KR20120007855A - A microorganism having enhanced l-ornithine productivity and a method of producing l-ornithine using the same - Google Patents
A microorganism having enhanced l-ornithine productivity and a method of producing l-ornithine using the same Download PDFInfo
- Publication number
- KR20120007855A KR20120007855A KR1020100068619A KR20100068619A KR20120007855A KR 20120007855 A KR20120007855 A KR 20120007855A KR 1020100068619 A KR1020100068619 A KR 1020100068619A KR 20100068619 A KR20100068619 A KR 20100068619A KR 20120007855 A KR20120007855 A KR 20120007855A
- Authority
- KR
- South Korea
- Prior art keywords
- microorganism
- ornithine
- activity
- ala
- leu
- Prior art date
Links
Images


Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P13/00—Preparation of nitrogen-containing organic compounds
- C12P13/04—Alpha- or beta- amino acids
- C12P13/10—Citrulline; Arginine; Ornithine
-
- A—HUMAN NECESSITIES
- A23—FOODS OR FOODSTUFFS; TREATMENT THEREOF, NOT COVERED BY OTHER CLASSES
- A23K—FODDER
- A23K20/00—Accessory food factors for animal feeding-stuffs
- A23K20/10—Organic substances
- A23K20/142—Amino acids; Derivatives thereof
-
- A—HUMAN NECESSITIES
- A23—FOODS OR FOODSTUFFS; TREATMENT THEREOF, NOT COVERED BY OTHER CLASSES
- A23L—FOODS, FOODSTUFFS, OR NON-ALCOHOLIC BEVERAGES, NOT COVERED BY SUBCLASSES A21D OR A23B-A23J; THEIR PREPARATION OR TREATMENT, e.g. COOKING, MODIFICATION OF NUTRITIVE QUALITIES, PHYSICAL TREATMENT; PRESERVATION OF FOODS OR FOODSTUFFS, IN GENERAL
- A23L33/00—Modifying nutritive qualities of foods; Dietetic products; Preparation or treatment thereof
- A23L33/10—Modifying nutritive qualities of foods; Dietetic products; Preparation or treatment thereof using additives
- A23L33/17—Amino acids, peptides or proteins
- A23L33/175—Amino acids
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/12—Transferases (2.) transferring phosphorus containing groups, e.g. kinases (2.7)
- C12N9/1205—Phosphotransferases with an alcohol group as acceptor (2.7.1), e.g. protein kinases
Landscapes
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Health & Medical Sciences (AREA)
- Engineering & Computer Science (AREA)
- Zoology (AREA)
- Organic Chemistry (AREA)
- Wood Science & Technology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Genetics & Genomics (AREA)
- Polymers & Plastics (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Food Science & Technology (AREA)
- Molecular Biology (AREA)
- Biotechnology (AREA)
- General Health & Medical Sciences (AREA)
- Microbiology (AREA)
- General Engineering & Computer Science (AREA)
- Biochemistry (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Animal Husbandry (AREA)
- General Chemical & Material Sciences (AREA)
- Mycology (AREA)
- Nutrition Science (AREA)
- Medicinal Chemistry (AREA)
- Biomedical Technology (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
Description
본 발명은 글루코네이트 키나아제(GntK) 활성이 내재적 활성에 비하여 약화된 L-오르니틴 생산 미생물에 관한 것이다. 또한, 본 발명은 상기 L-오르니틴 생산 미생물을 배양하는 단계 및 L-오르니틴을 회수하는 단계를 포함하는 L-오르니틴의 생산방법에 관한 것이다.
The present invention relates to L-ornithine producing microorganisms in which gluconate kinase (GntK) activity is attenuated compared to endogenous activity. The present invention also relates to a method for producing L-ornithine, comprising culturing the L-ornithine producing microorganism and recovering L-ornithine.
오르니틴은 생체 내의 요소 회로 상에 존재하는 아미노산 중의 하나이다. 혈중의 암모니아 농도가 높으면 간 손상을 일으키는데 오르니틴과 시트룰린은 혈중의 암모니아와 결합하여 각각 시트룰린과 아르기닌이 되어 혈중의 암모니아 농도를 감소시켜준다. L-오르니틴은 시트룰린, 프롤린과 같이 암모니아에 대한 해독 기능을 갖고 있어 간장 질환의 치료제로서 사용될 뿐만 아니라, 비만을 예방하는 식품소재로서도 이용되고 있으며, 주름살 개선과 같은 피부미용에의 효과도 밝혀지고 있다.Ornithine is one of the amino acids present on the urea cycle in vivo. High levels of ammonia in the blood cause liver damage. Ornithine and citrulline combine with ammonia in the blood to form citrulline and arginine, respectively, to reduce the level of ammonia in the blood. L-ornithine has a detoxifying function against ammonia, such as citrulline and proline, and is used not only as a therapeutic agent for liver disease but also as a food material for preventing obesity. have.
한편, 코리네박테리움 균주(Corynebacterium), 특히 코리네박테리움 글루타미쿰(Corynebacterium glutamicum)은 L-아미노산 생산에 많이 이용되고 있는 그람 양성의 미생물이다. 이와 같이 코리네박테리움 균주를 이용한 L-아미노산의 제법은 중요하기 때문에 그 제조방법을 개선하기 위해 많은 시도가 행해지고 있다. 구체적으로 오르니틴 생산에 이용되는 미생물로는 자외선 처리 등에 의한 시트룰린 또는 아르기닌 영양 요구성 돌연변이주인 마이크로코커스 글루타미쿠스(Micrococcus glutamicus)(교와하꼬공업(주), 일본, 미국등록특허 제2,988,489호), 브레비박테리움 락토페르멘툼(Brevibacterium lactofermentum), 브레비박테리움 카와사키(B. kawasaki), 브레비박테리움 플라붐(B. flavum)(아지노모토, 일본공고특허 제68-8712호, 제68-8714호 및 제69-26,910호), 브레비박테리움 케토글루타미쿰(B. ketoglutamicum), 아스로박터 파라피네우스(Arthrobacter paraffineus), 코리네박테리움 히드로카보클라스투스(Corynebacterium hydrocarboclastus)(교와하꼬공업(주), 일본, 미국등록특허 제3,574,061호) 등이 알려져 있다. 또한 아르기닌이나 시트룰린 영양 요구성 균주인 아스로박터 시트레우스(A. citreus), 브레비박테리움 락토페르멘툼(B. Lactofermentum), 또는 코리네박테리움 글루타미쿰(C. glutamicum)(아지노모토(주), 유럽공개특허 제0393708호)은 마이코페놀산(mycophenolic acid) 또는 오르니티놀(ornithinol)에 저항성을 갖는 영양 요구성 돌연변이주를 선별하여 오르니틴을 생산하는데 사용한다. 미국등록특허 제3,574,061호에서는 영양 요구성 돌연변이주인 브레비박테리움 케토글루타미쿰 ATCC 21092을 탄화수소, 예를 들어, 파라핀을 주요 탄소원으로 하는 배지에서 배양하여 L-오르니틴을 제조하는 방법을 개시하고 있으나, 이 방법에 의하면 오르니틴의 생산성은 4일간의 배양에서 8.6 mg/ml로 그다지 높지 않았다. 이처럼 L-오르니틴을 생산하는 방법에 관한 연구들이 보고되고 있으나 생산 수율이 높지 않다는 문제점이 있다. 그러나 L-오르니틴의 산업적 대량생산은 여전히 요구되고 있기 때문에 L-오르니틴의 고효율, 고수율적인 생산 방법을 개발할 필요성이 있다. On the other hand, Corynebacterium strain (Corynebacterium), in particular Corynebacterium glutamicum ( Corynebacterium glutamicum ) is a Gram-positive microorganism that is widely used for L-amino acid production. Thus, since the production of L-amino acid using the Corynebacterium strain is important, many attempts have been made to improve the production method thereof. Specifically, microorganisms used for ornithine production include micrococcus glutamicus , which is a citrulline or arginine nutritionally demanding mutant strain by ultraviolet light treatment, etc. (Kyowahako Co., Ltd., Japan, US Patent No. 2,988,489). , Brevibacterium lactofermentum (Brevibacterium lactofermentum), Brevibacterium Kawasaki (B. kawasaki), Brevibacterium Plastic boom (B. flavum) (Ajinomoto, Japan Patent Publication No. 68-8712, No. 68 8714 and No. 69-26910 call), Brevibacterium keto glutamicum (B. ketoglutamicum), bakteo as asparagus pinewooseu (Arthrobacter paraffineus), Corynebacterium hydrocarbyl carbonyl class tooth (Corynebacterium hydrocarboclastus ) (Kyowahako Industry Co., Ltd., Japan, US Patent No. 3,574,061) and the like are known. In addition, arginine or citrulline nutrient strains Asrobacter citruus ( A. citreus ), Brevibacterium lactofermentum ( B. Lactofermentum ), or Corynebacterium glutamicum ( C. glutamicum ) (Ajinomoto ( European Patent No. 0393708) is used to produce ornithine by selecting a nutritionally demanding mutant that is resistant to mycophenolic acid or ornithinol. US Patent No. 3,574,061 discloses a method for producing L-ornithine by culturing the nutritionally demanding mutant Brevibacterium ketoglutamcum ATCC 21092 in a medium containing hydrocarbons, for example, paraffin as the main carbon source. However, according to this method, the productivity of ornithine was not very high at 8.6 mg / ml in 4 days of culture. As such, studies on how to produce L-ornithine have been reported, but there is a problem that the production yield is not high. However, industrial mass production of L-ornithine is still required, and therefore, there is a need to develop a high-efficiency, high-yield production method of L-ornithine.
한편, L-오르니틴은 코리네박테리움 글루타미쿰에서 L-글루탐산으로부터 순환경로(cyclic pathway)를 통해 생합성 되고(Cunin et al., Microbiol Rev 50: 314-352, 1986), 상기 생합성 과정 중 세번째 단계는 argC 유전자에 의해 코딩된 NADPH 의존성 환원효소에 의해 촉매화된다. NADPH의 재생과 특정 대사체의 생합성 간의 직접적인 연관관계는 코리네박테리움 글루타미쿰에서의 L-라이신의 생합성을 통해 밝혀졌다(Wittmann and Heinzle, Microbiol 68:5843-5849, 2002; Marx et al., J Biotechnol 104:185-197, 2003; Ohnishi et al., Microbiol Lett 242:265-274, 2005). NADPH는 코리네박테리움 글루타미쿰에서 라이신을 생산하는 동안 오탄당 인산화 경로(pentose phosphate pathway)를 촉매하는 효소인 글루코오스-6-포스페이트 디하이드로게나제(glucose-6-phosphate dehydrogenase, G6PDH) 및 6-포스포글루코네이트 디하이드로게나제(6-phosphogluconate dehydrogenase, 6PGD)에 의한 산화과정에서 주로 생산된다(Marx et al., J Biotechnol 104:185-197, 2003; Wittmann and Heinzle, Microbiol 68:5843-5849, 2002). 일련의 연구들은 오탄당 인산화 경로로의 탄소 흐름의 전환이, NADPH 공급이 필요한 특정 대사물질의 생합성을 증대하기 위한 대사공학적 타겟이 될 수 있다는 점을 제시하였다. (Mascarenhas et al., Appl Environ Microbiol 57:2995-2999, 1991; Sauer et al., Appl Environ Microbiol 62:3687-3696, 1996; Marx et al., J Biotechnol 104:185-197, 2003; Ohnishi et al., Microbiol Lett 242:265-274, 2005).Meanwhile, L-ornithine is biosynthesized from L-glutamic acid via cyclic pathway in Corynebacterium glutamicum (Cunin et al., Microbiol Rev 50: 314-352, 1986), and during the biosynthesis process The third step is catalyzed by NADPH dependent reductase encoded by the argC gene. A direct link between the regeneration of NADPH and the biosynthesis of certain metabolites was revealed through the biosynthesis of L-lysine in Corynebacterium glutamicum (Wittmann and Heinzle, Microbiol 68: 5843-5849, 2002; Marx et al. , J Biotechnol 104: 185-197, 2003; Ohnishi et al., Microbiol Lett 242: 265-274, 2005). NADPH is an enzyme that catalyzes the pentose phosphate pathway during the production of lysine in Corynebacterium glutamicum (glucose-6-phosphate dehydrogenase (G6PDH) and 6- It is mainly produced during oxidation by 6-phosphogluconate dehydrogenase (6PGD) (Marx et al., J Biotechnol 104: 185-197, 2003; Wittmann and Heinzle, Microbiol 68: 5843-5849 , 2002). A series of studies suggested that the shift of carbon flow to the pentose phosphorylation pathway could be a metabolic target for increasing the biosynthesis of certain metabolites that require NADPH supply. (Mascarenhas et al., Appl Environ Microbiol 57: 2995-2999, 1991; Sauer et al., Appl Environ Microbiol 62: 3687-3696, 1996; Marx et al., J Biotechnol 104: 185-197, 2003; Ohnishi et al., Microbiol Lett 242: 265-274, 2005).
본 발명자들은 이전의 연구에서, 시트룰린과 프롤린 생합성 유전자가 결손되고 아르기닌 억제자(ArgR)에 의한 피드백 제어가 차단된 돌연변이 균주에서 argCJBD의 상동적 발현(homologous expression)으로 L-오르니틴의 생산량이 증가하였음을 보여주었다(Hwang et al., J Microbiol Biotechnol 18:704-710, 2008). 그러나 L-오르니틴의 생합성 경로로 최대의 카본 플럭스를 유지하기 위해서는 argCJBD의 과발현뿐만 아니라, 오탄당 인산화 경로로의 탄소 흐름의 전환에 따른 NADPH가 충분하게 공급되어야 한다.In a previous study, we found that the production of L-ornithine is increased by homologous expression of argCJBD in mutant strains that lack citrulline and proline biosynthesis genes and block feedback control by an arginine inhibitor ( ArgR ). (Hwang et al., J Microbiol Biotechnol 18: 704-710, 2008). However, in order to maintain the maximum carbon flux in the biosynthetic pathway of L-ornithine, NADPH must be supplied sufficiently as well as the overexpression of argCJBD as well as the conversion of carbon flow to the pentose phosphorylation pathway.
이러한 배경 하에서 본 발명자들은 L-오르니틴을 고효율, 고수율로 수득하기 위한 방법을 찾기 위해 예의 노력한 결과, 산화적 오탄당 인산화 경로의 대사적 활성을 유전적으로 변형시켜 오탄당 인산화 경로를 통해 NADPH 생산을 증가시킴으로서 결과적으로 L-오르니틴 생산량을 증가시킬 수 있음을 확인하여 본 발명을 완성하였다.
Against this background, the inventors have made diligent efforts to find a method for obtaining L-ornithine with high efficiency and high yield. As a result, we have genetically modified the metabolic activity of the oxidative pentane phosphorylation pathway to increase NADPH production through the pentose phosphorylation pathway. The present invention was completed by confirming that L-ornithine production can be increased as a result.
본 발명의 하나의 목적은 글루코네이트 키나아제(GntK) 활성이 내재적 활성에 비하여 약화된 L-오르니틴 생산 미생물을 제공하는 것이다. One object of the present invention is to provide L-ornithine producing microorganisms in which gluconate kinase (GntK) activity is attenuated compared to endogenous activity.
본 발명의 또 다른 목적은 1) 상기 L-오르니틴 생산 미생물을 배양하는 단계: 및 2) L-오르니틴을 회수하는 단계를 포함하는 L-오르니틴의 생산방법을 제공하는 것이다.
Still another object of the present invention is to provide a method for producing L-ornithine, comprising 1) culturing the L-ornithine producing microorganism: and 2) recovering L-ornithine.
하나의 양태로서 본 발명은, 글루코네이트 키나아제(gluconate kinase, GntK) 활성이 내재적 활성에 비하여 약화된 L-오르니틴 생산 미생물을 제공하는 것이다.In one embodiment, the present invention provides an L-ornithine producing microorganism in which gluconate kinase (GntK) activity is attenuated compared to endogenous activity.
본 발명에서 용어, "L-오르니틴"은 고등동물의 생체 내의 요소 회로 상에 존재하는 염기성 아미노산으로서, 간에 대해 독성이 강한 암모니아를 무독성의 요소로 변환, 오줌으로 배출함으로써 간을 보호하는 역할을 하는 아미노산을 말한다. L-오르니틴은 성장호르몬을 분비시킴으로써 근육합성을 증가시키고 기초대사를 촉진시켜 비만을 예방하는 식품소재로서 많이 이용되고 있다. 최근에는 주름살 개선과 같은 피부미용 작용 등의 새로운 기능이 있는 것으로 밝혀졌다. L-오르니틴은 L-오르니틴알파케토글루타릴산염(OKG: L-오르니틴과 알파케토글루타릴산이 2:1의 비율로 함유되어 있음)의 형태로 근육증강, 비만예방 및 면역증강소재로서도 폭넓게 이용되고 있다. L-오르니틴은 유럽에서는 간장장애를 개선하는 의약품으로서 이용되고 있고, 일본에서는 L-오르니틴산염의 형태로 식품소재로서 사용되고 있으며, 미국에서는 식품보조제로서 사용되고 있다. In the present invention, the term "L-ornithine" is a basic amino acid present in the urea circuit in a living body of a higher animal, and serves to protect the liver by converting ammonia, which is highly toxic to the liver, into a non-toxic urea and releasing it with urine. Refers to amino acids. L-ornithine is widely used as a food material for preventing obesity by increasing muscle synthesis and promoting basal metabolism by secreting growth hormone. Recently, it has been found that there are new functions such as skin beauty action such as wrinkle improvement. L-ornithine is in the form of L-ornithine alpha ketoglutarate (OKG: L-ornithine and alpha ketoglutaric acid in a 2: 1 ratio) for muscle enhancement, prevention of obesity and immunity. It is widely used as a material. L-ornithine is used as a medicine for improving liver disorders in Europe, in the form of L-ornithine salt in Japan, and as a food supplement in the United States.
본 발명에서 용어, "글루코네이트 키나아제(gluconate kinase, GntK)"란 미생물의 오탄당 인산화 경로의 산화과정 중간체인 6-포스포글루콘산의 생합성을 위한 GntK 경로에 관여하는 효소를 의미한다. 코리네박테리움 속 미생물은 오탄당 인산화 경로의 산화과정의 중간체인 6-포스포글루콘산의 생합성에 있어 2가지 대사경로, 즉 G6PDH 경로 및 GntK 경로를 지니고 있으며(도 1), 이것은 GntK 경로를 통해 적어도 글루코스의 일부를 대사할 수 있다는 것을 의미한다. 이에, 본 발명자들은 상기 GntK 경로의 차단을 통하여 6-포스포글루콘산으로의 탄소흐름을 차단함으로써 오탄당 인산화 경로의 산화과정으로 더 많은 탄소흐름이 진행되어, 6-포스포글루코네이트 디하드로게나제(6PGD)의 활성이 특이적으로 증가하여 NADPH 재생량이 증가할 것으로 예측하고, 결과적으로 L-오르니틴의 생산량이 증가할 수 있는 메커니즘을 제안하였다. 이에, 미생물 내에서 글루코네이트 키나아제(GntK) 활성을 갖는 것으로 추정되는 효소의 활성을 약화시키고자 하였다. 바람직하게 본 발명에서 글루코네이트 키나아제(GntK) 활성을 갖는 효소는 서열번호 1로 나타내는 NCgl2399 또는 서열번호 2로 나타내는 NCgl2905이다.As used herein, the term "gluconate kinase (GntK)" refers to an enzyme involved in the GntK pathway for biosynthesis of 6-phosphogluconic acid, which is an intermediate of the oxidation process of the pentose phosphorylation pathway of microorganisms. Microorganisms of Corynebacterium have two metabolic pathways, 6 G6PDH pathway and GntK pathway, for the biosynthesis of 6-phosphogluconic acid, an intermediate of the oxidation of the pentose phosphorylation pathway (Fig. 1). It means that it can metabolize at least part of glucose. Accordingly, the present inventors further block the carbon flow to 6-phosphogluconic acid through the blocking of the GntK pathway, and thus more carbon flow proceeds through oxidation of the pentose phosphorylation pathway, resulting in 6-phosphogluconate dehydrogena. It was predicted that NADPH regeneration was increased due to the specifically increased activity of (6PGD), and consequently, a mechanism for increasing the production of L-ornithine was proposed. Thus, it was intended to weaken the activity of enzymes that are believed to have gluconate kinase (GntK) activity in microorganisms. Preferably, the enzyme having gluconate kinase (GntK) activity in the present invention is NCgl2399 represented by SEQ ID NO: 1 or NCgl2905 represented by SEQ ID NO: 2.
본 발명에서 용어, "내재적 활성"이란 미생물이 천연의 상태로 가지고 있는 효소의 활성 상태를 의미하는 것으로, 본 발명에서는 미생물이 천연적으로 가지고 있는 GntK의 활성 상태을 의미한다. As used herein, the term "intrinsic activity" refers to an active state of an enzyme possessed by a microorganism in a natural state, and in the present invention, means an active state of GntK naturally possessed by a microorganism.
본 발명에서 "GntK 활성이 내재적 활성에 비해 약화" 되었다는 것은, GntK를 코딩하는 염색체상의 유전자의 결손 또는 돌연변이에 의해 GntK가 정상적으로 작용하지 않아 미생물이 천연의 상태로 가지고 있는 GntK 활성이 약해진 상태를 의미한다. 바람직하게는 NCgl2399 또는 NCgl2905를 암호화하는 유전자 또는 상기 유전자 모두가 결실, 치환 또는 삽입에 의하여 돌연변이 되거나 이의 발현 조절 서열이 돌연변이 되어, GntK가 정상적으로 작용하지 않는 상태를 의미한다. In the present invention, "GntK activity is weakened compared to the intrinsic activity" means that GntK is not normally acted by the deletion or mutation of the gene on the chromosome encoding GntK, so that the GntK activity in which the microorganism has a natural state is weakened. do. Preferably, the gene encoding NCgl2399 or NCgl2905 or all of the genes is mutated by deletion, substitution or insertion, or its expression control sequence is mutated to mean that GntK does not function normally.
본 발명에서는 글루코네이트 키나아제 유전자의 일부를 포함하는 재조합 벡터를 미생물에 도입하여 내재적 글루코네이트 키나아제 유전자를 결손 또는 돌연변이시킴으로써 GntK 내재적 활성이 약화된 미생물을 제조할 수 있다. 상기 유전자의 염색체 내로의 삽입은 당업계에 알려진 임의의 방법, 예를 들면 상동 재조합에 의하여 이루어질 수 있다.In the present invention, by introducing a recombinant vector containing a portion of the gluconate kinase gene into the microorganism, microorganisms with reduced GntK endogenous activity can be prepared by deleting or mutating the endogenous gluconate kinase gene. Insertion of the gene into the chromosome can be by any method known in the art, for example homologous recombination.
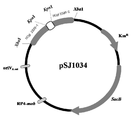
본 발명에서 용어, "재조합 벡터"란 적당한 숙주세포에서 목적 단백질 또는 목적 RNA를 발현할 수 있는 벡터로서, 유전자 삽입물이 발현되도록 작동가능하게 연결된 필수적인 조절요소를 포함하는 유전자 작제물을 말한다. 바람직하게 본 발명에서는 도 2 또는 도 3의 개열지도로 표시되는 재조합 벡터를 사용할 수 있다. As used herein, the term "recombinant vector" refers to a gene construct that is capable of expressing a target protein or RNA of interest in a suitable host cell, and includes a gene construct comprising essential regulatory elements operably linked to express the gene insert. In the present invention, a recombinant vector represented by the cleavage map of FIG. 2 or 3 may be used.
상기 재조합 벡터를 도입하는 모균주로는 L-오르니틴을 생산할 수 있는 미생물이라면 제한없이 사용할 수 있으나, 바람직하게는 코리네박테리움 속 또는 브레비박테리움 속 미생물을 사용할 수 있다. 예를 들면, 코리네박테리움 히드로카보클라스투스(Corynebacterium hydrocarboclastus), 코리네박테리움 글루타미쿰(C. glutamicum), 코리네박테리움 글루타미쿰 ATCC13032, 코리네박테리움 써모아미노게네스(thermoaminogenes) FERM BP-1539, 브레비박테리움 플라붐(Brevibacterium flavum) ATCC 14067, 브레비박테리움 락토페르멘툼(lactofermentum) ATCC 13869, 및 이들로부터 제조된 L-아미노산 생산 돌연변이체 또는 균주, 예를 들면, 코리네박테리움 글루타미쿰 KFCC10881, 코리네박테리움 글루타미쿰 KFCC11001, 코리네박테리움 글루타미쿰 KCCM10770P가 포함될 수 있으며, 바람직하게는, 코리네박테리움 글루타미쿰 SJC8039(C. glutamicum ATCC13032, argF△, argR△)일 수 있다. 본 발명의 구체적인 실시예에서는 코리네박테리움 글루타미쿰 SJC8039(C. glutamicum ATCC13032, argF△, argR△)이 사용되었으나, 이에 제한되는 것은 아니다.As a parent strain introducing the recombinant vector, any microorganism capable of producing L-ornithine may be used without limitation, and preferably, a microorganism of the genus Corynebacterium or Brevibacterium may be used. For example, Corynebacterium hydrocarbyl carbonyl class tooth (Corynebacterium hydrocarboclastus), Corynebacterium glutamicum (C. glutamicum), Corynebacterium glutamicum ATCC13032, Corynebacterium thermoaminogenes amino to Ness (thermoaminogenes) FERM BP-1539, Brevibacterium Plastic boom (Brevibacterium flavum ) ATCC 14067, Brevibacterium lactofermentum ATCC 13869, and L-amino acid producing mutants or strains prepared therefrom such as Corynebacterium glutamicum KFCC10881, Corynebacterium glutami Kum KFCC11001, Corynebacterium glutamicum KCCM10770P may be included, and preferably, Corynebacterium glutamicum SJC8039 ( C. glutamicum ATCC13032, argF Δ, argR Δ). In a specific embodiment of the present invention, Corynebacterium glutamicum SJC8039 ( C. glutamicum ATCC13032, argF △, argR △) was used, but is not limited thereto.
구체적인 일 구현예에 따르면, 코리네박테리움 글루타미쿰 내에서 복제되지 않고 표적 유전자의 마커-프리 결실(marker-free deletion)을 수행하는 기능이 있는 pK18mobsacB 벡터(Schafer et al., Gene 145: 69-73, 1994)를 이용함으로써 유전자 결실을 수행하였다. 즉, 상기 pK18mobsacB 벡터에 NCgl2399 또는 NCgl2905 를 암호화하는 유전자의 일부분과 항생제 마커 및 레반수크레아제 유전자(SacB)를 포함하도록 재조합 벡터를 제작한 후, 당업계에 알려진 임의의 방법으로 상기 재조합 벡터를 미생물에 형질전환 및 염색체 내 상동 재조합시킴으로써, GntK 활성이 약화된 L-오르니틴 생산용 코리네박테리움 미생물을 제작하였다. According to one specific embodiment, the pK18mobsacB vector (Schafer et al., Gene 145: 69), which does not replicate in Corynebacterium glutamicum but has a function of performing marker-free deletion of the target gene -73, 1994) gene deletion was performed. That is, a recombinant vector is prepared to include a portion of a gene encoding NCgl2399 or NCgl2905 in the pK18mobsacB vector and an antibiotic marker and a Levansucrease gene ( SacB ), and then the recombinant vector is transferred to a microorganism by any method known in the art. By transformation and homologous recombination in chromosomes, Corynebacterium microorganisms for L-ornithine production with reduced GntK activity were produced.
본 발명에서는 pK18mobsacB 벡터에서 NCgl2399 를 암호화하는 유전자의 일부를 포함하는 pK18mobsacB 유도체를 제조하여 pSJ1034 로 명명하였으며, 이를 형질전환시켜 NCgl2399 를 암호화하는 유전자가 파괴된 재조합 균주를 SJC8245 라고 명명하였다. 또한, pK18mobsacB 벡터에서 NCgl2905를 암호화하는 유전자의 일부를 포함하는 pK18mobsacB 유도체를 제조하여 pSJ1035 로 명명하였으며, 이를 상기 SJC8245 균주 내에 형질전환시켜 NCgl2905를 암호화하는 유전자가 추가로 파괴된 재조합 균주를 SJC8399로 명명하였다.In the present invention, a pK18mobsacB derivative including a part of the gene encoding NCgl2399 in the pK18mobsacB vector was prepared and named pSJ1034. The recombinant strain whose gene encoding NCgl2399 was destroyed by transforming it was named SJC8245. In addition, a pK18mobsacB derivative including a part of the gene encoding NCgl2905 in the pK18mobsacB vector was prepared and named as pSJ1035, which was transformed into the SJC8245 strain, and the recombinant strain which further destroyed the gene encoding NCgl2905 was named SJC8399. .
구체적인 일 구현예에 따르면, NCgl2399 유전자가 결손된 코리네박테리움 글루타미쿰 균주인 SJC8245를 배양한 결과, GntK의 특이적 활성이 41% 정도 남아있었고, NADPH의 농도는 23.5% 증가하였으며, L-오르니틴의 생산량은 24.8% 증가함을 확인하였다. 또한 상기 SJC8245 균주 내에서 글루코네이트 키나아제 활성을 갖는 효소의 또 다른 유전자로 추정되는 NCgl2905 유전자를 추가적으로 파괴한 코리네박테리움 글루타미쿰 균주인 SJC8399를 배양한 결과, SJC8245에 비하여 상기 GntK 경로의 활성이 더욱 감소되는 것을 확인하였고, NADPH의 농도는 51.8% 증가하였으며, L-오르니틴의 생산량은 63.8% 증가함을 확인하였다(표 3 및 4). 이러한 결과는 본 발명의 미생물에 의해 L-오르니틴 생산량이 증가되었음을 보여주며, 또한 과량의 NADPH가 L-오르니틴 생산량의 증가에 영향을 주는 요인임을 보여주었다.According to a specific embodiment, the culture of SJC8245, a Corynebacterium glutamicum strain lacking the NCgl2399 gene, resulted in a 41% specific activity of GntK and a 23.5% increase in NADPH concentration. The production of ornithine was found to increase by 24.8%. In addition, as a result of culturing SJC8399, a Corynebacterium glutamicum strain that additionally destroyed the NCgl2905 gene, which is assumed to be another gene of the enzyme having gluconate kinase activity in the SJC8245 strain, the activity of the GntK pathway was higher than that of SJC8245. It was confirmed that the decrease further, the concentration of NADPH was increased by 51.8%, the production of L- ornithine was increased by 63.8% (Tables 3 and 4). These results show that the L-ornithine production was increased by the microorganism of the present invention, and also showed that excessive NADPH is a factor influencing the increase of L-ornithine production.
결과적으로 본 발명의 미생물은 GntK의 활성이 약화되고, 6GDP 의 활성이 특이적으로 증가함으로써 오탄당 인산화 경로의 산화과정이 증대되었으며, 세포 내 NADPH의 농도가 유의적으로 증가 되었다(표 4). 코리네박테리움 속 미생물에서의 L-오르니틴의 생합성 경로는 NADPH 의존성 환원효소에 의해 촉매화되므로, 결과적으로 본 발명의 미생물이 L-오르니틴 생산량을 증가시킴을 알 수 있었다.As a result, the microorganism of the present invention attenuated the GntK activity, specifically increased the activity of 6GDP, increased the oxidation process of the pentose phosphorylation pathway, and significantly increased the concentration of NADPH in the cells (Table 4). Since the biosynthetic pathway of L-ornithine in the microorganism of the genus Corynebacterium is catalyzed by NADPH dependent reductase, it was found that the microorganism of the present invention increased L-ornithine production.
또한 본 발명에서는 L-오르니틴 제조량을 더욱 증가시키기 위해, L-오르니틴의 생합성에 관련된 공지의 효소들의 활성을 추가로 조절할 수 있다. 바람직하게 본 발명에서는 추가로 argCJBD 오페론 활성을 조절하여 argCJBD 유전자를 과발현시켜 L-오르니틴 생산량을 증대시킬 수 있다(Hwang et al., J Microbiol Biotechnol 18:704-710, 2008).
In addition, in the present invention, in order to further increase the L-ornithine production amount, the activity of known enzymes related to the biosynthesis of L-ornithine can be further controlled. Preferably, the present invention may further increase the L-ornithine production by overexpressing the argCJBD gene by regulating argCJBD operon activity (Hwang et al., J Microbiol Biotechnol 18: 704-710, 2008).
또 하나의 양태로서 본 발명은, 1) 상기 L-오르니틴 생산 미생물을 배양하는 단계; 및 2) L-오르니틴을 회수하는 단계를 포함하는 L-오르니틴의 생산방법을 제공하는 것이다. As another aspect, the present invention, 1) culturing the L- ornithine producing microorganism; And 2) to provide a method for producing L- ornithine comprising the step of recovering L- ornithine.
본 발명에서 용어 "배양"은 미생물을 적당히 인공적으로 조절한 환경조건에서 생육시키는 것을 의미한다. 본 발명에서 코리네박테리아를 이용하여 L-오르니틴을 배양하는 방법은 당업계에 널리 알려져 있는 방법을 이용하여 수행할 수 있다. 구체적으로 상기 배양은 배치 공정 또는 주입 배치 또는 반복 주입 배치 공정(fed batch or repeated fed batch process)에서 연속식으로 배양할 수 있으나, 이에 제한되는 것은 아니다.The term "culture" in the present invention means to grow microorganisms under environmental conditions that are appropriately artificially controlled. In the present invention, the method of culturing L-ornithine using Corynebacteria may be performed using a method well known in the art. Specifically, the culture may be continuously cultured in a batch process or in a fed batch or repeated fed batch process, but is not limited thereto.
배양에 사용되는 배지는 적절한 방식으로 특정 균주의 요건을 충족해야 한다. 코리네박테리아 균주에 대한 배양 배지는 공지되어 있다 (예를 들면, Manual of Methods for General Bacteriology. American Society for Bacteriology. Washington D.C., USA, 1981). 사용될 수 있는 당원으로는 글루코즈, 사카로즈, 락토즈, 프락토즈, 말토즈, 전분, 셀룰로즈와 같은 당 및 탄수화물, 대두유, 해바라기유, 피마자유, 코코넛유 등과 같은 오일 및 지방, 팔미트산, 스테아린산, 리놀레산과 같은 지방산, 글리세롤, 에탄올과 같은 알코올, 아세트산과 같은 유기산이 포함된다. 이들 물질은 개별적으로 또는 혼합물로서 사용될 수 있다. 사용될 수 있는 질소원으로는 펩톤, 효모 추출물, 육즙, 맥아 추출물, 옥수수 침지액, 대두밀 및 요소 또는 무기 화합물, 예를 들면 황산암모늄, 염화암모늄, 인산암모늄, 탄산암모늄 및 질산암모늄이 포함된다. 질소원 또한 개별적으로 또는 혼합물로서 사용할 수 있다. 사용될 수 있는 인원으로는 인산이수소칼륨 또는 인산수소이칼륨 또는 상응하는 나트륨-함유 염이 포함된다. 또한, 배양 배지는 성장에 필요한 황산마그네슘 또는 황산철과 같은 금속염을 함유해야 한다. 마지막으로, 상기 물질에 더하여 아미노산 및 비타민과 같은 필수 성장 물질이 사용될 수 있다. 또한, 배양 배지에 적절한 전구체들이 사용될 수 있다. 상기된 원료들은 배양과정에서 배양물에 적절한 방식에 의해 회분식으로 또는 연속식으로 첨가될 수 있다. The medium used for cultivation must meet the requirements of the particular strain in an appropriate manner. Culture media for Corynebacteria strains are known (eg, Manual of Methods for General Bacteriology. American Society for Bacteriology.Washington D.C., USA, 1981). Sugar sources that can be used include sugars and carbohydrates such as glucose, saccharose, lactose, fructose, maltose, starch, cellulose, oils and fats such as soybean oil, sunflower oil, castor oil, coconut oil, palmitic acid, stearic acid Fatty acids such as linoleic acid, alcohols such as glycerol, ethanol, and organic acids such as acetic acid. These materials can be used individually or as a mixture. Nitrogen sources that can be used include peptone, yeast extract, gravy, malt extract, corn steep liquor, soybean wheat and urea or inorganic compounds such as ammonium sulfate, ammonium chloride, ammonium phosphate, ammonium carbonate and ammonium nitrate. Nitrogen sources can also be used individually or as a mixture. Personnel that may be used include potassium dihydrogen phosphate or dipotassium hydrogen phosphate or the corresponding sodium-containing salts. In addition, the culture medium should contain metal salts such as magnesium sulfate or iron sulfate required for growth. Finally, in addition to the above substances, essential growth substances such as amino acids and vitamins can be used. In addition, suitable precursors to the culture medium may be used. The above-mentioned raw materials may be added batchwise or continuously by a suitable method for the culture during the culture.
수산화나트륨, 수산화칼륨, 암모니아와 같은 기초 화합물 또는 인산 또는 황산과 같은 산 화합물을 적절한 방식으로 사용하여 배양물의 pH를 조절할 수 있다. 또한, 지방산 폴리글리콜 에스테르와 같은 소포제를 사용하여 기포 생성을 억제할 수 있다. 호기 상태를 유지하기 위해 배양물 내로 산소 또는 산소-함유 기체 (예, 공기)를 주입한다. 배양물의 온도는 보통 20℃ 내지 45℃, 바람직하게는 25℃ 내지 40℃이다. 배양은 L-오르니틴의 생성량이 최대로 얻어질 때까지 계속한다. 이러한 목적으로 보통 10 내지 160 시간에서 달성된다. L-오르니틴은 배양 배지 중으로 배출되거나, 세포 중에 포함되어 있을 수 있다. Basic compounds such as sodium hydroxide, potassium hydroxide, ammonia or acid compounds such as phosphoric acid or sulfuric acid can be used in an appropriate manner to adjust the pH of the culture. In addition, antifoaming agents such as fatty acid polyglycol esters can be used to inhibit bubble generation. Inject oxygen or oxygen-containing gas (eg, air) into the culture to maintain aerobic conditions. The temperature of the culture is usually 20 ° C to 45 ° C, preferably 25 ° C to 40 ° C. Incubation is continued until the maximum amount of L-ornithine production is obtained. For this purpose it is usually achieved in 10 to 160 hours. L-ornithine may be excreted in culture medium or contained in cells.
본 발명의 L-오르니틴을 생산하는 방법은, 세포 또는 배양 배지로부터 오르니틴을 회수하는 단계를 포함한다. 세포 또는 배양 배지로부터 L-오르니틴을 회수하는 방법은 당업계에 널리 알려져 있다. 상기 L-오르니틴 회수 방법에는, 여과, 음이온 교환 크로마토그래피, 결정화 및 HPLC 등이 사용될 수 있으나, 이들 예에 한정되는 것은 아니다.
The method for producing L-ornithine of the present invention includes recovering ornithine from cells or culture medium. Methods of recovering L-ornithine from cells or culture media are well known in the art. Filtration, anion exchange chromatography, crystallization and HPLC may be used for the L-ornithine recovery method, but is not limited to these examples.
본 발명은 글루코네이트 키나아제(GntK) 활성이 내재적 활성에 비하여 약화된 L-오르니틴 생산 미생물을 제공함에 따라 고수율 및 고효율로 L-오르니틴을 생산할 수 있으며, 이와 같이 생성된 L-오르니틴은 인간 의약 및 약학산업, 식품산업, 사료산업, 화학산업에 유용하게 사용할 수 있다.
The present invention is capable of producing L-ornithine with high yield and high efficiency as the gluconate kinase (GntK) activity provides an attenuated L-ornithine producing microorganism compared to intrinsic activity, and thus the L-ornithine It can be usefully used in human medicine and pharmaceutical industry, food industry, feed industry and chemical industry.
도 1은 코리네박테리움 글루타미쿰의 오탄당 인산화 경로 대사과정의 개략도이다.
도 2는 코리네박테리움 염색체 내 NCgl2399 유전자의 마커-프리 결손용 벡터 pSJ1034를 나타내는 도이다.
도 3은 코리네박테리움 염색체 내 NCgl2905 유전자의 마커-프리 결손용 벡터 pSJ1035를 나타내는 도이다.1 is a schematic diagram of the metabolism of the pentose phosphorylation pathway of Corynebacterium glutamicum.
2 is a diagram showing a marker-free deletion vector pSJ1034 of the NCgl2399 gene in the Corynebacterium chromosome.
3 is a diagram showing a marker-free deletion vector pSJ1035 of the NCgl2905 gene in the Corynebacterium chromosome.
이하 본 발명을 하기의 실시예에 의해 상세히 설명한다. 단, 하기 실시예는 본 발명을 예시하는 것일 뿐, 본 발명의 내용이 하기 실시예에 의해 한정되는 것은 아니다. Hereinafter, the present invention will be described in detail by the following examples. However, the following examples are illustrative of the present invention, and the contents of the present invention are not limited by the following examples.
실시예Example 1: 부위-특이적 유전자가 결실된 변이주의 제조 1: Preparation of Mutant Wines Deleting Site-Specific Genes
본 발명에서는 모균주로 코리네박테리움 글루타미쿰 SJC8039(C. glutamicum ATCC13032, argF△, argR△) (Hwang et al., J Microbiol Biotechnol 18:704-710, 2008)을 사용하였다. 상기 모균주로부터 본 발명의 재조합 균주 SJC8399와 SJC8245를 제조하기 위하여 프라이머를 제작하였다. 우선, NCgl2905 또는 NCgl2399를 암호화하는 유전자의 불활성화 단편을 제작하기 위해 NCgl2399(서열번호 1) 및 NCgl2905(서열번호 2)의 서열을 근거로 각각 프라이머 2399F1, 2399F2, 2399R1, 2399R2, 2905F1, 2905F2, 2905R1, 2905R2를 제작하였다(표 1). 밑줄 친 서열은 괄호 안에 나타낸 바와 같은 제한효소에 대한 제한위치를 나타내며, 대문자는 미생물 유전자의 서열을 의미한다.
In the present invention, the parent strain Corynebacterium glutamicum SJC8039 ( C. glutamicum ATCC13032, argF △, argR △) (Hwang et al., J Microbiol Biotechnol 18: 704-710, 2008) was used. Primers were prepared to prepare recombinant strains SJC8399 and SJC8245 of the present invention from the parent strain. First, primers 2399F1, 2399F2, 2399R1, 2399R2, 2905F1, 2905F2 and 2905R1, respectively, based on the sequences of NCgl2399 (SEQ ID NO: 1) and NCgl2905 (SEQ ID NO: 2) to construct inactivated fragments of genes encoding NCgl2905 or NCgl2399. , 2905R2 was produced (Table 1). Underlined sequences indicate restriction positions for restriction enzymes as shown in parentheses, and uppercase letters indicate sequences of microbial genes.
부위-특이적 유전자 결실(site specific gene disruption)을 위하여 코리네박테리움 글루타미쿰 내에서 복제되지 않고 표적 유전자의 마커-프리 결실(marker-free deletion)을 수행하는 기능이 있는 pK18mobsacB 벡터(Schafer et al., Gene 145: 69-73, 1994)를 사용하여 수행하였으며, 유전자가 파괴된 변이주(gene-disrupted mutant strain)를 만들기 위하여 NCgl2399와 NCgl2905의 개방해독틀(open reading frame)이 내부적으로 결실된 pK18mobsacB 유도체를 만들었다. 상기 pK18mobsacB 유도체는 각각 pSJ1034와 pSJ1035로 명명하였다. PK18mobsacB vector (Schafer et al.) With the ability to perform marker-free deletion of the target gene without replication in Corynebacterium glutamicum for site specific gene disruption al., Gene 145: 69-73, 1994), in which the open reading frames of NCgl2399 and NCgl2905 were internally deleted to create gene-disrupted mutant strains. pK18mobsacB derivatives were made. The pK18mobsacB derivatives were named pSJ1034 and pSJ1035, respectively.
pSJ1034는 2,267 bp짜리 NCgl2399의 XbaI 말단을 포함하는 pK18mobsacB 유도체로 NCgl2399의 내부적 유전자 소실을 포함한다. 상기 NCgl2399 내부적 유전자 소실은 코리네박테리움 글루타미쿰 SJC8039 게놈 DNA를 주형으로 2399F1-2399R1 프라이머와 2399F2-2399R2 프라이머 쌍으로 교차 PCR을 하여 생성된 것이다. pSJ1035는 2,819 bp짜리 NCgl2905의 XbaI 말단이 있는 pK18mobsacB 유도체로 NCgl2905의 내부적 유전자 소실을 포함한다. 상기 NCgl2905 내부적 유전자 소실은 코리네박테리움 글루타미쿰 SJC8039 게놈 DNA를 주형으로 2905F1-2905R1 프라이머와 2905F2-2905R2 프라이머 쌍으로 교차 PCR을 하여 생성된 것이다. pSJ1034 is a pK18mobsacB derivative containing the Xba I terminus of 2,267 bp NCgl2399, which includes the internal gene loss of NCgl2399. The NCgl2399 internal gene loss was generated by cross-PCR using 2399F1-2399R1 primer and 2399F2-2399R2 primer pair as a template of Corynebacterium glutamicum SJC8039 genomic DNA. pSJ1035 is a pK18mobsacB derivative with the Xba I terminus of 2,819 bp NCgl2905, which includes the internal gene loss of NCgl2905. The NCgl2905 internal gene loss was generated by cross PCR with 2905F1-2905R1 primer pair and 2905F2-2905R2 primer pair as a template of Corynebacterium glutamicum SJC8039 genomic DNA.
상기의 재조합 플라스미드를 야생의 코리네박테리움 글루타미쿰에 전기천공법으로 형질전환시키고(van der Rest et al., Appl Microbiol Biotechnol 52:541-545, 1999), 상기 플라스미드는 1차 재조합(교차)에 의해 염색체에 도입하였다. 그 후 10%의 수크로오스를 포함하는 고체평판배지에서 2차 재조합(교차)을 통해 염색체로부터 플라스미드를 절제(excision)하였다. The recombinant plasmid was transformed into wild Corynebacterium glutamicum by electroporation (van der Rest et al., Appl Microbiol Biotechnol 52: 541-545, 1999), and the plasmid was first recombinant (crossed out). ) Into the chromosome. The plasmid was then excised from the chromosome via secondary recombination (crossing) in a solid plate medium containing 10% sucrose.
상기 2차 재조합이 완료된 코리네박테리움 글루타미쿰 형질전환주들을 대상으로 각 유전자 특이적 프라이머(gene-specific primer) 쌍인 2905F1-2905R2 및 2399F1-2399R2를 사용한 다이아그너스틱 PCR(diagnostic PCR)을 통해 각각 NCgl2905 및 NCgl2399 유전자의 결실을 확인하였다. 상기 NCgl2905, NCgl2399의 개방해독틀 DNA서열의 Genbank accession number는 NC_003450이다.
Corynebacterium glutamicum transformants having completed the second recombination through diagnostic PCR using pairs of gene-specific primers 2905F1-2905R2 and 2399F1-2399R2 The deletion of the NCgl2905 and NCgl2399 genes, respectively, was confirmed. The Genbank accession numbers of the open reading frame DNA sequences of NCgl2905 and NCgl2399 are NC_003450.
실시예Example 2: 2: NCgl2905NCgl2905 및/또는 And / or NCgl2399NCgl2399 유전자가 결실된 Gene deleted 코리네박테리움Corynebacterium 글루타미쿰의Glutamicum 배양 culture
L-오르니틴 생산균주인 코리네박테리움 글루타미쿰 SJC8399와 코리네박테리움 SJC8245를 L-오르니틴 생산을 위해 아래와 같은 방법으로 배양하였다. 종배지로 Recovery Glucose 배지(80g BHI, 20g 글루코오스, 60g 소르비톨/L)을 사용하여 24시간 배양하였다. 생산배지는 MMY 배지[0.8 g KH2PO4, 10 g (NH4)2SO4, 1 g MgSO4ㆍ7H2O, 1.2 g Na2HPO4, 20 mg MnSO4ㆍH2O, 20 mg FeSO4ㆍ7H2O, 10 mg,ZnSO4ㆍ7H2O, 10 g yeast extract, 및 60 g 글루코오스/L, pH 7.0]를 사용하였다. 종배지를 함유하는 100㎖ 배플 플라스크에 코리네박테리움 글루타미쿰 모균주 SJC8039와 본 발명의 균주 코리네박테리움 글루타미쿰 SJC8399 및 코리네박테리움 글루타미쿰 SJC8245를 각각 접종하고 진탕배양(200rpm)하였다. 종배지에서 배양된 상기 글루타미쿰을 분리하여 수확한 후 상기 글루타미쿰은 600nm에서 흡광도가 0.4~0.5일 때까지 상기 생산배지 10㎖을 함유하는 100㎖ 배플 플라스크에서 재현탁하였다. 그 후, 10㎖ 당 0.2 g의 멸균된 CaCO3를 첨가하고, 필요한 경우 카나마이신(kanamycin)은 50 μg/㎖ 의 최종농도까지 첨가하였다. 상기의 배양은 30℃의 200rpm 회전식 교반기에서 수행하였다.
Corynebacterium glutamicum SJC8399 and Corynebacterium SJC8245, L-ornithine producing strains, were cultured in the following manner for L-ornithine production. The seed medium was incubated for 24 hours using Recovery Glucose medium (80 g BHI, 20 g glucose, 60 g sorbitol / L). The production medium was MMY medium [0.8 g KH 2 PO 4 , 10 g (NH 4 ) 2 SO 4, 1 g MgSO 4 ㆍ 7H 2 O, 1.2 g Na 2 HPO 4 , 20 mg MnSO 4 ㆍ H 2 O, 20 mg FeSO 4 7H 2 O, 10 mg, ZnSO 4 7H 2 O, 10 g yeast extract, and 60 g glucose / L, pH 7.0] were used. A 100 ml baffle flask containing a seed medium was inoculated with Corynebacterium glutamicum mother strain SJC8039 and the strain Corynebacterium glutamicum SJC8399 and Corynebacterium glutamicum SJC8245, respectively, and shaken (200 rpm). ). After separating and harvesting the glutamicum cultured in the seed medium, the glutamicum was resuspended in a 100 ml baffle flask containing 10 ml of the production medium until the absorbance at 600 nm was 0.4-0.5. Thereafter, 0.2 g of sterile CaCO 3 per 10 ml was added, and kanamycin was added to a final concentration of 50 μg / ml if necessary. The incubation was carried out in a 200 rpm rotary stirrer at 30 ° C.
실시예Example 3: 효소 활성 및 이에 따른 L-오르니틴 생산량 측정 3: Determination of enzyme activity and thus L-ornithine production
상기 생산배지에서 배양된 코리네박테리움 글루타미쿰은 지수 성장기(exponetial phase)동안 원심분리법을 사용하여 의해 수확하였고 100mM Tris/HCl buffer(pH 7.5)으로 재현탁 하였다. 세포벽은 유리 비드(glass bead)를 사용하여 파괴하였고 세포 추출물을 얻기 위해 원심분리 하였다. 이 모든 처리는 4℃에서 이루어졌으며, 상층액은 특이적 효소 활성 분석을 위하여 채취하였다. Corynebacterium glutamicum cultured in the production medium was harvested by centrifugation during the expontial phase and resuspended in 100 mM Tris / HCl buffer (pH 7.5). Cell walls were broken down using glass beads and centrifuged to obtain cell extracts. All these treatments were performed at 4 ° C., and the supernatants were harvested for specific enzyme activity analysis.
효소분석을 위한 단백질의 양은 브래드포드법에 의해 결정하였다(Bradford 1976). G6PDH와 6PGD의 활성은 340nm에서 NADPH 분광학적 형성을 통해 측정하였다(Frunzke et al., Mol Microbiol 67:305-322, 2008). GntK 활성은 6PGD의 coupled enzymatic assay으로 측정하였다(Frunzke et al., Mol Microbiol 67:305-322, 2008). 600nm에서 분광법으로 셀 성장률을 측정하였고, L-오르니틴 농도는 닌하이드린을 사용한 비색법으로 결정하였다(Chinard et al., J Biol Chem 199: 91-95, 1952). 건조균체량은 600nm에서의 흡광도 값 1은 1L당 건조중량 0.3g에 해당한다는 점을 기초로 측정하였다. NADPH의 농도는 EnzyChromTMNADP+/NADPH 분석 키트를 사용한 enzymatic cycling reaction으로 결정하였다(BioAssay Systems, CA, USA). The amount of protein for enzymatic analysis was determined by the Bradford method (Bradford 1976). The activity of G6PDH and 6PGD was measured by NADPH spectroscopic formation at 340 nm (Frunzke et al., Mol Microbiol 67: 305-322, 2008). GntK activity was determined by coupled enzymatic assay of 6PGD (Frunzke et al., Mol Microbiol 67: 305-322, 2008). Cell growth rate was measured spectroscopically at 600 nm, and L-ornithine concentration was determined by colorimetry using ninhydrin (Chinard et al., J Biol Chem 199: 91-95, 1952). Dry cell mass was measured based on the absorbance value of 1 at 600 nm corresponds to a dry weight of 0.3g per 1L. The concentration of NADPH was determined by an enzymatic cycling reaction using the EnzyChrom ™ NADP + / NADPH Assay Kit (BioAssay Systems, CA, USA).
실험결과, 본 발명의 NCgl2399를 암호화하는 유전자가 결손된 코리네박테리움 글루타미쿰 균주인 SJC8245는 GntK의 특이적 활성이 41% 정도 남아있게 되는 것으로 나타났다(0.51 vs 0.21 U mg/protein). 또한 상기 SJC8245 균주 내에서 역시 글루코네이트 키나아제 (EC:2.7.1.12) 활성을 갖는 효소의 또 다른 유전자로 추정되는 NCgl2905를 추가적으로 파괴함으로써 SJC8399로 명명되는 코리네박테리움 글루타미쿰을 얻었고, 상기 SJC8399의 경우 상기 SJC8245에 비하여 상기 GntK 경로의 활성이 더욱 감소되는 것을 확인하였다(0.11 U mg/protein). As a result, SJC8245, a Corynebacterium glutamicum strain lacking the gene encoding NCgl2399 of the present invention, was found to retain 41% of GntK specific activity (0.51 vs 0.21 U mg / protein). In addition, by additionally destroying NCgl2905, another gene of the enzyme also possessing gluconate kinase (EC: 2.7.1.12) activity in the SJC8245 strain, Corynebacterium glutamicum named SJC8399 was obtained. It was confirmed that the activity of the GntK pathway was further reduced compared to the SJC8245 (0.11 U mg / protein).
한편, NCgl2399의 불활성화의 경우 NADPH의 농도는 23.5% 증가하였고, L-오르니틴의 생산량은 24.8% 증가하였다. 또한 NCgl2399 및 NCgl2905를 모두 불활성화시킨 경우에는 NADPH의 농도는 51.8% 증가하였고, L-오르니틴의 생산량은 63.8% 증가하였다. 이러한 결과는 과량의 NADPH는 코리네박테리움 글루타미쿰 내의 L-오르니틴의 농도를 증가시키기 위해 요구된다는 것을 나타낸다. In the case of inactivation of NCgl2399, NADPH concentration was increased by 23.5%, and L-ornithine production was increased by 24.8%. In addition, when both NCgl2399 and NCgl2905 were inactivated, NADPH concentration was increased by 51.8%, and L-ornithine production was increased by 63.8%. These results indicate that excess NADPH is required to increase the concentration of L-ornithine in Corynebacterium glutamicum.
표 2는 코리네박테리움 글루타미쿰의 성장률을, 표 3은 NADPH와 L-오르니틴의 축적농도를 나타내며, 표 4는 중심탄소대사과정의 핵심 효소의 특이적 활성도를 나타낸다. 표 2와 표 3에서 보는 바와 같이 SJC8245(NCgl2399이 불활성화된 것)과 SJC8399(NCgl2399 및 NCgl2905 모두가 불활성화된 것)는 SJC8039(모균주)에 비교하여, 성장률은 거의 비슷하거나 다소 감소하지만, 세포내 NADPH 농도와 L-오르니틴의 생산량은 현격히 증가하였다. Table 2 shows the growth rate of Corynebacterium glutamicum, Table 3 shows the accumulation concentration of NADPH and L-ornithine, and Table 4 shows the specific activity of the core enzymes of the core carbon metabolism process. As shown in Tables 2 and 3, SJC8245 (which NCgl2399 is inactivated) and SJC8399 (which both NCgl2399 and NCgl2905 are inactivated) have a similar or slightly reduced growth rate compared to SJC8039 (parent strain). Intracellular NADPH levels and L-ornithine production increased significantly.
구체적으로 G6PDH와 6PGD의 특이적 활성을 비교하면, 표 4에서 보는 것과 같이 6PGD의 특이적 활성은 SJC8039에 비해 SJC8245 및 SJC8399 내에서 현격히 증가하였다. 그러나 G6PDH 활성은 거의 변화가 없다는 것이 관측되었다.
Specifically, when comparing the specific activities of G6PDH and 6PGD, as shown in Table 4, the specific activity of 6PGD was significantly increased in SJC8245 and SJC8399 compared to SJC8039. However, little change was observed in G6PDH activity.
상기의 값은 적어도 3개의 독립적인 배양에서 얻어진 결과를 기초로 한 평균값을 나타내며 모든 경우에 있어서 표준편차가 10% 이하였다. The above values represent mean values based on the results obtained in at least three independent cultures and in all cases the standard deviation was less than 10%.
<110> SANGJI UNIVERSITY INDUSTRY ACADEMIC COOPERATION FOUNDATION <120> A microorganism having enhanced L-ornithine productivity and a method of producing L-ornithine using the same <130> PA100296/KR <160> 10 <170> KopatentIn 1.71 <210> 1 <211> 167 <212> PRT <213> Corynebacterium glutamicum ATCC 13032 <220> <221> PEPTIDE <222> (1)..(167) <223> NCgl2399 gluconate kinase <400> 1 Met Ser Ala Ala Glu Gly Leu His Ile Val Val Met Gly Val Ser Gly 1 5 10 15 Cys Gly Lys Ser Ser Val Gly Lys Ala Leu Ala Ala Glu Leu Gly Ile 20 25 30 Glu Tyr Lys Asp Gly Asp Glu Leu His Pro Gln Glu Asn Ile Asp Lys 35 40 45 Met Ala Ser Gly Gln Ala Leu Asp Asp Asp Asp Arg Ala Trp Trp Leu 50 55 60 Val Gln Val Gly Lys Trp Leu Arg Asp Arg Pro Ser Gly Val Ile Ala 65 70 75 80 Cys Ser Ala Leu Lys Arg Ser Tyr Arg Asp Leu Leu Arg Thr Lys Cys 85 90 95 Pro Gly Thr Val Phe Val His Leu His Gly Asp Tyr Asp Leu Leu Leu 100 105 110 Ser Arg Met Lys Ala Arg Glu Asp His Phe Met Pro Ser Thr Leu Leu 115 120 125 Asp Ser Gln Phe Ala Thr Leu Glu Pro Leu Glu Asp Asp Glu Asp Gly 130 135 140 Lys Val Phe Asp Val Ala His Thr Ile Ser Glu Leu Ala Ala Gln Ser 145 150 155 160 Ala Glu Trp Val Arg Asn Lys 165 <210> 2 <211> 494 <212> PRT <213> Corynebacterium glutamicum ATCC 13032 <220> <221> PEPTIDE <222> (1)..(494) <223> NCgl2905 gluconate kinase <400> 2 Met Gly Ser Ile Pro Thr Met Ser Ile Pro Phe Asp Asp Ser Arg Gly 1 5 10 15 Pro Tyr Val Leu Ala Met Asp Ile Gly Ser Thr Ala Ser Arg Gly Gly 20 25 30 Leu Tyr Asp Ala Ser Gly Cys Pro Ile Lys Gly Thr Lys Gln Arg Glu 35 40 45 Ser His Glu Phe Thr Thr Gly Glu Gly Val Ser Thr Ile Asp Ala Asp 50 55 60 Gln Val Val Ser Glu Ile Thr Ser Val Ile Asn Gly Ile Leu Asn Ala 65 70 75 80 Ala Asp His His Asn Ile Lys Asp Gln Ile Ala Ala Val Ala Leu Asp 85 90 95 Ser Phe Ala Ser Ser Leu Ile Leu Val Asp Gly Glu Gly Asn Ala Leu 100 105 110 Thr Pro Cys Ile Thr Tyr Ala Asp Ser Arg Ser Ala Gln Tyr Val Glu 115 120 125 Gln Leu Arg Ala Glu Ile Asp Glu Glu Ala Tyr His Gly Arg Thr Gly 130 135 140 Val Cys Leu His Thr Ser Tyr His Pro Ser Arg Leu Leu Trp Leu Lys 145 150 155 160 Thr Glu Phe Glu Glu Glu Phe Asn Lys Ala Lys Tyr Val Met Thr Ile 165 170 175 Gly Glu Tyr Val Tyr Phe Lys Leu Ala Gly Ile Thr Gly Met Ala Thr 180 185 190 Ser Ile Ala Ala Trp Ser Gly Ile Leu Asp Ala His Thr Gly Glu Leu 195 200 205 Asp Leu Thr Ile Leu Glu His Ile Gly Val Asp Pro Ala Leu Phe Gly 210 215 220 Glu Ile Arg Asn Pro Asp Glu Pro Ala Thr Asp Ala Lys Val Val Asp 225 230 235 240 Lys Lys Trp Lys His Leu Glu Glu Ile Pro Trp Phe His Ala Ile Pro 245 250 255 Asp Gly Trp Pro Ser Asn Ile Gly Pro Gly Ala Val Asp Ser Lys Thr 260 265 270 Val Ala Val Ala Ala Ala Thr Ser Gly Ala Met Arg Val Ile Leu Pro 275 280 285 Ser Val Pro Glu Gln Ile Pro Ser Gly Leu Trp Cys Tyr Arg Val Ser 290 295 300 Arg Asp Gln Cys Ile Val Gly Gly Ala Leu Asn Asp Val Gly Arg Ala 305 310 315 320 Val Thr Trp Leu Glu Arg Thr Ile Ile Lys Pro Glu Asn Leu Asp Glu 325 330 335 Val Leu Ile Arg Glu Pro Leu Glu Gly Thr Pro Ala Val Leu Pro Phe 340 345 350 Phe Ser Gly Glu Arg Ser Ile Gly Trp Ala Ala Ser Ala Gln Ala Thr 355 360 365 Ile Thr Asn Ile Gln Glu Gln Thr Gly Pro Glu His Leu Trp Arg Gly 370 375 380 Val Phe Glu Ala Leu Ala Leu Ser Tyr Gln Arg Val Trp Glu His Met 385 390 395 400 Gly Lys Ala Gly Ala Ala Pro Glu Arg Val Ile Ala Ser Gly Arg Val 405 410 415 Ser Thr Asp His Pro Glu Phe Leu Ala Met Leu Ser Asp Ala Leu Asp 420 425 430 Thr Pro Val Ile Pro Leu Glu Met Lys Arg Ala Thr Leu Arg Gly Thr 435 440 445 Ala Leu Ile Val Leu Glu Gln Leu Glu Pro Gly Gly Thr Arg Ala Thr 450 455 460 Pro Pro Phe Gly Thr Thr His Gln Pro Arg Phe Ala His His Tyr Ser 465 470 475 480 Lys Ala Arg Glu Leu Phe Asp Ala Leu Tyr Leu Lys Leu Val 485 490 <210> 3 <211> 26 <212> DNA <213> Artificial Sequence <220> <223> primer 2399F1 <400> 3 gctctagacc ccagaacatg ctgacg 26 <210> 4 <211> 26 <212> DNA <213> Artificial Sequence <220> <223> primer 2399R1 <400> 4 ggggtacccg ctgctagggc tttacc 26 <210> 5 <211> 27 <212> DNA <213> Artificial Sequence <220> <223> primer 2399F2 <400> 5 ggggtaccgg aaccgtcttc gtccacc 27 <210> 6 <211> 26 <212> DNA <213> Artificial Sequence <220> <223> primer 2399R2 <400> 6 gctctagagt caccgctggt acatcc 26 <210> 7 <211> 26 <212> DNA <213> Artificial Sequence <220> <223> primer 2905F1 <400> 7 gctctagaca ttggagcatc cgtagc 26 <210> 8 <211> 27 <212> DNA <213> Artificial Sequence <220> <223> primer 2905R1 <400> 8 tcccccgggg ccttcaccat cgaccaa 27 <210> 9 <211> 27 <212> DNA <213> Artificial Sequence <220> <223> primer 2905F2 <400> 9 tcccccgggg ttcccgaaca gatcccc 27 <210> 10 <211> 26 <212> DNA <213> Artificial Sequence <220> <223> primer 2905R2 <400> 10 gctctagacg ctctgacctg cctaac 26 <110> SANGJI UNIVERSITY INDUSTRY ACADEMIC COOPERATION FOUNDATION <120> A microorganism having enhanced L-ornithine productivity and a method of producing L-ornithine using the same <130> PA100296 / KR <160> 10 <170> KopatentIn 1.71 <210> 1 <211> 167 <212> PRT <213> Corynebacterium glutamicum ATCC 13032 <220> <221> PEPTIDE (222) (1) .. (167) <223> NCgl2399 gluconate kinase <400> 1 Met Ser Ala Ala Glu Gly Leu His Ile Val Val Met Gly Val Ser Gly 1 5 10 15 Cys Gly Lys Ser Ser Val Gly Lys Ala Leu Ala Ala Glu Leu Gly Ile 20 25 30 Glu Tyr Lys Asp Gly Asp Glu Leu His Pro Gln Glu Asn Ile Asp Lys 35 40 45 Met Ala Ser Gly Gln Ala Leu Asp Asp Asp Asp Arg Ala Trp Trp Leu 50 55 60 Val Gln Val Gly Lys Trp Leu Arg Asp Arg Pro Ser Gly Val Ile Ala 65 70 75 80 Cys Ser Ala Leu Lys Arg Ser Tyr Arg Asp Leu Leu Arg Thr Lys Cys 85 90 95 Pro Gly Thr Val Phe Val His Leu His Gly Asp Tyr Asp Leu Leu Leu 100 105 110 Ser Arg Met Lys Ala Arg Glu Asp His Phe Met Pro Ser Thr Leu Leu 115 120 125 Asp Ser Gln Phe Ala Thr Leu Glu Pro Leu Glu Asp Asp Glu Asp Gly 130 135 140 Lys Val Phe Asp Val Ala His Thr Ile Ser Glu Leu Ala Ala Gln Ser 145 150 155 160 Ala Glu Trp Val Arg Asn Lys 165 <210> 2 <211> 494 <212> PRT <213> Corynebacterium glutamicum ATCC 13032 <220> <221> PEPTIDE (222) (1) .. (494) <223> NCgl2905 gluconate kinase <400> 2 Met Gly Ser Ile Pro Thr Met Ser Ile Pro Phe Asp Asp Ser Arg Gly 1 5 10 15 Pro Tyr Val Leu Ala Met Asp Ile Gly Ser Thr Ala Ser Arg Gly Gly 20 25 30 Leu Tyr Asp Ala Ser Gly Cys Pro Ile Lys Gly Thr Lys Gln Arg Glu 35 40 45 Ser His Glu Phe Thr Thr Gly Glu Gly Val Ser Thr Ile Asp Ala Asp 50 55 60 Gln Val Val Ser Glu Ile Thr Ser Val Ile Asn Gly Ile Leu Asn Ala 65 70 75 80 Ala Asp His His Asn Ile Lys Asp Gln Ile Ala Ala Val Ala Leu Asp 85 90 95 Ser Phe Ala Ser Ser Leu Ile Leu Val Asp Gly Glu Gly Asn Ala Leu 100 105 110 Thr Pro Cys Ile Thr Tyr Ala Asp Ser Arg Ser Ala Gln Tyr Val Glu 115 120 125 Gln Leu Arg Ala Glu Ile Asp Glu Glu Ala Tyr His Gly Arg Thr Gly 130 135 140 Val Cys Leu His Thr Ser Tyr His Pro Ser Arg Leu Leu Trp Leu Lys 145 150 155 160 Thr Glu Phe Glu Glu Glu Phe Asn Lys Ala Lys Tyr Val Met Thr Ile 165 170 175 Gly Glu Tyr Val Tyr Phe Lys Leu Ala Gly Ile Thr Gly Met Ala Thr 180 185 190 Ser Ile Ala Ala Trp Ser Gly Ile Leu Asp Ala His Thr Gly Glu Leu 195 200 205 Asp Leu Thr Ile Leu Glu His Ile Gly Val Asp Pro Ala Leu Phe Gly 210 215 220 Glu Ile Arg Asn Pro Asp Glu Pro Ala Thr Asp Ala Lys Val Val Asp 225 230 235 240 Lys Lys Trp Lys His Leu Glu Glu Ile Pro Trp Phe His Ala Ile Pro 245 250 255 Asp Gly Trp Pro Ser Asn Ile Gly Pro Gly Ala Val Asp Ser Lys Thr 260 265 270 Val Ala Val Ala Ala Ala Thr Ser Gly Ala Met Arg Val Ile Leu Pro 275 280 285 Ser Val Pro Glu Gln Ile Pro Ser Gly Leu Trp Cys Tyr Arg Val Ser 290 295 300 Arg Asp Gln Cys Ile Val Gly Gly Ala Leu Asn Asp Val Gly Arg Ala 305 310 315 320 Val Thr Trp Leu Glu Arg Thr Ile Ile Lys Pro Glu Asn Leu Asp Glu 325 330 335 Val Leu Ile Arg Glu Pro Leu Glu Gly Thr Pro Ala Val Leu Pro Phe 340 345 350 Phe Ser Gly Glu Arg Ser Ile Gly Trp Ala Ala Ser Ala Gln Ala Thr 355 360 365 Ile Thr Asn Ile Gln Glu Gln Thr Gly Pro Glu His Leu Trp Arg Gly 370 375 380 Val Phe Glu Ala Leu Ala Leu Ser Tyr Gln Arg Val Trp Glu His Met 385 390 395 400 Gly Lys Ala Gly Ala Ala Pro Glu Arg Val Ile Ala Ser Gly Arg Val 405 410 415 Ser Thr Asp His Pro Glu Phe Leu Ala Met Leu Ser Asp Ala Leu Asp 420 425 430 Thr Pro Val Ile Pro Leu Glu Met Lys Arg Ala Thr Leu Arg Gly Thr 435 440 445 Ala Leu Ile Val Leu Glu Gln Leu Glu Pro Gly Gly Thr Arg Ala Thr 450 455 460 Pro Pro Phe Gly Thr Thr His Gln Pro Arg Phe Ala His His Tyr Ser 465 470 475 480 Lys Ala Arg Glu Leu Phe Asp Ala Leu Tyr Leu Lys Leu Val 485 490 <210> 3 <211> 26 <212> DNA <213> Artificial Sequence <220> <223> primer 2399F1 <400> 3 gctctagacc ccagaacatg ctgacg 26 <210> 4 <211> 26 <212> DNA <213> Artificial Sequence <220> <223> primer 2399R1 <400> 4 ggggtacccg ctgctagggc tttacc 26 <210> 5 <211> 27 <212> DNA <213> Artificial Sequence <220> <223> primer 2399F2 <400> 5 ggggtaccgg aaccgtcttc gtccacc 27 <210> 6 <211> 26 <212> DNA <213> Artificial Sequence <220> <223> primer 2399R2 <400> 6 gctctagagt caccgctggt acatcc 26 <210> 7 <211> 26 <212> DNA <213> Artificial Sequence <220> <223> primer 2905F1 <400> 7 gctctagaca ttggagcatc cgtagc 26 <210> 8 <211> 27 <212> DNA <213> Artificial Sequence <220> <223> primer 2905R1 <400> 8 tcccccgggg ccttcaccat cgaccaa 27 <210> 9 <211> 27 <212> DNA <213> Artificial Sequence <220> <223> primer 2905F2 <400> 9 tcccccgggg ttcccgaaca gatcccc 27 <210> 10 <211> 26 <212> DNA <213> Artificial Sequence <220> <223> primer 2905R2 <400> 10 gctctagacg ctctgacctg cctaac 26
Claims (10)
L-ornithine producing microorganism, wherein gluconate kinase (GntK) activity is attenuated relative to intrinsic activity.
The microorganism of claim 1, wherein the activity of at least one of a gluconate kinase having an amino acid sequence of SEQ ID NO: 1 and a gluconate kinase having an amino acid sequence of SEQ ID NO: 2 is attenuated relative to intrinsic activity.
The microorganism of claim 1, wherein the microorganism is a genus Corynebacterium.
The microorganism of claim 1, wherein the endogenous activity of the protein is attenuated by deletion, substitution, or insertion of a gene on a chromosome that encodes a protein having gluconate kinase activity.
2. The microorganism according to claim 1, wherein the expression control sequence of a gene on a chromosome encoding a protein having gluconate kinase activity is mutated to attenuate the endogenous activity of the protein.
The microorganism of claim 1, wherein a recombinant vector comprising a portion of the gluconate kinase is introduced to attenuate intrinsic activity.
The microorganism of claim 6, wherein the microorganism is introduced with at least one of the recombinant vector of FIG. 2 and the recombinant vector of FIG. 3 such that the activity of the gluconate kinase is weakened relative to the endogenous activity.
The microorganism of claim 1, wherein the microorganism further has increased activity of argCJBD operon.
The microorganism of claim 1, wherein the microorganism is a parent strain of Corynebacterium glutamicum ATCC13032.
Priority Applications (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| KR1020100068619A KR101300185B1 (en) | 2010-07-15 | 2010-07-15 | A microorganism having enhanced L-ornithine productivity and a method of producing L-ornithine using the same |
| PCT/KR2011/005251 WO2012008809A2 (en) | 2010-07-15 | 2011-07-15 | Microorganism having improved l-ornithine productivity, and method for producing l-ornithine using same |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| KR1020100068619A KR101300185B1 (en) | 2010-07-15 | 2010-07-15 | A microorganism having enhanced L-ornithine productivity and a method of producing L-ornithine using the same |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| KR20120007855A true KR20120007855A (en) | 2012-01-25 |
| KR101300185B1 KR101300185B1 (en) | 2013-08-26 |
Family
ID=45469966
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020100068619A KR101300185B1 (en) | 2010-07-15 | 2010-07-15 | A microorganism having enhanced L-ornithine productivity and a method of producing L-ornithine using the same |
Country Status (2)
| Country | Link |
|---|---|
| KR (1) | KR101300185B1 (en) |
| WO (1) | WO2012008809A2 (en) |
Cited By (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| KR101429815B1 (en) * | 2012-10-05 | 2014-08-12 | 상지대학교산학협력단 | Corynebacterium sp. microorganism having enhanced L-threonine productivity by regulation of GntK activity and a method of producing L-threonine using the same |
| KR101526047B1 (en) * | 2013-04-26 | 2015-06-04 | 상지대학교산학협력단 | Microorganism having improved L-ornithin production by increasing the aminotransferase activity and process for preparing the L-ornithin employing the same |
Families Citing this family (6)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP2430152B1 (en) * | 2010-07-15 | 2016-08-24 | CJ CheilJedang Corporation | Microorganism with enhanced l-lysine productivity and method for producing l-lysine using the same |
| CA2922849A1 (en) * | 2012-08-31 | 2014-03-06 | Ixchel Pharma, Llc | Agents useful for treating obesity, diabetes and related disorders |
| CN103794988B (en) | 2012-10-26 | 2017-07-28 | 施耐德电器工业公司 | Supply gas component |
| CN103014086B (en) * | 2012-12-12 | 2015-02-11 | 武汉圣佳化工有限公司 | Method for continuously producing L-ornithine composite salt by enzyme immobilization |
| WO2016144247A1 (en) | 2015-03-12 | 2016-09-15 | Biopetrolia Ab | L-ornithine production in eukaryotic cells |
| CN113604390B (en) * | 2021-08-16 | 2023-04-25 | 江苏澳创生物科技有限公司 | Corynebacterium glutamicum and application thereof in fermentation production of L-ornithine |
-
2010
- 2010-07-15 KR KR1020100068619A patent/KR101300185B1/en active IP Right Grant
-
2011
- 2011-07-15 WO PCT/KR2011/005251 patent/WO2012008809A2/en active Application Filing
Cited By (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| KR101429815B1 (en) * | 2012-10-05 | 2014-08-12 | 상지대학교산학협력단 | Corynebacterium sp. microorganism having enhanced L-threonine productivity by regulation of GntK activity and a method of producing L-threonine using the same |
| KR101526047B1 (en) * | 2013-04-26 | 2015-06-04 | 상지대학교산학협력단 | Microorganism having improved L-ornithin production by increasing the aminotransferase activity and process for preparing the L-ornithin employing the same |
Also Published As
| Publication number | Publication date |
|---|---|
| WO2012008809A2 (en) | 2012-01-19 |
| KR101300185B1 (en) | 2013-08-26 |
| WO2012008809A3 (en) | 2012-05-18 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| KR101300185B1 (en) | A microorganism having enhanced L-ornithine productivity and a method of producing L-ornithine using the same | |
| KR100838038B1 (en) | - - a microorganism of corynebacterium genus having enhanced l-lysine productivity and a method of producing l-lysine using the same | |
| KR101592140B1 (en) | Corynebacterium sp. having xylose availability and process for preparing L-lysine employing the same | |
| US7794990B2 (en) | Microorganism of corynebacterium genus having enhanced L-lysine production ability and method of producing L-lysine using the same | |
| KR100838035B1 (en) | - - a microorganism of corynebacterium genus having enhanced l-lysine productivity and a method of producing l-lysine using the same | |
| KR100789270B1 (en) | - - A microorganism of corynebacterium genus having enhanced L-lysine productivity and a method of producing L-lysine using the same | |
| RU2671106C1 (en) | Microorganism of corynebacterium genus for production of l-arginine and method of production of l-arginine using this microorganism | |
| KR100837844B1 (en) | - - Microoragnism of Corynbacterium genus with an enhanced L-glutamic acid productivity and method of producing L-glutamic acid using the microorganism | |
| JP2019115341A (en) | Corynebacterium genus microorganism for producing l-lysine and method for producing l-lysine by using the same | |
| EP1004671B1 (en) | Process for producing l-glutamic acid by fermentation method | |
| KR970001238B1 (en) | Process for l-tryptophan | |
| KR101285945B1 (en) | Corynebacterium sp. Having Improved L-lysine Production and Process for Preparing the L-lysine Employing the Same | |
| KR101300186B1 (en) | Corynebacterium sp. Having Improved L-ornithin Production and Process for Preparing the L-ornithin Employing the Same | |
| KR20120007965A (en) | A microorganism having enhanced l-lysine productivity and a method of producing l-lysine using the same | |
| JP2000201692A (en) | Production of l-glutamic acid by fermentation | |
| JPH0755155B2 (en) | Amino acid manufacturing method | |
| KR100789272B1 (en) | - - A microorganism of corynebacterium genus having enhanced L-lysine productivity and a method of producing L-lysine using the same | |
| KR101755349B1 (en) | Microorganism producing L-threonine and process for producing L-threonine using the same | |
| KR101622460B1 (en) | Corynebacterium glutamicum having improved L-ornithine production and method for preparing L-ornithine using the same | |
| KR101687474B1 (en) | Microorganism of the genus corynebacterium with enhanced l-arginine productivity and method for producing l-arginine using the same | |
| KR101526047B1 (en) | Microorganism having improved L-ornithin production by increasing the aminotransferase activity and process for preparing the L-ornithin employing the same | |
| KR20120098235A (en) | Microorganism producing unnatural amino acids and the method of producing unnatural amino acids using the microorganism | |
| KR102016050B1 (en) | Novel promoter and uses thereof |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A201 | Request for examination | ||
| N231 | Notification of change of applicant | ||
| E902 | Notification of reason for refusal | ||
| E902 | Notification of reason for refusal | ||
| E701 | Decision to grant or registration of patent right | ||
| GRNT | Written decision to grant | ||
| FPAY | Annual fee payment |
Payment date: 20160811 Year of fee payment: 4 |
|
| FPAY | Annual fee payment |
Payment date: 20170601 Year of fee payment: 5 |
|
| FPAY | Annual fee payment |
Payment date: 20180920 Year of fee payment: 6 |