterizado pelo fato que as células hospedeiras são células de vertebrados, preferivelmente células de mamíferos, mais preferivelmente células de humanos ou roedores, ainda mais preferivelmente células linfóides, ainda mais preferivelmente células B, ainda mais preferivelmente células B progenitoras ou células B precursoras, particularmente preferidas células B progenitoras ou células B precursoras transformadas de vírus de leucemia Abelson-Murina e células preB e proB humanas transformadas EBV precoces sem imunoglobulina.
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201261673144P | 2012-07-18 | 2012-07-18 | |
| US61/673,144 | 2012-07-18 | ||
| EP12178529.9A EP2692865B1 (en) | 2012-07-30 | 2012-07-30 | Transposition-mediated identification of specific binding or functional proteins |
| EP12178529-9 | 2012-07-30 | ||
| PCT/EP2013/065214 WO2014013026A1 (en) | 2012-07-18 | 2013-07-18 | Transposition-mediated identification of specific binding or functional proteins |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| BR112015000836A2 BR112015000836A2 (pt) | 2017-12-12 |
| BR112015000836B1 true BR112015000836B1 (pt) | 2021-12-21 |
Family
ID=
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US20190153433A1 (en) | Transposition-mediated identification of specific binding or functional proteins | |
| US10502745B2 (en) | Identification of antigen- or ligand-specific binding proteins | |
| BR112015000836B1 (pt) | Método para identificar um polipeptídeo tendo uma funcionalidade ou especificidade de ligação desejadas, biblioteca de moléculas de polinucleotídeo, método para gerar uma biblioteca de polinucleotídeos e método para gerar uma população de células hospedeiras | |
| AU2014274595B2 (en) | Identification of antigen- or ligand-specific binding proteins |