WO2011043834A1 - Combinatorial libraries based on c-type lectin domain - Google Patents
Combinatorial libraries based on c-type lectin domain Download PDFInfo
- Publication number
- WO2011043834A1 WO2011043834A1 PCT/US2010/023803 US2010023803W WO2011043834A1 WO 2011043834 A1 WO2011043834 A1 WO 2011043834A1 US 2010023803 W US2010023803 W US 2010023803W WO 2011043834 A1 WO2011043834 A1 WO 2011043834A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- loop
- amino acid
- ctld
- library
- polypeptide
- Prior art date
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/46—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates
- C07K14/47—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates from mammals
- C07K14/4701—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates from mammals not used
- C07K14/4726—Lectins
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/005—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies constructed by phage libraries
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/2866—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against receptors for cytokines, lymphokines, interferons
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/2878—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the NGF-receptor/TNF-receptor superfamily, e.g. CD27, CD30, CD40, CD95
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/10—Processes for the isolation, preparation or purification of DNA or RNA
- C12N15/1034—Isolating an individual clone by screening libraries
- C12N15/1037—Screening libraries presented on the surface of microorganisms, e.g. phage display, E. coli display
-
- C—CHEMISTRY; METALLURGY
- C40—COMBINATORIAL TECHNOLOGY
- C40B—COMBINATORIAL CHEMISTRY; LIBRARIES, e.g. CHEMICAL LIBRARIES
- C40B40/00—Libraries per se, e.g. arrays, mixtures
- C40B40/02—Libraries contained in or displayed by microorganisms, e.g. bacteria or animal cells; Libraries contained in or displayed by vectors, e.g. plasmids; Libraries containing only microorganisms or vectors
-
- C—CHEMISTRY; METALLURGY
- C40—COMBINATORIAL TECHNOLOGY
- C40B—COMBINATORIAL CHEMISTRY; LIBRARIES, e.g. CHEMICAL LIBRARIES
- C40B40/00—Libraries per se, e.g. arrays, mixtures
- C40B40/04—Libraries containing only organic compounds
- C40B40/06—Libraries containing nucleotides or polynucleotides, or derivatives thereof
- C40B40/08—Libraries containing RNA or DNA which encodes proteins, e.g. gene libraries
-
- C—CHEMISTRY; METALLURGY
- C40—COMBINATORIAL TECHNOLOGY
- C40B—COMBINATORIAL CHEMISTRY; LIBRARIES, e.g. CHEMICAL LIBRARIES
- C40B40/00—Libraries per se, e.g. arrays, mixtures
- C40B40/04—Libraries containing only organic compounds
- C40B40/10—Libraries containing peptides or polypeptides, or derivatives thereof
-
- C—CHEMISTRY; METALLURGY
- C40—COMBINATORIAL TECHNOLOGY
- C40B—COMBINATORIAL CHEMISTRY; LIBRARIES, e.g. CHEMICAL LIBRARIES
- C40B50/00—Methods of creating libraries, e.g. combinatorial synthesis
- C40B50/06—Biochemical methods, e.g. using enzymes or whole viable microorganisms
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/505—Medicinal preparations containing antigens or antibodies comprising antibodies
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/10—Immunoglobulins specific features characterized by their source of isolation or production
- C07K2317/14—Specific host cells or culture conditions, e.g. components, pH or temperature
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/30—Immunoglobulins specific features characterized by aspects of specificity or valency
- C07K2317/35—Valency
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/73—Inducing cell death, e.g. apoptosis, necrosis or inhibition of cell proliferation
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/75—Agonist effect on antigen
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/76—Antagonist effect on antigen, e.g. neutralization or inhibition of binding
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/90—Immunoglobulins specific features characterized by (pharmaco)kinetic aspects or by stability of the immunoglobulin
- C07K2317/92—Affinity (KD), association rate (Ka), dissociation rate (Kd) or EC50 value
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2318/00—Antibody mimetics or scaffolds
- C07K2318/20—Antigen-binding scaffold molecules wherein the scaffold is not an immunoglobulin variable region or antibody mimetics
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/10—Processes for the isolation, preparation or purification of DNA or RNA
- C12N15/102—Mutagenizing nucleic acids
- C12N15/1027—Mutagenizing nucleic acids by DNA shuffling, e.g. RSR, STEP, RPR
Definitions
- sequence listing is filed in this application in electronic format only and is incorporated by reference herein.
- sequence listing text file "09-493_Subst_SeqList.txt” was created on March 21 , 2010, and is 385 kilobytes in size.
- This invention relates to polypeptide libraries comprising polypeptides having a C-type lectin domain (CTLD) with a randomized loop region, as well as nucleic acid libraries comprising nucleic acid molecules encoding such polypeptides.
- CTLD C-type lectin domain
- the invention also relates to methods for generating the randomized polypeptides and the polypeptide libraries.
- the invention further relates to methods of screening the polypeptide and nucleic acid libraries based on the specific binding of the modified CTLDs to a target molecule of interest.
- the invention also relates to polypeptides derived from such libraries that bind to target molecules of interest.
- the C-type lectin-like domain is a protein motif that has been identified in a number of proteins isolated from a variety of animal species (reviewed in Drickamer and Taylor ( 993) and Drickamer (1999)). Initially, the CTLD domain was identified as a domain common to the so-called C-type lectins (calcium-dependent carbohydrate binding proteins) and named "Carbohydrate Recognition Domain” ("CRD").
- CTLD canonical domain
- the CTLD contains approximately 120 amino acid residues and, characteristically, contains two or three intra-chain disulfide bridges. Although the primary sequences of CTLDs from different proteins share relatively low amino acid sequence homology, the secondary and tertiary structures of a number of CTLDs are similar, resulting in a highly conserved three dimensional structure, in which the structural variability is essentially confined to the CTLD loop-region.
- the CTLD loop region which typically contains up to five loops, plays a role in ligand and calcium binding.
- Several CTLDs contain either one or two binding sites for calcium and most of the side chains which interact with calcium are located in the loop-region.
- the canonical CTLD is characterized by seven main secondary structure elements (five ⁇ -strands and two a-helices) sequentially appearing in the following order: ⁇ ; al ; a2; ⁇ 2; ⁇ 3; ⁇ 4; and ⁇ 5 (FIG. 1).
- the ⁇ -strands are arranged in two anti-parallel ⁇ -sheets, one composed of ⁇ 1 and ⁇ 5, the other composed of ⁇ 2, ⁇ 3 and ⁇ 4.
- An additional ⁇ -strand, ⁇ often precedes ⁇ in the sequence and, where present, forms an additional strand integrating with the ⁇ , ⁇ 5 sheet.
- two disulfide bridges one connecting al and ⁇ 5 (d-Crv, FIG. 1) and one connecting ⁇ 3 and the polypeptide segment connecting ⁇ 4 and ⁇ 5 (Cu-Cm, FIG. 1) are invariantly found in all CTLDs characterized so far.
- the conserved secondary and tertiary structural elements form a compact scaffold for a number of loops, which in the present context
- LSA loop segment A
- LSB loop segment B
- FIGs 4-6 A schematic of a CTLD, including the loop region, is shown in FIGs 4-6. Residues in LSA, together with single residues in ⁇ 4, have been shown to specify the Ca 2+ - and ligand-binding sites of several CTLDs, including that of tetranectin.
- Tetranectin is a trimeric glycoprotein (Holtet et al. (1 97), Nielsen et al. (1997)) which has been isolated from human plasma and found to be present in the extracellular matrix in certain tissues. Tetranectin is known to bind calcium, complex polysaccharides, plasminogen, fibrinogen/fibrin, and apolipoprotein (a). The interaction with plasminogen and apolipoprotein (a) is mediated by the kringle 4-protein domain therein. This interaction is known to be sensitive to calcium and to derivatives of the amino acid lysine (Graversen et al. (1998)).
- a human tetranectin gene has been characterized, and both human and murine tetranectin cDNA clones have been isolated.
- the mature protein of both the human and murine tetranectin comprises 181 amino acid residues. See US Patent Application Publication 2007/0154901, which is incorporated here in its entirety.
- the three dimensional structures of full length recombinant human tetranectin and of the isolated tetranectin CTLD have been determined independently in two separate studies (Nielsen et al. (1997) and Kastrup et al. (1998)).
- Tetranectin is a two- or possibly three-domain protein, i.e.
- the main part of the polypeptide chain comprises the CTLD (amino acid residues Gly53 to Vail 81), whereas the region Leu26 to Lys52 encodes an alpha- helix governing trimerization of the protein via the formation of a homotrimeric parallel coiled coil.
- the polypeptide segment Glul to Glu25 contains the binding site for complex
- a number of other proteins having CTLDs are known, including the following non- limiting examples: lithostatin, mouse macrophage galactose lectin, Kupffer cell receptor, chicken neurocan, perlucin, asialoglycoprotein receptor, cartilage proteoglycan core protein, IgE Fc receptor, pancreatitis-associated protein, mouse macrophage receptor, Natural Killer group, stem cell growth factor, factor IX/X binding protein, mannose binding protein, bovine conglutinin, bovine CL43, collectin liver 1 , surfactant protein A, surfactant protein D, e-selectin, tunicate c- type lectin, CD94 NK receptor domain, LY49A NK receptor domain, chicken hepatic lectin, trout c-type lectin, HIV gp 120-binding c-type lectin, dendritic cell immunoreceptor, and many snake venom proteins.
- CTLDs are therefore particularly well suited to serve as a basis for constructing new and useful protein products with desired binding properties to target molecules of interest.
- the CTLDs have advantages relative to antibody derivatives as each binding site in a CTLD-based protein product is harbored in a single structurally autonomous protein domain.
- the CTLD domains are resistant to proteolysis, and neither stability nor access to the ligand-binding site is compromised by the attachment of other protein domains to the N- or C-terminus of the CTLD.
- CTLD-based protein products are identical to the corresponding natural CTLD protein already present in the body, and are therefore expected to elicit minimal immunological response in the patient.
- Single CTLDs are about half the mass of an antibody and may in some applications be advantageous as it may provide better tissue penetration and distribution, as well as a shorter half-life in circulation.
- Multivalent formats of CTLD proteins may provide increased binding capacity and avidity and longer circulation half- life.
- the present invention provides combinatorial CTLD polypeptide libraries and methods for identifying and isolating CTLDs to serve as a basis for constructing new and useful protein products with desired binding properties to target molecules of interest.
- the invention provides a combinatorial polypeptide library comprising polypeptide members having a C-type lectin domain (CTLD) with a randomized loop region, in which the randomized loop region has been modified from the native sequence of the CTLD.
- CTLD C-type lectin domain
- the invention provides a combinatorial polypeptide library, and a library of nucleic acids encoding the library of polypeptides, comprising polypeptide members having a C-type lectin domain (CTLD) with a randomized loop region, wherein the loop region of the CTLD is randomized according to one of the following Schemes:
- amino acid modifications in at least one of the four loops in loop segment A (LSA) of the CTLD wherein the amino acid modifications comprise random substitution of at least five amino acids within Loop 1 and random substitution of at least three amino acids within Loop 2;
- amino acid modifications in at least one of the four loops in the loop segment A (LSA) of the CTLD wherein the amino acid modifications comprise at least one amino acid insertion in Loop 3 and random substitution of at least three amino acids within Loop 3;
- amino acid modifications in at least one of the four loops in the loop segment A (LSA) of the CTLD wherein the amino acid modifications comprise a modification that combines two loops into a single loop, wherein the two combined loops are Loop 3 and Loop 4;
- amino acid modifications in at least one of the four loops in the loop segment A (LSA) of the CTLD wherein the amino acid modifications comprise at least one amino acid insertion in Loop 4 and random substitution of at least three amino acids within Loop 4;
- amino acid modifications in at least one of the four loops in the loop segment A (LSA) of the CTLD wherein the amino acid modifications comprise random substitution of at least one amino acid and insertion of at least six amino acids in Loop 3;
- amino acid modifications in at least one of the four loops in the loop segment A (LSA) of the CTLD wherein the amino acid modifications comprise a mixture of (1) random
- amino acid modifications in at least one of the four loops in the loop segment A (LSA) of the CTLD wherein the amino acid modifications comprise at least four or more amino acid insertions in at least one of the four loops in the loop segment A (LSA) or loop 5 in loop segment B (LSB) of the CTLD.
- the CTLD of the polypeptides of the library have the following secondary structure: a. five ⁇ -strands and two a-helices sequentially appearing in the order ⁇ , ⁇ , ⁇ 2, ⁇ 2, ⁇ 3, ⁇ 4, and ⁇ 5, the ⁇ -strands being arranged in two anti-parallel ⁇ -sheets, one composed of ⁇ and ⁇ 5, the other composed of ⁇ 2, ⁇ 3 and ⁇ 4,
- the polypeptides of the library have a random substitution of the amino acid located adjacent the C-terminal end of Loop 2 in the C-terminal direction.
- the CTLD when the CTLD is from human tetranectin, the CTLD can further comprise random substitution of Arginine-130.
- the CTLD when the CTLD is from mouse tetranectin, the CTLD can further comprise random substitution of Leucine- 130.
- the CTLD when the CTLD is from human or mouse tetranectin, the CTLD can further comprise a random substitution of proline 144.
- the polypeptides of the library can have random substitution of one or more amino acids involved in calcium coordination and/or plasminogen binding.
- the CTLD when the CTLD is from tetranectin, the CTLD can further comprise substitution of Lysine- 148 to Alanine (in Loop 4).
- the amino acid modifications when the combinatorial library has the modified CTLD of Scheme (a), the amino acid modifications comprise two amino acid insertions in Loop 1 and random substitution of at least five amino acids within Loop 1. In other embodiments, when the combinatorial library has the modified CTLD of scheme (a) and the CTLD is from human tetranectin, the amino acid modifications comprise at least one amino acid insertion in Loop 1, random substitution of at least five amino acids within Loop 1 , and include a random substitution of Arginine 130.
- the amino acid modifications when the combinatorial library has the modified CTLD of scheme (a) and the CTLD is from human tetranectin, the amino acid modifications comprise two amino acid insertions in Loop 1, random substitution of five amino acids within Loop 1 , and a random substitution of Arginine 130.
- the amino acid modifications when the combinatorial library has the modified CTLD of scheme (a) and the CTLD is from mouse tetranectin, the amino acid modifications comprise two amino acid insertions in Loop 1 , random substitution of five amino acids within Loop 1, and a random substitution of Leucine 130.1n any of the embodiments for scheme (a), the amino acid modifications can further comprise a substitution of Lysine- 148 to Alanine.
- the amino acid modifications include random substitutions of at least five amino acids in Loop 1 , random substitution of at least three amino acids in Loop 2, and include a random substitution of Arginine 130.
- the amino acid modifications include random substitutions of five amino acids in Loop 1 , random substitution of three amino acids in Loop 2, and a random substitution of Arginine 130.
- the amino acid modifications include random substitutions of at least five amino acids in Loop 1 , random substitution of at least three amino acids in Loop 2, and include a random substitution of Leucine 130.
- the amino acid modifications include random substitutions of five amino acids in Loop 1, random substitution of three amino acids in Loop 2, and a random substitution of Leucine 130.
- the amino acid modifications can further comprise a substitution of Lysine- 148 to Alanine.
- individual members of the combinatorial library include loop regions including any or all of the polyeptpide sequences provided by Table 3 in the Examples below.
- the amino acid modifications optionally further comprise random substitution of at least two amino acids.
- the amino acid modifications when the combinatorial library has the modifications of Scheme (c), the amino acid modifications comprise three amino acid insertions within Loop 4 and optionally further comprise random substitution of at least two amino acids.
- the amino acid modifications comprise random substitution of at least seven amino acids within Loop 1 , at least three amino acid insertions in Loop 4, and random substitution of at least two amino acids within Loop 4.
- the amino acid modifications comprise random substitution of seven amino acids within Loop 1 , three amino acid insertions in Loop 4, and random substitution of two amino acids within Loop 4.
- the amino acid modifications comprise random substitution of seven amino acids within Loop 1 , three amino acid insertions in Loop 4, and random substitution of two amino acids within Loop 4.
- individual members of the combinatorial library include loop regions including any or all of the polyeptpide sequences provided by Table 3 in the Examples below.
- the amino acid modifications can further comprise at least one amino acid insertion in Loop 4, and can further comprise random substitution of at least three amino acids within Loop 4.
- the amino acid modifications can comprise three amino acid insertions in Loop 3.
- the amino acid modifications can comprise three amino acid insertions in Loop 4.
- the amino acid modifications comprise random substitution of at least three amino acids within Loop 3, random substitution of at least three amino acids within Loop 4, at least one amino acid insertion in Loop 3 and at least one amino acid insertion in Loop 4.
- the amino acid modifications comprise random substitution of at least three amino acids within Loop 3, random substitution of at least three amino acids within Loop 4, at least three amino acid insertions in Loop 3 and at least three amino acid insertions in Loop 4.
- the amino acid modifications comprise random substitution of three amino acids within Loop 3, random substitution of three amino acids within Loop 4, three amino acid insertions in Loop 3, and three amino acid insertions in Loop 4.
- individual members of the combinatorial library include loop regions including any or all of the polyeptpide sequences provided by Table 3 in the Examples below.
- the amino acid modifications comprise random substitution of at least six amino acids within Loop 3 and random substitution of at least four amino acids within Loop 4. In one specific embodiment, the amino acid modifications comprise random substitution of six amino acids within Loop 3 and random substitution of four amino acids within Loop 4. In any of the embodiments for scheme (e), when the CTLD is from human tetranectin, the amino acid modifications can further comprise random substitution of Proline- 144.
- the amino acid modifications comprise random substitution of six amino acids within Loop 3, random substitution of four amino acids within Loop 4, and a random substitution of proline 144, resulting in a combined Loop 3 and Loop 4 amino acid sequence, comprising, for example, NWEXXXXXXX XGGXXXN (SEQ ID NO: 578), wherein X is any amino acid and wherein the amino acid sequence of SEQ ID NO: 578 forms a single Loop region.
- individual members of the combinatorial library include loop regions including any or all of the polyeptpide sequences provided by Table 3 in the Examples below.
- the amino acid modifications when the combinatorial library has the modified CTLD of Scheme (f) , the amino acid modifications comprise four amino acid insertions in Loop 4. In one embodiment, when the combinatorial library has the modified CTLD of Scheme (f), the amino acid modifications comprise at least four amino acid insertions in Loop 4 and random
- amino acid substitutions comprise four amino acid insertions in Loop 4 and random substitution of three amino acids within Loop 4.
- individual members of the combinatorial library include loop regions including any or all of the polyeptpide sequences provided by Table 3 in the Examples below.
- the amino acid modifications can further comprise one or more amino acid modifications in Loop 4 that modulates plasminogen binding affinity of the CTLD, for example, the substitution of Lysine 148 to Alanine.
- the amino acid modifications when the CTLD is from human or mouse tetranectin, the amino acid modifications comprise random substitution of at least five amino acid residues in Loop 3, random substitution of at least three amino acid residues in Loop 5, and substitution of Lysine 148 to Alanine in Loop 4.
- the amino acid modifications comprises random substitution of five amino acid residue ' s in Loop 3 and random substitution of three amino acid residues in Loop 5, and, in another specific embodiment, when the CTLD is from human or mouse tetranectin, the amino acid modifications further comprise substitution of Lysine 148 to Alanine in Loop 4.
- individual members of the combinatorial library include loop regions including any or all of the polyeptpide sequences provided by Table 3 in the Examples below.
- the amino acid modifications can further comprise one or more amino acid modifications in Loop 4 that modulates plasminogen binding affinity of the CTLD, for example, the substitution of lysine 148 to Alanine.
- the members of the combinatorial library have random substitution of at least one amino acid and insertion of at least six amino acids in Loop 3, and substitution of Lysine 148 to Alanine in Loop 4.
- the amino acid modifications comprise random substitution of one amino acid and insertion of six amino acids in Loop 3.
- the members of the combinatorial library have random substitution of one amino acid and insertion of six amino acids in Loop 3, and substitution of lysine 148 to alanine in Loop 4. In any of these
- CTLD when the CTLD is from human or mouse tetranectin, one of the substitutions is the substitution of Isoleucine 140.
- individual members of the combinatorial library include loop regions including any or all of the polyeptpide sequences provided by Table 3 in the Examples below.
- the amino acid modifications comprise a mixture of random substitution of six amino acids in Loop 3, random substitution of six amino acids and one amino acid insertion in Loop 3, and random substitution of six amino acids and two amino acid insertions in Loop 3.
- the amino acid modifications when the CTLD is from tetranectin, the amino acid modifications further comprise a substitution of Lysine 148 to Alanine in Loop 4.
- the polypeptide members of the combinatorial polypeptide library have one or more amino acid modifications in any combination of two, three, four, or five of the loops in loop segment A (LSA) and loop segment B (LSB).
- the polypeptide members can also comprise a CTLD region having amino acid modifications in regions outside of the LSA and LSB.
- individual members of the combinatorial library include loop regions including any or all of the polyeptpide sequences provided by Table 17 in the Examples below.
- the combinatorial library is composed of polypeptide members having modified loop regions in the CTLD from human or murine tetranectin.
- the polypeptide members can also have an N-terminal extension and/or a C-terminal extension of the CTLD.
- the N-terminal extension and/or C- terminal extension can provide effector ftinction, enzyme function, further binding function, or multimerizing function.
- at least one of the N-terminal extension and the C- terminal extension includes the non-CTLD-portions of a native C-type lectin-Iike protein or C- type lectin or a C-type lectin lacking a functional transmembrane domain.
- the proteins are multimers of a moiety comprising the CTLD.
- the polypeptide members can have additional alterations in the loop regions, introduced by peptide grafting or identified by panning, that can provide effector function, enzyme function, further binding function, or multimerising function.
- the combinatorial library is composed of polypeptide members having modified loop regions in the CTLD region of a full-length human or murine tetranectin.
- the polypeptide members can have an N-terminal extension of the trimerization domain of tetranectin.
- the N-terminal extension can provide effector function, enzyme function, further binding function, or multimerizing function.
- the N- terminal extension is a peptide or a polypeptide with known function or a peptide identified by panning.
- the invention is directed to a library of nucleic acid molecules that encode any of the polypeptides described herein.
- the invention provides a library of nucleic acid molecules encoding polypeptides having a CTLD with a randomized loop region, wherein the loop region of the CTLD is randomized according to any of the Schemes (a)
- the invention provides a library of nucleic acid molecules encoding polypeptides having a CTLD randomized according to any of the Schemes (a) - (i) and having any of the further modifications or sequences described herein.
- the library of nucleic acid molecules can be expressed in a display system having an observable phenotype that represents at least one property of the displayed expression products and the corresponding genotypes.
- suitable display systems include a phage display system; a yeast display system; a viral display system; a cell-based display system; a ribosome- linked display system; or a plasmid-linked display system.
- the invention is directed to a method for generating a combinatorial library of any of the polypeptides described herein.
- the invention provides a method for generating a combinatorial library of polypeptides having a CTLD with a randomized loop region, wherein the loop region of the CTLD is randomized according to any of the Schemes (a)
- the method comprises generating at least one random mutation in at least one of the four loops in the LSA region of the CTLD.
- the method comprises generating at least one random mutation in at least one of the four loops in the LS A region and generating at least one random mutation in the loop in the LBA region of the CTLD.
- the random mutation can be created by oligonucleotide-directed randomization, DNA shuffling by random fragmentation, loop shuffling, loop walking, or error- prone PCR mutagenesis and other methods known in the art.
- the invention provides a method for generating a combinatorial library of polypeptides having a CTLD randomized according to any of the Schemes (a) - (j) and having any of the further modifications or sequences described herein.
- the invention is directed to a method for identifying and isolating a polypeptide having specific binding activity to a target molecule.
- the method comprises providing a combinatorial library of polypeptides having a CTLD wherein the loop region of the CTLD is randomized according to any of the Schemes (a) - (j), contacting the combinatorial polypeptide library with the target molecule under conditions that allow for binding between a polypeptide and the target molecule; and isolating a polypeptide that binds to the target molecule.
- the method comprises providing a combinatorial library of polypeptides having a CTLD randomized according to any of the Schemes (a) - (j) and any of the further modification or sequences described herein, contacting the combinatorial polypeptide library with the target molecule under conditions that allow for binding between a polypeptide and the target molecule; and isolating a polypeptide that binds to the target molecule.
- the method can further include a library of nucleic acid molecules encoding polypeptides of the combinatorial polypeptide library described herein, wherein the library of nucleic acids is expressed in a display system, wherein the display system comprises an observable phenotype that represents at least one property of the displayed expression products and the corresponding genotypes.
- the invention is also directed to a method for the identification and isolation of a polypeptide that specifically binds to a target using a library of nucleic acid molecules.
- the invention provides a method for the identification and isolation of a polypeptide capable of specifically binding to a target comprising the steps of: providing a library of nucleic acids encoding polypeptides having a CTLD with a randomized loop region, wherein the loop region of the CTLD is randomized according to any of Schemes (a) - (j), expressing the nucleic acid library in a display system to obtain an ensemble of polypeptides, in which the amino acid residues at one or more sequence positions differ between different members of said ensemble of polypeptides, contacting the ensemble of polypeptides with said target, and isolating a polypeptide that is capable of specifically binding to said target.
- the method comprises providing a library of nucleic acid molecules encoding polypeptides having a CTLD randomized according to any of the Schemes
- the invention provides a polypeptide having the scaffold structure of a C-type Lectin Like Domain (CTLD), wherein the polypeptide binds to a target other than a natural target for that CTLD and wherein the CTLD scaffold structure of the CTLD is modified according to any of the schemes (a) - (j).
- CTLD scaffold structure is modified according to any of the schemes (a) - (j) and further comprises any of the further modifications described herein, for example, modifications outside the CTLD loop region.
- the polypeptide has the scaffold structure of the CTLD from human or mouse tetranectin and binds to a target other than plasminogen.
- the polypeptide can be produced using a combinatorial library of polypeptides having a CTLD, wherein the loop region of the CTLD is randomized according to any of the Schemes (a) - (j), contacting the combinatorial polypeptide library with the target molecule under conditions that allow for binding between a polypeptide and the target molecule; and isolating a polypeptide that binds to the target molecule, wherein the the target molecule is not the natural target for that CTLD.
- the CTLD is human or mouse tetranectin.
- the CTLD is randomized according to any of the Schemes (a) - (j) and comprises any of the further modifications described herein, for example, modifications outside the CTLD loop region.
- Figure 1 depicts an alignment of the amino acid sequences of ten CTLDs of known three-dimensional structure.
- sequence locations of main secondary structural elements are indicated above each sequence and labeled in sequential numerical order wherein "aX" denotes an a-helix number X, and ⁇ denotes a ⁇ -strand number Y.
- the four cysteine residues involved in the formation of the two conserved disulfide bridges of the CTLDs are indicated and numbered as Ci, Cn, Cm, and Crv, where the disulfide bridges are formed by Ci-Crv and Cn- Cm.
- the various loop regions in the human tetranectin sequence are indicated by underlining.
- CTLDs include: "hTN” (human tetranectin, Nielsen et al., (1997)); “MBP” (mannose binding protein, Weis et al., (1991); Sheriff et al., (1994)); “SP-D” (surfactant protein D, Hakansson et al., (1999)); “LY49A” (N receptor LY49A, Tormo et al., (1999)); “Hl-ASR” (HI subunit of the asialoglycoprotein receptor, Meier et al., (2000)); “MMR-4" (macrophage mannose receptor domain 4, Feinberg et al., (2000)); "IX-A” and “IX-B” (coagulation factors IX/X-binding protein domain A and B, respectively, Mizuno et al., (1997); “Lit” (lithostatine, Bertrand et al., (1996)); and "TU14" (tunicate C
- Figure 2 depicts an alignment of the nucleotide and amino acid sequences of the coding regions of the mature forms of human and murine tetranectin with an indication of known secondary structural elements.
- Figure 3 depicts an alignment of several C-type lectin domains from tetranectins isolated from human (Swissprot P05452), mouse (Swissprot P43025), chicken (Swissprot Q9DDD4), bovine (Swissprot Q2 IS7), Atlantic salmon (Swissprot B5XCV4), frog (Swissprot Q5I0R9), zebrafish (GenBank XP 701303), and related CTLD homologues isolated from cartilage of cattle (Swissprot u22298) and reef shark (Swissprot p26258).
- Figure 4 depicts the three dimensional structure (ribbon format) for human tetranectin, depicting the secondary structural features of the protein. The structure was solved in the Ca - bound form.
- Figure 5A depicts the three dimensional overlay structures of the CTLDs for human tetranectin (HTN) and several tetranectin homologues, including human mannose binding protein (MBP), rat mannose binding protein-C (MBP-C), human surfactant protein D, rat mannose binding protein-A (MBP- A), and rat surfactant protein A.
- the CTLD overlay structures were generated using Swiss PDB Viewer DeepView v. 4.0.1 for Macintosh using the three-dimensional structure of human tetranectin as a template.
- Figure 5B shows the corresponding amino acid sequences of the CTLDS for human tetranectin and the tetranectin homologues depicted in Figure 5 A.
- 1 HUP human mannose binding protein
- 1 BV4A rat mannose binding protein
- 2GGUA human surfactant protein D
- 1 XO A rat mannose binding protein A
- 1R13 rat surfactant protein A.
- FIG. 6A depicts the three dimensional overlay structures of the CTLDs for human tetranectin (HTN) and several tetranectin homologues, including human pancreatitis-associated protein, human dendritic cell-specific ICAM-3 -grabbing non-integrin 2 (DC-SIGNR), rat aggrecan, mouse scavenger receptor, and human scavenger receptor.
- the CTLD overlay structures were generated using Swiss PDB Viewer Deep View v. 4.0.1 for Macintosh using the three-dimensional structure of human tetranectin as a template.
- Figure 6B shows the
- Figure 7 shows the PCR strategy for creating randomized loops in a CTLD.
- Figure 8 shows the DNA and amino acid sequence of the human tetranectin CTLD modified to contain restriction sites for cloning, indicating the Ca2+ binding sites. Restriction sites are underscored with solid lines. Loops are underlined with dashed lines. Calcium coordinating residues are in bold italics and include Site 1 : D 1 16, El 20, CI 47, El 50, Nl 51 ; Site 2: (?143, / 145, £"150, Z 165. The CTLD domain starts at amino acid A45 in bold (i.e.
- Figure 9 depicts a non-limiting strategy for lengthening and introducing randomization in a CTLD loop region.
- Figure 10 shows the results of experiments measuring cell death in the presence of five DR5 ATRIMERsTM: 4a8c, 2a la, la7b, 9b3d and 8b6b. H2122 lung adenocarnoma cells and A2780 ovarian carcinoma cells were incubated at lxl 0 4 cells/well with DR5 ATRIMERsTM (20 ⁇ g/mL) or TRAIL (0.2 ⁇ g mL). Data are expressed as percent cell death relative to the respective buffer control.
- Figure 11 shows the results of an experiment comparing binding of the polypeptides of the invention and native human IL-23 to human IL-23R.
- Figure 12 shows the results of an experiment comparing IL-23-induced IL-17 production in the presence of ATRIMERTM complex 4G8 of the invention, native human IL-23, and Ustekinumab.
- Figure 13 shows the results of an experiment comparing IL-23 induced IL-17 production in the presence of ATRIMERTM complex 1 A4 of the invention and Ustekinumab.
- Figure 14 shows the results of an experiment comparing IL-12-induced IFNy production in the presence of ATRIMERTM complex 4G8 of the invention, native human IL-23, and Ustekinumab.
- Figure 15 shows the results of an experiment comparing Stat-3 phosphorylation in NKL cell in in response to IL-23 and the the polypeptides of the invention.
- Figures 16A and 16B are tables showing experimental results associated with several ATRIMERTM polypeptide complexes of the invention.
- C-type lectin-like protein and “C-type lectin” are used to refer to any protein or polypeptide present in or encoded in the genomes of any eukaryotic species, wherein the protein or polypeptide contains one or more C-type lectin domains (CTLDs) or one or more domains belonging to any subgroup of CTLD, (e.g., the CRDs, which can bind carbohydrate ligands).
- CTLDs C-type lectin domains
- CRDs which can bind carbohydrate ligands
- the definition includes membrane attached C-type lectin-like proteins and C-type lectins, "soluble" C-type lectin-like proteins and C-type lectins lacking a functional transmembrane domain and variant C-type lectin-like proteins and C-type lectins in which one or more amino acid residues have been altered in vivo by glycosylation or any other post-synthetic modification, as well as any product that is obtained by chemical and enzymatic modification of C-type lectin-like proteins and C-type lectins.
- certain alterations can be defined with reference to particular amino acid residue numbers of a CTLD or a CTLD-containing protein. See, Essentials of Glycobiology, second edition. Edited by A. Varki, R.D. Cummings, J.D. Esko, HH. Freeze, P. Stanley, C.R. Bertozzi, G.W. Hart, M.E. Etzler. CHS Press.
- the CTLD consists of roughly 120 amino acid residues and, characteristically, contains two or three intra-chain disulfide bridges. Although the similarity at the amino acid sequence level between CTLDs from different proteins is relatively low, the three dimensional structures of a number of CTLDs have been found to be highly conserved, with the structural variability essentially confined to the loop-region, often defined by up to five loops. Several CTLDs contain either one or two binding sites for calcium and most of the side chains which interact with calcium are located in the loop-region.
- CTLDs for which three dimensional structural information is available, it has been inferred that the canonical CTLD is structurally characterized by seven main
- Figure 1 illustrates an alignment of the CTLDs of known three dimensional structures of ten C-type lectins.
- the ⁇ -strands are arranged in two anti-parallel ⁇ -sheets, one composed of ⁇ and ⁇ 5, the other composed of ⁇ 2, ⁇ 3 and ⁇ 4.
- An additional ⁇ -strand, ⁇ often precedes ⁇ in the sequence and, where present, forms an additional strand integrating with the ⁇ , ⁇ 5-8 ⁇ 66 ⁇ .
- the conserved secondary structure elements form a compact scaffold for a number of loops, which in the present context collectively are referred to as the "loop-region", protruding out from the core.
- these loops are organized in two segments, loop segment A, LSA, and loop segment B, LSB.
- LSA represents the long polypeptide segment connecting ⁇ 2 and ⁇ 3 that often lacks regular secondary structure and contains up to four loops.
- LSB represents the polypeptide segment connecting the ⁇ -strands ⁇ 3 and ⁇ 4.
- Residues in LSA together with single residues in ⁇ 4, have been shown to specify the Ca 2+ - and ligand-binding sites of several CTLDs, including that of tetranectin.
- mutagenesis studies involving substitution of one or a few residues, have shown that changes in binding specificity, Ca 2+ -sensitivity and/or affinity can be accommodated by CTLD domains
- tetranectin tetranectin
- lithostatin mouse macrophage galactose lectin
- upffer cell receptor chicken neurocan
- perlucin asialoglycoprotein receptor
- cartilage proteoglycan core protein IgE Fc receptor
- pancreatitis-associated protein mouse macrophage receptor
- Natural Killer group stem cell growth factor, factor IX X binding protein, mannose binding protein, bovine conglutinin, bovine CL43, collectin liver 1, surfactant protein A, surfactant protein D, e-selectin, tunicate c-type lectin, CD94 NK receptor domain, LY49A NK receptor domain, chicken hepatic lectin, trout c-type lectin, HIV gp 120-binding c-type lectin, and dendritic cell immunoreceptor. See U.S. 2007/0275393, which is incorporated
- amino acid refers to all naturally occurring L-amino acids, as well as non-naturally occurring amino acids. This definition is meant to include norleucine, ornithine, and homocysteine.
- the naturally occurring L-amino acids can be classified according to the chemical composition and properties of their side chains. They are broadly classified into two groups, charged and uncharged. Each of these groups is divided into subgroups to classify the amino acids more accurately: A. Charged Amino Acids - (A. l . Acidic Residues): Asp, Glu; (A.2. Basic Residues): Lys, Arg, His, Orn; B.
- Uncharged Amino Acids - (B.1. Hydrophilic Residues): Ser, Thr, Asn, Gin; (B.2. Aliphatic Residues): Gly, Ala, Val, Leu, He, Nle; (B.3. Non-polar Residues): Cys, Met, Pro, Hey; (B.4. Aromatic
- Residues Phe, Tyr, Trp.
- non-natural amino acid refers to an amino acid that is not one of the 20 common amino acids including, for example, amino acids that occur by modification (e.g. post-translational modifications) of a naturally encoded amino acid (including but not limited to, the 20 common amino acids or pyrolysine and selenocysteine) but are not themselves naturally incorporated into a growing polypeptide chain by the translation complex.
- non-naturally-occurring amino acids include, but are not limited to, N-acetylglucosaminyl-L-serine, N-acetylglucosaminyl-L-threonine, and O-phosphotyrosine.
- amino acid modification(s) and “modification(s)” refer to amino acid substitutions, deletions or insertions or any combinations thereof in an amino acid sequence relative to the native sequence.
- substitutional variants herein are those that have at least one amino acid residue in a native CTLD sequence removed and a different amino acid inserted in its place at the same position.
- the substitutions may be single, where only one amino acid in the molecule has been substituted, or they may be multiple, where two or more amino acids have been substituted in the same molecule.
- Specific reference to more than one amino acid substitution in a CTLD refers to multiple substitutions in which each individual amino acid substitution can occur at any amino acid position within the CTLD, including consecutive and non-consecutive amino acid positions.
- specific reference to more than one amino acid insertion or deletion in a CTLD refers to multiple insertions or deletions in which each individual amino acid insertion or deletion can occur at any amino acid position within the CTLD, including consecutive and non-consecutive amino acid positions.
- nucleic acid molecule encoding refers to the order or sequence of deoxyribonucleotides along a strand of
- deoxyribonucleic acid The order of these deoxyribonucleotides determines the order of amino acids along the polypeptide chain.
- the DNA sequence thus encodes the amino acid sequence.
- Randomize refers to ensembles of polypeptide or nucleic acid sequences or segments, in which the amino acid residue or nucleotide at one or more sequence positions may differ between different members of the ensemble of polypeptides or nucleic acids, such that the amino acid residue or nucleotide occurring at each such sequence position may belong to a set of amino acid residues or nucleotides that may include all possible amino acid residues or nucleotides or any restricted subset thereof.
- modulate or “modulating” when used with reference to either the binding affinity of a CTLD to plasminogen, metal (e.g., Mg 2+ , Ca 2+ , Zn 2+ , Mn 2+ , etc.) or any other target molecule refer to a change in the binding affinity of a modified CTLD polypeptide to either plasminogen or metal ion or target molecule relative to the binding affinity of the native (unmodified) CTLD polypeptide.
- metal e.g., Mg 2+ , Ca 2+ , Zn 2+ , Mn 2+ , etc.
- any other target molecule refer to a change in the binding affinity of a modified CTLD polypeptide to either plasminogen or metal ion or target molecule relative to the binding affinity of the native (unmodified) CTLD polypeptide.
- modulating includes increasing binding affinity, decreasing binding affinity, and/or abolishing or abrogating binding affinity (although not to the exclusion of the specific recitation of the terms “abolishing” or “abrogating” plasminogen, metal ion, or target molecule binding activity).
- binding pair such as ligand/receptor, antibody/antigen, or other binding pair
- binding is measured in a binding reaction which is determinative of the presence of a member of a binding pair in a heterogeneous population of another member of the binding pair.
- binding occurs when one member of the binding pair binds to another member of the binding pair in a heterologous population and does not bind in a significant amount to other proteins or polypeptides present in the sample.
- Specific binding can be measured using the methods described herein, including Biacore and ELISA.
- 1X-2 Library refers to a combinatorial polypeptide library comprising polypeptide members that have a C-type lectin domain (CTLD) comprising amino acid modifications in at least one of the four loops in the LSA of the CTLD, wherein the amino acid modifications comprise at least two amino acid insertions in Loop 1 and random substitution of at least five amino acids within Loop 1 of the CTLD.
- C-type lectin domain C-type lectin domain
- 1-2 library refers to a combinatorial polypeptide library comprising polypeptide members that have a C-type lectin domain (CTLD) comprising amino acid modifications in at least one of the four loops in the LSA of the CTLD, wherein the amino acid modifications comprise random substitution of at least five amino acids within Loop 1 and random substitution of at least three amino acids within Loop 2.
- C-type lectin domain C-type lectin domain
- 1-4 library refers to a combinatorial polypeptide library comprising polypeptide members that have a C-type lectin domain (CTLD) comprising amino acid modifications in at least one of the four loops in the LSA of the CTLD, wherein the amino acid modifications comprise random substitution of at least seven amino acids within Loop 1 , at least three amino acid insertions in Loop 4, and random substitution of at least two amino acids.
- C-type lectin domain C-type lectin domain
- 3X library refers to a combinatorial polypeptide library comprising polypeptide members that have a C-type lectin domain (CTLD) comprising amino acid modifications in at least one of the four loops in the LSA of the CTLD, wherein the amino acid modifications comprise a mixture of random substitution of at least six amino acids, random substitution of at least six amino acids and at least one amino acid substitution, and random substitution of at least six amino acids and at least two amino acid substitutions in Loop 3.
- C-type lectin domain C-type lectin domain
- 3-4X library refers to a combinatorial polypeptide library comprising polypeptide members that have a C-type lectin domain (CTLD) comprising amino acid modifications in at least one of the four loops in the LSA of the CTLD, wherein the amino acid modifications comprise at least three amino acid insertions in Loop 3 and random substitution of at least three amino acids within Loop 3 and comprise at least three amino acid insertions in Loop 4 and random substitution of at least three amino acids within Loop 4.
- C-type lectin domain C-type lectin domain
- 3-4 combo library refers to a combinatorial polypeptide library comprising polypeptide members that have a C-type lectin domain (CTLD) comprising amino acid modifications in at least one of the four loops in the LSA of the CTLD, wherein the amino acid modifications comprise a modification that combines two loops into a single loop, wherein the two combined loops are Loop 3 and Loop 4.
- C-type lectin domain C-type lectin domain
- 4 library refers to a combinatorial polypeptide library comprising polypeptide members that have a C-type lectin domain (CTLD) comprising amino acid modifications in at least one of the four loops in the LSA of the CTLD, wherein the amino acid modifications comprise at least four amino acid insertions in Loop 4 and random substitution of at least three amino acids within Loop 4.
- C-type lectin domain C-type lectin domain
- 3-5 library refers to a combinatorial polypeptide library comprising polypeptide members that have a C-type lectin domain (CTLD) comprising amino acid modifications in at least one of the four loops in the LSA of the CTLD, wherein the amino acid modifications comprise random substitution of at least five amino acids within Loop 3 and random substitution of at least three amino acids within Loop 5.
- C-type lectin domain C-type lectin domain
- Loop 3X loop library refers to a combinatorial polypeptide library comprising polypeptide members that have a C-type lectin domain (CTLD) comprising amino acid modifications in at least one of the four loops in the LSA of the CTLD, wherein the amino acid modifications comprise random substitution of at least one amino acid and at least six amino acid insertions.
- C-type lectin domain C-type lectin domain
- the invention relates generally to a combinatorial polypeptide library comprising polypeptide members having a C-type lectin domain (CTLD) with a randomized loop region, in which the randomized loop region has been modified from the native sequence of the CTLD.
- the randomized loop region of the CTLD can comprise one or more amino acid modifications in at least one of the four loops in the loop segment A (LSA) of the CTLD and can further comprise one or more amino acid modifications in the loop in Loop Segment B (LSB) (also known as loop 5).
- LSA loop segment A
- LSB loop Segment B
- the invention also relates to methods for generating and using the randomized combinatorial polypeptide libraries.
- the libraries and methods of the invention allow for the generation, screening, and identification of protein products that exhibit binding specificity to target molecules of interest.
- CTLDs are therefore particularly well suited to serve as a basis for constructing such new and useful protein products with desired binding properties.
- the invention relates to combinatorial polypeptide libraries comprising modifications to the loop region of the CTLD (LSA and LSB), other modifications to the general CTLD core structure (i.e., the ⁇ -strands and a-helices) can be made without affecting the utility of the libraries described herein.
- One of skill in the art can target particular modifications in the CTLD core structure that will retain CTLD functionality. For example, based on secondary and tertiary structures of various polypeptides comprising CTLDs, hydropathy, charge (ionic), and hydrogen bonding interactions can all be taken into
- variants such as deletion, insertion, or substitution variants in the region outside of the loop region of the CTLD
- percent identity can be as low as 50%.
- variants are at least 80% identical to any given CTLD sequence, or CTLD consensus sequence.
- such variants are at least 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, or 99%, identical to any CTLD sequence, or CTLD consensus sequence.
- CTLD used in the combinatorial libraries can be derived from any CTLD.
- suitable CTLDs are CTLDs described herein (i.e., Figures 1 -3) and in US 2007/0275393, which is incorporated by reference herein in its entirety (i.e., Figure 1 and Table 1) and CTLDs otherwise known in the art.
- the CTLD has the following secondary structure: five ⁇ -strands and two a-helices sequentially appearing in the order ⁇ , ⁇ , ⁇ 2, ⁇ 2, ⁇ 3, ⁇ 4, and ⁇ 5, the ⁇ - strands being arranged in two anti-parallel ⁇ -sheets, one composed of ⁇ and ⁇ 5, the other composed of ⁇ 2, ⁇ 3 and ⁇ 4, at least two disulfide bridges, one connecting al and ⁇ 5 and one connecting ⁇ 3 and the polypeptide segment connecting ⁇ 4 and ⁇ 5, and a loop region containing loop segment A (LSA) and loop segment B (LSB) in which LSA connects ⁇ 2 and ⁇ 3, and LSB connects ⁇ 3 and ⁇ 4.
- LSA loop segment A
- LSB loop segment B
- the CTLD sequence is a human or murine tetranectin CTLD sequence that is modified according to the invention.
- FIG. 2 shows the alignment of the nucleic acid and polypeptide sequences of human and mouse tetranectin CTLDs.
- the CTLD is from a variety of peptides, for example, those shown in FIG.
- the invention provides a polypeptide library comprising polypeptide members that comprise a C-type lectin domain (CTLD), wherein the CTLD comprises one or more amino acid modifications in at least one of the four loops in the loop segment A (LSA) of the CTLD, and/or in the loop in loop segment B (LSB) (Loop 5).
- CTLD comprises one or more amino acid modifications in at least one of the four loops in the loop segment A (LSA) of the CTLD, and/or in the loop in loop segment B (LSB) (Loop 5).
- LSA loop segment A
- LSB loop in loop segment B
- the polypeptide members have CTLDs in which one, two, three, four, or five of the CTLD loops have one or more amino acid modifications, wherein the one or more modifications include at least one amino acid insertion that extends the loop region beyond its original length.
- the one or more modifications include from 1 to about 30 amino acid insertions (e.g., 1 , 2, 3, 4, 5, 6, 7, 8, 9, 10, 1 1 , 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, or 30 amino acid insertions) in any single loop in the loop region (LSA and LSB). In certain of these
- the one or more modifications include at least one amino acid insertion in at least two of the five loops in the loop region (e.g. , two, three, or four loops in LSA or one, two, or three loops in LSA and one loop in LSB).
- the polypeptide libraries comprise polypeptide members that comprise a C-type lectin domain (CTLD), wherein the CTLD comprises one or more amino acid modifications in at least one of the five loops in the loop region (LSA and LSB), wherein certain Ca 2+ coordinating amino acids in the loop regions are retained.
- the polypeptide libraries comprise polypeptide members that comprise a C-type lectin domain (CTLD), wherein the CTLD comprises one or more amino acid modifications in at least one of the five loops in the loop region (LSA and LSB), wherein certain amino acid(s) involved with plasminogen binding activity are eliminated.
- the polypeptide library comprises polypeptide members that comprise a C-type lectin domain (CTLD), wherein the CTLD comprises one or more amino acid modifications in regions of the CTLD that fall outside of the LSA and LSB regions. Accordingly, such modifications can be designed or randomly generated in any one or more of the beta strand and/or alpha helical regions. An example of this is shown in Table 17.
- CTLD C-type lectin domain
- the loop region of any CTLD can be identified by using any variety of structural or sequence-based analysis using the existing sequence based information for any single structurally characterized CTLD or any combination of structurally characterized CTLDs.
- the loop regions are stretches of amino acids found between more ordered regions of the CTLD amino acid sequence (e.g., between the a-helices or ⁇ - strands), and typically have a more flexible conformation.
- Loop segment A (LSA) in a CTLD typically falls between the ⁇ 2 and ⁇ 3 strands of the canonical CTLD motif.
- the (LSA) contains smaller loop regions (loops 1, 2, 3, and 4), which are usually located between small beta sheet structures that provide a degree of order to the (LSA) (see, e.g. , FIG 4).
- CTLDs typically have a smaller loop structure (loop segment B, "LSB” or "loop 5") located between ⁇ 3 and ⁇ 4.
- the loop region of any CTLD can be identified using structural and/or sequence-based analyses based on the existing sequence information for any single structurally characterized CTLD or any combination of structurally characterized CTLDs.
- the location of the loop region of any uncharacterized CTLD can be identified by aligning a prospective CTLD sequence with the group of structure-characterized CTLDs presented in FIG. 1.
- the sequence alignments shown in FIG. 1 were strictly elucidated from actual three dimensional structure data. Given that the polypeptide segments of corresponding structural elements of the framework also exhibit strong amino acid sequence similarities, FIG. 1 provides a set of direct sequence-structure signatures, which can readily be inferred from the sequence alignment. As shown in FIG.
- the loop region (LSA and LSB) is flanked by segments corresponding to the ⁇ 2-, ⁇ 3-, and ⁇ 4-strands (loops 1-4 of LSA typically fall between the ⁇ 2 and ⁇ 3 strands of the canonical CTLD and loop 5 of LSB is typically located between ⁇ 3 and ⁇ 4 of the CTLD).
- the ⁇ 2-, ⁇ 3-, and ⁇ 4-strands can be identified by identification of their respective consensus sequences (published in US Patent Application Publication 2007/0275393).
- the loop region of the prospective CTLD can be identified by aligning the sequence of the prospective CTLD with the sequence shown in FIG.
- the loop regions of a prosective CTLD can be identified using known protein structure modeling programs, such as Swiss PDB Viewer DeepView v. 4.0.1 for Macintosh, by aligning the sequence of prospective CTLD with any of the CTLD sequences in FIG 1.
- protein modeling programs that can be used in the same manner are known in the art and available for public use, for example,
- the sequence-structure analyses demonstrate that CTLDs can be used as frameworks in the construction of new classes of CTLD libraries.
- the additional steps involved in preparing starting materials for the construction of a new class of CTLD library on the basis of a CTLD for which the precise three dimensional structure has not yet been determined includes the following: (1) alignment of the sequence of the new CTLD with the sequence shown in FIG. 1; and (2) assignment of approximate locations of framework structural elements as guided by the sequence alignment, observing any requirement for minor adjustment of the alignment to ensure precise alignment of the four canonical cysteine residues involved in the formation of the two conserved disulfide bridges (C
- the polypeptides comprising a CTLD used in the polypeptide libraries of the invention can be full-length proteins or partial proteins having a CTLD, for example, the full-length amino acid sequence or partial amino acid sequence of any of the proteins described herein and otherwise known.
- the polypeptides comprising a CTLD used in the polypeptide libraries of the invention can be polypeptides comprising only CTLD sequence, for example, the amino acid sequence of any of the CTLDs described herein and otherwise known.
- the polypeptides comprising CTLD sequence can have additional flanking C-terminal and/or N- terminal (non-CTLD) amino acid sequence.
- the invention provides a combinatorial peptide library, and a library of nucleic acid sequences encoding the polypeptides of the library, wherein the CTLDs of the polypeptides have been modified according to a number of schemes, which have been labeled for the purposes of identification only as Schemes (a)-(j). While each scheme is more particularly described herein, the modifications are at least as follows:
- amino acid modifications in at least one of four loops in loop segment A (LSA) of the CTLD wherein the amino acid modifications comprise an insertion of at least one amino acid in Loop 1 and random substitution of at least five amino acids within Loop 1 ;
- amino acid modifications in at least one of four loops in loop segment A (LSA) of the CTLD wherein the amino acid modifications comprise random substitution of at least five amino acids within Loop 1 and random substitution of at least three amino acids within Loop 2;
- amino acid modifications in at least one of four loops in loop segment A (LSA) of the CTLD wherein the amino acid modifications comprise random substitution of at least seven amino acids within Loop 1 and at least one amino acid insertion in Loop 4;
- amino acid modifications in at least one of four loops in loop segment A (LSA) of the CTLD wherein the amino acid modifications comprise at least one amino acid insertion in Loop
- amino acid modifications in at least one of four loops in loop segment A (LSA) of the CTLD wherein the amino acid modifications comprise a modification that combines two loops into a single loop, wherein the two combined loops are Loop 3 and Loop 4;
- amino acid modifications in at least one of four loops in loop segment A (LSA) of the CTLD wherein the amino acid modifications comprise at least one amino acid insertion in Loop
- amino acid modifications in at least one of the four loops in loop segment A (LSA) of the CTLD wherein the amino acid modifications comprise random substitution of at least one amino acid and insertion of at least six amino acids in Loop 3;
- amino acid modifications in at least one of the four loops in the loop segment A (LSA) of the CTLD wherein the amino acid modifications comprise a mixture of (1) random
- amino acid modifications in at least one of the four loops in the loop segment A (LSA) of the CTLD wherein the amino acid modifications comprise at least four or more amino acid insertions in at least one of the four loops in the loop segment A (LSA) or loop 5 in loop segment B (LSB) of the CTLD.
- the invention provides a combinatorial polypeptide library comprising polypeptide members having a randomized C-type lectin domain (CTLD), wherein the randomized CTLD includes amino acid modifications in at least one of the four loops in LSA or in the loop in LSB of the CTLD, wherein the amino acid modifications comprise at least one amino acid insertion in Loop 1 and random substitution of at least five amino acids within Loop 1.
- CTLD C-type lectin domain
- the CTLD when the CTLD is from human tetranectin, the CTLD also has a random substitution of Arginine-130.
- this peptide is located immediately adjacent to the C- terminal peptide of Loop 2 in the C-terminal direction.
- this peptide is Gly-130.
- the CTLD when the CTLD is from tetranectin, for example human or mouse tetranectin, the CTLD includes a substitution of Lysine- 148 to Alanine in Loop 4.
- the amino acid modifications when the combinatorial library has the modified CTLD of Scheme (a), the amino acid modifications comprise two amino acid insertions in Loop 1 and random substitution of at least five amino acids within Loop 1. In other embodiments, when the combinatorial library has the modified CTLD of scheme (a) and the CTLD is from human tetranectin, the amino acid modifications comprise at least one amino acid insertion in Loop 1 , random substitution of at least five amino acids within Loop 1 , and include a random substitution of Arginine 130.
- the amino acid modifications when the combinatorial library has the modified CTLD of scheme (a) and the CTLD is from human tetranectin, the amino acid modifications comprise two amino acid insertions in Loop 1 , random substitution of five amino acids within Loop 1 , and a random substitution of Arginine 130.
- the amino acid modifications when the combinatorial library has the modified CTLD of scheme (a) and the CTLD is from mouse tetranectin, the amino acid modifications comprise two amino acid insertions in Loop 1 , random substitution of five amino acids within Loop 1 , and a random substitution of Leucine 130.
- the amino acid modifications can further comprise a substitution of Lysine-148 to Alanine.
- the CTLD comprises two amino acid insertions in Loop 1 , random substitution of at least five amino acids within Loop 1 , random substitution of Arginine- 130 or other amino acid located outside and adjacent to loop 2 in the C-terminal direction, and a substitution of lysine-148 to alanine in Loop 4.
- the invention provides a combinatorial polypeptide library comprising polypeptide members having a randomized C-type lectin domain (CTLD), wherein the randomized CTLD comprises amino acid modifications in at least one of the four loops in the LSA of the CTLD, wherein the amino acid modifications comprise random substitution of at least five amino acids within Loop 1 and random substitution of at least three amino acids within Loop 2.
- CTLD C-type lectin domain
- the amino acid modifications comprise random substitution of at least five amino acids within Loop 1 , random substitution of at least three amino acids within Loop 2, and random substitution of Arginine- 130, or other amino acid located outside and adjacent to loop 2 in the C-terminal direction.
- the amino acid modifications include random substitutions of at least five amino acids in Loop 1 , random substitution of at least three amino acids in Loop 2, and include a random substitution of Arginine 130.
- the amino acid modifications include random substitutions of five amino acids in Loop 1 , random substitution of three amino acids in Loop 2, and a random substitution of Arginine 130.
- the amino acid modifications include random substitutions of at least five amino acids in Loop 1, random substitution of at least three amino acids in Loop 2, and include a random substitution of Leucine 130.
- the amino acid modifications include random substitutions of five amino acids in Loop 1, random substitution of three amino acids in Loop 2, and a random substitution of Leucine 130.
- the amino acid modifications can further comprise a substitution of Lysine- 148 to Alanine.
- the amino acid modifications comprise random substitution of at least five amino acids within Loop 1 , random substitution of at least three amino acids within Loop 2, and random substitution of Arginine-130, or other amino acid located outside and adjacent to loop 2 in the C-terminal direction and a substitution of Lysine- 148 to Alanine in Loop 4.
- the invention provides a combinatorial polypeptide library comprising polypeptide members that have a randomized C-type lectin domain (CTLD), wherein the randomized CTLD comprises amino acid modifications in at least one of the four loops in loop segment A (LS A) of the CTLD, wherein the amino acid modifications comprise random substitution of at least seven amino acids within Loop 1 and at least one amino acid insertion in Loop 4.
- CTLD C-type lectin domain
- the polypeptide members of the combinatorial library further comprise random substitution of at least two amino acids within Loop 4.
- the amino acid modifications comprise three amino acid insertions within Loop 4 and optionally further comprise random substitution of at least two amino acids.
- the amino acid modifications comprise random substitution of at least seven amino acids within Loop 1 , at least three amino acid insertions in Loop 4, and random substitution of at least two amino acids within Loop 4.
- the amino acid modifications comprise random substitution of seven amino acids within Loop 1 , three amino acid insertions in Loop 4, and random substitution of two amino acids within Loop 4.
- the invention provides a combinatorial polypeptide library comprising polypeptide members that have a randomized C-type lectin domain (CTLD), wherein the randomized CTLD comprises amino acid modifications in at least one of the four loops in the loop segment A (LSA) of the CTLD, wherein the amino acid modifications comprise at least one amino acid insertion in loop 3 and random substitution of at least three amino acids within Loop 3.
- CTLD C-type lectin domain
- the amino acid modifications when the combinatorial library has the modified CTLD of Scheme (d), can further comprise at least one amino acid insertion in Loop 4, and can further comprise random substitution of at least three amino acids within Loop 4.
- the amino acid modifications can comprise three amino acid insertions in Loop 3.
- the amino acid modifications can comprise three amino acid insertions in Loop 4.
- the amino acid modifications comprise random substitution of at least three amino acids within Loop 3, random substitution of at least three amino acids within Loop 4, at least one amino acid insertion in Loop 3 and at least one amino acid insertion in Loop 4.
- the amino acid modifications comprise random substitution of at least three amino acids within Loop 3, random substitution of at least three amino acids within Loop 4, at least three amino acid insertions in Loop 3 and at least three amino acid insertions in Loop 4.
- the amino acid modifications comprise random substitution of three amino acids within Loop 3, random substitution of three amino acids within Loop 4, three amino acid insertions in Loop 3, and three amino acid insertions in Loop 4.
- the amino acid modifications can further compr random substitution of Lysine- 148 to Alanine or in Loop 4.
- the invention provides a combinatorial polypeptide library comprising polypeptide members that have a randomized C-type lectin domain (CTLD), wherein the randomized CTLD comprises amino acid modifications in at least one of the four loops in the loop segment A (LSA) of the CTLD, wherein the amino acid modifications comprise a modification that combines two Loops into a single Loop, wherein the two combined Loops are Loop 3 and Loop 4.
- CTLD C-type lectin domain
- the amino acid modifications comprise random substitution of at least six amino acids within Loop 3 and random substitution of at least four amino acids within Loop 4.
- the amino acid modifications comprise random substitution of six amino acids within Loop 3 and random substitution of four amino acids within Loop 4.
- the amino acid modifications can further comprise random substitution of Proline- 144.
- the amino acid modifications comprise random substitution of six amino acids within Loop 3, random substitution of four amino acids within Loop 4, and a random substitution of proline 144, resulting in a combined Loop 3 and Loop 4 amino acid sequence, comprising, for example, NWEXXXXXXX XGGXXXN (SEQ ID NO: 578), wherein X is any amino acid and wherein the amino acid sequence of SEQ ID NO: 578 forms a single Loop region.
- polypeptide members of the combinatorial library comprise the sequence NWEXXXXXXX XGGXXXN (SEQ ID NO: 578), wherein X is any amino acid and wherein the amino acid sequence of SEQ ID NO: 578 forms a single loop from combined and modified Loop 3 and Loop 4.
- the invention provides a combinatorial polypeptide library comprising polypeptide members that have a randomized C-type lectin domain (CTLD), wherein the randomized CTLD comprises amino acid modifications in at least one of the four loops in the loop segment A (LSA) of the CTLD, wherein the amino acid modifications comprise at least one amino acid insertion in Loop 4 and random substitution of at least three amino acids within Loop 4.
- the amino acid modifications comprise four amino acid insertions in Loop 4.
- the amino acid modifications comprise at least four amino acid insertions in Loop 4 and random substitution of at least three amino acids within Loop 4.
- the amino acid substitutions comprise four amino acid insertions in Loop 4 and random substitution of three amino acids within Loop 4.
- the polypeptide members of the combinatorial library comprise a modified Loop 3 and a modified Loop 5, wherein the modified Loop 3 comprises randomization of five amino acid residues and the modified Loop 5 comprises randomization of three amino acid residues.
- the polypeptide members of the combinatorial library comprise a modified Loop 3, a modified Loop 5, and a modified Loop 4, wherein the modification to Loop 4 abrogates plasminogen binding.
- the amino acid modifications can further comprise one or more amino acid modifications in Loop 4 that modulates plasminogen binding affinity of the CTLD, for example, the substitution of Lysine 148 to Alanine.
- the amino acid modifications when the CTLD is from human or mouse tetranectin, comprise random substitution of at least five amino acid residues in Loop 3, random substitution of at least three amino acid residues in Loop 5, and substitution of Lysine 148 to Alanine in Loop 4.
- the amino acid modifications comprises random substitution of five amino acid residues in Loop 3 and random substitution of three amino acid residues in Loop 5, and, in another specific embodiment, when the CTLD is from human or mouse tetranectin, the amino acid modifications further comprise substitution of Lysine 148 to Alanine in Loop 4.
- the invention provides a combinatorial polypeptide library comprising polypeptide members that have a randomized C-type lectin domain (CTLD), wherein the randomized CTLD comprises amino acid modifications in at least one of the four loops in the loop segment A (LSA) of the-CTLD, wherein the amino acid modifications comprise random substitution of at least one amino acid and at least six amino acid insertions.
- CTLD C-type lectin domain
- the amino acid modifications can further comprise one or more amino acid modifications in Loop 4 that modulates plasminogen binding affinity of the CTLD, for example, the substitution of lysine 148 to Alanine.
- the members of the combinatorial library when the CTLD is from human or mouse tetranectin, have random substitution of at least one amino acid and insertion of at least six amino acids in Loop 3, and substitution of Lysine 148 to Alanine in Loop 4.
- the amino acid modifications comprise random substitution of one amino acid and insertion of six amino acids in Loop 3.
- the members of the combinatorial library when the CTLD is from human or mouse tetranectin, have random substitution of one amino acid and insertion of six amino acids in Loop 3, and substitution of lysine 148 to alanine in Loop 4.
- one of the substitutions is the substitution of Isoleucine 140.
- the invention provides a combinatorial polypeptide library comprising polypeptide members that have a randomized C-type lectin domain (CTLD), wherein the randomized CTLD comprises amino acid modifications in at least one of the four loops in the loop segment A (LSA) of the CTLD, wherein the amino acid modifications comprise a mixture of random substitution of six amino acids in Loop 3 and random substitution of six amino acids and one amino acid insertion in Loop 3.
- the mixture further comprises random substitution of six amino acids and two amino acid insertions in Loop 3.
- the amino acid modifications comprises a mixture of random substitution of six amino acids in Loop 3, random substitution of six amino acids and one amino acid insertion in Loop 3, and random substitution of six amino acids and two amino acid insertions in Loop 3.
- the amino acid modifications when the CTLD is from tetranectin, the amino acid modifications further comprise a substitution of Lysine 148 to Alanine in Loop 4.
- the invention provides a combinatorial polypeptide library comprising polypeptide members that have a randomized C-type lectin domain (CTLD), wherein the randomized CTLD comprises amino acid modifications in at least one of the four loops in the loop segment A (LSA) of the CTLD, wherein the amino acid modifications in at least one of the four loops in the loop segment A (LSA) of the CTLD, wherein the amino acid modifications comprise at least four or more amino acid insertions in at least one of the four loops in the loop segment A (LSA) or loop 5 in loop segment B (LSB) of the CTLD.
- CTLD randomized C-type lectin domain
- the combinatorial library comprises one or more amino acid modifications to the Loop 4 region (alone or in combination with modifications to other regions of the CTLD)
- certain of the modification(s) are designed to maintain, modulate, or abrogate the metal ion-binding affinity of the CTLD.
- Such modifications affect the plasminogen-binding activity of the CTLD (see, e.g., Nielbo, et al., Biochemistry, 2004, 43 (27), pp 8636-8643; or Graversen 1998).
- the polypeptide members of the libraries can comprise one or more amino acid modifications (e.g., by insertion, substitution, extension, or randomization) in any combination of the four LSA loops and the LSB loop (Loop 5) of the CTLD.
- the randomized CTLD can comprise one or more amino acid modifications in the loop of the LSB loop region (Loop 5), either alone, or in combination with one or more amino acid modifications in any one, two, three, or four loops of the LSA loop region (Loops 1-4).
- the invention provides a combinatorial polypeptide library comprising polypeptide members that have a randomized C-type lectin domain (CTLD), wherein the randomized CTLD comprises one or more amino acid modifications in at least one of the four loops in loop segment A (LSA) and one or more amino acid modifications in the loop in loop segment B (LSB) (Loop 5) of the CTLD, wherein the one or more amino acid modifications comprises randomization of the LSB amino acid residues.
- C-type lectin domain C-type lectin domain
- the polypeptide members of the combinatorial libraries can have one or more amino acid modifications in any two, three, four, or five loops in the loop region (LSA and LSB) of the CTLD (e.g., any random
- polypeptide members of the combinatorial libraries can further comprise additional amino acid modifications to regions of the CTLD outside of the loop region (LSA and LSB), such as in the a-helices or ⁇ -strands (see, e.g. , FIG. 1).
- CTLD loop regions can be extended beyond the exemplary constructs detailed in the non-limiting Examples below.
- the invention also provides a library of nucleic acid molecules encoding polypeptides of the combinatorial polypeptide library according to any one of the above- described aspects and embodiments.
- the invention provides a library of nucleic acid sequences encoding the polypeptides of the library, wherein the CTLDs of the polypeptides have been modified according to Schemes (a)-(j).
- the invention provides methods for generating a polypeptide library comprising polypeptide members that have a C-type lectin domain (CTLD), wherein the CTLD comprises one or more amino acid modifications in at least one of the four loops in loop segment A (LSA) and/or in the loop in loop segment B (LSB) (Loop 5) of the CTLD.
- CTLD comprises one or more amino acid modifications in at least one of the four loops in loop segment A (LSA) and/or in the loop in loop segment B (LSB) (Loop 5) of the CTLD.
- the method comprises generating at least one random mutation in at least one of the four loops in the LSA region and/or in the loop in the LSB region of the CTLD, wherein the at least one random mutation comprises (a) an insertion of one or more amino acids in the at least one loop; or (b) a substitution of one or more amino acids within or immediately adjacent to the at least one loop; or (c) a deletion of one or more amino acids within or immediately adjacent to the at least one loop; (d) a modification that combines two adjacent loops, or (e) any combination thereof.
- the method comprises generating random mutations in at least one of the four loops in the LSA region and/or in the loop in the LSB region of the CTLD in accordance with any of Schemes (a)-(j).
- the polypeptides of the recombinant CTLD libraries comprise modified CTLDs in which certain Ca 2+ coordinating amino acid(s) in the loop regions is retained and/or comprise modified CTLDs in which plasminogen binding activity is eliminated.
- the recombinant CTLD libraries can comprise polypeptides having modified CTLD regions, wherein the amino acid modifications fall outside of the loop region (LSA and LSB) of the CTLD. Accordingly, such modifications can be designed or randomly generated in any one or more of the beta strand and/or alpha helical regions.
- Generating randomized and optimized recombinant CTLD libraries to obtain protein products that can bind specifically to targets of interest can be performed by any technique known in the art such as, for example, oligonucleotide-directed randomization, error-prone PCR mutagenesis, DNA shuffling by random fragmentation, loop shuffling, loop walking, somatic hypermutation (see, e.g., US Patent Publication 2009/0075378, which is incorporated by reference), and other known methods in the art to create sequence diversity in order to generate molecules with optimal binding activity. (See, e.g., Stemmer, W.P., Proc Natl Acad Sci USA, (Oct.
- the generating and optimizing methods comprise an oligonucleotide-directed randomization (NN or NNS) strategy for mutagenizing the loops.
- NNS oligonucleotide-directed randomization
- Random amino acid sequences (generated via randomization, substitution, insertion, etc) can be introduced into one or more of these loops to create libraries from which CTLD domains with the desired binding properties can be selected. Construction of these libraries containing random peptides constrained within any or all of the five loops of the human tetranectin CTLD can be accomplished using either a NNK or NNS as described herein. These libraries can comprise further amino acid modifications that are introduced in regions of the CTLD that are outside of the LSA or LSB regions (e.g., the a-helices and/or ⁇ -strands). The following procedure describes a non-limiting, illustrative example of a method by which seven random peptides can be inserted into loop 1 of the hTN CTLD.
- PCR can be used to generate a first fragment (fragment A, see FIG. 7) using the following strategy.
- Oligo lXfor (SEQ ID NO: 137) can be annealed with the reverse oligo lXrev2 (5'-GGC GGT GAT CTC AGT TTC CCA GTT CTT GTA GGC GAT GCG GGC GCC AGT CAT ATC CAC CCA-3'; SEQ ID NO: 580).
- the two oligos are complementary across 21 nucleotides of their 3' ends.
- PCR is used to generate Fragment A (101 bp) from these two overlapping oligos.
- a Fragment B (see FIG.
- PCR can be created by performing PCR using forward oligo BstXl for (5'- ACT GGG AAA CTG AGA TCA CCG CCC AAC CTG ATG GCG GCG CAA CCG AGA ACT GCG CGG TCC TG-3'; SEQ ID NO: 139) and the reverse primer PstBssRevC (5'- CCC TGC AGC GCT TGT CGA ACC ACT TGC CGT TGG CGG CGC CAG ACA GGA CCG CGC AGT TCT-3'; SEQ ID NO: 140) to generate a 105 bp fragment.
- PCR can be performed using a high fidelity polymerase or tag blend and standard PCR thermocycling conditions.
- fragment A is complementary to the 5' end of fragment B.
- fragments can be gel isolated and subsequently combined for overlap extension PCR using outer primers Bglforl2 (SEQ ID NO: 141) and PstRev (SEQ ID NO: 142).
- the resulting 195 bp fragment can be gel isolated and then digested with the restriction enzymes Bgl II and Pst I, after which the final 185 bp fragment can be gel isolated and cloned into a phage display vector (such as CANTAB 5E) containing the restriction modified CTLD shown below fused to Gene III, which is similarly digested with Bgl II and Pst I for cloning.
- a phage display vector such as CANTAB 5E
- Modification of other loops by replacement with randomized amino acids can be similarly performed as described herein.
- the replacement of defined amino acids within a loop with randomized amino acids is not restricted to any specific loop, nor is it restricted to the original size of the loops. Likewise, total replacement of the loop is not required, partial replacement is possible for any of the loops. In some cases retention of some of the original amino acids within the loop, such as the calcium coordinating amino acids, may be desirable. In these cases, replacement with randomized amino acids may occur for either fewer of the amino acids within the loop to retain the calcium coordinating amino acids, or additional randomized amino acids may be added to the loop to increase the overall size of the loop yet still retain these calcium coordinating amino acids. Very large peptides can be accommodated and tested by combining loop regions, such as loops 1 and 2 or loops 3 and 4, into one larger replacement loop.
- the nucleic acid molecules can be obtained by ordinary methods for chemical synthesis of nucleic acids by directing the step- wise synthesis to add pre-defined combinations of pure nucleotide monomers or a mixture of any combination of nucleotide monomers at each step in the chemical synthesis of the nucleic acid fragment. In this way it is possible to generate any level of sequence degeneracy, from one unique nucleic acid sequence to the most complex mixture, which will represent a complete or incomplete representation of maximum number unique sequences of 4 N , where N is the number of nucleotides in the sequence.
- compositions comprising a plurality of nucleic acid fragments can,
- the crude mixtures of fragments, obtained in the initial cleavage step would typically be size- fractionated to obtain fragments of an approximate molecular mass range which would then typically be adjoined to a suitable pair of linker nucleic acids, designed to facilitate insertion of the linker-embedded mixtures of size-restricted oligonucleotide fragments into the receiving nucleic acid vector.
- Nucleic acid fragments can be inserted in specific locations into receiving nucleic acids by any common method of molecular cloning of nucleic acids, such as by appropriately designed PCR manipulations in which chemically synthesized nucleic acids are copy-edited into the receiving nucleic acid, in which case no endonuclease restriction sites are required for insertion.
- the insertion/excision of nucleic acid fragments may be facilitated by engineering appropriate combinations of endonuclease restriction sites into the target nucleic acid into which suitably designed oligonucleotide fragments may be inserted using standard methods of molecular cloning of nucleic acids.
- DNA is isolated from the specific phages, and the nucleotide sequence of the segments encoding the ligand-binding region determined, excised from the phagemid DNA and transferred to the appropriate derivative expression vector for heterologous production of the desired product.
- Heterologous production in a prokaryote can be used for the isolation of the desired product.
- restriction sites can be introduced into the CTLD.
- suitable restriction sites located in the vicinity of the nucleic acid sequences encoding ⁇ 2, ⁇ 3 and ⁇ 4 in both human and murine tetranectin were designed with minimal perturbation of the polypeptide sequence encoded by the altered sequences. It was found possible to establish a design strategy, as detailed below, by which identical endonuclease restriction sites could be introduced at corresponding locations in the two sequences, allowing interesting loop-region variants to be readily excised from a recombinant murine CTLD and inserted correctly into the CTLD framework of human tetranectin or vice versa.
- GGCGCC GGCGCC
- CTCAG restriction endonuclease sites
- CAATTG Mfe I
- nucleic acid constructs in the form of plasmids, vectors, transcription or expression cassettes which comprise at least one nucleic acid described herein are provided.
- Suitable vectors can be chosen or constructed, containing appropriate regulatory sequences, including promoter sequences, terminator sequences, polyadenylation sequences, enhancer sequences, marker genes and other sequences as appropriate.
- Vectors may be plasmids, viral e.g. phage, or phagemid, as appropriate.
- Molecular Cloning a Laboratory Manual: 2nd edition, Sambrook et al., 1989, Cold Spring Harbor Laboratory Press.
- the invention also provides a recombinant host cell which comprises one or more of the constructs as described herein.
- Suitable host cells include bacteria, mammalian cells, yeast, and baculovirus systems.
- Mammalian cell lines available in the art for expression of a heterologous polypeptide include Chinese hamster ovary cells, HeLa cells, baby hamster kidney cells, NSO mouse melanoma cells and many others.
- the host cell is HE 293 cells.
- the resulting recombinant CTLD libraries described herein can be displayed using a number of alternative techniques that are described herein and known in the art. Methods for expressing the nucleic acid molecule library in a display system are described in US Patent Application Publication 2007/0275393, which is incorporated by reference herein in its entirety.
- the display system comprises an observable phenotype that represents at least one property of the displayed expression products and the corresponding genotypes.
- suitable display systems include a phage display system; a yeast display system; a viral display system; a cell-based display system; a ribosome-linked display system; or a plasmid-linked display system; any combinations thereof, or any other suitable display system that is known in the art.
- the invention provides a display system comprising the
- the invention provides a display system comprising the combinatorial polypeptide library according to Schemes (a)-(i).
- the display system comprises a phage display system; a yeast display system; a viral display system; a cell-based display system; a ribosome- linked display system; or a plasmid-linked display system; any combinations thereof, or any other display system that is known in the art.
- phage display e.g., the filamentous phage fd (Dunn (1996); Griffiths and Duncan (1998); Marks et al. (1992)
- phage lambda display e.g., phage lambda display
- eukaryotic virus e.g., baculovirus (Ernst et al. (2000))
- cell display e.g., display on bacterial cells (Benhar et al. (2000))
- yeast cells e.g., yeast cells (Boder and Wittrup (1997)
- mammalian cells Whitehorn et al. (1995)
- ribosome linked display Schoaffitzel et al. (1 99)
- plasmid linked display Gates et al. (1996).
- a commonly used method for phenotype display and linking this to genotype is by phage display. This is accomplished by insertion of the reading frame encoding the scaffold protein or protein of interest to a surface exposed phage protein.
- the filamentous phage fd e.g. Ml 3 has proven useful for this purpose.
- US Patent Application Publication No: 2007/0275393 describes a procedure for accomplishing a display system for the generation of CTLD libraries.
- a method for generating a display system for the described CTLD libraries comprises: [0160] (1) identifying the location of the loop-region of a CTLD;
- the location of the loop region of a CTLD can be identified using the methods previously described herein. Briefly, the loop region can be identified by referring to the three dimensional structure of the CTLD of choice, if such information is available, or, if not, identifying the sequence locations of the ⁇ 2-, ⁇ 3- and by sequence alignment with the sequences shown in FIG. 1, as aided by the identification of sequence elements corresponding to the ⁇ 2 and ⁇ 3 consensus sequence elements and ⁇ 4-strand characteristics, and the conserved cysteine residues also disclosed herein in FIG. 1.
- the invention provides a method for identifying and isolating a polypeptide having specific binding activity to a target molecule, wherein the method comprises (a) providing a combinatorial polypeptide library of the invention; (b) contacting the polypeptides of the combinatorial polypeptide library with the target molecule under conditions that allow for binding between a polypeptide and the target molecule; and (c) isolating a polypeptide that binds to the target molecule.
- the target molecule can comprise any molecule associated with the surface of a cell (such as eukaryotic cells, tumor cells, immune cells, bacterial cells, protozoa, fungi and a cell infected with a virus); proteins (such as receptor proteins, soluble proteins, enzymes, or antibodies); polysaccharides; polymers; and small organic compounds.
- a cell such as eukaryotic cells, tumor cells, immune cells, bacterial cells, protozoa, fungi and a cell infected with a virus
- proteins such as receptor proteins, soluble proteins, enzymes, or antibodies
- polysaccharides polymers
- small organic compounds such as eukaryotic cells, tumor cells, immune cells, bacterial cells, protozoa, fungi and a cell infected with a virus
- proteins such as receptor proteins, soluble proteins, enzymes, or antibodies
- polysaccharides such as receptor proteins, soluble proteins, enzymes, or antibodies
- polymers such as receptor proteins, soluble proteins, enzymes,
- the invention provides a method for identifying and isolating a polypeptide having specific binding activity to a target molecule comprising the steps of: (a) providing a library of nucleic acid molecules encoding the polypeptide library of claim 1 ; (b) expressing the library of nucleic acid molecules in a display system to obtain an ensemble of polypeptides, in which the amino acid residues at one or more sequence positions differ between different members of said ensemble of polypeptides; (c) contacting the ensemble of polypeptides with said target molecule under conditions that allow for binding between a polypeptide and the target molecule; and (d) isolating a polypeptide that is capable of binding to said target molecule.
- the invention provides a method for identifying and isolating a polypeptide having specific binding activity to a target molecule, wherein the polypeptide has been modified in accordance with any of Schemes (a)-(i).
- a specific binding member for a target molecule of interest can be obtained from a random library of polypeptides by selection of members of the library that specifically bind to the target molecule.
- a number of systems for displaying phenotypes with putative ligand binding sites are known. These include: phage display (e.g. the filamentous phage fd [Dunn (1996), Griffiths and Duncan (1998), Marks et al. (1992)], phage lambda
- eukaryotic virus e.g. baculovirus [Ernst et al. (2000)]
- cell display e.g. display on bacterial cells [Benhar et al. (2000)], yeast cells [Boder and Wittrup (1997)], and mammalian cells [Whitehorn et al. (1995)], ribosome linked display [Schaffitzel et al. (1999)], and plasmid linked display [Gates et al. (1996)].
- libraries can be constructed and initially screened for binding to the target molecule as monomeric elements, either as single monomeric CTLD domains or individual peptides displayed on the surface of phage. Libraries can be constructed by randomizing the amino acids in one or more of the five different loops (or outside the loops) within the CTLD scaffold displayed on the surface of phage. Binding to the target molecules can be selected for by phage display panning.
- Random linear peptides and/or random peptides constructed as disulfide constrained loops can be individually displayed on the surface of phage particles and selected for binding to the desired target molecule through phage display "panning". After obtaining peptide clones with the desired binding activity, these peptides can be grafted on to the trimerization domain of human tetranectin or into loops of the CTLD domain followed by grafting on the trimerization domain and screened for agonist activity.
- Another strategy for construction of phage display libraries and trimerization domain constructs include obtaining CTLD derived binders. Libraries can be constructed by
- the CTLD scaffold i.e., of human tetranectin displayed on the surface of phage. Binding to the target molecule can be selected for through phage display panning. After obtaining CTLD clones with peptide loops demonstrating the desired binding activity, the CTLD clones can then be grafted on to the trimerization domain of human tetranectin and screened for agonist activity.
- Another strategy includes using peptide sequences with known binding capabilities to the target of interest and first improving their binding by creating new libraries with randomized amino acids flanking the peptide or/and randomized selected internal amino acids within the peptide, followed by selection for improved binding through phage display. After obtaining binders with improved affinity, the binders of these peptides can be fused to other functional protein domains such as, for example, the trimerization domain of human tetranectin (discussed herein below and discussed in detail in PCT US09/60271 and US. 2010/0028995, which are incorporated herein by reference in their entirety), and evaluated for desired activity.
- initial libraries can be constructed as either free peptides displayed on the surface of phage particles, as in the first strategy, or as constrained loops within the CTLD scaffold as in the second strategy discussed above.
- Peptide display library kits such as, but not limited to, the New England Biolabs Ph.D. Phage display Peptide Library Kits are sold commercially and can be purchased for use in selection of new and novel peptides with specific binding activity for a target molecule of interest.
- Ph.D.-7 Peptide Library Kit containing linear random peptides 7 amino acids in length, with a library size of 2.8x10 9 independent clones
- Ph.D.-C7C Disulfide Constrained Peptide Library Kit containing peptides constructed as disulfide constrained loops with random peptides 7 amino acids in length and a library size of 1.2xl0 9 independent clones
- Ph.D.- 12 Peptide Library Kit containing linear random peptides 12 amino acids in length, with a library size of 2.8 10 9 independent clones.
- NNK an equal mixture of the four nucleic acid bases A, C, G and T.
- the K represents an equal mixture of either G or T, and S represents and equal mixture of either G or C.
- S represents and equal mixture of either G or C.
- ribosome display libraries might be beneficial where larger/ longer random peptides are involved.
- a similar NN or NNS random nucleotide strategy can be used for disulfide constrained libraries. However, these random positions are flanked by cysteine amino acid residues, to allow for disulfide bridge formation. The N-terminal cysteine is often preceded by an additional amino acid such as alanine.
- a flexible linker made up of but not limited to several glycine residues may act as a spacer between the peptides and the gene III protein for any of the above random peptide libraries.
- the human tetranectin CTLD shown in FIGs. 1 and 4 contains five loops (four loops in LSA and one loop comprising LSB), which can be altered to confer binding of the CTLD to different protein targets. Random amino acid sequences can be placed in one or more of these loops to create libraries from which CTLD domains with the desired binding properties can be selected.
- any of the CTLD polypeptide libraries described herein can be used, i.e., polypeptides having CTLDs modified in accordance with any of Schemes (a)-(i). Construction these libraries containing random peptides constrained within any or all of the five loops of the human tetranectin CTLD can be accomplished (but is not limited to) using either a NNK. or NNS as described above in strategy 1 and also described in detail elsewhere herein.
- a strategy can be utilized in which these peptides can be cloned directly on to either the N- or C- terminal end of the trimerization domain of tetranectin as free linear peptides or as disulfide constrained loops using cysteines can be utilized.
- Single-chain antibodies or domain antibodies capable of binding to the target of interest can also be cloned on to either end of the trimerization domain.
- peptides with known binding properties can be cloned directly into any one of the loop regions of the TN CTLD.
- Peptides selected as disulfide constrained loops or as complementarity-determining regions of antibodies might be quite amenable to relocation into the loop regions of the CTLD of human tetranectin. Binding can be tested for all of these constructs in monomeric form, and binding and agonist activation can be tested in trimeric form, when the CTLD is fused with the trimerization domain
- combinatorial polypeptide libraries of the invention can be used to generate and identify polypeptides comprising CTLDs with desired binding properties to target molecules of interest.
- the invention provides a polypeptide having the scaffold structure of a C- type Lectin Like Domain (CTLD), wherein the polypeptide binds to a target other than a natural target for that CTLD and wherein the CTLD scaffold structure of the CTLD is modified according to any of the schemes (a) - (j).
- CTLD scaffold structure is modified according to any of the schemes (a) - (j) and further comprises any of the further modifications described herein, for example, modifications outside the CTLD loop region.
- the polypeptide has the scaffold structure of the CTLD from human or mouse tetranectin and binds to a target other than plasminogen.
- the CTLD polypeptide of the invention can be produced using any of the methods and combinatorial libraries described herein.
- the polypeptide can be produced using a combinatorial library of polypeptides having a CTLD, wherein the loop region of the CTLD is randomized according to any of the Schemes (a) - (j), contacting the combinatorial polypeptide library with the target molecule under conditions that allow for binding between a polypeptide and the target molecule; and isolating a polypeptide that binds to the target molecule, wherein the the target molecule is not the natural target for that CTLD.
- the CTLD is human or mouse tetranectin.
- the CTLD is randomized according to any of the Schemes (a) - (j) and comprises any of the further modifications described herein, for example, modifications outside the CTLD loop region.
- a non-natural target for a modified CTLD according to the invention can be any chemical compound in free or conjugated form which exhibits features of an immunological hapten, a hormone such as steroid hormones, or any biopolymer or fragment thereof, for example, a protein or protein domain; a peptide; an oligodeoxynucleotide; a nucleic acid;
- the target is a protein.
- the protein can be any globular soluble protein or a receptor protein, for example, a transmembrane protein involved in cell signaling, a component of the immune systems such as an MHC molecule or cell surface receptor that is indicative of a specific disease.
- the protein can be a post translationally modified protein having the addition of a biochemical functional group such as acetate, phosphate,and/or various lipids and carbohydrates, including but not limited to, glycosylation and myristoylation.
- the modified CTLD of the invention can also bind protein fragments.
- the CTLD can bind to a domain of a cell surface receptor, when it is part of the receptor anchored in the cell membrane as well as to the same domain in solution, if this domain can be produced as a soluble protein as well.
- the CTLDs can also have specific binding affinity to ligands of low(er) molecUlar weight such as biotin, fluorescein or digoxigenin.
- the CTLD polypeptide sequences that bind one or more target molecule(s) can have binding affinities that are about equal to the binding affinities of naturally occurring ligands for the one or more target molecule(s).
- the binding affinities that are about equal to the binding affinities of naturally occurring ligands for the one or more target molecule(s).
- polypeptides of the invention have a binding affinity for one or more target molecule(s) that is stronger than the binding affinity that a native ligand has for the same target molecule(s). Such polypeptides are useful, for example, for blocking the activity of binding members in some cases, or for more potently agonizing in other cases, e.g., in cases in which the modified CTLD binds to a receptor and is further selected to agonize the receptor. In other embodiments, the polypeptides of the invention have a binding affinity for one or more target molecule(s) that is weaker than the binding affinity that a native ligand has for the same target molecule(s).
- CTLD polypeptides having a weaker affinity for a target molecule(s) than a native ligand may have an improved ability to penetrate tumors or tissues and/or may be useful in cases where the desired goal is to dampen the activity of the target rather than completely block it.
- CTLDs with a lower binding affinity over a native ligand could also be desired, for example, in cases where the optimal selected activity is based on internalization into the cell following binding to the target.
- the modified CTLDs can also bind to one or more receptor(s) and act as agonists.
- the respective binding affinity of the agonists can be determined and compared to the binding properties of native ligands, or a portion thereof, by EL1SA, RIA, and/or BIAcore assays, as well as other assays known in the art.
- the receptor-selective agonists of the invention inhibit or induce a biological activity in at least one type of mammalian cell (e.g., a cancer cell), and such activity can be determined by known art methods.
- Examples of CTLDs identified using the methods provided herein that act as agonists are polypeptides that bind to TRAIL-R1 and TRAIL- R2.
- the modified CTLDs can bind to one or more receptor(s) or one or more ligand(s) having affinity for a receptor(s) and act as antagonists (receptor blockers).
- the respective binding affinity of the agonists can be determined and compared to the binding properties of native ligands, or a portion thereof, by ELISA, RIA, and/or BIAcore assays, as well as other assays known in the art.
- the antagonists of the invention inhibit or induce a biological activity in at least one type of mammalian cell (e.g., a cancer cell), and such activity can be determined by known art methods. Examples of CTLDs identified using the methods provided herein that act as antagonists are polypeptides that bind to IL-23R.
- Polypeptides comprising CTLDs that specifically bind to a target molecule of interest can comprise a "binding member", which includes all or a portion of the CTLD.
- binding member refers to a member of a pair of molecules which have binding specificity for one another.
- the members of a binding pair may be naturally derived or wholly or partially synthetically produced.
- One member of the pair of molecules has an area on its surface, or a cavity, which binds to and is therefore complementary to a particular spatial and polar organization of the other member of the pair of molecules.
- the members of the pair have the property of binding specifically to each other.
- CTLD-based protein products are derived from a
- mammalian tetranectin as exemplified herein with murine and human tetranectin, the structure is nearly identical with all other mammalian tetranectins.
- This species-conserved structure allows for straightforward swapping of polypeptide segments defining ligand-binding specificity between orthologs (e.g. murine and human tetranectin derivatives).
- this platform provides a particular advantage over the "humanization" of murine antibody derivatives, which can involve a number of complications.
- the invention provides a polypeptide having a multimerizing domain and comprises at least one CTLD polypeptide-binding member that binds to at least one target molecule.
- the term "multimerizing domain” means an amino acid sequence that comprises the functionality that can associate with two or more other amino acid sequences to form trimers or other multimeric complexes.
- the multimerizing domain is a dimerizing domain, a trimerizing domain, a tetramerizing domain, a pentamerizing domain, etc. These domains are capable of forming polypeptide complexes of two, three, four, five or more polypeptides of the invention.
- the polypeptide contains an amino acid sequence - a "trimerizing domain”— which forms a trimeric complex with two other trimerizing domains.
- a trimerizing domain can associate with other trimerizing domains of identical amino acid sequence (forming a homotrimer), or with trimerizing domains of different amino acid sequence (forming a heterotrimer).
- the interaction is of the type that produces trimeric proteins or polypeptides. Such an interaction may be caused by covalent bonds between the components of the trimerizing domains as well as by hydrogen bond forces, hydrophobic forces, van der Waals forces and salt bridges.
- trimerizing effect of trimerizing domain is caused by a coiled coil structure that interacts with the coiled coil structure of two other trimerizing domains to form a triple alpha helical coiled coil trimer that is stable even at relatively high temperatures.
- a trimerizing domain based upon a tetranectin structural element the complex is stable at least 60 °C, for example in some embodiments at least 70 °C.
- the multimerized polypeptide is a trimer, for example a tetranectin trimerizing module ⁇ see US 2007/0154901).
- a trimeric complex including a CTLD is referred to herein as an "atrimer.”
- An "ATRIMERTM” polypeptide complex refers to a trimeric complex of three trimerizing domains that also include CLTDs (Anaphore, Inc., San Diego, California).
- a binding member may either be linked to the N- or the C-terminal amino acid residue of the multimerizing domain. Also, in certain embodiments it may be advantageous to have a binding member at both the N-terminus and the C-terminus of the multimerizing domain of the monomer, thereby providing a multimeric polypeptide complex. For example, when the multimeric peptide forms trimers with like molecules, six binding members capable of binding a target molecule of interest can be associated with a single trimeric complex.
- a polypeptide that specifically binds to a target molecule of interest is contained in one or more loops in the loop region of a CTLD.
- the CTLD can be attached to any known trimerizing domain at the C-terminus of the trimerizing domain.
- a fusion protein of the invention can include a second CTLD domain, fused at the N-terminus of the trimerizing domain.
- the fusion protein includes a polypeptide that binds to a first target molecule at one of the termini of the trimerizing domain and a CTLD at the other of the termini.
- One, two or three such proteins can be part of a trimeric complex containing up to six specific CTLD binding members for one or more target molecules.
- the invention provides a multimeric complex of three proteins, each of the proteins comprising a multimerizing domain and at least one CTLD polypeptide that binds to at least one target molecule of interest.
- the multimeric complex comprises a fusion protein having a multimerizing domain selected from a tetranectin trimerizing structural element (tetranectin trimerizing module), a mannose binding protein (MBP) trimerizing domain, a collectin neck region, and other similar moieties.
- the multimeric complex can be comprised of multimerizing domains that are able to associate with each other to form a multimer.
- the multimeric complex is a homomultimeric complex comprised of proteins having the same amino acid sequences.
- the multimeric complex is a heteromultimeric complex comprised of proteins having different amino acid sequences such as, for example, different multimerizing domains, and/or different CTLD polypeptides that bind to a different target molecule.
- the CTLD the CTLD
- the multimeric complex comprises fusion proteins of the invention, wherein each of the fusion proteins comprise at least one CTLD polypeptide that binds to one target molecule, wherein the polypeptides can be the same or different, and/or at least one CTLD polypeptide that binds to a second target molecule, wherein the second target molecule-binding polypeptide can be the same or different.
- the trimerizing domain of a polypeptide of the invention can be derived from tetranectin as described in U.S. Patent Application Publication No. 2007/0154901 ('901 Application), which is incorporated by reference in its entirety.
- the mature human tetranectin single chain polypeptide sequence is provided herein as SEQ ID NO: 11.
- Examples of a tetranectin trimerizing domain include the amino acids 17 to 49, 17 to 50, 17 to 51 and 17-52 of SEQ ID NO: 40, which represent the amino acids encoded by exon 2 of the human tetranectin gene, and optionally the first one, two or three amino acids encoded by exon 3 of the gene.
- Other examples include amino acids 1 to 49, 1 to 50, 1 to 51 and 1 to 52, which represents all of exons 1 and 2, and optionally the first one, two or three amino acids encoded by exon 3 of the gene.
- the N-terminus of the trimerizing domain may begin at any of residues 1 , 2, 3, 4, 5, 6, 7, 8, 9, 10, 1 1 , 12, 13, 14, 15, 16 and 17 of SEQ ID NO: 40.
- the N terminus is 110 or VI 7 and the C-terminus is Q47, T48, V49, C(S)50, L51 or 52 (numbering according to SEQ ID NO: 40). See PCT US09/60271, which is incorporated by reference herein in its entirety.
- the trimerizing domain can be a tetranectin trimerizing structural element ("TTSE") having an amino acid sequence of SEQ ID NO: 40 which is a consensus sequence of the tetranectin family trimerizing structural element as more fully described in US 2007/00154901 , which is incorporated herein by reference in its entirety.
- the TTSE embraces variants of a naturally occurring member of the tetranectin family of proteins, and in particular variants that have been modified in the amino acid sequence without adversely affecting, to any substantial degree, the ability of the TTSE to form alpha helical coiled coil trimers.
- the trimeric polypeptide according to the invention includes a TTSE as a trimerizing domain having at least 66% amino acid sequence identity to the consensus sequence of SEQ ID NO: 49; for example at least 73%, at least 80%, at least 86% or at least 92% sequence identity to the consensus sequence of SEQ ID NO: 40 (counting only the defined (not X) residues). In other words, at least one, at least two, at least three, at least four, or at least five of the defined amino acids in SEQ ID NO: 40 may be substituted.
- cysteine at position 50 (C50) of SEQ ID NO: 40 can be advantageously mutagenized to serine, threonine, methionine or to any other amino acid residue in order to avoid formation of an unwanted inter-chain disulphide bridge, which can lead to unwanted multimerization.
- Other known variants include at least one amino acid residue selected from amino acid residue nos. 6, 21, 22, 24, 25, 27, 28, 31, 32, 35, 39, 41, and 42
- the TTSE has a repeated heptad having the formula a-b-c-d-e-f-g (N to C), wherein residues a and d (i.e., positions 26, 30, 33, 37, 40, 44, 47, and 51 may be any hydrophobic amino acid (numbering according to SEQ ID NO: 40).
- the TTSE trimerization domain can be modified by the incorporation of polyhistidine sequence and/or a protease cleavage site, e.g. Blood Coagulating Factor Xa or Gran2yme B (see US 2005/0199251 , which is incorporated herein by reference), and by including a C-terminal KG or KGS sequence. Also, to assist in purification, Proline at position 2 may be substituted with Glycine .
- a protease cleavage site e.g. Blood Coagulating Factor Xa or Gran2yme B (see US 2005/0199251 , which is incorporated herein by reference)
- Proline at position 2 may be substituted with Glycine .
- TTSE truncations and variants are shown in PCT US09/60271 (FIGs 3A-3D).
- Taeniopygia guttata SEQ ID NO: 75
- trimerizing domain is disclosed in US 6,190,886 (incorporated by reference herein in its entirety), which describes polypeptides comprising a collectin neck region. Trimers can then be made under appropriate conditions with three polypeptides comprising the collectin neck region amino acid sequence. A number of collectins are identified, including:
- the invention also provides for a general and simple procedure for reliable conversion of an initially selected protein derivative into a final protein product, which without further reformatting may be produced in bacteria (e.g. Escherichia coli) both in small and in large scale (International Patent Application Publication No. WO 94/18227 A2).
- bacteria e.g. Escherichia coli
- WO 94/18227 A2 e.g. Escherichia coli
- several identical or non-identical binding sites can be included in the same functional protein unit by simple and general means, enabling the exploitation even of weak affinities by means of avidity in the interaction, or the construction of bi- or hetero-functional molecular assemblies (International Patent Application Publication No. WO 98/56906, which is incorporated by reference in its entirety).
- binding can be modulated by the addition or removal of divalent metal ions (e.g. calcium ions) in combinational libraries with one or more preserved metal binding site(s) in the CTLDs.
- binding can be modulated by altering the pH.
- the combinatorial polypeptide libraries of the invention can be used to generate and identify CTLDs with desired binding properties to target molecules of interest for use in a number of applications including, for example, diagnostic or therapeutic applications in which antibody products are typically used as reagents, in biochemical assay systems, medical in vitro or in vivo diagnostic assay systems, or as active components in therapeutic compositions.
- the combinatorial polypeptide library comprises altered loop regions that allow for the generation of high affinity binding molecules to selected target moieties.
- CTLDs For use in vitro assay systems, the CTLDs (or CTLD-based protein products) have advantages relative to antibody derivatives as each binding site in a CTLD-based protein product is harbored in a single structurally autonomous protein domain. CTLD domains are resistant to proteolysis, and neither stability nor access to the ligand-binding site is compromised by the attachment of other protein domains to the N- or C-terminus of the CTLD.
- the CTLD binding module may readily be utilized as a building block for the construction of modular molecular assemblies (e.g., N- and/or C-terminal extensions), for example, harboring multiple CTLDs of identical or non-identical specificity, reporter molecules, enzymatic molecules (peroxidases, phosphatases), effector molecules, radioisotopes, or any other signaling molecule known in the art.
- modular molecular assemblies e.g., N- and/or C-terminal extensions
- CTLD-based protein products are virtually identical to the corresponding natural CTLD protein already present in the body, and are therefore expected to elicit minimal immunological response in the patient.
- Single CTLDs are about half the mass of the smallest functional antibody derivative, the single-chain Fv derivative, and this small size may in some applications be advantageous as it may provide better tissue penetration and distribution, as well as a shorter half-life in circulation.
- Multivalent formats of CTLD proteins such as those based on the complete tetranectin trimer or the further multimerized collectins, (e.g., mannose binding protein) provide increased binding capacity and avidity and longer circulation half-life.
- the vectors discussed in the following Examples are derived from vectors that have been previously described [see US 2007/0275393]. Certain vector sequences are provided in the Sequence Listing and one of skill will be able to derive vectors given the description provided herein.
- the pPhCPAB phage display vector (SEQ ID NO: 50) has the gill signal peptide coding region has been fused with a linker to the hTN sequence encoding ALQT (etc.). The C-terminal end of the CTLD region is fused via a linker to the remaining gill coding region.
- the murine TN CTLD phage display vectors are similarly designed. Another embodiment of these vectors is pANA27 (SEQ ID NO: 64) in which the gene III C-terminal region has been truncated and the suppressible stop codon at the end of the hTN coding sequence has been altered to encode glutamine.
- the murine vector pANA28 (SEQ ID NO: 65) was constructed in a similar fashion.
- FIGs. 1, 2 and 4 The sequences of human tetranectin and mouse tetranectin, and the positions of loops 1 , 2, 3, 4 (LSA) and 5 (LSB) are shown in FIGs. 1, 2 and 4.
- the coding sequences for Loop 1 were modified to encode the sequences shown in Table 3, where the five amino acids AAEGT (SEQ ID NO: 579; human) or AAEGA (SEQ ID NO: 581 ; mouse) were substituted with seven random amino acids encoded by the nucleotides NN NNK NN NN NN NNK (SEQ ID NO: 582); N denotes A, C, G, or T; K denotes G or T.
- the amino acid arginine immediately following Loop 2 was also fully randomized by using the nucleotides NNK in the coding strand. This amino acid was randomized because the arginine contacts amino acids in Loop 1 , and might constrain the configurations attainable by Loop 1 randomization.
- the coding sequence for Loop 4 was altered to encode an alanine (A) instead of Lysine 148 (K) in order to abrogate plasminogen binding, which has been shown to be dependent on the Loop 4 lysine (Graversen et al., 1998).
- the sequences of human tetranectin and mouse tetranectin, and the positions of Loops 1 , 2, 3, 4, and 5 are shown in FIG. 2.
- the human Loop 1 extended library was generated using overlap PCR in the following manner (primer sequences are shown in Table 4).
- Primers IXfor (SEQ ID NO: 137) and IXrev (SEQ ID NO: 138) were mixed and extended by PCR, and primers BstXl for (SEQ ID NO: 139) and PstBssRevC (SEQ ID NO: 140) were mixed and extended by PCR.
- the resulting fragments were purified from gels, and mixed and extended by PCR in the presence of the outer primers Bglforl2 (SEQ ID NO: 141) and PstRev (SEQ ID NO: 142).
- the resulting fragment was gel purified and cut with Bgl II and Pst I and cloned into a phage display vector pPhCPAB or pANA27.
- the phage display vector pPhCPAB was derived from pCANTAB (Pharmacia), and contained a portion of the human tetranectin CTLD fused to the Ml 3 gene III protein.
- the CTLD region was modified to include BglW and Pst ⁇ restriction enzyme sites flanking Loops 1- 4, and the 1 -4 region was altered to include stop codons, such that no functional gene III protein could be produced from the vector without ligation of an in-frame insert.
- pANA27 was derived from pPhCPAB by replacing the BamHl to Clal regions with the BamHl to Clal sequence of SEQ ID NO:64 (pANA27). This replaces the amber suppressible stop codon with a g!utamine codon and truncates the amino terminal region of gene III.
- Ligated material was transformed into electrocompetent XL I -Blue E. coli (Stratagene) and four to eight liters of cells were grown overnight and DN A isolated to generate a master library DNA stock for panning. A library size of 1.5 x 10 was obtained, and clones examined showed diversified sequence in the targeted regions.
- the mouse Loop 1 extended library was generated using overlap PCR in the following manner.
- Primers MulXfor (SEQ ID NO: 143) and MulXrev (SEQ ID NO: 144) were mixed and extended by PCR, and primers MulXSallfor (SEQ ID NO: 145) and MulXPstRev (SEQ ID NO: 146) were mixed and extended by PCR.
- the resulting fragments were purified from gels, mixed and extended by PCR in the presence of the outer primers BstBBssH (SEQ ID NO: 147) and Mu Pst (SEQ ID NO: 148).
- the resulting fragment was gel purified and cut with BssH II and Pst I and ligated into similarly digested phage display vector ⁇ or ANA28.
- Phage display vector pANA1 (SEQ ID NO: 63) was derived from pPhCPAB by replacing the human tetranectin CTLD with the mouse tetranectin CTLD.
- the mouse tetranectin CTLD included BstBl, BssHll, and Sail sites within the Loop 1 -4 region and a Pst site after the Loop 4 region similar to pPhCPAB in order to facilitate cloning.
- the region was altered to include stop codons as described above.
- Phage display vector pANA28 (SEQ ID NO:65) was derived from pANA16 (SEQ ID NO:63) by replacing the BamHl to Clal region with the BamHl to Clal sequence given in SEQ ID NO:65.
- Ligated material was transformed into electrocompetent XLl -Blue E. coli (Stratagene) and four to eight liters of cells were grown overnight and DNA isolated to generate a master library DNA stock for panning. A library size of 2.65 XI 0 10 was obtained, and clones examined showed diversified sequence in the targeted regions.
- M A or C
- N A, C, G, or T
- K G or T
- S G or C
- W A or T.
- Loop3AF2 GAGCGTGGGCAACGAGGCCGAGATCTGGCTGGGCCTCAACGACATGGCCGCCGA 187
- Loop3BF ATCGCCTACAAGAACTGGGAGACTGRGNNKNNKNNKNNKNNKNNKNNKNNKNNKNNKNNKNNKNNKACCGCG
- Loop30R GATCCCTGCAGCGCTTGTCGAACCACTTGCCGT 191 3X OR GCAGATGTAGGGCAACTGATCTCT 192 HuBgl f or GCCGAGATCTGGCTGGGCCTGA 193
- Loop 1-2 libraries of human and mouse tetranectin C-type lectin binding domains (“Human 1-2" and "Mouse 1-2,” respectively)
- the coding sequences for Loop 1 were modified to encode the sequences shown in Table 1 , where the five amino acids AAEGT (SEQ ID NO: 579; human) or AAEGA (SEQ ID NO: 581 ; mouse) were replaced with five random amino acids encoded by the nucleotides NNK NNK NNK NNK (SEQ ID NO: 583); N denotes A, C, G, or T; K denotes G or T).
- Loop 2 (including the neighboring arginine), the four amino acids TGAR (SEQ ID NO: 584) in human or TGGR (SEQ ID NO: 585) in mouse were replaced with four random amino acids encoded by the nucleotides NNK NNK NNK NNK (SEQ ID NO: 586).
- the coding sequence for Loop 4 was altered to encode an alanine (A) instead of the lysine (K) in the loop, in order to abrogate plasminogen binding, which has been shown to be dependent on the Loop 4 lysine (Graversen et al., 1998).
- the human 1-2 library was generated using overlap PCR in the following manner (primer sequences are shown in Table 4).
- Primers 1 -2 for (SEQ ID NO: 149) and 1-2 rev (SEQ ID NO: 150) were mixed and extended by PCR.
- the resulting fragment was purified from gels, mixed and extended by PCR in the presence of the outer primers Bglforl2 (SEQ ID NO: 141) and PstRevl2 (SEQ ID NO: 151).
- the resulting fragment was gel purified and cut with Bgl II and Pst I and cloned into similarly digested phage display vector pPhCPAB or pANA27, as described above.
- a library size of 4.86 x 10 was obtained, and clones examined showed diversified sequence in the targeted regions.
- the mouse Loop 1-2 library was generated using overlap PCR in the following manner.
- Primers MulXfor (SEQ ID NO: 143) and Mul2rev (SEQ ID NO: 152) were mixed and extended by PCR, and primers Mul234for (SEQ ID NO: 153) and MulXPstRev (SEQ ID NO: 146) were mixed and extended by PCR.
- the resulting fragments were purified from gels, mixed and extended by PCR in the presence of the outer primers BstBBssH (SEQ ID NO: 147) and Mu Pst (SEQ ID NO: 148).
- the resulting fragment was gel purified and cut with BssH II and Pst I and cloned into similarly digested phage display vector pANAl 6 or pANA28, as described above.
- a library size of 1.63 x 10 9 was obtained, and clones examined showed diversified sequence in the targeted regions.
- Loop 1 Library construction: mutation and extension of Loops 1 and 4
- the coding sequences for Loop 1 were modified to encode the sequences shown in Table 3, where the seven amino acids DMAAEGT (see SEQ ID NO: 587; human) or DMAAEGA (see SEQ ID NO: 588; mouse) were replaced with seven random amino acids encoded by the nucleotides NN NNK NNK NNK NNK NNK NNK NNK (SEQ ID NO: 582); N denotes A, C, G, or T; K denotes G or T).
- two amino acids KT in human or KA in mouse were replaced with five random amino acids encoded by the nucleotides NNK NNK NNK NNK NNK (SEQ ID NO: 583).
- the human 1-4 library was generated using overlap PCR in the following manner (primer sequences are shown in Table 4).
- Primers BglBssfor SEQ ID NO: 154) and BssBglrev (SEQ ID NO: 155) were mixed and extended by PCR, and primers BssPstfor (SEQ ID NO: 156) and PstBssRev (SEQ ID NO: 157) were mixed and extended by PCR.
- the resulting fragments were purified from gels, mixed and extended by PCR in the presence of the outer primers Bglfor (SEQ ID NO: 158) and PstRev (SEQ ID NO: 142).
- the resulting fragment was gel purified and cut with Bgl II and Pst I restriction enzymes, and cloned into similarly digested phage display vector pPhCPAB or pANA27, as described above.
- a library size of 2 x 10 9 was obtained, and 12 clones examined prior to panning showed diversified sequence in the targeted regions.
- the mouse 1-4 library was generated using overlap PCR in the following manner (primer sequences are shown in Table 4).
- the resulting fragments were purified from gels, mixed and extended by PCR in the presence of the outer primers Mu 1 -4 OF (SEQ ID NO: 205) and Mu 1-4 OR (SEQ ID NO: 206).
- the resulting fragment was gel purified and cut with BstB I and Pst I restriction enzymes, and cloned into similarly digested phage display vector pANA28, as described above.
- a library size of 4.7 x 10 9 was obtained, and >20 clones were examined prior to panning showed diversified sequence in the targeted regions.
- Loop 3-4 extended libraries of human and mouse tetranectin C-type lectin binding domains ("Human 3-4X” and "Mouse 3-4X,” respectively)
- the coding sequences for Loop 3 were modified to encode the sequences shown in Table 4, where the three amino acids EIT of human or mouse tetranectin were replaced with six random amino acids encoded by the nucleotides N K NNK NNK NNK NNK NN (SEQ ID NO: 589) in the coding strand (N denotes A, C, G, or T; denotes G or T).
- N denotes A, C, G, or T
- the three amino acids TE in human or KAE in mouse were replaced with six random amino acids encoded by the nucleotides NNK NNK NNK NNK NNK (SEQ ID NO: 589).
- the human 3-4 extended library was generated using overlap PCR in the following manner (primer sequences are shown in Table 4).
- Primers H Loop 1 -2-F (SEQ ID NO: 163) and H Loop 3-4 Ext-R (SEQ ID NO: 164) were mixed and extended by PCR, and primers H Loop 3- 4 Ext-F (SEQ ID NO: 165) and H Loop 5-R (SEQ ID NO: 166) were mixed and extended by PCR.
- the resulting fragments were purified from gels, and mixed and extended by PCR in the presence of additional H Loop 1-2-F (SEQ ID NO: 163) and H Loop 5-R (SEQ ID NO: 166).
- the resulting fragment was gel purified and cut with Bgl II and Pst I restriction enzymes, and cloned into similarly digested phage display vector pPhCPAB or pANA27, as described above.
- a library size of 7.9 x 10 8 was obtained, and clones examined showed diversified sequence in the targeted regions.
- the mouse 3-4 extended library was generated using overlap PCR in the following manner.
- Primers M SacII-F (SEQ ID NO: 167) and M Loop 3-4 Ext-R (SEQ ID NO: 168) were mixed and extended by PCR, and primers M Loop 3-4 Ext-F (SEQ ID NO: 1 9) and M Loop 5- R (SEQ ID NO: 170) were mixed and extended by PCR.
- the resulting fragments were purified from gels, and mixed and extended by PCR in the presence of additional M SacII-F (SEQ ID NO: 1 7) and M Loop 5-R (SEQ ID NO: 170).
- the resulting fragment was gel purified and cut with Sac II and Pst I restriction enzymes, and cloned into similarly digested phage display vector pANA16 or pANA28, as described above.
- a library size of 4.95 x 10 9 was obtained, and clones examined showed diversified sequence in the targeted regions.
- the human 3-4 combo library was generated using overlap PCR in the following manner (primer sequences are shown in Table 4).
- Primers H Loop 1-2-F (SEQ ID NO: 163) and H Loop 3-4 Combo-R (SEQ ID NO: 171) were mixed and extended by PCR and the resulting fragment was purified from gels and mixed and extended by PCR in the presence of additional H Loop 1- 2-F (SEQ ID NO: 163) and H loop 5-R (SEQ ID NO: 166).
- the resulting fragment was gel purified and cut with Bgl II and Pst I restriction enzymes, and cloned into similarly digested phage display vector pPhCPAB or ANA27, as described above.
- a library size of 4.95 x 10 9 was obtained, and clones examined showed diversified sequence in the targeted regions.
- the mouse 3-4 combo library was generated using overlap PCR in the following manner.
- Primers M SacII-F (SEQ ID NO: 167) and M Loop 3-4 Combo-R (SEQ ID NO: 172) were mixed and extended by PCR and the resulting fragment was purified from gels and mixed and extended by PCR in the presence of the outer primers M SacII-F (SEQ ID NO: 167) and M Loop 5-R (SEQ ID NO: 170).
- the resulting fragment was gel purified and cut with Sac II and Pst I restriction enzymes, and cloned into similarly digested phage display vector pANA16 or pANA28, as described above.
- a library size of 7.29 x 10 was obtained, and clones examined showed diversified sequence in the targeted regions.
- Loop 4 extended libraries of human and mouse tetranectin C-type lectin binding domains ("Human 4" and "Mouse 4," respectively)
- the coding sequences for Loop 4 were modified to encode the sequences shown in Table 3, where the three amino acids TE of human or AE of mouse tetranectin were replaced with seven random amino acids encoded by the nucleotides NN NNK NNK NNK NNK NNK NNK (SEQ ID NO: 582); N denotes A, C, G, or T; K denotes G or T).
- the human 4 extended library was generated using overlap PCR in the following manner (primer sequences are shown in Table 4).
- Primers H Loop 1-2-F (SEQ ID NO: 163) and H Loop 3-R (SEQ ID NO: 173) were mixed and extended by PCR, and primers H Loop 4 Ext-F (SEQ ID NO: 174) and H Loop 5-R (SEQ ID NO: 166) were mixed and extended by PCR.
- the resulting fragments were purified from gels, and mixed and extended by PCR in the presence of additional H Loop 1-2-F (SEQ ID NO: 163) and H Loop 5-R (SEQ ID NO: 166).
- the resulting fragment gel purified and was cut with Bgl II and Pst I restriction enzymes, and cloned into similarly digested phage display vector pPhCPAB or pANA27, as described above.
- a library size of 2.7 x 10 9 was obtained, and clones examined showed diversified sequence in the targeted regions.
- the mouse 4 extended library was generated using overlap PCR in the following manner.
- Primers M SacII-F (SEQ ID NO: 167) and M Loop 3-R (SEQ ID NO: 175) were mixed and extended by PCR
- primers M Loop 4 Ext-F (SEQ ID NO: 176) and M Loop 5-R (SEQ ID NO: 170) were mixed and extended by PCR.
- the resulting fragments were purified from gels, and mixed and extended by PCR in the presence of the additional M SacII-F (SEQ ID NO: 167) and M Loop 5-R (SEQ ID NO: 170).
- the resulting fragment was gel purified, digested with Sacll and Pstl restriction enzymes, and cloned into similarly digested phage display vector pANA16 or pANA28, as described above.
- Loop 3 altered libraries of human and mouse tetranectin C-type lectin binding domains
- the coding sequences for Loop 3 were modified to encode the sequences shown in Table 3, where the six amino acids ETEITA (SEQ ID NO: 593) of human or ETEITT (SEQ ID NO: 594) of mouse tetranectin were replaced with six, seven, or eight random amino acids encoded by the nucleotides NNK NNK NNK NNK NN NNK (SEQ ID NO: 583), NN NNK NN NNK NNK NNK NNK (SEQ ID NO: 582), and NNK NNK NNK NNK NNK NNK NNK NNK (SEQ ID NO: 595); N denotes A, C, G, or T; and K denotes G or T.
- Loop 4 the three amino acids KTE in human or KAE in mouse were replaced with six random amino acids encoded by the nucleotides NNK NNK NNK NNK NNK NNK NNK (SEQ ID NO: 589).
- the coding sequence for loop 4 was altered to encode an alanine (A) instead of the lysine (K) in the loop, in order to abrogate plasminogen binding, which has been shown to be dependent on the loop 4 lysine (Graversen et al., 1998).
- the human Loop 3 altered library was generated using overlap PCR in the following manner.
- Primers HLoop3F6, HLoop3F7, and HLoop3F8 (SEQ ID NOS: 177-179, respectively) were individually mixed with HLoop4R (SEQ ID NO: 180) and extended by PCR.
- the resulting fragments were purified from gels, and mixed and extended by PCR in the presence of oligos H Loop 1-2F (SEQ ID NO: 163), HuBglfor (SEQ ID NO: 193) and PstRev (SEQ ID NO: 142).
- the resulting fragments were gel purified, digested with Bgll and Pstl restriction enzymes, and cloned into similarly digested phage display vector pPhCPAB or pANA27, as above. After library generation, the three libraries were pooled for panning.
- the mouse Loop 3 altered library was generated using overlap PCR in the following manner.
- Primers MLoop3F 6, MLoop3F 7, and MLoop3F 8 (SEQ ID NOS: 181-183, respectively) were individually mixed with primer M SacII-F (SEQ ID NO: 167) and extended by PCR.
- primers MLoop4F (SEQ ID NO: 207) and M MfeR (SEQ ID NO: 208) were mixed and extended by PCR.
- the resulting fragments were purified from gels, mixed, and subjected to PCR in the presence of primers M 3X OF (SEQ ID NO: 184) and M 3X OR (SEQ ID NO: 192). Products were digested with Sal I (or Sac II) and Pstl restriction enzymes, and the purified fragments were cloned into similarly digested phage display vector pANA16 or pANA28, as described above.
- the human loop 3 loop library was generated using overlap PCR in the following manner.
- Primers Loop3AF2 (SEQ ID NO: 187) and Loop3AR2 (SEQ ID NO: 188) are mixed and extended by PCR, and primers Loop3BF (SEQ ID NO: 189) and Loop3BR (SEQ ID NO: 190) are mixed and extended by PCR.
- the resulting fragments are purified from gels, mixed, and subjected to PCR in the presence of primers Bgl for (SEQ ID NO: 158) and Loop30R (SEQ ID NO: 191).
- Products are digested with Bgl II and Pst I restriction enzymes, and the purified fragments are cloned into similarly digested phage display vector pPhCPAB or pANA27, as above.
- the coding sequence for loop 4 was altered to encode an alanine (A) instead of the lysine (K) in the loop, in order to abrogate plasminogen binding, which has been shown to be dependent on the loop 4 lysine (Graversen et al., 1998).
- a similar approach can be used to generate the corresponding mouse TN library.
- loop 3 and 5 altered libraries of human and mouse tetranectin C-type lectin binding domains
- the coding sequences for loops 3 and 5 were modified to encode the sequences shown in Table 3, where the five amino acids TEITA (SEQ ID NO: 596) of human or TEITT (SEQ ID NO: 597) of mouse tetranectin were replaced with five amino acids encoded by the nucleotides NNK NNK NNK NNK (SEQ ID NO: 583), and the three Loop 5 amino acids AAN of human or mouse were replaced with three amino acids encoded by the nucleotides NNK NNK NNK.
- the coding sequence for loop 4 was altered to encode an alanine (A) instead of the lysine (K) in the loop, in order to abrogate plasminogen binding, which has been shown to be dependent o n the loop 4 lysine (Graversen et al., 1998).
- the human loop 3 and 5 altered library was generated using overlap PCR in the following manner.
- Primers h3-5AF (SEQ ID NO: 213) and h3-5AR (SEQ ID NO: 214) were mixed and extended by PCR, and primers h3-5BF (SEQ ID NO: 215) and h3-5 BR (SEQ ID NO: 21 ) were mixed and extended by PCR.
- the resulting fragments were purified from gels, and mixed and extended by PCR in the presence of h3-5 OF (SEQ ID NO: 217) and PstRev (SEQ ID NO: 1 2).
- the resulting fragment was gel purified, digested with Bgl I and Pst I restriction enzymes, and cloned into similarly digested phage display vector pPhCPAB or pANA27 as described above.
- the mouse loop 3 and 5 altered library was generated using overlap PCR in the following manner.
- Primers m3-5 for (SEQ ID NO: 209) and m3-5 rev (SEQ ID NO: 210) were mixed and extended by PCR.
- the resulting fragment was purified from gels, and reamplified by PCR with primers m3-5 OF (SEQ ID NO: 21 1) and m3-5 OR (SEQ ID NO: 212).
- Products were digested with Sal I and Pst I restriction enzymes, and the purified fragments were cloned into similarly digested phage display vector pANA16 or pANA28 as described above.
- Examples 9-22 provide exemplary methods for isolating polypeptide sequences specific for TRAIL death receptors using the combinatorial polypeptide libraries of the invention.
- TRAIL tumor necrosis factor-related apoptosis-inducing ligand, also referred to in the literature as Apo2L and TNFSFIO, among other things
- TNF tumor necrosis factor
- TRAIL is expressed in cells of the immune system including NK cells, T cells, macrophages, and dendritic cells and is located in the cell membrane.
- TRAIL can be processed by cysteine proteases, generating a soluble form of the protein. Both the membrane-bound and soluble forms of TRAIL function as trimers and are able to trigger apoptosis via interaction with TRAIL receptors located on target cells.
- TRAIL receptors located on target cells.
- TRAIL-R1 DR4, TNFSFl Oa
- TRAIL-R2 DR5, TNFRSFl 0b
- DD death domain
- the death domain on these two receptor molecules is required for TRAIL-activation of the extrinsic apoptotic pathway upon the binding of TRAIL to the receptors.
- TRAIL-R3 DcRl, TNFRSFlOc
- TRAIL-R4 DcR2, TNFRSFlOd
- OPG circulating osteoprotegerin
- TRAIL TRA1L-R1 (DR4) or -R2 (DR5)
- the trimerized receptors recruit several cytosolic proteins that form the death-inducing signaling complex (DISC) which subsequently leads to activation of caspase-8 or caspase-10.
- DISC death-inducing signaling complex
- caspase-8 directly activates the effector caspases (caspases-3, -6, -7) leading to the disassembly of the cell, and the other route involving the caspase-8 dependent cleavage of the pro-death Bcl-2 family protein, Bid, and engaging the mitochondrial or intrinsic death pathway.
- CTLD polypeptide libraries of the invention were screen in an effort to identify and isolate CTLD-based polypeptides having specific binding activity to TRAILRl and TRAIL R2.
- Phage generated from human library 1-4 were panned on recombinant TRAIL Rl (DR4)/Fc chimera, and TRAIL R2 (DR5)/Fc chimera. Screening of these binding panels after three, four, and/or five rounds of panning using an ELISA plate assay identified receptor-specific binders in all cases.
- Phage libraries expressing linear or cyclized randomized peptides of varying lengths can be purchased commercially from manufacturers such as New England Biolabs (NEB).
- phage display libraries containing randomized peptides in loops of the C-type lectin domain (CTLD) of human tetranectin can be generated. Loops 1, 2, 3, and 4 of the LSA of CTLD are shown in Fig. 4. Amino acids within these loops can be randomized using an NNS or NNK overlapping PCR mutagenesis strategy. From one to seven codons in any one loop may be replaced by a mutagenic NNS or NNK codon to generate libraries for screening; alternatively, the number of mutagenized amino acids may exceed the number being replaced (two amino acids may be replaced by five, for example, to make larger randomized loops). In addition, more than one loop may be altered at the same time.
- C-type lectin domain C-type lectin domain
- the overlap PCR strategy can generate either a Kpn I site in the final DNA construct between loops 2 and 3, which alters one of the amino acids between the loops, exchanging a threonine for the original alanine.
- a BssH II site can be incorporated between loops 2 and 3 that does not alter the original amino acid sequence.
- Example 11 Selection and screening of agonists for TRAIL receptors DR4 and DR5
- Bacterial colonies expressing phage were generated by infection or transfection of bacteria such as E. coli TG-1 or XL-1 Blue using either glycerol phage stocks of phage libraries or library DNA, respectively. Fifty milliliters of infected/transfected bacteria at an O.D. 6 oo of 1.0 are grown for 15 min at room temperature (RT), after which time 40% of the final concentration of selectable drug marker is added to the culture and incubated for 1 h at 37°C. Following that incubation the remaining drug for selection is added and incubated for another hour at 37°C. Helper phage VCS M13 are added and incubated for 2 h.
- RT room temperature
- Phage libraries are titered in E. coli TG-1 , XLl-Blue, or other appropriate bacterial host.
- Phage are panned in rounds of positive selection against human DR4 and/or DR5.
- Human DR4 and DR5 (aka human TRAIL death receptors 1 and 2) are commercially available in a soluble form (Antigenix America, Cell Sciences, or as Fc (Genway Biotech, R&D Systems) or GST fusions (Novus Biologicals).
- Soluble DR4 or DR5 in PBS is bound directly to a solid support, such as the bottom of a microplate well (Immulon 2B plates) or to magnetic beads such as Dynabeads. About 250 ng to 500 ng of soluble DR4 or DR5 is bound to the solid substrate by incubation overnight in PBS at either 4°C or RT.
- the plates (or beads) are then washed three times in PBS/0.05% Tween 20, followed by addition of a blocking agent such as 1% BSA, 0.05% sodium azide in PBS and is incubated for at least 0.5 h at RT to prevent binding of material in future steps to non-specific surfaces.
- a blocking agent such as 1% BSA, 0.05% sodium azide in PBS and is incubated for at least 0.5 h at RT to prevent binding of material in future steps to non-specific surfaces.
- Blocking agents such as PBS with 3% non-fat dry milk or boiled casein can also be used.
- Phage are incubated for at least 2 h at RT, although to screen for different binding properties the incubation time and temperature can be varied. Wells are washed at least eight times with PBS/0.05% Tween 20, followed by PBS washes (8x). Wells can be washed in later rounds of selection with increasingly acidic buffers, such as 100 mM Tris pH 5.0, Tris pH 4.0, and Tris pH 3.0. Bound phages are eluted by trypsin digestion (100 ⁇ of 1 mg/mL trypsin in PBS for 30 min). Bound phages can also be eluted using 0.1 M glycine, pH 2.2.
- bound phages can be eluted using TRAIL (available commercially from AbD Serotec) to select for CTLDs or peptides that compete with TRAIL for binding to the death receptors. Further, bound phage can be eluted with compounds that are known to compete with TRAIL for death receptor binding.
- TRAIL available commercially from AbD Serotec
- Eluted phage are incubated for 15 min with 10 mL of freshly grown bacteria at an OD 6 oo of 0.8, and the infected bacteria are treated as above to generate phage for the second round of panning. Two or three additional rounds of positive panning are performed.
- DR4 and/or DR5 expressed endogenously by cancer cell lines or expressed by transfected cells such as 293 cells may be used in rounds of positive selection.
- transfection is performed two days prior to panning using the Qiagen AttracteneTM protocol, for example, and an appropriate expression plasmid such as pcDNA3.1, pCEP4, or pCEP5 bearing DR4 or DR5.
- Cells are dissociated in a non-trypsin dissociation buffer and 6 x 10 6 cells are resuspended in 2 mL IMDM buffer.
- Phage to be panned are dialyzed prior to being added to cells and incubated for 2 h, RT. Cells are washed by pelleting and resuspending multiple times in IMDM, and phage are eluted with glycine buffer.
- negative selection rounds or negative selection concomitant with positive selection are performed. Negative selection is done using the decoy receptors DcRl, DcR2, soluble DcR3, and/or osteoprotegerin (OPG, R&D systems). OPG and soluble DcR3 are commercially available(GeneTex, R&D systems), as are DcRl and DcR2 conjugated to Fcor GST (R&D Systems, Novus Biologicals). For negative selection rounds, decoy receptor is bound to plates or beads and blocked as described above for positive rounds of selection.
- Beads are more desirable as a larger surface area of negative selection molecules can be exposed to the library being panned.
- the primary library or the phage from other rounds of positive selection are incubated with the decoy receptors for 2 h at room temperature, or overnight at 4°C. Unbound phage are then removed and subjected to a positi ve round of selection.
- Positive selection is also performed simultaneously with negative selection.
- Wells or beads coated with soluble DR4 or DR5 are blocked and exposed to the primary library or phage from a selection round as described above, but a decoy receptor such as DcRl is included at a concentration of 10 ⁇ g mL.
- Incubation time may be extended from 2 h to several days at 4°C prior to elution in this strategy in order to obtain phage with greater specificity and affinity for DR4 or DR5.
- Negative selection using DR4, in order to obtain DR5-specific, or DR5, in order to obtain DR4-specific binders, can also be performed using the approaches detailed above.
- Negative selection can also be performed on cancerous or transfected cells that express one or more of the decoy receptors. Negative selection is performed similarly to positive selection as described above except that phage are recovered from the supernatant after spinning cells down after incubation and then used in a positive round of selection.
- trimeric TRAIL receptor agonists and trimeric CTLD-derived TRAIL receptor agonists from phage display or from peptide-grafted, peptide-trimerization domain (TD) fusions, peptide-TD-CTLD fusion, or their various combinations are sub-cloned into bacterial expression vectors (pT7 in house vector, or pET, NovaGen) and mammalian expression vectors (pCEP4, pcDNA3, Invitrogen) for small scale or large-scale production.
- bacterial expression vectors pT7 in house vector, or pET, NovaGen
- mammalian expression vectors pCEP4, pcDNA3, Invitrogen
- Primers are designed to PCR amplify DNA fragments of binders/agonists from various functional display vectors from Example 1.
- Primers for the 5 '-end are flanked with BamH I restriction sites and are in frame with the leader sequence in the vector pT7CIIH6.
- 5' primers also can be incorporated with a cleavage site for protease Granzyme B or Factor Xa.
- 3 ' primers are flanked with EcoRI restriction sites. PCR products are digested with BamHI/EcoRl, and then ligated into pT7CIIH6 digested with the same enzymes, to create bacterial expression vectors pT7CIIH6-TRAILa.
- the TRAIL receptor agonist DNAs can be sub-cloned into vector pT7CIIH6 or pET28a (NovoGen), without any leader sequences and 6XHis. 5' primers are flanked with Ndel restriction sites and 3' primers are flanked with EcoRI restriction sites. PCR products are digested with Ndel/EcoRI, and ligated into the vectors digested with the same enzymes, to create expression vectors pT7-TRAILa and pET-TRAILa.
- the TRAIL receptor agonist DNAs can be sub-cloned into vector pT7CIIH6 or pET28a (NovoGen), with a secretion signal peptide. Expressed proteins are exported into bacterial periplasm, and secretion signal peptide is removed during translocation. 5' primers are flanked with Ndel restriction sites and the primers are incorporated into a bacterial secretion signal peptide, PelB, OmpA or OmpT. 3' primers are flanked with EcoRIrestriction sites. A 6xHis tag coding sequence can optionally be incorporated into the 3' primers.
- PCR products are digested with Ndel/EcoRI, and ligated into vectors that are digested with the same enzymes, to create the expression vectors pT7-sTRAILa, pET-sTRAILa, pT7-sTRAILaHis, and pET-sTRAILHis.
- the TRAIL receptor agonist DNAs can also be sub-cloned into mammalian expression vector pCEP4 or pcDNA3.1 , along with a secretion signal peptide. Expressed proteins are secreted into the culture medium, and the secretion signal peptide is removed during the secretion processes.
- 5' primers are flanked with Nhel restriction sites and the primers are incorporated into a tetranectin secretion signal peptide, or another secretion signal peptide (e.g., Ig peptide).
- 3' primers are flanked with Xhol restriction sites.
- a 6xHis tag is optionally incorporated into the 3' primers.
- PCR products are digested with Nhel/Xhol, and ligated into the vectors that are digested with the same enzymes, to create expression vectors pCEP4-TRAILa, pcDNA-TRAILa, pCEP4-TRAILaHis, and pcDNA-TRAILaHis.
- Bacterial expression constructs are transformed into bacterial strain BL21(DE3) (Invitrogen). A single colony on a fresh plate is inoculated into 100 mL of 2xYT medium in a shaker flask. The flask is incubated in a shaker rotating at 250 rpm at 37°C for 12 h or overnight. Overnight culture (50 mL) is used to inoculate 1 L of 2xYT in a 4 L shaker flask. Bacteria are cultured in the flask to an OD 6 oo of about 0.7, at which time IPTG is added to the culture to a final concentration of 1 mM. After a 4 h induction, bacterial pellets are collected by
- Bacterial fermentation is performed under fed-batch conditions in a 10-liter fermentor.
- One liter of complex fermentation medium contains 5 g of yeast extract, 20 g of tryptone, 0.5 g of NaCl, 4.25 g of KH 2 P0 4 , 4.25 g of K 2 HP0 4 -3H 2 0, 8 g of glucose, 2 g of MgS0 4 7H 2 0, and 3 mL of trace metal solution (2.7% FeCl 3 -6H 2 O/0.2% ZnCl 2 -4H 2 O/0.2% CoCl 2 -6H 2 O/0.15% Na 2 MoO 4 -2H 2 O/0.1% CaCl 2 -2H 2 O/0.1% CuCl 2 /0.05% H 3 B0 3 /3.7% HC1).
- the fermentor is inoculated with an overnight culture (5% vol/vol) and grown at constant operating conditions at pH 6.9 (controlled with ammonium hydroxide and phosphoric acid) and at 30°C.
- the airflow rate and agitation are varied to maintain a minimum dissolved oxygen level of 40%.
- the feed (with 40% glucose) is initiated once the glucose level in the culture is below 1 g /L, and the glucose level is maintained at 0.5 g/L for the rest of the fermentation.
- IPTG is added into the culture to a final concentration of 0.05 mM.
- the cells are harvested.
- the bacterial pellet is obtained by centrifugation and stored at -80°C for subsequent protein purification.
- Expressed proteins that are soluble, secreted into the periplasm of the bacterial cell, and include an affinity tag are purified using standard chromatographic methods, such as metal chelation chromatography (e.g., Ni affinity column), anionic/cationic affinity chromatography, size exclusion chromatography, or any combination thereof, which are well known to one skilled in the art.
- affinity tag e.g., 6xHis tagged proteins
- Expressed proteins can form insoluble inclusion bodies in bacterial cells. These proteins are purified under denaturing conditions in initial purification steps and undergo a subsequent refolding procedure, which can be performed on a purification chromatography column.
- the bacterial pellets are suspended in a lysis buffer (0.5 M NaCl, lOmM Tris-HCl, pH 8, and lmM EDTA) and sonicated.
- the inclusion body is recovered by centrifugation, and subsequently dissolved in a binding buffer containing 6M guanidinium chloride, 50mM Tri-HCl, pH8, and 0.1M DTT. The solubilized portion is applied to a Ni affinitycolumn.
- the proteins are eluted with an elution buffer (6M guanidinium chloride, 50mM Tris-HCI pH8.0, 10mM 2-mercaptoethanol, 250mM imidazole). Isolated proteins are buffer exchanged into the binding buffer, and are re-applied to the Ni + column to remove the denaturing agent. Once loaded onto the column, the proteins are refolded by a linear gradient (0-0.5M NaCl) using 5 C.V. (column volumes) of a buffer that lacks the denaturant (50mM Tris-HCI pH8.0, lOmM 2-mercaptoethanol, plus 2mM CaCl 2 ).
- an elution buffer 6M guanidinium chloride, 50mM Tris-HCI pH8.0, 10mM 2-mercaptoethanol, 250mM imidazole.
- Isolated proteins are buffer exchanged into the binding buffer, and are re-applied to the Ni + column to remove the denaturing agent.
- the proteins are
- the proteins are eluted with a buffer containing 0.5M NaCl, 50mM Tris-HCI pH8.0, and 250mM imidazole.
- the fusion tags (6xHis, CII6His) are cleaved with Factor Xa or Granzyme B, and removed from protein samples by passage through a Ni + -NTA affinity column.
- the proteins are further purified by ion- exchange chromatography on Q-sepharose (GE) using linear gradients (0-0.5M NaCl) over 10 C.V. in a buffer (50mM Tris-HCI, pH8.0 and 2mM CaCl 2 ). Proteins are dialyzed into 1XPBS buffer. Optionally, endotoxin is removed by passing through a Mustang E filter (PALL).
- PALL Mustang E filter
- soluble extracts from bacterial cells for expressed proteins in the periplasm are suspended in a loading buffer (lOmM phosphate buffer pH6.0), and lysed using sonication (or alternatively a French press). After spinning down the insoluble portion in a centrifuge, the soluble extract is applied to an SP FF column (GE). Periplasmic extracts are also prepared by osmotic shock or "soft" sonication. Secreted soluble 6xHis tagged proteins are purified by Ni + -NTA column as described above. Crude extracts are buffer exchanged into an affinity column loading buffer, and then applied to an SP FF column. After washing with 4 C.V.
- the proteins are eluted using a 100% gradient over 8 C.V. with a high salt buffer (lOmM phosphate buffer, 0.5M NaCl, pH6.0). Eluate is filtered by passing through a Mustang E filter to remove endotoxin.
- the partially purified proteins are buffer exchanged into lOmM phosphate buffer, pH7.4, and then loaded to a Q FF column. After washing with 7 C.V. with lOmM phosphate buffer pH 6.0, the proteins are eluted using a 100% gradient over 8 C.V. with a high salt buffer (lOmM phosphate buffer, pH6.0, 0.5M NaCl). Once again endotoxin is removed by passing through a Mustang E filter.
- Example 14 Expression and purification of TRAIL receptor agonists from mammalian cells
- Plasmids for each expression construct are prepared using a Qiagen Endofree Maxi Prep Kit. Plasmids are used to transiently transfect HEK293-EBNA cells. Tissue culture supernatants are collected for protein purification 2-4 days after transfection.
- Potential binding regions or epitopes located on the DR4/DR5 extracellular domain are identified for the agonists using a combination of manual and molecular dynamics-based docking.
- the binding domains are further confirmed by performing binding assays using deletion or point mutations of DR4 DR5 extracellular domain(s) and the agonists.
- Amino acid residues (or sequences) that are involved in determining binding specificity are defined on both DR4/DR5 and TRAIL CTLD agonists.
- a combination of random mutations at various target positions is screened using structure-based computation to determine the compatibility with the structure template. Based on the analysis of apparent packing defects, residues are selected for mutagenesis to construct a library for phage display.
- the 3D models of TRAIL receptor agonist peptides and DR4/DR5 can be used as a reference to refine the peptide-grafted CTLD and DR4 DR5 modeling.
- TRAIL receptor agonist peptides When TRAIL receptor agonist peptides are grafted into CTLD loops, loop conformations are optimized and re-surfaced to match agonist peptides/DR4/DR5 binding by changing the flanking and surrounding amino acid residues using in silico modeling.
- Peptide grafted CTLD agonist homology models are built based on the known tetranectin 3D structures. Loop conformations of homology models of agonists are refined and optimized using LOOPER (DS2.1, Accelrys) and their related algorithms as described above. A combination of random mutations at various target positions is screened by structure-based computation for their compatibility with the structure
- amino acid residues flanking and surrounding peptides are selected for mutagenesis to construct a library for phage display.
- Human cancer cell lines expressing DR4 and/or DR5 such as COLO205 (colorectal adenocarcinoma), NCI-H2122 (non-small cell lung cancer), MIA PaCa-2 (pancreatic carcinoma), ACHN (renal cell carcinoma), WM793B (melanoma) and U266B1 (lymphoma) (all purchased from American Type Tissue Collection (Manassas, VA) are cultured under the appropriate condition for each cell line and seeded at cell densities of 5,000-20,000 cells/well (as determined appropriate by growth curve for each cancer cell line).
- DR4/5 agonistic molecules are added at concentrations ranging from 0.0001 -100 ⁇ g/mL.
- DR4/DR5 agonists are combined with therapeutic methods, including chemotherapeutics (e.g., bortezomib) or cells that are pre- sensitized by radiation, to generate a synergistic effect that upregulates DR4 or DR5 or alters caspase activity.
- chemotherapeutics e.g., bortezomib
- cells that are pre- sensitized by radiation to generate a synergistic effect that upregulates DR4 or DR5 or alters caspase activity.
- the number of viable cells is assessed after 24 and 48 h using "CellTiter 96 Q U eous One Solution Cell Proliferation Assay" (Promega) according to the manufacturer's instructions, and the IC50 concentrations for the DR4/DR5 agonists are determined.
- Human cancer cell lines expressing DR4 and/or DR5 such as COLO205 (colorectal adenocarcinoma), NCI-H2122 (non-small cell lung cancer), MIA PaCa-2 (pancreatic carcinoma), ACHN (renal cell carcinoma), WM793B (melanoma) and U266B1 (lymphoma) (all purchased from American Type Tissue Collection (Mannasas, VA)) are cultured under the appropriate condition for each cell line and seeded at cell densities of 5,000-20,000 cells/well (as determined appropriate by growth curve for each cancer cell line).
- DR4/5 agonistic molecules are added at concentrations ranging from 0.0001 -100 ⁇ g/mL.
- DR4 DR5 agonists can be combined with other therapies such as chemotherapeutics (e.g., bortezomib) or cells that are pre-sensitized by radiation to determine whether such a combination has a synergistic effect on up-regulation of DR4 or DR5 or altering caspase activity.
- chemotherapeutics e.g., bortezomib
- Caspase activity is determined at various timepoints using the " APO-ONE Caspase assay" (Promega) according to the manufacturers instruction.
- Cancer cell lines e.g. HCT-1 16, SW620, COLO205
- Tumor length and width is measured twice a week using a caliper. Once the tumor reaches 250 mm 3 in size, mice will be randomized and treated i.v. or s.c. with 10-100 mg/kg DR4 or DR5 agonist. Treatment can be combined with other therapeutics such as
- chemotherapeutics e.g. irinotecan, bortezomib, or 5FU
- Tumor size is observed for 30 days unless tumor size reaches 1500 mm 3 in which case mice have to be sacrificed.
- Panning was performed using the human Loop 1-4 library of human CTLDs on DR4/Fc antigen-coated (R&D Systems) wells prepared fresh the night before bound with 250 ng to 1 ⁇ g of the carrier free target antigen diluted in 100 of PBS per well. Antigen plates were incubated overnight at 4°C then for 1 hour at 37°C, washed twice with PBS/0.05% Tween 20 and twice with PBS, and then blocked with 1 % BSA/PBS for 1 hr at 37°C prior to panning.
- phage supernatant stocks contained 1 ⁇ g/mL soluble IgGl Fc acting as soluble competitor.
- phage supernatants were pre-bound to antigen wells with human IgGl Fc to remove Fc binders (no soluble IgGl Fc competitor was present during the pre-binding).
- the bacteria were spun down and resuspended in 500 mL SB containing 50 ⁇ g/mL carbenicillin and 100 ⁇ g/mL kanamycin and grown overnight at room temperature shaking at 250 rpm. The following day bacteria were spun out and the phage precipitated with a final concentration of 4% PEG/0.5 M NaCl on ice for 1 hr. Precipitated phage were then spun down at 10,500 rpm for 20 minutes at 4°C. Phage pellets were resuspended in 1% BSA/PBS containing the Roche EDTA free complete protease inhibitors. Resuspended phage were then spun in a microfuge for 10 minutes at 13,200 rpm and passed through a 0.2 ⁇ filter to remove residual bacteria.
- Target antigen bound phage were eluted with 60 ⁇ iL per well acid elution buffer (glycine pH 2) and then neutralized with 2M Tris 3.6 ⁇ ⁇ . Eluted phage were then used to infect TG-1 bacteria (2 mL at OD 6 oo of 0.8-1.0) for 15 minutes at room temperature.
- the culture volume was brought up to 10 mL in SB with 40 ⁇ g/mL carbenicillin and 2% glucose and grown for 1 hour at 37°C shaking at 250 rpm.
- Panning was performed using the TG-1 strain of bacteria for at least four rounds. At each round of panning sample titers were taken and plated on LB plates containing 50 ⁇ g mL carbenicillin and 2% glucose. To screen for specific binding of phagemid clones to the receptor target, individual colonies were picked from these titer plates from the later rounds of panning and grown up overnight at room temperature with shaking at 250 rpm in 250 ⁇ of 2xYT medium containing 2% glucose and 50 ⁇ g/mL carbenicillin in a polypropylene 96-well plate with an air-permeable membrane on top. The following day a replica plate was set up in a 96- deep-well plate by inoculating 500 ⁇ . of 2xYT containing 2% glucose and 50 g/mL
- kanamycin An air-permeable membrane was placed on the culture block and cells were grown overnight at room temperature with shaking at 250 rpm.
- Antigen plates were incubated overnight at 4°C then for 1 hour at 37°C, washed twice with PBS/0.05% Tween 20 and twice with PBS, and then blocked with 3% milk/PBS for 1 hr at 37°C prior to the ELISA. Blocked phage were bound to blocked antigen- bound plates for 1 hr then washed twice with 0.05% Tween 20/PBS and then twice more with PBS. A HRP-conjugated anti-M13 secondary antibody diluted in 3% milk/PBS was then applied, with binding for 1 hr and washing as described above. The ELISA signal was developed using 90 TMB substrate mix and then stopped with 90 xL 0.2 M sulfuric acid, then ELISA plates were read at 450 nM.
- Loop 1 contained seven randomized amino acids in the screened library, whereas Loop 4 had an insertion of 5 randomized amino acids in place of 2 native amino acids (underlined regions in Table 6).
- an amber-suppressible stop codon (TAG) encoded the glutamine, and this is indicated by a lower case "q”.
- TAG amber-suppressible stop codon
- loop region DNA fragments were released from DR4/DR5 binder DNA by double digestion with Bglll and Mfel restriction enzymes, and were ligated to bacterial expression vectors pANA4 (SEQ ID: 54), pANAlO (SEQ ID NO: 60) or pANA19 to produce secreted ATRIMERTM in E. coli.
- the expression constructs were transformed into E. coli strains BL21 (DE3), and the bacteria were plated on LB agar with ampicillin. Single colony on a fresh plate was inoculated into 2xYT medium with ampicillin. The cultures were incubated at 37°C in a shaker at 200 rpm until OD600 reached 0.5, then cooled to room temperature. Arabinosis was added to a final concentration of 0.002-0.02%. The induction was performed overnight at room temperature with shaking at 120-150 rpm, after which the bacteria were collected by centrifugation. The periplasmic proteins were extracted by osmotic shock or gentle sonication.
- the 6xHis-tagged ATRIMERsTM were purified by Ni + -NTA affinity chromatography. Briefly, periplasmic proteins were reconstituted in a His-binding buffer (100 mM HEPES, pH 8.0, 500 mM NaCl, 10 mM imidazole) and loaded onto a Ni + -NTA column pre-equivalent with His-binding buffer. The column was washed with 1 OX vol. of binding buffer. The proteins were eluted with an elution buffer (100 mM HEPES, pH 8.0, 500 mM NaCl, 500 mM imidazole). The purified proteins were dialyzed into PBS buffer and bacterial endotoxin was removed by anion exchange.
- a His-binding buffer 100 mM HEPES, pH 8.0, 500 mM NaCl, 10 mM imidazole
- periplasmic proteins were reconstituted in IX binding buffer (20 mM Tris-HCl, pH 8.5, 150 mM NaCl, 2 mM CaCl 2s 0.1% Triton X-100) and loaded onto a Strep- Tactin column pre-equivalent with binding buffer. The column was washed with 10X vol. of binding buffer. The proteins were eluted with an elution buffer (binding buffer with 2.5 mM desthiobiotin). The purified proteins were dialyzed into binding buffer and bacterial endotoxin was removed by anion exchange.
- the DNA fragments of loop region were sub-cloned into mammalian expression vectors pANA2 (SEQ ID NO: 52) and pANAl 1 (SEQ ID NO: 61) to produce ATRlMERsTM in a HE 293 transient expression system.
- the DNA fragments of the loop region were released from IL-23R binder DNA by double digestion with Bglll and Mfel restriction enzymes, and ligated to the expression vectors pANA2 and pANAl 1 , which were pre-digested with Bglll and Mfel.
- the expression plasmids were purified from bacteria by Qiagen HiSpeed Plasmid Maxi Kit
- HEK293 adhesion cells For HEK293 adhesion cells, the transient transfection was performed by Qiagen SuperFect Reagent (Qiagene) according to the manufacturer's protocol. The day after transfection, the medium was removed and changed to 293 Isopro serum-free medium (Irvine Scientific). Two days later, 20% glucose in 0.5M HEPES was added into the media to a final concentration of 1%. The tissue culture supernatant was collected 4-7 days after transfection for purification. For HEK293F suspension cells, the transient transfection was performed by Invitrogen's 293Fectin and its protocol. The next day, IX volume of fresh medium was added into the culture. The tissue culture supernatant was collected 4-7 days after transfection for purification. The His- or Strep II-tagged ATRIMERTM purification from mammalian tissue culture supernatant was performed as described above.
- pANA5 SEQ ID NO: 55
- pANA6 SEQ ID NO: 56
- pANA7 SEQ ID NO: 57
- pANA8 SEQ ID NO: 58
- pANA9 SEQ ID NO: 59
- pANA5 is a modified pCEP4 vector containing a C-terminal His-tag and a V49 deletion in human TN.
- pANA6 has a ⁇ 48 deletion
- pANA7 has T48 and V49 deletions.
- pANA8 has a CSO.CGO— >Sso,S6o double mutation to provide a more flexible CTLD than wildtype TN.
- pANA9 has E1-V17 deletions to remove the glycosylation site.
- the DNA fragments of loop region were released from IL-23R binder DNA by double digestion with Bglll and Mfel restriction enzymes, and were ligated to the expression vectors pANA5, pANA6, pANA7, pANA8 and pANA9, which were pre-digested with BgHI and Mfel.
- Apparent affinities of the trimeric DR4 and DR5 binders are provided in Tables 7 and 8, respectively. Immobilization of an anti-human IgG Fc antibody (Biacore) to the CMS chip (Biacore) was performed using standard amine coupling chemistry and this surface was used to capture recombinant human DR4 or DR5 receptor Fc fusion protein (R&D Systems).
- ATRIMERTM COMPLEX dilutions (1-500 nM) were injected over the IL-23 receptor surface at 30 ⁇ /min and kinetic constants were derived from the sensorgram data using the Biaevaluation software (version 3.1 , Biacore). Data collection was 3 minutes for the association and 5 minutes for dissociation. The anti-human IgG surface was regenerated with a 30s pulse of 3 M
- H2122 lung adenocamoma cells (ATCC# CRL-5985) and A2780 ovarian carcinoma cells (European Collection of Cell Culture, #931 12519) were incubated at lxl 0 4 cells/well with DR5 ATRIMERsTM (20 ⁇ / ⁇ ) or TRAIL (0.2 ⁇ / ⁇ , R&D Systems) in 10% FBS RMPI media (Invitrogen) in a 96-well white opaque plate (Costar).
- the control wells received media and the respective buffer: TBS for DR5 ATRIMERsTM and PBS for TRAIL. After 20 hours, cell viability was determined by ViaLight Plus (Lonza) and detected on a Glomax luminometer (Promega).
- Panning of peptide libraries was performed using the New England Biolabs (NEB) Ph.D. Phage Display Libraries. Panning was performed on DR5 Fc antigen-coated (R&D Systems) wells prepared fresh the night before bound with 3 ⁇ g of the carrier free target antigen diluted in 150 ⁇ - of 0.1 M NaHCC pH 8.6 per well. Duplicate wells were used in each round. Antigen plates were incubated overnight at 4°C then for 1 hour at 37°C. The antigen was removed and the well was then blocked with 0.5% boiled Casein in PBS pH 7.4 for 1 hr at 37°C prior to panning.
- Eluted phage were then removed from the wells and used to infect 20 mis of ER2738 bacteria at an OD 6 oonm of 0.05 to 0.1 , and grown shaking at 250 rpm at 37°C for 4.5 hrs. Bacteria were then spun out of the culture at 12 X G for 20 min at 4°C. Bacteria were transferred to a fresh tube and re-spun. The supernatant was again transferred to a fresh tube and the Phage were precipitated by adding l/6 th the volume of 20% PEG/2.5M NaCl. Phage were precipitated overnight at 4°C. The following day the precipitated phage were spun down at 12K X G for 20 min at 4°C.
- the supernatant was discarded and the phage pellet re-suspended in 1 ml of TBST(0.1% Tween). Residual bacteria were cleared by spinning in a micro fuge at 13.2 for 10 minutes at 4°C. The phage supernatant was then transferred to a new tube and re-precipitated by adding l/6 ,h the volume of 20% PEG/2.5M NaCl, and incubating at 4°C on ice for lhr. The precipitated phage were spun down in a microfuge at 13.2 for 10 minutes at 4°C. The supernatant was discarded and the phage pellet re-suspended in 200 ⁇ , of TBS.
- Panning was performed using the ER2738 strain of bacteria for at least four rounds. At each round of panning sample titers were taken and plated using top agar on LB/Xgal plates to obtain plaques. To screen for specific binding of phage clones to the receptor target, individual plaques were picked from these titer plates from the later rounds of panning and used to infect ER2738 bacteria at an OD 6 oonm of 0.05 to 0.1 , and grown shaking at 250 rpm at 37°C for 4.5 hrs. Then stored at 4°C overnight.
- Antigen plates were incubated overnight at 4°C then for 1 hour at 37°C, washed twice with PBS/0.05% Tween 20 and twice with PBS, and then blocked with 3% milk PBS for 1 hr at 37°C prior to the ELISA. Blocked phage were bound to blocked antigen-bound plates for 1 hr then washed twice with 0.05% Tween 20/PBS and then twice more with PBS. A HRP-conjugated anti-M13 secondary antibody diluted in 3% milk/PBS was then applied, with binding for 1 hr and washing as described above.
- the ELISA signal was developed using 90 ⁇ L ⁇ TMB substrate mix and then stopped with 90 )iL 0.2 M sulfuric acid, then ELISA plates were read at 450 nM. Secondary ELISA screens were performed on the positive binding clones identified, screening against additional TRAIL receptors and decoy receptors to test for specificity (DR4, DR5, DcRl and DcR2). Secondary ELISA screens were performed similarly to the protocol detailed above.
- Examples 23- 32 provide exemplary methods for identifying and isolating CTLD polypeptides that specifically bind IL-23 receptors using the combinatorial polypeptide libraries of the invention.
- IL-23 is an essential cytokine for generation and survival of Thl7 cells.
- Thl7 cells play a critical role in pathology of many autoimmune diseases, including rheumatoid arthritis, inflammatory bowel disease, psoriasis, systemic lupus erythematosus (SLE) and multiple sclerosis.
- IL-23R is a key target on Thl 7 cells.
- the IL-23 cytokine is composed of two subunits: pi 9 and p40, with the pi 9 subunit being unique to IL-23, and p40 shared with IL-12.
- the IL-23 receptor is a heterodimeric receptor that binds IL-23 and mediates activation of certain T cell subsets, NK cells and myeloid cells.
- the IL-23 heterodimeric receptor is composed of two subunits: IL-23R and IL-12Rpl , with IL-23R being the subunit unique to the IL-23 pathway.
- IL-12Rpi is shared with the IL-12 receptor and hence the IL-12 pathway.
- IL-23R has been associated with susceptibility to psoriasis and Crohn's disease and also has been implicated in susceptibility to ankylosing spondylitis, Vogt-Koyanagi-Harada disease, Systemic Sclerosis, Behcet's disease (BD), Primary Sjogren's Syndrome, Goodpasture disease. Also, importance of IL-23 in Graft Versus Host disease and chronic ulcers has been suggested, and IL-23 has been implicated in tumorigenesis.
- Blockade of the IL-23 pathway is efficacious in many preclinical models of autoimmune disease.
- the nature of shared ligand and receptor subunits between IL-23 and IL-12 pathways has led to more complex biology than previously appreciated, and separation of IL-23 blockade from IL-12 blockade appears to have important therapeutic implications regarding both efficacy and safety.
- Blockade of one or the other, or both can be done at the level of the cytokine subunits or the receptor subunits.
- Phage generated from human library 1-4 were panned on recombinant human IL-23R/Fc chimera (R&D Systems), and recombinant mouse IL-23R/Fc chimera (R&D Systems).
- the supernatant was incubated with one-fourth volume (usually 250 mL of supernatant/bottle + 62.5 mL PEG solution) of 20% PEG/2.5 M NaCl on ice for 30 min.
- the phage is pelleted by centrifugation at 15,000 g and 4°C for 20 min.
- the phage pellet was resuspended in 1% bovine serum albumin (BSA) in phosphate buffered saline (PBS) containing 0.1% sodium azide (BSA PBS/azide) and complete mini-EDTA-free protease inhibitors (Roche), prepared according to the manufacturer's instructions.
- phage was resuspended in Buffer D, containing 0.05% boiled cassein, 0.025% Tween-20, and protease inhibitors. Material was filter-sterilized using Whatman Puradisc 25 mm diameter, 0.2 ⁇ ⁇ pore size filters.
- Phage generated from human library 1-4 were panned on recombinant human IL-23R/Fc chimera (R&D Systems cat #1686-MR). Library panning was performed either using a plate or a bead format. For the plate format, six to eight wells of a 96-well Immulon HB2 ELISA plate were coated with 250 -1000 ng/well of carrier- free human IL-23R/Fc in Dulbecco's PBS.
- bound phage were washed once with either IX PBS/0.05% Tween or with Buffer D, and were eluted using glycine buffer, pH 2.2, containing 1 mg/mL BSA. After neutralization with 2 M Tris base (pH 1 1.5) the eluted phage were incubated for 15 minutes at room temperature with two to four milliliters of TGI (Stratagene), XLl -Blue (Stratagene), ER2738 (Lucigen or NEB), or SS320 (Lucigen) cells at an optical density of approximately 0.9 measured at 600 nm (OD 6 oo) in yeast extract-tryptone (YT) medium .
- Phage were prepared from this infection using the protocol above, but scaled down by about 20% (volume). Phage prepared from eluted phage were subjected to additional rounds of panning. At each round, titers of input and output phage were determined by plating on agar with appropriate antibiotics, and colonies from these plates were used later for screening for binders by ELIS A.
- human IL-23R was biotinylated and purified using a Sulfo- NHS micro biotinylation kit (Thermo-Scientific) according to the manufacturer's instructions. Phage were generated for panning from the master library as per the protocol above, except that the phage pellet was resuspended in a casein buffer containing 0.5% boiled casein, 0.025% Tween 20 in PBS with added EDTA-free protease inhibitors (Roche).
- streptavidin magnetic beads (2 tubes with 50 or 0.5 mg each of Myone Tl Dynabeads (Invitrogen)) were washed several times in 0.5% boiled casein, 1% Tween 20 to remove preservatives.
- a 150 xL aliquot of the phage prep was preincubated with one tube of beads for 30 min at 37°C to remove streptavidin binders.
- the phage prep was then removed from the beads and 1 ⁇ g of biotinylated 1L-23R was added along with 10 ⁇ of human Fc at 100 ⁇ g/mL and incubated for 2 h at 37°C with rotation.
- ELISA plates were prepared by incubating overnight at 4°C with 50-100 of PBS containing 75-100 ng/well of recombinant human IL-23R/Fc. A duplicate plate coated with human IgG Fc (R&D Systems) was used as a control. Plates were washed 3 times with PBS, blocked for 1 h at 37°C with 3% milk in IX PBS, and incubated for 1 hour with 100 uL/well of each milk-treated phage mixture.
- Binders to human IL-23R were identified from the third and fourth rounds of panning. Examples of the sequences from the randomized regions of Loops 1 and 4 from phage-displayed CTLD binders to human IL-23R Fc chimera are given in Table 11.
- Loop 4 the second and fifth amino acids were always glycine, the fourth amino acid was always one of the cyclic amino acids tryptophan or phenylalanine, the first amino acid was hydrophobic, and usually a cyclic amino acid, such as phenylalanine, tyrosine, or tryptophan, and the third amino acid was hydrophobic, and was usually valine.
- the Loop 1 region had less of a consensus, though glycine and serine appeared predominantly in the first and second positions, and valine was often in the seventh position.
- ELISA assays indicated that these binders did not cross-react with either human IgGl Fc or with recombinant mouse IL-23R.
- ELISA and Biacore binding assays indicated that purified monomelic CTLD or full-length trimers from candidate clones 001 -69.4G8 and other competed with IL-23 for binding to the human IL-23R.
- Competitive candidates have been identified that have nanomolar affinities.
- Example 24 [0364] Affinity maturation of binders to human IL-23R
- Loop 4 region of the human IL-23R appeared to be a relevant motif, a shuffling approach was developed preserving the diversity of Loop 4 regions already obtained by panning, but resorting them with all possible Loop 1 regions from the original naive library.
- DNA from the round 4 panning of human IL-23R was digested with EcoRI and Bss U restriction enzymes, which cut between the Loop 1 and Loop 4 regions, and a fragment of about 1.4 kb, containing the Loop 4 region, was isolated.
- the original human 1 -4 library DNA was digested with the same enzymes, and a fragment of about 3.5 kb, containing the Loop 1 region, was isolated.
- Loop 1 and 4 sequences from affinity-matured human Loop 1 -4 binders to human IL-23R
- a separate affinity maturation library was generated in which the diversity of the Loop 1 regions obtained in the initial panning round 4 was maintained, a limited selection of Loop 4 options was utilized, and Loop 3 was randomized in six positions. This was achieved by generating primers to amplify the Loop 1 region using DNA from the original panning round 4 of the human Loop 1-4 library as template, along with primers Bglfor (SEQ ID NO: 158) and H1-3-4R (SEQ ID NO: 185). This primer encodes the following amino acid sequence for loops 3 and 4:
- Affinity matured IL-23R binding sequences obtained from these libraries are provided in Table 14. Some of the binders obtained were altered by swapping more favorable loop 4 or loop 1 sequences for others to obtain additional affinity-matured binders, and these are included in Table 14.
- Table 15 shows some additional clones that were made with a primer similar to H1-3-4R (SEQ ID NO: 185), but having a coding sequences for the following loop modications.
- Another affinity maturation library was generated by limiting loop 4 to five amino acid sequences: FGVFG (SEQ ID NO: 381), WGVFG (SEQ ID NO: 404), FGYFG (SEQ ID NO: 389), WGYFG (SEQ ID NO: 413), and WGVWG (SEQ ID NO: 409), while maintaining the GlySer found at the beginning of loop 1 in IL-23R binders, and varying the subsequent five amino acids in loop 1 using an NN strategy.
- Primers GSXX (SEQ ID NO: 194) and 090827 BssBglrev (SEQ ID NO: 195) were mixed and extended using PCR, and primers FGVFGfor, FGYFGfor, WGVFGfor, WGYFGfor, and WGVWGfor (SEQ ID NOS: 196 to 200) were mixed individually with primer Pst Loop 4 rev (SEQ ID NO: 186) and extended using PCR.
- the resulting fragments were gel purified and mixed and extended by PCR in the presence of primers Bgl for (SEQ ID NO: 158) and Pst rev (SEQ ID NO: 142).
- the resulting fragments were digested with Bgl II and Pst I and inserted into vector pANA27 for phage display. Bead panning with successive target dilution was used to select affinity-matured candidates from the library. Sequences of the candidates obtained from this library are provided in Table 16.
- the DNA fragments encoding loop regions were obtained by restriction digestion with Bglll and Pstl (or Mfel) restriction enzymes, and ligated to the bacterial CTLD expression vectors pANAl (SEQ ID NO: 51), pANA3 (SEQ ID NO: 53), or pANA12 (SEQ ID NO: 62) that were pre-digested with Bglll and Pstl.
- pANAl is a T7 based expression vector designed to express C-terminal 6xHis-tagged human monomelic CTLD.
- the pelB signal peptide directs the proteins to the periplasm or growth medium.
- pANA3 is the C-terminal HA-His-tagged version of pANAl .
- pANA12 is the C-terminal HA-StrepII-tagged version of pANAl .
- the loop regions can be sub-cloned into ATRIMERTM polypeptide complex expression vectors pANA4 (SEQ ID NO: 54) or pANAlO (SEQ ID NO: 60)to produce secreted ATRIMERTM polypeptide complexes in E. coli.
- pANA4 is a pB AD based expression vector containing C-terminal His/Myc-tagged full length human TN with an ompA signal peptide to direct the proteins to periplasm or growth medium.
- pANAlO is the C-terminal HA-StrepII- tagged version of pANA4.
- the cultures were incubated at 37°C on a shaker at 200 rpm to an OD 60 o of 0.5, then cooled to room temperature.
- IPTG was added to a final concentration of 0.05 mM for pANAl and pANA12, while arabinosis was added to a final concentration of 0.002-0.02% for pANA4 and pANAlO.
- the induction was performed overnight at room temperature with shaking at 120-150 rpm, after which the bacteria were collected by centrifugation.
- the periplasmic proteins were extracted by osmotic shock or gentle sonication.
- periplasmic proteins were reconstituted in a His-binding buffer (100 mM HEPES, pH 8.0, 500 mM NaCl, 10 mM imidazole) and loaded onto a Ni + -NTA column pre-equilibrated with His-binding buffer. The column was washed with lOx volume of binding buffer. The bound proteins were eluted with an elution buffer (100 mM HEPES, pH 8.0, 500 mM NaCl, 500 mM imidazole). The purified proteins were dialyzed into IX PBS buffer and bacterial endotoxin was removed by anion exchange.
- His-binding buffer 100 mM HEPES, pH 8.0, 500 mM NaCl, 10 mM imidazole
- strep II-tagged monomelic CTLDs and ATRIMERTM polypeptide complexes were purified by Strep-Tactin affinity chromatography. Briefly, periplasmic proteins were reconstituted in IX PBS buffer and loaded onto a Strep-Tactin column pre-equivatent with IX PBS buffer. The column was washed with 10X volume of PBS buffer. The proteins were eluted with elution buffer (IX PBS with 2.5 mM desthiobiotin). The purified proteins were dialyzed into IX PBS buffer and bacterial endotoxin was removed by anion exchange.
- ATRIMERTM polypeptide complexes were produced by mammalian cells. DNA fragments encoding loop regions were sub-cloned into the mammalian expression vector pANA2 or pANAl 1 to produce ATRIMERTM polypeptide complexes in the HEK293 transient expression system.
- pANA2 is a modified pCEP4 vector containing a C- terminal His tag.
- pANAl 1 is the C-terminal HA-StrepII-tagged version of pANA2.
- the DNA fragments encoding loop region were obtained by double digestion with Bglll and Mfel and ligated into the expression vectors pANA2 and pANAl 1 pre-digested with Bglll and Mfel.
- the expression plasmids were purified from bacteria using a Qiagen HiSpeed Plasmid Maxi Kit (Qiagene).
- Qiagene Qiagen HiSpeed Plasmid Maxi Kit
- transient transfection was performed using Qiagen SuperFect Reagent according to the manufacturer's protocol. The day after transfection, the medium was removed and changed to 293 Isopro serum-free medium (Irvine Scientific). Two days later, glucose in 0.5 M HEPES buffer was added into the media to a final concentration of 1%.
- tissue culture supernatant was collected 4-7 days after transfection for purification.
- the transient transfection was performed by Invitrogen's 293Fectin according to the manufacturer's protocol. The next day, IX volume of fresh medium was added into the culture. The tissue culture supernatant was collected 4-7 days after transfection for purification.
- IL-23-containing solutions were prepared as follows. Human IL- 23 (eBioscience) was added at a concentration of 100 ng mL. Competitor was included at a final concentration of 1 ⁇ g/mL. After incubation, plates were washed as described and wells were incubated for 40 min at RT with 100 ⁇ ⁇ each of PBS containing a 1 :5000 dilution of
- FIG. 1 An example of the results of the competition assay (inhibiting IL-23/IL-23R interaction) using the ATRIMERTM polypeptide complexes from the initial panning is presented in Figure 1 1.
- ATRIMERTM polypeptide complexes to the left of the wild-type human tetranectin control (TN) were obtained from the third round of panning against human IL-23R using the human Loop 1-4 library (except for P 1D1).
- ATRIMERTM polypeptide complexes to the right of the tetranectin control were obtained from the human 1-4 shuffle library after 3-4 rounds of panning on decreasing quantities of IL-23R.
- ATRIMERTM polypeptide complexes were tested in competition ELISA more extensively to determine IC50 values. As shown in Table 19, ATRIMERTM polypeptide complexes displayed low to subnanomolar IC50s.
- ATRIMERTM polypeptide complexes to compete with IL-23 for binding to IL-23R.
- the ATRIMERTM polypeptide complex 056-53.H4E was chosen as a standard for comparison, and additional competition assays were performed with affinity-matured ATRIMERTM polypeptide complexes.
- Table 20 provides the ratio of the IC50 of tested ATRIMERTM polypeptide complexes to that of 056-53.H4E performed in the same assay, in order to better compare competition results among assays.
- ATRIMERTM polypeptide complex dilutions (1-500 nM) were injected over the IL-23R surface at 30 ⁇ /min and kinetic constants were derived from the sensorgram data using the Biaevaluation software (version 3.1 , GE Healthcare). Data collection was 3 minutes for the association and 5 minutes for dissociation. The anti-human IgG surface was regenerated with a 30s pulse of 3M magnesium chloride. All sensorgrams were double-referenced against an activated and blocked flow-cell as well as buffer injections.
- Affinities of full-length ATRIMERTM polypeptide complex IL-23R binders from the original and the first affinity-matured library refers to mammalian-cell produced material. All other material was produced in E. coli.
- ATRIMERTM complexes binding to IL-23R do not recognize IL-12Rpl or IL-12Rp2
- a Biacore 3000 biosensor (GE Healthcare) was used to evaluate the interaction of human IL-12Rpl/Fc or IL-12Rp2 Fc with IL-23R binding ATRIMERTM complexes. Immobilization of an anti-human IgG Fc antibody (GE Healthcare) to the CM5 chip (GE Healthcare) was performed using standard amine coupling chemistry, and this modified surface was used to capture recombinant human IL-12Rpi/Fc or IL-12RP2/Fc fusion protein (R&D Systems). A low-density receptor surface, less than 200 RU, was used for all of the analyses. ATRIMERTM complex dilutions (100 nM) were injected over the IL-12R surface at 30 ⁇ /min.
Landscapes
- Chemical & Material Sciences (AREA)
- Health & Medical Sciences (AREA)
- Organic Chemistry (AREA)
- Life Sciences & Earth Sciences (AREA)
- Biochemistry (AREA)
- Molecular Biology (AREA)
- Genetics & Genomics (AREA)
- Medicinal Chemistry (AREA)
- General Health & Medical Sciences (AREA)
- Immunology (AREA)
- Biophysics (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Engineering & Computer Science (AREA)
- Zoology (AREA)
- Microbiology (AREA)
- Wood Science & Technology (AREA)
- Biomedical Technology (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Virology (AREA)
- Physics & Mathematics (AREA)
- Plant Pathology (AREA)
- Crystallography & Structural Chemistry (AREA)
- Toxicology (AREA)
- Gastroenterology & Hepatology (AREA)
- Peptides Or Proteins (AREA)
Abstract
Description














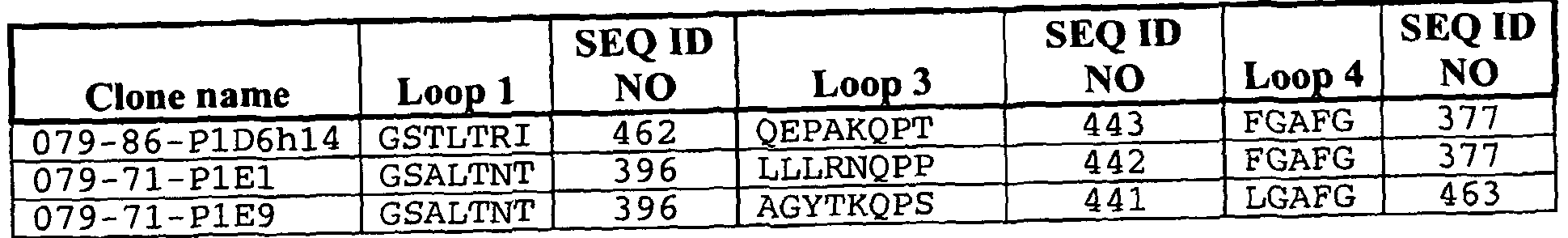











Claims
Priority Applications (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| AU2010303879A AU2010303879A1 (en) | 2009-10-09 | 2010-02-10 | Combinatorial libraries based on C-type lectin domain |
| EP10704873A EP2486132A1 (en) | 2009-10-09 | 2010-02-10 | Combinatorial libraries based on c-type lectin domain |
| CA2776954A CA2776954A1 (en) | 2009-10-09 | 2010-02-10 | Combinatorial libraries based on c-type lectin domain |
| CN2010800560215A CN102686727A (en) | 2009-10-09 | 2010-02-10 | Combinatorial libraries based on C-type lectin domain |
| JP2012533143A JP2013507123A (en) | 2009-10-09 | 2010-02-10 | Combinatorial library based on C-type lectin domain |
Applications Claiming Priority (4)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| USPCT/US2009/060271 | 2009-10-09 | ||
| US12/577,067 | 2009-10-09 | ||
| US12/577,067 US20100105620A1 (en) | 2008-10-10 | 2009-10-09 | Polypeptides that bind Trail-R1 and Trail-R2 |
| PCT/US2009/060271 WO2010042890A2 (en) | 2008-10-10 | 2009-10-09 | Polypeptides that bind trail-ri and trail-r2 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO2011043834A1 true WO2011043834A1 (en) | 2011-04-14 |
Family
ID=42103903
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/US2010/023803 WO2011043834A1 (en) | 2009-10-09 | 2010-02-10 | Combinatorial libraries based on c-type lectin domain |
Country Status (5)
| Country | Link |
|---|---|
| EP (1) | EP2486132A1 (en) |
| JP (1) | JP2013507123A (en) |
| CN (1) | CN102686727A (en) |
| CA (1) | CA2776954A1 (en) |
| WO (1) | WO2011043834A1 (en) |
Cited By (10)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US9388215B2 (en) | 2013-03-15 | 2016-07-12 | Shenzhen Hightide Biopharmaceutical, Ltd. | Compositions and methods of using islet neogenesis peptides and analogs thereof |
| US20170246281A1 (en) * | 2016-02-16 | 2017-08-31 | President And Fellows Of Harvard College | Pathogen vaccines and methods of producing and using the same |
| US10682400B2 (en) | 2014-04-30 | 2020-06-16 | President And Fellows Of Harvard College | Combination vaccine devices and methods of killing cancer cells |
| US11096997B2 (en) | 2005-12-13 | 2021-08-24 | President And Fellows Of Harvard College | Scaffolds for cell transplantation |
| US11150242B2 (en) | 2015-04-10 | 2021-10-19 | President And Fellows Of Harvard College | Immune cell trapping devices and methods for making and using the same |
| US11202759B2 (en) | 2010-10-06 | 2021-12-21 | President And Fellows Of Harvard College | Injectable, pore-forming hydrogels for materials-based cell therapies |
| US11278604B2 (en) | 2012-04-16 | 2022-03-22 | President And Fellows Of Harvard College | Mesoporous silica compositions comprising inflammatory cytokines comprising inflammatory cytokines for modulating immune responses |
| US11555177B2 (en) | 2016-07-13 | 2023-01-17 | President And Fellows Of Harvard College | Antigen-presenting cell-mimetic scaffolds and methods for making and using the same |
| US11752238B2 (en) | 2016-02-06 | 2023-09-12 | President And Fellows Of Harvard College | Recapitulating the hematopoietic niche to reconstitute immunity |
| US11786457B2 (en) | 2015-01-30 | 2023-10-17 | President And Fellows Of Harvard College | Peritumoral and intratumoral materials for cancer therapy |
Families Citing this family (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN104961833B (en) * | 2015-07-21 | 2018-08-03 | 中国科学技术大学 | A kind of people source NKp80-Fc fusion proteins and preparation method thereof and the application in oncotherapy |
Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2002048189A2 (en) * | 2000-12-13 | 2002-06-20 | Borean Pharma A/S | Combinatorial libraries of proteins having the scaffold structure of c-type lectin-like domains |
-
2010
- 2010-02-10 EP EP10704873A patent/EP2486132A1/en not_active Withdrawn
- 2010-02-10 WO PCT/US2010/023803 patent/WO2011043834A1/en active Application Filing
- 2010-02-10 CA CA2776954A patent/CA2776954A1/en not_active Abandoned
- 2010-02-10 JP JP2012533143A patent/JP2013507123A/en active Pending
- 2010-02-10 CN CN2010800560215A patent/CN102686727A/en active Pending
Patent Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2002048189A2 (en) * | 2000-12-13 | 2002-06-20 | Borean Pharma A/S | Combinatorial libraries of proteins having the scaffold structure of c-type lectin-like domains |
Non-Patent Citations (120)
| Title |
|---|
| "Organic Chemistry by Fessendon and Fessendon", 1982, WILLARD GRANT PRESS, BOSTON MASS |
| AGNEW, CHEM INTL. ED. ENGL., vol. 33, 1994, pages 183 - 186 |
| ASHKENAZI ET AL., J CLIN INVEST., vol. 104, no. 2, July 1999 (1999-07-01), pages 155 - 62 |
| AUSUBEL ET AL.,: "Current Protocols in Molecular Biology", 1994, GREEN PUBLISHERS INC. AND WILEY AND SONS 1994 |
| AZOULAY, M.; VILMONT, M.; FRAPPIER, F.: "Glutamine analogues as Potential Antimalarials", EUR. J. MED. CHEM., vol. 26, 1991, pages 201 - 5, XP023870208, DOI: doi:10.1016/0223-5234(91)90030-Q |
| BARTON ET AL.: "Synthesis of Novel a-Amino-Acids and Derivatives Using Radical Chemistry: Synthesis of L-and D-a-Amino-Adipic Acids, L-a-aminopimelic Acid and Appropriate Unsaturated Derivatives", TETRAHEDRON LETT., vol. 43, 1987, pages 4297 - 4308, XP001008711, DOI: doi:10.1016/S0040-4020(01)90305-9 |
| BASS, S.; GREENE, R.; WELLS, J. A.: "Hormone phage: an enrichment method for variant proteins with altered binding properties", PROTEINS, vol. 8, 1990, pages 309 - 314, XP000938404, DOI: doi:10.1002/prot.340080405 |
| BENHAR, I.; AZRIEL, R.; NAHARY, L.; SHAKY, S.; BERDICHEVSKY, Y.; TAMARKIN, A.; WELS, W.: "Highly efficient selection of phage antibodies mediated by display of antigen as Lpp-OmpA' fusions on live bacteria", J. MOL. BIOL., vol. 301, 2000, pages 893 - 904, XP004461742, DOI: doi:10.1006/jmbi.2000.4021 |
| BERGLUND, L.; PETERSEN, T. E.: "The gene structure of tetranectin, a plasminogen binding protein", FEBS LETTERS, vol. 309, 1992, pages 15 - 19, XP002077572, DOI: doi:10.1016/0014-5793(92)80729-Z |
| BERTRAND, J. A.; PIGNOL, D.; BERNARD, J-P.; VERDIER, J-M.; DAGORN, J-C.; FONTECILLA-CAMPS, J. C.: "Crystal structure of human lithostathine, the pancreatic inhibitor of stone formation", EMBO J., vol. 15, 1996, pages 2678 - 2684 |
| BETTLER, B.; TEXIDO, G.; RAGGINI, S.; RUEGG, D.; HOFSTETTER, H.: "Immunoglobulin E-binding site in Fc epsilon receptor (Fc epsilon RII/CD23) identified by homolog-scanning mutagenesis", J. BIOL. CHEM., vol. 267, 1992, pages 185 - 191 |
| BLANCK, O.; IOBST, S. T.; GABEL, C.; DRICKAMER, K.: "Introduction of selectin- like binding specificity into a homologous mannose-binding protein", J. BIOL. CHEM., vol. 271, 1996, pages 7289 - 7292 |
| BODER, E. T.; WITTRUP, K. D.: "Yeast surface display for screening combinatorial polypeptide libraries", NATURE BIOTECH., vol. 15, 1997, pages 553 - 557, XP002945515, DOI: doi:10.1038/nbt0697-553 |
| BURROWS L; IOBST S T; DRICKAMER K.: "Selective binding of N-acetylglucosamine to the chicken hepatic lectin", BIO-CHEM J., vol. 324, 1997, pages 673 - 680 |
| CAREY AND SUNDBERG: "Advanced Organic Chemistry", 1990, PLENUM PRESS, NEW YORK |
| CHA, JBIOL CHEM., vol. 275, no. 40, 6 October 2000 (2000-10-06), pages 31171 - 7 |
| CHIBA, H.; SANO, H.; SAITOH, M.; SOHMA, H.; VOELKER, D. R.; AKINO, T.; KUROKI, Y.: "Introduction of mannose binding protein-type phosphatidylinositol recognition into pulmonary surfactant protein A", BIOCHEMISTRY, vol. 38, 1999, pages 7321 - 7331 |
| CHRISTENSEN, J. H.; HANSEN, P. K.; LILLELUND, O.; THOGERSEN, H. C.: "Sequence- specific binding of the N-terminal three-finger fragment of Xenopus transcription factor IIIA to the internal control region of a 5S RNA gene", FEBS LETTERS, vol. 281, 1991, pages 181 - 184 |
| CHRISTIE, B. D.; RAPOPORT, H.: "Synthesis of Optically Pure Pipecolates from L-Asparagine. Application to the Total Synthesis of (+)-Apovincamine through Amino Acid Decarbonylation and Iminium Ion Cyclization", J. ORG. CHEM., 1985, pages 1859 - 1866 |
| CRAIG, J. C. ET AL.: "Absolute Configuration of the Enantiomers of 7-Chloro-4[[4-(diethylamino)-1- methylbutyl]amino]quinoline (Chloroquine)", J. ORG. CHEM., vol. 53, 1988, pages 1167 - 1170 |
| CYR, J. L.; HUDSPETH, A. J.: "A library of bacteriophage-displayed antibody fragments directed against proteins of the inner ear", PROC. NATL. ACAD. SCI, vol. 97, 2000, pages 2276 - 2281, XP002975242, DOI: doi:10.1073/pnas.030535797 |
| DEGLI-ESPOSTI ET AL., IMMUNITY, vol. 7, no. 6, December 1997 (1997-12-01), pages 813 - 820 |
| DEGLI-ESPOSTI ET AL., J EXP. MED., vol. 186, no. 7, 6 October 1997 (1997-10-06), pages 1165 - 1170 |
| DRICKAMER, K.: "C-type lectin-like domains", CURR. OPINION STRUC. BIOL., vol. 9, 1999, pages 585 - 590, XP002175480, DOI: doi:10.1016/S0959-440X(99)00009-3 |
| DRICKAMER, K.: "Engineering galactose-binding activity into a C-type mannose-binding protein", NATURE, vol. 360, 1992, pages 183 - 186 |
| DRICKAMER, K.; TAYLOR, M. E.: "Biology of animal lectins", ANNU. REV. CELL BIOL., vol. 9, 1993, pages 237 - 264, XP055018928, DOI: doi:10.1146/annurev.cellbio.9.1.237 |
| DUNN, I. S.: "Phage display of proteins", CURR. OPINION BIOTECH., vol. 7, 1996, pages 547 - 553, XP002092910, DOI: doi:10.1016/S0958-1669(96)80060-7 |
| ERNST, W. J.; SPENGER, A.; TOELLNER, L.; KATINGER, H.; GRABHERR, R. M.: "Expanding baculovirus surface display. Modification of the native coat protein gp64 of Autographa californica NPV", EUR. J. BIOCHEM., vol. 267, 2000, pages 4033 - 4039, XP002258435, DOI: doi:10.1046/j.1432-1327.2000.01439.x |
| EWART, K. V.; LI, Z.; YANG, D. S. C.; FLETCHER, G. L.; HEW, C. L.: "The ice- binding site of Atlantic herring antifreeze protein corresponds to the carbohydrate-binding site of C-type lectins", BIOCHEMISTRY, vol. 37, 1998, pages 4080 - 4085 |
| FEINBERG, H.; PARK-SNYDER, S.; KOLATKAR, A. R.; HEISE, C. T.; TAYLOR, M. E.; WEIS, W. 1.: "Structure of a C-type carbohydrate recognition domain from the macrophage mannose receptor", J. BIOL. CHEM., vol. 275, 2000, pages 21539 - 21548 |
| FIRTH, A.E.; PATRICK, W.M., BIOINFORMATICS, vol. 21, no. 15, 2005, pages 3314 - 3315 |
| FISER A; SALI A.: "Modeller: generation and refinement of homology-based protein structure models.", METHODS ENZYMOL., vol. 374, 2003, pages 461 - 91 |
| FRIEDMAN, O. M.; CHATTERRJI, R.: "Synthesis of Derivatives of Glutamine as Model Substrates for Anti-Tumor Agents", J. AM. CHEM. SOC., vol. 81, 1959, pages 3750 - 3752 |
| FUJII, I.; FUKUYAMA, S.; IWABUCHI, Y.; TANIMURA, R.: "Evolving catalytic antibodies in a phage-displayed combinatorial library", NATURE BIOTECH., vol. 16, 1998, pages 463 - 467, XP000783677, DOI: doi:10.1038/nbt0598-463 |
| GATES, C. M.; STEMMER, W. P. C.; KAPTEIN, R.; SCHATZ, P. J.: "Affinity selective isolation of ligands from peptide libraries through display on a lac repressor "headpiece dimer", J. MOL. BIOL., vol. 255, 1996, pages 373 - 386, XP002941960, DOI: doi:10.1006/jmbi.1996.0031 |
| GRAVERSEN, J. H.; JACOBSEN, C.; SIGURSKJOLD, B. W.; LORENTSEN, R. H.; MOESTRUP, S. K.; THOGERSEN, H. C.; ETZERODT, M.: "Mutational Analysis of Affinity and Selectivity of Kringle- Tetranectin Interaction. Grafting novel kringle affinity onto the tetranectin lectin scaffold", J. BIOL. CHEM., vol. 275, 2000, pages 37390 - 37396, XP002175478, DOI: doi:10.1074/jbc.M004873200 |
| GRAVERSEN, J. H.; LORENTSEN, R. H.; JACOBSEN, C.; MOESTRUP, S. K.; SIGURSKJOLD, B. W.; THOGERSEN, H. C.; ETZERODT, M.: "The plasminogen binding site of the C-type lectin tetranectin is located in the carbohydrate recognition domain, and binding is sensitive to both calcium and lysine", J. BIOL. CHEM., vol. 273, 1998, pages 29241 - 29246, XP002175479, DOI: doi:10.1074/jbc.273.44.29241 |
| GRIFFITHS, A. D.; DUNCAN, A. R.: "Strategies for selection of antibodies by phage display", CURR. OPINION BIOTECH., vol. 9, 1998, pages 102 - 108, XP002505900, DOI: doi:10.1016/S0958-1669(98)80092-X |
| HAKANSSON, K.; LIM, N. K.; HOPPE, H-J.; REID, K. B. M.: "Crystal structure of the trimeric alpha-helical coiled-coil and the three lectin domains of human lung surfactant protein D", STRUCTURE FOLDING AND DESIGN, vol. 7, 1999, pages 255 - 264, XP001157313, DOI: doi:10.1016/S0969-2126(99)80036-7 |
| HOLTET, T. L.; GRAVERSEN, J. H.; CLEMMENSEN, I.; THOGERSEN, H. C.; ETZERODT, M.: "Tetranectin, a trimeric plasminogen-binding C-type lectin", PROT. SCI., vol. 6, 1997, pages 1511 - 1515 |
| HONMA, T.; KUROKI, Y.; TZUNEZAWA, W.; OGASAWARA, Y.; SOHMA, H.; VOELKER, D. R.; AKINO, T.: "The mannose-binding protein A region of glutamic acid185-alanine221 can functionally replace the surfactant protein A region of glutamic acidl95-phenylalanine228 without loss of interaction with lipids and alveolar type II cells", BIOCHEMISTRY, vol. 36, 1997, pages 7176 - 7184 |
| HUANG, W.; ZHANG, Z.; PALZKILL, T.: "Design of potent beta-lactamase inhibitors by phage display of beta-lactamase inhibitory protein", J. BIOL. CHEM., vol. 275, 2000, pages 14964 - 14968 |
| HUFTON, S. E.; VAN NEER, N.; VAN DEN BEUKEN, T.; DESMET, J.; SABLON, E.; HOOGENBOOM, H. R.: "Development and application of cytotoxic T lymphocyte- associated antigen 4 as a protein scaffold for the generation of novel binding ligands", FEBS LETTERS, vol. 475, 2000, pages 225 - 231, XP004337267, DOI: doi:10.1016/S0014-5793(00)01701-4 |
| IOBST, S. T.; DRICKAMER, K.: "Binding of sugar ligands to Ca(2+)-dependent animal lectins. II. Generation of high-affinity galactose binding by site-directed mutagenesis", J. BIOL. CHEM., vol. 269, 1994, pages 15512 - 15519, XP002209824 |
| IOBST, S. T.; DRICKAMER, K.: "Selective sugar binding to the carbohydrate recognition domains of the rat hepatic and macrophage asialoglycoprotein receptors", J. BIOL. CHEM., vol. 271, 1996, pages 6686 - 6693, XP055182220, DOI: doi:10.1074/jbc.271.12.6686 |
| IOBST, S. T.; WORMALD, M. R.; WEIS, W. I.; DWEK, R. A.; DRICKAMER, K.: "Binding of sugar ligands to Ca(2+)-dependent animal lectins. I. Analysis of mannose binding by site-directed mutagenesis and NMR", J. BIOL. CHEM., vol. 269, 1994, pages 15505 - 15511, XP002209823 |
| JANEWAY, NATURE, vol. 341, no. 6242, 12 October 1989 (1989-10-12), pages 482 - 3 |
| JAQUINOD, M.; HOLTET, T. L.; ETZERODT, M.; CLEMMENSEN, I.; THOGERSEN, H. C.; ROEPSTORFF, P.: "Mass Spectrometric Characterisation of Post-Translational Modification and Genetic Variation in Human Tetranectin", BIOL. CHEM., vol. 380, 1999, pages 1307 - 1314, XP002260659, DOI: doi:10.1515/BC.1999.166 |
| JIN ET AL., CANCER RES., vol. 64, no. 14, 15 July 2004 (2004-07-15), pages 4900 - 5 |
| KASTRUP, J. S.; NIELSEN, B. B.; RASMUSSEN, H.; HOLTET, T. L.; GRAVERSEN, J. H.; ETZERODT, M.; THOGERSEN, H. C.; LARSEN, I. K.: "Structure of the C-type lectin carbohydrate recognition domain of human tetranectin", ACTA. CRYST. D, vol. 54, 1998, pages 757 - 766 |
| KING, F. E.; KIDD, D. A. A.: "A New Synthesis of Glutamine and of .gamma.-Dipeptides of Glutamic Acid from Phthylated Intermediates", J. CHEM. SOC., 1949, pages 3315 - 3319 |
| KOGAN, T. P.; REVELLE, B. M.; TAPP, S.; SCOTT, D.; BECK, P. J.: "A single amino acid residue can determine the ligand specificity of E-selectin", J. BIOL. CHEM., vol. 270, 1995, pages 14047 - 14055 |
| KOLATKAR, A. R.; LEUNG, A. K.; ISECKE, R.; BROSSMER, R.; DRICKAMER, K.; WEIS, W. I.: "Mechanism of N-acetylgalactosamine binding to a C-type animal lectin carbohydrate- recognition domain", J. BIOL. CHEM., vol. 273, 1998, pages 19502 - 19508 |
| KOSKINEN, A. M. P.; RAPOPORT, H.: "Synthesis of 4-Substituted Prolines as Conformationally Constrained Amino Acid Analogues", J. ORG. CHEM., vol. 54, 1989, pages 1859 - 1866 |
| LANGER ET AL., J BIOMED. MATER. RES., vol. 15, 1981, pages 167 - 277 |
| LANGER, CHEM. TECH., vol. 12, 1982, pages 98 - 105 |
| LORENTSEN, R. H.; GRAVERSEN, J. H.; CATERER, N. R.; THOGERSEN, H. C.; ETZERODT, M.: "The heparin-binding site in tetranectin is located in the N-terminal region and binding does not involve the carbohydrate recognition domain", BIOCHEM. J., vol. 347, 2000, pages 83 - 87, XP002260660, DOI: doi:10.1042/0264-6021:3470083 |
| LUTZ S.; PATRICK, W.M., CURR. OPIN. BIOTECHNOL., vol. 15, 2004, pages 291 - 297 |
| M. C. PERRY: "Chemotherapy Service", 1992, WILLIAMS & WILKINS, BALTIMORE, MD. |
| MANN K; WEISS I M; ANDRE S; GABIUS H J; FRITZ M.: "The amino-acid sequence of the abalone (Haliotis laevigata) nacre protein perlucin. Detection of a functional C-type lectin domain with galactose/mannose specificity", EUR. J. BIOCHEM., vol. 267, 2000, pages 5257 - 5264, XP002450404, DOI: doi:10.1046/j.1432-1327.2000.01602.x |
| MARCH: "Advanced Organic Chemistry", 1985, WILEY AND SONS, NEW YORK |
| MARKS, J. D.; HOOGENBOOM, H. R.; GRIFFITHS, A. D.; WINTER, G.: "Molecular evolution of proteins on filamentous phage. Mimicking the strategy of the immune system", J. BIOL. CHEM., vol. 267, 1992, pages 16007 - 16010, XP002025816 |
| MARSHAK, D. R.: "9th Meeting: Tech. Prot. Chem VII", 1996, ACADEMIC, SAN DIEGO, CALIF, article NEAME, P. J., ET AL.: "Protein Soc. Symposium", pages: 401 - 407 |
| MARSTERS ET AL., CURR. BIOL., vol. 7, 1997, pages 1003 - 1006 |
| MARTI-RENOM MA; STUART A; FISER A; SÁNCHEZ R; MELO F; SALI A.: "Comparative protein structure modeling of genes and genomes", ANNU. REV. BIOPHYS. BIOMOL. STRUCT., vol. 29, 2000, pages 291 - 325, XP001085249, DOI: doi:10.1146/annurev.biophys.29.1.291 |
| MATSOUKAS ET AL., J. MED. CHEM., vol. 38, 1995, pages 4660 - 4669 |
| MCCAFFERTY, J.; JACKSON, R. H.; CHISWELL, D. J.: "Phage-enzyrnes: expression and affinity chromatography of functional alkaline phosphatase on the surface of bacteriophage", PROT. ENG., vol. 4, 1991, pages 955 - 961, XP009005463 |
| MCCORMACK, F. X.; FESTA, A. L.; ANDREWS, R. P.; LINKE, M.; WALZER, P. D.: "The carbohydrate recognition domain of surfactant protein A mediates binding to the major surface glycoprotein of Pneumocystis carinii", BIOCHEMISTRY, vol. 36, 1997, pages 8092 - 8099 |
| MCCORMACK, F. X.; KUROKI, Y.; STEWART, J. J.; MASON, R. J.; VOELKER, D. R.: "Surfactant protein A amino acids Glu195 and Argl97 are essential for receptor binding, phospholipid aggregation, regulation of secretion, and the facilitated uptake of phospholipid by type II cells", J. BIOL. CHEM., vol. 269, 1994, pages 29801 - 29807 |
| MCFARLANE ET AL., J BIOL. CHEM., vol. 272, 1997, pages 25417 - 25420 |
| MEIER, M.; BIDER, M. D.; MALASHKEVICH, V. N.; SPIESS, M.; BURKHARD, P.: "Crystal structure of the carbohydrate recognition domain of the Hi subunit of the asialoglycoprotein receptor", J. MOL. BIOL., vol. 300, 2000, pages 857 - 865 |
| MENDELSOHN AND ISRAEL: "The Molecular Basis of Cancer", 15 January 1995, WB SAUNDERS, PHILADELPHIA, article MURAKAMI ET AL.,: "Cell cycle regulation, oncogenes, and antineoplastic drugs", pages: 13 |
| MIKAWA, Y. G.; MARUYAMA, I. N.; BRENNER, S.: "Surface display of proteins on bacteriophage lambda heads", J. MOL. BIOL., vol. 262, 1996, pages 21 - 30, XP002203952, DOI: doi:10.1006/jmbi.1996.0495 |
| MIO H; KAGAMI N; YOKOKAWA S; KAWAI H; NAKAGAWA S; TAKEUCHI K; SEKINE S; HIRAOKA A.: "Isolation and characterization of a cDNA for human mouse, and rat full-length stem cell growth factor, a new member of C-type lectin superfamily", BIOCHEM. BIOPHYS. RES. COMMUN., vol. 249, 1998, pages 124 - 130 |
| MIZUNO, H.; FUJIMOTO, Z.; KOIZUMI, M.; KANO, H.; ATODA, H.; MORITA, T.: "Structure of coagulation factors IX/X-binding protein, a heterodimer of C-type lectin domains", NAT. STRUC. BIOL., vol. 4, 1997, pages 438 - 441 |
| MONGKOLSAPAYA ET AL., J IMMUNOL., vol. 160, 1998, pages 3 - 6 |
| MORDENTI ET AL., PHARMACEUT. RES., vol. 8, 1991, pages 1351 |
| NEAME ET AL., PROTEIN SCI., vol. 1, no. 1, 1992, pages 161 - 8 |
| NG, K. K.; PARK-SNYDER, S.; WEIS, W. 1.: "Ca.sup.2+-dependent structural changes in C-type mannose-binding proteins", BIOCHEMISTRY, vol. 37, 1998, pages 17965 - 17976 |
| NG, K. K.; WEIS, W. I.: "Coupling of prolyl peptide bond isomerization and Ca2+ binding in a C-type mannose-binding protein", BIOCHEMISTRY, vol. 37, 1998, pages 17977 - 17989 |
| NIELBO ET AL., BIOCHEMISTRY, vol. 43, no. 27, 2004, pages 8636 - 8643 |
| NIELSEN, B. B.; KASTRUP, J. S.; RASMUSSEN, H.; HOLTET, T. L.; GRAVERSEN, J. H.; ETZERODT, M.; THOGERSEN, H. C.; LARSEN, I. K.: "Crystal structure of tetranectin, a trimeric plasminogen-binding protein with an alpha-helical coiled coi", FEBS LETTERS, vol. 412, 1997, pages 388 - 396, XP002260662, DOI: doi:10.1016/S0014-5793(97)00664-9 |
| NISSIM A.; HOOGENBOOM, H. R.; TOMLINSON, I. M.; FLYNN, G.; MIDGLEY, C.; LANE, D.; WINTER, G.: "Antibody fragments from a 'single pot' phage display library as immunochemical reagents", EMBO J., vol. 13, 1994, pages 692 - 698 |
| OFFNER ET AL., SCIENCE, vol. 251, 1991, pages 430 - 432 |
| OGASAWARA, Y.; VOELKER, D. R.: "Altered carbohydrate recognition specificity engineered into surfactant protein D reveals different binding mechanisms for phosphatidylinositol and glucosylceramide", J. BIOL. CHEM., vol. 270, 1995, pages 14725 - 14732 |
| OHTANI, K.; SUZUKI, Y.; EDA, S.; TAKAO, K.; KASE, T.; YAMAZAKI, H.; SHIMADA, T.; KESHI, H.; SAKAI, Y.; FUKUOH, A.: "Molecular cloning of a novel human collectin from liver (CL-L1)", J. BIOL. CHEM., vol. 274, 1999, pages 13681 - 13689, XP002242030, DOI: doi:10.1074/jbc.274.19.13681 |
| OSOL, A: "Remington's Pharmaceutical Sciences", 1980 |
| PAN ET AL., FEBS LETTERS, vol. 424, 1998, pages 41 - 45 |
| PAN ET AL., SCIENCE, vol. 276, 1997, pages 111 - 113 |
| PAN ET AL., SCIENCE, vol. 277, 1997, pages 815 - 818 |
| PATRICK, W.M.; FIRTH, A.E., BIOMOLECULAR ENGINEERING, vol. 22, 2005, pages 105 - 112 |
| PATTANAJITVILAI, S.; KUROKI, Y.; TSUNEZAWA, W.; MCCORMACK, F. X.; VOELKER, D. R.: "Mutational analysis of Argl97 of rat surfactant protein A. His197 creates specific lipid uptake defects", J. BIOL. CHEM., vol. 273, 1998, pages 5702 - 5707 |
| POGET, S. F.; LEGGE, G. B.; PROCTOR, M. R.; BUTLER, P. J.; BYCROFT, M.; WILLIAMS, R. L.: "The structure of a tunicate C-type lectin from Polyandrocarpa misakiensis complexed with D-galactose", J. MOL. BIOL., vol. 290, 1999, pages 867 - 879, XP004451660, DOI: doi:10.1006/jmbi.1999.2910 |
| REVELLE, B. M.; SCOTT, D.; KOGAN, T. P.; ZHENG, J.; BECK, P. J.: "Structure- function analysis of P-selectinsialyl LewisX binding interactions. Mutagenic alteration of ligand binding specificity", J. BIOL. CHEM., vol. 271, 1996, pages 4289 - 4297 |
| RUOSLAHTI, E.; YAMAGUCHI, Y.: "The C-type lectin domains of lecticans, a family of aggregating chondroitin sulfate proteoglycans, bind tenascin-R by protein-protein interactions independent of carbohydrate moiety", PROC. NATL. ACAD. SCI., vol. 94, 1997, pages 10116 - 10121 |
| S. G. HYMOWITZ, MOL CELL, vol. 4, no. 4, October 1999 (1999-10-01), pages 563 - 71 |
| SALI A; BLUNDELL TL.: "Comparative protein modelling by satisfaction of spatial restraints", J MOL. BIOL., vol. 234, 1993, pages 779 - 815, XP024008717, DOI: doi:10.1006/jmbi.1993.1626 |
| SAMBROOK, ET AL.: "Molecular Cloning: A Laboratory Manual", 1989, COLD SPRING HARBOR LABORATORY PRESS, COLD SPRING HARBOR, NY |
| SANO, H.; KUROKI, Y.; HONMA, T.; OGASAWARA, Y.; SOHMA, H.; VOELKER; D. R.; AKINO, T.: "Analysis of chimeric proteins identifies the regions in the carbohydrate recognition domains of rat lung collections that are essential for interactions with phospholipids, glycolipids, and alveolar type II cells", J. BIOL. CHEM., vol. 273, 1998, pages 4783 - 4789, XP002268026, DOI: doi:10.1074/jbc.273.8.4783 |
| SCHAFFITZEL, C.; HANES, J.; JERMUTUS, L.; PLUCKTUN, A.: "Ribosome display: an in vitro method for selection and evolution of antibodies from libraries", J. IMMUNOL. METHODS, vol. 231, 1999, pages 119 - 135, XP004186079, DOI: doi:10.1016/S0022-1759(99)00149-0 |
| SCHNEIDER ET AL., FEBS LETTERS, vol. 416, 1997, pages 329 - 334 |
| SCREATON ET AL., CURR. BIOL., vol. 7, 1997, pages 693 - 696 |
| SHERIDAN ET AL., SCIENCE, vol. 277, 1997, pages 818 - 821 |
| SHERIFF, S.; CHANG, C. Y.; EZEKOWITZ, R. A.: "Human mannose-binding protein carbohydrate recognition domain trimerizes through a triple alpha-helical coiled-coil", NAT. STRUC. BIOL., vol. 1, 1994, pages 789 - 794 |
| SIDMAN ET AL., BIOPOLYMERS, vol. 22, 1983, pages 547 - 556 |
| SORENSEN, C. B.; BERGLUND, L.; PETERSEN, T. E.: "Cloning of a CDNA encoding murine tetranectin", GENE, vol. 152, 1995, pages 243 - 245, XP004042684, DOI: doi:10.1016/0378-1119(94)00703-U |
| STEMMER, W.P., PROC NATL ACAD SCI USA, vol. 91, October 1994 (1994-10-01), pages 10747 - 751 |
| SUBASINGHE ET AL.: "Quisqualic acid analogues: synthesis of beta-heterocyclic 2-aminopropanoic acid derivatives and their activity at a novel quisqualate-sensitized site", J. MED. CHEM., vol. 35, 1992, pages 4602 - 7, XP002198373, DOI: doi:10.1021/jm00102a014 |
| TORGERSEN, D.; MULLIN, N. P.; DRICKAMER, K.: "Mechanism of ligand binding to E- and P-selectin analyzed using selectin/mannose-binding protein chimeras", J. BIOL. CHEM., vol. 273, 1998, pages 6254 - 6261, XP002209822, DOI: doi:10.1074/jbc.273.11.6254 |
| TORMO, J.; NATARAJAN, K.; MARGULIES, D. H.; MARIUZZA, R. A.: "Crystal structure of a lectin-like natural killer cell receptor bound to its MHC class I ligand", NATURE, vol. 402, 1999, pages 623 - 631 |
| TSUNEZAWA, W.; SANO, H.; SOHMA, H.; MCCORMACK, F. X.; VOELKER, D. R.; KUROKI, Y.: "Site-directed mutagenesis of surfactant protein A reveals dissociation of lipid aggregation and lipid uptake by alveolar type II cells", BIOCHIM. BIOPHYS. ACTA, vol. 1387, 1998, pages 433 - 446, XP004278498, DOI: doi:10.1016/S0167-4838(98)00159-9 |
| TURK B E ET AL: "Determination of protease cleavage site motifs using mixture-based oriented peptide libraries", NATURE BIOTECHNOLOGY, NATURE PUBLISHING GROUP, NEW YORK, NY, US LNKD- DOI:10.1038/90273, vol. 19, no. 7, 1 July 2001 (2001-07-01), pages 661 - 667, XP002254824, ISSN: 1087-0156 * |
| VARKI A., ET AL.: "Essentials of Glycobiology, second edition", CHS PRESS |
| WALCZAK ET AL., EMBO J., vol. 16, 1997, pages 5386 - 5387 |
| WEIS, W. I.; DRICKAMER, K.: "Structural basis of lectin-carbohydrate recognition", ANNU. REV. BIOCHEM., vol. 65, 1996, pages 441 - 473 |
| WEIS, W. I.; KAHN, R.; FOURME, R.; DRICKAMER, K.; HENDRICKSON, W. A.: "Structure of the calcium-dependent lectin domain from a rat mannose-binding protein determined by MAD phasing", SCIENCE, vol. 254, 1991, pages 1608 - 1615 |
| WHITEHORN, E.A.; TATE, E.; YANOFSKY, S. D., KOCHERSPERGER, L.; DAVIS A.; MORTENSEN, R. B.; YONKOVIC, S.; BELL, K.; DOWER, W. J.; B: "A generic method for expression and use of "tagged" soluble versions of cell surface receptors", BIO/TECHNOLOGY, vol. 13, 1995, pages 1215 - 1219 |
| WRAGG, S.; DRICKAMER, K.: "Identification of amino acid residues that determine pH dependence of ligand binding to the asialoglycoprotein receptor during endocytosis", J. BIOL. CHEM., vol. 274, 1999, pages 35400 - 35406 |
| WU ET AL., NATURE GENETICS, vol. 17, 1997, pages 141 - 143 |
| ZHANG, H.; ROBISON, B.; THORGAARD, G. H.; RISTOW, S. S.: "Cloning, mapping and genomic organization of a fish C-type lectin gene from homozygous clones of rainbow trout (Oncorhynchos Mykiss)", BIOCHIM. ET BIOPHYS. ACTA, vol. 1494, 2000, pages 14 - 22, XP004275787, DOI: doi:10.1016/S0167-4781(00)00198-6 |
Cited By (14)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US11096997B2 (en) | 2005-12-13 | 2021-08-24 | President And Fellows Of Harvard College | Scaffolds for cell transplantation |
| US11202759B2 (en) | 2010-10-06 | 2021-12-21 | President And Fellows Of Harvard College | Injectable, pore-forming hydrogels for materials-based cell therapies |
| US11278604B2 (en) | 2012-04-16 | 2022-03-22 | President And Fellows Of Harvard College | Mesoporous silica compositions comprising inflammatory cytokines comprising inflammatory cytokines for modulating immune responses |
| US10899815B2 (en) | 2013-03-15 | 2021-01-26 | Shenzhen Hightide Biopharmaceutical, Ltd. | Compositions and methods of using islet neogenesis peptides and analogs thereof |
| US9738695B2 (en) | 2013-03-15 | 2017-08-22 | Shenzhen Hightide Biopharmaceutical, Ltd. | Compositions and methods of using islet neogenesis peptides and analogs thereof |
| US9388215B2 (en) | 2013-03-15 | 2016-07-12 | Shenzhen Hightide Biopharmaceutical, Ltd. | Compositions and methods of using islet neogenesis peptides and analogs thereof |
| US10682400B2 (en) | 2014-04-30 | 2020-06-16 | President And Fellows Of Harvard College | Combination vaccine devices and methods of killing cancer cells |
| US11786457B2 (en) | 2015-01-30 | 2023-10-17 | President And Fellows Of Harvard College | Peritumoral and intratumoral materials for cancer therapy |
| US11150242B2 (en) | 2015-04-10 | 2021-10-19 | President And Fellows Of Harvard College | Immune cell trapping devices and methods for making and using the same |
| US11752238B2 (en) | 2016-02-06 | 2023-09-12 | President And Fellows Of Harvard College | Recapitulating the hematopoietic niche to reconstitute immunity |
| US10813988B2 (en) * | 2016-02-16 | 2020-10-27 | President And Fellows Of Harvard College | Pathogen vaccines and methods of producing and using the same |
| US20170246281A1 (en) * | 2016-02-16 | 2017-08-31 | President And Fellows Of Harvard College | Pathogen vaccines and methods of producing and using the same |
| US11638748B2 (en) | 2016-02-16 | 2023-05-02 | President And Fellows Of Harvard College | Pathogen vaccines and methods of producing and using the same |
| US11555177B2 (en) | 2016-07-13 | 2023-01-17 | President And Fellows Of Harvard College | Antigen-presenting cell-mimetic scaffolds and methods for making and using the same |
Also Published As
| Publication number | Publication date |
|---|---|
| CN102686727A (en) | 2012-09-19 |
| JP2013507123A (en) | 2013-03-04 |
| EP2486132A1 (en) | 2012-08-15 |
| CA2776954A1 (en) | 2011-04-14 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US20110086770A1 (en) | Combinatorial Libraries Based on C-type Lectin-like Domain | |
| WO2011043834A1 (en) | Combinatorial libraries based on c-type lectin domain | |
| EP2105503B1 (en) | Method for the identification and isolation of binding polypeptides from combinatorial libraries of proteins having the scaffold structure of C-type lectin-like domains | |
| US20110028403A1 (en) | HSP70-Based Treatment for Autoimmune Diseases and Cancer | |
| JP4907542B2 (en) | Protein complexes for use in therapy, diagnosis and chromatography | |
| AU2005265150B2 (en) | C-MET kinase binding proteins | |
| EP2284270A2 (en) | Method for the identification and isolation of binding polypeptides from combinatorial libraries of proteins having the scaffold structure of C-type lectin-like domains | |
| AU2002221568A1 (en) | Combinatorial libraries of proteins having the scaffold structure of C-type lectin-like domains | |
| JP2016053031A (en) | Collections of repeat proteins comprising repeat modules | |
| WO2012171541A1 (en) | Human fusion proteins comprising interferons and hetero-dimeric modified ubiquitin proteins | |
| KR20150016585A (en) | Non-natural consensus albumin binding domains | |
| JP2009501517A (en) | IL-6 binding protein | |
| WO2003091429A1 (en) | Antimicrobial polypeptide and utizliation thereof | |
| Eisenmesser et al. | Expression, purification, refolding, and characterization of recombinant human interleukin-13: utilization of intracellular processing | |
| AU2010303879A1 (en) | Combinatorial libraries based on C-type lectin domain | |
| US20110086806A1 (en) | Polypeptides that Bind IL-23R | |
| US20090203541A1 (en) | Msp and its domains as frameworks for novel binding molecules | |
| EP2486050A1 (en) | Polypeptides that bind il-23r | |
| CN113811542A (en) | Fusion proteins comprising a cytokine and a scaffold protein | |
| JP2839837B2 (en) | DNA encoding the ligand-binding domain protein of granulocyte colony-stimulating factor receptor | |
| Class et al. | Patent application title: HSP70-Based Treatment for Autoimmune Diseases and Cancer Inventors: Isabelle Caroline Le Poole (Downers Grove, IL, US) Josephus Dirk Nieland (Arhus C, DK) Thor Las Holtet (Ronde, DK) Thor Las Holtet (Ronde, DK) |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| WWE | Wipo information: entry into national phase |
Ref document number: 201080056021.5 Country of ref document: CN |
|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 10704873 Country of ref document: EP Kind code of ref document: A1 |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2776954 Country of ref document: CA |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2012533143 Country of ref document: JP |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2010303879 Country of ref document: AU |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2010704873 Country of ref document: EP |
|
| ENP | Entry into the national phase |
Ref document number: 2010303879 Country of ref document: AU Date of ref document: 20100210 Kind code of ref document: A |