WO2007076115A2 - Nitrogen-efficient monocot plants - Google Patents
Nitrogen-efficient monocot plants Download PDFInfo
- Publication number
- WO2007076115A2 WO2007076115A2 PCT/US2006/049241 US2006049241W WO2007076115A2 WO 2007076115 A2 WO2007076115 A2 WO 2007076115A2 US 2006049241 W US2006049241 W US 2006049241W WO 2007076115 A2 WO2007076115 A2 WO 2007076115A2
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- plant
- monocot
- transgenic
- rice
- maize
- Prior art date
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8241—Phenotypically and genetically modified plants via recombinant DNA technology
- C12N15/8261—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield
- C12N15/8262—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield involving plant development
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/1096—Transferases (2.) transferring nitrogenous groups (2.6)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8216—Methods for controlling, regulating or enhancing expression of transgenes in plant cells
- C12N15/8222—Developmentally regulated expression systems, tissue, organ specific, temporal or spatial regulation
- C12N15/8223—Vegetative tissue-specific promoters
- C12N15/8227—Root-specific
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8241—Phenotypically and genetically modified plants via recombinant DNA technology
- C12N15/8242—Phenotypically and genetically modified plants via recombinant DNA technology with non-agronomic quality (output) traits, e.g. for industrial processing; Value added, non-agronomic traits
- C12N15/8243—Phenotypically and genetically modified plants via recombinant DNA technology with non-agronomic quality (output) traits, e.g. for industrial processing; Value added, non-agronomic traits involving biosynthetic or metabolic pathways, i.e. metabolic engineering, e.g. nicotine, caffeine
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8241—Phenotypically and genetically modified plants via recombinant DNA technology
- C12N15/8261—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y02—TECHNOLOGIES OR APPLICATIONS FOR MITIGATION OR ADAPTATION AGAINST CLIMATE CHANGE
- Y02A—TECHNOLOGIES FOR ADAPTATION TO CLIMATE CHANGE
- Y02A40/00—Adaptation technologies in agriculture, forestry, livestock or agroalimentary production
- Y02A40/10—Adaptation technologies in agriculture, forestry, livestock or agroalimentary production in agriculture
- Y02A40/146—Genetically Modified [GMO] plants, e.g. transgenic plants
Definitions
- the invention relates to monocot plants having enhanced nitrogen utilization efficiency (NUE), to methods for enhancing NUE in monocot plants, and to methods of increasing biomass and seed yield in monocot plants grown under nitrogen limiting conditions.
- NUE nitrogen utilization efficiency
- This invention also relates to monocot antiquitin promoters.
- Rice is routinely used as the model crop for genetic and physiological studies in other monocot crops including maize, wheat, sugarcane, barley, sorghum, rye and grass. Because of its importance as a model crop, rice was the first crop plant to be sequenced.
- the International Rice Genome Sequencing Project a consortium of publicly funded laboratories, completed the sequencing of the rice genome in December 2004.
- Rice has a small, diploid genome that is well conserved and syntenic across monocots. It is easily transformed and transgenic studies have been performed in rice to study a number of phenotypic traits, including flowering, abiotic stress response, disease resistance, drought tolerance, and morphological development. [010] Because of the critical importance of nitrogen to plant growth, previous studies have attempted to increase the efficiency of nitrogen utilization in plants using a variety of means. These methods have included conventional breeding programs directed toward the development of plants that are more efficient at nitrogen utilization. Recombinant deoxyribonucleic acid (DNA) and transgenic plant methods have also been employed in an attempt to generate nitrogen efficient plants.
- DNA deoxyribonucleic acid
- transgenic plant methods have also been employed in an attempt to generate nitrogen efficient plants.
- AIaAT alanine aminotransferase
- Brassica turgor gene-26 also known as antiquitin
- the invention addresses the need for monocot plants with enhanced growth characteristics and nitrogen utilization efficiencies when grown under low nitrogen conditions by providing such plants and methods for generating transgenic monocot plants with elevated levels of AIaAT.
- the invention provides transgenic monocot plants including a recombinant DNA sequence encoding an AIaAT.
- the transgenic monocot plant may be barley, rice, sugar cane, maize, sorghum, rye, wheat, or grass. Grass includes lawn, turfgrass, forage and the like.
- the AIaAT is operably linked to a promoter, most preferably, a monocot antiquitin promoter. Seeds from the transgenic monocot plants are also provided.
- transgenic rice, maize, wheat, sorghum, barley, and sugar cane include a recombinant DNA sequence encoding an AIaAT and seeds therefrom.
- a method of producing a transgenic monocot plant including the steps of: (1) selecting a nucleic acid encoding an AIaAT, (2) selecting a promoter that is operable in a monocot plant, (3) coupling the selected nucleic acid to the selected promoter to form a genetic construct, (4) transforming a monocot plant cell with the genetic construct to form a transformed cell, and (5) growing a transgenic monocot plant from the transformed cell to produce a transgenic plant.
- overexpression of AlaAT causes at least a 5% to 7.5%, 7.5 to 10%, IO to 15% or 15 to 20%, or more increase in plant biomass and/or seed yield when expressed in a transgenic monocot plant compared to the plant biomass or seed yield of a comparable monocot plant not expressing this construct when the plants are grown under suboptimal nitrogen conditions.
- transgenic monocot plants are described wherein the transgenic monocot plant expresses a recombinant AlaAT and exhibits at least a 5% increase in plant biomass or seed yield compared to biomass or seed yield of a comparable plant lacking the recombinant AlaAT. Also described are seeds produced from the transgenic monocots.
- the monocots include but are not limited to, maize, wheat, rice, barley and rye.
- a method for increasing biomass of a monocot plant by contacting and introducing into a plant an AlaAT coding region in operative linkage with monocot antiquitin promoter is described. Similar methods for increasing seed yield of a plant and are also provided.
- the nucleic acids encoding AlaAT that are used in the genetic constructs of these inventions may be derived from any organism preferably a plant, and most preferably from a monocot plant including, but not limited to, barley, rice, sugar cane, rye, wheat, maize, or grass.
- the invention provides an isolated monocot antiquitin promoter sequence.
- the monocot promoter sequence may be from barley, rice, sugar cane, maize, sorghum, rye, wheat, or grass.
- it is a sorghum promoter that includes SEQ ID NO: 9 or an active fragment thereof.
- it is a maize promoter that includes SEQ ID NO: 10 or an active fragment thereof.
- the target gene encodes a nitrogen utilization protein, such as, for example, a high affinity nitrate transporter, a low affinity nitrate transporter, an ammonium transporter, an ammonia transporter, an amino acid transporter, alanine dehydrogenase, glutamine synthetase, asparagine synthetase, glutamate synthase, glutamate 2:oxogluturate amino transferase, asparaginase, glutamate dehydrogenase, nitrate reductase, aspartate aminotransferase, or AIaAT.
- a nitrogen utilization protein such as, for example, a high affinity nitrate transporter, a low affinity nitrate transporter, an ammonium transporter, an ammonia transporter, an amino acid transporter, alanine dehydrogenase, glutamine synthetase, asparagine synthetase, glutamate synthase, glutamate 2
- FIGURE 1 shows a schematic representation of the key steps in nitrogen utilization in a plant cell.
- Nitrate NCV
- NR nitrate reductase
- Nitrite is translocated from the cytoplasm to the chloroplast where it is reduced by nitrite reductase (NiR) to ammonium (NH 4 + ).
- Glutamine synthetase GS functions in assimilating or recycling ammonium.
- FIGURE 2 shows an alignment of the amino acid sequences (SEQ ID NO:s 29 to 45) of AIaAT from various organisms. Note that some of sequences used for these alignments are truncated sequences which contain less than the complete sequence of the cited AIaAT.
- FIGURE 3 shows an alignment of the amino acid sequences (SEQ ID NO:s 29 to 40) of AIaAT from various plant species. Note that some of sequences used for these alignments are truncated sequences that contain less than the complete sequence of the cited AIaAT. The alignment was performed using the methionine (M) of the barley AIaAT sequence as the reference first residue.
- FIGURE 4 shows the nucleotide sequence for the OS Ant 1 promoter of the invention (SEQ ID NO:!).
- the sequence was isolated using a blastn search of the National Center for Biotechnology Information (NCBI) database using the nucleotide sequence (366-3175 bp) of the Brassica btg26 gene (Stroeher et al., 1995, Plant MoI. Biol. 27:541-551) to identify the homologous rice nucleotide sequence (accession number AF323586).
- This sequence was then used in turn against the TIGR Oryza saliva sequencing project (see: tigr.org/tdb/e2kl/osal/), as set out in Example 1.
- the putative TATA box is shown in bold and the primers used in PCR amplifying the sequence from the rice genome are underlined.
- FIGURE 5 shows a schematic representation of the steps for producing the genetic construct OsAntlpro-Gus, using the reporter gene beta-glucuronidase (GUS) in accordance with the method described in Example 1.
- FIGURE 6 shows a schematic representation of the steps for producing the genetic construct OsAntlpro-AlaAT in accordance with the method described in Example 1.
- FIGURE 7 shows expression of the GUS reporter gene directed by the
- FIGURE 8 shows the average dry weight biomass (grams) of Oryza sativa plants transformed with the genetic construct OsAntl pro-AlaAT as shown in FIGURE 6 compared to the average dry weight biomass (grams) of control, wild-type Oryza sativa plants grown under the same growth conditions as given in Example 1.
- FIGURE 9 shows the average total seed weight (grams) of seeds collected from Oryza sativa plants transformed with the genetic construct OsAntl pro-AlaAT as shown in FIGURE 6 compared to the average total seed weight (grams) of seeds collected From control, wild-type Oryza sativa plants grown under the same growth conditions as given in Example 1.
- FIGURE 10 shows the relationship between dry weight biomass (grams) and total seed weight (grams) for each transgenic plant.
- FIGURE 11 shows the nucleotide sequence of the sorghum antiquitin promoter of the invention (SEQ ID NO:9). The sequence was derived from accession CW033386 as described in Example 5 and includes 443 nucleotides of sequence upstream of the ATG start codon of a sorghum antiquitin gene.
- FIGURE 12 shows the nucleotide sequence of a partial maize antiquitin promoter (SEQ ID NO: 10). The sequence was derived from accession BH215004 as described in Example 5 and contains 204-bp upstream of a maize antiquitin gene.
- Nitrogen assimilation and metabolism in plants occurs through the coordinated action of a variety of enzymes acting upon a variety of substrates ( Figure 1). Nitrogen assimilation occurs primarily through the activities of glutamine synthetase (GS) and glutamate synthase (GOGAT).
- glutamate is used as a nitrogen source to supply nitrogen for other required metabolic reactions.
- the metabolic flow of nitrogen is principally mediated by transamination reactions in which an amino group of glutamate is transferred to other carbon skeletons.
- the transfer of the amino group from glutamate to these other carbon skeletons results in the disposition of nitrogen in more readily usable forms such as other amino acids like aspartate or alanine.
- Examples of such enzymes are the aminotransferases.
- Figure 1 shows the reaction catalyzed by the enzyme AIaAT which catalyzes the transfer of an amino group from glutamate to pyruvate thus generating alanine.
- transgenic refers to a monocot plant that contains an exogenous nucleic acid molecule that can be derived from the same monocot plant species, from a heterologous plant species, or from a non-plant species.
- a “promoter” is a regulatory nucleic acid sequence, typically located upstream (5') of a gene or protein coding sequence that, in conjunction with various cellular proteins, is responsible for regulating the expression of the gene or protein coding sequence.
- Such promoters can be the full length promoter or active fragments thereof.
- active fragment is meant a fragment that has at least about 0.1%, preferably at least about 10%, and more preferably at least about 25% of the activity of a reference promoter sequence as tested via methods known to those of skill in the art for detecting promoter activity, e.g., measurement of GUS reporter gene levels.
- DNA sequences necessary for activity can be identified by synthesizing various fragments and testing for expression or introducing point mutations in certain regions and testing for loss of activity.
- coding region of interest or “target gene” includes any gene that is desirably expressed in one or more than one plant tissue.
- target protein refers to any protein that is desirably expressed in one or more than one plant tissue. Examples of a coding region of interest which may advantageously be utilized in conjunction with the methods described herein include nucleic acid sequences that encode one or more than one protein involved in nitrogen assimilation, nitrogen utilization, nitrogen uptake or a combination thereof.
- the term "elevated levels" of a protein of interest, as used herein in reference to protein levels in a transgenic monocot plant, means higher levels of protein as compared to the protein levels of a corresponding monocot plant variety lacking the transgene such as an over expressed nucleic acid molecule encoding an AIaAT.
- the gene constructs described herein can also include further enhancers, either translation or transcription enhancers, as may be required. These enhancer regions are well known to persons skilled in the art and can include the ATG initiation codon and adjacent sequences. The initiation codon must be in phase with the reading frame of the coding sequence to ensure translation of the entire sequence.
- the translation control signals and initiation codons can be from a variety of origins, both natural and synthetic. Translational initiation regions may be provided from the source of the transcriptional initiation region or from the structural gene. The sequence can also be derived from the promoter selected to express the gene and can be specifically modified to increase translation of the messenger ribonucleic acid (mRNA).
- the gene constructs of the invention can further include a 3'-untranslated (or terminator) region that contains a polyadenylation signal and other regulatory signals capable of effecting mRNA processing or gene expression.
- Nonlimiting examples of suitable 3 -regions are the 3'-transcribed non-translated regions containing a polyadenylation signal of Agrobacterium tumor-inducing (Ti) plasmid genes such as the nopaline synthase ⁇ Nos gene), plant genes such as the soybean storage protein genes, and the small subunit of the ribulose-1 , 5-bisphosphate carboxylase (ssRUBISCO) gene.
- Ti Agrobacterium tumor-inducing
- ssRUBISCO small subunit of the ribulose-1 , 5-bisphosphate carboxylase
- exogenous nucleic acid molecule means a nucleic acid molecule originating from outside the plant.
- An exogenous nucleic acid molecule can have a naturally occurring or non-naturally occurring nucleotide sequence.
- an exogenous nucleic acid molecule can be a heterologous nucleic acid molecule derived from the same plant species or a different plant species than the plant into which the nucleic acid molecule is introduced. Alternatively, it can be a nucleic acid molecule derived from a non-plant species such as fungi, yeast, bacteria or other non-plant organisms.
- AIaATs Overview of alanine aminotransferases
- aminotransferases are pyridoxal phosphate-dependent enzymes that catalyze reactions known as transamination reactions.
- the transamination reaction catalyzed by aminotransferases involves the transfer of an ⁇ - amino group from an amino acid to the ⁇ -keto position of an ⁇ -keto acid. In the process, the amino acid becomes an ⁇ -keto acid while the ⁇ -keto acid acceptor becomes an ⁇ - amino acid.
- AIaAT sequences for use in the invention is database searching. Because the genomes of a number of organisms have been seque ⁇ ced, computer-based database searching based on amino acid or nucleic acid homology will reveal sequences which are homologous to a known AIaAT that is used as the query sequence.
- One common tool for such computer database searching is the BLAST program available from the NCBl.
- the NCBl Basic Local Alignment Search Tool (BLAST) (Altschu! et al., J.
- MoL Biol 215(3):403-410, 1990 is available from several sources, including the National Center for Biotechnology Information (NCBl, Bethesda, MD) and on the Internet, for use in connection with the sequence analysis programs blastp, blastn, blastx, tblastn and tblastx. It can be accessed at the NCBI website. A description of how to determine sequence identity using this program is available at the NCBl website. An example of using a BLAST program to identify members of the AIaAT family is described in Example 7. The use of computer programs such as Softberry and PSORT can be used to determine the subcellular localization of these enzymes to exclude enzymes that are targeted to less optimal sites, i.e., to the peroxisome.
- NCBl National Center for Biotechnology Information
- PSORT can be used to determine the subcellular localization of these enzymes to exclude enzymes that are targeted to less optimal sites, i.e., to the peroxisome.
- homologous sequences identified using computer-based search methods such as those described above, can be reasonably suspected of encoding an AIaAT. Whether such a sequence actually encodes an AIaAT can be determined by a number of means. As a first indicator, the annotation to a GenBank entry is used. Many sequences have been previously identified and tested by investigators as corresponding to AIaAT activity and the annotation to such a GenBank entry would so indicate. [056] Alternatively, a sequence identified from a search can be tested experimentally to determine if it encodes an AIaAT activity. In the case of a nucleic acid sequence that has been identified, it can be isolated for testing using a variety of methods known in the art.
- the sequence of interest can be amplified by polymerase chain reaction (PCR) using primers that correspond to the 5' and 3' ends of the complementary DNA (cDNA).
- PCR polymerase chain reaction
- cDNA complementary DNA
- PCR methods are well known in the art and are disclosed in sources such as the laboratory manual PCR Protocols: A Guide to Methods and Applications by M. Innes, et ah, Academic Press, 1989.
- the desired sequence can be obtained by conventional hybridization screening using oligonucleotides corresponding to the known nucleic acid sequence to screen a cDNA library.
- the AIaAT coding region can be modified in any suitable way. For example, it can be modified to be transcribable and translatable in the plant system; for example, the nucleotide sequence encoding the AIaAT protein can be modified such that it contains all of the necessary poly-adenylation sequences, start sites and termination sites which allow the coding sequence to be transcribed to mRNA and the mRNA to be translated in the monocot plant.
- the coding region may be modified such that its codon usage is more similar to that of native genes of the monocot plant ⁇ i.e., plant optimized sequence may be used).
- plant optimized sequence may be used.
- nucleotide sequence modifications and the methods by which they may be made are well known to one of skill in the art.
- Many vectors for protein expression in E. coli, yeast, mammalian cells, or plants are commercially available. Expression of such a construct containing an AIaAT in an appropriate host cell, such as an E. coli, using a plamid such as pET vectors available from Novagen (www.Novagen.com), wilJ reveal if the plasmid encodes an AIaAT activity. Methods for assaying for AIaAT activity are well known in the art.
- AIaATs that may be used in the invention.
- high, medium, or low stringency hybridization methods can be used to isolate orthologues or homologues of known AIaATs that maybe used in the practice of this invention.
- Hybridization conditions are sequence dependent and vary according to the experimental parameters used. Generally, stringent hybridization conditions are selected to be about 5° C to 20° C lower than the thermal melting point (T m ) for the specific sequence at a defined ionic strength and pH. The T m is the temperature (under defined ionic strength and pH) at which 50% of the target sequence hybridizes to a perfectly matched probe. Conditions for nucleic acid hybridization and calculation of stringencies can be found in Sambrook el al.
- nucleic acid hybridization examples include: temperature, salt conditions, the presence of organic solvents in the hybridization mixtures, and the lengths and base compositions of the sequences to be hybridized and the extent of base mismatching.
- An example of high stringency conditions for hybridizing a probe to a filter-bound DNA is 5 X SSC, 2% sodium dodecyl sulfate
- SDS 100 ug/ml single stranded DNA at 55-65° C overnight, and washing twice in 0.1 X SSC and 0.1% SDS at 60-65° C for 20 minutes.
- Reduced stringency conditions can be used to isolate nucleic acid sequences that are related but have mismatches. Examples of such conditions include lowering the hybridization and wash temperatures or raising the salt concentrations of the wash solutions. Protocols for such medium and low stringency hybridization methods can be found in commonly used molecular biology manuals such as the aforementioned Sambrook, et al, and Ausubel, et al. references. [062] Other methods that can be used to isolate orthologues or homologues suitable for use in the invention include PCR cloning. Unique or degenerate primers can be designed to encode conserved regions in AIaAT nucleotide or amino acid sequences.
- Such conserved regions can be identified by aligning the sequences of known AIaATs using the alignments disclosed above.
- the PCR primers so designed can be used in PCR reactions to generate a portion of an AIaAT sequence from a species of interest which then can be used to isolate a full length cDNA by conventional library screening methods or by means of additional PCR methods such as Rapid Amplification of cDNA Ends (RACE).
- RACE Rapid Amplification of cDNA Ends
- An alternative strategy for identifying AIaATs for use in the invention entails the biochemical purification of AIaATs from a source of interest based on enzymatic activity. Because enzymatic assays for AIaAT activity are well known in the art, a skilled artisan would be able to fractionate a cell or tissue of interest and use conventional biochemical methods such as chromatography to purify an AIaAT to homogeneity. Such biochemical methods are available in sources such as Protein Purification: Principles and Practice by Robert K. Scopes, Springer Advanced Texts in Chemistry, 3rd edition, 1994; Guide to Protein Purification (Methods in Enzymology Series, Vol.
- AIaAT once purified to homogeneity, can be used to derive partial amino acid sequences, from which oligonucleotides can be designed to clone the corresponding cDNA by conventional molecular biological methods such as library screening or PCR as described above.
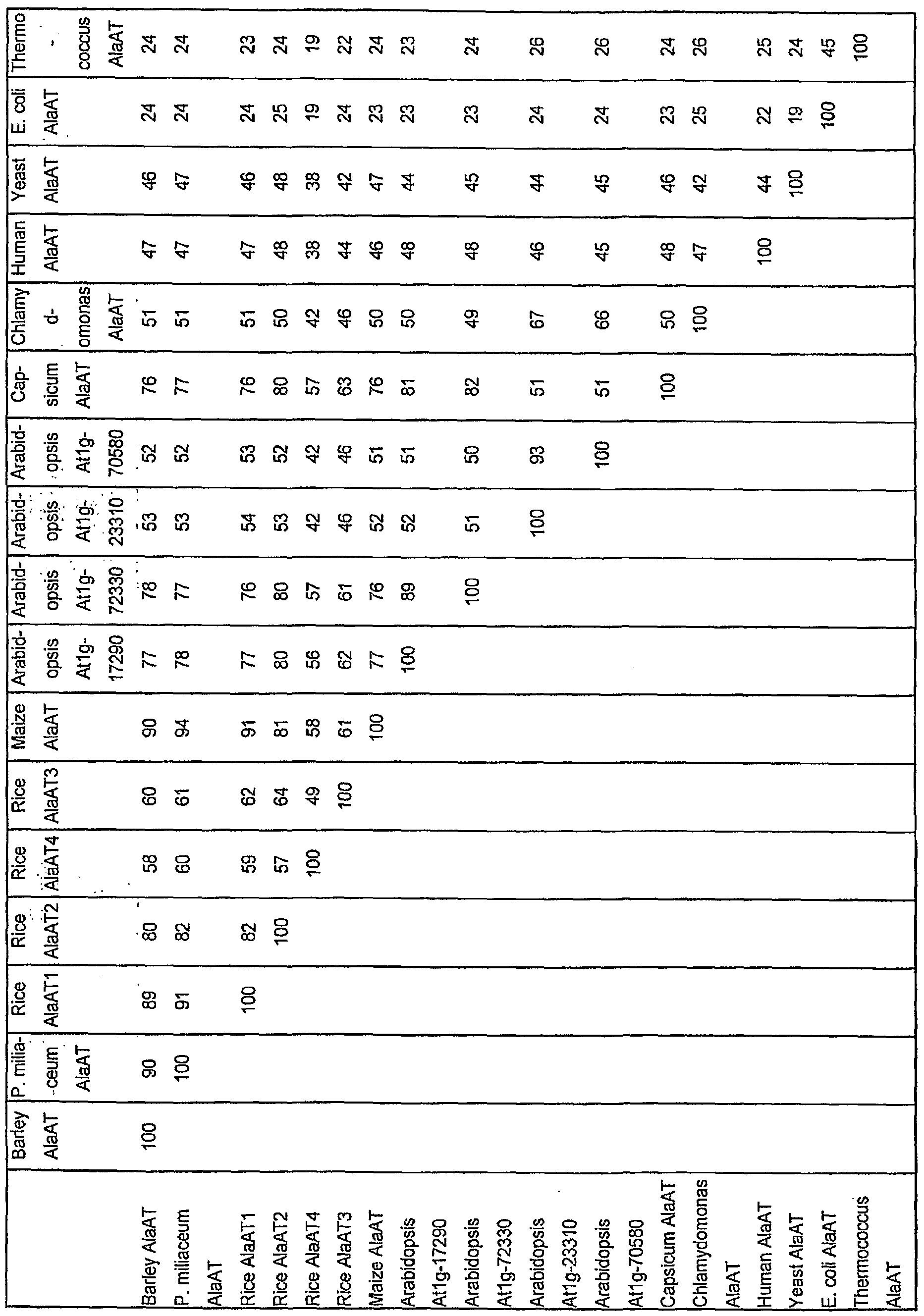
- Figures 2 and 3 and Tables 1 and 2 show alignments between AIaATs from a variety of species, ranging from E. coli to humans and including a number of plant species. The percent homologies range from over 90% to under 25% when the sequence of each AIaAT is compared with that of every other AIaAT as shown in Table 1.
- an AIaAT has been identified and verified as corresponding to a bona fide AIaAT, a construct for overexpression of the AIaAT in a monocot plant of interest is generated using methods well known in the art.
- a variety of plasmids are available for this purpose as disclosed below.
- a variety of promoters such as constitutive promoters, various inducible promoters, or tissue-specific promoters can be used for expression. Promoters
- the promoters suitable for use in the constructs of this invention are functional in monocot plants and in host organisms used for expressing the constructs described. Many plant promoters are publicly known and several examples are listed below. These include constitutive promoters, inducible promoters, tissue- and cell-specific promoters and developmentally regulated promoters. Methods are disclosed below for the selection of promoters that are suitable for use in practicing the invention. [067] Promoters can be isolated by procedures well known in the art of plant molecular biology.
- One type of promoter particularly useful for expression of a target gene such as AIaAT in a plant is a monocot antiquitin promoter.
- the rice antiquitin promoter is described in Example 1.
- Other antiquitin promoters are described in Example 5. Knowing the monocot antiquitin promoters disclosed in these Examples, one of skill could readily identify other monocot antiquitin promoters using methods similar to those described in Example 1 for identification of the rice antiquitin promoter using the btg 26 gene.
- the sequence can be subject to analysis with a promoter prediction software such as the TSSP plant promoter prediction software found at http://softberry.com to identify likely TATA box sequences and other promoter sequence elements and further analyzed for promoter motifs that may be recognition sites for transcription factors using Signal Scan Software (Prestridge, 1991 ; available at bimas.dcrt.nih.gov/molbio/signal).
- a promoter prediction software such as the TSSP plant promoter prediction software found at http://softberry.com to identify likely TATA box sequences and other promoter sequence elements and further analyzed for promoter motifs that may be recognition sites for transcription factors using Signal Scan Software (Prestridge, 1991 ; available at bimas.dcrt.nih.gov/molbio/signal).
- Sequences likely to encode promoters can be confirmed by synthesizing various fragments and testing for expression or introducing point mutations in certain regions and testing for loss of activity using any assay system known to those of skill in the art as being useful for measuring the promoter
- Reporter genes can be any polynucleotide the transcription of which under the control of a promoter sequence, the subsequent translation thereof, or both, can be readily detected by a skilled artisan.
- the reporter gene does not have to encode a full length protein. In some instances, the reporter gene can even be an oligonucleotide. Most commonly, the reporter gene encodes a protein with detectable activity.
- Common reporter genes include GUS, luciferase, GFP, beta-galactosidase, CAT, alkaline phosphatase, etc. In preferred embodiments, the reporter gene is GUS.
- the expression of the reporter gene can be measured at either the mRNA or protein level using any method known to those of skill in the art.
- mRNA levels can be detected using a cell-free transcription assay.
- protein levels can be measured by detecting enzyme activity, using antibodies specific for the protein, or a transcription-translation assay, which allows detection of both the mRNA level and the protein or peptide level.
- Promoters from genes that are regulated similarly to the a ⁇ tiquitin genes in plants might also find use in the invention. These genes could be turgor responsive genes that are expressed in root tissues and could be induced by ABA and/or under stress conditions such as drought and salt.
- transgenic plants of interest can be generated using transformation methods well known in the art and described herein as well as in the Examples below.
- An exogenous nucleic acid molecule can be introduced into a monocot plant for ectopic expression using a variety of transformation methodologies including Agrobacterium-mediated transformation and direct gene transfer methods such as electroporation and microprojectile-mediated transformation (see, generally, Wang et al. (eds), Transformation of Plants and Soil Microorganisms, Cambridge, UK: University Press, 1995, which is incorporated herein by reference).
- Transformation methods based upon the soil bacterium, Agrobacterium lumefaciens, are particularly useful for introducing an exogenous nucleic acid molecule into a seed plant.
- the wild-type form of Agrobacterium contains a Ti (tumor-inducing) plasmid that directs production of tumorigenic crown gall growth on host plants. Transfer of the tumor- inducing T-DNA region of the Ti plasmid to a plant genome requires the Ti plasmid- encoded virulence genes as well as T-DlMA borders, which are a set of direct DNA repeats that delineate the region to be transferred.
- An Agrobacterium-based vector is a modified form of a Ti plasmid, in which the tumor inducing functions are replaced by the nucleic acid sequence of interest to be introduced into the plant host.
- Agrob ⁇ cterium-m ⁇ d ' iated transformation generally employs cointegrate vectors or, preferably, binary vector systems, in which the components of the Ti plasmid are divided between a helper vector, which resides permanently in the Agrobacterium host and carries the virulence genes, and a shuttle vector, which contains the gene of interest bounded by T-DNA sequences.
- a variety of binary vectors are well known in the art and are commercially available, for example, from Clontech (Palo Alto, CA).
- Microprojectile-mediated transformation also can be used to produce a transgenic plant that ectopically expresses AIaAT. This method, first described by Klein et al. ⁇ Nature 327:70-73, 1987, which is incorporated herein by reference), relies on microprojectiles such as gold or tungsten that are coated with the desired nucleic acid molecule by precipitation with calcium chloride, spermidine or PEG. The microprojectile particles are accelerated at high speed into a plant tissue using a device such as the BIOLISTIC PD- 1000 (Biorad, Hercules, CA).
- Microprojectile-mediated delivery or "particle bombardment” is especially useful to transform plants that are difficult to transform or regenerate using other methods.
- Microprojectile-mediated transformation has been used, for example, to generate a variety of transgenic plant species, including cotton, tobacco, maize, hybrid poplar and papaya (see G lick and Thompson, supra, 1993) as well as cereal crops such as wheat, oat, barley, sorghum and rice (Duan et al, Nature Biotech. 14:494-498, 1996; Shimamoto, Curr. Opin. Biotech. 5:158-162, 1994; each of which is incorporated herein by reference).
- transgenic plant species including cotton, tobacco, maize, hybrid poplar and papaya (see G lick and Thompson, supra, 1993) as well as cereal crops such as wheat, oat, barley, sorghum and rice (Duan et al, Nature Biotech. 14:494-498, 1996; Shimamoto, Curr. Opin. Biotech. 5:158
- Agrobacterium-medlated or microprojectile-mediated transformation as disclosed herein, or other methods known in the art can be used to produce a transgenic seed plant of the invention.
- Alternative gene transfer and transformation methods useful in the invention include, but are not limited to, liposomes, electroporation or chemical-mediated uptake of free DNA, calcium phosphate co-precipitation techniques, and micro- or macro injection, direct DNA transformation, and may involve Ti plasmids, Ri plasmids, or plant virus vectors. Such transformation methods are well documented in the art. Growth and NUE assays [077] The resulting transgenic plant of interest are tested for expression of the
- AIaAT transgene and those plant lines that express the AIaAT transgene are tested for the effect of the expressed transgene on plant growth or nitrogen utilization.
- Suitable tests for monocot plant growth can include a variety of assays such as measuring plant height, seed weight, stem diameter, number of plant leaves, plant biomass as measured in fresh weight or dry weight of roots, leaves, shoots, buds, and flowers, to name but a few such measurement parameters.
- Tests for NUE can include growth of transgenic plants under different suboptimal nitrogen conditions. Tests may be Field test, greenhouse or growth chamber tests or in vitro tests. Plants may be grown hydroponically in PerliteTM, other commercially available growing material, soil, or in agar-based media.
- Use of monoeot antiquitin promoters to direct expression of other coding regions [078] Monoeot antiquitin promoters can also be used to direct expression of coding regions other than AIaAT.
- the coding region of interest, or target gene, operatively linked to the monoeot antiquitin promoter may be any nucleotide sequence that is desirably expressed within a plant.
- General classes of coding regions which may be advantageously employed in the methods and constructs of the invention include nucleotide sequences encoding structural proteins; proteins involved in the transport of nitrogen; proteins involved in the uptake of nitrogen; proteins involved in both the transport and uptake of nitrogen; enzymes and proteins involved in nitrogen utilization; proteins involved in plant resistance to pesticides or herbicides; proteins involved in plant resistance to nematodes, viruses, insects, or bacteria; proteins involved in plant resistance to stress, for example but not limited to osmotic, temperature, pH, or oxygen stress; proteins involved in stimulation or continuation of plant growth; proteins involved in phytoremediation; or proteins having pharmaceutical properties or encoding enzymes which produce compounds having pharmaceutical properties.
- the coding region of interest may encode a nitrogen utilization protein and, in particular, an enzyme that assimilates ammonia into amino acids or uses the formed amino acids in biosynthetic reactions.
- This protein may be selected from, but not limited to, a nitrate transporter (high or low affinity), an ammonium transporter, an ammonia transporter, an amino acid transporter, alanine dehydrogenase, glutamine synthetase (GS), asparagine synthetase (AS), glutamate synthase (also known as glutamate 2:oxogluturate amino transferase and GOGAT), asparaginase (ANS), glutamate dehydrogenase (GDH), nitrate reductase, aspartate aminotransferase (AspAT), AIaAT, and other known aminotransferases.
- a nitrate transporter high or low affinity
- an ammonium transporter an ammonia transporter
- the target gene or coding region of interest may be naturally expressed in the plant or it may be heterologous to the plant.
- the gene may originate from any source, including viral, bacterial, plant or animal sources.
- the coding region of interest is heterologous to the monocot antiquitin promoter sequence to which it is operatively linked, in that it is not from the gene the monocot antiquitin promoter sequence is naturally linked to.
- the coding region can be modified in any suitable way in order to engineer a gene with desirable properties.
- the coding region can be modified to be transcribable and translatable in the plant system; for example, the nucleotide sequence encoding the protein of interest can be modified such that it contains all of the necessary poly- adenylation sequences, start sites and termination sites which allow the coding sequence to be transcribed to mRNA (messenger ribonucleic acid) and the mRNA to be translated in the plant. Further, the coding region may be modified such that its codon usage is more similar to that of native genes of the plant (i.e., plant optimized sequence may be used). Such nucleotide sequence modifications and the methods by which they may be made are well known to one of skill in the art.
- Example 1 Demonstration of NUE in rice expressing barley AIaAT Identification and characterization of a rice antiquitin promoter COs ⁇ ntl ) [085] The nucleotide sequence (bp 366-3175) of the btg26 gene (Stroeher et al.,
- OsAntl gene was selected for further analysis. To determine if the sequence was likely to function as a promoter sequence, the sequence was analyzed using the TSSP plant promoter prediction software found at http://softberry.com/. The analysis predicted that the sequence was a plant promoter sequence. The most likely location of the TATA box (bold in Figure 4), as well as other promoter sequence elements, was determined.
- the OsAnt l sequence was compared to nucleic acid sequences of btg26 promoter sequences from Brassica napus and Arabidopsis using the ClustalW 1.8 multiple sequence alignment software on the BCM Search Launcher homepage (searchlauncher.bcm.tmc.edu/) and BOXSHADE server (ch.embnet.org/software/BOX_forrn.html). Inspection of conserved nucleotides revealed that the Brassica and Arabidopsis turgor gene-26 promoter sequences are more similar to each other than to the OsAntl sequence.
- a feature among all three promoter sequences is the polypyrimidine (CT) tracts evident within the nucleotide sequences. These tracts range from 20-22 bases and are found just upstream of the probable TATA boxes in all three promoter sequences. Furthermore, the OsAntl sequence has a second polypyrimidine tract just upstream of the ATG start codon.
- Rice genomic DNA was isolated from cv. Kitaake. The following PCR primers (positions underlined in Figure 4) corresponding to the OsAntl promoter region were selected: Primer 1 : AGGAAGTGATTTTTAGCGTAGCTG (SEQ ID NO:2);
- Primer 2 ATGGCAGAAGAGAGAGAGAGAGG (SEQ ID NO:3).
- Touch-down PCR was conducted using rice genomic DNA and the above primers. A 975-bp fragment was produced. The amplified PCR fragment was ligated into pCR ® II-TOPO vector (Invitrogen) and transformed into E. coli, TOP 10 cells. The resulting piasmid is designated pT-riceOsAntl pro.
- the beta-glucuronidase (GUS) reporter gene driven by OsAntl was produced using the steps shown schematically in Figure 5.
- the RiceOsAntl pro-GUS construct was produced by amplifying the pT-RiceOsAntlpro template using the following primers:
- Primer 3 EcoRI-OsAntl promoter sequence GGAATTCAGGAA GTGATTTTT
- Primer 4 NcoI-OsAnt 1 promoter sequence CATGCCATGGATGGCAGAAGA
- the resultant PCR fragments were ligated into the plant binary vector, pCAMBlA 1305.1 , digested with EcoRl and Ncol to produce a pCAMBlA 1305.1- riceOsAntlpro-GUS construct.
- the Ncol sequence (CCATGG) includes a Met codon, ATG, which is in frame with the GUS reporter gene and allows expression of the GUS reporter gene from the OsAntl promoter sequence.
- the barley AIaAT gene driven by OsAntl was produced using the steps shown schematically in Figure 6.
- the RiceOsAntl pro-AlaAT construct was produced by amplifying the pT-RiceOsAnt 1 pro template using the following primers:
- Primer 3 EcoRl-OsAntl promoter sequence GG AATTCAGG A AGTG ATTTTT
- Primer 5 Pstl-OsAntl promoter sequence AACTGCAGATGGCAGAAGA (SEQ ID NO.-6)
- An AIaAT DNA fragment was amplified by PCR using pAGOOl as a template.
- pAGOOl is described in U.S. Patent No. 6,084,153 where it is identified as pbtg26/AlaAT/nos. It contains the btg26 promoter linked to the barley AIaAT gene with a nopaline synthase terminator.
- the barley AlaAT/nos terminator sequences were amplified from pAGOOl using the following primers;
- Primer 7 HindlH-NOS terminator sequence CCCAAGCTTCCCGATCTAGTA
- R2-CS R2 major and minor salts, vitamins and iron source
- R2-AS R2 major and minor salts, vitamins and iron source
- R2S R2 major and minor salts, vitamins and iron source +
- Pre-regeneration medium mg/1 NAA + 50 mg/1 hygromycin + 100 mg/1
- the putatively transgenic, hygromycin-resistant calli were gently picked out, transferred, cultured on pre-regeneration medium (PRN, Table 3) and then incubated for a further week. All of the resistant calli originating from a single co-cultured embryogenic nodular unit were grouped in a sector of the PRN dish. Creamy-white, lobed calli with a smooth and dry appearance were individually transferred to regeneration medium (RN, Table 3), incubated for 2 days in the dark, then maintained for three weeks under a 12/12-h (day/night) photoperiod with light provided at an intensity of 55 ⁇ mo l/m per sec.
- Green shoots regenerating from a resistant callus were dissected and sub-cultured in test tube containing rooting medium (R, Table 3) for 1 -2 weeks to promote vigorous roots and tillers before being transferred to pots in growth rooms.
- Transgenic plants were grown to maturity in 16-cm pots containing soil-less potting mixture (Metromix 220). Plants were maintained in growth rooms set to 28° C and 14/10 hours day/night photoperiods. Fertilizer was applied twice a week starting two weeks after planting in pots.
- the fertilizer mix contained 225 g 20/20/20 fertilizer, 50 g of plant micronutrients, 6.1 g of CuSO 4 .5H 2 O, 140 g FeEDTA, 13.8 g ZnSO 4 .7H 2 0, 260 g
- Example 2 Demonstration of NUE in maize using OsAntl/barley AIaAT
- the OsAntl -pro-AlaAT construct can be incorporated into suitable plant binary vectors for use in Agrobacterium-mediated transformation of maize.
- Many methods for transformation of immature embryos of maize using a variety of selectable markers are known in the art (Ishida et ⁇ l, Nature Biotech. 14:745-750, 1996; Lupotto, Maydica 44:21 1-218, 1999; Zhao et al, Molec. Breeding 8:323-333, 2001; Frame et al, Plant Physiol 129: 13-22, 2002 and Miller et al., Transgenic Res. 1 1 :381 -396, 2002, U.S. Patent No. 5,591 ,616. Contract production of transgenic maize plants is also available through facilities such as the Plant Transformation Facility, Iowa State University, Ames, Iowa.
- the OsA ntlpro-A IaAT sequence can be used similarly in biolistic transformation methods for maize (Wright et al, Plant Cell Reports 20(5):429-436, 2001 ; Brettschneider et al, Theorel. Appl Genet. 94:737-748, 1997; Gordon-Kamm et al, Plant Cell 2(7):603-6l 8, 1990; Fromm et al, Biotechnology (N Y). 8(9):833-9. 1990).
- Maize plants can be tested forNUE by measurement of biomass and seed yield during growth under various nitrogen fertilizer regimes including limiting nitrogen. Plant biomass can be fresh weight or dry weight, total plant weight, leaf weight or root weight.
- Suboptimal nitrogen conditions are those conditions in which nitrogen concentrations limit growth. Under such conditions, addition of added nitrogen such as fertilizer will increase growth. For each of these tests, biomass and seed yield can be evaluated in growth chamber, greenhouse or field tests .
- Example 3 Demonstration of NUE in wheat using OsAntl/barley AIaAT [0120] Similar to maize, the OsAntl-pro-AlaAT construct can be used for particle-gun bombardment transformation methods of wheat (Pastori et al, J. Exp. Bot. 52(357):857- 863, 2001 ; Becker et al, Plant J.
- Example 4 Demonstration of NUE in sorghum using OsAntl/barley AIaAT [0122] ⁇ grobacte ⁇ um-mediated sorghum transformation of immature embryos with a binary vector containing any of the OsAnt promoter/AlaAT constructs can be achieved according to methods established in the art (Zhao et ⁇ l., Plant MoI. Biol. 44(6):789-98, 2000; Gao et al, Genome 48(2):321-33, 2005; Zhao, Z. Y., Methods MoI Biol 343:233- 44, 2006; Howe et al, Plant Cell Rep. 25(8):784-91, 2006).
- Sorghum plants can be tested for NUE by measurement of biomass and seed yield during growth under various nitrogen fertilizer regimes including limiting nitrogen.
- Plant biomass can be fresh weight or dry weight, total plant weight, leaf weight or root weight.
- Suboptimal nitrogen conditions are those conditions in which nitrogen concentrations limit growth. Under such conditions, addition of added nitrogen such as fertilizer will increase growth. For each of these tests, biomass and seed yield can be evaluated in growth chamber, greenhouse or field tests.
- Example 5 Identification of alternate (antiquitin) promoter sequences for use in NUE constructs
- nucleotide sequence (bp 366-3175) of the btg26 gene (Stroeher et al., Plant MoI. Biol. 27:541-551 , 1995; accession number S77096) is used to search the nucleotide database at NCBI using the blastn search tool.
- sorghum accession number U87982
- maize accession numbers AY103614 and BT017791
- cocoa Theobroma cacao; accession number DQ448866; and Curculigo latifolia, accession number X641 10
- ESTs for wheat, sugarcane and switchgrass can also be identified in databases using the identified rice antiquitin nucleotide or amino acid sequences using various search algorithms.
- a sorghum promoter sequence was identified by using the rice nucleotide sequence of the antiquitin clone (accession number AF323586) in a BLAST search of the sorghum sequences in the NCBl Genome Sequence Survey (gss) Database.
- Clone CW033386 was identified as containing 443 nucleotides of sequence upstream of the ATG start codon of a sorghum antiquitin gene (SEQ ID NO:9, Figure 1 1). This sequence can be used as a promoter sequence alone or methods to clone and sequence larger genomic fragments can be used to identify sequences further upstream.
- This sequence can be used as a promoter sequence alone or methods to clone and sequence larger genomic fragments can be used to identify sequences up to 1.5 kb upstream of this particular antiquitin gene. Sequences including the longer promoters could be used to design promoter/A IaAT gene constructs as described below.
- Promoter cassettes for expression of various genes are constructed by combining the promoter of interest with a nos terminator with convenient restriction sites in between the promoter and terminator for gene cloning. Other restriction sites flank the promoter and terminator to facilitate movement of the cassette to a binary vector for plant transformation.
- a base vector containing the nos terminator is constructed by PCR amplifying the nos region contained in the binary vectors described in U.S. Patent No. 6,084,153 with the primers NOSupper2: 5'-CCTAGGCCATGGTTCAAACATTTGGCAATAAAGTTT-
- NOSupper2 supplies Avrll and Ncol restriction sites at the 5'-end of the nos terminator and NOSlower supplies a Pacl site at the 3'end of the amplified fragment.
- PCR was performed using the BD AdvantageTM 2 PCR kit following manufacturer's instructions.
- the resulting 263 bp fragment is cloned into pCR ® 2.1 -TOPO ® vector using a TOPO TA
- Ncol site Deletion of the Ncol site may be verified by restriction analysis and growth of the E. coli on kanamycin. This resulting plasmid is Nos/pCR2.1 mut.
- OsAntl promoter An alternative expression cassette for expressing genes from the OsAntl promoter is made in the following manner.
- the OsAntl promoter is cloned from rice var. Nipponbare genomic DNA (made by manufacturer's recommendation, Sigma Extract-n-
- the forward primer incorporates Avr II, Pad and Pmel restriction sites and the reverse primer incorporates BspHl, Aat II and AsiSI and restriction sites to facilitate further cloning steps.
- the resulting 1.1 kb fragment (corresponding to nucleotides 101336-100216 of AP005570) is cloned into pCR®2.1-TOPO® vector using a TOPO
- the resulting plasmid is digested with restriction enzymes Avr II and BspH l and is cloned into Nos/pCR2.1 mut that has been digested with Avr 11 and Ncol .
- the resulting construct has an OsAntl promoter and a nos3'-region with unique AsiSI and AatII sites between them for cloning genes of interest.
- the expression cassette is flanked
- TTTAATTCATGAGACGTCTTTGCGATCGCGGCGCCGGCGGCGTTGGCAGGT- 3' can be used to amplify a 443-bp Ant promoter (SEQ ID NO:9) from sorghum genomic DNA as described above for the OsAntl promoter and rice DNA.
- the cloned promoter SEQ ID NO:9
- 25 fragment is flanked by Avrll, Pac 1 and Pme 1 restriction sites on the 5'-end and BspHl, Aat II and Asi SI sites on the 3'-end.
- the promoter fragment is digested with restriction enzymes Avr II and BspH l and is cloned into Nos/pCR2.tmut that has been digested with Avr II and Ncol .
- the resulting construct has a sorghum Ant promoter and a nos3'- region with unique AsiSI and Aat Il sites between them for cloning genes of interest.
- 30 expression cassette is flanked by Avr II, Pac I, and Pme I restriction sites on the 5'-end and a Pacl restriction site on the 3'-end to facilitate movement into a plant binary expression vector.
- An expression cassette utilizing a maize Ant promoter (see Example 5) is also designed in a similar manner to that described for the rice and sorghum. Promoter regions from other antiquitin genes can also be used as they are identified from genome sequencing projects and other technologies.
- Example 7 Identification and cloning of alternate alanine aminotransferase (AIaAT) genes for use in NUE constructs
- Aminotransferases are enzymes which catalyze the reversible transfer of amino groups from amino acids to oxo acids. They can be divided into four subgroups based on mutual structural relatedness (Mehta et ai, Eur. J. Biochem. 214(2):549-561, 1993). AIaAT enzymes catalyze the reversible interconversion of alanine and 2-oxoglutarate to pyruvate and glutamate and belong to subgroup 1. In addition to the barley alanine aminotransferase, other alanine aminotransferases are useful for conferring NUE in monocots.
- the barley AIaAT protein sequence (NCBl accession number CAA81231) was used as a query to search the " NCBI protein sequence database using the BLAST algorithm. Genes with a high degree of sequence homology to barley AIaAT were found in all major classes of eukaryotes. Related sequences were also found in bacteria. A tBlastn search of the NCBI EST database revealed that AIaAT homologs are widespread in plants, but because most of these sequences were not full length they were not analyzed further. As additional genomic sequences for monocots become available, additional homologs may be identified using these methods.
- Tissue for RNA isolation was prepared from maize (A 188) and rice (Nipponbare) in the following manner. Seeds were germinated in H 2 O on germination paper at 24°C in a sealed bag (maize, rice). After 7 days root tissue was collected and stored in RNAlater ® (Ambion) for RNA isolation. Seedlings of pepper ⁇ Capsicum annuum, Pepper Hot Asia, Santaka, Botanical Interests Broomf ⁇ eld, CO) were sterilized and germinated in half strength MS and whole seedlings were used. Leaves from soil-grown Arabidopsis plants (Columbia 0) were used.
- RNA was prepared from the plant tissues using the RNAqueousTM-4PCR kit (Ambion).
- cDN ⁇ was synthesized from purified RNA using the Superscript Hl platinum ® 2-step q-RT-PCR kit (Invitrogen) as per the manufacturer's instructions.
- PCR amplification of AIaAT [0139] AIaAT genes may be amplified by PCR from cDNA from many sources including maize ⁇ Zea mays), rice ⁇ Otyza sativa), Arabidopsis thaliana, or pepper ⁇ Capsicum annuum). The template for barley (Hordeum vulgar ⁇ L.
- AIaAT is plasmid pAGOOl (obtained from Allen Good, University of Alberta) which contains the barley AIaAT coding sequences as described in Muench and Good, 1994, GenBank accession CAA81231.
- PCR primers contain an AsiS I restriction site on the 5'-end and an Aat II restriction site at the 3 '-end to facilitate cloning into expression cassettes.
- AIaAT genes may be amplified using the BD AdvantageTM 2 PCR kit following manufacturer's instructions (Clontech, Mountain View, CA). The resulting PCR products may be purified using QIAquickTM Purification Kit (Qiagen®, Hilden,
- the AIaAT gene in each of the expression constructs is sequence verified for PCR fidelity and integrity of the ATG start codon.
- Example 8 Binary vector construction and plant transformation.
- Ant promoter/AlaAT gene/nos 3' expression cassettes are cloned into a binary vector for plant transformation by digestion with Pmel and Pacl and ligation with pARC 1 10 digested with the same enzymes.
- pARCl 10 is an Agrobacterium binary vector originally based on pZPl OO (Hajdukiewicz et al, Plant MoI. Biol. 25, 989-994,
- pARCl 10 utilizes a Basta selectable marker driven by a CaMV 35S promoter and a nos terminator.
- the selectable marker is located near the left border, and the unique restriction sites Xba I, Avr II, Pac 1, and Pst I have been engineered close to the RB for gene cloning.
- the chloramphenicol bacterial selectable marker in the backbone of pZPlOO was also replaced with the kanamycin resistance gene (nptl ⁇ l) from the pCAMBlA 1304 vector (found on the internet at the site cambia.org.au').
- the promoter/AlaAT/nos 3' gene binary vectors can be introduced into Agrobacte ⁇ um turnefaciens strains for Agrobacterium-mediate ⁇ transformation of monocot crop plants or vector DNA is used for particle gun bombardment methods of plant transformation.
- Example 9 Use of alternate antiquitin/AlaAT constructs in rice transformation using selection on bialophos [0154] Agrob ⁇ cteriurn-med ⁇ ated rice transformation with the OsAntl /AIaAT construct, or any alternate Ant/AlaAT construct, is achieved using a transformation method based on the method described in U.S. Patent No. 7,060,876 and European Patent No. 672752B1. A detailed description follows.
- Plasmids were transferred into Agrob ⁇ cterium strain EHA 105 (Hood el ⁇ l., Transgenic Res. 2: 208-218, 1993) by electroporation (Sambrook el at. in Molecular Cloning, A Laboratory Manual Cold Spring Harbor, N.Y.: Cold Spring Harbor
- Agrobacterium cells were plated on solid AB medium (Chilton el al, 1974) containing 50 mg/1 kanamycin and incubated at 28° C for 3 days. The bacteria were then collected with a flat spatula and resuspended in liquid co-cultivation medium (R2-CL, Table 4) by gentle vortexing prior to transforming the rice tissues.
- R2-CL liquid co-cultivation medium
- the seeds were dehusked and surface sterilized by dipping (1 min) in 70 % (v/v) ethanol followed by soaking in 50% bleach plus 0.1% Tween-20 for 10 min and then rinsing five times in sterile distilled water. Following sterilization, seeds were cultured on callus induction medium (N6C, Table 4) and incubated for three weeks in the dark at 26°C.
- callus induction medium N6C, Table 4
- Table 4 Medium used for callus induction, inoculation, co-culture, resting phase, selection, regeneration and rooting
- Liquid co-culture medium without sucrose (Ohira et al. (1973) Plant and Cell (filter sterilize) Physiol. 14:1 1 13-1 121) + 0.25 M glucose + 125 ⁇ M acetosyringone + 2.0 mg/1 2,4-D, pH 5.2
- N6 liquid medium containing 400 mg/1 Timentin for disinfection and placed for 4 hours on an orbital shaker (100 rpm) at 26°C in the dark. After dry blotting on sterile filter paper, calli were placed on N6 selection medium (N6S, Table 4) and kept at 26°C in dark. 10159] After 4 weeks of culture, uncontaminated embryogenic units had developed into large yellowish globular structures that were transferred onto fresh N6S medium and cultured for another 4-5 weeks at 26°C in dark.
- the globular structures had proliferated many round-shaped, compact and yellowish calli. These putatively transgenic, bialaphos-resistant calli were gently picked out, transferred and cultured on regeneration medium (RH, Table 4), incubated for 1 week in the dark, then maintained for 4-5 weeks under a 14/10 hours day/night photoperiod with light provided at an intensity of 70 ⁇ mol/m per sec. Green shoots regenerating from a resistant callus were dissected and sub-cultured in culture vessels containing rooting medium (R, Table 4) for 2 weeks to promote vigorous roots and tillers before being transferred to 2-inch pots filled with sterile Sunshine Mix #3.
- RH regeneration medium
- the transgenic plantlets were acclimated by maintaining them in growth rooms set to 26° C, 14/10 hours day/night photoperiod and high humidity. Fertilizer was applied three times a week starting two weeks after planting in pots.
- the fertilizer mix is Simmons Solution (San Joaquin Sulphur Co., Lodi, CA) with addition of calcium nitrate. Sixteen g of Simmons and 6Og of calcium nitrate are mixed for 40 gallons of fertilizer.
- Nitrogen efficient monocol plants including but not limited to maize, sorghum, barley, wheat, rye and grass can be developed using the methods outlined in the above examples.
- the invention has been described with regard to one or more embodiments.
Landscapes
- Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Chemical & Material Sciences (AREA)
- Zoology (AREA)
- Organic Chemistry (AREA)
- Biomedical Technology (AREA)
- Biotechnology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Wood Science & Technology (AREA)
- General Engineering & Computer Science (AREA)
- Molecular Biology (AREA)
- Microbiology (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Cell Biology (AREA)
- Physics & Mathematics (AREA)
- Biophysics (AREA)
- Plant Pathology (AREA)
- Medicinal Chemistry (AREA)
- Nutrition Science (AREA)
- Breeding Of Plants And Reproduction By Means Of Culturing (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Enzymes And Modification Thereof (AREA)
Abstract
Description

Claims
Priority Applications (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| AU2006330817A AU2006330817B2 (en) | 2005-12-23 | 2006-12-21 | Nitrogen-efficient monocot plants |
| CA2634925A CA2634925C (en) | 2005-12-23 | 2006-12-21 | Nitrogen-efficient monocot plants |
| BRPI0620312-4A BRPI0620312A2 (en) | 2005-12-23 | 2006-12-21 | monocotyledon plants with nitrogen efficiency |
| EP06848873A EP1983819A4 (en) | 2005-12-23 | 2006-12-21 | Nitrogen-efficient monocot plants |
| CN2006800487181A CN101378651B (en) | 2005-12-23 | 2006-12-21 | Nitrogen-efficient monocot plants |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US75381805P | 2005-12-23 | 2005-12-23 | |
| US60/753,818 | 2005-12-23 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| WO2007076115A2 true WO2007076115A2 (en) | 2007-07-05 |
| WO2007076115A3 WO2007076115A3 (en) | 2008-06-19 |
Family
ID=38218711
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/US2006/049241 WO2007076115A2 (en) | 2005-12-23 | 2006-12-21 | Nitrogen-efficient monocot plants |
Country Status (9)
| Country | Link |
|---|---|
| US (2) | US8288611B2 (en) |
| EP (1) | EP1983819A4 (en) |
| CN (1) | CN101378651B (en) |
| AR (2) | AR059119A1 (en) |
| AU (1) | AU2006330817B2 (en) |
| BR (1) | BRPI0620312A2 (en) |
| CA (1) | CA2634925C (en) |
| WO (1) | WO2007076115A2 (en) |
| ZA (1) | ZA200805007B (en) |
Cited By (31)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP1968372A2 (en) * | 2005-12-23 | 2008-09-17 | Arcadia Biosciences Inc. | Promoter sequence obtained from rice and methods of use |
| WO2009013225A2 (en) * | 2007-07-20 | 2009-01-29 | Cropdesign N.V. | Plants having enhanced yield-related traits and a method for making the same |
| WO2009149304A2 (en) * | 2008-06-04 | 2009-12-10 | Edenspace Systems Corporation | Plant gene regulatory elements |
| US7786343B2 (en) | 1996-02-14 | 2010-08-31 | The Governors Of The University Of Alberta | Transgenic plants expressing recombinant barley alanine aminotransferase |
| WO2010120862A1 (en) * | 2009-04-14 | 2010-10-21 | Pioneer Hi-Bred International, Inc. | Modulation of acc synthase improves plant yield under low nitrogen conditions |
| WO2010111568A3 (en) * | 2009-03-27 | 2010-11-18 | E. I. Du Pont De Nemours And Company | Plants having altered agronomic characteristics under nitrogen limiting conditions and related constructs and methods involving genes encoding snf2 domain-containing polypeptides |
| CN101942512A (en) * | 2010-08-25 | 2011-01-12 | 中国农业科学院作物科学研究所 | Development and application of molecule marker for corn with considerable number of kernels and excellent allele function in low-nitrogen adverse environment |
| WO2011022608A3 (en) * | 2009-08-20 | 2011-04-14 | Pioneer Hi-Bred International, Inc. | Functional expression of yeast nitrate transporter (ynt1) in maize to improve nitrate uptake |
| WO2012049663A1 (en) | 2010-10-15 | 2012-04-19 | Genoplante-Valor | Production of plants having improved water-deficit tolerance |
| WO2012049661A1 (en) | 2010-10-15 | 2012-04-19 | Genoplante-Valor | Production of plants with improved tolerance to water deficit |
| WO2012059497A1 (en) | 2010-11-02 | 2012-05-10 | Bayer Cropscience Ag | N-hetarylmethyl pyrazolylcarboxamides |
| WO2012089757A1 (en) | 2010-12-29 | 2012-07-05 | Bayer Cropscience Ag | Fungicide hydroximoyl-tetrazole derivatives |
| US8288611B2 (en) | 2005-12-23 | 2012-10-16 | Arcadia Biosciences, Inc. | Nitrogen-efficient monocot plants |
| EP2535417A1 (en) * | 2007-07-20 | 2012-12-19 | BASF Plant Science GmbH | Plants having increased yield-related traits and a method for making the same |
| WO2012176167A1 (en) | 2011-06-24 | 2012-12-27 | Genoplante-Valor | Obtaining plants with improved tolerance to water stress |
| WO2014033672A1 (en) | 2012-08-30 | 2014-03-06 | Institut National De La Recherche Agronomique | Use of a receptor kinase having lysm motifs in order to improve the response of plants to lipochitooligosaccharides |
| US8722072B2 (en) | 2010-01-22 | 2014-05-13 | Bayer Intellectual Property Gmbh | Acaricidal and/or insecticidal active ingredient combinations |
| EP2753698A1 (en) * | 2011-09-07 | 2014-07-16 | Bayer CropScience AG | Plants having enhanced nitrogen efficiency |
| US9206137B2 (en) | 2010-11-15 | 2015-12-08 | Bayer Intellectual Property Gmbh | N-Aryl pyrazole(thio)carboxamides |
| US9265252B2 (en) | 2011-08-10 | 2016-02-23 | Bayer Intellectual Property Gmbh | Active compound combinations comprising specific tetramic acid derivatives |
| WO2020035486A1 (en) | 2018-08-13 | 2020-02-20 | Aarhus Universitet | Genetically altered plants expressing heterologous receptors that recognize lipo-chitooligosaccharides |
| WO2020035488A1 (en) | 2018-08-13 | 2020-02-20 | Aarhus Universitet | Genetically altered lysm receptors with altered agonist specificity and affinity |
| WO2020115181A1 (en) | 2018-12-06 | 2020-06-11 | Wageningen Universiteit | Methods of genetically altering a plant nin-gene to be responsive to cytokinin |
| WO2020187995A1 (en) | 2019-03-21 | 2020-09-24 | University Of Essex Enterprises Limited | Methods of enhancing biomass in a plant through stimulation of rubp regeneration and electron transport |
| WO2021233904A1 (en) | 2020-05-19 | 2021-11-25 | Aarhus Universitet | Lysm receptor motifs |
| WO2022251428A2 (en) | 2021-05-26 | 2022-12-01 | The Board Of Trustees Of The University Of Illinois | C4 plants with increased photosynthetic efficiency |
| WO2023201230A1 (en) | 2022-04-11 | 2023-10-19 | The Regents Of The University Of California | Methods of screening for plant gain of function mutations and compositions therefor |
| WO2024126805A1 (en) | 2022-12-15 | 2024-06-20 | Aarhus Universitet | Synthetic activation of multimeric transmembrane receptors |
| WO2024161012A2 (en) | 2023-02-03 | 2024-08-08 | Aarhus Universitet | Enhancing nitrogen fixation with fun |
| WO2024189171A1 (en) | 2023-03-14 | 2024-09-19 | Aarhus Universitet | Genetically altered nfr5 receptor kinases |
| WO2024189170A1 (en) | 2023-03-14 | 2024-09-19 | Aarhus Universitet | Genetically altered nfr1 receptor kinases |
Families Citing this family (14)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP4977691B2 (en) | 2006-04-13 | 2012-07-18 | 中外製薬株式会社 | Taurine transporter gene |
| JP5635260B2 (en) | 2007-03-15 | 2014-12-03 | 中外製薬株式会社 | Method for producing polypeptide |
| JP5337033B2 (en) * | 2007-08-07 | 2013-11-06 | 中外製薬株式会社 | Method for producing heterologous protein |
| US20100281761A1 (en) * | 2007-09-21 | 2010-11-11 | Eric John Rey | Energy efficient and greenhouse gas efficient biofuel |
| BRPI0818039A2 (en) * | 2007-10-15 | 2014-10-14 | Chugai Pharmaceutical Co Ltd | METHOD FOR PRODUCING A CAPABLE OF HIGH-INCOME HYPEROPROTEIN PRODUCTION CELL. |
| DK2213746T3 (en) * | 2007-10-24 | 2015-08-03 | Chugai Pharmaceutical Co Ltd | CELL FOR USE IN THE PREPARATION OF exogenous protein, AND MANUFACTURING METHOD THAT USE CELL |
| BRPI0909285B1 (en) * | 2008-03-31 | 2020-01-28 | Ceres Inc | nucleic acid, vector construction, transcription conduction method, method of expression of an exogenous coding region in a plant, method of altering the expression of a gene in a plant, method of producing a transgenic plant |
| US20110004961A1 (en) * | 2008-08-29 | 2011-01-06 | Unkefer Pat J | Plant glutamine phenylpyruvate transaminase gene and transgenic plants carrying same |
| US20110030104A1 (en) | 2008-08-29 | 2011-02-03 | Los Alamos National Security, Llc | Nucleic acids encoding plant glutamine phenylpyruvate transaminase (GPT) and uses thereof |
| US20110030089A1 (en) | 2008-08-29 | 2011-02-03 | Los Alamos National Security, Llc | Transgenic plants with enhanced growth characteristics |
| EP2423309B1 (en) | 2009-04-22 | 2018-01-03 | Chugai Seiyaku Kabushiki Kaisha | A method for producing a cell capable of high-yield production of heteroproteins |
| AU2014224716A1 (en) * | 2013-03-06 | 2015-09-10 | Bayer Cropscience Lp | Plants having enhanced nitrogen use efficiency and methods of producing same |
| CN103392591B (en) * | 2013-07-05 | 2016-08-10 | 安徽省农业科学院水稻研究所 | A kind of breeding of new variety method that rice high yield and nitrogen efficiently utilize |
| CN109112138B (en) * | 2017-12-01 | 2020-12-08 | 华中农业大学 | Gene OsVAS1 for regulating and controlling ideal plant type of rice |
Family Cites Families (19)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| DE3719053A1 (en) | 1987-06-06 | 1988-12-15 | Hoechst Ag | IMPROVED USE OF PLANT-RECOVERABLE NITROGEN BY CULTURAL PLANTS WITH OVEREXPRESSION OF GLUTAMINE SYNTHETASE |
| US5955651A (en) | 1989-05-03 | 1999-09-21 | New York University | Transgenic plants that exhibit enhanced nitrogen assimilation |
| US5256558A (en) | 1989-05-03 | 1993-10-26 | The Trustees Of Rockefeller University | Gene encoding plant asparagine synthetase |
| JPH05500156A (en) | 1989-09-15 | 1993-01-21 | ザ・ジェネラル・ホスピタル・コーポレーション | Membrane-bound nitrate reductase activity |
| US5399680A (en) | 1991-05-22 | 1995-03-21 | The Salk Institute For Biological Studies | Rice chitinase promoter |
| JPH05199877A (en) | 1991-10-03 | 1993-08-10 | Sumitomo Chem Co Ltd | Controlling region for induction type plant protecting gene of potato and rice plant, its use and method for assay |
| DE69434715D1 (en) | 1993-10-06 | 2006-06-01 | Univ New York | METHOD FOR THE PRODUCTION OF TRANSGENIC PLANTS THAT SHOW AN INCREASED NITROGEN DETECTION |
| US5750399A (en) | 1994-11-14 | 1998-05-12 | The Samuel Roberts Noble Foundation, Inc. | Isoflavone reductase promoter |
| US6084153A (en) | 1996-02-14 | 2000-07-04 | The Governors Of The University Of Alberta | Plants having enhanced nitrogen assimilation/metabolism |
| US7390937B2 (en) | 1996-02-14 | 2008-06-24 | The Governors Of The University Of Alberta | Plants with enhanced levels of nitrogen utilization proteins in their root epidermis and uses thereof |
| WO1997030163A1 (en) * | 1996-02-14 | 1997-08-21 | The Governors Of The University Of Alberta | Plants having enhanced nitrogen assimilation/metabolism |
| US6080913A (en) | 1996-09-25 | 2000-06-27 | Pioneer Hi-Bred International, Inc. | Binary methods of increasing accumulation of essential amino acids in seeds |
| US20040116682A1 (en) | 1998-03-06 | 2004-06-17 | Nordine Cheikh | Nucleic acid molecules and other molecules associated with the carbon assimilation pathway |
| US7365185B2 (en) | 2000-07-19 | 2008-04-29 | Monsanto Technology Llc | Genomic plant sequences and uses thereof |
| ES2290152T3 (en) * | 2000-01-28 | 2008-02-16 | The Governors Of The University Of Alberta | SPECIFIC EXPRESSION IN THE ROOT OF DESIRED GENES IN PLANTS. |
| WO2003000898A1 (en) | 2001-06-22 | 2003-01-03 | Syngenta Participations Ag | Plant genes involved in defense against pathogens |
| AU2003279752A1 (en) | 2002-06-28 | 2004-01-19 | Paradigm Genetics, Inc. | Method for improving plant traits by altering the activity of plant g-protein alpha and beta subunits |
| AR059119A1 (en) | 2005-12-23 | 2008-03-12 | Arcadia Biosciences Inc | EFFECTIVE USE OF NITROGEN IN MONOCOTILEDONEAS PLANTS |
| PL1968372T3 (en) | 2005-12-23 | 2012-07-31 | Arcadia Biosciences Inc | Promoter sequence obtained from rice and methods of use |
-
2006
- 2006-12-21 AR ARP060105745A patent/AR059119A1/en not_active Application Discontinuation
- 2006-12-21 CA CA2634925A patent/CA2634925C/en not_active Expired - Fee Related
- 2006-12-21 CN CN2006800487181A patent/CN101378651B/en not_active Expired - Fee Related
- 2006-12-21 WO PCT/US2006/049241 patent/WO2007076115A2/en active Application Filing
- 2006-12-21 US US11/644,321 patent/US8288611B2/en active Active
- 2006-12-21 BR BRPI0620312-4A patent/BRPI0620312A2/en not_active Application Discontinuation
- 2006-12-21 AU AU2006330817A patent/AU2006330817B2/en not_active Ceased
- 2006-12-21 EP EP06848873A patent/EP1983819A4/en not_active Ceased
-
2008
- 2008-06-09 ZA ZA200805007A patent/ZA200805007B/en unknown
-
2012
- 2012-09-18 US US13/622,311 patent/US8642840B2/en not_active Expired - Fee Related
-
2013
- 2013-11-04 AR ARP130104024A patent/AR093346A2/en unknown
Non-Patent Citations (1)
| Title |
|---|
| See references of EP1983819A4 * |
Cited By (52)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US7786343B2 (en) | 1996-02-14 | 2010-08-31 | The Governors Of The University Of Alberta | Transgenic plants expressing recombinant barley alanine aminotransferase |
| US8115062B2 (en) | 1996-02-14 | 2012-02-14 | The Governors Of The University Of Alberta | Transgenic plants expressing recombinant barley alanine aminotransferase |
| EP1968372A4 (en) * | 2005-12-23 | 2009-09-09 | Arcadia Biosciences Inc | Promoter sequence obtained from rice and methods of use |
| US8288611B2 (en) | 2005-12-23 | 2012-10-16 | Arcadia Biosciences, Inc. | Nitrogen-efficient monocot plants |
| US8642840B2 (en) | 2005-12-23 | 2014-02-04 | Arcadia Biosciences, Inc. | Nitrogen-efficient monocot plants |
| US7982093B2 (en) | 2005-12-23 | 2011-07-19 | Arcadia Biosciences, Inc. | Promoter sequence obtained from rice and methods of use |
| EP1968372A2 (en) * | 2005-12-23 | 2008-09-17 | Arcadia Biosciences Inc. | Promoter sequence obtained from rice and methods of use |
| WO2009013225A3 (en) * | 2007-07-20 | 2009-03-26 | Cropdesign Nv | Plants having enhanced yield-related traits and a method for making the same |
| AU2008280200B2 (en) * | 2007-07-20 | 2014-11-06 | Cropdesign N.V. | Plants having enhanced yield-related traits and a method for making the same |
| US8697947B2 (en) | 2007-07-20 | 2014-04-15 | Basf Plant Science Gmbh | Plants having increased yield-related traits and a method for making the same |
| US8569576B2 (en) | 2007-07-20 | 2013-10-29 | Cropdesign N.V. | Plants having enhanced yield-related traits and a method for making the same |
| WO2009013225A2 (en) * | 2007-07-20 | 2009-01-29 | Cropdesign N.V. | Plants having enhanced yield-related traits and a method for making the same |
| EP2535417A1 (en) * | 2007-07-20 | 2012-12-19 | BASF Plant Science GmbH | Plants having increased yield-related traits and a method for making the same |
| WO2009149304A2 (en) * | 2008-06-04 | 2009-12-10 | Edenspace Systems Corporation | Plant gene regulatory elements |
| WO2009149304A3 (en) * | 2008-06-04 | 2010-08-19 | Edenspace Systems Corporation | Plant gene regulatory elements |
| CN102365365A (en) * | 2009-03-27 | 2012-02-29 | 纳幕尔杜邦公司 | Plants having altered agronomic characteristics under nitrogen limiting conditions and related constructs and methods involving genes encoding snf2 domain-containing polypeptides |
| US9085633B2 (en) | 2009-03-27 | 2015-07-21 | E I Du Pont De Nemours And Company | Plants having altered agronomic characteristics under nitrogen limiting conditions and related constructs and methods involving genes encoding SNF2 domain-containing polypeptides |
| EP2511374A1 (en) * | 2009-03-27 | 2012-10-17 | E. I. du Pont de Nemours and Company | Plants having altered agronomic characteristics under nitrogen limiting conditions and related constructs and methods involving genes encoding SNF2 domain-containing polypeptides |
| CN102365365B (en) * | 2009-03-27 | 2014-11-26 | 纳幕尔杜邦公司 | Plants having altered agronomic characteristics under nitrogen limiting conditions and related constructs and methods involving genes encoding SNF2 domain-containing polypeptides |
| WO2010111568A3 (en) * | 2009-03-27 | 2010-11-18 | E. I. Du Pont De Nemours And Company | Plants having altered agronomic characteristics under nitrogen limiting conditions and related constructs and methods involving genes encoding snf2 domain-containing polypeptides |
| CN102395674B (en) * | 2009-04-14 | 2015-07-29 | 先锋国际良种公司 | Plant biomass under regulating acc synthase to improve low nitrogen condition |
| US8987553B2 (en) | 2009-04-14 | 2015-03-24 | Pioneer Hi Bred International Inc | Modulation of ACC synthase improves plant yield under low nitrogen conditions |
| CN102395674A (en) * | 2009-04-14 | 2012-03-28 | 先锋国际良种公司 | Modulation of acc synthase improves plant yield under low nitrogen conditions |
| WO2010120862A1 (en) * | 2009-04-14 | 2010-10-21 | Pioneer Hi-Bred International, Inc. | Modulation of acc synthase improves plant yield under low nitrogen conditions |
| CN102549149A (en) * | 2009-08-20 | 2012-07-04 | 先锋国际良种公司 | Functional expression of yeast nitrate transporter (ynt1) in maize to improve nitrate uptake |
| US8975474B2 (en) | 2009-08-20 | 2015-03-10 | Pioneer Hi Bred International Inc | Functional expression of yeast nitrate transporter (YNT1)and a nitrate reductase in maize |
| WO2011022608A3 (en) * | 2009-08-20 | 2011-04-14 | Pioneer Hi-Bred International, Inc. | Functional expression of yeast nitrate transporter (ynt1) in maize to improve nitrate uptake |
| US8722072B2 (en) | 2010-01-22 | 2014-05-13 | Bayer Intellectual Property Gmbh | Acaricidal and/or insecticidal active ingredient combinations |
| CN101942512A (en) * | 2010-08-25 | 2011-01-12 | 中国农业科学院作物科学研究所 | Development and application of molecule marker for corn with considerable number of kernels and excellent allele function in low-nitrogen adverse environment |
| WO2012049661A1 (en) | 2010-10-15 | 2012-04-19 | Genoplante-Valor | Production of plants with improved tolerance to water deficit |
| WO2012049663A1 (en) | 2010-10-15 | 2012-04-19 | Genoplante-Valor | Production of plants having improved water-deficit tolerance |
| WO2012059497A1 (en) | 2010-11-02 | 2012-05-10 | Bayer Cropscience Ag | N-hetarylmethyl pyrazolylcarboxamides |
| US9206137B2 (en) | 2010-11-15 | 2015-12-08 | Bayer Intellectual Property Gmbh | N-Aryl pyrazole(thio)carboxamides |
| WO2012089757A1 (en) | 2010-12-29 | 2012-07-05 | Bayer Cropscience Ag | Fungicide hydroximoyl-tetrazole derivatives |
| WO2012176167A1 (en) | 2011-06-24 | 2012-12-27 | Genoplante-Valor | Obtaining plants with improved tolerance to water stress |
| US9265252B2 (en) | 2011-08-10 | 2016-02-23 | Bayer Intellectual Property Gmbh | Active compound combinations comprising specific tetramic acid derivatives |
| AU2012307006B2 (en) * | 2011-09-07 | 2017-08-03 | Bayer Cropscience Ag | Plants having enhanced nitrogen efficiency |
| EP2753698A4 (en) * | 2011-09-07 | 2015-02-11 | Bayer Cropscience Ag | Plants having enhanced nitrogen efficiency |
| EP2753698A1 (en) * | 2011-09-07 | 2014-07-16 | Bayer CropScience AG | Plants having enhanced nitrogen efficiency |
| US10246720B2 (en) | 2011-09-07 | 2019-04-02 | Bayer Intellectual Property Gmbh | Plants having enhanced nitrogen efficiency |
| WO2014033672A1 (en) | 2012-08-30 | 2014-03-06 | Institut National De La Recherche Agronomique | Use of a receptor kinase having lysm motifs in order to improve the response of plants to lipochitooligosaccharides |
| WO2020035486A1 (en) | 2018-08-13 | 2020-02-20 | Aarhus Universitet | Genetically altered plants expressing heterologous receptors that recognize lipo-chitooligosaccharides |
| WO2020035488A1 (en) | 2018-08-13 | 2020-02-20 | Aarhus Universitet | Genetically altered lysm receptors with altered agonist specificity and affinity |
| WO2020115181A1 (en) | 2018-12-06 | 2020-06-11 | Wageningen Universiteit | Methods of genetically altering a plant nin-gene to be responsive to cytokinin |
| WO2020187995A1 (en) | 2019-03-21 | 2020-09-24 | University Of Essex Enterprises Limited | Methods of enhancing biomass in a plant through stimulation of rubp regeneration and electron transport |
| WO2021233904A1 (en) | 2020-05-19 | 2021-11-25 | Aarhus Universitet | Lysm receptor motifs |
| WO2022251428A2 (en) | 2021-05-26 | 2022-12-01 | The Board Of Trustees Of The University Of Illinois | C4 plants with increased photosynthetic efficiency |
| WO2023201230A1 (en) | 2022-04-11 | 2023-10-19 | The Regents Of The University Of California | Methods of screening for plant gain of function mutations and compositions therefor |
| WO2024126805A1 (en) | 2022-12-15 | 2024-06-20 | Aarhus Universitet | Synthetic activation of multimeric transmembrane receptors |
| WO2024161012A2 (en) | 2023-02-03 | 2024-08-08 | Aarhus Universitet | Enhancing nitrogen fixation with fun |
| WO2024189171A1 (en) | 2023-03-14 | 2024-09-19 | Aarhus Universitet | Genetically altered nfr5 receptor kinases |
| WO2024189170A1 (en) | 2023-03-14 | 2024-09-19 | Aarhus Universitet | Genetically altered nfr1 receptor kinases |
Also Published As
| Publication number | Publication date |
|---|---|
| AR093346A2 (en) | 2015-06-03 |
| ZA200805007B (en) | 2009-08-26 |
| US8642840B2 (en) | 2014-02-04 |
| US8288611B2 (en) | 2012-10-16 |
| US20130097738A1 (en) | 2013-04-18 |
| CN101378651B (en) | 2012-03-14 |
| EP1983819A4 (en) | 2009-09-02 |
| AU2006330817A1 (en) | 2007-07-05 |
| CA2634925A1 (en) | 2007-07-05 |
| CN101378651A (en) | 2009-03-04 |
| BRPI0620312A2 (en) | 2011-11-08 |
| AR059119A1 (en) | 2008-03-12 |
| US20070162995A1 (en) | 2007-07-12 |
| EP1983819A2 (en) | 2008-10-29 |
| CA2634925C (en) | 2015-06-23 |
| WO2007076115A3 (en) | 2008-06-19 |
| AU2006330817B2 (en) | 2012-07-19 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US8642840B2 (en) | Nitrogen-efficient monocot plants | |
| EP1968372B1 (en) | Promoter sequence obtained from rice and methods of use | |
| US20210102218A1 (en) | Expression of transcription regulators that provide heat tolerance | |
| EP0590086A4 (en) | Novel organ-specific plant promoter sequences. | |
| AU2009287446B2 (en) | Transgenic plants with enhanced growth characteristics | |
| Erpen et al. | Isolation, characterization, and evaluation of three Citrus sinensis-derived constitutive gene promoters | |
| WO2010025465A1 (en) | Plant glutamine phenylpyruvate transaminase gene and transgenic plants carrying same | |
| US9458455B2 (en) | High efficiency plant expression promoter from Capsicum annuum serine hydroxymethyl transferase gene and uses thereof | |
| US20100263090A1 (en) | Plant Glutamine Phenylpyruvate Transaminase Gene and Transgenic Plants Carrying Same | |
| CN116768991B (en) | Soybean four-transmembrane-region protein GmTET270,270 related to lipid metabolism regulation and encoding gene and application thereof | |
| US10988771B2 (en) | Endosperm-specific promoter from the lipid transfer protein 1 gene of Coffea arabica | |
| Kim JaeYoon | Identification and functional analysis of S-AdenosylMethionine Synthetase (HvSAMS) genes in early maturing barley (Hordeum vulgare subsp. vulgare). | |
| Biswas | prospects, perspectives, and problems of plant genetic engineering |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| WWE | Wipo information: entry into national phase |
Ref document number: 200680048718.1 Country of ref document: CN |
|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application | ||
| WWE | Wipo information: entry into national phase |
Ref document number: MX/a/2008/008180 Country of ref document: MX Ref document number: 2006330817 Country of ref document: AU |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2634925 Country of ref document: CA |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |
|
| ENP | Entry into the national phase |
Ref document number: 2006330817 Country of ref document: AU Date of ref document: 20061221 Kind code of ref document: A |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2870/KOLNP/2008 Country of ref document: IN |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2006848873 Country of ref document: EP |
|
| ENP | Entry into the national phase |
Ref document number: PI0620312 Country of ref document: BR Kind code of ref document: A2 Effective date: 20080623 |