KR20160077750A - Mass production method of recombinant trans glutaminase - Google Patents
Mass production method of recombinant trans glutaminase Download PDFInfo
- Publication number
- KR20160077750A KR20160077750A KR1020140188039A KR20140188039A KR20160077750A KR 20160077750 A KR20160077750 A KR 20160077750A KR 1020140188039 A KR1020140188039 A KR 1020140188039A KR 20140188039 A KR20140188039 A KR 20140188039A KR 20160077750 A KR20160077750 A KR 20160077750A
- Authority
- KR
- South Korea
- Prior art keywords
- recombinant
- transglutaminase
- protein
- gene encoding
- expression vector
- Prior art date
Links
Images






Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P21/00—Preparation of peptides or proteins
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/70—Vectors or expression systems specially adapted for E. coli
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y02—TECHNOLOGIES OR APPLICATIONS FOR MITIGATION OR ADAPTATION AGAINST CLIMATE CHANGE
- Y02E—REDUCTION OF GREENHOUSE GAS [GHG] EMISSIONS, RELATED TO ENERGY GENERATION, TRANSMISSION OR DISTRIBUTION
- Y02E50/00—Technologies for the production of fuel of non-fossil origin
- Y02E50/10—Biofuels, e.g. bio-diesel
Abstract
Description
본 발명은 재조합 트랜스 글루타미나아제의 대량 생산을 위한 벡터 및 이를 이용한 재조합 트랜스글루타미나아제의 대량 생산 방법에 관한 것이다.
The present invention relates to a vector for mass production of a recombinant transglutaminase and a method for mass-production of the recombinant transglutaminase using the same.
트랜스 글루타미나아제(Transglutaminase, TGase; protein-glutamine γ-glutamyltransferase, EC2.3.2.13)는 펩티드 사슬의 글루타민 잔기(glutamine residue)의 γ-carboxyamide group과 lysine residue의 ε-amino group 사이의 아실 그룹 이동 반응(acyl group transfer reaction)을 촉매하는 효소로서 ε-(γ-glutamyl)lysine cross-linkage를 형성하는 효소이다.Transglutaminase (TGase; protein-glutamine? -Glutamyltransferase, EC2.3.2.13) is an acyl group between the? -Carboxyamide group of the glutamine residue of the peptide chain and the? -Amino group of the lysine residue It is an enzyme that catalyzes the acyl group transfer reaction and forms ε- (γ-glutamyl) lysine cross-linkage.
트랜스 글루타미나아제는 식품 및 어류가공 산업에서 다양하게 활용되고 있고, 비식품산업에서 그 용도가 다양하게 개발되어 의료 공학, 재료과학, 섬유, 피혁가공 등 많은 분야에서 응용되고 있다. Transglutaminase has been widely used in the food and fish processing industries and has been developed in various fields such as medical engineering, material science, textiles, leather processing and so on.
현재 트랜스 글루타미나아제는 대부분 스트렙토마이시스 모바렌시스(Streptomyces mobaraensis)로 부터 전통적 발효공정에 의해 생산하고 있다. 최근에는 유전공학기술을 이용하여, 대장균이나 코리네박테리움에서도 트랜스 글루타미나아제의 생산을 시도하고 있다. 그러나 현재의 트랜스 글루타미나아제 생산 기술은 생산 수율이 낮아 대량생산이 어려운바(대장균: 5 mg/l, S. lividans: 0.1 mg/l, Corynebacterium: 132 mg/l (native type); Shino Takehana, Hiroshi Takagi, Bioscience Biotechnology and Biochemistry, 1994 Jan;58(1):88~92; Kinya Washizu, Kazuyuki Takeuchi, Bioscience Biotechnology and Biochemistry, 1994 Jan;58(1):82~87; Masayo Date, Yoshimi Kikuchi, Appl Environ Microbiol. May 2003;69(5):3011~3014), 생산 수율을 획기적으로 향상 시킬 필요가 있는 실정이다. Currently, transglutaminase is produced mostly by the conventional fermentation process from Streptomyces mobaraensis . Recently, genetic engineering technology has been used to produce transglutaminase in E. coli and Corynebacterium. However, the current production technology of transglutaminase is difficult to mass-produce due to low production yield (E. coli: 5 mg / l, S. lividans: 0.1 mg / l, Corynebacterium: 132 mg / Masayo Date, Yoshimi Kikuchi, < / RTI >< RTI ID = 0.0 > 1, < / RTI > Appl Environ Microbiol. May 2003; 69 (5): 3011-3014), it is necessary to dramatically improve the production yield.
또한, 활성형의 트랜스 글루타미나아제는 세포 내에서 단백질의 가교(crosslinking)를 유도하기 때문에 toxic하여, 세포 내에서는 비활성형의 유지를 위해 성숙형(mature form)의 N-말단(N-terminal)에 활성 억제 서열인 pro-sequence가 붙여져 있다. pro가 붙여져 있는 형태의 트랜스 글루타미나아제 (pro-TGase)는 활성형을 띄지 않지만, 특정 단백질 분해효소에 의해서 pro 서열이 제거되면 활성형을 가지게 된다. 그러나 상기 활성 억제 서열의 제거를 위해 특정 서열을 인식하는 단백질 분해 효소는 고가인바 트랜스 글루타미나아제의 대량생산에 적합하지 않다. 또한 트랜스 글루타미나아제가 불용해성으로 발현되는 것은 복잡한 재접힘(refolding) 과정이 요구되기 때문에 대량생산에 부적합하다.In addition, since active transglutaminase induces cross-linking of proteins in cells, it is toxic, and in order to maintain an inactive form in a cell, a mature form of N-terminal ) Is attached to the pro-sequence, which is the activity inhibition sequence. The pro-TGase in which the pro is attached does not have an active form, but when the pro-sequence is removed by a specific protease, it has an active form. However, proteolytic enzymes that recognize a specific sequence for the removal of the activity-inhibiting sequence are not suitable for the mass production of expensive in vitro transglutaminase. In addition, the expression of transglutaminase in an insoluble state is not suitable for mass production because a complicated refolding process is required.
이에 본 발명자들은, 상기한 문제점을 해결하고 전량 수입에 의존하는 트랜스 글루타미나아제의 국내 자체 대량 생산을 위해, 분자유전학 기술을 이용해 트랜스 글루타미나아제의 대량 발현 방법 및 정제, 분리 공정을 확립함으로써 본 발명을 완성하기에 이르렀다.
Therefore, the present inventors have established a mass expression method, purification and separation process of transglutaminase by using molecular genetics technology for the domestic mass production of transglutaminase, which solves the above problems and relies on the whole amount of the transglutaminase. Thereby completing the present invention.
본 발명의 목적은 재조합 트랜스 글루타미나아제의 대량 생산 방법을 제공하는 것이다.It is an object of the present invention to provide a mass production method of recombinant transglutaminase.
본 발명의 다른 목적은 재조합 트랜스 글루타미나아제를 대량 생산하기 위한 재조합 발현 벡터를 제공하는 것이다.
It is another object of the present invention to provide a recombinant expression vector for mass production of recombinant transglutaminase.
상기 목적을 달성하기 위하여, 본 발명은 수용성 발현 유도용 단백질을 암호화하는 유전자, 재조합 트랜스 글루타미나아제의 세포 내 활성 억제 서열, 단백질 분해 효소 인식부위를 암호화하는 유전자 및 트랜스 글루타미아나제를 암호화하는 유전자를 포함하는 제1 재조합 발현 벡터; 및 수용성 발현 유도용 단백질을 암호화하는 유전자 및 단백질 분해 효소를 암호화하는 유전자를 포함하는 제2 재조합 발현 벡터;로 미생물을 형질전환하는 단계; 및In order to accomplish the above object, the present invention provides a gene encoding a protein for inducing water-soluble expression, a cell activation inhibition sequence of a recombinant transglutaminase, a gene encoding a protease recognition site and a transglutaminase A first recombinant expression vector comprising a gene encoding the gene of interest; And a second recombinant expression vector comprising a gene encoding a protein for inducing water-soluble expression and a gene encoding a proteolytic enzyme; And
상기 형질전환된 미생물을 배양하여 활성형의 재조합 트랜스 글루타미나아제를 생산하는 단계; 를 포함하는 재조합 트랜스 글루타미나아제의 생산 방법을 제공한다. Culturing the transformed microorganism to produce an active recombinant transglutaminase; And a method for producing the recombinant transglutaminase.
본 발명의 용어, "벡터"는 적합한 숙주내에서 DNA를 발현시킬 수 있는 적합한 조절 서열에 작동가능하게 연결된 외부 DNA 서열을 포함하는 DNA 작제물을 말한다. 본 발명의 벡터는 전형적으로 발현을 위한 벡터로서 구축될 수 있다. 바람직하게는, 본 발명의 벡터는 재조합 펩타이드 또는 단백질을 발현시키기 위한 벡터이다. 또한, 본 발명의 벡터는 원핵세포 또는 진핵세포를 숙주세포로 하여 구축될 수 있다. 본 발명의 재조합 발현 벡터는 예컨대, 박테리오파지 벡터, 코스미드 벡터, YAC(Yeast Artificial Chromosome) 벡터 등일 수 있다. 본 발명의 목적상, 플라스미드 벡터를 이용하는 것이 바람직하다. 그러한 목적에 사용될 수 있는 전형적인 플라스미드 벡터는 (a) 숙주세포당 수백 개의 플라스미드 벡터를 포함하도록 복제가 효율적으로 이루어지도록 하는 복제 개시점, (b) 플라스미드 벡터로 형질전환된 숙주세포가 선발될 수 있도록 하는 항생제 내성 유전자 및 (c) 외래 DNA 절편이 삽입될 수 있는 제한효소 절단부위를 포함하는 구조를 지니고 있다. 적절한 제한효소 절단부위가 존재하지 않을지라도 통상의 방법에 따른 합성 올리고뉴클레오타이드 어댑터(oligonucleotide adaptor) 또는 링커(linker)를 사용하면 벡터와 외래 DNA를 용이하게 라이게이션(ligation)할 수 있다. 본 발명에 이용되는 벡터는 당업계에 공지된 다양한 방법을 통해 구축될 수 있으며, 이에 대한 구체적인 방법은 Sambrook et al. Molecular Cloning. A Laboratory Manual. Cold Spring Harbor Laboratory Press(2001)에 개시되어 있으며, 이 문헌은 본 명세서에 참조로서 삽입된다.The term "vector" of the present invention refers to a DNA construct containing an exogenous DNA sequence operably linked to a suitable regulatory sequence capable of expressing DNA within the appropriate host. The vector of the present invention can typically be constructed as a vector for expression. Preferably, the vector of the present invention is a vector for expressing a recombinant peptide or protein. In addition, the vector of the present invention can be constructed by using prokaryotic cells or eukaryotic cells as host cells. The recombinant expression vector of the present invention may be, for example, a bacteriophage vector, a cosmid vector, a yeast artificial chromosome (YAC) vector, or the like. For the purpose of the present invention, it is preferable to use a plasmid vector. Typical plasmid vectors that can be used for such purposes include (a) a cloning start point that allows replication to be efficiently made to include several hundred plasmid vectors per host cell, (b) a host cell transformed with the plasmid vector And (c) a restriction enzyme cleavage site into which the foreign DNA fragment can be inserted. Even if an appropriate restriction enzyme cleavage site is not present, the vector and the foreign DNA can be easily ligated using a synthetic oligonucleotide adapter or a linker according to a conventional method. The vectors used in the present invention can be constructed through various methods known in the art, and specific methods for this can be found in Sambrook et al. Molecular Cloning. A Laboratory Manual. Cold Spring Harbor Laboratory Press (2001), which is incorporated herein by reference.
본 발명의 용어, "작동가능하게 연결"되어 있다는 것은 핵산 발현 조절 서열(예: 프로모터, 시그널 서열, 또는 전사조절인자 결합 위치의 어레이)과 다른 핵산 서열(예컨대, 트랜스 글루타미나아제-코딩 서열)사이의 기능적인 결합을 의미하며, 이에 의해 상기 조절 서열은 상기 다른 핵산 서열의 전사 및/또는 번역 과정을 조절하게 된다.
The term "operably linked" of the present invention means that a nucleic acid sequence other than a nucleic acid expression control sequence (e.g., an array of promoter, signal sequence or transcription factor binding site) (e.g., transglutaminase- ), Whereby the regulatory sequence regulates transcription and / or translation of the other nucleic acid sequences.
본 발명의 상기 제1 재조합 발현 벡터는 수용성 발현 유도용 단백질을 암호화하는 유전자, 재조합 트랜스 글루타미나아제의 세포 내 활성 억제 서열, 단백질 분해 효소 인식부위를 암호화하는 유전자 및 트랜스 글루타미아나제를 암호화하는 유전자를 포함할 수 있으며, 상기 유전자를 차례로 포함할 수 있으나 이에 제한되는 것은 아니다. The first recombinant expression vector of the present invention comprises a gene encoding a protein for inducing a water-soluble expression, an intracellular activity-inhibiting sequence of a recombinant transglutaminase, a gene encoding a protease recognition site, and a transglutaminase And may include, but is not limited to, the gene in turn.
본 발명의 용어, “수용성 발현 유도용 단백질”은 비용해성 단백질 형태의 발현이 대량생산에 부적합하다는 단점을 개선하기 위해 재조합 단백질을 수용성의 융합 단백질로 생산할 수 있는 단백질을 말한다. 재조합 단백질을 불용성 형태에서 수용성형태로 생산하기 위해, 보통의 경우 수용성 증진성 단백질의 결합형태인 융합단백질형태로 생산하는 방법을 사용하는데, 모든 단백질이 수용성 증진성 단백질을 부착한다고 하더라도 수용성형태로 생산되지 않을 수 있으며, 또한 융합단백질형태로 수용성 형태로 발현이 될 경우에는 목적단백질의 고유의 활성을 잃어버릴 수가 있으며, 더욱이, 목적단백질에서 수용성 증진성 태그를 제거해야하는 번거로운 일이 발생할 수 있다. 본 발명에서는 복잡한 재접힘(refolding) 과정이 불필요한 수용성 단백질로의 발현을 극대화하기 위하여, 베타-갈락토시다제(β-galactosidase), CAT(chloramphenicol acetyltransferase), GST(glutathione S transferase) 및 MBP(maltose binding protein) 등의 단백질을 사용할 수 있으나 이에 제한되는 것은 아니며, 가장 바람직하게는 MBP(maltose binding protein)를 사용할 수 있다. 본 발명의 구체예에서, 수용성 발현 유도용 단백질로 서열번호 1의 아미노산 서열로 이루어진 말토오스 결합 단백질을 결합하여, 융합단백질(Fusion protein) 형태로 발현시켜 수용성 단백질로의 발현을 유도하는 공정을 개발하였다. 본 발명의 구체예에서, 상기 수용성 단백질로의 발현은 플라스크 배양 및 5L 발효조를 통한 배양에서도 동일한 양상을 나타내었다.The term " protein for inducing water-soluble expression " of the present invention refers to a protein capable of producing a recombinant protein as a water-soluble fusion protein in order to improve the disadvantage that the expression of the insoluble protein form is not suitable for mass production. In order to produce a recombinant protein from an insoluble form to a water-soluble form, a method of producing a recombinant protein in the form of a fusion protein, usually a binding form of a water-soluble promoting protein, is used. In addition, when expressed as a water soluble form in the form of a fusion protein, the intrinsic activity of the target protein may be lost, and furthermore, the troublesome task of removing the water soluble enhancing tag from the target protein may occur. In the present invention, β-galactosidase, chloramphenicol acetyltransferase (GST), glutathione S transferase (GST) and maltose (MBP) are used in order to maximize the expression of a water-soluble protein which does not require a complicated refolding process. binding protein may be used, but the present invention is not limited thereto, and MBP (maltose binding protein) can be most preferably used. In the embodiment of the present invention, a process for binding a maltose binding protein consisting of the amino acid sequence of SEQ ID NO: 1 to a water-soluble expression inducing protein and expressing it in the form of a fusion protein to induce the expression of the soluble protein . In the embodiment of the present invention, the expression of the soluble protein was found to be the same in both flask culture and 5L fermentation.
본 발명의 “재조합 트랜스 글루타미나아제의 세포 내 활성 억제 서열”은 재조합 트랜스 글루타미나아제 발현 시, 활성형 트랜스 글루타미나아제 효소는 세포 내 기작에 영향을 미쳐 발현이 어려운 문제를 해결하고자 결합한 것이다. 구체적으로, 재조합 트랜스 글루타미나아제의 세포 내 활성 억제 서열(pro sequence)을 통해 트랜스 글루타미나아제를 발현시킨 후, 효소 정제 과정에서 추가적인 활성 억제 서열 제거 과정을 통해 상기한 세포 내 기작에 영향을 미치는 문제점을 해결할 수 있다. 본 발명의 구체예에서, 상기 재조합 트랜스 글루타미나아제의 세포 내 활성 억제 서열은 서열번호 2의 아미노산 서열로 이루어진 단백질 분해 저항성(proteolytic-resistant sequence, pro sequence) 서열일 수 있다. The " intracellular activity inhibition sequence of recombinant transglutaminase " of the present invention is intended to solve the problem that the active transglutaminase enzyme affects the intracellular mechanism and is difficult to express at the time of expression of the recombinant transglutaminase It is a combination. Specifically, the transglutaminase is expressed through the intracellular activity inhibition sequence (prosequence) of the recombinant transglutaminase, followed by further activity-inhibition sequencing in the enzyme purification step to affect the intracellular mechanism Can be solved. In an embodiment of the present invention, the intracellular activity inhibition sequence of the recombinant transglutaminase may be a proteolytic-resistant sequence (prosequence) sequence consisting of the amino acid sequence of SEQ ID NO: 2.
본 발명의 “단백질 분해 효소 인식부위”는 본 발명의 재조합 트랜스 글루타미나아제의 발현 후, 트랜스 글루타미나아제 활성형으로의 변환을 위하여, 활성 억제 서열 제거를 위해 단백질 분해 효소가 인식하는 아미노산 서열 부위를 말한다.
The " protease recognition site " of the present invention refers to an amino acid sequence of the amino acid sequence of the amino acid sequence of the amino acid sequence of the amino acid sequence of SEQ ID NO: It refers to the sequence region.
본 발명의 상기 제2 재조합 발현 벡터는 수용성 발현 유도용 단백질을 암호화하는 유전자 및 단백질 분해 효소를 암호화하는 유전자를 포함할 수 있으며, 상기 유전자를 차례로 포함할 수 있으나 이에 제한되는 것은 아니다. The second recombinant expression vector of the present invention may include a gene encoding a protein for inducing water-soluble expression and a gene encoding a protease, and the gene may be in turn included, but is not limited thereto.
상기 수용성 발현 유도용 단백질은 본 발명의 상기 제1 재조합 발현 벡터의 수용성 발현 유도용 단백질과 동일할 수 있으나 이에 제한되는 것은 아니며, 바람직하게는 베타-갈락토시다제(β-galactosidase), CAT(chloramphenicol acetyltransferase), GST(glutathione S transferase) 및 MBP(maltose binding protein) 등의 공지된 단백질을 사용할 수 있으며, 특히 MBP(maltose binding protein)를 사용할 수 있다. The water-soluble expression inducing protein may be the same as the water-soluble expression inducing protein of the first recombinant expression vector of the present invention, but is not limited thereto, preferably beta-galactosidase, CAT ( chloramphenicol acetyltransferase (GST), glutathione S transferase (GST), and maltose binding protein (MBP). In particular, maltose binding protein (MBP) can be used.
상기 제2 재조합 발현 벡터의 “단백질 분해 효소”는 종래 활성 억제 서열 제거를 위해 고가의 단백질 분해 효소 사용함으로써 대량 생산이 어려웠던 문제를 극복하고자, 특정 서열을 인식하는 단백질 분해효소를 자가 생산하기 위한 것이다. 상기 단백질 분해 효소는 바람직하게 트롬빈, TEV(tobacco etch virus)프로테아제, 프리시션(PreScission) 프로테아제, Factor Xa 및 엔테로키나아제(Enterokinase)를 사용할 수 있으며, 바람직하게 TEV(tobacco etch virus)프로테아제를 사용할 수 있으나 이에 제한되는 것은 아니다. 본 발명의 구체예에서는 재조합 트랜스 글루타미나아제의 발현 후, 트랜스 글루타미나아제를 활성형으로 변환시키기 위하여, 활성 억제 서열 제거를 위한 특정 아미노산 서열을 인식하는 단백질 분해 효소(Protease)로 Tabacco etch virus 유래의 서열번호 5의 아미노산 서열로 이루어진 TEV 단백질 분해 효소를 선정하였다. The "protease" of the second recombinant expression vector is used for self-production of a protease which recognizes a specific sequence in order to overcome the problem of mass production by using an expensive protease for removing the activity inhibition sequence . Preferably, the protease is selected from the group consisting of thrombin, tobacco etch virus (TEV) protease, PreScission protease, Factor Xa, and Enterokinase. Preferably, TEV (tobacco etch virus) But are not limited thereto. In a specific example of the present invention, a proteinase (Protease) recognizing a specific amino acid sequence for the removal of the activity-inhibiting sequence is transformed into Tabacco etch virus-derived TEV protease consisting of the amino acid sequence of SEQ ID NO: 5 was selected.
본 발명의 폴리뉴클레오티드들은 각각의 염기 서열과 각각 70% 이상, 더욱 바람직하게는 80% 이상, 더 더욱 바람직하게는 90% 이상, 가장 바람직하게는 95% 이상의 서열 상동성을 가지는 염기 서열을 포함할 수 있다. 폴리뉴클레오티드에 대한 "서열 상동성의 %"는 두 개의 최적으로 배열된 서열과 비교 영역을 비교함으로써 확인되며, 비교 영역에서의 폴리뉴클레오티드 서열의 일부는 두 서열의 최적 배열에 대한 참고 서열(추가 또는 삭제를 포함하지 않음)에 비해 추가 또는 삭제(즉, 갭)를 포함할 수 있다.The polynucleotides of the present invention comprise a nucleotide sequence having a sequence homology of at least 70%, more preferably at least 80%, even more preferably at least 90%, most preferably at least 95% . "% Of sequence homology to polynucleotides" is ascertained by comparing the comparison region with two optimally aligned sequences, and a portion of the polynucleotide sequence in the comparison region is the reference sequence for the optimal alignment of the two sequences (I. E., A gap) relative to the < / RTI >
본 발명의 상기 제1 재조합 발현 벡터는 도 1a로 표시되는 것일 수 있으며, 상기 제2 재조합 발현 벡터는 도 1c로 표시되는 것일 수 있다.
The first recombinant expression vector of the present invention may be represented by FIG. 1A, and the second recombinant expression vector may be represented by FIG. 1C.
본 발명의 상기 상기 제1 재조합 발현 벡터 및 제2 재조합 발현 벡터는 수용성 융합 단백질로 동시 발현되는 것일 수 있으며, 본 발명의 구체예에서, 말토오스 결합 단백질과 융합단백질 형태의 수용성 단백질로 발현되는 것일 수 있다.The first recombinant expression vector and the second recombinant expression vector of the present invention may be co-expressed as a water soluble fusion protein, and in the embodiment of the present invention may be expressed as a maltose binding protein and a water soluble protein in the form of a fusion protein have.
본 발명의 상기 제2 재조합 발현 벡터는 미생물에 형질전환되어 융합단백질 형태로 발현된 후, 발현 구조 내에 있는 단백질 분해 효소 절단 부위를 인식하여 수용성 발현 유도용 단백질과 단백질 분해 효소 사이를 자가 절단(self-cleavage)할 수 있다. The second recombinant expression vector of the present invention is transformed into a microorganism and expressed in the form of a fusion protein. Then, the second recombinant expression vector of the present invention recognizes a proteolytic enzyme cleavage site in the expression structure and then cleaves self-cleavage between the protein -cleavage).
또한 본 발명의 상기 제2 재조합 발현 벡터로 발현된 단백질 분해 효소는 상기 제1 재조합 발현 벡터로 발현된 수용성 융합 단백질의 활성 억제 부위를 제거하여 트랜스 글루타미나아제를 활성형으로 만들 수 있다. 본 발명의 구체예에서, 상기 발현된 단백질 분해 효소는 발현 구조 내에 있는 단백질 분해 효소 절단 부위를 인식하여 수용성 발현 유도용 단백질과 활성 억제 서열을 제거하는 것일 수 있다. In addition, the proteolytic enzyme expressed by the second recombinant expression vector of the present invention can be made into an active form by removing the inhibitory site of the soluble fusion protein expressed by the first recombinant expression vector. In an embodiment of the present invention, the expressed proteolytic enzyme can recognize the proteolytic enzyme cleavage site in the expression structure and remove the protein and the activity inhibition sequence for inducing the water-soluble expression.
본 발명의 구체예에서, 상기 형질전환 미생물의 배양은 유가식 배양방법으로 배양할 수있으나 이에 제한되는 것은 아니다. 배양 온도는 20 내지 40 ℃ 일 수 있으며, 본 발명의 상기 재조합 발현 벡터를 형질전환한 미생물은 28℃ 이상에서도 100% 수용성 융합단백질로 발현될 수 있으며, 37℃ 이상에서 50% 이상의 수용성 융합단백질로 발현될 수 있다.In the embodiment of the present invention, the culturing of the transforming microorganism can be carried out by a fed-batch culture method, but is not limited thereto. The microorganism transformed with the recombinant expression vector of the present invention may be expressed as a 100% water-soluble fusion protein even at 28 ° C or higher, and more than 50% of the water-soluble fusion protein at 37 ° C or higher Lt; / RTI >
본 발명의 구체예에서, 상기 형질전환된 미생물은 대장균일 수 있으나 이에 제한되는 것은 아니며, 바람직하게 E. coli MG1655 또는 BL21 일 수 있다. In an embodiment of the present invention, the transformed microorganism may be E. coli but is not limited thereto, and preferably E. coli MG1655 or BL21.
본 발명의 상기 재조합 트랜스 글루타미나아제의 생산 방법은 재조합 트랜스 글루타미나아제를 생산 및 분비하는 단계를 추가적으로 포함할 수 있으며, 상기 생산, 분비된 트랜스 글루타미나아제를 정제 및 분리하는 공정을 추가적으로 포함할 수 있다.
The production method of the recombinant transglutaminase of the present invention may further include a step of producing and secreting a recombinant transglutaminase, and the step of purifying and separating the produced and secreted transglutaminase may be performed May be included.
본 발명은 또한, The present invention also relates to
수용성 발현 유도용 단백질을 암호화하는 유전자, 재조합 트랜스 글루타미나아제의 세포 내 활성 억제 서열(proteolytic-resistant sequence), 단백질 분해 효소 인식부위를 암호화하는 유전자 및 트랜스 글루타미아나제를 암호화하는 유전자; 수용성 발현 유도용 단백질을 암호화하는 유전자 및 단백질 분해 효소를 암호화하는 유전자를 차례로 포함하는 재조합 트랜스 글루타미나아제 생산용 재조합 발현 벡터를 제공한다.A gene encoding a protein for inducing water-soluble expression, a proteolytic-resistant sequence of a recombinant transglutaminase, a gene encoding a protease recognition site, and a gene encoding a transglutaminase; There is provided a recombinant expression vector for the production of recombinant transglutaminase comprising in sequence a gene encoding a protein for inducing water-soluble expression and a gene encoding a protease.
본 발명의 구체예에서, 상기 재조합 발현 벡터는 도 9로 표시되는 것일 수 있으나 이에 제한되는 것은 아니다. In an embodiment of the present invention, the recombinant expression vector may be as shown in FIG. 9, but is not limited thereto.
상기 재조합 발현 벡터는 균체 내에서 동시에 트랜스 글루타미나아제와 단백질 분해 효소를 발현시킬 수 있다.
The recombinant expression vector can simultaneously express transglutaminase and proteolytic enzyme in the cells.
본 발명의 구체예에서는, pACYC MBP-proTGase::MBP-TEV 발현 벡터를 E. coli MG1655와 BL21 균주에 형질 전환하여 플라스크 배양한 결과, 세포 내에서 발현된 TEV 단백질이 수용성 억제서열 결합 융합 단백질 형태의 트랜스 글루타미나아제를 100% 분해하여 활성형의 트랜스글루타미나아제로 생성하는 것을 확인하였다.
In a specific example of the present invention, the pACYC MBP-proTGase :: MBP-TEV expression vector was transformed into E. coli MG1655 and BL21, and the cells were cultured in a flask. As a result, the TEV protein expressed in the cell was expressed as a water- Of the transglutaminase was decomposed at 100% and it was confirmed that it was produced as the active type transglutaminase.
본 발명에 따른 활성형 트랜스 글루타미나아제의 대량 생산 방법은 세포 내 기작에 영향을 주지 않아 활성형 효소의 대량 생산이 가능하다. 또한, 추가적인 재접힘(refolding) 과정이 불필요하여 생산 방법이 간단하다.
The mass production method of the active transglutaminase according to the present invention does not affect the intracellular mechanism, so that the active enzyme can be mass-produced. In addition, the production method is simple since an additional refolding process is unnecessary.
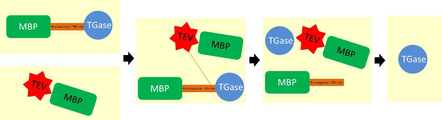
도 1은 본 발명의 융합 단백질 형태의 발현 벡터를 나타낸다(a: pKPM::MBP-pro-TEV site-TGase 벡터; b: pKPM::pro-TEV site-TGase 벡터; c: pET::MBP-TEV site-TEV 벡터; T7: T7 프로모터; MBP: 말토오스 결합 도메인(maltolse-binding domain); linker: proteolytic-resistant sequences, TGase: 트랜스 글루타미나아제 codon-optimized gene; H6: 6XHistidine tag; TEV: codon-optimized gene; T7t: T7 터미네이터; 화살표를 통해, 각각의 재조합 단백질 밴드를 나타냄).
도 2는 대장균에서 도 1의 새가지 벡터를 배양 온도별로 발현시킨 웨스턴 블롯 결과이다(a: pKPM::pro-TEV site-TGase 벡터 발현 결과, b: pKPM::MBP-pro-TEV site-TGase 벡터 발현 결과; c: pET::MBP-TEV site-TEV 벡터 발현 결과임).
도 3은 5L 발효조를 이용한 pKPM::MBP-pro-TEV site-TGase 벡터의 유가식 배양 결과이다.
도 4는 본 발명의 활성형의 트랜스 글루타미나아제의 대량 생산 방법의 모식도이다.
도 5는 in-vitro 조건에서 본 발명의 활성형 트랜스 글루타미나아제의 생산을 위한 벡터에 클로닝된 모든 효소들의 발현을 웨스턴 블롯으로 확인한 결과이다.
도 6은 in-vivo 시스템에서 본 발명의 활성형의 트랜스 글루타미나아제의 대량 생산하기 위한 방법의 모식도이다.
도 7은 in-vivo 시스템 적용하기 위한 pACYC MBP-proTGase::MBP-TEV 발현 벡터의 모식도이다.
도 8은 In-vivo 발현 조건에서 pACYC MBP-proTGase::MBP-TEV 발현 벡터를 E. coli MG1655와 BL21 균주에 형질 전환한 후의 활성형 트랜스 글루타미나아제의 생성을 확인한 단백질 발현 결과이다.
도 9는 본 발명의 활성형 트랜스 글루타미나아제를 Ni 컬럼을 이용해 정제한 결과이다(레인 1은 정제 전 시료이며, 레인 4~9(E1~E6)는 활성형 트랜스 글루타미나아제 정제 프랙션을 나타낸다).
도 10은 발효조를 이용한 in-vivo 시스템에서의 트랜스 글루타미나아제 발현 양상을 확인한 결과이다(숫자는 IPTG 발현 유도 후의 시간을 나타내며, 16I와 16S는 각각 16시간 후 불용해성 분획물과 용해성 분획물을 나타낸다).
Figure 1 shows the expression vector of the fusion protein form of the present invention (a: pKPM :: MBP-pro-TEV site-TGase vector; b: pKPM :: pro-TEV site-TGase vector; c: pET :: MBP- TEV site-TEV vector, T7: T7 promoter, MBP: maltolse-binding domain, linker: proteolytic-resistant sequences, TGase: transglutaminase codon-optimized gene, H6: 6XHistidine tag, TEV: codon -optimized gene; T7t: T7 terminator; through arrows, representing each recombinant protein band).
FIG. 2 is a Western blot result of Escherichia coli expressing the novel vector of FIG. 1 at different culture temperatures (a: pKPM :: pro-TEV site-TGase vector expression b: pKPM :: MBP-pro-TEV site-TGase Vector expression result; c: pET :: MBP-TEV site-TEV vector expression result).
FIG. 3 shows the results of a fed-batch culture of pKPM :: MBP-pro-TEV site-TGase vector using a 5 L fermenter.
4 is a schematic diagram of a mass production method of the active type transglutaminase of the present invention.
Figure 5 shows the results of western blot analysis of the expression of all the enzymes cloned in the vector for the production of the active transglutaminase of the present invention under in-vitro conditions.
Figure 6 is a schematic diagram of a method for mass production of the active form of transglutaminase of the present invention in an in-vivo system.
FIG. 7 is a schematic diagram of pACYC MBP-proTGase :: MBP-TEV expression vector for in vivo system application.
FIG. 8 shows the results of protein expression confirming the production of active transglutaminase after transformation of pACYC MBP-proTGase :: MBP-TEV expression vector into E. coli MG1655 and BL21 strain under in vivo expression conditions.
9 shows the result of purification of the active transglutaminase of the present invention by using a Ni column (
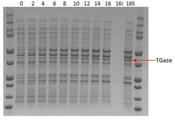
FIG. 10 shows the results of transglutaminase expression in an in-vivo system using a fermenter (the numbers represent the time after induction of IPTG expression, and 16I and 16S represent insoluble fractions and soluble fractions after 16 hours, respectively ).
이하, 본 발명의 이해를 돕기 위하여 실시예를 들어 상세하게 설명하기로 한다. 다만 하기의 실시예는 본 발명의 내용을 예시하는 것일 뿐 본 발명의 범위가 하기 실시예에 한정되는 것은 아니다. 본 발명의 실시예는 이 기술분야에서 평균적인 지식을 가진 자에게 본 발명을 보다 완전하게 설명하기 위해 제공되는 것이다.
BEST MODE FOR CARRYING OUT THE INVENTION Hereinafter, the present invention will be described in detail with reference to the following examples. However, the following examples are intended to illustrate the contents of the present invention, but the scope of the present invention is not limited to the following examples. Embodiments of the present invention are provided to more fully describe the present invention to those skilled in the art.
<< 실시예Example 1> 재조합 발현벡터의 제조 1> Preparation of Recombinant Expression Vector
트랜스 글루타미나아제(이하, TGase로 혼용됨)와 TEV protease를 코딩하는 유전자를 코돈 최적화하여 대장균에서 발현가능한 벡터를 도 1과 같이 제조하였다. A vector capable of expressing in Escherichia coli was prepared as shown in Fig. 1 by codon-optimizing a gene coding for transglutaminase (hereinafter referred to as TGase) and TEV protease.
알려진 유전정보를 기반으로 미국의 DNA2.0사에서 코돈 최적화를 수행하였다. 코돈 최적화된 글루타미나아제(CpTgase-optEc)는 5` 말단에 EcoR I, 3` 말단에 Xho I 제한효소 사이트를 삽입하여 pJ204 벡터에 삽입하여 제작되었으며, CpTgase-optEc-pJ204로 명명하였다. 유전자 서열 132번째 아미노산 phenylalanine(F)을 valine(V), 142번째 아미노산 alanine(A)을 aspartic acid(D), 255번째 아미노산 glycine(G)을 aspartic acid(D)로 변경하였다. 코돈 최적화된 TEV(CpTEVp-optEc)는 5` 말단에 EcoR I, 3` 말단에 Xho I 제한효소 사이트를 삽입하여 pJ204 벡터에 삽입하여 제작되었으며, CpTEVp-optEc-pJ204로 명명하였다. 유전자 서열 17번째 아미노산 threonine(T)을 serine(S), 56번째 아미노산 leucine(L)을 valine(V), 68번째 아미노산 asparagine(N)을 aspartic acid(D), 77번째 아미노산 isoleucine(I)을 valine(V), 135번째 아미노산 serine(S)을 glycine(G), 219번째 아미노산 serine(S)을 valine(V)로 변경하였다. Based on known genetic information, codon optimization was performed in DNA2.0 of USA. The codon-optimized glutaminase (CpTgase-optEc) was amplified by EcoR I, by inserting the Xho I restriction enzyme site on the 3` end was fabricated by inserting the pJ204 vector, was named CpTgase-optEc-pJ204. Gene sequence 132th amino acid phenylalanine (F) was changed to valine (V), 142th amino acid alanine (A) was changed to aspartic acid (D) and 255th amino acid glycine (G) was changed to aspartic acid (D). The codon optimized TEV (CpTEVp-optEc) is the 5` end EcoR I, at the 3 'end Xho I restriction enzyme site and inserted into pJ204 vector, and designated as CpTEVp-optEc-pJ204. The amino acid sequence of the 17th amino acid threonine (T) is serine (S), the 56th amino acid leucine (L) is valine (V), the 68th amino acid asparagine (N) is aspartic acid (D), the 77th amino acid isoleucine (I) the amino acid serine (S) at the 135th position was substituted with glycine (G) and the amino acid serine (S) at position 219 was replaced with valine (V).
최적화된 글루타미나아제와 TEV의 플라스미드를 EcoR I과 Xho I 제한 효소 처리 후, 같은 제한 효소로 분해된 pKPM::MBP 벡터와 pKPM 벡터에 도입되었다. 이 플라스미드는 pKPM::pro-TEVsite-TGase, pKPM::MBP-pro-TEVsite-TGase와 pKPM::MBP-TEV로 최종 명명하였다.
The optimized glutaminase and TEV plasmids were digested with EcoR I and Xho I restriction enzyme treatment, pKPM :: MBP vector and pKPM vector were digested with the same restriction enzymes. This plasmid was finally named pKPM :: pro-TEVsite-TGase, pKPM :: MBP-pro-TEVsite-TGase and pKPM :: MBP-TEV.
<< 실시예Example 2> 플라스크에서 단백질 발현결과 2> Result of protein expression in flask
상기 실시예 1에 따른 각각의 벡터로 대장균을 형질전환하여 발현을 유도하였다.E. coli was transformed with each vector according to Example 1 to induce expression.
이때 사용된 대장균은 B 계열 대장균 BL21(DE3) (E. coli B F- dcm opmT hsdS(rB- mB -a) gal λ,Novagen, USA)과 K 계열 대장균인 MG1655(DE3) [F- λ- ilvG- rfb -50 rph -1]를 효소 유전자 발현을 위한 숙주세포로 사용하였으며, 상기 형질 전환체는 50 μg/ml 의 kanamycin을 함유하는 auto induction 배지(표 1)를 이용하여 37°C, 28°C, 22℃에서 각각 약 16시간 동안 배양하였다. 발현시스템은 일반적으로 고가의 IPTG를 이용하여 T7 promotor를 작동시켜 단백질의 과발현을 유도하나 본 연구에서 사용된 auto induction 배지를 이용하여 이 IPTG 대신에 락토오즈를 유도 물질로 대체하여, 재조합 균주의 세포농도 증식과 단백질 과발현을 유도하였다. The Escherichia coli used was B-type E. coli BL21 (DE3) ( E. coli B F- dcm We used opmT hsdS (rB- mB -a) gal λ, Novagen, USA) and E. coli K series of MG1655 (DE3) [F- λ- ilvG- rfb -50 rph -1] as a host cell for gene expression , And the transformants were cultured at 37 ° C, 28 ° C and 22 ° C for about 16 hours using an auto induction medium containing 50 μg / ml kanamycin (Table 1). Expression system generally induces overexpression of protein by activating T7 promoter using expensive IPTG. However, instead of using this auto induction medium used in this study, lactoose was replaced with inducer in place of IPTG, and the recombinant strain cell Concentration and protein overexpression were induced.
그 결과, pKPM::pro-TEV site-TGase 벡터를 형질 전환한 균주는 37도 배양에서는 모두 불용성의 단백질로 발현 되었으며, 28도 배양에서는 50%의 단백질만이 수용성, 22도 배양에서는 100% 모두 수용성으로 발현되었다(도 2a). pKPM::MBP-pro-TEVsite-TGase 벡터를 형질 전환한 균주는 37도 배양에서 단백질 중 50%가 수용성으로 발현 되었으며, 28도와 22도 배양에서는 100% 수용성으로 발현되었다(도 2b). pET::MBP-TEV site-TEV 벡터를 형질 전환한 균주는 세포 내에서 TEV 단백질이 발현되어 발현 구조내에 있는 TEV site를 인식하여 MBP와 TEV 사이를 자가 절단(self-cleavage)하였다. 웨스턴 블롯 분석 결과, 37도 배양에서 수용성으로 발현된 MBP 단백질 밴드(42kDa)와 TEV(25kDa) 단백질 밴드를 확인하였다(도 2c).
As a result, the strain transformed with the pKPM :: pro-TEV site-TGase vector was expressed as an insoluble protein in all 37 ° cultures. In 28 ° culturing, only 50% of the protein was soluble and 100% Soluble (Fig. 2a). The strain transformed with pKPM :: MBP-pro-TEVsite-TGase vector was expressed in 50% of the proteins in a water-soluble state at 37 ° C and 100% in water at 28 ° C and 22 ° C (FIG. The strain transformed with the pET :: MBP-TEV site-TEV vector recognized the TEV site within the expression structure by expressing the TEV protein in the cell and self-cleavage between the MBP and the TEV. Western blot analysis revealed that the water soluble MBP protein band (42 kDa) and the TEV (25 kDa) protein band were detected in 37 ° culture (FIG. 2c).
<< 실시예Example 3> 발효조에서 단백질 발현결과 3> Result of Protein Expression in Fermenter
pKPM::MBP-pro-TEVsite-TGase 벡터를 형질 전환한 균주를 5L 발효조에서 배양하여, 단백질의 대량발현을 유도하였다. 발효조 배양을 위한 배지의 조성은 표 2와 같다. 초기의 포도당 농도를 15 g/L로 하고, 이 포도당이 다 고갈되어 pH, DO가 급격하게 증가하는 시점에서 대체 유도물질인 락토오즈를 15 g/L을 일시적으로 공급함과 동시에 대장균의 생존에 필요한 최소한의 탄소원 공급을 위해 600 g/L의 포도당을 이용하여 6 g/L/h로 지속적으로 공급하였다. 배양 결과, 균체는 155.2 OD 정도의 고농도로 성장하였으며, 배양 후 전기영동을 통한 단백질 발현 분석결과, 배양말기에 TGase 발현양은 총단백질의 45% 였으며, 모두 수용성으로 12.62 g/L가 발현된 것을 확인하였다.The strain transformed with the pKPM :: MBP-pro-TEVsite-TGase vector was cultured in a 5 L fermenter to induce a large amount of protein expression. Table 2 shows the composition of the medium for fermentation. At an initial glucose concentration of 15 g / L, when the pH and DO are rapidly increased due to depletion of glucose, an alternative inducing substance, lactose, is supplied at a rate of 15 g / L and at the same time necessary for the survival of E. coli For a minimum supply of carbon, 600 g / L of glucose was continuously fed at 6 g / L / h. As a result of the culture, the cells were grown at a high concentration of about 155.2 OD. After analysis of the protein expression by electrophoresis after culture, the amount of TGase expressed at the end of the culture was 45% of the total protein and all the water was 12.62 g / L Respectively.
(초기)g / L
(Early)
(첨가)g / L
(adding)
<< 실시예Example 4> 발효조에서 활성형의 4> In the fermentation tank, MBPMBP -- TGaseTGase 분리 결과 확인 Confirm separation result
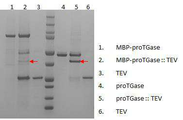
실시예 2에서 발현 확인된 세포를 소니케이션으로 파쇄하고 상층액으로부터 Ni-NTA agaros 레진을 이용하여 pro-TGase, MBP-pro-TGase, TEV 단백질을 정제하였다. 정제된 단백질은 50mM NaH2PO4 ㆍ2H2O, 250mM NaCl pH7.5 버퍼로 투석하였다. Pro-TGase와 TEV 단백질, MBP-pro-TGase와 TEV 단백질을 20ug씩 혼합하여 37℃에서 1시간 반응하였다. Cells expressed in Example 2 were disrupted by sonication, and pro-TGase, MBP-pro-TGase and TEV proteins were purified from the supernatant using Ni-NTA agarose resin. The purified protein was dialyzed with 50mM NaH 2 PO 4 and 2H 2 O, 250mM NaCl pH7.5 buffer. Pro-TGase and TEV proteins, MBP-pro-TGase and TEV proteins were mixed at 20 ug each and reacted at 37 ° C for 1 hour.
도 5의 레인 1과 4는 정제된 MBP-proTGase와 proTGase 단백질, 레인 3과 6은 정제된 TEV 단백질, 레인 2는 MBP-proTGase와 TEV 혼합 후 37℃에서 1시간 반응 후의 시료, 5는 proTGase와 TEV 혼합 후 37℃에서 1시간 반응 후의 시료이다. 즉, MBP-pro-TGase와 pro-TGase의 형태로 발현된 단백질들을 TEV에 의한 절단을 통해, 활성형 TGase를 분리하는 시도한 결과, TEV에 의해서 활성형 TGase가 분리되는 것을 확인하였다(도 5).
<< 실시예Example 5> 5> inin vivovivo 조건에서 트랜스 글루타미나아제의 활성화 확인Confirm activation of transglutaminase in conditions
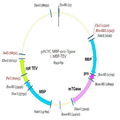
본 발명에서 활성형의 트랜스 글루타미나아제를 생성하기 위해서 확립한 방법은 기질로 이용되는 두 가지 형태의 TGase와 TEV를 모두 정제한 단백질을 이용한 것이다. 활성형 트랜스 글루타미나아제 분리 공정 단계를 최소화하고 TEV 단백질 분해 효소의 효율을 높이기 위해서 in-vivo에서 트랜스 글루타미나아제와 TEV 단백질을 동시 발현하는 벡터를 개발하였다(도 7). In the present invention, a method established for producing active-type transglutaminase is a protein in which both types of TGase and TEV used as a substrate are purified. To minimize the active transglutaminase separation process step and to increase the efficiency of TEV protease, a vector expressing transglutaminase and TEV protein in vivo was developed (Fig. 7).
활성형 TGase만을 획득하기 위하여 TGase 단백질의 C 말단에 his tag을 첨가하고 TEV 단백질에는 tag을 부착하지 않았다. 실시예 1에서 구축된 pKPM MBP-pro-TEV site-TGase 플라스미드의 T7 promotor와 T7 terminator를 포함하도록 프라이머를 디자인 하였으며 프라이머의 양 말단에 Hind III 제한 효소를 도입하였다. 증폭된 DNA는 In-fusion™ Advantage PCR cloning kit (Clontech, USA)를 이용하여 Hind III로 분해된 pACYC184에 삽입하였다. 구축된 플라스미드는 pACYC MBP-pro-TGase 로 명명하였다. To obtain only active TGase, the tag was added to the C terminal of the TGase protein and the tag was not attached to the TEV protein. Primers were designed to include the T7 promoter and T7 terminator of the pKPM MBP-pro-TEV site-TGase plasmid constructed in Example 1, and HindIII restriction enzymes were introduced at both ends of the primers. The amplified DNA was inserted into pACYC184 digested with Hind III using the In-fusion ™ Advantage PCR cloning kit (Clontech, USA). The constructed plasmid was named pACYC MBP-pro-TGase.
pKPM MBP-TEV의 T7 promotor와 T7 terminator를 포함하도록 양 말단에 Sal I 효소를 포함하는 프라이머를 이용하여 증폭하였다. PCR 산물은 In-fusion™ Advantage PCR cloning kit (Clontech, USA)를 이용하여 Sal I으로 분해된 pACYC184 MBP-pro-TGase에 각각 삽입하였다. 구축된 플라스미드는 pACYC MBP-pro-TGase :: MBP-TEV로 최종 명명하였다. 본 발명의 in-vivo 발현 시스템은 도 6으로 모식화하였다.
pKPM The T7 promoter of MBP-TEV and the T7 terminator at both ends And amplified with a primer containing Sal I enzyme. The PCR product was Sal using the In-fusion ™ Advantage PCR cloning kit (Clontech, USA) I < / RTI > digested pACYC184 MBP-pro-TGase. The constructed plasmid was finally named pACYC MBP-pro-TGase :: MBP-TEV. The in vivo expression system of the present invention is illustrated in FIG.
상기의 방법으로 제작된 발현벡터를 E. coli MG1655와 BL21 균주에 형질 전환하였다. 상기 형질 전환체는 25 μg/ml 의 chloramphenicol을 함유하는 LB 배지에서 37°C에서 3시간 배양 후(OD 0.4) 0.04mM, 0.4mM의 IPTG를 첨가하여 200rpm, 25°C에서 각각 약 16시간 동안 배양하였다. The expression vector prepared by the above method was transformed into E. coli MG1655 and BL21 strains. The transformants were cultured in LB medium containing 25 μg / ml of chloramphenicol for 3 hours at 37 ° C. (OD 0.4), after which 0.04 mM, 0.4 mM IPTG was added and incubated at 200 rpm and 25 ° C. for about 16 hours Lt; / RTI >
상기 형질 전환체의 플라스크 배양결과, MG1655 균주에서 TEV에 의해 융합 단백질이 100% 분해되어 활성형의 트랜스 글루타미나아제가 생성됨을 확인하였다(도 8). As a result of the flask culture of the transformant, it was confirmed that the fusion protein was 100% degraded by TEV in MG1655 strain to generate active transglutaminase (FIG. 8).
발현 확인된 세포는 소니케이션으로 파쇄하고 상층액으로부터 Ni-NTA agarose 레진을 이용하여 정제하여 순도 90%의 활성형 TGase를 분리하였다(도 9). The expression-confirmed cells were disrupted by sonication and purified from the supernatant using Ni-NTA agarose resin to isolate active TGase having a purity of 90% (FIG. 9).
분리된 활성형 트랜스 글루타미나아제는 한국기초과학연구소에 N-말단 분석을 의뢰하여 활성형 트랜스 글루타미나아제의 단백질 서열을 확인하였다. Microbial transglutaminase assay kit (Zedira)을 사용하여 단백질 활성을 확인한 결과, 31.17U/mg의 활성을 가짐을 확인하였다. The isolated active transglutaminase was subjected to N-terminal analysis by the Korea Basic Science Institute to confirm the protein sequence of the active transglutaminase. The protein activity was confirmed by using a microbial transglutaminase assay kit (Zedira), and it was confirmed that the activity was 31.17 U / mg.
또한, 유가식 배양을 통해서도 TGase가 in vivo에서 활성화가 되는 것을 확인하였다(도 10). 구체적으로, pACYC MBP-pro-TGase :: MBP-TEV 플라스미드를 형질 전환한 MG1655균주를 5L 발효조에서 배양하여, 단백질의 대량발현을 유도하였다. 발효조 배양을 위한 배지의 조성은 표 4와 같다. 초기의 포도당 농도를 15 g/L로 하고, 이 포도당이 다 고갈되어 pH, DO가 급격하게 증가하는 시점에서 대장균의 생존에 필요한 최소한의 탄소원 공급을 위해 600 g/L의 포도당을 이용하여 6 g/L/h로 지속적으로 공급하였다. 균체가 8 OD가 되었을 때 유도물질인 IPTG 0.04mM을 첨가하였다. 수용성의 단백질 발현을 유도하기 위해 배양 온도는 25℃로 낮추었다. 배양 결과, 균체는 26.48 OD 정도로 성장하였으며, 배양 후 전기영동을 통한 단백질 발현 분석결과, 배양말기에 TGase 발현양은 총단백질의 3.1% 였으며, 모두 수용성으로 156mg/L가 발현된 것을 확인하였다.
Also, it was confirmed that TGase was activated in vivo by fed-batch culture (FIG. 10). Specifically, the MG1655 strain transformed with the pACYC MBP-pro-TGase :: MBP-TEV plasmid was cultured in a 5 L fermenter to induce a large amount of protein expression. Table 4 shows the composition of the medium for fermentation. At an initial glucose concentration of 15 g / L, and at the point when the pH and DO increase rapidly due to depletion of glucose, 600 g / L of glucose was added to the solution to obtain the minimum amount of carbon source necessary for the survival of E. coli. / L / h. When the cells reached 8 OD, 0.04 mM IPTG as an inducing substance was added. The culture temperature was lowered to 25 ° C to induce water-soluble protein expression. As a result of the culture, the cells were grown to about 26.48 OD. After the culture, the expression of TGase was 3.1% of the total protein at the end of the culture, and 156 mg / L was expressed as water soluble.
(초기)g / L
(Early)
(첨가)g / L
(adding)
<110> Korea Research Institute of Bioscience & Biotechnology <120> MASS PRODUCTION METHOD OF RECOMBINANT TRANS GLUTAMINASE <130> p14r16d1756 <160> 14 <170> KopatentIn 2.0 <210> 1 <211> 367 <212> PRT <213> Artificial Sequence <220> <223> maltose binding protein <400> 1 Met Lys Ile Glu Glu Gly Lys Leu Val Ile Trp Ile Asn Gly Asp Lys 1 5 10 15 Gly Tyr Asn Gly Leu Ala Glu Val Gly Lys Lys Phe Glu Lys Asp Thr 20 25 30 Gly Ile Lys Val Thr Val Glu His Pro Asp Lys Leu Glu Glu Lys Phe 35 40 45 Pro Gln Val Ala Ala Thr Gly Asp Gly Pro Asp Ile Ile Phe Trp Ala 50 55 60 His Asp Arg Phe Gly Gly Tyr Ala Gln Ser Gly Leu Leu Ala Glu Ile 65 70 75 80 Thr Pro Asp Lys Ala Phe Gln Asp Lys Leu Tyr Pro Phe Thr Trp Asp 85 90 95 Ala Val Arg Tyr Asn Gly Lys Leu Ile Ala Tyr Pro Ile Ala Val Glu 100 105 110 Ala Leu Ser Leu Ile Tyr Asn Lys Asp Leu Leu Pro Asn Pro Pro Lys 115 120 125 Thr Trp Glu Glu Ile Pro Ala Leu Asp Lys Glu Leu Lys Ala Lys Gly 130 135 140 Lys Ser Ala Leu Met Phe Asn Leu Gln Glu Pro Tyr Phe Thr Trp Pro 145 150 155 160 Leu Ile Ala Ala Asp Gly Gly Tyr Ala Phe Lys Tyr Glu Asn Gly Lys 165 170 175 Tyr Asp Ile Lys Asp Val Gly Val Asp Asn Ala Gly Ala Lys Ala Gly 180 185 190 Leu Thr Phe Leu Val Asp Leu Ile Lys Asn Lys His Met Asn Ala Asp 195 200 205 Thr Asp Tyr Ser Ile Ala Glu Ala Ala Phe Asn Lys Gly Glu Thr Ala 210 215 220 Met Thr Ile Asn Gly Pro Trp Ala Trp Ser Asn Ile Asp Thr Ser Lys 225 230 235 240 Val Asn Tyr Gly Val Thr Val Leu Pro Thr Phe Lys Gly Gln Pro Ser 245 250 255 Lys Pro Phe Val Gly Val Leu Ser Ala Gly Ile Asn Ala Ala Ser Pro 260 265 270 Asn Lys Glu Leu Ala Lys Glu Phe Leu Glu Asn Tyr Leu Leu Thr Asp 275 280 285 Glu Gly Leu Glu Ala Val Asn Lys Asp Lys Pro Leu Gly Ala Val Ala 290 295 300 Leu Lys Ser Tyr Glu Glu Glu Leu Ala Lys Asp Pro Arg Ile Ala Ala 305 310 315 320 Thr Met Glu Asn Ala Gln Lys Gly Glu Ile Met Pro Asn Ile Pro Gln 325 330 335 Met Ser Ala Phe Trp Tyr Ala Val Arg Thr Ala Val Ile Asn Ala Ala 340 345 350 Ser Gly Arg Gln Thr Val Asp Glu Ala Leu Lys Asp Ala Gln Thr 355 360 365 <210> 2 <211> 45 <212> PRT <213> Artificial Sequence <220> <223> proteolytic-resistant sequence <400> 2 Asp Asn Gly Ala Gly Glu Glu Thr Lys Ser Tyr Ala Glu Thr Tyr Arg 1 5 10 15 Leu Thr Ala Asp Asp Val Ala Asn Ile Asn Ala Leu Asn Glu Ser Ala 20 25 30 Pro Ala Ala Ser Ser Ala Gly Pro Ser Phe Arg Ser Pro 35 40 45 <210> 3 <211> 7 <212> PRT <213> Artificial Sequence <220> <223> Tobacco etch virus cleavage site <400> 3 Glu Asn Leu Tyr Phe Gln Gly 1 5 <210> 4 <211> 339 <212> PRT <213> Artificial Sequence <220> <223> Transglutaminase <400> 4 Asp Ser Asp Asp Arg Val Thr Pro Pro Ala Glu Pro Leu Asp Arg Met 1 5 10 15 Pro Asp Pro Tyr Arg Pro Ser Tyr Gly Arg Ala Glu Thr Val Val Asn 20 25 30 Asn Tyr Ile Arg Lys Trp Gln Gln Val Tyr Ser His Arg Asp Gly Arg 35 40 45 Lys Gln Gln Met Thr Glu Glu Gln Arg Glu Trp Leu Ser Tyr Gly Cys 50 55 60 Val Gly Val Thr Trp Val Asn Ser Gly Gln Tyr Pro Thr Asn Arg Leu 65 70 75 80 Ala Phe Ala Ser Phe Asp Glu Asp Arg Phe Lys Asn Glu Leu Lys Asn 85 90 95 Gly Arg Pro Arg Ser Gly Glu Thr Arg Ala Glu Phe Glu Gly Arg Val 100 105 110 Ala Lys Glu Ser Phe Asp Glu Glu Lys Gly Phe Gln Arg Ala Arg Glu 115 120 125 Val Ala Ser Val Met Asn Arg Ala Leu Glu Asn Ala His Asp Glu Ser 130 135 140 Ala Tyr Leu Asp Asn Leu Lys Lys Glu Leu Ala Asn Gly Asn Asp Ala 145 150 155 160 Leu Arg Asn Glu Asp Ala Arg Ser Pro Phe Tyr Ser Ala Leu Arg Asn 165 170 175 Thr Pro Ser Phe Lys Glu Arg Asn Gly Gly Asn His Asp Pro Ser Arg 180 185 190 Met Lys Ala Val Ile Tyr Ser Lys His Phe Trp Ser Gly Gln Asp Arg 195 200 205 Ser Ser Ser Ala Asp Lys Arg Lys Tyr Gly Asp Pro Asp Ala Phe Arg 210 215 220 Pro Ala Pro Gly Thr Gly Leu Val Asp Met Ser Arg Asp Arg Asn Ile 225 230 235 240 Pro Arg Ser Pro Thr Ser Pro Gly Glu Gly Phe Val Asn Phe Asp Tyr 245 250 255 Gly Trp Phe Gly Ala Gln Thr Glu Ala Asp Ala Asp Lys Thr Val Trp 260 265 270 Thr His Gly Asn His Tyr His Ala Pro Asn Gly Ser Leu Gly Ala Met 275 280 285 His Val Tyr Glu Ser Lys Phe Arg Asn Trp Ser Glu Gly Tyr Ser Asp 290 295 300 Phe Asp Arg Gly Ala Tyr Val Ile Thr Phe Ile Pro Lys Ser Trp Asn 305 310 315 320 Thr Ala Pro Asp Lys Val Lys Gln Gly Trp Pro Leu Glu His His His 325 330 335 His His His <210> 5 <211> 241 <212> PRT <213> Artificial Sequence <220> <223> codon-optimized tobacco etch virus protease <400> 5 Gly Glu Ser Leu Phe Lys Gly Pro Arg Asp Tyr Asn Pro Ile Ser Ser 1 5 10 15 Ser Ile Cys His Leu Thr Asn Glu Ser Asp Gly His Thr Thr Ser Leu 20 25 30 Tyr Gly Ile Gly Phe Gly Pro Phe Ile Ile Thr Asn Lys His Leu Phe 35 40 45 Arg Arg Asn Asn Gly Thr Leu Val Val Gln Ser Leu His Gly Val Phe 50 55 60 Lys Val Lys Asp Thr Thr Thr Leu Gln Gln His Leu Val Asp Gly Arg 65 70 75 80 Asp Met Ile Ile Ile Arg Met Pro Lys Asp Phe Pro Pro Phe Pro Gln 85 90 95 Lys Leu Lys Phe Arg Glu Pro Gln Arg Glu Glu Arg Ile Cys Leu Val 100 105 110 Thr Thr Asn Phe Gln Thr Lys Ser Met Ser Ser Met Val Ser Asp Thr 115 120 125 Ser Cys Thr Phe Pro Ser Gly Asp Gly Ile Phe Trp Lys His Trp Ile 130 135 140 Gln Thr Lys Asp Gly Gln Cys Gly Ser Pro Leu Val Ser Thr Arg Asp 145 150 155 160 Gly Phe Ile Val Gly Ile His Ser Ala Ser Asn Phe Thr Asn Thr Asn 165 170 175 Asn Tyr Phe Thr Ser Val Pro Lys Asn Phe Met Glu Leu Leu Thr Asn 180 185 190 Gln Glu Ala Gln Gln Trp Val Ser Gly Trp Arg Leu Asn Ala Asp Ser 195 200 205 Val Leu Trp Gly Gly His Lys Val Phe Met Val Lys Pro Glu Glu Pro 210 215 220 Phe Gln Pro Val Lys Glu Ala Thr Gln Leu Met Asn Arg Arg Arg Arg 225 230 235 240 Arg <210> 6 <211> 1101 <212> DNA <213> Artificial Sequence <220> <223> maltose binding protein <400> 6 atgaaaatcg aagaaggtaa actggtaatc tggattaacg gcgataaagg ctataacggt 60 ctcgctgaag tcggtaagaa attcgagaaa gataccggaa ttaaagtcac cgttgagcat 120 ccggataaac tggaagagaa attcccacag gttgcggcaa ctggcgatgg ccctgacatt 180 atcttctggg cacacgaccg ctttggtggc tacgctcaat ctggcctgtt ggctgaaatc 240 accccggaca aagcgttcca ggacaagctg tatccgttta cctgggatgc cgtacgttac 300 aacggcaagc tgattgctta cccgatcgct gttgaagcgt tatcgctgat ttataacaaa 360 gatctgctgc cgaacccgcc aaaaacctgg gaagagatcc cggcgctgga taaagaactg 420 aaagcgaaag gtaagagcgc gctgatgttc aacctgcaag aaccgtactt cacctggccg 480 ctgattgctg ctgacggggg ttatgcgttc aagtatgaaa acggcaagta cgacattaaa 540 gacgtgggcg tggataacgc tggcgcgaaa gcgggtctga ccttcctggt tgacctgatt 600 aaaaacaaac acatgaatgc agacaccgat tactccatcg cagaagctgc ctttaataaa 660 ggcgaaacag cgatgaccat caacggcccg tgggcatggt ccaacatcga caccagcaaa 720 gtgaattatg gtgtaacggt actgccgacc ttcaagggtc aaccatccaa accgttcgtt 780 ggcgtgctga gcgcaggtat taacgccgcc agtccgaaca aagagctggc aaaagagttc 840 ctcgaaaact atctgctgac tgatgaaggt ctggaagcgg ttaataaaga caaaccgctg 900 ggtgccgtag cgctgaagtc ttacgaggaa gagttggcga aagatccacg tattgccgcc 960 actatggaaa acgcccagaa aggtgaaatc atgccgaaca tcccgcagat gtccgctttc 1020 tggtatgccg tgcgtactgc ggtgatcaac gccgccagcg gtcgtcagac tgtcgatgaa 1080 gccctgaaag acgcgcagac t 1101 <210> 7 <211> 135 <212> DNA <213> Artificial Sequence <220> <223> proteolytic-resistant sequence <400> 7 gacaatggcg cgggggaaga gacgaagtcc tacgccgaaa cctaccgcct cacggcggat 60 gacgtcgcga acatcaacgc gctcaacgaa agcgctccgg ccgcttcgag cgccggcccg 120 tcgttccggt ccccc 135 <210> 8 <211> 21 <212> DNA <213> Artificial Sequence <220> <223> tobacco etch virus cleavage site <400> 8 gagaatcttt attttcaggg c 21 <210> 9 <211> 1020 <212> DNA <213> Artificial Sequence <220> <223> codon-optimized trans glutaminase <400> 9 gattctgatg accgcgtaac cccaccagct gaaccactcg atcgcatgcc agacccttat 60 cgcccgtcct atggtcgcgc ggaaacggtc gttaacaact atattcgtaa atggcaacaa 120 gtttacagcc atcgtgacgg tcgtaagcag cagatgacgg aagagcagcg cgagtggctg 180 agctacggtt gcgtcggtgt tacctgggtc aatagcggcc agtacccgac gaaccgtctg 240 gcgtttgcga gcttcgatga ggatcgtttt aagaacgagc tgaagaacgg tcgtccgcgt 300 agcggtgaaa cccgtgcgga gtttgagggc cgtgtcgcaa aagagtcgtt cgatgaagaa 360 aaaggcttcc aacgcgctcg tgaagtggcc agcgtgatga atcgtgcgct ggagaacgca 420 cacgacgaaa gcgcgtacct ggacaatttg aagaaagagc tggcgaatgg caatgatgcg 480 ctgcgtaacg aggatgcgcg cagcccgttt tactctgcct tgcgtaacac cccgtccttc 540 aaagagcgta acggtggtaa tcatgatccg agccgcatga aagcggtgat ctatagcaag 600 catttttgga gcggtcaaga tcgtagcagc agcgcagaca agcgcaaata cggcgacccg 660 gacgcgttcc gccctgcacc gggtaccggc ctggtggata tgagccgcga ccgtaatatc 720 ccgcgtagcc cgacctctcc gggcgaaggt ttcgttaact ttgactacgg ctggttcggt 780 gcccagactg aggcggacgc tgataaaacc gtgtggaccc acggtaatca ctatcacgcc 840 ccgaacggca gcctgggcgc aatgcacgtt tacgaaagca agttccgtaa ttggtccgag 900 ggttacagcg acttcgaccg tggtgcctat gttattacgt ttatcccgaa gtcctggaat 960 acggcaccgg acaaggtgaa acagggttgg ccgctcgagc accaccacca ccaccactga 1020 1020 <210> 10 <211> 729 <212> DNA <213> Artificial Sequence <220> <223> tobacco etch virus protease <400> 10 ggtgaatcgc tgtttaaagg tccgcgcgac tataatccga ttagcagctc tatctgtcac 60 ctgacgaatg agagcgatgg tcacaccacc tctctgtacg gcatcggttt tggtccgttc 120 attatcacga ataaacactt gtttcgccgc aataatggca ccctggtcgt tcagagcctg 180 cacggtgtgt tcaaggttaa agacacgacc actctgcagc aacacttggt ggatggccgt 240 gatatgatta tcatccgtat gcctaaagat ttcccgccgt tcccgcagaa gctgaagttc 300 cgcgaaccac aacgtgaaga gcgtatctgc ctggttacga ccaattttca gaccaagagc 360 atgtccagca tggtgagcga cacgtcctgt accttcccga gcggtgatgg catcttttgg 420 aagcattgga ttcaaactaa ggacggtcag tgcggcagcc cgctggttag cacccgtgac 480 ggcttcattg tcggcattca ttcggcgagc aactttacga ataccaataa ctactttacc 540 agcgtcccga agaatttcat ggagttgctg acgaatcaag aggcacaaca atgggtaagc 600 ggttggcgtc tgaacgccga ctccgtgctg tggggtggtc ataaagtgtt catggttaaa 660 ccagaagagc cgtttcagcc ggtcaaagag gcgacccagc tgatgaatcg tcgtcgtcgc 720 cgttaataa 729 <210> 11 <211> 48 <212> DNA <213> Artificial Sequence <220> <223> Hind III Forward primer <400> 11 gtttgacagc ttatcatcga taagcttaat taatacgact cactatag 48 <210> 12 <211> 48 <212> DNA <213> Artificial Sequence <220> <223> Hind III Reverse primer <400> 12 ctgtgataaa ctaccgcatt aaagcttcaa aaaacccctc aagacccg 48 <210> 13 <211> 49 <212> DNA <213> Artificial Sequence <220> <223> Sal I Forward primer <400> 13 caggagtcgc ataagggaga gcgtcgacaa ttaatacgac tcactatag 49 <210> 14 <211> 47 <212> DNA <213> Artificial Sequence <220> <223> Sal I Reverse primer <400> 14 gaaggctctc aagggcatcg gtcgaccaaa aaacccctca agacccg 47 <110> Korea Research Institute of Bioscience & Biotechnology <120> MASS PRODUCTION METHOD OF RECOMBINANT TRANS GLUTAMINASE <130> p14r16d1756 <160> 14 <170> Kopatentin 2.0 <210> 1 <211> 367 <212> PRT <213> Artificial Sequence <220> <223> maltose binding protein <400> 1 Met Lys Ile Glu Glu Gly Lys Leu Val Ile Trp Ile Asn Gly Asp Lys 1 5 10 15 Gly Tyr Asn Gly Leu Ala Glu Val Gly Lys Lys Phe Glu Lys Asp Thr 20 25 30 Gly Ile Lys Val Thr Val Glu His Pro Asp Lys Leu Glu Glu Lys Phe 35 40 45 Pro Gln Val Ala Ala Thr Gly Asp Gly Pro Asp Ile Ile Phe Trp Ala 50 55 60 His Asp Arg Phe Gly Gly Tyr Ala Gln Ser Gly Leu Leu Ala Glu Ile 65 70 75 80 Thr Pro Asp Lys Ala Phe Gln Asp Lys Leu Tyr Pro Phe Thr Trp Asp 85 90 95 Ala Val Arg Tyr Asn Gly Lys Leu Ile Ala Tyr Pro Ile Ala Val Glu 100 105 110 Ala Leu Ser Leu Ile Tyr Asn Lys Asp Leu Leu Pro Asn Pro Pro Lys 115 120 125 Thr Trp Glu Glu Ile Pro Ala Leu Asp Lys Glu Leu Lys Ala Lys Gly 130 135 140 Lys Ser Ala Leu Met Phe Asn Leu Gln Glu Pro Tyr Phe Thr Trp Pro 145 150 155 160 Leu Ile Ala Ala Asp Gly Gly Tyr Ala Phe Lys Tyr Glu Asn Gly Lys 165 170 175 Tyr Asp Ile Lys Asp Val Gly Val Asp Asn Ala Gly Ala Lys Ala Gly 180 185 190 Leu Thr Phe Leu Val Asp Leu Ile Lys Asn Lys His Met Asn Ala Asp 195 200 205 Thr Asp Tyr Ser Ile Ala Glu Ala Ala Phe Asn Lys Gly Glu Thr Ala 210 215 220 Met Thr Ile Asn Gly Pro Trp Ala Trp Ser Asn Ile Asp Thr Ser Lys 225 230 235 240 Val Asn Tyr Gly Val Thr Val Leu Pro Thr Phe Lys Gly Gln Pro Ser 245 250 255 Lys Pro Phe Val Gly Val Leu Ser Ala Gly Ile Asn Ala Ala Ser Pro 260 265 270 Asn Lys Glu Leu Ala Lys Glu Phe Leu Glu Asn Tyr Leu Leu Thr Asp 275 280 285 Glu Gly Leu Glu Ala Val Asn Lys Asp Lys Pro Leu Gly Ala Val Ala 290 295 300 Leu Lys Ser Tyr Glu Glu Glu Leu Ala Lys Asp Pro Arg Ile Ala Ala 305 310 315 320 Thr Met Glu Asn Ala Gln Lys Gly Glu Ile Met Pro Asn Ile Pro Gln 325 330 335 Met Ser Ala Phe Trp Tyr Ala Val Arg Thr Ala Val Ile Asn Ala Ala 340 345 350 Ser Gly Arg Gln Thr Val Asp Glu Ala Leu Lys Asp Ala Gln Thr 355 360 365 <210> 2 <211> 45 <212> PRT <213> Artificial Sequence <220> <223> proteolytic-resistant sequence <400> 2 Asp Asn Gly Ala Gly Glu Glu Thr Lys Ser Tyr Ala Glu Thr Tyr Arg 1 5 10 15 Leu Thr Ala Asp Asp Val Ala Asn Ile Asn Ala Leu Asn Glu Ser Ala 20 25 30 Pro Ala Ala Ser Ser Ala Gly Ser Ser Phe Arg Ser Pro 35 40 45 <210> 3 <211> 7 <212> PRT <213> Artificial Sequence <220> <223> Tobacco etch virus cleavage site <400> 3 Glu Asn Leu Tyr Phe Gln Gly 1 5 <210> 4 <211> 339 <212> PRT <213> Artificial Sequence <220> <223> Transglutaminase <400> 4 Asp Ser Asp Asp Arg Val Thr Pro Pro Ala Glu Pro Leu Asp Arg Met 1 5 10 15 Pro Asp Pro Tyr Arg Pro Ser Tyr Gly Arg Ala Glu Thr Val Val Asn 20 25 30 Asn Tyr Ile Arg Lys Trp Gln Gln Val Tyr Ser His Arg Asp Gly Arg 35 40 45 Lys Gln Gln Met Thr Glu Glu Gln Arg Glu Trp Leu Ser Tyr Gly Cys 50 55 60 Val Gly Val Thr Trp Val Asn Ser Gly Gln Tyr Pro Thr Asn Arg Leu 65 70 75 80 Ala Phe Ala Ser Phe Asp Glu Asp Arg Phe Lys Asn Glu Leu Lys Asn 85 90 95 Gly Arg Pro Arg Ser Gly Glu Thr Arg Ala Glu Phe Glu Gly Arg Val 100 105 110 Ala Lys Glu Ser Phe Asp Glu Glu Lys Gly Phe Gln Arg Ala Arg Glu 115 120 125 Val Ala Ser Val Met Asn Arg Ala Leu Glu Asn Ala His Asp Glu Ser 130 135 140 Ala Tyr Leu Asp Asn Leu Lys Lys Glu Leu Ala Asn Gly Asn Asp Ala 145 150 155 160 Leu Arg Asn Glu Asp Ala Arg Ser Pro Phe Tyr Ser Ala Leu Arg Asn 165 170 175 Thr Pro Ser Phe Lys Glu Arg Asn Gly Gly Asn His Asp Pro Ser Arg 180 185 190 Met Lys Ala Val Ile Tyr Ser Lys His Phe Trp Ser Gly Gln Asp Arg 195 200 205 Ser Ser Ser Ala Asp Lys Arg Lys Tyr Gly Asp Pro Asp Ala Phe Arg 210 215 220 Pro Ala Pro Gly Thr Gly Leu Val Asp Met Ser Arg Asp Arg Asn Ile 225 230 235 240 Pro Arg Ser Pro Thr Ser Pro Gly Glu Gly Phe Val Asn Phe Asp Tyr 245 250 255 Gly Trp Phe Gly Ala Gln Thr Glu Ala Asp Ala Asp Lys Thr Val Trp 260 265 270 Thr His Gly Asn His Tyr His Ala Pro Asn Gly Ser Leu Gly Ala Met 275 280 285 His Val Tyr Glu Ser Lys Phe Arg Asn Trp Ser Glu Gly Tyr Ser Asp 290 295 300 Phe Asp Arg Gly Ala Tyr Val Ile Thr Phe Ile Pro Lys Ser Trp Asn 305 310 315 320 Thr Ala Pro Asp Lys Val Lys Gln Gly Trp Pro Leu Glu His His His 325 330 335 His His His <210> 5 <211> 241 <212> PRT <213> Artificial Sequence <220> <223> codon-optimized tobacco etch virus protease <400> 5 Gly Glu Ser Leu Phe Lys Gly Pro Arg Asp Tyr Asn Pro Ile Ser Ser 1 5 10 15 Ser Ile Cys His Leu Thr Asn Glu Ser Asp Gly His Thr Thr Ser Leu 20 25 30 Tyr Gly Ile Gly Phe Gly Pro Phe Ile Ile Thr Asn Lys His Leu Phe 35 40 45 Arg Arg Asn Asn Gly Thr Leu Val Val Gln Ser Leu His Gly Val Phe 50 55 60 Lys Val Lys Asp Thr Thr Thr Leu Gln Gln His Leu Val Asp Gly Arg 65 70 75 80 Asp Met Ile Ile Ile Arg Met Pro Lys Asp Phe Pro Pro Phe Pro Gln 85 90 95 Lys Leu Lys Phe Arg Glu Pro Gln Arg Glu Glu Arg Ile Cys Leu Val 100 105 110 Thr Asn Phe Gln Thr Lys Ser Met Ser Ser Met Val Ser Asp Thr 115 120 125 Ser Cys Thr Phe Pro Ser Gly Asp Gly Ile Phe Trp Lys His Trp Ile 130 135 140 Gln Thr Lys Asp Gly Gln Cys Gly Ser Pro Leu Val Ser Thr Arg Asp 145 150 155 160 Gly Phe Ile Val Gly Ile His Ser Ala Ser Asn Phe Thr Asn Thr Asn 165 170 175 Asn Tyr Phe Thr Ser Val Pro Lys Asn Phe Met Glu Leu Leu Thr Asn 180 185 190 Gln Glu Ala Gln Gln Trp Val Ser Gly Trp Arg Leu Asn Ala Asp Ser 195 200 205 Val Leu Trp Gly Gly His Lys Val Phe Met Val Lys Pro Glu Glu Pro 210 215 220 Phe Gln Pro Val Lys Glu Ala Thr Gln Leu Met Asn Arg Arg Arg Arg 225 230 235 240 Arg <210> 6 <211> 1101 <212> DNA <213> Artificial Sequence <220> <223> maltose binding protein <400> 6 atgaaaatcg aagaaggtaa actggtaatc tggattaacg gcgataaagg ctataacggt 60 ctcgctgaag tcggtaagaa attcgagaaa gataccggaa ttaaagtcac cgttgagcat 120 ccggataaac tggaagagaa attcccacag gttgcggcaa ctggcgatgg ccctgacatt 180 atcttctggg cacacgaccg ctttggtggc tacgctcaat ctggcctgtt ggctgaaatc 240 accccggaca aagcgttcca ggacaagctg tatccgttta cctgggatgc cgtacgttac 300 aacggcaagc tgattgctta cccgatcgct gttgaagcgt tatcgctgat ttataacaaa 360 gatctgctgc cgaacccgcc aaaaacctgg gaagagatcc cggcgctgga taaagaactg 420 aaagcgaaag gtaagagcgc gctgatgttc aacctgcaag aaccgtactt cacctggccg 480 ctgattgctg ctgacggggg ttatgcgttc aagtatgaaa acggcaagta cgacattaaa 540 gacgtgggcg tggataacgc tggcgcgaaa gcgggtctga ccttcctggt tgacctgatt 600 aaaaacaaac acatgaatgc agacaccgat tactccatcg cagaagctgc ctttaataaa 660 ggcgaaacag cgatgaccat caacggcccg tgggcatggt ccaacatcga caccagcaaa 720 gtgaattatg gtgtaacggt actgccgacc ttcaagggtc aaccatccaa accgttcgtt 780 ggcgtgctga gcgcaggtat taacgccgcc agtccgaaca aagagctggc aaaagagttc 840 ctcgaaaact atctgctgac tgatgaaggt ctggaagcgg ttaataaaga caaaccgctg 900 ggtgccgtag cgctgaagtc ttacgaggaa gagttggcga aagatccacg tattgccgcc 960 actatggaaa acgcccagaa aggtgaaatc atgccgaaca tcccgcagat gtccgctttc 1020 tggtatgccg tgcgtactgc ggtgatcaac gccgccagcg gtcgtcagac tgtcgatgaa 1080 gccctgaaag acgcgcagac t 1101 <210> 7 <211> 135 <212> DNA <213> Artificial Sequence <220> <223> proteolytic-resistant sequence <400> 7 gacaatggcg cgggggaaga gacgaagtcc tacgccgaaa cctaccgcct cacggcggat 60 gacgtcgcga acatcaacgc gctcaacgaa agcgctccgg ccgcttcgag cgccggcccg 120 tcgttccggt ccccc 135 <210> 8 <211> 21 <212> DNA <213> Artificial Sequence <220> <223> tobacco etch virus cleavage site <400> 8 gagaatcttt attttcaggg c 21 <210> 9 <211> 1020 <212> DNA <213> Artificial Sequence <220> <223> codon-optimized trans glutaminase <400> 9 gattctgatg accgcgtaac cccaccagct gaaccactcg atcgcatgcc agacccttat 60 cgcccgtcct atggtcgcgc ggaaacggtc gttaacaact atattcgtaa atggcaacaa 120 gtttacagcc atcgtgacgg tcgtaagcag cagatgacgg aagagcagcg cgagtggctg 180 agctacggtt gcgtcggtgt tacctgggtc aatagcggcc agtacccgac gaaccgtctg 240 gcgtttgcga gcttcgatga ggatcgtttt aagaacgagc tgaagaacgg tcgtccgcgt 300 agcggtgaaa cccgtgcgga gtttgagggc cgtgtcgcaa aagagtcgtt cgatgaagaa 360 aaaggcttcc aacgcgctcg tgaagtggcc agcgtgatga atcgtgcgct ggagaacgca 420 cacgacgaaa gcgcgtacct ggacaatttg aagaaagagc tggcgaatgg caatgatgcg 480 ctgcgtaacg aggatgcgcg cagcccgttt tactctgcct tgcgtaacac cccgtccttc 540 aaagagcgta acggtggtaa tcatgatccg agccgcatga aagcggtgat ctatagcaag 600 catttttgga gcggtcaaga tcgtagcagc agcgcagaca agcgcaaata cggcgacccg 660 gcgcgttcc gccctgcacc gggtaccggc ctggtggata tgagccgcga ccgtaatatc 720 ccgcgtagcc cgacctctcc gggcgaaggt ttcgttaact ttgactacgg ctggttcggt 780 gcccagactg aggcggacgc tgataaaacc gtgtggaccc acggtaatca ctatcacgcc 840 ccgaacggca gcctgggcgc aatgcacgtt tacgaaagca agttccgtaa ttggtccgag 900 ggttacagcg acttcgaccg tggtgcctat gttattacgt ttatcccgaa gtcctggaat 960 acggcaccgg acaaggtgaa acagggttgg ccgctcgagc accaccacca ccaccactga 1020 1020 <210> 10 <211> 729 <212> DNA <213> Artificial Sequence <220> <223> Tobacco etch virus protease <400> 10 ggtgaatcgc tgtttaaagg tccgcgcgac tataatccga ttagcagctc tatctgtcac 60 ctgacgaatg agagcgatgg tcacaccacc tctctgtacg gcatcggttt tggtccgttc 120 attatcacga ataaacactt gtttcgccgc aataatggca ccctggtcgt tcagagcctg 180 cacggtgtgt tcaaggttaa agacacgacc actctgcagc aacacttggt ggatggccgt 240 gatatgatta tcatccgtat gcctaaagat ttcccgccgt tcccgcagaa gctgaagttc 300 cgcgaaccac aacgtgaaga gcgtatctgc ctggttacga ccaattttca gaccaagagc 360 atgtccagca tggtgagcga cacgtcctgt accttcccga gcggtgatgg catcttttgg 420 aagcattgga ttcaaactaa ggacggtcag tgcggcagcc cgctggttag cacccgtgac 480 ggcttcattg tcggcattca ttcggcgagc aactttacga ataccaataa ctactttacc 540 agcgtcccga agaatttcat ggagttgctg acgaatcaag aggcacaaca atgggtaagc 600 ggttggcgtc tgaacgccga ctccgtgctg tggggtggtc ataaagtgtt catggttaaa 660 ccagaagagc cgtttcagcc ggtcaaagag gcgacccagc tgatgaatcg tcgtcgtcgc 720 cgttaataa 729 <210> 11 <211> 48 <212> DNA <213> Artificial Sequence <220> <223> Hind III Forward primer <400> 11 gtttgacagc ttatcatcga taagcttaat taatacgact cactatag 48 <210> 12 <211> 48 <212> DNA <213> Artificial Sequence <220> <223> Hind III Reverse primer <400> 12 ctgtgataaa ctaccgcatt aaagcttcaa aaaacccctc aagacccg 48 <210> 13 <211> 49 <212> DNA <213> Artificial Sequence <220> <223> Sal I Forward primer <400> 13 caggagtcgc ataagggaga gcgtcgacaa ttaatacgac tcactatag 49 <210> 14 <211> 47 <212> DNA <213> Artificial Sequence <220> <223> Sal I Reverse primer <400> 14 gaaggctctc aagggcatcg gtcgaccaaa aaacccctca agacccg 47
Claims (13)
상기 형질전환된 미생물을 배양하여 활성형의 재조합 트랜스 글루타미나아제를 생산하는 단계; 를 포함하는 재조합 트랜스 글루타미나아제의 생산 방법.A first recombinant expression vector comprising a gene encoding a protein for inducing water-soluble expression, an intracellular activity-inhibiting sequence of a recombinant transglutaminase, a gene encoding a protease recognition site, and a gene encoding a transglutaminase ; And a second recombinant expression vector comprising a gene encoding a protein for inducing water-soluble expression and a gene encoding a proteolytic enzyme; And
Culturing the transformed microorganism to produce an active recombinant transglutaminase; ≪ RTI ID = 0.0 > transglutaminase. ≪ / RTI >
13. The recombinant expression vector for production of recombinant transglutaminase according to claim 12, wherein said recombinant expression vector is as shown in Fig.
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| KR1020140188039A KR20160077750A (en) | 2014-12-24 | 2014-12-24 | Mass production method of recombinant trans glutaminase |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| KR1020140188039A KR20160077750A (en) | 2014-12-24 | 2014-12-24 | Mass production method of recombinant trans glutaminase |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| KR20160077750A true KR20160077750A (en) | 2016-07-04 |
Family
ID=56501156
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020140188039A KR20160077750A (en) | 2014-12-24 | 2014-12-24 | Mass production method of recombinant trans glutaminase |
Country Status (1)
| Country | Link |
|---|---|
| KR (1) | KR20160077750A (en) |
Cited By (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| KR20180089736A (en) * | 2017-02-01 | 2018-08-09 | (주)포바이오코리아 | Expression and purification method of soluble crm197 proteins |
| CN109055339A (en) * | 2018-09-19 | 2018-12-21 | 生工生物工程(上海)股份有限公司 | TEV protease mutant, gene, biomaterial, preparation method, reagent or kit and application |
Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO1996006931A1 (en) | 1994-08-26 | 1996-03-07 | Novo Nordisk A/S | Microbial transglutaminases, their production and use |
| KR20100029566A (en) | 2008-09-08 | 2010-03-17 | 주식회사 엘지생활건강 | Damaged hair recovering composition containing transglutaminase and extract of zanthoxylum bungeanum maxim |
-
2014
- 2014-12-24 KR KR1020140188039A patent/KR20160077750A/en active Search and Examination
Patent Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO1996006931A1 (en) | 1994-08-26 | 1996-03-07 | Novo Nordisk A/S | Microbial transglutaminases, their production and use |
| KR20100029566A (en) | 2008-09-08 | 2010-03-17 | 주식회사 엘지생활건강 | Damaged hair recovering composition containing transglutaminase and extract of zanthoxylum bungeanum maxim |
Cited By (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| KR20180089736A (en) * | 2017-02-01 | 2018-08-09 | (주)포바이오코리아 | Expression and purification method of soluble crm197 proteins |
| WO2018143648A3 (en) * | 2017-02-01 | 2019-04-11 | (주)포바이오코리아 | Method for expressing and isolating soluble protein of crm197 |
| US11124548B2 (en) | 2017-02-01 | 2021-09-21 | Forbiokorea Co., Ltd. | Method for expressing and purifying soluble protein of CRM197 |
| CN109055339A (en) * | 2018-09-19 | 2018-12-21 | 生工生物工程(上海)股份有限公司 | TEV protease mutant, gene, biomaterial, preparation method, reagent or kit and application |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| KR20170085129A (en) | Fusion partners for peptide production | |
| JP2012531198A (en) | Bacterial expression of artificial genes for production of CRM197 and its derivatives | |
| EP3289088B1 (en) | Uncoupling growth and protein production | |
| KR20100086717A (en) | Method for the secretory production of heterologous protein in escherichia coli | |
| KR20160077750A (en) | Mass production method of recombinant trans glutaminase | |
| CN109136209B (en) | Enterokinase light chain mutant and application thereof | |
| KR101677090B1 (en) | Polypeptide for purification of target protein and use thereof | |
| US8759065B2 (en) | Protein and DNA sequence encoding a cold adapted subtilisin-like activity | |
| KR20090039239A (en) | Method for over-expressing human epidermal growth factor as bioactive form in escherichia. coli | |
| US8530217B2 (en) | Processing of peptides and proteins | |
| US7109015B2 (en) | Removal of N-terminal methionine from proteins by engineered methionine aminopeptidase | |
| CN113201074B (en) | PKEK fusion protein and preparation method and application thereof | |
| RU2593172C2 (en) | RECOMBINANT PLASMID DNA pER-TA1 GyrA-AcSer CODING SERINE ACETYLTRANSFERASE CAPABLE OF in vivo ACETYLATION OF N-TERMINAL SERINE DEACETYL-THYMOSIN α1 AND HYBRID PROTEIN CAPABLE OF AUTOCATALYTIC BREAKDOWN TO FORM HUMAN THYMOSIN α1, STRAIN OF Eschrichia coli C3030/pER-TA1GyrA-AcSer PRODUCER OF SAID PROTEINS AND METHOD OF PRODUCING GENETICALLY ENGINEERED HUMAN THYMOSIN | |
| KR101778878B1 (en) | Highly active GABA-producing glutamate decarboxylase from Bacteroides sp. and use thereof | |
| RU2435863C2 (en) | Method for producing protein | |
| KR102014901B1 (en) | Method for producing virus like particle with enhanced purity and stability as nanocarrier for protein | |
| CA2549587A1 (en) | Processing of peptides and proteins | |
| Novikov et al. | The highly efficient expression of the aspartase gene (L-aspartate ammonia-lyase) in Escherichia coli cells | |
| RU2592860C2 (en) | RECOMBINANT PLASMID DNA pER-TB4GyrA-AcSer CODING SERINE ACETYLTRANSFERASE CAPABLE OF in vivo ACETYLATION OF N-TERMINAL SERINE DEACETYL-THYMOSIN β4 AND HYBRID PROTEIN CAPABLE OF AUTOCATALYTIC BREAKDOWN TO FORM HUMAN THYMOSIN β4, STRAIN OF Eschrichia coli C3030/pER-TB4GyrA-AcSer PRODUCER OF SAID PROTEINS AND METHOD OF PRODUCING GENETICALLY ENGINEERED HUMAN THYMOSIN | |
| KR101423713B1 (en) | Vector for Mass Producing of Recombinant Protein Using Barley Ribosome-inactivating Protein and Method for Mass Producing of Protein Using the Same | |
| JP6235815B2 (en) | Method for preparing recombinant human μ-calpain | |
| KR20220162226A (en) | A novel fusion tag system promising soluble expression and purification in Escherichia coli using CBM66 and levan, and their applications | |
| KR20230057844A (en) | Recombinant strain for extracellular secretion of PETase | |
| CN114381471A (en) | Application of auxiliary protein in recombinant protein production and fusion expression system | |
| JP2022544277A (en) | Caspase-2 variant |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A201 | Request for examination | ||
| E902 | Notification of reason for refusal | ||
| AMND | Amendment | ||
| E90F | Notification of reason for final refusal | ||
| AMND | Amendment | ||
| E601 | Decision to refuse application | ||
| E801 | Decision on dismissal of amendment | ||
| AMND | Amendment | ||
| E902 | Notification of reason for refusal | ||
| AMND | Amendment |