도 1은 본 발명에 따른 다양한 ENTER(Environmental transition guided cellular reprogramming) 세포로의 리프로그래밍 방법을 도시한 모식도이다.
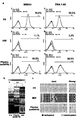
도 2a-b는 초음파 자극에 의한 세포막 손상 및 회복을 보여주는 결과로, 도 2a는 SEM으로 촬영된 세포 표면 이미지이고 도 2b는 live/dead kit 염색 이미지이다.
도 3a-c는 세포 내 Ca 유입 분석(a), ATP 반응 분석(b) 및 ATP 수용체 유전자 발현에 대한 RT-PCR 분석 결과(c)이다.
도 4a-c는 QD605을 이용하여 초음파 자극에 의한 세포 내 외부물질 유입 가능성을 보여주는 결과로, 도 4a는 usMC-S(ultrasound-exposed medium and cells) 스페로이드와 단일세포에서 QD605 유입 분석 결과이고, 도 4b 및 4c는 각각의 환경 조건별 usMC-S 스페로이드(b)와 단일세포(c) 내 QD605 유입 분석 및 전사 인자 발현 결과를 나타낸 것이다.
도 5는 초음파 처리된 배양액 내 엑소좀의 RNA를 RT-PCR 분석한 결과이다.
도 6a-c는 엑소좀에 의한 물질 전달을 보여주는 결과로, 도 6a는 엑소좀에 의해 QD605가 다른 세포로 이동되는 과정을 캡쳐한 이미지이고, 도 6b는 엑소좀 내 단백질 마커의 발현을 보여주며, 도 6c는 엑소좀에 의한 poly(A)27-Cy5.5 전달 실험 결과를 나타낸 것이다.
도 7a-b는 초음파 처리된 세포의 배양액 내 엑소좀과 초음파 처리되지 않은 세포의 공동배양에 따른 세포의 변화를 나타낸 것으로, 도 7a는 배양시간에 따른 세포 형태의 변화이고, 도 7b는 엑소좀과 6일간 배양된 세포내 Oct4의 발현을 보여주는 결과이다.
도 8은 본 발명에 따른 인간 진피 섬유아세포의 직접 분화 방법을 도시한 것이다.
도 9a-b는 지방세포 분화유도배지에서 초음파 처리된 인간 진피 섬유아세포의 지방세포로의 분화 결과를 나타낸 것으로, 도 9a는 분화 유도 후 20일째 세포의 변화와 지방세포의 oil red O 염색 결과이고, 도 9b는 분화 유도된 세포의 지방세포 마커 유전자 발현에 대한 RT-PCR 분석 결과이다.
도 10a-c는 신경줄기세포 분화유도배지와 초음파 처리된 인간 진피 섬유아세포의 신경전구세포로의 분화 결과를 나타낸 것으로, 도 10a는 분화 유도 후 3일째 세포의 변화, 도 10b는 분화 유도된 신경전구세포 마커 발현, 도 10c는 부착된 세포에서 신경전구세포 마커 발현을 분석한 결과이다.
도 11은 신경전구세포로 분화 유도된 세포(n/ENTER cells)에서 Pax6 및 Nestin의 발현을 분석한 플로우 사이토메트리 분석 결과이다.
도 12는 n/ENTER 세포에서의 증식 마커인 Ki67 발현을 나타낸 것이다.
도 13a는 n/ENTER 세포의 단일 스피어에서 세포가 증식되는 과정을, 도 13b는 증식된 세포에서의 신경전구세포의 속성 유지 결과를 보여준다.
도 14a-b는 n/ENTER 세포를 쥐의 뇌에 이식한 후 분화를 보여주는 결과로, 도 14a는 이식된 세포(HNA 염색된 세포)에서의 성상세포 마커(Gfap)의 발현, 도 14b는 이식 후 분화된 세포에서 시냅신(Syn1)의 분비를 보여준다.
도 15a-c는 신경줄기세포 분화유도배지와 초음파 자극에 의한 MEF의 신경전구세포로의 분화를 보여주는 결과로, 도 15a는 세포 형태 변화, 도 15b는 스피어의 신경전구세포 마커의 발현, 도 15c는 플로우 사이토메트리를 이용한 신경전구세포 마커의 발현 분석 결과를 나타낸다.
도 16은 MEF에서 분화 유도된 신경전구세포(mouse n/ENTER cells)의 분화 마커 발현 분석 결과이다.
도 17은 간세포로의 직접 분화를 보여주는 도식도이다.
도 18a-b는 간세포 분화유도배지와 초음파 처리된 HDF의 간세포로의 분화 결과로, 도 18a는 세포 형태 변화, 도 18b는 간세포 마커의 발현을 보여준다.
[규칙 제91조에 의한 정정 12.10.2016]
도 19a-c는 간세포 분화유도배지와 초음파 처리된 HeLa 세포의 간세포(HeLa h/ENTER)로의 분화 결과로, 도 19a는 세포 형태 변화, 도 19b는 qPCR을 이용한 간세포 마커의 발현 결과, 도 19c는 면역세포화학법을 이용한 간세포 마커의 발현 결과를 보여준다.
도 20a-b는 간세포 분화유도배지와 초음파 처리된 Hep3B 세포의 간세포(Hep3B h/ENTER cell)로의 분화 결과로, 도 20a는 세포 형태 변화, 도 20b는 간세포 마커의 발현을 보여준다.
도 21a-b는 인간 ES 배양배지와 초음파 처리된 HDF의 es/ENTER 세포로의 분화 결과로, 도 21a는 세포 형태 변화, 도 21b는 배양시간에 따른 Oct4 발현의 변화를 보여준다.
도 22a-b는 es/ENTER 세포에서의 다능성 마커 유전자(a) 및 단백질(b)의 발현을 나타낸 것이다.
도 23a-c는 es/ENTER 세포에서의 다능성 속성 분석 결과로, 도 23a는 플로우 사이토메트리를 이용한 다능성 마커 분석 결과, 도 23b는 마이크로어레이를 이용한 다능성 유전자 발현 패턴 분석 결과, 도 23c는 바이설파이트 시퀀싱을 이용한 Oct4 및 Nanog 프로모터의 메틸화 여부 분석 결과이다.
도 24a-b는 es/ENTER 세포에서의 3배엽 마커 유전자(a) 및 단백질(b)의 발현을 나타낸 것이다.
도 25는 배양시간 별 es/ENTER 세포에서의 Oct4 및 3배엽 마커 발현의 변화를 나타낸 것이다.
도 26a-b는 부착 배양된 es/ENTER 세포에서의 3배엽 마커 단백질의 발현(a) 및 바이설파이트 시퀀싱을 이용한 es/ENTER 세포의 3배엽 마커 DNA의 메틸화 분석(b) 결과를 나타낸 것이다.
도 27a-c는 es/ENTER 세포의 신경세포(a), 심근세포(b) 및 간세포(c)로의 인 비트로 분화 결과를 나타낸 것이다.
도 28a-c는 HDF에서 신경세포(a), 심근세포(b) 및 간세포(c) 마커의 발현 여부를 분석한 결과를 나타낸 것이다.
도 29a-c는 분화 유도된 es/ENTER 세포의 신경세포(a), 심근세포(b) 및 간세포(c) 분화 마커 유전자 발현에 대한 RT-PCR 분석 결과를 나타낸 것이다.
도 30은 es/ENTER 세포의 염색체 G-밴드 분석에 의한 핵형분석 결과를 나타낸 것이다.
도 31a-b는 es/ENTER 세포의 근육 이식 및 인 비보 분화 결과로, 도 31a는 이식된 세포의 골격 근육으로의 분화 결과를, 도 31b는 이식된 세포의 Oct4 발현 및 증식(Ki67) 유무를 보여준다.
도 32a-c는 es/ENTER 세포의 쥐의 뇌로의 이식 및 인 비보 분화 결과로, 도 32a는 세포가 이식된 좌표, 도 32b는 이식된 세포의 성상세포 분화 마커(Gfap)와 신경세포 기능성 마커(시냅신, Syn1 및 Vesicular Glutamate transpoter(vGlut1))의 분비, 도 32c는 이식된 세포의 Oct4 발현과 증식(Ki67) 유무를 보여준다.
도 33a-c는 인간 ES 배양배지와 초음파 자극에 의한 MEF의 mouse es/ENTER 세포로의 분화를 보여주는 결과로, 도 33a는 배양시간에 따른 세포 형태 및 Oct4-GFP 발현의 변화, 도 33b는 배양시간에 따른 스페로이드 형성율, 도 33c는 Oct4 발현의 변화를 보여준다.
도 34a-d는 mouse es/ENTER 세포에서의 다능성 속성 분석 결과로, 도 34a는 면역세포화학법에 의한 다능성 마커 단백질의 발현, 도 34b는 RT-PCR에 의한 다능성 마커 유전자의 발현, 도 34c는 플로우 사이토메트리에 의한 다능성 마커 유전자의 발현 비율 분석 결과, 도 34d는 mouse es/ENTER 세포의 알칼라인 포스파타아제(AP) 염색 결과를 보여준다.
도 35a-c는 mouse es/ENTER 세포에서 3배엽 특성을 분석한 결과로, 도 35a는 면역세포화학법에 의한 3배엽 마커의 발현, 도 35b 및 35c는 배양시간 별 3배엽 마커 유전자(b) 및 단백질(c)의 발현 패턴의 분석 결과를 보여준다.
도 36a-c는 mouse es/ENTER 세포의 신경세포(a), 심근세포(b)로의 인 비트로 분화 결과와, mouse es/ENTER 세포의 염색체 G-밴드 분석에 의한 핵형 분석 결과(c)를 나타낸 것이다.
도 37a-c는 인간 ES 배양배지와 초음파 자극에 의한 L132 세포의 L132 es/ENTER 세포로의 분화를 보여주는 결과로, 도 37a는 배양시간에 따른 세포 형태 변화, 도 37b 및 37c는 L132 es/ENTER 세포의 다능성(b) 및 3배엽(c) 속성을 보여준다.
도 38a-c는 인간 ES 배양배지와 초음파 자극에 의한 MSC의 MSC es/ENTER 세포로의 분화를 보여주는 결과로, 도 38a는 배양시간에 따른 세포 형태 변화, 도 38b 및 38c는 MSC es/ENTER 세포의 다능성(b) 및 3배엽(c) 속성을 보여준다.
도 39a-c는 인간 ES 배양배지와 초음파 자극에 의한 인간 피부 섬유아세포의 SF es/ENTER 세포로의 분화를 보여주는 결과로, 도 39a는 배양시간에 따른 세포 형태 변화, 도 39b 및 39c는 SF es/ENTER 세포의 다능성(b) 및 3배엽(c) 속성을 보여준다.
도 40a-c는 열 처리와 hES 배지를 이용한 HDF의 es/ENTER 세포로의 분화를 보여주는 결과로, 도 40a는 분화 유도된 HDF 스페로이드, 도 40b 및 40c는 es/ENTER 세포의 다능성(b) 및 3배엽(c) 속성을 보여준다.
도 41a-c는 레이저 자극과 hES 배지를 이용한 HDF의 es/ENTER 세포로의 분화를 보여주는 결과로, 도 41a는 분화 유도된 HDF 스페로이드, 도 41b 및 41c는 es/ENTER 세포의 다능성(b) 및 3배엽(c) 속성을 보여준다.
도 42는 정상 체세포로의 세포 밖 소포체(EVs)의 전달 여부를 확인한 결과이다.
도 43은 EVs에 의한 인간 섬유아세포의 리프로그래밍 유도 여부를 확인한 결과이다.
도 44는 es/ENTER 유도 시 회수한 EVs 처리된 인간 섬유아세포의 배양시간별 세포 형태의 변화를 나타낸 것이다.
도 45는 EVs 첨가량에 따른 6일 배양된 HDF의 Oct4 발현 여부를 확인한 Flow cytometry 분석 결과이다.
도 46a-b는 es/ENTER 유도 시 회수한 EVs 처리된 3일 배양된 HDF에서 다능성 마커 단백질(a: ICC 이미지) 및 유전자(b: qPCR 분석) 발현 여부를 확인한 결과이다.
도 47a-b는 n/ENTER 유도 시 회수한 EVs 처리된 3일 배양된 HDF에서 신경줄기세포 마커 단백질(a: ICC 이미지) 및 유전자(b: qPCR 분석) 발현 여부를 확인한 결과이다.
이하, 본 발명의 구성을 구체적으로 설명한다.
본 발명은 분화 또는 미분화 세포 및 배양 배지의 혼합물에 환경유입(environmental influx)을 촉진할 수 있는 물리적 자극을 제공하고, 상기 물리적 자극을 제공받은 혼합물을 일정 시간 배양하여 리프로그래밍된 세포를 수득하는 것을 포함하는, 세포의 리프로그래밍 방법에 관한 것이다.
본 발명은 분화 또는 미분화 세포에 초음파, 레이저 또는 열처리 등 환경 유입을 촉진할 수 있는 물리적 자극을 제공하면서 목적하는 리프로그래밍된 세포를 유도할 수 있는 임의의 배지에서 분화 또는 미분화 세포를 배양하여 다능성(pluripotency) 세포; 또는 상기 분화 또는 미분화된 세포와 표현형이 상이한 임의의 분화 세포, 예컨대, 간세포, 조골세포, 지방세포, 근육세포, 신경세포, 성상세포, 각질세포, 모근세포, 췌장 베타세포 또는 심근세포 등으로 세포의 리프로그래밍을 유도할 수 있는 것을 특징으로 한다.
일 예로, 리프로그래밍된 세포로 다능성 세포를 목적으로 하는 경우 분화된 세포와 줄기세포 배양 배지를 혼합하고, 물리적 자극을 제공하여 일정 시간 배양함으로써 분화된 세포는 다능성 세포로 리프로그래밍될 수 있다.
다른 예로, 리프로그래밍된 세포로 분화된 세포와 표현형이 상이한 임의의 분화 세포를 목적으로 하는 경우, 분화된 세포와 목적하는 분화 세포의 분화유도배지를 혼합하고, 물리적 자극을 제공하여 일정 시간 동안 배양함으로써 분화된 세포는 표현형이 상이한 임의의 분화 세포로 리프로그래밍될 수 있다.
또 다른 예로, 미분화 세포, 예컨대 유도만능줄기세포 또는 배아줄기세포와 목적하는 분화 세포의 분화유도배지를 혼합하고, 물리적 자극을 제공하여 일정 시간 동안 배양함으로써 기존 기술 대비 분화율이 개선된 목적하는 분화 세포로 리프로그래밍될 수 있다.
본 발명의 세포의 리프로그래밍 방법은 분화 또는 미분화 세포에의 물리적 자극을 통해 세포 외의 환경유입에 따라 분화 또는 미분화 세포의 리프로그래밍이 유도되는 것으로 보인다. 이러한 환경유입은 물리적 자극을 제공받은 분화된 세포에서 배출되는 유전물질, 화학물질, 소분자, 엑소좀, 또는 엑소좀 함유 세포 밖 소포체(extracellular vesicles); 또는 배양 배지 성분 등의 이웃하는 분화 또는 미분화 세포로의 유입을 의미한다.
본 발명의 세포의 리프로그래밍 방법에 따르면, 분화 또는 미분화 세포로의 환경유입은 다능성 마커와 3배엽 마커를 안정적으로 발현하는 다능성 세포와 상기 분화 또는 미분화 세포와 표현형이 상이한 분화 세포로의 리프로그래밍 방향성을 결정할 수 있는 것으로 보인다.
덧붙여, 리프로그래밍 방향성은 배양 배지의 종류에 의해 결정될 수 있는 것으로 보인다.
즉, 상술한 바와 같이 분화 또는 미분화 세포에서 다능성 세포로의 리프로그래밍은 분화된 세포와 줄기세포 배양 배지의 혼합물에 물리적 자극을 제공한 경우 유도될 수 있고, 분화된 세포에서 표현형이 상이한 임의의 분화 세포로의 리프로그래밍은 분화된 세포와 임의의 분화 세포의 분화유도배지의 혼합물에 물리적 자극을 제공한 경우 유도될 수 있으며, 미분화 세포와 임의의 분화 세포의 분화유도배지의 혼합물에 물리적 자극을 제공하는 경우 임의의 분화 세포로 리프로그래밍될 수 있다.
분화 또는 미분화 세포로의 환경유입과 관련하여, 본 발명자들은 특히 물리적 자극에 의한 세포막 손상과 세포 분비 물질(엑소좀 또는 엑소좀 함유 세포 밖 소포체)을 고려하였다. 즉, 초음파, 레이저, 또는 열 처리는 에너지에 의한 온도 상승, 초음파에 의해 생성되는 마이크로버블의 진동, 액체의 흐름 생성 유도 즉, 세포막을 따라 마이크로스트림 생성을 유도하여 이러한 효과로 인해 세포막에 미세한 손상을 가하고, 구멍 생성을 유도함으로써 외부 물질의 흡수가 증가하도록 하는데, 이는 세포 내 Ca2
+ 농도 변화 즉, 세포 내 Ca2
+ 농도 변화 분석은 세포막에 손상이나 막의 유동성이 증가될 경우 순간적으로 세포 내(cytosol) Ca2
+ 농도가 증가함을 반영하여 세포막 유동성의 증가를 확인하는 것으로, 본 발명의 일 구체 예에 따르면, 초음파 처리 직후 Ca2
+ 농도가 빠르게 증가하다가 점차 감소하여 초음파를 처리하지 않는 대조군의 수준으로 감소함을 통해 세포막의 손상이 유도된 후 회복됨을 알 수 있었다. 또한, 초음파로 인한 ATP 발생 및 증가가 다양한 세포성 스트레스에 대한 반응 및 세포막내 ATP 수용체와 반응하여 엔도사이토시스를 유도하는 것으로 알려져 있다. 즉 ATP 농도와 세포 손상 및 세포내 물질 유입과의 연관성이 있고, 이를 확인하기 위해, 초음파 처리 후 세포에서의 ATP 농도를 분석한 결과, 미처리 대조군에 비해 높은 수준으로 ATP를 농도가 높게 나타났다. 아울러, ATP 의해 영향을 받는 세포막내 이온성 P2X 수용체와 대사성 P2Y 수용체 발현 역시 초음파 처리된 세포에서 대조군 대비 활성화되었다. 이와 같은 결과는 초음파로 세포내 손상뿐만 아니라 세포 외부환경의 유입 가능성을 나타낸다.
한편, 엑소좀 또는 엑소좀 함유 세포 밖 소포체는 내부에 유전정보 물질(DNA, mRNA, microRNA, protein)을 포함하고 있는 것으로 알려져 있는데, 세포막 손상을 통해 세포막 밖으로 빠져 나온 엑소좀 또는 엑소좀 함유 세포 밖 소포체가 주변 다른 세포에 다시 들어가는 과정을 통해 엑소좀 또는 엑소좀 함유 세포 밖 소포체의 내부에 존재하는 유전정보 물질이 전달될 수 있다. 따라서, 초음파 처리에 의한 자극으로 인해 세포 내 저발현되거나 발현이 억제된 상태로 유지되었던 다능성 마커들, 3배엽 마커들, 또는 분화 세포 마커들의 발현이 유도 내지 촉진됨과 동시에 세포막에 손상이 생김에 따라, 상기 발현이 유도 내지 촉진된 다능성 마커들, 3배엽 마커들, 또는 분화 세포 마커들을 포함하는 세포 내부에 존재하는 엑소좀 또는 엑소좀 함유 세포 밖 소포체가 외부로 배출되어 주변 세포에 전달되는데 주변 세포 역시 세포막이 부분적으로 손상된 상태이기 때문에 세포막 유동성이 증가되어 세포 내부로 엑소좀 또는 엑소좀 함유 세포 밖 소포체가 들어가는 효율이 정상 상태에 비해 더 높을 것으로 추정되며, 이러한 엑소좀 또는 엑소좀 함유 세포 밖 소포체 내부에 존재하는 상기 발현 유도 내지 촉진된 다능성, 발생, 분화 관련 유전정보가 전달되어 다능성 세포 또는 임의의 분화 세포가 만들어지는 것으로 생각되었다. 본 발명의 일구체예에서 다능성 세포 유도 과정 중 배양 배지를 회수하고, 배지 내 엑소좀 또는 엑소좀 함유 세포 밖 소포체를 추출하여 이들 내부에 다능성 세포 관련 다능성 마커 내지는 분화 마커들이 존재하는지 확인한 결과, 알려진 다능성 마커들, 분화 마커들이 높은 발현 정도를 보이면서 확인되어 본 발명자들의 이러한 가설을 뒷받침하는 것으로 사료된다. 또한, 이러한 초음파, 레이저, 또는 열 처리에도 핵형 기형이 없이 정상인 것으로 나타났다.
이러한 가설은 세포막 손상에 의한 엑소좀 또는 엑소좀 함유 세포 밖 소포체의 방출 유도를 통해 다능성 세포, 또는 분화 세포를 제조할 수 있음을 가능케 한다.
상기 분화된 세포로, 포유류 유래의 진피섬유아세포, 피부섬유아세포 등을 포함하는 체세포; 자궁암세포(HeLa), 간암세포(Hep3B) 등을 포함하는 암세포; 또는 폐 상피세포(L132 cell)를 포함하는 기관 내 조직세포 등을 사용할 수 있다.
본 명세서에서, 용어 "체세포"란 성체를 구성하는 세포로서 분화능 및 자가생산능이 제한된 세포를 의미한다. 일 구현예에 따르면, 상기 체세포는 포유류의 피부, 모발, 지방을 구성하는 체세포일 수 있고, 바람직하게는 포유류 유래의 섬유아세포일 수 있으나, 이에 제한되지는 않는다.
본 명세서에서, 용어 “미분화 세포”란 분화능 및 자가생산능을 가진 세포를 의미한다. 예컨대, 유도만능줄기세포, 배아줄기세포, 전구세포 등을 예로 들 수 있다.
본 명세서에서, 용어 "다능성 세포"는 물리적 자극, 엄격한 의미에서 초음파, 레이저, 자기장, 플라즈마, 발광 다이오드(light-emitting diode), 전기적 자극, 화학적 노출, 열 처리, 또는 산 처리 후 다능성을 획득한 세포를 말한다. 본 명세서에서 상기 다능성은 포괄적으로 줄기세포에서 발현되는 다능성 마커를 안정적으로 발현하는 상태를 의미한다. 아울러, 세 종류의 내배엽, 외배엽 및 중배엽의 3배엽 마커를 발현하는 상태를 의미한다. 상기 다능성 세포는 "Embryonic stem cell media-based En vironmental t ransition-guided c e llular
r eprogramming(es/ENTER) 세포 사용될 수 있다.
본 발명에 따른 다능성 세포는 외부 환경에 따라 분화 유도가 잘되며, 줄기세포의 속성 대비 분화속성이 강한 전구세포(progenitor cell)의 속성이 더 강하다는 점에서 공지의 유도만능줄기세포와 차별화된 특징을 갖는다. 즉, 유도만능줄기세포와 같은 배아줄기세포를 세포 치료제로 사용하는 경우, 어느 정도 분화 과정을 거치는 준비 단계가 요구되며, 암으로 변할 수 있는 위험 요소를 내포하고, 역분화 유도인자 도입을 위해 바이러스 벡터를 사용함에 따른 안전성 문제가 대두되나, 본 발명의 다능성 세포는 유전자 변이를 위한 역분화 유도인자 또는 화학물질 등의 역분화 유도 물질을 별도로 도입하지 않고 유도되므로 다른 종류의 세포와 공동 배양을 통한 배양이 필요 없어 세포 오염(다른 세포가 섞이는 문제점) 문제점이 없고 인 비보 실험에서 암세포와 유사한 테라토마를 형성하지 않아 암 발생의 문제점이 없어 안전성이 확보되어 있다. 다시 말해, 본 발명의 다능성 세포는 유도 과정이 단순하고 짧아 자가세포를 처리하여 이식 때까지 시간을 획기적으로 단축할 수 있는 장점을 가진다.
상기 다능성 세포는 OCT3/4, SOX2, NANOG, c-MYC, KLF4, TDGF1, SSEA4, TRA-1-60, PAX6, Nestin, Brachyury, SMA, GATA4, 또는 AFP 중 어느 하나의 다능성 마커 또는 중배엽 및 내배엽으로 이루어진 3배엽 마커 유전자를 안정적으로 발현하는 것을 특징으로 한다.
본 명세서에서, 용어 "리프로그래밍(reprogramming)"은 분화능이 없는 세포 또는 일정 부분 분화능이 있는 세포 등 서로 다른 양태로 존재하는 분화된 세포로부터 최종적으로 새로운 유형의 세포 또는 새로운 유형의 분화잠재력을 갖는 상태로 복원 또는 전환될 수 있는 프로세스를 의미한다. 뿐만 아니라 분화능이 있는 세포로부터 최종적으로 새로운 유형의 세포로 전환될 수 있는 프로세스 역시 포함한다. 본 발명에 따르면, 분화된 세포는 환경유입을 촉진할 수 있는 물리적 자극을 받을 시 다능성 세포 또는 분화된 세포와 표현형이 상이한 목적하는 임의의 분화 세포로 리프로그래밍될 수 있다. 또한, 미분화 세포는 환경유입을 촉진할 수 있는 물리적 자극을 받을 시 현저히 우수한 분화율을 보이면서 임의의 분화 세포로 리프로그래밍될 수 있다.
상기 분화 세포는 예시로, PAX6, SOX1, SOX2, Nestin, MAP2, TuJ1, GFAP 또는 O4 중 어느 하나를 발현하는 신경세포("neuronal stem cell media-based ENTER, n/ENTER"라고 함); Desmin, Pax3, Actinin, SMA, GATA4 또는 NKX2-5 중 어느 하나를 발현하는 근육세포("muscle differentiation media-based ENTER, m/ENTER"라고 함); AFP, HNF1a, HNF4a, CK18 또는 ALB 중 어느 하나를 발현하는 간세포("hepatocyte differentiation media-based ENTER, h/ENTER"라고 함); 또는 Pparc2, C/ebpa, aP2 또는 Fabp4 중 어느 하나를 발현하는 지방세포("adipocyte differentiation media-based ENTER, a/ENTER"라고 함)를 포함할 수 있으나, 이에 제한되지는 않는다.
본 명세서에서, "배양 배지"는 포괄적 의미에서 인 비트로 세포 배양에 사용되는 배지로, 본 발명에서는 줄기세포 배양 배지 또는 분화유도배지를 의미하며, 상기 줄기세포 배양 배지는 더 구체적으로 배아줄기세포 배양 배지를 의미한다. 또한, "분화유도배지"는 통상의 줄기세포의 분화 세포로의 유도에 사용하는 배지로, 예를 들어, 다분화능 (multipotent)세포 분화유도배지, 간세포 분화유도배지, 골형성 분화유도배지, 지방세포 분화유도배지, 근육세포 분화유도배지, 성상세포 분화유도배지, 신경계세포 분화유도배지, 혈관내피세포 분화유도배지, 각질세포 분화유도배지, 췌장 베타세포 분화유도배지 또는 심근세포 분화유도배지 등을 사용할 수 있으나, 이에 제한되지는 않는다.
본 발명의 세포의 리프로그래밍 방법은 도 1을 참고하여 구체적으로 설명하면 다음과 같다.
우선, 배양 배지와 분화 또는 미분화 세포를 혼합하고, 상기의 혼합물에 물리적 자극을 제공한다.
분화 또는 미분화 세포를 포함하는 혼합물에 물리적 자극을 제공하기 전에 배양 배지에 물리적 자극을 제공하여 세포의 리프로그래밍 효율을 높일 수 있다.
상기 물리적 자극은 초음파, 레이저, 플라즈마, 발광 다이오드(light-emitting diode), 전기적 자극, 화학적 노출, 열 처리 또는 산 처리 중 어느 하나일 수 있다.
상기 배양 배지에 대한 초음파 처리는 출력강도 1W/cm2
내지 20W/cm2의 초음파를 1 내지 20 분, 구체적으로, 출력강도 2W/cm2
내지 10W/cm2의 초음파를 5 내지 15 분, 더 구체적으로, 출력강도 3W/cm2
내지 7W/cm2의 초음파를 7 내지 13 분 동안 동안 동안 수행하는 것일 수 있다.
배양 배지에 대한 레이저 처리는 300 내지 900 nm 파장 대역의 펄스형 레이저 빔을 1분 내지 20분, 더 구체적으로, 상기 파장 대역의 펄스형 레이저 빔을 3분 내지 10분, 보다 구체적으로, 상기 파장 대역의 펄스형 레이저 빔을 4분 내지 6분 동안 조사하는 것일 수 있다. 상기 파장 대역은 예를 들어 400 nm, 808 nm, 880 nm의 파장을 사용할 수 있다.
배양 배지에 대한 열 처리는 40 내지 50 ℃의 온도 조건에서 5분 내지 20분 동안 수행할 수 있다.
분화 또는 미분화 세포에 물리적 자극을 제공할 경우, 일정 세기로 노출시키는 것이 좋고, 이 범위를 벗어날 경우 세포 생존율이 감소할 수 있다.
따라서, 배양 배지와 분화 또는 미분화 세포의 혼합물에 대한 초음파 처리는 출력강도 0.5W/cm2 내지 3W/cm2로 1 내지 5초 동안, 더 구체적으로, 출력강도 0.7W/cm2 내지 2W/cm2로 1 내지 5초 동안, 보다 구체적으로, 출력강도 0.8W/cm2 내지 1.5W/cm2로 1 내지 5초 동안 수행하는 것일 수 있다.
배양 배지와 분화 또는 미분화 세포의 혼합물에 대한 레이저 처리는 300 내지 900 nm 파장 대역의 펄스형 레이저 빔을 1초 내지 20초, 더 구체적으로, 상기 파장 대역의 펄스형 레이저 빔을 3초 내지 10초, 보다 구체적으로, 상기 파장 대역의 펄스형 레이저 빔을 4초 내지 6초 동안 조사하는 것일 수 있다. 상기 파장 대역은 예를 들어 400 nm, 808 nm, 880 nm의 파장을 사용할 수 있다.
배양 배지와 분화 또는 미분화 세포의 혼합물에 대한 열 처리는 40 내지 50 ℃의 온도 조건에서 1분 내지 10분 동안 노출한 후 0℃ 내지 4℃의 온도 조건에서 5 내지 10초간 노출하여 수행하는 것일 수 있다.
다음으로, 물리적 자극이 제공된 혼합물을 일정 시간 동안 배양하여 리프로그래밍된 세포를 수득한다.
물리적 자극이 제공된 혼합물의 배양은 부유 배양(suspended culture) 또는 부착 배양(monolayer culture) 방식을 통해 다능성 마커 또는 분화 마커를 안정적으로 발현하는 스페로이드가 형성되는 기간, 즉, 2일 내지 10일 동안 수행하는 것일 수 있으나, 이에 특별히 제한하는 것은 아니다.
본 발명의 일 구체예에 따르면, 부유 배양은 부착 배양에 비해 더 높은 스페로이드 형성 효율을 나타낸다. 또한, 부유 배양은 부착 배양에 비해 스페로이드의 수와 크기가 더 크며, 일정한 크기 분포를 나타낸다.
본 발명의 일 구체예에 따르면, 초음파 또는 레이저 처리된 사람 피부 섬유아세포의 부유 배양 시 대략 3일째부터 다능성 마커 또는 분화 마커의 발현이 증가하거나 안정화되어 이 시기부터 리프로그래밍이 시작되는 것으로 보인다. 또한, 열 처리된 사람 피부 섬유아세포의 부유 배양 시 대략 8일째부터 다능성 마커의 발현이 증가하거나 안정화되어 이 시기부터 리프로그래밍이 시작되는 것으로 보인다.
상기 다능성 마커, 예컨대, OCT3/4, SOX2, NANOG, c-MYC, KLF4, TDGF1, SSEA4, TRA-1-60 등이 발현됨을 통해 스페로이드가 다능성을 가짐을 확인할 수 있다. 다능성 마커의 확인은 RT-PCR 또는 면역세포화학법을 통해 분석할 수 있으나, 이에 특별히 제한하는 것은 아니다.
또한, 본 발명의 다능성 세포는 3배엽 마커, 즉 외배엽(PAX6, Nestin), 중배엽(Brachyury, SMA), 내배엽(GATA4, AFP) 마커들을 높은 수준으로 발현하는 특징을 나타낸다.
다른 구체예에서, 분화유도배지에서 피부 섬유아세포에 대해 물리적 자극을 제공하는 경우, 배양 후 약 1일 내지 20일 사이에 스페로이드가 형성될 수 있다.
분화 마커는, 신경세포로 리프로그래밍되는 경우, PAX6, SOX1, SOX2, Nestin, MAP2, TuJ1, GFAP 또는 O4 중 하나 이상일 수 있다.
근육세포로 리프로그래밍되는 경우, Desmin, Actinin, Pax3, SMA, GATA4 또는 NKX2-5 중 하나 이상일 수 있다.
간세포로 리프로그래밍되는 경우, AFP, HNF1a, HNF4a, CK18 또는 ALB 중 하나 이상일 수 있다.
지방세포로 리프로그래밍되는 경우, oil red O 염색이 되며, Pparc2, C/ebpa, aP2 또는 Fabp4 중 어느 하나일 수 있다.
또한, 본 발명의 다능성 세포는 증식 마커 단백질인 Ki-67를 발현하여 증식능을 가지는 특징이 있다.
또한, 상기의 리프로그래밍된 다능성 세포를 영양세포와 공배양할 경우 다능성 세포의 증식이 증가될 수 있다.
또한, 본 발명의 세포의 리프로그래밍 방법은 상기 다능성 세포를 분화유도배지에서 배양하는 단계를 더 포함할 수 있다. 상기 분화유도배지의 종류에 따라 다능성 세포는 목적하는 분화 세포로 분화될 수 있다.
상기 분화유도배지로, 다분화능 (multipotent)세포 분화유도배지, 간세포 분화유도배지, 골형성 분화유도배지, 지방세포 분화유도배지, 근육세포 분화유도배지, 성상세포 분화유도배지, 신경계세포 분화유도배지, 혈관내피세포 분화유도배지, 각질세포 분화유도배지, 췌장 베타세포 분화유도배지 또는 심근세포 분화유도배지 등을 사용할 수 있으나, 이에 특별히 제한하는 것은 아니다.
본 발명은 또한 분화 또는 미분화 세포 및 배양 배지의 혼합물에 환경유입(environmental influx)을 촉진할 수 있는 물리적 자극을 제공하고,
상기 물리적 자극을 제공받은 혼합물을 1일 내지 6일 동안 배양하고,
상기의 배양물에서 분리한 엑소좀 함유 세포 밖 소포체(extracellular vesicles)와 분화 또는 미분화 세포를 혼합하여 일정 시간 동안 배양하여 리프로그래밍된 세포를 수득하는 것을 포함하는, 세포의 리프로그래밍 방법에 관한 것이다.
본 발명의 세포의 리프로그래밍 방법은 물리적 자극을 받은 분화 또는 미분화 세포에서 분리한 엑소좀 함유 세포 밖 소포체를 분화 또는 미분화 세포와 일정 시간 동안 배양하여 임의의 분화 세포로 리프로그래밍할 수 있는 것을 특징으로 한다.
상기 엑소좀 함유 세포 밖 소포체는 분화 또는 미분화 세포 및 배양 배지의 혼합물에 환경유입을 촉진할 수 있는 물리적 자극을 제공하고, 상기 물리적 자극을 제공받은 혼합물을 1일 내지 6일 동안 배양하여 원심분리를 통해 회수할 수 있다.
상기 분화 또는 미분화 세포의 물리적 자극은 상기에 기재되어 있어 중복 기재를 피하기 위해 생략한다.
상기 엑소좀 함유 세포 밖 소포체는 OCT3/4, SOX2, NANOG, c-MYC, KLF4, TDGF1, SSEA4, TRA-1-60, PAX6, Nestin, Brachyury, SMA, GATA4, 또는 AFP 중 어느 하나의 다능성 마커 또는 3배엽 마커; PAX6, Nestin, Sox1, Sox2, MAP2, TuJ1, GFAP 또는 O4 중 어느 하나의 신경세포 마커; Desmin, Pax3, Actinin, SMA, GATA4 또는 NKX2-5 중 어느 하나의 근육세포 마커; AFP, HNF1a, HNF4a, CK18 또는 ALB 중 어느 하나의 간세포 마커; 또는 oil red O 염색이 되며, Pparc2, C/ebpa, aP2 또는 Fabp4 중 어느 하나의 지방세포 마커를 발현하는 것일 수 있다.
예컨대, 상기 엑소좀 함유 세포 밖 소포체가 다능성 마커를 포함하는 경우, 분화 세포와 배양하면 상기 분화 세포는 다능성 세포로 리프로그래밍될 수 있다. 또한, 상기 엑소좀 함유 세포 밖 소포체가 분화 마커를 포함하는 경우, 분화 세포와 배양하면 표현형이 상이한 임의의 분화 세포로 리프로그래밍될 수 있다. 또한, 상기 엑소좀 함유 세포 밖 소포체가 분화 마커를 포함하는 경우, 미분화 세포와 배양하면 임의의 분화 세포로 리프로그래밍될 수 있다.
본 발명의 일 구체예에 따르면, es/ENTER 유도 시 회수한 엑소좀 마커인 CD63이 염색된 세포 밖 소포체(EVs)에서 다양한 다능성 마커의 발현이 확인되었고, 상기 EVs가 처리된 정상 인간 체세포에서 3일 배양 후 다능성 마커인 Oct4, Sox2 및 Nanog가 발현되어 세포의 리프로그래밍이 확인되었다.
또한, n/ENTER 유도 시 회수한 엑소좀 마커인 CD63이 염색된 세포 밖 소포체(EVs)에서 Pax6와 같은 신경줄기세포의 마커의 발현이 확인되었고, 상기 EVs가 처리된 정상 인간 체세포에서 3일 배양 후 신경줄기세포 마커인 Sox1, Sox2, Pax6, Nestin 발현이 확인되었다.
또한, m/ENTER 유도 시 회수한 엑소좀 마커인 CD63이 염색된 세포 밖 소포체(EVs)에서 Pax3와 같은 근육세포 마커의 발현이 확인되었고, h/ENTER 유도 시 회수한 엑소좀 마커인 CD63이 염색된 세포 밖 소포체(EVs)에서 HNF1a와 같은 간세포 마커의 발현이 확인되었다.
따라서, 물리적 자극을 받은 분화 또는 미분화 세포는 리프로그래밍 인자가 포함된 세포 밖 소포체를 분비함을 알 수 있고, 이들을 분화 또는 미분화 세포에 처리하여 부유 배양 또는 부착 배양 방식을 통해 1일 내지 20일 동안 배양하면 임의의 다능성 세포, 또는 분화 세포로 리프로그래밍될 수 있다.
본 발명의 세포의 리프로그래밍 방법을 통해 리프로그래밍될 수 있는 세포로 상술한 종류의 다능성 세포 또는 분화 세포일 수 있고, 중복 기재를 피하기 위해 기재를 생략한다.
이하, 본 발명을 실시예에 의해 상세히 설명한다. 단, 하기 실시예는 본 발명을 예시하는 것일 뿐, 본 발명의 내용이 하기 실시예에 한정되는 것은 아니다.
<실시예 1> 물리적 자극에 의한 세포 내 환경유입 증명 실험
본 실시예는 물리적 자극에 의한 세포 내로의 환경유입에 대한 증명 실험으로, 이를 위해, 세포는 invitrogen에서 구입한 primary HDF 세포를 10% FBS(Gibco)와 1% penicillin/streptomycin(Gibco)이 첨가된 DMEM에서 배양하였으며, 배양액에 대한 초음파 처리는 5W/cm2, 10분간 수행하고, 세포 처리는 1×106 세포에 1W/cm2, 5초간 처리 후 처리된 배양액과 함께 35mm 배양접시에 2×105 세포를 배양하였다.
SEM 이미지 분석을 위해 처리되지 않은 HDF 세포와 위와 같이 처리된 직후와 37℃, 5% CO2 인큐베이터에서 2시간 동안 배양한 세포를 4℃에서 4% 파라포름알데히드로 12시간 동안 고정한 후 0.1% 타닌산 용액에 1시간, 1% 오스뮴 테트록사이드 용액에서 2시간 처리 후 농도 단계별 아세톤으로 탈수시킨 후 액체 CO2로 세포를 건조하고, 금-팔라듐으로 코팅된 표면에 고정시켜 전자현미경(1555 VP-FESEM, Carl Zeiss)으로 세포를 관찰하였다.
Live/dead 이미지 분석을 위해 세포는 처리되지 않은 HDF와 초음파 처리 직 후와 37℃, 5% CO2 인큐베이터에서 2시간 동안 배양한 세포에 Live/dead viability/cytotoxicity assay kit(Molecular Probes, Eugene, OR, USA)을 사용하여 염색하였다. 염색 과정은 살아있는 세포에 2μM 칼세인(live cell staining dye) 및 4μM 에티디움 호모다이머-1(EthD-1, dead cell staining dye)을 세포 배양액에 첨가한 후 37℃, 5% CO2 인큐베이터에서 30분 동안 배양 후 형광현미경(IX3-ZDC, Olympus)으로 붉은색(EthD-1 염색, 죽거나 손상된 세포, excitation/emission, 528/617 nm)과 녹색 형광(calcein 염색, 살아있는 세포, excitation/emission, 494/517 nm)을 분석하였다.
도 2a에 나타난 바와 같이, 초음파에 의해 세포막이 손상되어 외부환경이 들어올 수 있는 구멍이 형성되며, 이와 같은 손상은 2시간 후 회복되었다.
또한, 초음파 자극 후 손상된 세포 및 세포의 회복을 확인하기 위해 세포 사멸을 분석하는데 이용되는 Live/dead kit를 이용하여 세포를 염색하였다.
도 2b에서와 같이, 초음파를 세포에 처리한 후 처리 직후 염색하고 2시간 지난 후 염색한 결과, 처리 직후 녹색 형광과 적색 형광이 같이 존재하는 세포가 관찰되었고 2시간 이후 적색 형광을 나타내는 세포의 수가 현저히 줄어들어도 2a와 같이 세포막이 회복됨으로 인해 적색 형광이 줄어드는 것으로 나타났다.
이는 초음파 자극으로 인해 세포손상이 일어나나 회복이 가능하며, 이렇게 자극에 의한 세포막 손상으로 인해 배지 환경유입이 가능할 수 있을 것이라 생각하고 세포 내 물질 유입에 의해 나타나는 현상을 분석하였다.
한편, 도 2에서 usMC(ultrasound-exposed medium and cells)은 세포 및 배양배지 각각에 초음파를 처리한 경우를 말하며, usMC-S는 부유 배양된 usMC를 의미한다.
<실시예 2> 물리적 자극에 의한 세포 내 외부 물질 유입에 대한 증명 실험
세포 내 외부 물질 유입에 민감한 세포 내 칼슘 농도의 변화를 측정하고, 초음파에 의한 ATP 발생에 따른 물질 유입 가능성을 확인하기 위해 세포 내 ATP 측정과 ATP 반응에 의해 세포막내 수용체가 세포막 외부물질 유입 통로를 개방하는 것으로 알려진 바 있는 ATP 수용체의 발현을 RT-PCR로 분석하였다.
칼슘 농도 분석은 Fluo-4 NW Calsium Assay Kit(Molecular Probes)를 이용하여 분석하였다. 처리되지 않은 HDF 세포와 초음파 (1W/cm2, 5초)로 직접 처리된 후 초음파 처리된 배지(5W/cm2, 10분)에 노출된 세포(usMC-S)를 각각 키트 내 구성 품 중 분석 버퍼와 섞어 주고 96-웰 플레이트의 웰당 3×104 세포를 분주한 후 웰당 50㎕의 Fluo-4 NW 시약과 섞은 후 Varioskan Flash Fluorescent Microplate Fluorometer(Thermo Fisher Scientific, Waltham, MA, USA)로 여기파장 494nm/방출파장 516nm 범위의 형광을 10초 간격으로 15분 동안 측정하였다.
ATP는 Adenosine 5'-triphosphate(ATP) Bioluminescent assay kit를 이용하여 측정하였다. 세포는 처리되지 않은 세포와 초음파(1W/cm2, 5초)로 직접 처리된 후 초음파 처리된 배지(5W/cm2, 10분)에 노출된 세포 (usMC-S)를 96-웰 플레이트의 웰당 3×104 세포를 분주한 후 웰당 100㎕의 ATP 분석 믹스와 ATP 표준물질을 분주하여 실온에서 3분 배양 후 Varioskan Flash Fluorescent Microplate Fluorometer(Thermo Fisher Scientific)로 발광 강도를 측정하였다.
ATP 수용체 발현 분석을 위한 RT-PCR은 처리된 세포를 RNeasy plus mini kit(Qiagen, Hilden, Germany)를 이용하여 RNA를 추출하였고, Super ScripⅡ kit(Invitrogen, Carlsbad CA, USA)으로 cDNA를 합성하였다. PCR은 PCR 프리믹스(Bioneer, Daejeon, Korea)에 cDNA와 프라이머를 섞은 후 Thermal cycler dice PCR machine(TP600, TAKARA, Otsu, Japan)을 이용하여 95℃ 5분 변성, 95℃로 30초, gradient(50-65℃) 30초 및 72℃로 1분을 35 사이클, 및 72℃ 15분 조건으로 수행되었다.
P2 수용체의 PCR 프라이머 리스트
| 유전자 부호 |
프라이머 서열(5'-3') |
| 정방향 |
역방향 |
|
P2X4
|
TCTCAACAGGCAGGTGCGTAGCTT |
GCTCAACGTCCCGTGTATCGAGG |
|
P2X7
|
CAGAAGGCCAAGAGCAGCGGTT |
GGACACGTTGGTGGTCTTGTCGTCA |
|
P2Y1
|
CTTGGTGCTGATTCTGGGCTG |
GCTCGGGAGAGTCTCCTTCTG |
|
P2Y2
|
CCGCTCGCTGGACCTCAGCTG |
CTCACTGCTGCCCAACACATC |
|
P2Y11
|
GAGGCCTGCATCAAGTGTCTG |
ACGTTGAGCACCCGCATGATG |
도 3에 나타난 바와 같이, 초음파 자극 후 세포 내 칼슘 유입이 60초까지 증가하고, 60분에 세포 내 ATP 농도가 최대로 증가하였으며, 세포막내 ATP 수용체 역시 1시간 및 4시간 경에 발현이 증가되는 것으로 나타났다. 이는 초음파 자극에 의해 초반 세포 내로 외부 물질이 유입됨을 칼슘의 농도 증가로 확인되었으며 초음파에 의해 ATP가 발생되고, 그로 인해 ATP 수용체가 반응하여 세포막 통로가 개방되어 외부물질 유입이 가능함을 알 수 있었다.
<실시예 3> QD605를 이용한 물리적 자극에 의한 세포 내 외부 물질 유입 증명 실험
외부 물질로 QD605로 정하고 QD605가 초음파에 의해 세포 내로 유입되는지를 확인하였다. QD605는 형광을 띠는 나노 물질로 살아있는 세포에서 침투성이 떨어지는 것으로 알려져 있어 QD605를 이용한 초음파에 의한 세포 내 외부 물질 유입을 확인하였다.
이를 위해, HDF에 실시예 1과 같이 초음파 자극을 준 후 QD605 100pmol을 처리하고, 24시간 후 단일 세포와 스페로이드 내 QD605의 존재를 확인하였다.
도 4에 나타난 바와 같이, 초음파 자극을 받지 않은 HDF에는 형광이 보이지 않았다. 그러나, 초음파 자극을 받은 HDF에는 형광이 관찰되었다.
또한, 외부 물질 유입에 따른 세포 변화에 대한 가능성을 확인하기 위해 각각의 배지 환경(ES, Neuroprogenitor, Hepatocyte, muscle)에서 초음파를 처리한 후 QD605 100pmol을 넣고 24시간 후에 각각의 분화과정상의 전사 인자(ES: Oct4, Neuroprogenitor: Pax6, Hepatocyte: HNF1a, Muscle: Pax3)의 발현을 ICC를 통해 확인하였다.
도 4b 및 4c에 나타난 바와 같이, 외부 물질(QD605)이 들어간 세포 및 스페로이드 각각에서 전사 인자가 관찰되었다. 이는 외부 물질 유입에 따른 세포의 리프로그래밍 가능성을 나타내는 결과이다.
실험과정에서 4가지 환경유입 샘플 중 es/ENTER와 n/ENTER만 스페로이드를 형성하였다. 그러나 m/ENTER와 h/ENTER는 스페로이드를 형성하지 않았다. 이는 세포의 특성과 배지 조성에 있다. ES와 신경전구세포의 경우 부유 배양 과정에서 스페로이드나 스피어 형성이 되는 세포이나, 근육세포와 간세포의 경우는 스페로이드를 형성하지 않는다. 이는 분화유도과정에서 코팅된 배양접시에서 부착시켜 배양하였기 때문이며, 특히 근육세포의 배지에 FBS가 들어 있는데 FBS는 세포 부착력을 높여 주기 때문에 스페로이드가 형성되지 못한 것으로 보인다.
<실시예 4> 물리적 자극을 받은 세포 배양액 내 엑소좀 분석
초음파에 의한 세포 자극은 모든 세포에 동일하게 자극되지 않으므로, 세포의 리프로그래밍은 일부 세포에서 일어날 수 있으며, 이렇게 변화된 세포와 그렇지 않은 세포의 세포 교류의 가능성을 생각해 보았다. 최근 엑소좀에 의한 세포간의 물질 교류에 대한 가능성을 참고로 하여 초음파 처리된 세포에서 배출된 배양 배지 내 엑소좀이 유전물질을 포함하고 있을 가능성이 있으며, 이는 리프로그래밍된 세포에서 분비되는 물질 중 리프로그래밍에 중요한 역할을 하는 유전물질을 포함하고 있을 가능성을 가지고 있어 초음파 처리 후 배양된 배양 배지내 엑소좀을 배양시간별 배지 교환 시 회수하여 배양액 내 엑소좀의 RNA를 Amicon Ultra-0.5 kit(Millipore)로 추출하였으며, cDNA 합성은 Super ScripⅡ kit(Invitrogen, Carlsbad CA, USA)로 하였다. PCR은 PCR 프리믹스(Bioneer, Daejeon, Korea)에 cDNA와 프라이머를 섞은 후 Thermal cycler dice PCR machine(TP600, TAKARA, Otsu, Japan)을 이용하여 95℃로 5분 변성, 95℃로 30초, gradient 30초 및 72℃로 1분을 35 사이클, 72℃로 15분 조건으로 RT-PCR 분석을 수행하였다(표 2).
도 5에 나타난 바와 같이, 엑소좀의 RNA 중 다능성 RNA의 발현이 확인되었다.
한편, 도 5에서 usMC은 세포 및 배양배지 각각에 초음파를 처리한 경우를 말하며, usMC-A는 부착 배양된 usMC을 의미한다.
다능성 마커 유전자의 PCR 프라이머 리스트
| 유전자 부호 |
프라이머 서열(5'-3') |
어닐링 온도(℃) |
| 정방향 |
역방항 |
|
Oct4
(POU5F1)
|
GACAGGGGGAGGGGAGGAGCTAGG |
CTTCCCTCCAACCAGTTGCCCCAAAC |
60 |
|
Sox2
|
GGGAAATGGGAGGGGTGCAAAAGAGG |
TTGCGTGAGTGTGGATGGGATTGGTG |
63 |
|
Nanog
|
CAGCCCCGATTCTTCCACCAGTCCC |
CGGAAGATTCCCAGTCGGGTTCACC |
64 |
|
Utf1
|
CCGTCGCTGAACACCGCCCTGCTG |
CGCGCTGCCCAGAATGAAGCCCAC |
65 |
|
Lin28a
|
AGCGCAGATCAAAAGGAGACA |
CCTCTCGAAAGTAGGTTGGCT |
50 |
|
Rex1
|
CAGATCCTAAACAGCTCGCAGAAT |
GCGTACGCAAATTAAAGTCCAGA |
52 |
|
Fgf4
|
CTACAACGCCTACGAGTCCTACA |
GTTGCACCAGAAAAGTCAGAGTTG |
55 |
|
Foxd3
|
AAGCTGGTCGAGCAAACTCA |
CTCCCATCCCCACGGTACTA |
50 |
|
Esg1
|
ATATCCCGCCGTGGGTGAAAGTTC |
ACTCAGCCATGGACTGGAGCATCC |
60 |
|
Tdgf1
|
CTGCTGCCTGAATGGGGGAACCTGC |
GCCACGAGGTGCTCATCCATCACAAGG |
65 |
|
c-
Myc
|
AATGAAAAGGCCCCCAAGGTAGTTATCC |
GTCGTTTCCGCAACAAGTCCTCTTC |
50 |
|
Klf4
|
CCCACATGAAGCGACTTCCC |
CAGGTCCAGGAGATCGTTGAA |
54 |
<실시예 5> 엑소좀에 의한 물질 전달 증명 실험
상기 실시예 4에서 초음파 처리된 세포의 배양액 내 엑소좀에서 다능성 마커의 발현이 확인되었으므로, 이러한 엑소좀에 의해 유전물질 및 단백질이 전달되는지를 확인하였다.
초음파 처리 후 QD605를 첨가하여 살아있는 세포의 이미지를 촬영한 결과, 도 6a에 나타난 바와 같이, 7시간 45분경에 QD605가 유입된 세포의 QD605가 세포질의 일부와 함께 떨어져 나와 다른 세포로 이동되는 현상을 확인할 수 있었다.
이렇게 떨어져 나온 세포질의 일부분이 엑소좀일 것으로 예상하고, usMC 처리된 세포를 다양한 배지 환경에 노출시킨 후 4% 파라포름알데히드에 10분간 고정시키고 0.1% 트리톤 X-100을 포함하는 PBS에 40분간 스며들게 하였다. 5%(v/v) 염소 혈청을 포함하는 PBS 용액으로 1시간 동안 블록킹하고 1차 항체로 엑소좀 마커인 CD63(1:100, Santa Cruz Biotechnology)과 각각의 분화유도배지에 의해 유도될 세포의 초기 발현 마커인 배아줄기세포(Oct4 1:200; Nanog 1:200; abcam), 신경줄기세포(Pax6, 1:200; abcam), 근육세포(Pax3, 1:200; abcam) 및 간세포(HNF1a, 1:200; Cell Signaling Technology) 등을 4℃에서 하룻밤 동안 염색하였다. 그리고 0.03% 트리톤 X-100을 포함하는 PBS 버퍼로 세척하고, 2차 항체, Alexa-488 또는 -594이 결합된 항-토끼, 항-마우스 항체(1:1000, Thermo, excitation/emission, 495/519 nm, excitation/emission, 590/617 nm)를 실온에서 1시간 반 정도 염색 후 0.03% 트리톤 X-100을 포함하는 PBS 버퍼로 세척하고, DAPI가 포함된 마운팅 졸(Vector Laboratories, Inc., Burlingame, CA, excitation/emission, 420/480 nm)으로 마운팅하여 공초점 레이져 형광현미경(LSM 700; Carl Zeiss)으로 이미지를 분석하였다.
도 6b에 나타난 바와 같이, 세포 주변에 나와 있던 세포질의 일부분으로 추측되는 것들이 CD63에 염색되었으며, es/ENTER 유도 시 엑소좀 마커인 CD63이 염색된 세포 밖 소포체(EVs)에서 Oct4, Nanog와 같은 다능성세포 마커의 발현이 확인되었고, n/ENTER 유도 시 엑소좀 마커인 CD63이 염색된 세포 밖 소포체(EVs)에서 Pax6와 같은 신경줄기세포의 마커의 발현이 확인되었고, m/ENTER 유도 시 엑소좀 마커인 CD63이 염색된 세포 밖 소포체(EVs)에서 Pax3와 같은 근육세포 마커의 발현이 확인되었고, h/ENTER유도 시 엑소좀 마커인 CD63이 염색된 세포 밖 소포체(EVs)에서 Hnf1a와 같은 간세포 마커의 발현이 확인되었다.
상기 결과로부터 엑소좀이 세포질에서 떨어져 나오고, 그것은 유전물질과 단백질들을 포함하고 있어 주변세포에 전달시켜 주변세포의 변화를 유도할 수 있다는 가설을 세웠다. 이를 입증하기 위해, 엑소좀을 추출하여 엑소좀을 CD63으로 염색(배양중인 세포에서도 엑소좀이 존재하기 때문에 새로 주입하는 엑소좀이라는 것을 구분하기 위해 염색함)한 후 그 엑소좀 내에 poly(A)27-Cy5.5를 수식한 유전물질을 엑소좀에 유입시킨 후 처리하지 않은 HDF와 같이 배양시키면 엑소좀에 의해 poly(A)27-Cy5.5가 전달되지 않을까 생각하였다.
도 6c에 나타난 바와 같이, CD63이 염색된 엑소좀이 세포 내에서 발견되었으며, CD63과 같이 cy5.5가 발현되는 것을 확인하였다. 이는 엑소좀에 의해 주입한 유전자(poly-A)가 전달되었음을 의미한다.
<실시예 7> 물리적 자극을 받은 세포와 정상 세포의 공동 배양
엑소좀에 의해 유전물질이 전달될 수 있으므로, 인간 ES 배지에서 배양된 세포에서 분비되는 엑소좀이 주변의 세포나 아니면 처리되지 않은 세포의 속성도 변화시킬 수 있다는 가설을 세웠고, 이를 증명하기 위해 인간 ES 배지환경에서 배양된 초음파 처리된 세포의 2일 배양된 배지에서 엑소좀을 추출하여 인간 ES 배지와 섬유아세포 배양 배지인 DMEM으로 처리되지 않은 세포를 배양하는 과정에서 엑소좀 추출물을 섞어서 6일간 배양하였다.
그 결과, 엑소좀이 첨가된 그룹에서 스페로이드가 생성되었으며(도 7a), 이 세포에서의 Oct4 발현을 확인한 결과 다능성 마커인 Oct4의 발현이 관찰되었다(도 7b). 이는 엑소좀에 의한 유전물질 전달이 세포 리프로그래밍을 유도할 수 있다는 의미이다.
<실시예 8> 섬유아세포의 직접 리프로그래밍
상기 실시예 1 내지 7에서 배지 환경의 변화에 의한 세포 리프로그래밍 가능성과 주변세포까지 리프로그래밍할 수 있음이 입증되었는바, 이를 바탕으로 다양한 배지 환경을 적용하여 세포의 리프로그래밍을 확인해 보았다.
이를 위해, 도 8과 같이 인간 섬유아세포를 1mL의 분화유도배지로 1×106 세포로 모은 후 초음파를 1W/cm2 강도로 5초간 처리한 후 35mm 배양접시 또는 6-웰 플레이트에 2×105/Well로 분주한 후 초음파를 10W/cm2 강도로 10분간 처리한 2mL의 분화유도배지에서 배양하였다.
분화유도배지의 종류에 따라 차이가 있으나, 배양 후 약 2일 내지 6일 사이에 스페로이드가 형성되었다.
또한, 지방세포 분화유도배지를 이용하여 usMC 처리 후 20일간 배양한 결과 세포 내 기포가 생성되는 것이 관찰되었으며, 이 기포에 대한 분석으로 지방세포 판별을 위한 지질 염색시약인 oil red O가 염색되는 것으로 나타났다. 이는 세포가 지방을 생산함을 나타내는 지표이다(도 9a).
그리고, 세포의 RNA를 추출하여 RT-PCR을 이용하여 지방세포 마커 유전자, Pparc2, C/ebpa, aP2, Fabp4의 발현을 확인한 결과 분화유도 후 발현이 증가되는 것으로 나타났다(도 9b).
또한, 신경줄기세포(신경전구세포) 분화유도배지와 초음파에 의한 HDF의 신경전구세포로의 분화를 확인하기 위해, 분화 유도 후 3일째 생성된 스페로이드와 부착된 세포에서 신경전구세포 마커, Oct4, Sox2, Pax6, Nestin의 발현을 면역세포화학적 방법으로 염색하여 확인하였다.
도 10a는 분화 유도된 세포의 모습이며, 도 10b는 스페로이드에서의 신경전구세포 마커를 나타낸 것으로 분화 유도되면 Oct4의 발현이 감소되며, Sox2와 Pax6 그리고 Nestin의 발현이 높게 나타나는 것을 확인하였다. 도 10c는 부착된 세포에서의 발현을 본 결과로, 역시 위와 같은 발현 패턴을 보였다. 이때, 신경전구세포나 신경줄기세포의 마커는 Sox2, Pax6, Nestin이며, Oct4는 다능성 마커로 성체줄기세포나 전구세포의 경우 Oct4의 발현이 떨어진다.
분화유도배지의 조성
| |
성분 |
함량 |
| 신경전구세포 분화유도 배지 |
DMEM F12 |
| bFGF |
20ng/mL |
| EGF |
20ng/mL |
| B27 supplement (×50) |
1/50 |
| N2 supplement (×100) |
1/100 |
도 11은 분화 유도 후 7일간 분화 유도된 세포에서의 Pax6/Nestin의 발현 패턴을 플로우 사이토메트리를 통해 분석한 결과로, 처리 후 1일째 Pax6와 Nestin의 발현이 상대적으로 50% 이상 발현되었으며, 3일째에 Pax6, Nestin의 발현이 제일 높게 나타났다.
도 12는 분화 유도 후 3일째 세포 중 신경전구세포 마커(Pax6/nestin)가 발현되는 세포에서 증식 유무를 확인하고자 ki67을 발현을 확인한 결과로, 하얀색 화살표가 가리키는 세포를 보면, Nestin이 발현되는 세포에서의 ki67 발현을 확인할 수 있었다. 이와 같은 결과는 분화 유도된 세포가 증식 능력을 가짐을 의미한다. 도 12에서 화살표는 Nestin이 염색된 세포가 증식하고 있음을 보여주는 표시이다.
도 13은 분화 유도된 세포(n/ENTER cells)의 자기재생(self-renewal)을 확인한 실험 결과로, 도 13a는 동영상으로 1개의 스페로이드로부터 증식되어 나온 세포들이 Pax6와 nestin을 발현함을 확인함으로써 이후 증식된 세포에 신경전구세포 속성이 그대로 전해짐을 알 수 있었다.
다음으로, 5주령된 쥐의 뇌에 분화 유도된 세포(n/ENTER cells)를 주입하여 4주 후에 뇌를 회수한 후 주입된 세포의 주변 세포로의 분화를 확인하고, 분화된 세포의 기능을 확인하였다.
도 14a에 나타난 바와 같이, 뇌에 주입된 세포(n/ENTER cells)를 Human Nuclear antigen(HNA)으로 염색하여 표시한 후 표시된 세포를 Gfap 항체를 염색한 결과 주입된 세포에서의 Gfap가 발현되는 것을 확인하였다.
그리고, Gfap가 발현되는 세포가 정상적인 기능을 하는지 확인하기 위해 synapsin을 분비하는지를 synapsin 1 항체(1:500, R&D system)로 염색하였다. 그 결과, HNA가 발현되는 세포 중 Gfap가 발현되는 세포에서 synapsin 1 발현이 관찰되었다(도 14b).
다음으로, 분화 유도 후 3일째 생성된 스페로이드에서 신경전구세포 마커(Oct4, Sox2, Pax6, Nestin)의 발현을 면역세포화학방법으로 염색하여 확인하였다.
이를 위해, 스페로이드와 부착된 세포는 4% 파라포름알데히드에 10분간 고정시키고 0.1% 트리톤 X-100을 포함하는 PBS에 40분간 스며들게 하였다. 5%(v/v) 염소 혈청을 포함하는 PBS로 1시간 동안 블록킹하고 1차 항체로 Oct4(1:200), Sox2(1:200), Pax6(1;200), Nestin(1:200, Cell Signaling Technology) 등을 4℃에서 하룻밤 동안 염색하였다. 그리고 0.03% 트리톤 X-100을 포함하는 PBS 버퍼로 세척하고, 2차 항체, Alexa-488 또는 -594가 결합된 항-토끼, 항-마우스 항체(1:1000, Thermo, excitation/emission, 495/519 nm, excitation/emission, 590/617 nm)를 실온에서 1시간 반 정도 염색 후 0.03% 트리톤 X-100을 포함하는 PBS 버퍼로 세척하고, DAPI가 포함된 마운팅 졸(Vector Laboratories, Inc., excitation/emission, 420/480 nm)으로 마운팅하여 공초점 레이져 형광현미경(LSM 700; Carl Zeiss)으로 이미지를 분석하였다.
도 15a에서 스페로이드 형성을 확인하였고, 스페로이드에서 신경전구세포 마커인 Pax6 및 Nestin의 발현이 높게 나타나는 것을 확인하였다(도 15b).
도 15c는 분화 유도 후 3일간 분화유도된 세포에서의 Pax6/Nestin의 발현 패턴을 플로우 사이토메트리를 통해 분석한 결과로, 처리 후 1일째 Pax6와 Nestin의 발현이 70% 이상 발현되었으며, 3일째에 Pax6, Nestin의 발현이 제일 높게 나타났다.
도 16은 분화 유도 후 20일된 세포에서 신경전구세포 마커(Sox2, Pax6, Nestin)의 발현을 면역세포화학 방법으로 염색하여 확인한 결과로, 상단 및 중간 사진은 Sox2와 Pax6 그리고 Nestin의 발현이 높게 나타남을 보여주며, 하단 사진은 희돌기교세포 마커의 발현을 본 결과로, 역시 위와 같은 발현 패턴을 보였다.
이 결과는 분화 유도된 세포가 신경전구세포 분화능력과 비슷한 분화 능력이 있다는 의미이다.
<실시예 9> 직접 간세포 분화
도 17의 도식도와 같이, HDF, HeLa 세포 및 Hep3B 세포를 1mL의 분화유도배지로 1×106 세포로 모은 후 초음파를 1W/cm2 강도로 5초간 처리한 후 35mm Laminin 코팅 배양접시에 2×105로 분주한 후 초음파를 10W/cm2 강도로 10분간 처리한 2mL의 간세포 분화유도배지에서 배양하였다. 간세포 분화유도배지를 이용하여 분화 유도된 세포를 h/ENTER 로 명명하였다.
도 18은 간세포 분화유도배지와 초음파 처리된 HDF의 간세포(h/ENTER)로의 분화를 유도하였다. HDF를 분화 유도 20일 후 세포 외형의 변화를 나타낸 것으로, HDF 분화 유도 20일 후 면역세포화학 방법을 통하여 간세포 마커(AFP, HNF4a, CK18, ALB)를 확인한 결과, h/ENTER 세포에서 간세포 마커의 발현이 확인되었다.
다음으로, 간세포 분화유도배지와 초음파 처리된 HeLa 세포의 간세포(HeLa h/ENTER)로의 분화를 유도하였다. HeLa 세포를 분화 유도 19일 후 세포(HeLa h/ENTER) 외형의 변화를 나타낸 것으로, HeLa 세포 분화 유도 20일 후 (HeLa h/ENTER) qPCR를 통하여 간세포 마커(ALB, HNF4a, CYP3A4F, CYP3A7F, AIAT, SOX7, GATA6)를 확인하였다.
도 19a에서와 같이, HeLa 세포와 비교하여 분화유도한 HeLa 세포에서 간세포 마커의 대부분이 증가한 것을 확인하였다.
또한, HeLa 세포 분화 유도 3주 후 면역세포화학법을 통하여 간세포 마커(HNF4a, CK18, ALB)를 확인한 결과, HeLa 세포와 비교하여 분화시킨 HeLa 세포(HeLa h/ENTER)에서 간세포 마커의 발현이 증가됨을 확인하였다(도 19b).
[규칙 제91조에 의한 정정 12.10.2016]
HeLa 세포 분화 유도 3주 후 면역세포화학법을 통하여 간세포 마커(HNF4a, CK18, ALB)를 확인한 결과, HeLa 세포와 비교하여 분화시킨 HeLa 세포(HeLa h/ENTER)에서 간세포 마커의 발현이 증가됨을 확인하였다(도 19c).
다음으로, 간세포 분화유도배지와 초음파 처리된 Hep3B 세포의 간세포(Hep3B h/ENTER cell)로의 분화를 유도하였다. Hep3B 세포를 분화 유도한 19일 후 세포 외형의 변화와 Hep3B 세포 분화 유도 3주 후 면역세포화학에 의한 간세포 마커(HNF4a, CK18, ALB)의 발현 여부를 확인하였다.
도 20b에 나타난 바와 같이, Hep3B 세포와 비교하여 분화시킨 Hep3B 세포(Hep3B h/ENTER)에서 간세포 마커의 발현이 증가됨을 확인하였다.
다음으로, 인간 ES 배양배지와 초음파 처리된 HDF의 es/ENTER 세포로의 분화를 유도하였다. 도 21a는 배양시간에 따른 세포 형태 변화를, 도 21b는 배양시간에 따른 Oct4 발현의 차이를 보여주는 결과로, 인간 ES 배지로 분화 유도된 후 1일에 스페로이드가 형성되었으며, 다능성 마커인 Oct4의 발현이 배양시간에 따라 증가되었다.
6일간 배양되어 형성된 스페로이드를 회수하여 다능성 마커 발현을 RT-PCR과 ICC를 통해 확인하였다. 그 결과, es/ENTER 세포에서 다능성 마커 유전자(a) 및 단백질(b)의 발현이 확인되었다.
상기 es/ENTER 세포에서 다능성 속성을 분석한 결과, 플로우 사이토메트리를 이용하여 es/ENTER 세포의 SSEA4와 TRA-1-60의 발현을 확인하였고(도 23a), 다능성 유전자 발현의 패턴을 마이크로어레이(affymetrix chip)로 분석한 결과 HDF에 비해 ES세포에 비슷한 패턴을 나타냈다(도 23b). 그리고, 바이설파이트 시퀀싱을 통해 발현되는 DNA 유전자에서의 합성이 시작되는 프로모터 부분의 메틸화가 풀려있는지 확인한 결과, 중요한 다능성 유전자 Oct4와 Nanog이 개방되어 있음을 확인하였다. 이와 같은 결과들로 보아 인간 ES 배지로 분화 유도된 es/ENTER는 다능성 속성을 가진다는 것을 알 수 있다(도 23c).
메틸화 분석을 위한 바이설파이트 시퀀싱 프라이머 리스트
| 유전자 부호 |
프라이머 서열(5'-3') |
어닐링 온도(℃) |
| 정방향 |
역방향 |
|
Oct4
|
CCAGGTTCAATGGATTCTCC |
GTATCCGACCAGGGTTAGGG |
58 |
|
Nanog
|
TTCTCTCCTCCTCCCTCTCC |
CTCCCAAAATGCTGGGATTA |
56 |
|
Tdgf1
|
1 |
GTGGGTCCTCTTCAGTGCAT |
GCTGCTGGAGAGGTGCTTAG |
60 |
| 2 |
GACCCTCGCCTTATCCTTTC |
CACTGCCCTACTGCTTGGTT |
60 |
| 3 |
GCACAGAGGGTGTCCATCTT |
CTGCCCCTCTCACTCATCTC |
60 |
|
Afp
|
CAGTCCAGCAACAAGCCTTT |
ACTGGAGTCACTGGGAGGAA |
58 |
|
Gata4
|
TAGGATGCCTGCTGGATTTC |
CATTCATTCGCCCTCTCTTC |
58 |
|
Acta2
|
GGAGCACTTGAGAAGCAAAGA |
CTCAGGAAAGCCTCCCTCTT |
60 |
|
Msx1
|
GTAGACGCGGTTTGTGGAAC |
TTGGGGCTCTGTTTTTAACG |
60 |
|
Pax6
|
GTTGCAGCTGGTGTGTTGAC |
GCATTGTTGTGAATGCTGCT |
60 |
|
Nestin
|
GGGTCAAGTGGACTTTCCTG |
CACCCTCCTTGTCACTCCTC |
60 |
다음으로, es/ENTER 세포에서 분화 마커를 확인하였다. 도 24는 es/ENTER 세포에서의 3배엽 마커 유전자의 발현 (a) 과 단백질 발현 (b)을 나타낸 것이고, 도 25는 배양시간별 es/ENTER 세포에서의 Oct4와 3배엽 마커 유전자 발현의 변화를 나타낸 것이며, 도 26은 부착 배양된 es/ENTER 세포에서의 3배엽 마커 단백질의 발현(a) 및 바이설파이트 시퀀싱을 이용한 es/ENTER 세포의 3배엽 마커 DNA 메틸화 분석(b) 결과이다.
분화 마커 유전자의 PCR 프라이머 리스트
| 유전자 부호 |
프라이머 서열 (5-'3') |
어닐링 온도(℃) |
| 정방향 |
역방향 |
| 내배엽 |
Afp
|
AGCAGCTTGGTGGTGGATGA |
CCTGAGCTTGGCACAGATCC |
63 |
|
Foxa2
|
TTCAGGCCCGGCTAACTCTG |
CCTTGCGTCTCTGCAACACC |
58 |
|
Gata6
|
TGTGCGTTCATGGAGAAGATCA |
TTTGATAAGAGACCTCATGAACCGACT |
60 |
| 외배엽 |
Nestin
|
GAAACAGCCATAGAGGGCAAA |
TGGTTTTCCAGAGTCTTCAGTGA |
50 |
|
Pax6
|
ACCCATTATCCAGATGTGTTTGCCCGAG |
ATGGTGAAGCTGGGCATAGGCGGCAG |
58 |
| 중배엽, 심근세포 |
Acta2
(a-SMA)
|
CTATGAGGGCTATGCCTTGCC |
GCTCAGCAGTAGTAACGAAGGA |
50 |
| 중배엽 |
Brachyury
(T)
|
GCCCTCTCCCTCCCCTCCACGCACAG |
CGGCGCCGTTGCTCACAGACCACAGG |
66 |
|
Msx1
|
CGAGAGGACCCCGTGGATGCAGAG |
GGCGGCCATCTTCAGCTTCTCCAG |
58 |
| 심근세포 |
Myl7
|
GGGCCCCATCAACTTCACCGTCTTCC |
TGTAGTCGATGTTCCCCGCCAGGTCC |
58 |
|
Nkx2
-5
|
CCAAGGACCCTAGAGCCGAA |
ATAGGCGGGGTAGGCGTTAT |
50 |
|
TnTc
|
ATGAGCGGGAGAAGGAGCGGCAGAAC |
TCAATGGCCAGCACCTTCCTCCTCTC |
63 |
| 신경세포 |
Map2
|
CAGGTGGCGGACGTGTGAAAATTGAGAGTG |
CACGCTGGATCTGCCTGGGGACTGTG |
66.5 |
|
TuJ1
|
GAGCGGATCAGCGTCTACTACAA |
GATACTCCTCACGCACCTTGCT |
52 |
|
Gfap
|
CCTCTCCCTGGCTCGAATG |
GGAAGCGAACCTTCTCGATGTA |
52 |
|
Vglut1
|
CGACGACAGCCTTTTGTGGT |
GCCGAGACGTAGAAAACAGAG |
50 |
|
Vmat2
|
CTTTGGAGTTGGTTTTGC |
GCAGTTGTGGTCCATGAG |
43 |
| 지방세포 |
Pparc2
|
ATTGACCCAGAAAGCGATTC |
CAAAGGAGTGGGAGTGGTCT |
52.7 |
|
C/
ebpa
|
GCAAACTCACCGCTCCAATG |
TTAGGTTCCAAGCCCCAAGTC |
56.7 |
|
aP2
|
AACCTTAGATGGGGGTGTCCTG |
TCGTGGAAGTGACGCCTTTC |
57.2 |
|
Fabp4
|
ACTGGGCCAGGAATTTGACG |
CTCGTGGAAGTGACGCCTT |
55 |
| 간세포 |
Alb
|
AGCTGTTATGGATGATTTCGCAG |
CCTCGGCAAAGCAGGTCTC |
60 |
|
Cyp3a4
|
GTGACTTTGCCCATTGTTTAGAAAG |
CAGGCGTGAGCCACTGTG |
60 |
|
Cyp3a7
|
GATTCTGTACGTGCATTGTGCTC |
ATTTGGTCATCTCCTCTATATTACCAAGT |
60 |
|
Tat
|
CCACACCCACACTCAGATCCT |
ATTAGTGAGTCACTCTAGCAGCGC |
60 |
|
A1AT
|
GGTCACAGAGGAGGCACCC |
AGTCCCTTTCTCGTCGATGGT |
60 |
|
Sox7
|
TGCCCACTTCATGCAACTCC |
AGGTACCCTGGGTCTTTGGTCA |
60 |
| 하우스키핑유전자 |
beta-actin
|
CATGTACGTTGCTATCCAGGC |
CTCCTTAATGTCACGCACGAT |
50 |
|
Gapdh
|
ATGGGGAAGGTGAAGGTCG |
GGGTCATTGATGGCAACAATATC |
60 |
es/ENTER 스페로이드에서 3배엽 마커의 발현이 RT-PCR과 ICC를 통해 확인되었으며, 이와 같은 결과는 es/ENTER 세포가 다분화 분화속성을 가짐은 나타내는 지표이다.
따라서, 배양시간에 따른 다능성 마커인 Oct4의 발현 패턴을 확인한 결과 6일 이후 다능성 속성인 Oct4의 발현이 감소되고 3배엽 마커의 발현들이 증가되는 것을 확인하였다. 이와 같은 결과는 es/ENTER 세포가 다능성 속성에서 분화가 진행됨을 나타낸다. 이에 es/ENTER 스페로이드를 부착하여 2일간 배양한 스페로이드에서 나온 세포들이 3배엽 마커를 발현함이 확인되었고, 대표적인 3배엽 마커 유전자의 DNA 메틸화 분석 결과, 3배엽 마커 유전자는 개방되어 있음을 확인하였다.
도 27은 es/ENTER 세포의 신경세포(a), 심근세포(b) 및 간세포(c)로의 인 비트로 분화 실험 결과로, es/ENTER 세포를 각각의 3배엽에서 분화되는 세포로 분화 유도하는 배지를 이용하여 4주간 분화 유도한 결과, 신경세포(외배엽), 심근세포(중배엽), 간세포(내배엽)로 분화 유도되었으며, 각각의 분화 마커들이 발현되었다.
도 28은 HDF에서 신경세포(a), 심근세포(b) 및 간세포(c)의 분화 마커의 발현을 확인한 결과로, HDF에서는 분화 마커가 발현되지 않았다.
도 29는 분화 유도된 es/ENTER 세포의 신경세포(a), 심근세포(b) 및 간세포(c)의 분화 마커 유전자의 발현을 보여주는 RT-PCR 분석 결과로, 분화 유도된 es/ENTER에서의 분화 마커 유전자들의 발현이 증가되었다. 이와 같은 결과들은 es/ENTER의 다분화 능력을 증명하는 결과들이다.
도 30은 es/ENTER 세포의 염색체 G-밴드 분석에 의한 핵형을 보여주는 결과로, es/ENTER 생성에 있어서 초음파 자극으로 인한 세포 염색체의 돌연변이 여부를 분석한 결과, 정상으로 나타났다.
다음으로, es/ENTER 세포를 5주령 SCID 마우스의 다리 근육에 이식 후 4주 후에 HNA를 이용하여 이식한 세포를 확인하였다.
도 31에 나타난 바와 같이, es/ENTER 세포는 골격 근육으로 분화되었음을 확인하였다. 그리고 이식된 세포에서의 Oct4가 발현 및 증식되지 않음을 확인하였다.
다음으로, es/ENTER 세포를 쥐의 뇌에 이식하여 인 비보 분화를 확인하였다. 이를 위해, es/ENTER 세포를 5주령 SCID 쥐의 뇌에 이식 후 4주 후에 HNA를 이용하여 이식한 세포를 확인하였다.
그 결과, es/ENTER 세포는 성상세포(Gfap)로 분화되었음을 확인하였고, 시냅신과 Vesicular Glutamate transpoter가 분비됨을 확인하였다. 이는 이식된 세포가 정상적으로 분화되어 기능을 수행하고 있음을 의미한다. 그리고 이식된 세포에서의 Oct4가 발현되지 않고 있으며, 증식되지 않음을 확인하였다(도 32).
다음으로, MEF(마우스 태아 섬유아세포)도 HDF와 같은 방법인 hES 배지로 mouse es/ENTER 세포로 분화 유도하였다. 이번 실험에 이용된 MEF는 OG2-MEF로 Oct4 프로모터를 벡터로 형질전환된 마우스의 태아 섬유아세포를 가지고 실시하였다. 이 세포의 경우 Oct4가 발현되면 GFP 형광이 발현되는 세포로 Oct4의 발현을 관찰하기 위해 사용하였다.
도 33에 나타난 바와 같이, 초음파 처리를 통해 유도된 세포는 시간이 지남에 따라 스페로이드의 수와 크기 그리고 GFP 발현이 증가되었다.
다음으로, 상기 mouse es/ENTER 세포에서 다능성 속성을 분석하였다.
도 34에 나타난 바와 같이, mouse es/ENTER의 ICC, RT-PCR, 플로우 사이토메트리, AP 염색 결과, 마우스 ES와 비슷한 경향을 나타냈다.
다음으로 상기 mouse es/ENTER 세포에서 3배엽 특성을 분석하였다.
실험 결과, 인간 es/ENTER에서 나타난 3배엽 속성이 mouse es/ENTER 세포에서도 나타났으며, 배양시간별 RT-PCR과 ICC 분석을 통해 시간이 지남에 따라 그 발현의 차이가 나타났다. 이와 같은 결과는 인간 es/ENTER의 결과와 같았다(도 35).
도 36은 mouse es/ENTER의 신경세포(a)과 심근세포(b)로의 인 비트로 분화를 보여주며, 도 36c는 mouse es/ENTER의 염색체 G 밴드 분석에 의한 핵형 분석 결과를 보여준다. 핵형 분석은 GTG banding chromosome analysis(GenDix, Inc. Seoul, Korea)를 사용하여 수행하였다.
실험 결과, mouse es/ENTER 세포에서 신경세포 및 심근세포 분화 마커를 확인하여 분화되었음을 확인하였고, 초음파에 의한 염색체 변이를 핵형 분석한 결과 변이가 일어나지 않았음을 확인하였다.
<실시예 10> 다른 세포를 이용한 es/ENTER의 분화 유도 실험
이와 같은 결과를 통해 HDF 뿐만 아니라 다른 개체의 세포에도 이 방법이 가능하다는 결론을 내리고, 다양한 세포 (L132, MSC, 환자 피부섬유아세포)에 적용해 보았다.
L132(폐 상피세포), MSC(mesenchymal stem cell, 간엽줄기세포) 및 Skin fibroblast(환자유래 피부섬유아세포) 등을 이용하여 es/ENTER와 동일한 방법으로 분화를 유도한 결과, 세포 스페로이드가 형성되었고, es/ENTER와 비슷하게 다능성 마커 및 3배엽 마커들이 발현되는 것으로 나타났다.
도 37은 인간 ES 배양배지와 초음파 자극에 의한 L132 세포의 L132 es/ENTER 세포로의 분화를 보여주는 결과로, 도 37a는 배양시간에 따른 세포 형태 변화, 도 37b 및 37c는 L132 es/ENTER 세포의 다능성(b) 및 3배엽(c) 속성을 보여준다.
도 38은 인간 ES 배양배지와 초음파 자극에 의한 MSC의 MSC es/ENTER 세포로의 분화를 보여주는 결과로, 도 38a는 배양시간에 따른 세포 형태 변화, 도 38b 및 38c는 MSC es/ENTER 세포의 다능성(b) 및 3배엽(c) 속성을 보여준다.
도 39는 인간 ES 배양배지와 초음파 자극에 의한 인간 피부 섬유아세포의 SF es/ENTER 세포로의 분화를 보여주는 결과로, 도 39a는 배양시간에 따른 세포 형태 변화, 도 39b 및 39c는 SF es/ENTER 세포의 다능성(b) 및 3배엽(c) 속성을 보여준다.
<실시예 11> 다른 물리적 지극을 이용한 es/ENTER 세포로의 분화 유도
인간 ES 배양액이라는 동일한 배지환경에 이번엔 분화 유도를 위한 물리적 자극으로 열 처리(Heat shock)와 레이저를 사용하였다.
우선, 열 처리(Heat shock)와 hES 배지를 이용하여 HDF의 es/ENTER 세포의 분화를 유도하였다. 열 처리를 위해, HDF를 42℃로 2분간 노출시킨 후 아이스에서 약 5초간 정치하였다. 도 40a는 분화 유도된 HDF 스페로이드, 도 40b 및 40c는 es/ENTER 세포의 다능성(b) 및 3배엽(c) 속성을 보여준다.
다음으로, 레이저 자극과 hES 배지를 이용한 HDF의 es/ENTER 세포의 분화를 유도하였다. 레이저 처리 조건은 Ocla 치료용 레이저(Ndlux)를 사용하여, 808nm의 레이저를 5초간 조사한 후 배양하였다. 도 41a는 분화 유도된 HDF 스페로이드, 도 41b 및 41c는 es/ENTER 세포의 다능성(b) 및 3배엽(c) 속성을 보여준다.
도 40 및 41에 나타난 바와 같이, 두 자극 모두 초음파에 의한 효과와 유사하게 세포 스페로이드가 형성되었으며, 다능성 및 3배엽 마커들이 발현되는 것을 확인하였다. 이와 같은 결과들은 배지 환경 유입으로 인한 세포 리프로그래밍이 다양한 세포에 적용이 가능하며, 배지환경과 더불어 환경유입을 위한 물리적인 자극 또한 다양한 방법이 가능함을 나타내며, 이전의 결과에서와 같이 환경에 의해 세포의 속성이 변할 수 있다.
<실시예 12> 세포 밖 소포체(extracellular vesicles, EVs)를 이용한 세포의 리프로그래밍
세포는 invitrogen에서 구입한 primary HDF를 10% FBS(Gibco)와 1% penicillin/streptomycin(Gibco)이 첨가된 DMEM에서 배양하였으며, 배양 배지에 대한 초음파 처리는 5W/cm2, 10분간 수행하고, 세포 처리는 1×106 HDF에 1W/cm2, 5초간 처리 후 상기에서 초음파 처리된 배양 배지와 함께 35mm 배양접시에 2×105 세포를 1일간 37℃, 5% CO2 조건으로 배양하였다. 배양액을 회수하여 Amicon Ultra centrifugal filter (Millipore)에 넣고 14000 rpm, 20분간 원심분리하여 배양액내 EVs를 필터로 걸러 회수하였다.
다음으로, HDF를 배양접시에 약 70-80% 채워지게 배양한 후, 배양액을 회수하여 D-PBS로 2회 세척한 다음, 배아줄기세포 배지 또는 신경줄기세포 분화배지(Gibco)에 각각 es/ENTER 와 n/ENTER 1일째 배양 배지로부터 회수된 10㎕/mL(v/v)의 농축 EVs를 첨가한 후 상기에서 세척한 HDF와 혼합하여 3일간 배양하였다.
<실시예 13> 정상 체세포(HDF)에서 EVs의 전달 실험
EVs를 이용한 체세포의 리프로그래밍을 확인하기 위해 상기 실시예 12와 같이 물리적 자극을 받은 세포의 1일 배양 후 얻은 EVs를 농축한 후 Did dye를 이용하여 EVs를 표지하고, 정상 체세포에 상기 EVs가 전달되고, 전달된 세포에서 각각의 다능성 마커인 Oct4와 신경줄기세포 마커인 Pax6의 발현을 확인하였다.
이를 위해, 상기 실시예 12에서 얻은 EVs 50㎕를 D-PBS 450㎕와 혼합하여 희석하고, 여기에 2.5㎕의 Vybrant DiD cell-labelling solution(molecular probe, excitation/emission, 644/667 nm)을 첨가하여 37℃에서 30분간 엑소좀을 염색하였다. 염색 후 다시 Amicon Ultra centrifugal filter(Millipore)로 14000 rpm, 20분 원심분리하여 Did 염색된 EVs를 농축 후 D-PBS로 희석하기를 2회 반복한 후 3mL의 HDF 배양액(5% FBS가 포함된 DMEM(Gibco) 배양액)에 첨가한 후 37℃, 5% CO2에서 24시간 배양하였다. 24시간 배양된 HDF를 4% paraformaldehyde로 10분간 고정하고, 0.2% triton X100 in PBS buffer로 10분간 투과시켰다. 이후 3% BSA in PBS buffer로 1시간 블록킹한 후 1차 항체인 토끼-항 Oct4(1:250, abcam)와 Pax6(1:200, abcam)로 4℃에서 하룻밤 동안 염색한 후 2차 항체인 항-토끼 컨쥬게이티드 Alexa-488(1:1000, Thermo, excitation/emission, 495/519 nm)을 1시간 동안 염색하였다. 2차 항체가 염색된 샘플을 DAPI(4′, 6-diamidino-2-phenylindole dihydrochloride, Vector Laboratories, excitation/emission, 420/480 nm)가 포함된 마운팅 용액을 사용하여 공초점 레이저현미경(Confocal laser scanning microscope, LSM 700; Carl Zeiss) 이미지를 분석하고 도 42에 결과를 나타내었다. 도면에서, 녹색은 Oct4, 적색은 Did dye로 염색된 EVs이다.
도 42에 나타난 바와 같이, 물리적 자극 후 세포에서 분비되는 EVs는 세포 배지 환경에 따라 다양한 다능성 마커의 유전자 및 단백질을 포함하고 있음을 확인하였고, 이러한 인자들은 EVs에 의해 인접하는 세포로 전달될 수 있음을 확인하였다. 상기 결과는 다양한 배지 환경에서 물리적 자극을 받은 세포에서 분비된 EVs는 정상 체세포의 리프로그래밍을 유도할 가능성이 있음을 시사한다.
<실시예 14> EVs에 의한 인간 섬유아세포의 리프로그래밍 효과
상기 실시예 13에서 다양한 배지 환경에서 물리적 자극을 받은 세포에서 분비된 EVs는 정상 체세포의 리프로그래밍을 유도할 가능성이 있으므로 이를 검증하기 위해 인간 섬유아세포 배양 배지인 DMEM 배지와 인간 배아줄기세포 또는 iPS 세포의 배양 배지인 hESC 배지를 사용하여 실험하였다. 대조군은 EVs가 첨가되지 않은 각 배지에서 3일간 배양하였고, 처리군은 각 배지에 10㎕/mL(v/v)의 EVs를 첨가하여 3일간 배양하였다.
배양된 세포는 상기 실시예 13과 같이 1차 항체로 토끼-항-Oct4(1:250, abcam), 2차 항체로 항-토끼 컨쥬게이티드 Alexa-488(1:1000, Thermo excitation/emission, 495/519nm)를 사용하여 염색하고, DAPI가 포함된 마운팅 용액으로 마운팅한 후 공초점 레이저현미경으로 이미지를 분석하여 도 43에 나타내었다. 도면에서, hESC는 human ESC 배지, DMEM은 섬유아세포 배양 배지, EVs는 es/ENTER 유도 시 회수한 EVs를 나타낸다.
도 43에 나타난 바와 같이, 대조군에서는 Oct4의 발현이 관찰되지 않았지만, 처리군에서는 Oct4의 발현과 세포가 스페로이드를 형성하는 현상이 관찰되었다. 상기 결과는 세포 리프로그래밍이 배양 배지에 의한 영향이 아닌 오직 EVs에 의해 유도되었음을 나타낸다.
<실시예 15> EVs 처리된 인간 섬유아세포의 배양시간별 세포 형태의 변화
es/ENTER 유도 시 회수한 10㎕/mL(v/v)의 EVs를 인간 섬유아세포에 처리한 후 6일 동안 배양하여 세포의 형태 변화를 관찰하였다.
도 44에 나타난 바와 같이, 배양 시간에 따라 세포의 형태가 달라지고 있으며, 3일차에서 스페로이드가 형성되는 것을 관찰하였다.
<실시예 16> es/ENTER 유도 시 회수한 EVs 첨가량에 따른 6일 배양된 HDF의 다능성 마커 발현 확인 실험
세포 리프로그래밍을 위한 적정 EVs의 농도를 알아보기 위해 EVs의 첨가량을 달리하여 HDF에 처리하고 6일 동안 배양하였다. 또한, es/ENTER 유도 시 회수한 EVs를 사용하므로 다능성 마커인 Oct4가 발현되는 세포를 Flow cytometry로 분석하였다.
이를 위해, es/ENTER 유도 시 회수한 EVs를 섬유아세포 배양 시 각각 0, 5, 12.5, 25, 50 및 100㎕/mL(v/v)의 농도로 첨가형 37℃, 5% CO2 조건에서 6일간 배양하였다. 배양된 세포는 상기 실시예 13과 같이 1차 항체로 토끼-항-Oct4(1:250, abcam), 2차 항체로 항-토끼 컨쥬게이티드 Alexa-488(1:1000, Thermo excitation/emission, 495/519nm)를 사용하여 염색하고, BD AccuriTM C6 Flow cytometry(BD biosciences)를 이용하여 분석하였다.
도 45에 나타난 바와 같이, 12.5㎕/mL(v/v)의 EVs를 처리하였을 때 가장 많은 84.6%의 세포에서 Oct4의 발현이 확인되었다.
<실시예 17> es/ENTER 유도 시 회수한 EVs 처리된 3일 배양된 HDF에서 다능성 마커 발현 확인 실험
es/ENTER 유도 시 회수한 10㎕/mL(v/v)의 EVs를 인간 섬유아세포에 처리한 후 3일 동안 배양하여 세포 리프로그래밍 효과를 확인하였다. 배양된 세포는 ICC 분석을 위해 상기 실시예 13과 같이 1차 항체로 토끼-항-Oct4(1:250, abcam), Sox2 (1:250, abcam) 및 Nanog (1:250, abcam), 2차 항체로 항-토끼 컨쥬게이티드 Alexa-488(1:1000, Thermo excitation/emission, 495/519nm)를 사용하고, DAPI가 포함된 마운팅 용액으로 마운팅한 후 공초점 레이저현미경으로 이미지를 분석하였다. qPCR 분석은 3일간 배양된 세포에서 Trizol(Takara)을 이용하여 전체 RNA를 회수한 후 Superscrip 2 kit(Invitrogen)로 cDNA를 합성하였다. PCR 분석은 다능성 마커인 Oct4, Sox2 및 Nanog에 대해 Real time PCR 기기(ab step one plus, AB)로 분석하였다.
도 46에 나타난 바와 같이, ICC 분석 결과 인간 섬유아세포 핵에서 다능성 마커인 Oct4, Sox2 및 Nanog의 발현이 관찰되었으며, 유전자 발현을 real time PCR로 qPCR 분석한 결과 Oct4, Sox2 및 Nanog 유전자는 EVs 처리되지 않은 정상 섬유아세포 대비 약 50 배 정도 과발현되었다.
<실시예 18> n/ENTER 유도 시 회수한 EVs 처리된 3일 배양된 HDF에서의 신경줄기세포 마커 발현 확인 실험
n/ENTER 유도 시 회수한 10㎕/mL(v/v)의 EVs를 인간 섬유아세포에 처리한 후 3일 동안 배양하여 세포 리프로그래밍 효과를 확인하였다. 배양된 세포는 ICC 분석을 위해 상기 실시예 13과 같이 1차 항체로 토끼-항-Sox1(1:200, abcam), Sox2(1:250, abcam), Pax6(1:200, abcam) 및 마우스-항-Nestin (1:250, Thermo Scientific), 2차 항체로 항-토끼 컨쥬게이티드 Alexa-488(1:1000, Thermo excitation/emission, 495/519nm) 및 항-마우스 컨쥬게이티드 Alexa-594(1:1000, Thermo, alexa 488 excitation/emission, 495/519 nm; alexa 594 excitation/emission, 590/617 nm)를 사용하고, DAPI가 포함된 마운팅 용액으로 마운팅한 후 후 공초점 레이저현미경으로 이미지를 분석하였다. qPCR 분석은 3일간 배양된 세포에서 Trizol(Takara)을 이용하여 전체 RNA를 회수한 후 Superscrip 2 kit(Invitrogen)로 cDNA를 합성하였다. PCR 분석은 신경줄기세포 마커인 Sox1, Sox2, Pax6 및 Nestin에 대해 Real time PCR 기기(ab step one plus, AB)로 분석하였다.
도 47에 나타난 바와 같이, ICC 분석을 통해 인간 섬유아세포 핵에서 신경줄기세포 마커인 Sox1, Sox2 및 Pax6의 발현이 관찰되었으며 세포질에서 Nestin의 발현이 관찰되었다. 유전자 발현을 real time PCR로 qPCR 분석한 결과 Sox1, Sox2, Pax6 및 Nestin 유전자는 EVs 처리되지 않은 정상 섬유아세포 대비 약 200 배 정도 과발현되었다.
qPCR 프라이머 리스트
| 유전자 부호 |
프라이머 서열 (5'-3') |
| 정방향 |
역방향 |
| 다능성마커 |
Oct4
|
GGGTTTTTGGGATTAAGTTCTTCA |
GCCCCCACCCTTTGTGTT |
|
Sox2
|
CAAAAATGGCCATGCAGGTT |
AGTTGGGATCGAACAAAAGCTATT |
|
Nanog
|
ACAACTGGCCGAAGAATAGCA |
GGTTCCCAGTCGGGTTCAC |
| 신경 줄기세포 마커 |
Sox1
|
TCTGTTAACTCACCGGGACC |
ACTCCAGGGTACACACAGGG |
|
Sox2
|
GGAGTGCAATAGGGCGGAAT |
CCAGTTGTAGACACGCACCT |
|
Pax6
|
GTCCATCTTTGCTTGGGAAA |
TAGCCAGGTTGCGAAGAACT |
|
Nestin
|
CTCCAGAAACTCAAGCACC |
TCCTGATTCTCCTCTTCCA |
|
Gapdh
|
ATGGGGAAGGTGAAGGTCG |
GGGTCATTGATGGCAACAATATC |