JP2008546775A - Modulation of negative immune regulators and application for immunotherapy - Google Patents
Modulation of negative immune regulators and application for immunotherapy Download PDFInfo
- Publication number
- JP2008546775A JP2008546775A JP2008518123A JP2008518123A JP2008546775A JP 2008546775 A JP2008546775 A JP 2008546775A JP 2008518123 A JP2008518123 A JP 2008518123A JP 2008518123 A JP2008518123 A JP 2008518123A JP 2008546775 A JP2008546775 A JP 2008546775A
- Authority
- JP
- Japan
- Prior art keywords
- antigen
- cell
- composition
- sirna
- cells
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
- 229940124644 immune regulator Drugs 0.000 title claims abstract description 101
- 238000009169 immunotherapy Methods 0.000 title description 6
- 210000004027 cell Anatomy 0.000 claims abstract description 313
- 108091007433 antigens Proteins 0.000 claims abstract description 243
- 239000000427 antigen Substances 0.000 claims abstract description 241
- 102000036639 antigens Human genes 0.000 claims abstract description 239
- 239000000203 mixture Substances 0.000 claims abstract description 146
- 238000000034 method Methods 0.000 claims abstract description 121
- 206010028980 Neoplasm Diseases 0.000 claims abstract description 64
- 210000002865 immune cell Anatomy 0.000 claims abstract description 44
- 230000005764 inhibitory process Effects 0.000 claims abstract description 20
- 230000002708 enhancing effect Effects 0.000 claims abstract description 17
- 108090000623 proteins and genes Proteins 0.000 claims description 206
- 108020004459 Small interfering RNA Proteins 0.000 claims description 175
- 102000004169 proteins and genes Human genes 0.000 claims description 119
- 239000004055 small Interfering RNA Substances 0.000 claims description 110
- 239000003112 inhibitor Substances 0.000 claims description 105
- 230000014509 gene expression Effects 0.000 claims description 101
- 102000040430 polynucleotide Human genes 0.000 claims description 101
- 108091033319 polynucleotide Proteins 0.000 claims description 101
- 239000002157 polynucleotide Substances 0.000 claims description 101
- 108090000765 processed proteins & peptides Proteins 0.000 claims description 98
- 239000013598 vector Substances 0.000 claims description 92
- 230000011664 signaling Effects 0.000 claims description 87
- 150000007523 nucleic acids Chemical class 0.000 claims description 79
- 230000028993 immune response Effects 0.000 claims description 67
- 102000039446 nucleic acids Human genes 0.000 claims description 66
- 108020004707 nucleic acids Proteins 0.000 claims description 66
- 102000004196 processed proteins & peptides Human genes 0.000 claims description 66
- 102000004127 Cytokines Human genes 0.000 claims description 58
- 210000004443 dendritic cell Anatomy 0.000 claims description 58
- 108090000695 Cytokines Proteins 0.000 claims description 57
- 229920001184 polypeptide Polymers 0.000 claims description 52
- 239000002502 liposome Substances 0.000 claims description 51
- 238000013518 transcription Methods 0.000 claims description 46
- 230000035897 transcription Effects 0.000 claims description 46
- 101000979342 Homo sapiens Nuclear factor NF-kappa-B p105 subunit Proteins 0.000 claims description 44
- 102100023050 Nuclear factor NF-kappa-B p105 subunit Human genes 0.000 claims description 44
- 239000013604 expression vector Substances 0.000 claims description 43
- 241000124008 Mammalia Species 0.000 claims description 42
- 210000001744 T-lymphocyte Anatomy 0.000 claims description 40
- 108020004414 DNA Proteins 0.000 claims description 37
- 241000700605 Viruses Species 0.000 claims description 34
- 238000001727 in vivo Methods 0.000 claims description 33
- 230000000692 anti-sense effect Effects 0.000 claims description 29
- 101150108366 Foxj1 gene Proteins 0.000 claims description 28
- 101000832631 Homo sapiens Small ubiquitin-related modifier 3 Proteins 0.000 claims description 27
- 102100024534 Small ubiquitin-related modifier 3 Human genes 0.000 claims description 27
- 102100026940 Small ubiquitin-related modifier 1 Human genes 0.000 claims description 25
- 238000000338 in vitro Methods 0.000 claims description 25
- 101000628899 Homo sapiens Small ubiquitin-related modifier 1 Proteins 0.000 claims description 24
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 claims description 23
- 108091034117 Oligonucleotide Proteins 0.000 claims description 22
- 201000011510 cancer Diseases 0.000 claims description 22
- 102000053642 Catalytic RNA Human genes 0.000 claims description 21
- 108090000994 Catalytic RNA Proteins 0.000 claims description 21
- 101000832643 Homo sapiens Small ubiquitin-related modifier 4 Proteins 0.000 claims description 21
- 102100024535 Small ubiquitin-related modifier 4 Human genes 0.000 claims description 21
- 102000040945 Transcription factor Human genes 0.000 claims description 21
- 108091023040 Transcription factor Proteins 0.000 claims description 21
- 239000002955 immunomodulating agent Substances 0.000 claims description 21
- 229940121354 immunomodulator Drugs 0.000 claims description 21
- 230000003834 intracellular effect Effects 0.000 claims description 21
- 108091092562 ribozyme Proteins 0.000 claims description 21
- 101000832685 Homo sapiens Small ubiquitin-related modifier 2 Proteins 0.000 claims description 20
- 102100024542 Small ubiquitin-related modifier 2 Human genes 0.000 claims description 20
- -1 TRP Proteins 0.000 claims description 20
- 210000003719 b-lymphocyte Anatomy 0.000 claims description 19
- 230000036039 immunity Effects 0.000 claims description 17
- 201000010099 disease Diseases 0.000 claims description 16
- 241000701022 Cytomegalovirus Species 0.000 claims description 15
- 210000001151 cytotoxic T lymphocyte Anatomy 0.000 claims description 15
- 239000013603 viral vector Substances 0.000 claims description 15
- 241000725303 Human immunodeficiency virus Species 0.000 claims description 14
- 241000282414 Homo sapiens Species 0.000 claims description 12
- 108700005077 Viral Genes Proteins 0.000 claims description 12
- 210000002443 helper t lymphocyte Anatomy 0.000 claims description 12
- 230000002584 immunomodulator Effects 0.000 claims description 12
- 208000015181 infectious disease Diseases 0.000 claims description 12
- 108060008682 Tumor Necrosis Factor Proteins 0.000 claims description 11
- 230000001580 bacterial effect Effects 0.000 claims description 11
- 230000030279 gene silencing Effects 0.000 claims description 11
- 210000002540 macrophage Anatomy 0.000 claims description 11
- 239000013612 plasmid Substances 0.000 claims description 11
- 208000035473 Communicable disease Diseases 0.000 claims description 9
- 241000700721 Hepatitis B virus Species 0.000 claims description 9
- 102000000589 Interleukin-1 Human genes 0.000 claims description 9
- 108010002352 Interleukin-1 Proteins 0.000 claims description 9
- 238000004519 manufacturing process Methods 0.000 claims description 9
- 208000023275 Autoimmune disease Diseases 0.000 claims description 8
- 102000004889 Interleukin-6 Human genes 0.000 claims description 8
- 108090001005 Interleukin-6 Proteins 0.000 claims description 8
- 230000005867 T cell response Effects 0.000 claims description 8
- 241000894006 Bacteria Species 0.000 claims description 7
- 101001012157 Homo sapiens Receptor tyrosine-protein kinase erbB-2 Proteins 0.000 claims description 7
- 108700011259 MicroRNAs Proteins 0.000 claims description 7
- 108010072866 Prostate-Specific Antigen Proteins 0.000 claims description 7
- 239000002679 microRNA Substances 0.000 claims description 7
- 150000003384 small molecules Chemical class 0.000 claims description 7
- 241001529453 unidentified herpesvirus Species 0.000 claims description 7
- 101000654734 Homo sapiens Septin-4 Proteins 0.000 claims description 6
- 241001631646 Papillomaviridae Species 0.000 claims description 6
- 102100030086 Receptor tyrosine-protein kinase erbB-2 Human genes 0.000 claims description 6
- 102100032743 Septin-4 Human genes 0.000 claims description 6
- 241000700584 Simplexvirus Species 0.000 claims description 6
- 241000233866 Fungi Species 0.000 claims description 5
- 102000003812 Interleukin-15 Human genes 0.000 claims description 5
- 108090000172 Interleukin-15 Proteins 0.000 claims description 5
- 108010002350 Interleukin-2 Proteins 0.000 claims description 5
- 102000000588 Interleukin-2 Human genes 0.000 claims description 5
- 108010002586 Interleukin-7 Proteins 0.000 claims description 5
- 206010060862 Prostate cancer Diseases 0.000 claims description 5
- 208000000236 Prostatic Neoplasms Diseases 0.000 claims description 5
- 102000000852 Tumor Necrosis Factor-alpha Human genes 0.000 claims description 5
- 102000003425 Tyrosinase Human genes 0.000 claims description 5
- 108060008724 Tyrosinase Proteins 0.000 claims description 5
- 102100024222 B-lymphocyte antigen CD19 Human genes 0.000 claims description 4
- 102100022005 B-lymphocyte antigen CD20 Human genes 0.000 claims description 4
- 206010006187 Breast cancer Diseases 0.000 claims description 4
- 208000026310 Breast neoplasm Diseases 0.000 claims description 4
- 206010008342 Cervix carcinoma Diseases 0.000 claims description 4
- 101000980825 Homo sapiens B-lymphocyte antigen CD19 Proteins 0.000 claims description 4
- 101000897405 Homo sapiens B-lymphocyte antigen CD20 Proteins 0.000 claims description 4
- 102100030703 Interleukin-22 Human genes 0.000 claims description 4
- 208000008839 Kidney Neoplasms Diseases 0.000 claims description 4
- 206010038389 Renal cancer Diseases 0.000 claims description 4
- 208000006105 Uterine Cervical Neoplasms Diseases 0.000 claims description 4
- 201000010881 cervical cancer Diseases 0.000 claims description 4
- 230000010354 integration Effects 0.000 claims description 4
- 108010074108 interleukin-21 Proteins 0.000 claims description 4
- 201000010982 kidney cancer Diseases 0.000 claims description 4
- 210000001616 monocyte Anatomy 0.000 claims description 4
- 108010014186 ras Proteins Proteins 0.000 claims description 4
- 208000011231 Crohn disease Diseases 0.000 claims description 3
- 241000701044 Human gammaherpesvirus 4 Species 0.000 claims description 3
- 102000013264 Interleukin-23 Human genes 0.000 claims description 3
- 108010065637 Interleukin-23 Proteins 0.000 claims description 3
- 102100022430 Melanocyte protein PMEL Human genes 0.000 claims description 3
- 201000004681 Psoriasis Diseases 0.000 claims description 3
- 101800001271 Surface protein Proteins 0.000 claims description 3
- 229940123384 Toll-like receptor (TLR) agonist Drugs 0.000 claims description 3
- 108010057085 cytokine receptors Proteins 0.000 claims description 3
- 201000001441 melanoma Diseases 0.000 claims description 3
- 201000006417 multiple sclerosis Diseases 0.000 claims description 3
- 210000003289 regulatory T cell Anatomy 0.000 claims description 3
- 206010039073 rheumatoid arthritis Diseases 0.000 claims description 3
- 201000000596 systemic lupus erythematosus Diseases 0.000 claims description 3
- WEVYNIUIFUYDGI-UHFFFAOYSA-N 3-[6-[4-(trifluoromethoxy)anilino]-4-pyrimidinyl]benzamide Chemical compound NC(=O)C1=CC=CC(C=2N=CN=C(NC=3C=CC(OC(F)(F)F)=CC=3)C=2)=C1 WEVYNIUIFUYDGI-UHFFFAOYSA-N 0.000 claims description 2
- 102000006306 Antigen Receptors Human genes 0.000 claims description 2
- 108010083359 Antigen Receptors Proteins 0.000 claims description 2
- 102100035526 B melanoma antigen 1 Human genes 0.000 claims description 2
- 108091007914 CDKs Proteins 0.000 claims description 2
- 101710132601 Capsid protein Proteins 0.000 claims description 2
- 101150013359 E7 gene Proteins 0.000 claims description 2
- 241000711549 Hepacivirus C Species 0.000 claims description 2
- 101000874316 Homo sapiens B melanoma antigen 1 Proteins 0.000 claims description 2
- 101000859758 Homo sapiens Cartilage-associated protein Proteins 0.000 claims description 2
- 101000721661 Homo sapiens Cellular tumor antigen p53 Proteins 0.000 claims description 2
- 101000916686 Homo sapiens Cytohesin-interacting protein Proteins 0.000 claims description 2
- 101000726740 Homo sapiens Homeobox protein cut-like 1 Proteins 0.000 claims description 2
- 101001136592 Homo sapiens Prostate stem cell antigen Proteins 0.000 claims description 2
- 101000761460 Homo sapiens Protein CASP Proteins 0.000 claims description 2
- 241000701041 Human betaherpesvirus 7 Species 0.000 claims description 2
- 241000701027 Human herpesvirus 6 Species 0.000 claims description 2
- 102000000440 Melanoma-associated antigen Human genes 0.000 claims description 2
- 108050008953 Melanoma-associated antigen Proteins 0.000 claims description 2
- 101000761459 Mesocricetus auratus Calcium-dependent serine proteinase Proteins 0.000 claims description 2
- 102100036735 Prostate stem cell antigen Human genes 0.000 claims description 2
- 108010076504 Protein Sorting Signals Proteins 0.000 claims description 2
- 108010002687 Survivin Proteins 0.000 claims description 2
- 108010017842 Telomerase Proteins 0.000 claims description 2
- 230000036755 cellular response Effects 0.000 claims description 2
- 102000052116 epidermal growth factor receptor activity proteins Human genes 0.000 claims description 2
- 108700015053 epidermal growth factor receptor activity proteins Proteins 0.000 claims description 2
- 230000002519 immonomodulatory effect Effects 0.000 claims description 2
- 238000012737 microarray-based gene expression Methods 0.000 claims description 2
- 244000000010 microbial pathogen Species 0.000 claims description 2
- 238000012243 multiplex automated genomic engineering Methods 0.000 claims description 2
- YOHYSYJDKVYCJI-UHFFFAOYSA-N n-[3-[[6-[3-(trifluoromethyl)anilino]pyrimidin-4-yl]amino]phenyl]cyclopropanecarboxamide Chemical compound FC(F)(F)C1=CC=CC(NC=2N=CN=C(NC=3C=C(NC(=O)C4CC4)C=CC=3)C=2)=C1 YOHYSYJDKVYCJI-UHFFFAOYSA-N 0.000 claims description 2
- 108700004028 nef Genes Proteins 0.000 claims description 2
- 101150023385 nef gene Proteins 0.000 claims description 2
- 208000025189 neoplasm of testis Diseases 0.000 claims description 2
- 102000000704 Interleukin-7 Human genes 0.000 claims 2
- 101000617738 Homo sapiens Survival motor neuron protein Proteins 0.000 claims 1
- XQFRJNBWHJMXHO-RRKCRQDMSA-N IDUR Chemical compound C1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C(I)=C1 XQFRJNBWHJMXHO-RRKCRQDMSA-N 0.000 claims 1
- 102100021768 Phosphoserine aminotransferase Human genes 0.000 claims 1
- 102100024933 Protein CASP Human genes 0.000 claims 1
- 102100021947 Survival motor neuron protein Human genes 0.000 claims 1
- 102000000763 Survivin Human genes 0.000 claims 1
- 208000024313 Testicular Neoplasms Diseases 0.000 claims 1
- 108040000983 polyphosphate:AMP phosphotransferase activity proteins Proteins 0.000 claims 1
- 201000003120 testicular cancer Diseases 0.000 claims 1
- 229960005486 vaccine Drugs 0.000 abstract description 29
- 230000007246 mechanism Effects 0.000 abstract description 14
- 230000030741 antigen processing and presentation Effects 0.000 abstract description 9
- 238000002255 vaccination Methods 0.000 abstract description 6
- 238000002560 therapeutic procedure Methods 0.000 abstract description 5
- 235000018102 proteins Nutrition 0.000 description 101
- 210000000612 antigen-presenting cell Anatomy 0.000 description 74
- 230000002401 inhibitory effect Effects 0.000 description 74
- 108700027336 Suppressor of Cytokine Signaling 1 Proteins 0.000 description 64
- 101150045565 Socs1 gene Proteins 0.000 description 63
- 102000058018 Suppressor of Cytokine Signaling 1 Human genes 0.000 description 63
- 239000002773 nucleotide Substances 0.000 description 60
- 125000003729 nucleotide group Chemical group 0.000 description 60
- 102000002689 Toll-like receptor Human genes 0.000 description 57
- 108020000411 Toll-like receptor Proteins 0.000 description 57
- 150000002632 lipids Chemical class 0.000 description 44
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 39
- 230000000694 effects Effects 0.000 description 34
- 101000652220 Homo sapiens Suppressor of cytokine signaling 4 Proteins 0.000 description 31
- 101000652226 Homo sapiens Suppressor of cytokine signaling 6 Proteins 0.000 description 31
- 102100030524 Suppressor of cytokine signaling 4 Human genes 0.000 description 31
- 108020004999 messenger RNA Proteins 0.000 description 24
- 239000000047 product Substances 0.000 description 24
- 108010057466 NF-kappa B Proteins 0.000 description 23
- 102000003945 NF-kappa B Human genes 0.000 description 23
- 210000001519 tissue Anatomy 0.000 description 23
- 108091028043 Nucleic acid sequence Proteins 0.000 description 22
- 230000001105 regulatory effect Effects 0.000 description 20
- 102100034922 T-cell surface glycoprotein CD8 alpha chain Human genes 0.000 description 19
- 230000004913 activation Effects 0.000 description 19
- 230000019491 signal transduction Effects 0.000 description 19
- 239000000243 solution Substances 0.000 description 19
- 101000879203 Caenorhabditis elegans Small ubiquitin-related modifier Proteins 0.000 description 18
- 101710132484 Cytokine-inducible SH2-containing protein Proteins 0.000 description 18
- 102100032218 Cytokine-inducible SH2-containing protein Human genes 0.000 description 18
- 102000051619 SUMO-1 Human genes 0.000 description 18
- 239000002158 endotoxin Substances 0.000 description 18
- 229920006008 lipopolysaccharide Polymers 0.000 description 18
- 101000652229 Homo sapiens Suppressor of cytokine signaling 7 Proteins 0.000 description 17
- 102100030529 Suppressor of cytokine signaling 7 Human genes 0.000 description 17
- 102100030987 E3 SUMO-protein ligase PIAS4 Human genes 0.000 description 16
- 101710191253 E3 SUMO-protein ligase PIAS4 Proteins 0.000 description 16
- 101000924577 Homo sapiens Adenomatous polyposis coli protein Proteins 0.000 description 16
- 101150086423 PIAS2 gene Proteins 0.000 description 16
- 230000000295 complement effect Effects 0.000 description 16
- 239000003814 drug Substances 0.000 description 16
- 230000001225 therapeutic effect Effects 0.000 description 16
- 102100034540 Adenomatous polyposis coli protein Human genes 0.000 description 15
- 102100035989 E3 SUMO-protein ligase PIAS1 Human genes 0.000 description 15
- 102100030837 E3 SUMO-protein ligase PIAS3 Human genes 0.000 description 15
- 101000583444 Homo sapiens E3 SUMO-protein ligase PIAS3 Proteins 0.000 description 15
- 241000699666 Mus <mouse, genus> Species 0.000 description 15
- 108010038241 Protein Inhibitors of Activated STAT Proteins 0.000 description 15
- 101150043341 Socs3 gene Proteins 0.000 description 15
- 108700027337 Suppressor of Cytokine Signaling 3 Proteins 0.000 description 15
- 102100024283 Suppressor of cytokine signaling 3 Human genes 0.000 description 15
- 102100033019 Tyrosine-protein phosphatase non-receptor type 11 Human genes 0.000 description 15
- 101710116241 Tyrosine-protein phosphatase non-receptor type 11 Proteins 0.000 description 15
- 102100021657 Tyrosine-protein phosphatase non-receptor type 6 Human genes 0.000 description 15
- 101710128901 Tyrosine-protein phosphatase non-receptor type 6 Proteins 0.000 description 15
- 210000001185 bone marrow Anatomy 0.000 description 15
- 230000006870 function Effects 0.000 description 15
- 101000687808 Homo sapiens Suppressor of cytokine signaling 2 Proteins 0.000 description 14
- 101000652224 Homo sapiens Suppressor of cytokine signaling 5 Proteins 0.000 description 14
- 108700018351 Major Histocompatibility Complex Proteins 0.000 description 14
- 102100024784 Suppressor of cytokine signaling 2 Human genes 0.000 description 14
- 102100030523 Suppressor of cytokine signaling 5 Human genes 0.000 description 14
- 238000003752 polymerase chain reaction Methods 0.000 description 14
- 238000013519 translation Methods 0.000 description 14
- 102100036342 Interleukin-1 receptor-associated kinase 1 Human genes 0.000 description 13
- 241000699670 Mus sp. Species 0.000 description 13
- 101150037166 Twist2 gene Proteins 0.000 description 13
- 230000033228 biological regulation Effects 0.000 description 13
- 239000003795 chemical substances by application Substances 0.000 description 13
- 150000001875 compounds Chemical group 0.000 description 13
- 230000000875 corresponding effect Effects 0.000 description 13
- ONCZDRURRATYFI-QTCHDTBASA-N methyl (2z)-2-methoxyimino-2-[2-[[(e)-1-[3-(trifluoromethyl)phenyl]ethylideneamino]oxymethyl]phenyl]acetate Chemical compound CO\N=C(/C(=O)OC)C1=CC=CC=C1CO\N=C(/C)C1=CC=CC(C(F)(F)F)=C1 ONCZDRURRATYFI-QTCHDTBASA-N 0.000 description 13
- 108060003951 Immunoglobulin Proteins 0.000 description 12
- 108010029485 Protein Isoforms Proteins 0.000 description 12
- 102000001708 Protein Isoforms Human genes 0.000 description 12
- 239000004480 active ingredient Substances 0.000 description 12
- 230000015572 biosynthetic process Effects 0.000 description 12
- 239000002299 complementary DNA Substances 0.000 description 12
- 230000003053 immunization Effects 0.000 description 12
- 238000002649 immunization Methods 0.000 description 12
- 102000018358 immunoglobulin Human genes 0.000 description 12
- 230000001717 pathogenic effect Effects 0.000 description 12
- 230000004044 response Effects 0.000 description 12
- 239000000523 sample Substances 0.000 description 12
- 230000020382 suppression by virus of host antigen processing and presentation of peptide antigen via MHC class I Effects 0.000 description 12
- 238000003786 synthesis reaction Methods 0.000 description 12
- 229940124597 therapeutic agent Drugs 0.000 description 12
- 238000012546 transfer Methods 0.000 description 12
- 101001125026 Homo sapiens Nucleotide-binding oligomerization domain-containing protein 2 Proteins 0.000 description 11
- 102100029441 Nucleotide-binding oligomerization domain-containing protein 2 Human genes 0.000 description 11
- 239000003623 enhancer Substances 0.000 description 11
- 239000012634 fragment Substances 0.000 description 11
- 230000037361 pathway Effects 0.000 description 11
- 230000000770 proinflammatory effect Effects 0.000 description 11
- 102000005962 receptors Human genes 0.000 description 11
- 108020003175 receptors Proteins 0.000 description 11
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 11
- 108010058846 Ovalbumin Proteins 0.000 description 10
- 230000008901 benefit Effects 0.000 description 10
- 239000002552 dosage form Substances 0.000 description 10
- 238000009472 formulation Methods 0.000 description 10
- 230000035800 maturation Effects 0.000 description 10
- 244000052769 pathogen Species 0.000 description 10
- 239000008194 pharmaceutical composition Substances 0.000 description 10
- 230000034190 positive regulation of NF-kappaB transcription factor activity Effects 0.000 description 10
- 238000001890 transfection Methods 0.000 description 10
- 239000012980 RPMI-1640 medium Substances 0.000 description 9
- 108700008625 Reporter Genes Proteins 0.000 description 9
- 238000003556 assay Methods 0.000 description 9
- 230000003308 immunostimulating effect Effects 0.000 description 9
- 239000000463 material Substances 0.000 description 9
- 230000001404 mediated effect Effects 0.000 description 9
- 229940092253 ovalbumin Drugs 0.000 description 9
- 150000003904 phospholipids Chemical class 0.000 description 9
- 125000006850 spacer group Chemical group 0.000 description 9
- 238000012384 transportation and delivery Methods 0.000 description 9
- 101000914484 Homo sapiens T-lymphocyte activation antigen CD80 Proteins 0.000 description 8
- 108010024121 Janus Kinases Proteins 0.000 description 8
- 102000015617 Janus Kinases Human genes 0.000 description 8
- 102100027222 T-lymphocyte activation antigen CD80 Human genes 0.000 description 8
- 230000007423 decrease Effects 0.000 description 8
- 238000009396 hybridization Methods 0.000 description 8
- 239000003550 marker Substances 0.000 description 8
- 239000002904 solvent Substances 0.000 description 8
- 230000000638 stimulation Effects 0.000 description 8
- 239000000126 substance Substances 0.000 description 8
- 108010043121 Green Fluorescent Proteins Proteins 0.000 description 7
- 101710199015 Interleukin-1 receptor-associated kinase 1 Proteins 0.000 description 7
- 108010077432 Myeloid Differentiation Factor 88 Proteins 0.000 description 7
- 102000010168 Myeloid Differentiation Factor 88 Human genes 0.000 description 7
- 108020004511 Recombinant DNA Proteins 0.000 description 7
- 108060008683 Tumor Necrosis Factor Receptor Proteins 0.000 description 7
- 239000002671 adjuvant Substances 0.000 description 7
- 235000001014 amino acid Nutrition 0.000 description 7
- 238000006243 chemical reaction Methods 0.000 description 7
- 208000035475 disorder Diseases 0.000 description 7
- 238000002474 experimental method Methods 0.000 description 7
- 239000005090 green fluorescent protein Substances 0.000 description 7
- 230000001965 increasing effect Effects 0.000 description 7
- 230000001939 inductive effect Effects 0.000 description 7
- 230000004048 modification Effects 0.000 description 7
- 238000012986 modification Methods 0.000 description 7
- 238000012360 testing method Methods 0.000 description 7
- 238000011282 treatment Methods 0.000 description 7
- 102000003298 tumor necrosis factor receptor Human genes 0.000 description 7
- 239000003981 vehicle Substances 0.000 description 7
- 102000040650 (ribonucleotides)n+m Human genes 0.000 description 6
- 102000003916 Arrestin Human genes 0.000 description 6
- 108090000328 Arrestin Proteins 0.000 description 6
- HEDRZPFGACZZDS-UHFFFAOYSA-N Chloroform Chemical compound ClC(Cl)Cl HEDRZPFGACZZDS-UHFFFAOYSA-N 0.000 description 6
- 108091033380 Coding strand Proteins 0.000 description 6
- 108010038537 Dual Specificity Phosphatase 1 Proteins 0.000 description 6
- 102000010778 Dual Specificity Phosphatase 1 Human genes 0.000 description 6
- 102000004144 Green Fluorescent Proteins Human genes 0.000 description 6
- 101000852483 Homo sapiens Interleukin-1 receptor-associated kinase 1 Proteins 0.000 description 6
- 101000852968 Homo sapiens Interleukin-1 receptor-like 1 Proteins 0.000 description 6
- 101000585365 Homo sapiens Sulfotransferase 2A1 Proteins 0.000 description 6
- 102000014150 Interferons Human genes 0.000 description 6
- 108010050904 Interferons Proteins 0.000 description 6
- 102100036706 Interleukin-1 receptor-like 1 Human genes 0.000 description 6
- 102000015696 Interleukins Human genes 0.000 description 6
- 108010063738 Interleukins Proteins 0.000 description 6
- 108091054455 MAP kinase family Proteins 0.000 description 6
- 102000043136 MAP kinase family Human genes 0.000 description 6
- OKKJLVBELUTLKV-UHFFFAOYSA-N Methanol Chemical compound OC OKKJLVBELUTLKV-UHFFFAOYSA-N 0.000 description 6
- 238000010240 RT-PCR analysis Methods 0.000 description 6
- 102100040247 Tumor necrosis factor Human genes 0.000 description 6
- 238000007792 addition Methods 0.000 description 6
- 125000003275 alpha amino acid group Chemical group 0.000 description 6
- 150000001413 amino acids Chemical class 0.000 description 6
- 230000003321 amplification Effects 0.000 description 6
- 230000000890 antigenic effect Effects 0.000 description 6
- 230000027455 binding Effects 0.000 description 6
- 239000003153 chemical reaction reagent Substances 0.000 description 6
- 238000012217 deletion Methods 0.000 description 6
- 230000037430 deletion Effects 0.000 description 6
- 230000002163 immunogen Effects 0.000 description 6
- 230000006698 induction Effects 0.000 description 6
- 229940079322 interferon Drugs 0.000 description 6
- 239000012528 membrane Substances 0.000 description 6
- 210000004379 membrane Anatomy 0.000 description 6
- 239000000693 micelle Substances 0.000 description 6
- 238000003199 nucleic acid amplification method Methods 0.000 description 6
- 239000002245 particle Substances 0.000 description 6
- 229920000642 polymer Polymers 0.000 description 6
- 238000012545 processing Methods 0.000 description 6
- 230000002441 reversible effect Effects 0.000 description 6
- 230000028327 secretion Effects 0.000 description 6
- 241000894007 species Species 0.000 description 6
- 210000000130 stem cell Anatomy 0.000 description 6
- 238000006467 substitution reaction Methods 0.000 description 6
- 238000010361 transduction Methods 0.000 description 6
- 230000026683 transduction Effects 0.000 description 6
- 102000007469 Actins Human genes 0.000 description 5
- 108010085238 Actins Proteins 0.000 description 5
- 102100024220 CD180 antigen Human genes 0.000 description 5
- 101150013553 CD40 gene Proteins 0.000 description 5
- 108091026890 Coding region Proteins 0.000 description 5
- 102000053602 DNA Human genes 0.000 description 5
- 102100031573 Hematopoietic progenitor cell antigen CD34 Human genes 0.000 description 5
- 101000980829 Homo sapiens CD180 antigen Proteins 0.000 description 5
- 101000777663 Homo sapiens Hematopoietic progenitor cell antigen CD34 Proteins 0.000 description 5
- 101000852255 Homo sapiens Interleukin-1 receptor-associated kinase-like 2 Proteins 0.000 description 5
- 101000652846 Homo sapiens Single Ig IL-1-related receptor Proteins 0.000 description 5
- 101000634866 Homo sapiens TRAF-type zinc finger domain-containing protein 1 Proteins 0.000 description 5
- 108010074328 Interferon-gamma Proteins 0.000 description 5
- 102100036433 Interleukin-1 receptor-associated kinase-like 2 Human genes 0.000 description 5
- 241000713666 Lentivirus Species 0.000 description 5
- 239000000232 Lipid Bilayer Substances 0.000 description 5
- 108091007960 PI3Ks Proteins 0.000 description 5
- 102000038030 PI3Ks Human genes 0.000 description 5
- 108010033276 Peptide Fragments Proteins 0.000 description 5
- 102000007079 Peptide Fragments Human genes 0.000 description 5
- 102000004160 Phosphoric Monoester Hydrolases Human genes 0.000 description 5
- 108090000608 Phosphoric Monoester Hydrolases Proteins 0.000 description 5
- 102000007066 Prostate-Specific Antigen Human genes 0.000 description 5
- 102100030929 Single Ig IL-1-related receptor Human genes 0.000 description 5
- 102100029451 TRAF-type zinc finger domain-containing protein 1 Human genes 0.000 description 5
- 102100024584 Tumor necrosis factor ligand superfamily member 12 Human genes 0.000 description 5
- 101710097155 Tumor necrosis factor ligand superfamily member 12 Proteins 0.000 description 5
- 102100040245 Tumor necrosis factor receptor superfamily member 5 Human genes 0.000 description 5
- 229940030156 cell vaccine Drugs 0.000 description 5
- 238000003501 co-culture Methods 0.000 description 5
- 239000003085 diluting agent Substances 0.000 description 5
- 239000003937 drug carrier Substances 0.000 description 5
- 239000000839 emulsion Substances 0.000 description 5
- 238000001943 fluorescence-activated cell sorting Methods 0.000 description 5
- 238000010369 molecular cloning Methods 0.000 description 5
- 230000036961 partial effect Effects 0.000 description 5
- 239000002953 phosphate buffered saline Substances 0.000 description 5
- 230000026731 phosphorylation Effects 0.000 description 5
- 238000006366 phosphorylation reaction Methods 0.000 description 5
- 235000020004 porter Nutrition 0.000 description 5
- 230000008569 process Effects 0.000 description 5
- 230000004936 stimulating effect Effects 0.000 description 5
- 230000001629 suppression Effects 0.000 description 5
- 230000008685 targeting Effects 0.000 description 5
- 238000011144 upstream manufacturing Methods 0.000 description 5
- 238000001262 western blot Methods 0.000 description 5
- 108010022366 Carcinoembryonic Antigen Proteins 0.000 description 4
- 102100025475 Carcinoembryonic antigen-related cell adhesion molecule 5 Human genes 0.000 description 4
- 108010086291 Deubiquitinating Enzyme CYLD Proteins 0.000 description 4
- 102100037830 Docking protein 2 Human genes 0.000 description 4
- 108091029865 Exogenous DNA Proteins 0.000 description 4
- 101000805166 Homo sapiens Docking protein 2 Proteins 0.000 description 4
- 101000669447 Homo sapiens Toll-like receptor 4 Proteins 0.000 description 4
- 102000008379 I-kappa B Proteins Human genes 0.000 description 4
- 108010021699 I-kappa B Proteins Proteins 0.000 description 4
- 102100030126 Interferon regulatory factor 4 Human genes 0.000 description 4
- 102000004182 Mitogen-Activated Protein Kinase Phosphatases Human genes 0.000 description 4
- 108010082747 Mitogen-Activated Protein Kinase Phosphatases Proteins 0.000 description 4
- 241000194017 Streptococcus Species 0.000 description 4
- IQFYYKKMVGJFEH-XLPZGREQSA-N Thymidine Chemical compound O=C1NC(=O)C(C)=CN1[C@@H]1O[C@H](CO)[C@@H](O)C1 IQFYYKKMVGJFEH-XLPZGREQSA-N 0.000 description 4
- 102100039360 Toll-like receptor 4 Human genes 0.000 description 4
- 102100024250 Ubiquitin carboxyl-terminal hydrolase CYLD Human genes 0.000 description 4
- DRTQHJPVMGBUCF-XVFCMESISA-N Uridine Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C=C1 DRTQHJPVMGBUCF-XVFCMESISA-N 0.000 description 4
- 230000003213 activating effect Effects 0.000 description 4
- 230000004721 adaptive immunity Effects 0.000 description 4
- 238000013459 approach Methods 0.000 description 4
- 230000010261 cell growth Effects 0.000 description 4
- 238000010367 cloning Methods 0.000 description 4
- 230000000139 costimulatory effect Effects 0.000 description 4
- 210000000805 cytoplasm Anatomy 0.000 description 4
- 230000001086 cytosolic effect Effects 0.000 description 4
- 230000003828 downregulation Effects 0.000 description 4
- 229940079593 drug Drugs 0.000 description 4
- 210000003958 hematopoietic stem cell Anatomy 0.000 description 4
- 230000002209 hydrophobic effect Effects 0.000 description 4
- 230000003463 hyperproliferative effect Effects 0.000 description 4
- 230000001900 immune effect Effects 0.000 description 4
- 230000002458 infectious effect Effects 0.000 description 4
- 238000001802 infusion Methods 0.000 description 4
- 238000003780 insertion Methods 0.000 description 4
- 230000037431 insertion Effects 0.000 description 4
- 230000002452 interceptive effect Effects 0.000 description 4
- 108010051920 interferon regulatory factor-4 Proteins 0.000 description 4
- 239000003446 ligand Substances 0.000 description 4
- 230000000670 limiting effect Effects 0.000 description 4
- 238000011068 loading method Methods 0.000 description 4
- BSOQXXWZTUDTEL-ZUYCGGNHSA-N muramyl dipeptide Chemical compound OC(=O)CC[C@H](C(N)=O)NC(=O)[C@H](C)NC(=O)[C@@H](C)O[C@H]1[C@H](O)[C@@H](CO)O[C@@H](O)[C@@H]1NC(C)=O BSOQXXWZTUDTEL-ZUYCGGNHSA-N 0.000 description 4
- 210000000822 natural killer cell Anatomy 0.000 description 4
- 239000012071 phase Substances 0.000 description 4
- 230000008741 proinflammatory signaling process Effects 0.000 description 4
- 229940121649 protein inhibitor Drugs 0.000 description 4
- 239000012268 protein inhibitor Substances 0.000 description 4
- 230000002829 reductive effect Effects 0.000 description 4
- 230000027404 regulation of phosphorylation Effects 0.000 description 4
- 230000010076 replication Effects 0.000 description 4
- 238000011160 research Methods 0.000 description 4
- 239000007787 solid Substances 0.000 description 4
- 235000000346 sugar Nutrition 0.000 description 4
- 230000010741 sumoylation Effects 0.000 description 4
- 239000006228 supernatant Substances 0.000 description 4
- 239000000725 suspension Substances 0.000 description 4
- 231100000331 toxic Toxicity 0.000 description 4
- 230000002588 toxic effect Effects 0.000 description 4
- 230000003612 virological effect Effects 0.000 description 4
- 238000005406 washing Methods 0.000 description 4
- 108091005957 yellow fluorescent proteins Proteins 0.000 description 4
- 108010042708 Acetylmuramyl-Alanyl-Isoglutamine Proteins 0.000 description 3
- 101100508533 Drosophila melanogaster IKKbeta gene Proteins 0.000 description 3
- 102100037569 Dual specificity protein phosphatase 10 Human genes 0.000 description 3
- 102100037567 Dual specificity protein phosphatase 14 Human genes 0.000 description 3
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 3
- 238000002965 ELISA Methods 0.000 description 3
- 241000282412 Homo Species 0.000 description 3
- 101000881127 Homo sapiens Dual specificity protein phosphatase 10 Proteins 0.000 description 3
- 101000881116 Homo sapiens Dual specificity protein phosphatase 14 Proteins 0.000 description 3
- 101000830560 Homo sapiens Toll-interacting protein Proteins 0.000 description 3
- 101000831567 Homo sapiens Toll-like receptor 2 Proteins 0.000 description 3
- 102100037850 Interferon gamma Human genes 0.000 description 3
- 102000004388 Interleukin-4 Human genes 0.000 description 3
- 108090000978 Interleukin-4 Proteins 0.000 description 3
- 102100021592 Interleukin-7 Human genes 0.000 description 3
- 108091092195 Intron Proteins 0.000 description 3
- 241000829100 Macaca mulatta polyomavirus 1 Species 0.000 description 3
- 241001465754 Metazoa Species 0.000 description 3
- 101100257268 Mus musculus Socs1 gene Proteins 0.000 description 3
- 108091000080 Phosphotransferase Proteins 0.000 description 3
- DNIAPMSPPWPWGF-UHFFFAOYSA-N Propylene glycol Chemical compound CC(O)CO DNIAPMSPPWPWGF-UHFFFAOYSA-N 0.000 description 3
- 238000011529 RT qPCR Methods 0.000 description 3
- 241000700159 Rattus Species 0.000 description 3
- 102000014400 SH2 domains Human genes 0.000 description 3
- 108050003452 SH2 domains Proteins 0.000 description 3
- 241000251131 Sphyrna Species 0.000 description 3
- 241000223892 Tetrahymena Species 0.000 description 3
- 102100024652 Toll-interacting protein Human genes 0.000 description 3
- 102100024333 Toll-like receptor 2 Human genes 0.000 description 3
- 102000000887 Transcription factor STAT Human genes 0.000 description 3
- 108050007918 Transcription factor STAT Proteins 0.000 description 3
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Chemical class Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 3
- 230000002159 abnormal effect Effects 0.000 description 3
- 239000012190 activator Substances 0.000 description 3
- 239000000556 agonist Substances 0.000 description 3
- 238000004458 analytical method Methods 0.000 description 3
- 239000005557 antagonist Substances 0.000 description 3
- 230000006907 apoptotic process Effects 0.000 description 3
- 239000007864 aqueous solution Substances 0.000 description 3
- 239000012472 biological sample Substances 0.000 description 3
- 230000000903 blocking effect Effects 0.000 description 3
- 239000001506 calcium phosphate Substances 0.000 description 3
- 229910000389 calcium phosphate Inorganic materials 0.000 description 3
- 235000011010 calcium phosphates Nutrition 0.000 description 3
- 238000004422 calculation algorithm Methods 0.000 description 3
- 230000015556 catabolic process Effects 0.000 description 3
- 239000011651 chromium Substances 0.000 description 3
- KRKNYBCHXYNGOX-UHFFFAOYSA-N citric acid Chemical compound OC(=O)CC(O)(C(O)=O)CC(O)=O KRKNYBCHXYNGOX-UHFFFAOYSA-N 0.000 description 3
- 238000012761 co-transfection Methods 0.000 description 3
- 230000001276 controlling effect Effects 0.000 description 3
- 238000006731 degradation reaction Methods 0.000 description 3
- 238000001514 detection method Methods 0.000 description 3
- 230000009504 deubiquitination Effects 0.000 description 3
- 239000012636 effector Substances 0.000 description 3
- 210000003162 effector t lymphocyte Anatomy 0.000 description 3
- 238000004520 electroporation Methods 0.000 description 3
- 230000004927 fusion Effects 0.000 description 3
- 210000000987 immune system Anatomy 0.000 description 3
- 230000001976 improved effect Effects 0.000 description 3
- 238000011534 incubation Methods 0.000 description 3
- 239000004615 ingredient Substances 0.000 description 3
- 230000003993 interaction Effects 0.000 description 3
- 229940028885 interleukin-4 Drugs 0.000 description 3
- 238000007918 intramuscular administration Methods 0.000 description 3
- 238000001990 intravenous administration Methods 0.000 description 3
- 150000002500 ions Chemical class 0.000 description 3
- 238000002955 isolation Methods 0.000 description 3
- 229920002521 macromolecule Polymers 0.000 description 3
- 238000002826 magnetic-activated cell sorting Methods 0.000 description 3
- 210000004962 mammalian cell Anatomy 0.000 description 3
- 238000002844 melting Methods 0.000 description 3
- 230000008018 melting Effects 0.000 description 3
- 239000011325 microbead Substances 0.000 description 3
- 230000035772 mutation Effects 0.000 description 3
- 238000007911 parenteral administration Methods 0.000 description 3
- 230000035699 permeability Effects 0.000 description 3
- 102000020233 phosphotransferase Human genes 0.000 description 3
- 238000002360 preparation method Methods 0.000 description 3
- 230000035755 proliferation Effects 0.000 description 3
- 150000003180 prostaglandins Chemical class 0.000 description 3
- 108091008146 restriction endonucleases Proteins 0.000 description 3
- 230000001177 retroviral effect Effects 0.000 description 3
- 150000003839 salts Chemical class 0.000 description 3
- 210000004989 spleen cell Anatomy 0.000 description 3
- 238000010186 staining Methods 0.000 description 3
- 229940104230 thymidine Drugs 0.000 description 3
- 230000007704 transition Effects 0.000 description 3
- QORWJWZARLRLPR-UHFFFAOYSA-H tricalcium bis(phosphate) Chemical compound [Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O QORWJWZARLRLPR-UHFFFAOYSA-H 0.000 description 3
- 238000010798 ubiquitination Methods 0.000 description 3
- 230000034512 ubiquitination Effects 0.000 description 3
- 241000701161 unidentified adenovirus Species 0.000 description 3
- 229930024421 Adenine Natural products 0.000 description 2
- GFFGJBXGBJISGV-UHFFFAOYSA-N Adenine Chemical compound NC1=NC=NC2=C1N=CN2 GFFGJBXGBJISGV-UHFFFAOYSA-N 0.000 description 2
- NLXLAEXVIDQMFP-UHFFFAOYSA-N Ammonia chloride Chemical compound [NH4+].[Cl-] NLXLAEXVIDQMFP-UHFFFAOYSA-N 0.000 description 2
- CIWBSHSKHKDKBQ-JLAZNSOCSA-N Ascorbic acid Chemical compound OC[C@H](O)[C@H]1OC(=O)C(O)=C1O CIWBSHSKHKDKBQ-JLAZNSOCSA-N 0.000 description 2
- 208000003950 B-cell lymphoma Diseases 0.000 description 2
- 241000606125 Bacteroides Species 0.000 description 2
- DWRXFEITVBNRMK-UHFFFAOYSA-N Beta-D-1-Arabinofuranosylthymine Natural products O=C1NC(=O)C(C)=CN1C1C(O)C(O)C(CO)O1 DWRXFEITVBNRMK-UHFFFAOYSA-N 0.000 description 2
- 102000008203 CTLA-4 Antigen Human genes 0.000 description 2
- 108010021064 CTLA-4 Antigen Proteins 0.000 description 2
- 206010009944 Colon cancer Diseases 0.000 description 2
- 108020004635 Complementary DNA Proteins 0.000 description 2
- 241000711573 Coronaviridae Species 0.000 description 2
- 238000001712 DNA sequencing Methods 0.000 description 2
- 102000004163 DNA-directed RNA polymerases Human genes 0.000 description 2
- 108090000626 DNA-directed RNA polymerases Proteins 0.000 description 2
- 241000702421 Dependoparvovirus Species 0.000 description 2
- 241000196324 Embryophyta Species 0.000 description 2
- 241000709661 Enterovirus Species 0.000 description 2
- 102000004190 Enzymes Human genes 0.000 description 2
- 108090000790 Enzymes Proteins 0.000 description 2
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 2
- 108010017213 Granulocyte-Macrophage Colony-Stimulating Factor Proteins 0.000 description 2
- 102100039620 Granulocyte-macrophage colony-stimulating factor Human genes 0.000 description 2
- NYHBQMYGNKIUIF-UUOKFMHZSA-N Guanosine Chemical compound C1=NC=2C(=O)NC(N)=NC=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O NYHBQMYGNKIUIF-UUOKFMHZSA-N 0.000 description 2
- 229920000209 Hexadimethrine bromide Polymers 0.000 description 2
- 102100037907 High mobility group protein B1 Human genes 0.000 description 2
- 101710168537 High mobility group protein B1 Proteins 0.000 description 2
- 101000977771 Homo sapiens Interleukin-1 receptor-associated kinase 4 Proteins 0.000 description 2
- 241000701085 Human alphaherpesvirus 3 Species 0.000 description 2
- 241000713772 Human immunodeficiency virus 1 Species 0.000 description 2
- 206010061218 Inflammation Diseases 0.000 description 2
- 102100022297 Integrin alpha-X Human genes 0.000 description 2
- 102000008070 Interferon-gamma Human genes 0.000 description 2
- 102000019223 Interleukin-1 receptor Human genes 0.000 description 2
- 108050006617 Interleukin-1 receptor Proteins 0.000 description 2
- 102100023533 Interleukin-1 receptor-associated kinase 4 Human genes 0.000 description 2
- 102100039897 Interleukin-5 Human genes 0.000 description 2
- 108010002616 Interleukin-5 Proteins 0.000 description 2
- 230000004163 JAK-STAT signaling pathway Effects 0.000 description 2
- 241000588915 Klebsiella aerogenes Species 0.000 description 2
- 241000588747 Klebsiella pneumoniae Species 0.000 description 2
- JVTAAEKCZFNVCJ-UHFFFAOYSA-M Lactate Chemical compound CC(O)C([O-])=O JVTAAEKCZFNVCJ-UHFFFAOYSA-M 0.000 description 2
- 206010058467 Lung neoplasm malignant Diseases 0.000 description 2
- 206010025323 Lymphomas Diseases 0.000 description 2
- 102000043131 MHC class II family Human genes 0.000 description 2
- 108091054438 MHC class II family Proteins 0.000 description 2
- 101100503253 Mus musculus Foxj1 gene Proteins 0.000 description 2
- 101100369989 Mus musculus Tnfaip3 gene Proteins 0.000 description 2
- 206010033128 Ovarian cancer Diseases 0.000 description 2
- 206010061535 Ovarian neoplasm Diseases 0.000 description 2
- 238000010222 PCR analysis Methods 0.000 description 2
- 206010061902 Pancreatic neoplasm Diseases 0.000 description 2
- 241000701945 Parvoviridae Species 0.000 description 2
- 239000004698 Polyethylene Substances 0.000 description 2
- 241001505332 Polyomavirus sp. Species 0.000 description 2
- RJKFOVLPORLFTN-LEKSSAKUSA-N Progesterone Chemical compound C1CC2=CC(=O)CC[C@]2(C)[C@@H]2[C@@H]1[C@@H]1CC[C@H](C(=O)C)[C@@]1(C)CC2 RJKFOVLPORLFTN-LEKSSAKUSA-N 0.000 description 2
- 102000001253 Protein Kinase Human genes 0.000 description 2
- 238000012228 RNA interference-mediated gene silencing Methods 0.000 description 2
- 241000725643 Respiratory syncytial virus Species 0.000 description 2
- 241000710799 Rubella virus Species 0.000 description 2
- 241000193985 Streptococcus agalactiae Species 0.000 description 2
- 230000006044 T cell activation Effects 0.000 description 2
- 108091008874 T cell receptors Proteins 0.000 description 2
- 102000016266 T-Cell Antigen Receptors Human genes 0.000 description 2
- 108700026226 TATA Box Proteins 0.000 description 2
- 102000006601 Thymidine Kinase Human genes 0.000 description 2
- 108020004440 Thymidine kinase Proteins 0.000 description 2
- 102000044159 Ubiquitin Human genes 0.000 description 2
- 108090000848 Ubiquitin Proteins 0.000 description 2
- 241000711975 Vesicular stomatitis virus Species 0.000 description 2
- 108010067390 Viral Proteins Proteins 0.000 description 2
- 239000013543 active substance Substances 0.000 description 2
- 229960000643 adenine Drugs 0.000 description 2
- OIRDTQYFTABQOQ-KQYNXXCUSA-N adenosine Chemical compound C1=NC=2C(N)=NC=NC=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O OIRDTQYFTABQOQ-KQYNXXCUSA-N 0.000 description 2
- 239000000443 aerosol Substances 0.000 description 2
- 230000000735 allogeneic effect Effects 0.000 description 2
- 230000005809 anti-tumor immunity Effects 0.000 description 2
- 230000005875 antibody response Effects 0.000 description 2
- 239000012736 aqueous medium Substances 0.000 description 2
- 239000008346 aqueous phase Substances 0.000 description 2
- 230000002238 attenuated effect Effects 0.000 description 2
- 230000001363 autoimmune Effects 0.000 description 2
- 230000006472 autoimmune response Effects 0.000 description 2
- 239000011324 bead Substances 0.000 description 2
- 230000009286 beneficial effect Effects 0.000 description 2
- IQFYYKKMVGJFEH-UHFFFAOYSA-N beta-L-thymidine Natural products O=C1NC(=O)C(C)=CN1C1OC(CO)C(O)C1 IQFYYKKMVGJFEH-UHFFFAOYSA-N 0.000 description 2
- DRTQHJPVMGBUCF-PSQAKQOGSA-N beta-L-uridine Natural products O[C@H]1[C@@H](O)[C@H](CO)O[C@@H]1N1C(=O)NC(=O)C=C1 DRTQHJPVMGBUCF-PSQAKQOGSA-N 0.000 description 2
- 230000004071 biological effect Effects 0.000 description 2
- 210000000988 bone and bone Anatomy 0.000 description 2
- 210000004556 brain Anatomy 0.000 description 2
- 239000000969 carrier Substances 0.000 description 2
- 239000003054 catalyst Substances 0.000 description 2
- 238000004113 cell culture Methods 0.000 description 2
- 210000000170 cell membrane Anatomy 0.000 description 2
- 230000007969 cellular immunity Effects 0.000 description 2
- OSASVXMJTNOKOY-UHFFFAOYSA-N chlorobutanol Chemical compound CC(C)(O)C(Cl)(Cl)Cl OSASVXMJTNOKOY-UHFFFAOYSA-N 0.000 description 2
- HVYWMOMLDIMFJA-DPAQBDIFSA-N cholesterol Chemical compound C1C=C2C[C@@H](O)CC[C@]2(C)[C@@H]2[C@@H]1[C@@H]1CC[C@H]([C@H](C)CCCC(C)C)[C@@]1(C)CC2 HVYWMOMLDIMFJA-DPAQBDIFSA-N 0.000 description 2
- 230000004186 co-expression Effects 0.000 description 2
- 238000001246 colloidal dispersion Methods 0.000 description 2
- CVSVTCORWBXHQV-UHFFFAOYSA-N creatine Chemical compound NC(=[NH2+])N(C)CC([O-])=O CVSVTCORWBXHQV-UHFFFAOYSA-N 0.000 description 2
- YPHMISFOHDHNIV-FSZOTQKASA-N cycloheximide Chemical compound C1[C@@H](C)C[C@H](C)C(=O)[C@@H]1[C@H](O)CC1CC(=O)NC(=O)C1 YPHMISFOHDHNIV-FSZOTQKASA-N 0.000 description 2
- 102000003675 cytokine receptors Human genes 0.000 description 2
- 230000010250 cytokine signaling pathway Effects 0.000 description 2
- 230000001461 cytolytic effect Effects 0.000 description 2
- OPTASPLRGRRNAP-UHFFFAOYSA-N cytosine Chemical compound NC=1C=CNC(=O)N=1 OPTASPLRGRRNAP-UHFFFAOYSA-N 0.000 description 2
- 231100000433 cytotoxic Toxicity 0.000 description 2
- 230000001472 cytotoxic effect Effects 0.000 description 2
- 230000034994 death Effects 0.000 description 2
- 230000003247 decreasing effect Effects 0.000 description 2
- 238000013461 design Methods 0.000 description 2
- RNPXCFINMKSQPQ-UHFFFAOYSA-N dicetyl hydrogen phosphate Chemical compound CCCCCCCCCCCCCCCCOP(O)(=O)OCCCCCCCCCCCCCCCC RNPXCFINMKSQPQ-UHFFFAOYSA-N 0.000 description 2
- PSLWZOIUBRXAQW-UHFFFAOYSA-M dimethyl(dioctadecyl)azanium;bromide Chemical compound [Br-].CCCCCCCCCCCCCCCCCC[N+](C)(C)CCCCCCCCCCCCCCCCCC PSLWZOIUBRXAQW-UHFFFAOYSA-M 0.000 description 2
- 229940092559 enterobacter aerogenes Drugs 0.000 description 2
- 238000011124 ex vivo culture Methods 0.000 description 2
- 239000010408 film Substances 0.000 description 2
- 238000000684 flow cytometry Methods 0.000 description 2
- 239000012530 fluid Substances 0.000 description 2
- 239000012737 fresh medium Substances 0.000 description 2
- 230000009368 gene silencing by RNA Effects 0.000 description 2
- 238000001415 gene therapy Methods 0.000 description 2
- UYTPUPDQBNUYGX-UHFFFAOYSA-N guanine Chemical compound O=C1NC(N)=NC2=C1N=CN2 UYTPUPDQBNUYGX-UHFFFAOYSA-N 0.000 description 2
- 230000016784 immunoglobulin production Effects 0.000 description 2
- 239000012678 infectious agent Substances 0.000 description 2
- 230000004054 inflammatory process Effects 0.000 description 2
- 230000000977 initiatory effect Effects 0.000 description 2
- 238000002347 injection Methods 0.000 description 2
- 239000007924 injection Substances 0.000 description 2
- 230000015788 innate immune response Effects 0.000 description 2
- 229940100602 interleukin-5 Drugs 0.000 description 2
- 229940047122 interleukins Drugs 0.000 description 2
- 239000000543 intermediate Substances 0.000 description 2
- 208000032839 leukemia Diseases 0.000 description 2
- 230000021633 leukocyte mediated immunity Effects 0.000 description 2
- 238000001638 lipofection Methods 0.000 description 2
- 239000007788 liquid Substances 0.000 description 2
- 201000007270 liver cancer Diseases 0.000 description 2
- 208000014018 liver neoplasm Diseases 0.000 description 2
- 201000005202 lung cancer Diseases 0.000 description 2
- 208000020816 lung neoplasm Diseases 0.000 description 2
- 230000014759 maintenance of location Effects 0.000 description 2
- 208000015486 malignant pancreatic neoplasm Diseases 0.000 description 2
- 230000000873 masking effect Effects 0.000 description 2
- 238000000520 microinjection Methods 0.000 description 2
- 239000004005 microsphere Substances 0.000 description 2
- 238000002703 mutagenesis Methods 0.000 description 2
- 231100000350 mutagenesis Toxicity 0.000 description 2
- 239000002088 nanocapsule Substances 0.000 description 2
- 230000007935 neutral effect Effects 0.000 description 2
- 239000003921 oil Substances 0.000 description 2
- 210000003463 organelle Anatomy 0.000 description 2
- 201000002528 pancreatic cancer Diseases 0.000 description 2
- 208000008443 pancreatic carcinoma Diseases 0.000 description 2
- 239000003182 parenteral nutrition solution Substances 0.000 description 2
- 239000008188 pellet Substances 0.000 description 2
- 239000000546 pharmaceutical excipient Substances 0.000 description 2
- WTJKGGKOPKCXLL-RRHRGVEJSA-N phosphatidylcholine Chemical compound CCCCCCCCCCCCCCCC(=O)OC[C@H](COP([O-])(=O)OCC[N+](C)(C)C)OC(=O)CCCCCCCC=CCCCCCCCC WTJKGGKOPKCXLL-RRHRGVEJSA-N 0.000 description 2
- 150000004713 phosphodiesters Chemical class 0.000 description 2
- 210000004180 plasmocyte Anatomy 0.000 description 2
- 229920001308 poly(aminoacid) Polymers 0.000 description 2
- 229920001200 poly(ethylene-vinyl acetate) Polymers 0.000 description 2
- 229920000728 polyester Polymers 0.000 description 2
- 239000000843 powder Substances 0.000 description 2
- 238000001556 precipitation Methods 0.000 description 2
- 239000003755 preservative agent Substances 0.000 description 2
- QELSKZZBTMNZEB-UHFFFAOYSA-N propylparaben Chemical compound CCCOC(=O)C1=CC=C(O)C=C1 QELSKZZBTMNZEB-UHFFFAOYSA-N 0.000 description 2
- 108060006633 protein kinase Proteins 0.000 description 2
- 102000016914 ras Proteins Human genes 0.000 description 2
- 230000011663 regulation of signaling Effects 0.000 description 2
- 230000003248 secreting effect Effects 0.000 description 2
- DAEPDZWVDSPTHF-UHFFFAOYSA-M sodium pyruvate Chemical compound [Na+].CC(=O)C([O-])=O DAEPDZWVDSPTHF-UHFFFAOYSA-M 0.000 description 2
- GEHJYWRUCIMESM-UHFFFAOYSA-L sodium sulfite Chemical compound [Na+].[Na+].[O-]S([O-])=O GEHJYWRUCIMESM-UHFFFAOYSA-L 0.000 description 2
- 210000000952 spleen Anatomy 0.000 description 2
- 210000004988 splenocyte Anatomy 0.000 description 2
- 239000003381 stabilizer Substances 0.000 description 2
- 238000007619 statistical method Methods 0.000 description 2
- UCSJYZPVAKXKNQ-HZYVHMACSA-N streptomycin Chemical compound CN[C@H]1[C@H](O)[C@@H](O)[C@H](CO)O[C@H]1O[C@@H]1[C@](C=O)(O)[C@H](C)O[C@H]1O[C@@H]1[C@@H](NC(N)=N)[C@H](O)[C@@H](NC(N)=N)[C@H](O)[C@H]1O UCSJYZPVAKXKNQ-HZYVHMACSA-N 0.000 description 2
- 238000007920 subcutaneous administration Methods 0.000 description 2
- 150000008163 sugars Chemical class 0.000 description 2
- 230000009885 systemic effect Effects 0.000 description 2
- RWQNBRDOKXIBIV-UHFFFAOYSA-N thymine Chemical compound CC1=CNC(=O)NC1=O RWQNBRDOKXIBIV-UHFFFAOYSA-N 0.000 description 2
- 238000011200 topical administration Methods 0.000 description 2
- 230000002103 transcriptional effect Effects 0.000 description 2
- 210000004881 tumor cell Anatomy 0.000 description 2
- 210000003171 tumor-infiltrating lymphocyte Anatomy 0.000 description 2
- 241000712461 unidentified influenza virus Species 0.000 description 2
- DRTQHJPVMGBUCF-UHFFFAOYSA-N uracil arabinoside Natural products OC1C(O)C(CO)OC1N1C(=O)NC(=O)C=C1 DRTQHJPVMGBUCF-UHFFFAOYSA-N 0.000 description 2
- 229940045145 uridine Drugs 0.000 description 2
- OZFAFGSSMRRTDW-UHFFFAOYSA-N (2,4-dichlorophenyl) benzenesulfonate Chemical compound ClC1=CC(Cl)=CC=C1OS(=O)(=O)C1=CC=CC=C1 OZFAFGSSMRRTDW-UHFFFAOYSA-N 0.000 description 1
- LNAZSHAWQACDHT-XIYTZBAFSA-N (2r,3r,4s,5r,6s)-4,5-dimethoxy-2-(methoxymethyl)-3-[(2s,3r,4s,5r,6r)-3,4,5-trimethoxy-6-(methoxymethyl)oxan-2-yl]oxy-6-[(2r,3r,4s,5r,6r)-4,5,6-trimethoxy-2-(methoxymethyl)oxan-3-yl]oxyoxane Chemical compound CO[C@@H]1[C@@H](OC)[C@H](OC)[C@@H](COC)O[C@H]1O[C@H]1[C@H](OC)[C@@H](OC)[C@H](O[C@H]2[C@@H]([C@@H](OC)[C@H](OC)O[C@@H]2COC)OC)O[C@@H]1COC LNAZSHAWQACDHT-XIYTZBAFSA-N 0.000 description 1
- JVJGCCBAOOWGEO-RUTPOYCXSA-N (2s)-2-[[(2s)-2-[[(2s)-2-[[(2s)-2-[[(2s)-4-amino-2-[[(2s,3s)-2-[[(2s,3s)-2-[[(2s)-2-azaniumyl-3-hydroxypropanoyl]amino]-3-methylpentanoyl]amino]-3-methylpentanoyl]amino]-4-oxobutanoyl]amino]-3-phenylpropanoyl]amino]-4-carboxylatobutanoyl]amino]-6-azaniumy Chemical compound OC[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@H](C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(O)=O)CC1=CC=CC=C1 JVJGCCBAOOWGEO-RUTPOYCXSA-N 0.000 description 1
- MZOFCQQQCNRIBI-VMXHOPILSA-N (3s)-4-[[(2s)-1-[[(2s)-1-[[(1s)-1-carboxy-2-hydroxyethyl]amino]-4-methyl-1-oxopentan-2-yl]amino]-5-(diaminomethylideneamino)-1-oxopentan-2-yl]amino]-3-[[2-[[(2s)-2,6-diaminohexanoyl]amino]acetyl]amino]-4-oxobutanoic acid Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CC(O)=O)NC(=O)CNC(=O)[C@@H](N)CCCCN MZOFCQQQCNRIBI-VMXHOPILSA-N 0.000 description 1
- YYGNTYWPHWGJRM-UHFFFAOYSA-N (6E,10E,14E,18E)-2,6,10,15,19,23-hexamethyltetracosa-2,6,10,14,18,22-hexaene Chemical compound CC(C)=CCCC(C)=CCCC(C)=CCCC=C(C)CCC=C(C)CCC=C(C)C YYGNTYWPHWGJRM-UHFFFAOYSA-N 0.000 description 1
- PORPENFLTBBHSG-MGBGTMOVSA-N 1,2-dihexadecanoyl-sn-glycerol-3-phosphate Chemical compound CCCCCCCCCCCCCCCC(=O)OC[C@H](COP(O)(O)=O)OC(=O)CCCCCCCCCCCCCCC PORPENFLTBBHSG-MGBGTMOVSA-N 0.000 description 1
- UHDGCWIWMRVCDJ-UHFFFAOYSA-N 1-beta-D-Xylofuranosyl-NH-Cytosine Natural products O=C1N=C(N)C=CN1C1C(O)C(O)C(CO)O1 UHDGCWIWMRVCDJ-UHFFFAOYSA-N 0.000 description 1
- GZCWLCBFPRFLKL-UHFFFAOYSA-N 1-prop-2-ynoxypropan-2-ol Chemical compound CC(O)COCC#C GZCWLCBFPRFLKL-UHFFFAOYSA-N 0.000 description 1
- 108020004463 18S ribosomal RNA Proteins 0.000 description 1
- JKMHFZQWWAIEOD-UHFFFAOYSA-N 2-[4-(2-hydroxyethyl)piperazin-1-yl]ethanesulfonic acid Chemical compound OCC[NH+]1CCN(CCS([O-])(=O)=O)CC1 JKMHFZQWWAIEOD-UHFFFAOYSA-N 0.000 description 1
- 238000010600 3H thymidine incorporation assay Methods 0.000 description 1
- 108020005029 5' Flanking Region Proteins 0.000 description 1
- 102000013563 Acid Phosphatase Human genes 0.000 description 1
- 108010051457 Acid Phosphatase Proteins 0.000 description 1
- 241000186046 Actinomyces Species 0.000 description 1
- 208000026872 Addison Disease Diseases 0.000 description 1
- 241000701242 Adenoviridae Species 0.000 description 1
- 102000002260 Alkaline Phosphatase Human genes 0.000 description 1
- 108020004774 Alkaline Phosphatase Proteins 0.000 description 1
- 206010002556 Ankylosing Spondylitis Diseases 0.000 description 1
- 241000712892 Arenaviridae Species 0.000 description 1
- 206010003827 Autoimmune hepatitis Diseases 0.000 description 1
- 241000714230 Avian leukemia virus Species 0.000 description 1
- 102000019260 B-Cell Antigen Receptors Human genes 0.000 description 1
- 108010012919 B-Cell Antigen Receptors Proteins 0.000 description 1
- 230000003844 B-cell-activation Effects 0.000 description 1
- 241000193830 Bacillus <bacterium> Species 0.000 description 1
- 241000193738 Bacillus anthracis Species 0.000 description 1
- 108020000946 Bacterial DNA Proteins 0.000 description 1
- 102100021663 Baculoviral IAP repeat-containing protein 5 Human genes 0.000 description 1
- 208000023328 Basedow disease Diseases 0.000 description 1
- 102100026189 Beta-galactosidase Human genes 0.000 description 1
- 241000702628 Birnaviridae Species 0.000 description 1
- 206010005003 Bladder cancer Diseases 0.000 description 1
- 241000335423 Blastomyces Species 0.000 description 1
- 241000589969 Borreliella burgdorferi Species 0.000 description 1
- 208000003174 Brain Neoplasms Diseases 0.000 description 1
- 239000002126 C01EB10 - Adenosine Substances 0.000 description 1
- 108010029697 CD40 Ligand Proteins 0.000 description 1
- 102100032937 CD40 ligand Human genes 0.000 description 1
- 101150108228 CYLD gene Proteins 0.000 description 1
- 101100289995 Caenorhabditis elegans mac-1 gene Proteins 0.000 description 1
- 101100257262 Caenorhabditis elegans soc-1 gene Proteins 0.000 description 1
- 241000222122 Candida albicans Species 0.000 description 1
- 229920002134 Carboxymethyl cellulose Polymers 0.000 description 1
- 102000014914 Carrier Proteins Human genes 0.000 description 1
- 108010078791 Carrier Proteins Proteins 0.000 description 1
- 241000606153 Chlamydia trachomatis Species 0.000 description 1
- 108010035563 Chloramphenicol O-acetyltransferase Proteins 0.000 description 1
- VYZAMTAEIAYCRO-UHFFFAOYSA-N Chromium Chemical compound [Cr] VYZAMTAEIAYCRO-UHFFFAOYSA-N 0.000 description 1
- 241000193449 Clostridium tetani Species 0.000 description 1
- 241000223203 Coccidioides Species 0.000 description 1
- 108020004705 Codon Proteins 0.000 description 1
- 108700010070 Codon Usage Proteins 0.000 description 1
- 101100332459 Coffea arabica DXMT1 gene Proteins 0.000 description 1
- 206010009900 Colitis ulcerative Diseases 0.000 description 1
- 208000001333 Colorectal Neoplasms Diseases 0.000 description 1
- 241000186216 Corynebacterium Species 0.000 description 1
- 241000186227 Corynebacterium diphtheriae Species 0.000 description 1
- 241000709687 Coxsackievirus Species 0.000 description 1
- MIKUYHXYGGJMLM-GIMIYPNGSA-N Crotonoside Natural products C1=NC2=C(N)NC(=O)N=C2N1[C@H]1O[C@@H](CO)[C@H](O)[C@@H]1O MIKUYHXYGGJMLM-GIMIYPNGSA-N 0.000 description 1
- 201000007336 Cryptococcosis Diseases 0.000 description 1
- 241000221204 Cryptococcus neoformans Species 0.000 description 1
- UHDGCWIWMRVCDJ-CCXZUQQUSA-N Cytarabine Chemical compound O=C1N=C(N)C=CN1[C@H]1[C@@H](O)[C@H](O)[C@@H](CO)O1 UHDGCWIWMRVCDJ-CCXZUQQUSA-N 0.000 description 1
- UHDGCWIWMRVCDJ-PSQAKQOGSA-N Cytidine Natural products O=C1N=C(N)C=CN1[C@@H]1[C@@H](O)[C@@H](O)[C@H](CO)O1 UHDGCWIWMRVCDJ-PSQAKQOGSA-N 0.000 description 1
- 102000000311 Cytosine Deaminase Human genes 0.000 description 1
- 108010080611 Cytosine Deaminase Proteins 0.000 description 1
- NYHBQMYGNKIUIF-UHFFFAOYSA-N D-guanosine Natural products C1=2NC(N)=NC(=O)C=2N=CN1C1OC(CO)C(O)C1O NYHBQMYGNKIUIF-UHFFFAOYSA-N 0.000 description 1
- 108010008286 DNA nucleotidylexotransferase Proteins 0.000 description 1
- 230000007023 DNA restriction-modification system Effects 0.000 description 1
- 241000725619 Dengue virus Species 0.000 description 1
- 102100029588 Deoxycytidine kinase Human genes 0.000 description 1
- 108010033174 Deoxycytidine kinase Proteins 0.000 description 1
- GZDFHIJNHHMENY-UHFFFAOYSA-N Dimethyl dicarbonate Chemical compound COC(=O)OC(=O)OC GZDFHIJNHHMENY-UHFFFAOYSA-N 0.000 description 1
- 239000012591 Dulbecco’s Phosphate Buffered Saline Substances 0.000 description 1
- 208000010975 Dystrophic epidermolysis bullosa Diseases 0.000 description 1
- 108091035710 E-box Proteins 0.000 description 1
- 101150029707 ERBB2 gene Proteins 0.000 description 1
- 241001115402 Ebolavirus Species 0.000 description 1
- 241001466953 Echovirus Species 0.000 description 1
- 108010009900 Endothelial Protein C Receptor Proteins 0.000 description 1
- 102100030024 Endothelial protein C receptor Human genes 0.000 description 1
- 241000194033 Enterococcus Species 0.000 description 1
- 241000194032 Enterococcus faecalis Species 0.000 description 1
- 241000991587 Enterovirus C Species 0.000 description 1
- 208000000832 Equine Encephalomyelitis Diseases 0.000 description 1
- 206010051841 Exposure to allergen Diseases 0.000 description 1
- 241000710781 Flaviviridae Species 0.000 description 1
- 102000004315 Forkhead Transcription Factors Human genes 0.000 description 1
- 108090000852 Forkhead Transcription Factors Proteins 0.000 description 1
- 241000605909 Fusobacterium Species 0.000 description 1
- 241000605986 Fusobacterium nucleatum Species 0.000 description 1
- 241000287828 Gallus gallus Species 0.000 description 1
- 208000005577 Gastroenteritis Diseases 0.000 description 1
- 108700028146 Genetic Enhancer Elements Proteins 0.000 description 1
- 108700039691 Genetic Promoter Regions Proteins 0.000 description 1
- 108700007698 Genetic Terminator Regions Proteins 0.000 description 1
- 206010018364 Glomerulonephritis Diseases 0.000 description 1
- 102100041003 Glutamate carboxypeptidase 2 Human genes 0.000 description 1
- JZNWSCPGTDBMEW-UHFFFAOYSA-N Glycerophosphorylethanolamin Natural products NCCOP(O)(=O)OCC(O)CO JZNWSCPGTDBMEW-UHFFFAOYSA-N 0.000 description 1
- 208000015023 Graves' disease Diseases 0.000 description 1
- 208000035895 Guillain-Barré syndrome Diseases 0.000 description 1
- 239000007995 HEPES buffer Substances 0.000 description 1
- 208000031886 HIV Infections Diseases 0.000 description 1
- 208000037357 HIV infectious disease Diseases 0.000 description 1
- 108010058597 HLA-DR Antigens Proteins 0.000 description 1
- 102000006354 HLA-DR Antigens Human genes 0.000 description 1
- 241000606768 Haemophilus influenzae Species 0.000 description 1
- 206010061192 Haemorrhagic fever Diseases 0.000 description 1
- 208000001204 Hashimoto Disease Diseases 0.000 description 1
- 208000030836 Hashimoto thyroiditis Diseases 0.000 description 1
- 241000590002 Helicobacter pylori Species 0.000 description 1
- 102000001554 Hemoglobins Human genes 0.000 description 1
- 108010054147 Hemoglobins Proteins 0.000 description 1
- 241000700739 Hepadnaviridae Species 0.000 description 1
- 208000005176 Hepatitis C Diseases 0.000 description 1
- 241000709721 Hepatovirus A Species 0.000 description 1
- 208000009889 Herpes Simplex Diseases 0.000 description 1
- 241000700586 Herpesviridae Species 0.000 description 1
- 241000238631 Hexapoda Species 0.000 description 1
- 102100022130 High mobility group protein B3 Human genes 0.000 description 1
- 241000228402 Histoplasma Species 0.000 description 1
- 208000017604 Hodgkin disease Diseases 0.000 description 1
- 208000021519 Hodgkin lymphoma Diseases 0.000 description 1
- 208000010747 Hodgkins lymphoma Diseases 0.000 description 1
- 102100030234 Homeobox protein cut-like 1 Human genes 0.000 description 1
- 101000892862 Homo sapiens Glutamate carboxypeptidase 2 Proteins 0.000 description 1
- 101001006375 Homo sapiens High mobility group nucleosome-binding domain-containing protein 4 Proteins 0.000 description 1
- 101001045794 Homo sapiens High mobility group protein B3 Proteins 0.000 description 1
- 101001046686 Homo sapiens Integrin alpha-M Proteins 0.000 description 1
- 101000777628 Homo sapiens Leukocyte antigen CD37 Proteins 0.000 description 1
- 101100495232 Homo sapiens MS4A1 gene Proteins 0.000 description 1
- 101000578784 Homo sapiens Melanoma antigen recognized by T-cells 1 Proteins 0.000 description 1
- 101000763579 Homo sapiens Toll-like receptor 1 Proteins 0.000 description 1
- 108060006678 I-kappa-B kinase Proteins 0.000 description 1
- 102000001284 I-kappa-B kinase Human genes 0.000 description 1
- 101150103227 IFN gene Proteins 0.000 description 1
- 101150007193 IFNB1 gene Proteins 0.000 description 1
- 101150106931 IFNG gene Proteins 0.000 description 1
- 102000043138 IRF family Human genes 0.000 description 1
- 108091054729 IRF family Proteins 0.000 description 1
- 208000001718 Immediate Hypersensitivity Diseases 0.000 description 1
- 229930010555 Inosine Natural products 0.000 description 1
- UGQMRVRMYYASKQ-KQYNXXCUSA-N Inosine Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C2=NC=NC(O)=C2N=C1 UGQMRVRMYYASKQ-KQYNXXCUSA-N 0.000 description 1
- 102100022338 Integrin alpha-M Human genes 0.000 description 1
- 102100023530 Interleukin-1 receptor-associated kinase 3 Human genes 0.000 description 1
- 101710199012 Interleukin-1 receptor-associated kinase 3 Proteins 0.000 description 1
- 108010002386 Interleukin-3 Proteins 0.000 description 1
- 241000701377 Iridoviridae Species 0.000 description 1
- 102000042838 JAK family Human genes 0.000 description 1
- 108091082332 JAK family Proteins 0.000 description 1
- 102000019145 JUN kinase activity proteins Human genes 0.000 description 1
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 1
- 229930182816 L-glutamine Natural products 0.000 description 1
- 241000446313 Lamella Species 0.000 description 1
- 241000589248 Legionella Species 0.000 description 1
- 208000007764 Legionnaires' Disease Diseases 0.000 description 1
- 241000589902 Leptospira Species 0.000 description 1
- 102100031586 Leukocyte antigen CD37 Human genes 0.000 description 1
- 241000186779 Listeria monocytogenes Species 0.000 description 1
- 108060001084 Luciferase Proteins 0.000 description 1
- 239000005089 Luciferase Substances 0.000 description 1
- 241000283923 Marmota monax Species 0.000 description 1
- 102100027754 Mast/stem cell growth factor receptor Kit Human genes 0.000 description 1
- 241000712079 Measles morbillivirus Species 0.000 description 1
- 102100028389 Melanoma antigen recognized by T-cells 1 Human genes 0.000 description 1
- 102000018697 Membrane Proteins Human genes 0.000 description 1
- 108010052285 Membrane Proteins Proteins 0.000 description 1
- 206010027480 Metastatic malignant melanoma Diseases 0.000 description 1
- 206010049567 Miller Fisher syndrome Diseases 0.000 description 1
- 241000713333 Mouse mammary tumor virus Species 0.000 description 1
- 241000711386 Mumps virus Species 0.000 description 1
- 241000711408 Murine respirovirus Species 0.000 description 1
- 101100335081 Mus musculus Flt3 gene Proteins 0.000 description 1
- 101000981253 Mus musculus GPI-linked NAD(P)(+)-arginine ADP-ribosyltransferase 1 Proteins 0.000 description 1
- 101001043827 Mus musculus Interleukin-2 Proteins 0.000 description 1
- 101001002703 Mus musculus Interleukin-4 Proteins 0.000 description 1
- 101000628985 Mus musculus Small ubiquitin-related modifier 1 Proteins 0.000 description 1
- 101100204386 Mus musculus Sumo1 gene Proteins 0.000 description 1
- 101100481579 Mus musculus Tlr11 gene Proteins 0.000 description 1
- 101100481580 Mus musculus Tlr12 gene Proteins 0.000 description 1
- 241000186359 Mycobacterium Species 0.000 description 1
- 241000187484 Mycobacterium gordonae Species 0.000 description 1
- 241000186364 Mycobacterium intracellulare Species 0.000 description 1
- 102000003505 Myosin Human genes 0.000 description 1
- 108060008487 Myosin Proteins 0.000 description 1
- 206010028665 Myxoedema Diseases 0.000 description 1
- 102000018745 NF-KappaB Inhibitor alpha Human genes 0.000 description 1
- 108010052419 NF-KappaB Inhibitor alpha Proteins 0.000 description 1
- 102100033457 NF-kappa-B inhibitor beta Human genes 0.000 description 1
- 101710204094 NF-kappa-B inhibitor beta Proteins 0.000 description 1
- 102100033104 NF-kappa-B inhibitor epsilon Human genes 0.000 description 1
- 101710093997 NF-kappa-B inhibitor epsilon Proteins 0.000 description 1
- 241000588652 Neisseria gonorrhoeae Species 0.000 description 1
- 241000588650 Neisseria meningitidis Species 0.000 description 1
- 208000015914 Non-Hodgkin lymphomas Diseases 0.000 description 1
- 238000000636 Northern blotting Methods 0.000 description 1
- 239000004677 Nylon Substances 0.000 description 1
- REYJJPSVUYRZGE-UHFFFAOYSA-N Octadecylamine Chemical compound CCCCCCCCCCCCCCCCCCN REYJJPSVUYRZGE-UHFFFAOYSA-N 0.000 description 1
- 206010048685 Oral infection Diseases 0.000 description 1
- 241000702259 Orbivirus Species 0.000 description 1
- 241000150218 Orthonairovirus Species 0.000 description 1
- 241000702244 Orthoreovirus Species 0.000 description 1
- 102000004316 Oxidoreductases Human genes 0.000 description 1
- 108090000854 Oxidoreductases Proteins 0.000 description 1
- 241000711504 Paramyxoviridae Species 0.000 description 1
- 208000002606 Paramyxoviridae Infections Diseases 0.000 description 1
- 206010034038 Parotitis Diseases 0.000 description 1
- 241000606860 Pasteurella Species 0.000 description 1
- 201000011152 Pemphigus Diseases 0.000 description 1
- 229930182555 Penicillin Natural products 0.000 description 1
- JGSARLDLIJGVTE-MBNYWOFBSA-N Penicillin G Chemical compound N([C@H]1[C@H]2SC([C@@H](N2C1=O)C(O)=O)(C)C)C(=O)CC1=CC=CC=C1 JGSARLDLIJGVTE-MBNYWOFBSA-N 0.000 description 1
- 208000031845 Pernicious anaemia Diseases 0.000 description 1
- 208000037581 Persistent Infection Diseases 0.000 description 1
- 241000223960 Plasmodium falciparum Species 0.000 description 1
- 239000002202 Polyethylene glycol Substances 0.000 description 1
- 108010039918 Polylysine Proteins 0.000 description 1
- 241000700625 Poxviridae Species 0.000 description 1
- 208000024777 Prion disease Diseases 0.000 description 1
- 108010007568 Protamines Proteins 0.000 description 1
- 102000007327 Protamines Human genes 0.000 description 1
- 102000004245 Proteasome Endopeptidase Complex Human genes 0.000 description 1
- 108090000708 Proteasome Endopeptidase Complex Proteins 0.000 description 1
- 101710149951 Protein Tat Proteins 0.000 description 1
- 108010001859 Proto-Oncogene Proteins c-rel Proteins 0.000 description 1
- 102000000850 Proto-Oncogene Proteins c-rel Human genes 0.000 description 1
- 235000014443 Pyrus communis Nutrition 0.000 description 1
- 230000006819 RNA synthesis Effects 0.000 description 1
- 241000711798 Rabies lyssavirus Species 0.000 description 1
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 description 1
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 description 1
- 241000702247 Reoviridae Species 0.000 description 1
- 241000712907 Retroviridae Species 0.000 description 1
- 241000711931 Rhabdoviridae Species 0.000 description 1
- 108091028664 Ribonucleotide Proteins 0.000 description 1
- PYMYPHUHKUWMLA-LMVFSUKVSA-N Ribose Natural products OC[C@@H](O)[C@@H](O)[C@@H](O)C=O PYMYPHUHKUWMLA-LMVFSUKVSA-N 0.000 description 1
- 239000006146 Roswell Park Memorial Institute medium Substances 0.000 description 1
- 241000702670 Rotavirus Species 0.000 description 1
- 241000714474 Rous sarcoma virus Species 0.000 description 1
- 108010044012 STAT1 Transcription Factor Proteins 0.000 description 1
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 1
- 206010039491 Sarcoma Diseases 0.000 description 1
- 206010039710 Scleroderma Diseases 0.000 description 1
- 206010040070 Septic Shock Diseases 0.000 description 1
- 102100029904 Signal transducer and activator of transcription 1-alpha/beta Human genes 0.000 description 1
- 208000021386 Sjogren Syndrome Diseases 0.000 description 1
- 208000000453 Skin Neoplasms Diseases 0.000 description 1
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 1
- 238000002105 Southern blotting Methods 0.000 description 1
- 241000191940 Staphylococcus Species 0.000 description 1
- 241000191967 Staphylococcus aureus Species 0.000 description 1
- 241001478880 Streptobacillus moniliformis Species 0.000 description 1
- 241000194049 Streptococcus equinus Species 0.000 description 1
- 241000193996 Streptococcus pyogenes Species 0.000 description 1
- 241001505901 Streptococcus sp. 'group A' Species 0.000 description 1
- 241000187747 Streptomyces Species 0.000 description 1
- 238000000692 Student's t-test Methods 0.000 description 1
- 102000008036 Suppressor of Cytokine Signaling Proteins Human genes 0.000 description 1
- 108010075383 Suppressor of Cytokine Signaling Proteins Proteins 0.000 description 1
- 102100024779 Suppressor of cytokine signaling 1 Human genes 0.000 description 1
- 101150052863 THY1 gene Proteins 0.000 description 1
- 108700012920 TNF Proteins 0.000 description 1
- 239000004098 Tetracycline Substances 0.000 description 1
- 101001081186 Tetrahymena pyriformis High mobility group protein Proteins 0.000 description 1
- BHEOSNUKNHRBNM-UHFFFAOYSA-N Tetramethylsqualene Natural products CC(=C)C(C)CCC(=C)C(C)CCC(C)=CCCC=C(C)CCC(C)C(=C)CCC(C)C(C)=C BHEOSNUKNHRBNM-UHFFFAOYSA-N 0.000 description 1
- RYYWUUFWQRZTIU-UHFFFAOYSA-N Thiophosphoric acid Chemical class OP(O)(S)=O RYYWUUFWQRZTIU-UHFFFAOYSA-N 0.000 description 1
- 108010046722 Thrombospondin 1 Proteins 0.000 description 1
- 102100036034 Thrombospondin-1 Human genes 0.000 description 1
- 102100037357 Thymidylate kinase Human genes 0.000 description 1
- 241000710924 Togaviridae Species 0.000 description 1
- 102100027010 Toll-like receptor 1 Human genes 0.000 description 1
- 241000223997 Toxoplasma gondii Species 0.000 description 1
- 102100035100 Transcription factor p65 Human genes 0.000 description 1
- 241000589886 Treponema Species 0.000 description 1
- 206010045240 Type I hypersensitivity Diseases 0.000 description 1
- 201000006704 Ulcerative Colitis Diseases 0.000 description 1
- 208000007097 Urinary Bladder Neoplasms Diseases 0.000 description 1
- 208000002495 Uterine Neoplasms Diseases 0.000 description 1
- 241000700618 Vaccinia virus Species 0.000 description 1
- 241000700647 Variola virus Species 0.000 description 1
- 206010047115 Vasculitis Diseases 0.000 description 1
- 206010047642 Vitiligo Diseases 0.000 description 1
- 108091093126 WHP Posttrascriptional Response Element Proteins 0.000 description 1
- 241000710772 Yellow fever virus Species 0.000 description 1
- ATBOMIWRCZXYSZ-XZBBILGWSA-N [1-[2,3-dihydroxypropoxy(hydroxy)phosphoryl]oxy-3-hexadecanoyloxypropan-2-yl] (9e,12e)-octadeca-9,12-dienoate Chemical compound CCCCCCCCCCCCCCCC(=O)OCC(COP(O)(=O)OCC(O)CO)OC(=O)CCCCCCC\C=C\C\C=C\CCCCC ATBOMIWRCZXYSZ-XZBBILGWSA-N 0.000 description 1
- XLLNINGEDIOQGQ-UHFFFAOYSA-N [acetyloxy(hydroxy)phosphoryl] acetate Chemical compound CC(=O)OP(O)(=O)OC(C)=O XLLNINGEDIOQGQ-UHFFFAOYSA-N 0.000 description 1
- 229960004150 aciclovir Drugs 0.000 description 1
- MKUXAQIIEYXACX-UHFFFAOYSA-N aciclovir Chemical compound N1C(N)=NC(=O)C2=C1N(COCCO)C=N2 MKUXAQIIEYXACX-UHFFFAOYSA-N 0.000 description 1
- 230000009471 action Effects 0.000 description 1
- 108010042591 activated protein C receptor Proteins 0.000 description 1
- 230000013564 activation of immune response Effects 0.000 description 1
- 208000009956 adenocarcinoma Diseases 0.000 description 1
- 229960005305 adenosine Drugs 0.000 description 1
- 108700010877 adenoviridae proteins Proteins 0.000 description 1
- 230000001464 adherent effect Effects 0.000 description 1
- 230000004520 agglutination Effects 0.000 description 1
- 150000001298 alcohols Chemical class 0.000 description 1
- 150000001299 aldehydes Chemical class 0.000 description 1
- 150000001338 aliphatic hydrocarbons Chemical class 0.000 description 1
- 230000000961 alloantigen Effects 0.000 description 1
- 108010004469 allophycocyanin Proteins 0.000 description 1
- 208000004631 alopecia areata Diseases 0.000 description 1
- HMFHBZSHGGEWLO-UHFFFAOYSA-N alpha-D-Furanose-Ribose Natural products OCC1OC(O)C(O)C1O HMFHBZSHGGEWLO-UHFFFAOYSA-N 0.000 description 1
- AWUCVROLDVIAJX-UHFFFAOYSA-N alpha-glycerophosphate Natural products OCC(O)COP(O)(O)=O AWUCVROLDVIAJX-UHFFFAOYSA-N 0.000 description 1
- 229940037003 alum Drugs 0.000 description 1
- 150000001412 amines Chemical class 0.000 description 1
- 125000000539 amino acid group Chemical group 0.000 description 1
- 150000001414 amino alcohols Chemical class 0.000 description 1
- 235000019270 ammonium chloride Nutrition 0.000 description 1
- 238000010171 animal model Methods 0.000 description 1
- 238000000137 annealing Methods 0.000 description 1
- 230000001745 anti-biotin effect Effects 0.000 description 1
- 230000000259 anti-tumor effect Effects 0.000 description 1
- 239000003963 antioxidant agent Substances 0.000 description 1
- 235000006708 antioxidants Nutrition 0.000 description 1
- 239000007900 aqueous suspension Substances 0.000 description 1
- 239000008135 aqueous vehicle Substances 0.000 description 1
- 239000000823 artificial membrane Substances 0.000 description 1
- 235000010323 ascorbic acid Nutrition 0.000 description 1
- 239000011668 ascorbic acid Substances 0.000 description 1
- 229960005070 ascorbic acid Drugs 0.000 description 1
- 244000309743 astrovirus Species 0.000 description 1
- 208000010216 atopic IgE responsiveness Diseases 0.000 description 1
- 229940065181 bacillus anthracis Drugs 0.000 description 1
- 210000003651 basophil Anatomy 0.000 description 1
- 230000006399 behavior Effects 0.000 description 1
- 229960000686 benzalkonium chloride Drugs 0.000 description 1
- CADWTSSKOVRVJC-UHFFFAOYSA-N benzyl(dimethyl)azanium;chloride Chemical compound [Cl-].C[NH+](C)CC1=CC=CC=C1 CADWTSSKOVRVJC-UHFFFAOYSA-N 0.000 description 1
- 108010005774 beta-Galactosidase Proteins 0.000 description 1
- 239000003012 bilayer membrane Substances 0.000 description 1
- 108091008324 binding proteins Proteins 0.000 description 1
- 230000003115 biocidal effect Effects 0.000 description 1
- 238000010170 biological method Methods 0.000 description 1
- 230000031018 biological processes and functions Effects 0.000 description 1
- 229930189065 blasticidin Natural products 0.000 description 1
- 229920001400 block copolymer Polymers 0.000 description 1
- 210000003103 bodily secretion Anatomy 0.000 description 1
- 210000001124 body fluid Anatomy 0.000 description 1
- 239000010839 body fluid Substances 0.000 description 1
- 210000002798 bone marrow cell Anatomy 0.000 description 1
- 239000000872 buffer Substances 0.000 description 1
- 239000007975 buffered saline Substances 0.000 description 1
- DQXBYHZEEUGOBF-UHFFFAOYSA-N but-3-enoic acid;ethene Chemical compound C=C.OC(=O)CC=C DQXBYHZEEUGOBF-UHFFFAOYSA-N 0.000 description 1
- 238000004364 calculation method Methods 0.000 description 1
- 229940095731 candida albicans Drugs 0.000 description 1
- 239000001768 carboxy methyl cellulose Substances 0.000 description 1
- 235000010948 carboxy methyl cellulose Nutrition 0.000 description 1
- 239000008112 carboxymethyl-cellulose Substances 0.000 description 1
- 230000003197 catalytic effect Effects 0.000 description 1
- 150000001768 cations Chemical class 0.000 description 1
- 230000011712 cell development Effects 0.000 description 1
- 230000024245 cell differentiation Effects 0.000 description 1
- 230000003915 cell function Effects 0.000 description 1
- 230000011748 cell maturation Effects 0.000 description 1
- 239000006285 cell suspension Substances 0.000 description 1
- 238000002659 cell therapy Methods 0.000 description 1
- 230000001413 cellular effect Effects 0.000 description 1
- 238000005119 centrifugation Methods 0.000 description 1
- 230000008859 change Effects 0.000 description 1
- 238000007385 chemical modification Methods 0.000 description 1
- 229940038705 chlamydia trachomatis Drugs 0.000 description 1
- 229960004926 chlorobutanol Drugs 0.000 description 1
- 210000003763 chloroplast Anatomy 0.000 description 1
- 235000012000 cholesterol Nutrition 0.000 description 1
- 229910052804 chromium Inorganic materials 0.000 description 1
- 230000001684 chronic effect Effects 0.000 description 1
- 208000025302 chronic primary adrenal insufficiency Diseases 0.000 description 1
- 238000003776 cleavage reaction Methods 0.000 description 1
- 238000012411 cloning technique Methods 0.000 description 1
- 239000000084 colloidal system Substances 0.000 description 1
- 230000000052 comparative effect Effects 0.000 description 1
- 230000006957 competitive inhibition Effects 0.000 description 1
- 238000004590 computer program Methods 0.000 description 1
- 230000001268 conjugating effect Effects 0.000 description 1
- 230000004940 costimulation Effects 0.000 description 1
- 229960003624 creatine Drugs 0.000 description 1
- 239000006046 creatine Substances 0.000 description 1
- 239000012228 culture supernatant Substances 0.000 description 1
- 210000004748 cultured cell Anatomy 0.000 description 1
- 238000012258 culturing Methods 0.000 description 1
- UHDGCWIWMRVCDJ-ZAKLUEHWSA-N cytidine Chemical compound O=C1N=C(N)C=CN1[C@H]1[C@H](O)[C@@H](O)[C@H](CO)O1 UHDGCWIWMRVCDJ-ZAKLUEHWSA-N 0.000 description 1
- 230000016396 cytokine production Effects 0.000 description 1
- 230000009089 cytolysis Effects 0.000 description 1
- 229940104302 cytosine Drugs 0.000 description 1
- 230000003013 cytotoxicity Effects 0.000 description 1
- 231100000135 cytotoxicity Toxicity 0.000 description 1
- 108010000742 dTMP kinase Proteins 0.000 description 1
- 230000006378 damage Effects 0.000 description 1
- 230000007123 defense Effects 0.000 description 1
- 230000002950 deficient Effects 0.000 description 1
- 239000003405 delayed action preparation Substances 0.000 description 1
- 230000002939 deleterious effect Effects 0.000 description 1
- 238000002716 delivery method Methods 0.000 description 1
- 239000005547 deoxyribonucleotide Substances 0.000 description 1
- 125000002637 deoxyribonucleotide group Chemical group 0.000 description 1
- 238000011161 development Methods 0.000 description 1
- 230000018109 developmental process Effects 0.000 description 1
- 239000008121 dextrose Substances 0.000 description 1
- 206010012601 diabetes mellitus Diseases 0.000 description 1
- 229940093541 dicetylphosphate Drugs 0.000 description 1
- 235000014113 dietary fatty acids Nutrition 0.000 description 1
- 230000029087 digestion Effects 0.000 description 1
- 238000010790 dilution Methods 0.000 description 1
- 239000012895 dilution Substances 0.000 description 1
- BPHQZTVXXXJVHI-UHFFFAOYSA-N dimyristoyl phosphatidylglycerol Chemical compound CCCCCCCCCCCCCC(=O)OCC(COP(O)(=O)OCC(O)CO)OC(=O)CCCCCCCCCCCCC BPHQZTVXXXJVHI-UHFFFAOYSA-N 0.000 description 1
- LOKCTEFSRHRXRJ-UHFFFAOYSA-I dipotassium trisodium dihydrogen phosphate hydrogen phosphate dichloride Chemical compound P(=O)(O)(O)[O-].[K+].P(=O)(O)([O-])[O-].[Na+].[Na+].[Cl-].[K+].[Cl-].[Na+] LOKCTEFSRHRXRJ-UHFFFAOYSA-I 0.000 description 1
- ZGSPNIOCEDOHGS-UHFFFAOYSA-L disodium [3-[2,3-di(octadeca-9,12-dienoyloxy)propoxy-oxidophosphoryl]oxy-2-hydroxypropyl] 2,3-di(octadeca-9,12-dienoyloxy)propyl phosphate Chemical compound [Na+].[Na+].CCCCCC=CCC=CCCCCCCCC(=O)OCC(OC(=O)CCCCCCCC=CCC=CCCCCC)COP([O-])(=O)OCC(O)COP([O-])(=O)OCC(OC(=O)CCCCCCCC=CCC=CCCCCC)COC(=O)CCCCCCCC=CCC=CCCCCC ZGSPNIOCEDOHGS-UHFFFAOYSA-L 0.000 description 1
- 239000002270 dispersing agent Substances 0.000 description 1
- 239000006185 dispersion Substances 0.000 description 1
- 238000004090 dissolution Methods 0.000 description 1
- NAGJZTKCGNOGPW-UHFFFAOYSA-N dithiophosphoric acid Chemical class OP(O)(S)=S NAGJZTKCGNOGPW-UHFFFAOYSA-N 0.000 description 1
- PRAKJMSDJKAYCZ-UHFFFAOYSA-N dodecahydrosqualene Natural products CC(C)CCCC(C)CCCC(C)CCCCC(C)CCCC(C)CCCC(C)C PRAKJMSDJKAYCZ-UHFFFAOYSA-N 0.000 description 1
- 230000008030 elimination Effects 0.000 description 1
- 238000003379 elimination reaction Methods 0.000 description 1
- 210000002257 embryonic structure Anatomy 0.000 description 1
- 239000003995 emulsifying agent Substances 0.000 description 1
- 206010014599 encephalitis Diseases 0.000 description 1
- 230000012202 endocytosis Effects 0.000 description 1
- 210000001163 endosome Anatomy 0.000 description 1
- 238000005516 engineering process Methods 0.000 description 1
- 210000001842 enterocyte Anatomy 0.000 description 1
- 230000002255 enzymatic effect Effects 0.000 description 1
- 238000001976 enzyme digestion Methods 0.000 description 1
- 238000003114 enzyme-linked immunosorbent spot assay Methods 0.000 description 1
- 208000004298 epidermolysis bullosa dystrophica Diseases 0.000 description 1
- 210000003743 erythrocyte Anatomy 0.000 description 1
- 239000005038 ethylene vinyl acetate Substances 0.000 description 1
- 210000003527 eukaryotic cell Anatomy 0.000 description 1
- 238000001704 evaporation Methods 0.000 description 1
- 230000001747 exhibiting effect Effects 0.000 description 1
- 229930195729 fatty acid Natural products 0.000 description 1
- 239000000194 fatty acid Substances 0.000 description 1
- 150000004665 fatty acids Chemical class 0.000 description 1
- XRECTZIEBJDKEO-UHFFFAOYSA-N flucytosine Chemical compound NC1=NC(=O)NC=C1F XRECTZIEBJDKEO-UHFFFAOYSA-N 0.000 description 1
- 229960004413 flucytosine Drugs 0.000 description 1
- MHMNJMPURVTYEJ-UHFFFAOYSA-N fluorescein-5-isothiocyanate Chemical compound O1C(=O)C2=CC(N=C=S)=CC=C2C21C1=CC=C(O)C=C1OC1=CC(O)=CC=C21 MHMNJMPURVTYEJ-UHFFFAOYSA-N 0.000 description 1
- 238000013467 fragmentation Methods 0.000 description 1
- 238000006062 fragmentation reaction Methods 0.000 description 1
- 238000004108 freeze drying Methods 0.000 description 1
- 230000002538 fungal effect Effects 0.000 description 1
- 229940044627 gamma-interferon Drugs 0.000 description 1
- 229960002963 ganciclovir Drugs 0.000 description 1
- IRSCQMHQWWYFCW-UHFFFAOYSA-N ganciclovir Chemical compound O=C1NC(N)=NC2=C1N=CN2COC(CO)CO IRSCQMHQWWYFCW-UHFFFAOYSA-N 0.000 description 1
- 230000002496 gastric effect Effects 0.000 description 1
- 239000000499 gel Substances 0.000 description 1
- 108091008053 gene clusters Proteins 0.000 description 1
- 238000010363 gene targeting Methods 0.000 description 1
- 230000002068 genetic effect Effects 0.000 description 1
- 239000011521 glass Substances 0.000 description 1
- 239000003862 glucocorticoid Substances 0.000 description 1
- 239000008103 glucose Substances 0.000 description 1
- 150000002334 glycols Chemical class 0.000 description 1
- 239000008187 granular material Substances 0.000 description 1
- 210000003714 granulocyte Anatomy 0.000 description 1
- 230000012010 growth Effects 0.000 description 1
- 239000003102 growth factor Substances 0.000 description 1
- 229940029575 guanosine Drugs 0.000 description 1
- 229940047650 haemophilus influenzae Drugs 0.000 description 1
- 238000003306 harvesting Methods 0.000 description 1
- 229940037467 helicobacter pylori Drugs 0.000 description 1
- 208000007475 hemolytic anemia Diseases 0.000 description 1
- 208000006454 hepatitis Diseases 0.000 description 1
- 231100000283 hepatitis Toxicity 0.000 description 1
- 208000002672 hepatitis B Diseases 0.000 description 1
- 206010073071 hepatocellular carcinoma Diseases 0.000 description 1
- 238000004128 high performance liquid chromatography Methods 0.000 description 1
- 210000003630 histaminocyte Anatomy 0.000 description 1
- 230000013632 homeostatic process Effects 0.000 description 1
- 239000000710 homodimer Substances 0.000 description 1
- 102000055691 human APC Human genes 0.000 description 1
- 208000033519 human immunodeficiency virus infectious disease Diseases 0.000 description 1
- 235000020256 human milk Nutrition 0.000 description 1
- 210000004251 human milk Anatomy 0.000 description 1
- 239000000017 hydrogel Substances 0.000 description 1
- 230000008004 immune attack Effects 0.000 description 1
- 238000003018 immunoassay Methods 0.000 description 1
- 230000009851 immunogenic response Effects 0.000 description 1
- 229940072221 immunoglobulins Drugs 0.000 description 1
- 210000000428 immunological synapse Anatomy 0.000 description 1
- 230000006872 improvement Effects 0.000 description 1
- 238000000099 in vitro assay Methods 0.000 description 1
- 238000011065 in-situ storage Methods 0.000 description 1
- 230000000415 inactivating effect Effects 0.000 description 1
- 238000010348 incorporation Methods 0.000 description 1
- 239000000411 inducer Substances 0.000 description 1
- 230000002757 inflammatory effect Effects 0.000 description 1
- 238000011081 inoculation Methods 0.000 description 1
- 239000002054 inoculum Substances 0.000 description 1
- 229960003786 inosine Drugs 0.000 description 1
- 229960003130 interferon gamma Drugs 0.000 description 1
- 230000000968 intestinal effect Effects 0.000 description 1
- 210000004020 intracellular membrane Anatomy 0.000 description 1
- 230000031146 intracellular signal transduction Effects 0.000 description 1
- 238000007912 intraperitoneal administration Methods 0.000 description 1
- 238000010253 intravenous injection Methods 0.000 description 1
- 238000005304 joining Methods 0.000 description 1
- 230000002147 killing effect Effects 0.000 description 1
- 238000002372 labelling Methods 0.000 description 1
- 238000011005 laboratory method Methods 0.000 description 1
- 210000001821 langerhans cell Anatomy 0.000 description 1
- 238000011031 large-scale manufacturing process Methods 0.000 description 1
- GZQKNULLWNGMCW-PWQABINMSA-N lipid A (E. coli) Chemical compound O1[C@H](CO)[C@@H](OP(O)(O)=O)[C@H](OC(=O)C[C@@H](CCCCCCCCCCC)OC(=O)CCCCCCCCCCCCC)[C@@H](NC(=O)C[C@@H](CCCCCCCCCCC)OC(=O)CCCCCCCCCCC)[C@@H]1OC[C@@H]1[C@@H](O)[C@H](OC(=O)C[C@H](O)CCCCCCCCCCC)[C@@H](NC(=O)C[C@H](O)CCCCCCCCCCC)[C@@H](OP(O)(O)=O)O1 GZQKNULLWNGMCW-PWQABINMSA-N 0.000 description 1
- 210000001165 lymph node Anatomy 0.000 description 1
- 210000004324 lymphatic system Anatomy 0.000 description 1
- 210000004698 lymphocyte Anatomy 0.000 description 1
- 210000003563 lymphoid tissue Anatomy 0.000 description 1
- 239000006166 lysate Substances 0.000 description 1
- 230000002132 lysosomal effect Effects 0.000 description 1
- 210000003712 lysosome Anatomy 0.000 description 1
- 230000001868 lysosomic effect Effects 0.000 description 1
- 230000003211 malignant effect Effects 0.000 description 1
- 239000011159 matrix material Substances 0.000 description 1
- 239000002609 medium Substances 0.000 description 1
- 210000004779 membrane envelope Anatomy 0.000 description 1
- 230000002503 metabolic effect Effects 0.000 description 1
- 230000004060 metabolic process Effects 0.000 description 1
- 208000021039 metastatic melanoma Diseases 0.000 description 1
- 229920000609 methyl cellulose Polymers 0.000 description 1
- 125000002496 methyl group Chemical group [H]C([H])([H])* 0.000 description 1
- 235000010270 methyl p-hydroxybenzoate Nutrition 0.000 description 1
- 239000004292 methyl p-hydroxybenzoate Substances 0.000 description 1
- 239000001923 methylcellulose Substances 0.000 description 1
- 229960002216 methylparaben Drugs 0.000 description 1
- YACKEPLHDIMKIO-UHFFFAOYSA-N methylphosphonic acid Chemical class CP(O)(O)=O YACKEPLHDIMKIO-UHFFFAOYSA-N 0.000 description 1
- 239000013586 microbial product Substances 0.000 description 1
- 239000003094 microcapsule Substances 0.000 description 1
- 210000003470 mitochondria Anatomy 0.000 description 1
- 238000002156 mixing Methods 0.000 description 1
- 238000000329 molecular dynamics simulation Methods 0.000 description 1
- 238000012544 monitoring process Methods 0.000 description 1
- 239000000178 monomer Substances 0.000 description 1
- 210000000865 mononuclear phagocyte system Anatomy 0.000 description 1
- 210000003205 muscle Anatomy 0.000 description 1
- 206010028417 myasthenia gravis Diseases 0.000 description 1
- 208000003786 myxedema Diseases 0.000 description 1
- DAZSWUUAFHBCGE-KRWDZBQOSA-N n-[(2s)-3-methyl-1-oxo-1-pyrrolidin-1-ylbutan-2-yl]-3-phenylpropanamide Chemical compound N([C@@H](C(C)C)C(=O)N1CCCC1)C(=O)CCC1=CC=CC=C1 DAZSWUUAFHBCGE-KRWDZBQOSA-N 0.000 description 1
- 210000000581 natural killer T-cell Anatomy 0.000 description 1
- 239000013642 negative control Substances 0.000 description 1
- 230000017128 negative regulation of NF-kappaB transcription factor activity Effects 0.000 description 1
- 210000000440 neutrophil Anatomy 0.000 description 1
- 239000002736 nonionic surfactant Substances 0.000 description 1
- 238000010899 nucleation Methods 0.000 description 1
- 239000002777 nucleoside Substances 0.000 description 1
- 125000003835 nucleoside group Chemical group 0.000 description 1
- 230000030648 nucleus localization Effects 0.000 description 1
- 229920001778 nylon Polymers 0.000 description 1
- 229940046166 oligodeoxynucleotide Drugs 0.000 description 1
- 229940124276 oligodeoxyribonucleotide Drugs 0.000 description 1
- 238000006384 oligomerization reaction Methods 0.000 description 1
- 210000000056 organ Anatomy 0.000 description 1
- 229940037201 oris Drugs 0.000 description 1
- LXCFILQKKLGQFO-UHFFFAOYSA-N p-hydroxybenzoic acid methyl ester Natural products COC(=O)C1=CC=C(O)C=C1 LXCFILQKKLGQFO-UHFFFAOYSA-N 0.000 description 1
- 238000004806 packaging method and process Methods 0.000 description 1
- 230000007170 pathology Effects 0.000 description 1
- 102000007863 pattern recognition receptors Human genes 0.000 description 1
- 108010089193 pattern recognition receptors Proteins 0.000 description 1
- 201000001976 pemphigus vulgaris Diseases 0.000 description 1
- 229940049954 penicillin Drugs 0.000 description 1
- 108010042234 peptide SVYDFFVWL Proteins 0.000 description 1
- 238000010647 peptide synthesis reaction Methods 0.000 description 1
- 210000005259 peripheral blood Anatomy 0.000 description 1
- 239000011886 peripheral blood Substances 0.000 description 1
- 210000001539 phagocyte Anatomy 0.000 description 1
- 239000008177 pharmaceutical agent Substances 0.000 description 1
- 230000003285 pharmacodynamic effect Effects 0.000 description 1
- 150000008104 phosphatidylethanolamines Chemical class 0.000 description 1
- 150000003905 phosphatidylinositols Chemical class 0.000 description 1
- 150000008298 phosphoramidates Chemical class 0.000 description 1
- 150000003014 phosphoric acid esters Chemical class 0.000 description 1
- 238000000053 physical method Methods 0.000 description 1
- 230000004962 physiological condition Effects 0.000 description 1
- 239000013600 plasmid vector Substances 0.000 description 1
- 229920001983 poloxamer Polymers 0.000 description 1
- 229920000747 poly(lactic acid) Polymers 0.000 description 1
- 229920001481 poly(stearyl methacrylate) Polymers 0.000 description 1
- 229920001223 polyethylene glycol Polymers 0.000 description 1
- 229920000656 polylysine Polymers 0.000 description 1
- 239000002861 polymer material Substances 0.000 description 1
- 230000004481 post-translational protein modification Effects 0.000 description 1
- 230000001124 posttranscriptional effect Effects 0.000 description 1
- 230000032361 posttranscriptional gene silencing Effects 0.000 description 1
- 239000002243 precursor Substances 0.000 description 1
- 230000002265 prevention Effects 0.000 description 1
- 229940002612 prodrug Drugs 0.000 description 1
- 239000000651 prodrug Substances 0.000 description 1
- 229960003387 progesterone Drugs 0.000 description 1
- 239000000186 progesterone Substances 0.000 description 1
- 230000001737 promoting effect Effects 0.000 description 1
- 235000010232 propyl p-hydroxybenzoate Nutrition 0.000 description 1
- 239000004405 propyl p-hydroxybenzoate Substances 0.000 description 1
- 229960003415 propylparaben Drugs 0.000 description 1
- 210000002307 prostate Anatomy 0.000 description 1
- 229950008679 protamine sulfate Drugs 0.000 description 1
- 230000001681 protective effect Effects 0.000 description 1
- 238000000159 protein binding assay Methods 0.000 description 1
- 230000017854 proteolysis Effects 0.000 description 1
- 230000004063 proteosomal degradation Effects 0.000 description 1
- 238000000746 purification Methods 0.000 description 1
- HNJBEVLQSNELDL-UHFFFAOYSA-N pyrrolidin-2-one Chemical compound O=C1CCCN1 HNJBEVLQSNELDL-UHFFFAOYSA-N 0.000 description 1
- QQXQGKSPIMGUIZ-AEZJAUAXSA-N queuosine Chemical compound C1=2C(=O)NC(N)=NC=2N([C@H]2[C@@H]([C@H](O)[C@@H](CO)O2)O)C=C1CN[C@H]1C=C[C@H](O)[C@@H]1O QQXQGKSPIMGUIZ-AEZJAUAXSA-N 0.000 description 1
- 230000009467 reduction Effects 0.000 description 1
- 230000022532 regulation of transcription, DNA-dependent Effects 0.000 description 1
- 102000037983 regulatory factors Human genes 0.000 description 1
- 108091008025 regulatory factors Proteins 0.000 description 1
- 230000008844 regulatory mechanism Effects 0.000 description 1
- 210000002345 respiratory system Anatomy 0.000 description 1
- 238000012340 reverse transcriptase PCR Methods 0.000 description 1
- 201000003068 rheumatic fever Diseases 0.000 description 1
- 239000002336 ribonucleotide Substances 0.000 description 1
- 125000002652 ribonucleotide group Chemical group 0.000 description 1
- 229920002477 rna polymer Polymers 0.000 description 1
- 210000003296 saliva Anatomy 0.000 description 1
- 201000000306 sarcoidosis Diseases 0.000 description 1
- 230000007017 scission Effects 0.000 description 1
- 238000000926 separation method Methods 0.000 description 1
- 230000036303 septic shock Effects 0.000 description 1
- 238000012163 sequencing technique Methods 0.000 description 1
- 238000013207 serial dilution Methods 0.000 description 1
- 210000002966 serum Anatomy 0.000 description 1
- 238000007493 shaping process Methods 0.000 description 1
- 230000035939 shock Effects 0.000 description 1
- 108091006024 signal transducing proteins Proteins 0.000 description 1
- 102000034285 signal transducing proteins Human genes 0.000 description 1
- 102000035025 signaling receptors Human genes 0.000 description 1
- 108091005475 signaling receptors Proteins 0.000 description 1
- 238000002741 site-directed mutagenesis Methods 0.000 description 1
- 210000003491 skin Anatomy 0.000 description 1
- 201000000849 skin cancer Diseases 0.000 description 1
- WBHQBSYUUJJSRZ-UHFFFAOYSA-M sodium bisulfate Chemical compound [Na+].OS([O-])(=O)=O WBHQBSYUUJJSRZ-UHFFFAOYSA-M 0.000 description 1
- 229910000342 sodium bisulfate Inorganic materials 0.000 description 1
- 239000011780 sodium chloride Substances 0.000 description 1
- PXLIDIMHPNPGMH-UHFFFAOYSA-N sodium chromate Chemical compound [Na+].[Na+].[O-][Cr]([O-])(=O)=O PXLIDIMHPNPGMH-UHFFFAOYSA-N 0.000 description 1
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 1
- 229940054269 sodium pyruvate Drugs 0.000 description 1
- 235000010265 sodium sulphite Nutrition 0.000 description 1
- 239000007790 solid phase Substances 0.000 description 1
- 238000010532 solid phase synthesis reaction Methods 0.000 description 1
- 238000001179 sorption measurement Methods 0.000 description 1
- 230000003393 splenic effect Effects 0.000 description 1
- 201000005671 spondyloarthropathy Diseases 0.000 description 1
- 229940031439 squalene Drugs 0.000 description 1
- TUHBEKDERLKLEC-UHFFFAOYSA-N squalene Natural products CC(=CCCC(=CCCC(=CCCC=C(/C)CCC=C(/C)CC=C(C)C)C)C)C TUHBEKDERLKLEC-UHFFFAOYSA-N 0.000 description 1
- 230000000087 stabilizing effect Effects 0.000 description 1
- 238000010561 standard procedure Methods 0.000 description 1
- 239000011550 stock solution Substances 0.000 description 1
- 229960005322 streptomycin Drugs 0.000 description 1
- 150000003457 sulfones Chemical class 0.000 description 1
- 150000003467 sulfuric acid derivatives Chemical class 0.000 description 1
- 230000008093 supporting effect Effects 0.000 description 1
- 239000000829 suppository Substances 0.000 description 1
- 239000000375 suspending agent Substances 0.000 description 1
- 238000013268 sustained release Methods 0.000 description 1
- 239000012730 sustained-release form Substances 0.000 description 1
- 208000011580 syndromic disease Diseases 0.000 description 1
- 238000001308 synthesis method Methods 0.000 description 1
- 238000007910 systemic administration Methods 0.000 description 1
- 238000012385 systemic delivery Methods 0.000 description 1
- 210000001138 tear Anatomy 0.000 description 1
- 210000001550 testis Anatomy 0.000 description 1
- 229930101283 tetracycline Natural products 0.000 description 1
- 229960002180 tetracycline Drugs 0.000 description 1
- 235000019364 tetracycline Nutrition 0.000 description 1
- 150000003522 tetracyclines Chemical class 0.000 description 1
- 229940021747 therapeutic vaccine Drugs 0.000 description 1
- RYYWUUFWQRZTIU-UHFFFAOYSA-K thiophosphate Chemical compound [O-]P([O-])([O-])=S RYYWUUFWQRZTIU-UHFFFAOYSA-K 0.000 description 1
- 229940113082 thymine Drugs 0.000 description 1
- 208000008732 thymoma Diseases 0.000 description 1
- 210000001541 thymus gland Anatomy 0.000 description 1
- 206010043778 thyroiditis Diseases 0.000 description 1
- 230000005026 transcription initiation Effects 0.000 description 1
- 230000009466 transformation Effects 0.000 description 1
- 230000001052 transient effect Effects 0.000 description 1
- 230000005945 translocation Effects 0.000 description 1
- 102000035160 transmembrane proteins Human genes 0.000 description 1
- 108091005703 transmembrane proteins Proteins 0.000 description 1
- XETCRXVKJHBPMK-MJSODCSWSA-N trehalose 6,6'-dimycolate Chemical compound C([C@@H]1[C@H]([C@H](O)[C@@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](O)[C@H](O)[C@@H](COC(=O)C(CCCCCCCCCCC3C(C3)CCCCCCCCCCCCCCCCCC)C(O)CCCCCCCCCCCCCCCCCCCCCCCCC)O2)O)O1)O)OC(=O)C(C(O)CCCCCCCCCCCCCCCCCCCCCCCCC)CCCCCCCCCCC1CC1CCCCCCCCCCCCCCCCCC XETCRXVKJHBPMK-MJSODCSWSA-N 0.000 description 1
- 201000008827 tuberculosis Diseases 0.000 description 1
- 238000005199 ultracentrifugation Methods 0.000 description 1
- 241000724775 unclassified viruses Species 0.000 description 1
- 230000004222 uncontrolled growth Effects 0.000 description 1
- 241001515965 unidentified phage Species 0.000 description 1
- 241001430294 unidentified retrovirus Species 0.000 description 1
- 201000005112 urinary bladder cancer Diseases 0.000 description 1
- 206010046766 uterine cancer Diseases 0.000 description 1
- 229920002554 vinyl polymer Polymers 0.000 description 1
- QAOHCFGKCWTBGC-QHOAOGIMSA-N wybutosine Chemical compound C1=NC=2C(=O)N3C(CC[C@H](NC(=O)OC)C(=O)OC)=C(C)N=C3N(C)C=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O QAOHCFGKCWTBGC-QHOAOGIMSA-N 0.000 description 1
- QAOHCFGKCWTBGC-UHFFFAOYSA-N wybutosine Natural products C1=NC=2C(=O)N3C(CCC(NC(=O)OC)C(=O)OC)=C(C)N=C3N(C)C=2N1C1OC(CO)C(O)C1O QAOHCFGKCWTBGC-UHFFFAOYSA-N 0.000 description 1
- 229940051021 yellow-fever virus Drugs 0.000 description 1
Images































































































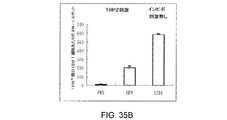





















Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/395—Antibodies; Immunoglobulins; Immune serum, e.g. antilymphocytic serum
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/46—Cellular immunotherapy
- A61K39/461—Cellular immunotherapy characterised by the cell type used
- A61K39/4615—Dendritic cells
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/46—Cellular immunotherapy
- A61K39/462—Cellular immunotherapy characterized by the effect or the function of the cells
- A61K39/4622—Antigen presenting cells
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/46—Cellular immunotherapy
- A61K39/464—Cellular immunotherapy characterised by the antigen targeted or presented
- A61K39/4643—Vertebrate antigens
- A61K39/4644—Cancer antigens
- A61K39/464454—Enzymes
- A61K39/464456—Tyrosinase or tyrosinase related proteinases [TRP-1 or TRP-2]
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K48/00—Medicinal preparations containing genetic material which is inserted into cells of the living body to treat genetic diseases; Gene therapy
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P1/00—Drugs for disorders of the alimentary tract or the digestive system
- A61P1/04—Drugs for disorders of the alimentary tract or the digestive system for ulcers, gastritis or reflux esophagitis, e.g. antacids, inhibitors of acid secretion, mucosal protectants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P13/00—Drugs for disorders of the urinary system
- A61P13/12—Drugs for disorders of the urinary system of the kidneys
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P15/00—Drugs for genital or sexual disorders; Contraceptives
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P17/00—Drugs for dermatological disorders
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P17/00—Drugs for dermatological disorders
- A61P17/06—Antipsoriatics
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P17/00—Drugs for dermatological disorders
- A61P17/14—Drugs for dermatological disorders for baldness or alopecia
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P19/00—Drugs for skeletal disorders
- A61P19/02—Drugs for skeletal disorders for joint disorders, e.g. arthritis, arthrosis
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P21/00—Drugs for disorders of the muscular or neuromuscular system
- A61P21/04—Drugs for disorders of the muscular or neuromuscular system for myasthenia gravis
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P29/00—Non-central analgesic, antipyretic or antiinflammatory agents, e.g. antirheumatic agents; Non-steroidal antiinflammatory drugs [NSAID]
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P3/00—Drugs for disorders of the metabolism
- A61P3/08—Drugs for disorders of the metabolism for glucose homeostasis
- A61P3/10—Drugs for disorders of the metabolism for glucose homeostasis for hyperglycaemia, e.g. antidiabetics
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
- A61P37/04—Immunostimulants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/08—Antiallergic agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P5/00—Drugs for disorders of the endocrine system
- A61P5/14—Drugs for disorders of the endocrine system of the thyroid hormones, e.g. T3, T4
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P7/00—Drugs for disorders of the blood or the extracellular fluid
- A61P7/06—Antianaemics
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/113—Non-coding nucleic acids modulating the expression of genes, e.g. antisense oligonucleotides; Antisense DNA or RNA; Triplex- forming oligonucleotides; Catalytic nucleic acids, e.g. ribozymes; Nucleic acids used in co-suppression or gene silencing
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/06—Animal cells or tissues; Human cells or tissues
- C12N5/0602—Vertebrate cells
- C12N5/0634—Cells from the blood or the immune system
- C12N5/0639—Dendritic cells, e.g. Langherhans cells in the epidermis
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/10—Type of nucleic acid
- C12N2310/14—Type of nucleic acid interfering N.A.
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/60—Transcription factors
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Medicinal Chemistry (AREA)
- Pharmacology & Pharmacy (AREA)
- Veterinary Medicine (AREA)
- Public Health (AREA)
- Animal Behavior & Ethology (AREA)
- Organic Chemistry (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Immunology (AREA)
- Cell Biology (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Microbiology (AREA)
- Biomedical Technology (AREA)
- Genetics & Genomics (AREA)
- Epidemiology (AREA)
- Mycology (AREA)
- Biotechnology (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Hematology (AREA)
- Diabetes (AREA)
- General Engineering & Computer Science (AREA)
- Biochemistry (AREA)
- Molecular Biology (AREA)
- Oncology (AREA)
- Endocrinology (AREA)
- Dermatology (AREA)
- Neurology (AREA)
- Orthopedic Medicine & Surgery (AREA)
- Rheumatology (AREA)
- Physical Education & Sports Medicine (AREA)
- Urology & Nephrology (AREA)
- Physics & Mathematics (AREA)
- Pulmonology (AREA)
Abstract
本発明は、細胞内での負の免疫調節因子の阻害を用いて免疫細胞の免疫能力を増強させるための組成物及び方法を含む。本発明は、負の免疫調節因子の阻害を通して抗原提示が増強されるワクチン及び療法を提供する。本発明はまた、自己腫瘍抗原の提示に依存する腫瘍ワクチン接種方法における自己寛容を破壊するための機序をも提供している。 The present invention includes compositions and methods for enhancing immune capacity of immune cells using inhibition of negative immune modulators within the cells. The present invention provides vaccines and therapies in which antigen presentation is enhanced through inhibition of negative immune regulators. The present invention also provides a mechanism for breaking self-tolerance in tumor vaccination methods that rely on presentation of self-tumor antigens.
Description
ヒトにおける先天及び適応免疫における不適切な抗原提示及び活性化は結果として、数多くの病原性感染及び悪性細胞成長を制御し一掃するヒト免疫系に障害をもたらす。慢性感染症及び癌に対する有効な治療用ワクチン及び免疫療法は、その病理学に関連する攻撃的抗原を制御し一掃する能力をもつ活発な免疫応答を誘発する効率のよい手段に対する新しいアプローチの開発に依存している。 Inappropriate antigen presentation and activation in innate and adaptive immunity in humans results in damage to the human immune system that controls and eliminates many pathogenic infections and malignant cell growth. Effective therapeutic vaccines and immunotherapy for chronic infections and cancers will develop new approaches to efficient means of eliciting an active immune response with the ability to control and clear off aggressive antigens associated with its pathology It depends.
抗原を認識するT細胞の能力は、主要組織適合複合体(MHC)I又はMHCIIタンパク質のいずれかと抗原の会合に左右される。例えば、細胞傷害性T細胞は、MHC−Iタンパク質と会合した状態で提示される抗原に応答する。かくしてウイルスの感染した細胞を殺すべき細胞傷害性T細胞は、その細胞が適切なMHC−Iタンパク質を発現しない場合その細胞を殺さなくなる。ヘルパーT細胞はMHC−IIを認識する。ヘルパーT細胞の活性は、一般に、抗原提示細胞上の抗原の認識及び「自己」MHC−IIタンパク質のこれらの細胞上の存在の両方に左右される。自己MHCタンパク質と会合状態にある抗原の認識に対する必要条件は、MHC制限と呼ばれている。MHC−Iタンパク質は、ほぼ全ての有核細胞の表面上に見出される。MHC−IIタンパク質は、皮膚のランゲルハンス細胞及び脾臓の樹状細胞、B細胞及びマクロファージを含めた或る種の細胞の表面上に見出される。 The ability of a T cell to recognize an antigen depends on the association of the antigen with either the major histocompatibility complex (MHC) I or MHCII protein. For example, cytotoxic T cells respond to antigens that are presented in association with MHC-I protein. Thus, cytotoxic T cells that should kill virus-infected cells will not kill the cells if they do not express the appropriate MHC-I protein. Helper T cells recognize MHC-II. Helper T cell activity generally depends on both antigen recognition on antigen presenting cells and the presence of “self” MHC-II protein on these cells. The prerequisite for recognition of antigens associated with self MHC proteins is called MHC restriction. MHC-I protein is found on the surface of almost all nucleated cells. MHC-II protein is found on the surface of certain cells, including Langerhans cells of the skin and splenic dendritic cells, B cells and macrophages.
哺乳動物内で免疫応答を開始させる上での決定的な工程は、MHC−II制限された外因性抗原を認識するCD4+ヘルパーT細胞の活性化である。これらの抗原は、樹状細胞(DC)などの抗原提示細胞内の細胞エンドソーム経路の中で捕捉されプロセッシングされる。エンドソーム及びリソソーム内では、抗原は、抗原−MHC−II複合体を形成するべくゴルジ画分内のMHC−II上で複合化される小抗原ペプチドへとプロセシングされる。これらの複合体は細胞表面上で発現され、この発現は、CD4+T細胞の活性化を誘発する。 A critical step in initiating an immune response in a mammal is the activation of CD4 + helper T cells that recognize MHC-II restricted exogenous antigens. These antigens are captured and processed in the cellular endosomal pathway within antigen-presenting cells such as dendritic cells (DC). Within endosomes and lysosomes, the antigen is processed into small antigen peptides that are complexed on MHC-II in the Golgi fraction to form antigen-MHC-II complexes. These complexes are expressed on the cell surface and this expression triggers activation of CD4 + T cells.
哺乳動物内の有効な免疫応答の誘発におけるその他の決定的な事象には、CD8+T細胞及びB細胞の活性化が関与する。CD8+細胞は、MHC−I抗原と複合化されるプロセッシング済みタンパク質として細胞表面上に提示されるような形で細胞を通して所望のタンパク質が送られた時点で活性化される。B細胞は、MHCタンパク質を必要とせずに表面免疫グロブリン(IgM及びIgD)を介して抗原と相互作用できる。しかしながら、CD4+T細胞の活性化は、免疫系の全てのアームを刺激する。活性化時点でCD4+T細胞(ヘルパーT細胞)はインターロイキンを産生する。これらのインターロイキンは免疫系のその他の腕を活性化するのを助ける。例えば、ヘルパーT細胞は、B細胞が抗体を産生するのを助けるインターロイキン−4(IL−4)及びインターロイキン−5(IL−5);CD4+及びCD8+T細胞を活性させるインターロイキン−2(IL−2);及びマクロファージを活性化させるガンマインタフェロンを産生する。MHC−II制限を受けた抗原を認識するヘルパーT細胞は、細胞傷害性T細胞、マクロファージ、ナチュラルキラー細胞及びB細胞の活性化及びクローン拡張において中心的な役割を果たすことから、抗原に応答してヘルパーT細胞を活性化させるという初期事象は、その抗原に対抗して向けられた有効な免疫応答の誘発にとって決定的である。リソソーム膜貫通タンパク質から誘導された配列を用いてヘルパーT細胞活性を刺激するための試みが報告されている。しかしながら、これらの試みは、テスト対象の哺乳動物の体内でCD8+T細胞及びB細胞に関する有効な免疫応答の誘発を結果としてもたらさなかった。 Other critical events in the induction of an effective immune response in mammals involve CD8 + T cell and B cell activation. CD8 + cells are activated when the desired protein is delivered through the cell in such a way that it is presented on the cell surface as a processed protein complexed with the MHC-I antigen. B cells can interact with antigen via surface immunoglobulin (IgM and IgD) without the need for MHC proteins. However, activation of CD4 + T cells stimulates all arms of the immune system. At the time of activation, CD4 + T cells (helper T cells) produce interleukins. These interleukins help activate the other arms of the immune system. For example, helper T cells are interleukin-4 (IL-4) and interleukin-5 (IL-5) that help B cells produce antibodies; interleukin-2 (IL that activates CD4 + and CD8 + T cells. -2); and produces gamma interferon that activates macrophages. Helper T cells that recognize MHC-II restricted antigens respond to antigens because they play a central role in the activation and clonal expansion of cytotoxic T cells, macrophages, natural killer cells and B cells. The initial event of activating helper T cells is crucial for the induction of an effective immune response directed against the antigen. Attempts have been reported to stimulate helper T cell activity using sequences derived from lysosomal transmembrane proteins. However, these attempts did not result in the induction of an effective immune response for CD8 + T cells and B cells in the mammal being tested.
免疫応答においてT細胞が果たす決定的な役割に加えて、DCも同等に重要である。D
Cは、自己抗原に対する寛容の維持及び先天免疫及び適応免疫の活性化において主要な調節の役目を果たす専門の抗原提示細胞である(非特許文献1、非特許文献2)。DCが微生物産物などの前炎症性刺激に遭遇した場合、細胞の成熟プロセスは、細胞表面で発現された抗原ペプチド負荷されたMHC分子及び共同刺激分子をアップレギュレートすることによって開始される。成熟及び局所リンパ節への帰還の後、DCは免疫学的シナプスを形成することによりT細胞との接触を確立し、ここでT細胞レセプタ(TCR)及び共同刺激分子は接着分子がとり囲む中心部域内に集合する(非特許文献3)。ひとたび活性化された時点で、CD8+T細胞は数世代にわたり自律的に増殖し、さらなる抗原刺激無く細胞傷害機能を獲得する(非特許文献4、非特許文献5)。従って、DCにより提供されるペプチドMHC複合体(シグナル1)及び共同刺激分子(シグナル2)のレベル及び持続時間は、抗原特異的T細胞応答の規模及び運命を判定するために不可欠である(非特許文献6、非特許文献7)。
In addition to the critical role played by T cells in the immune response, DC is equally important. D
C is a specialized antigen-presenting cell that plays a major regulatory role in maintaining tolerance to self-antigens and activating innate and adaptive immunity (Non-patent
樹状細胞(DC)及びマクロファージなどの抗原提示細胞(APC)は、先天及び適合免疫の活性化ならびに免疫学的寛容の維持において重要な役割を果たす。APCが病原菌を検知し免疫応答を開始させる機序は、過去数年間にわたり充分に研究されてきた。APCは、リポ多糖類(LPS)、未メチル化細菌DNA(CpG)及びRNAなどの保存された病原菌構造を認識するために、トール様レセプタ(TLR)などのパターン認識レセプタを使用する。TLRは、TIR(トール/インターロイキン−1(IL−1)レセプタ)スーパーファミリに属し、このスーパーファミリは2つの主要亜群すなわちIL−1レセプタ及びTLRに細分される。TLRは11のメンバー(TLR1〜TLR11)からなる。このスーパーファミリ−の全てのメンバーは、共通のシグナリング経路、特に転写因子核因子−κB(NFκB)及びストレス活性化タンパク質キナーゼの活性化を導くシグナリング経路を活性化する保存されたサイトゾルTIRドメインの存在に起因して同様にシグナリングする。NF−κB活性化は、腫瘍壊因子(TNF)、IFN、インターロイキン1(IL−1)、IL−6、及びIL−12などの前炎症性サイトカインを分泌することによってか又はCD80、CD86及びCD40などの共同刺激分子を発現することによって、免疫応答の触媒として作用する。TNFα及びCD40LなどのTNFファミリーのメンバーは、そのレセプタ又はリガンドと相互作用し、同様にNF−κBを活性化させる。 Antigen presenting cells (APC) such as dendritic cells (DC) and macrophages play an important role in the activation of innate and adaptive immunity and in maintaining immunological tolerance. The mechanism by which APC detects pathogens and initiates an immune response has been well studied over the past few years. APC uses pattern recognition receptors such as Toll-like receptors (TLRs) to recognize conserved pathogen structures such as lipopolysaccharide (LPS), unmethylated bacterial DNA (CpG) and RNA. TLRs belong to the TIR (Toll / Interleukin-1 (IL-1) receptor) superfamily, which is subdivided into two major subgroups: the IL-1 receptor and the TLR. TLR consists of 11 members (TLR1 to TLR11). All members of this superfamily are members of a conserved cytosolic TIR domain that activates common signaling pathways, particularly signaling pathways that lead to activation of transcription factor nuclear factor-κB (NFκB) and stress-activated protein kinases. Signaling similarly due to presence. NF-κB activation is by secreting pro-inflammatory cytokines such as tumor necrosis factor (TNF), IFN, interleukin 1 (IL-1), IL-6, and IL-12 or CD80, CD86 and By expressing a costimulatory molecule such as CD40, it acts as a catalyst for the immune response. TNF family members such as TNFα and CD40L interact with their receptors or ligands and activate NF-κB as well.
腫瘍ワクチンを開発する大きな研究努力が、抗腫瘍免疫を増強させる一手段としてDCの成熟及び共同刺激を促進する目的で試みられてきた。しかしながら、自己腫瘍関連抗原(TAA)に対抗する免疫の誘発は、固有の阻害機序により制限されており、これらの機序の多くがいまだに定義づけされていない。DC又はその他の細胞上のB7ファミリー分子との細胞−細胞接触を介してエフェクタT細胞活性の規模を制御するべくT細胞上の細胞傷害性Tリンパ球抗原4(CTLA4)及びそれに関連する分子により、既知の阻害機序が利用されている。DC成熟は、自己寛容の維持から免疫誘発への臨界スイッチとして役立つ。しかしながら、成熟点を超えて適合免疫の規模及び持続時間を制御できるようにするような負の免疫調節機序を成熟抗原提示DCが有するか否かはいまだに不明である。 Great research efforts to develop tumor vaccines have been attempted with the aim of promoting DC maturation and costimulation as a means to enhance anti-tumor immunity. However, induction of immunity against self-tumor associated antigens (TAA) is limited by inherent inhibition mechanisms, and many of these mechanisms have not yet been defined. By cytotoxic T lymphocyte antigen 4 (CTLA4) and related molecules on T cells to control the magnitude of effector T cell activity via cell-cell contact with B7 family molecules on DCs or other cells Known inhibition mechanisms are utilized. DC maturation serves as a critical switch from maintaining self tolerance to immune induction. However, it remains unclear whether mature antigen-presenting DCs have a negative immune regulatory mechanism that allows the size and duration of adaptive immunity to be controlled beyond the maturity point.
前炎症性シグナリングについては多くの注目が集められてきたが、炎症を抑制しかつ消散させる機序についてはほとんど知られていない。TLRが開始させる免疫応答の規模及び持続時間は、前炎症性シグナリングの強度と持続時間及びシグナル伝達経路の調節によって決定づけられる。転写因子NF−κBのTLR誘発型活性化は、多数の前炎症性遺伝子の転写にとって不可欠であることから、敗血性ショックなどの過度の免疫応答から宿主を保護するため多数のレベルでTLRシグナリングを負に調節するため、及び腸内微環境などの慢性的病原菌曝露の状態で免疫ホメオスタシスを維持するために、数多くの機序が利用されている。TLRシグナル経路の負の免疫調節因子としては、IRAK−M、MyD88s、PI3K、TOLLIP、A20、TRIAD3A、NOD2、可溶性TLR
2/4及びSIGRR及びST2などのTIRドメインを含有する膜結合分子が含まれる(非特許文献8)。
Although much attention has been focused on pro-inflammatory signaling, little is known about the mechanisms that suppress and resolve inflammation. The magnitude and duration of the immune response initiated by the TLR is determined by the intensity and duration of pro-inflammatory signaling and modulation of signaling pathways. Since TLR-induced activation of the transcription factor NF-κB is essential for transcription of many pro-inflammatory genes, TLR signaling at multiple levels is protected to protect the host from excessive immune responses such as septic shock. A number of mechanisms have been utilized to negatively regulate and maintain immune homeostasis in conditions of chronic pathogen exposure such as the intestinal microenvironment. Negative immune regulators of the TLR signaling pathway include IRAK-M, MyD88s, PI3K, TOLLIP, A20, TRIAD3A, NOD2, and soluble TLR
Membrane-bound molecules containing TIR domains such as 2/4 and SIGRR and ST2 are included (Non-patent Document 8).
サイトカインは、多数の免疫細胞機能の調節に決定的に関与している(非特許文献9、非特許文献10)。DCは、核因子−κB(NF−κB)シグナリング経路を活性化することによりDC成熟を促進するため、リポ多糖類(LPS)などの保存された病原菌構造を認識するトール様レセプタ(TLR)を使用する(非特許文献11)。NF−κBメンバーはこのときIL−12などの前炎症性サイトカインの発現を媒介する(非特許文献11、非特許文献12、非特許文献13)。DC成熟の後、サイトカイン産生及び細胞内シグナリング経路は、過度の自己免疫活性化を制限する一方で外来性抗原に対する有益な免疫応答を促進するように綿密に調節されると思われる。しかしながら、これらの経路についての特異的フィードバック阻害機序の重要性及びその結果としての自己抗原特異的免疫応答の制御は、いまだにほとんど定義づけされていないままである。
Cytokines are critically involved in the regulation of many immune cell functions (Non-patent
SOCS1は、インターフェロン(IFN)−g、インターロイキン(IL)−2、IL−6、IL−7、IL−12及びIL−15を含めたさまざまなサイトカインによるシグナリングの誘発可能な負のフィードバック調節因子である(非特許文献14、非特許文献15)。SOCS1は、偽基質インヒビターとして上流側ヤーヌスキナーゼ(JAK)の活性化ループ結合しかつ/又はプロテアソーム分解についてJAKをターゲティングすることによって転写(STAT)シグナリング経路の多重シグナルトランスデューサ及び活性化因子を抑制する(非特許文献14、非特許文献15)。SOCS1は同様に、そのSOCSボックス領域を通ってユビキチン媒介型タンパク質分解についてp65タンパク質をターゲティングすることによりNF−κBシグナリングを遮断する(非特許文献16)。SOCS1欠損(−/−)マウスは、主として抑制のきかないサイトカインシグナリングの結果として、T及びNKT細胞の異常活性化及び重大な全身的炎症を伴って、新生児として死亡する(非特許文献17、非特許文献18、非特許文献19)。DCにおけるSOCS1の機能についてはほとんど知られていないが、最近の研究から、抗原提示細胞(APC)内でのシグナリングを制御する上でのSOCS1の役割が示唆されている(非特許文献14、非特許文献20)。マクロファージ内でのSOCS1の発現はLPS又はCpG−DNAの刺激によって誘発され、SOCS1−/−マウスは、その野生型同腹子に比べてよりLPS誘発型ショックに対し感受性が高い(非特許文献21、非特許文献22、非特許文献23、非特許文献24)。その上SOCS1発現がSOCS1−/−バックグラウンド上のT及びB細胞内で回復されているマウスに由来するSOCS1−/−DCは、IFNγ及びLPSに対し過反応性であり、同種異型細胞拡張を開始させ、B細胞の異常拡張及び自己反応性抗体の産生を誘発する(非特許文献20)。
SOCS1 is a negative feedback regulator capable of inducing signaling by various cytokines including interferon (IFN) -g, interleukin (IL) -2, IL-6, IL-7, IL-12 and IL-15 (Non-Patent
DC内でのSOCS1の役割についてはほとんど知られていないが、最近の研究から、サイトカインシグナル伝達経路を調節する上でのSOCS1の役目が実証された。例えば、SOCS1−/−DCがより成熟した表現型を示し、シグナリングのためにトール様レセプタ(TLR)4と相互作用するリポ多糖類(LPS)に過反応性を有することが観察された、ということが実証されている。同様に観察されたのは、SOCS1−/−DCが自己反応性抗体産生を誘発するということであった。これらの観察事実は、場合によってJAK/STAT経路及びTLR/NF−κB経路を制御することによるDCの負の調節におけるSOCS1の考えられる役割の可能性を示唆していた。 Little is known about the role of SOCS1 in DCs, but recent studies have demonstrated the role of SOCS1 in regulating cytokine signaling pathways. For example, it was observed that SOCS1-/-DC showed a more mature phenotype and was overreactive with lipopolysaccharide (LPS) interacting with Toll-like receptor (TLR) 4 for signaling It has been proven. Similar observations were that SOCS1-/-DC induced autoreactive antibody production. These observations suggested a possible role for SOCS1 in the negative regulation of DCs, possibly by controlling the JAK / STAT pathway and TLR / NF-κB pathway.
哺乳動物の体内で免疫応答を刺激するべく免疫療法においてDCを使用するための数多くの試みが行なわれてきた。これらの研究努力においては、DCは、抗原をそれに負荷しエクスビボ状況下でそれらを成熟させかくしてそれが癌患者の体内で抗腫瘍免疫を刺激するようにすることによって操作された。ヒト免疫不全ウイルス(HIV)を撲滅するための免疫療法の使用に関しては、有効なヒト免疫不全ウイルス(HIV)ワクチンはまだ全
く現われていない。従って、当該技術分野では、哺乳動物において疾病の治療のための免疫応答を惹起する効率のよい直接的な手段に対するニーズが長年にわたって認識されている。本発明はこのニーズを満たすものである。
Numerous attempts have been made to use DC in immunotherapy to stimulate an immune response in the mammalian body. In these research efforts, DCs have been engineered by loading it with antigen and allowing them to mature under ex vivo conditions, thus stimulating antitumor immunity in the body of cancer patients. Regarding the use of immunotherapy to eradicate human immunodeficiency virus (HIV), no effective human immunodeficiency virus (HIV) vaccine has yet emerged. Thus, the art has long recognized a need for an efficient and direct means of eliciting an immune response for the treatment of disease in mammals. The present invention satisfies this need.
本発明は、免疫細胞の免疫能力を増強させるための組成物を包含する。好ましくは、本組成物は、負の免疫調節因子のインヒビターを含んでなる。一態様においては、負の免疫調節因子は、ユビキチン化、脱ユビキチン化及びスモ化(sumoylation)により分子安定性に関与するタンパク質及びNFκBのインヒビターの発現を誘発するか又はNFκB標的遺伝子の転写を抑制する転写因子、又はその任意の組合せよりなる群から選択される。 The present invention includes a composition for enhancing the immune capacity of immune cells. Preferably, the composition comprises an inhibitor of a negative immune modulator. In one aspect, the negative immunomodulator induces the expression of proteins and NFκB inhibitors involved in molecular stability by ubiquitination, deubiquitination and sumoylation or represses transcription of NFκB target genes Selected from the group consisting of transcription factors, or any combination thereof.
一態様においては、ユビキチン化、脱ユビキチン化及びスモ化により分子安定性に関与する負の免疫調節因子はA20及びSUMO(SUMO1、SUMO2、SUMO3、SUMO4)よりなる群から選択される。もう1つの態様においては、NFκB標的遺伝子の転写を抑制するか又はNFκBのインヒビターの発現を誘発しNFκB標的遺伝子の転写を抑制する転写因子のファミリーの一部を成す負の免疫調節因子は、Twist−1、Twist−2、Foxj1、Foxo3a及びそれらの変異体よりなる群から選択される。 In one aspect, the negative immune regulator involved in molecular stability by ubiquitination, deubiquitination and sumoylation is selected from the group consisting of A20 and SUMO (SUMO1, SUMO2, SUMO3, SUMO4). In another embodiment, the negative immune regulator that forms part of a family of transcription factors that repress transcription of NFκB target genes or induce expression of inhibitors of NFκB and repress transcription of NFκB target genes is Twist -1, Twist-2, Foxj1, Foxo3a, and variants thereof.
本発明はまた、負の免疫調節因子が(SOCS)をシグナリングするサイトカインのサプレッサであり、さらに、SOCSはSOCS1、SOCS2、SOCS3、SOCS4、SOCS5、SOCS6、SOCS7及びサイトカイン誘発性SH2−ドメイン含有タンパク質(CIS)よりなる群から選択される、負の免疫調節因子のインヒビターをも含む。 The present invention is also a suppressor of cytokines in which negative immune regulators signal (SOCS), and SOCS is SOCS1, SOCS2, SOCS3, SOCS4, SOCS5, SOCS6, SOCS7 and cytokine-induced SH2-domain-containing proteins ( Also included are inhibitors of negative immune modulators selected from the group consisting of (CIS).
もう1つの態様においては、本発明はSH2含有ホスファターゼ(SHP)のインヒビターであって、SHPがSHP−1及びSHP−2よりなる群から選択されるインヒビターを含む。 In another aspect, the invention includes an inhibitor of SH2-containing phosphatase (SHP), wherein the SHP is selected from the group consisting of SHP-1 and SHP-2.
さらなる態様においては、本発明は、活性化されたSTAT(PIAS)のタンパク質インヒビターであって、PIASが、PIAS1、PIAS3、PIASx及びPIASyよりなる群から選択されるインヒビターを含む。 In a further aspect, the present invention comprises a protein inhibitor of activated STAT (PIAS), wherein PIAS is selected from the group consisting of PIAS1, PIAS3, PIASx and PIASy.
一態様においては、負の免疫調節因子は、A20、SUMO(SUMO1、SUMO2、SUMO3、及びSUMO4)、Foxj1、Foxo3a、TWIST(Twist1、Twist2)、SOCS(SOCS1、SOCS2、SOCS3、SOCS4、SOCS5、SOCS6、SOCS7、及びCIS)、PIAS(PIAS1、PIAS3、PIASx及びPIASy)、SHP(SHP−1及びSHP−2)などよりなる群から選択される。 In one aspect, the negative immune modulator is A20, SUMO (SUMO1, SUMO2, SUMO3, and SUMO4), Foxj1, Foxo3a, TWIST (Twist1, Twist2), SOCS (SOCS1, SOCS2, SOCS3, SOCS4, SOCS5, SOCS5, SOCS5, SOCS5 , SOCS7, and CIS), PIAS (PIAS1, PIAS3, PIASx and PIASy), SHP (SHP-1 and SHP-2), and the like.
特定の一実施形態においては、インヒビターは、小分子干渉RNA(siRNA)、マイクロRNA、アンチセンス核酸、リボザイム、トランス優性負突然変異体をコードする発現ベクター、細胞内抗体、ペプチド及び小分子よりなる群から選択される。好ましくは、インヒビターはsiRNAである。 In one particular embodiment, the inhibitor consists of small interfering RNA (siRNA), microRNA, antisense nucleic acid, ribozyme, expression vector encoding transdominant negative mutant, intracellular antibody, peptide and small molecule. Selected from the group. Preferably, the inhibitor is siRNA.
さらなる態様においては、siRNAが2本鎖オリゴヌクレオチド、1本鎖オリゴヌクレオチド及びポリヌクレオチドよりなる群から選択される。 In a further aspect, the siRNA is selected from the group consisting of a double stranded oligonucleotide, a single stranded oligonucleotide and a polynucleotide.
さらにもう1つの態様においては、siRNAは化学的に合成されている。 In yet another embodiment, the siRNA is chemically synthesized.
本発明のもう1つの実施形態は、負の免疫調節因子のインヒビターを含む組成物であって、生理学的に許容可能な担体をさらに含んでなる組成物を含む。好ましくは、生理学的に許容可能な担体は、リポソームである。 Another embodiment of the invention includes a composition comprising an inhibitor of a negative immune modulator, further comprising a physiologically acceptable carrier. Preferably, the physiologically acceptable carrier is a liposome.
もう1つの実施形態においては、負の免疫調節因子のインヒビターは、発現ベクター内にクローニングされた単離ポリヌクレオチドによりコードされる。発現ベクターは、プラスミドDNA、ウイルスベクター、細菌ベクター及び哺乳動物ベクターよりなる群から選択される。もう1つの態様においては、発現ベクターは、宿主細胞のゲノム内への単離ポリヌクレオチドの組込みを容易にする組込みシグナル配列をさらに含んでなる。 In another embodiment, the negative immune modulator inhibitor is encoded by an isolated polynucleotide cloned into an expression vector. The expression vector is selected from the group consisting of plasmid DNA, viral vector, bacterial vector and mammalian vector. In another embodiment, the expression vector further comprises an integration signal sequence that facilitates integration of the isolated polynucleotide into the genome of the host cell.
本発明はまた、免疫細胞の免疫能力を増強するための組成物であって、少なくとも1つのエピトープをさらに含んでなる組成物をも含む。好ましくは、エピトープは哺乳動物の体内で免疫応答を惹起する能力をもつ。もう1つの態様においては、エピトープは、哺乳動物の体内でCD4+T細胞応答を誘発する。さらにもう1つの態様においては、エピトープは、哺乳動物の体内でCD8+T細胞応答を誘発する。さらなる態様では、エピトープは哺乳動物の体内でB細胞応答を誘発する。 The present invention also includes a composition for enhancing the immunity of immune cells, further comprising at least one epitope. Preferably, the epitope is capable of eliciting an immune response in the mammalian body. In another aspect, the epitope elicits a CD4 + T cell response in the mammalian body. In yet another aspect, the epitope elicits a CD8 + T cell response in the mammalian body. In a further aspect, the epitope elicits a B cell response in the mammalian body.
もう1つの実施形態においては、抗原は発現ベクターにより発現される。さらなる態様においては、抗原は単離ポリペプチドである。 In another embodiment, the antigen is expressed by an expression vector. In a further aspect, the antigen is an isolated polypeptide.
さらにもう1つの実施形態においては、抗原は1つの疾患に関連している。好ましくは、疾患は、感染性疾患、癌及び自己免疫疾患よりなる群から選択される。 In yet another embodiment, the antigen is associated with one disease. Preferably, the disease is selected from the group consisting of infectious diseases, cancer and autoimmune diseases.
一態様においては、感染性疾患はウイルス、細菌、真菌及び原生動物よりなる群から選択された病原性微生物によってひき起こされる。 In one aspect, the infectious disease is caused by a pathogenic microorganism selected from the group consisting of viruses, bacteria, fungi and protozoa.
もう1つの実施形態においては、抗原はウイルス遺伝子によりコードされる。好ましくは、ウイルス遺伝子は、B型肝炎ウイルス、C型肝炎ウイルス、ヒト免疫不全ウイルス、乳頭腫ウイルス及びヘルペスウイルスよりなる群から選択されたウイルスから誘導されている。 In another embodiment, the antigen is encoded by a viral gene. Preferably, the viral gene is derived from a virus selected from the group consisting of hepatitis B virus, hepatitis C virus, human immunodeficiency virus, papilloma virus and herpes virus.
一態様においては、抗原は、B型肝炎ウイルスe抗原遺伝子、B型肝炎ウイルス表面抗原遺伝子及びB型肝炎ウイルスコア抗原遺伝子よりなる群から選択されたウイルス遺伝子によりコードされている。 In one aspect, the antigen is encoded by a viral gene selected from the group consisting of hepatitis B virus e antigen gene, hepatitis B virus surface antigen gene and hepatitis B virus core antigen gene.
もう1つの態様においては、抗原は、ヒト免疫不全ウイルスEnv gp160遺伝子、Gag遺伝子、Pol遺伝子、Rev遺伝子、Tat遺伝子、Vif遺伝子及びNef遺伝子よりなる群から選択されたウイルス遺伝子によりコードされている。 In another embodiment, the antigen is encoded by a viral gene selected from the group consisting of human immunodeficiency virus Env gp160 gene, Gag gene, Pol gene, Rev gene, Tat gene, Vif gene and Nef gene.
さらなる態様においては、抗原は、乳頭腫ウイルスE7遺伝子及び乳頭腫ウイルスE6よりなる群から選択されたウイルス遺伝子によりコードされている。 In a further aspect, the antigen is encoded by a viral gene selected from the group consisting of papilloma virus E7 gene and papilloma virus E6.
さらにもう1つの態様においては、抗原は、1型単純ヘルペスウイルス、2型単純ヘルペスウイルス、エプスタイン・バーウイルス、サイトメガロウイルス、ヒトヘルペスウイルス6型、ヒトヘルペスウイルス7型及びヒトヘルペスウイルス8型よりなる群から選択されたヘルペスウイルスから誘導されたウイルス遺伝子によりコードされる。
In yet another embodiment, the antigen is from
1つの実施形態においては、抗原は癌に関連しており、ここで、癌は、乳癌、子宮頸癌、黒色腫、腎臓癌及び前立腺癌よりなる群から選択される。 In one embodiment, the antigen is associated with cancer, wherein the cancer is selected from the group consisting of breast cancer, cervical cancer, melanoma, kidney cancer and prostate cancer.
さらなる態様においては、腫瘍関連抗原は、過剰発現された腫瘍関連抗原、睾丸−腫瘍抗原、突然変異腫瘍関連抗原、分化腫瘍関連抗原チロシナーゼ、MART、trp、MAGE−1、MAGE−2、MAGE−3、gp100、HER−2、Ras及びPSAよりなる群から選択される。 In a further aspect, the tumor associated antigen is an overexpressed tumor associated antigen, testis-tumor antigen, mutant tumor associated antigen, differentiated tumor associated antigen tyrosinase, MART, trp, MAGE-1, MAGE-2, MAGE-3 , Gp100, HER-2, Ras and PSA.
さらにもう1つの態様においては、腫瘍関連抗原は、BCR−ABL、CASP、CDK、Ras、p53、HER−2/neu、CEA、MUC、TW1、PAP、スルビビン、テロメラーゼ、EGFR、PSMA、PSA、PSCA、チロシナーゼ、MART、TRP、gp100、MART、MAGE、BAGE、GAGE、LAGE/NY−ESO、RAGE、SSX−2、CD19及びCD20よりなる群から選択される。 In yet another embodiment, the tumor associated antigen is BCR-ABL, CASP, CDK, Ras, p53, HER-2 / neu, CEA, MUC, TW1, PAP, survivin, telomerase, EGFR, PSMA, PSA, PSCA , Tyrosinase, MART, TRP, gp100, MART, MAGE, BAGE, GAGE, LAGE / NY-ESO, RAGE, SSX-2, CD19 and CD20.
もう1つの実施形態においては、抗原は、関節リウマチ、全身性エリテマトーデス、多発性硬化症、乾癬及びクローン病よりなる群から選択される疾患に関連している。 In another embodiment, the antigen is associated with a disease selected from the group consisting of rheumatoid arthritis, systemic lupus erythematosus, multiple sclerosis, psoriasis and Crohn's disease.
本発明はまた、免疫応答を惹起する能力をもつ少なくとも1つのエピトープを有する抗原及び負の免疫調節因子のインヒビターを含んでなり、しかもさらにサイトカインを含んでなる組成物をも含む。好ましくは、サイトカインは、単離されたポリペプチドである。もう1つの態様においては、サイトカインは、IL−12、TNFα、IFNα、IFNβ、IFNγ、IL−7、IL−1、IL−2、IL−6、IL−15、IL−21及びIL−23よりなる群から選択される。 The present invention also includes a composition comprising an antigen having at least one epitope capable of eliciting an immune response and an inhibitor of a negative immune modulator, and further comprising a cytokine. Preferably, the cytokine is an isolated polypeptide. In another aspect, the cytokine is from IL-12, TNFα, IFNα, IFNβ, IFNγ, IL-7, IL-1, IL-2, IL-6, IL-15, IL-21 and IL-23. Selected from the group consisting of
本発明はまた、免疫応答を惹起する能力をもつ少なくとも1つのエピトープを有する抗原及び負の免疫調節因子のインヒビターを含んでなり、かつさらにトール様レセプタ(TLR)アゴニストを含んでなる組成物をも含む。一態様においては、TLRアゴニストは発現ベクターにより発現される。好ましくは、TLRアゴニストは単離ポリペプチドである。 The present invention also includes a composition comprising an antigen having at least one epitope capable of eliciting an immune response and an inhibitor of a negative immune modulator and further comprising a toll-like receptor (TLR) agonist. Including. In one aspect, the TLR agonist is expressed by an expression vector. Preferably, the TLR agonist is an isolated polypeptide.
本発明はまた、負の免疫調節因子のインヒビターを含んでなる組成物であって、インヒビターがヤーヌスキナーゼ(JAK)シグナリングの阻害を抑制する、組成物をも含む。 The invention also includes a composition comprising an inhibitor of a negative immunomodulator, wherein the inhibitor suppresses Janus kinase (JAK) signaling inhibition.
もう1つの実施形態においては、負の免疫調節因子のインヒビターを含んでなる組成物は、トール様レセプタ(TLR)シグナリングの阻害を抑制する。 In another embodiment, a composition comprising an inhibitor of a negative immune regulator suppresses inhibition of Toll-like receptor (TLR) signaling.
さらにもう1つの実施形態においては、負の免疫調節因子のインヒビターを含んでなる組成物はNF−κBシグナリングの阻害を抑制する。 In yet another embodiment, a composition comprising an inhibitor of a negative immune modulator inhibits inhibition of NF-κB signaling.
本発明は、細胞の免疫能力を増強させるための組成物であって、前記細胞内の負の免疫調節因子を阻害するインヒビターをコードする第1のポリヌクレオチド及び哺乳動物の体内で免疫応答を誘発する少なくとも1つのエピトープを有する抗原をコードする第2のポリヌクレオチドを含んでなる組成物をも含む。 The present invention relates to a composition for enhancing the immunity of a cell, the first polynucleotide encoding an inhibitor that inhibits a negative immune regulator in the cell, and an immune response in a mammalian body. And a composition comprising a second polynucleotide encoding an antigen having at least one epitope.
本発明はまた、細胞の免疫能力を増強させるための組成物であって、前記細胞内の負のIRMを阻害するインヒビターをコードする第1のポリヌクレオチド及びサイトカインをコードする第2のポリヌクレオチドを含んでなるベクターを含んでなる組成物をも含む。好ましくは、サイトカインをコードする第2のポリヌクレオチドは、IL−12、TNFα、IFNα、IFNβ、IFNγ、IL−7、IL−2、IL−6、IL−15、IL−21及びIL−23よりなる群から選択される。 The present invention also provides a composition for enhancing the immunity of a cell, comprising: a first polynucleotide encoding an inhibitor that inhibits negative IRM in the cell; and a second polynucleotide encoding a cytokine. Also included is a composition comprising a vector comprising. Preferably, the second polynucleotide encoding a cytokine is from IL-12, TNFα, IFNα, IFNβ, IFNγ, IL-7, IL-2, IL-6, IL-15, IL-21 and IL-23 Selected from the group consisting of
本発明はまた、負の免疫調節因子のインヒビターを含んでなる細胞をも含む。好ましく
は、細胞はAPC、樹状細胞、単球/マクロファージ、T細胞及びB細胞よりなる群から選択された免疫細胞である。
The invention also includes a cell comprising an inhibitor of a negative immune regulator. Preferably, the cell is an immune cell selected from the group consisting of APC, dendritic cells, monocytes / macrophages, T cells and B cells.
もう1つの態様においては、細胞はさらに、少なくとも1つのエピトープを有する抗原を含んでなり、ここで少なくとも1つのエピトープは、哺乳動物の体内で免疫応答を惹起する能力をもつ。 In another embodiment, the cell further comprises an antigen having at least one epitope, wherein the at least one epitope is capable of eliciting an immune response in the mammalian body.
さらにもう1つの態様においては、細胞はさらに、サイトカインをコードするポリヌクレオチドを含んでなる発現ベクターを含んでなる。 In yet another embodiment, the cell further comprises an expression vector comprising a polynucleotide encoding a cytokine.
本発明はまた、負の免疫調節因子のインヒビターと細胞を接触させる工程を含んでなるサイレンシング化された細胞を生成する方法をも含む。 The invention also includes a method of generating a silenced cell comprising contacting the cell with an inhibitor of a negative immune modulator.
もう1つの実施形態においては、本発明は、サイレンシング化及びパルスを受けた細胞を生成する方法であって、細胞を負の免疫調節因子と接触させる工程及びさらに細胞を、哺乳動物の体内で免疫応答を惹起する能力をもつ少なくとも1つのエピトープを有する抗原と接触させる工程を含んでなる方法を含む。 In another embodiment, the present invention provides a method of producing a cell that has been silenced and pulsed, wherein the step of contacting the cell with a negative immunomodulator and further comprising the cell in the body of a mammal. Contacting with an antigen having at least one epitope capable of eliciting an immune response.
本発明はまた、哺乳動物の体内で免疫応答を惹起する方法であって、それを必要としている哺乳動物の体内に負の免疫調節因子のインヒビターを含んでなる組成物を投与する工程を含んでなる方法をも含む。 The present invention also includes a method of eliciting an immune response in a mammalian body, comprising the step of administering a composition comprising an inhibitor of a negative immune modulator to the body of the mammal in need thereof. Including the method.
本発明のもう1つの実施形態は、哺乳動物の体内で免疫応答を惹起する方法であって、それを必要としている哺乳動物の体内にサイレンシング化された細胞を含んでなる組成物を投与する工程を含んでなる方法であって、サイレンシング化された細胞が負の免疫調節因子のインヒビターを含んでなる方法をも含む。好ましくは、サイレンシング化された細胞は、それを必要とする哺乳動物の体内にサイレンシング化された細胞を投与する前にインビトロで抗原と接触させられる。もう1つの態様においては、それを必要としている哺乳動物内へのサイレンシング化された細胞の投与の後にインビボで抗原とサイレンシング化された細胞接触させることもできる。 Another embodiment of the invention is a method of eliciting an immune response in a mammalian body, comprising administering a composition comprising silenced cells in the mammalian body in need thereof Also included is a method comprising a step, wherein the silenced cell comprises an inhibitor of a negative immune modulator. Preferably, the silenced cell is contacted with the antigen in vitro prior to administering the silenced cell into the body of a mammal in need thereof. In another embodiment, the silenced cells can be contacted with the antigen in vivo after administration of the silenced cells into the mammal in need thereof.
本発明を例示する目的で、図面中には、本発明のいくつかの実施形態が描かれている。しかしながら、本発明は、図面中に描かれている実施形態の精確な配置及び手段に限定されるわけではない。 For the purpose of illustrating the invention, there are shown in the drawings several embodiments of the invention. However, the invention is not limited to the precise arrangement and means of the embodiments depicted in the drawings.
本発明は、免疫細胞内の負の免疫調節因子を変調(modulating)させることによる免疫細胞の免疫能力の増強に関する。本発明は、負の免疫調節因子を変調させることにより免疫細胞内の抗原提示を変調させるための組成物及び方法を提供する。負の免疫調節因子は、シグナル伝達に関与するタンパク質の阻害性相同体(例えば可溶性デコイTLR、阻害性TIR相同体、阻害性シグナリング分子イソ型、阻害性サイトカイン相同体など)、分子安定性に関与するタンパク質、シグナリング分子複合体の阻害性構成要素、シグナリング分子リン酸化反応の調節に関与するタンパク質、NFκB標的遺伝子の転写を抑制する転写因子、RNA翻訳及び安定性の調節に関与するタンパク質、サイトカインシグナリング調節因子などを含む(ただしこれらに限定されるわけではない)異なるタンパク質ファミリーに属することができる。 The present invention relates to enhancing the immune capacity of immune cells by modulating negative immune regulators in immune cells. The present invention provides compositions and methods for modulating antigen presentation in immune cells by modulating negative immune regulators. Negative immune regulators are involved in inhibitory homologues of proteins involved in signal transduction (eg soluble decoy TLR, inhibitory TIR homologues, inhibitory signaling molecule isoforms, inhibitory cytokine homologues, etc.), molecular stability Protein, inhibitory component of signaling molecule complex, protein involved in regulation of signaling molecule phosphorylation, transcription factor that suppresses transcription of NFκB target gene, protein involved in regulation of RNA translation and stability, cytokine signaling It can belong to different protein families, including but not limited to regulatory factors.
本発明は、負の免疫調節因子の変調によって免疫細胞の免疫能力が増強されているワクチン及び療法を提供する。さらに、本発明は、同様に腫瘍ワクチン接腫における自己寛容を破るための機序をも提供している。従って本発明は、細胞の免疫刺激能力を増強するこ
とにより免疫細胞内でTLR−、TNFR−媒介型シグナリング及びサイトカインレセプタ媒介型JAK/STATシグナリングなどの前炎症性シグナル伝達経路の負の免疫調節因子に干渉することのもつ治療上の恩恵を提供している。
The present invention provides vaccines and therapies in which the immune capacity of immune cells is enhanced by modulation of negative immune regulators. Furthermore, the present invention also provides a mechanism for breaking self-tolerance in tumor vaccine contact. Thus, the present invention relates to negative immune regulators of proinflammatory signaling pathways such as TLR-, TNFR-mediated signaling and cytokine receptor-mediated JAK / STAT signaling within immune cells by enhancing the immune stimulatory capacity of the cells Provides the therapeutic benefits of interfering with
定義
本書で使用される通り、以下の用語の各々は、本節で関連づけされているその意味を有している。
Definitions As used in this document, each of the following terms has its meaning associated with it in this section.
冠詞「a」及び「an」は、本書では、その物品の文法的目的語の1つ又は2つ以上(すなわち少なくとも1つ)を意味するように用いられている。一例としては、「an element(要素)」というのは1要素又は2つ以上の要素を意味する。 The articles “a” and “an” are used herein to mean one or more (ie at least one) of the grammatical objects of the article. By way of example, “an element” means one element or more than one element.
「約」という用語は、当業者により理解されるものであり、それが用いられる文脈に応じて幾分か変動することになる。 The term “about” is understood by one of ordinary skill in the art and will vary somewhat depending on the context in which it is used.
「同種異系」という用語は、同じ種の異なる動物に由来する移植片を意味する。 The term “homologous” means grafts from different animals of the same species.
「アロ抗原」というのは、レシピエントにより発現される抗原とは異なる抗原である。 An “alloantigen” is an antigen that is different from the antigen expressed by the recipient.
本書で使用される「抗原」という用語は、抗原上で特定のエピトープに特異的に結合することのできる免疫グロブリン分子を意味する。抗体は、天然供給源又は組換え型供給源に由来する無傷の免疫グロブリンであり得、無傷の免疫グロブリンの免疫活性部分であり得る。抗体は標準的には免疫グロブリン分子の四量体である。本発明における抗体は、例えばポリクローナル抗体、モノクローナル抗体、Fv、Fab及びF(ab)2ならびに1本鎖抗体及びヒト化抗体を含めたさまざまな形態で存在し得る(ハーロウ(Harlow)ら、1988年;ハウストン(Houston)ら、1988年;バード(Bird)ら、1988年)。 As used herein, the term “antigen” refers to an immunoglobulin molecule capable of specifically binding to a particular epitope on an antigen. The antibody can be an intact immunoglobulin derived from a natural source or a recombinant source, and can be an immunoactive portion of an intact immunoglobulin. An antibody is typically a tetramer of immunoglobulin molecules. The antibodies in the present invention may exist in various forms including, for example, polyclonal antibodies, monoclonal antibodies, Fv, Fab and F (ab) 2 and single chain antibodies and humanized antibodies (Harlow et al., 1988). Houston et al., 1988; Bird et al., 1988).
本書で使用される「抗原」又は「Ag」という用語は、免疫応答をひき起こす分子として定義される。この免疫応答には抗体産生及び免疫学的にコンピテントな細胞又はその両方が関与する。当業者であれば、ほぼ全てのタンパク質又はペプチドを含むあらゆる巨大分子が抗原として役立ち得るということを理解するであろう。さらに、抗原は組換え型又はゲノムDNAから誘導され得る。当業者であれば、かくして免疫応答を惹起するタンパク質をコードするヌクレオチド配列又は部分的ヌクレオチド配列を含んでなるあらゆるDNAが、本書で使用される通りの「抗原」をコードするということを理解することだろう。さらに、当業者は、抗原が遺伝子の全長ヌクレオチド配列のみによりコードされる必要がないということも理解するであろう。本発明が2つ以上の遺伝子の部分的ヌクレオチド配列の使用を含むがこれに限定されるわけではないということ、そしてこれらのヌクレオチド配列が所望の免疫応答を惹起するためにさまざまな組合せで配置されていることは、直ちに明らかである。その上、当業者であれば、抗原が「遺伝子」によりコードされる必要が全くないことを理解するだろう。抗原を合成した形で生成することができ、そうでなければ生体試料から誘導することもできるということは直ちに明らかである。このような生体試料としては、組織試料、腫瘍試料、細胞又は体液が含まれ得るがこれに限定されるわけではない。 The term “antigen” or “Ag” as used herein is defined as a molecule that provokes an immune response. This immune response involves antibody production and immunologically competent cells or both. One skilled in the art will understand that any macromolecule, including almost any protein or peptide, can serve as an antigen. Furthermore, the antigen can be derived from recombinant or genomic DNA. Those skilled in the art will understand that any DNA comprising a nucleotide sequence or partial nucleotide sequence encoding a protein that elicits an immune response thus encodes an “antigen” as used herein. right. Furthermore, those skilled in the art will also understand that the antigen need not be encoded solely by the full-length nucleotide sequence of the gene. The present invention includes, but is not limited to, the use of partial nucleotide sequences of two or more genes, and these nucleotide sequences are arranged in various combinations to elicit the desired immune response. It is immediately obvious. Moreover, those skilled in the art will appreciate that the antigen need not be encoded by a “gene” at all. It will be readily apparent that the antigen can be produced in synthetic form or otherwise derived from a biological sample. Such biological samples can include, but are not limited to, tissue samples, tumor samples, cells or body fluids.
「抗原提示細胞」(APC)は、T細胞を活性化する能力をもつ細胞であり、単球/マクロファージ、B細胞及び樹状細胞(DC)を含むがこれに限定されるわけではない。 An “antigen-presenting cell” (APC) is a cell that has the ability to activate T cells and includes, but is not limited to, monocytes / macrophages, B cells and dendritic cells (DC).
「樹状細胞」又は「DC」という用語は、リンパ系組織又は非リンパ系組織の中に発見される形態学的に類似の細胞型のさまざまな集団の任意のメンバーを意味する。これらの
細胞は、その独特の形態、高レベルの表面MHC−クラスII発現によって特徴づけされる。DCは、数多くの組織供給源から単離され得る。DCは、MHC制限されたT細胞を感作させるための高い能力を有し、インサイチュでT細胞に対し抗原を提示する上できわめて有効である。抗原は、T細胞の発達及び寛容中に発現される自己抗原及び、通常の免疫プロセス中に存在する外来性抗原であり得る。
The term “dendritic cell” or “DC” means any member of various populations of morphologically similar cell types found in lymphoid or non-lymphoid tissues. These cells are characterized by their unique morphology, high levels of surface MHC-class II expression. DC can be isolated from a number of tissue sources. DC has a high ability to sensitize MHC-restricted T cells and is very effective at presenting antigens to T cells in situ. Antigens can be self-antigens expressed during T cell development and tolerance and foreign antigens present during normal immune processes.
本書で使用されているように、「活性化されたDC」は、抗原でのパルスを受け免疫細胞を活性化させる能力をもつDCである。 As used herein, an “activated DC” is a DC that has the ability to activate immune cells in response to a pulse with an antigen.
本書で使用されている「成熟DC」という用語は、高レベルのMHCクラスII、CD80(B7.1)及びCD86(B7.2)分子を発現する樹状細胞として定義される。これとは対照的に、未成熟樹状細胞は低レベルのMHCクラスII、CD80(B7.1)及びCD86(B7.2)分子を発現するが、抗原を取り込む大きな能力を有する。 The term “mature DC” as used herein is defined as dendritic cells that express high levels of MHC class II, CD80 (B7.1) and CD86 (B7.2) molecules. In contrast, immature dendritic cells express low levels of MHC class II, CD80 (B7.1) and CD86 (B7.2) molecules, but have a great ability to take up antigens.
「抗原負荷を受けたAPC」又は「抗原パルスを受けたAPC」には、抗原に曝露され抗原により活性化されたAPCが含まれる。例えばAPCは、例えば抗原の存在下での培養中にインビトロでAg負荷を受けた状態となり得る。APCは同様に、抗原に対する曝露によりインビボで負荷され得る。 “APC under antigen loading” or “APC under antigen pulse” includes APCs that have been exposed to and activated by an antigen. For example, APC can become Ag loaded in vitro, for example during culture in the presence of antigen. APC can also be loaded in vivo by exposure to an antigen.
「抗原を負荷されたAPC」は、従来次の2つの方法のうちの1つにおいて調製される:(1)抗原ペプチドとして知られる小ペプチドフラグメントが、APCの外側に直接「パルスされる;又は(2)APCが全タンパク質又はタンパク質粒子とインキュベートされ、これらのタンパク質がその後APCにより取り込まれる。これらのタンパク質はAPCにより小ペプチドフラグメントへと消化され、場合によってはAPC表面上に提示される。さらに、抗原を負荷されたAPCは、細胞内に抗原をコードするポリヌクレオチドによっても生成され得る。 An “antigen-loaded APC” is conventionally prepared in one of two ways: (1) A small peptide fragment known as an antigenic peptide is “pulsed” directly outside the APC; or (2) APC is incubated with total protein or protein particles, and these proteins are then taken up by APC, which are digested by APC into small peptide fragments and possibly presented on the APC surface. An APC loaded with an antigen can also be produced by a polynucleotide encoding the antigen in a cell.
本書で使用する「サイレンシング化されたAPC」又は「サイレンシング化されたDC」という用語は、(別途負の免疫調節因子を阻害するものとして知られている)負の免疫調節因子のインヒビターに曝露され、かくして負の免疫調節因子が免疫応答の調節に関連することになるAPC又はDCをそれぞれ意味している。インヒビターはTLR、RP105、SIGIRR、ST2、NOD2、MyD88s、IRAK(IRAKM、IRAK1、IRAK2)、IRF−4、FLN29、TWEAK、TRIAD3A、CYLD、Cbl、A20、SUMO(SUMO1、SUMO2、SUMO3、及びSUMO4)、IkBタンパク質、MKPs、Foxj1、Foxo3a、TWIST(Twist1、Twist2)、Roquin、Dok(Dok−1、Dok−2)、PI3K、プロスタグランジン、TRAIL−R、アレスチン、TOLLIP、SOCS(SOCS1、SOCS2、SOCS3、SOCS4、SOCS5、SOCS6、SOCS7及びCIS)、PIAS(PIAS1、PIAS3、PIASx及びPIASy)、SHP(SHP−1及びSHP−2)などを含む(ただしこれに限定されるわけではない)本書で開示されている任意の負の免疫調節因子を阻害する能力をもつ。サイレンシング化されたAPCは、負の免疫調節因子のインヒビターで処理されていないその他の点では同一のAPCに比べて増強された免疫能力を有する。 As used herein, the term “silenced APC” or “silenced DC” refers to inhibitors of negative immune regulators (otherwise known as inhibiting negative immune regulators). It means APC or DC, respectively, that is exposed and thus negative immune modulators will be associated with modulation of the immune response. Inhibitors are TLR, RP105, SIGIRR, ST2, NOD2, MyD88s, IRAK (IRAKM, IRAK1, IRAK2), IRF-4, FLN29, TWEAK, TRIAD3A, CYLD, Cbl, A20, SUMO (SUMO1, SUMO4, SUMO3, SUMO3, SUMO3, SUMO3, SUMO3, SUMO3, SUMO3 , IkB protein, MKPs, Foxj1, Foxo3a, TWIST (Twist1, Twist2), Roquin, Dok (Dok-1, Dok-2), PI3K, prostaglandin, TRAIL-R, arrestin, TOLLIP, SOCS (SOCS1, SOCS2, SOCS1, SOCS1) SOCS3, SOCS4, SOCS5, SOCS6, SOCS7 and CIS), PIAS (PIAS1, PIAS3, PIASx and PIASy), SHP (SHP -1 and SHP-2) and the like, and has the ability to inhibit any negative immunomodulator disclosed herein. Silenced APCs have enhanced immune capacity compared to otherwise identical APCs that have not been treated with inhibitors of negative immune regulators.
「アンチセンス」というのは特に、ポリペプチドをコードする2本鎖DNA分子の非コーディング鎖の核酸配列、又は非コーディング鎖と実質的に相同である配列を意味する。本書で定義されている通り、アンチセンス配列は、ポリペプチドをコードする2本鎖DNA分子の配列に相補的である。アンチセンス配列はDNA分子のコーディング鎖のコーディング部分のみに相補的である必要はない。アンチセンス配列は、コーディング配列の発現を制御する、ポリペプチドをコードするDNA分子のコーディング鎖上で特定された調節配列に相補的である。 “Antisense” specifically refers to the nucleic acid sequence of the non-coding strand of a double-stranded DNA molecule encoding a polypeptide, or a sequence that is substantially homologous to the non-coding strand. As defined herein, an antisense sequence is complementary to the sequence of a double-stranded DNA molecule that encodes a polypeptide. The antisense sequence need not be complementary only to the coding portion of the coding strand of the DNA molecule. The antisense sequence is complementary to the regulatory sequences identified on the coding strand of the DNA molecule encoding the polypeptide that control the expression of the coding sequence.
本書で使用される「自己免疫疾患」という用語は、自己免疫応答の結果としてもたらされる障害として定義される。自己免疫疾患は、自己抗原に対する不適切な及び過剰な応答の結果である。自己免疫疾患の例としては、アジソン病、円形脱毛症、強直性脊椎炎、自己免疫性肝炎、自己免疫性耳下腺炎、クローン病、糖尿病(I型)、栄養障害性表皮水泡症、精巣上体炎、糸球体腎炎、グレーブス病、ギラン・バレー症候群、橋本病、溶血性貧血、全身性エリテマトーデス、多発性硬化症、重症筋無力症、尋常性天疱瘡、乾癬、リウマチ熱、関節リウマチ、サルコイドーシス、強皮症、シェーグレン症候群、脊椎関節症、甲状腺炎、脈管炎、白斑、粘液水腫、悪性貧血、潰瘍性大腸炎などが含まれるがこれに限定されるわけではない。 The term “autoimmune disease” as used herein is defined as a disorder resulting from an autoimmune response. Autoimmune diseases are the result of inappropriate and excessive responses to self antigens. Examples of autoimmune diseases include Addison's disease, alopecia areata, ankylosing spondylitis, autoimmune hepatitis, autoimmune parotitis, Crohn's disease, diabetes (type I), dystrophic epidermolysis bullosa, testis Epidermitis, glomerulonephritis, Graves' disease, Guillain-Barre syndrome, Hashimoto's disease, hemolytic anemia, systemic lupus erythematosus, multiple sclerosis, myasthenia gravis, pemphigus vulgaris, psoriasis, rheumatic fever, rheumatoid arthritis, Examples include, but are not limited to, sarcoidosis, scleroderma, Sjogren's syndrome, spondyloarthritis, thyroiditis, vasculitis, vitiligo, myxedema, pernicious anemia, and ulcerative colitis.
本書で使用される「自己移植性(autologous)」という用語は、後にその個体に再導入されるべき同じ個体に由来するあらゆる材料を意味するものとされている。 As used herein, the term “autologous” is intended to mean any material derived from the same individual that is to be subsequently reintroduced into the individual.
「癌」という用語は、本書では、異常細胞の急速な及び無制御の成長により特徴づけされる疾患として定義される。癌細胞は、局所的に又は血流及びリンパ系を通して、体の他の部分に蔓延し得る。さまざまな癌の例としては乳癌、前立腺癌、卵巣癌、子宮頚癌、皮膚癌、膵臓癌、直腸結腸癌、腎臓癌、肝臓癌、脳腫瘍、リンパ腫、白血病、肺癌などが含まれるが、これに限定されるわけではない。 The term “cancer” is defined herein as a disease characterized by rapid and uncontrolled growth of abnormal cells. Cancer cells can spread to other parts of the body locally or through the bloodstream and lymphatic system. Examples of various cancers include breast cancer, prostate cancer, ovarian cancer, cervical cancer, skin cancer, pancreatic cancer, colorectal cancer, kidney cancer, liver cancer, brain tumor, lymphoma, leukemia, lung cancer, etc. It is not limited.
「サイトカインシグナリング調節因子」又は「サイトカインシグナリングの調節因子」又は「サイトカインシグナル伝達の調節因子」という語は、1つの細胞内でサイトカインシグナリング伝達経路を負に調節する能力をもつタンパク質を意味する。サイトカインシグナル伝達の調節因子は、サイトカインシグナル伝達のサプレッサ(SOCS1−SOCS7、サイトカイン誘発可能SH2−ドメイン含有タンパク質(CIS))、SHS含有ホスファターゼ(SHP)、及び活性化されたSTATのタンパク質インヒビター(PIAS)を含むが、これに限定されるわけではない。 The term “cytokine signaling regulator” or “regulator of cytokine signaling” or “regulator of cytokine signaling” means a protein that has the ability to negatively regulate the cytokine signaling pathway in one cell. Modulators of cytokine signaling include suppressors of cytokine signaling (SOCS1-SOCS7, cytokine-inducible SH2-domain containing protein (CIS)), SHS-containing phosphatase (SHP), and activated STAT protein inhibitors (PIAS) Including, but not limited to.
本書で使用する「DNA」という用語は、デオキシリボ核酸として定義される。 The term “DNA” as used herein is defined as deoxyribonucleic acid.
「ドナー抗原」というのは、レシピエント内に移植すべきドナー組織により発現される抗原を意味する。 By “donor antigen” is meant an antigen expressed by donor tissue to be transplanted into a recipient.
「レシピエント抗原」という用語は、ドナー抗原に対する免疫応答のための標的を意味する。 The term “recipient antigen” means a target for an immune response against a donor antigen.
本書で使用する「エフェクタ細胞」というのは、抗原に対抗する免疫応答を媒介する細胞を意味する。エフェクタ細胞の一例としてはT細胞及びB細胞が含まれるが、これに限定されるわけではない。 As used herein, “effector cell” refers to a cell that mediates an immune response against an antigen. Examples of effector cells include, but are not limited to, T cells and B cells.
「コードする」という用語は、遺伝子、cDNA又はmRNAなどのポリヌクレオチド内のヌクレオチドの特定の配列が有する、定義されたヌクレオチド配列(すなわちrRNA、tRNA及びmRNA)又は定義されたアミノ酸配列のいずれかを有するその他のポリマー及び巨大分子の生物学的プロセス中の合成のために鋳型として役立つという固有の特性、及びその結果としてもたらされる生物学的特性を意味する。かくして、1つの遺伝子は、その遺伝子に対応するmRNAの転写及び翻訳が細胞又はその他の生体系の中でタンパク質を産生する場合に、そのタンパク質をコードする。mRNA配列と同一でかつ通常配列のリスト中に提供されているヌクレオチド配列を有するコーディング鎖及び遺伝子又はcDNAの転写のための鋳型として使用される非コーディング鎖の両方共が、その遺
伝子又はcDNAのタンパク質又はその他の産物をコードするものとして言及され得る。
The term “encode” refers to either a defined nucleotide sequence (ie, rRNA, tRNA and mRNA) or a defined amino acid sequence possessed by a particular sequence of nucleotides within a polynucleotide, such as a gene, cDNA or mRNA. It refers to the unique properties of serving as a template for the synthesis of other polymers and macromolecules in the biological process and the resulting biological properties. Thus, a gene encodes a protein when transcription and translation of mRNA corresponding to that gene produces the protein in a cell or other biological system. Both the coding strand having the nucleotide sequence identical to the mRNA sequence and usually provided in the list of sequences and the non-coding strand used as a template for transcription of the gene or cDNA are proteins of that gene or cDNA Or it may be mentioned as encoding other products.
本書で使用する「内因性」というのは、生体、細胞、組織又は系由来の又はそれらの内部で産生されるあらゆる材料を意味する。 As used herein, “endogenous” refers to any material derived from or produced within a living organism, cell, tissue or system.
本書で使用する「外因性」という用語は、生体、細胞、組織又は系の外部から導入される又はそれらの外部で産生されるあらゆる材料を意味する。 As used herein, the term “exogenous” refers to any material that is introduced from or produced outside a living organism, cell, tissue, or system.
本書で使用する「エピトープ」という用語は、免疫応答を惹起して、B及び/又はT細胞応答を誘発することのできる抗原上の小化学分子として定義される。抗原は1つもしくはそれ以上のエピトープを有することができる。大部分の抗原は数多くのエピトープを有する。すなわち多価である。一般に、エピトープは、サイズ的におおよそアミノ酸5個及び/又は糖である。当業者は、一般に分子の特定的線形配列よりもむしろ全体的に3次元の構造が抗原特異性の主たる基準であり、従ってこれがエピトープ同士を区別するということを了解している。 The term “epitope” as used herein is defined as a small chemical molecule on an antigen that can elicit an immune response and elicit a B and / or T cell response. An antigen can have one or more epitopes. Most antigens have many epitopes. That is, it is multivalent. In general, an epitope is approximately 5 amino acids and / or sugars in size. One skilled in the art understands that generally an overall three-dimensional structure, rather than a specific linear array of molecules, is the primary measure of antigen specificity and thus distinguishes epitopes.
本書で使用する「発現」という用語は、そのプロモータにより駆動される特定のヌクレオチド配列の転写及び/又は翻訳として定義される。 The term “expression” as used herein is defined as the transcription and / or translation of a particular nucleotide sequence driven by its promoter.
本書で使用する「発現ベクター」という用語は、転写可能な遺伝子産物の少なくとも一部分についてコードする核酸配列を含有するベクターを意味している。場合によっては、RNA分子は次にタンパク質、ポリペプチド又はペプチドへと翻訳される。その他の場合には、これらの配列は、例えばアンチセンス分子、siRNA、リボザイムなどの産生において翻訳されない。発現ベクターは、特定の宿主生体内での作動的に連結されたコーディング配列の転写及び場合によってはその翻訳にとって必要な核酸配列を意味するさまざまな制御配列を含み得る。転写及び翻訳を支配する制御配列に加えて、ベクター及び発現ベクターは、その他の機能にも役立つ核酸配列を含有し得る。 As used herein, the term “expression vector” refers to a vector that contains a nucleic acid sequence that encodes at least a portion of a transcribable gene product. In some cases, the RNA molecule is then translated into a protein, polypeptide or peptide. In other cases, these sequences are not translated, for example, in the production of antisense molecules, siRNA, ribozymes and the like. An expression vector may contain various control sequences that refer to the nucleic acid sequences necessary for transcription and optionally translation of the operably linked coding sequence in a particular host organism. In addition to control sequences that govern transcription and translation, vectors and expression vectors may contain nucleic acid sequences that serve other functions as well.
本書で用いられる「ヘルパーT細胞」という用語は、その他のB及びTリンパ球及び/又はマクロファージの活性化及び機能を促進することを主たる機能とするエフェクタT細胞として定義される。ほとんどのヘルパーT細胞はCD4T細胞である。 The term “helper T cells” as used herein is defined as effector T cells whose primary function is to promote the activation and function of other B and T lymphocytes and / or macrophages. Most helper T cells are CD4 T cells.
本書で用いられる「非相同な」という用語は、異なる種から誘導されるDNA又はRNA配列又はタンパク質として定義される。 The term “heterologous” as used herein is defined as a DNA or RNA sequence or protein derived from a different species.
本書で用いられる「相同な」という語は、2つのポリマー分子間例えば2つの核酸分子、例えば2つのDNA分子又は2つのRNA分子の間又は2つのポリペプチド分子の間のサブユニット配列類似性を意味する。2つの分子の両方においてサブユニット位置が同じモノマーサブユニットにより占有されている場合、例えば2つのDNA分子の各々の中の1つの位置がアデニンにより占有されている場合、それらはその位置において相同である。2つの配列の間の相同性は、整合する又は相同な位置の数に正比例し、例えば2つの化合物配列内の位置の半分(例えば長さ10サブユニットのポリマー内の5位置)が相同である場合、2つの配列は50%相同であり、90%の位置、例えば10中9位が整合しているか相同である場合、その2つの配列は90%の相同性を共有する。例を挙げると、DNA配列3’ATTGCC5’と3’TATGGCは50%の相同性を共有する。 As used herein, the term “homologous” refers to subunit sequence similarity between two polymer molecules, eg, two nucleic acid molecules, eg, between two DNA molecules or two RNA molecules or between two polypeptide molecules. means. If the subunit position is occupied by the same monomer subunit in both two molecules, for example if one position in each of the two DNA molecules is occupied by adenine, they are homologous at that position. is there. Homology between two sequences is directly proportional to the number of matching or homologous positions, eg, half of the positions in the two compound sequences (eg, 5 positions in a 10 subunit long polymer) are homologous In some cases, two sequences are 50% homologous, and if 90% of the positions, eg, 9 out of 10, are matched or homologous, the two sequences share 90% homology. As an example, the DNA sequences 3'ATTGCC5 'and 3'TATGGC share 50% homology.
本書で使用されている「相同性な」というのは、「同一性」と同義で用いられる。 As used herein, “homology” is used interchangeably with “identity”.
本書で使用されている「免疫原」というのは、哺乳動物の体内で液性抗体及び/又は細胞媒介型免疫応答を刺激又は誘発することのできる物質を意味する。 As used herein, “immunogen” means a substance capable of stimulating or inducing a humoral antibody and / or a cell-mediated immune response in a mammalian body.
本書で使用されている「免疫グロブリン」又は「Ig」という用語は、抗体として機能する1種類のタンパク質として定義される。このタンパク質クラスの中に含まれている5つのメンバーは、IgA、IgG、IgM、IgD及びIgEである。IgAは、唾液、涙液、母乳、胃腸分泌物及び呼吸管及び尿生殖路の粘腋分泌などの身体の分泌物の中に存在する一次抗体である。IgGは、最も一般的な循環抗体である。IgMは、大部分の哺乳動物の体内で一次免疫応答において産生される主要な免疫グロブリンである。それは、凝集反応、補体固定及びその他の抗体応答において最も効率の良い免疫グロブリンであり、細菌及びウイルスに対する防御において重要である。IgDは、既知の抗体機能をもたないものの抗原レセプタとして役立ち得る免疫グロブリンである。IgEは、アレルゲンに対する暴露の時点でマスト細胞及び好塩基球からメディエータを放出させることにより即時型過敏性を媒介する免疫グロブリンである。 As used herein, the term “immunoglobulin” or “Ig” is defined as a type of protein that functions as an antibody. The five members included in this protein class are IgA, IgG, IgM, IgD and IgE. IgA is the primary antibody present in bodily secretions such as saliva, tears, breast milk, gastrointestinal secretions, and mucous secretions of the respiratory tract and urogenital tract. IgG is the most common circulating antibody. IgM is the major immunoglobulin produced in the primary immune response in most mammalian bodies. It is the most efficient immunoglobulin in agglutination, complement fixation and other antibody responses and is important in defense against bacteria and viruses. IgD is an immunoglobulin that has no known antibody function but can serve as an antigen receptor. IgE is an immunoglobulin that mediates immediate hypersensitivity by releasing mediators from mast cells and basophils upon exposure to allergens.
本書で使用されている「負の免疫調節因子」又は「負の調節因子」又は「免疫応答の負の調節因子」というのは、細胞中の免疫シグナリング形質導入経路を負に調節する能力をもつタンパク質を意味する。好ましくは、負の免疫調節因子は、前炎症性シグナル伝達経路を負に調節する。本書で開示されている負の免疫調節因子には、sTLR、RP105、SIGIRR、ST2、NOD2、MyD88s、IRAK(IRAKM、IRAK1、IRAK2)、IRF−4、FLN29、TWEAK、TRIAD3A、CYLD、Cbl、A20、SUMO(SUMO1、SUMO2、SUMO3及びSUMO4)、IkBタンパク質、MKPs、Foxj1、Foxo3a、TWIST(Twist1、Twist2)、Roquin、Dok(Dok−1、Dok−2)、PI3K、プロスタグランジン、TRAIL−R、アレスチン、TOLLIP、SOCS(SOCS1、SOCS2、SOCS3、SOCS4、SOCS5、SOCS6、SOCS7及びCIS)、PIAS(PIAS1、PIAS3、PIASx及びPIASy)、SHP(SHP−1及びSHP−2)などが含まれるが、これに限定されるわけではない。負の免疫調節因子は、シグナル伝達に関与するタンパク質の阻害性相同体(例えば可溶性デコイTLR、阻害性TIR相同体、阻害性シグナリング分子イソ型、阻害性サイトカイン相同体など)、分子安定性に関与するタンパク質、シグナリング分子複合体の阻害性構成要素、シグナリング分子のリン酸化反応の調節に関与するタンパク質、NFκB標的遺伝子の転写を抑制する転写因子、RNA翻訳及び安定性の調節に関与するタンパク質、サイトカインシグナリング調節因子などを含む(ただしこれに限定されるわけではない)異なるタンパク質ファミリーに属し得る。一態様においては、免疫細胞内の1つもしくはそれ以上の負の免疫調節因子を阻害することが、細胞の免疫能力を増強させるのに役立つ。 As used herein, “negative immune regulator” or “negative regulator” or “negative regulator of immune response” has the ability to negatively regulate immune signaling transduction pathways in cells It means protein. Preferably, the negative immune regulator negatively regulates the proinflammatory signaling pathway. Negative immunomodulators disclosed herein include sTLR, RP105, SIGIRR, ST2, NOD2, MyD88s, IRAK (IRAKM, IRAK1, IRAK2), IRF-4, FLN29, TWEAK, TRIAD3A, CYLD, Cbl, A20. , SUMO (SUMO1, SUMO2, SUMO3 and SUMO4), IkB protein, MKPs, Foxj1, Foxo3a, TWIST (Twist1, Twist2), Roquin, Dok (Dok-1, Dok-2), PI3K, Prostagland R, Arrestin, TOLLIP, SOCS (SOCS1, SOCS2, SOCS3, SOCS4, SOCS5, SOCS6, SOCS7 and CIS), PIAS (PIAS1, PIAS3, PIASx and PIASy), SHP (SHP-1 and SHP-2), and the like, but are not limited thereto. Negative immune regulators are involved in inhibitory homologues of proteins involved in signal transduction (eg soluble decoy TLR, inhibitory TIR homologues, inhibitory signaling molecule isoforms, inhibitory cytokine homologues, etc.), molecular stability Protein, inhibitory component of signaling molecule complex, protein involved in regulation of phosphorylation of signaling molecule, transcription factor that suppresses transcription of NFκB target gene, protein involved in regulation of RNA translation and stability, cytokine It may belong to different protein families including but not limited to signaling regulators and the like. In one aspect, inhibiting one or more negative immune modulators in immune cells serves to enhance the immune capacity of the cell.
「単離核酸」というのは、天然に発生する状態でそれがフランキングする配列から分離された核酸セグメント又はフラグメントすなわち通常そのフラグメントに隣接している配列つまりそれが内部で天然に発生するゲノム中でそのフラグメントに隣接している配列から除去されたDNAフラグメントを意味する。用語は同様に、天然に核酸に随伴しているその他の構成要素すなわち細胞内で天然にそれに随伴しているRNA又はDNA又はタンパク質から実質的に精製された核酸にもあてはまる。従ってこの用語は、例えば、ベクター内、自己複製プラスミド又はウイルス内、又は原核生物又は真核生物のゲノムDNA内に取込まれる組換え型DNA、又はその他の配列とは独立して別の分子として(すなわちPCR又は制限酵素消化により産生されるcDNA又はゲノム又はcDNAフラグメントとして)存在する組換え型DNAを含む。これには又、付加的なポリペプチド配列をコードするハイブリッド遺伝子の一部である組換え型DNAも含まれる。 An "isolated nucleic acid" is a nucleic acid segment or fragment that is separated from the sequence that it flanks in a naturally occurring state, i.e. a sequence that is usually adjacent to that fragment, i.e., in the genome in which it occurs naturally. Means a DNA fragment removed from the sequence adjacent to the fragment. The term also applies to nucleic acids that have been substantially purified from other components that naturally accompany the nucleic acid, ie, RNA or DNA or proteins that naturally accompany it in the cell. Thus, the term is used as a separate molecule independent of, for example, recombinant DNA or other sequences incorporated into vectors, self-replicating plasmids or viruses, or prokaryotic or eukaryotic genomic DNA. Including recombinant DNA present (ie, as cDNA or genomic or cDNA fragments produced by PCR or restriction enzyme digestion). This also includes recombinant DNA that is part of a hybrid gene encoding additional polypeptide sequences.
本発明に関連して、一般に発生する核酸塩基について以下の略号が用いられる。すなわち「A」はアデノシンを意味し、「C」はシトシンを意味し、「G」はグアノシンを意味し、「T」はチミジンを意味しかつ「U」はウリジンを意味する。 In connection with the present invention, the following abbreviations are used for commonly occurring nucleobases. That is, “A” means adenosine, “C” means cytosine, “G” means guanosine, “T” means thymidine and “U” means uridine.
本書で使用されている「主要組織適合複合体」又は「MHC」という用語は、その多くが、組織適合性の最も重要な決定因子の中に入る抗原提示に関与する進化的に関連性ある細胞表面タンパク質をコードするものである、特定の遺伝子クラスタとして定義されている。クラスIMHC、又はMHC−Iは主としてCD8Tリンパ球に対する抗原提示において機能する。クラスIIMHC、又はMHC−IIは主としてCD4Tリンパ球に対する抗原提示において機能する。 As used herein, the term “major histocompatibility complex” or “MHC” refers to evolutionarily relevant cells, many of which are involved in antigen presentation that falls among the most important determinants of histocompatibility. It is defined as a specific gene cluster that encodes a surface protein. Class IMHC, or MHC-I, functions primarily in antigen presentation to CD8 T lymphocytes. Class II MHC, or MHC-II functions primarily in antigen presentation to CD4 T lymphocytes.
本書で使用されている「変調する」という用語は、生物学的状態の何らかの変化すなわち増加、減少などを意味するものとされている。例えば、「変調する」という用語は、所望の負の免疫調節因子mRNAの転写、所望の負の免疫調節因子mRNAの安定性、所望の負の免疫調節因子mRNAの翻訳、所望の負の免疫調節因子ポリペプチドの安定性、所望の負の免疫調節因子の翻訳後修飾又はそれらの任意の組合せを含む(ただしこれに限定されるわけではない)負の免疫調節因子の発現又は活性を正又は負に調節する能力を意味する。さらに、「変調する」という用語は、樹状細胞の免疫能力に関連している所望の負の免疫調節因子の活性を含む(ただしこれに限定されるわけではない)活性の増大、減少、マスキング、改変、オーバーライド又は復元を意味するものとして使用することができる。「変調する」という用語は、sTLR、RP105、SIGIRR、ST2、NOD2、MyD88s、IRAK(IRAKM、IRAK1、IRAK2)、IRF−4、FLN29、TWEAK、TRIAD3A、CYLD、Cbl、A20、SUMO(SUMO1、SUMO2、SUMO3及びSUMO4)、IkBタンパク質、MKPs、Foxj1、Foxo3a、TWIST(Twist1、Twist2)、Roquin、Dok(Dok−1、Dok−2)、PI3K、プロスタグランジン、TRAIL−R、アレスチン、TOLLIP、SOCS(SOCS1、SOCS2、SOCS3、SOCS4、SOCS5、SOCS6、SOCS7及びCIS)、PIAS(PIAS1、PIAS3、PIASx及びPIASy)、SHP(SHP−1及びSHP−2)などを含めた(ただしこれに限定されるわけではない)本書に開示されているあらゆる負の免疫調節因子にあてはまる。相反する規定のないかぎり、「アミノ酸配列をコードするヌクレオチド配列」には、互いの縮重バージョンであり同じアミノ酸配列をコードする全てのヌクレオチド配列が含まれる。タンパク質又はRNAをコードするヌクレオチド配列という語句は同様に、タンパク質が一部のバージョンでイントロンを含有し得るかぎりにおいて、イントロンをも含み得る。 As used herein, the term “modulate” is intended to mean any change in biological state, ie, increase, decrease, etc. For example, the term “modulate” refers to transcription of the desired negative immune regulator mRNA, stability of the desired negative immune regulator mRNA, translation of the desired negative immune regulator mRNA, desired negative immune regulator. Positive or negative expression or activity of negative immune modulators including, but not limited to, stability of the factor polypeptide, post-translational modification of the desired negative immune regulator or any combination thereof Means ability to adjust to. In addition, the term “modulate” includes, but is not limited to, the activity of a desired negative immune modulator associated with the immune capacity of a dendritic cell, increasing, decreasing, masking. Can be used to mean altering, overriding or restoring. The term “modulate” is sTLR, RP105, SIGIRR, ST2, NOD2, MyD88s, IRAK (IRAKM, IRAK1, IRAK2), IRF-4, FLN29, TWEAK, TRIAD3A, CYLD, Cbl, A20, SUMO (SUMO1, SUMO2 , SUMO3 and SUMO4), IkB protein, MKPs, Foxj1, Foxo3a, TWIST (Twist1, Twist2), Roquin, Dok (Dok-1, Dok-2), PI3K, prostaglandin, TRAIL-R, SOLIP, COLIP (SOCS1, SOCS2, SOCS3, SOCS4, SOCS5, SOCS6, SOCS7 and CIS), PIAS (PIAS1, PIAS3, PIASx and PIASy), S This applies to any negative immune regulator disclosed herein, including but not limited to HP (SHP-1 and SHP-2) and the like. Unless otherwise specified, a “nucleotide sequence encoding an amino acid sequence” includes all nucleotide sequences that are degenerate versions of each other and that encode the same amino acid sequence. The phrase nucleotide sequence encoding a protein or RNA can also include introns, as long as the protein can contain introns in some versions.
本書で使用されている「ポリヌクレオチド」という用語は、ヌクレオチドの鎖として定義される。さらに、核酸はヌクレオチドのポリマーである。かくして、本書で使用されている核酸及びポリヌクレオチドは互換性がある。当業者は、核酸がモノマー「ヌクレオチド」へと加水分解され得るポリヌクレオチドであるという一般的知識を有している。モノマーヌクレオチドはヌクレオシド内に加水分解され得る。本書で使用されているポリヌクレオチドには、組換え体手段、すなわち通常のクローニング技術及びPCRTMなどを用いた組換え体ライブラリ又はセルゲノムからの核酸配列のクローニング、及び合成手段を限定的意味なく含めた当該技術分野において利用可能なあらゆる手段によって得られる全ての核酸配列が含まれるが、これに限定されるわけではない。 The term “polynucleotide” as used herein is defined as a chain of nucleotides. In addition, nucleic acids are nucleotide polymers. Thus, the nucleic acids and polynucleotides used herein are interchangeable. One skilled in the art has the general knowledge that nucleic acids are polynucleotides that can be hydrolyzed into monomeric “nucleotides”. Monomeric nucleotides can be hydrolyzed into nucleosides. Polynucleotides used in this document include, without limitation, recombinant means, ie cloning of nucleic acid sequences from recombinant libraries or cell genomes using conventional cloning techniques and PCR ™ , and synthetic means, without limitation. All nucleic acid sequences obtained by any means available in the art are included, but are not limited to such.
本書で使用されている「ポリペプチド」という用語は、通常は確定した配列を有するアミノ酸残基の鎖として定義づけされる。本書で使用されているポリペプチドという用語は、「ペプチド」及び「タンパク質」という用語と相互に包含し合う。 The term “polypeptide” as used herein is usually defined as a chain of amino acid residues having a defined sequence. As used herein, the term polypeptide is mutually inclusive of the terms “peptide” and “protein”.
「増殖」というのはここでは、例えば細胞の増幅といった類似の形態の実体の複製又は繁殖を意味する。すなわち、増殖には、より多くの細胞の産生が包含され、これは、なかでも、単に細胞数を計数すること、細胞内への3H−チミジンの取り込みを測定すること
などによって測定され得る。
“Proliferation” as used herein means the replication or propagation of a similar form of entity, eg, amplification of cells. That is, proliferation includes the production of more cells, which can be measured, among other things, by simply counting the number of cells, measuring 3 H-thymidine incorporation into the cells, and the like.
本書で使用されている「プロモータ」という用語は、細胞の合成機構により認識されるか、又は合成機構により導入される、ポリヌクレオチド配列の特異的転写を開始するのに必要とされるDNA配列として定義される。 As used herein, the term “promoter” refers to the DNA sequence required to initiate specific transcription of a polynucleotide sequence that is recognized by or introduced by a synthetic machinery of a cell. Defined.
本書で使用されている「プロモータ/調節配列」というのは、プロモータ/調節因子配列に対して作動的に連結された遺伝子産物の発現に必要とされる核酸配列を意味する。場合によっては、この配列はコアプロモータ配列であり得、その他の場合には、この配列は、遺伝子産物の発現のために必要とされるエンハンサ配列及びその他の調節要素をも内含し得る。プロモータ/調節配列は、例えば、組織特異的に遺伝子産物を発現するものであり得る。 As used herein, “promoter / regulatory sequence” refers to a nucleic acid sequence required for expression of a gene product operably linked to a promoter / regulator sequence. In some cases, this sequence may be a core promoter sequence; in other cases, this sequence may also include enhancer sequences and other regulatory elements required for expression of the gene product. A promoter / regulatory sequence can be, for example, one that expresses a gene product in a tissue-specific manner.
「構成性」プロモータは、遺伝子産物をコードするか又は特定するポリヌクレオチドと作動的に連結された時点で、遺伝子産物を細胞の大部分の又は全ての生理学的条件下でその細胞内で産生させるヌクレオチド配列である。 A “constitutive” promoter, when operably linked to a polynucleotide that encodes or identifies a gene product, causes the gene product to be produced in the cell under most or all physiological conditions of the cell. Nucleotide sequence.
「誘発性」プロモータは、遺伝子産物をコードするか又は特定するポリヌクレオチドと作動的に連結された時点で、プロモータに対応するインデューサが細胞内部に存在する場合にのみ実質的に細胞内で遺伝子産物を産生させるヌクレオチド配列である。 An “inducible” promoter is a gene that is substantially intracellularly only when an inducer corresponding to the promoter is present inside the cell when operably linked to a polynucleotide that encodes or identifies the gene product. A nucleotide sequence that produces a product.
「組織特異的」プロモータは、遺伝子産物をコードするか又は特定するポリヌクレオチドと作動的に連結された時点で、その細胞がプロモータに対応するタイプの組織型の細胞である場合にのみ実質的に細胞内で遺伝子産物を産生させるヌクレオチド配列である。 A “tissue specific” promoter is substantially only when the cell is a cell of a tissue type of the type corresponding to the promoter when operably linked to a polynucleotide encoding or identifying the gene product. A nucleotide sequence that produces a gene product in a cell.
本書で使用されている「RNA」という用語はリボ核酸として定義される。 As used herein, the term “RNA” is defined as ribonucleic acid.
本書で使用されている「組換え型DNA」という用語は、異なる供給源からのDNAの小片を接合させることにより産生されるDNAとして定義される。 As used herein, the term “recombinant DNA” is defined as DNA that is produced by joining pieces of DNA from different sources.
本書で使用されている「組換え型ポリペプチド」は、組換え型DNA方法を用いることによって産生されたポリペプチドとして定義される。 A “recombinant polypeptide” as used herein is defined as a polypeptide produced by using recombinant DNA methods.
本書で使用されている「自己抗原」という用語は、宿主細胞又は組織によって発現される抗原として定義される。自己抗原は腫瘍抗原であるが、一部の実施形態においては、正常な細胞及び腫瘍細胞の両方において発現される。当業者であれば、自己抗原が細胞内で過剰発現され得るということを容易に理解するものと思われる。 The term “self antigen” as used herein is defined as an antigen expressed by a host cell or tissue. The autoantigen is a tumor antigen, but in some embodiments it is expressed on both normal and tumor cells. One skilled in the art will readily appreciate that self-antigens can be overexpressed in cells.
本書で使用されている「実質的に精製された」細胞というのは、基本的にその他の細胞型を含まない細胞である。実質的に精製された細胞は同様に、それがその天然に発生する状態において通常会合されるその他の細胞型から分離された細胞をも意味する。場合によっては、実質的に精製された細胞の集団というのは、相同な細胞集団を意味する。その他の場合には、この用語は単純に、それがその天然の状態で天然に会合されている細胞から分離された細胞のことを意味している。一部の実施形態では、細胞はインビトロ培養である。その他の実施形態では、細胞はインビトロで培養されない。 As used herein, a “substantially purified” cell is a cell that is essentially free of other cell types. Substantially purified cells also mean cells that are separated from other cell types with which they are normally associated in their naturally occurring state. In some cases, a substantially purified population of cells means a homogenous population of cells. In other cases, the term simply refers to a cell that has been separated from a cell with which it is naturally associated in its natural state. In some embodiments, the cell is an in vitro culture. In other embodiments, the cells are not cultured in vitro.
本書で使用されている「T細胞」という用語は、さまざまな細胞媒介型免疫反応に参加する胸腺由来の細胞として定義される。 The term “T cell” as used herein is defined as a thymus-derived cell that participates in various cell-mediated immune responses.
本書で使用されている「B細胞」という用語は、骨髄及び/又は脾臓に由来する細胞と
して定義される。B細胞は、抗体を産生するプラズマ細胞へと発達し得る。
The term “B cell” as used herein is defined as a cell derived from the bone marrow and / or spleen. B cells can develop into plasma cells that produce antibodies.
本書で使用されている「治療上有効な量」というのは、組成物が投与される哺乳動物に対し有益な効果をもたらすのに充分な治療用組成物の量である。 As used herein, a “therapeutically effective amount” is an amount of a therapeutic composition sufficient to provide a beneficial effect to the mammal to which the composition is administered.
本書で使用されている「トランスフェクションを受けた」又は「形質転換を受けた」又は「形質導入を受けた」という用語は、外因性核酸を宿主細胞内に転移又は導入させるプロセスを意味する。「トランスフェクションを受けた」又は「形質転換を受けた」又は「形質導入を受けた」細胞は、外因性核酸でのトランスフェクション、形質転換又は形質導入を受けた細胞である。細胞には、一次対象細胞及びその後代が含まれる。 As used herein, the term “transfected” or “transformed” or “transduced” refers to the process of transferring or introducing exogenous nucleic acid into a host cell. A “transfected” or “transformed” or “transduced” cell is a cell that has been transfected, transformed or transduced with an exogenous nucleic acid. Cells include primary target cells and their progeny.
本書で使用される「転写制御下の」又は「作動的に連結された」という語句は、プロモータが、RNAポリメラーゼによる転写及びポリヌクレオチドの発現の開始を制御するべくポリヌクレオチドとの関係において正しい場所及び配向にあることを意味する。 As used herein, the phrase “under transcriptional control” or “operably linked” means that the promoter is the right place in relation to the polynucleotide to control transcription by RNA polymerase and initiation of expression of the polynucleotide. And in an orientation.
本書で使用されている「ワクチン」という用語は、哺乳動物に対して材料を投与した後に免疫応答をひき起こすのに用いられる材料として定義される。 The term “vaccine” as used herein is defined as a material used to raise an immune response after the material has been administered to a mammal.
本書で使用されている「変異体」という用語は、それぞれ基準核酸配列又はペプチド配列と配列が異なっているものの基準分子の基本的特性を保持する核酸配列又はペプチド配列である。核酸変異体の配列の変化は、基準核酸によりコードされるペプチドのアミノ酸配列を改変しないかもしれず、又そうでなければ、アミノ酸の置換、付加、欠失、融合及び切断を結果としてもたらす可能性がある。ペプチド変異体の配列の変化は、標準的には制限されるか又は保存的であり、かくして、基準ペプチド及び変異体の配列は、全体的に類似しており、かつ数多くの領域で同一である。変異体及び基準ペプチドは、任意の組合せでの1つもしくはそれ以上の置換、付加、欠失によりアミノ酸配列において異なっている可能性がある。核酸又はペプチドの変異体は、対立遺伝子変異体などの天然に発生するものであり得、そうでなければ天然に発生するものとして知られていない変異体であり得る。核酸及びペプチドの天然に発生しない変異体は、突然変異誘発技術又は直接合成により作ることができる。 As used herein, the term “variant” is a nucleic acid or peptide sequence that retains the basic properties of a reference molecule, although the sequence differs from the reference nucleic acid sequence or peptide sequence, respectively. Changes in the sequence of the nucleic acid variant may not alter the amino acid sequence of the peptide encoded by the reference nucleic acid, or may otherwise result in amino acid substitutions, additions, deletions, fusions and truncations. is there. Peptide variant sequence changes are typically limited or conservative, thus the reference peptide and variant sequences are generally similar and identical in many regions. . A variant and reference peptide may differ in amino acid sequence by one or more substitutions, additions, deletions in any combination. Nucleic acid or peptide variants can be naturally occurring, such as allelic variants, and can otherwise be variants not known to occur naturally. Non-naturally occurring variants of nucleic acids and peptides can be made by mutagenesis techniques or direct synthesis.
「ベクター」は、単離核酸を含みかつ細胞の内部に単離核酸を送達するために使用可能である物質の組成物である。線形ポリヌクレオチド、イオン又は両親媒性化合物と会合されたポリヌクレオチド、プラスミド及びウイルスを含めた(ただしこれに限定されるわけではない)数多くのベクターが当該技術分野において知られている。かくして、「ベクター」という用語には、自己複製プラスミド又はウイルスが含まれている。用語は同様に、例えばポリリシン化合物、リポソームなどの細胞内への核酸の移入を容易にする非プラスミド及び非ウイルス化合物を含むものとみなされるべきである。ウイルスベクターの例としては、アデノウイルスベクター、アデノ関連ウイルスベクター、レトロウイルスベクターなどが含まれる。 A “vector” is a composition of matter that contains an isolated nucleic acid and can be used to deliver the isolated nucleic acid to the interior of a cell. Numerous vectors are known in the art, including but not limited to linear polynucleotides, polynucleotides associated with ions or amphiphiles, plasmids and viruses. Thus, the term “vector” includes self-replicating plasmids or viruses. The term should also be taken to include non-plasmid and non-viral compounds that facilitate the transfer of nucleic acids into cells such as, for example, polylysine compounds, liposomes. Examples of viral vectors include adenoviral vectors, adeno-associated viral vectors, retroviral vectors and the like.
本書で使用されている「ウイルス」という用語は、全細胞内で複製する能力をもつ、外部脂質エンベロープを伴う又は伴わないタンパク膜内に封入された核酸(RNA又はDNA)からなる粒子として定義される。 The term “virus” as used herein is defined as a particle consisting of nucleic acid (RNA or DNA) encapsulated in a protein membrane with or without an external lipid envelope that has the ability to replicate in whole cells. The
「異種の」というのは、異なる種の動物に由来する移植片を意味する。 By “heterologous” is meant a graft derived from a different species of animal.
記述
哺乳動物の体内の免疫応答に応答した無制御のシグナル伝達は、生物学的に破滅的な結果をもたらし得る。従って、異なるレベルでシグナリング経路が綿密に制御される。さま
ざまなタイプの免疫応答調節因子が存在する。免疫応答を調節する1つの方法は、負の免疫調節因子を通したものである。従って、本発明は、細胞の免疫能力を増強するべく免疫細胞内の負の免疫調節因子を阻害することに関する。
DESCRIPTION Uncontrolled signaling in response to an immune response in a mammalian body can have biologically devastating consequences. Therefore, the signaling path is closely controlled at different levels. There are various types of immune response modulators. One way to modulate the immune response is through negative immune regulators. Accordingly, the present invention relates to inhibiting negative immune regulators in immune cells to enhance the immune capacity of the cells.
負の免疫調節因子は、シグナル伝達に関与するタンパク質の阻害性相同体(例えば可溶性デコイTLR、阻害性TIR相同体、阻害性シグナリング分子イソ型、阻害性サイトカイン相同体など)、分子安定性に関与するタンパク質、シグナリング分子複合体の阻害性構成要素、シグナリング分子のリン酸化反応の調節に関与するタンパク質、NFκB標的遺伝子の転写を抑制する転写因子、RNA翻訳及び安定性の調節に関与するタンパク質、サイトカインシグナリング調節因子などを含む(ただしこれに限定されるわけではない)異なるタンパク質ファミリーに属し得る。負の免疫調節因子の例としてはsTLR、RP105、SIGIRR、ST2、NOD2、MyD88s、IRAK(IRAKM、IRAK1、IRAK2)、IRF−4、FLN29、TWEAK、TRIAD3A、CYLD、Cbl、A20、SUMO(SUMO1、SUMO2、SUMO3、及び SUMO4)、IkBタンパク質、MKPs、Foxj1、Foxo3a、TWIST(Twist1、Twist2)、Roquin、Dok(Dok−1、Dok−2)、PI3K、プロスタグランジン、TRAIL−R、アレスチン、TOLLIP、SOCS(SOCS1、SOCS2、SOCS3、SOCS4、SOCS5、SOCS6、SOCS7及びCIS)、PIAS(PIAS1、PIAS3、PIASx及びPIASy)、SHP(SHP−1及びSHP−2)などが含まれるが、これに限定されるわけではない。 Negative immune regulators are involved in inhibitory homologues of proteins involved in signal transduction (eg soluble decoy TLR, inhibitory TIR homologues, inhibitory signaling molecule isoforms, inhibitory cytokine homologues, etc.), molecular stability Protein, inhibitory component of signaling molecule complex, protein involved in regulation of phosphorylation of signaling molecule, transcription factor suppressing transcription of NFκB target gene, protein involved in regulation of RNA translation and stability, cytokine It can belong to different protein families including but not limited to signaling regulators and the like. Examples of negative immune regulators include sTLR, RP105, SIGIRR, ST2, NOD2, MyD88s, IRAK (IRAKM, IRAK1, IRAK2), IRF-4, FLN29, TWEAK, TRIAD3A, CYLD, Cbl, A20, SUMO (SUMO1, SUMO2, SUMO3, and SUMO4), IkB protein, MKPs, Foxj1, Foxo3a, TWIST (Twist1, Twist2), Roquin, Dok (Dok-1, Dok-2), PI3K, prostaglandin, TRAIL-TOL, Lestin L , SOCS (SOCS1, SOCS2, SOCS3, SOCS4, SOCS5, SOCS6, SOCS7 and CIS), PIAS (PIAS1, PIAS3, PIASx and PIASy) Although the like SHP (SHP-1 and SHP-2), but is not limited thereto.
一態様においては、負の免疫調節因子は、サイトカインシグナル伝達のサプレッサ(SOCS)、SH2含有ホスファターゼ(SHP)及び活性化されたSTATのタンパク質インヒビター(PIAS)を含むサイトカインシグナリングの調節因子であり、これらを(ただしこれに限定されるわけではない)含む。 In one aspect, the negative immune regulator is a regulator of cytokine signaling comprising a suppressor of cytokine signaling (SOCS), an SH2-containing phosphatase (SHP) and an activated STAT protein inhibitor (PIAS), (But not limited to).
サイトカインシグナリングの誘発性インヒビターは、サイトカインシグナリング(SOCS)タンパク質のサプレッサであり、その中にはSOCS1〜SOCS7及びサイトカイン誘発性SH2ドメイン含有タンパク質(CIS)という8つのファミリーメンバーが存在する。SOCSタンパク質は、サイトカインレセプタ又は付随するJAKを認識し、シグナリングでの直接的干渉及びユビキチン媒介型プロテアソーム分解についてレセプタ複合体をターゲティングすることの両方によって、シグナル伝達を減衰させる。 Inducible inhibitors of cytokine signaling are suppressors of cytokine signaling (SOCS) proteins, among which there are eight family members: SOCS1-SOCS7 and cytokine-induced SH2 domain-containing protein (CIS). SOCS proteins recognize cytokine receptors or associated JAKs and attenuate signal transduction both by direct interference in signaling and targeting the receptor complex for ubiquitin-mediated proteasome degradation.
(SHP−1及びSHP−2)を含む(ただしこれに限定されるわけではない)SHPタンパク質は、構成的に発現され、ヤーヌスキナーゼ(JAK)及びそのレセプタなどの脱リン酸化シグナリング中間体によりサイトカインシグナル伝達を減衰させることができる。PIAS1、PIAS3、PIASx及びPIASyを含むもののこれに限定されるわけではない活性化されたSTATのタンパク質インヒビター(PIAS)ファミリーのメンバーも又、構成的に発現され、STAT活性を抑止することによりシグナル伝達を減衰させる。スモ化プロセスは、STAT活性のPIAS媒介型抑止に関与してきた。 SHP proteins including (but not limited to) (SHP-1 and SHP-2) are constitutively expressed and cytokines by dephosphorylated signaling intermediates such as Janus kinase (JAK) and its receptors Signal transduction can be attenuated. Members of the activated STAT protein inhibitor (PIAS) family, including but not limited to PIAS1, PIAS3, PIASx and PIASy, are also constitutively expressed and signaling by inhibiting STAT activity. Is attenuated. The sumoylation process has been implicated in PIAS-mediated suppression of STAT activity.
本発明は、負の免疫調節因子の阻害が哺乳動物に対して治療上の利益を提供するという発見に関係する。すなわち、免疫細胞内の負の免疫調節因子の阻害は、細胞内の免疫能力と結びつけられたシグナリング経路の増強に役立つ。一態様においては、シグナル伝達に関与するタンパク質の阻害性相同体(例えば可溶性デコイTLR、阻害性TIR相同体、阻害性シグナリング分子イソ型、阻害性サイトカイン相同体など)、分子安定性に関与するタンパク質、シグナリング分子複合体の阻害性構成要素、シグナリング分子のリン酸化反応の調節に関与するタンパク質、NFκB標的遺伝子の転写を抑制する転写因子、RNA翻訳及び安定性の調節に関与するタンパク質、サイトカインシグナリング調節因子又はそれらの任意の組合せの阻害が、哺乳動物に対して治療上の利益を提供する。 The present invention relates to the discovery that inhibition of negative immune modulators provides a therapeutic benefit to mammals. That is, inhibition of negative immune regulators in immune cells serves to enhance signaling pathways associated with intracellular immune capacity. In one aspect, inhibitory homologues of proteins involved in signal transduction (eg, soluble decoy TLR, inhibitory TIR homologues, inhibitory signaling molecule isoforms, inhibitory cytokine homologues, etc.), proteins involved in molecular stability , Inhibitory components of signaling molecule complexes, proteins involved in regulation of phosphorylation of signaling molecules, transcription factors that suppress transcription of NFκB target genes, proteins involved in regulation of RNA translation and stability, cytokine signaling regulation Inhibition of the factors or any combination thereof provides a therapeutic benefit to the mammal.
かくして、本発明は、免疫細胞内の負の免疫調節因子を阻害して免疫細胞の免疫能力を増強するための組成物及び方法を含んでなる。好ましくはA20、SUMO(SUMO1、SUMO2、SUMO3及びSUMO4)、Foxj1、Foxo3a、TWIST(Twist1、Twist2)、SOCS(SOCS1、SOCS2、SOCS3、SOCS4、SOCS5、SOCS6、SOCS7及びCIS)、PIAS(PIAS1、PIAS3、PIASx及びPIASy)、SHP(SHP−1及びSHP−2)などよりなる群から選択された1つもしくはそれ以上の負の免疫調節因子を阻害することで、治療上の利益が得られる。 Thus, the present invention comprises compositions and methods for inhibiting negative immune regulators in immune cells to enhance immune capacity of immune cells. Preferably, A20, SUMO (SUMO1, SUMO2, SUMO3 and SUMO4), Foxj1, Foxo3a, TWIST (Twist1, Twist2), SOCS (SOCS1, SOCS2, SOCS3, SOCS4, SOCS6, SOCS6, SOCS3 and PISI, SIS7, CIS, PISC3, PISI, SPI, PI , PIASx and PIASy), SHP (SHP-1 and SHP-2), etc., to inhibit one or more negative immune modulators selected from the group consisting of therapeutic benefits.
さらなる態様においては、サイトカインシグナル調節因子からの任意の1つもしくはそれ以上のメンバーの阻害が、治療上の利益を提供する。かくして本発明は、免疫細胞内のサイトカインシグナリング調節因子を変調させて免疫細胞内の免疫能力を増強するための組成物及び方法を含んでなる。 In a further aspect, inhibition of any one or more members from a cytokine signal modulator provides a therapeutic benefit. Thus, the present invention comprises compositions and methods for modulating cytokine signaling regulators within immune cells to enhance immune capacity within immune cells.
阻害すべき所望の負の免疫調節因子の如何に関わらず、細胞の免疫能力を増強する本発明の組成物は、負の免疫調節因子のインヒビター(例えばインヒビターは所望の負の免疫調節因子に対応するsiRNAである)、抗原、サイレンシング化された免疫細胞、パルスを受けた細胞、抗原でのパルスを受けたサイレンシング化された細胞、サイトカインなどのうちの少なくとも1つもしくはそれ以上のものの任意の組合せを含む。組成物は、インビボ免疫化及び/又はエクスビボ療法のためのワクチンであり得る。 Regardless of the desired negative immune modulator to be inhibited, the compositions of the present invention that enhance the immunity of a cell can be an inhibitor of a negative immune modulator (eg, the inhibitor corresponds to the desired negative immune modulator). Any of at least one or more of: an antigen, a silenced immune cell, a pulsed cell, a silenced cell pulsed with an antigen, a cytokine, etc. Including a combination of The composition can be a vaccine for in vivo immunization and / or ex vivo therapy.
本発明は、APC内の臨界制御点を無効化することによりワクチンの効能を増強するための包括的手段としてのサイレンシング化されたAPCを提供する。本発明のサイレンシング化されたAPC又は負の免疫調節因子のインヒビターでのワクチン接種は、負の免疫調節因子のサイレンシング化により抗原提示免疫原性APCがインビボで持続的に抗原特異的T細胞を刺激できるようになることから、抗原特異的免疫を増強する。本発明の一実施形態においては、サイレンシング化されたAPCは、調節T細胞抑制を阻害する前炎症性サイトカインの産生及びAPCの成熟を増強することによって、調節T細胞をオフ切換えする能力をもつ。 The present invention provides silenced APC as a comprehensive means to enhance vaccine efficacy by overriding critical control points within APC. Vaccination with the silenced APCs or inhibitors of negative immune regulators of the present invention can result in antigen-presenting immunogenic APCs persistently antigen-specific T cells in vivo by silencing negative immune regulators Can enhance antigen-specific immunity. In one embodiment of the invention, the silenced APC has the ability to switch off regulatory T cells by enhancing the production of proinflammatory cytokines that inhibit regulatory T cell suppression and APC maturation. .
サイレンシング化されたAPCを生成することに加えて、本発明は、同様に、サイレンシング化された細胞傷害性Tリンパ球(CTL)をも含む。本開示は、本発明の方法を用いてサイレンシング化されたCTLが増強した細胞溶解活性を示すことを実証している。増強した細胞溶解活性は、細胞療法及び/又はワクチン接種において治療上の利益を提供する。 In addition to generating silenced APCs, the present invention also includes silenced cytotoxic T lymphocytes (CTLs). The present disclosure demonstrates that CTLs silenced using the methods of the invention exhibit enhanced cytolytic activity. Enhanced cytolytic activity provides a therapeutic benefit in cell therapy and / or vaccination.
負の免疫調節因子の阻害:
本書で提示されている開示に基づき、本発明は、負の免疫調節因子を阻害しかくして負の免疫調節因子が免疫応答の調節と関係しているという包括的概念を包含している。負の免疫調節因子は異なるタンパク質ファミリーに属し得る。負の免疫調節因子の各ファミリーは、免疫応答を調節するための異なる戦略を有する。異なるファミリーには、シグナル伝達に関与するタンパク質の阻害性相同体(例えば可溶性デコイTLR、阻害性TIR相同体、阻害性シグナリング分子イソ型、阻害性サイトカイン相同体など)、分子安定性に関与するタンパク質、シグナリング分子複合体の阻害性構成要素、シグナリング分子のリン酸化反応の調節に関与するタンパク質、NFκB標的遺伝子の転写を抑制する転写因子、RNA翻訳及び安定性の調節に関与するタンパク質、サイトカインシグナリング調節因子などが含まれるが、これに限定されるわけではない。本書で提示されている開示に基づくと、免疫細胞内の負の免疫調節因子を阻害することでこの細胞中の免疫能力は増強される。
Inhibition of negative immune regulators:
Based on the disclosure presented herein, the present invention encompasses the generic concept that negative immune regulators are thus inhibited and thus negative immune regulators are associated with modulation of the immune response. Negative immune modulators can belong to different protein families. Each family of negative immune regulators has a different strategy for modulating the immune response. Different families include inhibitory homologues of proteins involved in signal transduction (eg soluble decoy TLR, inhibitory TIR homologues, inhibitory signaling molecule isoforms, inhibitory cytokine homologues, etc.), proteins involved in molecular stability , Inhibitory components of signaling molecule complexes, proteins involved in regulation of phosphorylation of signaling molecules, transcription factors that suppress transcription of NFκB target genes, proteins involved in regulation of RNA translation and stability, cytokine signaling regulation Including, but not limited to, factors. Based on the disclosure presented herein, the immune capacity in the cell is enhanced by inhibiting negative immune regulators in the immune cell.
一部の態様においては、免疫細胞内の負の免疫調節因子を阻害することで、細胞内のトール様レセプタ(TLR)及び腫瘍壊死因子レセプタ(TNFR)は増強される。細胞内のTLR及びTNFRを増強した結果として、その細胞の免疫能力は増大する。 In some embodiments, inhibition of negative immune regulators in immune cells enhances intracellular Toll-like receptor (TLR) and tumor necrosis factor receptor (TNFR). As a result of enhancing intracellular TLR and TNFR, the immune capacity of the cell increases.
シグナル伝達に関与するタンパク質の阻害性相同体
有効な免疫応答は、病原性感染に応答したDCの活性化に起因する。この活性化には、TLRなどの病原体認識レセプタを通したシグナリングが含まれる。TLRのメンバーは、共通のシグナリング経路特にNF−κB及びストレス活性化を受けたタンパク質キナーゼの活性化を導く経路を活性化する保存されたサイトゾルトール/インターロイキン−1レセプタ(TIR)ドメインの存在に起因して、同様にシグナリングする。NF−κB活性化は、TNF、IFN、インターロイキン1(IL−1)、IL−6、及びIL−12などの前炎症性サイトカインを分泌することによって、及びCD80、CD86及びCD40などの共同刺激分子を発現することにより、免疫応答の触媒として作用する。
Inhibitory homologues of proteins involved in signal transduction Effective immune responses result from activation of DCs in response to pathogenic infections. This activation includes signaling through a pathogen recognition receptor such as TLR. Members of TLRs are present in a conserved cytosolitol / interleukin-1 receptor (TIR) domain that activates common signaling pathways, particularly those that lead to activation of NF-κB and stress-activated protein kinases. Signaling in the same way. NF-κB activation is secreted by secreting proinflammatory cytokines such as TNF, IFN, interleukin 1 (IL-1), IL-6, and IL-12, and co-stimulation such as CD80, CD86 and CD40 By expressing the molecule, it acts as a catalyst for the immune response.
細胞内のTLRシグナリングに基づき免疫応答を調節するための頻繁に使用されている戦略は、TLRの細胞外部分で刺激活性をもたないその非機能的相同体又はイソ型によって主要なシグナリング分子を競合的に阻害することである。負の免疫調節因子のこのファミリー内のタンパク質としては、可溶性デコイTLR(sTLR)が含まれるが、これに限定されるわけではない。sTLRは、病原性産物(例えば病原菌産物)とTLRの相互作用を抑制する。例えば、可溶性デコイTLR2は、対応する病原菌リガンドとの相互作用についてTLR2と競合する。別の場合には、可溶性デコイTLR4はMD2と相互作用し、MD2−TLR4複合体の形成を阻害し、かくしてTLR4によるLPS媒介型シグナリングを遮断する。 A frequently used strategy for modulating the immune response based on intracellular TLR signaling is to identify the major signaling molecule by its non-functional homologue or isoform that has no stimulatory activity in the extracellular portion of the TLR. Competitive inhibition. Proteins within this family of negative immune regulators include, but are not limited to, soluble decoy TLR (sTLR). sTLR suppresses the interaction between a pathogenic product (eg, a pathogen product) and TLR. For example, soluble decoy TLR2 competes with TLR2 for interaction with the corresponding pathogen ligand. In another case, soluble decoy TLR4 interacts with MD2 and inhibits the formation of the MD2-TLR4 complex, thus blocking LPS-mediated signaling by TLR4.
本開示に基づくと、当業者であれば本発明が負の免疫調節因子を阻害するための組成物及び方法であって、負の免疫調節因子がシグナリング分子の非機能的相同体である組成物及び方法を含むことを認識するものと思われる。好ましくは、シグナリング分子の非機能的相同体は可溶性デコイTLRである。シグナリング分子の非機能的相同体を阻害すること(又はsTLRをその他の形で阻害すること)は、シグナリング分子上の非機能的相同体の阻害効果を阻害するのに役立つ。 Based on the present disclosure, those skilled in the art will understand that the present invention is a composition and method for inhibiting negative immune regulators, wherein the negative immune regulator is a non-functional homologue of a signaling molecule. And would include a method. Preferably, the non-functional homologue of the signaling molecule is a soluble decoy TLR. Inhibiting a non-functional homologue of a signaling molecule (or otherwise inhibiting sTLR) serves to inhibit the inhibitory effect of the non-functional homologue on the signaling molecule.
可溶性デコイTLRを阻害することに加えて、TLRシグナリングを増強するもう1つの戦略は、TLRの細胞質領域で非機能的TIR相同体又はイソ型を含有するタンパク質を阻害することにある。これらのタンパク質は、いかなる刺激活性ももたないTIRドメインを含有する膜結合非機能的TIR相同体(別途非機能的レセプタとしても知られているもの)を共有するタンパク質のファミリーのメンバーである。かかるタンパク質には、STGIRR(単一免疫グロブリンIL−IR−関連分子)、ST2及びRP105が含まれるが、これらに限定されるわけではない。 In addition to inhibiting soluble decoy TLRs, another strategy to enhance TLR signaling is to inhibit proteins containing non-functional TIR homologues or isoforms in the cytoplasmic region of TLRs. These proteins are members of a family of proteins that share membrane-bound non-functional TIR homologs (otherwise known as non-functional receptors) that contain TIR domains without any stimulating activity. Such proteins include, but are not limited to, STGIRR (single immunoglobulin IL-IR-related molecule), ST2 and RP105.
いずれかの特定の理論により束縛されることは望まないが、これらのタンパク質は、TLRがその対応するシグナリング分子に結合するのを阻害するか又は封鎖することにより、TLRシグナリングを抑制する。例えば、SIGIRRは、シグナリング分子と相互作用するべくTLR4と競合することによりTLR4シグナリングを減衰させる。かくして、本発明は、負の免疫調節因子を阻害する組成物及び方法であって、負の免疫調節因子が免疫細胞内のTLRシグナリングを増強するための膜結合非機能的TIR相同体である組成物及び方法を含む。この戦略は、免疫細胞の免疫能力を増強するべく負の免疫調節因子を阻害するという一般的概念に対する裏づけを提供する。 While not wishing to be bound by any particular theory, these proteins suppress TLR signaling by inhibiting or blocking the TLR from binding to its corresponding signaling molecule. For example, SIGIRR attenuates TLR4 signaling by competing with TLR4 to interact with signaling molecules. Thus, the present invention provides compositions and methods for inhibiting negative immune regulators, wherein the negative immune regulator is a membrane bound non-functional TIR homologue for enhancing TLR signaling in immune cells. Products and methods. This strategy provides support for the general concept of inhibiting negative immune regulators to enhance the immune capacity of immune cells.
TLRシグナリングは、同様に、NOD2によっても調節され得る。NOD2は、細菌
産物ムラミルジペプチド(MDP)を認識できるヌクレオチド結合オリゴマー化ドメインファミリの一員である。NOD2はTLR2シグナリングの負の調節因子である。本書に提示される本開示に基づくと、免疫細胞内でNOD2を阻害することにより、細胞内のTLRシグナリングに対するNOD2の阻害効果は抑制される。従って、免疫者の体内でNOD2を阻害すると、その細胞の免疫能力は増強される。
TLR signaling can be regulated by NOD2 as well. NOD2 is a member of a family of nucleotide-binding oligomerization domains that can recognize the bacterial product muramyl dipeptide (MDP). NOD2 is a negative regulator of TLR2 signaling. Based on the present disclosure presented herein, inhibiting NOD2 in immune cells suppresses the inhibitory effect of NOD2 on intracellular TLR signaling. Therefore, when NOD2 is inhibited in the body of an immunizer, the immunity of the cell is enhanced.
TLRシグナリングを調節するもう1つの戦略は、MyD88及びIRAKなどの細胞質シグナリングタンパク質を調節することにある。細胞質シグナリング分子を阻害する負の免疫調節因子は、TLRシグナリングにおける主要なシグナリング分子の非機能性イソ型(又は別途シグナリング分子イソ型としても知られている)であるものとみなされる。非機能的イソ型であるとみなされる負の免疫調節因子の一例としては、中間ドメインが欠如したMyD88の交互にスプライシングされた短い変異体がある。MyD88と表示されるMyD88のこのスプライス変異体は、TLRシグナリングを抑制することがわかっている。例えば、MyD88は、LPS誘発されたNF−κB活性を阻害する。MyD88は、IRAK4に結合しIRAK1リン酸化を促進する能力をもたないことから、LPS誘発型NF−κB活性を阻害するものと考えられている。本書に提示されている開示に基づくと、1つの細胞内の非機能的イソ型(例えばMyD88)である負の免疫調節因子を阻害することで、細胞内のTLRシグナリングを増強しかくしてその細胞の免疫能力を増強するための手段が提供される。 Another strategy to regulate TLR signaling is to regulate cytoplasmic signaling proteins such as MyD88 and IRAK. Negative immunomodulators that inhibit cytoplasmic signaling molecules are considered to be non-functional isoforms of the major signaling molecules in TLR signaling (or alternatively known as signaling molecule isoforms). An example of a negative immune regulator that is considered to be a non-functional isoform is a short alternatively spliced variant of MyD88 that lacks an intermediate domain. This splice variant of MyD88, designated MyD88, has been found to suppress TLR signaling. For example, MyD88 inhibits LPS-induced NF-κB activity. MyD88 is thought to inhibit LPS-induced NF-κB activity because it does not have the ability to bind to IRAK4 and promote IRAK1 phosphorylation. Based on the disclosure presented herein, inhibiting a negative immunomodulator that is a non-functional isoform in one cell (eg, MyD88) enhances intracellular TLR signaling, thus Means are provided for enhancing immune capacity.
シグナル伝達に関与するタンパク質の阻害性相同体のファミリーに属するタンパク質のもう1つの例はIRAKである。キナーゼのIRAKファミリーは4つのメンバーすなわちIRAK1、IRAK2、IRAK4及びIRAKMを含んでなる。さらに、IRAKMにはキナーゼ活性が欠如しており、それは細胞内のTLRシグナリングの包括的な負の免疫調節因子として機能する。従って、IRAKMなどの非機能的イソ型である負の免疫調節因子を阻害することにより、免疫細胞内の負の免疫調節因子を阻害して細胞の免疫能力を増強するという一般的概念に対する裏づけが提供される。 Another example of a protein belonging to a family of inhibitory homologues of proteins involved in signal transduction is IRAK. The IRAK family of kinases comprises four members: IRAK1, IRAK2, IRAK4 and IRAKM. Furthermore, IRAKM lacks kinase activity, which functions as a global negative immune regulator of intracellular TLR signaling. Thus, supporting the general concept of inhibiting negative immune regulators that are non-functional isoforms, such as IRAKM, by inhibiting negative immune regulators in immune cells to enhance the immune capacity of the cells. Provided.
TLRシグナリングのその他の負の免疫調節因子には、転写因子のIFN調節因子(IRF)ファミリーのメンバー、FLN29(新規インターフェロン及びLPS誘発型遺伝子)及びTWEAKが含まれるがこれに限定されるわけではない。従って、これらの負の免疫調節因子のうちの1つもしくはそれ以上のものを阻害することにより、免疫応答に対する各々の負の免疫調節因子の阻害効果を妨げることができる。従って、これらの負の免疫調節因子のうちのいずれか1つもしくはそれ以上のものを阻害することにより、細胞内でTLRシグナリングを増強しかくして細胞の免疫能力を増強するための手段が提供される。 Other negative immune regulators of TLR signaling include, but are not limited to, members of the IFN regulator (IRF) family of transcription factors, FLN29 (a novel interferon and LPS-inducible gene) and TWEAK. . Thus, inhibiting one or more of these negative immune regulators can prevent the inhibitory effect of each negative immune regulator on the immune response. Thus, inhibiting any one or more of these negative immunomodulators provides a means to enhance TLR signaling and thus enhance the cell's immune capacity within the cell. .
分子安定性:
負の免疫調節因子の1つのファミリーは、シグナリング分子の安定性を調節する戦略を利用する。この戦略は、ユビキチン化/脱ユビキチン化により主要シグナリング分子の安定性を調節すること、そしてスモ化及びその他の機序によりシグナリング分子複合体の阻害構成要素の安定性を増大することと結びつけられている。TRIAD3A、Cyld、Cbl、A20及びSUMOなどのこれらの負の免疫調節因子の多くは、標的TLR及びシグナリング分子を修飾しその分解を促進してTLRシグナル伝達を減衰させる2ビキチン修飾酵素である。場合によっては、これらの負の免疫調節因子は、同様にTNFRシグナル伝達をも減衰できる。従って、本書に提示されている開示に基づくと、当業者であれば、免疫細胞内のこれらの負の免疫調節因子のうちの1つもしくはそれ以上のものを阻害することにより細胞内のTLR及び/又はTNFRシグナリングが増強されるということを認識するであろう。細胞内のTLRの増強及び/又はTNFRシグナリングの結果は、細胞の免疫能力の増大である。
Molecular stability:
One family of negative immune regulators utilizes strategies that modulate the stability of signaling molecules. This strategy is coupled with regulating the stability of the major signaling molecules by ubiquitination / deubiquitination and increasing the stability of the inhibitory component of the signaling molecule complex by sumoylation and other mechanisms. Yes. Many of these negative immune regulators such as TRIAD3A, Cyld, Cbl, A20 and SUMO are 2-biquitin modifying enzymes that modify target TLRs and signaling molecules and promote their degradation to attenuate TLR signaling. In some cases, these negative immune regulators can attenuate TNFR signaling as well. Thus, based on the disclosure presented herein, one of ordinary skill in the art will recognize the intracellular TLR and the intracellular TLR by inhibiting one or more of these negative immune modulators in the immune cell. It will be appreciated that / or TNFR signaling is enhanced. The result of enhancement of intracellular TLR and / or TNFR signaling is an increase in the immune capacity of the cell.
分子安定性を調節することに関与するその他の負の免疫調節因子としては、アレスチンファミリーが含まれる。本開示に基づくと、当業者であれば、免疫細胞内のアレスチンファミリーメンバーを阻害することにより、NF−κBの活性化に対するアレスチンファミリーメンバーの阻害効果が抑制されるということを認識するであろう。従ってアレスチンファミリーメンバーの阻害は細胞の免疫能力を増大させる。 Other negative immune regulators involved in regulating molecular stability include the arrestin family. Based on this disclosure, one skilled in the art will recognize that inhibiting the arrestin family member in immune cells suppresses the inhibitory effect of the arrestin family member on NF-κB activation. . Thus, inhibition of arrestin family members increases the immune capacity of the cell.
分子複合体をシグナリングする阻害性構成要素
シグナリング分子複合体の阻害性構成要素のファミリーのメンバーである負の免疫調節因子には、IκBタンパク質が含まれる。IκBα、IκBβ及びIκBεを含むIκBタンパク質は、細胞質内のNF−κBタンパク質を封鎖しかくして、NF−κBをその正常な機能から阻害する。IκBタンパク質は、NF−κBサブユニット上の核局在化配列(NLS)をマスキングすることによって細胞質内のNF−κBを保持する。
Inhibitory components that signal molecular complexes Negative immunomodulators that are members of the family of inhibitory components of signaling molecule complexes include the IκB protein. IκB proteins, including IκBα, IκBβ and IκBε, sequester NF-κB proteins in the cytoplasm and inhibit NF-κB from its normal function. The IκB protein retains NF-κB in the cytoplasm by masking the nuclear localization sequence (NLS) on the NF-κB subunit.
NF−κBの活性化における重要な事象は、IκBキナーゼ(IKK)複合体によるIκBのリン酸化である。IKK複合体は、TLR、リガンド及びTNFを含めたさまざまな刺激によるNF−κBの活性化のための合流点である。IKK複合体は、2つの触媒サブユニットIKKα及びIKKβを含有し、NF−κB転写因子の活性化を制御する。IKKβは、前炎症性サイトカイン及び病原菌産物に応答したNF−κB活性化を媒介する。NF−κBの活性化を制御する上でのIKKαについての負の調節という役目が観察されてきた。IKKαは、NF−κBサブユニットRelA及びcRelの分解及び前炎症性遺伝子プロモータからのRelA及びc−Relの除去の両方を加速することによりNF−κB活性の抑制に寄与する。 An important event in the activation of NF-κB is the phosphorylation of IκB by the IκB kinase (IKK) complex. The IKK complex is a confluence for the activation of NF-κB by various stimuli including TLR, ligand and TNF. The IKK complex contains two catalytic subunits, IKKα and IKKβ, and controls the activation of the NF-κB transcription factor. IKKβ mediates NF-κB activation in response to proinflammatory cytokines and pathogenic products. The role of negative regulation for IKKα in controlling the activation of NF-κB has been observed. IKKα contributes to the suppression of NF-κB activity by accelerating both the degradation of the NF-κB subunits RelA and cRel and the removal of RelA and c-Rel from the pro-inflammatory gene promoter.
本書に提示されている開示に基づくと、NF−κBを負に調節するか又はその他の形でNF−κBの活性化を防止するタンパク質は、細胞の免疫能力を増強するべく本書に開示されている方法を用いるターゲティングされた阻害のための候補である。当業者であれば、IκBタンパク質IKKα、IKKβ又は任意の組合せを阻害することにより、NF−κB活性化に対するこれらのタンパク質の阻害効果を抑制することができるということを認識するであろう。 Based on the disclosure presented herein, proteins that negatively regulate NF-κB or otherwise prevent activation of NF-κB are disclosed herein to enhance cellular immunity. Are candidates for targeted inhibition using the method. One skilled in the art will recognize that inhibiting the IκB proteins IKKα, IKKβ, or any combination can suppress the inhibitory effect of these proteins on NF-κB activation.
シグナリング分子のリン酸化の調節
MAPKシグナリング経路は、免疫応答の活性化と結びつけられる。MAPK経路は、ホスファターゼがMAPK経路の活性化の後に誘発されることから、MAPKホスファターゼ(MKP)によるフィードバック阻害を受ける。本書で提示されている開示に基づくと、MKP(例えばMKP5及びMKP6)を阻害することは、その対応するMAPK経路に対するMKPの阻害効果を抑制するのに役立つ。MKP5及びMKP6は、そのそれぞれのMAPK標的の活性を阻害することによりT細胞活性化を負に調節することが示されてきた。
Regulation of signaling molecule phosphorylation The MAPK signaling pathway is linked to the activation of immune responses. The MAPK pathway is subject to feedback inhibition by MAPK phosphatase (MKP) because phosphatase is induced after activation of the MAPK pathway. Based on the disclosure presented herein, inhibiting MKP (eg, MKP5 and MKP6) helps to suppress the inhibitory effect of MKP on its corresponding MAPK pathway. MKP5 and MKP6 have been shown to negatively regulate T cell activation by inhibiting the activity of their respective MAPK targets.
本書で提示されている開示に基づくと、当業者であれば、MKP(例えばMKP5及びMKP6)を阻害することによりMAPKシグナル経路に対するホスファターゼの阻害効果を抑制できるということを認識することだろう。免疫細胞内のMKPを阻害することにより、細胞の免疫能力を増強することが可能である。 Based on the disclosure presented herein, one of ordinary skill in the art will recognize that inhibiting MKP (eg, MKP5 and MKP6) can suppress the inhibitory effect of phosphatases on the MAPK signaling pathway. By inhibiting MKP in immune cells, the immune capacity of the cells can be enhanced.
標的遺伝子転写の調節
免疫応答を調節するためのもう1つの戦略は、NFκB標的遺伝子の転写を抑制するか又はシグナリング分子複合体中の負の構成要素の転写を増強することにある。かかる戦略には、Twist−1、Twist−2、Foxj1、及びFoxo3aを含む(これに限定されるわけではない)負の免疫調節因子を抑制することが包含されている。本書で提
示されている開示は、これらのタンパク質のうちの少なくとも1つのタンパク質のレベルを抑制することで細胞の免疫能力を増強することができるということを実証している。
Regulation of target gene transcription Another strategy for regulating immune responses is to repress transcription of NFκB target genes or enhance transcription of negative components in signaling molecule complexes. Such strategies include the suppression of negative immune modulators including but not limited to Twist-1, Twist-2, Foxj1, and Foxo3a. The disclosure presented herein demonstrates that suppression of the level of at least one of these proteins can enhance the immune capacity of the cell.
Foxo3a及びFoxj1は、NF−κBの負の調節に活発に関与するフォークヘッド転写因子ファミリーのメンバーである。Twistは、NF−κBのもう1つの負の免疫調節因子である。Twistは、サイトカインプロモータの中のEボックスに結合し、隣接するκB部位に結合されたNF−κBの活性を阻害する。 Foxo3a and Foxj1 are members of the forkhead transcription factor family that are actively involved in the negative regulation of NF-κB. Twist is another negative immune regulator of NF-κB. Twist binds to the E box in the cytokine promoter and inhibits the activity of NF-κB bound to the adjacent κB site.
負の免疫調節因子を阻害する方法
本書の開示に基づくと、本発明は、細胞の免疫能力を増強するべくその細胞内の負の免疫調節因子を阻害するための包括的概念を含む。好ましくは、本発明は、A20、SUMO(SUMO1、SUMO2、SUMO3及びSUMO4)、Foxj1、Foxo3a、TWIST(Twist1、Twist2)、SOCS(SOCS1、SOCS2、SOCS3、SOCS4、SOCS5、SOCS6、SOCS7及びCIS)、PIAS(PIAS1、PIAS3、PIASx及びPIASy)、SHP(SHP−1及びSHP−2)などよりなる群から選択された1つもしくはそれ以上の負の免疫調節因子を阻害する工程を含む。
Methods of Inhibiting Negative Immunomodulators Based on the disclosure herein, the present invention includes a generic concept for inhibiting negative immune regulators in a cell to enhance the immune capacity of the cell. Preferably, the present invention includes A20, SUMO (SUMO1, SUMO2, SUMO3, and SUMO4), Foxj1, Foxo3a, TWIST (Twist1, Twist2), SOCS (SOCS1, SOCS2, SOCS3, SOCS4, SOCS5, SOCS6, SOCS7, and SOCS7, SOCS7, and SOCS7). Inhibiting one or more negative immune regulators selected from the group consisting of PIAS (PIAS1, PIAS3, PIASx and PIASy), SHP (SHP-1 and SHP-2) and the like.
一実施形態においては、本発明は免疫細胞の免疫能力を増強するための組成物を含んでなる。好ましくは、組成物は、A20、SUMO(SUMO1、SUMO2、SUMO3及びSUMO4)、Foxj1、Foxo3a、TWIST(Twist1、Twist2)、SOCS(SOCS1、SOCS2、SOCS3、SOCS4、SOCS5、SOCS6、SOCS7及びCIS)、PIAS(PIAS1、PIAS3、PIASx及びPIASy)、SHP(SHP−1及びSHP−2)などよりなる群から選択された1つもしくはそれ以上の負の免疫調節因子を阻害するインヒビターを含んでなる。 In one embodiment, the present invention comprises a composition for enhancing the immune capacity of immune cells. Preferably, the composition comprises A20, SUMO (SUMO1, SUMO2, SUMO3 and SUMO4), Foxj1, Foxo3a, TWIST (Twist1, Twist2), SOCS (SOCS1, SOCS2, SOCS3, SOCS4, SOCS5, SOCS6, SOCS7, SOCS7, SOCS7, An inhibitor that inhibits one or more negative immune modulators selected from the group consisting of PIAS (PIAS1, PIAS3, PIASx and PIASy), SHP (SHP-1 and SHP-2), and the like.
負の免疫調節因子のインヒビターを含んでなる組成物は、小分子干渉RNA(siRNA)、microRNA、アンチセンス核酸、リボザイム、トランス優性遺伝子負突然変異体、細胞内抗体、ペプチド及び小分子よりなる群から選択される。 A composition comprising an inhibitor of a negative immune regulator comprises a group consisting of small interfering RNA (siRNA), microRNA, antisense nucleic acid, ribozyme, transdominant gene negative mutant, intracellular antibody, peptide and small molecule Selected from.
当業者であれば、本書で提供されている開示に基づき、1つの細胞内のサイトカインシグナリング調節因子などの負の免疫調節因子のmRNA及び/又はタンパク質レベルを減少させるための1つの方法が、調節因子をコードする核酸の発現を削減又は阻害することによるものである、ということを認識するであろう。かくして、細胞内の負の免疫調節因子のタンパク質レベルは、例えばアンチセンス分子又はリボザイムなどの遺伝子発現を阻害するか又は削減する分子又は化合物を用いても低減させることができる。 One skilled in the art, based on the disclosure provided herein, is one method for reducing mRNA and / or protein levels of negative immune regulators such as cytokine signaling regulators in one cell. It will be appreciated that this is due to reducing or inhibiting the expression of the nucleic acid encoding the factor. Thus, the protein levels of negative immune regulators in cells can also be reduced using molecules or compounds that inhibit or reduce gene expression, such as antisense molecules or ribozymes.
好ましい一実施形態においては、変調配列は、プラスミドベクターにより発現されるアンチセンス核酸配列である。アンチセンス発現ベクターは、哺乳動物細胞又は哺乳動物自体をトランスフェクトし、かくして所望の負の免疫調節因子の削減された内因性発現をひき起こすために使用される。しかしながら、本発明は、アンチセンス分子での細胞のトランスフェクションにより負の免疫調節因子の発現を阻害することに限定されるものとして解釈されてはならない。むしろ本発明は、リボザイムの使用、非機能的負免疫調節因子(すなわちトランス優性負突然変異体)の発現及び細胞内抗体の使用を含めた(ただしこれに限定されるわけではない)細胞内のタンパク質の発現又は活性を阻害するための当該技術分野において既知のその他の方法を包含している。 In a preferred embodiment, the modulation sequence is an antisense nucleic acid sequence expressed by a plasmid vector. Antisense expression vectors are used to transfect mammalian cells or the mammal itself, thus causing reduced endogenous expression of the desired negative immune regulator. However, the present invention should not be construed as being limited to inhibiting the expression of negative immune regulators by transfection of cells with antisense molecules. Rather, the present invention includes the use of ribozymes, the expression of non-functional negative immune regulators (ie trans dominant negative mutants) and the use of intracellular antibodies (but not limited to) Other methods known in the art for inhibiting protein expression or activity are included.
アンチセンス分子及び遺伝子発現を阻害するためのそれらの使用は、当該技術分野において周知である(例えば、コーエン(Cohen)、1989年、「Oligodeoxyribonucleotides、Antisense Inhibitors of
Gene Expression」、CRC Press中を参照のこと)。アンチセンス核酸は、その用語が本書の他の場所で定義されている通り、少なくとも特異的mRNA分子の一部分に相補的であるDNA又はRNA分子である(ワイントラウブ(Weintraub)、1990年、Scientific American第262号:40頁)。細胞内では、アンチセンス核酸は対応するmRNAに対しハイブリッド形成し、2本鎖分子を形成して遺伝子の翻訳を阻害する。
Antisense molecules and their use to inhibit gene expression are well known in the art (see, for example, Cohen, 1989, “Oligodeoxyribonucleotides, Antisense Inhibitors of
See “Gene Expression”, in CRC Press). An antisense nucleic acid is a DNA or RNA molecule that is complementary to at least a portion of a specific mRNA molecule, as that term is defined elsewhere in this document (Weintraub, 1990, Scientific American No. 262: 40). Within the cell, antisense nucleic acids hybridize to the corresponding mRNA, forming double-stranded molecules and inhibiting gene translation.
遺伝子の翻訳を阻害するためのアンチセンス方法の使用は、当該技術分野において既知であり、例えばマーカス−サクラ(Marcus−Sakura)(1988年、Anal.Biochem.第172号:289頁)の中で記述されている。このようなアンチセンス分子は、イノウエ(Inoue、1993年、米国特許第5,190,931号明細書)により教示されている通りアンチセンス分子をコードするDNAを用いて遺伝子発現を介して細胞に提供され得る。 The use of antisense methods to inhibit gene translation is known in the art, for example in Marcus-Sakura (1988, Anal. Biochem. 172: 289). is described. Such antisense molecules can be transferred to cells via gene expression using DNA encoding the antisense molecule as taught by Inoue (1993, US Pat. No. 5,190,931). Can be provided.
あるいは、本発明のアンチセンス分子は合成的に作製しその後細胞に提供され得る。約10〜約30、より好ましくは約15個のヌクレオチドのアンチセンスオリゴマーは、容易に合成され標的細胞に導入されることから好まれる。本発明により考慮されている合成アンチセンス分子には、未修飾オリゴヌクレオチドに比べて改善された生物活性を有する当該技術分野において既知のオリゴヌクレオチド誘導体が含まれる(米国特許第5,023,243号明細書参照)。 Alternatively, antisense molecules of the invention can be made synthetically and then provided to cells. Antisense oligomers of about 10 to about 30, more preferably about 15 nucleotides are preferred because they are readily synthesized and introduced into target cells. Synthetic antisense molecules contemplated by the present invention include oligonucleotide derivatives known in the art that have improved biological activity compared to unmodified oligonucleotides (US Pat. No. 5,023,243). See the description).
遺伝子発現を阻害するためのリボザイム及びそれらの使用も同様に当該技術分野において周知である(例えば、チェフ(Cech)ら、1992年、J.Biol.Chem.第267号:17479〜17482頁;ハンペル(Hampel)ら、1989年、Biochemistry第28号:4929〜4933頁;エックスタイン(Eckstein)ら、国際公開第WO92/07065号パンフレット;アルトマン(Altman)ら、米国特許第5,168,053号明細書を参照のこと)。リボザイムは、DNA制限エンドヌクレアーゼと同様にその他の1本鎖RNAを特異的に分割する能力を有するRNA分子である。これらのRNAをコードするヌクレオチド配列の修飾を通して、RNA分子内の特異的ヌクレオチド配列を認識しそれを分割するように分子を工学処理することができる(チェフ、1988年、J.Amer.Med.Assn.第260号:3030頁)。このアプローチの主たる利点は、リボザイムが配列特異的である、という事実にある。 Ribozymes and their use to inhibit gene expression are also well known in the art (eg, Chech et al., 1992, J. Biol. Chem. 267: 17479-17482; Hampel (Hampel et al., 1989, Biochemistry 28: 4929-4933; Eckstein et al., WO 92/07005; Altman et al., US Pat. No. 5,168,053). See the book). Ribozymes are RNA molecules that have the ability to specifically split other single-stranded RNAs, similar to DNA restriction endonucleases. Through modification of the nucleotide sequences encoding these RNAs, the molecules can be engineered to recognize and split specific nucleotide sequences within the RNA molecule (Chef, 1988, J. Amer. Med. Assn. 260: 3030). The main advantage of this approach lies in the fact that the ribozyme is sequence specific.
リボザイムには2つの基本型、すなわちテトラヒメナ型(ハッセルホッフ(Hasselhoff)、1988年、Nature第334号:585頁)及びハンマーヘッド型がある。テトラヒメナ型リボザイムは、4塩基の長さをもつ配列を認識し、一方ハンマーヘッド型リボザイムは、長さが11〜18塩基の塩基配列を認識する。配列が長くなるほど、配列が標的mRNA種内で専ら発生することになる確率は大きくなる。その結果として、特異的mRNA種を不活性化するためには、テトラヒメナ型リボザイムよりもハンマーヘッド型リボザイムの方が好ましく、さまざまな無関係のmRNA分子の内部で無作為に発生し得る比較的短い認識配列よりも、18塩基の認識配列が好ましい。 There are two basic types of ribozymes, the Tetrahymena type (Hasselhoff, 1988, Nature 334: 585) and the hammerhead type. Tetrahymena ribozymes recognize sequences with a length of 4 bases, while hammerhead ribozymes recognize base sequences with a length of 11-18 bases. The longer the sequence, the greater the probability that the sequence will occur exclusively within the target mRNA species. As a result, hammerhead ribozymes are preferred over tetrahymena ribozymes to inactivate specific mRNA species and are relatively short recognitions that can occur randomly within various unrelated mRNA molecules. An 18-base recognition sequence is preferred over the sequence.
負の免疫調節因子の発現を阻害するのに有用なリボザイムは、所望の負の免疫調節因子のmRNA配列に相補的である基本的リボザイム構造の中に標的配列を取込むことにより設計可能である。所望の負の免疫調節因子をターゲティングするリボザイムは、市販の試薬を用いて合成され得(アプライド・バイオシステムズ社(Applied Biosystems,Inc.)、カリフォルニア州フォスターシティー(Foster City,CA))、そうでなければ、それらをコードするDNAから遺伝学的に発現され得る。 Ribozymes useful for inhibiting the expression of negative immune regulators can be designed by incorporating the target sequence into a basic ribozyme structure that is complementary to the mRNA sequence of the desired negative immune regulator. . Ribozymes targeting the desired negative immunomodulator can be synthesized using commercially available reagents (Applied Biosystems, Inc., Foster City, CA), and so on. Otherwise, it can be genetically expressed from the DNA that encodes them.
本発明のもう1つの態様においては、所望の負の免疫調節因子を不活性化及び/又は封鎖することにより、負の免疫調節因子を阻害することができる。従って、トランス優性負突然変異体を使用することにより、負の免疫調節因子の効果の阻害を達成することができる。あるいは、その他の点では負の免疫調節因子に対するアンタゴニストとして知られている所望の負の免疫調節因子に特異的な細胞内抗体を使用することが可能である。一実施形態においては、アンタゴニストは、負の免疫調節因子の結合パートナーと相互作用しかくして対応する野生型負免疫調節因子と競合するという所望の特性をもつタンパク質及び/又は化合物である。もう1つの実施形態においては、アンタゴニストは、負の免疫調節因子と相互作用しかくして負の免疫調節因子を封鎖するという所望の特性を有するタンパク質及び/又は化合物である。 In another aspect of the invention, negative immunomodulators can be inhibited by inactivating and / or sequestering the desired negative immune regulator. Thus, by using a transdominant negative mutant, inhibition of the effects of negative immune modulators can be achieved. Alternatively, intracellular antibodies specific for the desired negative immune modulator, otherwise known as antagonists to the negative immune regulator, can be used. In one embodiment, the antagonist is a protein and / or compound that has the desired property of interacting with the binding partner of the negative immune regulator and thus competing with the corresponding wild-type negative immune regulator. In another embodiment, the antagonist is a protein and / or compound that has the desired property of interacting with a negative immune regulator and thus sequestering the negative immune regulator.
小分子干渉RNA(siRNA)
小分子干渉RNA(siRNA)は、問題の遺伝子又はポリヌクレオチドに対しターゲティングされている1組のヌクレオチドを含んでなるRNA分子である。本書で使用される通り、「siRNA」という用語は、(i)2本鎖RNAポリヌクレオチド、(ii)1本鎖ポリヌクレオチド及び(iii)1、2、3、4個以上のヌクレオチド改変又は置換を内部に有する(i)又は(ii)のいずれかのポリヌクレオチドを含めた(ただしこれに限定されるわけではない)全ての形態を包含する。
Small interfering RNA (siRNA)
Small interfering RNA (siRNA) is an RNA molecule that comprises a set of nucleotides that are targeted to a gene or polynucleotide of interest. As used herein, the term “siRNA” refers to (i) a double stranded RNA polynucleotide, (ii) a single stranded polynucleotide and (iii) 1, 2, 3, 4 or more nucleotide modifications or substitutions. All forms including (but not limited to) the polynucleotide of either (i) or (ii) are included.
2本鎖ポリヌクレオチドの形態をしたsiRNAは、長さ約18塩基対、約19塩基対、約20塩基対、約21塩基対、約22塩基対、約23塩基対、約24塩基対、約25塩基対、約26塩基対、約27塩基対、約28塩基対、約29塩基対又は約30塩基を含んでなる。2本鎖siRNAは、負の免疫調節因子の発現及び/又は活性に干渉する能力をもつ。 SiRNAs in the form of double-stranded polynucleotides are about 18 base pairs, about 19 base pairs, about 20 base pairs, about 21 base pairs, about 22 base pairs, about 23 base pairs, about 24 base pairs, about 25 base pairs, about 26 base pairs, about 27 base pairs, about 28 base pairs, about 29 base pairs or about 30 bases. Double-stranded siRNA has the ability to interfere with the expression and / or activity of negative immune regulators.
1本鎖siRNAは、問題の遺伝子又はポリヌクレオチドにターゲティングされているRNAポリヌクレオチド配列の一部分を含んでなる。1本鎖siRNAは長さがヌクレオチド約18個、ヌクレオチド約19個、ヌクレオチド約20個、ヌクレオチド約21個、ヌクレオチド約22個、ヌクレオチド約23個、ヌクレオチド約24個、ヌクレオチド約25個、ヌクレオチド約26個、ヌクレオチド約27個、ヌクレオチド約28個、ヌクレオチド約29個又はヌクレオチド約30個のポリヌクレオチドを含んでなる。1本鎖siRNAは、A20、SUMO(SUMO1、SUMO2、SUMO3、及びSUMO4)、Foxj1、Foxo3a、TWIST(Twist1、Twist2)、SOCS(SOCS1、SOCS2、SOCS3、SOCS4、SOCS5、SOCS6、SOCS7及びCIS)、PIAS(PIAS1、PIAS3、PIASx及びPIASy)、SHP(SHP−1及びSHP−2)などの標的ポリヌクレオチドの発現及び/又は活性に干渉する能力をもつ。1本鎖siRNAは同様に、相補的配列にアニールして負の免疫調節因子の発現及び/又は活性に干渉する能力をもつdsRNAをもたらす能力も有している。 Single-stranded siRNA comprises a portion of an RNA polynucleotide sequence that is targeted to a gene or polynucleotide of interest. A single-stranded siRNA is about 18 nucleotides in length, about 19 nucleotides, about 20 nucleotides, about 21 nucleotides, about 22 nucleotides, about 23 nucleotides, about 24 nucleotides, about 25 nucleotides, about 25 nucleotides A polynucleotide comprising 26, about 27 nucleotides, about 28 nucleotides, about 29 nucleotides or about 30 nucleotides. Single-stranded siRNAs are A20, SUMO (SUMO1, SUMO2, SUMO3, and SUMO4), Foxj1, Foxo3a, TWIST (Twist1, Twist2), SOCS (SOCS1, SOCS2, SOCS3, SOCS4, SOCS6, SOCS6, It has the ability to interfere with the expression and / or activity of target polynucleotides such as PIAS (PIAS1, PIAS3, PIASx and PIASy), SHP (SHP-1 and SHP-2). Single-stranded siRNAs also have the ability to anneal to complementary sequences to yield dsRNAs that have the ability to interfere with the expression and / or activity of negative immune regulators.
さらにもう1つの態様においては、siRNAは、2本鎖又は1本鎖のいずれかのポリヌクレオチドを含んでなるポリヌクレオチドを含んでなり、ここでsiRNAは内部に1、2、3、4個以上のヌクレオチド改変又は置換を有している。 In yet another embodiment, the siRNA comprises a polynucleotide comprising a polynucleotide that is either double-stranded or single-stranded, wherein the siRNA has 1, 2, 3, 4 or more therein. Of nucleotide modifications or substitutions.
siRNAポリヌクレオチドは、一般に転写後の遺伝子サイレンシング化機序を介して発生すると考えられているRNA活性に干渉するRNA核酸分子である。siRNAポリヌクレオチドは好ましくは、2本鎖RNA(dsRNA)を含んでなるが、そのように限定されることを意図されておらず、1本鎖RNAを含んでなっていてよい(例えば、マルチネス(Martinez)ら、2002年、Cell第110号:563〜74頁を参
照のこと)。本発明中に含まれるsiRNAポリヌクレオチドは、本書で提供されているようなヌクレオチド(リボヌクレオチド又はデオキシリボヌクレオチド又は両方の組合せ)及び/又はヌクレオチド類似体(例えば、標準的に5’〜3’中のリン酸ジエステル連結中のオリゴヌクレオチド又はポリヌクレオチドなど)のその他の天然に発生する、組換え型、又は合成1本鎖又は2本鎖ポリマーをも含んでなり得る。従って、siRNAポリヌクレオチドの転写を導く能力をもつDNA配列などの本書に開示されているいくつかの配列例は同様に、相補性ヌクレオチド塩基対合の充分に立証済みの原理を考慮して、対応するRNA配列及びその補体を記述するためにも意図されているということがわかるだろう。
siRNA polynucleotides are RNA nucleic acid molecules that interfere with RNA activity that is generally thought to occur through post-transcriptional gene silencing mechanisms. The siRNA polynucleotide preferably comprises double stranded RNA (dsRNA), but is not intended to be so limited and may comprise single stranded RNA (eg, multiness ( Martinez) et al., 2002, Cell 110: 563-74). SiRNA polynucleotides included in the present invention may comprise nucleotides (ribonucleotides or deoxyribonucleotides or a combination of both) and / or nucleotide analogs (eg, typically in 5 ′ to 3 ′) as provided herein. Other naturally occurring, recombinant or synthetic single or double stranded polymers (such as oligonucleotides or polynucleotides in phosphodiester linkages) may also be included. Thus, several example sequences disclosed herein, such as DNA sequences capable of directing transcription of siRNA polynucleotides, are similarly addressed in view of the well-proven principle of complementary nucleotide base pairing. It will be appreciated that it is also intended to describe the RNA sequence to be complemented and its complement.
RNAポリメラーゼプロモータのためのプロモータを含有するDNA(ゲノム、cDNA、又は合成)を鋳型として用いて、siRNAを転写することができる。例えば、プロモータはU6プロモータ又はHIRNAポリメラーゼIIIプロモータであり得る。あるいは、siRNAは合成的に誘導されたRNA分子であり得る。一部の実施形態においては、siRNAポリヌクレオチドは平滑末端を有し得る。いくつかのその他の実施形態においては、siRNAポリヌクレオチドの少なくとも1つのストランドが、siRNAポリヌクレオチドのいずれかのストランドの3’末端において「オーバーハングし」(すなわち相対するストランドの中の相補的塩基と塩基対合しない)少なくとも1つ、好ましくは2つのヌクレオチドを有している。本発明の好ましい実施形態においては、siRNAポリヌクレオチド2重鎖の各ストランドは、3’末端で2ヌクレオチドオーバーハングを有する。2−ヌクレオチドオーバーハングは好ましくはチミジンジヌクレオチド(TT)であるが、同様にその他の塩基例えばTCジヌクレオチド又はTGジヌクレオチド又はその他のあらゆるジヌクレオチドを含んでなり得る。オーバーハングジヌクレオチドは同様に、干渉のためにターゲティングされているポリヌクレオチドの配列の5’末端にある2つのヌクレオチドに対しても相補的であり得る。siRNAポリヌクレオチドの3’末端の論述については、例えば国際公開第01/75164号パンフレットなどを参照のこと。 SiRNA can be transcribed using DNA (genome, cDNA, or synthesis) containing a promoter for the RNA polymerase promoter as a template. For example, the promoter can be a U6 promoter or a HIRNA polymerase III promoter. Alternatively, the siRNA can be a synthetically derived RNA molecule. In some embodiments, siRNA polynucleotides can have blunt ends. In some other embodiments, at least one strand of the siRNA polynucleotide “overhangs” at the 3 ′ end of either strand of the siRNA polynucleotide (ie, with a complementary base in the opposite strand). It has at least one, preferably two nucleotides (not base-paired). In a preferred embodiment of the invention, each strand of the siRNA polynucleotide duplex has a 2 nucleotide overhang at the 3 'end. The 2-nucleotide overhang is preferably a thymidine dinucleotide (TT), but may similarly comprise other bases such as TC dinucleotides or TG dinucleotides or any other dinucleotide. The overhanging dinucleotide may also be complementary to the two nucleotides at the 5 'end of the polynucleotide sequence targeted for interference. For a discussion of the 3 'end of siRNA polynucleotides, see eg WO 01/75164.
好ましいsiRNAポリヌクレオチドは、約18〜30ヌクレオチド塩基対、好ましくは約18、約19、約20、約21、約22、約23、約24、約25、約26又は約27塩基対の2本鎖ポリヌクレオチドを含んでなり、その他の好ましい実施形態においては、約19、約20、約21、約22又は約23塩基対、或いは約27塩基対を含んでなり、かくして、「約」という用語の使用は、一部の実施形態において及び一部の条件下で、選択されたポリペプチドの発現に干渉する能力をもつ機能的siRNAポリヌクレオチドを発生させ得る前進する連続的分割工程が絶対的に効果的でない可能性がある、ということを標示している。従って、siRNAポリヌクレオチドは、siRNAのプロセッシング、生合成又は人工的合成における可変性の帰結として、長さが1、2、3、4塩基対以上異なる(例えばヌクレオチド挿入又は欠失による)1つもしくはそれ以上のsiRNAポリヌクレオチド分子を内含し得る。本発明のsiRNAポリヌクレオチドは同様に、特定の配列からの1、2、3、又は4個のヌクレオチドにおいて異なることによって(例えば塩基転位又は塩基転換を含めたヌクレオチド置換による)可変性を示すポリヌクレオチド配列を含んでなることもできる。これらの差は、2本鎖ポリヌクレオチドのセンス又はアンチセンスストランドのいずれの中にあるかに関わらず、分子の長さに応じて特定のsiRNAポリヌクレオチド配列のヌクレオチド位置のいずれかで発生可能である。ヌクレオチド差は、2本鎖ポリヌクレオチドの1本のストランド上に発見され得、ここで代替ヌクレオチドと共に水素結合塩基対合を標準的に形成することになる相補的ヌクレオチドは必ずしも対応する形で置換され得ない。好ましい実施形態においては、siRNAポリヌクレオチドは、特異的ヌクレオチド配列との関係において相同である。 Preferred siRNA polynucleotides are two of about 18-30 nucleotide base pairs, preferably about 18, about 19, about 20, about 21, about 22, about 23, about 24, about 25, about 26 or about 27 base pairs. In other preferred embodiments comprising about 19, about 20, about 21, about 22 or about 23 base pairs, or about 27 base pairs, thus the term "about" In some embodiments and under certain conditions, an absolute sequential cleavage step that can generate a functional siRNA polynucleotide capable of interfering with the expression of a selected polypeptide is absolutely necessary. Indicates that it may not be effective. Thus, siRNA polynucleotides can vary in length by 1, 2, 3, 4 base pairs or more (eg, due to nucleotide insertions or deletions) as a consequence of variability in siRNA processing, biosynthesis or artificial synthesis, or Further siRNA polynucleotide molecules may be included. The siRNA polynucleotides of the present invention are also polynucleotides that exhibit variability by differing in 1, 2, 3, or 4 nucleotides from a particular sequence (eg, by nucleotide substitutions including translocations or transversions) It can also comprise an array. These differences can occur at any of the nucleotide positions of a particular siRNA polynucleotide sequence, depending on the length of the molecule, whether in the sense or antisense strand of the double-stranded polynucleotide. is there. Nucleotide differences can be found on one strand of a double-stranded polynucleotide, where complementary nucleotides that would normally form hydrogen-bonded base pairings with alternative nucleotides are not necessarily replaced in a corresponding manner. I don't get it. In preferred embodiments, siRNA polynucleotides are homologous in relation to a specific nucleotide sequence.
一部の実施形態では、本発明のsiRNAポリヌクレオチドを含んでなるポリヌクレオ
チドは、標準的にはスペーサ配列により分離された、1本鎖オリゴヌクレオチドフラグメント(例えば約18〜30ヌクレオチドの)及びその逆補体を含んでなる1本鎖ポリヌクレオチドから誘導され得る。或る種のこのような実施形態に従うと、スペーサの分割が1本鎖オリゴヌクレオチドフラグメント及びその逆補体を提供し、かくして、これらはアニールして、場合によりいずれかの又は両方のストランドの3’末端及び/又は5’末端からの1、2、3又は4個以上のヌクレオチドの付加又は除去を結果としてもたらすさらなるプロセッシング工程を伴って、本発明の2本鎖siRNAポリヌクレオチドを形成することができる。一部の実施形態においては、スペーサは、その分割に先立ってかつ場合により、いずれか又は両方のストランドの3’末端及び/又は5’末端からの1、2、3、4個以上のヌクレオチドの付加又は除去を結果としてもたらし得る後続するプロセッシング工程の前に、フラグメント及びその逆補体がアニールし2本鎖構造(例えばヘアピンポリヌクレオチドのようなもの)を形成できるようにする長さを有している。従って、スペーサ配列は、2本鎖核酸内にアニールされた時点でsiRNAポリヌクレオチドを結果としてもたらす2つの相補的ポリヌクレオチド配列領域の間にある本書で提供されている通りのあらゆるポリヌクレオチド配列であり得る。好ましくは、スペーサ配列は少なくとも4つのヌクレオチドを含んでなる。一部の実施形態においては、スペーサは5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21〜25、26〜30、31〜40、41〜50、51〜70、71〜90、91〜110、111〜150、151〜200個又はそれ以上のヌクレオチドを含んでなり得る。1つのスペーサにより分離された2つの相補的ヌクレオチド配列を含んでなる単一のヌクレオチドストランドから誘導されるsiRNAポリヌクレオチドの例が記述されてきた(例えば、ブルンメルカンプ(Brummelkamp)ら、2002年、Science第296号:550頁;パディソン(Paddison)ら、2002年、Genes Develop.第16号:948頁;ポール(Paul)ら、2002年、Nat.Biotechnol.第20号:505〜508頁;グラバレク(Grabarek)ら、2003年、BioTechniques第34号:734〜44頁)。
In some embodiments, a polynucleotide comprising a siRNA polynucleotide of the invention is a single stranded oligonucleotide fragment (eg, of about 18-30 nucleotides) and vice versa, typically separated by a spacer sequence. It can be derived from a single stranded polynucleotide comprising complement. According to certain such embodiments, spacer splitting provides single stranded oligonucleotide fragments and their reverse complements, thus they anneal and optionally 3 of either or both strands. Forming a double-stranded siRNA polynucleotide of the present invention with further processing steps resulting in the addition or removal of 1, 2, 3 or 4 or more nucleotides from the 'end and / or 5' end. it can. In some embodiments, the spacer is one, two, three, four or more nucleotides from the 3 ′ end and / or 5 ′ end of either or both strands prior to and optionally in the splitting. Has a length that allows the fragment and its reverse complement to anneal to form a double stranded structure (such as a hairpin polynucleotide) prior to subsequent processing steps that may result in addition or removal. ing. Thus, a spacer sequence is any polynucleotide sequence as provided herein that is between two complementary polynucleotide sequence regions that result in an siRNA polynucleotide when annealed into a double stranded nucleic acid. obtain. Preferably, the spacer sequence comprises at least 4 nucleotides. In some embodiments, the spacer is 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21-25, 26-30, It can comprise 31-40, 41-50, 51-70, 71-90, 91-110, 111-150, 151-200 or more nucleotides. Examples of siRNA polynucleotides derived from a single nucleotide strand comprising two complementary nucleotide sequences separated by one spacer have been described (e.g., Brunmelkamp et al., 2002, Science 296: 550; Paddison et al., 2002, Genes Develop. 16: 948; Paul et al., 2002, Nat. Biotechnol. 20: 505-508; (Grabarek) et al., 2003, BioTechniques 34: 734-44).
ポリヌクレオチド変異体は、siRNAポリヌクレオチドの活性が実質的に減少しないような形で1つもしくはそれ以上の置換、付加、欠失及び/又は挿入を含有し得る。siRNAポリヌクレオチドの活性に対してヌクレオチド含有量の何らかのこのような改変が有する効果は一般に、本書の他の箇所で記述されているように査定可能である。変異体は、好ましくは未変性A20、SUMO(SUMO1、SUMO2、SUMO3及びSUMO4)、Foxj1、Foxo3a、TWIST(Twist1、Twist2)、SOCS(SOCS1、SOCS2、SOCS3、SOCS4、SOCS5、SOCS6、SOCS7及びCIS)、PIAS(PIAS1、PIAS3、PIASx及びPIASy)、SHP(SHP−1及びSHP−2)などをコードするポリヌクレオチド配列の一部分に対し、好ましくは少なくとも約75%、78%、80%、85%、87%、88%又は89%の同一性、そしてより好ましくは少なくとも約90%、92%、95%、96%又は97%の同一性を示す。同一性百分率は、当業者にとっては周知のコンピュータアルゴリズムを用いることを含めた任意の方法を用いて、標的ポリヌクレオチドの対応する部分とポリヌクレオチドの配列を比較することによって容易に判定可能である。これらのアルゴリズムとしては、Align又はBLASTアルゴリズムが含まれる(アルトシュール(Altschul)、1991年、J.Mol.Biol.第219号:555〜565頁;ヘニコフ(Henikoff)及びヘニコフ、1992年、Proc.Natl.Acad.Sci.USA第89号:10915〜10919頁)。 A polynucleotide variant may contain one or more substitutions, additions, deletions and / or insertions such that the activity of the siRNA polynucleotide is not substantially reduced. The effect of any such modification of nucleotide content on the activity of siRNA polynucleotides can generally be assessed as described elsewhere in this document. The variants are preferably native A20, SUMO (SUMO1, SUMO2, SUMO3 and SUMO4), Foxj1, Foxo3a, TWIST (Twist1, Twist2), SOCS (SOCS1, SOCS2, SOCS3, SOCS4, SOCS6, and SOCS6, SOCS6, CS7). , Preferably at least about 75%, 78%, 80%, 85% to a portion of the polynucleotide sequence encoding PIAS (PIAS1, PIAS3, PIASx and PIASy), SHP (SHP-1 and SHP-2), etc. It exhibits 87%, 88% or 89% identity, and more preferably at least about 90%, 92%, 95%, 96% or 97% identity. The percent identity can be readily determined by comparing the sequence of the polynucleotide with the corresponding portion of the target polynucleotide using any method, including using computer algorithms well known to those skilled in the art. These algorithms include the Align or BLAST algorithm (Altschul, 1991, J. Mol. Biol. 219: 555-565; Henikoff and Henikov, 1992, Proc. Natl. Acad. Sci. USA 89: 10915-10919).
一部のsiRNAポリヌクレオチド変異体は、標的ポリヌクレオチドをコードするポリヌクレオチドの一部分と実質的に相同であり得る。これらのポリヌクレオチド変異体から誘導された1本鎖ポリヌクレオチドは、中程度にストリンジェントな条件下で、標的ポリペプチドをコードする天然に発生するDNA又はRNA配列に対しハイブリッド形成する
能力を有する。中程度にストリンジェントな条件下で標的ポリペプチドをコードするポリヌクレオチド配列に対し検出可能な形でハイブリッド形成するsiRNAポリヌクレオチドが、特定の標的ポリヌクレオチドに対し相補的である少なくとも10個の連続するヌクレオチド配列、より好ましくは11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29または30個の連続するヌクレオチドを有し得る。一部の好ましい実施形態においては、このようなsiRNA配列(又はその補体)は、発現との干渉が望まれている標的ポリペプチドをコードする単一の特定のポリヌクレオチドに固有のものとなり得る。一部のその他の実施形態においては、配列(又はその補体)は、ポリペプチド発現との干渉が望まれている標的ポリペプチドをコードする2つ以上の関係するポリヌクレオチドによって共有され得る。
Some siRNA polynucleotide variants can be substantially homologous to a portion of the polynucleotide encoding the target polynucleotide. Single stranded polynucleotides derived from these polynucleotide variants have the ability to hybridize to naturally occurring DNA or RNA sequences encoding the target polypeptide under moderately stringent conditions. At least 10 consecutive siRNA polynucleotides that hybridize in a detectable manner to a polynucleotide sequence encoding a target polypeptide under moderately stringent conditions are complementary to a particular target polynucleotide Nucleotide sequence, more preferably 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 or 30 consecutive nucleotides Can have. In some preferred embodiments, such siRNA sequences (or their complements) can be unique to a single specific polynucleotide that encodes a target polypeptide for which interference with expression is desired. . In some other embodiments, the sequence (or its complement) may be shared by two or more related polynucleotides that encode target polypeptides that are desired to interfere with polypeptide expression.
適切な中程度にストリンジェントな条件としては、例えば、5×SSC、0.5%SDS、1.0mMEDTA(pH8.0)の溶液中でポリヌクレオチドを予備洗浄すること;50℃〜70℃で、5×SSCで1〜16時間(例えば一晩中)ポリヌクレオチドをハイブリッド形成すること;そしてその後に、各々0.05〜0.1%のSDSを含有する2×、0.5×及び0.2×のSSCのうちの1つもしくはそれ以上のもので20〜40分間22〜65℃で1回又は2回ポリヌクレオチドを洗浄することが含まれる。さらなるストリンジェンシーについては、ハイブリダイゼーション条件は、15〜40分間50〜60℃で0.1×SSC及び0.1%SDS中での付加的な洗浄を含む。当業者であれば、予備ハイブリダイゼーション、ハイブリダイゼーション及び洗浄工程のために用いられる溶液の濃度及び/又は温度、時間を改変することによってハイブリダイゼーション条件のストリンジェンシーの変動を達成できるということを理解することだろう。適切な条件は同様に、使用されるプローブ及びブロッティングされたプロバンド核酸試料の特定の核酸配列によっても一部左右され得る。従って、或る種のその他の発端者配列にはハイブリッド形成しない一方で1つもしくはそれ以上の或る種のプロバンド配列にはハイブリッド形成するその能力に基づいて、ポリヌクレオチドの所望の選択性が同定された時点で、過度に実験せずに、適切にストリンジェントな条件を容易に選択することができるということがわかるだろう。 Suitable moderately stringent conditions include, for example, pre-washing the polynucleotide in a solution of 5 × SSC, 0.5% SDS, 1.0 mM EDTA (pH 8.0); Hybridize polynucleotides at 5 × SSC for 1-16 hours (eg overnight); and then 2 ×, 0.5 × and 0 each containing 0.05-0.1% SDS Washing the polynucleotide once or twice with one or more of the 2X SSCs for 20-40 minutes at 22-65 ° C. For further stringency, hybridization conditions include additional washing in 0.1 × SSC and 0.1% SDS at 50-60 ° C. for 15-40 minutes. Those skilled in the art will appreciate that variations in the stringency of hybridization conditions can be achieved by modifying the concentration and / or temperature and time of the solution used for the prehybridization, hybridization and washing steps. That would be true. Appropriate conditions may also depend in part on the probe used and the specific nucleic acid sequence of the blotted proband nucleic acid sample. Thus, the desired selectivity of the polynucleotide is based on its ability to hybridize to one or more certain proband sequences while not hybridizing to certain other proband sequences. Once identified, it will be appreciated that appropriately stringent conditions can be readily selected without undue experimentation.
本発明の配列特異的siRNAポリヌクレオチドは、複数の基準のうちの1つもしくはそれ以上のものを用いて設計可能である。例えば、問題のポリペプチドをコードする配列と同一の約21個の連続するヌクレオチドを有するsiRNAポリヌクレオチドを設計するためには、以下の特徴のうちの1つもしくはそれ以上のものを有する約21の塩基配列という長さにわたりポリヌクレオチド配列の読取り枠を走査することができる:すなわち(1)約1:1、ただし2:1又は1:2以下のA+T/G+C比;(2)5’末端にあるAAジヌクレオチドもしくはCAジヌクレオチド;(3)55℃未満の内部ヘアピンループ融解温度;(4)37℃未満のホモ二量体融解温度((3)及び(4)で記述されている融解温度計算は、当業者にとっては既知のコンピュータソフトウェアを用いて決定可能である);(5)その他の何らかの既知のポリヌクレオチド配列内に存在しているものとして同定されていない少なくとも16個の連続するヌクレオチドの配列。あるいは、siRNAポリヌクレオチド配列は、例えば、オリゴエンジン(OligoEngine)TM(ワシントン州シアトル(Seattle,Wash.));ダーマコン社(Dharmacon,Inc.)(コロラド州ラファイエット(Lafayette,Colo.));アンビオン社(Ambion Inc.)(テキサス州オースティン(Austin,Tex.));及びキアゲン社(QIAGEN,Inc.)(カリフォルニア州バレンシア(Valencia,Calif.))などのさまざまな供給メーカーから市販されているコンピュータソフトウェアを用いて設計及び選択され得る。エルバシール(Elbashir)ら、2000年、Genes & Development第15号:188〜200頁;エルバシールら、2001年、Nature第411号:494〜98頁も同じく参照のこと。このときsiRNAポリヌクレオチドは、当該技術分野におい
て既知であり本書の他の場所で記述されている方法に従って、標的ポリペプチドの発現に干渉する能力についてテストされ得る。siRNAポリヌクレオチドの有効性の判定には、標的ポリペプチドの発現に干渉するその能力の考慮のみならずsiRNAポリヌクレオチドが宿主細胞に対して毒性をもつか否かが含まれる。例えば、所望のsiRNAはRNA干渉活性を示すと同時に、望ましくない生物学的帰結を示さないと思われる。望ましくない生物学的帰結の例としては、宿主細胞内へのsiRNAの導入の結果としてのその細胞死が望まれていない細胞のアポトーシスがある。
The sequence-specific siRNA polynucleotides of the present invention can be designed using one or more of a plurality of criteria. For example, to design a siRNA polynucleotide having about 21 contiguous nucleotides identical to a sequence encoding the polypeptide of interest, about 21 having one or more of the following characteristics: The reading frame of the polynucleotide sequence can be scanned over the length of the base sequence: (1) A + T / G + C ratio of about 1: 1, but not more than 2: 1 or 1: 2; (2) at the 5 ′ end An AA dinucleotide or CA dinucleotide; (3) an internal hairpin loop melting temperature below 55 ° C .; (4) a homodimer melting temperature below 37 ° C. (the melting temperature described in (3) and (4) Calculations can be determined by those skilled in the art using known computer software); (5) exist within some other known polynucleotide sequence. A sequence of at least 16 contiguous nucleotides not identified as present. Alternatively, siRNA polynucleotide sequences can be obtained from, for example, OligoEngine ™ (Seattle, Wash.); Dharmacon, Inc. (Lafayette, Colo.); Ambion Commercially available from a variety of suppliers, such as Ambion Inc. (Austin, Tex.); And QIAGEN, Inc. (Valencia, Calif.). It can be designed and selected using software. See also Elbashir et al., 2000, Genes & Development 15: 188-200; Elbashir et al., 2001, Nature 411: 494-98. The siRNA polynucleotide can then be tested for the ability to interfere with expression of the target polypeptide according to methods known in the art and described elsewhere herein. Determining the effectiveness of a siRNA polynucleotide includes not only considering its ability to interfere with expression of the target polypeptide, but also whether the siRNA polynucleotide is toxic to the host cell. For example, the desired siRNA appears to exhibit RNA interference activity while not exhibiting undesirable biological consequences. An example of an undesirable biological consequence is apoptosis of a cell whose death as a result of introduction of siRNA into the host cell is not desired.
本開示に基づくと、本発明のsiRNAが異なる度合で標的ポリペプチド発現のサイレンシング化をもたらし得ることがわかるはずである。かくしてsiRNAはまずその有効性についてテストされなくてはならない。一定の与えられたsiRNAがもつ標的ポリペプチドの発現に干渉するか又はこれを変調する能力に基づいてそこからsiRNAが選択される。従って、所望の標的ポリペプチドの発現に干渉する能力をもつ特異的siRNAポリヌクレオチド配列の同定には、各siRNAの産生及び試験が必要である。本発明において使用するための適切なsiRNAの選択及び各siRNAの試験のための方法は、本書の実施例中で充分に記されている。タンパク質発現に干渉する全てのsiRNAが生理学的に重要な効果を有することにはならないことから、本開示は同様に、本発明のsiRNAを用いた標的タンパク質発現との干渉レベルが臨床的に関連する意義を有するか否かを判定するためのさまざまな生理学的に関連する検定についても説明している。 Based on this disclosure, it should be understood that the siRNAs of the invention can result in silencing of target polypeptide expression to different degrees. Thus, siRNA must first be tested for its effectiveness. An siRNA is selected therefrom based on its ability to interfere with or modulate the expression of a target polypeptide possessed by a given siRNA. Accordingly, the production and testing of each siRNA is necessary to identify specific siRNA polynucleotide sequences that have the ability to interfere with the expression of the desired target polypeptide. Selection of appropriate siRNA for use in the present invention and methods for testing each siRNA are fully described in the examples herein. The present disclosure is also clinically relevant to the level of interference with target protein expression using the siRNA of the invention, since not all siRNAs that interfere with protein expression will have a physiologically significant effect. It also describes various physiologically relevant tests for determining whether they are significant.
当業者であれば、遺伝コードの縮重の結果として、数多くの異なるヌクレオチド配列が同じポリペプチドをコードし得るということを容易に認識することになる。すなわち、アミノ酸は、複数の異なるコドンの1つによってコードされ得、当業者であれば、1つの特定のヌクレオチド配列は互いに異なり得るもののポリヌクレオチドは実際同一のアミノ酸配列をもつポリペプチドをコードすることができるということを容易に判定できる。従って、本発明では、コドン使用法が異なっているために変動するポリヌクレオチドが特定的に考慮されている。 One skilled in the art will readily recognize that as a result of the degeneracy of the genetic code, many different nucleotide sequences can encode the same polypeptide. That is, an amino acid can be encoded by one of a plurality of different codons, and those skilled in the art will encode a polypeptide that actually has the same amino acid sequence, although one specific nucleotide sequence may differ from each other. Can be easily determined. Accordingly, the present invention specifically considers polynucleotides that vary due to different codon usage.
siRNAのポリヌクレオチドは、特定的に望まれるsiRNAポリヌクレオチドの調製のために有用であるさまざまな技術のうちのいずれかを用いて調製可能である。例えば、適切な細胞又は組織型から調製されたcDNAからポリヌクレオチドを増幅することができる。かかるポリヌクレオチドはポリメラーゼ連鎖反応(PCR)を介して増幅可能である。このアプローチを用いて、本書に提供されている配列に基づき配列特異的プライマが設計され、これは購入することもできるし又直接合成することもできる。プライマの増幅された部分を用いて、周知の技術を使用して適切なDNAライブラリから全長遺伝子又はその所望の一部分を単離することが可能である。ライブラリ(cDNA又はゲノム)が、増幅に適した1つもしくはそれ以上のポリヌクレオチドプローブ又はプライマを用いてスクリーニングされる。好ましくは、ライブラリは、より大きいポリヌクレオチド配列を含むようにサイズ選択される。遺伝子の5’及びその他の上流領域を同定するためにもランダムプライミングされたライブラリが好まれる可能性がある。イントロンを獲得し5’配列を拡張するためには、ゲノムライブラリが好ましい。本発明で考慮されているsiRNAポリヌクレオチドを同様に、siRNAポリヌクレオチド配列のライブラリから選択することも可能である。 siRNA polynucleotides can be prepared using any of a variety of techniques that are useful for the preparation of specifically desired siRNA polynucleotides. For example, a polynucleotide can be amplified from cDNA prepared from an appropriate cell or tissue type. Such polynucleotides can be amplified via the polymerase chain reaction (PCR). Using this approach, sequence-specific primers are designed based on the sequences provided herein, which can be purchased or synthesized directly. The amplified portion of the primer can be used to isolate the full length gene or a desired portion thereof from an appropriate DNA library using well known techniques. The library (cDNA or genome) is screened using one or more polynucleotide probes or primers suitable for amplification. Preferably, the library is sized to include larger polynucleotide sequences. Randomly primed libraries may also be preferred to identify 5 'and other upstream regions of genes. Genomic libraries are preferred for obtaining introns and extending 5 'sequences. The siRNA polynucleotides contemplated by the present invention can also be selected from a library of siRNA polynucleotide sequences.
ハイブリダイゼーション技術のためには、周知の技術を用いて、部分的ポリヌクレオチド配列を(例えばニック翻訳又は32Pでの末端標識により)標識することができる。このとき、標識されたプローブを伴う変性細菌コロニーを含有するフィルタ(又はファージプラークを含有するローン)に対するハイブリダイゼーションにより、細菌又はバクテリオファージライブラリをスクリーニングすることができる(例えば、サンブルック(Sambrook)ら、「Molecular Cloning:A Laboratory
Manual」、コールド・スプリング・ハーバー・ラボラトリーズ(Cold Spring Harbor Laboratories)、ニューヨーク州コールド・スプリング・ハーバー(Cold Spring Harbor,N.Y.)、2001年を参照)。ハイブリッド形成用コロニー又はプラークが選択され拡張され、DNAはさらなる分析を目的として単離される。
For hybridization techniques, partial polynucleotide sequences can be labeled (eg, by nick translation or end labeling with 32 P) using well-known techniques. At this time, bacteria or bacteriophage libraries can be screened (eg, Sambrook et al.) By hybridization to a filter containing denatured bacterial colonies with labeled probes (or a lawn containing phage plaques). , “Molecular Cloning: A Laboratory
Manual ", Cold Spring Harbor Laboratories, Cold Spring Harbor, NY (see 2001), Cold Spring Harbor Laboratories, Cold Spring Harbor Laboratories, NY. Hybridizing colonies or plaques are selected and expanded, and the DNA is isolated for further analysis.
あるいは、部分的cDNA配列から全長コーディング配列を獲得するため、数多くの増幅技術が当該技術分野において知られている。このような技術の範囲内で、一般にPCRを介して増幅が実施される。このような1つの技術は、「cDNA末端の高速増幅」又はRACEとして知られている(例えば、サンブルックら、「Molecular Cloning:A Laboratory Manual」、コールド・スプリング・ハーバー・ラボラトリーズ、ニューヨーク州コールド・スプリング・ハーバー、2001年を参照)。 Alternatively, a number of amplification techniques are known in the art for obtaining a full-length coding sequence from a partial cDNA sequence. Within the scope of such techniques, amplification is generally performed via PCR. One such technique is known as “fast amplification of cDNA ends” or RACE (eg, Sunbrook et al., “Molecular Cloning: A Laboratory Manual”, Cold Spring Harbor Laboratories, Cold, NY). (See Spring Harbor, 2001).
標的ポリペプチド発現に干渉するために有用な一定数の特定的siRNAポリヌクレオチド配列が本書中の実施例、図面及び配列リストの中で提示されている。siRNAポリヌクレオチドは一般に、例えば固相化学合成などを含めた当該技術分野において既知のあらゆる方法によって調製可能である。ポリヌクレオチド配列内の修飾も同様に、オリゴヌクレオチド指向の部位特異的突然変異誘発などの標準的な突然変異誘発技術を用いて導入され得る。さらに、siRNAをその他の分子で化学的に修飾するか又はそれと接合させて、その安定性及び/又は送達特性を改善させることもできる。本発明の一態様として含まれているのは、本書で記述されている通りの、1つもしくはそれ以上のリボース糖が除去されたsiRNAである。 A number of specific siRNA polynucleotide sequences useful for interfering with target polypeptide expression are presented in the examples, drawings and sequence listings herein. siRNA polynucleotides can generally be prepared by any method known in the art, including, for example, solid phase chemical synthesis. Modifications in the polynucleotide sequence can also be introduced using standard mutagenesis techniques such as oligonucleotide-directed site-directed mutagenesis. In addition, siRNA can be chemically modified or conjugated with other molecules to improve its stability and / or delivery properties. Included as an aspect of the present invention is an siRNA from which one or more ribose sugars have been removed, as described herein.
あるいは、そのDNAが適切なRNAポリメラーゼプロモータ(例えばT7、U6、H1又はSP6、ただしその他のプロモータも同等に有用であり得る)と共にベクター内に取込まれることを条件として、適切なDNA配列(例えば標的ポリペプチドをコードするポリヌクレオチド配列又はその所望の一部分)のインビトロ又はインビボ転写により、siRNAポリヌクレオチド分子を生成することができる。さらに、所望のsiRNAがインビボで生成されるような形で転写(及び場合により該当するプロセッシング工程)を裏づけるDNA配列(例えば本書で提供されているような組換え型核酸構成体)がそうであり得るように、siRNAポリヌクレオチドを哺乳動物に投与することが可能である。 Alternatively, the appropriate DNA sequence (eg, T7, U6, H1 or SP6, although other promoters may be equally useful), as long as the DNA is incorporated into the vector (eg, SiRNA polynucleotide molecules can be generated by in vitro or in vivo transcription of a polynucleotide sequence encoding a target polypeptide or a desired portion thereof. In addition, there are DNA sequences (eg, recombinant nucleic acid constructs as provided herein) that support transcription (and possibly applicable processing steps) in such a way that the desired siRNA is generated in vivo. As obtained, siRNA polynucleotides can be administered to a mammal.
一実施形態においては、標的ポリペプチドの発現に干渉する能力をもつsiRNAポリヌクレオチドを、サイレンシング化された細胞を生成する目的で使用することができる。一定の時間生物学的供給源と接触させられた時点で標的ポリペプチドの発現の大幅な減少を結果としてもたらすあらゆるsiRNAポリヌクレオチドが本発明に含まれる。好ましくは、減少は、siRNAの不在下で検出される標的ポリペプチドの発現レベルとの関係において約10%超、より好ましくは約20%超、より好ましくは約30%超、より好ましくは約40%、約50%、約60%、約70%、約75%、約80%、約85%、約90%、約95%又は約98%超である。好ましくは、細胞内のsiRNAポリヌクレオチドの存在により、例えば内部でのアポトーシスがRNA干渉の所望の効果ではない1つの細胞の死又はアポトーシスなどの何らかの望ましくない毒性効果が結果としてもたらされること又はひき起こされることはない。 In one embodiment, siRNA polynucleotides capable of interfering with target polypeptide expression can be used to generate silenced cells. Any siRNA polynucleotide that results in a significant decrease in expression of the target polypeptide when contacted with a biological source for a period of time is included in the invention. Preferably, the decrease is greater than about 10%, more preferably greater than about 20%, more preferably greater than about 30%, more preferably about 40% relative to the expression level of the target polypeptide detected in the absence of siRNA. %, About 50%, about 60%, about 70%, about 75%, about 80%, about 85%, about 90%, about 95% or more than about 98%. Preferably, the presence of siRNA polynucleotides in the cell results in or causes some undesirable toxic effect, such as death of one cell or apoptosis where internal apoptosis is not the desired effect of RNA interference. It will never be.
もう1つの実施形態においては、siRNAポリヌクレオチドは、一定の時間生物学的供給源と接触させられた時点で標的ポリペプチドの発現の大幅な減少を結果としてもたらす。好ましくは、減少は、siRNAの不在下で検出される標的ポリペプチドの発現レベルとの関係において約10%〜20%、より好ましくは約20%〜30%、より好ましくは約30%〜40%、より好ましくは約40%〜50%、より好ましくは約50%〜60
%、より好ましくは約60%〜70%、より好ましくは約70%〜80%、より好ましくは約80%〜90%、より好ましくは約90%〜95%、より好ましくは約95%〜98%である。好ましくは、細胞内のsiRNAポリヌクレオチドの存在により望ましくない何らかの毒性効果が結果としてもたらされるか又はひき起こされるわけではない。
In another embodiment, the siRNA polynucleotide results in a significant decrease in expression of the target polypeptide when contacted with the biological source for a period of time. Preferably, the decrease is about 10% to 20%, more preferably about 20% to 30%, more preferably about 30% to 40% in relation to the expression level of the target polypeptide detected in the absence of siRNA. , More preferably about 40% to 50%, more preferably about 50% to 60%.
%, More preferably about 60% to 70%, more preferably about 70% to 80%, more preferably about 80% to 90%, more preferably about 90% to 95%, more preferably about 95% to 98%. %. Preferably, the presence of intracellular siRNA polynucleotides does not result in or cause any undesirable toxic effects.
さらにもう1つの実施形態においては、siRNAポリヌクレオチドは、一定の時間生物学的供給源と接触させられた時点で標的ポリペプチドの発現の大幅な減少を結果としてもたらす。好ましくは、減少は、siRNAの不在下で検出される標的ポリペプチドの発現レベルとの関係において約10%以上、より好ましくは約20%以上、より好ましくは約30%以上、より好ましくは約40%以上、より好ましくは約50%以上、より好ましくは約60%以上、より好ましくは約70%以上、より好ましくは約80%以上、より好ましくは約90%以上、より好ましくは約95%以上、より好ましくは約98%以上である。好ましくは、細胞内のsiRNAポリヌクレオチドの存在により望ましくない何らかの毒性効果が結果としてもたらされるか又はひき起こされるわけではない。 In yet another embodiment, the siRNA polynucleotide results in a significant decrease in the expression of the target polypeptide when contacted with the biological source for a period of time. Preferably, the decrease is about 10% or more, more preferably about 20% or more, more preferably about 30% or more, more preferably about 40% relative to the expression level of the target polypeptide detected in the absence of siRNA. % Or more, more preferably about 50% or more, more preferably about 60% or more, more preferably about 70% or more, more preferably about 80% or more, more preferably about 90% or more, more preferably about 95% or more. More preferably, it is about 98% or more. Preferably, the presence of intracellular siRNA polynucleotides does not result in or cause any undesirable toxic effects.
従って、本発明は配列番号1〜3、21〜23及び28〜57の中で例示されている通りのsiRNAのようなsiRNAポリヌクレオチドを含む。配列番号1〜3及び21〜23は、それぞれSOCS1のためのマウス及びヒトsiRNA候補配列の配列である。配列番号28〜33、34〜39、40〜45、46〜51及び52〜57はそれぞれPIAS1、PIAS3、PIASx、PIASy及びSHP−1のためのヒトsiRNA候補配列の配列である。siRNAの配列は、図30の中で描かれている。 Accordingly, the present invention includes siRNA polynucleotides such as siRNA as exemplified in SEQ ID NOs: 1-3, 21-23 and 28-57. SEQ ID NOs: 1-3 and 21-23 are sequences of mouse and human siRNA candidate sequences for SOCS1, respectively. SEQ ID NOs: 28-33, 34-39, 40-45, 46-51 and 52-57 are the sequences of human siRNA candidate sequences for PIAS1, PIAS3, PIASx, PIASy and SHP-1, respectively. The sequence of siRNA is depicted in FIG.
本発明に包含されているその他のsiRNA配列としては、マウスA20 siRNA:5’−CAAAGCACUUAUUGACAGA−3’、配列番号58;マウスSUMO−1 siRNA:5’−GAUGUGAUUGAAGUUUAUC−3’、配列番号59;マウスFoxj1 siRNA:5’−AGAUCACUCUGUCGGCCAU−3’、配列番号60;及びマウスTwist−2 siRNA:5’−GCGACGAGAUGGACAAUAA−3’、配列番号61(Twist−2siRNA1)及び5’−CAAGAAAUCGAGCGAAGAU−3’、配列番号62(Twist−2siRNA2)が含まれる。 Other siRNA sequences encompassed by the present invention include mouse A20 siRNA: 5′-CAAAGCACUUAUUGAGAGA-3 ′, SEQ ID NO: 58; mouse SUMO-1 siRNA: 5′-GAUGUGAUGAGAGUUUAUC-3 ′, SEQ ID NO: 59; mouse Foxj1 siRNA: 5′-AGAUCACUCUGUCGGCCAU-3 ′, SEQ ID NO: 60; and mouse Twist-2 siRNA: 5′-GCGACGGAGAUGGACAAUAA-3 ′, SEQ ID NO: 61 (Twist-2 siRNA1) and 5′-CAAGAAAUCGAGCGAAGAU-3 ′ Twist-2 siRNA 2) is included.
もう1つの態様においては、本発明は所望の負の免疫調節因子にターゲティングされたsiRNAポリヌクレオチドを含んでなる。好ましくは、siRNAはA20、SUMO(SUMO1、SUMO2、SUMO3、SUMO4)、Twist−1、Twist−2、Foxj1、Foxo3a及びそれらの変異体よりなる群から選択された負の免疫調節因子にターゲティングされる。A20、SUMO1、SUMO2、SUMO3、SUMO4、Twist−1、Twist−2、Foxj1及びFoxo3aのための標的配列の例はそれぞれ配列番号63〜84、85〜92、93〜105、106〜112、113〜128、129〜136、137〜149、150〜161及び162〜185内で例示されている。これらのsiRNA標的配列の配列は図53に描かれている。 In another aspect, the invention comprises siRNA polynucleotides targeted to a desired negative immune regulator. Preferably, the siRNA is targeted to a negative immune modulator selected from the group consisting of A20, SUMO (SUMO1, SUMO2, SUMO3, SUMO4), Twist-1, Twist-2, Foxj1, Foxo3a and variants thereof. . Examples of target sequences for A20, SUMO1, SUMO2, SUMO3, SUMO4, Twist-1, Twist-2, Foxj1 and Foxo3a are SEQ ID NOS: 63-84, 85-92, 93-105, 106-112, 113-, respectively. 128, 129-136, 137-149, 150-161 and 162-185. The sequences of these siRNA target sequences are depicted in FIG.
さまざまな負の免疫調節因子のためのポリヌクレオチド及びポリペプチド配列は、当業者にとって既知のコンピュータ化されたデータベースで見出すことができる。このような1つのデータベースは、米国立バイオテクノロジー情報センター(National Center for Biotechnology Information)のGenbank及びGenPeptデータベースである。これらの既知の遺伝子についての核酸配列は、増幅され、本書に開示されている配列と組み合わされ(例えばライゲートされる)、かつ/又は本書で開示されている技術を用いて又は当業者にとって既知であると思われるあらゆる技術によって発現され得る(例えばサンブルックら、2001年)。核酸はインビトロ発現系の中で発現され得るが、好ましい実施形態においては、核酸はインビボ
複製及び/又は発現のためのベクターを含んでなる。
Polynucleotide and polypeptide sequences for various negative immune regulators can be found in computerized databases known to those skilled in the art. One such database is the Genbank and GenPept databases of the National Center for Biotechnology Information. Nucleic acid sequences for these known genes are amplified, combined (eg, ligated) with sequences disclosed herein, and / or using techniques disclosed herein or known to those of skill in the art. It can be expressed by any technique that appears to be present (eg, Sunbrook et al., 2001). Although the nucleic acid can be expressed in an in vitro expression system, in preferred embodiments, the nucleic acid comprises a vector for in vivo replication and / or expression.
siRNAの修飾
本発明のsiRNAポリヌクレオチドの生成の後、当業者であれば、siRNAポリヌクレオチドが治療用組成物としてsiRNAを改善するように修飾され得る或る種の特徴を有することになるということを理解するだろう。従って、siRNAポリヌクレオチドはさらに、ホスホロチオアート又はその他の連結、メチルホスホナート、スルホン、スルファート、ケチル、ホスホロジチオアート、ホスホルアミダート、リン酸エステルなどを含むようにそれを修飾することによって分解に耐えるように設計され得る(例えば、アグルワル(Agrwal)ら、1987年、Tetrahedron Lett.第28号:3539〜3542頁;ステック(Stec)ら、1985年、Tetrahedron Lett.第26号:2191〜2194頁; ムーディ(Moody)ら、1989年、Nucleic Acids Res.第12号:4769〜4782頁;エックスタイン、1989年、Trends Biol.Sci.第14号:97〜100頁;スタイン(Stein)、「Oligodeoxynucleotides:Antisense Inhibitors of Gene Expression」中、コーエン編、Macmillan Press、London、97〜117頁(1989年)参照のこと)。
Modification of siRNA After production of the siRNA polynucleotides of the present invention, one of ordinary skill in the art will have certain characteristics that siRNA polynucleotides can be modified to improve siRNA as therapeutic compositions. Will understand. Thus, the siRNA polynucleotide may be further modified to include phosphorothioate or other linkages, methylphosphonates, sulfones, sulfates, ketyls, phosphorodithioates, phosphoramidates, phosphate esters, and the like. (Eg, Agrwal et al., 1987, Tetrahedron Lett. 28: 3539-3542; Stec et al., 1985, Tetrahedron Lett. 26: 2191). Moody et al., 1989, Nucleic Acids Res. 12: 4769-4782; Xtine, 1989, Trends Biol. Sci. 14: 97-100; Tine (Stein), "Oligodeoxynucleotides: Antisense Inhibitors of Gene Expression" in, Cohen eds., Macmillan Press, London, pp. 97-117 (1989) see).
本発明の任意のポリヌクレオチドをさらに修飾してインビボでのその安定性を増大させることができる。考えられる修飾としては、5’及び/又は3’末端でのフランキング配列の添加;主鎖の中でのホスホジエステルではなくホスホロチオアート又は2’O−メチルの使用;及び/又はイノシン、キューオシン及びワイブトシンなどといった非伝統的塩基ならびにアセチル−、メチル−、チオ−及びその他の修飾された形態のアデニン、シチジン、グアニン、チミン及びウリジンが含まれるが、これに限定されるわけではない。 Any polynucleotide of the invention can be further modified to increase its stability in vivo. Possible modifications include the addition of flanking sequences at the 5 ′ and / or 3 ′ ends; the use of phosphorothioates or 2′O-methyl rather than phosphodiester in the main chain; and / or inosine, Non-traditional bases such as queuosine and wybutosine and acetyl-, methyl-, thio- and other modified forms of adenine, cytidine, guanine, thymine and uridine are included but are not limited thereto.
ベクター
その他の関連する態様では、本発明は、インヒビターをコードする単離され核酸において、好ましくは核酸によりコードされるタンパク質の発現を導く能力を核酸がもつような形でプロモータ/調節配列を含む核酸に作動的に連結された負の免疫調節因子をインヒビター好ましくはsiRNAが阻害する、単離核酸を含む。かくして、本発明は、サンブルックら、(2001年、「Molecular Cloning:A Laboratory Manual」、コールド・スプリング・ハーバー・ラボラトリー、ニューヨーク)中及びアウスベル(Ausubel)ら、(1997年、「Current Protocols in Molecular Biology」、ジョン・ウィリー・アンド・サンズ(John Wiley & Sons)、ニューヨーク)の中で記述されているもののような細胞内の外因性DNAの同時発現を伴う細胞内への外因性DNAの導入のための発現ベクター及び方法を含む。
Vectors In other related aspects, the invention provides a nucleic acid comprising a promoter / regulatory sequence in an isolated nucleic acid encoding an inhibitor, preferably in such a way that the nucleic acid has the ability to direct expression of the protein encoded by the nucleic acid. An isolated nucleic acid that inhibits a negative immunomodulator operably linked to an inhibitor, preferably an siRNA. Thus, the present invention is disclosed in Sunbrook et al. (2001, “Molecular Cloning: A Laboratory Manual”, Cold Spring Harbor Laboratory, New York) and Ausubel et al. (1997, “Current Protocols in Molecular. Biology ", introduction of exogenous DNA into cells with co-expression of exogenous DNA in cells such as those described in John Wiley & Sons, New York) Expression vectors and methods for
もう1つの態様においては、本発明はsiRNAポリヌクレオチドを含んでなるベクターを含む。好ましくは、siRNAポリヌクレオチドは、標的ポリペプチドの発現を阻害する能力をもち、ここで標的ポリペプチドはA20、SUMO(SUMO1、SUMO2、SUMO3及びSUMO4)、Foxj1、Foxo3a、TWIST(Twist1、Twist2)、SOCS(SOCS1、SOCS2、SOCS3、SOCS4、SOCS5、SOCS6、SOCS7及びCIS)、PIAS(PIAS1、PIAS3、PIASx及びPIASy)、SHP(SHP−1及びSHP−2)またはそれらの任意の組合せよりなる群から選択される。ベクター内への所望のポリヌクレオチドの取込み及びベクターの選択は例えばサンブルックら(前掲書)及びアウスベルら(前掲書)の中で記述されている通り、当該技術分野において周知である。 In another aspect, the invention includes a vector comprising an siRNA polynucleotide. Preferably, the siRNA polynucleotide has the ability to inhibit the expression of the target polypeptide, wherein the target polypeptide is A20, SUMO (SUMO1, SUMO2, SUMO3 and SUMO4), Foxj1, Foxo3a, TWIST (Twist1, Twist2), From the group consisting of SOCS (SOCS1, SOCS2, SOCS3, SOCS4, SOCS5, SOCS6, SOCS7 and CIS), PIAS (PIAS1, PIAS3, PIASx and PIASy), SHP (SHP-1 and SHP-2) or any combination thereof Selected. Incorporation of the desired polynucleotide into the vector and selection of the vector is well known in the art, as described, for example, in Sunbrook et al. (Supra) and Ausvel et al. (Supra).
siRNAポリヌクレオチドは数多くのタイプのベクターの中にクローニングされ得る。しかしながら、本発明は、いずれかの特定のベクターに限定されるものとみなされるべきではない。むしろ本発明は、当該技術分野において容易に利用可能でありかつ/又は周知である多数のベクターを包含するものと解釈されるべきである。例えば、本発明のsiRNAポリヌクレオチドは、プラスミド、ファージミド、ファージ誘導体、動物ウイルス及びコスミドを含めた(ただしこれに限定されるわけではない)ベクター内にクローニングされ得る。特に有利なベクターとしては、発現ベクター、複製ベクター、プローブ生成ベクター及び配列決定ベクターが含まれる。 siRNA polynucleotides can be cloned into many types of vectors. However, the present invention should not be regarded as limited to any particular vector. Rather, the present invention should be construed to include a number of vectors that are readily available and / or well known in the art. For example, the siRNA polynucleotides of the present invention can be cloned into vectors, including but not limited to plasmids, phagemids, phage derivatives, animal viruses and cosmids. Particularly advantageous vectors include expression vectors, replication vectors, probe generation vectors and sequencing vectors.
特定の実施形態においては、発現ベクターは、ウイルスベクター、細菌ベクター及び哺乳動物細胞ベクターよりなる群から選択される。上述の組成物の少なくとも一部分又は全てを含む数多くの発現ベクター系が存在する。ポリヌクレオチド又はその同族ポリペプチドを産生するのに本発明と共に使用する目的で、原核生物及び/又は真核生物ベクターをベースとした系を利用することができる。数多くのこのような系が市販され広く利用可能である。 In certain embodiments, the expression vector is selected from the group consisting of a viral vector, a bacterial vector, and a mammalian cell vector. There are numerous expression vector systems that include at least a portion or all of the compositions described above. Prokaryotic and / or eukaryotic vector based systems may be utilized for use with the present invention to produce polynucleotides or their cognate polypeptides. Many such systems are commercially available and widely available.
さらに、ウイルスベクターの形で細胞に対し発現ベクターを提供することができる。ウイルスベクター技術は当該技術分野において周知であり、例えばサンブルックら、(2001年)中、及びアウスベルら(1997年)中及びその他のウイルス学及び分子生物学マニュアルの中で記述されている。ベクターとして有用であるウイルスとしては、レトロウイルス、アデノウイルス、アデノ関連ウイルス、ヘルペスウイルス及びレンチウイルスが含まれるが、これに限定されるわけではない。一般に、適切なベクターは、少なくとも1つの生体の中で機能する複製起点、プロモータ配列、便利な制限エンドヌクレアーゼ部位及び1つもしくはそれ以上の選択可能なマーカーを含有する(例えば、国際公開第01/96584号パンフレット;国際公開第01/29058号パンフレット;及び米国特許第6,326,193号明細書参照)。 Furthermore, expression vectors can be provided to cells in the form of viral vectors. Viral vector technology is well known in the art and is described, for example, in Sunbrook et al. (2001), and in Außbel et al. (1997) and in other virology and molecular biology manuals. Viruses that are useful as vectors include, but are not limited to, retroviruses, adenoviruses, adeno-associated viruses, herpes viruses, and lentiviruses. In general, suitable vectors contain an origin of replication that functions in at least one organism, a promoter sequence, a convenient restriction endonuclease site, and one or more selectable markers (eg, WO01 / 96584 pamphlet; WO 01/29058 pamphlet; and US Pat. No. 6,326,193).
siRNAの発現については、各プロモータ内の少なくとも1つのモジュールが、RNA合成のための開始部位を位置づけするように機能する。これの最高の既知の例はTATAボックスであるが、哺乳動物末端デオキシヌクレオチジルトランスフェラーゼ遺伝子のためのプロモータ及びSV40遺伝子のためのプロモータなどの、TATAボックスが欠如している一部のプロモータ内では、開始部位自体の上にある離散的要素が、開始場所を定める働きをする。 For siRNA expression, at least one module within each promoter functions to position the start site for RNA synthesis. The best known example of this is the TATA box, but within some promoters lacking the TATA box, such as the promoter for the mammalian terminal deoxynucleotidyl transferase gene and the promoter for the SV40 gene, The discrete elements above the start site itself serve to define the start location.
付加的なプロモータ要素すなわちエンハンサは、転写開始頻度を調節する。標準的には、これらは開始部位の上流側30〜110bpの領域に場所設定されるが、数多くのプロモータが開始部位の下流側にも機能的要素を含有することが近年示されてきた。プロモータ要素間の間隔どりは往々にして融通性があり、従って、要素が互いとの関係において逆転されるか又は移動された時点でプロモータ機能が保たれるようになっている。チミジンキナーゼ(tk)プロモータにおいては、プロモータ要素間の間隔どりは、活性が衰え始める前に50bp離隔するまで増大させることができる。プロモータに応じて、転写を活性化させるために個々の要素が協働的に又は独立して機能できると思われる。 Additional promoter elements or enhancers regulate the frequency of transcription initiation. Typically these are located in the region 30-110 bp upstream of the start site, but it has recently been shown that many promoters also contain functional elements downstream of the start site. The spacing between the promoter elements is often flexible so that the promoter function is maintained when the elements are reversed or moved in relation to each other. In the thymidine kinase (tk) promoter, the spacing between the promoter elements can be increased until they are separated by 50 bp before activity begins to decline. Depending on the promoter, it appears that individual elements can function cooperatively or independently to activate transcription.
プロモータは、コーディングセグメント及び/又はエキソンの上流側にある5’非コーディング配列を単離することにより得ることができるような、遺伝子又はポリヌクレオチド配列と天然に会合されたものであり得る。このようなプロモータは「内因性」プロモータと呼ぶことができる。同様にして、エンハンサは、その配列の下流側又は上流側のいずれかに位置設定されたポリヌクレオチド配列と天然に会合されたものであり得る。あるいは、通常はその天然の環境においてポリヌクレオチド配列と会合されていないプロモータを意味する組換え型又は非相同プロモータの制御下にコーディングポリヌクレオチドセグ
メントを位置づけすることによって、一部の利点が得られることになる。組換え型又は非相同エンハンサは同様に、通常はその天然の環境においてポリヌクレオチド配列と会合されていないエンハンサを意味する。このようなプロモータ又はエンハンサとしては、その他の遺伝子のプロモータ及びエンハンサ、及びその他のあらゆる原核生物、ウイルス又は真核生物細胞から単離されたプロモータ又はエンハンサ、及び「天然発生の」ものでない、すなわち異なる転写調節領域の異なる要素及び/又は発現を改変させる突然変異を含有するプロモータ又はエンハンサが含まれ得る。プロモータ及びエンハンサの核酸配列を合成的に産生させることに加えて、本書で開示されている組成物と関連させてPCRTMを含めた組換え体クローニング及び/又は核酸増幅技術を用いて配列を産生させることも可能である(米国特許第4,683,202号明細書、米国特許第5,928,906号明細書)。さらに、ミトコンドリア、葉緑体などといった非核オルガネラの内部での配列の転写及び/又は発現を導く制御配列も同様に利用可能であると考えられている。
A promoter can be one that is naturally associated with a gene or polynucleotide sequence, such as can be obtained by isolating a coding segment and / or a 5 ′ non-coding sequence upstream of an exon. Such promoters can be referred to as “endogenous” promoters. Similarly, an enhancer can be one that is naturally associated with a polynucleotide sequence located either downstream or upstream of the sequence. Alternatively, some advantage may be gained by positioning the coding polynucleotide segment under the control of a recombinant or heterologous promoter that usually means a promoter not associated with the polynucleotide sequence in its natural environment. become. A recombinant or heterologous enhancer also refers to an enhancer that is not normally associated with a polynucleotide sequence in its natural environment. Such promoters or enhancers include promoters and enhancers of other genes, and promoters or enhancers isolated from any other prokaryotic, viral or eukaryotic cell, and are not “naturally occurring”, ie different Promoters or enhancers containing mutations that alter different elements of the transcriptional regulatory region and / or expression may be included. In addition to synthetically producing promoter and enhancer nucleic acid sequences, the sequences are produced using recombinant cloning and / or nucleic acid amplification techniques, including PCR ™ , in conjunction with the compositions disclosed herein. (US Pat. No. 4,683,202, US Pat. No. 5,928,906). In addition, it is believed that regulatory sequences leading to transcription and / or expression of sequences within non-nuclear organelles such as mitochondria and chloroplasts can be used as well.
当然のことながら、発現のために選ばれた細胞型、オルガネラ及び生体内でDNAセグメントの発現を有効に導くプロモータ及び/又はエンハンサを利用することが重要になる。分子生物学の当業者は一般に、タンパク質の発現のためのプロモータ、エンハンサ及び細胞型の組合せをいかに使用するかを知っている。例えばサンブルックら、(2001年)を参照のこと。利用されるプロモータは、組換え型タンパク質及び/又はペプチドの大規模産生において有利であるような、導入されたDNAセグメントの高レベル発現を導くために適切な条件下で有用でありかつ/又は構成性、組織特異的、誘発型であり得る。プロモータは非相同的又は内因性であり得る。 Of course, it will be important to utilize a promoter and / or enhancer that effectively directs the expression of the DNA segment in the cell type, organelle and organism chosen for expression. Those skilled in molecular biology generally know how to use a combination of promoters, enhancers and cell types for the expression of proteins. See, for example, Sunbrook et al. (2001). The promoter utilized is useful and / or configured under appropriate conditions to direct high level expression of the introduced DNA segment, which is advantageous in large scale production of recombinant proteins and / or peptides. It can be sex, tissue specific, or induced. Promoters can be heterologous or endogenous.
本書に提示されている実験例の中で例示されているプロモータ配列は、初前期サイトメガロウイルス(CMV)プロモータ配列である。このプロモータ配列は、作動的に連結されたあらゆるポリヌクレオチド配列の高レベルの発現を駆動する能力をもつ強い構成性プロモータ配列である。しかしながら、シミアンウイルス40(SV40)早期プロモータ、マウス乳癌ウイルス(MMTV)、ヒト免疫不全ウイルス(HIV)長末端反復(LTR)プロモータ、モロニーウイルスプロモータ、トリ白血病ウイルスプロモータ、エプスタイン・バーウイルス初前期プロモータ、ラウス肉腫ウイルスプロモータならびにヒト遺伝子プロモータ例えば(ただしこれに限定されるわけではない)アクチンプロモータ、ミオシンプロモータ、ヘモグロビンプロモータ及び筋肉クレアチンプロモータを含めた(ただしこれに限定されるわけではない)その他の構成性プロモータ配列も同様に使用可能である。さらに、本発明は、構成性プロモータの使用に限定されるべきではない。本発明の一部分として誘発性プロモータも同様に考慮されている。本発明における誘発性プロモータの使用は、発現が望まれる場合に作動的に連結されているポリヌクレオチド配列の発現をオンに切換え、発現が望ましくない場合発現をオフに切換えることのできる分子スイッチを提供する。誘発性プロモータの例としては、メタロチオニンプロモータ、グルココルチコイドプロモータ、プロゲストロンプロモータ及びテトラサイクリンプロモータが含まれるが、これに限定されるわけではない。さらに、本発明は、所望の組織内のみで活性である組織特異的プロモータの使用を含む。組織特異的プロモータは、当該技術分野において周知であり、HER−2プロモータ及びPSA関連プロモータ配列がこれに含まれるが、これに限定されるわけではない。 The promoter sequence illustrated in the experimental examples presented herein is the early early cytomegalovirus (CMV) promoter sequence. This promoter sequence is a strong constitutive promoter sequence with the ability to drive high level expression of any operably linked polynucleotide sequence. However, Simian virus 40 (SV40) early promoter, mouse mammary tumor virus (MMTV), human immunodeficiency virus (HIV) long terminal repeat (LTR) promoter, Moloney virus promoter, avian leukemia virus promoter, Epstein-Barr virus early promoter, Other constitutive properties including (but not limited to) the rous sarcoma virus promoter and the human gene promoter including but not limited to the actin promoter, myosin promoter, hemoglobin promoter and muscle creatine promoter. Promoter sequences can be used as well. Furthermore, the present invention should not be limited to the use of constitutive promoters. Inducible promoters are considered as part of the present invention. The use of an inducible promoter in the present invention provides a molecular switch that can turn on expression of a polynucleotide sequence that is operably linked when expression is desired, and turn off expression when expression is not desired. To do. Examples of inducible promoters include, but are not limited to, metallothionine promoter, glucocorticoid promoter, progesterone promoter and tetracycline promoter. Furthermore, the present invention includes the use of tissue specific promoters that are active only in the desired tissue. Tissue specific promoters are well known in the art and include, but are not limited to, the HER-2 promoter and PSA-related promoter sequences.
siRNAの発現を査定するため、1つの細胞中に導入すべき発現ベクターは同様に、ウイルスベクターの同定及び選択を容易にするべく選択可能なマーカー遺伝子又はリポータ遺伝子のいずれか又はその両方をも含有することができる。その他の実施形態においては、選択可能なマーカーは、別のDNA片上に担持され、同時トランスフェクション手順内で使用され得る。選択可能なマーカー及びリポータ遺伝子の両方共が、宿主細胞内での発現を可能にするため適切な調節配列でフランキングされ得る。当該技術分野においては有用な選択可能なマーカーが知られており、例えばneoなどといった抗生物質耐性遺伝
子が含まれる。
To assess siRNA expression, the expression vector to be introduced into one cell also contains either a selectable marker gene or reporter gene or both to facilitate identification and selection of viral vectors. can do. In other embodiments, the selectable marker can be carried on a separate piece of DNA and used in a co-transfection procedure. Both the selectable marker and the reporter gene can be flanked with appropriate regulatory sequences to allow expression in the host cell. Useful selectable markers are known in the art and include antibiotic resistance genes such as neo.
潜在的にトランスフェクションを受けた細胞を同定するため及び調節配列の機能性を評価するために、リポータ遺伝子が使用される。容易に検定可能なタンパク質についてコードするリポータ遺伝子は当該技術分野において周知である。一般に、リポータ遺伝子は、レシピエント生体又は組織の中に存在しないか又はそれらにより発現されず、例えば酵素活性などのいくつかの容易に検出可能な特性によってその発現が明らかになるタンパク質をコードする遺伝子である。リポータ遺伝子の発現は、DNAがレシピエント細胞内に導入されてから適切な時間後に検定される。 Reporter genes are used to identify potentially transfected cells and to evaluate the functionality of regulatory sequences. Reporter genes encoding for readily assayable proteins are well known in the art. In general, a reporter gene is a gene that encodes a protein that is not present in or expressed by the recipient organism or tissue, and whose expression is revealed by some readily detectable property, such as enzymatic activity, for example. It is. Reporter gene expression is assayed an appropriate time after the DNA is introduced into the recipient cell.
適切なリポータ遺伝子としては、ルシフェラーゼ、ベーターガラクトシダーゼ、クロラムフェニコールアセチルトランスフェラーゼ、分泌されたアルカリホスファターゼをコードする遺伝子又は緑色螢光タンパク質遺伝子が含まれ得る(例えば、ウイ・テイ(Ui−Tei)ら、2000年、FEBS Lett.第479号:79〜82頁参照)。適切な発現系は周知であり、周知の技術を用いて調製されるか又は商業的に入手可能である。内部欠失構成体は、固有内部制限部位を用いて、又は非固有制限部位の部分的消化より生成され得る。構成体はこのとき、高レベルのsiRNAポリヌクレオチド及び/又はポリペプチド発現を標示する細胞内にトランスフェクトされ得る。一般に、リポータ遺伝子の最高レベルの発現を示す最小限の5’フランキング領域を伴う構成体は、プロモータとして同定される。このようなプロモータ領域はリポータ遺伝子に連結され、プロモータ駆動型転写を変調させる能力について作用物質を評価するのに用いられ得る。 Suitable reporter genes may include luciferase, beta-galactosidase, chloramphenicol acetyltransferase, a gene encoding secreted alkaline phosphatase or a green fluorescent protein gene (eg, Ui-Tei et al. 2000, FEBS Lett. No. 479: 79-82). Suitable expression systems are well known and are prepared using well known techniques or are commercially available. Internal deletion constructs can be generated using unique internal restriction sites or by partial digestion of non-unique restriction sites. The construct can then be transfected into cells that display high levels of siRNA polynucleotide and / or polypeptide expression. In general, a construct with a minimal 5 'flanking region that exhibits the highest level of expression of a reporter gene is identified as a promoter. Such a promoter region is linked to a reporter gene and can be used to evaluate an agent for its ability to modulate promoter-driven transcription.
サイレンシング化された免疫細胞の生成
一実施形態においては、本発明は、細胞内に負の免疫調節因子のインヒビターを発現させるための細胞ベースの系を提供している。細胞ベースの系は、「サイレンシング化された細胞」を意味し、細胞とインヒビターを発現するための発現ベクターを含んでなる。しかしながら、本発明は、発現ベクターを含んでなる細胞に限定されず、むしろ本発明のサイレンシング化された細胞は、本発明のあらゆるタイプのインヒビター、すなわち化学合成されたsiRNAで修飾された細胞を含むものとして解釈されるべきである。いずれにせよ、インヒビターを含んでなるサイレンシング化された細胞は、その他の点では同一であるもののこのようにインヒビターでのサイレンシング化を受けていない細胞に比べて高い免疫能力を有している。サイレンシング化された細胞は、単独で又はその他の療法と組合せた形で、哺乳動物のレシピエントに投与するのに適している。
Generating Silenced Immune Cells In one embodiment, the present invention provides a cell-based system for expressing inhibitors of negative immune modulators in cells. A cell-based system means a “silenced cell” and comprises an expression vector for expressing the cell and the inhibitor. However, the present invention is not limited to cells comprising an expression vector; rather, the silenced cells of the present invention include any type of inhibitor of the present invention, ie cells modified with chemically synthesized siRNA. Should be construed as including. In any case, a silenced cell comprising an inhibitor has a higher immunity compared to a cell that is otherwise identical but not yet silenced with an inhibitor. . The silenced cells are suitable for administration to a mammalian recipient, either alone or in combination with other therapies.
本発明は、負の免疫調節因子のインヒビターを含む細胞を含む。一態様においては、インヒビターは、A20、SUMO(SUMO1、SUMO2、SUMO3及びSUMO4)、Foxj1、Foxo3a、TWIST(Twist1、Twist2)、SOCS(SOCS1、SOCS2、SOCS3、SOCS4、SOCS5、SOCS6、SOCS7及びCIS)、PIAS(PIAS1、PIAS3、PIASx及びPIASy)、SHP(SHP−1及びSHP−2)などのうちの少なくとも1つを阻害する能力を有する。一態様においては、細胞は、インヒビターをコードするポリヌクレオチドを含んでなるベクターでのトランスフェクションを受けることができる。ポリヌクレオチドは細胞内に組込まれている必要はない。もう1つの態様においては、細胞がベクターによるトランスフェクションを受けている必要は全くなく、むしろ、細胞はベクターから発現されないインヒビターに曝露される。かかるインヒビターの一例は、化学的に合成されたsiRNAである。 The present invention includes cells comprising inhibitors of negative immune modulators. In one embodiment, the inhibitor is A20, SUMO (SUMO1, SUMO2, SUMO3 and SUMO4), Foxj1, Foxo3a, TWIST (Twist1, Twist2), SOCS (SOCS1, SOCS2, SOCS3, SOCS4, SOCS5, SOCS6, SOCS7, SOCS7). , Have the ability to inhibit at least one of PIAS (PIAS1, PIAS3, PIASx and PIASy), SHP (SHP-1 and SHP-2), and the like. In one aspect, the cells can be transfected with a vector comprising a polynucleotide encoding the inhibitor. The polynucleotide need not be incorporated into the cell. In another embodiment, the cell need not have been transfected with the vector, but rather, the cell is exposed to an inhibitor that is not expressed from the vector. An example of such an inhibitor is a chemically synthesized siRNA.
発現ベクターに関連して、ベクターは、当該技術分野のいずれかの方法により、例えば哺乳動物、細菌、酵母又は昆虫の細胞といった宿主細胞内に容易に導入され得る。例えば発現ベクターは、物理的、化学的又は生物学的手段により宿主細胞内に移入され得る。本
発明のポリヌクレオチドを含んでなる発現ベクターの導入が、負の免疫調節因子との関係においてサイレンシング化された細胞を生み出すということは容易に理解できる。
In the context of an expression vector, the vector can be easily introduced into a host cell, such as a mammalian, bacterial, yeast or insect cell, by any method in the art. For example, an expression vector can be transferred into a host cell by physical, chemical or biological means. It can be readily appreciated that the introduction of an expression vector comprising a polynucleotide of the present invention produces a cell that is silenced in the context of a negative immune regulator.
宿主細胞内にポリヌクレオチドを導入するための物理的方法としては、リン酸カルシウム沈降、リポフェクション、粒子衝突、マイクロインジェクション、電気穿孔などが含まれる。ベクター及び/又は外因性核酸を含んでなる細胞を産生するための方法は、当該技術分野において周知である。例えばサンブルックら、(2001年、「Molecular Cloning:A Laboratory Manual」、コールド・スプリング・ハーバー・ラボラトリー、ニューヨーク)及びアウスベルら、(1997年、「Current Protocols in Molecular Biology」、ジョン・ウィリー・アンド・サンズ、ニューヨーク)中を参照のこと。 Physical methods for introducing polynucleotides into host cells include calcium phosphate precipitation, lipofection, particle bombardment, microinjection, electroporation, and the like. Methods for producing cells comprising vectors and / or exogenous nucleic acids are well known in the art. For example, Sunbrook et al. (2001, “Molecular Cloning: A Laboratory Manual”, Cold Spring Harbor Laboratory, New York) and Ausbell et al. (1997, “Current Protocols in Molecular Biology”, John Willy and. (Sands, New York).
宿主細胞内に有利なポリヌクレオチドを導入するための生物学的方法としては、DNA及びRNAベクターの使用が含まれる。ウイルスベクター特にレトロウイルスベクターは、哺乳動物例えばヒトの細胞内に遺伝子を挿入するための最も広く用いられている方法となっている。その他のウイルスベクターは、レンチウイルス、ポックスウイルス、単純ヘルペスウイルスI、アデノウイルス及びアデノ関連ウイルスなどから誘導可能である。例えば、米国特許第5,350,674号明細書及び第5,585,362号明細書を参照のこと。 Biological methods for introducing advantageous polynucleotides into host cells include the use of DNA and RNA vectors. Viral vectors, particularly retroviral vectors, have become the most widely used method for inserting genes into mammalian cells such as humans. Other viral vectors can be derived from lentiviruses, poxviruses, herpes simplex virus I, adenoviruses and adeno-associated viruses. See, for example, US Pat. Nos. 5,350,674 and 5,585,362.
宿主細胞内にポリヌクレオチドを導入するための化学的手段には、巨大分子複合体、ナノカプセル、ミクロスフェア、ビーズ、及び水中油エマルジョン、ミセル、混合ミセル及びリポソームを含めた脂質ベースの系などのコロイド分散系が含まれる。インビトロ及びインビボでの送達ビヒクルとして使用するための好ましいコロイド系が、リポソーム(すなわち人工膜小胞)である。かかる系の調製及び用途は、当該技術分野において周知である。 Chemical means for introducing polynucleotides into host cells include macromolecular complexes, nanocapsules, microspheres, beads, and lipid-based systems including oil-in-water emulsions, micelles, mixed micelles and liposomes. Colloidal dispersions are included. A preferred colloidal system for use as a delivery vehicle in vitro and in vivo is a liposome (ie, an artificial membrane vesicle). The preparation and use of such systems is well known in the art.
宿主細胞内へ外因性核酸を導入するか又はその他の形で細胞を本発明のインヒビターに曝露して宿主細胞内の組換え型DNA配列の存在を確認するのに用いられる方法の如何に関わらず、さまざまな検定を実施することができる。かかる検定には、例えば、サザン及びノーザンブロット法、RT−PCR及びPCRなどの当業者にとって周知の「分子生物学」検定;例えば本発明の範囲内に入る作用物質を同定するための本書で記述された検定によってか又は免疫学的手段(ELISA及びウェスタンブロット)によって特定のペプチドの有無を検出することなどの「生化学的」検定が含まれる。 Regardless of the method used to introduce exogenous nucleic acid into the host cell or otherwise expose the cell to the inhibitors of the present invention to confirm the presence of the recombinant DNA sequence in the host cell. Various tests can be performed. Such assays include, for example, “molecular biology” assays well known to those skilled in the art, such as Southern and Northern blots, RT-PCR and PCR; described herein for identifying agents that fall within the scope of the invention Included are “biochemical” assays such as detecting the presence or absence of a particular peptide by an assay performed or by immunological means (ELISA and Western blot).
サイレンシング化された細胞を生成するためには、インビトロ又はインビボで免疫細胞まで所望のsiRNAポリヌクレオチドを移送するべく、任意のDNAベクター又は送達ビヒクルを利用することができる。非ウイルス送達系が利用される場合、好ましい送達ビヒクルはリポソームである。従って、上述の送達系及びプロトコルは、「Gene Targeting Protocols」、第2版、1〜35頁(2002年)及び「Gene Transfer and Expression Protocols」、第7巻、マリー(Murray)編、81〜89頁(1991年)の中に見出すことができる。 To generate a silenced cell, any DNA vector or delivery vehicle can be utilized to transfer the desired siRNA polynucleotide to the immune cell in vitro or in vivo. When non-viral delivery systems are utilized, the preferred delivery vehicle is a liposome. Thus, the delivery systems and protocols described above are described in “Gene Targeting Protocols”, 2nd edition, pages 1-35 (2002) and “Gene Transfer and Expression Protocols”, Vol. 7, edited by Murray, 81-89. Can be found in the page (1991).
宿主細胞内に(インビトロ、エクスビボ又はインビボで)本発明の負の免疫調節因子のインヒビターを導入するために、脂質処方物の使用が考慮されている。本発明の特定の実施形態においては、インヒビターを脂質と会合させることが可能である。脂質と会合したインヒビターは、リポソームの水性内部の中にカプセル化されるか、リポソームの脂質2重層内に散在させられるか、リポソーム及びオリゴヌクレオチドの両方と会合させられた連結用分子を介してリポソームに付着させられるか、リポソーム内に取り込まれるか、リ
ポソームと複合されるか、脂質を含有する溶液中に分散されるか、脂質と混合されるか、脂質と組み合わされるか、脂質内に懸濁物として含有されるか、ミセルと共に含有されるか複合されるか複合されるか又はその他の形で脂質と会合させられ得る。本発明の脂質、脂質/siRNA又は脂質/発現ベクター会合組成物は、溶解状態の任意の特定の構造に限定されない。例えば、2重層構造内で、ミセルとして、又は「崩壊した」構造を伴って存在し得る。これらは又単に溶液中に散在させられ、場合によってはサイズ的又は形状的に均一でない凝集体を形成し得る。
The use of lipid formulations is contemplated to introduce inhibitors of the negative immunomodulators of the present invention into host cells (in vitro, ex vivo or in vivo). In certain embodiments of the invention, the inhibitor can be associated with a lipid. Inhibitors associated with lipids are either encapsulated within the aqueous interior of the liposome, dispersed within the lipid bilayer of the liposome, or via a linking molecule associated with both the liposome and the oligonucleotide. Attached to, incorporated into liposomes, complexed with liposomes, dispersed in a solution containing lipids, mixed with lipids, combined with lipids, suspended in lipids It can be contained as a product, contained with micelles, complexed, complexed or otherwise associated with lipids. The lipid, lipid / siRNA or lipid / expression vector association composition of the present invention is not limited to any particular structure in solution. For example, within a bilayer structure, it may exist as micelles or with a “collapsed” structure. They can also be simply scattered in the solution and in some cases form aggregates that are not uniform in size or shape.
脂質は、天然発生の又は合成の脂質であり得る脂肪性物質である。例えば、脂質には、細胞質内に天然に発生する脂肪性液滴ならびに、長鎖脂肪族炭化水素及びその誘導体を含有する、当業者にとって周知の化合物の種類、例えば脂肪酸、アルコール、アミン、アミノアルコール及びアルデヒトが含まれる。 Lipids are fatty substances that can be naturally occurring or synthetic lipids. For example, lipids include naturally occurring fatty droplets in the cytoplasm, as well as types of compounds well known to those skilled in the art, including long chain aliphatic hydrocarbons and derivatives thereof, such as fatty acids, alcohols, amines, amino alcohols. And aldehyde.
本発明に係るリポソームを調製するために、リン脂質を使用することができ、これは、正味の正電荷、負電荷又は中性電荷を担持し得る。リポソーム上に負電荷を付与するためにはリン酸ジアセチルを利用することができ、リポソーム上に正電荷を付与するためには、ステアリルアミンを使用することができる。リポソームは、1つもしくはそれ以上のリン脂質からできている可能性がある。 Phospholipids can be used to prepare the liposomes according to the present invention, which can carry a net positive charge, a negative charge or a neutral charge. Diacetyl phosphate can be used to impart a negative charge on the liposome, and stearylamine can be used to impart a positive charge on the liposome. Liposomes can be made of one or more phospholipids.
中性荷電脂質は、無電荷の脂質、実質的に非荷電の脂質又は正と負の電荷を同数有する脂質混合物を含んでなり得る。適切なリン脂質には、当業者にとって周知であるホスファチジルコリン及びその他は含まれる。 Neutral charged lipids can comprise uncharged lipids, substantially uncharged lipids or lipid mixtures having the same number of positive and negative charges. Suitable phospholipids include phosphatidylcholine and others well known to those skilled in the art.
本発明に従って用途に適した脂質は、商業的供給源から入手可能である。例えば、ジミリスチルホスファチジルコリン(「DMPC」)はシグマ・ケミカル・カンパニー(Sigma Chemical Co.)(ミズーリ州セントルイス(St.Louis,MO))から入手可能であり、リン酸ジセチル(「DCP」)はKアンドKラボラトリーズ(K & K Laboratories)(ニューヨーク州プレインビュー(Plainview,NY))から入手され;コレステロール(「Chol」)はカルビオケム・ベーリング(Calbiochem−Behring)から入手され;ジミリスチルホスファチジルグリセロール(「DMPG」)及びその他の脂質はアヴァンティ・ポーラー・リピズ社(Avanti Polar Lipids,Inc.)(アラバマ州バーミングハム(Birmingham,AL))から入手され得る。クロロホルム又はクロロホルム/メタノール中の脂質の原液は約−20℃で貯蔵できる。好ましくは、メタノールよりも容易に蒸発することから、クロロホルムが唯一の溶媒として用いられる。 Lipids suitable for use according to the present invention are available from commercial sources. For example, dimyristylphosphatidylcholine (“DMPC”) is available from Sigma Chemical Co. (St. Louis, Mo.) and dicetyl phosphate (“DCP”) is available from K And K Laboratories (Plainview, NY); cholesterol (“Chol”) is obtained from Calbiochem-Behring; dimyristyl phosphatidylglycerol (“DMPG”) )) And other lipids are available from Avanti Polar Lipids, Inc. (Birmi, Alabama). Gham, may be obtained from the AL)). Stock solutions of lipids in chloroform or chloroform / methanol can be stored at about -20 ° C. Preferably, chloroform is used as the only solvent because it evaporates more easily than methanol.
卵又は大豆ホスファチジルコリン、脳ホスファチジン酸、脳又は植物ホスファチジルイノシトール、心臓カルジオリピン及び植物又は細菌ホスファチジルエタノールアミンなどの天然供給源由来のリン脂質は、結果として得られるリポソームが不安定であり漏出しやすいことから、一次ホスファチドすなわち全ホスファチド組成物の50%以上を構成するホスファチドとしては好ましくは使用されない。 Phospholipids from natural sources such as egg or soy phosphatidylcholine, brain phosphatidic acid, brain or plant phosphatidylinositol, heart cardiolipin and plant or bacterial phosphatidylethanolamine can result in unstable and prone to leakage of the resulting liposomes It is preferably not used as the primary phosphatide, ie the phosphatide constituting more than 50% of the total phosphatide composition.
「リポソーム」というのは、封入された脂質2重層又は凝集体の生成によって形成されるさまざまな単一及び多重膜脂質ビヒクルを包含する包括的用語である。リポソームは、リン脂質2重層膜及び内部水性媒質を伴う小胞状構造をもつものとして特徴づけされ得る。多重膜リポソームは、水性媒質により分離された多重脂質層を有する。これらは、リン脂質が過剰な水溶液中に懸濁させられている場合に自然に形成する。脂質構成要素は、閉鎖構造の形成前に自己再配置を受け、脂質2重層の間に水及び溶解した溶質をカプセル化する(ゴッシュ(Ghosh)及びバチャワット(Bachhawat)、1991年)。しかしながら、本発明はまた、通常の小胞状構造と異なる溶解状態の構造を有する組成
物をも包含する。例えば、脂質は、ミセル構造をとることもでき、又単に脂質分子の不均一な凝集体として存在することもできる。同様に考慮されているのは、リポフェクタミン−核酸複合体である。
“Liposome” is a generic term encompassing various single and multilamellar lipid vehicles formed by the formation of encapsulated lipid bilayers or aggregates. Liposomes can be characterized as having a vesicular structure with a phospholipid bilayer membrane and an internal aqueous medium. Multilamellar liposomes have multiple lipid layers separated by an aqueous medium. These form spontaneously when phospholipids are suspended in an excess of aqueous solution. The lipid component undergoes self-rearrangement prior to the formation of a closed structure, encapsulating water and dissolved solutes between the lipid bilayers (Ghosh and Bachhawat, 1991). However, the present invention also includes a composition having a dissolved structure different from a normal vesicular structure. For example, lipids can have a micellar structure or can simply exist as heterogeneous aggregates of lipid molecules. Also considered are lipofectamine-nucleic acid complexes.
リン脂質は、脂質対水のモル比に応じて、水中に分散された場合にリポソーム以外のさまざまな構造を形成することができる。低い比率では、リポソームは好ましい構造である。リポソームの物理的特徴は、pH、イオン強度及び/又は2価のカチオンの存在に左右される。リポソームはイオン及び/又は極性物質に対する低い浸透性を示し得るが、高温では、その浸透性を著しく改変する相転移を受ける。相転移には、ゲル状態として知られる密に詰まった秩序構造から流体状態として知られる緩く詰まったあまり秩序立っていない構造までが関与する。これは、特徴的な相転移温度で起こり、かつ/又はイオン、糖及び/又は薬物への浸透性の増加を結果としてもたらす。 Phospholipids can form various structures other than liposomes when dispersed in water, depending on the molar ratio of lipid to water. At low ratios, liposomes are the preferred structure. The physical characteristics of liposomes depend on pH, ionic strength and / or the presence of divalent cations. Liposomes may exhibit low permeability to ions and / or polar substances, but at high temperatures undergo a phase transition that significantly modifies their permeability. Phase transitions involve from tightly packed ordered structures known as gel states to loosely packed and less ordered structures known as fluid states. This occurs at a characteristic phase transition temperature and / or results in increased permeability to ions, sugars and / or drugs.
リポソームは、4つの異なる機序すなわちマクロファージ及び/又は好中球などの細網内皮系の食細胞によるエンドサイトーシス;非特異的弱疎水性及び/又は静電気力、及び/又は細胞−表面構成要素との特異的相互作用のいずれかによる細胞表面への吸着;細胞質内へのリポソームの中味の同時放出を伴う、原形質膜内へのリポソームの脂質2重層の挿入によるプラズマ細胞膜との融合;及び/又はリポソームの中味の会合無しの、細胞及び/又は細胞内膜へのリポソーム脂質の移送及び/又はその逆によるもの、といった機序を介して細胞と相互作用する。リポソーム処方を変動させることにより、どの機序が作動状態にあるかを改変させることができるが、2つ以上が同時に作動することもできる。 Liposomes have four different mechanisms: endocytosis by phagocytic cells of the reticuloendothelial system such as macrophages and / or neutrophils; nonspecific weakly hydrophobic and / or electrostatic forces, and / or cell-surface components. Adsorption to the cell surface by either specific interaction with the plasma; fusion with the plasma cell membrane by insertion of a lipid bilayer of the liposome into the plasma membrane with simultaneous release of the contents of the liposome into the cytoplasm; and It interacts with cells via mechanisms such as by transfer of liposomal lipids to cells and / or intracellular membranes and / or vice versa, without association of liposome contents. Varying the liposome formulation can alter which mechanism is in operation, but two or more can also operate simultaneously.
インビトロでの外来性DNAの発現及びリポソーム媒介型オリゴヌクレオチド送達はきわめて好首尾であった。ウォン(Wong)ら、(1980年)は、培養されたニワトリ胚、HeLa及びヘパトーマ細胞内での外来性DNAの発現及びリポソーム媒介型送達の実現可能性を実証した。ニコラウ(Nicolau)ら、(1987年)は、静脈注射後にラット内でリポソーム媒介型遺伝子移入の成功を収めた。 In vitro expression of exogenous DNA and liposome-mediated oligonucleotide delivery have been very successful. Wong et al. (1980) demonstrated the feasibility of exogenous DNA expression and liposome-mediated delivery in cultured chicken embryos, HeLa and hepatoma cells. Nicolau et al. (1987) have had successful liposome-mediated gene transfer in rats after intravenous injection.
本発明の一部の実施形態においては、脂質は、センダイウイルス(HVJ)と会合され得る。これは、細胞膜との融合を容易にし、リポソームカプセル化DNAの細胞進入を促進するということが示されてきた(カネダ(Kaneda)ら、1989年)。その他の実施形態においては、脂質は、核非ヒストン染色体タンパク質(HMG−1)と複合又は併用され得る(カトー(Kato)ら、1991年)。さらにもう1つの実施形態においては、脂質は、HVJ及びHMG−1の両方と複合されるか又は併用され得る。このような発現ベクターがインビトロ及びインビボでのオリゴヌクレオチドの移入及び発現においてうまく利用されてきたことから、今度はそれらを本発明のために応用することができる。細菌プロモータがDNA構成体において利用される場合には、リポソーム内部に適切な細菌ポリメラーゼを含み入れることも望ましくなる。 In some embodiments of the invention, the lipid may be associated with Sendai virus (HVJ). This has been shown to facilitate fusion with the cell membrane and promote cell entry of liposome-encapsulated DNA (Kaneda et al., 1989). In other embodiments, the lipid can be complexed or combined with nuclear non-histone chromosomal protein (HMG-1) (Kato et al., 1991). In yet another embodiment, the lipid can be complexed or combined with both HVJ and HMG-1. Since such expression vectors have been successfully used in oligonucleotide transfer and expression in vitro and in vivo, they can now be applied for the present invention. If a bacterial promoter is utilized in the DNA construct, it may also be desirable to include an appropriate bacterial polymerase inside the liposome.
本発明に従って用いられるリポソームは、異なる方法で作ることができる。リポソームのサイズは、合成方法によって変動する。水溶液中に懸濁されたリポソームは一般に、脂質2重層分子の1つもしくはそれ以上の同心層を有する球形小胞の形をしている。各層は、Xが親水性部分でありYが疎水性部分であるものとして構造式XYで表わされる分子の平行な列からなる。水性懸濁液中では、同心層は、親水性部分が水相と接触した状態にとどまる傾向をもちかつ疎水性領域が自己会合する傾向をもつように配置されている。例えば、リポソーム内部及びリポソーム限外の両方で水相が存在する場合、脂質分子は、XY−YXという配置のラメラとして知られる2重層を形成し得る。2つ以上の脂質分子の親水性及び疎水性部分が互いに会合状態となった時点で、脂質凝集体が形成し得る。これらの凝集体のサイズ及び形状は、溶媒の性質及び溶液中のその他の化合物の存在などの数多くの異なる変数により左右されることになる。 Liposomes used according to the present invention can be made in different ways. The size of the liposome varies depending on the synthesis method. Liposomes suspended in an aqueous solution are generally in the form of spherical vesicles having one or more concentric layers of lipid bilayer molecules. Each layer consists of parallel rows of molecules represented by structural formula XY where X is a hydrophilic moiety and Y is a hydrophobic moiety. In aqueous suspension, the concentric layers are arranged so that the hydrophilic portion tends to stay in contact with the aqueous phase and the hydrophobic regions tend to self-associate. For example, when an aqueous phase is present both inside and outside the liposome, lipid molecules can form a bilayer known as a lamella with the configuration XY-YX. Lipid aggregates can form when the hydrophilic and hydrophobic portions of two or more lipid molecules become associated with each other. The size and shape of these aggregates will depend on many different variables such as the nature of the solvent and the presence of other compounds in the solution.
本発明の範囲内に入るリポソームは、既知の実験室技術に従って調製可能である。1つの好ましい実施形態においては、例えばガラス、梨形フラスコといったコンテナの中で溶媒中にリポソーム脂質を混合することにより、リポソームが調製される。コンテナは、予想されるリポソーム懸濁液の体積より10倍大きい体積を有するべきである。回転蒸発器を用いて、負の圧力下にて約40℃で溶媒が除去される。溶媒は通常、リポソームの所望の体積に応じて約5分乃至2時間以内に除去される。組成物は、さらに真空下のデシケータ内で乾燥され得る。乾燥された脂質は一般に、経時的に劣化する傾向があることから、約一週間後に処分される。 Liposomes falling within the scope of the invention can be prepared according to known laboratory techniques. In one preferred embodiment, liposomes are prepared by mixing liposomal lipids in a solvent in a container such as a glass, pear flask. The container should have a volume that is 10 times greater than the expected volume of the liposome suspension. The solvent is removed at about 40 ° C. under negative pressure using a rotary evaporator. The solvent is usually removed within about 5 minutes to 2 hours, depending on the desired volume of the liposomes. The composition can be further dried in a desiccator under vacuum. Dried lipids are generally disposed after about a week because they tend to degrade over time.
乾燥した脂質は、全ての脂質薄膜が再懸濁させられるまで振とうすることにより、無菌で発熱物質を含まない水中で約25〜50mMのリン脂質にて水和させることができる。このとき、リポソーム水をアリコートに分離し、各々バイアル内に置き、凍結乾燥させ、真空下で密封することができる。 The dried lipid can be hydrated with about 25-50 mM phospholipid in sterile, pyrogen-free water by shaking until all lipid film is resuspended. At this time, the liposomal water can be separated into aliquots, each placed in a vial, lyophilized, and sealed under vacuum.
代替案としては、リポソームをその他の既知の実験室手順、すなわちその内容が本発明に参照により援用されているバンガム(Bangham)ら、(1965年)の方法;その内容が本発明に参照により援用されている「Drug Carriers in Biology and Medicine」、G.グレゴリアディス(Gregoriadis)編、(1979年)、287〜341頁に記述されている通りのグレゴリアディスの方法;その内容が本発明に参照により援用されている「the method of Deamer and Uster」、1983年;及びスゾカ(Szoka)及びパパハジョポウロス(Papahadjopoulos)、1978年により記述されている通りの逆相蒸発方法に従って調製することができる。上述の方法は、水性材料を封入するそのそれぞれの能力及びそのそれぞれの水空間対脂質比に関して異なっている。 Alternatively, liposomes may be used in other known laboratory procedures, ie, the method of Bangham et al. (1965), the contents of which are hereby incorporated by reference; the contents of which are incorporated herein by reference. "Drug Carriers in Biology and Medicine", G. Gregoriadis, Ed. (1979) Gregoriadis method as described on pages 287-341; “the method of Deamer and Uster”, 1983, the contents of which are incorporated by reference in the present invention. And the reverse phase evaporation method as described by Szoka and Papahadjopoulos, 1978. The methods described above differ with respect to their respective ability to encapsulate aqueous materials and their respective water space to lipid ratios.
上述の通りに調製された凍結乾燥されたリポソーム又は乾燥した脂質は、脱水され阻害性ペプチドの溶液中で戻され、適当な溶媒、例えばDPBSで適切な濃度まで希釈され得る。次に、混合物は、渦流ミキサー内で激しく振とうされる。未封入の核酸は、29,000×gでの遠心分離により除去され、リポソームペレットは、洗浄される。洗浄されたリポソームは、例えば約50〜200mMといった適切な総リン脂質濃度で再懸濁される。封入された核酸の量は、標準的な方法に従って決定可能である。リポソーム調製物中に封入された核酸の量を判定した後、リポソームを適切な濃度まで希釈し、使用まで4℃で貯蔵することができる。 Lyophilized liposomes or dried lipids prepared as described above can be dehydrated and reconstituted in a solution of inhibitory peptides and diluted to a suitable concentration with a suitable solvent, such as DPBS. The mixture is then vigorously shaken in a vortex mixer. Unencapsulated nucleic acid is removed by centrifugation at 29,000 × g and the liposome pellet is washed. The washed liposomes are resuspended at an appropriate total phospholipid concentration, eg, about 50-200 mM. The amount of nucleic acid encapsulated can be determined according to standard methods. After determining the amount of nucleic acid encapsulated in the liposome preparation, the liposomes can be diluted to an appropriate concentration and stored at 4 ° C. until use.
活性化(パルス)を受けた免疫細胞の生成
本発明は、曝露されるか又はその他の形で抗原での「パルスを受け」、抗原により活性化された細胞を含む。例えば、APCは抗原の存在下でのエクスビボ培養によってインビトロで又は抗原に対する曝露によりインビボでAg負荷された状態となり得る。
Generation of activated (pulsed) immune cells The invention includes cells that have been exposed or otherwise “pulsed” with an antigen and activated by the antigen. For example, APC can be Ag-loaded in vitro by ex vivo culture in the presence of the antigen or in vivo by exposure to the antigen.
当業者であれば同様に、APCの表面上でのその抗原の提示を促進するのに充分な時間APCを抗原に曝露するような形でAPCを「パルス」することができる、ということも容易に理解するものと思われる。例えば、APCは、抗原ペプチドとして知られる小ペプチドフラグメントが直接APCの外側に「パルス」されるような形で抗原に曝露され得(メヘタ・ダマニ(Mehta−Damani)ら、1994年);そうでなければAPCは、後でAPCにより摂取されることになる全タンパク質又はタンパク質粒子と共にインキュベートされ得る。これらの全タンパク質は、APCにより小ペプチドフラグメント内に消化され、場合によってはAPC表面まで搬送されその上に提示される(コーエンら、1994年)。ペプチド形態の抗原が、本書で記述されている標準的「パルス」技術によって細胞に対し曝露され得る。 It is also easy for those skilled in the art to “pulse” APC in such a way that APC is exposed to the antigen for a time sufficient to promote presentation of that antigen on the surface of the APC. Seems to understand. For example, APC can be exposed to an antigen such that a small peptide fragment known as an antigenic peptide is “pulsed” directly outside the APC (Mehta-Damani et al., 1994); Otherwise, APC can be incubated with whole protein or protein particles that will later be ingested by APC. All these proteins are digested into small peptide fragments by APC and are sometimes transported to and presented on the APC surface (Cohen et al., 1994). Peptide forms of antigen can be exposed to cells by the standard “pulse” technique described herein.
特定の理論により束縛されることは望まないが、外来性の抗原又は自己抗原の形をした抗原は、抗原の免疫原性形態を保持する目的で、本発明のAPCによりプロセッシングされる。抗原の免疫原性形態は、例えばT細胞といった免疫細胞により認識され得この細胞を刺激する抗原の一形態を産生する目的でのフラグメント化を通した抗原のプロセッシングを暗に意味している。好ましくは、かかる外来性又は自己抗原は、APCによりペプチドへとプロセッシングされるタンパク質である。APCにより産生される関連性あるペプチドは、免疫原性組成物として使用するために抽出され精製され得る。APCによりプロセッシングされたペプチドは同様に、APCによりプロセッシングされたタンパク質に対する寛容を誘発するのに使用することもできる。 While not wishing to be bound by any particular theory, foreign antigens or antigens in the form of autoantigens are processed by the APCs of the present invention in order to retain the immunogenic form of the antigen. An immunogenic form of an antigen implies the processing of the antigen through fragmentation in order to produce a form of the antigen that can be recognized by and stimulates immune cells such as T cells. Preferably, such foreign or autoantigen is a protein that is processed into a peptide by APC. Relevant peptides produced by APC can be extracted and purified for use as an immunogenic composition. Peptides processed by APC can also be used to induce tolerance to proteins processed by APC.
自己免疫疾患は、自己抗原として別途知られている「自己タンパク質」すなわち、1つの個体の中に存在する又はその中で内因性である自己抗原に対し免疫応答が向けられていることの結果としてもたらされると考えられている。自己免疫応答において、これらの「自己タンパク質」はT細胞に提示され、こうしてT細胞は「自己反応性」を付与される。本発明の方法に従うと、APCは、関連性ある「自己ペプチド」を産生するべく抗原でのパルスを受ける。関連性ある自己ペプチドは、MHC産物がきわめて多型であり各々の個々のMHC分子が異なるペプチドフラグメントと結合する可能性があることから、各個体について異なっている。このとき「自己ペプチド」及びサイトカインシグナリングのインヒビターのアゴニストを用いて、競合するペプチドを設計するか又は治療を必要とする個体の中で自己タンパク質に対する寛容を誘発することができる。 Autoimmune diseases are the result of an immune response directed against a “self-protein” otherwise known as an autoantigen, ie, an autoantigen present in or endogenous to one individual. It is thought to be brought about. In an autoimmune response, these “self proteins” are presented to T cells, thus conferring “self-reactivity”. According to the method of the invention, APCs are pulsed with an antigen to produce the relevant “self-peptide”. Relevant self-peptides are different for each individual because the MHC product is highly polymorphic and each individual MHC molecule can bind to a different peptide fragment. “Self peptides” and agonists of inhibitors of cytokine signaling can then be used to design competing peptides or induce tolerance to self proteins in individuals in need of treatment.
本発明の「パルスを受けたAPC」としても別途知られている抗原活性化されたAPCは、インビトロ又はインビボのいずれかで抗原に対してAPCを曝露することによって産生される。APCがインビトロでパルスを受ける場合、APCは、培養皿上で平板固定され、抗原がAPCに結合できるのに充分な時間、充分な量で抗原に曝露される。APCに対する抗原の結合を達成するのに必要な量及び時間は、当該技術分野において既知の又は本書中にその他の形で開示されている方法を使用することにより判定され得る。抗原に対する曝露の後のAPC上の抗原の存在を検出するために、例えば免疫検定又は結合検定といった当業者にとって既知のその他の方法を使用することも可能である。 Antigen activated APC, otherwise known as “pulsed APC” of the present invention, is produced by exposing APC to an antigen either in vitro or in vivo. When APC is pulsed in vitro, it is plated on a culture dish and exposed to the antigen for a sufficient amount of time for the antigen to bind to the APC. The amount and time required to achieve antigen binding to APC can be determined by using methods known in the art or otherwise disclosed herein. Other methods known to those skilled in the art can be used to detect the presence of the antigen on the APC after exposure to the antigen, such as an immunoassay or binding assay.
本発明のさらなる実施形態においては、APCによる特異的タンパク質に発現を可能にするベクターでAPCをトランスフェクトすることができる。このとき、APCにより発現されるタンパク質をプロセッシングし、MHCレセプタ上の細胞表面上で提示することができる。トランスフェクションを受けたAPCはこのとき、ベクターによりコードされたタンパク質に対する免疫応答を生成するための免疫原性組成物として使用することができる。 In a further embodiment of the invention, APC can be transfected with a vector that allows expression by a specific protein by APC. At this time, the protein expressed by APC can be processed and presented on the cell surface on the MHC receptor. The transfected APC can then be used as an immunogenic composition to generate an immune response against the protein encoded by the vector.
本書のその他の場所で論述されているように、ベクターは、免疫原性応答が望まれるタンパク質をコードし発現する特異的ポリヌクレオチドを含むように調製され得る。好ましくは、細胞を感染させるために、レトロウイルスベクターが使用される。より好ましくは、細胞を感染させるためにアデノウイルスベクターが使用される。 As discussed elsewhere in this document, vectors can be prepared to contain specific polynucleotides that encode and express proteins for which an immunogenic response is desired. Preferably, retroviral vectors are used to infect cells. More preferably, an adenoviral vector is used to infect cells.
本発明のもう1つの実施形態においては、APC上のレセプタにより認識されるタンパク質又はその一部分をコードするべくウイルスベクターを修飾することによってベクターをAPCにターゲティングすることができ、かくしてベクターによるAPCレセプタの占有はウイルスベクターの核酸によりコードされる抗原のプロセッシング及び提示を可能にすることになる。ウイルスにより送達される核酸は、APC上にて発現された時点でウイルスタンパク質をコードしこれらのウイルスタンパク質が次にプロセッシングされAPCのMHCレセプタ上で提示されることになるようなウイルスに天然のものである。 In another embodiment of the invention, the vector can be targeted to APC by modifying the viral vector to encode a protein or portion thereof recognized by a receptor on the APC, thus the vector-based APC receptor. Occupancy will allow the processing and presentation of the antigen encoded by the viral vector nucleic acid. The nucleic acids delivered by the virus are natural to the virus when encoded on the APC, which encodes the viral proteins and these viral proteins are then processed and presented on the APC MHC receptor. It is.
本書の他の箇所で論述した通り、宿主細胞内にポリヌクレオチドをトランスフェクトするためにさまざまな方法を使用することができる。本方法には、リン酸カルシウム沈殿、リポフェクション、粒子衝突、マイクロインジェクション、電気穿孔、コロイド分散系(すなわち、巨大分子複合体、ナノカプセル、ミクロスフェア、ビーズ及び水中油型エマルジョン、ミセル、混合ミセル及びリポソームを含めた脂質ベース系)が含まれるがこれに限定されるわけではない。 As discussed elsewhere in this document, various methods can be used to transfect a polynucleotide into a host cell. The method includes calcium phosphate precipitation, lipofection, particle bombardment, microinjection, electroporation, colloidal dispersion (ie, macromolecular complexes, nanocapsules, microspheres, beads and oil-in-water emulsions, micelles, mixed micelles and liposomes. Including, but not limited to, lipid-based systems).
もう1つの態様においては、抗原をコードするポリヌクレオチドを発現ベクター内にクローニングでき、ベクターをAPC内に導入してその他の形で活性化APCを生成することができる。細胞内に核酸を導入するさまざまなタイプのベクター及び方法が、本書の中の他の箇所で論述されている。例えば、当該技術分野におけるあらゆる方法により宿主細胞内に、抗原をコードするベクターを導入することができる。例えば、物理的、化学的又は生物学的手段により宿主細胞内に発現ベクターを移入することができる。例えばサンブルックら、(2001年、「Molecular Cloning:A Laboratory Manual」、コールド・スプリング・ハーバー・ラボラトリー、ニューヨーク)、及びアウスベルら、(1997年、「Current Protocols in
Molecular Biology」、ジョン・ウィリー・アンド・サンズ、ニューヨーク)中を参照のこと。抗原をコードするポリヌクレオチドを含んでなる発現ベクターの導入が、パルスを受けた細胞を生み出す、ということは容易に理解される。
In another embodiment, the polynucleotide encoding the antigen can be cloned into an expression vector, and the vector can be introduced into APC to otherwise produce activated APC. Various types of vectors and methods for introducing nucleic acids into cells are discussed elsewhere in this document. For example, a vector encoding an antigen can be introduced into a host cell by any method in the art. For example, the expression vector can be transferred into the host cell by physical, chemical or biological means. For example, Sanbrook et al. (2001, “Molecular Cloning: A Laboratory Manual”, Cold Spring Harbor Laboratory, New York), and Ausbell et al. (1997, “Current Protocols in
See "Molecular Biology", John Willy and Sons, New York). It will be readily appreciated that the introduction of an expression vector comprising a polynucleotide encoding an antigen produces a pulsed cell.
本発明は、タンパク質、cDNA又はcRNAの形をした全抗原をAPCに負荷することを含めた(ただしこれに限定されるわけではない)、APCをパルスするためのさまざまな方法を含む。しかしながら、本発明は、APCをパルスするために用いられる抗原の特定の形態に限定されるものとして解釈されるべきではない。むしろ、本発明は、抗原負荷されたAPCを生成するための当該技術分野において既知のその他の方法をも包含している。好ましくは、APCは、規定された抗原をコードするmRNAでのトランスフェクションを受ける。その配列が既知である遺伝子産物に対応するmRNAは、適切なプライマ及び転写反応と合わせた逆転写酵素−ポリメラーゼ連鎖反応(RT−PCR)を用いてインビトロで急速に生成することが可能である。mRNAでのAPCのトランスフェクションは、パルスを受けたAPCを生成するためのその他の抗原負荷技術に比べて1つの利点を提供する。例えば、微細な量の組織すなわち腫瘍組織からRNAを増幅する能力は、数多くの患者に対するワクチン接種向けにAPCの用途を拡大する。 The present invention includes various methods for pulsing APC, including (but not limited to) loading APC with a whole antigen in the form of a protein, cDNA or cRNA. However, the present invention should not be construed as limited to the particular form of antigen used to pulse APC. Rather, the present invention also encompasses other methods known in the art for generating antigen-loaded APCs. Preferably, the APC undergoes transfection with mRNA encoding a defined antigen. MRNA corresponding to a gene product whose sequence is known can be rapidly generated in vitro using reverse transcriptase-polymerase chain reaction (RT-PCR) combined with appropriate primers and transcription reactions. Transfection of APC with mRNA offers one advantage over other antigen loading techniques for generating pulsed APC. For example, the ability to amplify RNA from minute amounts of tissue, ie tumor tissue, expands the use of APC for vaccination against a large number of patients.
抗原は、ウイルス、真菌又は細菌に由来するものであり得る。抗原は、感染性疾患、癌、自己免疫疾患よりなる群から選択された疾病に結びつけられた抗原又は自己抗原であり得る。 Antigens can be derived from viruses, fungi or bacteria. The antigen may be an antigen or autoantigen associated with a disease selected from the group consisting of infectious diseases, cancer, autoimmune diseases.
1つの抗原組成物がワクチンとして有用であるためには、その抗原組成物は、細胞、組織又は哺乳動物(例えばヒト)の中で抗原に対する免疫応答を誘発しなければならない。本書で使用される「免疫学的組成物」は、抗原(例えばペプチド又はポリペプチド)、抗原をコードする核酸(例えば抗原発現ベクター)、抗原又は細胞組成物を発現又は提示する細胞を含んでなる。特定の実施形態においては、抗原組成物は、本書に記述されているあらゆる抗原の全て又は一部分又はその免疫学的機能的等価物を含んでなるか又はコードする。その他の実施形態においては、抗原組成物は、付加的な免疫刺激性作用物質又はかかる作用物質をコードする核酸を含んでなる混合物の中にある。免疫刺激性作用物質には、付加的な抗原、免疫調節薬、抗原提示細胞又はアジュバンドが含まれるが、これに限定されるわけではない。その他の実施形態においては、付加的な作用物質のうちの1つもしくはそれ以上のものは、あらゆる組合せで抗原又は免疫刺激性作用物質に共有結合されている。一部の実施形態においては、抗原組成物はHLAアンカーモチーフアミノ酸に接合
されるか又はこれを含んでなる。
In order for one antigen composition to be useful as a vaccine, the antigen composition must elicit an immune response against the antigen in a cell, tissue or mammal (eg, a human). An “immunological composition” as used herein comprises an antigen (eg, a peptide or polypeptide), a nucleic acid encoding the antigen (eg, an antigen expression vector), a cell that expresses or presents the antigen or cell composition. . In certain embodiments, the antigen composition comprises or encodes all or a portion of any antigen described herein or an immunological functional equivalent thereof. In other embodiments, the antigen composition is in a mixture comprising additional immunostimulatory agents or nucleic acids encoding such agents. Immunostimulatory agents include, but are not limited to, additional antigens, immunomodulators, antigen presenting cells or adjuvants. In other embodiments, one or more of the additional agents are covalently bound to the antigen or immunostimulatory agent in any combination. In some embodiments, the antigen composition is conjugated to or comprises an HLA anchor motif amino acid.
本発明のワクチンは、その核酸及び/又は細胞組成物の組成において変動し得る。限定的意味のない例においては、抗原をコードする核酸を、アジュバンドと共に処方することもできる。当然のことながら、本書で記述するさまざまな組成物がさらに付加的な組成物を含み得るということも理解できるだろう。例えば、脂質又はリポゾームの中には、1つもしくはそれ以上のワクチン組成物が含まれ得る。もう1つの限定的な意味のない例では、ワクチンが1つもしくはそれ以上のアジュバンドを含んでなる可能性がある。本発明のワクチン及びそのさまざまな構成要素は、本書で開示されているあらゆる方法によって、又は本開示に照らして当業者にとって既知のものとなるように、調製及び/又は投与可能である。 The vaccine of the present invention may vary in the composition of its nucleic acid and / or cell composition. In a non-limiting example, the nucleic acid encoding the antigen can be formulated with an adjuvant. Of course, it will also be understood that the various compositions described herein may include additional compositions. For example, one or more vaccine compositions can be included in a lipid or liposome. In another non-limiting example, the vaccine may comprise one or more adjuvants. The vaccine of the invention and its various components can be prepared and / or administered by any method disclosed herein or as would be known to one of ordinary skill in the art in light of this disclosure.
本発明の抗原組成物は、固相合成による化学合成及びHPLCによる化学反応のその他の生成物からの精製、又はインビトロ翻訳系内又は生きた細胞内での本発明の抗原を含むペプチド又はポリペプチドをコードする核酸配列(例えばDNA配列)の発現による産生を含めた(ただしこれに限定されるわけではない)当該技術分野において周知の方法により作ることができるということがわかっている。さらに、抗原組成物は、生体試料から単離された細胞組成物を含んでなり得る。好ましくは、抗原組成物は、単離され1つもしくはそれ以上の望ましくない低分子量分子を除去するべく大規模に透析されかつ/又はより容易に所望のビヒクルへと処方されるよう凍結乾燥される。さらに、ワクチン組成物中で作られる(行なわれる)付加的なアミノ酸、突然変異、化学的修飾などがある場合、これらは、好ましくはエピトープ配列の抗体認識と実質的に干渉しないということもわかっている。 The antigen composition of the present invention is a peptide or polypeptide comprising the antigen of the present invention in a chemical synthesis by solid phase synthesis and purification from other products of chemical reaction by HPLC, or in an in vitro translation system or in a living cell. It has been found that can be made by methods well known in the art, including but not limited to production by expression of a nucleic acid sequence encoding (eg, a DNA sequence). Further, the antigen composition can comprise a cell composition isolated from a biological sample. Preferably, the antigen composition is isolated and lyophilized to be extensively dialyzed and / or more easily formulated into the desired vehicle to remove one or more undesirable low molecular weight molecules. . Furthermore, it is also understood that if there are additional amino acids, mutations, chemical modifications, etc. made (performed) in the vaccine composition, these preferably do not substantially interfere with antibody recognition of the epitope sequence. Yes.
本発明の1つもしくはそれ以上の抗原決定因子に対応するペプチド又はポリペプチドは、一般に長さが少なくとも5個又は6個のアミノ酸でなければならず、最高約10、約15、約20、約25、約30、約35、約40、約45又は約50個などの残基を含有し得る。ペプチド配列は、例えばアプライド・バイオシステムズ社(カリフォルニア州フォスターシティー)から入手可能なものなどの自動ペプチド合成機を用いたペプチド合成などの当業者にとっては既知の方法によって合成可能である。 Peptides or polypeptides corresponding to one or more antigenic determinants of the invention generally must be at least 5 or 6 amino acids in length, up to about 10, about 15, about 20, about It may contain 25, about 30, about 35, about 40, about 45, or about 50 residues. Peptide sequences can be synthesized by methods known to those skilled in the art, such as peptide synthesis using an automated peptide synthesizer such as those available from Applied Biosystems (Foster City, Calif.).
組換え型手段などにより、さらに長いペプチド又はポリペプチドを調製することもできる。一部の実施形態においては、例えば、本発明のさまざまな組成物及び方法のためにインビトロ又はインビボで抗原組成物を生産する目的で、本書に記述されている抗原組成物及び/又は構成要素をコードする核酸を使用することが可能である。例えば、一部の実施形態では、抗原をコードする核酸が、例えば組換え型細胞内のベクター内に含まれる。抗原配列を含んでなるペプチド又はポリペプチドを産生するために核酸を発現することが可能である。ペプチド又はポリペプチドは細胞から分泌され得るか又は細胞の一部として又は細胞の内部に含まれ得る。 Longer peptides or polypeptides can also be prepared, such as by recombinant means. In some embodiments, the antigen compositions and / or components described herein are used, for example, for the purpose of producing the antigen compositions in vitro or in vivo for the various compositions and methods of the invention. It is possible to use encoding nucleic acids. For example, in some embodiments, the nucleic acid encoding the antigen is contained within a vector, eg, in a recombinant cell. Nucleic acids can be expressed to produce peptides or polypeptides that comprise an antigen sequence. The peptide or polypeptide can be secreted from the cell or can be contained as part of or within the cell.
一部の実施形態においては、抗原をコードする核酸で哺乳動物をトランスフェクトするか又は接種することにより、免疫応答を促進することができる。このとき、標的哺乳動物の内部に含まれた1つもしくはそれ以上の細胞が、哺乳動物に対する核酸の投与後核酸によりコードされる配列を発現する。ワクチンは同様に、例えば1つの抗原のペプチド又はポリペプチド配列の全て又は一部分をコードする核酸(例えばcDNA又はRNA)の形をしていてもよい。核酸によるインビボでの発現は例えば、プラスミド型ベクター、ウイルスベクター又はウイルス/プラスミド構成体ベクターによるものであり得る。 In some embodiments, an immune response can be promoted by transfecting or inoculating a mammal with a nucleic acid encoding an antigen. At this time, one or more cells contained within the target mammal express a sequence encoded by the nucleic acid after administration of the nucleic acid to the mammal. A vaccine may also be in the form of a nucleic acid (eg, cDNA or RNA) encoding, for example, all or part of a peptide or polypeptide sequence of one antigen. In vivo expression by a nucleic acid can be, for example, by a plasmid-type vector, a viral vector, or a virus / plasmid construct vector.
好ましい態様では、核酸は、適切な抗原をコードする配列の全て又は一部分をコードするコーディング領域又はその免疫学的に機能的な等価物を含んでなる。当然のことながら
、核酸は、1つもしくはそれ以上の免疫調節薬又はアジュバンドを含んでなるものを含めた(ただしこれに限定されるわけではない)付加的な配列を含んでなるかつ/又はコードすることができる。
In a preferred embodiment, the nucleic acid comprises a coding region that encodes all or a portion of the sequence encoding the appropriate antigen, or an immunologically functional equivalent thereof. It will be appreciated that the nucleic acid comprises additional sequences, including but not limited to those comprising one or more immunomodulatory agents or adjuvants, and / or Can be coded.
腫瘍関連抗原
本発明に関連して「腫瘍抗原」又は「過剰増殖性障害抗原」又は「過剰増殖性障害と結びつけられる抗原」という用語は、特定の過剰増殖性障害に共通の抗原を意味する。一部の態様においては、本発明の過剰増殖性障害抗原は、原発性又は転移性悪性黒色腫、胸腺腫、リンパ腫、肉腫、肺癌、肝臓癌、非ホジキンリンパ腫、ホジキンリンパ腫、白血病、子宮癌、子宮頚癌、膀胱癌、腎臓癌及び腺癌、例えば乳癌、前立腺癌、卵巣癌、膵臓癌などを含めた(ただしこれに限定されるわけではない)癌に由来する。
Tumor-associated antigen The term “tumor antigen” or “hyperproliferative disorder antigen” or “antigen associated with a hyperproliferative disorder” in the context of the present invention means an antigen common to a particular hyperproliferative disorder. In some aspects, the hyperproliferative disorder antigen of the invention comprises primary or metastatic malignant melanoma, thymoma, lymphoma, sarcoma, lung cancer, liver cancer, non-Hodgkin lymphoma, Hodgkin lymphoma, leukemia, uterine cancer, Derived from cancers including, but not limited to, cervical cancer, bladder cancer, kidney cancer and adenocarcinoma such as breast cancer, prostate cancer, ovarian cancer, pancreatic cancer and the like.
一実施形態においては、本発明の腫瘍抗原は、哺乳動物の癌腫瘍に由来する腫瘍浸潤性リンパ球(TIL)によって免疫学的に認識される1つもしくはそれ以上の抗原癌エピトープを含んでなる。 In one embodiment, the tumor antigens of the invention comprise one or more antigenic cancer epitopes that are immunologically recognized by tumor infiltrating lymphocytes (TILs) derived from mammalian cancer tumors. .
悪性腫瘍は、免疫攻撃のための標的抗原として役立ち得る一定数のタンパク質を発現する。これらの分子は、黒色腫におけるMART−1、チロシナーゼ及びGP−100及び前立腺癌における前立腺酸性ホスファターゼ(PAP)及び前立腺特異的抗原(PSA)などの組織特異的抗原を含むが、これに限定されるわけではない。その他の標的分子は、癌遺伝子HER−2/Neu/ErbB−2などの形質転換関連分子のグループに属する。さらにもう1つの標的抗原グループは、癌胎児性抗原(CEA)などの腫瘍胎児抗原である。B細胞リンパ腫においては、腫瘍特異的イディオタイプ免疫グロブリンは個々の腫瘍に固有の真に腫瘍特異的な免疫グロブリン抗原を構成する。CD19、CD20及びCD37などのB細胞分化抗原は、B細胞リンパ腫における標的抗原のためのその他の候補である。これらの抗原のいくつか(CEA、HER−2、CD19、CD20、イディオタイプ)が、モノクローナル抗体での受動免疫療法のための標的として使用され、或る程度の成功を収めた。 Malignant tumors express a certain number of proteins that can serve as target antigens for immune attack. These molecules include, but are not limited to, tissue specific antigens such as MART-1, tyrosinase and GP-100 in melanoma and prostate acid phosphatase (PAP) and prostate specific antigen (PSA) in prostate cancer. Do not mean. Other target molecules belong to the group of transformation-related molecules such as the oncogene HER-2 / Neu / ErbB-2. Yet another target antigen group is oncofetal antigens such as carcinoembryonic antigen (CEA). In B-cell lymphoma, tumor-specific idiotype immunoglobulins constitute truly tumor-specific immunoglobulin antigens that are unique to individual tumors. B cell differentiation antigens such as CD19, CD20 and CD37 are other candidates for target antigens in B cell lymphomas. Some of these antigens (CEA, HER-2, CD19, CD20, idiotype) have been used with some success as targets for passive immunotherapy with monoclonal antibodies.
腫瘍抗原及びその抗原癌エピトープは、一次臨床分離株、細胞系列などといった天然の供給源から精製及び単離され得る。癌ペプチド及びその抗原エピトープは、当該技術分野において既知の化学合成又は組換え型DNA技術によっても得られ得る。化学合成のための技術はスチュワード(Steward)ら、(1969年);ボダンスキー(Bodansky)ら、(1976年);マイエンホッファー(Meienhofer)(1983年);及びシュローダー(Schroder)ら、(1965年)の中で記述されている。さらに、レンクヴィスト(Renkvist)ら、(2001年)の中で記述されているように、当該技術分野においては数多くの抗原が知られている。以下の表は、腫瘍抗原によってコードされるT細胞で確定されたエピトープを記述しており、T細胞により認識される腫瘍抗原(細胞傷害性CD8+又はヘルパーCD4+のいずれか)のみが列挙されている。類似体又は人工的に修飾されたエピトープは列挙されていないものの、当業者であれば、当該技術分野における標準的な手段によりこれらをいかに獲得又は生成するかを認識する。抗体により同定されSerex法(サヒン(Sahin)ら、(1997年)及びチェン(Chen)ら、(2000年)参照))により検出されるその他の抗原は、ルードゥイング・インスティチュート・フォー・キャンサー・リサーチ(Ludwig
Institute for Cancer Research)のデータベース内で同定される。
Tumor antigens and their antigenic cancer epitopes can be purified and isolated from natural sources such as primary clinical isolates, cell lines, and the like. Cancer peptides and their antigenic epitopes can also be obtained by chemical synthesis or recombinant DNA techniques known in the art. Techniques for chemical synthesis are: Steward et al. (1969); Bodansky et al. (1976); Meienhofer (1983); and Schroder et al. (1965). ). In addition, a number of antigens are known in the art, as described in Renkvist et al. (2001). The table below describes the epitopes identified in T cells encoded by tumor antigens, listing only those tumor antigens (either cytotoxic CD8 + or helper CD4 +) that are recognized by T cells. . Although analogs or artificially modified epitopes are not listed, those skilled in the art will recognize how to obtain or generate them by standard means in the art. Other antigens identified by antibodies and detected by the Serex method (see Sahin et al., (1997) and Chen et al., (2000)) include the Ludouing Institute for Cancer. Research (Ludwig)
Identified in the Institute for Cancer Research database.
病原菌抗原
病原菌抗原は、ウイルス、細菌又は真菌由来であり得る。感染性ウイルスの例としては、レトロウイルス科(例えばヒト免疫不全ウイルス例えばHIV−1(HTLV−III
、LAV又はHTLV−III/LAV、又はHIV−III)とも呼ばれる:及びその他の分離株例えばHIV−LP;ピコルナウイルス科(Picornaviridae)(例えばポリオウイルス(polio virus)類、肝炎Aウイルス(hepatitis A virus);エンテロウイルス(enterovirus)類;ヒトコクサッキーウイルス(human coxsackie virus)類、ライノウイルス(rhinovirus)類、エコーウイルス(echovirus)類);カルシウイルス科(Calciviridae)(例えば胃腸炎を起こす株);トガウイルス科(Togaviridae)(例えばウマ脳炎ウイルス(equine encephalitis virus)類、風疹ウイルス(rubella virus)類);フラビウイルス科(Flaviviridae)(例えば、デング熱ウイルス(dengue virus)類、脳炎ウイルス(encephalitis virus)類、黄熱病ウイルス(yellow fever virus)類);コロナウイルス科(Coronaviridae)(例えば、コロナウイルス(coronavirus)類);ラブドウイルス科(Rhabdoviridae)(例えば、水疱性口内炎ウイルス(vesicular stomatitis virus)類、狂犬病ウイルス(rabies virus)類);フィロウイルス科(Filoviridae)(例えばエボラウイルス(ebolavirus)類);パラミクソウイルス科(Paramyxoviridae)(例えば、パラインフルエンザウイルス(parainfluenza virus)類、流行性耳下腺炎ウイルス(mumps virus)、麻疹ウイルス(measles virus)、呼吸器合抱体(RS)ウイルス(respiratory syncytial virus));オルソミクソウイルス科(Orthomyxoviridae)(例えば、インフルエンザウイルス(influenza virus)類);ブンガウイルス科(Bungaviridae)(例えば、ハンターンウイルス(Hantaan virus)類、ブンガウイルス(bunga virus)類、フレボウイルス(phlebovirus)類およびナイロウイルス(Nairo virus)類);アレナウイルス科(Arena viridae)(出血性熱ウイルス(hemorrhagic fever virus)類);レオウイルス科(Reoviridae)(例えば、レオウイルス(reovirus)類、オルビウイルス(orbivirus)類およびロタウイルス(rotavirus)類);ビルナウイルス科(Birnaviridae);ヘパドナウイルス科(Hepadnaviridae)(B型肝炎ウイルス(Hepatitis B virus));パルボウイルス科(Parvovlridae)(パルボウイルス(parvovirus)類);パポバウイルス科(Papovaviridae)(乳頭腫ウイルス(papilloma virus)類、ポリオーマウイルス(polyomavirus)類);アデノウイルス科(Adenoviridae)(大部分のアデノウイルス(adenovirus)類);ヘルペスウイルス科(Herpesviridae)(単純ヘルペスウイルス(herpes simplex virus (HSV))1および2、水痘帯状疱疹ウイルス(varicella zoster virus)、サイトメガロウイルス(cytomegalovirus(CMV))、ヘルペスウイルス(herpes virus)類);ポックスウイルス科(Poxviridae)(痘瘡ウイルス(variola virus)類、ワクシニアウイルス(vaccinia virus)類、ポックスウイルス(pox
virus)類);およびイリドウイルス科(Iridoviridae)(例えばブタコレラウイルス(African swine fever virus));未分類ウイルス(例えば、海綿状脳症(Spongiform encephalopathy)の病因病原体、デルタ肝炎の病原体(B型肝炎ウイルスの不完全付随体と考えられているもの)、非A、非B型肝炎の病原体(クラス1=経口感染、クラス2=非経口感染(すなわち、C型肝炎));ノーウォーク(Norwalk)および関連ウイルス、およびアストロウイルス(astrovirus)類)が含まれる。
Pathogen antigens Pathogen antigens can be derived from viruses, bacteria or fungi. Examples of infectious viruses include retroviridae (eg, human immunodeficiency virus such as HIV-1 (HTLV-III
, LAV or HTLV-III / LAV, or HIV-III): and other isolates such as HIV-LP; Picoraviridae (eg, polioviruses, hepatitis A virus) enteroviruses; human coxsackie viruses, rhinoviruses, echoviruses; calciviridae (for example, gastroenteritis) Togaviridae (eg equine encephalitis virus, rubella virus) (Rubella virus)); Flaviviridae (eg, dengue virus, encephalitis virus, yellow fever virus) or coronaviridae For example, coronaviruses); Rhabdoviridae (eg, vesicular stomatitis viruses, rabies viruses (Firoviridae), for example, Firoviridae (Firoviridae) ebolavirus)); Paramyxoviridae idae) (eg parainfluenza viruses, mumps virus, measles virus, respiratory syncytial virus (RS) virus) Viridae (for example, influenza virus (influenza virus) class); Nairoviruses); Arenaviridae (Arena) viridae (hemorrhagic fever viruses); Reoviridae (eg, reoviruses, orbiviruses and rotaviruses); (Birnaviridae); Hepadnaviridae (Hepatitis B virus); Parvoviridae (Parvoviridae); ), Polyomavirus (Polyomavirus)); Adenoviridae Adenoviridae (most adenoviruses); Herpesviridae (Herpes simplex virus (HSV) 1 and 2, varicella zoster virus (Varicella zoster virus) cytomegalovirus (CMV)), herpes viruses); Poxviridae (variola viruses), vaccinia viruses, poxvirus (pox)
viruses)); and Iridoviridae (eg, African swinever virus); Unclassified viruses (eg, Spongiform encephalopathy pathogens, hepatitis B What is considered an incomplete incidental virus), non-A, non-B hepatitis pathogens (
感染性細菌の例としては、ヘリコバクターピロリ(Helicobacter pyl
oris)、ボレリア・ブルグドルフェリBorelia burgdorferi、レジオネラ・ニューモフィリア(Legionella pneumophilia)、ミコバクテリウム(Mycobacteria)種(例えば結核菌(M.tuberculosis)、トリ結核菌(M.avium)、M.イントラセルラレ(M.intracellulare)、M.カンサイ(M.kansaii)、M.ゴルドナイ(M.gordonae))、黄色ブドウ球菌(Staphylococcus aureus)、淋菌(Neisseria gonorrhoeae)、髄膜炎菌(Neisseria
meningitidis)、リステリア菌(Listeria monocytogenes)、化膿性連鎖球菌(Streptococcus pyogenes)(A群連鎖球菌)、ストレプトコックス・アガラクティアイ(Streptococcus agalactiae)(B群連鎖球菌)、連鎖球菌(Streptococcus)(ヴィリダンス群(viridans group))、糞便連鎖球菌(Streptococcus faecalis)、ストレプトコックス・ボビス(Streptococcus bovis)、連鎖球菌(Streptococcus)(嫌気性種)、肺炎連鎖球菌(Streptococcus pneumoniae)、病原性カンピロバクター(Campylobacter)種、腸球菌(Enterococcus)種、インフルエンザ菌(Haemophilus influenzae)、炭疽菌(Bacillus antracis)、ジフテリア菌(Corynebacterium diphtheriae)、コリネバクテリウム(Corynebacterium)種、ブタ丹毒菌(Erysipelothrix rhusiopathiae)、ウェルシュ菌(Clostridium perfringens)、破傷風菌(Clostridium
tetani)、エンテロバクター・アエロゲネス(Enterobacter aerogenes)、肺炎桿菌(Klebsiella pneumoniae)、パスツレラ・ムルトチダ(Pasturella multocida)、バクテロイド(Bacteroides)種、フゾバクテリウム・ヌタレアツム(Fusobacterium nucleatum)、ストレプトバチラス・モニリフォルミス(Streptobacillus moniliformis)、トレポネーマ(Treponema)、フランベジアトレポネーマ(Treponema pertenue)、レプトスピラ属(Leptospira)およびイスラエル放線菌(Actinomyces israelli)が含まれる。
Examples of infectious bacteria include Helicobacter pylori
oris), Borrelia burgdorferi Borelia burgdorferi, Legionella pneumophilia, Mycobacterium species (for example, M. tuberculosis, M. tuberculum (M. M. intracellulare, M. kansai, M. gordonae, Staphylococcus aureus, Neisseria gonorrhoeae, Neisseria meningitidis
meningitidis), Listeria monocytogenes, Streptococcus pyogenes (Group A Streptococcus), Streptococcus agalactiae (Streptococcus agalactiae) (Streptococcus staphylococcus p Group (viridans group), Streptococcus faecalis, Streptococcus bovis, Streptococcus species, Streptococcus spp., Streptococcus spp. , Enterocytes Enterococcus spp., Haemophilus influenzae, Bacillus anthracis, Corynebacterium diphtheriae, Corynebacterium spp. Clostridium
tetani), Enterobacter aerogenes (Enterobacter aerogenes), Klebsiella pneumoniae (Klebsiella pneumoniae), Pasteurella Murutochida (Pasturella multocida), Bacteroides (Bacteroides) species, Fusobacterium Nutareatsumu (Fusobacterium nucleatum), Streptomyces Bacillus Las Moniriforumisu (Streptobacillus moniliformis), Treponema, Tremone pertenue, Leptospira and Actinomyces israelli are included.
感染性真菌の例としては、クリプトコックス・ネオフォルマンス(Cryptococcus neoformans)、ヒストプラズマ・カプスラーツム(Histoplasma capsulatum)、コクシジオイデス・イミティス(Coccidioides immitis)、ブラストミセス・デルマティティディス(Blastomyces dermatitidis)、トラコーマ病原体(Chlamydia trachomatis)、口瘡カンジダ(Candida albicans)が含まれる。その他の感染性生体(すなわち原生生物)には、熱帯性マラリア原虫(Plasmodium falciparum)およびトキソプラズマ・ゴンジ(Toxoplasma gondii)が含まれる。 Examples of infectious fungi include Cryptococcus neoformans, Histoplasma capsultatum, Coccidioides imtis, Blastomyces dermatoid B. Chlamydia trachomatis), Candida albicans. Other infectious organisms (ie, protists) include tropical Plasmodium falciparum and Toxoplasma gondii.
サイレンシング化された及びパルスを受けた免疫細胞
もう1つの実施形態においては、細胞を培養、組織、臓器又は生体から単離し、細胞ワクチンとして哺乳動物に投与することが可能である。かくして、本発明は「細胞ワクチン」を考慮している。当然のことながら、細胞は、免疫調節薬又はアジュバンドなどの1つもしくはそれ以上の付加的なワクチン構成要素を発現することもできる。ワクチンは、細胞の全て又は一部を含んでなっていてよい。好ましい実施形態においては、本発明の細胞ワクチンは、ヒトAPCを含んでなり、さらに好ましい実施形態においては、APCはDCである。
Silenced and pulsed immune cells In another embodiment, cells can be isolated from cultures, tissues, organs or organisms and administered to a mammal as a cellular vaccine. Thus, the present invention contemplates “cellular vaccines”. Of course, the cells can also express one or more additional vaccine components, such as immunomodulators or adjuvants. The vaccine may comprise all or part of the cells. In a preferred embodiment, the cellular vaccine of the present invention comprises human APC, and in a more preferred embodiment, the APC is DC.
細胞ワクチンは、その免疫能力を増強するため本発明に従ってサイレンシング化されたAPCを含んでなり得る。サイレンシング化されたAPCはこのとき、抗原負荷された細胞を生成するべく抗原をコードする核酸でのトランスフェクションを受けることができる。もう1つの態様においては、抗原負荷細胞を生成するべく抗原を含む免疫刺激タンパク質で、サイレンシング化されたAPCをパルスすることができる。本開示に基づくと、サイレンシング化されたAPCは、抗原を負荷するためにあらゆるタイプの抗原を利用してあらゆる方法でパルス可能である。さらに、本発明のインヒビターでのAPCのサイレンシング化の前、又はそれと同時に又はその後に、任意の方法によりAPCをパルスすることができる。 A cellular vaccine may comprise APC that has been silenced according to the present invention to enhance its immune capacity. The silenced APC can then be transfected with the nucleic acid encoding the antigen to produce antigen-loaded cells. In another embodiment, the silenced APC can be pulsed with an immunostimulatory protein containing the antigen to generate antigen-loaded cells. Based on the present disclosure, the silenced APC can be pulsed in any way utilizing any type of antigen to load the antigen. Furthermore, APC can be pulsed by any method prior to, simultaneously with, or after silencing of APC with the inhibitors of the present invention.
本書の他の箇所で開示されているように、さまざまな方法を用いて抗原で細胞をパルスすることができる。本発明の抗原は、少なくとも1つのエピトープを含有し、ここで前記エピトープは哺乳動物内で免疫応答を惹起する能力を有する。一実施形態においては、抗原は発現ベクターにより発現される。もう1つの実施形態においては、抗原は単離されたポリペプチドである。好ましくは、抗原は、感染性疾患、癌及び自己免疫疾患よりなる群から選択された1疾患と結びつけられる。本発明の細胞をパルスする上で有用な一定数の好ましい抗原が本書の他の箇所で開示されている。抗原は、腫瘍溶解物、タンパク質、ペプチド、mRNA、ベクターから発現されたDNA、リポソームなどのうちの少なくとも1つもしくはそれ以上の形をとり得る。 As disclosed elsewhere herein, a variety of methods can be used to pulse cells with antigen. The antigen of the present invention contains at least one epitope, wherein said epitope is capable of eliciting an immune response in a mammal. In one embodiment, the antigen is expressed by an expression vector. In another embodiment, the antigen is an isolated polypeptide. Preferably, the antigen is associated with one disease selected from the group consisting of infectious diseases, cancer and autoimmune diseases. A number of preferred antigens useful in pulsing the cells of the invention are disclosed elsewhere in this document. The antigen can take the form of at least one or more of tumor lysates, proteins, peptides, mRNA, DNA expressed from vectors, liposomes, and the like.
負の免疫調節因子のインヒビターでのサイレンシング化されたAPCは、増大した免疫能力を有し、従って増強された免疫応答すなわち抗原を提示しそれに対する免疫応答を活性化させる増強された能力を惹起する。本発明に従ってサイレンシング化及びパルスを受けたAPCは、エフェクタT細胞を刺激し、サイレンシング化を受けていないその他の点では同一であるAPCに比べて、それに対する抗原に対して改善された免疫応答を惹起する。 Silenced APCs with inhibitors of negative immune regulators have an increased immune capacity, thus eliciting an enhanced immune response, ie an enhanced ability to present and activate an immune response thereto To do. APCs that have been silenced and pulsed in accordance with the present invention stimulate effector T cells and have improved immunity against antigens compared to otherwise identical APCs that have not been silenced. Invoke a response.
治療的応用
本発明は、APCなどの免疫細胞の免疫能力を増強するのに有用な組成物を含む。APCにより提示される抗原に対する応答は、当該技術分野において周知の方法を用いて細胞溶解性T細胞応答、ヘルパーT細胞応答及び/又は抗原に対する抗体応答の誘発を監視することによって測定可能である。
Therapeutic Applications The present invention includes compositions useful for enhancing the immune capacity of immune cells such as APC. The response to an antigen presented by APC can be measured by monitoring the induction of a cytolytic T cell response, a helper T cell response and / or an antibody response to the antigen using methods well known in the art.
本発明は、抗原組成物と1つもしくはそれ以上のリンパ球を接触させる工程を含んでなる哺乳動物の体内の免疫応答を増強する方法において、抗原がAPCなどの免疫細胞により提示される方法を含む。本開示に基づくと、APCは、負の免疫調節因子のインヒビターに対する曝露によってサイレンシング化され得、かくしてインヒビターに対する曝露はAPCの免疫能力を増強する。APCは、その他の形でAPCをパルスするべく、抗原組成物にAPCを曝露する前、又はそれと同時又はその後に本書で開示されている方法を用いてサイレンシング化され得る。 The present invention relates to a method for enhancing an immune response in a mammalian body comprising the step of contacting an antigen composition with one or more lymphocytes, wherein the antigen is presented by immune cells such as APC. Including. Based on the present disclosure, APC can be silenced by exposure to inhibitors of negative immune modulators, thus exposure to inhibitors enhances the immune capacity of APC. The APC can be silenced using the methods disclosed herein prior to, simultaneously with, or after exposing the APC to the antigen composition to otherwise pulse the APC.
増強された免疫応答は、能動又は受動免疫応答であり得る。応答は、樹状細胞、B細胞又は単球/マクロファージといったAPCが哺乳動物(例えば患者)から得られ、その後(その他の形で免疫細胞をサイレンシング化するべく負の免疫調節因子のインヒビターに対し細胞を曝露する前、又はそれと同時又はその後に)抗原組成物を含んでなる組成物でパルスされ、次に必要としている哺乳動物に対しAPCを投与する養子免疫療法アプローチの一部であり得る。 The enhanced immune response can be an active or passive immune response. The response is that APCs such as dendritic cells, B cells or monocytes / macrophages are obtained from mammals (eg patients) and then (against negative immunomodulator inhibitors to silence immune cells in other ways). It may be part of an adoptive immunotherapy approach in which APC is administered to a mammal in need thereof, pulsed with a composition comprising an antigen composition (before, simultaneously with, or after cell exposure).
組成物は、負の免疫調節因子、抗原、サイレンシング化された免疫細胞、パルスを受けた細胞、そして同じく抗原でのパルスも受けたサイレンシング化された免疫細胞のうちの
少なくとも1つもしくはそれ以上のもののあらゆる組合せを含む。組成物は、哺乳動物内のエクスビボ免疫化及び/又はインビボ療法のためのワクチンであり得る。好ましくは、哺乳動物はヒトである。
The composition comprises at least one or more of a negative immune modulator, an antigen, a silenced immune cell, a pulsed cell, and a silenced immune cell that is also pulsed with an antigen. Includes any combination of the above. The composition can be a vaccine for ex vivo immunization and / or in vivo therapy in a mammal. Preferably the mammal is a human.
エクスビボ免疫化に関連して、哺乳動物内に細胞を投与する前にインビトロで以下のもののうちの少なくとも1つが起こる。すなわち、i)細胞のサイレンシング化、ii)細胞のパルス又は iii)細胞のサイレンシング化及びパルス。本発明の免疫細胞を、免疫細胞をパルスするべく抗原でAPCを処理する前、それと同時又はその後に本書の他の箇所で開示された方法を用いて、本発明の免疫細胞(すなわちAPC)をサイレンシング化することができる、ということを充分に理解すべきである。 In connection with ex vivo immunization, at least one of the following occurs in vitro prior to administering the cells into the mammal. I) cell silencing, ii) cell pulse or iii) cell silencing and pulse. The immune cells of the present invention can be treated using the methods disclosed elsewhere herein before, simultaneously with, or after treatment of the APC with an antigen to pulse the immune cells. It should be fully understood that it can be silenced.
もう1つの実施形態においてはサイレンシング化されたAPCを、抗原に予めインビトロで曝露することなく、必要としている患者に対し投与することが可能である。すなわち、本発明は、患者の体内でインビボで細胞のパルスが発生する、患者に対するサイレンシング化されたAPCの投与を包含している。 In another embodiment, the silenced APC can be administered to a patient in need without prior in vitro exposure to the antigen. That is, the present invention encompasses the administration of a silenced APC to a patient in which a pulse of cells occurs in vivo within the patient's body.
さらにもう1つの実施形態においては、パルスを受けたAPCを、負の免疫調節因子のインヒビターに対する細胞の事前のインビトロ曝露無く、それを必要とする患者に対して投与することができる。すなわち、本発明は、細胞のサイレンシング化が患者の体内においてインビボで起こる、患者に対するパルスを受けたAPCの投与を包含している。 In yet another embodiment, pulsed APCs can be administered to patients in need thereof without prior in vitro exposure of cells to inhibitors of negative immune modulators. That is, the present invention encompasses the administration of pulsed APCs to a patient, where cell silencing occurs in vivo in the patient's body.
エクスビボ手順は当該技術分野において周知であり、以下でさらに詳述されている。簡単に言うと、細胞が哺乳動物(好ましくはヒト)から単離され、負の免疫調節因子のインヒビターを発現するベクター又は本書で開示されている任意のその他の形態の負の免疫調節因子(すなわち化学的に合成されたsiRNA)でサイレンシング化(すなわちインビトロで形質導入又はトランスフェクション)される。サイレンシング化された細胞は、治療上の利益を提供するべく哺乳動物レシピエントに投与され得る。哺乳動物レシピエントはヒトであり得、このようにサイレンシング化された細胞はレシピエントとの関係において自己移植性であり得る。あるいは、細胞はレシピエントとの関係において同種異系、同系又は異種のものであり得る。 Ex vivo procedures are well known in the art and are described in further detail below. Briefly, a cell is isolated from a mammal (preferably a human) and a vector expressing an inhibitor of a negative immune regulator or any other form of negative immune regulator disclosed herein (ie, Chemically synthesized siRNA) is silenced (ie transduced or transfected in vitro). The silenced cells can be administered to a mammalian recipient to provide a therapeutic benefit. The mammalian recipient can be a human and the cells thus silenced can be autograft in the context of the recipient. Alternatively, the cells can be allogeneic, syngeneic or xenogeneic in relation to the recipient.
本発明に参照により援用されている米国特許第5,199,942号明細書の中で記述された造血幹及び前駆細胞のエクスビボ拡張のための手順を本発明の細胞に適用することができる。その他の適切な方法も当該技術分野において既知であり、従って本発明は細胞のエクスビボ拡張のいずれか特定の方法に限定されるわけではない。簡単に言うと、エクスビボ培養及びDCの拡張には、(1)末梢血の収穫物又は骨髄外植体から哺乳動物由来のCD34+造血幹及び前駆細胞を収集する工程及び(2)かかる細胞をエクスビボで拡張させる工程が含まれる。米国特許第5,199,942号明細書に記述されている細胞成長因子に加えて、flt3−L、TL−1、IL−3及びC−kitリガントなどのその他の因子を、細胞の培養及び拡張のために使用することができる。 The procedure for ex vivo expansion of hematopoietic stem and progenitor cells described in US Pat. No. 5,199,942, incorporated herein by reference, can be applied to the cells of the invention. Other suitable methods are also known in the art, and thus the present invention is not limited to any particular method of ex vivo expansion of cells. Briefly, ex vivo culture and DC expansion include (1) collecting mammalian derived CD34 + hematopoietic stem and progenitor cells from a peripheral blood harvest or bone marrow explant and (2) ex vivo such cells. The process of expanding by is included. In addition to the cell growth factors described in U.S. Pat. No. 5,199,942, other factors such as flt3-L, TL-1, IL-3 and C-kit ligand can be added to cell culture and Can be used for expansion.
細胞集団からCD34+造血幹又は前駆細胞を同定しかつ分離するためのさまざまな細胞選択技術が知られている。例えば、幹及び前駆細胞上に見られるマーカータンパク質又は表面抗原タンパク質に結合するように、モノクローナル抗体(又はその他の特異的細胞結合タンパク質)を使用することができる。造血幹細胞のためのいくつかのかかるマーカーまたは細胞表面抗原(すなわち、flt−3、CD34、My−10およびThy−1)は、当該技術分野において既知である。 Various cell selection techniques are known for identifying and separating CD34 + hematopoietic stem or progenitor cells from a cell population. For example, monoclonal antibodies (or other specific cell binding proteins) can be used to bind to marker proteins or surface antigen proteins found on stem and progenitor cells. Several such markers or cell surface antigens (ie flt-3, CD34, My-10 and Thy-1) for hematopoietic stem cells are known in the art.
収集されたCD34+細胞は適切なサイトカインと共に培養される。その後CD34+細胞は、分化して樹状系統の細胞に対応することができるようになる。これらの細胞は次
に、CD1a、HLA DR、CD80及び/又はCD86などの樹状細胞に特徴的であるマーカーを用いて、フローサイトメトリ又は類似の手段によってさらに精製される。DCの培養の単離後、細胞を本発明の方法に従って修飾することができる。あるいは、DC様の細胞に分化される前に前駆細胞を修飾することができる。
Collected CD34 + cells are cultured with appropriate cytokines. CD34 + cells can then differentiate to accommodate dendritic lineage cells. These cells are then further purified by flow cytometry or similar means using markers characteristic of dendritic cells such as CD1a, HLA DR, CD80 and / or CD86. After isolation of the DC culture, the cells can be modified according to the methods of the invention. Alternatively, progenitor cells can be modified before being differentiated into DC-like cells.
エクスビボ免疫化に関して細胞ベースのワクチンを使用することに加えて、本発明はまた、患者の体内の抗原に対し向けられた免疫応答を惹起するためのインビボ免疫化用の組成物及び方法をも提供している。 In addition to using cell-based vaccines for ex vivo immunization, the present invention also provides compositions and methods for in vivo immunization to elicit an immune response directed against antigens in a patient's body is doing.
インビボ免疫化に関し、本発明は、APC内の臨界制御点を無効にすることによりワクチンの効能を増強するための包括的手段としての負の免疫調節因子のインヒビターの使用を提供している。従って、インビボ免疫化にとって有用なワクチンは、少なくとも負の免疫調節因子を阻害することのできるインヒビター構成要素を含んでなる。もう1つの態様では、ワクチンは、インヒビター構成要素と抗原構成要素の両方を含んでなり、抗原構成要素は哺乳動物の体内で免疫応答を惹起する能力をもつ。 With respect to in vivo immunization, the present invention provides the use of inhibitors of negative immune modulators as a comprehensive means to enhance vaccine efficacy by overriding critical control points within APC. Thus, a vaccine useful for in vivo immunization comprises an inhibitor component capable of inhibiting at least negative immune modulators. In another aspect, the vaccine comprises both an inhibitor component and an antigen component, the antigen component having the ability to elicit an immune response in the mammalian body.
インビボ免疫化に関しては、患者から得られた細胞は、サイレンシング化された細胞をその他の形で生成するべくインビボでトランスフェクト又は形質導入される。細胞はインビボで、サイトカインレギュレータのインヒビターを発現するベクターでサイレンシング化される。あるいは、細胞は、(化学的に合成されたsiRNAである)ベクターにより発現されない本書で開示された負の免疫調節因子のその他の任意の形のインヒビターを用いてサイレンシング化される。インビボでサイレンシング化された細胞を生成する方法については、本書のその他の箇所で論述されている。 For in vivo immunization, cells obtained from a patient are transfected or transduced in vivo to generate silenced cells in other ways. Cells are silenced in vivo with vectors that express inhibitors of cytokine regulators. Alternatively, the cells are silenced using any other form of inhibitor of the negative immunomodulator disclosed herein that is not expressed by the vector (which is a chemically synthesized siRNA). Methods for generating silencing cells in vivo are discussed elsewhere in this document.
ワクチンのもう1つの態様は、インビボで細胞をパルスするのに有用な抗原構成要素を含む。本発明の負の免疫調節因子のインヒビターと組み合わせて、任意の抗原を投与することができる。インヒビターを含んでなるワクチンでの細胞のサイレンシング化の前に、又はそれと同時に又はその後に、本書のその他の箇所で論述されている通りのあらゆる方法を用いて細胞をパルスすることができる。細胞を同時にパルスしサイレンシング化すべきである場合には、インヒビター及び抗原を両方共含んでなる単一のワクチンで哺乳動物を免疫化できるということが容易にわかる。あるいは、インヒビターを含んでなるもの及び抗原を含んでなる第2のワクチンという2つの別々のワクチンで哺乳動物を免疫化することが可能である。 Another aspect of the vaccine includes an antigen component useful for pulsing cells in vivo. Any antigen can be administered in combination with the negative immunomodulator inhibitors of the present invention. Cells can be pulsed using any method as discussed elsewhere herein prior to, simultaneously with, or after silencing of cells with a vaccine comprising an inhibitor. It can easily be seen that if cells are to be pulsed and silenced simultaneously, a mammal can be immunized with a single vaccine comprising both inhibitor and antigen. Alternatively, it is possible to immunize a mammal with two separate vaccines, one comprising an inhibitor and a second vaccine comprising an antigen.
本発明は、癌及び感染性疾患のためのインビボ免疫化を包含する。一実施形態においては、患者の体内の抗原に対抗する免疫応答を生成するべく抗原と組合せた形で又は単独でsiRNAをインビボで投与することにより、障害又は疾患を治療することができる。本開示に基づくと、負の免疫調節因子のインヒビター(例えばA20、SUMO(SUMO1、SUMO2、SUMO3及びSUMO4)、Foxj1、Foxo3a、TWIST(Twist1、Twist2)、SOCS(SOCS1、SOCS2、SOCS3、SOCS4、SOCS5、SOCS6、SOCS7及びCIS)、PIAS(PIAS1、PIAS3、PIASx及びPIASy)、SHP(SHP−1及びSHP−2のためのsiRNA又はそのいずれかの組合せ)を抗原処方物と組合せた形で投与することにより、負の免疫調節因子のインヒビターを使用することなくその他の点では同一であるワクチン接種プロトコルの効能が増強される。何らかの特定の理論により束縛されることを望んではいないが、患者の体内の抗原に対する免疫応答は、(1)投与されたsiRNA組成物、(2)投与の持続時間、用量及び頻度、(3)患者の全身状態、そして該当する場合には(4)投与される抗原組成物により左右されると考えられている。 The present invention includes in vivo immunization for cancer and infectious diseases. In one embodiment, a disorder or disease can be treated by administering siRNA in vivo in combination with an antigen or alone to generate an immune response against the antigen in the patient's body. Based on this disclosure, inhibitors of negative immune modulators (eg A20, SUMO (SUMO1, SUMO2, SUMO3 and SUMO4), Foxj1, Foxo3a, TWIST (Twist1, Twist2), SOCS (SOCS1, SOCS2, SOCS3, SOCS4, SOCS5) , SOCS6, SOCS7 and CIS), PIAS (PIAS1, PIAS3, PIASx and PIASy), SHP (siRNA for SHP-1 and SHP-2 or any combination thereof) in combination with the antigen formulation This enhances the efficacy of vaccination protocols that are otherwise identical without the use of inhibitors of negative immune regulators, although not wishing to be bound by any particular theory, The immune response to the antigen is: (1) the administered siRNA composition, (2) the duration, dose and frequency of administration, (3) the general condition of the patient, and (4) the administered antigen composition, if applicable. It is thought to be influenced by things.
一実施形態においては、哺乳動物は、腫瘍特異的抗原を発現するタイプの癌を有してい
る。本発明に従うと、腫瘍特異的抗原配列構成要素を含んでなる免疫刺激タンパク質を作ることができる。このような場合、負の免疫調節因子のインヒビターは、それを必要とする患者に対し免疫刺激タンパク質との組合せの形で投与され、その結果、例えば腫瘍特異的抗原を発現する中実腫瘍又は癌細胞の成長の減速又は減少或いは癌細胞の総数又は総全身腫瘍組織量の低減によって証明される、患者にとっての治療成果の改善がもたらされる。
In one embodiment, the mammal has a type of cancer that expresses a tumor-specific antigen. According to the present invention, an immunostimulatory protein can be made comprising a tumor specific antigen sequence component. In such cases, the inhibitor of the negative immune modulator is administered to a patient in need thereof in combination with an immunostimulatory protein, resulting in, for example, a solid tumor or cancer that expresses a tumor-specific antigen Improvement in therapeutic outcome for the patient is evidenced by slowing or reducing cell growth or reducing the total number of cancer cells or total systemic tumor tissue volume.
関連する実施形態においては、患者は、例えばウイルス抗原といった特定の抗原の発現と結びつけられるウイルス、細菌、真菌又はその他のタイプの感染を有するとの診断を受けている。本発明に従うと、例えばHIV特異的抗原などの抗原からなる配列構成要素を含んでなる免疫刺激タンパク質を作ることができる。このような場合には、負の免疫調節因子のインヒビターが、それを必要とする患者に対して免疫刺激タンパク質と組合せた形で投与され、その結果、患者の体内の原因感染物質の成長の減速及び/又は特定の感染性疾患に標準的に付随する検出可能な症候群の低減又は除去により証明されるような患者にとっての治療成果の改善がもたらされる。 In related embodiments, the patient has been diagnosed as having a virus, bacterial, fungal or other type of infection that is associated with the expression of a particular antigen, eg, a viral antigen. In accordance with the present invention, an immunostimulatory protein can be made comprising a sequence component consisting of an antigen, for example an HIV specific antigen. In such cases, an inhibitor of a negative immune modulator is administered to a patient in need thereof in combination with an immunostimulatory protein, resulting in a slowing of the growth of the causative infectious agent in the patient's body. And / or results in improved therapeutic outcomes for patients as evidenced by the reduction or elimination of detectable syndromes typically associated with certain infectious diseases.
いずれの状況下でも、障害又は疾患は、それを必要とする患者に対する抗原と組合せた形での負の免疫調節因子のインヒビターの投与により治療され得る。本発明は、患者の体内で抗原に対する防御的DC誘発型免疫応答を生成するための手段を提供する。本開示に基づいて、当業者であれば、負の免疫調節因子ワクチンのインヒビターの効能を増強するため、本書に開示された治療計画に対し炎症性サイトカイン(すなわち、IL−12、TNFa、IFNa、IFNb、IFNgなど)を添加することができるということがわかるだろう。 Under any circumstances, a disorder or disease can be treated by administration of an inhibitor of a negative immune modulator in combination with an antigen to a patient in need thereof. The present invention provides a means for generating a protective DC-induced immune response to an antigen in a patient. Based on this disclosure, one of ordinary skill in the art would recognize inflammatory cytokines (ie, IL-12, TNFa, IFNa, etc.) against the treatment regimes disclosed herein to enhance the efficacy of inhibitors of negative immunomodulator vaccines. It will be appreciated that IFNb, IFNg, etc.) can be added.
投薬量及び処方(薬学組成物)
本発明は、治療薬例えばsiRNAの投与により哺乳動物の体内のHIV感染、癌などの疾患を治療することを想定している。本発明に係る治療薬の投与は、例えば患者の生理学的身体条件、投与の目的が治療か予防か、そして専門の医師にとって既知であるその他の要因に応じて、連続的又は間欠的であってよい。本発明の作用物質の投与は、基本的に、予め選択された期間にわたり連続的であり得、そうでなければ一連の間隔どりされた用量の形をとり得る。局所的及び全身的投与の両方が考慮される。投与される量は、選択された組成物、特定の疾患、哺乳動物の体重、身体条件及び年令、及び予防又は治療のいずれを達成すべきかを含めた(ただしこれに限定されるわけではない)さまざまな要因によって変動することになる。かかる要因は、当該技術分野において周知である動物モデル又はその他のテスト系を利用する臨床医により、容易に判定可能である。
Dosage and formulation (pharmaceutical composition)
The present invention envisages treating diseases such as HIV infection and cancer in the body of mammals by administration of therapeutic agents such as siRNA. Administration of the therapeutic agent according to the present invention may be continuous or intermittent, depending on, for example, the patient's physiological body condition, whether the purpose of the treatment is treatment or prevention, and other factors known to the specialist physician. Good. Administration of the agents of the present invention can be essentially continuous over a preselected period of time, otherwise it can take the form of a series of spaced doses. Both local and systemic administration is considered. The amount administered includes (but is not limited to) the selected composition, the particular disease, the mammal's weight, physical condition and age, and whether to prevent or treat. ) It will vary depending on various factors. Such factors can be readily determined by clinicians utilizing animal models or other test systems that are well known in the art.
siRNAの投与は、siRNAをコードする核酸分子の投与を通して達成され得る(例えば、フェルグナー(Felgner)ら、米国特許第5,580,859号明細書、パルドール(Pardoll)ら、1995年;スティーブンソン(Stevenson)ら、1995年;モーリング(Molling)1997年;ドネリー(Donnelly)ら、1995年;ヤン(Yang)ら、II;アブダラ(Abdallah)ら、1995年を参照)。例えば、フェルグナーら(前掲書)の中などで、核酸の薬学処方物、投薬量及び投与経路が一般的に開示されている。 Administration of siRNA can be accomplished through administration of a nucleic acid molecule encoding siRNA (eg, Felgner et al., US Pat. No. 5,580,859, Pardoll et al., 1995; Stevenson). ) Et al. 1995; Molling 1997; Donnelly et al. 1995; Yang et al. II; Abdallah et al. 1995). For example, in Fergner et al. (Supra), nucleic acid pharmaceutical formulations, dosages and routes of administration are generally disclosed.
以下で論述されているように場合により持続的放出のために処方され得る(例えばマイクロカプセル化を用いたもの、その開示が本発明に参照により援用されている国際公開第94/07529号パンフレット及び米国特許第4,962,091号明細書を参照)本発明の治療薬を有する1つもしくはそれ以上の適切な単位剤形を、静脈内及び筋肉を含めた非経口経路ならびに罹患組織内への直接注入を含むさまざまな経路で投与することができる。例えば、治療薬を直接腫瘍内に注射することが可能である。処方物は、該当する場
合、個別の単位剤形で便利な体裁をとり得、薬学では周知の方法のいずれかにより調製可能である。かかる方法は、液体担体、固体マトリクス、半固体担体、細分した固体担体又はそれらの組合せと治療薬を会合させ、その後必要であらば製品を所望の送達系内に導入又は整形する工程を含み得る。
May be optionally formulated for sustained release as discussed below (e.g., using microencapsulation, WO 94/07529, the disclosure of which is incorporated herein by reference) and (See US Pat. No. 4,962,091) One or more suitable unit dosage forms having the therapeutic agents of the invention can be administered into parenteral routes, including intravenous and intramuscular routes, and into affected tissues. It can be administered by a variety of routes including direct injection. For example, the therapeutic agent can be injected directly into the tumor. Formulations may take any convenient form in individual unit dosage forms, where applicable, and may be prepared by any of the methods well known in pharmacy. Such methods can include the step of associating the therapeutic agent with a liquid carrier, solid matrix, semi-solid carrier, finely divided solid carrier, or combinations thereof, and then introducing or shaping the product into the desired delivery system, if necessary. .
本発明の治療薬が投与のために調製される場合、これらは好ましくは、薬学処方物又は単位剤形を形成するため薬学的に許容可能な担体、希釈剤又は賦形剤と組み合わされる。かかる処方物中の総活性成分は、0.1〜99.9重量%の処方物を含む。「薬学的に許容可能な」というのは、処方物のその他の成分と適合可能でありそのレシピエントに有害でない担体、希釈剤、賦形剤及び/又は塩である。投与のための活性成分は、粉末又は顆粒として、溶液、懸液又はエマルジョンとして存在し得る。 When the therapeutic agents of the present invention are prepared for administration, these are preferably combined with a pharmaceutically acceptable carrier, diluent or excipient to form a pharmaceutical formulation or unit dosage form. The total active ingredient in such a formulation comprises 0.1 to 99.9% by weight of the formulation. “Pharmaceutically acceptable” is a carrier, diluent, excipient and / or salt that is compatible with the other ingredients of the formulation and not deleterious to the recipient thereof. The active ingredient for administration may be present as a powder or granule, as a solution, suspension or emulsion.
本発明の治療薬を含有する薬学的処方物は、周知の及び容易に入手可能な成分を用いて当該技術分野において既知の手順により調製可能である。本発明の治療薬は、非経口投与、例えば筋肉、皮下又は静脈内経路による投与に適した溶液としても処方可能である。 Pharmaceutical formulations containing the therapeutic agents of the invention can be prepared by procedures known in the art using well-known and readily available ingredients. The therapeutic agents of the invention can also be formulated as solutions suitable for parenteral administration, eg, by intramuscular, subcutaneous or intravenous routes.
本発明の治療薬の薬学処方物は同様に、水溶液又は無水溶液又は分散の形、あるいはエマルジョン又は懸濁液の形をとることもできる。 The pharmaceutical formulation of the therapeutic agent of the present invention can also take the form of an aqueous solution or no solution or dispersion, or an emulsion or suspension.
かくして、治療薬は、非経口投与(例えば注入、例えばボーラス注入又は持続注入による投与)のために処方され得、アンプル、薬剤充填済み注射器、少量注入器又は防腐剤が添加された多回用量コンテナに入った単回剤形の体裁をとり得る。活性成分は、油性又は水性ビヒクル中の懸濁液、溶液又はエマルジョンの形をとり得、懸濁、安定化及び/又は分散剤などの処方用作用物質を含有し得る。あるいは、活性成分は、使用前に滅菌で発熱物質を含まない水といった適切なビヒクルで構成するように、溶液からの凍結乾燥により又は滅菌固体の無菌単離によって得られる粉末形態であってもよい。 Thus, the therapeutic agent can be formulated for parenteral administration (eg, infusion, eg, by bolus injection or continuous infusion), ampoules, drug-filled syringes, small dose infusions or multi-dose containers with preservatives added. Can take the form of a single dosage form. The active ingredient may take the form of a suspension, solution or emulsion in an oily or aqueous vehicle and may contain formulatory agents such as suspending, stabilizing and / or dispersing agents. Alternatively, the active ingredient may be in powder form, obtained by lyophilization from solution or by aseptic isolation of sterile solids, such that it is composed of a suitable vehicle, such as sterile, pyrogen-free water prior to use. .
各剤形の個々のエアロゾル用量中に含有されている活性成分の単位含有量は、複数の剤形の投与によって必要な有効量を達成することが可能であるため、それ自体で特定の適応症又は疾病を治療するための1有効量を構成する必要はない、ということがわかるだろう。その上、剤形中の用量より少ない量を個別に又は一連の投与の中で用いることにより、有効量を達成することも可能である。 The unit content of the active ingredient contained in the individual aerosol dose of each dosage form is itself capable of achieving the required effective amount by administration of multiple dosage forms and as such is a specific indication. Alternatively, it will be appreciated that it is not necessary to constitute one effective amount for treating a disease. Moreover, effective amounts can be achieved by using less than the dose in the dosage form individually or in a series of administrations.
本発明の薬学処方物は、任意の成分として、薬学的に許容可能な担体、希釈剤、可溶化剤又は乳化剤、及び当該技術分野において周知であるタイプの塩を内含し得る。本発明の薬学処方物において有用である担体及び/又は希釈剤の限定的な意味のない特定の例としては、水及び生理学的に許容可能な緩衝生理食塩溶液、例えばpH7.0〜8.0のリン酸緩衝生理食塩溶液が含まれる。 The pharmaceutical formulations of the present invention can include, as optional ingredients, pharmaceutically acceptable carriers, diluents, solubilizers or emulsifiers, and salts of the type well known in the art. Specific, non-limiting examples of carriers and / or diluents useful in the pharmaceutical formulations of the present invention include water and physiologically acceptable buffered saline solutions, such as pH 7.0-8.0. A phosphate buffered saline solution.
本発明の発現ベクター、形質導入された細胞、ポリヌクレオチド及びポリペプチド(活性成分)は、生体の体内でその作用物質の作用部位と活性成分の接触を作り出すあらゆる手段によりさまざまな疾病状態を治療するように処方及び投与可能である。これらは、個々の治療用活性成分としてか又は治療用活性成分の組合せの中で、調合薬と併用する利用可能なあらゆる従来の手段によって投与され得る。これらは単独で投与され得るが、一般に、標準的な薬学的実践方法及び選択された投与経路に基づいて、薬学的担体と共に投与される。 The expression vectors, transduced cells, polynucleotides and polypeptides (active ingredients) of the present invention treat a variety of disease states by any means that create contact of the active ingredient with the active site in the body. Can be formulated and administered. They can be administered by any conventional means available in combination with pharmaceutical agents, either as individual therapeutic active ingredients or in a combination of therapeutic active ingredients. They can be administered alone, but are generally administered with a pharmaceutical carrier based on standard pharmaceutical practice and the chosen route of administration.
一般に、非経口溶液のためには、水、適切な油、食塩水、水性デキストロース(グルコース)、及び関連する糖溶液及びプロピレングリコール又はポリエチレングリコールといったグリコールが適切な担体である。非経口投与のための溶液は、活性成分、適切な可溶
化剤、そして必要に応じて緩衝物質を含有する。重硫酸ナトリウム、亜硫酸ナトリウム又はアスコルビン酸などの酸化防止剤が、単独であれ、組み合わされた形であれ、適切な安定化剤である。同様に使用されるのはクエン酸及びその塩、そしてエチレンジアミン四酢酸(EDTA)である。さらに、非経口溶液は、塩化ベンザルコニウム、メチル又はプロピルパラベン及びクロロブタノールなどの防腐剤を含有し得る。適切な薬学的担体が、この分野での標準的な参考書である「Remington’s Pharmacentical Sciences」の中で記述されている。
In general, for parenteral solutions, water, suitable oils, saline, aqueous dextrose (glucose), and related sugar solutions and glycols such as propylene glycol or polyethylene glycol are suitable carriers. Solutions for parenteral administration contain the active ingredient, suitable solubilizers and, if necessary, buffer substances. Antioxidants such as sodium bisulfate, sodium sulfite or ascorbic acid are suitable stabilizers, either alone or in combination. Also used are citric acid and its salts, and ethylenediaminetetraacetic acid (EDTA). In addition, parenteral solutions can contain preservatives, such as benzalkonium chloride, methyl or propylparaben, and chlorobutanol. Suitable pharmaceutical carriers are described in “Remington's Pharmaceutical Sciences”, a standard reference book in the field.
本発明の活性成分は、哺乳動物特にヒトにおいて使用するのに適した薬学的に許容可能な組成物の中で懸濁されるように処方可能である。このような処方物は、米国特許第4,082,735号明細書;同第4,082,736号明細書;同第4,101,536号明細書;同第4,185,089号明細書;同第4,235,771号明細書;及び同第4,406,890号明細書の中で記述されているムラミルジペプチド誘導体(MDP)又は類似体などのアジュバンドの使用を含む。有用なその他のアジュバンドには、ミョウバン(ピアス・ケミカル・カンパニー(Pierce Chemical Co.))、脂質A、トレハロースジミコレート及び臭化ジメチルジオクタデシルアンモニウム(DDA)、フロインドアジュバンド及びIL−12が含まれる。その他の構成要素としては、ポリオキシプロピレン−ポリオキシレンブロックコポリマー(プルロニック(Pluronic)(登録商標))、非イオン性界面活性剤及びスクアレンなどの代謝性油(米国特許第4,606,918号明細書)が含まれ得る。 The active ingredients of the present invention can be formulated to be suspended in a pharmaceutically acceptable composition suitable for use in mammals, particularly humans. Such formulations are described in U.S. Pat. Nos. 4,082,735; 4,082,736; 4,101,536; 4,185,089. No. 4,235,771; and 4,406,890, including the use of adjuvants such as the muramyl dipeptide derivatives (MDP) or analogs described therein. Other useful adjuvants include alum (Pierce Chemical Co.), lipid A, trehalose dimycolate and dimethyldioctadecylammonium bromide (DDA), Freundajuband and IL-12. It is. Other components include polyoxypropylene-polyoxylene block copolymers (Pluronic®), nonionic surfactants and metabolic oils such as squalene (US Pat. No. 4,606,918). Letter).
さらに、標準的な薬学的方法を利用して作用持続時間を制御することが可能である。これらの方法は、当該技術分野において周知であり、制御放出調製物を内含し、例えばポリマー、ポリエステル、ポリアミノ酸、ポリビニル、ピロリドン、エチレンビニルアセタート、メチルセルロース、カルボキシメチルセルロース又は硫酸プロタミンといった適切な巨大分子を含み得る。巨大分子の濃度ならびに取込み方法は、放出を制御するために調整可能である。さらに、ポリエステル、ポリアミノ酸、ヒドロゲル、ポリ(乳酸)又はエチレン酢酸ビニルコポリマーなどの作用物質をポリマー材料の粒子中に取込むことができる。取込まれることに加えて、これらの作用物質は、マイクロカプセル内に化合物を捕捉するのにも使用可能である。 Furthermore, the duration of action can be controlled using standard pharmaceutical methods. These methods are well known in the art and include controlled release preparations such as polymers, polyesters, polyamino acids, polyvinyls, pyrrolidone, ethylene vinyl acetate, methylcellulose, carboxymethylcellulose or protamine sulfate. It can contain molecules. The concentration of macromolecules as well as the uptake method can be adjusted to control the release. In addition, agents such as polyesters, polyamino acids, hydrogels, poly (lactic acid) or ethylene vinyl acetate copolymers can be incorporated into the particles of polymer material. In addition to being incorporated, these agents can also be used to entrap compounds within the microcapsules.
従って、本発明の薬学組成物は、特定の効果を達成するべく哺乳動物の体内のさまざまな部位に対しさまざまな経路を介して送達可能である(例えば、ローゼンフェルド(Rosenfeld)ら、1991年;ローゼンフェルドら、1991年a;ジャフェ(Jaffe)ら、上掲書;バークナー(Berkner)、上掲書を参照)。当業者であれば、投与のために2つ以上の経路を用いることができるものの、特定の経路が、もう一方の経路に比べより即刻のかつより効果的な反応を提供する可能性がある、ということを認識するだろう。体腔内への処方物の点滴又は塗布、エアロゾルの吸入又は吹送又は、筋肉、静脈内、腹腔内、皮下、皮内ならびに局所的投与を含んでなる非経口導入を含んでなる投与により、局所的又は全身的送達を達成することが可能である。 Accordingly, the pharmaceutical compositions of the invention can be delivered via a variety of routes to various sites within a mammal to achieve a particular effect (eg, Rosenfeld et al., 1991; Rosenfeld et al., 1991a; Jaffe et al., Supra; see Berkner, supra). Although one of skill in the art can use more than one route for administration, a particular route may provide a more immediate and more effective response than the other route. You will recognize that. By topical administration by infusion or application of the formulation into body cavities, inhalation or insufflation of aerosols or parenteral introduction comprising intramuscular, intravenous, intraperitoneal, subcutaneous, intradermal as well as topical administration Alternatively, systemic delivery can be achieved.
本発明の活性成分は、例えば茶さじ一杯、錠剤、溶液又は座薬などの各投薬量単位が、単独で又はその他の活性作用物質と適切に組合せた形で、予め定められた量の組成物を含有している、単位剤形の形で提供され得る。本書で使用する「単位剤形」という用語は、ヒト及び哺乳動物の対象のための単一の投薬量として適切でかつ、各々該当する場合薬学的に許容可能な希釈剤、担体又はビヒクルと会合した状態で、所望の効果を生み出すのに充分な量で計算された、その他の活性作用物質と組合せた形又は単独の予め定められた数量の本発明の組成物を含有している、標準的に個別の単位を意味する。本発明の単位剤形のための仕様は、達成すべき特別な効果及び特定の宿主の中で薬学組成物に付随する特別な薬物力学によって左右される。 The active ingredient of the present invention comprises a predetermined amount of a composition in which each dosage unit, for example, a teaspoon, a tablet, a solution or a suppository, alone or in appropriate combination with other active agents. It can be provided in a unit dosage form containing. As used herein, the term “unit dosage form” is appropriate as a single dosage for human and mammalian subjects, and is associated with a pharmaceutically acceptable diluent, carrier or vehicle, as applicable. A standard amount containing the composition of the present invention alone or in combination with other active agents, in an amount sufficient to produce the desired effect. Means an individual unit. The specifications for the unit dosage form of the present invention depend on the particular effects to be achieved and the particular pharmacodynamics associated with the pharmaceutical composition within the particular host.
本書で記述されているこれらの方法は、決して包括的ではなく、当業者には特定の利用分野に適したさらなる方法が明らかになることだろう。その上、組成物の有効量は、所望の効果を及ぼすものとして知られている化合物に対する類推を通してさらに近似され得る。 These methods described in this document are in no way comprehensive, and those skilled in the art will find additional methods suitable for a particular field of application. Moreover, the effective amount of the composition can be further approximated through analogy to compounds known to have the desired effect.
遺伝子療法の投与
当業者であれば、細胞内にベクターを投与するために異なる送達方法を利用できることを認識する。例としては、(1)物理的手段、例えば電気穿孔(電気)、遺伝子統(物理的力)又は大量の液体(圧力)の付加を用いた方法、及び(2)リポソーム、凝集タンパク質又はトランスポータ分子などのもう1つの実体に対して前記ベクターを複合させる方法、が含まれる。
Gene Therapy Administration The skilled artisan will recognize that different delivery methods are available for administering the vector into the cell. Examples include (1) methods using physical means such as electroporation (electro), genetics (physical force) or application of large amounts of liquid (pressure), and (2) liposomes, aggregated proteins or transporters A method of conjugating the vector to another entity, such as a molecule.
さらに、実際の用量及び計画は、その組成物がその他の薬学組成物と組合せて投与されているか否か又は薬物動態、薬物挙動及び代謝における個体間差に応じて変動し得る。同様に、量はインビトロ利用分野において、利用される特定の細胞系列に応じて(例えば細胞表面上に存在するベクターレセプタの数、又はその細胞系列内で複製する遺伝子移送に利用された特定のベクターの能力に基づく)変動し得る。さらに、細胞1個あたりの付加すべきベクターの量は、ベクター内に挿入された治療用遺伝子の長さ及び安定性ならびに配列の性質に応じて変動する確率が高く、特に経験的に判定される必要のあるパラメータであり、本発明の方法に固有の因子(例えば合成に付随するコスト)に起因して改変され得る。当業者であれば、特定の状況の要件に従って、任意の必要な調整を容易に行なうことができる。 Furthermore, actual dosages and regimens can vary depending on whether the composition is being administered in combination with other pharmaceutical compositions or on individual differences in pharmacokinetics, drug behavior and metabolism. Similarly, the amount depends on the particular cell line utilized in the in vitro field (eg, the number of vector receptors present on the cell surface, or the particular vector utilized for gene transfer that replicates within that cell line. Based on the ability of Furthermore, the amount of vector to be added per cell is likely to vary depending on the length and stability of the therapeutic gene inserted into the vector and the nature of the sequence, and is determined especially empirically. It is a necessary parameter and can be modified due to factors inherent in the method of the invention (eg, costs associated with synthesis). A person skilled in the art can easily make any necessary adjustments according to the requirements of the particular situation.
治療薬を含有する細胞は同様に、自殺遺伝子すなわち細胞を破壊するのに使用可能な産物をコードする遺伝子をも含有し得る。数多くの遺伝子療法の状況下で、宿主細胞内で治療目的で遺伝子を発現できることのみならず意のままに宿主細胞を破壊する能力をもつことができることも望ましい。治療薬は、アクチベータ化合物の不在下でその発現が活性化されない自殺遺伝子に連結され得る。作用物質及び自殺遺伝子の両方が導入された細胞の死滅が望まれる場合、アクチベータ化合物は、その細胞に対し投与され、かくして自殺遺伝子の発現を活性化し、細胞を死滅させる。使用可能な自殺遺伝子/プロドラッグの組合せの例としては、単純ヘルペスウイルス−チミジンキナーゼ(HSV−tk)及びガンシクロビル、アシクロビル;オキシドレダクターゼ、及びシクロヘキシミド;シトシンデアミナーゼ及び5−フルオロシトシン;チミジンキナーゼチミジラートキナーゼ(Tdk::Tmk)及びAZT;及びデオキシシチジンキナーゼ及びシトシンアラビノシドがある。 Cells containing therapeutic agents can also contain genes that code for suicide genes, ie products that can be used to destroy the cells. In many gene therapy situations, it is desirable not only to be able to express a gene for therapeutic purposes in a host cell, but also to have the ability to destroy host cells at will. The therapeutic agent can be linked to a suicide gene whose expression is not activated in the absence of the activator compound. If it is desired to kill a cell into which both the agent and the suicide gene have been introduced, the activator compound is administered to the cell, thus activating the expression of the suicide gene and killing the cell. Examples of suicide gene / prodrug combinations that can be used include herpes simplex virus-thymidine kinase (HSV-tk) and ganciclovir, acyclovir; oxidoreductase, and cycloheximide; cytosine deaminase and 5-fluorocytosine; thymidine kinase thymidylate kinase (Tdk :: Tmk) and AZT; and deoxycytidine kinase and cytosine arabinoside.
本書で記述されているこれらの方法は、決して包括的ではなく、特定の利用分野に適したさらなる方法が当業者には明らかとなるであろう。その上、所望の効果を及ぼすものとして知られている化合物に対する類推を通して、組成物の有効量をさらに近似することが可能である。 These methods described herein are by no means exhaustive and additional methods suitable for a particular field of application will be apparent to those skilled in the art. Moreover, it is possible to further approximate the effective amount of the composition through analogy to compounds known to have the desired effect.
本発明についてここで以下の実施例を参考にしながら記述する。これらの実施例は、あくまで例示を目的として提供されており、本発明はこれらの実施例に限定されず、むしろ本書で提供されている教示の結果として明白である全ての変形形態を包含する。 The invention will now be described with reference to the following examples. These examples are provided for illustrative purposes only, and the invention is not limited to these examples, but rather encompasses all variations that are apparent as a result of the teachings provided herein.
本書で開示されている実験は、さまざまな癌及び感染性病原体に対する有効なワクチンを開発する目的でDCを活用するためDCによる抗原提示の調節を探索するべく実施された。本書で開示されている結果は、免疫細胞内での負の免疫調節因子の阻害として別途知
られている負の免疫調節経路との干渉が、その免疫刺激能力を増強する、ということを実証している。細胞の免疫能力を強化するために負の免疫調節因子を阻害するという概念は、より効果的なワクチンを開発する新規の方法として役立つ。
The experiments disclosed herein were conducted to explore the regulation of antigen presentation by DCs to exploit DCs for the purpose of developing effective vaccines against various cancers and infectious agents. The results disclosed herein demonstrate that interference with negative immune regulatory pathways, otherwise known as inhibition of negative immune regulators within immune cells, enhances its immunostimulatory capacity. ing. The concept of inhibiting negative immune regulators to enhance cellular immunity serves as a new way to develop more effective vaccines.
本書で開示されている実験において利用される材料及び方法についてここで記述する。 The materials and methods utilized in the experiments disclosed herein are now described.
siRNAオリゴによるDCのトランスフェクション
21、26又は同等の塩基対siRNAオリゴスクレオチドで、骨髄DCをトランスフェクトした。例えば、メーカーのプロトコルに従いジーンポーター(GenePorter)(ゲンランティス(Genlantis)、カリフォルニア州サンディエゴ(San
Diego,CA))を用いて、DCをSOCS1−siRNA3(5’−CTACCTGAGTTCCTTCCCCTT−3’;配列番号3)でトランスフェクトした。ただし、本書の他の箇所で論述されているいずれのsiRNAオリゴヌクレオチドでも、本書で論述されている方法を用いてDC内にトランスフェクト可能である。簡単に言うと、20μMのオリゴヌクレオチドの溶液3μlを3μlのジーンポーター試薬及び94μlの無血清RPMI1640に添加した。30分間25℃で混合物をインキュベートし、その後、100μlのジーンポーター/オリゴヌクレオチドの混合物を骨髄−DCの各ウェルに添加し、37℃で4時間インキュベートした。インキュベーションの後、20%のFBSで捕足された500μl/ウェルのRPMI1640を骨髄DCに添加した。
Transfection of DCs with siRNA oligos Bone marrow DCs were transfected with 21, 26 or equivalent base pair siRNA oligos. For example, according to the manufacturer's protocol, GenePorter (Genlantis, San Diego, CA)
Diego, CA)) were used to transfect DCs with SOCS1-siRNA3 (5′-CTACCTGAGTTCCTTCCCCTT-3 ′; SEQ ID NO: 3). However, any siRNA oligonucleotide discussed elsewhere in this document can be transfected into DC using the methods discussed herein. Briefly, 3 μl of a 20 μM oligonucleotide solution was added to 3 μl Gene Porter reagent and 94 μl serum-free RPMI 1640. The mixture was incubated for 30 minutes at 25 ° C., after which 100 μl of the gene porter / oligonucleotide mixture was added to each well of the bone marrow-DC and incubated at 37 ° C. for 4 hours. After incubation, 500 μl / well RPMI 1640 supplemented with 20% FBS was added to bone marrow DC.
骨髄由来のDCのレンチウイルスベクターによる形質導入
マウスの骨髄由来のDCを、当該技術分野において既知の方法を用いて調製した。簡単に言うと、マウス骨髄を四肢から洗い流し、ナイロンメッシュを通し、塩化アンモニウムで赤血球を枯渇させた。RPMI−1640での広範な洗浄の後、細胞を、10%のFBS、mGM−CSF/ml(20ng/ml)及び組換え型マウスIL−4(20ng/ml:ペプロテック社(PeproTech.Inc.)ニュージャージー州ロッキーヒル(Rocky Hill,NJ))で補足された2.5mlのRPMI−1640と共に培養した。培養2日目と4日目に、上清を除去し、20ng/mlのrmGM−CSF及び20ng/mlのrmIL−4を含有する新鮮な培地と交換した。5%加湿したCO2中で37℃で全ての培養をインキュベートした。48時間の培養の後、非接着性顆粒球を除去し、新鮮培地を添加した。7日間の培養後、80%を超える細胞が、FACSにより決定される通り、特徴的なDC特異的マーカーを発現した。マウス骨髄由来のDCの形質導入(培養5日目〜7日目)を、5μg/mlのポリブレン(Polybrene)(シグマ、ミズーリ州セントルイス)の添加により、24ウェルのプレート上で実施した。DCを洗浄し、400μlの無血清RPMI1640中2×105細胞/ウェルの濃度で24ウェルのプレート内にプレート固定した。5×105細胞/mlという細胞密度で、異なる感染多重度(MOI)をもつレンチウイルスベクターに細胞を曝露した。8時間の形質導入の後、細胞をPBSで洗浄し、さらに新鮮な組織培地内でインキュベートした。
Transduction of bone marrow-derived DCs with lentiviral vectors Mouse bone marrow-derived DCs were prepared using methods known in the art. Briefly, mouse bone marrow was washed from the extremities, passed through a nylon mesh, and red blood cells were depleted with ammonium chloride. After extensive washing with RPMI-1640, cells were treated with 10% FBS, mGM-CSF / ml (20 ng / ml) and recombinant mouse IL-4 (20 ng / ml: PeproTech. Inc.). Incubated with 2.5 ml RPMI-1640 supplemented with Rocky Hill, NJ. On
サイトカイン及びウェスタンブロット法
メーカーの指示に従って、ELISA分析(BDバイオサイエンシーズ(Biosciences)、ニュージャージー州リンカーンパーク(Lincoln Park,NJ))を用いて細胞培養の上清を使用してさまざまなサイトカインレベルを定量化した。ウェスタンブロット分析のためには、SOCS1、A20、Foxj1、SUMO及びTwist2−siRNA又は対照siRNA(例えばGFP−siRNA)を含めた(ただしこれに限定されるわけではない)所望のsiRNAを発現するpSUPERベクター及び対応するタグ付き発現ベクターで、293T細胞を同時トランスフェクトした。
Cytokines and Western blots Quantify various cytokine levels using cell culture supernatants using ELISA analysis (BD Biosciences, Lincoln Park, NJ) according to manufacturer's instructions Turned into. For Western blot analysis, pSUPER vectors expressing desired siRNA, including but not limited to SOCS1, A20, Foxj1, SUMO and Twist2-siRNA or control siRNA (eg, GFP-siRNA) And 293T cells were co-transfected with the corresponding tagged expression vector.
例えば、293T細胞を、10:1の比のマウスSOCS1−siRNA又は無関係のGFP−siRNAとFLAGタグ付きSOCS1ベクターで同時トランスフェクトした
。細胞を48時間後に収穫し、SDS−PAGEに付した。Hybond−P膜(アマシャム(Amersham)、イリノイ州アーリントンハイツ(Arlington Heights,IL))への移送の後、試料を、抗Flag(シグマ、ミズーリ州セントルイス)又はアクチン((サンタクルス・バイオテクノロジー社(Santa Cruz Biotechnology,Inc.)、カリフォルニア州サンタクルス(Santa
Cruz,CA)))抗体でのウェスタンブロット法とそれに続くECL−Plus試薬(アマシャム、イリノイ州アーリントンハイツ)での検出により分析した。フィルムをデンシトメータ(Densitometer)SIで走査し、SOCS−1/アクチンバンドをイメージ・クアント(ImageQuant)ソフトウエア(モレキュラー・ダイナミクス(Molecular Dynamics)、ニュージャージー州ピスカタウェイ(Piscataway,NJ))で定量した。SOCS1バンドの強度をベータアクチンバンドの強度に正規化した。
For example, 293T cells were co-transfected with a 10: 1 ratio of mouse SOCS1-siRNA or irrelevant GFP-siRNA and a FLAG-tagged SOCS1 vector. Cells were harvested 48 hours later and subjected to SDS-PAGE. After transfer to Hybond-P membrane (Amersham, Arlington Heights, Ill.), The samples were either anti-Flag (Sigma, St. Louis, MO) or actin ((Santa Cruz Biotechnology ( Santa Cruz Biotechnology, Inc., Santa Cruz, CA
Cruz, CA))) was analyzed by Western blotting with antibodies followed by detection with ECL-Plus reagent (Amersham, Arlington Heights, IL). Films were scanned with a Densitometer SI and SOCS-1 / actin bands were quantified with ImageQuant software (Molecular Dynamics, Piscataway, NJ). The intensity of the SOCS1 band was normalized to the intensity of the beta actin band.
SOCS1の定量RT−PCR分析
トランスフェクションを受けたマウスBM−DC内のSOCS1の相対的発現を、定量的実時間PCRにより評価した。トリゾール試薬(インビトロジェン、カリフォルニア州カールズバッド(Carlsbad,CA))を用いて3.5〜5×105BM−DCから、全RNAを抽出した。各試料についての全RNA1.0μgをランダム六量体プライマ及びSuperScript First−Strand Synthesisキット(インビトロジェン、カリフォルニア州カールズバッド)を用いて逆転写した。鋳型として反応一回につき5ngの出発RNA材料の当量を用い20μlの四重反応中のABI7900HTシーケンス・デテクション・システム(Sequence Detection System)(アプライド・バイオシステムズ社、カリフォルニア州フォスターシティー)上で、実時間5’−ヌクレアーゼ螢光性PCR分析を実施した。マウスSOCS1(6FAM)及び18Sリボソーム対照(VIC)のための予め開発されたプライマ/プローブセットをアプライド・バイオシステムズ社(カリフォルニア州フォスターシティー)(SOCS1のためのプライマ、5’−ACCTTCTTGGTGCGCGAC−3’;配列番号12及び5’−AAGCCATCTTCACGCTGAGC−3’;配列番号13及びハイブリダイゼーションプローブ、6FAM−TCGCCAACGGAACTGCTTCTTCG−TAMRA;配列番号14)から購入した。PCRパラメータは、SOCS1及び18S反応を別々の管で実施して、タクマン・ユニバーサルPCRマスター・ミックス・キット(TaqMan Universal PCR Master Mix kit)(アプライド・バイオシステムズ社、カリフォルニア州フォスターシティー)のために推奨された通りであった。SOCS1レベルを18SrRNAに正規化した。モックトランスフェクション及び刺激を受けたBM−DCの対照値との関係におけるSOCS1発現を、コンパラティブ(Comparative)Ct方法(ライバック(Livak)ら、2001年、Methods第25号:402〜408頁)を用いて計算した。
Quantitative RT-PCR analysis of SOCS1 Relative expression of SOCS1 in transfected mouse BM-DC was assessed by quantitative real-time PCR. Total RNA was extracted from 3.5-5 × 10 5 BM-DC using Trizol reagent (Invitrogen, Carlsbad, Calif.). 1.0 μg of total RNA for each sample was reverse transcribed using a random hexamer primer and SuperScript First-Strand Synthesis kit (Invitrogen, Carlsbad, Calif.). Real time on ABI 7900HT Sequence Detection System (Applied Biosystems, Foster City, Calif.) In a 20 μl quadruplicate reaction using an equivalent of 5 ng starting RNA material per reaction as template. 5'-nuclease fluorescent PCR analysis was performed. A pre-developed primer / probe set for mouse SOCS1 (6FAM) and 18S ribosome control (VIC) was applied to Applied Biosystems, Inc. (Foster City, Calif.) (Primer for SOCS1, 5′-ACCTCTTTGTGTGCGCGAC-3 ′; SEQ ID NO: 12 and 5′-AAGCCATCTTCACCGCTGAGC-3 ′; SEQ ID NO: 13 and hybridization probe, 6FAM-TCGCCAACGGGAACTGCTTTCTTCG-TAMRA; SEQ ID NO: 14). PCR parameters are recommended for the TaqMan Universal PCR Master Mix kit (Applied Biosystems, Foster City, Calif.) With SOCS1 and 18S reactions performed in separate tubes As it was. SOCS1 levels were normalized to 18S rRNA. SOCS1 expression in relation to control values for mock transfection and stimulated BM-DC was determined using the Comparative Ct method (Livak et al., 2001, Methods 25: 402-408). Used to calculate.
A20、Foxj1及びSUMO1の定量的RT−PCR分析
定量的実時間PCRにより、トランスフェクションを受けたマウスBM−DC内のA20、Foxj1及びSUMO1の相対的発現を評価した。全RNAを、トリゾール試薬(インビトロジェン、カリフォルニア州カールズバッド)を用いて3.5〜5×105BM−DCから抽出した。各試料についての全RNA1.0μgをランダム六量体プライマ及びSuperScript First−Strand Synthesisキット(インビトロジェン、カリフォルニア州カールズバッド)を用いて逆転写した。鋳型として反応一回につき5ngの出発RNA材料の当量を用い20μlの四重反応中のABI7900HT Sequence Detection System(アプライド・バイオシステムズ社、カリフォルニア州フォスターシティー)上で、実時間5’−ヌクレアーゼ螢光性PCR分析を実施した。マウスA20、Foxj1及びSUMO1及び18Sリボソ
ーム対照(VIC)のための予め開発されたプライマ/プローブセットをアプライド・バイオシステムズ社(カリフォルニア州フォスターシティー)から購入した(A20のためのプライマ、5’−GCATCTGCAGTACCTGTTTC’−3;配列番号186及び5’−GACAGGAGGCAGGGATA−3’;配列番号187及びハイブリダイゼーションプローブ:5’−ACTACAGCAGAGCCCAGCTCCAGCC−3’;配列番号188;Foxj1のためのプライマ、5’−GCCATGCAAGCCAGCAA−3’;配列番号189及び5’−GCAGAAGTTGTCCGTGATCCA−3’;配列番号190;及びハイブリダイゼーションプローブ:5’−AAGATCACTCTGTCGGCCATCTACAA−3’;配列番号191;SUMO1のためのプライマ、5’−GAAGGTCAGAGAATTGCTGATAATCAT−3’;配列番号192及び5’−CCCCGTTTGTTCCTGATAAACT−3’;配列番号193、及びハイブリダイゼーションプローブ:5’−TCCGAAAGAACTGGGAATGGAGGAAGAA−3’;配列番号194)。PCRパラメータは、本書の他の箇所で論述されたものに従った。
Quantitative RT-PCR analysis of A20, Foxj1 and SUMO1 The relative expression of A20, Foxj1 and SUMO1 in transfected mouse BM-DC was assessed by quantitative real-time PCR. Total RNA was extracted from 3.5-5 × 10 5 BM-DC using Trizol reagent (Invitrogen, Carlsbad, Calif.). 1.0 μg of total RNA for each sample was reverse transcribed using a random hexamer primer and SuperScript First-Strand Synthesis kit (Invitrogen, Carlsbad, Calif.). Real-
OT−I細胞のインビトロ検定
OT−Iマウスから脾臓を収穫し、プールし、単一の細胞懸濁液を得るべく分断した。MACSCD8+T細胞単離キット(ミルテニー・バイオテク社(Miltenyi Biotec Inc.)、カリフォルニア州オウバーン(Auburn,CA))を用いて負の選択によりCD8+OT−IT細胞を収集した。簡単に言うと、細胞を、CD4(L3T4)、CD45R(B220)、DX5、CD11b(Mac−1)及びTer−119に特異的なビオチン標識された抗体でコーティングした。抗ビオチン磁気ミクロビーズ(MicroBeads)(ミルテニー・バイオテク社、カリフォルニア州オウバーン)を細胞に添加し、これらを、MACS磁石に取付けられた分離カラム全体に通過させた。カラムに結合しなかった細胞を収集し、それはFACSにより判定されるとき95%超のCD8+であった。10%のFCS、4mMのL−グルタミン、1mMのピルビン酸ナトリウム、100U/mlのペニシリン及びストレプトマイシン、10mMのHEPES及び5μMの2−MEで補足された200μlのRPMI1640培地の中で丸底96ウェルのマイクロタイタープレートの各ウェル内に、合計5×104個の精製済みCD8+OT−IT細胞及び5×103の未熟なDCを入れた。最後の8時間の培養中、1ウェルあたり1μのCi[3H]TdRを添加することにより2日後に増殖を測定した。3重判定を行ない、これは3通りの実験に相当する。OT−I/DC同時培養内のサイトカイン分泌を、指示されたサイトカインについてELISA分析を用いて判定した(BDバイオサイエンシーズ、カリフォルニア州サンノゼ(San Jose,CA))。
In vitro assay of OT-I cells Spleens from OT-I mice were harvested, pooled and split to obtain a single cell suspension. CD8 + OT-IT cells were collected by negative selection using a MACSCD8 + T cell isolation kit (Miltenyi Biotec Inc., Auburn, Calif.). Briefly, cells were coated with biotinylated antibodies specific for CD4 (L3T4), CD45R (B220), DX5, CD11b (Mac-1) and Ter-119. Anti-biotin magnetic microbeads (MicroBeads) (Miltenyi Biotech, Auburn, Calif.) Were added to the cells and they were passed through a separation column attached to a MACS magnet. Cells that did not bind to the column were collected and were > 95% CD8 + as judged by FACS. Round bottom 96-well in 200 μl RPMI 1640 medium supplemented with 10% FCS, 4 mM L-glutamine, 1 mM sodium pyruvate, 100 U / ml penicillin and streptomycin, 10 mM HEPES and 5 μM 2-ME. A total of 5 × 10 4 purified CD8 + OT-IT cells and 5 × 10 3 immature DCs were placed in each well of the microtiter plate. During the last 8 hours of culture, proliferation was measured after 2 days by adding 1 μCi [ 3 H] TdR per well. A triple determination is made, which corresponds to three experiments. Cytokine secretion within the OT-I / DC co-culture was determined using ELISA analysis for the indicated cytokine (BD Biosciences, San Jose, Calif.).
フローサイトメトリ分析
FCγレセプタを予め遮断した後、0.1%のNaN3及び2%のFCSを含有するPBS中のFITC、PE、アロフィコシアニン(APC)又はPerCP接合型mAbsで細胞を染色した。ラットCD4(RM4−5)、CD8(53−6.7)、CD11c(HL3)、CD40(3/23)、CD80(16−10A1)、CD86(GL1)に特異的なラットmAbs及び整合したイソタイプ対照をBDバイオサイエンシーズ(カリフォルニア州サンノゼ)から購入した。染色した細胞をFACSCalibur(ベクトン・ディッキンソン(Becton Dickinson)、ニュージャージー州リンカーンパーク)フローサイトメーター及びセルクエスト(CELLQuest)ソフトウエア上で分析した。
Flow cytometric analysis After pre-blocking the FC gamma receptor, cells were stained with FITC, PE, allophycocyanin (APC) or PerCP conjugated mAbs in PBS containing 0.1% NaN 3 and 2% FCS. . Rat mAbs specific for rat CD4 (RM4-5), CD8 (53-6.7), CD11c (HL3), CD40 (3/23), CD80 (16-10A1), CD86 (GL1) and matched isotypes Controls were purchased from BD Biosciences (San Jose, Calif.). Stained cells were analyzed on a FACSCalibur (Becton Dickinson, Lincoln Park, NJ) flow cytometer and CELLQuest software.
四量体染色
オボアルブミン特異的CD8+T細胞を検出するためにH2−Kb/オボアルブミン四量体検定を使用した。免疫化したマウスに由来する脾細胞又はT細胞を、DC免疫化から異なる日数後に抗CD8α−FITC及びH2−Kb/オボアルブミン(SIINFEK
L)−PE四量体;配列番号11(ベックマン・カルター・イムノミクス(Beckman Coulter Immunomics)、カリフォルニア州サンディエゴ)で2重染色した。メーカーの指示に従って、細胞106個あたり1μgの抗CD8α及び10μlのオボアルブミン四量体で1時間4℃で四量体染色を行なった。
Tetramer staining The H2-K b / ovalbumin tetramer assay was used to detect ovalbumin-specific CD8 + T cells. Spleen cells or T cells derived from immunized mice were treated with anti-CD8α-FITC and H2-K b / ovalbumin (SIINFEK) after different days after DC immunization.
L) -PE tetramer; double stained with SEQ ID NO: 11 (Beckman Coulter Immunos, San Diego, CA). Tetramer staining was performed with 1 μg anti-CD8α and 10 μl ovalbumin tetramer per 10 6 cells for 1 hour at 4 ° C. according to the manufacturer's instructions.
酵素連結型イムノスポット(ELISPOT)
CTLペプチドをCD8+T細胞刺激のために使用した。負の対照として、ヒトCD20分子からの無関係なペプチドも使用した。CD8+T細胞を、MACS CD4(L3T4)又はMACS CD8(Ly−2)マイクロビーズ(ミルテニー・バイオテク社、カリフォルニア州オウバーン)
を用いることで脾細胞から単離した。
Enzyme-linked immunospot (ELISPOT)
CTL peptide was used for CD8 + T cell stimulation. An irrelevant peptide from the human CD20 molecule was also used as a negative control. CD8 + T cells were transformed into MACS CD4 (L3T4) or MACS CD8 (Ly-2) microbeads (Miltenyi Biotech, Auburn, Calif.).
Was isolated from spleen cells.
CTL及びNK検定
標的細胞を溶解するインビトロ再刺激された脾細胞の能力を測定する標準クロム放出検定でCD8+CTL応答を評価した。免疫化されたマウスからプールした脾細胞を、4〜6日間ペプチドを含有するRPMI−1640内でインビボで再刺激した。標的細胞及び対照細胞を90分間、51Crクロム酸ナトリウム溶液で標識した。異なる数のエフェクタ細胞を、37℃で3時間96ウェルのV字底プレート(200μl/ウェル)の中で一定数の標的細胞(1×104/ウェル)と共にインキュベートした。三重培養からの上清(100μl)を収集した。(実験的放出−自然放出)/(最大放出−自然放出)×100として溶解百分率を計算した。500U/mlの組換え型マウスIL−2で1×106細胞/mlを培養することにより、マウスの脾細胞からNK細胞を生成した。NK細胞による溶解をきわめて受けやすいYAC−1細胞を37℃で1時間51Crでインキュベートし、洗浄し、105細胞/mlで再懸濁した。異なるE:T細胞比を得るため、標的細胞に対し三重にNK細胞を添加した。インキュベーションの後、プレートを遠心分離に付し、ガンマ計数器で上清流体中の放射能を計数した(ベックマン・カルター社(Beckman Coulter,Inc.)、カリフォルニア州フラートン(Fullerton,CA))。
CTL and NK assays CD8 + CTL responses were assessed with a standard chromium release assay that measures the ability of in vitro restimulated splenocytes to lyse target cells. Pooled splenocytes from immunized mice were restimulated in vivo in RPMI-1640 containing peptides for 4-6 days. Target and control cells were labeled with 51 Cr sodium chromate solution for 90 minutes. Different numbers of effector cells were incubated with a fixed number of target cells (1 × 10 4 / well) in 96-well V-bottom plates (200 μl / well) at 37 ° C. for 3 hours. Supernatants (100 μl) from triplicate cultures were collected. Percent dissolution was calculated as (experimental release-spontaneous release) / (maximum release-spontaneous release) x 100. NK cells were generated from mouse spleen cells by culturing 1 × 10 6 cells / ml with 500 U / ml recombinant mouse IL-2. YAC-1 cells highly susceptible to lysis by NK cells were incubated with 51 Cr for 1 hour at 37 ° C., washed and resuspended at 10 5 cells / ml. To obtain different E: T cell ratios, NK cells were added in triplicate to the target cells. After incubation, the plates were centrifuged and the radioactivity in the supernatant fluid was counted with a gamma counter (Beckman Coulter, Inc., Fullerton, Calif.).
DC免疫化及び腫瘍モデル
SOCS1、A20、Foxj1、Foxo3a、SUMO、Twist−1及びTwist−2−siRNAを含む(ただしこれに限定されるわけではない)所望のsiRNAを発現するレンチウイルスで、骨髄由来のDC(骨髄培養5日目)を形質導入した。例えば、骨髄由来のDCを、5のMOIでLV−SOCS1−siRNA又はLV−GFP−siRNAで形質導入した。その後DCを8時間TRP2ペプチド又はオボアルブミンタンパク質でパルスし、PBSで3回洗浄し、培養中で、さらに36時間後、免疫化のために使用した。一部の実験については、抗原パルスを受けたDCを24時間LPS(100ng/ml、シグマ、ミズーリ州セントルイス)で刺激し、PBSで洗浄し、その後C57BL/6マウス(ジャクソン・ラボラトリー)に足蹠から注入した。治療モデルにおいて、EG7又はB16腫瘍細胞(2.5〜5×105)を同系C57BL/6マウスの右脇腹内に皮下(s.c.)注射した。腫瘍接種から異なる日数後で、マウスをランダムにグループに分け、50μlの抗原パルスを受け形質導入されたDC又はPBC対照を注入した。一部のマウスにおいて、ワクチン接種から指示された日数後にLPSを腹腔内(i.p.)投与した。キャリパで一週間に2〜3回、腫瘍体積を測定した。
DC immunization and tumor model Lentivirus expressing the desired siRNA, including but not limited to SOCS1, A20, Foxj1, Foxo3a, SUMO, Twist-1 and Twist-2-siRNA, derived from bone marrow DC (bone marrow culture day 5) were transduced. For example, bone marrow-derived DCs were transduced with LV-SOCS1-siRNA or LV-GFP-siRNA at an MOI of 5. DCs were then pulsed with TRP2 peptide or ovalbumin protein for 8 hours, washed 3 times with PBS, and used for immunization in culture after an additional 36 hours. For some experiments, antigen-pulsed DCs were stimulated with LPS (100 ng / ml, Sigma, St. Louis, MO) for 24 hours, washed with PBS, and then footpadd into C57BL / 6 mice (Jackson Laboratory). Injected from. In the treatment model, EG7 or B16 tumor cells (2.5-5 × 10 5 ) were injected subcutaneously (sc) into the right flank of syngeneic C57BL / 6 mice. Different days after tumor inoculation, mice were randomly grouped and injected with DC or PBC controls transduced with 50 μl of antigen pulses. In some mice, LPS was administered intraperitoneally (ip) after the number of days indicated after vaccination. Tumor volumes were measured with calipers 2-3 times a week.
統計学的分析
統計学的分析のためには、スチューデントt検定が使用され、P<0.05と定義づけされる95%の信頼性限界が有意なものとされた。結果は標準的に、平均±標準誤差として提示される。
Statistical analysis For statistical analysis, Student's t-test was used and a 95% confidence limit defined as P <0.05 was considered significant. Results are typically presented as mean ± standard error.
本実施例で提示されている実験の結果について、ここで記述する。 The results of the experiment presented in this example will now be described.
実施例1:マウスSOC−1siRNAの同定と分析
SiRNA配列ターゲティングマウスSOCS1:SOCS1−siRNA1(CCTTCCGCTCCCACTCCGA;配列番号1)、SOCS1−siRNA2(CAGTCGCCAACGGAACTGC;配列番号2)及びSOCS1−siRNA3(CTACCTGAGTTCCTTCCCCTT;配列番号3)を選択するためにコンピュータプログラムを使用した。その他の既知の遺伝子に対する相同性の欠如を確認するために、全ての標的配列をNCBI Blastクエリに付した。9ntのスペーサで分離されたセンス及びアンチセンス19nt標的配列をコードするために順方向及び逆方向オリゴを設計した。このコアsiRNA配列は、H1RNA転写開始及び5Tターミネータ配列によってフランキングされ、アニーリングの時点で5’Bg1II及び3’HindIII相溶性オーバーハングを取込んだ。DNAベースのsiRNA発現ベクターpSUPER(ブルンメルカンプら、2002年、Science第296号:550〜553頁)は、siRNAのデノボ合成を導くためにH1−RNAプロモータを使用する。合成されアニールされたオリゴヌクレオチド対を、Bg1II/HindIIIで消化されたpSUPERベクター内にクローニングした。制限消化物により正のクローンを同定し、DNA配列決定により確認した。
Example 1: Identification and analysis of mouse SOC-1 siRNA SiRNA sequence targeting mouse SOCS1: SOCS1-siRNA1 (CCTCTCCCTCCCACTACTGA); Used a computer program to select. All target sequences were subjected to NCBI Blast queries to confirm the lack of homology to other known genes. Forward and reverse oligos were designed to encode sense and
マウスSOCS1−siRNAのためのレンチウイルスベクターの生成及び産生
本研究において使用されたHIV移入ベクターは、内部サイトメガロウイルス(CMV)プロモータを含んでなり、かつ転写活性のある配列を除去する3’長末端反復(LTR)のU3領域内に400bpの欠失をもつ自己不活性化ベクター(SINベクター)である、pTRIP△U3 CMV eGFPであった。レンチウイルス移入ベクターpTRIP△U3 CMV GFPは、もとのpHR’主鎖の固有のClaI部位の中に中央ポリプリントラクト(cPPT)及び中央終結配列(CTS)を包含する178bpのフラグメントを含有する。pTRIPΔU3 CMV GFPは、H1 RNAプロモータからのsiRNAの発現及びビシストロンブラスチシジン耐性/eYEP選択マーカーの同時発現のために修飾された。クローニングに対処し未変性CMVプロモータを除去するために、クローニングのための5’−EcoRI及び3’−BamHI部位を挿入するべくプライマ(5’−GATCGAATTCACAAATGGC−3’;配列番号4及び5’−CTAGGGATCCATCGCCCCAAAGTGG−3’;配列番号5)を用いて、pTRIPベクターの中央ポリプリントラクト/中央終結配列(cPPT/CTS)をPCR増幅した。その後、cPPT−CTSPCR産物をEcoRI/BamHIで消化し、EcoRI/BamHI消化済みpTRIPΔU3 CMV GFPベクターの中に再挿入した。ウッドチャック転写後調節要素(wPRE)配列を、プライマ(5’−GATCCTCGAGGTCGACAATCAACCTCTGGA−3’;配列番号6及び5’−GATCGGTACCCAGGCGGGGAGG−3’;配列番号7)を用いてpBS−SK−WPREからPCR増幅して、5’−XhoI/SalI及び3’KpnI部位を添加した。その後、XhoI/KpnIでWPREフラグメントを消化し、修飾済みのpTRIPΔU3 CMV GFP主鎖の中に挿入してpTRIP−Wを生成した。siRNA及びビシストロン選択マーカーカセットをまず、pTRIP−W内への移送のためにpSUPER主鎖内で組立てた。ビシストロン選択マーカーCMV−BlastiR−IRES−eYFP(BY)をプライマ(5’−CAGTATCGATTTAATTAATCAATATTGGCCATTAG−3’;配列番号8及び5’−CAGTGTCGACTTAATTAAGTGGCCGCTTTACTTG−3’;配列番号9)を用いてプラスミドPYAP6からPCR増幅して、5’−ClaI及び3’−SalI部位を取込んだ。PCR産物をClaI及びSalIで消化し、次にClaI/SalI消化したpSUPERベクター内にライゲートさせて、H1−RNAプロモータ及びpSUPERMCSの3’末端でBYマーカーを添加し、pSUPER−BYを生成させた。その後、
pSUPER−BYをBamHI/SalI消化し、pTRIP−W主鎖内にライゲートさせて、pTRIP−H1−BY−Wを生成させた。
Generation and production of lentiviral vectors for mouse SOCS1-siRNA The HIV transfer vector used in this study comprises an internal cytomegalovirus (CMV) promoter and is 3 ′ long to remove transcriptionally active sequences. PTRIPΔU3 CMV eGFP, a self-inactivating vector (SIN vector) with a 400 bp deletion in the U3 region of the terminal repeat (LTR). The lentiviral transfer vector pTRIPΔU3 CMV GFP contains a 178 bp fragment containing a central polyprinted lactate (cPPT) and a central termination sequence (CTS) within the unique ClaI site of the original pHR ′ backbone. pTRIPΔU3 CMV GFP was modified for expression of siRNA from the H1 RNA promoter and co-expression of the bicistronic blasticidin resistance / eYEP selectable marker. Primers (5′-GATCGAATTCACAAATGGC-3 ′; SEQ ID NO: 4 and 5′-CTAGGGATCCCATCGCCCCAAAGTGGG) to insert the 5′-EcoRI and 3′-BamHI sites for cloning in order to deal with cloning and remove the native CMV promoter -3 ′; SEQ ID NO: 5) was used to PCR amplify the central polyprinted lactate / central termination sequence (cPPT / CTS) of the pTRIP vector. The cPPT-CTS PCR product was then digested with EcoRI / BamHI and reinserted into the EcoRI / BamHI digested pTRIPΔU3 CMV GFP vector. The woodchuck post-transcriptional regulatory element (wPRE) sequence was PCR amplified from pBS-SK-WPRE using primers (5'-GATCCTCGAGGGTCGACAATCAACCCTTGGA-3 '; SEQ ID NO: 6 and 5'-GATCGGTACCCAGGCGGGAGG-3'; SEQ ID NO: 7). 5'-XhoI / SalI and 3'KpnI sites were added. The WPRE fragment was then digested with XhoI / KpnI and inserted into the modified pTRIPΔU3 CMV GFP backbone to generate pTRIP-W. siRNA and bicistronic selectable marker cassettes were first assembled in the pSUPER backbone for transfer into pTRIP-W. Bicistron selection marker CMV-Blasti R -IRES-eYFP (BY) was amplified using primer (5′-CAGTATCGATTTAATTAATCAATATTGGCCCATTAG-3 ′; SEQ ID NO: 8 and 5′-CAGTGTCGACTTAATTAAGTGGCCCGCTTTTACTTG-3 ′; SEQ ID NO: 9) 5'-ClaI and 3'-SalI sites were incorporated. The PCR product was digested with ClaI and SalI and then ligated into the CSUP / SalI digested pSUPER vector, and a BY marker was added at the 3 'end of the H1-RNA promoter and pSUPERMCS to generate pSUPER-BY. afterwards,
pSUPER-BY was digested with BamHI / SalI and ligated into the pTRIP-W backbone to generate pTRIP-H1-BY-W.
hrGFP又はSOCS1(3)siRNAヘアピン配列を含有する後続するpSUPERベクターを、pTRIP−H1−BY−W内への挿入のためにBstB1及びClaIで消化して、pTRIP−hrGFP−siRNA−BY−W(GFP−siRNA)、及びpTRIP−SOCS1−siRNA−BY−W(SOCS1−siRNA)を生成させた。最後の390bpのスペーサーフラグメントを、最終ベクターのClaI部位で挿入して、CMVプロモータの最初からsiRNAの終結配列を離隔させた。全てのベクターをDNA配列決定によって確認した(ローン・スター・ラブス(Lone Star Labs)、米国テキサス州ヒューストン(Houston,TX))。 Subsequent pSUPER vectors containing hrGFP or SOCS1 (3) siRNA hairpin sequences were digested with BstB1 and ClaI for insertion into pTRIP-H1-BY-W to obtain pTRIP-hrGFP-siRNA-BY-W ( GFP-siRNA), and pTRIP-SOCS1-siRNA-BY-W (SOCS1-siRNA). The last 390 bp spacer fragment was inserted at the ClaI site of the final vector to separate the siRNA termination sequence from the beginning of the CMV promoter. All vectors were confirmed by DNA sequencing (Lone Star Labs, Houston, TX, USA).
293T細胞内への3つのプラスミドの同時トランスフェクションにより、組換え型シュードタイプレンチウイルスベクターを生成した。HIV由来のパッケージング構成体pCMV△R8.9は、HIV−1gag、及びpol前駆物質ならびに調節タンパク質tat及びrevをコードする。水疱性口内炎ウイルスの糖タンパク質G(VSV−G)が、プラスミドpMD.Gから発現された。pCMV△R8.9、pMD.G及びレンチウイルスpTRIPsiRNA移入ベクターでの293T細胞の過渡的リン酸カルシウム同時トランスフェクションによりシュードタイプレンチウイルスを産生させた。トランスフェクションから60〜72時間後に、4℃で2時間50、000×gで超遠心分離により上清を濃縮させた。ウイルスペレットをRPMI内で再懸濁し、さらなる研究のため−80℃で凍結させた。濃縮ウイルスの段階希釈物及び8μg/mlのポリブレンと共に6時間293T細胞をインキュベートしその後、72〜96時間後にeYEP−陽性細胞を判定するべく蛍光活性化細胞分類(FACS)分析を行なうことによって、ウイルス力価を決定した。ベクター力価を次のように計算した:力価=F×2×C0/V×D(なお式中Dはウイルス希釈因子、Vは接種材料の体積、FはeYFP陽性293T細胞の頻度、そしてC0は播種の時点での標的細胞の数である)。 Recombinant pseudotyped lentiviral vector was generated by co-transfection of three plasmids into 293T cells. The HIV-derived packaging construct pCMVΔR8.9 encodes HIV-1 gag and pol precursors and regulatory proteins tat and rev. Vesicular stomatitis virus glycoprotein G (VSV-G) is expressed in plasmid pMD. Expressed from G. pCMVΔR8.9, pMD. Pseudotype lentivirus was generated by transient calcium phosphate co-transfection of 293T cells with G and lentiviral pTRIP siRNA transfer vectors. 60-72 hours after transfection, the supernatant was concentrated by ultracentrifugation at 50,000 × g for 2 hours at 4 ° C. The virus pellet was resuspended in RPMI and frozen at −80 ° C. for further study. By incubating 293T cells with serial dilutions of concentrated virus and 8 μg / ml polybrene for 6 hours followed by fluorescence activated cell sorting (FACS) analysis to determine eYEP-positive cells after 72-96 hours. The titer was determined. Vector titers were calculated as follows: titer = F × 2 × C 0 / V × D (where D is the virus dilution factor, V is the volume of the inoculum, F is the frequency of eYFP positive 293T cells, And C 0 is the number of target cells at the time of seeding).
実施例2:ジーンポーターでのSOCS1siRNAを伴うマウスBM−DCのトランスフェクション及びSOCS1mRNAダウンレギュレーション
DCによる抗原提示のSOCS1調節を調査するために、特異的にSOCS1をダウンレギュレートする短鎖干渉RNA(siRNA)を以下で記述する通りに同定した。
Example 2: Transfection of mouse BM-DC with SOCS1 siRNA at GenePorter and SOCS1 mRNA downregulation To investigate the SOCS1 regulation of antigen presentation by DC, short interfering RNA (siRNA) that specifically downregulates SOCS1 ) Was identified as described below.
合成siRNAオリゴ2重鎖を、83%のトランスフェクション効率でジーンポーターにより、顆粒球マクロファージ・コロニー刺激因子(GM−CSF)及びIL−4の存在下でエクスビボでマウス骨髄細胞に由来するDCの中に効率良くトランスフェクトさせた。簡単に言うと、メーカーのプロトコルに従ってジーンポーターを用いて21の塩基対siRNAオリゴヌクレオチド(5’−CTACCTGAGTTCCTTCCCCTT−3’;配列番号3)で骨髄DCをトランスフェクトした。3μlのジーンポーター試薬及び94μlの無血清RPMI1640に対し3μlの20μMオリゴヌクレオチドを添加し、30分間25℃でインキュベートし、その後100μlのジーンポーター/オリゴヌクレオチド混合物を骨髄−DCの各ウェルに添加して37℃で4時間インキュベートした。インキュベーションの後、20%のFBSで補足された500μl/ウェルのRPMI1640を骨髄DCに添加した。本開示に基づくと、生理学的に許容可能な担体という状況下で合成siRNAオリゴが細胞に送達され得る。許容可能な担体の例としてはリポソームがある。
In a DC derived from mouse bone marrow cells ex vivo in the presence of granulocyte-macrophage colony stimulating factor (GM-CSF) and IL-4 by a gene porter with 83% transfection efficiency at a synthetic siRNA oligo duplex Efficiently transfected. Briefly, bone marrow DCs were transfected with 21 base pair siRNA oligonucleotides (5'-CTACCTGAGTTCCTTCCCCTT-3 '; SEQ ID NO: 3) using a gene porter according to the manufacturer's protocol. Add 3
図1Aは、SOCS1−siRNAを発現するベクターで形質導入された細胞の中でSOCS1がダウンレギュレートされることを立証している。簡単に言うと、ジーンポーターを用いて10:1の比でマウスSOCS1−siRNA又は無関係なGFP−siRN
Aを発現するpSUPER(pSUR)ベクター及びFLAGタグ付きmSOCS1発現ベクターで293T細胞を同時トランスフェクトし、48時間後にウェスタンブロット法に付した。SOCS1バンドの強度をベータアクチンバンドのものに正規化し、相対的強度(比)を示している。SOCS−siRNAと呼ばれるSOCS1−siRNA3(5’−CTACCTGAGTTCCTTCCCCTT−3’;配列番号3)を後続する研究で使用した。
FIG. 1A demonstrates that SOCS1 is down-regulated in cells transduced with a vector expressing SOCS1-siRNA. Briefly, mouse SOCS1-siRNA or irrelevant GFP-siRN at a 10: 1 ratio using a gene porter.
293T cells were co-transfected with a pSUPER (pSUR) vector expressing A and a FLAG-tagged mSOCS1 expression vector and subjected to
定量的RT−PCR検定によって確認される通り、SOCS1siRNAでのトランスフェクションを受けた全DC集団中のSOCS1mRNAのレベルは、SOCS1(図1B)をダウンレギュレートできないSOCS1siRNA突然変異体でのトランスフェクションを受けたDC中のレベルと比べて約60%だけ特異的に減少した。さらに、インビトロでの骨髄のDC培養中及び成熟の後、SOCS1発現はさらに高いものであることが観察された。 As confirmed by quantitative RT-PCR assay, the level of SOCS1 mRNA in the total DC population transfected with SOCS1 siRNA is subject to transfection with a SOCS1 siRNA mutant that cannot down-regulate SOCS1 (FIG. 1B). It was specifically reduced by about 60% compared to the level in DC. Moreover, SOCS1 expression was observed to be even higher during bone marrow DC culture in vitro and after maturation.
同様に、刺激の時点でのSTAT1、I−κB及びJNKのリン酸化の増強及びIL−6及びTNF−α(図2)などの前炎症性サイトカインの分泌の増強によって示されるように、siRNA突然変異体を伴うDCよりもSOCS1siRNAでのトランスフェクションを受けたDCの方がLPS又はIFN−γに対しより高い応答性を有するということも観察された。図1Bは、3つの独立した実験の1つからの、24時間LPS(100ng/ml)又はIFN−γ(10ng/ml)に応答してsiRNAオリゴ−又はモック−トランスフェクションを受けたDCが分泌するサイトカインのレベルを描いている。SiRNA突然変異体(5’−ACTATCTAAGTTACTACCCCTT−3’;配列番号10)は、SOCS1siRNA3配列中に4つの突然変異を含有する。 Similarly, siRNA abruptity, as shown by enhanced phosphorylation of STAT1, I-κB and JNK at the time of stimulation and secretion of proinflammatory cytokines such as IL-6 and TNF-α (FIG. 2) It was also observed that DCs transfected with SOCS1 siRNA were more responsive to LPS or IFN-γ than DCs with mutants. FIG. 1B shows the secretion of siRNA oligo- or mock-transfected DCs in response to 24-hour LPS (100 ng / ml) or IFN-γ (10 ng / ml) from one of three independent experiments. Depicts the level of cytokines to be. The SiRNA mutant (5'-ACTATCTAAGTTACTACCCCT-3 '; SEQ ID NO: 10) contains 4 mutations in the SOCS1 siRNA3 sequence.
これらのデータは、JAK/STAT経路及びTLR/NF−κB経路の調節にSOCS1が関与するという報告と一致している(ハナダら、2003年、Immunity第19号:437〜450頁;チョン(Chong)ら、2003年、Immunity第18号:475〜487頁)。SOCS1siRNAでのトランスフェクションを受けたDCは、IFN−γ及びLPS刺激の前後いずれかでのsiRNA−DC突然変異体よりもわずかに成熟度の高い表現型を示した。トランスフェクションを受けた両方のDC共、モックトランスフェクションを受けたDCよりも成熟度が高かったが、このことはsiRNAによるIFN遺伝子の非特異的活性化の効果を反映しているかもしれない。 These data are consistent with reports that SOCS1 is involved in regulation of the JAK / STAT pathway and the TLR / NF-κB pathway (Hanada et al., 2003, Immunity 19: 437-450; Chong ) Et al., 2003, Immunity 18: 475-487). DCs transfected with SOCS1 siRNA showed a slightly more mature phenotype than siRNA-DC mutants either before or after IFN-γ and LPS stimulation. Both transfected DCs were more mature than mock transfected DCs, which may reflect the effect of non-specific activation of the IFN gene by siRNA.
実施例3:ウイルスベクターでのマウスBM−DCのトランスフェクション
SOCS1がインビボでDC抗原提示を負に調節するか否かを評価するために、DCを安定した形で形質導入する能力をもつレンチウイルスベクター(LV)内に、SOCS1siRNA又は対照緑色蛍光タンパク質(GFP)siRNAをクローニングして(ルビンソンら、2003年、Nat.Genet.第33号:401〜406頁;スクロアーら、2004年、Methods Mol.Biol.第246号:451〜459頁)、SOCS1サイレンシング化の効果をより高い信頼性で査定できるようにした。本書の他の箇所で記述された方法に従って黄色螢光タンパク質(YFP)マーカー(図3)を共に含有するLV−SOCS1−siRNA及びLV−GFP−siRNAという2つの構成体を生成した。LV−SOCS1−siRNA又はLV−GFP−siRNAベクターのいずれかでの骨髄由来のDCの形質導入(80%超のCD11c+)は、日常的に、YFPについて陽性の培養細胞の58〜63%を生成した。siRNAオリゴトランスフェクションを受けたDCに関する先行する観察と一致して、LV−GFP−siRNA−DCと比べLV−SOCS1−siRNA−DCの刺激の時点で、形質導入を受けた全DC集団内のSOCS1mRNAのレベルの低下及び前炎症性サイトカインの分泌の増強が観察された。形質導入を受けたDC中のSOCS1mRNAレベルを判定するために、螢光活性化細胞分類(FACS)を用いてYFP+での形質導入を受けたDCを単離し、その
後、実時間定量的PCRにより、SOCS1mRNAの相対的発現を判定した。YEP+LV−SOCS1−siRNA−DC集団中のSOCS1mRNAのレベルは、モック形質導入されたDC内のレベルと比べて約90%低いことが観察された。LPS刺激を伴う又はこれを伴わないLV−SOCS1−siRNA及びLV−GFP−siRNAでの形質導入を受けたDCは、比敵するレベルのCD86及びCD40発現を示した。いずれかの特定の理論により束縛されることは望まないが、LV−GFP−siRNA−DCが結果としてモックDCよりも高いレベルのCD86及びCD40をもたらしたという観察事実は、siRNA及びレンチウイルス形質導入による非特異的活性化の効果に起因するものである確率が高いと考えられている。
Example 3: Transfection of mouse BM-DC with viral vectors To assess whether SOCS1 negatively regulates DC antigen presentation in vivo, a lentivirus with the ability to transduce DCs in a stable fashion SOCS1 siRNA or control green fluorescent protein (GFP) siRNA was cloned into the vector (LV) (Rubinson et al., 2003, Nat. Genet. 33: 401-406; Scroar et al., 2004, Mols Mol. Biol.246: 451-459), the effect of silencing the SOCS1 can be evaluated with higher reliability. Two constructs, LV-SOCS1-siRNA and LV-GFP-siRNA, containing both yellow fluorescent protein (YFP) markers (FIG. 3) were generated according to the methods described elsewhere in this document. Transduction of bone marrow-derived DCs with either LV-SOCS1-siRNA or LV-GFP-siRNA vectors ( > 80% CD11c + ) routinely results in 58-63% of cultured cells positive for YFP. Generated. Consistent with previous observations on siRNA oligo-transfected DCs, SOCS1 mRNA in the total DC population transduced at the time of stimulation of LV-SOCS1-siRNA-DC compared to LV-GFP-siRNA-DC Decreased levels and increased secretion of pro-inflammatory cytokines was observed. To determine SOCS1 mRNA levels in transduced DCs, fluorescently activated cell sorting (FACS) was used to isolate DCs transduced with YFP + , followed by real-time quantitative PCR. The relative expression of SOCS1 mRNA was determined. It was observed that the level of SOCS1 mRNA in the YEP + LV-SOCS1-siRNA-DC population was approximately 90% lower than the level in mock-transduced DCs. DCs transduced with LV-SOCS1-siRNA and LV-GFP-siRNA with or without LPS stimulation showed comparable levels of CD86 and CD40 expression. While not wishing to be bound by any particular theory, the observation that LV-GFP-siRNA-DC resulted in higher levels of CD86 and CD40 than mock DC was the result of siRNA and lentiviral transduction. It is thought that there is a high probability that it is due to the effect of non-specific activation by.
実施例4:マウスSOCS1siRNA DCにより誘発されるOVA特異的CTL及び抗腫瘍活性
特異的細胞傷害性Tリンパ球(CTL)のDC刺激がSOCS1により調節されるか否かをテストするために、次の一連の実験を実施した。成熟に至るまでさらに刺激されなかった未成熟のSOSC1−siRNA−DC又はsiRNA突然変異体DCをオボアルブミンI−ペプチド(SIINFEKL;配列番号11)でパルスされ、オボアルブミン特異的TCRT細胞(OT−I)と同時培養させた時点で、OT−I細胞は、siRNA−DC突然変異体同時培養中よりもSOCS1−siRNA−DC同時培養中で多く増殖した(図4A)。これらのデータと一致して、SOCS1−siRNA−DC同時培養中で、より高いレベルの前炎症性サイトカインが分泌された(図4B)。さらに、これらの細胞のCTL検定は、SOCS1−siRNA−DCとの同時培養の後オボアルブミン+同系EG7細胞に対するより活性な細胞傷害性を示し、このことは、SOCS1が抗原特異的T細胞のDC刺激の調節に貢献することを実証している。
Example 4: OVA-specific CTL and anti-tumor activity elicited by mouse SOCS1 siRNA DC To test whether DC stimulation of specific cytotoxic T lymphocytes (CTL) is regulated by SOCS1, the following A series of experiments was performed. Immature SOSC1-siRNA-DC or siRNA mutant DC, which was not further stimulated until maturity, was pulsed with ovalbumin I-peptide (SIINFEKL; SEQ ID NO: 11) to produce ovalbumin-specific TCRT cells (OT-I). OT-I cells proliferated more in SOCS1-siRNA-DC co-culture than in siRNA-DC mutant co-culture (FIG. 4A). Consistent with these data, higher levels of proinflammatory cytokines were secreted in SOCS1-siRNA-DC co-cultures (FIG. 4B). Furthermore, CTL assays of these cells showed more active cytotoxicity against ovalbumin + syngeneic EG7 cells after co-culture with SOCS1-siRNA-DC, indicating that SOCS1 is a DC of antigen-specific T cells. It has been demonstrated to contribute to the regulation of stimuli.
インビボで抗原特異的応答をプライミングされるSOCS1サイレンシング化されたDCの能力を次に、エクスビボでの成熟の不在下でオボアルブミンでのパルスを受けた形質導入されたDCでマウスを直接免疫化することによって、テストした。四量体染色は、LV−GFP−siRNA−DC又はモックDCで免疫化されたマウスにおいてはそれぞれ0.64%及び0.43%にすぎなかった(図5A)のに比べ、LV−SOCS1−siRNA−DCで免疫化されたマウスでは、全CD8+T細胞の2.3%がオボアルブミン−四量体について陽性であることを示した。LV−SOCS1−siRNA−DC又はLV−GFPsiRNA−DC間では表面成熟マーカーにわずかしか差異がなかったことから、これらのデータは、増大したDC成熟だけがLV−SOCS1−siRNA−DCの機能的効能に貢献する要因ではなかったということを示唆している。免疫化されたマウスの体内のCD8+T細胞の機能的状態を、インターフェロンγ(IFNγ)ELISPOT検定を用いてさらに評価した。未成熟なLV−SOCS1−siRNA−DCで免疫化されたマウスは、それぞれに未成熟モックDC又はLV−GFP−siRNA−DCを与えられたものにおける1及び18スポットに比べて、2×105のCD8+T細胞あたり68のIFNγ+スポットを有していた(図5B)。これらの結果は、未成熟LV−SOCS1−siRNA−DCが与えられたマウスからの脾細胞のオボアルブミン+標的細胞に対するより強力な細胞傷害性を示しているCTL検定と一致していた(図5C)。未成熟LV−SOC1−siRNA−DCでの免疫化は同様に、観察可能な強力な抗原特異的CD4+T−ヘルパー応答をも誘発した。かくして、SOCS1サイレンシング化は、未成熟抗原提示DCが、インビボで抗原特異的CD8+CTL応答をプライミングさせる能力をもつ免疫原性状態に達することができるようにした。何らかの特定の理論により束縛されることは望まないが、SOCS1サイレンシング化されたDCは先行する成熟無く適応免疫をプライミングすることから、SOCS1は、DCの寛容原性状態を維持する上で調節の役目を果たすと考えられる。 The ability of SOCS1-silenced DCs to be primed with an antigen-specific response in vivo, then directly immunize mice with transduced DCs pulsed with ovalbumin in the absence of ex vivo maturation By testing. Tetramer staining was only 0.64% and 0.43% in mice immunized with LV-GFP-siRNA-DC or mock DC, respectively (Figure 5A), compared to LV-SOCS1- In mice immunized with siRNA-DC, 2.3% of total CD8 + T cells showed positive for ovalbumin-tetramer. Since there were only slight differences in surface maturation markers between LV-SOCS1-siRNA-DC or LV-GFP siRNA-DC, these data indicate that only increased DC maturation is functional efficacy of LV-SOCS1-siRNA-DC. This suggests that it was not a factor contributing to The functional status of CD8 + T cells in immunized mice was further evaluated using an interferon gamma (IFNγ) ELISPOT assay. Mice immunized with immature LV-SOCS1-siRNA-DC were 2 × 10 5 compared to 1 and 18 spots in those given immature mock DC or LV-GFP-siRNA-DC, respectively. Had 68 IFNγ + spots per CD8 + T cell (FIG. 5B). These results were consistent with a CTL assay showing more potent cytotoxicity of splenocytes from mice given immature LV-SOCS1-siRNA-DC to ovalbumin + target cells (FIG. 5C). ). Immunization with immature LV-SOC1-siRNA-DC also elicited an observable strong antigen-specific CD4 + T-helper response. Thus, SOCS1 silencing allowed immature antigen-presenting DCs to reach an immunogenic state with the ability to prime antigen-specific CD8 + CTL responses in vivo. While not wishing to be bound by any particular theory, since SOCS1-silenced DCs prime adaptive immunity without prior maturity, SOCS1 regulates in maintaining the tolerogenic state of DCs It is thought to play a role.
インビボでの成熟DCのSOCS1媒介型調節
以下の実験は、SOCS1サイレンシング化されたDCによるCTL応答のプライミングが、内因性の環境刺激に応答したDCの増強された成熟の結果としてもたらされたか否かを調査するために実施された。24時間エクスビボでLPSでオボアルブミンパルスを受けたLV−SOCS1−siRNA−DCを最初に成熟させ、次にマウスの体内にそれらを投与する前に3回洗浄した。LPS−成熟LV−SOCS1−siRNA−DCは、成熟LV−GFP−siRNA−DCのものに匹敵する成熟表現型を示し、エクスビボ成熟の不在下でのDCに比べ、両DC共、CTLをより強力にプライミングした。しかしながら、成熟LV−SOCS1−siRNA−DCはなお、オボアルブミン四量体染色により実証される通り(図5A)、抗原特異的CTLを誘発する上で明らかに成熟LV−GFP−siRNA−DCよりも優れていた。IFN−γELISPOT検定は同様に、LV−SOCS1−siRNA−DCマウスにおける増強されたオボアルブミン特異的CD8+T細胞応答をも示し、SOCS1サイレンシング化が内因性環境刺激に対する成熟DCのより大きな応答性を可能にし、CTL応答の増大を導くことを示唆している。
SOCS1-mediated regulation of mature DCs in vivo. The following experiments have demonstrated that priming the CTL response by SOCS1-silenced DCs resulted from enhanced maturation of DCs in response to endogenous environmental stimuli Conducted to investigate whether or not. LV-SOCS1-siRNA-DCs that received an ovalbumin pulse with LPS ex vivo for 24 hours were first matured and then washed 3 times before administering them into the body of mice. LPS-mature LV-SOCS1-siRNA-DC exhibits a mature phenotype comparable to that of mature LV-GFP-siRNA-DC, both DCs being more potent than CTL in the absence of ex vivo maturation Primed to. However, mature LV-SOCS1-siRNA-DC is still clearly superior to mature LV-GFP-siRNA-DC in inducing antigen-specific CTL, as demonstrated by ovalbumin tetramer staining (FIG. 5A). It was excellent. The IFN-γ ELISPOT assay also shows an enhanced ovalbumin-specific CD8 + T cell response in LV-SOCS1-siRNA-DC mice, where SOCS1 silencing is greater responsiveness of mature DC to endogenous environmental stimuli Suggesting that it may lead to an increase in CTL response.
さらなるインビボ試験においては、1日前に未成熟LV−SOCS1−siRNA−DCで免疫化されたマウスに一回LPSを注射した。この刺激は、未成熟LV−SOCS1−siRNA−DCマウス内でのCTL応答を有意に(P<0.01)高めた(CD8+T細胞内で7.87%のオボアルブミン四量体+)が、未成熟DC−GFP−siRNAマウス内ではその効果は低いものであった(CD8+T細胞内で0.64%のオボアルブミン−四量体+)(図6A)。インビボLPS刺激後の未成熟LV−SOCS1−siRNA−DCマウス中のオボアルブミン特異的CTL応答の増強は、IFN−γELISPOT検定によって確認された(図6B)。 In a further in vivo study, mice immunized with immature LV-SOCS1-siRNA-DC one day before were injected with LPS once. This stimulation, a CTL response in the immature LV-SOCS1-siRNA-DC mice significantly (P <0.01) elevated (CD8 + T cells in a 7.87% ovalbumin tetramer +) but immature DC-GFP-siRNA its effect in mice was low (CD8 + T cells in 0.64% of the ovalbumin - tetramer +) (Fig. 6A). Enhancement of the ovalbumin-specific CTL response in immature LV-SOCS1-siRNA-DC mice after in vivo LPS stimulation was confirmed by the IFN-γ ELISPOT assay (FIG. 6B).
環境刺激に応答した成熟DCのシグナリングがT細胞プライミングにおいて役割を果たすか否かをテストするために、一日前にエクスビボ成熟LV−SOCS1−siRNA−DCで免疫化されたマウスに一度又は反復的にLPSを注射した。LPS注射は、成熟LV−SOCS1−siRNA−DCマウスにおけるCTL応答を有意に(P<0.01)高め、T細胞応答をプライミングするための成熟DCのシグナリングの重要性を示した(図6C及び6D)。その上、LPS注射は、成熟LV−SOCS1−siRNA−DCで免疫化したマウスにおけるオボアルブミン特異的CTL応答を高める上でさらに有効であった。 To test whether signaling of mature DCs in response to environmental stimuli plays a role in T cell priming, mice immunized with ex vivo matured LV-SOCS1-siRNA-DCs once or repeatedly one day ago LPS was injected. LPS injection significantly (P <0.01) enhanced the CTL response in mature LV-SOCS1-siRNA-DC mice, indicating the importance of mature DC signaling to prime the T cell response (FIGS. 6C and 6D). Moreover, LPS injection was further effective in enhancing ovalbumin-specific CTL responses in mice immunized with mature LV-SOCS1-siRNA-DC.
反復されたLPS刺激に対するDC−SOCS1−siRNAの応答性を直接調べるため、インビトロで内毒素寛容を発達させるSOCS1−siRNA−DC及びGFP−siRNA−DCの能力を評価した。高レベルの前炎症性サイトカインを産生することによる反復的LPS刺激に対しSOCS1−siRNA−DCはなおも強く応答したがGFP−siRNA−DCは強く応答せず、このことは、SOCS1サイレンシング化されたDCが刺激に対し連続して応答することを示唆している。さらに、天然の危険な分子として機能する熱ショックタンパク質(HSP)などの内因性刺激に対するDC応答性の閾値は、SOCS1サイレンシング化によって低減された。 To directly examine the responsiveness of DC-SOCS1-siRNA to repeated LPS stimulation, the ability of SOCS1-siRNA-DC and GFP-siRNA-DC to develop endotoxin tolerance in vitro was evaluated. SOCS1-siRNA-DC still responded strongly to repetitive LPS stimulation by producing high levels of pro-inflammatory cytokines, but GFP-siRNA-DC did not respond strongly, which was silenced in SOCS1 This suggests that DC responds continuously to stimuli. Furthermore, the threshold for DC responsiveness to endogenous stimuli such as heat shock proteins (HSP) that function as natural dangerous molecules was reduced by SOCS1 silencing.
合わせて考慮するとこれらのデータは、DC内のSOCS1のサイレンシング化が、DC応答性の閾値を低減させる確率が高く、未成熟及び成熟抗原提示DCが内因性刺激に対して連続的に応答し、結果として抗原特異的CTL応答をもたらすことを可能にする、ということを表わしている。この機序は、成熟DCによる抗原提示の範囲、ひいては適応免疫の規模の制御におけるSOCS1のきわめて重要な役割を強調するものである。 Taken together, these data indicate that silencing of SOCS1 in DCs is likely to reduce the DC responsiveness threshold, and immature and mature antigen-presenting DCs respond continuously to endogenous stimuli. , Which can result in an antigen-specific CTL response. This mechanism highlights the critical role of SOCS1 in controlling the extent of antigen presentation by mature DCs and thus the scale of adaptive immunity.
インビボ刺激によるSOCS1サイレンシング化されたDCの免疫化の増強
サイトカイン又はトール様レセプタ(TLR)アゴニストでのインビボ刺激がSOCS1サイレンシング化されたDCの効力をさらに増強できるか否かを調査するために、OV
Aパルスを受けたDC−LV−SOCS1−siRNAで免疫化されたマウスに、連続3日間一日一回腹腔内でLPS(30μg/マウス)、CpG(60μg/マウス)、PolyI:C(50μg/マウス)、抗CD40(100μg/マウス)又はIFN−g(1μg/マウス)を注射した。これらの刺激がDC−LV−SOCS1−siRNAマウスにより誘発されたCTL応答を優先的に高めるということが観察された(図6E)。これらの結果は、数多くの刺激がLPSに加えてさらに、SOCS1サイレンシング化されたDCの免疫化の効力を増強し得るということを表わしている。
Enhancement of immunization of SOCS1-silenced DCs by in vivo stimulation To investigate whether in vivo stimulation with cytokines or toll-like receptor (TLR) agonists can further enhance the potency of SOCS1-silenced DCs , OV
Mice immunized with A-pulsed DC-LV-SOCS1-siRNA were injected intraperitoneally once daily for 3 consecutive days with LPS (30 μg / mouse), CpG (60 μg / mouse), PolyI: C (50 μg / Mice), anti-CD40 (100 μg / mouse) or IFN-g (1 μg / mouse). It was observed that these stimuli preferentially enhance the CTL response elicited by DC-LV-SOCS1-siRNA mice (FIG. 6E). These results indicate that numerous stimuli can further enhance the efficacy of immunization of SOCS1-silenced DCs in addition to LPS.
DC内のSOCS1サイレンシング化による抗腫瘍免疫の増強
DC抗原提示におけるSOCS1の調節機能が観察されたことで、SOCS1サイレンシング化されたDCがより強力な抗原特異的抗腫瘍免疫を誘発し、かくして事前準備された腫瘍成長の制御を導くか否かの調査が促されることになった。エクスビボ成熟の不在下でのオボアルブミンでパルスされたLV−SOCS1−siRNA−DCでの免疫化は、エクスビボ成熟の不在下でオボアルブミンパルスを受けたLV−GFP−siRNA−DC又はモックDCを与えられたマウスにおける腫瘍成長の控えめな減少とは対照的に、テスト対象の全てのマウスにおいて事前準備されたオボアルブミン+EG7腫瘍の成長を完全に遮断した(図7A)。インビボ抗体遮断実験は、抗CD4ではなく抗CD8抗体遮断が、オボアルブミンパルスを受けたLV−SOCS1−siRNA−DCにより誘発される抗腫瘍活性を無効にする(図7B)ことを立証しており、このことは、抗腫瘍応答におけるCD8+CTLの極めて重要な役割を示している。
Enhancement of anti-tumor immunity by SOCS1 silencing in DC The observation of the regulatory function of SOCS1 in DC antigen presentation allows SOCS1-silenced DCs to induce stronger antigen-specific anti-tumor immunity, thus An investigation will be encouraged whether it will lead to pre-prepared control of tumor growth. Immunization with LV-SOCS1-siRNA-DC pulsed with ovalbumin in the absence of ex vivo maturation gives LV-GFP-siRNA-DC or mock DC that received ovalbumin pulse in the absence of ex vivo maturation. In contrast to the modest decrease in tumor growth in the generated mice, the pre-prepared ovalbumin + EG7 tumor growth was completely blocked in all tested mice (FIG. 7A). In vivo antibody blocking experiments have demonstrated that anti-CD8 antibody blocking but not anti-CD4 abolishes the anti-tumor activity elicited by ovalbumin pulsed LV-SOCS1-siRNA-DC (FIG. 7B). This indicates a critical role for CD8 + CTL in anti-tumor responses.
SOCS1siRNAのオリゴ2重鎖トランスフェクションを受けたDCが、抗腫瘍活性の増強を示した否かをテストするため、DCをSOCS1siRNAオリゴ2重鎖又は対照オリゴ2重鎖でトランスフェクトした。その後マウスのグループを、OVA又はTC−1腫瘍リゼイトでパルスされた、トランスフェクションを受けたDCを用いて免疫化した。パルスされたDCでのマウスの免疫化の後、マウスをインビボで3回LPS(30μg/マウス)で刺激した。DC免疫化から2週間後に、OVA+EG7腫瘍又はTC−1腫瘍を用いて、免疫化済みマウスに抗原投与した。EG7及びTC−1の両方の腫瘍モデル(図7C及び7D)の中で、抗腫瘍活性の増強が観察された(図7C及び7D)。その上IFN−γELISPOT検定は、TC−1腫瘍リゼイトでパルスされたSOCS1siRNAオリゴ−DCで免疫化されたマウスにおける腫瘍特異的CTL応答の増強を示した(図7E)。 To test whether DCs that had undergone oligo duplex transfection of SOCS1 siRNA showed enhanced anti-tumor activity, DCs were transfected with SOCS1 siRNA oligo duplex or control oligo duplex. Groups of mice were then immunized with transfected DCs pulsed with OVA or TC-1 tumor lysates. After immunization of mice with pulsed DC, mice were stimulated with LPS (30 μg / mouse) three times in vivo. Two weeks after DC immunization, immunized mice were challenged with OVA + EG7 or TC-1 tumors. In both EG7 and TC-1 tumor models (FIGS. 7C and 7D), enhanced anti-tumor activity was observed (FIGS. 7C and 7D). Moreover, the IFN-γ ELISPOT assay showed enhanced tumor-specific CTL responses in mice immunized with SOCS1 siRNA oligo-DC pulsed with TC-1 tumor lysate (FIG. 7E).
SOCS1サイレンシング化されたDCが自己腫瘍関連抗原に対する免疫応答を増強できるか否かがさらにテストされた。これらの実験のためには、弱免疫原性のB16黒色腫細胞の中で天然に発現されるマウス黒色腫分化抗原チロシナーゼ関連タンパク質(TRP)2を使用した。C57BL/6マウスにB16腫瘍細胞を接腫し、3日後にLPSでエクスビボで刺激された、TRP2ペプチドでのパルスを受けた成熟LV−SOCS1−siRNA−DC又はLV−GFP−siRNA−DCを用いて一回これらのマウスを処置した。成熟LV−SOCS1−siRNA−DCは、事前準備されたB16腫瘍の成長を有効に遮断し、一方成熟LV−GFP−siRNA−DCはいかなる阻害効果も有していなかった(図8A)。増強された抗腫瘍活性は、IFN−γELISPOT及びCTL検定により検出される通り(図8B及び8C)、LV−SOCS1−siRNA−DCマウスにおける強力なTRP2特異的CTL応答と相関した。成熟LV−SOCS1−siRNA−DCが与えられたマウスにおいてのみ、活発なNK活性が検出された。オボアルブミン+EG7腫瘍での結果とは対照的に、未成熟TRP2パルスを受けたLV−SOCS1−siRNA−DCでの免疫化は、B16腫瘍に対する有意な(P>0.05)阻害効果を生み出すことができず、このことは、有効な抗腫瘍免疫を生成するためにはDCの完全成熟及び成熟後の連続的シグナリングが必要とされる、ということを示唆している。外来性抗原(オボアルブミン)に対するCTL応答に及ぼすGFPsiRNA形質導入の非
特異的な刺激効果は一貫して観察された。しかしながら、非特異的刺激効果は、自己抗原(TRP2)に対するCTL応答を増強するのに不充分であった。
It was further tested whether SOCS1-silenced DCs could enhance the immune response against self tumor associated antigens. For these experiments, mouse melanoma differentiation antigen tyrosinase-related protein (TRP) 2 naturally expressed in weakly immunogenic B16 melanoma cells was used. Using mature LV-SOCS1-siRNA-DC or LV-GFP-siRNA-DC pulsed with TRP2 peptide stimulated ex vivo with
LV−SOCS1−siRNA−DCでの免疫化により誘発されると考えられる不利な自己免疫病変を検討した。TRP2でパルスされたSOCS1サイレンシング化されたDCで免疫化したマウスの脱色(白斑)が観察された。しかしながら、免疫化から3ヵ月目までは、オボアルブミン又はTRP2でパルスされたLV−SOCS1−siRNA−DCで免疫化された200匹超のマウスにおいて、その他の明らかな毒性は全く観察されなかった。免疫化を受けたマウスの主要な臓器及び組織の全ての組織学的分析は、病理学的炎症を全く明らかにせず、免疫組織化学的染色は、腎臓内のIgG又はIgM被着物を全く示さなかった。IgG(IgG1、IgG2a)及び抗−dsDNAのレベルは、LV−SOCS1−siRNA−DC及びモックDCマウスにおいても匹敵するものであった。これらのデータは、LV−SOCS1−siRNA−DC免疫化がマウスの体内で病的炎症をひき起こさないことを示唆している。いずれかの特定の理論により束縛されることは望まないものの、SOCS1サイレンシング化されたDCは、事前準備された弱免疫原性の腫瘍の成長を遮断することのできる有効で抗原特異的な抗腫瘍免疫を誘発する大幅に増強された能力を有するものと考えられる。 Disadvantageous autoimmune lesions that were thought to be induced by immunization with LV-SOCS1-siRNA-DC were examined. Decolorization (white spots) was observed in mice immunized with SOCS1-silenced DC pulsed with TRP2. However, up to 3 months after immunization, no other apparent toxicity was observed in over 200 mice immunized with LV-SOCS1-siRNA-DC pulsed with ovalbumin or TRP2. All histological analyzes of the major organs and tissues of the immunized mice reveal no pathological inflammation and immunohistochemical staining does not show any IgG or IgM deposits in the kidney It was. The levels of IgG (IgG1, IgG2a) and anti-dsDNA were comparable in LV-SOCS1-siRNA-DC and mock DC mice. These data suggest that LV-SOCS1-siRNA-DC immunization does not cause pathological inflammation in the mouse body. While not wishing to be bound by any particular theory, SOCS1-silenced DCs are effective and antigen-specific anti-antigens that can block the growth of pre-prepared weakly immunogenic tumors. It is believed to have a greatly enhanced ability to elicit tumor immunity.
SOC1のサイレンシング化が、DCによる抗原提示及び抗原特異的抗腫瘍免疫を増強させる
本書で提示されている結果は、DCの刺激能力及び適応免疫の規模がDC中のSOCS1により決定的に調節されるということを実証している。抗原提示DC内でSOCS1をサイレンシング化することは、抗原特異的抗腫瘍免疫を大幅に増強する。SOCS1は、DCによる抗原提示及び適応免疫の規模を定性的及び定量的に制御するための阻害的機序を表わす。
SOC1 silencing enhances antigen presentation by DC and antigen-specific anti-tumor immunity. The results presented here indicate that the stimulatory capacity of DC and the magnitude of adaptive immunity are critically regulated by SOCS1 in DC. This proves that Silencing SOCS1 within the antigen-presenting DC greatly enhances antigen-specific anti-tumor immunity. SOCS1 represents an inhibitory mechanism for qualitatively and quantitatively controlling the magnitude of antigen presentation by DC and adaptive immunity.
本開示は、成熟したDCによる抗原提示の範囲を調節する上でのSOCS1の決定的な役割を実証し、かくして適応免疫の規模及び持続時間をDCが制御できるようにする調節機序を実証している。DCの寛容原性状態を維持する上でのSOCS1の重要性は、本書において、野生型DCとは対照的に、SOCS1サイレンシング化されたDCがエクスビボ成熟の必要性が無い状態でインビボでT細胞応答をプライミングするための刺激性抗原提示能力に恵まれている、という点で例示されている。何らかの特定の理論により束縛されることは望まないが、DC内のSOCS1が適応免疫の規模を制御する精確な機序には、サイトカイン、病原菌産物そして又恐らくは細胞と細胞の接触での刺激に応答した抗原性ペプチド提示、共同刺激/同時阻害及びサイトカイン産生に関する成熟DCのシグナリング及び産出量の調節が関与する、と考えられている。 This disclosure demonstrates the critical role of SOCS1 in regulating the extent of antigen presentation by mature DCs, thus demonstrating regulatory mechanisms that allow DCs to control the magnitude and duration of adaptive immunity. ing. The importance of SOCS1 in maintaining the tolerogenic state of DC is demonstrated here in contrast to wild-type DC, in which SOCS1-silenced DCs are in vivo without the need for ex vivo maturation. It is exemplified in that it is endowed with the ability to present stimulating antigens to prime cellular responses. While not wishing to be bound by any particular theory, the precise mechanism by which SOCS1 in DCs controls the magnitude of adaptive immunity is in response to cytokines, pathogenic products, and possibly also stimuli at cell-cell contacts. It is thought to be involved in mature DC signaling and output regulation with respect to antigenic peptide presentation, costimulation / co-inhibition and cytokine production.
成熟DCは一般に、限られた数の研究に基づいて、寿命が短かい。しかしながら、信頼性の高い遺伝的方法を用いた最近の研究は、成熟抗原提示DCの寿命が以前に推定されたものよりもはるかに長く、インビボで2週間持続するということを実証しており、成熟DCにより抗原提示の範囲を調節することの必要性及び重要性を裏づけている。 Mature DCs generally have a short lifetime based on a limited number of studies. However, recent studies using reliable genetic methods have demonstrated that the life span of mature antigen-presenting DCs is much longer than previously estimated and lasts 2 weeks in vivo, Supporting the need and importance of regulating the extent of antigen presentation by mature DCs.
本発明は、DC内の決定的な抑制を無効にすることにより、腫瘍ワクチンの効力を増強するための遺伝的手段としてDC内でSOCS1をサイレンシング化する新規の原理に関する。SOCS1サイレンシング化されたDCでのワクチン接種は、SOCS1のサイレンシング化により抗原提示免疫原性DCがインビボで抗原特異的T細胞を持続的に刺激できるようになるため、抗原特異的抗腫瘍免疫を大幅に増強する。SOCS1サイレンシング化されたDCは、調節T細胞抑制を阻害するIL−6などの前炎症性サイトカインの産生及びDC成熟を増強することにより、調節T細胞をオフに切換えることができる。 The present invention relates to a novel principle of silencing SOCS1 in DC as a genetic means to enhance the efficacy of tumor vaccines by overriding critical suppression in DC. Vaccination with SOCS1-silenced DCs allows antigen-specific anti-tumor immunity because silencing of SOCS1 allows antigen-presenting immunogenic DCs to continuously stimulate antigen-specific T cells in vivo. Will be greatly enhanced. SOCS1-silenced DCs can switch off regulatory T cells by enhancing the production of proinflammatory cytokines such as IL-6 that inhibit regulatory T cell suppression and DC maturation.
T細胞上のCTLA−4の遮断は、寛容性を有効に破るが、患者の体内で重大な非特異的自己免疫炎症をひき起こす。抗原提示レベルでDC内でSOCS1をターゲティングすることにより、より抗原特異的な抗腫瘍応答を達成することが可能である。第1に、腫瘍関連抗原が豊富に負荷されているSOCS1サイレンシング化されたDCでの免疫化は、正常な腫瘍組織に対する自己反応性T細胞を不可避的に活性化させるアプローチである、エフェクタCTL上でのCTLA−4のターゲティングとは対照的に、抗原特異的免疫を誘発することになる。第2に、残留SOCS1レベルを伴う部分的にSOCS1サイレンシング化されたDCの使用は、ヘテロ接合SOCS1+/−マウスが全く又は穏やかにしか自己免疫炎症の徴候を示さないことから、重大な自己免疫炎症をひき起こさない可能性があると思われる。さらに、SOCS1−/−マウスにおいて見られる重大な自己免疫炎症は、DC内のみならずT及びNKT細胞などのその他の免疫細胞系統においてもSOCS1の完全な欠損を必要とする。本書で提示されている結果は、抗原提示及び適応免疫の定性的及び定量的調節を理解するためのみならず、DCの刺激潜在力を増強することによる癌及び感染性疾患に対する有効なワクチンの開発のためにも、見識を提供する。 Blocking CTLA-4 on T cells effectively breaks tolerance but causes significant non-specific autoimmune inflammation in the patient's body. By targeting SOCS1 in DC at the antigen presentation level, it is possible to achieve a more antigen-specific anti-tumor response. First, immunization with SOCS1-silenced DCs rich in tumor-associated antigens is an approach that unavoidably activates autoreactive T cells against normal tumor tissue, effector CTL In contrast to CTLA-4 targeting above, it will elicit antigen-specific immunity. Secondly, the use of partially SOCS1-silenced DC with residual SOCS1 levels results in significant self-immunity since heterozygous SOCS1 +/− mice show no or only mild signs of autoimmune inflammation. It may not cause immune inflammation. Furthermore, the significant autoimmune inflammation seen in SOCS1 − / − mice requires a complete deficiency of SOCS1 not only in DC but also in other immune cell lines such as T and NKT cells. The results presented in this document are not only for understanding qualitative and quantitative regulation of antigen presentation and adaptive immunity, but also for the development of effective vaccines against cancer and infectious diseases by enhancing the stimulation potential of DC Also provide insight for.
実施例5:マウスSOCS1−siRNA−DCにより誘発されるTRP2特異的CTL及び抗腫瘍活性
本開示は、成熟DCにおいてSOCS1siRNAで低減されたSOCS1発現を発現するレンチウイルスベクターが、自己抗原特異的CTL応答の規模を増大できるということを実証している。この研究のためには、モデル自己抗原として、マウスメラニン細胞分化抗原であるチロシナーゼ関連タンパク質2(TRP2)が使用された。正常なメラニン細胞及び弱免疫原性のB16黒色腫細胞の両方の中で天然に発現されていること、そしてTRP2内で数多くのクラスIMHCエピトープが同定されていることから、TRP2が使用された(ファン・エルサス(van Elsas)ら、2001年、J.Exp.Med.第194号:481〜9頁)。
Example 5: TRP2-specific CTL and anti-tumor activity elicited by mouse SOCS1-siRNA-DC The present disclosure shows that lentiviral vectors expressing reduced SOCS1 expression with SOCS1 siRNA in mature DCs are self-antigen-specific CTL responses It is demonstrating that the scale of can be increased. For this study, mouse melanocyte differentiation antigen tyrosinase-related protein 2 (TRP2) was used as a model autoantigen. TRP2 was used because it is naturally expressed in both normal melanocytes and weakly immunogenic B16 melanoma cells, and a number of class IMHC epitopes have been identified within TRP2 ( Van Elsas et al., 2001, J. Exp. Med. 194: 481-9).
この実施例中で提示されている実験において用いられる材料及び方法についてここで記述する。 The materials and methods used in the experiments presented in this example are now described.
マウス/動物モデル
4〜6週齢の雌のC57BL/6、CD4KO、CD8KO又はp35(IL−12)KOマウスをジャクソン・ラボラトリーズ(米国メーン州バーハーバー(Bar Harbor,Main))から購入し、機関の指針に従ってベイラー・カレッジ・オブ・メディシン(Baylor College of Medicine)(米国テキサス州ヒューストン)で無菌のマウス施設の中に維持した。
Mice / Animal Model 4-6 week old female C57BL / 6, CD4KO, CD8KO or p35 (IL-12) KO mice were purchased from Jackson Laboratories (Bar Harbor, Maine, USA) And maintained in a sterile mouse facility at Baylor College of Medicine (Houston, Texas, USA).
ペプチド
H2−Kb制限されたTRP2a(VYDFFVWL;配列番号15)及びTRP2b(SVYDFFVWL;配列番号16)(ファン・エルサスら、2001年、J.Exp.Med.第194号:481〜9頁)及び対照H2−Kb制限されたOVA−I(SIINFEKL;配列番号11)を合成し、ジーンメッド・シンセシス社(Genemed
Synthesis Inc.)(米国カリフォルニア州サウスサンフランシスコ(South San Francisco,CA))により95%超の純度までHPLCにより精製した。内毒素を含まないPBS(シグマ、ミズーリ州セントルイス)中での最終希釈の前にDMSO中で全てのペプチドを溶解させた。
Peptides H2- Kb restricted TRP2a (VYDFFVWL; SEQ ID NO: 15) and TRP2b (SVYDFFVWL; SEQ ID NO: 16) (Van Elsus et al., 2001, J. Exp. Med. 194: 481-9) and A control H2- Kb restricted OVA-I (SIINFEKL; SEQ ID NO: 11) was synthesized and generated by Genemed Synthesis.
Synthesis Inc. ) (South San Francisco, Calif.) To a purity of greater than 95%. All peptides were dissolved in DMSO prior to final dilution in PBS without endotoxin (Sigma, St. Louis, MO).
レンチウイルスベクターでのBM由来のDCの形質導入
組換え型レンチウイルスベクター(LV−SOCS1−siRNA及びLV−GFP−siRNA)を、本書の他の箇所で記述されている通りに産生し、滴定しDCを形質導入するのに使用した。
Transduction of BM-derived DCs with lentiviral vectors Recombinant lentiviral vectors (LV-SOCS1-siRNA and LV-GFP-siRNA) are produced and titrated as described elsewhere in this document. Used to transduce DC.
サイトカインELISA及び酵素連結型イムノスポット(ELISPOT)検定
提示された時点で指示された刺激を用いてメーカーの指示に従って、ELISA分析用のDC培養の上清(BDバイオサイエンシーズ、ニュージャージー州リンカーンパーク)を用いて様々な前炎症性サイトカインのレベルを定量した。フアン(Huang)ら、2003年、Cancer Res.第63号:7321〜9頁の中で記述されているように、単離されたCD4+又はCD8+T細胞のELISPOT検定を実施した。マウスCD8+T細胞刺激のためには、H2−Kb/TRP2クラスIペプチドを使用した。OVAからの無関係のペプチドも、負の対象として使用した。MACS CD8(Ly−2)マイクロビーズ(ミルテニー・バイオテク社、カリフォルニア州オウバーン)を用いることにより、脾細胞からCD8+T細胞を単離した。
Cytokine ELISA and enzyme linked immunospot (ELISPOT) assay DC culture supernatants for ELISA analysis (BD Biosciences, Lincoln Park, NJ) using the indicated stimuli according to the manufacturer's instructions. Used to quantify the levels of various proinflammatory cytokines. Huang et al., 2003, Cancer Res. No. 63: ELISPOT assay of isolated CD4 + or CD8 + T cells was performed as described in pages 7321-9. H2-K b / TRP2 class I peptides were used for mouse CD8 + T cell stimulation. An irrelevant peptide from OVA was also used as a negative subject. CD8 + T cells were isolated from splenocytes by using MACS CD8 (Ly-2) microbeads (Miltenyi Biotech, Auburn, CA).
フローサイトメトリ分析
細胞を、0.1%のNaN3及び2%のFCSを含有するPBS中のFITC又はPEmAbsで染色した。マウスのCD8(53−6.7)、CD11c(HL3)、CD40(3/23)、CD80(16−10A1)、CD86(GL1)、OX40L(RM134L)又はPDL1(MIH5)及び整合したイソタイプ対照に特異的な抗体を、BDファーミンゲン(Pharmingen)(ニュージャージー州フランクリンレイクス(Franklin Lakes,NJ))又はイーバイオサイエンス(eBioscience)(カリフォルニア州サンディエゴ)から購入した。染色した細胞をFACS Cslibur(ベクトン・ディッキンソン、ニュージャージー州リンカーンパーク、ニュージャージー州フランクリンレイクス)フローサイトメトリ上で分析した。
Flow cytometry analysis Cells were stained with FITC or PE mAbs in PBS containing 0.1% NaN 3 and 2% FCS. Mouse CD8 (53-6.7), CD11c (HL3), CD40 (3/23), CD80 (16-10A1), CD86 (GL1), OX40L (RM134L) or PDL1 (MIH5) and matched isotype controls Specific antibodies were purchased from BD Pharmingen (Franklin Lakes, NJ) or eBioscience (San Diego, Calif.). Stained cells were analyzed on FACS Cslibur (Becton Dickinson, Lincoln Park, NJ, Franklin Lakes, NJ) flow cytometry.
四量体染色
TRP2特異的マウスCD8+T細胞を検出するために、H2−Kb/TRP2−PE四量体検定を使用した。べイラー・カレッジ・オブ・メディシン、テトラマー・コアー・ファシリティー(Tetramer Core Facility)(米国、テキサス州ヒューストン)でTRP2−四量体を合成した。免疫化したマウス由来の脾臓を抗−CD8a−FITC/抗−CD3−PerCP及びH2−Kb/TRP2−PEで同時染色した。四量体染色は、メーカーの指示に従い、106個の細胞につきTRP2−PE四量体の1:100希釈物及び抗−CD8α−Fitcの1μgを用いて1時間4℃で行なった。
Tetramer staining The H2- Kb / TRP2-PE tetramer assay was used to detect TRP2-specific mouse CD8 + T cells. TRP2-tetramers were synthesized at Bayer College of Medicine, Tetramer Core Facility (Houston, Texas, USA). Spleens from immunized mice were co-stained with anti-CD8a-FITC / anti-CD3-PerCP and H2- Kb / TRP2-PE. Tetramer staining was performed at 4 ° C. for 1 hour using a 1: 100 dilution of TRP2-PE tetramer and 1 μg of anti-CD8α-Fitc per 10 6 cells per manufacturer's instructions.
DC免疫化及び腫瘍マウス研究
本書の他の個所で記述されている通り、5というMOIでSOCS1−siRNA又はGFP−siRNAを用いてBM由来のDC(BM培養4〜5日目)を形質導入した。簡単に言うと、DCを20時間ペプチドでパルスし、PBSで3回洗浄し、24時間LPS(100ng/ml、シグマ、ミズーリ州セントルイス)又はTNFa(500ng/ml、アールアンドディー・システムズ(R&D Systems)、ミネソタ州ミネアポリス(Minneapolis,MN))で刺激し、PBSで3回洗浄し、次にC57BL/6、CD8KO、CD4KO又はp35KOマウスの体内に後足蹠から注射した。治療モデルにおいて、B16腫瘍細胞(2.5×105)を、同系マウスの右脇腹に皮下(s.c)注射して、腫瘍モデルを確立した。腫瘍接種の後3日目に、マウスをランダムにグループ分けし、30μlのペプチドパルスされたDC(50μg/ml)、形質導入されたDC(1.5×106)又はPBS対照を注射した。一部のマウスにおいては、DCワクチン接種から指示された日数後に、LPS(30μg/マウス)又は組換え型マウスIL−12タンパク質(1mg/マウス、ぺプロテック、ニュージャージー州ロッキーヒル)を腹腔内(i.p.)投与した。実験が完了するまで2日に一度、キャリパで腫瘍体積を測定した。
DC Immunization and Tumor Mouse Studies Transduced BM-derived DCs (BM cultures 4-5 days) with SOCS1-siRNA or GFP-siRNA at MOI of 5 as described elsewhere in this document . Briefly, DC were pulsed with peptide for 20 hours, washed 3 times with PBS, and 24 hours LPS (100 ng / ml, Sigma, St. Louis, MO) or TNFa (500 ng / ml, R & D Systems (R & D Systems). ), Minneapolis, MN), washed 3 times with PBS, then injected into the body of C57BL / 6, CD8KO, CD4KO or p35KO mice from the hind footpad. In the treatment model, B16 tumor cells (2.5 × 10 5 ) were injected subcutaneously (sc) into the right flank of syngeneic mice to establish the tumor model. On
CTL検定
標的細胞を溶解するインビトロ再刺激された脾細胞の能力を測定する標準クロム放出検定で、CD8+CTL応答を査定した(ホアンら、2003年、Cancer Res.第63号:7321〜9頁)。2〜3匹の免疫化したマウスからプールした脾細胞を、4〜6日間、H2−Kb/TRP2ペプチドを含有するRPMI−1640中でインビトロで再刺激した。TRP2+標的B16細胞(H2−Kb)及び対照EG7細胞(ATCC、バージニア州マナサス(Manassas,VA))を、振とうさせながら37℃で、90分間51Crクロム酸ナトリウム溶液で標識した。異なる数のエフェクタ細胞を37℃で4時間、96ウェルのU字底平板(200μl/ウェル)内で、一定数の標的細胞(5×104/ウェル)と共にインキュベートした。3重培養から上清を収集し、分析した。(実験的放出−自然放出)/(最大放出−自然放出)×100として溶解百分率を計算した。
CTL Assay CD8 + CTL responses were assessed with a standard chromium release assay that measures the ability of in vitro restimulated splenocytes to lyse target cells (Hoan et al., 2003, Cancer Res. 63: 7321-9). Pooled splenocytes from 2-3 immunized mice were restimulated in vitro in RPMI-1640 containing H2- Kb / TRP2 peptide for 4-6 days. TRP2 + target B16 cells (H2-K b ) and control EG7 cells (ATCC, Manassas, Va.) Were labeled with 51 Cr sodium chromate solution for 90 minutes at 37 ° C. with shaking. Different numbers of effector cells were incubated with a fixed number of target cells (5 × 10 4 / well) in 96-well U-bottom plates (200 μl / well) for 4 hours at 37 ° C. Supernatants from triplicate cultures were collected and analyzed. Percent dissolution was calculated as (experimental release-spontaneous release) / (maximum release-spontaneous release) x 100.
統計学的分析
統計学的分析のためには、スチューデントt検定が使用され、P<0.05と定義づけされる95%の信頼性限界が有意なものとされた。結果は標準的に、平均±標準誤差として提示される。
Statistical analysis For statistical analysis, Student's t-test was used and a 95% confidence limit defined as P <0.05 was considered significant. Results are typically presented as mean ± standard error.
本実施例で提示されている実験の結果について、ここで記述する。 The results of the experiment presented in this example will now be described.
SOCS1により制限された成熟DC内のシグナリングが特異的CTL応答及び寛容を制御する
LPSで、エクスビボで成熟させられたTRP2ペプチドパルスを受けた形質導入されたDCを用いて、C57BL/6マウスを免疫化した。免疫化したマウスを次にLPSの存在下又は不在下で1回又は3回インビボで刺激し、四量体分析を用いてTRP2特異的CTL応答を測定した。誘発する前炎症性サイトカインの数が多く、その多くがSOCS1により調節されていること、ならびに、NF−κB(p65)シグナリングの直接的調節におけるSOCS1の役割が立証されていることを理由として、インビボ刺激のためにはLPSが選択された(リョーら、2003年、Mol.Cell.第12号:1413〜26頁)。インビボLPS刺激の不在下で、GFP−siRNA−DCで免疫化されたマウス内では3.1%にすぎなかったのに比べ、SOCS1−siRNA−DCで免疫化されたマウスにおいては、TRP2−四量体について総CD8+T細胞の5.1%が陽性であった(図9A)。1回又は3回のインビボLPS刺激で、TRP2−四量体について陽性であるCD8+T細胞の百分率は、SOCS1−siRNA−DCで免疫化されたマウスにおいて実質的に増大し(それぞれ9.7%及び19.4%)、GFP−siRNA−DCで免疫化されたマウスにおいては概して変化がなかった(それぞれ3.0%及び4.0%)(図9A)。これと一致して、CTL検定(図9C)及びインターフェロンγ(IFNγ)ELISPOTは類似の結果を示した。その上、免疫化から3ヵ月後に、LPSが同時注入され(図9B)、TRP2ペプチドパルスを受けたSOCS1−siRNA
DCで免疫化されたマウスの大部分に、白斑(外被の明色化、脱色及び/又は脱毛)がはっきりと見え、このことは、宿主メラニン細胞内で通常発現されるTRP2に対する自己寛容の破壊を表わしていた。これとは対照的に、反復的にインビボでLPSを投与した場合でさえGFP−siRNA DCで免疫化されたマウスのいずれにも全く白斑が観察されず、これは、自己抗原に対する寛容を維持するためのDC内のSOCS1の決定的な役割を示唆していた。これらの結果は、成熟抗原提示DC内でのシグナリングがCTL応答及び自己寛容の規模を制御すること、そして成熟DCのシグナリングがSOCS1により厳格に制限されることを実証している。
Signaling in mature DCs restricted by SOCS1 regulates specific CTL responses and tolerance Immunizing C57BL / 6 mice with transduced DCs receiving LPS and ex vivo matured TRP2 peptide pulses Turned into. Immunized mice were then stimulated in vivo once or three times in the presence or absence of LPS, and TRP2-specific CTL responses were measured using tetramer analysis. Because of the high number of pro-inflammatory cytokines to induce, many of which are regulated by SOCS1, and the role of SOCS1 in the direct regulation of NF-κB (p65) signaling has been demonstrated in vivo LPS was selected for stimulation (Ryo et al., 2003, Mol. Cell. 12: 1413-26). In mice immunized with SOCS1-siRNA-DC, in the absence of in vivo LPS stimulation, only 3.1% in mice immunized with GFP-siRNA-DC, TRP2-4 5.1% of total CD8 + T cells were positive for the mer (FIG. 9A). Upon one or three in vivo LPS stimulation, the percentage of CD8 + T cells that are positive for TRP2-tetramer is substantially increased in mice immunized with SOCS1-siRNA-DC (9.7% and 19.4%), there was generally no change in mice immunized with GFP-siRNA-DC (3.0% and 4.0%, respectively) (FIG. 9A). Consistent with this, the CTL test (FIG. 9C) and interferon gamma (IFNγ) ELISPOT showed similar results. In addition, 3 months after immunization, SOCS1-siRNA co-injected with LPS (FIG. 9B) and received a TRP2 peptide pulse.
In most of the mice immunized with DC, vitiligo (lightening, depigmentation and / or hair loss) of the coat is clearly visible, indicating self-tolerance to TRP2 normally expressed in host melanocytes. It represents destruction. In contrast, no vitiligo was observed in any of the mice immunized with GFP-siRNA DC even when repeatedly administered LPS in vivo, which maintains tolerance to self-antigens. Suggested a critical role for SOCS1 in DCs. These results demonstrate that signaling within mature antigen presenting DCs controls the magnitude of CTL responses and self-tolerance, and that mature DC signaling is severely restricted by SOCS1.
SOCS1−制限されたシグナリングが、自己寛容を破断し有効な抗腫瘍免疫を誘発するDCの能力を制御する
腫瘍ワクチン接種の第1の目的は、腫瘍細胞上で優先的に発現される自己抗原に対する強い適応免疫応答を誘発することにより自己寛容を破断することにある。自己抗原特異的CTL応答及び自己寛容の規模を調節する上でのSOCS1の役割が観察されたことから、有効な抗腫瘍免疫を誘発する成熟DCの能力がSOCS1発現により制御されるか否かの調査が促されることになった。これをテストするために、C57BL/6マウスをB16腫瘍細胞で皮下接種し、3日後に、LPSによりエクスビボで成熟させたTRP2パルスされ形質導入されたDCで一回免疫化した。図10Aは、SOCS1−siRNADC免疫化だけで、GFP−siRNA−DC免疫化と比べたB16腫瘍の成長を著しく阻害することができた(P<0.01)ということを示している。しかしながら、SOCS1−siRNA−DCで免疫化したマウスの50%は、腫瘍接種から30日後に1,500mm3を上回る全身腫瘍組織量のため最終的に死亡した。前炎症性シグナルを増強することによりSOCS1−siRNA−DCで免疫化したマウスにおけるマウス生存率を改善できるか否かを判定するために、マウスをDC免疫化から一日後にLPSによりインビボで一回刺激した。免疫化プロトコルに対するLPS刺激の付加は、SOCS1−siRNA−DCで免疫化したマウスにおけるB16腫瘍の成長を実質的に遮断した(図10B)。このことは、LPS刺激を受けていないマウスに比べて全身腫瘍組織量の減少を全く示さなかったGFP−siRNA−DC及びモック形質導入を受けたDC対照と好対照を成していた(図10Aと10Bを比較のこと)。SOCS1−siRNA−DCでの免疫化とLPS抗原投与の組合せは同様に、マウスの生存率を60日超の間100%まで大幅に増大させた(図10C)。抗腫瘍活性の増強は、SOCS1−siRNA−DCで免疫化されたマウスの体内の強力なTRP2特異的CTL活性と相関された(図10D)。SOCS1−siRNA−DCでCD4及びCD8ノックアウト(KO)マウスを免疫化することにより、免疫化されたCD4KOマウスにおいては弱い抗腫瘍活性しか観察されなかったものの、抗腫瘍活性がCD8+及びCD4+細胞の両方を必要とすることがさらに実証された(図10B−10D)。集合的に、これらの結果は、成熟DCにおけるSOCS1で制限されたシグナリングが、寛容を破断し有効な抗腫瘍免疫を誘発するそれらの能力を制御すること、そして通常SOCS1により調節される付加的な前炎症性シグナルが、事前準備された全身腫瘍組織量を制御する誘発された抗腫瘍免疫応答の能力をさらに増大し得ることを表わしている。
SOCS1-restricted signaling controls DC's ability to break self-tolerance and elicit effective anti-tumor immunity The primary objective of tumor vaccination is against self-antigens preferentially expressed on tumor cells To break self-tolerance by inducing a strong adaptive immune response. Since the role of SOCS1 in regulating the magnitude of autoantigen-specific CTL responses and self-tolerance has been observed, whether the ability of mature DCs to induce effective anti-tumor immunity is controlled by SOCS1 expression Investigation was encouraged. To test this, C57BL / 6 mice were inoculated subcutaneously with B16 tumor cells and 3 days later immunized once with TRP2 pulsed transduced DCs matured ex vivo with LPS. FIG. 10A shows that SOCS1-siRNADC immunization alone could significantly inhibit B16 tumor growth compared to GFP-siRNA-DC immunization (P <0.01). However, 50% of the mice immunized with SOCS1-siRNA-DC eventually died due to systemic tumor tissue volume exceeding 1500
自己寛容及び抗腫瘍免疫の制御におけるSOCS1で制限されたシグナル3の決定的役割
SOCS1は、抗原ペプチド/MHC提示/共同刺激及び/又はサイトカインシグナリング及び分泌を調節することにより、成熟DCのシグナリング及び産出量に影響を及ぼす確率が高い。以下の実験は、自己抗原特異的CTL応答及び抗腫瘍免疫の制御のためにこれらの3つのシグナルのうちのどれが主としてSOCS1により調節されるかを調査するために着手された。
The critical role of SOCS1-restricted
まず第1に、SOCS1サイレンシング化が、共同刺激分子の発現に影響を及ぼすか否かがテストされた(シグナル2)。フローサイトメトリ検定により、LPS誘発型成熟の前後両方で、GFP−siRNA−DC上に比べSOCS1−siRNA−DC上には検出不能な又はわずかにしか増強されていない共同刺激/阻害分子(B7.1、B7.2、OX40L、CD40又はPDL1)表面レベルが存在する、ということが一貫して観察された(図11A)。匹敵するレベルのMHC−I及びII分子がSOCS1−siRNA−DC及びGFP−siRNA−DC上でも検出された。 First, it was tested whether SOCS1 silencing affects the expression of costimulatory molecules (signal 2). Co-stimulatory / inhibitory molecules (B7.) That were undetectable or slightly enhanced on SOCS1-siRNA-DC compared to GFP-siRNA-DC both before and after LPS-induced maturation by flow cytometry assay. 1, B7.2, OX40L, CD40 or PDL1) It was consistently observed that surface levels were present (FIG. 11A). Comparable levels of MHC-I and II molecules were also detected on SOCS1-siRNA-DC and GFP-siRNA-DC.
抗腫瘍免疫を誘発する上でのCD8+T細胞の重要性のため、MHC−I制限型ペプチド免疫原性(TCR親和性)がインビボでのSOCS1制限型抗原提示において重要な役割を果たすか否かがさらに調査された。高親和性形態(TRP2b)及び低親和性形態(TRP2a)のTRP2CTLペプチドが以前に同定された(ファン・エルサス(van
Elsas)ら、2001年、J.Exp.Med.第194号:481〜9頁)こと
から、抗腫瘍免疫応答を誘発する形質導入済みDCの能力に対しシグナル1の強度が影響を及ぼし得るか否かをテストするために、両方のTRP2ペプチドを使用した。図11Bは、TRP2bペプチドで負荷されたGFP−siRNA−DCはほんの少しの抗腫瘍活性(統計学的には重要でない)しか示さなかったものの低(TRP2a)又は高(TRP2b)親和性ペプチドのいずれかが負荷された成熟GFP−siRNA−DCはインビボLPS刺激でB16腫瘍の退行を誘発できなかったということを示している。これとは対照的に、低又は高親和性TRP2ペプチドのいずれかで負荷されたSOCS1−siRNA−DCは両方共、腫瘍の成長を有効に遮断した。免疫化されたマウスにおけるTRP2特異的CTL活性も同様に、IFNγELISPOT検定を用いて調査された。図11Cは、高親和性ペプチドで負荷されたGFP−siRNA−DCが低親和性ペプチドで負荷されたGFP−siRNA−DCよりも強いIFNγ応答を誘発したことを示している。しかしながら、低又は高親和性ペプチドのいずれかで負荷された両方のSOCS1−siRNA−DCグループが、高親和性ペプチドで負荷されたGFP−siRNA−DCよりもはるかに強いIFNγ応答を誘発しており(P<0.01)、このことは観察された抗腫瘍活性と一致している(図11B)。さらに、低又は高親和性ペプチドで負荷されたSOCS1−siRNA−DCが類似のIFNγ応答を誘発した(図11C)。
Because of the importance of CD8 + T cells in inducing anti-tumor immunity, whether MHC-I restricted peptide immunogenicity (TCR affinity) plays an important role in SOCS1-restricted antigen presentation in vivo Further investigation was made. A high affinity form (TRP2b) and a low affinity form (TRP2a) of the TRP2CTL peptide have been previously identified (van
Elsas) et al., 2001, J. MoI. Exp. Med. 194: 481-9) to test whether the intensity of
これらの実験の結果は、SOCS1サイレンシング化が、成熟の存在下又は不在下でDC上の共同刺激/阻害性分子(シグナル2)及びMHC−I及びII分子の発現に対し有意な影響を及ぼさないこと、そしてSOCS1がサイレンシング化されないかぎり、高親和性又は低親和性TRP2ペプチド(シグナル1)で負荷された成熟DCが強力なIFNγ応答を誘発する上で有効でないこと、を実証している。 The results of these experiments indicate that SOCS1 silencing has a significant effect on the expression of costimulatory / inhibitory molecules (signal 2) and MHC-I and II molecules on DCs in the presence or absence of maturation. And, unless SOCS1 is silenced, demonstrates that mature DCs loaded with high or low affinity TRP2 peptide (signal 1) are not effective in inducing a strong IFNγ response .
実施例6:マウスのSOCS1−siRNA−DCにより誘発されたCTL及び抗腫瘍活性のインビボIL−12増強
樹状細胞(DC)による細胞傷害性T細胞(CTL)応答の開始は充分に研究されてきているものの、自己寛容の維持又は破断を調節するための機序はなおも充分に定義されていないままである。この実施例では、成熟野生型DCではなく、サイトカインシグナリング(SOCS)1のサプレッサが内部でシグナリングされた成熟DCが、特に病原菌産物又はIL−12によりインビボで刺激された場合に、自己寛容を破断する上で有効である、ということが実証されている。本書で開示される実験は、抗原提示DCにより提供される(抗原親和性(シグナル1)及び共同刺激分子レベル(シグナル2)ではなく)SOCS1制限されたシグナル3(IL−12)が自己寛容を制御することを実証している。さらに、本開示は、SOCS1サイレンシング化されたDCが自己抗原に対する強力な免疫応答を誘発し、事前準備されたB16腫瘍の成長を遮断することを実証している。その上、ヒトのSOCS1サイレンシング化されたDCは、溶解エフェクタ機能で自己抗原特異的ヒトCTLを完全に活性化するさらに優れた能力を有し、このSOCS1サイレンシング化アプローチの翻訳上の潜在性を暗示している。
Example 6: Murine SOCS1-siRNA-DC-induced CTL and in vivo IL-12 enhancement of antitumor activity Initiation of cytotoxic T cell (CTL) responses by dendritic cells (DC) has been well studied. However, the mechanisms for adjusting self-tolerance or regulating breakage still remain poorly defined. In this example, mature DCs, in which the suppressor of cytokine signaling (SOCS) 1 is signaled, but not mature wild-type DCs, break self-tolerance, especially when stimulated in vivo by pathogenic products or IL-12 It has been proven to be effective in doing so. Experiments disclosed herein show that SOCS1-restricted signal 3 (IL-12) provided by antigen-presenting DC (rather than antigen affinity (signal 1) and costimulatory molecule level (signal 2)) is self-tolerant. Demonstrates control. Furthermore, the present disclosure demonstrates that SOCS1-silenced DC elicits a strong immune response against self-antigens and blocks the growth of pre-prepared B16 tumors. Moreover, human SOCS1-silenced DCs have a better ability to fully activate autoantigen-specific human CTLs with lytic effector function, and the translational potential of this SOCS1 silencing approach Is implied.
自己抗原特異的CTL活性化及び寛容の制御においてSOCS1で制限されたシグナル3が重要である確率が高いことから、成熟DCにおいてSOCS1により調節される主要なサイトカインの同定が促された。CTL活性化に影響を及ぼすものとして知られている複数の前炎症性サイトカイン候補の重要性は、当初、SOCS1サイレンシング化と組合せた形でのDCワクチン接種のために遺伝子的にホモ接合性の異なるKOマウスに由来するDCを用いることによってテストされた。IL−12(p35−/−)KOマウス由来のDCが免疫化のために使用された場合、TRP2ペプチドで負荷されたp35−/−SOCS1−siRNA−DCは、もはやB16腫瘍を阻害しないことが観察され(図12A−12B)、このことは、腫瘍の退行のためにSOCS1制限されDC産生されたIL−12の決定的役割を示唆していた。SOCS1制限されたDCの機能におけるIL−12の役割をさらに判定するため、p35−/−SOCS1−siRNA−DCにより誘発
されたCTL応答がさらに査定された。IFNγELISPOT及びCTL検定を用いて、野生型SOCS1−siRNA−DCに比べ、p35−/−SOCS1−siRNA−DCのTRP2特異的CTL応答を誘発する能力が著しく減少していたことが観察された(図12C及び12D)。さらにLPSでのインビボ刺激は、TRP2−四量体分析により測定される通り、p35−/−SOCS1−siRNA DCにより誘発されるCTL応答を高めることができなかった(図9A)。興味深いことに、抗原提示DCによって産生されたIL−12の基本的な役割とは対照的に、野生型SOCS1−siRNA−DCで免疫化されたIL−12(p35−/−)KOマウスは、活発な抗腫瘍免疫及びCTL応答をも誘発することが観察され、このことは、自己抗原特異的CTL応答の誘発のために常在宿主細胞により産生されたIL−12は必要とされないということを示していた(図12A−12B)。合わせて考えた場合これらの結果は、抗原提示DCにより産生されたSOCS1制限型IL−12が、強力なTRP2特異的CTL応答及びB16腫瘍の根絶を誘発する上で重要であることを表わしている。
The high probability that signal 3 restricted with SOCS1 is important in the control of autoantigen-specific CTL activation and tolerance prompted the identification of the major cytokines regulated by SOCS1 in mature DCs. The importance of multiple pro-inflammatory cytokines known to affect CTL activation is initially genetically homozygous for DC vaccination in combination with SOCS1 silencing Tested by using DCs from different KO mice. When DCs from IL-12 (p35 − / − ) KO mice were used for immunization, p35 − / − SOCS1-siRNA-DCs loaded with TRP2 peptide no longer inhibit B16 tumors Observed (FIGS. 12A-12B), suggesting a critical role for SOCS1 restricted and DC produced IL-12 for tumor regression. To further determine the role of IL-12 in SOCS1-restricted DC function, the CTL response elicited by p35 − / − SOCS1-siRNA-DC was further assessed. Using IFNγ ELISPOT and CTL assay, it was observed that the ability of p35 − / − SOCS1-siRNA-DC to elicit a TRP2-specific CTL response was significantly reduced compared to wild-type SOCS1-siRNA-DC (FIG. 12C and 12D). Furthermore, in vivo stimulation with LPS failed to enhance the CTL response elicited by p35 − / − SOCS1-siRNA DC as measured by TRP2-tetramer analysis (FIG. 9A). Interestingly, in contrast to the basic role of IL-12 produced by antigen-presenting DCs, IL-12 (p35 − / − ) KO mice immunized with wild type SOCS1-siRNA-DC are It has also been observed to elicit active anti-tumor immunity and CTL responses, indicating that IL-12 produced by resident host cells is not required for the induction of autoantigen-specific CTL responses. (FIGS. 12A-12B). Taken together, these results indicate that SOCS1-restricted IL-12 produced by antigen-presenting DCs is important in inducing a potent TRP2-specific CTL response and B16 tumor eradication. .
SOCS1サイレンシング化されたDCによるIL−12及びIL−12誘発型サイトカインの持続的かつ増強された産生とwtDCによる過渡的で低い産生の関係
DCは、LPS及びCD40ライゲーションなどの病原菌産物に応答して有意な量のIL−12を産生する(シュルツ(Schulz)ら、2000年、Immunity第13号:453〜62頁)。DCによるIL−12の産生は、成熟の誘発後の短い期間(8〜16時間)に綿密に制限され(ランゲンカンプ(Langenkamp)ら、2000年、Nat.Immunol.第1号:311〜6頁)、SOCS1により調節される(アイルズ(Eyles)ら、2002年、J.Biol.Chem.第277号:43735〜40頁)。適応免疫の調節におけるSOCS1制限型IL−12の重要な役割が識別されたことから、免疫寛容を破断しTRP2に対する有効な抗腫瘍免疫応答を誘発するSOCS1−siRNA−DCの能力に密に関係し得るDCによるIL−12産生の強度及び持続時間に対するSOCS1サイレンシング化の効果が検討された。
Relationship between sustained and enhanced production of IL-12 and IL-12-induced cytokines by SOCS1-silenced DCs and transient and low production by wtDC DCs respond to pathogenic products such as LPS and CD40 ligation Significant amounts of IL-12 (Schulz et al., 2000, Immunity 13: 453-62). The production of IL-12 by DC is tightly limited to a short period (8-16 hours) after induction of maturation (Langenkamp et al., 2000, Nat. Immunol. 1: 311-6). ), Regulated by SOCS1 (Eyles et al., 2002, J. Biol. Chem. 277: 43735-40). Since the important role of SOCS1-restricted IL-12 in the regulation of adaptive immunity has been identified, it is closely related to the ability of SOCS1-siRNA-DC to break immune tolerance and elicit an effective anti-tumor immune response against TRP2. The effect of SOCS1 silencing on the intensity and duration of IL-12 production by the resulting DCs was investigated.
図13Aは、GFP−siRNA−DC及びモック形質導入を受けたDCに比べて、72時間にわたるLPS及び抗CD40mAbでの連続的刺激に応答してSOCS1−siRNA−DCによって著しく増大したレベルのIL−12(p70)が産生されたということを示している。次に、24時間にわたりLPS/抗−CD40で刺激し次にLPSを含まない新鮮な培地の中のDCを新しい培養平板に移すことにより、もとの刺激の除去にも関わらずIL−12レベルを維持するSOCS1−siRNA−DCの能力を検討した。図13Bは、GFP−siRNA−DC及びモック形質導入を受けたDCが刺激時点で過渡的にのみIL−12を産生し、一方SOCS1−siRNA−DCは刺激の除去に関わらずはるかに高いレベルのIL−12を持続的に産生したことを示している。何らかの特定の理論により束縛されることを望まないが、LPS/抗CD40の除去の後のSOCS1−siRNA−DCによるIL−12の長時間にわたる増強した産生は、IL−12又はその他のDCが分泌した前炎症性サイトカインによる場合によって自己分泌/傍分泌の刺激及び/又はもとの刺激によって誘発されるシグナリング経路の長時間にわたる活性化に起因する可能性がある。これらの結果は、SOCS1サイレンシング化によりDCが刺激に応答して持続的な増加したレベルのIL−12を産生でき、これが、寛容を破断し事前準備された腫瘍を根絶するSOCS1サイレンシング化されたDCの能力の原因であり得る、ということを示している。 FIG. 13A shows significantly increased levels of IL-by SOCS1-siRNA-DC in response to continuous stimulation with LPS and anti-CD40 mAb over 72 hours compared to GFP-siRNA-DC and mock-transduced DC. 12 (p70) was produced. Next, IL-12 levels were achieved despite the removal of the original stimulus by stimulating with LPS / anti-CD40 for 24 hours and then transferring DCs in fresh medium without LPS to a new culture plate. The ability of SOCS1-siRNA-DC to maintain the FIG. 13B shows that GFP-siRNA-DC and mock-transduced DC produce IL-12 only transiently at the time of stimulation, while SOCS1-siRNA-DC shows much higher levels regardless of stimulation removal. It shows that IL-12 was produced continuously. While not wishing to be bound by any particular theory, prolonged production of IL-12 by SOCS1-siRNA-DC after removal of LPS / anti-CD40 is secreted by IL-12 or other DCs. May be due to prolonged activation of signaling pathways triggered by possibly autocrine / paracrine stimuli and / or original stimuli by selected pro-inflammatory cytokines. These results indicate that SOCS1 silencing allows DCs to produce sustained increased levels of IL-12 in response to stimulation, which is SOCS1 silenced that breaks tolerance and eradicates pre-prepared tumors. This indicates that it may be the cause of DC capability.
SOCS1は、IL−12及びその他のサイトカインのシグナリングを媒介するJak/Stat経路の決定的調節因子であることから(クボら、2003年、Nat.Immunol.第4号:1169〜76頁;アレキサンダーら、2004年、Annu.Rev.Immunol.第22号:503〜29頁)、互いの間及び場合によってはその他の近傍のDCの間でのサイトカインフィードバックループの発達を通してDC内のSOC
S1サイレンシング化がサイトカインの産生を増大させるか否かをテストするために、次の実験セットに着手した。これに対応するため、SOCS1−siRNA−DCによる腫瘍壊死因子(TNF)α及びIL−6の産生を測定した。かかるサイトカインはIL−12刺激により誘発されるものとして知られていることから、(TNF)α及びIL−6をテストした(トリンキエリ(Trinchieri)ら、2003年、Nat.Rev.Immunol.第3号:133〜46頁)。
SOCS1 is a critical regulator of the Jak / Stat pathway that mediates IL-12 and other cytokine signaling (Kubo et al., 2003, Nat. Immunol. 4: 1169-76; Alexander et al. 2004, Annu.Rev.Immunol.22: 503-29), SOC in DCs through the development of cytokine feedback loops between each other and possibly other nearby DCs.
In order to test whether S1 silencing increases cytokine production, the next set of experiments was undertaken. In response to this, the production of tumor necrosis factor (TNF) α and IL-6 by SOCS1-siRNA-DC was measured. Since such cytokines are known to be induced by IL-12 stimulation, (TNF) α and IL-6 were tested (Trinchieri et al., 2003, Nat. Rev. Immunol. No. 3). : 133-46).
図13Cは、過渡的に低レベルのTNFα及びIL−6を産生したにすぎないGFP−siRNA−DC及びモック形質導入を受けたDCとは異なり、SOCS1−siRNA−DCがもとの刺激の除去後も持続的により高いレベルのTNFα及びIL−6を産生したことを示している。フィードバックループの発達のためのIL−12の重要性をさらに判定するために、p35−/−SOCS1−siRNA−DC及びwtSOCS1−siRNA−DCによるTNFα及びIL−6の産生を比較した。図13Dは、p35−/−SOCS1−siRNA−DCかもはや増大した長時間にわたる量のTNFα及びIL−6を産生することができなかったということを示しており、これは、IL−12のフィードバックがSOCS1−siRNA−DCによるTNFα及びIL−6産生の主要な誘発因子であることを表わしている。これらのデータは、SOCS1サイレンシング化がDC中の決定的なシグナリング抑制を無効化し、従ってこれらが増強されたフィードバックループを介して連続的にIL−12のみならずIL−12誘導型前炎症性サイトカインに対しても応答しこれを産生することができるようにしている、ということを表わしている。本書で開示されている結果は、白斑及びB16腫瘍細胞に対する有効な抗腫瘍免疫の両方を誘発するTRP2負荷されたSOCS1−siRNA−DCの能力を説明する確立の高い機序を提供している。 FIG. 13C shows that, unlike GFP-siRNA-DC and mock-transduced DC that only produced transiently low levels of TNFα and IL-6, SOCS1-siRNA-DC removed the original stimulus. It was later shown that sustained higher levels of TNFα and IL-6 were produced. To further determine the importance of IL-12 for feedback loop development, the production of TNFα and IL-6 by p35 − / − SOCS1-siRNA-DC and wtSOCS1-siRNA-DC was compared. FIG. 13D shows that p35 − / − SOCS1-siRNA-DC could no longer produce increased amounts of TNFα and IL-6 over time, indicating that IL-12 feedback Are the major inducers of TNFα and IL-6 production by SOCS1-siRNA-DC. These data show that SOCS1 silencing abolishes critical signaling repression in DC, so they are not only IL-12 but also IL-12-induced pro-inflammatory through continuous feedback loops It also means that it is able to respond to and produce cytokines. The results disclosed herein provide a well-established mechanism that accounts for the ability of TRP2-loaded SOCS1-siRNA-DC to elicit both vitiligo and effective anti-tumor immunity against B16 tumor cells.
自己寛容を破断するDCの能力を制御するためのSOCS1制限型IL−12シグナリングの重要性
自己腫瘍関連抗原に対する強力なCTL応答を誘発するSOCS1−siRNA−DCの能力は、SOCS1サイレンシング化戦略の治療適応用を示唆する。患者の体内での刺激としてのLPSの臨床的使用には過度に毒性があるため、SOCS1によりシグナリングが調節されているIL−12がSOCS1−siRNA−DCの効能を高める上でも有効であるか否かを査定した。
The importance of SOCS1-restricted IL-12 signaling to control the ability of DC to break self-tolerance The ability of SOCS1-siRNA-DC to elicit a strong CTL response against self-tumor-associated antigens is Suggest for therapeutic use. Whether or not IL-12, whose signaling is regulated by SOCS1, is also effective in enhancing the efficacy of SOCS1-siRNA-DC because it is too toxic for clinical use of LPS as a stimulus in the patient's body Was assessed.
C57BL/6マウスにB16腫瘍細胞を接種し、3日後に、組換え型マウスTNFαによりエクスビボで成熟させられたTRP2パルスを受け形質導入されたDCで一度、マウスを免疫化した。DC免疫化の後、レシピエントマウスをインビボで3回、低用量の組換え型マウスIL−12(マウス一匹あたり1μg)で刺激した。図14Aは、SOCS1−siRNA−DCで免疫化されたマウス内のB16の腫瘍の成長が効率よく遮断されたことを示している。これとは対照的に、IL−12を用いたインビボ投与を伴うGFP−siRNA−DCでの免疫化は、PBS対照に比べて腫瘍の効果に対しほとんど効果をもたらさなかった。抗腫瘍活性は、IFNγELISPOT検定により示される通り、SOCS1−siRNA−DCで免疫化されたマウスにおけるTRP2特異的CTL活性の増大と相関された(図14B)。先の観察事実(図10A)と一致して、インビボでのIL−12刺激の不在下でのTRP2パルス、TNFα成熟を受けたSOCS1−siRNA−DCでの免疫化は、同様に、GFP−siRNA−DC対照に比べて増大した抗腫瘍活性をも示した。 C57BL / 6 mice were inoculated with B16 tumor cells and 3 days later, mice were immunized once with DCs transduced with TRP2 pulses matured ex vivo with recombinant mouse TNFα. Following DC immunization, recipient mice were stimulated 3 times in vivo with low doses of recombinant mouse IL-12 (1 μg per mouse). FIG. 14A shows that B16 tumor growth in mice immunized with SOCS1-siRNA-DC was efficiently blocked. In contrast, immunization with GFP-siRNA-DC with in vivo administration with IL-12 had little effect on tumor effect compared to PBS control. Anti-tumor activity was correlated with increased TRP2-specific CTL activity in mice immunized with SOCS1-siRNA-DC as shown by the IFNγ ELISPOT assay (FIG. 14B). Consistent with previous observations (FIG. 10A), immunization with SOCS1-siRNA-DC undergoing TRP2 pulse, TNFα maturation in the absence of IL-12 stimulation in vivo is also similar to GFP-siRNA. -Also showed increased anti-tumor activity compared to DC control.
これらの結果は、おそらくはIL−12及びIL−12誘発型サイトカインの増強されたシグナリングに由来して、IL−12のインビボ投与は、SOCS1サイレンシング化されたDCの免疫刺激能力を著しく増強するものの野生型DCのものは増強しないということを示している。これらの結果はさらに、IL−12などのサイトカインの全身的濃度
ではなく抗原提示DC内のSOCS1制限型サイトカインシグナリングが、自己腫瘍関連抗原に対する有効な抗腫瘍免疫を誘発するために重要である、ということを暗示している。
These results probably come from enhanced signaling of IL-12 and IL-12-induced cytokines, although in vivo administration of IL-12 significantly enhances the immunostimulatory capacity of SOCS1-silenced DCs It shows that the wild type DC does not enhance. These results further indicate that SOCS1-restricted cytokine signaling in antigen-presenting DCs, but not systemic concentrations of cytokines such as IL-12, is important for inducing effective anti-tumor immunity against self-tumor associated antigens. It implies.
白斑以外のいかなる明白な毒性も、免疫化から6ヵ月までは、LPS又はIL−12のいずれかが同時注射されたTRP2パルスを受けたSOCS1−siRNA−DCマウスにおいて観察されなかった。免疫化されたマウスの全ての主要な器官及び組織の組織学的分析は、いかなる病的炎症も明らかにしなかった。IgG及び抗dsDNAのレベルは、SOCS1−siRNA−DC及びモックDCマウスにおいて比較可能なものであった。これらのデータは、TRP2パルスを受けたSOCS1−siRNA−DCでの免疫化が、マウスにおいて病的炎症をひき起こさないということを示唆している。 No obvious toxicity other than vitiligo was observed in SOCS1-siRNA-DC mice receiving TRP2 pulses co-injected with either LPS or IL-12 up to 6 months after immunization. Histological analysis of all major organs and tissues of immunized mice did not reveal any pathological inflammation. IgG and anti-dsDNA levels were comparable in SOCS1-siRNA-DC and mock DC mice. These data suggest that immunization with SOCS1-siRNA-DC receiving TRP2 pulses does not cause pathological inflammation in mice.
SOCS1制限型シグナル3の強度は、CTL活性化、寛容及び抗腫瘍免疫を制御する
本書に開示されている結果は、腫瘍ワクチンの開発に深いかかわりを有するはずである成熟DCによるCTL応答の調節への新たな見識を提供する。成熟は未成熟の寛容原性状態から成熟した免疫原性状態へのDC遷移のための制御点であることが知られている(バンシュローら、1998年、Nature第392号:245〜52頁;スタインマンら、2003年、Annu.Rev.Immunol.第21号:685〜711頁)。抗原提示DC及びT細胞の間の初期接触(シグナル1及び2)の強度は、成熟DCが限られた数の研究に基づいて短命であると考えられていることから、CTL応答の規模及び命運を決定すると考えられている(ポルガドール(Porgador)ら、1998年、Journal of Experimental Medicine第188号:1075〜82頁;インガリ(Ingulli)ら、1997年、J.Exp.Med.第185号:2133〜41頁;リュードル(Ruedl)ら、2000年、J.Immunol.第165号:4910〜6頁)。しかしながら、成熟抗原提示DCの寿命は以前に推定されたものよりもはるかに長く、インビボで2週間持続するということが、信頼性の高い遺伝子方法を用いたさらに近年の研究により実証されており(ガルグ(Garg)ら、2003年、Nat.Immunol.第4号:907〜12頁)、このことはT細胞との初期係合の後に成熟DCが調節の役割を果たすことを示唆している。
The intensity of SOCS1-restricted
この結果は、SOCS1により綿密に制限される成熟DC内での前炎症性シグナリングが、自己抗原特異的CTL反応の規模を決定的に制御することを実証している。このことは、CTL応答が少なくとも2つのレベル、すなわち、SOCS1発現によりその規模が調節される、CTL反応の開始に必要なDC成熟及び自らと及びCTLとの成熟DCの進行中のサイトカインシグナリングというレベルでDCにより制御されるということを表わしている。当該結果は、さらに、さまざまな免疫細胞のDC及びその周囲環境とサイトカイン及び病原菌産物の組成及び濃度の間の動的相互作用を暗示しており、これが集合的に、自己寛容の維持又は破断、ひいては自己抗原特異的CTL反応の命運を決定している。 This result demonstrates that pro-inflammatory signaling in mature DCs that is tightly restricted by SOCS1 critically controls the magnitude of autoantigen-specific CTL responses. This means that the CTL response is at least two levels: the DC maturation required for initiation of the CTL response and the ongoing cytokine signaling of the mature DC with itself and the CTL, the magnitude of which is regulated by SOCS1 expression This means that it is controlled by DC. The results further imply a dynamic interaction between the DC of various immune cells and their surrounding environment and the composition and concentration of cytokines and pathogen products, which collectively maintain or break self-tolerance, As a result, the fate of the autoantigen-specific CTL reaction is determined.
本書で開示されている結果は、自己寛容の維持又は破断を調節するための新規の機序を明らかにしている。DCにより提供される(ペプチド親和性及び共同刺激分子レベルではなく)SOCS1制限型シグナル3(IL−12)が自己寛容を決定的に制御することが発見された。自己免疫疾患(バンシュローら、2004年、Immunity第20号:539〜50頁)及びその他のモデル(カートシンガーら、2003年、J.Exp.Med.第197号:1141〜51頁;バレンズエラら、2002年、J.Immunol.第169号:6842〜9頁)の数多くの研究において、T細胞のDC媒介型活性化又は過剰活性化のためのサイトカインの重要性が暗示されてきた。 The results disclosed herein reveal a novel mechanism for adjusting self-tolerance maintenance or breakage. It was discovered that SOCS1-restricted signal 3 (IL-12) provided by DC (rather than peptide affinity and costimulatory molecule levels) controls self-tolerance critically. Autoimmune disease (Banschlo et al., 2004, Immunity 20: 539-50) and other models (Kartsinger et al., 2003, J. Exp. Med. 197: 1141-51; Valenzuela et al., (2002, J. Immunol. 169: 6842-9), the importance of cytokines for DC-mediated activation or overactivation of T cells has been implied.
wtDCによるIL−12といったサイトカインの産生は、過渡的であり、本書で及び他の研究(ランゲカンプら、2000年、Nat.Immunol.第1号:311〜6
頁)内で実証されている通り、刺激時点でSOCS1により阻害される。wtDCとは対照的に、SOCS1サイレンシング化されたDCは、この経路の誘発可能なフィードバックインヒビターを無効化した後、DC内のJak/Statシグナリング経路を通って入り組んだ自己分泌(そして場合によっては傍分泌も)シグナリングループを形成することにより、初期刺激に応答して、著しく増強したIL−12及びIL−12誘発型サイトカインレベルを連続的に産生することができるということが観察された。これと一致して、低用量のIL−12でのインビボ投与は、自己寛容を破断する(wtDCではなく)SOCS1サイレンシング化されたDCの能力を有効に増強することが発見された。
The production of cytokines such as IL-12 by wtDC is transient and is described in this and other studies (Langecamp et al., 2000, Nat. Immunol. 1: 311-6.
As demonstrated in page), it is inhibited by SOCS1 at the time of stimulation. In contrast to wtDC, SOCS1-silenced DCs have intruded autocrine (and possibly in some cases) through the Jak / Stat signaling pathway within the DC after disabling the inducible feedback inhibitor of this pathway. It was observed that by forming a signaling loop (also paracrine), it was possible to continuously produce significantly enhanced IL-12 and IL-12-induced cytokine levels in response to initial stimulation. Consistent with this, in vivo administration with low doses of IL-12 was found to effectively enhance the ability of SOCS1-silenced DCs to break self-tolerance (rather than wtDC).
集合的にこれらの結果は、SOCS1サイレンシング化されたDCにより産生されたIL−12及びIL−12誘発型サイトカインの連続的かつ増強された産生及びシグナリングが、自己寛容の破断及び抗腫瘍CTL反応の増強において主要な役割を果たす確率が高い、ということを表わしている。これらの結果はさらに、DC内のSOCS1による刺激性シグナリングの細胞内阻害が自己寛容の維持に寄与することを示唆している。SOCS1は、そのSOCSボックス領域を通してのユビキチン媒介型タンパク質分解のためにp65タンパク質をターゲティングすることによりNF−κBシグナリングを直接遮断することが発見された(リョーら、2003年、Mol.Cell.第12号:1413〜26頁)が、ギングラらは、SOCS1が、I型IFNのシグナリングを阻害することによってLPS誘発型毒性についてのマクロファージ内のTLR シグナリングを間接的に調節することを報告した(ギングラ(Gingras)ら、2004年、J.Biol.Chem.第279号:54702〜7頁)。I型IFNシグナリングを通したLPS誘発型毒性とは異なり、本書中の結果は、SOCS1サイレンシング化されたDCにより誘発されたCTL応答がIL−12により主として媒介されることを実証している。本書中の結果は同様に、LPSに応答したSOCS1サイレンシング化されたDCによるTNF−α、IL−6及びIL−12などのさまざまなサイトカインの産生の増強をも実証している。 Collectively, these results indicate that continuous and enhanced production and signaling of IL-12 and IL-12-induced cytokines produced by SOCS1-silenced DCs are disrupted by self-tolerance and anti-tumor CTL responses This means that there is a high probability of playing a major role in augmenting These results further suggest that intracellular inhibition of stimulatory signaling by SOCS1 in DCs contributes to maintaining self-tolerance. SOCS1 was found to directly block NF-κB signaling by targeting the p65 protein for ubiquitin-mediated proteolysis through its SOCS box region (Ryo et al., 2003, Mol. Cell. 12th). No. 1413-26) reported that SOCS1 indirectly regulates TLR signaling in macrophages for LPS-induced toxicity by inhibiting type I IFN signaling (Gingla ( Gingras) et al., 2004, J. Biol. Chem. 279: 54702-7). Unlike LPS-induced toxicity through type I IFN signaling, the results herein demonstrate that the CTL response elicited by SOCS1-silenced DCs is mediated primarily by IL-12. The results in this document also demonstrate enhanced production of various cytokines such as TNF-α, IL-6 and IL-12 by SOCS1-silenced DC in response to LPS.
本研究は、自己腫瘍関連抗原に対する有効な抗腫瘍免疫を誘発するためにDC内でSOCS1をサイレンシング化することの必要性を実証している。DCワクチンは、近年において1000人超の患者が関与した98件の公表されたDCワクチン治験において証明されているように、腫瘍ワクチン接種のための最も見込みある戦略の1つと考えられている(ローゼンバーグ(Rosenberg)ら、2004年、Nat.Med.第10号:909〜15頁)。主として多様なアプローチによりDCの成熟及び抗原提示を促進することを目的としたこれらの試みは、目標の臨床反応奏功率がきわめて低く、大いに失望させるものである(ローゼンバーグら、2004年、Nat.Med.第10号:909〜15頁)。本書中のデータは、高親和性ペプチドで負荷されたwtDCが、LPS又はIL−12でのインビボ刺激の後でさえ自己寛容を破断することがなおもできなかったということを実証している。従って、本書中の結果は、現在記述されている腫瘍ワクチン(ローゼンバーグら、2004年、Nat.Med.第10号:909〜15頁)の全般的無効性を説明することができ、DC抗原の提示及び成熟を促進する現行の戦略と組合せた形でSOCS1といった決定的なシグナリングインヒビターをサイレンシング化させることを介してさらに有効な腫瘍ワクチンを開発するための新たな途を提供する(ギルボア(Gilboa)、2004年、Nat.Rev.Cancer第4号:401〜11頁;ユー(You)ら、2000年、J.Immunol.第165号:4581〜4592頁;ソイファー(Soiffer)ら、2003年、J.Clin.Oncol.第21号:3343〜50頁;パルドール(Pardoll)、2002年、Nat.Rev.Immunol.第2号:227〜38頁)。 This study demonstrates the need for silencing SOCS1 within DCs to elicit effective anti-tumor immunity against self-tumor associated antigens. DC vaccines are considered one of the most promising strategies for tumor vaccination, as evidenced in 98 published DC vaccine trials involving more than 1000 patients in recent years (Rosen (Rosenberg et al., 2004, Nat. Med. 10: 909-15). These attempts, primarily aimed at promoting DC maturation and antigen presentation through a variety of approaches, are extremely disappointing and highly disappointing with the targeted clinical response success (Rosenberg et al., 2004, Nat. Med. 10: 909-15). The data in this document demonstrates that wtDC loaded with high affinity peptides was still unable to break self-tolerance even after in vivo stimulation with LPS or IL-12. Thus, the results in this document can explain the general ineffectiveness of the currently described tumor vaccines (Rosenberg et al., 2004, Nat. Med. 10: 909-15) and the DC antigen Provides a new way to develop more effective tumor vaccines by silencing critical signaling inhibitors such as SOCS1 in combination with current strategies that promote the presentation and maturation of (Gilboa ( Gilboa), 2004, Nat. Rev. Cancer 4: 401-11; You et al., 2000, J. Immunol. 165: 4581-4592; Soiffer et al., 2003. J. Clin. Oncol 21: 3343-50; 2002, Nat.Rev.Immunol No. 2:. 227-38, pp.).
抗原負荷されたDCにより誘発される抗原特異的免疫応答を特異的に増強する能力をもつこのSOCS1サイレンシング化アプローチは同様に、主要組織又は器官に対するもの
を含めた自己反応性T細胞を差別なく過剰活性化する、エフェクタT細胞上でCTLA4を全身的に遮断するアプローチよりもさらに魅力的なものでもある(ホッジ(Hodi)ら、2003年、Proc.Natl.Acad.Sci.USA第100号:4712〜7頁;ファン(Phan)ら、2003年、Proc.Natl.Acad.Sci.USA第100号:8372〜7頁)。要約すると、腫瘍ワクチンを増強するための集約的な努力が抗原親和性/用量及び共同刺激の改善に焦点を合わされてきたことから、本研究の中で見出されるSOCS1サイレンシング化されたDCの増強された免疫刺激能力及び自己寛容を調節するための新しい機序は、腫瘍に対する自己寛容制限を破断する一般的に応用可能な新規のワクチン接種戦略を提供するはずである。
This SOCS1 silencing approach, which has the ability to specifically enhance antigen-specific immune responses elicited by antigen-loaded DCs, also differentiates autoreactive T cells, including those against major tissues or organs, without discrimination. It is also more attractive than the approach of systemically blocking CTLA4 on effector T cells that are overactivated (Hodi et al., 2003, Proc. Natl. Acad. Sci. USA 100: 4712-7; Phan et al., 2003, Proc. Natl. Acad. Sci. USA 100: 8372-7). In summary, since the intensive efforts to enhance tumor vaccines have been focused on improving antigen affinity / dose and costimulation, the enhancement of SOCS1 silenced DCs found in this study New mechanisms for modulating the immunostimulatory capacity and self-tolerance should provide a generally applicable new vaccination strategy that breaks the self-tolerance limit on tumors.
実施例7:マウスSOCS1−siRNA−DCにより誘発されるHIV、特異的抗体及びCTL応答
この例は、宿主の天然免疫インヒビターを阻害することにより抗HIV免疫応答を誘発するための代替的戦略を実証している。本研究は、DC内のJAK/STAT経路の負の免疫調節因子であるSOCS1がHIV特異的細胞傷害性Tリンパ球(CTL)のみならず抗体応答をも制御することを実証している。SOCS1サイレンシング化されたDCは、HIVエンベロープ媒介型抑制に対する耐性を有し、マウスの体内でバランスのとれたメモリーHIVエンベロープ特異的抗体及びCTL応答を有効に誘発する。本開示は、宿主の免疫インヒビターを阻害することによりHIV特異的抗体及びCTL応答を惹起する最初の試みである。
Example 7: HIV, specific antibodies and CTL responses elicited by murine SOCS1-siRNA-DC This example demonstrates an alternative strategy for inducing an anti-HIV immune response by inhibiting the host's natural immune inhibitors is doing. This study demonstrates that SOCS1, a negative immune regulator of the JAK / STAT pathway in DC, regulates antibody responses as well as HIV-specific cytotoxic T lymphocytes (CTLs). SOCS1 silenced DCs are resistant to HIV envelope mediated suppression and effectively induce a balanced memory HIV envelope specific antibody and CTL response in the mouse body. The present disclosure is the first attempt to elicit HIV-specific antibodies and CTL responses by inhibiting host immune inhibitors.
この実施例中で提示される実験において使用される材料及び方法についてここで記述する。 The materials and methods used in the experiments presented in this example are now described.
レンチウイルスベクターでのBM由来のDCの形質導入
組換え型レンチウイルスベクター、LV−SOCS1−siRNA及びLV−GFP−siRNAを、本書の他の箇所で記述されている通りに産生し、滴定しDCを形質導入するのに使用した。
Transduction of BM-derived DCs with lentiviral vectors Recombinant lentiviral vectors, LV-SOCS1-siRNA and LV-GFP-siRNA were produced, titrated and DCs as described elsewhere in this document. Was used to transduce.
サイトカイン及び抗体ELISA検定
細胞培養上清中のサイトカインレベルを、メーカーの指示に従ってELISA分析(BDバイオサイエンシーズ、ニュージャージー州リンカーンパーク)により定量化した。Gp120特異的抗体及びサブクラスの力価を判定するためgp120タンパク質(炭酸塩緩衝液中5μg/ml[pH9.6])を4℃で一晩コーティングし、室温で1時間、壁にPBS−5%FBS中の血清の12倍階段希釈物を添加した。8回の洗浄後、ビオチニル化した抗マウス抗体(抗マウスIgM、IgG、IgG1、IgG2a、IgG2b又はIgG3)を室温で1時間ウェルに添加した。ペルオキシダーゼ基質としてストレプトアビジン−HRPを使用した。50μlの2MのH2SO4を添加することにより、反応を停止させた。バイオアッセイリーダー(Bio Assay Reader)上にて450nmで光学密度を読み取った。χ軸目盛がその対数であるものとして、χ軸に希釈物−1をy軸に光学密度(OD)値をとった散布図から判定した相互終点力価として、結果を表現している。データをプロットした後、個々の希釈系列の各々に対し対数曲線フィットを適用し、曲線フィットが正負カットオフ値と交差する点を決定した。対照マウスの血清由来の全ての希釈物の平均(+3SD)として各抗体イソタイプについてカットオフ値を計算した。各実験中でテストした全ての試料を同時に検定した。
Cytokine and antibody ELISA assays Cytokine levels in cell culture supernatants were quantified by ELISA analysis (BD Biosciences, Lincoln Park, NJ) according to the manufacturer's instructions. To determine the titer of Gp120-specific antibodies and subclasses, gp120 protein (5 μg / ml in carbonate buffer [pH 9.6]) was coated overnight at 4 ° C., 1 hour at room temperature, PBS-5% on the wall A 12-fold serial dilution of serum in FBS was added. After 8 washes, biotinylated anti-mouse antibody (anti-mouse IgM, IgG, IgG1, IgG2a, IgG2b or IgG3) was added to the wells for 1 hour at room temperature. Streptavidin-HRP was used as the peroxidase substrate. The reaction was stopped by adding 50 μl of 2M H 2 SO 4 . Optical density was read at 450 nm on a bioassay reader. Assuming that the χ axis scale is the logarithm, the result is expressed as a mutual end point titer determined from a scatter diagram in which the
T細胞酵素連結型イムノスポット(ELISPOT)検定
単離されたCD4+又はCD8+T細胞のELISPOT検定を、本書の他の個所で記述されている通りに実施した。
T Cell Enzyme Linked Immunospot (ELISPOT) Assay An ELISPOT assay of isolated CD4 + or CD8 + T cells was performed as described elsewhere in this document.
B細胞単離及びgp120抗体産生B細胞ELISPOT検定
完全RPMI1640培地内で脾臓から調製された単細胞懸濁液を、5%のCO2中で、37℃で1時間プラスチック皿の上で平板固定して、接着性マクロファージを除去した。非接着性細胞を抗Thg1.2及びウサギ補体で37℃で45分間処置してT細胞を溶解させた。残りのB細胞の純度は通常90%を超えていた。前述のような修正された方法により、B細胞ELISPOT検定を実施した((ル・ボン(Le Bon)ら、2001年、Immunity第14号:461〜7023頁))。簡単に言うと、96ウェルのニトロセルロースベース平板(ミリポアマルチスクリーンPI)を一晩PBS中のgp120でコーティングした。平板を6回PBSで洗浄し、2時間37℃で10%のFBSを含有するRPMI1640で遮断した。単離したB細胞をウェル内に播種し(5×105細胞/ウェル)、5%のCO2中で、37℃で20時間インキュベートした。その後、PBS0.5%トゥイーン(Tween)20(シグマ、ミズーリ州セントルイス)で6回洗浄して細胞を除去した。0.5%のFBSを含むPBS内で1μg/mlまで希釈させたビオチニル化抗マウスIgG(BDファーミンゲン)を添加し、混合物を2時間37℃でインキュベートした。アビジン:ビオチニル化酵素複合体(ABC、ベクター・ラボラトリーズ社(Vector Laboratories,Inc.)、カリフォルニア州バーリンゲーム(Burlingame,CA))をさらに1時間添加した。AEC(3−アミノ−9−エチルカバゾール;シグマ、ミズーリ州セントルイス)で4分間反応させた後、抗gp120IgGを検出した。KSELISPOT4.3ソフトウエアを用いて、自動ELISPOT読取り機システム(カール・ツァイス社(Carl Zeiss,Inc.)、ニューヨーク州ソーンウッド(Thornwood,NY))で、ゼルネット・コンサルティング社(ZellNet Consulting Inc.)(ニューヨーク州ニューヨーク(New York,NY))が結果を評価した。
B cell isolation and single cell suspensions prepared from spleen gp120 antibody producing B cells ELISPOT assays in complete RPMI1640 medium, in 5% CO 2, and a flat plate fixed on the 1 hour plastic dishes at 37 ° C. Adhesive macrophages were removed. Non-adherent cells were treated with anti-Thg1.2 and rabbit complement at 37 ° C. for 45 minutes to lyse T cells. The purity of the remaining B cells was usually above 90%. A B cell ELISPOT assay was performed by the modified method as described above ((Le Bon et al., 2001, Immunity No. 14: 461-7023)). Briefly, 96 well nitrocellulose based plates (Millipore Multiscreen PI) were coated overnight with gp120 in PBS. The plates were washed 6 times with PBS and blocked with RPMI 1640 containing 10% FBS at 37 ° C. for 2 hours. Isolated B cells were seeded in wells (5 × 10 5 cells / well) and incubated for 20 hours at 37 ° C. in 5% CO 2 . The cells were then removed by washing 6 times with PBS 0.5% Tween 20 (Sigma, St. Louis, MO). Biotinylated anti-mouse IgG (BD Farmingen) diluted to 1 μg / ml in PBS containing 0.5% FBS was added and the mixture was incubated for 2 hours at 37 ° C. Avidin: biotinylase complex (ABC, Vector Laboratories, Inc., Burlingame, Calif.)) Was added for an additional hour. After reacting for 4 minutes with AEC (3-amino-9-ethylcarbazole; Sigma, St. Louis, MO), anti-gp120 IgG was detected. Using the KSELISPOT 4.3 software, an automated ELISPOT reader system (Carl Zeiss, Inc., Thornwood, NY) at ZellNet Consulting Inc. ( New York, NY evaluated the results.
BAFF及びAPRILの定量的RT−PCR分析
トランスフェクションを受けたマウスBM−DC中のSOCS1の相対的発現を定量的実時間PCRによって評価した。トリゾール試薬(インビトロジェン、カリフォルニア州カールズバッド)を用いて、全RNAをDCから抽出し、ランダム六量体プライマ及びスーパー・スクリプト・ファースト・ストランド・シンセシス・キット(SuperScript First Strand Synthesis Kit)(インビトロジェン、カリフォルニア州カールズバッド)を用いて、各試料について1.0μgの全RNAを逆転写させた。鋳型として1回の反応につき5ngの出発RNA材料の当量を用いて20μlの4重反応でABI7900HT シーケンス・デテクション・システム(Sequence Detection System)(アプライド・バイオシステムズ社、カリフォルニア州フォスターシティー)上で、実時間5’−ヌクレアーゼ蛍光発生PCR分析を実施した。BAFT及びAPRILのためには次のプライマを使用した:BAFF、センス5’−TGCTATGGGTCATGTCATCCA−3’(配列番号17)及びアンチセンス5’−GGCAGTGTTTTGGGCATATTC−3’(配列番号18);APRIL、センス5’−TCACAATGGGTCAGGTGGTATC−3’(配列番号19)及びアンチセンス5’−TGTAAATGAAAGACACCTGCACTGT−3’(配列番号:20)。18Sのための順方向及び逆方向プライマであるTag Manプローブを、タックマン・ローデント(Taqman Rodent)18S対照試薬(アプライド・バイオシステムズ社、カリフォルニア州フォスターシティー)から得た。PCRパラメータは、タックマン・ユニバーサル・PCR・マスター・ミックス・キット(TaqMan Universal PCR Master Mix kit)(アプライド・バイオシステムズ社、カリフォルニア州フォスターシティー)のために推奨されたものであり、BAFF、APRIL及び18S反応は別々の管内で行なった。BAFF及びAPRILのレベルを18SrRNAに正規化し、一方BAFF又はAPRIL発現(モックトランスフェクションを受けたシミュレーションされたDCの対照値との関係におけるもの)は、コンパラティブCt方法(ライバックら、2001年、Met
hods第25号:402〜8頁)により計算した。
Quantitative RT-PCR analysis of BAFF and APRIL The relative expression of SOCS1 in transfected mouse BM-DC was assessed by quantitative real-time PCR. Total RNA was extracted from DCs using Trizol reagent (Invitrogen, Carlsbad, Calif.) And random hexamer primers and SuperScript First Strand Synthesis Kit (Invitrogen, CA) Carlsbad) was used to reverse transcribe 1.0 μg of total RNA for each sample. On an ABI 7900HT Sequence Detection System (Applied Biosystems, Foster City, Calif.) In a 20 μl quadruplicate reaction using an equivalent of 5 ng starting RNA material per reaction as a template. Time 5'-nuclease fluorogenic PCR analysis was performed. The following primers were used for BAFT and APRIL: BAFF,
hods No. 25: pages 402-8).
CTL検定
標的細胞を溶解するインビトロ再刺激された脾細胞の能力を測定する本書の他の個所で記述されている通りの標準クロム放出検定で、CD8+CTL応答を査定した。免疫化したマウスからプールした脾細胞を、4〜6日間、gp120タンパク質(20μg/ml)を含有するRPMI−1640中でインビトロで再刺激した。一晩20μg/mlのgp120タンパク質でパルスした標的細胞を90分間51Crクロム酸ナトリウム溶液で標識した。異なる数のエフェクタ細胞を37℃で3時間、96ウェルのV字底平板(200μl/ウェル)内で、一定数の標的細胞(1×104/ウェル)と共にインキュベートした。3重培養から上清(100μl)を収集した。(実験的放出−自然放出)/(最大放出−自然放出)×100として細胞溶解百分率を計算した。
CTL Assay CD8 + CTL response was assessed with a standard chromium release assay as described elsewhere in this document that measures the ability of in vitro restimulated splenocytes to lyse target cells. Splenocytes pooled from immunized mice were restimulated in vitro in RPMI-1640 containing gp120 protein (20 μg / ml) for 4-6 days. Target cells pulsed with 20 μg / ml gp120 protein overnight were labeled with 51 Cr sodium chromate solution for 90 minutes. Different numbers of effector cells were incubated with a fixed number of target cells (1 × 10 4 / well) in 96-well V-bottom plates (200 μl / well) at 37 ° C. for 3 hours. Supernatants (100 μl) were collected from triplicate cultures. The percentage of cell lysis was calculated as (experimental release-spontaneous release) / (maximum release-spontaneous release) x 100.
T及びB細胞増殖検定
本書の他の個所で記述されている通りに単離したCD4+又はCD8+T細胞(1ウェルにつき1×106個)及びB細胞(1ウェルにつき1×105個)を、さまざまな刺激の存在下又は不在下で96ウェル平板の3重ウェルの中で完全培地内で培養した。培養4日目に、ウェルを16時間、1μのCi[3H]−チミジンでパルスした。その後平板を収獲し、ミクロ・ベータ(Micro Beta)シンチレーションカウンタ(トップ・カウント(Top Count)NXT、パッカード(Packard))を用いて、取込まれた[3H]−チミジンを測定した。
T and B cell proliferation assays CD4 + or CD8 + T cells (1 x 10 6 per well) and B cells (1 x 10 5 per well) isolated as described elsewhere in this document Cultured in complete medium in triplicate wells of 96-well plates in the presence or absence of various stimuli. On
DC免疫化
本書の他の個所で記述されている通りの5のMOIで、骨髄由来のDC(BM培養5日目)をLV−SOCS1−siRNA又はLV−GFP−siRNAで形質導入した。
DC Immunization Bone marrow derived DCs (BM culture day 5) were transduced with LV-SOCS1-siRNA or LV-GFP-siRNA at 5 MOIs as described elsewhere in this document.
この実施例で提示されている実験の結果についてここで記述する。 The results of the experiment presented in this example will now be described.
DC中のSOCS1のサイレンシング化が、HIVEnv特異的抗体応答を増強する
抗HIV抗体を誘発するDCの能力に対するSOCS1サイレンシング化の効果についてまず調査した。HIVEnvは、細胞及び中和抗体の両方の応答を誘発できることから、この研究ではこれを使用した。本書の他の個所で記述されている通り、トランスフェクションを受けた細胞内でSOCS1mRNAの約90%をダウンレギュレートする能力をもつSOCS1siRNAを発現する組換え型レンチウイルスベクター(LV−SOCS1−siRNA)及び対照ベクター(LV−GFP−siRNA)を生成した。マウス骨髄(BM)由来のDCをLV−SOCS1−siRNA又はLV−GFP−siRNAで形質導入し、組換え型HIVgp120タンパク質を負荷し、LPSによりエクスビボで成熟させた。マウスのグループを次に週間隔で2回、形質導入されたDCで免疫化し、続いて各DC免疫化の後にインビボでLPS刺激を行なった。本書の他の個所で記述されている通り、SOCS1サイレンシング化されたDCによって誘発された腫瘍関連抗原に対するCTL応答をさらに増強することができるという観察事実に基づき、インビボ刺激が使用された。図15Aは、LV−SOCS1−siRNA−DCが、対照のLV−GFP−siRNA−DCよりも著しく頑強なgp120特異的IgM及びIgG反応を惹起したということを示している。
Silencing of SOCS1 in DC enhances HIV Env-specific antibody response We first investigated the effect of SOCS1 silencing on the ability of DC to elicit anti-HIV antibodies. HIV Env was used in this study because it can elicit both cellular and neutralizing antibody responses. Recombinant lentiviral vector (LV-SOCS1-siRNA) expressing SOCS1 siRNA with the ability to down-regulate about 90% of SOCS1 mRNA in transfected cells as described elsewhere in this document And a control vector (LV-GFP-siRNA) was generated. Mouse bone marrow (BM) -derived DCs were transduced with LV-SOCS1-siRNA or LV-GFP-siRNA, loaded with recombinant HIV gp120 protein, and matured ex vivo with LPS. Groups of mice were then immunized twice weekly with transduced DCs, followed by LPS stimulation in vivo after each DC immunization. In vivo stimulation was used based on the observation that CTL responses to tumor-associated antigens elicited by SOCS1-silenced DCs could be further enhanced as described elsewhere in this document. FIG. 15A shows that LV-SOCS1-siRNA-DC elicited a gp120-specific IgM and IgG response that was significantly more robust than the control LV-GFP-siRNA-DC.
そのプロファイルが全く異なる免疫学的状態を表わしている異なる抗体サブクラスの産生は、CD4+T−ヘルパー(Th)1及びTh2−分極サイトカイン及びTh細胞の機能によって左右される(アーレン(Allen)ら、1997年、Immunol.Today第18号:387〜92頁)。図15Bは、LV−GFP−siRNA−DCマウス内の対応するサブクラスに比べた、LV−SOCS1−siRNA−DCで免疫化されたマウスにおける全てのIgGサブクラス内のHIVEnv特異的抗体力価の大幅な増加
を示す。Env特異的抗体サブクラスのプロファイルは、Th2応答の産物であるIgG1とTh1応答と結びつけられたサブクラスであるIgG2aの混合応答を標示し、これは、Th1−及びTh2−依存性免疫応答の両方がLV−SOCS1−siRNA−DCにより誘発されたということを表わす。反復実験においても、類似の結果が得られた。マウスはHIV中和抗体の信頼性の高い試験のために適した種でないことから、中和検定は実施しなかった((バートン(Burton)ら、2004年、Nat.Immunol.第5号:233〜6頁))。さらに、SOCS1サイレンシング化が、HIVEnvタンパク質のその他の株及びオボアルブミン(OVA)といった抗原に対する抗体反応を増強することが観察された。これらの結果は、全てのIgGサブクラスを包含するHIVEnv特異的抗体応答が、DC内のSOCS1のサイレンシング化によって大幅に増強されることを実証しており、このことは、抗原特異的抗体応答を制御する上でのDC内のSOCS1の決定的役割を暗示している。
The production of different antibody subclasses whose profiles represent completely different immunological states depends on the function of CD4 + T-helper (Th) 1 and Th2-polarized cytokines and Th cells (Allen et al., 1997 Immunol.Today 18: 387-92). FIG. 15B shows a significant increase in HIV Env-specific antibody titers in all IgG subclasses in mice immunized with LV-SOCS1-siRNA-DC compared to the corresponding subclass in LV-GFP-siRNA-DC mice. Shows an increase. The profile of the Env-specific antibody subclass shows a mixed response of the product of the Th2 response, IgG1 and the subclass IgG2a associated with the Th1 response, indicating that both Th1- and Th2-dependent immune responses are LV -Indicates that it was induced by SOCS1-siRNA-DC. Similar results were obtained in repeated experiments. Since mice are not suitable species for reliable testing of HIV neutralizing antibodies, neutralization assays were not performed ((Burton et al., 2004, Nat. Immunol. 5: 233). -6 pages)). Furthermore, SOCS1 silencing was observed to enhance antibody responses to other strains of HIV Env protein and antigens such as ovalbumin (OVA). These results demonstrate that the HIV Env-specific antibody response, encompassing all IgG subclasses, is greatly enhanced by silencing of SOCS1 in DCs, indicating that the antigen-specific antibody response is The critical role of SOCS1 in the DC in controlling is implied.
DC内のSOCS1のサイレンシング化はHIVgp120特異的CTL応答を増強する
以下の実験は、SOCS1サイレンシング化がHIVEnv特異的CTL応答を増強できるか否かを査定するために実施された。免疫化されたマウスの体内のCD8+T細胞の機能的状況をテストするために、IFNγELISPOT、細胞内サイトカイン染色及びCTL検定を使用した。LV−SOCS1−siRNA−DCマウス体内でのgp120パルスされた標的細胞に対するCTL活性は、LV−GFP−siRNA−DCマウスにおけるものと比べはるかに強力であった(P<0.01)(図15C)。これらの検定において検出されたCTL活性は、LV−SOCS1−siRNA−DCマウス由来の脾細胞に、gp120パルスを受けていない標的細胞に対する明白なあらゆるCTL活性が欠如していたことから、gp120特異的であった。これと一致して、LV−SOCS1−siRNA−DCで免疫化されたマウスにおいては、それぞれ、LV−GFP−siRNA−DCマウスにおける191スポットに比べ、5×105のCD8+T細胞あたり363のIFNγ+スポットが検出された(図15D)。IFN−γでの脾細胞の細胞内染色は同様に、LV−SOCS1−siRNA−DCマウス内のIFN−γ+T細胞のさらに高い百分率を示した。これらの結果を合わせて考慮すると、SOCS1サイレンシング化されたDCで免疫化されたマウスにおけるHIVEnvに対するバランスのとれた増強された抗体及びCTL応答が実証され、このことは、DC中のSOCS1が液性免疫及び細胞性免疫の両方を決定的に調節することを示唆している。
Silencing of SOCS1 in DC enhances HIV gp120-specific CTL response The following experiment was performed to assess whether SOCS1 silencing can enhance a HIV Env-specific CTL response. To test the functional status of CD8 + T cells in immunized mice, IFNγ ELISPOT, intracellular cytokine staining and CTL assay were used. CTL activity against gp120-pulsed target cells in LV-SOCS1-siRNA-DC mice was much more potent than in LV-GFP-siRNA-DC mice (P <0.01) (FIG. 15C). ). The CTL activity detected in these assays was gp120 specific because spleen cells from LV-SOCS1-siRNA-DC mice lacked any apparent CTL activity against target cells not receiving gp120 pulses. Met. Consistent with this, in mice immunized with LV-SOCS1-siRNA-DC, 363 IFNγ + spots per 5 × 10 5 CD8 + T cells, respectively, compared to 191 spots in LV-GFP-siRNA-DC mice. Was detected (FIG. 15D). Intracellular staining of splenocytes with IFN-γ also showed a higher percentage of IFN-γ + T cells in LV-SOCS1-siRNA-DC mice. Taken together, these results demonstrate a balanced and enhanced antibody and CTL response to HIVEnv in mice immunized with SOCS1-silenced DCs, indicating that SOCS1 in DCs is fluid It suggests critical regulation of both sexual and cellular immunity.
SOCS1サイレンシング化されたDCにより誘発された混合型の増強されたTh1及びTh2応答
何らかの特定の理論により束縛されることは望まないが、Th1対Th2応答をプログラミングする上でのサイトカインの役目からみて(マクドナルド(MacDonald)ら、2002年、J.Immunol.第168号:3127〜30頁;ゴール(Gor)ら、2003年、Nat.Immunol.第4号:503〜5頁)、SOCS1サイレンシング化が、DCによるサイトカインの産生を調節することによってCTL及び抗体応答に影響を及ぼす可能性があると考えられている。図16Aは、LPSでの刺激の後のGFP−siRNA−DCと比べた、LV−SOCS1−siRNA−DCにより産生されたTh1分極された応答を促進するIL−12、IFN−γ及びTNFαのレベルの著しい増大を実証している。興味深いことに、SOCS1サイレンシング化されたDC内でも、Th2分極された応答を促進するIL−4、IL−6及びIL−10の著しい応答が見られた(P<0.01)。SOCS1サイレンシング化されたDCにより産生されたTh1及びTh2促進サイトカインの両方のより高いレベルによって、HIV Env特異的CTL及び抗体応答の両方を誘発するSOCS1サイレンシング化されたDCの能力増強を説明することができる。
Mixed enhanced Th1 and Th2 responses elicited by SOCS1-silenced DCs While not wishing to be bound by any particular theory, in view of the role of cytokines in programming Th1 versus Th2 responses (MacDonald et al., 2002, J. Immunol. 168: 3127-30; Gor et al., 2003, Nat. Immunol. 4: 503-5), SOCS1 silencing Are thought to have the potential to affect CTL and antibody responses by modulating the production of cytokines by DCs. FIG. 16A shows IL-12, IFN-γ and TNFα levels that promote the Th1 polarized response produced by LV-SOCS1-siRNA-DC compared to GFP-siRNA-DC after stimulation with LPS. Has demonstrated a significant increase in Interestingly, significant responses of IL-4, IL-6, and IL-10 that promote a Th2-polarized response were also found in SOCS1-silenced DCs (P <0.01). Higher levels of both Th1 and Th2-promoting cytokines produced by SOCS1-silenced DCs explain the enhanced ability of SOCS1-silenced DCs to elicit both HIV Env-specific CTL and antibody responses be able to.
これらの結果に基づくと、DCの中のSOCS1サイレンシング化は、抗体及びCTL
の応答を明らかに促進した。以下の実験は、抗体及びCTL応答の誘発に密接に関与するHIVEnv特異的CD4+Th応答がSOCS1サイレンシング化によっても増強されるか否かに注目している。CD4+T細胞は、CD4+マイクロビーズを用いて、免疫化されたマウスから単離され、さまざまな検定を用いて分析された。図16Bで描かれているように、gp120特異的CD4+T細胞の頻度は、LV−GFP−siRNA−DCマウス内よりもLV−SOCS1−siRNA−DCマウス内で著しく高いものであった。3H−チミジン取込み検定は、LV−SOCS1−siRNA−DC由来のCD4+T細胞が、gp120パルスを受けたDCでの刺激に応答してLV−GFP−siRNA−DCマウスからのものに比べより活発に増殖することを示した(図16C)。gp120パルスを受けたDCでの刺激の後にLV−SOCS1−siRNA−DCから単離されたCD4+T細胞が生成したサイトカインプロファイルの分析は、Th1分極(IFN−γ、IL−2及びTNFα)及びTh2分極(IL−4及びIL−10)の両方のサイトカインの増大したレベルを明らかにした(図16D)。これらの結果は、SOCS1サイレンシング化されたDCが、HIVEnvに対する増強した混合型Th1及びTh2応答を誘発することを示しており、これは、図15B中の混合型gp120特異的IgGサブクラスプロファイルと一致している。
Based on these results, SOCS1 silencing in DCs is due to antibody and CTL
Clearly promoted the response. The following experiments focus on whether the HIVEnv-specific CD4 + Th response, which is closely involved in the induction of antibody and CTL responses, is also enhanced by SOCS1 silencing. CD4 + T cells were isolated from immunized mice using CD4 + microbeads and analyzed using various assays. As depicted in FIG. 16B, the frequency of gp120-specific CD4 + T cells was significantly higher in LV-SOCS1-siRNA-DC mice than in LV-GFP-siRNA-DC mice. The 3 H-thymidine incorporation assay shows that LV-SOCS1-siRNA-DC-derived CD4 + T cells are more active than those from LV-GFP-siRNA-DC mice in response to stimulation with DCs receiving gp120 pulses. It was shown to proliferate (FIG. 16C). Analysis of the cytokine profile produced by CD4 + T cells isolated from LV-SOCS1-siRNA-DC after stimulation with DCs receiving gp120 pulses was analyzed for Th1 polarization (IFN-γ, IL-2 and TNFα) and Th2 polarization. Increased levels of both (IL-4 and IL-10) cytokines were revealed (FIG. 16D). These results indicate that SOCS1-silenced DC induces an enhanced mixed Th1 and Th2 response to HIV Env, which is consistent with the mixed gp120-specific IgG subclass profile in FIG. 15B. I'm doing it.
SOCS1サイレンシング化されたDCによる増強されたgp120特異的B細胞の活性化
DCは、TNFスーパーファミリのメンバーであるAPRIL(増殖誘発リガンド)及びBAFF(BLySとしても知られているB−リンパ球刺激因子)を産生することによりB細胞増殖、成熟及びクラススイッチ組換えを直接誘発することが示されてきた(バラーシ(Balazs)ら、2002年、Immunity第17号:341〜52頁;リチンスキー(Litinskiy)ら、2002年、Nat.Immunol.第3号:822〜9頁;マクレナン(MacLennan)ら、2002年、Immunity第17号:235〜8頁)。従って実時間RT−PCRを用いたDCによるAPRIL及びBAFFの産生に対するSOCS1サイレンシング化の効果を査定した。LV−SOCS1−siRNA−DCは、SOCS1−/−DCにおけるBAFF及びAPRILの発現の増加と一致して、LV−GFP−siRNA−DCよりも高いレベルのAPRIL及びBAFFmRNAをLPS刺激時点で発現した(図17A)(ハナダら、2003年、Immunity第19号:437〜50頁)。
Enhanced gp120-specific B cell activation by SOCS1-silenced DCs DCs stimulate B-lymphocytes, also known as APRIL (proliferation-inducing ligand) and BAFF (also known as BLyS), members of the TNF superfamily Have been shown to directly induce B cell proliferation, maturation and class switch recombination (Balass et al., 2002, Immunity 17: 341-52; Litinsky). ) Et al., 2002, Nat. Immunol. 3: 822-9; MacLennan et al., 2002, Immunity 17: 235-8). Therefore, the effect of SOCS1 silencing on the production of APRIL and BAFF by DC using real-time RT-PCR was assessed. LV-SOCS1-siRNA-DC expressed higher levels of APRIL and BAFF mRNA at the time of LPS stimulation than LV-GFP-siRNA-DC, consistent with increased expression of BAFF and APRIL in SOCS1 − / − DC ( FIG. 17A) (Hanada et al., 2003, Immunity 19: 437-50).
gp120特異的B細胞の活性化を増強するべくSOCS1サイレンシング化されたDCの能力をテストするために、免疫化されたマウスの体内の抗gp120IgG産生B細胞の頻度を直接検討するべく、抗gp120IgG特異的B細胞Elispot検定を使用した。抗gp120IgG産生B細胞の頻度は、LV−GFP−siRNA−DCマウスよりもLV−SOCS1−siRNA−DCマウスにおいて著しく高いものであった(P<0.01)(図17B)。LV−GFP−siRNA−DCマウス由来のB細胞と比べてより高い百分率のB細胞が、LV−SOCS1−siRNA−DCマウスにおいて高レベルのCD69、CD40及びCD86で特徴づけされる活性化済み表現型を示した。さらに、免疫化されたマウスの脾臓に由来するB細胞を精製し、さまざまな刺激物で刺激させた。図17Cは、LV−SOCS1−siRNA−DCマウス由来のB細胞が、LV−GFP−siRNA−DCマウス由来のB細胞に比べ、抗CD40及びIL−4で同時刺激した際にさらに激しく増殖したことを示している。興味深いことに、LV−GFP−siRNA−DCマウス由来のものではなくLV−SOCS1−siRNA−DCマウス由来のB細胞がIL−4又は抗−CD40のみに対し強く応答しており、このことは、LV−SOCS1−siRNA−DCでの免疫化により増大した数のB細胞がインビボですでに活性化されたということを示唆している。同様に、LV−SOCS1−siRNA−DCマウスからのB細胞がさまざまな刺激に応答してIL−6、IL−2及びTNF−αを含めたさまざまなサイトカインをより高いレベルで産生することも観察された(図17
D)。集合的に、これらの結果は、SOCS1サイレンシング化されたDCが、増強したレベルのB−リンパ球刺激因子(BAFF及びAPRIL)及びTh2分極サイトカインを産生し、HIVEnv特異的B細胞及びTh細胞のより効果的な活性化を導くということを示唆している。
In order to directly test the frequency of anti-gp120 IgG producing B cells in the immunized mice to test the ability of SOCS1-silenced DCs to enhance the activation of gp120-specific B cells, anti-gp120 IgG A specific B cell Elispot assay was used. The frequency of anti-gp120 IgG producing B cells was significantly higher in LV-SOCS1-siRNA-DC mice than in LV-GFP-siRNA-DC mice (P <0.01) (FIG. 17B). An activated phenotype characterized by a higher percentage of B cells compared to B cells from LV-GFP-siRNA-DC mice is characterized by high levels of CD69, CD40 and CD86 in LV-SOCS1-siRNA-DC mice showed that. In addition, B cells derived from the spleens of immunized mice were purified and stimulated with various stimuli. FIG. 17C shows that B cells derived from LV-SOCS1-siRNA-DC mice proliferated more vigorously when co-stimulated with anti-CD40 and IL-4 than B cells derived from LV-GFP-siRNA-DC mice. Is shown. Interestingly, B cells derived from LV-SOCS1-siRNA-DC mice but not from LV-GFP-siRNA-DC mice responded strongly to IL-4 or anti-CD40 only, This suggests that an increased number of B cells were already activated in vivo by immunization with LV-SOCS1-siRNA-DC. Similarly, it has been observed that B cells from LV-SOCS1-siRNA-DC mice produce higher levels of various cytokines including IL-6, IL-2 and TNF-α in response to various stimuli. (Fig. 17
D). Collectively, these results indicate that SOCS1-silenced DCs produce enhanced levels of B-lymphocyte stimulating factors (BAFF and APRIL) and Th2 polarized cytokines, and that of HIV Env-specific B cells and Th cells. It suggests that it leads to more effective activation.
SOCS1サイレンシング化されたDCにより誘発された長期HIVEnv特異的CTL及び抗体応答
DC内でのSOCS1サイレンシング化は一次的HIVEnv特異的CTL及び抗体応答を増強するということを示してきたが、SOCS1サイレンシング化されたDCがメモリーHIV特異的CTL及び抗体応答を誘発することになるか否かということもさらに査定した。図18Aは、LV−GFP−siRNA−DCで免疫化されたマウスが免疫化から6カ月後にきわめて低レベルのgp120特異的抗体を有し、一方LV−SOCS1−siRNA−DCマウスはなおもその血清中にgp120特異的IgG1及びIgG2抗体を有意な力価で保持していたということを示している。追加免疫化から1週間目に、LV−SOCS1−siRNA−DCマウスは強いリコール抗体応答を示し、抗gp120IgG1の平均力価は2×105及び抗gp120IgG2の平均力価は1×105であり、一方、LV−GFP−siRNA−DCマウスは低いリコール抗体応答を示し、IgG1の平均力価は3×103、IgG2は4×102であった。これらのデータは、SOCS1サイレンシング化されたDCがGFP−siRNA−DCに比べてそれぞれIgG1及びIgG2aの抗体の力価の約64倍及び255倍の増加を示すことを表わしている。
Long-term HIV Env-specific CTL and antibody responses elicited by SOCS1-silenced DCs Although SOCS1 silencing within DCs has been shown to enhance primary HIV Env-specific CTL and antibody responses, SOCS1 silencing It was further assessed whether the singed DC would elicit a memory HIV specific CTL and antibody response. FIG. 18A shows that mice immunized with LV-GFP-siRNA-DC have very low levels of gp120-
メモリーHIV特異的CTL及びThの維持を、IFNγ−ELISPOT検定でgp120特異的CD8+及びCD4+T細胞応答を検査することによって査定した。図18Bは、免疫化から6カ月目に、LV−GFP−siRNA−DCマウスではなくLV−SOCS1−siRNA−DCマウスの体内で強いgp120特異的CTL応答が検出されたということを示している((LV−SOCS1−siRNA−DCマウスの体内でCD8+T細胞5×105個あたりIFNγスポット249個対LV−GFP−siRNA−DCマウスの体内でIFNγスポット3個))。LV−SOCS1−siRNA−DCマウスにおける勢いのよいgp120特異的CTL応答が追加免疫化によって急速に誘発されたが、LV−GFP−siRNA−DCマウス内では誘発されなかった((追加免疫後7日目にLV−SOCS1−siRNA−DCマウスの体内でCD8+T細胞5×105個あたりIFNγスポット446個対LV−GFP−siRNA−DCマウスの体内でIFNγスポット16個)(図18B)。CD8+T細胞の表面CD44メモリーマーカー及び細胞内IFN−γの同時染色も又、免疫化から6カ月後にLV−GFP−siRNA−DCマウスと比べて、LV−SOCS1−siRNA−DCマウスの体内でより高い百分率のCD44hi及びIFNg+CD8+T細胞を示した(図18C)。同様にして、免疫化から6カ月目に、LV−SOCS1−siRNA−DCマウス内でgp120特異的CD4+Th応答が維持され急速に誘発された(追加免疫後7日目にLV−SOCS1−siRNA−DCマウスの体内でCD4+T細胞5×105個あたりIFNγスポット391個対LV−GFP−siRNA−DCマウスの体内でIFNγスポット37個)(図18D)。かくして、SOCS1サイレンシング化されたDCでの免疫化は、長期HIV Env特異的CTL、Th及び抗体応答を有効に誘発する。
Maintenance of memory HIV-specific CTL and Th was assessed by examining gp120-specific CD8 + and CD4 + T cell responses with an IFNγ-ELISPOT assay. FIG. 18B shows that a strong gp120-specific CTL response was detected in the body of LV-SOCS1-siRNA-DC mice but not LV-GFP-siRNA-
免疫化から7カ月目までは、gp120でパルスされたLV−SOCS1−siRNA−DCで免疫化されたマウスの体内には明白な毒性は全く観察されなかった。免疫化されたマウスの全ての主要器官及び組織の組織学的分析が、病的炎症を全く、明らかにしなかった。IgG及び抗dsDNAのレベルは、DC−LV−SOCS1−siRNA及びモックDCマウス内で匹敵するものであった。これらのデータは、gp120パルスを受けたLV−SOCS1−siRNA−DCの免疫化がマウスの体内で病的炎症をひき起こさ
ないということを示唆している。
From the 7th month after immunization, no obvious toxicity was observed in the body of mice immunized with LV-SOCS1-siRNA-DC pulsed with gp120. Histological analysis of all major organs and tissues of immunized mice revealed no pathological inflammation. IgG and anti-dsDNA levels were comparable in DC-LV-SOCS1-siRNA and mock DC mice. These data suggest that immunization of LV-SOCS1-siRNA-DC that received gp120 pulses does not cause pathological inflammation in the mouse body.
HIVEnv媒介型免疫抑制に対するSOCS1サイレンシング化されたDCの耐性
gp120タンパク質を含むHIVウイルスは、前炎症性サイトカインを産生しT細胞を刺激するDCの能力を抑制することができる(ファントゥッジ(Fantuzzi)ら、2004年、J.Virol.第78号:9763〜72頁;グラネッリ・ピペルノ(Granelli−Piperno)ら、2004年、Proc.Natl.Acad.Sci.USA.第101号:7669〜74頁;バロン(Barron)ら、2003年、J.Infect.Dis.第187号:26〜37頁;パカノウスキー(Pacanowski)ら、2001年、Blood第98号:3016〜21頁)。SOCS1サイレンシング化によるDCの増強された活性化がDCのサイトカイン産生及び免疫刺激能力に対するgp120タンパク質の阻害効果を克服できるか否かを査定するために、以下の実験に着手した。DC由来のIL−12は、Th1の発達を駆動することならびに液性応答を発生させるためにB細胞を直接シグナリングすることという2重の役割を果たすことが発見されたことから、これらの実験のための代表的サイトカインとしてIL−12が選択された(デュボア(Dubois)ら、1998年、J.Immunol.第161号:2223〜31頁;デュボアら、1997年、J.Exp.Med.第185号:941〜51頁;スコック(Skok)ら、1999年、J.Immunol.第163号:4284〜91頁)。
Resistance of SOCS1-silenced DCs to HIV Env-mediated immunosuppression HIV viruses containing the gp120 protein can suppress the ability of DCs to produce pro-inflammatory cytokines and stimulate T cells (Fantuzzi et al. 2004, J. Virol. 78: 9763-72; Granelli-Pipeno et al., 2004, Proc.Natl.Acad.Sci.USA.101: 7669-74; (Barron et al., 2003, J. Infect. Dis. 187: 26-37; Pacanoski et al., 2001, Blood 98: 3016-21). In order to assess whether enhanced activation of DC by SOCS1 silencing could overcome the inhibitory effect of gp120 protein on DC cytokine production and immunostimulatory capacity, the following experiment was undertaken. DC-derived IL-12 was found to play a dual role in driving Th1 development and signaling B cells directly to generate a humoral response. IL-12 was selected as a representative cytokine for (Dubois et al., 1998, J. Immunol. 161: 2223-31; Duboa et al., 1997, J. Exp. Med. 185. No .: 941-51; Skok et al., 1999, J. Immunol. 163: 4284-91).
図19Aに示されているように、gp120タンパク質の存在下でのLV−SOCS1−siRNA−DCは、LPSに応答する能力を保持していた。これとは対照的に、LPS刺激に対するLV−GFP−siRNA−DCの応答は、gp120タンパク質の存在によりひどく低下した。gp120媒介型抑制に対するSOCS1サイレンシング化されたDCの感受性をさらにインビボで調査した。gp120タンパク質のエクスビボでの予備処置を伴って又は伴わずに、OVAでのパルスを受けた形質導入されたDCでマウスを免疫化した。gp120タンパク質に対する予備曝露は、OVA特異的抗体応答を誘発するLV−SOCS1−siRNA−DCの能力に対する明白な効果をもたず(図19B及び19C)LV−SOCS1−siRNA−DCにより誘発されるOVA特異的CD8+CTL及びCD4+Th応答を低下させることもなかった(P>0.05)(図19D及び19E)。しかしながら、このような予備処置は、OVA特異的抗体及びCTL応答を誘発するLV−GFP−siRNA−DCの能力を著しく低減させた(P<0.05)(図19B〜19E)。これらの結果は、SOCS1サイレンシング化が、おそらくは増強されたサイトカイン産生及びSOCS1サイレンシング化されたDCの過剰活性化状態に起因して、DCにHIVgp120媒介型抑制に対する耐性を付与する、ということを表わしている(ハナダら、2003年、Immunity、第19号:437〜50頁)。 As shown in FIG. 19A, LV-SOCS1-siRNA-DC in the presence of gp120 protein retained the ability to respond to LPS. In contrast, the response of LV-GFP-siRNA-DC to LPS stimulation was severely reduced by the presence of gp120 protein. The sensitivity of SOCS1-silenced DCs to gp120-mediated suppression was further investigated in vivo. Mice were immunized with transduced DCs pulsed with OVA with or without ex vivo pretreatment of gp120 protein. Pre-exposure to gp120 protein has no obvious effect on the ability of LV-SOCS1-siRNA-DC to elicit an OVA-specific antibody response (FIGS. 19B and 19C) OVA induced by LV-SOCS1-siRNA-DC It did not reduce specific CD8 + CTL and CD4 + Th responses (P> 0.05) (FIGS. 19D and 19E). However, such pretreatment significantly reduced the ability of LV-GFP-siRNA-DC to elicit OVA specific antibodies and CTL responses (P <0.05) (FIGS. 19B-19E). These results indicate that SOCS1 silencing confers resistance to HIV gp120-mediated suppression, presumably due to enhanced cytokine production and the over-activated state of SOCS1-silenced DCs. (Hanada et al., 2003, Immunity, 19: 437-50).
実施例8:HIV特異的抗体及びCTL応答を増強するためのインビボDNAワクチン接種
本実施例は、SOCS1siRNA発現DNAでの同時免疫化によりHIVDNAワクチン接種の効力が著しく増強されるということを実証している。この研究は、宿主の免疫インヒビターを阻害することによりHIV−特異的抗体及びCTL応答を惹起する最初の試みを表わしており、これは、より有効なHIVワクチンを開発するための新たな途を提示している。
Example 8: In vivo DNA vaccination to enhance HIV-specific antibodies and CTL responses This example demonstrates that co-immunization with SOCS1 siRNA expressing DNA significantly enhances the efficacy of HIV DNA vaccination. Yes. This study represents the first attempt to elicit HIV-specific antibodies and CTL responses by inhibiting host immune inhibitors, which presents a new path to developing more effective HIV vaccines. is doing.
本実施例の中で提示されている実験において使用される材料及び方法についてここで記述する。 The materials and methods used in the experiments presented in this example are now described.
DNAワクチン接種
先に本書の他の個所で記述されている通りに、pSuper−SOCS1−siRNA
発現ベクターを生成した。HIVEnvレトロゲン発現ベクターを生成するためには、HIVEnvの分泌を促進するべくHIVgp160のgp120/gp41切断部位及びgp41の融合ドメイン(コドン使用頻度最適化されたJRFL)を欠失させることにより、gp140CFプラスミドをまず構築した。結果として得たpCMV/R−gp140CF−Fcレトロゲンベクタは、CMVプロモータの制御下でIgGFcフラグメントに融合されたgp140CF遺伝子を含有している。CHO細胞から組換え型gp120(JFRL)タンパク質が産生され、これは、NIHAIDS研究参照プログラムによって提供された。キアゲン(Qiagen)キットで内毒素を含まないDNAを調製し、1μg/μlの最終濃度で、内毒素を含まないPBS(シグマ、ミズーリ州セントルイス;アルドリッチ社、ミズーリ州セントルイス)の中に再懸濁させ、注入のために使用するまで−20℃で保管した。ワクチン接種予定日に、50μgのgp140CF−FcDNA又は、gp140CF−FcDNA(50μg)とpSuper−SOCS1−siRNAエクスプレッサDNA(150μg)の混合物200μgを各マウスの大腿四頭筋内に筋内注射した(ハウザー(Hauser)ら、2004年、Gene Ther.第11号:924〜32頁;ユーら、2001年、Cancer Research第61号:3704〜11頁)。免疫化したマウスをその後、各々のDNA免疫化の後1、3及び5日目に3回LPS(30μg/マウス)(IP)で処置した。
DNA vaccination As previously described elsewhere in this document, pSuper-SOCS1-siRNA
An expression vector was generated. To generate a HIV Env retrogen expression vector, the gp140CF plasmid is deleted by deleting the gp120 / gp41 cleavage site of HIV gp160 and the fusion domain of gp41 (JRFL with codon usage optimization) to promote secretion of HIV Env. First built. The resulting pCMV / R-gp140CF-Fc retrogen vector contains the gp140CF gene fused to an IgG Fc fragment under the control of a CMV promoter. Recombinant gp120 (JFRL) protein was produced from CHO cells and was provided by the NIHAIDS research reference program. Endotoxin-free DNA is prepared with the Qiagen kit and resuspended in endotoxin-free PBS (Sigma, St. Louis, MO; Aldrich, St. Louis, MO) at a final concentration of 1 μg / μl. And stored at −20 ° C. until used for injection. On the scheduled day of vaccination, 50 μg of gp140CF-FcDNA or a mixture of gp140CF-FcDNA (50 μg) and pSuper-SOCS1-siRNA expressor DNA (150 μg) was injected intramuscularly into the quadriceps of each mouse (Houser (Hauser et al., 2004, Gene Ther. No. 11: 924-32; You et al., 2001, Cancer Research 61: 3704-11). Immunized mice were then treated 3 times with LPS (30 μg / mouse) (IP) 1, 3, and 5 days after each DNA immunization.
本実施例中に提示された実験の結果についてここで記述する。 The experimental results presented in this example will now be described.
SOCS1 siRNA DNAでの同時免疫化により増強されたHIVDNAワクチンの効能
HIVEnv特異的CTL及び抗体の両方の応答を増強するSOCS1サイレンシング化されたDCの能力は、HIVDNAワクチン接種の効能を改善する上でこのSOCS1サイレンシング化アプローチが有用であり得るということを示唆していた。抗原のDCターゲティング及びMHC提示を増強するためにレセプタ媒介型エンドサイトーシスを用いる「レトロゲン」免疫化戦略を、ハウザー(Hauser)ら、2004年、Gene Ther.第11号:924〜32頁及びユーら、2001年、Cancer Research第61号:3704〜11頁に従って使用した。簡単に言うと、gp120/gp41分割部位及びgp41の融合ドメインが欠失されているgp140CF遺伝子に対してIgGFcフラグメントを枠内融合させることによってgp140CFレトロゲンを産生させた。結果として得られたgp140CF−Fc融合タンパク質を発現させ、gp140CF−Fcベクターでのトランスフェクションを受けた細胞から分泌させた。
Efficacy of HIV DNA vaccines enhanced by co-immunization with SOCS1 siRNA DNA The ability of SOCS1-silenced DCs to enhance both HIV Env-specific CTL and antibody responses may improve the efficacy of HIV DNA vaccination. It has been suggested that this SOCS1 silencing approach may be useful. A “retrogen” immunization strategy that uses receptor-mediated endocytosis to enhance DC targeting and MHC presentation of antigens has been described by Hauser et al., 2004, Gene Ther. 11: 924-32 and You et al., 2001, Cancer Research 61: 3704-11. Briefly, gp140CF retrogen was produced by in-frame fusion of an IgG Fc fragment to the gp140CF gene lacking the gp120 / gp41 split site and the gp41 fusion domain. The resulting gp140CF-Fc fusion protein was expressed and secreted from cells that had been transfected with the gp140CF-Fc vector.
DNAワクチン接種の時点でのSOCS1 siRNAの効果をテストするために、gp140CF−FcDNAのみで又はgp140CF−FcDNAとp Super−SOCS1−siRNA発現DNAの混合物を一週間に一回3週間にわたりマウスに注射し、次に一週間後のHIVEnv特異的免疫応答についてマウスを監視した。HIVEnv特異的抗体力価の増強は、pSuper−SOCS1−siRNA DNAで同時免疫化されたマウスにおいて明白であった(図20A)。CTL及びELISPOT検定(図20B及び20C)により実証されているように、pSuper−SOCS1−siRNA
DNAの同時注射により、HIVEnv特異的CTL応答を著しく増強させた。その上、SOCS1−siRNADNAの同時注入により、HIVEnc特異的CD4+Th応答を増強させた(図20D)。サイトカインでの細胞内染色も同様に、pSuper−SOCS1 siRNA DNAで同時免疫化されたマウス内のgp120特異的CD4+T細胞応答の増強を示した。これらの結果は、免疫化されたマウスの体内での同時トランスフェクションを受けた抗原提示細胞の免疫刺激能力の増強に起因して、pSuper−SOCS1 siRNA DNAの同時免疫化がHIVDNAワクチン接種の効能を増強することを表わしている。かくして、本書で提示されているSOCS1サイレンシング化戦略は、エクスビボDCベースの及びインビボワクチン接種の設定に応用可能である。
To test the effect of SOCS1 siRNA at the time of DNA vaccination, mice were injected with gp140CF-FcDNA alone or a mixture of gp140CF-FcDNA and pSuper-SOCS1-siRNA expressing DNA once a week for 3 weeks. The mice were then monitored for HIV Env-specific immune responses one week later. Enhancement of HIV Env-specific antibody titers was evident in mice co-immunized with pSuper-SOCS1-siRNA DNA (FIG. 20A). PSuper-SOCS1-siRNA as demonstrated by CTL and ELISPOT assays (FIGS. 20B and 20C)
Simultaneous injection of DNA significantly enhanced the HIV Env-specific CTL response. Moreover, co-injection of SOCS1-siRNA DNA enhanced the HIVEn specific CD4 + Th response (FIG. 20D). Intracellular staining with cytokines also showed enhanced gp120-specific CD4 + T cell responses in mice co-immunized with pSuper-SOCS1 siRNA DNA. These results indicate that co-immunization of pSuper-SOCS1 siRNA DNA demonstrates the efficacy of HIV DNA vaccination due to the enhanced immunostimulatory capacity of antigen-presenting cells that have been co-transfected in the immunized mouse. It means to strengthen. Thus, the SOCS1 silencing strategy presented herein is applicable to ex vivo DC-based and in vivo vaccination settings.
DC内の天然免疫インヒビターの阻害に基づく代替的かつ有効なHIVワクチン接種アプローチ
本開示は、DC中の負のシグナリング調節因子SOCS1のサイレンシング化が、マウスの体内のHIV Env−特異的抗体及びCTL応答の両方の大幅な増強を結果としてもたらすということを実証している。SOCS1サイレンシング化されたDCにより誘発されたHIV Env−特異的抗体及びCTL応答の両方が長く持続することが観察された。さらに、結果は、SOCS1 siRNA DNAでの同時免疫化が、HIVDNAワクチン接種の効能を著しく増強するということを実証した。かくして、HIVに対するバランスのとれたメモリー液性及び細胞応答がSOCS1サイレンシング化されたDCで誘発され得、このSOCS1サイレンシング化戦略は、治療的及び予防的HIVワクチン接種という両方の設定に応用可能である。
Alternative and Effective HIV Vaccination Approach Based on Inhibition of Natural Immune Inhibitors in DC The present disclosure shows that silencing of the negative signaling regulator SOCS1 in DCs causes HIV Env-specific antibodies and CTLs in the mouse body It demonstrates that it results in a significant enhancement of both responses. It was observed that both HIV Env-specific antibodies and CTL responses elicited by SOCS1-silenced DCs last long. Furthermore, the results demonstrated that co-immunization with SOCS1 siRNA DNA significantly enhances the efficacy of HIV DNA vaccination. Thus, a balanced memory humoral and cellular response to HIV can be induced in SOCS1-silenced DCs, and this SOCS1 silencing strategy is applicable to both therapeutic and prophylactic HIV vaccination settings It is.
液性応答の誘発におけるDCの役割は伝統的に、T細胞とB細胞の間の同族相互作用のためのCD4+Thプライミングの当然の結果と考えられてきた。しかしながら、液性応答の刺激におけるDCの直接的役割は、インビトロ及びインビボで実証されてきた(デュボアら、1997年、J.Exp.Med.第185号:941〜5138頁;イナバ(Inaba)ら、1983年、Proc.Natl.Acad.Sci.USA第80号:6041〜5頁)。特に、DCは、CD40で活性化されたB細胞の増殖及び抗体産生の両方を強く増強することが発見された(デュボアら、1997年、J.Exp.Med.第185号:941〜51頁)。抗原で負荷されたDCでの免疫化は、防御的液性応答を誘発することができる(フラマンド(Flamand)ら、1994、Eur.J.Immunol.第24号:605〜10頁))。本書の結果は、SOCS1サイレンシング化されたDCが、SOCS1サイレンシング化されたDCで免疫化されたマウスの中で見られる増強されたTh及びB細胞活性化の原因である確率の高い、Th2分極サイトカインならびにB−リンパ球刺激性サイトカイン(BAFF及びAPRIL)の産生を増強する、ということを実証している。この知見は、SOCS1−/−DCがB細胞の異常な拡張及び自己反応性抗体産生を誘発するという以前の報告((ハナダら、2003年、Immunity第19号:437〜50頁))により裏づけされている。従って、本研究は、HIV特異的抗体応答を制御する上でのDC中のSOCS1の決定的役割を実証しており、SOCS1のサイレンシング化を包括的に用いてHIVEnv以外の抗原に対する抗体応答を高めることができるということを暗示している。 The role of DCs in inducing humoral responses has traditionally been considered a natural consequence of CD4 + Th priming for cognate interactions between T and B cells. However, the direct role of DCs in stimulating humoral responses has been demonstrated in vitro and in vivo (Duboa et al., 1997, J. Exp. Med. 185: 941-5138; Inaba et al. 1983, Proc. Natl. Acad. Sci. USA 80: 6041-5). In particular, DC was found to strongly enhance both proliferation and antibody production of CD40 activated B cells (Duboa et al., 1997, J. Exp. Med. 185: 941-51). ). Immunization with DCs loaded with antigen can elicit a protective humoral response (Flamand et al., 1994, Eur. J. Immunol. 24: 605-10)). The results in this document indicate that ThCS1-silenced DC is likely to be responsible for the enhanced Th and B cell activation found in mice immunized with SOCS1-silenced DC. It has been demonstrated to enhance the production of polarized cytokines and B-lymphocyte stimulating cytokines (BAFF and APRIL). This finding is supported by a previous report that (SOCS1 − / − DC induces abnormal expansion of B cells and autoreactive antibody production (Hanada et al., 2003, Immunity 19: 437-50)). Has been. Thus, this study demonstrates the critical role of SOCS1 in DCs in controlling HIV-specific antibody responses, and comprehensively uses SOCS1 silencing to develop antibody responses against antigens other than HIV Env. It implies that it can be increased.
本研究の重要な知見は、SOCS1サイレンシング化されたDCが、HIV感染を予防又は制御するために望ましい可能性のあるバランスのとれたメモリーHIV−Env−特異的抗体及びCTL応答で誘発するということにある((バートン(Burton)ら、2004年、Nat.Immunol.第5号:233〜6頁;マクマイケル(McMichael)ら、2003年、Nat.Med.第9号:874〜80頁;Nabel、2001年、Nature第410号:1002〜7頁;レトビン(Letvin)ら、2002年、Annu.Rev.Immunol.第20号:73〜99頁;Zolla−Pazner、2004年、NatRev.Immunol.第4号:199〜210頁;イマイ(Imami)ら、2002年、J.Virol.第76号:9011〜23頁;レトビンら、2003年、Nat.Med.第9号:861〜6頁))。何らかの特定の理論により束縛されることは望まないが、SOCS1サイレンシング化がバランスのとれたメモリー液性及び細胞性応答を誘発する機序には、SOCS1サイレンシング化されたDC及びgp120特異的CD4+T細胞によるTh1及びTh2分極サイトカインの混合パターンの産生が関与し得る。これらの結果は、ウイルスなどの数多くの病原体に対して天然に生成される混合型抗体及びCTL応答と一貫性をもつものであり(アーレン(Allen)ら、1997年、Immunol.Today第18号:387〜92頁)、Th1及びTh2分極が互いに排他的でないことを表わしている(ゴール(Gor)
ら、2003年、Nat.Immunol.第4号:503〜5頁;コロナ(Colonna)、2001年、Nat.Immunol.第2号:899〜900頁))。
An important finding of this study is that SOCS1-silenced DCs elicit with a balanced memory HIV-Env-specific antibody and CTL response that may be desirable to prevent or control HIV infection (Burton et al., 2004, Nat. Immunol. 5: 233-6; McMichael et al., 2003, Nat. Med. 9: 874-80; Nabel, 2001, Nature 410: 1002-7; Letvin et al., 2002, Annu. Rev. Immunol. 20: 73-99; Zolla-Pazner, 2004, Nat Rev. Immunol. No. 4: pp. 199-210; Imai et al. 002 years, J.Virol No. 76:.. 9011-23, pp; Retobin et al., 2003, Nat.Med No. 9: 861-6 pages)). While not wishing to be bound by any particular theory, the mechanisms by which SOCS1 silencing induces a balanced memory humoral and cellular response include SOCS1-silenced DC and gp120-specific CD4 + T Production of a mixed pattern of Th1 and Th2 polarized cytokines by the cell may be involved. These results are consistent with naturally occurring mixed antibodies and CTL responses against a number of pathogens such as viruses (Allen et al., 1997, Immunol. Today 18: Pp. 387-92), indicating that Th1 and Th2 polarizations are not mutually exclusive (Gor)
Et al., Nat. Immunol. 4: 503-5; Colonna, 2001, Nat. Immunol. No. 2: pages 899-900)).
SOCS1は、さまざまなサイトカインにより用いられるJAK/STATシグナリング経路のフィードバックインヒビターとして機能し、直接的又は間接的にTLRシグナリング経路を調節することに関与している(ベッツ(Baetz)ら、2004年、J.Biol.Chem.第279号:54708〜15頁;ギングラら、2004年、J.Biol.Chem.第279号:54702〜7頁)。本書中の結果は、LPSに応答した、SOCS1サイレンシング化されたDCによるTNF−α、IL−6及びIL−12などのさまざまなサイトカインの産生の増強を、一貫して標示している。LPS−TLRシグナリングは、自己分泌及び傍分泌に機能することのできる数多くの炎症性サイトカインを含めた豊富なNF−κβ応答性遺伝子を活性化する(ベッツら、2004年、J.Biol.Chem.第279号:54708〜15頁;グローマン(Grohmann)ら、1998年、Immunity第9号:315〜23頁;パン(Pan)ら、2004年、Immunol.Lett.第94号:141〜51頁)。SOCS1がTLRSGLを間接的に減衰させるのに関与しているという事実を考慮すると、JAK/STAT経路の臨界的抑制を無効化にすることでサイトカインが自己分泌又は/及び傍分泌刺激ループを確立できるようになるはずであり、こうしてサイトカイン産生の削減ではなく増強が導かれることになる。サイトカイン及びサイトカインレセプタノックアウトマウスの使用が関与する本書に開示されている結果は、自己分泌サイトカイン刺激ループがSOCS1サイレンシング化されたDCによるサイトカインの過剰産生に貢献することを示唆している。 SOCS1 functions as a feedback inhibitor of the JAK / STAT signaling pathway used by various cytokines and is involved in directly or indirectly regulating the TLR signaling pathway (Baetz et al., 2004, J Biol. Chem. 279: 54708-15; Gingla et al., 2004, J. Biol. Chem. 279: 54702-7). The results in this document consistently indicate enhanced production of various cytokines such as TNF-α, IL-6 and IL-12 by SOCS1-silenced DCs in response to LPS. LPS-TLR signaling activates abundant NF-κβ responsive genes, including numerous inflammatory cytokines that can function in autocrine and paracrine (Bets et al., 2004, J. Biol. Chem. 279: 54708-15; Grohmann et al., 1998, Immunity 9: 315-23; Pan et al., 2004, Immunol. Lett. 94: 141-51) . Given the fact that SOCS1 is involved in indirectly attenuated TLRSGL, cytokines can establish autocrine or / and paracrine stimulation loops by overriding critical suppression of the JAK / STAT pathway This should lead to an enhancement rather than a reduction in cytokine production. The results disclosed herein involving the use of cytokines and cytokine receptor knockout mice suggest that the autocrine cytokine stimulation loop contributes to cytokine overproduction by SOCS1-silenced DCs.
HIV感染した個体においては、DCの機能的欠陥及び枯渇が一般的であり、漸進的な免疫不全に寄与する確率が高い。HIVgp120タンパク質は、前炎症性サイトカインを産生しT細胞を刺激するDCの能力を抑制することができる(ファントゥッジら、2004年、J.Virol.第78号:9763〜72頁;カルボネイル(Carbonneil)ら、2004年、J.Immunol.第172号:7832〜40頁)。SOCS1サイレンシング化されたDCは、前炎症性サイトカインの産生の増強及びSOCS1サイレンシング化されたDCの過剰活性化状態のため、HIVgp120媒介型抑制に耐える、ということが実証された((ハナダら、2003年、Immunity第19号:437〜50頁))。この知見は、免疫抑制されたHIV感染個体の中で使用されると思われる治療用HIVワクチンの開発と特に関連性が高い(リュー(Lu)ら、2004年、Nat.Med.第10号:1359〜1365頁)。 In HIV-infected individuals, DC functional defects and depletion are common and have a high probability of contributing to progressive immune deficiency. The HIV gp120 protein can suppress the ability of DC to produce pro-inflammatory cytokines and stimulate T cells (Fantuge et al., 2004, J. Virol. 78: 9763-72; Carbonneil). Et al., 2004, J. Immunol., 172: 7832-40). It has been demonstrated that SOCS1-silenced DCs are resistant to HIV gp120-mediated suppression because of the enhanced production of pro-inflammatory cytokines and the over-activated state of SOCS1-silenced DCs ((Hanada et al. 2003, Immunity 19: 437-50)). This finding is particularly relevant to the development of therapeutic HIV vaccines that would be used in immunosuppressed HIV-infected individuals (Lu et al., 2004, Nat. Med. No. 10: 1359-1365).
ここで記述されているワクチン接種戦略は、DC内で宿主の免疫インヒビターを阻害することにより抗HIV免疫応答を増強するための最初の努力である。自然免疫はHIV−1感染を制御するのに有効でないことから、宿主の免疫インヒビターを無効化することは、有効な抗HIV免疫応答を生成するのに決定的であり得る。しかしながら、HIV特異的免疫応答の増強だけでは防御的HIV抗体及びCTL応答の誘発を導き得ない。この点において、当該戦略は、DNAワクチンとSOCS1siRNADNAの同時免疫により実証されるように、現在利用可能なワクチンとの組合せ免疫化の機会を提供する。改良されたHIV免疫原と送達系((バートンら、2004年、Nat.Immunol.第5号:233〜6頁;ヤン(Yang)ら、2002年、J.Virol.第76号:4634〜42頁))と共に使用された場合、このワクチン接種アプローチは、弱防御免疫応答を増強するか又は優性エピトープに対してのみならず弱免疫原性又は潜在性でしかも防御的なエピトープに対してもより広くより強い応答を生成する新たな途を提供し得る。要約すると、本開示は、HIV特異的抗体及びCTL応答の両方を増強するべくDC内の宿主のシグナリングインヒビターを阻害するという原則を実証しサルそして究極的にはヒトにおいてこの戦略により防御的抗HIV応答を誘発できるか否かを判定するためにさらな
る調査を要請している。さらにこのSOCS1サイレンシング化戦略は、その他の病原体に対する免疫応答を増強するために使用することができると思われる。
The vaccination strategy described here is the first effort to enhance the anti-HIV immune response by inhibiting host immune inhibitors in DCs. Since innate immunity is not effective in controlling HIV-1 infection, disabling host immune inhibitors can be crucial in generating an effective anti-HIV immune response. However, enhancing HIV-specific immune responses alone cannot lead to the induction of protective HIV antibodies and CTL responses. In this regard, the strategy provides an opportunity for combined immunization with currently available vaccines, as demonstrated by co-immunization of DNA vaccines and SOCS1 siRNA DNA. Improved HIV immunogens and delivery systems ((Burton et al., 2004, Nat. Immunol. 5: 233-6; Yang et al., 2002, J. Virol. 76: 4634-42 This vaccination approach, when used in conjunction with))) enhances the weak protective immune response or not only against a dominant epitope but also against a weak immunogenic or latent and protective epitope It can provide a new way to generate broader and stronger responses. In summary, this disclosure demonstrates the principle of inhibiting host signaling inhibitors in DCs to enhance both HIV-specific antibodies and CTL responses, and this strategy protects anti-HIV in monkeys and ultimately humans. Requests further investigation to determine if a response can be triggered. Furthermore, this SOCS1 silencing strategy could be used to enhance the immune response against other pathogens.
実施例9:ヒトSOCS1siRNAの同定及び分析
ヒトSOCS1:hSOCS1−siRNA1(CACGCACUUCCGCACAUUC.dT.dT;配列番号21)、hSOCS1−siRNA2(UUCCGUUCGCACGCCGAUU.dT.dT;配列番号22)及びhSOCS1−siRNA3(GAGCUUCGACUGCCUCUUC.dT.dT;配列番号23)をターゲティングするsiRNA配列を選択するためにはコンピュータプログラムを使用した。全ての標的配列は、その他の既知の遺伝子に対する相同性の欠如を確認するためにNCBIBlastクエリに付した。「.dT.dT」という呼称は、siRNA標的配列のすぐ下流側のポリdT配列を意味する。
Example 9: Identification and analysis of human SOCS1 siRNA Human SOCS1: hSOCS1-siRNA1 (CACGCACUUCCCCACAUC.UC). Computer program was used to select siRNA sequences targeting .dT; SEQ ID NO: 23). All target sequences were subjected to NCBIBlast queries to confirm the lack of homology to other known genes. The designation “.dT.dT” means a poly dT sequence immediately downstream of the siRNA target sequence.
実施例10:ジーンポーターでのヒト単球−DCのトランスフェクション
ヒトDCの調節におけるヒトSOCS1の役目を調査するために、ヒトSOCS1を特異的にダウンレギュレートする能力をもつ小型干渉RNA(siRNA)がまず同定された。ヒトSOCS1及び293T細胞をターゲティングするsiRNA配列を選択するために、コンピュータプログラムが使用された。その後、各々の合成ヒトSOCS−1−siRNAオリゴヌクレオチド2重鎖を、293T細胞へとジーンポータートランスフェクション試薬を用いて10:1の比でフラグでタグ付けされたヒトSOCS1発現と共に同時トランスフェクトさせた。トランスフェクションから48時間後に、細胞を採取し、本書の他の個所で記述されている通りに、ウェスタンブロット法により分析した。
Example 10: Human Monocyte-DC Transfection with Gene Porter To investigate the role of human SOCS1 in the regulation of human DC, a small interfering RNA (siRNA) with the ability to specifically down-regulate human SOCS1 Was first identified. A computer program was used to select siRNA sequences targeting human SOCS1 and 293T cells. Each synthetic human SOCS-1-siRNA oligonucleotide duplex is then co-transfected into 293T cells with human SOCS1 expression flagged at a 10: 1 ratio using Gene Porter transfection reagent. It was. Forty-eight hours after transfection, cells were harvested and analyzed by Western blot as described elsewhere in this document.
図21Aは、ヒトSOCS1siRNA1がヒトSOCS1発現を効率良くダウンレギュレートしたことを示している。siRNAによるヒトSOCS1mRNAダウンレギュレーションの特異性は、スクランブルされたsiRNA1オリゴヌクレオチドがSOCS1mRNAをダウンレギュレートすることができないことによって確認された。従って、ヒトSOCS1siRNA1が、さらなる調査のために選択された。合成siRNA2重鎖がジーンポーターにより、85.5%のトランスフェクション効率で、ヒト単球に由来するDC内にトランスフェクトされた(図21B)。定量的RT−PCR検定によって確認される通り、hSOCS1siRNA2重鎖でのトランスフェクションを受けた全DC集団内のhSOCS1mRNAのレベルは、モックトランスフェクションを受けたDC内でのレベルと比べて約60%だけ特異的に減少した(図21C、P<0.01)。siRNAの効率及びSOCS1RNAの減少は、全マウス骨髄由来DC集団内のマウスSOCS1をターゲティングする合成siRNA2重鎖を用いた実験において観察されたものと類似している。 FIG. 21A shows that human SOCS1 siRNA1 efficiently downregulated human SOCS1 expression. The specificity of human SOCS1 mRNA downregulation by siRNA was confirmed by the inability of scrambled siRNA1 oligonucleotides to downregulate SOCS1 mRNA. Therefore, human SOCS1 siRNA1 was selected for further investigation. Synthetic siRNA duplexes were transfected into DCs derived from human monocytes by Gene Porter with a transfection efficiency of 85.5% (FIG. 21B). As confirmed by quantitative RT-PCR assay, the level of hSOCS1 mRNA in the total DC population transfected with hSOCS1 siRNA duplex was only about 60% compared to the level in mock-transfected DCs. There was a specific decrease (FIG. 21C, P <0.01). The siRNA efficiency and SOCS1 RNA reduction is similar to that observed in experiments with synthetic siRNA duplexes targeting mouse SOCS1 within the total mouse bone marrow derived DC population.
ヒトDCにおけるヒトSOCS1の相対的発現を、本書の他の個所で記述されている通りに定量的実時間RT−PCRによって評価した。カリフォルニア州フォスターシティーのアプライド・バイオシステムズ社製のヒトSOCS1用予め開発されたプライマ/プローブセット(プライマ、5’−TTTTTCGCCCTTAGCGGGAA−3’;配列番号24及び5’−CTGCCATCCAGGTGAAAGC−3’;配列番号25、及びプローブ、6FAM−ATGGCCTCGGGACCCACGAG−TAMRA;配列番号26)を使用した。 The relative expression of human SOCS1 in human DCs was assessed by quantitative real-time RT-PCR as described elsewhere in this document. A pre-developed primer / probe set for human SOCS1 (Primer, 5′-TTTTTCGCCCCTTAGCGGGAA-3 ′; SEQ ID NO: 24 and 5′-CTGCCCATCCAGGTGAAAGC-3 ′; SEQ ID NO: 25, manufactured by Applied Biosystems of Foster City, Calif. And the probe, 6FAM-ATGGCCCTGGGACCCACGAG-TAMRA; SEQ ID NO: 26).
ヒトのSOCS1サイレンシング化されたDCの特徴づけ
ヒトSOCS1がフローサイトメトリ分析によってDC上の共同刺激分子の発現を調節するか否かを査定するために、次の実験セットを実施した。本書の他の個所で記述されている通りに、マウスSOCS1を査定する上で使用されたものに従って、ヒトSOCS1のためのフローサイトメトリ分析を進めた。マウスDCにおける観察事実と一致して、h
SOCS1−siRNAでのトランスフェクションを受けたDC及び対照のhSOCS1−siRNA突然変異体でのトランスフェクションを受けたDCが、LPS誘発型成熟の前後に代表的な同時刺激性分子のそれらの発現における差異をわずかしか示さないということが観察された(図22A)。hSOCS1−siRNADC及び突然変異体−siRNADC上では、比較可能なレベルのMHC−I及びII分子も検出された。これとは対照的に、IL−12、IL−6及びTNF−αなどの前炎症性サイトカインの大幅に増強された分泌によって示されているように、hSOCS1siRNAでのトランスフェクションを受けたDCは、siRNA突然変異体でのトランスフェクションを受けたヒトDCよりもLPSでの刺激に対する応答性が高いということが観察された(図22B及び22C)。
Characterization of human SOCS1 silenced DCs To assess whether human SOCS1 regulates the expression of costimulatory molecules on DCs by flow cytometry analysis, the following set of experiments was performed. Flow cytometric analysis for human SOCS1 proceeded as described in assessing mouse SOCS1 as described elsewhere in this document. Consistent with observations in mouse DC, h
DCs transfected with SOCS1-siRNA and DCs transfected with control hSOCS1-siRNA mutant differ in their expression of representative costimulatory molecules before and after LPS-induced maturation Was observed to show only a small amount (FIG. 22A). Comparable levels of MHC-I and II molecules were also detected on hSOCS1-siRNADC and mutant-siRNADC. In contrast, DCs transfected with hSOCS1 siRNA, as shown by greatly enhanced secretion of pro-inflammatory cytokines such as IL-12, IL-6 and TNF-α, It was observed that responsiveness to stimulation with LPS was higher than human DCs that had been transfected with siRNA mutants (FIGS. 22B and 22C).
ヒトSOCS1siRNAを発現する組換え型アデノウイルスベクターの生成
アドイージ・システム(AdEasy system)(E1及びE3を削除したAd(5);カンタム・バイオテクノロジーズ社(Quantum Biotechnologies Inc.)、カリフォルニア州パロアルト(Palo Alto,CA))を用いて、複製欠損アデノウイルスを構築し生成した。アドイージーベクターの中にH1−ヒトSOCS1−siRNADNAフラグメントを挿入することによってシャトルベクターAd−hSOCS1−siRNAを、構築した(図28)。ヒトSOCS1−siRNAの挿入をDNA配列決定によって確認した。その後、メーカー(カンタム・バイオテクノロジーズ社、カリフォルニア州パロアルト)の指示に従って、組換え型アデノウイルスAd−hSOCS1−siRNAを生成した。メーカー(カンタム・バイオテクノロジーズ社、カリフォルニア州パロアルト)の指示に従って、組換え型アデノウイルスを293個の細胞中で産生させ滴定した。組換え型Ad(5)がヒト単球由来のDCをトランスフェクトできるということが観察された。
Generation of recombinant adenoviral vectors expressing human SOCS1 siRNA AdEasy system (Ad (5) with E1 and E3 deleted; Quantum Biotechnologies Inc., Palo Alto, Calif.) , CA)) was used to construct and generate a replication defective adenovirus. The shuttle vector Ad-hSOCS1-siRNA was constructed by inserting the H1-human SOCS1-siRNA DNA fragment into the adeasy vector (FIG. 28). The insertion of human SOCS1-siRNA was confirmed by DNA sequencing. Subsequently, recombinant adenovirus Ad-hSOCS1-siRNA was generated according to the manufacturer's instructions (Quantum Biotechnology, Palo Alto, Calif.). Recombinant adenovirus was produced and titrated in 293 cells according to the manufacturer's instructions (Quantum Biotechnology, Palo Alto, Calif.). It was observed that recombinant Ad (5) can transfect DCs derived from human monocytes.
実施例11:ヒトSOCS1−siRNADCによりプライミングされたMAGE3特異的CTL応答
ヒトDCの免疫刺激能力を調節する上でのヒトSOCS1の役割を調査するために以下の実験に着手した。この実施例で提示されている結果は、ヒトのSOCS1サイレンシング化されたDCが病原菌産物での刺激に対し過剰反応し、自己抗原特異的ヒト細胞傷害性Tリンパ球(CTL)をプライミングする増強された、刺激能力を有することを実証している。重要なことに、野生型DCではなく、ヒトのSOCS1サイレンシング化されたDCは、天然の抗原発現ヒト腫瘍細胞に対する活発な溶解活性を有するCTLを完全に活性化する能力をもつ。同様に、CTLをプライミングするヒトのSOCS1サイレンシング化されたDCの能力は、IL−12産生及びシグナリングのSOCS1制限によって制御される確率が高いと考えられている。これらの結果は、ヒトDCを負に調節する上でのヒトSOCS1の決定的な役割を標示し、ヒトの患者のためのより有効な腫瘍ワクチンを開発するためのこのSOCS1サイレンシング化アプローチの翻訳潜在能力が関係するとみなしている。
Example 11: MAGE3-specific CTL response primed with human SOCS1-siRNADC The following experiment was undertaken to investigate the role of human SOCS1 in modulating the immunostimulatory capacity of human DC. The results presented in this example show that human SOCS1 silenced DCs overreact to stimulation with pathogen products and prime autoantigen-specific human cytotoxic T lymphocytes (CTLs) It has been demonstrated that it has the ability to stimulate. Importantly, human SOCS1 silenced DCs, but not wild type DCs, have the ability to fully activate CTLs with active lytic activity against native antigen-expressing human tumor cells. Similarly, it is believed that the ability of human SOCS1-silenced DCs to prime CTL is likely to be controlled by SOCS1 restriction of IL-12 production and signaling. These results demonstrate the critical role of human SOCS1 in negatively regulating human DCs and translation of this SOCS1 silencing approach to develop more effective tumor vaccines for human patients We consider that potential is involved.
本書で開示されている実験において利用される材料及び方法についてここで記述する。 The materials and methods utilized in the experiments disclosed herein are now described.
ペプチド
ジーンメッド・シンセシス社(米国カリフォルニア州サウスサンフランシスコ)により、HPLCを用いて95%超の純度までHLA−A2−制限されたMAGE3 CTLペプチド(FLWGPRALV;配列番号27)(ファン・デル・ブルゲン(van der Bruggen)ら、1994年、European Journal of Immunology第24号:3038〜43頁)及び対照H−2Kb−制限されたOVA−I(SIINFEKL;配列番号11)が合成され精製された。内毒素を含まないPBS中での最終希釈の前に、ペプチドをDMSO中で溶解させた(シグマ、ミズーリ州セン
トルイス)。
The peptide MAGE3 CTL peptide (FLWGPRALV; SEQ ID NO: 27) (van der burgen), which was HLA-A2-restricted to> 95% purity using HPLC by the peptide GeneMed Synthesis (South San Francisco, Calif., USA) Brugen) et al., 1994, European Journal of Immunology 24: 3038-43) and control H-2K b -restricted OVA-I (SIINFEKL; SEQ ID NO: 11) was synthesized and purified. Peptides were dissolved in DMSO (Sigma, St. Louis, MO) prior to final dilution in PBS without endotoxin.
ヒトSOCS1発現のウェスタンブロット分析
293T細胞を、本書の他の個所で記述されている通りにジーンポーター試薬を用いて10:1の比で合成ヒトSOCS−1−siRNAオリゴヌクレオチド2重鎖(21bp)又は無関連オリゴ2重鎖及びフラグタグ付きヒトSOCS1発現ベクター(pCMV−hSOCS1)と同時トランスフェクトした。トランスフェクションから48時間後に、細胞を採取し、SDS−PAGEに付した。ハイボンド(Hybond)−P膜(アマシャム、イリノイ州アーリントンハイツ)への移送の後、抗−Flag(シグマ、ミズーリ州セントルイス)又はアクチン(サンタクルス・バイオテクノロジー社、カリフォルニア州サンタクルス)抗体でのウェスタンブロット法に続けて、ECL−Plus試薬(アマシャム、イリノイ州アーリントンハイツ)での検出により試料を分析した。薄膜を密度計SIで走査し、SOCS−1/アクチンバンドをイメージ・クアントソウトウェア(モレキュラー・ダイナミクス、ニュージャージー州ピスカタウェイ)で定量した。SOCS1バンドの強度をベータアクチンバンドの強度に正規化した。
Western Blot Analysis of Human SOCS1 Expression 293T cells were synthesized at a 10: 1 ratio using GenePorter reagent as described elsewhere in this document, and a human SOCS-1-siRNA oligonucleotide duplex (21 bp) Or co-transfected with an irrelevant oligo duplex and flag-tagged human SOCS1 expression vector (pCMV-hSOCS1). 48 hours after transfection, cells were harvested and subjected to SDS-PAGE. Western with anti-Flag (Sigma, St. Louis, MO) or actin (Santa Cruz Biotechnology, Santa Cruz, CA) antibodies after transfer to Hybond-P membranes (Amersham, Arlington Heights, Ill.) Following blotting, samples were analyzed by detection with ECL-Plus reagent (Amersham, Arlington Heights, Ill.). Thin films were scanned with a densitometer SI and SOCS-1 / actin bands were quantified with Image Quant software (Molecular Dynamics, Piscataway, NJ). The intensity of the SOCS1 band was normalized to the intensity of the beta actin band.
ヒトSOCS1発現の定量的RT−PCR分析
ヒトDC中のヒトSOCS1の相対的発現を、本書の他の個所で記述されている通り、定量的実時間RT−PCRにより評価した。
Quantitative RT-PCR analysis of human SOCS1 expression Relative expression of human SOCS1 in human DCs was assessed by quantitative real-time RT-PCR as described elsewhere in this document.
ヒト単球由来のDCのトランスフェクション及びヒトT細胞のインビトロプライミング
スクロアース(Schroers)ら、2003年、Clinical Can.Res.第9号:4743〜4755頁;及びスクロアースら、2004年、Methods
Mol.Biol.第246号:451〜9頁に記述されている通りに、PBMCに由来するヒトDCを生成し培養した。HLA−A2+の健康なボランティアからヘパリン化された血液を収集した。PCR−SSP−DNAベースの手順によりHLA分類を実施した(メソジスト・ホスピタル(The Methodist Hospital)、テキサス州ヒューストン)。無血清DC培地(セルジェニックス(CellGenix)、イリノイ州アンティアック(Antioch,Il))の中にPBMCを再懸濁させ、加湿した5%CO2中で37℃でインキュベートした。プラスチックに接着した細胞画分を、1000IU/mlの組換え型ヒトGM−CSF(rhGM−CSF;アールアンドディー・システムズ社、ミネソタ州ミネアポリス)及び1000IU/ml rhIL−4(アールアンドディー・システムズ社、ミネソタ州ミネアポリス)を伴う無血清DC培地内で培養させた。5日目又は6日目に、メーカーの指示に従ってジーンポーターを用いて120nMのsiRNAオリゴヌクレオチドで単球由来のDCをトランスフェクトした。その後、トランスフェクションを受けたDCを一晩MAGE3ペプチド(20μg/ml)でパルスした。24ウェルの平板の1ウェルあたり合計1×106のヒトT細胞を、5%ABヒト血清、rhIL−2(50U/ml)、及びTNFa(10ng/ml、アールアンドディー・システムズ社、ミネソタ州ミネアポリス)で補足された0.5mlのRPMI−1640中で、5×104個のMAGE3パルスされたトランスフェクションを受けたDCと同時培養させた(20:1)。同時培養したT細胞を、同時培養7日目に、自己移植性MAGE3パルスされたトランスフェクションを受けたDCで一回再刺激した。一部の実験については、3日に一度DCとT細胞の同時培養の中に抗−ヒトIL−12(p70)抗体(20mg/ml、アールアンドディー・システムズ社、ミネソタ州ミネアポリス)を添加した。2週間の同時培養の後、T細胞を免疫検定のために使用した。
Transfection of DCs derived from human monocytes and in vitro priming of human T cells Schroers et al., 2003, Clinical Can. Res. 9: 4743-4755; and Scroars et al., 2004, Methods.
Mol. Biol. No. 246: Human DCs derived from PBMC were generated and cultured as described on pages 451-9. Heparinized blood was collected from healthy volunteers with HLA-A2 +. HLA classification was performed by PCR-SSP-DNA based procedure (The Methodist Hospital, Houston, TX). PBMC were resuspended in serum-free DC medium (CellGenix, Antioch, Ill.) And incubated at 37 ° C. in humidified 5% CO 2 . The cell fraction adhered to the plastic was treated with 1000 IU / ml recombinant human GM-CSF (rhGM-CSF; R & D Systems, Minneapolis, MN) and 1000 IU / ml rhIL-4 (R & D Systems). And cultured in serum-free DC medium with Minneapolis, Minnesota). On
サイトカインELISA及び酵素連結型イムノスポット(ELISPOT)検定
メーカーの指示に従い、指示された時点で指示された刺激物で、ELISA分析(BDバイオサイエンシーズ、ニュージャージー州リンカーンパーク)のためのDC培養の上清を用いてさまざまな前炎症性サイトカインのレベルを定量化した。ヒト末梢リンパ球のE
LISPOT検定を、本書の他の個所で記述されている通りに実施した。
Cytokine ELISA and Enzyme Linked Immunospot (ELISPOT) Assay DC culture supernatant for ELISA analysis (BD Biosciences, Lincoln Park, NJ) at the indicated time points according to the manufacturer's instructions Was used to quantify the levels of various pro-inflammatory cytokines. E of human peripheral lymphocytes
The LISPOT test was performed as described elsewhere in this document.
フローサイトメトリ分析
0.1%のNaN3及び2%のFCSを含有するPBS中のFITC又はPEmAbsを用いて、細胞を染色した。ヒトCD40、CD80及びCD86に特異的な抗体及び整合したイソタイプ対照をBDバイオサイエンシーズ(カリフォルニア州サンノゼ)から購入した。染色した細胞をFACSCalibur(ベクトン・ディッキンソン、ニュージャージー州リンカーンパーク)フローサイトメーター上で分析した。
Flow cytometry analysis Cells were stained with FITC or PE mAbs in PBS containing 0.1% NaN 3 and 2% FCS. Antibodies specific for human CD40, CD80 and CD86 and matched isotype controls were purchased from BD Biosciences (San Jose, Calif.). Stained cells were analyzed on a FACSCalibur (Becton Dickinson, Lincoln Park, NJ) flow cytometer.
四量体染色
ベイラー・カレッジ・オブ・メディシン・テトラマー・コア・ファシリティー(米国テキサス州ヒューストン)において、ヒトMAGE3/HLA−A2四量体が合成された。同時培養中のリンパ球又はヒト末梢血リンパ球を抗−hCD8a−FITC/抗−hCD3−PerCP及びMAGE3−PE三量体と同時染色した。四量体染色を4℃で1時間、メーカーの指示に従い、106個の細胞あたり1μlの抗CD8α及びMAGE3−PE四量体の1:100希釈物を用いて行なった。
Tetramer Staining Human MAGE3 / HLA-A2 tetramer was synthesized at Baylor College of Medicine Tetramer Core Facility (Houston, Texas, USA). Lymphocytes in co-culture or human peripheral blood lymphocytes were co-stained with anti-hCD8a-FITC / anti-hCD3-PerCP and MAGE3-PE trimers. Tetramer staining was performed at 4 ° C. for 1 hour according to the manufacturer's instructions using 1 μl of anti-CD8α and MAGE3-PE tetramer 1: 100 dilution per 10 6 cells.
HLA−A2遺伝子導入マウスのDC免疫化
4〜6週令の雌のHLA−A2.1遺伝子導入マウスをジャクソン・ラボラトリー(米国メーン州)から購入し、制度的指針に従ってベイラー・カレッジ・オブ・メディシン(米国テキサス州ヒューストン)において無菌のマウス施設内に維持した。本書の他の個所で記述されている通りマウスBM由来のDCを、HLA−A2.1遺伝子導入マウスから調製し、5のMOIで組換え型レトロウイルスベクターLV−SOCS1−siRNA又はLV−GFP−siRNAで形質導入した。その後DCを20時間、MAGE3ペプチドでパルスし、PBSで3回洗浄し、TNFαで24時間刺激した(500ng/ml、アールアンドディー・システムズ社、ミネソタ州ミネアポリス)。その後DCを、後足蹠を介してHLA−A2遺伝子導入マウス内に注入した。一部のマウスにおいて、DCワクチン接種から指示された日数後に、LPS(30μg/マウス)又は組換え型マウスIL−12(1μg/マウス、ペプロテック社、ニュージャージー州ロッキーヒル)を腹腔内(i.p.)投与した。
DC immunization of HLA-A2 transgenic mice 4-6 week old female HLA-A2.1 transgenic mice were purchased from Jackson Laboratory (Maine, USA) and in accordance with institutional guidelines Baylor College of Medicine Maintained in a sterile mouse facility (Houston, Texas, USA). Mouse BM-derived DCs were prepared from HLA-A2.1 transgenic mice as described elsewhere in this document and recombinant retroviral vectors LV-SOCS1-siRNA or LV-GFP- at a MOI of 5 Transduced with siRNA. DC were then pulsed with MAGE3 peptide for 20 hours, washed 3 times with PBS, and stimulated with TNFα for 24 hours (500 ng / ml, R & D Systems, Minneapolis, MN). Thereafter, DC was injected into the HLA-A2 transgenic mouse via the hind footpad. In some mice, LPS (30 μg / mouse) or recombinant mouse IL-12 (1 μg / mouse, Peprotech, Rocky Hill, NJ) was injected intraperitoneally (ip) after the days indicated after DC vaccination. .) Was administered.
CTL検定
本書の他の個所で記述されている通り標的細胞を溶解するインビトロ再刺激された脾細胞の能力を測定する標準クロム放出検定で、CD8+CTL応答を査定した(フアンら、2003年、Cancer Res.第63号:7321〜9頁)。2〜3匹の免疫化したマウスからプールした脾細胞を、4〜6日間、MAGE3ペプチドを含有するRPMI−1640中でインビトロで再刺激した。ヒトMAGE3+、HLA−A2+黒色腫細胞(SK−Mel−37)及び対照ヒトMAGE3+、HLA−A2−黒色腫細胞(NA−6−Mel)を37℃で90分間51Crクロム酸ナトリウム溶液で標識した。異なる数のエフェクタ細胞を37℃で4時間、96ウェルのU字底平板(200μl/ウェル)内で、一定数の標的細胞(5×104/ウェル)と共にインキュベートした。3重培養から上清を収集し、分析した。(実験的放出−自然放出)/(最大放出−自然放出)×100として溶解百分率を計算した。
CTL Assay A CD8 + CTL response was assessed in a standard chromium release assay that measures the ability of in vitro restimulated splenocytes to lyse target cells as described elsewhere in this document (Huan et al., 2003, Cancer Res. 63: 7321-9). Pooled splenocytes from 2-3 immunized mice were restimulated in vitro in RPMI-1640 containing MAGE3 peptide for 4-6 days. Human MAGE3 +, HLA-A2 + melanoma cells (SK-Mel-37) and control human MAGE3 +, HLA-A2 - melanoma cells (NA-6-Mel)
本実施例で提示されている実験の結果について、ここで記述する。 The results of the experiment presented in this example will now be described.
抗原特異的CTL応答をプライミングするためのヒトのSOCS1サイレンシング化されたDCの増強された免疫刺激性効能
サイレンシング化されたヒトSOCS1が自己抗原特異的CTLをプライミングする上でヒトDCの刺激効能を増強し得るか否かを査定するために、次の実験セットに着手した
。成人のヒトの睾丸及び黒色腫細胞の中で発現されるものとして知られている胚芽腫抗原であるヒトMAGE3由来のHLA−A2制限されたペプチド(ファン・デル・ブルゲンら、1994年、European Journal of Immunology第24号:3038〜43頁)を、モデルヒト自己抗原として用いた。HLA−A2+の健康なボランティアからのヒト単球由来DCをhSOCS1siRNAオリゴヌクレオチドでトランスフェクトし、次に一晩MAGE3ペプチド(20μg/ml)でパルスした。TNFα(成熟刺激物)(10ng/ml、アールアンドディー・システムズ社、ミネソタ州ミネアポリス)の存在下で5×104個のMAGE3パルスされたトランスフェクションを受けたDCと、1ウェルあたり合計1×106個の自己移植性ヒトT細胞を同時培養した(20:1)。同時培養したT細胞を、同時培養7日目に、自己移植性MAGE−3パルスされたトランスフェクションを受けたDCで一回再刺激した。2週間の同時培養後、免疫検定のためにT細胞を用いた。MAGE3ペプチドでパルスされたhSOCSsiRNAでのトランスフェクションを受けたDCとの同時培養中では、MAGE3でパルスされたmut−siRNADC又はモックDCとの同時培養中でそれぞれ5.4%及び4.3%にすぎなかったのに比べて、13.9%のCD8+T細胞がMAGE3−四量体について陽性であった(図23A)。同じドナーからの実験未使用の(刺激を受けていない)一次ヒトリンパ球の四量体染色は、低レベルの陽性MAGE3四量体染色を示した(0.5%のCD8+T細胞)。これと一致して、細胞内IFNγ染色(図23B)は、MAGE3ペプチドパルスを受けたmut−siRNADC(6.9%のIFNγ+CD8+T細胞)又はモックDC(5.7%のIFNγ+CD8+T細胞)に比べて、hSOCS1−siRNADCがMAGE3特異的CTL応答を実質的に改善する(11.18%のIFNγ+CD8T細胞)ことを示した。さらに、IFNγELISPOT検定(図23C)は、hSOCS1−siRNADCにより、増大した数のMAGE3特異的CTLが活性化されることを示した。HLA−A2+ドナーからの繰り返しの実験は同様の結果を示した。一次ヒトT細胞の大部分は、抗原でパルスされていないDCとの2週間の同時培養後死滅した。集合的に、これらの結果は、ヒトのSOCS1サイレンシング化されたDCが自己抗原特異的CTLをプライミングする増強された免疫刺激能力を有することを表わしている。
Enhanced immunostimulatory efficacy of human SOCS1-silenced DC for priming antigen-specific CTL responses Silenced human SOCS1 stimulates human DC to prime autoantigen-specific CTL The next set of experiments was undertaken to assess whether it could be enhanced. HLA-A2 restricted peptide from human MAGE3, a germinoma antigen known to be expressed in adult human testicular and melanoma cells (Van der Burgen et al., 1994, European Journal of Immunology 24: 3038-43) was used as a model human autoantigen. Human monocyte-derived DCs from HLA-A2 + healthy volunteers were transfected with hSOCS1 siRNA oligonucleotides and then pulsed overnight with MAGE3 peptide (20 μg / ml). 5 × 10 4 MAGE3 pulsed transfected DCs in the presence of TNFα (maturation stimulant) (10 ng / ml, R & D Systems, Minneapolis, Minn.) And a total of 1 × per well 10 6 autologous human T cells were co-cultured (20: 1). Co-cultured T cells were restimulated once on
CTLプライミングにおけるSOCS1制限されたIL−12の決定的役割
SOCS1サイレンシング化されたDCは、病原菌産物での刺激に応答してCTL応答の活性化における主要なサイトカインであるIL−12を増強された量だけ産生すること(トリンキエリ、2003年、Nat.Rev.Immunol.第3号:133〜46頁)そしてIL−12シグナリングがSOCS1により制限されていること(アイルズら、2002年、J.Biol.Chem.第277号:43735〜40頁))から、ヒトのSOCS1サイレンシング化されたDCによりCTLをプライミングする上でのIL−12の役割を検査した。従って、T細胞とMAGE3でパルスされたトランスフェクションを受けたDCの同時培養に対して抗ヒトIL−12抗体を3日に1度添加した。図23A〜23Cは、抗IL12(p70)抗体を用いたIL−12の阻害が、四量体染色、細胞内IFN−γ染色及びELISPOT検定により実証される通り、MAGE3特異的CTLを刺激するhSOCS1−siRNADCの増強した能力を無効にしたということを示している。
The critical role of SOCS1-restricted IL-12 in CTL priming SOCS1-silenced DCs augmented IL-12, a major cytokine in the activation of CTL responses in response to stimulation with pathogenic products Only in quantities (Trinquieri, 2003, Nat. Rev. Immunol. 3: 133-46) and that IL-12 signaling is restricted by SOCS1 (Ayles et al., 2002, J. Biol. Chem., 277: 43735-40)), the role of IL-12 in priming CTLs with human SOCS1 silenced DCs was examined. Therefore, anti-human IL-12 antibody was added once every 3 days to cocultures of DCs transfected with T cells and pulsed with MAGE3. FIGS. 23A-23C show hSOCS1 that stimulates MAGE3-specific CTL as inhibition of IL-12 using anti-IL12 (p70) antibody is demonstrated by tetramer staining, intracellular IFN-γ staining and ELISPOT assay. -Indicates that the enhanced ability of siRNADC has been negated.
ヒト化されたHLA−A2.1遺伝子導入マウスを用いて、SOCS1サイレンシング化されたDCにより誘発された増強されたCTL応答におけるIL−12の役割をさらにテストした。マウスSOCS1−siRNAを発現する組換え型レンチウイルスベクター(LV−mSOC1siRNA)又は対照のベクターLV−GFPsiRNAで、HLA−A2.1遺伝子導入マウスBM由来DCを形質導入し、A2制限MAGE3ペプチドでパルスした。TNFαでの成熟の後、一週間に一度の間隔で、HLA−A2.1遺伝子導入マウスの体内に2回足蹠から形質導入されたDCを投与した。各々のDC免疫化の後、
マウスを、LPSか又は低用量の組換え型TL−12サイトカインで、3回インビボで刺激した。LPSは多数の前炎症性サイトカインを誘発し、その多くがSOCS1により調節されていること、そしてNF−κB(p65)シグナリングの調節におけるSOCS1の直接的役割が可能であることから、LPSが使用された(リョーら、2003年、Mol.Cell、第12号:1413〜26頁)。インビボのIL−12刺激で、MAGE3でのパルスを受けたGFP−siRNA−DCで免疫化されたマウスの体内では2×105個のT細胞あたりわずか10IFNγ+スポットであるのに比べ、MAGE3パルスを受けたSOCS1−siRNA−DCで免疫化されたマウスにおいては、2×105個のT細胞あたり239のIFNγ+スポットが検出された(図24)。インビボでのLPS刺激は同様に、SOCS1−siRNA DCs(SOCS1−siRNA−DCマウス中T細胞2×105個あたりIFNγ+スポット63個対GFP−siRNA−DCマウスの体内でT細胞2×105個あたりIFNγ+スポット24個)により誘発されたCTL応答を優先的に増強した(図24)。しかしながらIL−12刺激は、SOCS1−siRNA−DCで免疫化されたマウスにおけるMAGE3特異的CTL応答を高めるために、LPS刺激よりもさらに有効であった(P<0.01)。IL−12のこのさらに優れた刺激能力は、Jak/Stat経路を通してIL−12刺激を直接的に調節するSOCS1の能力、ならびに、SOCS1−siRNA−DCで免疫化されたマウスの体内で活性化されたCTLに対するIL−12の直接的効果((トリンキエリ、2003年、Nat.Rev.Immunol.第3号:133〜46頁))に起因している確率が高い。合わせて考慮すると、これらの結果は、SOCS1が抗原提示細胞内でIL−12のシグナリングを制限することのさらなる証拠を提供する。これらの結果は同様に、サイトカインベースの腫瘍療法の効き目を判定する上でのサイトカインシグナリングの重要性をも強調している。
Humanized HLA-A2.1 transgenic mice were used to further test the role of IL-12 in the enhanced CTL response induced by SOCS1-silenced DCs. Transduced HLA-A2.1 transgenic mouse BM-derived DCs with a recombinant lentiviral vector (LV-mSOC1 siRNA) expressing mouse SOCS1-siRNA or a control vector LV-GFP siRNA and pulsed with A2 restricted MAGE3 peptide . After maturation with TNFα, DC transduced from the footpad were administered twice into the body of the HLA-A2.1 transgenic mice at an interval of once a week. After each DC immunization,
Mice were stimulated in vivo three times with LPS or low doses of recombinant TL-12 cytokines. LPS is used because it induces a number of pro-inflammatory cytokines, many of which are regulated by SOCS1 and allows a direct role for SOCS1 in the regulation of NF-κB (p65) signaling. (Ryo et al., 2003, Mol. Cell, 12: 1413-26). In vivo IL-12 stimulation, MAGE3 pulses were compared to only 10 IFNγ + spots per 2 × 10 5 T cells in mice immunized with GFP-siRNA-DC pulsed with MAGE3. In mice immunized with the received SOCS1-siRNA-DC, 239 IFNγ + spots were detected per 2 × 10 5 T cells (FIG. 24). LPS stimulation in vivo is also similar to SOCS1-siRNA DCs (63 IFNγ + spots per 2 × 10 5 T cells in SOCS1-siRNA-DC mice versus 2 × 10 5 T cells in GFP-siRNA-DC mice. CTL responses elicited by IFNγ + 24 spots per) were preferentially enhanced (FIG. 24). However, IL-12 stimulation was even more effective than LPS stimulation to enhance the MAGE3-specific CTL response in mice immunized with SOCS1-siRNA-DC (P <0.01). This even greater stimulation ability of IL-12 is activated in the body of mice immunized with SOCS1-siRNA-DC as well as the ability of SOCS1 to directly modulate IL-12 stimulation through the Jak / Stat pathway. There is a high probability that it is attributed to the direct effect of IL-12 on CTL ((Trinkieri, 2003, Nat. Rev. Immunol. No. 3: 133-46)). Taken together, these results provide further evidence that SOCS1 limits IL-12 signaling in antigen-presenting cells. These results also highlight the importance of cytokine signaling in determining the efficacy of cytokine-based tumor therapy.
野生型DCによってではなく、ヒトのSOCS1サイレンシング化されたDCにより活性化されたヒトCTLは、腫瘍溶解エフェクタ機能を有する
自己腫瘍関連抗原MAGE3に特異的な活性化されたT細胞が腫瘍溶解エフェクタ機能を有するか否かを判定するため、CTL検定のための標的細胞として、天然MAGE3+ヒト腫瘍細胞を使用した。MAGE3パルスを受けたSOCS1−siRNA−DC又はmut−siRNADCによって活性化されたヒトT細胞は、MAGE3ペプチドパルスされたMAGE3+HLA−A2+黒色腫細胞(SK−Mel−37)を容易に殺した。しかしながら、MAGE3パルスを受けたmut−siRNADCは、MAGE3でパルスされていない天然SK−Mel−37細胞に対しては弱い細胞溶解活性しか示さなかった(図25)。これとは対照的に、MAGE3−パルスを受けたSOCS1−siRNA−DCによって活性化されたこれらのT細胞は、MAGE3でパルスされていない天然のSK−Mel−37細胞に対し強い細胞溶解活性をなおも有していた(図25)。hSOCS1−siRNA−DCとの同時培養中でのT細胞の腫瘍溶解活性は、抗hIL−12抗体処置により著しく低下させられた。MAGE3パルスを受けたSOCS1−siRNA−DCにより活性化されたヒトT細胞のみがHLA−A2−陰性、MAGE3+黒色腫細胞(NA−6−Mel)に対する背景細胞溶解活性を有していたことから、腫瘍細胞溶解活性はCTLにより特異的に媒介された。異なるドナーからの反復実験が、類似の結果を示した。
Human CTLs activated by human SOCS1 silenced DCs, but not by wild type DCs, have oncolytic effector functions. Activated T cells specific for the autologous tumor-associated antigen MAGE3 have oncolytic effectors. In order to determine whether it has function, natural MAGE3 + human tumor cells were used as target cells for the CTL assay. Human T cells activated by SOCS1-siRNA-DC or mut-siRNADC that received MAGE3 pulses readily killed MAGE3 peptide-pulsed MAGE3 + HLA-A2 + melanoma cells (SK-Mel-37). However, mut-siRNADCs that received MAGE3 pulse showed only weak cytolytic activity against native SK-Mel-37 cells that were not pulsed with MAGE3 (FIG. 25). In contrast, these T cells activated by SOCS1-siRNA-DCs that received MAGE3-pulse showed strong cytolytic activity against native SK-Mel-37 cells that were not pulsed with MAGE3. It still had (FIG. 25). The oncolytic activity of T cells in co-culture with hSOCS1-siRNA-DC was significantly reduced by anti-hIL-12 antibody treatment. Since only human T cells activated by SOCS1-siRNA-DC that received MAGE3 pulses had background cytolytic activity against HLA-A2-negative, MAGE3 + melanoma cells (NA-6-Mel), Tumor cytolytic activity was specifically mediated by CTL. Repeated experiments from different donors showed similar results.
以上の観察事実を確認するために、MAGE3パルスされたSOCS1−siRNA−DC又はGFP−siRNA−DCで免疫化されたHLA−A2.1遺伝子導入マウス由来のT細胞の腫瘍溶解活性をテストした。図26は、MAGE3パルスされたmSOCS1−siRNA−DCで免疫化された遺伝子導入マウスに由来するT細胞が、天然のMAGE3+HLA−A2陽性黒色腫細胞SK−Mel−37に対する活発な細胞溶解活性を有していたことを示している。これとは対照的に、MAGE3でのパルスを受けたGFP−siRNA−DCで免疫化された遺伝子導入マウス由来のT細胞のみが、図25に示さ
れた結果と一致して、天然黒色腫細胞に対する弱い細胞溶解活性を有していた。さらに、低用量の組換え型IL−12でのインビボ刺激が、GFP−siRNA−DCではなくSOCS1−siRNA−DCにより誘発されたCTL応答を著しく増強することも観察された(図26)。合わせて考えると、本書中の結果は、おそらくは抗原提示細胞によるIL−12の増強された産生及びシグナリングに起因して、天然腫瘍細胞に対する活発な溶解エフェクタ機能を有するCTLを完全に活性化させる固有の能力を、ヒトのSOCS1サイレンシング化されたDCが有することを示している。
In order to confirm the above observation facts, the oncolytic activity of T cells derived from HLA-A2.1 transgenic mice immunized with MAGE3-pulsed SOCS1-siRNA-DC or GFP-siRNA-DC was tested. FIG. 26 shows that T cells derived from transgenic mice immunized with MAGE3-pulsed mSOCS1-siRNA-DC have active cytolytic activity against native MAGE3 + HLA-A2-positive melanoma cells SK-Mel-37. It shows that he was doing. In contrast, only T cells from transgenic mice immunized with GFP-siRNA-DC pulsed with MAGE3 were found in natural melanoma cells, consistent with the results shown in FIG. Had weak cytolytic activity against. Furthermore, it was also observed that in vivo stimulation with low doses of recombinant IL-12 significantly enhanced the CTL response elicited by SOCS1-siRNA-DC but not GFP-siRNA-DC (FIG. 26). Taken together, the results herein are unique to fully activate CTL with active lytic effector function on native tumor cells, presumably due to enhanced production and signaling of IL-12 by antigen presenting cells It shows that human SOCS1-silenced DCs have the ability of
ヒトのSOCS−サイレンシング化されたDCによる自己抗原特異的ヒトCTLの完全な活性化
本開示は、ヒトDC中でのヒトSOCS1の調節の役割を実証しており、JAK/STATシグナリング経路のインヒビターをサイレンシング化することによりヒトDCの免疫刺激効能を増強する代替的戦略を提供している。本書で開示している結果は、抗原特異的CTLをプライミングするヒトDCの免疫刺激能力を負に調節する上でのヒトSOCS1の決定的な役割を実証している。ヒトのSOCS1サイレンシング化されたDCは、天然の抗原発現腫瘍細胞に対する頑強な溶解機能を有するヒトCTLを完全に活性化する固有の能力を有する。CTLをプライミングするヒトのSOCS1サイレンシング化されたDCの能力は、IL−12の産生及びシグナリングのSOCS1制限により制御される確率が高い。かくして、本開示は、より有効な腫瘍ワクチンを開発するためのこの一般的に応用可能なSOCS1シグナリングアプローチの翻訳潜在能力を実証している。
Complete activation of autoantigen-specific human CTLs by human SOCS-silenced DCs This disclosure demonstrates a regulatory role for human SOCS1 in human DCs and is an inhibitor of the JAK / STAT signaling pathway Provides an alternative strategy to enhance the immunostimulatory efficacy of human DCs. The results disclosed herein demonstrate the critical role of human SOCS1 in negatively regulating the immunostimulatory ability of human DCs that prime antigen-specific CTL. Human SOCS1 silenced DCs have the inherent ability to fully activate human CTLs with robust lytic function against native antigen-expressing tumor cells. The ability of human SOCS1-silenced DCs to prime CTLs is likely to be controlled by the production of IL-12 and SOCS1 restriction of signaling. Thus, the present disclosure demonstrates the translational potential of this generally applicable SOCS1 signaling approach to develop more effective tumor vaccines.
本発明のSOCS1シグナリングアプローチは、腫瘍関連抗原が負荷されたDCにより誘発された抗原特異的免疫応答を増強する能力を有する。過去10年間に、腫瘍免疫学における主要な進歩は、ヒト腫瘍特異的又は関連抗原を数多く同定し認証したということにあった((ファン・デン・アインデ(Van den Eynde)ら、1997年、Current Opinion in Immunology第9号:684〜93頁))。かくして、腫瘍関連抗原で負荷されたSOCS1サイレンシング化されたDCでのワクチン接種は、抗原特異的抗腫瘍応答を誘発するのに、CTL上でのCTLA4の遮断よりもさらに魅力的なものであると思われる。ヘテロ接合のSOCS1+/−マウスが自己免疫炎症の徴候を全く又は軽度にしか示さないことから、SOCS1サイレンシング化されたDCは重大な自己免疫炎症をひき起こさないかもしれないという点で、SOCS1サイレンシング化されたDC免疫化の使用は、付加的な治療上の利点を提供する。その上、SOCS1−/−マウスの体内にみられる重大な自己免疫炎症には、DC中のみならずT及びNKT細胞といった免疫細胞のその他の系統においてもSOCS1の完全な欠損が必要である(クボら、2003年、Nat.Immunol.第4号:1169〜76頁;アレキサンダーら、2004年、Annu.Rev.Immunol.第22号:503〜29頁;メトカーフ(Metcalf)ら、2003年、Proc.Natl.Acad.Sci.USA第100号「8436〜41頁;チョン(Chong)ら、2003年、Immunity第18号:475〜87頁;ハナダら、2003年、Immunity第19号:437〜50頁;キンジョーら、2002年、Immunity第17号:583〜91頁)。
The SOCS1 signaling approach of the present invention has the ability to enhance antigen-specific immune responses elicited by DCs loaded with tumor-associated antigens. Over the past decade, a major advance in tumor immunology has been the identification and authentication of a number of human tumor-specific or related antigens ((Van den Einde et al., 1997, Current Opinion in Immunology 9: 684-93)). Thus, vaccination with SOCS1-silenced DCs loaded with tumor-associated antigens is more attractive than blocking CTLA4 on CTLs to elicit antigen-specific anti-tumor responses I think that the. SOCS1 silenced DCs may not cause significant autoimmune inflammation since heterozygous SOCS1 +/− mice show no or only mild signs of autoimmune inflammation. The use of silenced DC immunization provides additional therapeutic benefits. Moreover, the serious autoimmune inflammation seen in the body of SOCS1 − / − mice requires a complete deficiency of SOCS1 not only in DC but also in other strains of immune cells such as T and NKT cells. 2003, Nat.Immunol.4: 1169-76; Alexander et al., 2004, Annu.Rev.Immunol.22: 503-29; Metcalf et al., 2003, Proc. Natl.
腫瘍ワクチンの効能を増強する大規模な研究努力は、腫瘍関連抗原の親和性/用量(シグナル1)及び共同刺激分子媒介型シグナル2のレベルの改善に焦点があてられてきた(ギルボア、2004年、Nat.Rev.Cancer第4号:401〜11頁;ローゼンバーグら、2004年、Nat.Med.第10号:909〜15頁)。本開示は、抗原特異的CTLをプライミングするヒトのSOCS1サイレンシング化されたDCのより優れた能力が、IL−12の増強された産生及びシグナリングに起因する確率が高い(シグナル3)ということを示している。この結論は、以下の観察事実に基づいている。すなわち、1)ヒトのSOCS1サイレンシング化されたDCは、病原菌産物での刺激に応答
して、増強されたレベルのIL−12を産生する;2)IL−12の抗体遮断は、ヒトのSOCS1サイレンシング化されたDCの免疫刺激能力を低下させる、そして3)低用量のIL−12のインビボ投与は、野生型DCではなく、SOCS1サイレンシング化されたDCにより誘発された抗原特異的CTL応答を大幅に増強する。これらの結果は、アイルズ(Eyles)J.L、ら(アイルズら、2002年、J.Biol.Chem.第277号;43735〜40頁)によるIL−12シグナリングのマウスSOCS1調節の知見及び本書の他の箇所で開示されている結果によって裏づけされている。
Large-scale research efforts to enhance the efficacy of tumor vaccines have focused on improving the affinity / dose (signal 1) and costimulatory molecule-mediated
サイトカインは、CTLを活性化するためDCにより提供される第3のシグナルとして提案されてきた(カートシンガーら、2003年、J.Exp.Med.第197号:1141〜51頁)。サイトカインの産生及びシグナリングは、過度の自己免疫活性化を制限する一方で外因性抗原に対する免疫応答を活性化するために綿密に調節される(ダーネル(Darnell)ら、1994年、Science第264号:1415〜21頁)。サイトカインは一般にJAKを活性化し、これは次にサイトカインレセプタの細胞質ドメインをリン酸化して、シグナリング物質及び転写活性化剤(STAT)のメンバーのドッキング部位を作り上げる(アレキサンダーら、2004年、Annu.Rev.Immunol.第22号:503〜29頁)。サイトカインは同様に、フィードバックインヒビターとしてSOCS1の発現をアップレギュレートし、このことが次にDCによる前炎症性サイトカインの産生及びシグナリングをオフ切換えし、かくして進行中の免疫応答を減衰させ自己寛容を維持する(アレキサンダーら、2004年、Annu.Rev.Immunol.第22号:503〜29頁)。SOCS1は、そのSH2ドメインを介して偽基質インヒビターとしてJAK活性ループに特異的に結合すること及びユビキチン依存性タンパク質分解のためのJAK2をターゲティングすることにより、STATを抑制する(クボら、2003年、Nat.Immunol.第4号:1169〜76頁; アレキサンダーら、2004年、Annu.Rev.Immunol.第22号:503〜29頁)。合わせて考えると、本書で開示されている結果は、CTL応答の規模を決定する上で抗原提示細胞内のSOCS1により制限されるIL−12の産生及びシグナリングの決定的重要性を表わしている。 Cytokines have been proposed as a third signal provided by DCs to activate CTL (Kartsinger et al., 2003, J. Exp. Med. 197: 1141-51). Cytokine production and signaling are closely regulated to activate an immune response against exogenous antigens while limiting excessive autoimmune activation (Darnell et al., 1994, Science 264: 1415-21). Cytokines generally activate JAK, which in turn phosphorylates the cytoplasmic domain of the cytokine receptor to create docking sites for signaling agents and transcription activator (STAT) members (Alexander et al., 2004, Annu. Rev. Immunol.22: 503-29). Cytokines likewise up-regulate the expression of SOCS1 as feedback inhibitors, which in turn switches off proinflammatory cytokine production and signaling by DCs, thus attenuating the ongoing immune response and maintaining self-tolerance (Alexander et al., 2004, Annu. Rev. Immunol. 22: 503-29). SOCS1 represses STAT by specifically binding to the JAK active loop through its SH2 domain as a pseudosubstrate inhibitor and targeting JAK2 for ubiquitin-dependent proteolysis (Kubo et al., 2003, Nat. Immunol. 4: 1169-76; Alexander et al., 2004, Annu. Rev. Immunol. 22: 503-29). Taken together, the results disclosed herein represent the critical importance of IL-12 production and signaling limited by SOCS1 in antigen presenting cells in determining the magnitude of the CTL response.
本研究の1つの有意な知見は、ヒトのSOCS1サイレンシング化されたDCが活発な溶解エフェクタ機能で自己反応性T細胞を完全に活性化させる固有の能力を有するということにある。臨床及び実験室研究において、四量体染色及びELISPOT検定などのさまざまな免疫検定によって決定されるように、自己抗原特異的T細胞がDCワクチン接種又はインビトロ感作によって活性化され得るということが、頻繁に観察されてきた。しかしながら、このような活性化されたT細胞は、自己抗原を発現するように遺伝子修飾された人工的な抗原パルスを受けた腫瘍細胞を有効に殺すことができるものの、通常、天然腫瘍細胞に対する弱い細胞溶解活性を示し、これが現行の腫瘍ワクチンの効き目の低さの主たる理由である(ザックス(Zaks)ら、1998年、Cancer Research第58号:4902〜8頁;ユー(Yu)ら、2002年、J.Clin.Invest.第110号:289〜94頁)。本書で開示されている結果は、SOCS1サイレンシング化されたDCにより提供される持続的かつ増強された抗原提示/刺激が、自己反応性の低親和性T細胞を完全に活性化するため及び天然の腫瘍細胞に対する活発な溶解エフェクタ機能をそれらに与えるために必要とされ得るということを示唆している。 One significant finding of this study is that human SOCS1-silenced DCs have the unique ability to fully activate autoreactive T cells with active lytic effector function. In clinical and laboratory studies, autoantigen-specific T cells can be activated by DC vaccination or in vitro sensitization, as determined by various immunoassays such as tetramer staining and ELISPOT assays. Has been observed frequently. However, although such activated T cells can effectively kill tumor cells that have received artificial antigen pulses that have been genetically modified to express self-antigens, they are usually weak against natural tumor cells. It exhibits cytolytic activity, which is the main reason for the low efficacy of current tumor vaccines (Zaks et al., 1998, Cancer Research 58: 4902-8; Yu et al., 2002) J. Clin. Invest. 110: 289-94). The results disclosed herein show that the sustained and enhanced antigen presentation / stimulation provided by SOCS1-silenced DCs fully activates autoreactive, low affinity T cells and naturally Suggests that they may be required to give them active lytic effector function on tumor cells.
実施例12:SOCS1 siRNAオリゴ2重鎖がタンパク質免疫化を増強する
SOCS1 siRNAオリゴ2重鎖でのインビボ免疫化がタンパク質免疫化を増強できるか否かをテストするために、1)BCGを伴うOVAタンパク質での免疫化により誘発されるCTL応答を、2)BCGを伴うOVAタンパク質及びSOCS1−siRNAオリゴ2重鎖リポソームでの同時免疫化、と比較した。マウスのグループを、一週間間隔で2回足蹠を介して、OVAタンパク質とBCGの混合物、SOCS1−siRNAオリ
ゴ2重鎖−リポソーム及びOVAタンパク質とBCGの混合物で免疫化させるか又は、OVAタンパク質とBCG及びSOCS1 siRNAオリゴ2重鎖−リポソームの混合物を同時注入した。2回目の免疫化から2週間後に、細胞内サイトカイン染色及びELISPOT検定(IFNγ)から、SOCS1 siRNAオリゴ2重鎖で同時免疫化されたマウスの中により強力なOVA特異的CTL応答が誘発されることがわかる、ということが観察された(図27A及び図27B)。
Example 12: SOCS1 siRNA oligo duplex enhances protein immunization To test whether in vivo immunization with SOCS1 siRNA oligo duplex can enhance protein immunization 1) OVA with BCG The CTL response elicited by protein immunization was compared to 2) co-immunization with OVA protein with BCG and SOCS1-siRNA oligo duplex liposomes. Groups of mice are immunized with a mixture of OVA protein and BCG, SOCS1-siRNA oligo duplex-liposomes and a mixture of OVA protein and BCG, twice a week at a footpad, or with OVA protein A mixture of BCG and SOCS1 siRNA oligo duplex-liposomes was co-injected. Two weeks after the second immunization, intracellular cytokine staining and ELISPOT assay (IFNγ) induce a more potent OVA-specific CTL response in mice co-immunized with SOCS1 siRNA oligo duplex Was observed (FIGS. 27A and 27B).
実施例13:SOCS1サイレンシング化されたCTLは増強された細胞溶解活性を有する
T細胞内のSOCS1がCTL活性を調節する上で1つの役割を果たすか否かを調査するために、OT−I遺伝子導入マウスから単離されたOVAエピトープに特異的な遺伝子導入TCRを有するCD8+OT−I細胞(ジャクソン・ラボラトリー、メーン州バーハーバー)を、ジーンポーターを用いてSOCS1又は突然変異体siRNAオリゴでトランスフェクトした。トランスフェクションを受けたOT−I細胞を、さらなる刺激無しでCTL検定のために使用した。SOCS1−siRNAオリゴのトランスフェクションを受けたOT−Iは、突然変異体siRNA−オリゴでのトランスフェクションを受けたOT−I細胞と比較して同系のOVA陽性EG7細胞に対する増強した細胞溶解活性を有することが観察された(図29)。この結果は、T細胞中のSOCS1シグナリングがそれらの細胞溶解活性を増強することを示している。
Example 13: SOCS1 Silenced CTL Has Enhanced Cytolytic Activity To investigate whether SOCS1 in T cells plays a role in regulating CTL activity, OT-I CD8 + OT-I cells (Jackson Laboratory, Bar Harbor, Maine) with a transgenic TCR specific for the OVA epitope isolated from transgenic mice were transfected with SOCS1 or mutant siRNA oligos using a gene porter. did. Transfected OT-I cells were used for CTL assays without further stimulation. OT-I transfected with SOCS1-siRNA oligo has enhanced cytolytic activity against syngeneic OVA-positive EG7 cells compared to OT-I cells transfected with mutant siRNA-oligo Was observed (FIG. 29). This result indicates that SOCS1 signaling in T cells enhances their cytolytic activity.
実施例14:A20を変調させることによる免疫細胞の免疫能力の増強
A20は、多数の細胞型によって発現されるジンクフィンガータンパク質である。A20は、TLR4リガンド及びTNFによって急速に誘発される。A20は、NF−κBシグナリングに決定的に関与するシグナリングタンパク質のユビキチン化と脱ユビキチン化の調節において2重の機能を有している。A20は、タンパク質中に見出される脱ユビキチン化酵素ドメインを通して脱ユビキチン化に参加する。このドメインは、TNFレセプタ複合体の重要な構成要素であるレセプタ相互作用性タンパク質(RIP)からK63連結型ユビキチン鎖を除去することができる。A20によるRIP上でのK63連結型ユビキチン鎖の除去は、RIP分解を導く。その上、A20のカルボキシル末端にあるジンクフィンガーは、プロテアソーム分解のためにRIPとK48連結型ユビキチン鎖を接合させることによってユビキチンリガーゼとして役立つ。従って、RIPからのK63ユビキチン鎖の除去及びA20によるK48連結型ユビキチン鎖の付加が、RIPの分解に導く。
Example 14: Enhancement of immune capacity of immune cells by modulating A20 A20 is a zinc finger protein expressed by numerous cell types. A20 is rapidly induced by TLR4 ligand and TNF. A20 has a dual function in the regulation of ubiquitination and deubiquitination of signaling proteins critically involved in NF-κB signaling. A20 participates in deubiquitination through the deubiquitinase domain found in proteins. This domain can remove the K63-linked ubiquitin chain from the receptor-interacting protein (RIP), an important component of the TNF receptor complex. Removal of the K63-linked ubiquitin chain on RIP by A20 leads to RIP degradation. Moreover, the zinc finger at the carboxyl terminus of A20 serves as a ubiquitin ligase by joining RIP and K48-linked ubiquitin chains for proteasome degradation. Therefore, removal of K63 ubiquitin chain from RIP and addition of K48-linked ubiquitin chain by A20 leads to degradation of RIP.
さらに、A20は同様に、TRAF6中でK63連結型ユビキチン鎖を脱ユビキチン化し、これがTRAF6の分解という結果をもたらす。TRAF6がTLRファミリーの全メンバーによって共有されている共通のシグナル構成要素であるという事実を考えると、A20は、MyD88依存性及びMyD88非依存性TLRシグナリング経路の両方を抑制することができる固有の負の免疫調節因子である。 Furthermore, A20 similarly deubiquitinates the K63-linked ubiquitin chain in TRAF6, which results in the degradation of TRAF6. Given the fact that TRAF6 is a common signal component shared by all members of the TLR family, A20 is a unique negative that can suppress both MyD88-dependent and MyD88-independent TLR signaling pathways. Is an immunoregulatory factor.
以下の実験及び結果の組合せが、免疫細胞中でA20を変調させることで細胞の免疫能力がもたらされることを実証している。 The following experiment and combination of results demonstrate that modulating A20 in immune cells results in cellular immunity.
ジーンポーターを用いたA20siRNAオリゴでのマウスBM−DCのA20mRNAダウンレギュレーション
DCによる抗原提示のA20調節を調査するため、A20を特定的にダウンレギュレートする小さな干渉性RNA(siRNA)を最初に以下に記述する通りに同定した。
A20 mRNA down-regulation of mouse BM-DC with A20 siRNA oligos using gene porters To investigate the A20 regulation of antigen presentation by DC, a small interfering RNA (siRNA) that specifically down-regulates A20 is first described below. Identified as described.
合成siRNAオリゴ2重鎖を、約83%のトランスフェクション効率でジーンポーターを用い、顆粒球・マクロファージ・コロニー刺激因子(GM−CSF)及びIL−4の
存在下で、エクスビボでマウス骨髄細胞由来のDC内にトランスフェクトした。簡単に言うと、ジーンポーターを用い、メーカーのプロトコルに従って、21塩基対siRNAオリゴヌクレオチドで骨髄DCをトラスフェクトした。3μlの20μMオリゴヌクレオチドを3μlのジーンポーター試薬及び94μlの無血清RPMI1640に添加し、30分間25℃でインキュベートし、その後100μlのジーンポーター/オリゴヌクレオチド混合物を骨髄−DCの各ウェルに添加して、37°Cで4時間インキュベートした。インキュベーション後、20%FBSで補足された1ウェルあたり500μlのRPMI1640を骨髄DCに添加した。本開示に基づき、合成siRNAオリゴは、生理学的に許容可能な担体という状況下で細胞に対して送達することが可能である。許容可能な担体の一例としては、リポソームがある。
Synthetic siRNA oligo duplexes derived from mouse bone marrow cells ex vivo in the presence of granulocytes, macrophages, colony stimulating factor (GM-CSF) and IL-4 using a gene porter with a transfection efficiency of about 83% Transfected into DC. Briefly, bone marrow DCs were transfected with 21 base pair siRNA oligonucleotides using a gene porter and following the manufacturer's protocol. Add 3 μl of 20 μM oligonucleotide to 3 μl of Gene Porter reagent and 94 μl of serum-free RPMI 1640, incubate for 30 minutes at 25 ° C., then add 100 μl of Gene Porter / oligonucleotide mixture to each well of bone marrow-DC, Incubated for 4 hours at 37 ° C. After incubation, 500 μl RPMI 1640 per well supplemented with 20% FBS was added to bone marrow DC. Based on the present disclosure, synthetic siRNA oligos can be delivered to cells in the context of a physiologically acceptable carrier. An example of an acceptable carrier is a liposome.
図31は、A20がA20−siRNAオリゴで形質導入を受けた骨髄−DC中でダウンレギュレートされたことを実証している。定量的RT−PCR検定により判定された通り、20siRNA−2オリゴでのトランスフェクションを受けた全DC集団中のA20mRNAレベルは、A20をダウンレギュレートすることができないA20siRNA突然変異体でのトランスフェクションを受けた骨髄−DC中のレベルと比較しておよそ70%だけ減少した。マウスA20siRNA配列は、5’−CAAAGCACUUAUUGACAGA−3’;配列番号58であった。 FIG. 31 demonstrates that A20 was downregulated in bone marrow-DC transduced with A20-siRNA oligos. As determined by quantitative RT-PCR assay, A20 mRNA levels in the total DC population transfected with the 20 siRNA-2 oligos indicate transfection with an A20 siRNA mutant that is unable to down-regulate A20. There was a reduction of approximately 70% compared to the level in bone marrow-DC received. The mouse A20 siRNA sequence was 5'-CAAAGCACUUAUUGACAGA-3 '; SEQ ID NO: 58.
マウスA20siRNAオリゴDCによって誘発された、増強された抗原特異的T細胞応答
抗原特異的細胞傷害性Tリンパ球(CTL)及びCD4+TヘルパーのDC刺激がA20によって調節されたか否かをテストするために一連の実験を実施した。A20siRNAオリゴ2重鎖でのトランスフェクションを受けたDCが、抗原特異的T細胞応答を誘発する増強された効能を有する否かをテストするために、A20siRNA−2オリゴ2重鎖又は対照オリゴ2重鎖でDCをトランスフェクトした。次に、マウスのグループを、OVAでパルスされたトランスフェクトを受けたDCで免疫化した。パルスを受けたDCを用いたマウスの免疫化の後、インビボで3回PolyI:C(50μg/マウス)を伴って又は伴わずにマウスを刺激した。DC免疫化から2週間後に、免疫化されたマウス体内の免疫応答を検査した。四量体染色及びIFNγELISPOT検定は、インビボPolyI:C刺激を伴って又は伴わずにA20siRNAオリゴ−DCで免疫化されたマウスの体内で増強されたOVA特異的CD8+及びCD4+T細胞応答を示した(図32〜34を参照)。これと一致して、IFNγELISPOT検定は、インビボPolyI:C刺激を伴って又は伴わずにA20siRNAオリゴ−DCで免疫化されたマウスの体内で増強されたTRP2−特異的CD8+T細胞応答を示した(図35)。
Enhanced antigen-specific T cell response elicited by mouse A20 siRNA oligo DC A series of tests to test whether DC stimulation of antigen-specific cytotoxic T lymphocytes (CTLs) and CD4 + T helper was regulated by A20 The experiment was conducted. To test whether DCs transfected with A20 siRNA oligo duplex have an enhanced efficacy in eliciting an antigen-specific T cell response, A20 siRNA-2 oligo duplex or control oligo duplex DCs were transfected with the strands. Next, groups of mice were immunized with DCs transfected with OVA pulsed. After immunization of mice with pulsed DC, mice were stimulated in vivo with or without PolyI: C (50 μg / mouse) three times. Two weeks after DC immunization, the immune response in the immunized mice was examined. Tetramer staining and IFNγ ELISPOT assay showed enhanced OVA-specific CD8 + and CD4 + T cell responses in mice immunized with A20 siRNA oligo-DC with or without in vivo PolyI: C stimulation (FIG. 32-34). Consistent with this, the IFNγ ELISPOT assay showed enhanced TRP2-specific CD8 + T cell responses in mice immunized with A20 siRNA oligo-DC with or without in vivo PolyI: C stimulation (FIG. 35).
マウスA20siRNAオリゴDCにより誘発された増強された抗腫瘍応答
抗腫瘍免疫を誘発するその能力に関してDC中においてA20を阻害することの影響をテストするために、DCをA20siRNA−2オリゴ2重鎖又は対照オリゴ2重鎖でトランスフェクトした。マウスのグループを、OVA抗原でパルスされたトランスフェクトを受けたDCで免疫化した。パルスを受けたDCを用いたマウスの免疫化の後、インビボで3回CpG(50μg/マウス)を伴って又は伴わずにマウスを刺激した。DC免疫化から2週間後に、免疫化されたマウスにOVA+EG7腫瘍で抗原投与した。インビボ刺激を伴って又は伴わずに免疫化されたマウスの体内で、増強された抗腫瘍活性が観察された(図36及び37)。siA20オリゴ−トランスフェクトしたDCを受けた免疫化が、インビボで事前準備されたEG.7腫瘍を根絶する観察された。
Enhanced anti-tumor response elicited by murine A20 siRNA oligo DC To test the effect of inhibiting A20 in DC with respect to its ability to elicit anti-tumor immunity, DC was tested with A20 siRNA-2 oligo duplex or control Transfected with oligo duplex. Groups of mice were immunized with transfected DCs pulsed with OVA antigen. After immunization of mice with pulsed DC, mice were stimulated in vivo with or without CpG (50 μg / mouse) three times. Two weeks after DC immunization, the immunized mice were challenged with OVA + EG7 tumors. Enhanced anti-tumor activity was observed in immunized mice with or without in vivo stimulation (FIGS. 36 and 37). Immunizations that received siA20 oligo-transfected DC were pre-prepared in vivo in EG. 7 tumors were observed to eradicate.
マウスA20siRNAオリゴDCによって誘発された増強された抗体応答
A20siRNAオリゴ2重鎖トランスフェクションを受けたDCが、抗原特異的抗体応答を誘発する増強された効能を有するか否かをテストするために以下の実験を設計した
。簡単に言うと、DCをA20siRNA−2オリゴ2重鎖又は対照オリゴ2重鎖を用いてトランスフェクトした。次にマウスのグループを、OVAでパルスされたトランスフェクションを受けたDCで免疫化した。パルスを受けたDCを用いたマウスの免疫化の後、マウスを、インビボで3回PolyI:C(50μg/マウス)で刺激した。DC免疫化から2週間後に、免疫化されたマウスの体内の抗体応答を検査した。ELISA検定は、A20siRNAオリゴ−DCで免疫化したマウスの体内の増強されたOVA特異的抗体応答を示した(図38)。さらに、SUMO1siRNAオリゴ−DC及びFoxj1 siRNAオリゴ−DCも同様に、免疫化されたマウスの体内でOVA−特異的抗体応答を誘発する増強された効能を示した(図38)。
Enhanced antibody response elicited by murine A20 siRNA oligo DCs To test whether DCs that received A20 siRNA oligo duplex transfections have an enhanced efficacy to elicit an antigen-specific antibody response: The experiment was designed. Briefly, DCs were transfected with A20 siRNA-2 oligo duplex or control oligo duplex. A group of mice was then immunized with transfected DCs pulsed with OVA. After immunization of mice with pulsed DCs, mice were stimulated with PolyI: C (50 μg / mouse) three times in vivo. Two weeks after DC immunization, antibody responses in the immunized mice were examined. The ELISA assay showed an enhanced OVA specific antibody response in the body of mice immunized with A20 siRNA oligo-DC (FIG. 38). Furthermore, SUMO1 siRNA oligo-DC and Foxj1 siRNA oligo-DC also showed enhanced efficacy in inducing OVA-specific antibody responses in immunized mice (FIG. 38).
ウイルスベクターでの形質導入を受けたマウスBM−DCの増強された免疫刺激効能
A20がインビボでのDC抗原提示を負に調節するか否かを評価するために以下の実験を設計した。簡単に言うと、A20siRNA又は対照緑色蛍光タンパク質(GFP)siRNAをレンチウイルスベクター(LV)中にクローニングした。レンチウイルスベクターは、DCを安定した形で形質導入する能力をもち(ルビンソン(Rubinson)ら、2003年、Nat.Genet.第33号:401〜406頁;スクロアースら、2004年、Methods Mol.Biol.第246号:451〜459頁)、そのためA20サイレンシング化の効果をより高い信頼性で評価し得ると考えられることから選択された。本書中の他の個所で記述されている方法に従い、共に黄色蛍光タンパク質(YFP)マーカーを含むLV−A20−siRNA及びLV−GFP−siRNAという2つの構成体が生成された(図33)。LV−A20−siRNA又はLV−GFP−siRNAベクターのいずれかを用いた骨髄由来のDCの形質導入(>80%CD11c+)は、YFPについて陽性である約58〜63%の培養細胞を日常的に生成した。LV−GFP−siRNA−DCの場合と比較してLV−A20−siRNA−DCの刺激時点で、形質導入を受けた全DC集団中のより低いレベルのA20mRNA及び増強された前炎症性サイトカインの分泌が観察された。
Enhanced immunostimulatory efficacy of mouse BM-DCs transduced with viral vectors The following experiment was designed to evaluate whether A20 negatively regulates DC antigen presentation in vivo. Briefly, A20 siRNA or control green fluorescent protein (GFP) siRNA was cloned into a lentiviral vector (LV). Lentiviral vectors have the ability to transduce DCs in a stable fashion (Rubinson et al., 2003, Nat. Genet. 33: 401-406; Scroars et al., 2004, Methods Mol. Biol. 246: 451-459), and therefore, it was selected because it is considered that the effect of silencing A20 can be evaluated with higher reliability. According to the methods described elsewhere in this document, two constructs were generated, LV-A20-siRNA and LV-GFP-siRNA, both containing a yellow fluorescent protein (YFP) marker (FIG. 33). Transduction of bone marrow-derived DCs using either LV-A20-siRNA or LV-GFP-siRNA vectors (> 80% CD11c + ) routinely resulted in approximately 58-63% cultured cells positive for YFP Generated. Secretion of lower levels of A20 mRNA and enhanced pro-inflammatory cytokines in the total transduced DC population at the time of stimulation of LV-A20-siRNA-DC compared to LV-GFP-siRNA-DC Was observed.
次の実験セットは、LV−A20−siRNA−DCが増強された免疫応答誘発能力を有するか否かをテストするように設計された。マウスのグループを、OVA又はTRP2でパルスを受けたLV−A20−siRNA−DCで免疫化した。パルスを受けたDCを用いたマウスの免疫化の後に、インビボ刺激の不在下でマウスを刺激した。DC免疫化から2週間後に、免疫化されたマウス体内の免疫応答を検査した。細胞内IFNγ染色は、LV−A20−siRNA−DCで免疫化されたマウスにおいて、増強されたTRP2特異的CD8+T細胞又はOVA特異的CD4+T細胞応答を示した(図39及び40)。 The next set of experiments was designed to test whether LV-A20-siRNA-DC has an enhanced ability to elicit an immune response. Groups of mice were immunized with LV-A20-siRNA-DC pulsed with OVA or TRP2. Following immunization of mice with pulsed DC, mice were stimulated in the absence of in vivo stimulation. Two weeks after DC immunization, the immune response in the immunized mice was examined. Intracellular IFNγ staining showed an enhanced TRP2-specific CD8 + T cell or OVA-specific CD4 + T cell response in mice immunized with LV-A20-siRNA-DC (FIGS. 39 and 40).
抗腫瘍応答を誘発するLV−A20−siRNA−DCの能力をテストするために、C57BL/6マウスにB16−OVA腫瘍細胞を接種した。3日後、マウスを、LPSでエクスビボ刺激されたOVA−パルスを受けた成熟LV−A20−siRNA−DC又はLV−GFP−siRNA−DCで1回処置した。LV−A20−siRNA−DCが事前準備されたB16−OVA腫瘍の成長を効果的に遮断することが観察された(図41及び42)。LV−Foxj1−siRNA−DCが抗腫瘍応答の誘発において効果的であることも同様に観察された。 To test the ability of LV-A20-siRNA-DC to elicit an anti-tumor response, C57BL / 6 mice were inoculated with B16-OVA tumor cells. Three days later, mice were treated once with mature LV-A20-siRNA-DC or LV-GFP-siRNA-DC that received LPS ex vivo stimulated OVA-pulse. LV-A20-siRNA-DC was observed to effectively block the growth of pre-prepared B16-OVA tumors (FIGS. 41 and 42). It was also observed that LV-Foxj1-siRNA-DC was effective in inducing an anti-tumor response.
LV−A20−siRNA DCにより誘発される、増強されたHIV特異的T細胞応答
A20siRNAオリゴ2重鎖トランスフェクションを受けたDCが、HIVに対する免疫応答を誘発する増強された効能を有するか否かをテストするために以下の実験を設計した。簡単に言うと、DCを、LV−A20siRNA−2又は対照でトランスフェクトした。次にマウスのグループを、HIVgp120でパルスを受けた形質導入されたDCで免疫化した。パルスを受けたDCを用いてマウスを免疫化した後、マウスを、インビボで3回PolyI:C(50μg/マウス)で刺激した。DC免疫化から2週間後に、免
疫化されたマウス体内の免疫応答を検査した。細胞内IFNγ染色は、A20siRNAオリゴ−DCで免疫化したマウスにおいて、増強されたHIVgp120−特異的CD8+T細胞応答を示した(図43)。
Enhanced HIV-specific T cell response elicited by LV-A20-siRNA DCs Whether DCs undergoing A20 siRNA oligo duplex transfection have an enhanced efficacy to elicit an immune response against HIV The following experiments were designed to test. Briefly, DCs were transfected with LV-A20 siRNA-2 or control. Groups of mice were then immunized with transduced DCs pulsed with HIV gp120. After immunization of mice with pulsed DC, mice were stimulated with PolyI: C (50 μg / mouse) three times in vivo. Two weeks after DC immunization, the immune response in the immunized mice was examined. Intracellular IFNγ staining showed an enhanced HIV gp120-specific CD8 + T cell response in mice immunized with A20 siRNA oligo-DC (FIG. 43).
実施例15:Foxj1−サイレンシング化されたDCの増強された免疫刺激効能
何らかの特定の理論に束縛されることは望まないが、IκB発現の誘発は、DCの免疫刺激効能を調節すると考えられている。IκBの発現は動的制御下にあり、IκBは活性化シグナルが無くても定常代謝回転を受け、新しいIκB分子の合成を通して恒常的に補充されて、NF−κBが活性化されないようにしている。Foxo3a及びFoxj1は、IκB転写の活性化因子として近年同定されたフォークヘッド転写因子ファミリーのメンバーである。Foxj1及びFoxo3aは、NF−κBの負の調節に積極的に寄与する。foxj1遺伝子のノックアウトは、子宮内早期死亡に導く。RAG-/-バックグラウンド中にfoxj1-/-リンパ系を伴うキメラ動物は、多臓器全身性炎症、TH1サイトカイン産生の上昇及びT細胞過剰増殖を示すことが分かった。これはNF−κBシグナリング経路の調節異常が原因であると考えられている。
Example 15: Enhanced immunostimulatory efficacy of Foxj1-silenced DC While not wishing to be bound by any particular theory, it is believed that induction of IκB expression modulates the immune stimulatory efficacy of DC Yes. IκB expression is under dynamic control, and IκB undergoes constant turnover without an activation signal and is constantly recruited through the synthesis of new IκB molecules to prevent NF-κB from being activated. . Foxo3a and Foxj1 are members of the forkhead transcription factor family recently identified as activators of IκB transcription. Foxj1 and Foxo3a actively contribute to the negative regulation of NF-κB. The knockout of the foxj1 gene leads to early in utero death. Chimeric animals with foxj1 − / − lymphatic system in the RAG − / − background were found to show multi-organ systemic inflammation, increased TH1 cytokine production and T cell hyperproliferation. This is believed to be due to dysregulation of the NF-κB signaling pathway.
IκBの1つの主要なイソ型であるIκBβは、foxj1欠損T細胞中には不在であり、このことが、休止状態のfoxj1−/−T細胞中のNF−κBの過剰活性化及びTCR刺激に対するこれらの細胞の過敏性を説明している。Foxo3a−欠損T細胞は、foxj1−/−T細胞中の場合のようなIκBβ、そしてIκBεという2つのIκBイソ型の発現の欠落のためにNF−κB活性の高い基底レベルを有し、このことはFoxo3aが、Foxj1と同様に、休止状態のT細胞中におけるIκBの産生に求められることを示している。Foxo3a欠損T細胞は、TCRシグナリング後に過剰増殖とTH1サイトカイン及びTH2サイトカインの両方の実質的上昇を示した。Foxj1及びFoxo3aは、重複するものの、完全に冗長ではない機能を有するように思われる。IκBαの存在は、foxj1−/−及びfoxo3a−/−マウスの両方におけるNF−κBアップレギュレーションに対して観測可能な影響は及ぼさず、このことはNF−κB活性の基底レベル制御において3つのIκBタンパク質中に機能的冗長性が欠落していることを示している。 IκBβ, one major isoform of IκB, is absent in foxj1-deficient T cells, which is responsible for overactivation of NF-κB and TCR stimulation in dormant foxj1 − / − T cells. It explains the hypersensitivity of these cells. Foxo3a-deficient T cells have a high basal level of NF-κB activity due to the lack of expression of two IκB isoforms, IκBβ and IκBε, as in foxj1 − / − T cells, Indicates that Foxo3a is required for production of IκB in resting T cells, similar to Foxj1. Foxo3a deficient T cells showed hyperproliferation and substantial elevation of both TH1 and TH2 cytokines after TCR signaling. Foxj1 and Foxo3a appear to have overlapping but not fully redundant functions. The presence of IκBα has no observable effect on NF-κB up-regulation in both foxj1 − / − and foxo3a − / − mice, indicating that three IκB proteins in basal level regulation of NF-κB activity. It shows that there is a lack of functional redundancy.
抗原提示の調節におけるFoxj1の役割を検査するために以下の実験セットを設計した。図44は、Foxj1が、Foxj1siRNAオリゴ(5’−AGAUCACUCUGUCGGCCAU−3’;配列番号60)でトランスフェクトされた細胞中でダウンレギュレートされたことを実証している。簡単に言うと、ジーンポーターを用いて10:1の比で、Foxj1 siRNAオリゴとFLAG−タグ付きmFoxj1発現ベクターで、293T細胞を同時トランスフェクトした。トランスフェクションから48時間後に、細胞を処理し、対応する細胞溶解物をウェスタンブロット法に付した。ウェスタンブロット法は、ベータアクチンバンドの強度に正規化されたFoxj1バンドの強度を示している。(後続の研究ではFoxj1−siRNAと呼称されている)Foxj1−siRNA2オリゴでのトランスフェクションを受けたDCが、例えばIL−6のような、前炎症性サイトカインの分泌の増強が示す通り、siRNA突然変異体でのトランスフェクトを受けたDCと比較して、IL−6LPSに対する応答性がより高いことが観察された(図45)。 The following experimental set was designed to examine the role of Foxj1 in regulating antigen presentation. FIG. 44 demonstrates that Foxj1 was down-regulated in cells transfected with Foxj1 siRNA oligo (5′-AGAUCACUCUGUCGCCAU-3 ′; SEQ ID NO: 60). Briefly, 293T cells were co-transfected with Foxj1 siRNA oligo and FLAG-tagged mFoxj1 expression vector in a 10: 1 ratio using a gene porter. 48 hours after transfection, cells were treated and the corresponding cell lysates were subjected to Western blotting. Western blot shows the intensity of the Foxj1 band normalized to the intensity of the beta actin band. DCs transfected with Foxj1-siRNA2 oligos (referred to as Foxj1-siRNA in subsequent studies) showed siRNA mutations as shown by enhanced secretion of pro-inflammatory cytokines such as IL-6. Higher responsiveness to IL-6LPS was observed compared to DCs transfected with the mutant (FIG. 45).
Foxj1−siRNAオリゴがIL−6の分泌の増強において効果的であったという事実に基づき、形質導入を受けたDCの免疫刺激効能を増強させるLV−Foxj1−siRNAの能力を査定するために組換え型LV−Foxj1−siRNAを生成した。LV−Foxj1−siRNA又はLV−GFP−siRNAベクターのいずれかを用いたDC形質導入(CD11c+80%超)は、YFPについて陽性である60%の培養細胞を日常的に生成した。形質導入を受けたDCを、LV−A20−siRNA−DCが増強
された免疫応答誘発能力を有するか否かをテストするために用いた。
Based on the fact that Foxj1-siRNA oligos were effective in enhancing IL-6 secretion, recombination to assess the ability of LV-Foxj1-siRNA to enhance the immunostimulatory efficacy of transduced DCs Type LV-Foxj1-siRNA was generated. DC transduction (greater than CD11c + 80%) with either LV-Foxj1-siRNA or LV-GFP-siRNA vectors routinely produced 60% cultured cells that were positive for YFP. Transduced DCs were used to test whether LV-A20-siRNA-DC had an enhanced ability to elicit an immune response.
次にマウスのグループを、例えばTRP2又はHIVgp120といった所望の抗原でのパルスを受けたLV−Foxj1−siRNA−DCで免疫化した。パルスを受けたDCを用いたマウスの免疫化の後、マウスをインビボ刺激無しで刺激した。DC免疫化から2週間後に、免疫化されたマウス体内の免疫応答を検査した。細胞内IFNγ染色は、LV−Foxj1−siRNA−DCで免疫化したマウスの体内で増強されたTRP2特異的CD8+T細胞応答を示した(図46)。さらに、細胞内IFNγ染色は、LV−Foxj1−siRNA−DCで免疫化したマウスで、増強されたHIVgp120特異的CD8+及びCD4+T細胞応答を示した(図47)。 A group of mice was then immunized with LV-Foxj1-siRNA-DC pulsed with the desired antigen, eg, TRP2 or HIV gp120. After immunization of mice with pulsed DC, mice were stimulated without in vivo stimulation. Two weeks after DC immunization, the immune response in the immunized mice was examined. Intracellular IFNγ staining showed enhanced TRP2-specific CD8 + T cell responses in mice immunized with LV-Foxj1-siRNA-DC (FIG. 46). Furthermore, intracellular IFNγ staining showed enhanced HIV gp120-specific CD8 + and CD4 + T cell responses in mice immunized with LV-Foxj1-siRNA-DC (FIG. 47).
実施例16:Twist2−サイレンシング化されたDCの増強された免疫刺激効能
何らかの特定の理論に束縛されることは望まないが、NF−κB標的遺伝子発現の阻害は、DC免疫刺激効能をもたらすと考えられている。Twistは、NF−κBシグナリングの負の免疫調節因子である。哺乳動物中にはTwist−1とTwist−2の2つのtwistタンパク質が存在し、広範囲の生後組織中において低レベルで発現される。twist−2遺伝子の欠損は、結果としての悪液質及び成長障害を伴う多組織中の増強された前炎症性サイトカイン遺伝子発現という結果をもたらす。この表現型は、twist−1及びtwist−2複合ヘテロ接合体中で反復され、サイトカイン発現の阻害におけるこれらの遺伝子の冗長性及び投薬量依存性を反映している。twist−2ノックアウトマウスの表現型は、Twistが通常はNF−κBのp65と結びつきサイトカインプロモータを抑制する負のフィードバックループの喪失を反映しているように思われる。Twistは、サイトカインプロモータ中のEボックスに結合し、隣接するNF−κB部位に結合したNF−κBの活性を阻害する。Twistは、p65とのタンパク質−タンパク質コンタクトを通して、DNA結合とは独立して標的プロモータのNF−κB−媒介型活性化も同様に阻害することができる。
Example 16: Enhanced immunostimulatory efficacy of Twist2-silenced DC Although not wishing to be bound by any particular theory, inhibition of NF-κB target gene expression results in DC immunostimulatory efficacy It is considered. Twist is a negative immune regulator of NF-κB signaling. In mammals, there are two twist proteins, Twist-1 and Twist-2, which are expressed at low levels in a wide range of postnatal tissues. Twist-2 gene deficiency results in enhanced pro-inflammatory cytokine gene expression in multiple tissues with resulting cachexia and growth disorders. This phenotype is repeated in twist-1 and twist-2 complex heterozygotes, reflecting the redundancy and dose dependence of these genes in inhibiting cytokine expression. The phenotype of twist-2 knockout mice appears to reflect the loss of a negative feedback loop that Twist normally associates with p65 of NF-κB and suppresses the cytokine promoter. Twist binds to the E box in the cytokine promoter and inhibits the activity of NF-κB bound to the adjacent NF-κB site. Twist can also inhibit NF-κB-mediated activation of target promoters independently of DNA binding through protein-protein contacts with p65.
抗原提示の調節におけるTwist2の役割を検査するために以下の実験セットを設計した。図48は、Twist2が、Twist2siRNAオリゴでトランスフェクトされた細胞中においてダウンレギュレートされることを実証している。簡単に言うと、ジーンポーターを用いて10:1の比で、Twist2siRNAオリゴ及びFLAG−タグ付きTwist2発現ベクターで293T細胞を同時にトランスフェクトした。トランスフェクションから48時間後に、細胞を処理し、対応する細胞溶解物をウェスタンブロットに付した。ベータアクチンバンドの強度にTwist2バンドの強度を正規化し、相対的強度(比)を示す(図48)。Twist発現は、Twist−2siRNAオリゴ(5’−GCGACGAGAUGGACAAUAA−3’;配列番号61及び5’−CAAGAAAUCGAGCGAAGAU−3’;配列番号62)によってダウンレギュレートされた。 The following experimental set was designed to examine the role of Twist2 in regulating antigen presentation. FIG. 48 demonstrates that Twist2 is downregulated in cells transfected with Twist2 siRNA oligos. Briefly, 293T cells were co-transfected with Twist2 siRNA oligo and FLAG-tagged Twist2 expression vector at a 10: 1 ratio using a gene porter. 48 hours after transfection, the cells were treated and the corresponding cell lysates were subjected to Western blot. The intensity of the Twist2 band is normalized to the intensity of the beta actin band, and the relative intensity (ratio) is shown (FIG. 48). Twist expression was downregulated by Twist-2 siRNA oligos (5'-GCGACGAGAUGGACAAUAA-3 '; SEQ ID NO: 61 and 5'-CAAGAAAUCGAGGCGAAGAU-3'; SEQ ID NO: 62).
Twist2siRNAオリゴ2重鎖でのトランスフェクションを受けたDCが、抗原特異的T細胞応答を誘発する増強された効能を有するか否かをテストするために、Twist2siRNA−2オリゴ2重鎖又は対照オリゴ2重鎖をDCにトランスフェクトした。次にマウスのグループを、OVAでのパルスを受けたトランスフェクトされたDCで免疫化した。パルスを受けたDCを用いたマウスの免疫化の後に、インビボで3回PolyI:C(50μg/マウス)を用いてマウスを刺激した。DC免疫化から2週間後に、免疫化されたマウス体内の免疫応答を検査した。四量体染色及びIFNγELISPOT検定は、インビボPolyI:C刺激を伴うTwist2siRNAオリゴ−DCでの免疫化を受けたマウスの体内で、増強されたOVA−特異的CD8+T細胞応答を示した(図49及び50)。
To test whether DCs transfected with Twist2 siRNA oligo duplex have enhanced potency to elicit an antigen-specific T cell response, Twist2 siRNA-2 oligo duplex or
実施例17:SUMO1サイレンシング化されたDCの増強された免疫刺激効能
SUMOタンパク質は、その輸送、位置づけ及び安定性(別途SUMO化(SUMOylation)としても知られている)を改変することによってその結合パートナーを負に調節する。SUMOは、脊椎動物種中においてSUMO1、SUMO2、SUMO3及びSUMO4という4つのメンバーを有している。SUMO1は、ユビキチンリガーゼによってK48−連結型ユビキチン鎖と接合させることのできる部位であるK21上でIκBαをSUMO化することによってNF−κBシグナリングを負に調節する。SUMOによるIκBαの修飾は、IκBαをユビキチン媒介型分解から保護し、かつその結果としてNF−κBの活性化を阻害する。さらに、保存されたメチオニン(Met55)が内部でバリン残基(M55V)によって置換されているSUMO4の機能的変異体が、NF−κB活性化に対するSUMO4の阻害効果を低減させかつI型糖尿病に関連することが発見された。
Example 17: Enhanced immunostimulatory efficacy of SUMO1-silenced DCs SUMO proteins bind by altering their transport, positioning and stability (also known as SUMOylation). Adjust partner negatively. SUMO has four members in vertebrate species: SUMO1, SUMO2, SUMO3 and SUMO4. SUMO1 negatively regulates NF-κB signaling by sumoylating IκBα on K21, a site that can be joined to a K48-linked ubiquitin chain by ubiquitin ligase. Modification of IκBα by SUMO protects IκBα from ubiquitin-mediated degradation and consequently inhibits NF-κB activation. Furthermore, a functional variant of SUMO4 in which the conserved methionine (Met55) is internally replaced by a valine residue (M55V) reduces the inhibitory effect of SUMO4 on NF-κB activation and is associated with type I diabetes It was discovered that
抗原提示の調節におけるSUMO1の役割を検査するために以下の実験セットを設計した。SUMO1siRNAオリゴ2重鎖でのトランスフェクションを受けたCが、抗原特異的T細胞応答を誘発する増強された効能を有するか否かをテストするために、DCを所望のSUMO1siRNAオリゴ2重鎖(5’−GAUGUGAUUGAAGUUUAUC−3’;配列番号59)でトランスフェクトした。次にマウスのグループを、OVAでのパルスを受けたトランスフェクトされたDCで免疫化した。パルスを受けたDCでのマウスの免疫化の後に、インビボで3回LPS(30μg/マウス)によりマウスを刺激した。DC免疫化から2週間後に、免疫化されたマウス体内の免疫応答を検査した。IFNγELISPOT検定は、インビボPolyI:C刺激を伴うSUMO1siRNAオリゴ(2及び3)−DCでの免疫化を受けたマウスの体内で、増強されたOVA特異的CD8+T細胞応答を示した(図51)。さらに、100ng/mlLPSでの刺激の時点でSUMO1siRNAオリゴ(3)でのトランスフェクションを受けたDC中に観測可能なIKβαのリン酸化反応の増大が存在した(図52)。 The following experimental set was designed to examine the role of SUMO1 in regulating antigen presentation. In order to test whether C transfected with the SUMO1 siRNA oligo duplex has an enhanced efficacy in eliciting an antigen-specific T cell response, the DC is transformed into the desired SUMO1 siRNA oligo duplex (5 '-GAUGUGAUUGAAGUUUAUC-3'; SEQ ID NO: 59). A group of mice was then immunized with transfected DCs pulsed with OVA. Following immunization of mice with pulsed DCs, mice were stimulated with LPS (30 μg / mouse) three times in vivo. Two weeks after DC immunization, the immune response in the immunized mice was examined. The IFNγ ELISPOT assay showed enhanced OVA-specific CD8 + T cell responses in mice immunized with SUMO1 siRNA oligos (2 and 3) -DC with in vivo PolyI: C stimulation (FIG. 51). Furthermore, there was an observable increase in phosphorylation of IKβα in DCs transfected with SUMO1 siRNA oligo (3) at the time of stimulation with 100 ng / ml LPS (FIG. 52).
実施例18:分子安定性の調節に関連する遺伝子の阻害
様々な負の免疫調節因子によって頻繁に利用される戦略は、ユビキチン化/脱ユビキチン化によって主要なシグナリング分子の安定性を調節すること及びSUMO化及びその他の機序によってシグナリング分子複合体の阻害構成要素の安定性を増大させることである。これらの負の免疫調節因子の多くのもの、例えばTRIAD3A、Cyld、SUMO、Clb及びA20などは、標的Toll様レセプタ(TLR)及びシグナリング分子を修飾し、かつそれらの分解を促進してTLR及び腫瘍壊死因子レセプタ(TNFR)シグナル伝達を減衰させるユビキチン修飾酵素である。
Example 18: Inhibition of genes involved in the regulation of molecular stability Strategies frequently utilized by various negative immune regulators regulate the stability of key signaling molecules by ubiquitination / deubiquitination and By increasing the stability of the inhibitory component of the signaling molecule complex by sumoylation and other mechanisms. Many of these negative immunomodulators, such as TRIAD3A, Cyld, SUMO, Clb and A20, modify target Toll-like receptors (TLR) and signaling molecules and promote their degradation to promote TLR and tumors. A ubiquitin-modifying enzyme that attenuates necrosis factor receptor (TNFR) signaling.
TRIAD3Aは、TRIAD3ファミリーのRINGフィンガーE3ユビキチンリガーゼである。TRIAD3Aは、TLR4とTLR9の細胞質ドメインを結合させかつユビキチル化することができ、これがそれらの分解という結果をもたらす。TRIAD3Aの過剰発現は、LPS及びCpGDNAに応答した、TLR4及びTLR9媒介型のNF−κBの活性化を低減させる。従って、本開示に基づくと、本書中に開示されている方法を用いたTRIAD3A発現レベルのダウンレギュレーションは、TLR発現の増強及びインビトロでのLPS及びCpGに対する応答の増大を導き得る。従って、本発明は、TRIAD3Aを阻害することよって免疫細胞の免疫能力を増強させることを含む。 TRIAD3A is a TRIAD3 family RING finger E3 ubiquitin ligase. TRIAD3A can bind and ubiquitylate the cytoplasmic domains of TLR4 and TLR9, which results in their degradation. TRIAD3A overexpression reduces TLR4 and TLR9 mediated activation of NF-κB in response to LPS and CpG DNA. Thus, based on the present disclosure, down-regulation of TRIAD3A expression levels using the methods disclosed herein can lead to enhanced TLR expression and increased responses to LPS and CpG in vitro. Thus, the present invention includes enhancing the immune capacity of immune cells by inhibiting TRIAD3A.
タンパク質の安定性調節に関与するもう一つの遺伝子はCYLDである。CYLDは、956個のアミノ酸でタンパク質をコードする腫瘍サプレッサ遺伝子である。早期終結又はフレームシフト改変を含むCYLD遺伝子の突然変異は、多数の良性皮膚付属器腫瘍に付随する疾病である円柱腫症中で発見された。NF−κBシグナリングカスケード中において、TRAF2及びIKKγは、リジン63(K63)連結型ポリユビキチン鎖と接合
される。プロテアソーム分解のためのK48ポリユビキチン鎖によるタンパク質の通常のユビキチン化とは異なり、TRAF2、IKKγ及び場合によってはTRAF6のK63ユビキチン化は、IKK複合体の組立て及び活性化のために必要である。CYLDは、TRAF2及びIKKγ上で、K63ポリユビキチン鎖を脱ユビキチン化する能力をもち、IKK複合体の分解及びNF−κB活性化の阻害という結果をもたらす。突然変異CYLDの脱ユビキチン化活性の喪失は、NF−κBの持続的活性化と相関している。従って、本書中に開示されている方法を用いた免疫細胞中でのCYLDの阻害は、細胞の免疫能力を増強する。
Another gene involved in protein stability regulation is CYLD. CYLD is a tumor suppressor gene that encodes a protein with 956 amino acids. Mutations in the CYLD gene, including premature termination or frameshift alterations, have been found in columnomatosis, a disease associated with many benign skin appendage tumors. In the NF-κB signaling cascade, TRAF2 and IKKγ are joined to lysine 63 (K63) -linked polyubiquitin chains. Unlike normal ubiquitination of proteins with K48 polyubiquitin chains for proteasome degradation, K63 ubiquitination of TRAF2, IKKγ and possibly TRAF6 is required for assembly and activation of the IKK complex. CYLD has the ability to deubiquitinate the K63 polyubiquitin chain on TRAF2 and IKKγ, resulting in degradation of the IKK complex and inhibition of NF-κB activation. Loss of mutated CYLD deubiquitination activity correlates with sustained activation of NF-κB. Thus, inhibition of CYLD in immune cells using the methods disclosed herein enhances the cell's immunity.
分子の安定性に関与するもう一つの遺伝子は、Cbl(Casitas Bリンパ腫に相当)である。Cblタンパク質ファミリーは3つのメンバー、すなわちCbl、Cbl−b及びCbl−cを有している。Cblタンパク質ファミリーのシグネチュアモチーフは、チロシンキナーゼ結合(TKB)ドメイン及びユビキチン接合酵素(E2)と相互作用するRINGフィンガードメインという、2つの高度に保存されたN−末端ドメインである。これら2つのドメインは、合わさって、活性化されたチロシンキナーゼに向けられたユビキチンリガーゼであるCblタンパク質の基本的機能ユニットを画定している。本書中に提示されている開示に基づくと、Cblを阻害することは、活性化されたチロシンキナーゼ、好ましくは免疫応答の増強に関連するチロシンキナーゼが分解されるのを予防するのに役立つ。従って、本書中に開示されている方法を用いて免疫細胞中のCblを阻害することによって、細胞の免疫能力は増強される。 Another gene involved in molecular stability is Cbl (corresponding to Casitas B lymphoma). The Cbl protein family has three members: Cbl, Cbl-b and Cbl-c. The signature motif of the Cbl protein family is two highly conserved N-terminal domains, the tyrosine kinase binding (TKB) domain and the RING finger domain that interacts with the ubiquitin conjugating enzyme (E2). Together, these two domains define the basic functional unit of the Cbl protein, a ubiquitin ligase directed at activated tyrosine kinases. Based on the disclosure presented herein, inhibiting Cbl helps to prevent degradation of activated tyrosine kinases, preferably tyrosine kinases associated with enhanced immune responses. Thus, by inhibiting Cbl in immune cells using the methods disclosed herein, the immune capacity of the cells is enhanced.
アレスチンファミリーは、分子安定性の調節に関連するタンパク質のもう一つのファミリーである。アレスチンファミリーは、4つのメンバー、すなわちβ−アレスチン1、β−アレスチン2、x−アレスチン及びs−アレスチンを含む。β−アレスチン1及びβ−アレスチン2の1つの機能は、β2−アドレナリン作動性レセプタにより媒介されるセカンドメッセンジャーシグナリングを脱感作することにある。β−アレスチン1及びβ−アレスチン2のもう一つの機能は、IκBαリン酸化反応/分解に対する対策に由来する、NF−κBのシグナル誘発型活性化を消滅させることにあると考えられている。β−アレスチンは、IκBαのカルボキシ末端と結合し、IKKはIκBαのPESTドメインにアクセスできないことになり、PESTドメインのリン酸化反応がIκBα分解を促進することから、IκBαタンパク質の安定化が導かれる。従って、本書中に開示されている方法を用いて免疫細胞中でアレスチンファミリーのメンバーを阻害することによって、細胞の免疫能力は増強される。
The arrestin family is another family of proteins involved in the regulation of molecular stability. The arrestin family includes four members: β-
TOLLIP(Toll相互作用性タンパク質)は、IRAK1のユビキチン化及び後続するその分解を容易にする。TOLLIPはIRAK1と相互作用し、IRAK1の自己リン酸化反応のレベルは、TOLLIPの存在下で減少する。TOLLIPの過剰発現は、TLR2及びTLR4媒介型NF−κB活性化の阻害という結果をもたらす。本書中に提示されている本開示に基づくと、本書中に開示されている方法を用いて免疫細胞中のTOOLIPを阻害することで、細胞の免疫能力は増強される。 TOLLIP (Toll interacting protein) facilitates the ubiquitination of IRAK1 and its subsequent degradation. TOLLIP interacts with IRAK1 and the level of IRAK1 autophosphorylation is reduced in the presence of TOLLIP. TOLLIP overexpression results in inhibition of TLR2 and TLR4 mediated NF-κB activation. Based on the present disclosure presented herein, the immunity of cells is enhanced by inhibiting TOOLIP in immune cells using the methods disclosed herein.
実施例19:その非機能的相同体によるシグナリング分子の阻害
Toll様レセプタ(TLR)及び腫瘍壊死因子レセプタ(TNFR)のシグナリングを調節するためのもう一つの戦略は、刺激活性を全く有していない非機能的相同体又はイソ型によって主要シグナリング分子を競合的に阻害することにある。RP105、ST2、SIGIRR、IRAK−2、IRAK−M、sMyD88及びsTLRといった負の免疫調節因子がこの戦略を例示している。
Example 19: Inhibition of signaling molecules by their non-functional homologues Another strategy for modulating Toll-like receptor (TLR) and tumor necrosis factor receptor (TNFR) signaling has no stimulatory activity Competitive inhibition of major signaling molecules by non-functional homologues or isoforms. Negative immune modulators such as RP105, ST2, SIGIRR, IRAK-2, IRAK-M, sMyD88 and sTLR illustrate this strategy.
可溶性デコイTLR(sTLR)は、TLRと病原菌産物の相互作用を抑制する。可溶性TLR2は、病原菌リガンドとの相互作用についてTLR2と競合することができる。
可溶性TLR4はMD2と相互作用し、MD2−TLR4複合体の形成を阻害することができ、かくしてTLR4によるLPS媒介型シグナリングを遮断する。
Soluble decoy TLR (sTLR) suppresses the interaction of TLR and pathogenic products. Soluble TLR2 can compete with TLR2 for interaction with pathogenic ligands.
Soluble TLR4 can interact with MD2 and inhibit the formation of the MD2-TLR4 complex, thus blocking LPS-mediated signaling by TLR4.
何らかの特定の理論に束縛されることは望まないが、本書中に開示されている方法を用いて免疫細胞中で、RP105、ST2、SIGIRR、IRAK−2、IRAK−M、sMyD88及びsTLRといった負の免疫調節因子を阻害することにより、細胞の免疫能力が増強されると考えられている。 Although not wishing to be bound by any particular theory, negative such as RP105, ST2, SIGIRR, IRAK-2, IRAK-M, sMyD88 and sTLR in immune cells using the methods disclosed herein It is believed that inhibiting immune regulators enhances the immunity of cells.
非機能的TIR相同体
免疫細胞の免疫刺激効能を増強するためのもう一つの戦略は、SIGIRR、ST2及びRP105といった、TIRドメインを含有しかつ刺激活性を有していない膜結合非機能的TIR相同体と結びつけられたタンパク質を阻害することにある。これらのタンパク質は、TLRシグナリングを抑制することができる。RP105はTLR4の相同体であり、APC上で発現される。RP105はMD−1と複合体を形成し、これがTLR4−MD−2と直接相互作用し、LPSに対するTLR4−MD−2複合体の結合を阻害する。RP105は、一次樹状細胞におけるTLR4シグナリング及びインビボでの内毒素に対する応答の生理学的調節因子である。
Non-functional TIR homologs Another strategy for enhancing the immune stimulatory efficacy of immune cells is a membrane-bound non-functional TIR homologue that contains a TIR domain and has no stimulating activity, such as SIGIRR, ST2 and RP105. It is to inhibit proteins associated with the body. These proteins can suppress TLR signaling. RP105 is a homologue of TLR4 and is expressed on APC. RP105 forms a complex with MD-1, which directly interacts with TLR4-MD-2 and inhibits binding of the TLR4-MD-2 complex to LPS. RP105 is a physiological regulator of TLR4 signaling in primary dendritic cells and the response to endotoxin in vivo.
SIGIRR欠損マウスは、LPSにより誘発される内毒素ショックに対して非常に感受性が高いことが分かった。SIGIRRは、シグナリング分子と相互作用するべくTLR4と競合することによってTLR4シグナリングを減衰させる。同様にして、ST2欠損マウスは、LPSに応答した炎症性サイトカインの産生の増大とLPS寛容の誘発不良を示した。ST2がMyD88及びTIRAPと会合しこれらを封鎖したことから、ST2の過剰発現は、NF−κBの活性化を阻害すると考えられている。 SIGIRR-deficient mice were found to be very sensitive to endotoxin shock induced by LPS. SIGIRR attenuates TLR4 signaling by competing with TLR4 to interact with signaling molecules. Similarly, ST2-deficient mice showed increased production of inflammatory cytokines in response to LPS and poor induction of LPS tolerance. Since ST2 associates with MyD88 and TIRAP and sequesters them, overexpression of ST2 is thought to inhibit NF-κB activation.
シグナリング分子イソ型
MyD88及びIRAKといった主要シグナリング分子のいくつかの非機能的イソ型は、TLRシグナリングを抑制することが分かっている。中間ドメインが欠如しているMyD88の代替的にスプライシングされた短い変異体であるMyD88sは、LPSで刺激された場合に単球内で誘発される。MyD88sは、IRAK4に対しての結合しIRAK1のリン酸化反応を促進することができないことから、LPS誘発型NF−κB活性化を阻害する。
Signaling molecule isoforms Several non-functional isoforms of major signaling molecules such as MyD88 and IRAK have been found to suppress TLR signaling. MyD88s, an alternatively spliced short variant of MyD88 that lacks an intermediate domain, is induced in monocytes when stimulated with LPS. MyD88s inhibits LPS-induced NF-κB activation because it cannot bind to IRAK4 and promote IRAK1 phosphorylation.
IRAK−Mはキナーゼ活性が欠如し、APCs中においてTLRシグナリングの包括的な負の免疫調節因子として機能する。キナーゼのIRAKファミリーは、IRAK1、IRAK2、IRAK4及びIRAKMという4つのメンバーを含んでなる。偏在的に発現されるその他のIRAKと異なり、非刺激性IRAK−Mの発現は、単球及びマクロファージに制限され、かつTLRリガンドでの刺激の後に誘発可能な形で発現される。IRAK−M−欠損マウスは、TLRリガンドに応答した炎症性サイトカイン産生の増大及びLPS寛容誘発の欠損を示した。IRAK−Mは、IRAK1及びIRAK4のリン酸化反応を阻害すること、又はTLR−MyD88−IRAK4複合体を安定化させることのいずれかによって、IRAK1−IRAK4複合体のMyD88からの解離を予防し、かくしてIRAK1−TRAF6複合体の形成を予防する。 IRAK-M lacks kinase activity and functions as a global negative immune regulator of TLR signaling in APCs. The IRAK family of kinases comprises four members: IRAK1, IRAK2, IRAK4 and IRAKM. Unlike other ubiquitously expressed IRAKs, unstimulated IRAK-M expression is restricted to monocytes and macrophages and is expressed in an inducible form after stimulation with TLR ligands. IRAK-M-deficient mice showed an increase in inflammatory cytokine production in response to TLR ligands and a deficiency in LPS tolerance induction. IRAK-M prevents dissociation of the IRAK1-IRAK4 complex from MyD88, either by inhibiting the phosphorylation of IRAK1 and IRAK4, or by stabilizing the TLR-MyD88-IRAK4 complex, thus Prevents formation of IRAK1-TRAF6 complex.
IRAK2は、マウスの体内で、IRAK2a、IRAK2b、IRAK2c及びIRAK2dという4つのスプライス変異体を有する。IRAK2c及びIRAK2dには、全長IRAK2中及びIRAK2a及びIRAK2bイソ型中に見出される死ドメインが欠如し、かつLPS刺激の後にマクロファージ中において誘発可能な形で発現される。IRAK2c及びIRAK2dの過剰発現が、LPSで誘発されたNF−κBの活性化を、用量非依存的に阻害し、これにはシグナリングに対する負のフィードバック効果が関与す
ることが示された。
IRAK2 has four splice variants in the mouse body: IRAK2a, IRAK2b, IRAK2c and IRAK2d. IRAK2c and IRAK2d lack the death domain found in full-length IRAK2 and in IRAK2a and IRAK2b isoforms and are expressed in an inducible form in macrophages after LPS stimulation. Overexpression of IRAK2c and IRAK2d was shown to inhibit LPS-induced NF-κB activation in a dose-independent manner, involving a negative feedback effect on signaling.
転写因子のIFN調節因子(IRF)ファミリーの2つのメンバーであるIRF−5及びIRF−7は、MyD88と相互作用し、それぞれ前炎症性サイトカイン及びI型IFNsを誘発させる。しかしながら、IRF−4は同様にMyD88とも相互作用し、TLRシグナリングの負の免疫調節因子として作用する。IRF−4mRNAはTLR活性化によって誘発され、IRF−4はMyD88相互作用のためにIRF−5と競合する。前炎症性サイトカインのTLR依存性誘発は、Irf4遺伝子欠損マウス由来の腹腔マクロファージ中において顕著に増強され、一方、誘発はマクロファージ細胞系統中IRF−4の異所性発現によって阻害される。インビボでのTLRシグナリングにおけるIRF−4の機能の決定的な機能は、Irf4−欠損マウスが、高い血清前炎症性サイトカインレベルを伴って、DNA誘発性のショックに対して過敏性を示すという観察事実によって強調されている。 Two members of the IFN regulator (IRF) family of transcription factors, IRF-5 and IRF-7, interact with MyD88 and induce proinflammatory cytokines and type I IFNs, respectively. However, IRF-4 also interacts with MyD88 and acts as a negative immune regulator of TLR signaling. IRF-4 mRNA is induced by TLR activation and IRF-4 competes with IRF-5 for MyD88 interaction. TLR-dependent induction of pro-inflammatory cytokines is markedly enhanced in peritoneal macrophages from Irf4 gene-deficient mice, while induction is inhibited by ectopic expression of IRF-4 in macrophage cell lines. The critical function of IRF-4 function in TLR signaling in vivo is the observation that Irf4-deficient mice are hypersensitive to DNA-induced shock with high serum proinflammatory cytokine levels Is emphasized by.
FLN29は、TRAF6関連ジンクフィンガーモチーフ及びTANK(TRAFファミリーのメンバー関連NFカッパB活性化因子)関連配列を含有する新規のインターフェロン及びLPS誘発性遺伝子である。マクロファージ様RAW細胞中でのFLN29の発現は、TLR媒介型NF−カッパB及びMAPキナーゼ活性化の抑制という結果をもたらし、一方小分子干渉性RNAによるFLN29の発現の低減は、LPSシグナリングのダウンレギュレーションを部分的に無効にした。さらにTRAF6及びTAB2によって誘発されたNF−カッパB活性化は、FLN29の同時発現によって損なわれたことが実証されており、このことは、FLN29がTRAF6の下流側で調節し得ることを表している。FLN29はTLRシグナリングの負のフィードバック調節因子であると考えられている。 FLN29 is a novel interferon and LPS-inducible gene containing a TRAF6-related zinc finger motif and a TANK (TRAF family member-related NF kappa B activator) -related sequence. Expression of FLN29 in macrophage-like RAW cells results in suppression of TLR-mediated NF-kappa B and MAP kinase activation, while reduction of FLN29 expression by small interfering RNA is a down-regulation of LPS signaling Was partially disabled. Furthermore, it has been demonstrated that NF-kappa B activation induced by TRAF6 and TAB2 was impaired by coexpression of FLN29, indicating that FLN29 can be regulated downstream of TRAF6 . FLN29 is believed to be a negative feedback regulator of TLR signaling.
シグナリング分子複合体の負の構成要素
NF−κBタンパク質は、IκBα、IκBβ及びIκBεを含むIκBタンパク質とのその会合の結果として不活性形態で細胞質中に封鎖されている。IκBタンパク質は、NF−κBサブユニット上の核局在化配列(NLS)をマスキングすることによって、又は核及び細胞質の間の核内輸送プロセスよりも核外輸送プロセスの方をより効果的に促進することによって、細胞質中にNF−κBを保持している。IκBαは、過渡的NF−κB活性化を調節し、IκBβは、持続的なNF−κB活性化を維持する。IκBαは、刺激物に応答して急速に分解され、かつそのプロモータ中にNF−κB応答要素が存在するために急速に再合成される。これとは対照的に、IκBβはIκBαに比べて刺激物誘発型分解に対する反応性が低い。遺伝学研究は、NF−κB活性化を制御する上でのIκBα、IκBβ及びIκBεの全く異なる機能及び冗長性のある機能の両方を示した。IκBαの欠損は、造血組織内における高いNF−κB活性という結果をもたらす。
Negative component of signaling molecule complex The NF-κB protein is sequestered in the cytoplasm in an inactive form as a result of its association with IκB proteins including IκBα, IκBβ and IκBε. The IκB protein promotes the nuclear export process more effectively by masking the nuclear localization sequence (NLS) on the NF-κB subunit or than the nuclear transport process between the nucleus and the cytoplasm By doing so, NF-κB is retained in the cytoplasm. IκBα regulates transient NF-κB activation, and IκBβ maintains sustained NF-κB activation. IκBα is rapidly degraded in response to stimuli and is rapidly re-synthesized due to the presence of the NF-κB response element in its promoter. In contrast, IκBβ is less responsive to irritant-induced degradation than IκBα. Genetic studies have shown both completely different and redundant functions of IκBα, IκBβ and IκBε in controlling NF-κB activation. IκBα deficiency results in high NF-κB activity in hematopoietic tissues.
NF−κBの活性化における決定的な事象は、IKK複合体により媒介されるIκBsのリン酸化反応である。IKK複合体は、TLRリガンド及びTNFを含む様々な刺激物によるNF−κB活性化の合流点である。IκBキナーゼ(IKK)複合体は、2つの触媒サブユニットIKKα及びIKKβを含有し、NF−κB転写因子の活性化を制御する。IKKβは、前炎症性サイトカイン及び病原菌産物に応答してNF−κB活性化を媒介する。 A critical event in the activation of NF-κB is the phosphorylation of IκBs mediated by the IKK complex. The IKK complex is the confluence of NF-κB activation by various stimuli including TLR ligands and TNF. The IκB kinase (IKK) complex contains two catalytic subunits, IKKα and IKKβ, and controls the activation of the NF-κB transcription factor. IKKβ mediates NF-κB activation in response to proinflammatory cytokines and pathogenic products.
IKKαは、NF−κB活性化の制御において負の免疫調節の役割に関与することが実証された。IKKαは、NF−κBサブユニットRelA及びc−Relの分解並びに前炎症性遺伝子プロモータからのRelA及びc−Relの除去の両方を加速することによって、NF−κB活性の抑制に寄与する。マウスの体内でのIKKαの不活性化は、炎症及び細菌クリアランスを増強する(Lawrenceら、2005年Nature第43
4号:1138頁)。
IKKα has been demonstrated to be involved in a negative immune regulatory role in the control of NF-κB activation. IKKα contributes to the suppression of NF-κB activity by accelerating both the degradation of the NF-κB subunits RelA and c-Rel and the removal of RelA and c-Rel from the pro-inflammatory gene promoter. Inactivation of IKKα in the mouse body enhances inflammation and bacterial clearance (Lawrence et al., 2005 Nature 43
4: pp. 1138).
何らかの特定の理論によって拘束されることは望まないが、本書中で論述されている何らかの遺伝子又はそれらの組合せを阻害することは、インヒビターを阻害する方法として役立つ。本開示に基づくと、免疫者の体内で負の免疫調節因子を阻害することによって細胞の免疫能力を増強することができる。 While not wishing to be bound by any particular theory, inhibiting any gene or combination thereof discussed herein serves as a method of inhibiting the inhibitor. Based on the present disclosure, the immune capacity of a cell can be enhanced by inhibiting negative immune regulators in the body of the immunizer.
実施例20:シグナリング分子リン酸化反応の調節
MAPK経路は、MAPKホスファターゼ(MKPs)によるフィードバック阻害を受ける。MKPsは、MAPK経路の活性化後に誘発される。MKPs、MKP5及びMKP6のうちの2つは、そのそれぞれのMAPK標的の活性を阻害することによってT細胞活性化を負に調節する。
Example 20: Modulation of signaling molecule phosphorylation The MAPK pathway undergoes feedback inhibition by MAPK phosphatases (MKPs). MKPs are induced after activation of the MAPK pathway. Two of MKPs, MKP5 and MKP6 negatively regulate T cell activation by inhibiting the activity of their respective MAPK targets.
MKP5は当初、JNK及びp38上で優先的に作用するホスファターゼとして特徴づけされた。Mkp5ノックアウト動物は、JNK活性のアップレギュレーション及びその結果としての先天免疫及び適応免疫の両方の応答の調節異常を示した。MKP6は、CD28コレセプタの細胞質テールと相互作用するタンパク質として同定された。 MKP5 was initially characterized as a phosphatase that acts preferentially on JNK and p38. Mkp5 knockout animals showed upregulation of JNK activity and the resulting dysregulation of both innate and adaptive immunity responses. MKP6 has been identified as a protein that interacts with the cytoplasmic tail of the CD28 coreceptor.
MKP6はCD28が一次ヒトT細胞を同時刺激した際に誘発される。MKP6の優性阻害形態のレトロウイルス駆動型異所性発現は、TCR及びCD28の同時刺激に応答してIL−2の発現を増大させる。IL−2産生の増大は同様に、MKP6ともはや相互作用することができない突然変異体CD28コレセプタを用いることによって得られた。合わせて考えれば、MKP6が、CD28同時刺激に応答したMAPK経路の負の調節に関与するということをこれらの観察が実証している。 MKP6 is induced when CD28 co-stimulates primary human T cells. Retrovirus-driven ectopic expression of a dominant negative form of MKP6 increases IL-2 expression in response to TCR and CD28 costimulation. Increased IL-2 production was also obtained by using a mutant CD28 coreceptor that can no longer interact with MKP6. Taken together, these observations demonstrate that MKP6 is involved in the negative regulation of the MAPK pathway in response to CD28 costimulation.
何らかの特定の理論によって拘束されることは望まないが、ホスファターゼが免疫応答シグナル伝達経路に関与するキナーゼの阻害に関連している1細胞の中でホスファターゼを阻害することは、細胞の免疫能力を増強する1つの方法として役立つ。 While not wishing to be bound by any particular theory, inhibiting phosphatase in one cell where phosphatase is associated with inhibition of kinases involved in the immune response signaling pathway enhances the cell's immune capacity It serves as a way to do this.
実施例21:その他の調節因子機序
Dok−1及びDok−2
Dok−1及びDok−2が元来多数のタンパク質チロシンキナーゼ(PTK)の基質として同定された(シノハラ(Shinohara)ら、2005年、J Exp Med.第3号:333−339)。Dok−1及びDok−2は、PTKの下流側でRas−Erkシグナリングを負に調節するアダプタータンパク質である。LPSは、Dok−1及びDok−2のアダプター機能及びチロシンリン酸化反応を急速に誘発した。Dok−1又はDok−2が欠如したマウス由来のマクロファージのLPSによる刺激は、高いErk活性化を誘発したが、その他のMAPキナーゼ又はNF−κBを誘発せず、TNF−α及び酸化窒素の過剰産生という結果をもたらした。さらに、突然変異体マウスは、TNF−αの過剰産生とLPSに対する過敏性を示した。マクロファージ中のDok−1又はDok−2何らかの強制発現は、LPS−誘発型Erk活性化及びTNF−α産生を阻害した。従って、Dok−1及びDok−2は、TLR4シグナリングの負の免疫調節因子とみなされている。
Example 21: Other Regulator Mechanisms Dok-1 and Dok-2
Dok-1 and Dok-2 were originally identified as substrates for a number of protein tyrosine kinases (PTKs) (Shinohara et al., 2005, J Exp Med. 3: 333-339). Dok-1 and Dok-2 are adapter proteins that negatively regulate Ras-Erk signaling downstream of PTK. LPS rapidly induced adapter function and tyrosine phosphorylation of Dok-1 and Dok-2. Stimulation by LPS of macrophages from mice lacking Dok-1 or Dok-2 induced high Erk activation, but not other MAP kinases or NF-κB, and an excess of TNF-α and nitric oxide The result was production. Furthermore, the mutant mice showed overproduction of TNF-α and hypersensitivity to LPS. Any forced expression of Dok-1 or Dok-2 in macrophages inhibited LPS-induced Erk activation and TNF-α production. Thus, Dok-1 and Dok-2 are considered negative immune regulators of TLR4 signaling.
NOD2
NOD2は、細菌産物ムラミルジペプチド(MDP)を認識することのできるヌクレオチド結合オリゴマー化ドメインファミリのメンバーである。NOD2欠損マウス由来の脾細胞は、TLR2アゴニスへの暴露に応答して高いTH1細胞応答を発生させたが、その他のTLRリガンドに応答してはこれを発生させなかった。従って、NOD2は、TLR2シグナリングの負の免疫調節因子であるとみなされる。これと一致して、クローン病を
患う患者は、NOD2遺伝子ロイシンリッチ反復領域中に突然変異を担持し、これがMDP認識障害という結果をもたらす。さらに、NOD2機能障害は、TLR2刺激での単核細胞の刺激後に前炎症性サイトカインのバイアスという結果をもたらす。
NOD2
NOD2 is a member of the nucleotide-binding oligomerization domain family that can recognize the bacterial product muramyl dipeptide (MDP). Spleen cells from NOD2-deficient mice generated a
PI3K
PI3Kは、p85調節サブユニット及びp110触媒鎖で構成されるヘテロ二量体である。PI3Kは、大半の細胞よって構成的に発現され、数多くの細胞内事象中で早期シグナルとして機能する。p85欠損マウスは、増強されたTLRシグナリング及び優性なTH1細胞応答を示した。PI3K欠損マウス由来の樹状細胞によるIL−12合成は、それぞれTLR4、TLR2及びTLR9リガンドLPS、ペプチドグリカン及びCpGDNAに応答して顕著に増大した。インビボで、感受性の高いBALB/cバックグラウンドのPI3K欠損マウスは、プロトタイプのTH2媒介型寄生虫病であるリーシュマニア・メジャーによる感染に対して野生型マウスよりも耐性が高かった。これらの結果は、PI3KがTLRシグナリングの有効な負の免疫調節因子であり、それがIL−12合成の阻害という結果をもたらし、かつTH1応答の過剰発現を予防することを示している。何らかの特定の理論に束縛されることは望まないが、PI3KがTLRシグナリングを阻害する機序には、p38、JNK、ERK1/ERK2及びNF−κBの抑制が関与していると考えられている。
PI3K
PI3K is a heterodimer composed of a p85 regulatory subunit and a p110 catalytic chain. PI3K is constitutively expressed by most cells and functions as an early signal in many intracellular events. p85-deficient mice showed enhanced TLR signaling and a dominant TH1 cell response. IL-12 synthesis by dendritic cells from PI3K-deficient mice was significantly increased in response to TLR4, TLR2 and TLR9 ligands LPS, peptidoglycan and CpG DNA, respectively. In vivo, susceptible BALB / c background PI3K-deficient mice were more resistant to infection by Leishmania major, a prototype TH2-mediated parasitic disease, than wild-type mice. These results, PI3K is a valid negative immune regulator of TLR signaling, indicating that it resulted in inhibition of IL-12 synthesis and prevent over-expression of T H 1 responses. While not wishing to be bound by any particular theory, it is believed that the mechanism by which PI3K inhibits TLR signaling involves the suppression of p38, JNK, ERK1 / ERK2 and NF-κB.
プロスタグランジン
cyPG15−デオキシ−D12,14−プロスタグランジンJ2(15d−PGJ2)は、天然のリガンド及びシクロペンテノンプロスタグランジンのアゴニストである。15d−PGJ2は、シクロオキシゲナーゼ代謝産物プロスタグランジンD2の脱水及び異性化から誘発される。ペルオキソーム増殖因子活性化レセプタγ(PPARγ)の活性化は、NF−κBの転写活性に拮抗する可能性がある。15d−PGJ2は、IKKβのキナーゼ活性の阻害を通してIBαのリン酸化反応並びに分解を阻害する。15d−PGJ2及びその代謝産物の特徴的な構造は、求電子反応性α,β不飽和カルボニル基を含有するシクロペンテノン環にある。15d−PGJ2のシクロペンタノン環構造によって同様に、NF−κBのDNA結合活性を損なうp50及びp65サブユニット内のシステイン残基の共有結合による修飾を説明することもできる。
Prostaglandin cyPG15-deoxy-D 12,14 -prostaglandin J 2 (15d-PGJ 2 ) is a natural ligand and an agonist of cyclopentenone prostaglandin. 15d-PGJ 2 is induced from the dehydration and isomerization of cyclooxygenase metabolite prostaglandin D 2. Activation of peroxome growth factor activated receptor γ (PPARγ) may antagonize the transcriptional activity of NF-κB. 15d-PGJ 2 inhibits phosphorylation and degradation of IBα through inhibition of the kinase activity of IKK [beta]. The characteristic structure of 15d-PGJ 2 and its metabolites is in the cyclopentenone ring containing electrophilic reactive α, β unsaturated carbonyl groups. Similarly, the cyclopentanone ring structure of 15d-PGJ 2 can explain the covalent modification of cysteine residues in the p50 and p65 subunits that impair the DNA binding activity of NF-κB.
TRAIL−R
TRAILに対するレセプタであるTRAIL−Rは、TIRドメインを有しておらず、TNFスーパーファミリに属している。TRAIL欠損及びTRAILR欠損マウスは、マウスサイトメガロウイルス感染のクリアランスの増強を示し、これはIL−12、IFN−β及びIFN−γのレベルの増大と相関関係を有していた。さらに、TLR2、TLR3及びTLR4リガンドによるマクロファージの刺激は、TRAILR欠損細胞中においてより高レベルのTRAILアップレギュレーションと増強されたサイトカイン産生という結果をもたらした。これらの結果は、TRAILRシグナリングが、TLRの選択的な負の免疫調節因子であることを示している。
TRAIL-R
TRAIL-R, which is a receptor for TRAIL, does not have a TIR domain and belongs to the TNF superfamily. TRAIL-deficient and TRAILR-deficient mice showed enhanced clearance of mouse cytomegalovirus infection, which correlated with increased levels of IL-12, IFN-β and IFN-γ. Furthermore, stimulation of macrophages with TLR2, TLR3 and TLR4 ligands resulted in higher levels of TRAIL up-regulation and enhanced cytokine production in TRAILR-deficient cells. These results indicate that TRAILR signaling is a selective negative immune regulator of TLR.
ロキン(Roquin)(Rc3h1)
ロキンは、RING型E3ユビキチンリガーゼタンパク質ファミリーの高度に保存されたメンバーである。ロキンタンパク質は、RNA−結合タンパク質中に見出されるCCCHジンクフィンガーの存在及びそのシトゾールRNA顆粒への局在化によって特徴づけされる。ロキンは、メッセンジャーRNAの翻訳と安定性の調節に関与していると考えられている。何らかの特定の理論に束縛されることは望まないが、成熟T細胞内のロキン遺伝子の突然変異(そうでなければロキンの阻害)は、過剰な数の濾胞性ヘルパーT細胞及び胚中心を形成させる原因であると考えられている。本開示に基づくと、ロキンを阻害することは、免疫細胞の免疫能力を増強するのに役立ち得る。
Roquin (Rc3h1)
Rokin is a highly conserved member of the RING-type E3 ubiquitin ligase protein family. Rokin proteins are characterized by the presence of CCCH zinc fingers found in RNA-binding proteins and their localization to cytosolic RNA granules. Rokin is thought to be involved in the regulation of messenger RNA translation and stability. Without wishing to be bound by any particular theory, mutations in the rokin gene in mature T cells (otherwise inhibition of rokin) cause an excessive number of follicular helper T cells and germinal centers to form. It is thought to be the cause. Based on the present disclosure, inhibiting rokin may help to enhance the immune capacity of immune cells.
本書中に引用されている各々及び全ての特許、特許出願及び公開の開示は、その全体が本明細書に参照により援用されている。 The disclosures of each and every patent, patent application, and publication cited herein are hereby incorporated by reference in their entirety.
特定の実施形態を参照しながら本発明を開示してきたが、当業者であれば本発明の真の趣旨及び範囲から逸脱することなく、本発明のその他の実施形態及び変形形態も考案し得ることは明白である。添付の特許請求の範囲は、かかる全ての実施形態及び等価の変形形態を包含するように解釈されるものである。 While the invention has been disclosed with reference to specific embodiments, those skilled in the art can devise other embodiments and variations of the invention without departing from the true spirit and scope of the invention. Is obvious. The appended claims are to be construed to include all such embodiments and equivalent variations.
Claims (66)
SMA、PSA、PSCA、チロシナーゼ、MART、TRP、gp100、MART、MAGE、BAGE、GAGE、LAGE/NY−ESO、RAGE、SSX−2、CD19及びCD20よりなる群から選択される請求項31に記載の組成物。 The tumor-associated antigen is BCR-ABL, CASP, CDK, Ras, p53, HER-2 / neu, CEA, MUC, TW1, PAP, survivin, telomerase, EGFR, P
The SMA, PSA, PSCA, tyrosinase, MART, TRP, gp100, MART, MAGE, BAGE, GAGE, LAGE / NY-ESO, RAGE, SSX-2, CD19 and CD20. Composition.
、SUMO3、SUMO4)よりなる群から選択される請求項43に記載の組成物。 The proteins involved in molecular stability are A20 and SUMO (SUMO1, SUMO2
44. The composition of claim 43, selected from the group consisting of: SUMO3, SUMO4).
SUMO3、SUMO4)及びその任意の変異体よりなる群から選択される請求項56に記載の方法。 The protein involved in molecular stability is A20, SUMO (SUMO1, SUMO2,
58. The method of claim 56, selected from the group consisting of SUMO3, SUMO4) and any variant thereof.
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| PCT/US2005/022396 WO2006112869A2 (en) | 2004-07-19 | 2005-06-23 | Modulation of cytokine signaling regulators and applications for immunotherapy |
| PCT/US2006/001751 WO2007001487A2 (en) | 2005-06-23 | 2006-01-19 | Modulation of negative immune regulators and applications for immunotherapy |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2008546775A true JP2008546775A (en) | 2008-12-25 |
| JP2008546775A5 JP2008546775A5 (en) | 2009-02-12 |
Family
ID=37595624
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2008518123A Pending JP2008546775A (en) | 2005-06-23 | 2006-01-19 | Modulation of negative immune regulators and application for immunotherapy |
Country Status (6)
| Country | Link |
|---|---|
| EP (1) | EP1917038A4 (en) |
| JP (1) | JP2008546775A (en) |
| KR (1) | KR20080052549A (en) |
| AU (1) | AU2006262917A1 (en) |
| CA (1) | CA2613166A1 (en) |
| WO (1) | WO2007001487A2 (en) |
Families Citing this family (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2012175735A1 (en) * | 2011-06-23 | 2012-12-27 | Vib Vzw | A20 inhibitors for the treatment of respiratory viral infections |
| CN104830786A (en) * | 2015-05-05 | 2015-08-12 | 杨光华 | DC cell based on HER-2/NEU antigen, targeting immune cell population, preparation method and applications thereof |
| CN116875520B (en) * | 2023-07-12 | 2024-06-14 | 吉林农业大学 | Lactic acid bacteria expressing ribosome inactivating protein and application thereof in resisting rotavirus |
Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2003518507A (en) * | 1999-12-24 | 2003-06-10 | シノヴィス・リミテッド | Activation and inhibition of the immune system |
Family Cites Families (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US5710129A (en) * | 1995-02-23 | 1998-01-20 | Ariad Pharmaceuticals, Inc. | Inhibitors of SH2-mediated processes |
| KR100522483B1 (en) * | 2001-03-01 | 2005-10-18 | 캐논 가부시끼가이샤 | Novel polyhydroxyalkanoate containing unit with phenylsulfanyl structure in the side chain, process for its production, charge control agent, toner binder and toner which contain novel polyhydroxyalkanoate, and image-forming method and image-forming apparatus which make use of the toner |
| US20030171253A1 (en) * | 2001-04-19 | 2003-09-11 | Averil Ma | Methods and compositions relating to modulation of A20 |
-
2006
- 2006-01-19 WO PCT/US2006/001751 patent/WO2007001487A2/en active Application Filing
- 2006-01-19 CA CA002613166A patent/CA2613166A1/en not_active Abandoned
- 2006-01-19 JP JP2008518123A patent/JP2008546775A/en active Pending
- 2006-01-19 KR KR1020087001727A patent/KR20080052549A/en not_active Application Discontinuation
- 2006-01-19 EP EP06718772A patent/EP1917038A4/en not_active Withdrawn
- 2006-01-19 AU AU2006262917A patent/AU2006262917A1/en not_active Abandoned
Patent Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2003518507A (en) * | 1999-12-24 | 2003-06-10 | シノヴィス・リミテッド | Activation and inhibition of the immune system |
Also Published As
| Publication number | Publication date |
|---|---|
| WO2007001487A3 (en) | 2009-04-23 |
| EP1917038A2 (en) | 2008-05-07 |
| CA2613166A1 (en) | 2007-01-04 |
| AU2006262917A1 (en) | 2007-01-04 |
| EP1917038A4 (en) | 2010-01-13 |
| WO2007001487A2 (en) | 2007-01-04 |
| KR20080052549A (en) | 2008-06-11 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US7868159B2 (en) | Modulation of negative immune regulators and applications for immunotherapy | |
| US7868158B2 (en) | Modulation of cytokine signaling regulators and applications for immunotherapy | |
| US8324369B2 (en) | Dendritic cell vaccine compositions and uses of same | |
| Song et al. | An alternative and effective HIV vaccination approach based on inhibition of antigen presentation attenuators in dendritic cells | |
| JP6118491B2 (en) | Methods for increasing immune response | |
| JP2008521410A (en) | Inhibition of Ii expression in mammalian cells | |
| US20090123501A1 (en) | Inhibition of the sh2-domain containing protein tyr-phosphatase, shp-1, to enhance vaccines | |
| WO2007097370A1 (en) | IMMUNOTHERAPY BY USING CELL CAPABLE OF CO-EXPRESSING TARGET ANTIGEN AND CD1d AND PULSED WITH CD1d LIGAND | |
| Jiang | Blockade of B7-H1 enhances dendritic cell-mediated T cell response and antiviral immunity in HBV transgenic mice | |
| WO2018137295A1 (en) | Transgenic lymphocyte co-expressing anti-msln chimeric antigen receptor and immune checkpoint inhibitor molecule and use thereof | |
| US20090220528A1 (en) | Stimulation of Toll-Like Receptors on T Cells | |
| US20090028875A1 (en) | Use of gilz protein expressed in dendritic cells to modulate an antigen-specific immune response | |
| Li et al. | Induction of RNA interference in dendritic cells | |
| JP2008546775A (en) | Modulation of negative immune regulators and application for immunotherapy | |
| US8314076B2 (en) | Method for inhibiting scavenger receptor-A and increasing immune Response to antigens | |
| US20140105885A1 (en) | Method for Inhibiting Scavenger Receptor-A and Increasing Immune Response to Antigens | |
| Chen | DNA-based vaccines against cancer |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| RD02 | Notification of acceptance of power of attorney |
Free format text: JAPANESE INTERMEDIATE CODE: A7422 Effective date: 20081027 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A821 Effective date: 20081027 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20081219 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20081219 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20110823 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20111122 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20111130 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20111226 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20120410 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20120809 |
|
| A911 | Transfer to examiner for re-examination before appeal (zenchi) |
Free format text: JAPANESE INTERMEDIATE CODE: A911 Effective date: 20120824 |
|
| A912 | Re-examination (zenchi) completed and case transferred to appeal board |
Free format text: JAPANESE INTERMEDIATE CODE: A912 Effective date: 20120914 |