RU2692932C2 - BIOCATALYST FOR PRODUCING 11α-ACETOXYPROGESTERONE - Google Patents
BIOCATALYST FOR PRODUCING 11α-ACETOXYPROGESTERONE Download PDFInfo
- Publication number
- RU2692932C2 RU2692932C2 RU2017138536A RU2017138536A RU2692932C2 RU 2692932 C2 RU2692932 C2 RU 2692932C2 RU 2017138536 A RU2017138536 A RU 2017138536A RU 2017138536 A RU2017138536 A RU 2017138536A RU 2692932 C2 RU2692932 C2 RU 2692932C2
- Authority
- RU
- Russia
- Prior art keywords
- progesterone
- hydroxyprogesterone
- nidulans
- acetoxyprogesterone
- strain
- Prior art date
Links
- IWRPVTXREVYBHT-ZQEATNLPSA-N [(8s,9s,10r,11r,13s,14s,17s)-17-acetyl-10,13-dimethyl-3-oxo-1,2,6,7,8,9,11,12,14,15,16,17-dodecahydrocyclopenta[a]phenanthren-11-yl] acetate Chemical compound O=C1CC[C@]2(C)[C@H]3[C@H](OC(=O)C)C[C@]4(C)[C@@H](C(C)=O)CC[C@H]4[C@@H]3CCC2=C1 IWRPVTXREVYBHT-ZQEATNLPSA-N 0.000 title claims abstract description 41
- 108090000790 Enzymes Proteins 0.000 title claims description 6
- 102000004190 Enzymes Human genes 0.000 title claims description 6
- 239000011942 biocatalyst Substances 0.000 title claims description 3
- 241000351920 Aspergillus nidulans Species 0.000 claims abstract description 67
- 244000005700 microbiome Species 0.000 claims abstract description 11
- 101100295959 Halobacterium salinarum (strain ATCC 700922 / JCM 11081 / NRC-1) arcB gene Proteins 0.000 claims description 19
- 101150008194 argB gene Proteins 0.000 claims description 19
- 101150039489 lysZ gene Proteins 0.000 claims description 19
- 101150054232 pyrG gene Proteins 0.000 claims description 19
- RJKFOVLPORLFTN-LEKSSAKUSA-N Progesterone Chemical compound C1CC2=CC(=O)CC[C@]2(C)[C@@H]2[C@@H]1[C@@H]1CC[C@H](C(=O)C)[C@@]1(C)CC2 RJKFOVLPORLFTN-LEKSSAKUSA-N 0.000 abstract description 166
- 239000000186 progesterone Substances 0.000 abstract description 82
- 229960003387 progesterone Drugs 0.000 abstract description 82
- BFZHCUBIASXHPK-QJSKAATBSA-N 11alpha-hydroxyprogesterone Chemical compound C1CC2=CC(=O)CC[C@]2(C)[C@@H]2[C@@H]1[C@@H]1CC[C@H](C(=O)C)[C@@]1(C)C[C@H]2O BFZHCUBIASXHPK-QJSKAATBSA-N 0.000 abstract description 68
- 238000005805 hydroxylation reaction Methods 0.000 abstract description 24
- 230000036983 biotransformation Effects 0.000 abstract description 23
- 239000000203 mixture Substances 0.000 abstract description 23
- 150000003431 steroids Chemical class 0.000 abstract description 23
- 230000000694 effects Effects 0.000 abstract description 19
- 239000000126 substance Substances 0.000 abstract description 15
- 238000005481 NMR spectroscopy Methods 0.000 abstract description 13
- 239000000758 substrate Substances 0.000 abstract description 13
- 238000003786 synthesis reaction Methods 0.000 abstract description 9
- 241000233866 Fungi Species 0.000 abstract description 8
- 238000005160 1H NMR spectroscopy Methods 0.000 abstract description 5
- 238000001644 13C nuclear magnetic resonance spectroscopy Methods 0.000 abstract description 4
- 238000004949 mass spectrometry Methods 0.000 abstract description 4
- 238000005084 2D-nuclear magnetic resonance Methods 0.000 abstract description 2
- 238000009614 chemical analysis method Methods 0.000 abstract 1
- 150000002894 organic compounds Chemical class 0.000 abstract 1
- 238000004940 physical analysis method Methods 0.000 abstract 1
- YMWUJEATGCHHMB-UHFFFAOYSA-N Dichloromethane Chemical compound ClCCl YMWUJEATGCHHMB-UHFFFAOYSA-N 0.000 description 48
- 230000009466 transformation Effects 0.000 description 40
- 238000000034 method Methods 0.000 description 31
- 230000021736 acetylation Effects 0.000 description 24
- 238000006640 acetylation reaction Methods 0.000 description 24
- 230000015572 biosynthetic process Effects 0.000 description 22
- OKKJLVBELUTLKV-UHFFFAOYSA-N Methanol Chemical compound OC OKKJLVBELUTLKV-UHFFFAOYSA-N 0.000 description 21
- 239000002609 medium Substances 0.000 description 20
- CSCPPACGZOOCGX-UHFFFAOYSA-N Acetone Chemical compound CC(C)=O CSCPPACGZOOCGX-UHFFFAOYSA-N 0.000 description 18
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 18
- 238000006243 chemical reaction Methods 0.000 description 14
- 239000000047 product Substances 0.000 description 14
- 238000004809 thin layer chromatography Methods 0.000 description 14
- WFDIJRYMOXRFFG-UHFFFAOYSA-N Acetic anhydride Chemical compound CC(=O)OC(C)=O WFDIJRYMOXRFFG-UHFFFAOYSA-N 0.000 description 12
- 238000001228 spectrum Methods 0.000 description 12
- 230000008569 process Effects 0.000 description 10
- KDHBHXVDYRGRDL-KDZLLLCRSA-N 6beta,11alpha-Dihydroxyprogesterone Chemical compound C1([C@H](O)C2)=CC(=O)CC[C@]1(C)[C@@H]1[C@@H]2[C@@H]2CC[C@H](C(=O)C)[C@@]2(C)C[C@H]1O KDHBHXVDYRGRDL-KDZLLLCRSA-N 0.000 description 9
- XEKOWRVHYACXOJ-UHFFFAOYSA-N Ethyl acetate Chemical compound CCOC(C)=O XEKOWRVHYACXOJ-UHFFFAOYSA-N 0.000 description 9
- 150000001875 compounds Chemical class 0.000 description 9
- 239000002904 solvent Substances 0.000 description 9
- ISAKRJDGNUQOIC-UHFFFAOYSA-N Uracil Chemical compound O=C1C=CNC(=O)N1 ISAKRJDGNUQOIC-UHFFFAOYSA-N 0.000 description 8
- 239000001963 growth medium Substances 0.000 description 8
- 239000000243 solution Substances 0.000 description 8
- 230000002210 biocatalytic effect Effects 0.000 description 7
- 238000007429 general method Methods 0.000 description 7
- 125000002887 hydroxy group Chemical group [H]O* 0.000 description 7
- WEVYAHXRMPXWCK-UHFFFAOYSA-N Acetonitrile Chemical compound CC#N WEVYAHXRMPXWCK-UHFFFAOYSA-N 0.000 description 6
- OKTJSMMVPCPJKN-UHFFFAOYSA-N Carbon Chemical compound [C] OKTJSMMVPCPJKN-UHFFFAOYSA-N 0.000 description 6
- IAZDPXIOMUYVGZ-UHFFFAOYSA-N Dimethylsulphoxide Chemical compound CS(C)=O IAZDPXIOMUYVGZ-UHFFFAOYSA-N 0.000 description 6
- DRTQHJPVMGBUCF-XVFCMESISA-N Uridine Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C=C1 DRTQHJPVMGBUCF-XVFCMESISA-N 0.000 description 6
- 230000033444 hydroxylation Effects 0.000 description 6
- 241000894007 species Species 0.000 description 6
- -1 steroid compounds Chemical class 0.000 description 6
- 229940035893 uracil Drugs 0.000 description 6
- 229920001817 Agar Polymers 0.000 description 5
- 239000008272 agar Substances 0.000 description 5
- 239000012531 culture fluid Substances 0.000 description 5
- 239000003814 drug Substances 0.000 description 5
- 230000032050 esterification Effects 0.000 description 5
- 238000005886 esterification reaction Methods 0.000 description 5
- 239000000284 extract Substances 0.000 description 5
- 230000002906 microbiologic effect Effects 0.000 description 5
- VLTRZXGMWDSKGL-UHFFFAOYSA-N perchloric acid Chemical compound OCl(=O)(=O)=O VLTRZXGMWDSKGL-UHFFFAOYSA-N 0.000 description 5
- 238000000844 transformation Methods 0.000 description 5
- WKAVAGKRWFGIEA-DADBAOPHSA-N 11-Ketoprogesterone Chemical compound C1CC2=CC(=O)CC[C@]2(C)[C@@H]2[C@@H]1[C@@H]1CC[C@H](C(=O)C)[C@@]1(C)CC2=O WKAVAGKRWFGIEA-DADBAOPHSA-N 0.000 description 4
- ZESRJSPZRDMNHY-YFWFAHHUSA-N 11-deoxycorticosterone Chemical compound O=C1CC[C@]2(C)[C@H]3CC[C@](C)([C@H](CC4)C(=O)CO)[C@@H]4[C@@H]3CCC2=C1 ZESRJSPZRDMNHY-YFWFAHHUSA-N 0.000 description 4
- 241000228212 Aspergillus Species 0.000 description 4
- RTZKZFJDLAIYFH-UHFFFAOYSA-N Diethyl ether Chemical compound CCOCC RTZKZFJDLAIYFH-UHFFFAOYSA-N 0.000 description 4
- VYPSYNLAJGMNEJ-UHFFFAOYSA-N Silicium dioxide Chemical compound O=[Si]=O VYPSYNLAJGMNEJ-UHFFFAOYSA-N 0.000 description 4
- 230000000397 acetylating effect Effects 0.000 description 4
- 238000004458 analytical method Methods 0.000 description 4
- ZESRJSPZRDMNHY-UHFFFAOYSA-N de-oxy corticosterone Natural products O=C1CCC2(C)C3CCC(C)(C(CC4)C(=O)CO)C4C3CCC2=C1 ZESRJSPZRDMNHY-UHFFFAOYSA-N 0.000 description 4
- 238000010828 elution Methods 0.000 description 4
- 238000001704 evaporation Methods 0.000 description 4
- 230000008020 evaporation Effects 0.000 description 4
- 238000004519 manufacturing process Methods 0.000 description 4
- 238000001819 mass spectrum Methods 0.000 description 4
- 230000000813 microbial effect Effects 0.000 description 4
- 239000000741 silica gel Substances 0.000 description 4
- 229910002027 silica gel Inorganic materials 0.000 description 4
- 230000028070 sporulation Effects 0.000 description 4
- 239000007858 starting material Substances 0.000 description 4
- 239000000725 suspension Substances 0.000 description 4
- BFZHCUBIASXHPK-UHFFFAOYSA-N 11beta-hydroxy-progesterone Natural products C1CC2=CC(=O)CCC2(C)C2C1C1CCC(C(=O)C)C1(C)CC2O BFZHCUBIASXHPK-UHFFFAOYSA-N 0.000 description 3
- BFZHCUBIASXHPK-ATWVFEABSA-N 11beta-hydroxyprogesterone Chemical compound C1CC2=CC(=O)CC[C@]2(C)[C@@H]2[C@@H]1[C@@H]1CC[C@H](C(=O)C)[C@@]1(C)C[C@@H]2O BFZHCUBIASXHPK-ATWVFEABSA-N 0.000 description 3
- 239000004475 Arginine Substances 0.000 description 3
- 241000122824 Aspergillus ochraceus Species 0.000 description 3
- 241000203233 Aspergillus versicolor Species 0.000 description 3
- LFQSCWFLJHTTHZ-UHFFFAOYSA-N Ethanol Chemical compound CCO LFQSCWFLJHTTHZ-UHFFFAOYSA-N 0.000 description 3
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 3
- DRTQHJPVMGBUCF-PSQAKQOGSA-N beta-L-uridine Natural products O[C@H]1[C@@H](O)[C@H](CO)O[C@@H]1N1C(=O)NC(=O)C=C1 DRTQHJPVMGBUCF-PSQAKQOGSA-N 0.000 description 3
- IJKVHSBPTUYDLN-UHFFFAOYSA-N dihydroxy(oxo)silane Chemical compound O[Si](O)=O IJKVHSBPTUYDLN-UHFFFAOYSA-N 0.000 description 3
- 229940079593 drug Drugs 0.000 description 3
- 239000003480 eluent Substances 0.000 description 3
- 238000011534 incubation Methods 0.000 description 3
- 238000002955 isolation Methods 0.000 description 3
- 239000007788 liquid Substances 0.000 description 3
- 238000002803 maceration Methods 0.000 description 3
- 230000035772 mutation Effects 0.000 description 3
- 238000002360 preparation method Methods 0.000 description 3
- 150000003839 salts Chemical class 0.000 description 3
- 238000003756 stirring Methods 0.000 description 3
- DRTQHJPVMGBUCF-UHFFFAOYSA-N uracil arabinoside Natural products OC1C(O)C(CO)OC1N1C(=O)NC(=O)C=C1 DRTQHJPVMGBUCF-UHFFFAOYSA-N 0.000 description 3
- 229940045145 uridine Drugs 0.000 description 3
- WQZGKKKJIJFFOK-SVZMEOIVSA-N (+)-Galactose Chemical compound OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@H]1O WQZGKKKJIJFFOK-SVZMEOIVSA-N 0.000 description 2
- IWRPVTXREVYBHT-UHFFFAOYSA-N (17-acetyl-10,13-dimethyl-3-oxo-1,2,6,7,8,9,11,12,14,15,16,17-dodecahydrocyclopenta[a]phenanthren-11-yl) acetate Chemical compound O=C1CCC2(C)C3C(OC(=O)C)CC4(C)C(C(C)=O)CCC4C3CCC2=C1 IWRPVTXREVYBHT-UHFFFAOYSA-N 0.000 description 2
- WKAVAGKRWFGIEA-UHFFFAOYSA-N 11-Ketoprogesterone Natural products C1CC2=CC(=O)CCC2(C)C2C1C1CCC(C(=O)C)C1(C)CC2=O WKAVAGKRWFGIEA-UHFFFAOYSA-N 0.000 description 2
- 102000004277 11-beta-hydroxysteroid dehydrogenases Human genes 0.000 description 2
- 108090000874 11-beta-hydroxysteroid dehydrogenases Proteins 0.000 description 2
- FUFLCEKSBBHCMO-UHFFFAOYSA-N 11-dehydrocorticosterone Natural products O=C1CCC2(C)C3C(=O)CC(C)(C(CC4)C(=O)CO)C4C3CCC2=C1 FUFLCEKSBBHCMO-UHFFFAOYSA-N 0.000 description 2
- QZLYKIGBANMMBK-UGCZWRCOSA-N 5α-Androstane Chemical compound C([C@@H]1CC2)CCC[C@]1(C)[C@@H]1[C@@H]2[C@@H]2CCC[C@@]2(C)CC1 QZLYKIGBANMMBK-UGCZWRCOSA-N 0.000 description 2
- 201000004384 Alopecia Diseases 0.000 description 2
- 241000228197 Aspergillus flavus Species 0.000 description 2
- 241000134719 Aspergillus tamarii Species 0.000 description 2
- 241001465318 Aspergillus terreus Species 0.000 description 2
- IJGRMHOSHXDMSA-UHFFFAOYSA-N Atomic nitrogen Chemical compound N#N IJGRMHOSHXDMSA-UHFFFAOYSA-N 0.000 description 2
- MFYSYFVPBJMHGN-ZPOLXVRWSA-N Cortisone Chemical compound O=C1CC[C@]2(C)[C@H]3C(=O)C[C@](C)([C@@](CC4)(O)C(=O)CO)[C@@H]4[C@@H]3CCC2=C1 MFYSYFVPBJMHGN-ZPOLXVRWSA-N 0.000 description 2
- MFYSYFVPBJMHGN-UHFFFAOYSA-N Cortisone Natural products O=C1CCC2(C)C3C(=O)CC(C)(C(CC4)(O)C(=O)CO)C4C3CCC2=C1 MFYSYFVPBJMHGN-UHFFFAOYSA-N 0.000 description 2
- 102000002004 Cytochrome P-450 Enzyme System Human genes 0.000 description 2
- 108010015742 Cytochrome P-450 Enzyme System Proteins 0.000 description 2
- DCXYFEDJOCDNAF-REOHCLBHSA-N L-asparagine Chemical compound OC(=O)[C@@H](N)CC(N)=O DCXYFEDJOCDNAF-REOHCLBHSA-N 0.000 description 2
- 239000001888 Peptone Substances 0.000 description 2
- 108010080698 Peptones Proteins 0.000 description 2
- JUJWROOIHBZHMG-UHFFFAOYSA-N Pyridine Chemical compound C1=CC=NC=C1 JUJWROOIHBZHMG-UHFFFAOYSA-N 0.000 description 2
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 2
- MUMGGOZAMZWBJJ-DYKIIFRCSA-N Testostosterone Chemical compound O=C1CC[C@]2(C)[C@H]3CC[C@](C)([C@H](CC4)O)[C@@H]4[C@@H]3CCC2=C1 MUMGGOZAMZWBJJ-DYKIIFRCSA-N 0.000 description 2
- HEDRZPFGACZZDS-MICDWDOJSA-N Trichloro(2H)methane Chemical compound [2H]C(Cl)(Cl)Cl HEDRZPFGACZZDS-MICDWDOJSA-N 0.000 description 2
- 239000002253 acid Substances 0.000 description 2
- 150000007513 acids Chemical class 0.000 description 2
- 230000009471 action Effects 0.000 description 2
- 239000007864 aqueous solution Substances 0.000 description 2
- 125000004429 atom Chemical group 0.000 description 2
- QVGXLLKOCUKJST-UHFFFAOYSA-N atomic oxygen Chemical group [O] QVGXLLKOCUKJST-UHFFFAOYSA-N 0.000 description 2
- 229940041514 candida albicans extract Drugs 0.000 description 2
- OKTJSMMVPCPJKN-BJUDXGSMSA-N carbon-11 Chemical compound [11C] OKTJSMMVPCPJKN-BJUDXGSMSA-N 0.000 description 2
- 230000015556 catabolic process Effects 0.000 description 2
- 239000003153 chemical reaction reagent Substances 0.000 description 2
- 229960004544 cortisone Drugs 0.000 description 2
- 239000013078 crystal Substances 0.000 description 2
- 238000006731 degradation reaction Methods 0.000 description 2
- FDSGHYHRLSWSLQ-UHFFFAOYSA-N dichloromethane;propan-2-one Chemical compound ClCCl.CC(C)=O FDSGHYHRLSWSLQ-UHFFFAOYSA-N 0.000 description 2
- 238000002518 distortionless enhancement with polarization transfer Methods 0.000 description 2
- 238000000132 electrospray ionisation Methods 0.000 description 2
- 238000005516 engineering process Methods 0.000 description 2
- 210000000416 exudates and transudate Anatomy 0.000 description 2
- 239000000417 fungicide Substances 0.000 description 2
- 238000003919 heteronuclear multiple bond coherence Methods 0.000 description 2
- 238000004128 high performance liquid chromatography Methods 0.000 description 2
- JYGXADMDTFJGBT-VWUMJDOOSA-N hydrocortisone Chemical compound O=C1CC[C@]2(C)[C@H]3[C@@H](O)C[C@](C)([C@@](CC4)(O)C(=O)CO)[C@@H]4[C@@H]3CCC2=C1 JYGXADMDTFJGBT-VWUMJDOOSA-N 0.000 description 2
- 229960002899 hydroxyprogesterone Drugs 0.000 description 2
- 239000003112 inhibitor Substances 0.000 description 2
- 230000002401 inhibitory effect Effects 0.000 description 2
- 229910052500 inorganic mineral Inorganic materials 0.000 description 2
- 238000002844 melting Methods 0.000 description 2
- 230000008018 melting Effects 0.000 description 2
- 239000011707 mineral Substances 0.000 description 2
- 230000004048 modification Effects 0.000 description 2
- 238000012986 modification Methods 0.000 description 2
- 230000007935 neutral effect Effects 0.000 description 2
- 235000019319 peptone Nutrition 0.000 description 2
- 239000002244 precipitate Substances 0.000 description 2
- 150000003145 progesterone derivatives Chemical class 0.000 description 2
- 238000000425 proton nuclear magnetic resonance spectrum Methods 0.000 description 2
- 238000011160 research Methods 0.000 description 2
- 230000002441 reversible effect Effects 0.000 description 2
- 210000004761 scalp Anatomy 0.000 description 2
- 238000000926 separation method Methods 0.000 description 2
- 238000003797 solvolysis reaction Methods 0.000 description 2
- 239000008399 tap water Substances 0.000 description 2
- 235000020679 tap water Nutrition 0.000 description 2
- CZDYPVPMEAXLPK-UHFFFAOYSA-N tetramethylsilane Chemical compound C[Si](C)(C)C CZDYPVPMEAXLPK-UHFFFAOYSA-N 0.000 description 2
- 239000012138 yeast extract Substances 0.000 description 2
- DUHUCHOQIDJXAT-OLVMNOGESA-N 3-hydroxy-(3-α,5-α)-Pregnane-11,20-dione Chemical compound C([C@@H]1CC2)[C@H](O)CC[C@]1(C)[C@@H]1[C@@H]2[C@@H]2CC[C@H](C(=O)C)[C@@]2(C)CC1=O DUHUCHOQIDJXAT-OLVMNOGESA-N 0.000 description 1
- PWCLWZOSAFOXFL-CXICGXRGSA-N 6beta-hydroxyprogesterone Chemical compound C1([C@H](O)C2)=CC(=O)CC[C@]1(C)[C@@H]1[C@@H]2[C@@H]2CC[C@H](C(=O)C)[C@@]2(C)CC1 PWCLWZOSAFOXFL-CXICGXRGSA-N 0.000 description 1
- 241000235389 Absidia Species 0.000 description 1
- QTBSBXVTEAMEQO-UHFFFAOYSA-M Acetate Chemical compound CC([O-])=O QTBSBXVTEAMEQO-UHFFFAOYSA-M 0.000 description 1
- 208000002874 Acne Vulgaris Diseases 0.000 description 1
- NLXLAEXVIDQMFP-UHFFFAOYSA-N Ammonium chloride Substances [NH4+].[Cl-] NLXLAEXVIDQMFP-UHFFFAOYSA-N 0.000 description 1
- VHUUQVKOLVNVRT-UHFFFAOYSA-N Ammonium hydroxide Chemical compound [NH4+].[OH-] VHUUQVKOLVNVRT-UHFFFAOYSA-N 0.000 description 1
- 241000131314 Aspergillus candidus Species 0.000 description 1
- 238000006220 Baeyer-Villiger oxidation reaction Methods 0.000 description 1
- UXVMQQNJUSDDNG-UHFFFAOYSA-L Calcium chloride Chemical compound [Cl-].[Cl-].[Ca+2] UXVMQQNJUSDDNG-UHFFFAOYSA-L 0.000 description 1
- 241000222120 Candida <Saccharomycetales> Species 0.000 description 1
- OMFXVFTZEKFJBZ-UHFFFAOYSA-N Corticosterone Natural products O=C1CCC2(C)C3C(O)CC(C)(C(CC4)C(=O)CO)C4C3CCC2=C1 OMFXVFTZEKFJBZ-UHFFFAOYSA-N 0.000 description 1
- ITRJWOMZKQRYTA-RFZYENFJSA-N Cortisone acetate Chemical compound C1CC2=CC(=O)CC[C@]2(C)[C@@H]2[C@@H]1[C@@H]1CC[C@@](C(=O)COC(=O)C)(O)[C@@]1(C)CC2=O ITRJWOMZKQRYTA-RFZYENFJSA-N 0.000 description 1
- IAZDPXIOMUYVGZ-WFGJKAKNSA-N Dimethyl sulfoxide Chemical compound [2H]C([2H])([2H])S(=O)C([2H])([2H])[2H] IAZDPXIOMUYVGZ-WFGJKAKNSA-N 0.000 description 1
- 206010059866 Drug resistance Diseases 0.000 description 1
- LLQPHQFNMLZJMP-UHFFFAOYSA-N Fentrazamide Chemical compound N1=NN(C=2C(=CC=CC=2)Cl)C(=O)N1C(=O)N(CC)C1CCCCC1 LLQPHQFNMLZJMP-UHFFFAOYSA-N 0.000 description 1
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 1
- 244000285963 Kluyveromyces fragilis Species 0.000 description 1
- 235000014663 Kluyveromyces fragilis Nutrition 0.000 description 1
- 102000004317 Lyases Human genes 0.000 description 1
- 108090000856 Lyases Proteins 0.000 description 1
- 101000745728 Mesocricetus auratus Lithocholate 6-beta-hydroxylase Proteins 0.000 description 1
- 102000008109 Mixed Function Oxygenases Human genes 0.000 description 1
- 108010074633 Mixed Function Oxygenases Proteins 0.000 description 1
- 102000004316 Oxidoreductases Human genes 0.000 description 1
- 108090000854 Oxidoreductases Proteins 0.000 description 1
- 208000005107 Premature Birth Diseases 0.000 description 1
- 206010036590 Premature baby Diseases 0.000 description 1
- 108010029485 Protein Isoforms Proteins 0.000 description 1
- 102000001708 Protein Isoforms Human genes 0.000 description 1
- 244000253911 Saccharomyces fragilis Species 0.000 description 1
- 235000018368 Saccharomyces fragilis Nutrition 0.000 description 1
- 206010039792 Seborrhoea Diseases 0.000 description 1
- 229930182558 Sterol Natural products 0.000 description 1
- 244000057717 Streptococcus lactis Species 0.000 description 1
- APQHKWPGGHMYKJ-UHFFFAOYSA-N Tributyltin oxide Chemical compound CCCC[Sn](CCCC)(CCCC)O[Sn](CCCC)(CCCC)CCCC APQHKWPGGHMYKJ-UHFFFAOYSA-N 0.000 description 1
- 238000006137 acetoxylation reaction Methods 0.000 description 1
- WETWJCDKMRHUPV-UHFFFAOYSA-N acetyl chloride Chemical compound CC(Cl)=O WETWJCDKMRHUPV-UHFFFAOYSA-N 0.000 description 1
- 239000012345 acetylating agent Substances 0.000 description 1
- 206010000496 acne Diseases 0.000 description 1
- 239000004480 active ingredient Substances 0.000 description 1
- 239000008186 active pharmaceutical agent Substances 0.000 description 1
- 210000004100 adrenal gland Anatomy 0.000 description 1
- GZCGUPFRVQAUEE-SLPGGIOYSA-N aldehydo-D-glucose Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)[C@@H](O)C=O GZCGUPFRVQAUEE-SLPGGIOYSA-N 0.000 description 1
- 229960003305 alfaxalone Drugs 0.000 description 1
- 231100000360 alopecia Toxicity 0.000 description 1
- 235000011114 ammonium hydroxide Nutrition 0.000 description 1
- 230000003444 anaesthetic effect Effects 0.000 description 1
- 150000001441 androstanes Chemical class 0.000 description 1
- 239000000051 antiandrogen Substances 0.000 description 1
- 229960001230 asparagine Drugs 0.000 description 1
- 239000003613 bile acid Substances 0.000 description 1
- 230000004071 biological effect Effects 0.000 description 1
- 238000011138 biotechnological process Methods 0.000 description 1
- 239000006227 byproduct Substances 0.000 description 1
- 239000001110 calcium chloride Substances 0.000 description 1
- 229910001628 calcium chloride Inorganic materials 0.000 description 1
- 238000001460 carbon-13 nuclear magnetic resonance spectrum Methods 0.000 description 1
- 239000003054 catalyst Substances 0.000 description 1
- 238000006555 catalytic reaction Methods 0.000 description 1
- 230000022131 cell cycle Effects 0.000 description 1
- 230000008859 change Effects 0.000 description 1
- 238000003889 chemical engineering Methods 0.000 description 1
- 239000003795 chemical substances by application Substances 0.000 description 1
- 238000004587 chromatography analysis Methods 0.000 description 1
- 238000003776 cleavage reaction Methods 0.000 description 1
- 238000004440 column chromatography Methods 0.000 description 1
- 230000002860 competitive effect Effects 0.000 description 1
- 238000001816 cooling Methods 0.000 description 1
- 210000004246 corpus luteum Anatomy 0.000 description 1
- 239000003246 corticosteroid Substances 0.000 description 1
- 229960001334 corticosteroids Drugs 0.000 description 1
- OMFXVFTZEKFJBZ-HJTSIMOOSA-N corticosterone Chemical compound O=C1CC[C@]2(C)[C@H]3[C@@H](O)C[C@](C)([C@H](CC4)C(=O)CO)[C@@H]4[C@@H]3CCC2=C1 OMFXVFTZEKFJBZ-HJTSIMOOSA-N 0.000 description 1
- 229960003290 cortisone acetate Drugs 0.000 description 1
- 239000002537 cosmetic Substances 0.000 description 1
- 238000002425 crystallisation Methods 0.000 description 1
- 230000008025 crystallization Effects 0.000 description 1
- 230000001419 dependent effect Effects 0.000 description 1
- 238000010511 deprotection reaction Methods 0.000 description 1
- 201000010099 disease Diseases 0.000 description 1
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 1
- 239000012153 distilled water Substances 0.000 description 1
- 230000035622 drinking Effects 0.000 description 1
- 238000001035 drying Methods 0.000 description 1
- 239000003792 electrolyte Substances 0.000 description 1
- 238000000921 elemental analysis Methods 0.000 description 1
- 230000008030 elimination Effects 0.000 description 1
- 238000003379 elimination reaction Methods 0.000 description 1
- 150000002085 enols Chemical class 0.000 description 1
- 230000008686 ergosterol biosynthesis Effects 0.000 description 1
- 239000003687 estradiol congener Substances 0.000 description 1
- 238000011156 evaluation Methods 0.000 description 1
- 238000002474 experimental method Methods 0.000 description 1
- 230000002349 favourable effect Effects 0.000 description 1
- 238000001914 filtration Methods 0.000 description 1
- 230000002538 fungal effect Effects 0.000 description 1
- 239000007789 gas Substances 0.000 description 1
- 230000002068 genetic effect Effects 0.000 description 1
- 238000010353 genetic engineering Methods 0.000 description 1
- 239000008103 glucose Substances 0.000 description 1
- 208000024963 hair loss Diseases 0.000 description 1
- 230000003676 hair loss Effects 0.000 description 1
- 125000005843 halogen group Chemical group 0.000 description 1
- 230000003054 hormonal effect Effects 0.000 description 1
- 229960000890 hydrocortisone Drugs 0.000 description 1
- 238000011065 in-situ storage Methods 0.000 description 1
- 230000006698 induction Effects 0.000 description 1
- 238000011090 industrial biotechnology method and process Methods 0.000 description 1
- 239000011261 inert gas Substances 0.000 description 1
- 150000002500 ions Chemical class 0.000 description 1
- 239000003446 ligand Substances 0.000 description 1
- 238000004895 liquid chromatography mass spectrometry Methods 0.000 description 1
- 230000014759 maintenance of location Effects 0.000 description 1
- 239000000463 material Substances 0.000 description 1
- 238000005259 measurement Methods 0.000 description 1
- 239000002395 mineralocorticoid Substances 0.000 description 1
- 229910052757 nitrogen Inorganic materials 0.000 description 1
- 238000000655 nuclear magnetic resonance spectrum Methods 0.000 description 1
- 229940127234 oral contraceptive Drugs 0.000 description 1
- 239000003539 oral contraceptive agent Substances 0.000 description 1
- 239000003960 organic solvent Substances 0.000 description 1
- 210000001672 ovary Anatomy 0.000 description 1
- 239000001301 oxygen Substances 0.000 description 1
- 229910052760 oxygen Inorganic materials 0.000 description 1
- 238000006213 oxygenation reaction Methods 0.000 description 1
- 230000007918 pathogenicity Effects 0.000 description 1
- 239000000575 pesticide Substances 0.000 description 1
- 230000000144 pharmacologic effect Effects 0.000 description 1
- 239000007981 phosphate-citrate buffer Substances 0.000 description 1
- 230000001766 physiological effect Effects 0.000 description 1
- 230000035479 physiological effects, processes and functions Effects 0.000 description 1
- 210000002826 placenta Anatomy 0.000 description 1
- 230000003389 potentiating effect Effects 0.000 description 1
- 239000002243 precursor Substances 0.000 description 1
- JWMFYGXQPXQEEM-WZBAXQLOSA-N pregnane Chemical compound C1CC2CCCC[C@]2(C)[C@@H]2[C@@H]1[C@@H]1CC[C@H](CC)[C@@]1(C)CC2 JWMFYGXQPXQEEM-WZBAXQLOSA-N 0.000 description 1
- 238000004237 preparative chromatography Methods 0.000 description 1
- 230000019525 primary metabolic process Effects 0.000 description 1
- 230000000757 progestagenic effect Effects 0.000 description 1
- RJKFOVLPORLFTN-UHFFFAOYSA-N progesterone acetate Natural products C1CC2=CC(=O)CCC2(C)C2C1C1CCC(C(=O)C)C1(C)CC2 RJKFOVLPORLFTN-UHFFFAOYSA-N 0.000 description 1
- 239000000583 progesterone congener Substances 0.000 description 1
- 238000000746 purification Methods 0.000 description 1
- UMJSCPRVCHMLSP-UHFFFAOYSA-N pyridine Natural products COC1=CC=CN=C1 UMJSCPRVCHMLSP-UHFFFAOYSA-N 0.000 description 1
- 238000012797 qualification Methods 0.000 description 1
- 230000001105 regulatory effect Effects 0.000 description 1
- 230000008844 regulatory mechanism Effects 0.000 description 1
- 230000004044 response Effects 0.000 description 1
- 230000007017 scission Effects 0.000 description 1
- 208000008742 seborrheic dermatitis Diseases 0.000 description 1
- 230000024053 secondary metabolic process Effects 0.000 description 1
- 230000028327 secretion Effects 0.000 description 1
- 210000001625 seminal vesicle Anatomy 0.000 description 1
- 208000017520 skin disease Diseases 0.000 description 1
- 239000003270 steroid hormone Substances 0.000 description 1
- 230000003637 steroidlike Effects 0.000 description 1
- 150000003432 sterols Chemical class 0.000 description 1
- 235000003702 sterols Nutrition 0.000 description 1
- 125000001424 substituent group Chemical group 0.000 description 1
- 229960003604 testosterone Drugs 0.000 description 1
- 230000007704 transition Effects 0.000 description 1
- 229910021642 ultra pure water Inorganic materials 0.000 description 1
- 239000012498 ultrapure water Substances 0.000 description 1
- 238000001291 vacuum drying Methods 0.000 description 1
- MWOOGOJBHIARFG-UHFFFAOYSA-N vanillin Chemical compound COC1=CC(C=O)=CC=C1O MWOOGOJBHIARFG-UHFFFAOYSA-N 0.000 description 1
- FGQOOHJZONJGDT-UHFFFAOYSA-N vanillin Natural products COC1=CC(O)=CC(C=O)=C1 FGQOOHJZONJGDT-UHFFFAOYSA-N 0.000 description 1
- 235000012141 vanillin Nutrition 0.000 description 1
- 201000010653 vesiculitis Diseases 0.000 description 1
Images





















Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P33/00—Preparation of steroids
- C12P33/06—Hydroxylating
- C12P33/08—Hydroxylating at 11 position
- C12P33/10—Hydroxylating at 11 position at 11 alpha-position
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K31/00—Medicinal preparations containing organic active ingredients
- A61K31/56—Compounds containing cyclopenta[a]hydrophenanthrene ring systems; Derivatives thereof, e.g. steroids
- A61K31/57—Compounds containing cyclopenta[a]hydrophenanthrene ring systems; Derivatives thereof, e.g. steroids substituted in position 17 beta by a chain of two carbon atoms, e.g. pregnane or progesterone
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N1/00—Microorganisms, e.g. protozoa; Compositions thereof; Processes of propagating, maintaining or preserving microorganisms or compositions thereof; Processes of preparing or isolating a composition containing a microorganism; Culture media therefor
- C12N1/14—Fungi; Culture media therefor
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Organic Chemistry (AREA)
- Engineering & Computer Science (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Genetics & Genomics (AREA)
- General Health & Medical Sciences (AREA)
- Biotechnology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Medicinal Chemistry (AREA)
- Biochemistry (AREA)
- Microbiology (AREA)
- General Engineering & Computer Science (AREA)
- Pharmacology & Pharmacy (AREA)
- Animal Behavior & Ethology (AREA)
- Virology (AREA)
- Mycology (AREA)
- Botany (AREA)
- Biomedical Technology (AREA)
- Epidemiology (AREA)
- Tropical Medicine & Parasitology (AREA)
- Public Health (AREA)
- Veterinary Medicine (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Steroid Compounds (AREA)
Abstract
Description
Область техникиTechnical field
Настоящее изобретение относится к области биотехнологии и касается применения известных штаммов мицелиальных грибов для гидроксилирования и ацетилирования стероидных соединений, в частности, соединений ряда прегнана (производных прогестерона) и может быть использовано в промышленной биотехнологии, а также в фармацевтической промышленности, для производства стероидных медицинских препаратов.The present invention relates to the field of biotechnology and relates to the application of known strains of filamentous fungi for hydroxylation and acetylation of steroid compounds, in particular compounds of several pregnan (derivatives of progesterone) and can be used in industrial biotechnology, as well as in the pharmaceutical industry, for the production of steroid medical drugs.
Уровень техникиThe level of technology
Введение гидроксильной группы в положение С11 стероидной молекулы имеет большое промышленное значение, так как не только обеспечивает изменение биологической активности исходной молекулы в желаемом направлении, но и создает возможность для дальнейшей модификации молекулы, например, введения атома галогена в положение С9, с целью создания фармакологически более активных производных. Осуществление 11(α/(β)-гидроксилирования стероидов микроорганизмами, особенно прогестерона, является экономически важным для производства кортикостероидов, так как одной из основных проблем их синтеза является введение кислородной функции в 11 положение, трудно осуществимое химическими методами. Так, например, химический синтез кортизона из желчных кислот включает -30 стадий и общий выход составляет несколько десятых долей процента [Л. Физер, М. Физер. Стероиды. Пер. с англ. Под ред. д.х.н. Н.Н Суворова и д.х.н. И.В. Торгова. М: Мир, 1964, с. 673]. Напротив, синтез ацетата кортизона из 11α-гидроксипрогестерона состоит из 10 стадий и общий выход достигает 14,6% [SU 106380, 1956].The introduction of a hydroxyl group in the C 11 position of the steroid molecule is of great industrial importance, since it not only provides a change in the biological activity of the parent molecule in the desired direction, but also creates an opportunity for further modification of the molecule, for example, introducing a halogen atom in the C 9 position pharmacologically more active derivatives. Implementation of 11 (α / (β) -hydroxylation of steroids by microorganisms, especially progesterone, is economically important for the production of corticosteroids, since one of the main problems of their synthesis is the introduction of oxygen function in the 11 position, which is difficult to implement by chemical methods. For example, chemical synthesis cortisone from bile acids includes -30 stages and the total yield is several tenths of a percent [L. Fizer, M. Fizer. Steroids. Translated from English. Edited by Dr. Kh.N.N. Suvorov and D.h. N.V.Torgovaya, Moscow: Mir, 1964, p. 673]. in the synthesis of cortisone acetate 11α-hydroxyprogesterone is composed of 10 stages and the overall yield reaches 14,6% [SU 106380, 1956].
Известно, что 11-гидроксилирование стероидов в одну стадию с высокой степенью селективности возможно только биотехнологически с применением ферментной системы микроорганизмов, в основном, мицелиальных грибов, одним из которых является родIt is known that 11-hydroxylation of steroids in one stage with a high degree of selectivity is possible only by biotechnology using the enzyme system of microorganisms, mainly filamentous fungi, one of which is the genus
Aspergillus. Однако этому роду присуща видовая специфичность трансформации стероидов ряда прегнана. Так, трансформация прогестерона у ряда видов (например, А. terreus [K.Yildirim, A.Uzuner, and E.Y. Gulcuoglu. Biotransformation of some steroids by Aspergillus terreus MRC 200365. Collect. Czech. Chem. Commun. 2010, Vol. 75(6), 665-673], A.tamarii [K.Yildirim, A.Uzuner and E.Y. Gulcuoglu. Baeyer-Villiger oxidation of some steroids by Aspergillus tamarii MRC 72400. Collect. Czech. Chem. Commun. 2011, Vol. 76(6), 743-754], A. versicolor [H.Y. Yang, H.L. Su, G. Du, G.J. Shen, J.X. Sun, and H.Y. Chen. Progesterone side-chain cleavage by Aspergillus versicolor. Advanced Materials Research. Advances in Chemical Engineering III. 2013.Vols. 781-784, 1164-1167; Abed Nosrat M. Side chain degradation of progesterone by Aspergillus versicolor 79. Pakistan Journal of Biochemistry, 1972, Vol. 5(1), 5-7], A. flavus [M.E. Mostafa, and A.A.Zohri. Progesterone side-chain degradation by some species of Aspergillus flavus group.Folia Microbiol (Praha). 2000; Vol. 45(3), 243-7]) может протекать не в направлении гидроксилирования, а в направлении элиминирования боковой прегнановой цепи с образованием соединений ряда андростана. Другие штаммы рода Aspergillus способны гидроксилировать прогестерон, вводя гидроксильную группу в различные положения молекулы, в том числе в 11α-положение. При этом направление гидроксилирования может существенно зависеть не только от видовой принадлежности штамма, но и от условий проведения трансформации.Aspergillus. However, this genus is inherent in the species specificity of transformation of steroids of the series pregnan. Thus, the transformation of progesterone in a number of species (for example, A. terreus [K. Yildirim, A. Uzuner, and EY Gulcuoglu. Biotransformation of some steroids by Aspergillus terreus MRC 200365. Collect. Czech. Chem. Commun. 2010, Vol. 75 ( 6), 665-673], A.tamarii [K.Yildirim, A.Uzuner and EY Gulcuoglu. Baeyer-Villiger oxidation of Aspergillus tamarii MRC 72400. Collect. Czech. Chem. Commun. 2011, Vol. 76 ( 6), 743-754], A. versicolor [HY Yang, HL Su, G. Du, GJ Shen, JX Sun, and HY Chen. Progesterone side-chain cleavage by Aspergillus versicolor. Advanced Materials Research. Advances in Chemical Engineering III 2013.Vols. 781-784, 1164-1167; Abed Nosrat M. Side chain degradation of progesterone by Aspergillus versicolor 79. Pakistan Journal of Biochemistry, 1972, Vol. 5 (1), 5-7], A. flavus [ ME Mostafa, AAZohri Progesterone side chain degradation of some species of Aspergillus flavus group.Folia Microbiol (Praha) 2000; Vol .45 (3), 243-7]) can proceed not in the direction of hydroxylation, but in the direction of elimination of the side of the pregnanic chain with the formation of compounds of the Androstane series. Other strains of the genus Aspergillus are able to hydroxylate progesterone by introducing a hydroxyl group into various positions of the molecule, including the 11α position. The direction of hydroxylation may significantly depend not only on the species of the strain, but also on the conditions of the transformation.
Биотрансформация прогестерона с образованием 11α-гидроксипрогестерона наиболее изучена с применением грибов Aspergillus ochraceus, селективность 11α-гидроксилирования у которых может достигать 90% и более [Т.К. Dutta, and Т.В. Samanta. Bioconversion of progesterone by the activated immobilized conidia of Aspergillus ochraceus TS. Curr. Microbiol. 1999; Vol. 39(6), 309-312; P.Somal, and C.L. Chopra. Microbial conversion of steroids. III: 11α-hydroxylation by fungal mycelium. Applied Microbiology and Biotechnology, 1985, Vol. 21(5), 267-269]. Штаммы A. ochraceus нашли применение в качестве промышленных культур.Biotransformation of progesterone with the formation of 11α-hydroxyprogesterone is the most studied using Aspergillus ochraceus fungi, the selectivity of 11α-hydroxylation in which can reach 90% or more [T.K. Dutta, and T.V. Samanta. Bioconversion of progesterone by the activated immobilized conidia of Aspergillus ochraceus TS. Curr. Microbiol. 1999; Vol. 39 (6), 309-312; P.Somal, and C.L. Chopra. Microbial conversion of steroids. III: 11α-hydroxylation by fungal mycelium. Applied Microbiology and Biotechnology, 1985, Vol. 21 (5), 267-269]. The strains of A. ochraceus have been used as industrial crops.
Мицеллиальный гриб Aspergillus nidulans является одной из наиболее известных эукариотических генетических систем и широко используется в качестве модельной системы для расшифровки биологии клеточного цикла, патогенности, лекарственной устойчивости, болезней человека, первичного и вторичного метаболизма других микроорганизмов. Несмотря на это, штаммы гриба A. nidulans как стероид-трансформирующие микроорганизмы мало изучены. Однако имеются сообщения, что этот вид обладает 11α-монооксигеназной активностью и способен трансформировать прогестерон с образованием преимущественно 11α-гидрокси-производного. Так, M.J. Henry и H.D. Sisler [M.J. Henry, and H.D. Sisler. Effects of sterol biosynthesis-inhibiting (SBI) fungicides on cytochrome P-450 oxygenations in fungi. Pesticide Biochemistry and Physiology, 1984, Vol. 22(3),262-275], изучая влияние фунгицидов, ингибирующих биосинтез эргостерина в грибах, на цитохором-Р450-зависимое гидроксилирование прогестерона, сообщили, что гриб A. nidulans (Eidam) Winter (штамм 003) гидроксилирует прогестерон при начальной концентрации 100 мкг/мл с образованием смеси, содержащей 11α-гидроксипрогестерон (50%), 6β-гидроксипрогестерон (5%), 6β,11α-дигидроксипрогестерон (14%) и неконвертированный прогестерон (31%). При этом лиазная активность (расщепление боковой цепи по связи С17-С20) у этого штамма не наблюдалась.The mycelial fungus Aspergillus nidulans is one of the most well-known eukaryotic genetic systems and is widely used as a model system for deciphering cell cycle biology, pathogenicity, drug resistance, human diseases, primary and secondary metabolism of other microorganisms. Despite this, the strains of the fungus A. nidulans as steroid-transforming microorganisms have been little studied. However, there are reports that this species has 11α-monooxygenase activity and is able to transform progesterone with the formation of predominantly 11α-hydroxy-derivative. So, MJ Henry and HD Sisler [MJ Henry, and HD Sisler. Effects of sterol biosynthesis-inhibiting (SBI) fungicides on cytochrome P-450 oxygenations in fungi. Pesticide Biochemistry and Physiology, 1984, Vol. 22 (3), 262-275], studying the effect of fungicides that inhibit ergosterol biosynthesis in fungi on cytochore-P450-dependent progesterone hydroxylation, reported that A. nidulans (Eidam) Winter fungus (strain 003) hydroxylates progesterone at an initial concentration of 100 µg / ml to form a mixture containing 11α-hydroxyprogesterone (50%), 6β-hydroxyprogesterone (5%), 6β, 11α-dihydroxyprogesterone (14%) and unconverted progesterone (31%). At the same time, lyase activity (splitting of the side chain by the C 17 –C 20 bond) was not observed in this strain.
Также известно, что культура гриба A. nidulans (из коллекции Центра культур лаборатории микробиологической химии, National Research Centre, Каир, местная среда обитания), обладая стероид-11α-гидроксилирующей активностью, может трансформировать прогестерон не только в 11α -гидроксипрогестерон, но и в 21-гидроксипрогестерон (он же 11-дезоксикортикостерон) [А.Н. El-Refai, and К.М. Ghanem. Some physiological relations of progesterone conversion by Aspergillus nidulans. Egyptian Journal of Microbiology, 1987, Vol. 22(2), 327-338; A.H. El-Refai, and K.M. Ghanem. Microbial response to steroids. Egyptian Journal of Microbiology, 1989, Vol. 23(1), 1-11]. Авторы вносили прогестерон в ростовую среду в растворе 96% этанола с финальной концентрацией растворителя 1%. Трансформацию прогестерона проводили с нагрузкой 1 г/л при температуре 30±2°С на качалке (200 об/мин, амплитуда 7 см) в течение 48 ч, при этом наблюдалось образование дигидроксилированного производного - 6β,11α-дигидроксипрогестерона. Был сделан вывод, что при значениях рН среды, близких к нейтральным, 6β-гидроксилирование является вторичным процессом и 6β,11α-дигидроксипрогестерон образуется из первично образованного 11α-гидроксипрогестерона [А.Н. El-Refai, and К.М. Ghanem. Some physiological relations of progesterone conversion by Aspergillus nidulans. Egyptian Journal of Microbiology, 1987, Vol. 22(2), 327-338].It is also known that the culture of the fungus A. nidulans (from the collection of the Crop Center for Microbiological Chemistry Laboratories, National Research Center, Cairo, local habitat), possessing steroid-11α-hydroxylation activity, can transform progesterone not only into 11α-hydroxyprogesterone, but also 21-hydroxyprogesterone (aka 11-deoxycorticosterone) [A.N. El-Refai, and KM Ghanem. Some physiological relations of progesterone conversion by Aspergillus nidulans. Egyptian Journal of Microbiology, 1987, Vol. 22 (2), 327-338; A.H. El-Refai, and K.M. Ghanem. Microbial response to steroids. Egyptian Journal of Microbiology, 1989, Vol. 23 (1), 1-11]. The authors applied progesterone to the growth medium in a solution of 96% ethanol with a final solvent concentration of 1%. Transformation of progesterone was performed with a load of 1 g / l at a temperature of 30 ± 2 ° C on a rocking chair (200 rpm,
Известно, что микроорганизмы способны ацетилировать стероидные вторичные спирты. Так, известна способность некоторых видов дрожжей и дрожжеподобных организмов {Saccharomyces fragilis, S. lactis, Candida pseudotropicalis и Torulopsis sphaerica) ацетилировать тестостерон [![]()
![]()
![]()
![]()
Из уровня техники известно, что ацетилирование 11α-гидроксипрогестерона осуществляют химически, используя в качестве ацетилирующего агента уксусный ангидрид. Так, известным химическим методом [D.H. Peterson, Н.С. Murray, S.H. Eppstein, L.M. Reineke, A. Weintraub, P.D. Meister, and H.M. Leigh. Microbiological Transformations of Steroids. I. Introduction of Oxygen at Carbon-11 of Progesterone. J. Am. Chem. Soc, 1952, Vol. 74 (23), 5933-5936] получают 11α-ацетоксипрогестерон, ацетилируя 11α-гидроксипрогестерон (20 мг) действием уксусного ангидрида (0,6 мл) в среде пиридина (0,6 мл). Смесь выдерживают в течение 16 ч при комнатной температуре. По окончании реакции реакционную массу разбавляют 25 мл воды, выдерживают в течение 1 ч, охлаждают для начала кристаллизации. Кристаллы отделяют фильтрацией, промывают водой и сушат. Получают 16,1 мг 11α-ацетоксипрогестерона с т. пл. 176-177°С.It is known in the art that acetylation of 11α-hydroxyprogesterone is carried out chemically using acetic anhydride as the acetylating agent. Thus, the well-known chemical method [D.H. Peterson, N.S. Murray, S.H. Eppstein, L.M. Reineke, A. Weintraub, P.D. Meister, and H.M. Leigh. Microbiological Transformations of Steroids. I. Introduction of Oxygen at Carbon-11 of Progesterone. J. Am. Chem. Soc, 1952, Vol. 74 (23), 5933-5936], 11α-acetoxyprogesterone is obtained by acetylating 11α-hydroxyprogesterone (20 mg) by the action of acetic anhydride (0.6 ml) in pyridine (0.6 ml). The mixture was incubated for 16 hours at room temperature. Upon completion of the reaction, the reaction mass is diluted with 25 ml of water, incubated for 1 h, cooled to start crystallization. The crystals are separated by filtration, washed with water and dried. 16.1 mg of 11α-acetoxyprogesterone with m.p. 176-177 ° C.
Таким образом, из уровня техники следует, что образование 11α-ацетоксипрогестерона при трансформации прогестерона культурами мицеллиальных грибов вообще, в частности, рода Aspergillus, конкретно Aspergillus nidulans, ранее не наблюдалось.Thus, it follows from the prior art that the formation of 11α-acetoxyprogesterone during transformation of progesterone by cultures of micellar fungi in general, in particular, the genus Aspergillus, specifically Aspergillus nidulans, has not been previously observed.
11α-Гидроксипрогестерон (11α-гидроксипрегн-4-ен-3,20-дион, СAS №80-75-1) является физиологически активным производным прогестерона (прегн-4-ен-3, 20-диона, CAS №57-83-0) - нативного стероидного гормона, синтезируемого желтым телом яичника, корковым веществом (корой) надпочечников, семенными пузырьками и плацентой. Препараты на основе прогестерона применяются в качестве лекарственных средств в медицине и ветеринарии.11α-Hydroxyprogesterone (11α-hydroxypregn-4-ene-3,20-dione, CAS No. 80-75-1) is a physiologically active progesterone derivative (Pregn-4-en-3, 20-dione, CAS No. 57-83- 0) - a native steroid hormone synthesized by the corpus luteum of the ovary, the cortex (cortex) of the adrenal glands, the seminal vesicles and the placenta. Progesterone-based preparations are used as medicines in medicine and veterinary medicine.
11α-Гидроксипрогестерон, как и его 11β-эпимер, обладает ингибирующей активностью в отношении фермента 11β-гидроксистероид-дегидрогеназы, который окисляет 11-гидрокси-группу в 11-кето-группу, таким образом играя регулятивную роль в поддержании электролитного баланса [G.W. Souness, S.A. Latif, J.L.Laurenzo, D.J. Morris. 11 alpha- and 11 beta-hydroxyprogesterone, potent inhibitors of 11 beta-hydroxysteroid dehydrogenase (isoforms 1 and 2), confer marked mineralocorticoid activity on corticosterone in the ADX rat. Endocrinology. 1995, Vol. 136(4), 1809-1812].11α-hydroxyprogesterone, like its 11β-epimer, has inhibitory activity against the enzyme 11β-hydroxysteroid dehydrogenase, which oxidizes the 11-hydroxy group to the 11-keto group, thus playing a regulatory role in maintaining electrolyte balance [G.W. Souness, S.A. Latif, J.L.Laurenzo, D.J. Morris. 11 alpha- and 11 beta-hydroxyprogesterone, potent inhibitors of 11 beta-hydroxysteroid dehydrogenase (
11α-Ацетоксипрогестерон (11α-гидроксипрогестерона ацетат, CAS №2268-98-6) обладает физиологической активностью, является активным фармацевтическим ингредиентом и относится к средствам, снижающим риск преждевременных родов [http://www.imcopharma.cz/ru/api/11 -alfa-gidroksiprogesteron-acetat].11α-Acetoxyprogesterone (11α-hydroxyprogesterone acetate, CAS No. 2268-98-6) has a physiological activity, is an active pharmaceutical ingredient and refers to the means that reduce the risk of premature birth [http://www.imcopharma.cz/ru/api/11 -alfa-gidroksiprogesteron-acetat].
11α-Ацетоксипрогестерон и 11α-гидроксипрогестерон, являясь производными прогестерона, обладающими гестагенной активностью, могут быть использованы в комбинации с природными или синтетическими эстрогенами в качестве активного ингредиента как в препаратах для пероральной контрацепции, так и в препаратах для гормональной заместительной терапии женщин в постклимактерический период.11α-Acetoxyprogesterone and 11α-hydroxyprogesterone, being progesterone derivatives with progestogen activity, can be used in combination with natural or synthetic estrogens as an active ingredient in both oral contraceptive preparations and in post-menopausal women.
Кроме того, 11α-гидроксипрогестерон и его ацетат могут быть использованы не только в качестве прогестагенных средств, как указано выше, но и в качестве антиандрогенных агентов для снижения сальных выделений у пациентов, страдающих себореей, аллопецией и другими заболеваниями кожи, связанными с гиперандрогенизацией [RU 2432952, 2011], также известны косметические композиции для ухода за волосами и кожей головы, содержащие 11α-гидроксипрогестерон или 11α-ацетоксипрогестерон [DE 2757024, 1979], которые могут применяться для лечения выпадения волос, чрезмерной засаленности волос и кожи головы, вульгарных угрей и т.п.При этом отмечено, что у этих соединений при местном применении гормональные побочные эффекты отсутствуют.In addition, 11α-hydroxyprogesterone and its acetate can be used not only as progestogenic agents, as mentioned above, but also as antiandrogenic agents to reduce sebaceous secretions in patients with seborrhea, alopecia and other skin diseases associated with hyperandrogenization [RU 2432952, 2011], also known cosmetic compositions for hair and scalp, containing 11α-hydroxyprogesterone or 11α-acetoxyprogesterone [DE 2757024, 1979], which can be used to treat hair loss, is excessive oh greasy hair and scalp, acne vulgaris and t.p.Pri it noted that these compounds, when applied topically hormonal side effects are absent.
11α-Гидроксипрогестерон и 11α-ацетоксипрогестерон могут быть использованы как исходные продукты в синтезе других лекарственных средств для медицинского применения. Например, 11α-гидроксипрогестерон может быть использован в синтезе 11-кетопрогестерона (кетогестина, CAS №516-15-4), который далее превращают в кортизон или 11β-гидроксипрогестерон - предшественник нативного гидрокортизона - методами, известными из уровня техники. Химический метод перехода от 11α-гидроксипрогестерона к 11-кетопрогестерону с выходом более 90% и далее в 11β-гидроксипрогестерон описан в патенте [US 2015376225, 2015, примеры 1 и 7 соответственно]. Кроме того, 11α-гидроксипрогестерон может быть использован в качестве исходного соединения в синтезе перспективных аналогов нейростероидного анестетика альфаксолона [P.Y. Savechenkov, D.C. Chiara, R. Desai, A.T. Stern, X.Zhou, A.M. Ziemba, A.L. Szabo, Y.Zhang, J.B. Cohen, S.A. Forman, K.W. Miller, K.S. Bruzik. Synthesis and pharmacological evaluation of neurosteroid photoaffinity ligands. European Journal of Medicinal Chemistry. 2017. Vol. 136, P. 334-347].11α-hydroxyprogesterone and 11α-acetoxyprogesterone can be used as starting materials in the synthesis of other medicines for medical use. For example, 11α-hydroxyprogesterone can be used in the synthesis of 11-ketoprogesterone (ketogestin, CAS No. 516-15-4), which is further converted into cortisone or 11β-hydroxyprogesterone, the precursor of native hydrocortisone, by methods known from the prior art. The chemical method of transition from 11α-hydroxyprogesterone to 11-ketoprogesterone with a yield of more than 90% and further to 11β-hydroxyprogesterone is described in the patent [US 2015376225, 2015, examples 1 and 7, respectively]. In addition, 11α-hydroxyprogesterone can be used as a starting compound in the synthesis of promising analogues of the neurosteroid anesthetic alfaxolone [P.Y. Savechenkov, D.C. Chiara, R. Desai, A.T. Stern, X.Zhou, A.M. Ziemba, A.L. Szabo, Y.Zhang, J.B. Cohen, S.A. Forman, K.W. Miller, K.S. Bruzik. Synthesis and pharmacological evaluation of neurosteroid photoaffinity ligands. European Journal of Medicinal Chemistry. 2017. Vol. 136, p. 334-347].
Однако о применении грибов A. nidulans, обладающих стероид-11α-гидроксилирующей активностью и способных конвертировать прогестерон в 11α-гидроксипрогестерон, для получения 11α-ацетоксипрогестерона из уровня техники не известно.However, the use of fungi A. nidulans with steroid-11α-hydroxylation activity and able to convert progesterone to 11α-hydroxyprogesterone, to obtain 11α-acetoxyprogesterone in the prior art is not known.
Раскрытие изобретенияDISCLOSURE OF INVENTION
Технической проблемой, решаемой с помощью настоящего изобретения, является расширение номенклатуры микроорганизмов, способных проводить 11α-гидроксилирование прогестерона, а именно, применение известных штаммов аскомицета A. nidulans для 11Α-гидроксилирования прогестерона, а также расширение ассортимента методов ацетилирования вторичной гидроксильной группы при атоме С11 молекулы прогестерона, благодаря наличию способности штаммов по настоящему изобретению ацетилировать вторичную гидроксильную группу при атоме С11 молекулы прогестерона, как альтернативы химическому методу ацетилирования.The technical problem to be solved with the help of the present invention is the expansion of the range of microorganisms capable of conducting 11α-hydroxylation of progesterone, namely, the use of known strains of A. nidulans ascomycete for 11Α-hydroxylation of progesterone, as well as the expansion of the range of methods for acetylation of the secondary hydroxyl group at the C 11 atom progesterone molecules due to the ability of the strains of the present invention to acetylate the secondary hydroxyl group at the C 11 atom of the progesterone molecule as an alternative to the chemical method of acetylation.
Техническая проблема решалась путем использования следующих штаммов:The technical problem was solved by using the following strains:
1. Штамм дикого типа Aspergillus nidulans, депонированный во Всероссийской Коллекции Промышленных Микроорганизмов (ВКПМ) под регистрационным номером F-1069 (synonym FGSC А4; АТСС 3863, 12996, 26451; CBS 112.46; NRRL 194).1. Wild-type strain Aspergillus nidulans, deposited in the All-Russian Collection of Industrial Microorganisms (VKPM) under registration number F-1069 (synonym FGSC A4; ATCC 3863, 12996, 26451; CBS 112.46; NRRL 194).
2. Штамм A. nidulans 031 (argB-; pyrG-) (synonym FP-308.1) - ауксотроф штамма Aspergillus nidulans ВКПМ F-1069, несущий мутации argB2 и pyrG89 [U. Christensen, B.S. Gruben, S. Madrid, H. Mulder, I. Nikolaev, and R. P. de Vries. Unique Regulatory Mechanism for D-Galactose Utilization in Aspergillus nidulans. Appl Environ Microbiol. 2011, Vol. 77(19), 7084-7087].2. Strain A. nidulans 031 (argB - ; pyrG - ) (synonym FP-308.1) - auxotroph of the strain Aspergillus nidulans VKPM F-1069, carrying the mutations argB2 and pyrG89 [U. Christensen, BS Gruben, S. Madrid, H. Mulder, I. Nikolaev, and RP de Vries. Unique Regulatory Mechanism for D-Galactose Utilization in Aspergillus nidulans. Appl Environ Microbiol. 2011 Vol. 77 (19), 7084-7087].
Штамм A. nidulans ВКПМ F-1069Strain A. nidulans VKPM F-1069
Штамм A. nidulans ВКПМ F-1069 ранее для биотрасформации прогестерона не применялся, его стероид-11α-гидроксилирующая активность обнаружена впервые. Этот штамм впервые применен для биотрасформации прогестерона. Установлена способность этого штамма конвертировать прогестерон в 11α-гидроксипрогестерон и 11α-ацетоксипрогестерон.The A. nidulans VKPM F-1069 strain was not previously used for progesterone biotransformation, its steroid-11α-hydroxylation activity was detected for the first time. This strain was first used for progesterone biotraformation. The ability of this strain to convert progesterone to 11α-hydroxyprogesterone and 11α-acetoxyprogesterone has been established.
Штамм A. nidulans 031 (argB-: pyrG-)Strain A. nidulans 031 (argB - : pyrG - )
Штамм A. nidulans 031 (argB-; pyrG-) - ауксотроф штамма Aspergillus nidulans ВКПМ F-1069, несущий мутации argB2 (требующий аргинин) и pyrG89 (требующий уридин и урацил), ранее для биотрасформации прогестерона не применялся, его стероид-11α-гидроксилирующая активность обнаружена впервые. Этот штамм впервые применен для биотрасформации прогестерона. Установлена способность этого штамма конвертировать прогестерон в 11α-гидроксипрогестерон и 11α-ацетоксипрогестерон.The strain A. nidulans 031 (argB - ; pyrG - ) - auxotroph of the Aspergillus nidulans strain VKPM F-1069, carrying the argB2 (requiring arginine) and pyrG89 (requiring uridine and uracil) mutations, had not previously been used for progesterone biotransformation, it did not use its steroid-11, uterine-uracil, it did not use its steroid-11 for the progesterone, its steroid-11, uridine-uracil and uracil didn’t use the progesterone biotransformation. hydroxylation activity is detected for the first time. This strain was first used for progesterone biotraformation. The ability of this strain to convert progesterone to 11α-hydroxyprogesterone and 11α-acetoxyprogesterone has been established.
Было обнаружено, что наличие мутаций argB2 и pyrG89 у штамма A. nidulans ВКПМ F-1069 не изменяет его цитохром Р450-зависимую монооксигеназную способность и направление гидроксилирования молекулы прогестерона, а также не приводит к изменению его ацетилирующей способности - компонентный состав продуктов одинаков.It was found that the presence of argB2 and pyrG89 mutations in A. nidulans VKPM F-1069 strain does not alter its cytochrome P450-dependent monooxygenase ability and hydroxylation direction of the progesterone molecule, and does not alter its acetylating ability - the component composition of the products is the same.
Техническим результатом является биокаталитическое получение стероидных спиртов и их этерификация без применения химических методов. Получение 11α-ацетоксипрогестерона биотрансформацией прогестерона в одну стадию имеет преимущества перед известным из уровня техники двухстадийным процессом, включающим следующие этапы: 1) микробиологическое 11α-гидроксилирование прогестерона и 2) химическое ацетилирование вторичной 11α-гидроксильной группы. Биотехологическое 11α-гидроксилирование прогестерона и ацетилирование образованного 11α-гидроксипрогестерона с выделением или без выделения его из реакционной массы является экономически эффективным процессом, преимущества которого перед химическим методом ацетилирования состоят в следующем:The technical result is the biocatalytic production of steroid alcohols and their esterification without the use of chemical methods. Obtaining 11α-acetoxyprogesterone by biotransformation of progesterone in one stage has advantages over the two-stage process known from the prior art, including the following steps: 1) microbiological 11α-hydroxylation of progesterone and 2) chemical acetylation of the secondary 11α-hydroxyl group. Biotechnological 11α-hydroxylation of progesterone and acetylation of the formed 11α-hydroxyprogesterone, with or without isolation from the reaction mass, is a cost-effective process, the advantages of which over the chemical method of acetylation are as follows:
- процесс проводится без использования агрессивных химических реагентов, таких как уксусный ангидрид или хлорангидрид уксусной кислоты, без использования сильных минеральных кислот в качестве катализаторов ацетилирования, таких как 60% хлорная кислота;- the process is carried out without the use of aggressive chemical reagents, such as acetic anhydride or acetic acid chloride, without using strong mineral acids as acetylation catalysts, such as 60% perchloric acid;
-биокаталитическая этерификация 11α-гидроксипрогестерона является альтернативой химическому методу ацетилирования, является экологически более чистым и безопасным методом;-biocatalytic esterification of 11α-hydroxyprogesterone is an alternative to the chemical method of acetylation, it is an environmentally cleaner and safer method;
- биокаталитическое ацетилирование протекает регионаправленно без образования побочного продукта енолацетилирования Δ4-3-кетогруппы молекулы 11α-гидроксипрогестерона - 3, 11α-диацетоксипрегна-3,5-диен-20-она, содержание которого при использовании химического метода ацетилирования может сотавлять 20-30%, что существенно осложняет очистку целевого 11α-ацетоксипрогестерона и приводит к потерям его выхода.- biocatalytic acetylation proceeds regionally without the formation of a byproduct of enol acetylation of the Δ 4 -3-keto group of the 11α-hydroxyprogesterone molecule — 3, 11α-diacetoxypregna-3,5-diene-20-one, the content of which can be 20–30% when using the chemical method of acetylation , which significantly complicates the purification of the target 11α-acetoxyprogesterone and leads to a loss of its release.
Для достижения указанного выше технического результата по настоящему изобретению предлагается использовать известные штаммы Aspergillus nidulans в качестве биокатализаторов для биотехнологических процессов 11α-гидроксилирования и 11α-ацетоксилирования прогестерона, а также биокаталитической этерификации 11α-гидроксипрогестерона.To achieve the above technical result of the present invention, it is proposed to use known strains of Aspergillus nidulans as biocatalysts for biotechnological processes 11α-hydroxylation and 11α-acetoxylation of progesterone, as well as biocatalytic esterification of 11α-hydroxyprogesterone.
11α-Ацетоксипрогестерон при необходимости может быть гидролизован любым удобным способом с образованием 11α-гидроксипрогестерона, известным из уровня техники. Так, например, известен химический метод получения 11α-гидроксипрогестерона сольволизом 11α-ацетоксипрогестерона действием (бистрибутилолово)оксида [M.G. Perez, and M.S. Maier. Mild deprotection of steroid esters by bis(tributyltin)oxide. Tetrahedron Letters, 1995. Vol. 36(19), 3311-3314] или традиционным способом химического сольволиза в условиях основного катализа: в среде метанола в присутствии КОН [Т. Kubota, and F. Hayashi. Studies on A-norsteroids - VI. Directing effects of С11 substituents on the addition of osmium tetroxide to steroidal Δ1,4-3-ketones. Tetrahedron, 1967. Vol. 23, 995-1006].11α-Acetoxyprogesterone, if necessary, can be hydrolyzed in any convenient way with the formation of 11α-hydroxyprogesterone, known from the prior art. For example, a chemical method is known for producing 11α-hydroxyprogesterone by solvolysis of 11α-acetoxyprogesterone by the action of (bi-tributyltin) oxide [MG Perez, and MS Maier. Mild deprotection of steroid esters by bis (tributyltin) oxide. Tetrahedron Letters, 1995. Vol. 36 (19), 3311-3314] or the traditional method of chemical solvolysis under basic catalysis conditions: in methanol in the presence of KOH [T. Kubota, and F. Hayashi. Studies on A-norsteroids - VI. Directing effects of C 11 substituents of steroidal Δ 1,4 -3-ketones. Tetrahedron, 1967. Vol. 23, 995-1006].
Культуралыго-морфологические особенности заявляемых штаммов.Culture-morphological features of the claimed strains.
Штамм A. nidulans ВКПМ F1069Strain A. nidulans VKPM F1069
На агаризованной среде образует круглые колонии диаметром 30-35 мм через 7 суток роста. Поверхность ровная, выпуклая, пушистая. Текстура средней плотности. Край колоний плотный, ровный. Цвет колоний в зоне спороношения зеленовато-белый. Обратная сторона палево-коричневая. Эксудат отсутствует. Характеризуется интенсивным спороношением. Цвет спор зеленый.On agar medium forms round colonies with a diameter of 30-35 mm after 7 days of growth. The surface is flat, convex, fluffy. Medium density texture. The edge of the colonies tight, smooth. The color of the colonies in the area of sporulation greenish-white. The reverse side is pale brown. Exudate is absent. It is characterized by intense sporulation. The color of the dispute is green.
Штамм A. nidulans 031 (argB-: pyrG-)Strain A. nidulans 031 (argB - : pyrG - )
На агаризованной среде образует круглые колонии диаметром 30-35 мм через 10 суток роста (растет медленнее). Поверхность ровная, выпуклая, пушистая. Текстура средней плотности. Край колоний плотный, ровный. Цвет колоний в зоне спороношения зеленовато-белый. Обратная сторона палево-коричневая. Эксудат отсутствует. Характеризуется умеренным спороношением. Цвет спор зеленый.On agar medium forms round colonies with a diameter of 30-35 mm after 10 days of growth (grows slower). The surface is flat, convex, fluffy. Medium density texture. The edge of the colonies tight, smooth. The color of the colonies in the area of sporulation greenish-white. The reverse side is pale brown. Exudate is absent. It is characterized by moderate sporulation. The color of the dispute is green.
Основным свойством штаммов по настоящему изобретению является наличие стероид-11α-гидроксилирующей активности и способности конвертировать прогестерон в 11α-ацетоксипрогестерон, а также ацетилирующей активности и способности этерифицировать 11α-гидроксипрогестерон.The main property of the strains of the present invention is the presence of steroid-11α-hydroxylation activity and the ability to convert progesterone to 11α-acetoxyprogesterone, as well as acetylation activity and the ability to esterify 11α-hydroxyprogesterone.
На фиг. 1 представлена схема трансформации прогестерона известными штаммами A. nidulans ВКПМ F-1069 и A. nidulans 031 (argB-; pyrG-).FIG. 1 shows the progesterone transformation scheme by known strains of A. nidulans VKPM F-1069 and A. nidulans 031 (argB - ; pyrG - ).
Трансформация прогестерона протекает с первичным образованием 11α-гидроксипрогестерона. Процессы ацетилирования гидроксильной группы образованного 11α-гидроксипрогестерона и его 6β-гидроксилирования с образованием 6β,11α-дигидроксипрогестерона являются вторичными, конкурентными процессами трансформации, причем процесс биокаталитической этерификации имеет преимущества. Этот вывод подтвержден экспериментально. При продолжительности трансформации 14 ч штаммами A. nidulans ВКПМ F-1069 и A. nidulans 031 (argB-; pyrG-) в культуральной среде определено наличие только 11α-ацетоксипрогестерона и нетрансформированного исходного субстрата. Кроме того, используя 11α-гидроксипрогестерон вместо прогестерона в качестве исходного субстрата в аналогичных условиях трансформации штаммами A. nidulans ВКПМ F-1069 и A. nidulans 031 (argB-; pyrG-), было отмечено, что в течение 23 ч имеет место образование исключительно 11α-ацетоксипрогестерона.Transformation of progesterone proceeds with the initial formation of 11α-hydroxyprogesterone. The processes of acetylation of the hydroxyl group of the formed 11α-hydroxyprogesterone and its 6β-hydroxylation to form 6β, 11α-dihydroxyprogesterone are secondary, competitive transformation processes, and the process of biocatalytic esterification has advantages. This conclusion is confirmed experimentally. When the duration of transformation was 14 h, A. nidulans VKPM F-1069 and A. nidulans 031 (argB - ; pyrG - ) strains in the culture medium determined the presence of only 11α-acetoxyprogesterone and the untransformed initial substrate. In addition, using 11α-hydroxyprogesterone instead of progesterone as a starting substrate under similar transformation conditions with A. nidulans VKPM F-1069 and A. nidulans 031 strains (argB - ; pyrG - ), it was noted that within 23 hours 11α-acetoxyprogesterone.
Таким образом, сущность заявленного изобретения заключается в применении известных штаммов Aspergillus nidulans ВКПМ F-1069 и A. nidulans 031 (argB-; pyrG-) по новому назначению, а именно для 11α-гидроксилирования прогестерона и ацетилирования lla-гидроксипрогестерона с выделением или без выделения из культуральной жидкости.Thus, the essence of the claimed invention is the application of known strains of Aspergillus nidulans VKPM F-1069 and A. nidulans 031 (argB - ; pyrG - ) for a new purpose, namely for 11α-hydroxylation of progesterone and acetylation of lla-hydroxyprogesterone with or without isolation from the culture fluid.
Преимущества применения штаммов по настоящему изобретению состоят в способности конвертировать прогестерон в 11α-гидроксипрогестерон и ацетилировать гидроксильную группу в молекуле 11α-гидроксипрогестерона с образованием 11α-ацетоксипрогестерона как с выделением 11α-гидроксипрогестерона из культуральной среды, так и без выделения, in situ.The advantages of using the strains of the present invention are the ability to convert progesterone to 11α-hydroxyprogesterone and acetylate the hydroxyl group in the 11α-hydroxyprogesterone molecule with the formation of 11α-acetoxyprogesterone with the release of 11α-hydroxyprogesterone from the culture medium, and without isolation, in situ.
Краткое описание чертежейBrief Description of the Drawings
На фиг. 1 представлена схема трансформации прогестерона штаммами A. nidulans ВКПМ F-1069 и A. nidulans 031 (argB-; pyrG-).FIG. Figure 1 shows the progesterone transformation scheme by A. nidulans VKPM F-1069 and A. nidulans 031 strains (argB - ; pyrG - ).
На фиг. 2, 5 и 13 представлены хроматограммы хромато-масс-спектрометрического анализа 11α-гидроксипрогестерона, 11α-ацетоксипрогестерона и 6β, 11α-дигидроксипрогестерона соответственно.FIG. 2, 5, and 13 are chromatography-mass spectrometry analyzes of 11α-hydroxyprogesterone, 11α-acetoxyprogesterone, and 6β, 11α-dihydroxyprogesterone, respectively.
На фиг. 3 и 4 представлены 1Н ЯМР-спектры 11α-гидроксипрогестерона.FIG. 3 and 4 show 1 H NMR spectra of 11α-hydroxyprogesterone.
На фиг. 6-12 представлены 1Н ЯМР-, 13С ЯМР-, DEPT-ЯМР-, HSQC-ЯМР- и НМВС-ЯМР- спектры 11α-ацетоксипрогестерона.FIG. 6-12 are 1 H NMR, 13 C NMR, DEPT-NMR, HSQC-NMR and HMBC-NMR spectra of 11α-acetoxyprogesterone.
На фиг. 14-21 представлены 1Н-ЯМР-, 13С-ЯМР-, DEPT-ЯМР-, HSQC-ЯМР- и НМВС-ЯМР - спектры 6β,11α-дигидроксипрогестерона.FIG. 14-21 are 1 H-NMR, 13 C-NMR, DEPT-NMR, HSQC-NMR and HMBC-NMR spectra of 6β, 11α-dihydroxyprogesterone.
Осуществление изобретенияThe implementation of the invention
Штамм дикого типа Aspergillus nidulans ВКПМ F-1069 (synonym FGSC A4; АТСС 3863, 12996, 26451; CBS 112.46; NRRL 194) был получен из Всероссийской Коллекции Промышленных Микроорганизмов (ВКПМ).The wild-type strain Aspergillus nidulans VKPM F-1069 (synonym FGSC A4; ATCC 3863, 12996, 26451; CBS 112.46; NRRL 194) was obtained from the All-Russian Collection of Industrial Microorganisms (VKPM).
Прогестерон (I) (CAS №57-83-0, С21Н30О2), 11α-гидроксипрогестерон (II) (CAS №80-75-1, С21Н30О3) и 11α-ацетоксипрогестерон (С23Н32О4, M.w. 372.5 являются коммерчески доступными и могут быть приобретены, например, у компании Steraloids Inc. (USA) или у других производителей.Progesterone (I) (CAS No. 57-83-0, C 21 H 30 O 2 ), 11α-hydroxyprogesterone (II) (CAS No. 80-75-1, C 21 H 30 O 3 ) and 11α-acetoxyprogesterone (C 23 H 32 O 4 , Mw 372.5 are commercially available and can be purchased, for example, from Steraloids Inc. (USA) or from other manufacturers.
Неорганические соли были приобретены у компании Fluka (Germany). Дрожжевой экстракт и агар - Difco Becton Dickinson and company (Sparks, USA).Inorganic salts were purchased from Fluka (Germany). Yeast Extract and Agar - Difco Becton Dickinson and company (Sparks, USA).
Другие реагенты, растворители, и инертные газы являются коммерчески доступными, были приобретены у российских производителей.Other reagents, solvents, and inert gases are commercially available, were purchased from Russian manufacturers.
Для приготовления сред для культивирования и трансформации, водных растворов кислот, солей и щелочей использовали дистиллированную воду. Для промывки экстрактов в органических растворителях использовали питьевую водопроводную воду, если не оговорено особо.For the preparation of media for cultivation and transformation, aqueous solutions of acids, salts and alkalis used distilled water. Drinking tap water was used to rinse the extracts in organic solvents, unless otherwise noted.
Все процедуры, если не оговорено особо, осуществляли при комнатной температуре или температуре окружающей среды, то есть в диапазоне от 20 до 25°С.Для процессов, требующих более низкие температуры, чем комнатная, охлаждение обеспечивали холодной водопроводной водой (в диапазоне от 10 до 20°С), или смесью колотого льда и холодной воды (в диапазоне от 5 до 10°С), или смесью колотого льда и хлорида кальция (при температуре ниже 5°С).All procedures, unless otherwise specified, were carried out at room or ambient temperature, that is, in the range from 20 to 25 ° C. For processes requiring lower temperatures than room temperature, cooling was provided with cold tap water (in the range from 10 to 20 ° C), or a mixture of crushed ice and cold water (in the range from 5 to 10 ° C), or a mixture of crushed ice and calcium chloride (at a temperature below 5 ° C).
Культивирование микроорганизмов осуществляли на качалке New Brunswick™ Innova® 44/44R в термостатированном помещении при температуре 37°С (240-250 об/мин., амплитуда 5 см).The cultivation of microorganisms was carried out on a New Brunswick ™ Innova® 44 / 44R shaker in a thermostatically controlled room at 37 ° C (240–250 rpm,
Упаривание растворителей в вакууме осуществляли с использованием ротационного вакуумного испарителя Rotavapor ![]()
![]()
Высушивание кристаллов продуктов до постоянного веса осуществляли при температуре 35-45°С при атмосферном давлении или с использованием вакуум-сушильного шкафа при остаточном давлении 0,35±0,05 кгс/см2 (35±5 кПа).Drying of product crystals to a constant weight was carried out at a temperature of 35-45 ° C at atmospheric pressure or using a vacuum drying cabinet at a residual pressure of 0.35 ± 0.05 kgf / cm 2 (35 ± 5 kPa).
Для определения рН промывных вод использовали универсальную индикаторную бумагу с диапазоном значений от 0 до 12 (Лахема, Чехия).To determine the pH of the wash water used universal indicator paper with a range of values from 0 to 12 (Lahema, Czech Republic).
Колоночную хроматографию осуществляли на колонке (16×650 мм), используя силикагель марки Silica gel 60 (0.040-0.063 mm) (Merck, Germany).Column chromatography was carried out on a column (16 × 650 mm) using silica gel of the brand Silica gel 60 (0.040-0.063 mm) (Merck, Germany).
Контроль за ходом элюирования осуществляли методом тонкослойной хроматографии (ТСХ), используя пластины Silica gel 60 F254 (Merck, Germany) и хроматографические системы растворителей: дихлорметан-ацетон 9:1 (v/v) или дихлорметан-ацетон 4:1 (v/v). Пластинки просматривали в УФ-свете при длине волны 254 нм, затем опрыскивали 1% раствором ванилина в 10% водном растворе НСlO4, проявляли при температуре 100-120°С.The control of the elution was carried out by thin layer chromatography (TLC) using
Структуру и чистоту всех выделенных соединений подтверждали, по меньшей мере, одним из следующих методов: ТСХ (пластины для ТСХ Silica gel 60 F254 (Merck, Germany)), масс-спектрометрия, элементный анализ, ядерный магнитный резонанс (ЯМР), высокоэффективная жидкостная хроматография (ВЭЖХ).The structure and purity of all isolated compounds was confirmed by at least one of the following methods: TLC (plates for
Температуру плавления выделенных соединений определяли на приборе для определения точки плавления М-565 ![]()
![]()
1Н- и 13С- ЯМР спектры были определены на спектрометре Bruker Avance-400 (Bruker BioSpin GmbH) с рабочей частотой 400 МГц и 100.6 МГц соответственно, используя дейтерированный хлороформ (99,8% D, Sigma-Aldrich), или дейтерированный диметилсульфоксид (99,9% D, Sigma-Aldrich) в качестве растворителя относительно тетраметилсилана (TMS NMR grade ≥99,9%, Sigma-Aldrich) в качестве внутреннего стандарта, в миллионных долях (м.д.). 1 H- and 13 C-NMR spectra were determined on a Bruker Avance-400 spectrometer (Bruker BioSpin GmbH) with an operating frequency of 400 MHz and 100.6 MHz, respectively, using deuterated chloroform (99.8% D, Sigma-Aldrich), or deuterated dimethyl sulfoxide (99.9% D, Sigma-Aldrich) as a solvent relative to tetramethylsilane (TMS NMR grade ≥99.9%, Sigma-Aldrich) as an internal standard, in ppm (ppm).
Масс-спектры высокого разрешения (HRMS) регистрировали на приборе Bruker Daltonics micrOTOF-Q II, используя электрораспылительную ионизацию (ESI). Измерения были получены в режиме положительных ионов со следующими параметрами: граничное напряжение капилляра 4500 В; диапазон масс от m/z 50 до 3000; внешняя калибровка (Electricpray Calibrant Solution, Fluka); давление распылителя 0,4 бар; скорость потока 3 мкл/мин; азот применяли в виде сухого газа (6 л/мин); температура интерфейса была установлена при 180°С. Образец вводили в камеру масс-спектрометра из системы HPLC Agilent 1260, снабженной колонкой Agilent Poroshell 120 ЕС-С18 (3.0×50 μm; 2,7 urn) и защитой, используя автосамплер. Образец вводили в растворе 50% ацетонитрила (квалификация LC-MS) в воде (ультрачистая вода MilliQ, Merck Millipore KGaA, Germany), и колонку элюировали смесью ацетонитрила (А) и воды (В) в градиенте концентраций со скоростью потока 400 мкл/мин в следующих параметрах градиента: 0-15% А в течение 6 мин, 15% -85% А в течение 1,5 мин, 85% -0% А в течение 0,1 мин, 0% А в течение 2,4 мин. Время удерживания было следующим: 11α-гидроксипрогестерон - 5,2 мин; прогестерон - 6,7 мин; 11α-ацетоксипрогестерон - 6,0 мин; 6β, 11α дигидроксипрогестерон - 4,2 мин.High-resolution mass spectra (HRMS) were recorded on a Bruker Daltonics micrOTOF-Q II instrument using electrospray ionization (ESI). The measurements were obtained in the positive ion mode with the following parameters: capillary boundary voltage of 4500 V; mass range from m /
Образование 11α-ацетоксипрогестерона в результате трансформации прогестерона или lla-гидроксипрогестерона культурами A. nidulans ВКПМ F-1069 и A. nidulans 031 (argB-; pyrG-) подтверждено данными спектров 1Н ЯМР, 13С ЯМР, 2DЯМР, хромато-масс-спектрометрии высокого разрешения. На фиг. №№5,6,7-14 и 16-21 приведены спектры и на фиг.№№4,7 и 15 - хроматограммы полученных соединений.The formation of 11α-acetoxyprogesterone as a result of the transformation of progesterone or lla-hydroxyprogesterone by the cultures of A. nidulans VKPM F-1069 and A. nidulans 031 (argB - ; pyrG - ) was confirmed by 1 H NMR, 13 C NMR, 2D NMR, and chromatography mass spectrometry high resolution. FIG. Nos. 5, 6, 7-14 and 16-21 shows the spectra and in Fig. Nos. 4.7 and 15 are chromatograms of the compounds obtained.
Для подтверждения образования 11α-ацетоксипрогестерона в процессе трансформации прогестерона или 11α-гидроксипрогестерона культурами A. nidulans 11α-ацетоксипрогестерон был получен химическим синтезом из 11α-гидроксипрогестерона с применением метода ацетилирования.To confirm the formation of 11α-acetoxyprogesterone during the transformation of progesterone or 11α-hydroxyprogesterone by cultures of A. nidulans 11α-acetoxyprogesterone was obtained by chemical synthesis from 11α-hydroxyprogesterone using the acetylation method.
При осуществлении изобретения помимо методов, подробно раскрытых в нижеследующих примерах, использовали хорошо известные специалистам методики, описанные в руководствах по молекулярной биологии и генетической инженерии [J. Sambrook, Т. Maniatis, and E.F. Fritsch. Molecular cloning a laboratory manual (Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.; 1989; F.M. Ausubel, R. Brent, R.E. Kingston, D.D. Moore, J.G. Seidman, J.A. Smith, and K. Struhl. Current Protocols in Molecular Biology, John Wiley and Sons, N.Y., 1997].In carrying out the invention, in addition to the methods described in detail in the following examples, the techniques described in the manuals on molecular biology and genetic engineering [J. Sambrook, T. Maniatis, and E.F. Fritsch. Molecular cloning a laboratory manual (Cold Spring Harbor Laboratory, Cold Spring Harbor, NY; 1989; FM Ausubel, R. Brent, RE Kingston, DD Moore, JG Seidman, JA Smith, and K. Struhl. Current Protocols in Molecular Biology, John Wiley and Sons, NY, 1997].
Выходы 11α-гидроксипрогестерона и 11α-ацетоксипрогестерона, полученные биотрансформацией прогестерона и 11α-гидроксипрогестерона штаммами по настоящему изобретению и приведенные в примерах заявляемого изобретения, безусловно, ниже тех, которые необходимы для эффективного промышленного процесса. Однако примеры даны лишь для иллюстрации биокаталитической способности заявляемых штаммов конвертировать прогестерон не только в 11α-гидроксипрогестерон, но и в ацетат 11α-гидроксипрогестерона. Следует отметить, что указанная ацетилирующая способность выявлена впервые. Выходы могут быть значительно улучшены за счет оптимизации условий проведения биотрансформации с целью минимизации образования 6β, 11α-дигидроксипрогестерона и сдвига направления вторичной модификации 11α-гидроксипрогестерона в сторону ацетилирования. Известно, что состав продуктов зависит от состава среды, рН среды, возраста культуры, от индукции исходным субстратом или продуктом [А.Н. El-Refai, and K.M.Ghanem Some physiological relations of progesterone conversion by Aspergillus nidulans. Egyptian Journal of Microbiology, 1987. Vol. 22(2), 327-338]. Поэтому для снижения активности вторичного энзима 6β-гидроксилазы и уменьшения количества, образованного 6β, 11α-дигидроксипрогестерона необходимо оптимизировать параметры проведения процесса трансформации, например, значение рН среды, возраст культуры, способ внесения субстрата, состав среды для трансформации, температурный режим и другие, известные из уровня техники. Для примера в таблице 1 приведено влияние состава среды на конверсию прогестерона заявляемыми штаммами и на их 11α-монооксигенаазную активность (суммарно), а в таблице 2 - на относительную селективность образования 11α-гидроксипрогестерона и 11α-ацетоксипрогестерона, при прочих одинаковых условиях.The yields of 11α-hydroxyprogesterone and 11α-acetoxyprogesterone obtained by biotransformation of progesterone and 11α-hydroxyprogesterone by the strains of the present invention and given in the examples of the claimed invention are, of course, lower than those necessary for an effective industrial process. However, examples are given only to illustrate the biocatalytic ability of the claimed strains to convert progesterone not only into 11α-hydroxyprogesterone, but also into 11α-hydroxyprogesterone acetate. It should be noted that this acetylating ability was revealed for the first time. The yields can be significantly improved by optimizing the biotransformation conditions in order to minimize the formation of 6β, 11α-dihydroxyprogesterone and shift the direction of the secondary modification of 11α-hydroxyprogesterone towards acetylation. It is known that the composition of the products depends on the composition of the medium, the pH of the medium, the age of the culture, on the induction of the initial substrate or product [A.N. El-Refai, and K.M.Ghanem Some physiological relations of Astergillus nidulans. Egyptian Journal of Microbiology, 1987. Vol. 22 (2), 327-338]. Therefore, to reduce the activity of the secondary enzyme 6β-hydroxylase and reduce the amount formed by 6β, 11α-dihydroxyprogesterone, it is necessary to optimize the parameters of the transformation process, for example, the pH value of the medium, the age of the culture, the method of introducing the substrate, the composition of the medium for transformation, temperature, and other known from the level of technology. For example, Table 1 shows the effect of the composition of the medium on the conversion of progesterone by the claimed strains and on their 11α-monooxygenase activity (total), and in Table 2 on the relative selectivity of the formation of 11α-hydroxyprogesterone and 11α-acetoxyprogesterone, all other conditions being the same.
Для трансформации мы использовали 2 варианта сред следующего состава: 1 - среда СМ, использованная ![]()
![]()
![]()
![]()
2 - минимальная синтетическая среда ММ. Состав среды, г/л: глюкоза - 20; MgS04×7Н2О2 - minimal synthetic medium MM. The composition of the medium, g / l: glucose - 20; MgS0 4 × 7H 2 O
- 0,52; КН2РО4 - 1,52; NaNO3 - 0,85; CuSO4×5Н2O - 0,13; СаСl2 - 0,11; KСl - 0,52, Н3 ВО3 -5×10-6; MnSO4×5Н2O - 1×10-3; ZnSO4×7Н2O - 0,8×10-3; Na2MoO4×4Н2O - 0,8×10-3; FeSO4×5H2О - 0,8×10-3; 0.5 М фосфатно-цитратный буфер (рН 6.6) - 200 мл/л.- 0,52; KN 2 PO 4 - 1.52; NaNO 3 - 0.85; CuSO 4 × 5H 2 O — 0.13; L 2 = 0.11; KCl - 0.52, H 3 BO 3 -5 × 10 -6 ; MnSO 4 × 5H 2 O - 1 × 10 -3 ; ZnSO 4 × 7H 2 O — 0.8 × 10 -3 ; Na 2 MoO 4 × 4H 2 O — 0.8 × 10 -3 ; FeSO 4 × 5H 2 O — 0.8 × 10 -3 ; 0.5 M phosphate-citrate buffer (pH 6.6) - 200 ml / l.
При культивировании штамма A. nidulans 031 (argB-; pyrG-) в среды ММ и СМ добавляли аргинин - 0,21 г/л, уридин - 1,1 г/л и урацил - 1,2 г/л.When A. nidulans 031 (argB - ; pyrG - ) was cultivated, arginine — 0.21 g / l, uridine — 1.1 g / l, and uracil — 1.2 g / l were added to the MM and CM media.
Известно, что минеральные соли, добавленные в среду для культивирования, в частности, CuSO4, могут быть ингибиторами стероид-С21-монооксигеназы (ЕС №1.14.99.10 [Springer Handbook of Enzymes, Vol. 27, Class 1. Oxidoreductases XII. EC 1.14.15-1.97. Second edition. Springer. 2006, p.302]). Однако результаты показывают, что на среде СМ образование 21-гидроксипрогестерона как продукта трансформации не наблюдается. Следует отметить, что 21-гидроксипрогестерон наряду с 11α-гидроксипрогестероном является одним из продуктов трансформации прогестерона в этой среде штаммом А. nidulans известным способом [А.Н. El-Refai, and K.M.Ghanem. Some physiological relations of progesterone conversion by Aspergillus nidulans. Egyptian Journal of Microbiology, 1987, Vol. 22(2), 327-338].It is known that the mineral salts added to the culture medium, in particular CuSO 4 , can be steroid-C21 monooxygenase inhibitors (EC No. 1.1.14.99.10 [Springer Handbook of Enzymes, Vol. 27,
Заявленное изобретение иллюстрируется следующими примерами.The claimed invention is illustrated by the following examples.
Пример 1. Общий метод биотрансформации прогестерона штаммами Aspergillus nidulans на средах ММ и СМ в течение 66 чExample 1. General method of biotransformation of progesterone by Aspergillus nidulans strains on MM and CM media for 66 h
Для получения споровой суспензии штаммы выращивали при температуре 37°С в течение 7-10 дней на агаризованной среде МПА (мальт-экстракт - 3 г/л, пептон -1 г/л, агар - 20 г/л). При культивировании штамма A. nidulans 031 (argB-; pyrG-) в среду добавляли аргинин - 0,21 г/л, уридин - 1,1 г/л и урацил - 1,2 г/л. Споровую суспензию высевали в качалочные колбы вместимостью 750 мл, содержащие по 100 мл ростовой среды (ММ или СМ), выращивали на качалке (240-250 об/мин, амплитуда 5 см) при температуре 37°С в течение 4 дней. Затем в ростовую среду вносили прогестерон в виде раствора в диметилсульфоксиде до финальной концентрации 1 г/л, при этом концентрация растворителя не превышала 6% (об.). Трансформацию проводили в течение 66 ч в тех же условиях, параллельно культивируя штаммы без прогестерона в качестве контроля. Каждый эксперимент выполняли в трех повторностях.To obtain a spore suspension, the strains were grown at 37 ° C for 7–10 days on MPA agar medium (malt extract — 3 g / l, peptone -1 g / l, agar — 20 g / l). When A. nidulans 031 (argB - ; pyrG - ) was cultivated , arginine was added to the medium - 0.21 g / l, uridine - 1.1 g / l and uracil - 1.2 g / l. The spore suspension was sown in 750 ml rocking flasks containing 100 ml growth medium (MM or CM), grown on a rocking chair (240-250 rpm,
Для извлечения продуктов трансформации прогестерона по окончании инкубации мицелий отфильтровывали, промывали дихлорметаном на фильтре. Культуральную жидкость экстрагировали трижды порциями дихлорметана равного объема. Стероиды, адсорбированные на мицелии, извлекали ре-мацерацией (трижды), используя метанол и выдерживая без перемешивания в течение 4 ч. Метанол упаривали, остаток, содержащий воду, экстрагировали дихлорметаном трижды. Объединенные экстракты культуральной жидкости и мицелия промывали водой, сушили Na2SO4, осветляли активированным углем и упаривали до прекращения погона. Остаток растворяли в 5 мл смеси этилацетата и дихлорметана (1:2) и хроматографировали на колонке, используя 30-кратное количество силикагеля к весу остатка после упаривания (фиг. 2). В качестве элюэнта использовали смесь дихлорметана и ацетона (от 0 до 25 об.% ацетона), для контроля элюции использовали ТСХ. Хроматографическую чистоту соединений подтверждали с помощью ТСХ и ЯМР-спектроскопии.To extract progesterone transformation products at the end of incubation, the mycelium was filtered, washed with dichloromethane on the filter. The culture fluid was extracted three times with equal volumes of dichloromethane. Steroids adsorbed on the mycelium were recovered by re-maceration (three times) using methanol and kept without stirring for 4 hours. Methanol was evaporated, the residue containing water was extracted with dichloromethane three times. The combined extracts of the culture liquid and mycelium were washed with water, dried with Na 2 SO 4 , lightened with activated charcoal and evaporated until cessation stopped. The residue was dissolved in 5 ml of a mixture of ethyl acetate and dichloromethane (1: 2) and chromatographed on a column using a 30-fold amount of silica gel to the weight of the residue after evaporation (Fig. 2). A mixture of dichloromethane and acetone (from 0 to 25 vol.% Of acetone) was used as eluent, TLC was used to control the elution. Chromatographic purity of the compounds was confirmed by TLC and NMR spectroscopy.
Результаты биотрансформации прогестерона штаммами A. nidulans по настоящему изобретению приведены в таблицах 1 и 2.The results of progesterone biotransformation by A. nidulans strains of the present invention are shown in Tables 1 and 2.
Пример 2. Биотрансформация прогестерона штаммами A. nidulans в течение 14 чExample 2. Biotransformation of progesterone by A. nidulans strains for 14 h
Вариант 1. Споровую суспензию штамма A. nidulans ВКПМ F-1069 высевали в 4 качалочные колбы вместимостью 750 мл, содержащие по 100 мл среды ММ для культивирования, выращивали на качалке (240-250 об/мин, амплитуда 5 см) при температуре 37°С в течение 4 дней. Затем в ростовую среду вносили прогестерон в растворе диметилсульфоксида до финальной концентрации 1 г/л, при этом концентрация растворителя составляла 4% (об.). Трансформацию проводили в течение 14 ч в тех же условиях.
Для извлечения продуктов трансформации прогестерона по окончании инкубации мицелий отфильтровывали, промывали дихлорметаном на фильтре. Культуральную жидкость экстрагировали трижды порциями дихлорметана равного объема. Стероиды, адсорбированные на мицелии, извлекали ре-мацерацией (трижды), используя метанол и выдерживая без перемешивания в течение 4 ч. Метанол упаривали, остаток, содержащий воду, экстрагировали дихлорметаном трижды. Объединенные экстракты культуральной жидкости и мицелия промывали водой, сушили Na2SO4, осветляли активированным углем и упаривали до прекращения погона. Остаток растворяли в 5 мл смеси этилацетата и дихлорметана (1:2) и хроматографировали на колонке, используя 30-кратное количество силикагеля к весу остатка после упаривания. В качестве элюэнта использовали смесь дихлорметана и ацетона (от 0 до 25 об.% ацетона), для контроля элюции использовали ТСХ. Одинаковые по составу стероидов фракции элюата объединяли и упаривали досуха. Кристаллизующийся остаток растирали с эфиром, осадок отфильтровывали, сушили до постоянного веса в вакуум-сушильном шкафу. Хроматографическую чистоту соединений подтверждали с помощью ТСХ и ЯМР-спектроскопии.To extract progesterone transformation products at the end of incubation, the mycelium was filtered, washed with dichloromethane on the filter. The culture fluid was extracted three times with equal volumes of dichloromethane. Steroids adsorbed on the mycelium were recovered by re-maceration (three times) using methanol and kept without stirring for 4 hours. Methanol was evaporated, the residue containing water was extracted with dichloromethane three times. The combined extracts of the culture liquid and mycelium were washed with water, dried with Na 2 SO 4 , lightened with activated charcoal and evaporated until cessation stopped. The residue was dissolved in 5 ml of a mixture of ethyl acetate and dichloromethane (1: 2) and chromatographed on a column using a 30-fold amount of silica gel to the weight of the residue after evaporation. A mixture of dichloromethane and acetone (from 0 to 25 vol.% Of acetone) was used as eluent, TLC was used to control the elution. Fractions of the eluate of the same composition of steroids were combined and evaporated to dryness. The residue crystallized was triturated with ether, the precipitate was filtered off, dried to constant weight in a vacuum oven. Chromatographic purity of the compounds was confirmed by TLC and NMR spectroscopy.
Из 400 мг прогестерона, загруженного на трансформацию, получили 394 мг прогестерона (98,5%) и 3,8 мг 11α-ацетоксипрогестерона с выходом 0,8% на загруженный субстрат (53,5% на конвертированный субстрат). По данным ТСХ анализа другие продукты трансформации отсутствуют.Of the 400 mg of progesterone loaded on the transformation, 394 mg of progesterone (98.5%) and 3.8 mg of 11α-acetoxyprogesterone were obtained in a yield of 0.8% on the loaded substrate (53.5% on the converted substrate). According to TLC analysis, there are no other transformation products.
Вариант 2. Биотрансформацию прогестерона штаммом A. nidulans 031 (argB-; pyrG-) проводили в условиях варианта 1.
Из 400 мг прогестерона, загруженного на трансформацию, получили 390 мг прогестерона (97,5%) и 7,4 мг 11α-ацетоксипрогестерона с выходом 1,6% на загруженный субстрат (62,5% на конвертированный субстрат). По данным ТСХ анализа другие продукты трансформации отсутствуют.Of the 400 mg of progesterone loaded on the transformation, 390 mg of progesterone (97.5%) and 7.4 mg of 11α-acetoxyprogesterone were obtained with a yield of 1.6% per loaded substrate (62.5% per converted substrate). According to TLC analysis, there are no other transformation products.
Пример 3. Биокаталитическое ацетилирование 11α-гидроксипрогестерона штаммами A. nidulansExample 3. Biocatalytic acetylation of 11α-hydroxyprogesterone with A. nidulans strains
Вариант 1. Споровую суспензию штамма A. nidulans ВКПМ F-1069 высевали в 4 качалочные колбы вместимостью 750 мл, содержащие по 100 мл среды ММ для культивирования, выращивали на качалке (240-250 об/мин, амплитуда 5 см) при температуре 37°С в течение 4 дней. Затем в ростовую среду вносили 11α-гидроксипрогестерон в растворе диметилсульфоксида до финальной концентрации 0,5 г/л, при этом концентрация растворителя составляла 2% (об.). Трансформацию проводили в течение 23 ч в тех же условиях.
Для извлечения продуктов трансформации 11α-гидроксипрогестерона по окончании инкубации мицелий отфильтровывали, промывали дихлорметаном на фильтре. Культуральную жидкость экстрагировали трижды порциями дихлорметана равного объема. Стероиды, адсорбированные на мицелии, извлекали ре-мацерацией (трижды), используя метанол и выдерживая без перемешивания в течение 4 ч. Метанол упаривали, остаток, содержащий воду, экстрагировали дихлорметаном трижды. Объединенные экстракты культуральной жидкости и мицелия промывали водой, сушили Na2SO4, осветляли активированным углем и упаривали до прекращения погона. Остаток растворяли в 5 мл смеси этилацетата и дихлорметана (1:2) и хроматографировали на колонке, используя 30-кратное количество силикагеля к весу остатка после упаривания. В качестве элюэнта использовали смесь дихлорметана и ацетона (от 0 до 20 об.% ацетона), для контроля элюции использовали ТСХ. Одинаковые по составу стероидов фракции элюата объединяли и упаривали досуха. Кристаллизующийся остаток растирали с эфиром, осадок отфильтровывали, сушили до постоянного веса в вакуум-сушильном шкафу. Хроматографическую чистоту соединений подтверждали с помощью ТСХ и ЯМР-спектроскопии.To extract the transformation products of 11α-hydroxyprogesterone at the end of the incubation, the mycelium was filtered, washed with dichloromethane on the filter. The culture fluid was extracted three times with equal volumes of dichloromethane. Steroids adsorbed on the mycelium were recovered by re-maceration (three times) using methanol and kept without stirring for 4 hours. Methanol was evaporated, the residue containing water was extracted with dichloromethane three times. The combined extracts of the culture liquid and mycelium were washed with water, dried with Na 2 SO 4 , lightened with activated charcoal and evaporated until cessation stopped. The residue was dissolved in 5 ml of a mixture of ethyl acetate and dichloromethane (1: 2) and chromatographed on a column using a 30-fold amount of silica gel to the weight of the residue after evaporation. A mixture of dichloromethane and acetone (from 0 to 20 vol.% Of acetone) was used as eluent, TLC was used to control the elution. Fractions of the eluate of the same composition of steroids were combined and evaporated to dryness. The residue crystallized was triturated with ether, the precipitate was filtered off, dried to constant weight in a vacuum oven. Chromatographic purity of the compounds was confirmed by TLC and NMR spectroscopy.
Из 200 мг 11α-гидроксипрогестерона, загруженного на трансформацию, получили 178 мг 11α-гидроксипрогестерона (88%) и 19,75 мг 11α-ацетоксипрогестерона с выходом 8,76% на загруженный субстрат (79,64% на конвертированный субстрат). По данным ТСХ анализа другие продукты трансформации отсутствуют.Out of 200 mg of 11α-hydroxyprogesterone loaded for transformation, 178 mg of 11α-hydroxyprogesterone (88%) and 19.75 mg of 11α-acetoxyprogesterone were obtained with a yield of 8.76% on the loaded substrate (79.64% on the converted substrate). According to TLC analysis, there are no other transformation products.
Вариант 2. Битрансформацию 11α-гидроксипрогестерона штаммом A. nidulans 031 (argB-, pyrG-) проводили в условиях варианта 1.
Из 200 мг 11α-гидроксипрогестерона, загруженного на трансформацию, получили 170 мг 11α-гидроксипрогестерона (85%) и 31,6 мг 11α-ацетоксипрогестерона с выходом 14,02% на загруженный субстрат (93,45% на конвертированный субстрат). По данным ТСХ анализа другие продукты трансформации отсутствуют.Of the 200 mg of 11α-hydroxyprogesterone loaded on the transformation, 170 mg of 11α-hydroxyprogesterone (85%) and 31.6 mg of 11α-acetoxyprogesterone were obtained with a yield of 14.02% on the loaded substrate (93.45% on the converted substrate). According to TLC analysis, there are no other transformation products.
ЯМР-спектры полученных образцов 11α-ацетоксипрогестерона идентичны спектрам стандартного образца и спектрам образца, полученного химическим методом ацетилирования 11α-гидроксипрогестерона.The NMR spectra of the obtained samples of 11α-acetoxyprogesterone are identical to the spectra of the standard sample and the spectra of the sample obtained by the chemical method of acetylation of 11α-hydroxyprogesterone.
Пример 4. Получение 11α-ацетоксипрогестерона химическим методомExample 4. Obtaining 11α-acetoxyprogesterone by chemical method
К раствору 34 мг 11α-гидроксипрогестерона в 0,2 мл уксусного ангидрида добавили 1 каплю 60% хлорной кислоты. Реакционную массу выдерживали при комнатной температуре в течение 1 ч. По окончании реакции реакционную массу добавляли по каплям в 5 мл воды, содержащей 1 мл 25% раствора аммиака (рН ~7.5). Перемешивали 30 мин., затем реакционную массу экстрагировали дихлорметаном трижды, экстракт промыли водой до нейтральной реакции, упарили досуха. Получили 39 мг остатка, из которого препаративной хроматографией извлекли 27 мг 11α-ацетоксипрогестерона с выходом 70,45%. Т.пл. 173-174°С (лит.Т.пл. 176-177°С [D.H. Peterson, Н.С. Murray, S.H. Eppstein, L.M. Reineke, A. Weintraub, P.D. Meister, and H.M. Leigh. Microbiological Transformations of Steroids. I. Introduction of Oxygen at Carbon-11 of Progesterone. J. Am. Chem. Soc, 1952, Vol. 74 (23), 5933-5936]To a solution of 34 mg of 11α-hydroxyprogesterone in 0.2 ml of acetic anhydride was added 1 drop of 60% perchloric acid. The reaction mass was kept at room temperature for 1 hour. At the end of the reaction, the reaction mass was added dropwise to 5 ml of water containing 1 ml of 25% ammonia solution (pH ~ 7.5). Stirred for 30 minutes, then the reaction mass was extracted with dichloromethane three times, the extract was washed with water until neutral, evaporated to dryness. 39 mg of residue was obtained, from which 27 mg of 11α-acetoxyprogesterone was recovered by preparative chromatography in 70.45% yield. M.p. 173-174 ° C (lit. T.p. 176-177 ° C [DH Peterson, NS Murray, SH Eppstein, LM Reineke, A. Weintraub, PD Meister, and HM Leigh. Microbiological Transformations of Steroids. I Carbon-11 of Progesterone, J. Am. Chem. Soc., 1952, Vol. 74 (23), 5933-5936]
Характеристика продуктов биотрансформации 11α-Гидроксипрогестерон, полученный в условиях общего метода биотрансформации прогестерона штаммом Aspergillus nidulans ВКПМ F-1069 Т.пл. 164-166°С (лит.Т.пл. 164-165°С [K. Yildirim, and A. Kuru. Biotransformation of some steroids by Aspergillus candidus. Journal of Chemical Research, 2015. Vol. 39(9), 546-549]). M.w. 330.46.The characteristics of the products of biotransformation 11α-Hydroxyprogesterone, obtained in terms of the general method of biotransformation of progesterone by the strain Aspergillus nidulans VKPM F-1069 m. 164-166 ° C (lit. T.pl. 164-165 ° C [K. Yildirim, and A. Kuru. Biotransformation of some steroids by Aspergillus candidus. Journal of Chemical Research, 2015. Vol. 39 (9), 546 -549]). M.w. 330.46.
Масс-спектр высокого разрешения C21H30O3 (m/z): рассчитано для [М+Н]+331.2268, найдено 331.2264; рассчитано для [M+Na]+353.2087, найдено 353.2088.High resolution mass spectrum C 21 H 30 O 3 (m / z): calculated for [M + H] + 331.2268, found 331.2264; calculated for [M + Na] + 353.2087, found 353.2088.
1Н-ЯМР (400 МГц, CDCl3): 5.73 (с, 1Н СН-4), 4.04 (ддд (псевдо-дт), 1Н, 3J11,9=10.3 Гц, 3J11,12α 4.8 Гц, 3J11,12β 4.6 Гц, СН-11), 2.66 (дт, 1H, 2J1β,1α 13.7 Гц, 3J1β, 2 4.4 Гц, СН-1β), 2,55 (дд (псевдо-т), 1H, 3J17, 16α 8.7 Гц, 3J17, 16β 9.1 Гц, СН-17), 2.48-2.26 (м, 5Н, СН2-2 & СН2-6 & СН-12β), 2.22-2.10 (м, 1Н, СН-16β), 2.13 (с, 3Н, СН3-21), 2.02 (тд, 1H, 2J1a, 1b 13.7 Гц, 3J1α, 2, 4.5 Гц, СН-1α), 1.87-1.81 (м, 1H, СН-7β), 1.78-1.64 (м, 3Н, СН-8 & СН-15α & СН-16α), 1.58-1.47 (2Н (т, 1Н, 3J12α, 12β 11.3 Гц, СН-12α), 1.31 (с, 3Н, СН3-19), 1.29-1.20 (м, 2Н, СН-12α & СН-14), 1.14 (т, 1Н, 3J9, 8, 10.3 Гц, СН-9), 1.14-1.04 (2×ддд, 2Н, 3J7α, 7β 13.1 Гц, 3J7,6 13.0 Гц, 3J7α, 8 3.8 Гц, СН2-7α), 0.69 (с, 3Н, СН3-18) (фиг. 3). 1 H-NMR (400 MHz, CDCl 3 ): 5.73 (s, 1H CH-4), 4.04 (ddd (pseudo-dt), 1H, 3 J 11.9 = 10.3 Hz, 3 J 11.12 α 4.8 Hz, 3 J 11.12β 4.6 Hz, CH-11), 2.66 (dt, 1H, 2 J 1β , 1α 13.7 Hz, 3 J 1β, 2 4.4 Hz, CH-1β), 2.55 (dd (pseudo-t) , 1H, 3 J 17, 16α 8.7 Hz, 3 J 17, 16β 9.1 Hz, CH-17), 2.48-2.26 (m, 5H, CH 2 -2 & CH 2 -6 & CH-12β), 2.22-2.10 (m, 1H, CH-16β), 2.13 (s, 3H, CH 3 -21), 2.02 (td, 1H, 2 J 1a, 1b 13.7 Hz, 3 J 1α, 2 , 4.5 Hz, CH-1α), 1.87-1.81 (m, 1H, CH-7β), 1.78-1.64 (m, 3H, CH-8 & CH-15α & CH-16α), 1.58-1.47 (2H (t, 1H, 3 J 12α, 12β 11.3 Hz, CH-12α), 1.31 (s, 3H, CH 3 -19), 1.29-1.20 (m, 2H, CH-12α & CH-14), 1.14 (t, 1H, 3 J 9, 8 , 10.3 Hz , CH-9), 1.14-1.04 (2 × ddd, 2H, 3 J 7α, 7β 13.1 Hz, 3 J 7 , 6 13.0 Hz, 3 J 7α, 8 3.8 Hz, CH 2 -7α), 0.69 (s, 3H, CH 3 -18) (Fig. 3).
Спектры идентичны спектрам стандартного образца и образца, полученного в условиях общего метода биотрансформации прогестерона штаммом A. nidulans 031 (argB-; pyrG-).The spectra are identical to the spectra of the standard sample and the sample obtained under the conditions of the general method of biotransformation of progesterone by the strain A. nidulans 031 (argB - ; pyrG - ).
11α-Ацетоксипрогестерон. полученный в условиях общего метода биотрансформации прогестерона штаммом Aspergillus nidulans ВКПМ F-1069 Т.пл. 172-174°С (лит.Т.пл. 176-177°С [D.H. Peterson, Н.С.Murray, S.H. Eppstein, L.M. Reineke, A. Weintraub, P.D. Meister, and H.M. Leigh. Microbiological Transformations of Steroids. I. Introduction of Oxygen at Carbon-11 of Progesterone. J. Am. Chem. Soc, 1952, Vol. 74 (23), 5933-5936]. M.w. 372.5;11α-Acetoxyprogesterone. obtained in terms of the general method of biotransformation of progesterone by a strain of Aspergillus nidulans VKPM F-1069 So pl. 172-174 ° C (lit. T.pl. 176-177 ° C [DH Peterson, N.S. Murray, SH Eppstein, LM Reineke, A. Weintraub, PD Meister, and HM Leigh. Microbiological Transformations of Steroids. I .Program of Progesterone, J. Am. Chem. Soc., 1952, Vol. 74 (23), 5933-5936]. Mw 372.5;
Масс-спектр высокого разрешения C23H32O4 (m/z): рассчитано для [М+Н]+373.2373, найдено 373.2361; рассчитано для [M+Na]+395.2193, найдено 395.2183.High resolution mass spectrum C 23 H 32 O 4 (m / z): calculated for [M + H] + 373.2373, found 373.2361; calculated for [M + Na] + 395.2193, found 395.2183.
1Н-ЯМР (300 МГц, DMSO-d6): 5.67 (с, 1Н СН-4), 5.14 (дт, 1Н, 3J11, 9=10.6 Гц, 3J11, 10 5.0 Гц, СН-11), 2,62 (дд (псевдо-т), 1Н, 3J17, 16α 8.9 Гц, 3J17,16β 9.1 Гц, СН-17), 2.47-2.35 (м, 2Н, СН-2β & СН-6β),2.29-2.12 (м, 3Н, СН-6α & СН-12β & СН-2α), 2.10-1.96 (м, 1Н, СН-16β), 2.05 (с, 3Н, СН3-21), 2.01 (с, 3Н, СН3-23), 1.91-1.78 (м, 3Н, СН2-1 & СН-7α), 1.72-1.60 (м, 3Н, СН-16α & СН-15α & СН-8), 1.50 (т, 1Н, 3J12α, 12β 11.6 Гц, СН-12α), 1.44 (т, 1H, 3J9, 8 10.6 Гц, СН-9), 1.39-1.29 (ддд, 1Н, 3J14, 8 10.7 Гц, 3J14, 15β 6.4 Гц, 3J14, 15α 5.6 Гц, СН-14) 1.29 (уш.с, 4Н, СН3-19 & СН-15β), 1.17-1.00 (2×ддд, 2Н, 3J7α, 7β 12.6 Гц, 3J7, 6α~3J7, 6β 12.2 Гц, 3J7α,8 3.1 Гц, СН2-7), 0.64 (с, 3Н, СН3-18). 1 H-NMR (300 MHz, DMSO-d 6 ): 5.67 (s, 1H CH-4), 5.14 (dt, 1H, 3 J 11, 9 = 10.6 Hz, 3 J 11, 10 5.0 Hz, CH-11 ), 2.62 (dd (pseudo-t), 1H, 3 J 17 , 16α 8.9 Hz, 3 J 17.16β 9.1 Hz, CH-17), 2.47-2.35 (m, 2H, CH-2β & CH- 6β), 2.29-2.12 (m, 3H, CH-6α & CH-12β & CH-2α), 2.10-1.96 (m, 1H, CH-16β), 2.05 (s, 3H, CH 3 -21), 2.01 (s, 3H, CH 3 -23), 1.91-1.78 (m, 3H, CH 2 -1 & CH-7α), 1.72-1.60 (m, 3H, CH-16α & CH-15α & CH-8), 1.50 (t, 1H, 3 J 12α, 12β 11.6 Hz, CH-12α), 1.44 (t, 1H, 3 J 9, 8 10.6 Hz, CH-9), 1.39-1.29 (ddd, 1H, 3 J 14, 8 10.7 Hz, 3 J 14, 15β 6.4 Hz, 3 J 14, 15α 5.6 Hz, CH-14) 1.29 (broad s, 4H, CH 3 -19 & CH-15β), 1.17-1.00 (2 × ddd, 2H, 3 J 7α, 7β 12.6 Hz, 3 J 7 , 6α ~ 3 J 7, 6β 12.2 Hz, 3 J 7α, 8 3.1 Hz, CH 2 -7), 0.64 (s, 3H, CH 3 -18).
13С-ЯМР (300 МГц, DMSO-d6): 208.61 (с, СО-20), 199.40 (с, СО-3), 170.14 (с, С-5), 170.10 (с, СО-22) 124.48 (с, СН-4), 70.58 (с, СНОН-11), 62.30 (с, СН-17), 55.34 (с, СН-9), 54.39 (с, СН-14), 45.02 (с, СН2-12), 43.54 (с, С-13), 39.75 (с, С-10), 36.79 (с, СН2-1), 34.72 (с, СН-8) 34.19 (с, СН2-2), 33.03 (с, СH2-6), 31.67 (с, СН2-7), 31.37 (с, СН3-21) 30.92 (с, СН3-21), 24.16 (с, СH2-15), 22.96 (с, СН2-16), 22.02 (с, СН3-21), 18.20 (с, СН3-19), 14.24 (с, СН3-18). 13 C-NMR (300 MHz, DMSO-d 6 ): 208.61 (s, CO-20), 199.40 (s, CO-3), 170.14 (s, C-5), 170.10 (s, CO-22) 124.48 (s, CH-4), 70.58 (s, CHON-11), 62.30 (s, CH-17), 55.34 (s, CH-9), 54.39 (s, CH-14), 45.02 (s, CH 2 -12), 43.54 (s, C-13), 39.75 (s, C-10), 36.79 (s, CH 2 -1), 34.72 (s, CH-8) 34.19 (s, CH 2 -2), 33.03 (s, CH 2 -6), 31.67 (s, CH 2 -7), 31.37 (s, CH 3 -21) 30.92 (s, CH 3 -21), 24.16 (s, CH 2 -15), 22.96 (s, CH 2 -16), 22.02 (s, CH 3 -21), 18.20 (s, CH 3 -19), 14.24 (s, CH 3 -18).
Спектры идентичны спектрам стандартного образца, образца, полученного в условиях общего метода биотрансформации прогестерона штаммом A. nidulans 031 (argB-; pyrG-), а также спектрам образцов, полученных методом биокаталитического ацетилирования 11α-гидроксипрогестерона штаммом A. nidulans 031 (argB-; pyrG-) и химическим методом ацетилирования 11α-гидроксипрогестерона.The spectra are identical to the spectra of a standard sample, a sample obtained under the general method of biotransformation of progesterone with A. nidulans 031 strain (argB - ; pyrG - ), and also the spectra of samples obtained by biocatalytic acetylation of 11α-hydroxyprogesterone with A. nidulans 031 strain (argB - ; pyrG - ) and by the chemical method of acetylation of 11α-hydroxyprogesterone.
6β, 11α-Дигидроксипрогестерон, полученный в условиях общего метода биотрансформации прогестерона штаммом Aspergillus nidulans ВКПМ F-1069 Т.пл. 242-244°С (лит.т.пл. 244-246°С [Z. Habibi, M.Yousefi, Sh. Ghanian, M. Mohammadi, S. Ghasemi. Biotransformation of progesterone by Absidia griseolla var. igachii and Rhizomucor pusillus. Steroids, 2012, Vol. 77, p.1446-1449].M.w. 346.46;6β, 11α-Dihydroxyprogesterone, obtained under the general method of biotransformation of progesterone by the strain Aspergillus nidulans VKPM F-1069 m.p. 242-244 ° С (lit.t.pl. 244-246 ° С [Z. Habibi, M. Yousefi, Sh. Ghanian, M. Mohammadi, S. Ghasemi. Biotransformation of progesterone by Absidia griseolla var. Igachii and Rhizomucorus pusillus .Steroids, 2012, Vol. 77, p. 1446-1449] .Mw 346.46;
Масс-спектр высокого разрешения C23H32O4 (m/z): рассчитано для [М+Н]+373.2373, найдено 373.2361; рассчитано для [M+Na]+395.2193, найдено 395.2183.High resolution mass spectrum C 23 H 32 O 4 (m / z): calculated for [M + H] + 373.2373, found 373.2361; calculated for [M + Na] + 395.2193, found 395.2183.
1Н-ЯМР (300 МГц, DMSO-d6): 5.67 (с, 1Н СН-4), 5.14 (дт, 1H, 3J11, 9=10.6 Гц, 3J11, 10 5.0 Гц, СН-11), 2,62 (дд (псевдо-т), 1Н, 3J17, 16α 8.9 Гц, 3J17, 16β 9.1 Гц, СН-17), 2.47-2.35 (м, 2Н, СН-2β & СН-6β),2.29-2.12 (м, 3Н, СН-6α & СН-12β & СН-2α), 2.10-1.96 (м, 1H, СН-16β), 2.05 (с, 3Н, СН3-21), 2.01 (с, 3Н, СН3-23), 1.91-1.78 (м, 3Н, СН2-1 & СН-7α), 1.72-1.60 (м, 3Н, СН-16α & СН-15α & СН-8), 1.50 (т, 1H, 3J12α, 12β П.6 Гц, СН-12α), 1.44 (т, 1H, 3J9, 8 10.6 Гц, СН-9), 1.39-1.29 (ддд, 1Н, 3J14, 8 10.7 Гц, 3J14, 15β 6.4 Гц, 3J14, 15α 5.6 Гц, СН-14) 1.29 (уш.с, 4Н, СН3-19 & СН-15β), 1.17-1.00 (2×ддд, 2Н, 3J7a, 7β 12.6 Гц, 3J7, 6α~3J7, 6β 12.2 Гц, 3J7α, 8 3.1 Гц, СН2-7), 0.64 (с, 3Н, СН3-18). 1 H-NMR (300 MHz, DMSO-d 6 ): 5.67 (s, 1H CH-4), 5.14 (dt, 1H, 3 J 11 , 9 = 10.6 Hz, 3 J 11, 10 5.0 Hz, CH-11 ), 2.62 (dd (pseudo-t), 1H, 3 J 17, 16α 8.9 Hz, 3 J 17, 16β 9.1 Hz, CH-17), 2.47-2.35 (m, 2H, CH-2β & CH- 6β), 2.29-2.12 (m, 3H, CH-6α & CH-12β & CH-2α), 2.10-1.96 (m, 1H, CH-16β), 2.05 (s, 3H, CH 3 -21), 2.01 (s, 3H, CH 3 -23), 1.91-1.78 (m, 3H, CH 2 -1 & CH-7α), 1.72-1.60 (m, 3H, CH-16α & CH-15α & CH-8), 1.50 (t, 1H, 3 J 12α, 12β P.6 Hz, CH-12α), 1.44 (t, 1H, 3 J 9, 8 10.6 Hz, CH-9), 1.39-1.29 (ddd, 1H, 3 J 14, 8 10.7 Hz, 3 J 14, 15β 6.4 Hz, 3 J 14, 15α 5.6 Hz, CH-14) 1.29 (broad s, 4H, CH 3 -19 & CH-15β), 1.17-1.00 (2 × ddd, 2H, 3 J 7a, 7β 12.6 Hz, 3 J 7, 6α ~ 3 J 7, 6β 12.2 Hz, 3 J 7α, 8 3.1 Hz, CH 2 -7), 0.64 (s, 3H, CH 3 -18 ).
13С-ЯМР (300 МГц, DMSO-d6): 208.61 (с, СО-20), 199.40 (с, СО-3), 170.14 (с, С-5), 170.10 (с, СО-22) 124.48 (с, СН-4), 70.58 (с, СНОН-11), 62.30 (с, СН-17), 55.34 (с, СН-9), 54.39 (с, СН-14), 45.02 (с, СН2-12), 43.54 (с, С-13), 39.75 (с, С-10), 36.79 (с, СН2-1), 34.72 (с, СН-8) 34.19 (с, СН2-2), 33.03 (с, СН2-6), 31.67 (с, СН2-7), 31.37 (с, СН3-21) 30.92 (с, СН3-21), 24.16 (с, СН2-15), 22.96 (с, СН2-16), 22.02 (с, СН3-21), 18.20 (с, СН3-19), 14.24 (с, СН3-18). 13 C-NMR (300 MHz, DMSO-d 6 ): 208.61 (s, CO-20), 199.40 (s, CO-3), 170.14 (s, C-5), 170.10 (s, CO-22) 124.48 (s, CH-4), 70.58 (s, CHON-11), 62.30 (s, CH-17), 55.34 (s, CH-9), 54.39 (s, CH-14), 45.02 (s, CH 2 -12), 43.54 (s, C-13), 39.75 (s, C-10), 36.79 (s, CH 2 -1), 34.72 (s, CH-8) 34.19 (s, CH 2 -2), 33.03 (s, CH 2 -6), 31.67 (s, CH 2 -7), 31.37 (s, CH 3 -21) 30.92 (s, CH 3 -21), 24.16 (s, CH 2 -15), 22.96 (s, CH 2 -16), 22.02 (s, CH 3 -21), 18.20 (s, CH 3 -19), 14.24 (s, CH 3 -18).
Спектры идентичны спектрам образцов, полученных в условиях общего метода биотрансформации прогестерона штаммом A. nidulans 031 (argB-; pyrG-).The spectra are identical to the spectra of samples obtained under the conditions of the general method of biotransformation of progesterone by the strain A. nidulans 031 (argB - ; pyrG - ).
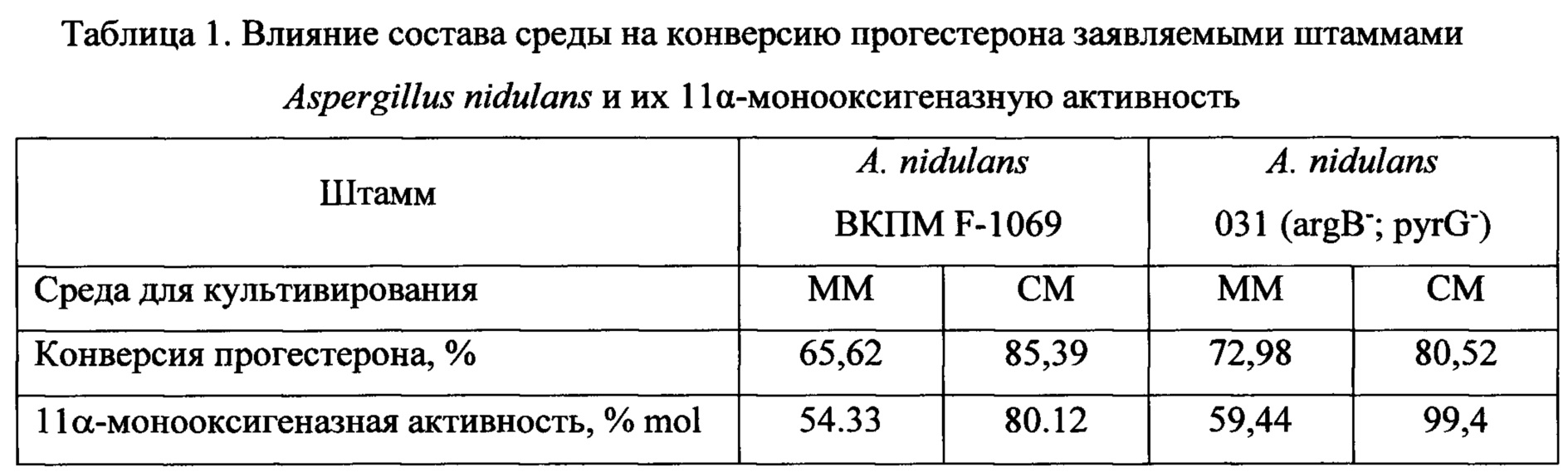
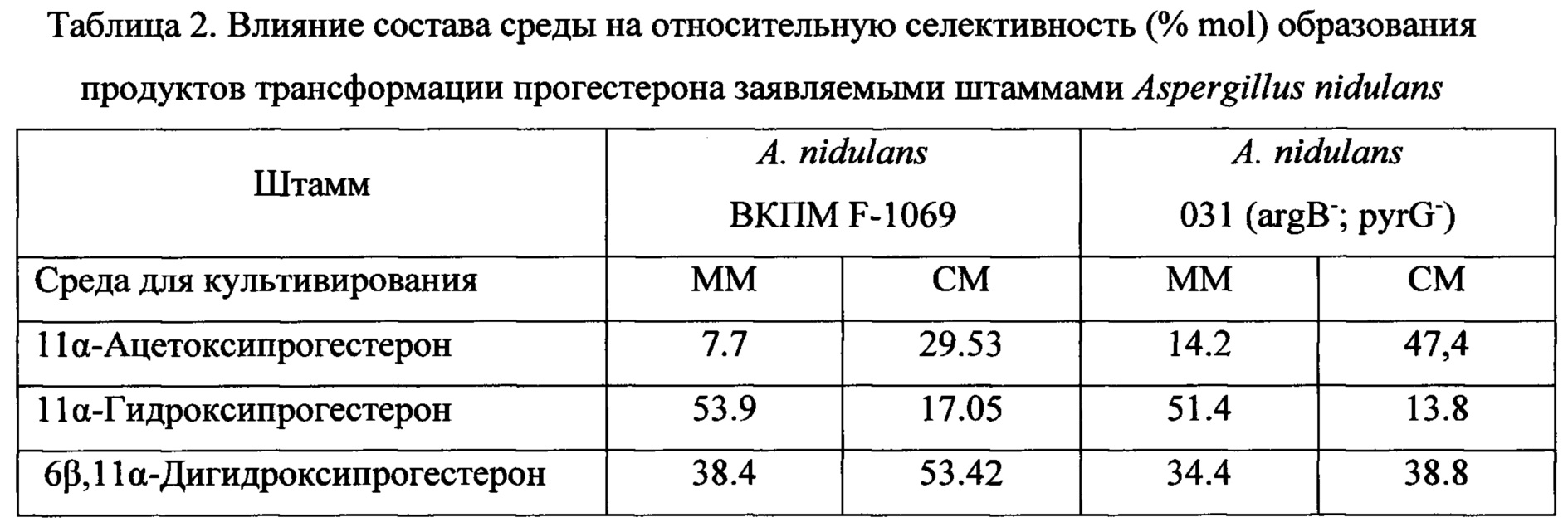
Таким образом, раскрыто применение грибов A. nidulans, обладающих стероид-11α-гидроксилирующей активностью и способных конвертировать прогестерон в 11α-гидроксипрогестерон, для получения 11α-ацетоксипрогестерона из уровня техники не известно. При этом этерификация 11α-гидроксипрогестерона грибом A. nidulans с образованием 11α-ацетоксипрогестерона является вторым этапом процесса трансформиции прогестерона и может быть осуществлена как с выделением, так и без выделения 11α-гидроксипрогестерона из культуральной жидкости, образованного на первом этапе биотрансформации.Thus, the use of A. nidulans fungi with steroid-11α-hydroxylation activity and capable of converting progesterone to 11α-hydroxyprogesterone to produce 11α-acetoxyprogesterone is not known in the prior art. At the same time, esterification of 11α-hydroxyprogesterone by the fungus A. nidulans with the formation of 11α-acetoxyprogesterone is the second stage of the progesterone transformation process and can be carried out with or without 11α-hydroxyprogesterone from the culture fluid formed in the first stage of biotransformation.
Claims (1)
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| RU2017138536A RU2692932C2 (en) | 2017-11-07 | 2017-11-07 | BIOCATALYST FOR PRODUCING 11α-ACETOXYPROGESTERONE |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| RU2017138536A RU2692932C2 (en) | 2017-11-07 | 2017-11-07 | BIOCATALYST FOR PRODUCING 11α-ACETOXYPROGESTERONE |
Publications (3)
| Publication Number | Publication Date |
|---|---|
| RU2017138536A RU2017138536A (en) | 2019-05-08 |
| RU2017138536A3 RU2017138536A3 (en) | 2019-05-08 |
| RU2692932C2 true RU2692932C2 (en) | 2019-06-28 |
Family
ID=66430123
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| RU2017138536A RU2692932C2 (en) | 2017-11-07 | 2017-11-07 | BIOCATALYST FOR PRODUCING 11α-ACETOXYPROGESTERONE |
Country Status (1)
| Country | Link |
|---|---|
| RU (1) | RU2692932C2 (en) |
Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| RU2524617C2 (en) * | 2007-02-06 | 2014-07-27 | Коламбиа Лэборетериз (Бермуда) Лимитед | Progesteron for treatment and prevention of spontaneous preterm birth |
-
2017
- 2017-11-07 RU RU2017138536A patent/RU2692932C2/en active
Patent Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| RU2524617C2 (en) * | 2007-02-06 | 2014-07-27 | Коламбиа Лэборетериз (Бермуда) Лимитед | Progesteron for treatment and prevention of spontaneous preterm birth |
Non-Patent Citations (5)
| Title |
|---|
| A.H. EI-RIFAI, K.M. CHANEM, Some physiological relations of progesterone conversion by Aspergillus nidulans, Egyptian journal of microbiology, 1987, v.22(2), p. 327-338. * |
| A.H. EI-RIFAI, K.M. CHANEM, Some physiological relations of progesterone conversion by Aspergillus nidulans, Egyptian journal of microbiology, 1987, v.22(2), p. 327-338. T.K. DUTTA, T.B. SAMANTA, Bioconversion of progesterone by the activated immobilized conidia of Aspergillus ochraceus TS, Current Microbilogy, 1999, v.39 (6), p. 309-312. M.J. HENRY, H.D. SISLER, Effects of sterol biosynthesis - inhibiting (SBI) fungicides on cytochrome P-450 oxygenations in fungi, Pesticide Biochemistry and physiology, 1984, v.22(3), p. 262-275. P.SOMAL, C.L. CHOPRA, Microbial conversion of steroids. III: 11a- hydroxylation by fungal mycelium, Applied Microbiology, 1985, v. 21(5), p. 267-269. * |
| M.J. HENRY, H.D. SISLER, Effects of sterol biosynthesis - inhibiting (SBI) fungicides on cytochrome P-450 oxygenations in fungi, Pesticide Biochemistry and physiology, 1984, v.22(3), p. 262-275. * |
| P.SOMAL, C.L. CHOPRA, Microbial conversion of steroids. III: 11a- hydroxylation by fungal mycelium, Applied Microbiology, 1985, v. 21(5), p. 267-269. * |
| T.K. DUTTA, T.B. SAMANTA, Bioconversion of progesterone by the activated immobilized conidia of Aspergillus ochraceus TS, Current Microbilogy, 1999, v.39 (6), p. 309-312. * |
Also Published As
| Publication number | Publication date |
|---|---|
| RU2017138536A (en) | 2019-05-08 |
| RU2017138536A3 (en) | 2019-05-08 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US4435327A (en) | 3β,7β,15α-Trihydroxy-5-androsten-17-one, its 3,15-dipivalate, and their preparation | |
| Bartmańska et al. | Steroids’ transformations in Penicillium notatum culture | |
| Wilson et al. | Steroid transformations with Fusarium oxysporum var. cubense and Colletotrichum musae | |
| Savinova et al. | Biotransformation of progesterone by Aspergillus nidulans VKPM F-1069 (wild type) | |
| Wu et al. | Efficient hydroxylation of functionalized steroids by Colletotrichum lini ST-1 | |
| US5834452A (en) | 1- or 6-hydroxylated steroids | |
| Faramarzi et al. | Formation of hydroxysteroid derivatives from androst-4-en-3, 17-dione by the filamentous fungus Mucor racemosus | |
| Faramarzi et al. | Metabolism of androst-4-en-3, 17-dione by the filamentous fungus Neurospora crassa | |
| Bartmańska et al. | Transformation of steroids by Trichoderma hamatum | |
| Brzezowska et al. | Biotransformation XXXIX. Metabolism of testosterone, androstenedione, progesterone and testosterone derivatives in Absidia coerulea culture | |
| GB2123833A (en) | Steroid 1,2-dehydrogenation using dried microbial cells | |
| RU2692932C2 (en) | BIOCATALYST FOR PRODUCING 11α-ACETOXYPROGESTERONE | |
| AKAO et al. | 3β-Hydroxysterod Dehydrogenase of Ruminococcus sp. from Human Intestinal Bacteria | |
| Lacroix et al. | Microbial models of drug metabolism: microbial transformations of trimegestone®(RU27987), a 3-Keto-Δ4, 9 (10)-19-norsteroid drug | |
| Zhang et al. | Transformation of prednisolone to a 20β-hydroxy prednisolone compound by Streptomyces roseochromogenes TS79 | |
| Madyastha et al. | Transformation of dehydroepiandrosterone and pregnenolone by Mucor piriformis | |
| Huszcza et al. | Transformations of testosterone and related steroids in Absidia glauca culture | |
| HU194318B (en) | Process for the delta 1-dehydrogenation of steroids in the presence of materials relieving from toxic kinds of oxygen | |
| RU2677332C1 (en) | Recombinant strain of filamentous fungus aspergillus nidulans and its use for progesterone 11α-hydroxylation and for acetylation of 11α-hydroxyprogesterone | |
| IE42168B1 (en) | New 1-hydroxy-steroids | |
| Savinova et al. | Biotransformation of progesterone by the ascomycete Aspergillus niger N402 | |
| RU2731712C2 (en) | Microbiological method of producing 11α-acetoxyprogesterone | |
| Ambrus et al. | Microbial transformation of β-sitosterol and stigmasterol into 26-oxygenated derivatives | |
| Dovbnya et al. | Microbial side-chain degradation of ergosterol and its 3-substituted derivatives: a new route for obtaining of deltanoids | |
| Madyastha et al. | Transformation of 16-dehydroprogesterone and 17α-hydroxyprogesterone by Mucor piriformis |