JP4514708B2 - Identification and utilization of chondroitin polymerization factor - Google Patents
Identification and utilization of chondroitin polymerization factor Download PDFInfo
- Publication number
- JP4514708B2 JP4514708B2 JP2005503149A JP2005503149A JP4514708B2 JP 4514708 B2 JP4514708 B2 JP 4514708B2 JP 2005503149 A JP2005503149 A JP 2005503149A JP 2005503149 A JP2005503149 A JP 2005503149A JP 4514708 B2 JP4514708 B2 JP 4514708B2
- Authority
- JP
- Japan
- Prior art keywords
- chpf
- chondroitin
- chsy
- protein
- amino acid
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Expired - Fee Related
Links
Images





Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P19/00—Preparation of compounds containing saccharide radicals
- C12P19/26—Preparation of nitrogen-containing carbohydrates
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/1048—Glycosyltransferases (2.4)
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/68—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving proteins, peptides or amino acids
- G01N33/6887—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving proteins, peptides or amino acids from muscle, cartilage or connective tissue
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; CARE OF BIRDS, FISHES, INSECTS; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K2217/00—Genetically modified animals
- A01K2217/05—Animals comprising random inserted nucleic acids (transgenic)
Description
本発明は、コンドロイチン合成酵素によるコンドロイチン鎖の重合反応を促進する重合化因子(重合促進因子)の同定と、同因子を利用したコンドロイチン糖鎖の合成方法などに関するものである。 The present invention relates to identification of a polymerization factor (polymerization promoting factor) that promotes a polymerization reaction of a chondroitin chain by a chondroitin synthase, a method for synthesizing a chondroitin sugar chain using the same factor, and the like.
コンドロイチン硫酸は、細胞表面や細胞外マトリックスに広く分布する硫酸化グリコサミノグリカンの一種であり、細胞間相互作用、免疫における細胞認識機構、リンパ球の特異的ホーミング現象、がん細胞の接着と転移等、多方面にわたる生物学的現象に関与すると考えられ、その多彩な生理的機能から、医薬品、化粧品、食品等に広く利用されている。また、コンドロイチン硫酸は、ヘパラン硫酸と同様に、脊椎動物等における脳の神経ネットワーク形成に重要な役割を果たすことが最近の研究から明らかになってきている。
コンドロイチン硫酸の生合成は、コアタンパク質の特定のセリン(Ser)残基にキシロース(Xyl)が転移されることにより始まり、続いて2つのガラクトース(Gal)およびグルクロン酸(GlcA)が単糖単位で転移され、四糖結合領域(GlcAβ1−3Galβ1−3Galβ1−4Xylβ1−O−Ser)が合成される。その後、結合領域にN−アセチルガラクトサミン(GalNAc)とグルクロン酸(GlcA)とが交互に繰り返し転移されることによりコンドロイチン鎖の重合反応が起こり、コンドロイチン硫酸の二糖繰り返し領域が形成される。そして、硫酸基転移酵素によって構成糖が硫酸化されてコンドロイチン硫酸となる。
コンドロイチン硫酸を取得する従来方法としては、クジラ軟骨等の天然物から調製する方法が知られていた。しかしながら、クジラ軟骨等から調製できるコンドロイチン硫酸の量は限られている。また、近年、クジラは動物愛護の観点から捕鯨は制限されており、クジラの調達自体が困難となっている。
そこで、コンドロイチン糖鎖の合成技術の開発、さらに得られたコンドロイチン糖鎖を硫酸化してコンドロイチン硫酸を製造する方法の開発が望まれている。コンドロイチン糖鎖の合成については、次のような報告がある。即ち、ヒトの悪性黒色腫G361細胞の培養上清を酵素源とし、四糖結合領域(GlcAβ1−3Galβ1−3Galβ1−4Xyl)で糖鎖の伸長が停止しているパートタイムプロテオグリカンであるα−トロンボモジュリン(α−TM)を糖受容体基質として用いると、GalNAcとGlcAの二糖が交互に繰り返し重合し、重合化した糖鎖は天然のコンドロイチンと同程度の長さにまで伸長することが報告されている(Nadanaka,S.,Kitagawa,H.,Goto,F.,Tamura,J.,Neumann,K.W.,Ogawa,T.,and Sugahara,K.(1999)Biochem.J.340,353−357)。
他方、コンドロイチンの重合化に関与する酵素のcDNAクローニングは遅れていたが、最近、コンドロイチン鎖の二糖繰り返し領域の合成に関与するN−アセチルガラクトサミン転移酵素−II(GalNAcT−II)およびグルクロン酸転移酵素−II(GlcAT−II)の両活性をもつコンドロイチン合成酵素(ChSy)のcDNAがクローニングされた(Kitagawa,H.,Uyama,T.,and Sugahara,K.(2001)J.Biol.Chem.276,38721−38726)。しかしながら、in vitroにおいて組換え型タンパク質として発現させたChSyは、GalNAcやGlcAをそれぞれ転移させる活性は示したが、α−TM上にコンドロイチン鎖を重合させることができず、上記のG361細胞の培養上清中に検出されたコンドロイチン鎖の重合反応を再現することができなかった。Chondroitin sulfate is a type of sulfated glycosaminoglycan that is widely distributed on the cell surface and extracellular matrix. It is a cell-cell interaction, cell recognition mechanism in immunity, specific homing phenomenon of lymphocytes, cancer cell adhesion and It is considered to be involved in various biological phenomena such as metastasis, and is widely used in pharmaceuticals, cosmetics, foods, etc. due to its various physiological functions. Also, chondroitin sulfate, like heparan sulfate, has been revealed from recent studies to play an important role in the formation of neural networks in the brain in vertebrates and the like.
Chondroitin sulfate biosynthesis begins with the transfer of xylose (Xyl) to a specific serine (Ser) residue of the core protein, followed by two galactose (Gal) and glucuronic acid (GlcA) in monosaccharide units. The tetrasaccharide binding region (GlcAβ1-3Galβ1-3Galβ1-4Xylβ1-O-Ser) is synthesized. Thereafter, N-acetylgalactosamine (GalNAc) and glucuronic acid (GlcA) are alternately and repeatedly transferred to the binding region, whereby a polymerization reaction of chondroitin chains occurs, and a disaccharide repeating region of chondroitin sulfate is formed. Then, the constituent sugars are sulfated by sulfate group transferase to become chondroitin sulfate.
As a conventional method for obtaining chondroitin sulfate, a method of preparing from a natural product such as whale cartilage has been known. However, the amount of chondroitin sulfate that can be prepared from whale cartilage or the like is limited. In recent years, whales are restricted from whaling from the viewpoint of animal welfare, and it is difficult to procure whales themselves.
Accordingly, development of a technique for synthesizing chondroitin sugar chains and development of a method for producing chondroitin sulfate by sulfating the obtained chondroitin sugar chains are desired. There are following reports on the synthesis of chondroitin sugar chains. That is, α-thrombomodulin, which is a part-time proteoglycan in which the growth of a sugar chain is stopped in a tetrasaccharide-binding region (GlcAβ1-3Galβ1-3Galβ1-4Xyl) using the culture supernatant of human malignant melanoma G361 cells as an enzyme source. When α-TM) is used as a sugar acceptor substrate, GalNAc and GlcA disaccharides are alternately and repeatedly polymerized, and the polymerized sugar chain has been reported to extend to the same length as natural chondroitin. (Nadanaka, S., Kitagawa, H., Goto, F., Tamura, J., Neumann, K.W., Ogawa, T., and Sugawara, K. (1999) Biochem. J. 340, 353-. 357).
On the other hand, cDNA cloning of enzymes involved in the polymerization of chondroitin was delayed, but recently, N-acetylgalactosamine transferase-II (GalNAcT-II) and glucuronic acid transfer involved in the synthesis of the disaccharide repeat region of chondroitin chain The cDNA of chondroitin synthase (ChSy) having both activities of enzyme-II (GlcAT-II) has been cloned (Kitagawa, H., Uyama, T., and Sugahara, K. (2001) J. Biol. Chem. 276, 38721-38726). However, ChSy expressed as a recombinant protein in vitro showed activity to transfer GalNAc and GlcA, respectively, but could not polymerize chondroitin chains on α-TM, and culture of the above G361 cells. The polymerization reaction of chondroitin chains detected in the supernatant could not be reproduced.
本発明者は、上記の知見から、ChSyと共同してコンドロイチン鎖を重合化させるために必須の、未だ同定されていないタンパク性因子が存在するのではないかと予想し、同分子の探索を行った。
本発明の目的は、コンドロイチン鎖の重合化に関与する新規分子を同定すると共に、同分子を利用したコンドロイチン糖鎖の合成方法、さらにはコンドロイチン硫酸の製造方法などを提供することにある。
ごく最近、コンドロイチン硫酸の生合成に関与する糖転移酵素としてChSyを含む5つの糖転移酵素が同定された。これらはすべてChSyと高い相同性を示すので、コンドロイチン鎖の重合化に関与する因子もChSyと相同性を示すのではないかと考え、ChSyのアミノ酸配列をもとにデータベースを検索した。得られたデータをもとに解析を進めた結果、775アミノ酸からなり、アミノ酸レベルでヒトChSyと23%の相同性を示すヒト新規遺伝子の存在を同定した。さらに機能解析の結果、この遺伝子産物がChSyと共同してin vitroでコンドロイチン鎖の重合化を促進する作用を有すること等を見出し、本発明を完成させるに至った。
即ち、本発明は、産業上有用な方法・物として、下記A)〜P)の発明を含むものである。
A) 下記(a)〜(d)の何れかのアミノ酸配列からなるタンパク質をコードするポリヌクレオチド。
(a)配列番号2に示されるアミノ酸配列、
(b)配列番号4に示されるアミノ酸配列、
(c)配列番号2又は4に示されるアミノ酸配列を含むアミノ酸配列、
(d)上記(a)〜(c)の何れかのアミノ酸配列において、1又は数個のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列。
B) 配列番号1又は3に示される塩基配列を有する上記A)記載のポリヌクレオチド。
C) 上記A)又はB)記載のポリヌクレオチドを含む組換え発現ベクター。組換え発現ベクターの作製には、プラスミド、ファージ、又はコスミドなどを用いることができるが、特に限定されるものではない。
D) 上記A)又はB)記載のポリヌクレオチドが導入された形質転換体。ここで、「ポリヌクレオチドが導入された」とは、公知の遺伝子工学的手法(遺伝子操作技術)により、対象細胞(宿主細胞)内にポリヌクレオチド(遺伝子)が発現可能に導入されることを意味する。また、上記「形質転換体」とは、細胞・組織・器官のみならず、動物個体(線虫、キイロショウジョウバエ、マウス、ラット、ウサギ、ブタ、サルなどの形質転換動物)を含む意味である。
E) 下記(a)〜(d)の何れかのアミノ酸配列からなるタンパク質。
(a)配列番号2に示されるアミノ酸配列、
(b)配列番号4に示されるアミノ酸配列、
(c)配列番号2又は4に示されるアミノ酸配列を含むアミノ酸配列、
(d)上記(a)〜(c)の何れかのアミノ酸配列において、1又は数個のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列。
F) 上記E)記載のタンパク質を含むコンドロイチン重合促進剤。
G) 上記E)記載のタンパク質とコンドロイチン合成酵素とを共発現させることにより得られた複合体。
H) 上記E)記載のタンパク質に対する抗体。抗体は、タンパク質の全長又はその部分ペプチドを抗原として、公知の方法によりポリクローナル抗体またはモノクローナル抗体として得ることができる。
I) 上記E)記載のタンパク質を用いてコンドロイチン鎖の重合化を促進する方法。
J) 上記E)記載のタンパク質とコンドロイチン合成酵素とを共発現させることにより得られた複合体を使用することを特徴とする上記I)記載のコンドロイチン鎖重合化方法。
K) 四糖が結合したα−トロンボモジュリン等のコアタンパク質上にコンドロイチン鎖を合成することを特徴とする上記I)又はJ)記載のコンドロイチン鎖重合化方法。
L) 上記I)〜K)の何れかに記載の方法を用いて製造されたコンドロイチン鎖。
M) 上記I)〜K)の何れかに記載の方法を用いて製造されたコンドロイチン硫酸。
N) 上記I)〜K)の何れかに記載の方法を用いて製造されたデルマタン硫酸。
O) 配列番号2に示されるアミノ酸配列からなるタンパク質をコードする遺伝子上の変異を検出することにより遺伝子病の有無を判定する方法。同タンパク質は、後述のように、ChSyと共同してコンドロイチン鎖の重合化に重要な役割を果たすヒト由来コンドロイチン重合化因子(ChPF)であり、その変異によりコンドロイチン硫酸の生合成に異常を生じ、疾患の原因となっている可能性がある。このような疾患については、ChPF遺伝子上の変異を検出することにより当該遺伝子病の有無を判定することができる。
P) ゲノム中のコンドロイチン重合化因子(ChPF)遺伝子配列の全部または一部を改変することにより得られ、相同染色体上の一方又は双方のChPF遺伝子が破壊されたことを特徴とするノックアウト非ヒト動物。
本発明のさらに他の目的、特徴、および優れた点は、以下に示す記載によって十分わかるであろう。また、本発明の利益は、添付図面を参照した次の説明で明白になるであろう。Based on the above findings, the present inventor predicted that there may be an unidentified protein factor that is essential for polymerizing chondroitin chains in cooperation with ChSy, and searches for the same molecule. It was.
An object of the present invention is to identify a novel molecule involved in polymerization of a chondroitin chain, and to provide a method for synthesizing a chondroitin sugar chain using the same molecule, a method for producing chondroitin sulfate, and the like.
Most recently, five glycosyltransferases, including ChSy, have been identified as glycosyltransferases involved in the biosynthesis of chondroitin sulfate. Since these all show high homology with ChSy, it was considered that factors involved in the polymerization of the chondroitin chain also show homology with ChSy, and the database was searched based on the amino acid sequence of ChSy. As a result of further analysis based on the obtained data, the existence of a novel human gene consisting of 775 amino acids and showing 23% homology with human ChSy at the amino acid level was identified. Furthermore, as a result of functional analysis, it was found that this gene product has an action of promoting the polymerization of chondroitin chains in vitro in cooperation with ChSy, and the present invention has been completed.
That is, the present invention includes the following inventions A) to P) as industrially useful methods and products.
A) A polynucleotide encoding a protein comprising the amino acid sequence of any one of (a) to (d) below.
(A) the amino acid sequence shown in SEQ ID NO: 2,
(B) the amino acid sequence shown in SEQ ID NO: 4,
(C) an amino acid sequence comprising the amino acid sequence shown in SEQ ID NO: 2 or 4,
(D) An amino acid sequence in which one or several amino acids are substituted, deleted, inserted and / or added in any one of the amino acid sequences of (a) to (c) above.
B) The polynucleotide according to A), which has the base sequence represented by SEQ ID NO: 1 or 3.
C) A recombinant expression vector comprising the polynucleotide described in A) or B) above. For the production of the recombinant expression vector, a plasmid, phage, cosmid or the like can be used, but it is not particularly limited.
D) A transformant in which the polynucleotide described in A) or B) is introduced. Here, “the polynucleotide has been introduced” means that the polynucleotide (gene) is introduced into the target cell (host cell) so as to be expressed by a known genetic engineering technique (gene manipulation technique). To do. The above-mentioned “transformant” means not only cells / tissues / organs but also animals (transformed animals such as nematodes, Drosophila melanogaster, mice, rats, rabbits, pigs, monkeys).
E) A protein comprising the amino acid sequence of any one of (a) to (d) below.
(A) the amino acid sequence shown in SEQ ID NO: 2,
(B) the amino acid sequence shown in SEQ ID NO: 4,
(C) an amino acid sequence comprising the amino acid sequence shown in SEQ ID NO: 2 or 4,
(D) An amino acid sequence in which one or several amino acids are substituted, deleted, inserted and / or added in any one of the amino acid sequences of (a) to (c) above.
F) A chondroitin polymerization accelerator comprising the protein described in E) above.
G) A complex obtained by co-expressing the protein described in E) above and chondroitin synthase.
H) An antibody against the protein described in E) above. The antibody can be obtained as a polyclonal antibody or a monoclonal antibody by a known method using the full-length protein or a partial peptide thereof as an antigen.
I) A method for promoting polymerization of chondroitin chains using the protein described in E) above.
J) A chondroitin chain polymerization method as described in I) above, wherein a complex obtained by co-expressing the protein as described in E) and chondroitin synthase is used.
K) The chondroitin chain polymerization method as described in I) or J) above, wherein a chondroitin chain is synthesized on a core protein such as α-thrombomodulin to which a tetrasaccharide is bound.
L) A chondroitin chain produced using the method according to any one of I) to K) above.
M) Chondroitin sulfate produced using the method according to any one of I) to K) above.
N) Dermatan sulfate produced using the method according to any one of I) to K) above.
O) A method for determining the presence or absence of a genetic disease by detecting a mutation on a gene encoding a protein consisting of the amino acid sequence represented by SEQ ID NO: 2. As described later, this protein is a human-derived chondroitin polymerization factor (ChPF) that plays an important role in the polymerization of chondroitin chains in cooperation with ChSy, and its mutation causes an abnormality in the biosynthesis of chondroitin sulfate, It may be the cause of the disease. About such a disease, the presence or absence of the genetic disease can be determined by detecting a mutation on the ChPF gene.
P) A knockout non-human animal obtained by modifying all or part of the chondroitin polymerization factor (ChPF) gene sequence in the genome, wherein one or both of the ChPF genes on the homologous chromosome are disrupted .
Other objects, features, and advantages of the present invention will be fully understood from the following description. The benefits of the present invention will become apparent from the following description with reference to the accompanying drawings.
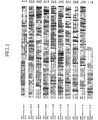
図1は、本発明に係るヒト由来コンドロイチン重合化因子(ChPF)の配列・構造を、ヒト由来コンドロイチン合成酵素(ChSy)およびヒト由来コンドロイチン・グルクロン酸転移酵素(Chondroitin GlcAT)と比較して示す図である。図中、ボックスは膜貫通ドメインを示し、ChPFの推定上のN−glycosylation siteは★で示される。
図2は、上記ヒト由来コンドロイチン重合化因子(ChPF)のゲノム構造を、コンドロイチン・グルクロン酸転移酵素(Chondroitin GlcAT)と比較して示す図である。図中、ボックスはエキソン、バーはイントロンを示し、白いボックスは非翻訳領域、黒いボックスは翻訳領域を表す。ATGは開始コドン、TGA・TAGは終止コドンを示す。
図3のA・Bは、結合領域オリゴ糖、人工基質およびα−トロンボモジュリン(α−TM)をそれぞれ受容体基質として重合反応を行い、得られたコンドロイチン鎖のサイズを比較検討した実験結果を示すグラフである。
図4のA〜Cは、(GlcAβ1−3GalNAc)3やコンドロイチンポリマーを受容体基質として重合反応を行い、得られたコンドロイチン鎖のサイズを比較検討した実験結果を示すグラフである。
図5のA・Bは、α−TM上に合成されたコンドロイチン糖鎖に由来する糖−タンパク質結合領域の六糖の分析結果を示すグラフである。
図6は、pull−down法を用いたChPFとChSyとの相互作用解析の結果を示す図である。
図7は、ChPFおよびChSyのノーザンブロット分析の結果を示す図である。FIG. 1 is a diagram showing the sequence and structure of human-derived chondroitin polymerization factor (ChPF) according to the present invention in comparison with human-derived chondroitin synthase (ChSy) and human-derived chondroitin glucuronyltransferase (Chandroitin GlcAT). It is. In the figure, the box indicates the transmembrane domain, and the putative N-glycosylation site of ChPF is indicated by *.
FIG. 2 is a diagram showing the genome structure of the human-derived chondroitin polymerization factor (ChPF) in comparison with chondroitin glucuronyltransferase (Chondroitin GlcAT). In the figure, boxes represent exons, bars represent introns, white boxes represent untranslated regions, and black boxes represent translated regions. ATG represents a start codon and TGA • TAG represents a stop codon.
3A and 3B show experimental results of conducting a polymerization reaction using a binding domain oligosaccharide, an artificial substrate and α-thrombomodulin (α-TM) as a receptor substrate, and comparing the sizes of the obtained chondroitin chains. It is a graph.
4A to 4C are graphs showing experimental results of conducting a polymerization reaction using (GlcAβ1-3GalNAc) 3 or a chondroitin polymer as a receptor substrate and comparing the sizes of the obtained chondroitin chains.
5A and 5B are graphs showing the analysis results of the hexasaccharide in the sugar-protein binding region derived from the chondroitin sugar chain synthesized on α-TM.
FIG. 6 is a diagram showing the results of an interaction analysis between ChPF and ChSy using the pull-down method.
FIG. 7 shows the results of Northern blot analysis of ChPF and ChSy.
本発明は、コンドロイチン合成酵素(ChSy)と共同して、コンドロイチン鎖の重合化を促進する重合化因子およびこれをコードする遺伝子を同定すると共に、その機能を詳細に解析し、この重合化因子を利用したコンドロイチン鎖の合成方法などを提案するものである。そこで以下では、このコンドロイチン重合化因子とその遺伝子(ポリヌクレオチド)の配列、構造について説明し、次いでその機能、利用方法等について説明することにする。
(1)本発明に係るコンドロイチン重合化因子とその遺伝子の配列、構造
ヒトChSyのアミノ酸配列をもとにBLASTサーチを行った。その結果、ChSyと相同性の高い遺伝子情報(アクセッション番号:BC021223)を見出した。しかし、この情報に開示される推定アミノ酸配列は522残基からなる短いものであった。本発明者は、得られたデータをもとに解析を進めた結果、775アミノ酸からなり、ヒトChSyと23%の相同性を示し、ヒトChondroitin GlcATとは57%の相同性を示す新規タンパク質(ChPF)の存在を同定した(図1参照)。
後述のように、この新規タンパク質には、糖供与体であるUDP−糖の結合モチーフであるDXDモチーフが存在しなかった。このことより、この遺伝子産物はコンドロイチン硫酸の生合成に関与しているが、糖転移活性を保持していないことが予想された。機能解析の結果からも、この遺伝子産物は糖転移酵素ではなく、ChSyと共同してコンドロイチンを重合化させる因子であることが判明したので、本発明者は、この新規タンパク質をコンドロイチン重合化因子(chondroitin polymerizing factor(ChPF))と名付けた。
配列表の配列番号1には、ヒトChPF遺伝子のcDNA配列が示され、配列番号2には、同遺伝子によってコードされるヒトChPFのアミノ酸配列が示される。また、配列番号4には、上記ChPFの推定膜貫通ドメインを含むN末端側から62個のアミノ酸を他の配列と組換えた可溶性のChPF組換え型タンパク質のアミノ酸配列が示され、配列番号3には、そのcDNA配列が示される。後述のように、ChPFの機能解析は、この可溶性ChPFを発現させることにより行った。
ゲノム情報を検索し解析した結果、図2に示すように、ChPFは第2番染色体上で約5kbに広がって存在し、翻訳領域の遺伝子は4つのエキソンから構成されていた。このゲノム構造は、Chondroitin GlcAT遺伝子と類似するものであった。
本発明に係るタンパク質、即ちコンドロイチン重合化因子は、(a)配列番号2に示されるアミノ酸配列からなるタンパク質、(b)配列番号4に示されるアミノ酸配列からなる可溶性タンパク質、(c)配列番号2又は4に示されるアミノ酸配列を含むアミノ酸配列からなるタンパク質(例えば、融合タンパク質)、(d)上記(a)〜(c)の何れかのアミノ酸配列において、1又は数個のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列からなるタンパク質(コンドロイチン重合化因子として機能するもの)、のいずれであってもよい。また、ヒト由来に限らず、他の生物由来のコンドロイチン重合化因子であってもよい。
上記「1又は数個のアミノ酸が置換、欠失、挿入、及び/又は付加」とは、点変異導入法等の公知の変異タンパク質作製法により置換、欠失、挿入、及び/又は付加できる程度の数のアミノ酸が置換、欠失、挿入、及び/又は付加されることを意味する。このように、上記(d)のタンパク質は、換言すれば、上記(a)〜(c)の変異タンパク質であり、ここにいう「変異」は、主として公知の変異タンパク質作製法により人為的に導入された変異を意味するが、天然に存在する同様の変異タンパク質を単離精製したものであってもよい。
本発明に係るタンパク質は、タンパク質の精製や検出等を容易に行うために、公知のHAやFlag等の付加配列を末端に含ませてもよいし、融合タンパク質としてもよい。また、公知の各種修飾を施してもよいし、二量体以上の多量体を形成するものであってもよい。
本発明の「ポリヌクレオチド(遺伝子)」は、上記(a)〜(d)のいずれかのタンパク質をコードする配列を有するものであればよく、さらに非翻訳領域(UTR)の配列などを含むものであってもよい。
尚、本発明の「ポリヌクレオチド(遺伝子)」には、RNAおよびDNAが含まれるものとする。RNAにはmRNAやsiRNAなどが含まれ、DNAには、cDNAやゲノムDNAなどが含まれる。また、DNAは二本鎖でも一本鎖でもよく、一本鎖DNAは、センス鎖となるコードDNAでもよく、アンチセンス鎖となるアンチコード鎖でもよい。アンチセンス鎖は、プローブとして又はアンチセンス薬物として利用できる。また、siRNAは、RNAiとして遺伝子発現抑制などに利用できる。
(2)コンドロイチン重合化因子の機能とその利用方法
機能解析の結果、上記コンドロイチン重合化因子(ChPF)には以下の機能が見出されている(詳細は、後述の実施例参照)。1)GalNAcT−IIおよびGlcAT−II活性を保持するChSyとChPFを同一細胞内で共発現させると、ChSy単独発現時と比べて、GalNAcT−IIおよびGlcAT−IIの活性が20倍以上に上昇した。2)α−TMを受容体基質、ChSyとChPFを共発現させた分泌型タンパク質を酵素源として用い、UDP−GalNAcとUDP−GlcA共存下で酵素反応を行った結果、天然のコンドロイチン鎖と同程度の長さにまで重合化を起こすことに成功した。3)ChPFを用いたコンドロイチン鎖の重合反応には、受容体基質としてα−TM等のコアタンパク質を用いることが好ましい。4)分泌型タンパク質として共発現させたChSyとChPFとは複合体を形成し、相互作用していると考えられる。5)ChPFは調べた殆どの組織で発現しており、その発現パターンはChSyと類似していた。このことより生体内で両者が共同して、コンドロイチン硫酸の生合成に関与していると考えられる。
したがって、上記コンドロイチン重合化因子(ChPF)およびその遺伝子は、生理活性を有する硫酸化グリコサミノグリカン(コンドロイチン硫酸、デルマタン硫酸)の糖鎖骨格の人工合成に利用することができる。例えば、コンドロイチンはこれまでクジラの軟骨などから調製されたコンドロイチン硫酸を化学的に脱硫酸化することによって調製されていたが、ChSyとChPFとを共発現させ、これを酵素源として用いることにより、in vitroでコンドロイチン糖鎖を安定かつ容易に大量生産することが可能となる。
上記コンドロイチン重合化因子(ChPF)を用いた糖鎖合成において、α−トロンボモジュリン(α−TM)等のコアタンパク質上にコンドロイチン糖鎖を合成することは好ましい。
さらに、特異性の異なる種々の硫酸化転移酵素を用いることによって、生理活性を有する種々のコンドロイチン硫酸やその異性体であるデルマタン硫酸の合成に利用できる。
その他にも、本発明のコンドロイチン重合化因子およびその遺伝子は、研究用試薬や検査用試薬として利用可能であるほか、コンドロイチン重合化因子の異常に起因する疾患や、コンドロイチン硫酸、デルマタン硫酸等の硫酸化グリコサミノグリカンが関係する疾患の病態解析、並びにその病気への治療薬、治療法、診断薬、および診断法等の開発に利用可能である。
(3)本発明に係るタンパク質、遺伝子の取得方法
本発明に係るChPF遺伝子の取得方法は、特に限定されるものではなく、前述の開示された配列情報等に基づいて種々の方法により、上記遺伝子配列を含むDNA断片を単離し、クローニングすることができる。
例えば、上記ChPF遺伝子のcDNAの一部配列と特異的にハイブリダイズするプローブを調製し、ヒトゲノムDNAライブラリーやcDNAライブラリーをスクリーニングすればよい。このようなプローブとしては、上記ChPF遺伝子のcDNA配列又はその相補配列の少なくとも一部に特異的にハイブリダイズするプローブであれば、いずれの配列・長さのものを用いてもよい。また、インシリコ・スクリーニングを行ってもよい。
上記スクリーニングによって得られたクローンは、制限酵素地図の作成及びその塩基配列決定(シークエンシング)によって、さらに詳しく解析することができる。これらの解析によって、本発明に係る遺伝子配列を含むDNA断片を取得したか容易に確認することができる。
また、上記プローブの配列を、上記ChPF遺伝子の特異性の高い領域の中から選択し、ヒト・マウスやその他の生物のゲノムDNA(又はcDNA)ライブラリーをスクリーニングすれば、上記ChPFタンパク質と同様の機能を有する相同分子や類縁分子をコードする遺伝子を単離しクローニングできる可能性が高い。
本発明に係る遺伝子(ポリヌクレオチド)を取得する方法は、上記スクリーニング法以外にも、PCR等の増幅手段を用いる方法がある。例えば、上記ChPFのcDNA配列のうち、5’側及び3’側の非翻訳領域の配列(又はその相補配列)の中からそれぞれプライマーを調製し、これらプライマーを用いてヒトのゲノムDNA(又はcDNA)等を鋳型にしてPCR等を行い、両プライマー間に挟まれるDNA領域を増幅することで、本発明のポリヌクレオチドを含むDNA断片を大量に取得できる。
本発明に係る上記ChPFタンパク質等を取得する方法についても、特に限定されるものではなく、例えば、上述のようにして取得された遺伝子(上記ChPFタンパク質又はその相同分子等をコードするcDNA等)を導入した組換え発現ベクターを作製し、周知の方法により大腸菌や酵母等の微生物又は動物細胞などに組み入れて形質転換体として、そのcDNAがコードするタンパク質を発現させ精製することで、本発明に係るタンパク質を容易に取得することができる。
上記ChPFタンパク質の変異タンパク質を作製する方法についても特に限定されるものではなく、例えば、点変異導入法などの公知の変異タンパク質作製法を用いて、上述のようにして取得された遺伝子の塩基配列において、1又はそれ以上の塩基が置換、欠失、挿入、及び/又は付加されるように改変を加えることによって作製することができる。また、変異タンパク質の作製には、市販のキットを利用してもよい。
(4)本発明のChPFノックアウト動物
本発明のChPFノックアウト動物、即ち、ゲノム中のChPF遺伝子配列の全部または一部を改変することにより得られ、相同染色体上の一方または双方のChPF遺伝子が破壊された非ヒト動物は、遺伝子ターゲッティング法を用いて作製することができる。
ここで「ChPF遺伝子が破壊された」とは、換言すれば、ChPF本来の機能を持った蛋白質が発現されないことを意味する。したがって、ChPF本来の活性を持たないChPF蛋白の一部断片(ポリペプチド)が発現されるものであってもよい。
遺伝子ターゲッティング法は、通常の方法にしたがって行えばよい。一例を挙げれば、まず、相同組換えのためのターゲッティングベクター(ターゲッティングコンストラクト)を構築する。ターゲッティングベクターは、公知の方法により作製することができ、大略、ChPFゲノムDNAクローン、市販のプラスミド、ポジティブセレクション用のマーカー(PGK−neoカセット等)、およびネガティブセレクション用のマーカー(DTA遺伝子、HSV−tk遺伝子等)などの各フラグメントを適切に連結することにより作製することができる。このとき、目的とする制限酵素切断部位が適切な位置に配されるようターゲッティングベクターを設計するとよい。また、ターゲッティングの効率は相同領域の長さに依存するので、相同領域はできるだけ長いほうが好ましい。さらに、ターゲッティングベクターは環状より直鎖状のほうが好ましいので、直鎖状化のため相同領域以外の部分に一カ所適当な制限酵素切断部位を設けておくとよい。
上記方法により作製したChPF遺伝子ターゲッティングベクターを、ES細胞等の対象動物由来の全能性細胞にエレクトロポレーション法等により導入し、その後、目的とする相同組換えが起こった細胞を選別する。選別は、ポジティブ−ネガティブ選択法により薬剤を用いて効率よくスクリーニングできる。選別後、目的とする相同組換えが起こった細胞を、サザンブロットやPCR法などによって確認する。最終的に相同組換えが確認された全能性細胞を、妊娠中の子宮から採取された胚盤胞(ブラストシスト)に導入する。胚盤胞への細胞の導入は、マイクロインジェクション法等により行うことができるが、これに限定されるものではない。
上記胚盤胞を常法に従い仮親に移植する。仮親から生まれた生殖系列キメラ動物(好ましくは雄)と、野生型ChPF遺伝子をホモで持つ野生型動物(好ましくは雌)とを交配させることにより、第1世代(F1)として、相同染色体上の一方のChPF遺伝子が相同組換えにより破壊されたヘテロ接合体を得ることができる。さらに、これらヘテロ接合体同士を交配させることにより、第2世代(F2)として、相同染色体上の双方のChPF遺伝子が破壊されたホモ接合体を得ることができる。ホモ接合体の同定は、体の一部(例えば尻尾)を切断し、DNAを抽出してサザンブロットやPCR法などによって遺伝子型を調べればよい。また、第2世代(F2)として、ChPFノックアウト動物と同腹の野生型動物(野生型遺伝子をホモで持つ)を得ることができるが、この野生型動物は対照実験に好適に用いることができる。
以上説明した遺伝子ターゲッティング法は、あくまでその一例を示すものであって、公知の種々の変更が可能であることはいうまでもない。
ChPFノックアウト動物の対象となる動物は、特に限定されるものではないが、ウシ、ブタ、ヒツジ、ヤギ、ウサギ、イヌ、ネコ、モルモット、ハムスター、マウス、ラットなどの哺乳動物が例示される。これらのうち、実験動物として用いるには、ウサギ、イヌ、ネコ、モルモット、ハムスター、マウス、ラットが好ましく、なかでも齧歯目がさらに好ましく、近交系が多数作出されており、受精卵の培養、体外受精等の技術が整っているマウスが、特に好ましい。また、全能性細胞としては、受精卵や初期胚のほか、多分化能を有するES細胞、体性幹細胞のような培養細胞を対象とすることができる。
本発明のChPFノックアウト動物は、ChPFの機能解析などに有用である。
以下、実施例により本発明をより具体的に説明するが、本発明はこれら実施例により何ら限定されるものではない。尚、各実施例における詳しい実験方法については、最後の項においてまとめて説明する。
〔実施例1:ChPFのクローニング〕
ヒトのChSyのアミノ酸配列を用いたBLASTサーチの結果、ChSyと相同性のある新規遺伝子が存在することが判明した。この遺伝子は236bpの5’非翻訳領域、775個のアミノ酸をコードする2325bpの翻訳領域(図1および配列番号1・2)と、約500bpの3’非翻訳領域から構成されていた。その翻訳開始領域と予想される配列はKozak consensus配列に従っていた。また、この遺伝子から翻訳されるタンパク質は、Kyte−Doolittle hydropathy analysisより、N末端側に23個のアミノ酸からなる膜貫通ドメインを1つ持つ、ゴルジ体に局在する糖転移酵素に特徴的なII型膜蛋白質であることが同定され、さらに3カ所のN−glycosylation siteを保持していた。この遺伝子はChSyとアミノ酸配列レベルで23%の相同性を示し、ヒトChondroitin GlcATとは57%の相同性を示した(図1)。しかしながら、糖供与体であるUDP−糖の結合モチーフであるDXDモチーフが存在しなかった。このことより、この遺伝子産物は、コンドロイチン硫酸の生合成に関与しているが、糖転移活性を保持していないことが予想された。機能解析の結果からも、この遺伝子産物は糖転移酵素ではなく、ChSyと共同してコンドロイチン(以下、略して「Chn」という。)を重合化させる因子であることが判明したので、本発明者は、この新規タンパク質をコンドロイチン重合化因子(chondroitin polymerizing factor(ChPF))と名付けた。
解析の結果、Caenorhabditis elegansやDrosophila melanogasterにおいても上記ChPFのorthologが存在することがわかり、それぞれ27および25%の相同性があった。
〔実施例2:ChPFの遺伝子構造と染色体マッピング〕
図2に示すように、ChPFは第2番染色体上で約5kbに広がって存在し、翻訳領域の遺伝子は4つのエキソンから構成されていた。それぞれのイントロン/エキソンの境界はGT/AG ruleに適合していた。
〔実施例3:ChPFの発現と機能解析〕
ChPFの機能を解析するために、ChPFの膜貫通ドメインを含むN末端側から62個のアミノ酸を、細胞外分泌シグナルであるinsulin leaderおよびprotein AのIgG−結合ドメインのシークエンスと組換えることにより、COS−1細胞でprotein Aが融合した分泌型タンパク質を発現させた。このChPF組換え型タンパク質を酵素源に用い、Chnを糖受容体基質にUDP−GalNAcあるいはUDP−GlcAを糖供与体として糖転移活性を測定したが、糖転移活性はほとんど検出できなかった(下記表1)。
そこで、ChPFは糖転移酵素ではなく重合化因子であると考え、Chn鎖の二糖繰り返し領域を合成する糖転移活性をもつChSyをChPFと同一細胞内で共発現させた。その結果、ChPFをChSyと共発現させたものを酵素源に用いた場合では、ChSy単独時よりもGalNAcT−IIおよびGlcAT−II活性は20倍以上に上昇した(表1)。また、ChSyとChPFをそれぞれ単独に発現させた組換え型タンパク質を混合したものでは、糖転移活性の上昇は見られなかった。このことより、ChSyとChPFが細胞内で複合体を形成することが示唆された。さらに、複合体は単独で発現させたときには形成されず、同一細胞内でChSyとChPFを共発現させることにより、複合体が形成されると考えられた。

前述のように、組換え型タンパク質として発現させたChSyはGalNAcとGlcAのそれぞれを単糖で転移させるGalNAcT−IIおよびGlcAT−II両活性を保持しているにもかかわらず、α−TM上にChn鎖を重合させる活性は保持していなかった。
そこで、ChPFとChSyを共発現させたものに糖転移活性の上昇が見られたことより、ChPFとChSyを共発現させたものがChn鎖を重合させる活性を保持しているのではないかと考えた。この点について解析するため、ChSyとChPFを共発現させた組換え型タンパク質を酵素源として、α−TM、GlcAβ1−3Galβ1−3Galβ1−4Xylβ1−O−Ser、四糖結合領域にα−TM由来のペプチドが結合したGlcAβ1−3Galβ1−3Galβ1−4Xylβ1−O−(Gly)Ser−Gly−Glu、四糖結合領域にβ−glycanとよばれるプロテオグリカン(PG)由来のペプチドが結合したGlcAβ1−3Galβ1−3Galβ1−4Xylβ1−O−Ser−Gly−Trp−Pro−Asp−Glyおよび人工基質であるGlcAβ1−3Galβ1−O−C2H4NH−benzyloxycarbonylを糖受容体基質として用い、重合活性を測定した。その結果が図3のグラフA、Bに示される。
図3の説明:A.Chn鎖の重合反応の基質としてGlcAβ1−3Galβ1−3Galβ1−4Xylβ1−O−Ser(●)、GlcAβ1−3Galβ1−3Galβ1−4Xylβ1−O−(Gly)Ser−Gly−Glu(×)、GlcAβ1−3Galβ1−3Galβ1−4Xylβ1−O−Ser−Gly−Trp−Pro−Asp−Gly(△)、GlcAβ1−3Galβ1−O−C2H4NH−benzyloxycarbonyl(◇)、α−TM(○)を用いた。[3H]標識重合反応生成物をSuperdex peptideカラムを用いてゲルろ過クロマトグラフィーを行うことにより分析した。α−TMを受容体基質に用いた場合には、[3H]標識重合反応生成物をアルカリ還元処理した後、分析に用いた。矢印はChn由来八糖〜十糖の溶出位置を示す。Vtはフラクション(Fr.)60付近である。B.重合反応によりα−TM上で合成された糖鎖(○)をSuperdex 75カラム(1.6×60cm)を用いて分析した。ゲルろ過クロマトグラフィー後の溶出液の放射活性を測定した。分子量スタンダードとしてChnポリマーについても分析し、溶出液の210nmにおける紫外吸収を測定した(●)。
図3に示すように、用いた様々な基質上にChn鎖の重合が起ったが、α−TM上には最も効率よく、より長くChn鎖が伸長した。
また、既に糖鎖が伸長したChn鎖にもChSyとChPFを共発現させた融合タンパク質により、Chn鎖が伸長されるかを確かめるため、Chn由来の六糖(GlcAβ1−3GalNAc)3およびChnポリマーを受容体基質として、Chn鎖の重合反応を行った。その結果が図4のグラフA〜Cに示される。
図4の説明:A.Chn鎖の重合反応の受容体基質としてChn由来のオリゴ糖(GlcAβ1−3GalNAc)3を用いた。[3H]標識重合反応生成物をSuperdex peptideカラムを用いてゲルろ過クロマトグラフィーにより分離し、溶出液の放射活性を測定した(●)。矢印はChn由来八糖〜十糖の溶出位置を示す。VtはFr.60付近である。B.Chnポリマーを受容体基質としてChPFとChSyを共発現させたもの(○)およびChSy単独のもの(●)を酵素源に用い重合反応を行い、[3H]重合反応生成産物をゲルろ過クロマトグラフィーにより単離した。単離した[3H]標識重合反応生成物をSuperdex 200カラム(1.6×60cm)を用いたゲルろ過クロマトグラフィーを行い、溶出液の放射活性を測定した。矢印は分子量マーカーとして用いたデキストランにより求めたそれぞれの分子量における溶出位置を示している。C.上記Bで単離した反応生成物をCSaseACII消化し、消化後Superdex Peptideカラムを用いてゲルろ過クロマトグラフィーを行うことによって分離し、液体シンチレーションカウンターで測定した。ChPFとChSyを共発現させたものを酵素源に用いたときの反応生成物のCSase消化後(○)およびChSy単独で発現させたものを酵素源に用いたときの反応生成物のCSase消化後(●)のクロマトグラムを示す。矢印はChn由来の二糖およびGalNAcの溶出位置を示す。
図4に示すように、Chn由来の六糖(GlcAβ1−3GalNAc)3を受容体基質として使用した場合は八〜十糖の位置にピークが検出されたことより、二糖〜四糖程度の糖鎖の伸長が起ったと予想された(同図A)。Chnポリマーを受容体基質とした場合、伸長した糖鎖の長さは、controlとしてChSyのみを酵素源として得られた生成物と比べて、ゲルろ過クロマトグラフィーによる分析で大きな差異は見られなかった(同図B)。しかし、CSaseACII消化の結果、ChSyを用いて調製した反応生成物からは単糖の位置のみに放射活性のあるピークが検出されたのに対して、ChPFとChSyを共発現させたものを酵素源に用いた場合の反応生成物からは、二糖の位置にも顕著なピークが検出された(同図C)。このことより、ChSy単独時よりも、Chnポリマー基質上に少なくとも数糖程度の糖鎖の伸長反応が起ったことが明らかになった。
〔実施例5:α−TMの四糖結合領域に重合したChn鎖の分析〕
次に、α−TMの四糖結合領域をプライマーとしてChn鎖の伸長反応が起ったかを確認した。その結果が図5のグラフA、Bに示される。
図5の説明:A.α−TMを受容体基質として重合反応を行い、[3H]標識重合反応生成物をゲルろ過により単離した。単離した[3H]標識重合反応生成物をアルカリ処理した後、2−aminobenzamide(AB)により蛍光標識した。蛍光標識した糖鎖をCSaseABCで完全に消化し、不飽和二糖標準品としてΔDi−0Sを添加し、Superdex peptideカラムを用いたゲルろ過クロマトグラフィーにより分画し、溶出液の放射活性(●)を測定した。矢印は不飽和二糖(ΔHex α 1−3GalNAc)の溶出位置を示す。B.結合領域を含む放射標識された画分(グラフAのFr.1)を回収し、陰イオン交換カラムを用いたHPLCで分析した。Fr.1のchondro−glycuronidaseによる消化前(○)および消化後(●)のクロマトグラムを示す。溶出液の放射活性を測定した。矢印は2−ABで標識された結合領域五糖と六糖の溶出位置を示す。波線はNaH2PO4の濃度勾配を示す。
図5Aに示すように、放射標識された糖鎖がCSaseABCにより消化され、結合領域を含むFr.1および二糖を含むFr.2を得た。Fr.1とFr.2の放射活性の比は1:47であるため、合成されたChn鎖は平均約百糖から成っていると予想された。
その後、Fr.1を陰イオン交換HPLCで分析したところ、放射活性をもった1本のピーク(図5Bの矢印2)が検出され、その溶出位置は、結合領域六糖標準品ΔHexA α 1−3GalNAcβ1−4GlcAβ1−3Galβ1−3Galβ1−4Xyl−2−ABのものと一致した。また、このピークはchondro−glycuronidaseによる消化によって非還元末端のΔHexAが消化され、GalNAcβ1−4GlcAβ1−3Galβ1−3Galβ1−4Xyl−2−ABの溶出位置(図5Bの矢印1)へ移動した。これらの結果より、糖鎖の伸長はα−TMの結合領域四糖上で起ったことが示された。
興味深いことに、ChSyとChPFの複合体がGalNAcT−II、GlcAT−II活性だけでなく、結合領域に最初にGalNAcを転移するGalNAcT−I活性をも保持していることが明らかとなった。この結果は、以前報告された、ヒトの悪性黒色腫G361細胞の培養上清中に見出した重合活性と同様の結果であった。
〔実施例6:ChPFとChSyの相互作用解析〕
以上の結果より、ChSyとChPFを同じ細胞内で共発現させると、糖転移活性の上昇およびChn鎖の重合が起こることが明らかとなった。さらに、それぞれを単独に発現させた融合タンパク質を混合したものでは、糖転移活性の上昇は見られなかった。このことより、単独で発現させたときには複合体は形成されず、同じ細胞内でChSyとChPFを共発現させることにより、複合体が形成されることが示唆された。そこで、ChSyとChPFが実際に複合体を形成しているかを確認するため、ChPFおよびChSyを6×Hisあるいはprotein Aとの分泌型融合タンパク質として同一細胞内で共発現させ、pull−down法によりChPFおよびChSy間の相互作用を調べた。その結果が図6に示される。
図6の説明:まず、ChPFおよびChSyを6×Hisあるいはprotein Aとの分泌型融合タンパク質として単独および同一細胞内で発現させ、Ni−NTA agaroseを用いたpull−down assayを行った。その後、SDS−PAGEにより分離し、protein Aとの融合タンパク質をIgG抗体を用いたwestern blottingにより検出した。Lane1ではprotein A−ChPFの大きさと予想される約110kDaのバンドが検出された。Lane4ではprotein A−ChSyの大きさと予想される約120kDaのバンドが検出された。各レーンのサンプルは以下のとおりである。Lane1:protein A−ChPF/His−ChSy、lane2:His−ChSy、lane3:protein A−ChSy+His−ChPF(単独に発現させたタンパク質を混合したもの)、lane4:protein A−ChSy/His−ChPF、lane5:His−ChPF。
このように、protein A−ChPF/His−ChSyおよびprotein A−ChSy/His−ChPFの組合せでChPFとChSyを共発現させたものでは、それぞれprotein A−ChPF(約110kDa)およびprotein A−ChSy(約120kDa)と予想されるバンドが検出された。しかし、単独で発現させたものや、単独で発現させそれぞれのタンパク質を混合したものでは、バンドは検出されなかった。このことより、ChPFおよびChSyは同一細胞内で相互作用していることが示された。
〔実施例7:Northern blotによるChPFの発現パターン解析〕
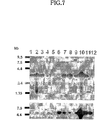
ChPFの発現パターンを調べるため、northern blot分析を行った。その結果が図7に示される。
図7の説明:ヒトの様々な組織から抽出したmRNAを用いて、ChPF(上段)およびChSy(下段)の発現を調べた。各レーンのサンプルは以下のとおりである。
Lane1,brain;lane2,heart;lane3,skeletal muscle;lane4,colon;lane5,thymus;lane6,spleen;lane7,kidney;lane8,liver;lane9,small intestine;lane10,placenta;lane11,lung;lane12,peripheral blood leukocytes。
図7に示すように、mRNAのサイズは約3.4kbであり、末梢白血球を除き調べた組織全てに発現が認められた。このことは、コンドロイチン硫酸がほとんどすべての組織に発現しているという結果と一致した。しかし、組識により発現レベルは異なり、胎盤および心臓では発現がもっとも強く、続いて脳、骨格筋、腎臓および肝臓で強い発現が見られた。さらに、ChPFはChSyのmRNAの発現パターンと非常に類似していた。このことより、両者が共同してコンドロイチン硫酸の生合成に関与していることが示された。
〔実験方法〕
(1)ChPFの機能解析のためのsoluble formの発現プラスミドの作製
内在性の糖転移酵素の混入をさけるために、培養上清中に可溶性の組換え型タンパク質としてChPFを発現させるための発現プラスミドを作製した。ChPFのN末端側から膜貫通ドメインと予想される最初の62アミノ酸を除いた配列をPCR法によって増幅させた。5’−Primerは5’末端にBamHI site(下線)を含む5’−CGGGATCCAACTCGGTGCAGC−3’を、3’−primerは5’末端にBamHI site(下線)を含む5’−CGGGATCCGCTCTGGTTTTGGGGGAGAAG−3’を用いた。PCRはKOD DNA polymeraseを使い、鋳型としてヒト悪性黒色腫細胞株G361(ATCC CRL−1424)由来cDNA library 50ngを使用した。反応は、5%DMSO存在下で94℃,30sec;55℃,30sec;68℃,180secを32cycle行った。増幅したfragmentをBamHIで消化し、pEF−BOS/IPベクターに組込み発現プラスミドとした(pEF−BOS/IP−ChPF)。
(2)培養細胞への遺伝子の導入と組換えタンパク質の精製
上記発現プラスミドpEF−BOS/IP−ChPF 6.0μgをFuGENETM6transfection reagentを用いてアフリカミドリザル腎臓由来COS−1細胞に遺伝子導入した。共発現の場合には、pEF−BOS/IP−ChPFとpEF−BOS/IP−ChSyを3.0μgずつ同様にFuGENETM6transfection reagentを用いて、COS−1細胞に両方のプラスミドを導入した。Transfection後、DMEM/10% FBS培地で2日間、37℃/5% CO2の条件でincubationした後、培養上清をIgG−Sepharoseと4℃で1時間混合し、protein Aとの融合タンパク質を結合させた。その後、1,000rpmで2min遠心し上清を除き、沈殿した樹脂を0.15M NaCl/0.02% Tween−20/50mM Tris−HCl(pH7.5)で洗浄した。これを3回繰り返し、精製された融合タンパク質/IgG−Sepharoseを酵素源とした。
(3)糖転移酵素活性の測定条件
(3−1)GalNAcT−II活性の測定
最終濃度として、50mM MES−NaOH(pH6.5)、10mM MnCl2、8.57μM UDP−[3H]GalNAc(3.60x105dpm)を加えた条件で171μgのChnを糖受容体基質として加えた。酵素源は上記方法(2)で精製したものを使用した。この反応溶液を37℃で2時間incubationした。シリンジカラムを用いて反応産物から遊離のアイソトープを除去し、溶出液の放射活性を液体シンチレーションカウンターで測定した。
(3−2)GlcAT−II活性の測定
最終濃度として、50mM sodium acetate buffer(pH5.6)、10mM MnCl2、14.3μM UDP−[14C]GlcA(1.46x105dpm)を加えた条件で171μgのChnを糖受容体基質として加えた。酵素源は上記方法(2)で精製したものを使用した。この反応溶液を37℃で2時間incubationした。シリンジカラムを用いて反応産物から遊離のアイソトープを除去し、溶出液の放射活性を液体シンチレーションカウンターで測定した。
(3−3)Polymerization活性の測定
最終濃度として100mM MES−NaOH(pH6.5)、10mM MnCl2、250μM UDP−[3H]GalNAc(50.0x105dpm)、250μM UDP−GlcAを加えた条件で1nmolのα−TM、GlcAβ1−3Galβ1−3Galβ1−4Xylβ1−O−Ser、GlcAβ1−3Galβ1−3Galβ1−4Xylβ1−O−(Gly)Ser−Gly−Glu、GlcAβ1−3Galβ1−3Galβ1−4Xylβ1−O−Ser−Gly−Trp−Pro−Asp−Gly、100nmol GlcAβ1−3Galβ1−O−C2H4NH−benzyloxycarbonyl、10nmol(GlcAβ1−3GalNAc)3または171μg Chnをそれぞれ糖受容体基質として加えた。酵素源は上記方法(2)で精製したものを使用した。この反応溶液を37℃で12時間incubationした。
(4)Polymerization reaction productの同定
(4−1)α−TMを糖受容体基質に用いた際のproductの同定
α−TMの重合反応後の精製物を、シリンジカラムを用いて、遊離のアイソトープを除去することにより単離した。α−TM上の[3H]GalNAcで標識された糖鎖を0.5M LiOHを用いたアルカリ処理によりコアタンパク質より切り出し、4分の1をSuperdex75カラムを用いたゲルろ過クロマトグラフィーにかけ、分画した画分の放射活性を液体シンチレーションカウンターで測定した。アルカリ処理した残りの4分の3を2−aminobenzamide(AB)により蛍光標識した。得られた蛍光標識された糖鎖を50mIUのCSaseABCを用いて1時間、完全に消化した。このCSaseABCによる消化物をSuperdex Peptideカラムを用いたゲルろ過クロマトグラフィーにかけ、2−ABで標識された結合領域六糖と考えられる画分を分取し、ポリアミン結合シリカカラム(PA−03)を用いたHPLCで分析した。また、chondro−glycuronidaseによる消化を行い、HPLCで分析した。反応生成物の同定は2−ABで標識された結合領域六糖標準品および結合領域五糖標準品とのco−クロマトグラフィーにより行った。
(4−2)糖鎖または糖ペプチドを受容体基質に用いた際のproductの同定
重合反応後の生成物をSuperdex Peptideカラムを用いたゲルろ過クロマトグラフィーによって分離し、分画した溶出液の放射活性を液体シンチレーションカウンターで測定した。
(4−3)Chnポリマーを受容体基質に用いた際のproductの同定
重合反応後の生成物を、シリンジカラムを用いて遊離のアイソトープを除去することにより単離した。その後、精製物をそのままSuperdex 200カラムを用いたゲルろ過クロマトグラフィーで分析した。また、単離した精製物をCSaseACII消化し、消化後Superdex Peptideカラムを用いたゲルろ過クロマトグラフィーを行うことによって分離し、分画した画分の放射活性を液体シンチレーションカウンターで測定した。
(5)ChPFとChSyの相互作用解析
(5−1)相互作用解析用のHis−tag融合タンパク質発現プラスミドの作製
ChPFとChSyとの相互作用を確認する際、内在性の糖転移酵素の影響を除去するため、培養上清中にHis−tagされたChPFおよびChSyを分泌型組換えタンパク質として発現させるための発現プラスミドを作製した。ChPFおよびChSyの膜貫通ドメイン以降の配列をPCR法によって以下のように増幅した。
a)ChPF:5’−Primerは5’末端にEcoRI site(下線)を含む5’−CGGAATTCAACTCGGTGCAGCCCGGAGC−3’を、3’−primerは5’末端にBamHI site(下線)を含む5’−CGGGATCCGCTCTGGTTTTGGGGGAGAAG−3’を用い、上記方法(1)と同条件でPCRを行った。増幅したfragmentをEcoRIとBamHIで消化し、pcDNA3(Ins.leader+6×His)ベクターに組込み発現プラスミドとした{pcDNA3(Ins.leader+6×His)−ChPF}。
b)ChSy:5’−Primerは5’末端にXbaI site(下線)を含む5’−GCTCTAGAGGCTGCCGGTCCGGGCAG−3’を、3’−primerは5’末端にXbaI site(下線)を含む5’−GCTCTAGACAATCTTAAAGGAGTCCTATGTA−3’を用い、上記方法(1)と同条件でPCRを行った。増幅したfragmentをXbaIで消化し、pcDNA3(Ins.leader+6×His)に組込み発現プラスミドとした{pcDNA3(Ins.leader+6×His)−ChSy}。
(5−2)培養細胞への遺伝子の導入と組換えタンパク質の精製
上記a)およびb)で調製した発現プラスミド6.0μgをFuGENETM6transfection reagentを用いてCOS−1細胞に遺伝子導入した。共発現の場合には、pcDNA3(Ins.leader+6×His)−ChSyとpEF−BOS/IP−ChPFあるいはpcDNA3(Ins.leader+6×His)−ChPFとpEF−BOS/IP−ChSyを3.0μgずつ同様にFuGENETM6transfection reagentを用いて、COS−1細胞に両方のプラスミドを導入した。Transfection後、DMEM/10% FBS培地で2日間、37℃/5% CO2の条件でincubationした後、培養上清をNi−NTA agaroseと4℃で1時間混合し、His−tag融合タンパク質を結合させた。その後、1,000rpmで2min遠心し上清を除き、沈殿した樹脂を0.15M NaCl/0.02% Tween−20/50mM Tris−HCl(pH7.5)で洗浄した。
(5−3)SDS−PAGEおよびwestern blot分析によるタンパク質相互作用の確認
上記方法で得たHis−tag融合タンパク質/Ni−NTA agaroseを1.3mM dithiothreitol(DTT)の存在下で94℃、5分間incubationした。その後、5,000rpmで5min遠心し、上清をサンプルとした。そのサンプルをSDS−PAGEによって分離し、分離されたタンパク質を電気的にPVDF膜に移行させた。1次抗体にはprotein Aと高親和性を持つマウスモノクローナル抗体IgG3(MSW113)、2次抗体にはhorseradish peroxidase(HRP)が結合したanti−mouse IgGを用い、ECL Advance Detection Reagents kitのプロトコールに従い、目的タンパク質を検出した。
(6)ChPFのnorthern blot解析
メンブランはHuman12−Lane MTNTM blot用い、probeは、ChPFが組込まれたプラスミドを鋳型として、PCR法により増幅し、増幅したフラグメントをBglII消化することにより、ChPFに特異的な840bpのフラグメントを精製したものを用いた。上記方法(1)と同条件でPCRを行い、5’−primerは5’−CGGGATCCACTCCTCTGGCTGCTCTGG−3’を、3’−primerは5’−CGGGATCCGCTCTGGTTTTGGGGGAGAAG−3’を用いた。Probeの標識は[α−32P]dCTPを使用し、ランダムプライム法に基づくoligolabelling kitを用いて行った。次に、メンブランをプレハイブリダイゼーションし、32P標識probeを加え、68℃で24時間ハイブリダイゼーションを行った。メンブランを洗浄した後、BIOMAXTM MA(Kodak社)に1週間露光し、検出した。
(7)酵素活性の測定方法
(7−1)Superdex Peptide、Supredex 75、Superdex 200の各種カラムを用いたゲルろ過クロマトグラフィー
酵素反応後のサンプルを遠心する事によって夾雑物を取り除き、予め0.2M NH4HCO3で平衡化しておいたカラムにアプライした。0.4ml/minの流速でゲルろ過クロマトグラフィーを行い、0.4mlずつ分画した画分の放射活性を、液体シンチレーションカウンターで測定した。
(7−2)シリンジカラムアッセイ
1mlのシリンジ(0.45x6.8cm)に0.2M NH4HCO3で平衡化したSephadex G−25(superfine)を充填し、150gx5min、続いて900gx5min遠心して、ゲル内の溶媒を除去した。0.2M NH4HCO3を30μl加え、全量50μlとしたサンプルをこの樹脂に添加して900gx5minで遠心し、さらに0.2M NH4HCO3を50μl加えてもう1度、900gx5minで遠心し、未反応のUDP−糖を取り除き、その溶出液の放射活性を液体シンチレーションカウンターで測定した。
〔実験結果の考察〕
ChPFとChSyを共発現させることにより、in vitroでChn鎖の重合化が可能になることを明らかにした。ChPF自身はGalNAcT−IIおよびGlcAT−II活性をほとんど保持していないにも関わらず、ChPFは今までに同定されている他の5つのコンドロイチン硫酸(CS)の生合成に関与している糖転移酵素と高い相同性を示す。それゆえ、ChPFはCSの生合成に関与する6番目の遺伝子ファミリー(ChSy family)のmemberであると考えられる。
以前、in vitroでヒトの悪性黒色腫G361細胞培養上清を酵素源に用い、α−TM上にChn鎖の重合化がおこることが報告されている。さらに基質阻害実験により、この推定されるchondroitin polymeraseは3つの糖転移活性を保持していることが予想された。1つはCS合成の開始に関与する四糖結合領域に最初のGalNAcを転移するGalNAcT−I活性、2つ目および3つ目はCS鎖を伸長させるGalNAcT−IIおよびGlcAT−II活性である。今回、ChPFとChSyを共発現した組換えタンパク質を酵素源とした際に、α−TMの四糖結合領域上にChn鎖の重合化が起こったことより、Chnの重合化は単独のタンパク質により担われるのではなく、ChPFとChSyの複合体により担われていることが示唆された。Pull−down法による相互作用解析からも、ChPFとChSyが同一細胞内で複合体を形成していることが強く示唆された(図6)。さらに、G361細胞の培養上清から部分精製した酵素活性の分子量は160kDaであり、今回クローニングしたChPFおよびChSyの分泌型のタンパク質の分子量は、それぞれ約80kDaであり、両者が1:1で複合体を形成していることが示唆された。
以前の報告と同様に、ChSyはChnポリマーやChn由来のオリゴ糖に対して低い糖転移活性しか示さなかった(表1)。しかし、ChSyとChPFを可溶性の融合タンパク質として共発現させると、ChSy単独時よりもGalNAcT−IIおよびGlcAT−II両活性は20倍以上に上昇した(表1)。このことから、ChSyが十分な活性を発揮するためにはChPFが必要であると考えられる。
さらに、Chn鎖の重合化にはコアタンパク質が重要な役割を果たしていることが示唆された。GlcAβ1−3Galβ1−3Galβ1−4Xylβ1−O−Ser、GlcAβ1−3Galβ1−3Galβ1−4Xylβ1−O−(Gly)Ser−Gly−Glu、GlcAβ1−3Galβ1−3Galβ1−4Xylβ1−O−Ser−Gly−Trp−Pro−Asp−Gly、GlcAβ1−3Galβ1−O−C2H4NH−benzyloxycarbonyl、(GlcAβ1−3GalNAc)3、およびChnポリマーを受容体基質に用いたときと比較し、α−TMを受容体基質に用いたときの方がより長く効率よくChn鎖の重合化が起った(図3)。これに関連して、以前、PGであるdecorinのコアタンパク質の様々な欠失変異体では短いGAG鎖しか合成されず、その理由としてコアタンパク質と糖転移酵素間の親和性が低下したためであるとの報告がある。それ故、今回の結果はChn鎖の長さが、受容体となるコアタンパク質によって影響を受け、ChSyとChPFの複合体がコアタンパク質と相互作用し、GAG鎖の長さを調節していることが示唆された。
動物細胞においてβ−D−xylosideをプライマーとしてGAGが合成されることが知られており、このことより疎水的な性質を保持しているxylosideのアグリコン部分はコアタンパク質を模倣することが示唆されている。実際、ChPFとChSyの組換え型タンパク質を同一細胞内で共発現させたものを酵素源とし、GlcAβ1−3Galβ1−O−C2H4NH−benzyloxycarbonylを受容体とした際、Chn鎖の重合反応が起った(図3)。しかし、α−TMを受容体に用いたときよりも短いChn鎖しか重合しなかった(図3)。このことは疎水的なアグリコンがGAG鎖の付加するコアタンパク質の最小限の性質を保持しており、ChSyとChPFの複合体と相互作用することが可能であることを示唆している。
これまでの研究では、通常、重合化反応と硫酸化修飾は同じGolgi区画で起り、硫酸化修飾と糖鎖の伸長および停止との間には密接な相関があると考えられている。伸長しているCS鎖の硫酸化は糖鎖の重合化に影響を与えるといわれているが、今回の結果よりin vitroではChn鎖の重合化には硫酸化修飾は必須ではないことが示された。
Northern blot解析により、ChPFは末梢白血球を除いて調べた組織ほとんど全てに発現が認められた。また、ChPFはChSyのmRNAの発現パターンと非常に類似していたことより、両者が共同してCSの生合成に関与していることが強く示唆された(図7)。
今回、組換え型タンパク質として共発現させたChSyとChPFの複合体を用いてin vitroでChn鎖の重合化を起こすことができた。この知見は、CSの生合成機構の解明およびその機能解析にも役立つと考えられる。
尚、発明を実施するための最良の形態の項においてなした具体的な実施態様または実施例は、あくまでも、本発明の技術内容を明らかにするものであって、そのような具体例にのみ限定して狭義に解釈されるべきものではなく、次に記載する特許請求の範囲内で、様々に変更して実施することができる。 The present invention, in collaboration with chondroitin synthase (ChSy), identifies a polymerization factor that promotes the polymerization of chondroitin chains and a gene that encodes the same, and analyzes the function in detail. We propose a method for synthesizing chondroitin chains. Therefore, in the following, the sequence and structure of this chondroitin polymerization factor and its gene (polynucleotide) will be described, and then its function, usage method, etc. will be described.
(1) Chondroitin polymerization factor and gene sequence and structure according to the present invention
A BLAST search was performed based on the amino acid sequence of human ChSy. As a result, gene information (accession number: BC021223) having high homology with ChSy was found. However, the deduced amino acid sequence disclosed in this information was a short one consisting of 522 residues. As a result of proceeding with the analysis based on the obtained data, the present inventor has a novel protein comprising 775 amino acids, showing 23% homology with human ChSy and 57% homology with human Chondroitin GlcAT ( The presence of ChPF) was identified (see FIG. 1).
As will be described later, this novel protein did not have a DXD motif which is a binding motif of UDP-sugar which is a sugar donor. This suggests that this gene product is involved in the biosynthesis of chondroitin sulfate, but does not retain glycosyl transfer activity. The results of functional analysis also revealed that this gene product is not a glycosyltransferase but a factor that polymerizes chondroitin in cooperation with ChSy. Therefore, the present inventor has identified this novel protein as a chondroitin polymerization factor ( It was named chondroitin polymerizing factor (ChPF)).
SEQ ID NO: 1 in the sequence listing shows the cDNA sequence of the human ChPF gene, and SEQ ID NO: 2 shows the amino acid sequence of human ChPF encoded by the gene. SEQ ID NO: 4 shows the amino acid sequence of a soluble ChPF recombinant protein obtained by recombining 62 amino acids from the N-terminal side containing the predicted transmembrane domain of ChPF with other sequences. SEQ ID NO: 3 Shows the cDNA sequence. As will be described later, functional analysis of ChPF was performed by expressing this soluble ChPF.
As a result of searching and analyzing the genome information, as shown in FIG. 2, ChPF was present on the second chromosome in a spread of about 5 kb, and the gene in the translation region was composed of four exons. This genomic structure was similar to the Chondroitin GlcAT gene.
The protein according to the present invention, that is, chondroitin polymerization factor is (a) a protein consisting of the amino acid sequence shown in SEQ ID NO: 2, (b) a soluble protein consisting of the amino acid sequence shown in SEQ ID NO: 4, (c) SEQ ID NO: 2 Or a protein consisting of an amino acid sequence comprising the amino acid sequence shown in 4 (for example, a fusion protein), (d) in the amino acid sequence of any one of (a) to (c) above, one or several amino acids are substituted or missing It may be any of proteins consisting of lost, inserted and / or added amino acid sequences (functioning as a chondroitin polymerization factor). Moreover, not only human origin but chondroitin polymerization factor derived from other organisms may be sufficient.
The above “one or several amino acids are substituted, deleted, inserted, and / or added” is a degree that can be replaced, deleted, inserted, and / or added by a known mutant protein production method such as a point mutagenesis method. Means that a certain number of amino acids are substituted, deleted, inserted, and / or added. Thus, in other words, the protein of (d) above is the mutant protein of (a) to (c) above, and the “mutation” mentioned here is artificially introduced mainly by a known mutant protein production method. The same mutant protein that exists in nature may be isolated and purified.
The protein according to the present invention may contain a known additional sequence such as HA or Flag at the end or may be a fusion protein in order to easily purify and detect the protein. Moreover, you may give various well-known modifications and may form a multimer more than a dimer.
The “polynucleotide (gene)” of the present invention only needs to have a sequence encoding the protein of any one of the above (a) to (d), and further includes a sequence of an untranslated region (UTR). It may be.
The “polynucleotide (gene)” of the present invention includes RNA and DNA. RNA includes mRNA, siRNA, and the like, and DNA includes cDNA, genomic DNA, and the like. The DNA may be double-stranded or single-stranded, and the single-stranded DNA may be a coding DNA serving as a sense strand or an anti-coding strand serving as an antisense strand. The antisense strand can be used as a probe or as an antisense drug. Moreover, siRNA can be utilized for gene expression suppression etc. as RNAi.
(2) Function of chondroitin polymerization factor and its utilization method
As a result of functional analysis, the following functions have been found in the chondroitin polymerization factor (ChPF) (for details, refer to Examples described later). 1) When ChSy and ChPF retaining GalNAcT-II and GlcAT-II activities were co-expressed in the same cell, the activities of GalNAcT-II and GlcAT-II were increased by 20 times or more compared to the ChSy single expression. . 2) As a result of enzymatic reaction in the presence of UDP-GalNAc and UDP-GlcA using α-TM as a receptor substrate and secreted protein coexpressed with ChSy and ChPF as an enzyme source, the result is the same as that of natural chondroitin chain We succeeded in causing polymerization to a certain length. 3) In the polymerization reaction of chondroitin chains using ChPF, it is preferable to use a core protein such as α-TM as a receptor substrate. 4) It is considered that ChSy co-expressed as a secretory protein and ChPF form a complex and interact with each other. 5) ChPF was expressed in most tissues examined, and its expression pattern was similar to ChSy. From this, it is considered that both of them are involved in the biosynthesis of chondroitin sulfate in vivo.
Therefore, the chondroitin polymerization factor (ChPF) and its gene can be used for artificial synthesis of a sugar chain skeleton of a sulfated glycosaminoglycan (chondroitin sulfate, dermatan sulfate) having physiological activity. For example, chondroitin has been prepared by chemically desulfating chondroitin sulfate prepared from whale cartilage and the like, but ChSy and ChPF are co-expressed and used as an enzyme source. It becomes possible to stably and easily mass-produce chondroitin sugar chains in vitro.
In the sugar chain synthesis using the chondroitin polymerization factor (ChPF), it is preferable to synthesize a chondroitin sugar chain on a core protein such as α-thrombomodulin (α-TM).
Furthermore, by using various sulfated transferases having different specificities, it can be used for the synthesis of various chondroitin sulfates having physiological activity and dermatan sulfate, which is an isomer thereof.
In addition, the chondroitin polymerization factor and gene thereof of the present invention can be used as a research reagent and a test reagent, as well as diseases caused by abnormal chondroitin polymerization factor, sulfates such as chondroitin sulfate and dermatan sulfate. The present invention can be used for pathological analysis of diseases related to glycated glycosaminoglycan and development of therapeutic agents, therapeutic methods, diagnostic agents, diagnostic methods and the like for the diseases.
(3) Protein and gene acquisition method according to the present invention
The method for obtaining the ChPF gene according to the present invention is not particularly limited, and DNA fragments containing the gene sequence can be isolated and cloned by various methods based on the above-described disclosed sequence information and the like. it can.
For example, a probe that specifically hybridizes with a partial sequence of the cDNA of the ChPF gene may be prepared, and a human genomic DNA library or cDNA library may be screened. As such a probe, any probe of any sequence / length may be used as long as it specifically hybridizes to at least part of the cDNA sequence of the ChPF gene or its complementary sequence. In silico screening may also be performed.
The clone obtained by the above screening can be analyzed in more detail by preparing a restriction enzyme map and determining its nucleotide sequence (sequencing). From these analyses, it can be easily confirmed whether a DNA fragment containing the gene sequence according to the present invention has been obtained.
Further, if the sequence of the probe is selected from the highly specific region of the ChPF gene and a genomic DNA (or cDNA) library of human, mouse or other organisms is screened, the same sequence as the ChPF protein is obtained. There is a high possibility that a gene encoding a homologous molecule or a related molecule having a function can be isolated and cloned.
The method for obtaining the gene (polynucleotide) according to the present invention includes a method using an amplification means such as PCR in addition to the above screening method. For example, among the above-mentioned cDNA sequences of ChPF, primers are prepared from sequences of untranslated regions on the 5 ′ side and 3 ′ side (or their complementary sequences), and human genomic DNA (or cDNA) is prepared using these primers. ) And the like as a template, and amplifying a DNA region sandwiched between both primers, a large amount of DNA fragments containing the polynucleotide of the present invention can be obtained.
The method for obtaining the ChPF protein or the like according to the present invention is not particularly limited. For example, the gene obtained as described above (cDNA or the like encoding the ChPF protein or a homologous molecule thereof) is used. According to the present invention, the introduced recombinant expression vector is prepared and incorporated into microorganisms such as Escherichia coli and yeast or animal cells by a well-known method, and the protein encoded by the cDNA is expressed and purified as a transformant. Protein can be easily obtained.
The method for producing the mutant protein of the ChPF protein is not particularly limited. For example, the base sequence of the gene obtained as described above using a known mutant protein production method such as a point mutation introduction method. In which one or more bases can be modified by substitutions, deletions, insertions and / or additions. Moreover, you may utilize a commercially available kit for preparation of a mutein.
(4) ChPF knockout animal of the present invention
The ChPF knockout animal of the present invention, that is, a non-human animal obtained by modifying all or part of the ChPF gene sequence in the genome and having one or both ChPF genes on the homologous chromosome disrupted, is a gene targeting method. Can be used.
Here, the phrase “ChPF gene has been destroyed” means that a protein having the original function of ChPF is not expressed. Therefore, a partial fragment (polypeptide) of the ChPF protein having no ChPF intrinsic activity may be expressed.
The gene targeting method may be performed according to a normal method. For example, first, a targeting vector (targeting construct) for homologous recombination is constructed. The targeting vector can be prepared by a known method. Generally, a ChPF genomic DNA clone, a commercially available plasmid, a marker for positive selection (eg, PGK-neo cassette), and a marker for negative selection (DTA gene, HSV- Each fragment such as a tk gene) can be appropriately ligated. At this time, the targeting vector may be designed so that the target restriction enzyme cleavage site is located at an appropriate position. Moreover, since the efficiency of targeting depends on the length of the homologous region, it is preferable that the homologous region is as long as possible. Furthermore, since the targeting vector is preferably linear rather than circular, an appropriate restriction enzyme cleavage site may be provided in a portion other than the homologous region for linearization.
The ChPF gene targeting vector prepared by the above method is introduced into a totipotent cell derived from a target animal such as an ES cell by electroporation or the like, and then a cell in which the desired homologous recombination has occurred is selected. The screening can be efficiently performed using a drug by a positive-negative selection method. After selection, the cells in which the desired homologous recombination has occurred are confirmed by Southern blotting or PCR. The totipotent cells finally confirmed to have homologous recombination are introduced into blastocysts (blast cysts) collected from the uterus during pregnancy. The introduction of cells into the blastocyst can be performed by a microinjection method or the like, but is not limited thereto.
The blastocyst is transplanted to a temporary parent according to a conventional method. By crossing a germline chimeric animal (preferably male) born from a foster parent with a wild-type animal (preferably female) having a wild-type ChPF gene homozygously, as a first generation (F1), A heterozygote in which one ChPF gene is disrupted by homologous recombination can be obtained. Furthermore, by mating these heterozygotes, a homozygote in which both ChPF genes on the homologous chromosome are disrupted can be obtained as the second generation (F2). The homozygote can be identified by cutting a part of the body (eg, tail), extracting the DNA, and examining the genotype by Southern blotting or PCR. Further, as a second generation (F2), a wild-type animal (having a wild-type gene homozygous) that is the same as a ChPF knockout animal can be obtained, and this wild-type animal can be suitably used for a control experiment.
The gene targeting method described above is merely an example, and it goes without saying that various known modifications can be made.
The animal that is the subject of the ChPF knockout animal is not particularly limited, and examples thereof include mammals such as cows, pigs, sheep, goats, rabbits, dogs, cats, guinea pigs, hamsters, mice, and rats. Of these, rabbits, dogs, cats, guinea pigs, hamsters, mice, and rats are preferred for use as experimental animals, of which rodents are more preferred, and many inbred strains have been produced. Particularly preferred are mice that are equipped with techniques such as in vitro fertilization. As totipotent cells, in addition to fertilized eggs and early embryos, pluripotent ES cells and cultured cells such as somatic stem cells can be targeted.
The ChPF knockout animal of the present invention is useful for functional analysis of ChPF.
EXAMPLES Hereinafter, although an Example demonstrates this invention more concretely, this invention is not limited at all by these Examples. Detailed experimental methods in each example will be described together in the last section.
[Example 1: Cloning of ChPF]
As a result of a BLAST search using the amino acid sequence of human ChSy, it was found that a novel gene having homology with ChSy exists. This gene was composed of a 236 bp 5 'untranslated region, a 2325 bp translated region encoding 775 amino acids (Figure 1 and SEQ ID NOs: 1 and 2), and an approximately 500 bp 3' untranslated region. The sequence predicted to be the translation initiation region followed the Kozak consensus sequence. Moreover, the protein translated from this gene is a characteristic II of a glycosyltransferase localized in the Golgi apparatus having one transmembrane domain consisting of 23 amino acids on the N-terminal side from Kyte-Doolittle hydropathy analysis. It was identified as a type membrane protein and further retained three N-glycosylation sites. This gene showed 23% homology with ChSy at the amino acid sequence level and 57% homology with human Chondroitin GlcAT (FIG. 1). However, there was no DXD motif, which is a binding motif of UDP-sugar, which is a sugar donor. From this, it was predicted that this gene product is involved in the biosynthesis of chondroitin sulfate but does not retain transglycosylation activity. The results of functional analysis also revealed that this gene product is not a glycosyltransferase but a factor that polymerizes chondroitin (hereinafter referred to as “Chn” for short) in cooperation with ChSy. Named this novel protein a chondroitin polymerizing factor (ChPF).
As a result of the analysis, it was found that the above-mentioned ChPF ortholog was also present in Caenorhabditis elegans and Drosophila melanogaster, with 27 and 25% homology, respectively.
[Example 2: ChPF gene structure and chromosome mapping]
As shown in FIG. 2, ChPF was present on the second chromosome extending over about 5 kb, and the gene in the translation region was composed of four exons. Each intron / exon boundary was GT / AG rule compatible.
[Example 3: ChPF expression and functional analysis]
In order to analyze the function of ChPF, 62 amino acids from the N-terminal side including the transmembrane domain of ChPF were recombined with a sequence of the IgG-binding domain of insulin leader and protein A, which are extracellular secretion signals. -1 cells expressed secreted protein fused with protein A. This ChPF recombinant protein was used as an enzyme source, and transglycosylation activity was measured using Chn as a sugar acceptor substrate and UDP-GalNAc or UDP-GlcA as a sugar donor. Table 1).
Therefore, ChPF was considered to be a polymerization factor rather than a glycosyltransferase, and ChSy having glycosyltransferase activity that synthesizes the disaccharide repeat region of the Chn chain was coexpressed with ChPF in the same cell. As a result, when the co-expression of ChPF with ChSy was used as the enzyme source, the GalNAcT-II and GlcAT-II activities increased more than 20 times compared to ChSy alone (Table 1). In addition, no increase in transglycosylation activity was observed in a mixture of recombinant proteins in which ChSy and ChPF were individually expressed. This suggested that ChSy and ChPF form a complex in the cell. Furthermore, it was considered that the complex was not formed when expressed alone, but was formed by co-expressing ChSy and ChPF in the same cell.
As described above, ChSy expressed as a recombinant protein has both GalNAcT-II and GlcAT-II activities that transfer GalNAc and GlcA with monosaccharides, but is not present on α-TM. The activity of polymerizing the Chn chain was not retained.
Therefore, since the increase in transglycosylation activity was observed in the co-expressed ChPF and ChSy, the co-expressed ChPF and ChSy may have the activity of polymerizing the Chn chain. It was. In order to analyze this point, α-TM, GlcAβ1-3Galβ1-3Galβ1-4Xylβ1-O-Ser, and α-TM derived from the tetrasaccharide-binding region were obtained using the recombinant protein coexpressed with ChSy and ChPF as the enzyme source. GlcAβ1-3Galβ1-3Galβ1-GlcAβ1-3Galβ1-3Galβ1-Peptide-bound GlcAβ1-3Galβ1-3Galβ1-O- (Gly) Ser-Gly-Glu; 4Xylβ1-O-Ser-Gly-Trp-Pro-Asp-Gly and the artificial substrate GlcAβ1-3Galβ1-OC2H4Polymerization activity was measured using NH-benzoxycarbonyl as a sugar acceptor substrate. The results are shown in graphs A and B in FIG.
Explanation of FIG. GlcAβ1-3Galβ1-3Galβ1-4Xylβ1-O-Ser (●), GlcAβ1-3Galβ1-3Galβ1-4Xylβ1-O- (Gly) Ser-Gly-Glu (×), GlcAβ1-3Galβ1-3Gal1 -4Xylβ1-O-Ser-Gly-Trp-Pro-Asp-Gly (Δ), GlcAβ1-3Galβ1-OC2H4NH-benzoxycarbonyl (() and α-TM (◯) were used. [3H] -labeled polymerization reaction products were analyzed by gel filtration chromatography using a Superdex peptide column. When α-TM is used as a receptor substrate, [3The H] labeled polymerization reaction product was subjected to alkali reduction treatment and then used for analysis. The arrow indicates the elution position of Chn-derived octasaccharide to decasaccharide. Vt is around the fraction (Fr.) 60. B. The sugar chain (◯) synthesized on α-TM by the polymerization reaction was analyzed using a Superdex 75 column (1.6 × 60 cm). The radioactivity of the eluate after gel filtration chromatography was measured. Chn polymer was also analyzed as a molecular weight standard, and ultraviolet absorption at 210 nm of the eluate was measured (●).
As shown in FIG. 3, polymerization of the Chn chain occurred on the various substrates used, but the Chn chain was elongated most efficiently on α-TM.
In addition, Chn-derived hexasaccharide (GlcAβ1-3GalNAc) is used to confirm whether the Chn chain is extended by the fusion protein in which ChSy and ChPF are co-expressed in the already extended Chn chain.3Then, a Chn chain polymerization reaction was carried out using the Chn polymer as an acceptor substrate. The results are shown in graphs AC in FIG.
Explanation of FIG. Chn-derived oligosaccharide (GlcAβ1-3GalNAc) as an acceptor substrate for polymerization reaction of Chn chain3Was used. [3H] The labeled polymerization reaction product was separated by gel filtration chromatography using a Superdex peptide column, and the radioactivity of the eluate was measured (●). The arrow indicates the elution position of Chn-derived octasaccharide to decasaccharide. Vt is Fr. Around 60. B. Polymerization reaction was carried out using Chn polymer as an acceptor substrate and co-expression of ChPF and ChSy (◯) and ChSy alone (●) as an enzyme source, [3H] The product of the polymerization reaction was isolated by gel filtration chromatography. Isolated [3H] The labeled polymerization reaction product was subjected to gel filtration chromatography using a Superdex 200 column (1.6 × 60 cm), and the radioactivity of the eluate was measured. Arrows indicate the elution positions at the respective molecular weights determined by dextran used as molecular weight markers. C. The reaction product isolated in B above was digested with CSaseACII, and after digestion, the reaction product was separated by gel filtration chromatography using a Superdex Peptide column, and measured with a liquid scintillation counter. After CSase digestion of reaction product when using ChPF and ChSy co-expressed as enzyme source (○) and after CSase digestion of reaction product when using ChSy alone expressed as enzyme source The chromatogram of (●) is shown. The arrow indicates the elution position of Chn-derived disaccharide and GalNAc.
As shown in FIG. 4, Chn-derived hexasaccharide (GlcAβ1-3GalNAc)3When was used as a receptor substrate, it was predicted that the sugar chain was elongated from about disaccharide to tetrasaccharide because the peak was detected at the position of octadecasaccharide (Fig. A). When Chn polymer was used as the acceptor substrate, the length of the extended sugar chain was not significantly different from the product obtained by using only ChSy as the control as the enzyme source and analyzed by gel filtration chromatography. (Fig. B). However, as a result of CSaseACII digestion, a radioactive peak was detected only at the position of the monosaccharide from the reaction product prepared using ChSy, whereas the enzyme source was obtained by co-expressing ChPF and ChSy. From the reaction product used in the above, a prominent peak was also detected at the position of the disaccharide (Fig. C). From this, it was clarified that a sugar chain elongation reaction of at least several sugars occurred on the Chn polymer substrate compared with ChSy alone.
[Example 5: Analysis of Chn chain polymerized to tetrasaccharide binding region of α-TM]
Next, it was confirmed whether the elongation reaction of the Chn chain occurred using the tetrasaccharide binding region of α-TM as a primer. The results are shown in graphs A and B in FIG.
Explanation of FIG. Perform polymerization reaction using α-TM as the acceptor substrate, [3The H] labeled polymerization reaction product was isolated by gel filtration. Isolated [3H] The labeled polymerization reaction product was alkali-treated and then fluorescently labeled with 2-aminobenzamide (AB). The fluorescently labeled sugar chain is completely digested with CSaseABC, ΔDi-0S is added as an unsaturated disaccharide standard, fractionated by gel filtration chromatography using a Superdex peptide column, and the radioactivity of the eluate (●) Was measured. The arrow indicates the elution position of unsaturated disaccharide (ΔHex α 1-3GalNAc). B. The radiolabeled fraction containing the binding region (Fr. 1 in graph A) was collected and analyzed by HPLC using an anion exchange column. Fr. 1 shows chromatograms before digestion (◯) and after digestion (●) with 1 chondro-glycuronidase. The radioactivity of the eluate was measured. The arrows indicate the elution positions of the binding region pentasaccharide and hexasaccharide labeled with 2-AB. The wavy line is NaH2PO4The concentration gradient is shown.
As shown in FIG. 5A, the radiolabeled sugar chain was digested with CSaseABC, and Fr. Fr. containing 1 and disaccharides. 2 was obtained. Fr. 1 and Fr. Since the ratio of the radioactivity of 2 was 1:47, the synthesized Chn chain was expected to consist of an average of about a hundred sugars.
Thereafter, Fr. When 1 was analyzed by anion exchange HPLC, one radioactive peak (
Interestingly, it was revealed that the complex of ChSy and ChPF retains not only GalNAcT-II and GlcAT-II activity but also GalNAcT-I activity that first transfers GalNAc to the binding region. This result was similar to the previously reported polymerization activity found in the culture supernatant of human malignant melanoma G361 cells.
[Example 6: Analysis of interaction between ChPF and ChSy]
From the above results, it has been clarified that when ChSy and ChPF are co-expressed in the same cell, an increase in sugar transfer activity and polymerization of the Chn chain occur. Furthermore, no increase in transglycosylation activity was observed in a mixture of fusion proteins in which each was expressed alone. This suggests that no complex was formed when expressed alone, but a complex was formed by co-expressing ChSy and ChPF in the same cell. Therefore, in order to confirm whether ChSy and ChPF actually form a complex, ChPF and ChSy are co-expressed as a secreted fusion protein with 6 × His or protein A in the same cell, and the pull-down method is used. The interaction between ChPF and ChSy was examined. The result is shown in FIG.
Explanation of FIG. 6: First, ChPF and ChSy were expressed alone and in the same cell as a secreted fusion protein with 6 × His or protein A, and then pull-down assay using Ni-NTA agarose was performed. Then, it isolate | separated by SDS-PAGE and the fusion protein with protein A was detected by Western blotting using IgG antibody. In
Thus, protein A-ChPF (about 110 kDa) and protein A-ChSy (about 110 kDa) and protein A-ChPF (about 110 kDa) and protein A-ChPF / His-ChSy and protein A-ChSy / His-ChPF co-expressed with ChPF and ChSy, respectively, A band expected to be about 120 kDa) was detected. However, no band was detected in those expressed alone or in the case where each protein was expressed alone and mixed with each protein. This indicates that ChPF and ChSy interact in the same cell.
[Example 7: Expression pattern analysis of ChPF by Northern blot]
In order to examine the expression pattern of ChPF, a northern blot analysis was performed. The result is shown in FIG.
Explanation of FIG. 7: Expression of ChPF (upper) and ChSy (lower) was examined using mRNA extracted from various human tissues. Samples for each lane are as follows.
As shown in FIG. 7, the size of mRNA was about 3.4 kb, and expression was observed in all the tissues examined except for peripheral leukocytes. This was consistent with the result that chondroitin sulfate was expressed in almost all tissues. However, expression levels differed depending on the tissue, with the strongest expression in the placenta and heart followed by strong expression in the brain, skeletal muscle, kidney and liver. Furthermore, ChPF was very similar to the expression pattern of ChSy mRNA. From this, it was shown that both were jointly involved in the biosynthesis of chondroitin sulfate.
〔experimental method〕
(1) Preparation of a soluble form expression plasmid for functional analysis of ChPF
In order to avoid contamination with endogenous glycosyltransferases, an expression plasmid for expressing ChPF as a soluble recombinant protein in the culture supernatant was prepared. A sequence excluding the first 62 amino acids expected to be a transmembrane domain from the N-terminal side of ChPF was amplified by PCR. 5'-Primer contains 5'-CG containing a BamHI site (underlined) at the 5 'endGGATCCAACTCGGTGCAGC-3 ', 3'-primer contains 5'-CG containing BamHI site (underlined) at the 5' endGGATCCGCTCTGGTTTTGGGGGAGAAG-3 'was used. For PCR, KOD DNA polymerase was used and 50 ng of cDNA library derived from human malignant melanoma cell line G361 (ATCC CRL-1424) was used as a template. The reaction was carried out in the presence of 5% DMSO for 32 cycles at 94 ° C., 30 sec; 55 ° C., 30 sec; 68 ° C., 180 sec. The amplified fragment was digested with BamHI and incorporated into a pEF-BOS / IP vector to form an expression plasmid (pEF-BOS / IP-ChPF).
(2) Introduction of genes into cultured cells and purification of recombinant proteins
6.0 μg of the expression plasmid pEF-BOS / IP-ChPF was added to FuGENE.TMThe gene was introduced into COS-1 cells derived from African green monkey kidney using 6-translation reagent. In the case of co-expression, FuGENE is similarly used for 3.0 μg each of pEF-BOS / IP-ChPF and pEF-BOS / IP-ChSy.TMBoth plasmids were introduced into COS-1 cells using 6 transfection reagent. After Transfection, DMEM / 10% FBS medium for 2 days at 37 ° C / 5% CO2After incubation under the conditions described above, the culture supernatant was mixed with IgG-Sepharose at 4 ° C. for 1 hour to bind the protein A fusion protein. Thereafter, the mixture was centrifuged at 1,000 rpm for 2 minutes, the supernatant was removed, and the precipitated resin was washed with 0.15 M NaCl / 0.02% Tween-20 / 50 mM Tris-HCl (pH 7.5). This was repeated three times, and purified fusion protein / IgG-Sepharose was used as the enzyme source.
(3) Glycosyltransferase activity measurement conditions
(3-1) Measurement of GalNAcT-II activity
As final concentration, 50 mM MES-NaOH (pH 6.5), 10 mM MnCl28.57 μM UDP- [3H] GalNAc (3.60x105171 μg of Chn was added as a sugar receptor substrate under the condition of adding dpm). The enzyme source used was purified by the above method (2). This reaction solution was incubated at 37 ° C. for 2 hours. The free isotope was removed from the reaction product using a syringe column, and the radioactivity of the eluate was measured with a liquid scintillation counter.
(3-2) Measurement of GlcAT-II activity
As final concentration, 50 mM sodium acetate buffer (pH 5.6), 10 mM MnCl2, 14.3 μM UDP- [14C] GlcA (1.46 × 105171 μg of Chn was added as a sugar receptor substrate under the condition of adding dpm). The enzyme source used was purified by the above method (2). This reaction solution was incubated at 37 ° C. for 2 hours. The free isotope was removed from the reaction product using a syringe column, and the radioactivity of the eluate was measured with a liquid scintillation counter.
(3-3) Measurement of Polymerization activity
Final concentration of 100 mM MES-NaOH (pH 6.5), 10 mM MnCl2, 250 μM UDP- [3H] GalNAc (50.0x105dpm), 1 nmol α-TM, GlcAβ1-3Galβ1-3Galβ1-4Xylβ1-O-Ser, GlcAβ1-3Galβ1-3Galβ1-4Xylβ1-O- (Gly) Ser-Gly-GluGlu, with 250 μM UDP-GlcA added -3Galβ1-3Galβ1-4Xylβ1-O-Ser-Gly-Trp-Pro-Asp-Gly, 100 nmol GlcAβ1-3Galβ1-O-C2H4NH-benzoxycarbonyl, 10 nmol (GlcAβ1-3GalNAc)3Alternatively, 171 μg Chn was added as a sugar acceptor substrate. The enzyme source used was purified by the above method (2). This reaction solution was incubated at 37 ° C. for 12 hours.
(4) Identification of Polymerization reaction product
(4-1) Identification of product when α-TM is used as a sugar receptor substrate
The purified product after the polymerization reaction of α-TM was isolated by removing a free isotope using a syringe column. On α-TM3H] GalNAc-labeled sugar chain was excised from the core protein by alkali treatment with 0.5M LiOH, and a quarter was subjected to gel filtration chromatography using a Superdex 75 column, and the radioactivity of the fractionated fraction was determined. Measured with a liquid scintillation counter. The remaining three quarters treated with alkali were fluorescently labeled with 2-aminobenzamide (AB). The resulting fluorescently labeled sugar chain was completely digested with 50 mIU CSaseABC for 1 hour. The digested product of CSaseABC was subjected to gel filtration chromatography using a Superdex Peptide column, and a fraction considered to be a binding region hexasaccharide labeled with 2-AB was fractionated, and a polyamine-bound silica column (PA-03) was used. Was analyzed by HPLC. In addition, digestion with chondro-glycuronidase was performed and analyzed by HPLC. The reaction product was identified by co-chromatography with a binding region hexasaccharide standard and a binding region pentasaccharide standard labeled with 2-AB.
(4-2) Identification of product when sugar chain or glycopeptide is used as receptor substrate
The product after the polymerization reaction was separated by gel filtration chromatography using a Superdex Peptide column, and the radioactivity of the fractionated eluate was measured with a liquid scintillation counter.
(4-3) Identification of product when Chn polymer is used as receptor substrate
The product after the polymerization reaction was isolated by removing free isotopes using a syringe column. Thereafter, the purified product was directly analyzed by gel filtration chromatography using a Superdex 200 column. In addition, the isolated purified product was digested with CSaseACII, separated by gel filtration chromatography using a Superdex Peptide column after digestion, and the radioactivity of the fractionated fraction was measured with a liquid scintillation counter.
(5) Analysis of interaction between ChPF and ChSy
(5-1) Preparation of His-tag fusion protein expression plasmid for interaction analysis
When confirming the interaction between ChPF and ChSy, in order to remove the influence of endogenous glycosyltransferase, expression for expressing HisPF-labeled ChPF and ChSy as secretory recombinant proteins in the culture supernatant A plasmid was prepared. Sequences after the transmembrane domain of ChPF and ChSy were amplified by PCR as follows.
a) ChPF: 5'-Primer contains an EcoRI site (underlined) at the 5 'end 5'-CGGAATTCAACTCGGTGCAGCCCGGAGC-3 ', 3'-primer contains 5'-CG containing BamHI site (underlined) at the 5' endGGATCCPCR was performed using GCTCTGGTTTGGGGGAGAAG-3 'under the same conditions as in the above method (1). The amplified fragment was digested with EcoRI and BamHI, and incorporated into a pcDNA3 (Ins.leader + 6 × His) vector as an expression plasmid {pcDNA3 (Ins.leader + 6 × His) -ChPF}.
b) ChSy: 5'-Primer contains 5'-GC containing an XbaI site (underlined) at the 5 'endTCTAGAGCGTCGCCGTCGCGGGCAG-3 ', 3'-primer contains 5'-GC containing XbaI site (underlined) at the 5' endTCTAGAPCR was performed using CAATCTTAAAGGAGCTCTATGTA-3 'under the same conditions as in the above method (1). The amplified fragment was digested with XbaI and incorporated into pcDNA3 (Ins.leader + 6 × His) as an expression plasmid {pcDNA3 (Ins.leader + 6 × His) -ChSy}.
(5-2) Introduction of genes into cultured cells and purification of recombinant proteins
6.0 μg of the expression plasmid prepared in the above a) and b) was added to FuGENE.TMThe gene was introduced into COS-1 cells using 6-transfecting reagent. In the case of co-expression, pcDNA3 (Ins.leader + 6 × His) -ChSy and pEF-BOS / IP-ChPF or pcDNA3 (Ins.leader + 6 × His) -ChPF and pEF-BOS / IP-ChSy in the same amount of 3.0 μg FuGENETMBoth plasmids were introduced into COS-1 cells using 6 transfection reagent. After Transfection, DMEM / 10% FBS medium for 2 days at 37 ° C / 5% CO2After incubation under the conditions, the culture supernatant was mixed with Ni-NTA agarose at 4 ° C. for 1 hour to bind the His-tag fusion protein. Thereafter, the mixture was centrifuged at 1,000 rpm for 2 minutes, the supernatant was removed, and the precipitated resin was washed with 0.15 M NaCl / 0.02% Tween-20 / 50 mM Tris-HCl (pH 7.5).
(5-3) Confirmation of protein interaction by SDS-PAGE and Western blot analysis
The His-tag fusion protein / Ni-NTA agarose obtained by the above method was incubated at 94 ° C. for 5 minutes in the presence of 1.3 mM dithiothreitol (DTT). Thereafter, the mixture was centrifuged at 5,000 rpm for 5 minutes, and the supernatant was used as a sample. The sample was separated by SDS-PAGE, and the separated protein was electrically transferred to a PVDF membrane. Mouse monoclonal antibody IgG with high affinity for protein A as the primary antibody3(MSW113) Anti-mouse IgG conjugated with horseradish peroxidase (HRP) was used as the secondary antibody, and the target protein was detected according to the protocol of ECL Advance Detection Reagents kit.
(6) Northern blot analysis of ChPF
The membrane is Human12-Lane MTNTM Blot was used and probe was obtained by amplifying by PCR using a plasmid incorporating ChPF as a template, and digesting the amplified fragment with BglII to purify a 840 bp fragment specific for ChPF. PCR was performed under the same conditions as in the above method (1), and 5'-primer was 5'-CGGGATCCACTCCCTCTGGCTGCCTCTGG-3 ', and 3'-primer was 5'-CGGGATCCGCTCTGGTTTTGGGGGGAGAAG-3'. The label of Probe is [α-32P] dCTP was used, and an oligobolating kit based on the random prime method was used. Next, prehybridize the membrane,32P-labeled probe was added and hybridization was performed at 68 ° C. for 24 hours. After washing the membrane, BIOMAXTM MA (Kodak) was exposed for 1 week and detected.
(7) Measuring method of enzyme activity
(7-1) Gel filtration chromatography using various columns of Superdex Peptide, Superdex 75, Superdex 200
The sample after the enzyme reaction is centrifuged to remove impurities, and 0.2M NH in advance.4HCO3And applied to the column that had been equilibrated with. Gel filtration chromatography was performed at a flow rate of 0.4 ml / min, and the radioactivity of fractions fractioned by 0.4 ml was measured with a liquid scintillation counter.
(7-2) Syringe column assay
0.2 ml NH in a 1 ml syringe (0.45 x 6.8 cm)4HCO3Sephadex G-25 (superfine) equilibrated in (1) was filled, followed by centrifugation at 150 g × 5 min and then 900 g × 5 min to remove the solvent in the gel. 0.2M NH4HCO3Was added to this resin, centrifuged at 900 g × 5 min, and further 0.2 M NH4HCO3Was added, and centrifuged again at 900 g × 5 min to remove unreacted UDP-sugar, and the radioactivity of the eluate was measured with a liquid scintillation counter.
[Consideration of experimental results]
It was clarified that Chn chains can be polymerized in vitro by co-expressing ChPF and ChSy. Although ChPF itself retains little GalNAcT-II and GlcAT-II activity, ChPF is involved in the biosynthesis of the other five chondroitin sulfates (CS) that have been identified so far. High homology with the enzyme. Therefore, ChPF is considered to be a member of the sixth gene family (ChSy family) involved in CS biosynthesis.
Previously, it has been reported that Chn chain polymerization occurs on α-TM using human malignant melanoma G361 cell culture supernatant in vitro as an enzyme source. Furthermore, it was predicted from the substrate inhibition experiment that this estimated chondroitin polymerase retained three glycosyltransferase activities. One is GalNAcT-I activity that transfers the first GalNAc to the tetrasaccharide binding region involved in the initiation of CS synthesis, and the second and third are GalNAcT-II and GlcAT-II activities that extend the CS chain. In this study, when a recombinant protein coexpressed with ChPF and ChSy was used as the enzyme source, the polymerization of Chn occurred on the tetrasaccharide binding region of α-TM. It was suggested that it was carried by a complex of ChPF and ChSy rather than being carried. The interaction analysis by the Pull-down method also strongly suggested that ChPF and ChSy formed a complex in the same cell (FIG. 6). Furthermore, the molecular weight of the enzyme activity partially purified from the culture supernatant of G361 cells is 160 kDa, and the molecular weights of the secreted proteins of ChPF and ChSy cloned this time are about 80 kDa, both of which are 1: 1 and complex. It was suggested that
Similar to previous reports, ChSy showed only low transglycosylation activity for Chn polymers and Chn-derived oligosaccharides (Table 1). However, when ChSy and ChPF were co-expressed as a soluble fusion protein, both GalNAcT-II and GlcAT-II activities increased more than 20 times compared to ChSy alone (Table 1). From this, it is considered that ChPF is necessary for ChSy to exhibit sufficient activity.
Furthermore, it was suggested that the core protein plays an important role in the polymerization of the Chn chain. GlcAβ1-3Galβ1-3Galβ1-4Xylβ1-O-Ser, GlcAβ1-3Galβ1-3Galβ1-4Xylβ1-O- (Gly) Ser-Gly-Glu, GlcAβ1-3Galβ1-3Galβ1-4Xylβ1-O-Srp-Gly-β -Gly, GlcAβ1-3Galβ1-OC2H4NH-benzoxycarbonyl, (GlcAβ1-3GalNAc)3As compared with the case where the Chn polymer was used as the receptor substrate, polymerization of the Chn chain occurred longer and more efficiently when α-TM was used as the receptor substrate (FIG. 3). In this connection, various deletion mutants of the core protein of decorin, PG, previously synthesized only a short GAG chain because the affinity between the core protein and glycosyltransferase was reduced. There is a report. Therefore, the result of this time is that the length of the Chn chain is affected by the core protein as a receptor, and the complex of ChSy and ChPF interacts with the core protein and regulates the length of the GAG chain. Was suggested.
It is known that GAG is synthesized by using β-D-xyloside as a primer in animal cells. This suggests that the aglycon part of xyloside that retains hydrophobic properties mimics the core protein. Yes. In fact, the enzyme source is a co-expressed recombinant protein of ChPF and ChSy in the same cell, and GlcAβ1-3Galβ1-OC2H4When NH-benzoylcarbonyl was used as an acceptor, a Chn chain polymerization reaction occurred (FIG. 3). However, only a shorter Chn chain polymerized than when α-TM was used for the receptor (FIG. 3). This suggests that the hydrophobic aglycon retains the minimal properties of the core protein to which the GAG chain is added and can interact with the complex of ChSy and ChPF.
In previous studies, polymerization reaction and sulfation modification usually occur in the same Golgi compartment, and it is considered that there is a close correlation between sulfation modification and sugar chain elongation and termination. Although it is said that the sulfation of the extending CS chain affects the polymerization of sugar chains, the present results indicate that sulfation modification is not essential for polymerization of Chn chains in vitro. It was.
According to the Northern blot analysis, ChPF was found to be expressed in almost all tissues examined except for peripheral leukocytes. Moreover, since ChPF was very similar to the expression pattern of mRNA of ChSy, it was strongly suggested that both were involved in the biosynthesis of CS (FIG. 7).
This time, the Chn chain could be polymerized in vitro using a complex of ChSy and ChPF co-expressed as a recombinant protein. This finding may be useful for elucidating the biosynthetic mechanism of CS and analyzing its function.
It should be noted that the specific embodiments or examples made in the best mode for carrying out the invention are merely to clarify the technical contents of the present invention, and are limited to such specific examples. Therefore, the present invention should not be interpreted in a narrow sense, and various modifications can be made within the scope of the following claims.
以上のように、本発明のコンドロイチン重合化因子(ChPF)は、コンドロイチン合成酵素(ChSy)と複合体を形成し、コンドロイチン糖鎖の重合化を促進する作用を有するので、前述したとおり、コンドロイチン硫酸やデルマタン硫酸といった有用生理活性を有する硫酸化グリコサミノグリカンの糖鎖骨格の人工合成などに利用することができるほか、種々の有用性を有するものである。 As described above, the chondroitin polymerization factor (ChPF) of the present invention forms a complex with chondroitin synthase (ChSy) and promotes the polymerization of chondroitin sugar chains. It can be used for artificial synthesis of the sugar chain skeleton of sulfated glycosaminoglycans having useful physiological activities such as sulfa and dermatan sulfate, and has various usefulness.
Claims (3)
(a)配列番号2に示されるアミノ酸配列、
(b)配列番号4に示されるアミノ酸配列、
(c)配列番号2又は4に示されるアミノ酸配列を含むアミノ酸配列、
(d)上記(a)又は(b)に示されるアミノ酸配列において、1又は数個のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列
(但し(c)又は(d)はコンドロイチン合成酵素と共同してコンドロイチン鎖の重合化作用を有する)。 A complex obtained by co-expressing a protein comprising any one of the following amino acid sequences (a) to (d) and chondroitin synthase :
(A) the amino acid sequence shown in SEQ ID NO: 2,
(B) the amino acid sequence shown in SEQ ID NO: 4,
(C) an amino acid sequence comprising the amino acid sequence shown in SEQ ID NO: 2 or 4,
(D) Amino acid sequence in which one or several amino acids are substituted, deleted, inserted, and / or added in the amino acid sequence shown in (a) or (b) above
(However, (c) or (d) has a polymerization effect of chondroitin chains in cooperation with chondroitin synthase).
(a)配列番号2に示されるアミノ酸配列、
(b)配列番号4に示されるアミノ酸配列、
(c)配列番号2又は4に示されるアミノ酸配列を含むアミノ酸配列、
(d)上記(a)又は(b)に示されるアミノ酸配列において、1又は数個のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列を含む
(但し(c)又は(d)はコンドロイチン合成酵素と共同してコンドロイチン鎖の重合化作用を有する)。 The chondroitin chain polymerization method as described in the above, wherein a complex obtained by co-expressing a protein comprising any one of the following amino acid sequences (a) to (d) and chondroitin synthase :
(A) the amino acid sequence shown in SEQ ID NO: 2,
(B) the amino acid sequence shown in SEQ ID NO: 4,
(C) an amino acid sequence comprising the amino acid sequence shown in SEQ ID NO: 2 or 4,
(D) In the amino acid sequence shown in (a) or (b) above, an amino acid sequence in which one or several amino acids are substituted, deleted, inserted, and / or added is included.
(However, (c) or (d) has a polymerization effect of chondroitin chains in cooperation with chondroitin synthase).
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US45342503P | 2003-03-06 | 2003-03-06 | |
| US60/453,425 | 2003-03-06 | ||
| PCT/JP2004/002893 WO2004078969A1 (en) | 2003-03-06 | 2004-03-05 | Identification and utilization of chondroitin polymerization factor |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JPWO2004078969A1 JPWO2004078969A1 (en) | 2006-06-08 |
| JP4514708B2 true JP4514708B2 (en) | 2010-07-28 |
Family
ID=32962762
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2005503149A Expired - Fee Related JP4514708B2 (en) | 2003-03-06 | 2004-03-05 | Identification and utilization of chondroitin polymerization factor |
Country Status (2)
| Country | Link |
|---|---|
| JP (1) | JP4514708B2 (en) |
| WO (1) | WO2004078969A1 (en) |
Families Citing this family (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP5885136B2 (en) * | 2011-01-31 | 2016-03-15 | 学校法人 愛知医科大学 | Method for synthesizing highly sulfated chondroitin sulfates, highly sulfated chondroitin sulfates, and analytical reagents |
Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO1999063088A2 (en) * | 1998-06-02 | 1999-12-09 | Genentech, Inc. | Membrane-bound proteins and nucleic acids encoding the same |
| JP2003102194A (en) * | 2001-09-25 | 2003-04-04 | Hitachi Ltd | Drive device for brushless motor |
Family Cites Families (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| AU2003241687A1 (en) * | 2002-05-31 | 2003-12-19 | Amersham Biosciences K.K. | Chondroitin synthetase and dna coding for the enzyme |
-
2004
- 2004-03-05 WO PCT/JP2004/002893 patent/WO2004078969A1/en active Application Filing
- 2004-03-05 JP JP2005503149A patent/JP4514708B2/en not_active Expired - Fee Related
Patent Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO1999063088A2 (en) * | 1998-06-02 | 1999-12-09 | Genentech, Inc. | Membrane-bound proteins and nucleic acids encoding the same |
| JP2003102194A (en) * | 2001-09-25 | 2003-04-04 | Hitachi Ltd | Drive device for brushless motor |
Also Published As
| Publication number | Publication date |
|---|---|
| WO2004078969A1 (en) | 2004-09-16 |
| JPWO2004078969A1 (en) | 2006-06-08 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP2002507409A (en) | Glycosyl sulfotransferase-3 | |
| JP4642301B2 (en) | UDP-N-acetylglucosamine: galactose-β1,3-N-acetyl-galactosamine-α-R / (GlcNAc: GalNAc) β1,6-N-acetylglucosaminyltransferase, C2GnT3 | |
| US7476730B2 (en) | Sulfotransferase and DNA encoding the enzyme | |
| DeBose-Boyd et al. | Molecular cloning and characterization of an α1, 3 fucosyltransferase, CEFT-1, from Caenorhabditis elegans | |
| JP4702819B2 (en) | Chondroitin synthase | |
| JP4482233B2 (en) | Novel polypeptide | |
| US6461835B1 (en) | Fucosyltransferases, polynucleotides encoding fucosyltransferases, and transgenic mammal incorporating same | |
| JP4514708B2 (en) | Identification and utilization of chondroitin polymerization factor | |
| JP3964982B2 (en) | Heparan sulfate 6-O sulfotransferase polypeptide and DNA encoding the same | |
| JP3802570B2 (en) | DNA sequence encoding mammalian glucuronyl C5-epimerase and method for producing the same | |
| US20020115839A1 (en) | 8797, a novel human galactosyltransferase and uses thereof | |
| JP4451036B2 (en) | New chondroitin synthase | |
| US20060141463A1 (en) | Method of detecting bone paget's disease | |
| US6849437B2 (en) | 47153, a human glycosyltransferase family member and uses therefor | |
| JP2004033091A (en) | New chondroitin synthase and gene encoding the same | |
| US20020142426A1 (en) | 33945, a human glycosyltransferase family member and uses therefor | |
| JP2004166702A (en) | Method for detecting bone paget's disease | |
| US20020068823A1 (en) | 33877 and 47179, novel human glycosyltransferase family members and uses thereof | |
| JP2004033064A (en) | New glucronic acid transferase and gene thereof | |
| CA2514461A1 (en) | Glycosyltransferase, nucleic acid encoding the glycosyltransferase and method of testing canceration using the nucleic acid | |
| US20020155499A1 (en) | 32624, a novel human UDP-glucuronosyl and glycosyl transferase family member and uses thereof | |
| EP1266907A1 (en) | 67108, a Human phospholipid transporter family member and uses therefor | |
| Olson | Chondroitin proteoglycans in Caenorhabditis elegans development | |
| JP2000312587A (en) | Galactose transferase and dna coding for the same | |
| JP2005058162A (en) | Knockout animal |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20070302 |
|
| RD03 | Notification of appointment of power of attorney |
Free format text: JAPANESE INTERMEDIATE CODE: A7423 Effective date: 20080307 |
|
| RD04 | Notification of resignation of power of attorney |
Free format text: JAPANESE INTERMEDIATE CODE: A7424 Effective date: 20080321 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20091216 |
|
| A521 | Written amendment |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20100212 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20100421 |
|
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20100511 |
|
| R150 | Certificate of patent or registration of utility model |
Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20130521 Year of fee payment: 3 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20130521 Year of fee payment: 3 |
|
| S533 | Written request for registration of change of name |
Free format text: JAPANESE INTERMEDIATE CODE: R313533 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20130521 Year of fee payment: 3 |
|
| R350 | Written notification of registration of transfer |
Free format text: JAPANESE INTERMEDIATE CODE: R350 |
|
| LAPS | Cancellation because of no payment of annual fees |